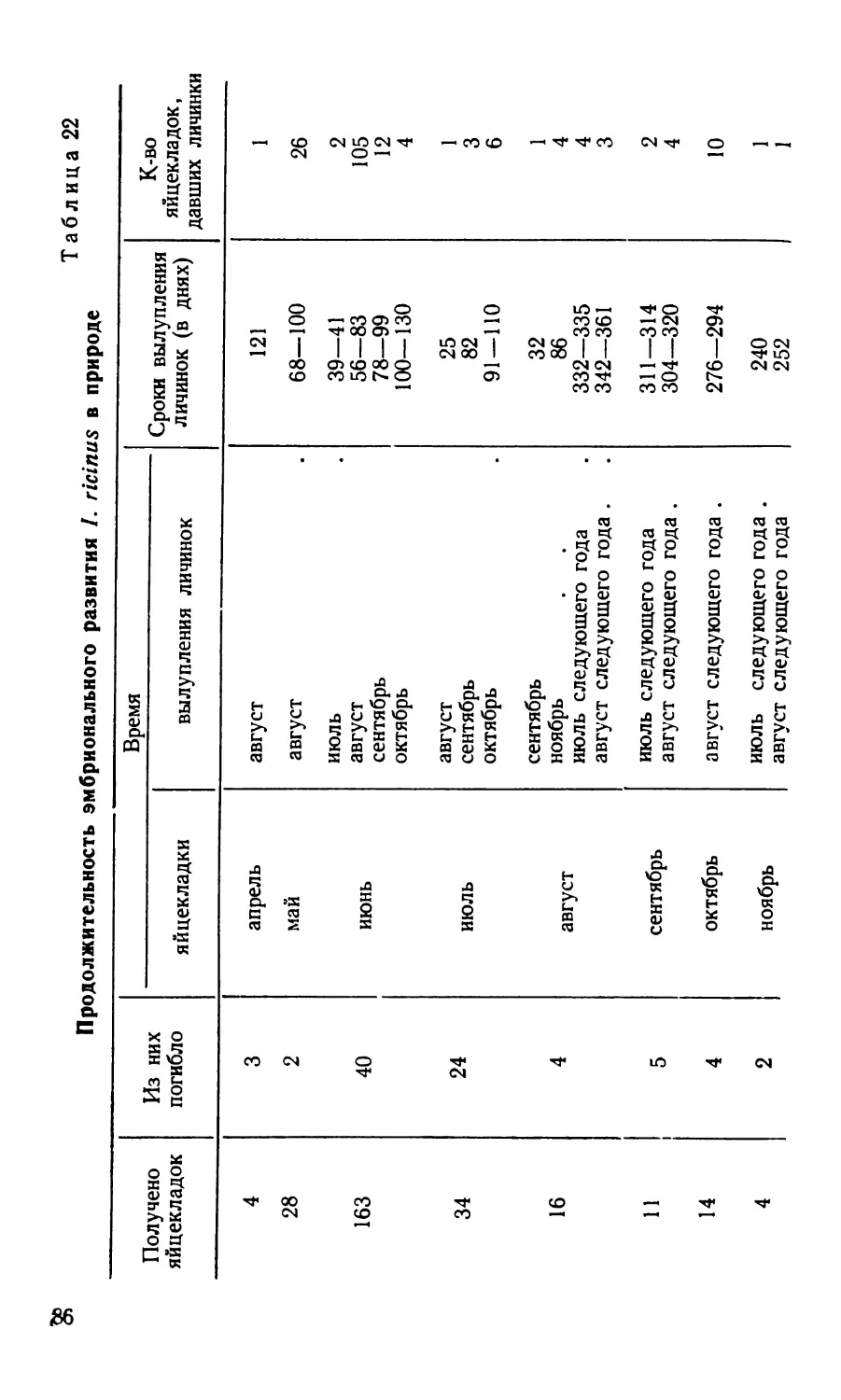
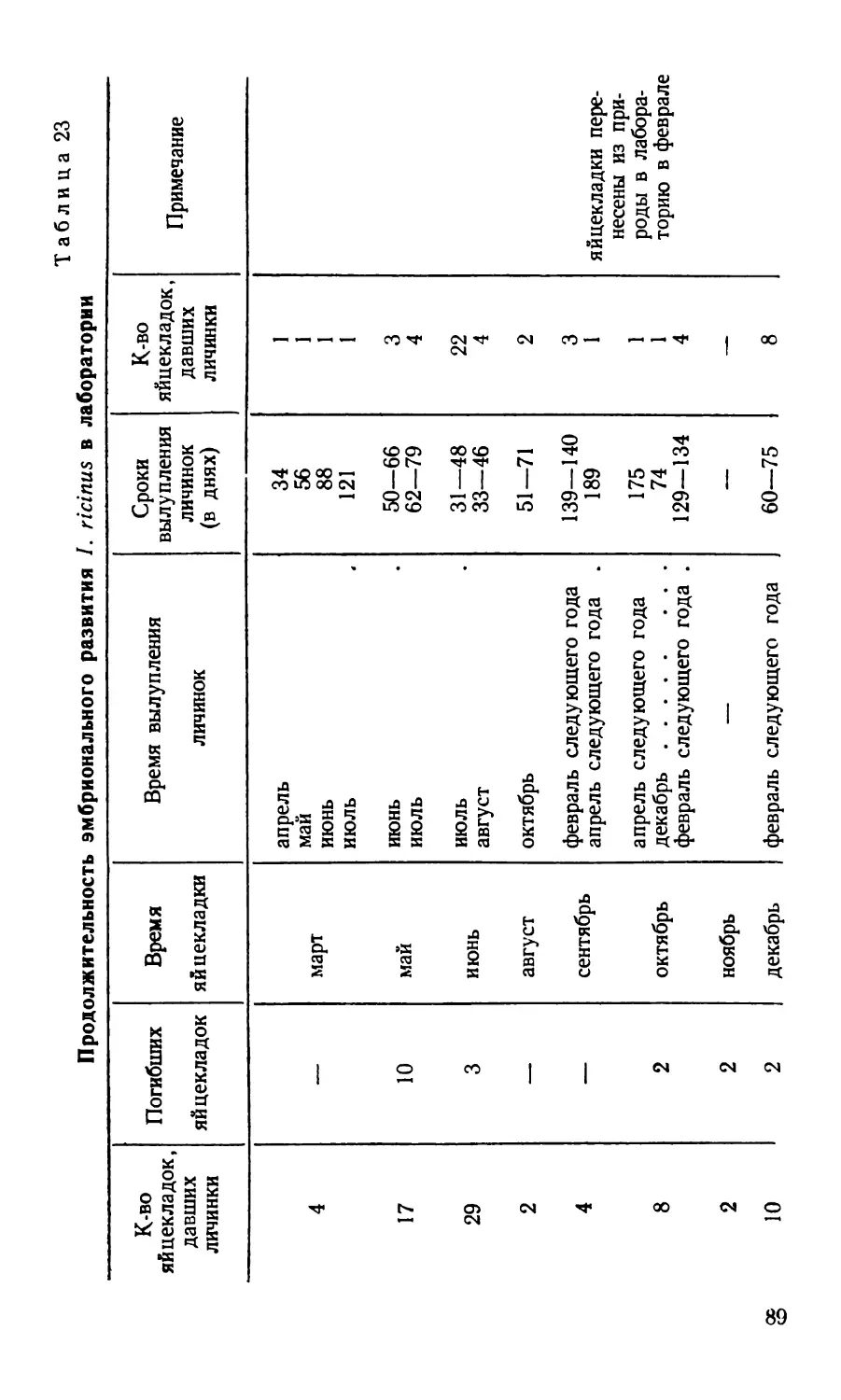
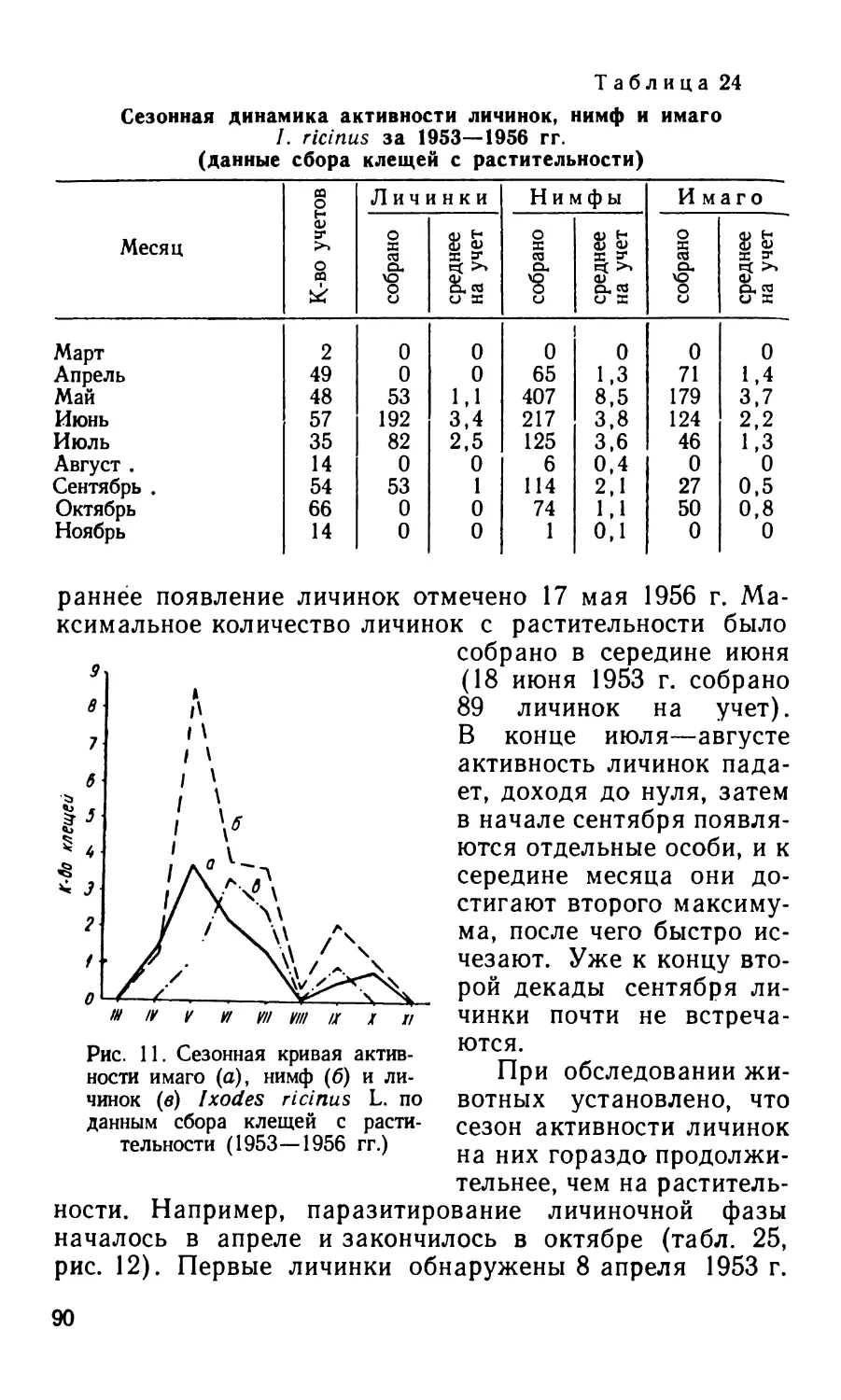
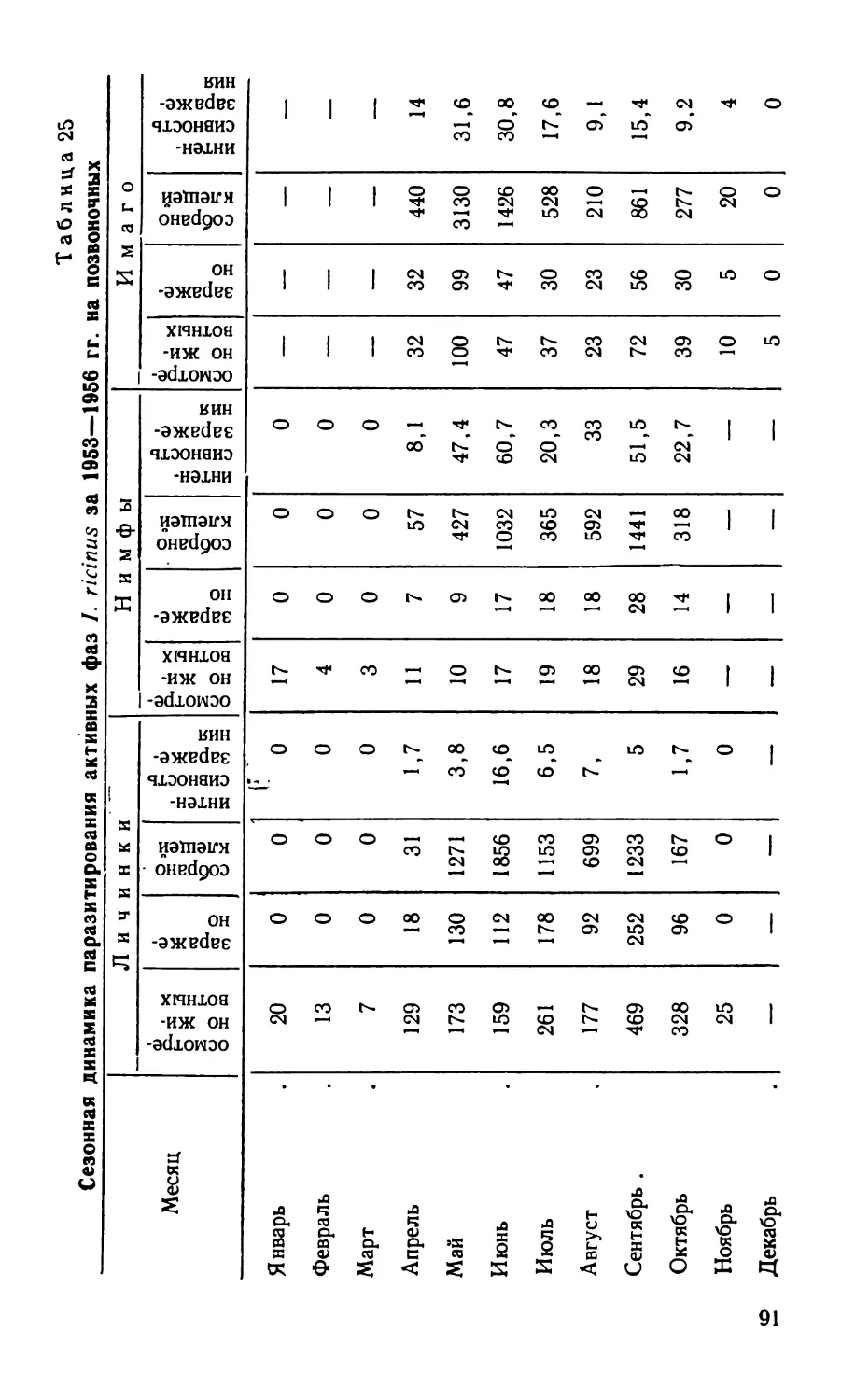
/

















































































































Author: Арзамасов И.Т.
Tags: медицина биология насекомые зоология ветеринария мир насекомых
Year: 1961




Text
И.Т. АРЗАМАСОВ
АКАДЕМИЯ НАУК БЕЛОРУССКОЙ ССР
ОТДЕЛ ЗООЛОГИИ И ПАРАЗИТОЛОГИИ
И. Т. Арзамасов
ИКСОДОВЫЕ
КЛЕЩИ
Издательство Академии наук Белорусской ССР
Минск 1961
Редактор проф.
И. Н. СЕРЖАНИН
В работе приводятся сведения о географическом распростране-
нии иксодовых клещей в Белоруссии, о распределении иксодид в
различных биотопах, указываются круг хозяев и степень заражен-
ности их паразитами, цикл развития наиболее распространенных в
республике иксодовых клещей. Большое внимание уделяется мерам
борьбы с иксодовыми клещами, применение которых позволит зна-
чительно сократить заболеваемость домашних животных и человека.
Книга рассчитана на медицинских и ветеринарных врачей, зоо-
техников и зоологов.
ВВЕДЕНИЕ
Теория природной очаговости академика Е. Н. Пав-
ловского способствовала выявлению переносчиков и хра-
нителей возбудителей трансмиссивных болезней. Соглас-
но этому учению, в природе имеются очаги инфекции,
где происходит циркуляция возбудителя заболевания
между эктопаразитами и их хозяевами. Учение о перенос-
чиках возбудителей вирусных, бактериальных и прото-
зойных болезней является одним из важнейших звеньев
паразитологии. Изучение биологии переносчиков, в осо-
бенности специфических, является такой же важной за-
дачей, как и изучение самого возбудителя болезни (Е. Н.
Павловский, 1940). Природным очагам болезни соответ-
ствуют определенные участки ландшафта, ибо возбуди-
тель болезни, его переносчик, животные-доноры и реци-
пиенты являются сочленами биоценоза, эволюционно
сложившегося на определенных участках географическо-
го ландшафта со свойственной ему флорой (Е. Н. Пав-
ловский, 1955). О важности проблемы природной очаго-
вости говорит увеличивающееся число выявляемых
болезней с природной очаговостью. На территории Совет-
ского Союза уже обнаружено около 15 таких болезней
(Е. Н. Павловский, 1957).
Из эктопаразитов—переносчиков заболеваний наибо-
лее вредны иксодовые клещи. Они передают человеку
клещевой энцефалит, туляремию, клещевые риккетсиозы.
Некоторые виды непосредственно сохраняют вирус чумы
и другие заболевания (Е. Н. Павловский, 1948). Многие
являются специфическими переносчиками пироплазмоз-
иых заболеваний домашних и диких животных, вызывае-
3
мых простейшими эндоглобулярными паразитами. Эти
заболевания, поражая крупный рогатый скот, лошадей,
овец, коз, свиней, собак, а также домашних северных
оленей, наносят большой урон животноводству (В. Л.
Якимов, 1931).
В Белоруссии отмечено заболевание овец и человека
шотландским энцефалитом—Leaping ill (Л. А. Зильбер,
1945; С. С. Фолитарек и др., 1949), заболевание человека
клещевым весенне-летним энцефалитом (П. П. Чумаков,
1941; Г. А. Найденова, 1941), клещевым нейротропным
энцефалитом (Л. В. Федорчук, 1955), сезонным ви-
русным менингоэнцефалитом (В. И. Вотяков, 1957;
Е. И. Смирнова и др., 1957; К. П. Госмер, 1957; А. Н. Фи-
липпович, 1957), туляремией (Ф. Г. Рубанова, 1957), пе-
редаваемых клещом Ixodes ricinus L. Отмечено значи-
тельное распространение гемоспоридиозов крупного ро-
гатого скота (Babesiella bovis, Francaiella occidentalis,
F, caucasica), передаваемых клещами /. ricinus, пиро-
плазмоза (Piroplasma caballi) и нутталлиоза (Nuttallia
equi) лошадей, передаваемых клещами Dermacentor
pictus Herm. и D. marginatus Sulz. (В. И. Бурцев, 1925;
H. И. Барцевич, 1927; В. Л. Якимов и В. И. Бурцев,
1928; В. Л. Якимов и др., 1929; 3. 3. Сергеева, 1932;
М. Н. Судзиловский и Н. В. Щербинин, 1936; С. Ф. Сузь-
ко, 1949, 1953; В. Ф. Гусев, 1954; Н. И. Дылько, 1953,
1954).
Большой вред наносят иксодовые клещи как эктопа-
разиты-гематофаги. Результатом их паразитирования
является истощение животных и уменьшение удоев. На-
пример, в Белоруссии при массовом нападении клещей
на коров отмечено уменьшение удоев в среднем на 2—
3 литра (Н. И. Дылько, 1953).
Ранее при исследовании иксодовых клещей на тер-
ритории Белоруссии основное внимание уделялось гео-
графическому распространению их, степени заражения
ими домашних животных в зависимости от сезона года
и некоторым моментам экологии—влиянию затопления
на выживаемость клещей, распределению по различным
биотопам и др. Мало внимания уделялось исследованию
роли дикой фауны в развитии клещей, что отразилось
на полноте изученности видового состава иксодовых кле-
щей и круга их хозяев. Совсем не изучались циклы раз-
вития.
4
Нашей задачей являлось более углубленное изучение
видового состава и биологии иксодовых клещей в усло-
виях Белоруссии, необходимых для обоснования систе-
мы противоклещевых мер борьбы и профилактики.
Основное внимание нами уделялось выявлению видо-
вого состава иксодовых клещей, изучению стадиального
распространения и круга хозяев отдельных видов, опре-
делению продолжительности цикла развития у двух мас-
совых видов—/. ricinus и D. pictus—основных перенос-
чиков клещевого энцефалита, бабезиеллоза, пироплаз-
моза и нутталлиоза в БССР.
Считаю своим долгом выразить глубокую благодар-
ность академику АН СССР Е. Н. Павловскому и канди-
дату биологических наук Г В. Сердюковой за консуль-
тации, критические замечания и методическую помощь
в постановке полевых исследований.
ПРИРОДНЫЕ УСЛОВИЯ БССР
Территория Белоруссии является продолжением
Средне-Русской равнины и расположена в зоне средних
широт с мягким климатом. Это наиболее увлажняемая
часть Европейской территории. Среднегодовая темпера-
тура воздуха 4,4—7,3°. Среднегодовое количество осад-
ков 540—700 мм. Наибольшее их количество выпадает
летом (в июле) и меньше зимой. Зима длится 130—
160 дней. Промерзание почвы невелико. Снеговой покров
составляет 20—40 см. Наиболее холодный месяц январь.
Минимальная температура доходит до —30, —44° Зима
богата оттепелями. Для весны характерна частая смена
теплых и холодных дней с заморозками. Лето умеренно
теплое и влажное, продолжительность его 100—120 дней.
Максимальная температура воздуха достигает 38°. Выпа-
дение осадков летом иногда значительное (до 280 мм в
месяц}. Осень теплая и сухая (А. И. Кайгородов, 1928,
1934).
Почвы Белоруссии в основном дерново-подзолистые,
они составляют 58% всей площади республики. Широко
распространены дерновые и торфяно-болотные (23,8%) и
дерново-подзолистые заболачиваемые (12,2 %) почвы.
Редко встречаются аллювиально-луговые (1,7%) и дер-
новые, близкие к черноземным (0,5%) почвы (П. П. Ро-
говой и др., 1952).
Территория Белоруссии находится в лесной зоне, в
подзоне смешанных, или хвойно-широколиственных лесов
(Л. С. Берг, 1937; В. Б. Сачава, 1953). По рельефу, поч-
венно-гидрологическим и климатическим условиям БССР
подразделяется на шесть лесорастительных районов.
6
Северный район еловых лесов занимает крайнюю
северную часть БССР с южной границей по 55° с. ш.
Для этого района характерны еловые насаждения с не-
значительной примесью широколиственных пород, глав-
ным образом дуба, клена, реже вяза и липы. Имеется
много озер.
Центральный переходный район смешанных елово-
широколиственных лесов располагается примерно меж-
ду 54 и 55° с. ш. В этом районе наблюдается увеличе-
ние сосновых насаждений до 62,8% и уменьшение ели
до 19,3%. Сосновые насаждения больше распространены
в его западной части. Березовые, осиновые и ольховые
составляют более 15%. В древостоях отсутствуют граб
и берест.
Западный район елово-широколиственных лесов (не-
манский) охватывает крайнюю западную часть террито-
рии БССР с границами между 53 и 54° с. ш. и 28° в. д.
По структуре и составу насаждений этот район сложен
и многообразен. Сосновые насаждения составляют до
65%, еловые—20%. В юго-западной части его встреча-
ется граб, в северной—ольха серая. Удельный вес бере-
зы, осины и ольхи резко снижается по сравнению с
Центральным районом.
Восточный район елово-дубовых лесов (межречный)
расположен в восточной части БССР между 54 и
53° с. ш. и 28—32°30' в. д. На почвах с глубоким залега-
нием морены произрастают дубравы с примесью ели,
которые по мере увеличения песчаного наноса сменяются
сосновыми лесами. Процент древостоя с преобладанием
сосны наименьший (около 45%) по сравнению с други-
ми районами; участие ели уменьшается до 3%; заметно
увеличивается количество березы, ольхи, составляющих
до 26—27% лесных площадей. Граб почти отсутствует.
Юго-западный район ольхово-широколиственных ле-
сов (водораздельно-бугскоприпятский) занимает край-
нюю юго-западную часть территории республики между
53 и 52° с. ш. и 24 и 28° в. д. Район характеризуется сме-
шанными насаждениями, в которых сложные дубравы
составляют 11,9% общего числа лесов. Северная грани-
ца района является южной границей распространения
ели. Значительно распространены граб, березовые
(14,7%) и ольховые (17,1%) древостои. Сосновые на-
саждения занимают 58,9% лесных площадей.
7
Юго-восточный район дубово-грабовых лесов (при-
пятский) расположен между 52 и 53° с. ш. и 28 и
32° в. д. и охватывает центральную часть Припятского и
Дисненского Полесья. Территория его представляет со-
бой низину с большим количеством низменных и верхо-
вых сфагновых болот. Плоские понижения с близкими
грунтовыми водами и небольшой их проточностью заняты
ольхово-ясеневыми и дубово-ясенево-ольховыми насажде-
ниями. Невысокие водораздельные плато между реками
покрыты сложными насаждениями дубрав с грабом, дуб-
рав с сосной (В. И. Переход, 1954; Н. И. Костюкевич,
1954).
В лесах БССР преобладает сосна (57,2%), широко
представлена ель (13,2%), значительно распространены
береза (10,8%), ольха (8,1%), дуб (5,3%), осина (4,4%),
очень мало ясеня (0,2%), клена (0,1 %), граба (0,5%),
липы (0,1%) и прочих мягколиственных пород (И. Д.
Юркевич, 1948).
Фауна позвоночных Белоруссии неоднородна. Основ-
ную группу составляют виды, свойственные европейскому
широколиственному лесу: обыкновенный еж, малая куто-
ра, сони, желтогорлая мышь, лесная куница, черный
хорек, малый подорлик, клинтух, горлинка, средний пест-
рый дятел, безногая ящерица, болотная черепаха, квак-
ша и др. Вторую группу образуют таежные виды: трех-
палый дятел, снегирь, кедровка, белобровый дрозд и др.;
третью группу—лесостепные и степные виды: обыкновен-
ный хомяк, крапчатый суслик, заяц-русак, дрофа, поле-
вой и хохлатый жаворонки, серая куропатка и др.; чет-
вертую—лесные виды: обыкновенная и малая бурозубки,
обыкновенная белка, водяная полевка, полевка-эконом-
ка, рысь, лось, глухарь, тетерев и др.; пятую—интерзо-
нальные виды: волк, лисица, барсук, горностай, ласка,
выдра и др. (И. Н. Сержанин, 1957).
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Для обоснования системы противоклещевых меро-
приятий необходимы рекогносцировочные и стационар-
ные обследования с целью выявления фауны клещей,
распространения их в связи с экологическими особенно-
стями местности, установления круга хозяев и изучения
биологии клещей. Эти вопросы выяснялись путем сбора
клещей в Лунинецком (д. Крестуново), Ивацевичском
(д. Козики), Малоритском (д. Ласки, д. Ляховичи) рай-
онах Брестской области, в Брагинском (с. Брагин,
д. Глуховичи, д. Ясени, д. Ильичи, д. Лубино, д. Рыж-
кова), Хойникском (д. Аврамовская) районах Гомель-
ской области, в Борисовском (д. Погодина), Дзержинском
(д. Каменка), Вилейском (д. Седыца) районах Минской
области, в Витебском (с. Старосельск), Сиротинском
(д. Полтево), Лепельском (д. Кветча) районах Витеб-
ской области, а также в заповеднике «Беловежская
пуща». Для стационарных исследований наиболее инте-
ресным оказался заповедник «Беловежская пуща». На
его территории встречаются почти все типы леса, харак-
терные для Белоруссии.
За 1953—1958 гг. проведено 397 учетов пропашником
и обследовано 4699 животных, относящихся к 86 видам
(млекопитающих 47, птиц 38, рептилий 1 вид). Обсле-
довано 7 нор барсуков и лисиц и 93 гнезда птиц, сонь и
белок. Всего собрано и определено 29 052 иксодовых
клеща.
Количественные подекадные учеты клещей с расти-
тельности производились пропашником (Г. В. Сердюко-
ва и Б. И. Померанцев, 1940, 1948). Это отрезок фанеры
9
50X^0 см, обтянутый вафельной тканью. Пропашник
протаскивали по растительности в вертикальном поло-
жении так, чтобы его нижняя грань касалась почвы. За
один учет принималось расстояние в 500 м. Через
каждые 10 м пропашник осматривался. Мелкие грызуны
и насекомоядные для учета клещей отлавливались ло-
вушками «Геро». При подекадном учете в стации стави-
лось по 20 ловушек через 5 м в одну линию на 3—
5 дней. Производился также отстрел оленей, белок, ли-
сиц, зайцев и птиц. Отловленные мелкие животные по-
мещались в белые матерчатые мешочки, которые пере-
вязывались для предотвращения расползания эктопара-
зитов до момента осмотра животных. Животные
обрабатывались в лаборатории в белой кювете или на
белой бумаге. Сначала осматривалась внутренняя сто-
рона мешочка, а затем тщательно обследовалось живот-
ное. Содержимое нор и гнезд обрабатывалось вручную,
но чаще с помощью термоэклектора (С. О. Высоцкая,
1953). Кроме того, подекадно учитывались клещи на до-
машних животных, главным образом на крупном рога-
том скоте.
Наблюдения над развитием клещей /. ricinus и
D. pictus велись одновременно в естественных условиях
и в лаборатории. Сытые самки этих видов сажались в
малые металлические садочки (рис. 1), которые поме-
Рис. 1. Маленькие металлические садочки
щались в толще подстилки в стациях обитания клещей.
Садочки регулярно просматривались, отмечалась дата
начала откладки яиц и вылупления личинок. Для опре-
деления срока физиологического доразвития (Г В. Сер-
дюкова, 1952) садочки переносились в большие полевые
садки (рис. 2) размером 40X20X20 см, имевшие откид-
ную крышку и обтянутые со всех сторон густой метал-
10
лической сеткой, непроходимой для личинок клещей.
Садки устанавливались в местах обитания клещей в
грунте на глубине 5 см. Дно садка выстилалось слоем
дерна толщиной 3—4 см. Внутри на расстоянии 5 см от
края часть садка отгораживалась крупной металличе-
Рис. 2. Полевой садок с ежом
ской сеткой для изоляции животного-хозяина от клещей.
В большое отделение садка сажался еж, а в малое по-
мещался садочек с приоткрытой крышечкой для свобод-
ного выхода личинок. По окончании развития личинки
могли выползать из садочков и проникать через пере-
городку к ежу. Животное содержалось в полевом садке
в течение суток, затем пересаживалось в специальный
садок для сбора сытых клещей. Этот садок из крупно-
ячеистой металлической сетки помещался над тазом с
Н
водой (рис. 3). Собранные в воде сытые клещи снова
помещались в маленькие металлические садочки, кото-
рые закладывались в толще лесной подстилки. Подсад-
ка животного в полевой садок с оставшимися голодными
Рис. 3. Садок из крупноячеистой металлической сетки для сбора
сытых клещей
личинками повторялась примерно через 10 дней до пол-
ного их исчерпания. Свежеперелинявшие нимфы и взрос-
лые особи также переносились в маленьких садочках в
полевые садки, где по мере доразвития нападали на жи-
вотное. Наблюдение велось за потомством каждой
самки отдельно, что позволяло наиболее точно устано-
вить начало откладки яиц каждой генерации, сроки раз-
вития сытых личинок и нимф, а также сроки физиоло-
гического доразвития вылупившихся личинок и перели-
нявших клещей в природе.
В лабораторных условиях клещи содержались при
температуре 18—20° в стеклянных пробирках и корми-
лись на кроликах под колпачками, которые приклеива-
лись на ушах. Чтобы предотвратить сдирание колпачков
с ушей, на шею кролика одевался фанерный круг диа-
метром 25 см (рис. 4).
12
Для контроля с отстрелянных и отловленных диких
животных собирались сытые личинки и нимфы, которые
также помещались в маленькие садочки в естественных
условиях с целью наблюдения за сроками их линьки.
Рис 4. Кормление клещей под колпач-
ками на ушах кролика
Длительность паразитирования всех фаз клещей
определялась путем маркирования клещей на крупном
рогатом скоте и ежедневного подсчета отпавших сытых
особей с ежей и кроликов. Для определения клещей
использовалась отечественная литература (Б. И. Поме-
ранцев, 1946, 1950; Г. В. Сердюкова, 1955; Н. И. Джапа-
ридзе, 1950; Н. А. Филиппова, 1954).
Правильность результатов определения иксодовых
клещей проверена Г В. Сердюковой (лаборатория ЗИН
АН СССР).
ВИДОВОЙ СОСТАВ
И ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
ИКСОДОВЫХ КЛЕЩЕЙ В БССР
В Белоруссии были хорошо известны три вида иксодо-
вых клещей: Ixodes ricinus L. и Dermacentor pictus
Herm., встречающиеся почти повсеместно, и Dermacen-
tor marginatus Sulz. из юго-восточных районов (М. Н. Су-
дзиловский и Н. В. Щербинин, 1936; С. Ф. Сузько,
1949, 1953; В. Ф. Гусев, 1954, 1955 и др.). За последние
годы обнаружены единичные экземпляры переносчика
клещевого энцефалита человека—Ixodes persulcatus Sch.
(Е. Е. Вапник и др., 1956), Haeniaphysalis concinna
Koch. (Г. В. Сердюкова, 1956; Е. Е. Вапник и др., 1957) и
Haemaphysatis punctata Can. et Fanz. (В. Ф. Гусев, 1952,
1954).
Нами подтверждено широкое распространение /. rici-
nus и D. pictus и выявлены не известные ранее для БССР
Ixodes trianguliceps Bir., I. apronophorus Sch. и /. crenu-
latus Koch. (И. T. Арзамасов, 1955). Позднее /. triangu-
liceps и /. apronophorus найдены также в центральной,
западной и юго-западной частях БССР (X. М. Модель
и др., 1956; Е. Е. Вапник и др., 1956, 1957).
Численность иксодовых клещей в сильной степени за-
висит от ландшафта и климатических особенностей.
В северном районе еловых лесов обнаружены /. ricinus,
I. trianguliceps, I. apronophorus и D. pictus, причем пер-
вый вид встречается повсеместно. Ареал D. pictus про-
стирается полосой по долине Западной Двины, главным
образом к северу от нее, заходя отдельными вкрапле-
ниями на левобережье в районах Витебска и Полоцка.
/. trianguliceps и /. apronophorus обнаружены в Сиро-
тинском районе на правобережье Западной Двины.
14
В Центральном переходном районе смешанных ело-
во-широколиственных лесов почти повсеместно распро-
странен I. ricinus. Он отсутствует только в нескольких
административных районах севернее Могилева и в рай-
оне Молодечно, что связано, очевидно, с освоением бо-
лее 50% территории под пашни. D. pictus распространен
в основном по территории бассейна Березины. В северной
части этого района ареал D. pictus сужается, ограничи-
ваясь с запада и востока Минской и Оршанской возвы-
шенностями, а к югу расширяется, распространяясь по
всей Центрально-Березинской равнине. В Центральном
районе довольно часто встречаются /. trianguliceps (Ви-
лейский, Лепельский, Борисовский и Крупский районы),
/. apronophorus (Лепельский, Борисовский и Крупский
районы) и единично I. persulcatus (Борисовский район).
В Западном районе елово-широколиственных лесов
почти повсеместно распространен /. ricinus, не обнару-
жен он в нескольких административных районах, что,
очевидно, связано с недостаточной изученностью этих
пунктов. D. pictus обнаружен в западной и северной
частях этого района, а также в районах, примыкающих
непосредственно к Полесью. Кроме того, в северной
(б. Юратишский район) и юго-западной (Свислочский
район) частях обнаружен /. trianguliceps.
В Восточном районе елово-дубовых лесов повсемест-
но распространены /. ricinus (за исключением единич-
ных пунктов в восточной части) и D. pictus. В юго-во-
сточной части его (Ветковский и Уваровичский районы)
обнаружен Н. concinna.
В Юго-западном районе ольхово-широколиственных
лесов распространены I. ricinus и D. pictus. Довольно
часто встречаются /. trianguliceps (Лунинецкий, Иваце-
вичский, Малоритский, Каменецкий и Пружанский
районы) и /. apronophorus (Ивацевичский, Малоритский,
Каменецкий и Пружанский районы). Имеются единичные
находки /. crenulatus (Каменецкий и Пружанский
районы).
Юго-восточный район дубово-грабовых лесов харак-
терен повсеместным распространением /. ricinus и D. pic-
tus. В северной части его (Копаткевичский и Терехов-
ский районы) имеются единичные находки Н. concinna,
в юго-восточной (Хойникский, Наровлянский, Калинко-
вичский и Комаринский районы) встречается D. margi-
15
natus. В Калинковичском районе зарегистрированы еди-
ничные находки Н. punctata.
На территории Белоруссии с широколиственно-хвои-
ными лесами широко распространены характерные для
этой зоны /. ricinus и D. pictus. Свойственный лесным
стациям /. ricinus встречается почти во всех районах
БССР. Клеща нет там, где исчезли его естественные
биотопы вследствие сплошных распашек или они заняты
большими массивами болот, экологически не свойствен-
ных этому виду. Возможно также, что в результате сла-
бой изученности некоторых местностей в отношении за-
селения их клещами имеются районы, как бы свободные
от клещей. Массовый вид D. pictus—обитатель луговых
формаций по долинам рек—распространен главным
образом в южных и центральных районах республики.
В северных районах он встречается реже, так как здесь
появляются леса таежной подзоны, не характерной для
него.
Юго-восточные районы Белоруссии сходны с лесо-
степью. Здесь в открытых биотопах встречается степной
вид D. marginatus. Таежный клещ /. persulcatus найден
в центральной части БССР, занятой хвойными и широ-
колиственно-хвойными лесами, где, видимо, имеются
экологически близкие к таежным стации, свойственные
этому виду.
В южной части Белоруссии, расположенной в зоне
европейских широколиственных лесов, найден характер-
ный для этих лесов вид Я. concinna.
Частые находки в БССР /. trianguliceps, который
свойствен для лесных формаций, и норового вида
I. apronophorus, имеющего вместе со своим основным
хозяином—водяной полевкой широкий ареал, позволяют
предположить, что эти два вида в Белоруссии встреча'
ются повсеместно.
Судить о степени распространения в БССР норового
вида /. crenulatus по единичным находкам пока невоз-
можно.
Единичные находки Н. punctata можно объяснить
заносом клещей птицами из области распространения
вида, лежащей южнее БССР.
Таким образом, к настоящему времени в БССР обна-
ружены девять видов иксодовых клещей: Ixodes ricinus
L, /. persulcatus Sch. /. trianguliceps Bir., /. apronopho-
16
rus Sch., I. crenulatus Koch., Haetnaphysails concinna
Koch., H. punctata Can. et Fanz., Dermacentor pictus
Herm. и D. marginatus Sulz.
Возможно, что при дальнейших исследованиях юго-
восточных районов будет найден известный переносчик
туляремии степной вид Ixodes laguri 01.
2. Иксодовые клещи
СТАЦИАЛЬНОЕ РАСПРЕДЕЛЕНИЕ
ИКСОДОВЫХ КЛЕЩЕЙ
Стадиальное распределение иксодовых клещей нами
выявлялось путем сбора паразитов пропашником и с от-
ловленных мелких зверьков в основных растительных
формациях БССР (боры, субори, ельники, дубравы,
ольсы, березовые и осиновые леса), а также на выруб-
ках, лесных полянах, лугах, болотах, в прибрежных зо-
нах водоемов, на целинных участках, культурных полях,
огородах, садах, в домашних постройках и скотных дво-
рах. При этом наблюдалась неодинаковая заселенность
их клещами.
СОСНОВЫЕ ЛЕСА
Бор ы-з еленомошники (Pineta hylocomiosa) —
самая распространенная группа сосновых лесов. Из них
нами обследованы боры мшистые (Pinetum pleurozio-
sum) и бор-брусничник (Pinetum vacciniosum).
Боры мшистые обследовались в Беловежской
пуще, Лунинецком (д. Крестуново), Малоритском
(д. Ласки) и Сиротинском (д. Полтево) районах. Из
мелких грызунов в них отловлены лесная соня, европей-
ская рыжая полевка, желтогорлая мышь, полевая мышь,
лесная мышь; из насекомоядных—обыкновенная буро-
зубка, малая бурозубка; отстреляны обыкновенная бел-
ка и заяц-русак. Заселенность боров мшистых мелкими
грызунами и насекомоядными была низкая, не превы-
шающая 8 зверьков на 100 ловушко-суток.
Обследования в Беловежской пуще показали, что от-
18
носительный запас клещей1 /. ricinus в данной стации
незначителен: 0—4 (весной) и 10—11 (конец лета—
осень). Интенсивность заражения зверьков также невы-
сокая (0—1 весной, до 2 осенью). Некоторое повышение
к осени как относительного запаса клещей, так и интен-
сивности заражения грызунов, вероятно, происходит за
счет увеличения отлова зверьков, мигрировавших из
других типов леса, более насыщенных клещами и их хо-
зяевами.
Сборы клещей с растительности были также немно-
гочисленны. Например, в мае 1956 г. в среднем на один
учет приходилось 2—3 нимфы и одна самка.
Обследование боров мшистых в Лунинецком, Мало-
ритском и Сиротинском районах дало отрицательный
результат. На растительности и мелких грызунах /. rici-
nus не обнаружен (табл. 1 и 2).
Обследованием более крупных зверьков, добытых в
борах мшистых, установлена несколько иная заклещев-
ленность. Так, интенсивность заражения белок и зайцев
преимагинальными фазами /. ricinus оказалась почти
такая же, как в ельнике дубняковом, и немного меньше,
чем в ольшанике. По-видимому, зараженность их клеща-
ми не может служить показателем зараженности для
стации, где они добыты, так как эти зверьки в поисках
корма передвигаются на значительные расстояния и в
течение суток успевают побывать в различных расти-
тельных формациях, где могут подвергаться нападению
клещей. Данные же сборов клещей с мелких грызунов и
насекомоядных, имеющих малую площадь деятельности,
а также сборы клещей с растительности указывают на
малую заселенность боров мшистых клещами /. ricinus.
Очень редко в этом типе леса встречаются /. triangu-
liceps (с мышевидных грызунов снято 11 особей) и
D. pictus (снято 7 личинок), еще реже—/. crenulatus (най-
дено 3 самца в норе лисицы).
Бор брусничников ый обследован нами в Бере-
зинском заповеднике (Лепельский район, окрестности
д. Кветча). Здесь добыты желтогорлая мышь, лесная
мышь и обыкновенная бурозубка. На них иксодовых
клещей не обнаружено.
1 Величина относительного запаса клещей получается от перемно-
жения процента отлова грызунов на среднюю зараженность их клещами.
19
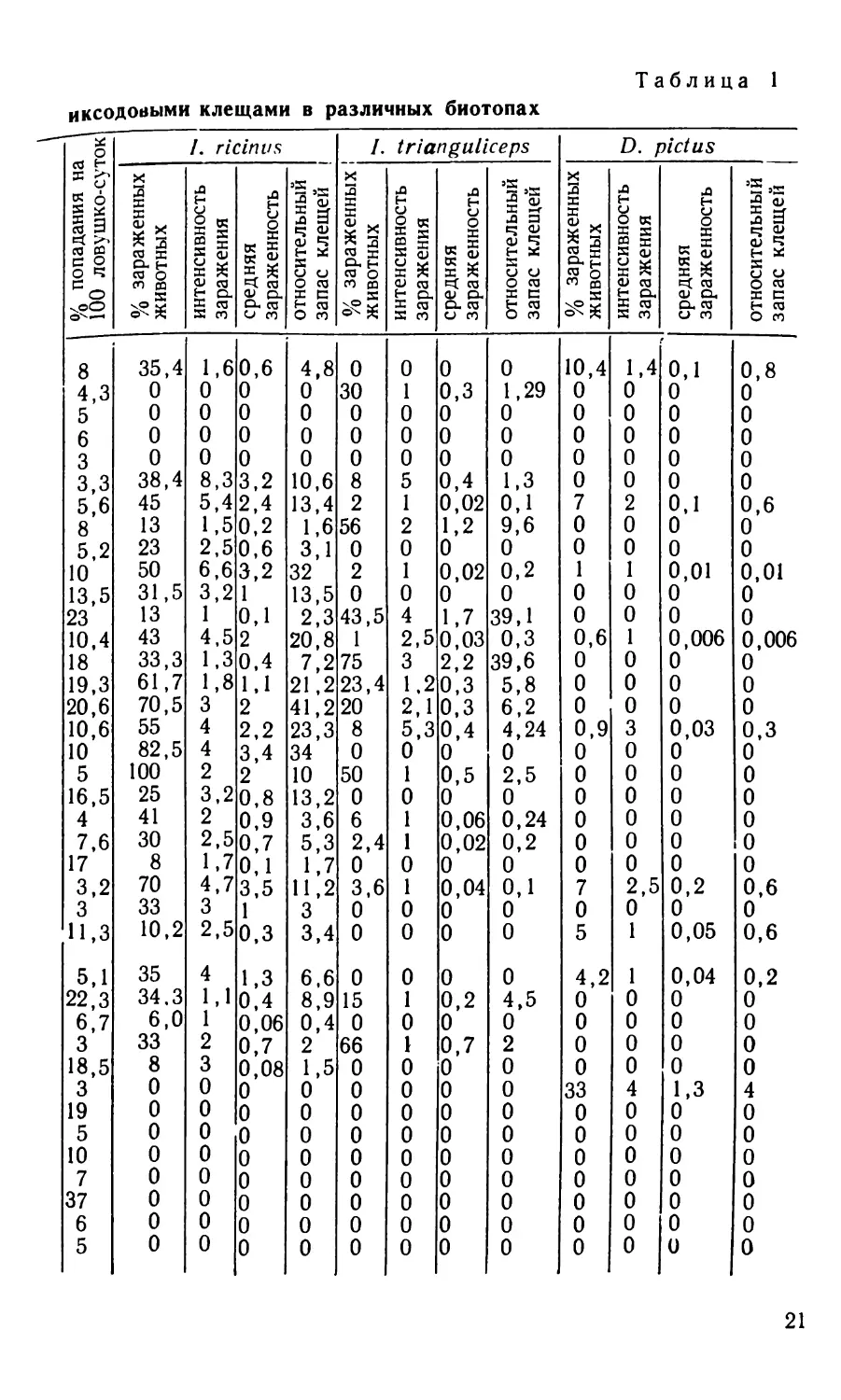
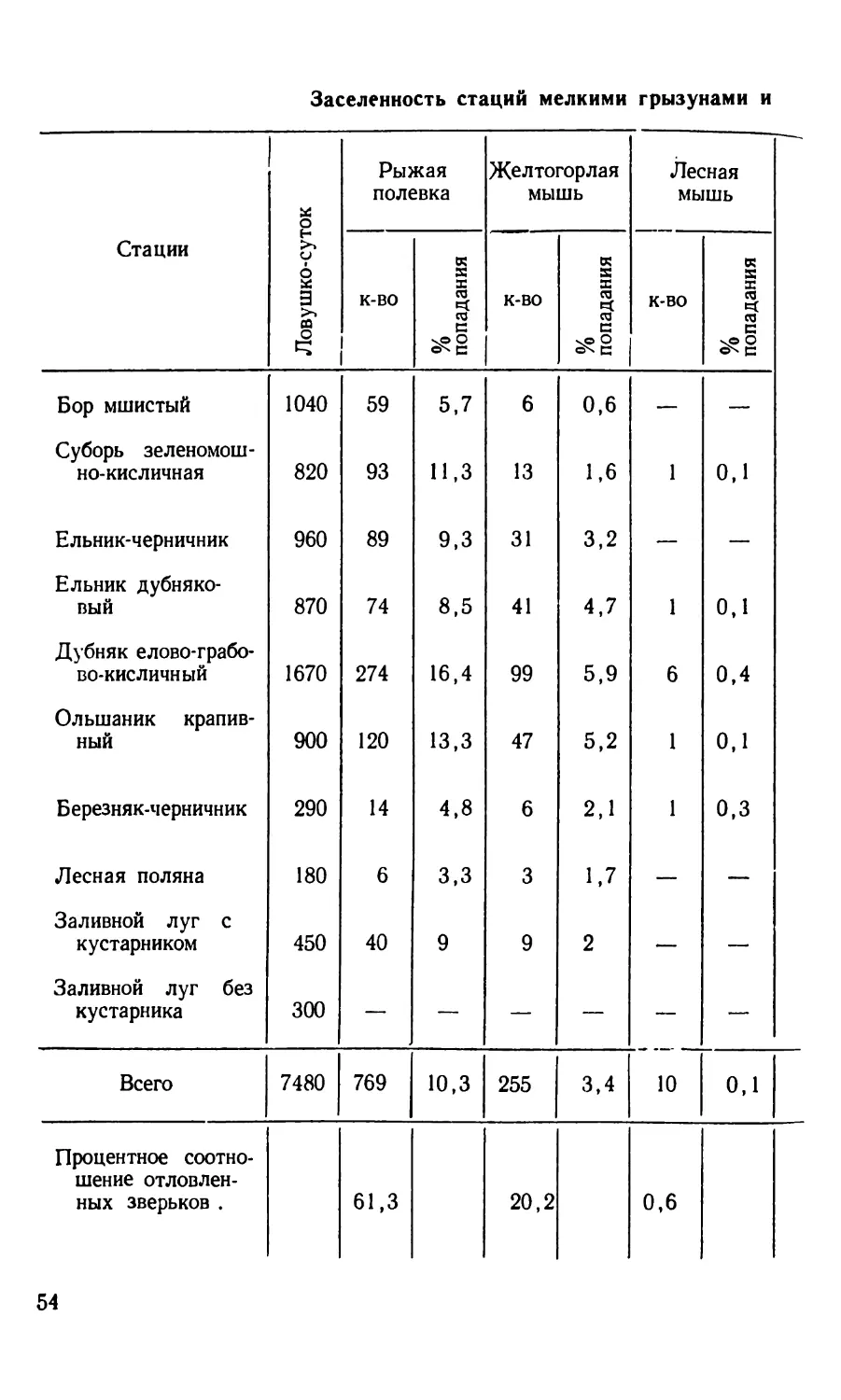
Зараженность мелких грызунов и насекомоядных
Биотоп Пункт обследован ия
Бор мшистый » » » » » » Бор брусничниковый Суборь зеленомошно-кисличная Ельник-черничник » » » » Ельник дубняковый » лещиновый » приручьево-травяной Дубняк елово-грабово-кисличный » елово-грабово-черничный Дубово-сосновый лес Грабняк кисличниковый Ольшаник крапивный » кочедыжниковый » крапивный » снытевый Березняк-черничник Осинник кисличниковый » » Смешанный молодняк Вырубка Лесная поляна Заросли кустарника в прибрежной зоне водоемов » » » » » » Низинное болото Скотомогильник Культурные поля Фруктовый сад Огороды Жилые дома и скотные дворы » » » » » » » » » Беловежская пуща Лунинецкий район Малоритский район Сиротинский район Березинский заповедник Беловежская пуща » . . Березинский заповедник Сиротинский район Беловежская пуща Сиротинский район . . Березинский заповедник Беловежская пуща Лунинецкий район » » Беловежская пуща Лунинецкий район . . Березинский заповедник Сиротинский район Беловежская пуща » Сиротинский район Беловежская пуща . Березинский заповедник Беловежская пуща » Лунинецкий район Сиротинский район . Березинский заповедник Лунинецкий район Брагинский район » » Беловежская пуща Малоритский район Брагинский район . . . Березинский заповедник Сиротинский район
20
nd
СО — — — ND сло-^-^ослсооооооооюсл СП -4J со >— — — — — — ND — — — ND —— — GOGO^J-^^OOlOOOOOOOGOCOOCnOOCnGOCOOCn^Qo GO ND О СП О О GO hU СП ND О GO GO % попадания на I 100 ловушко-суток
00 00 00 0000000000000^01 О 00 — GO GO 4*- ND О 00 СП -q О GO 4*. — GO СЛ ND — 4^ GO GO OGOOOOO — CnONOCnO-GOGOGO — OG0G0Cn00OOOOCn ND СП СП GO СП 4*> 4*- % зараженных животных
4 1,1 1 2 3 0 0 0 0 0 0 0 0 ND GO 4^ — NDNDGOND4^4^GO — — 4^ — GO О ND — СЛОООООО — СП СП ND 00 GO СП ND О СП СП 4*. GO О интенсивность заражения /. г и
1,3 0,4 0,06 0,7 0,08 0 0 0 0 0 0 0 0 О — G0OOOONDG0NDND — ONDO — Q0OONDG0OOOOO co СП — о 00 4^ ND — 4^ — ND О ND 4^ ND О средняя зараженность пЛпиь
ОООООООО — ND О 00 О СП 4^ о о — — — GO ND 4^ ND ND -GO — — GO GO — — CnG0G0O4^G0 — — -<1 О ND GO ND GO — GO О О О О О 4^ 4ь. ND •<! GO О ND GO ND ND ND 00 GO СП — О 4* О 00 относительный запас клещей I
0 15 0 66 0 0 0 0 0 0 0 0 0 СП ND ND 4ь. СП GO OOGOONDOOOOOOOGOCn-GoONDOONDODOOOOO О 4*- 4^ СП % зараженных 1 животных
000000000—0—0 OO — О — — О — О СП ND — GO ND 4^ О — О ND — СпООО — О GO — ND СП интенсивность заражения tria
0 0,2 0 0,7 0 0 0 0 0 0 0 0 0 0 0,3 0 0 0 0,4 0,02 1,2 0 0,02 0 1,7 0,03 2,2 0,3 0,3 0,4 0,5 0 0,06 0,02 0 0,04 0 0 средняя зараженность nguli
0 4,5 0 2 0 0 0 0 0 0 0 0 0 GO GO OOOOOOONDO4xOCHCDOCDOOOCDO — ООО — О — ND ND СП ND ND 00 О GO — ND О — GO ND 4^ 4^ О относительный запас клещей ceps I
4,2 0 0 0 0 33 0 0 0 0 0 0 0 OiO-JOOOOOOOOOOOOO-OO-JOOOOOO О о 4^ % зараженных животных
0000000^0000— 1,4 0 0 0 0 0 2 0 0 1 0 0 1 0 0 0 3 0 0 0 0 0 0 2,5 0 1 интенсивность заражения о
0,04 0 0 0 0 1,3 0 0 0 0 0 0 0 0,1 0 0 0 0 0 0,1 0 0 0,01 0 0 0,006 0 0 0 0,03 0 0 0 0 0 0 0,2 0 0,05 средняя зараженность lictus
0,2 0 0 0 0 4 0 0 0 0 0 0 0 0,8 0 0 0 0 0 0,6 0 0 0,01 0 0 0,006 0 0 0 0,3 0 0 0 0 0 0 0,6 0 0,6 относительный запас клещей
иксодовыми клещами в различных биотопах
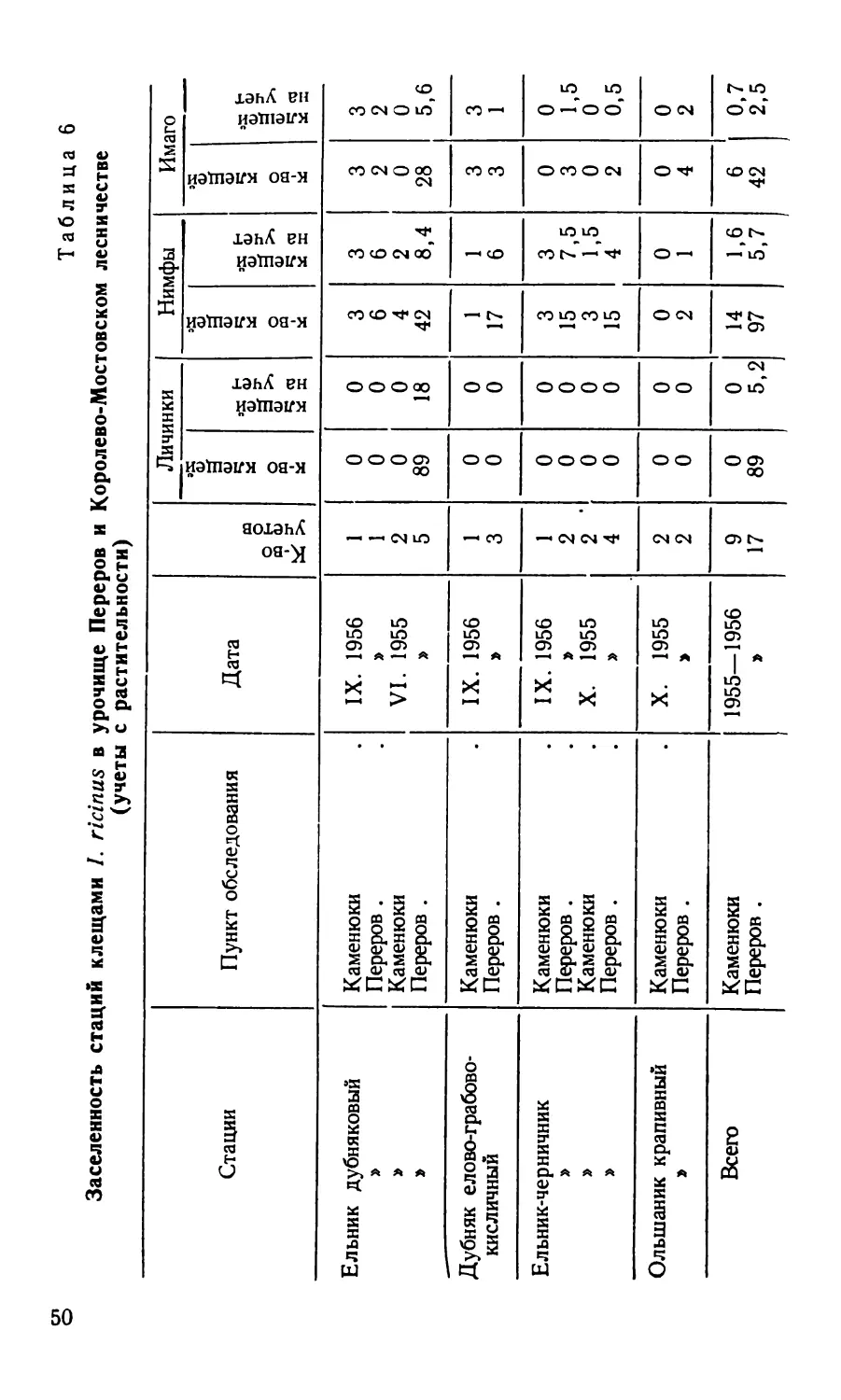
Таблица 2
Заселенность различных биотопов иксодовыми клещами
(по данным сбора с растительности)
Биотоп Пункт обследования К-во клещей на один учет
/. ri- cinus D. pic- tus
Бор мшистый Беловежская пуща 2—3 0
» Малоритский район 0 0
Суборь зеленомошно-кис- Беловежская пуща 4—5 0
личная
Ельник-черничник 9—10 0
» дубняковый 13—14 0
Дубняк елово-грабово-кис- 8 0
личный
» елово-грабово-чернич- Лунинецкий район 4 0
ный
Дубово-сосновый лес » » 2-3 0
Грабняк кисличниковый » » 2—3 0
» Малоритский район 14—15 0
Ольшаник крапивный Беловежская пуща 7—8 0-1
» Малоритский район 15 0
Березняк-черничник Беловежская пуща 4—5 0—1
Лесная поляна » » 0—1 0
» Малоритский район 0 0
Пойменные заросли кустар- Беловежская пуща 1—2 6-7
ника
Заливной луг без кустар- 0 0—1
ника
Лунинецкий район 0 0
Малоритский район 0 0
Из сказанного можно сделать заключение о незначи-
тельной заселенности сосновых лесов /. ricinus, I. trian-
guliceps, I. crenulatus и D. pictus, которые, очевидно,
периодически заносятся туда животными из других био-
топов. Об этом говорит и увеличение зараженных зверь-
ков в сосняках к осени—в период расселения грызунов
из других биотопов, более богатых в фаунистическом
отношении. Г. А. Найденова (1941), Б. П. Савицкий
(1957) в Белоруссии, В. В. Судаченков (1941) в Ленин-
градской области и А. К. Вильбасте (1956) в Эстонии
также пришли к заключению о чрезвычайно малой за-
раженности сосняков /. ricinus. В. В. Судаченков объяс-
22
няет это чрезмерной разреженностью сосняков, подтвер-
ждая фактами различной зараженности в одних и тех же
стациях. Например, клещи встречаются в основном на
затененных склонах, на ярко освещенных их в 5—6 раз
меньше. По нашему мнению, вполне возможно, что
здесь сказывается более низкая относительная влаж-
ность по сравнению с другими типами леса. Бедность
подстилки и редкий травяной покров, очевидно, также
способствуют ухудшению условий для развития клещей.
Слабая зараженность сосняков клещами зависит и от
незначительной плотности заселения этой стации мелки-
ми млекопитающими, что препятствует успешному раз-
множению иксодовых клещей.
СОСНОВО-ЕЛОВЫЕ ЛЕСА
Субори зеленомошные (Pineta piceeta hylo-
comiosa)—наиболее широко распространенная группа из
сосново-еловых лесов БССР. В Беловежской пуще нами
обследована суборь зеленомошно-кисличниковая (Pine-
lum piceeto-hylocomioso-oxalidosum). Здесь часто встре-
чаются европейская рыжая полевка, желтогорлая мышь,
редко лесная мышь и полевая мышь, соня-полчок, лес-
ная соня; из насекомоядных редко—обыкновенная буро-
зубка; отстреляны благородный олень, заяц-русак и
обыкновенная белка. Процент попадания мелких млеко-
питающих почти такой же, как и в борах брусничнико-
вых. Относительный запас /. ricinus колебался как по се-
зонам, так и в различные годы. Но в среднем субори
более насыщены клещами этого вида, чем боры (табл. 1).
Например, в 1955 г. относительный запас клещей в бо-
рах достигал 11, а в субори—74. Интенсивность зара-
жения в суборях была в 7 раз, а процент зараженных
зверьков в 1,5—2 раза выше. На растительности собрано
клещей в среднем в два раза больше, чем в бору мши-
стом (табл. 2), а в отдельные периоды значительно боль-
ше. Так, в мае 1956 г. в среднем на учет было 10 взрос-
лых, 5 нимф и 1 личинка, что превышало сборы в этот
же период в сосняках в 4—5 раз. Благородный олень,
заяц-русак, обыкновенная белка были также сильно за-
ражены /. ricinus.
Таким образом, вследствие более разнообразного
флористического и фаунистического состава сосново-
23
еловый лес является более благоприятной стацией оби-
тания /. ricinus, чем сосняки. В данном биотопе с убито-
го оленя сняты 2 самки D. piebus. В сборах на раститель-
ности и с отловленных мелких позвоночных этот вид не
обнаружен. Очевидно, крупные животные (как олень)
могут заносить его с открытых стаций (луга, лесные по-
ляны) в лесные. Из 141 отловленного здесь мелкого
зверька на 10 найдено 19 клещей I. trianguliceps.
ЕЛОВЫЕ ЛЕСА
Ельник и-зеленомошники (Piceeta hyloco-
miosa)—наиболее широко распространенная группа из
еловых лесов. Нами обследован ельник-черничник (Pi-
ceetum myrtillosum) в Беловежской пуще (рис. 5), Бере-
Рис 5. Ельник-черничник. Биотоп Ixodes ricinus L,
зинском заповеднике и в Сиротинском районе. В ельнике
очень часты европейская рыжая полевка, реже желто-
горлая мышь, соня-полчок и лесная соня, очень редко
мышь-малютка; часто обыкновенная бурозубка, реже
крот. Встречаются благородный олень, заяц-русак,
обыкновенная белка. Частота попадания в ловушки
мелких млекопитающих и процент заражения их не-
сколько выше, чем в субори (табл. 1). На отловленных
24
животных часто обнаруживается /. ricinus, значительно
реже—/. trianguliceps, D. pictus и единично—/, aprono-
phorus.
В Беловежской пуще отловленные в июне Г955 г.
зверьки были заражены /. ricinus на 89—100%. Интен-
сивность заражения, относительный запас клещей и
данные учетов на растительности выше по сравнению с
предыдущими стациями. На пропашник в мае 1956 г.
было собрано в среднем на учет 10 взрослых и 20 нимф
/. ricinus. Степень же заражения крупных животных
(благородный олень) была меньше, чем в субори, а
обыкновенной белки и зайца-русака—примерно одина-
кова с предыдущей стацией.
Зараженность животных в южных, центральных и се-
верных областях различная. Так, интенсивность зара-
жения /. ricinus мелких зверьков, добытых в Лепельском
и Сиротинском районах, значительно ниже (в централь-
ных областях в 12 раз, в северных в 4 раза), чем на юго-
западе республики (Беловежская пуща). Такая же зако-
номерность отмечается и по величине относительного
запаса клещей в этих пунктах (табл. 1). Таким образом,
интенсивность заражения мелких млекопитающих в ель-
нике-черничнике на юге несколько выше, чем в предыду-
щих стациях, процент заражения животных и плотность
их заселения также больше. Заселенность растительно-
сти клещами в ельнике-черничнике в 2 раза больше, чем
в сосново-еловом лесу.
Следовательно, ельники-черничники более заражены
/. ricinus, чем сосново-еловые леса и сосняки-зеленомош-
ники. Вероятно, здесь положительно сказывается более
мощная подстилка, создающая благоприятные условия
для прохождения метаморфоза /. ricinus, а также более
густой травяной и кустарниковый покров.
Интересен факт изменения соотношения численности
некоторых видов иксодовых клещей в ельниках чернич-
никовых. На юге республики на обследованных животных
в этих биотопах преобладающим оказался /. ricinus, в
центральных областях (Березинский заповедник)—
/. trianguliceps. Процент заражения животных послед-
ним был в 7 раз, а относительный запас клещей в 6 раз
выше, чем /. ricinus.
Ельники сложные (Piceeta composita). Из этой
группы в Беловежской пуще обследован ельник дубня-
25
ковый (Piceet'um quercetum), а в Сиротинском районе
(д. Полтево)—ельник лещиновый (Piceetum corylostun).
В ельнике дубняковом (рис. 6) часто встре-
чаются европейская рыжая полевка, желтогорлая мышь,
реже лесная и полевая мыши, лесная соня, часто попа-
дается обыкновенная белка; из насекомоядных—обыкно-
венная бурозубка и крот. Плотность заселения его мел-
кими грызунами и насекомоядными выше, чем в ельни-
ках черничниковых.
Рис 6. Елово-широколиственный лес. Биотоп Ixodes ricinus L.
В этом типе леса преобладает /. ricinus, единично
встречается D. pictus (обнаружена одна нимфа на евро-
пейской рыжей полевке) и редко—/. trianguliceps (с 150
зверьков собрано 35 особей этого вида клеща). Зара-
женность зверьков клещами I, ricinus, как и в ельнике-
черничнике на юге, в среднем около 50%, а в отдельные
периоды (июнь—сентябрь) до 100%. Интенсивность за-
ражения животных в ельнике дубняковом несколько
выше (6,6), чем в ельнике-черничнике (5,4). Относитель-
ный запас клещей в ельнике дубняковом очень большой
(в среднем 32), превосходит все обследованные нами
стации, кроме грабняков и ольшаников. Сборы на расти-
тельности показали также значительную заселенность
26
ельников дубняковых I. ricinus (табл. 2). Например, в
мае 1956 г. в среднем на один учет собрано 7 взрослых,
22 нимфы и 17 личинок этого вида.
В ельнике лещиновом часто встречается ры-
жая полевка, несколько реже лесная мышь; из насеко-
моядных—обыкновенная бурозубка. Процент отлова
мелких зверьков в среднем составил 13,5%, что на 3,5%
превышает показатели предыдущей стации, но процент
зараженных животных /. ricinus был меньше, чем в ель-
нике дубняковом (соответственно 31,5 и 50%), хотя в
отдельные периоды (в мае 1958 г.) составлял 63,6.
Интенсивность заражения в 2 раза меньше (в ельнике
дубняковом 6,6, в ельнике лещиновом 3,2). Разница в
относительном запасе клещей еще значительнее —
в 2,5 раза (табл. 1). Относительный запас клещей /. ri-
cinus в ельнике лещиновом примерно такой же, как в
ельнике-черничнике Беловежской пущи.
При сравнении заселенности I. ricinus ельника дуб-
някового и ельника лещинового, относящихся к группе
сложных ельников, вскрывается такое же соотношение
между югом и севером, как мы наблюдали в ельниках-
черничниках. В частности, /. ricinus гораздо интенсивнее
распространен в южных областях республики, чем в се-
верных. Но в общем ельники сложные заражены этим
видом в большей степени, чем субори и ельники-зелено-
мошники, не говоря уже о сосновых лесах. Это подтвер-
ждают и данные сборов клещей с растительности на тер-
ритории питомника, расположенного в районе сложных
ельников, где содержатся зубры и олени. В начале мая
1956 г. с растительности там собрана масса нимф: на
учет приходилось 52 нимфы. Такого большого количе-
ства /. ricinus с растительности нами не было собрано ни
в одной стации. В мае 1956 г. в загоне с одной более
спокойной оленицы собрано также большое количество
нимф (76). В октябре 1955 г. с убитой в районе питом-
ника белки снято 44 нимфы и 2 личинки /. ricinus. Таким
образом, ельники сложные при достаточной базе пита-
ния для /. ricinus (в данном случае наличие питомника)
могут быть мощным резервуаром их размножения.
Ельники травяные (Piceeta herbosa) зани-
мают небольшие узкие полоски возле речек и ручьев.
В Березинском заповеднике обследован ельник (рис. 7)
приручьево-травяной (Piceetum fontinale-herbo^um), где
27
наиболее часты европейская рыжая полевка, из насеко-
моядных—обыкновенная бурозубка, редко обыкновенная
кутора. Попадание указанных зверьков довольно частое
Рис. 7. Ельник приручьево-травяной. Биотоп Ixodes
trianguliceps Bir.
(в 1958 г. в среднем 23%), и превышает отлов многих
обследованных стаций.
В этой стации обнаружены /. ricinus (7% всех сбо-
ров) и /. trianguliceps. Зараженных 1. ricinus животных
было 13%, а /. trianguliceps—43,5% всех добытых зверь-
ков. Разница в интенсивности заражения животных
28
между этими видами клещей также значительная (/. ri-
cinus один на зараженного зверька, а /. trianguliceps 4).
Относительный запас клещей /. ricinus 2, 3, а /. triangu-
liceps 39,1. Общий же запас иксодовых клещей (41,4 без
подразделения на виды) в этом типе леса намного пре-
восходит все ранее рассмотренные биотопы.
ДУБОВЫЕ ЛЕСА
Из елово-грабовых дубрав (Querceta picee-
ta carpinosa) нами обследованы дубняк елово-грабово-
кисличный (Quercetum piceeto-carpineto-oxalidosum) в
Беловежской пуще и дубняк елово-грабово-черничный
(Quercetum piceeto-carpineto-myrtillosum) в Лунинецком
районе.
В дубняке елово-г рабов о-к неличном
отмечено много следов благородного оленя, при отлове
часто попадались европейская рыжая полевка, желто-
горлая мышь, лесная соня, реже лесная мышь, обыкно-
венная, пашенная и кустарниковая полевки, соня-полчок,
встречались обыкновенная белка, обыкновенная буро-
зубка и кутора малая. Заселенность мелкими грызунами
и насекомоядными (10,4%) почти такая же, как в ельни-
ке дубняковом. В отдельные периоды, например в октяб-
ре 1955 г., попадание мелких зверьков достигало 34%.
В этом типе леса встречался преимущественно I. ri-
cinus, значительно реже—I. trianguliceps (с 527 зверьков
собрано 77 особей) и очень редко—D. pictus (найдены
4 личинки). Зараженных I. ricinus животных было на
26% больше, чем в ельнике дубняковом. В июне—августе
зараженность животных достигала 100%. Интенсивность
же заражения позвоночных за весь теплый сезон была,
наоборот, на 2,1 меньше. В период наибольшей актив-
ности преимагинальных фаз (в июне—июле) интенсив-
ность заражения здесь была 18,3, а в ельнике дубняко-
вом—26. Относительный запас клещей /. ricinus также
был меньше (соответственно 20,8 и 32). При сборах кле-
щей на растительности оказалось, что в среднем на учет
/. ricinus было почти столько же, сколько в ельнике-
черничнике, но в 1,5 раза меньше, чем в ельнике дубня-
ковом (табл. 2). В апреле 1955 г. на пропашник собрано
5 взрослых особей и 25 нимф (на учет). Таким обра-
зом, дубравы елово-грабово-кисличные наиболее зараже-
29
ны I. ricinus и уступают в этом отношении только ельни-
кам дубняковым.
В дубняке елово-грабов о-ч ерничном
часто встречаются европейская рыжая полевка, полевая
мышь, желтогорлая мышь, редко лесная мышь и мышь-
малютка; из насекомоядных—обыкновенная бурозубка.
Отлов (18%) мышевидных значительно больший, чем в
дубняке елово-грабово-кисличном, но уступает в этом
отношении ельнику приручьево-травяному.
В этой стации распространены /. ricinus и /. triangu-
liceps. Зараженность животных первым видом ниже, чем
в дубняке елово-грабово-кисличном (соответственно 33,3
и 43%). Относительный запас клещей (7,2) также почти
в 3 раза меньший, чем в предыдущей стации. При сборе
/. ricinus на растительности оказалось 4 клеща на учет
(табл. 2). Но в этой стации больше всего обнаружено
/. trianguliceps (84% всех сборов клещей), которым за-
ражено 75% животных. Относительный запас (39,6) это-
го вида клеща в дубняке елово-грабово-черничном пре-
восходит все обследованные биотопы (табл. 1). Таким
образом, мы опять (как в ельнике-черничнике и ельнике
приручьево-травяном в Березинском заповеднике) на-
блюдаем изменение соотношений в пользу /. triangu-
liceps.
Дубово-сосновый лес обследован в Лунинец-
ком районе в окрестности д. Крестуново. В нем встре-
чаются желтогорлая мышь, европейская рыжая полевка,
полевая мышь, лесная мышь, обыкновенная бурозубка.
Отлов зверьков большой (19,3%), но меньше, чем в ель-
нике приручьево-травяном (табл. 1).
Из иксодовых клещей часто встречается /. ricinus,
реже /. trianguliceps. Первый составляет 80% сбора
клещей. Им заражено в среднем 50% животных, что в
4 раза выше, чем в дубняке елово-грабово-черничном в
этом же районе. Также выше интенсивность заражения
(1,8) и относительный запас клещей. Но в этой стации
уменьшилось количество клещей /. trianguliceps по
сравнению с предыдущей. Однако при обследовании ду-
бово-соснового леса в Лунинецком районе в начале апре-
ля 1956 г. вследствие низкой температуры (местами еще
лежал снег) на пропашник клещей попадалось мало:
2—3 на учет.
Грабняк кисличников ы й обследован в Лу-
30
нинецком и Малоритском районах. Из грызунов здесь
часто встречаются желтогорлая мышь, европейская ры-
жая полевка, редко лесная мышь, полевая мышь; из на-
секомоядных часто—обыкновенная бурозубка. Попа-
дание зверьков еще выше (20,6%), чем в дубово-сосно-
вом лесу.
В этой стации очень часто встречается /. ricinus
(86%) и значительно реже—/. trianguliceps (14%).
Первым животные заражены в среднем на 93%. Относи-
тельный запас клещей 41,2, что превышает все обследо-
ванные стации. Сбор с растительности в Малоритском
районе показал также самые высокие результаты (14—
15 клещей на учет). При сборе же клещей пропашником
в Лунинецком районе в начале апреля при низкой тем-
пературе на учет приходилось 2—3 особи /. ricinus.
ОЛЬХОВЫЕ ЛЕСА
В Беловежской пуще, Березинском заповеднике и
Малоритском районе обследован ольшаник крапивный
(Alnetum urticosum), в Лунинецком районе — ольшаник
кочедыжниковый (Alnetum filicosum) и в Сиротинском
районе—сероолыпаник снытевый (Jncano-Alnetum aego~
podiosum). В ольшанике часты европейская рыжая по-
левка, желтогорлая мышь, реже полевая и лесная мы-
ши, пашенная полевка, степная мышовка, мышь-малют-
ка, водяная крыса, попадается соня-полчок, лесная
соня и обыкновенная белка; из насекомоядных часто—
обыкновенная бурозубка, редко малая бурозубка и ма-
лая кутора. Наибольшая плотность мелких зверьков в
ольшанике наблюдалась в Сиротинском районе (16,5%),
несколько меньшая—в Беловежской пуще (10,6%) и Лу-
нинецком районе (10%) и совсем незначительная в Бе-
резинском заповеднике (5%). Но в отдельные периоды,
например в августе 1955 г., в Беловежской пуще попа-
дание зверьков на 100 ловушко-суток достигало 33%.
В ольшаниках очень часто встречаются /. ricinus и
trianguliceps, единично обнаружен D. pictus. В мае
1955 г. (Беловежская пуща) зараженных зверьков вто-
рым видом было 31 %, а интенсивность заражения достиг-
ла 1,8. С пойманного в этой стации в июле 1956 г.
обыкновенного ежа снята нимфа /. crenulatus.
31
Зараженность животных /. ricinus, за исключением
северной части Белоруссии (Сиротинский район), была
высока (82,5—100%). Высок и относительный запас кле-
щей (в Лунинецком районе в среднем равен 34, в Бело-
вежской пуще—23,3), который уступал только грабняку
кисличниковому.
В Сиротинском районе в центральной части респуб-
лики (Березинский заповедник) относительный запас
/. ricinus был значительно меньше, чем на юге в подоб-
ных стациях (табл. 1). Так, /. ricinus в ольшанике Ма-
лоритского района на один учет в среднем приходилось
15, т. е. столько же, сколько в грабняке кисличниковом.
Большая заселенность клещами /. ricinus черно-
ольшаников на юге, дубово-грабовых и хвойно-широко-
лиственных лесов является следствием их достаточной
влажности, оптимальной температуры, благоприятного
растительного и древесного покрова, богатой подстилки
и большой плотности заселения хозяевами клещей. Ши-
роколиственные породы и черноольшаники создают
сомкнутую крону, под которую редко проникают солнеч-
ные лучи, а густой подлесок и толстая подстилка способ-
ствуют поддержанию более постоянной температуры и
повышенной влажности. Все это создает благоприятные
условия для успешного развития /. ricinus.
В сходных лесных формациях в условиях Белоруссии
зараженность /. ricinus наблюдали также Г. А. Найде-
нова (1941), В. И. Бурцев (1925), Ф. А. Закович (1957),
С. С. Фолитарек и др. (1949). Н. О. Оленев (1927) для
Новгородской области считает наиболее благоприятны-
ми для I. ricinus стации сырые, сплошь покрытые зарос-
лями, главным образом ольхи, пастбища. В. В. Суда-
ченков (1941) в Ленинградской области к наиболее зара-
женным относит стации из серой ольхи, березы, рябины,
осины с небольшой примесью хвойных пород. Б. И. По-
меранцев (1935) указывает также на благоприятные
стации обитания клещей в густых зарослях белой ольхи.
Н. И. Алфеев (1935) в районе Череменецкого озера
наибольшее количество клещей находил на лесных паст-
бищах, относительно возвышенных, но достаточно влаж-
ных и покрытых древесной растительностью. М. П. Мар-
ков (1927) в условиях Украины указывает на повсемест-
ное распространение /. ricinus, но более частое в сыро-
ватых кустарниковых зарослях орешника, ольхи и дуба.
32
д. Н. Засухин и другие (1936) на юго-востоке РСФСР
отмечают наиболее благоприятные для I. ricinus стации
с сильно пониженным рельефом и повышенной влаж-
ностью. Г. С. Первомайский (1948) в Иране обнаружил
/. ricinus в лесу с преобладанием клена, граба, липы,
дуба. А. С. Лутта и другие (1953) в условиях Карель-
ской АССР указывают как основной биотоп для I. rici-
nus мелколиственный лес, где в основном преобладают
ольха, береза, осина. А. К. Вильбасте (1956) в условиях
Эстонской ССР к наиболее благоприятным стациям от-
носит ольшаники. Kossel и другие (1903) встречали
/. ricinus в Германии главным образом на влажной поч-
ве с густой растительностью. К. Samson (1909) чаще все-
го встречал I. ricinus вокруг Берлина на возвышенных
пастбищах вдоль тропинок во влажных лесах. В. Rosicky
и V. Cerni (1954) в Чехословакии наблюдали наибольшее
заражение мелких млекопитающих личинками I. ricinus
в низменном пойменном лесу.
Эти данные говорят о наибольшей зараженности
I. ricinus стаций, достаточно увлажненных, с наличием
в древесном ярусе ольхи и широколиственных пород.
При этом, по нашему мнению, следует учитывать пер-
востепенный фактор—питание. Ольшаники же дубово-
грабовые и хвойно-широколиственные леса и являются
такими стациями, где наряду с оптимальными темпера-
турными условиями и влажностью имеется большая
плотность заселения различными мелкими грызунами
и насекомоядными—хозяевами преимагинальных фаз
клещей /. ricinus. И. Н. Сержанин (1955) указывает, что
эти стации наиболее богаты и разнообразны в фаунисти-
ческом отношении. Наряду с большой плотностью мелких
зверьков здесь часто встречаются и такие крупные живот-
ные, как европейская косуля, лось, благородный олень,
уссурийская енотовидная собака, зайцы—хозяева взрос-
лых клещей /. ricinus. Таким образом, удачное сочетание
климатического и трофического факторов, безусловно,
создает благоприятные условия для успешного развития
всех фаз /. ricinus в указанных типах леса.
БЕРЕЗОВЫЕ ЛЕСА
В березняке-черничнике (Betuletum myrtillo-
sum) из мышевидных грызунов встречаются редко евро-
пейская рыжая полевка, желтогорлая, лесная и полевая
3- Иксодовые клещи
33
мыши, лесная соня; из насекомоядных—обыкновенная и
малая бурозубки. Плотность заселения их в 2—4 раза
ниже, чем в ольшаниках, и почти такая же, как в сосно-
вых борах. Наибольшее попадание (17,5%) наблюдалось
в октябре 1955 г.
В березняках с мышевидных грызунов были собраны
клещи I. ricinus (94%) и единичные экземпляры /. trian-
guliceps. Редко попадались на растительности D. pictus.
Например, на границе с лесной поляной на пропашник в
мае 1956 г. попались 2 самки и самец. По количеству за-
раженных (41%) и интенсивности заражения (2) зверь-
ков /. ricinus березняки значительно уступают ольша-
никам. Относительный запас /. ricinus в березовом лесу
почти в 7 раз меньше, чем в ольшаниках (сравнение
по Беловежской пуще). Сбор клещей с растительности в
среднем на учет такой же, как в сосново-еловом лесу
(табл. 2). В мае 1956 г. попадались 3 взрослых и 7 нимф
/. ricinus. По степени зараженности березняки можно
приравнивать к сосновым лесам.
ОСИНОВЫЕ ЛЕСА
В осиннике кисличниковом (Tremuletum
oxalidosum) встречаются европейская рыжая полевка,
желтогорлая и лесная мыши, обыкновенная бурозубка.
Попадание мелких зверьков незначительно (7,6—17%).
Здесь обнаружены /. ricinus и единично I. trianguliceps,
причем второй вид найден только в Беловежской
пуще.
Заселенность данной стации /. ricinus неравномерна
в различных областях. Количество зараженных живот-
ных и относительный запас клещей'на севере (Сиротин-
ский район) в 3 раза меньше, чем на юге (Беловежская
пуща). По интенсивности заражения (2,5) и относитель-
ному запасу клещей (5,3) осинники занимают промежу-
точное положение между березняками и суборями
(сравнение по Беловежской пуще).
СМЕШАННЫЕ МОЛОДНЯКИ
Нами в Беловежской пуще обследован участок мо-
лодняка в возрасте 15 лет. Высота древостоя 6 м, пол-
нота 0,8. Древостой смешанный, представлен в основном
34
осиной, березой, ольхой, елью, сосной, дубом. Из кустар-
ников много малины, можжевельника, бересклета. По-
кров составляют черника, зеленые мхи, злаки, ветреница,
осоки. Из мышевидных грызунов часто встречаются
европейская рыжая полевка, реже желтогорлая мышь;
из насекомоядных—обыкновенная бурозубка. Попадание
зверьков (3,2) по сравнению с другими стациями не-
большое. Самый высокий отлов (10%) наблюдался в
августе 1957 г. Из иксодовых клещей очень часто встре-
чается /. ricinus и единично—/. trianguliceps и D. pictus.
Зараженность животных клещами /. ricinus в этой
стации довольно высокая (в среднем до 70%). Интенсив-
ность заражения значительная (4,7), не уступающая
даже широколиственным лесам. В связи с незначитель-
ным процентом отлова мелких грызунов относительный
запас клещей (11,2) снизился, поэтому смешанные мр-
лодняки по запасу клещей /. ricinus могут занять проме-
жуточное положение между еловыми лесами и суборями.
ВЫРУБКИ
Вырубка 1000X500 м обследована в Березинском за-
поведнике. Тип условий местопроизрастания ясенево-
крапивный. Вырубка сплошь покрыта зарослями осины
высотой 1,5—2 м. Изредка встречаются березки, елочки,
ясень высотой 5—10 м, очень часто кустики черемухи,
рябины, малины, ежевики. Покров густой—много кра-
пивы, черники, брусники, земляники, встречаются папо-
ротник, вероника лекарственная, осоки, поповник, клевер
белый, майник, лютик едкий. Из мелких зверьков отлов-
лены европейская рыжая полевка и обыкновенная буро-
зубка. Отлов их очень незначительный (3%), даже мень-
ший, чем в березняках.
Иксодовые клещи встречались редко. Кроме I. ricinus,
с европейской рыжей полевки снята одна нимфа /. apro-
nophorus. Зараженность животных (3%) и относитель-
ный запас (3) /. ricinus очень низки.
ЛЕСНЫЕ ПОЛЯНЫ
В Беловежской пуще обследована лесная поляна
300ХЮ0 м, обработанная в 1952 г. под посадку сосны.
На поляне редко разбросаны небольшие кустики березы.
35
Почва супесчаная свежая. Подстилка почти отсутствует.
Из мышевидных грызунов часто встречаются обыкновен-
ная полевка, редко европейская рыжая полевка и желто-
горлая мышь. Заселенность мелкими грызунами (в сред-
нем 11,3%) была довольно значительная (в августе
1955 г. попадание достигало 27%), но зараженные зверь-
ки составляли всего 10,2%. Чрезвычайно мал также и
относительный запас (3,4) клещей /. ricinus. Такие же
результаты получены и при сборе клещей на раститель-
ности (табл.2).
При сборе клещей на пропашник на одной из лесных
полян в Малоритском районе (д. Ласки) клещей не
обнаружено. Таким образом, заселенность лесной поля-
ны /. ricinus низка по сравнению с другими стациями.
Возможно, что более высокая температура и низкая
влажность открытых растительных формаций создают
неблагоприятную среду для обитания /. ricinus.
В. В. Судаченков (1941) при обследовании открытых
пастбищ Ленинградской области также пришел к заклю-
чению о полной незараженности их /. ricinus. Б. И. По-
меранцев указывает, что наличие древесной раститель-
ности нужно признать обязательным для клещевых ста-
ций /. ricinus. Д. Н. Засухин и другие (1936) в условиях
юго-востока РСФСР (Саратовская область) на выгонах
и залежах также не встречали I. ricinus. К такому же
выводу пришли А. С. Лутта и другие в условиях Карель-
ской АССР.
Следовательно, лесные поляны не являются местом
обитания клещей I. ricinus. В этой стации с грызунов
собраны лишь единичные экземпляры D. pictus.
ПРИБРЕЖНАЯ ЗОНА ВОДОЕМОВ
Заросли кустарника около водоемов обследованы в
Беловежской пуще, Березинском заповеднике, Лунинец-
ком и Сиротинском районах.
В Беловежской пуще на заливных лугах в пойме
р. Лесной обследованы заросли кустарников (рис. 8) из
ольхи, березы, ивы, крушины ломкой, калины, черемухи,
малины, смородины черной и красной. Покров в кустар-
никах составляют осоки, лабазник, крапива, хвощи,
белокрыльник болотный, вахта. Часто встречаются толс-
тые пни—остатки старого ольхового леса. Почва илова-
36
то-торфянистая. Подстилка толщиной до 3 см. В кустар-
никах обитает много мелких млекопитающих. Из мыше-
видных грызунов часто попадаются европейская ры-
жая полевка, полевая мышь, реже желтогорлая мышь,
обыкновенная и пашенная полевки; из насекомоядных—
обыкновенная бурозубка, реже—малая бурозубка, малая
кутора и обыкновенный еж. Плотность заселения мелкими
грызунами и насекомоядными довольно большая. На-
Рис. 8. Река Лесная. Биотоп Dermacentor pictus Herm
пример, процент попадания их даже несколько выше,
чем в ольшаниках (в октябре 1955 г. 38,1%). Заражен-
ность зверьков клещами /. ricinus на 4% больше, чем
в осиннике, а относительный запас клещей (6,6) примерно
такой же (табл. 1). Зверьков, зараженных D. pictus (в
июле 1955 г. 70%), а также интенсивность заражения их
преимагинальными фазами (в июле 1955 г. 3,4 клеща на
одного зверька) в этой стации были гораздо больше, чем
I. ricinus. В результате сбора пропашником в мае 1956 г.
в среднем клещей /. ricinus на учет оказалось 1,7 взрос-
лого и 0,7 нимфы, a D. pictus—29.
При обследовании кустарников на заливных лугах
Хойникского района (д. Аврамовская) установлено, что
интенсивность заражения зверьков I. ricinus была 1,5, а
37
D. pictus—26. Такие показатели для D. pictus на залив-
ных лугах в кустарниковых биотопах являются наиболь-
шими по сравнению со всеми другими обследованными
стациями. Таким образом, подтверждается приурочен-
ность D. pictus к луговым формациям с наличием кустар-
ников, в то время как /. ricinus доминирует в лесу.
Здесь также обнаружены /. apronophorus (с обыкновен-
ной полевки и водяной крысы сняты 3 самки и одна ним-
фа) и /. crenulatus (с обыкновенного ежа в июле 1955 г.
снята одна самка).
В Лунинецком районе обследованы кустарниковые
заросли по берегам осушительных канав. Здесь часто
встречаются полевая мышь, европейская рыжая полевка,
обыкновенная полевка, реже водяная крыса, полевка-
экономка, мышь-малютка, северная мышовка. Попада-
ние их очень велико (22,3), превышает почти все обсле-
дованные стации.
Зараженность животных клещами /. ricinus и отно-
сительный запас их почти такие же, как в сходных ста-
циях в Беловежской пуще.
Обнаружен и /. trianguliceps. Зараженность им зверь-
ков и относительный запас клещей в 2 раза меньше, чем
I. ricinus.
В Березинском заповеднике обследован берег канала,
проходящего среди хвойно-широколиственного леса и
сплошь заросшего ивняком и ольхой. Здесь редко встре-
чается водяная крыса (попадание ее не превышает 3%)-
Зараженность зверьков иксодовыми клещами, как и от-
носительный запас их, незначительна (табл. 1). Здесь
/. ricinus и I. trianguliceps обнаружены в одинаковом ко-
личестве.
В Сиротинском районе обследован берег озера Го-
роднянского, где редко разбросаны кустики серой ольхи
и березы. Покров густой, преимущественно из мужского
папоротника. У самой воды заросли тростника. Из мел-
ких грызунов часто встречаются европейская рыжая по-
левка, реже лесная мышь и водяная полевка; из насе-
комоядных—обыкновенная бурозубка и обыкновенная
кутора. Попадание зверьков незначительно (6,7%).
Здесь редко встречается /. ricinus и единично—/. apro-
nophorus. 24 июня 1958 г. с обыкновенной куторы сняты
2 нимфы второго вида. Относительный запас клещей
/. ricinus в этой стации наиболее беден.
38
НИЗИННОЕ БОЛОТО
Низинное болото обследовано в Лунинецком районе.
Из мышевидных грызунов здесь чаще попадаются поле-
вая мышь, реже европейская рыжая полевка, полевка-
экономка и единично—черная крыса, мышь-малютка,
домовая мышь. Попадание указанных зверьков доволь-
но значительное (18,5%).
Из иксодовых клещей обнаружен только /. ricinus,
зараженность которым зверьков (8%) и относительный
запас (1,5) очень незначительны. По плотности заселе-
ния клещами эта стация уступает даже прибрежным
кустарниковым биотопам.
ЗАЛИВНЫЕ ЛУГА ОКОЛО ВОДОЕМОВ
В пойме р. Лесной (Беловежская пуща) нами обсле-
довалась также полоса (100—300 м шириной) заливных
лугов без кустарников. Покров густой из злаковых, осо-
ки, хвоща, лютика едкого. Почва иловато-торфянистая,
мокрая. Мышевидные грызуны и насекомоядные в этой
стации в ловушки не попадались. Пойманные вручную
кроты и обыкновенные ежи оказались заражены /. rici-
nus и D. pictus в очень слабой степени. При одновремен-
ном осмотре в 1955 г. коров в д. Селищи, пасущихся на
чистых лугах, и в д. Каменюки, пасущихся в лесу, обна-
ружена большая разница в степени заражения клещами
/. ricinus. Например, с третьей декады апреля до второй
декады мая зараженность коров в д. Селищи была равна
нулю, а в д. Каменюки—17,5; в третьей декаде мая
интенсивность заражения в д. Селищи—0,3, а в д. Ка-
менюки—36; в течение всего июня в д. Селищи интен-
сивность заражения коров—нуль, а в д. Каменюки—от
29,5 до 41.
Отловленные на заливном лугу в Лунинецком районе
(д. Крестуново) мелкие грызуны оказались незаражен-
ными. Почти то же наблюдалось и в Малоритском райо-
не при обследовании в мае 1956 г. коров, пасущихся в
лесу (д. Ляховичи) и на чистом лугу (д. Ласки). У пер-
вых интенсивность заражения 11,8, у вторых—0,2. При
сборе на пропашник на чистом лугу найдены единич-
ные экземпляры D. pictus, а /. ricinus на заливных лугах
без кустарников на пропашник вообще не попадались.
39
В опытах по влиянию затопления на жизненный цикл
/. ricinus и D. pictus (С. Ф. Сузько, 1949, 1953; В. Ф. Гу-
сев и С. Ф. Сузько, 1950) доказано, что погружение яиц,
голодных личинок и нимф /. ricinus в воду на 20—30 су-
ток вызывало их гибель на 99,5—100%. Напитавшиеся
личинки, нимфы и голодные имаго погибали через 12—
15 суток; упитанные самки погибали полностью через
четверо суток затопления, а напитавшиеся самки, яйца
и голодные личинки D. pictus—на 3—5-е сутки, напитав-
шиеся личинки—на 5-е сутки, голодные нимфы—на 1 —
5-е сутки. Напитавшиеся нимфы погибали только на
55-е сутки, а голодные имаго D. pictus выживали до
30 суток.
По данным Н. Г. Олсуфьева (1953), при затоплении
водой гибель взрослых клещей D. pictus при температуре
18—21° началась с 5-го и закончилась на 10-й день, а
при температуре 10—14° гибель наступила только на
15—20-й день. Поскольку весенние паводки рек Белорус-
сии продолжительны, то можно предположить, что все
фазы развития /. ricinus и D. pictus, за исключением
сытых нимф и взрослых D. pictus, обитающие в поймах
рек, очевидно, погибнут. Например, в 1953 г. паводок на
р. Лесной начался во второй половине февраля и про-
должался до апреля. Высокий уровень воды продержал-
ся весь март, причем средняя температура воздуха была
1,2°, а максимальная достигла 17°. В 1954 г. паводок
начался 5 марта и продолжался до 26 апреля (длитель-
ность паводка 52 дня). Средняя температура в марте 1°
(максимальная 11,7°), в апреле 4,3° (максимальная до
18°). В 1955 г. весенний паводок был непродолжителен
(с 24 марта до 9 апреля, всего 16 дней). Затем был вто-
рой паводок с 8 по 22 июля при среднемесячной темпера-
туре воздуха 18° (максимальная 28,3°). Таким образом,
в условиях БССР весенние паводки, как правило, затяги-
ваются и действуют, вероятно, губительно на все фазы
развития /. ricinus.
На заливных лугах клещи выживают только в остров-
ках кустарников с остатками старых пней и высоких ко-
чек. На отсутствие клещей /. ricinus на полосе, ежегодно
затапливаемой полыми водами, указывает и Н. И. Дыль-
ко (1953, 1954, 1958), проводивший исследования в пой-
ме р. Горыни. В. В. Судаченков (1941) при обследовании
пастбищ в Ленинградской области пришел также к за-
40
ключению, что на моховых и осоковых болотах, а также
в пойменных лугах /. ricinus отсутствует. Б. И. Поме-
ранцев (1935) указывает на слабое заражение /. ricinus
в западной части Лычковского района (Ленинградская
область), характеризующегося отрицательными форма-
ми рельефа и обширными пространствами, занятыми бо-
лотами. Н. И. Алфеев (1935), изучая экологию /. rici-
nus в районе Череменецкого озера, отмечает отсутствие
клещей в низких заболоченных местах, по берегам рек
и озер. А. С. Лутта и другие (1953) при изучении ста-
диального распространения иксодовых клещей в Карель-
ской АССР пришли к заключению, что в болотистых
лесах, а также в пойме реки /. ricinus отсутствовал со-
вершенно или встречался единично. Наоборот, в низмен-
ном лесу на берегу озера, но без застаивающихся по-
верхностных вод клещи встречались часто.
Таким образом, клещи /. ricinus избегают слишком
сырых заболоченных стаций с плохим или затяжным сто-
ком вод. Очевидно, здесь сказывается чрезмерная влаж-
ность, а главное—продолжительный застой вод, губитель-
но действующие на все фазы развития, особенно опасные
для сытых самок /. ricinus.
D. pictus был широко распространен в пойменных лу-
гах в кустарниковых биотопах. Очевидно, весеннее поло-
водье на взрослых клещей этого вида влияет менее отри-
цательно, чем на /. ricinus. К тому же концентрация мел-
ких млекопитающих в районе кустарников создает более
благоприятные условия для развития этих клещей, чем
на открытых и чистых от кустарников заливных лугах.
Кроме того, в кустарниковых биотопах более благо-
приятные условия для перезимовки клещей и их хозяев,
так как снеговой покров в открытом поле достигает
30 см, а в кустарниках увеличивается на 50%. В поле
почва промерзает на глубину 50 см, а под кустарниками
до 20 см (данные по Беловежской пуще).
На лесных полянах клещи D. pictus встречались очень
редко. Отдельные находки их были в лесных стациях:
сосняке, ельнике-черничнике, елово-широколиственных
лесах, ольшанике, смешанном молодняке и березняках.
Н. И. Дылько (1953, 1954, 1958) в долине р. Горыни
также обнаруживал D. pictus, но не на сплошных масси-
вах лугов, а очагово, в местах произрастания хотя бы
незначительного количества древесной растительности и
41
наличия небольших, не затопляемых половодьем возвы-
шений. Н. Г. Олсуфьев (1941) основной стацией D. pictus
в Московской области считает луга среднего увлажне-
ния. В меньшем количестве клещи населяют мелкие и
разреженные кустарники и лесные опушки. Клещи со-
вершенно не встречались в высокоствольном лесу.
Н. Г. Олсуфьев (1947) отмечает, что D. pictus в Запад-
ной Сибири обитает главным образом на суходольных
лугах, а в условиях средней полосы РСФСР—на кочко-
ватых суходольных лугах, вырубках, лесных опушках.
В поймах же крупных рек этот вид не встречается, так
как не выдерживает длительных затоплений. В долинах
мелких речушек, и особенно их верховьях, он может
встречаться в тех случаях, когда весенний паводок на них
кратковременный. На основании обследований поймен-
ных лугов в долине р. Оки Н. Г. Олсуфьев (1953) при-
ходит к выводу о большой зараженности кочковатых
пойменных лугов с кустарниками
же лесных стаций вблизи опушек
численность клещей. На вырубках
ивы. Обследование
указало на низкую
клещи встречаются
при условии, если последние были произведены больше
года назад и не слишком заросли молодой порослью.
Н. И. Алфеев и С. М. Кулагин (1953), изучая экологию
D. pictus в Сибири, указывают на большое количество
клещей на луговых участках в мелколесье на понижен-
ных местах. В Орловской области (Н. И. Алфеев, 1939)
D, pictus распространен на полянах, покрытых березо-
вым и осиновым кустарником, в дубовых молодняках,
очень редко в биотопах высокоствольных лесов и совер-
шенно отсутствует на болотах.
О. Macicka и другие (1956) при изучении экологии
D. pictus в Чехословакии установили, что ыа пастбищах
возле речек, заливаемых весенними паводками, клещи
имеются, а в пойме речек с летним наводнением они от-
сутствуют из-за гибели личинок от воды. Зараженными,
по их данным, являются поросшие кустарниками паст-
бища в поймах речек. Клещи совершенно отсутствуют
на чистых лугах и в лесах. В. Rosicky и О. Havlic (1952)
отмечают в условиях Чехословакии нахождение D. pictus
на лесных лугах. В. Rosicky и В. Rysavy (1955) пришли к
заключению, что кусты сохраняют более определенную
температуру и влажность, в зимний период под ними и
вокруг них образуются снежные заносы, предохраняющие
42
от резких колебаний температуры, вследствие чего созда-
ются благоприятные условия для зимовки грызунов и
клещей. Личинки D. pictus не выносят прямых солнечных
лучей, и поэтому куст для них является хорошим убежи-
щем, а хозяев клещей защищает от хищников. Концент-
рации клещей и их хозяев около кустов, по мнению авто-
ров, способствуют также отсутствие вытаптывания почвы
скотом под кустами и более разнообразный травяной
покров под ними.
ПРИУСАДЕБНЫЕ УЧАСТКИ И ЖИЛЫЕ ПОСТРОЙКИ
При рекогносцировочном обследовании приусадебных
участков под огородами, а также скотных дворов и жи-
лых построек в Беловежской пуще и Березинском запо-
веднике, Брагинском, Малоритском, Сиротинском райо-
нах в апреле—июле 1956 г. установлено отсутствие зара-
женности клещами мелких грызунов и насекомоядных.
Исключением являлось урочище Переров (Беловежская
пуща), где при обследовании огорода и жилого дома
лесника в смешанном хвойно-широколиственном лесу
обнаружена незначительная зараженность зверьков /. ri-
cinus, что, вероятно, связано с заносами клещей из леса.
При обследовании фруктового сада в июле 1956 г.
(д. Глуховичи, Брагинский район) зараженности зверь-
ков не обнаружено.
Отсутствие клещей в жилых домах, скотных дворах,
приусадебных участках под огородами и садами можно,
очевидно, объяснить тем, что выживаемость заносимых
с пастбищ или из близлежащего леса напитавшихся осо-
бей в несвойственных им экологических условиях ни-
чтожна или вообще отсутствует. Возможно, влияет так-
же и суточная ритмичность отпадения напитавшихся
клещей (Б. И. Померанцев и Н. И. Алфеев, 1935;
Г. В. Сердюкова, 1945 и Ю. С. Балашов, 1954).
культурные поля
При обследовании отловленных на культурных полях
(рожь и клевер с тимофеевкой) в июле 1956 г. в Брагин-
ском районе (деревни Глуховичи, Ясени, Ильичи) мыше-
видных грызунов клещей не обнаружено (табл. 1).
43
Н. И. Алфеев (1935) указывает на незначительное
заражение клещами I. ricinus культурных пастбищ, за-
сеянных клевером в районе Череменецкого озера.
Д. Н. Засухин и другие (1936) на юго-востоке РСФСР
(Саратовская область) на посевах культурных трав I. ri-
cinus также не встречали. В Московской области
Ц. Г. Олсуфьев (1941, 1953) изредка находил D. pictus
на старых клеверных участках, но совершенно не встре-
чал на свежих пашнях, так как клещи не переносят за-
пашки почвы. О. Macicka и другие (1956) в условиях
Чехословакии не обнаружили D. pictus на культурных
полях. А. В. Федюшин (1949) наблюдал некоторое на-
копление D. pictus на многолетних сеяных пастбищах
в Омской лесостепи. Даже на 4—5-й год хозяйственного
использования этих пастбищ клещей на них в 10—30 раз
меньше, чем на естественных пастбищах. Очевидно,
большое влияние оказывает распашка почвы, при кото-
рой гибнут как клещи, так и их хозяева.
ЦЕЛИННЫЕ ЗЕМЛИ
При обследовании скотомогильника—небольшого
участка целины (50X50 м), густо заросшего бурьяном и
ивняком и со всех сторон окруженного рожью, около
д. Глуховичи (Брагинский район) в июле 1956 г. на
обыкновенной полевке обнаружены личинки D. pictus.
Интересно, что этот заброшенный участок оказался рас-
садником клещей, тогда как на окружающих массивах
культурных полей их нет.
Н. Г. Олсуфьев (1941, 1953) в Московской области и
в пойме р. Оки изредка находил D. pictus на межах и
обочинах дорог. Он указывает, что при превращении
пашни в многолетние залежи возможно заселение их
клещами D. pictus. Н. И. Алфеев и С. М. Кулагин (1953)
в Сибири также обнаружили много клещей на целинных
степных участках и на прошлогодней сухой полыни.
Следовательно, сильно заражены /. ricinus чернооль-
шаники, хвойно-широколиственные и грабовые леса,
несколько менее—ельники, смешанные молодняки, сосно-
во-еловые леса, осинники, кустарники у водоемов, очень
слабо—сосняки, березняки, лесные поляны, вырубки. За-
ливные луга без кустарников, болото, открытые целин-
ные земли, культурные поля, а также жилые дома, скот-
44
ные дворы и приусадебные участки можно считать не
заселенными клещами.
Таким образом, /. ricinus—довольно пластичный вид
по заселению различных стаций. Однако в массе клещи
наблюдались в умеренно увлажненных стациях (черно-
ольшаники, хвойно-широколиственные и грабовые леса),
где встречаются в заметных количествах и их хозяева—
млекопитающие, и нет их в слишком сырых (заливные
луга, болото) и чрезмерно сухих (лесная поляна, сосня-
ки, вырубки, целинные участки) стациях, даже при зна-
чительном наличии их хозяев (лесная поляна, низинное
болото).
D. pictus широко распространен в пойменных лугах в
кустарниковых биотопах. Встречается и на целинных
участках, заросших бурьяном и кустарником. На откры-
тых и чистых от кустарников заливных лугах и на лес-
ных полянах, а также в жилых домах и скотных по-
стройках, на культурных полях эти клещи не встречают-
ся. Очень редки они в лесных стациях.
/. trianguliceps наиболее часто встречается в ельни-
ках, хвойно-широколиственных и грабовых лесах, значи-
тельно реже в ольшаниках, сосново-еловых лесах, борах,
прибрежных кустарниках. Единичные находки обнару-
жены в березняке, осиннике и смешанных молодняках.
Этот вид, как и I. ricinus, придерживается лесных ста-
ций. В открытых биотопах не встречен. Литературные
данные по экологии и биологии /. trianguliceps незначи-
тельны. С. О. Высоцкая (1951), А. С. Лутта и другие
(1956) также указывают на распространение этого вида
в лесных стациях.
ХОЗЯЕВА КЛЕЩЕЙ
Возникновение очагов массового размножения кле-
щей связывают с фактором питания (Е. Н. Павловский,
1935; Б. И. Померанцев, 1935; Г. В. Сердюкова, 1945).
Поэтому мы обратили особое внимание на обследование
позвоночных животных с целью выяснения среди них
хозяев клещей.
Так как личинки и нимфы /. ricinus и D. pictus напа-
дают на мелких зверьков, а взрослые—на крупных мле-
копитающих, то мы считаем целесообразным круг хозяев
этих видов рассматривать отдельно для каждой фазы.
Как известно, /. ricinus свойствен широкий круг хо-
зяев (Н. И. Алфеев, 1935, 1939; Б. И. Померанцев, 1935,
1950; А. И. Щеглова, 1939; Н. А. Золотарев, 1947;
А. С. Лутта и др., 1953). В СССР он паразитирует на 115
видах позвоночных животных (Г. В. Сердюкова, 1956).
Нами выявлено 35 видов (табл. 3—5) позвоночных
животных—хозяев /. ricinus, из них 7 оказались новыми:
обыкновенный канюк (Buteo buteo L.), хохлатая синица
(Parus cristatus L./ зорянка (Erithaous rubecula L.), ма-
лая бурозубка (Sorecs minutus L.), малая кутора (Neo-
mys anomalus Cabr.), пашенная полевка (Microtus agres-
tis L.) и зубр (Bison bonasus L.).
При сравнении зараженности отдельных видов жи-
вотных взрослыми клещами установлено, что на лесных
пастбищах они заражают преимущественно крупный ро-
гатый скот. В период наибольшей активности взрослых
клещей заражение отдельных животных достигало зна-
чительной цифры. Например, в начале мая 1953 г. на
одном животном обнаружено 350 клещей. Значительно
46
заражаются взрослыми клещами собаки, менее—лоша-
ди, овцы, кошки (табл. 3). На свиньях клещей совсем
не обнаружено. Как отмечалось выше, интенсивность за-
ражения животных клещами зависит от характера вы-
паса. Например, интенсивность заражения крупного ро-
Таблица 3
Хозяева взрослых клещей /. ricinus
Животные-хозяева Осмотре- но живот- ных Собрано клещей Интенсив- ность заражения Макси- мальное зараже- ние
Обыкновенный еж 48 27 2,5 7
Заяц-русак . . 7 69 17,2 29
Обыкновенная белка 108 6 1,2 2
Лесная соня . 96 1 1 1
Собака 14 124 9 33
Лисица 6 39 7,8 23
Кошка 4 6 1,5 2
Лошадь 50 36 3 4
Благородный олень 11 73 9,1 37
Овца домашняя . . 58 28 2,6 3
Крупный рогатый скот 538 11075 21,7 350
Зубр замечено много клещей
гатого скота на лесных пастбищах во много раз больше,
чем на чистых лугах. Однако степень зараженности по-
звоночных клещами I. ricinus зависит не только от ха-
рактера пастбища, а, очевидно, и от индивидуальных ви-
довых особенностей животного. Например, при осмотре
в октябре 1955 г. в урочище Переров пасшихся в сме-
шанном хвойно-широколиственном лесу коров обнару-
жено 103 клеща, у лошади—всего 4, свиньи не были за-
ражены. Таким образом, крупный рогатый скот можно
считать основным или главным хозяином взрослой фазы
/. ricinus. Наши данные подтверждают вывод Б. И. Поме-
ранцева (1935) о том, что из сельскохозяйственных жи-
вотных I. ricinus паразитирует преимущественно на круп-
ном рогатом скоте. Овцы, лошади и свиньи не имеют су-
щественного значения в прокорме клещей. К таким же
выводам приходят М. П. Марков (1927) в условиях
Украины и В. Ф. Гусев (1955) в условиях Белоруссии.
Паразитирование взрослых I. ricinus отмечено на ди-
ких млекопитающих: благородном олене, лисице, зайце-
русаке, обыкновенной белке, лесной соне и обыкновен-
47
Таблица 4
Хозяева нимф I. ricinus
Животные-хозяева Осмотре- но живот- ных Собрано клещей Интенсив- ность за- ражения Макси- мальное зараже- ние
Ящерица прыткая . 6 3 3 3
Обыкновенный канюк . 4 1 1 1
Рябчик 4 10 3,3 4
Сойка . . 14 2 1 1
Черный дрозд 1 7 7 7
Лесной конек . . 4 10 3,3 3
Обыкновенный еж 48 1137 31,6 258
Крот 37 1 1 1
Обыкновенная бурозубка 170 4 1 1
Заяц-русак .... 7 76 19 47
Обыкновенная белка 108 3033 36,5 199
Соня-полчок . 11 49 4,9 17
Лесная соня . 96 108 2,5 7
Полевая мышь 29 1 1 1
Лесная мышь . . 18 1 1 1
Желтогорлая мышь 354 103 1,5 8
Мышь-малютка . . . 6 4 4 4
Европейская рыжая полевка 940 39 1 1
Водяная полевка . . . 4 4 1,3 2
Обыкновенная полевка 59 2 1 1
Лисица 6 1 1 1
Кошка . ... 4 1 1 1
Благородный олень 11 85 17 76
Крупный рогатый скот 534 194 1,15 18
ном еже. При визуальном осмотре в паху у зубра было
замечено много напитавшихся самок иксодовых клещей.
Вероятно, это был /. ricinus, так как при сборе на рас-
тительности и с оленей в загоне, где жил зубр, других
клещей не обнаружено. Таким образом, мы пришли к
заключению, что главными хозяевами I. ricinus являются
благородный олень, лисица, заяц-русак, которые по
интенсивности заражения часто не уступают крупному
рогатому скоту. Например, при осмотре убитого в сен-
тябре 1953 г. благородного оленя было снято 37 клещей,
тогда как в этот же период у коров, пасущихся в лесу
(д. Каменюки), средняя зараженность была 1—3 клеща
на голову. У зайца-русака в мае 1954 г. снято 29 имаго,
а интенсивность заражения коров в этот период была
7—17. Б. И. Померанцев (1935) отмечает сильную за-
раженность зайцев в Ленинградской области, а Н. А. Зо-
48
Таблица 5
Хозяева личинок /. ricinus
-——— Животные-хозяева Осмотре- но живот- ных Собрано клещей Интенсив- ность за- ражения Макси- мальное зараже- ние
Рябчик .... 4 3 1,5 2
Хохлатая синица 1 1 1 1
Лесной конек 4 8 4 7
Зяблик 4 1 1 1
Зорянка ... 4 2 1 1
Обыкновенный еж 48 1251 37 284
Крот .... 37 11 1,8 5
Малая бурозубка . . . 15 3 1,5 2
Обыкновенная бурозубка 170 219 3,3 20
Малая кутора 6 2 2 2
Заяц-русак .... 7 83 28 68
Обыкновенная белка 108 1697 25 132
Соня-полчок 11 24 4 10
Лесная соня . 96 328 5,2 32
Полевая мышь 29 23 3 14
Лесная мышь . . 18 50 3,5 10
Желтогорлая мышь 354 979 4,2 50
Мышь-малютка 6 10 5 7
Европейская рыжая полевка 940 1775 4,4 52
Обыкновенная полевка 59 14 2,3 9
Пашенная полевка . 5 3 1,5 2
Благородный олень 11 1 1 1
лотарев (1947)—в Дагестанской АССР. С лисицы в сен-
тябре 1955 г. снято 23 клеща при средней зараженности
коров в это время 3—5. О том, что дикие животные могут
играть значительную роль в прокормлении взрослых
клещей I. ricinus, говорит и такой пример. Урочище
Переров—массив леса, мало затронутый рубками, рас-
положен почти в центре заповедника, где происходит
концентрация диких животных, в том числе и оленей.
Населенных пунктов нет, за исключением кордона, где
имеется 7 голов крупного рогатого скота. Королево-Мос-
товское лесничество занимает крайнюю юго-западную
часть заповедника, окруженного с южной стороны на-
селенными пунктами. Концентрация диких животных
здесь значительно меньшая, но в то же время скота па-
сется больше. При сравнении результатов обследований
растительности (табл. 6) и диких животных (табл. 7)
оказалось, что в урочище Переров клещей больше, чем
Иксодовые клещи 49
Таблица 6
Заселенность стаций клещами /. ricinus в урочище Переров и Королево-Мостовском лесничестве
(учеты с растительности)
Стации Пункт обследования Дата К-во учетов Личинки Нимфы Имаго
к-во клещей клещей на учет к-во клещей клещей на учет к-во клещей клещей на учет
Ельник дубняковый Каменюки IX. 1956 1 0 0 3 3 3 3
» Переров . » 1 0 0 6 6 2 2
Каменюки VI. 1955 2 0 0 4 2 0 0
» Переров . 5 89 18 42 8,4 28 5,6
Дубняк елово-грабово- Каменюки IX. 1956 1 0 0 1 1 3 3
кисличный Переров . » 3 0 0 17 6 3 1
Ельник-черничник Каменюки IX. 1956 1 0 0 3 3 0 0
Переров . » 2 0 0 15 7,5 3 1,5
» Каменюки X. 1955 2 ‘ 0 0 3 1,5 0 0
» Переров . 4 0 0 15 4 2 0,5
Ольшаник крапивный Каменюки X. 1955 2 0 0 0 0 0 0
» Переров . > 2 0 0 2 1 4 2
Всего Каменюки 1955—1956 9 0 0 14 1,6 6 0,7
Переров . » 17 89 5,2 97 5,7 42 2,5
Таблица 7
Зараженность мелких млекопитающих личинками и нимфами
/. ricinus в различных стациях в урочище Переров и в Королево-
Мостовском лесничестве
Стации Пункт обследования Дата Животных Собрано клещей Интенсивность заражения
отловлено заражено
Ельник-чернич- Каменюки X. 1955 16 3 5 1,7
ник Переров . » 65 18 25 1,4
Каменюки VIII. 1956 1 0 0 0
Переров » 15 15 135 9
Дубняк елово- Каменюки VII. 1955 41 19 114 6
грабово-кис- Переров » 9 8 37 4,6
личный Каменюки VI. 1956 5 4 10 2.5
Переров » 2 2 19 9,5
Ельник-дубняко- Каменюки VII. 1955 20 8 16 2
вый Переров . . . » 4 4 21 5,3
Ольшаник Каменюки VII. 1955 18 16 71 4,5
крапивный Переров . » 6 3 16 5,3
Всего Каменюки 1955— 101 50 216 4,3
Переров 1956 101 49 253 5,1
в Королево-Мостовском лесничестве. Поэтому наряду с
домашними животными дикие, как олени, лисицы, зайцы,
имеют не меньшее, а в некоторых случаях даже реша-
ющее значение в питании взрослых /. ricinus. Другие
дикие позвоночные, например ежи, в условиях заповед-
ника мало заражены имаго. Последние очень редко
встречаются на белках и сонях (табл. 3). Например, из
108 обследованных белок зараженными оказались 5, а
из 96 лесных сонь—одна. Довольно часты случаи напа-
дения клещей /. ricinus на человека.
Таким образом, главными хозяевами для взрослых
клещей /. ricinus из домашних животных оказались
крупный рогатый скот, собака, из диких—благородный
51
олень, лисица, заяц-русак, а локально для Беловежской
пущи и зубр. К второстепенным хозяевам можно отнести
лошадь, овцу, кошку, ежа. На мелких млекопитающих
взрослые клещи встретились только на нескольких бел-
ках и лесной соне.
В прокормлении нимфальной фазы I. ricinus значи-
тельную роль играют более мелкие позвоночные, особен-
но ежи и белки (табл. 6). Максимальное заражение от-
дельных ежей, пойманных в кустарнике в июне 1956 г.,
достигало 258, а белок, отстрелянных в сосново-еловом
лесу в сентябре 1956 г.,—199 нимф. М. П. Марков (1927)
в условиях Украины также отмечает чрезвычайную зара-
женность белок и ежей. Б. И. Померанцев (1935) отво-
дит ежам значительное место в прокормлении нимф. То
же отмечают Д. Н. Засухин и другие (1936) в условиях
Саратовской области. В Белоруссии Г. А. Найденова
(1941) установила сильную зараженность белок, а
В. Ф. Гусев (1955)—ежей.
Менее зараженными оказались заяц-русак и благо-
родный олень. С зайца-русака в июне 1953 г. собрано
47 нимф, с благородного оленя в мае 1956 г.—76. Круп-
ный рогатый скот в питании нимфальной фазы играет
менее значительную роль. Мышевидные грызуны и мел-
кие насекомоядные нимфами заражены очень редко. Из
мелких грызунов по интенсивности заражения несколько
выделяются сони. Из птиц наиболее часто зараженными
нимфами оказались черный дрозд, рябчик, лесной конек.
Единичные нимфы отмечены на обыкновенном канюке и
сойке (табл. 4). Но поскольку птиц обработано мало
(93 птицы 36 видов), то сделать определенные выводы
относительно их роли в клещеносительстве еще нельзя.
Казалось бы, что такие виды, как рябчик, лесной конек,
дрозд, связанные экологически с травяным покровом,
должны быть сильно заражены клещами. Однако с чер-
ного дрозда в сентябре 1955 г. снято 7, с рябчика 4 и с
лесного конька (июнь, 1955 г.) 3 нимфы, тогда как интен-
сивность заражения нимфами белок была во много раз
выше: в июне 61, в сентябре 52. Поэтому мы склонны
придерживаться мнения Е. Н. Павловского (1935) и
Б. И. Померанцева (1935) относительно незначительной
зараженности птиц клещами /. ricinus. Из рептилий об-
следовано 6 прытких ящериц, из которых зараженной
оказалась только одна.
52
Таким образом, мы считаем, что главными хозяевами
нимф можно считать обыкновенную белку и обыкновен-
ного ежа. Меньшую роль в прокормлении нимф играют
благородный олень, заяц-русак, черный дрозд, соня-пол-
чок, мышь-малютка, рябчик, лесной конек, ящерица прыт-
кая, лесная соня, желтогорлая мышь, водяная полевка,
крупный рогатый скот, обыкновенный канюк, кошка, ли-
сица, сойка, лесная мышь, полевая мышь, европейская
рыжая полевка, обыкновенная полевка, крот, обыкновен-
ная бурозубка. Нимфы могут нападать и на человека.
Как и нимфы, личинки наиболее часто паразитируют
на ежах и белках. Интенсивность заражения их очень
велика (табл. 5). Максимальное заражение отдельных
животных доходило до нескольких сотен личинок. На-
пример, с ежа, пойманного в кустарнике в мае 1956 г.,
собрано 284 личинки, а с белки, отстрелянной в сосново-
еловом лесу (сентябрь 1956 г.) —132. На крупных млеко-
питающих (например, олень) личинки встречаются чрез-
вычайно редко. Интенсивность заражения мелких гры-
зунов незначительная, причем самая высокая была у
лесной сони, желтогорлой мыши и европейской рыжей
полевки. Но поскольку сони встречаются очень редко, то
они не играют большой роли в прокорме личинок. То же
можно сказать и о других мелких грызунах, за исключе-
нием европейской рыжей полевки и желтогорлой мыши,
которые являются фоновыми видами в лесных форма-
циях. Европейская рыжая полевка и желтогорлая мышь,
по нашим данным, составили 81,5% общего числа отлов-
ленных зверьков (табл. 8), а по данным Е. П. Пивова-
ровой (1955, 1956) —87%. В силу своей многочисленно-
сти именно эти два вида мышевидных грызунов, обитаю-
щих почти во всех стациях, могут иметь не меньшее
значение в прокормлении личинок, чем, например, менее
многочисленные ежи и белки. Мелкие насекомоядные по
интенсивности заражения несколько уступают мышевид-
ным грызунам. К тому же они составляли 12% общего
количества отловленных зверьков. На птицах личинки
паразитировали редко, наибольшее количество клещей
было собрано только с лесного конька и рябчика.
В ряде работ (А. И. Щеглова, 1939; А. С. Вильбасте,
1956) указывается на большое значение птиц как доми-
нирующих хозяев преимагинальных фаз /. ricinus. Осо-
бенно детально изучили этот вопрос В. Rosicky и F. Ва-
53
Заселенность стаций мелкими грызунами и
Стации
Рыжая
полевка
Желтогорлая
мышь
Лесная
мышь
попадания
Бор мшистый 1040 59 5,7 6 0,6 — —
Суборь зеленомош- но-кисличная 820 93 11,3 13 1,6 1 о,1
Ельник-черничник 960 89 9,3 31 3,2 — —
Ельник дубняко- вый 870 74 8,5 41 4,7 1 0,1
Дубняк елово-грабо- во-кисличный 1670 274 16,4 99 5,9 6 0,4
Ольшаник крапив- ный 900 120 13,3 47 5,2 1 0,1
Березняк-черничник 290 14 4,8 6 2,1 1 0,3
Лесная поляна 180 6 3,3 3 1,7 — —
Заливной луг с кустарником 450 40 9 9 2 — —
Заливной луг без кустарника 300 — — — — — —
Всего 7480 769 10,3 255 3,4 10 о,1
Процентное соотно-
шение отловлен-
ных зверьков .
61,3
20,2
0,6
54
Таблица 8
насекомоядными за апрель—октябрь 1955 г.
Полевая
мышь
Обыкно-
венная
полевка
Пашенная
полевка
Кустарни-
ковая
полевка
Насекомо-
ядные
Всего
к-во
попадания
1 0,1 2 0,2 — —
1 0,1 — — — —
— — — — — —
— — — — — —
— — 3 0,2 2 0,1
1 о,1 — — — —
1 0,3 — — — —
— — 45 25 — —
12 2,7 3 0,7 1 0,2
— — 10 1 78 7,5
— — 9 1,1 117 14,1
— — 20 2,1 140 15,6
— -- 5 0,6 121 14
1 0,1 42 2,5 427 25,6
— — 37 4,1 206 22,9
— — 12 4,1 34 11,4
— — — — 54 30
15 3,3 80 17,8
1257 16,8
16 0,2 53 0,7
0,04 1 0,01 150
55
lat (1954). Эти авторы отмечают высокую зараженность
птиц, связанных с травяным покровом, и не нашли клещей
на птицах, живущих преимущественно в ветвях деревьев,
а также на полях и лугах. Авторы анализируют значе-
ние птиц как возможных переносчиков клещей в другие
районы при перелетах, причем первостепенное значение
в образовании вторичных очагов заражения придают
осенним медленным перелетам. Оседлые же птицы яв-
ляются мощным фактором в поддержании первичных
очагов.
Таким образом, главными хозяевами личинок /. rici-
nus являются обыкновенный еж, обыкновенная белка,
желтогорлая мышь и европейская рыжая полевка. К вто-
ростепенным хозяевам относятся заяц-русак, лесная со-
ня, лесная мышь, соня-полчок, лесной конек, мышь-ма-
лютка, обыкновенная бурозубка, полевая мышь, обыкно-
венная полевка, малая кутора, крот, малая бурозубка,
рябчик, зорянка, зяблик, хохлатая синица. Очень редко
личинки встречаются на крупных млекопитающих, на-
пример на благородном олене.
Интересен факт резкой депрессии численности всех
видов мышевидных грызунов в Беловежской пуще в
1956 г. по сравнению с 1955 г. В 1955 г. попадание евро-
пейской рыжей полевки было почти в 10 раз больше,
чем в 1956 г. (соответственно 10,2 и 1,1%). У желтогор-
лой мыши эта разница еще больше—в 17 раз (соответ-
ственно 3,4 и 0,2%). Процент же заражения европейской
рыжей полевки клещами в течение 2 лет был почти оди-
наков (41,3 и 41,4), у желтогорлой мыши несколько по-
высился (с 58 до 69). Среднее количество клещей на од-
ного зверька у европейской рыжей полевки увеличилось
менее чем в 2 раза (с 1,6 до 2,8), у желтогорлой мыши в
4,5 раза (с 1,9 до 8,5).
На основании выявленных различий в степени зара-
жения европейской рыжей полевки и желтогорлой мыши
личинками клещей можно предположить, что общее ко-
личество личинок, получивших возможность питаться на
этих двух видах хозяев, было гораздо больше в 1955 г.,
в год их массового размножения, чем в 1956 г., в год де-
прессии их численности. Казалось бы, что запас голод-
ных личинок, не имея главных хозяев, перейдет на вто-
ростепенных, например на лесную мышь, полевую мышь
и мелких насекомоядных (крот, обыкновенная бурозуб-
56
ка, малая бурозубка, малая кутора). Но оказывается,
интенсивность заражения у последних изменяется еще в
меньшей мере, чем у главных хозяев. Так, у лесной мы-
ши при резком снижении численности в 1956 г. средняя
зараженность осталась та же, что и в предыдущем году.
У полевой мыши, обыкновенной полевки и насекомояд-
ных одновременно с депрессией их численности несколь-
ко снизились количество зараженных особей и интенсив-
ность заражения.
Таким образом, в годы глубокой депрессии числен-
ности мышевидных грызунов интенсивность заражения
клещами изменяется главным образом среди основных
хозяев личиночной фазы. Заражение второстепенных хо-
зяев остается почти неизмененным или изменяется очень
мало. При этом интенсивность заражения главных хо-
зяев клещей в годы депрессии возрастает, но не настоль-
ко, по-видимому, чтобы можно было бы предполагать,
что весь запас личинок, существующий в природе, напи-
тался. Значит, часть голодных личинок в год депрессии
численности основных хозяев клещей остается голодной,
в связи с чем на следующий год имеется вероятность
снижения количества нимф, а на 3-й год — взрослых.
Если это предположение оправдается, то при условии
ежегодных наблюдений за численностью главных хозяев
личинок клещей можно будет давать прогноз за несколь-
ко лет вперед о возможном резком увеличении численно-
сти взрослых клещей и предпринять против подобных
вспышек соответствующие меры. Кроме того, можно сде-
лать заключение, что /. ricinus, хотя и является чрезвы-
чайно пластичным видом в отношении хозяев, но пред-
почитает до некоторой степени определенные виды жи-
вотных, к которым он приспособился в процессе своего
эволюционного развития.
Для D. pictus известен 51 вид животных-хозяев
(Г. В. Сердюкова, 1956). Нами обнаружено 16 видов, из
которых лесная соня (Dyromys nitedula Pall.) в качестве
хозяина D. pictus ранее не была еще отмечена.
Крупные домашние животные (лошади, овцы, круп-
ный рогатый скот), выпасающиеся на лугах, поросших
кустарником, в весенний и осенний периоды, как прави-
ло, были заражены взрослыми клещами D. pictus. Наи-
более часто клещи встречались на коровах и овцах, на
которых и интенсивность заражения была наибольшая,
57
реже—на лошадях (табл. 9). Существенную роль в про-
кормлении взрослых клещей D. pictus могут играть соба-
ки. В ряде работ (Н. И. Алфеев, 1939; Н. Г. Олсуфьев,
1947, 1953; Н. И. Дылько, 1954; В. Ф. Гусев, 1955) ука-
зывается на крупных домашних млекопитающих как на
главных хозяев взрослой фазы D. pictus. На свиньях
имаго встречались очень редко. Н. И. Алфеев (1939) в
условиях Унечского района Орловской области также
отмечает редкую зараженность свиней имаго, связывая
данный факт с характером выпаса. Из диких млекопитаю-
щих, хотя и редко, взрослые клещи встречались на бла-
городном олене, что известно и для Чехословакии (В. Ro-
sicky и О. Havlik, 1952; О. Macicka и др., 1956). На бо-
лее мелких диких животные имаго встречается как
исключение. Например, при осмотре 48 ежей, отловлен-
ных преимущественно на лугах в кустарниковых биото-
пах, найдена всего одна самка D. pictus. Н. А. Золота-
рев (1947) в условиях Дагестанской АССР при осмотре
ежей обнаружил только одну самку и одного самца
D. pictus.
Из изложенного следует, что главными хозяевами
для взрослых клещей D. pictus являются крупный рога-
тый скот и овцы, существенную роль могут играть так-
же лошади и собаки. Очень редко клещи встречались на
благородном олене и обыкновенном еже.
Очень часто клещи D. pictus прицепляются к одежде
человека, но случаев присасывания не наблюдалось.
В литературе же есть указания о единичных случаях на-
падения этого вида и на человека (И. Г. Галузо, 1948;
Н. И. Алфеев и С. М. Кулагин, 1953; Н. Г. Олсуфьев,
1953; В. Ф. Гусев, 1955).
Нимфы D. pictus в наших сборах попадались редко.
Они были найдены на еже обыкновенном и европейской
рыжей полевке, отловленных на лугах с кустарником,
обыкновенной белке, отстрелянной в ольшанике. Интен-
сивность заражения нимфами этих зверьков очень мала.
Наибольшее количество личинок собрано с мелких
млекопитающих, обитающих главным образом в луго-
вых стациях с кустарником. Наибольшая интенсивность
заражения наблюдалась на водяной полевке. Несколько
менее была заражена обыкновенная полевка. Как по
интенсивности, так и по максимальному заражению эти
зверьки во много раз превосходят другие виды. Значи-
58
69
Обыкновенный еж Крот ..... Малая бурозубка . Обыкновенная белка Лесная соня Полевая мышь Желтогорлая мышь . Европейская рыжая по- левка . Водяная полевка . Обыкновенная полевка Собака . Лошадь Свиньи .... Благородный олень Овца . . Крупный рогатый сыт Животные-хозяева
СП С© 00 ND — GO СЛ — — СЛ — СЛ 4^ СЛ СО С© О *—00 4* 4» <© О 00 СЛ 00 Осмотрено жи- вотных
Й nd — СЛ^ totouiro ООО OOOOOOW собрано X
сичю-слсоосю оооооо—* интенсивность заражения м а г ।
00 — <© с© ьэ — о о о о о оооооо — максимальное заражение о
ООООООООЮ ОООГОООСП собрано | Нимфы
1,67 0 0 1 0 0 0 1,2 0 0 0 0 0 0 0 0 интенсивность заражения
ООООООООЮ ООО — О О ND максимальное заражение
ND ND О — — — 0000000000 о — — — 4-— О собрано
3,1 1 4 L 1,4 2,7 26 5,7 0 0 0 0 0 0 интенсивность заражения и ч и н к
— ND О О О О О О-4 О <© 00 ОС — — 4^ — С© максимальное заражение S
Хозяева клещей D. pictus
тельная интенсивность заражения наблюдалась и у
ежей.
Европейская рыжая полевка по интенсивности зара-
жения уступала указанным выше хозяевам личинок, но
из-за ее многочисленности в кустарниковых луговых ста-
циях она может иметь существенное значение в про-
кормлении молодых фаз D. pictus. Что касается других
мышевидных грызунов и мелких насекомоядных, то в
силу своей малочисленности в стациях, наиболее зара-
женных D. pictus, они играют менее существенную роль
в питании личинок. Случайны единичные находки личи-
нок D. pictus на обыкновенной белке и лесной соне, до-
бытых в лесных стациях. На птицах клещей D. pictus не
обнаружено. Не обнаружены преимагинальные фазы и
на более крупных позвоночных, например на зайцах. Но
в ряде работ имеются указания на нахождение нимф и
даже личинок на зайцах (Н. И. Алфеев, 1939; Н. А. Зо-
лотарев, 1947; О. Macicka и др., 1956).
Следовательно, главными хозяевами преимагинальных
фаз D. pictus являются мелкие грызуны (водяная, обык-
новенная и европейская рыжая полевки) и обыкновен-
ный еж. К второстепенным хозяевам можно отнести ма-
лую бурозубку, полевую мышь, желтогорлую мышь, кро-
та, лесную соню, обыкновенную белку.
Среди хозяев клещей /. trianguliceps известно 15 ви-
дов мелких млекопитающих (Г. В. Сердюкова, 1956).
Мы обнаружили /. trianguliceps на 5 видах, из которых
полевая мышь (Apodemus agrarius Pall.) является но-
вым хозяином. Интенсивность заражения зверьков неве-
лика (табл. 10). Несколько выделялась европейская ры-
жая полевка—максимальное заражение ее достигало
9 личинок. Все активные фазы клеща /. trianguliceps
встречались только на европейской рыжей полевке и
желтогорлой мыши; нимфы и личинки попадались на
лесной мыши и обыкновенной бурозубке; на полевой
мыши обнаружены только личинки. Все особи этого вида
клещей паразитировали на зверьках, отловленных в лес-
ных стациях. По данным С. О. Высоцкой (1951),
А. С. Лутта и Р. Е. Шульман-Альбова (1956), все особи
этого клеща также паразитируют только на мышевид-
ных грызунах и мелких насекомоядных.
Относительно распределения по хозяевам клешей
/. crenulatus и /. apronophorus ничего определенного
60
Таблица 10
Хозяева клещей 1. trianguliceps
Животные-хозяева
Обыкновенная буро- зубка 170 0
Полевая мышь 29 0
Лесная мышь 18 0
Европейская рыжая полевка 940 15
Желтогорлая мышь . 354 2
0 0 5 1 1 8 1,1 2
0 0 0 0 0 1 1 1
0 0 1 1 1 1 1 1
1 2 27 1,5 3 102 2 9
1 1 2 1 1 25 1,3 3
сказать пока нельзя, так как эти клещи встречались в
единичных случаях. Две самки и нимфа /. crenulatus
сняты с ежей, 3 самца обнаружены при обследовании
нор лисиц. 4 самки и 4 нимфы /. apronophorus сняты с
европейской рыжей полевки, водяной крысы, обыкновен-
ной полевки и обыкновенной куторы. По литературным
данным (Г. В. Сердюкова, 1956), для /. crenulatus извест-
но 46 видов хозяев, для /. apronophorus 9. Причем клещ
/. apronophorus как паразит европейской рыжей полевки
(Cletrionomys glareolus Schreb.) и обыкновенной куто-
ры (Neomys fodiens Schr.) еще не отмечался.
В наших обследованиях свободными от иксодовых
клещей оказались: из птиц ястреб-тетеревятник, голубь-
вяхирь, курочка малая, коростель, белый аист, кулик-
черныш, седой дятел, зеленый дятел, трехпалый дятел,
большой пестрый дятел, средний пестрый дятел, ворон,
большая синица, болотная гаичка, серый сорокопут,
обыкновенный поползень, дрозд деряба, белая трясогуз-
ка, обыкновенный скворец, дубонос, домовой воробей,
обыкновенная овсянка, полевой жаворонок, жулан, вара-
кушка, деревенская ласточка, пеночка-трещотка, обыкно-
венная сизоворонка; из рукокрылых водяная ночница, ле-
61
тучая мышь Наттерера, ушан, европейская широкоушка,
рыжая вечерница, двухцветный кожан, поздний кожан,
усатая ночница; из грызунов орешниковая соня, степная
мышовка, серая крыса, черная крыса, домовая мышь, ку-
старниковая полевка, ондатра; из хищных ласка, рысь.
Все виды перечисленных птиц добыты с апреля по сен-
тябрь, рукокрылые—с июня по август, млекопитающие-
с апреля по октябрь.
При обследовании гнезд открыто гнездящихся птиц в
древесном ярусе и дуплогнездников (зяблика, дрозда
певчего, большой синицы, мухоловки-пеструшки, во-
робья, поползня, деревенской ласточки), а также гнезд
белки и лесной сони иксодовые клещи не обнаружены.
Очевидно, это зависит от таких неблагоприятных для
клещей факторов, как температура и влажность, так
как гнезда данных птиц и млекопитающих не связаны с
почвой.
Клещи чаще всего встречаются на таких животных,
которые ведут экологически сходный с ними образ
жизни.
/. ricinus и /. trianguliceps паразитируют преимуще-
ственно на животных, обитающих в лесных стациях. На-
оборот, D pictus чаще встречается на млекопитающих,
живущих в открытых местах. Дикие позвоночные, при-
держивающиеся поселений человека и культурных по-
лей, клещами не заражены. Птицы, связанные по образу
жизни с травяным покровом, т. е. когда возможен непо-
средственный контакт клещей с их хозяевами, часто за-
ражены /. ricinus. Единичные случаи заражения /. rici-
nus наблюдались у хищных птиц (обыкновенный канюк)
при попадании на них клещей с жертвы. Птицы, обитаю-
щие главным образом в верхнем древесном ярусе, а
также водоплавающие, болотно-луговые и связанные
преимущественно с культурными полями клещами не
заражены. То же можно сказать и о летучих мышах, ко-
торые большую часть своей активной жизни проводят в
воздухе.
ЦИКЛ РАЗВИТИЯ D. PICTUS
СРОКИ ОТКЛАДКИ яиц
Наблюдения в природе за развитием самок D. pictus,
питавшихся в апреле—мае, показали, что они приступа-
ли к откладке яиц в июне и только частично в июле то-
го же года (табл. 11). От момента насыщения самок до
начала откладки ими яиц проходило 7—68 дней, в зави-
симости от времени питания. Так, у самок, питавшихся
в апреле, этот период продолжался до 2 и более меся-
цев, тогда как у самок, насытившихся в мае, за исклю-
чением единичных особей,—до месяца и меньше. Одна
самка, насосавшаяся в осенний период (ноябрь), при-
ступила к откладке яиц в этом же сезоне с непродолжи-
тельным промежутком (через 15 дней от момента насы-
щения). Две самки, напитавшиеся в октябре—'Ноябре,
приступили к откладке яиц только на следующий год,
через 210—235 дней. Однако не все яйцекладки оказа-
лись жизнеспособными. Яйца, полученные в июле от са-
мок, питавшихся в мае, погибли, как и отложенные в
ноябре самкой, напитавшейся в том же месяце. Следова-
тельно, основная масса самок, питавшихся в весенний и
осенний периоды, приступает к откладке яиц в июне.
При этом самки, питавшиеся осенью, перезимовывают.
Н. И. Алфеев (1939), наблюдая в Орловской области
за развитием D. pictus в различных биотопах, отметил,
что у самок, напитавшихся в апреле, начало откладки
яиц колеблется от 32 до 60 дней, а у самок, напитавших-
ся в мае,—через 14—30 дней.
В лаборатории самки D. pictus, содержавшиеся при
температуре 18—20°, значительно сокращают сроки раз-
вития по сравнению с самками, содержавшимися в при-
63
Таблица 11
СП
Сроки откладки яиц самками D. pictus
Время питания самок К-во сытых самок Начало откладки Через сколь- ко суток на- чалась от- кладка Число кладок Условия содержания
взятых ДЛЯ опыта погиб- ших получен- ных погиб- ших
Апрель 1 2 июнь 66-68 1 2 1
Май 79 5 » 7—45 74 3
» . 7 — июль 41-58 7 5
Октябрь 2 1 июнь следующего
года 235 1 — 8. S
Ноябрь 1 — ноябрь . . . 15 1 1 &
» 1 — июнь следующего ш
года 210 1 1 —
Апрель 1 май 12 1 1
Май 25 — » 3—10 25 1
3 — июнь 9—13 3 3
Июль 1 — июль 3 1 — S S
1 — август 16 1 — О'
Август 52 3 сентябрь 20—36 45 4 й
» 13 » *« октябрь . 39—62 12 1 Q. О
Сентябрь 7 1 сентябрь 14—25 6 — Ю со
5 — октябрь . 30—37 5 — Ч
Октябрь 9 1 декабрь . 41—49 7 1 ш
роде. Например, самки, насосавшиеся в апреле—июле,
приступили к откладке яиц в том же месяце или в нача-
ле следующего. Срок до начала яйцекладки не превы-
шал 16 дней. Основная масса самок приступила к от-
кладке яиц через 5—6 дней, а единичные особи—через
3 дня после насыщения. Несколько иная картина наблю-
далась в осенних опытах. Большая часть самок, насо-
савшихся в августе—октябре, развивалась медленнее,
чем весенние. Кладка наступала через один-два месяца
после насыщения. И только в сентябре у некоторых са-
мок не наблюдалось такой сильной задержки в развитии
(Табл. 11). В лабораторных условиях Н. Г. Олсуфьев
(1953) получал от самок яйца на 7—12-й день после их
насыщения при температуре 18—22°, а О. Macicka и дру-
гие (1956) при температуре 27°—в пределах 1 —14 дней.
Н. И. Алфеев же (1948), следивший в лаборатории за
самками, насосавшимися в позднеосенний сезон, наблю-
дал задержку в развитии сытых самок до 69—83 дней.
Подобные результаты были получены П. И. Нецким и
Н. А. Ольшевской (1950), наблюдавшими яйцекладку от
самок весенне-летнего питания при 15—24° через 5 —
25 дней, а от самок осенне-зимнего кормления при тех
же условиях через 41—-133 (самки питались в августе) и
21 —152 дня (самки питались с августа по март).
Исходя из наших данных и выводов указанных авто-
ров, можно сделать следующее заключение. Преобла-
дающая масса сытых самок, напитавшихся весной, при-
ступает к откладке яиц в июне. В июне появляются яйца
и от перезимовавших самок, питавшихся осенью. За-
держка в развитии яиц у самок, насосавшихся осенью,
независимо от условий содержания (в природе или лабо-
ратории), говорит о наличии зимней диапаузы у сытых
самок осеннего кормления.
ПРОДОЛЖИТЕЛЬНОСТЬ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ
Вылупление личинок от летних кладок происходило
на протяжении лета (июль—август) и даже в начале
сентября (табл. 12). При этом сроки от момента начала
кладки до начала вылупления личинок были очень рас-
тянуты (от 39 до 97 дней). Основная масса личинок вы-
луплялась в июле—августе, через 45—67 дней. Н. И. Ал-
феев (1939) в различных биотопах Орловской области
5. Иксодовые клещи
65
Таблица 12
Продолжительность эмбрионального развития D. pictus
Время появления яиц К-во яйце- кладок Время вылуп- ления ли- чинок Через сколько суток после яйцекладки вышли личинки К-во кладок, давших личинки Условия содер- жания
полученных погибших |
июль 40—50 1 24
Июнь . 75 2 август 39-84 47 О)
сентябрь 95-97 2 а
Июль 7 5 август 48-52 2
Ноябрь 1 i 1 1 । — с 1 со
Май 1 26 2 июнь 17—28 24
Июнь . 3 3 — — — s
Июль . 1 — август 20 1 S
Август 1 — » 14 1 с
Сентябрь 51 4 октябрь 20-28 47 ОЗ
ноябрь . 20—22 10 \8
Октябрь 17 1 декабрь 22—26 6 ОЗ
Декабрь 7 2 » 15-20 6 а
наблюдал выход личинок через 28—56 дней, т. е. в зна-
чительно более короткие сроки, что можно объяснить
очень ранней и теплой весной 1934 г. в Унечском районе
(место его стационарных наблюдений). На месте наших
наблюдений весна (1955—1956 гг.) была очень холодной
и поздней. Так, по сведениям метеостанции заповедника,
средневесенняя температура воздуха в 1953 г. была 6,9,
в 1955 г.—3,5 и в 1956 г.—5,9°. Причем последние замо-
розки в 1953 г. наблюдались до 14 мая, а в 1955 и
1956 гг.—до 28 и 23 мая. Средняя температура за летний
период (июнь—август), когда происходило вылупление
основной массы личинок, в 1953 г. была 18, в 1955 г. —
17 и 1956 г.—16,6°. Максимальная температура в 1953 г.
достигала за летний период 32,3°, а в 1955—1956 гг. со-
ответственно—28,6 и 23,7° Такие низкие температуры,
очевидно, не могли не отразиться на развитии клещей, в
частности на сроках вылупления личинок.
Наблюдения за сроком вылупления личинок в лабо-
ратории при комнатной температуре велись с мая по де-
66
кабрь. Резких различий в сроках отрождения личинок за
этот период не наблюдалось. Личинки вылуплялись в
более короткие сроки, чем в природе, в пределах 14 —
28 дней. Основная масса их вылуплялась на 18—25-й
день. Н. Г. Олсуфьев (1953) в лабораторных условиях
при температуре 18—22° наблюдал вылупление личинок
примерно в те же сроки—на 21—27-й день. О. Macicka и
другие (1956) при 27°, т. е. при более повышенной тем-
пературе (в наших опытах 18—20°), получали личинки в
еще более короткие сроки—за 12—18 дней.
Таким образом, сроки созревания личинок D. pictus
не зависят от сезона откладки яиц и задержки в разви-
тии последних не наблюдалось, как это было с осенними
сытыми самками. Но сроки созревания личинок в силь-
ной степени зависят от температурных условий. Значи-
тельно удлиняются сроки в естественных условиях, осо-
бенно в затяжную холодную весну и в прохладное лето,
и резко сокращаются сроки в лабораторных условиях
при более высоких, чем в природе, температурах.
РАЗВИТИЕ ЛИЧИНОК
Сезонная динамика паразитирования. На обследо-
ванных мелких млекопитающих, добытых с апреля по
октябрь в стациях, наиболее зараженных клещами D. pi-
ctus (на лугах с кустарниковым биотопом), личинки на-
чинали попадаться с начала июня (1 июня 1956 г. снята
с европейской рыжей полевки) до конца второй декады
августа (последняя личинка снята с европейской рыжей
полевки 19 августа 1955 г.). Наибольшая интенсивность
заражения наблюдалась в июле (табл. 13). Максималь-
ное заражение зверька (9 личинок на европейской ры-
жей полевке) отмечено 20 июля 1955 г. Такие же сроки
паразитирования личинок указываются в ряде работ,
проведенных в Орловской (Н. И. Алфеев, 1939) и Мо-
сковской (Н. Г. Олсуфьев, 1953) областях, в Казахстане
(И. Г. Галузо, 1948) и в Чехословакии (В. Rosicky и
О. Havlik, 1954), а в Алтайском крае отмечены единич-
ные находки личинок даже в начале сентября (Н. И. Ал-
феев и С. М. Кулагин, 1953) и в первой декаде октября
(И. Г. Галузо, 1948).
Развитие непитавшихся личинок. В связи с неактивно-
стью свежевылупившихся личинок /. ricinus (Г. В. Сердю-
67
89
а о О > S S £ >
g О\ я ГР я CD § 3 я аз я« я та ГР
? О\ та а о\ 3 СТ 0* ь tr
£
cd
о
»
J=
4^ Go оо м в—в осмотрено
о сл CD оо ю о животных
о о о о 00 СП 00 о о заражено
Го собрано кле-
о о о со СО 00 о о щей
о о о , 1^ о о интенсивность
СП заражения
NS У . . осмотрено
о 00 о О> о животных
о о о О о о о заражено
собрано кле-
о о о 00 о о о щей
о о о . о о о интенсивность
00 00 заражения
осмотрено
о 00 со ьо ьэ 00 00 о о оо N3 животных
СО Го 00 о О 00 СП О заражено
оо собрано кле-
о о Го оо а> ьо СП щей
N3 СО ьо интенсивность
G0 4^ — о о 00 05 СП заражения
Сезонная динамика паразитирования активных фаз
D. pictus (1953—1956 гг.)
Личинки | Нимфы | Имаго
S
J5
аз
Go
кова, 1952) были поставлены опыты по выяснению
сроков развития непитавшихся личинок D. pictus. Опы-
ты проводились в полевых садках с подсадкой ежей. Ли-
чинки начинают нападать на хозяина с 3-дневного воз-
раста, но таких особей мало (табл. 14). С возрастом
увеличивается и число личинок, напавших на хозяина,
достигая максимума в возрасте 16—20 дней. Затем ко-
личество особей, способных к присасыванию, резко со-
Т а б л и ц а 14
Взаимосвязь между возрастом и количеством напитавшихся
личинок и нимф D. pictus
Личинки Нимфы
напитавшихся >S S напитавшихся
Е _ CU особей Е ГГ СП * особей
Q.
& t=( 5 х u 2 н «3 ж о , <ъ> 2 £3 t- S о ® Е Е X _ g Н н х о к СП ж о . 6 g г/ 2 х s <ъ> К Е
§ к 2 со 8 СХ _ сП О -й Е СХ Ш К X 2* со с <Ъ> сх СП О S Е СХ
л СО Е О СО со О) et ' СО Е О со <Ъ> К(
X со о со О СО Е О Е Ешо СО С- со Ш Е О Е
3 3—5 251 84 5 3-5 6 1
21 6—10 4572 218 11 6—10 260 24
15 11 — 15 3092 208 13 11 — 15 951 73
11 16-20 5287 481 5 16-20 726 145
16 21—25 3544 222 2 21—25 271 136
7 26—30 471 67 2 36—40 73 37
6 31—35 160 27 1 41—45 5 5
1 36—40 27 27 — — — —
2 41—45 112 56 — — — —
1 46—50 23 23 — — — —
1 58 4 4 — — — —
1 59 0 0 — — — —
кращается и уже в возрасте 58 дней нападают единич-
ные особи, а после 59 дней нападение прекращается.
Данное явление можно объяснить тем, что в первый пе-
риод (до 20-дневного возраста) свежевылупившиеся ли-
чинки физиологически доразвиваются, как это наблюда-
лось у личинок /. ricinus (Г. В. Сердюкова, 1952), а в
дальнейшем количество особей, способных к нападению
на хозяина, уменьшается, очевидно, в связи с истоще-
нием и гибелью от голода, так как известно, что личинки
Dermacentor не способны к длительному голоданию.
В полевых садках (опыт с 23 генерациями) гибель от-
дельных личинок наступала на 22 и 98-е сутки после вы-
69
лупления, но основная масса их гибла через 50—89 дней.
К зиме все личинки погибли. В лабораторных условиях
при комнатной температуре гибель личинок наступала
в основном через 35—64 дня (наблюдения над 14 генера-
циями), но отдельные особи оставались живыми до
86 дней. По литературным данным, в лабораторных усло-
виях гибель личинок Dermacentor наблюдалась через
30—65 дней (Е. А. Москачева, 1948; Н. И. Алфеев и
С. М. Кулагин, 1953; Н. Г. Олсуфьев, 1953).
Сроки паразитирования. Основная масса личинок па-
разитирует 3—4 дня. Отдельные особи питались в тече-
ние 12 дней. Питание в летний период (июнь—август)
продолжалось примерно 3—5 суток, а ближе к осени
срок его удлинялся. Так, уже в сентябре питание основ-
ной массы личинок длилось 3—8 дней (табл. 15). В ли-
Таблица 15
Сроки паразитирования личинок и нимф D. pictus
Личинки Нимфы
время паразитиро- вания к-во генераций, посаженных на хозяина срок паразити- рования (в днях) напитавшихся особей время паразитиро- вания к-во генераций, посаженных на хозяина срок паразити- рования (в днях) напитавшихся особей
Июнь 20 3— 8 9894 июль 17 3-15 1987
Июль 5 4—10 288 сентябрь 23 3—15 311
Август 32 3- 7 6347 октябрь г 7 1
Сентябрь 35 3—12 2299 — — — —
тературе имеются сведения, что сверх 5 суток личинки
не питаются, а в массе паразитируют около 3 суток
(Н. И. Алфеев, 1939). Н. Г. Олсуфьев (1953) длитель-
ность паразитирования связывает с видовой особенно-
стью хозяина. Например, на полевках личинки парази-
тируют 2,5—3,5 суток, в то время как на морских свин-
ках—3,5—4,5, а на ежах—4,5—5,5 суток.
Развитие сытых личинок. Для определения сроков
развития сытых личинок с учетом влияния сезона их
кормили в течение августа—октября. Линька личинок,
насосавшихся в августе, наблюдалась в том же месяце
и в сентябре, через 7—23 дня, и частично в октябре, спу-
70
стя 49 дней после насыщения. В этой серии опытов было
наибольшее количество личинок, превратившихся в
нимф. Личинки, насосавшиеся в сентябре, также пере-
линяли на нимф в сентябре—октябре, но значительная
часть их погибла. Все личинки, насосавшиеся в октябре,
погибли. Наблюдения проводились также над сытыми ли-
чинками, собранными с диких животных. Почти все ли-
чинки (из 67 погибло 9), добытые в июле, перелиняли на
нимф в августе, через 18—30 суток. Таким образом,
линька личинок на нимф в естественных условиях проис-
ходит в основном в августе—сентябре и совсем незначи-
тельная часть их линяет в октябре (табл. 16). При этом
Таблица 16
Развитие сытых личинок D. pictus
- . СО _ СО Н
ШОК, ЛИЧИНКИ ераций х после личино: Время питания Время линьки ли- иньки ( • момен личино: да ним< яераций нимф содер-
ВО КЛс вших . X X нм гч U ХО X S Е S личинок чинок на нимф ч о к g о к н _ £ х X g 1 к ш к О X 5 §
X § ' о X Е Е X X О О Е X § >> я
август 4-26 10 О)
32 2 август сентябрь 4—34 18 X
октябрь 49 2
24 20 сентябрь сентябрь октябрь 11 30—47 1 3 Е СО
1 1 октябрь — — —
15 — июнь июнь июль 6—8 6—7 10 5 СО is СО сх>
5 — июль » 12—16 5 со
наиболее жизнеспособными оказались особи, питающиеся
в июле—августе. У личинок, снятых с хозяина в более
поздние сроки, линька, как правило, не наступала. В усло-
виях полевого опыта в Орловской области линька проис-
ходила через 10—26 суток после кормления личинок
(Н. И. Алфеев, 1939), т. е. метаморфоз личинок проходил
в основном примерно в те же сроки, что и в наших опытах.
В лабораторных ^условиях при температуре 18 —
20° сроки линьки ока^лись значительно короче. Основ-
ная масса личинок линяла спустя 7—8 дней и частично
71
через 12—16 дней после их насыщения. Литературные
данные подтверждают эти наблюдения. Так, Н. Г Ол-
суфьев (1953) в лабораторных условиях при температу-
ре 18—22° линьку на нимф наблюдал на 10—12-й день.
Таким образом, температурные условия в значитель-
ной степени влияют на скорость развития сытых личи-
нок. В связи с этим при затяжной весне, а также в про-
хладное лето развитие личиночной фазы D. pictus может
в сильной степени затянуться.
РАЗВИТИЕ НИМФ
Сезонная динамика паразитирования. С мелких мле-
копитающих нами были добыты только единичные ним-
фы. Первая находка на еже была отмечена 10 июля
(табл. 13). Затем изредка отдельные особи встречались
на ежах и мышевидных грызунах в июле—августе. По-
следняя нимфа снята 30 августа с европейской рыжей
полевки. В ряде работ (Н. И. Алфеев, 1939; Н. Г. Ол-
суфьев, 1947, 1953; В. Rosicky и О. Havlik, 1952) указы-
вается на паразитирование основной массы нимф в
июле—августе. Имеются также случаи (в условиях теп-
лой весны) раннего появления нимф—в конце июня
(Н. Г. Олсуфьев, 1953) и окончания активности в начале
сентября (Н. И. Алфеев и С. М. Кулагин, 1953; Н. Г. Ол-
суфьев, 1953).
Развитие непитавшихся нимф. Опыты по выяснению
сроков развития непитавшихся нимф D. pictus проводи-
лись в полевых садках с подсадкой ежей. Отдельные
нимфы нападали на хозяина уже на 3-й день после вы-
хода. По мере увеличения возраста повышалось и число
нимф, напавших на хозяина, достигая своего максимума
в возрасте от 16 до 20 дней (табл. 14). По истечении
этого возраста количество присасывающихся нимф со-
кращается. Например, из всей генерации на 44-е сутки
напали на хозяина 5 нимф. Следовательно, как и непи-
тавшиеся личинки, нимфы в первый период (до 20-днев-
ного возраста) физиологически доразвиваются (Г. В. Се-
рдюкова, 1952), а в дальнейшем спад нападающих на
хозяина особей связан, по-видимому, с истощением и ги-
белью от голода. В наших опытах в естественных усло-
виях 3 генерации погибли на 12—50-е сутки, а в лабора-
тории 3 генерации погибли через 37—60 дней. В лите-
ратуре отмечается гибель нимф в природе через 16—
72
45 суток (Н. И. Алфеев, 1939; Н. И. Алфеев и С. М. Ку-
лагин, 1953). Н. Г Олсуфьев (1953) наблюдал начало
гибели с 3-й недели, причем массовое отмирание насту-
пало через месяц. Отдельные нимфы прожили 40 дней.
Таким образом, результаты опытов и литературные
данные указывают на неспособность нимф D. pictus к
длительному голоданию. Следовательно, оправдывается
предположение о снижении количества нападающих на
хозяина особей во второй период развития непитавшихся
нимф (после 20-дневного возраста) в связи с истоще-
нием и гибелью от голода. В соответствии с этим можно
сделать заключение, что вследствие неспособности к
длительному голоданию все нимфы должны погибнуть
до наступления зимнего периода.
Сроки паразитирования. В июле и сентябре — октяб-
ре нимфы кормились на ежах и кроликах для установ-
ления сроков паразитирования в различные сезоны года.
Оказалось, что продолжительность питания нимф D. pic-
tus может варьировать в пределах 3—15 дней и не за-
висит от сезона года (табл. 15). Как в июле, так и в сен-
тябре—октябре особых изменений в сроках паразитиро-
вания не наблюдалось. Основная маса нимф питалась в
течение 4—6 дней. Продолжительный срок паразитиро-
вания (до 15 дней), возможно, зависит от видовой осо-
бенности хозяина, так как, по литературным данным, в
зависимости от вида хозяина изменяются и сроки крово-
сосания. Например, на полевках паразитирование про-
должается 4—5, а на ежах и морских свинках—5—7 су-
ток (Н. Г. Олсуфьев, 1953).
Развитие сытых нимф. Сроки развития сытых нимф
с учетом влияния сезона определялись путем кормления
их в осенний период (сентябрь—октябрь), а также ис-
пользовались сытые нимфы, снятые в июле с диких жи-
вотных. Последние перелиняли на имаго в августе через
39 дней. Нимфы же, накормленные в сентябре—октябре
(табл. 17), не успели закончить метаморфоз и погибли
во время зимнего периода. Из 17 генераций, взятых для
опыта (14 генераций напитались в сентябре, 3 — в октяб-
ре), все сытые нимфы погибли при зимовке. Н. И. Алфеев
(1939) в естественных условиях Орловской области на-
блюдал линьку нимф, напитавшихся в июле, на имаго
через 26—54 дня, т. е. примерно в такие же сроки, что
и особи, снятые нами с диких животных.
73
Таблица 17
Развитие сытых нимф D. pictus
К-во генераций, взятых для опыта К-во генераций, погибших после питания нимф Время питания нимф Время линьки нимф на имаго Сроки линьки (в днях) от момента питания нимф до выхода имаго К-во генераций, давших имаго Условия содержа- ния
14 3 14 3 сентябрь октябрь — — — в при- роде
12 5 — июль август август » 14-18 13—15 12 5 в ла- бора- тории
В лабораторных условиях при комнатной температу-
ре (18—20°) развитие сытых нимф шло гораздо успеш-
нее. Нимфы, напитавшиеся в июле—августе, все пере-
линяли на имаго в августе, спустя 13—18 суток после
отпадения. Такие же результаты получены в лаборато-
рии Н. Г. Олсуфьева (1953), который наблюдал выход
имаго через 15—16 дней.
РАЗВИТИЕ ИМАГО
Сезонная динамика паразитирования. Активность
взрослых клещей D. pictus начиналась уже в первой де-
каде апреля, когда в лесу местами еще лежал снег. Пер-
вые сборы клещей были с растительности 4 апреля
1955 г. и 10 апреля 1956 г. на заливном лугу в кустар-
никах тотчас же после спада воды. Для Орловской обла-
сти первая находка имаго отмечена 25 марта (Н. И. Ал-
феев, 1939), а в Казахской ССР (И. Г. Галузо, 1948)
активность начинается даже с начала марта. В наших
условиях обследование в марте провести было невоз-
можно, так как заливные луга, где преимущественно
распространен D. pictus, в это время были еще залиты
полыми водами. На протяжении всего апреля кривая
активности взрослых клещей в природе при сборах на
пропашник резко возрастала, достигнув максимума в
74
Таблица 18
Сезонная динамика активности имаго D. pictus за 1955—1956 гг.
по данным сбора клещей с растительности
Месяц К-во учетов Собрано клещей В среднем кле- щей на учет
Апрель 7 95 13—14
Май 7 171 24—25
Июнь 9 2 0—1
Июль 4 0 0
Август 2 4 2
Сентябрь 6 18 3
Октябрь 5 1 0—1
Ноябрь 2 0 0
мае (табл. 18, рис. 9). Максимальное количество клещей с
растительности собрано 2 мая 1955 г.
(42 клеща на учет) и 5 мая 1956 г.
(60 клещей на учет). Затем начинается
быстрый спад численности клещей.
В конце мая и начале июня были обна-
ружены только единичные особи
(16 июня 1955 г. один клещ на учет).
В течение последних двух декад июня,
в июле и первой половине августа кле-
щи в природе не встречались. Только к
концу августа имаго появляются вновь,
образуя второй, осенний, пик активно-
сти в сентябре, но значительно мень-
ший, чем весенний (табл. 18). Послед-
няя находка клещей с растительности—
9 октября 1955 г. (один клещ на учет).
Для сравнения результатов учета
клещей в природе был обследован
крупный рогатый скот, на который сра-
зу же после выгона на пастбище
(14 апреля 1955 г.) нападали клещи.
В это время заражение животных еще
слабое (примерно по одному клещу на
животное). Затем, как и на раститель-
ности, зараженность скота стала резко
возрастать, достигнув максимума в мае
(табл. 18, рис. 9). 8 мая 1956 г.
v * п юти / г
Рис. 9. Сезонная
кривая активности
имаго Dermacentor
pictus Herm.
(1953—1956 гг.):
а—сбор клещей с жи-
вотных; б—с раститель-
ности
с отдельных животных собрано максимум 89 клещей.
В конце первой декады июня численнось клещей на ско-
75
те сходит на нет, и в течение почти всего летнего сезона
(с июня по август) имаго на скоте не встречались. В пер-
вых числах сентября клещи на скоте появились снова и
максимума достигли к концу сентября (табл. 13). Клещи
нападали до середины ноября (обнаружены 17 октября
1955 г.).
Таким образом, сезонная кривая паразитирования
взрослых клещей D. pictus как по данным сбора с расти-
тельности, так и в результате осмотра скота имеет две
вершины—весной (начало мая) и осенью (сентябрь).
При этом осенний пик примерно в 5—8 раз меньше ве-
сеннего.
Наши данные согласуются с данными ряда исследо-
ваний в Орловской (Н. И. Алфеев, 1939, 1947), Москов-
ской (Н. Г. Олсуфьев, 1953) областях, в Западном По-
лесье УССР (Е. М. Емчук, 1954), Чехословакии (В. Ro-
sicky и О. Havlik, 1952), а также в БССР (Н. И. Дылько,
1953, 1954; В. Ф. Гусев, 1955; Б. П. Савицкий, 1957). Не-
которые расхождения в сроках нападения D. pictus отме-
чены в Новосибирской области (Н. Г. Олсуфьев, 1947) и
Алтайском крае (Н. И. Алфеев и С. М. Кулагин, 1953).
где замечена активность имаго D. pictus, хотя и незначи-
тельная, в летний период (в июле), что нами не подтвер-
дилось. Кроме того, как отмечают Н. И. Алфеев и
С. М. Кулагин (1953), Ф. А. Закович (1957) и Б. П. Са-
вицкий (1959), бывают случаи паразитирования взрос-
лых клещей и в зимний период. Данный факт, по нашему
мнению, вполне возможен, так как при определении про-
должительности сроков питания имаго в различные се-
зоны года мы наблюдали чрезвычайную продолжитель-
ность паразитирования клещей на животных (отдель-
ные самцы пребывали на хозяине более 2 месяцев).
Сроки паразитирования взрослых клещей. Сроки па-
разитирования половозрелых особей D. pictus определя-
лись в различные сезоны года (табл. 19). Для опыта
были взяты клещи, собранные с апреля по июль 1956 г.
на растительности, т. е. опыты производились с перези-
мовавшими клещами. Затем опыт с кормлением был
проведен в сентябре с клещами, часть которых была
собрана на растительности весной, а часть—этой же
осенью (в сентябре). Самки насыщались за 1—2 недели.
Самцы находились на кроликах от нескольких дней до
нескольких месяцев. Некоторые самцы, очевидно, могли
76
Таблица 19
Сроки паразитирования взрослых клещей D. pictus на кроликах в 1956 г.
Время паразитирования Посажено на животное Сроки паразитиро- вания (в днях) К-ВО напитавшихся % напитав- шихся Примечание
X о S 3 о zj та о самок 1 самцов самок 1 Q О та самок самцов
Апрель 5 5 9—11 10—19 5 5 — — клещи собраны с
Май 80 80 5-12 5-30 72 70 90 88 растительности в апреле 1956 г.
Июль 10 10 8—15 6-30 8 5 — —
Сентябрь 3 3 12—17 51 3 3 — —
клещи собраны с
Сентябрь . 3 3 9-11 45 3 3 — — растительности в сентябре*1956 г.
Август 89 80 8-24 8-45 82 80 92 100 клещи выведены в лаборатории в
Сентябрь . 100 100 9-22 5^ 35 34 35 августе 1956 г
Октябрь 60 60 10—19 15-71 25 31 42 52
Декабрь 10 10 7—8 67 2 4 — —
бы еще дольше пребывать на хозяине, если бы не сор-
вались наклейки. При сравнении количества посажен
ных на кролика клещей с количеством напитавшихся
(сравнение берется по самкам) в разных сезонах наблю-
даются некоторые различия. Например, самки, собран-
ные на растительности в апреле и посаженные на кро-
лика в том же месяце, напитались нормально. При по-
садке клещей на кролика в мае получился отход (до
10%) ненапитавшихся самок. При кормлении в июле
отход ненапитавшихся особей был еще больше—до 20%.
При кормлении в сентябре все особи (как собранные на
растительности весной, так и осенью) напитались нор-
мально. Таким образом, ранней весной и осенью все
клещи, как правило, оканчивают питание благополучно.
При питании ближе к лету, и особенно в летний период,
процент отхода ненапитавшихся самок увеличивается.
Проведены также опыты по кормлению с клещами,
полученными в лаборатории. Клещи подсаживались -на
животное в августе, сентябре, октябре и декабре через
4—8, 32—41, 47—65, 145 дней после выхода имаго
(табл. 19). Сроки питания у самок варьировали от 1 до
2 недель. У единичных особей, особенно у самок, выли-
нявших на 4—8-й день, сроки питания растягивались до
24 дней. У самцов период пребывания на животном
длился от 8 до 71 дня. Самки, посаженные на кролика
в августе, через 4—8 дней после отрождения, напитались
почти все нормально (отход 8%). Клещи, посаженные на
хозяина в сентябре—октябре, через 32—65 дней после
отрождения, большей частью (в среднем до 60%) к пи-
танию не приступают, а в декабре из 10 самок в возрасте
145 дней, посаженных на кролика, напиталось только 2.
Н. Г. Олсуфьев (1953) при кормлении самок D. pictus
на морских свинках при температуре 17—18° получал
сытых особей через 9—12 суток. Н. И. Алфеев (1939) от-
мечает сроки паразитирования имаго на скоте от 3 до 10
(наиболее нормальный срок 4—6 суток) и на ежах от
10 до 15 суток. Н. И. Алфеев и С. М. Кулагин (1953)
при кормлении взрослых клещей на кроликах зимой
установили состояние длительного сцепления с хозяином:
самок до 75 дней, самцов до 36. Кроме того, эти авторы
указывают факты зимовки клещей на скоте.
Таким образом, длительность паразитирования у са-
мок от одной до двух недель. Значительного (до 75 дней)
78
удлинения срока паразитирования отдельных самок, как
указывают Н. И. Алфеев и С. М. Кулагин, в наших опы-
тах не наблюдалось. Но, очевидно, такое состояние в
природе бывает, если судить по сведениям, полученным
этими же авторами о зимовке клещей D. pictus на скоте,
а также о нахождении взрослых на диких животных
(волк) в Беловежской пуще (Ф. А. Закович, 1957). Что
касается чрезмерной длительности паразитирования сам-
цов D. pictus, то здесь, очевидно, наблюдается вполне
закономерное явление, связанное, как доказывает
Ю. С. Балашов (1956, 1957), с ходом сперматогенеза.
Заметно влияет на срок паразитирования и время напа-
дения на хозяина. Например, в наших опытах питание
проходило в более сжатые сроки весной (апрель—май) и
осенью (сентябрь), т. е. в период сезонной активности
взрослых клещей в природе. На длительность паразити-
рования, по нашему мнению, влияет и возраст клещей.
При посадке на кролика в сентябре клещей, отловлен-
ных в природе той же осенью, наблюдалось интенсивное
питание в более короткие и сжатые сроки. Посаженные
же на животное в сентябре, но из весеннего сбора в
природе, клещи питались вяло и в более растянутые
сроки, что зависело, вероятно, от истощения при длитель-
ном голодании (Ю. С. Балашов, 1957). На срок парази-
тирования влияют также и особенности того или иного
хозяина, как об этом указывают Н. И. Алфеев (1939) и
Ю. С. Балашов (1957). Действительно, при наблюдении
за питанием взрослых клещей D. pictus на коровах было
замечено, что весной (май) и осенью (сентябрь) срок
паразитирования длился в среднем 6—7 дней, тогда как
на кроликах процесс питания, как правило, удлинялся
у основной массы клещей до 9—10 дней.
ПРОДОЛЖИТЕЛЬНОСТЬ ЦИКЛА РАЗВИТИЯ
Активность взрослых клещей в природе начинается
с первой декады апреля и продолжается до начала
июня. В этот период они нападают на хозяина. Длитель-
ность паразитирования самок на скоте продолжается в
основном 7 дней. Сытые особи появляются с середины
апреля до середины июня. Затем происходит откладка
яиц, преимущественно в июне. Соответственно с этим
сроки созревания яиц у самок, напитавшихся в апреле,
79
удлиняются до 68 дней, а у самок, напитавшихся в
мае — июне, сокращаются до 7 дней. Небольшая часть
самок может откладывать яйца и в июле, но, как прави-
ло, такие кладки зачастую погибают. Продолжитель-
ность эмбрионального развития у основной массы яйце-
кладок длится 45—67 дней, и личинки вылупляются в
июле—августе. В зависимости от благоприятных кли-
матических условий (теплое и сухое лето) и стадиаль-
ного распространения личинки могут появляться и рань-
ше, т. е. с начала июня. В 3-дневном возрасте, и особенно
в возрасте 16—20 суток, личинки начинают нападать на
хозяина и паразитируют в течение 3—4 дней. Метамор-
фоз сытых личинок длится примерно от 7 до 23 дней, и
выход нимф в основной массе наблюдается в августе и
частично в сентябре. Но, исходя из данных сбора клещей
с диких животных, нимфы могут появляться и значи-
тельно раньше—в июле. Большинство личинок, напитав-
шихся в более поздние сроки (сентябре—октябре), поги-
бает. Как и личинки, нимфы нападают на хозяина с 3-
дневного, но большей частью 16—20-дневного возраста.
Паразитирование в основном продолжается 4—6 дней.
Метаморфоз нимф, напитавшихся в июле, длится 39 дней,
и имаго появляются к концу августа. Это наиболее ран-
нее появление имаго нового поколения. Нимфы, напи-
тавшиеся в более позднее время (сентябрь—октябрь), в
первый теплый сезон не успевают завершить свой мета-
морфоз и все погибают во время зимнего периода.
Имаго, появляющиеся в первый теплый сезон (ав-
густ—сентябрь), могут успеть насосаться и даже отло-
жить яйца до наступления зимы. Но яйца зимовки не
выдерживают и погибают. Что касается сытых самок
осеннего питания, то они в большинстве случаев благо-
получно перезимовывают, т. е. вступают в зимнюю диа-
паузу и в начале летнего сезона следующего года при-
ступают к откладке яиц. Взрослые клещи, которые не
успели напитаться в первый теплый сезон, зимуют и пи-
таются уже с наступлением второго теплого сезона, при-
ступая к яйцекладке так же, как и зимовавшие сытые
самки, в июне. Принимая во внимание сезонную кривую
активности имаго, можно предполагать, что большая
часть взрослых клещей зимует в голодном состоянии,
так как осенний пик в 5—8 раз меньше весеннего. Сле-
80
довательно, основная масса клещей питается с наступле-
нием теплого сезона на второй год.
Таким образом, развитие D. pictus в условиях Бело-
руссии протекает по одногодичному циклу (рис. 10). За
один теплый период развиваются последовательно все
фазы. Личинки и нимфы в голодном и сытом состоянии
Рис. 10. Схема развития Dermacentor pictus Herm. по од-
ногодичному циклу:
/г—голодные имаго; 1с—сытые самки; Ov—яйца; La—голодные ли-
чинки; Lc—сытые личинки; Wa—голодные нимфы; Nc—сытые нимфы
не зимуют: голодные погибают от истощения еще до на-
чала зимнего периода, а сытые в начале его, по-видимо-
му, от действия низких температур. Сытые самки впа-
дают в зимнюю диапаузу.
Иксодовые клещи
ЦИКЛ РАЗВИТИЯ I. RICINUS
СРОКИ ОТКЛАДКИ яиц
Наблюдения за сроками яйцекладок клеща в природе
проводились в весенне-летне-осенний период. Самки, на-
сосавшиеся весной (апрель—май), приступают к отклад-
ке яиц в основном в мае—июне и частично в июле и
даже в августе (табл. 20). Развитие некоторых из них
продолжалось до 120 дней. У преобладающего большин-
ства самок, накормленных в апреле—мае и приступаю-
щих к откладке в мае—июне, этот срок значительно ко-
роче—от 14 до 77 дней. Самки, насосавшиеся в летний
период (июнь—июль), частично приступают к яйцеклад-
ке в месяц кормления, а некоторые в августе. Сроки на-
чала яйцекладки у этих самок несколько сокращаются
по сравнению с весенними. Подобная закономерность
наблюдалась также в Карелии (Е. М. Хейсин и др.,
1954). Характерно, что большая часть яйцекладок, полу-
ченных в июле от весенних и летних самок, погибает.
Самки, насосавшиеся в августе—сентябре, приступают
к откладке яиц осенью (сентябрь—ноябрь). Однако не-
значительная часть самок, накормленных в сентябре,
приступила к яйцекладке только в апреле следующего
года, через 190 дней. Большинство самок, насосавшихся
в октябре, погибает во время зимовки, а небольшая
часть выживших приступает к яйцекладке только в ап-
реле—июне следующего года, через 172—227 дней. Боль-
шинство яйцекладок от таких самок получено в мае. Две
самки, насосавшиеся в ноябре, зимовку не перенесли и
погибли. В условиях Карельского перешейка (Г. В. Се-
рдюкова, 1951) основная масса яиц была получена в
июле—августе, т. е. несколько позднее, чем в условиях
82
Таблица 20
Сроки откладки яиц самками /. ricinus в природе
К-ВО сытых самок Из них погибло Время пи- тания самок Начало откладки яиц Сроки от- кладки яиц (в днях) К-во яйцекладок Из них погибло
май 27—49 19 2
72 1 апрель июнь июль 35-77 69-84 47 4 11 4
август 120 1 —
май 13 . 2
138 24 май июнь 14—41 109 26
июль 42 1 1
июнь 11—25 6 3
63 17 июнь июль 11—44 29 19
август 58-65 11 4
5 1 июль август 14—25 4
10 2 август сентябрь 19—37 8 4
сентябрь 18—27 3
27 3 сентябрь октябрь ноябрь 25—50 62 14 4 3 2
апрель следующего года 190 3 1
апрель следующего года 172—178 2 2
32 20 октябрь май следующего года 194—215 7 —
июнь следующего года 227 1 —
8 2 2 ноябрь —— — —
Белоруссии. Однако большинство самок, напитавшихся
осенью, созревает только к следующему году. Сходные с
нашими результаты получены Л. В. Бабенко (1956).
В Московской области яйцекладки от самок, насосав-
шихся в мае—начале сентября, получены в том же году,
а сытые самки, насосавшиеся в конце сентября—октября,
приступили к яйцекладке только весной (май) следую-
щего года.
В лабораторных условиях наблюдения за сроками
яйцекладок с марта по ноябрь дали несколько иные ре-
зультаты (табл. 21). Яйцекладка обычно наступала в
том же месяце, когда производилось кормление самок,
с интервалом 7—18 дней от момента насыщения или же
на следующий месяц, через 43 дня, и редко—на 3-й месяц,
через 70 дней. Каких-либо задержек в сроках развития
сытых самок и приуроченности яйцекладок к определен-
ному периоду в лабораторных условиях не наблюдалось.
Например, самки, питавшиеся поздней осенью (октяб-
ре—ноябре), не претерпевают такой длительной задерж-
ки в развитии, как в природе, а приступают к яйце-
кладке, как и в весенне-летний период, в сравнительно
короткие сроки. Исходя из этого, можно предположить,
что в природе сытые самки /. ricinus во время зимнего
периода не вступают в диапаузу, а претерпевают просто
холодовое оцепенение. Изучая развитие сытых самок
I. ricinus в лабораторных условиях, Q. Nuttall и С. War-
burton (1911) наблюдали начало яйцекладки через 8—
27 дней, Н. О. Оленев (1927)—через 4—24 (в основном
через 2 недели), Н. К. Ельманов (1930)—через 6—40,
Е. М. Хейсин (1953)—через 4—75 дней. Следовательно,
вполне подтверждаются наши предположения и выска-
зывания других исследователей (Е. М. Хейсин и
М. А. Лебешева, 1954) о зависимости сроков начала
яйцекладок от температурного фактора.
ПРОДОЛЖИТЕЛЬНОСТЬ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ
Изучение продолжительности эмбрионального разви-
тия в природе проводилось над яйцекладками, получен-
ными с апреля по ноябрь. Характерно, что основная
масса личинок в природе вылупляется в августе в год
откладки яиц или в этот же период на следующий год
(табл. 22). Например, из яйцекладок, полученных в
84
Таблица 21
Сроки откладки яиц самками /. ricinus в лаборатории
К-ВО сытых самок Из них погибло Время питания самок Начало откладки яиц Сроки откладки яиц (в днях) Получено яйцекладок Из них погибло
6 — март март май 15-17 46—61 4 2 2 2
17 — апрель май июнь 12—43 66—70 15 2 8 1
22 —- май июнь 7—14 22 2
11 1 июнь июнь июль 11—18 8—11 5 5 1
2 —- июль август 20 2 —
1 — август сентябрь 16 1 —
8 — сентябрь сентябрь октябрь 13 15-21 2 6 2
3 — октябрь октябрь ноябрь 15 17—18 1 2 2
8? 1 — ноябрь декабрь 37 1 —
Таблица 22
Продолжительность эмбрионального развития /. ricinus в природе
Получено яйцекладок Из них погибло Время Сроки вылупления личинок (в днях) К-во яйцекладок, давших личинки
яйцекладки вылупления личинок
4 3 апрель август 121 1
28 2 май август 68—100 26
163 40 июнь июль август сентябрь октябрь 39—41 56—83 78—99 100—130 2 105 12 4
34 24 июль август сентябрь октябрь 25 82 91—110 1 3 6
16 4 август сентябрь ноябрь июль следующего года август следующего года . 32 86 332—335 342—361 1 4 4 3
И 5 сентябрь июль следующего года август следующего года . 311—314 304—320 2 4
14 4 октябрь август следующего года . 276—294 10
4 2 ноябрь июль следующего года . август следующего года 240 252 1 1
апреле—мае, все личинки вылупились в августе в том
же году. Июньские и июльские яйцекладки дали личинки
в основном в августе и частично осенью (сентябрь—
октябрь), и только из двух июньских яйцекладок личин-
ки вылупились в июле. Августовские яйцекладки дают
личинки частично в том же году осенью (сентябрь—
ноябрь), а частично в июле—августе следующего года.
Из осенних (сентябрь—ноябрь) яйцекладок личинки вы-
ходят только на следующий год, но бпять-таки в июле—
августе. Таким образом, основная масса личинок вылуп-
ляется в августе этого же года от яйцекладок, получен-
ных в апреле—июле; часть яйцекладок, полученных в
июне—августе, дает личинки в том же году, но осенью,
а часть осенних кладок (конец августа—ноябрь) зимует
и дает личинки только на следующий год, но также в
конце теплого периода—в июле—августе. В связи с этим
соответственно меняются и сроки вылупления личинок.
Те яйцекладки, которые не зимуют, дают личинки через
25—130 дней (основная масса через 65—79 дней), и из
зимующих яйцекладок личинки выходят через 240—
361 день. Н. О. Оленевым (1924) в Новгородской обла-
сти установлено, что в природе личинки вылупляются
через 8 недель, а М. П. Марковым (1927) на Украине —
через 26—46 дней. Впоследствии при более детальной
разработке данного вопроса в условиях Карельского пе-
решейка (Г. В. Сердюкова, 1951, 1952) установлена зави-
симость между сроками вылупления личинок и временем
откладки яиц, т. е. основная масса личинок вылупляется
только на следующий год в начале июля—начале сен-
тября, и только часть весенних кладок дает личинки в
год яйцекладки, но также в конце теплого периода.
П. К. Кузнецов (1955, 1957) в Воронежской области
установил, что весенние кладки дают личинки в конце
теплого сезона в том же году, а из яйцекладок, получен-
ных во второй половине теплого сезона, личинки вылуп-
ляются только на следующий год, но также в конце теп-
лого сезона. Л. В. Бабенко (1956) в Московской области
также наблюдала сходные явления. Таким образом, вы-
ход личинок /. ricinus происходит в довольно определен-
ный период, т. е. в конце теплого сезона года.
В более суровых климатических условиях (Карель-
ский перешеек) большая часть кладок зимует и основ-
ная масса личинок вылупляется только на следующий
87
год, тогда как в более южных широтах, например в юго-
западной части Белоруссии (Беловежская пуща), боль-
шая часть яиц успевает развиться в том же году.
Параллельно с марта по декабрь велись опыты в ла-
боратории. Q. Nuttall и С. Warburton (1911) указывают
на вылупление личинок через 42—252 дня после яйце-
кладки. Н. О. Оленев (1924, 1927)—через 3 недели при
температуре 25—30° и через 50—67 дней при 16°, а в от-
дельных случаях через 400 дней. По М. П. Маркову
(1927), при комнатной температуре—через 30 дней, по
Н. К. Ельманову (1930)—через 15—128 дней, по
Е. М. Хейсину (1953) —через 33 дня при температуре
22,3° и через 3 месяца при температуре 14,5°. В наших
опытах сроки начала вылупления личинок колебались в
пределах 31 —189 дней (табл. 23), причем у части яйце-
кладок наблюдалась довольно определенная закономер-
ность. Например, у яйцекладок, полученных с марта по
август, задержки в развитии не было. Наоборот, у яйце-
кладок, полученных в сентябре—декабре, намечается
тенденция к задержке развития, и момент вылупления
личинок приурочивается к определенному периоду—
февралю.
Таким образом, яйца, полученные в осенний период и
остающиеся в природе на зимовку, вступают в диапаузу,
которая продолжается до февраля включительно. Затем
развитие продолжается в соответствии с внешними усло-
виями. В лаборатории после февраля наступает быстрое
развитие, а в природе яйца претерпевают холодовое
оцепенение, пока не наступят оптимальные температур-
ные условия. Например, из яйцекладок, полученных в
сентябре—октябре и зимовавших в естественных усло-
виях, а затем (в феврале) перенесенных в лабораторию,
вышли личинки не в конце теплого периода, как это на-
блюдается в природе, а значительно раньше—в апреле.
В противоположность нашим данным в Карельской
АССР (Е. М. Хейсин и Т. К. Кузнецова, 1956) диапауза
у зимующих яиц не наблюдалась.
РАЗВИТИЕ ЛИЧИНОК
Сезонная динамика паразитирования. Активность ли-
чинок, по данным сбора клещей с растительности, прояв-
лялась с мая по сентябрь (табл. 24, рис. 11). Наиболее
88
Таблица 23
Продолжительность эмбрионального развития I. ricinus в лаборатории
К-во яйцекладок, давших личинки Погибших яйцекладок Время яйцекладки Время вылупления личинок Сроки вылупления личинок (в днях) К-во яйцекладок, давших личинки Примечание
апрель 34 1
май 56 1
4 — март июнь 88 1
июль 121 1
июнь 50-66 3
17 10 май июль 62—79 4
29 июнь июль 31—48 22
3 август 33-46 4
2 — август октябрь 51—71 2
4 — сентябрь февраль следующего года апрель следующего года . 139—140 189 3 1 яйцекладки пере-
175 несены из при-
апрель следующего года 1 роды в лабора-
8 2 октябрь декабрь 74 1 торию в феврале
февраль следующего года . 129—134 4
2 2 ноябрь — — —
10 2 декабрь февраль следующего года 60—75 8
Таблица 24
Сезонная динамика активности личинок, нимф и имаго
/. ricinus за 1953—1956 гг.
(данные сбора клещей с растительности)
Месяц К-во учетов Личинки Нимфы Имаго
собрано среднее на учет собрано среднее на учет собрано среднее на учет
Март 2 0 0 0 0 0 0
Апрель 49 0 0 65 1,3 71 1,4
Май 48 53 1,1 407 8,5 179 3,7
Июнь 57 192 3,4 217 3,8 124 2,2
Июль 35 82 2,5 125 3,6 46 1,3
Август . 14 0 0 6 0,4 0 0
Сентябрь . 54 53 1 114 2,1 27 0,5
Октябрь 66 0 0 74 1,1 50 0,8
Ноябрь 14 0 0 1 0,1 0 0
раннее появление личинок отмечено 17 мая 1956 г. Ма-
ксимальное количество личинок с растительности было
Рис. 11. Сезонная кривая актив-
ности имаго (а), нимф (6) и ли-
чинок (в) Ixodes ricinus L. по
данным сбора клещей с расти-
тельности (1953—1956 гг.)
собрано в середине июня
(18 июня 1953 г. собрано
89 личинок на учет).
В конце июля—августе
активность личинок пада-
ет, доходя до нуля, затем
в начале сентября появля-
ются отдельные особи, и к
середине месяца они до-
стигают второго максиму-
ма, после чего быстро ис-
чезают. Уже к концу вто-
рой декады сентября ли-
чинки почти не встреча-
ЮТСЯ.
При обследовании жи-
вотных установлено, что
сезон активности личинок
на них гораздо продолжи-
тельнее, чем на раститель-
ности. Например, паразитирование личиночной фазы
началось в апреле и закончилось в октябре (табл. 25,
рис. 12). Первые личинки обнаружены 8 апреля 1953 г.
90
Таблица 25
Сезонная динамика паразитирования активных фаз /. ricinus за 1953—1956 гг. на позвоночных
Месяц Личинки Нимфы Имаго
осмотре- но жи- вотных зараже- но собрано клещей интен- сивность зараже- ния осмотре- но жи- вотных зараже- но собрано клещей интен- сивность зараже- ния осмотре- но жи- вотных зараже- но собрано клещей интен- сивность зараже- ния
Январь Февраль Март Апрель Май Июнь Июль Август Сентябрь . Октябрь Ноябрь 2 Декабрь 20 13 7 129 173 159 261 177 469 328 25 0 0 0 18 130 112 178 92 252 96 0 0 0 0 31 1271 1856 1153 699 1233 167 0 1: 0 0 0 1,7 3,8 16,6 6,5 7, 5 1,7 0 17 4 3 11 10 17 19 18 29 16 0 0 0 7 9 17 18 18 28 14 0 0 0 57 427 1032 365 592 1441 318 0 0 0 8,1 47,4 60,7 20,3 33 51,5 22,7 32 100 47 37 23 72 39 10 5 32 99 47 30 23 56 30 5 0 440 3130 1426 528 210 861 277 20 0 14 31,6 30,8 17,6 9,1 15,4 9,2 4 0
на желтогорлой мыши (4 личинки на двух зверьках)
при среднедекадной температуре воздуха 6,8° и относи-
тельной влажности 75,9 и одна личинка на белке 17 апре-
ля 1953 г. при среднедекадной температуре воздуха 6е
и относительной влажности 68,8. Максимум личинок
наблюдался в июне. Наибольшее заражение зверьков
отмечено 9 июня 1956 г. на еже (140 личинок), 21 июня
на белке (98 личинок) и 28 июня на европейской рыжей
полевке (24 личинки). Сред-
7Z71 немесячная температура воз-
Рис. 12. Сезонная кривая зара-
жения животных клещами /хо-
des ricinus L. (1953—1956 гг.):
а—имаго; б—нимфы; в—личинки
духа в этот период была
18,3°, относительная влаж-
ность 73. В июле наблюда-
лось понижение активности,
а затем снова подъем в ав-
густе (10 августа 1956 г. с
белки собрано 59 личинок и с
желтогорлой мыши 16 авгу-
ста 42 личинки). После вто-
рого пика начинается мед-
ленный спад до конца октяб-
ря. Последние личинки обна-
ружены 26 октября 1956 г.
на белке (1 личинка), евро-
пейской рыжей полевке (1 личинка) и землеройке обык-
новенной (1 личинка). Средняя температура воздуха в
последней декаде октября 6,3°, относительная влаж-
ность 86,7.
Таким образом, активность личинок наблюдается в
течение всего теплого периода, начиная с апреля и
кончая в октябре. Имеется два пика активности—в июне
и августе—сентябре. В зимний (январь—февраль) и ран-
невесенний (март) периоды личинки неактивны. Литера-
турные сведения по данному вопросу довольно разноре-
чивы. Например, в Ленинградской (Б. И. Померанцев,
1935) и Саратовской (Д. Н. Засухин и др., 1936) обла-
стях активность отмечается с мая по август—начало
сентября. В. Rosicky (1954), В. Rosicky и V. Сегпу (1954)
указывают на активность с апреля по октябрь и макси-
мум—с конца мая и июль—август, что наиболее близко
к нашим данным. Л. В. Бабенко (1956) в Московской
области период активности отмечает с середины мая до
середины августа с двумя максимумами: первый—ко-
92
иен мая и начало июня и второй—начало—конец июля.
В условиях Эстонии (А. К. Вильбасте, 1956) максимум
активности личинок отмечен в мае, затем он постепенно
понижается до октября. В условиях Белоруссии Б. П. Са-
вицкий (1957) встречал личинок только в июне—июле с
максимумом в последней декаде июня.
Такие несколько разноречивые данные объясняются,
вероятно, несколько различными сроками развития личи-
нок в различных климатических условиях, а также зави-
симостью от сезонной динамики размножения самих хо-
зяев. Доминирующими хозяевами личиночной фазы 1. ri-
cinus являются в основном мышевидные грызуны,
которые вследствие быстрого нарастания (в 5—6 раз)
численности с весны до осени (Е. П. Пивоварова, 1956)
несколько снижают интенсивность заражения клещами.
К тому же резкие колебания численности зверьков в раз-
личные годы, зависящие от метеорологических условий и
различного рода массовых заболеваний, в той или иной
степени отражаются на интенсивности заражения их ли-
чинками I. ricinus.
Развитие голодных личи-
нок. У личинок /. ricinus,
как и у личинок D. pictus,
наблюдается зависимость
количества способных к пи-
танию особей от их возраста
(табл. 26). Личинки способ-
ны приступать к питанию в
возрасте 7 дней, но таких
особей еще мало. Наиболь-
шее число личинок питается
в возрасте 20—60 дней, до-
стигая максимума примерно
на 24-й день. После 60-днев-
ного возраста количество ли-
чинок, способных питаться,
падает. Например, на 109-е
сутки питаются уже единич-
ные особи. Подобное перво-
Таблица 26
Взаимосвязь между количе-
ством напитавшихся личинок
/. ricinus и длительностью
их голодания
Возраст
личинок
(в днях)
Напитавшихся
личинок в одной
генерации
7 71
9 227
14 966
20 1054
24 2050
60 1094
66 585
82 398
97 109
109 24
начальное повышение, затем снижение количества осо-
бей, способных питаться в зависимости от возраста, явля-
ется, как уже говорилось, результатом физиологического
доразвития и истощения от голода. Следовательно, физи-
93
ологическое доразвитие у голодных особей данной фазы
заканчивается в среднем через 1—2 месяца, после чего
начинается истощение и гибель личинок, в связи с чем
количество питающихся особей падает. Таким образом,
основная масса личинок дважды зимовать не способна
и погибает в течение одного теплого сезона года. В усло-
виях северных широт (Е. М. Хейсин, 1954) сроки физио-
логического доразвития удлиняются до 2—3 месяцев.
Сроки паразитирования личинок. Кормление личинок
проводилось на протяжении всех сезонов года. Парази-
тирование продолжалось в зависимости от сезона в пре-
делах 2—14 дней. Основная масса личинок питалась 3-
4 дня. Особенное затягивание сроков питания наблюда-
лось весной (май) и поздней осенью (октябрь), что яв-
ляется, очевидно, результатом влияния более понижен-
ной температуры. В зимний период (декабрь—март) при
перенесении опытов в лабораторию с комнатной темпе-
ратурой питание проходит в еще более короткие сроки,
чем летом (табл. 27). При этом личинки быстро перехо-
Таблица 27
Сроки паразитирования личинок /. ricinus
Время паразитирования Сроки пара зитирования (в днях) Напитав- шихся личинок Средне- месячная температура воздуха, °C Примечание
Май 2—9 7092 10,4
Июнь . 2—7 13424 15
Июль . 2—7 2239 18,8
Август 2—5 8923 18,2
Сентябрь 2—8 3667 13,4
Октябрь 3-14 4702 7,3
Декабрь 3-6 2046 18-20 личинки кор-
Февраль 2—4 123 18-20 мились в
Март 2—4 483 18-20 лаборато- рии при комнатной температу- ре
дят из оцепенения в активное состояние, т. е. у голодных
личинок /. ricinus зимней диапаузы не наблюдается. По
всей вероятности, зимой в природе они находятся в хо-
94
ледовом оцепенении. Q. Nuttall и С. Warburton (1911),
Kossel и другие (1903) и Н. О. Оленев (1927) отмечают
колебания срока питания от 3 до 6, но чаще 4—5 дней. По
Н. К. Ельманову (1930), личинки питаются на взрослых
мышах 3—4 дня, на голых мышатах—4—6, на теленке —
7, на корове—5—6 дней.
Развитие сытых личинок. В развитии сытых личинок
в естественных условиях наблюдается довольно опреде-
ленная закономерность (табл. 28). Личинки, насосав-
Таблица 28
Развитие сытых личинок /. ricinus в природе
эХ X gs Время Время линьки иньки от мо- дтания до нимф iepa- нимф X X 3 ю X »х
К-во гене ВЗЯТЫХ J опыта питания личинок Сроки л (в днях) мента ш личинок выхода ] 2 g я m X * § К-во по] генерапр
12 июль 50-62 3
май август 68-79 9
август . 55—89 21
23 июнь сентябрь август следующего 102 1 —
года 422 1
4 июль октябрь 75-78 4 —
июль следующего
8 сентябрь года август следующего 312—324 5 —
года 319—331 3
июль следующего
26 октябрь года . август следующего 271—289 4 18
года 265—301 4
4 ноябрь — — — А
2 декабрь — — — и
шиеся в мае—июне, как правило, линяют на нимф в
июле—августе того же года. Часть личинок, питавшихся
в июне, и личинки, насосавшиеся в июле, частично ли-
няют осенью того же года (в сентябре—октябре) или в
августе на следующий год. Наконец, личинки осеннего
95
питания (сентябрь—октябрь) линяют на нимф, как пра-
вило, только на следующий год в июле—августе. При
этом большинство личинок октябрьского кормления по-
гибает во время зимовки, т. е. особи этого периода пи-
тания как бы не приспособлены к зимовке. Об этом гово-
рит и тот факт, что при кормлении в еще более поздние
сроки (ноябрь—декабрь) личинки не линяют на нимф
совсем, так как погибают в зимний период. При наблю-
дении развития сытых личинок, снятых с диких живот-
ных, получается такая же картина (табл. 29). Сытые ли-
Таблица 29
Развитие сытых личинок /. ricinus, снятых с диких животных
Время снятия сытых личинок Снято личинок Время линьки Сроки линьки (в днях) К-во нимф
Апрель 4 сентябрь 129—163 3
Май 410 август 72-93 380
Июнь 693 август 70 410
октябрь 123 203
Июль 83 сентябрь 48-81 80
Август 13 апрель следующего года 221 1
август следующего года 360 8
Октябрь 40 август следующего года . 301 2
чинки, добытые с апреля по июль, линяют на нимф в
основном в августе и частично в сентябре—октябре того
же года. Особи, питавшиеся на диких животных в авгу-
сте—октябре, линяют также в августе на следующий
год. При этом большинство особей октябрьского корм-
ления погибает в зимний период, за исключением одного
случая, когда одна личинка, снятая с белки в августе, пе-
релиняла на нимфу не в обычные летне-осенние сроки, а
в начале весны—в апреле следующего года.
Таким образом, все личинки, накормленные в различ-
ные сезоны года, линяют на нимф, как правило, в опре-
деленный период, т. е. в конце теплого и начале осенне-
го сезона. При этом особи, питавшиеся весной и летом,
96
линяют в том же году через 50—163 дня (основная мас-
са спустя 64—79 суток после кормления). Личинки осен-
него кормления—на следующий год через 221—422 дня
(основная масса на 312—331-е сутки). Личинки, насосав-
шиеся в слишком поздний период (октябрь—декабрь),
не приспособлены к зимовке и большей частью гибнут.
В условиях Карельского перешейка (Г. В. Сердюкова,
1952) также наблюдалось появление нимф как из лет-
них, так и из перезимовавших сытых личинок в опреде-
ленный сезон года—с конца июля по сентябрь. То же
наблюдали П. К. Кузнецов (1955, 1957) в Воронежской
и Л. В. Бабенко (1956) в Московской областях.
В лаборатории срок развития сытых личинок значи-
тельно сократился по сравнению с особями, находящими-
ся в природе (табл. 30). У некоторых генераций наблю-
далась определенная закономерность в выходе нимф.
Например, личинки, насосавшиеся в период февраля—
сентября, линяли на нимф без каких-либо задержек в
развитии в довольно короткие сроки (28—43 дня). Но
часть личинок, питавшихся в сентябре, и все личинки,
насосавшиеся с октября по январь, дают выход нимф
только в определенный период, а именно в феврале. При
этом сроки развития, особенно у особей, питавшихся
осенью (сентябрь — октябрь), удлиняются до 124—
161-го дня. По мере приближения момента питания к
февралю сроки линьки постепенно сокращаются. Q. Nut-
tall и С. Warburton (1911) наблюдали линьку личинок
на нимф через 28—140 дней. Н. О. Оленев (1927) и
Н. К. Ельманов (1930) указывают на еще большую
разбежку в сроках развития сытых личинок (первый от
54 до 426 дней, второй—45—286 суток). Безусловно, эти
исследователи наблюдали задержку в развитии у части
особей, несмотря на содержание клещей в оптимальных
лабораторных условиях. Н. О. Оленев, например, приво-
дит случай задержки развития личинок, напитавшихся в
сентябре, как и в наших опытах, до февраля. А. И. Хода-
ковский (1947), наблюдавший подобную задержку в раз-
витии сытых личинок и нимф у клещей /. persulcatus при
содержании в оптимальных лабораторных условиях,
объясняет данное явление наличием у молодых фаз зим-
ней диапаузы. Подобную задержку в развитии молодых
фаз I. ricinus наблюдали Н. И. Алфеев (1948) и
Е. М. Хейсин (1954), объясняя ее так же, как явление
7. Иксодовые клещи
97
Таблица 30
Развитие сытых личинок /. ricinus в лаборатории
К-во кладок, давших личинки Время питания личинок Время линьки Сроки линьки (в днях) . х W S О. Я g й £ S Примечание
О CQ X ЦИЙ, J нимф
1 январь февраль 42 1
1 январь февраль 47 1 голодные ли- чинки в ян-
варе пере- несены из
природы в лаборато- рию
1 февраль март 30 1
4 июль август 28—32 4
10 август сентябрь 25-43 10
5 сентябрь октябрь .... февраль следую- щего года . 29—39 161 4 1
4 8 1 октябрь ноябрь декабрь февраль следую- щего года . . февраль следую- щего года . февраль следую- щего года 124—129 97—101 46 4 8 1 сытые личин- ки в январе перенесены из природы в лаборато- рию
диапаузы. Мы склонны присоединиться к данному объяс-
нению.
Строгая приуроченность линьки к определенному пе-
риоду у особей, питавшихся в осенне-зимний период, не-
смотря на содержание их в оптимальных условиях, гово-
рит о наличии у них зимней диапаузы, не обязательной
для особей, питавшихся весной. Об этом говорит и тот
факт, что голодные личинки, содержавшиеся в природе, а
затем перенесенные в январе в лабораторию и накормлен-
98
ные в том же месяце, а также сытые личинки, питавшиеся
в октябре и содержавшиеся сначала в природе, а затем
перенесенные в январе в лабораторию, все перелиняли
на нимф наравне с лабораторными особями в феврале.
Следовательно, личинки, питавшиеся в осенне-зимний
период, вступают в зимнюю диапаузу до февраля, неза-
висимо от содержания их в лаборатории или в природе.
По истечении этого срока личинки, содержащиеся в лабо-
ратории, начинают быстро развиваться, а в природе на-
ходятся в состоянии холодового оцепенения до момента
наступления оптимальных температурных условий.
РАЗВИТИЕ НИМФ
Сезонная динамика паразитирования. Активность
нимф /. ricinus начинается с апреля и продолжается весь
теплый сезон до ноября включительно (табл. 24, рис. 11).
При сборе клещей с растительности первые нимфы обна-
ружены в 1956 г. 18 апреля (2 нимфы на учет), затем
активность быстро возрастает, достигая максимума в на-
чале мая (52 нимфы на учет 7 мая 1956 г.). В июне—
июле активность постепенно снижается, достигая мини-
мума в августе (2 августа 1955 г. было 2 нимфы на
учет). Осенью активность нимф снова возрастает и до-
стигает подъема в сентябре (6 сентября 1955 г. собрано
7 нимф на учет). Последние нимфы обнаружены 2 нояб-
ря 1955 г. При обследовании грызунов и насекомоядных
паразитирование нимф впервые отмечено раньше, чем на
растительности. Например, 4 апреля 1956 г. снята нимфа
с европейской рыжей полевки (среднедекадная темпера-
тура воздуха в этот период 0,7°, относительная влаж-
ность 80,1), а 10 апреля 1955 г.—4 нимфы с белки (средне-
декадная температура воздуха 2,2°, относительная влаж-
ность 74,9). Максимум паразитирующих нимф на млеко-
питающих приходится не на май, как на растительности,
а на июнь (табл. 25, рис. 12). Так, наибольшее количе-
ство нимф снято 9 июня 1956 г. (среднедекадная темпе-
ратура воздуха 20,9°, относительная влажность 62,2) с
ежа (258 нимф), а 18 июня 1953 г. с белки снято 120 нимф
(среднедекадная температура воздуха 18,9°, относитель-
ная влажность 74). В июле наблюдалось резкое сниже-
ние паразитирующих особей. Затем происходит повыше-
ние до сентября включительно (максимальное количе-
99
ство нимф собрано 6 сентября 1956 г. с белки—199 нимф
и 28 сентября 1955 г. с ежа—90 нимф). Последний слу-
чай паразитирования нимф отмечен 18 октября 1956 г.
на белке (7 нимф) и 26 октября 1956 г. на европейской ры-
жей полевке (1 нимфа). Средняя температура воздуха
за последнюю декаду октября 6,3°, относительная влаж-
ность 86,7. В зимний (январь—февраль) и ранневесен-
ний (март) периоды зараженности зверьков нимфами нс
наблюдалось (табл. 25).
Таким образом, кривая сезонной динамики активно-
сти нимф имеет две резко выраженные вершины с ма-
ксимумом весной (май—июнь) и осенью (сентябрь).
Наши наблюдения в некоторой степени согласуются
с данными, полученными Л. В. Бабенко (1956) в Мо-
сковской области, за исключением периода осеннего пи-
ка. Автор указывает, что осенний пик наступает в авгу-
сте, тогда как по нашим данным—в сентябре. В Ленин-
градской (Б. И. Померанцев, 1935) и Саратовской
(Д. Н. Засухин и др., 1936) областях авторы наблюдали
активность нимф: первый с мая до августа с максимумом
в июне, второй с мая по сентябрь с максимумом в мае
и августе. В условиях Эстонии (А. К. Вильбасте, 1956)
Таблица 31 максимальная активность нимф наблюдается в мае с
Взаимосвязь количества напи- постепенным понижением
тавшихся нимф /. ricinus с
Длительностью их голодания без второго пика вплоть до
— октября.
Возраст нимф (в днях) Напитавшихся Развитие голодных нимф.
нимф в одной Нимфы могут приступать к
генерации питанию даже на 9-е сутки
после отрождения. Но таких
9 136 особей мало (табл. 31).
16 ол 298 л со С увеличением возраста чис-
20 21 708 ло нимф, приступающих к пи-
47 501 танию, возрастает. В наших
52 477 опытах преобладающее чис-
57 1 ол 129 ло особей кормилось в воз-
144 оо 32 расте от 20 до 52 дней, при
268 21 этом наибольшее количество
нимф питалось в возрасте
21 дня. Но резкого падения,
как это наблюдалось у D. pictus, количества нимф, спо-
собных питаться, не происходит. Единичные особи могут
100
еще кормиться в возрасте 268 дней, что указывает на
способность отдельных нимф /. ricinus к длительному
голоданию. Однако, как и личинки, дважды зимовать
они, очевидно, не способны, а погибают в течение первого
теплого сезона года.
Сроки паразитирования нимф. Длительность питания
нимф I. ricinus определялась на протяжении февраля —
декабря. Кормление клещей в зимний период (декабрь—
март) проводилось в лаборатории. Общий срок парази-
тирования отмечен от 2 до 15 суток. Такая растянутость
зависит от температурного фактора (табл. 32). В наших
Таблица 32
Сроки паразитирования нимф /. ricinus
Время паразитирования Сроки пара- зитирования (в днях) Напитав- шихся нимф Среднемесяч- ная темпера- тура возду- ха, °C Примечание
Апрель 4-12 12 3,3
Май . 2-11 72 10,4
Август . 3-8 916 18,2
Сентябрь 3—15 1829 13,4
Октябрь 4—14 2038 7,3
Декабрь 3-6 25 18-20 нимфы корми-
Февраль 3—7 191 18—20 лись в ла-
Март 3-6 56 18—20 боратории при комнат- ной темпе- ратуре
опытах при кормлении в естественных условиях в апре-
ле—мае и сентябре—октябре, когда температура воз-
духа, особенно в ночное время, падает довольно низко,
паразитирование затягивалось зачастую до 14—15 су-
ток. После перенесения подопытных животных с клеща-
ми в лабораторные условия длительность питания резко
сокращается. Основная масса нимф в летний сезон
(август), как правило, заканчивала питание на 3—6-е
сутки. Q. Nuttall и С. Warburton (1911) определяют срок
питания нимф до 3—5 дней, Н. О. Оленев (1927)—до
4—6 суток, Н. К. Ельманов (1930)—6 дней, R. Totze
(1933), правда, в необычных условиях кормления (на
101
искусственной мембране) получал сытых нимф через
два с лишним дня.
Развитие сытых нимф. В развитии сытых нимф в при-
роде наблюдались такие же закономерности выхода
имаго, как и при вылуплении личинок и при линьке сы-
тых личинок на нимф. Нимфы, питавшиеся весной (май),
линяли на имаго в том же году в августе—октябре, через
78—156 дней. Особи, питавшиеся осенью (сентябрь —
октябрь), перелиняли только на следующий год также в
конце теплого сезона—в августе, через 285—306 дней
(табл. 33). Сытые нимфы, собранные с диких животных
Таблица 33
Развитие сытых нимф 7. ricinus в природе
4 май
4 сентябрь
10 октябрь
Время
питания
Время линьки
август
октябрь
Сроки
линьки
(в днях)
2
1
78 1
156 1
август следующего года 330—352 4
август следующего года 285—306
в апреле—июле, в основном перелиняли на имаго в кон-
це теплого сезона того же года и незначительная часть
в следующем году в этот же период. Нимфы же, собран-
ные в августе—октябре, перелиняли на имаго только по-
сле зимовки в конце теплого сезона (табл. 34). Такую же
картину появления имаго в природе отмечали Г. В.
Сердюкова (1952), П. К. Кузнецов (1955, 1957), Л.В. Ба-
бенко (1956).
В лабораторных условиях Q. Nuttall и С. Warburton
(1911) наблюдали выход имаго через 56—360 дней.
Н. О. Оленев (1927)—спустя 159—335 и Н. К. Ельманов
(1930)—через 243 дня после насыщения нимф. В наших
опытах при лабораторном содержании нимфы, питав-
102
Таблица 34
Развитие сытых нимф /. ricinus, снятых с диких животных
Время снятия Снято нимф Время линьки нимф Сроки линьки нимф (в днях) К-во имаго
август 99—106 2
Апрель 8 сентябрь 155 1
октябрь 175 1
август 82—117 242
Май 300 сентябрь 124—130 2
октябрь ... 149 1
август следующего года 471 2
август 66-79 16
Июнь 538 сентябрь 72—104 393
октябрь .... 103 1
август следующего года 444 1
Июль 19 сентябрь 59—64 5
Август 30 август следующего года 365—368 2
Г"* рмтпИпк 174 август следующего года 342—355 153
1 ziUUD сентябрь следующего года 371 1
Октябрь И август следующего года 325 1
шиеся в феврале—мае, развивались без задержки и да-
вали имаго в основном на 32—40-й день (табл. 35). На-
против, все нимфы, питавшиеся с августа по ноябрь,
включая содержавшихся в природе и затем перенесен-
ных в январе в лабораторию, перелиняли на имаго в од-
но и то же время—в феврале.
Таким образом, у нимф, питавшихся осенью, наблю-
дается задержка в развитии, как и у сытых личинок, до
определенного момента, а именно до февраля, несмотря
на содержание их в оптимальных условиях. Нимфы же,
питавшиеся в июле и содержавшиеся в природе, а затем
перенесенные в марте в лабораторные условия, быстро
перелиняли на имаго в том же месяце и частично в мае.
Следовательно, у нимф, напитавшихся в весенний пе-
риод, задержки в развитии нет, тогда как у особей осен-
него кормления даже в лабораторных условиях наблю-
103
Таблица 35
Развитие сытых нимф I. ricinus в лаборатории
К-во генераций, давших нимф Время питания К-во генераций с погибшими нимфами Время линьки Сроки линьки (в днях) К-во генераций, давших имаго Примечание
3 февраль — март апрель 34—40 40 2 1
4 март — июль 108—123 4
1 май — июнь 32 1
2 июль — март следую- щего года май следую- щего года 231 276 1 1 сытые нимфы перенесены в марте из природы в лаборато- рию
6 август февраль сле- дующего года . 162—180 6 4 генерации сытых нимф в январе перенесены из природы в лаборато- рию
5 сентябрь — февраль сле- дующего года 134-168 5
1 о ктябрь февраль сле- дующего года 114 1 сытые нимфы перенесены в январе из природы в лаборато- рию
9 ноябрь — февраль сле- дующего года 95—102 9
дается перерыв вплоть до февраля. Подобную задержку
в развитии у сытых нимф I. persulcatus в лабораторных
104
условиях наблюдали А. И. Ходаковский (1947), а
Н. И. Алфеев (1948) и Е. М. Хейсин (1954)—у I. ricinus.
Эти авторы объясняют данное явление наступлением
диапаузы. На основании сопоставления данных о разви-
тии в лаборатории и природе мы также склонны считать
вполне закономерным у нимф, питавшихся в осенний пе-
риод, наличие зимней диапаузы, которая отсутствует у
нимф весеннего кормления и частично у особей, питав-
шихся в летний период. После февраля нимфы, содержа-
щиеся в природе, находятся в холодовом оцепенении до
наступления благоприятных температурных условий.
РАЗВИТИЕ ИМАГО
Сезонная динамика паразитирования имаго. Актив-
ность взрослых клещей /. ricinus начинается, как и има-
го D, pictus, в первой декаде апреля при появлении про-
талин в лесу. Первая находка имаго в природе была
5 апреля 1955 г. (среднедекадная температура воздуха
2,2°, относительная влажность 74,9). Затем активность
повышалась, достигнув максимума в мае (табл. 24,
рис. 11). При этом возрастание активности у /. ricinus
идет медленнее, чем у D. pictus. Так, при сборе клещей
с растительности максимальная активность имаго D. pi-
ctus отмечена в начале мая, а у /. ricinus только к концу
мая (28 мая 1956 г. собрано 22 клеща на учет). Средне-
декадная температура воздуха в этот период была 20°,
относительная влажность 62,4. После подъема начинает-
ся не быстрый, как у D. pictus, а постепенный спад
активности вплоть до начала августа (минимум актив-
ности наблюдался 31 июля 1956 г.). В течение августа
взрослых клещей /. ricinus на растительности мы не об-
наружили. Клещи снова появились только 1 сентября
1955 г., после чего наблюдалось незначительное возра-
стание активности до начала октября (осенний пик от-
мечен 8 октября 1955 г.—5 клещей на учет) с последую-
щим падением кривой активности до середины октября
(последние клещи на растительности сняты 14 октября
1956 г.). Температура воздуха во второй декаде октября
6,5°, относительная влажность 84,5.
При учете сезонной динамики паразитирования кле-
щей на животных мы взяли за основу данные сбора кле-
щей за пастбищный сезон с каменюцкого стада крупно-
105
го рогатого скота. Выпас пролегал через пойменный
луг, ольшаник, ельник, сосняк, елово-широколиственный
и дубово-грабовый лес. Общая протяженность его до
5 км. С началом выпаса скота (апрель) сразу же наблю-
дается паразитирование клещей (первые сборы были
15 апреля 1955 г.), затем медленное возрастание количе-
ства паразитирующих особей до конца мая (максимум
отмечен 23 мая 1956 г.—на отдельных животных 256 кле-
щей). В июне интенсивность заражения снижается, но
очень незначительно (табл. 25, рис. 12). Постепенный
спад активности продолжается до начала августа (наи-
меньшая интенсивность заражения отмечена 5 августа
1955 г.), но совсем клещи не пропадают. В середине сен-
тября наблюдается второй пик активности (18 сентября
1955 г. на отдельных животных 67 клещей). Паразитиро-
вание на скоте продолжается до начала ноября (послед-
ние клещи сняты 5 ноября 1955 г.). Среднедекадная тем-
пература воздуха в этот период 3,9°, относительная
влажность 89,4.
Таким образом, активность взрослых клещей /. rici-
nus начинается с начала апреля и продолжается до на-
чала ноября, причем характер сезонной динамики актив-
ности имеет две вершины—в конце мая—начале июня и
в середине сентября—начале октября.
В ряде работ, кроме двух пиков активности (весной
и осенью), указывается на многовершинность кривой се-
зонной динамики активности взрослых клещей /. ricinus,
зависящей от смены пастбищ и в связи с этим от напа-
дения на скот запаса свежих клещей (Д. И. Благовещен-
ский, 1930; Е. Н. Павловский и др., 1932; Б. И. Померан-
цев, 1935). Сезонную динамику активности имаго /. rici-
nus изучали и многие другие исследователи (М. П. Мар-
ков, 1927; Н. О. Оленев, 1927, 1935; Д. Н. Засухин и др.,
1936; Н. И. Алфеев, 1939; В. Rosicky, 1954; Л. В. Бабен-
ко, 1956; А. К. Вильбасте, 1956; А. С. Лутта, 1957), ко-
торые пришли к выводу о наличии двух пиков активно-
сти весной (май) и осенью (сентябрь). В более южных
широтах активность клещей может длиться более про-
должительный период. Например, в условиях Крыма от-
мечается активность клещей с марта по декабрь
(В. И. Курчатов, 1940). На севере (Е. М. Хейсин и др.,
1954) период активности имаго сокращается с конца
апреля до сентября с максимумом в июне. В Белоруссии
106
(Н. И. Дылько, 1953, 1954; Л. В. Федорчук, 1955;
Б. П. Савицкий, 1957) сезонная кривая активности кле-
щей двухвершинная с максимумом в мае и сентябре.
В. Ф. Гусев (1955) на основании богатого фактического
материала из южных и северных районов БССР указы-
вает, что в северных районах республики максимум
активности клещей наступает на две недели позже по
сравнению с южными, т. е. в июне и сентябре, но может
иметь и одну вершину в июне—июле. В южных районах
имаго появляются в апреле и паразитируют до конца
октября. Пик активности приходится на конец мая и ко-
нец сентября—начало октября, что полностью согласует-
ся с нашими данными.
Сроки паразитирования взрослых клещей. На кроли-
ках продолжительность питания клещей в весеннее
(май) и осеннее (сентябрь) время варьировала у преоб-
ладающей массы особей от 7 до 8 дней. В летнее
(июль) время паразитирование значительно удлинялось
(табл. 36). Интересно, что срок паразитирования также
Таблица 36
Сроки паразитирования самок /. ricinus
Посажено клещей на животных Время паразитиро- вания Сроки паразити- рования (в днях) К-во насосав- шихся % сытых особей Среднемесячная температура воздуха, °C Примечание
14 апрель 7—11 14 100 3,3
74 май 6-8 66 89 10,4
15 июль 9-15 6 40 18,8
3 август 10—11 3 — 18,2
137 сентябрь 5—9 85 62 13,4
34 декабрь 6-18 23 68 18—20 клещи корми-
32 февраль 10-21 8 25 18—20 лись в лабо-
73 март 6—22 54 74 18-20 ратории при комнатной температуре
увеличивается у большинства особей в феврале—марте
и декабре даже при содержании подопытных животных
в лаборатории. Некоторые клещи пребывали на хозяине
до 22 суток (март 1956 г.).
107
В питании клещей на коровах в пастбищный сезон
также наблюдалось некоторое различие. Например, в
мае паразитирование продолжалось 6—7 дней, в сентяб-
ре—7—9, а в июле—8—10 дней. Таким образом, как и у
клещей D. pictus, наблюдается некоторая закономер-
ность в длительности паразитирования. Наиболее корот-
кие и сжатые сроки питания проходят в сезон наиболь-
шей активности клещей в природе. При этом они короче
на крупном рогатом скоте и продолжительнее на кроли-
ках. По Р. Bertkan (1881), Kossel и др. (1903), К. Sam-
son (1909), паразитирование взрослых длится 8—
14 дней. Н. О. Оленев (1927) и Ю. С. Балашов (1956) в
лабораторных условиях наблюдали длительность пита-
ния в 6—7 дней. R. Totze (1933) в лабораторных усло-
виях при питании клещей на искусственной мембране
отметил наиболее короткие сроки питания самок /. rici-
nus—4 дня, объясняя это отсутствием затраты времени
клещами на отыскание места присасывания на хозяине.
Д. И. Благовещенский и Б. И. Померанцев (1930) на
большом материале в естественных условиях делают
заключение о паразитизме основной массы клещей в те-
чение 6—7 дней, хотя были отклонения от 4 до 22 дней.
Нападение на хозяина свежеперелинявших самок мо-
жет происходить на 9-й день. По Wheler, этот срок длил-
ся 10 дней (по Q. Nuttall и С. Warburton, 1911).
В оптимальных температурных условиях имаго спо-
собны паразитировать и в зимнее время. При перенесе-
нии в декабре—феврале из естественных условий в ла-
бораторные клещи из оцепенения быстро приходили в
активное состояние и при посадке на кролика приступа-
ли к питанию. Отсюда можно сделать вывод, что имаго
/. ricinus в зимний период находятся не в состоянии диа-
паузы, а в состоянии холодового оцепенения.
ПРОДОЛЖИТЕЛЬНОСТЬ ЦИКЛА РАЗВИТИЯ
По литературным данным, на Карельском перешейке
(Г. В. Сердюкова, 1952, 1956), в Московской (Л. В. Ба-
бенко, 1956) и Воронежской (П. К. Кузнецов, 1957) об-
ластях развитие /. ricinus продолжается 3—4 года, в Ле-
нинградской области (Н. И. Алфеев, 1947) —от 3 до
6 лет, а в Карельской АССР—от 3 до 7 лет (Е. М. Хей-
син, 1954).
108
Цикл развития в наших опытах в природных усло-
виях прослежен у 18 генераций. Одна генерация
(табл. 37, № 1) с момента отпадения сытой самки дове-
дена до выхода дочернего поколения имаго. Имаго от
генерации № 1 получены на 3-й год. У 16 генераций пол-
ный цикл развития, если считать завершение его с мо-
мента отпадения сытых самок до насыщения дочернего
поколения взрослых (№ 2—17) или от фазы яйца до сле-
дующей фазы яйца (№ 13—17), был закончен также на
3-й год. И только одна генерация (№ 18) развивалась
по 4-годичному циклу.
Суммируя наши данные экспериментальных опытов в
природе, можно весь цикл развития /. ricinus в условиях
Белоруссии представить следующим образом. Самки,
напитавшиеся в весенне-летний период (апрель—июль)
в основном приступают к откладке яиц в этом же году
в мае—июне, а часть—в июле—августе. Самки, питав-
шиеся в летне-осенний период (август—ноябрь), присту-
пают к яйцекладке в том же году осенью (сентябрь—
ноябрь) и частично в апреле—июне следующего года.
Зимующие сытые самки в диапаузу не вступают, а на-
ходятся в состоянии холодового оцепенения. Яйца, отло-
женные в апреле—июле, созревают в том же году, и ли-
чинки в основном вылупляются в августе и частично в
сентябре—октябре. Из яиц, полученных в августе—нояб-
ре, личинки вылупляются частично в сентябре—ноябре
в том же году (из августовских яйцекладок), но боль-
шей частью—в июле—августе на следующий год.
Таким образом, из значительно большей части яиц,
полученных в весенне-летний период, личинки вылуп-
ляются в августе и частично в сентябре—октябре того
же года, а из яиц, полученных в осенний период, они вы-
лупляются также в конце теплого сезона, но уже на бу-
дущий год, т. е. осенняя группа яйцекладок отстает в
своем развитии от весенне-летней на календарный год.
В дальнейшем личинки, вылупившиеся в первый год в
конце теплого сезона, могут прийти в активное состоя-
ние в том же году и напитаться, что вполне возможно,
так как период физиологического доразвития у них за-
канчивается на 24-й, а у некоторых даже на 7-й день. За
счет этих личинок, пришедших в активное состояние, по-
лучается осенний пик сезонной активности, который мы
наблюдаем в природе. Напитавшиеся осенью личинки
109
Таблица 37
Развитие I. ricinus в природе
№ генера- ции Питание имаго Яйце- кладка Вылупление личинок Питание личинок Линька личинок на нимф Питание нимф Линька нимф на имаго Питание имаго Яйцекладка
1 IV.1953 VI.1953 VIII.1953 VI.1954 VIII.1954 V.1955 Х.1955
2 IV. 1954 V.1954 VIII.1954 VI.1955 VIII.1955 X » VIII.1956 Х.1956 —
3 IV » V » VIII » VI » VIII » V.1956 VIII » X » —
4 IV » V » VIII » VI » VIII » Х.1955 VIII » IX » —
5 IV » V » VIII » V » VIII » X » VIII » IX » —
6 IV » V » VIII » VI » VIII » IX » VIII » IX » —
7 IV » V » VIII » VI » VIII » X » IX » X —
8 IV » V » VIII » VI » VIII » V.1956 IX » X —
9 IV » V » VIII » VI » VIII » Х.1955 VIII IX —
10 VI » VII » X » VI » VIII » V.1956 VIII X —
И VI » VII » X » VI » VIII » Х.1955 VIII X —
12 VI » VII » IX » VI » VIII » X » VIII X —
13 IV » V » VIII.1955 VI » VIII » X » VIII IX Х.1956
14 IV » V » VIII.1954 VI » VIII » IX » VIII IX X »
15 IV » VI » VIII » VI » VIII » X » VIII IX X »
16 V » VI » VII » VI » VIII » IX » VIII IX X »
17 IV » VI » VIII » VI » VIII » X » VIII IX X »
18 Х.1953 V VIII » VI » VIII » IX » VIII IX IV. 1957
вступают в зимнюю диапаузу и линяют на нимф на вто-
рой год в конце теплого сезона, а личинки, которые не
успели напитаться осенью того же года (из-за отсутствия
хозяина, неблагоприятных температурных условий или
недоразвития), зимуют в состоянии холодового оцепене-
ния и имеют возможность напитаться только с наступле-
нием теплого периода следующего года. За счет перези-
мовавших голодных личинок наблюдается весенний пик
активности в природе. Напитавшиеся в весенне-летний
период личинки не вступают в диапаузу, а линяют на
нимф в конце теплого сезона одновременно с перезимо-
вавшими сытыми особями.
Таким образом, как перезимовавшие сытые, так и пе-
резимовавшие голодные и напитавшиеся в весенне-лет-
ний период личинки линяют на нимф одновременно на
второй год в конце теплого сезона (август—сентябрь)..
Из второй группы личинок, которые вылупились от осен-
них кладок в конце теплого сезона на второй год, нимфы
появятся уже на третий год. Затем нимфы первой груп-
пы, т. е. особи, которые появились в конце теплого сезо-
на на второй год развития генерации, как и личинки,
могут напитаться в год своего появления, так как часть
нимф заканчивает физиологическое доразвитие уже на
21-й день. Напитавшиеся осенью нимфы впадают в зим-
нюю диапаузу и линяют на имаго в конце теплого сезо-
на на следующий год, т. е. на третий год развития гене-
рации (рис. 13). За счет пришедших осенью в активное
1-и год 2-и год в 3-й год
Зинойка Зимоока Зимоока
Рис. 13. Схема развития Ixodes ricinus L. по трехгодичному
циклу:
/г—голодные имаго; 1с—сытые самки; Ov—яйца; Lг—голодные личинки;
Lc— сытые личинки; Ns—голодные нимфы; Nc—сытые нимфы
состояние особей наблюдается в природе осенний
подъем активности нимф. Особи, не успевшие напитать-
ся осенью, зимуют голодными в состоянии холодового
оцепенения, питаясь после зимовки в весенне-летний пе-
111
риод (за счет этих особей наблюдается весенний пик
активности в природе), и линяют на имаго так же, как и
зимовавшие сытые, одновременно с последними в конце
теплого периода на третий год. Соответственно нимфы
второй группы генераций перелиняют на имаго на чет-
вертый год (рис. 14). Появившиеся имаго могут частич-
но напитаться в том же году, так как некоторые особи
способны нападать на хозяина уже на 9—10-й день. За
f-й год 2-й год 3-й год 4-й год
ЗимоЬка Зинобка Зинобко ЗимоЬса
Qv
Рис. 14. Схема развития Ixodes ricinus L. по четырехгодичному
циклу:
/г-голодные имаго; /с—сытые самки; Ои—яйца; La—голодные личинки;
Lr—сытые личинки; Na—голодные нимфы; Nc—сытые нимфы
счет этих особей наблюдается осенний пик активности
взрослых клещей. Напитавшиеся самки вскоре после
линьки могут успеть отложить яйца в сезон их появле-
ния до зимовки, закончив свой цикл развития на третий
год. Голодные взрослые, перезимовавшие в состоянии
холодового оцепенения, питаются и откладывают яйца
после зимовки, закончив свой цикл развития на четвер-
тый год. За счет перезимовавших голодных особей на-
блюдается весенний пик активности. Соответственно
имаго второй группы могут напитаться и отложить яйца
осенью на четвертый или весной на пятый календар-
ный год.
Следовательно, для завершения метаморфоза каждой
активной фазы /. ricinus требуется один год или один
теплый сезон. Вылупление личинок, линька их на нимф
и выход имаго происходят в определенный период, а
именно в конце теплого сезона года. Яйцекладка может
произойти в весенне-летний период, и личинки могут вы-
лупиться в тот же сезон; тогда развитие пойдет по 3-го-
дичному циклу. При откладке яиц в конце лета или
осенью личинки вылупляются уже на следующий год и
развитие пойдет по 4-годичному циклу.
СИСТЕМА ПРОТИВОКЛЕЩЕВЫХ МЕРОПРИЯТИЙ
Для проведения борьбы с тем или иным паразитом в
определенных условиях местности должна быть осу-
ществлена система наблюдений и опытов, дающих ключ
к построению планов профилактических мероприятий.
Е. Н. Павловский (1932, 1935) и Д. И. Благовещенский
(1957) выдвинули основные задачи практического изу-
чения иксодовых клещей, решение которых является био-
логическим обоснованием борьбы с этими паразитами.
Задачи эти заключаются: 1) в выявлении фауны клещей,
так как рациональное исследование должно быть осно-
вано на точной диагностике видов; 2) в определении
стадиального распределения, т. е. приуроченности тех
или иных видов паразитов к определенным растительным
группировкам; 3) в изучении круговорота клещей в при-
роде, что сводится к выяснению круга хозяев и на осно-
вании экстенсивности и интенсивности их заклещевения
к выявлению среди них доминирующих хозяев, в отно-
шении которых должны быть организованы противокле-
щевые мероприятия; 4) в изучении сезонности заклеще-
вения животных, что в свою очередь служит показате-
лем сезона практических мероприятий; 5) в определении
продолжительности цикла развития клещей как показа-
теля срока исчерпаемости их запаса; 6) в изучении про-
должительности питания клещей для определения часто-
ты обработки скота; 7) в определении концентрации
апробированных химических противоклещевых средств
особо для каждой типичной местности.
Данные, полученные нами в результате изучения био-
логии Ixodes ricinus и Dermacentor pictus — наиболее
8. Иксодовые клещи
из
массовых видов и переносчиков различных заболеваний
человека и домашних животных, дают возможность
предложить систему противоклещевых мероприятий. При
изучении стадиального распределения клещей нами от-
мечена сильная зараженность /. ricinus черноольшани-
ков, хвойно-широколиственных и грабовых лесов. Не-
сколько менее заражены ельники, смешанные молодня-
ки, сосново-еловые леса, осинники, кустарники у во-
доемов. Слабо заражены сосняки, вырубки, березняки;
незараженными можно считать открытые целинные
земли. D. pictus широко распространен в пойменных лу-
гах в кустарниковых биотопах. Встречается и на целин-
ных участках, заросших бурьяном и кустарником. Оба
вида не встречаются на открытых и чистых от кустарни-
ков заливных лугах, на лесных полянах и культурных
полях.
Учитывая неравномерное распространение клещей по
различным стациям, самыми эффективными мероприя-
тиями против клещевых очагов нужно считать агротех-
нические—уничтожение кустарников на естественных лу-
гах и целинных участках, а еще лучше создание на месте
естественных лугов культурных пастбищ. Осуществле-
ние этих мероприятий создаст неблагоприятные экологи-
ческие условия как для мелких зверьков-хозяев преима-
гинальных фаз, так и для самих клещей.
Как указывает А. В. Федюшин (1949), накопление
D. pictus на многолетних сеяных пастбищах хотя и воз-
можно, но даже в конце срока хозяйственного использо-
вания таких пастбищ, т. е. на 4—5-й год, запас клещей
на них в 10—30 раз меньше, чем на естественных. На
однолетних же травяных полях после осенней вспашки
клещи погибают полностью в результате глубокого за-
сыпания землей и вымерзания зимой. Для поддержания
искусственных пастбищ в свободном от клещей состоя-
нии и в последующие годы необходимо по возможности
изолировать их от непосредственного соседства с лесны-
ми и целинными участками, особенно от заклещевелых
старых выгонов, полосами зерновых, пропашных или
технических культур; не допускать прогон скота к паст-
бищу по заклещевленным стациям.
Наглядные и убедительные примеры нерентабельно-
сти использования естественных пастбищ приводят
И. С. Авессаломов и др. (1957). От недостатка кормов и
114
изнуряющих переходов при пастьбе в кустарниковых за-
рослях и лесах сильно понижается сопротивляемость ор-
ганизма животных к заболеваниям. Нападение же кро-
вососущих паразитов, многочисленных на естественных
пастбищах, приводит к частым заболеваниям животных.
По данным этих авторов, заболеваемость бабезиеллозом
скота, выпасающегося на культурных пастбищах, отсут-
ствовала. На кустарниковых пастбищах заболеваемость
составляла 12—15%. Ушибов и ранений скота на куль-
турных пастбищах в 5—7 раз меньше. Вследствие ука-
занных причин молочная продуктивность и привес жи-
вотных на культурных пастбищах в 3 раза больше.
Таким образом, для ограничения развития клещевых
очагов и предупреждения домашних животных от зара-
жения следует по возможности свести к минимуму пасть-
бу скота в стациях с древесной и кустарниковой расти-
тельностью, особенно в ольшаниках, хвойно-широколи-
ственных и грабовых лесах, заселенных /. ricinus, и лу-
гах с кустарниковыми зарослями, заселенных D. pictus.
Следует отметить неравномерное распределение /. ri-
cinus в различных дубовых ассоциациях, обнаруженное
в Крымском заповеднике. Клещей много в дубняке с
подростом липы и травяным покровом из злаков и осок.
Очень мало или отсутствуют они в дубовых ассоциациях
с травяным покровом из пиретрума, чабреца и шалфея.
Нет клещей и на плантациях лаванды. Опыты показали,
что в пробирках со свежесрезанными листьями липы,
осоки и злаков клещи чувствовали себя нормально, в
пробирках с лавандой они полностью погибали через
4—5 часов, а в пробирках с чабрецом, шалфеем, пи-
ретрумом через 9—12 часов погибали на 60 — 90%
(И. М. Распопов, 1953). Следовательно, имеется возмож-
ность оздоровить дубравы и, очевидно, другие раститель-
ные ассоциации путем искусственного подсева отмечен-
ных трав. В литературе указывается на активный метод
борьбы с клещами в природных условиях путем выжига-
ния травы (А. И. Шпрингольц-Шмидт, 1936; Н. И. Аг-
ринский, 1941; С. П. Пионтковская, 1947). Однако такое
мероприятие, по нашему мнению, непрактично, так как
может привести к лесным пожарам и прочим нежела-
тельным последствиям.
Ряд исследователей рекомендует уничтожение кле-
щей непосредственно на местности. Например, для унич-
115
тожения I. persulcatus предлагается обработать мест-
ность ранней весной после схода снега и поздней осенью
под снег препаратами гексахлорана или ДДТ. Система-
тическая обработка в течение 2—3 лет приводит к пол-
ному уничтожению клещей.
Обработка может производиться моторным опылите-
лем «Серна», разбрасыванием акарицидов совками и
опрыскиванием из аппарата ДРП 13%-ной эмульсией
гексахлорана с дозировкой 0,3—0,5 г технического пре-
парата на 1 м2. Но производительность аппарата ДРП
мала (менее 1 га на человеко-день), а при отсутствии есте-
ственных водоемов на обрабатываемой территории со-
здается неудобство с доставкой воды для приготовления
эмульсии. Опылителем «Серна» при наличии лошади с
телегой и двух рабочих удается обработать 15—15,5 га
в день. Дозировка: 0,3 г технического гексахлорана на
1 м2. Недостаток аппарата—малая проходимость в лесу.
Ручная обработка устраняет этот недостаток, но при ней
очень трудно равномерно распределить ядоматериал по
поверхности. В результате получается большой расход
акарицида (не менее 0,5 г технического препарата на
1 м2) и небольшая производительность—2 —2,5 га в день
(С. П. Карпов и др., 1956).
В целях охраны людей от /. persulcatus на ограничен-
ных участках леса предлагается однократная за сезон
обработка лесной подстилки и нижнего яруса леса на
уровне 1 м высоты дустом, эмульсиями и суспензиями
ДДТ и ГХЦГ при нормах расхода 0,5—0,6 г техническо-
го вещества на 1 м2, что практически дает полное истреб-
ление половозрелых и значительное уменьшение преима-
гинальных фаз. При этом нападение клещей на человека
прекращалось на 2-й день после обработки (Н. Н. Гор-
чаковская, 1955; Ю. И. Гадалин и др., 1956).
Положительные результаты по уничтожению личинок
D. marginatus в природных условиях получены при опры-
скивании пастбищ водной суспензией ГХЦГ из расчета
200 мл жидкости на 1 м2 в 1%-ной концентрации, т. е.
10 кг 10%-ного дуста на 1 га (Е. И. Покровская, 1956).
Способы уничтожения клещей непосредственно в приро-
де при помощи акарицидов имеют большую перспекти-
ву, особенно при обработке небольших площадей на ме-
сте лесоразработок, различного рода лагерей и т. д. Но
116
в широких масштабах применение этого метода затруд-
нительно из-за несовершенства аппаратуры.
Применение всех этих химических способов обработ-
ки местности очень заманчиво, однако необходимо учи-
тывать, что применение различного рода акарицидов
действует не только на клещей, но и на все организмы в
данном биоценозе. Поэтому применять акарициды на
местности нужно только в исключительных случаях или
на ограниченных участках.
Поскольку основная масса половозрелых /. ricinus и
D. pictus выкармливается на крупном рогатом скоте, то
тщательное уничтожение клещей на животных значи-
тельно понизит численность паразитов. При незначитель-
ном заражении животных можно рекомендовать ручной
сбор клещей. Когда же заклещевение достигает значи-
тельных размеров, необходимо обрабатывать скот хими-
ческими средствами, истребляющими клещей. Например,
значительное снижение зараженности дает обработка
животных 10%-ным дустом ДДТ заводского приготов-
ления. Препарат втирается в шерсть животного. Норма
расхода на одно крупное животное 50—70 г дуста ДДТ
(С. П. Карпов и др., 1956). Хорошие результаты полу-
чены при обработке скота 5- и 2,5%-ными растворами
ДДТ в кондиционном соляровом масле и водными сус-
пензиями 10%-ного дуста ГХЦГ. Незначительное сни-
жение заклещевения при обработке скота водными рас-
творами объясняется быстрым высыханием шерсти, в
связи с чем дуст осыпается. Нормы расхода на одно жи-
вотное: масляных растворов 100—150 мл, водной суспен-
зии 0,5—1,5 л (в зависимости от возраста). Обработка
ведется вручную (Е. И. Покровская, 1956).
Значительных результатов можно достичь путем ку-
пания скота в ваннах с раствором мышьяковистокислого
натра (Н. О. Оленев, 1924; В. Л. Якимов, 1932). В Да-
гестанской АССР успешно применены мышьяковистые
ванны повышенной концентрации — до 0,21—0,23%
(Ю. Г. Егоров, 1955). В Западной Германии с успехом
применяется токсафен в виде 0,5—0,75%-ной эмульсии,
которой опрыскивают или купают в ней животных. Токса-
фен очень стоек, и действие его мало зависит от погоды.
Применяется в виде эмульсии, суспензии и порошка.
Норма расхода от 1 до 2 л 1%-ной эмульсии (0,5% дей-
ствующего начала) на животное. Даже особо устойчи-
117
вне клещи погибли на животных через 2 недели после
обработки. Для уничтожения более чувствительных экто-
паразитов достаточно одного опрыскивания в 3 недели
(I. Crittner и F. Caselitz, 1953).
В Белоруссии против /. ricinus и D. pictus предложен
ряд акарицидных препаратов контактного действия: СК-9
в виде 3- и 5%-ных водных эмульсий; 10°/о-ного дуста
пентахлорина; 12%-ного дуста гексахлорана; 2,5—5,0—
7,5- и 1О°/о-ных водных эмульсий из 20-и 25%-ного кон-
центрата гексахлорана; 3- и 5%-ных линиментов пента-
хлорина или гексахлорана на кондиционном соляровом
масле, предложенных М. Г. Хатиным; 30%-ного раство-
римого пентахлорина в порошке в виде водной эмульсии
с содержанием 0,5—1% чистого препарата; 30%-ного
минерально-масляного концентрата (препараты № 93 и
94) в виде водной эмульсии с содержанием 0,5—1,0—1,5%
чистого препарата; 40%-ного концентрата гексахлорана
(препарат № 100 )в виде водной эмульсии с содержа-
нием 0,5—4,0—1,5% чистого препарата; 30%-ного кон-
центрата пентахлорина и гексахлорана в равных соотно-
шениях (препарат № 99) в виде водной эмульсии с со-
держанием 0,5—1,0% чистого препарата, а также водно-
го раствора мышьяковистокислого натрия с содержанием
0,18—0,22 % мышьяковистого ангидрида; 2,5—5,0 % -ных
водных эмульсий концентрата, состоящего из 1 части
пентахлорина, 1 части гексахлорана и 8 частей ихтиоло-
вой пасты; 2,5%-ной водной эмульсии концентрата, со-
стоящего из 2 частей пентахлорина и 8 частей ихтиоло-
вой пасты; 5,0%-ной водной эмульсии концентрата, со-
стоящего из 30,0% пентахлорина и гексахлорана в рав-
ных соотношениях (препарат № 109); 2,5%-ной водной
эмульсии концентрата, состоящего из 1 части пентахло-
рина и 9 частей ихтиоловой пасты (В. Ф. Гусев, 1954).
Период активности — наиболее важный момент для
массового уничтожения клещей. Учитывая изменение се-
зонной активности взрослых /. ricinus и D. pictus, обра-
ботку животных ядохимикатами следует проводить в ве-
сенний (с начала апреля до начала июня) и осенний
(с начала сентября до середины ноября) периоды. Но
поскольку взрослые /. ricinus, хотя и в незначительном
количестве, нападают на скот и в летний период, то в
зависимости от заклещевения обработка животных мо-
жет быть рекомендована и в летние (июнь—август) ме-
118
сяцы. Особое внимание обработке следует уделять в пе-
риоды максимальной зараженности скота. Наиболее от-
ветственным моментом в профилактике и уничтожении
обоих видов клещей является май. Обработку скота в
соответствии с длительностью паразитирования имаго
следует проводить с интервалами: против /. ricinus че-
рез 6—7 дней весной, 7—9 дней осенью, 8—10 дней ле-
том и против D. pictus через 6—7 дней весной и осенью.
По литературным данным, наиболее удобным для об-
работки пораженного клещами скота следует считать ве-
чер (при дневном выпасе), так как большая часть насо-
савшихся клещей отпадает в дневное время на пастби-
ще. При вечерней обработке почти все клещи (96,1%)
подвергнутся действию яда на протяжении всей ночи,
что уже гарантирует значительную гибель паразитов
(Б. И. Померанцев и Н. И. Алфеев, 1935).
Дикие позвоночные—олень, лисица, заяц, белка, еж,
мышевидные грызуны также играют значительную роль
в развитии клещей, особенно преимагинальных фаз. По-
этому для ликвидации клещевых очагов нужно уничто-
жить диких животных—главных хозяев клещей, особен-
но в местах частого пребывания скота. Существенными
мероприятиями в этом отношении являются стимулиро-
вание заготовок пушнины таких видов, как лисица, заяц,
белка, истребление ежей и мышевидных грызунов, охра-
на полезных хищных птиц (канюк, малый подорлик, по-
левой лунь, пустельга, сова) — истребителей мышевид-
ных грызунов. Уничтожение грызунов в скирдах и сто-
гах—«стациях переживания» их в осенне-зимне-весенний
период имеет весьма существенное значение. По литера-
турным данным, плотность мышей в таких стациях ино-
гда достигает 70 экз. на 1 м3 соломы (Н. Г. Милютин,
1956). Эффективны при отлове грызунов приманки. В ча-
стности, хорошие результаты получены от приманок с
сахаром, приманок из запаренного и осоложенного теста
и пророщенных зерен (Л. В. Павленко, 1956). Еще более
действенным методом борьбы с грызунами является опы-
ливание нор грызунов порошковидными ядами, которые
применяются в виде смеси, приготовленной на крысиде
и фосфиде цинка с наполнителями из дорожной пыли и
талька. Смесь готовится из расчета 1 г крысида или
0,5—1 г фосфида цинка с 1—2 г наполнителя (М. И. Со-
ветова, 1956).
119
В литературе указывается, что одним из эффектив-
ных методов борьбы с клещами может быть смена паст-
бищ или исключение их из пользования на длительное
время (Н. О. Оленев, 1924; В. Л. Якимов, 1932; Е. И. По-
кровская, 1956). По нашему мнению, такой метод не со-
всем оправдывает себя, так как цикл развития /. ricinus
продолжается 3—4 года, a D. pictus—1 год и плюс к это-
му продолжительность жизни половозрелых клещей это-
го вида в природе до 2—3 лет и более. Следовательно,
изоляция пастбищ, зараженных как /. ricinus, так и
D. pictus, должна длиться минимум 3—4 года, что эко-
номически невыгодно. К тому же не исключена возмож-
ность заноса преимагинальных фаз клещей на изолиро-
ванные участки дикими позвоночными в связи с их
большой подвижностью и миграционной активностью
(В. Б. Дубинин и 3. М. Жмаева, 1947; В. Б. Дубинин,
1948; Д. И. Благовещенский, 1948). В результате изоля-
ция пастбищ может не дать ожидаемых результатов.
Таким образом, в конкретных условиях местности
клещевые очаги /. ricinus и D. pictus можно уничтожить
не отдельными мероприятиями, а применением комплек-
са противоклещевых мероприятий. Это — уничтожение
кустарников на естественных лугах и целинных участ-
ках, ограничение пастьбы скота на сильно зараженных
I. ricinus лесных угодьях, непосредственное уничтожение
клещей на домашних животных в периоды наибольшей
зараженности ими, уничтожение клещей вместе с дики-
ми животными — доминирующими хозяевами клещей.
Эти мероприятия должны проводиться систематически
на протяжении 3—4 лет с учетом цикла развития и дли-
тельности голодания иксодовых клещей.
ЛИТЕРАТУРА
Авессаломов И. С., Свирская С. А. и Мез др ин М. Д
Культурные пастбища как мера профилактики заболеваний сельскохо-
зяйственных животных. Бюллетень научно-технич. информации Ленинград-
ского научно-исслед. ветеринарного ин-та, вып. 3, 1957.
Агринский Н. И. Методы борьбы с клешами-переносчиками нут -
таллиоза и пироплазмоза лошадей в Средней Азии. Тезисы докладов
третьего совещания по паразитологическим проблемам, 1941.
Алфеев Н. И. О распространении клеща /. ricinus L. в районе
Черемецкого озера и наблюдения над его биологией и экологией. Вре-
дители животноводства, Изд. АН СССР, 1935.
Алфеев Н. И. Сравнительно-экологические особенности клещей
/. ricinus L. и D, marginatus Su\z. Зоологический журнал, т. XVIII, вып.
1, 1939.
А л ф е е в Н. И. Биология и экология D. marginatus Sulz. в усло-
виях Орловской области. Тр. Ленинградской пироплазмозной станции,
вып. 1, 1939.
Алфеев Н. И. К вопросу о сезонной активности D. marginatus
Sulz. в условиях Орловской области и о различных этапах в развитии
этого явления. Тр. Ленинградской пироплазмозной станции, вып. 1, 1939.
Алфеев Н. И. К вопросу о значении изоляции и смены пастбиш
в борьбе с клещом D. marginatus Sulz. Тр. Ленинградской пироплаз-
мозной станции, вып. 1, 1939.
Алфеев Н. И. О коррелятивной связи длительного голодания
взрослых D. pictus Негт. в природе с сезонной активностью их. Зооло-
гический журнал, т. XXVI, вып. 3, 1947.
Алфеев Н. И. О диапаузе у иксодовых клещей. Тр. Военно-
медицинской акад. им. С. М. Кирова, т. XLIV, 1948.
Алфеев Н. И. Затяжной цикл развития / ricinus L. в природ-
ных условиях Ленинградской области. «Ветеринария» № 7, 1947.
Алфеев Н. И. и Кулагин С. М. К изучению клещей D. margi-
natus Sulz. и D. pictus Негт. как переносчиков кле евого риккетсио-
за. Вопросы краевой, об ей и экспериментальной паразитологии и ме-
дицинской зоологии, т. VIII, 1953.
121
Арзамасов И. Т. Новые для БССР виды иксодовых клещей
«Изв. АН БССР» № 6, 1955.
Арзамасов И. Т. Развитие /. ricinus L. в условиях Белоруссии.
Рефераты научно-исслед. работ Ин-та биологии АН БССР, 1956.
Арзамасов И. Т. Стациальное распределение и круг хозяев
клеща /. ricinus L. в условиях Белоруссии. Тезисы докладов научно-
практич. конференции по заболеваниям с природной очаговостью. Бел.
ин-т эпидемиологии, микробиологии и гигиены, 1957.
Арзамасау I. Т. В1давы састау i распаусюджанне 1ксодавых
кляшчоу у Беларуси «Becni АН БССР» № 1, серыя б!ялапчных
навук, 1957.
Арзамасов И.Т. Иксодовые клещи БССР. Автореферат. Ин-т
биологии АН БССР, 1957.
Арзамасов И. Т. Круг хозяев иксодовых клещей. Бюллетень
научно-исслед. работ Ин-та биологии АН БССР, 1957.
Арзамасов И. Т. Взаимосвязь депрессии численности мышевид-
ных грызунов с интенсивностью заражения их личинками /. ricinus L.
Бюллетень научно-исслед. работ Ин-та биологии АН БССР, 1957.
Арзамасов И. Т. Клещи семейства Ixodidae в БССР. Тезисы
докладов первой зоологической конференции Белорусской ССР, 1958.
Арзамасов И. Т. К фауне иксодовых клешей БССР. Тезисы
докладов десятого совещания по паразитологическим проблемам и при-
родноочаговым болезням, вып. 2, 1959.
Бабенко Л. В. К вопросу о сезонных явлениях в жизни кле-
щей I. ricinus L. и /. persulcatus Sch. «Медицинская паразитология и
паразитарные болезни» № 4, 1956.
Балашов Ю. С. Особенности суточного ритма отпадения насо-
савшихся самок /. persulcatus Sch. с коров. ДАН СССР, т. XCVI1I,
№ 2, 1954.
Балашов Ю. С. Изменение веса скотского клеша /. ricinus L.
при кровососании. Зоологический журнал, т. XXXV, вып. 1, 1956.
Балашов Ю. С. Питание и ход сперматогенеза у самцов иксо-
довых кле ей. ДАН СССР, т. ПО, № 6, 1956.
Балашов Ю. С. Гонотрофические отношения у иксодовых кле-
шей. Тезисы докладов девятого совещания по паразитологическим про-
блемам, Изд. АН СССР, 1957.
Балашов Ю. С. Особенности питания иксодовых клешей. Тезисы
докладов девятого совещания по паразитологическим проблемам, Изд.
АН СССР, 1957.
Барцевич Н. И. Пироплазмоз крупного рогатого скота в Бе-
лоруссии и возможность борьбы с ним в белорусских условиях. ВС В,
№ 20, 1927.
Берг Л. С. Ландшафтные зоны Союза. Животный мир СССР, Изд.
АН СССР, т. I, 1937.
Благовещенский Д. И. и Померанцев Б. И. К биологии
клеща /. ricinus L.—переносчика пироплазмоза крупного рогатого скота.
«Практическая ветеринария» № 8—9, 1930.
Благовещенский Д. И. Биологические обоснования борьбы
с иксодовыми клещами. Энтомологическое обозрение, т. XXXVI, вып.
1. 1957.
122
Благовещенский Д. И. Клещи сем. Ixodidae и кровососущие
насекомые низовьев Амура. Паразитологический сборник Зоологии, ин-та
АН СССР, IX, 1948.
Бурцев В. И. Некоторые данные о пироплазмозе в БССР. «Бе-
лорусская ветеринария» № 3—4, 1925.
Вапник Е. Е. иСенчук Т. Т. Значение кле’па I. ricinus L.
в поддержании инфекции в туляремийном очаге пойменно-болотного ти-
па. Тезисы докладов научно-практич. конф, по заболеваниям с природ-
ной очаговостью. Бел. ин-т эпидемиологии, микробиологии и гигиены, 1956.
Вапник Е. Е., Савицкий Б. П. и Модель X. М. Распро-
странение иксодовых кле ”ей и заболевания с природной очаговостью
на территории БССР. Тезисы докладов научно-практич. конф, пс заболе-
ваниям с природной очаговостью. Бел. ин-т эпидемиологии, микро-
биологии и гигиены, 1957.
Вильбасте А. К. Иксодовые клещи Эстонской ССР. Авторефе-
рат, 1956.
Вотяков В. И. Некоторые итоги и задачи в изучении сезонных
вирусных менингоэнцефалитов в БССР. Тезисы докладов научно-практич.
конф, по заболеваниям с природной очаговостью. Бел. ин-т эпидемиоло-
гии, микробиологии и гигиены, 1957.
Высоцкая С. О. О биологии иксодового кле ца I. trianguliceps
Bir. Паразитологический сборник Зоологич. ин-та АН СССР, т.
XIII, 1951.
Высоцкая С. О. Методы сбора обитателей гнезд грызунов, Изд.
АН СССР, 1953.
ГадалинЮ. И., Гершкович Н. Л.,ГорчаковскаяН. Н.
и Левит А. Б. Опыт истребления переносчика кле евого энцефалита
/. persulcatus Sch. в природных условиях. Бюллетень Московского об-ва
испытателей природы, отд. биологии, т. LXI, вып. 3, 1956.
Галузо И. Г Кровососущие кле и Казахстана, Изд. АН
КазССР, т. 1, 1946; т. II, 1948; т. III, 1950.
Горчаковская Н. Н. Испытание метода прямого истребления
клеи ей /. persulcatus Sch. в природе. Природная очаговость болезней
человека и краевая эпидемиология, 1955.
Г о с м е р К- П. О заболеваниях вирусным менингоэнцефалитом в
Брестской области. Тезисы докладов научно-практич. конф, по заболе-
ваниям с природной очаговостью Бел. ин-т эпидемиологии, микроби-
ологии и гигиены, 1957.
Гусев В. Ф. и Сузько С. Ф. Влияние затопления на жизнен-
ный цикл /. ricinus L. и D. pictus Негт. Сб. научных трудов Ленин-
градского ин-та усовершенствования ветврачей, 1950.
Гусев В. Ф. Разработка мер борьбы с гемоспоридиозами сельско-
хозяйственных животных. Отчет Ин-та животноводства АН БССР, 1952.
Г у с е в В Ф Новые методы борьбы с кле ами / ricinus L. и
D. pictus Негт. в условиях БССР. Тезисы докладов на 1 Всес. конф,
по проблемам ветеринарной дерматологии, арахнологии и энтомоло-
гии, 1954.
Гусев В. Ф. Фенология и круг хозяев кле ей I ricinus L. и
D. pictus Негт. Сб. научных трудов Ленинградского ин-та усовершен-
ствования ветврачей, вып. X, 1955.
123
Джапаридзе Н. И. Описание молодых стадий клещей Ixodidae
Сб. АН ГрузССР, т. II, № 3, 1950.
Дубинин В. Б. иЖмаеваЗ.М. Кормовая миграция маньчжур-
ской белки и ее значение в расселении иксодовых клещей. Новости
медицины, вып. 5, 1947.
Дубинин В. Б. Значение миграций животных в распростране•
нии заболеваний. «Изв. АН Казахской ССР», серия паразитолог.,
вып. 5, № 43, 1948.
ДылькоН И. Материалы по бабезиеллозу крупного рогатого
скота (Горынского), распространенного вдоль нижнего течения р. Горы-
ни. Автореферат. Ленинградский ветеринарный ин-т, 1953.
ДылькоН И. Видовой состав и распространение иксодовых
клешей и гемоспоридий крупного рогатого скота по нижнему течению
р. Горыни. «Изв. АН БССР» № 2, 1954.
Д ы л ь к о Н. И. Микростации кле цей Ixodes ricinus L. и Dermacen-
tor pictus Негт. в Полесской низменности. Научные труды Белорус-
ского ин-та животноводства, т. I, 1958.
Егоров Ю. Г. Противоклещевые обработки мелкого рогатого
скота растворами мышьяка с повышенной концентрацией. Сб. научных
трудов Ленинградского ин-та усовершенствования ветврачей, вып.
X, 1955.
Ельманов Н. К- К биологии /. ricinus L. «Практическая ве-
теринария» № 5—6, 1930.
Емчук Е. М. Материалы к фауне иксодовых клещей УССР,
Изд. АН УССР, 1954.
За ков и4*! Ф. А. Об изучении иксодовых клецей на территории
госзаповедника «Беловежская пу.ла». Тезисы докладов научно-практич.
конф по заболеваниям с природной очаговостью. Бел. ин-т эпидемиоло-
гии. микробиологии и гигиены, 1957.
ЗасухинД Н. Материалы к познанию пироплазмозов диких
млекопитаю их на юго-востоке. Тр. Саратовского научно исслед. ве-
тевинарн. ин-та, т. III, 1936.
ЗасухинД. Н., Л о н зингер Г. К- иОкрокверцхо-
ва Л. А. Материалы к изучению кле лей I. ricinus L. на юго-востоке.
Тр. научно-исслед. ветеринарн. ин-та, т. III, 1936.
Захар ченков Л. В. Роль диких позвоночных в прокормле-
нии различных фаз развития /. ricinus L. и /. persulcatus Sch. в
К.-Ф.ССР. Сб. научных работ студентов КФГУ, № 2, 1954.
Зильбер Л. А. Эпидемические энцефалиты. Медгиз, 1945.
Золотарев Н. А. Значение диких млекопитающих в развитии
кле ей надсем Ixodoidea. Тр. 1 научной сессии Дагестанской научно-
йсслед базы АН СССР, 1947.
КарповС. П., Попов В М., Шипова А. А., Я в ь я А. Р.
и Лаптева К. Я. Организация мероприятий по оздоровлению очага
кле евого энцефалита и их эффективность. Тр. Томского научно-ис-
след. ин-та вакцин и сывороток Министерства здравоохранения СССР,
т. VIII, 1956.
Карцев Г. Беловежская пуша. СПб , 1903.
КайгородовА И. Краткий очерк сельскохозяйственных кли-
матических условий. Белгосиздат, 1928.
124
Кайгородов А. И. Климат БССР, Западной Белоруссии и
соседних стран. Минск, т. II, 1934.
Костюкевич Н. И. К порайонной характеристике лесорастительных
условий БССР. Леса БССР, Изд. АН БССР, 1954.
Кузнецов П. К. К вопросу о перезимовке клещей /. ricinus L.
в природных условиях Воронежской области. Зоологический журнал,
т. 34, № 2, 1955.
Кузнецов П. К. Некоторые данные о развитии клеща /. ricinus
L. по наблюдениям в природных условиях Воронежской области. Тези-
сы докладов девятого совещания по паразитологическим проблемам,
Изд. АН СССР, 1957.
Курчатов В. И. Обзор фауны кровососущих клещей сем. Ixodi-
dae в Крыму. «Сов. ветеринария» № 1, 1940.
Лутта А. С., X е й с и н Е. М. и Шульма н-А л ь б о в а Р. Е.
Распространение и экология /. ricinus L. и I. persulcatus Sch. в
К-ФССР Уч. зап. КФГУ, т. V, вып. 3, 1953.
Лутта А С и Шульма н-А льбова Р. Е. К распростра-
нению и экологии /. trianguliceps Bi г. в К.-Ф.ССР. Тр. К.-Ф. филиала
АН СССР, вып. 4, 1956.
Лутта А. С. Биологическое обоснование мер борьбы с гнусом
и иксодовыми клещами в Карельской АССР. Тезисы докладов девято-
го совещания по паразитологическим проблемам, Изд. АН СССР, 1957.
Марков М. П. Клещи—переносчики пироплазмоза на Украине,
их распространение и биология. Тр. II съезда научн. и практич. ве-
теринарн. работников Украины, 1927.
Марков М. П. Клетевая нейроинфекция (энцефаломиелит) в
Белоруссии. «Советская психоневрология» № 3, 1941.
Милютин Н. Г. Размножение мышевидных грызунов в скирдах
лесостепной зоны УССР и его значение в эпизоотологии и эпидеми-
ологии туляремии. Проблемы паразитологии. Тр. II научной конфе-
ренции паразитологов УССР, Изд. АН УССР, 1956.
Модель X. М., За кович Ф. А. и Поплевко И. А. Изу-
чение видового состава иксодовых клещей в некоторых очагах сезон-
ного менингоэнцефалита в БССР. Тезисы докладов научно-практич.
конф. Бел. ин-та эпидемиологии, микробиологии и гигиены, 1956.
Москачева Е. А. К вопросу о возможности зимовки голодных
личинок и нимф клецей D marginatus Sulz.—переносчика пироплаз-
моза лошадей. Тр. Бел. с.-х. ин-та, т. XIII, вып. 2, 1948.
Найденова Г. А. О клещах сем. Ixodidae в Белоруссии. Тезисы
докладов на третьем совещании по паразитологическим проблемам
АН СССР, 1941.
Нецкий П. И. и Ольшевская Н. А. Об осеннем торможе-
нии созревания и развития яиц клеща D. pictus Негт. Медицинская
паразитология, т. XIX, вып. 6, 1950.
Олсуфьев Н. Г. К методике лабораторного разведения иксодо-
вых клещей. Медицинская паразитология, т. X, вып. 3—4, 1941.
Олсуфьев Н. Г. Новое в изучении экологии и патогенной
роли D. pictus Негт. Тезисы докладов третьего совещания по пара-
зитологическим проблемам, 1941.
125
Олсуфьев Н. Е Заметка о животных-хозяевах молодых стадий
клещей D. pictus Негт. в условиях Западной Сибири. Зоологический
журнал, т. XXVI, вып. 3, 1947.
Олсуфьев Н. Г. О ландшафтных типах туляремийных очагов
средней полосы РСФСР. Зоологический журнал, т. XXVI, вып.З, 1947.
Олсуфьев Н. Г. К экологии лугового клеща D. pictus Негт.,
о происхождении его очагов и путях их ликвидации в средней поло-
се Европейской части РСФСР. Вопросы краевой общей и экспери-
ментальной паразитологии и медицинской зоологии, т. VIII, 1953.
Оленев Н. О. К биологии скотского клеща I. ricinus L. в ус-
ловиях Новгородской губернии. «Защита растений» № 1—2, 1924.
Оленев Н. О. К биологии /. ricinus L. в Новгородской области.
Зашита растений, т. IV, вып. 2, 1927.
Оленев Н. О. К систематике и географическому распростране-
нию клешей Ixodoidea. ДАН СССР, 1927.
Осадчий П. и Баврин В. Беловежская пуща. Минск, 1950.
Павленко Л. В. О вкусовых и обонятельных реакциях некото.
рых видов грызунов на пишевые приманки. Проблемы паразитологии-
Тр. II научн. конф, паразитологов УССР, Изд. АН УССР, 1956.
Павловский Е. Н. Наставление к собранию и исследованию
клешей (Ixndoidea)\ Наставления для собирания зоологич. коллекций,
т. XVI, 1928.
Павловский Е. Н. Метод хозяйственного и экологического
изучения клещей. Бюллетень VII Всес. съезда по защите растений,
№ 8, 1932.
Павловский Е. Н., Благовещенский Д. И. иАлфе-
е в Н. И. Основные задачи практического изучения клещей в целях
организации борьбы с ними. Изв. Ленинградского ин-та борьбы с вреди-
телями в сельском и лесном хозяйстве, вып. 2, 1932.
Павловский Е. Н. Изучение динамики клещей в природе как
обоснование мер борьбы с ними. Вредители животноводства, 1935.
Павловский Е. Н. Современное учение о переносчиках возбу-
дителей заболеваний и задачи советского здравоохранения. Медгиз, 1940.
Павловский Е. Н. Руководство по паразитологии человека,
т. II, 1948.
Павловский Е. Н. Состояние учения о природной очаговости
болезней человека. Кн. «Природная очаговость болезней человека и кра-
евая эпидемиология», 1955.
Павловский Е. Н. Дальнейшее развитие учения о природной
очаговости болезней человека и животных. Тезисы докладов девятого
совещания по паразитологическим проблемам, Изд. АН СССР, 1957.
Первомайский Г. С. К фауне клещей сем. Ixodidae, Ирана.
Тр. Военно-медицинской акад. им. С. М. Кирова, т. XLIV, 1948.
Переход В. И. Краткая характеристика и районирование лесов
БССР. Леса БССР, Изд. АН БССР, 1954.
Пивоварова Е. П. О размещении и численности грызунов в
заповеднике «Беловежская пуща». Уч. зап. Московского городск. пед.
ин-та им. В. П. Потемкина, т. XXXVIII, 1955.
Пивоварова Е. П. Основные черты экологии мышевидных гры-
126
зунов Беловежской пущи и их лесохозяйственное значение. Автореферат.
Московский городск. пед. ин-т им. В. П. Потемкина, 1956.
Пионтковская С. П. Материалы по биологии и экологии кле-
щей Hyalomma marginatum marginatum Koch, в северо-западном
очаге Крымской геморрагической лихорадки. Новости медицины, вып. V,
1947.
ПионтковскаяС П., Жмаева 3. М. и Корш у нова О. С.
Иксодовые клещи—переносчики риккетсиозов, Изд. Акад. мед. наук
СССР, 1952.
Покровская Е. И. Материалы к экологии клеща D. margina-
tus Sulz., а также о его патогенности и борьбе с ним по данным изу-
чения на юго-востоке черноземного центра. Автореферат. Московский ун-т
им. Ломоносова, 1956.
Померанцев Б. И. К вопросу о происхождении клещевых
очагов в Ленинградской области. Вредители животноводства, 1935.
Померанцев Б. И. и Алфеев Н. И. К вопросу о действии
мышьяковистых составов на клеща /. ricinus L. Вредители животновод-
ства, 1935.
Померанцев Б. И. Клещи (сем. Ixodidae) СССР и сопредель-
ных стран, Изд. АН СССР, 1946.
Померанцев Б. И. и Сердюкова Г. В. Экологические на-
блюдения над клещами сем. Ixodoidae—переносчиками весенне-летнего
энцефалита на Дальнем Востоке. Паразитологии, сб. Зоол. ин-та АН СССР,
IX, 1948.
Померанцев Б. И. Географическое распространение клещей
Ixodoidae и состав их фауны в Палеарктической области. Тр. Зоол. ин-та
АН СССР, т. VII, вып. 3, 1948.
Померанцев Б. И. Иксодовые клещи (Ixodidae). Фауна СССР,
Паукообразные, т. IV, вып. 2, 1950.
Распопов И. М. Растения и клеши. «Природа» № 11, 1953.
Роговой П. П., Медведев А. Г., Булгаков Н. П., Пиль-
ко В. М. и Ч е т в е р и к о в В. Н. Почвы БССР, Изд. АН БССР,
Минск, 1952.
Рубанова Ф. Г Ландшафтная типизация природных очагов ту-
ляремии в Белоруссии и меры профилактики в них. Тезисы докладов
научно-практнч. конф, по заболеваниям с природной очаговостью. Бел.
ин-т эпидемиологии, микробиологии и гигиены, 1957.
Савицкий Б. П. Изучение фенологии иксодовых клещей в оча-
гах сезонного менингоэнцефалита и организация борьбы с ними. Тезисы
докладов научно-практич. конф, по заболеваниям с природной очаговостью.
Бел. ин-т эпидемиологии, микробиологии и гигиены, 1957.
Савицкий Б. П. О зимовке клещей Ixodes ricinus L. и Derma-
centor pictus Негт. в условиях Белоруссии. Зоологический журнал,
т. XXXVIII, вып. 9, 1959.
С а ч а в а В. Б. Растительность лесной зоны. Животный мир СССР,
Изд. АН СССР, т. IV, 1953.
Сердюкова Г. В. Методика кормления эктопаразитов на лабо-
раторных животных. Сб. изобрет. и рацион, предложений. Изд. Военно-
мидицинской акад. им. С. М. Кирова, № 1, 1940.
Сердюкова Г. В. и Померанцев Б. И. Методика экологи-
127
ческих исследований в тайге над клещами сем. Ixodidae—переносчиками
весенне-летнего энцефалита и др. болезней. Сб. изобрет. и рацион, пред-
ложений, Изд. Военно-медицинской акад. им. С. М. Кирова, № 1, 1940.
Сердюкова Г. В. Случаи локального массового размножения
клещей Hyalomma anatolicum anatolicum Koch, в Таджикистане и их
причины. «Изв. Таджик, филиала АН СССР» № 6, 1945.
Сердюкова Г. В. Массовое размножение клещей и меры борь-
бы с ними в условиях колхозных животноводческих ферм Таджикистана.
Отчетная сессия Зоологич. ин-та АН СССР. Тезисы докладов, 1945.
Сердюкова Г. В. О цикле развития клеща Hyalomma anatolicum
anatolicum Koch. «Изв. АН СССР» № 2—3, 1946.
Сердюкова Г. В. Цикл развития клещей сем. Ixodidae. Рефера-
ты научно-исслед. работ за 1945 г., отдел, биологич., Изд. АН СССР,
1947.
Сердюкова Г. В. Метод определения продолжительности цикла
развития у клещей сем. Ixodidae. Рефераты научно-исслед. работ за
1945 г., отдел, биологич., Изд. АН СССР, 1947.
Сердюкова Г. В. Метод определения продолжительности цикла
развития у клещей сем. Ixodidae. Паразитологический сборник Зоологич
ин-та АН СССР, т. X, 1948.
Сердюкова Г. В. Зимовка яиц I. ricinus L. в условиях Карель-
ского перешейка. ДАН СССР, т LXXXI, № 6, 1951.
Сердюкова Г. В. Новые данные о развитии личинок и нимф I.
ricinus L. в природных условиях. ДАН СССР, т. XXXIII, № 5, 1952.
Сердюкова Г. В. К вопросу о дифференциальных признаках ли-
чинок и нимф иксодовых клещей. Зоологический журнал, т. XXXIV,
вып. 5, 1955.
Сердюкова Г. В. Иксодовые клещи. Клещи грызунов фауны
СССР, Изд. АН СССР, 1955.
Сердюкова Г. В. «Распределение иксодовых клещей в зональных
типах растительности. Тезисы докладов девятого совещания по парази-
тологии. проблемам. Л., ЗИН АН СССР, 1955.
СердюковаГ В. Иксодовые клещи фауны СССР, Изд. АН СССР,
1956.
Сержанин И. Н. Млекопитающие Белорусской ССР, Изд.
АН БССР, 1955.
Сержанин И. Н. Особенности распространения наземных позво-
ночных животных (гл. обр. млекопитающих) в Белоруссии. Тезисы до-
кладов научно-практич. конф, по заболеваниям с природной очаговостью.
Бел. ин-т эпидемиологии, микробиологии и гигиены, 1957.
Смирнова Е. И., Андриенко Г. В. иЧайкоГ У. Забо-
леваемость сезонным вирусным менингоэнцефалитом в Бегомльском .рай-
оне Минской области. Тезисы докладов научно-практич. конф, по за-
болеваниям с природной очаговостью. Бел. ин-т эпидемиологии, микро
биологии и гигиены, 1957
Смирнова Е. И., И н д м а н А. Я. и П о п о в В. Е. Особенности
эпидемиологии сезонных менингоэнцефалитов в Стародорожском районе.
Тезисы докладов научно-практич. конф, по заболеваниям с природной
очаговостью. Бел. ин-т эпидемиологии, микробиологии и гигиены, 1957.
Советова М. И. Опыт применения метода опыливания нор гры-
128
зунов в населенных пунктах при сплошной дератизации и на изолирован^
ном участке. Проблемы паразитологии. Тр. II научной конф, паразите-
логов УССР, Изд. АН УССР, 1956.
Судаченков В. В. Причины мозаичности заклещевения пастбищ
Ленинградской области и их значение для эпизоотологии бабезиеллоза
крупного рогатого скота. Тезисы докладов третьего совещания по пара-
зитологическим проблемам, Изд. АН СССР, 1941.
С у д з и л о в с к и й М. Н. и Щербинин Н. В. Географическое
распространение пироплазм и клещей—переносчиков пироплазмоза крупно-
го рогатого скота и лошадей БССР Уч. зап. Витебского ветеринарного
зоотехнич. ин-та, т. IV, 1936,
Сузь ко С. Ф. Изучение иксодовых клещей в целях предупрежде-
ния гемоспоридиозов сельскохозяйственных животных в Белоруссии.
Автореферат. Ленинградский ин-т усовершенствования ветврачей, 1949.
Сузько С. Ф. Изучение иксодовых клещей в целях предупрежде-
ния гемоспоридиозов сельскохозяйственных животных. Сб. научных трудов
Ленинградского ин-та усовершенствования ветврачей, вып. IX, 1953.
Сяргеева 3. 3. Шкоднш буйнай рагатай жывёлы i барацьба з
iMi. MiHCK, 1932.
Сяргеева 3. 3. Шкоднпо буйнай рагатай жывёлы Паганянска-
га балотнага мас!ву. 36. прац, ч. Ill, М1нск, 1933.
Сяргеева 3. 3. Х1м1я у барацьбе са шкодшкам! сельскагаспа-
дарчых раслш i жывёл. Мшск, 1933.
Федорчук Л. В.Материалы к этиологии и эпидемиологии клеще-
вого энцефалита в БССР. Автореферат. Отдел вирусологии Ин-та экспе-
риментальной медицины Акад. мед. наук СССР, 1955.
Федюшин А. В. Роль травопольной системы земледелия в борь-
бе с пастбищными клещами и клещевыми болезнями человека и сельско-
хозяйственных животных. Зоологический журнал, т. XXVIII, вып. 6,
1949.
Филиппова Н. А. К диагностике клеща Ixodes trianguliceps
Bir. по личинкам и нимфам. Зоологический журнал, т. XXXIII,
вып. 5, 1954.
Филиппова Н. А. К диагностике некоторых видов иксодовых
клещей рода Ixodes Latr. по личинкам и нимфам. Зоологический журнал,
т. XXXIII, вып. 1, 1954.
Филиппович А. Н. Острый сезонный серозный менингит (двух-
волновый вирусный менингоэнцефалит). Тезисы докладов научно-практич.
конф, по заболеваниям с природной очаговостью. Бел. ин-т эпидемиоло-
гии, микробиологии и гигиены, 1957.
ФолитарекС. С., Кв ит н и ц кая Г. В. и Саблина Т. Б.
Условия существования и методы уничтожения природных очагов шо-
тландского энцефалита в лесах западных районов СССР. Вопросы ме-
дицинской вирусологии, вып. 2, 1949.
Хейсин Е. М. Наблюдения над развитием скотского и таежного
клеща в лабораторных условиях. Уч. зап. КФГУ, т. V, вып. 3, 1953.
Хейсин Е. М. и Лебешева М. А. Яйцекладка и развитие
/. ricinus L. и /. persulcatus Sch. при разной температуре и влажности
окружающей среды. Тр. КФГУ, т. VI, 1954.
X е й с и н Е. М. Продолжительность развития личинок и нимф
9. Иксодовые клещи. 129
/. ricinus L. и /. persulcatus Sch. в разные сезоны года. Тр. КФГУ,
т. VI, 1954.
ХейсинЕ. М., Бочкарева К., ЛаврененкоЛ. и Ми-
хайлова Т. Яйцекладка и развитие /. ricinus L. в природных
условиях К.-Ф.ССР. Тр. КФГУ, т. VI, 1954.
Хейсин Е. М., Бочкарева К. и ЛаврененкоЛ. К вопро-
су о сезонной активности взрослых I. ricinus L. в К.-Ф.ССР. Тр. КФГУ,
Т: VI, 1954.
Хейсин Е. М. Продолжительность цикла развития I.ricinus L.
в природных условиях К-Ф.ССР. Тр. КФГУ, т. VI, 1954.
ХейсинЕ. М и Кузнецова Т. К. Холодостойкость яиц,
личинок и взрослых клещей /. ricinus L. и I.persulcatus Sch. Тр. К-Ф.
филиала АН СССР, вып. 4, 1956.
Ходаковский А. И. Клещевые очаги /. persulcatus Sch. та-
ежной полосы Европейской части СССР. Паразитологический сб. Зоологич.
ин-та АН СССР, т. IX, 1948.
Чумаков П. П. Дальнейшее изучение ареала распространения
и особенностей эпидемиологии клещевого энцефалита человека в Евро-
пейской части СССР. Тезисы докладов третьего совещания по паразито-
логии. проблемам, Изд. АН СССР, 1941.
Шпринголь ц-Ш м и д т А. И. Материалы по экологии и биологии
D.silvarum 01. на Дальнем Востоке. Вестник дальневосточного филиа-
ла АН СССР, № 16, 1936.
Щеглова А. И. О роли диких позвоночных на лесных пастби-
щах в качестве промежуточных хозяев клещей. «Вопросы экологии и
биоценологии» № 5—6, 1939.
Юркевич И. Д. Типы лесов Белорусской ССР. Госиздат
БССР, 1948.
Юркевич И. Д. О классификации типов леса Беловежской пу-
щи. Бюллетень МОИП, отд. биологии, т. IV (3), 1951.
Якимов В. Л. и Бурцев И. В. Пироплазмоз рогатого скота и
его возбудитель в Белоруссии. «Белорусская ветеринария» № 78, 1928.
Якимов В. Л., Судзиловский М. Н. и Растегае-
в а Е. Ф. К вопросу о видах пироплазм крупного рогатого скота в Бе-
лоруссии. «Белорусская ветеринария» № 6—7, 1929.
Якимов В. Л. Болезни домашних животных, вызываемые про-
стейшими (Protozoa). Сельхозгиз, 1931.
Якимов В. Л. Одна из актуальных проблем современного жи-
вотноводства в СССР. «Природа» № 1, 1932.
Balat F. a Rosicky В. Ptaci Nizinneho lesa, jejich pohyb a
vyznam v prirodnim ohnisku. Ceskoslovenska parasitologic, 1, 1954. t
В e г t k a n P. Aus der Lebens und Fortpflanzungageschichte unserer
Zecke I. ricinus. Verh. naturhist. Ver preuss. Rheinlande u Westfa-
len, 1881.
С r i t n n e r I. und C a s e 1 i t z F. H. Toxaphen—ein Insectizid mit
langanhalten der Wirkung. Tierarztl. Umschau. № 13—14, 1953.
Cerny V. К rozliseni larev a nymf Ixodes apronophorus P. Sch.
Casopis Ceskoslovenske spolecnosti entomologicke. Roc. 54, Cis. 4, Ri-
jen 1957.
130
Cerny V. Morfologicke rozdily larev a nymf stredoevropskycb
clenu rodu Dermacentor Koch. Zoologicke listy folia zoologica, rocnik
vi (xx), cislo 1, 1957.
Cerny V. Sezonni dinamika klistete Ixodes ricinus v divokem
miste zaklisteni. Ceskoslovenska parasitologie iv, 1957.
Cerny V. Vyvojovy cyklus klistete Ixodes ricinus L. v smisenem
lese strednich cech. Ceskoslovenska parasitologie v—1, 1958.
Kossel, Schutz, We her und M i e s s n e r. Uber die Haemog-
lobinurie der Binder in Deutschland. Arbeiten aus dem Kais. Gesundheit-
samte, 20, 1903.
Macicka О. О vyskovom rozvrstveni kliesta obycajneho (I. ricinus)
vo vysokych Totrach. Zoologicke a entomologicke listy folia zoologica et
entomologica Rocnik iv (xviii), cislo 4, 1955.
Macicka O., Nosek I. a Rosicky B. Poznamky к bionomii,
vyvoju a hospodarskemu vuznamu pijaka luzneho (D. pictus) v strednej
Europe. Biologicke prace. Edicia sekcie biolokgickych a lekarskyc vied
slovenskej akademie vied zvazok ii, Zosit 12, 1956.
Nuttall G. und Wua r b u r t о n C. Ticks. A monograph, of the
Ixodoidea pt. ii, 1911.
Rosicky B. a Havlik O. Nebenpecny parasit drubeze, Argas
persicus Oken., v ceskoslovensku. Zvlastni otisk z casopicu Zoologicke.
A entomologicke Listy Folia zoologica. et entomologica, cislo 2, roc-
nik 1 (xv), 1952.
Rosicky B. Bionomicko-faunisticky nastin klistat (Ixodidae) z
uzeml CSR. Zool. a entomol. listy №2, 1953.
Rosicky B. Poznamky к ekologii klistete I. ricinus ve stredni
Evrope se zretelem na prirodni ohniska nakaz. Doslo 24, 8, 1953.
Rfos'icky B. Klistata vdecny predmet studia. Ziva, № 2, 1954.
Rosicky B. Poznamky к ekologii klistete I. ricinus ve stredni Evrope
se zretelem na prirodni ohniska nakaz. Vest. Ceskosl., spolec, zool., 18,
№ 1, 1954.
Rosicky B., a Ba 1 at F. Kliste I. ricinus jako cizopasnik ptacuv
prirodnim ohnisku. Ceskoslovaska parasitologie № 1, 1954.
Rfosicky B. a Cerny V. Drobni stredoevropsti ssavci jaco hos-
titele klistete I. ricinus. Zool. a entomol. listy, 3, № 1, 1954.
Rosicky B. a R у s a v у В. Ekologicky vyzmankere pro udrzovani
vysoke pocetnosti nekterych cizopasnlku na pastvinach. Ceskosl. biol.,
№ 8, 1955.
Samson K. Zur anatomie und Biologie von I. ricinus. Zeitschr. f-
wiss. Zool., 93, 1909.
T о tze R. Beitrage zur Sinnesphysiologie der Zecken, Z. vergl.
Physiol., XIX, № 1, 1933.
131
СОДЕРЖАНИЕ
Введение .... 3
Природные условия БССР 6
Материал и методы исследования 9
Видовой состав и географическое распространение иксодовых
клещей в БССР 14
Стадиальное распределение иксодовых клещей 18
Хозяева клещей . . 46
Цикл развития D. pictus 63
Цикл развития /. ricinus 82
Система проги вок лещевых мероприятий 113
Литература 121
Редактор Издательства Т. Зайцева Корректор Н. ЛЬмако
Технический редактор Г Широкова
Утверждено РИСО АН БССР
АТ 03351. Сдано в набор 7/ХП 1960 г. Подписано к печати 7/11 1961 г.
Тираж 750 экз. Бумага 84ХЮ81/з2- Печ. л- 4,125. Усл. л. 6,7.
Уч.-изд. л. 7,4. Изд. заказ 109. Тип. заказ 41. Цена 37 коп.
Типография Издательства АН БССР, Минск, проспект Сталина, 68