/











































































































































































































Text
А.М. ПЕКЛО
МУХОЛОВКИ
ФАУНЫ
СССР
WWW.ZOOMET.RU
АКАДЕМИЯ НАУК УКРАИНСКОЙ ССР
ИНСТИТУТ ЗООЛОГИИ ИМ.И.И.ШМАЛЬГАУЗЕНА
А. М. ПЕКЛО
МУХОЛОВКИ
ФАУНЫ
СССР
WWW.ZOOIV1ET.RU
Киев Наукова думка 1987
УДК 598.813(47 + 57)
МОНОГРАФИЯ
Мухоловки фауны СССР / Пекло А.М. - Киев : Наук, думка, 1987. - 180 с.
В монографии описаны виды райских и настоящих мухоловок фауны СССР. Приведены детальная морфологическая характеристика возрастных групп каждого вида с указанием особенностей проявления сезонной и индивидуальной изменчивости, гнездовой ареал, места зимовок, пролета и точек залета. Рассматривается внутривидовая систематика, дан анализ географической изменчивости. Описаны биотопическое распределение, численность, размножение, линька, сроки и закономерности сезонных миграций на территории СССР. Характеризуется практическое значение и даются рекомендации по охране редких видов.
Для орнитологов-систематиков, фаунистов, экологов, специалистов по охране природы, преподавателей и студентов вузов.
Ил. 45. Табл. 14. Библиогр.: с.165—179.
Александр Михайлович Пекло
МУХОЛОВКИ ФАУНЫ СССР
Утверждено к печати ученым советом
Института зоологии им. И. И. Шмальгаузена АН УССР
Ответственный редактор Н.Н.Щербак
Рецензенты М.А.Воинственский, В.В.Серебряков
Редакторы В.И.Зубаток, Т.П.Горбань Художественный редактор Л.А.Комяхова Оформление художника М.Н. Усова Технический редактор И.Ф.Михалкина Оператор Т.Н.Шевченко
Корректоры Е.И.Мазниченко, А.Ф.Коровниченко, М.Е.Ролинская
Редакция общей биологии
п 2005000000-242 __
11 —------------ Jj /-о /
М221 (04)-87
ИБ№ 8532
Подп.впеч. 13.02.87.БФ24135. Формат70x108/16. Бум. офс. № 1. Гарн. Пресс Роман. Офс. печ. Уел. печ.л. 17,50. Усл.кр.-отт. 22,40. Уч.-изд. л. 22,16. Тираж 1150 зкз. Заказ. 1-189 Цена 3 р. 50 к.
Издательство "Наукова думка”. 252601 Киев 1, ул. Репина.З.
Киевская книжная типография научной книги.
252004 Киев 4, ул. Репина, 4.
© Издательство ’’Наукова думка”, 1987
ПРЕДИСЛОВИЕ
Мухоловковые птицы до настоящего времени остаются одним из наименее изученных семейств отряда воробьинообразных. В этой книге на основании собственных полевых исследований, а также изучения коллекционных материалов по данной группе и анализа литературных сведений автором дается описание всех видов мухоловок фауны СССР.
Настоящая работа выполнялась в Зоологическом музее и отделе эволюционной морфологии животных Института зоологии им.И.И.Шмальгаузена АН УССР в 1976-1985 гг. За этот период автором проведены полевые экспедиционные исследования на территории Украины, Белоруссии, Кавказа, Средней Азии и Приморского края. Стационары и пункты кратковременных работ располагались в наиболее характерных гнездовых биотопах и в местах массовых весенне-осенних миграций видов изучаемой группы. Полевыми исследованиями было охвачено большинство видов мухоловок фауны СССР, кроме черной райской, японской желтоспинной и сибирской.
Численность птиц определялась маршрутным методом по общепринятым методикам. Для некоторых видов на стационарах проводились абсолютные учеты всех гнездовых пар на контрольных участках.
За время полевых работ автором обследовано 56 гнезд 9 видов мухоловок. Кроме этого, обработано 185 гнезд с кладками, хранящихся в Зоологическом музее Московского государственного университета им.М.В.Ломоносова (ЗМ МГУ), на кафедре биогеографии МГУ (КБ МГУ), в Биологическом институте СО АН СССР (Новосибирск, БИН СО АН), Биолого-почвенном институте ДВНЦ АН СССР (Владивосток, БПиДВНЦ), Зоологическом музее Института зоологии им.И.И.Шмальгаузена АН УССР (Киев, ИЗАНУ), во Львовском природоведческом музее АН УССР (ЛПМ), в Киевском государственном университете им.Т.Г.Шевченко (КГУ), Харьковском государственном университете (ХГУ), Туркменском сельскохозяйственном институте им.М.И.Калинина (Ашхабад, Турк. СХИ),
а также в частных коллекциях А.П.Кузякина, В.В.Леоновича, С.Д.Кустановича (Москва), М.И.Лобановского (Минск), С.И.Пархоменко и Ю.В.Файделя (Киев). Ряд неопубликованных материалов по гнездам представлены В.В.Леоновичем, А.А.Виноку-ровым (Москва), В.П.Беликом, Б.А.Коза-ковым, НХЛомадзе (Ростов-на-Дону), М.И.Головушкиным (Киев), О.В.Митро-польским (Ташкент), П.А.Тильба (Сочи), Г.З.Гусаном (Кишинев). В общей сложности были собраны сведения о 310 гнездах с кладками и птенцами 12 видов мухоловок фауны СССР. Каждое гнездо подвергалось морфометрической обработке, описывалась его форма, расположение и строительный материал постройки. Лишайники из гнезд определены И Л.Навроцкой (Институт ботаники им.Н.Г.Холодного АН УССР), мхи - М.Ф.Бойко (Херсонский педагогический институт).
Окраска яиц описана по схеме, предложенной Ю.В.Костиным (1977), их форма - по В.Макачу (Makatsch, 1974). Для характеристики формы яйца использован также коэффициент удлиненности (К = L/D, где L - длина яйца, D - ширина). Пуховые птенцы описаны по И.А.Ней-фельдт (1970).
Линька оперения изучалась на коллекционном шкурковом и фиксированном материале, а для белобрюхой райской мухоловки - и на живых птицах при клеточном содержании.
Для изучения питания взрослых птиц и птенцов 12 видов мухоловок исследовано содержимое 515 желудков и пищевых проб от птенцов, полученных по методике наложения шейных лигатур. Кроме этого в работе использованы и визуальные наблюдения за питанием птиц и их способами добычи корма. Пищевые компоненты животного происхождения определены В.М.Ермоленко, Ю.А.Костюком, Ю.П.Некрутенко, В.Н.Логвиненко, А.А.Петрусенко, А.А.Пет-ренко, В.М .Бровдием, В.Г.Пучковым,
В.Г.Долиным и В.И.Вакаренко (ИЗАНУ), а также Б.А.Каротяевым (Зоологический институт АН СССР, Ленинград, ЗИН) и А.В.Богачевым (Кубанский сельскохозяйственный институт).
За время полевых работ автором по данной группе собрано 268 шкурок и фиксированных птиц. Обработаны также фондовые коллекции мухоловок ЗИН, БПиДВНЦ, ИЗАНУ, Зоологического музея Института зоологии и паразитологии АН ТаджССР (Душанбе, ИЗиП АН ТаджССР),
3
ЗМ МГУ, КБ МГУ, КГУ, ХГУ, Ростовского государственного университета (РГУ), а также частная коллекция птиц А.П.Кузя-кина, собранная в Китае. Даурские желтоспинные мухоловки с п-ова Малакка получены из Малайзийского университета (Куала-Лампур) от доктора Д.Р.Уэллса (D.R.Wells). Всего обработано более 2350 экз. мухоловок 17 видов, добытых в разные сезоны года на территории СССР, Албании, Турции, Ирана, Индии, Китая, Японии и Индонезии. Для анализа географической изменчивости морфологической обработке были подвержены только те птицы, гнездовая территориальная принадлежность которых не вызывала сомнений. Весь этот материал дифференцирован по полу и возрасту, обработан более чем по 24 морфологическим признакам (морфометрия, окраска, рисунок оперения). Для каждого морфометрического признака определены пределы (Нт) его вариабельности, средняя величина с ошибкой (М + т) и коэффициент вариации (CV, %). Достоверность различий между формами определялась по критерию Стьюдента (t). Вся статистическая обработка исходных данных проведена на вычислительной машине ’’Электроника БЗ-21”. Названия отдельных частей тела птиц и партий их оперения (за редкими исключениями) приняты по Н.В.Ви-ноградовой и др. (1976). Нумерация маховых перьев сквозная, от дистального конца крыла к телу, рулевых — от центра хвоста к его краям. Под шириной крыла понимается расстояние между кистевым суставом и вершиной 11-го махового (первого второстепенного) при сложенном крыле. Описание возрастных нарядов в книге дается в следующем порядке: характеристика окраски взрослой птицы, описание гнездового и далее первого послегнездового нарядов. Сведения о пуховых птенцах вынесены в разделы, в которых описано размножение. Объем семейства принят в соответствии с классификацией А.Уэтмора (Wetmore, 1960). Порядок подсемейств, родов и последовательность видовых единиц соответствуют принятым в сводке Ч.Вори (Vaurie, 1959).
Цветные рисунки в книге и большинство черно-белых иллюстраций являются оригинальными и выполнены автором.
Сведения о распространении мухоловок на территории СССР автору любезно предоставили также С.Д.Кустанович, В.В.Леоно-вич, С.М.Смиренский, В.Ю.Ильяшенко, А. А. Винокуров (Москва), С.В.Винтер, А.М.Волох (Мелитополь), Г.З.Гусан (Кишинев) , М.И.Головушкин, М.А.Осипова, Н.Л.Клестов (Киев), С.С.Москвитин (Томск), А.А.Естафьев (Сыктывкар), В.Д.Сонин (Иркутск), В.А.Нечаев (Владивосток), А.С.Вобленко, М.Ф.Самофалов (Чернигов), В.В.Ветров (Ворошиловград), В.И.Стригунов (Кривой Рог), Т.Б.Ардамац-кая (Голая Пристань), В.Г.Сулик (Луганский заповедник), М.М.Бескоровайный (Карадагский заповедник), А.Н.Гудина (Запорожье), Р.Г.Жордания (Тбилиси), АЛ.Тильба (Сочи) и Г.Б.Бахтадзе (Ростов-на-Дону) .
Настоящая работа выполнялась под руководством Н.Н.Щербака и С.Ф.Манзия. Ценные советы, консультации и замечания в процессе проведения данного исследования и при подготовке рукописи к печати автотором получены от К.А.Юдина, М.А.Во-инственского, В.П.Жежерина, В.М.Лоскота, И.А.Нейфельд, В.А.Нечаева, С.В.Винтера, Ю.А.Костюка, Ю.П.Некрутенко и О.А.Михай-левича. Всем перечисленным выше коллегам автор выражает искреннюю благодарность.
СПИСОК СОКРАЩЕНИЙ
д. - деревня
к. - кишлак
кл. - ключ
колл. - коллекция
с. - село, селение
сообщ. - сообщение
ст. - станция
стан. - станица
ур. - урочище
ущ. — ущелье
хр. - хребет
ц.кольц. - центр кольцевания
ОТРЯД PASSERIFORMES -ВОРОБЬИНООБРАЗНЫЕ
ПОДОТРЯД PASSERES -ПЕВЧИЕ ВОРОБЬИНЫЕ
СЕМЕЙСТВО MUSCICAPIDAE -МУХОЛОВКОВЫЕ
Большая группа певчих воробьиных птиц, насчитывающая более 300 видов1. Распространены в Старом Свете от умеренных широт Южного полушария к северу до 68—70° с.ш. почти повсеместно при наличии древесной и реже кустарниковой растительности. В СССР встречается 14 видов 4 родов; гнездится 13 видов.
Мелкие птицы. Телосложение довольно плотное. Клюв короткий или средней длины, у основания широкий и обычно заметно уплощенный. Хребет надклювья четко выражен. На конце надклювья часто имеются небольшой крючочек и выемка перед ним. Ноздри располагаются непосредственно перед лбом ближе к боковым краям надклювья и у большинства видов в разной степени прикрыты перышками, направленными вперед и обладающими волосовидными удлинениями стержней. Щетинки у основания клюва хорошо развиты. Ноги слабые. Цевка обычно тонкая и чаще всего короткая. Пальцы тонкие с маленькими когтями. Крылья относительно длинные, с заостренными или округлыми вершинами. Первостепенных маховых десять; 1-е часто сильно укорочено. Формула крыла, а также форма и длина хвоста в пределах семейства сильно варьируют. Рулевых обычно двенадцать; центральные у некоторых видов сильно удлинены и превышают длину крыла в 3—4 раза. На голове иногда имеется хохолок из удлиненных перьев. Оперение мягкое и приглаженное. Окраска его очень разнообразна: от буроватосерой до очень яркой, часто с металлическим блеском. У многих видов резко выражен половой диморфизм. Молодые в гнездовом наряде обычно имеют на перьях пятнистый или чешуйчатый рисунок; у некоторых видов он отсутствует.
Населяют различные типы хвойных, смешанных хвойно-лиственных и главным образом лиственных лесов на равнинах, в предгорьях и горах, а также саванны, кустарниковые заросли, сады и парки. Некоторые виды могут гнездится в населенных пунктах, в том числе и в крупных городах. В лесах обычно предпочитают разреженные
1 Объем семейства окончательно не выяснен.
участки, опушки и другие осветленные места, но иногда встречаются и в густых зарослях. Ведут дневной образ жизни. Обитатели умеренных широт - перелетные птицы, все тропические виды оседлы.
Моногамы, однако у некоторых видов отмечены случаи полигинии. Песня самцов чаще всего довольно однообразна, иногда бывает звучной и мелодичной. Гнезда строят в самых разнообразных местах, как открыто на ветвях деревьев и кустарников, так и в различных укрытиях: за отставшей корой, в дуплах, расщелинах, прогнивших пнях, нишах скальных выходов и береговых обрывов. Некоторые виды охотно гнездятся в искусственных гнездовьях, а также на постройках человека. В кладке обычно 4—6, у некоторых видов до 11 яиц. Повторные кладки (взамен утраченных) чаще всего меньших размеров и могут состоять из 1-3 яиц. У одних видов кладку насиживают оба партнера пары, у других -только самка. Инкубация длится Ills дней, птенцы вылупляются покрытые пухом и около двух недель находятся в гнезде. У открыто гнездящихся видов они покидают гнездо, еще не умея летать, у дуплогнездников — сравнительно хорошо перепархивающими. Выкармливают птенцов оба партнера. Обычно в месячном возрасте молодые становятся полностью самостоятельными. Вне сезона размножения большинство видов держится поодиночке, реже — небольшими группами.
Взрослые птицы имеют в году одну (полная послебрачная) или две линьки (полная послебрачная и частичная, или полная предбрачная). У некоторых видов самцы одевают окончательный наряд лишь на втором и даже третьем году жизни, но в размножении участвуют в возрасте около года.
Питаются преимущественно насекомыми, в меньшей степени другими беспозвоночными; иногда поедают мелкие сочные плоды некоторых растений. Гастролиты заглатывают редко. Для многих видов основным способом охоты является подкарауливание летающих насекомых с какой-либо присады (чаще всего ветвь дерева) с последующей ловлей их в воздухе на лету в стремительном броске. Преследуя добычу, проявляют большую маневренность Полета. Склевывают также мелких беспозвоночных с растений и .поверхности земли.
Поедая большое количество насекомых, многие из которых являются вредителями леса и садовых культур, а также
5
переносчиками возбудителей ряда заболеваний человека и животных, приносят несомненную пользу и заслуживают повсеместной всесторонней охраны.
ПОДСЕМЕЙСТВО MONARCHINAE -ДЛИННОХВОСТЫЕ МУХОЛОВКИ
Род Terpsiphone Gloger, 1827 — райские мухоловки
Terpsiphone Gloger, 1827, Froriep’s Notizen, 16: col. 278.
Типовой вид Corvus paradisi Linnaeus.
Род включает 10 видов (Wolters, 1979), 5 из которых распространены в Эфиопской, 3 — в Мадагаскарской и 2 — в Индо-Малайской и Палеарктической зоогеографических областях, причем 1 из двух последних проникает и в Австралийскую область. В СССР встречается 2 вида, 1 из которых гнездится.
Очень своеобразные длиннохвостые птицы, резко отличающиеся от настоящих мухоловок. Размеры средние и крупные. Большой клюв сильно уплощен и очень широк в основной своей половине. На конце надклювья расположен хорошо развитый крючочек и выемка перед ним. Ноздри овальные. Щетинки в углах рта очень хорошо развиты и резко превышают по длине лобные, прикрывающие сверху ноздри и осндвание клюва. Глаз окружен ярко окрашенным голым кольцом. Удлиненные перья образуют на голове хохол, достигающий наибольших размеров у взрослых самцов. Ноги довольно слабые. Цевка относительно короткая. Хвост клиновидноступенчатый. Центральная пара рулевых у взрослых самцов сильно удлинена и обычно превышает длину крыла в 3—4 раза. Крылья с округлыми вершинами. Первое маховое сильно выступает за вершины кроющих кисти и составляет около половины длины второго. Густое и мягкое оперение яркое, часто с металлическим блеском. Половой диморфизм выражен в окраске и в размерах отдельных частей тела. Некоторым видам свойственно явление морфизма: взрослые самцы имеют несколько цветовых морф. У молодых в гнездовом наряде на перьях отсутствует пятнистый или чешуйчатый рисунок.
Населяют главным образом различные типы тропических и субтропических лесов. В северной части ареала рода обитают в лиственных и значительно реже в смешанных хвойно-лиственных лесах равнин, пред
горий и гор, а также в парках и садах. Высоко в горы (до 2400 м н.у.м. и даже выше) поднимаются только некоторые представители рода. Два вида (T.atrocau-data и T.paradisi) являются частично перелетными, остальные ведут оседлый образ жизни.
Образование пар у райских мухоловок сопровождается пением самцов и их токовым демонстративным поведением. Чашевидные, очень искусные гнезда строят открыто на тонких ветвях деревьев и реже кустарников, обычно располагая их невысоко от поверхности земли. Некоторые формы могут использовать гнезда своего вида в течение нескольких лет, ежегодно достраивая новое на прошлогоднем. Яйца пятнистые. Длительность инкубации около 14 дней.
Питаются главным образом насекомыми разных групп и стадий развития, реже поедают других мелких беспозвоночных. Изредка заглатывают гастролиты. Для многих видов основным способом охоты является подкарауливание летающих насекомых с какой-либо присады с последующей ловлей их в воздухе на лету в стремительном броске. Склевывают также беспозвоночных с растений и поверхности земли.
Черная райская мухоловка — Terpsiphone atrocaudata (Eyton, 1839)
Muscipeta atrocaudata Eyton, 1839, Proc. Zool. Soc. London: 102, ’’Малайя” - ошибочно; Япония.
Син. (русск.): темноспинная длиннохвостая Мухоловка, черная длиннохвостая мухоловка, чернохвостая райская мухоловка.
Описание. Составлено по 9 экз. (8 — с Японских о-вов, 1 — из Южного Приморья).
Вершину крыла образуют 4-е и 5-е первостепенные маховые, причем 4-е всегда больше 5-го. Сильно выступает за вершины кроющих кисти 1-е маховое и составляет около половины длины 2-го; 2-е короче 3-го. У большинства особей наружные опахала 4—6-го первостепенных маховых в вершинной части заметно сужены. У взрослых самцов третья пара рулевых (от центра) значительно короче второй (рис. I)1. Расстояние между вершинами 2-го и 3-го рулевых составляет 25-40 мм (в среднем -31,5). Количество щетинок в углах рта (с каждой стороны головы) у основания
1 Рисунки 1,2,4-8, 10,11, 13, 15, 18, 22, 23, 25, 26, 30-32, 34 помешены на вклейку.
6
надклювья — 5 (14,3 %) — 6 (85,7 %), у основания подклювья — 3 (66,7 %) — 4 (33,3 %). Длина наибольшей щитинки у основания надклювья (п = 7) : 10,6— 14,1 мм (в среднем - 12,3). Количество щитков подотеки на передней части цевки варьирует от .7 до 9. Половой диморфизм резко выражен в окраске оперения и менее сильно в размерах отдельных частей тела (табл.1).
Распространение. Япония — от о-ва Хонсю на юг до о-вов Рюкю, о-в Чеч-жудо, о-ва Данной и Хошаодао восточнее о-ва Тайвань, о-ва Батан севернее о-ва Лусон. Характер пребывания на о-ве Тайвань точно не выяснен. В СССР изредка залетает в южную часть Приморского края. На большей части ареала птицы этого вида являются перелетными и зимуют главным образом в Южном Китае, на п-ове Индокитай и о-вах Хайнань и Суматра. Мухоловки южной части гнездового ареала оседлы.
Систематика. Таксономический статус этой формы до настоящего времени оставался спорным. Большинство исследователей (Bianchi, 1907; Hartert, 1907; Hartert, Steinbacher, 1934; Jahn, 1942; Спангенберг, 1954; Wynne, 1954; Иванов, 1976; Степанян, 1978; и др.) считают ее самостоятельным политипическим видом, образующим 2—3 подвида, однако некоторые зоологи, в том числе и Л.А.Портенко (1960), рассматривают форму atrocaudata как подвид T.paradisi.
Для решения этого вопроса Нами был применен метод анализа степени морфологических различий между интерградирую-щими подвидами внутри широко распространенных видов одного рода, предложенный Э.Майром (1971). Этот метод позволил установить, насколько сильные морфологические различия могут возникнуть в процессе эволюции у разных, в том числе и сильно разошедшихся подвидов T.paradisi, не сопровождаясь развитием репродуктивной изоляции. Изучение серии черных райских мухоловок, проведенное нами после этого, позволило выявить у них ряд морфологических особенностей, не свойственных ни одному из подвидов T.paradisi, в том числе и островным. У всех подвидов Т paradisi оперение верхней части головы, включая хохол, блестяще-темно-изумруднозеленое, в то время как у формы atrocaudata оно черное с фиолетовым отливом. По этому признаку черные райские мухоловки (как самцы, так и самки) во всех возрастных нарядах, начиная с первого после-
гнездового, четко отличаются от T.paradisi. Взрослые самцы формы atrocaudata в отличие от таковых T.paradisi имеют и несколько иную форму хвоста (см. рис. 1): третья пара рулевых (от центра) у них значительно короче второй пары. Идентичная форма хвоста не встречена ни у одного подвида T.paradisi. Белая цветовая морфа среди взрослых самцов, свойственная в разной степени проявления всем подвидам T.paradisi, у черной райской мухоловки отсутствует. Различия между этими аллопатрическими формами имеются также в окраске других участков оперения, в размерах отдельных частей тела (табл. 2), и в некоторых особенностях их образа жизни. На основании всего сказанного выше мы считаем T.atrocaudata самостоятельным видом.
Образует 3 подвида (Vaurie, 1959), 1 из которых — номинативный изредка встречается на территории СССР в южной части Приморского края.
Японская черная райская мухоловка — Terpsiphone atrocaudata atrocaudata (Eyton, 1839)
Muscipeta atrocaudata Eyton, 1839, Proc. Zool. Soc. London: 102, ’’Малайя” - ошибочно; Япония.
Описание. Составлено пр 9 экз. (8 — с Японских о-вов, 1 — из Южного Приморья).
У взрослого самца все оперение головы и шеи черное с фиолетовым отливом. Спина, надхвостье и большая часть верхних кроющих крыла блестяще-темно-лиловые (темно-красно-фиолетовые). Верхние кроющие хвоста и рулевые черные со слабым сине-фиолетовым отливом. Грудь или почти полностью черная, или черная только в верхней своей части. Бока тела от темнобурого до каштаново-бурого. Брюшко и нижние кроющие хвоста сливочно-белые. Маховые черные. Наружные опахала первостепенных маховых по краю отливают темно-сине-фиолетовым, а на второстепенных и третьестепенных имеют темно-каштановофиолетовые канты. Верхние кроющие первостепенных маховых черные со слабым металлическим отливом. Широкое голое кольцо вокруг глаза ярко-голубое.
У взрослой самки оперение верхней части головы черное, с фиолетовым, как и у самца, отливом. Бока головы и шеи, кроме передней ее части, темно-серые. В свежем пере на боках шеи имеется каштановорыжий налет, образованный узкими вер-
7
00
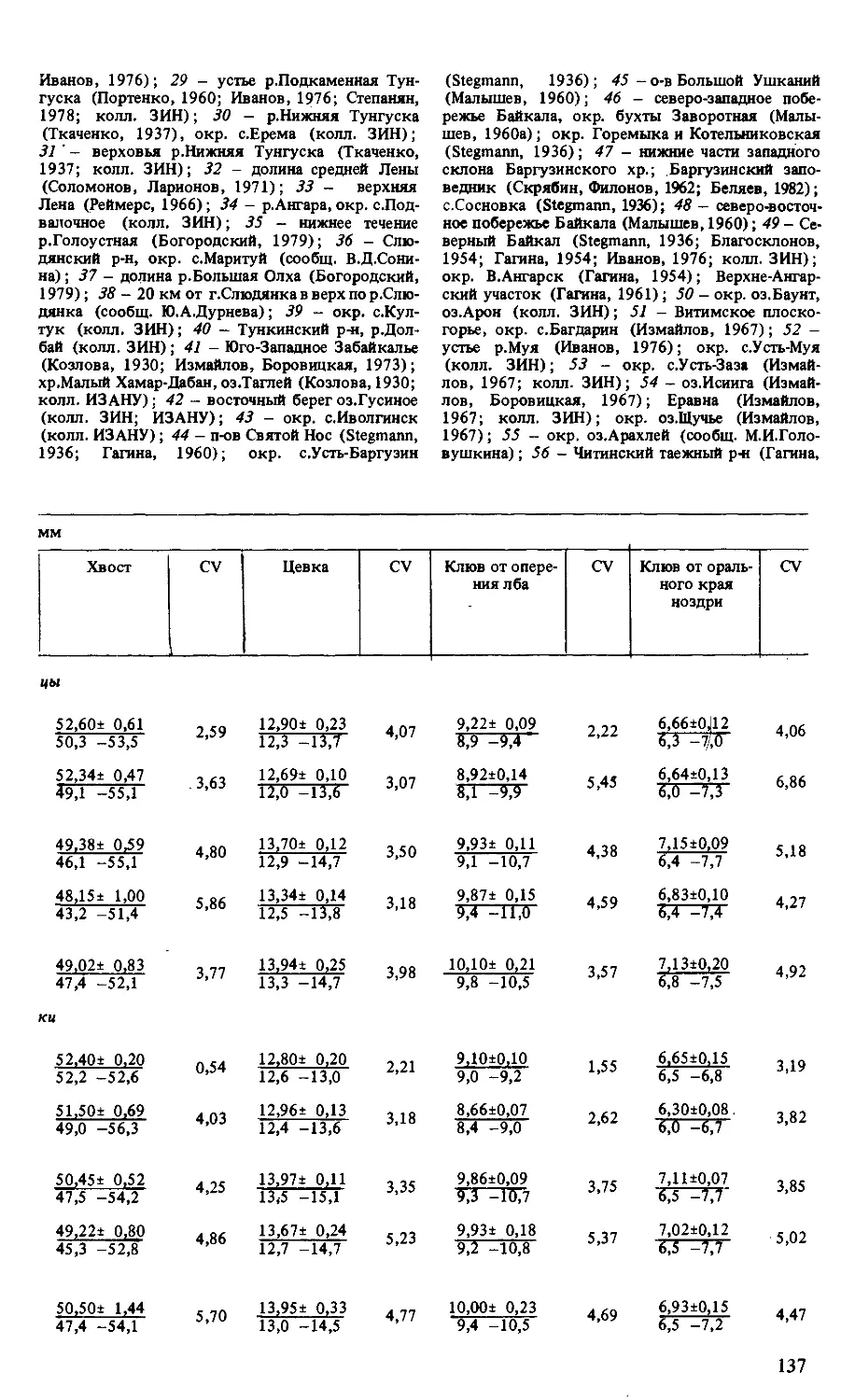
Таблица 1. Размеры 1 *?) и коэффициенты вариации (CV, %) райских мухоловок
Вид,подвид п Длина, мм
Крыло CV Центральная пара рулевых CV Хвост (без учета центральных рулевых) CV Цевка CV Клюв от оперения лба CV
Самцы первогодки
T.paradisi leucogaster 8 92,361 1,02 88,0 - 96,5 3,11 101,941 2,96 93,8 -115,7 7,68 97,54+ 1,81 5,24 17,611 0,12 17,2 -18,1 1,98 18,541 0,40 16,7 -19,8 5,69
91,0 -104,8
T.p.incei 3 90,571 0,30 90,0 - 91,0 0,57 87,201 0,20 87,0 - 87,4 0,32 86,651 0,35 86,3 - 87,0 0,57 15,401 0,20 15,0 -15,6 2,25 16,031 0,15 15,8 -16,3 1,57.
T.atrocaudata atrocaudata 1 - - - - - - 16,0 - 14,5
T.paradisi leucogaster 7 95,011 0,54 92,7 -96,9 1,51 Самцы второгодки 122,271 3,92 — 7,85 112,6 -138,0 (взрослые) 103,331 1,04 99,8 -106,3 2,47 17,671 0,18 17,0 -18,5 2,69 18,481 0,26 17,8 -19,0 3,19
Самцы старше двух лет (взрослые)
T.paradisi leucogaster 31 97,451 0,25 95,0 -100,0 1,42 236,181 Id,20 105,5 -305,0 23,70 111,631 1,09 100,6-128,0 5,14 17,381 0,08- 16,6 -18,1 2,54 18,50t 0,15 17,2 -20,1 4,30
T.p.incei 14 93,481 0,27 92,0 - 95,2 1,00 261,751 9,93 206,0 -321,0 13,15 106,051 3,27 89,6 -123,0 11,13 15,2310,12 14,3 -15,8 2,83 16,57t 0,37 14,7 -18,6 7,67
T.atrocaudata atrocaudata 5 94,081 0,64 92,8 - 96,5 1,52 311,751 15,34 279,0 -353,0 9,84 120,881 7,68 111,0-143,5 12,70 15,55t 0,25 15,3 -15,8 2,27 15,15t 0,20 14,6 -15,5 2,67
Самки
T.paradisi leucogaster 36 91,751 0,36 87,0 -96,4 2,34 104,501 1,33 90,7 -118,5 6,50 96,431 0,61 90,0 -103,8 3,45 17,261 0,08 15,7 -18,0 2,88 17,251 0,16 15,3 -18,9 5,29
T.p.incei 3 88,071 1,70 85,1 -91,0 3,35 86,401 4,10 Й2,3 - 90,5 6,71 85,951 4,55 81,4 - 90,5 7,49 15.301 0,12 15,1 -15,5 1,31 15,771 0,78 14,5 -17,2 8,61
T.atrocaudata atrocaudata 3 86,671 1,28 84,3 - 88,7 2,56 91,33t 5,75 84,8 -102,8 10,91 81,831 0,90 80,3 - 83,4 1,89 14,871 0,23 14,4 -15,1 2,72 14,30+ 0,20 13,9 -14,5 2,42
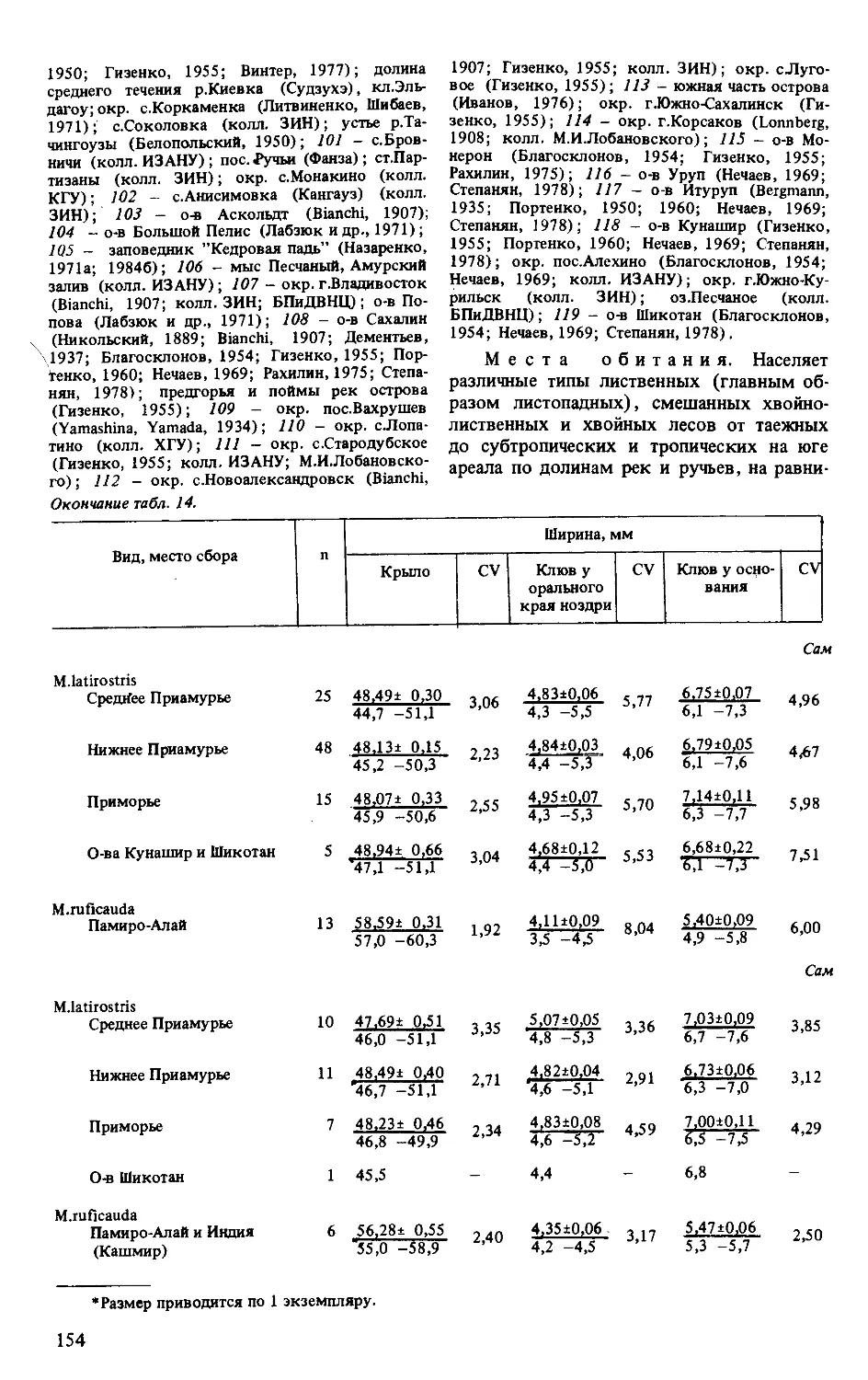
Вид,подвид п Длина, мм Ширина, мм Отношение шири-ны клюва у осно-вания к его длине от оперения лба, % CV Отношение длины хвоста к длине крыла, % CV
Клюв от орального края ноздри CV Клюв у орального края ноздри CV Клюв у основания CV
Самцы первогодки
T.paradisi leucogaster 8 13,75 + 0,27 13,0 -14,9 5,64 6,78+0,07 6,4 -7,0 2,82 8,09±0,06 7,8 -8,3 2,14 43,64± 0,93 40,00-47,31 5,63 105,55+ 0,95 102,28-109,74 2,54
T.p.incei 3 11,47+ 0,33 11,0 -12,1 4,96 6,03+ 0,27 5,5 -6,4 7,83 8,23t 0,67 8,1 -8,3 1,40 51,37± 0.86 49,69-52,53 2,90 95.91+ 0.76 95,15- 96,67 1,12
T.atrocaudata atrocaudata 1 10,6 — 6,2 — 7,7 — 53,10 — —
Самцы втерогодки (взрослые )
T.paradisi leucogaster 7 13,88+ 0,23 3,76 7,23+ 0,09 3,27 8,60± 0,07 2,12 46,34± 0,45 110,241 1,47 107,22-118,64 3,52
13,1 -14,3 6,8 -7,4 8,3 -8,8 44,74- 47,49
Самцы старше двух лет (взрослые)
T.paradisi leucogaster 31 13,86+ 0,13 12,8 -15,1 4,71 7,ОЗ1 0,07 6,4 -7,8 5,17 8,47+ 0,08 7,6 -9,3 5,16 45,80+ 0,50 40,00-51,10 5,63 115,06± 0,97 103,71-130,61 4,39
T.p.incei . 14 12,21± 0,11 3,19 6,51 ± 0,15 8,83 8,34± 0,13 5,90 50,31+ 0,99 6,85 115,90± 3,63 10,39
11,4 - 12,8 5,6 -8,2 7,5 -9,2 43,10-55,10 96,86-129,98
T.atrocaudata atrocaudata 5 11,38± 0,32 6,26 6,44± 0,07 2,35 8,46± 0,08 2,15 55,47± 0,83 2,98 128,29± 7,04 10,97
10,9 -12,6 6,2 -6,6 8,3 -8,7 53,90- 56,95 118,34-148,70
Самки
T.paradisi leucogaster 36 13,05 + 0,16 6,78 6,63± 0,06 5,50 8,06± 0,08 5,78 46,61 ± 0,63 7,65 105,17± 0,42 2,18
11,0 - 14,9 6,0 - 7,4 7,3 -8,9 41,71-57,52 101,09-111,97
T.p.incei 3 U,67± 0,23 3,46 6,30± 0,15 4,20 8,40± 0,10 2,06 53,54± 2,71 8,77 99,19± 3,53 5,04
11,3 - 12,1 6,0 -6,5 8,3 - 8,6 48,26-57,24 95,65-102,72
T.atrocaudata atrocaudata 3 10,57± 0,32 5,21 6,25 ± 0,25 5,66 7,60± 0,40 ' 7,44 53,49± 1,69 4,46 94,43± 0,41 0,76
10,2 -11,2 6,0 -6,5 7,2 -8,0 51,80-55,17 94,02- 95,26
Примечание. Птицы, размеры которых приводятся в таблице, собраны: T.paradisi leucogaster - Памиро-Алай, Западный Тянь-Шань; T.paradisi incei — Приморье, Приамурье, Китай; Т.atrocaudata atrocaudata - Южное Приморье и Япония.
Таблица 2. Достоверность отличий (Z) размеров Terpsiphone atrocaudata atrocaudata от T.paradisi
шинными каемками на перьях. Горло, зоб и верхняя часть груди шиферно-серые. Бока тела серые с охристым налетом. Нижняя часть груди, брюшко и нижние кроющие хвоста сливочно-белые. Маховые и их большие верхние кроющие темно-бурые. На наружных опахалах последних, а на третьестепенных маховых и на внутренних опахалах, расположены каштаново-рыжие каемки. Спина, надхвостье, верхние кроющие хвоста, рулевые, а также малые верхние кроющие крыла каштаново-рыжие.
Первый послегнездовой наряд идентичен наряду взрослой самки, но черные перья, образующие хохол, только на вершинах имеют фиолетовый отлив.
Размеры отдельных частей тела японской черной райской мухоловки приведены в табл. 1.
Распространение. Гнездится на островах Хонсю, Сикоку, Кюсю, Цусима, Якусима и Чечжудо. Характер пребывания на о-ве Тайвань точно не выяснен. К местам зимовок в Южный Китай, на п-ов Индокитай и о-ва Хайнань и Суматра мигрирует главным образом через Восточный Китай и о-в Тайвань. Зарегистрированы залеты в Северо-Восточный Китай, Корею, на о-в Хоккайдо (Chec-list of Japan. Birds, 1974) и на территорию СССР в южную часть Приморского края (см. рис. 9).
В фауну СССР этот вид включен по экземпляру (взрослый самец), добытому Т.Б.Саблиной в конце июня 1941 г. в бухте Находка (рис. 9, J)1 и доставленному А.Г.Банниковым в Зоологический музей МГУ (Воробьев, 1954; Спангенберг, 1954). По данным опроса, полученным К.А.Во-робьевым (1954) от местных жителей, птицы предположительно этого вида наблюдались и были добыты летом 1934 г. близ с.Славянка (рис. 9, 2) в Хасанском р-не и летом 1946 г. у государственной границы с КНДР (рис. 9, 5). 29.V 1964 г. на опушке мелколесья о-ва Большой Пелис (о-ва Римского-Корсакова в заливе Петра Великого) (рис. 9, 4) был добыт взрослый самец черной райской мухоловки, а 16— 17.VI 1964 г. на о-ве Стенина наблюдался и был добыт еще один самец (размеры семенников 6,0 х 4,0 мм). Он держался на просеке, густо поросшей кустарниками и высокотравьем (Лабзюк, Назаров, 1967).
11~4 - на рис. 9 номера географических пунктов.
10
Райская мухоловка — Terpsiphone paradisi (Linnaeus, 1758)
Corvus paradisi Linnaeus, 1758, Syst. Nat., ed. 10: 107, Индия.
Син. (русск.): длиннохвостая мухоловка.
Описание. Составлено по 109 экз. (85 - из Памиро-Алтая и Западного Тянь-Шаня, 13 - из Приамурья и Приморья, 11 - из Китая).
Вершину крыла образуют 4-е и 5-е первостепенные маховые (4=5 - 47,0 %; 4 > 5 - 31,8 %; 4 < 5 - 21,2 %). За вершины кроющих кисти сильно выступает 1-е маховое и составляет около половины длины 2-го; 2-е короче 3-го. У большинства особей наружные опахала 3-го - 6-го первостепенных маховых в вершинной части заметно сужены. У взрослых самцов третья пара рулевых (от центра) ненамного короче второй (рис. 1). Количество щитинок в углах рта (с каждой стороны головы) у основания надклювья — 5-7 (5 - 5,6 %; 6 -90,0 %; 7 - 4,4 %), у основания подклю-вья - 3-5 (3 - 77,5 %; 4 - 16,9 % 5 -5,6 %). Длина наибольшей щетинки у основания надклювья (п = 82): 12,2-19,1 мм (в среднем - 15,4). Количество щитков подотеки на передней части цевки варьирует от 6 до 9. Половой диморфизм резко выражен в окраске и менее сильно - в размерах отдельных частей тела (табл. 1).
Виду свойственно явление морфизма. Взрослые самцы райской мухолоНки имеют 3 цветовые морфы: темноокрашенную, белую и переходную (характеризуется частичным асимметричным развитием белой окраски). В различных частях ареала птицы одевают белый наряд в разном возрасте (Owen, 1963). Числовое соотношение цветовых морф в популяциях вида различно. На юго-востоке ареала (о-ва Калимантан, Сумба и др.) белый тип окраски имеют до 100 % взрослых длиннохвостых самцов; в Китае, Таиланде и на о-ве Шри Ланка -34-36 %; в Индии, Бирме и Гималаях -61-92 % (Owen, 1963). На территории СССР в Приморье и Приамурье белая морфа составляет около 25 % всей популяции (Степанян, 1978), а в Памиро-Алае и Западном Тянь-Шане не встречена. Птицы переходного типа окраски встречаются в основном в Индии и Китае, а также на территории СССР в Памиро-Алае и Западном Тянь-Шане.
Распространение. От южного склона Паропамиза, Средне-Афганских гор и Белуджистана к востоку до побережья
Японского, Желтого, Восточно-Китайского и Южно-Китайского морей. К северу до Паропамиза, Западного Гиндукуша, Западного Памиро-Алая, Западного Тянь-Шаня и южного склона Гималаев. От восточной оконечности Гималаев северная граница, становясь на этом участке ареала северо-западной, протягивается к северной оконечности Сино-Тибетских гор и далее проходит по хребту Алашань, откуда тянется к среднему течению Ляохэ, отсюда по долине Сунгари до устья последней и далее по долине Амура ниже устья Сунгари примерно до 49° с.ш. К югу до океанического побережья Южной и Юго-Восточной Азии и южной оконечности п-ова Малакка. О-ва Шри Ланка, Андаманские, Никобарские, Большие Зондские (предположительно), Малые Зондские к востоку до о-ва Алор (Степанян, 1978). Птицы северной части гнездового ареала являются перелетными и зимуют на юге ареала.
С и с т е матика. Географическая изменчивость проявляется в варьировании окраски оперения у самок и самцов темно-окрашенной морфы, в числовых соотношениях цветовых морф, в варьировании общих размеров и длины отдельных частей тела.
Выделяют обьино до 16 подвидов (Wynne, 1954; Ali, Ripley, 1972; Степанян, 1978), 2 из которых - T.p.leucogaster (Swainson, 1838) и T.p.incei (Gould, 1852) — встречаются и гнездятся на территории СССР. Мухоловки названных подвидов четко различаются по размерам отдельных частей тела (табл. 3; рис. 2), окраске оперения и числовому соотношению цветовых морф среди взрослых самцов.
Белобрюхая райская мухоловка — Terpsiphone paradisi leucogaster (Swainson, 1838)
Muscipeta leucogaster Swainson, 1838, Nat. Libr., 21, Flycatchers: 205, pl. 24, Индия.
Описание. Составлено по 85 экз. из Памиро-Алая и Западного Тянь-Шаня.
У взрослого самца старше двух лет темноокрашенной морфы все оперение головы, включая хохол, блестяще-темно-изумрудно-зеленое. Грудь, брюшко, бока тела и нижние кроющие хвоста белые. Перья, покрывающие голень и нижние кроющие крыла, светло-охристые. Верхние кроющие второстепенных маховых, спина, надхвостье, верхние кроющие хвоста и рулевые ярко-охристо-рыжие. Маховые черно-бурые. На наружных опахалах первостепен-
11
Таблица 3. Достоверность отличий (О размеров Terpsiphone paradisi leucogaster от T.paradisi incei
ных, а у некоторых особей и второстепенных маховых, имеются продольные белые канты, ширина которых увеличивается от второго первостепенного махового к второстепенным. У отдельных самцов белые канты проходят и по краю внутренних опахал маховых перьев. На наружных опахалах проксимальных второстепенных маховых канты охристо-рыжие, а на третьестепенных становятся очень широкими и опоясывают каждое перо со всех сторон, порою вытесняя основную темно-бурую окраску полностью. Верхние кроющие первостепенных маховых черно-бурые. В популяции Пами-ро-Алая и Западного Тянь-Шаня темноок-рашенная морфа среди взрослых самцов этого подвида составляет около 93 %. Самцы переходной морфы, составляющие 7 %, отличаются от описанных выше наличием и асимметричным расположением в хвосте и на крыльях отдельных белых перьев с черным стержнем и тонкой черной окантовкой по краю опахал. На некоторых белых перьях сохраняются рыжие пятна основной окраски. Самцы второгодки обладают такой же окраской, как и самцы старше двух лет темно-окрашенной морфы, но в отличие от последних канты на наружных опахалах первостепенных маховых у них охристо-рыжие. Кроме этого центральная пара рулевых перьев у них значительно короче, чем у самцов старше двух лет (табл. 1). У взрослых самцов голое кольцо вокруг глаза ярко-синее.Клюв сине-голубой. По краям надклювья и подклювья от вершины клюва до ноздри тянется узкий черный кант. Полость рта зеленовато-желтая, под языком — желто-зеленая. Ноги серо-голубые. Когти темно-бурые со светло-бурой внутренней полосой. Радужина глаза темно-бурая.
У взрослой самки хохол на голове более короткий, чем у самца. Верхняя часть g головы и тела, а также рулевые имеют ° такую же окраску, как и у взрослого сам-ца. Бока головы и зашеек темно-серые. * Горло, и область зоба светло-пепельно-о серые. Серый цвет заходит на верхнюю ° часть груди, а у некоторых особей распро-страняется и на бока тела. Нижняя часть $ груди, брюшко и нижние кроющие хвоста g белые. У некоторых особей светло-пепель-но-серые перья зоба и верхней части груди v имеют примесь охристо-рыжего. Маховые темно-берые с охристо-рыжими кантами по « краям наружных и внутренних опахал, начиная со второго первостепенного. Ширина кантов увеличивается от первостепенных маховых к третьестепенным и на последних
12
они полностью вытесняют основную окраску. Нижние кроющие крыла от молочно-белого до бледно-охристого. Клюв серовато-голубой, а голое кольцо вокруг глаза — с примесью серо-голубых тонов.
Молодые в гнездовом наряде сверху охристо-рыжие, с более темным лбом и теменем. Шея и передняя часть груди серо-вато-охристые, остальная часть нижней стороны тела белая. Клюв и ноги бурые. Крючочек на вершине надклювья отсутствует.
Первый послегнездовой наряд идентичен наряду взрослой самки, но перья, образующие хохол, в основаниях матовочерные и только на вершинах блестяще-темно-изумрудно-зеленые.
Размеры отдельных частей тела белобрюхой райской мухоловки приведены в табл.1.
Масса тела, г: самец первогодок (4.VI, Таджикистан, Варзобское ущ.) - 16,99; самцы второгодки (п = 3) (4.VI - 18.VI, Варзобское и Рамитское ущелья; окр. к. Косторош) — 18,50—19,42, в среднем -19,09; самцы старше двух лет (п = 6) (31.V—18.VI, Варзобское и Ратмитское ущелья) - 18,85—20,95, в среднем — 19,84; самки (п = 10) (31.V—7.VI, Варзобское ущ.) - 17,62-19,89, в среднем - 18,73.
Распространение. Гималаи от Восточного Непала до Кашмира, Восточный Афганистан и далее на юго-запад до Северного Белуджистана (Vaurie, 1959). Северная часть гнездового ареала заходит на территорию СССР в Памиро-Алай и Западный Тянь-Шань (рис. 3). Мухоловки северных популяций являются перелетными и зимуют на большей части п-ова Индостан, главным образом в его юго-западной части (Юго-Восточный Бомбей, Карнатака, Керала), а также на равнинах Восточного Пакистана (Рашид) (Vaurie, 1959; АН, Ripley, 1972). Более детально распространение белобрюхой райской мухоловки на территории СССР показано на карте (рис. 3).
Кадастр к рис. 3.
1 - хр. Кугитанг (Иванов, 1969; Степанян, 1978); окр. с.Свинцовый Рудник, ущ. Дарайдере (колл. КГУ; Турк. СХИ); Ходжипиль (колл. КГУ); 2 - южная оконечность хр. Кугитанг (Степанян, 1978); с.Кугитанг (Салихбаев, Остапенко, 1964; колл. ИЗАНУ); 3 - Зеравшанский хр., ур. Аман-Кутан, 50 км южнее г.Самарканд (Сагитов, 1961; колл. ИЗАНУ; А.П.Кузякина); лесхоз; Майдан-сай (Сагитов, 1961); 4 - окр. г.Педжикент (Bianchi, 1907; Абдусалямов,1973); Зебон (Абдусалямов, 1973); 5 - к. Ери (Лебедев, 1976); 6 - долина р. Шинк (Иванов, 1940; Абдусалямов, 1973); к. Шинг (Иванов, 1940;
колл. ЗИН; ИЗАНУ); р.Магиандарья у к.Шинг (колл. ЗИН); окр. к. Косторош (колл. ИЗАНУ);
7 - Рамитское ущ., с.Зайрон, лесистый склон с ручьем на левом берегу р.Кафирниган (колл. ИЗАНУ); заповедник ’’Рамит” (наши данные); Рамитское лесничество (Попов, 1959); долина Сарбо (Абдусалямов, 1973); окр. к. Сарбо (колл. ИЗАНУ); верховья р.Кафирниган, р.Сор-бух (Иванов, 1940; 1969); 8 - южные склоны Гиссарского хр.: долина р.Варзоб (Иванов, 1940; 1969; Попов, 1959; Воробьев, 1968; Абдусалямов, 1973; колл. ИЗАНУ); с.Варзоб, Альплагерь (колл. КБ МГУ; ИЗиП АН ТаджССР);
3 км южнее с.Варзоб (колл. ИЗАНУ); ур. Тан-гинур, 30 км севернее г.Душанбе (колл. ИЗиП АН ТаджССР); ур. Обисафед (Иванов, 1969; колл. КБ МГУ); ущ. Кондара (Попов, 1959; Воробьев, 1968; колл. ЗИН; БИН СО АН; ИЗАНУ; С.Д.Кустановича, А.П.Кузякина); левый берег р.Кондаринка (колл. ИЗАНУ); ущ. Джеринот, 1 км южнее к. Кондара; окр. к. Гажни (колл. ИЗАНУ); 9 - окр. с.Такоб, левый берег р.Та-кобка (колл. ИЗАНУ); ущ. Дей-Боло у устья р.Такобка (колл. ЗИН); 10 - ленточные леса по южному склону Туркестанского и северному склону Зеравшанского хребтов, между к.Суджино и 20 км не доезжая г.Айни (Лебедев, 1976); долина р.Кштут (Сагитов, 1961; Абдусалямов, 1973); к.Кштут (Иванов, 1940); восточнее к.Кштут (Bianchi, 1907); 11 - долина р.Сар-даи-Миеон (Иванов, 1940; 1969; Абдусалямов, 1973); 12 - долина Ханака; Каратаг (Абдусалямов, 1973); 13 - верховья р.Сурхан, вдоль ее притоков: р.Тупаланг и р.'Сангардакдарья (Иванов, 1969; Абдусалямов, 1973); р.Тупаланг (Салихбаев, Остапенко, 1964); 14 - хр.Аруктау, ур. Ак-Су (Абдусалямов, 1973); 15 - сан хр.
Рис. 3. Распространение белобрюхой райской мухоловки (Terpsiphone paradisi leucogaster) на территории СССР:
а — гнездится, б - отмечена в гнездовой период. См. кадастр, стр. 13, 14,
13
Хазрати-Шох, г.Муминабад, кишл. Ала-Ака (Абдусалямов, 1973); 16 - хр. Каратегин, горы Санглок (Иванов, 1969); окр. кишл. Санглок, ущ. Сары-Чашма (Попов, 1959; колл. ЗИН; ИЗиП АН ТаджССР); 17 - южные склоны Дар-вазского хр! (Абдусалямов, 1973); долина р.Пяндж до к.Калаи-Хумб и к.Тогмай (Иванов, 1940; 1969; Абдусалямов, 1973); долина
р. Пяндж ниже к.Калаи-Хумб (Степанян, 1978); к-Калаи-Хумб (Иванов, 1940); к.Тогмай (Иванов, 1976); окр. к.Хочильер (Попов, 1959); 18 -г.Гиссар (Ахмедов, 1957); 19 - окр. г.Исфара (Зарудный, 1915; Иванов, 1969; 1976); 20 -окр. к.Ворух; ур.Самаркандек (Зарудный, 1915; Иванов, 1969); 21 - хр.Каратау (Абдусалямов, 1973); 22 - хр. Нуратау, Нуратинский заповедник, ур. Хаят-сай (сообщ. О.В.Митропольского); 23 - западная часть Чаткальского хр. (Тараненко, 1974; Степанян, 1978); долина р.Аксак-Ата (сообщ. О.В.Митропольского); долина р.Угам; Башкизыл-сай; ур. Богучелпек, выше устья Та-наберды-сая; левый берег выше впадения Алтын-куль-сая; ореховая роща возле полевой базы Чаткальского заповедника; Шавази-Колон-сай; Кизыл-алма-сай; Кознакок-сай; Каракузы-сай (Тараненко, 1974); Пустунлик-сай (Тараненко, 1974; колл. РГУ); к.Брич-мулла у впадения р.Коксу в р.Чаткал (Зарудный, 1915; колл. КГУ); выше указанного кишлака (Зарудный, 1915); долина р.Ангрен и ее притока р.Аб-Яз; окр. к. Аблык (Зарудный, 1915); 24 - Киргизия, окр. орехосовхоза ’’Афлатунь” в 70 км от г.Наманган (колл. С.Д.Кустановича); 25 - Са маркандская обл., ущ. Ак-сай (Богданов, 1956; Сагитов, 1961; колл. БИН СО АН); 26 — г. Куляб; ур. Гамидара на р.Пяндж (Зарудный, 1915; Иванов, 1940, 1969; колл. ЗИН); 27 - г.Самар-канд (Зарудный, 1915; Дементьев, 1937; Спан-генберг, 1954; Портенко, 1960; Иванов, 1969; Степанян, 1978); 28 - ст. Яккобаг (Зарудный, 1915; Спангенберг, 1954; Иванов, 1969); 29 — горы Байсунтау (Сангенберг, 1954; Степанян, 1978); к. Байсун (Зарудный, 1915; Иванов, 1969); 30 - ур. Арчал на р.Ак-Бура в 30 км южнее г.Ош (Зарудный, 1915); 31 - бассейн р.Чирчик (Bianchi, 1907; Зарудный, 1915); к.Ходжикент; нижнее течение р.Угам (Зарудный, 1915); 32 - окр. г.Комсомолобад (Иванов, 1969); 33 - Западный Дарваз (Зарудный, 1926; Иванов, 1940; 1976); Юго-Западный Дарваз (Попов, 1959); р.Обиниоу, кЛянгар (Зарудный, 1915; 1926; Иванов, 1940; Абдусалямов, 1973); 34 - г.Ош (Дементьев, 1937; Спангенберг, 1954); 35 - Таласский Алатау (Зарудный, 1915; Дементьев, 1937; Степанян, 1978); заповедник ”Аксу-Джабаглы” (Шевченко, 1948; Ковшарь, 1962, 1966, 1970); каньон р.Аксу, ур. Кши-Каин-ды, окр. с.Новониколаевка (Шевченко, 1948; Ковшарь, 1962, 1966, 1970); окр. кордона Джа-баглы; ущ. р.Талдыбулак (Иващенко, 1982); 36,- западные отроги Каратау (Дементьев, 1937; Спангенберг, 1954); отроги восточной части хр.Каратау (Зарудный, 1915); южные склоны хр. (Портенко, 1960) и юго-восточная часть Каратау (Степанян, 1978); южная часть Каратау (Боролдай), долина р.Боролдай от среднего течения до верховьев; северные склоны хребта, истоки р.Арысь (Ковшарь, 1970); долина р.Боролдай и западная оконечность хребта (Степанян, 1978); р.Арысь (Hartert, 1907; Зарудный, 1915; Степанян, 1978); 37 - долина рЛскем (Ков
шарь, 1970); долина р.Пскем на север до Таласского Алатау; место Ланкат; сай Камыш-ат окр. кЛскем; Кара-булак (Зарудный, 1915); 38 -долина р.Угам (Ковшарь, 1970; колл. ЗИН); к.Хумсан (Bianchi, 1907; Зарудный, 1915; Тараненко, 1974; колл. ЗИН; КБ МГУ); восточные склоны Каржантау (Зарудный, 1915); 39 - западная часть Сусамырского хр. (Зарудный, 1915; Степанян, 1978); 40 - г.Чимкент (Зарудный, 1915; Спангенберг, 1954; Степанян, 1978); 41 -г.Джизак (Зарудный, 1915; Спангенберг, 1954);
42 - г.Ташкент (Степанян, 1978; Мекленбур-цев, 1982); окр. г.Ташкента место Капланбек; Кара-камыш; по Салару и Босу (Зарудный, 1915); ст.Келес (Зарудный, 1915; Спангенберг, 1954);43 - устье р.Хингоу, р.Вахш (колл. ЗИН); к.Помбачи на р.Вахш; р.Сарбог, приток р.Сур-хоб (Иванов, 1940; сборы Лидского, Абдусалямов, 1973; колл. ЗИН); р.Сарбог между кишлаками Шинглич и Гориф (колл. ЗИН); 44 - долина р.Пяндж: Сары-Гур; Парвар (Зарудный, 1915; Иванов, 1940); Шуроб-дара; Новачун; Мильол; Хирманэтау (Зарудный, 1915); 45 - восточная часть Ферганской долины (Дементьев, 1937; Спангенберг, 1954; Степанян, 1978); Андижанская обл., близ места Арсланбоб в бассейне р.Ка-радарья (Зарудный, 1915; Дементьев, 1937; Спангенберг, 1954); 46 — г.Наманган (Дементьев, 1937; Спангенберг, 1954); 47 — г.Чиназ; устье р.Чирчик (Зарудный, 1915); 48 - окр. ст. Бек-лярбек, северо-западнее к.Ходжикент (Зарудный, 1915); 49 - восточные склоны возвышенности, составляющей водораздел между р.Угам и р.Пскем между к.Хумсан на первом и кишлаками Багустан и Нанай на втором; к.Багустан; к.На-най; Кумтукай и далее к северу до перевала Май-дантал-ашу (Зарудный, 1915); 50 - с.Сосновка в предгорьях центральной части Киргизского Алатау (Спангенберг, 1954; Иванов, 1976); ущ.Ка-рабалты (Шнитников, 1949; Спангенберг, 1954); 51 - северо-восточная оконечность Пскемского хр. (Степанян, 1978); 52 - восточная часть Алайского хр. (Степанян, 1978); 53 - восточная часть Дарвазского хр. (Степанян, 1978); 54 - ур. Арал на р.Итокар, бассейн Нижнего Нарына (Зарудный, 1915).
Места обитания. На территории СССР в границах ареала населяет главным образом различные типы лиственных лесов, рощи, сады и значительно реже — смешанные лиственно-арчевые леса и заросли высоких кустарников с редкими деревьями в предгорьях и горах до 2000 м н.у.м. Изредка проникает на окраины населенных пунктов, в том числе и крупных городов (Ташкент),где поселяется в садах и парках, предпочитая участки, наименее посещаемые людьми. Может гнездиться в небольших куртинах старых развесистых деревьев и даже на одиночных деревьях, незначительно удаленных от ближайшего леса или зарослей высоких кустарников. При выборе места гнездования необходимыми для райской мухоловки являются два условия: чередование тенистых участков леса или сада с ярко освещенными полянами, а так
14
же наличие на гнездовом участке или поблизости от него воды (горных речек, ручьев или арыков).
В Западном Тянь-Шане на Пскемском, Угамском и Чаткальском хребтах предпочитает ореховые леса (Ковшарь, 1970; Тараненко, 1974), но там, где их нет, поселяется также в тополевых, ивовых и как исключение - березовых рощах (Ковшарь, 1970). В заповеднике ”Аксу-Джабаглы” гнездится в яблочнике с примесью арчи на северном склоне р.Аксу, в узкой полоске березово-тополевого галерейного леса по руслу этой реки, а также в маленькой березовой рощице, окруженной высокоствольными арчевниками и суходольными лугами в ур.Кши-Каинды (Ковшарь, 1970). В Боролдае (южная гряда Каратау) обитает в фрагментарных галерейных лесах из ясеня, шелковицы, магалепской вишни и каркаса. Верхняя граница гнездования в Таласском Алатау проходит на высоте около 1800мн.у.м. (Ковшарь, 1970).
В Памиро-Алае гнездовой ареал белобрюхой райской мухоловки в значительной мере совпадает с областью распространения горных мезофильных лесов, т.е. прежде всего орехово-кленовых насаждений. Вертикальное распространение этого вида определяется здесь высотами от 1000 до 2000 м н.у.м. (Иванов, 1969). В Кугитанге райская мухоловка найдена нами на гнездовании в островном участке чилондвого леса (рис. 4), а на северном склоне Зеравшан-ского хр. в Аман-Кутанской лесной даче -в разреженном лесу из грецкого ореха с примесью ивы, дикой сливы и боярышника (рис. 5). В Таджикистане на южном склоне Туркестанского и северном склоне Зе-равшанского хребтов (между к.Суджино и г.Айни) гнездится в смешанных ленточных лесах из арчи, миндаля, клена, фисташки и рябины с незначительной примесью тополя, алычи и винограда на высотах 1500—2000 м н.у.м. (Лебедев, 1976). На южных склонах Гиссарского хр. в Варзобском и Рамитском ущельях населяет разреженные лиственные леса из ореха, туркестанского клена, каркаса, платана, ясеня и шелковицы с примесью ивы, алычи и винограда, а также небольшие куртины грецкого ореха (3-5 старых деревьев). В ущ. Джеринот (окр. кишл. Кондара) райская мухоловка найдена нами на гнездовании в кустарниковых зарослях из гиссарского кизильника с редкими деревьями. Отсутствует в чистых фисташниках. Избегает арчевников, однако в мае во вре
мя образования пар изредка залетает выше своего верхнего предела распространения в зону ксерофильиых кустарников и даже к снежникам (Попов, 1959).
В СССР во всех частях гнездового ареала охотно поселяется в старых, запущенных, редко посещаемых людьми садах (главным образом ореховых); значительно реже в непосредственной близости от жилых построек человека (усадьбы).
В Кашмире чаще всего гнездится в рощах из ив или шелковиц у озер и горных потоков, а также на одиночных платанах и в садах до 1800 м и значительно реже до 2400 м н.у.м. В Западных Гималаях (Ла-хул) зарегистрирована на высоте 3100 м (Ali, Ripley, 1972).
Численность. Изучена недостаточно. В Западном Тянь-Шане райская мухоловка считается обычной только в каньоне р.Аксу (Таласский Алатау) и в долине р.Боролдай (Каратау) (Ковшарь, 1978).
В Памиро-Алае, по нашим данным, обычна во многих местах гнездового ареала, в характерных излюбленных биотопах бывает многочисленна. На южном склоне Гиссарского хр. в ущ. к.Хочильер в июне 1950 г. зарегистрированы 4 гнездящиеся пары н? 1 га платанового леса (Попов, 1959). Мнение И.А.Абдусалямова (1973) о повсеместной невысокой численности райской мухоловки в Таджикистане, на наш взгляд, не соответствует действительности, а фактические данные, приводимые им (1—2 пары на 1 км2 характерного биотипа), очень сильно занижены. 31.V и 1.VI 1978 г. нами был проведен абсолютный учет этого вида в одном из боковых урочищ Варзобского ущ. в 3 км южнее к.Вар-зоб. В ленточном лиственном лесу с ручьем на площади 1,5 га отмечено 8 гнездящихся пар (530 пар/км2). Следовательно, для райской мухоловки характерна не низкая численность, а крайне неравномерное распределение по территории. Последнее объясняется мозаичным расположением гнездовых биотопов, отвечающих всем требованиям данного вида.
К основным причинам, влияющим на снижение численности белобрюхой райской мухоловки, следует отнести сокращение в ряде мест площадей ее характерных гнездовых биотопов, степень обеспеченности пищей (в том числе на местах зимовок и миграционных путях), а также фактор частого беспокойства со стороны человека в период размножения. Большой урон
15
гнездам могут наносить лесные сони, многочисленные в совместных местах обитания (Ковшарь, 1978).
Прилет и весенний пролет на территории СССР. В Средней Азии проходит в III декаде апреля и в I—II декадах мая.
В Юго-Восточной Туркмении — хр.Ку-гитанг (окр. с.Свинцовый Рудник, с.Куги-танг) несколько птиц добыто О.З.Яценей (колл. КГУ; ИЗАНУ) - 4.V, 8.V, 9.V 1961 и 14.V 1964 гг.
В Узбекистан на Заревшанский хр. в ур. Аман-Кутан (4 года)1 первые мухоловки прилетают 25.IV—27.IV-30.IV—5.V (Сагитов, 1961). Высказывания А.К.Саги-това (1961) о изменившихся сроках прилета райской мухоловки в наши дни по сравнению с началом текущего столетия (Зарудный, 1915) и вывод на основании этого о похолодании весны в Узбекистане ошибочны. Даты, приводимые Н.А.Заруд-ным, автором не переведены на новый стиль.
В Таджикистане сроки прилета и пролета следующие: в Гиссарской долине самая ранняя птица добыта А.И.Ивановым (1940, 1969) - 24.IV 1934 г., а К.Р.Ахме-дов (1957) в другие годы отметил здесь пролет 26.IV-14.V. В долине р.Кафирни-ган (пос.Гулистон) (10 лет) пролет проходит 22.IV-26.IV - 5.V—6.V (Абдусалямов, 1973). В ущ. р.Варзоб (4 года) мухоловки прилетают 30.IV (Иванов, 1969), 5.V и 7.V (Попов, 1959), 4.V (Беме,Сытов, 1963). 10.V 1961 г. РЛ.Беме (колл. БПиДВНЦ) добыл в этом ущелье у альпинистского лагеря ’’Варзоб” взрослую самку, a 10.V 1966 г. Й.А.Абдусалямовым (колл. КБ МГУ) был добыт взрослый самец. Последние пролетные птицы в Варзобском ущ. отмечены 18.V (Воробьев, 1968). РЛ.Потапов (Иванов, 1969) встретил одиночных мухоловок 4.V-6.V в заповеднике ’’Тигровая балка”, а Р.Н.Мек-ленбурцев (Иванов, 1940) отметил ее 1.V 1936 г. на р.Пяндж в к.Барроу. 9.V 1878 г. райская мухоловка добыта В.Рус-совыму г.Чиназ (Pleske, 1888; колл. ЗИН).
По данным Н.А.Зарудного (1915), в окр. г.Ташкента (4 года) первые птицы прилетают 29.IV—1.V — 3.V—5.V, а по данным А.Ф.Ковшаря (1970), они появляются здесь 25.IV-5.V. В районе слияния рек Боролдай и Кашкараты пролет проходит 15.V-18.V (Ковшарь, 1970). В Чаткаль-
1 Здесь и далее в скобках - продолжительность наблюдений.
ский заповедник (3 года) райские мухоловки прилетают 27.IV—2.V-5.V (Тараненко, 1974).
Летят птицы поодиночке, значительно реже по две, придерживаясь низинных об-лесненных участков, как днем, так и ночью (Зарудный, 1915; Ali, Ripley, 1972). Стай не образуют и только на кормежках и отдыхе могут скапливаться небольшими группами.
Размножение. На места гнездования первыми прилетают самцы и только через несколько дней после их прилета появляются самки. В этот период птицы ведут себя особенно оживленно и подвижно. Постоянно перепархивают с дерева на дерево, перед посадкой на ветку обычно взмывают круто вверх, распуская при этом хвост. Песня самца, напоминающая приглушенную песню иволги, тиха и мелодична. Отмечен случай копирования райской мухоловкой песни обыкновенной чечевицы (Ковшарь, 1983). Призывный крик - тихое ’’тереуэ”, крик тревоги -резкий отрывистый ”четр”. Образование пар сопровождается интенсивным пением самцов и их токовым демонстративным поведением (рис. 18). Сведения об образовании пар райскими мухоловками на пролете (Сдангенберг, 1954) нуждаются в проверке.
Гнездовые участки пар невелики. При высокой плотности гнездования расстояние между гнездами соседних пар составляет (п = 9) 12-70 м (в среднем - 31,1) (Тараненко, 1974; наши данные). Гнезда райские мухоловки строят в нижних частях крон деревьев и кустарников (грецкий орех, каркас, клен, алыча, боярышник, ива, платан, ясень, гиссарский кизильник, яблоня, береза, шелковица, чилон, тополь, ильм и др.), редко — в их средних частях, на тонких (диаметр 3,5-20 мм, в среднем 9; п = 25), главным образом сухих и значительно реже живых ветвях. Высота расположения гнезд от поверхности земли варьирует от 0,5 до 11,0 м (Спангенберг, 1954; Сагитов, 1961; Ковшарь, 1970), однако чаще всего они строятся птицами на высоте 0,8—2,5 м.
В сборе строительных материалов и строительстве гнезда принимают участие самец и самка. Строительство гнезда обычно длится 6—11 дней (Сагитов, 1961; Ковшарь, 1978). Некоторые пары ремонтируют старые гнезда своего вида или достраивают новые на прошлогодних.
Гнезда представляют собой плотные, чашевидные очень аккуратные постройки
16
(рис. 7), которые обладают большой прочностью и выдерживают без разрушений, по данным И.А.Абдусалямова (1973), ураганные ветры. Основная часть гнезда состоит главным образом из тонких полосок луба древесных и кустарниковых растений (орех, виноград, чилон и др.), а также из тонких корешков, стебельков и листьев трав, колосков и метелок злаков: Alopecurus sp., Festuca rupicola (наши данные); Agrostis alba (Тараненко, 1974), соцветий ивы, маленьких кусочков коры и листьев древесных растений. Весь этот строительный материал скрепляется паутиной, шерстью или липким лубом травянистых растений (Тараненко, 1974). В толщу боковых стенок и снаружи к гнезду приплетаются маленькие пучочки шерсти, мха, хлопка, растительного пуха, отдельные перья и очень редко кусочки сухого помета копытных животных. Снаружи все гнездо густо оплетается паутиной, шерстью и растительными волокнами. Лоток выстилаемся полосками луба деревьев и кустарников (шириной до 4 мм), растительными волокнами, стебельками и листьями злаков и реже других трав. Размеры гнезд (п = 18), мм: диаметр 72—100, в среднем - 85,2; высота 50-130, в среднем - 77,3; диаметр лотка 57-70, в среднем - 63,8; глубина лотка 28-42, в среднем - 36,9 (наши данные).
В сезон размножения на территории СССР белобрюхая райская мухоловка имеет один репродуктивный цикл. Количество яиц в полной кладке 3-5 (3 - 20,8 %; 4 — 72,9 %, 5 — 6,3 %; п = 48) (Зарудный, 1915; Попов, 1959; Сагитов, 1961; Иванов, 1969; Ковшарь, 1970; Абдусалямов, 1973; Тараненко, 1974; наши данные). Полная кладка из 2 яиц, найденная А.К.Са-гитовым (1961), очевидно, является повторной взамен утраченной.
Яйца овальные и остроовальные, бледно-охристо-розовые с мелкими четкими крапинками (d до 1 мм) и точками (внутренние — светло-лиловые, наружные — красно-коричневые). Крапинки сгущаются на тупом полюсе яйца, образуя на нем "шапочку” (густота рисунка 20—30 %) или ’’венчик”. К средней части яйца их количество резко уменьшается, а на остром полюсе они единичны. Скорлупа полумато-вая. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
Яйца самка откладывает ежедневно (Ковшарь, 1970; 1978). Сведения об откладке яиц через день нуждаются в проверке. Насиживание начинается с момента отклад
ки последнего яйца. Длительность инкубации - 12-14 дней (Сагитов, 1961; Иванов, 1969; Ковшарь, 1978). Участие партнеров в насиживании кладки у разных пар неодинаково. По данным А.И.Иванова (1969), А.Ф.Ковшаря (1978) и нашим (окр. к.Вар-зоб), в этом процессе принимают участие оба партнера пары, сменяя попеременно друг друга на гнезде. Однако А.К.Сагитов (1961) приводит наблюдения (Аман-Кутан-ская лесная дача) за парой, у которой на протяжении всей инкубации насиживала только самка. Самец даже не приближался к гнезду и участвовал только в его строительстве и в выкармливании птенцов. Подобное явление, очевидно, имеет место также при гибели самца. Птенцы вылупляются почти одновременно, в редких случаях последний на 1 день позже (Ковшарь, 1970).
Пуховой птенец белобрюхой райской мухоловки имеет пух на надглазничных, затылочной, спинной, плечевых, локтевых, бедренных, брюшных, голенных, грудных, копчиковой и кистевых птерилиях. Пушинки на всех птерилиях, кроме кистевых, хорошо развиты и имеют следующую окраску и длину (мм): на первых четырех птерилиях они каштаново-рыжие от 3,3 (затылочная) до 5,2 (плечевые) ; на 5-6-й, 9-10-й - светло-охристо-рыжие от 2,0 (копчиковая) и 3,0 (грудные) до 4,0 (остальные) ; на 7—8-й - бледно-охристые от 2,0' (голенные) до 3,5 (брюшные). Сведения об отсутствии у пуховых птенцов райской мухоловки из Индии грудных, копчиковой и кистевых птерилий (Ti-cehurst, 1926. - Цит. по: Нейфельдт, 1970) нуждаются в проверке. Кожа у птенца на вентральной стороне тела светло-телесно-розовая, на дорсальной - более темная. Клюв бледно-охристый. Ротовая полость и язык желтые, складки в углах рта бледно-желтые.
Обогревают птенцов в гнезде оба партнера, но большее время самка. Она не покидает гнездо с яйцами и птенцами во время сильных ветров и ливневых дождей (Абдусалямов, 1973). Через 10-13 дней (Сагитов, 1961; Иванов, 1969; Ковшарь, 1970) птенцы с недоросшими рулевыми и маховыми перьями покидают гнездо. Вылетевшие слетки долгое время держатся около гнезда. Даже научившись летать и начав линьку, они появляются в его ближайших окрестностях (Ковшарь, 1970).
При появлении человека на гнездовом участке мухоловка незаметно и беззвучно слетает., с гнезда и возвращается на него
17
1 n i I
оо
Таблица 4. Размеры (limМ ± т).коэффициенты вариации (С V, %) и индексы удлиненности яиц мухоловок фауны СССР
Длина (L), мм
Диаметр (D), мм
Удлиненность яйца (К = Ь)
Вид,подвид п
Min
Мах
СУ
М ± т
СУ
М ± т
Min
Мах
СУ
Min
Мах
М ± т
Terpsiphone paradisi leucogastcr 92(25)* 18,0 23,0 20,8±0,10 4,56 14,0 16,8 15,2±0,05 3,15 1,26 1,47 l,36±0,01 3,37
T.p.incei 42(11) 18,7 21,5 20,0±0,ll 3,45 14,2 16,6 15,1 + 0,07 3,05 1,17 1,44 1,33+0,01 3,66
Ficedula hypoleuca hypoleuca 158(25) 15,9 19,5 17,7±0,05 3,81 12,3 14,4 13,3±0,03 2,88 1,18 1,44 l,33±0,01 3,91
F.albicollis albicollis 55 (11) 15,3 20,7 17,8+0,14 5,86 12,1 14,0 13,2±0,07 3,88 1,26 1,56 1,35+0,01 . 3,96
F.a.semitorquata 122(26) 15,9 19,8 17,6±0,06 3,90 12,0 14,0 13,3+0,03 2,80 1,18 1,49 l,33±0,01 '4,25
F.zanthopygia 93(18) 16,0 18,8 17,7+0,06 3,01 12,0 14,0 13,2+0,04 2,98 1,21 1,48 l,34±0,01 3,76
F.narcissina narcissina 28(5) 16,5 20,0 18,5+0,21 6,16 12,2 16,0 14,3+0,16 6,11 1,24 1,39 l,30±0,01 2,98
F.mugimaki 30(7) 16,0 18,5 17,2±0,13 4,04 12,2 14,2 13',0+0,10 4,33 1,23 1,46 1,33+0,01 4,19
F.parva parva 48(10) 15,5 17,4 16,6+0,06 2,48 12,2 13,3 12,7±0,04 2,14 1,23 1,38 l,31±0,01 2,48
F.p.albicilla 60(8) 16,0 19,0 17,3±0,09 4,23 12,1 13,8 13,0+0,04 2,54 1,23 1,53 l,34±0,01 4,72
Cyanoptila cyanomelana 22(5) 20,3 21,9 21,1+0,09 1,92 14,4 16,3 15,5+0,10 2,99 1,26 1,46 1,36+0,01 3,17
Muscicapa striata striata 111 (23) 17,0 20,8 18,6±0,08 4,60 13,0 15,0 14,1 ±0,05 3,42 1,21 1,53 1,32+0,01 4,98
M.s.neumanni 58(15) 16,0 21,0 18,9±0,ll 4,63 12,8 15,1 14,0+0,07 3,98 1,07 1,52 1,35 + 0,01 5,77
M.sibirica sibirica 24(7) 15,5 18,6 17,3+0,19 5,24 12,3 13,6 13,0±0,06 2,44 1,25 1,41 1,33+0,01 3,63
M.griseisticta 4(1) 16,1 17,8 16,9±0,35 4,18 12,6 12,9 12,8+0,07 1,10 1,28 1,38 l,32±0,02 3,43
M.latirostris latirostris 76(18) 14,6 17,9 16,1+0,10 5,58 11,9 13,6 12,7±0,05 3,33 1,14 1,40 l,27±0,01 5,02
M.ruficauda 4(1) 17,2 17,7 17,5±0,12 1,40 13,3 14,0 13,8+0,17 2,40 1,23 1,33 l,27±0,02 3,34
* Цифра перед скобками - количество промеренных яиц, в скобках - количество кладок, из которых они взяты.
Примечание. В таблице помимо собственных материалов автора использованы также сведения о размерах яиц мухоловок, содержащиеся в следующих литературных источниках: T.p.leucogastcr (Зарудный, 1915; Сагитов, 1961; Ковшарь, 1962), T.p.incei (Кисленко, 1965а; Нечаев, 1974; 1981), F.a.albicollis (Сомов, 1897), F.zanthopygia (Taczanowski, 1891; Нечаев, 1963), F.n.narcissina (Нечаев, 1969; 1984а), F.mugimaki (Taczanowski, 1891; Нечаев, 19846), F.p.parva (Сомов, 1897), F.p.albicilla (Воробьев, 1963), M.s.striata (Сомов, 1897), M.s.neumanni (Taczanowski, 1891; Благосклонов, 1954; Янушевич и др., 1960; Сагитов, 1961), M.s.sibirica (Taczanowski, 1891; Hartert, 1907), M.l.latirostris (Taczanowski, 1891; Dybowski - Цит. no: Hartert, 1907; Гизенко, 1955; Сонин, Липин, 1969; Винтер, 1977), M.ruficauda (Леонович, 1962).
только после его ухода. При частых беспокойствах птицы оставляют гнездо (Тараненко, 1974). При появлении у гнезда с птенцами истинного или ложного врага — синей птицы (Иванов, 1969) и др. —оба партнера проявляют сильное беспокойство. С тревожным цирканьем, но стараясь быть менее заметными, они перелетают по гнездовому участку, наблюдая за передвижением врага, или агрессивно пытаются его отогнать. Птенцы в это время затаиваются в гнезде.
Сезон размножения белобрюхой райской мухоловки на территории СССР длится с I декады мая по III декаду июля включительно. В Западном Тянь-Шане он проходит с конца III декады мая по III декаду июля, но основная масса пар начинает откладку яиц в I декаде июня, а их птенцы покидают гнезда в I половине июля.
На Пскемском и Угамском хребтах и частично в окр. г .Ташкент Н.А.Зарудный (1915) находил свежие кладки с 31.V по 23.VI; гнездо с сильно насиженными яйцами около к.Хумсан на р.Угам — 10.VII. 7.VI 1964 г. здесь же встречена полная свежая кладка, 14.VI яйца в ней были сильно насижены. В этот же день найдено еще 1 гнездо с сильно насиженной кладкой (Тараненко, 1974). Первые оперяющиеся птенцы зарегистрированы в долине р.Угам 24.VI 1957 г. (Тараненко, 1974), 23.VI -2 слетка добыты Н.А.Северцовым (колл.
ЗИН), a 7.VII отмечен выводок хорошо летающих молодых (Зарудный, 1915). В долине р.Пскем у Седжак Д.И.Чекменев 26.VI обнаружил гнездо с 3 насиженными яйцами, a 2.VII здесь же найдено гнездо с 2 оперившимися птенцами (Корелов, 1956. - Цит. по: Ковшарь, 1970). Самые ранние слетки отмечены М.Н.Кореловым в этой долине 29 .VI, но массовый вылет птенцов в том же году проходил около середины июля (Ковшарь, 1970).
На северных склонах Таласского Алатау в заповеднике ”Аксу-Джабаглы” 8.VI 1960 г. добыта самка, начавшая кладку, а 23 .VI в другом урочище найдено гнездо с полной насиженной кладкой, 12.VII в нем было 4 оперившихся птенца, а 19.VII выводок держался уже недалеко от гнезда (Ковшарь, 1962, 1966, 1970). Хорошо летающие молодые встречены в каньоне р.Аксу - 20.VII. В Каратау (долина Бо-ролдая) М.Н.Корелов (Ковшарь, 1970) добыл слетка 12.VII, а 20.VII обнаружил пустое гнездо, около которого держался выводок.
На хр.Нуратау (Нуратинский заповедник) в урочище Хаятсай 4.VI 1978 г. Е.Н.Коршуновой найдены 2 гнезда с 3 и 4 яйцами (степень насиженности неизвестна) (сообщ. О.В.Митропольского).
На Чаткальском хр. в долине р.Ак-сак-ата гнездо с сильно насиженной кладкой найдено 30.V 1978 г. (сообщ. О.В.Митро-
Таблица 5. Масса (lim, М ± m- CV, %) иеиасиженных яиц мухоловок фауны СССР
Вид,подвид n Масса, г Cl'
Min Max M ± m
Terpsiphone paradisi leucogaster 16(4)* 2,20 2,70 2,51 ±0,04 6,70
T.p.incei 15(4) 2,02 2,50 2,2 7 ±0,05 7,84
Ficedula hypoleuca hypoleuca 25 (4) 1,42 1,85 l,66±0,02 7,13
F.albicollis albicollis 11(2) 1,42 1,90 l,60±0,05 9,89
F.a.semitorquata 57(14) 1,40 1,95 l,63±0,02 7,16
F.zanthopygia 26(5) 1,55 1,80 1,67±O,O1 4,01
F.narcissina narcissina 6(1) 1,72 1,98 1,85 ±0,04 5,54
F.mugimaki 10(2) 1,30 1,61 I,43±0,04 8,63
F.parva parva 10(2) 1,31 " 1,55 l,45±0,02 5,00
F.p.albicilla 16(2) 1,25 1,70 1,45 ±0,02 6,86
Cyanoptila cyanomelana 6(1) 2,75 3,05 2,89 ±0,04 3,53
Muscicapa striata striata 29(6) 1,70 2,21 l,99±0,02 6,20
M.s.neumanni 23(5) 1,60 2,15 l,92±0,03 6,98
M.latirostris latirostris 26 (5) 1,12 1,45 1,31 ±0,02 7,44
M.ruficauda 4(1) 1,68 1,87 1,77 ±0,04 4,48
* Цифра перед скобками - количество яиц, в скобках - количество кладок, из которых они взяты.
Примечание. В таблице помимо собственных материалов автора использованы также сведения о массе яиц мухоловок, содержащиеся в следующих литературных источниках: T.p.leucogaster (Сагитов, 1961), T.p.incei (Кисленко, 1965а; Нечаев, 1981), F.n.narcissina (Нечаев, 1984а), F.mugimaki (Нечаев, 19846), M.ruficauda (Леонович, 1962).
19
польского). В Чаткальском заповеднике, по данным Л.И.Тараненко (1974), 2 гнезда с только что законченными кладками и 1 с неполной кладкой из 3 яиц найдены 30.V 1966 г. в низовьях Башкизыл-сая. 11-14.VI 1967 г. здесь же найдены 2 гнезда с сильно насиженными кладками, 1 - с надклюнувшимися яйцами и 3 - с недавно появившимися птенцами. Одна пара, запоздавшая с гнездованием, в это время имела в гнезде 2 яйца. 27.VI большинство гнезд птенцы покинули, но у одной из пар, бросившей первую кладку, в этот день отмечено новое гнездо с 2 свежими яйцами. 6.VII 1968 г. у впадения в Башкизыл-сай Кознакок-сая обнаружено гнездо с 4 оперившимися птенцами, 1 из которых при осмотре гнезда слетел на землю.
В окр. орехосовхоза "АфлатунЬ” (Киргизская ССР) в 70 км северо-восточнее г.Наманган гнездо с неполной кладкой из 2 яиц найдено 16.VI 1956 г. (колл. С.Д.Кустановича).
В Западном и Центральном Памиро-Алае сезон размножения длится с I декады мая по II декаду июля.
На хр. Кугитанг 2 свежепостроенных гнезда найдены 16.V 1964 г. (колл. ИЗАНУ). 26-30.V (другие годы) зарегистрировано строительство гнезд, а 5-12.VI - откладка яиц (в найденных гнездах было 1-2 яйца) (Салихбаев, Остапенко, 1964).
На северных склонах Зеравшанского хр. в Аман-Кутанской лесной даче, по данным А.К.Сагитова (1961), 18-29N 1958 г. отмечено строительство гнезда, a 30.V самка отложила в него первое яйцо. В другом гнезде откладка яиц продолжалась 21-27.V 1958 г., а вылупление птенцов — 8-9.VI. Самое позднее окончание кладки в этом году зарегистрировано здесь 23.VI. 8.VI 1958 г. найдено гнездо с полной ненасиженной кладкой (колл. А-ПКузяки-на). По нашим данным, собранным в 1976 г., массовое токование в этом месте проходило 1 .VI, но в этот день 1 пара строила гнездо, а у другой - самка утром отложила первое яйцо. В ущ. Ак-сай 6.VI 1950 г. найдена полная слабо насиженная кладка (колл. БИН СО АН). В ТаджССР в окр. к.Косторош 2 гнезда с полными кладками средней насиженности найдены Нами 11.VI 1976 г., a 12.VI 1976 г. у к.Шинг был добыт слеток.
На южных склонах Гиссарского хр. сроки размножения следующие: в Варзобском ущ. массовое токование отмечено 13—21.V 1954 г., 27.V птицы встречались 20
только парами (Попов, 1959). 4.VI найдена полная свежая кладка, а 18.VI - кладка, которую насиживала взрослая птица (Абдусалямов, 1973). Из 9 гнезд, найденных нами 31.V—1.VI 1978 г. у к.Варзоб: в 1 кладка еще не началась, в 2 были сильно насиженные кладки, в 6 — разновозрастные птенцы. Здесь же 26.VI 1964 г. были добыты слеток и молодая птица, перели-нивающая в первый послегнездовой наряд (колл. КБ МГУ). В ур.Обисафед (Иванов, 1969) первое яйцо в 1 гнезде было отложено 23.VI, а 26.VI в нем было 3 яйца. 25.VI найдено гнездо с 3 яйцами. В окр. к.Хочильер (Попов, 1959) 29.V 1950 найдено 1 гнездо с неполной кладкой из 3 яиц и 1 гнездо, кладка в котором еще не началась (5.VI в нем была полная кладка). 2-5.VI 1950 г. А.В.Попов (1959) нашел 4 гнезда с полными кладками (степень насиженности неизвестна). 18.VI в одном из найденных гнезд вылупились птенцы. Самый поздний слеток встречен здесь 18.VII, а 13.VII - полностью доросшая молодая птица в гнездовом наряде. В ущ. Кондара (Иванов, 1969; Абдусалямов, 1973) 31.V в 1 гнезде отмечена откладка первого яйца, 15-16.VI - вылупление, 26.VI — вылет. 3.VI найдено гнездо (взрослые насиживают); 14.VI — 2 гнезда, в 1-м полная свежая кладка, во 2-м — 3 недавно вылупившихся птенца, которые, за исключением одного, покинули гнездо к 22.VI;
15.VI - гнездо с 3 свежими яйцами. 17.VI зарегистрированы 2 гнезда: в 1-м - 3 яйца, во 2-м - 3 яйца и 1 вылупившийся птенец. 29.VI найдено гнездо с 1 птенцом перед вылетом. 1 и 3.VII найдены 2 гнезда: в 1-м — 2 птенца и 1 яйцо, во 2-м — 3 птенца в 3—4-дневном возрасте. 18 и 23.V 1971 г. две полные ненасиженные кладки найдены в ущ. Кондара Р.И.Малышевским (колл. ИЗАНУ; С.Д.Кустановича), а 2 и 4.VI 1959 г. — В.ВЛеоновичем (колл. БИН СО АН; А.П.Кузякина). Самый ранний слеток отмечен 13.VI 1949 г. (колл. ЗИН). Встречены они здесь и 3.VII (Воробьев, 1968). В ущ. Джеринот нами 4 и 5.VI 1978 г. обнаружены 2 гнезда с разновозрастными птенцами, а 6.VI 1978 г. у к.Гажни - 1 гнездо с 2 птенцами. В верховьях Варзобского ущ. у с.Такоб 7.VI 1978 г. нами найдено гнездо с 4 почти оперившимися птенцами и отмечена 1 пара, строящая гнездо. В Ра-митском ущ. полная слегка насиженная кладка найдена 29.V 1953 г. (Попов 1959), a 18.VI 1978 г. нами найдено гнездо с 3 7—8-дневными птенцами. В ущ. Сары-Шашма в горах Санглок гнездо с 2 не-
насиженными яйцами обнаружено 18.VI 1949 г. (Попов, 1959).
. Линька. Смена мелкого оперения гнездового наряда начинается у молодых еще при окончательно недоросших маховых и рулевых. У 2 молодых птиц, которых автор содержал в лаборатории, в первую очередь перелиняло оперение верхней части головы. Основная масса молодых птиц одевает первый послегнездовой наряд в конце июля - I декаде августа (Зарудный, 1915; Иванов, 1969; Абдусалямов, 1973; наши данные). К началу осенней миграции эта частичная линька полностью заканчивается. Через год в первую полную линьку самцы одевают наряд второгодка, а окончательный наряд приобретают на 3-м году жизни. Взрослые птицы имеют в году одну полную линьку. Она проходит на местах гнездования с конца I декады июня по I декаду сентября (Абдусалямов, 1973; наши данные). Большинство взрослых птиц, как и молодые, заканчивают смену оперения к началу осенней миграции, но у некоторых особей эта линька заканчивается на местах зимовок (Абдусалямов, 1973).
Питание. Основу пищи составляют насекомые разных групп. Редко белобрюхая райская мухоловка поедает паукообразных и мелких моллюсков. Взрослые птицы одинаково успешно используют различные способы охоты: от ловли насекомых в воздухе на лету в стремительном броске с присады (основной способ) до активного поиска с последующим склевыванием беспозвоночных со стволов, ветвей, стеблей, листьев и в редких случаях с открытых корней растений и поверхности земли. Активны весь световой день. Большую часть непереваренных пищевых остатков (главным образом хитин насекомых) взрослые птицы и птенцы отрыгивают в виде погадок (рис. 6). В неволе при естественных кормах много и охотно пьют воду. Поедание взрослыми белобрюхими райскими мухоловками гастролитов и скармливание их птенцам установлено нами впервые. Раковины съеденных моллюсков выполняют, очевидно, также функцию гастролитов.
В выкармливании птенцов принимают участие оба партнера пары, но в первые дни после вылупления птенцов, когда самка большую часть времени остается в гнезде, согревая птенцов, корм чаще приносит самец. По данным А.Ф.Ковшаря (1978), каждый птенец в среднем за 1 ч получает от родителей пищу 4-5 раз. Крупным на
секомым (некоторые виды бабочек) взрослые птицы предварительно перед скармливанием птенцам отрывают крылья. После того как птенцы покинут гнездо, родители продолжают кормить слетков и подкармливать летных молодых, поэтому выводки зачастую не распадаются довольно долго. Известен случай (Зарудный, 1915), когда взрослые кормили молодых перелиниваю-щих в первый послегнездовой наряд.
Отлет и осенний пролет на территории СССР. Проходит в августе и сентябре.
В северной части гнездового ареала — в горах Таласского Алатау (ур. Кши-Каинды) - самая поздняя встреча этого вида произошла 15.VIII 1961 г. (Ковшарь, 1966). У подножий северного склона — с.Новониколаевка (заповедник ”Аксу-Джа-баглы”) (6 лет) отлет начинается со II, редко III и еще реже с конца I (10.VIII) декады августа. В сентябре райские мухоловки здесь уже редки, самая поздняя встреча -27.IX (Ковшарь, 1962, 1966, 1970). В юго-западных отрогах Таласского Алатау, на Пскемском и Угамском хребтах, миграция начинается в конце августа. К середине сентября птиц становится мало. Последние отмечены 21.IX (Ковшарь, 1970). В окр. г.Ташкента отлет и пролет проходит с середины августа до середины сентября, самая поздняя встреча птиц 18.IX (Зарудный, 1915; Ковшарь, 1970). Наиболее интенсивно он выражен здесь 20.VIII-4.IX (Зарудный, 1915; Спангенберг, 1954). В Чаткаль-ском заповеднике (2 года) начало осенней миграции приходится на конец августа (Тараненко, 1974). На хр. Кугитанг, по данным Х.С.Салихбаева и М.М.Остапенко (1964), численность мухоловок заметно сокращается к 15-20.VIII. Последняя зарегистрирована 2.IX. В окр. г.Самарканд самая поздняя встреча отмечена А.К.Саги-товым (1961) 4.IX 1957 г., а на Зеравшан-ском хр. в ур. Аман-Кутан (3 года) 22.VIII, 2.IX и 8.IX. В.Руссов (Pleske, 1888; колл. ЗИН) добыл 2 птиц в к.Кштут 2.IX 1878 г., а Д.К.Глазунов (колл. ЗИН) добыл взрослого самца 13.IX 1892 г. в г.Пенджикент, где А.И.Иванов (1940) наблюдал пролет 1.IX. Т.И.Барей (Stolzmann, 1897) добыл райскую мухоловку 8.IX 1892 г. в к.Зар-дали на р. Сох. На Гиссарском хр. (лесничество Сильбурсай) самая поздняя встреча этого вида — 31.VIII 1951 г. (Попов, 1959). Ю.М.Павловым (колл. ИЗиП АН ТаджССР) эти птицы добывались 15.VIII 1959 г. в окр. Мулло-Коны, а Г.Н.Сапожниковым
21
(колл. ИЗиП АН ТаджССР) 22 и 23.VIII 1960 г. на хр.Хозретиши в урДечандон. На юге ТаджССР в г.Куляб самая поздняя райская мухоловка добыта 8.IX (Иванов, 1940, 1969).
По данным А.К.Сагитова (1961), молодые и взрослые особи летят раздельно. Первые склонны образовывать, стайки (возможно, выводки), вторые — движутся одиночно. Зарегистрирован единичный случай (Зарудный, 1915), когда райские мухоловки летели стаей около 30 особей. Указание Н.М.Маслова (1947) на зимовку этого вида в низовьях Зеравшана, несомненно, ошибочно.
Практическое значение и мероприятия по охране. Уничтожает насекомых — вредителей леса и садовых культур, а также переносчиков возбудителей ряда заболеваний человека и животных. Внесена в Красные книги Казахской и Киргизской ССР как редкая, малоизученная птица республик. В настоящее время этот подвид райской мухоловки охраняется на территории СССР в государственных заповедниках ”Аксу-Джабаглы”, ’’Рамит”, Нуратинском горно-орехоплодном и Чаткальском горно-лесном. Являясь одним из красивейших представителей орнитофауны СССР, белобрюхая райская мухоловка заслуживает повсеместной, всесторонней охраны.
Китайская 'райская мухоловка — Terpsip-honeparadisi incei (Gould, 1852)
Muscipeta incei Gould, 1852, Birds of Asia, 2: pl. 19, Шанхай.
Описание. Составлено по 24 экз. (13 — из Приамурья и Приморья, 11 — из Китая).
У взрослого самца старше двух лет темно-окрашенной морфы все оперение головы, включая хохол, блестяще-темно-изумрудно-зеленое. Спина, надхвостье и верхние кроющие крыла каштаново-лиловые. Область зоба, грудь и бока тела пепельно-серые. Серый цвет светлеет на верхней части брюшка. Нижняя часть брюшка и нижние кроющие хвоста белые. Рулевые каштановые. Маховые темно-бурые. На наружных опахалах маховых начиная со 2-го первостепенного расположены каштановые канты. Третьестепенные маховые от каштаново-рыжего до каштанового с темно-бурым полем вокруг стержня каждого пера. Нижние кроющие крыла бледносероватые. У взрослых самцов белой морфы оперение головы такой же окраски,
как и у темно-окрашенных. Маховые черно-бурые. На наружных опахалах первостепенных маховых, а на второстепенных маховых и на внутренних опахалах имеются белые канты, ширина которых увеличивается от первостепенных к третьестепенным и на последних черно-бурый цвет сохраняется только на стержнях перьев. Кроющие кисти черно-бурые с белыми каемками на наружных опахалах и вершинах перьев. Рулевые белые с черными стержнями и черной окантовкой по краям опахал. Все остальное оперение белое. Спина с продольными черно-бурыми пестринами по центру перьев.
Самец второгодок такой же окраски, как и самец старше двух лет темноокра-шенной морфы, но, в отличие от последнего, у него центральная пара рулевых перьев значительно короче (табл. 1). У взрослых самцов голое кольцо вокруг глаза яркосинее. Клюв голубой. Полость рта зеленовато-желтая, вершина языка светло-желтая, а его основная половина зеленая. Ноги серовато-голубые. Когти черные. Радужина глаза бурая.
У взрослой самки хохол на голове более короткий. Оперение верхней части головы такой же окраски, как и у взрослых самцов. Бока головы и зашеек черные. Горло и область зоба матово-черные. Черный цвет светлеет на верхней части груди и боках тела и переходит в пепельно-серый. Брюшко и нижние кроющие хвоста белые. Маховые темно-бурые с охристо-рыжими каемками иа наружных опахалах, начиная со 2-го первостепенного. Клюв серовато-голубой, а голое кольцо вокруг глаза с примесью серо-голубых тонов.
Гнездовой наряд китайской райской мухоловки отличается от такового предыдущего подвида более темной окраской оперения верхней части тела, маховых и рулевых. Ноги бледно-голубоватые. Когти бледно-серые.
Первый послегнездовой наряд идентичен наряду взрослой самки, но перья, образующие хохол, в основаниях матовочерные и только на вершинах блестяще-темно-изумрудно-зеленые.
Размеры отдельных частей тела китайской райской мухоловки приведены в табл. 1.
Масса тела, г: самец второгодок (4.VII, Хабаровский край, Вяземский р-н) — 19,87, самцы старше двух лет (п = 3) (4 — 14.VII, там же) - 19,75-21,17, в среднем — 20,49 (колл. ЗМ МГУ).
Распространение. Китай от се-
22
веро-восточной части на юг через восточные и центральные провинции до Гуанси-Чжу-анского автономного района и Южного Юньнаня, Северная Корея (предположительно). На территории СССР — юг Хабаровского края и Западное Приморье (рис. 9). К местам зимовок на п-ов Малакка, о-в Суматра и соседние острова мигрирует через Китай; Таиланд, Лаос и Кампучию (Vaurie, 1959). Более детально распространение китайской райской мухоловки на территории СССР показано на карте (рис. 9).
Кадастр к рис. 9.
1 - южный склон хр.Хехцир (Кулешова, Матюшкин, Кузнецов, 1965; Казаринов, 1973); 2 - Хабаровский р-н (Росляков, 1981); окр. оз .Петропавловское в 70 км северо-восточнее г.Хабаровска (Басов, Нечаев, 1961; Яхонтов, 1963; Нечаев, 1974); берег залива Мурашки; долина ключа Горбатого в 12 км от с.Петропав-ловка (Нечаев, 1974); окр. с.Петропавловка (Кисленко, 1965а); 3 - Вяземский р-н (Росляков, 1981; окр. с.Венюково, р.Аван (Кисленко, 1965а; колл. ЗМ МГУ); 4 - Бикинский р-н (Росляков, 1981); низовья р.Бикин (сообщ. Ю.Б.Шибнева и С.В.Винтера); 5 - низовья р.Боль-шая Уссурка (Иман); с.Вербовка (Спангенберг, 1954; 1965); о-ва по р.Большая Уссурка (Спангенберг, 1940; 1954); б - Приуссурийская низменность в р-не г.Лесозаводск (Дубинин, Гор-чаковская, 1949); 7 - пойменные заросли некоторых рек бассейна оз.Ханка (Шибнев, Глущенко, 1981), окр. с.Гайворон, р.Спасовка (Нечаев, 1981; колл. ИЗАНУ); 8 - окр. с.Раковка вблизи г.Уссурийск (Спангенберг, 1940, 1954; Дементьев, 1941; Воробьев, 1954; колл. ЗМ МГУ); 9 - верховья р.Борисовка (Куренцов, 1961); 10 - долина среднего течения р.Борисовка (Нечаев, 1981); 11 - долина р.Амур у устья р.Сун-гари (Степанян, 1978).
Места обитания. В северной части гнездового ареала населяет смешанные хвойно-лиственные и лиственные леса, кустарниковые заросли и реже сады. На юге — субтропические и тропические леса разных типов.
В Северной Маньчжурии, по данным А.СЛукашкина (Воробьев, 1954), встречается в разреженных широколиственных лесах отрогов Тачин-Шаня, где тяготеет к зарослям маньчжурского ореха, реже гнездится в садах.
На территории СССР в Хабаровском крае у оз.Петропавловское населяет разреженные кедрово-широколиственные леса (Нечаев, 1974), в том числе гнездится и в переплетенных виноградом зарослях лещины с редкими деревьями корейского кедра, амурской липы, мелколистного клена и амурского бархата. Отмечена также на старой гари в осиново-лещиновом лесу. На хр.Хехцир гнездится в кедрово-широколи
ственных лесах южного склона (Казаринов, 1973). В окр. с.Венюково (Вяземский р-н) обитает, по данным Г.С.Кисленко (1965а), в смешанных широколиственных лесах, где предпочитает густые заросли (высота до 2-2,5 м), состоящие из разнолистной лещины, маньчжурского ясеня и других деревьев и кустарников, растущих у подножий сопок. Гнездится также в приречных зарослях из лещины, амурской сирени с примесью маньчжурского ясеня. Реже встречается в островных лесах, где придерживается участков с более густым подлеском. На сопках гнездится только в густых зарослях по распадкам. На вершинах сопок, где подлесок почти отсутствует или представлен в основном зарослями леспедецы, — отсутствует.
В Приморье, в районе г.Лесозаводск встречена в гнездовой период в густых зарослях кустарников (Дубинин, Горчаков-ская, 1949). Заселяет пойменные заросли некоторых рек бассейна оз.Ханка (Шибнев, Глущенко, 1981). В окр. с.Гайворон гнездится в приречных ивняках (сообщение М.И.Головушкина). В долине среднего те-
Рис. 9. Распространение китайской райской мухоловки (Terpsiphone paradisi incei) (а, б) и точки залетов чернохвостой райской мухоловки (T.atrocaudata atrocaudata) (в) на территории СССР:
а - гнездится, б - отмечена в гнездовой период; в - залет. См. кадастр, стр. 23.
23
чения р.Борисовка, по данным В.А.Нечае-ва (1981), китайская райская мухоловка обитает в пойменных ольхово-черемухоивовых лесах с чозенйей, ильмом, тополем и боярышником, которое граничат с разреженными и низкорослыми дубовыми и липово-кленово-дубовыми лесами, покрывающими склоны долины. Кустарниковый ярус представлен здесь густыми зарослями из молодых ив, клена, ольхи, бархата, дуба и кустарников: лещины, крушины, леспедецы, розы - буйно перевитых виноградом и ломоносом. В среднем течении р.Раздольная (окр. с.Новогеоргиевка) гнездится также в пойменных лесах из ивы и ольхи с отдельными молодыми дубами и кленами, под пологом которых разрастаются кустарники. Здесь зарегистрирована (Нечаев, 1981) на гнездовье в молодом дубовом лесу на крутом склоне в 30 м от ивняка, а также в густых зарослях из жимолости Маака и акантопанакса на песчаной гриве. В высокоствольных и разреженных пойменных лесах не гнездится.
Предпочитает тенистые участки в пойменных лесах вблизи полян, по берегам ручьев, заростающих озер и стариц, где даже в летний зной всегда сыро и прохладно (Нечаев, 1981).
Численность. В СССР немногочисленна и неравномерно распределена на территории гнездового ареала.
В долине р.Уссури (окр. с.Венюково), по данным Г.С.Кисленко (1965а), численность птиц этого подвида составляет 9,4— 19,2 пары на 1 км2, а в местах массовых гнездовых локализаций на этом участке -2-3 пары на 1 га. В Южном Приморье (Нечаев, 1981, 1983) в долине р.Борисовка в 1971-72 гг. гнездилось лишь по 1 паре на протяжении 3 км пойменных лесов, а в долине среднего течения р.Раздольная (окр. с.Новогеоргиевка) в 1972—73 гг. зарегистрированы 3 пары на расстоянии 7 км.
К основным причинам, влияющим на снижение численности китайской райской мухоловки, как и у предыдущего подвида, следует отнести сокращение в ряде мест площадей характерных гнездовых биотопов в результате уничтожения приречных зарослей при пожарах (Шибнев, Глущенко, 1981), рубках, раскорчевках и других агролесомелиоративных работах. К не менее важным причинам относится также и фактор беспокойства со стороны человека в гнездовой период. Зарегистрированы случаи гибели гнезд и птенцов от града.
Прилет и весенний пролет
на территории СССР. Сроки выяснены недостаточно. Исходя из времени появления кладок и отрывочных литературных сведений, в Приморье и южной части Хабаровского края проходит в конце III декады мая и в I декаде июня. Во время пролета мухоловки ведут себя скрытно и молчаливо.
В Южном Приморье в долинах рек Бо-рисовка и Раздольная (окр. с.Новогеоргиевка) на местах гнездования птицы появляются в конце мая — I декаде июня (Нечаев, 1981). В долине р.Спасов-ка (окр. с. Гайворон) взрослый самец добыт Н.Л.Клестовым (колл. ИЗАНУ) 3.VI 1976 г., а самка предположительно этого вида была встречена на островах в низовьях р.Большая Уссурка (Иман) 30.V 1938 г. (Спангенберг, 1940). На юге Хабаровского края (окр. с.Венюково) первые птицы в 1963 г. отмечены 9.VI (Кисленко, 1965а).
Размножение. В большей части протекает так же, как и у описанного выше подвида.
На гнездовых участках, по данным В.А.Нечаева (1981), китайские райские мухоловки, как правило, держатся открыто и шумно. Самец регулярно облетает гнездовую территорию. После образования пары до начала насиживания самец и самка обычно держатся вместе. В период выкармливания птенцов облеты гнездового участка прекращаются. В это время самцы становятся скрытными и осторожными. В размножении принимают участие как самцы в возрасте двух лет и старше, так и первогодки (Нечаев, 1981).
Площадь гнездового участка пары 10000—50000 м2 (Нечаев, 1981), минимальное расстояние между гнездами соседних пар 15—75 м (Кисленко, 1965а; Нечаев, 1981). Новые гнезда птицы строят невдалеке от старых (не далее 40—100 м) и никогда не достраивают их на прошлогодних. Чаще всего китайские райские мухоловки строят их в развилках ветвей молодых деревьев и кустарников на высоте 1,1—2,35 м, чаще 1,5—1,8 (Кисленко, 1965а; Нечаев, 1981). Из 16 гнезд, найденных на юге Хабаровского края (10 — Кисленко, 1965а; 1 — Нечаев, 1974; 1 - колл.СД.Кус-тановича) и в Южном Приморье (4 - Нечаев, 1981), 5 были построены на сирени (Syringa amurensis), 4 - на лещине (Corylus sp.; C.heterophylla), по 2 — на ясене (Fraxinus mandshurica) и клене (Acer sp.; A.ginnala), по 1 — на дубе (Quercus
24
sp.), бархате (Phellodendron amurense) и крушине (Rhamnus sp.). Нижняя часть гнезда упирается в развилку, а боковые стенки гнезда прикрепляются к 2-3 (в редких случаях к 1) вертикальным ветвям.
Гнезда в отличие от описанного выше подвида менее плотные, с тонкими, а иногда даже с просвечивающимися стенками, чашевидной, конической или округлой формы снизу. Строительные материалы в основной массе такие же, как и у белобрюхой райской мухоловки, но используются также узкие полоски луба жимолости, липы (Нечаев, 1981), ясеня (Кисленко, 1965а), растительный пух (летучек семян тополя, чозении, ивы), перья птиц (голубой сороки, восточной горлицы ), зеленые и сухие листья (спиреи, ивы) (Нечаев, 1981), кусочки бересты (Кисленко, 1965а), талломов лишайника (Parmelia sinuosa) и пучочки мха (Anomodon thraustus). Снаружи, как и у предыдущего подвида, гнездо густо оплетается паутиной. Последней вместе с лубяными волокнами оно прикрепляется и к ветвям. Масса гнезда 8—14, в среднем (п = 6) 11 г (Кисленко, 1965а). Размеры гнезд (п = 20), мм: диаметр 74,0-90,0; высота 65,0—92,0; диаметр лотка 50,0— 64,0; глубина лотка 32,0-41,0 (Кисленко, 1965а; Нечаев, 1974; 1981; наши данные).
В сезон размножения на территории СССР имеет один репродуктивный цикл. Количество яиц в полной кладке 3—5 (3 -10,0 %, 4 - 80,0 %, 5 - 10,0 %; п = 10) (Кисленко, 1965а; Нечаев, 1974, 1981; колл. С.Д.Кустановича). Различия в форме и окраске яиц китайской и белобрюхой райских мухоловок выражены очень незначительно. Скорлупа яиц описываемого подвида блестящая. Наибольшие крапинки на ее поверхности диаметром до 2 мм. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5. К насиживанию птицы приступают после откладки последнего яйца. В этом процессе принимают участие оба партнера пары. Инкубация длится 13 дней (Кисленко, 1965 а).
Пуховой птенец китайской райской мухоловки имеет следующие 12 птерилий: надглазничные (длина пушинок 1—1,5 мм), затылочную (2—3), спинную (4—5), плечевые (6—7), локтевые (4,5), кистевые (1-2), на первом пальце (1, 5), грудные (2-3), брюшные (4—5), бедренные (3-4), голенные (1,5—2) и копчиковую (1). Кроме перечисленных пуховых птерилий у птенцов встречаются две обособленные парные пушинки длиной 2 мм за глазом (между
глазом и ушным отверстием) и по одной парной пушинке длиной 1 мм под глазом, в углах рта. Редкий пух на брюшных птерилиях серовато-белый, а на всех остальных — черный. Кожа светло-розовая. Ротовая полость и язык желтоватые. Клюв желтовато-розовый с серой вершиной, яйцевой зуб серый. Ноздри круглые. Цевка и пальцы светло-серые, когти белые (Нечаев, 1981).
Обогревают птенцов в гнезде оба партнера, но большее время самка. На 9—10 день после вылупления они покидают гнездо с недоросшими маховыми и рулевыми перьями (Кисленко, 1965а).
При появлении на гнездовом участке пары в период откладки яиц, насиживания и выкармливания птенцов человека или какого-либо врага мухоловки ведут себя так же, как и птицы описанного выше подвида.
Сезон размножения китайской райской мухоловки на территории СССР длится с I декады июня по III декаду июля включительно. У отдельных пар при повторных кладках взамен утраченных - до конца II декады августа.
В Хабаровском крае (окр. оз.Пегро-павловское) 17.VI 1958 г. найдено гнездо с насиженной кладкой, 26.VI в нем вылупились птенцы, 4,УПпри осмотре гнезда 1 птенец пытался выпрыгнуть наружу. Через 2 дня гнездо опустело. В этот же день в 100 м от гнезда был встречен слеток (Нечаев, 1974). В окр. с.Венюково, по данным Г.С.Кисленко (1965а), массовое токование в 1963 г. проходило 11-20.VI. У самца, добытого здесь И.VI, семенники достигали 10 и 8 мм. 24.VI найдено гнездо с полной кладкой из 5 свежих яиц, а 27.VI — с полной сильно насиженной кладкой. 28.VI обнаружены 2 гнезда. В одном незаконченная кладка из 2 яиц, в другом - сильно насиженная кладка. 1 .VII и 15.VII обнаружены 2 гнезда с сильно насиженными кладками, а 9.VII в одном из гнезд - 3 1-дневных птенца и 1 болтун, а 10.VII в другом гнезде 2 только что вылупившихся птенца; 10.VII в одном гнезде взрослые выкармливают птенцов; 15.VII в другом гнезде отмечено 3 птенца 1-2-дневного возраста. 17.VII наблюдались первые выводки слетков по 2-3 особи. 20.VII зарегистрированы 2 выводка слетков и 1 пара, собирающая строительный материал для гнезда. 4.VIII отмечены 1 выводок слетков и несколько летных молодых. 14.VIII встречены неполностью оперившие
25
ся молодые. Г.С. Кисленко (1965а) 16.VI 1968 г. здесь же нашел гнездо с полной кладкой из 4 слабо насиженных яиц (колл. С.Д.Кустановича).
В Южном Приморье у с.Гайворон 11.VII 1978 г. М.И.Головушкиным (личное сообщение) найдены 2 гнезда с 3 и 4 8— 10-дневными птенцами. В среднем течении рек Борисовка и Раздольная (Нечаев, 1981) строительство гнезд проходит в конце I — во II декадах июня. 2 гнезда, найденные здесь 15.VI, были полностью построены; в одном из них 18.VI 1972 г. обнаружена полная кладка. Пара птиц, поздно приступившая к размножению, 24.VI 1972 г. строила гнездо, но через 2 дня, будучи потревоженной, бросила его и принялась за строительство нового; 29.VI гнездо закончено, a 3.VII в нем была полная кладка из 4 яиц. В одном из гнезд птенцы вылупились 4.VII 1972 г. Выводки из 4 молодых зарегистрированы 27.VII 1971 г. в долине р.Борисовка и 11.VIII 1973 г. в окр. с.Но-вогеоргиевка.
Линька. Последовательность смены нарядов такая же, как и у белобрюхой райской мухоловки. Взрослые птицы имеют одну полную послебрачную линьку на местах гнездовий. Она протекает с конца июня по конец августа, у отдельных особей заканчивается в I декаде сентября. Смена первостепенных маховых у взрослых мухоловок идет от 10-го к 1-му, второстепенных - от' 1 -го к 5-му и от 9-го (или 8-го) к 7-му, рулевых - от 1-го к 6-му. У самца на крыле, по данным В.А.Нечаева (1981), сначала выпадают внутренние маховые, а в середине июля — центральная пара рулевых, затем линька охватывает все остальное оперение. У птиц наблюдается незначительная асинхронность в последовательности смены маховых на разных крыльях.
Питание. Основу пищи составляют насекомые разных групп. Значительно реже китайская райская мухоловка поедает паукообразных. Способы охоты такие же, как и у предыдущего подвида.
Отлет и осенний пролет на территории СССР. Проходит с III декады августа по II декаду сентября. Как исключение одиночная птица зарегистрирована во II декаде октября.
В южной части Хабаровского края -в западных подножьях хр.Хехцир мигрирующие мухоловки встречены 3.IX 1963 г. (Кулешова, Матюшкин, Кузнецов, 1965); из них одна птица добыта (колл. КБ МГУ).
В Южном Приморье пролетная взрослая самка добыта В.Д.Яхонтовым (колл. ИЗАНУ) — 29.VIII 1968 г. у оз.Ханка на п-ове Рябоконь. На о-ве Большой Пелис (залив Петра Великого) пролетные птицы отмечены: 20.IX 1965 г. добыт самец и 2.IX, 12.IX 1966 г. встречены 2 самца и самка (Лабзюк, Назаров, Нечаев, 1971). Самая поздняя встреча в Приморье зарегистрирована 14.Х 1948 г. на территории Лазовского заповедника им.Л.Г.Капланова в долине р.Сяухэ (Воробьев, 1954; Спангенберг, 1954).
Практическое значение и мероприятия по охране. Такое же, как и у белобрюхой райской мухоловки. Внесена в Красную книгу РСФСР как редкая птица республики. В настоящее время этот подвид райской мухоловки охраняется на территории СССР в Большехех-цирском государственном заповеднике. К необходимым мероприятиям по охране китайской райской мухоловки на территории СССР следует отнести сохранение ее гнездовых биотопов в Хабаровском и Приморском краях, а также создание в долине нижнего течения р.Аван (Вяземский р-н Хабаровского края) заказника республиканского или краевого подчинения.
ПОДСЕМЕЙСТВО MUSCICAPINAE -НАСТОЯЩИЕ МУХОЛОВКИ
Род Ficedula Brisscn, 1760 — пестрые мухоловки
Ficedula Brisson, 1760, Ornithologia, 3: 369.
Типовой вид Ficedulahypoleuca Pallas.
Род включает 26 видов (Vaurie, 1953) большая часть которых распространена в Индо-Малайской и меньшая — в Палеарктической и Австралийской зоогеографических областях. В фауне СССР представлен 6 гнездящимися видами.
Небольших размеров настоящие мухоловки. Клюв в отличие от такового представителей рода Muscicapa более конусовидный, менее уплощенный и чаще всего не такой широкий у основания. Боковые поверхности надклювья несколько выпуклы. Щетинки у основания надклювья наиболее хорошо развиты в углах рта. Их количество здесь (с каждой стороны) обычно равно 3-5. У многих видов они тонкие и не очень длинные. Хвост сравнительно короткий или умеренной длины с неглубокой выемкой на вершине или без нее. Цевка тонкая и по сравнению с серыми мухоловками (Muscicapa) относительно более длинная. Ее длина составляет
26
19-31 % длины крыла (Vaurie, 1953). Формула крыла довольно сильно варьирует. Вершина крыла у перелетных видов пестрых мухоловок по сравнению с таковыми рода Muscicapa более округлая. Первое первостепенное маховое очень мало и обычно только на несколько миллиметров выступает за вершины кроющих кисти. Половой диморфизм резко выражен в окраске и незначительно (у некоторых видов) -в размерах отдельных частей тела. Отдельным видам свойственно явление морфизма: взрослые самцы имеют несколько цветовых морф. У молодых в гнездовом наряде перья с пятнистым или чешуйчатым рисунком. Окраска оперения и особенности его рисунка у взрослых самок в основном однотипны, у взрослых самцов - очень разнообразны. Но при комплексном подходе к оценке признаков можно отметить много общего в закономерностях распределения цветов в разных частях оперения. Так, у самцов пестрых мухоловок почти всех видов фауны СССР большая часть оперения верха тела черная (разной степени интенсивности). Если оперение серовато-бурое (малая мухоловка и некоторые цветовые морфы пеструшки), то черный цвет сохраняется на верхних кроющих хвоста. Над глазом у самцов имеется светлая бровь (белая или желтая), которая тянется у желтоспинных мухоловок от лба, у таежной представлена белым пятном за глазом, у пеструшки и белошейки — полосой или двумя пятнами на лбу, и лишь у малой мухоловки полностью отсутствует. На верхней части крыла у большинства видов находится белое ’’зеркальце”. Если оно отсутствует (малая мухоловка), то белый цвет обязательно представлен в основании рулевых. Оперение низа тела светлое: у пеструшки и белошейки — белое, у желтоспинных — золотисто-желтое, у таежной и малой — охристо-ржавчато-рыжее в верхней части и бледно-охристое в нижней. Белый рисунок на рулевых убывает в ряду видов: у белошейки и пеструшки имеется как у самцов, так и у самок, однако у некоторых самцов пеструшки и европейской белошейки иногда полностью отсутствует; у малой мухоловки хорошо выражен у обоих полов; у таежной присутствует только у самцов; у желтоспинной даурской как реликтовый признак проявляется только у очень ограниченного количества самцов в виде маленьких асимметричных пятен, у желтоспинной японской полностью отсутствует у
обоих полов. Резкие различия в цвете надхвостья у самцов таежной и желтоспинных мухоловок, а также исчезновение у последних белого рисунка в основании рулевых связаны с формированием видовых опознавательных признаков у этих симпат-рических, несомненно, очень близкородственных видов (Пекло, 1977).
Населяют главным образом различные типы лиственных, а также смешанных хвойно-лиственных и хвойных лесов. Некоторые виды поселяются в рощах, парках и садах, часто в непосредственной близости от жилья человека. Все виды фауны СССР являются перелетными, один вид (F.strophiata) -частично перелетный, остальные представители рода ведут оседлый образ жизни.
Образование пар у пестрых мухоловок сопровождается интенсивным, довольно мелодичным пением самцов и их токовым демонстративным поведением. Гнезда строят как открыто на ветвях деревьев и реже кустарников, так и в различных укрытиях: дуплах, расщелинах и разнообразных нишах. Некоторые виды гнездятся исключительно в дуплах, в том числе и в искусственных гнездовьях (различные дуплянки). Подавляющее большинство пестрых мухоловок на территории СССР имеет в сезон размножения 1 репродуктивный цикл. У некоторых видов (пеструшка) в ряде мест отмечено 2 цикла. Количество яиц в первой полной кладке обычно 3-7, у отдельных видов до 11. Окраска яиц очень изменчива: от однотонно голубой до белой с коричневыми и лиловыми крапинками и точками. Длительность инкубации обычно 12-15 дней.
Питаются главным образом насекомыми разных групп и стадий развития, реже поедают других беспозвоночных (паукообразные, моллюски и др.), а также мелкие сочные плоды некоторых растений. Изредка заглатывают гастролиты. Способы охоты в отличие от серых мухоловок самые разнообразные.
У взрослых пестрых мухоловок фауны СССР в году обычно 2 линьки (полная послебрачная и частичная или полная предбрачная) .
Мухоловка-пеструшка — Ficedula hypoleuca (Pallas, 1764)
Motacilla hypoleuca Pallas, 1764, Vroeg’s, Cat. Adumbratiunculae: 3, Голландия.
Син. (русск.): пестрая мухоловка.
Описание. Составлено по 81 экз. из европейской части СССР.
27
Вершину крыла образуют 3-е и 4-е первостепенные маховые (3 < 4 — 4,2 %; 3=4 — 15,5 %; 3 > 4 — 80,3 %); 2-е и 5-е немного их короче (2 < 5 — 78,1 %; 2 = 5 — 17,8 %; 2 > 5 — 4,1 %). 2-е всегда больше 6-го. 1-е первостепенное маховое очень мало и только на несколько миллиметров выступает за вершины кроющих кисти или равно им (табл. 6). У большинства особей наружные опахала 3—5-го первостепенных маховых в вершинной части заметно сужены. Количество щетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3-4 (3 - 89,9 %; 4 - 10,1 %). Длина наибольшей щетинки (п = 41) 4,0 — 6,5 мм (в среднем - 5,2). На вершине хвоста (в центре) расположена небольшая выемка. Половой диморфизм выражен в окраске оперения и незначительно — в размерах отдельных частей тела (крыло) (табл. 6). Виду свойственно явление морфизма: взрослые самцы имеют до 7 типов окраски оперения (рис. 8). Числовое соотношение цветовых морф среди взрослых самцов в популяциях разных частей ареала вида различно.
Распространение. Европейская часть СССР и Сибирь на восток до долины Енисея (см. рис. 12), Западная Европа (к северу в Норвегии до 70° с.ш., а в Финляндии до 65° с.ш., на запад до Атлантического побережья и к югу до Средиземноморского побережья Испании и Франции, 43° с.ш. на Апеннинском п-ове, Южной Югославии и Западной Болгарии), Северо-Западная Африка (в Марокко — на юг до Среднего Атласа, в Тунисе — до 35— 36° с.ш.), острова Великобритания и Соловецкие (Степанян, 1978). Пеструшки европейской и азиатской частей гнездового ареала являются перелетными и зимуют главным образом в саваннах тропической Африки.
Систематика. Политипический вид, образующий 3 подвида, один из которых — номинативный встречается и гнездится на территории СССР.
Африканскую часть гнездового ареала вида населяет подвид F.h.speculigera (Bonaparte, 1850). На Пиренейском п-ове распространена форма — F.h.iberiae (Wit-herby, 1928), которую некоторые исследователи (Vaurie, 1959) объединяют с номинативным подвидом. Однако Е.Курио (Curio, 1960), изучив пеструшек из Испании, доказал, что подвид F.h.iberiae реально существует, а его представители по деталям окраски четко отличаются от таковых
номинативного подвида и стоят ближе к северо-африканской форме.
Таксономический статус пеструшек восточных (сибирских) популяций, описанных в качестве самостоятельного подвида — F.h.sibirica (Chachlov, 1915), до настоящего времени окончательно не выяснен. Одни исследователи (Vaurie, 1959; Портенко, 1960; Степанян, 1978; и др.) признают реальное существование этой формы, в то время как другие (Дементьев, 1937; Благосклонов, 1954; и др.) относят сибирских пеструшек к номинативному подвиду. Основными отличительными признаками формы F.h.sibirica от номинативной, по данным В.А.Хахлова (описание сделано по 3 самкам), является более светло-серая общая окраска оперения, а по данным Л.С.Степаняна (1978) — и значительно меньшая встречаемость черной морфы среди самцов в популяциях этой формы.
Детальное изучение окраски оперения взрослых и молодых птиц, проведенное нами на серийных материалах с учетом возрастной, сезонной и индивидуальной изменчивости, позволяет выступить против выделения формы F.h.sibirica. Цветовые диагностические признаки последней очень незначительны и полностью укладываются в пределы индивидуальной изменчивости номинативного подвида, а выделение сибирского подвида только на основании различий в числовом соотношении цветовых морф среди самцов мы считаем неоправданным, тем более что эти различия выявляются главным образом на коллекционных материалах и допустимость ошибки очень велика.
Северная мухоловка-пеструшка — Ficedula hypoleuca hypoleuca (Pallas, 1764)
Motacilla hypoleuca Pallas, 1764, Vroeg’s, Cat. Adum-bratiunculae: 3, Голландия.
Описание. Составлено по 81 экз. из европейской части СССР.
У взрослых самцов окраска оперения очень изменчива. Насчитывается до 7 цветовых морф (рис. 8). У самца крайней черной морфы верхняя часть и бока головы (кроме лба), шеи, а также спина и верхние кроющие хвоста бархатно-черные. На лбу небольшое белое пятно, форма и размеры которого сильно варьируют. Некоторые особи имеют 2 маленьких пятна. Нижняя часть спины и надхвостье — от серого до серовато-бурого. Оперение нижней части тела белое. Нижние кроющие крыла белые
28
Таблица б. Размеры + —) и коэффициенты вариации (CV,%) мухоловки-пеструшки lim
Место сбора п Длина, мм
Крыло CV Расстояние, на которое 1-е маховое выступает за вершины кроющих кисти CV Хвост CV Клюв от оперения лба CV Клюв от орального края ноздри CV Цевка CV
Европейская часть СССР 56 79,38± 0,24 75,8 -83,9 2,19 2,80Ю,14 0,6 -5,6
То же 21 77,611 0,39 73,6 -80,9 2,31 3,0510,24 0,0 -4,7
36,57 Самцы 52,731 0,22 47,9 -56,3 3,06 9,351 0,05 8,7 -10,1 3,69 6,7610,04 6,2 -7,5 4,16 17,071 0,08 15,0 -18,4 3,48
36,61 Самки 51,811 0,32 49,0 -54,4 2,86 9,181 0,08 8,7 - 9,8 3,62 6,5610,08 5,9 -7,1 5,19 17,211 0,13 15,5 -17,7 3,28
Место сбора п Ширина, мм Высота, мм Отношение ши-рины клюва у основания к его длине от оперения лба, % CV Отношение длины хвоста к длине крыла, % CV Отношение ширины крыла к его длине, % CV
Крыло CV Клюв у орального края ноздри CV Клюв у основания CV Клюв у основания CV
Самцы
\о
Европейская часть СССР 56 54,561 0,15 51,7 -57,2 2,09 3,8410,03 3,4 -4,3 5,98 5,6610,05 4,9 -6,5 6 3,6710,05 3,3 -4,8 7,57 60,661 0,59 50,00-71,26 7,09 66,461 0,23 61,10-70,86 2,42 68,731 0,18 66,50-71,94 1,96
Самки
То же 21 53,621 0,30 ,50,6 -55,7 2,57 3,8810,04 3,7 -4,3 4,69 5,7710,09 5,3 -6,4 6,38- 2^910,06 3,5 -4,0 4,90 62,851 0,84 57,61-68,82 5,68 66,761 0,25 64,83-68,86 1,71 69,091 0,26 66,25-70,79 1,74
в вершинной половине каждого пера и бурые (разной интенсивности) в основной. Хвост от однотонно-бархатно-черного до черного с белыми (часто асимметричными) пятнами и полосами на крайних рулевых (рис. 10). Маховые черные. В основании последних (п = 54), начиная с 5—8-го первостепенного махового пера (с 5 — 5,5 %; с 6 - 88,9 %, с 7 - 3,7 %; с 8 - 1,9 %) появляются белые пятна, которые при сложенном крыле закрыты кроющими кисти или только незначительно выступают за их вершины. На верхней части крыла расположено белое ’’зеркальце”, образованное пятнами в основании второстепенных маховых, на наружных опахалах третьестепенных маховых, а также на больших и средних верхних кроющих второстепенных маховых. Самцы крайней серой морфы (рис. 8) по окраске почти не отличимы от самок. Оперение верхней части тела у них от буровато-серого до коричневатобурого. Белое лобное пятно развито очень слабо или полностью отсутствует. Оперение низа от белого до бледно-буровато-охристого. Верхние кроющие хвоста и рулевые черные. На наружных опахалах трех крайних пар последних имеются белые пятна или полосы. ’’Зеркальце” на крыле значительно меньшего размера, с бледно-охристым оттенком. Между этими крайними противоположными цветовыми морфами имеются все переходные типы (рис. 8). Клюв черный. Ноги черно-бурые. Радужина глаза темно-коричневая. Сезонная изменчивость проявляется в частичном побурении черных перьев и в посветлении общей окраски в результате обноса и выцветания оперения.
У взрослой самки оперение верхней части тела коричневато-бурое. Бока головы и лоб более светлые. Горло, зоб, грудь и верхняя часть брюха от бледно-буроватого до светло-буровато-охристого. Нижняя часть брюшка и нижние кроющие хвоста белые. Верхние кроющие хвоста бурые. Рулевые темно-бурые. На трех крайних парах имеются бледно-охристые пятна и полосы. Маховые темно-бурые. В основании последних (п = 21), начиная с 6-го первостепенного махового пера (100 %), появляются бледноохристые пятна. ’’Зеркальце” на верхней части крыла от бледно-охристого до белого.
Молодые в гнездовом наряде сверху желтовато-буровато-коричневые с охристыми пятнами, окантованными темно-бурыми каемками. Низ от белого до бледно
желтовато-охристого с буровато-коричневым волнистым рисунком, который наименее выражен на брюшке. Нижние кроющие хвоста белые. Маховые и рулевые темно-бурые. По краю наружных опахал третьестепенных маховых и больших верхних кроющих второстепенных маховых имеются желтовато-охристые каемки.
Первый послегнездовой наряд идентичен наряду взрослой самки, но большие верхние кроющие второстепенных маховых (не перелинявшие партии перьев) и третьестепенные маховые с желтоватоохристыми каймами.
Размеры отдельных частей тела северной мухоловки-пеструшки приведены в табл. 6.
Масса тела, г: самцы (п = 2) (V, р.Урал) -13,2—13,5, в среднем — 13,4; самки (п = = 3) (IV-V, Западный Казахстан) - 12,1 — 12,5, в среднем - 12,3 (Ковшарь, 1970).
Распространение. Почти весь гнездовой ареал вида, кроме Пиренейского п-ова и Северо-Западной Африки. В СССР: европейская часть и Сибирь на восток до долины Енисея. На всем ареале пеструшки этого подвида являются перелетными и зимуют в Юго-Западной Европе и саваннах Африки к северу от экватора. Указания Н.А.Зарудного и С.И.Билькевича (1918) на зимовку этой мухоловки в верхнем течении р. Амударья маловероятны. В лучшем случае авторы имели дело с залетным экземпляром. Известен залет пеструшки в Западное Прибайкалье (Хамар-Дабан) (Васильченко, 1982). Более детально распространение сибирской мухоловки-пеструшки на территории СССР показано на карте (рис. 12).
Кадастр к рис. 12.
1 - вся территория Карельской АССР (Зимин, 1972; колл. ЗИН); 2 - оз.Имандра (Иванов, 1976; колл. ЗИН); 3 - Лапландский заповедник (Владимирская, 1948); 4 - г.Канда-лакша (Благосклонов, 1954; Иванов, 1976); 5 - Хибины (Шибанов, 1927); б - окр. ст.Поя-конда (Давыдова, 1970); 7 - о-ва Кандалакшского залива (Шутова, 1981); 8 - Кемский р-н, 8 км от устья р.Воньга (колл. ИЗАНУ); 9 - Соловецкие о-ва (Портенко, 1960; Степанян, 1978); 10 - заповедник ’’Кивач” (Ивантер, 1962); 11 -Заонежье, о-в Большой Клименецкий (Хохлова, 1977); 12 - Южная Карелия, окр. пос.Матросы (Нейфельдт, 1958); 13 - юго-восточный берег Ладожского озера, Гумбарицы (Головань, 1983); Свирская губа (Мальчевский, Пукинский, 1983); 14 - долина р. Золотица (Спангенберг, Леонович, 1958, 1960) ; 15 - г. Архангельск (Паровщиков, 19416; Благосклонов, 1954); Карпогорский лесхоз в среднем течении р.Пинега (Севастьянов, 1962); 16 - с.Семжа в устье р.Мезень (Спангенберг, Леонович, 1958; 1960); 17 - Ленинград-
30
екая обл., в том числе и Приозерский р-н (Мальчевский, 1959); 18 - г.Ленинград и его окрестности; парки Петродворца, Пушкина, Павловска (Мальчевский, Пукинский, 1983); окр. с.При-бытково; ст.Ушаки (колл.ЗИН); Лисинский лесной массив (Поспелов, 1957); 19 - южные р-ны Ленинградской обл. (Мальчевский, Пукинский, 1983); 20 — окр. г.Новгород (Благосклонов, 1954); 21 - Псковская обл. (Зарудный. - Цит. по: Благосклонов, 1954; Мальчевский, Пукинский, 1983; колл.ЗИН); р.Череха; Хорламова гора (колл.ЗИН); 22 - Эстония; Ванаярве; Лайксааре; Выйду; Тиммканая (Кумари, 1955);
23 - Нигула (Кумари, 1955); лес Рийтскулли в окр. г.Килинги-Нымме (Лейвитс, 1982); 24 -Матсалуский заповедник (Ренно, 1968); 25 -Латвия (Благосклонов, 1954); Варенский лесхоз (Езерскас, 1959); 26 - Лудзенский р-н, оз.Греб-люкалнс (Столбов, 1981); 27 — Валкский р-н (материалы ц.кольц.); 28 - окр. с.Цесис (материалы ц.кольц.); 29 - окр. г.Рига (Благосклонов, 1954); Рижский лесхоз (Михельсон и др., 1956) окр. с.Ропажа (материалы ц.кольц.); 30 - г.Рига -Юрмалский лесхоз; Стренчский лесхоз (Михельсон и др., 1956); Талсинский р-н (материалы ц.кольц.); 31 - Кульдичский р-н (материалы ц.кольц.); 32 - Литва (Валюс и др., 1977); г.Каунас и его окрестности (Навасайтис, Курла-вичюс, 1977); 33 - заповедник ’’Каманос” (Тиюшас, 1982); 34 -заповедник ’’Жувинтас” (Иванаускас, 1968); юго-запад Балтийской воз
вышенности (Валюс, Кигуолене, 1981); 35 -Калининградская обл. (Благосклонов, 1954);
36 - Белоруссия; Гродненская обл., лес ’’Пышки” (Сямашка, 1956); 37 - заповедник ’’Беловежская пуща” (Федюшин, Долбик, 1967);
38 - Березинский заповедник (Кожевникова, 1965); 39 - Оршанский р-н, окр. с.Тарасинка, Белозоры (Бианки, 1914); 40 - Лунинецкий и Сталинский р-ны (Федюшин, Долбик, 1967);
41 - Припятский заповедник (Клакоцкий, 1983); р.Припять (Федюшин, 1928. - Цит. по: Долбик, 1959); 42 - Лепельский р-н (Федюшин, До:^к, 1967); 43 - Бобруйский р-н (Шнитников, 1913);
44 - Ленинский р-н (Долбик, 1959); 45 - окр. пгт.Шацк (сообщ. М.И.Головушкина); 46 - Волынь (Страутман, 1963); окр. с.Сураж (колл. ЗИН); окр. ВИР, окр. Белого оз., Репкинский р-н (колл. ИЗАНУ); окр. с.Маневичи (сообщ. М.И.Головушкина); 47 - окр. с.Цумань (сообщ. М.И.Головушкина); 48 - Ровенская обл.; Владимирский р-н (колл. ИЗАНУ); окр. г.Ровно; окр. с.Корец (сообщ. М.И.Головушкина); 49 -окр. с.Быстричи (сообщ. М.И.Головушкина);
50 — окр. г.Олевска; окр. с.Городец и Усово (наши данные); 51 - Подолье (Страутман, 1963); окр. г.Кременец (сообщ. М.И.Головушкина) ; 52 - долина р.Уж в окр. пгт Перечни (Пор-тенко, 1950); 53 - Мукачевский лесокомбинат (Гвоздак, Симочко, 1977); 54 — окр. с.Гукли-вое, вблизи Воловецкого перевала (Страутман, 1954; 1963); 55 - окр. г.Житомир; Райгородец-
Рис. 12. Распространение мухоловки-пеструшки (Ficedula hypoleuca) на территории СССР: а - гнездится. См. кадастр, стр. 30-33.
31
кая дача в 10 км юго-западнее г.Бердичева (сообщ. А.С.Вобленко); окр. сел Яроповичи и Вче-райше; окр. г.Казатин (сообщ. М.И.Головушки-на); 56 - окр. пос. Малин (сообщ. М.И.Головуш-кина); 57 - окр. ст.Спартак; окр. с.Россоха (сообщ. М.И.Головушкина); 58 - окр. г.Киев (колл. ИЗАНУ); бор ’’Пуща Водица” (колл. ИЗАНУ); окр. ст.Боярка (колл. ИЗАНУ); окр. с.Бортничи; окр. г.Ирпень; окр. пгт Клавдиево-Тарасово (сообщ. М.И.Головушкина); 59 - окр. с.ЕаВовцы (сообщ. М.И.Головушкина); 60 -окр; с.Хотяновка; окр. с.Лебедевка (сообщ. М.И.Головушкина); окр. с.Тужар (сообщ. М.Ф.Самофалова); окр. с.Волчок и Самойловка (наши данные); 61 - Шосткинский и Ямпольский р-ны; г.Шостка (сообщ. В.П.Белика); г.Собычи; окр. с.Старая Гута (колл. ХГУ);
62 - окр. с.Козелец (Гавриленко, 1929); Грабовский лес (колл. ИЗАНУ); окр. г.Нежин (сообщ. М.Ф.Самофалова и А.С.Вобленко); окр. с.Вертиевка (сообщ. М.Ф.Самофалова); окр. с.Ядуты (сообщ. А.С.Вобленко); 63 - окр. с.Тихоновичи; окр. с.Олешня и окр. с.Кучиновка (сообщ. М.Ф.Самофалова); 64 - Полтавская обл. (Сомов, 1897); 65 - северная часть Харьковской обл. (Благосклонов, 1954;' Степанян, 1978); окр. г.Чугуев, окр. с.Гуты (колл. ХГУ); окр. г.Волчанска (Сомов, 1897); Коробовское лесничество (Ковалев, Присада, 1983); 66 -окр. г.Богуслав; Каневский заповедник (сообщ. М.И.Головушкина); 67 - г.Черкассы (Орлов. -Цит. no: ЕЬег, 1960в); Черкасский р-н, окр. с.Яснозерье (сообщ. А.М.Волоха); 68 - окр. г.Фастов; окр. с.Ракитное (сообщ. М.И.Головушкина) ; окр. с.Потнеевка (сообщ. С.И.Пархомен-ко); южная часть Киевской обл. (Благосклонов, 1954; Степанян, 1978); 69 - Кировоградская обл., Черный лес (Волчанецкий, 1959); 70 -территория, затопленная Днепродзержинским водохранилищем (Булахов, 1968); 71 - Гуров-ское лесничество, окр. с.Гуровка (сообщ. В.И.Стри-гунова); 72 - Черный лес (Шевченко. - Цит. по: ЕЬег, 1960в); 73 - Ворошиловградская обл., окр. г.Кременная (сообщ. В.В.Ветрова); 74 -Центральная Молдавия; заповедник ’’Кодры” (Иванов, 1976); 75 - Сорокский лес; окр. сел Бужеровка и Егоровка (Ганя, 1965); 76 - Лозовский лесхоз (Ганя, Литвак, 1961; Аверин, Ганя, 1970); юг республики (Степанян, 1978).; окр. с.Чобручи (колл. М.И.Лобановского); 77 -
окр. с.Литин (Герхнер. - Цит. по: ЕЬег, 1960в);
78 - окр. с.Нелидово (материалы ц.кольц.); 79 - Старо-Русский р-н; окр. с.Вылеги (колл. ЗИН); 80 — Калининская обл. (Зиновьев, Шапошников, 1978); 81 - окр. с.Берново (колл. ЗИН);
82 — Дарвинский заповедник (Благосклонов, 1954); 83 - Вологодская обл., окр. оз.Кубен-ское (Беме, 1962); 84 - г.Ярославль (Белоусов, 1984); 85 - северная часть Ивановской обл. (Шептуховский, 1981); 86 — Костромская обл. (Кирпичников, 1915); с.Писцово (колл. ЗИН);
87 - Георгиевский лес в 6 км юго-западнее г.Вла-димира; Собинский р-н, окр. г.Лакинска; Камеш-ковский р-н: леса к западу от с.Хохлово и к востоку от пос. Пенкино (Михлин,Сальников, 1981); Пенкинский лес (Измайлов, 1981); 88 - окр. г.Клин (колл. ЗМ МГУ); 89 - окр. ст.Челюс-кинская (колл. ЗМ МГУ); 90 - Московская обл.; зеленые насаждения г.Москва и его окрестностей (Строков, 1962; колл. ЗИН); ст.Лобио (колл. БИН СО АН); Красногорский р-н
(Иноземцев, 1963; 1978); Дмитровский и
Талдомский р-ны (Птушенко, Иноземцев, 1968); окр. с.Петровско-Разумовское (материалы ц.кольц.); Химкинский р-н; окр. с.Мель-кисарово (колл. С.Д.Кустановича); Орехово-Зуевский р-н (Дерим-Оглу, 1964); 91 - Звенигородская биологическая станция МГУ (Благосклонов, 1962; Птушенко, Иноземцев, 1968); 92 -Приокско-Террасный заповедник (Благосклонов, 1954; Лихачев, 1967; 1978; Анорова, 1976, колл. ЗМ МГУ); окр. с.Десна (колл. ЗМ МГУ); 93 -Тульская обл. (Птушенко, Иноземцев, 1968); Тульские засеки (Благосклонов, 1954); окр. с.Бобровка (колл. ЗИН)) 94 - истоки р.Ока (Ефимов, 1915); Орловская обл. (колл. ЗИН); 95 - Калужская обл. (Филатов, 1915); г.Калуга (Воронин, Акментьева, 1979); 96 - Смоленская обл. (Благосклонов, 1954); 97 - Карачаевский р-н (Суслова, 1937); окр. с.Пронкино (колл. ХГУ); 98 - Центрально-черноземный заповедник (Поливанов, 1956); 99 - Белгородская обл. (Будниченко, Козлов, 1980); 100 - Горьковская обл. (Пузанов и др., 1942); г.Горький, окр. с.Фроловское (колл. ЗИН); 101 - лесные участки Пустынской биостанции Горьковского университета (Воронцов и др., 1962); 702 -Мордовский заповедник (Семенов, 1979); Чер-новско-Выселская и Селищенская лесные дачи Краснослободского лесхоза Мордовской АССР (Щербаков, 1960; 1967); Симкинское лесничество (Луговой, 1975); 103 - окр. с.Саранск (Майхрук, Кузнецов, 1977); 104 - Присурье (Луговой, 1981); 105 - г.Пенза (Благосклонов, 1954; Степанян, 1978); 106 - Переделецкое лесничество (Френкина, 1981); 107 - Рязанская обл. (Хомяков, 1900; Птушенко, Иноземцев, 1968); Окский заповедник (Теплое, 1956; Птушенко, Иноземцев, 1968); окр. с.Ягодное (колл. ЗИН); 108 - Саратовская обл. (Иванов, 1976); 109 -Тамбовская обл.: Мичуринский, Знаменский и Моршанский р-ны (Щеголев, 1978); г.Моршанск (Резцов. - Цит. по: ЕЬег, 1960); ПО - Воронежский заповедник (Семенов, 1956); 111 - г.Во-ронеж (Благосклонов, 1954; Степанян, 1978): Савальское (Кадочников, 1960) и Усманское лесничества (Иванов, 1976); 112 - южная часть Воронежской обл. (Благосклонов, 1954); Графское лесничество (Благосклонов, 1954); 113 -Теллермановский лес; берег р.Хопер (Королькова, 1963); 114 — Синегорье (Плесский, 1927); 115 - г.Сыктывкар и его окрестности (Коганов, 1983; сообщ. А.А.Естафьева); 116 — среднее течение р.Вычегда (Гурьев, 1982); 117 - окр. с.Усть-Кулом (сообщ. А.А.Естафьева); 118 - верховья р.Мезень (сообщ. А.А.Естафьева); 119 -среднее течение р.Кедва (сообщение А.А.Естафьева) ; 120 - окр. г.Печора (сообщ. А.А.Естафьева) ; 121 - верховья р.Илыч (Естафьев, 1969); 722 - низовья р. Северная Мылва (Естафьев, 1969); 123 - Печоро-Ильгчский заповедник (Иванов, 1976); 124 - г.Казань (колл. ЗИН); биостанция Казанского университета (сообщ. Ю.В.Мищенко); 725 - Татарская АССР, Раифский лес (Ушакова, Ушаков, 1976); 126 -район г.Ульяновск (Благосклонов, 1954); 127 -Алексеевский р-н (Сербин, 1978); 128 - р.Ик (Благосклонов, 1954; Портенко, 1960); 729 -г.Куйбышев (Благосклонов, 1963); Жигулевский заповедник (Белянина, Беляиин, 1981); 130 -долина р.Самара (Благосклонов, 1954; Степанян, 1978); 131 - окр. с.Бузулук (Иванов, 1976;
32
колл. ЗИН); окр. пос.Смолачково (колл. С.Д.Кустановича) ; 132 - низовья р.Илек (Степанян, 1978); 133- район г.Оренбург (Степанян, 1978); 134 - Сакмара (Портенко, 1960); 135 - Шайтан-Тау (Кириков, 1952); 136 - Южный Урал (Благосклонов, 1954; Иванов, 1976); верховья р.Белая (Благосклонов, 1954); р.М.Кудаш, р.М.Сы-ростал (колл. ЗИН); 137 - Башкирия (Ильичев, 1960); Башкирский заповедник (Благосклонов, 1954; Филонов, 1965); 138 - г.Уфа (Благосклонов, 1954) ; Жилинская дача; окр. с.Ногаевой, с.Кустаревская (колл. ЗИН); 139 - р.Коса (приток р.Кама) (колл. ЗИН); 140 - окр. селений Кунгур и Серебрянка (колл. ЗИН); 141 - Добрянский р-н (Хазиева и др., 1975); 142 - Лысьвенский р-н (Шилова и др., 1963); 143 - Пермская обл. (Симкин, 1974) ; Александровский р-н (Шилова и др., 1963); 144 - Северное Прикамье (Воронцов. - Цит. по: Eber, 1960в); 145 - окр. с.Мужи на р.Обь (Брауде, 1970); 146 — с.Бере-зово (Благосклонов, 1954; Портенко, 1960; Иванов, 1976; колл. ЗИН); 147 — заповедник ’’Денежкин камень” (Кузнецов, 1958); 148 -р.Каква (приток р.Сосьва) (Резцов, 1904); 149 - окр. г.Свердловск (Благосклонов, 1954);
150 - окр. г.Тобольск (колл. ЗИН); Тобольский р-н (Тарунин, 1928); 151 - Тюменский р-н (Сушкин, 1938); с.Падерное (колл. ЗИН); среднее течение р.Конда (Тюменская обл.) на участке Леуши - Большой Тап (Пантелеев, 19726);
152 - Ишимский р-н (Сушкин, 1938); 153 -окр. ^Петропавловска (Портенко, 1960; Ковшарь. 1970); 154 - окр. г.Сургут (Портенко, 1960; Иванов, 1976; колл. ЗИН); 155 — окр. с.Ларьяк на р.Вах (Гордеев, 1965); 156 - р.Ва-сюган (Иванов, 1976); 157 - окр. г.Омск (Иванов, 1976; Гынгазов, Миловидов, 1977; Степанян, 1978); 158 - окр. оз.Чаны (Сушкин, 1938); оз.Сартлан (Гынгазов, Миловидов, 1977); о.Пе-рекопный (Юрлов и др., 1977); 159 - ленточные боры Кулунды (Юрлов, 1974); 160 - долина р.Тым (Степанян, 1978); окр. с.Напас (Москви-тин, 1968; 1972; Гынгазов, Миловидов, 1977); 161 - Томская обл. Чаинский р-н (Пантелеев, 1972а); 162 - р.Кеть (Гынгазов, Москвитин, 1965) и ее притоки (Гынгазов, Миловидов, 1977); окр. с.Красная Курья, окр. с.Карагасок (Гынгазов, Миловидов, 1977); 163 - устье р.Сисим (Сушкин, 1914, 1938); 164 - г.Томск и его окрестности (Благосклонов, 1954; Портенко, 1960; Иванов, 1976; Гынгазов, Миловидов, 1977); 165 - окр. г.Новосибирск (Гынгазов, Миловидов, 1977); Академгородок (Цыбу-лин, 1985); 166 - окр. г.Кемерово (Гынгазов, Миловидов, 1977); 167 - горно-таежные леса Салаирского кряжа в междуречье среднего течения рек Кара-Чумыш и Томь-Чумыш (Чунихин, 1965); 168 - г.Барнаул (Благосклонов, 1954; Портенко, 1960; Иванов, 1976; Гынгазов, Миловидов, 1977; Степанян, 1978); Петровская лесная дача (Велижанин, Велижанин, 1929);
169 - окр. г.Бийск (Гынгазов,Миловидов, 1977); 170 - р.Стерляжья (Москвитин и др., 1977);
171 - окр. с.Б Кемчуг; леса холмистого правобережья р.Большой Кемчуг (Наумов, 1960); 172 - окр. г.Красноярск (Сушкин, 1938; Юдин, 1952; Портенко, 1960; Степанян, 1978); 173 -оз.Синара (Волчанецкий. - Цит. по: Eber, 1960в); 174 - окр. г.Ачинск (Тугаринов, 1927); 175 - р.Чумыш (Дунаевский. - Цит. по: Eber, 1960в).
Места обитания. Населяют самые разнообразные типы лиственных, смешанных хвойно-лиственных и хвойных лесов, главным образом на равнинах, а также в предгорьях и горах до 1350 м н.у.м. В горные леса проникает главным образом по долинам рек. Тяготеет к старым разреженным и осветленным участкам леса, гарям, но гнездится и в сплошных густых массивах. Поселяется в старых парках, скверах, садах, в зеленых древесных насаждениях усадьб и улиц населенных пунктов, в том числе и крупных городов (Москва, Ленинград). Как исключение гнездится на постройках человека в значительном удалении от ближайших деревьев. При наличии искусственных гнездовий охотно заселяет разнообразные молодые леса и посадки.
В Карелии основными гнездовыми стациями пеструшки являются лиственные леса (старые осинники, березняки, серо-ольшанники). В хвойных насаждениях, особенно в лишайниковых борах и в различных ельниках без примеси лиственных пород гнездится спорадично (Зимин, 1972). Излюбленными местами обитания на юге республики (Нейфельдт, 1958) являются разреженные участки смешанных и сосновых лесов, опушки, вырубки и. гари с подходящими для устройства гнезд дуплистыми стволами деревьев. В Лапландском заповеднике гнездится как в сосновых, так и в еловых лесах (Владимирская, 1948). При наличии искусственных гнездовий (дуплянок) поселяется в самых разнообразных типах леса (в которых раньше практически не гнездилась): молодняках, 10—15-летних культурах сосны и других (Зимин, 1972). В Ленинградской и Псковской областях гнездится в лиственных и смешанных лесах. В чистых хвойных массивах, бедных дуплами — редка и если встречается, то держится обычно на разреженных участках. При наличии искусственных гнездовий заселяет почти все типы леса, а также снегозащитные лесополосы. Гнездится в парках, садах, скверах населенных пунктов (Мальчевский, Пукинский, 1983).
В Эстонии встречается в сплошных елово-широколиственных лесах, а также в отдельных его куртинах на островах среди болот, на опушках леса и в березовых рощах (Кумари, 1955). В Латвии (Бауманис и др., 1983) гнездится в парках, садах, лиственных и смешанных лесах, а при наличии дуплистых деревьев или вывешенных искусственных гнездовий - также
33
в еловых и сосновых лесах. Обычно является доминирующим видом птиц при заселении искусственных гнездовий в лесах.
На северо-восточном побережье Белого моря (Спангенберг, Леонович, 1960) населяет область лесов и проникает в самые южные части криволесья. В долине р.Зо-лотица встречается в лесах по ее берегам и полностью отсутствует в глухих участках тайги. В Вологодской обл. у оз.Кубен-ского гнездится в смешанных лесах из ели, осины, березы, сосны и различных кустарников (Беме, 1962).
В Московской обл. предпочитает разнообразные старые, осветленные, богатые дуплистыми деревьями леса и совершенно отсутствует в густоствольных молодняках, лишенных дупел или искусственных гнездовий (Птушенко, Иноземцев, 1968). Гнездится в дубравах, чистых мелколиственных и смешанных елово-сосново-мелколиственных лесах, сосняках, разнообразных ельниках, елово-сосновых лесах, на вырубках (Иноземцев, 1963), заболоченных участках, поросших осиной и черной ольхой (Михеев, 1953), в парках и на улицах населенных пунктов. В Приокско-Террасном заповеднике поселяется в смешанно-лиственных лесах с преобладанием березы, осины, липы с подлеском из бересклета, можжевельника и рябины, в сосняках, ельниках (Милованова, 1956), а также в дубово-сосновоберезовых лесах (Анорова, 1976). В Калуге зарегистрирована на гнездовании в городском сосновом бору с примесью осины, дуба, березы и липы с подлеском из лещины, крушины и бузины (Воронин, Ак-метьева, 1979). В Горьковской обл. на территории Пустынского заказника встречается в елово-сосновых, елово-сосново-липовых и липово-осиновых лесах (Хохлова, Олигер, 1982). Во Владимирской обл. гнездится в сосновых (Измайлов, 1981) и смешанных лесах (Михлин, Сальников, 1981), а в пойме р.Клязьма зарегистрирована в дубравно-кустарниковых комплексах (Измайлов, Сальников, 1978). В Северо-Западном Закамье (Татарская АССР) заселяет дубово-березово-осиново-липовые и чисто дубовые леса (Сербин, 1978). В Присурье встречается в островных лиственных (Луговой и др., 1978) и сосновых лесах (Луговой, 1981). В Пермской обл. гнездится в различных типах смешанных хвойно-лиственных, хвойных и лиственных лесов, в том числе в березовоосиновом старолесье, липняках, долгомош-но-зеленомошных ельниках (Симкин,
1974) и елово-пихтовых лесах-зеленомош-никах (Шилова и др., 1963). На Южном Урале в нагорно-лесных ландшафтах обычна на гнездовье в широколиственных лесах и довольно редка — в сосново-березовых (Кириков, 1952). В Башкирском заповеднике населяет нагорные сосново-березово-лиственничные и широколиственные леса (Филонов, 1965) и изредка встречается в куртинных ельниках с примесью березы среди высокотравных субальпийских лугов на высоте 1100—1350 м н.у.м. (Снигирев-ский. — Цит. по: Благосклонов, 1954). На южной оконечности нагорной широколиственной лесостепи (Шайтан-Тау) гнездится главным образом в дубово-липовых лесах и уреме (Кириков, 1952). В Тамбовской обл. (Щеголев, 1978) зарегистрирована на гнездовании в широколиственных, ольховых, березовых и осиновых лесах, а также в старых сосновых борах.
В Белоруссии пеструшка тяготеет к смешанным елово-дубово-грабовым и елово-ольховым лесам, дубравам, ольшан-никам и осинникам с большим количеством дуплистых деревьев. При наличии дуплянок поселяется и в сосновых лесах (Федюшин, Долбик, 1967). В Могилевской обл. зарегистрирована на гнездовье в березовой роще (Бианки, 1914).
На Украине заселяет также самые разнообразные леса. В Киевской обл. (Фас-товский р-н) гнездится в дубово-березовых лесах с примесью вяза и липы, с подлеском из лещины, а также на старых дуплистых деревьях усадьб населенных пунктов. Вокр. г.Киев встречается в сосновом бору ’’Пуща-Водица”. В Житомирской обл. нами найдена на гнездовье в широколиственнососновых лесах, а также в березняках с примесью груши. В Сумской обл. (Шост-кинский р-н) встречается также в старых сосновых и приспевающих дубово-березовых лесах (сообщ. В.П.Белика). В западных областях УССР (Страутман, 1963) гнездится в лесах, парках и садах на равнине и в небольшом количестве в предгорьях. По речным долинам проникает далеко в горы, где поселяется на высотах до 850— 900 м н.у.м.
В Молдавии придерживается высокоствольных разреженных насаждений с большим количеством дуплистых деревьев (Ганя, Литвак, 1961; Аверин, Ганя, 1970).
В Тюменской обл. (участок Леуши — Большой Тап) зарегистрирована в сосновомелколистных, темнохвойно-мелколистных лесах и сосновых борах (Пантелеев, 19726),
34
а в Нижневартовском р-не отмечена в ленточных рощах из березы, осины и ивы с хорошо развитым подлеском (Гордеев, 1965). В Томской обл. обитает в смешанных высокоствольных лесах, парковых березняках, сосновых борах, а также в городских скверах и парках (Гынгазов, Миловидов, 1977). В долине р.Кеть приурочена главным образом к пойменным лесам (Москвитин и др., 1977). По р.Обь заселяет самые разнообразные типы лесов от ивняков и осинников, а также смешанных пойменных березово-осиновых лесов до темнохвойных таежных (Равкин, 1978). На р.Тым (окр. с.Напас) гнездится исключительно в пойменных затопляемых лесах или прирусловых валах, но значительно реже (Москвитин, 1968; 1972). На Сала-ирском кряже встречается в горно-таежных хвойно-лиственных лесах и в ивняках по р.Томь-Чумыш (Чунихин, 1965). В окр. г .Новосибирск (Академгородок) населяет осиново-березовые леса, сады (Цыбулин, 1985) и сосново-березовые насаждения (Смирнов, 1980), а в окр. г.Красноярск зарегистрирована на гнездовании в старом осиннике (Юдин, 1952).
Численность. На территории СССР в большинстве мест западной и центральной части гнездового ареала обычна, в излюбленных биотопах - многочисленна. У границ ареала чаще всего редка. При развешивании искусственных гнездовий в характерных биотопах численность пеструшки на этих участках может быть поднята до 900-3800 пар/км2 (Птушенко, Иноземцев, 1968).
Прилег и весенний пролет на территории СССР. Проходит с I декады апреля по. I декаду июня включительно. В отдаленных северных районах гнездового ареала (Приобье) прилет длится вплоть до конца июня.
На юге Азербайджана (Талыш) в 1937 г. первые птицы появились 4.IV, а массовый пролет проходил 8—14.IV (Тугаринов, 1950).
В Краснодарском крае пеструшка редкий пролетный вид, а не залетный, как указывает А.И.Иванов (1976). На левобережье р.Кубань пролетные мухоловки отмечены В.С.Очаповским 4.IV 1953 г. и 1965 г., а на Черноморском побережье края (Анапский р-н) одиночные птицы зарегистрированы 24.IV 1964 г. у с.Джемете и 2.V 1975 г. на Кизилташской косе.
В Крыму (Костин, 1983) начало пролета в середине апреля (16-19.IV; разные годы). Разгар пролета приходится на конец
апреля и начало мая. Наиболее поздняя встреча у с.Портовое 12.V. В Измаильском р-не Одесской обл. пролетные пеструшки добыты 3.V 1948 г. (колл. КГУ). В Молдавию мухоловки прилетают (8 лет) 5—11 и 14.IV (Ганя, Литвак, 1961); Аверин, Ганя, 1970). В Киевской обл. первые птицы появляются (9 лет) 10-26.IV (Данилович. - Цит. по: Благосклонов, 1954; наши данные). На Волыно-Подолье прилетают во второй половине или в конце апреля - начале мая. В окр. с.Поторица 2 мухоловки добыты 16 и 24.IV , а в Краковце в 1962 г. первая птица зарегистрирована 7.V (Страутман, 1963; колл. ЛПМ). В окр. г.Львов появляется (6 лет) 19, 26, 30.IV, 1 и 3.V (Miczynski, 1962).
На территорию Белоруссии пеструшка прилетает во II декаде апреля - I декаде мая. В Беловежской пуще первые птицы появляются (5 лет) 15, 23, 24.IV и 10.V (Федюшин, Долбик, 1967), в Пинском р-не (2 года) - 21 и 22.IV (Шнитников, 1913), Столинском (1 год) - 15.IV, Гродненском (2 года) — 12 и 16.IV, Минском (1 год) — 1.V, Пуховичском (1 год) - 25.IV, Лепель-ском (2 года) - 25 и 26.IV, Витебском (3 года) — 25, 28, 30.IV (Федюшин, Долбик, 1967).
В Воронежской обл. (Барабаш-Никифоров, Семаго, 1963) прилет самцов проходит в конце апреля — начале мая. Пролетные самки встречаются до 20.V. В Орловскую обл. (истоки р.Ока) первые мухоловки прилетают (10 лет) 18-25 .IV. В отдельные годы пролет длится до 5-11.V (Ефимов, 1915). В Карачаевском р-не в 1933 г. первые птицы зарегистрированы 4-5.V (Суслова, 1937). В Тульскую обл. (Тульские засеки) пеструшка прилетает (15 лет) 27—28.IV (Благосклонов, 1954). В Рязанской обл. (Татарская дача) первое появление в 1898 г. отмечено 28.IV (Хомяков, 1900), а в Калужской в 1911 г. -21.IV (Филатов, 1915). В Подмосковье первые птицы прилетают (23 года) 13.IV-3.V (Карпов, Паровщиков, 1941; Паров-щиков, 1941а), а в Москву (3 года) -23-25.IV, 28-30.IV, 2.V (Строков, 1962).
В юго-западной части Литвы пеструшка появляется (14 лет) 18-28.IV (Алекно-нис, 1977), а в г.Вильнюс и его окрестностях (8 лет) - 14.IV-4.V, в среднем -25.IV (Идзелис, 1981). В Эстонию прилетает (15 лет) 24.IV— 17.V (Тамм. — Цит. по: Благосклонов, 1954); в г.Новгород (25 лет) - 28.IV-14.V (Мантейфель. - Цит. по: Благосклонов, 1954). В Псковской обл.,
35
а также на юге Ленинградской (Мальчевский, Пукинский, 1983) появляется также в последних числах апреля — начале мая. Наиболее ранняя дата прилета в Ленинградскую обл. — 23.IV. Несколько позднее (1-6.V) прилетает в северные и восточные р-ны области. Пролет здесь идет в течение всего мая и частично даже в июне. Под г.Архангельск первое появление (1 год) отмечено 4.V (Паровщиков, 19416).
В Южной Карелии появляется (8 лет) 3-11 .V (Зимин, 1972), а в Лапландском заповеднике (Владимирская, 1948) средняя дата прилета (6 лет) - 23.V (17.V 1937 -28.V 1933). В отдельные годы первые птицы на Кольском п-ове регистрируются даже в начале июня (Портенко, I960).
В Казахстане слабо выраженный пролет проходит с I декады апреля iio середину III декады мая. На п-ове Мангышлак пеструшка в небольшом количестве летит 9.IV-4.V. В нижнем течении р.Урал (с.Яманка) одиночный самец зарегистрирован 22.IV, севернее у с.Тельнова начало пролета отмечено 28.IV, а у с.Фурманово — 8.IV. Позже 23.V пеструшка здесь не отмечалась. 26.V одиночная пролетная птица встречена на ст.Арысь, а 21.V добыта в г.Це-линоград (Ковшарь, 1970). В низовьях р.Волги появляется, по данным В.Н.Бос-танжогло (1911), в начале мая, а под г.Оренбург слабый пролет проходит с начала апреля до середины мая (Ковшарь, 1970). Севернее г.Петропавловск пеструшка появляется в начале мая, у с.Суворовка пролет отмечен 7.V (Ковшарь, 1970).
В Куйбышевскую обл. (Жигулевский заповедник) мухоловки прилетают (4 года) 4—6.IV и 24.IV (Белянина, Белянин, 1981). У устья р.Кама первые птицы появляются (4 года) 16—30.IV (Птицы Волжско-Камского края, 1978), а в Свердловскую обл. (23 года) — 24.IV—19.V (Батманов. — Цит. по: Благосклонов, 1954). В окр. г.Новосибирск первые прилетевшие птицы отмечены (4 года) 1—9.V (Цыбулин, 1985). У г.Томск пеструшка появляется (больше 5 лет) 30.IV—13.V (Гынгазов, Миловидов, 1977; Дубовик и др., 1977), на р.Кеть (2 года) - 8 и 9.V (Гынгазов, Миловидов, 1977; Москвитин и др., 1977), на р.Малая Сосьва (2 года) — 12 и 16.V (Тарунин, 1959; Вартапетов, 1984), у с.Нарын (2 года) — 8, 16.V (Дубовик и др., 1977). В Приобье и Прииртышье пролет идет во второй половине мая — начале июня. В северных районах — до конца июня (Равкин, 1978).
Известен случай залета пеструшки в период весенней миграции в Западное Прибайкалье. На хр. Хамар-Дабан в устье р.Переемная 16.V 1974 г. добыт самец из стайки в 6 особей (Васильченко, 1982).
Летят пеструшки в период миграции как днем, так и ночью - поодиночке и небольшими стайками, стараясь придерживаться облесенных мест. Первыми на места гнездовий прилетают старые самцы и только 3—10 дней спустя самки и передовые годовалые особи.
Размножение. В большинстве районов СССР прилетевшие на места гнездования пеструшки приступают к размножению не сразу, а только через некоторое время. На юге гнездового ареала — в Молдавии этот период составляет 20—25 дней (Ганя, Литвак, 1961), в Белоруссии — 15—16 (Федюшин, Долбик, 1967), в Московской обл. — 10—16 (Благосклонов, 1954; Птушенко, Иноземцев, 1968), в Волжско-Камском крае - 5-8 (Птицы Волжско-Камского края, 1978), в Латвии приступают к гнездованию сразу после прилета (Тауриньш, Тима. - Цит. по: Благосклонов, 1954). Ухудшение погодных условий задерживает сроки начала гнездования и сказывается на ходе размножения в целом.
До начала образования пар самцы занимают гнездовые участки и ведут себя в это время очень оживленно. Петь они начинают еще на пролете, но наиболее активно токуют у гнездовий. Мелодичная и громкая песня самца состоит из коротких и отрывистых строф и сильно подвержена индивидуальной изменчивости. По данным А.С.Мальчевского (1959), пеструшка включает в нее также отдельные звуки и даже строфы из криков и песен других птиц: клеста, лесного конька, большой синицы, певчего дрозда, зарянки. Образование пар сопровождается, помимо интенсивного пения самцов, их токовым демонстративным поведением. Для большинства особей характерна сезонная моногамия. Однако известны случаи и полигинии (Щербаков, 1967; Анорова, 1976; Шутова, 1981), при которой некоторые самцы за сезон размножения спариваются с двумя и даже тремя самками. Это происходит как в период откладки яиц (в этом случае самец одновременно и кормит птенцов в двух или трех гнездах), так и в конце периода насиживания яиц первой самкой. Площадь гнездовых участков пар 210-220-950 м2 (Благосклонов, 1962).
36
Являясь типичными дуплогнездниками, пеструшки чаще всего строят гнезда в естественных или в выдолбленных другими птицами (дятлы, синицы) дуплах, моро-зобойных трещинах и глубоких расщелинах стволов, ветвей и пней деревьев (сосна, береза, липа, рябина, вяз, ольха, осина, груша, дуб), а также в искусственных гнездовьях (разнообразные дуплянки), а в парках и садах населенных пунктов — и в щелях построек человека. Высота расположения гнезд от поверхности земли сильно варьирует. Чаще всего они помещаются на высоте (п = 30) 0,3—7,0 м, в среднем - 2,3 (наши данные; сообщ. В.П.Белика и С.И.Пархоменко), реже - до 15,0 (Зимин, 1972) и как исключение -до 20,0 (Дергунов. — Цит. по: Благосклонов, 1954) или — на одном уровне с поверхностью земли (Зарудный, 1910). Интересен факт гнездования пеструшки в искусственных гнездовьях (дуплянки), закопанных в землю (Вилкс, 1965). Строительство гнезда длится 4—11 дней (Зимин, 1972). Строит его самка. Иногда пара начинает строить гнезда сразу в нескольких местах, но полностью достраивает, как правило, только одно, в которое и производится откладка яиц. Сведения А. В. Михеева (1953) о гнездовании пеструшки в дуплах, не имеющих внутри никакого гнезда, кроме трухи от древесины, сомнительны и нуждаются в проверке.
Гнездо представляет собой рыхлую, довольно массивную пострс/йку, форма и размеры которой зависят главным образом от таковых нижней части гнездового дупла. В качестве строительных материалов для гнезда птицы используют тонкие чешуи коры (береза, сосна), сухие листья древесных и травянистых растений, тонкие веточки деревьев и кустарников, стебельки и корешки трав, соцветия диких злаков, полоски луба, пучочки мхов, хвою сосны, а по данным Е.С.Птушенко, А.А.Инозем-цева (1968) — паклю, обрывки тряпочек и нитки. Лоток или не вымащивается, или выстилается тоненькими стебельками и листьями трав, жилками листьев, полосками луба и другими растительными волокнами, а также волосом крупных млекопитающих, тончайшими чешуями коры и отдельными хвоинками. В редких случаях, по нашим данным, мухоловки используют для этой цели также спорогонии мха (Polytrichum commune), одиночные мелкие перья, кусочки талломов лишайников, а в городах даже кусочки бумаги (Благо
склонов, 1954). Размеры гнезд (п = 26), мм: диаметр 70—120 х 75—130, в среднем — 95 х 106; высота 30-83, в среднем - 54; диаметр лотка 45—68 х 43—90, в среднем — 55 х 61; глубина лотка 20—45, в среднем — 31.
В сезон размножения, на территории СССР северная мухоловка-пеструшка имеет один, в редких случаях два репродуктивных цикла (Анорова, 1976). Количество яиц в полной кладке 3-11 (Портенко, 1960; Лихачев, 1967; Анорова, 1976; 1984; Мальчевский, Пукинский, 1983; наши данные), чаще 6-8. Повторные кладки содержат 1—6 яиц (Мальчевский, Пукинский, 1983). Количество яиц в кладке зависит от многих факторов и подвержено хронографической, сезонной, возрастной (Анорова, 1976, 1984) и географической изменчивости (Berndt, Winkel, 1967). Обычно мухоловка-пеструшка откладывает яйца ежедневно. Однако этот процесс может быть прерван неблагоприятными погодными условиями (резкое похолодание). Лишь при наступлении теплой погоды откладка яиц возобновляется. В результате самка продолжает откладку яиц с перерывом в 1—4, даже 6 дней. Очень часто в результате похолодания или обилия осадков кладка яиц резко снижается, а у отдельных особей прекращается полностью (Анорова, 1976; 1984).
Яйца укороченно-остроовальные, овальные, остроовальные, реже эллипсовидные, однотонные (от бледно- до светло-голубых со слабым зеленоватым оттенком). Скорлупа блестящая. Размеры, индекс удлиненности И масса яиц северной мухоловки-пеструшки приведены в таблицах 4 и 5. Насиживание кладки начинается с откладки последнего яйца. Длительность инкубации 10—17, чаще 12-13 дней (Портенко, 1960; Ганя, Литвак, 1961; Федюшин, Долбик, 1967; Мальчевский, Пукинский, 1983). Самец в насиживании яиц участия не принимает. В этот период он обычно держится поблизости от гнезда и подкармливает насиживающую самку. В большинстве гнезд птенцы вылупляются в 1 день.
Пуховой птенец северной мухоловки-пеструшки имеет короткий, редкий серый пух на надглазничных, затылочной, спинной, плечевых и глазных птерилиях. На последних он рудиментарный и бывает не всегда. Клюв высокий, довольно короткий и массивный, несколько вздутый в области ноздрей. Ноздри круглые. Когти темные. Ротовая полость однотонно-желтая с оран
37
жевым оттенком. Широкие складки в углах рта желтовато-белые (Мальчевский, 1959). Масса вылупившегося птенца 1,3-1,6 г.
Рост и развитие птенцов протекают быстро и на 13-18-й день (Портенко, 1960; Птушенко, Иноземцев, 1968), чаще 15-16-й (Мальчевский, Пукинский, 1983) они покидают гнездо. Несмотря на относительно хорошие летные способности по сравнению со слетками малой и серой мухоловок, слетки пеструшки после вылета из гнезда большую часть времени сидят неподвижно, периодически издавая призывные крики, по которым их находят взрослые птицы. В месячном возрасте молодые мухоловки переходят к самостоятельной жизни и держатся отдельно от родителей (Головань, 1982). В это время выводки распадаются и в дальнейшем молодые птицы держатся небольшими группами и поодиночке.
Охрану гнездового участка в период откладки яиц, насиживания кладки и выкармливания птенцов выполняет самец. Хотя расположенные в дуплах гнезда пеструшек защищены значительно лучше, чем гнезда открытогнездящихся видов птиц, тем не менее гибель яиц и птенцов в отдельные годы, по данным А.С.Маль-чевского и Ю.Б.Пукинского (1983), достигает значительных размеров (до 43 %). Основными причинами отхода яиц и птенцов являются неблагоприятные погодные условия, хищническая деятельность со стороны некоторых видов птиц и млекопитающих (хищные, грызуны), наличие в кладках неоплодотворенных яиц, а в населенных пунктах и их окрестностях и беспокойство со стороны человека.
Сезон размножения северной мухоловки-пеструшки на территории СССР длится с III декады апреля по II декаду августа включительно. Сроки начала размножения у отдельных пар варьируют и зависят главным образом от погодных условий и возраста партнеров. Первыми приступают к гнездованию 2-летние самки, а не более старые птицы или первогодки (Анорова, 1976).
В Молдавии (Лазовский р-н) начало кладки зарегистрировано (3 года): 5, 8 и 10.V (Ганя, Литвак, 1961). У с.Чобручи гнездо с полной ненасиженной кладкой найдено 12.V 1959 г. Первое вылупление птенцов на территории республики отмечено (2 года) 21 и 23.V (Ганя, Литвак, 1961).
В западных областях Украины откладка яиц проходит в середине мая (Страут
ман, 1963). В долине р.Уж самец, кормивший птенцов, встречен в середине июля 1947 г., а молодая птица с недоросшими маховыми добыта 13.VII (Портенко, 1950). В Черкасской обл. (окр. с.Яснозерье) гнездо с полной кладкой обнаружено 24.VI 1974 г. (сообщ. А.М.Волоха). В Киевской обл. в окр. г.Киев (сосновый бор ”Пуща-Водица”) начало кладки зарегистрировано (2 года) 10 и 17.V. Полные свежие или слабо насиженные кладки встречены здесь (4 года) 15, 16, 18, 24.V и 12.VI (колл. ИЗАНУ; КГУ; наши данные). У ст. Боярка 2 кладки средней насиженно-сти найдены (2 года) 24 и 29.V, а в Фас-товском р-не полные кладки встречены (2 года) 19 и 28.V. 3.VI 1968 г. здесь обнаружено гнездо с 4 птенцами и 1 яйцом (сообщ. С.И.Пархоменко). В Житомирской обл. (окр. с.Городец) откладка первого яйца отмечена 9.V 1970 г. В Сумской обл. (Шосткинский р-н) пеструшки занимают гнездовые участки (2 года) 22 и 27.IV, а к строительству гнезд приступают (3 года) 25.IV и 4.V, некоторые пары даже 3.VI. Начало кладки зарегистрировано здесь (3 года) 10, 15, 29. V и 6.VI, а гнезда с птенцами найдены 8, 15, 16 и 23.VI. В г.Шостке выводок слетков встречен 11 .VI 1966 г. (сообщ. В.П.Белика).
На территории Белоруссии (Федюшин, Долбик, 1967) полные и незаконченные кладки встречаются 10-26.V, насиженные -в конце III декады мая — I декаде июня. Вылет птенцов проходит в середине и второй половине июня.
В Окском заповеднике полная кладка из 6 яиц найдена 30.V 1962 г. (колл. ЗМ МГУ). В Брянской обл. (Карачаевский р-н) летные молодые зарегистрированы 20 и 28.VI 1932 г. (Суслова, 1937; колл. ЗИН). В Тульской обл. (Тульские засеки) начало кладки проходит 3.V—23.VI, конец кладки - 10.V-27.VI и 1.VII, вылупление птенцов - 24.V-7.V1I, а их вылет - 10.VI-23.VII (Благосклонов, 1954). В Московской обл. (многолетние материалы) откладка яиц начинается 10.V-24.V—15.VI; полные ненасиженные кладки встречаются 13.V—19.VI; вылупление птенцов проходит 30.V- 12.VI—4.VII; вылет птенцов - 13.VI-28.VI-19.VII (в большинстве гнезд -20.VI-4.VII), а появление самостоятельных молодых - 21.VI—5.VII-25.VII (Благосклонов, 1954; Птушенко, Иноземцев, 1968; колл. БИН СО АН; ЗМ МГУ). В Красногорском р-не вторые кладки найдены 6, 11 и 13.VII 1959 г., 12.VII в двух из них
38
вывелись птенцы и 26-29.VI1 вылетели из гнезд (Птушенко, Иноземцев, 1968). В Приокско-Террасном заповеднике начало откладки яиц проходит с середины I декады мая до конца июня, в большинстве гнезд -со II декады до конца мая (Лихачева, 1967, 1978). Полные кладки разной наси-женности найдены здесь 28.V 1960 г. (колл. ЗМ МГУ). В г.Калуга смешанная кладка пеструшки и большой синицы обнаружена 14.VI 1977 г. (Воронин, Акментьева, 1979).
В лесах Литвы (Езерскас, 1962) в ранние весны (1961 г.) яйцекладка у пеструшки начинается 10-15.V; в поздние (1955— 56 гг.) - 17-25.V; при своевременной весне (1960 г.) - 15-20.V. В населенных пунктах откладка яиц начинается на 3— 5 дней раньше, чем в лесных биотопах. В Латвии (окр. г.Рига) первые птенцы появляются в гнездах в III декаде мая -I декаде июня (Тауриньш, Тима. - Цит. по: Благосклонов, 1954). Гнезда с птенцами находили здесь также 17.VI 1962 г.; в Кульдичском р-не - 27.VI 1954 г., а в Валкском - 13.VII 1955 г. (материалы ц.кольц.).
В окр. г.Новгород птенцы в большинстве гнезд вылупляются 1-15.VI,а их вылет проходит 17-29.VI (Мантейфель. -Цит. по: Благосклонову, 1954). В Псковской обл. гнезда со свежими кладками отмечаются в первой половине июня, реже — во II половине этого месяца (Зарудный, 1910). У Харламовой горы молодая пеструшка добыта 4.VII 1884 г. (колл. ЗИН). В Ленинградской обл. (Мальчевский, Пу-кинский, 1983) откладка яиц начинается во II половине мая. Разница в сроках появления первых кладок в разные годы в районе Свирской губы (12 лет) составляет 12 дней (15-27.V). При резких длительных похолоданиях птицы приступают к гнездованию в конце III декады мая. Все кладки, появляющиеся во II и III декадах июня, являются повторными, возникшими в результате разорения первого гнезда. Наиболее поздняя такая кладка обнаружена в районе Свирской губы 29.VI 1976 г. При повторном размножении наблюдается совмещение гнездостроения и откладки яиц. Новая кладка появляется обычно на 3—8-й день после разорения гнезда. При этом в большинстве случаев образуются новые пары. Вылет птенцов из гнезд проходит с III декады июня по конец июля.
В Карелии (Зимин, 1972; Хохлова, 1976) к строительству гнезд пеструшки приступают в конце I — начале III декады
мая. Откладка яиц начинается с середины II декады этого месяца и длится до I декады июля включительно; у большинства пар заканчивается к 10. VI. Массовый вылет птенцов проходит в конце июня - начале июля, в то время как самые поздние гнезда птенцы покидают лишь в I декаде августа. В Архангельской обл. у с.Семжа кладка из 5 ненасиженных яиц найдена 21.VI 1956 г. (Спангенберг, Леонович, 1958,1960).
В Волжско-Камском крае образование пар отмечено 26.IV. Откладка яиц у большинства пар проходит здесь в III декаде мая, а массовый вылет птенцов (2 года) зарегистрирован 25, 28-30. VI (Птицы Волжско-Камского края, 1978). В Раиф-ском лесу смешанные кладки пеструшки с большой синицей и обыкновенной горихвосткой обнаружены 23.VI и 14.VI11969 г. (Ушакова, Ушаков, 1976).
В Пермской обл. (Добрянский р-н) гнездо с полной ненасиженной кладкой в 1972 г. найдено 10.VI, а с птенцами -26.VI (Хазиева и др., 1982). В заповеднике ’’Денежкин камень” откладка яиц проходит в I декаде июня (Кузнецов, 1958). В окр. г.Свердловска наиболее ранний вылет птенцов (14 лет) зарегистрирован 20.VI, а наиболее поздний - 10.VI1 (Батманов. - Цит. по: Благосклонов, 1954). В Сургутском р-не Тюменской обл. молодые птицы добыты 27.VII и 8.VIII 1947 г. (колл. ЗИН). В Новосибирской обл. на о-ве Перекопном (оз. Большой Чан) гнездо с полной ненасиженной кладкой обнаружено 25.VI 1975 г. (Юрлов и др., 1977). Слетки у Новосибирского водохранилища встречены 27.VII 1959 г., а у г.Бийск — 24.VII 1972 г. (Гынгазов, Миловидов, 1977). В окр. г.Красноярск (Гремячин-ская сопка) пара пеструшек, занимающаяся строительством гнезда, зарегистрирована 28.V 1934 г., a 17.VI это гнездо содержало 3 свежих яйца (Юдин, 1952). У г.Томск (несколько лет) строительство гнезд отмечено 16.V— 21.VI, а у с.Светлая Протока на р.Обь - 2.VI. Полные кладки в окр. г.Томск (21 год) встречены 18.V-12.VI, 7.VII, а вылет птенцов зарегистрирован 7.VII, 2—14.VIII (Иоганзен, 1914; Гынгазов, Миловидов, 1977). На р.Кеть у с.Крас-ная Курья самка с наседным пятном добыта 9.VI 1963 г. Полные кладки найдены у с.Белый Яр - 20.V 1963 v (Москвитин и др., 1977) и в окр. с.Карагасок - 31.V 1934 г. (Гынгазов, Миловидов, 1977).
Линька. Через некоторое время
39
после того, как маховые и рулевые окончательно закончат свой рост, молодые пеструшки начинают смену гнездового наряда на первый послегнездовой. Эта частичная линька охватывает почти все мелкое оперение, кроме некоторых больших верхних кроющих второстепенных маховых. На местах зимовок у первогодков проходит частичная предбрачная линька, в ходе которой сменяются рулевые и часть мелких перьев. Отдельные партии последних линяют уже на местах гнездования (Виноградова и др., 1976). Взрослые птицы имеют в году 2 линьки оперения: полную послебрачную на местах гнездования и частичную предбрачную на местах зимовок. Послебрачная линька обычно начинается у взрослых пеструшек лишь на поздних этапах гнездования (вскоре после того, как в гнездах появляются птенцы). Самцы приступают к смене оперения несколько раньше самок. Замена оперения у них, вероятно, в меньшей степени затормаживается процессами, связанными с размножением. При позднем гнездовании самцы нередко не участвуют в выкармливании птенцов, видимо, вследствие интенсивной линьки в это время. Самки реже совмещают смену оперения с размножением. Линька у них начинается на более поздних стадиях гнездования, она менее интенсивна и чаще идет асимметрично (Артемьев, 1981). Однако некоторые самки, гнездящиеся повторно, могут совмещать линьку с насиживанием (Мальчевский, Пукинский, 1983). Темпы смены оперения у еще размножающихся особей заметно медленнее, чем у птиц, закончивших гнездование. Частичная предбрачная линька у взрослых птиц охватывает только мелкое оперение и протекает с большими индивидуальными различиями.
Питание. Основу пищи составляют насекомые разных групп и стадий развития, а также паукообразные. Значительно реже пеструшки поедают многоножек, моллюсков, кольчатых червей, а во второй половине лета и осенью на пролете - мелкие сочные плоды и даже семена некоторых растений. Способы сбора корма у этого вида разнообразны, а сами птицы очень пластичны в их применении. В выкармливании птенцов принимают участие оба партнера пары. По данным В.И.Головань (1982), в течение первых 3—4 дней после вылета из гнезда родители подлетают к слеткам и кормят их. В 20-дневном возрасте молодые начинают перелетать за взрослыми птицами, настойчиво выпрашивая пищу. Первые по
пытки самостоятельного поиска и схватывания добычи отмечаются у молодых пеструшек в конце I — начале II недели вне-гнездовой жизни, в возрасте 22-24 дня. Летающие насекомые уже доступны 24-25-дневным молодым.
Отлет и осенний пролет на территории СССР. Проходит с середины III декады июля по I декаду октября включительно. Отдельные особи задерживаются до начала III декады октября.
В Карелии передвижение пеструшек становится заметным со второй половины августа и заканчивается в начале I декады сентября. В Лапландском заповеднике последние птицы в 1938 г. отмечены 1.IX (Владимирская, 1948), а в районе Пряжи (Южная Карелия, другой год) - 29.VIII (Нейфельдт, 1958). В Ленинградской обл. отлет проходит главным образом в первой половине августа, хотя отдельные особи и встречаются до середины сентября (Мальчевский, Пукинский, 1983). В районе г.Пскова миграция протекает с середины II декады августа до середины, реже до двадцатых чисел сентября (Зарудный. -Цит. по: Благосклонов, 1954), а у г.Новгород (11 лет) - 20.VIII-11.IX (Мантейфель. — Цит. по: Благосклонов, 1954).
В Волжско-Камском крае заметное уменьшение численности мухоловок-пеструшек отмечается уже в начале августа. На севере края отлет начинается в конце III декады июля. В сентябре птицы встречаются уже редко и, как правило, одиночными особями. Самая поздняя регистрация - 9.IX (Птицы Волжско-Камского
края, 1978). В Мордовии отлет начинается также в начале августа. В середине этого месяца окольцованных здесь пеструшек отлавливали уже в Италии (Луговой, 1975).
В Подмосковье отлет части популяций начинается уже 24—30.VII, 25.VII—4.VIII он усиливается и 1—10.VIII основная масса местных птиц исчезает, однако еще продолжается пролет мухоловок более северных популяций, которые иногда образуют небольшие скопления, отмеченные (2 года) 8 и 11.VIII (Птушенко, Иноземцев, 1968). Последние птицы задерживаются до 21.Х (Карпов, Паровщиков, 1941). У истоков р.Ока в Орловской обл. последние встречи мухоловок зарегистрированы (5 лет) 17.VIII, 2,7 и 22.IX (Ефимов, 1915). В Воронежской обл. пролетные птицы появляются в середине августа, а миграция здесь длится до середины сентября. Максимум пролета -в конце августа (Огнев, Воробьев, 1923).
40
В Белоруссии основная часть пеструшек отлетает в августе. В отдельные годы задерживаются до I декады сентября (Федюшин, Долбик, 1967). В Пинском Полесье стайка пролетных птиц отмечена 6.IX 1905 г. (Шнитников, 1913).
В западных областях Украины (Портенко, 1950; Страутман, 1963) и в Молдавии (Ганя, Литвак, 1961; Аверин, Ганя, 1970) миграция протекает в сентябре — I декаде октября. В окр. гЛьвов последние мухоловки в 1916 г. отмечены 25 .IX (Miczynski, 1962). На пролете в Крыму (Костин, 1983) пеструшки встречаются в разные годы между 9 и 30.VIII, но чаще в III декаде августа. Явно задерживавшаяся птица добыта на Тарханкуте 25.IX. За последние 20 лет позднее 1 .IX не встречались.
На Кубани пролетная птица в 1909 г. добыта 1.VIII (Моламусов, 1967; колл. ЗИН).
Под г.Оренбург пролет протекает с конца августа до конца I декады октября включительно, а в Северо-Западном Казахстане проходит с I декады августа по II декаду сентября (Ковшарь, 1970).
В Верхнем Приобье миграция пеструшки начинается в начале августа (Юрлов, 1959). У г.Томск (17 лет) пролет заканчивается 1-3.IX (Гынгазов, Миловидов, 1977), а в окр. г.Новосибирск (1 год) -19.IX (Цыбулин, 1985).
Практической значение. Уничтожает многих насекомых — вредителей леса и садовых культур, а также переносчиков возбудителей ряда заболеваний человека и животных.
Пластичность этой мухоловки в питании, выборе местообитаний вместе с возможностями быстрого увеличения ее численности на любом участке леса при развешивании искусственных гнездовий характеризует этот вид как очень перспективный для широкого использования в борьбе с вредителями леса, особенно в очагах массового размножения последних, а также в монокультурах хвойных пород северной тайги, где общая численность насекомоядных птиц низка.
Мухоловка-белошейка — Ficedula albicollis (Temminck, 1815)
Muscicapa albicollis Temminck, 1815, Manuel d’Ornith.. ed. 1: 100, новое имя для Muscicapa collaris Bechstein, 1795, Gemeinn. Naturgesch. Deutschlands, 4 : 495, Европа и Германия, преоккупировано Muscicapa collaris Latham, 1790, Index om., 2 : 471.
Сии. (русск.) : белошейная мухоловка.
Описание. Составлено по 162 экз. (116 - из европейской части СССР, 36 — с Кавказа, 3 - из Турции, 6 - из Ирана, 1 - из Северной Албании).
Вершину крыла образуют 3-е и 4-е первостепенные маховые (3 < 4 — 0,7 %; 3 = 4 - 6,1 %; 3 > 4 - 93,2 %. 2-е и 5-е немного их короче (2 < 5 — 15,0 %; 2 = = 5 — 13,6 %; 2 > 5 — 71,4 %); 2-е всегда больше 6-го. Первое первостепенное маховое очень мало (табл. 7). У большинства особей (78,0 %) наружные опахала 3-4-го первостепенных маховых в вершинной части заметно сужены. У меньшего количества особей (22,0 %) наружное опахало в вершинной части сужено и на 5-м маховом. Количество щетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3-4 (3 - 87,8 %; 4 - 12,2 %); длина наибольшей (п = 42) 4,3-6,3 мм (в среднем — 5,2). Количество щитков подотеки на передней части цевки варьирует от 3 до 6. На вершине хвоста (в центре) имеется небольшая выемка. Половой диморфизм выражен в окраске оперения и незначительно в размерах отдельных частей тела (крыло, хвост) (табл. 7).
Распространение. Западная Европа на запад до Восточной Франции и к северу до 51—52° с.ш. К югу до южных оконечностей Апеннинского и Балканского полуостровов и северо-западного побережья Черного моря. От западных частей Малой Азии к востоку до Хорасанских гор. К северу до южного побережья Черного моря, Кавказа, Хорасанских гор и предположительно Западного Копетдага. К югу до южного побережья Малой Азии, Армянского (Восточного) Тавра, гор Загрос. Острова Сицилия и Готланд (Степанян, 1978). В СССР: юго-запад европейской части и Кавказ (см. рис. 14, 16). На всем гнездовом ареале птицы этого вида являются перелетными и зимуют главным образом в саваннах тропической Африки.
Систематика. Одни исследователи (Благосклонов, 1954; Vaurie, 1959; Портенко, 1960; Ctowacinski, 1974; Makatsch, 1976; Степанян, 1978; и др.) считают • мухолов ку-белошейку политипиче-ским видом, образующим 2 подвида: F.a.al-bicollis и F.a.semitorquata, другие (Curio, 1959; Mauersberger, Portenko, 1960; Мат-Bejee, 1976; Иванов, 1976; и др.) - возводят форму semitorquata в ранг самостоятельного вида или рассматривают ее как подвид мухоловки-пеструшки - F.hypo-leuca (Hartert, 1907; Пузанов, 1938; и др.).
41
Таблица 7. Размеры и коэффициенты вариации (CV, %) мухоловки белошейки
Пт
Подвид, место сбора п Длина, мм -
Крыло CV Расстояние, на которое 1-е маховое выступает за вершины кроющих кисти CV Хвост CV Клюв от оперения лба CV Клюв от орального края ноздри CV Цевка CV
Самцы первогодки
F.albicollis albicollis, 56,93 50,221 0,24 "47,0 -52,4 2,86 9,121 0,06 8,5 - 9,9 4,00 6,4610,06 5,7 -7,5 5,58 16,911 0,10 15,7 -18,3 3,46
европейская часть СССР 37 80,76± 0,29 76,3 -84,5 , 2,6810,26 ’ 0,0 -7,6
F.a.semitorquata,
Кавкази сопре- 11 81,671 0,57 э по 90±0,20 32,12 52,241 0,37 2,02 9,461 0,21 6,31 6,5510,09 4,08 16,721 0,19 3,71
дельные районы Ирана и Турции 79,4 -84,6 ’ 1,0 -2,7 50,2 -53,3 8,6 -10,5 6,2 -7,1 15,8 -17,7
Самцы взрослые (старше одного года)
F.albicollis albicollis.
европейская часть СССР 59 81,661 0,21 77,8 -85,3 . qq 1,3610,19 ’ -2,4 -4,6 100,00 51,231 0,23 47,3 -55,6 3,35 9,361 0,07 8,3 -10,5 5,25 6,5510,05 5,6 -7,5 5,42 16,931 0,06 15,9 -18,1 2,62
F.a.semitorquata,
Кавкази сопредельные 20 81,641 0,30 79,6 -84,0 1 ас 0.8810.23 1>65 -0,3 -2,9 100,00 52,511 0,29 50,0 -54,6 2,50 9,021 0,10 8,4 - 9,7 4,48 6,4910,08 5,9 -7,3 5,12 16,871 0,10 16,0 -17,7 2,69
районы Ирана и Турции Самки
F.albicollis albicollis,
европейская часть СССР 18 79,491 0,45 "773 -Й5,6' , „ 1,8710,28 232 TO~-4,6 63,94 49,591 0,48 46,5 —52,6 3,78 9,151 0,15 7,$ -10,7 6,6.2 6,4410,09 5,7 -7,3 5,64 16,841 0.15 15,6 -17,7 3,66
F.a.semitorquata,
Кавкази со- 12 78,601 0,58 2 46 2-05±0-52 84,71 50,341 0,52 3,40 9,121 0,16 5,74 6,3010,09 4,49 16,631 0,12 2,59
предельные районы Ирана и Турции 75,8 -81,8 ’ -0,9 -4,8 46,7 -53,0 Т,5 -10,1 5,8 -6,7 16,0 -17,7"
Подвид, место сбора п Ширина, мм Высота, мм Отношение ши-рины клюва у основания к его длине от оперения лба,% CV Отношение длины хвоста к длине крыла, % CV Отношение ширины крыла к его длине, % CV
Крыло CV Клюв у орального края нозДри CV Клюв у основания CV Клюв у основания CV
Самцы первогодки
F.albicollis albicollis,
европейская часть СССР 37 54,231 0,21 51,5 -56,4 2,33 3,8510,03 3,4 -4,3 5,25 5,8010,07 5,2 -6,7 6,77 3,6210,06 5,0 -4,1 7,30 63,451 0,62 5'7,14-72,64 5,53 62,201 0,23 59,32-64,83 2,18 67.151 0,20 64,54-69,70 1,72
F.a.semitorquata, Кавказ и сопредельные районы Ирана и Турции 11 54,О5± 1,22 Лб,1 -56,7 6,37 3,6410,08 з’Г-4,0 7,71 6,71 3,5010,06 3,1 -3,7 5,07 62,741 1,95 52,38—70,93 8,78 63,741 0,40 62,05 -65,04 1,66 66,391 1Л5 56,22-69,79 6,62
F.albicollis albicollis, европейская часть СССР 59' 54.231 0.16 51,5 -56,7 2,28 3,87 *0,Q? 3,4 -4,6 6,39 Самцы взрослые (старше одного года) 5,8110,05 „g 3,6210.04 , 5,2 -6,7 3,3 -4,1 ’ 62,241 0,72 51,43-75,90 8,53 62,751 0,24 59,71-69,32 2,84 66,431 0,12 63,51-68,38 1,36
F.a.semitorquata, Кавказ и сопредельные районы Ирана и Турции 20 55,631 0,28 54,1 -59,1 2,26 3,8610,06 3,5 -4,3 6,59 5,8610,08 5,1 -6,4 6,21 3,5310,05 3,3 -3,9 5,38 64,821 0,96 57,89-71,76 6,27 64,321 0,31 61,76-67,79 2,13 68,151 0,26 66,30—70,78 1,73
F.albicollis albicollis, европейская часть СССР 18 53,051 0,35 51,0 -56,0 2,72 3,8810,04 3,7 -4,2 -4,33 5,7310,10 5,1 -6,3 7,08 Самки 3,4410,13 3,1 -3,9 8,62 62,411 1,26 52,34-72,09 8,05 62,541 0,39 60,00-65,22 2,35 66,741 0,36 64,48-69,01 2,22
F.a.semitorquata, Кавказ и сопредельные районы Ирана и Турции 12 53,541 0,26 52,0 -55,1 1,71 3,7610,12 3,2 -4,7 1,06 5,6110,13 5,0 -6,3 7,83 3,5510,09 3,2 -4,1 7,53 62,541 1,43 56,82-69,41 7,23 64.161 0,55 61,61-66,97 2,71 68,171 0,40 “65,51-70,21 1,94
Анализ морфометрических показателей, изучение окраски и рисунка оперения всех возрастных нарядов у этих форм, а также их экологии и распространения, проведенные нами, позволяют считать возведение формы semitorquata в ранг вида преждевременным, а отнесение ее на правах самостоятельного подвида к F.hypoleuca ошибочным. По целому комплексу таксономически весомых признаков форма semitorquata стоит значительно ближе к F.albicollis нежели к F.hypoleuca. Действительным является и то, что эта форма приобрела уже значительные морфологические и экологические отличия от формы albicollis. Однако между ними до настоящего времени еще происходит гибридизация в некоторых районах их гнездовых ареалов, о чем свидетельствуют встречи гибридных особей с полным комплексом переходных морфологических признаков (колл. ИЗАНУ; РГУ). На основании сказанного мы считаем, что форма semitorquata еще не достигла полной репродуктивной изоляции с формой albicollis и в настоящее время они должны рассматриваться как подвиды одного политипи-ческого вида F.albicollis.
Оба подвида мухоловки-белошейки встречаются и гнездятся на территории СССР.
Европейская мухоловка-белошейка — Ficedula albicollis albicollis (Temminck, 1815)
Muscicapa albicollis Temminck, 1815, Manuel d’Or-nith., ed. 1: 100, новое имя для Muscicapa collaris Bechstein, 1795, Gemeinn. Naturgesch. Deuts-ehlands, 4: 495, Европа и Германия, преоккупиро-вано Muscicapa collaris Latham, 1790. Index orn., 2: 471.
Описание. Составлено по 117 экз. (116 - из европейской части СССР, 1 - из Северной Албании).
У взрослого самца верхняя часть и бока головы (кроме лба), спина и верхние кроющие хвоста блестяще-бархатно-черные. На лбу расположено большое (до 11,0 мм) белое пятно, которое у отдельных особей нередко заходит на переднюю часть темени. Шею опоясывает широкая белая перевязь. Нижняя часть спины и надхвостье светло-белесо-серые с шиферно-серым налетом на вершинах перьев. Оперение нижней части тела от чисто-белого до белого с сероватым оттенком. Нижние кроющие крыла бурые с широкими белыми окаймлениями. Рулевые от однотонно-бархатночерных до черных с белыми (часто асим
метричными) пятнами и полосами на двух крайних парах (рис. И). Маховые бархатно-черные. В основаниях последних (п = 60) на наружных опахалах начиная с 1-го - 4-го первостепенного махового (с 1 - 26,6 %; со 2 - 1,7; с 3 - 70,0; с 4 - 1,7 %) и на внутренних опахалах со 2-3-го (со 2-98,3 %; с 3 - 1,7 %) появляются белые пятна, которые при сложенном крыле образуют на нем полосу. На верхней части крыла кроме этого имеется белое ’’зеркальце”, образованное пятнами в основаниях второстепенных маховых, на вершинах больших верхних кроющих последних и на третьестепенных маховых. Клюв черный. Ноги черно-бурые. Радужина глаза темно-коричневая. Индивидуальная изменчивость проявляется в варьировании величины и формы белого ’’зеркальца”, лобного пятна, пятен в основании первостепенных маховых и на двух крайних парах рулевых, а также ширины шейного кольца и густоте черного цвета оперения верхней части тела. Сезонная изменчивость проявляется в частичном исчезновении блеска и в побурении черных перьев в результате их обноса и выцветания.
Взрослая самка почти не отличима по общей окраске от таковой мухоловки-пеструшки, но, в отличие от последней, белые пятна в основаниях первостепенных маховых (п = 17) появляются у нее на наружных опахалах с 3-5-го пера (с 3 - 23,5 %; с 4 — 64,7 %; с 5 — 11,8 %), а на внутренних — со 2—3-го (со 2 — 66,7 %; с 3 — 33,3 %).
Молодые в гнездовом наряде сверху коричнево-серовато-оливковые, с округлыми, нерезкими охристыми пятнами и темнобурыми вершинными каемками. Оперение нижней части тела бледно-охристое с бурыми окантовками по краю опахал верхней части каждого пера, образующими волнистый рисунок. Последний становится менее выраженным на брюшке. Нижние кроющие хвоста белые. Маховые и рулевые такой же окраски, как и у взрослых самок в свежем пере, но ’’зеркальце” на крыле светло-охристое и меньшего размера.
, Первый послегнездовой наряд идентичен наряду взрослой самки, но общий тон окраски оперения сверху более серовато-коричневый. Самцы в этом наряде уже имеют черный цвет в оперении. Белые пятна в основаниях первостепенных маховых появляются у них (п = 35) на наружных опахалах с 1-5-го пера (с 1-го - 2,9 %;
44
со 2-го — 2,9 %; с 3-го — 45,7 %; с 4-го — 37,1 %; с 5-го — 11,4 %), а на внутренних со 2-го — 3-го (со 2-го - 91,4 %; с 3-го — 8,6%).
Размеры отдельных частей тела европейской мухоловки-белошейки приведены в табл. 7.
Масса тела, г: самец (V. долина р.Урал, с.Чапаево) - 12,0 (Ковшарь, 1970).
Распространение. Европейская часть ареала вида, кроме Кавказа и южной части Балканского п-ова. В Центральной Европе за последние пять десятилетий отмечено расширение гнездового ареала на север (Gfowacirfski, 1974). Осваивает новые территории скачкообразно, путем образования островных популяций. Зимует главным образом в саваннах тропической Африки. Известны залеты птиц этого подвида в Латвию и Эстонию. Детально распространение европейской мухоловки-белошейки на территории СССР показано на карте (рис. 14).
Кадастр к рис. 14.
1 - Литва (изолированное гнездование) (Степанян, 1978); 2 - Белоруссия, Гродненский р-н (Иванов, 1976); окр. с.Кашеневка - лес "Панямунь” (Сямашка, 1956; Федюшин, Долбик, 1967); 3 - заповедник "Беловежская пуща” (Цедлиц. - Цит. по: Долбик, 1959; Портенко, 1960); 4 - Хойникский р-н, с.Загалье (Долбик, 1959); Загальское лесничество (Федюшин, Долбик, 1967); 5 - Сталинский р-н (Федюшин, Долбик, 1967); окр. г.Столин (Долбик, 1959); 6 - Припятский заповедник (Клакоцкий, 1983); 7 - с.Мозырь (Портенко, 1960); 8 - Василе-вичский р-н (Долбик, 1959); Василевичское лесничество (Федюшин, Долбик, 1967); 9 - окр. г.Речица (Иванов, 1976); 10 - Макановичское и Великоборское лесничества (Федюшин, Долбик, 1967); И - Украина, Волынская обл. (Дементьев, 1937); Андрушевское лесничество (колл. ИЗАНУ); 12 - окр. с.Сопачев, хут. Шоков, р.Стыр (колл. ИЗАНУ); 13 - Костопольский лес (колл. ИЗАНУ); 14 - Коростеньский р-н, с.Ушомир (колл. ИЗАНУ); 15 - Северное Подолье (Дементьев, 1937; Марисова, Холина, 1959; колл. ИЗАНУ); Кременецкий р-н, г.Креме-нец и его окрестности (Марисова, Холина, 1959; колл. ЗИН); окр. с.Бережцы; окр. с.Лишня; Велико-Дедеркальский р-н, с.Вилия; Почаевский р-н, г.Почаев (Марисова, Холина, 1959; колл. ЗИН); 16 - Шумский р-н, окр. с.Сураж (Марисова, Холина, 1959; колл. ЗИН); Волынское лесничество, окр. с.Стожок (наши данные); 17 -Летичевский р-н, окр. сЛысогорка; окр.с.Голов-чинцы (колл. ЗИН); 18 - окр. с.Новая Рудня (сообщ. А.С.Вобленко); 19 - окр. г .Житомир, с.Дениши (колл. ИЗАНУ; наши данные);20 -окр. г.Коростышев (сообщ. А.С.Вобленко); 21 -с.Розважев (сообщ. М.И.Головушкина); 22 -окр. с.Залесье (сообщ. М.И.Головушкина); 23 -окр. с.Горенка (сообщ. М.И.Головушкина); 24 -г.Ю1ев и его окрестности (наши данные); 25 -ст.Ирпень и ее окрестности (сообщ. М.И.Голо
вушкина) ; 26 - окр. пгт Клавдиево-Тарасово (сообщ. М.И.Головушкина); 27 - г.Фастов и его окрестности; окр. с.Скригалевка (колл. ИЗАНУ); 28 - с.Корнин (сообщ. М.И.Головушкина);
29 - окр. с.Яроповичи (сообщ. М.И.Головушкина) ; 30 - окр. с.Ивница (сообщение М.И.Головушкина); 31 - окр. с.Вчерайше (сообщ. М.И.Головушкина) ; 32 - окр. с.Ржищев (сообщ. М.И.Головушкина); 33 - окр. г.Канев (сообщ. М.И.Головушкина); 34 - 40 км от г.Чернигов вверх по р.Десне (сообщ. А.С.Вобленко); 35 -окр. с.Сосница (Портенко, 1960); 36 - Кировоградская обл., Черный лес (Шевченко, 1940; Волчанецкий, 1959); 37 - истоки р.Ингул (Портенко, 1960); 38 - Одесская обл., Савранский лес (сообщ. Т.Б.Ардамацкой); 39 - Балтийский р-н, окр. г.Балта (Портенко, 1960; колл. ЗИН); Кищевский лес (Волчанецкий, 1959); 40,- Черноморский заповедник, участок Волыжин лес (сообщ. Т.Б.Ардамацкой); 41 - с.Новая Ушица (Михайловский. - Цит. no:Mauersberger,Portenko, 1960); 42 - Глыбокский р-н, с.Турянка (колл. ИЗАНУ); 43 - Раховский р-н, окр. Апшенсцкой полонины (колл. ХГУ); окр. г.Рахов (Страутман, 1954; колл. ХГУ); 44 - Лумшор, Велико-Уголек, окр. с.Квасы (Кистяковский, 1950);
45 - пойма р.Боржавы, окр. с.Кушница (Страутман, 1954; 1963); 46 - лесничество Мукачевско-го лесокомбината (Гвоздак, Симочко, 1977); окр. пгт Чинадиево (наши данные); 47 - г.Пе-речин (Портенко, 1960; колл. ЗИН); 48 - окр. пгт Моршин (колл. С.Д.Кустановича); окр. с.Пе-няки и с.Поторицы (Страутман, 1963); 49 - окр. пгт Бурштын (сообщ. Ю.В.Мищенко); 50 -г.Львов и его окрестности; буковые и смешанные леса Голоско, Брюховичи, Винников, Сихов (Miczynski, 1962); 51 - г.Павлоград (Портенко, 1960); 52 - р.Ворскла, с.Богачка; Руднянская дача; г.Полтава, Монастырский лес (колл. ХГУ);
53 - окр. с.Диканька, уроч.Братерское и Дубове (колл. ХГУ); 54 - окр. с.Тростянец, Краснянский и Маковский леса (колл. ХГУ); 55 -Богодуховский р-н, окр. с.Гуты (колл. ХГУ); 56 - окр. с.Просяное (колл. ХГУ); 57 - окр. г.Волчанск, овраг Паренский (Сомов, 1897);
58 - окр. г.Чугуев (колл. ХГУ); Чугуево-Баб-чанское лесничество (Сомов, 1897); 59 - окр. г.Готвальд (Змиев) (Портенко, 1960); с.Гай-дары-Беспаловка; окр. с.Лебяжье; Змиев ское лесничество (Сомов, 1897); с.Кочеток (колл. ЗИН); Коробовское лесничество (Ковалев, При-сада, 1983); окр. с.Мохнач на правом берегу р.Северский Донец (колл. ИЗАНУ); 60 - Изюм-ский р-н (колл. ХГУ); 61 - окр. г.Кременная (сообщ. В.В.Ветрова); 62 - окр. г.Счастье (сообщ. В.В.Ветрова); 63 - Луганский заповедник (сообщ. В.В.Ветрова); 64 - нижнее течение р.Деркул: окр. ст.Н.Ильенко (сообщ. В.В.Ветрова) ; 65 - окр. с.Станично-Луганское (сообщ. В.В.Ветрова); 66 - Молдавия, Оргеевский лесхоз у с.Морозены (Аверин, Ганя, 1970; Иванов, 1976); 67 - нагорные леса Приднестровья (сообщ. Г.З.Гусана); 68 - заповедник "Кодры” (сообщ. Г.З.Гусана); 69 - Слободзенский и Суворовский р-ны (сообщ. Г.З.Гусана); 70 - Ростовская обл., окр. стан. Нижнекундрюченская (колл. РГУ); 71 - Азовский р-н, Ленинский лесхоз в 7 км восточнее ст.Александровка (колл. РГУ; сообщ. Б.А.Казакова); 72 - Белгородская обл. (Будниченко, Козлов, 1980); окр. г.Белго
45
род (Мальчевский, 1959); 73 - заповедник ’’Лес на Ворскле” (Мальчевский, 1959); 74 - г.Пав-ловск (Огнев, Воробьев, 1923; Портенко, 1960);
75 - окр. г.Воронеж (Давыдова, 1970); Саваль-ское лесничество (Мальчевский, 1959; Будни-ченко, 1968); Усманский лес (Барабаш-Никифоров, Семаго, 1963); лесополосы Воронежской обл. (Будниченко, 1968); 76 - Воронежский заповедник (Барабаш-Никифоров, Павловский, 1947); 77 - истоки р.Ока (Иванов, 1976); Черномошный лес (Ефимов, 1915); г.Орел (Портенко, 1960); 78 - окр. г.Тула (Портенко, 1960; колл. ЗИН); Ефремовский р-н, с.Прудки (колл. ЗИН); 79 - Крапивницкий р-н, Тульские засеки (колл. БПиДВНЦ); 80 - окр. г.Москва, окр. с.Черепково (Lorenz, 1894; колл. ЗИН);
81 - окр. с.Камкино в долине р.Пахра (Каминский, 1916; Иванов, 1976); окр. г.Подольск (Поляков, 1924; Портенко, 1960); 82 - Калининская обл. (Зиновьев, Шапошников, 1978); 83 -- Хоперский заповедник (Измайлов, 1940; Иванов, 1976); Теллермановский лесной массив (Королькова, 1963); 84 - г.Саратов (Портенко, 1960; Иванов, 1976); 85 - Пензенская обл. (Федорович по Луговому, 1975); окр. г.Пенза (колл. КБ МГУ); 86 - Южная Мордовия; р.Синяш, Сабаевское лесничество (Луговой, 1975); 87 - Западная Мордовия (Луговой, 1975); Мордовский заповедник (Бородин, 1967);
88 - окр. сЛустынь, Арзамасский р-н Горьковской обл. (Хохлова, 1965; Иванов, 1976); 89 -Лукояновский р-н (Хохлова, 1965; Иванов, 1976); 90 - Чувашия, окр. с.Ядрин (Луговой, 1975); 91 - северо-западная часть Татарской АССР (Птицы Волжско-Камского края, 1978); 92 - г.Казань (Портенко, 1960; Иванов, 1976);
93 - перестойные кайбицкие дубравы на западе Татарии (Птицы Волжско-Камского края, 1978);
94 - дельта р.Кама (Птицы Волжско-Камского края, 1978); 95 - дельта р.Вятка (правый берег)
(Птицы Волжско-Камского края, 1978); 96 -окр. с.Сл. Черемуховая (Птицы Волжско-Камского края, 1978); 97 - Жигулевский заповедник (Белянина, Белянин, 1981); 98 - 20-22 км северо-западнее г.Белебея (Фомин, 1977).
Места обитания. Населяет главным образом различные типы лиственных, реже смешанных хвойно-лиственных и хвойных лесов на равнинах, в предгорьях и по склонам гор, поднимаясь вверх до 1700 м н.у.м. Тяготеет к наиболее старым, часто разреженным участкам леса, с большим количеством дуплистых деревьев, по поймам рек, у опушек, вдоль просек и лесных дорог. Поселяется в старых рощах, парках, садах населенных пунктов и их окрестностей. Как исключение гнездится на отдельно стоящих старых, дуплистых деревьях, растущих на значительном удалении от массивов леса, и постройках человека.
В Белорусском Полесье (Василевич-ское лесничество) гнездится как на опушках и вдоль лесных дорог, так и в глубине дубово-грабовых лесов с примесью ясеня, клена, береста, с подлеском из лещины, бересклета, смородины, ивы, а также в дубово-ольховых лесах. Зарегистрирована на опушках сосново-дубового леса и белях с примесью ольхи и ивы. На берегу р.Льва встречена в старом пойменном дубняке (Федюшин, Долбик, 1967).
На Украине в окр. г.Житомир найдена нами на гнездовании в разреженном участке
Рис. 14. Распространение европейской мухоловки-бслошейки (Ficedula albicollis albicollis) на территории СССР:
а - гнездится, б - отмечена в гнездовой период. См. кадастр, стр. 45-46.
46
смешанного леса с преобладанием дуба. В Киевской обл. предпочитает различные типы старых дубовых лесов с примесью березы, груши и осины, с подлеском из боярышника и лещины, с густым травостоем и папоротниками в нижнем ярусе, но гнездится также в дубово-березовых лесах без подлеска, на границах последних с ельниками, в липовых с незначительной примесью березы (сообщ. С.И.Пархоменко; наши данные), а также в грабово-дубовых лесах (Орлов. — Цит. по: Благосклонов, 1954). В г.Киев обычна в парках, скверах и ботаническом саду АН УССР. В Северном Подолье (Тернопольская обл.) мухоловка-белошейка является одной из характерных многочисленных птиц смешанных лесов и парковых насаждений, в том числе и городских (г.Кременец). Обычна также в грабово-дубовых и нередко в сосновых высокоствольных лесах (Марисова, Холина, 1959; колл. А.П.Кузякина). В Харьковской обл. населяет также лиственные леса разных типов с подлеском. Отмечена в борах (Сомов, 1897; Благосклонов, 1954; колл. ИЗАНУ). В западных областях Украины поселяется по речным долинам северных и в буковых лесах южных и юго-западных склонов Украинских Карпат, поднимаясь вверх до 1000 м и выше. Гнездится спорадически в дубравах, в лиственных лесах с преобладанием граба и в смешанных лесах равнин, в парках, садах и в пойменных зарослях из ольхи и ивы (Кистя-ковский, 1950; Страутман, 1954, 1963; наши данные). •
В Молдавии (сообщ. Г.3.Гусана) населяет смешанные лиственные леса из граба, дуба, ясеня и липы, а также буково-дубовые и реже липово-ясенево-дубовые леса. При наличии искусственных гнездовий охотно поселяется в неспелых лесах с редким подлеском. На юго-западе республики (Слободзейский и Суворовский районы) в гнездовой период отмечена в высокоствольных лесополосах из дуба, вяза, липы и акации.
В Северо-Западном Предкавказье (Ростовская обл., Ленинский лесхоз) гнездится в старых смешанных, лиственных лесах с преобладанием дуба (сообщ. Б.А.Каза-кова и Н.Х.Ломадзе).
В Воронежской обл. у южной границы лесостепи (Савальский и Теллермановский леса) встречается главным образом в старых дубравах, но отмечена также в полосах старолесья среди осветленного жердняка (Королькова, 1963), а также в сме
шанных и чисто сосновых насаждениях (Мальчевский, 1959). Обычна в сосновых лесах Хоперского заповедника, где ежегодно заселяет большую часть искусственных гнездовий (Барабаш-Никифоров, Семаго, 1963) и зарегистрирована в пойменном широколиственном лесу по р.Хопер (Измайлов, 1940). В Воронежском заповеднике европейская белошейка отмечена в старой дубраве и в ольшаннике поймы р.Усманка (Барабаш-Никифоров, Павловский, 1947). В Мордовии также поселяется в лиственных и сосновых лесах при наличии дуплистых деревьев или искусственных гнездовий. Вдоль р.Синяш гнездится в дубово-осиновых лесах (Луговой, 1975). В Арзамасском р-не Горьковской обл. (окр. с.Пустынь) гнездится на участках старого разреженного сосново-елового леса с подлеском из липы, а в Лукояновском р-не — на приопушечных участках леса с примесью липы и березы (Хохлова, 1965). В Подмосковье на р.Пахра (окр. с.Кам-кино) отмечена на гнездовании в лиственном лесу (Каминский, 1916).
В Волжско-Камском крае предпочитает, как и в большинстве других мест гнездового ареала, высокоствольные леса с большим количеством дуплистых деревьев. В молодняках, на вырубках и в полезащитных лесополосах не отмечена (Птицы Волжско-Камского края, 1978). В Башкирии (Белебеевский р-н) гнездится на участках старого высокоствольного леса из липы, осины, березы и клена, с густым подлеском (Фомин, 1977).
Численность. В большинстве мест гнездового ареала на территории СССР обычна, в излюбленных оптимальных биотопах, где может образовывать плотные поселения — многочисленна. В Литве, Подмосковье, Башкирии и Северо-Западном Предкавказье — редка. Ввиду мозаичного распределения на территории ареала этой формы характерных гнездовых биотопов (старых лесов с большим количеством дуплистых деревьев или искусственных гнездовий) численность европейской мухоловки-белошейки неравномерна, а ее распространение в некоторых местах часто носит спорадичный характер.
Прилет и весенний пролет на территории СССР. Проходит с I декады апреля по III декаду мая включительно. На юге европейской части СССР в отдельные годы первые птицы появляются уже в III декаде марта.
На Северном Кавказе у г.Орджоникид
47
зе одиночная европейская белошейка добыта 21.IV 1935 г. (Моламусов, 1967; колл. ЗИН).
В Крыму самая ранняя птица зарегистрирована А.И.Дулицким 21.III 1974 г. в 12 км от г.Алушта, а основной пролет проходит на полуострове (многолетние наблюдения) 3.IV - 19.V (Костин, 1983). Нами в 1981 г. пролетные мухоловки отмечены в центральной части Арабатской Стрелки 6.V, а в Карадагском заповеднике — 9 и 11 .V. Самая поздняя птица добыта в низовьях р.Салгир 27.V 1866 г. (Костин, 1983; колл. ЗМ МГУ). На юге Одесской обл. несколько пролетных самцов в 1948 г. добыты 25 и 27.IV (колл. КГУ).
В Молдавии самый ранний прилет зарегистрирован Г.З.Гусаном 27.III 1983 г. Обычно первые птицы появляются здесь в I декаде апреля, а основной прилет и пролет протекают во II—III декадах этого месяца (Ганя, Литвак, 1961; Аверин, Ганя, 1970), Сроки и продолжительность пролета зависят главным образом от погодных условий. В годы с холодными веснаьш пролет затягивается до начала II декады мая (сообщ. Г.З.Гусана).
В Луганском заповеднике белошейки летят (9 лет) 18-28.IV (Сулик, Енакиев, 1978). В Харьковской обл. прилети пролет проходит (9 дет) со II декады апреля по конец Г декады: 19.IV-17.V; 24—29.IV; 24.IV-2.V; 27-29.IV; 29.IV; 30.IV; 2.V; 2—7.V; 10.V (Сомов, 1897). Кроме этого 2 мухоловки добыты здесь 21.IV 1950 г. и 27.IV 1924 г. (колл. ХГУ). В Полтавской обл. (3 года) птицы добывались 12 и 27.IV (колл. ХГУ).
В Украинских Карпатах европейская белошейка отмечалась: под полониной Ква-совский Менчул — 26.IV 1957 г. (Страутман, 1963); у Чорногоры — 27.IV 1959 г., у с-Лумшоры - 28 и 30.IV 1948 г. (колл. ИЗАНУ), а также 3.V 1950 г. у г.Рахов (Страутман, 1954, 1963). В окр. гЛьвов прилетает 18—25 .IV, реже - в начале I декады мая (Miczynski, 1962), а в Тернопольской обл. (4 года) появляется 20— 23 и 26.IV (Марисова, Холина, 1959).
В Днепропетровской обл. у с.Мыс Доброй Надежды самец добыт 18.IV 1952 г. (колл. ИЗАНУ), в Александровском р-не Кировоградской обл. — 30.IV 1907 г. (колл. ЗИН), а в Черкасской обл. несколько птиц встречены 20, 22, 24 и 27.IV 1922 г. (колл. ЗИН). В Киевской обл. в окр. г.Фастов (3 года) первые белошейки появляются 12, 13 и 16.IV, а в окр. г.Киев
(9 лет) - 17-26.IV (Данилович. - Цит. по: Благосклонов, 1954). В Николаевском лесничестве Сумской обл. самец добыт 29.IV 1961 г. (колл. ИЗАНУ).
В Воронежской обл. (Теллерманов-ский лес) прилет и пролет длятся с III декады апреля (26—27.IV 1952) до начала I декады мая включительно (Благосклонов, 1954; Королькова, 1963). У истоков р.Ока (5 лет) — 22.IV—13.V (Ефимов, 1915). В Мордовии пролет также проходит в конце апреля — первых числах мая (Луговой, L975). В Мордовском заповеднике 2 птицы встречены (2 года) 10.V (Бородин, 1967). В Подмосковье у д.Че-репково самец белошейки пойман 16.V 1890 г. (Lorenz, 1894).
В районе устья р.Кама появляется во II декаде апреля - начале мая. Первые птицы здесь отмечены (5 лет) 16, 24, 30.IV и 2.V (Птицы Волжско-Камского края, 1978).
В Западном Казахстане пролет выражен очень слабо. Две пролетные особи, ранее считавшиеся залетными, встречены в долине р.Урал: 28.IV - в садах Александров Гай и 12.V (тот же год) у хут. Данилка (Чапаевский р-н Уральской обл.) (Ков-шарь, 1970).
На места гнездования первыми прилетают самцы и только через 4-5 дней самки. Сразу после прилета самцы часто держатся неплотными группами до 10—15 особей.
Размножение. После появления самок самцы начинают вести себя очень оживленно и много поют. Пение сходно с пением пеструшки, но громче. Образование пар сопровождается наиболее интенсивным пением самцов и их токовым демонстративным поведением.
Гнездовые участки пар невелики, площадью 150—200 м2. В местах плотных поселений гнезда в среднем располагаются друг от друга на расстоянии 37,5 м, редко -96,5 (Марисова, Холина, 1959). Строят их белошейки в глубоких трещинах, нишах и чаще всего в естественных или выдолбленных другими птицами (малый и большой пестр&е дятлы, обыкновенный поползень) дуплах стволов и ветвей деревьев (дуб, клен, ива, граб, ясень, береза, липа, осина, вяз) на высоте (п = 8) 1,24-12,0 м от поверхности земли, в среднем - 3,52, а также в дуплах выгнивших пней на высоте (п = 4) 0,0—0,5, в среднем — 0,21 м (Каминский, 1916; Марисова, Холина, 1959; Птицы Волжско-Камского края, 1978; колл. ИЗАНУ и С.Д.Кустановича; сообщ.
48
С.И.Пархоменко; наши данные). Размеры гнездового дупла и летка сильно варьируют. Птицы этого вида охотно заселяют и искусственные гнездовья (разнообразные дуплянки). Интересен факт гнездования мухоловки-белошейки в окр. г.Готвальда три года подряд в трубе турника (Харченко и др., 1984). В сборе строительных материалов и строительстве гнезда, которое длится 7-12 дней (Марисова, Холина, 1959), участвуют оба партнера пары, однако самка более часто залетает в дупло и укладывает там материал.
Гнезда представляют собой очень рыхлые постройки, форма и размеры которых зависят от таковых нижцей части дупла. В качестве строительных материалов мухоловки используют сухие листья деревьев и кустарников, полоски луба, кусочки коры, веточки, а также тонкие корешки, стебли и листья трав (главным образом дикорастущих злаков). Лоток выстилается тонкими полосками луба, корешками трав, черешками, жилками и кусочками листьев и реже конским волосом. Отдельные постройки состоят исключительно из луба деревьев и кустарников. Размеры гнезд (п = 6) (Сомов, 1897; наши данные), мм: диаметр 90-120, в среднем - 106; высота 30—55, в среднем - 46; диаметр лотка 50-72, в среднем - 61; глубина лотка 23—35, в среднем -31.
Для европейской мухоловки-белошейки характерна сезонная моногамия, однако известны случаи и полигинии (Ковалев, Присада, 1983), при которой отдельные самцы за сезон размножения спариваются сразу с двумя самками. В сезон размножения птицы имеют один репродуктивный цикл. Повторные кладки возникают в результате гибели первых. Количество яиц в первой полной кладке 5—8, как исключение — 4. Повторные кладки обычно меньше и могут состоять даже из 3 яиц.
Яйца укороченно-остроовальные, овальные, остроовальные, в редких случаях -удлиненно-остроовальные, однотонные от бледно- до светло-голубых со слабым зеленоватым оттенком). Как исключение нами встречена 1 кладка, яйца в которой по светло-голубому фону имели редкие (густота рисунка менее 10 %) четкие и размытые крапинки (d до 0,6 мм) и точки светло-рыжевато-охристого цвета. Последние распределялись по всей поверхности блестящей скорлупы, но наибольшее их количество локализовалось вокруг тупого полюса яйца. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
Насиживает кладку самка. Самец кормит ее на гнезде и редко (не у всех пар) на незначительное время садится насиживать кладку, подменяя слетевшую кормиться самку. Инкубация длится 13-15 дней. Начиная с 7-8 дня родители сидят на гнезде очень плотно и покидают его только в случае сильного беспокойства. Вылупление птенцов обычно проходит в течение суток, в гнездах с большими кладками — в течение полутора-двух (сообщ. Г.З.Гусана).
Пуховой птенец европейской мухоловки-белошейки (Мальчевский, 1959) имеет короткий, редкий, светло-серый пух на надглазничных, затылочной, спинной, плечевых и глазных птерилиях. На последних он рудиментарный и бывает не всегда. Кожа буровато-розовая. Клюв светлый с незначительным потемнением на конце. Яйцевой зуб и когти серые. Ротовая полость желтая. Складки в углах рта хорошо развиты, белые.
Птенцы очень подвижны. Растут и развиваются быстро и на 13—16 день (Мальчевский, 1959; Марисова, Холина, 1959; сообщ. Г.З.Гусана) с недоросшими рулевыми. и маховыми перьями, покидают гнезда. Первые 2-3 дня после вылета слетки держатся на гнездовых участках, а затем начинают кочевать с родителями за их пределами.
Основные причины отхода яиц и птенцов у европейской мухоловки-белошейки такие же, как и у пеструшки. На юго-западе европейской части СССР большой урон гнездам в совместных местах обитания на территории Украины и Молдавии (Гвоздак, Симочко, 1977; сообщ. Г.З.Гусана) могут наносить сони орешниковая (Muscardinus avellanarius) и лесная (Dryomys nitedula), а в Черноморском заповеднике на участке Волыжин лес (сообщ. Т.Б.Ардамацкой) — четырехполо-сый полоз (Elaphe quatuorlineata).
Сезон размножения европейской мухоловки-белошейки на территории СССР длится с I декады мая по II декаду июля включительно. В годы с ранними веснами отдельные пары приступают к размножению в III декаде апреля.
В Азовском районе Ростовской обл. (Ленинский лесхоз) 29.V 1977 г. найдено 1 гнездо с сильно насиженной кладкой из 7 яиц и 1 гнездо, кладка в котором только началась (отложено 1 яйцо). 11.VI в последнем была полная насиженная кладка из 5 яиц, а 21 .VI 3 птенца 5-6-дневного возраста и 2 болтуна. В 1978 г. строящееся
49
гнездо обнаружено 11.V, а 4.VI в нем находилась сильно насиженная кладка из 7 яиц. Во втором гнезде, найденном 4.VI, были 2 только что вылупившихся птенца, 1 яйцо перед вылуплением и 3 болтуна (сообщ. Б.А.Казакова и Н.ХЛомадзе).
В Молдавии сроки откладки яиц растянуты почти на месяц. Первые отложенные яйца отмечены здесь 3.V, а последние кладки зарегистрированы 27 .V (сообщ. Г.З.Гусана). В Оргеевском лесхозе гнездо с 4 яйцами обнаружено 15.V 1957 г. (Ганя, Литвак, 1961; Аверин, Ганя, 1970).
Во Львовской обл. полные кладки встречаются (6 лет) 16.V—5.VI (Страутман, 1963; колл. ЛПМ). У Моршин-здроя (колл. С.Д.Кустановича) полная ненаси-женная кладка найдена 5.VI 1966 г., а 7.VI здесь обнаружено второе гнездо с 3 яйцами (очевидно, повторная кладка взамен утраченной). В западных областях УССР, по данным Ф.И.Страутмана (1963), летные молодые появляются во второй половине июня — начале июля. В Мукачевском р-не у пгт Чинадиево летная молодая птица с взрослой самкой встречены нами 24.VI 1983 г. В долине р.Уж выводок летных молодых отмечен 8.VII 1947 г. (Портенко, 1950). В окр. пгт Перечни на р.Вильшава молодая белошейка добыта 25.VII 1947 г. (колл. ЗИН), а в Раховском р-не у Аншенс-цкой полонины — 27.VII 1956 г. (колл. ХГУ). В Тернопольской обл. (4 года) самое раннее строительство гнезда отмечено в окр. сЛишня 27.IV 1955 г. Свежие кладки в окр. г.Кременец встречаются 8— 28 .V, а насиженные — во второй половине мая и I декаде июня (2, 6.VI) (Марисова, Холина, 1959; колл. А.П.Кузякина). Птенцы появляются в конце мая — первой половине июня. В окр. г.Почаев гнездо с 2-дневными птенцами найдено 29.V 1955 г. (Марисова, Холина, 1959). В Шумском р-не у с.Сураж молодая птица добыта 7.VII 1889 г. (колл. ЗИН). В Киевской обл. в окр. г.Фастов (сообщ. С.И.Пархоменко) гнездо с начавшейся кладкой (отложено 1 яйцо) обнаружено 7.V 1966 г., 9.V в нем было 3 яйца. В 1969 г. полные кладки встречены здесь 19—25.V, а в 1970 г. — 16.V. В окр. г.Киев гнездо с 3—4-дневными птенцами найдено 30.V 1968 г. В Харьковской обл. (3 года) полные свежие кладки появляются во второй половине мая и как исключение в I—II декадах июня: 17.V -гнездо с 4 слабо насиженными яйцами; 21.V — кладка такой же насиженности; 1.VI - поздняя ненасиженная кладка из
6 яиц; 19.VI - слабо насиженная (очевидно, повторная взамен утраченной) кладка из 5 яиц (Сомов, 1897; колл. ИЗАНУ).
В Хоперском заповеднике (Барабаш-Никифоров, Семаго, 1963) в 4 гнездах бе-лошейки, обнаруженных 3.VI 1957 г., зарегистрировано: в одном - 4 двухдневных птенца, в другом — 2 ненасиженных яйца, а в остальных двух - кладки из 4 и 6 сильно насиженных яиц, из которых уже начали вылупляться птенцы. 21.VI также было найдено в одном гнезде 2 пятидневных птенца, а в двух других — 2 и 5 ненасиженных яиц. В Савальском лесничестве Воронежской обл. (3 года) гнезда с птенцами встречены 18.VI-7.VII (материалы ц.кольц.). В Тел-лермановском лесу вылет птенцов наблюдается во второй половине июня (Королькова. — Цит. по: Барабаш-Никифоров, Семаго, 1963).
В окр. г.Пенза гнездо с 1 яйцом найдено 17.V 1968 г. (колл. КБ МГУ), а в Подмосковье у с.Камкино на р.Пахра гнездо с 6 неоперенными птенцами обнаружено 10.VI 1914 г. В Башкирии (Белебеевский р-н) 19.VI 1975 г. у гнезда добыт самец с кормом, a 22.VI встречен выводок слетков (Фомин, 1977).
Линька. Через некоторое время после того, как маховые и рулевые окончательно закончат свой рост, молодые начинают смену гнездового наряда на первый послегнездовой. Эта частичная линька охватывает почти все мелкое оперение, кроме части больших, средних и малых верхних кроющих второстепенных маховых (большие индивидуальные различия). На местах зимовок у первогодков проходит частичная предбрачная линька, включающая у большинства птиц мелкое оперение, кроме нескольких кроющих крыла. У некоторых особей линяет почти все оперение, в том числе маховые и рулевые, но при этом остаются старыми, контрастирующими по цвету, 1-2 пера из больших, средних и малых верхних кроющих второстепенных маховых, а иногда и все верхние кроющие крыла (Виноградова и др., 1976). Взрослые птицы имеют в году две полные линьки: послебрачную на местах гнездований и предбрачную на местах зимовок.
Питание. Основу пищи составляют насекомые разных групп и стадий развития, а также паукообразные. Значительно реже поедают мелких моллюсков и как исключение кивсяков, мокриц и малощетинковых червей. При добывании корма белошейка пользуется такими же способами
50
охоты, как и пеструшка. Птенцов выкармливают оба партнера пары, но в первые дни после их вылупления, когда самка большую часть времени проводит в гнезде, согревая птенцов, корм чаще приносит самец.
Отлет и осенний пролет на территории СССР. Проходит очень растянуто с конца III декады июля по III декаду сентября включительно. Отдельные особи задерживаются до середины октября и встречаются даже на юге в I декаде ноября.
В Волжско-Камском крае отлет начинается уже в конце июля, а последняя встреча европейской белошейки зарегистрирована здесь в 1961 г. — 28.VIII (Птицы Волжско-Камского края, 1978). В Рязанской обл. одиночный взрослый самец добыт в окр. с.Рубецкое 27.VIII 1959 г. (Птушенко, 1968). В Воронежской обл. отлет проходит в августе, отдельные птицы задерживаются до середины сентября (Барабаш-Никифоров, Семаго, 1963).
На Украине в Подолье отлет протекает с середины II декады августа до середины второй половины сентября, затягиваясь иногда до начала октября (Марисова, Холина, 1959; Страутман, 1963), однако в отдельные годы заканчивается к началу II декады сентября (в* окр. г.Кременец в 1957 г. исчезла после И.IX) (Марисова, Холина, 1959). В Харьковской обл. (6 лет) отлет и пролет белошеек начинается с середины августа и особенно интенсивно проходит в конце III декады этого месяца и в первой половине сентября: 24.VIII, 27.VIII, 27.VIII-15.IX, 27.VIII-23.IX, 1 .IX, 4.IX (Сомов, 1897). В 1889-1890 гг. мухоловки исчезли уже 14 и 17.VIII, а в 1886 г. — последняя птица была отмечена 23.IX. В Полтавской обл. 3 белошейки добыты 25.VIII 1913 г. (колл. ЗИН), 21.VIII и 26.IX 1928 г. (колл. ХГУ), а в Николаевской обл. (окр. г.Очаков) 1 мухоловка -в октябре (колл. ЗИН). В Крыму (4 года) пролетные птицы зарегистрированы 13, 21-22, 24, 28.VIII и 4.IX (Костин, 1983; колл. ЗИН; ХГУ).
В Молдавии отлет и пролет белошеек длится с августа по конец сентября (Ганя, Литвак, 1961; Аверин, Ганя, 1970). Отдельные особи задерживаются на территории республики до середины октября, а в теплые осени — даже до I декады ноября (4—6.XI 1983 г.) (сообщ. Г.З.Гусана).
Практическое значение. Уничтожает многих насекомых — вредите
лей леса и переносчиков возбудителей ряда заболеваний человека и животных. Учитывая возможность быстрого увеличения ее численности во многих типах леса, а также в парковых зонах населенных пунктов путем развешивания искусственных гнездовий, этот вид может быть широко использован в лесном хозяйстве для борьбы с вышеуказанными вредителями, в том числе и в очагах их массового размножения.
Кавказская мухоловка-белошейка — Ficedula albicollis semitorquata (Homeyer, 1885)
Muscicapa semitorquata Homeyer, 1885, Zeitschr. gesamm. Omith., 2: 185, t. 10, Кавказ.
Син. (русск.): закаспийская черная мухоловка, полуошейниковая мухоловка.
Описание. Составлено по 45 экз. (36 - с Кавказа, 3 - из Турции, 6 - из Ирана).
Взрослый самец в отличие от такового номинативного подвида в большинстве случаев (74,1 %) имеет значительно меньшее белое пятно на лбу, реже (18,5 %) оно представлено в виде двух (часто асимметричных) точек, которые у некоторых особей (7,4 %) сливаются между собой. Полная перевязь вокруг шеи отсутствует и белая полоса заходит только на бока шеи, имея полный разрыв в задней ее части. В основании первостепенных маховых белый цвет появляется (п = 20) на наружных опахалах с 3-го пера (100 %), на внутренних - со 2-го или 4-го (со 2-го - 95,0 %; с 4-го — 5,0 %), а по краям хвоста (4—6-я пары рулевых от центра) он развит значительно сильнее (рис. 13). У некоторых особей белые асимметричные пятна несет даже 3-я пара рулевых, а на черных перьях верха тела Имеется значительная примесь буровато-серого. Надхвостье у большинства особей значительно темнее. Индивидуальная и сезонная изменчивость проявляется так же, как и у номинативного подвида.
Взрослая самка такой же окраски, как и у предыдущего подвида, но в области лба у некоторых особей намечается незначительное посветление. Белый или светлоохристый цвет в основании первостепенных маховых появляется (п = 12) на наружных опахалах с 3—5-го пера (с 3-го - 25,0 %; с 4-го — 58,3 %; с 5-го - 16,7 %), а на внутренних — со 2, 3-го или 5-го (со 2-го — 40,0 %; с 3-го — 40,0 %; с 5-го — 20,0 %).
Гнездовой и первый послегнездовой наряды идентичны таковым европейской белошейки. У самцов белый цвет в осно
51
вании маховых появляется (п = 5) на наружных опахалах с 4—5-го пера (с 4-го — 20,0 %; с 5-го - 80,0 %), а на внутренних -со 2-3-го (со 2-го - 40,0 %; с 3-го - 60,0 %).
Размеры отдельных частей тела кавказской мухоловки-белошейки приведены в табл. 7.
Масса тела, г: самец первогодок (21.IV. Краснодарский край, окр. пос. Никель) — 13,28; взрослый самец (21.IV., там же) — 14,40; взрослая самка (12.VII., окр. г.Крас-нодар) — 13,90.
Распространение. Азиатская часть ареала вида и южная половина Балканского п-ова. В СССР: Кавказ к северу до северных подножий Скалистого хребта и г.Краснодар, к югу до государственной границы. Зарегистрирована в весенний период в Крыму и Юго-Западной Туркмении, но гнездование в этих местах не доказано. Известен залет в окр. г.Оренбург (колл.
ЗИН). На всем ареале птицы этого подвида являются перелетными и зимуют главным образом в Восточной Африке. Более детально распространение кавказской мухоловки-белошейки на территории СССР показано на карте (рис. 16).
Кадастр к рис. 16.
1 - г.Краснодар и его окрестности (колл. ИЗАНУ; наши данные); пойменный лес Кут на берегу р.Кубань (наши данные); 2 - Адыгейская АО, окр. аула Октябрьский (колл. РГУ); 3 - окр. г.Горячий Ключ (колл. РГУ; наши данные) ; 4 - окр. стан. Крепостная (колл. ХГУ); 5 - окр. стан. Убинская (наши данные); 6 -р.Дюрсо (Пузанов, 1938); 7 - окр. г.Геленд-жик (колл. ХГУ); 8 - с.Михайловский Перевал (колл. РГУ); окр. с.Пшада (Пузанов, 1938); Мезыбь (Пузанов, 1938); 9 - Туапсинский р-н, г.Индюк (колл. ХГУ); 10 -окр. с.Аше (колл. ЗИН); 11 - окр. пос. Лазаревское (колл. БИН СО АН; БПиДВНЦ); 12 - с.Вардане (Кудашев, 1917; колл. ЗИН); 13 - г.Сочи (Иванов, 1976); р.Агура (Кудашев, 1917; колл. ЗИН); 14 - окр. пос. Кудепста
Рис. 16. Распространение кавказской мухоловки-белошейки (Ficedula albicollis semitorquata) на территории СССР:
а - гнездится, б - отмечена в гнездовой период, в - место встречи гибридной особи (F.a.albicollis х х F.a.semitorquata). См. кадастр, стр. 52, 53.
52
(колл. ЗИН); р.Кудепста (Кудашев, 1917);
15 — окр. пос. Адлер, нижнее течение р.Мэым-та (наши'данные); 16 - окр. с.Красная Поляна (Сушкин, 1914; Аверин, Насимович, 1938; колл. ЗИН); с.Эстонка (Кудашев, 1917); 17 -окр. пос. Никель, р.Белая (колл. РГУ; наши данные); ручей Сюг (наши данные); 18 - Кавказский заповедник (Иванов, 1976); Абаго (Аверин, Насимович, 1938); 19 - р.Киша (Аверин, Насимович, 1938); участок между поляной ’’Сенной” и лагерем ’’Туровым” (наши данные);
20 - 16 км шоссе г.Армавир - стан. Отрадная, пойменный лес по берегу р.Уруп (колл. РГУ);
21 - северная часть Тебердинского заповедника (Ткаченко, 1966); 22 - широколиственные высокоствольные леса Кабардино-Балкарии (Чуни-хин, 1962); 23 - окр. г.Орджоникидэе (Беме, 1958); 24 - Дагестан, Дылымский р-н (колл. ХГУ); 25 - дельта р.Самур (Дроздов, 1965);
26 - Абхазия (Nordmann по Сатунину и др., 1913); 27 - Верхняя Аджария (Вильконский, 1897; Иванов, 1976); 28 - северная часть Лих-ского хребта (Жордания, Гогилашвили, 1969);
29 - Сурамский перевал (Иванов, 1976); 30 -Сурамская равнина (Радде, 1884); 31 - долина р.Арагви (Иванов, 1976); 32 - окр. г.Тбилиси (Радде, 1884); 33 - окр. г.Тетри-Цкаро (Цветков, 1901; колл. ЗИН); 34 - окр. г.Ахалкалаки (Чхиквишвили, 1933); 35 - окр. гЛагодехи (колл. ЗИН); 36 - Армения, окр. с.Амасия, ущелье р.Ахурьян (Ляйстер, Соснин, 1942);
37 — Дилижанский заповедник, ущелье р.Агстев (Адамян, Абрамян, 1973; Адамян, 1975); 38 -окр. с.Дилижан (колл. ЗИН; В.ВЛеоновича);
39 — долина р.Мармарик, склоны хр.Цахкуняц до 1980 м (Даль, 1954); 40 - сДрзакенд и его окрестности (Ляйстер, Соснии, 1942); 41 -окр. с.Элар (Ляйстер, Соснин, 1942); 42 - окр. с.Паракар; с.Зейва (Ляйстер, Соснин, 1942);
43 — с-Инаклу; с.Казнафар (Ляйстер, Соснин, 1942); 44 - склоны г.Арагац до 1700 м (Даль, 1954; Иванов, 1976); 45 - Баргушатский хр. (Даль, 1954); 46 - Айоцдзорский хр. (Даль, 1954); 47 - окр. г.Ордубад (Иванов, 1976); с.Аза (Ляйстер, Соснин, 1942); 48 - лесная зона Главного хребта Малого Кавказа (в границах Азербайджана) (Гамбаров, 1941); 49 - Лерик-ский р-н (Иванов, 1976); окр. сЛерик (Сату-нин, 1907); окр. с.Визазамин (колл. ИЗАНУ);
50 - Талышские горы (Гамбаров, 1941).
Места обитания. Населяет различные типы лиственных, реже смешанных хвойно-лиственных и хвойных лесов в предгорьях и по склонам гор, поднимаясь вверх до 2000 м н.у.м. На равнинах поселяется в пойменных лиственных лесах, как правило, недалеко от предгорной зоны. Встречается в старых рощах, садах, парках населенных пунктов и их окрестностей.
У северной границы ареала, по нашим данным, населяет островные старые лиственные леса поймы р.Кубань и ее притоков, а также зарегистрирована в парках, рощах и садах г.Краснодар и его окрестностей. В предгорной зоне Краснодарского края встречается в широколиственных лесах,
но отмечена на гнездовании и в дубовых с примесью сосны в окр. г.Горячий Ключ. В Кавказском заповеднике и его окрестностях гнездится в широколиственных горных лесах, в том числе и по долинам рек (Белая, Киша), изредка поднимаясь в зону пихтовых лесов (Аверин, Насимович, 1938; наши данные). Охотно заселяет здесь искусственные гнездовья (дуплянки) . В северной части Тебердинского заповедника распространена как в широколиственных, так и в буково-мелколистных лесах подножий горных склонов (Ткаченко, 1966), а в Кабардино-Балкарии населяет горные высокоствольные леса (в основном буковые) на высотах 500—1600 м (Чунихин, 1962).
На Малом Кавказе в Грузии встречается главным образом в лиственных и реже в хвойных лесах и рощах (Жордания, 1962). В Азербайджане на Главном хребте Малого Кавказа и в Талышских горах зарегистрирована в лесах с редким подлеском и в садах (Гамбаров, 1941), а в дельте р.Самур - в низовом лиственном лесу с зарастающими вырубками (Дроздов, 1965).
В Армении обитает в садах и парках населенных пунктов и их окрестностей, галерейных лесах южной части республики, а также в редколесье северных лесных районов, поднимаясь в горы до 1980 м (Ляйстер, Соснин, 1942; Даль, 1954). Обычна в дубовых и грабовых лесах по ущельям. Изредка встречается в сосняках (Благосклонов, 1954). В Дилижанском заповеднике кроме этого гнездится в лесах из дуба, ясеня, граба, бука, березы (Адамян, 1975) и плодовом саду из яблонь и груш, охотно заселяя искусственные гнездовья (Адамян, Абрамян, 1973).
Численность. В большинстве мест гнездового ареала на территории СССР редка и распространена спорадически. В Майкопском р-не Адыгейской АО у пос.Никель и в Дилижанском заповеднике, где привлекается искусственными гнездовьями — обычна.
Прилет и весенний пролет на территории СССР. Проходит с середины III декады марта по конец апреля включительно. В Крыму птицы этого подвида отмечены в мае.
В окр. г.Ереван (2 года) первые птицы появляются 25.III и 27.IV (Адамян, 1974). В ущ. р.Занга (теперь Раздан) 2 самки добыты 6.IV 1928 г. (Ляйстер, Соснин, 1942). В Дилижанском заповеднике (3 го
53
да) прилет зарегистрирован 8, 10 и 13.IV (Адамян, 1975). В Закатальском округе Закавказья пролетная мухоловка отмечена 10.IV 1899 г. (Серебровский, 1925). В Южном Азербайджане самец добыт 24.III 1880 г., а на о-ве Сара в Каспийском море 2 птицы —4 и 8.IV 1937 г. (колл. ЗИН). В Азербайджане пролет длится по II декаду апреля включительно (Гамбаров, 1941).
В Грузии прилет и пролет кавказской белошейки проходит в эти же сроки. На Лихском хр. появляется в конце марта (Жордания, Гогилашвили, 1969). В Лаго-дехском заповеднике добыта 11.IV 1955 г., в окр. г.Тбилиси - 15.IV 1910 г. (колл. ЗИН), а в уреме р.Арагви близ г.Мцхета пролетные птицы встречены 14—17.IV 1910 г. (Баньковский,1913).
В центральной части Северного Кавказа (10 лет) прилет зарегистрирован в I декаде апреля: 3-6, 8 и 10.IV (Моламу-сов, 1967). В окр. г.Орджоникидзе мухоловки добыты (3 года): 6-7.IV (Беме, 1958) и 10.IV (колл. БПиДВНЦ).
В Кавказском заповеднике белошейка появляется в начале апреля (Аверин, Наси-мович, 1938). В Майкопском р-не Адыгейской АО у пос.Никель несколько птиц добыто 7.IV 1977 г. и 27.IV 1975 г. (колл. РГУ), а в Сочинском р-не Краснодарского края — 23 и 30.IV 1914 г. (колл. ЗИН).
В Крыму отмечена на г.Опук (Иванов, 1976), a 8.V 1966 г. два самца добыты у перевала Кебит-Богаз (Костин, 1983). Кроме этого одиночные взрослые самцы предположительно кавказской белошейки встречены Ю.В.Костиным (1983) 9.V 1963 г. в верховьях Сухой Альмы и 30.V 1964 г. -на склоне горы Большая Чучель.
В Юго-Западной Туркмении 2 самца в 1911 г. добыты 29.111 в окр. с.Багир и ЗО.Ш около с.Махтум-кала (Зарудный, Билькевич, 1918а). В 1942 г. в Кызыл-Ат-реке на территории питомника станции субтропических растений 31. III отмечены 2 стаи этих птиц (более 100 и около 30 особей) , a 2.IV была встречена еще одна большая стая мухоловок, пролетевшая в юго-западном направлении (Дементьев и др., 1955). В Гасан-Кули птицы зарегистрированы 5 и 9.IV 1939 г., у Чикишляра 2 особи добыты в начале второй половины апреля 1938 г. (Исаков, Воробьев, 1940), а на хр.Кюрендаг у с.Даната одиночный самец — 27.IV 1982 г. (Мищенко, 1983). У Беумба-ша встречена в мае (Radde, Walter, 1889).
Залетный взрослый самец этого подвида добыт 2.IV 1882 г. у г.Оренбург (колл. ЗИН).
54
Размножение. В основном проходит так же, как и у номинативного подвида. Как и европейская белошейка, кавказская гнездится исключительно в дуплах стволов и ветвей деревьев, а также в искусственных гнездовьях. В местах плотных поселений гнезда соседних пар располагаются на расстоянии 50—150 м, а при гнездовании в дуплянках это расстояние может быть еще меньше.
В качестве строительных материалов для гнезда птицы используют пучочки мхов, листья деревьев и кустарников, их черешки, а также стебли, листья и тончайшие корешки трав. Лоток выстилается лубяными волокнами, сухими стебельками, листьями и корешками трав (главным образом дикорастущих злаков), реже волосками шерсти (олень, кабан), конским волосом, отдельными перьями и пучочками ваты. Строительство гнезда длится 5-16 дней. Размеры гнезд (п = 2), мм: диаметр 72-80, в среднем - 76; высота 53, диаметр лотка - 60; глубина лотка - 40.
Откладка первого яйца производится самкой обычно через 1—2 дня после окончания гнездостроеиия. В сезон размножения кавказская белошейка имеет один репродуктивный цикл. Количество яиц в полной кладке 4—7, чаще 5-6. Кладку из 3 яиц, найденную М.С.Адамяном (1975), очевидно, следует относить к повторной взамен утраченной. Яйца такой же однотонной окраски, как и у номинативного подвида, но голубой цвет более насыщен, с синим оттенком. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4, 5.
Птенцы вылупляются на 13—14 день насиживания (Адамян, 1975).
Сезон размножения на Кавказе длится со II декады апреля до середины июля включительно.
На территории Армении в Дилижан-ском заповеднике (Адамян, 1975) начало откладки яиц в 1969 г. отмечено 12-21.V (10 гнезд), а в 1970 г. - 19.IV-23.V (16 гнезд) и 13.VI (1 гнездо). Полная не-насиженная кладка из 5 яиц найдена у с.Дилижан 13.V 1968 г. (колл. В.ВЛеоновича). Молодые птицы добыты в окр. с.Инаклу 18.VII 1924 г. и у с.Казнафар - 24.VII 1924 г. (Ляйстер, Соснин, 1942).
В Грузии на Лихском хребте откладка яиц начинается в конце апреля (Жордания, Гогилашвили, 1969).
В Краснодарском крае у пос. Никель (Майкопский р-н) (сообщ. Б.А.Казакова и Н.Х.Ломадзе) брачные игры у кавказ
ской белошейки отмечены 4—5 .IV 1977 г. Строительство гнезд (2 года) у большинства пар проходит во П-Ш декадах апреля. Полные кладки разной степени насижен-ности (3 года) встречаются 20.IV—18.V. Вылупление птенцов в большинстве гнезд происходит в I-II декадах мая, а выводки перепархивающих слетков (3 года) встречены 6—13 и 16.VI. Летная молодая бело-шейка добыта здесь 26.VI 1974 г. (колл. РГУ). В окр. г.Горячий Ключ 2 слетка отмечены В.С.Очаповским 4.VI 1964 г. В Сочинском р-не у с.Вардане молодая птица добыта 18.VI 1913 г. (Кудашев, 1917; колл. ЗИН), а в окр. г.Геленджик - 26.VI 1960 г. (колл. ХГУ).
В окр. г.Орджоникидзе на склоне г.Ил пара мухоловок, носивших корм птенцам, встречена 13.VI 1947 г. (Беме, 1958), а у с.Аргудан 2 слетка отмечены 15.VI 1960 г. (Моламусов, 1967).
Линька. Изучена недостаточно. Проходит в основном так же, как и у номинативного подвида.
Питание. Основу пищи кавказской мухоловки-белошейки составляют насекомые разных групп и стадий развития. Изредка поедает паукообразных. Способы сбора корма такие же, как и у номинативного подвида. В выкармливании птенцов принимают участие оба партнера пары.
Отлет и осенний пролет на территории СССР. Материалов недостаточно. Проходит в августе-сентябре.
В центральной части Северного Кавказа (8 лет) отлетает 4, 10, 15, 19, 20, 22, 25.IX (Моламусов, 1967).
Практическое значение. Уничтожает некоторых насекомых — вредителей леса и садовых культур.
Даурская желтоспинная мухоловка — Ficedula zanthopygia (Hay, 1845)
Muscicapa zanthopygia Hay, 1845, Madras Journ. Lit. Sci., 13, pt. 2: 162, Малакка.
Син. (русск.): белобровая мухоловка, желтая мухоловка, желтоспинная мухоловка, белобровая желтобрюхая мухоловка.
Описание. Составлено по 330 экз. (322 — из Амурской обл., Хабаровского и Приморского краев, 3 — из Читинской обл., 5 - из Китая).
Вершину крыла образуют 3-е и 4-е первостепенные маховые (3 < 4 — 4,3 %; 3=4 — 33,4; 3 > 4 — 62,3 %); 5-е немного их короче. Первое первостепенное маховое очень мало и только на несколько миллиметров выступает за вершины кроющих
кисти (табл. 8); 2-е всегда меньше 5-го и больше 7-го (2 < 6 — 19,1 %; 2 = 6 — 29,1; 2 > 6 - 51,8 %). Наружные опахала 3-го -5-го первостепенных маховых в вершинной части заметно сужены. Количество щетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3-5 (3 -4,9 %; 4 — 88,5; 5 — 6,6 %). Длина наибольшей (п = 186): 5,3—8,9 мм (в среднем — 7,1). Количество щитков подотеки на передней части цевки варьирует от 3 до 6. У взрослых особей вершина хвоста имеет прямой обрез или небольшую выемку в центре. Половой диморфизм выражен в окраске оперения и рамфотеки.
У взрослого самца в свежем пере горло, зоб и верхняя часть груди от ярко-зо-лотисто-желтого до оранжево-желтого. Нижняя часть груди, бока тела и верхняя часть брюшка ярко-желтые. Нижняя часть брюшка и нижние кроющие хвоста белые. От лба и далее над глазом к затылку тянется широкая белая бровь. У некоторых особей (20,0 %) на ней имеются очень редкие, неясные светло-желтые ’’искорки”, а у 4,5 % особей — асимметричная примесь желтого, охватывающая 5—10 % поверхности всей брови. Последнее мы объясняем проявлением возрастной и индивидуальной изменчивости. Просмотрев большую серию самцов-первогодков в весеннем наряде, нами не обнаружено ни одного экземпляра с примесью желтой окраски (5-10 %) на бровях, а особи с незначительно выраженными ’’искорками” составили только 3,4 %. Верхняя часть и бока головы, зашеек, спина, верхние кроющие хвоста и рулевые — блестяще-бархатно-черные. У некоторых особей в основании последних расположены белые пятна разной степени выраженности (табл. 8). Нижняя часть спины и надхвостье ярко-золотисто-желтые. Нижние кроющие крыла белые. Маховые и большинство верхних кроющих крыла черные. На верхней части крыла имеется белое ’’зеркальце”, образованное белыми проксимальными партиями больших, средних и малых верхних кроющих второстепенных маховых, а также широкими белыми каймами на наружных опахалах третьестепенных маховых и пятнами на проксимальных второстепенных маховых. Очень редко у взрослых самцов небольшие белые пятна находятся и в основаниях некоторых первостепенных маховых. Клюв черный. Ноги голубовато-серые. Радужина глаза темно-коричневая. Индивидуальная изменчивость
55
Таблица 8. Размеры (---) и коэффициенты вариации (CV,%) желтоспинных мухоловок и наличие hm
Вид, подвид, место сбора л Длина, мм
Крыло CV Расстояние, на которое 1 маховое выступает за вершины кроющих кисти CV
Самцы
F.narcissina narcissina,
Кунашир, Сахалин 22 76,13± 0,54
и Японские острова 70,5 -79,5
F.zanthopygia,
Среднее Приамурье 12 70,21 ± 0,34 68,8 -72,5
Нижнее Приамурье 47 69,93± 0,19 67,5 -72,4
Приморье 22 69,92± 0,23 67,7 -72,1
3,23 2,98t0,37 0,5 -6,4 56,04
1,62 4,79±0,31 3,0 -6,9 22,15
1,80 4,18±0,18 1,2 -6,9 29,00
1,57 4,53±0,29 2,4 -7,3 29,89
F.narcissina narcissina, Куиашир, Сахалин и Японские острова 16 78,09± 0,52 74,4 -81,5
F.zanthopygia, Среднее Приамурье 18 71,77± 0,28 69,8 -74,3
Нижнее Приамурье 77 71,7О± 0,15 68,5 -75,4
Приморье 36 71,23± 0,27 68,3 -74,4
F.narcissina narcissina,
Кунашир, Сахалин и Японские острова 15 73,84± 0,51 69,7 -77,2
F.zanthopygia, Среднее Приамурье 6 68,27± 0,77 65,7 -70,8
Нижнее Приамурье 44 68,13± 0,23 65,3 -71,6
Приморье 13 67,23± 0,49 64,8 -70,2
Самцы взрослые
2,59 2,77±0,41 0,4 -6,3 55,71
1,68 3,93±0,34 1,4 -7,1 36,37
1,85 3,54±0,14 1,4 -6,6 35,06
2,26 3,44±0,19 0,8 -5,5 33,54
Сам
2,57 4,19±0,28 2,3 -5,7 25,88
2,75 4,00±0,31 2,7 -5,0 19,10
2,20 3,73±О,21 0,0 -6,8 36,70
2,51 3,80±0,18 2,8 -5,2 16,88
проявляется в окраске брови, незначительном варьировании величины белого ’’зеркальца”, наличии или отсутствии белых пятен в основаниях рулевых и первостепенных маховых, а также черных поперечных штрихов (вершинные каемки) на некоторых желтых перьях надхвостья. Сезонная изменчивость проявляется в частичном исчезновении блеска и в побурении черных перьев в результате их обноса и выцветания.
Взрослая самка сверху оливково-серая с примесью зеленого на нижней части спины. Надхвостье светло-желтое. Бока головы и
шеи светло-оливково-серые. Оперение нижней части тела бледно-желтовато-лимонно-белое со слабым серым чешуйчатым рисунком на горле, зобе и груди. Рулевые, маховые и большинство верхних кроющих крыла темно-бурые. На верхней части крыла имеется белое ’’зеркальце”, но размеры его меньше, чем у самцов. Клюв черно-бурый.
Молодые в гнездовом наряде сверху коричнево-бурые, с мелкими охристыми пятнами на вершинах отдельных перьев и слабо выраженной темно-бурой окантовкой. Надхвостье светло-желтое. Рулевые,
56
у них белых пятен в основании рулевых
Длина, мм
Хвост CV Клюв от оперения лба CV Клюв от орального края ноздри CV Цевка CV
первогодки
50.351 0,41 46,5 -53,1 3,74 10,201 0,12 9,2 -11,4 5,47 7,2710,07 6,8 -8,0 4,46 16,331 0,10 13,4 —17,3 2,78
47,491 0,45 3,27 10,191 0,18 5,86 7,5410,10 4,61 16,431 0,20 3,99
44,9 -50,1 8,9 -10,9 6,8 - 8,0 14,8 -17,1
47,771 0,23 3,16 10,121 0,08 5,09 7,6110,05 4,61 16,501 0,07 2,94
44,9 -50,9 8,7 -11,4 d,9 -8,3 15,3 -17,3
46,041 0,34 42,4 -48,4 3,36 10,121 0,08 9,2 -10,7 3,68 7,3510,07 6,7 -7,9 4,32 16,471 0,09 13'4 -17,1 2,58
(старше одного года )
51,641 0,48 3,73 10,911 0,17 6,19 7,6610,08 4,29 16,241 0,10 2,46
49,0 -55,6 10,1 -12,0 7,3 -8,3 15,3 —16,8
48,491 0,28 2,47 10,341 0,16 6,19 7,5610,11 6,14 16.641 0,15 3,77
46,8 -50,9 9,5 -11,6 6,9 -8,6 15,6 -18,1
48,471 0,17 3,06 10,341 0,05 ' 4,26 7,7510,04 4,03 16,6410,06 3,28
44,3 -52,4 9,3 —11,3 1,2 -8,4 15,0 -18,1
46,851 0,25 3,14 10,241 0,10 5,59 7,4410,06 4,69 16.351 0,07 2,73
44,0 -50,44 8,8 -11,3 6,7 -8,3 15,3 -17,4
ки
46,671 0,27 2,25 10,751 0,09 3,17 7,4810,10 4,95 16,421 0,08 1,67
45,0 -47,4 10,2 -11,3 6,8 -8,1 15,8 -16,8
45,581 0,62 3,06 10,231 0,14 3,37 7,3510,15 4,92 16,621 0,25 3,39
44,6 -48,0 9,9 -10,9 6,9 -7,7 15,8 -17,1
45.4910.21 3,05 3,92 7,5310,05 4,25 16.501 0,07 2,90
41,6 -47,9 9,2 -10,9 6,9 -8,2 15,4 -17,5
44,111 0,39 3,04 10,451 0,14 4,42 7,3910,13 5,82 16,2710,18 3,87
41,7 -46,2 9,8 -11,2 6,8 -8,1 15,1 -17,2
маховые, верхние кроющие хвоста и крыла маховых несут охристые каемки. Раз-
темно-бурые с коричнево-охристыми кан- меры отдельных частей тела даурской
тами по краям наружных опахал маховых желтоспинной мухоловки приведены в
и с каемками, а также вершинными пятна- табл. 8.
ми (такого же цвета) на верхних кроющих Масса тела, г: слетки (п = 2) (27.VI
крыла. Оперение нижней части и боков и 1.VII, окр. Уссурийского заповедника) -
тела белое со слабым темно-бурым чешуй- 11,40-11,88, в среднем - 11,64; взрослые
чатым рисунком. самцы (п = 11) (17.VI—1.VII, Южное При-
Первый послегнездовой наряд иденти- морье) — 11,62—14,32, в среднем - 12,62;
чен наряду взрослой самки, но белое ”зер- взрослые самцы (г = 27) (Нижнее При-
кальце” на крыле' отсутствует, общий тон амурье) - 12 ,20—14,00, в среднем - 13,20
окраски оперения сверху более коричнево- (Кистяковский, Смогоржевский, 1973);
оливковый, а вершины больших и сред- взрослая самка (27.VI, окр.Уссурийского
них верхних кроющих второстепенных заповедника) — 12,16; взрослые самки
57
Окончание табл. 8
Вид, подвид, место сбора п Ширина, мм
Крыло CV Клюв у орального края ноздри CV Клюв у основания CV
Самцы
F.narcissina narcissina, Кунашир, Сахалин и Японские острова 22 54,72± 0,40 51,5 -57,9 3,23 3,8210,05 3,5 -4,3 5,65 5,7510,08. 5,1 -6,6 6,37
F.zanthopygia, Среднее Приамурье 12 50,991 0,45 48,5 -52,9 2,90 3,9710,09 3,4 -4,3 7,22 5,6410,09 5,2 -6,1 5,28
Нижнее Приамурье 47 51,301 0,19 48,8 -53,7 2,45 4,0910,04 3,6 -4,7 6,21 5,6910,04 5,1 -6,4 4,99
Приморье 22 50,561 0,31 47,6 -53,7 2,88 4,1510,07 3,5 -4,7 8,09 5,9310,08 5,1 -6,8 6,40
Самцы взрослые
F.narcissina narcissina, Кунашир, Сахалин и Японские острова 16 55,651 0,63 49,0 -58,0 4,38 3,8810,08 3,3 -4,3 7,74 5,7810,10 5,0 -6,4’ 6,85
F.zanthopygia, Среднее Приамурье 18 51.981 0,26 49,4 -53,8 2,10 4,0810,08 3,5 -4,6 7,73 5,7210,07 5,2 -6,2 5,03
Нижнее Приамурье 77 52,161 0,12 49,6 -55,4 2,08 4,0410,03 3,3 -4,6 5,43 5,7610,04 5,0 -6,6 5,53
Приморье 36 51,261 0,25 48,6 -54,7 2,76 4,0310,05 3,1 -4,6 7,68 5,7810,05 4,8 -6,3 5,67
Сам
F.narcissina narcissina, Кунашир, Сахалин и Японские острова 15 52,891 0,43 50,5 -56,3 2,79 4,0910,09 3,4 -4,8 8,07 6,0610,16 5,5 -7,7 9,92
F.zanthopygia, Среднее Приамурье 6 49,831 0,59 47,7 -51,6 2,92 4,2010,07 4,1 -4,5 3,98 6,1510,09 5,8 -6,4 3,53
Нижнее Приамурье 44 49,921 0,19 47,5 -53,0 2,56 4,0910,04 3,6 -4,7 5,86 6,0210,06 5,3 -6,9 6,36
Приморье 13 49,55+ 0,26 48,4 -51,1 1,86 4,1710,08 3,7 -4,5 6,53 5,8510,07 5,4 -6,1 4,00
* Цифра перед скобками - количество особей из выборки, имеющих в основании рулевых белые
(п = 21) (Нижнее Приамурье) — 12,20— 15,90, в среднем - 13,9 (Кистяковский, Смогоржевский, 1973).
Распространение. Восточная Монголия, Северо-Восточный и Центральный Китай (к югу до нижнего течения и устья р.Янцзы), Корейский п-ов. На территории СССР гнездится от долины р.Ар-гунь на восток до устья р.Амур, от подножий юго-восточной части хр.Тукурингра, верхнего течения р.Селемджа и оз.Орель на юг, включая Среднее и Нижнее Приамурье, Приморье, а также о-ва Аскольд
58
и Римского-Корсакова. На всем гнездовом ареале птицы этого вида являются перелетными и мигрируют к местам зимовок на п-ов Малакка и о-ва Андаманские, Суматра и Ява через Приморье, Маньчжурию, Корею, о.Чечжудо, Юго-Восточный и Южный Китай и Индокитай. Зарегистрированы залеты на п-ов Сахалин (Vaurie, 1959; Иванов, 1976) и в Японию на о-ва Хонсю и Кюсю (Checklist of Jap. Birds, 1974). Более детально распространение даурской желтоспинной мухоловки на территории СССР показано на карте (рис. 17).
> Высота, мм Отношение ши-рины клюва у основания к его длине от оперения лба, % см Отношение длины хвоста к длине крыла, % CV Отношение ширины крыла к его длине, % CV Белые пятна в основании рулевых
Клюв у основания CV
первогодки
3.7210,05 3,4 -4,4 5,42 56.441 0,86 50,00-66,67 6,85 66.141 0.41 62,67-69,60 2,85 71,771 0,29 69,10-73,33 1,80 Отсутствуют
3.7ОЮ.О5 3,4 -3,9 4,05 55.441 1,12 48,60-62,92 6,72 67.661 0,55 65,09-70,95 2,69 72.881 0.54 70,23-75,18 2,34 2(16,7%)»
3,8610,05 3,4 -4,8 6,99 56.501 0,56 49,12-64,37 6,60 68,311 0,27 64,33-72,51 2,60 73.371 0.23 70,43-76,78 2,09 К 2,3%)
3.91 ±0,06 3,5 -4,4 5,36 58,671 0,81 47,66-65,38 6,50 65,791 0,35 62,05-68,71 2,43 72,311 0,37 67,90-76,04 2,43 Отсутствуют
(старше одного года)
3.7710.04 3,5 -4,1 4,53 5-3.14* 1.23 42,50-59,80 9,27 66.341 0.37 63,54-68,89 2,17 71.251 0,51 65,86-74,26 2,77
3.8Н0.08 3,5 -4,2 5,68 55.481 0,97 50,86-63,16 7,19 67.571 0.26 66,12-69,44 1,61 72.431 0,32 69,43-74,65 1,85 2(11,1 %)
3.7610.03 3,3 -4,1 5,01 55.731 0,40 46,30-63,46 6,21 67.601 0.19 63,39-71,59 2,44 72.761 0.14 69,00-76,47 1,68 4( 5,2%)
3.9910.04 3,5 -4,4 5,36 56.361 0,80 45,71-65,93 8,13 65,771 0,33 62,77-69,59 2,92 71,931 0,25 68,43-74,93 2,02 1( 2,8%)
ки
3,76±О,О7 3,5 -4,1 5,30 56,421 1,79 50,45-73,33 11,44 63J81 0,29 бТД^-65,28 1.73 71.871 0,27 70,66-72,51 1,32 Отсутствуют
4,08±0,09 3,9 -4,3 4,19 60,131 0,91 56,88-62,75 3,69 67,2610,55 65,89-68,87 1,82 73.001 0.30 72,13-74,03 1,00
2.73*0,02 3,6 -3,9 2,48 59.491 0,71 51,38-71,13 7,67 66,751 0,26 63,04 -70,64 2,56 73,291 0,24 70,46-78,64 2,19
3,96±0,10 3,5 -4,3 6,50 56,001 0,64 52,94-61,00 3,78 65,19± 0,42 61,97-66,47 2,11 73,801 0,56 71,37-78,24 2,61 »♦
пятна (разной степени выраженности), цифры в скобках - процент общего количества особей в выборке.
Кадастр к рис. 17.
1 - Приморский край, окр. ст.Хасан: сопка Змеиная у оз.Тальми (Панов, 1973; колл. ИЗАНУ); Голубиный утес (кол. ИЗАНУ); 2 -п-ов Краббе (колл. ИЗАНУ; наши данные); п-ов Новгородский (колл. ЗИН); пгт Ново-киевск в заливе Посьета (колл. ЗИН); территория между ст.Краскино и пгт Посьет (Панов, 1973); 3 - заповедник ’’Кедровая падь” (Назаренко, 1971а; Поливанов, 1981); среднее течение р.Пойма (Адами), уремы рек Барабашевка (Большой Мангугай) и Панчунэ (Панов, 1973); 4 - о-ва Римского Корсакова в заливе Петра Великого (Лабзюк и др., 1971); 5 - о-в Попова (колл. БПиДВНЦ); 6 ' - окр. г.Владивосток (колл. ЗМ МГУ); р.Седанка (колл. ЗИН;
ИЗАНУ); с.Сидеми, окр.Сад-города (колл. ЗИН); 7 - о-в Аскольд (Hartert, 1907; Воробьев, 1954; колл. ЗМ МГУ); 8 - р.Суворовка (Пейшула) (колл. ЗИН); р.Артемовна (Майхе) (колл. ЗМ МГУ); 9 - Уссурийский заповедник имЗЛ.Комарова (Иванов, 1952); 10 - нижиее течение р.Каменка, долина р.Комаровка (Супу-тинка) (Иванов, 1952); окр. пос.Комарово-зало-ведное.окр.пос.Каймановка (колл. ИЗАНУ); 11 -окр. ст.Екатериновка (колл. БПиДВНЦ); 12 -Партизанский р-н: с.Бровничи, с.Хлыстовка (колл. ИЗАНУ); ст.Сица, «.Партизаны, пос. Ручьи (Фанза) (колл. ЗИН); 13 - Лазовский заповедник им.Л.Г.Капланова (Белопольский, 1950; Литвиненко, Шибаев, 1971; Поливанов, Поливанова, 1971) ; пойма р.Тачингоуза, база заповед-
59
ника, подножья сопки Горалья (Белопольский, 1950); 14 - р.Киевка (Судзухе), с.Киевка (Литвиненко, Шибаев, 1971); о-в Петрова (Воробьев. -Цит. по: Белопольский, 1950); 15 - бухта Моряк-Рыболов, с.Маргаритово (колл. ЗИН); 16 - с.Щербаковка (колл. ЗИН); 17 - с.Сива-ковка (колл. ЗИН; ИЗАНУ); 18 - устье р.Илис-тая (Лефу) (колл. ЗИН); 19 - Спасский р-н (Кушнарев, 1984); окр. с.Татьяновка, с.Зеленов-ка, с.Одаровка (Иогансен, 1927); Горный хутор (колл. ИЗАНУ); 20 - верховья р.Уссури, с.Шмаков ка (колл. ИЗАНУ; С.Д.Кустановича); 21 -сопки на р.Большая Уссурка (Иман) в 45-50 км от устья (Спангенберг, 1940; колл. БПиДВНЦ; КБ МГУ; А.П.Кузякина); окр. с.Саровка (Спангенберг, 1940; колл. БИН СО АН); окр. с.Вер-бовка (Спангенберг, 1940; 1965; колл. БИН СО АН; БПиДВНЦ; В.В.Леоновича); окр. пос.Островной (Санчихеза) (Спангенберг, 1965); с.Евсеевка (колл. ЗИН); 22 - широколиственные и смешанные леса островов и сопок на всем протяжении р.Большая Уссурка (Спангенберг, 1965);
23 - р.Черемуховая (Большая Синанча) (колл. ЗИН); 24 - нижнее течение р.Рудная (Теттохе) (Черский, 1915а); 25 - пгт Терней (Благосклонов, 1954; Воробьев, 1954; Иванов, 1976); 26 -Сихотэ-Алинский заповедник (Елсуков, 1982); окр. кардона Усть-Серебряный, правый берег р.Серебрянка (наши данные); пойма р.Серебрян-ка (Санхобэ) (Воробьев, 1954); 27 - р.Бикин (Поливанов, 1981); 28 - низовья р.Г.умин (приток р.Бикии) в 10-15 км от г.Бикин (колл. А.П.Кузякина); 29 - Хабаровский край; пойма р.Аван в окр. с.Венюково (колл. М.И.Лобанов-ского); 30 — окр. пос. Кия в нижнем течении р.Хор (Кисленко, 19656); с.Полетное (колл. ЗМ МГУ; ИЗАНУ); 31 - р.Уссури, 48° с.ш. (колл. ЗИН); 32 - хр.Хехцир (Кулешова и др.,
1965; Казаринов, 1973); окр. с.Корсаково (колл. ЗИН); окр. с.Казакевичево (колл. ЗИН); 33 -с.Троицкое, с.Славянка (колл. ИЗАНУ); 34 -с.Болонь (Кистяковский, Смогоржевский, 1973; колл. КГУ); берег оз.Болонь (колл. ЗМ МГУ); с.Верхний Нерген (колл. КГУ) ; 35 - тальники по берегам р.Харпи (Мищенко, Смиренский, 1981);
36 - с.Омми (колл. КГУ); 37 - окр. г.Комсо-мольска-на-Амуре (Кистяковский, Смогоржевский, 1973; колл. КГУ); с.Хурба (колл. КГУ); 38 - нижнее течение р.Горюн (Воробьев, 1938; 1954; Портенко, 1960); Комсомольский заповедник (Штильмарк, 1973); 39 - уремные леса по р.Гур (Хунгари) (Штильмарк, 1973); 40 - р.Эвур в 50 км от устья (Нечаев, 1963); 41 - оз.Эворон, устье р.Холодной (Нечаев, 1963); 42 - г.Амурск (Кистяковский, Смогоржевский, 1973); 43 -р.Бичи вверх от устья (колл. ЗИН); 44 - с.Ки-селевка (Кистяковский, Смогоржевский, 1973; колл. КГУ); 45 - окр. с.Циммермановки (колл. КГУ); 46 - с.Софийск (колл. КГУ); 47 - окр. с.Мариинское, Щучья гора (колл. ЗМ МГУ);
48 - с.Савинское (колл. КГУ); 49 - с.Нижняя Гавань (колл. КГУ); р.Тучка близ с.Богород-ское (Иванов, 1976; колл. КГУ); 50 ~ с.Суса-нино (Кистяковский, Смогоржевский, 1973; колл. КГУ); 51 - долина р.Амур у 53° с.ш. (Степанян, 1978); 52 - с.Маго (колл. КГУ); 53 -устье р.Амур (Иванов, Штегман, 1978; Степанян, 1978); окр. г.Николаевск-на-Амуре (Кузякин, Второв, 1963; Кистяковский, Смогоржевский, 1973); 54 ~ окр. пос.Орель - Чля; восточный берег оз.Орель (Бабенко, 1984); 55 - пос. Чля (Бабенко, 1984); 56 - Малый Хинган, урочище ’’Тигровая падь” (колл. КГУ); 57 - окр. с.Радде (колл. ЗИН); 58 - окр. с.Пашково (Лавренченко, 1983; 1984а); 59 - Амурская обл.; окр. с.Северное (колл. ЗИН); 60 - р.Бурея в 5 км
Рис. 17. Распространение даурской (Ficedula zanthopygia) (1) и японской (F.narcissina narcissina) (2) желтоспинных мухоловок на территории СССР.
а - гнездятся, б - отмечены в гнездовой период. См. кадастр, стр. 59-61, 68.
60
на юго-запад от с.Украинка; окр. оз .Крив ун (сообщ. С.В.Винтера); 61 — окр. сДжелунда (колл. КГУ); 62 - окр. пгт Экимчан (Назаренко, 1984а); 63 - устье р.Селемджа (Иванов, 1976; колл. КГУ); окр. сЛебедино (колл. КГУ); 64 - окр. с.Климоуцы, окр. с.Зиговка, междуречье рек Малая Пера и Большой Эргель (колл. ЗИН); 65 - устье р.Гулик (сообщ. В-Ю.Ильяшенко); 66 - устье р.Алгая (в настоящий период территория затоплена Зейским водохранилищем) (сообщ. В.Ю.Ильяшенко); 67 -окр. с.Кумара (Благосклонов, 1954; колл.ЗИН);
68 - окр. с.Симоново (колл. ЗИН); 69 - с.Джа-линда (Благосклонов, 1954; Иванов, 1976; колл. ЗИН); 70 - Юго-Восточное Забайкалье (Портенко, 1960); Даурская степь (Гагина, 1960а); о-в Торей-Нор (Дементьев, 1973; Благо
склонов, 1954); с.Кулусутай (Иванов, 1976); сопка Кухадан у о.Зун-Торей (колл. ИЗАНУ); 71 - окр. ст.Мациевская (Иванов, 1976; колл. ЗИН); 72 - р.Аргунь (Дементьев, 1937; Благосклонов, 1954; Степанян, 1978); Приаргунье (Гагина, 1960а).
Систематика. Таксономический статус даурской желтоспинной мухоловки до настоящего времени оставался спорным. Одни исследователи (Bianchi, 1907; Wynne, 1954; Vaurie, 1959; Степанян, 1971; 1978; Иванов, 1976; и др.), считают ее монотипическим видом, в то время как другие (Harter, 1907; Hartert, Steinbacher, 1934; Дементьев, 1937; Благосклонов, 1954;
Таблица 9. Достоверность отличий (t) размеров Ficedula narcissina narcissina от F .zanthopygya
Ficedula zanthopygya Длина
Крыло Хвост Клюв от оперения лба Клюв от орального края ноздри Цевка
Самцы первогодки
Среднее Приамурье 9,28*** 4,70*** 0,05 2,21* 0,45
Нижнее Приамурье 10,83*** 5,49*** 0,55 3,95*** 1,39
Приморье 10,58*** 8,09*** 0,55 0,81 .1,04
Самцы взрослые (старше одного года)
Среднее Приамурье 10,70*** 5,67*** 2,44* 0,74 2,22*
Нижнее Приамурье 11,81*** 6,23*** 3,22** 1,01 3,43***
Приморье 11,71*** 8,85*** 3,40** 2,20* 0,90
Самки
Среднее Приамурье 6,03*** 1,61 3,12** 0,72 0,76
Нижнее Приамурье 10,21*** 3,45** 6,01*** 0,45 0,75
Приморье 9,35*** 5,40*** 1,80 0,55 0,76
Ficedula zanthopygya Ширина Высота Отношение
Крыло Клюв Клюв у ос-нования ширины клюва у основания к его длине от оперения лба длины хвое* та к длине крыла ширины крыла к его длине
у орального края ноздри у основания
Сял<цы первогодки
Среднее Приамурье 6,20*** 1,46 0,91 0,28 0,71 2,22* 1,81
Нижнее Приамурье 7,72*** 4 22*** 0,67 1,98 0,06 4 42*** 4,32***
Приморье 8,22*** 3,84*** 1,59 2,43* 1,89 Самцы взрослые (старше одного года) 0,65 1,15
Среднее Приамурье 5,38*** 1,77 0,49 0,45 1,49 2,72** 1,96
Нижнее'Приамурье 5,44*** 1,87 0,19 0,20 2,90* 3,03** 2,86**
Приморье 6,48*** 1,59 0,00 Самки 3,89*** 2,19* 1,15 1,20
Среднее Приамурье 4,19*** 0,96 0,49 2,81* 1,85 6,56*** 2,80*
Нижнее Приамурье 6,32*** 0,00 0,23 0,41 1,59 g 22*** 3,93***
Приморье 6,65*** 0,66 1,20 1,64 0,22 3 94*** 3,10**
*** Р < 0,001. ** Р < 0,01. * Р < 0,05.
61
Портенко, 1960; Нейфельдт, 1970; Виноградова и др., 1976; Поливанов, 1981; и др.) - рассматривают как материковый подвид японской желтоспинной мухоловки - Ficedula narcissina.
Статистический анализ морфометрических показателей, исследование окраски и рисунка оперения желтоспинных мухоловок, проведенные нами на большом серийном материале, а также изучение экологических особенностей и географического распространения этих форм позволяют говорить об их видовой самостоятельности. Во всех возрастных нарядах, включая и гнездовой, они четко различаются окраской оперения и размерами отдельных частей тела (табл. 8, 9). Ранее одним из основных диагностических признаков между этими формами являлся цвет брови взрослых самцов (белый у даурской и желтый у японской). Этот признак в популяциях даурской желтоспинной мухоловки подвержен, по нашим данным, возрастной и индивидуальной изменчивости (см. описание). В связи с этим указание Е.Н.Панова (1973) на некоторую интерградацию этих форм (по данному признаку) на о-ве Аскольд маловероятно. Взрослые самцы даурской желтоспинной мухоловки с примесью желтого на белых бровях, по нашим данным, характерны не только для птиц с 6-ва Аскольд, но и для материковых популяций, в том числе и сильно удаленных от берега моря.
Исходя из сказанного выше, мы считаем, что даурская и японская желтоспинные мухоловки несомненно уже достигли видовой самостоятельности и должны рассматриваться как монотипический (первая) и политипический (вторая) виды. О правильности наших выводов говорит и карио-анализ, проведенный для них. Так, кариотип Ficedula narcissina равен 2п = 80 (Udagawa, 1954), в то время как у F.zanthopygia - 2п = 76 при наличии ряда различий в морфологии хромосом (Крюков, 1977). Если даже предположить, что Т.Удагава ошибся (кариоанализ проводился им по старым методикам), и принять, что японская желтоспинная мухоловка имеет такой же кариотип (2п = 76), то различия в морфологии хромосом также являются важным критерием видовой самостоятельности этих форм.
Места обитания. Населяет различные типы лиственных и реже смешанных хвойно-лиственных лесов, главным образом по поймам рек, долинам ключей и берегам
озер, придерживаясь низинных участков. В небольшом количестве проникает в горные долинные леса. Избегает сплошных лесных массивов и поселяется в них только по окраинам или не далее 5—7 км от ближайших полян и вырубок. Обитаете рощах, отдельных куртинах лиственного леса среди открытых пространств недалеко от воды и реже в кустарниковых зарослях островов больших рек. В хвойных лесах на территории СССР не встречается.
В Нижнем Приамурье излюбленными биотопами даурской желтоспинной мухоловки являются уремные леса по берегам рек Амур (Кистяковский, Смогоржевский, 1973) и Гур, довольно часто встречается в кустарниковых зарослях по островам Амура в окр. г.Комсомольск (Штильмарк, 1973), в тальниках по берегам р.Хар-пи (Мищенко, Смиренский, 1981), гнездится в прибрежных ивняках нижнего течения р.Эвур и в островных осиновых лесах среди обширных моховых болот в устье р.Холодной (Нечаев, 1963). Встречена на гнездовании в старых ивняках в окр. пос.Орель — Чля, а также отмечена в мелколиственных насаждениях из березы, ольхи и ивы по восточному берегу о.Орель на 15—20 км к северу (Бабенко, 1984). На хр.Хехцир поселяется главным образом в перелесках предгорной зоны (Казаринов, 1973) в разреженных участках леса, на вырубках и в поймах, но гнездится и в широколиственных лесах с примесью кедра (Кулешова и др., 1964). В нижнем течении р.Хор встречается в темнохвой-но-широколиственных лесах (Кисленко, 19656).
В Среднем Приамурье у с.Пашково гнездится в густом пойменном лесу и разреженных ивняках (Лавренченко, 1984а). В нижнем течении рек Аван, Бурея и по берегам оз.Кривун также населяет пойменные ивняки (сообщ.С.В.Винтера; колл. С.Д.Кус-тановича), а в устье р.Гулик (приток р.Зея) встречается в пойменном лесу из ивы и ольхи (сообщ. В.Ю.Ильяшенко). В верхнем течении р.Селемджа у пгт Экимчан отмечена в высокоствольных пойменных чозениево-тополевых лесах (Назаренко, 1984а).
В Приморье по р.Большая Уссурка гнездится в смешанных широколиственных лесах береговых сопок и островов реки, тяготея к ивовым зарослям, но поселяется и в низинных перелесках, окруженных открытыми участками (Спангенберг, 1940; 1965). В Сихотэ-Алинском заповеднике,
62
по нашим данным, обычна в старых ивняках по берегам рек, но найдена на гнездовании и в лиственном лесу с преобладанием березы на берегу р.Серебрянка. В Уссурийском заповеднике им.В.Л.Комарова излюбленными биотопами являются также пойменные широколиственные леса по р.Комаровка, но найдена нами на гнездовании и в разреженных смешанных широколиственных лесах с примесью хвойных деревьев недалеко от воды. В Лазовском заповеднике имЛ.Г.Капланова населяет сходные биотопы, но отмечена также в дубняках (Белопольский, 1950) и в кедрово-широколиственных лесах по долинам ключей (Литвиненко, Шибаев, 1971). В Южном Приморье этот вид является характерным для комплекса галерейных широколиственных лесов речных долин. В липняках распространена только в придолинных участках (Назаренко, 19716). Значительно реже населяет фрагментарные молодые дубняки с полянами, заросшими лещиной и леспедецей, по нижним частям склонов сопок (Панов, 1973) и в небольшом количестве проникает в горные долинные леса в заповеднике ’’Кедровая падь” (Назаренко, 1971а). В куртинных участках лиственного лес? с примесью кустарников среди открытых участков отмечена нами в гнездовой период в Хасанском р-не у подножья сопки ’’Голубиный утес” и на п-ове Краббе по дну распадка к Новгородской бухте (залив Посьета).
На рХалкин-Гол поселяется в ильмовых рощах (Благосклонов, 1954).
Численность. На территории СССР в большинстве мест Приморья, Среднего и Нижнего Приамурья обычна, в излюбленных биотопах - многочисленна. У северо-западной границы гнездового ареала - в долине р.Аргунь - редка. На некоторых участках ареала численность подвержена годовым колебаниям (Белопольский, 1950; Назаренко, 1984а).
Прилет и весенний пролет на территории СССР. Проходит с конца III декады апреля по III декаду мая включительно. Однако есть указания В.К.Ра-хилина (1975) о начале весеннего прилета и пролета в середине апреля.
На острова в северо-западной части залива Петра Великого мухоловки прилетают в конце III декады апреля и в I декаде мая (Лабзюк и др., 1971). На о-ве Путятин взрослый самец добыт А.И.Черским 15.V 1914 г. (колл. ЗИН).
В Южном Приморье, по данным К.А.Во
робьева (1954) (несколько лет), прилет и пролет проходят 10—20.V, а, по данным Е.Н.Панова (1973) (2 года), первые самцы зарегистрированы 8 и 10.V, но пролет в устье р.Илистая еще проходил 29 и 31.V 1962 г. В окр. г.Владивосток самцы были добыты 16.V 1912 г. (Черский, 1915а) и 18.V 1909 г. (колл. ЗИН). В верхнем течении р.Одарка у с.Нововладимировка (южный берег оз. Ханка) взрослый самец в 1911 г. добыт 19.V, а самка — 20.V (Черский, 1915а; 19156). В Лазовский заповедник (4 года) первые самцы прилетают 14, 16, 17 и 18.V (Белопольский, 1950; Литвиненко, Шибаев, 1971). В Партизанском р-не пролетный самец добыт у ст.Си-ца 19.V 1926 г., а самка у ст.Мучная -19.V 1928 г. (колл. ЗИН). В низовьях р.Большая Уссурка (2 года) первые птицы появляются 15 и 18.V (Спангенберг, 1965).
В Приамурье первые мухоловки появляются уже в I декаде мая, но основная масса птиц прилетает в Ш декаде этого месяца. Так, самец в окр. с.Кумары добыт 7.V 1928 г.; в окр. г.Николаевск - 10.V 1921 г., а самка — 19.V 1922 г. (колл. ЗИН). На нижнем Амуре (несколько лет) начало прилета отмечено: у с.Даур - 19.V, у сел Омми, Гавань и г.Комсомольск-на-Амуре — 23.V, в окр. с.Гячи — 28.V (Раш-кевич, 1965; Кистяковский, Смогоржевский, 1973).
В Борзинском р-не Читинской обл. у ст.Мациевская самец добыт 20.V 1939 г. (колл. ЗИН).
Обычно первыми на места гнездования прилегают са^щы и только через несколько дней после них появляются самки.
Размножение. Гнездовые участки самцы занимают еще до прилета самок. Их площадь 1350-5000 м2, в среднем (п = 12) - 2613 (Лавренченко, 1983). В этот период самцы много и охотно поют, часто сидя или перелетая в кронах лиственных деревьев. Песня даурской желтоспинной мухоловки звонка и мелодична. После прилета самок начинается разбивка на пары, сопровождающаяся интенсивным пением самцов и их токовым демонстративным поведением (рис. 22). Сбор самкой строительных материалов для гнезда, спаривание и сбор корма при выкармливании птенцов происходят в пределах гнездового участка (Лавренченко, 1983).
По характеру гнездования даурская желтоспинная мухоловка является пассивным облигатным дуплогнездником. Из 103 гнезд этого вида, найденных в При
63
морье, Среднем и Нижнем Приамурье (Белопольский, 1950; Нечаев, 1963; Росляков, 1965; Литвиненко, Шибаев, 1971; Панов, 1973; Поливанов, 1981; колл. ИЗАНУ; ЗМ МГУ; КБ МГУ; БИН СО АН; БПиДВНЦ; КГУ; В.В.Леоновича; А.П.Ку-зякина; С.Д.Кустановича; М.И.Лобанов-ского; наши данные), 72 были построены в дуплах стволов, сучьев и пней живых, сухих и гниющих деревьев (ива, осина, липа, дуб, бархат, чозения, ясень, береза, ольха, черемуха, жимолость), в том числе и в дуплах дятлов: малого острокрылого (11), малого пестрого (12) и белосПин-ного (1); 12 — в полудуплах стволов, ветвей и пней деревьев (ива, дуб, ольха); 1 - в расщелине ствола ивы; 17 — в искусственных гнездовьях (дуплянки) и 1 — в жестяной консервной банке, подвешенной на дереве на берегу р.Амур. Высота расположения гнезд от поверхности земли 0,3— 7,0 м, в среднем — 2,7. Обычно мухоловки занимают одно и то же гнездо в течение нескольких лет или строят новые гнезда недалеко от прошлогодних (Поливанов, 1981). Размеры гнездовых дупел, мм: диаметр летка 25—70 х 20-85, в среднем — 40 х 46; глубина дупла 75-350, в среднем — 128; внутренний диаметр — 60—75 х х 70-75, в среднем — 67 х 72.
Гнезда птицы чаще всего строят в глубине дупла или полудупла и значительно реже в обрез с нижним краем летка. Е.С .Спангенберг (1965) находил также гнезда в нишах между древесиной и отставшей корой ствола дерева. В трухлявых деревьях мухоловки иногда расширяют и углубляют дупла, выщипывая клювом кусочки мягкой древесины (сообщ. С.В.Винтера) . При строительстве гнезда птицы безразлично относятся к содержимому дупла или дуплянки и если в них есть прошлогодние гнезда птиц или мелких млекопитающих (бурундук), то свое гнездо они выстраивают на них. В Лазовском заповеднике отмечено строительство двух гнезд даурскими желтоспинными мухоловками на гнездах полевого воробья, в которых были яйца второй кладки, и четырех — на гнездах большой синицы, построенных в этом же году (Литвиненко, Шибаев, 1971).
Гнезда представляют собой очень рыхлые постройки неправильной формы. В качестве строительных материалов птицы используют полоски луба (шириной до 9 мм), тонкие веточки и прошлогодние листья древесных и кустарниковых растений, стебельки, листья, соцветия и корешки
трав (главным образом дикорастущих злаков), чешуи цветочных почек, кусочки коры, листья ивы, пучочки мхов (Sanionia uncinata, Hypnum lindbergii), а по данным Н.М.Литвиненко, Ю.В.Шибаева (1971), веточки с колосками метелок тростника и кусочки его листьев. Лоток выстилается тонкими полосками луба, корешками, стебельками (часто рассученными) и листьями трав (большей частью злаков), растительными волокнами, небольшим количеством пучочков мха и очень редко одиночными пуховыми перьями, волосками шерсти крупных млекопитающих, спорогониями бокоплодных мхов, а по данным Л.Тача-новского (Taczanowski, 1981) - и тонкими усиками вьющихся растений. В глубоких дуплах в основание гнезда птицы часто приносят большое количество мха, чем в той или иной степени уменьшают глубину гнездового дупла (Шибнев. - Цит. по: Поливанов, 1981). Гнездо самка строит 2—4 дня, иногда несколько дольше (Поливанов, 1981). Размеры гнезд (п = 18) (Taczanowski, 1891; Нечаев, 1963; наши данные), мм: диаметр 70-115, в среднем -89; высота — 34-75, в среднем - 51; диаметр лотка 45-65, в среднем — 56; глубина лотка 20—50, в среднем -31.
В сезон размножения на территории СССР даурская желтоспинная мухоловка имеет один репродуктивный цикл. Количество яиц в полной кладке 4—8, чаще 5—6. Овальные, укороченно-овальные, укороченно-остроовальные и значительно реже — эллипсовидные и остроовальные яйца имеют следующую окраску: фон от молочно-белого до белого со слабым розоватым или бледно-охристым оттенком. По нему разбросаны мелкие светло-красновато-бурые и светло-лиловые крапинки и точки. Распределение их по поверхности скорлупы различно. На большинстве яиц количество крапинок увеличивается к тупому полюсу, образуя вокруг последнего ’’венчик” или ’’шапочку” (густота рисунка 20—40 %), но встречаются также яйца, на которых рисунок сгущается к экваториальной части яйца, образуя вокруг последней ’’поясок” или образует ’’венчик” вокруг острого полюса. Как исключение встречаются яйца, на которых большая часть крапинок локализуется на скорлупе с одной стороны — в центре яйца в виде пятна (густота рисунка -60 %). Скорлупа матовая или полумато-вая. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
Яйца самка откладывает ежедневно.
64
Как исключение зарегистрирован случай, когда самка за сутки отложила 2 яйца (Литвиненко, Шибаев, 1971). Насиживание начинается чаще всего после откладки последнего яйца, а при больших кладках (7 яиц) и после 5-го (Литвиненко, Шибаев, 1971). Инкубация длится 12-15 дней (Росляков, 1965; Литвиненко, Шибаев, 1971; Поливанов, 1981) . Самец в насиживании кладки участия не принимает. В этот период он держится поблизости от гнездового дерева и много поет. Самка на гнезде очень осторожна и, почувствовав малейшую опасность, сразу его покидает. В светлое время суток она от 1 до 5 раз в час вылетает кормиться, отсутствуя от 2 до 20 мин, в среднем 5—7 мин (Поливанов, 1981). В очень редких случаях самец подкармливает самку.
При вторжении на гнездовой участок близко к гнезду других птиц (малый пестрый дятел, Панов, 1973; 3-длиннохвостых синиц, наши данные) самец агрессивно с шипением атакует их и отгоняет. Самка в это время слетает с гнезда. Эта же участь постигает и других самцов, подлетевших близко к гнезду.
Вылупление птенцов обычно проходит 1—2 дня.
Пуховой птенец даурской желтоспинной мухоловки имеет негустой светлосерый пух (длина 5—6 мм) на надглазничных, затылочной, спинной, плечевых и локтевых птерилиях. На последних пушинки рудиментарны и у некоторых особей отсутствуют. Ротовая полость и язык — желтые, клювные валики - бледно-желтоватые (Нейфельд, 1970).
В выкармливании птенцов принимают участие оба партнера пары, но в первые дни после вылупления корм чаще приносит самец, а самка большее время проводит в гнезде, обогревая птенцов. Растут птенцы быстро и на 12-1441 день покидают гнездо. Родители продолжают кормить слетков, легко отыскивая их в зарослях по характерному писку. По данным Л.А.Лавренченко (1983), характерной чертой послегнездовой жизни даурских желтоспинных мухоловок является разделение выводка на две части, каждая из которых сопровождается одним из партнеров. Эти группы перемещаются независимо и в дальнейшем не объединяются как друг с другом, так и с другими выводками. В это время птицы привязаны к местам гнездования. Максимальное расстояние, на которое удаляется выводок от гнезда, — 700 м.
Основными причинами отхода яиц и
птенцов у этого вида являются хищническая деятельность со стороны некоторых видов рептилий (амурский полоз - Elaphe schrencki), птиц (голубая сорока и некоторые др.) и млекопитающих, неблагоприятные погодные условия и наличие в кладках неоплодотворенных яиц.
Сезон размножения даурской желтоспинной мухоловки на территории СССР длится со II декады мая по конец июля.
В Южном Приморье начало весеннего пения самцов в 1962 г. отмечено 10.V, 16.V отмечена первая пара, которая 21.V занималась выбором гнездового дупла, а 5 .VI найдено гнездо, в котором самка насиживала полную кладку (Панов, 1973). В окр. г .Владивосток гнездо с полной свежей кладкой из 6 яиц обнаружено 8.VI 1967 г. (колл. ЗМ МГУ). Самка, строящая гнездо, встречена 9.VI 1962 г., три гнезда с полными кладками найдены в уремах рек Бараба-шевка (Мангугай) и Панчунэ 9—13.VI 1960 г., почти достроенное гнездо в среднем течении р.Пойма (Адими) отмечено 14.VI, пара, выкармливающая птенцов, зарегистрирована 26.VI 1963 г., а выводок хорошо летающих молодых обнаружен 4.VII (Панов, 1973). В Лазовском заповеднике начало весеннего токования в 1945 г. отмечено 16.V (Белопольский, 1950), а начало откладки яиц в р-не Киевки (2 года) зарегистрировано 25, 27, 28, 30.V и 12 и 18.VI (Литвиненко, Шибаев, 1971). Гнездо с полной кладкой найдено в заповеднике 21.VI 1945 г. (Белопольский, 1950). В Партизанском р-не гнездо с 4 свежими яйцами обнаружено 12.VI 1930 г. (Благосклонов, 1954), а в окр. с.Екатериновка слабо насиженная кладка из 5 яиц встречена 25 .VI 1975 г. (колл. БП и ДВНЦ). В Ольгинском р-не у с.Щербаковка молодые птицы добыты 6 и 18.VII 1927 г. (колл. ЗИН). В окр. первого кордона Уссурийского заповедника 2 слетка добыты нами 27.VI и 1 .VII 1977 г.
На р.Уссури у с.Шмаковка 8.VI 1964 г. найдены 4 гнезда: 1 - законченная постройка (кладка не началась), в 1 - полная свежая и 2 - насиженные кладки (колл. ИЗАНУ; С.Д.Кустановича). Здесь же 12.VI 1964 г. обнаружено гнездо с 4 оперившимися птенцами, 16.VI — с 3 оперившимися птенцами и 2 яйцами (болтуны), a 18.VI — с полной сильно насиженной кладкой (материалы С.Д.Кустановича).
В Сихотэ-Алинском заповеднике (3 года) в долине р.Серебрянка пустое гнездо найдено 26.V, с полной свёжей кладкой из
65
7 яиц - 5.VI, с незаконченной кладкой из 3 яиц - 10.VI, строящаяся постройка и гнездо с 6 птенцами — 27 .VI, 2 гнезда с 4 и 7 птенцами (стадия раскрытия опахал) -30.VI и 7.VII. В окр. пгт Терней 2 гнезда с полными кладками отмечены 15.VI, а в окр. кордона Усть-Серебряный 16.VI зарегистрирована пара у гнезда (картотека заповедника; наши данные).
На р.Большая Уссурка в окр. пос.Ост-ровной (Санчихеза) и с.Вербовка 2 гнезда с полными свежими кладками найдены 9 и 11.VI 1950 г. (Спангенберг, 1965; колл. КБ МГУ). Здесь же в окр. с.Саровка 11.VI 1954 г. встречено гнездо с 4 ненасиженными яйцами (колл. БИН СО АН), а в окр. с.Вер-бовка 17.VI 1953 г. — с полной кладкой средней насиженности (колл. БИН СО АН), 21 .VI - с 4 оперившимися птенцами, а 27.VI поймана молодая летная птица (Спангенберг, 1965). В сопках в 45 км южнее устья р.Большая Уссурка 16.VI 1953 г. найдено гнездо с полной кладкой из 6 яиц, а 21 .VI -с сильно насиженной кладкой из 5 яиц (колл. БП и ДВНЦ; А.П.Кузякина).
На р.Бикин (Поливанов, 1981) самое раннее начало откладки яиц зарегистрировано 21.V, массовая откладка яиц проходит 25.V—5.VI. В наиболее поздних гнездах первые яйца обнаружены 26.VI. На р.Гумин (в 10-15 км от г.Бикин) полная кладка средней насиженности из 6 яиц отмечена 7.VI 1953 г. (колл. А.П.Кузякина).
В Вяземском р-не Хабаровского края у с.Венюково гнездо с полной кладкой средней насиженности обнаружено 12.VI 1963 г. (колл. М.И.Лобановского). В Нижнем Приамурье откладка яиц зарегистрирована: 1.VI - с.Болонь, 4.VI — с.Киселевка и 10.VI — с.Софийск, а полные кладки разной насиженности отмечены (3 года): 9.VI — окр. г.Амурск, 13.VI — окр. г.Комсомольск, 22.VI - устье р.Холодная, 25 и 26.VI - окр. с.Сусанино (Нечаев, 1963; Кистяковский, Смогоржевский, 1973; колл. КГУ). Молодые птицы на нижнем Амуре встречены 7—18.VII (Рашкевич, 1965).
В Свободненском р-не Амурской обл. гнездо с пуховыми птенцами найдено 24.VI 1961 г. (колл. ЗИН). В низовьях р.Бурея (окр. с.Северное) 28.VI 1975 г. встречены 2 слетка, a 29.VII 1974 г. добыта летная мог-_ лодая птица (колл. ЗИН). Вокр. с.Зиговка летные молодые добыты 19.VII 1957 г. и 10.VIII 1958 г., а в окр.с.Симоново (Шиманский р-н) - 4.VIII 1959 г. (колл. ЗИН). У устья р.Гулик (приток р.Зея) самка, кормившая 2 молодых, отмечена 1.VII 1982 г. (сообщ. В.Ю.Ильяшенко).
66
Линька. После того как маховые и рулевые окончательно закончат рост, молодые птицы начинают смену гнездового наряда на первый послегнездовой. Эта частичная линька охватывает почти все мелкое оперение, кроме некоторых кроющих крыла. У большинства мухоловок она заканчивается к началу осенней миграции. На местах зимовок у первогодков начинается частичная предбрачная линька оперения, которая протекает с большими индивидуальными вариациями. Перепиливают большая часть мелкого оперения, третьестепенные маховые и рулевые перья. В первую очередь сменяются участки оперения (бровь, горло, спина, надхвостье, грудь, участок белого ’’зеркальца” на крыле, рулевые), несущие у самцов видовые опознавательные признаки. Большинство особей к началу весенней миграции полностью заканчивает линьку и приступает к размножению в свежем пере, однако маховые и некоторые верхние кроющие крыла остаются старые и сменяются в послебрачную линьку на местах гнездования. Характер линьки взрослых птиц из-за недостаточного количества материала и противоречивых литературных сведений окончательно не выяснен.
Питание. Основу пищи составляют насекомые разных групп и стадий развития, а также пауки. Значительно реже мухоловки поедают мелких брюхоногих моллюсков, многоножек и плоды некоторых растений. Отыскивают пищу птицы в кронах средних и невысоких деревьев и в нижнем ярусе леса - в древесно-кустарниковой поросли и травяном покрове. Мухоловки этого вида очень подвижны и одинаково успешно пользуются разными способами сбора корма: чаще всего от активного поиска, выпугивания и ловли, а также склевывания беспозвоночных с растений и поверхности земли, до ловли насекомых в воздухе на лету в стремительном броске с присады (Пекло, Смогоржевский, 1980).
Отлет и осенний пролет на территории СССР. Проходит с середины августа по I декаду сентября включительно.
На нижнем Амуре отлет начинается в августе. Последняя птица в 1962 г. добыта 1.IX (Рашкейич, 1965).
В Южном Приморье (2 года) последние самцы отмечены 21 и 26.VIII (Панов, 1973). В Партизанском р-не у с.Бровничи самец добыт 15.VIII 1928 г. (колл. ИЗАНУ), а в окр. г.Владивосток самка добыта 25 .VIII 1903 г. и молодой самец в долине р.Седан-ка— 27.VIII 1912 г. (Черский, 1915а).
Практическое значение. Уничтожает многих насекомых — вредителей леса, а также переносчиков возбудителей некоторых заболеваний человека и животных. Эта подвижная красивая птица является украшением орнитофауны Дальнего Востока СССР и заслуживает повсеместной всесторонней охраны.
Японская желтоспинная мухоловка — Ficedula narcissina (Temminck, 1835)
Muscicapa narcissina Temminck, 1835, Temminck et Laugier, Planches color., livr. 97: pl. 577, fig. 1, Япония.
Син. (русск.): желтоспинная мухоловка, японская мухоловка, желтобровая мухоловка.
Описание. Составлено по 55 экз. (13 — с о-ва Сахалин, 23 — с о-ва Кунашир, 14 — с Японских о-вов, 5 — из Китая).
Вершину крыла образуют 3-е и 4-е первостепенные маховые (3 < 4 — 9,5 %; 3 = = 4 — 21,5; 3 > 4 — 69,0 %). Пятое немного их короче (2 < 5 — 96,8 %; 2 > 5 — 3,2; 2<6-6,5; 2>6 - 93,5 %). Наружные опахала 3-го — 5-го первостепенных маховых в вершинной части заметно сужены. Количество щетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3—4 (3 - 42,9 %; 4 - 57,1 %). Длина наибольшей (п = 39) 6,1 — 9,3 мм (в среднем — 7,7). Количество щитков подотеки на передней части цевки варьирует от 3 до 6. Половой диморфизм выражен в окраске и незначительно в размерах отдельных частей тела (табл. 8).
Распространение. Горная область Восточного Китая, лежащая между верховьями рЛяохэ и побережьем Ляодунского залива и Япония от о-ва Хоккайдо на юг до о-вов Рюкю. На территории СССР гнездится на Сахалине, южных Курильских о-вах и о-ве Монерон. Птицы северной части гнездового ареала являются перелетными и зимуют на о-вах Хайнань, Калимантан (северная часть) и Филиппинских (Vaurie, 1959). Мухоловки южных популяций, очевидно, оседлы.
Систематика. Политипический вид, образующий 3 подвида, один из которых (номинативный) встречается и гнездится на территории СССР.
Географическая изменчивость проявляется в варьировании общей окраски оперения у самцов и незначительно у самок, а также в размерах отдельных частей тела (крыло). На большей части гнездового ареала она носит клинальный характер. Клина идет с северных островов Японии на юг через
о-ва Рюкю и проявляется в замене черной окраски в оперении взрослых самцов на зеленую и в дальнейшем ее посветлении, а также в уменьшении длины крыла (Vaurie, 1959).
Японская желтоспинная мухоловка — Ficedula narcissina narcissina (Temminck, 1835)
Muscicapa narcissina Temminck, 1835, Temminck et Laugier, color., livr. 97 : pl. 577, fig. 1, Япония.
Описание. Составлено по 50 экз. (13 — с о-ва Сахалин, 23 - с о-ва Кунашир, 14 — с Японских о-вов.
У взрослого самца в свежем пере горло, область зоба и верхняя часть груди от желто-оранжевого до ярко-оранжевого у старых птиц. На груди и боках тела этот цвет светлеет и переходит в светло-желтый. Брюшко и нижние кроющие хвоста белые. От лба и далее над глазом к затылку тянется широкая желтая бровь. Верхняя часть и бока головы, зашеек и верхняя часть спины черные. Черный цвет с боков шеи заходит на бока груди. У многих птиц черное оперение верха тела с примесью зеленого разной интенсивности. Существует мнение (Нечаев, 1969), что это проявление возрастной изменчивости. Нижняя часть спины и надхвостье золотисто-желтые. Бока тела серые со слабой примесью светло-желтого на вершинах перьев. Нижние кроющие крыла белые. Маховые, рулевые, а также большая часть верхних кроющих крыла черные. На верхней части крыла расположено белое ’’зеркальце”, образованное белыми проксимальными партиями больших, средних и малых верхних кроющих второстепенных маховых, а также в незначительной степени белыми каемками и пятнами на наружных опахалах третьестепенных маховых. Клюв черно-бурый. Ноги бурые. Радужина глаза темно-коричневая. Индивидуальная изменчивость проявляется в незначительном варьировании величины белого ’’зеркальца” и в появлении у некоторых особей (очень редко) черных пятен на вершинах нижних кроющих хвоста. Сезонная изменчивость проявляется в некотором побурении черных перьев в результате их обноса и выцветания.
Взрослая самка сверху от зеленовато- . оливкового цвета до оливково-серого. Надхвостье такое же, но к верхним кроющим хвоста появляется примесь ржавчато-рыжего. Рулевые темно-роговые. Маховые и верхние кроющие крыла бурые. Первые по краю наружных опахал имеют оливково
67
охристые канты, вторые — каемки и вершины такого же цвета. Нижняя часть тела белая с соловым налетом и слабым серым чешуйчатым рисунком. У некоторых Особей на горле находится слабо выраженное свет-ло-желто-охристое пятно. Клюв темно-рогового цвета.
Молодые в гнездовом наряде сверху коричнево-оливковые со светло-охристыми большими пятнами на перьях и черно-бурыми узкими окантовками по краю опахал. Надхвостье со значительной примесью охристо-рыжего. Рулевые ржавчато-коричневые. Маховые и верхние кроющие крыла бурые со светлоохристыми кантами й каемками на наружных опахалах, а для последних — и на вершинах. Горло, зоб и верхняя часть груди бледно-охристые со слабым бурым волнистым рисунком. Брюшко и нижние кроющие хвоста белые.
Первый послегнездовой наряд идентичен наряду взрослой самки в осеннем пере, но каемки на наружных опахалах и вершинах верхних кроющих маховых шире, а нижняя сторона тела более желтоватая.
Размеры отдельных частей тела японской желтоспинной мухоловки приведены в табл. 8.
Масса тела, г: молодые птицы в гнездовом наряде (п = 3) (18 и 22.VII, 15.IX, Кунашир) - 13,8-15,0, в среднем - 14,4 (колл. ЗИН); молодые птицы в гнездовом наряде (п = 4) (о-в Сахалин) - 13,3—15,2, в среднем — 14,1 (Нечаев, 1984а); молодые птицы в первом послегнездовом наряде (п = 3) (о-в Сахалин) - 13,2—15,0, в среднем - 14,2 (Нечаев, 1984а); самцы первогодки (п = 3) (21.V, 13.VI, 20.VII, о-в Кунашир) — 13,2—13,9, в среднем — 13,6 (колл. ЗИН; БПиДВНЦ; КБ МГУ); самцы первогодки (п = 10) (о-в Сахалин) — 12,2-14,8, в среднем 13,6 (Нечаев, 1984а); самцы старше 1 года (п = 3) (15.V, 28.VI, 15.IX, о-в Кунашир) — 13,5—15,4, в среднем — 14,6 (колл. ЗИН, БПиДВНЦ); самцы старше 1 года (п = 15) (о-в Сахалин) — 13,3-16,5, в среднем - 14,7 (Нечаев, 1984а); взрослые самки (п = 2) (2.VII, 15.IX, о-в Кунашир) — 13,0—13,8, в среднем — 13,4 (колл. ЗИН); взрослые самки (п = 2) (о-в Сахалин) - 14,7-17,5, в среднем — 16,1 (Нечаев, 1984а; колл. ЗМ МГУ).
Распространение. Япония (о-ва Хоккайдо, Хонсю, Сикоку, предположительно Кюсю и Цусима). На территории СССР гнездится на островах Сахалин и Монерон и южных Курильских о-вах. В гнездовой период зарегистрированы залеты японской
желтоспинной мухоловки в Приморье [летом 1926 г. в Шкотовском р-не близ пос.На-речное (ст.Сица) встречена залетная птица (Шульпин, 19276), а 4.VI 1974 г. в окр. пгт Терней отмечен активно поющий самец, 7.VI с ним была самка (гнездо не обнаружено) (Елсуков, 1977)], в Хабаровский край [13.VII 1980 г. в долине р.Амур у с.Пашково встречен одиючный самец (Лавренченко, 19846)] и на о-в Тюлений [в I декаде июня 1962 г. добыт самец (колл. БПиДВНЦ)]. К местам зимовок на о-ва Хайнань, Калимантан (северная часть) и Филиппинские большинство птиц мигрирует через Японские о-ва и о-в Тайвань, меньшая — по окраине материка через Корейский п-ов и Восточный Китай (Checklist of Jap. Birds, 1974). Более детально распространение японской желтоспинной мухоловки на территории СССР показано на карте (рис. 17).
Кадастр к рис. 17.
1 - о-в Сахалин (Hartert, 1907; Дементьев, 1937; Благосклонов, 1954; Гизенко, 1955; Vaurie, 1959; Портенко, 1960; Нечаев, 1969; 1984а; Иванов, 1976; Степанян, 1978); северные отроги Западно-Сахалинских (Камышовый хр.) гор (Нечаев, 1984а); окр. г.Александровка (Гизенко, 1955); долина р.Александровка (Никольский, 1889); 2 - северные отроги Восточно-Сахалинских (Набильский хр.) гор (Нечаев, 1984а); долина верхнего течения р.Набиль (Нечаев, 1984а); 3 — долина р.Ныш (Нечаев, 1984а); 4 — бассейн р.Тымь (Yanukai, Matsuki, 1925; Нечаев, 1984а); 5 - долина р.Поронай (Нечаев, 1984а); 6 - окр. г.Южно-Сахалинск, окр. г.Новоалександ-ровск, окр. с.Луговое, окр. базы АН СССР (Гизенко, 1955); 7 - г.Корсаков (Корсаковский пост) (колл. ЗИН); 8 - южная часть острова (Иванов, Штегман, 1978; колл. ЗИН); Крильон-ский п-ов: склоны и долины рек Кузнецовка и Ше-бунинка (Нечаев, 1984а); 9 - долина р.Айнская (Нечаев, 1984а); 10 - о-в Монерон (Благосклонов, 1954; Гизенко, 1955; Нечаев, 1975; Иванов, 1976); 11 - о-в Кунашир (Bergmann, 1935; Austin, Kuroda, 1953; Благосклонов, 1954; Гизенко, 1955; Портенко, 1960; Коротких и др., 1963; Нечаев, 1969; Иванов, 1976; Степанян, 1978); с.Горячий Пляж (колл. БПиДВНЦ); с.Алехино (Нечаев, 1969; 1971; колл. ЗИН; БПиДВНЦ; КБ МГУ); окр. оз. Горячее (Нечаев, 1969; колл. ЗИН); сопка Вишневая, окр. оз.Пес-чаное (Нечаев, 1969); 12 - долина р.Тятина (Нечаев, 1969); 13 - о-в Шикотан (Гизенко, 1955; Степанян, 1978; Иванов, Штегман, 1978).
Места обитания. Населяет различные типы смешанных лиственных, хвойно-лиственных и реже хвойных лесов по долинам, поймам и террасам рек и озер, в том числе и вулканических, а также разреженные высокоствольные леса с опушками и вырубками по склонам гор и водоразделам. Избегает глухих хвойных лесов
68
и как исключение поселяется в них только по окраинам (о-в Хоккайдо). На о-ве Сикоку обитает в субтропических лесах. В горы поднимается др высоты 1800 м н.у.м. (Jahn, 1942).
На о-ве Сахалин (Нечаев, 1984а) в южных и центральных районах острова японская желтоспинная мухоловка гнездится как в разнообразных долинных лесах, так и в лесах горных склонов до 700—800 м н.у.м. На склонах населяет хвойно-березовые и разреженные вследствие старых рубок елово-пихтовые леса. Однако наиболее оптимальные условия находит в долинных тополево-ольхово-ивовых лесах с хорошо развитым подлеском. Встречается в ольхово-лиственничных заболоченных лесах. У северной границы гнездового ареала на острове зарегистрирована только в долинных лесах. На о-ве Монерон населяет каменноберезовые колки с примесью бархата, ольхи и клена (Гизенко, 1955), а также ольховые леса (Нечаев, 1975).
На о-вах Кунашир и Шикотан гнездится в смешанных широколиственных лесах из клена, ильма и дуба с примесью вишни, рябины, калопанакса, лиан и высоко-травьем в нижнем ярусе, а также в смешанных пихтово-дубовых с примесью ели, тиса, вяза, бархата, березы и значительно реже в заболоченных елово-пихтовых лесах (Гизенко, 1955; Нечаев, 1969).
Численность. В большинстве мест гнездового ареала на территории СССР — обычна.
На о-ве Сахалин, по данным В.А.Нечае-ва (1984а), наиболее высокая численность - до 10 пар на 1 км учетного маршрута — отмечена в долинных лесах, в частности в долинах рек Поронай и Тымь. На Крильонском п-ове на склонах и в долинах рек Кузнецовка и Шебунинка она составляет 6—8 пар на 1 км, а у северной границы ареала в долине р.Тымь ниже устья р.Ныш и в долине последней не превышает 1—2 пары на 1 км учетного маршрута. На о-ве Монерон редка. Общее количество гнездящихся мухоловок на острове составляет 2—3 пары (Нечаев, 1975).
В южной части о-ва Кунащир по склону долины р.Алехина на маршруте протяженностью 1 км зарегистрировано 8—10 гнездящихся пар, а в пойменных лесах р.Тяти-на — 2-3 пары на 1 км (Нечаев, 1969).
Прилет и весенний пролет на территории СССР. Проходит в мае, однако первые птицы появляются даже в конце 111 декады апреля.
На о-вах Большой Пелис и Матвеева в заливе Петра Великого (3 года) пролет отмечен 13—18.V (Лабзюк, Назаров, 1967); 15 и 27.V, 8—29.V (Лабзюк и др., 1971).
На о-ве Кунашир в окр. с.Алехино в 1963 г. первые самцы появились 14.V, а первые самки в III декаде этого месяца (Нечаев, 1969; 1971).
На о-ве Монерон в 1973 г. пролетные мухоловки отмечены во второй половине мая (Нечаев, 1975), а на о-ве Сахалин первые птицы появляются во II декаде этого месяца (Гизенко, 1955; Нечаев, 1984а).
В период весеннего пролета японская желтоспинная мухоловка зарегистрирована и на материке: 30.IV 1974 г. в Приморье в пгт Терней найден мертвый самец (Ел-суков,1977).
Первыми на места гнездования прилетают самцы и только через несколько дней после них появляются самки.
Размножение. Прилетевшие самцы ведут себя оживленно и много поют. Песня японской желтоспинной мухоловки, по данным В.А.Нечаева (1984а), — длительно звучащая громкая переливчатая трель. Она состоит как из мелодичных, флейтового характера строф, так и из скрежещущих звуков и сильно подвержена индивидуальной изменчивости. Очень типична имитация песен и криков других птиц: соловья-красношейки, черного дятла, канюка и др.
После занятия гнездовых участков и разбивки на пары мухоловки приступают к строительству гнезд. Из 27 гнезд этого вида, найденных на о-вах Сахалин и Кунашир (Нечаев, 1969; 1984а), 24 были построены в дуплах и полудуплах живых и сухостойных деревьев (ольха, ива, бархат, пихта, бузина, клен), а также их пней, в том числе и в старых, несколько разрушенных дуплах карликового дятла и пухляка (3): 1 — в углублении ствола пихты, выдолбленном черным дятлом; 1 — в изломе ствола ольхи и 1 — под коньком крыши брошенной постройки человека. Высота расположения гнезд от поверхности земли 1,2-9,5 м, чаще 3-4. Размеры гнездовых дупел, мм: диаметр летка (п = 10) - 32-70 х 40-80, в среднем - 47 х 63; глубина дупла (п = 11) - 55-165, в среднем — 103; внутренний диаметр на уровне гнезда (п = 8) — 65-130, в среднем - 84 (Нечаев, 1984а). Наиболее охотно заселяются достаточно просторные дупла с широким входом, но когда их недостаточно птицы поселяются и в очень тесных. В последнем
69
случае клюв, а иногда и голова сидящей на гнезде самки видны снаружи.
Гнездо представляет собой рыхлую постройку, форма и размеры которой зависят главным образом от таковых нижней части гнездового дупла. В качестве строительных материалов для гнезда мухоловки используют лубяные волокна древесных и кустарниковых растений, кусочки бересты, талломы лишайника-бородача, зеленые мхи, тонкие корешки, стебельки и листья осок и злаков, кусочки листовых пластинок, жилки и черешки листьев древесных растений (ильм, дуб, береза, клен, ива, ольха), плодоножки рябины, кусочки побегов хвоща. Лоток выстилается тонкими стебельками и листьями трав (в том числе злаков и осок), корешками папоротников, лубяными волокнами, а также ножками спорогониев мхов. В некоторых гнездах лоток оплетается по краю листьями ильма (Гизенко, 1955; Нечаев, 1969; 1984а). Одно из найденных гнезд было построено исключительно из талломов лишайников (Alectoria sp., Usnea sp. и др.), в лотке другого выстилка состояла на 50 % из волоса шерсти енотовидной собаки (Нечаев, 1984а). Гнездо строит самка. Размеры гнезд, мм: диаметр (п = 16) — 50—125 х х 50—130, в среднем — 82 х 85; высота гнезда (п = 8) - 50—80, в среднем — 64; диаметр лотка (п = 12) — 45—67, в среднем - 55; глубина лотка (п = 13) — 20-60, в среднем - 41 (Нечаев, 1984а).
В сезон размножения на территории СССР японская желтоспинная мухоловка имеет один репродуктивный цикл. Количество яиц в полной кладке 5—6, в одной повторной, взамен утраченной, было 4 яйца. (Нечаев, 1984а). По данным В.А.Нечаева (1984а), правильно-яйцевидные и реже укороченно-округлые яйца бывают голубоватозелеными со светло-бурыми пятнами, сконцентрированными на тупой половине яйца, либо образующими ’’венчик” у тупого полюса, и розовато-белыми с коричневыми пятнами, разбросанными более или менее равномерно по всей поверхности скорлупы. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
К насиживанию самка приступает после откладки последнего, реже — предпоследнего яйца. Инкубация (2 гнезда) длится 11 дней. Самец в насиживании кладки участия не принимает. Он изредка подкармливает самку, сидящую на яйцах. Птенцы вылупляются в течение суток (Нечаев, 1984а).
70
Пуховой птенец японской желтоспинной мухоловки (Нечаев, 1983) имеет серый пух на надглазничных (длина пушинок 5—7 мм), затылочной (5—8), плечевых (5-8), локтевых (2-3) и спинной (5-8) птерилиях, а у некоторых особей он отмечен и на веках (до 3). Кожа розовато-желтая. Ноги розовые, когти бледно-розовые. Клюв серовато-розовый. Яйцевой зуб серый. Складки в углах рта бледно-желтые. Ротовая полость и язык желтые.
В выкармливании птенцов принимают участие оба партнера пары. На 12-й день с недоросшими маховыми и рулевыми птенцы покидают гнездо. При отсутствии беспокойства они находятся в нем до 14— 16 дней (Тамануки, 1942; Нечаев, 1984а). Родители продолжают кормить слетков. В августе выводки распадаются, молодые и взрослые держатся в этот период поодиночке или группами из 2—3 особей. Иногда их можно наблюдать в смешанных стаях синиц, корольковых пеночек и ширококлювых мухоловок (Нечаев, 1984а).
По данным В.А.Нечаева (1984а), отмечается значительная гибель гнезд японской желтоспинной мухоловки. Так, на о-ве Сахалин лишь в 6 (30 %) из 18 гнезд, находившихся под наблюдением, птенцы благополучно вылетели. В период откладки яиц и насиживания было разорено 5 гнезд, 3 брошено с законченными кладками, 1 с птенцами — разорено, 1 — оставлено в период строительства. В 1 гнезде (находящемся в 3 м от поверхности земли) птенцы были покусаны землеройкой-бурозубкой и погибли. Как считает В.А.Нечаев (1984а), судьба найденных гнезд усугубилась тем, что они находились под наблюдением. Основными врагами этого вида на острове являются большеклювая и черная вороны.
Сезон размножения японской желтоспинной мухоловки на территории СССР длится с конца III декады мая до середины августа.
На о-ве Кунашир (Нечаев, 1969) массовое строительство гнезд проходит в первой половине июня: в окр. пос.Алехине в 1963 г. законченная постройка найдена 9.VI, а строящееся гнездо — 14.VI. К откладке яиц и насиживанию птицы приступают здесь во второй половине этого месяца. В окр. оз.Песчаное гнездо с полной кладкой из 5 яиц (степень насиженности 1—2 дня) обнаружено 25.VI 1963 г. На сопке Вишневая 2.VII 1963 г. и 6.VH 1962 г. встречены 2 гнезда с полностью оперившимися птенцами (по 4 особи в каждом): готовых к выле
ту в первом и вылетевших при сотрясении ствола гнездового дерева во втором. Первые выводки молодых появляются в I декаде июля (2.VII 1963 г. добыт слеток в окр. пос.Алехино (колл. БПиДВНЦ) и значительно чаще — в конце июля — начале августа. Самостоятельно кормившиеся летные молодые, державшиеся без взрослых, отмечены в лесу на берегу оз.Горячее 18.VII 1962г. (Нечаев, 1969).Вокр.пос.Алехино молодые в гнездовом наряде добыты 22.VII 1962 г. (колл. ЗИН) и 7-8.VIII 1965 г. (колл. ИЗАНУ).
На о-ве Монерон в 1973 г. самцы уже держались на гнездовых участках в начале I декады июня (Нечаев, 1975). На о-ве Сахалин разбивка на пары отмечена А.И.Ги-зенко (1955) в конце мая. По данным В.А.Нечаева (1984а), полностью законченные гнезда без яиц найдены на острове 12 и 17.VI; полные ненасиженные кладки — 23 и 26.VI, повторная — 7.VII; гнезда с птенцами перед вылетом — 7.VII 1980 г., а первые слетки (2 года) - 14 и 17.VII; выводки слетков встречены также (3 года) 26.VII, 1 и 3.VIII, а взрослые птицы с кормом (2 года) - 23, 27, 28.VII и 1, 12.VIII.
Линька. У большинства молодых частичная послегнездовая линька, в ходе которой сменяется только мелкое оперение гнездового наряда, начинается в конце июля — начале августа и заканчивается в первой половине сентября. У некоторых особей из поздних выводков она начинается в сентябре. Так, мухоловки, добытые 12 и 15.IX на о-ве Кунашир (колл. ЗИН), закончили линьку и были в свежем пере первого послегнездового наряда, в то время как 1 птица, добытая здесь 15.IX (колл. ЗИН), только начала смену гнездового наряда. На местах зимовок у первогодков проходит частичная предбрачная линька, которая протекает с некоторыми индивидуальными вариациями. Она охватывает в первую очередь участки оперения (бровь, горло, спина, надхвостье, бока груди, участок белого ’’зеркальца” на крыле), несущие у самцов видовые опознавательные признаки. У некоторых особей происходит частичная или полная смена рулевых, а также некоторых второстепенных маховых. В комбинированном наряде, имея на теле мелкие перья от 2 генераций (Нечаев, 1984а; наши данные), первогодки прилетают на места гнездования и приступают к размножению. После завершения гнездового периода у них проходит полная послебрачная линька, в ходе которой в первую
очередь сменяются первостепенные и второстепенные маховые (Нечаев, 1984а). Взрослые птицы имеют в году полную послебрачную и частичную предбрачную линьки. Полная линька проходит у них на местах гнездования с конца июля по первую половину сентября (Нечаев, 1984а), частичная предбрачная — на местах зимовок. В ходе последней меняется мелкое оперение, а также по-видимому, проксимальные второстепенные маховые и некоторые рулевые.
Питание. Основу пищи составляют насекомые разных групп и стадий развития, а также пауки. Очень редко поедает мелких брюхоногих моллюсков, многоножек и некоторых других беспозвоночных. В конце лета и осенью питается также сочными плодами некоторых растений. По данным В.А.Нечаева (1984а), птицы этого вида охотятся главным образом на находящихся на субстрате и слабо летающих насекомых, а также на пауков. Сравнительно редко они ловят насекомых в воздухе на лету, подобно другим видам мухоловок. Иногда весной кормятся в несвойственных им местах обитания: на лугах, болотах, в кустарниково-травянистых зарослях, вблизи построек человека и скал на морском берегу.
Отлет и осенний пролет на территории СССР. Проходит с середины августа по I декаду октября включительно. Одиночные птицы задерживаются до середины II декады октября и как исключение зарегистрированы в ноябре.
Остров Сахалин мухоловки покидают во второй половине августа и в сентябре, хотя одиночные особи еще встречаются здесь до середины октября (Нечаев, 1984а). Самая поздняя встреча японской желтоспинной мухоловки на острове - 16.Х 1926 г. В этот день была добыта птица у пос.Вахрушев (Yamashina, 1927).
На о-ве Кунашир осенняя миграция проходит в сентябре. За период 10—28.IX 1948 г, в окр. пос.Серноводск и г.Южно-Курильск добыто 19 птиц (Гизенко, 1955; колл. ЗИН). В окр. г.Южно-Курильск Л.А.Портенко (Нечаев, 1969) наблюдал пролетных птиц 24.IX, а Ю.Н.Назаров 17.IX 1963 г. добывал мухоловок в окр. пос.Алехино.
Самая поздняя встреча японской желтоспинной мухоловки на территории СССР зарегистрирована А.А.Назаренко в заповеднике ’’Кедровая падь”, где 21.XI 1959 г. в уреме р.Кедровой он добыл сильно упи-
71
Таблица 10. Размеры и коэффициенты вариации (CV,%) мухоловок малой и таежной
Вид,подвид, место сбора п Длина, мм
Крыло CV Расстояние, на которое 1-е маховое выступает за вершины кроющих кисти CV Хвост CV
Самцы пер-
F. parva parva
европейская часть СССР 29 66.551 0.18 64,8 -68,3 1,46 4,7410,21 2,8 -7,1 23,88 50,181 0,25 47,4 -53,2 2,63
Кавказ 5 66,601 0,73 65,0 -68,5 2,44 5,8010,46 4,8 -6,8 15,74 50,221 0,36 49,5 -51,5 1,59
F.p.albicilla Восточная Сибирь 1 68,0 — 3,7 — 52,0 —
F.mugimaki Южндо Сибирь и Дальний восток СССР к югу от Якутии 46 71,691 0,23 67,6 -74,0 2,08 4,3010,17 1,9 -6,7 25,94 49.6210,19 46,6 -53,1 2,58
Самцы взрослые
F.parva parva европейская часть СССР 37 67,801 0,19 65,6 -69,9 1,67 4,3910,23 1,7 -7,5 30,16 43,7 -54,9 3,61
Кавказ 13 66.741 0.32 64,0 -68,0 1,74 5.0710.32 3,5 -7,6 23,07 51,111 0,33 49,1 -52,7 2,22
F.p.albicilla, юг и юго-восток Сибири и север Дальнего Востока СССР 30 68.401 0.27 65,3 -70,5 2,09 4.4010,23 1,6 -6,4 27,75 50,211 0,36 46,0 -53,4 3,88
F.mugimaki Южная Сибирь и Дальний Восток СССР к югу от Якутии 30 74.101 0.27 71,5 -77,3 2,02 3.61Ю.24 -0,5 -6,1 35,95 51,361 0,27 48,5 -54,5 2,81
Сам
F.parva parva европейская часть СССР 15 65.531 0,44 63,4 -69,9 2,61 4,8210,34 3,1 -7,2 27,68 49,431 0,45 40 -4J.J 3,55
Кавказ 7 64.601 0.48 62,8 -67,0 1,95 4,9810,65 3,2 -6,3 26,02 49,121 0,63 47,2 -51,0 3,16
F.p.albicilla юг и юго-восток Сибири и север Дальнего Востока СССР 11 66,731 0,55 64,6 -70,0 2,74 5,5410,53 3,1 -8,8 30,37 49,381 0,45 47,8 -52,8 3,05
F.mugimaki
Южная Сибирь и Дальний Восток 28 70.371 0.32 67,2 -73,4 2,30 4.3910,24 2,1 -6,6 29,18 47,561 0,35 43,1 -50,3 3,79
СССР к югу от Якутии
72
Длина, мм
Белое поле в основании рулевых CV Цевка CV Клюв от оперения лба CV Клюв от орального края ноздри CV
вогодки
Ш4* Q.33 29,8 -37,8 5,21 16.821 0.08 15,9 -17,4 2,40 9,531 0,11 8,5 -10,7 5,99 6.6310,05 6,1 -7,1 4,13
36.301 0.52 35,0 -38,0 3,19 17.181 0.21 16,6 -17,8 2,74 9.201 0,24 8,5 - 9,6 5,25 7,0310,26 6*3 -У,3 7,48
36,8 - 17,6 - 9,4 - 7,0 -
-26.441 0Л0 19,5 -29,8 10,14 15,641 0,06 15,0 -16,6 2,72 8.701 0.06 7,8 -9,6 4,76 6,3610,05 ^4 -У,2 5,13
(старше одного года )
37.06* 30,3 -40,2 5,12 17.091 0.09 15,0 -18,2 3,22 9.5410,07 8,8 -10,3 4,42 6,8010.06 6,2 -7,5 4,88
36.761 0Л5 35,1 -40,1 4,74 17,081 0,13 16,6^-18,0 2,69 9,221 0,08 8,9 - 9,7 2,96 7,0110,05 Т7 -7,3 2,54
Ш5* 0.32 31,8 -38,2 5,10 16.981 0,12 15,1 -18,3 3,69 9,431 0,05 8,9 - 9,9 3,04 6.8010.05 6,2 -7,3 4,19
ЗД.14* ОЛб 26,7 —33,5 6,76 13.31* ОДД. 14,3 -16,5 3,11 8-951 0.10 8,1 -10,2 6,17 6.57Ю.07 5,8 -7,7 5,70
ки
.33,26* 0,44 32,2 -38/) 4,83 17.03* 013 15,9 -17,6 2,95 9.491 0.13 8,5 -10,2 5,28 6.6110.08 6,1 -7,2 4,66
35.081 0.60 33,9 -37,3 3,82 1Й661 0,22 15,5 -17,2 3,55 9.411 0.18 8,9 -10,3 5,15 6.8910,10 6,5 -7,3 3,70
32,941 048 3д!3- 34,6 4,63 16,851 0,12 16,2 -1У,4 2,27 9,111 0,14 -1(£Г 5,02 6,4210,06 6,1 -6,8 3,25
Отсутствует - 15.491 0.10 14,0 -16,6 3,38 8,441 0,08 L4 - 5,07 6,1910,05 & “ 4,48
73
Окончание табл. 10
Вид, подвид место сбора п Ширина, мм Высота, мм
Крыло CV Клюв у орального края ноздри CV Клюв у основания CV Клюв у основания CV
Самцы перво
F. parva parva европейская часть СССР 29 49.91* 0,19 48,1 -51,6 1,98 3.70*0,04 3,1 -4,2 5,87 5,40*0,05 5,0 -5,9 4,73 3,35*0.04 3,1 -3,7 5,50
Кавказ 5 50,44* 0,37 49,4 -51,3 1,66 3,98*0,09 3% -4 Д 4,30 5.38*0,27 4,7 -6,0 10,00 3,1* -
F.p.albicilla Восточная Сибирь 1 50,0 — 3,6 — 5,4 — 3,5 —
F.mugimaki Южная Сибирь и Дальний Восток СССР к югу от Якутии 46 51,57* 0,17 49,3 -54,0 2,24 3,29*0,03 2,8 —3,5 5,15 5,00*0,05 4,3 -6,1 7,22 3,20*0,04 3,0 -3,6 6,36
Самцы взрослые
F.parva parva европейская часть СССР 37 49,62* 0,19 46,3 -51,2 2,20 3,78*0,04 3,0 -4,6 7,02 5,42*0,06 4,6 -6,1 6,25 3,41*0,05 3,0 -3,9 6,88
Кавказ 13 50,16* 0,35 48,5 -52,4 •2,42 3,83*0,06 3,5 -4,1 5,48 ^23*0.0? 4,5 -5,6 5,66 3.46*0.15 3.F -3,8 9,50
F.p.albicilla, юг и юго-восток Сибири и север Дальнего Востока СССР 30 50,58* 0,33 <3 -ЗМ' 3,48 3,0 -3,9 5,89 5.30*0.07 4,6 -6,1 7,30 3,42*0,05 3*1 -5,5 5,98
F.mugimaki Южная Сибирь и Дальний Восток СССР к югу от Якутии 30 52,57* 0,16 50,7 —54,2 •1,70 3,36*0,04 Тб -3,6 • 6,63 5.01*0.06 4,5 -5,5 6,16 3,39*0.06 5,1 -5,9 6,75 Сам
F.parva parva европейская часть СССР 15 48.55* 0.30 46,4 -50,8 2,41 3,68*0.04 3,4 -3,9 4,00 5.35*0,11 4,4 -6,2 8,15 3,38*0,08 5,1 -3,8 6,75
Кавказ 7 48.67* 0.32 47,6 -49,5 1,63 3,88*0,08 3,6 -4,1 4,10 5,40*0,12 5,1 -5,8 5,37 3,40*0,17 3*5 -<9 9,90
F.p.albicilla
юг и юго-восток Сибири и север Дальнего Востока СССР 11 49,75* 0,47 47,8 —52,5 3,li 3,48*0,08 3,1 -5,9 7,69 5.32*0.09 3*0 -5,8 5,07 13^.2*0.14 3,2 -4,0 8,62
F.mugimaki
Южная Сибирь и Дальний Восток СССР к югу от Якутии 28 49.95* 0,25 47,8 -53,0 2,51 3.36*0.04 2,8 -3,7 6,14 4,87*0,09 3,8 -5,7 9,20 3,26*0,04 3,0 -3,6 5,21
* Размер дается по 1 экз.
74
Отношение ширины клюва у основания к его длине от оперения лба, % CV Отношение длины хвоста к длине крыла, % CV Отношение ширины крыла к его длине, % CV
годки
56.87* 0.79 49,53-64,13 7,38 75,37* 0,39 71,60—79,58 2,72 74,98* 0,25 72,86—77,76 1,77
58.35* 1.88 55,29-63,83 6,44 75,45* 1,06 72,73-78,27 3,15 75,79* 1,32 72,43-78,92 3,88
57,45 - 76,47 - 73,53 -
57,57* 0,68 48,86-69,77 7,96 69.22* 0,25 64,45-72,78 2,40 71.85* 0.17 69,18-73,75 1,53
(старше одного года)
57,12* 0,62 51,46-64,13 6,29 75,47* 0,46 64,64-79,42 3,53 73,17* 0,19 69,00-74,96 1,54
56.72* 0,99 49,45-60,44 6,02 76,70* 0Л2 73,95-80,06 2,35 75,26* 0,29 73,99-77,74 1,34
56,31* 0,77 48,94-64,21 7,25 73.56* 0.43 68,91-77,28 3,11 73,95* 0.38 68,90-76,47 2,74
56,07* 0,61 50,00-61,11 5,80 69,33* 0,38 65,98-73,65 2,91 70,96* 0,24 68,18-73,43 1,85
ки
.56,49*. 1.43 48,35-72,94 9,81 75.42* 0.35 72,97-77,84 1,81 74,10* 0,34 71,92-76,81 1,79
56.91* 1.33 53,13-61,05 5,72 75.93* 0.54 74,84-78,46 1,73 75.43* 0.43 73,88—76,74 1,39
58.64* 1.48 49,50-65,88 7,99 74.02* 0,52 71,54-76,63 2,31 74,57* 0.45 71,54-76,69 2,02
57,9Q* U6 45,78-68,75 10,41 67,83* 0,39 62,19-71,26 2,86 70,99* 0,29 67,72-73,26 2,05
75
тайную молодую самку (Назаренко, 1971а; Панов, 1973; колл. ЗИН).
Практическое значение. Такое же, как и у даурской желтоспинной мухоловки.
Таежная мухоловка — Ficedula mugimaki (Temminck, 1835)
Muscicapa mugimaki Temminck, 1835, Temminck et Laugier, Planches color., livr. 97 : pl. 577, fig. 2, Япония.
Син. (русск.): мухоловка Мугимаки, рыжая мухоловка, японский лоцманчик, рыжебрюхая мухоловка.
Описание. Составлено по 102 экз. (30 — из Южной Сибири, 71 — с Дальнего Востока СССР к югу от Якутии, 1 — с о-ва Сахалин).
Вершину крыла образуют 3-е и 4-е первостепенные маховые (3 < 4 — 24,2 %; 3=4 — 40,3 %; 3 > 4 — 35,5 %). У большинства особей (98,4 %) 5-е немного их короче, но у некоторых экземпляров (1,6 %) оно равно 3-му или 4-му. Первое первостепенное маховое очень мало и только на несколько миллиметров выступает за вершины кроющих кисти (табл. 10); 2-е всегда меньше 5-го и больше 7-го (2 < 6 — 23,8 %; 2 = 6 — 25,0 %; 2 > 6 — 51,2 %). У большинства особей (92,5 %) наружные опахала 3—5-го первостепенных маховых в вершинной части заметно сужены. У некоторых экземпляров (7,5 %) сужено в вершинной части и наружное опахало 6-го махового. Количество щетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3-4 (3 — 78,5 %; 4 - 21,5 %). Длина наибольшей (п = 69) 4,0—7,0 мм (в среднем 5,7). Количество щитков подотеки на передней части цевки варьирует от 3 до 6. У взрослых птиц на вершине хвоста (в центре) имеется небольшая выемка. Половой диморфизм выражен в окраске оперения и рамфотеки клюва.
У взрослого самца (старше 1 года) в свежем пере горло, зоб и грудь ярко оранжево-рыжие. Последний цвет незначительно светлеет на верхней части брюшка и реже на боках тела. Нижняя часть брюшка белая. Нижние кроющие хвоста от белого до светло-рыжевато-охристого. От верхней части глаза к затылку тянется широкая белая бровь. У некоторых особей она имеет вид пятна, расположенного за глазом. Верхняя часть и бока головы, зашеек, спина и верхние кроющие хвоста от черно-серого до черного. Надхвостье темно-серое. Маховые, большинство верхних кроющих
крыла и рулевые черные. В основании рулевых находится белое поле (табл. 10), размеры которого увеличиваются от крайней пары к центральной и на последней оно очень мало (отдельные, часто асимметричные пятна) или отсутствует. Нижние кроющие крыла от светло-охристо-желтого до светло-желтовато-рыжего. Черные третьестепенные маховые по краю наружных опахал несут широкие белые каймы. На верхней части крыла расположено белое ’’зеркальце”, образованное белыми проксимальными партиями больших, средних и в незначительной степени малыми верхними кроющими второстепенных маховых. Очень редко у некоторых самцов белые канты находятся на наружных опахалах проксимальных второстепенных маховых. Клюв черный. Ноги бурые. Радужина глаза темно-коричневая. Индивидуальная изменчивость проявляется в варьировании величины бровей и ’’зеркальца” на крыле, в появлении у некоторых особей черного надхвостья и узкой охристой поперечной полосы между надхвостьем и верхними кроющими хвоста, а также, как исключение, в полной замене черного цвета в оперении верхней части тела на темно-пепельно-серый. Сезонная изменчивость проявляется в побурении маховых, частично верхних кроющих крыла и черных частей рулевых в результате их обноса и выцветания.
У взрослой самки оперение верхней части тела от светло-серовато-оливкового до оливково-серого. На верхних кроющих хвоста появляется примесь светло-буровато-коричневого. Маховые, верхние кроющие крыла и рулевые темно-бурые. В отличие от самцов, белое поле в основании рулевых и белое ’’зеркальце” на крыле ' отсутствуют. По краю наружные опахала маховых несут рыжевато-охристые канты. Горло, зоб, грудь и верхняя часть брюшка от светло-рыжевато-охристого до бледно-оранжево-рыжего. Нижняя часть брюшка и нижние кроющие хвоста — белые. Нижние кроющие крыла бледно-охристые. Клюв темно-коричнево-бурый. Сезонная изменчивость проявляется в варьировании оттенков основной окраски верхней части тела и в общем посветлении оперения в результате его обноса и выцветания.
Молодые в гнездовом наряде сверху орехово-бурые с редкими продолговатыми желтовато-белыми или охристыми пятнами на перьях. Оперение нижней части тела бледно-желтовато-охристое со слабым черновато-серым чешуйчатым рисунком. На
76
наружных опахалах и вершинах больших кроющих второстепенных маховых имеются широкие светлогжелтовато-охристые каемки, которые при сложенном крыле образуют на нем поперечную полосу. Уже в этом наряде самцы четко отличаются от самок по наличию белого поля в основании рулевых.
Первый послегнездовой наряд похож на наряд взрослой самки, но оперение верхней части тела со значительной примесью зеленовато-оливкового цвета. У самцов горло, зоб и грудь оранжево-рыжие, но в отличие от старых птиц этот цвет более светлый, далеко на брюшко не заходит или полностью на нем отсутствует. У самок эти места — от светло-охристого до охристо-желтого.
Размеры отдельных частей тела таежной мухоловки приведены в табл. 10.
Масса тела, г: самцы (п = 17) (Нижнее Приамурье) — 10,9—13,1, в среднем - 12,1 (Кистяковский, Смогоржевский, 1973); самец 28.IX (о-в Рикорда) - 13,3 (колл. ЗИН); самка (Нижнее Приамурье) - 13,8 (Кистяковский, Смогоржевский, 1973); самка (29.V. Читинская обл.) — 11,7 (Павлов, 1948).
Распространение. Северо-Восточный Китай. На территории СССР гнездится в Южной Сибири, в южной части Дальнего Востока и на о-ве Сахалин. На всем гнездовом ареале мухоловки этого вида являются перелетными и к местам
зимовок в Южный Китай, на п-ов Индокитай и о-ва Хайнань (Check-list of Jap. Birds, 1974) Суматра, Ява и Калимантан (северная часть) мигрируют через Южную Сибирь, Приамурье, Приморье, Восточный Китай, Корейский п-ов, о-ва Японии, Тайвань, Хайнань и Индокитай. Детально распространение таежной мухоловки на территории СССР показано на карте (рис. 19).
Кадастр к рис. 19.
1 - о-в Сахалин (Дементьев, 1937; Гизенко, 1955; Портенко, 1960; Степанян, 1978; и др.); п-ов Шмидта (Нечаев, 19846); 2 - побережье залива Помрь (Нечаев, 19846); 3 - побережье оз.Сладкое (Нечаев, 19846); 4 - долина р.Пшпг тун (Нечаев, 19846); 5 - долина р.Вал (Нечаев, 19846); 6 - окр. мыса Погиби (Нечаев, 1984); 7 - долина р. Джим дан (Нечаев, 19846); 8 -побережье залива Терпения (Нечаев, 19846); 9 - долина р.Набиль; окр. пос. Ныш на р.Тымь (Нечаев, 19846); 10 - Долина р.Тымь (Нечаев, 19846); 11 - долина рЛоронай (Нечаев, 19846); 12 - пос. Гастелло (Yamashina, 1928; Благосклонов, 1954; Гизенко, 1955); 13 - Углегорский р-н (Благосклонов, 1954; Гизенко, 1955); 14 - побережье оз.Айнское и долина р.Айнская (Нечаев, 19845); 15 - подножие г Лопатина, долина р.Хребтовая (Нечаев, 19846); 16 - долина р.Сусун (Гизенко, 1955); окр. с.Третъя падь (Lbnnberg,1908); 17 - долина рЛютога (Нечаев, 19846); окр. с.Троицкое, окр. с.Пятиречье (Гизенко, 1955); 18 - Приморский край; окр. г.Владивосток, р.Нарва (Сидеми) (Bianchi, 1907; колл. ЗИН); 19 - р.Раздольная (Суйфун) (Bianchi,1907); 20 - бассейн истоков р.Уссури, ур.Мута, гора Облачная (Назаренко, 19846); 21 - Уссурийский заповедник им.В.Л.Комаро-ва: ельники по Егерскому ключу и Аникиной пади (Иванов, 1952); Даубихе - Майхинское
Рис. 19. Распространение таежной мухоловки (Ficedula mugimaki) на территории СССР: а — гнездится, б — отмечена в гнездовой период. См. кадастр, стр. 77, 78.
77
плато (Воробьев, 1954; колл. БПиДВНЦ); верховья рек Арсеньевка и Артемовна (Иванов, 1976); 22 — хвойные леса вершин гор Партизанского р-на (Шульпин, 19276; Воробьев, 1954); верховья р.Партизанская (Иванов, 1976); окр. пос.Ручьи (Фанза) (колл. ЗИН); 23 - Сихотэ-Алинский заповедник (Кулешова, 1976; наши данные); 24 - бассейн истоков р.Большая Уссур-ка, Краснореченское лесничество (Назаренко, 19846); 25 - среднее и верхнее течение р.Большая Уссурка (Флинт и др., 1959; Спангенберг, 1965); перевал Большая Синанча (колл. ЗИН); 26 - сопки и острова низовьев р.Болыпая Уссурка (Спангенберг, 1940); окр. с.Вербовка (Спангенберг, 1965); 27 - хр.Сихотэ-Алинь (Иванов, 1976); перевал Эдельштейна, реки Самарга и Сукпай (колл. ИЗАНУ); 28 - р.Ботчи в 60 км от устья (Емельянов, 1929); 29 - хр. Хехцир (Кулешова и др., 1965); 30 - пос. Омми (колл. КГУ); 31 - тайга по р.Гур (Хунгари) (Благосклонов, 1954; Воробьев, 1954); Комсомольский заповедник (Штильмарк, 1973); 32 - с.Гячи (Кистяковский, Смогоржевский, 1973); 33 -оз.Удыль (Яхонтов, 1963; колл. ЗИН); верховья р.Кенжи (колл. ЗИН); с.Богородское (Иванов, 1976; колл. ЗИН); с.Дуди (колл. КГУ); 34 -с.Покровка (Кистяковский, Смогоржевский, 1973; колл. КГУ); с.Новая ферма; с.Тальники (колл. КГУ); 35 - нижний Амур, с.Романовка (колл. КГУ); пойма р.Амур (Кистяковский, Смогоржевский, 1973); 36 - окр. оз.Орель (Бабенко, 1984); 37 - р.Амгунь, посЛаки (колл. ЗИН); 38 - хр.Малый Хинган, окр. с.Радде и с.Башурово (сообщ. С.М.Смиренского); 39 -с.Помпеевка; Хинганский заповедник, ст.Кундур (сообщ. С.М.Смиренского); 40 - устье р.Мель-гин (Иванов, 1976) ; устье р.Талая (колл. КГУ); 41 - окр. пгт Экимчан (колл. В.В.Леоновича); 42 - р.Селемджа, окр. пос.Дагмара (колл. КГУ); окр. с.Куравинское (колл. КГУ); 43 - окр. Инарогды возле ключа Бекельдеуль (Спаский и др., 1962); 44 - восточная часть хр.Тукурингра (сообщение Ю.В.Ильяшенко); 45 - Становой хр. в верховьях р.Зея (Иванов, 1976); 46 - хр.Джугд-жур в верховьях р.Мая (Иванов, 1976); 47 -р.Мая (колл. ЗИН); 48 — с.Устъ-Лантарь (колл. ЗИН); 49 - рЛантарь (Портенко, 1960; Иванов, 1976); р.Улейкан (колл. ЗИН); 50 - долина рЛена, 62° с.ш. (Степанян, 1978); 51 - долина р.Вилюй, 63° с.ш. (Степанян, 1978); 52 — окр. г.Олекминск (Сушкин, 1938; Воробьев, 1963; Иванов, 1976; колл. ЗМ МГУ); 53 - р.Токко (Воробьев, 1963); 54 - Олекмо-Чарское нагорье; тайга по островам и берегам р.Тяня (Воробьев, 1963); окр. пос. Тяня (колл. ЗИН);
55 - Приаргунье (Гагина, 1960); 56 - юго-восточная часть Витимского плоскогорья (Павлов, 1948); окр. г.Чита (Измайлов, 1967; Иванов, 1976); 57 - Яблоновый хр. (колл. ЗИН); 58 — окр. оз.Исинга (Измайлов, 1967); 59 — Муйская долина (Измайлов, 1959; 1967; Измайлов, Боровицкая, 1967); 60 - Баунтовская котловина (Измайлов, 1967); 61 - долина р.Томпуда (Малышев, 1960); 62 - Баргузинский заповедник (Скрябин, Филонов, 1962); 63 - оз.Байкал, губа Половинная (колл. ЗИН) ; о-в Большой Ушканий (колл. ЗИН); 64 - Байкальский заповедник (Баскаков, 1977); 65 - пгт Култук (Bianchi, 1907; колл. ЗИН); 66 - Тункинская долина, окр. с.Кырен (Измайлов, Боровицкая, 1973);
р.Долбай (колл. ЗИН); 67 - верховья р.Иркут (Иванов, 1976); 68 - истоки р.Ангара (сообщ. А.А.Винокурова); окр. г.Иркутск (Благосклонов, 1954); 69 - верхняя Лена (Реймерс, 1966); 70 - Киренский р-н, окр. с.Подвалочная; р.Кон-долка, тайга у оз.Мажарское (колл. ЗИН); 71 -окр. г.Братск (Иванов, 1976); 72 - верхнее течение р.Нижняя Тунгуска (от истоков до с.Ер-богачен) (Ткаченко, 1937); 73 - р.Нижняя Тунгуска (от с.Ербогачен, до 62° с.ш.) (Ткаченко, 1937); 74 — низовья р.Чона (Ткаченко, 1929; Иванов, 1976); 75 - р.Енисей, окр. с.Осипово (колл. БПиДВНЦ); 76 - окр. пос.Ворогово (Готфрид, 1982); 77 - р.Кеть (Гынгазов, Москвитин, 1965; Гынгазов, Миловидов, 1977); окр. с.Новоникольское (Гынгазов, Москвитин, 1965); окр. с.Красная Курья (Москвитин и др., 1977); 78 - низовья р.Кеть (Гынгазов, Мило-
видов, 1977); 79 - низовья р.Чулым (Гынгазов, Миловидов, 1977; Степанян, 1978); 80 - окр. пос.Карагай Бакчарского р-на (Фомин и др., 1972; Иванов, 1976); 81 - долина р.Обь в р-не устья р.Томь (Степанян, 1978); 50 км северо-западнее г.Томск (Гынгазов, 1962а); 82 - г.Томск (Гынгазов, Миловидов, 1977); 83 - с.Черная речка в 50 км северо-восточнее г.Томск (Гынгазов, 1962); Гынгазов, Москвитин, 1965); 84 -Тисульский р-н (Гуреев, 1979); 85 - горнотаежные леса Кузнецкого Алатау (Тугаринов, 1927; Гуреев, 1979); 86 - окр. с.Большой Кемчуг на одноименной реке (Наумов, 1960); 87 — р.Ка-ча (колл. ЗИН); 88 - окр. г.Красноярск, р.Ба-заиха (Юдин, 1952); р.Слизнева (колл. ЗИН); 89 - р.Мана (Иванов, 1976); 90 - с.Выезжий Лог (колл. ЗИН); 91 - восточная часть Минусинского края (Bianchi, 1907; Благосклонов, 1954); Минусинский р-н, устье Собачьего Ключа (Иванов, 1976; колл. ЗИН); 92 - окр. г.Туран и с.Усть-Уюк (Реймерс, 1960); 93 — северные склоны Западного Саяна (Сушкин, 1938; Благосклонов, 1954; Степанян, 1978); 94 - Северо-Восточный Алтай (Дементьев, 1937; Сушкин, 1938; Степанян, 1978); Березовый Ключ, приток р.Абакан; верховья р.Ижи; ключ Мойнак-су, приток р.Ада; нижнее течение р.Кайзас; р.Конису близ оз.Бу-лан-куль (Сушкин, 1938); 95 - Телецкое оз., р.Егач (Иванов, 1976; колл. ЗИН); 96 - верховья р.Бия (Равкин, 1973); 97 - северные предгорья Алтая: р.Клык, вверх от Узун-ока (Сушкин, 1938).
Систематика. Монотипический вид. Места обитания. Характерный представитель высокоствольной хвойной тайги, но населяет также хвойные леса с незначительной примесью лиственных деревьев и кустарников, а также смешанные хвойно-лиственные леса по низинным участкам, долинам рек, сопкам, склонам и гребням гор (избегая очень крутых склонов и ущелий) до высоты 1800 м н.у.м. В разреженных лесах чаще держится в среднем и нижнем ярусе, в густых — поднимается в кроны деревьев — в более освещенные и прогреваемые места.
На юго-востоке Западно-Сибирской равнины в Томской обл. гнездится на участках
78
заболоченной темнохвойной тайги со значительной примесью кедра (Гынгазов, Миловидов, 1977). В Красноярском крае населяет приречные ельники, тяготея к местам, где последние чередуются с заливными лугами и зарослями кустарников (Юдин, 1952), а на Среднем Енисее у с.Ворогово (61° с.ш.) отмечена в высокоствольном кедровнике (Готфрид, 1982). В Кузнецком Алатау таежная мухоловка поселяется в горно-таежных кедрово-пихтовых лесах (Тугаринов, 1927; Гуреев, 1979), а на Северо-Восточном Алтае — в кедровниках или в лесах со значительной примесью кедра, поднимаясь в горы до 1500 м (Сушкин, 1938).
На верхней Лене обитает в приручье-вых ельниках (Реймерс, 1966). У истоков р.Ангара отмечена в гнездовой период в смешанном елово-березовом лесу (сообщ. А.А.Винокурова).
В Баргузинском заповеднике гнездится главным образом в приручьевых смешанных лесах с примесью кедра, пихты и ели, реже — в сосновых и лиственных лесах, тяготея к равнинной части заповедника (Скрябин, Филонов, 1962). На юге Витимского плоскогорья (Исинга) и в Становом нагорье (Муйская долина) встречается в лиственных лесах (Измайлов, 1967; Измайлов, Боровицкая, 1967). На Олекмо-Чар-ском нагорье по рекам Тяня и Токко населяет как смешанные хвойные леса из ели, сосны, лиственницы, кедра, так и светлую лиственничную тайгу, а также ельники по берегам и островам этих рек (Воробьев, 1963).
В Хабаровском крае на хр.Хехцир отмечена в елово-пихтовой тайге основного водораздельного гребня (Кулешова и др., 1965), а в Нижнем Приамурье гнездится главным образом в темнохвойных таежных лесах, в том числе и в прилегающих к пойме р.Амур. Здесь эта мухоловка наиболее охотно поселяется в долинных ельниках, но гнездится и в смешанных лесах (Яхонтов, 1963; Штильмарк, 1971; Кистя-ковский, Смогоржевский, 1973).
В Приморье на восточных склонах Среднего Сихотз-Алиня таежная мухоловка населяет смешанные хвойные и хвойнолиственные леса по высоким надпойменным террасам, гребням и склонам гор, а также в верховьях рек и ключей (Кулешова, 1976). В южной части этого хребта она обычна на гнездовании в хвойных лесах вершин гор Партизанского р-на (Шуль-пин, 19276). В Уссурийском заповеднике
гнездится в чернопихтарниках (Коренкова, 1969), ельниках (Иванов, 1952) и хвойно-широколиственных лесах (Назаренко, 19846), а на Даубихэ-Майхинском плато -в глухой елово-пихтовой тайге с примесью березы и кедра на высотах 700-780 м (Воробьев, 1954).
На о-ве Сахалин (Нечаев, 19846) населяет разреженные высокоствольные хвойные, реже смешанные (хвойно-березовые) леса с обширными полянами и окраины лесных массивов, граничащих с долинными лесами и заболоченными участками. На побережье оз.Айнское и в долине р.Айнская гнездится на опушках елово-пихтовых лесов с одиночными березами, местами разреженных в результате старых рубок и пожаров, а также в заболоченных лесах. На побережье залива Терпения и в долине р.Набиль — в разреженных приречных лиственнично-еловых и елово-пихтовых лесах. В долинах рек Поронай и Тымь предпочитает окраины елово-пихтовых лесов и долинные смешанные леса. На северо-западном и северо-восточном побережье острова — разреженные и заболоченные лиственничные леса, а также березово-лиственничные с кедровым и ольховым стлаником. На п-ове Шмидта населяет елово-лиственничные леса с белой березой, рябиной и кедровым стлаником.
В Северо-Восточном Китае на хр.Чан-байшань отмечена в горных хвойных лесах на высоте 1200—1800 м (Чень Пзн, 1963).
Численность. Неравномерна. На территории СССР в большинстве мест гнездового ареала в характерных биотопах обычна, в излюбленных бывает даже многочисленна. В Северном Заангарье; южной части Витимского плоскогорья, Становом нагорье, Читинской обл. на о-ве Сахалин, а также у северной границы распространения на материке — редка. На некоторых участках ареала (верхняя Лена, Комсомольский заповедник и др.) численность подвержена годовым колебаниям (Реймерс, 1966; Штильмарк, 1973).
Прилет и весенний пролет на территории СССР. Проходит с I декады мая по II декаду июня включительно. Наиболее интенсивно протекает в мае. Одиночные птицы на юге Приморья как исключение появляются в апреле.
На островах в северо-западной части залива Петра Великого первые пролетные мухоловки отмечены в начале мая, а массовый их пролет проходит здесь во П декаде этого месяца (Лабзюк и др., 1971).
79
На о-ве Большой Пелис пролетная самка добыта 25.V 1964 г. (колл. БПиДВНЦ).
На о-в Сахалин (Нечаев, 19846) таежные мухоловки прилетают в середине мая. Наиболее интенсивно пролет протекает во второй половине этого месяца, однако отдельные особи продолжают лететь и в I декаде июня. На о-ве Монерон одиночные пролетные птицы отмечены 22.V и 4.VI 1973 г. (Нечаев, 1975).
В Южном Приморье у оз.Хасан самая ранняя для территории СССР таежная мухоловка добыта 16.IV 1964 г, (Панов, 1973). В заповеднике ’’Кедровая падь” пролет проходит (3 года): 7 - 23.V, 12-28.V, 18—23.V (Панов, 1973). У юго-восточного и южного берегов оз.Ханка пролетные птицы (4 года) добывались: 12, 18, 22, 23 и 26.V (Черский, 1915а; 19156; колл. ЗИН). В низовьях р.Большая Уссурка в 1938 г. первая мухоловка у с.Вербовка отмечена 23 .V; пролетные птицы встречены здесь также 29.V и 17.VI (Спангенберг, 1940; 1965).
На нижнем Амуре начало весеннего прилета в 1961 г. у с.Гавань зарегистрировано 23.V (Кистяковский, Смогоржевский, 1973; колл. КГУ), а на р.Бурее в устье р.Талой 3 самца добыты 25 и 26.V 1962 г. (колл. КГУ).
В Баргузинский заповедник прилетает в первой половине мая. В 1959 г. первое появйение отмечено 9.V (Скрябин, Филонов, 1962). В Южном Прибайкалье пролет проходит во второй половине мая; под г.Иркутск — 30.V—9.VI (Малышев. — Цит. по: Реймерс, 1966). На верхней Лене первые птицы в 1960 г. встречены 29.V (Реймерс, 1966).
В окр. г.Красноярск в тайге предгорьев Саян (2 года) первое появление отмечено 26 и 29.V (Юдин, 1952). На Северо-Восточный Алтай таежные мухоловки прилетают во II декаде июня (16.VI) (Сушкин, 1938).
В Томской обл. первые птицы зарегистрированы на Среднем Чулыме 12.VI 1969 г., а на р.Кеть у с .Красная Курья -16.VI 1963 г. (Гынгазов, Миловидов, 1977).
Размножение. Первыми на места гнездования прилетают самцы. Петь они начинают сразу после прилета. В период пролета поют и мигрирующие птицы (Юдин, 1952; Нечаев, 19846). Песня таежной мухоловки — однообразная торопливая трель, состоящая из звонких и мелодичных звуков (Нечаев, 19846). Ее длительность и структура подвержены возрастной изменчивости.
После прилета самок птицы разбиваются на пары и приступают к строительству гнезд. Гнездовые участки (в характерных биотопах) невелики и расстояние между гнездами соседних пар составляет 150-200 м (Юдин, 1952). Строит гнездо главным образом самка. Гнезда помещаются на ветвях и реже в развилках стволов хвойных деревьев (лиственница, ель, пихта) на высоте (п = 7) 1,0—8,0 м, в среднем — 4,5 от поверхности зёмли (Воробьев, 1963; Нечаев, 19846; наши данные). На ветвях они располагаются или в непосредственной близости со стволом гнездового дерева, или на некотором удалении (0,15—4,0 м, в среднем — 1,8, п = 5) от него (Воробьев, 1963; Нечаев, 19846; наши данные).
Гнезда неправильной чашевидной формы, очень аккуратные. В качестве строительных материалов птицы используют сухие веточки лиственницы (длина 70—90 мм, d до 3 мм), ели, тонкие полоски и волокна луба, рассученную кору жимолости, пучочки мхов (в том числе и Neckera pennata), тонкие стебельки трав, корешки, сухие листья, отдельные пучочки шерсти, хвою и кусочки талломов лишайников. Лоток выстилается растительными волокнами, тонкими корешками, волосом шерсти млекопитающих (изюбрь), а по данным Л.Тачановского (Taczanowski, 1891) — мягкими мхами и сухими листьями злаков. Снаружи к некоторым гнездам паутиной прикрепляются кусочки талломов лишайников и коконы науков (Нечаев, 19846). В.А.Нечаев (19846) находил на Сахалине гнезда этого вида, на 80—90 % состоящие из зеленых мхов (Pleurozium schreberi, Habrodon leucotrichus, Brachythecium ve-lutinum, Amblystegium serpens, Rauiella fuijsana, Rhytidiodelphus triquetrus и др.) и лишайников (Usnea sp., Alectoria sp. и др.). Размеры гнезд (n = 5; Нечаев, 19846), мм: диаметр - 85—110 х 80—125, в среднем - 97 х 108; высота - 55—73, в среднем - 61; диаметр лотка - 45-70 х х 52-70, в среднем 56 х 58; глубина лотка - 34—42, в среднем - 37.
В сезон размножения на территории СССР таежная мухоловка имеет один репродуктивный цикл. Количество яиц в полной кладке обычно 5—6, реже — 4, а по данным К.Н.Благосклонова (1954) и Л.АЛор-тенко (1960) - может быть 7 и даже 8. Яйца остроовальные насыщенно сероватобирюзовые (но не темные), с мельчайшими ржавчато-охристыми точками, которые сгущаются к тупому полюсу яйца. На отдельных яйцах в некоторых местах поверх
80
ности скорлупы этот рисунок сливается и воспринимается как налет по основному фону. Встречаются также яйца, на которых мелкие светло-бурые пятнышки образуют ’’венчик” или ’’шапочку” на тупом и реже -на остром полюсе яйца или покрывают более или менее равномерно всю поверхность скорлупы (Нечаев, 19846). Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
В период насиживания кладки самка сидит на гнезде очень плотно. Птенцы вылупляются в течение суток.
Пуховой птенец таежной мухоловки (Нечаев, 1983; 19846) имеет редкий черный пух на надглазничных (длина пуха 8-10 мм), затылочной (12—15), спинной (10-12) и плечевых (8-10) птерилиях. У некоторых особей он находится и на глазных (1-2). Отмечены также рудиментарные локтевые и копчиковая птерилии. Кожа желтовато-розовая. Ноги розоватожелтые, когти желтые. Клюв сероватожелтый с потемнением на вершине, яйцевой зуб серый. Ротовая полость и язык желтые, складки в углах рта светло-желтые.
В выкармливании птенцов принимают участие оба партнера пары. После того как птенцы покинут гнездо, родители продолжают кормить слетков. Подкармливают они первое время и летных молодых. В этот период птицы держатся выводками.
Сезон размножения таежной мухоловки на территории СССР растянут и длится с конца III декады мая по конец июля включительно. Выводки и самостоятельные летные молодые встречаются до конца августа.
На о-ве Сахалин (Нечаев, 19846) строительство гнезд начинается в конце I — начале II декад июня и у разных пар растягивается до I декады июля. В долине р.Айн-ская в 1978 г. строительство 1 гнезда было закончено 20.VI, другое 2.VII только строилось. Откладка яиц начинается в середине июня; у отдельных пар растягивается до середины июля. Полные кладки обнаружены на острове: 24,25 VI (2 гнезда) и 1LVII 1978 г. Первые птенцы появляются в середине III декады июня, массовое вылупление проходит в первой половине июля, а из поздних кладок — во второй половине этого месяца. Гнезда с 1-2-дневны-ми птенцами в 1978 г. найдены 27 и 29.VI, а гнездо с 7—8-дневными - 3.VII. Первые слетки зарегистрированы в середине июля. В долине р.Хребтовая они встречены 19.VII 1977 г., а в окр. пос.Ныш на р.Тымь -
24.VII 1980 г. Выводки в разных точках острова (4 года) отмечены: 3, 4, 10, 14 и 26.VIII.
В Сихотэ-Алинском заповеднике гнездо таежной мухоловки с 1 яйцом (начало кладки) обнаружено 8.VI 1973 г. (картотека заповедника).
На нижнем Амуре в окр. с.Гячи занятие гнездовых участков отмечено 27.V, а 18.VI добыта взрослая самка снаседным пятном в окр. с.Покровка (Кистяковский, Смогоржевский, 1973). Летные молодые птицы добыты: у с.Нижняя Гавань - 18.VIII 1967 г. (колл. БПиДВНЦ) и у оз.Кизи на р.Таба - 31.VIII 1928 г. (колл. ЗИН). На хр.Хехцир выводок таежной мухоловки встречен 28.VII 1963 г. (Кулешова и др., 1965).
В Амурской обл. у пгт Экимчан гнездо с полной ненасиженной кладкой из 5 яиц найдено 21.VI 1971 г. (колл. В.ВЛеонови-ча), а на Охотском побережье (р.Улейкан) птенец добыт 23.VII 1936 г. (колл. ЗИН).
На Олекмо-Чарском нагорье готовый к размножению самец добыт 13.VT 1956 г., a 27.VI здесь на р.Тяня найдено гнездо с 6-ю 2-дневными птенцами (Воробьев, 1963). В Читинской обл. самка с сильно увеличенными фолликулами яичника добыта 29.V (Павлов, 1948), а в Бурятии на рДолбай молодая летная птица - 30.VII 1912 г. (колл. ЗИН). В Иркутской обл. у с.Подвалочное 3 молодых мухоловки добыты 17, 26 и 27.VIII 1914 г. (колл. ЗИН).
В Красноярском крае (Юдин, 1952) разбивка на пары проходит в I декаде июня. Самка с наседным пятном добыта у гнезда близ г.Красноярск на р.Базаиха 10.VII 1941 г., а несколько выводков молодых с недоросшими рулевыми перьями встречены здесь 13.VIII 1941 г. На Енисее у пос.Ворогово выводок отмечен 18.VII 1976 г. (Готфрид, 1982). В Томской обл. на р.Кеть самец с увеличенными семенниками добыт 16.VI 1963 г. (Гынгазов, Москвитин, 1965).
Линька. После того как маховые и рулевые закончат рост, у молодых начинается частичная послегнездовая линька. Она охватывает почти все мелкое оперение, кроме некоторых кроющих маховых и рулевых и у большинства особей заканчивается к началу осенней миграции. У молодых из поздних выводков эта линька может прерываться осенней миграцией и заканчивается на зимовках (Виноградова и др., 1976). На местах зимовок у первогодков в частичную предбрачную линьку
81
сменяются рулевые (Виноградова и др., 1976). Полная послебрачная линька оперения проходит у них на местах гнездования после окончания периода размножения. У взрослых птиц в году отмечено 2 линьки: полная послебрачная на местах гнездования, которая начинается в конце июля - начале августа и заканчивается к началу осенней миграции, и полная предбрачная на зимовках.
Питацие. Основу пищи составляют насекомые разных групп и стадий развития, реже — пауки. В конце лета и осенью на пролете мухоловки также поедают мелкие сочные плоды некоторых растений.
Отлет и осенний пролет на территории СССР. Проходит с III декады августа по II декаду октября включительно. Наиболее интенсивно протекает в сентябре. Одиночные птицы задерживаются до середины III декады октября.
В районе Красноярска пролет проходит в первой половине сентября (Юдин, 1952). Самец сеголеток добыт здесь 15.IX 1931 г. (колл. ЗИН). В Южной тайге Средней Сибири осенняя миграция протекает в конце августа - начале сентября. Самая поздняя встреча таежной мухоловки зарегистрирована под г.Иркутск 23.Х 1959 г. (Реймерс, 1966).
В Баргузинском заповеднике пролет заканчивается в начале сентября (Скрябин, Филонов, 1962). В Читинской обл. на берегу р.Чита пролетная птица добыта 6.IX 1941 г. (Гагина, 1960), у пос.Кулусутай (Торейские озера) мухоловки добыты 10 и 11.IX 1856 г. (колл. ЗИН), а у сопки Кухадан - 16 и 18.IX 1980 г. (колл. ИЗАНУ).
На нижнем Амуре у г .Николаевск молодой самец добыт 7.IX 1854 г. (колл. ЗИН).
В Приморье на территории Уссурийского заповедника пролетные птицы отмечены 28 и 30.IX 1948 г. (колл. БПиДВНЦ); у устья р.Раздольная (Суйфун) (2 года) -27.IX и 2.Х (колл. ЗИН), а в Шкотовском р-не у пос. Анисимовка — 3.IX 1975 г. (колл. ИЗАНУ). У южного берега оз.Хан-ка (с.Нововладимировка) 2 самца добыты 15 и 27.IX 1911, а у Амурского залива на п-ове Янковского 1 птица - 1.Х 1912 г. (Черский, 1915а). На п-ове Де-Фриза небольшие стайки и одиночные пролетные мухоловки (2 года) встречены: 24—26, 29, 30.IX; 2-5.X (Омелько, 1979). В заповеднике ’’Кедровая падь” (3 года) пролет проходит: 16.IX-2.X; 30.IX-17.X; 2 и 82
18.Х (Панов, 1973), а в 1945 г. пролетный самец добыт здесь 15.IX (колл. БПиДВНЦ).
На о-ве Сахалин (Нечаев, 19846) отлет проходит в течение сентября. Последние птицы встречаются здесь до середины октября, отдельные особи задерживаются на острове до первых снегопадов, которые отмечаются обычно в I декаде октября.
На островах в северо-западной части залива Петра Великого первые пролетные мухоловки появляются 15.IX (Назаров и др., 1978) и 23.IX, но наиболее интенсивно пролет проходит в первой половине октября (Лабзюк и др., 1971). На о-ве Ри-корда взрослый самец добыт 28.IX 1965 г. (колл. БПиДВНЦ).
Практическое значение. Уничтожает некоторых насекомых-вредителей леса и переносчиков возбудителей некоторых заболеваний человека и животных. Переваривая только мякоть сочных плодов и оставляя при этом семена растений нетронутыми, птицы этого вида в какой-то степени участвуют в распространении последних на территории Южного Забайкалья, в Приморье и на о-ве Сахалин.
Малая мухоловка — Ficedula parva (Bechs-tein, 1794)
Muscicapa parva Bechstein, 1794, Latham’s AUg. Ubersicht Vogel, 2: 356, изображение на титульной странице 3 тома, хр.Тюрингенский Лес, Германия.
Син. (русск.): лоцманчик, малый мухолов.
Описание. Составлено по 190 экз. (148 - из европейской части СССР и Кавказа, 42 — из Сибири и Дальнего Востока СССР).
Вершину крыла образуют 3-е и 4-е первостепенные маховые (3 < 4 — 4,3 %; 3 = 4 — 37,1 %; 3 > 4 - 58,6 %). У большинства особей (97,4 %) 5-е немного их короче, но у некоторых экземпляров (2,6 %) равно по длине 3-му и 4-му. Первое первостепенное маховое очень мало и обычно только на несколько миллиметров выступает за вершины кроющих кисти (табл. 10); 2-е всегда меньше 5-го (2 < 6 -32,5 %; 2 = 6- 35,0 %; 2 > 6 - 32,5 %). У большинства особей (80,5 %) наружные опахала 3—5-го первостепенных маховых в вершинной части заметно сужены. У отдельных экземпляров (19,5 %) сужено в вершинной части и наружное опахало 6-го махового. Количество щетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3—5 (3 — 27,8 %; 4 — 70,4%; 5 — 1,8 %); длина наибольшей (п = 113): 4,6-7,3 мм (в среднем - 6,3). Количество
щитков подотеки на передней части цевки варьирует от 4 до 6. У взрослых птиц на вершине хвоста (в центре) имеется небольшая выемка, размеры которой варьируют. Половой диморфизм выражен в окраске оперения.
Распространение. Евразия от низовьев Эльбы, Среднего Рейна и верховьев Дуная к востоку до верхнего течения Анадыря, до Камчатки и побережья Охотского моря. В Европе (за пределами СССР) распространена к северу до Дании, Южной Швеции (предположительно) и Южной Финляндии, а к югу до восточной половины Альп и между Адриатическим и Черным морями до 41—42° с.ш. Северо-Западный Иран, Северная Монголия и Северо-Западные Гималаи от Кашмира к востоку до Гарвала (Иванов, 1976; Степанян, 1978). На территории СССР гнездится в европейской части, на Кавказе, в Сибири, на Дальнем Востоке, Шантарских о-вах и предположительно на о-ве Парамушир (см. рис. 20). На всем гнездовом ареале птицы этого вида являются перелетными и зимуют на юге Азии.
Систематика. Политипический вид, образующий 3 подвида, 2 из которых F.p.parva и F.p.albicilla встречаются и гнездятся на территории СССР. Таксономический статус формы subrubra, заселяющей изолированный участок гнездового ареала в Гималаях, окончательно не выяснен и остается спорным. Не исключено, что она представляет собой самостоятельный вид F.subrubra.
Кавказские малые мухоловки, описанные в 1911 г. Б.А.Домбровским в качестве самостоятельной формы F.p.colchica, по нашим данным, не отличаются от таковых из европейской части СССР как по морфометрическим показателям и окраске оперения, так и по основным чертам их биологии и, несомненно, должны рассматриваться в составе номинативного подвида.
Европейская малая мухоловка — Ficedula parva parva (Bechstein, 1794)
Muscicapa parva Bechstein, 1794, Latham’s Allg. Ubersicht Vogel, 2: 356, изображение на титульной странице 3 тома, хр.Тюрингенский Лес, Германия.
Описание. Составлено по 148 экз. из европейской части СССР и Кавказа.
У старого самца верхняя часть и бока головы, а также шеи от буровато-серого до темно-серого. Спина и надхвостье сероватобурые. Верхние кроющие хвоста бархатно
черные. На горле, зобе и верхней части груди располагается большое пятно от охристорыжего до ярко-ржавчато-рыжего цвета. Его величина и интенсивность окраски зависят главным образом от возраста птицы. У более старых самцов оно крупнее и ярче. Серая окраска с шеи распространяется на бока груди и окантовывает по краям (кроме низа) рыжее пятно. Нижняя часть груди, брюшко и нижние кроющие хвоста белые. Оперение на боках тела от светло-буровато-охристого до рыжевато-охристого. Нижние кроющие крыла от белого до светло-желтовато-охристого. Маховые и верхние кроющие крыла бурые. Центральные рулевые черные, остальные — двухцветные: белые в основаниях и черные на вершинах. Клюв коричнево-бурый с посветлением в основании подклювья. Ноги черно-бурые. Радужина глаза бурая. Индивидуальная изменчивость проявляется в варьировании величины белого поля в основании рулевых; сезонная - в общем посветлении окраски оперения в результате его обноса и выцветания.
У взрослой самки оперение верхней части тела от серовато-бурого до буроватокоричневого. Верхние кроющие хвоста и рулевые имеют такую же окраску, как и у самца. Бока головы светлее ее верхней части. Циз тела от белого с бледно-буровато-охристым оттенком до светло-охристого. Нижние кроющие хвоста белые. Маховые и верхние кроющие крыла бурые с незначительной примесью коричневого.
Молодые в гнездовом наряде сверху буровато-коричневые с светло-желтовато-охристыми пятнами на перьях, каждое из которых сверху окантовано бурым. Горло, зоб и верхняя часть груди бледно-буроватоохристые с бурым чешуйчатым рисунком, интенсивность которого уменьшается в верхней части брюшка. На нижней части брюшка этот рисунок отсутствует. Нижние кроющие хвоста белые.
Первый послегнездовой наряд идентичен наряду взрослой самки, но на второстепенных и третьестепенных маховых и верхних кроющих второстепенных маховых охристые каемки развиты сильнее.
Размеры отдельных частей тела европейской малой мухоловки приведены в табл. 10.
Масса тела, г: самцы первогодки (п = = 10) (13-18.IV, Южная Туркмения) -8,15—11,20, в среднем — 9,77; самцы старше 1 года (и =23) (13-18.IV, южная Турк
83
мения) - 8,90-11,70, в среднем - 10,32; самка (Тебердинский заповедник) — 9,5 (Ткаченко, 1966).
Рас пространение. Европейская часть гнездового ареала вида, Кавказ и Северо;3ападный Иран. На всем гнездовом ареале мухоловки этого подвида являются перелетными и зимуют главным образом на юге Азии, в том числе и на п-ове Индостан. Более детально распространение европейской малой мухоловки на территории СССР показано на карте (рис. 20).
Кадастр к рис. 20.
1 - Эстония; Матсалуский заповедник (Рен-но, 1968); 2 - Нигула; Лайксааре (Кумари,
1955); 3 - Латвия; Тукумский р-н (Меднис, 1965); 4 - окр. г.Елгава (Villes, 1943); 5 - Литва; заказник Биржулис (Гражулявичюс, 1977); 6 -г.Каунас и его окрестности (Алекнавичус, 1965); 7 - заповедник ’’Жувинтас” (Иванаускас, 1968); юго-запад Балтийской возвышенности (Валюс, Кигуолене, 1981); 8 - Белоруссия; Витебская обл.; Тиосто, Городище, Полтево (Дорофеев, 1969); 9 - Березинский заповедник (Кожевникова, 1965; Федюшин, Долбик,1967); 10-Оршанский р-н, окр. с.Протасово (Бианки, 1914); 11 -Кричевский р-н, с.Клины (Федюшин, Долбик, 1967); 12 -Минская обл., Пуховичский р-н (Федюшин, Долбик, 1967); 13 - Бобруйский р-н, Поток-ское лесничество (Федюшин по Долбику, 1959); 14 - Брестский р-н; заповедник ’’Беловежская пуща” (Федюшин, Долбик, 1967); 13 - Хойник-ский р-н, Великоборское лесничество (Долбик, 1959); 16 - Припятские болота; с.Таховичи; д.Переров; оз.Червоное (Федюшин, Долбик,
Рис.20. Распространение малой мухоловки (Ficedula parva) на территории СССР:
1 - F.p.parva, 2 - F.p.albicilla; а - гнездится, б - отмечена в гнездовой период. См. кадастр стр. 84-
84
1967); 17 - Лельчицкнй р-н; Марковское и Озе-ранское лесничества; окр. с.Лельчицы (Федюшин, Долбик, 1967); 18 - Василевичский р-н; Василевичское и Макановичское лесничества (Долбик, 1959); 19 - Украина; Волынская обл.; окр. оз.Бужня (Шацкие озера) (сообщ. М.И.Головушкина) ; 20 - Андрушевское лесничество (колл. ЗИН); 21 - окр. г.Нововолынск (сообщ. М.И.Головушкина); 22 - Тернопольская обл.; с.Бережаны; с.Холгоче (колл. ЛПМ); 23 - Шумский р-н, Волынское лесничество, окр. с.Стожок (наши данные); 24 - Львовская обл.; с.Пеняки (колл. ЛПМ); с-Подгайцы (Страутман, 1963); с.Тулиголово (колл. ЛПМ); 25 - северные и южные склоны Украинских Карпат; гора Круглая, Воловецкий перевал; Торуньский перевал; г.Во-дичное; долина р.Опор; окр. с.Микуличин (Страутман, 1954; 1963; колл. ИЗАНУ); 26 — долина р.Уж (Портенко, 1950); с.Каменица; пгт Пере
чни (колл. ЗИН); 27 - Закарпатская равнина (Страутман, 1963); 28 - долина р. Белый Чере-мош; с.Гринява (Страутман, 1954; 1963; колл. ИЗАНУ); 29 - долина р.Прут (Страутман, 1954; 1963); 30 - Хмельницкая обл., Летичевский р-н, с.Лысогорка (колл. ЗИН); 31 - Одесская обл., Балтийский р-н (колл. ЗИН); 32 - Савранский лес (сообщ. Т.Б.Ардамацкой); 33 -с.Песчаная, с.Бандурово (Mauersberger, Portenko, 1967); 34 - Кировоградская обл.; Черный лес (Шевченко, 1940); истоки рек Ингул и Ингулец (Иванов, 1976); 35 - Житомирская обл.; окр. г.Житомир (колл. ИЗАНУ); 36 - окр. с.Корнин; окр. с.Яроповичи (сообщ. М.И.Головушкина); 37 - Белоцерковский р-н (Кирпиченко, 1932); 38 - окр. ст. Спартак; окр. с.Розважев (сообщ. М.И.Головушкина); 39 - Киевская обл.; окр. ст. Боярка (колл. Ю.В.Файделя); г.Киев и его окрестности (колл. ИЗАНУ; КГУ): лесничество
87, 94-96.
85
Яковец (колл. ИЗАНУ); окр. пос.Ирпень; окр. с.Мостище (сообщ. М.И.Головушкина); 40 - Черкасская обл., окр. г.Канев (сообщ. М.И.Головушкина) ; 41 - окр. с.Чапаевка (сообщ. В.И.Стригунова); 42 — Черниговская обл., Черниговский р-н,*с.Неданчичи (сообщ. М.Ф.Самофалова) ; Грабовский лес (колл. ИЗАНУ); 43 -окр. сЛипов Рог; окр. с.Вертиевка (сообщ. М.Ф.Самофалова); 44 - Сумская обл., Шосткин-ский р-н (сообщ. В.П.Белика); 45 - с.Тростянец (колл. ХГУ); 46 - Полтавская обл. (Гавриленко, 1929); р.Ворскла; с.Парасоцкое; Бретерское; с.Диканька; с.Дубове (колл. ХГУ); 47 - окр. с.Гута (колл. ХГУ); р-н с.Краснокутск (колл. ХГУ); 48 - Харьковская обл. (Сомов, 1897; Степанян, 1978; колл. ХГУ); Волчанский р-н, долина верхнего течения р.Орчик (Сомов, 1897); 49 - окр. г.Харьков (колл. ИЗАНУ); 50 - Гот-вальдовский р-н (Сомов, 1897); окр. с.Гайдары (сообщ. А.Н.Гудины); г.Чугуев (колл. ХГУ); 51 - р-н г.Днепропетровск (Mauersberger, Рог-tenko, 1967); 52 - Донецкая обл., г .Волноваха (Будниченко. - Цит. по: Mauersberger, Portenko, 1967); 53 - Великоанадольское лесничество (колл. ХГУ); 54 - Ворошиловградская обл., окр. г.Кременная (сообщ. В.В.Ветрова); 55 -Луганский заповедник (сообщ. В.Г.Сулика); 56 - нижнее течение р.Деркул; окр. ст. Н.Ильен-ко (сообщ. В.В.Ветрова); 57 - Молдавия; заповедник ’’Кодры” (Ганя, Литвак, 1976); Садовский и Реденский леса (колл. ХГУ); с.Бужевка, с.Егоровка (Ганя, 1965); Трифауцкий, Рубленец-кий, Копанский, Кошницкий леса; окр. с.Моро-зены; окр. с.Ватич; Кицканская дача (Ганя, Литвак, 1961; Ганя, 1965); 58 - окр. г.Лазовск (Ганя, Литвак, 1976); 59 - Сорокский лес (Ганя, Литвак, 1961; Ганя, 1965); 60 - Калининградская обл. (Портенко, 1960); 61 - Карелия, 63° с.ш. (Степанян, 1978); р.Суна (Портенко, 1960); Черный лес (колл. КГУ); 62 — заповедник ’’Кивач” (Нейфельдг, 1958; Ивантер, 1962); 63 — северный берег Онежского озера (Благосклонов, 1954); 64 - Заонежье (Хохлова, 1977); 65 - Пряжинский р-н (Нейфельдт, 1958; колл. ЗИН); 66 — северо-восточное побережье Ладожского озера (Головань, 1982); 67 - Ленинградская обл. (Мальчевский, Пукинский, 1983); окр. г.Выборг (Иванов, 1976); 68 - г Ленинград и его окрестности (колл. ЗИН); Тосненский р-н, Лисинский лесной массив (Поспелов, 1957); 69 - с Лебяжье (колл. ЗИН); 70 - южные р-ны Ленинградской обл. (Мальчевский, Пукинский, 1983); 71 - Псковская обл. (Зарудный, 1910; Мальчевский, Пукинский, 1983); Харламова гора (колл. ЗИН); 72 - окр. Псковско-Чудского водоема (Мешков, 1961); 73 - Новгородская обл., ’’Княжий двор” (колл. КБ МГУ); с.Вылеги (колл. ЗИН); 74 - Калининская обл. (Зиновьев, Шапошников, 1978); 75 - Дарвинский заповедник (Спангенберг, Олигер, 1949); 76 - окр. оз.Кубенское (Беме, 1962); 77 - устье р.Онега (Иванов, 1976); 78 - г Архангельск (Иванов, 1976); юго-восточное побережье Белого моря (Степанян, 1978); 79 — Вологодская обл., Велико-Устюгский р-н (Второв, Дроздов, 1960); 80 - Тотемский р-н (Второв, Дроздов, 1960); 81 - территория между низовьями р.Вычегда и рЛинега (Паровщиков, 1959); 82 - Кировоградская обл. (Портенко, 1960); 83 - северная часть области, сЛальск (Благосклонов, 1954);
84 — верховья р.Мезень (сообщ. А.А.Естафьева) ;
85 - окр. г.Сыктывкар (сообщ. А.А.Естафьева); среднее течение р.Вычегда (Гурьев, 1982); 86 -устье рЛокчим (сообщ. А.А.Естафьева); 87 -Ярославская обл. (Портенко, 1960); 88 - Владимирская обл.: окр. г.Владимир, Пенкинский лес (Измайлов, 1981); 89 - Камешковский р-н, Знаменский лесной массив к востоку от пос.Пен-кино (Михлин, Сальников, 1981); 90 - Собин-ский р-н, леса юго-западиее г.Лакинск (Михлин, Сальников, 1981); 91 - Московская обл. (Пту-шенко, Иноземцев, 1968), г.Москва и ее окрестности (Флинт, Кривошеев, 1962; Птушенко, 1976; колл. ЗИН); 92 - Загорский р-н (колл. БИН СО АН); 93 - Дмитровский р-н (колл. В.В.Леоновича); 94 — г.Клин (колл. ЗМ МГУ);
95 - Звенигородская биостанция МГУ (Птушен-ко, Иноземцев, 1968); 96 - Можайский р-н (колл. КБ МГУ); 97 - Брянская обл. (колл. ХГУ); Карачаевский р-н (Суслова, 1937; колл. ЗИН); 98 - пос. Трубчевск (Суслова, 1937; колл. ЗИН); 99 - истоки р.Ока; Черномош-ный и Хуторской леса (Ефимов, 1915); 100 -Тульская обл., с.Бобровка; Карницкая дача; ст.Козловка; с.Никольское (колл. ЗИН); 101 -Рязанская обл. (Хомяков, 1900; колл. ЗИН); Переделецкое лесничество (Ежова, 1979; Френ-кина, 1981); 102 - Михайловский р-н, с.Нюхо-вец (колл. ЗИН); 103 - Белгородская обл. (Будниченко, Козлов, 1980); Борисовский р-н (Новиков и др., 1963); 104 - Тамбовская обл., Мичуринский р-н (Щеголев, 1978); 105 - Горьковская обл.; Пустынская биостанция Горьковского университета (Воронцов и др., 1962);
106 - Мордовия; Зуббво-Полянский р-н (Пред-теченский. - Цит. по: Луговой, 1975) ; 107 - Мордовский заповедник им.П.Г.Смидовича (Бородин, 1967); Темниковский р-н (Птушенко, 1938);
108 - Восточная Мордовия; Заводское лесничество в Приалатырских лесах (Луговой, 1975);
109 - Воронежская обл., окр. г.Воронеж; пойма р.Воронеж (Барабаш-Никифоров, Семаго, 1963);
110 - Воронежский заповедник (Барабаш-Никифоров, Павловский, 1947); 111 — г.Бобров (Иванов, 1976); с.Алексеевское (колл. ЗИН); 112 - Савальское лесничество (Кадочников. -Цит. по: Mauersberger, Portenko, 1967); 113 -Теллермановский лес (Благосклонов, 1954; Королькова, 1963); 114 - Кантемировский р-н (сообщ. В.П.Белика); 115 - пойма р.Хопер (сообщ. В.П.Белика); 116 - Ростовская обл., окр. ст.Мигулинская, ур. ’’Липняги” (Петров, 1965); Вешенский р-н, пойма р.Дон (сообщ. В.П.Белика); 117 - окр. стан.Нижнекундрючен-ская, ур. ’’Большой стан” (Петров, 1965); 118 -Цимлянский р-н; Сальская лесная дача (сообщ. В.П.Белика); 119 - г.Саратов (Иванов, 1976); 120 - устье р.Коса (приток р.Кама) (колл. ЗИН); 121 - с.Бисерово (Благосклонов, 1954); 122 -северная часть Удмуртской АССР (Птицы Волжско-Камского края, 1978); 123 - Пермская обл.: южные р-ны (Симкин, 1974); 124 - Лысьвенский р-н (Шилова и др., 1963); 125 - Александровский р-н (Шилова и др., 1963); 126 - Татарская АССР, Алексеевский р-н (Сербин, 1978); 127 -устье р.Свияга (Птицы Волжско-Камского края, 1978); 128 - Жигулевский заповедник (Белянина, Белянин, 1981) ; г.Куйбышев (Птицы Волжско-Камского края, 1978); 129 - р-н г.Бугурус-лана (Mauersberger, Portenko, 1967); 130 - верховья р.Самара (Благосклонов; 1954); 131 -г.Оренбург (Иванов, 1976); реки Сакмара и Боль
86
шой Ик (Зарудный по Кирикову, 1952); 132 -Башкирия (Ильичев, Фомин, 1979); г.Уфа (Птицы Волжско-Камского края, 1978); 133 - г.Ка-тав-Ивановск (Mauersberger, Portenko, 1967);
134 - г.Миасс (Снигиревский. - Цит. по: Mauersberger, Portenko, 1967); 135 - с.Верхняя Сы-серть (Данилов. - Цит. по: Mauersberger, Portenko, 1967); 136 - г.Тюмень (Портенко, 1960; Mauersberger, Portenko, 1967; Иванов, 1976); 137 -Краснодарский край, окр. г.Славянск-на-Кубани, Красный лес и лес Круглик (Кистяковский, 1932); 138 - Северский р-н, ст.Крепостная; с.Хребтовое (колл. ХГУ); 139 - с.Михайловский Перевал, Плисецкая щель (Пекло и др., 1978; колл. РГУ); гора Абляго, с.Пшада, Авраамова щель (Пузанов, 1938); 140 - окр. с-Джубга (колл. ХГУ); 141 - посЛазаревское (колл. БПиДВНЦ); с.Аше (колл. ЗИН); окр. г.Сочи, гора Ахун (Пекло и др., 1978); села Вардане, Эстонка, пос.Хоста, Псеашхо (Кудашев, 1917); верховья рЛсешва (Волчанецкий и др., 1962); окр. с.Красная Поляна (Сушкин, 1914; Кудашев, 1917); 142 - Кавказский заповедник (Аверин, Насимович, 1938); окр. пос.Никель, окр. пос.Гу-зерипль(Пекло и др., 1978); 143 - г .Ставрополь (Будниченко. - Цит. по: Mauersberger, Portenko, 1967); 144 - Ставропольская возвышенность; верховья р.Томузловка, Томузловский лес в 90 км юго-восточнее г.Ставрополь (Лиховид, 1977); 145 - Лопатинский лес на горе Недре-маной в 30 км южнее г.Ставрополь (Лиховид, 1977); 146 - Карачаево-Черкесская АО, с.Ар-хыз (колл. ХГУ); 147 - Тебердинский заповедник (Ткаченко, 1966); 148 - Северная Осетия; окр. пос.Редант; с.Дзаурикау (Беме, 1926); окр. г.Орджоникидзе (колл. КГУ); 149 - Грузия; Абхазия, Пицундский заповедник (Бернацкий, 1958); 150 - Аджария, Чуруксу (колл. ИЗАНУ); 151 - Лихский хребет (Жордания, Го-гилашвили, 1969); 152 - окр. г.Боржоми; с.Ахалдаба; Зекарское ущелье (Жордания, 1962); с.Сурами (колл. ЗИН); 153 - окр. г.Тбилиси (Сатунин, 1907); окр. г.Мцхета (Жордания, 1962); 154 - г.Тетри-Цкаро (Цветков, 1901; колл. ЗИН); 755 - г.Телави (Mauersberger, Portenko, 1967); 156 - г.Лагодехи (Mauersberger, Portenko, 1967); 157 - Армения; с.Шамлуг; с.Узунтала (Ляйстер, Соснин, 1942; Даль, 1954);
158 - Азербайджан (Гамбаров, 1941); 159 -Закатальский заповедник (Дроздов, 1965); 160 -хр.Муровдаг, оз.Гек-Гель (Дроздов, 1965); 161 - Талыш (Сатунин, 1907; Гамбаров, 1941); предгорья Талышских гор (Дроздов, 1965); с.Бутусар, окр. сЛерик (Сатунин, 1907); хр.Уля-сы (Дроздов, 1965).
Места обитания. Населяет самые разнообразные типы хвойных, смешанных хвойно-лиственных и лиственных лесов от северных таежных до южных широколиственных и субтропических вечнозеленых, на равнинах, в предгорьях и горах до 2350 м н.у.м. Встречается в пойменных лесах и зарослях, парках, старых фруктовых садах и как исключение на виноградниках. Тяготеет к среднему и нижнему древесно-кустарниковому ярусам леса. Любит участки с повышенной влажностью.
В Архангельской обл. местообитаниями малой мухоловки являются сухие высокоствольные ельники, близ осветленных мест (Паровшиков, 1959). В Карелии (заповедник ’’Кивач”) встречается как в хвойных (еловые) и смешанных хвойно-лиственных, так и в лиственных лесах (Ивантер, 1962). В Ленинградской обл. и на сопредельных территориях (Мальчевский, Пукинский, 1983) тяготеет к елово-лиственным лесам с хорошо развитым подростом и подлеском. Охотно поселяется также в березовоольховых молодняках, где встречаются куртины елочек или где ель выходит в первый ярус. Реже заселяет здесь зрелые высокоствольные ельники.
В Эстонии гнездится главным образом в елово-широколиственных лесах с хорошо развитым подлеском из липы, лешины, ели и других деревьев и кустарников (Кумари, 1955). В Латвии обитает в тенистых смешанных и еловых лесах (Бауманис и др., 1983), а в Литве гнездится локализовано в старых смешанных заболоченных лесах (Валюс и др., 1977).
В Вологодской обл. встречается в ельниках среди смешанного леса (Беме, 1962), а в северной части Волжско-Камского края — в зоне темнохвойных лесов (Птицы Волжско-Камского края, 1978). В Пермской обл. населяет елово-пихтовые леса и сложные ельники (Шилова и др., 1963), березово-осиновое старолесье, липняки, а в южных районах области — самые разнообразные смешанные леса (Симкин, 1974). В Алексеевском р-не Татарской АССР зарегистрирована в дубово-березово-осино-во-липовых и в дубово-липовых лесах, а также в чистых дубравах и на зарастающих вырубках (Сербин, 1978).
В Белоруссии предпочитает тенистые высокоствольные и влажные участки леса — главным образом различные ельники, елово-лиственные леса и субори, значительно реже поселяется в старых дубравах, смешанных дубово-сосново-осиновых лесах, а в Поозерье - также в осинниках и оль-шанниках (Федюшин, Долбик, 1967; Дорофеев, 1969). В Березинском заповеднике отмечена в молодых лишайниковых сосняках (Кожевникова, 1965).
В Московской обл. встречается в самых разнообразных лесах. В Можайском р-не найдена на гнездовании в еловом лесу — зеленомошнике (колл. КБ МГУ), у г.Дмитров — в густом сосняке с примесью березы (колл. В.В. Леоновича), в г .Москве — в парках (Измайловский и др.) (Флинт,
87
Кривошеев, 1962). В Орловской обл. гнездится в глубинных участках высокоствольных старых лесов с подлеском из кустарников (Ефимов, 1915), а в Рязанской (Переделецкое лесничество) у оз.Ур-жинского — в березовых лесах (Ежова, 1979). В Воронежском заповеднике придерживается темных лиственных и смешанных лесов.
В Киевской обл. встречается в смешанных лесах, а в Черниговской, по данным М.Ф.Самофалова, - и в старых сосняках у дорог. В Харьковской обл. отмечена в больших светлых лесах с подлеском, главным образом в дубовых (Сомов, 1897; колл. ИЗАНУ). В западных областях УССР на равнине Волыно-Подолья и Закарпатской низменности населяет сосново-буковые, буково-грабовые и буково-дубовые леса, парки, фруктовые сады и виноградники (Страутман, 1963). В Карпатах гнездится как на северных, так и на южных склонах. Здесь встречается повсюду в буковых лесах и поднимается в горы до их верхней границы распространения (1150— 1200 м). Кроме этого на северных склонах населяет также смешанные леса, а по речным долинам — и пойменные лиственные, по которым проникает глубоко в горы (Страутман, 1954; 1963).
В Молдавии гнездится в больших светлых дубово-грабовых и дубовых лесах с подлеском (Аверин, Ганя, 1970), а также в садах (Ганя, Литвак, 1961).
В Ростовской обл. отмечена в вязов-никах у стан.Нижнекундрюченской, в бай-рачной дубраве урочиша ’’Липняги” у х.Вяжа (Петров, 1965), а также гнездится в пойменных вязово-фруктовых лесах Вешен-ского р-на (сообш. В.П.Белика). На Северо-Западном Кавказе в гнездовой период малая мухоловка обычна на северных и более многочисленна на южных склонах Главного Кавказского хребта и его отрогов, от пояса широколиственных до верхней границы темнохвойных лесов, изредка поднимается даже до криволесья и субальпики (Аверин, Насимович, 1938). По нашим данным (Пекло и др., 1978), в окр.Кав-казского заповедника гнездится в лиственных с преобладанием бука (пос. Никель) и в смешанных хвойно-лиственных лесах (окр. пос.Гузерипль). В Тебердинском заповеднике излюбленными местами обитания являются долинные темнохвойные и смешанные темнохвойные Леса по берегам рек и ручьев. Поселяется здесь и в нагорных пихтарниках, но значительно реже
(Ткаченко, 1966). На Черноморском побережье встречается в самшитовых участках леса в верховьях р.Псешва (Волча-нецкий, и др., 1962), а нами (Пекло и др., 1978) найдена на гнездовании в тисо-самшитовой роще на склоне г.Ахун у г.Сочи.
В Азербайджане (Дроздов, 1965) в За-катальском заповеднике (Катехский участок) отмечена в буково-грабовых (с подлеском и без него), буковых и парковых кленово-дубовых лесах, а на хр.Муров-даг у оз .Гек-Гель - в грабово-буковых лесах. В Талышских горах населяет предгорные широколиственные с пышным подлеском и дубово-железняковые леса, а на склоне хр.Улясы - буково-грабовые. В Армении поднимается в горы до 2350 м (Ляй-стер, Соснин, 1942).
Численность. На территории СССР неравномерна. В ряде мест гнездового ареала распространена спорадично.
Прилет и весенний пролет на территорииСССР. Проходит с III'декады марта по конец мая. Самые ранние одиночные птицы на Кавказе появляются даже в I и II декадах марта. Во время пролета мухоловки летят поодиночке и небольшими стайками, часто образуя на местах отдыха и кормежек большие скопления. На пролете одинаково часто встречаются как в облесенных местностях, так и в открытых биотопах - степях и пустынях.
На Кавказе в Восточной Грузии (р.Ку-ра), по данным К.А.Сатунина (1907), первые одиночные птицы появляются (3 года) 4 и 14.Ш, а также 12, 14 и 17.IV. Наиболее интенсивно пролет протекает 20.IV. У г.Тетри-Цкаро самка добыта 5.V 1897 г., а у с.Сурами самец - 6.V 1878 г. (колл. ЗИН). На о-ве Сара в Каспийском море первая мухоловка в 1937 г. отмечена 8.IV, а валовый пролет зарегистрирован 5—7.V (Тугаринов, 1950). У г .Орджоникидзе (2 года) пролетные птицы наблюдались 10 и 24.IV (Беме, 1958).
На Черноморском побережье в окр. г.Сочи первые особи отмечены нами в середине апреля, основной прилет и пролет мухоловок проходит в III декаде апреля — I декаде мая. На юге Таманского п-ова (Кизилташская коса) пролет в 1975 г. отмечен нами 2.V. Пролетные птицы держались в полезащитных лесополосах (Пекло и др., 1978). В Тебердинском заповеднике (Ткаченко, 1966) сроки миграции те же.
88
В Крыму (многолетние наблюдения) пролет протекает 23.IV—25.V, наиболее интенсивно - в первой половине мая (Костин, 1983). Нами пролетные мухоловки встречены в центре Арабатской Стрелки 6 и 19.V 1981 г.
В Восточном Приазовье одиночная особь отмечена А.А.Комаровым 13.V 1960 г. на Безымянном лимане. В Ростовской обл. у пос.Рогожкино пролетные птицы добыты М.А.Воинственским (колл. ИЗАНУ) 29.IV, 2 и 6.V 1951, а в Донском лесхозе встречены В.П.Беликом (2 года) 23 и 24.IV.
В Измаильском р-не (окр. с.Вилково) Одесской обл. пролетная мухоловка добыта 7.V 1948 г. (колл. ИЗАНУ), а в Херсонской обл. (с.Знаменка) — 1.V 1907 г. (колл. ЗИН).
В Молдавию прилетают в конце апреля - начале мая: Кицканская дача - 27— 28.IV; окр. с.Суклея - 30.IV; Дубоссар-ский р-н (2 года) - 28.IV и 5.V; Лазовское лесничество (2 года) - 27 и 29.IV (Ганя, Литвак, 1961; Аверин, Ганя, 1970). В Закарпатье и на Волыно-Подолье (Miczynski, 1962; Страутман, 1963; колл. ХГУ; ЛПМ) сроки прилета и пролета те же.
В Донецкой обл. в Великоанадольском лесничестве пролетные птицы добыты 1-8.V 1949 г. (колл. ИЗАНУ), в г.Запо-рожье - 11.V 1948 г. (колл. ХГУ), а в Черкасской обл. (Лысянский р-н) — 14 и 15.V 1922 г. (колл. ЗИН). В окр. г.Киев малые мухоловки появляются (4 года) 25.IV-1.V (Данилович.-Цит. по: Благосклонов, 1954), в 1928 г. взрослый самец добыт 3.V (колл. ИЗАНУ). На Полтавщине самые ранние птицы зарегистрированы 16.IV 1924 г. (Гавриленко, 1929). В Харьковской обл. прилет и пролет (11 лет) проходят 30.IV—17.V (Сомов, 1897), ауг.Нежин на Черниговщине самец добыт 9.V 1929 г. (колл. ИЗАНУ).
В Воронежской обл. (Барабаш-Никифоров, Семаго, 1963) пролет начинается в конце апреля - начале мая. В 1956 г. он проходил 7-31.V, основная масса птиц пролетела 10—17.V. В Бобровском р-не мухоловки добыты 2 и 16.V 1893 г. (колл. ЗИН). У истоков р.Ока в Орловской обл. появляются также в мае (Ефимов, 1915).
В Белоруссию прилетают в I—II декадах мая: в Гродненский р-н — 3.V; в Пинский — 11.V; в Березинский заповедник — 12-17.V, реже 22V (Шнитников, 1913; Сямашка, 1956; Федюшин, Долбик; 1967; Дорофеев, 1969). В Литве (окр. сЛеке-
чяй) появляются (11 лет) 6—13.V (Алек-нонис, 1977). В Калининградскую обл. прилетают также в мае (Портенко, 1960).
В Московской обл. старые самцы появляются 21—30.IV—7.V. Незначительный пролет самцов и самок всех возрастов начинается 3—9—17.V и заканчивается к 9—22.V (Паровшиков, 1941а; Птушенко, Иноземцев, 1968). В Тульской обл. у Кар-ницкой дачи мухоловка добыта 7.V 1889 г. (колл. ЗИН). В Псковскую и Ленинградскую области птицы прилетают в первой половине мая (Зарудный, 1910; Мальчевский, Пукинский, 1983). Даже в позднюю весну 1980 г. первые особи на побережье Свирской губы встречены 14.V (Мальчевский, Пукинский, 1983). Наиболее раннее появление малой мухоловки в Лужском р-не зарегистрировано 24.IV 1970 г. (Гавлюк. - Цит. по: Мальчевскому, Пукин-скому, 1983). В Карелию прилетают в середине мая (Марвин,1951).
В дельте р.Волга пролет проходит в I декаде мая (Воробьев, 1936). Одиночная птица у с.Эльтон добыта 3.V 1949 г. (колл. ХГУ). В район устья р.Кама прилетают (5 лет) — 27—29.IV, а в Райфе’(Татарская АССР) в 1974 г. появились 26.IV (Птицы Волжско-Камского края, 1978). В Марийской АССР 2 птицы добыты 5 и 10.V 1927 г. (колл. ЗИН).
В Южной Туркмении весенний пролет проходит с конца марта до середины мая, валовый — в апреле (Исаков, Воробьев, 1940; Благосклонов, 1954; Дементьев и др., 1955; Гептнер, 1956; Шукуров, 1962; Полозов, 1980; наши данные).
У г.Ташкент малые мухоловки пролетают (3 года) 2-27.IV-3-9.V; в Южных Кызылкумах (оазис Тамды) - 22.IV-5.V; на Средней Сырдарье - 8.IV-4.V; у г .Аральск — с 22.IV; на п-ове Мангышлак -с конца апреля до начала мая включительно (в 1947 г. слабый пролет отмечен 8— 20.V); в Северном Прикаспии (Джаман-сор) - 9.V; в низовьях р.Урал - 17-21.V; севернее с.Фурманово — 20.IV-28.V (Ковшарь, 1970).
В Памиро-Алае первые пролетные птицы появляются (многолетние наблюдения) 3, 4, 10 и 12.IV (Иванов, 1969). В заповеднике ’’Тигровая балка” пролет проходит с 5.IV по 14.V, наиболее интенсивно — 6—8 и 11 —16.IV (Попов, 1959; Иванов, 1969). В окр. пгт Гиссар мухоловки добывались А.И.Ивановым (Попов, 1959) 10-26.IV 1934 г. Указание И.А.Абдусаля-
89
мова (1973) о том, что на пролете в Таджикистане встречается только подвид F.p.albicilla не соответствует действительности. Изучение серии малых мухоловок (добытых в разных точках республики), проведенное нами, показало, что все они, несомненно, принадлежат к номинативному подвиду.
Размножение. В большинстве мест гнездового ареала первыми на места гнездования прилетают самцы и только через несколько дней после них появляются самки. Птицы разбиваются на пары и приступают к строительству гнезд. Образование пар сопровождается наиболее интенсивным пением самцов и их токовым демонстративным поведением. Продолжительность песни довольно изменчива и, по данным В.Г.Гептнера (1958), колеблется в пределах 3—6,6 с. Размеры гнездовых участков сильно варьируют. В оптимальных гнездовых биотопах их площадь обычно составляет 5000—7000 м2, а в островных ельниках Белорусского Поозерья - от 100— 120 до 15 000-20000 м2 (Дорофеев, 1969). В выборе места для гнезда принимают участие оба партнера пары. Строит гнездо главным образом самка.
Гнезда птицы помешают на живых, частично гниющих и сухостойных деревьях (ольха, береза, дуб, граб, бук, ива, груша, ель, пихта, сосна, самшит, слива и др.), а также на их пнях: в естественных и выдолбленных другими птицами (дятлы) дуплах, полудуплах, нишах на месте выпавших сучьев, выбоинах, подолбах, различных расщелинах, пустотах стволов, за стволовой порослью и отставшей корой, в развилках стволов и боковых ветвей, на вершинах пней и стволовых сломов, а также (как исключение) — в местах соприкосновения 2 деревьев на высоте (п = 18) 0,9— 7,0 м от поверхности земли, в среднем — 2,6 (Ткаченко, 1966; Федюшин, Долбик, 1967; Дорофеев, 1969; Пекло и др., 1978; Птицы Волжско-Камского края, 1978; Мальчевский, Пукинский, 1983; наши данные). В редких случаях гнезда располагаются на высоте 0,6 и даже 12,0 м (Птушенко, Иноземцев, 1968).
Гнезда представляк£г собой довольно рыхлые, часто массивные постройки округлой (если построены открыто) или неправильной формы (в дуплах, полудуплах и щелях). В качестве строительных материалов мухоловки используют пучки мхов (в том числе: Pleurozium schreberi — в Подмосковье; Hypnum sp.,
Mnium sp., — на Северо-Западном Кавказе), тонкие сухие веточки хвойных растений, пленки березовой коры, корешки, прошлогодние листья деревьев и кустарников, хвою (ель, сосна), полоски луба, стебли и листья трав, кусочки талломов лишайников, остевой волос шерсти крупных млекопитающих, одиночные перья, а по данным Е.С.Птушенко, А.А.Иноземце-ва (1968) — также паутину и коконы пауков. Лоток обычно выстилается тонкими стебельками трав, растительными волокнами, волосками шерсти млекопитающих, пучочками мха (Mnium sp.), жилками и черешками листьев древесных растений, отдельными перьями и хвоинками (ель, пихта). В редких случаях лоток или не выстилается вообще, или для этой цели птицы используют спорогонии мхов (Вгуасеае). Размеры гнезд (п = 14) (Сомов, 1897; наши данные), мм: диаметр 55-105 х 55—130 мм, в среднем — 78 х 95; высота 30—100, в среднем — 57; диаметр лотка 40-68 х 45-72, в среднем - 52 х 54, глубина лотка 22—45, в среднем — 36.
В сезон размножения на территории СССР европейская малая мухоловка имеет один, предположительно два репродуктивных цикла. Повторные кладки часто возникают в результате гибели первых. Количество яиц в первой полной кладке 4—7, чаше — 5. Средние размеры кладки увеличиваются с юга на север гнездового ареала. Количество яиц в повторной кладке может быть 3 и даже 2.
Укороченно-остроовальные, остроовальные, овальные и как исключение эллипсовидные яйца имеют следующую окраску: фон от бледно-зеленоватого до молочно-белого со слабым голубовато-зеленоватым оттенком. По нему густо разбросаны мелкие размытые охристо-рыжевато-розоватые и светло-лиловые крапинки и точки. На одних яйцах они настолько густо покрывают поверхность скорлупы, что яйцо кажется однотонно бледно-рыжевато-бежевым, а на других — концентрируются главным образом у тупого полюса и вокруг центральной части яйца. Скорлупа матовая или полуматовая. Форма и окраска яиц варьируют как внутри одной, так и между разными кладками. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
Насиживание начинается после снесения последнего яйца или за 1—2 дня до его откладки. Длительность инкубации 12-15, чаше — 12—13 дней (Ганя, Литвак, 1961;
90
Дерим-Оглу, 1961; Птушенко, Иноземцев, 1968; Дорофеев, 1969; Мальчевский, Пукинский, 1983; наши данные). Насиживает кладку самка. Самец в этот период держится поблизости от гнезда и периодически ее подкармливает.
Пуховой птенец европейской малой мухоловки (Новиков и др., 1963) имеет редкий, но довольно длинный пух на надглазничных, затылочной, спинной, плечевых и глазных птерилиях. Ротовая полость желтая, клювные валики белые. Масса тела вылупившегося птенца 1,17—1,51 г, в среднем — 1,35.
В выкармливании птенцов принимают участие оба партнера пары, но в первые дни, когда самка большую часть времени проводит в гнезде, согревая птенцов, корм в основном приносит самец. Растут и развиваются птенцы быстро и через 11—15 дней, чаше 12-13 покидают гнездо (Птушенко, Иноземцев, 1968; Дорофеев, 1969; Пекло и др., 1978; Головань, 1982). Родители держатся рядом со слетками и продолжают их кормить. В течение 2-3 дней выводки держатся у земли, а затем поднимаются в верхние ярусы леса. На 4—5-й день нахождения вне гнезда молодые следуют за взрослыми птицами, настойчиво выпрашивая у них корм. Переход к самостоятельному поиску и схватыванию кормовых объектов отмечается у них на 7—8-й день после вылета из гнезда, а в 27—29-дневном возрасте молодые мухоловки встречаются уже без родителей (Головань, 1982). Вплоть до отлета старые и молодые птицы обитают в районе гнездования, иногда удаляясь от гнезда на несколько километров.
Основной причиной отхода яиц и гибели птенцов малой мухоловки является хищническая деятельность со стороны некоторых млекопитающих (куньи, обыкновенная белка) и птиц (врановые) (Мальчевский, Пукинский,1983).
Сезон размножения европейской малой мухоловки на территории СССР сильно растянут и длится с III декады апреля по I декаду августа включительно.
На Северо-Западном Кавказе в Тебер-динском заповеднике строительство гнезда отмечено в середине мая (Ткаченко, 1966). В Майкопском р-не Краснодарского края (пос.Никель) полная кладка найдена 2.VI 1976 г., птенцы вылупились 11.VI, а вылетели 24.VI. В другом гнезде, обнаруженном 3.VI 1975 г„ вылупление проходило 5—6.VI, а вылет птенцов — 17.VI (Пекло и др., 1978). В окр. г.Сочи гнездо с 4 оперивши
мися птенцами найдено 1.VI 1977 г., последние покинули его 2.VI. Во втором гнезде, обнаруженном 29.VI, были птенцы 8—9-дневного возраста, которые вылетели 4.VII (Пекло и др., 1978). Слетки в Сочинском р-не у пос .Лазаревское встречены 30.V 1966 г. (колл. БПиДВНЦ), а у с.Крас-ная Поляна — 8.VI 1935 г. (Волчанецкий и др., 1962). Летные молодые добыты здесь (3 года): 13, 18.VII и 29.VIII (Кудашев, 1917; колл. ЗИН), а в Геленджикском р-не на Михайловском перевале (Плисецкая шель) зарегистрированы В.С.Оча-повским 2—6.VIII 1962 г. В Северной Осетии у с.Дзаурикау слетки добыты 31.VII 1925 г. (Беме, 1926).
В Вешенском р-не Ростовской обл. гнездо с 5 птенцами 2—3-дневного возраста найдено 17.VI 1982 г., а 11.VII здесь обнаружено второе гнездо с кладкой из 4 ненасиженных яиц (сообш. В.П.Белика). В окр. г.Воронеж гнездо с 6 слабо насиженными яйцами найдено в начале июня 1945 г., а выводок слетков в пойме р.Воронеж отмечен 9.VII 1959 г. (Барабаш-Никифоров, Семаго, 1963).
В Молдавии (Ганя, Литвак, 1961) свежие кладки встречаются с конца мая до первой половины июня: в Оргеевском лес-ходе у с.Морозены гнездо с 3 ненасиженны-ми яйцами найдено 26.V 1956 г.; на Киц-канской даче с 4 свежими - 29.V 1955 г., а в Сорокском лесу ненасиженные кладки отмечены 30.V и 3.VI 1957 г. В большинстве гнезд птенцы появляются 2—14.VI.
В западных областях Украины откладка яиц проходит в основном в III декаде мая — начале июня. Во Львовской обл. у селений Пеняки и Подгайцы полные и незаконченные кладки находили 24.V—3.VI— 5.VII (Страутман, 1963; колл. ЛПМ). В долине р.Уж Л.А.Портенко (1950) наблюдал выкармливание птенцов в середине июля 1947 г. Летные молодые встречаются в начале июля (Страутман, 1963; колл. ЛПМ).
В окр. г.Харьков гнездо с 5 яйцами средней насиженности обнаружено 22.VI 1938 г. (колл. ИЗАНУ). В Киевской обл. у ст.Боярка кладка из 3 яиц средней насиженности отмечена 20.VI 1961 г. (колл. ИЗАНУ), а в Черниговской обл. у с.Недан-чичи гнездо с 4 яйцами найдено 13.VI 1977 г. (сообщ. М.Ф.Самофалова). В Шост-кинском р-не Сумской обл. полная свежая кладка обнаружена В.П.Беликом 11.VI 1966 г.
В Белоруссии у с.Лельчицы гнездо с пол-
91
ной сильно насиженной кладкой встречено 5.VI 1965 г., птенцы вылупились в нем 6.VI (Федюшин, Долбик, 1967). В Поозерье основная масса полных свежих кладок отмечается в I декаде июня (Дорофеев, 1969).
В Московской обл. к строительству гнезд приступают 20.V—15.VI, чаще -1—7 .VI; первое яйцо откладывают 25 .V— 18.VI; полные кладки встречаются 30.V— 24.VI, а вылупление птенцов проходит 13—24.VI. Из яиц повторных кладок (очевидно, взамен утраченных) птенцы вылупляются в июле (14.VII 1957 г.). Вылет птенцов — 25.VI—4.VII—18.VII (Птушенко, Иноземцев, 1968). В Мордовии (Заводское лесничество) слеток добыт 29.VII 1967 г. (Луговой,1975).
В Волжско-Камском крае малые мухоловки приступают к гнездованию в III декаде мая - начале июня. В гнезде, найденном 10.VI 1958 г. в устье р.Свитяга, было 6 сильно насиженных яиц, а в обнаруженном 1-VII 1958 г. на севере Удмуртской АССР — 3 свежих яйца (Птицы Волжско-Камского края, 1978).
В Псковской обл. самые ранние оперившиеся птенцы зарегистрированы 3.VII, в это время в большинстве других гнезд находятся ненасиженные кладки (Зарудный, 1910). В Лениградской обл. (Мальчевский, Пукинский, 1983) строить гнезда малые мухоловки начинают во второй половине мая, а уже в середине III декады этого месяца встречаются неполные кладки. Период размножения сильно растянут: свежие кладки отмечаются до конца июня — первых чисел июля. Наиболее ранняя зарегистрированная кладка начата 25.V 1981 г., а самая поздняя (судя по срокам вылупления птенцов) — в начале I декады июля. В Южной Карелии выводок летных молодых с недоросшими рулевыми встречен 9.VII 1955 г. (Нейфельдт, 1958).
Линька. Уже в середине июня молодые птицы из ранних выводков начинают смену гнездового наряда на первый послегнездовой. Эта частичная линька охватывает почти все мелкое оперение, кроме наружных больших верхних кроющих второстепенных маховых. У молодых из поздних выводков послегнездовая линька обычно заканчивается в конце августа — сентябре. На местах зимовок у некоторых первогодков перелинивают отдельные перья на горле. Взрослые птицы имеют в году две линьки: полную послебрачную на местах гнездования и частичную предбрачную на
зимовках. Послебрачная линька обычно проходит с июля по сентябрь и у некоторых особей совмещается с выкармливанием птенцов, находящихся еще в гнездах. Частичная предбрачная протекает в феврале-марте. Происходит смена всех или части рулевых, некоторых больших верхних кроющих второстепенных маховых и оперения горла (Виноградова и др., 1976).
Питание. Основу пиши составляют насекомые разных групп и стадий развития, а также пауки. Реже склевывает мелких моллюсков, а во второй половине лета и осенью поедает также и сочные плоды некоторых растений (бузина, ежевика). Самый пластичный вид мухоловок нашей фауны по способам добывания корма. Очень подвижная и постоянно меняет способы охоты. Кормится главным образом в среднем — древесно-кустарниковом ярусе леса, реже — в нижних частях крон высоких деревьев, травяном покрове и на поверхности земли.
Отлет и осенний пролет на территории СССР. Сильно растянут и проходит со II (в редких случаях с I) декады августа до конца ноября. Одиночные задержавшиеся птицы встречаются в южных районах в декабре и даже в январе.
В Ленинградской обл. (Мальчевский, Пукинский, 1983) отлет проходит, видимо, с середины августа до конца сентября. Подавляющее большинство птиц покидает места гнездования уже в конце августа. В Калининградской обл. на Куршской косе пролетная особь добыта 6.IX 1960 г., а в окр. г Ленинград — 26.IX 1898 г. (колл. ЗИН). В Псковской обл., по данным Н.А.За-рудного (1910), миграция начинается в I—II декадах сентября и протекает до 26.IX.
В Московской обл. (Птушенко, Иноземцев, 1968) к 25—30.VIII основная масса мухоловок постепенно отлетает, 20— 30.VIII наблюдается слабо выраженный пролет, который заканчивается к 5—7.IX. Последние птицы в Подмосковье отмечены 18 и 21 .IX 1956 г. В Рязанской обл. пролет проходит с конца августа по конец сентября (Хомяков, 1900), последняя встреча зарегистрирована 5.Х (Павлов, 1879). В Тульской обл. (Ефремовский и Богородский р-ны) пролетные мухоловки добывались (2 года) 8, 13, 14, 25 и 28.IX (колл. ЗИН). В Калужской обл. (Козелецкое лесничество) отмечены 30.VIII и 1.IX 1910 г. (Филатов, 1915). В Орловской обл. миграция проходит (7 лет) с 23 .VIII до се
92
редины октября, одиночные особи задерживаются до 12.XI (Ефимов, 1915). У г.Горь-кий зарегистрирована 21.IX 1954 г„ а в окр. г.Казань — 26.IX 1959 г. Отдельные задержавшиеся птицы в Волжско-Камском крае встречаются до 26.Х (Птицы Волжско-Камского края, 1978). В Мордовии (Тем-никовский р-н) последние мухоловки в 1936l г. отмечены 18 и 19.IX (Птушенко, 1938), а в Жигулевском заповеднике -в конце октября (Белянина, Белянин, 1981).
В Белоруссии основная часть птиц отлетает во второй половине августа — первой половине сентября (Федюшин, Долбик, 1967). В Поозерье миграция полностью заканчивается во II декаде сентября (Дорофеев, 1969).
В Воронежской обл. малые мухоловки исчезают в конце сентября — первой половине октября. В Усманском лесу последняя встреча зарегистрирована 22.Х 1950 г. (Семенов. — Цит. по: Барабаш-Никифоров, Семаго, 1963), а в Бобровском р-не птица добыта 23.Х (колл. ЗИН).
На Украине в Киевской обл. пролетные особи отмечены у с.Гребени — 1 .IX 1925 г. (колл. ЗИН) и в окр. г.Фастов — 5.Х 1968 г. (колл. ИЗАНУ). В Харьковской обл. миграция начинается в конце августа (иногда 11 и 14.VIII) и продолжается до середины сентября, реже — до начала октября. Одиночные птицы задерживаются до 15.Х (Сомов, 1897). В Изюмском р-не (х.Гадки) мухоловка добыта 28.IX 1960 г., а в окр. г.Харьков — 10.Х 1946 г. (колл. ХГУ). На Полтавщине самые поздние особи встречены (3 года) 12, 15.Х и 1.XI (Гавриленко, 1929). В Запорожской обл. пролетные мухоловки добыты: в Богатырском лесничестве — 2.Х 1959 г. (колл. ИЗАНУ), у с.Водяное — 18.Х 1951 г. (колл. ХГУ) и у г.Бердянск — ЗО.Х 1939 г. (колл. ИЗАНУ), а в Донецкой обл. в Великоана-дольском лесничестве — 20.X 1937 г. (колл. ХГУ). В западных областях УССР отлетают в середине сентября — второй его половине (Страутман, 1963). В долине р.Уж Л.А.Портенко (1950) последний раз в 1947 г. встретил этот вид 18.IX.
В Молдавии миграция начинается во второй половине августа и продолжается до конца октября (Ганя, Литвак 1961; Аверин, Ганя, 1970). В Одесской обл. пролетные особи добыты: в Александровском лесничестве — 23, 25 и 29.IX 1937 г. (колл. ХГУ), а в окр. г.Одесса — 4, 28.IX и 7.Х 1902 г. (колл. ЗИН).
В Крыму (Костин, 1983) наиболее ранняя послегнездовая встреча у с.Портовое зарегистрирована 18.VIII, а в III декаде августа в некоторые годы наблюдается первая волна массового пролета. С перерывами пролет идет в сентябре и октябре, но наиболее многочисленны малые мухоловки в сентябре (в течение 10—12 дней подряд). Наиболее поздняя осенняя встреча отмечена 31.X 1973 г. На о-ве Бирючий вазовском море пролетные птицы добыты 22.VIII, 16.1Хи4.Х 1932 г. (колл. ХГУ).
В дельте р.Волга пролет протекает (6 лет) с конца августа до 3.XI (Воробьев, 1936). В Оренбургской обл. (Бугурусланский р-н) мухоловки добыты (2 года) 28.1Хи 1.XI (колл. ЗИН).
На Черноморском побережье Кавказа пролетные птицы зарегистрированы: у г.Сочи (Хоста) — 6 и 7.IX 1914 г., а в окр. с Ла-заревское — 8.IX 1968 г. (колл. ЗИН). В Ставропольском крае они отмечены: у г.Кисловодск — 6.IX 1898 г. (колл. ЗИН) и в окр. г.Ставрополь (Грачинская балка) — 17.IX 1951 г. (колл. ХГУ). В Северной Осетии у г.Орджоникидзе пролет зарегистрирован (2 года) 10—11.Х и 7.Х—2.XI (Беме,1926,1958).
В Грузии пролетные мухоловки добыты: у с-Лагодехи - 19.IX 1953 г., на Су-рамском перевале — 9.Х 1894 г., в окр. г.Тбилиси — 1, 5, 18 и 22.Х 1911 п, а у г.Мцхета - 26.Х 1909 г. (колл. ЗИН). В Азербайджане они встречены: у с.Скобе-левка — 14.IX 1915 г., а на о-ве Сара в Каспийском море - 1 .XI, ЗО.Х и даже 21.XII 1935 г. (колл. ЗИН). По данным К.А.Са-тунина (1907), в тугаях по р.Кура малые мухоловки держатся до конца октября, а одиночные особи встречаются и в ноябре (15и24.Х1).
В Казахстане на северо-западе республики (Ковшарь, 1970) появляются в середине, а в некоторые годы даже в начале августа: в Тельнове - 19—28.VIII, а близ Яманки - даже 1 .VIII. Массовый пролет у с.Фурманово проходит около середины сентября, у Яманки — во второй половине сентября, но иногда с 17-21.VIII. Последние птицы здесь зарегистрированы: в Тель-ново - 17.Х, в Фурманово - 19.Х, в песках южнее Новой Казанки — 17.Х, в Урде — 18.Х, у Краснояра — 24.Х, а в Яманке — 10.Х. На п-ове Мангышлак пролетают с 11.IX до конца октября (Ковшарь, 1970). В Восточном Казахстане на р.Каратал пролетные мухоловки встречены (2 года) 28.IX и 5.Х (Шнитнйков, 1949), у г.Кара
93
ганда - 8.Х, а у г.Алма-Ата — 26.Х (Ковшарь, 1970); у г .Чимкент добыта 25.Х (колл. ЗМ МГУ). В окр. г.Ташкент миграция идет в III декаде сентября — первой половине октября (Ковшарь, 1970).
В Киргизии в восточной части хр.Тер-скей-Алатоо (уш.Джиланды) одиночная пролетная особь добыта 23.Х 1952 г. (Януше-вич и др., 1960). В Таджикистане птицы зарегистрированы: в средней части долины Шах-Дары (2800 м) — 2,6 и 11.XI 1955 г. (Попов, 1959), в Рамитском уш. — 2.XI 1955 г. (колл. ИЗиПТаджССР), в заповеднике ’’Тигровая балка” — 30.VIII, 15 и 20.Х, а также 5 .XII (Иванов, 1969).
В Юго-Западной Туркмении пролет проходит с конца августа по конец ноября, отдельные задержавшиеся особи встречены в Юго-Западном Копетдаге 7.XII 1947 г. (Дементьев и др., 1955), 28.XII 1978 г. и даже - 1 и 14.1 1979 г. (Полозов, 1980). В р-не Гасан-Кули (Исаков, Воробьев, 1940) малые мухоловки пролетают в октябре — первой половине ноября. Последние птицы в 1937 г. у Чикишляра зарегистрированы 20.XI. В других точках Туркмении пролетные особи отмечены: у г.Ашхабад (2 года) - 15, 25.VIII, 2.IX и 12.Х; в Восточном Копетдаге — 6.IX 1889 г.; у г.Серахс — 6JX и 7.Х 1935 г. (колл. ЗИН); в Бадхызе - в октябре 1973 г. (Второв, 1976); у г.Кушка - 11 .IX и 5-18.Х (Благосклонов, 1954); в арчевниках Кугитан-га — 3 и 9.Х (Иванов, 1969).
Практическое значение. Уничтожает многих насекомых — вредителей леса и садовых культур, а также — переносчиков возбудителей некоторых заболеваний человека и животных.
Сибирская малая мухоловка — Ficedula parva albiciIla (Pallas, 1811)
Muscicapa albicilla Pallas, 1811, Zoogr. Rosso-Asiat., 1: 462, fig.l, близ р.Онон, Даурия.
Описание. Составлено по 42 экз. из Сибири и Дальнего Востока СССР.
Старый самец, в отличие от такового номинативного подвида, имеет значительно меньшее рыжее горловое пятно, которое не заходит на грудь. По бокам и снизу оно окантовано широким серым полем. Цвет последнего светлеет на груди. Брюшко и нижние кроющие хвоста бледно-охристые. Бока тела светло-охристо-серые. Верхняя часть головы, как и спина, однотонно серовато-бурые. Индивидуальная и сезонная
изменчивость проявляется так же, как и у предыдущего подвида.
Взрослая самка окрашена так же, как и у номинативного подвида.
Молодые в гнездовом наряде сверху более темные, без коричневых тонов в окраске. Бледно-охристые пятна на перьях сверху окантованы черно-бурым. Оперение низа тела чисто-белое или белое с бледнобуроватым оттенком. Волнистый рисунок наиболее хорошо выражен на зобе и груди и менее — на горле и верхней части брюшка.
Первый послегнездовой наряд идентичен таковому номинативного подвида.
Размеры отдельных частей тела сибирской малой мухоловки приведены в табл. 10.
Масса тела, г: самец (26.VII, р.Вилюй) — 11,1 (Андреев, 1974); самец (17.VIII, Читинская обл., оз.Зун-Торей) — 9,65; самки (п = 2) (20 и 22.VIII, северо-западное побережье оз. Байкал) - 10,0—11,3, в среднем — 10,65 (колл. КБ МГУ); самка (16.VI, Южный Ямал) - 10,2 (Данилов и др., 1984).
Распространение. СССР: восточная часть гнездового ареала вида на запад до западных подножий Среднего Урала. Северная Монголия. Ведет перелетный образ жизни и зимует в Южной и Юго-Восточной Азии. Детально распространение сибирской малой мухоловки на территории СССР показано на карте (рис. 20).
Кадастр к рис. 20.
1 - окр. г.Киров, р.Вятка (Иоганзен. - Цит. по: Mauersberger, Portenko, 1967); 2 - г.Пермь (Иоганзен по Mauersberger, Portenko, 1967); 3 -с.Ивдель (Никитоивдель) (Резцов, 1904); 4 -Уральский хр., 65° с.ш. (Портенко, 1960; Степанян, 1978); долина р.Маньи в р-не горы Сальур-нэ (Портенко, 1937); 5 - окр. с.Мужи (Брауде, 1970); 6 - с.Октябрьский (Данилов и др., 1984); 7 - Ласточкин берег на р.Хадытаяха (Данилов и др., 1984); 8 - р.Тромъеган (Иванов, 1976); 9 -р.Тым (Иванов, 1976; Гынгазов, Миловидов, 1977); окр. с.Напас (Гынгазов, Миловидов, 1977); 10 - левые притоки р.Обь (Гынгазов, Миловидов, 1977); 11 - Чаинский р-н, пойма р.Обь (Пантелеев, 1972а); 12 — р.Орловка (Гынгазов, Миловидов, 1977); 13 - среднее течение р.Кеть; окр. с.Красная Курья (Гынгазов, Миловидов, 1977); Верхнекетский р-н, д.Тайное (сообщ. С.С.Москвитина); 14 - верховья р.Таз (Скалой, Слудский, 1941); 75 — р.Елогуй (Скалой, Слудский, 1941; Юдин, 1952: Рогачева, 1962; колл. ЗИН); 16 - р.Енисей, 68° с.ш. (Степанян, 1978); 17 - истоки р.Рыбная (Кречмар, 1963); оз.Конон (колл. ЗИН); 18 - западная часть плато Путорана (Кречмар, 1962); окр. оз.Капчук (Морозов, 1984); 19 - устье р.Ниж-няя Тунгуска (Сушкин, 1914; Портенко, 1960; Иванов, 1976); окр. г.Туруханск (Сушкин, 1914; Благосклонов, 1954); 20 -с.Тура (Ткаченко,
94
1937); 21 - средняя треть р.Нижняя Тунгуска (Ткаченко, 1937); 22 - р.Нижняя Тунгуска от истоков до с.Ербогачен (Ткаченко, 1937); 23 -истоки р.Вилюй (Андреев, 1974); 24 - р.Катанга (колл. ЗИН); 25 - р.Чадобец (Реймерс, 1966); 26 - Удерейский р-н (колл. КБ МГУ) ; 27 - окр. г.Енисейск (Сушкии, 1914); 28 - Средний Чулым (Гынгазов, Миловидов, 1977); 29 - р.Большой Кемчуг; с.Большой Кемчуг; Козульская равнина (Наумов, 1960); 30 - окр. г.Красноярск (Сушкин, 1938; колл. ЗИН); предгорья Восточного Саяна у города (Юдин, 1952); р.Базаиха (колл. ЗИН); 31 - р.Бирюса (колл. ЗИН); 32 - Нижне-Ингашский р-н (Равкин и др., 1983); 33 - окр. с.Выезжий Лог (колл. ЗИН); 34 - Шушенский р-н, окр. с.Ермаковское (Земкова, Штильмарк, 1965);
35 - Усинская котловина (Янушевич, Юрлов, 1950); р.Ус от Казачьего станка до с.Усинское; р.Шираспа (Сушкин, 1914); 36 - окр. с.Монок (Губарь, 1929); 37 — Салаирский кряж, р.Томь-Чумыш (Чунихин, 1965; колл. КБ МГУ); 38 -южная часть Кемеровской обл. (Горная Шория) (Сушкин, 1938); 39 - Алтай (Благосклонов, 1954; Портенко, 1960; Иванов, 1976; Степанян, 1978); Телецкое оз.; устья рек Колдора и Камма (Сушкин, 1938; колл. ЗИН); 40 - гора Монгун-Тайга (сообщ. М.И.Головушкина); 41 - р.Хем-чик между Ужеданом и Джергаком (Сушкин, 1914); 42 - хр.Танну-Ола (Янушевич, 1948); 43 - Тувинская котловина (Янушевич, 1948); 44 - Восточно-Тувинское нагорье (Янушевич, 1948); 45 - Саяны (Благосклонов, 1954); 46 -Тулунский р-н, окр. с.Гадалей; г.Тулун, долина р.Ия (сообщ. В.Д.Сонина); 47 - Приангарье на юге Иркутской обл. (Зонов и др., 1967); г.Иркутск и его окр. (Пшеничников, 1959); 48 - Усть-Удинский р-н, пос. Майск (сообщ. В.Д.Сонина); 49 - Заларинский р-н, д.Кольче-жино (колл. ЗМ МГУ; сообщ. В.Д.Сонина); 50-Тункинский р-н, с.Ахалик (сообщ. В.Д.Сонина); р.Мойгот (колл. ЗИН); 51 - с. Култук (колл. ЗИН); 52 - хр.Хамар-Дабан (Доржиев, 1982); южные склоны хр. (колл. ЗИН); 53 - Слюдянский р-н, ст.Выдрино (сообщ. В.Д.Сонина); 54 -западный берег оз.Щучье и долина горного ручья, впадающего в озеро (Гусиноозерская котловина) (Боровицкая, Измайлов, 1978); окр. г.Гусино-озерск (сообщ. М.И.Головушкина); 55 - Селен-гинский р-н (Измайлов, Боровицкая, 1973); р.Селенга; с.Ильинка (колл. ИЗАНУ); 56 - окр. г.Петровск-Забайкальский (сообщ. М.И.Голо-вушкина); 57 - р.Менза (Линдеман, 1976);
58 - р.Бобровая (верхнее течение р.Чикой) (Леонтьев, Павлов, 1963); 59 - окр. пос.Кыра (сообщ. М.И.Головушкина); 60 - Агинская лесостепь (Гагина, 1960); 61 - окр. с.Цаган-Олуй (сообщ. М.И.Головушкина); 62 - окр. пос.Клич-ка (сообщ. М.И.Головушкина); 63 - окр. пос.Бырка (сообщ. М.И.Головушкина); 64 -Александрозаводской подтаежный р-н (Гагина, 1960); 65 - Приаргунье (Гагина, 1960); 66 -долина р.Шилка (Иванов, 1976); с.Шилкинский завод (Маак. - Цит. по: Mauersberger, Portenko, 1967); 67 - верхнее течение р.Чита (Павлов, 1948); 68 - район озер Тасей, Арахлей и Иван-озера (Павлов, 1948); 69 - окр. оз.Исинга (Измайлов, Боровицкая, 1967); 70 - окр. оз.Тур-хел; падь Киро (Измайлов, 1967; Измайлов, Боровицкая, 1967); 71 - п-ов Святой Нос (Гагина, 1960); 72 - северо-восточный берег Байкала;
Баргузинский заповедник (Скрябин, Филонов, 1962); 73 - Баргузинский хр. (Федоров, 1982); 74 - р.Витим (Измайлов,. 1967); 75 - с.Багда-рин (Измалой, Боровицкая, 1967; колл. ЗИН); 76 - Муйская долина (Измайлов, Ханхасаев, 1959); 77 - Икатский хр. (Измайлов, 1967); 78 - берег оз.Иркан (Гагина, 1954); 79 - окр. бухты Заворотная (Малышев, 1960а); 80 -верхняя Лена (Реймерс, 1966); 81 - Северное Заангарье (Гибет, Артамошин, 1977); 82 - р.Чо-на у впадения в р.Вилюй (колл. ЗИН); 83 - окр. с.Сунтар (колл. ЗИН); 84 - окр. с.Эльгяй (Андреев, 1974; колл. ЗИН); 85 - окр. с.Шея (Андреев, 1974); 86 - долина средней Лены (Соломонов, Ларионов, 1971); 87 - окр. г.Якутск (Иванов, 1929); междуречье рек Лена и Кенгке-ме (Шубникова, Морозов, 1959); 88 - левый берег р.Лена, ур.Тюбе Баха; Хомурган (Иванов, 1929); 89 - р.Лунгха (Маак. - Цит. по: Mauersberger, Portenko, 1967); 90 - устье р.Вилюй (Андреев, 1974); 91 - р.Лена, 68° с.ш. (Степанян, 1978); 92 - хр.Орулган, р.Тара-Сала (Воробьев, 1963); 93 - среднее течение р.Бытантай; пос. Кустур (Воробьев, 1963; колл, ЗИН); 94 -долина р.Ойун-Урях (левый приток р.Яна) (Наумов, Лабутии, 1961); 95 - р.Яна, 68° с.ш. (Сушкин, 1938; Портенко, 1960; Воробьев, 1963); р.Кумах на водоразделе рек Яна и Адыча (Ткаченко, 1932; Сушкин, 1938; колл. ЗИН); 96 -долина р.Нельгесе (Кривошеев, 1960); 97 -долина ручья ’’Соревнование” в 40 км ниже устья Неньделге (Кривошеев, 1960); 98 - устье р.Ал-дан (Иванов, 1929); 99 - р.Амга (колл. ЗИН); 100 - Ленско-Амгинское плато (Иванов, 1929);
101 - Олемо-Чарское нагорье; р.Тяня (Воробьев, 1963; колл. ЗИН); 102 - верхнее течение р.Алдан (Благосклонов, 1954); 103 - устье р.Мая (Иванов, 1929); 104 - р.Мая, перек. Оленькю, перек. Перочта (Иванов, 1929); 105 - р.Инди-гирка, 68° с.ш. (Сушкин, 1938; Портенко, 1960; Иванов, 1976; Степанян, 1978); устье р.Уяндина (Сушкин, 1938); 106 - хр. Черского; долина р.Нера (Воробьев, 1959); 107 - пойма р.Омолон; 30 км ниже устья р.Олойчан (Кречмар и др., 1978); 108 - верхнее течение р.Анадырь (Портенко, 1960; Степанян, 1978); 109 - среднее течение р.Анадырь (Портенко. - Цит. по: Кищин-скому, 1980); р.Марковка; урочище Ягодное (колл. ЗИН); окр. с.Марково (Иванов, 1976; колл. А.П.Кузякина); 110 - Корякское нагорье, р.Кай-Тэклаваям (среднее течение р.Ачайваям) (Кищинский, 1980); р.Ачайваям (Иванов, 1976; колл. ЗИН); верховья реки (Кищинский, 1980); 111 - Пенжинский р-н (Яхонтов, 1979); 112 -Парапольский дол (Лобков по Кищинскому, 1980); ИЗ - окр. с.Омсукчан (колл. В.В.Леоно-вича); 114 - нижнее течение р.Детрин близ побережья залива Бабушкина (Кищинский, 1968); 115 - поймы рек Кухтуй и его крупных притоков (Кузякин, Второв, 1963); Ц6 - поймы рек Охота и ее крупных притоков (Кузякин, Второв, 1963); 117 - р.Кетанда (Линдгольм. - Цит. по: Mauersberger, Portenko, 1967); 118 - Охотское побережье (Иванов, 1976; Степанян, 1978); р.Урак (Линдгольм. - Цит. по: Mauersberger, Portenko, 1967); 119 - Алдано-Учурский хр. (Воробьев, 1963); 120 - с.Бомнак (Штегман. -Цит. по: Mauersberger, Portenko, 1967); 121 -восточная часть хр.Тукурингра; Зейский заповедник; побережье Зейского водохранилища (сообщ.
95
В.Ю.Ильяшенко); 1.22 - р.Зея, ключ Теплый (колл. БПиДВНЦ); 123 - с.Кумара (Иванов, 1976); 124 - Свободненский р-н, с.Клнмоуцы; междуречье Малой Перы и Большого Эргеля (колл. ЗИН); 125 - окр. с-Дагмара на р.Селемд-жа (колл. КГУ); 126 - с.Экимчан (колл. В.В.Ле-оновича); 127 - хр.Малый Хинган (Яхонтов, 1973); 128 - с.Коидон (южнее оз.Эворон) (Федотов, Брунов, 1977); 129 - с.Киселевка (Кистяковский, Смогоржевский, 1973; колл. КГУ); 130 — окр. пос.Софнйск, о-в Бычий; долина кл.Юдинка (Нечаев, 1960; колл. ЗИН); 131 -с.Дуди (Кистяковский, Смогоржевский, 1973; колл. КГУ); 132 - устье р.Амур (Гизенко, 1955; Портенко, 1960; Степанян, 1978); с.Талышки (Кистяковский, Смогоржевский, 1973; колл. КГУ); с.Сусанино (колл. КГУ); 133 - р-н оз.Орель (Бабенко, 1984); 134 - Шантарские о-ва (Благосклонов, 1954; Гизенко, 1955; Степанян, 1978); о-в Большой Шантар (Яхонтов, 1977); 135 - о-в Феклистова (Яхонтов, 1977); 136 -о-в Малый Шантар (Яхонтов, 1977); '137 - Камчатка (Гизенко, 1955; Портенко, 1960; Иванов, 1976; Степанян, 1978); долина среднего течения р.Камчатка, долины рек Кунч н Яковская (Вронский, 1977); 138 - окр. г.Козыревск (колл. ЗИН); 139 - с.Ключи (Bergman - Цит. по: Mauersberger, Portenko, 1967); 140 - Кроноцкий заповедник (Аверин, 1948; Лобков, 1978); 141 - окр. сЛущино (колл. БПиДВНЦ; КБ МГУ); 142 - окр. г.Петропавловск-Камчатский; Авачинская бухта; р.Авача (колл. ЗИН); 143 — о-в Парамушир (Yamashina, 1931); 144 — долина р.Большой Чивыркуй (Малышев, 1960); р.Том-пуда, протока Тихая (колл. КБ МГУ).
Места обитания. Населяет различные типы хвойных, смешанных хвойнолиственных и лиственных лесов.
На Северо-Восточном Алтае предпочитает частично заболоченные березово-еловые долинные леса среднегорья (Равкин, 1984). У северного берега Телецкого озера встречается поблизости от воды в березняках низин и по краю тайги (Сушкин, 1938). В горы поднимается до 500 м (Портенко, I960). В Тувинской котловине отмечена в лиственных и смешанных уремах (Сушкин, 1938). На Салаирском кряже населяет исключительно приречные ивняки по р.Томь-Чумыш (Чунихин, 1965).
В юго-восточной части Западно-Сибирской равнины гнездится в темнохвойных лесах со значительной примесью спелого осинника (Гынгазов, Миловидов, 1977). На р.Кеть встречается во всех хвойных лесах (Москвитин и др., 1977), а на р.Обь в Чаинском р-не Томской обл. отмечена в пойменных уремных лесах (Пантелеев, 1972а).
В р-не Красноярска (Юдин, 1952) гнездится в светлых смешанных лесах из сосны, березы и осины. Тяготеет к участкам леса, прилежащим к старым гарям и лесосекам. В Нижне-Ингашском р-не отмечена
в елово-пихтовых лесах (Равкин и др., 1983). В Приенисейской тайге на Нижнем Елогуе населяет сосновые и сосново-лиственные леса, предпочитая участки около полян, а также зарегистрирована на вырубках по берегам реки (Рогачева, 1962). В р-не Норильских озер встречается в высокоствольных смешанных лесах северотаежного типа из сибирской лиственницы, ели и березы (Кречмар, 1962). У оз.Капчук отмечена только на участках густо захламленного елово-лиственничного леса с густым подлеском из ольховника (Морозов, 1984).
В южной части п-ова Ямал найдена на гнездовании в редком елово-лиственничном лесу в окр. Ласточкинового берега на р.Хадытаяха (Данилов и др., 1984).
В горах юга Средней Сибири (Реймерс, 1966) населяет придолинную полосу лесов с высоким участием лиственных деревьев. Нарушенные приречные и приручьевые ельники или эти же ельники, перемежающиеся с кустарниками, являются здесь исконными стациями сибирской малой мухоловки. Предпочитают лес с хорошо развитым подлеском, у истоков ручьев, гари при-ручьевых ельников и приручьевым участкам темнохвойных лесов водоразделов. Менее охотно поселяются в сосняках.
В Иркутской обл. гнездится в сосновоберезовых лесах, на старых вырубках соснового леса, в ельниках (сообщ. В.Д.Со-нина) и на гарях среди последних (колл. КБ МГУ). В Приангарье предпочитает лесоболотные комплексы (Равкин, 1984). В Северном Заангарье населяет таежные хвойные леса (Гибет, Артамошин, 1977). На северо-восточном побережье оз.Байкал отмечена на опушках смешанного леса с примесью березы (Скрябин, Филонов, 1962), а на берегу оз.Иркан добыта в сосновом лесу (Гагина, 1954). На Баргузин-ском хр. зарегистрирована на гнездовании в условиях субвысокогорья на высоте свыше 900 м (Федоров, 1982). В Юго-Западном Забайкалье (Измайлов, Боровицкая, 1973) встречается в приключевых смешанных лесах, в горной темнохвойной тайге по долинам горных рек и ручьев; реже — в сосновых лесах и приречных кустарниковых зарослях. В верхнем течении р.Чикой найдена в ивняках по р.Бобровая (Леонтьев, Павлов, 1963). На Витимском плоскогорье населяет высокоствольные лиственничные леса и лиственничные редколесья, а также березово-лиственничные и лиственнично-сосновые леса (Измайлов, 1967). В юго-восточной части плоскогорья у озер
96
Тасей, Иван и Арахлей обитает в редколесье смешанного леса, но не избегает и открытых пространств, поросших кустарниковой березой и ивой с островками лиственйич-ного леса, а в верхнем течении р.Чита гнездится в ивняках по речным долинам и в лиственничном лесу, особенно по опушкам (Павлов, 1948).
В Верхне-Чарской котловине в гнездовой период встречается в рододендроновобрусничниковых лиственничниках, а также в лиственничниках ерниково-голубичнико-вых с заболоченными лесными полянами, в пойменных лесах и в предгольцовом редколесье (Павлов, Парыгин,1969).
В Амурской обл. в восточной части хр.Тукурингра населяет преимущественно разреженные белоберезовые леса, обычно по краю опушек, но гнездится также в лиственнично-белоберезовых лесах и черноберезовых с густым подлеском из монгольского дуба (сообщ. В.Ю.Ильяшенко). В Хабаровском крае у с.Кондон (южнее оз.Эво-рон) отмечена в березово-лиственничных и лиственничных лесах со вторым ярусом из дуба (Федотов, Брунов, 1977). На нижнем Амуре у с.Софийск обычна в ивняках о-ва Бычий и в смешанных лесах долины кл.Юдинка (Нечаев, 1960), а в р-не оз.Орель встречается в лиственничных лесах (Бабенко, 1984).
В Южной Якутии поселяется в смешанных лесах из лиственницы, ели, сосны и березы, с подлеском из рябины и боярышника (Воробьев, 1963). На р.Алдан излюбленными местами обитания являются сырые высокоствольные ельники с густым подседом и кучами бурелома. Гораздо реже встречается здесь в сухих лиственничниках и в зарослях кустарников по долинам таежных рек (Иванов, 1929). По р.Вилюй придерживается глухих крупноствольных лиственничных и смешанных лесов (Андреев, 1974), а в междуречье рек Лена и Кенгке-ме, помимо лиственничников, гнездится и в березняках (Шубникова, Морозов, 1959). В долине средней Лены населяет ельники, смешанные сосново-лиственничные леса и кустарниковые заросли (Соломонов, Ларионов, 1971).
В р-не г.Охотск встречается в пойменных лесах из ольхи, тополя, ивы и черемухи, с подлеском из розы в осветленных местах (Кузякин, Второв, 1963). В Магаданской обл. у с.Омсукчан также найдена на гнездовании в приречных лесах (колл. В.В. Леоновича).
На хр.Черского и в Верхоянье населяет
речные поймы и острова, поросшие тополем, ивой и чозенией, однако не избегает и часто лиственничной тайги (Воробьев, 1959; 1963).
В р-не р.Омолон и у устья р.Олойчан встречается в горной и пойменной тайге (Кречмар и др., 1978), а в пойме р.Ана-дырь у с.Марково гнездится в старых ивняках (колл. А.П.Кузякина).
В Корякском нагорье (Кишинский, 1980) населяет высокоствольные тополь-ники. Так, найдена на гнездовании в тополевой роше по р.Кай-Тэклаваям. К западу довольно обычна в высокоствольных пойменных лесах Парапольского дола (Лобков. — Цит. по: Кишинскому, 1980), а в Пенжинском р-не отмечена в светлохвойной тайге, по поймам рек и в лесотундре (Яхонтов, 1979).
На Камчатке (Вронский, 1977) в долине верхнего течения р.Камчатка встречается в пойменных многоярусных лесах из душистого тополя, чозении, ивы и ольхи с подлеском и кустарниковым ярусом из ивы и бузины, а в центральной части долины этой реки — в смешанных лесах из лиственницы и белой березы. По склонам горных цепей, увалов и всхолмлений населяет каменноберезовые леса. В Кроноцком заповеднике также гнездится в каменноберезовых лесах (Лобков, 1978).
Численность. Очень неравномерна. На Камчатке и в Нижнем Приамурье в характерных биотопах обычна. В ряде других мест гнездового ареала редка и распространена спорадично.
Прилет и весенний пролет на территории СССР. Проходит с конца I декады мая по конец II декады июня включительно. Летит в светлое время суток низко над землей на высоте 0,5— 2,0 м (Беляев, 1979).
В Южном Приморье слабо выраженный пролет проходит во II—III декадах мая. На о-ве Большой Пелис в заливе Петра Великого пролетные малые мухоловки зарегистрированы в 1968 г. в середине мая (Лабзюк и др., 1971). В Партизанском р-не (ст.Мучная) — 19.V 1928 г. (колл. ЗИН), в Лазовском заповеднике - 25.V 1945 г. (Белопольский, 1950), а у оз.Ханка (3 года) — 22, 23, 29 и 31.V (Черский, 1915а; Панов, 1973; колл. БПиДВНЦ).
В Среднем Приамурье взрослые самцы добыты: нар.Селемджа (окр.пгтЭкимчан) — 14.V 1963 г., на р.Бурея (с.Чекунда) -16 и 17.V 1962 г. (колл. КГУ) и в междуречье рек Малая Пера и Большой Эргель —
97
19.V 1958 г. (колл. ЗИН). Первые птицы в г.Зея встречены 15.V, а массовый пролет зарегистрирован здесь 25—27.V (сообш. В.Ю.Ильяшенко). На нижнем Амуре пролет отмечен: у с.Ченки — 19.V, у с.Гячи — 26.V (интенсивный) (Кистяковский, Смогоржевский, 1973) ив р-не оз.Орель - до 1.VI 1981 г. (Бабенко, 1984),
На юге Читинской обл. пролетные мухоловки добыты: у оз.Зун-Торей — 10.V 1975 г. (колл. ИЗАНУ) и в окр. с.Кулу-сутай — 17, 18, 21 и 28.V 1856 г. (колл. ЗИН). В юго-восточной части Витимского плоскогорья в р-не озер Тасей, Иван и Арах-лей самцы на весеннем пролете отмечены 21.V, самки появляются позднее — 26 .V, а в верхнем течении р.Чита первое появление малых мухоловок зарегистрировано 17.V (Павлов, 1948). В Юго-Западном Забайкалье первые птицы встречены (2 года) 18 и 21.V (Измайлов, Боровицкая, 1973), а в северной части Витимского плоскогорья прилет и пролет проходят 24—25 .V (Измайлов, 1967). В Баргузинском заповеднике миграция протекает (3 года) 22.V—4.VI, средняя дата — 27 .V (Филонов, 1967). В Верхнечарской котловине появляются с III декады мая, первый самец отмечен 21.V (Павлов, Парыгин, 1969).
На верхней Лене в 1961 г. появление зарегистрировано 1.VI, а на р.Чадобец в 1962 г. лишь в середине июня (Реймерс, 1966). У г.Красноярск прилет отмечен (4 года) 26-28 и 30.V (Юдин, 1952).
В Якутию прилетают во второй половине мая: на р.Чара (Олекминский р-н) птицы в 1956 г. появились 22.V (Воробьев, 1963), на р.Нера (хр.Черского) в 1958 г. — 25.V (Воробьев, 1959; 1963), а в Верхоянье на р.Бытантай в 1959 г. — 17.V (Воробьев, 1963). На северо-востоке СССР в р-не р.Омолон в 1973 г. отмечена 20.V (Кречмар и др., 1978), а на р.Анадырь прилетает в I декаде июня (Портенко, 1960).
На Камчатке в Кроноцком заповеднике первые мухоловки появляются (многолетние данные) в конце мая (Лобков, 1978) -первой половине июня (4—6, 10, 13.VI) (Аверин, 1948). Пролет длится до 20.VI (Аверин, 1948). На севере полуострова в пгт Корф пролетная птица отмечена 5.VI 1976 г. (Кишинский, 1980).
Размножение. Проходит в общих чертах так же, как и у номинативного подвида. Гнезда сибирские малые мухоловки строят в подавляющем большинстве случаев в дуплах и редко — в полудуплах пней, стволов и ветвей живых, гниющих и сухо
стойных деревьев (лиственница, береза, осина, ива, тополь, чозения и др.) на высоте (п = 28): 0,6-17 м от поверхности земли, в среднем — 4,0 (Юдин, 1952; Воробьев, 1963; Зонов и др., 1967; Измайлов, 1967; Измайлов, Боровицкая, 1973; Кишинский, 1980; Данилов и др., 1984; Морозов, 1984; колл. В.ВЛеоновича; А.П.Кузякина; сооб. В.Д.Сонина и В.Ю.Ильяшенко). Помимо естественно образовавшихся, охотно заселяют дупла, выдолбленные дятлами (трехпалый, малый и большой пестрые). Зарегистрированы одиночные случаи гнездования в искусственных гнездовьях (различные дуплянки) (Андреев, 1974; Измайлов, Боровицкая, 1973). Нарастание дуплогнез-ности проходит с юга на север и у северной границы ареала птицы этого подвида гнездятся исключительно в дуплах.
В отличие от номинативного подвида в качестве строительных материалов для гнезд используют главным образом полоски луба деревьев и кустарников (шириной до 14 мм) и значительно реже кусочки мелких листьев и стебельки трав (в том числе и злаков), сухие листья древесных и кустарниковых растений, кусочки коры, а по данным других исследователей (Шубникова, Морозов, 1959); Воробьев, 1963; Данилов и др., 1984) — волос шерсти млекопитающих и талломы лишайциков. Лоток чаше всего не выстилается, в редких случаях птицы используют для этой цели мягкие полоски луба, растительные волокна и тончайшие пленки бересты. Размеры гнезд (п = 4), мм: диаметр 75—108 х 80—135, в среднем — 86 х 106; высота 48—60, в среднем — 54; диаметр лотка 50—62 х х 60-62, в среднем - 58 х 61; глубина лотка 24-40, в среднем - 35.
В сезон размножения на территории СССР сибирская малая мухоловка имеет один репродуктивный цикл. Полная кладка состоит из 5—9 яиц. Фоновая окраска яиц от почти белого цвета до бледно-голубого. Расположение рыжевато-охристых мелких крапинок и точек (четких и размытых) по поверхности скорлупы различно. На одних яйцах их очень мало, на других они почти полностью покрывают всю поверхность скорлупы или концентрируются у тупого полюса. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
Пуховой птенец сибирской малой мухоловки (Нейфельд, 1970) имеет дымчатосерый, довольно редкий и короткий пух на тех же самых птерилиях, что и птенец номинативного подвида. Пигментация по
98
кровов и другие птенцовые признаки совершенно идентичны у обеих форм.
Сезон размножения сибирской малой мухоловки на территории СССР длится с начала III декады мая по I декаду августа включительно.
В р-не г.Красноярск (Юдин, 1952) образование пар и токовые полеты самцов отмечены 1-2.VI 1943 г. у Гремячинской сопки кладка из 9 слабо насиженных яиц найдена 14.VI 1945 г., а слетки добыты (2 года) 12 и 15.VII. На р.Кеть у протоки Анга (д.Карбино) гнездо с 3 яйцами встречено 31.V 1968 г., а у с.Красная Курья полная кладка обнаружена 12.VI 1963 г. На р.Орловка выводки с взрослыми птицами зарегистрированы 26-29.VI 1967 г., в верховьях р.Кеть - 18.VII, на р.Тым (окр. с.Напас) - 27-29.VII 1967 г., а на среднем Чулыме слетки отмечены 2-4.VII1969 г. (Гынгазов, Миловидов, 1977; Москвитин и др., 1977). В бассейне р.Елогуй несколько выводков встречены 10.VII 1932 г. (Юдин, 1952).
В Тункинском р-не Бурятской АССР (сообш. В.Д.Сонина) самка с готовым яйцом в яйцеводе добыта 30.V 1970 г., а 17.VI 1970 г. найдена сильно насиженная кладка из 9 яиц. В Иркутской обл. (сообш. В.Д.Сонина) у пос.Майск (Усть-Удинский р-н) гнездо с 2 яйцами обнаружено 2.VI 1966 г., а у ст.Выдрино (Слюдянский р-н) с 3 яйцами — 15.VI 1965 г.; в Тулунском р-не у с.Гадалей полная слабо насиженная кладка из 7_ яиц зарегистрирована 12.VI 1964 г., a 26.VI 1964 г. в пойме р.Ия у г.Ту-лун найдено 2 гнезда малой мухоловки: в одном находилось 4 свежих яйца, в другом — 8 птенцов, которые вылетели при осмотре гнезда. В окр. с.Кударейка (Эхи-рит-Булагатский р-н) сильно насиженная кладка из 8 яиц найдена 16.VI 1974 г.
В Юго-Западном Забайкалье гнездо с 8 сильно насиженными яйцами обнаружено 23.VI 1962 г., птенцы вылупились в нем 28.VI (Измайлов, Боровицкая, 1973). Слетки отмечены здесь (2 года) 3 и 28 .VII (Измайлов, Боровицкая, 1973; Линдеман, 1976). На Витимском плоскогорье (Измайлов, 1967) подавляющее большинство пар строит гнезда и откладывает яйца в I декаде июня, во II декаде этого месяца встречаются насиженные кладки, а в III декаде июня — начале июля в гнездах появляются птенцы. В верхнем течении р.Чита слетки зарегистрированы с 8.VII (Павлов, 1948), а на северо-восточном побережье оз.Байкал — 11.VII 1958 г. (Скрябин, Филонов, 1962).
В Зейском заповеднике образование пар и токовые полеты самцов отмечены В.Ю.Ильяшенко 23.V 1980 г., a 23.VII 1980 г. на побережье Зейского водохранилища им найдено гнездо, в котором взрослые птицы выкармливали птенцов. В верховьях р.Селемджа у пгт Экимчан полная кладка из 8 яиц обнаружена В.В.Ле-онтовичем 16.VI 1971 г. На нижнем Амуре токование зарегистрировано у с.Киселев-ка - 3.VI, у с.Дуди - 15.VI и у с.Тальни-ки - 20.VI (Кистяковский, Смогоржевский, 1973). В Якутии на р.Вилюй (Андреев, 1974) в окр. с.Шея полная кладка из 8 яиц найдена 5.VI. У с.Эльгяй 30.VI 1968 г. обнаружено гнездо с 7 птенцами, которые вылетели из него 5.VII. 14.VII ранее пустовавшее рядом дупло было занято парой малой мухоловок. В Олекминском р-не на р.Тяня гнездо с полной ненасиженной кладкой найдено 16.VI 1956 г. (Воробьев, 1963). В междуречье рек Лена и Кенгкеме первая полная кладка отмечена 21.VI, первый выводок — 2.VII, а последняя кладка найдена 11.VII (Шубникова, Морозов, 1959). В Хомургане слетки только что покинувшие гнездо встречены 2.VII, а летная молодая птица - 15.VII (Иванов, 1929). В Верхоянье на р.Тара-Сала и в долине р.Нельгесе выводки зарегистрированы 5.VII (Кривошеев, 1960; Воробьев, 1963).
На Южном Ямале (р.Хадытаяха) гнездо с 7 птенцами в возрасте 3—4 дней и 1 не-оплодотворенным яйцом обнаружено 8.VII 1980 г. (Данилов и др., 1984). В р-не Норильских озер у оз.Капчук (Морозов, 1984) начало строительства гнезда в 1980 г. отмечено 5.VI. Законченная постройка найдена 27.VI, 30.VI в ней было 3 яйца, а 3.VII - 5 яиц. Выводок летных молодых встречен здесь 31.VII.
В Магаданской обл. (нижнее течение р.Детрин) поющий самец на гнездовом участке зарегистрирован 9.VI 1964 г. (Ки-шинский, 1968). Полная слабо насиженная кладка обнаружена у с.Омсукчан 24.VI 1972 г. (колл. В.ВЛеоновича), a 11.VIII 1964 г. у залива Бабушкина добыта летная молодая птица (Кишинский, 1968). В пойме р.Анадырь (с.Марково) полная ненаси-женная кладка найдена 25.VI 1960 г. (колл. А.П.Кузякина). В Корякском нагорье (Кишинский, 1980) на р.Кай-Тэклаваям, в среднем течении р.Ачайваям гнездо с 5 птенцами, которые при осмотре вылетели из него, обнаружено 26.VII 1960 г. Летная молодая мухоловка встречена в верховьях р.Ачайваям — 14.VIII 1960 г.
99
Линька.- Проходит в основном так же, как и у номинативного подвида. Большинство самцов первогодков на местах зимовок в предбрачную линьку приобретают рыжие перья на горле.
Питание. Основу пиши составляют насекомые разных групп и стадий развития. Реже поедает пауков.
Отлет и осенний пролет на территории СССР. Проходит с конца I декады августа по III декаду октября включительно.
На северо-востоке СССР покидает места гнездования в начале I декады сентября. У устья р.Пятьковенде последняя птица в 1974 г. отмечена 5.IX (Кречмар и др., 1978), а на Охотском побережье у устья р.Немуй мухоловка добыта 17.1Х 1935 г. (колл. ЗИН).
На Камчатке в Кроноцком заповеднике осенняя миграция начинается (в разные годы) в конце I, во II или в III декадах августа (Аверин, 1948; Лобков, 1978). В 1941 г. на побережье после летнего отсутствия взрослые и молодые птицы появились 9 .VIII, а в 1945 г. интенсивный пролет в долине Гейзерной проходил 2-7.IX (Аверин, 1948).
На Амуре в г.Хабаровск одиночная пролетная особь встречена 18.IX (Воробьев, 1938), а в окр. г.Николаевск — 7.IX 1854 г. (колл. ЗИН). В Приморье пролет проходит во второй половине сентября - октябре: у оз.Ханка (с.Ново-Владимировка) мухоловки добыты 15 и 29.IX 1911 г. (Черский, 1915а; 19156); в окр. с.Троицкое — 21.IX 1909 г. (Черский, 1915а); на р.Раздольная (Суйфун) — 2.Х 1882 г. (колл. ЗИН); в заповеднике ’’Кедровая падь” встречены 9 и 16.Х 1960 г., на п-ове Де-Фриза - 25.Х 1961 г. (Панов, 1973), а на о-ве Большой Пелис в заливе Петра Великого - 16 и 24.IX 1977 г. (Назаров и др., 1978).
На Витимском плоскогорье покидает места гнездования в конце августа (Измайлов, 1967). В Баргузинском заповеднике миграция проходит в конце августа - сентябре, последняя встреча — 29.IX (Скрябин, Филонов, 1962). На юге Читинской обл. у с.Кулусутай мухоловки добыты Г.Радде 11 и 13.IX 1856 г. (колл. ЗИН), а в Борзинском р-не пролет проходит (5 лет) 16.VIII-22.IX (сообш. М.А.Оси-повой). На верхней Лене в 1961 г. последняя птица зарегистрирована 8.IX (Реймерс, 1966), а в окр. г.Иркутск пролетные мухоловки добыты 6, 8 и И.IX 1908 г. (колл. ЗИН).
В окр. г.Томск встречены на пролете (3 года) 17, 27.VIII и 5.IX (Гынгазов, Миловидов, 1977), а в р-не г.Красноярск (с.Торгашино) - 9.IX 1912 г. (колл. ЗИН).
Практическое значение. Такое же, как и у номинативного подвида. Род Cyanoptila Blyth, 1847 - синяя мухоловка
Cyanoptila Blyth, 1847, Jour. Asiat. Soc. Bengal, 16: 124.
Типовой вид Muscicapa cyanomelana Temminck
Род включает 1 вид: синюю мухоловку - Cyanoptila cyanomelana (Temm.), которая распространена на Дальнем Востоке, в том числе и на территории СССР.
Средних размеров настоящие мухоловки. Клюв широкий и толстый. Боковые поверхности надклювья заметно выпуклы. На вершине надклювья имеются крючочек и выемка перед ним. Лобные шетинки и шетинки в углах рта тонкие, умеренной длины. Их количество в углах рта у основания надклювья (с каждой стороны головы) равно 3-5. Хвост умеренной длины с очень маленькой выемкой на вершине в центре. Ноги относительно сильные. Заостренную вершину крыла образуют 3-е и 4-е первостепенные маховые; 2-е и 5-е немного их короче. Первое первостепенное маховое очень мало. Оперение густое и мягкое. Его окраска определяется как наличием пигментов, так и своеобразной микроструктурой пера, по-разному преломляющей световые лучи. Половой диморфизм резко выражен в окраске уже с гнездового наряда. У молодых в гнездовом наряде перья с пятнистым рисунком.
Населяют различные типы лиственных, смешанных хвойно-лиственных и хвойных лесов. Единственный представитель рода является перелетной птицей.
Самцы интенсивно и звучно поют. Сравнительно большие плотные гнезда синие мухоловки строят почти полностью из мха, помещая их как открыто — на уступах скал, так и в каменных нишах, а также расщелинах стволов и толстых ветвей деревьев. На территории СССР в сезон размножения имеют один репродуктивный цикл. Яйца белые с редкими мелкими бледно-коричневатыми крапинками.
Питаются главным образом насекомыми разных групп и стадий развития, реже поедают других беспозвоночных (паукообразные, многоножки, моллюски), а также мелкие сочные плоды некоторых растений.
100
Взрослые птицы имеют в году две линьки: полную предбрачную на местах зимовок (Виноградова и др., 1976) и полную послебрачную на местах гнездования.
Синяя мухоловка — Су anopt ila cyanomelana (Temminck, 1829)
Muscicapa cyanomelana Temminck, 1829, Temminck et Laugier, Planches color., livr. 79, pl. 470, Япония.
Описание. Составлено по 128 экз. (124 - из Приамурья, Приморья, с о-вов Кунашир и Монерон, 1 — из Японии, 3 -из Китая).
Вершину крыла образуют 3-е и 4-е первостепенные маховые (3 < 4 - 17,3 %; 3=4- 39,5 %; 3 > 4 - 43,2 %); 2-е и 5-е немного их короче (2 < 5 — 98,7 %; 2 = = 5 — 1,3 %); 2-е всегда больше 6-го. У большинства особей 1 -е первостепенное маховое очень мало и только на несколько миллиметров выступает за вершины кроющих кисти, но значительно реже бывает равно им или немного короче (табл. 11). Наружные опахала 3-5-го первостепенных маховых в вершинной части заметно сужены. Количество шетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3-5 (3 - 30,3 %; 4 - 67,1; 5 - 2,6 %). Длина наибольшей (п = 50) 5,0-9,0 мм (в среднем — 7,3). Количество шитков подотеки на передней части цевки варьирует от 2 до 5. Половой диморфизм выражен в окраске оперения, рамфотеки, подотеки, а также в размерах отдельных частей тела (табл. 11).
У старого самца в свежем пере верхняя часть головы ярко-блестяше-сине-голубая, с усилением синего на затылке. Верхняя часть шеи, частично ее бока и спина яркосиние. Синий цвет светлеет на надхвостье и верхних кроющих хвоста. Перья на боках головы, горле, зобе и верхней части груди матово-черные с сине-голубыми вершинными каемками шириной до 3 мм. Нижняя часть груди, брюшко и нижние кроюшие хвоста белые. Бока тела темносерые с примесью синего на вершинах отдельных перьев. Маховые черно-бурые. Их наружные опахала, начиная с 3-го первостепенного махового, несут по краю ярко-голубоватосиние канты, ширина которых увеличивается к третьестепенным маховым. Большие верхние кроюшие маховых черно-бурые с синими каемками на наружных опахалах. Остальные верхние кроюшие крыла черно-бурые в основаниях и ярко-блестяще-синие на вершинах. Синий цвет на них усиливается
к кистевому сгибу. Нижние кроющие крыла от бурого до черно-бурого с незначительной примесью синего и зеленовато-голубого по краям наружных опахал и на вершинах перьев. Центральная пара рулевых сине-голубая. Остальные рулевые в верхней части каждого пера имеют сине-голубые наружные опахала и черно-бурые внутренние. Основания рулевых белые. Клюв и ноги черные. Радужина глаза темно-бурая. Индивидуальная изменчивость проявляется в незначительном варьировании размеров белого поля в основании рулевых и в появлении у некоторых особей узких продольных черных пестрин на спине, надхвостье и верхних кроющих хвоста; сезонная — в незначительном посветлении окраски и в появлении на верхней части тела примеси зеленовато-голубых тонов в результате частичного обноса и выцветания оперения. Верхняя часть головы приобретает блестяше-ярко-голубой цвет с бирюзовым оттенком. Синие вершинные каемки на черных перьях горла, зоба и верхней части груди частично или полностью обнашиваются.
Взрослая самка в свежем пере сверху однотонно-буровато-оливковая со слабым блеском. Надхвостье и верхние кроющие хвоста с примесью коричнево-охристого. Оперение верхней части головы со слабо выраженным волнистым рисунком. Бока головы, шеи, зоб и верхняя часть груди светло-буровато-оливковые. Оливковый цвет светлеет на боках тела и нижней части груди. Брюшко, нижние кроюшие хвоста и небольшое пятно на горле бледно-охристые. Маховые и их большие верхние кроюшие бурые с коричневоохристыми каемками по краю наружных опахал, а на последних — и на вершинах перьев. Рулевые однотонно-буровато-коричневые. Клюв и ноги бурые. Сезонная изменчивость проявляется в общем посветлении окраски оперения, в побурении его на верхней части тела, а также в частичной или полной утрате боковых и вершинных каемок на маховых и верхних кроющих крыла в результате их обноса и выцветания.
Молодые в гнездовом наряде сверху коричнево-бурые с желтовато-охристыми пятнами и узкими черными каемками на вершинах перьев. На боках шеи, а также на горле и груди светло-желтовато-охристые перья окантованы узкими черно-бурыми каемками. Уже в этом наряде самцы резко отличаются по окраске от самок. На наружных опахалах маховых и верхних кроюших первостепенных маховых у них
101
Таблица 11. Размеры и коэффициенты вариации (CV,%) синей мухоловки
Место сбора п Длина, мм
Крыло CV Расстояние, на которое 1-е маховое выходит за вершины кроющих кисти CV Хвост CV Клюв от оперения лба CV Клюв от орального края ноздри CV Цевка CV
Самцы первогодки
Нижнее 19 92,261 0,38 1,78 235Ю30 85,47 63,081 0,41 2,77 11,121 0,12 4,44 8,0410,08 4,07 16,341 0,11 2,93
Приамурье 89 Л -953 -1,8 -7,2 59,4—66,4 10,3 —12,0 73 -8,6 15,4 -17,1
Приморье 15 92,06± 031 2,14 2,7510,33 4634 61371 0,64 4,00 11,231 0,14 4,69 8,1510,11 532 16,041 0,14 3,35
89,4 -953 03 -4,8 56,3 —65,0 10,3 -12,2 73 -9,0 15,1 -17,1
Самцы взрослые (старше одного года)
Нижнее 8 93,191 0,53 139 1,731039 97 33 64381 0,74 3,26 11,071 0,14 3,06 7,7010,12 3,67 16,211 1,69 2,96
Приамурье 91,0 -95,8 0,0 -3,8 62,0 -68,8 10,4 -11,3 7,3 -8,0 153 -16,8
Приморье 26 92,291 0,39 2,14 13510,25 77,70 63,101 032 2,43 11,311 0,11 4,42 8,4010,08 4,38 16,161 0,09 2,98
87,0 -953 -03 -4,1 60,3 -65,1 10,2 -12,1 7,7 -9,1 15,3 -17,0
Остров 2 883* - 3,0011,00 47,14 58,651 0,35 0,84 12,001 0,00 0,00 9,0010,00 0,00 16,451 0,05 0,43
Кунашир 2,0 -4,0 58,3 -59,0 12,0 9,0 16,4 -16,5
Самки
Нижнее 2 89,901 0,10 0,16 0,6511,65 35,90 61,651 1,95 4,47 11,001 0,60 7,71 8,0010,80 14,14 16,1010,00 0,00
Приамурье 89,8 -90,0 -1,0 -2,3 59,7 -63,6 40,4 -11,6 7,2 -8,8 16,1
Приморье И 89,031 0,69 239 2,6810,33 38,89 593U 0.72 4,02 11,041 0,15 4,25 8,2410,11 4,25 16,491 0,14 2,76
85,0 -923 1,1 -4,0 543 -62,0 10,3 -11,7 7,7 -8,8 15,7 -17,2
Место сбора п Ширина, мм Высота, мм Отношение ши-рины клюва у основания к его длине от оперения лба, % CV Отношение длины хвоста к длине крыла, % CV Отношение ширины крыла к его длине, % CV
Крыло CV Клюв у орального края ноздри CV Клюв у основания CV Клюв у основания CV
Самцы первогодки
Нижнее Приамурье 19 63,841 0,35 -59,4 -66,1 2,32 4,6810,07 4,1 -5,1 6,30 7,1810,09 6,4 -7,7 5,74 4,5410,14 4,3 -5,3 7,92 64,811 0,51 61,26-69,16 3,35 68,281 0,40 65,27 -71,47 2,46 69,181 0,28 66,00-71,51 1,74
Приморье 15 63,921 0,41 61,4 -67,1 2,46 4,6610,06 4,3 -5,0 5,05 7,1110,11 6,4 -7,9 6,26 4,5310,17 4,2 -5,3 9,32 63,281 0,79 58,26-68,81 4,86 67,301 0,53 62,98-70,36 3,05 69,441 0,35 66,88-71,57 1,93
Самцы взрослые (старше одного года )
Нижнее Приамурье 8 64,051 0,53 62,2 -67,2 2,36 4,4910,10 4,1 -4,7 5,96 6,7410,15 6,0 -7,1 5,98 4,5510,06 4,4 -4,7 2,84 62,131 1,50 57,52-68,27 5,92 69,071 0,57 66,84-71,82 2,35 68,731 0,26 67,75-70,15 1,07
Приморье 26 63,771 0,33 60,0 -67,3 2,62 4,75 10,06 4,2 —5,5 6,43 6,9810,09 5,6 -7,9 6,42 4,5110,09 4,0 -5,3 7,38 61,781 0,76 54,90-68,27 5,60 68,141 0,24 66,12-69,95 1,66 69,051 0,25 67,03-70,89 1,74
Остров Кунашир 2 61,9511,45 60,5 -63,4 3,31 5.4010,50 4,9 -5,9 13,09 7.35Ю.95 6,4 —8,3 18,28 4,3 61,251 7,92 55,33-69,17 18,29 65,88 - 68,36 -
Самки
Нижнее Приамурье 2 63,451 0,15 63,3 -63,6 0,33 4,8510,25 4,6 -5,1 • 7,29 6,8510,25 6,6 -7,1 5,16 4,8 - 62,341 1,13 61,21-63,46 2,55 68,581 2,10 66,48-70,67 4,32 70,581 0,09 70,49-70,67 0,18
Приморье 11 61,721 0.53 59,1 -64,1 2,57 4,8910,08 4,6 -5,3 4,96 7,2410,11 6,8 -7,7 4,66 4,4410,06 4,3 -4,6 3,02 65,601 0,65 63,39-70,64 3,11 66,501 0,56 62,92-68,87 2,79 68,781 0,36 67,53—71,06 1,56
о
* Размер дается по 1 экз.
расположены зеленовато-голубые канты и каемки. Рулевые окрашены так же, как и у взрослых самцов, но синий цвет на них менее яркий с примесью зеленовато-серых тонов. У самок маховые и их кроюшие бурые с оливково:коричневыми кантами и каемками. Рулевые рыжевато-охристые.
В первом послегнездовом наряде у самца нижняя часть спины, надхвостье и верхние кроюшие хвоста зеленовато-сине-голу-бые. Маховые и рулевые остаются те же. Верхние кроюшие крыла с сине-голубым цветом разной яркости. Остальные участки оперения такой же окраски, как и у взрослой самки. Первый послегнездовой и взрослый наряды у самое практически идентичны.
Размеры отдельных частей тела синей мухоловки приведены в табл. 11.
Масса тела, г: молодой самец (о-в Кунашир) - 23,2 (Нечаев, 1965); молодой самец (1.VII, окр. Уссурийского заповедника) - 21,7; взрослые самцы (п = 2) (16.V и 18.VII о.Кунашир) - 22,5-24,8, в среднем - 23,6 (колл. БПиДВНЦ); взрослые самцы (п = 21) (25.VI—1.VII,
Рис. 21. Распространение синей мухоловки (Cyanoptila cyanomelana) иа территории СССР: а - гнездится, б — отмечена в гнездовой период. См. кадастр, стр. 104, 105.
окр. Уссурийского заповедника) — 19,63— 23,58, в среднем — 21,82; молодая самка (28.VI, там же) — 23, 15; взрослые самки (и = 2) (25 и 28.У1,там же) - 21,30-22,15, в среднем — 21,73; взрослая самка (30.VI, Партизанский р-н, р.Тигровая) — 25,1 (колл. ЗМ МГУ); взрослая самка (29.V, о-в Монерон) - 26,0 (колл. ЗМ МГУ).
Распространение. Япония (о-ва: Хоккайдо, Хонсю, Сикоку, Кюсю и Цусима), п-ов Корея, Северо-Восточный Китай (от Маньчжурии на юг до провинции Хубэй). На территории СССР гнездится в Среднем и Нижнем Приамурье (Хабаровский край), Приморье, на о-вах Кунашир, Монерон и предположительно на Южном Сахалине. Зарегистрирован залет синей мухоловки на юг Читинской обл. К местам зимовок на п-ов Индокитай, Малакка и о-ва Тайвань, Хайнань, Филиппинские и Калимантан (северная часть) мигрирует через Приморье, Восточный Китай, п-ов Корея и Японские о-ва (Vaurie, 1959). Детально распространение синей мухоловки на территории СССР показано на карте (рис. 21).
Кадастр к рис. 21.
1 — Хабаровский край, устье р.Горюн (Степанян, 1978); 2 - с.Хурба, окр. г.Комсомольск-на-Амуре (Кистяковский, Смогоржевский, 1973; Иванов, 1976); 3 - окр. пос.Пивань (Штиль-марк, 1973); 4 - с.Малмыж (Яхонтов, 1963); 5 - с.Болонь (копл. КГУ); 6 - с.Джуен (Яхонтов, 1963; колл. ИЗАНУ); 7 - с.Троицкое (Яхонтов, 1963); с.Славянка (колл. КГУ); 8 - с.Ела-буга (Яхонтов, 1963); 9 - оз.Петропавловское, Кухаринский залив (Нечаев, 1963); 10 - верховья р.Кия, 13-15 км от с.Кия (колл. ЗМ МГУ); окр. с.Кия (Кисленко, 19656); 11 - с.Веню-ково (колл. С.Д.Кустановича); 12 - хр.Хехцир (Нечаев, 1963; Кулешова и др., 1965; Казаринов, 1973); Соснинский ключ (Казаринов, 1973); 13 - устье р.Уссури (Портенко, 1960; колл. КБ МГУ); 14 - долина р.Амур ниже устья р.Сун-гари (Степанян, 1978); 75 - хр.Малый Хинган, нижние течения рек, впадающих в р.Амур (Сми-ренский, Смиренская, 1980); с.Радде, с.Башурово (сообщ. С.М.Смиренского); 16 - с.Пашково (сообщ. С.М.Смиренского); 17 - Хинганский заповедник, ст.Кундур, с.Помпеевка (сообщ. С.М.Смиренского); 18 - Приморский край, долина р.Уссури у устья р.Бикин (колл. ЗМ МГУ); 19 - р.Бикин (Поливанова, Ходков, 1975); 20 - нижнее течение р.Большая Уссурка (Иван), окр. сЛукьяновка, сопки и острова по реке между селениями Гончаровка и Ново-покровка (Спангенберг, 1940; 1965); окр. с.Вер-бовка (колл. БПиДВНЦ); 27 - с.Вострецово' (Карту н) (колл. КБ МГУ); 22 - среднее течение р.Болыпая Уссурка (Спангенберг, 1965; колл. ЗМ МГУ); с.Дерсу (Лаулю) (колл. КБ МГУ); 23 - поймы рек верховьев р.Большая Уссурка (Флинт и др., 1959); 24 - бухта Терней (Воробьев, 1951); пгт Терней (Воробьев, 1954); 25 - Сихотэ-Алинский заповедник (колл. КБ МГУ; наши данные); 26 - восточные склоны
104
Сихотэ-Алиня, р.Рудная (Тетюхе) (колл.ЗИН); 21 - бухта (залив) Ольги (Лабзюк, 1975; колл. БПиДВНЦ); 28 — бассейн истоков р.Уссури, урочище ’’Мута” (Назаренко, 19846); р.Маргаритов-ка (Пфусунг), с.Щербаковка (Щульпин, 1928; колл. ЗИН; ИЗАНУ); 29 - Лазовский заповедник им Л.Г.Капланова (Белопольский, 1950; колл. ЗИН); р.Киевка (Судзухе) (Поливанова, Ходков, 1975); 30 км к западу от р.Киевка (колл. ЗМ МГУ); 30 - окр. с.Киевка (колл. ЗИН); 31 - с.Монакино (колл. КГУ); тайга по р.Тачингоуза (Воробьев, 1954; колл. БПиДВНЦ) ; 32 - г.Партизанск (Сучан) (Щульпин, 19276; 1928; копп. ЗИН; ИЗАНУ); р.Сергеевка (Мала-за) (Воробьев, 1951; 1954); р.Тигровая (Сица) (Воробьев, 1951; колл. ЗМ МГУ); Сучанский рудник, Каменка, с.Бровничи (колл. ИЗАНУ); пос.Ручьи (Фанза), ст.Партизаны (колл. ЗИН); 33 - р.Суворовка (Пейшула) (колл. ЗИН); верховья реки, с.Анисимовка (Кангауз) (Воробьев, 1951; 1954); 34 - окр. пос.Большой Камень, р.Пинканка (наши данные); 35 - побережье Уссурийского залива (колл. ЗМ МГУ); 36 - п-ов Муравьева-Амурского (колл. ЗИН); окр. г.Владивосток: долина р .Л анчин (колл. БИН СО АН); долина р.Седанка (Черский, 1915а; колл. ИЗАНУ); окр. Сад-города (колл. ЗИН); 37 - заповедник ’’Кедровая падь” (Neufeldt, 1968; Назаренко, 1971а; 19846; Панов, 1973; Поливанова, Ходков, 1975); среднее течение р.Барабашевка (Большой Мангугай) (колл. ИЗАНУ); 38 - мыс Песчаный (сообщ. Н.Л.Клес-това); 39 - среднее течение р.Нежинка (Сан-дуга) (Панов, 1973); 40 - ст.Кипарисово, р.Па-чихеза (колл. ЗИН); 41 - Уссурийский заповедник им.В. Л.Комарова (Иванов, 1952; Панов, 1973; наши данные); окр. пос.Комарово-запо-ведное, пос.Каймановка (наши данные); верховья Бателянзы (колл. БПиДВНЦ); 42 - окр. пос Лесной Кордон (Пейшула), верхнее течение р.Артемовка (Майхэ) (Neufeldt, 1968; колл. ЗИН); верховья р.Комаровка (Супутинка) (колл. ЗИН); 43 - Горнотаежная станция ДВНЦ, окр. г.Уссурийск (Панов, 1973); 44 - с.Барабаш-Левада, долина р.Комиссаровка (Синтуха) (колл. ЗИН); 45 - окр. г.Камень-Рыболов, окр. с.Троиц-кое (Черский, 1915а); 46 — с.Сиваковка (колл. ЗИН); 47 - с.Черниговка (колл. ИЗАНУ); 48 - с.Евсеевка (колл. ЗИН); 49 - Спасский р-н (Кушнарев, 1984); верхнее течение р.Одарка близ с.Нововладимировка (Черский, 1915а; 19156); 50 - с.Гайворон (колл. ИЗАНУ); 51 -о-в Сахалин; южная часть острова (сообщ. В.А.Нечаева) ; окр. пос.Новоалександровск, западные склоны Сусунайского хр. (сообщ. В.Г.Вороно-ва); 52 - о-в Монерон (Степанян, 1978); 53 -о-в Кунашир; вулкан Головнина, северный берег оз.Горячее, окр. с.Алехино (Нечаев, 1965; 1969; 1971; колл.ЗИН; БПиДВНЦ).
Систематика. Ряд исследователей (Hartert, Steinbacher, 1934; Благосклонов, 1954; Vaurie, 1959; Портенко, 1960; Степанян, 1978; и др.) считают вид Суапор-tila cyanomelana политипическим, образующим два подвида: C.c.cyanomelana (Temminck, 1829), распространенный на Японских о-вах, п-ове Корея; о-вах Кунашир, Монерон и предположительно на Южном
Сахалине, и C.c.cumatilis (Thayer et Bangs, 1909), заселяющий материковую часть гнездового ареала вида. Основные отличия формы cumatilis от номинативной заключаются по мнению этих авторов, в следующем: самцы cumatilis имеют светло-синюю с зеленоватым оттенком окраску в оперении в противоположность насышенно-чисто-си-ней самцов cyanomelana; у самок верхняя сторона тела серовато-оливковая, а не охристо-оливковая, как у номинативной формы, рулевые коричнево-бурые, а не коричневые, как у самок cyanomelana. Г.П.Де-ментьев (1937) приводит в работе этотвид как политипический, но вместе с тем отмечает, что различия между указанными выше формами не резки и этот вид, вероятно, лучше считать монотипическим. Последней точки зрения придерживается и ряд других исследователей (Neufeldt, 1968; Нечаев, 1969; Назаренко, 1971а; Поливанова, Ходков, 1975; Виноградова и др., 1976; идр.), считающих синюю мухоловку монотипическим видом. И.А.Нейфельд (Neufeldt, 1968) доказала зависимость интенсивности синей окраски в оперении самцов от их возраста. Самцы первогодки (в весеннем наряде) имеют голубовато-зеленый оттенок в синей окраске и только в начале второго года жизни после полной линьки на местах гнездования приобретают насыщенно-чистосиний наряд. В.А.Нечаев (1969), проведя сравнение самцов курильских синих мухоловок с материковыми и японскими, не нашел между ними резких отличий. Как среди материковых, так и среди островных птиц встречены экземпляры с чисто синей и с зеленоватым оттенком спиной.
Изучение окраски синих мухоловок, приведенное нами на большом серийном коллекционном материале с учетом возрастной, сезонной и индивидуальной изменчивости склоняет нас к мнению исследователей, считающих вид C.cyanomelana монотипическим. Все перечисленные выше различия в окраске между формами cyanomelana и cumatilis полностью укладываются в пределы их возрастной и сезонной изменчивости. Не найдены нами также отличия между мухоловками материковых и островных популяций и по другим морфологическим признакам. К сожалению, нам так и не удалось получить для обработки птиц с южных о-вов Японии, а просмотренные серии островных популяций невелики.
Места обитания. Населяет различные типы смешанных хвойно-лиственных, лиственных и как исключение окраи
105
ны хвойных лесов по долинам рек и ручьев, распадкам, сопкам, а также по невысоким скалистым склонам предгорий и значительно реже морских побережий. Типичными местами обитания в гнездовой период являются участки леса с небольшими скальными выходами обычно по обрывистым берегам рек, ручьев и ключей недалеко от воды. Микроклимат здесь характеризуется очень высокой влажностью и на камнях в обилии произрастают зеленые мхи. При недостатке характерных мест гнездования поселяется в лесах по склонам сопок со скальными выходами, отличающимися значительно бблыпей сухостью и меньшим развитием наскальной растительности, а также на участках леса, имеющих земляные обрывы, выворотни (поваленные деревья с вывернутыми из земли корнями), пни и старые деревья с глубокими расщелинами, нишами и полудуплами. В сплошных лесных массивах предпочитает пади с высокой влажностью. Стволы деревьев здесь в изобилии покрыты зелеными мхами, имеется много пней и выворотней. В некоторых районах проникает в окультуренный ландшафт, где порою гнездится даже на заброшенных или редко посещаемых постройках человека (Панов, 1973). Избегает безлесые склоны сопок и предгорий, обширные скальные выходы и высокие скалы. Верхний предел распространения на территории СССР 600—700 м н.у.м.
В Хабаровском крае на хр.Хехцир обитает в широколиственных (Кулешова и др., 1965) и кедрово-широколиственных лесах (Нечаев, 1963). У оз .Петропавловское найдена на гнездовании в дубоволиповом лесу с примесью бархата и клена, с подлеском из чубушника, лешины и жимолости (Нечаев, 1963), а в нижнем течении р.Хор гнездится в темнохвойно-широ-колиственных лесах (Кисленко, 19656). В Нижнем Приамурье населяет как кедрово-широколиственные (Штильмарк, 1973), так и старые лиственные леса, в том числе и дубравы (Яхонтов, 1963) по сопкам и обрывистым берегам р.Амур (Кистяков-ский, Смогоржевский, 1973).
В Приморье на р.БоЛьшая Уссурка гнездится в широколиственных лесах речных островов, а также в лиственных, хвойно-лиственных и по окраинам разреженных лиственничных лесов береговых сопок, предпочитая высокоствольные участки с обилием сухих вершин поблизости от речных каменистых обрывов (Спангенберг, 1965). На Среднем Сихотэ-Алине нами за
регистрирована в сходных биотопах, но в окр. пгт Терней изредка гнездится и на облесенных скалах морского побережья (Воробьев, 1954), а также на заброшенных постройках человека. В бассейне истоков р.Уссури встречена в переходных от смешанных к темнохвойным (экотонных) лесах урочища ”Мута” на высоте 600 м (Назаренко, 19846). В Лазовском заповеднике предпочитает дубравы, а также леса с густым подлеском из кустарников с примесью лимонника в верховьях рек и ручьев (Белопольский, 1950).
В заповедниках ’’Кедровая падь” и Уссурийском излюбленными местами обитания этого вида являются высокоствольные смешанные лиственные и хвойно-лиственные леса по долинам рек и ручьев. По их берегам имеются скальные выходы, густо поросшие мхами, а также травянистыми и реже кустарниковыми растениями. Верхний ярус в этих лесах образуют главным образом ясень, ильм, клен, липа, береза, бархат, маньчжурский орех, пихта и сосна. Подлесок состоит из сирени, черемухи, граба, бересклета, чубушника, элеутерококка и акантопанакса. Деревья и кустарники здесь густо оплетены актинидией и амурским виноградом. В нижнем ярусе много папоротников и травянистых растений. На стволах упавших и гниюших деревьев в массе произрастают зеленые мхи, а из расщелин скал и камней вытекают многочисленные ручейки. В таких местах обычно сумрачно и очень влажно. Однако для гнездования синяя мухоловка выискивает довольно освещенные участки, обычно вблизи воды (Neufeldt, 1968; наши данные). Верхний предел распространения этого вида в заповеднике ’’Кедровая падь” определяется высотами 600—700 м (Назаренко, 1971а). В среднем течении р.Нежинка (Сандуга) населяет леса с заметным участием белокорой пихты, а в окр. г.Уссурийск (Горнотаежная станция) отмечены случаи гнездования на постройках человека (Панов, 1973).
На о-ве Кунашир гнездится в темно-хвойно-широколиственных лесах по долинам горных рек южной части острова и в ольхово-березовом лесу на берегу оз.Горя-чее (Нечаев, 1965,1969).
Численность. На территории СССР в Приморье и на о-ве Кунашир в характерных биотипах обычна. У северной границы гнездового ареала в Среднем и Нижнем Приамурье встречается значительно реже. На хр.Малый Хинган распростра
ни
йена спорадично (Смиренский, Смирен-ская, 1980). На Южном Сахалине редка. В ряде мест Приморья зарегистрированы периодические годовые колебания численности этого вида (Поливанова, Ходков, 1975; Назаренко, 19846).
Прилет и весенний пролет на территории СССР. Проходит в I и II декадах мая. В отдельные годы одиночные первые птицы появляются уже во II декаде апреля. Первыми на места гнездования прилетают самцы и только через несколько дней после их прилета появляются самки.
В Южном Приморье у оз.Тальми (Ха-санский р-н) пролетные самцы в 1947 г. зарегистрированы 7.V (Воробьев, 1954). Здесь же у ст .Хасан мертвый самец найден 12.V 1971 г. (колл. БПиДВНЦ). На островах в северо-западной части залива Петра Великого первые пролетные мухоловки появляются в начале I декады мая, а массовый пролет проходит здесь во II декаде этого месяца (Лабзюк и др., 1971). Нао^ве Аскольд самец добыт 7.V 1931 г. (колл. ЗМ МГУ).
В заповеднике ’’Кедровая падь” (Панов, 1973; колл. КБ МГУ) первые самцы отмечены (2 года) 3 и 4.V, массовый пролет проходит (4 года) 5, 7 и 9.V, а первые самки появляются (2 года) 8 и 10.V.
В период весенней миграции самцы синей мухоловки добыты: в окр. г.Влади-восток на р.Седанка - 10.V 1910 г. (Черский, 1915а); на р.Суворовка (Пейшу-ла) — 7.V 1946 г. (колл. ЗМ МГУ); в Партизанском р-не (3 года) - 12.IV, 5-6.V (колл. ЗИН; ИЗАНУ); нарЛачихеза (2 года) - 3 и 10.V (колл. ЗИН; БПиДВНЦ); в Спасском р-не у с.Нововладимировка -1 и 6.V 1911 (Черский, 1915а; 19156).
В Сихотэ-Алинском заповеднике первое появление в 1977 г. зарегистрировано 19.IV (кордон Усть-Серебряный) (картотека заповедника). На р.Большая Уссурка у с.Вострецово (Картун) самец добыт ВЛ.Флинтом 12.V 1957 г. (колл. КБ МГУ).
На нижнем Амуре (2 года) у с.Вятское прилет отмечен 9.V (Кистяковский, Смогоржевский, 1973), а в окр. селений Елабуга, Троицкое и Малмыж - 11-14.V (Яхонтов, 1963),.
На о-ве Кунашир появляется (1 год) 16.V (Нечаев, 1965; 1969; 1971).
Зарегистрирован случай залета синей мухоловки в период весенней миграции на юг Читинской обл.: 21.IV 1951 г. одиночная птица этого вида поймана в с.Кай-ластуй (Гагина, 1967).
Размножение. Сразу после прилета на места гнездования самцы не проявляют антагонизма по отношению друг к другу. В это время большинство из них начинает петь только вполголоса. Интенсивность пения резко возрастает с момента занятия самцами гнездовых участков, на границах которых между ними начинают возникать антагонистические столкновения, сопровождающиеся элементами демонстративного поведения. Песня самца синей мухоловки очень звучна и мелодична. Ее основная часть состоит из 3-х многократно повторяющихся колен, между которыми некоторые самцы могут вставлять отдельные звуки и даже строфы из песен других птиц (Neufeldt, 1968). Таким образом, индивидуальная часть песни очень изменчива, в то время как основная — постоянна. Поют самцы на протяжении почти всего периода размножения, но во время выкармливания птенцов интенсивность пения заметно падает. После прилета самок происходит образование пар также сопровождающееся элементами демонстративного поведения (рис. 15), и птицы приступают к строительству гнезд.
Размеры гнездового участка пары 200— 250 х 300—400 м (Neufeldt, 19(>8).
Из 75 гнезд синей мухоловки, найденных в Приморье и в Приамурье (Воробьев, 1954; Нечаев, 1963; Neufeldt, 1968; Панов, 1973; Поливанова, Ходков, 1975; колл. ЗМ МГУ; БИН СО АН; С.ДКустановича; наши данные), 18 были построены в нишах береговых скал или крупных камней; 18 — на уступах скал и береговых обрывов с козырьком над гнездовой площадкой; 12 — на выступах и карнизах без козырька на высоте 1,8-7,5 м; 2 — в выворотнях больших деревьев; 8 — в расщелинах деревьев (осина, ильм, тополь, клен) на высоте 0,7—9,0, в среднем - 2,7; 4 - в трещинах коры (клен, береза); 2 — в полудуплах деревьев (мелколистный клен, .кедр); 1 — в выдолбе крупного дятла; 1 — в нише полусгнившего пня; 4 - в разросшихся корнях деревьев и кустарников и в корнях под камнями; 5 - на заброшенных или малопосешаемых постройках человека (между крышей и карнизом, на козырьке окна, на угловом выступе и на перекладине). Кроме этого, по данным Е.Н.Панова (1973), гнезда сравнительно часто устраиваются в углублениях земляных обрывов. На довольно больших скалах птицы выбирают для гнезд обычно те участки, которые в большей степени подверглись эрозии и заросли мхами, травой, кустами
107
и деревьями. Обычно возле свежепостроен-ного гнезда, в соседних нишах можно найти несколько (до 6—7) прошлогодних гнезд, это указывает на то обстоятельство, что одни и те же скалы регулярно занимаются птицами из года в год (Панов, 1973). Н.Н.Поливанова, Г.Н.Ходков (1975) считают, что популяции приморских синих мухоловок распадаются на 2 группы: для первой выходы скал являются необходимым условием при постройке гнезд и нормального размножения, у второй связь со скалами отсутствует или значительно ослаблена.
Сбором строительного материала и строительством гнезда занимается самка. Участие самца в строительстве гнезда носит скорее ритуальный характер (Панов, 1973). Гнезда представляют собой сравнительно большие, плотные постройки чашевидной или неправильной формы, сделанные исключительно из пучков зеленых мхов, в том числе Anomodon thraustus, A.minor, Hete-rophyllium haldanianum. Изредка среди мха (в основании гнезда и снаружи на его боковых стенках) попадаются маленькие кусочки коры и отдельные сухие листья древесных растений, а также тонкие стебельки трав. Лоток птицы или не выстилают или используют для этой цели спорогонии бо-коплодных мхов. Размеры гнезд (п =6), мм: диаметр 130-146, в среднем - 135; высота 57-90, в среднем — 72; диаметр лотка 60—84, в среднем — 71; глубина лотка 31—50, в среднем — 39.
На территории СССР в сезон размножения синяя мухоловка имеет один репродуктивный цикл. Количество яиц в полной кладке 5-6, значительно реже - 7 (Neufeldt, 1986; Панов, 1973) и как исключение - 4 (Поливанова, Ходков, 1975). Овальные, остроовальные и очень редко укороченно-остроовальные яйца по белому или бледно-палево-розоватому фону имеют мелкие размытые крапинки и точки светло-терракотового и бледно-телесно-розового цвета. На большинстве яиц они сгуша-ются к тупому полюсу и образуют вокруг него ’’венчик” (густота рисунка 10 %). Однако встречаются и однотонные яйца без рисунка, а также очень редко с ’’венчиком” вокруг острого полюса. Скорлупа полуматовая. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
При 4—6 яйцах в кладке насиживание начинается с откладки последнего яйца, а при 7 — с предпоследнего (Neufeldt, 1968). Инкубация длится 13 дней. Наси
живает кладку только самка. Самец в это время держится недалеко от гнезда и много поет. Самка сидит на яйцах очень крепко и подпускает к себе часто вплотную. Потревоженная человеком она слетает с гнезда и, устроившись неподалеку, остается в полной неподвижности, как бы находясь в оцепенении (Панов, 1973). Периодически она слетает с гнезда и на кормежку, отсутствуя при этом в течение нескольких минут, после чего вновь садится насиживать кладку. Вылупление птенцов длится от 4—5 ч до 2 дней (Neufeldt, 1968; Поливанова, Ходков, 1975).
Пуховой птенец синей мухоловки (Ней-фельд, 1970) имеет длинный (до 19 мм) темно-серый, почти черный пух на надглазничных, затылочной, спинной и плечевых птерилиях. Кожа телесно-розовая с характерной желтизной, возрастающей по мере роста птенца. Ротовая полость и язык желтые, складки в углах рта бледно-желтые. Клюв розовато-желтый, яйцевой зуб и кончик надклювья темные, когти светлые.
Выкармливают птенцов оба партнера пары (рис. 25), но в первые дни после вылупления, когда самка большую часть времени проводит в гнезде, согревая птенцов, корм приносит в основном самец. В обогреве птенцов он не участвует. Растут птенцы быстро и обычно в возрасте 11— 12 дней с недоросшими маховыми и рулевыми покидают гнездо (Neufeldt, 1968; Поливанова, Ходков, 1975). Летать в этом возрасте слетки еше не могут. Поэтому, покинув гнездо, они прячутся в кронах деревьев, кустарниках, различных нишах и среди камней, периодически издавая громкую и резкую позывку, которая слышна на 50—70 м. По ней их находят родители и продолжают кормить. В возрасте 13— 14 дней слетки способны перепархивать на короткие расстояния, а на 16-17 день молодые начинают нормально летать (Neufeldt, 1968).
Установлено, что синяя мухоловка является одним из хозяев ширококрылой (Hierococcyx fugas; Jahn, 1942; Neufeldt, 1968), глухой (Cuculus saturatus; Jahn, 1942) и предположительно обыкновенной (C.canorus; Поливанова, Ходков, 1975) кукушек, которые изредка подбрасывают свои яйца к ней в гнезда.
Сезон размножения этого вида на территории СССР длится со II декады мая по II декаду августа включительно. Большая растянутость его определяется как погодными условиями, так и гибелью у отдель
108
ных пар первых гнезд, кладок и реже птенцов, а по мнению Н.Н.Поливановой, Г.И.Ходкова (1975), и асинхронностью биологических ритмов у различных особей одной популяции.
На о-ве Кунашир самцы с готовыми к размножению семенниками добывались 16.V, 16.VI и 19.VIL Молодой летный самец добыт здесь 10.VII, а выводок из 3 перепархивающих слетков отмечен 2. VIII (Нечаев, 1965; 1969).
В южном Приморье занятие гнездовых участков синими мухоловками зарегистрировано И.А.Нейфельдт (Neufeldt, 1968) 7—10.V, а строительство гнезд — 19—20.V. Откладка первого яйца в наиболее ранней кладке отмечена здесь Н.Н.Поливановой, Г.И.Ходковым (1975) 21.V, а в одной из поздних - 11.VII. В заповеднике ’’Кедровая падь” начало токования в 1961 г. зарегистрировано Е.Н.Пановым (1973) 13.V. По его наблюдениям за 1 парой 27.V 1963 г. самка собирала строительный материал и строила гнездо, 5.VI в нем была полная кладка из 6 яиц, a 28.VI - птенцы незадолго перед вылетом. Гнездо с полной кладкой из 5 свежих яиц найдено в заповеднике 28.V 1963 г. (Панов, 1973; колл. ЗММГУ); законченная постройка без яиц — 19.VI (другой год) (Neufeldt, 1968); гнезда со слепыми птенцами - 1 .VII 1960 г. (Панов, 1973) и 7.VII 1966 г. (колл. ЗИН), а с повторной кладкой из 3 яиц - 2.VII 1961 г. (Панов, 1973). В Надеждинском р-не на р.Нежинка (Сандуга) слеток встречен 23.VII 1965 г. (Панов, 1973). В окр. г.Вла-дивосток (долина рЛанчин) гнездо с полной ненасиженной кладкой обнаружено 1 .VII 1956 г. (колл. БИН СО АН). В Шко-товском р-не у с.Анисимовка (Кангауз) массовое строительство гнезд отмечено 22.V (Воробьев, 1951; 1954), а у пос.Лес-ной Кордон (Пейшула) гнездо с пуховыми птенцами найдено И.А.Нейфельд (колл. ЗИН) 11.VI 1967 г. На р.Тачингоуза самка с яйцом без скорлупы в яйцеводе добыта 26.VI (Воробьев, 1954). В Партизанском р-не на берегу р.Сергеевка (Малаза) гнездо с 4 птенцами 6—7-дневного возраста встречено 7.VIII 1949 г. (Воробьев, 1951; 1954). В этом же районе молодые птицы в разные годы добывались: 20, 22.VI, 3.VII (Панов, 1973; колл. ЗМ МГУ), 11 и 21.VIII (Белопольский, 1950; колл. ЗИН; КГУ). В Уссурийском р-не (окр. Горнотаежной станции) по наблюдениям Е.Н.Панова (1973) за 1 парой: 15.V 1963 г. проходило токование, 8.VI — в гнезде полная кладка
из 6 яиц, а 11.VI вылупление птенцов. В другом гнезде, обнаруженном 11.VI 1963 г., было 7 птенцов недельного возраста, а в гнезде, найденном здесь 21 .VI 1963 г., находилась кладка из 6 яиц (Панов, 1973). В Уссурийском заповеднике гнездо с 6 маленькими птенцами встречено 14.VI (Панов, 1973). У с.Каймановка (окрестности заповедника) гнездо с 1 яйцом найдено нами 26.VI 1977 г., a 30.VI в нем было 4 яйца. Летные молодые в гнездовом наряде добыты нами здесь 28.VI, 1 и 9.VII, а молодые с признаками послегнездовой линьки добыты в заповеднике 5 и 19.VII 1940 г. (Иванов, 1952). В окр. пос.Гайво-рон (Спасский р-н) гнездо с 1 яйцом обнаружено 27.V 1978 г., a 5.VI в нем была полная кладка из 5 яиц (сообщ. М.И.Головушкина) .
В Сихотэ-Алинском заповеднике у кордона Усть-Серебряный в 1977 г. спаривание отмечено 19.V, строительство гнезда — 24.V, начало кладки — 31.V. В окр. пгт Терней 2 гнезда с полными насиженными кладками найдены 10.VI 1976 г. и 17.VII 1968 г. Во втором птенцы вылупились 19. VII (картотека заповедника).
В Хабаровском крае у с.Венюково (Вяземский р-н) гнездо с 2 яйцами обнаружено С.Д.Кустановичем 26.V 1968 г., 31.V -в нем была полная кладка из 6 яиц. В окр. оз .Петропавловское гнездо с птенцами найдено 28.VI 1957 г. (Нечаев, 1963), а на хр.Хехцир с полной кладкой из 6 яиц (степень насиженности не определена) -17.VII 1964 г. (колл. А.П.Кузякина).
Линька. Частичная линька из гнездового пера начинается у молодых синих мухоловок в возрасте 27—29 дней (Neufeldt, 1968). Она не затрагивает маховые, рулевые, а также большие верхние кроюшие второстепенных маховых и, как правило, заканчивается у большинства особей к концу августа — середине сентября. Молодые птицы, добытые в Южном Приморье 21 .VIII, 2 и 5.IX (колл. ЗИН; ИЗАНУ), уже закончили линьку и были в 1-м послегнездовом наряде. На местах зимовок у первогодков (по крайней мере у самцов) проходит частичная предбрачная линька оперения, в ходе которой самцы приобретают синие перья на верхней части тела, черные — на боках головы, горле, зобе и верхней части груди и белые - на нижней части груди и брюшке. Старыми (с гнездового наряда) остаются маховые, рулевые и некоторые большие верхние кроющие второстепенных маховых-. Синий цвет оперения верхней
109
части тела самцов первогодков с зеленовато-голубым оттенком. В этом наряде они возвращаются весной на места гнездования и приступают к размножению. С конца июля по начало сентября, одновременно со взрослыми птицами, у первогодков проходит полная послебрачная линька, в ходе которой самцы одевают насышенно-чисто-синий наряд.
Взрослые птицы имеют в году две линьки: полную предбрачную на местах зимовок (Виноградова и др., 1976) и полную послебрачную на местах гнездования.
Питание. Основу пиши составляют насекомые разных групп и стадий развития. Значительно реже синие мухоловки поедают паукообразных, многоножек, мелких моллюсков, а во второй половине лета — и сочные плоды некоторых растений. Кормятся птицы чаше всего в нижнем ярусе леса (древесно-кустарниковая поросль и травяной покров) и на земле, напоминая по способам добычи пиши скорее некоторых мелких дроздовых, чем мухоловок. Реже беспозвоночных склевывают с листьев, ветвей и стволов деревьев. Активная ловля насекомых в воздухе на лету для этого вида не характерна. Этим способом охоты синие мухоловки пользуются очень редко. По данным Н.Н.Поливановой, Г.Н.Ходкова (1975), ведущую роль среди насекомых в питании птенцов играют прямокрылые (кобылки, кузнечики) и гусеницы. Остальные насекомые служат дополнительной или случайной пишей. Гусеницы в периоды их массового появления становятся основным объектом питания взрослых птиц и составляют до 100 % пиши, приносимой родителями птенцам.
Отлет и осенний пролет на территории СССР. Проходит с середины II декады августа по I декаду октября включительно.
В Южном Приморье в Партизанском (Сучанском) р-не молодой самец добыт 2.IX 1926 г. (Белопольский, 1950; колл. ЗИН). В Шкотовском р-не у с.Анисимовка Н.Л.Клестов отметил самца 4.IX, a 5.IX встретил 3 птицы в зарослях жимолости, одна из которых (самка) была добыта (колл. ИЗАНУ). Район заповедника ’’Кедровая падь” синие мухоловки покидают в конце августа — начале сентября (Поливанова, Ходков, 1975). Последний раз в 1960 г. птицы этого вида встречены в Южном Приморье (точно место не указано) 12.IX (Панов, 1973).
На островах в северо-западной части
110
залива Петра Великого пролет проходит с середины августа до первых чисел октября, наиболее интенсивно — в первой половине сентября (Лабзюк и др., 1971). На о-ве Верховского молодой самец добыт 16.VIII 1966 г., а на о-ве Большой Пелис самка встречена 26.IX 1964 г. (колл. БПиДВНЦ).
Практическое значение. Уничтожает многих насекомых — вредителей леса. Благодаря великолепному пению и красивой окраске эта птица является украшением орнитофауны юга Дальнего Востока СССР и заслуживает всесторонней охраны.
Род Muscicapa Brisson, 1760 — серые мухоловки
Muscicapa Brisson, 1760, Omithologia, 1: 32.
Типовой вид Muscicapa striata Brisson
Род включает 211 вид, 13 из которых распространены в Эфиопской, 1 — в Австралийской (о-в Сумба) и 7 — в Палеарктической и Индо-Малайской зоогеографических областях. В фауне СССР представлен 5 гнездящимися видами.
Средних и небольших размеров настоящие мухоловки. Клюв в отличие от такового представителей родов Ficedula и Cyanoptila наиболее приспособлен к ловле насекомых на лету. Его форма и длина изменчивы, однако у большинства видов он значительно уплошен и очень широк у основания. Щетинки у основания надклювья наиболее хорошо развиты в углах рта. Их количество здесь (с каждой стороны) обычно равно 3-6. Хвост сравнительно короткий или умеренной длины, у многих видов с неглубокой выемкой на вершине. Ноги относительно слабые. Цевка короткая и чаше всего тонкая. Ее длина составляет 13-20 % длины крыла. Исключением является M.griseigularis (Африка), у которой этот показатель равен 25 % (Vaurie, 1953). Формула крыла сильно варьирует. Перелетные и частично перелетные виды имеют относительно длинные крылья с заостренными вершинами, оседлые — более короткие и округлые. Первое первостепенное маховое очень мало. Самец и самка окрашены идентично. Окраска оперения не яркая. Верхняя часть тела различных тонов бурого, коричневого и серого цветов, иногда с примесью оливкового и рыжего. Исключение составляет M.ferruginea (Южная
1 К данному роду мы склонны относить только виды номинативного подрода в понимании Ч.Вори (Vaurie, 1953).
и Юго-Восточная Азия), обладающая хорошо развитой рыжей окраской. Оперение низа светлое, чаше всего беловатое, однако у некоторых представителей рода на отдельных участках оно может быть бурым, серовато-коричневым или серым разной интенсивности. У некоторых видов на перьях верхней части головы и нижней части тела расположены четкие или размытые темные продольные пестрины. Рулевые однотонные. Молодые в гнездовом наряде имеют на перьях пятнистый или чешуйчатый рисунок.
Населяют различные типы хвойных, смешанных хвойно-лиственных и главным образом лиственных лесов на равнинах, в предгорьях и горах, саванны, кустарниковые заросли, сады и парки. Некоторые виды могут гнездиться в населенных пунктах (в том числе и в крупных городах) и даже в открытых ландшафтах — на отдельно стоящих деревьях и как исключение — среди травянистой растительности. Все палеарктические и индо-малайские виды являются перелетными или частично перелетными и зимуют в Южной и Юго-Восточной Азии и Африке. Остальные представители рода ведут оседлый образ жизни.
В отличие от представителей родов Ficedula и Cyanoptila песня самцов серых мухоловок менее звучна, мало мелодична и у большинства видов довольно примитивна. Гнезда строят на ветвях деревьев и кустарников, в развилках стволов и на их пнях, в полудуплах, расшелинах, разнообразных нишах (в том числе и скальных выходов), в старых гнездах других видов птиц и на постройках человека. Высота расположения гнезд от поверхности земли сильно варьирует. Одни виды строят их главным образом в кронах высоких деревьев, в то время как другие — на поверхности земли в кустиках травянистых растений (Волчанецкий, 1972). В кладке обычно 3—6 яиц. Длительность инкубации 11-15 дней. Птенцы находятся в гнезде около 2 недель и покидают его с недоросшими рулевыми и маховыми перьями.
Питаются главным образом насекомыми разных групп, реже паукообразными, мелкими моллюсками и другими беспозвоночными, а также сочными плодами некоторых растений. Для большинства видов основным способом охоты является подкарауливание летающих насекомых с какой-либо присады (чаще всего ветви дерева) с последующей их ловлей в воздухе на лету в стремительном броске.
Взрослые серые мухоловки фауны СССР имеют в году одну линьку (полная
послебрачная), которая у некоторых видов бывает сильно растянута.
Серая мухоловка — Muscicapa striata (Pallas, 1764}
Motacilla striata Pallas, 1764, Vroeg, Cat. Adumbra-tiunculae: 3, Голландия.
Описание. Составлено по 220 экз. (95 — из центральной европейской части СССР, 22 — из Крыма, 10 - с Кавказа, 71 — из Туркмении, Узбекистана, Таджикистана и Киргизии, 22 - из Тувы и Бурятии).
Вершину крыла образуют 3-е и 4-е первостепенные маховые (3 < 4 — 0,6 %; 3=4- 9,5; 3 > 4 - 89,9 %); 2-е и 5-е немного их короче (2 < 5 - 8,3 %; 2=5-4,4; 2 > 5 — 87,3 %); 2-е всегда длиннее 6-го. У большинства особей 1-е первостепенное маховое очень мало и только на несколько миллиметров выступает за вершины кроющих кисти (табл. 12). У большей части особей (54,5 %) наружные опахала 3-5-го первостепенных маховых в вершинной части заметно сужены: у меньшей части (45,5 %) суженные наружные опахала находятся только на 3-м и 4-м маховых. Количество щетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3-6 (3 - 0,7 %; 4 - 85,4; 5 - 13,2; 6 - 0,7 %), длина наибольшей (п = 133) -5,4-9,3 мм (в среднем - 7,4). На вершине хвоста в центре имеется небольшая выемка. Самец и самка окрашены одинаково.
Распространение. Северо-Западная Африка от Марокко к востоку до Туниса, к северу до побережья Средиземного моря и к югу до Высокого и Сахарского Атласа; в Тунисе на юг до 35° с.ш., Западная Евразия от Скандинавии и атлантического побережья к востоку в Забайкалье до 115“ в.д., до Хэнтэя; в Монгольском Алтае до 96° в.д., до Тарбагатая; в Тянь-Шане к востоку до хр.Джумголтау, долины Кокомерена, хр.Ферганского, до Алайского, восточной части Гиссарского, Дарвазского хребтов; в Северо-Западных Гималаях до Гилгита и северной части Белуджистана. К северу в Скандинавии до 69-70“ с.ш., на Кольском п-ове до Печенги и Пялицы, до южной части п-ова Канин, между Канином и Уралом (рис. 24) до 65“ с,ш., между Уралом и Енисеем до 62° с.ш., от нижнего течения Подкаменной Тунгуски северная граница ареала протягивается к верховьям Лены, в Забайкалье к северу предположительно до 52“ с.ш. К югу до побережья Средиземного моря, южного побережья Малой Азии, по восточ
111
ному побережью Средиземного моря, к югу до его юго-восточного угла, восточнее к югу до Армянского (Восточного) Тавра, северного побережья Персидского залива, до
южной оконечности Восточно-Иранских гор, до Средне-Афганских гор. Восточнее южная граница ареала протягивается к северо-востоку в указанных выше пределах и при
Рис. 24. Распространение серой мухоловки (Muscicapa striata) на территории СССР:
а - гнездится, б - отмечена в гнездовой период. См. кадастр, стр. 114-120, 128-130.
112
обретает юго-восточный характер. Острова Британские, Балеарские, Корейка, Сардиния, Крит, Кипр (Степанян, 1978; наши данные). Птицы этого вида являются пе-
1 - M.s.striata, 2 - M.s.neumanni;
релетными и зимуют главным образом в Африке, в Южной и Юго-Западной Азии. Систематика. Географическая изменчивость проявляется в варьировании
113
оттенков и интенсивности окраски оперения, в степени развития пестрин на нижней стороне тела и в размерах. Общий характер изменчивости клинальный. Вид образует несколько подвидов (Vaurie, 1959), 2 из которых - Muscicapa striata striata (Pallas, 1764) и M.s.neumanni (Poche, 1904) -обитают на территории СССР.
Кроме названных подвидов, некоторые исследователи (Vaurie, 1959; Портенко, 1960; Волчанецкий, 1972; и др.) указывают для территории СССР еше 3 формы этого вида: M.s.inexpectata Dementiev, 1932-Крым; M.s.sarudnyi Snigirewski, 1928 -самые южные части Туркмении; M.s.mon-gola Portenko, 1955 — Южный Алтай, южная оконечность Байкала и Забайкалье. Однако, как показали наши исследования,'серые мухоловки из Крыма по окраске оперения, особенностям его рисунка и морфометрическим показателям практически не отличаются от таковых из центральной европейской части СССР и, несомненно, должны рассматриваться в составе номинативного подвида. Что касается птиц из Южной Туркмении, Южного Алтая и Забайкалья, то мы склонны относить их к подвиду M.s.neumanni, как это делает Л.С.Степа-нян (1978).
Европейская серая мухоловка — Muscicapa striata striata (Pallas, 1764)
Motacilla striata Pallas, 1764, Vroeg, Cat. Adumbra-tiunculae: 3, Голландия.
Описание. Составлено по 127 экз. (95 - из центральной европейской части СССР, 22 — из Крыма, 10 — с Кавказа).
У взрослой птицы (рис. 26) оперение верхней части тела бурое. Лоб и передняя часть темени бледно-буроватые. На лбу и темени имеются темно-бурые стержневые пятна. Оперение нижней части тела от бледно-буроватого до бледно-охристого с продольными темными пестринами вокруг горла, поперек груди и на боках. Маховые и рулевые темно-бурые. На наружных опахалах второстепенных маховых, а на третьестепенных и больших верхних кроющих второстепенных маховых и на вершинах имеются светло-буроватые канты и каемки. Нижние кроюшие крыла светлобуроватые. Надклювье темно-рогового цвета. Подклювье телесно-бурое в основании и темно-роговое в вершинной части. Ноги темно-серые. Когти темные. Радужина коричнево-бурая. Сезонная изменчивость проявляется в обшем посветлении окраски оперения и в частичном или полном исчез
новении светлых каемок на маховых и больших верхних кроющих второстепенных маховых в результате их обноса и выцветания.
Молодые в гнездовом наряде (рис. 30) сверху беловато-охристые с бурыми каемками на перьях. Оперение нижней части тела белое с незначительной примесью бледно-бурого. По нему разбросаны размытые пестрины, локализующиеся главным образом на груди и боках тела. Рулевые и маховые темно-бурые. По краю наружных опахал второстепенных маховых, на третьестепенных и больших верхних кроющих второстепенных маховых и на вершинах расположены широкие светлые каемки и канты, наиболее развитые на третьестепенных маховых.
Первый послегнездовой наряд идентичен наряду взрослой птицы, но на вершинах самых длинных верхних кроющих хвоста сохраняются охристо-беловатые пятна, а на третьестепенных маховых и больших верхних кроющих второстепенных маховых — широкие светло-охристые каемки на вершинах и по краю опахал этих перьев.
Размеры отдельных частей тела европейской серой мухоловки приведены в табл. 12.
Масса тела, г: взрослый самец (побережье Белого моря) — 16,0 (Малышевский, 1962); самец (2.Х, Тебердинский заповедник) — 14,3 (Ткаченко, 1966).
Распространение. В СССР (рис. 24): вся европейская часть ареала вида, Кавказ, Северный Казахстан и Западная Сибирь к востоку до долины р.Обь и западного подножья Алтая. У восточных пределов распространения интерградирует с M.s.neumanni (Степанян, 1978). На всем ареале серые мухоловки этого подвида являются перелетными и зимуют главным образом в Африке.
Кадастр к рис. 24.
1 — Эстония: Ванаярве, Лайксааре, Выйду, Толкусе (Кумари, 1955); д.Кадана (материалы ц.кольц.); 2 - Матсалуский заповедник (Ренно, 1968); 3 - Нигула (Кумари, 1955); 4 - г.Тарту (Мазинг, 1960); 5 - окр. г.Нарва (колл. ЗИН); 6 — Латвия; г.Рига (Греве, 1915); 7 - заповедник Слитере (Бауманис и др., 1983); 8 - Литва (Валюс и др., 1976; Алекнонис, 1977а); г.Клайпеда (Гражулявичюс, 1984); 9 - с.Неринга (материалы ц.кольц.); 10 - г.Каунас и его окрестности (Навасайтис, Курлавичюс, 1977); с.Ионава (материалы ц.кольц.); 11 - юго-западная дуга Балтийской возвышенности (Валюс, Кигуолене, 1981); заповедник ’’Жувинтас” (Иванаускас, 1968); 12 - Рокишкский р-н, д.Шилейкяй (Ба-раускас, 1981); 13 - Молетский лесхоз (мате
114
риалы ц.кольц.); 14 - Варенский лесхоз (материалы ц.кольц.); 15 - Белоруссия: Минская обл. с.Юревичи (колл. ИЗАНУ); 16 - Витебская обл., Лепельский р-н, с.Крайцы (Федюшин, Долбик, 1967); 17 - Городокский р-н (Федюшин, Долбик, 1967); 18 - Оршанский р-н, окр. с.Тарасинка, окр. с.Схоляны, с.Загорье (Бианки, 1914); 19 -Гродненская обл. (Сямашка, 1956); г.Гродно (колл. ЗИН); 20 - Брестская обл. заповедник ’’Беловежская пуща”, Меднянское лесничество (Федюшин, Долбик, 1967); 21 - Пинский р-н (Шнитников, 1913; Федюшин, Долбик, 1967); 22 - Лунинецкий р-н (Долбик, 1959; Федюшин, Долбик, 1967); 23 - Столинский р-н (Долбик, 1959; Федюшин, Долбик, 1967); 24 - Припятский заповедник (Клакоцкий, 1983); 25 - Ленинский р-н (Долбик, 1959); 26 - с.Копаткевичи, с.Лучицы (Штамм, 1923); 27 - Гомельская обл.; Мозырьский р-н, Макановичское лесничество (Федюшин, Долбик, 1967); 28 - Украина: Волынская обл. (наши данные); с.Маневичи (наши данные); 29 - г.Шацк (колл. ЛПМ); 30 - с.Ново-волынск (сообщ. М.И.Головушкина); 31 -г.Луцк, с.Цумань (сообщ. М.И.Головушкина);
32 - Ровенская обл., Владимирецкий р-н, р.Стырь (колл. ИЗАНУ); с.Быстричи (сообщ. М.И.Головушкина) ; Константиновское лесничество (колл. ИЗАНУ); Березновский р-н (колл. ИЗАНУ);
33 — окр. оз.Белое (сообщ. М.И.Головушкина);
34 - г.Ровно и его окрестности (сообщ. М.И.Головушкина) ; окр. с.Сусь, левый берег р.Горынь (наши данные); окр. г.Костополь (колл. ИЗАНУ); 35 - Житомирская обл.: г.Житомир и его окрестности (сообщ. М.И.Головушкина); долина р.Случь, окр. с.Вирля (сообщ. А.С.Вобленко); пгт Мирополь (колл. ЗИН); 15 км северо-восточнее г.Коростышев (сообщ. А.С.Вобленко) ; 36 - г.Олевск, с.Перга, с.Копище (сообщ. М.И.Головушкина); Словечанский р-н (колл. ИЗАНУ); р.Словечна (сообщ. М.И.Головушкина) ; 37 - пгт Попельня и с.Вчерайше (сообщ. М.И.Головушкина); 38 - Киевская обл. (Вла-дышевский, 1972; наши данные): г.Киев и его окрестности (наши данные); Броварской р-н (колл. ИЗАНУ; М.И.Лобановского), г. Б ров ары (сообщ. М.И.Головушкина); Киево-Святошин-ский р-н (колл. ИЗАНУ); с.Ходосовка (сообщ. М.И.Головушкина); села Бортничи, Гнедин, Пеники (сообщ. М.И.Головушкина); 39 - г.Иванков, с.Розважев (сообщ. М.И.Головушкина); 40 -г.Ирпень, пгт Клавднево-Тарасово (наши данные) ; 41 — Фастовский р-н, г.Фастов и его окрестности (колл. ЗИН; ИЗАНУ); 42 - Белоцерковский р-н (Кирпиченко, 1932; колл. ЗИН); селения Ракитное и Синява (сообщ. М.И.Головушкина) ; 43 - окр. с.Масловка (сообщ. Ю.В.Ми-щенко); г.Кагарлык (сообщ. М.И.Головушкина); 44 - с.Григоровка, с.Великий Букрин (сообщ. М.И.Головушкина); Переяслав-Хмельницкий р-н; с.Комаровка (колл. М.И.Лобановского); с.Ер-ковцы (сообщ. М.И.Головушкина); 45 - с.Се-миполки, сЛебедевка (сообщ. М.И.Головушкина); р-н с.Высшая Дубечня (колл. ИЗАНУ);
46 - Черниговская обл.: г.Чернигов (сообщ. М.Ф.Самофалова); окр. с.Михайло-Коцюбинское (наши данные); окр. с.Неданчичи, окр. с.Тараса Шевченко, - окр. с.Кези, окр. с.Кувечичи, окр. с.Любеч (сообщ. М.Ф.Самофалова); 47 - окр. с.Жеведь, окр. с.Днепровское, окр. с.Тужар, г.Остер и его окрестности (сообщ. М.Ф.Самофалова) ; окр. с.Самойловка (наши данные); 48 -
окр. пгт Репки, окр. г.Городня, окр. с.Перепись (сообщ. М.Ф.Самофалова); 49 - окр. г.Щорс, окр. с.Тихоновичи (сообщ. М.Ф.Самофалова); 50 - окр. с.Семеновка (сообщ. М.Ф.Самофалова); 51 - окр. пос.Куликовка, окр. с.Локиистое, окр. с.Березна (сообщ. М.Ф.Самофалова); 52 -окр. с.Ольшаное, окр. с.Батурин, окр. г.Бахмач, окр. с.Стрельники (сообщ. М.Ф.Самофалова); окр. с.Ядуты (сообщ. А.С.Вобленко); 53 - Сумская обл., с.Старая Гута (колл. ХГУ); 54 - Шост-кинский р-н (сообщ. В.П.Белика); Собичский бор (колл. ХГУ); 55 - г.Путивль, г.Конотоп (сообщ. М.И.Головушкина); 56 - окр. с,Тростянец (колл. ХГУ); 57 - Черкасская обл.: г.Черкассы и его окрестности (наши данные); 58 - р.Ирдынка, окр. с.Свидивок (наши данные); 59 - Каневский р-н, окр. с.Селище (колл. М.И.Лобановского); г.Канев (сообщ. М.И.Головушкина); Каневский заповедник (наши данные); 60 - окр. с.Чапаевка (сообщ. В.И.Стригунова); 61 - Полтавская обл. (Гавриленко, 1929): г.Полтава, р.Ворскла, с.Яковцы, с.Диканька, Монастырский лес (колл. ХГУ); 62 - Харьковская обд. (колл. ХГУ), окр. г.Готвальд (Харченко и др., 1984); Готваль-довский р-н, окр. с.Гайдары (сообщ. А.Н.Гудины) ; хут.Коробов (колл. ЗИН; М.И.Лобановского); 63 - р.Орчик, с.Бабаях (Сомов, 1897);
64 - г.Чугуев (колл. ХГУ); 65 - Купянский р-н (колл. ХГУ); 66 - районы населенных пунктов Изюм, Балаклея, Петровское (колл. ХГУ); 67 — Днепропетровская обл.: Самарский лес, с.В.Тарасовка (колл. ИЗАНУ); 68 - территория, затопленная Днепродзержинским водохранилищем (Булахов, 1968); 69 - среднее течение р.Орель (Апостолов и др., 1968); 70 - пос.За-чепиловка (сообщ. А.Н.Гудины); 71 - Донецкая обл. (Уварова, 1959), окр. г.Славянск (сообщ. С.В.Винтера); 72 -Ворошиловградская обл.: окр. г.Ворошиловград, окр. с.Счастье (сообщ. В.В.Ветров а); 73 - окр. г.Кременная (сообщ. В.В.Вет-рова); 74 - окр. с.Городище, ботанический заказник (сообщ. В.В.Ветрова); 75 - Луганский заповедник, окр. с.Станично-Луганское, нижнее течение р.Деркул, правый берег р.Северский Донец, юго-восточнее с.Станично-Луганское (сообщ. В.В.Ветрова); 76 - пгт Краснодон (сообщ. В.В.Ветрова); 77 - г.Запорожье и его окрестности (колл. ХГУ; наши данные); 78 - юго-западная часть Запорожской обл., г.Мелитополь и его окрестности (Филонов, 1967); Старобердянское лесничество (сообщ. А.М.Волоха); 79 - с.Ал-тагир; Богатырское лесничество (сообщ. А.М.Волоха) ; 80 - г.Кривой Рог (Боченко, 1969); 81 - Кировоградская обл.: Аникиевское лесничество (колл. ИЗАНУ); 82 - Херсонская обл., ст.Новоалексеевка (сообщ. М.И.Головушкина);
83 - г.Голая Пристань (сообщ. Т.Б.Ардамацкой); с.Новая Збурьевка, г.Херсон (сообщ. М.И.Головушкина) ; 84 - Черноморский заповедник (Ардамацкая, Семенов, 1977); 85 - Одесская обл.: г.Одесса (сообщ. М.И.Головушкина); 86 -Беляевский р-н, с.Яськи (сообщ. Л.Ф.Назарен-ко); с.Беляевка, с.Маяки (сообщ. М.И.Головушкина) ; 87 - Балтийский р-н (колл. ХГУ); 88 -г.Винница (сообщ. М.И.Головушкина); 89 -г.Казатин (сообщ. М.И.Головушкина); 90 -сЛитин (сообщ. М.И.Головушкина); 91 - Хмельницкая обл.: г.Хмельницкий и его окрестности (сообщ. М.И.Головушкина); Летичевский р-н, сЛысогорка (колл. ЗИН); 92 - г.Каменец-По-дольский (сообщ. М.И.Головушкина); 93 -
115
г.Черновцы (сообщ. М:И.Головушкина); 94 - долина р.Белый Черемош (Страутман, 1954); 95 - Северные и южные склоны Украинских Карпат (Страутман, 1954; 1963); ст.Сияки, Торуньский перевал, окр. сЛопушное, с.Воло-вец, Воловецкий перевал (Страутман, 1954); окр. с.Синевир (сообщ. М.И.Головушкина); 96 -г. Рахов (Луговой, 1984); 97 - с.Ясиня (сообщ. М.И.Головушкина); 98 - Закарпатская равнина (Страутман, 1963); г.Мукачево (сообщ. М.И.Головушкина) ; 99 - г.Ужгород и его окрестности (наши данные); долина р.Уж, с.Невицкое (Портенко, 1950); пгт Перечни (колл. ЗИН); 100 - Ужокский перевал (Страутман, 1954);
101 - окр. пгт Буршгын (сообщ. Ю.В.Мищен-ко); 102 - г.Тернополь и его окрестности (сообщ. М.И.Головушкина); 103 - Кременецкий р-н (колл. ЗИН); г.Кременец и гЛочаев (сообщ. М.И.Головушкина); 104 - Львовская обл. (Страутман, 1963), г.Львов и его окрестности (Страутман, 1963; Горбань и др., 1984); села Поторица, Пеняки, Заречье (Страутман, 1963; колл. ЛПМ); 105 - Крым (Портенко, 1960; Волчанецкий, 1962; колл. ИЗАНУ; ХГУ); Евпаторийский р-н (колл. ИЗАНУ); 106 - Белогорский р-н (колл. ХГУ); 107 - Судакский р-н (колл. ХГУ); Судакское лесничество (колл. ЗИН); 108 - г.Феодосия (сообщ. М.И.Головушкина); 109 - южные склоны Главной гряды Крымских гор, окр. Большой Чучели (Костин,
1983); Никитский ботанический сад (сообщ. М.М.Бескоровайного); г. Орлиный Залет, окр. с.Соколиное (сообщ. А.М.Волоха); г.Ялта (колл. ХГУ); южные склоны Ай-Петри (колл. ИЗАНУ);
110 - р.Кача, с.Машино (колл. ИЗАНУ); 111 -Молдавия: г.Кишинев (Аверин, Ганя, 1970); Трифауцкий, Рублеиецкий, Копанский, Кошниц-кий леса; окр. с.Морозены, окр. с.Ватич (Ганяг Литвак, 1961); Малачуиский лес (Аверин, Ганя, 1970); 112 - Сорокский лес (Ганя, Литвак, 1961); 113 - Тираспольский р-н, Кицканский лес, биостанция Тернопольского пединститута, с.Бужевка, с.Егоровка (Ганя, 1965); 114 -РСФСР: с.Печеига (Иванов, 1976); 115 - Лапландский заповедник, берег Маврагубы, северный берег Чуиозера (Владимирская, 1948);
116 - средняя часть Кольского п-ова (Портенко, 1960; Степанян, 1978; колл. ЗИН); Хибины, южный склон Поач-вум-чорра (Новиков, 1952а);
117 - г.Кандалакша (Благосклонов, 1954; Мальчевский, 1959; колл. ЗИН); 118 - побережье Кандалакшского залива (Бианки, Флеров, 1960); Кандалакшский заповедник (Благосклонов, 1960); о-ва в Кандалакшском заливе (Шутова, 1981а); 119 - сЛялица (Иванов, 1976); 120 -вся территория Карельской АССР (Марвин, 1951; Нейфельдт, 1958); 121 - заповедник ’’Кивач” (Ивантер, 1962); 122 - северо-восточ-ное побережье Ладожского оз. (Головань, 1982);
123 - о-в Большой Клименецкий (Хохлова, 1977);
М ± m lim
Таблица 12. Размеры
-) и коэффициенты вариации (CV, %) серой мухоловки
Подвиды, место сбора п Дли
Крыло CV Расстояние между вершинами 1-го махового и кроющих кисти CV
Сам
М. striata striata европейская часть СССР 44 87,59± 0,27 83,4 -90,2 2,00 1,50±0,17 -1,0 -3,5 7-7,47
Крым 8 87,28± 0,74 84,1 -90,0 2,39 1,18±0,39 0,0 -2,7 93,20
M.s.neumanni Средняя Азия 33 86,50± 0,39 Й2,1 -92,2 2,45 1,95±О,22 0,0 -4,2 61,84
Тува и Бурятия 8 86,51± 0,59 84,6 -89,3 1,92 2,16±0,29 0,6 -3,1 37,81
Сам
M.striata striata европейская часть СССР 24 86,99± 0,31 83,8 -89,8 1.77 1,52±0,28 -1,3 -3,6 89,76
Крым 5 86,38± 0,69 84,8 -83,1 1,59 1,30*0,53 Ъ,о -з,1 91,83
M.s.neumanni Средняя Азия 27 85,1 It 0,48 81,2 -90,4 2,93 2,25±0,23 0,0 -4,6 51,21
Тува и Бурятия 9 84,78± 0,57 81,9 -87,1 1,91 2,12±0,44 0,0 -4,9 61,70
(
116
124 - Ленинградская обл. (Мальчевский, 1959; Мальчевский, Пукинский, 1983; колл. ЗИН; ХГУ), г Ленинград и его окрестности (Мартынов, 1977; Мальчевский, Пукинский, 1983); Петродворец (колл. ЗИН); 125 — Лужский р-н, д.Мерево (Мальчевский, Пукинский, 1983);
126 - р-н с.Рощино, окр. пос. Смолячково (колл. С.Д.Кустановича); 127 - г.Выборг (колл. ЗИН);
128 - окр. гЛриозерск (материалы ц.кольц.);
129 - острова в западной части Ладожского оз. (Мальчевский, Пукинский, 1983); 130 - Юго-Западное Приладожье (Мальчевский, Пукинский, 1983); 131 - Юго-Восточное Приладожье (Мальчевский, Пукинский, 1983); 132 - Свирская губа (Мальчевский, Пукинский, 1983); 133 -Новгородская обл. (колл. ЗИН); г.Новгород (Мантейфель. - Цит. по: Благосклонов, 1954);
134 - Псковская обл. (Зарудный, 1910), пос.Елизарово (материалы ц.кольц.); Харламова гора (колл. ЗИН); 135 - верховья р.Пскова (Мальчевский, Пукинский, 1983); 136 - окр. Псковско-Чудского водоема (Мешков, 1961);
137 - Куршская коса, пос.Рьгбачий (материалы ц.кольц.); 138 - Вологодская обл., Велико-Устюгский р-н (Второв, Дроздов, 1960); 139 -р-н с.Тотьма (Второв, Дроздов, 1960); 140 -окр. оз.Кубенское (Беме, 1962); 141 — Дарвинский заповедник (Спангенберг, Олигер, 1949);
142 - г.Архангельск (Паровщиков, 19416;
Асоскова, 1984); 143 - долина р.Золотица (Спан
генберг, Леонович, 1958; 1960); 144 - г.Пинега (Портенко, 1960; Севастьянов, 1962; Карногор-ский лесхоз (Севастьянов, 1962); 145 - устье р.Мезень (Иванов, 1976); 146 - Конушинский берег между селами Каменка и Семжа, окр. с.Сем-жа, в том числе и в 3-х км севернее (Спангенберг, Леонович, 1958; 1960); 147 - южная часть п-ова Канин (Степанян, 1978); лайда р.Несь (Зубцовский, Рябицев, 1976); 148 - Соловецкие о-ва (Goebel. - Цит. по: Eber, 1960а); Беломорские о-ва (Благосклонов, 1954; колл. ЗИН);
149 - Коми АССР, окр. г.Сыктывкар (Гурьев, 1982; сообщ. А.А.Естафьева); 150 - окр. с.Усть-Кулом (сообщ. А.А.Естафьева); 151 - верховья р.Мезень (сообщ. А.А.Естафьева) ; 152 - среднее течение р.Кедва (приток р.Ижма) (сообщ. А.А.Естафьева) ; 153 - окр. с.Кожва на р.Печора (Портенко, 1960; Иванов, 1976); окр. г.Печора (сообщ. А.А.Естафьева); 154 - окр. с.Аранец (Дмо-ховский, 1933); 155 - низовья р.Северная Мылва (Естафьев, 1969); 156 - верховья р.Илыч (Естафьев, 1969); 157 - д.Порог (Дмоховский, 1933); 158 - Печоро-Илычский заповедник (Теп-лова, 1957); 159 - Калининская обл. (Зиновьев, Шапошников, 1978); Старицкий р-н (колл. ЗИН);
160 - Брянская обл., г.Трубчевск (колл. ЗИН);
161 - Карачаевский р-н, окр. г.Карачаев (Суслова, 1937; колл. ЗИН); 162 - Калужская обл., г.Калуга (материалы ц.кольц.); 163 - Московская обл. (Портенко, Иноземцев, 1968; колл.
на, мм
Хвост CV Цевка CV Клюв от оперения лба CV Клюв от орального края ноздри CV
ЦЫ
60,91* 0,22 57,1 -64,4 2,38 14,75* 0,08 13,8 -15,7 3,22 11,82* 0,09 10,6 -13,6 4,78 8,75*0,07 7,9 -9,9 5,45
61,65* 0,47 59,5 -63,3 2,14 14,81* 0,13 14,2 -15,4 2,56 11,87* 0,12 11,3 -12,3 2,78 8,61*0,18 7,9 -9,1 5,46
62,53* 0,35 59,5 -66,5 3,06 14,78* 0,08 13,7 -15,9 3,16 12,24* 0,09 11,2 -13,6 4,25 9,01*0,06 8,3 -9,7 4,11
64,13* 0,48 62,1 -66,2 2,13 15,16* 0,15 14,8 -16,0 2,86 12,61* 0,18 12,0 -13,4 3,96 9,25*0,12 8,7 -9,6 3,74
KU
60,26* 0,40 57,1 -64,6 3,24 15,06* 0,10 14,2 -16,2 3,25 11,64* 0,10 10,$ -13,1 3,99 8,67*0,08 7,8 -9,Г 4,45
59,72* 0,70 57,7 —61,5 2,62 15,12* 0,23 14,4 -15,7 3,42 11,68*0,24 11,1 -12,2 4,10 8,25*0,31 7,7 -9,1 7,50
62,16* 0,43 58,9 -68,2 3,60 14,83* 0,08 14,1 -15,6 2,93 12,13* 0,12 11,2 -13,2 4,79 8,99* 0,10 8,1 -10,2 5,37
62,70± 0,63 58,7 -64,8 3,02 15,41* 0,12 15,0 -16,0 2,42 12,50* 0,28 11,5 -13,3 5,47 9,05* 0,21 8,1 - 9,6 5,62
117
ЗИН; ЗМ МГУ, КБ МГУ), г.Москва и ее окрестности (Строков, 1962; Флинт, Кривошеев, 1962; Птушенко, 1976; колл. ЗИН); 164 — Звенигородская биостанция МГУ (Птушенко, Иноземцев, 1968); 165 - окр. г.Крюково (Птушенко, Иноземцев, 1968); 166 - Талдомский р-н (Птушенко, Иноземцев, 1968); 167 - Дмитровский р-н (Птушенко, Иноземцев, 1968); 168 - Орехово-Зуевский р-н (Дерим-Оглу, 1964); 169 - окр. с.Горы (Степанян, 1962); 170 - Приокско-Террасный заповедник (Александрова, 1956); 171 - г.Яро-славль (Белоусов, 1984); 172 - Владимирская обл. (Благосклонов, 1954); пойма р.Клязьма (Измайлов, Сальников, 1978); окр. г.Владимир, Пенкинский лес (Измайлов, 1981); Георгиевский лес (Михлин, Сальников, 1981); 173 - Ивановская обл., Шуйский р-н (Шептуховский, 1981); 174 - Южский р-н (Шептуховский, 1981); 175 -Рязанская обл., Окский заповедник (колл. ЗМ МГУ); Мещерская низина, Переделецкое лесничество (Ежова, 1979; Френкина, 1981); Липецкая засека (Хомяков, 1900); 176 - окр. с.Гусь-Железный (колл. БИН СО АН); 177 -Тульская обл., окр. с.Никольское (колл. ЗИН); 178 - Орловская обл., истоки р.Ока (Ефимов, 1915); 179 - Курская обл., р.Сейм (Макаров, 1980); 180 - Белгородская обл. (Будничен-ко, Козлов, 1980), окр. г.Белтород (Новиков и др., 1963); 181 - заповедник ’’Лес на Ворскле” (Новиков и др., 1963а); 182 - Липецкая обл.,
Данковский р-н, с.Ягодное (колл. ЗИН); 183 -Тамбовская обл. Мичуринский р-н (Щеголев, 1978); 184 - Моршанский р-н (Щеголев, 1978); 185' - Воронежская обл., Савальское лесничество (Мальчевский, 1959; колл. ЗИН); Каменная степь (Барабаш-Никифоров, Семаго, 1963); 186 - Воронежский заповедник (Барабаш-Никифоров, Павловский, 1947); 187 - Эртильские лесополосы (Барабаш-Никифоров,Семаго, 1963);
188 — Бобровский р-н (колл. ЗИН); 189 — Хоперский заповедник (Измайлов, 1940); 190 - Тел-лермановский лес (Королькова, 196-3); 191 -Ростовская обл., г.Ростов-на-Дону и его окрестности (сообщ. В.П.Белика); 192 — г.Каменолом-ни (сообщ. В.П.Белика); 193 - Красносулинский р-н, Донской лесхоз (сообщ. В.П.Белика);
194 - Каменский р-н, пойма р.Северский Донец (сообщ. В.П.Белика); 195 - Вешенский р-н, стан.Вешенская (сообщ. В.П.Белика); 196 -
хут.Терновский (сообщ. В.П.Белика); 197 -г.Цимлянск, г.Волгодонск (сообщ. В.П.Белика);
198 - г.Константиновск (сообщ. В.П.Белика); 199 - Усть-Донецкий р-н, ст.Роздорская, окр. стан.Нижнекундрюченская (сообщ. В.П.Белика); Багаевский р-н (колл. РГУ); 200 - Сальский р-н, с.Гигант (сообщ. В.П.Белика); 201 - Азовский р-н, Ленинский лесхоз (сообщ. В.П.Белика) ; 202 - г.Таганрог (сообщ. В.П.Белика);
203 - Матвеево-Курганское лесничество (колл. РГУ); 204 - г.Саратов и его окрестности (По
Окончание табл. 12
Подвиды, место сбора п Ширина, мм
Крыло CV Клюв у орального края ноздри CV Клюв у основания CV
Сам
М.striata striata
европейская часть СССР 44 59,151 0,20 56,7 —61,5 2,25 4,5310,04 4,0 -5,3 5,75 6,7710,06 5,8 -7,8 6,15
Крым 8 59,691 0,65 57,2 -63,0 3,10 4,5810,08 4,3 -4,9 5,19 6,83Ю,13 6,3 -7,3 5,18
M.s.neumanni Средняя Азия 33 60,391 0,33 56,0 -64,4 2,89 4,67Ю,05 4,1 -5,1 6,23 6,5410,06 6,1 -7,3 5,50
Тува и Бурятия 8 60,61± 0,38 59,5 -62,9 1,75 4,5810,09 4,1 -4,9 5,45 6,7510,19 5,7 -7,3 8,04
M.striata striata европейская часть СССР 24 58,59± 0,29 55,6 -60,9 2,43 4,6310,05 4,1 -5,0 4,87 6,7410,06 6,2 -7,2 Сам 4,31
Крым 5 59,321 0,89 57,4 -62,3 3,34 4,4210,02 4,4 -4,5 1,01 6,4610,08 6,3 -6,7 2,81
M.s.neumanni Средняя Азия 27 59,221 0,31 56,0 -61,9 2,66 4,65Ю,06 4,1 -5,3 6,72 6,7010,07 6,1 -7,3 5,31
Тува и Бурятия 9 60,281 0,61 56,7 -62,0 2,85 4,65Ю,11 4,1 -5,0 6,60 7,0010,19 6,2 —7,9 8,18
118
дольский, Харин, 1984); 205 - Жигулевский заповедник (Белянина, Белянин, 1981); 206 -Пензенская обл., Присурье (Луговой, 1981); 207 - Мордовская АССР: Сабаевское, Краснослободское и Симкинское лесничества (Луговой, 1975); 208 - Мордовия, Присурье (Луговой, 1981); 209 - Мордовский заповедник им.П.Г.Смидовича (Птушенко, 1938; Бородин, 1967); 210 - Горьковская обл. (Пузанов и др., 1942), окр. поселков Колосовой Алешино (Аникин, 1970); с.Фроловское (колл. ЗИН); 211 -Арзамасский р-н Горьковской обл., окр. с.Ста-рая Пустынь (Аникин, 1970; Хохлова, Олигер, 1982); Пустынская биостанция (Воронцов и др., 1962); 47 км шоссе г.Горький - Арзамас (Аникин, 1970); 212 - Чувашская АССР, Присурье (Луговой, 1981); 213 — Тойзинское лесничество (колл. ЗИН); 214 - Татарская АССР (Благосклонов, 1954); г.Казань (Птицы Волжско-Камского края, 1978; колл. ЗИН); 215 - окр. ст.Обсерваторная, биостанция Казанского университета (сообщ. Ю.В.Мищенко); 216 - устье р.Свияга (сообщ. Ю.В.Мищенко); 217 - Кировская обл. (Птицы Волжско-Камского края, 1978); 218 - Пермская обл., пойма р.Колва у г.Чердынь (Симкин, 1974); окр. г.Чердынь (Резцов, 1904); 219 - пойма р.Кама у с.Гайны, с.Верхняя Старица (Симкин, 1974) ; 220 - с.Усть-Черная (Симкии, 1974); 221 - Александровский р-н (Шилова и др., 1963); 222 - Лысьвен-
ский р-н (Шилова и др., 1963); 223 - г.Кунгур (колл. ЗИН); 224 - южные р-ны Пермской обл. (Симкии, 1974);' 225 - устье р.Умпин на р.Лозь-ва (Портенко, 1960); 226 - заповедник ’’Денежкин камень” (Кузнецов, 1958); 227 - Свердловская обл., Сысертский р-н, окр. с.Черданцево (Данилов, 1959); 228 - Челябинская обл., с.Тур-гояк (колл. ЗИН); Ильменский заповедник им.В.И.Ленина (колл. ЗИН); 229 - с.Каштым (колл. ХГУ); 230 - окр. г.Златоуст (колл. ЗИН); 231 — Башкирская АССР (Ильичев, 1960; Ильичев, Фомин, 1979); Жилинская дача (колл. ЗИН); 232 - Башкирский заповедник (Кириков, 1952; Филонов, 1965); д.Сосновка (Кириков, 1952); 233 - р.Белая (колл. ЗИН); 234 - окр. с.Кананикольское (Кириков, 1952); 235 - окр. с.Зилаир, окр. пос.Крепостной Зилаир (Кириков, 1952); 236 - р.Илек (Благосклонов, 1954); 237 - Курганская обл., окр. с.Зырянка (Ларионов, 1927); 238 - г.Курган (Благосклонов, 1954); 239 - г.Тюмень (Портенко, 1960; колл. ЗИН); 240 - Тобольский р-н (Тарунин, 1928); г.Тобольск (Портенко, 1960; колл. ЗИН); 241 -г.Сургут (Портенко, 1960; Гынгазов, Москвитин, 1965; Иванов, 1976); 242 - г.Ханты-Ман-сийск (Самарово) (Иоганезен. - Цит. по: ЕЬег, 1960а); 243 — 62° с.ш. в бассейне р.Обь (Степанян, 1978); 244 - р.Ларьеган (Гынгазов, Миловидов, 1977); 245 - окр. г.Омск (Гынгазов, Миловидов, 1977); 246 - окр. оз.Сартлан (Иоганзен,
Высота, мм Отношение шири-яы клюва у осно-вания к его длине от оперения лба, % CV Отношение длины хвоста к длине крыла, % CV Отношение ширины крыла к его длине, % CV 1
Клюв у основания CV
4,00*0,05 3,6 -4,7 6,20 57,10* 0,61 49,15-67,54 6,73 69,53* 0,24 65,41-73,21 2,24 67,49* 0,16 65,76-69,87 1,57
3,90*0,07 5,6 -tl 4,68 57,20* 1,27 51,22-61^4 5,86 70,65* 0,43 69,26-73,25 1,74 68,38* 0,37 66,67-70,00 1,54
4,03*0,04 i,6 -4,4 4,71 53,47* 0,48 49,59-60,00 5,10 72,20* 0,41 67,23-75,12 2,99 69,78* 0,38 66,21-72,84 2,87
4,20*0,14 3,9 -4,7 7,34 53,51* 1,28 47,11-57,94 6,76 74,13* 0,47 72,13-76,27 1,79 70,07* 0,25 68,70-71,04 1,02
KU
3,94*0,06 34 -4,3 4,89 57,96* 042 53,85-61,61 4,25 69,28* 0,38 64,74-72,79 2,70 67,36* 0,30 65,08-70,76 2,18
3,90*0,23 34 -4,3 10,26 55,75* 1,40 51,64-57,66 5,02 68,92* 0,80 66,63-70,23 2,33 68,38* 0,80 67,09-70,72 2,35
4,16*0,08 3,6 -4,9 8,13 55,70* 0,84 48,80-63,48 7,07 73,08* 047 '68,25-82,17 4Д6 69,77* 0,40 34,99-74,13 2,91
4,13*0,15 3,7 -4,8 9,01 56,70* 2,69 50,82-68,70 11,64 73,75* 048 71,67-75,87 2,22 71,09* 0,31 69,23-72,01 1,23
119
1923) ; 247 - г.Ставрополь (сообщ. В.П.Белика); 248 - Ставропольский край, Левокумский р-н, с.Владимировка (колл. ХГУ); 249 - Георгиевский р-н (колл. РГУ); 250 - с.Архыз (колл. ХГУ); 251 - Кабардино-Балкария, окр. г.Наль-чик (Радищев, 1926; Моламусов, 1967); с.Мур-тазово (Радищев. - Цит. по: Моламусову, 1967); 252 - Северо-Осетинская АССР (Беме, 1926); 253 - Чечено-Ингушская АССР (Рашкевич, 1980) ;
254 - г.Краснодар и его окрестности (наши данные) ; 255 - Каневский р-н, окр. стан.Стародере-вянковская (колл. РГУ); Челбасское лесничество (наши данные); 256 - окр. г.Славянск-на-Кубани (Кистяковский, 1932; наши данные); 257 - Северский р-н (Волчаиецкий и др., 1962); стан.Крепостная (колл. ХГУ); 258 - Апшеронский р-н, с.Хребтовое (колл. ХГУ); 259 - Майкопский р-н, окр. пос.Никель, стан.Даховская (колл. РГУ; наши данные); 260 - Кавказский заповедник, Лаго-FI аки, р.Киша (колл. РГУ; ХГУ); окр. пос.Гу-зерипль (наши данные); сЛсебай (колл. ЗИН); 261 - г.Сочи и его окрестности (Сушкин, 1914; наши данные); пос.Адлер и его окрестности (Кудашев, 1917; наши данные); с.ВардаНе (Кудашев, 1917); с.Красная Поляна и его окрестности (Сушкин, 1914; Кудашев, 1917; колл. ЗИН); 262 - Аше (колл. ЗИН); 263 - с.Джубга (колл. ХГУ); 264 - Геленджикский р-н, г.Геленджик и его окрестности (наши данные); Михайловский перевал (колл. РГУ); с.Пшада (Пузанов, 1938; колл. ХГУ); 265 - Дюрсо (Пузанов, 1938; колл. ИЗАНУ); берег Навагир (Пузанов, 1938);
266 - г.Абинск и его окрестности, пос.Нижне-баканский (наши данные); 267 - Грузия; Абхазия, г.Сухуми (Jordania, 1975); 268 - Пицундский заповедник (Бернацкий. 1958); 269 -с.Алхино (Jordania, 1975); 270 - с.кжцл (Jordania, 1975); 271 - Нижняя Сванетия, с.Зе-мо-Карши (Жордания, Гогилашвили, 1976);
272 - Верхняя Сванетия, бассейн р.Ингури (Кузнецов, 1983); 273 - Аджария (Вильконский, 1897; колл. ЗИН); г.Батуми (Jordania, 1975); 274 — Лихский хр. (Жордания, Гогилашвили, 1969); окр. с.Сурами (Михайловский, 1880); 275 - г.Тетри-Цкаро (Цветков, 1901; колл. ЗИН); Атепское ущ. (Чхиквишвили, 1933); 276 - окр. г.Тбилиси, г.Мцхета (Жордания, 1962; колл. ЗМ МГУ); 277 - долина р.Алазани (БанГуковский, 1913); 278 - с.Лагодехи (Jordania, 1975; колл. ЗИН); 279 - Азербайджан, г.Баку (наши данные); 280 - Закатальский заповедник; долина р.Катех-чай (Дроздов, 1965); 281 - долина р.Кура (Гамбаров, 1941); 282 - долина р.Кура у впадения в последнюю р.Аракс (Гамбаров, 1941); 283 - долина р.Аракс (Гамбаров, 1941); 284 - Талышские горы, хр.Улясы (Дроздов, 1965); предгорья Талышских гор (Сатунин, .1907); 285 - Армения (Ляйстер, Соснин, 1942; Даль, 1954); 286 - Казахстан, форт Шевченко (Ковшарь, 1970); 287 - с.Фурманово (Ковшарь, 1970); 288 - 60 км севернее г.Уральск (Батыршина, 1977); 289 - 60 км северо-восточнее г.Уральск (Левин, 1982); 290 - Чапаевский р-н (Ковшарь, 1970); 291 — верховья р.Уил, лес Кокжар (Ковшарь, 1970); 292 - верхнее течение р.Эмба (Портенко, 1960); 293 - окр. г.Петро-павловск (Резцов, 1904); 294 - Кокчетавские боры, Дубрава, Аиртау, Бойсары, оз.Бобровое (Ковшарь, 1970); 295 - оз.Щучье (Ковшарь, 1970; колл. ЗИН); 296 - с.Зеренда (Ковшарь, 1970);
297 - с.Имантау (Ковшарь, 1970); 298 - с.Сан-
дыктау (Ковшарь, 1970); 299 - окр. с.Ямышево (Ковшарь, 1970) ; 300 - пойма р.Иртыш от г.Се-мипалатинск до с.Ямышево (Ковшарь, 1970); 301 - окр. ^Семипалатинск (Ковшарь, 1970); 302 - низовья р.Малая Ульба, г.Усть-Каменогорск (Ковшарь, 1970); 303 - западные подножья Алтая (Ковшарь, 1970; Степанян, 1978).
Места обитания. Населяет самые разнообразные типы лиственных, смешанных хвойно-лиственных и хвойных лесов на равнинах, в предгорьях и горах до 2000 м н.у.м. У северных границ распространения в ограниченном количестве проникает в самые южные части лесотундры. Обитает в полезащитных лесополосах, рощах, парках, садах и населенных пунктах (в том числе и в больших городах). Есть указания В.Н.Бостанжогло (1911) на гнездование этого вида в открытых местах с кустиками полыни по р.Урал и в прикаспийских степях, однако эти данные, на наш взгляд, нуждаются в проверке.
На Кольском п-ове в Хибинах отмечена в смешанном лесу на южном склоне Поач-вум-чорра (Новиков, 1952а), а на Терском берегу Белого моря у р.Усть-Пялке — в березовом криволесье (Малышевский, 1962). В Лапландском заповеднике найдена на гнездовании в елово-сосновом лесу (Владимирская, 1948), а в Архангельской обл. на Конушинском берегу Белого моря в окр. с.Семжа - среди заболоченного мелколесья (Спангенберг, Леонович, 1958). В Карелии встречается по опушкам сосновых, смешанных и лиственных лесов, в садах и парках городов (Марвин, 1951). В южной части республики обычна в негустых сосновых лесах на вырубках и гарях, а также в смешанных насаждениях и в небольших лесных поселках (Нейфельдт, 1958). В заповеднике ’’Кивач” гнездится в лиственных, смешанных и сосновых лесах (Ивантер, 1962).
В Ленинградской обл. (Мальчевский, Пукинский, 1983) населяет самые разнообразные биотопы: парки, населенные пункты и различные типы лесов. Предпочитает светлые средневозрастные или спелые смешанные и лиственные леса, а также сосняки. Наиболее многочисленна на лесных опушках и в старых пригородных парках. В Вологодской обл. у оз.Кубенское обитает в смешанных лесах из ели, осины, березы, сосны и различных кустарников (Беме, 1962). В Дарвинском заповеднике гнездится главным образом в сухих сосновых борах, реже - в сырых сосняках и смешанных хвойно-лиственных лесах (Спангенберг, Олигер, 1949).
120
В Эстонии встречается в широколиственных лесах, тяготея к разреженным участкам, и в сосновых борах (Кумари, 1955). В Латвии обитает в парках, садах, населенных пунктах, а также в смешанных и сосновых редколесьях (Бауманис и др., 1983). В районе юго-западной дуги Балтийской возвышенности отмечена в старых садах и других насаждениях (Валюс, Кигуоле-не, 1981).
В Белоруссии встречается на разреженных участках елово-лиственных и сосноводубовых лесов, в дубравах, ольшанниках, борах, парках, садах (Федюшин, Долбик, 1967) и населенных пунктах. В Оршанском р-не Витебской обл. найдена на гнездовании в березовой роше (Бианки, 1914).
В Московской обл. населяет разнообразные леса, занимая осветленные участки, окраины опушек, полян, вырубок, просек и лесных дорог, а также лесные поселки,, парки, сады, а в открытых ландшафтах — населенные пункты, имеющие большое количество старых раскидистых деревьев (Птушенко, Иноземцев, 1968). Гнездится на территории г.Москва. Во Владимирской обл. обитает в различных сосновых лесах, в том числе и в высокоствольных с примесью лиственных деревьев и с хорошо развитым подлеском во многих местах (Измайлов, 1981). В пойме р.Клязьма - редкий вид дубравно-кустарникового комплекса (Измайлов, Сальников, 1978). В Горьковской обл. предпочитает светлые сосняки, встречается по окраинам полян в смешанных лесах и очень часто в непосредственной близости от жилья человека — в садах и парках (Пузанов и др., 1942). Гнездится в злаково-разнотравных дубравах и березняках (Аникин, 1970), в еловососновых, липово-осиновых и елово-сосново-липовых лесах (Хохлова, Олигер, 1982). В Рязанской обл. у оз.Уржинское обитает в разреженных березняках, в том числе и с примесью сосны (Ежова, 1979), а в Пе-ределецком лесничестве — как в чисто лиственных насаждениях (береза, осина), так и на участках смешанного леса, где к названным лиственным деревьям прибавляется также сосна (Френкина, 1981).
В Мордовии населяет разновозрастные хвойные и лиственные леса вблизи опушек, полян, дорог и населенных пунктов (Луговой, 1975). В Мордовском заповеднике приурочена к окраинам обширных вырубок, полян, к лесным поселкам и окраинам самого заповедника (Птушенко, 1938).
В Моршанском р-не Тамбовской обл. гнездится в старых борах, а в Мичуринском
р-не — в широколиственных, березовых и осиновых лесах (Щеголев, 1978). В Орловской обл. также населяет разнообразные леса и сады, часто вблизи от жилья человека (Ефимов, 1915). В Воронежской обл. встречается почти повсеместно: в лесах различного типа, садах, парках и населенных пунктах (Барабаш-Никифоров, Семаго, 1963). В Хоперском заповеднике приурочена к опушкам пойменного и нагорного лесов, а также к мелколесью. Часто гнездится в садах и рядом с жильем человека (Измайлов, 1940).
В Пермской обл. обитает в борах, ельниках, елово-пихтовом старолесье (Шилова и др., 1963), припойменных сложных лесах с лиственницей и сосной, в березовоосиновых, липовых и смешанных хвойнолиственных лесах (Симкин, 1974).
В Волжско-Камском крае отмечена в темнохвойных лесах с примесью сосны и лиственных деревьев, в сосново-широко-лиственных, осиновых и березовых лесах, в хвойно-лиственных и широколиственных молодняках, а также в пойменных лесах и полезащитных лесополосах (Птицы Волжско-Камского края, 1978).
На Среднем Урале (окр. с.Черданцево) гнездится в чистых сосновых борах (Данилов, 1959), а в районе’ южной оконечности Урала — в нагорных сосново-лиственничных лесах (Кириков, 1952). В Башкирском заповеднике обитает в нагорных сосново-березово-лиственничных и широколиственных лесах (Филонов, 1965).
В Тюменской обл. найдена на гнездовании в сосновых лесах (Ларионов, 1927), а также в ивняках Нижне-Вартовского р-на (Гордеев, 1965).
В Казахстане на Кокчетавском поднятии многочисленна в сосновых борах по опушкам и гарям, а также обычна в березовых колках (Ковшарь, 1970).
На Украине населяет разнообразные леса (березняки, осинники, боры) и встречается в населенных пунктах. В Киевской обл. гнездится в основном в сосняках, смешанных сосново-дубовых лесах, дубравах, парках и скверах населенных пунктов. В Украинских Карпатах обитает на северных и южных склонах во фруктовых садах селений, на деревьях по берегам рек, опушкам, полянам среди леса и особенно лесосекам (Страутман, 1954). По речным долинам проникает до водоразделов и встречается даже на перевалах. В Ворошиповград-ской обл. (сообщ. В.В.Ветрова) гнездится в пойменных лесах по р.Северский Донец (где охотно поселяется на территориях
121
пионерских лагерей и баз отдыха и тд.), а также в старых сосняках нижнего течения рДеркул. В Крыму обычна лишь местами в сосновых лесах южного склона Главной гряды Крымских гор (Костин, 1983). В окр. г.Ялта гнездится в Никитском ботаническом саду.
На Кавказе населяет смешанные лиственные, хвойно-лиственные и реже хвойные леса на равнинах, в предгорьях и горах, поднимаясь в Азербайджане до 1500мн.у.м. (Гамбаров, 1941), а в Армении — до 1980 м (Даль, 1954). Гнездится в пойменных лесах и зарослях, садах и населенных пунктах. В Чечено-Ингушетии обитает в лесополосах, тугаях, парках, светлых лесах степной и лесостепной зон, а также в' горных лесах (Рашкевич, 1980). Обычна по всему Черноморскому побережью Кавказа, где встречается в различных лесах подножий и нижнего пояса гор, в садах и населенных пунктах. В Закатальском заповеднике и Та-лышских горах отмечена в буково-грабовых лесах (Дроздов,1965).
Численность. В СССР на большей части гнездового ареала обычна; в характерных, излюбленных биотопах — многочисленна. У северных, восточных и юго-восточных границ ареала — чаше всего редка. На территории СССР в ряде районов численность подвержена годовым колебаниям.
Прилет и весенний пролет на территории СССР. Проходит с III декады марта по I декаду июня включительно. Во время миграции серые мухоловки движутся как поодиночке, так и небольшими стайками, придерживаясь облесенных мест, но довольно часто в этот период встречаются и в открытых биотопах.
В окр. г.Ереван (2 года) прилетает 12 и 28.IV (Адамян, 1974). В Азербайджане первые одиночные птицы появляются в Г декаде апреля (9.IV), а наиболее'интенсивно пролет проходит здесь в I декаде мая (Гамбаров, 1941; Тугаринов, 1950). Нами пролетная серая мухоловка добыта в г.Баку 26.IV 1976 г. В Грузию прилетает в конце марта — начале апреля (Жордания, 1962), а в центральной части Северного Кавказа (10 лет) появляется 29.Ш-7. V (Моламусов, 1967). В Восточном Приазовье в 1959 г. отмечена на пролете 4.V.
В Крыму, по данным Ю.В.Костина (1983), весенний пролет этого вида выражен слабее осеннего и численность птиц весной невелика. Наиболее ранняя встреча зарегистрирована в сЛортовое 24.IV 1972 г.,
а 28-29.IV 1972 г. мухоловки отмечены на Тарханкутском п-ове. Обычно первые особи появляются в степном Крыму (нескблько лет) 5—8.V. Миграция продолжается в течение всего мая, а в некоторые годы и в начале июня. Разгар пролета наблюдается иногда в III декаде мая. Так, у сЛортовое серые мухоловки были относительно обычны 20.V 1971 г. и 22.V 1975 г., а на юге Керченского п-ова — 22—25.V Ф.А.Киселев по дороге из с.Оленевка (Тарханкутский п-ов) в г.Евпатория отметил их 2.VI, а в 1977 г, явно пролетные группы из 4—6 особей встречены в г.Алушта 3, 4 и 12.VI (Костин, 1983). Нами в 1981 г, птицы этого вида зарегистрированы в Карадагском заповеднике 1, 9 и ll.V, a 6.V одиночная самка добыта в центральной части Арабатской Стрелки.
В Молдавию прилетает в середине апреля. Первые птицы (8 лет) появляются здесь 15-25.IV (Ганя, Литвак, 1961). На Украине в Волыно-Подолье прилетает в конце апреля — начале мая (Страутман, 1963), а в Харьковскую обл. (8 лет) — 19.IV-10.V (Сомов, 1897).
В Белоруссии появляется в III декаде апреля — I декаде мая: в заповеднике ’’Беловежская пуща” (5 лет) — 15.IV, 4, 8, 11 и 14.V, в Сталинском р-не (2 года) - 5 и 7.V, в Лепельском р-не (2 года) — 3 и 7.V (Федюшин, Долбик, 1967); в Пинском р-не. (3 года) — 2 ,4 и 5.V (Шнитников, 1913); в Гродненской обл. (3 года) -23 и 25.IV (Сямашка, 1956).
В юго-восточную часть Литвы прилетает (13 лет) 30.IV—12.V (Алекнонис, 1976); в г.Вильнюс и его окрестности (8 лет) — 9-12.V, в среднем — 10.V (Идзелис, 1981); а в Эстонию (15 лет) — 6—27.V (Тамм. -Цит. по: Благосклонов, 1954).
В Воронежской обл. в Хоперском заповеднике первое появление в 1937 г. отмечено 5.V (Измайлов, 1940), а в Воронежском заповеднике в 1938 г. — 14.V (Барабаш-Никифоров, Павловский, 1947).
В Белгородскую обл. в заповедник ’’Лес на Ворскле” прилетает во второй половине апреля - начале мая (5 лет): 19, 30.IV; 5, 8.V (Новиков и др., 1963). В Орловской обл. появляется (9 лет) 20.IV— 31.V (Ефимов, 1915), а в Карачаевском р-не Брянской обл. первые птицы в 1933 г. зарегистрированы 5—6.V (Суслова, 1937). В Рязанскую обл. прилетает в I—II декадах мая (Хомяков, 1900), а в Подмосковье (20 лет) — 1—10—19.V (Птушенко, Иноземцев, 1968).
122
В Мордовии появляется в III декаде апреля (Луговой, 1975), а в Горьковской обл. — в середине мая (Пузанов и др., 1942).
В окр. г.Новгород первые серые мухоловки прилетают (24 года) 30.IV— 22.V (Мантейфель. - Цит. по: Благосклонов, 1954). В эти же сроки прилет проходит в Псковской обл. (Зарудный, 1910). В Ленинградской обл. (Мальчевский, Пукинский, 1983) появляется, как правило, в конце I - начале II декады мая. Исключительно ранний прилет (28.IV) отмечен В.Л.Бианки (Цит. по: Мальчевский, Пукинский, 1983) у пос Лебяжье. В позднюю, холодную весну 1981 г. серые мухоловки в р-не Свирской губы впервые появились только 26.V. В Карелию прилетает во второй половине мая (Марвин, 1951). Первые птицы в Пряжинском р-не в 1955 г. зарегистрированы 15.V (Нейфельд, 1958), а на о-ве Большой Клименецкий (Онежское озеро) в 1975 г. — 17.V (Хохлова, 1976). В Лапландском заповеднике первое появление в 1938 г. отмечено 1.VI (Владимирская, 1948).
В дельте Волги пролет проходит с конца III декады апреля до конца мая (Воробьев, 1936). В Волжско-Камский край прилетает в конце апреля — начале мая. У устья р.Кама (несколько лет) наиболее раннее появление зарегистрировано 17.IV, а наиболее позднее - 17.V (Птицы Волжско-Камского края, 1978).
В Пермской обл. в р-не Княснинского оз. прилет в 1896 г. отмечен 11.VI, а у с.Чердынь в 1897 г. - 28.V (Резцов, 1904). На Средний Урал в окр. с.Черданцево (3 года) прилетает 28-31.V (Данилов, 1959).
Размножение. Прилетев на места гнездования, птицы вскоре приступают к размножению. Занятие гнездовых участков и образование пар сопровождается пением самцов и их токовым, демонстративным поведением. Песня серой мухоловки довольно примитивна и состоит из отрывистых одиночных или сдвоенных циканий и скрипов, которые следуют друг за другом иногда довольно долго. Гнездовые участки невелики и при высокой плотности гнездования расстояние между гнездами соседних пар составляет обычно 50-150 м (Михеев, 1953), в редких случаях - 25 м (Горбань, 1982) и как исключение — 1 м (Федюшин, Долбик, 1967). По наблюдениям А.В.Ми-хеева (1953), птицы охраняют только центральную часть гнездового участка в радиусе 20—30 м от гнезда, однако зареги
стрирован случай, когда гнездо серой мухоловки располагалось вплотную к жилому гнезду дрозда-рябинника, причем птицы в обоих гнездах насиживали кладки (Подольский, Харин, 1984).
Гнезда серые мухоловки строят главным образом на лиственных или хвойных деревьях и кустарниках (дуб, липа, ильм, клен, ясень, береза, ольха, тополь, осина, ива, яблоня, груша, рябина, белая акация, сирень, сосна, пихта, ель, можжевельник): в полудуплах; различных расщелинах; в неглубоких дуплах; в углублениях, образовавшихся в местах выпадения сучьев; в изъянах стволов, поврежденных дятлами; за отставшей корой стволов и ветвей; в их развилках; на горизонтальных ветвях; за стволовой порослью; между стволами вплотную растущих деревьев; на сломах и пнях; в корневых выворотнях; в металлических и глиняных чашках для сбора смолы (живицы), закрепленных на стволах сосен. Гнездятся в старых гнездах других видов птиц: ласточек деревенской (Барабаш-Никифоров, Семаго, 1963; Харченко и др., 1984) и городской (Stegmann, 1980), кольчатой горлицы, щегла (Горбань, 1982), зяблика, дроздов (Мальчевский, Пукинский, 1983), а также в искусственных гнездовьях ящичного типа. На островах в западной части Ладожского оз. иногда устраивают гнезда в трещинах и на карнизах скал (Мальчевский, Пукинский, 1983). В населенных пунктах часто строят их на карнизах домов, за наличниками окон, в нишах стен, на балконах, подоконниках, под крышами и на чердаках сараев. Изредка поселяются внутри заброшенных домов, проникая в них через приоткрытые двери или окна (Мальчевский, Пукинский, 1983). Кроме этого гнезда серых мухоловок находили также внутри уличных фонарей, на почтовых яшиках, на рукоятке приставленной к стене лопаты (Мальчевский, Пукинский, 1983), в консервной банке, оставленной под крышей (Федюшин, Долбик, 1967), в поленницах дров и в других самых разнообразных местах. Высота расположения гнезд от поверхности земли сильно варьирует. В одних случаях (на гнилых пнях и в изъянах стволов) они помешаются у самой земли или прямо на ней (Мальчевский, Пукинский, 1983), в других — в 10—15 м от ее поверхности (Новиков и др., 1963), чаше - на высоте (п = 26) 1,0-7,0 м, в среднем - 2,6. Строят гнездо оба партнера пары в течение 5— 10 дней.
123
Гнезда в зависимости от места их расположения могут иметь правильную чашевидную (на ветвях деревьев) или неправильную форму (в расщелинах и полудуплах). В качестве строительных материалов птицы используют полоски луба, тонкие веточки деревьев и кустарников, их листья, корешки, кусочки кбры, пленки бересты, хвою (сосна), соцветия ивы, пучочки шерсти, ваты, мха, листья, стебли и соцветия трав (в том числе и дикорастущих злаков), чешуи почек, коконы пауков, нитки, кусочки бумаги, талломы лишайников, одиночные перья. Лоток выстилается мягкими полосками луба, растительными волокнами, шерстью, конским волосом, талломами лишайников, мелкими контурными и пуховыми перьями, а в редких случаях и спорогониями мхов. Размеры гнезд (Сомов, 1897; наши данные) (п = = 24), мм: диаметр 65—125 х 65—130 мм, в среднем - 96 х 106; высота 40—70, в среднем - 53; диаметр лотка 48—74 х х 50-74, в среднем — 57 х 60; глубина лотка 25-47, в среднем - 36.
На территории СССР в сезон размножения европейская серая мухоловка имеет один, предположительно два репродуктивных цикла. Указания на полицикличность у этой формы (наличие 2 и даже 3 кладок), имеющиеся в литературе (Благосклонов, 1954; Портенко, 1960; Федюшин, Долбик, 1967; и др.), лишены прямых доказательств и нуждаются в проверке. К повторному гнездованию птицы приступают чаше всего вынужденно в результате гибели первых гнезд и кладок. Первые полные кладки обычно содержат 4-5, реже — 6 яиц. Повторные имеют чаще всего меньшие размеры и могут состоять из 1—3 яиц.
Яйца остроовальные или укороченно-овальные. Реже встречаются овальные и укороченно-остроовальные. Фон скорлупы от бледно-серого с голубоватым оттенком до бледно-голубовато-зеленоватого. По нему разбросаны четкие и размытые пятна, крапинки и точки (густота рисунка 20— 50 %) охристо-рыжего, розовато-лилового, светло-коричневого и красно-бурого цветов. На одних яйцах они локализуются главным образом на тупом полушарии, часто образуя вокруг полюса ’’венчик”, на других — равномерно распределяются по всей поверхности скорлупы. Форма и окраска яиц сильно варьируют как внутри одной, так и между разными кладками. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
В норме откладка яиц производится
самкой ежедневно. Насиживание начинается с откладки последнего яйца, в редких случаях (при максимальных размерах кладки) - с предпоследнего и как исключение (при атипическом размножении) - с первого (Степанян, 1962). Как показали наблюдения за мечеными птицами, насиживает кладку только самка (Мальчевский, Пукинский, 1983). Самец в зто время держится недалеко от гнезда и периодически приносит ей корм. Инкубация длится 11— 15, обычно 12—14 дней (Сямашка, 1956; Данилов, 1959; Долбик, 1959; Мальчевский, 1959; Птушенко, Иноземцев, 1968; Рашкевич, 1980; наши данные).
Пуховой птенец европейской серой мухоловки имеет серый пух на глазных, надглазничных, затылочной, спинной, плечевых, локтевых и бедренных птерилиях. Кожа желтоватая. Клюв серый. Яйцевой зуб бурый. Ротовая полость желтая.
В выкармливании птенцов участвуют оба партнера пары, но в первые дни после вылупления, когда самка почти все время проводит в гнезде, согревая птенцов, корм приносит в основном самец. В возрасте 11-14 дней птенцы с недоросшими маховыми и рулевыми покидают гнездо. Родители продолжают кормить слетков. Первые попытки самостоятельного схватывания насекомых слетки делают в возрасте 26-28 дней, а на 32—34 день молодые становятся полностью самостоятельными (Го-ловань, 1962).
Европейская серая мухоловка является одним из хозяев обыкновенной кукушки, которая изредка подбрасывает свои яйца к ней в гнезда.
Отмечены случаи разорения гнезд серой мухоловки врановыми птицами, большим пестрым дятлом и мелкими куньими. При гнездовании в чашках для сбора смолы (живицы) кладки и птенцы иногда гибнут от скопления в гнездах смолы или воды (Мальчевский, Пукинский, 1983).
Сезон размножения европейской серой мухоловки на территории СССР длится с III декады апреля по II декаду августа включительно.
В Грузии на Лихском хр. откладка яиц в большинстве гнезд проходит в конце апреля — начале мая (Жордания, Гогилашвили, 1969). В эти же сроки птицы приступают к размножению и в центральной части Северного Кавказа. Гнезда с полными кладками встречаются здесь с I декады мая (Моламусов, 1967). В окр. г.Нальчик самка с готовым к откладке яйцом добыта 13.V 1925 г. (Радищев, 1926). В Красно
124
царском крае у г.Краснодар гнездо с полной кладкой обнаружено 7.VI 1964 г., а в окр. пос.Нижнебаканский с 4 птенцами - 26.VII 1961 г. В Северском р-не выводок встречен в конце мая (Волчанецкий и др., 1962), а у г.Краснодар — 4.VII 1964 г.
В Ростовской обл., по данным В.П.Бе-лика, 2 гнезда (строящееся и с полной кладкой) найдены в г.Таганрог 4.VI 1972 г. Несколько выводков и перепархивающих слеток зарегистрированы здесь 25 .VI 1967 г. В г.Ростов свежепостроенное гнездо обнаружено 9.VI 1972 г., а выводок летных молодых, которых изредка подкармливали взрослые птицы, встречен 8.VIII 1979 г. В Вешенском р-не в 1982 г. насиженная кладка из 5 яиц найдена 17.VI, строящееся гнездо — 24.VI, а слетки отмечены 10 и 13. VII.
В Южном Крыму у горы Большая Чу-чель пара, отыскивающая место для гнезда, зарегистрирована 17.V 1965 г., 15.VI здесь обнаружно гнездо с насиженной кладкой, 20.VI в нем вылупились птенцы, a 3.VII вылетели из гнезда (Костин, 1983). У г.Ял-та в Никитском ботаническом саду гнездо с птенцами найдено М.М.Бескоровайным 23.VI 1980 г., а в Бахчисарайском р-не у с.Соколиное погибший слеток серой мухоловки подобран А.М.Волохом 29.VI 1984 г.
В Молдавии самая ранняя откладка первого яйца зарегистрирована 15 .V, самая поздняя - 28.VI. Однако в большинстве гнезд первое яйцо птицы откладывают во II—III декадах мая (Ганя, Литвак, 1961).
В западных областях Украины (Страутман, 1963; колл. ЛПМ) первые кладки появляются в конце мая - начале июня, повторные — в конце июня — начале июля. Летные молодые встречаются с конца июня. В долине р.Уж Молодые птицы отмечены Л.А.Портенко (1950) 18.VIII.
В Черкасской обл. у с.Селище полная слегка насиженная кладка найдена 8.VI 1956 г. (колл. М.И.Лобановского), а у с.Свидивок на р.Ирдынка - 18.VI 1982 г. (наши данные). В Харьковской обл. самая ранняя кладка зарегистрирована 23.V 1886 г. (Сомов, 1897). В Готвальдовском р-не гнездо с 3 свежими яйцами обнаружено 15.VI 1938 г. (колл. М.И.Лобановского). В Киевской обл. кладки разной степени насиженности отмечены: в г.Киеве и его окрестностях - 26.V 1975 г., 30.V 1973 г. и 14.VI 1951 г., в Макаровском р-не у с.Ко-маровка — 29.V 1955 г., у ст.Боярка — 3.VI 1961 г. и 10.VI 1959 г;; в Броварском
р-не - 6.VI 1953 г., в Вышгородском р-не на р.Десна — 31.V 1955 г. (колл. ИЗАНУ; М.ЙЛобановского; Ю.В.Файделя; наши данные), в Фастовском р-не (4 года) -31.V-28.VI. Вылупление птенцов в 1 гнезде у г.Фастов зарегистрировано С.И.Пархомен-ко 3.VI 1968 г. В Черниговской обл. у г.Нежин гнездо с 4 яйцами найдено 8.VI 1976 г., а в Борзнянском р-не — 1 .VI (сообщ. М.Ф.Самофалова).
В Белоруссии свежие кладки встречаются во второй половине мая, насиженные — в первой половине июня, а птенцы вылетают из гнезд в конце июня — I декаде июля (Шнитников, 1913; Федюшин, Долбик, 1967). В Гродненской обл. откладка яиц (3 года) заканчивается 16, 17 и 21.V (Сямашка, 1956). В Оршанском р-не Витебской обл. в 1913 г. гнездо с оперяющимися птенцами обнаружено у д.Тарасинки -21.VI, а с пуховыми птенцами в Загорье — 26.VI (Бианки, 1914).
В Белгородской обл. в заповеднике ’’Лес на Ворскле” (Новиков и др., 1963) строящиеся и готовые пустые гнезда находили с середины мая до середины июня. Полные кладки обычно встречаются здесь (6 лет) во второй половине мая - первой половине июня. Гнезда с птенцами отмечены в III декаде мая — I декаде июля. Основная масса птенцов вылетает из гнезд в конце июня - начале июля. Поздние гнезда с кладками найдены на территории заповедника 4.VII 1950 г., 17.VII 1962 г., а самые поздние выводки слетков зарегистрированы 22.VII 1946 г. и 23.VII 1962 г. В Саваль-ском лесхозе Воронежской обл. самая ранняя кладка обнаружена 25 .V, самая поздняя - 3.VII (Мальчевский, 1959). В Воронежском заповеднике (Барабаш-Никифоров, Павловский, 1947) в 1938 г. начало откладки яиц в 1 гнезде отмечено 29.V вылет птенцов — 27 .VI. В 1942 г. летные молодые встречены 26.VII, а в 1945 г. массовый вылет птенцов из гнезд зарегистрирован 5-7.VII. В Курской обл. у берега р.Сейм гнездо с 5 яйцами найдено 5.VI (Макаров, 1980). В Орловской обл. у истоков р.Ока полные свежие кладки обнаружены 2 и 3.VI, а гнезда с птенцами встречены с 12.VI до конца июня (Ефимов, 1915). В Мордовии (Луговой, 1975) гнезда с кладками (2 года) найдены 9, 13, 16 и 26.VI, в двух из них птенцы вылупились 21 и 23 .VI. Летные молодые появляются здесь с I декады июля. В Московской обл. (Птушенко, Иноземцев, 1968; колл. ЗМ МГУ; КБ МГУ) строительство гнезд (мно
125
голетние наблюдения) отмечено 19—31.V— 20.VI, первое яйцо мухоловки откладывают 28.V—9.VI—26.VI, полные свежие кладки встречаются 1—29.VI, птенцы вылупляются 14—25.VI—11.VII, их вылет из гнезд проходит 26.VI-9.VII-24.VII. Повторные кладки встречаются в конце июня — I декаде июля. Слетки в 1956 г. отмечены также 31.VII— 3.VIII.
В Дарвинском заповеднике в 1947 г. гнезда со свежими кладками найдены в р-не Борка 19 и 28.VII (Спангенберг, Оли-гер, 1949).
В г.Новгород в большинстве гнезд первое яйцо птицы откладывают (18 лет) 22.V-14.VI, а вылет птенцов проходит здесь 14.VI-6.VII (Мантейфель. - Цит. по: Благосклонов, 1954). В Ленинградской и Псковской областях (Мальчевский, Пукинский, 1983) откладка яиц начинается в III декаде мая - I декаде июля, обычно -в I и II декадах июня. В Лужском р-не самое раннее появление первого яйца в одном из гнезд отмечено 23.V 1966 г., а в Юго-Восточном Приладожье (9 лет) -30.V 1977 г. Вылет птенцов в Ленинградской обл. проходит в конце июня — I декаде августа. Наиболее ранний вылет зарегистрирован в р-не Раковых озер 24— 25.VI 1966 г., а наиболее поздний - в д.Ме-рёво 6.VIII 1977 г. В Южной Карелии самое раннее начало гнездования отмечено 28 .V (Ивантер, Зимин. — Цит. по: Мальчевский, Пукинский, 1983). По данным И.А.Ней-фельд (1958), в 1954 г. птицы, строящие гнездо, встречены 9.VI, 16.VI в гнезде было 3, a 18.VI - 5 яиц. В этом же году откладка первого яйца в 2 гнездах зарегистрирована 9 и 10.VI, а вылет птенцов (также в 2 гнездах) - 10 и 13.VII. В 1955 г. кладка из 5 сильно насиженных яиц найдена 19.VI; вылупление в одном из гнезд отмечено 6.VII, а вылет птенцов (5 гнезд) — 13, 16 и 18.VII (Нейфельдт, 1958). В Архангельской обл. на Конушинском берегу Белого моря у с.Семжа 2 гнезда с кладками из 5 свежих яиц обнаружены 21.VI 1956 г. (Спангенберг, Леонович, 1958; 1960). В Лапландском заповеднике в 1937 г. гнездо с 4 яйцами найдено 16.VII, а выводок летных молодых встречен 4.VIII (Владимирская, 1948). В Хибинах на южном склоне Поач-вум-чорра выводок отмечен 24.VII 1931 г. (Новиков, 1952а).
В Волжско-Камском крае массовое строительство гнезд проходит во II декаде мая. Самое позднее строящееся гнездо обнаружено 22.VI. Из 14 гнезд, найденных в июне, 2 строились, в 4 были полные клад
ки, в 8 птенцы. 2 гнезда с птенцами встречены в июле. Наиболее ранний вылет птенцов зарегистрирован 22.VI, самый поздний - 22.VII (Птицы Волжско-Камского края, 1978).
В среднем течении р.Урал (Левин, 1982) откладка яиц в большинстве гнезд начинается во II декаде мая — II декаде июня, однако в 4 найденных гнездах первое яйцо было отложено в III декаде июня, а в 1 гнезде - в I декаде июля.
В Кокчетавской обл. Казахстана (Ковшарь, 1970) спаривание отмечено 19.V, гнездо с насиженной кладкой - 16.VI, а с 5 оперившимися птенцами - 19.VI. В окр. с.Лобаново полная насиженная кладка найдена 21.VI. Слетки в селах Зеренда и Дубрава встречены 12.VII, а у озер Аше-куль и Щучье — 10.VII.
В Западной Сибири у г.Омск кладка из 5 свежих яиц найдена 12.VI 1972 г. (Гынгазов, Миловидов, 1977).
Линька. Изучена недостаточно. Сведения, имеющиеся в литературе, противоречивы. В возрасте 23-26 дней (Мальчевский, Пукинский, 1983) у молодых серых мухоловок начинается послегнездо-вая линька, в ходе которой происходит смена- почти всего оперения гнездового наряда, кроме маховых, рулевых и части верхних кроющих второстепенных маховых. У некоторых особей перелинивают также (полностью или частично) и третьестепенные маховые (Виноградова и др., 1976). Эта смена оперения у большинства мухоловок заканчивается к середине августа, однако еше и в середине сентября у птиц из поздних выводков обнаруживаются следы линьки (Мальчевский, Пукинский, 1983). На зимовках, по данным Н.В.Вино-градовой и др. (1976), у первогодков проходит полная смена оперения, но у некоторых особей к весне вторично уже не меняются линявшие летом-осенью третьестепенные маховые и большие верхние кроющие второстепенных маховых. Весной в свежем пере птицы прилетают на места гнездования и приступают к размножению. У взрослых серых мухоловок обычно с середины июля начинается послебрачная линька, которая, по данным Н.В.Виноградовой и др. (1976), растягивается до февраля-марта. В Ленинградской обл. у птиц, отловленных у гнезд с птенцами во второй половине июля — начале августа, сменялись третьестепенные маховые и кроющие рулевых, а также перья на голове, шее, спине и нижней стороне тела (Мальчевский, Пукинский,1983).
126
Питание. Основу пищи европейской серой мухоловки составляют насекомые разных групп, главным образом двукрылые, жесткокрылые, перепончатокрылые и чешуекрылые. Изредка поедает паукообразных (в основном пауков), мелких наземных моллюсков и как исключение - мокриц и даже, по данным Л.А.Портенко (1960), — дождевых червей. Во второй половине лета и осенью в пише, помимо беспозвоночных животных, иногда встречаются и сочные плоды некоторых растений. Основным способом охоты серой мухоловки является подкарауливание летающих насекомых с какой-либо присады (чаше всего ветвь дерева) с последующей ловлей их в воздухе на лету в стремительном броске. Значительно реже птицы склевывают беспозвоночных с растений и поверхности земли. Последний способ становится основным при похолоданиях и в дождливую погоду, когда лет насекомых резко снижается.
Отлет и осенний пролет на территории СССР. Проходит с I декады августа по III декаду октября. Есть указания Е.С.Птушенко, А.А.Инозем-цева (1968) на начало миграции даже со II декады июля. Наиболее интенсивно в большинстве районов европейской части СССР протекает во второй половине августа - первой половине сентября. Одиночные сильно задержавшиеся особи в ряде мест изредка встречаются даже в ноябре.
В Лапландском заповеднике последние птицы в 1937 г. наблюдались 13.VIII (Владимирская, 1948). В Южной Карелии в заповеднике ’’Кивач” начавшееся передвижение серых мухоловок в 1955 г. отмечено И.А.Нейфельдт (1958) 16-17.VIII. По ее данным, в Пряжинском р-не в этом году почти все мухоловки исчезли к 22 .VIII, одиночные особи попадались до 8.IX.
В Ленинградской обл. (Мальчевский, Пукинский, 1983) осенняя миграция начинается с начала или с середины августа. Самые поздние поимки местных птиц приходятся на II декаду августа. Незначительная часть особей, вероятно, задерживается и до начала сентября. Редкие встречи пролетных серых мухоловок возможны даже в октябре. Так, одиночные птицы зарегистрированы в г.Лениград 10.Х 1979 г. и 22.Х 1968 г. В окр. г.Новгород (14 лет) массовый отлет наблюдается 3—20.IX, одиночные особи задерживаются до 1.Х (Мантейфель. — Цит. по: Благосклонов, 1954). В Дарвинском заповеднике миграция начи
нается в середине августа и, вероятно, заканчивается к концу этого месяца (Спангенберг, Олигер, 1949).
В Подмосковье (Птушенко, Иноземцев, 1968) отлет начинается 10-20.VII. 25-30.VIII изредка встречаются одиночные местные птицы, окончательно исчезающие к 31.VIII-5.IX. В августе здесь наблюдается и незначительный пролет мухоловок более северных популяций, который заканчивается к 6—10.IX. Последние пролетные птицы отмечены 14.IX 1957 г. и 15.IX 1956 г. В Орловской обл. и на сопредельных территориях (6 лет) миграция проходит 15.VIII-30.IX (Ефимов, 1915; Суслова, 1937).
В Воронежской обл. отлет и пролет серых мухоловок протекает в конце августа — начале сентября. В Воронежском заповеднике последние особи в 1938 г. отмечены 19.1Х, а в 1942 г. - 7.IX (Барабаш-Никифоров, Павловский, 1947). Самые поздние регистрации мухоловок этого вида 7.Х 1947 г. и 11.XI 1951 г. (Семенов. -Цит. по: Барабаш-Никифоров, Семаго, 1963).
В Волжско-Камском крае основная масса птиц отлетает в сентябре. Одиночные особи задерживаются до октября. Самая поздняя встреча 23 .X (Птицы Волжско-Камского края, 1978). В Мордовии миграция начинается в середине августа и заканчивается ко II декаде сентября (Луговой, 1975). В Жигулевском заповеднике последние особи отмечены в 1974 г. - 5.Х, а в 1975 г. - 19.VIII (Белянина, Белянин, 1981).
В Белоруссии серые мухоловки исчезают постепенно в августе. Самые поздние встречи здесь (2 года) у г.Минск 26.VIII и 3.IX (Федюшин, Долбик, 1967).
На Украине в Харьковской обл. (3 года) миграция проходит с 14.V1II по 24.IX (Сомов, 1897). На Полтавщине птица этого вида добыта 12.Х 1926 г. (Гавриленко, 1929), а в Киевской обл. (Вышгородский р-н) у сЛебедевка - 9.Х 1978 г. (наши данные). В западных областях УССР серые мухоловки отлетают во второй половине сентября. Одиночные особи встречаются в начале октября (Страутман, 1963). В Николаевской обл. наиболее интенсивно пролет проходит во II декаде сентября, но и в I декаде октября еше регистрируются одиночные птицы (Хохлова, 1960).
В Молдавии в середине августа численность серых мухоловок увеличивается за счет появления пролетных особей популяций северной части гнездового ареала (Ганя, Литвак, 1976). Наиболее интенсивно
127
миграция на территории республики проходит в I декаде сентября — II декаде октября (Ганя, Литвак, 1961).
Осенний пролет этого вида через Крым (Костин, 1983) выражен хорошо и протекает отдельными волнами с начала августа до середины октября. Первое появление в сЛортовое отмечено в разные годы 8-19.VIII. Исчезают обычно в начале октября. Самые поздние встречи в г.Алушта (2 года) 17 и 18.Х.
В Восточном Приазовье (Краснодарский край) пролетные птицы появляются в конце августа (22.VHI — Приморско-Ахтарский р-н). У г.Майкоп одиночная особь встречена 14.Х. В центральной части Северного Кавказа (8 лет) серые мухоловки исчезают 20.IX-2.X (Моламусов, 1967), В Тебердинском заповеднике птица этого вида добыта 2Х 1959 г. (Ткаченко, 1966).
В Грузии отлетают в августе-сентябре (Жордания, 1962). Так, у г.Тбилиси серая мухоловка встречена на пролете 11.IX (Сатунин, 1907), а в северной части Лих-ского хр. добыта 20.IX 1964 г. (Жордания, Гогилащвили, 1969). В Азербайджане пролет проходит в середине сентября (Гамбаров, 1941).
В дельте Волги пролет начинается в середине августа и продолжается весь сентябрь. В 1930 г. серые мухоловки отмечены здесь даже 20.Х (Воробьев, 1936). В Западном Казахстане по р.Урал и в Северном Прикаспии миграция проходит с 11.VII по 18.Х, наиболее интенсивно во второй половине августа и в начале сентября (Ковшарь, 1970). На Юго-Восточном Каспии пролет длится с конца III декады августа и весь сентябрь. Одиночные птицы встречаются здесь и в октябре (Исаков, Воробьев, 1940).
Практическое значение. Уничтожает многих насекомых — вредителей леса и садовых культур, а также переносчиков возбудителей ряда заболеваний человека и животных. Заслуживает повсеместной всесторонней охраны.
Восточная серая мухоловка — Muscicapa striata neumanni (Poche, 1904)
Muscicapa grisola neumanni Poche , 1904, Omith. Monatsber. ,12: 26 , новое имя для Muscicapa grisola sibiiica Neumann, 1900, Jour. Omith.,48: 259, земля Массаев, Северная Танганьика, преокку-пировано Muscicapa sibirica Gmelin ,1789.
Син. (русск.): сибирская серая мухоловка.
Описание. Составлено по 93 экз. (71 - из Туркмении, Узбекистана, Тад
жикистана и Киргизии, 22 - из Тувы и Бурятии).
У взрослой птицы общий тон окраски оперения более светлый, чем у номинативного подвида. Оперение верхней части тела, а также боков головы и шеи светло-бурое. Низ от белого до бледно-охристо-буровато-го. Пестрины на груди развиты слабо. Рулевые и маховые черно-бурые. Каемки на больших верхних кроюших второстепенных маховых и на третьестепенных маховых почти белые. Молодые в гнездовом наряде практически не отличимы от таковых номинативного подвида, но общий тон окраски их оперения несколько светлее.
Размеры отдельных частей тела восточной серой мухоловки приведены в табл. 12.
Масса тела, г: взрослый самец (22.IV, Копетдаг, с.Дурдыхан) - 15,30; взрослый самец (11.V, окр. г.Теджен) - 16,62; взрослые самцы (п = 6) (1—11.VI, Таджикистан, Варзобское ущ. и его окрестности) - 12,90-14,18, в среднем — 13,48; взрослые самки (п = 2) (6, 17.VI, Варзобское и Рамитское ущ.) — 15,45.
Распространение. В СССР: от долины рек Обь, верхний Иртыш и западных подножий Алтая к востоку в Забайкалье до 115° в.д., Тарбагатай, Тянь-Шань (к западу до Каратау и западного подножья и к востоку до хр.Джумголтау, долины Кеке-Мерена и Ферганского хр.), Алайская система (к востоку до хребтов Алайского, восточной части Гиссарского, Дарвазского и к западу до западных склонов хребтов Туркестанского, Зеравшанского, Гиссарского, Байсунтау и Кугитанга), Копетдаг, Большой Б ал хан, Бадхыз, долина Мурга-ба и долина Теджена. В долине Оби и западного подножья Алтая интерградирует с M.s.striata (Степанян, 1978). На всем ареале птицы этого подвида являются перелетными. Детально распространение восточной серой мухоловки на территории СССР показано на карте (рис. 24).
Кадастр к рис. 24.
1 - Большой Балхан (Зарудный, Билькевич, 1918; Степанян, 1978); родники Баш-Мугур, Кекилик-Сув, колодец Коша-Гуйджик (Шукуров, 1962); 2 - Западный Копетдаг (Дементьев и др., 1955); Кара-Калинский^н окр. с.Дурдыхан (наши данные); долина левого притока р.Сумбар (Полозов, 1980); 3 - Копетдаг (Иванов, 1976; Степанян, 1978); окр. с.Гермаб (сообщ. Ю.В.Мищенко) ; Фирюзинское ущ. (колл. ИЗАНУ); 4 - окр. г.Ашхабад (колл. ЗИН); 5 - долина р.Теджен (Степанян, 1978); 6 - Бадхыз (Степанян, 1978); 7 - долина р.Мургаб (Степанян, 1978); 8 - окр. с.Карамет - Нияз (колл. ИЗАНУ); 9 - хр.Кугитанг (Салихбаев, Остапенко, 1969; Абдусалямов, 1973; Степанян, 1978);
128
10 - хр.Байсунтау (Иванов, 1969); Степанян, 1978; колл. ЗИН); 11 - хр.Бабатаг (Иванов, 1969; Абдусалямов, 1973); 12 - междуречье Кафирнигана н Вахша, склоны хр.Актау (Абдусалямов, 1973); р.Вахш (колл. ЗИН); 13 - западная часть Зеравшанскцго хр. (Степанян, 1978); Аман-Кутанская лесная дача (колл. ИЗАНУ; наши данные); 14 - Зеравшанский хр. и долина р.Зеравшан (Сагитов, 1961; Иванов, 1969; Абдусалямов, 1973); селения Палдарак, Худгиф, Лянглиф, Шинг, Артуч, Косторош, Зебон, Магиян, Кштут (Иванов, 1969; Абдусалямов, 1973; наши данные); 15 - западная часть Гиссарского хр. (Степанян, 1978); Ак - кишлак на р.Катта-Уру-дарья, селения Лянгар и Хитай, ручей Кызылсу, р.Кашкадарья (Мекленбурцев, 1958); 16 - Гис-сарский хр. (Степанян, 1978); Варзобское ущ. (Абдусалямов, 1973; колл. ЗИН; ИЗАНУ); ущ. Кондара (Попов, 1959; Иванов, 1969; наши данные); ур.Обисафед (колл. ИЗиП АН Тадж.); окр. с.Варзоб (наши данные); окр. с.Гажни (колл. ЗИН; наши данные); уш.Джеринот (наши данные); ур.Квак (колл. ЗИН; наши данные); ущ.Такоб (колл. ЗИН; БИН СО АН; наши данные) ; Рамитское ущ. (наши данные); 17 -с.Вознаут (Л аз дин. - Цит. по: Попов, 1959); горы Санглок, с.Хочильер (Попов, 1959); Шуроб-дара (Иванов, 1969); р.Сарбо, Сардан-Миён, Ха-нак, Каратаг, Тупаланг (Абдусалямов, 1973); с.Гарм (Иванов, 1969); Каратегинский хр. (Иванов, 1940; Попов, 1959); 18 - Туркестанский хр. (Степанян, 1978; Абдусалямов, 1973); селения Акдаршай, Баляндчакыр (Иванов, 1969); с.Ур-митан (Абдусалямов, 1973); 19 - бассейн р.Ис-фара; селения Ворух и Матча; р.Нурлоу; Кше-мыш; Чичак (Иванов, 1969; Абдусалямов, 1973); р.Ходжабакирган (Абдусалямов, 1973);
20 - Алайский хр. (Иванов, 1969; Степанян, 1978); северный склон хребта (Янушевич и др., 1960); с.Шахимардан (Иванов, 1969; колл. ЗИН); 21 - горы восточнее г.Куляб (Иванов, 1969); окр. г.Куляб (колл. ЗИН); 22 - Дар-вазский хр. (Степанян, 1978); р.Обихингоу у с.Товиль-Дора (Иванов, 1969); 23 - с.Рушан (Зарудный. - Цит. по: Eber, 1960а); 24 - Ферганская долина (колл. БИН СО АН); 25 - Ку-раминский хр.; Нодак-Сай, Караул-Хона (Абдусалямов, 1973); 26 - Чаткальский заповедник (Железняков, Колесников, 1958); 27 - хр.Кар-жантау (Ковшарь, 1970); 28 - заповедник ”Аксу-Джабаглы”; Угамский хр. (Ковшарь, 1970);
29 - Пскемский хр. (Ковшарь, 1970); 30 - Чаткальский хр. (Янушевич и др., 1960); 31 - окр, оз.Сары-Челек (колл. БИН СО АН); 32 - хр.Та-ласский Алатау (Янушевич и др., 1960); 33 -Киргизский хребет; ущелье Ала-Арча (Кузнецов, 1962); 34 - окр. г.Фрунзе (Благосклонов, 1954; Янушевич и др., 1960); 35 - Иссык-Атинская лесная дача, ущелье Туюк (Янушевич и др., 1960); 36 - хр.Джумгалтау (Степанян, 1978); бассейн р.Джумгал (Янушевич и др., 1960);
37 - р.Кеке-Мерен (Янушевич и др., 1960; Степанян, 1978); 38 - Ферганский хр. (Янушевич и др., 1960; Степанян, 1978); 39 - хр.Молдо-Тоо (Шнитников, 1949); 40 - Алма-Атинский заповедник' (Щульпин, 1939) на хр.Заилийский Алатау; 41 - хр.Каратау (Степанян, 1978); 42 -оз.Ащиколь (Ковшарь, 1970); 43 - восточная оконечность хр.Джунгарский Алатау (Шнитников, 1949); 44 - хр.Тарбагатай (Шнитников7 1949; Степанян, 1978); 45 - р.Черный Иртыш (Хох
лов. - Цит. по: Eber, 1960а); 46 - верховья р.Иртыш (Степанян, 1978); 47 - Южный Алтай (Сушкин, 1938; Портенко, 1960; Ковшарь, 1970); окр. с.Катон-Карагай (Сушкин, 1938; Ковшарь, 1970); окр. оз.Маркаколь (Сушкин, 1938); окр. с.Рахмановские Ключи (Ковшарь, 1970); 48 - Западный Алтай (Сушкин, 1938; Ковшарь, 1970; Степанян, 1978); 49 — Центральный Алтай (Сушкин, 1938); 50 - Курай-ская степь (Сушкин, 1938); 51 - Северо-Восточный Алтай (Сушкин, 1938; Равкин, 1967); Алтайский заповедник (Фолитарек, Дементьев, 1938); окр. Телецкого оз. (Стахеев, 1981; колл. ЗИН); 52 - Турочакский р-н, окр. с.Кебезень (Дроздова и др., 1960); р.Бия (Равкин, 1972);
53 - окр. с.Чемал (Хахлов, 1927); 54 - Северо-Западный Алтай (Сушкин, 1938); 55 - бассейн верхнего течения р.Алей и его правых притоков в р-не Змеиногорска (Селевин, 1928); 56 -г.Бийск (колл. ЗИН); 57 - р-н г.Барнаул (Велижанин, Велижанин, 1929); 58 - Кузнецкая степь (Хахлов, 1937); 59 - Салаирский кряж (Чунихин, 1965); 60 - г.Новосибирск и его окрестности (Гынгазов, Миловидов, 1977; Цыбу-лин, 1985); 61 - окр. с.Боровое (Гынгазов, Миловидов, 1977); 62 - окр. г.Томск (Гынгазов, Миловидов, 1977; Вартапетов, 1984); 63 -долина р.Кеть (Гынгазов, Миловидов, 1977); селения Макзырь, Комаровка, Тархово (Москвитин и др., 1977); 64 - Чаинский р-н (Пантелеев, 1972а); 65 - р.Васюган (Гынгазов, Миловидов, 1977); 66 - р.Тым, окр. с.Напас (Гынгазов, Миловидов, 1977); 67 - с.Ларьяк (Портенко, 1960; Иванов, 1976; Гордеев, 1977); 68 - р.Ени-сей, 62° с.ш. (Степанян, 1978); с.Комса и его окрестности (Сыроечковский, 1960; Иванов, 1976); 69 - нижнее течение р.Подкаменная Тунгуска (Портенко, 1960; Степанян, 1978); 70 -г.Енисейск (Благосклонов, 1954); 71 - г.Крас-ноярск (Благосклонов, 1954) и его окрестности (Юдин, 1952); 72 - Козульская равнина; окр. с.Большой Кемчуг, р.Большой Кемчуг (Наумов, 1960; колл. С.Д.Кустановича); 73 - окр. с.Ужур (Сушкин, 1914); 74 - р.Базаиха (колл. ЗИН);
75 - окр. с.Выезжий Лог (колл. ЗИН); 76 -г.Минусинск (Сушкин, 1914; колл. ЗИН); села Означенное, Ермаковское (колл. ЗИН); 77 - окр. г.Абакан (сообщ. М.И.Головушкина); 78 - р. А бакан (Сушкин, 1914; колл. ЗИН); 79 - р.Можарка, окр. с.Можарка, Можарские озера (Сушкин, 1914; колл. ЗИН); 80 - окр. с.Гри-горьевка (Сушкин, 1914); 81 - р.Мирская выше с.Ус (Сушкин, 1914); 82 - р.Хемчик (Сушкин, 1914); 83 - Тува (Благосклонов, 1954); Тувинская котловина (Янушевич, 1948); окр. г.Кы зыл (сообщ. М.И.Головушкина); 84 - окр г.Туран и с.Усть-Уюк (Реймерс, 1960); 85 - окр с.Тоджа (сообщ. М.И.Головушкина); 86 — Вос точно-Тувинское нагорье (Янушевич, 1948); 87 -окр. оз.Чагытай; окр. с.Самагалтай (сообщ. М.И.Головушкина); 88 - с.Эрзин (колл. В.В.Лео-новича); ^ -- окр. заставы Цаган-Тологой (сообщ. М.И.Толовушкина); 90 - хр.Танну-Ола (Сушкин, 1914; Янушевич, 1948); северные подножья хребта (Сушкин, 1914); 91 - окр. пос. Хандагайты (сообщ. М.И.Головушкина); 92 - окр. с.Мугур-Аксы (сообщ. М.И.Головушкина) ; 93 - Тункинский р-н, р.Долбай (колл. ЗИН); 94 - р.Чадобец (Реймерс, 1966); с.Чадо-бец (Иванов, 1976); 95 - Северное Заангарье (Гнбет, Артамошин, 1977); 96 — окр. г.Усолье-
129
Сибирское (Taczanowski, 1891); 97 - г.Иркутск, (Благосклонов, 1954); р.Ангара (колл. ЗИН); 98 - окр. сЛиствянка (сообщ. М.И.Головушкина) ; 99 - с.Култук (колл. ЗИН); 100 - верховья р.Лена (Портенко, 1960; Иванов, 1976; Степанян, 1978); 101 - с.Красиоярово (колл. ЗИН); 102 - Баргузинская долина, ур.Зармат (р-н Ба-рагхана); берег р.Баргузин; по обоим Куйтунам; восточные склоны Баргузинского хр. (Гагина, 1960; Гусев, 1965); 103 - западные склоны Икатского хр. (Гусев, 1965); 104 - окр. г.Улан-Удэ (Левин, 1979); р.Селенга (колл. ЗИН; ИЗАНУ); окр. ст.Тольцы (Левин, 1979); 105 -р.Чикой (Портенко, 1960; колл. ИЗАНУ); окр. с.Мурочи (колл. ИЗАНУ); 106 - окр. г.Кяхта (ЕЬег, 1960а); окр. пос. Киран (сообщ. М.И.Головушкина); 107 - окр. оз. Гусиное (сообщ. М.И.Головушкина); окр. оз.Щучье (Измайлов, Боровицкая, 1973); 108 - окр. с.Улзар (колл. ЗИН); 109 - Южный Байкал (Благосклонов, 1954; Портенко, 1960); НО - г.Чита (Портенко, 1960; Иванов, 1976; колл. ЗИН); Читинский таежный р-н (Гагина, 1960); 111 - Ононский р-н, окр. с.Нижний Цасучей (сообщ. М.И.Головушкина); 112 - с.Большетархово (Гордеев, 1965).
Места обитания. Населяет различные типы лиственных, смешанных хвойно-лиственных и хвойных лесов на равнинах, в предгорьях и горах до 2300 м н.у.м. Гнездится в рошах, парках, садах, в том числе и в населенных пунктах.
В Западной Туркмении найдена на гнездовании в арчевниках и по густым кустарникам ушелий хр. Большой Балхан (Зарудный, Билькевич, 1918; Шукуров, 1962). В Копетдаге также встречается в арчевниках, а по р.Мургаб обитает в тугайных лесах с преобладанием тополя.
В Памиро-Алае гнездится в садах у населенных пунктов, в березняках и в других самых разнообразных типах лиственных и смешанных хвойно-лиственных лесов на высотах главным образом 1000—2100 м н.у.м. Проникает в фисташники и арчев-ники. В Узбекистане на северных склонах Зеравшанского хр. (Аман-Кутанская лесная дача) найдена нами в большом количестве в лесах из грецкого ореха с примесью других лиственных деревьев и кустарников (см. рис. 5). В Таджикистане обитает в лиственных, смешанных хвойно-лиственных и реже в чисто арчовых лесах, а также в горных садах на высотах 800—2300 м н.у.м. (оптимальные высоты 1200-1600 м). В горах на юго-западе республики поселяется в фисташниках и в смешанных лесах из арчи, клена, боярышника и миндаля.
В Западном Тянь-Шане предпочитает лиственные леса (ясеневые, орехоплодные), в том числе и прирусловые галерейные, но в заповеднике ”Аксу-Джабаглы” гнездится и в старых высокоствольных арчевниках до 2000 м н.у.м. (Ковшарь, 1970).
В Киргизии населяет орехоплодные леса, пойменные заросли, сады и парки (Янушевич и др., 1960), а на Киргизском хр. встречена А.А.Кузнецовым (1962) и в ельниках.
На Алтае обитает в сосновых борах, но гнездится также по опушкам и в лиственнично-пихтовой тайге, а на южном берегу оз.Маркакуль местами многочисленна в березняках по кочкарнику (Ковшарь, 1970). Встречается в лиственничниках (Сушкин, 1938) и в сосново-березовых лесах (Равкин, 1972), На Северо-Восточном Алтае в гнездовой период предпочитает предгорные осинники, сосново-березовые леса и залесенные низинные болота (Равкин, 1984). В горы поднимается до 1600 м н.у.м. (Портенко, 1960). На Салаирском кряже в междуречье среднего течения рек Кара- и Томь-Чумыш населяет хвойно-лиственные, кедрово-елово-пихтовые леса и приречные ивняки (Чунихин, 1965).
В окр. г.Новосибирск обитает в сосново-березовых лесах (Цыбулин, 1985), а также отмечена в смешанных лесах и кустарниках по р.Зырянке (Смирнов, 1980). В Томской обл. в долине р.Обь гнездится во вторичных осиново-березовых лесах с незначительной примесью ели, а также в заболоченных смешанных притеррасной части реки и в уремах поймы (Пантелеев, 1972а). В северной тайге Приобья многочисленна в смешанных полузаболоченных лесах и обычна в темнохвойной тайге (Равкин,1978).
В Красноярском крае у с.Большой Кемчуг отмечена в лиственных и сосноволиственных лесах (Наумов, 1960), а в окр. г.Красноярск — и в сосняках (Юдин, 1952). В Тувинской АССР многочисленна в парковых лиственничных лесах и в лесах Тувинской котловины по долинам рек (Благосклонов, 1954).
В зоне южной тайги Средней Сибири встречается почти исключительно в при-долинных и водораздельных сосняках, как в чистых, так и с примесью мелколиственных деревьев (Реймерс, 1966).
В Забайкалье (Баргузинская долина) гнездится в сосновой лесостепи ур.Зармат и в отдельных островках соснового леса по берегам р.Баргузин и обоим Куйтунам, а также в сосняках восточных склонов Баргузинского и западных Икатского хр., нигде не поднимаясь высоко в горы (Гусев, 1965). В разреженном сосновом лесу отмечена на гнездовании в 25 км восточнее г.Улан-Удэ (Левин, 1969), а также добыта в гнездовой период в сосняке у оз.Щучье (Измайлов, Боровицкая, 1973).
130
Численность. В СССР на большей части гнездового ареала обычна в ряде мест в характерных биотопах — многочисленна. У северо-восточной границы ареала редка.
Прилет и весенний пролет на территории СССР. Проходит с I декады апреля по I декаду июня включительно. В Южной Туркмении первые одиночные птицы как исключение иногда появляются даже в марте.
В Туркмении на р.Мургаб весенняя миграция зарегистрирована 1—17.IV (Таш-лиев по Благосклонову, 1954). Нами интенсивный пролет серых мухоловок на возвышенности Карабиль наблюдался 11-17.V 1979 г.
На территории Памиро-Алая в разные годы птицы этого вида на пролете отмечены: в пойме Аму-Дарьи у кишл. Окузбу-лак - 19.V, а у кишл. Карлюк - 20.V (Иванов, 1969); в низовьях р.Сурхан -25-28.IV (Иванов, 1969); на хр.Кугитанг -в начале мая (Салихбаев, Остапенко, 1964); в ушельях рек Варзоб и Кафирниган — 5-8.V (Попов, 1959; Беме, Сытов, 1963; Иванов, 1969); в Гиссарской долине -20.V — I декаде июня, в верховьях р.Зе-равшан - 17-23.V, в долине этой реки на территории Таджикистана — 1—10.V (Иванов, 1969; Абдусалямов, 1973), в низовьях реки на территории Бухарской обл. — 26.IV (Бакаев, 1978); на хр.Нуратау — 10.V-5.VI (Иванов, 1969).
В Казахстане (Ковшарь, 1970) миграция проходит со второй половины апреля по конец мая. Отставшие одиночные особи встречаются в начале июня. В Киргизию серые мухоловки прилетают в конце апреля - в мае (Янушевич и др., 1960).
На Алтае в р-не Телецкого оз. (8 лет) первое появление отмечено 11—23.V, в среднем - 17.V (Стахеев, 1981), а у Онгудая -26.V (Сушкин, 1938). В окр.г.Омск (1 год) прилетает 13.V (Гынгазов, Миловидов, 1977), в окр. г.Новосибирск (3 года) — 6, 16, 17.V (Гынгазов, Миловидов, 1977; Цыбулин, 1985), в окр. г.Томск - 12— 21.V, а на р.Малая Сосьва — 16.V (Гынгазов, Миловидов, 1977).
В Красноярском крае у г.Минусинск серая мухоловка отмечена 30.V (Сушкин, 1914), в окр. г.Красноярск птицы появляются в начале II декады мая (Юдин, 1952), а на р.Чадобец (правый приток р.Ангара) прилет в 1962 г. зарегистрирован 28—29.V (Реймерс, 1966).
Размножение. Проходит в основном так же, как и у номинативного подвида. Гнезда восточная серая мухоловка стро
ит главным образом на деревьях и кустарниках (слива, грецкий орех, ива, сосна, береза, алыча, боярышник, тополь, платан, арча, миндаль и др.) и значительно реже на постройках человека и выступах скал на высоте 0,0—10,0 м от поверхности земли (Юдин, 1952; Ковшарь, 1970; Абдусалямов, 1973; Левин, 1979; наши данные), чаше (п =7; Ковшарь, 1970; наши данные): 1,1-5,5, в среднем - 3,3. Строительство гнезда длится 4-7 дней. При высокой плотности гнездования гнезда соседних пар обычно располагаются на расстоянии 30— 300 м друг от друга (Абдусалямов, 1973; Левин, 1979; наши данные).
В качестве строительных материалов для гнезда птицы используют полоски луба деревьев и кустарников, стебли, листья, тонкие корешки и соцветия трав (в том числе и дикорастущих злаков), растительные волокна, конский волос, нитки, пучочки хлопка, кусочки листьев и цветы древесных растений. Некоторые постройки снаружи оплетаются паутиной. Лоток выстилается волосками шерсти и мягкими растительными волокнами, однако в некоторых гнездах выстилку лотка птицы не делают. Размеры гнезд (п =8), мм: диаметр 72-120 х 72—130, в среднем — 90 х 98; высота 32—80, в среднем - 56; диаметр лотка 33—60 х 33—60, в среднем - 51 ,х 52; глубина лотка 15-44, в среднем — 30.
В сезон размножения на территории СССР восточная серая мухоловка имеет 1—2 репродуктивных цикла. Количество яиц в полной первой кладке обычно 3—6. Повторные кладки могут быть меньших размеров. Форма и окраска яиц укладываются в пределы изменчивости таковых номинативного подвида. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5. Длительность инкубации 11-14 дней. Птенцы находятся в гнезде обычно 13—14 дней. В выкармливании птенцов участвуют оба партнера пары, но в первые дни после вылупления, когда самка большую часть времени проводит в гнезде, согревая птенцов, корм приносит в основном самец.
Основными причинами отхода яиц и птенцов являются неблагоприятные погодные условия, хищническая деятельность со стороны некоторых видов млекопитающих, птиц и рептилий (разноцветный полоз -Coluber ravergieri), наличие в кладках неоплодотворенных яиц, а в населенных пунктах — и частое беспокойство со стороны человека.
Сезон размножения восточной серой
131
мухоловки на территории СССР длится с III декады апреля по II декаду августа включительно.
В Западной Туркмении на хр.Большой Бал хан у родника Кекелик-Сув гнездо с 4 еше не оперившимися птенцами найдено 13.V 1949 г., а у колодца Коша-Гуйджик с 5 птенцами перед вылетом — 29.V 1948 г. (Шукуров, 1962).
В Памиро-Алае на хр.Кугитанг строительство гнезд отмечено 17-25.V, насиженные кладки обнаружены 10—20.VI, а вылет птенцов зарегистрирован 25—30.VI (Салихбаев, Остапенко, 1964). На южных склонах Гиссарского хр. в уш.Кондара (Попов, 1959; Иванов, 1969; наши данные) строящиеся гнезда отмечены 29. V и 4.VI; 7 .VI найдены 2 гнезда, в каждом из которых было по 3 яйца; 8.VI - с 4 яйцами (птенцы вылупились 17.VI, а вылетели из гнезда 29.VI); 10.VI — также с 4 яйцами (птенцы вылетели 27 .VI); 14.VI найдены 2 гнезда: в одном — 4 яйца (птенцы вылетели — 27 .VI), в другом — 4 птенца перед вылетом; 17.VI отмечены 2 гнезда: одно — незаконченная постройка, во втором самка насиживала кладку; 27.VI встречены взрослые птицы, выкармливающие птенцов, и 1 гнездо с 3 яйцами (птенцы вылупились 2-4 .VII). В Варзобской долине (Абдусалямов, 1973) гнезда с полной кладкой и только что вылупившимися птенцами найдены 18 и 22.VI, а с разновозрастными птенцами - 24.VI-5.VII, однако 28.VI обнаружено гнездо и с сильно насиженной кладкой. У Сагардака строительство гнезд отмечено 4—7.VI (Салихбаев, Остапенко, 1964). В горах Санглок гнезда с кладками встречены 18-20.VI, а вылет птенцов зарегистрирован здесь 2.VII (Попов, 1959). В окр. Шуробдара гнездо с 2 слабо насиженными яйцами найдено 26 .VI (Алексеев по Иванову, 1969). На р.Катта-Уру-дарья у Ак-кишлака выводок молодых с недоросшими рулевыми отмечен 1.VII 1955 г. (Мекленбурцев, 1958). На северном склоне Зеравшанского хр. в ур.Аман-Кутан строящиеся гнезда обнаружены нами в 1976 г. 31.V и 1.VI, в одном из них 1.VI самка отложила первое яйцо.
В Киргизии самое раннее гнездование отмечено у г,Фрунзе (Якушевич и др., 1960): 23.IV 1939 г. одна пара начала строить гнездо, 28.IV гнездо было готово, 29.IV самка отложила первое яйцо, a 3.V в гнезде была полная кладка. В июне эта пара загнездилась вторично: 20.VI — подправка гнезда, 29.VI-1.VII — откладка яиц, 132
15.VII - вылупление птенцов, 31.VII -вылет их из гнезда.
В Западном Тянь-Шане на юге Казахстана спаривание и строительство гнезд проходит во второй половине мая - начале июня (Ковшарь, 1970). На хр.Каржантау М.Н.Корелов наблюдал строительство гнезд 31.V, а в южных отрогах Чаткальского хр. (р.Аксак-Ата) 17.VI им найдено гнездо, с 3 яйцами (Ковшарь, 1970). В Чаткаль-ском заповеднике в 1937 г. выводки начали встречаться со второй половины июля, а в 1949 г. в пойме Шавас-сая слетки отмечены в первых числах августа (Железняков, Колесников, 1958). Ряд гнезд серой мухоловки найден в заповеднике ”Аксу-Джа-баглы” (Ковшарь, 1970): 30.V— с 3 яйцами; 6.VI — почти готовое, но пустое; 7.VI — пустое, но птица сидит в гнезде; 10.VI — с первым яйцом, 4 птенца вылупились 28.VI; 11.VI - также с первым яйцом; 14.VI - с 5 яйцами; 18.VI - с 3 яйцами; 24.VI — с 4 оперяющимися птенцами; a 2.VII - с 2 пуховыми птенцами и 2 ’’болтунами” (Шульпин. - Цит. по: Ковшарь, 1970). Покинувших гнезда птенцов, которых еше кормили взрослые птицы, Л.М.Шулышн (Цит. по: Ковшарь, 1970) встречал в заповеднике 2.VII—13.VIII, а М.Н.Корелов на Пскемском и Угамском хребтах — 18.VII— 21.VIII. В долине Бо-ролдая (Каратау) гнездо с 3 свежими яйцами найдено М.Н.Кореловым 11 .VII (Ковшарь, 1970).
На Алтае у Телецкого оз. начало откладки яиц отмечено в 9 гнездах в III декаде мая, в 20 — в I декаде июня и в 6 -во II декаде июня (Стахеев, 1981).
У Обского водохранилища (с.Боровое) строительство гнезда зарегистрировано 12.V 1959 г., готовая пустая постройка обнаружена 5.VI, а гнезда со свежими кладками найдены 9.VI (Гынгазов, Миловидов, 1977). У г.Томск гнезда с полными кладками встречены (4 года) 5 - 21.VI, а с оперившимися птенцами (6 лет) - 25-29.VI; слетки добыты 11.VII 1970 г. (Гынгазов, Миловидов, 1977). На р.Кеть у пос .Мозырь гнездо с 4 яйцами найдено в конце мая, а у с.Комаровка 3.VII 1968 г. отмечено выкармливание птёнцов (Москвитин и др., 1977). На р.Тым в окр. с.Напас насиженная кладка обнаружена 9.VI 1967 г., а слетки в бассейнах рек Кеть, Васюган и Тым встречены 20.VII-5.VIII (Гынгазов, Миловидов,1977).
В Туве у с.Эрзин 2 гнезда со свежими кладками найдены 12.VI 1973 г. (колл.
В.ВЛеоновича). В районе г .Красноярск (Юдин, 1952) готовое, но еше пустое гнездо обнаружено в 1944 г, 4.VI, с 5 свежими яйцами — 8.VI и с 5 слепыми птенцами -20.VI; в 1934 г. кладка из 5 сильно насиженных яиц найдена 17.VI; птенцы, недавно покинувшие гнездо, отмечены 3-5.VII 1942 г., а вполне доросшие молодые в гнездовом наряде добыты 9.VII 1940 г. На р.Ча-добец (правый приток Ангары) первые молодые в 1962 г. добыты 15.VII (Реймерс, 1966).
В Юго-Западном Забайкалье у ст.Толь-цы (25 км восточнее г.Улан-Удэ) (Левин, 1979) в 1974 г. строительство гнезд зарегистрировано в последних числах мая -начале июня. 3.VI найдены 2 гнезда: одно -начатая постройка, второе - почти готово. Во втором гнезде кладка началась 11.VI, первое гнездо 9.VI было еше пустым, а 15.VI содержало полную кладку из 6 яиц. Строящееся гнездо найдено также 13.VI.
Линька. В основном проходит так же, как и номинативного подвида.
Пи т а н и е. Основу пиши взрослых птиц составляют насекомые разных групп, главным образом жесткокрылые, перепончатокрылые, двукрылые и чешуекрылые. Значительно реже поедает пауков, моллюсков и мелкие сочные плоды некоторых растений. Изредка заглатывает гастролиты и иногда скармливает их даже птенцам. Основным способом охоты является ловля насекомых на лету в стремительном броске с присады. Активный роиск беспозвоночных с последующим склевыванием их с растений и поверхности земли восточной серой мухоловке не характерен и пользуется она им редко, главным образом в ненастную, дождливую погоду, когда массовый лет большинства насекомых резко снижается. В населенных пунктах вечером взрослые птицы иногда охотятся у зажженных уличных фонарей и ламп, поедая насекомых, привлеченных на свет. Кроме этого на южных склонах Гиссарского хр. в ущ.Кон-дара мы неоднократно наблюдали, как взрослые серые мухоловки поедали спелые, сочные плоды белой шелковицы (Morus alba). Это происходило даже в ясную солнечную погоду, когда лет насекомых был интенсивным (Пекло, 1982). Основу пиши птенцов составляют также насекомые разных групп, главным образом двукрылые, чешуекрылые и стрекозы. В редких случаях взрослые птицы скармливают им пауков и мелких моллюсков. Поймав крупное насекомое (большие стрекозы, цикады,
бабочки), мухоловка умертвляет его, отрывает крылья и другие жесткие хитиновые части тела и только после этого несет его птенцам. За все время исследований нами ни разу не отмечено скармливание птенцам сочных плодов растений.
В Западном Тянь-Шане в пище серой мухоловки зарегистрированы: жесткокрылые (щелкуны, пластинчатоусые, листоеды), двукрылые, перепончатокрылые, полужесткокрылые, стрекозы, а также богомолы, саранчовые, уховертки и пауки (Ковшарь, 1970).
В 17 желудках птиц этого вида, добытых в мае—июле 1958 г. в Тувинской АССР (окр. г.Турана и с.Усть-Уюк), встречены: стрекозы - в 1 желудке, полужесткокрылые — в 1, нарывники (Mylabris sp.) — в 1, усач (Monochamus sp.) — 3, яйца этого усача — 1, усачи ближе не опред. — 1, листоеды - 1, долгоносики - 1, жесткокрылые ближе не опред. — 10, наездники (Ich-neumonidae) — 1, муравьи древоточцы (Camponotus sp.) - 3, муравей (Formica rufa L.) — 2, комары - 1, ктыри — 2, ручейники - 1, дневные чешуекрылые - 2, совки — 2, коконопряд (Dendrolimus sibi-ricus) - 1, яйца этого коконопряда - 2, насекомые ближе не опред. — 2, пауки - 2 и гастролиты (мелкие камешки) — 2 (Реймерс, 1960 (1961).
По нашим данным (Пекло, 1982), в Памиро-Алае взрослые серые мухоловки поедают главным образом перепончато-крылых (пилильщиков, наездников -Ichneumonidae, ос-блестянок, пчел, муравьев) , жесткокрылых (жужелиц, ста-филин, пластинчатоусых, щелкунов, златок, усачей, коровок, долгоносиков), а также чешуекрылых и двукрылых насекомых. Основу пищи птенцов составляют двукрылые, стрекозы и чешуекрылые (имаго). Изредка взрослые птицы приносят и скармливают им прямокрылых, цикад, жесткокрылых, пауков и мелких моллюсков.
В 2 желудках взрослых мухоловок, добытых М.И.Головушкиным 26.VII 1985 г. в Ононском р-не Читинской обл. у сЛиж-ний Цасучей, обнаружены: жужелицы (Harpalus sp. и Anisodactylus signatus) -3 особи (в 1 желудке), златка (Agrilus sp.) - 2(1), усач (Plagionotus sp.) - 1(1), коровка (Halyzia sp.) — 1 (1), долгоносик (Pissodes sp.) — 1 (1), совка (Noctuidae) — 1(1), белянка (Pieridae) - 1(1), журчалка (Syrphus sp.) - 3(1) и пчеловидка (Eristalis sp.) - 1 (1).
133
Отлет и осенний пролет на территории СССР. Проходит с I декады августа до начала II декады октября включительно.
В окр. г.Томск (16 лет) осенняя миграция протекает 20.VIII-10.IX (Гынгазов, Миловидов, 1977), а в окр. г.Новосибирск наиболее интенсивно проходит в первой половине сентября (Цыбулин, 1985). Последние птицы у г.Новосибирск в 1959 г. и 1978 г. отмечены 6.IX (Гынгазов, Миловидов, 1977; Цыбулин, 1985).
В Красноярском крае на р.Чадобец наиболее поздняя серая мухоловка в 1962 г. добыта 14.VIII (Реймерс, 1966). Под г .Красноярск пролет проходит в I и II декадах августа (Юдин, 1952), а в окр. г.Мину-синск — во второй половине этого месяца (Сушкин, 1914). Последние птицы на р.Хемчик зарегистрированы (1 год) 25.VIII (Сушкин, 1914).
На Алтае начало миграции приходится на первую половину августа, а в Зайсан-ской котловине пролетные мухоловки появляются 15- 19. VIII. Заканчивается пролет 2—28.IX (Ковшарь, 1970). Последние птицы у с.Рахмановские Ключи встречены 2.1Х, а у Катон-Карагая — 3-5.IX (Сушкин, 1938).
В окр. г. Алма-Ата (Благосклонов, 1954) пролет начинается (2 года) 12 и 26.VIII. Наиболее интенсивно протекает в конце III декады августа и обычно заканчивается в середине или в конце II декады сентября (Благосклонов, 1954).
В Киргизии миграция проходит в конце августа - начале сентября, последние птицы у г.Фрунзе в 1955 г. зарегистрированы 21.IX (Янушевич и др., 1960). В окр. г.Ташкент пролет заканчивается в конце сентября — начале октября, наиболее поздняя встреча мухоловок здесь 11 .X (Ковшарь, 1970).
В долине р.Кашкадарья у устья Танха-са пролетные серые мухоловки в 1938 г. появились 7-8 .VIII, а в 1939 г. отмечены в долине этой реки (в 6—7 км ниже устья Тан-хаса) 20—26.IX (Мекленбурцев, 1958).
В Таджикистане основной пролет проходит с 15-18.VIII по 20-25.IX (Абдусалямов, 1973). Самые поздние одиночные особи у оз.Искандер-Куль встречены 28.IX (Попов, 1959). В р-не Гарма пролетные птицы отмечаются с 26.VHI до начала октября. В эти же сроки осенняя миграция протекает и в Гиссарской долине у г.Ку-ляб (Иванов, 1969).
На р.Мургаб и в Юго-Западной Туркмении пролет проходит с середины августа 134
до конца сентября (Ташлиев. — Цит. по: Благосклонов, 1954; Дементьев и др., 1955). На Кушке серая мухоловка добыта 18.IX 1945 г. (Гептнер, 1956).
Практическое значение. Такое же, как и у номинативного подвида.
Сибирская мухоловка — Mucsicapa sibirica Gmelin, 1789
Muscicapa sibirica Gmelin, 1789, Syst. Nat., 1: 936, Байкал.
Син. (русск.): мухоловка-касатка.
Описание. Составлено по 36 экз. (7 - из южной части Восточной Сибири, 27 - из Амурской обл., Хабаровского и Приморского краев, 2 — с Курильских о-вов).
Вершину крыла образуют 2—4-е первостепенные маховые (2 < 3 — 100%; 2 < 4 -71,0, 2 = 4- 25,8, 2 > 4 - 3,2; 3 > 4 -100 %); 5-е немного их короче; 2-е маховое всегда больше 5 и 6-го. Первое первостепенное маховое очень мало и не доходит до вершин кроющих кисти (табл. 13). Наружные опахала 3-4-го первостепенных маховых в вершинной части заметно сужены. У некоторых особей сужено в вершинной части и наружное опахало 5-го махового. Количество шетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3—4 (3 — 86,2 %; 4 - 13,8). Длина наибольшей шетинки (п = 21): 4,9—7,4 мм (в среднем — 6,1). Количество щитков подотеки на передней части цевки варьирует от 3 до 6. На вершине хвоста (в центре) имеется небольшая выемка. Самец и самка окрашены одинаково.
Распространение. Южная Сибирь к западу до долины Оби и Западного Алтая и Дальний Восток СССР к северу до Пенжинского р-на Камчатской обл. (рис. 27). Далее к югу за пределами СССР до Хангая, Хэнтэя, долины Керулена, южной оконечности^ Большого Хингана, среднего течения Ляохэ и Корейского п-ова. Изолированный участок гнездового ареала охватывает хр.Сафедкох (Спингар) в Восточном Афганистане, Гималаи, Юго-Восточный Тибет, Юго-Западный Сикан, Северо-Восточную Бирму и Северную Юньнань до Северной Сычуани (Степанян, 1978), острова: Шантарские, Сахалин, Курильские (рис. 27), Хоккайдо и Хонсю. Зимуют в Южной и Юго-Восточной Азии.
Систематика. Географическая изменчивость проявляется в особенностях окраски и рисунка оперения на нижней стороне тела и в общих размерах. Вид образует
4 подвида, 1 из которых - номинативный -встречается и гнездится на территории СССР.
Сибирские мухоловки из Приморского края, описанные Л.М.Шульпиным (19276) в качестве самостоятельной формы M.s.opaca (Shulpin, 1927), по нашим данным, практически. не отличаются от таковых номинативного подвида и, несомненно, должны рассматриваться в его составе.
Северная сибирская мухоловка — Muscicapa sibirica sibirica Gmelin, 1789
Muscicapa sibirica Gmelin, 1789, Syst. Nat., 1 : 936, Байкал.
Описание. Составлено по 36 экз. (7 — из южной части Восточной Сибири, 27 - из Амурской обл., Хабаровского и Приморского краев, 2 — с Курильских о-вов).
У взрослой птицы оперение верхней части тела и боков головы от чисто-бурого до бурого с незначительной примесью серого цвета. Оперение верхней части головы с чешуйчатым рисунком, образованным двухцветными перьями (темные в центре и более светлые по краям опахал). На горле имеется светлое пятно от белого до бледно-охристого цвета. Бока горла, тела, а также верхняя часть груди от светло-серовато-бурого до охристо-бурого. На центральной части груди и брюшке охристо-бурый цвет светлеет и локализуется на перьях в виде сильно размытых пестрин. Нижние кроющие хвоста бурые с широкими бледно-буровато-белыми вершинными и боковыми каемками. Нижние кроющие крыла бурые с узкими ржавчатоохристыми каймами по краям их наружных опахал. Рулевые, маховые и верхние кроющие крыла темно-бурые. По краям наружных опахал проксимальных второстепенных маховых, а на третьестепенных маховых и больших верхних кроющих второстепенных маховых также и на вершинах расположены узкие светло-буровато-охристые каемки. Подклювье темно-коричнево-бурое. Надклювье желтовато-охристое в основании и темно-коричнево-бурое в вершинной части. Ноги тёмно-бурые. Радужина глаза темно-коричневая. Сезонная изменчивость проявляется в общем посветлении окраски оперения, частичном исчезновении чешуйчатого рисунка на верхней части головы, а также в исчезновении светлых каемок на маховых и верхних кроющих крыла в результате их обноса и выцветания.
Молодые в гнездовом наряде сверху
темно-бурые с мелкими заостренными пятнами от чисто-белого до бледно-охристого цвета. На горле и зобе имеется треугольное пятно такого же цвета. Грудь, верхняя часть брюшка и бока тела с чешуйчатым рисунком (бледно-охристая или белая центральная часть каждого пера окантована бурыми каймами). Интенсивность его проявления уменьшается к нижней части боков тела. Центральная часть брюшка и нижние кроющие хвоста белые. Нижние кроющие крыла охристо-рыжие. Маховые и рулевые черно-бурые. У некоторых особей на вершинах рулевых находятся мелкие бледно-охристые пятна. Проксимальные второстепенные маховые, третьестепенные маховые, большие и средние верхние кроющие второстепенных маховых имеют светло-охристые каемки по краю наружных опахал и на вершинах этих перьев.
Первый послегнездовой наряд идентичен наряду взрослой птицы, но на третьестепенных маховых и больших верхних кроюших второстепенных маховых сохраняются бледно-охристые каемки и вершинные пятна.
Размеры отдельных частей тела северной сибирской мухоловки приведены в табл.13.
Масса тела, г: самец (10.VI, Алтай, с.Ке-безень) - 14,0 (колл. БПиДВНЦ); самец (8.VI, Командорские о-ва, о-в Топорок) -12,0 (Мараков, 1962); самец (23.IX, Курильские о-ва, о-в Юрий) — 9,6 (колл. БПиДВНЦ).
Распространение. Весь ареал вида на территории СССР, а также Северная Монголия, Северо-Восточный Китай, п-ов Корея и о-ва Хоккайдо и Хонсю. На всем гнездовом ареале^ северные сибирские мухоловки являютёя перелетными и к местам зимовок в Южный Китай, на п-ов Индокитай, и о-ва Хайнань, Калимантан, Суматра и Ява мигрируют через Восточный Китай, Японию, Корею и о-в Тайвань. Залетные птицы встречены на Командорских о-вах (Беринга, Медный, Топорок) (Благосклонов, 1954; Мараков, 1962). Детально распространение северной сибирской мухоловки на территории СССР показано на карте (рис. 27).
Кадастр к рис. 27.
1 - р.Кеть, окр. с.Белый Яр (Гынгазов, Миловидов, 1977); окр. с.Комаровка (Москви-тии и др., 1977); 2 - устье р.Томь (Степанян, 1978); окр. г.Томск (Гынгазов, Москвитин, 1965); окр. с.Конинино (Гынгазов, 1962); окр. с.Туган (Гынгазов, Миловидов, 1977); 3 - окр. ст.Итатка (Гынгазов, 1962; колл. ЗМ МГУ); окр. с.Мелетьево (Гынгазов, 1962); 4 - 50 км
135
северо-западнее г.Томск (Гынгазов, 1962а); 5 -Бакчарский р-н, окр. с.Карагай (Гынгазов, Миловидов, 1977); 6 - Шегарский р-н, окр. с.Поздня-ково (сообщ. С.С.Москвитина); 7 - среднее течение р.Чулым (Гынгазов, Миловидов, 1977); 8 - Ачинский р-н, Божье оз. (Сушкин, 1938); 9 - Кузнецкий Алатау (Чунихин, 1965); 10 -Салаирский кряж в междуречье среднего течения рек Кара-Чумыш и Томь-Чумыш (Чунихин, 1965); 11 - окр. г.Бийск (колл. БИН СО АН); 12 - Центральный Алтай (Дементьев, 1937; Сушкин, 1938; Иванов, 1976); Теректайский хр., на пути между селениями Теньга и Уймон (Силантьев. - Цит. по: Сушкин, 1938; колл. ЗИН); 13 - Чулышманское нагорье (Иванов, 1976); верховья р.Чулышман, устье притока р.Еланду (колл. ЗИН); 14 - окр.Телецкого оз. (колл. ЗИН); северный берег (Сушкин, 1938; колл. ЗИН); перевал из долины р.Колдор к оз.Булан-куль (Сушкнн, 1938); р.Егач (колл. ЗИН); верховья р.Чалышкар (Сушкин, 1938);
15 - р.Клык от Узун-ока до верховьев (Сушкин,
1938; колл. ЗИН); верховья кл. Дадайчак, приток р.Байгол (Сушкин, 1938); окр. с.Кебезень (Доброхотов, 1962; колл. БПиДВНЦ); 16 - Северо-Восточный Алтай (Дементьев, 1937; Сушкин, 1938; Иванов, 1976); между селениями Сея и Матур, окрестности этих селений, р.Кони-су (Сушкин, 1938; колл. ЗИН); 17 — окр. г.Крас-ноярск (Благосклонов, 1954); р.Кача (колл. ЗИН); 18 - окр. с.Выезжий Лог (колл. ЗИН); 19 - Можарские озера (Сушкин, 1914; колл. ЗИН); 20 - Шушенский р-н, Ермаковский стационар Ин-та леса и древесины СО АН СССР (Земкова, Штильмарк, 1965); 21 - подножье Араданского хр., долина Каменного ключа (Сушкин, 1914; колл. ЗИН); 22 - окр. г.Туран (колл. ЗИН); 23 - хр. Танну-Ола (Янушевич, 1948; Иванов, 1976); 24 - Восточно-Тувинское нагорье (Янушевич, 1948); . 25 - Тоджинская впадина, окр. оз. Тоджа (колл. ИЗАНУ); 26 -р.Бирюса, окр. с.Бирюса (колл. ЗИН); 27 -р.Мура (Сушкин, 1914); 28 - водораздел рек Ангара и Подкаменная Тунгуска (Реймерс, 1966;
Таблица 13. Размеры " коэффициенты вариации (CV, %) сибирской и пестрогрудой мухоловок
Вид, подвид, место сбора п Длина,
Крыло CV Расстояние, на которое кроющие кисти выступают за вершину 1-го махового CV
Сам
M.sibrica sibrica южная часть Восточной Сибири 5 80,16* 0,22 79,5 -80,8 0,60 3,62*0,25 3,0 -4,2 15,31
Среднее и Нижнее Приамурье, Приморье 16 79,98* 0,55 76,5 -84,6 2,75 4,06*0,34 1,6 -6,7’ / 6,01*0,34 3,7 -8,8 33,56
M.griseisticta griseisticta, Среднее и Нижнее Приамурье 17 84,39* 0,60 81,1 -91,3 2,95 22,67
Приморье 9 82,58* 0,96 76,5 -86,0 3,48 5,86*0,77 1,5 -9,1 39,42
M.g.pallens Камчатка 5 84,40* 1,36 82,2 -88,3 3,22 5,32*0,63 4,3 -7,8 26,56
Сам
M.sibirica sibirica южная часть Восточной Сибири 2 79,65* 1,15 78,5 -80,8 2,04 3,75*0,35 3,4 -4,1 13,20
Среднее и Нижнее Приамурье, Приморье 10 79,17* 0,61 76,7 -83,1 2,45 2,62*0,30 0,7 -3,8 36,16
M.griseisticta griseisticta Среднее и Нижнее Приамурье 19 85,09* 0,47 81,1 -89,8 2,42 6,06*0,33 3,6 -9,5 23,16
Приморье 9 83,49* 1,13 77,3 -86,6 4,07 6,39*0,44 3,8 —8,3 20,76
M.g.pallens Камчатка 4 85,65* 1,79 81,5 -90,1 4,17 6,45*0,57 5,6 -8,0 17,61
136
Иванов, 1976); 29 - устье р.Подкаменная Тунгуска (Портенко, 1960; Иванов, 1976; Степанян, 1978; колл. ЗИН); 30 - р.Нижняя Тунгуска (Ткаченко, 1937), окр. с.Ерема (колл. ЗИН); 31 ' - верховья р.Нижняя Тунгуска (Ткаченко, 1937; колл. ЗИН); 32 - долина средней Лены (Соломонов, Ларионов, 1971); 33 - верхняя Лена (Реймерс, 1966); 34 - р.Ангара, окр. с.Под-валочное (колл. ЗИН); 35 - нижнее течение р.Голоустная (Богородский, 1979); 36 - Слюдянский р-н, окр. с.Маритуй (сообщ. В.Д.Сони-на); 37 - долина р.Большая Олха (Богородский, 1979); 38 - 20 км от г.Слюдянка в верх по р.Слю-дянка (сообщ. Ю.А.Дурнева); 39 - окр. с.Кул-тук (колл. ЗИН); 40 - Тункинский р-н, р.Дол-бай (колл. ЗИН); 41 - Юго-Западное Забайкалье (Козлова, 1930; Измайлов, Боровицкая, 1973); хр.Малый Хамар-Дабан, оз.Таглей (Козлова, 1930; колл. ИЗАНУ); 42 - восточный берегоз.Гусиное (колл. ЗИН; ИЗАНУ); 43 — окр. с.Иволгинск (колл. ИЗАНУ); 44 - п-ов Святой Hoc (Stegmann, 1936; Гагина, 1960); окр. с.Усть-Баргузин
(Stegmann, 1936); 45 - о-в Большой Ушканий (Малышев, 1960); 46 - северо-западное побережье Байкала, окр. бухты Заворотная (Малышев, 1960а); окр. Горемыка и Котельниковская (Stegmann, 1936); 47 - нижние части западного склона Баргузинского хр.; Баргузинский заповедник (Скрябин, Филонов, 1962; Беляев, 1982); с.Сосновка (Stegmann, 1936); 48 - северо-восточное побережье Байкала (Малышев, 1960); 49 - Северный Байкал (Stegmann, 1936; Благосклонов, 1954; Гагина, 1954; Иванов, 1976; колл. ЗИН); окр. В.Ангарск (Гагина, 1954); Верхне-Ангарский участок (Гагина, 1961); 50 - окр. оз.Баунт, оз.Арон (колл. ЗИН); 51 - Витимское плоскогорье, окр. с.Багдарин (Измайлов, 1967); 52 -устье р.Муя (Иванов, 1976); окр. с.Усть-Муя (колл. ЗИН); 53 - окр. с.Усть-Заза (Измайлов, 1967; колл. ЗИН); 54 - оз.Исиига (Измайлов, Боровицкая, 1967); Еравна (Измайлов, 1967; колл. ЗИН); окр. оз.Щучье (Измайлов, 1967); 55 - окр. оз.Арахлей (сообщ. М.И.Головушкина) ; 56 - Читинский таежный р-н (Гагина,
мм
Хвост CV Цевка CV Клюв от оперения лба CV Клюв от орального края ноздри CV
ЦЫ
52,601 0,61 50,3 -53,5 2,59 12,901 0,23 12,3 -13,7 4,07 9,221 0,09 8,9 -9,4' 2,22 6.6610J12 6,3 -7/,б' 4,06
52,341 0,47 49,1 -55,1 3,63 12,691 0,10 12,6 —13,6 3,07 8,9210,14 8,1 -5,9 5,45 6,6410,13 6,6 -7,3 6,86
49,381 0,59 46,1 -55,1 4,80 13,701 0,12 12,9 -14,7 3,50 9,931 0,11 9,1 -10,7 4,38 7,1510,09 6,4 -7,7 5,18
48,151 1,00 43,2 -51,4 5,86 13,341 0,14 12,5 -13,8 3,18 9,871 0,15 9,4 -11,0 4,59 6,8310,10 6,4 —7,4 4,27
49,021 0,83 47,4 -52,1 3,77 13,941 0,25 13,3 -14,7 3,98 10,101 0,21 9,8 -10,5 3,57 7,1310,20 6,8 -7,5 4,92
ки
52,401 0,20 52,2 -52,6 0,54 12,801 0,20 12,6 -13,0 2,21 9,1010,10 9,0 -9,2 1,55 6,6510,15 6,5 -6,8 3,19
51,501 0,69 49,0 -56,3 4,03 12,961 0,13 12,4 -13,6 3,18 8,6610,07 8,4 -9,0 2,62 6,3010,08. 6,0 -6,7 3,82
50,451 0,52 47,5 -54,2 4,25 13,971 0,11 13,5 -15,1 3,35 9,8610,09 9,3 -10,7 3,75 7,1110,07 6,3 -7,7’ 3,85
49,221 0,80 45,3 -52,8 4,86 13,671 0,24 12,7 -14,7 5,23 9,931 0,18 9,2 -10,8 5,37 7,0210,12 6,3 -7,7 5,02
50,501 1,44 47,4 -54,1 5,70 13,951 0,33 13,0 -14,5 4,77 10,001 0,23 9,4 -10,5 4,69 6,9310,15 6,5 -7,2 4,47
137
1960а); окр. г.Чита (сообщ. М.И.Головушкина); 57 - верхнее течение р.Чита (Павлов, 1948); 58 - Агинская лесостепь (Гагина, 1960а); 59 -р.Онои (Благосклонов, 1954); 60 - Хэнтэй-ЧикоЙское нагорье, бассейн верхнего течения р.Бальджа (Назаренко, 1978); 61 — окр. с.Ца-ган-Олуй (колл. ЗИН); 62 - окр. с.КайластуЙ (Скалой, 1935); 63 - окр. с.Кличка (сообщ. М.И.Головушкина); 64 - Александрозаводский подтаежный р-н (Гагина, 1960а); 65 - Приар-гунье (Гагина, 1960а; 1961); 66 - хр.Удокан (Гагина, 19606); 67 - долина р.Чара; окр. с.Кюсть-Кенды; верховья р.Икабия (Гагина, 19606); 68 - Олекмо-Чарское нагорье, р.Тяня (колл. ЗИН); 69 - Становой хр., верховья р.Тимптон (Иванов, 1976); окр. с.Нагорный (колл. ЗИН); 70 - среднее течение р.Или (Воробьев, 1963); 71 - р.Мая (Иванов, 1929, 1976) у переката Оленькю (Иванов, 1929); 72 - р.Уп-тар в 45 км к северу от г.Магадан (Васьков-ский, 1956); 73 - окр. г.Магадан (Васьковский, 1956; Портенко, 1960; Воробьев, 1963); 74 -
залив Бабушкина (Иванов, 1976); 75 - Пенжинский p-и (Яхонтов, 1979); 76 - верхнее течение р.Таймень (колл. ЗИН); 77 — окр. посЛан-тарь (колл. ЗИН); 78 — Шантарские о-ва (Яхонтов, 1977; Степанян, 1978); 79 - южные склоны Станового хребта, р.Самурай (колл. ЗИН);
80 - с.Бомнак (колл. ЗИН); 81 — Верхнезейская равнина, побережье Зейского водохранилища вблизи устья р.Тымна (сообщ. В.Ю.Ильяшенко);
82 - 30 км от устья р.Мотовая (сообщ. В.Ю.Илья-шеико); 83 - Свободненский р-н, окр. с.Кли-моуцы (колл. ЗИН); междуречье Малой Перы и Большого Эргеля (колл. ЗИН); 84 - окр. с.Эки-мчан (сообщ. В.В.Леоновича); 85 - верховья р.Амгуиь против устья р.Баджал (Федотов, Брунов, 1977); 86 - хр.Малый Хииган (Яхонтов, 1973); 87 - устье р.Сунгари (Благосклонов, 1954); 88 - устье р.Уссури (Благосклонов, 1954); 89 - хр. Хехцир (Кулешова и др., 1965);
90 - окр. г.Комсомольск-на-Амуре (Рашкевич, 1965); 91 - окр. с.Калииовка (колл. КГУ);
92 - окр. оз.Кизи (Рашкевич, 1965); 93 - окр.
Окончание табл. 13.
Вид, подвид, место сбора п Ширина, мм
Крыло CV Клюв у орального края ноздри CV Клюв у основания CV
Сам
M.sibrica sibrica южная часть Восточной Сибири 5 52,18* 0.41 51,3 -53,6 1,74 4,06* 0,08 • 4,47 6,62*0,19 6,0 -7,0 6,26
Среднее и Нижнее Приамурье, Приморье 16 51,57* 0,31 48,7 -53,9 2,44 4,40*0,09 4,0 -5,0 7,24 6,83*0,09 6,1 -7,2 4,97
M.griseisticta griseisticta Среднее и Нижнее Приамурье 17 53,06* 0,49 48,7 -57,3 3,85 4,29*0,06 3,9 -4,8 5,79 6,51*0,06 6,0 -6,9 3,91
Приморье 9 51,80* 0,77 47,7 -54,7 4,46 4,28*0,11 3,8 -4,7 7,91 6,67*0,09 6,4 -7,3 4,24
M.g.pallens Камчатка 5 52,86* 0,87 51,1 -56,2 3,68 4,55*0,13 4,3 -4,9 5,81 6,62*0,16 6,1 -7,0 5,49
Сам
M.sibirica sibirica южная часть Восточной Сибири 2 51,85* 0,25 51,6 -52,1 0,68 4,5* - 7,2* -
Среднее и Нижнее Приамурье, Приморье 10 51,23* 0,30 50,0 -53,0 1,85 4,29*0,04 4,1 -4,5 2,79 6,68*0,14 6,2 -7,8 6,69
M.griseisticta griseisticta Среднее и Нижнее Приамурье 19 53,79* 0.41 51,4 -57,7 3,33 4,38*0,06 3,9 -4,8 5,49 6.68*0,10 6,2 -7,8 6,58
Приморье 9 52,48* 0,67 48,0 -54,8 3,84 4,42*0,08 4,0 -4,8 5,40 6,63*0,13 6,3 -7,4 5,94
M.g.pallens Камчатка 4 53,35* 0,44 52,5 -54,6 1,67 4,25*0,14 4,0 -4,5 6,79 6,83*0,27 6,1 -7,4 7,97
* Размер приводится по 1 экз.
138
с.Савинское (колл. КГУ); 94 - окр. г.Никола-евск-на-Амуре (Благосклонов, 1954; Воробьев, 1954); 95 - долина р.Ботчи (Емельянов, 1929; Воробьев, 1954); 96 - р.Сукпай, метеостанция Тагему (колл. ИЗАНУ); 97 - Сихотэ-Алинский заповедник (Елсуков, 1982); окр. кл. Южного (картотека заповедника); 98 - истоки р.Большая Уссурка (Назаренко, 1971в; 19846; колл. ЗИН); верховья Красной Речки (Назаренко, 1971в); 99 - бассейн истоков р.Уссури; ур. ’’Мута”, гора Облачная (Назаренко, 19846); 100 - верховья р.Партизанская (Иванов, 1976; колл. ЗИН); окр. ст.Партизаны (Шульпин, 19276; Воробьев, 1954); окр. пос.Ручьи (Фанза) (колл. ЗИН); 101 - Майхэ-Даубихинское плато (Назаренко, 1971в; колл. ЗИН); ур.”Ларчеиково болото” (Назаренко, 1971в); 102 - о-в Сахалин (Дементьев, 1937; Благосклонов, 1954; Иванов, 1976; Степанян, 1978); побережье залива Терпения (Yamashina, 1927); окр. пос.Гастелло (Гизенко, 1955); 103 - п-ов Терпения близ пос.Китового (Гизенко, 1955); 104 - окр. г.Южно-Сахалииск
(Иванов, 1976); 105 - окр. г.Корсаков (Lonnberg, 1908); 106 — лагуна Буссе (Иванов, 1976);
107 - о-в Кунашир, подножия вулкана Менделеева (Стапанян, 1980); 108 - о-в Симушир (Иванов, 1976); 109 - о-в Парамушир (Иванов, 1976); 110 - о-в Шумшу (Иванов, 1976);
111 - Камчатка (Дементьев, 1937; Благосклонов, 1954; Иванов, 1976; Степанян, 1978; колл. ЗИН); окр. с.Ключи (Портенко, 1960; Иванов, 1976); 112 - окр. с.Козыревск (Вронский, 1977); 113 - Кроноцкий заповедник (Лобков, 1978); 114 - верхнее течение р.Камчатка, окр. с.Пущино (Вронский, 1977); 115 - р.Авача (колл. ЗИН).
Места обитания. На территории СССР населяет различные типы хвойных, смешанных хвойно-лиственных и реже лиственных лесов, поднимаяясь в горы до 1900 м н.у.м.
В юго-восточной части Западно-Сибир
Высота, мм Отношение шири-ны клюва у осно-вания к его длине от оперения лба, % CV Отношение длины хвоста к длине крыла, % CV Отношение ширины крыла к его длине, % CV
Клюв у основания CV
3.5810.09 3,4 -3,9 5,72 71,831 2.13 63,83-76,09 6,62 65,621 0,76 62,88-67,17 2,59 65.101 0,47 63,81-66,34 1,61
3.2410.05 3,1 -3,5 4,87 76,241 1,19 70,11-83,95 5,63 65,441 0,39 61,38-67,98 2,38 64,501 0,39 61,35-67,06 2,41
4.02Ю.18 3,5 -4,6 9,86 65,501 0,87 60,19-73,63 5,16 58,541 0,35 55,74-60,92 2,41 62,881 0,43 59,03-65,64 2,85
3,7110,10 3,3 -4,1 6,85 67,731 1,56 58,18-75,26 6,92 58,561 0,64 56,21-60,97 3,08 62,721 0,52 60,48-65,24 2,49
3.8310,28 3,5 -4,4 12,87 68,061 1,93 64,76-71,43 4,90 58,221 0,32 57,66-59,00 1,11 62,851 0,34 62,17-63,65 1,07
ки
3,5* - 78,26 — 65,801 0,70 65,10-66,50 1,50 65,111 0,63 64,48-65,73 1,36
3,3810,09 3,1 -3,7 6,85 77,161 1,60 72,09-88,64 6,57 65,181 0,56 62,18-67,75 2,60 64,721 0,32 63,35-66,50 1,54
- — 67,671 0,95 60,19-74,74 5,81 59,38^1 0,37 56,84-61,10 2,59 63,221 0,28 61,55-65,69 1,91
3.9310.09 3,7 -4,3 5,49 66,821 0,95 60,95-71,15 4,26 58,941 0.31 57,86-60,97 1,60 62,881 0,50 60,68-65,98 2,39
3.6310,15 3,4 -3,9 6,93 68,311 2.74 61,62-74,47 8,02 58.931 0,48 58,09-60,04 1,64 62.33* 1.06 60,60-65,15 3,39
139
ской равнины обитает в смешанной тайге на осветленных участках (Гынгазов, Миловидов, 1977). На р.Кеть (с.Комаровка) встречена в заболоченном смешанном лесу (Москвитин и др., 1977), а на Салаирском кряже в междуречье среднего течения рек Кара-Чумыш и Томь-Чумыш зарегистрирована в горнотаежных лесах: смешанных хвойно-лиственных и кедрово-елово-пихто-вых (Чунихин,1965).
На Северо-Восточном Алтае поднимается вверх до зоны редколесья (1900 м) и держится у просветов елово-пихтовой тайги поблизости от воды, в борах по окраинам тайги в густых лиственничниках и кедровниках, а также в естественно нарушенных пихтовых лесах. К западу от Те-лецкого оз. поселяется в глубоких долинах в высокой зоне тайги, поднимаясь с выводками недоросших молодых даже в пояс редколесья (Сушкин, 1938).
В долине средней Лены встречается в смешанных сосново-лиственничных лесах (Соломонов, Ларионов, 1971). В р-не северо-западного побережья Байкала обычна в сосняках и лиственничниках (Малышев, 1960а). На северо-восточном побережье озера обитает также в светлых сос-
Рис. 27. Распространение сибирской мухоловки (Muscicapa sibirica) иа территории СССР:
а - гнездится, б - отмечена в гнездовой период. См. кадастр, стр. 135-139.
новых и лиственничных лесах и на лесных опушках (Малышев, 1960), а в долине р.Баргузин отмечена на лесостепных участках в старых сосняках (Гагина, 1960). На Витимском плоскогорье зарегистрирована в высокоствольном лиственничном лесу и в лиственнично-березовых рошах у с.Баг-дарин и у оз.Щучье (Измайлов, 1967), а в Юго-Западном Забайкалье населяет сосновые, приключевые смешанные, пойменные леса и горную тайгу (Измайлов, Боровицкая, 1973). На Хэнтой-Чикойском нагорье встречается в долинных и придолин-ных лиственничных и лиственнично-кедровых лесах, а также в приручьевых кедровниках, предпочитая участки, где среди березово-лиственничного молодняка имеются редкие куртины из старых лиственниц и места, где высокоствольные участки леса чередуются с полянами и просветами у дорог (Назаренко, 1978).
В окр. г.Магадан и далее к северу почти до Охотско-Колымского водораздела обитает на вырубках й захламленных участках лиственничного леса, а также в тополевых лесах (Васьковский, 1956).
В верховьях р.Амгуньна водоразделах населяет лиственничные травяно-брусничномоховые леса с подлеском из рододендрона, а в долинах — багульниково-моховые мари с ельниками и отдельными лиственницами (Федотов, Брунов, 1977). На Малом Хингане гнездится в смешанных лесах
140
с кустарниками и перелесками (Яхонтов, 1973), а на хр.Хехцир в гнездовой период отмечена в широколиственных лесах по основному водораздельному гребню (Кулешова и др., 1965).
В Приморье на Южном Сихотэ-Алине приурочена к поясам хвойно-широколиственных и частично пихтово-еловых лесов. Здесь сибирская мухоловка повсеместно населяет лесные участки по узким долинам горных ключей и рек, поднимаясь до 1400 м н.у.м., а на Майхэ-Даубихинском плато (800 м) как исключение найдена в двух массивах лиственничного леса (Назаренко, 1971в; 19846).
В Пенжинском р-не Камчатской обл. встречается в светло-хвойной тайге, по поймам рек и даже проникает в лесотундру (Яхонтов, 1979). На Камчатке тяготеет к пойменным высокоствольным, многоярусным лесам из душистого тополя, чо-зении, ивы и ольхи с подлеском и кустарниковым ярусом из ивы и бузины, но поселяется также в равнинных каменноберезовых и смешанных лесах из лиственницы, белой березы и осины в центральной части долины р.Камчатка (Вронский, 1977), а также отмечена в пихтовых рошах (Лобков, 1978). На о-ве Кунашир в гнездовой период зарегистрирована в смешанном лесу у подножья вулкана Менделеева (Степанян, 1980).
Численность. Неравномерна. На Салаирском кряже в междуречье среднего течения рек Кара-Чумыш и Томь-Чумыш на глубинных участках хвойно-лиственного и кедрово-елово-пихтового лесов численность сибирской мухоловки составляет 0,1 особей/км2 (Чунихин, 1965). В Хабаровском крае в верховьях р.Амгунь в лиственничных травяно-бруснично-моховых лесах на водоразделах, а также на багульниково-моховых марях с ельниками и отдельными лиственницами этот показатель равен 13,0 особей/км2 (Федотов, Брунов, 1977). В Приморье в бассейне истоков р.Уссури численность этого вида в переходных (экотонных) лесах ур. ”Мута” составляет 1,2 пар/км2, в ’’высокогорных” зеленомошных пихтово-еловых лесах горы Облачная - до 1,0, а в пихтово-еловых лесах бассейна истоков р.Большая Уссурка - 1,0— 5,0 пар/км2 (Назаренко, 19846). На Камчатке в пойменных многоярусных лесах долины верхнего течения р.Камчатки численность сибирской мухоловки составляет 32,0 особи/км2, во вторичных смешанных лесах центральной части долины этой ре
ки — 4,0, а в равнинных каменноберезовых лесах - 8,0 особи/км2 (Вронский, 1977).
Прилет и весенний пролет на территории СССР. Проходит со II декады мая до середины II декады июня включительно. Во время миграции летят чаще всего на высоте 2-3,5 м, иногда -0,5—10 (Беляев, 1979).
На островах в северо-западной части залива Петра Великого обычна на пролете во второй половине мая (Лабзюк и др., 1971). На о-ве Монерон в 1973 г. одиночки и стайки пролетных птиц отмечены 26.V— 4.VI (Нечаев, 1975), а на о-ве Шумшу (Курилы) в 1952 г. первые мухоловки появились 13.VI (Благосклонов, 1954).
В Приморье пролет протекает со II декады мая до конца этого месяца: в окр. оз.Ханка 9 пролетных мухоловок добыты между 21 и 30.V, а основная часть птиц -28—29.V (Панов, 1973), у с.Нововладими-ровка пролетные мухоловки добыты 13 и 24.V 191.1 г., у с.Камень-Рыболов - 27.V 1908 г., у с.Троицкое - 31.V 1909 г. (Черский, 1915а; 19156); в Партизанском р-не (ст.Партизаны) - 28.V 1926 г. (Шуль-пин, 19276; колл. ЗИН), а на р.Большая Уссурка у с.Вербовка — 27.V 1938 г. (Спангенберг, 1964).
На нижнем Амуре (Кистяковский, Смогоржевский, 1973) начало пролета отмечено у с.Кисилевка 3.VI и у с.Омми - 4.VI. Пролет протекает интенсивно и в короткий срок. Птицы держатся стайками иногда совместно с пестрогрудными мухоловками.
На Камчатку в Кроноцкий заповедник прилетают также в I декаде июня (Лобков, 1978). В июне в период весенней миграции часто залетают на Командорские о-ва (Беринга, Медный, Топорок) (Мараков, 1962).
В окр. г.Улан-Удэ прилет первых птиц отмечен в 1963 г. - 26.V, а в 1964 г. -1 .VI; на Витимском плоскогорье: в 1961 г. около с.Багдарин — 2.VI, а в 1960 г. на Усть-Зазе — 7.VI (Измайлов, Боровицкая, 1973). На северо-восточном побережье Байкала пролет длится всю первую половину июня (Малышев, 1960). На верхней Лене в 1961 г. первые мухоловки появились 2.VI (Реймерс, 1966). На р.Иртыш у г.Тару прилет зарегистрирован 20.V (Ушаков. -Цит. по: Благосклонов, 1954); на Среднем Чулыме - 23.V 1969, у г.Томск (4 года) 25.V—2.VI (Гынгазов, Миловидов, 1977), а на р.Енисей — в начале и даже в середине июня (Портенко, 1960). На Алтае у Сеи первые птицы встречены П.П.Сушкиным (1938) - 12.VI.
141
Размножение. Изучено недостаточно. Прилетев на места гнездования, мухоловки разбиваются на пары и приступают к строительству гнезд. Последние птицы строят открыто в развилках Стволов и на боковых ветвях деревьев (береза, ива, лиственница и др.) на высоте 1,0—8,0 м от поверхности земли. В сборе строительных материалов принимают участие оба партнера, но строит гнездо главным образом самка (Портенко, 1960).
Гнезда представляют собой сравнительно рыхлые постройки чашевидной формы. В качестве строительных материалов мухоловки используют стебли и листья трав, чешуи и кусочки коры, отдельные тонкие веточки деревьев и кустарников, хвою сосны, одиночные перья и большое количество талломов лишайников. Последние вплетаются как в толшу, так и в наружные стенки гнезда и хорошо его маскируют. Лоток выстилается хвоей (лиственница) и другими мягкими материалами (Taczanowski, 1891; Благосклонов, 1954; Портенко, 1960; наши данные). Размеры гнезд, мм: диаметр 70-120; высота 50—60; диаметр лотка - 43—60; глубина лотка 16—25 (Taczanowski, 1891; Благосклонов, 1954; наши данные).
На территории СССР в сезон размножения сибирская мухоловка имеет 1 репродуктивный цикл. Количество яиц в полной кладке обычно 4—5. Овальные яйца по светло-голубовато-зеленому (бирюзовому) или светло-голубому фону с охристо-бурыми и коричневыми пятнышками, крапинками и точками, большинство из которых размыты. Густота рисунка на тупом полюсе обычно меньше, чем на других участках скорлупы. ’’Венчик”, как правило, не выражен. Размеры и индекс удлиненности яиц приведены в табл. 4.
Пуховой птенец сибирской мухоловки (Нейфельдт, 1970) имеет недлинный (5-6 мм) серый пух на надглазничных, затылочной, спинной, плечевых, бедренных и глазных птерилиях. Кожа желтовато-розовая. Клюв, складки в углах рта, язык и ротовая полость желтые.
В выкармливании птенцов принимают участие оба партнера пары. Первое время после вылета из гнезда слетки держатся очень скрытно. Родители продолжают их кормить. В дальнейшем выводки начинают кочевать, собираясь порой на небольшой территории большими группами. Так, 18.VII 1969 г. в верховьях Красной Речки (истоки р.Большая Уссурка) в разреженном берез
няке с лиственницей на плошади около 1 га А.А.Назаренко (1971 в) встретил не менее 10 выводков этого вида.
Сезон размножения сибирской мухоловки на территории СССР длится с I декады июня по конец июля.
У северо-западной границы гнездового ареала у устья р.Подкаменная Тунгуска молодая птица добыта 31.VII 1929 г. (колл. ЗИН). На р.Кеть выводки встречены у с.Комаровка 15.VII 1968 г. (Москвитин и др., 1977) и у с.Белый Яр - 30.VII 1975 г. (Гынгазов, Миловидов, 1977), а на Среднем Чулыме слетки зарегистрированы 8.VII 1969 г. (Гынгазов, Миловидов, 1977). В окр. г.Томск (с.Туган) 17.VI 1960 г. найдено гнездо с полной ненасиженной кладкой из 4 яиц, а у с.Черная речка 4 выводка отмечены 9-15.VII (Гынгазов, Миловидов, 1977). На Алтае в окр. г.Бийск гнездо с полной кладкой из 4 яиц обнаружено 10.VI 1933 г. (колл. БИН СО АН). На Северо-Восточном Алтае первые слетки в 1960 г. встречены 10—12.VII (Доброхотов, 1962), в верховьях р.Чалышнар несколько молодых особей добыты 21.VII 1912 г. (Сушкин, 1938), а у подножия Саяна выводок недоросших мухоловок отмечен 12.VII (Сушкин, 1914).
На северо-восточном побережье Байкала в Баргузинском заповеднике гнездо с птенцами перед вылетом найдено 14.VII 1960 г. (Скрябин, Филонов, 1962). Первый вылет птенцов Л.М.Малышев (1960) в 1954 г. наблюдал 8.VII. В долине р.Баргу-зин несколько выводков отмечено 14-19.VII (Гагина,. 1960), а у Усть-Баргузина молодые только что покинувшие гнездо зарегистрированы 16.VII (Stegmann, 1936). На Витимском плоскогорье у с.Усть-Заза 8-20.VI 1960 г. встречались пары, самцы пели на гнездовых участках; у 3 добытых птиц семенники были сильно увеличены (Измайлов, 1967).
В Якутии в среднем течении р.Илли разновозрастные выводки отмечены 28.VII и 1.VIII (Воробьев, 1963), а на р.Тимп-тон молодая птица добыта 18.VII 1928 (колл. ЗИН).
В Амурской обл. в 30 км от устья р.Гилюй (долина р.Мотовая) самостоятельная молодая птица зарегистрирована 31 .VII 1981 г. (сообш. В.Ю.Ильяшенко); а в верхнем течении р.Селемджа у пгт Экимчан гнездо с 3 птенцами и 1 болтуном обнаружено 28.VI 1971 г., здесь же второе гнездо с птенцами найдено 2.VII 1971 г. (сообш. В.В.Лео-новича).
142
В Приморье на территории Сихотэ-Алинского заповедника слеток сибирской мухоловки пойман в долине ключа Южного 11.VII 1973 г. (картотека заповедника), а в верховьях Красной Речки (истоки р.Большая Уссурка) около 10 разновозрастных выводков встречены 18.VII 1969 г. (Назаренко, 1971 в). На Майхэ-Даубихин-ском плато в урочише ’’Ларченково болото” в 1965 г. массовый вылет птенцов зарегистрирован 3—10.VII, a 5.VII здесь было найдено гнездо с 3 птенцами перед вылетом (Назаренко, 1971в).
На о-ве Кунашир самец с увеличенными семенниками добыт 8.VI (Степанян, 1980).
Линька. После того как маховые и рулевые закончат рост, у молодых птиц начинается частичная послегнездовая линька, в ходе которой происходит смена почти всего мелкого оперения, кроме больших верхних кроюших второстепенных маховых. На местах зимовок у первогодков проходит повторная полная линька с некоторыми индивидуальными отклонениями (Виноградова и др., 1976). В приобретенном наряде мухоловки прилетают на места гнездования и приступают к размножению. Взрослые птицы имеют в году одну полную послебрачную линьку оперения, которая протекает на местах гнездования с конца июля.
Питание. Питается насекомыми разных групп. Основной способ охоты — ловля насекомых в воздухе на лету в стремительном броске с присады, как между деревьями, так и над лесом. По данным А.А.Назаренко (1971в), характерной особенностью кормового поведения этого вида является длительное использование одной присады, на которую мухоловка возвращается после каждого очередного броска.
Отлет и осенний пролет на территории СССР. Проходит с III декады июля по I декаду октября включительно. В большинстве р-нов наиболее интенсивно миграция протекает в августе и сентябре. Самая поздняя встреча сибирской мухоловки на р.Лена осенью 1960 г.- 12.IX (Реймерс, 1966).
На северо-западном и северо-восточном побережье Байкала отлет заканчивается во II декаде августа (Малышев, 1960; 1960а), а в Юго-Западном Забайкалье — в конце августа — начале сентября (Измайлов, Боровицкая, 1973).
На Камчатке в Кроноцком заповеднике отлетает в I декаде сентября (Лобков, 1978). На южных Курильских о-вах пролет длится весь сентябрь. Здесь мухоловки
наблюдались и были добыты на о-вах: Кунашир — 2.IX 1948 г. (колл. ЗИН); Юрий — 22 и 23.IX 1962 г., Зеленый - 25.IX 1962 г. (Нечаев, 1969; колл. БПиДВНЦ).
В Южном Приморье пролет протекает с III декады августа по I декаду октября включительно: в нижнем течении р.Рудная (Тетюхе) пролетные птицы добыты 20 и 25.1Х 1909 г. (Черский, 1915а), у ст.Пачи-хеза — 28.IX 1973 г. (колл. БПиДВНЦ), на п-ове Де-Фриза — 20 .VIII, 8 и 12.IX 1955 г. (Омелько, 1963), в заповеднике ’’Кедровая падь” — 10.IX 1968 г. (колл. БПиДВНЦ) и 15.IX 1945 г. (Воробьев, 1954; колл. БПиДВНЦ). Самая поздняя встреча сибирской мухоловки в Приморье — 10.Х, птица отмечена у Горнотаежной станции (окр. г.Уссурийск) (Назаренко, 1971в).
На о-вах в северо-западной части залива Петра Великого осенняя миграция наиболее интенсивно проходит в середине сентября и заканчивается в I декаде октября (Лаб-зюк и др., 1971).
Практическое значение. Уничтожает многих насекомых — вредителей леса и переносчиков возбудителей ряда заболеваний человека и животных. Заслуживает повсеместной всесторонней охраны.
Пестрогрудая мухоловка - Muscicapa gri-seisticta (Swinhoe, 1861)
Hemichelidon griseisticta Swinhoe, 1861,-Ibis, 3: 330, Амой и Таку, Восточный Китай.
Син. (русск.): полосатая мухоловка-касатка, дальневосточная мухоловка.
Описание. Составлено по 77 экз. (3 — из Якутии, 37 - из Хабаровского края и Амурской обл., 28 - из Приморья и 9 -с Камчатки).
Вершину крыла образуют 2—4-е первостепенные маховые (2 < 3 - 93,7 %, 2 = = 3 - 4,8, 2>3- 1,5; 2<4- 17,7; 2 = 4-37,1, 2 > 4 - 45,2; 3 > 4 - 100 %); 5-е маховое немного их короче; 2-е всегда больше 6-го. 1-е первостепенное маховое очень мало и не выступает за вершины кроюших кисти (табл. 13). У большинства особей наружные опахала 3—4-го первостепенных маховых в вершинной части заметно сужены. Количество шетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3-4 (3 - 72,9 %; 4 - 27,1 %). Длина наибольшей (п = 47): 3,5—7,0 мм (в среднем - 5,5). Количество щитков подотеки на передней части цевки варьирует от 3 до 7. На вершине хвоста (в центре) находится небольшая выемка. Самец и самка окрашены одинаково.
143
Распространение. Восточная Азия от долины р.Лена в р-не г.Якутск, бассейна р.Зея, долины Кумары и Большого Хингана к востоку до побережья Охотского и Японского морей. К северу предположительно до 62° с.ш. К югу в области Большого Хингана и в бассейне Сунгари до 45° с.ш. и до Южного Приморья, Камчатка к северу до 56-57° с.ш. Предположительно о-ва Сахалин и Курильские (Степанян, 1978). На всем гнездовом ареале птицы этого вида являются перелетными и к местам зимовок на о-ва Филиппинские, Каролинские (Палау), Сулавеси, Сула, Молуккские и Новая Гвинея (северо-западная часть) мигрируют через Приамурье, Приморье, Восточный Китай, Корейский п-ов, о-ва Японии и о-в Тайвань. Отмечены залеты в окр. г.Иркутск и на о-в Беринга.
Систематика. Географическая изменчивость проявляется в интенсивности обшей окраски оперения и в размерах отдельных частей тела. Вид образует 2 подвида: M.g.griseisticta (Swinhoe, 1861) и M.g.pallens (Smejneger, 1887). Оба подвида встречаются и гнездятся на территории СССР. Однако некоторые исследователи (Hertert, 1907; Дементьев, 1937; Благосклонов, 1954; Wynne, 1954; Vaurie, 1959; Назаренко, 1971а; Степанян, 1978; и др.) считают этот вид монотипическим. Приморская пестрогрудая мухоловка — Muscicapagriseistictagriseisticta (Swinhoe, 1861)
Hemichelidon griseisticta Swinhoe, 1861, Ibis, 3: 330, Амой и Таку, Восточный Китай.
Описание. Составлено по 68 экз. (3 — из Якутии, 37 — из Хабаровского края и Амурской обл. и 28 - из Приморья).
У взрослой птицы оперение верхней части тела бурое. Этот цвет светлеет на боках головы и шеи. Перья на верхней части головы несут чешуйчатый рисунок или слабо выраженные продольные пестрины более темного цвета. Оперение нижней части тела сливочно-белое. На боках горла и тела, а также на зобе и груди имеются широкие размытые продольные светло-бурые пестрины. У некоторых особей они заходят и на брюшко. Нижние кроющие хвоста белые. Нижние кроющие крыла светло-бурые с узкими рыжевато-охристыми каймами на вершинах и по краям наружных опахал этих перьев. Маховые и рулевые темно-бурые. По краю наружных опахал второстепенных маховых, а на третьестепенных маховых, больших и средних верхних кроющих второстепенных маховых и на вершинах расположены бледно-буроватые каемки разной
интенсивности. Надклювье и ноги черно-бурые. Подклювье коричнево-роговое в основании и черно-бурое в вершинной части. Радужина глаза темно-коричневая. Сезонная изменчивость проявляется в общем посвет-лении оперения и в частичном исчезновении светлых каемок на маховых и кроющих крыла в результате их обноса и выцветания.
У молодых в гнездовом наряде оперение верхней части тела темно-бурое с каплевидными бледно-охристыми пятнами. На наружных опахалах проксимальных второстепенных маховых, третьестепенных маховых и верхних кроюших крыла находятся светло-охристые каемки и вершинные пятна. Оперение нижней части тела белое с темнобурым чешуйчатым рисунком (кроме нижних кроюших хвоста).
Первый послегнездовой наряд идентичен наряду взрослой птицы, но на третьестепенных маховых и больших верхних кроюших второстепенных маховых сохраняются бледно-охристые каемки и вершинные пятна.
Размеры отдельных частей тела приморской пестрогрудой мухоловки приведены в табл.13.
Масса тела, г: взрослые самцы (п = = 10) (Нижнее Приамурье) — 14,7—21,2 (Кистяковский, Смогоржевский, 1973); взрослый истощенный самец (8.VI. Лазовский заповедник) — 13,75 (Литвиненко, Шибаев, 1971); взрослые самки (п = 10) (Нижнее Приамурье) - 19,0—22,3 (Кистяковский, Смогоржевский, 1973).
Распространение. Весь ареал вида, кроме Камчатки и Курильских о-вов. Детально распространение приморской пестрогрудой мухоловки на территории СССР показано на карте (рис. 28).
Кадастр к рис. 28.
1 - долина рЛена в 35 км севернее г.Якутск (Шубникова, Морозов, 1959; Воробьев, 1963; Нейфельдт, 1971; Степанян, 1978; колл. ЗМ МГУ); 2 - р-н р.Ола, р.Сиглан (Портенко, 1960; Иванов, 1976); 3 - побережье Охотского моря (Воробьев, 1954; Гизенко, 1955; Степанян, 1978); 4 - Шантарские о-ва (Яхонтов, 1977); 5 - 50 км от устья р.Гилюй; берег Зейского водохранилища в 10 км ниже устья р.Гилюй (сообщ. В.Ю.Илья-шенко); 6 - р.Зея (Степанян, 1978; сообщ. С.М.Смиренского); 7 - долина р.Ушумун (Воробьев, 1963; Иванов, 1976; колл. ЗИН); 8 -южная часть Амуро-Зейского междуречья (Нейфельдт, 1971); Свободненский р-н, окр. с.Кли-моуцы; междуречье Малой Перы и Большого Эр-геля (колл. ЗИН) ; 9 - р.Селемджа, окр. с.Ново-российка (колл. КГУ); 10 - окр. с.Дагмара (колл. КГУ); 11 - окр. пгт Экимчан (колл. В.ВЛео-иовича); 12 - окр. с.Тальники (Кистяковский, Смогоржевский, 1973; колл. КГУ); 13 - окр. с-Дуди (колл. КГУ); 14 - окр. с.Калиновка
144
(колл. КГУ); 15 - окр. с.Кисилевка (колл. КГУ); 16 - окр. с.Манома, р.Манома (Нечаев, 1963; колл. БПиДВНЦ); 17 - окр. оз .Петропавловское, Свечинская коса (Нечаев,'1963); 18 - темно-хвойно-широколиственные леса нижнего течения р.Хор (Кисленко, 19656); 19 - Красноармейский р-н (колл. КБ МГУ); 20 - р.Наице в бассейне верхнего течения р.Большая Уссурка (Флинт и др., 1959; колл. ЗИН); 21 - p-и истоков р.Большая Уссурка, бассейн Красной Речки (Назаренко, 1971 г.; колл. ЗИН); 22 - восточные склоны хр.Сихотэ-Алинь (Shulpin, 1931); Сихотэ-Алин-ский заповедник (Елсуков, 1982); верховья р.Джигитовка (Хантахеза) (картотека заповедника) ; 23 - верховья р.Черемуховая (Большая Синанча) (Назаренко, 1971 г.; колл. ЗИН); 24 - заповедник ’’Кедровая падь”, среднее течение
р.Кедровая (Назаренко, 1971 г.; Нейфельдт, 1971; колл. БПиДВНЦ); 25 - восточные склоны Южного Сихотэ-Алиня, долина р.Маргаритовка (Пфусунг) (Шульпин, 1927а); окр. с.Щербаковка (колл. ЗИН; ИЗАНУ); 26 - р-н истоков р.Уссури (Улахе, Ян-Муть-Хоуза) (Назаренко, 1971 г.); ур.”Мута” (Назаренко, 19846); водораздел между р.Милоградовка (Ванчин) и р.Уссури (Ян-Муть-Хоуза) (Назаренко, 1971 г.); 27 - бассейн р.Милоградовка (Назаренко, 1971 г); 28 - Лазовский заповедник имЛ.Г.Капланова, долина р.Киевка (Судзухэ); окр. Звездочки (Литвиненко, Шибаев, 1971; колл. ЗИН); 29 - о-в Сахалин, окр. пос.Гастелло (Yamashina, 1928).
Места обитания. Населяет различные типы хвойных (главным образом
Рис. 28. Распространение пестрогрудой мухоловки (Muscicapa griseisticta) на территории СССР:
1 - M.g.griseisticta, 2 - M.g.pallens; а - гнездитс’я, б - отмечена в гнездовой период. См. кадастр, стр. 144, 149.
145
лиственничных), смешанных хвойно-лиственных и как исключение лиственных лесов, а также старые гари и вырубки по долинам рек, склонам сопок (в том числе и круто спускающимся в широкие ложбины) и вершинам возвышенных плато, поднимаясь вверх до 700 м н.у.м.
В Якутии на р.Лена (35 км севернее г.Якутск) встречена в чистых лиственничниках как с большим количеством сухостоя без подлеска, так и в толокнянковых с обильным подлеском из березы и ольхи (Шубникова, Морозов, 1959).
В восточной части Амурской обл. у пгт Экимчан гнездится также в лиственничной тайге с примесью березы (колл. В.В.Леоновича). На юге Амуро-Зейского междуречья поселяется на выровненных поверхностях возвышенных плато и по крутым склонам сопок в сильно нарушенных рубками и пожарами дубово-лиственничных лесах с подлеском из леспедецы, рододендрона и спиреи (Нейфельдт, 1971). На о-вах р.Амур отмечена в сосновых лесах (Портенко, 1960), В Хабаровском крае в нижнем течении р.Хор (кор. пос. Кия) населяет темнохвойно-широколиственные леса (Кисленко, 19656), а у оз.Петропавловское в конце августа 1959 г. (очевидно, уже на пролете) встречена в леспедецевой дубраве с ивой Гмелина (Нечаев, 1963).
В Южном Приморье, по данным А.А.Назаренко (1971 г.), этот вид связан главным образом с различными типами лиственничных лесов. Так, у истоков р.Улахе пестрогрудая мухоловка с высокой плотностью населяет лиственничные редколесья на высоте 600 м н.у.м., где гнездится в высокоствольных участках, на марях и старых лиственничных гарях. Вместе с тем этот вид найден здесь и в березово-елово-осиновом редколесье, а также в приречной полосе долинных хвойно-широколиственных лесов. В верхнем течении р.Большая Уссурка (600—700 м н.у.м.) обитает исключительно в лиственничниках, на старых гарях и вырубках, возникших на месте этих лесов. Пихтово-еловые, еловые, хвойно-широколиственные и березовые леса в этом р-не пестрогрудая мухоловка избегает. В южной части гнездового ареала в заповеднике ’’Кедровая падь” гнездится в разреженной опушечной части высокоствольного долинного широколиственного леса с примесью одиночных высоких пихт и с густым подлеском (Нейфельдт, 1971), а на р.Марга-ритовка (Пфусунг) как исключение — в дубовых лесах (Щульпин, 1927а).
Численность. На территории СССР в границах гнездового ареала повсеместно невысока. Наибольшая плотность гнездования — в различных типах лиственничных лесов, отвечающих всем требованиям данного вида.
Прилет и весенний пролет на территории СССР. Проходит с I декады мая по I декаду июня включительно. Во время миграции летят поодиночке и стайками по 3—4 особи, часто образуя на местах отдыха и кормежки скопления до 25 экз.
На островах в северо-западной части залива Петра Великого пролет начинается 5-12.V, а наиболее интенсивно протекает во II половине этого месяца (Лабзюк и др., 1971). На о-ве Большой Пелис пролетная самка добыта 9.V, а на о.Аскольд - 30.V (Назаренко, 1971 г.).
В Южном Приморье прилет и пролет проходит с I декады мая по I декаду июня включительно. В 1960 г. отмечен 6—20.V (Панов, 1973). Пролетные птицы добывались здесь также: в окр. ст.Хасан — 12.V 1964 г., на р.Лянчихэ — 19.V 1931 г., у с.Новая Деревня - 20.V, на р.Нарва (Сидими) -27.V 1886 г. и 2.VI 1885 г., в заповеднике ’’Кедровая падь” — 27.V 1945 г., в окр. оз.Тальми — 30.V 1977 г., в окр. с.Крас-кино — 4.VI (Назаренко, 1971 г.; Панов, 1973; колл. ЗИН; БПиДВНЦ; наши данные) . . В Пограничном р-не у с.Барабаш-Левада пролетная особь добыта 12.V 1972 г. (колл. БПиДВНЦ), а на р.Средняя Синтуха 3 мухоловки встречены 30.V (Назаренко, 1971 г.). В окр. оз.Ханка птицы добывались: 13 и 24 .V 1911 — в верхнем течении р.Одарка у с.Нововладимировка, 29, 30.V 1927 г. и 26.V 1928 г. — у с.Сиваковка, 27.V 1908 г. — в окр. с.Камень-Рыболов (Черский, 1915а; 19156; колл. ЗИН). В нижнем и среднем течении р.Большая Уссурка мухоловки добывались 27.V и 1.VI 1955 г. и 1954 г. (Спангенберг, 1965; Назаренко, 1971 г.).
На нижнем Амуре начало весеннего пролета (несколько лет) отмечено: в с.Вятское — 14.V, в окр.г.Комсомольск — 29.V, в с.Киселевка — 3.VI (Кистяковский, Смогоржевский, 1973; колл. КГУ) и в окр. с.Мано-ма 31.V (Нечаев, 1963). На р.Амур в окр. с.Радде (Малый Хинган) пролетный самец добыт 27.V 1928 г. (колл. ЗИН), а у с.Кли-моуцы в 1959 г. первое появление отмечено 28.V (Нейфельдт, 1971). 22.V 1914 г. найдена В.ЧДорогостайским в окр. ур.Бомнак нар.Зея (Воробьев, 1963).
146
На Сахалине пролетная птица встречена в окр. г.Корсаков 20.V 1947 г. (Гизенко, 1955), а также добывалась на юге о-ва с 20.V по 3.VI (Иванов, 1976). На о-в Мо-нерон 2 птицы зарегистрированы 4.VI 1973 г. (Нечаев, 1975).
Размножение. А.А.Назаренко (1971 г.) на основании находок ранних кладок считает, что у пестрогрудых мухоловок большинства популяций пары образуются еще на зимовках или на пролете и, прилетев на места гнездования, птицы сразу приступают к размножению. На наш взгляд, эти сведения нуждаются в проверке. Против них свидетельствует и такой факт, что в период весенней миграции мухоловки летят чаще всего поодиночке или небольшими стайками.
Песня самца очень похожа на таковую сибирской и ширококлювой мухоловок.
Гнезда птицы строят открыто в развилках и на боковых ветвях деревьев, как вплотную к стволу, так и на некотором удалении (1-2 м) от него. В редких случаях гнезда помещаются в пучках лишайника (колл. В.В.Леоновича) или на старых гнездах других птиц (большая горлица) (Нейфельдт, 1971). Над гнездами, построенными на лиственницах, имеется обычно редкий навес из хвойных веточек, который маскирует их сверху и защищает'в некоторой степени от прямых лучей солнца. Из 13 гнезд приморской пестрогрудой мухоловки, найденных в Приморском крае и Амурской обл. (Назаренко, 1971 г; Нейфельдт, 1971; колл. В.В.Леоновича), 10 были построены на лиственницах и по одному на пихте, дубе и березе на высоте 1,8-15,0 м от поверхности земли (в среднем - 8,0). В строительстве гнезда участвуют оба партнера пары.
Гнезда представляют собой очень аккуратные, плотные, чашевидные постройки, диаметр нижней части которых в большинстве случаев превышает таковой верхней. В качестве строительных материалов птицы используют талломы лишайников (Ever-nia mesomorpha; Usnea longissima; U.an-nulata; Ramalina sp.; Bryopogon sp.) (Назаренко, 1971 г; наши данные), значительно реже также веточки лиственницы, голубики и леспедецы, сухие стебли травянистых растений, лубяные волокна, пучочки мхов и сухую хвою (сосна, кедр, лиственница). Снаружи стенки гнезда облицовываются кусочками талломов лишайников, часто растущих по-соседству на ветвях деревьев, что делает гнездо хорошо замаскирован
ным. Лоток выстилается главным образом сухими хвоинками лиственницы и значительно реже тонкими корешками и усиками вьющихся трав (Назаренко, 1971 г; Нейфельдт, 1971; наши данные). Размеры гнезд (п = 11), мм: нижний диаметр 110-120; верхний диаметр — 60-94; высота 54; диаметр лотка 50—58, глубина лотка 28—40.
На территории СССР в сезон размножения приморская пестрогрудая мухоловка имеет один репродуктивный цикл. Количество яиц в полной кладке 3—5 (3 — 12,5 %, 4 - 75,0, 5 - 12,5 %; п = 8) (Назаренко, 1971 г; Нейфельдт, 1971; наши данные). Яйца остроовальные, реже - укороченно-остроовальные. Фон скорлупы голубовато-зеленовато-бирюзовый. По нему равномерно густо распределены размытые мелкие ржавчато-охристо-бурые крапинки. Кроме описанного типа окраски встречаются яйца (Назаренко, 1971 г), на которых по бледно-серому, со слабым голубоватым или зеленоватым оттенком фону, разбросаны мельчайшие крапинки табачно-бурого цвета. Расположение рисунка на поверхности скорлупы этих яиц не одинаково. Крапинки или равномерно покрывают всю поверхность, или образуют сгущения в виде ’’венчиков” вокруг тупого или острого полюсов. Размеры и индекс удлиненности яиц приведены в табл. 4.
После откладки последнего яйца самка приступает к насиживанию. Длительность инкубации 13 дней (Нейфельдт, 1971). Самец, по-видимому, не участвует в насиживании кладки, но иногда подкармливает самку на гнезде (Назаренко, 1971 г).
Пуховой птенец пестрогрудой мухоловки имеет такую же топографию опущения и окраску тела и пуха, как и птенец сибирской мухоловки. В выкармливании птенцов участвуют оба партнера пары. После того как птенцы покинут гнездо, родители первое время продолжают кормить слетков, а несколько дней спустя выводки начинают кочевать. При появлении птенцов и особенно перед их вылетом взрослые птицы совершенно утрачивают территориальность (Назаренко, 1971 г).
Сезон размножения приморской пестрогрудой мухоловки на территории СССР длится с конца II декады мая по III декаду июля включительно.
На юге Приморья в заповеднике ’’Кедровая падь” активное пение самцов на своих гнездовых участках в V966 г. отмечено 15.V, а 24.V найдено почти готовое гнездо, самка в котором в начале июня начала наси-
147
живать полную кладку (Нейфельдт, 1971). У истоков р.Большая Уссурка самец с увеличенными семенниками добыт на гнездовом участке 1.VI 1966 г. (колл. ЗИН). В р-не истоков р.Улахе 4 гнезда с полными кладками средней насиженности найдены 8, 10 и 12.VI 1967 г., a 12.VI было обнаружено еше 1 гнездо с 3 сильно насиженными яйцами (Назаренко, 1971 г); Массовый вылет птенцов в 1964 г. проходил 30.VI—4.VII, а в 1969 г. с 4 по 11 .VII первый слеток был зарегистрирован только 10.VII. В гнездах, осмотренных в этот период, птенцы находились на следующих этапах развития: в гнезде от 7.VII - еше слепые, покрытые пеньками перьев; в гнезде от 8.VII — начали покрываться перьями, глаза почти открылись; в гнезде от 9.VII — полностью оперились, реакция на наблюдателя — затаивание. При попытке осмотреть гнездо от 11 .VII птенцы из него вылетели (Назаренко, 1971г). На р.Маргаритовка Л.М.Шульпин (колл. ЗИН) добыл полностью доросшую молодую птицу 4.VII, а 5.VII — почти доросшую. Чуть позже — 9.VII — им был добыт слеток (колл. ЗИН). У с.Щербаковка слеток добыт также 11 .VII 1927 г. (колл. ИЗАНУ). В Сихотэ-Алин-ском заповеднике выводок из 6 молодых встречен в верховьях р.Джигитовка 1 .VIII 1970 г', (картотека заповедника).
На нижнем Амуре в окр. с.Манома самцы, гоняющиеся за самками и друг за другом, отмечены 31.V 1958 г.,a 1.V добыт самец с созревшими семенниками (длина 5,5 мм) (Нечаев, 1963). В окр. с.Тальники самка, закончившая откладку яиц, добыта 20.VI (Кистяковский, Смогоржевский, 1973). В Амурской обл. на Амуро-Зейском плато 1.VI 1962 г. найдено свежепостроен-ное гнездо, в котором 6.VI после откладки 4-го яйца, самка приступила к насиживанию; 5.VI обнаружено строящееся гнездо, 7.VI птицы возвели стенки гнезда и начали вымащивать лоток, 11.VI самка отложила 1-е, a 14.VI - 4-е яйцо и начала насиживать кладку. Все птенцы вылупились 26.VI, а покинули гнездо 9.VII (Нейфельдт, 1971; колл. ЗИН). В 1958 г. самые ранние короткохвостые слетки встречены 7.VII, а молодые из других выводков 27 .VII и 31.VII по размерам и летным способностям не отличались от родителей (Нейфельдт, 1971; колл. ЗИН). В окр. пгт Экимчан гнездо с полной кладкой из 4 яиц семидневной насиженности найдено 20.VI 1971 г. (колл. В.В.Леоновича). У с.Натальино молодая птица, на 2/3 сменившая гнездовой наряд, добыта 29.VIII 1971 г. (колл. КБ МГУ), 148
a 28.VIII 1959 г. в Хабаровском крае у оз.Петропавловское встречен выводок из 5—7 мухоловок (Нечаев, 1963).
В Якутии (35 км севернее г.Якутск) 18,VII добыта взрослая самка и слеток из выводка, a 6.VI1I — здесь был встречен еше 1 выводок молодых (Шубникова, Морозов, 1959).
Линька. После того как маховь!е и рулевые окончательно закончат рост, у молодых начинается послегнездовая линька, которая охватывает почти все оперение кроме маховых и рулевых. На местах зимовок у первогодков проходит повторная, но уже полная линька оперения и на места гнездования они прилетают в свежем пере (Виноградова и др., 1976). Взрослые птицы имеют в году одну полную послебрачную линьку, которая проходит на местах гнездования с июля. По данным И.А.Нейфельдт (1971), первостепенные маховые сменяются строго центробежно от 10-го к 1-му, а рулевые от центральных к крайним (на каждой половине хвоста).
Питание. Основу пиши составляют насекомые разных групп, реже поедает пауков и как исключение мелкие сочные плоды некоторых растений. Во время кормления старается держаться на внешних частях древесных крон, чаше всего на вершинах деревьев (в особенности сухих). Схватывает главным образом летающих насекомых среди деревьев и высоко над лесом. В кронах деревьев охотится редко. Кроме этого (обычно в дождливую погоду и на пролете) склевывает беспозвоночных также с древесных, кустарниковых и травянистых растений и даже поверхности земли (Назаренко, 1971 г; Нейфельдт, 1971; наши данные)
Отлет и осенний пролет на территории СССР. Проходит с конца II декады августа до середины II декады октября.
В Приморье самые ранние пролетные птицы добыты 16.VIII 1911 г. у южного берега оз.Ханка в окр. с.Нововладимиров-ка (Черский, 1915а; 19156). В Партизанском р-не 2 пролетные особи добыты 28 и 29.VIII (колл. ЗИН), а в Дальнегорском р-не (нижнее течение р.Рудная) - 20 и 25.IX 1909 г. (Черский, 1915а). В Лазовском заповеднике у устья р.Тачингоуза 2 особи, неверно определенные Л.О.Белопольским как M.sibirica, были добыты 23.VIII 1944 г. (Белопольский, 1950; Воробьев, 1963), а 26.VIII1960 г. 2 пролетных птицы наблюдались на морском побережье у вершины ropbi Туманная (Литвиненко, Ши-
баев, 1971). В Хасанском р-не пролетные мухоловки были отмечены и добыты: 23.VIII — у с.Новая Деревня (Назаренко, 1971 г.); 30 и 31.VIII 1960 г. — в заповеднике ’’Кедровая падь” (Назаренко, 1971 г; Панов, 1973); 7.IX 1926 г. на п-ове Новгородский у залива Посьета (колл. ЗИН); 10.IX 1968 г. — в низовьях р.Кедровая (колл. БПиДВНЦ); 19 и 20.IX 1968 г. - в окр. ст .Приморская (колл. БПиДВНЦ). Кроме этого одиночные птицы в 1960 г. зарегистрированы в Южном Приморье 1, 6, 12 и 22.IX (Панов, 1973).
На островах в северо-западной части залива Петра Великого осенняя миграция протекает с середины августа до первых чисел октября. Максимум пролетав первой половине сентября (Лабзюк и др„ 1971). На о-в Большой Пелис пестрогрудые мухоловки добывались 12.1Х 1964 г. (колл. БПиДВНЦ) и 21.IX (Назаренко, 1971 г).
На Сахалине в окр. пос .Ново-Александровск одиночные пролетные птицы, задержавшиеся с отлетом, отмечены в сильный снегопад 12 и 13.Х 1968 г. (Нечаев, 1974а).
Практическое значение. Уничтожает многих насекомых - вредителей леса и переносчиков возбудителей ряда заболеваний человека и животных. Заслуживает повсеместной всесторонней охраны. Камчатская пестрогрудая мухоловка — Muscicapa griseisticta pa I lens (Smejneger. 1887)
Hemichelidon pollens Smejneger, 1887, Proc. U.StNat. Mus.. 10: 143, ”o.Беринга”-ошибочно; Камчатка.
Описание. Составлено по 9 экз.
с Камчатки.
У взрослой птицы общий тон окраски оперения более светлый, чем у номинативного подвида. У большинства особей пестрины на нижней части тела более размыты и покрывают только бока груди и тела, а на зобе образуют подобие узкого ’’ожерелья”. Все остальное оперение низа белое. Изредка встречаются экземпляры с более густым расположением пестрин, полностью покрывающих весь участок от зоба до верхней части брюшка включительно.
Размеры отдельных частей тела камчатской пестрогрудой мухоловки приведены в табл. 13.
Распространение. Камчатка и предположительно некоторые Курильские о-ва (рис. 28).
Кадастр к рис. 28.
1 - п-ов Камчатка к северу до 56-57° с.ш. (Степанян, 1978); с.Ключи (Портенко, 1960; колл. ЗИН); 2 - Тигильский р-н, близ мыса Бабушки (Остапенко и др., 1977); 3 - окр. г.Ко-
зыревск (Вронский, 1977; колл. ЗИН); 4 - окр. пос.Атласово (колл. КБ МГУ); 5 - окр. с.Пущи-но (Вронский, 1977; колл. КБ МГУ); 6 - Восточная Камчатка (Аверин, 1948); Кроноцкий заповедник, р-н Горячего Ключа (Лобков, 1977; 1978); 7 - Семлячикское лесничество Кроноц-кого заповедника (Лобков, 1977); 8 - окр. г.Петропавловск-Камчатский (колл. ЗИН); 9 -о-в Парамушир (Yamashina, 1931); 10 - о-в Итуруп (Yamashina, 1931).
Места обитания. В отличие от номинативного подвида населяет главным образом различные типы лиственных и смешанных хвойно-лиственных лесов на равнинах, а также в горных долинах и ушельях рек.
В верхнем течении р.Камчатка (Вронский, 1977) гнездится в долинных, многоярусных пойменных лесах из душистого тополя, чозении, ивы, ольхи, с подлеском из ивы и бузины, тяготея к среднему ярусу этого леса. Кроме этого поселяется на участках, где пойменный лес чередуется с большими полянами и в равнинных каменноберезовых лесах с обилием небольших полян и хорошо развитым подлеском из рябины, розы, можжевельника и спиреи. В центральной части долины этой реки населяет вторичные смешанные леса из лиственницы, белой березы и ольхи с подростом из тех же видов и хорошо развитым подлеском из жимолости, можжевельника, спиреи и розы. В горных долинах и ущельях рек Кунч и Яковская отмечена в березняках на высоте 500 м н.у.м. (Вронский, 1977), а в Кро-ноцком заповеднике обычна на гнездовании в каменноберезовых лесах приморской зоны (Лобков, 1977).
Численность. Обычна, местами многочисленна.
Прилет и весенний пролет на территории СССР. На Камчатку прилетает в I декаде июня.
В Кроноцком заповеднике появляется в первых числах июня (Лобков, 1978). В окр. пос.Жупаново в 1972 г. первые мухоловки отмечены 4.VI (Лобков, 1977). 6.VI 1909 г. птицы добыта у с.Ключи (колл. ЗИН). Одиночная птица этого подвида 27.V 1927 г. добыта в Приморье у южного берега оз .Ханка в окр. с.Сиваковка (колл. ЗИН).
Размножение. Единственное гнездо, описанное Е.Г.Лобковым (1977), было построено на березе в развилке толстой ветви в 2 м от ствола дерева на высоте 4,0 м от поверхности земли. Постройка была сделана из сухих стебелькоЬ и спорогониев мха, а снаружи облицована белыми перьями куропатки, которые заметно его демаскировали. Лоток был выстлан сте
149
бельками и спорогониями мха. Размеры, мм: диаметр — 97, высота — 48; диаметр лотка - 52; глубина лотка - 35. 19.VI 1972 г. в гнезде было 2 желтовато-зеленоватых яйца со слабо размытыми коричневыми пятнышками. Впоследствии кладка погибла.
Сезон размножения этого подвида на Камчатке длится со II декады июня по конец августа. У мыса Бабушки самка с увеличенным яичником добыта 10.VI 1974,г. (Остапенко и др., 1977). Южнее Горячих Юпочей 27 .VI наблюдалась пара, строивШая гнездо, а в 18 км севернее с.Жупаново 17.VI 1972 г. найдено уже готовое гнездо. 19.VI в нем было 2 яйца, а при осмотре его 21.VI яиц в гнезде не оказалось (Лобков, 1977). Слетки появляются с начала I декады августа, в III декаде этого месяца их численность достигает максимума (Лобков, 1977).
Линька. Проходит так же, как и у номинативного подвида.
Питание. Не изучено. В 2 желудках пролетных еамцов, добытых 2.IX 1948 г. на о-ве Кунашир, обнаружены: цикадки (Sinophora submacula), комар и семена плодов бузины Микеля (Нечаев, 1969).
Отлет и осенний пролет на территории СССР. Покидает места гнездования на Камчатке в I декаде сентября.
В Кроноцком заповеднике отлетает в начале сентября (Лобков, 1978). В окр. пос.Жупаново в 1972 г. последние птицы отмечены 2.IX (Лобков, 1977). Нао-ве Кунашир пролетные птицы добыты 2.IX 1948 г. (Нечаев, 1969) и 11.IX 1974 г. (Остапенко и др., 1977). Пролетные мухоловки, добытые С.И.Снигиревским (Воробьев, 1963) 31.VIII и 17.IX 1935 г. в р-не Аяна на Охотском побережье, очевидно, принадлежат также к этому подвиду.
Практическое значение. Такое же, как и у номинативного подвида. Ширококлювая мухоловка — Muscicapa latirostris Raffles, 1822
Muscicapa latirostris Raffles, 1822, Trans. Linn. Soc. London, 13: 312, Суматра.
Описание. Составлено по 158 экз. (150 — из Южной Сибири и Дальнего Востока СССР и 8 — с о-вов Кунашир, Шикотан и Итуруп).
Вершину крыла образуют 3-е и 4-е первостепенные маховые (3 > 4 — 63,5 %, 3=4- 33,9, 3 < 4 - 2,6 %); 5-е маховое немного их короче (2 < 5 — 94,6 %, 2 = 5 — 5,4 %); 2-е всегда больше 6-го. 1-е перво-150
степенное маховое очень мало и лишь незначительно выступает за вершины кроющих кисти (табл. 14). У большинства особей наружные опахала 3—5-го первостепенных маховых в вершинной части заметно сужены. Количество шетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3-5 (3 — 0,9 %; 4 - 77,9; 5 - 21,2 %). Длина наибольшей (п = 66) 5,3—8,3 мм (в среднем — 7,0). Количество щитков подотеки на передней части цевки варьирует от 4 до 5. На вершине хвоста (в центре) имеется небольшая выемка. Самец и самка окрашены одинаково.
Распространение. Восточная и Южная Азия. Ареал разобщен. СССР: Южная Сибирь (к западу до правых притоков верхнего течения р.Обь), Дальний Восток (к югу от Якутии), о-ва Сахалин и южные Курильские (рис. 29). Северная Монголия, Северо-Восточный Китай и северная часть Корейского п-ова. Южная Азия к западу в Гималаях до 76° в.д., на п-ове Индостан до 75° в.д. К востоку до 110° в.д. К северу до южного склона Гималаев, Ассама, восточнее Сино-Тибетских гор к северу до’ 32° с.ш. (Степанян, 1978). О-ва Хоккайдо, Хонсю, Сикоку, Кюсю, Цусима, Лусон и Негрос. Зимует в Южной и Юго-Восточной Азии. Отмечен залет в Англию.
Систематика. Географическая изменчивость проявляется в варьировании интенсивности окраски оперения, в размерах отдельных частей тела, в форме клюва и формуле крыла. Вид образует 2 подвида, 1 из которых - номинативный - встречается и гнездится на территории СССР. Номинативная форма населяет почти весь ареал вида, кроме Филиппинских о-вов, на которых обитает форма M.l.randi (Amadon et Pont, 1970).
Палеарктическая ширококлювая мухоловка — Muscicapa latirostris latirostris Raffles, 1822
Muscicapa latirostris Raffles, 1822, Trans. Linn. Soc. London, 13: 312, Суматра.
Описание. Составлено по 158 экз. (150 — из Южной Сибири и Дальнего Востока СССР и 8 — с о-вов Кунашир, Шикотан и Итуруп).
У взрослой птицы все оперение верхней части тела (от лба до верхних кроющих хвоста включительно), а также на боках головы и шеи светло-серовато-бурое. Верхняя часть головы более темная, чем спина, и в свежем пере имеет чешуйчатый рисунок, образованный двухцветными перьями (темные в центре и более светлые по краю опахал). Грудь и бока тела бледно-серовато
бурые. Уздечка, горло, кольцо вокруг глаза, брюшко и нижние кроющие хвоста белые с бледным буровато-серым оттенком. Нижние кроюшие крыла от белого до бледно-серовато-бурого. Рулевые, маховые и верхние кроющце крыла темно-бурые. По краю наружных опахал второстепенных маховых, а на третьестепенных маховых и больших верхних кроющих второстепенных маховых и на вершинах расположены узкие светло-буровато-охристые каемки. Надклювье темно-коричнево-бурое. Подклювье желтовато-охристое в основании и темно-коричнево-бурое в вершинной части. Ноги темно-бурые. Радужина глаза коричневая. Сезонная изменчивость проявляется в общем посветлении окраски оперения, частичном исчезновении чешуйчатого рисунка на верхней части головы, а также в исчезновении светлых каемок на маховых и кроющих крыла в результате их обноса и выцветания.
У молодых в гнездовом наряде оперение на верхней части тела и боках головы темно-коричнево-бурое с округлыми светло-охристыми пятнами на перьях. Горло зоб и бока груди бледно-охристо-буроватые с бурым чешуйчатым рисунком. Центральная часть груди, брюшко и нижние кроющие хвоста белые. Маховые и рулевые темно-бурые. Заостренные вершины последних изредка с примесью охристого оттенка. По краю наружных опахал и на вершинах второстепенных и третьестепенных маховых, а также больших и средних верхних кроющих второстепенных маховых расположены охристые каемки разной степени выраженности.
Первый послегнездовой наряд идентичен наряду взрослой птицы, но на больших верхних кроющих второстепенных маховых, третьестепенных маховых и некоторых верхних кроющих хвоста (не перелинявшие партии перьев) сохраняются охристые окаймления и пятна.
Размеры отдельных частей тела ширококлювой мухоловки приведены в табл. 14.
Масса тела, г: слетки (п = 8) (28— 30.VI, окр. Уссурийского заповедника) — 9^60—10,95, в среднем - 10,47; молодые самцы (п = 2) (южные Курильские о-ва) — 10,80—12,00, в среднем — 11,40 (Нечаев, 1969); взрослые самцы (п = 2) (там же) -10,80—11,80, в среднем - 11,30 (Нечаев, 1969); взрослые самцы (п = 5) (28 .VI-1 .VII, окр. Уссурийского заповедника) — 10,20-12,00, в среднем - 11,09; взрослые самцы (п = 17) (Нижнее Приамурье) -9,00-12,10, в среднем - 10,90 (Кистяков
ский, Смогоржевский, 1973); самки (п = = 3) (29 и 30.VI, окр. Уссурийского заповедника) — 10,57-10,89, в среднем -10,76; самка (южные Курильские о-ва) -11,20 (Нечаев, 1969).
Распространение. Весь ареал вида за исключением Филиппинских о-вов. Указание Н.А.Зарудного (Дементьев, 1937) на гнездование ширококлювой мухоловки в Южной Фергане у р.Нурлоу является, очевидно, ошибочным. Это сообщение не подтверждено коллекционными материалами. Не найден здесь этот вид на гнездовании и последующими исследователями. Детально распространение ширококлювой мухоловки на территории СССР показано на карте (рис. 29).
Кадастр к рис. 29.
1 - р.Тым, окр. с.Напас (Москвитин, 1973);
2 - р.Парабель, окр. с.Щука (Миловидов, Москвитин, 1973); 3 - бассейн р.Кеть (Гынгазов, Миловидов, 1977); 4 — окр. г.Томск (Москвитин, 1972; 1973); 5 - Шегарский р-н, окр. сЛоздня-ково на р.Обь (Гынгазов, Миловидов, 1977; колл. ЗИН); 6 - среднее течение р.Четь (Москвитин, 1972; 1973); окр. с.Четь-Конторка (Москвитин, 1973; Гынгазов, Миловидов, 1977); 7 -среднее течение р.Тонгул (Москвитин, 1972; 1973); 8 - бассейн р.Чулым (Гынгазов, Миловидов, 1977); 9 - окр. г.Енисейск (Bianchi, 1907; Сушкин, 1914); 10 - окр. г.Красноярск (Bianchi, 1907; Юдин, 1952); р.Собакина (Юдин, 1952); р.Бугач; гора Абатак близ с.Торгашино (колл. ЗИН); 11 - окр. с.Частоостровское (колл. ЗИН); 12 - р.Базаиха (колл. ЗИН); 13 - ручей Муртук, впадающий в р.Колбу (Портенко, 1950; колл. ЗИН); 14 - р.Мана, с.Выезжий Лог (Портенко, 1950; колл. ЗИН); 15 - р.Можарка (Сушкин, 1914; колл. ЗИН); окр. с.Бараксан (Сушкин, 1914; колл. ЗИН); 16 - окр. пос.Майна (колл. КБ МГУ); долина Уя (колл. С.Д.Куста-новича); 17 - хр.Танну-Ола (Степанян, 1978); 18 - Саяны (Bianchi, 1907); 19 - р.Бирюса (Благосклонов, 1954; колл. ЗИН); окр. с.Би-рюса (Bianchi, 1907; Юдин, 1952; колл. ЗИН); 20 - р.Чадобец (Реймерс, 1966); 21 - нижнее течение р.Ангара (Портенко, 1960); 22 - окр. с.Богучаны на р.Ангара (Иванов, 1976; колл. ЗИН); 23 - среднее течение р.Ангара (Реймерс, 1966); 24 - Тулунский р-н, окр. с.Гадалей (Сонин, Липин, 1969; колл. ЗМ МГУ); 25 - Тун-кинские Гольцы (колл. ЗИН; ХГУ); 26 - г.Ир-кутск и его окрестности (Bianchi, 1907; Сушкин, 1914; Пшеничников, 1959); 27 - нижнее течение р.Голоустная (Богородский, 1979); 28 -долина р.Большая Олха (Богородский, 1979); 29 - долина р.Иркут (Гагина, 1958; 1961); 30 -побережье Южного Байкала (Гагина, 1958); 31 - хр.Хамар-Дабан (Степанян, 1978; колл. ЗИН); 32 - Юго-Западное Забайкалье (Измайлов, Боровицкая, 1973); р-н оз.Гусиное (колл. ЗИН); окр. оз.Щучье (Измайлов, Боровицкая, 1973);
33 - долина р.Селенга (Гагина, 1958); 34 - п-ов Святой Нос (Гагина, 1960); 35 - долина р.Баргузин (Гагина, 1960; Лямкин, 1979); 36 - Баргузинский заповедник; Баргузинский хр. (Бе
151
ляев, 1982); устье р.Давша (Скрябин, Филонов, 1962); 37 - Верхне Ангарский участок (Гагина, 1961); 38 -• Западно-Прибайкальский участок (Гагина, 1961); 39 - Лено-Киренгский участок (Гагина, 1961); 40 - верхнее течение р.Лена (Гагина, 1958; Реймерс, 1966); окр. с.Жигалово (Реймерс, 1966) ; 41 - р.Тутура (Реймерс, 1966) ;
42 - Витимское плоскогорье, оз.Баунт (Иванов, 1976; колл, ЗИН); 43 - рощи Еравны (Измайлов, 1967); 44 - р.Чикой (колл. ЗИН); Красно-Чикойский р-н, окр. с.Большая Речка (колл. ИЗАНУ); 45 - верхнее течение р.Чикой, долины рек Ямаровка и Бабровая (Леонтьев, Павлов, 1963); 46 - р.Онон (Bianchi, 1907; Благосклонов, 1954); 47 - Читинский таежный р-н (Гагина, 1960а); окр. г.Чита (Шкатулова, 1965);
48 - с.Сретенск (Портенко, 1950; колл. ЗИН);
49 - окр. с.Цаган-Олуй (колл. ЗИН) ; 50 - Алек-сандрозаводской подтаежный р-н (Гагина, 1960а) ;
51 - Приаргунье (Гагина, 1960а); 52 - восточная часть хр.Тукурингра; 50 км выше устья по р.Гилюй (сообщ. В.Ю.Ильяшенко); 53 - окр. с.Бомнак (Иванов, 1976; колл. ЗИН); 54 — окр. пгт Экимчан (Назаренко, 1984а); 55 -
Таблица 14. Размеры (^^^7
окр. с.Усть-Норск, окр. пос.Дагмара (колл. КГУ); 56 - окр. с.Новороссийка (колл. КГУ);
57 - междуречье Малой Перы и Большого Эрге-ля (колл. ЗИН); окр. с.Климоуцы (колл. ЗИН; БПиДВНЦ); 58 - окр. с.Натальино (Спасский и др., 1962); 59 - устье р.Мальмальта (колл. КГУ); 60 - р.Бурея, устье р.Талая (колл. КГУ) ; с.Островское; устье р.Верхний Мельгин (Кистяковский, Смогоржевский, 1964); 61 - устье р.Тырма, устье р.Нижний Мельгин (колл. КГУ);
62 - окр. с.Чеугда (колл. КГУ); 63 - хр.Малый Хинган, долина р.Дичук (колл. ЗИН); ур. "Тигровая падь” (колл. КГУ); 64 - окр. с.Радде (колл. ЗИН); 65 - устье р.Сунгари (Bianchi, 1907; колл. ЗИН); 66 - р-н оз.Орель (Бабенко, 1984; колл. ИЗАНУ); 67 — устье р.Амур (Воробьев, 1938); окр. г.Николаевск-на-Амуре (Bianchi, 1907); 68 - с.Романовка (колл. КГУ);
69 - с .Тальники (Кистяковский, Смогоржевский, 1973; колл. КГУ); с.Новая Ферма (колл. КГУ); 70 — с.Богородское (Иванов, 1976; колл. ЗИН); 71 - окр. с.Дуди (колл. КГУ); с.Покров-ка (Кистяковский, Смогоржевский, 1973; колл. КГУ); 72 - оз.Кизи (Штильмарк, 1973; колл.
и коэффициенты вариации (CV, %) ширококлювой и рыжехвостой мухо
Вид, место сбора п Длина,
Крыло CV Расстояние, на которое 1-е маховое выступает за вершины кроющих кисти CV
Сам
M.latirostris
Среднее Приамурье 25 69.011 0.39 65,1 -73,0 2,74 2,3910,18 0,7 -3,8 37,03
Нижнее Приамурье 48 68,671 0,20 66,1 -71,8 2,07 2,6910,14 0,5 -5,0 36,43
Приморье 15 68,08± 0,50 64,8 -72,5 2,84 2,9910.20 1,8 -4,6 24,96
О-ва Кунашир и Шикотан 5 68,461 0,49 67,1 -70,1 1,62 3.92Ю.35 2,8 -4,6 19,71
M.ruficauda Памиро-Алай 13 77,241 0,47 74,2 -79,7 2,21 5,1410,34 4,0 -6,1 14,98
Сам
M.latirostris Среднее Приамурье 10 68.341 0,70 65,1 -73,0 3,09 2,8410,36 0,9 -4,2 39,71
Нижнее Приамурье 11 69,011 0,43 66,7 -71,0 2,09 2.6510,27 1,1 -3,9 33,70
Приморье . 7 68,351 0,56 66,1 -69,9 2,02 2,5310,43 1,1 -3,9 4130
О-в Шикотан 1 66,1 - 3,0 -
M.ruficauda Памиро-Алай и Индия (Кашмир) 6 73,Р.72 71,2 -76,2 2,40 5,0* -
152
ЗИН); р.Таба (колл. ЗИН); 73 - оз.Удыль (колл. ЗИН); 74 - с.Софийск (Штильмарк, 1973; колл. ЗИН; БПиДВНЦ); окр. с.Калинов-ка (колл. КГУ); 75 - с.Киселевка (Кистяков-ский, Смогоржевский,. 1973; колл. КГУ); 76 -с.Гячи (колл. КГУ); 77 - р.Горюн (Воробьев, 1938); 78 - Комсомольский заповедник (Штильмарк, 1973); окр. г.Комсомольск-на-Амуре (колл. КГУ); 79 - с.Болонь (Кистяковский, Смогоржевский, 1973; колл. КГУ); с.Омми, с .Верхний Нерген, с.Раздольное (колл. КГУ); 80 - г.Хабаровск (Черных, 1967); 81 - хр.Хехцир (Кулешова и др., 1965); южный склон хребта (Казаринов, 1973); 82 - окр. с.Кия в р-не нижнего течения р.Хор (Кисленко, 19656);
83 - 60 км от устья р.Ботчи вверх по реке (Емельянов, 1929); 84 - окр. устья р.Ботчи (Емельянов, 1929); 85 - Сихотэ-Алинский заповедник (Елсуков, 1982; наши данные); р.Серебрянка (Сида) (колл. БПиДВНЦ); окр. пос.Терней (колл. ИЗАНУ; наши данные); 86 - р.Рудная (Тетюхе) (колл. ЗИН); 87 - леса островов и сопок низовьев р.Большая Уссурка (Иман) (Спангенберг, 1940); 88 - среднее течение
р.Большая Уссурка между селениями Вахумбе и Санчихеза (Спангенберг, 1965); 89 - верхнее течение р.Большая Уссурка между р.Колумбе и р.Нанце (Спангенберг, 1965); 90 - бассейн истоков р.Большая Уссурка (Назаренко, 19846); верховья реки (Флинт и др., 1959) ; 91 - с.Михай-ловка (колл. БПиДВНЦ); 92 — р.Маргаритовка, окр. с.Щербаковка (колл. ЗИН; ИЗАНУ); 93 -бассейн истоков р.Уссури, ур. ’’Мута” (Назаренко, 19846); 94 - окр. с.Нововладимировка (Черский, 1915а; 19156); 95 — Спасский р-н, склоны хр.Синего (Кушнарев, 1984); 96 - южный берег оз.Ханка, окр. с.Сиваковка (колл. ЗИН; ИЗАНУ); 97 - окр. г.Уссурийск (колл. ЗМ МГУ); Горнотаежная станция ДВНЦ (Панов, 1973); 98 - Уссурийский заповедник им.В.Л.Ко-марова (Иванов, 1952; Коренкова, 1970; Назаренко, 19846; наши данные) ; окр. пос.Комарово-заповедное, пос.Каймановка (колл. ИЗАНУ); Егерский ключ (Иванов, 1952) ; р.Малая Каменка (Иванов, 1952); р.Комаровка (Супутинка) (Иванов, 1952; колл. ЗИН); 99 - верховья р.Суво-ровка (Пейшула) (колл. ЗИН); 100 - Лазовский заповедник им.Л.Г.Капланова (Белопольский,
ловок
мм
Хвост CV Цевка CV • Клюв от оперения лба CV Клюв от орального края ноздри CV
цы
47,47± 0,36 43,5 -50,2 3,67 12,971 0,09 12,1 -13,7 3,36 10,201 0,08 9,5 -ii,i 3,96 7,4610,06 7,0 -8,1 3,75
47.311 0.19 44,7 -50,1 2,72 12,901 0,05 11,9 -13,8 2,94 10,231 0,07 9,3 -1'1,4 4,78 7,35Ю,04 6,8 -8,0 4,19
46,051 0,23 44,i -4.7,Г 1,92 13,131 0,07 12,7 -13,6 2,20 10,291 0,15 9,3 -10,8 5,68 7,53Ю,10 6,7 - 8,0 0,05
47,001 0,57 46,1 -49,1 2,71 12,801 0,23 12,0 -13,3 4,02 9,781 0.14 9,5 -10,1 2,82 7,3310,06 7,2 -7,5 1,72
58,661 0,59 54,3 -61,3 3,66 15,791 0,16 14,8 -16,4 3,59 10,631 0,20 10,0 -12,1 6,86 8,4110,10 7,9 -8,9 4,19
ки
46,761 0,84 44,3 -52,0 5,10 13,071 0,13 12,2 -13,5 3,08 10.271 0.11 9,5 -10,6 3,30 7.5610,09 7,0 -7,9 3,57
47,701 0,54 44,2 -51,0 3,76 12.951 0.08 12,5 -13,4 2,07 10,201 0,12 9,6 -10,8 3,80 7,5510,08 7,0 -8,0 3,71
47,071 0,57 44,4 -4^Г 2,98 12,961 0,15 12,2 -13,4 3,05 10,341 0,19 9,7 -11,0 4,86 7,4010,04 7,3 -7,5 1,35
47,0 13,0 - - - -
55,351 0,41 53,6 -56,5 1,83 16,131 0.21 15,5 -16,8 3,20 10,431 0,15 9,9 -11,0 3,41 7,5 -8,5. 4,24
153
1950; Гизенко, 1955; Винтер, 1977); долина среднего течения р.Киевка (Судзухэ), кл.Эль-дагоу; окр. с.Коркаменка (Литвиненко, Шибаев, 1971); с.Соколовка (колл. ЗИН); устье р.Та-чингоузы (Белопольский, 1950); 101 - с.Бров-ничи (колл. ИЗАНУ) ; пос. ^учьи (Фанза); ст.Пар-тизаны (колл. ЗИН); окр. с.Монакино (колл. КГУ); 102 - с.Анисимовка (Кангауз) (колл. ЗИН); 103 - о-9 Аскольдт (Bianchi, 1907); 104 - о-в Большой Пелис (Лабзюк и др., 1971); 105 - заповедник ’’Кедровая падь” (Назаренко, 1971а; 19846); 106 - мыс Песчаный, Амурский залив (колл. ИЗАНУ); 107 - окр. г.Владивосток (Bianchi, 1907; колл. ЗИН; БПиДВНЦ); о-в Попова (Лабзюк и др., 1971); 108 - о-в Сахалин (Никольский, 1889; Bianchi, 1907; Дементьев, \1937; Благосклонов, 1954; Гизенко, 1955; Пор-тенко, 1960; Нечаев, 1969; Рахилин, 1975; Степанян, 1978); предгорья и поймы рек острова (Гизенко, 1955); 109 - окр. пос.Вахрушев (Yamashina, Yamada, 1934); НО - окр. с.Лопа-тино (колл. ХГУ); 111 - окр. с.Стародубское (Гизенко, 1955; колл. ИЗАНУ; М.И.Лобановского) ; 112 - окр. с.Новоалександровск (Bianchi, Окончание табл. 14.
1907; Гизенко, 1955; колл. ЗИН); окр. сЛуго-вое (Гизенко, 1955); 113 - южная часть острова (Иванов, 1976); окр. г.Южно-Сахалинск (Гизенко, 1955); 114 — окр. г.Корсаков (Lonnberg, 1908; колл. М.И.Лобановского); 115 - о-в Монерон (Благосклонов, 1954; Гизенко, 1955; Рахилин, 1975); 116 - о-в Уруп (Нечаев, 1969; Степанян, 1978); 117 - о-в Итуруп (Bergmann, 1935; Портенко, 1950; 1960; Нечаев, 1969; Степанян, 1978); 118 - о-в Кунашир (Гизенко, 1955; Портенко, 1960; Нечаев, 1969; Степанян, 1978); окр. пос.Алехино (Благосклонов, 1954; Нечаев, 1969; колл. ИЗАНУ); окр. г.Южно-Ку-рильск (колл. ЗИН); оз.Песчаное (колл. БПиДВНЦ); 119 — о-в Шикотан (Благосклонов, 1954; Нечаев, 1969; Степанян, 1978).
Места обитания. Населяет различные типы лиственных (главным образом листопадных), смешанных хвойнолиственных и хвойных лесов от таежных до субтропических и тропических на юге ареала по долинам рек и ручьев, на равни-
Вид, место сбора п Ширина, мм
Крыло CV Клюв у орального края ноздри CV Клюв у основания CV
Сам
M.latirostris Среднее Приамурье 25 48,49± 0,30 44,7 -51,1 3,06 4,8310,06 < -- 4,3 -5,5 ’ 6,7510,07 6,1 -7,3 4,96
Нижнее Приамурье 48 48,131 0,15 45,2 -50,3 2,23 4,8410,03 pg 4,4 -5,3 ’ 6,7910,05 6,1 -7,6 4,67
Приморье 15 48,071 0,33 45,9 -50,6 2,55 4,95 ±0,07 < in 4,3 -5,3 ’ U 7,14Ю,11 6,3 -7,7 5,98
О-ва Кунашир и Шикотан 5 48,941 0,66 '47,1 -51,1 3,04 4,6810,12 , # 5,53 6,6810,22 '?,1 -7,3 7,51
M.ruficauda Памиро-Алай 13 58,591 0,31 57,0 -60,3 1,92 8,04 3,5 -4,5 5,4010,09 4,9 -5,8 6,00
Сам
M.latirostris Среднее Приамурье 10 47,691 0,51 46,0 -51,1 3,35 5,0710,05 , Т§"-5,3 3,36 7,0310,09 6,7 -7,6 3,85
Нижнее Приамурье 11 48,491 0,40 '46,7 -51,1 2,71 4,8210,04 Тб -5,1 ’ 6,73Ю,06 6,3 -7,0 3,12
Приморье 7 48,231 0,46 46,8 -49,9 2,34 ±£310=08 4,59 4,6 -5,2 7,0010,11 4,5 -7,5 4,29
О-в Шикотан 1 45,5 - 4,4 6,8 -
M.ruficauda Памиро-Алай и Индия (Кашмир) 6 56,281 0,55 ‘55,0 -58,9 2,40 4,35±0,06 о я-? 4,2 -4,5 5,47Ю,06 5,3 -5,7 2,50
•Размер приводится по 1 экземпляру.
154
нах, в предгорьях, а также по склонам и гребням невысоких гор, поднимаясь вверх до 1900 м н.у.м. Поселяется в рошах, садах и парках, часто в непосредственной близости от жилья человека, а на юге ареала также в бамбуковых зарослях и на плантациях кофейного и тикового деревьев.
На территории СССР в северо-западной части гнездового ареала (Томская обл. й Красноярский край) гнездится в таежных смешанных березово-осиновых и светлых сосновых лесах с примесью березы ЛЪсины и лиственницы в поймах рек, низинах и предгорьях, тяготея к разреженным и хорошо освешенным местам в том числе и по окраинам полян, а также у дорог (Юдин, 1952; Наумов, 1960; Гынгазов, Миловидов, 1977). На лесостепных участках встречается только в приречных кус
тарниковых зарослях, избегая колков (Юдин, 1952). В Саянах верхняя граница обитания этого вида проходит в зоне рододендронов и ивовых кустарников (Tacza-nowski, 1891). В Среднем Приангарье обычна в лиственнично-сосновых с хорошо развитым подлеском и мелколиственных лесах, а также в кустарниковых зарослях на вырубках и гарях (Шведов, 1962).
В Иркутской обл. у с.Гадолей гнездится в сосново-еловых лесах с подлеском из розы, ивы и таволги (Сонин, Липин, 1969; колл. ЗМ МГУ), а на верхней Лене населяет леса долинной полосы, не проникая на водоразделы (Реймерс, 1966). В Забайкалье зарегистрирована в горной тайге и в лесостепи долины р. Баргузин и на п-ове Святой Нос (Гагина, 1960). На Витимском плоскогорье поселяется в лиственнич-
Высота, мм Отношение ширины клюва у основания к его длине от оперения лба, % CV Отношение длины хвоста к длине крыла, % CV Отношение ширины крыла к его длине, % CV
Клюв у основания CV
цы
3,4310,04 3,2 -3,8 4,60 66,19+ 0,70 60,95-72,92 5,16 68,70± 0,34 65,34-71,33 2,39 70,271 0,30 67,97-74,52 2,12
13,4610,04 3,1 -3,8 6,02 66,501 0,62 57,80-75,53 6,30 68,811 0,19 66,03-71,05 1,87 70,101 0,20 66,86-72,31 1,96
3,35 ±0,05 3,1 -3,7 5,24 69,531 1.15 61,17-77,00 6,40 67,68+ 0,51 64,28-70,72 2,91 70,551 0,51 65,52-73,33 2,71
3,32+0,12 3,0 -3,7 7,80 68,961 3,82 60,40-76,04 11,09 68,67+ 0,95 67,20-72,31 3,08 71,47+ 0,47 70,19-72,90 1,48
3,57+0,03 3,5 -3,6 1,62 50,91+ 0,91 46,28-57,00’ 6,45 75,931 0,42 73,18-77,89 1,98 75,88+ 0,37 73,79-77,97 1,78
ки
3.55±<Ш, 3,5 -3,6 1,63 68,871 0,92 66,02-73,68 3,99 68,771 0,56 67,06-71,23 2,14 70,06± 0,34 68,37-71,35 1,44
3,39+0,08 3,2 -3,8 6,59 65,991 0,55 • 63,11-68,32 . 2,75 69,11± 0,54 66,07-71,83 2,59 70,271 0,34 68,65-72,59 1,58
3,6010,12 3,4 -3,8 5,56 67,781 1,35 62,73-73,53 5,28 68,851 0,41 67,17-70,15 1,47 70,58+ 0,68 68,70-72,77 2,37
- - - - 71,10 - 68,84 -
4,3* 52,44+ 0,73 50,00-54,55 3,41 75,031 0,79 73,12-77,53 2,58 76,281 0,52 73,87-77,30 1,66
155
но-березовых рощах, разреженных лиственничных и сосновых лесах, а также в зарослях ивы по речным долинам (Измайлов, 1967).
В Амурской обл. в низовьях р.Бурея гнездится в пойменных ивняках и дубравах (сообщ. С.В.Винтера), а на р.Селемджа в окр. пгт Экимчан — в смешанных лесах из ели, пихты, березы и лиственницы (сообш. В.В.Леоновича). В Хабаровском крае в верховьях р.Амгунь встречается в осиноволиственничных лесах с подлеском из рододендрона (Федотов, Брунов, 1977), а в бассейнах рек Охота и Урак отмечена в пойменных лесах из чозении и тополя (Лемд-гольм, 1959). В нижнем течении р.Хор населяет темнохвойно-широколиственные леса (Кисленко, 19656), а на хр.Хехцир -широколиственные леса с примесью кедра (Кулешова и др., 1965; Казаринов, 1973). В Комсомольском заповеднике обитает в разнообразных хвойных и смешанных лесах, избегая глухой тайги и открытых ландшафтов (Штильмарк, 1973).
В Приморье на Среднем Сихотэ-Алине гнездится в различных лиственных, смешанных и хвойных лесах по долинам рек, ручьев, пойменным террасам, склонам и гребням гор (Кулешова, 1976; наши данные). В Уссурийском заповеднике населяет дубняки (колл. ИЗАНУ), ильмовники (Коренкова, '1969), ельники и кедровопихтовые массивы (Иванов, 1952), а нами найдена на гнездовании в смешанных широколиственных лесах с незначительной примесью кедра. В Лазовском заповеднике встречается в дубовых рощах (Белопольский, 1950), липово-широколиственных лесах (Винтер, 1977), а также гнездится в зарослях ивы и ольхи у кл.Эльдагоу (Литвинен
ко, Шибаев, 1971). В Южном Приморье поднимается по склонам сопок до вершинных сухих рододендроновых дубняков, камен-ноберезников и пихтарников (Панов, 1973).
На о-ве Сахалин населяет различные леса почти по всем поймам рек и предгорьям до верхнего предела распространения лиственного леса. На о-в Монерон гнездится в каменноберезовых лесах с примесью бархата, ольхи, бузины и других кустарников (Гизенко, 1955). На южных Курильских о-вах встречается в широколиственных, смешанных и хвойных лесах, предпочитая редколесья, лесные опушки, вырубки и долинные группировки с покосами и полями (Нечаев, 1969).
Численность. В СССР на оольшей части гнездового ареала обычна, в излюбленных биотопах - многочисленна. Однако в большинстве мест Южной Сибири по сравнению с Дальним Востоком более редка.
Прилет и весенний пролет на территории СССР. Проходит с I декады мая по I декаду июня включительно.
На южные Курильские оюва прилетает в середине II декады мая. Так, в 1963 г. первое появление на о-в Кунашир отмечено 19.V, но одиночные особи и стайки пролетных птиц отмечались до конца мая (Нечаев, 1969; 1971). На о-в Монерон пролетные мухоловки в 1973 г. встречались 17.V-8.V1 (Нечаев, 1975). В южной части Сахалина появляется во второй половине мая, а в северной — в I декаде июня (Гизенко, 1955). На о-вах в северо-западной части залива Петра Великого первые особи зарегистрированы 8.V, а основной пролет проходит здесь до середины мая (Лабзюк и др., 1971).
Рис. 29. Распространение ширококлювой мухоловки (Muscicapa latirostris) на территории СССР: а ~ гнездится, б - отмечена в гнездовой период. См. кадастр, стр. 151-154.
156
В Южном Приморье весенняя миграция проходит с 1 декады мая до начала I декады июня включительно. Е.НЛановым (1973) начало прилета (2 года) зарегистрировано 5 и 10.V, но пролетные птицы в заповеднике ’’Кедровая падь” отмечались также 22.V 1960 г., а в устье р.Илистая (Лефу) — даже 2.VI 1962 г. Нами одиночная пролетная мухоловка добыта 30.V 1977 г. на сопке Змеиная у оз.Тальми (окр. ст.Хасан). В окр. г.Владивосток на р.Седанка самец добыт 14.V 1911 г., а в верхнем течении р.Одарка у с.Нововладимировка 4 птицы добыты 7, 12, 14 и 15.V 1911 г. (Черский, 1915а; 19156). В Шкотовском р-не на р.Суворовка мухоловки добыты 14.V 1967, в Партизанском р-не — 15, 19, 20 и 23.V 1926 г., у ст.Мучная (с.Алтыновка) — 9.V 1928 г., а у ст.Дормидонтовка - 15.V 1928 г. (колл. ЗИН).
На нижнем Амуре пролет протекает с конца I декады мая до конца этого месяца. В р-не Пивании прилет первых птиц отмечен в 10-х числах мая (Штильмарк, 1973), у с.Даур - 19.V, у с.Омми — 23.V, интенсивный пролет у с.Раздольное наблюдался 24—25.V, а у с.Гячи - 28.V (Кистя-ковский, Смогоржевский, 1973).
В Амурской обл. в междуречье Малой Перы и Большого Эргеля самец добыт И.А.Нейфельдт 22.V 1958 г. (колл. ЗИН).
В Читинской обл. у с.Кулусутай самец добыт 19.V 1856 г. (колл. ЗИН). В Приангарье у с.Жигалова первое появление в 1961 г. отмечено 4.VI (Реймерс, 1966). В Красноярском крае ширококлювая мухоловка появляется в I декаде июня (Юдин, 1952).
Размножение. Песня самца ширококлювой мухоловки негромкая, но очень мелодичная. Занятие гнездовых участков и образование пар сопровождаются наиболее интенсивным пением самцов и их токовым демонстративным поведением (рис. 23). Размеры гнездовых участков невелики. Расстояние между гнездами соседних пар часто составляет 40-50 м (Винтер, 1977). В выборе места для гнезда принимают участие оба партнера. В этот период самцы часто продолжают еше токовать и ведут себя очень возбужденно.
Гнезда птицы строят открыто в развилках стволов, боковых горизонтальных и наклонных ветвей, а также на ветвях у сучьев и неровностей коры или вплотную к стволу деревьев. Из 21 гнезда ширококлювой мухоловки, найденных в Южной Сибири, на Дальнем Востоке СССР и на Са
халине (Гизенко, 1955; Шкатулова, 1965; Соснин, Липин, 1969; Панов, 1973; Винтер, 1977; колл. ЗМ МГУ; КБ МГУ; ИЗАНУ; В.В.Леоновича; М.И.Лобановско-го; С.Д.Кустановича; наши данные) 7 были построены на березах, 5 — на дубах (в том числе и на монгольских), 2 — на ивах, 2 — на маньчжурских липах (рис. 31, 32) и по одному на черемухе, мелколистном клене, сосне, ели и лиственнице, на высоте 2,0—9,0 м от поверхности земли, в среднем - 4,9. В единичных случаях гнезда помешаются на высоте 15,0 м (Nisbet, Neufeldt, 1975). В случае расположения гнезда на боковых ветвях расстояние от него до ствола дерева составляет 0,8— 3,6 м, в среднем — 2,2. В сборе строительных материалов для постройки принимают участие оба партнера пары, но строит гнездо главным образом самка. Строительство длится около 5 дней (Nisbet, Neufeldt, 1975).
Гнезда представляют собой очень аккуратные, плотные, чашевидные постройки (рис. 31, 32), диаметр нижней части которых часто превышает таковой верхней. В качестве строительных материалов птицы используют пучочки мхов (Thuidium philibertii, Platygyrium repens, Pylaisia polyantha, Leske palycarpa, Hypnum cup-ressiforme, Anomodon thrbustus), кусочки талломов лишайников (Hypogymnia pley-sodes, Parmelia sulcata, P.sinuosa), коры (в том числе чешуи коры сосны и пленки бересты), тонкие веточки, стебельки и листья дикорастущих злаков и других трав, сухую хвою, полоски луба и тонкие корешки. Лоток выстилается сухой хвоей (кедр, ель, пихта, лиственница), тончайшими корешками и полосками луба, а также стебельками и листьями трав, кусочками талломов лишайника (Bryopogon sp.), волосками шерсти млекопитающих, растительными волокнами, одиночными перьями, а по данным Я.Нисбета и И.А.Нейфельдт (Nisbet, Neufeldt, 1975), черешками, соцветиями и листьями леспедецы, цветами некоторых зонтичных и спорогониями мхов (Bryurn sp., Polytrichum sp.). Снаружи гнездо тщательно маскируется кусочками талломов лишайников, пленками березовой коры, пучочками мхов и чешуями цветочных почек. Все перечисленные материалы приплетаются к стенкам тонким слоем паутины и в редких случаях — шерстью млекопитающих. Размеры гнезд (п = 16), мм: диаметр 80—118, в среднем - 97; высота 37—80, в среднем - 52; диаметр лот
157
ка 45—60, в среднем — 53; глубина лотка 25-45, в среднем - 34,
На территории СССР в сезон размножения ширококлювая мухоловка имеет один репродуктивный цикл. Указания на поли-циклИчность у этого вида в СССР (Воробьев, 1954; Рашкевич, 1965) нуждаются в проверке. Полная кладка состоит из 5—6, значительно реже из 4 и как исключение из 3 яиц. Укороченно-остроовальные и значительно реже укороченно-овальные й остроовальные яйца по бледно-зеленовато-голубоватому или бледно-зеленовато-серому фону имеют бледно-лилово-розовый или светло-охристый налет. На одних яйцах он сгущается к тупому полюсу, образуя вокруг последнего слабо выраженный ’’венчик”, в то время как на других — равномерно покрывает всю поверхность полуматовой скорлупы, так, что яйцо выглядит Светло-бежевым. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
Насиживание начинается с откладки предпоследнего яйца (Винтер, 1977). Длительность инкубации 11—15 дней (Nisbet, Neufeldt, 1975; Винтер, 1977). Самец в насиживании кладки не участвует. В этот период он держится поблизости от гнезда и много поет. У большинства пар на протяжении всего периода насиживания самец кормит самку на гнезде или на ветке поблизости от него (рис. 30), приглашая ее характерной позывкой. Редко у некоторых пар самец этого не делает (Винтер, 1977). Вылупление птенцов проходит в течение суток.
Пуховой птенец ширококлювой мухоловки-(Нейфельдт, 1970) имеет недлинный, густой, двухцветный (желтовато-белый в базальной и темносерый в терминальной частях) пух на надглазничных, затылочной, спинной, плечевых, локтевых, бедренных и глазных птерилиях. Кожа дорсальной стороны тела темно-серая, вентральной - телесно-розовая. Ротовая полость и язык желтые. Складки в углах рта, клюв и когти бледно-желтые. Ноздри с темной окантовкой. В выкармливании птенцов принимают участие оба партнера пары, но в первые 7 дней после их вылупления корм чаше приносит самец, а самка большую часть времени проводит в гнезде, согревая птенцов. На 14 день с недоросшими маховыми и рулевыми птенцы покидают гнездо (Винтер, 1977). Взрослые продолжает кормить слетков и держатся рядом с ними. В дальнейшем выводки начинают кочевать. В некоторых случаях они не распадаются даже до начала осенней миграции.
Сезон размножения ширококлювой мухоловки на территории СССР длится с середины мая по II декаду августа включительно.
На о-ве Кунашир птицы с увеличенными гонадами встречены 11 и 22.VI, а выводки плохо летающих молодых отмечены на острове 27.VII и 2.VIII (Нечаев, 1969).
На Сахалине в окр. с.Стародубское гнездо с полной кладкой из 5 ненасиженных яиц найдено 23.VI 1951 г., а с кладкой незадолго до вылупления птенцов — 7.VII 1953 г. (Гизенко, 1955; колл. М.ИЛоба-новского). В окр. г.Южно-Сахалинск плохо летающие молодые в 1947 г. зарегистрированы 1.VIII (Гизенко, 1955), а в окр. Александровска молодая птица добыта 27.VII 1881 г. (колл. ЗИН).
На юге Приморского края начало токования (2 года) зарегистрировано 19 и 31.V; пара, выбирающая место для гнезда, отмечена 21.V 1962 г., a 31.V птицы держались уже у почти выстроенного гнезда (Панов, 1973). Самая ранняя откладка яиц зарегистрирована здесь 23.V (Nisbet, Neufeldt, 1975), а хорошо летающие молодые в заповеднике ’’Кедровая падь” встречены 11.VII (Панов, 1973). В Шкотовском р-не гнездо с пуховыми птенцами обнаружено 10.VI 1967 г„ a 19.VI добыта молодая самка (колл. ЗИН).
В Уссурийском заповеднике самка, закончившая откладку яиц, добыта на Егерском ключе 28.VI 1940 г. Самка и разновозрастные молодые встречены в заповеднике А.И.Ивановым (1952) 26.VII 1940 г., а нами в районе первого кордона — 28— 30.VI 1977 г. В окр. г.Уссурийск на территории горнотаежной станции кладка из 6 сильно насиженных яиц найдена 9.VI 1963 г. (Панов, 1973). В верхнем течении р.Одарка у с.Нововладимировка (Спасский р-н) молодые птицы добыты 25, 26 и 29.VII 1911 г. (Черский, 1915а, 19156), а на р.Маргаритовка у с.Щербаковка (Оль-гинский р-н) — 7,9 и 18.VII 1927 г. (колл. ЗИН). В Лазовском заповеднике несколько самок с готовыми к размножению яичниками и яйцеводами добыты 24.V и 2.VI 1945 г. (Белопольский, 1950), a 12.VI 1962 г. у кл.Эльдагоу зарегистрирована пара мухоловок, уже выкармливающая птенцов (Литвиненко, Шибаев, 1971). Откладка яиц в гнездах, найденных С.В .Винтером (1977) в 1971 г., началась 13.VI (в двух гнездах) и 21.VI (в одном), последующие яйца откладывались с интервалом в сутки; птенцы вылупились 28, 29.VI и 5.VII, а вылетели из всех гнезд на 14-й день жизни.
158
В окр. с.Коркаменка гнездо с 5 птенцами обнаружено 21.VI 1962 г., покинули его птенцы 22^1 (Литвиненко, Шибаев, 1971). Первые вывЬдки в заповеднике отмечены 13 и 17.VII 1971 г., а 7 и 20.VII отмечались слетки с недоросшими рулевыми (Белопольский, 1950; Винтер, 1977; колл. ЗИН). Две молодые птицы добыты 26.VII (Белопольский, 1950).
В Сихотэ-Алинском заповеднике гнезда с полными кладками находили 18.VI 1977 г. и 12.VII 1973 г., а молодой самец на р.Се-ребрянка (Сица) добыт К.А.Воробьевым 18.VII 1948 г. (колл. БПиДВНЦ). Самый поздний слеток на территории СССР встречен и добыт в Приморье 7.VIII (Воробьев, 1954; Портенко, I960).
В Хабаровском крае на нижнем Амуре токующий самец и строительство гнезда отмечены у с.Киселевка 3.VI, а начало откладки яиц зарегистрировано: у с.Болонь -1.VI, у с.Покровка — 17.VI и у с.Тальни-ки — 20.VI (Кистяковский, Смогоржевский, 1973). В окр. с.Тальники кроме этого 21.VI 1961 г. обнаружено также гнездо с уже законченной полной кладкой. В окр. оз.Кизи молодые птицы добывались 31.VII 1928 г. (колл. ЗИН), а на юго-востоке края - в 19 км от устья р.Б^тчи - 19.VII 1924г. (Емельянов, 1929).
В Амурской обл. самая ранняя откладка яиц отмечена 9.VI (Nisbet, Neufeldt, 1975). В окр. пгт ЭкимчаН гнездо с 4 яйцами найдено 13.VI 1971 г., a 16.VI в нем была полная кладка из 6 яиц (колл. В.В.Леоновича). 15 и 20.VI здесь были обнаружены еше 3 гнезда с полными слабо насиженными кладками (сообщ. В.ВЛеоно-вича). На р.Зея у с.Натальино 2 молодые птицы добыты 12.VII 1958 г. (Спасский и др., 1962), а в междуречье Малой Перы и Большого Эргеля - 19.VII 1958 г. (колл. ЗИН).
В Тулунском р-не Иркутской обл. у с.Гадолей начало откладки яиц в одном гнезде отмечено 8.VI 1964 г., a 13.VI в нем была полная кладка из 6 яиц (Сонин, Липин, 1969). На р.Зангисан молодая птица добыта 1.VIII 1912 г. (колл. ЗИН). В Приангарье у с.Жигалово слетки зарегистрированы 19.VII 1961 г. (Реймерс, 1966).
В Красноярском крае (Хакасская АО) у пос. Майна гнездо с 1 неоплодотворенным яйцом (самка насиживает) найдено 22.VI 1968 г. (колл. КБ МГУ). В 1964 г. строящаяся постройка обнаружена 26.VI, а гнездо с полной кладкой из 3 яиц средней насижениости — 7.VII (колл. С.Д.Кустано-вича). Самка с наседным пятном на р.Со
бакиной добыта 8.VII 1927 г., а молодые мухоловки с недоросшими рулевыми встречены у селений Отаровая и Бирюса на р.Ени-сей 18 и 20.VII 1931 г. (Юдин, 1952) и в окр. г.Красноярск на горе Абатак близ с.Торгашино — 23.VII 1910 г. (колл. ЗИН).
В юго-восточной части Западно-Сибирской равнины в бассейне р.Чулым у с.Четь-Конторка спаривание отмечено 7-12.VI, a 12.VI 1966 г. добыта самка, в яйцеводе которой обнаружено почти полностью сформировавшееся яйцо. 14.VI 1967 г. здесь найдено гнездо с незаконченной кладкой из 2 яиц. Выводок из 5 молодых, которых еше кормили родители, встречен 21.VI 1966 г., а 31.VII зарегистрированы самостоятельно кормившиеся молодые (Гынгазов, Миловидов, 1977). В долине р.Кеть летные молодые, которых кормили взрослые птицы, отмечены 15.VII, 5 и 19.VIII 1968 г. (Москвитин и др., 1977). В окр. г.Томск взрослые птицы с хорошо развитыми наседными пятнами добывались 28.VI и 16.VII, a 16.VII 1975 г. было найдено гнездо с 5 птенцами перед вылетом. Слетки отмечены 27.VII 1963 г., 25.VII 1973 г. и 30.VII 1974 г. (Гынгазов, Миловидов, 1977).
Линька. После того как маховые и рулевые закончат рост, у молодых начинается частичная послегнездовая линька, которая охватывает почти все мелкое оперение, кроме больших верхних кроюших второстепенных маховых и части кроюших хвоста. У большинства особей она заканчивается к началу осенней миграции. С февраля по апрель у первогодков проходит повторная частичная линька оперения, которая протекает с большими индивидуальными вариациями. По данным Н.В.Виногра-довой и др. (1976), у большинства особей появляются новые рулевые, перья лба и темени, а у некоторых и маховые. У части птиц сменяются только центральные рулевые, перья лба и часть больших верхних кроющих второстепенных маховых. Иногда все рулевые остаются старыми. В наряде, состоящем из свежих и обношенных прошлогодних перьев, первогодки прилетают на места гнездования и приступают к размножению. Взрослые птицы имеют в году 1 линьку (полная послебрачная), которая протекает на местах гнездования.
Питание. Основу пищи составляют насекомые разных групп.. Реже мухоловки поедают паукообразных (сенокосцы, пауки) и как исключение — мелкие сочные плоды некоторых растений. Чаше всего птицы охотятся за летающими насекомыми,
159
схватывая их в воздухе во время стремительного броска с присады. Реже склевывают кормовые объекты с растений, а по данным некоторых исследователей (Портенко, 1960; Nisbet, Neufeldt, 1975) -с ловчих паутинных сетей и даже поверхности земли. Несмотря на это, свою активность мухоловки ограничивают главным образом внутренними частями крон деревьев и очень редко присаживаются на их вершины. Птенцов выкармливают насекомыми разных стадий развития и пауками. В Лазовском заповеднике основную массу корма, приносимого родителями птенцам, составляют двукрылые и другие мелкие летающие насекомые (около 90 %). Бабочки и гусеницы чаше скармливались птенцам в последние дни пребывания их в гнезде. Крылья у бабочек перед скармливанием за редким исключением родители не отрывают (Винтер, 1977).
Отлет и осенний пролет на территории СССР. Проходит с III декады августа по I декаду октября включительно. Массовый пролет идет в сентябре. Во время осенней миграции мухоловки летят поодиночке и небольшими стайками.
В окр. г.Иркутск пролетная самка добыта 4.IX 1912 г. (колл. ЗИН).
На нижнем Амуре отлет проходит в I декаде сентября (Рашкевич, 1965).
В Приморье миграция проходит в сентябре и I декаде октября. В нижнем течении р.Рудная мухоловка добыта 2.Х 1909 г. (Черский, 1915а); в верховьях р.Одарка у с.Нововладимировка - 29.IX 1911 г. (Черский, 1915а; 19156), а у устья р.Илис-тая (Лефу) - 25.IX 1926 г. (колл. ЗИН). В Уссурийском заповеднике на р.Комаров-ка молодая самка добыта 22.IX 1921 г. (колл. ЗИН). На юге Приморья пролет начинается в I (2.IX 1961 г.) и в начале II (11.IX) декад сентября (Панов, 1973). Последние осенние встречи этого вида здесь (4 года): 18, 28, 30.IX и 7.Х (Панов, 1973; Омелько, 1979).
На островах в северо-западной части залива Петра Великого миграция проходит с середины августа до начала I декады октября включительно. Максимум пролета в первой половине сентября (Лабзюк и др., 1971). Здесь на о.Болыпой Пелис 12.IX 1962 г. добыт самец (колл. БПиДВНЦ).
На Сахалине отлет и осенний пролет основной массы птиц проходит с середины II декады сентября до конца этого месяца. Отдельные мухоловки (в долине р.Су-
сун) задерживаются до первых чисел октября (Гизенко,1955).
На о.Онекотан (Курильские о-ва) пролетные птицы отмечаются до середины сентября (Гизенко, 1955); на о-ве Уруп мухоловка зарегистрирована 14.IX 1947 г. (Гизенко, 1955); на о.Шикотан последняя птица (1 год) встречена 17.IX (Нечаев, 1969), а на о-в Кунашир пролетные особи наблюдались 2.IX—10.X (Гизенко, 1955, колл. ЗИН).
Известен случай залета ширококлювой мухоловки в период осенней миграции в Среднюю Азию: 10.IX 1892 г. в Памиро-Алае на р.Сох у к.Зардалы добыта одиночная птица (Stolzmann, 1897).
Практическое значение. Уничтожает многих насекомых - вредителей леса и переносчиков возбудителей ряда заболеваний человека и животных. Заслуживает повсеместной, всесторонней охраны.
Рыжехвостая мухоловка — Muscicapa rufi-cauda Swainson, 1838
Muscicapa ruficauda Swainson, 1838, Nat. Libr., 21, Flycatchers: 251, Кашмир, Индия.
Описание. Составлено по 23 экз. (22 - из Памиро-Алая и 1 - из Индии).
У птиц из Памиро-Алая вершину крыла образуют 3—5-е первостепенные маховые (3=4- 13,3 %, 4 - 80,0 %, 4 = 5 - 6,7 %). У просмотренного 1 экз. из Кашмира (Индия) вершина крыла образована 4—6-м первостепенными маховыми (4 = 5 =6). 2-е маховое всегда меньше 5-го; 1-е первостепенное маховое мало и лишь незначительно выступает за вершины кроющих кисти (табл. 14). У большинства особей наружные опахала 3—6-го первостепенных маховых в вершинной части заметно сужены. Количество шетинок в углах рта у основания надклювья (с каждой стороны головы) равно 3. Длина наибольшей (п = = 19): 6,0—8,5 мм (в среднем — 7,3). Количество щитков подотеки на передней части цевки варьирует от 4 до 5. Половой диморфизм незначительно выражен в размерах отдельных частей тела (табл. 14). Самец и самка окрашены одинаково.
У взрослой птицы все оперение верхней части тела от лба до надхвостья включительно оливково-бурое. На надхвостье по основному фону появляется незначительная примесь рыжего. У некоторых самцов (28,6 %) и самок (33,3 %) на верхней части головы (от лба до темени) просматри
160
ваются слабо выраженные продольные пестрины, образованные темно-бурыми стержневыми пятнами на перьях. Верхние кроющие хвоста и рулевые ржавчато-рыжие. Центральная пара последних несет примесь бурого на внутренних и наружных опахалах. На верхней трети этих перьев ржавчаторыжий основной цвет почти полностью вытесняется буроватым и сохраняется только на стержнях. Бока головы светло-оливково-бурые. Уздечка, перья, образующие кольцо вокруг глаза, и горло от грязно-белого до бледно-буровато-серого цвета. Нижняя часть тела от области зоба до брюшка включительно бледно-серовато-бурова-тая. Этот цвет незначительно темнеет на боках тела. Нижние кроющие хвоста и нижние кроющие крыла бледно-охристые. Маховые и верхние кроющие крыла темнобурые. Вершины больших верхних кроюших второстепенных маховых несут коричневато-охристые каемки, а по краям наружных опахал маховых (начиная с 3-го первостепенного) имеются узкие коричневатоохристые канты. Последние подвержены в значительной степени обносу. Надклювье коричневато-рогового цвета с более темной вершиной. Подклювье желтовато-телесное. Ноги бурые. Нижняя поверхность пальцев светло-желтовато-бурая. Когти темно-бурые с светло-бурой внутренней полосой.
Молодые в гнездовом наряде сверху оливкОвр-бурые с округлыми охристыми пестринами. Нижняя сторона бледно-охристая с темно-бурыми отметинами. Большие верхние кроющие второстепенных маховых на вершинах несут коричневато-желтые каемки, а маховые по краям наружных опахал имеют узкие коричневато-желтые канты. Верхние кроющие хвоста и рулевые такой же окраски, как и у взрослых птиц. Первый послегнездовой наряд практически идентичен наряду взрослой птицы.
Размеры отдельных частей тела рыжехвостой мухоловки приведены в табл. 14.
Масса тела, г: взрослые самцы (п = = 2) (11.VI. Таджикистан, ущ.Кондара, ур!Свак) — 11,55-12,05,в среднем- 11,80.
Распространение. В СССР (рис. 33) — Памиро-Алай, где этот вид гнездится к северу до северного склона Туркестанского хр. Однако одиночные (очевидно, залетные) птицы отмечены и значительно севернее — в Западном Тянь-Шане. К западу гнездовой ареал протягивается до хребтов Байсунтау (предположительно) и Кугитанга, а к востоку до Дар-вазского хр. (предположительно). За пре
делами СССР - Северо-Западные Гималаи к западу до Гиндукуша и хр.Сафедкох (Спингар) и к востоку от центральных частей Непала (Vaurie, 1959; Ali, Ripley, 1972; Степанян, 1978). К местам зимовок мигрирует через Северо-Восточный Афганистан, Пакистан, Северную Индию (Кашмир), вдоль подножий Гималаев и далее через Центральную Индию в ее юго-западную часть (штаты Карнатака и Керала), где зимует в Западных Гатах, поднимаясь вверх до 1000 м н.у.м. (Ali, Ripley, 1972). Детально распространение рыжехвостой мухоловки на территории СССР показано на карте (рис. 33).
Кадастр крио-. 33.
1 - Западный Тянь-Шань, заповедник ”Аксу-Джабаглы”; каньон Бала-Балдабрек (Шульпин. -Цит. по: Ковшарь, 1966; 1970); 2 - р.Пскем, бассейн р.Чирчик (Зарудный. - Цит. по: Иванов, 1940; Портенко, 1960); ур.Майдантал (Дементьев, 1937; Благосклонов, 1954; Ковшарь, 1966); 3 - Чаткальский хр., верховья р.Падыша-Ата (Зарудный по Иванову, 1940; Дементьев, 1937; Благосклонов, 1954; Янушевич и др., 1960); 4 -Туркестанский хр. (Портенко, 1960; Иванов, 1976; Степанян, 1978); ур.Гуралашсай в верховьях р.ЗааМинсу (Иванов, 1940; 1'969; Янушевич и др., 1960; Абдусалямов, 1973; колл. ЗИН); 5 - окр. оз.Каракулькатта в верховьях р.Исфара (Зарудный. - Цит. по: Иванов, 1969); 6 - Гиссарский хр., р.Сангардак (Иванов, 1940; 1969; Абдусалямов, 1973; колл. ЗИН); 7 -южные склоны хребта (Иванов, 1976); ущ. р.Варзоб (Леонович, 1962; Воробьев, 1968; Иванов, 1969; Абдусалямов, 1973); ущ.Кондара
Рис. 33. Распространение рыжехвостой мухоловки (Muscicapa ruficauda) на территории СССР:
а - гнездится, б - встречена в гнездовой период, в - предположительно гнездится
161
(Попов, 1959; Леонович, 1962; Воробьев, 1968; Иванов, 1969; Беме, 1975); ур. Квак (Леонович, 1962; Воробьев, 1968; Иванов, 1969; колл. ИЗАНУ); ур. Обисафед (Иванов, 1969; колл. ЗИН; ИЗиПАНТадж.); ущедья рек Зирч, Ровот, Джойрихонага (Иванов, 1969); 8 - устье р.Та-коб, ущ.Дейвбол? (Иванов, 1969; колл. ЗИН); 9 - долина р.Кафирниган (Абдусалямов, 1973); 10 - верховья рек Кафирниган и Сорбух (Иванов, 1969); 11 - Дарвазский хр. (Портенко, 1960; Иванов, 1976; Иванов, Штегман, 1978; Степанян, 1978); районы Ховалинга и Сарыхосо-ра (Иванов, 1969); 12 - хр.Байсунтау (Степанян, 1978); 13 - хр.Кугитанг (Дементьев, 1937; Благосклонов, 1954; Портенко, 1960; Ковшарь, 1970; Иванов, 1976); восточный склон хребта к западу от к.Кампыртеке (Зарудный. - Цит. по: Иванов, 1969).
Систематика. Монотипический вид.
Места обитания. Населяет хвойные, смешанные хвойно-лиственные и лиственные леса гор на высотах 1100— 3600 м н.у.м. В Гималаях гнездится в смешанных хвойно-лиственных и кедровых лесах на высотах 2100—3600 м (Ali, Ripley, 1972). В Кашмире обитает в сосновых, кедровых, еловых и смешанных лесах на высотах 1500-3300 м (Bates, Lowther, 1952).
На территории СССР в Памиро-Алае населяет различные типы горных лесов на высотах 1100—3200 м, чаше — 1800—1900 м. На южных склонах Гиссарского хр. гнездится в глухих лесах из туркестанского клена с примесью грецкого ореха, яблони и одиночных деревьев арчи в узких тенистых и сырых ушельях (Иванов, 1969), а также в орехово-плодовых лесах (Беме, 1975). В уш.Кондара отмечена на гнездовании в разреженном лесу из туркестанского клена с примесью арчи в ур.Квак (рис. 34). Обычно придерживается древесно-кустарниковых зарослей вдоль горных рек и ручьев. В лесах по безводным склонам гор в гнездовой период на территории СССР почти не встречается. В характерных гнездовых биотопах этого вида произрастают: клен, арча, грецкий орех, береза, миндаль, жимолость, роза, дикая вишня, дикий виноград и другие древесные и кустарниковые растения (Абдусалямов, 1973). В ар-чевниках отмечена Н.А.Зарудным (Ковшарь, 1970) на хр.Кугитанг, Туркестанском хр. у оз.Каракулькатта и на Ферганском хр.
Численность. На территории СССР повсеместно низка. А.И.Иванов (1969) считает рыжехвостую мухоловку немногочисленной, но не представляющей редкости птицей в орехово-кленовых насаждениях на южных склонах Гиссарского хр. в ущ. р.Варзоб. В некоторых пунктах этого ущелья им отмечено по 2—3 пары. В ур.Квак в 1959 г. плотность гнездования этого вида, по дан-162
ным В.ВЛеоновича (1962), составила 4-5 пар/км2. В долине р.Ишак-талды на 2 км приречных зарослей (18.VI) отмечены 2 гнездящиеся пары (Абдусалямов, 1973). Какие-либо данные о численности этого вида в других районах гнездового ареала на территории СССР полностью отсутствуют.
Проведенные нами в 1976 г. специальные поиски этого вида в гнездовой период в соответствующих биотопах во многих точках Туркестанского и Зеравшанского хребтов, а также хр.Кугитанг не дали положительных результатов. Рыжехвостая мухоловка обнаружена не была. Ю.А.Лебедев, в течение нескольких лет изучающий структуру населения птиц Центрального Таджикистана и проводящий постоянные учеты птиц, зарегистрировал только единичные встречи с этим видом и то главным образом в негнездовой период. В июне 1978 г. нами были проведены специальные поиски и повторный учет рыжехвостой мухоловки в ур.Квак (рис. 34). На плошади 1 км2 отмечены только 3 гнездящиеся пары. Поиски этого вида в соседних ушельях в таких же биотопах успехом не увенчались. На чрезвычайную редкость этой птицы указывают также А.В.Попов (1959) и некоторые другие исследователи.
На основании сказанного выше мы считаем рыжехвостую мухоловку редким, спорадично распространенным видом Памиро-Алая, численность которого подвержена годовым колебаниям и, очевидно, сокращается.
Прилет и весенний пролет на территории СССР. В Памиро-Алае проходит со II декады марта по I декаду мая включительно.
В районе северной оконечности Вахшс-кого хр. на стационаре ИЗиП АНТадж ССР Гирдоб 26.111 1976 г. отловлена и окольцована 1 птица.
На Гиссарском хр. в Варзобском ущ. самая ранняя одиночная пролетная особь отмечена 16.III (Абдусалямов, 1973). Здесь же в 1946—1947 гг. с 30.IV по 6 V А.И.Иванов (1969) встречал уже мухоловок на местах гнездования. 21.IV 1946 г. и 30.IV 1947 г. в ур.Обисафед он добыл 2 взрослых самца, a 4.V 1947 г. 2 взрослых самца добыты им на р.Зырч (приток р.Варзоб) (колл. ЗИН). Массовый прилет и пролет рыжехвостых мухоловок в Варзобском уш. в р-не альп. лагеря ’’Варзоб” отмечен 4-5.V 1961 г. (Беме, Сытов, 1963; Беме, 1975). На небольшом участке ореховой рощи встречейо более 20 птиц. Здесь же
5.V 1961 г. РЛ:Беме добыл взрослую самку (колл. БПиДВНЦ), a 4.V 1967 г. — взрослого самца (колл. КБ МГУ).
В одном из садов окраины г.Ташкент пролетный самец добыт 9.V 1908 г. (Иванов, 1940), а в г.Чиназ одиночная самка добыта та 23.V 1878 г. (Иванов, 1940; колл. ЗИН).
Размножение. Изучено недостаточно. После прилета на места гнездования птицы занимают гнездовые участки, разбиваются на пары и приступают к размножению. В этот период самцы активно поют, но самый разгар пения начинается с III декады мая. В ур.Квак в этот период В.ВЛео-нович (1962) регистрировал до 2—3 одновременно поюших самцов. Нами здесь же 11.VI 1978 г. отмечено одновременное пение 3 самцов. Песня по набору звуковых^ строф и их тембру довольно примитивна и отдаленно напоминает песню обыкновенной горихвостки. Поющий самец очень осторожен и держится скрытно в кронах деревьев, постоянно перелетая по гнездовому участку. Птица старается садиться на ветви среди листвы, которая ее полностью или почти полностью закрывает. Если же мухоловка садится на открытую сухую ветвь, то она, как правило, находится в центре кроны и со всех сторон окружена соседними ветвями. Во время пения самец'обычно сидит неподвижно с приподнятой головой.
С территории СССР известны только 2 гнезда этого вида, найденные В.В Леоновичем (1962) 25.V и 1.VI 1959 г. в ур.Квак. Первое было построено на ветке арчи на высоте чуть более 2 м от поверхности земли, второе — в развилке ствола тонкого клена на высоте 6 м.
Гнезда представляют собой аккуратные чашевидные, хорошо замаскированные постройки. Их наружный слой птицы строят из большого количества мха (Brachythe-cium sp.), кусочков талломов лишайников, тонких сухих стебельков и листьев трав; внутренний — из мягких лубяных и других растительных волокон. Последними выстилается и лоток гнезда с добавлением небольшого количества мягких перьев и волоса. Снаружи вся постройка оплетается паутиной. Размеры 1 гнезда, мм: диаметр 70 х 80; высота 65; диаметр лотка 58; глубина лотка 40.
На территории СССР в сезон размножения рыжехвостая мухоловка имеет 1 репродуктивный цикл. Полная кладка обычно состоит из 3-4 яиц (Леонович, 1962; АН, Ripley, 1972). Яйца бледнр-серовато-бирю-зовые с мелкими размытыми красновато
ржавчатыми крапинками и точками, образующими вокруг тупого полюса ’’венчик” шириной до 3,5 мм. Размеры, индекс удлиненности и масса яиц приведены в таблицах 4 и 5.
Сезон размножения этого вида на территории СССР длится со II декады мая по I декаду июля. В Кашмире гнезда с кладками находили и в июле (Bates, Lowther, 1952).
В 1 гнезде, найденном в ур.Квак 25.V 1959, находилась скорлупа разбитого яйца, в другом, обнаруженном здесь же 1.VI 1959, — полная кладка из 4 ненасиженных яиц (Леонович, 1962). К середине июня в большинстве гнезд появляются птенцы. В ур.Квак В.ВЛеонович (1962) 13.VI 1959 г. наблюдал 2 пары мухоловок, очевидно, уже у гнезд с вылупившимися птенцами. В III декаде июня и в июле И.А.Аб-дусалямов (1973) отмечал взрослых птиц, выкармливающих птенцов. С конца II декады июня начинают встречаться слетки. 18.VI 1948 г. А.И.Иванов добыл в ур.Оби-сафед (Варзобское ущ.) слетка с недоросшими рулевыми и маховыми- перьями (колл. ЗИН). 28.VI выводок из 2-х слетков, только что покинувших гнездо, встречен РЛ.Беме (1975).
Линька. Изучена недостаточно. Две молодые птицы (колл. ЗИН), добытые 28.VI 1948 г. в ур.Обисафед, полностью сменили оперение гнездового наряда на верхней части головы. Молодая самка, добытая 3.VIII 1885 г. в уш.Сангардак (колл. ЗИН), находилась в состоянии интенсивной линьки: оперение зашейка и спины сменилось наполовину, а перья верхней части головы заканчивали линьку. У слетка, пойманного 28.VI, послегнездовая линька полностью закончилась к сентябрю (Беме, 1975).
Взрослые птицы имеют в году 1 линьку оперения (полная послебрачная), которая протекает на местах гнездования. Взрослая самка, добытая 24.VIII 1915 г. в верховьях р.Яхсу у кишл.Обизелолак, закончила линьку и была в свежем пере (Зарудный, 1926).
Питание. Изучено недостаточно. Основу пиши составляют насекомые разных групп. Одинаково успешно пользуется различными способами охоты: от ловли насекомых в воздухе на лету в стремительном броске с присады до склевывания беспозвоночных с поверхности растений.
В 2 желудках взрослых самцов, добытых 11.VI 1978 г. в ур.Квак, нами обнаружены: полужесткокрылые - 1 особь
163
(в 1 желудке); жесткокрылые — 15(2): щелкун - 1(1), златка (Buprestidae) -1(1), долгоносики - 6(2); верблюдка (Raphidioptera) — 1(1); чешуекрылые — 5(2): голые гусеницы (длина 15-20 мм) -4(2), имаго - 1(1); перепончатокрылые - 12/2): муравьи - 4(2) (Пекло, 1982).
Отлет и осенний пролет на территори СССР. Данных недостаточно. В Таджикистане начинается со II декады августа (Абдусалямов, 1973). В верховьях р.Яхсу у кишл. Обизелолак одиночная самка добыта 24.VIII 1915 г. (Зарудный, 1926; Иванов, 1940; 1969). Во II декаде сентября рыжехвостые мухоловки полностью исчезают с мест гнездования. На южных склонах Гиссарско-го хр. в долине р.Варзоб самые поздние особи отмечены 13—14.IX (Абдусалямов, 1973).
Практическое значение и мероприятия по охране. Уничтожает многих насекомых — вредителей ле
са и переносчиков возбудителей ряда за-) болеваний человека и животных.
В связи с тем что рыжехвостая мухоловка является редким видом фауны СССР, а основные места ее гнездования на южных склонах Гиссарского хр. находятся за пределами заповедных территорий Таджикской ССР, мы предлагаем организовать полную охрану этого вида на территории СССР, а также мест его гнездования в ущелье р.Варзоб. Для этого необходимо внесение рыжехвостой мухоловки в Красные книги СССР и Таджикской ССР, а также создание заказника в ущ.Кондара, тем более что в нем сохранился и ряд других видов животных и растений, также нуждающихся в полной охране. Хозяйственные работы на этом участке, посещение ущелья людьми должны быть сведены до минимума и взяты под строгий контроль руководством биологической станции и Института ботаники АН ТаджССР, в ведении которых в настоящее время находится уш.Кондара.
164
СПИСОК ЛИТЕРАТУРЫ
Абдусалямов ИЛ. Фауна Таджикской ССР. -Душанбе, 1973. - Т.19, - ч.2. Птицы. - 404 с.
Аверин Ю.В. Наземные позвоночные Восточной Камчатки. - М., 1948. - 224 с. - (Тр. Кроной. заповедника; Вып. 1).
Аверин Ю.В,, Насимович А.А. Птицы горной части Северо-Западного Кавказа // Тр. Кавк, заповедника. - 1938. - Вып. 1. - С.5-56.
Аверин Ю.В., Ганя И.М, Птицы Молдавии. -Кишинев, 1970. - Т.1. - 240 с.
Адамян М.С. Среда обитания и некоторые экологические особенности птиц окрестностей Еревана // Фауна позвоноч. животных АрмССР. -1974.- Вып. 16.- С.5-30.
Адамян М.С. Экология кавказской мухоловки белошейки // Материалы конф. "Фауна и ее охрана в республиках Закавказья”. - Ереван : АН АрмССР, 1975. - С.14-16.
Адамян М.С., Абрамян А.А. Результаты привлечения насекомоядных птиц в Дилижанском заповеднике // Биол. журн. Армении. - 1973. -26, № 3. - С.45-54.
Алекнавичус А. Материалы по воробьиным птицам зеленых насаждений некоторых городов Литвы // Новости орнитологии: Материалы IV Все-союз. орнитол. конф. - Алма-Ата, 1965. - С.8-9.
Алекнонис А. Величина кладок лесных птиц Литвы // Экология птиц Литовской ССР. I. Материалы орнитологических исследований в Литве. -Вильнюс, 1977а. - С.107-113.
Алекнонис А. Весенний прилет птиц в окрестности Лекечяй в 1959-1974 годах // Там же. -19776. - С.114-120.
Александрова ИВ. Опыт привлечения серой мухоловки и изучение ее питания // Пути и методы использования птиц в борьбе с вредными насекомыми. - М., 1956. - С.57-61.
Андреев Б.Н. Птицы Вилюйского бассейна. -Якутск, 1974. - 312 с.
Аникин В.И. Об особенностях пространственной структуры населений птиц в различных местообитаниях // Материалы IV науч. конф, зоологов пед. ин-тов. - Горький, 1970. -С.327-329.
Анорова Н.С. Размножение популяции мухоловки-пеструшки в зависимости от возраста птиц // Орнитология. - 1976. - Вып. 12. - С.77-86.
Анорова Н.С. Факторы, определяющие успех размножения мухоловки-пеструшки // Там же. -1984. - Вып. 19. - С.100-112.
Апостолов Л.Г., Булахов В.Л., Губкин А.А. Характеристика питания птенцов воробьиных птиц в условиях лесов среднего течения р.Орель // Тр. комплекс, экспедиции Днепропетр. ун-та. -1968. - Вып. 1. - С.156-163.
Ардамацкая Т.Б., Семенов С.М. Экологофаунистический очерк птиц района Черноморского заповедника // Вести, зоологии. - 1977. -№2.-С.18-43.
Артемьев А.В. Совмещение линьки и гнездования у мухоловки-пеструшки // Биологические проблемы Севера: 9-й симпоз. "Экология жи
вотных, физиология и биохимия человека и животных, адаптация человека к Северу, охрана природы”: Тез. докл. - Сыктывкар, 1981. -Т.2.-С.5.
Асоскова Н.И. Особенности населения птиц города Архангельска // Птицы и урбанизированный ландшафт. - Каунас, 1984. - С.17-18.
Ахмедов К.Р, Птицы населенных пунктов юго-западного Таджикистана // Учен. зап. Ста-линобад. жен. пед. ин-та. - 1957. - 1. - с.27-100.
Бабенко В.Г Материалы к авифауне района озера Орель, Нижнее Приамурье // Фаунистика и биология птиц юга Дальнего Востока. - Владивосток, 1984. - С.18-27.
Бакаев С. Сроки весеннего пролета птиц в низовьях р.Зеравшана // Вторая Всесоюз. конф, по миграциям птиц: Тез. сообщ. - Алма-Ата: Наука, 1978. - 4.1. - С.82-83.
Баньковский В.Б. К орнитофауне Закавказья, преимущественно Тифлисской губернии // Изв. Кавк, музея. - 1913. - 7, вып. 3/4. -С.205-286.
Барабаш-Никифоров И.И.. Павловский Н.К. Фауна наземных позвоночных Воронежского гос. заповедника // Тр. Воронеж, заповедника. -1947.-Вып. 2.-С.7-128.
Барабаш-Никифоров И.И., Семаго Л.Л. Птицы юго-востока Черноземного центра. - Воронеж: Изд-во Воронеж, ун-та, 1963. - 212 с.
Бараускас Р. Плотность гнездящихся птиц и их распределение по биотопам в деревне Ши-лейкяй (Рокишский р-н, Северо-Восточная Литва) // Тез. докл. X Прибалт, орнитол. конф. -Рига, 1981.-Т.1.-С.8-9.
Баскаков В.В. К авифауне хребта Хамар-Дабан (Прибайкалье) // Экология и охрана птиц: Тез. VIII Всесоюз. орнитол. конф. - Кишинев : Штиинца, 1981. - С. 19.
Басов Ю.С., Нечаев В.А. Гнездовая жизнь птиц: (Обзор ст.) // Природа. - 1961. - № 5. -С.75-78.
Батыршина Н.К. К вопросу о зараженности кокцидиями птенцов некоторых видов воробьиных птиц // VII Всесоюз. орнитол. конф. - Киев : Наук, думка, 1977. - 4.2. - С.67.
Бауманис Я., Блум П., Виксне Я. и др. Птицы Латвии: Террит. размещение и численность. -Рига : Зинатне, 1983. - 224 с.
Белопольский Л.О. Птицы Судзухинского заповедника (воробьиные и ракшеобразные) // Сборник Памяти академика П.П.Сушкина. -М. ; Л. : Изд-во АН СССР, 1950. - С.360-406.
Белоусов Ю.А. Адаптация птиц к урбанизированному ландшафту // Птицы и урбанизированный ландшафт. - Каунас, 1984. - С.23-24.
Беляев К. Г. К характеристике миграций воробьиных птиц на северо-восточном побережье оз.Байкал // Миграции и экология птиц Сибири: Тез. докл. орнитол. конф. - Якутск, 1979. -С.8-10.
Беляев К.Г. Летнее население птиц лесного пояса Баргузинского хребта // Размещение и численность позвоночных Сибири. - Новосибирск : Наука, 1982. - С.217-227.
Белянина И.С., Белянин В.Н. Птицы Жигулевского заповедника // Эколого-фаунистические исследования в заповедниках. - М., 1981. -С.103-119.
Бернацкий Г.И. Птицы Пицундского заповедника: (Предвар. обзор) // Тр. Абхаз, музея. -1958.-Вып. 3.-С.31-81.
Беме Л.Б. Птицы Северной Осетии и Ингу-
165
шии (с прилежащими районами) // Учен. Зап. Сев.-Кавк. Ин-та краеведения. - 1926. - 1. -С.175-274.
Беме Р.Л. Птицы Центрального Кавказа // Учен. зап. Сев.-Осет. лед. ин-та. - 1958. - 23, выл. 1. - С.111-183.
Беме Р.Л. К познанию орнитофауны района Кубенского озера Вологодской области // Орнитология. - 1962. - Вып. 5. - С.92-95.
Беме Р.Л. Птицы гор южной Палеарктики. -М. : Изд-во МГУ, 1975. - 182 с.
Беме Р.Л., Сытое Н.А. Наблюдения над весенним пролетом птиц в Гиссарском хребте // Изв. Отд. биол. наук АН ТаджССР. - 1963. - 3. -С.108-112.
Бианки В.Л. Птицы, наблюдавшиеся в июне 1913 года в Оршанском уезде Могилевской губернии // Орнитол. вести. - 1914. - № 4. -С.260-266.
Бианки В.В., Флеров А.И. Список птиц Кандалакшского залива и его побережий // Тр. Кан-далакш. заповедника. - 1960. - Вып. 2. -С.105-112.
Благосклонов К.Н. Семейство мухоловковые - Muscicapidae // Птицы Советского Союза. -М. : Сов. наука, 1954. - Т.6. - С.73-118.
Благосклонов К.Н. Птицы Кандалакшского заповедника и окрестностей Беломорской биолог. станции Московского университета // Тр. Кандалакш. заповедника. - 1960. - Вып. 2. -С.5-106.
Благосклонов К.Н. Экспериментальное изучение гнездовых участков у мелких птиц // Проблемы орнитологии: Тр. Ill Всесоюз.орнитол.конф. -Львов, 1962. - С.168-173.
Богданов А.Н. Птицы бассейна реки Зерав-шан // Тр. Ин-та зоологии и паразитологии АН УзССР. - 1956. - 5 - С.107-163.
Богородский Ю.В. Численность лесных птиц южного Предбайкалья // Миграции и экология птиц Сибири: Тез. докл. орнитол. конф. - Якутск, 1979. - С.63-64.
Боровицкая Г.К, Измайлов ИВ. Изменения в структуре населения лесных птиц под влиянием деятельности человека // География и экология назем, позвоночных. - 1978. - Вып. 3. - С.8-10.
Бородин Л-П. Дополнения к фауне Мордовского заповедника по материалам И.Д.Щербакова // Тр. Мордов. заповедника. - 1967.- Вып.4. -С.58-71.
Бостанжогло В.Н. Орнитологическая фауна Арало-Каспийских степей. - М., 1911. - 410 с.
Боченко В.Е. Изменения в составе фауны птиц городских парков // Орнитология в СССР: Тез. V Всесоюз. орнитол. конф. - Ашхабад, 1969. - Кн.2. - С.78-80.
Брауде М.И, О северном пределе распространения некоторых птиц нижней Оби // Продуктивность биоценозов Субарктики. - Свердловск, 1970. - С.24-31.
Будниченко А.С. Птицы искусственных лесонасаждений степного ландшафта и их питание. -Воронеж, 1968. - 4.2. - 264 с.
Будниченко А.С., Козлов П.С. О составе и структуре авифауны Белгородской области // Науч. тр. Курск, пед. ин-та. - 1980. - 202. -С.64-82.
Булахов В.Л. Формирование орнитофауны Днепродзержинского водохранилища // Орнитология. - 1968. - Вып. 9. - С.178-187.
Валюс М., Калиндрене Э., Пятрайтис А., Скурдис В. Исторические и современные материалы по орнитологии Литовской ССР // Эколо-166
гия птиц Литовской ССР. Сообщ. 1. Материалы орнитологических исследований в Литве. - Вильнюс, 1977. - С.5-32.
Валюс М., Кигуолене В. Птицы холмистого агроландшафта в юго-западе Балтийской1* возвышенности // Там же. Сообщ. 2. Птицы агроландшафта. - Вильнюс, 1981. - С.60-91.
Вартапетов Л.Г. Птицы таежных междуречий Западной Сибири. - Новосибирск : Наука, 1984. -242 с.
Васильченко АЛ. Новые данные по орнитофауне Хамар-Дабана // Орнитология. - 1982. -Вып. 17. - С.130-134.
Васьковский А.Н. Новые орнитологические находки на северном побережье Охотского моря // Зоол. журн. - 1956. - 35, вып. 7. -С.1051-1058.
Велижанин А.П., Велижанин Г.А. Список птиц Барнаульского округа // Uragus. - 1929. -№ 1. - С.5-14.
Вилкс Е.К Удачный опыт направленного изменения стереотипа гнездования у мухоловки-пеструшки // Новости орнитологии: Материалы IV Всесоюз. орнитол. конф. - Алма-Ата: Наука, 1965. - С.65-67.
Вильконский Ф.В. Орнитологическая фауна Аджарии, Гурии и северо-восточного Лазистана. -М., 1897. - 115 с. - (Материалы к познанию флоры и фауны Российс. империи; Вып. 3).
Виноградова Н.В., Дольник В.Р., Ефремов В.Д., Паевский В.А. Определение пола и возраста воробьиных птиц фауны ССР: Справочник. -М. : Наука, 1976.- 192 с.
Винтер С.В. Гнездование ширококлювой мухоловки в Южном Приморье // Орнитология. -1977. -Вып. 13. -С.74-78.
, Владимирская М.И. Птицы Лапландского заповедника // Тр. Лапланд. заповедника. - 1948. -Вып. 3. - С.171-245.
Владышевский Д.В. Население птиц сосновых лесов Киевщины // Орнитология. - 1972. -Вып. 10. - С.130-138.
Волчанецкий И. Б. Материалы по орнитофауне юга Правобережной Украины и Молдавии // Учен, зап. Харьк. ун-та. Тр. НИИ биологии и биол. факультета. - 1959. - 28. - С.75-99.
Волчанецкий И.Б. Эндемики крымской орнитофауны с точки зрения закономерностей географической изменчивости // Там же. - 1962. - 32. -С.73-87.
Волчанецкий И.Б. К изучению географической изменчивости рисунка и окраски оперения птиц // Проблемы эволюции. - Новосибирск : Наука, 1972. - Т.2. - С.198-211.
Волчанецкий И.Б., Пузанов И.И., Петров В.С. Материалы по орнитофауне Северо-Западного Кавказа // Учен. зап. Харьк. ун-та. Тр. НИИ биологии и биол. факультета. - 1962. - 32. - С.7-72.
Воробьев К.А. Материалы к орнитологической фауне дельты Волги и прилежащих степей // Науч. тр. заповедников. Сер. 1. Астрахан. заповедник. - 1936. - Вып. 1. - С.3-52.
Воробьев КА. К орнитофауне Дальнего Востока // Тр. Сихотэ-Алин. заповедника. - 1938. -Вып. 2. — С.27-56.
Воробьев К.А. Новые сведения по орнитофауне Уссурийского края // Докл. АН СССР. -1951. - 77, № 1. - С.141-144.
Воробьев К.А. Птицы Уссурийского края. -М. : Изд-во АН СССР, 1954. - 359 с.
Воробьев К.А. Результаты орнитологических исследований хребта Черского // Орнитология. -1959. - Вып. 2. - С.115-121.
Воробьев К.А. Птицы Якутии. - М.: Изд-во АН СССР, 1963.- 336 с.
Воробьев КА. Некоторые результаты орнитологических исследований в Гиссарском хребте Ц Орнитология. - 1968. — Выл. 9. -С.164-168.
Воронин А.А., Акментьева В.Р. Размножение птиц в искусственных гнездовьях в условиях Калужского городского бора // Новые проблемы зоологической науки и их отражение в вузовском преподавании. - Ставрополь, 1979. - 4.2. -С.226-227.
Воронцов Е.М., Орлова Ю.Я., Творогова М.М., Хохлова Н.А. Изменения орнитофауны лесного участка Пустынской биостанции Горьковского университета // Орнитология. - 1962. - Вып. 4. -С.117-121.
Вронский Н.В. Размещение и количественная характеристика населения птиц долины реки Камчатки // Там же. - 1977. - Вып. 13. - С.12-21.
Второв П.П. Осенние группировки птиц фисташников Бадхыза // Там же. - 1976. - Вып. 12. -С.224-226.
Второв П.П., Дроздов Н.Н, Летне-осеннее население птиц таежных ландшафтов Вологодской области // Там же. - 1960. - Вып. 3. - С.131-138.
Гавриленко Н.И. Птицы Полтавщины. - Полтава: Полтав. союз охотников, 1929. - 142 с.
Гагина Т.Н. К фауне птиц Северного Байкала // Изв. Вост.-Сиб. отд. Геогр. о-ва СССР. -1954. - Т.58. - С.59-85.
Гагина Т.Н. Птицы Байкала и Прибайкалья // Зап. Иркут, обл. краевед, музея. - Иркутск, 1958. - С. 39-54.
Гагина Т.Н. Птицы Юго-Восточного Забайкалья // Биол. сб. Вост.-Сиб. отд. Геогр. о-ва СССР / Противочум. ин^г Сибири и Дал. Востока. -Иркутск, 1960а (1961). - С.179-213.
Гагина Т.Н. К фауне птиц Витимо-Олекмин-ской горной страны // Изв. Иркут, с.-х. ин<а. -19606. - Вып. 18. - С.211-240.
Гагина Т.Н. Птицы бассейна реки Баргузина // Тр. Баргуз. заповедника. - 1960в. - Вып. 2. -С.115-126.
Гагина Т.Н. Птицы Восточной Сибири (Список и распространение) // Там же. - 1961. -Вып. 3.- С.99-123.
Гагина Т.Н. Дальнейшие замечания и дополнения к списку птиц Восточной Сибири // Там же. - 1967. - Вып. 5. - С.52-64.
Гамборов К.М. Каталог птиц Азербайджана: Отряд Воробьиные - Passeriformes. - Баку : Фан, 1941. - 94 с.
Ганя И.М. Количественная характеристика орнитофауны садов в Приднестровье Молдавии // Орнитология. - 1965. - Вып. 7. - С.290-308.
Ганя И.М., Литвак МД. Экология и хозяйственное значение мухоловок в условиях Молдавии // Вопросы экологии и хозяйственного значения наземной фауны. - Кишинев : Штиинца, 1961.- С.60-74.
Ганя И.М., Литвак МД. Птицы - истребители вредных насекомых. - Кишинев : Штиинца, 1976,- 176 с.
Гвоздок АЛ., Симочко МД. Соня орешниковая (Muscardinus avellanarius L.) конкурент мелких дуплогнездных птиц // Веста, зоологии. -1977.-№3.-С.88-89.
Гептнер В.Г. Фауна позвоночных животных Бадхыза (Южный Туркменистан). - Ашхабад, 1956. - 336 с.
Гептнер В.Г. Интенсивность пения некоторых птиц // Учен. зап. Моск, ун-та. Орнитология. -1958. - Вып. 197. - С.131-143.
Гибет Л.А., Артамошин А.С. Птицы тайги Северного Заангарья // Фауна и систематика позвоночных Сибири. - Новосибирск : Наука, 1977. -С.286-292. - (Тр. Биол. ин-та; Вып.31).
Гизенко А.И. Птицы Сахалинской области. -М. : Изд-во АН СССР, 1955. - 328 с.
Головань В.И. О послегнездовой жизни некоторых представителей семейства мухоловковых // Вести. Ленигр. ун-та. - 1982. - № 9. - С.106-109.
Головань В.И. Поведение молодых мухоловок-пеструшек (Ficedula hypoleuca L.) в послегнездовой период // Сообщ. Прибалт, комис. по изуч. миграций птиц АН ЭССР. - 1983. - № 14. -С.113-118.
Горбань М.И. Гнездование серой мухоловки в старых гнездах других птиц // Орнитология. -1982.-Вып. 17.-С.180-181.
Горбань И.М., Давыдович Л.И., Сребродольская Е.Б. Об урбанизированной орнитофауне Львова и Львовской области // Птицы и урбанизированный ландшафт. - Каунас, 1984. -С.46-47.
Гордеев Ю.И. К орнитофауне восточной части Ханты-Мансийского национального округа (Тюменская область) // Орнитология. - 1965. -Вып. 7. - С.466-467.
Гордеев Ю.И. Материалы по распространению птиц в Ханты-Мансийском округе // Там же. -1977.-Вып. 13. -С.33-39.
Готфрид А.Б. Новые орнитологические находки в енисейской средней тайге // Там же. -1982. -Вып. 17. —С.163.
Гражулявичус Г.Б. Птицы заказника Бир-жулис // Экология птиц Литовской ССР. - Вильнюс, 1976. - С.96-99.
Гражулявичус ГБ. Видовой состав птиц города Клайпеда // Птицы и урбанизированный ландшафт. - Каунас, 1984. - С.47-49.
Греве КА. Птицы, живущие на воле в Рижском зоологическом саду // Орнитол. веста. -1915.-№ 1. - С.78-80.
Губарь В.В. К орнитофауне Минусинского края Ц Uragus. - 1929. - № 2. - С.12-14.
Гуреев С.П. Состав и структура летнего населения птиц северной часта Кузнецкого Алатау // Миграции и экология птиц Сибири: Тез. докл. конф. - Якутск, 1979. - С.71-73.
Гурьев В.Н. Видовой состав, численность и биомасса птиц среднего течения р.Вычегды // Фауна и экология птиц и млекопитающих европейского Северо-Востока СССР. - Сыктывкар, 1982.-С.10-20.
Гусев О.К. Новые данные по орнитофауне Прибайкалья // Орнитология. - 1965. - Вып. 7. -С.87-91.
Гынгазов А.М. Об изменении ареалов птиц в Западной Сибири в результате колебаний климата и воздействия человека // Материалы III Все-союз. орнитол. конф. - Львов : Изд-во Львов, ун-та, 1962а. - Кн.1. - С.119-120.
Гынгазов А.М. Новые данные о распространении птиц в Западной Сибири // Орнитология. -19626. - Вып. 4. - С.154-159.
Гынгазов AM., Москвитин С.С. О распространении некоторых птиц в таежной зоне западной Сибири // Там же. - 1965. - Вып. 7. - С.71-75.
Гынгазов А.М., Миловидов С.П. Орнитофауна Западно-Сибирской равнины. - Томск : Изд-во Томск, ун-та, 1977. - 351 с.
167
Давыдова А.С. Географическая изменчивость суточной активности пения мухоловковых // Материалы IV науч. конф, зоологов пед. ин-тов. -Горький, 1970. - С.342 343.
Даль С.К. Животный мир Армянской ССР. -Ереван: Изд-во АН АрмССР, 1954. - T.I Позвоночные. - 416 с.
Данилов Н.Н. Наблюдения над гнездованием серой мухоловки на Среднем Урале // Тр. III Прибалт, орнитол. конф. - Вильнюс, 1959. -С.81-93.
Данилов Н.Н., Рыжановский В.Н., Ряби-цев В.К. Птицы Ямала. - М. : Наука, 1984. - 334 с.
Дементьев Г.П. Полный определитель птиц СССР С.А.Бутурлина и Г.П.Дементьева. - М. ; Л. : КОИЗ, 1937. - Т.4. - 335 с.
Дементьев Г.П. Полный определитель птиц СССР С.А.Бутурлина и Г.П.Дементьева. - М. ; Л. : КОИЗ, 1941. - Т.5. - 384 с.
Дементьев Г.П., Караев М.К., Карташев Н.Н. Птицы Юго-Западной Туркмении // Учен. зап. Моск, ун-та. - 1955. - Вып. 171. - С.53-172.
Дерим-Оглу Е.Н. Наблюдения над гнездованием малой мухоловки (Muscicapa parva Bechst.) // Тр. по экологии и систематике животных Орехово-Зуев. пед. ин-та. - 1961. - Вып.2. -С.80-86.
Дерим-Оглу Е.Н. Гнездовая территория и территориальное поведение птиц леса // Экология и систематика животных. - М. : Орехово-Зуев. пед. ин-т, 1964. - С.119-157. - (Тр. Орехово-Зуев. пед. ин-та; Вып. 3).
Дмоховский А,В. Птицы Средней и Нижней Печоры // Бюл. Моск, о-ва испытателей природы. Отд. биол. - 1933. - Т.42. - С.214-241.
Доброхотов Б.П. Особенности применения метода линейного трансекта при учете птиц в лесных ландшафтах // Орнитология. - 1962. -Вып. 5. - с.379-385.
Долбик М.С. Птицы Белорусского Полесья. -Минск : Изд-во АН БССР, 1959. - 268 с.
Доржиев Ц.З. Роль большого пестрого дятла для птиц дуплогнездников на хребте Хамар-Дабан // Проблемы экологии Прибайкалья: Тез. докл. Всесоюз. научн. конф. IV. Экологический контроль наземных экосистем. - Иркутск, 1982. -С.85.
Дорофеев А.М. Экология малой мухоловки в Белорусском Поозерье // Орнитология в СССР: Тез. V Всесоюз. орнитол. конф. - Ашхабад, 1969. - Кн.2. - С.207-211.
Дроздов Н.Н. География летнего населения птиц в избранных ландшафтах Азербайджана // Орнитология. - 1965. - Вып. 7- - С.166-199.
Дроздова Ю.В., Таскаева Е.З., Доброхотов Б.П. Материалы по заклещевению птиц в горно-лесных ландшафтах северо-восточного Алтая // Там же. - 1960. - Вып. 3. - С.190-199.
Дубинин В.Б., Горчаковская Н.Н. Некоторые интересные находки птиц в Советском Приморье // Охрана природы. - 1949. - № 7. - С.100-103.
Дубовик А.Д., Миловидов СП, Стрелков В.Е. Фенология весеннего прилета птиц в Томской области // Миграция птиц в Азии. - Новосибирск : Наука, 1977. - С.108-115. - Тр. БИН СО АН СССР.
Ежова С.А. Размещение птичьих гнезд на участках леса с разным уровнем антропогенного воздействия // Новые проблемы зоологической науки и их отражение в вузовском преподавании: Тез. докл. науч.конф.зоологов пед.ин-тов. -Ставрополь, 1979. - 4.2. - С.251-252.
168
Езерскас Л, И. Об использовании искусствен-i ных гнездовий, как средств привлечения птиц-дуплогнездников в леса Литовской ССР // Тр. 111 Прибалт, орнитол. конф. - Вильнюс, 1959. -С.105-109.
Езерскас ЛИ. О биологии птиц-дуплогнез-дников семейств синицевых и мухоловковых в Литовской ССР // Экология и миграция птиц Прибалтики: Тр. IV Прибалт, орнитол. конф. -Рига, 1961. - С.115-122.
Езерскас Л.И. О цикле размножения скворца, мухоловок-пеструшек и большой синицы в лесах Литвы // Материалы 111 Всесоюз. орнитол. конф. -Львов. - 1962. - Кн.1. - С.141-142.
Елсуков С.В. О редких птицах северо-востока Приморья // Тез. VII Всесоюз. орнитол. конф. -Киев : Наук, думка, 1977. - 4.1. - С.58-59.
Елсуков С.В. Птицы // Растительный и животный мир Сихотэ-Алинского заповедника. - М. : Наука, 1982. - С.195-217.
Емельянов А.А. Сборы птиц лета 1924 года по рр.Ботчи и Копи и Северного Сихотэ-Алиня, Хабаровского округа ДВК // Зап. Владивост. отд. рус. геогр. о-ва. - 1929. - Т.4. - С.267-279.
Естафьев А.А. Орнитологическая характеристика бассейна верхней Печоры // Экология и физиология животных. - Сыктывкар, 1969. -С.101-108.
Ефимов А.Я. Птицы истоков Оки. - Орел; Киев: О-во для исслед. природы Орлов, губернии, 1915. - 100 с. - (Материалы к познанию природы Орлов, губернии; № 20).
Железняков Д.Ф., Колесников И.И. Фауна позвоночных Горно-Лесного заповедника // Тр. Горно-Лесн. заповедника. - 1958. - Вып. 1. -С.94-117.
Жордания Р.Г. Орнитофауна Малого Кавказа (в границах Грузинской ССР). - Тбилиси : Изд-во АН ГССР, 1962. - 290 с.
Жордания Р.Г., Гогилашвили ГС. Птицы Лихского хребта и сопредельных мест. - Тбилиси : Мецниереба, 1969. - 72 с.
Жордания Р.Г, Гогилашвили Г.С. Птицы Сванетии (ГрузССР) // Acta ornithol. - 1976. -15, № 6.-С.323-338.
Зарудный Н.А. Птицы Псковской губернии // Зап. Акад, наук по физ.-мат. отд. Сер. 8. -1910. - 25,№ 2. - С.1-181.
Зарудный Н.А. Индийская райская мухоловка в Туркестане (Tchitrea paradisi turkestanica Zar. et Harms) // Материалы к познанию фауны и флоры Рос. империи. - Отд. зоол. - 1915. -Вып. 14. - С.150-170.
Зарудный Н.А. Материалы к познанию орнитофауны Памира и Припамирья // Бюл. Моск, о-ва испытателей природы. Отд. биол. Н.С. -1926. - 35, вып. 3/4. - С.233-290.
Зарудный Н., Билькевич С. Закаспийская черная мухоловка (Muscicapa semitorquata trans-caspia subsp.n.) // Изв. Закасп. музея. - 1918а. -Кн.1. - С.17-18,
Зарудный Н.А., Билькевич СИ. Список птиц Закаспийской области и распределение их по зоологическим участкам страны // Там же. -19186. - Кн.1. - С.19-48.
Земкова Р.И., Штильмарк Ф.Р. Питание некоторых птиц в кедровых лесах Западного Саяна // Фауна кедровых лесов Сибири и ее использование. - М. : Наука, 1965. - С.103-120.
Зимин В.Б, Экология размножения мухоловки-пеструшки в Южной Карелии // Экология, - 1972. - № 5. - С.23-29.
Зиновьев В,И., Шапошников Л.В. Материалы по орнитофауне Калининской области // География и экология назем, позвоночных. - 1978. -Вып. 3. - С.40-53.
Зонов Г.Б., Липин С,И., Сонин В.Д. Особенности использования дупел некоторыми птицами в очагах клещевого энцефалита Приангарья // Изв. Иркут, н.-и. противочум. ин-та Сибири и Дал. Востока. - 1967. - 27. - С.178-182.
Зубцовский Н.Е., Рябицев В.К. Новые данные о птицах полуострова Канин // Орнитология. -1976. - Вып. 12. - С.228-231.
Иванаускас Т. Птицы заповедника Жувин-тас // Заповедник Жувинтас. - Вильнюс : Минтис, 1968. - С.320-337.
Иванов А.И. Птицы Якутского округа. -Л., 1929. - 206 с. - (Материалы комис. по изуч. Якут. авт. сов. соц. республики; Вып. 25).
Иванов А.И. Птицы Таджикистана. - М. ; Л. : Изд-во АН СССР, 1940. - 300 с. - (Тр. Тадж. базы; Т.10. Зоология и паразитология).
Иванов А.И. Летняя орнитофауна Супутии-ского заповедника // Тр. ЗИН АН СССР. - 1952. -9, вып. 4. - С.1081-1099.
Иванов А.И. Птицы Памиро-Алая. - Л. Наука, 1969. - 448 с.
Иванов А.И. Каталог птиц СССР. - Л. : Наука, 1976. - 276 с.
Иванов А.И., Штегман Б.К. Краткий определитель птиц СССР. - Л.: Наука, 1978. - 560 с.
Ивантер Э.В. Птицы заповедника Кивач // Орнитология. - 1962. - Вып. 5. - С.68-85.
Иващенко А.А. О гнездовании редких видов птиц в заповеднике Аксу-Джабаглы // Экологические исследования и охрана птиц Прибалтийских республик. - Каунас, 1982. - С.35-38.
Идзелис Р.Ф. Весенний прилет птиц в г.Виль-нюс и его окрестности // X Прибалт, орнитол. конф.: Тез. докл. - Рига, 1981.-Т.1.-С.124-128.
Измайлов И.В. Фауна птиц и млекопитающих Хоперского государственного заповедника // Тр. Хопер, заповедника. - 1940. - Вып. 1. -С.89-173.
Измайлов И.В. О некоторых особенностях состава птиц Муйской долины // II Всесоюз. орнитол. конф.: Тез. докл. - М.: Изд-во МГУ, 1959. -Т.З.-С.77-78.
Измайлов И.В. Птицы Витимского плоскогорья. -Улан-Удэ: Бурят.кн.изд-во, 1967. - 305 с.
Измайлов ИВ., Ханхасаев В.К. О птицах Муйской долины БурАССР // Краевед, сб. Бурят, фил. геогр. о-ва СССР. - 1959. - Вып. 4. -С.112-128.
Измайлов И.В. Многолетние изменения населения птиц в Пенкинском сосновом лесу (Владимирская область) // География и экология наземных позвоночных Нечерноземья: (Птицы). - Владимир, 1981. - С.5-21.
Измайлов И.В., Боровицкая ГК Характеристика населения птиц лиственничной тайги Витимского плоскогорья // Орнитология. - 1967. -Вып. 8. - С,192-197.
Измайлов И.В., Боровицкая Г-К. Птицы Юго-Западного Забайкалья. - Владимир, 1973. -316 с.
Измайлов И.В., Сальников Г.М. О некоторых географических и экологических закономерностях размещения птиц в пойме р.Клязьмы // География и экология назем, позвоночных. -1978. - Вып. 3.-С.54-65.
Ильичев В.Д. К орнитогеографии Башкир
ского Урала и Лредуралья Ц Орнитология. -1960.-Вып. 3.-С.180-189.
Ильичев В.Д., Фомин В.Е. Орнитофауна Башкирии и ее изменения в XX веке I/ Там же. -1979.-Вып. 14. -С.83-96.
Иноземцев А.А. Элективность питания птиц и некоторые причины ее изменчивости // Там же. - 1963. - Вып. 6. - С.424-450.
Иноземцев А.А. Роль насекомоядных птиц в лесных биоценозах. - Л. : Изд-во ЛГУ, 1978. -264 с.
Иоганзен В.Г. Наблюдения над гнездованием мухоловки-пеструшки (Muscicapa atricapilla Linn.) близ Томска // Орнитол. вести. - 1914. - № 4. -С.293-295.
Иоганзен В.Г. Описание яиц и гнезд птиц Сибири и прилегающих к ней стран // Изв. Том. ун-та, - 1923.- 72. - С.1-31.
Иогансен Г.Х. Материалы по орнитофауне Южно-Уссурийского края // Uragus. - 1927. -№3.-С.19-29.
Исаков Ю.А., Воробьев К.А. Обзор зимовок и пролета птиц на южном Каспии // Тр. Всесоюз. орнитол. заповедника Гасан-Кули. - 1940. -Вып. 1. - С.5-159.
Кадочников Н.П. Материалы по питанию гнездовых птенцов насекомоядных птиц Саваль-ского лесничества Воронежской области и оценка их с точки зрения лесохозяйственного значения птиц в гнездовой период // Тр. Всесоюз. ин-та защиты растений. - 1960. - Вып. 15. - С.225-316.
Казаринов А.П, Фауна позвоночных Больше-хехцирского заповедника // Вопр. географии Дал. Востока. - 1973. - 11. - С.3-29.
Каминский А.М. Наблюдения над редкими и малоизученными птицами Московской губернии Ц Орнитол. вести. - 1916. - № 4. -С.242-246.
Карпов А., Паровщиков В. Пеструшка или пестрая мухоловка // Природа и соц. хоз-во. -1941. - Сб.8,4.2. - С.391-401.
Кириков С.В. Птицы и млекопитающие в условиях ландшафтов южной оконечности Урала. - М.: Изд-во АН СССР, 1952. - 412 с.
Кирпиченко М. Матер1али до орштофауни Бшоцеркгвщини // Журн. бюзоол. циклу ВУАН. -1932,-№4.-С.91-104.
Кирпичников Б.Д. Материалы к познанию птиц Костромской губ // Материалы к познанию фауны и флоры. Отд. зоол. - 1915. - Вып. 14. -С.380-435.
Кисленко Г.С. Длиннохвостая мухоловка в Уссурийском крае // Орнитология. - 1965а. -Вып. 7. - С.108-114.
Кисленко Г.С. О численности птиц в нижнем течении р.Хор (Уссурийский край) Ц Там же. -19656. - Вып. 7. - С.472-473.
Кистяковский А.Б. Птицы садов низовьев Кубани Ц Тр. ин-та защиты растений. Сер. 2. -1932.- 4. -С.111-140.
Юстякгвський О.Б. Птахи ЗакарпатськоГ облает! Ц Труди 1н-ту зоологи АН УРСР. - 1950. -4. - С.3-77.
Кистяковский А.Б., Смогоржевский Л.А. О границе китайского орнитогеографического комплекса на р.Бурее // Науч. докл. высш. шк. Биол. науки. - 1964. - N® 3. - С.26-29.
Кистяковский А.Б., Смогоржевский Л.А. Материалы по фауне птиц Нижнего Амура // Вопросы географии ДальнегоВостока. - Хабаровск, 1973. -Сб.П.-С.182-224.
169
Кищинский А.А. Птицы Колымского нагорья. - М. : Наука, 1968. - 192 с.
Кищинский А.А. Птицы Корякского нагорья. - М. : Наука, 1980. - 336 с.
Клакоцкий В.П. Птицы Припятского ландшафтно-гидрологического заповедника // Заповедники Белоруссии. - 1983. - № 7. - С.98-106.
Ковалев В.А., Присада И.А. Поведение мухоловки-пеструшки и мухоловки-белошейки, гнездящихся на одной территории // Поведение животного в сообществах: Материалы 3 Всесоюз. конф, по поведению животных. - 1983. - Т.2. -С.169-170.
Ковшарь А.Ф. К экологии райской мухоловки (Таласский Алатау) // Орнитология. - 1962. -Вып. 4.-С.234-236.
Ковшарь А.Ф. Птицы Таласского Алатау. -Алма-Ата : Кайнар, 1966. - 436 с. - (Тр. заповедника ”Аксу-Джабаглы”; Вып. 3).
Ковшарь А.Ф. Семейство Мухоловковые -Muscicapidae // Птицы Казахстана. - Алма-Ата : Наука, 1970. - Т.З. - С.424-452.
Ковшарь А.Ф. Райская мухоловка // Красная книга Казахской ССР: Редкие и находящ. под угрозой исчезновения виды животных и растений. -Алма-Ата : Кайнар, 1978. - 4.1. Позвоноч. животные. - С.162-164.
Ковшарь А.Ф. Певчие птицы. - Алма-Ата : Кайнар, 1983. - 280 с.
Коганов С.К. Структура населения птиц г.Сыктывкара // Структура населения птиц европейского Северо-Востока СССР. - Сыктывкар, 1983.-С.50-56.
Кожевникова Р.К. Птицы сосновых насаждений Березинского заповедника // Орнитология. -1965.-Вып. 7. -С.55-62.
Козлова Е.В. Птицы Юго-Западного Забайкалья, Северной Монголии и Центральной Гоби. - Л. : Изд-во АН СССР, 1930. - 396 с. -(Материалы комис. по исслед. Монгол, и Тувин. нар. республик и Бурят-Монгол. АССР; Вып. 12).
Коренкова Р.И. О структуре населения птиц Супутинского заповедника (Приморский край) // Фауна, экология и география животных. - М., 1969. - С.145-151. (Учен. зап. Моск. пед. ин-та им Ленина; №362).
Коренкова Р.И. О структуре населения птиц смешанных лесов Южного Приморья // Материалы IV науч. конф, зоологов пед. ин-тов. - Горький, 1970. - С.351-353.
Королькова Г.Е. Влияние птиц на численность вредных насекомых: (По исслед. в лесостеп. дубравах). - М. : Изд-во АН СССР, 1963. - 126 с.
Коротких А.И., Кузякин А.П., Сучеленко-ва Л.М. О лесных фоновых птицах и млекопитающих о-ва Кунашир // Учен. зап. Моск. обл. пед, ин-та. - М., 1963. - 126; Вып. 6. - С.67-75.
Костин Ю.В. О методике ооморфопогических исследований и унификации описаний оологических материалов // Методики исследования продуктивности и структуры видов птиц в пределах их ареалов. - Вильнюс : Мокслас, 1977. - 4.1. -С.14-22.
Костин Ю.В. Птицы Крыма. - М. : Наука, 1983. - 240 с.
Кречмар А.В. О ландшафтном распределении птиц Юго-Западного Таймыра // Орнитология. -1962. - Вып. 4. - С.29-39.
Кречмар А.В. О сезонных явлениях в жизни птиц района Норильских озер // Там же. - 1963. -Вып.6. - С.37-48.
Кречмар А.В., Андреев А.В., Кондратьев А.Я. Экология и распространение птиц на Северо-Востоке СССР. - М. : Наука, 1978. - 196 с.
Кривошеев В.Г. Новые материалы по авифауне бассейна Яны // Орнитология. - 1960. -Вып. 3.-С.98-105.
Крюков А.П. Необычный для воробьиных птиц акроцентрический кариотип даурской желтоспинной мухоловки (Muscicapa zanthopygia) // Зоол. журн. - 1977. - 56, № 10. - С.1581-1582.
Кудашев А.Е. Предварительный список птиц, наблюдавшихся мною в Сочинском округе, 4ер-номорской губернии // Орнитол. вести. - 1917. -№ 1. - С.20-36.
Кузнецов А.А. Список птиц. Верхней Сване-тии // Распространение и систематика птиц: Исслед. по фауне Сов. Союза. - М.: Изд-во МГУ, 1983.-С.186-190.
Кузнецов А.А. Состав, численность и размещение авифауны высокогорья Киргизского хребта // Орнитология. - 1962. - Вып. 4. - С.237-255.
Кузнецов Н.И. Птицы заповедника ’’Денежкин камень” // Тр. заповедника ’’Денежкин камень”. - 1958. - Вы.1. - С.114-168.
Кузякин А.П., Второв П.П. К ландшафтной орнитогеографии Охотской тайги // Орнитология. - 1963. - Вып, 6. - С.184-194.
Кулешова Л.В. Закономерности обособления типов населения птиц в лесах Среднего Сихотэ-Алиня // Там же. - 1976. - Вып. 12. - С.26-54.
Кулешова Л.В., Матюшкин Е.Н., Кузнецов Г.В. Орнитогеографический очерк хребта Хехцир (Приамурье)//Тамже.- 1965.-Вып.7. -С.97-107.
Кумари Э.В. Фауна птиц природных ландшафтов юго-западной Эстонии // Тр. ЗИН АН СССР. - 1955. - 17. - С.266-294.
Куренцов А.И. В убежищах уссурийских реликтов. - Владивосток, 1961. - 182 с.
Кушнарев Е.Л. Антропогенные сукцессии ор-нитосообществ и территориальные связи местообитаний Западного Сихотэ-Алиня // Фаунистика и биология птиц юга Дальнего Востока. - Владивосток, 1984. - С.71-78.
Лабзюк В.И. Летняя авифауна морского побережья в районе залива Ольги // Орнитологические исследования на Дальнем Востоке. - Владивосток, 1975. - С.279-284, - (Бр. БПиДВНЦ. Н.С.; Т.29).
Лабзюк В.И., Назаров Ю.Н, О редких и новых птицах Южного Приморья // Орнитология. -1967. - Вып. 8. - С.363-364.
Лабзюк В.И, Назаров Ю.Н., Нечаев В.А. Птицы островов северо-западной части залива Петра Великого // Орнитологические исследования на юге Дальнего Востока. - Владивосток, 1971. - С.52-78. - (Тр. БПиДВНЦ. Н.С.; Т.6).
Лавренченко Л.А. Поведение желтоспинной мухоловки в период размножения // Птицы Сибири: Тез. докл. Второй сиб. орнитол. конф. -Горно-Алтайск, 1983. - С.191-192.
Лавренченко Л.А. Факторы, влияющие на успешность гнездования даурской желтоспинной мухоловки // Орнитология. - 1984а. - Вып. 19. -С.204-205.
Лавренченко Л.А. О залете японской желтоспинной мухоловки на Средний Амур // Там же. -19846. - Вып. 19. - С.181-182.
Ларионов В.Ф. К распределению птиц по станциям в окр. с.Зырянки Тюменского округа // Uragus. - 1927. - № 1. - С.10-14.
170
Лебедев Ю.А. Структура населения птиц Центрального Таджикистана: Автореф. дис. ... канд. биол. наук. - Душанбе, 1976. - 24 с.
Левин А.С. О гнездовании серой мухоловки в Юго-Западном Забайкалье // Миграции и экология птиц Сибири: Тез. докл. орнитол. конф. -Якутск, 1979. - С.156-157.
Левин А.С. О сроках гнездования некоторых воробьиных птиц в среднем течении Урала // Животный мир Казахстана и проблемы его охраны. -Алма-Ата : Наука, 1982. - С. 108-110.
Лейвитс А. Формирование местного населения мухоловки-пеструшки // Экологические исследования и охрана птиц Прибалтийских республик. - Каунас, 1982. - С.59-60.
Лемдгольм В.А. Летняя орнитофауна бассейнов рек Охоты, Кетанды и Урака // Сообщ. Дальневост. фил. СО АН СССР. - 1959. - Вып.Н. -С.24-26.
Леонович В.В. К биологии малоизученных птиц Таджикистана // Бюл. Моск, о-ва испытателей природы. Отд. биол. - 1962. - 67, № 2. -С.121-124.
Леонтьев А.Н., Павлов Е.И. Орнитологические наблюдения в долине Чикоя (Читинская область) // Орнитология. - 1963. - Вып. 6. -С.165-172.
Линдеман Г.В. Материалы по орнитофауне лесов Юго-Западного Забайкалья // Там же. -1976. -Вып.12. - С.236-238.
Литвиненко Н.М., Шибаев Ю.В. К орнитофауне Судзухинского заповедника и долины реки Судзухе // Экология и фауна птиц юга Дальнего Востока. - Владивосток, 1971. - С.127-186. -(Тр. заповедника "Кедровая падь”: Вып.11).
Лихачев Г.Н. О величине кладки некоторых птиц в центре Европейской части СССР // Орнитология. - 1967. - Вып. 8. - С.165-1 74.
Лихачев Г.Н. Размножение и численность мухоловки-пеструшки на юге Московской обл. // Птицы и пресмыкающиеся: Исслед. по фауне Сов. Союза. - М. : Изд-во МГУ, 1978. - С. 119-140.
Лиховид А.И. Летнее население птиц лесов Ставропольской возвышенности // Фауна Ставрополья. - Ставрополь, 1977. - Вып. 2. - С.25-37.
Лобков Е.Г. К биологии пестрогрудой мухоловки на Камчатке // Орнитология. - 1977. -Вып. 13. - С.207-208.
Лобков Е.Г. Краткий обзор птиц центральных районов Восточной Камчатки // Птицы и пресмыкающиеся: Исслед. по фауне Сов. Союза. -М. : Изд-во МГУ, 1978. - С.52-72. - (Сб. тр. Зоол. музея Моск, ун-та; Т.17).
Луговой А.Е. Птицы Мордовии. - Горький, 1975. - 300 с.
Луговой А.Е. Структура населения птиц хвойных лесов Присурья // География и экология наземных позвоночных Нечерноземья: (Птицы) . - Владимир, 1981. - С.49-60.
Луговой А.Е. Особенности городских орни-токомплексов в горах (Украинские Карпаты) // Птицы и урбанизированный ландшафт. - Каунас, 1984. - С.91-93.
Луговой А.Е., Кяжин И.С., Чаиркин А.С. Структура поселения птиц островных лиственных лесов Присурья // География и экология назем, позвоночных. - 1978. - Вып. 3. - С.75-85.
Ляйстер А.Ф., Соснин Г.В. Материалы по орнитофауне Армянской ССР (Omis Armenia-са). - Ереван, 1942. - 402 с.
Лямкин В.Ф. Картографирование населения птиц Баргузинской котловины (Северо-Западное
Забайкалье) // Миграции и экология птиц Сибири: Тез. докл. орнитол. конф. - Якутск, 1979. -С.89-91.
Мазинг В.В. О подборе древесных и кустарниковых пород для привлечения открытогнездя-щихся птиц в городах // Орнитология. - 1960. -Вып.З. - С.425-429.
Майр Э. Принципы зоологической систематики. - М. : Мир, 1971. - 456 с.
Майхрук М.И., Кузнецов Н.П. О привлечении птиц-дуплогнездников в пригородные леса Саранска // Тез. докл. VII Всесоюз. орнитол. конф. -Киев : Наук, думка, 1977. - 4.2. - С.152-153.
Макаров В.В. Материалы по биологии птиц Курской области в гнездовой период // Охрана фауны позвоночных животных лесостепной и степной зон европейской части СССР. - Курск, 1980. -С.49-57. - (Науч. тр. Курск, пед. инн'а; Т.202).
Мальчевский А.С. Гнездовая жизнь певчих птиц: (Размножение и постэмбрион, развитие лесных воробьиных птиц Европ. части СССР). - Л. : Изд-во ЛГУ, 1959. - 282 с.
Мальчевский А.С., Пукинский Ю.Б. Птицы Ленинградской области и сопредельных территорий: История, биология, охрана. - Л.: Изд-во ЛГУ, 1983. - Т.2. - Певчие птицы. - 504 с.
Малышев Л.И. Материалы к орнитофауне северо-западного побережья Байкала // Тр.Вост.-Сиб. фил. Сиб. отд. АН СССР. Сер. биол. - 1960а. -Вып. 23. - С.53-68.
Малышев Л.И. Птицы Северо-Восточного побережья Байкала // Тр. пробл. и темат. совещ. ЗИН АН СССР. - 19606. - Вып. 9. - С.81-91.
Малышевский Р.И. Летние наблюдения над птицами Терского берега Белого моря // Орнитология. - 1962. - Вып. 5. - С.13-27.
Мараков С.В. Редкие и новые птицы Командорских островов // Там же. - с.166-167.
Марвин М.Я. Животный мир Кйрепо-Финской ССР. - Петрозаводск, 1951. - 196 с.
Марисова И.В., Холина НМ. К биологии мухоловки-белошейки (Muscicapa albicollis Teinm.) в западных областях Украины // Фауна и животный мир Советских Карпат. - Ужгород, 1959. -С.75-81. (Науч. зап. Ужгор. ун-та; Т.40).
Мартынов Е.Н. Синантропность птиц на примере Ленинграда // VII Всесоюз. орнитол. конф. -Киев : Наук, думка, 1977. - 4.2. - С.154-155.
Маслов Н.М. Птицы Бухарской области // Сборник научных трудов Бухарского педагогического института. - Бухара, 1947. - С.3-26.
Матвеев С.Д. Методы и некоторые результаты ревизии систематических категорий птиц Балканского полуострова // Новости орнитологии: Материалы IV Всесоюз. орнитол. конф. - Алма-Ата : Наука, 1965. - С.234-237.
Матюшкин Е.Н. О населении птиц лесостепи Казахстана // Орнитология. - 1967. - Вып. 8. -С.198-210.
Меднис А.А. Результаты проверки маршрутного метода учета лесных воробьиных птиц в Латвийской ССР // Новости орнитологии: Материалы IV Всесоюз. орнитол. коиф. - Алма-Ата : Наука, 1965. - С.237-238.
Мекленбурцев Р.Н. Материалы по наземным позвоночным бассейна реки Кашка-дарья. - Ташкент: САГУ, 1958. - 141 с. - (Тр. Среднеаз. ун-та. Н.С.; Вып. 130. Биол. науки, кн. 30).
Мекленбурцев Р.Н. Население гнездящихся птиц Ташкента и многолетние изменения его состава // Бюл. Моск, о-ва испытателей природы. Отд. биол. - 19Й2. - 87, вып.4. - С.36-44.
171
Мешков М.М. Осенний пролет воробьиных в районе Псковско-Чудского водоема // Экология и миграция птиц Прибалтики: Тр. IV Прибалт, орнитол. конф. - Рига, 1961. - С.199-206.
Милованова Г.А. Материалы по питанию мухоловки-пеструшки и большой синицы // Пути и методы использования птиц в борьбе с вредными насекомыми. - М., 1956. - С.21-37.
Миловидов С.П., Москвитин С.С. Новые сведения по распространению птиц в Западной Сибири // Вопросы ботаники, зоологии и почвоведения: Сб. работ молодых ученых. - Томск : Изд-во Том. ун-та, 1973. - Вып. 1. - С.112-116.
Михаловский И. Орнитологические наблюдения в Закавказье летом 1878 года // Тр. Санкт-Петербург. о-ва естествоиспытателей. — 1880. -11, вып. 1. - С. 64-87.
Михеев А.В. О плотности населения насекомоядных птиц в связи с привлечением их в лесные насаждения. - М., 1953. - С.147-160. - (Учен, зап. МГПИ им Ленина ; Т.74. Вып. 2).
Михельсон Г.А., Чаун М.Г., Виксне Я.А. О расселении мухоловки-пеструшки на гнездование по данным кольцевания // Пути и методы использования птиц в борьбе с вредными насекомыми. - М., 1956. - С.144-152.
Михлин В.Е., Сальников ГМ. К изучению широтной изменчивости структуры Орнитонаселе-ния смешанного леса // География и экология наземных позвоночных Нечерноземья: (Птицы). -Владимир, 1981. - С.67-74.
Мищенко А.Л., Смиренский С.М. Птицы озера Болонь // Компоненты биоты и их роль в природе и народном хозяйстве: Докл. Моск, о-ва испытателей природы. Зоология и ботаника. - 1978. -М„ 1981. - С.40-42.
Мищенко Ю.В. Новые материалы по орнитофауне Копетдага // Вести, зоологии. - 1983. -№5. - С.87-88.
Моламусов Х.Т. Птицы центральной части Северного Кавказа. - Нальчик: Кабард.-Балкар. кн. изд-во, 1967. - 100 с.
Морозов В.В. Орнитофауна окрестностей озера Капчук, плато Путорана // Орнитология. -1984. - Вып. 19. - С.30-40.
Москвитин С.С. К авифауне лесной зоны Западной Сибири // Там же. - 1968. - Вып. 9. -С.360-361.
Москвитин С.С. К авифауне лесной зоны Западной Сибири // Там же. - 1972. - Вып. 10. -С.372-373.
Москвитин С.С. Материалы по распространению и образу жизни некоторых птиц Сибири // Фауна Сибири. - Новосибирск : Наука, 1973. -С.263-268. - (Тр. БИН СО АН СССР; 4.2. Вып.16 ).
Москвитин С.С., Дубовик А.Д., Горд Б.Я. Птицы долины р.Кеть // Фауна и систематика позвоночных Сибири. - Новосибирск : Наука, 1977. - С.245-279. - (Гр. БИН СО АН СССР; Вып. 31).
Навасайтис А., Курлавичюс П. Орнитофауна насаждений города Каунас и его окрестностей в 1970—1974 годах // Экология птиц Литовской ССР. I. Материалы орнитол. исслед. в Литве. -Вильнюс, 1977. - С.135-157.
Назаренко А.А. Краткий обзор птиц заповедника ’’Кедровая падь” // Орнитологические исследования на юге Дальнего Востока. - Владивосток, 1971а. - С.12-51. - (Тр. БПиДВНЦ. Н.С.; Т.6).
Назаренко А.А. Птицы вторичных широколиственных лесов Южного Приморья и некоторые аспекты формирования природных сообществ // Там же. - 19716. - С.79-97.
Назаренко А.А. О распространении некоторых птиц в Южном Приморье //Там же. - 1971в. -С.172-179.
Назарет.ко А.А. К распространению и биологии пестрогрудой мухоловки - Muscicapa griseisticta (Swinh.) в Южном Приморье // Там же. -1971г.-С.180-187.
Назаренко А А. К орнитофауне Хэнтэй-Чи-койского нагорья, Южное Забайкалье // Экология и зоогеография некоторых позвоночных суши Дальнего Востока. - Владивосток : ДВНЦ АН СССР, 1978.-С.40-56.
Назаренко А.А. О птицах окрестностей поселка Экимчан, крайний восток Амурской области, 1981-1983 гг. // Фаунистика и биология птиц юга Дальнего Востока. - Владивосток, 1984а. — С.28-33.
Назаренко А.А. Птичье население смешанных и темнохвойных лесов Южного Приморья, 1962-1971 гг. // Там же. - 19846. - С.60-70.
Назаров Ю.Н., Казыханова М.Г, Курин-ный В.Н. Пролет птиц на о-ве Большой Пелис в сентябре 1977 г. // Вторая Всесоюз. конф, по миграциям птиц: Тез. сообщ. — Алма-Ата : Наука, 1978. - 4.1.-С.148-150.
Наумов Р.Л. Фауна и распределение птиц окрестностей села Б.Кемчуг (Красноярский край) // Орнитология. - 1960. - Вып. 3. -С.200-211.
Наумов С.П., Лабутин Ю.В. Материалы по авифауне Верхоянской складчатой страны. I. Состав и некоторые особенности распространения видов авифауны Западного Верхоянья // Бюл. Моск, о-ва испытателей природы. Отд. биол. - 1961. -66, № 6. - С.116-125.
Нейфельдт И.А. Об орнитофауне южной Карелии // Тр. ЗИН АН СССР. - 1958. - 25. -С.183-254.
Нейфельдт И.А. Питание воробьиных птиц в Южной Карелии // Зоол. журн. - 1961. - 40, вып. 3. - С.416-426.
Нейфельдт И.А. Пуховые птенцы некоторых азиатских птиц // Орнитологический сборник к 100-летию со дня рождения академика П.П.Суш-кина. - Л.: Наука, 1970. - С.111-181. (Тр. ЗИН АН СССР; Т.47).
Нейфельдт ИА. Размножение и линька Не-michelidon griseisticta (Muscicapinae. Aves) // Докл. АН СССР. - 1971. - 200, № 2. - С.485-487.
Нечаев В.А. Орнитофауна основных биотопов Северного Сихотэ-Алиня // Вопросы географ'ии Дальнего Востока. - Хабаровск, 1960. - Сб. 4. -С.264-266.
Нечаев В.А. Новые данные о птицах Нижнего Амура // Орнитология. - 1963. - Вып. 6. -С.177-183.
Нечаев В.А. Орнитологические находки на южных Курилах // Там же. - 1965. - Вып. 7. -С.122-129.
Нечаев В.А., Назаров Ю.Н. О питании некоторых птиц Южного Приморья // Экология млекопитающих и птиц. - М. : Наука, 1967. - С.316-320.
Нечаев В.А. Птицы южных Курильских островов. — Л. : Наука, 1969. - 245 с.
Нечаев В.А. Периодические явления в жизни птиц острова Кунашир // Экология и фауна птиц юга Дальнего Востока. — Владивосток, 1971. -
172
С.69-83. - (Тр. заповедника 'Кедровая падь”; Вып.11).
Нечаев В.А. О новых и редких птицах Южного Сахалина // Орнитология. - 1974а. - Вып.И. -С.396-397.
Нечаев В.А. К распространению и биологии некоторых птиц Нижнего Амура //Фауна и экология наземных позвоночных юга Дальнего Востока СССР. — Владивосток, 19746. - С.145—154. -(Тр. БПиДВНЦ; Н.С.; Т.17).
Нечаев В.А. Птицы острова Монерон // Орнитологические исследования на Дальнем Востоке. - Владивосток, 1975. - С.5-25. - (Тр. БПиДВНЦ; Н.С.; Т.29).
Нечаев ВЛ. Новые данные о райской мухоловке — Terpsiphone paradisi incei (Gould) в Южном Приморье // Редкие птицы Дальнего Востока. - Владивосток, 1981. - С.74-79.
Нечаев ВА. Новые сведения о пуховых птенцах воробьиных птиц II Тр. ЗИН АН СССР. -1983.- 116. - С.58-60.
Нечаев В.А. Японская мухоловка - Ficedula narcissina Temm. на острове Сахалин (распространение и биология) // Фаунистика и биология птиц юга Дальнего Востока. - Владивосток, 1984а.-С.87-95.
Нечаев В.А, К биологии таежной мухоловки на острове Сахалин // Вести, зоологии. - 19846. — №4.-С.50-53.
Нечаев В.А., Назаров Ю.Н. Материалы по питанию некоторых птиц Южного Приморья // Науч. докл. высш. шк. Биол. науки. - 1968. -№6.-С.ЗО-ЗЗ.
Никольский А.М. Остров Сахалин и его фауна позвоночных животных. - Спб., 1889. - 334 с. -(Прил. к 9-му т. Зап. Императ. Акад, наук; Т.9. №5).
Новиков Г.А. Экология птиц Хибинских гор (Кольский полуостров) // Тр. ЗИН АН СССР. -1952а. -9, вып. 4. - С.1133-1154.
Новиков Г.А. Материалы по питанию лесных птиц Кольского полуострова // Там же. - 19526. — 9,вып.4. - С.1155-1198.
Новиков ГА., Мальчевский А.С., Овчинникова Н.П., Иванова Н.С. Птицы ’’Леса на Ворскле” и его окрестностей // Вопр. экологии и биоценологии. - 1963. - Вып. 8. - С.9-118.
Огнев С.И., Воробьев КА. Фауна позвоночных Воронежской губернии. - М. : Новая деревня, 1923. - 11. - 255 с.
Омелько М.А. Новые материалы к орнитофауне Южного Приморья // Сообщ. Дал. фил. Сиб. отд-ния АН СССР. - 1963. - Вып. 18. -С.119-121.
Омелько МА. О птицах-орнитохорах Южного Приморья // Орнитология. - 1979. - Вып.14. -С.219-221.
Осмоловская В.И., Формозов А.Н. Очерки экологии некоторых полезных птиц леса // Птицы и вредители леса. - М. : МОИП, 1950. - С.34-142.
Остапенко В.А,, ГавриловВ.М.,Ефремов В.Д., Голубева Т.Н, О некоторых интересных находках птиц на Камчатке и о-ве Кунашир // Орнитология. - 1977. - Вып. 13. - С.192-193.
Павлов Е. Птицы и звери Читинской области. - Чита: Чит. обл. изд-во, 1948. - 151 с. -(Материалы к изучению птиц и млекопитающих Забайкалья),
Павлов Е.И., Пары г ин Ю.А. Орнитологические наблюдения в Верхне-Чарской котловине //
Изв. Забайк. фил. геогр. о-ва СССР. - 1969. -5, вып. 5. - С.85-108.
Павлов П. Орнитологические наблюдения в Рязанской губернии. - СПб., 1879: - 102 с. -(Тр. Санкт-Петербург, о-ва естествоиспытателей; Т.10).
Панов Е.Н. Птицы Южного Приморья (фауна, биология и поведение). - Новосибирск : Наука, 1973. - 412 с.
Пантелеев П.А. О птичьем населении долины Оби и прилежащих ландшафтов в Нарымском крае // Орнитология. - 1972а. - Вып. 10. -С.161-172.
Пантелеев П.А. Материалы к количественной характеристике авифауны южной тайги Зауралья // Там же. - 19726. - Вып. 10. - С.374-377.
Паровщиков В.Я. Весенний прилет и пролет птиц в Москве // Природа и соц. хоз-во. - 1941а. -Кн.8,ч.2.-С.326-333.
Паровщиков В.Я. Систематический список птиц г.Архангельска н его окрестностей // Там же. - 19416. - Кн.8,ч.2.-С.355-366.
Паровщиков В.Я. Новые данные по воробьиным птицам Архангельского севера // Орнитология. - 1959. - Вып. 2. - С.102-103.
Пекло А.М. О родственных связях мухоловок желтоспинной даурской (Ficedula zanthopygia (Нау.) и мугимаки (F.mugimaki (Temm.) // Тез. докл. VII Всесоюз. орнитол. конф. - Киев : Наук, думка, 1977. - 4.1. - С.21-22.
Пекло А.М. Материалы по питанию некоторых воробьиных птиц Памиро-Алая //Вести, зоологии. - 1982. - № 5. - С.59-65.
Пекло А.М., Ломадзе Н.Х., Бахтадзе ГБ. и др. Экология мухоловки малой - Ficedula parva parva (Bechst.) (Aves, Muscicapidae) на Северо-Западном Кавказе // Там же. - 1978. - №5. т С.21-27.
Пекло А.М., Смогоржевский Л.А. Материалы по питанию мухоловок (Passeriformes, Muscicapidae) юга Советского Дальнего Востока // Там же. - 1980. - № 2. - С.17-27.
Петров В.С. Новые виды в летней орнитофауне Ростовской области // Орнитология. - 1965. -Вып. 7. -С.484-485.
Плесский П.В. Орнитологические исследования в Вятском крае // Тр. Вят. музея. - 1927. -1. - С.23-44.
Подольский А.Л., Харин В.Л. Некоторые случаи нетипичного гнездования птиц в Саратовской и Воронежской областях // Орнитология. -1984. - Вып.19. - С.209-210.
Поливанов В.М. Опыт массового переселения мухоловки-пеструшки в степные дубравы // Пути и методы использования птиц в борьбе с вредными насекомыми. - М., 1956. - С.137-138.
Поливанов В.М, Экология птиц-дуплогнездников Приморья. - М. : Наука, 1981. - 172 с.
Поливанов В.М., Поливанова НМ. К вопросу о соотношении внутривидовой специализации и экологической пластичности у птиц // Экология и фауна птиц юга Дальнего Востока- - Владивосток, 1971. - С.7-29. - (Тр. заповедника ’’Кедровая падь” ; вып. 11).
Поливанова Н.Н. Питание птенцов некоторых видов полезных насекомоядных птиц в Дарвинском заповеднике // Тр. Дарвин, заповедника. - 1957. - Вып.4. - С.157-244.
Поливанова Н.Н., Ходков ГН. О биологии синей мухоловки - Niltava cyanomelana (Temm.)
173
Орнитологические исследования на Дальнем Востоке. - Владивосток, 1975, - С.83-92. - (Тр. БПиДВНЦ; Н.С. ; Т.29).
Полозов С.А. Материалы по авифауне юго-западного Копетдага. - Люберцы, 1980. - 66 с. -Рукопись деп. в ВИНИТИ, - № 538-80.
Поляков Г.И. Птицы Богородского уезда с параллельным списком птиц остальной части Московской губернии // Фауна Богород. уезда Моск. губ. - М., 1924. - Вып. 1. - С.1-90.
Попов А.В. Птицы Гиссаро-Каратегина. Эко-лого-географический очерк. - Сталинабад: АН ТаджССР, 1959. - 184 с.
Портенко Л.А. Фауна птиц внеполярной части Северного Урала. - М. : Изд-во АН СССР, 1937- - 240 с.
Портенко Л.А. Очерк фауны птиц Западного Закарпатья // Сборник статей памяти академика П.П.Сушкина. - М. ; Л. : Изд-во АН СССР, 1950а. - С. 39-46.
Портенко ЛА. Новые и редкие находки птиц на крайнем юго-востоке СССР // Докл. АН СССР. -19506. - 70, № 2. - С.319-322.
Портенко ЛА. Новые подвиды воробьиных птиц (Aves, Passeriformes) // Тр. ЗИН АН СССР. -1955. - 18. - С.493-507.
Портенко Л.А. Птицы СССР. 4.IV. - М. ; Л. : Изд-во АН СССР, 1960. - 416 с. - (Определители по фауне СССР ЗИН АН СССР; Т.69).
Поспелов С.М. Птицы и млекопитающие ельника-черничника разных возрастов //' Зоол. журн. - 1957.,- 36, вып.4. - С.603-607.
Птицы Волжско-Камского края (воробьиные) . - М. : Наука, 1978. - 248 с.
Птушенко Е.С. Материалы к познанию фауны птиц Мордовского заповедника // Фауна Мордовского государственного заповедника им.П.Г.Сми-довича. - М„ 1938, - С.41-106.
Птушенко Е.С. Изменение авифауны Ленинских гор и их окрестностей // Орнитология. -1976, -Вып.12. - С.3-9.
Птушенко Е.С., Иноземцев АА. Биология и хозяйственное значение птиц Московской области и сопредельных территорий. - М. : Изд-во МГУ, 1968. - 462 с.
Пузанов И.И. Орнитофауна Северо-Западной Черкессии и некоторые соображения о ее происхождении и связях // Тр. зоол. сектора АН СССР. Грузинский филиал. - М. ; Л., 1938. -С.125-180.
Пузанов И.И., Кипарисов Г.П, Козлов В.И. Звери, птицы, гады и рыбы Горьковской области. - Горький: ОГИЗ, 1942. - 452 с.
Пшеничников Л.Н. К вопросу охраны и привлечения птиц в г.Иркутске н его окрестностях // Охрана природы Сибири: Материалы I Всесоюз. Сиб. конф., 1958. - Иркутск, 1959. - С.68-69.
Равкин Ю.С. Структурные особенности населения птиц Северо-Восточного Алтая // Орнитология. - 1967. - Вып.8. - С.175-191.
Равкин Ю.С. К характеристике весеннего населения птиц Северо-Восточного Алтая // Там же. - 1972. - Вып.10. -С.384-387.
Равкин Ю.С. Птицы лесной зоны Приобья: (Пространств, орг. лет. населения). - Новосибирск : Наука, 1978. - 288 с.
Равкин Ю.С. Пространственная организация населения птиц лесной зоны (Западная и Средняя Сибирь). - Новосибирск : Наука, 1984. - 264 с.
Равкин Ю.С., Глейх И.И., Черников О.А. О населении птиц юга Красноярского края //
Птицы Сибири: Тез, докл. Второй Сиб. орнитол. конф. - Горно-Алтайск, 1983. - С.50-52.
Радде Г.И. Орнитологическая фауна Кавказа (Ornis Caucasica). Систематическое и биологогеографическое описание кавказских птиц. -Тифлис, 1884. - 451 с.
Радищев А.М. Материалы к познанию авифауны Кабарды и Балкарии (отряды Passeriformes, и Coraciiformes) // Учен. зап.Сёв.-Кавк. ин^га краеведения. - 1926. - 1. - С.119-146.
Рахилин В,К. Пути пролета мухоловок Дальнего Востока // Материалы IX симпоз. "Экология вирусов”. - Душанбе : Дониш, 1975. -С.152-154.
Рашкевич Н.А. Материалы по экологии птиц-Нижнего Амура // Вопр. географии Дал. Востока. - 1965. - Сб.7. - С.283-291.
Рашкевич Н.А. Мир пернатых: (Птицы Чечено-Ингушетии) . - Грозный : Чечено-Ингуш. кн. изд-во, 1980. - 208 с.
Резцова С.А. Птицы Пермской губернии: (Сев. р-н: уезды Верхотур, и Чердын). - М., 1904. - 185 с.
Реймерс Н.Ф. Материалы к познанию орнитофауны лесных участков лесостепной зоны Тувинской автономной области // Биологический сборник Восточно-Сибирского отделения Географического общества и Противочумного института Сибири и Дальнего Востока. - Иркутск, 1960 (1961). - С.239-250.
Реймерс Н.Ф. Птицы и млекопитающие южной тайги Средней Сибири. - М. ; Л. : Наука, 1966. - 420 с.
Ренцо О.Я. Список птиц Матсалуского госзаповедника // Тр. заповедников Эстонской ССР. - Таллии : Валгус, 1968. - С.139-150.
Роачева Э.В. -Численность и размножение птиц, нижнего Елогуя: (ПриенИс. тайга) // Орнитология.-1962. - Вып.5. - С.118-134.
Росляков Г.Е. Гнездование некоторых Нижнеамурских птиц // Вопр. географии Дал. Востока. - 1965. - Сб.7. - С.292-296.
Росляков Г.Е. Редкие птицы Хабаровского края, нуждающиеся в особой охране // Редкие и исчезающие животные суши Дальнего Востока СССР: Материалы конф. - Владивосток, 1981. -С.141-144.
Сагитов А.К. К биологии мухоловок Зерав-шанской долины // Тр. Самарканд, унта. -1961. -109. - С.25-36.
Салихбаев Х.С., Остапенко М.М, Птицы // Экология и хозяйственное значение позвоночных животных юга Узбекистана. - Ташкент, 1964. -С.72-144.
Сатунин КА. Материалы к познанию птиц Кавказского края. - Тифлис, 1907. - 144 с.
Сатунин К.А., Берг Л.С., Кириченко А.Н., Муралевич В.С. Фауна Черноморского побережья Кавказа (Fauna Littons Orientalis Ponti Euxini) -Спб., 1913. -248c. - (Тр. о-ва изуч. Черномор, побережья; т.2).
Севастьянов Г.Н. О распределении гнезд-дупел в Архангельской тайге // Орнитология. -1962. - Вып.5. - С.86-91.
Селевин ВА. О птицах окрестностей Змеино-горска // Uragus. - 1928. - № 3/4. - С.14-18.
Семенов Н.Н. К экологии и этологии гнездования мухоловок-пеструшек // Новые проблемы зоологической науки и их отражение в вузовском преподавании. - Ставрополь, 1979. - 4.2. -С.334-335.
174
Семенов С.М. Материалы по питанию мухоловки-пеструшки в гнездовой период // Пути и методы использования птиц в борьбе с вредными насекомыми. - М., 1956. - С.38-39.
Сербин В.А. Структура населения птиц лесостепных дубрав Северо-Западного Закамья // География и экология назем, позвоночных. -1978. - Вып. 3. - С.97-103.
Серебровский П,В. Результаты орнитологических наблюдений в Закатальском округе Закавказья в 1916 г. - М., 1925. - 89 с. - (Новые мемуары Моск, о-ва испытателей природы; Т.18. Вып. 2).
Симкин Г.Н. Биогеоценозы таежного леса: (На примере Перм. обл.). - М. : Изд-во МГУ, 1974. - 176 с.
Скалой В.Н. Некоторые зоологические находки в Юго-Восточном Забайкалье // Изв. Иркут. Противочум. ин-та Сибири и Дальне-Вост. края. -1935. - 1. - С.83-101.
Скалой В.Н., Слудский А.А. Птицы Елогуй-ско-Тазовского бассейна // Природа и соц. хоз-во. - 1941. - Сб.8, ч.4. - С.421-434.
Скрябин Н.Г., Филонов К.П. Материалы к фауне птиц северо-восточного побережья Байкала // Тр. Баргуз. заповедника. - 1962. - Вып.4. -С.32-46.
Смиренский С.М., Смиренская Е.М. О некоторых редких и малоизученных птицах Еврейской автономной области (Хабаровский край) // Орнитология. - 1980. - Вып.15. - С.205-206.
Смирнов В.М. Материалы к количественной характеристике населения птиц приобских лесов в окрестностях Новосибирского научного центра // Проблемы зоогеографии и истории фауны. -Новосибирск : Наука, 1980. - С.166-169. -(Тр.БИН СО АН СССР; Вып.40).
Соломонов Н.Г., Ларионов Г.П. Фауна наземных позвоночных долины Средней Лены и ее изменение под влиянием деятельности человека // Охрана природы Якутии: Материалы V Респ. совещ. по охране природы Якутии. - Иркутск, 1971. - С.183-190.
Сомов Н.Н. Орнитологическая фауна Харьковской губернии. - Харьков, 1897. - 680 с.
Сонин В.Д., Липин С.И. Гнезда некоторых редких птиц в Прибайкалье // Орнитология в СССР. - Ашхабад, 1969. - Кн.2. - С.606-609.
Спангенберг Е.П. Наблюдения над распространением и биологией птиц в низовьях реки Имана // Тр. Моск, зоопарка. - 1940. - 1. -С.77-136.
Спангенберг Е.П. Длиннохвостые мухоловки - род Terpsiphone Gloger // Птицы Советского Союза. - М.: Сов.Наука, 1954,- Т.6. - С.118-126.
Спангенберг Е.П. Птицы бассейна реки Имана // Сб. тр. зоол. музея Моск, ун-та : Исслед. по фауне Сов. Союза: (Птицы). - 1965. - 9. -С.98-202.
Спангенберг ЕЛ., Леонович В.В. Новые сведения по географическому распространению и биологии птиц на восточном побережье Белого моря // Пробл. севера. - 1958. - Вып.2. -С.194-202.
Спангенберг Е.П., Леонович В.В. Птицы северо-восточного побережья Белого моря // Тр. Кандалакш. заповедника. - 1960. - Вып. 2. -С.213-336.
Спангенберг Е.П., Олигер И.М. Орнитологические исследования в Дарвинском заповеднике в 1946 и 1947 годах // Тр. Дарвин, заповед
ника на Рыбин, водохранилище. - 1949,- Вьш.1. -С.245—302/
Спасский А.А., Сонин ВД„ Парамонов Г.В. К орнитофауне Верхнего Приамурья // Орнитология. - 1962. - Вып.5. - С.161-163.
Стахеев В А. Фенологические аспекты весенней миграции и размножения перелетных воробьиных птиц в Северо-Восточном Алтае // Экология и биоценотические связи перелетных птиц Западной Сибири. - Новосибирск : Наука, 1981. - С.166-176.
Степанян Л.С. Случаи атипического размножения у некоторых воробьиных птиц // Бюл. Моск, о-ва испытателей природы. Отд. бнол. -1962. - 67, вып.1. - С.119-120.
Степанян Л.С. Систематический список видов птиц фауны СССР / Зоология позвоночных 1969 (Итоги ВИНИТИ АН СССР). - 1971. - С.136-175.
Степанян Л.С. Результаты поездки с орнитологическими целями на Южный Сахалин // Фауна и экология животных. - М., 1974. - С.148-173.
Степанян Л.С. Состав и распределение птиц фауны СССР: Воробьинообразные - Passeriformes. - М.: Наука, 1978. - 392 с.
Степанян Л.С. Заметки о птицах о-ва Кунашир (Курильские острова) // Орнитология. -1980. - Вып.15. - С. 16-25.
Столбов ИА. Птицы оза Креблюкапнс н его ближайших окрестностей // X Прибалт, орнитол. конф.: Тез. докл. - Рига, 1981. - Т.1. - С.62-64.
Страутман Ф.И. Птицы Советских' Карпат. -Киев : Изд-во АН УССР, 1954. - 332 с.
Страутман Ф.И. Птицы западных областей УССР, - Львов : Изд-во Льв. ун-та, 1963. - Т.2. -183 с.
Строков В.В. Насекомоядные птицы в Москве // Орнитология. — 1962. - Вып.4. - С.305-315.
Сулик В.Г, Енакиев С.В. Пролет птиц в Луганском гос. заповеднике // II Всесоюз. орнитол. конф, по миграциям Птиц: Тез. сообщ. - Алма-Ата, 1-978. - 4.1. - С.165.
Суслова П.В. Материалы по птицам пограничной полосы лесов в пределах Западной области // Сборник Памяти академика М.А.Мензбира. - М. ; Л. : Изд-во АН СССР, 1937. -С.503-550.
Сушкин П.П. Заметки о кавказских птицах // Орнитол. вести. - 1914. - № 1. - С.3-43.
Сушкин П.П. Птицы Минусинского края, Западного Саяна и Урянхайской земли. - М., 1914. - 551 с. - (Материалы к познанию фауны и флоры Росснйс. империи. Отд. зоол. ; Вып.13).
Сушкин П.П. Птицы Советского Алтая и прилежащих частей Северо-Западной Монголии. -М. ; Л. : Изд-во АН СССР, 1938. - Т.2. -436 с.
Сыроечковский Е.Е. Изменение ареалов птиц в Средней Сибири в результате потепления климата и воздействия человека // Орнитология. -1960. - Вып.З. - С.212-218.
Сямашка В-В. Птушю Гродзенскай воблас-ui. - Мшск : Вид-во АН БССР, 1956. - 88 с.
Тамануки К. Заметки о птицах // Карафуто сенринкай, Кайхо. - 1942. - № 53. - С.112-118 (на яп. яз.).
Тараненко Л.И. Новые материалы по экологии райской мухоловки // Орнитология. - 1974. -Вып.11.- С.268-273.
Таращук В.И. Птицы полезащитных насаждений. - Киев : Изд-во АН УССР, 1953. - 124 с.
Тарунин М.И Предварительный перечень
175
птиц Тобольского р-на // Uragus. - 1928. -№ 3/4. - С.6-14.
Тарунин М.П. Птицы реки Малой Сосьвы // Ежегодник Тюцен. обл. краеведч. музея. - 1959. -Вып.1. - С.124-148.
Теплое В.П. Работа по привлечению птиц в Окском заповеднике // Пути и методы использования птиц в борьбе с вредными насекомыми. -М., 1956. - С.121-122.
Теплова Е.Н. Птицы Печоро-Илычского заповедника. - Сыктывкар, 1957. - 94 с. - (Тр. Печоро-Илыч. заповедника; Вып.6).
Тиюшас Э. Опыт по привлечению мухоловки-пеструшки и большой синицы в заповеднике Каманос Литовской ССР // Экологические исследования и охрана птиц Прибалтийских республик. - Каунас, 1982. - С.25-26.
Ткаченко В.И. Птицы Тебердинского заповедника I/Тр. Теберд. заповедника. - 1966. - Вып.6. -С.145-230.
Ткаченко М.И. Предварительный отчет о работах Зоологического отряда в Вилюйском округе в 1926 г. // Материалы Комис, по изуч. Якут. АССР. - 1929. - Вып.Ю. - С.281-296.
Ткаченко М.И. Путевой дневник Верхоянского зоологического отряда Якутской экспедиции АН СССР 1927 года // Тр. Совета по изучению производит, сил АН СССР. Сер. Якут. -1932. - Вып.5. - С.5-75.
Ткаченко М.И. Птицы реки Нижней Тунгуски // Изв. Иркут, науч, музея. - 1937. — 2(57). -С.152-162.
Треус В.Д., Успенский Г.А. Опыты по переселению мухоловки-пеструшки в парки ’’Аскания-Нова” // Пути и методы использования птиц в борьбе с вредными насекомыми. - М., 1956. -С.139-143.
Тугаринов А.Я. Птицы Приенисейской Сибири, список и распространение. - Красноярск, 1927. - 43 с. - (Зап. Средне-Сиб. отд. Рус. Геогр. о-ва, Сер.2; Т.1, Вып.1).
Тугаринов А.Я. Весенний пролет птиц у берегов Талыша // Сборник Памяти академика А.А.Сушкина. - М. ; Л.: Изд-во АН СССР, 1950. -С.9-59.
Уварова В.Я. К вопросу о формировании орнитофауны полезащитных насаждений Сталинградской области // Материалы III Прибалт, орнитол. конф. - Вильнюс, 1959. - С.305-310.
Ушакова М.М., Ушаков В.А. О совместном гнездовании некоторых видов птиц-дуплогнездников // Орнитология. - 1976. - Вып.12. - С.248.
Федоров А.В. О гибели гнезд некоторых видов воробьиных в субвысокогорье Баргузинского хребта // Проблемы экологии Прибайкалья: Тез. докл. Всесоюз. науч. конф. — Иркутск, 1982. - 4. - С.104.
Федотов М.П., Брунов В.В. Население птиц двух ключевых участков восточной части трассы БАМа // Тез. VII Всесоюз. орнитол. конф. -Киев : Наук, думка, 1977. - 4.1. - С.107-108.
Федюшин А.В., Долбик М.С. Птицы Белоруссии. - Минск : Изд-во АН БССР, 1967. -520 с.
Филатов В.А. Птицы Калужской губ // Мате
риалы к познанию фауны и флоры Российск. империи. Отд. зоол. - 1915. - Вып. 14. - С.194-379.
Филонов К.П. Опыт количественной характеристики летней орнитофауны Башкирского заповедника // Орнитология. - 1965. - Вып.7. -С.63-66.
Филонов К.П. Перелеты птиц в Баргузинском заповеднике - свидетельство сезонных ритмов в природе // Тр. Баргуз. заповедника. - 1967. -Вып.5. - С.30-51.
Флинт В.Е., Земская А.А., Сидоров В.И. Роль экологических группировок птиц в прокормлении клеща Ixodes persulcatus Ц Зоол. жури. - 1959. - 38, вып. 3. - С.476-480.
Флинт В.Е., Кривошеев В.Г. Изменения орнитофауны Измайловского парка за двадцать пять лет // Орнитология. - 1962. - Вып.5. -С.300-308.
Фолитарек С.С., Дементьев Г.П. Птицы Алтайского государственного заповедника // Тр. Алтайс. заповедника. - 1938. - Вып.1. - С.7-91.
Фомин Б.И., Вартапетов Л.Г., Равкин Е.С. Новые данные о географическом распространении птиц в Западной Сибири // Зоологические проблемы Сибири. - Новосибирск: Наука, 1972. - С.
Фомин В.Е. К орнитофауне Башкирии // Орнитология. - 1977. - Вып.13. - С.198-199.
Френкина Г. И. О трофических отношениях близких видов мухоловок в лесах Рязанской области // Влияние антропогенных факторов на структуру и функцию биогеоценозов. - Калинин, 1981. - С.21-31.
Хазиева С.М., Болотников А.М., Каменский Ю.Н., Никольская В.И Материалы о гнездящихся птицах правобережья реки Камы Добрянского района Пермской области // Гнездовая жизнь птиц. - Пермь, 1975. - С.117-127. (Учен, зап. Перм. пед. ин-та; Т. 146).
Харченко Л.П., Надточий А.С, Зиоменко С.К. О специфике гнездования некоторых воробьиных птиц в урбанизированном ландшафте // Птицы и урбанизированный ландшафт. - Каунас, 1984. -С.136-137.
Хохлов В.А. Чемал. (Алтай) // Uragus. -1927. - № 3. - С.10-17.
Хохлов В.А. Кузнецкая степь и Салаир. (Птицы) // Учен. зап. Перм. пед. ин-та. - 1937. -Вьш.1. - С.1-243.
Хомяков М.М. Птицы Рязанской губ. - М., 1900. - 101 с.
Хохлова П.А. Материалы по питанию пролетных птиц в лесополосах юга Украины // Орнитология. - 1960. - Вып.З. - С.259-269.
Хохлова Н.А. Мухоловка-белошейка в Горьковской области // Там же. - 1965. - Вып.7. -С.496.
Хохлова Н.А., Олигер Т.Н. Многолетние изменения структуры населения н численности птиц лесного участка Пустынского заказника // Назем, и вод. экосистемы. - 1982. - № 5. - С.25-30.
Хохлова Т.Ю. Фенология прилета и размножения птнц в условиях ранней весны 1975 г. // Экология птиц и млекопитающих Северо-Запада СССР. - Петрозаводск, 1976. - С.38-44.
176
Хохлова Т.Ю. Эколого-фаунистическая характеристика гнездовой орнитофауны Заонежья // Вести. Ленингр. ун-та. - 1977. — № 15. - С.22-30.
Цветков Е.В. Орнитологические наблюдения в окрестностях Белого Ключа // Материалы к познанию фауны и флоры Рос. империи. Отд. зоол. - 1901. - Вып.5. - С.62-102.
Цыбулин С.М. Птицы диффузного города на примере новосибирского Академгородка. - Новосибирск : Наука, 1985. - 169 с.
Чень Пэн. Птицы хребтов Чанбайшаньи и их вертикальное распространение // Acta Zool. Sinica. - 1963. - 15, № 4. - С.648-664 (на кит. яз.).
Черных П.А. Материалы к орнитофауне г.Ха-баровска // Изв. Иркут, н.-и. противочум. ин-та Сибири и Дал. Востока. - 1967. - 27. - С.175-177.
Черский А.И. Орнитологическая коллекция музея о-ва изучения Амурского края во Владивостоке // Зап. о-ва изуч. Амур, края, Владивосток. Отд. Императ, рус. Геогр. о-ва. - 1915а. -14. - С.143-276.
Черский А.И. Орнитологические сборы с 8 марта по 20 октября 1911 года в долине верхнего течения речки Одарки, близ д.Ново-Владими-ровки Иманского уезда Приморской области // Там же. - 19156. - 14. - С.79-141.
Чунихин С.П. Вертикальная поясность в распределении птиц Кабардино-Балкарии // Орнитология. — 1962. - Вып.5. - С. 186-192.
Чунихин С.П. Фауна и распределение птиц горнотаежных лесов Салаирского кряжа // Там же. - 1965. - Вып.7. - С.76-82.
Чхиквишвили И.Д. Материалы по орнитофауне Джавахетии // Джавахетия, материалы по изучению природных ресурсов Ахалкалакскогд нагорья. - Тифлис, 1933. - С.
Шведов А.П. К распространению птиц Среднего Приангарья: (Брат, и Заяр. р-ны Иркут, обл.) // Орнитология. - 1962. - Вып.4. - С.169-175.
Шевченко В.В. К вопросу о заселении птицами искусственных лесонасаждений юга Украины // Пращ н.-д. зоол.-б1ол. 1н-ту Хар. ун-ту. - 1940. -8/9. - С.123-137.
Шевченко В.В. Птицы государственного заповедника Аксу-Джабаглы // Тр. заповедника Аксу-Джабаглы. - 1948. - Вып.1. - С.
Шептуховский М.В. О структуре населения птиц на участках смешанного леса различного возраста (Ивановская область) // География и экология наземных позвоночных Нечерноземья: (Птицы). - Владимир, 1981, - С.93-100.
Шибанов Н.В. К орнитологической фауне Русской Лапландии // Мемуары Зоол. отд-ния О-ва любителей естествознания, антропологии и этнографии. - 1927. - Вып.З. - С.1-29.
Шибнев Ю.Б., Глущенко Ю.Н. Редкие птицы Приморья, нуждающиеся в особой охране // Редкие и исчезающие животные суши Дальнего Востока СССР: Материалы конф. — Владивосток, 1981. - С.58-60.
Шилова С.А., Чабовский В.И, Морозов Ю.В. и др. Эпизоотологическое значение птиц в очагах клещевого энцефалита на Среднем Урале // Орнитология. - 1963. - Вып.6. - С. 126-139.
Шкатулова А.П. К экологии сибирской ширококлювой мухоловки // Изв. Забайкал. отд. Геогр. о-ва СССР. - 1965. - 1, № 2. -.50-55.
Шнитников В.Н. Птицы Минской губернии. -1913. - С.1-475. - (Материалы к познанию фауны и флоры Рос. империи. Отд. зоол.; Вып.12).
Шнитников В.Н. Птицы Семиречья. - М. ; Л. : Изд-во АН СССР, 1949. - 667 с.
Штамм А.Р. Материалы для познания фауны зверей и птиц Полесья // Нар. хоз-во Белоруссии. - 1923. - № 6. - С.324-326.
Штильмарк Ф.Р. Наземные позвоночные Ком-сомольского-на-Амуре заповедника и прилежащих территорий // Вопр. географии Дал. Востока. -1973. - Сб.И. - С.30-124.
Шубникова О.Н., Морозов Ю.В. Некоторые орнитологические наблюдения в Центральной Якутии // Бюл. Моск, о-ва испытателей природы. Отд. биол. Н.С. - 1959. - 64, вып.5. - С. 142-144.
Шукуров Г.Ш. Фауна позвоночных животных гор Большие Балканы: (Юго-Зап. Туркменистан) . - Ашхабад : Изд-во АН ТССР, 1962. - 158 с.
Шульпин Л.М. К распространению птиц в Южно-Уссурийском крае // Докл. АН СССР. Сер.А. - 1927а. - С.351-352.
Шульпин Л.М. Новые данные по распространению птиц в Южно-Уссурийском крае и описание новых форм // Ежегодник Зоол. музея АН СССР. - 19276. - 28, вып.З. - С.398-406.
Шульпин Л.М. Орнитологические исследования в Южно-Уссурийском крае в 1926-1927 годах и новые фаунистические данные Ц Тр. Третьего Всероссийс. съезда зоологов, анатомов и гистологов. - Л., 1928. - С.97-99.
Шульпин Л.М. Экологический очерк птиц Алма-Атинского государственного заповедника. (По наблюдениям в августе-сентябре 1932 г. и в мае 1933 г.). - Тр. Алма-Атин. заповедника. -1939. - Вып.1. - С. 1-120.
Шутова В.В. Динамика численности и биото-пическое размещение воробьиных на островах Белого моря //X Прибалт, орнитол. конф.: Тез. докл. - Рига, 1981а. - Т.1. - С.78-82.
Шутова Е.В. Полигамия и негнездование самцов - обычные явления у горихвостки-лысушки и мухоловки-пеструшки // Экология и охрана птиц: Тез. VIII Всесоюз. орнитол. конф. - Кишинев : Штиинца, 19816. - С.248-249.
Щеголев В.И. Население птиц Тамбовской области и его динамика // География и экология назем, позвоночных. - 1978. - Вып.З. - С.107-128.
Щербаков И.Д. Опыт активного заселения мухоловкой-пеструшкой и большой синицей островных лесов Мордовской АССР // Тр. пробл. и темат. совещ. - ЗИН АН СССР: I Всесоюз. орнитол. конф. - 1960. - Вып.9. — С.351-361.
Щербаков И.Д. Материалы по образованию местной популяции дуплогнездников в островных лесах Ц Тр. Мордов. заповедника. - 1967. -Вып.4. - С.8-52.
Юдин К.А. Наблюдения над распространением и биологией птиц Красноярского края // Тр. ЗИН АН СССР. - 1952. - 9, вып.4. - С. 1029-1099.
Юрлов КТ. Летняя авифауна Кулунды И Эпизоотологическая характеристика очагов Ом
177
ской геморрагической лихорадки Западной Сибири. - Наука, 1974. - С.76-112.
Юрлов К.Т., Чернышов В.М., Кошелев А.И. и др. Новые и редкие птицы в районе озера Чаны // Миграции птиц в Азии. - Новосибирск : Наука, 1977. - С.205-209.
Янушевич А.И. Материалы по позвоночным Тувинской области // Изв. Зап.-Сиб. фил. АН СССР. Сер. биол. - 1948. - Вып.2. - С.3-27.
Янушевич А.И., Юрлов КТ. Вертикальное распространение млекопитающих и птиц в Западном Саяне // Там же. - 1950(1949). - 3, вып.2. -С.З-
Янушевич А.И., Тюрин П.С., Яковлева И.Д. и др. Птицы Киргизии. - Фрунзе : АН КиргССР, 1960. - Т.2. - 274 с.
Яхонтов В.Д. Новые данные к орнитофауне Нижнего Амура // Вопр. географии Дал. Востока. - 1963. - Сб.6. - С.215-223.
Яхонтов В.Д. Орнитологический комплекс Малого Хингана // Там же. - 1973. - Сб.П. -С.225-235.
Яхонтов В.Д. Птицы Шантарских островов: некоторые вопросы экологии // Биогеография Приамурья: Вопр. географии Дал. Востока. -1977. - Сб.П. - С.150-171.
Яхонтов В.Д. Птицы Пенжинского района // Птицы Северо-Востока Азии. - Владивосток, 1979. - С.135-162.
AU S., Ripley S.D. Handbook of the Birds of India and Pakistan. Together with those of Nepal, Bangladesh, Sikkim, Bhutan, Sri Lanka. - Oxford : Univ, press, 1972. - Vol.7. - 236 p.
Bates R.S., Lowther E.H.N. Breeding birds of Kashmir. - London, 1952. - 367 p.
Bergmann S. Zur Kenntnis nordostasiatischer Vogel. Ein Beitrag zur Systematik, Biologie und Verbreitung der Vogel Kamtschatkas und der Kuriles. - Stokholm : Bonniers, 1935. - 268 S.
Berndt R., Winkel W. Die Gelegegrosse des Trauerschnapper (Ficedula hypoleuca) in Beziehung zu Ort, Zeit, Biotop und Alter // Vogelwelt. -1967. -88, H.4/5.-S.97-136.
Bianchi V. Preliminary review of the palaearctic and himalo-chinese species of the Muscicapidae or family of Flycatchers // Ежегодник Зоол. музея Императ. Акад. Наук. - 1907. - 12, № 1. - с. 13-85.
Check-list of Japanese Birds. - Tokyo : Orni-thol. Soc. Jap. Gakken Co., Ltd., 1974. - 364 p. (5th ed.).
Gtowacinski Z. Ekspancja muchotowki biafos-zyjej, Ficedula albicollis (Temm.) w Europie srod-kowej // Prz. zool. - 1974. - N4. - S.471-484.
Curio E. Beobachtungen am Halbringschnapper, Ficedula semitorquata, im mazedonischen Brutge-biet // J.Ornithol. - 1959. - N2. - S.176-209.
Curio E. Die systematische Stellung des spa-nischen Trauerschnappers // Vogelwelt. - 1960. -N4. - S.113-121.
Eber C. Muscicapa striata (Pallas) // Atlas der Verbreitung palaearktischer Vogel / Eds E.Strese-mann, L.A.Portenko. - Berlin : Akad. Verl., 1960a. -Bd.I.
Eber G. Muscicapa hypoleuca Pallas // Atlas
der Verbreitung palaearktischer Vogel / Eds E.Stre-semann, L.A.Portenko. - Berlin : Akad. Verl., 1960b. - Bd.I.
Hartert E. Die VSgel der palaarktischen Fauna (Systematische ubersicht der in Europa, Nord-Asien und der Mittelmeerregion vorkommenden Vogel). -Berlin, 1907. - H.4. - S.385-512.
Hartert E., Steinbacher F. Die VSgel der palltark-tischen Fauna (Systematische iibersicht der in Europa, Nord-Asien und der Mittelmeerregion vorkommenden Vogel.). - Berlin, 1934. - H.3. - S.193-288.
Jahn H. Zur Okologie und Biologie der Vogel Japans// J.Ornithol. - 1942. - 83, N 1/2. - S.l-138.
Jordania R. Die Vogelwelt Georgiens nach dem Material des Staatlichen Museums Georgiens // Beitr. Vogelk. - 1975. - 21, Nl/2. - S.132-139.
Lorenz Th. Die Vogel des Moskauer Gouver-nements // Bull. Soc. Nat. Moscou. N.S. - 1894. -8, H.3. - S.325-350.
Lonnberg E. Contributions to the Ornis of Saghalin // J.Coll. Sci., Imp. Univ. Tokyo. - 1908. -23, art.14. - P.1-69.
Makatsch W. Die Eier der Vogel Europas. Eine Darstellung der Brutbiologie alter in Europa briitenden Vogelarten. - Berlin : Neumann Verl., 1974. -Bd.I. - 468 S.
Makatsch W. Die Eier der Vogel Europas. Eine Darstellung der Brutbiologie aller in Europa brii-tenden Vogelarten. - Berlin : Neumann Verl., 1976. -Bd.2. - 460 S.
Mareejee С.Д. Преглед фауне птица Балкан-ског полуострва. I Део ДетлиАи и птице певачице (Piciformes et Passeriformes). - Београд, 1976. -365 с.
Mauersberger G., Portenko L.A. Muscicapa albicollis Temminck und Muscicapa semitorquata Homeyer // Atlas der Verbreitung palaearktischer Vogel / Eds E.Stresemann, L.A.Portenko. - Berlin : Akad. Verl., 1960. - Bd.I.
Mauersberger G., Portenko L.A. Muscicapa parva Bechstein // Atlas der Verbreitung palaearktischer Vogel. - Berlin : Akad. Verl., 1967. - Bd.2. Miczytfski K. Ptaki Dublan (Ukrainska SRR) // Acta Ornithol. - 1962. - 6, N10.
Nisbet Ian C.T., Neufeldt I. Studies of less familiar birds 175 Brown Flycatcher. Plate 8-10 // Brit. Birds. - 1975. - 68, N2. - P.68-75.
Neufeldt LA. Der Blaue Fliegenschnapper im Siiden von Primorje (UdSSR) // Falke. - 1968. -Nil. - S.364-371.
Owen D.F. The rufous and white forms of an Asiatic paradise flycatcher, Terpsiphone paradisi // Ardea. - 1963. - 51, N 2/4. - P.230-236.
Piechocki R., Bolod A. Beitrage zur Avifauna der Mongolei. Teil 2. Passeriformes // Mitt. Zool. Mus. Berlin. - 1972. - 48, H.l. - S.41-175.
Pleske Th. Revision der turkestanischen Ornis. Nach Sammlungen des verstorbenen Konservators V.Russow. - St. - Pdtersb., 1888. - 58 p. - (Mem. Acad. sci. St. - Petersb. Ser.7, t.36, N3).
Radde G., Walter A. Die Vogel Transcaspiens // Ornis. - 1889. - Jg.5, S.l-128, 165-279.
Shulpin L. Ubersicht der Verbreitung der an den Wald gebundenen VSgel aus dem Gebiete des Sichota-
178
Alin Bergriickens (Kiistengebiet der UdSSR) // Zool. Anz. - 1931. - 93, N 3/4. S.65-77.
Stegmann B. Die Vogel des nordlichen Baikal // J.Ornithol. - 1936. - 84, H.l. - S.50-139.
Stegmann K.-D. Zum Neststandort des Grausch-nappers, Muscicapa striata Pall. // Beitr. Vogelk. -1980. - 26, N 3/4. - S.227-228.
Stolzmann J. Oiseaux de la Ferghana d’apres les recherches faites par M.Thomas Barey (1892-1895) // Bull. Soc. nat. Moscou. N.S. - 1897. -11. - P.54-81.
Taczanowski L. Faune ornithologique de la Sibe'rie orientale. - St. - Petersb., 1891. - 684 p. -(Mem. Acad. sci. St.-P£tersb. Ser. 7, t.39).
Tauriqs E., Michelsons H. Maksligu ligzdosanas vietu ietekme uz sila putnu faunas kvaiitativo un kvantitativo sastavu Latvijas PSR apstaklos // Latv PSR ZinSt. akad. Mezsaimnicc. probl. inst. raksti. -1950. - 2. - 1pp.199-216.
Ticehurst C.B. On the down-plumage of some Indian birds // J.Bombay Natur. Hist. Soc. - 1926. -31, N 2. - P.268-378.
Udagawa T. Karyogram studies in birds. IV. The chromosomes of four passers and two columbine species // Ann. Zool. Jap. - 1954. - 27. - P.208-214.
Vaurie C. A generic revision of flycatchers of the tribe Muscicapini // Bull. Amer. Mus. Natur. Hist. - 1953. - Vol. 100, articl. 4. - P.1-538.
Vaurie C. The Birds of the Palearctic fauna.
A systematic reference. Order Passeriformes. - London : Witherby, 1959. - 762 p.
Vilks K. Avifauna aus vier Gegenden Lettlands // Fol. Zool. Hydrobiol. - 1943. - 12, N 1. - S.247-265.
Wetmore A. A classification for the birds on the world // Smithsonian Miscellaneous Collections. -Washington, 1960. - Vol. 139, N 11, P.1-37.
Wolters H.E. Die Vogelarten der Erde. Eine systematische Liste mit Verbreitungsangaden sowie deutschen und englischen Namen. - Hamburg ; Berlin : Parey, 1979. - Lief.4, Bogen 16-20. -S.241-320.
Wynne O. Key-list of the Palaearctic and Oriental Passerine birds. - 1954. - P.2. The North Western Naturalist new series. - P.123-137.
Yamashina Y. Notes on a collection of birds from southern Sakhalin // Tori. - 1927. - 5, N 24. -P.333-364.
Yamashina Y. Notes on a collection of birds from southern Sakhalin // Ibid. - 1928. - 5, N 25. -P.419-453.
Yamashina Y. Die Vogel der Kurilen // J.Ornithol. - 1931. - 79, H. 4. - S. 491-541.
Yamashina Y., Yamada S. On a collection of some birds from Saghalien // Tori. - 1934. - 8, N 39. - P.304-325.
Yanukai T., Matsuki T. On the birds of Northern Saghalien // J.Soc. Agric. - 1925. - N 71. -P. 120.
Рис. 1. Хвосты (вид снизу) взрослых самцов райских мухоловок:
а — Terpsiphone paradisi, б — Т. atrocaudata
Рис. 2. Клювы взрослых самцов двух подвидов райской мухоловки: а — Terpsiphone paradisi leucogaster, б — Т. р. incei
а
б
Рис. 4. Место гнездования белобрюхой райской мухоловки. Островной участок чилонового леса на хр. Кугитанг (Восточная Туркмения)
Рис. 5. Характерное местообитание белобрюхой райской и восточной серой мухоловок. Разреженный лиственный лес с преобладанием грецкого ореха по дну ущелья с ручьем на северном склоне Зеравшанского хр. (Узбекистан, Аман-Кутанская лесная дача)
Рис. 6. Погадки слетков белобрюхой райской мухоловки
Рис. 7. Форма и типы расположения гнезд белобрюхой райской мухоловки:
1—9 — обычные типы гнездовых построек, 10—12 — на основе прошлогодних гнезд; а, б — годовые надстройки
Рис. 8. Семь цветовых морф самцов мухоловки-пеструшки (по Портенко, I960 с изменениями)
Рис. 10. Изменчивость окраски трех крайних рулевых перьев у самцов мухоловки-пеструшки
Ill III III III
III III III III
5 6 7 8
9 10 » '2
III III III II*
Рис. 11. Изменчивость окраски трех крайних рулевых перьев у европейской мухоловки-белошейки:
1—14 — самцы, 15—-16 — самки
III III III III
1 2 3 4
III III IK IK
5 6 3 8
III III III III
9 10 11 12
13 14 15 16
Рис. 13. Изменчивость окраски трех крайних рулевых перьев у кавказской мухоловки-белошейки:
1—13 — самцы, 15— 16 — самки, 14 — гибридный самец (F. а. albicollisXF. a. semi-torquata)
/ 2 3 4
МШИ
8
5
1
6
II1 III II1 III
9 10 11 12
13 14 15 16
III III III III
Рис. 15. Элементы демонстративного поведения синей мухоловки (по Панову, 1973): а, б — последовательные позы самца, ухаживающего за самкой. Последняя настроена агрессивно, она пытается ударить клювом самца, севшего слишком близко (в); г, д-—демонстративный токовой полет самца и поза, принимаемая им по окончании полета. Демонстрируются белые пятна в основании хвоста, обычно скрытые
г
Рис. 18. Элементы демонстративного поведения белобрюхой райской мухоловки: а, б — позы самца, токующего перед самкой в период образования пар. Поющий самец разворачивает веером хвост и, быстро трепеща им и полураскрытыми крыльями, перелетает вокруг самки; в — самка настроена агрессивно, она пытается клюнуть самца, севшего слишком близко; г — летящий самец во время преследования самки
Рис. 22. Элементы демонстративного поведения даурской желтоспинной мухоловки (по Панову, 1973): а, б — последовательность поз самца, принимаемых им перед самкой или перед соперником; в — демонстративный замедленный полет; г — поведение самца в начале гнездостроения, когда самка носит материал для гнезда
Рис. 23. Элементы демонстративного поведения ширококлювой мухоловки (по Панову, 1973): а — самец (слева) токует перед самкой, выбирая для этого такую развилку ствола, где обычно помещается гнездо. Самка настроена агрессивно, она делает в сторону самца клюющие движения (первый период после образования пары); б — самец кормит самку, когда она на время покидает кладку в период насиживания
Рис. 25. Самка синей мухоловки у гнезда с птенцами (Лазовский заповедник им. Л. Г. Капланова). Фото С. В. Винтера
Рис. 26. Взрослая серая мухоловка (Донецкая обл.).
Фото С. В. Винтера
Рис. 30. Слеток европейской серой мухоловки
Рис. 31. Гнездо ширококлювой мухоловки, построенное на горизонтальной ветви липы (Лазовский заповедник им. Л. Г. Капланова). Фото С. В. Винтера
Рис. 32. Взрослая ширококлювая мухоловка у гнезда с птенцами (Лазовский заповедник им. Л. Г. Капланова). Фото С. В. Винтера
и
Рис. 34. Характерное местообитание рыжехвостой мухоловки.
Разреженный лес из туркестанского клена и арчи на южном склоне Гиссарского хребта (Таджикистан, урочище Квак, в ущелье Кондара)
ТабяЛЯИ^аЯМ7Х0Л0В
Чер Р trocauda-
“erp»PMm *
?***iS:“*
^взрослая самка
Таблица II.
Белобрюхая райская мухоловка (Terpsiphone paradisi leucogaster): 1 — самец второгодок, 2 — взрослый самец старше двух лет темноокрашенной морфы,
3 — взрослый самец старше двух лет переходной морфы,
4 —первогодок в весеннем наряде, 5 — взрослая самка, 6 — слеток
Таблица III.
1 — взрослый самец белой морфы китайской райской мухоловки (Terpsiphone paradisi incei) у гнезда (Приморье) (фото Ю. Б. Шибнева);
2 — полная кладка белобрюхой райской мухоловки
(Т. р. leucogaster) (Тадж. ССР, ущелье Кондара);
3 , 4 — слетки белобрюхой райской мухоловки у гнезда (Тадж. ССР, Рамитское ущелье)
Таблица IV.
Европейская мухоловка-белошейка (Ficedula albicollis albicollis):
1 — взрослый самец, 2 — взрослая самка, 3 — молодая птица
в гнездовом наряде; кавказская мухоловка-белошейка (F. a. semitorquata):
4 — взрослый самец; северная мухоловка-пеструшка (F. hypoleuca hypoleuca):
5 — взрослый самец, 6 — взрослая самка; европейская серая мухоловка (Muscicapa striata striata): 7 — взрослая птица, 9 — молодая птица в гнездовом наряде; восточная серая мухоловка (М. s. neumanni): 8 — взрослая птица
Таблица V.
Даурская желтоспинная мухоловка (Ficedula zanthopygia): 1 — взрослый самец, 2 — самец первогодок в весеннем наряде, 3—взрослая самка, 7 — молодая птица в гнездовом наряде;
японская желтоспинная мухоловка (F. narcissina narcissina):
4 — взрослый самец, 5 — самец первогодок в весеннем наряде, 6 — взрослая самка, 8 — молодая птица в гнездовом наряде
Таблица VI. Таежная мухоловка (Ficedula mugimaki): 1—3 — взрослые самцы, 4 — самец первогодок в весеннем наряде, 5, 6 — взрослые самки, 7 — молодая птица в гнездовом наряде, 8 — рулевые перья самца, 9 — рулевые перья самки
Таблица VII.
I — кладка северной мухоловки-пеструшки (Ficedula hypoleuca hypoleuca) (окр. г. Киев); 2 — кладка европейской мухоловки-белошейки (F. albicollis albicollis) (Харьковская обл.);
3 — яйца даурской желтоспинной мухоловки (F. zanthopygia) (Приморье);
4 — взрослый самец даурской желтоспинной мухоловки (Приморье) (фото Ю. Б. Шибнева)
Таблица VIII.
Европейская малая
мухоловка (Ficedula parva parva):
1 — взрослый самец,
2 — самец первогодок в весеннем наряде, 3 — взрослая самка,
7 — молодая птица в гнездовом наряде,
8 — рулевые перья самца,
9 — рулевые перья самки; сибирская малая мухоловка (F. р. albicilla):
4 — взрослый самец,
5 — самец первогодок,
6 —• взрослая самка
Таблица IX. Синяя мухоловка (Cyanoptila cyanomelana): 1, 2 — взрослые самцы, 3 — самец в первом послегнездовой наряде, 4, 5 — взрослые самки, 6 — молодой самец в гнездовом наряде, 7 — рулевые перья самца, 8 — рулевые перья самки
Таблица X.
Северная сибирская мухоловка (Muscicapa sibirica sibirica):
1 ,2 — взрослые птицы,
3 — молодая птица в гнездовом наряде;
палеарктическая ширококлювая мухоловка (М. latirostris latirostris):
4 — взрослая птица,
5 — молодая птица в гнездовом наряде; пестрогрудая мухоловка (М. griseisticta):
6 — взрослая птица,
7 — молодая птица в гнездовом наряде;
рыжехвостая мухоловка (М. ruficauda):
8 — взрослая птица,
9 — молодая в гнездовом наряде
Таблица XI. 1,2 — кладки европейской серой мухоловки (Muscicapa striata striata) (Киевская обл.);
3 — кладка ширококлювой мухоловки (М. latirostris latirostris) (Сахалин);
4 — яйца европейской малой
мухоловки (Ficedula parva parva)
(окр. г. Харьков)
СОДЕРЖАНИЕ
3
5 6
6 И 26 27 41 55 67 76 82
100 101 ПО 111 134 143 150 160
165
ПРЕДИСЛОВИЕ
СЕМЕЙСТВО Muscicapidae - Мухоловковые
Род Terpsiphone Gloger - Райские мухоловки
Черная райская мухоловка
Райская мухоловка
Род Ficedula Brisson - Пестрые мухоловки
Мухоловка-пеструшка
Мухоловка-белошейка
Даурская желтоспинная мухоловка
Японская желтоспинная мухоловка
Таежная мухоловка
Малая мухоловка
Род Cyanoptila Blyth - Синяя мухоловка
Синяя мухоловка
Род Muscicapa Brisson - Серые мухоловки
Серая мухоловка
Сибирская мухоловка
Пестрогрудая мухоловка
Ширококлювая мухоловка
Рыжехвостая мухоловка
СПИСОК ЛИТЕРАТУРЫ
www.zoomet.ru