














































































































































































































































































Author: Кучерук В.В.
Tags: математика естественные науки медицина млекопитающие инфекции териология грызуны хищные рукокрылые
ISBN: 5-02-005274-4
Year: 1989




МЕДИЦИНСКАЯ
ТЕРИОЛОГИЯ
ГРЫЗУНЫ
ХИЩНЫЕ
РУКОКРЫЛЫЕ
АКАДЕМИЯ НАУК СССР
ВСЕСОЮЗНОЕ ТЕРИОЛОГИЧЕСКОЕ ОБЩЕСТВО
ВОПРОСЫ ТЕРИОЛОГИИ
МЕДИЦИНСКАЯ
ТЕРИОЛОГИЯ
ГРЫЗУНЫ,
ХИЩНЫЕ,
РУКОКРЫЛЫЕ
Ответственный редактор
доктор биологических наук
В. В. КУЧЕРУК
в
МОСКВА «НАУКА» 1989
УДК 5Ы.67
Academy of Sciences USSR
All-Union Theriological Society
Medical Theriology
Rodents, carnivors, bats
Медицинская териология: Грызуны, хищные, рукокрылые.
М.: Наука, 1989.— 272 с.
ISBN 5-02-005274-4
Книга представляет собой сводку мировых данных о ме-
дицинском значении млекопитающих 3 отрядов, подготовлен-
ную ведущими специалистами медицинской териологии. Рас-
смотрена роль разных видов как хозяев возбудителей болез-
ней, общих для людей и животных. Показаны формы контакта
хозяина, переносчика и возбудителя и пути передачи инфек-
ции от млекопитающих к человеку. Обсуждены основные
принципы профилактики этих болезней. Книга является
продолжением издания «Медицинская териология» (М.: Наука,
1979), посвященного отряду грызунов.
Для зоологов, паразитологов, специалистов по профи-
лактической медицине, защите растений, охотоведов.
Главный редактор серии
академик В. Е. СОКОЛОВ
Редколлегия:
члены-корреспонденты АН СССР: И. А. ШИЛОВ, А. В. ЯБЛОКОВ;
доктора биологических наук: Н. К. ВЕРЕЩАГИН, Н. Н. ВОРОНЦОВ,
И. М. ГРОМОВ, |А. П. КУЗЯКИН], В. В. КУЧЕРУК,
В. Н. ОРЛОВ, П. А. ПАНТЕЛЕЕВ;
кандидаты биологических наук:
Г. В. КУЗНЕЦОВ (ответственный секретарь),
Т. Б. САБЛИНА
Рецензенты:
доктора биологических наук Е. В. КАРАСЕВА и Э. И. КОРЕНБЕРГ
.. 1907000000-294 ОА /гт . „ „ ' 1АОА
М—А.о,АГ1, оа— 165-89 (Доп.) © Издательство «Наука», 1989
U42 (UzJ-оУ
ISBN 5-02-005274-4
ПРЕДИСЛОВИЕ
Первый выпуск книги «Медицинская териология» (1979) содер-
жал главы, посвященные биоценотической характеристике зональ-
ных ландшафтов и их оценке как среды функционирования возбу-
дителей зоонозов (тундр, лесных ландшафтов, степей) и система-
тическому обзору млекопитающих — хозяев возбудителей при-
родно-очаговых болезней человека. Подробно рассмотрено медицин-
ское значение представителей отряда зайцеобразных — семейств
заячьих и пищух. Большую часть текста занимало описание ме-
дицинского значения грызунов — самого многочисленного, ши-
рокораспространенного и хозяйственно значимого отряда млеко-
питающих. В первую очередь были рассмотрены обильные видами
и имеющие важное медицинское значение семейства хомякообраз-
ных, мышиных, беличьих, а также более мелкие семейства: бобро-
вые, летяговые, слепышовые,. сониевые, селевинии, колючие сони,
мышовки, гунди, нутриевые. В заключение была дана подробная
таблица естественного носительства возбудителей болезней чело-
века представителями отрядов зайцеобразных и грызунов фауны
СССР.
Книга моментально разошлась и стала библиографической
редкостью, практически не появлялась в букинистических мага-
зинах. Она вызвала большой интерес и появление рецензий (Ол-
суфьев, 1980; Савицкий, 1980; Cerny, 1980). Во всех рецензиях
была дана весьма положительная общая оценка книги и выска-
зано пожелание о скорейшем издании следующего тома «Медицин-
ской териологии», посвященного другим отрядам млекопитающих.
Одновременно был высказан ряд критических замечаний и поже-
ланий. Рецензенты считали нецелесообразным описание тех си-
стематических групп млекопитающих, медицинское значение кото-
рых не установлено. Также предлагалось сократить текст, посвя-
щенный описанию внешнего вида и морфологии рассматриваемых
видов.
В предлагаемом вниманию читателей томе продолжено описание
грызунов, полностью описаны отряды хищных и рукокрылых
(крылановых).
В отряде грызунов продолжено описание представителей се-
мейства беличьих (суслики, бурундуки, тонкопалый суслик) и
семейств гоферовых, мешотчатых прыгунов, долгоноговых, тушкан-
чиковых, дикобразовых, американских дикобразов, свинковых,
агутиевых, колючих шиншилл. В связи с отсутствием сведений о
медицинском значении не рассматриваются следующие семейства:
1 *
4
Предисловие
аплодонтовые (Aplodontidae), шипохвостые (Anomaluridae), бам-
буковые крысы (Rhizomyidae), пакарновые (Dinomyidae), паковые
(Agoutidae), хутии (Capromyidae), восьмизубовые (Octodontidae),
шиншиллокрысовые (Abrocomidae), скальные крысы (Petromyidae),
тростниковые крысы (Thryonomyidae).
Весьма ограниченное медицинское значение имеет ряд семейств
грызунов, поэтому мы ограничимся лишь краткими упоминаниями
о них.
Водосвинковые (Hydrochaeridae) представлены одним видом;
среди них наблюдается регулярная гибель от американского три-
паносомоза (Schaller, Crawshaw, 1981).
Шиншилловые (Chinchillidae) — 3 рода с 7 видами. Заражен-
ные чумой особи равнинной вискаши (Lagostomus maximus) были
обнаружены в Аргентине. Они случайно вовлекаются в эпизоотии
и не имеют значения в передаче инфекции людям (Plague in the
Americas, 1965). От шиншиллы (Chinchilla laniger) выделен воз-
будитель псевдотуберкулеза (Davis et al., 1970).
Тукотуковые (Ctenomyidae) — 1 род с 27 видами. Зараженные
чумой зверьки (Ctenomys sp.?) зарегистрированы в Аргентине.
Случайно вовлекаются в эпизоотии, текущие среди других видов,
эпидемиологического значения не имеют (Pollitzer, 1954).
Землекоповые (Bathyergidae) — 5 родов с 14 видами. Есть
указание на выделение возбудителя чумы от- одного из видов пе-
скороев (Cryptomys sp.?) в Анголе (Pollitzer, 1954).
Картосхемы ареалов тушканчиков приведены в недавно вышед-
ших: популярной книге И. М. Фокина (1978) и специальной пуб-
ликации И. Л. Кулик (19806). Это позволяет не давать их в настоя-
щей книге, ограничившись одной картосхемой ареала всего
семейства.
Представители отряда хищных относятся к числу наиболее
изученных млекопитающих СССР. Опубликовано много видовых
монографий, большое внимание хищникам уделено в региональных
фаунистических сводках (Млекопитающие Якутии, 1971; Млеко-
питающие Киргизии, 1972; Млекопитающие Казахстана, 1981,
1982, и т. д.). Весьма полная характеристика хищных СССР дана
в коллективной монографии «Млекопитающие Советского Союза»,
т. 2 (1967, 1972). В ней приведены подробные карты ареалов се-
мейств, родов и видов, обитающих на территории нашей страны,
что позволяет не дублировать эти сведения в настоящей книге,
ограничившись лишь картой распространения семейства Viver-
ridae.
В то же время роль хищных млекопитающих как хозяев воз-
будителей зоонозов и их эпидемиологическое значение освещены
совершенно недостаточно. Особенно это касается бешенства. Ев-
ропейские и североамериканские ученые последние десятилетия
весьма активно изучали эпизоотии бешенства среди млекопитающих
и меры по их подавлению. Проведенные исследования не получили
Предисловие
5
должного отражения в советской териологической литературе,
а медицинские сводки (Селимов, 1978; Черкасский, 1985) не вос-
полнили этого пробела. Учитывая создавшееся положение, по
видам, имеющим наибольшее медицинское значение, представлены
развернутые видовые очерки (лисица, волк, енотовидная собака,
песец), а для менее значимых групп описание дается по родам или
семействам.
В классе млекопитающих отряд рукокрылых (крылановых)
второй после грызунов по числу входящих в него видов — 875.
Он характеризуется всемирным распространением (на всех мате-
риках, кроме Антарктиды, и на большинстве океанических остро-
вов). Для представителей отряда характерны скопления особей в
местах отдыха или на зимовках. У некоторых видов такие скопле-
ния могут насчитывать сотни и тысячи экземпляров. Нередко места
отдыха рукокрылых приурочены к постройкам человека как в
сельской местности, так и в городах.
Рукокрылые представляют собой четко очерченную группу
специфических и близких между собою жизненных форм. Среди
них есть потребители нектара, фруктов и плодов. Большое число
насекомоядных форм, ловящих добычу в полете или собирающих
ее с растений. Есть хищники, поедающие мелких наземных позво-
ночных, рыбоядные формы и кровососущие виды, нападающие
преимущественно на стадных копытных. Из-за способности к
полету и скопления в постоянных местах отдыха рукокрылым
свойственны весьма специфичные эктопаразиты, с трудом перехо-
дящие на других животных. Тем не менее среди них есть виды,
охотно кусающие людей, а некоторые клопы — исходные пара-
зиты рукокрылых — перешли к обитанию в человеческом жилище.
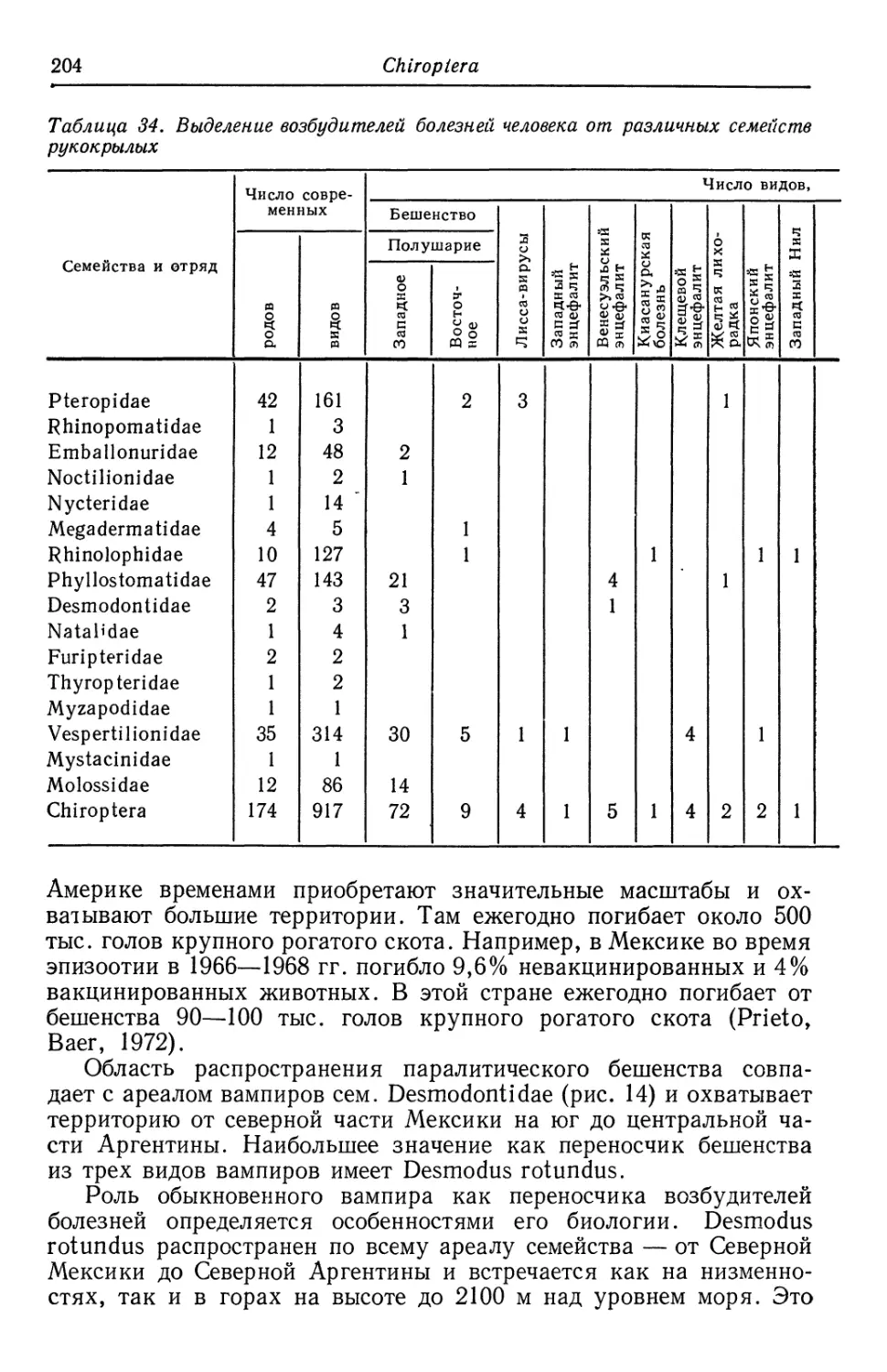
Рукокрылые относятся к важнейшим хозяевам бешенства.
Предполагается, что вирус бешенства был первичной инфекцией
рукокрылых, а уже затем перешел к паразитированию на хищных.
Массовое увлечение спелеологией привело к появлению случаев
аэрогенного заражения людей бешенством и вспышкам гистоплаз-
моза в результате посещения пещер, населенных летучими мышами.
Мировая литература по рукокрылым весьма обширна. На
большинстве европейских языков изданы монографии или попу-
лярные книги, посвященные этой группе млекопитающих. На рус-
ском языке обобщающие сведения по рукокрылым немногочисленны.
Единственная монография, посвященная фауне рукокрылых СССР,
издана более 35 лет тому назад (Кузякин, 1950). В шестом томе
«Жизни животных» (1971) на рукокрылых мира отведено 22 стра-
ницы. В. Е. Соколов (1973) в книге по систематике млекопитающих
не рассматривает общих проблем экологии, распространения и
происхождения рукокрылых. Практически нет обобщающих пуб-
ликаций по паразитам рукокрылых. Мы попытались в какой-то
мере восполнить отмеченные выше пробелы.
Разделы по грызунам подготовлены сотрудниками Института
G
Предисловие
эпидемиологии и микробиологии им. Н. Ф. Гамалеи АМН СССР
Н. А. Никитиной, И. Л. Кулик, Ю. А. Дубровским и сотрудником
Института вирусологии им. Д. И. Ивановского АМН СССР Г. А. Си-
доровой. Отряд хищных написан сотрудником НИИЭМ им. Н. Ф. Га-
малеи Т. Н. Дунаевой и сотрудником Института эволюционной
морфологии и экологии животных им. А. Н. Северцова АН СССР
Д. И. Бибиковым. Раздел по рукокрылым подготовлен сотрудником
ИЭМЭЖ им. А. Н. Северцова В. В. Кучеруком и И. Л. Кулик.
Общая тематика книги, а также схема составления отдельных
очерков разработаны В. В. Кучеруком. Им же доработаны отдель-
ные разделы и объединены в единую монографию.
Подготовила книгу к печати И. Л. Кулик, ею же совместно
с В. В. Кучеруком составлены карты распространения отдельных
таксонов.
Авторский коллектив надеется подготовить и третью книгу
«Медицинской териологии», основу которой составят отряды насе-
комоядных, парнопалых и приматов.
ОТРЯД RODENT1A BOWDICH, 1821 —
ГРЫЗУНЫ
В первом выпуске «Медицинской териологии» (1979) были рас-
смотрены наиболее важные в медицинском отношении группы
грызунов. Здесь мы продолжаем описание этого отряда. Семейство
беличьих (Sciuridae) частично вошло в первый том (подсемейство
Sciurinae и триба Marmotini подсемейства Marmotinae). Остальные
трибы подсемейства Marmotinae представлены в настоящей книге.
Триба Citellini I. Gromov, 1965 — суслики
Триба Citellini объединяет 5 родов (Громов и др., 1965)
Род Citellus Oken, 1816 (20 видов).
Подроды: Urocitellus (2 вида) — Сибирь, Аляска, Канада,
Citellus (7 видов) — Евразия,
Colobotis1 (11 видов) — Евразия, Северная Америка.
Род Otospermophilus Brandt, 1814 (8 видов) — запад Северной
Америки.
Род Ammospermophilus Merriam, 1892 (5 видов) — юго-запад
Северной Америки.
Род Xerospermophilus Merriam, 1892 (2 вида) — юго-запад Север-
ной Америки.
Род Ictidomys Allen, 1877 (5 видов) —Северная Америка.
В последней американской сводке (Honacki et al., 1982) все
перечисленные роды объединены в один род Spermophilus F. Cu-
vier, 1825 (36 видов), включающий 5 подродов: Otospermophilus,
Poliocitellus, Ictidomys, Callospermophilus.
Видовая систематика сусликов установилась относительно не-
давно. В старых работах нередко объединялись в один вид С. un-
dulatus и С. parryi, а также С. major (С. rufescens) и С. erythrogenys;
к С. citellus в качестве подвидов относили С. relictus, С. dauricus,
С. xanthoprymnus, а к С. pygmaeus — С. musicus (Бобринский и
др., 1965, и др.).
Зоологи, работавшие в Казахстане, видимо, справедливо вы-
деляют из С. erythrogenys отдельный вид С. intremedius (Млеко-
питающие Казахстана, 1969).
В сводке, изданной Британским музеем (Corbet, 1978), в один
вид объединены С. citellus и С. xanthoprymnus, С. dauricus и С. ala-
1 Иммуносистематика не дает оснований для выделения подрода Colobotis
(Воронцов и др., 1981).
8 Rodent i a
schanicus, C. pygmaeus и C. musicus, к C. major отнесены C. erythro-
genys и C. intermedins.
Проведенный сравнительный анализ сывороточных белков раз-
ных видов сусликов подтвердил самостоятельность видов С. undu-
latus и С. parryi, а также С. major, С. erythrogenys и С. fulvus
(Воронцов и др., 1981).
Суслики — зверьки средних и мелких размеров (длина тела
11—16 и 18—40 см), обитающие в открытых биотопах (лесостепь,
степь, полупустыня, пустыня и горно-степной пояс). Все суслики
впадают в зимнюю спячку, а многие виды и в летнюю, без перерыва
переходящую в зимнюю. Растительноядные зверьки, большое
значение в их питании имеют луковичные растения и эфемеры,
в их рацион включаются и животные корма, особенно в засуш-
ливый сезон. Активность всех сусликов дневная. Равнинные сус-
лики роют глубокие норы, горные виды часто для убежищ
используют пустоты в скалах, под камнями.
Род Citellus Oken, 1816 — настоящие суслики
В роде 20 видов, на территории СССР обитает 12 видов.
Citellus (Urocitellus) undulatus Pall., 1778 (2n=32) — азиат-
ский длиннохвостый суслик, или суслик Эверсманна, заселяет
степные и лугостепные ландшафты в предгорных и горных рай-
онах — от северных хребтов Тянь-Шаня и гор Южной Сибири до
Приамурья, Северного и Северо-Восточного Китая.
С. (Urocitellus) parryi Rich., 1825 (2n=34) — американский
длиннохвостый, или Берингийский, суслик занимает луговые и
луго-степные участки, а также субальпийские и альпийские луга
в Северо-Восточной Сибири (к востоку от Лены), на побережье
Чукотского и Охотского морей, на Камчатке, на Аляске и в Канаде.
С. (Citellus) relictus Kasch., 1923 (2n=36) — реликтовый сус-
лик занимает изолированные участки горных степей на Тянь-
Шане и Памиро-Алае, подымаясь до высоты 3000 м, наиболее рас-
пространен в Прииссыккульской котловине.
С. (Citellus) dauricus Brandt, 1843 (2n=36)—даурский суслик
распространен в ксерофитных степях Юго-Восточного Забайкалья,
Восточной Монголии и Северо-Восточного Китая.
С. (Citellus) musicus Menetrie, 1832 (2n=36) — горный суслик
занимает горные луга и степи центральной части Главного Кав-
казского хребта до высоты 3100 м над уровнем моря.
С. (Citellus) pygmaeus Pall., 1778 (2n=36) — малый суслик
распространен в степях и полупустынях — от Нижнего Днепра
до центральных районов Казахстана.
С. (Citellus) suslicus Giild., 1770 (2n=34—36) — крапчатый
суслик населяет разнотравные и ковыльные степи и южное лесо-
степье Европейской равнины от восточной части центральной
Польши до Волги.
Грызуны
9
С. (Citellus) citellus L., 1776 (2n=40) — европейский суслик
населяет равнинную и предгорную лесостепь Центральной и Юго-
Восточной Европы, от Эгейского моря (район Солоник) до север-
ной части Пруто-Днестровского междуречья.
С. (Citellus) xanthoprymnus Bennet, 1835 (2n=42) — малоазий-
ский суслик распространен по луговым, полупустынным и пустын-
ным степям в горах Закавказья и прилежащих районов Турции.
С. (Colobotis) major (rufescens) Pall., 1778 (2n=36) — большой,
или рыжеватый, суслик занимает лесостепи, злаковые и разно-
травные степи — от левобережья Камско-Куйбышевского Повол-
жья До Иртыша. По долинам рек и озерным котловинам прони-
кает в полупустынную зону.
С. (Colobotis) fulvus Licht., 1823 (2n=36) — желтый суслик,
или песчаник, распространен по песчаным и лессовым пустыням
и полупустыням равнин и предгорий — от Нижнего Заволжья
до Центрального и Юго-Восточного Казахстана, к югу идет до
Северного Ирана и Афганистана.
С. (Colobotis) erythrogenys Brandt, 1843 (2n=36) — красноще-
кий суслик заселяет ковыльно-типчаковые степи, лесостепи, по-
лупустыни и пустыни; распространен в Алтайском крае, Казах-
стане — от Кустаная до р. Томь на востоке, в Монголии, Северном
и Северо-Западном Китае (Каталог млекопитающих СССР, 1981).
Один вид этого рода обитает в Центральной Азии.
С. (Citellus) alaschanicus Buch., 1888 (2n=38) — алашанский
суслик заселяет горные ртепи Монголии (Гобийский Алтай) и За-
падного Китая (Алашань, Нань-Шань). Поднимается до высоты
3500—3800 м.
7 видов рода распространены в Северной Америке:
С. (Colobotis) columbianus Ord, 1815 (2n=32),
C. (Colobotis) richardsoni Sabine, 1822 (2n=36),
C. (Colobotis) beldingi Merriam, 1888 (2n=30),
C. (Colobotis) armatus Kennicott, 1863 (2n=34),
C. (Colobotis) townsendii Bachman, 1839 (2n=36—38—46),
C. (Colobotis) waschingtoni Howell, 1938 (2n=36),
C. (Colobotis) brunneus Howell, 1928 (2n=38).
БИОТОПЫ И ЧИСЛЕННОСТЬ
Все виды евразиатских сусликов, за исключением желтого,
обитают на плотных почвах, удобных для рытья глубоких нор.
Желтый суслик в северной части ареала заселяет песчаные био-
топы, в южной — лёссовые эфемеровые пустыни. Все суслики
обычно придерживаются мест с низкорослой негустой раститель-
ностью, дающей возможность большого обзора, чтобы вовремя
заметить опасность и скрыться в нору. Только большой суслик
селится на участках с богатой злаково-разнотравной раститель-
10
Rode nt i a
ностью, видимо не мешающей ему осматривать местность благо-
даря его большому размеру.
Суслики предпочитают целинные участки. При сельскохозяй-
ственном и промышленном освоении территории зверьки оттесня-
ются на сохранившиеся участки целины. Большая плотность их
населения наблюдается по обочинам дорог, на выгонах, склонах
балок, утоптанных участках возле кошар и ферм. Заселяют они
насыпи железных и шоссейных дорог. На пахотных землях посе-
ляются редко и только по их краям, чаще живут на полях много-
летних трав. В районах полезащитного лесоразведения поселяются
по опушкам лесных полос. На орошаемых землях сохраняются
по склонам оросительных каналов и на межполосных валиках
(Мамонтов, 1937; Лисецкий, 1959; Grulich, 1960; и др.).
По интразональным биотопам суслики проникают в соседние
зоны. Вырубка леса и остепнение луговых участков способствуют
продвижению их на север. Таким путем малый суслик появился в
Куйбышевской области, большой суслик проник по долине Камы
в Марийскую АССР и южную часть Кировской области. По участ-
кам с уплотненной почвой — «ашикам» — малый суслик прони-
кает в Волго-Уральские пески, по глинистым же участкам захо-
дит далеко в пустынную зону, доходя до побережья Аральско-
го моря.
Виды, обитающие в горах, поднимаются в субальпийскую и
альпийскую зоны, длиннохвостый суслик — до высоты 3400 м.
Этот же вид по степным склонам спускается в зону пустынных
степей в Монголии. Американский длиннохвостый суслик заходит
в тундру.
Живут суслики и по опушкам леса, если к ним примыкают
степные участки, а норы длиннохвостого суслика встречаются и
в светлых разреженных лиственничных и сосновых лесах.
Характер поселений сусликов связан с разнообразием ланд-
шафтов, в которых они обитают, и сельскохозяйственным освое-
нием территории. Кружевной характер носят поселения длинно-
хвостых сусликов в горах и предгорьях, он определяется распро-
странением там пригодных для них биотопов. Более равномерны
поселения даурского суслика, занимающего степи Забайкалья и
Монголии.
В зоне интенсивного сельского хозяйства распространение
сусликов спорадично. Так, пятнами распространены европейский
и крапчатый суслики, оттесненные на небольшие по площади не-
удобные земли. Также разрежены теперь поселения малого сус-
лика на Украине.
Поселения этого вида были сплошными еще в предвоенные и
послевоенные годы на юго-востоке РСФСР: в Ставропольском
крае, Волгоградской и Астраханской областях и в Калмыкии.
На этих территориях сусликов усиленно истребляли, а неблаго-
приятные метеорологические условия ряда лет и сельскохозяй-
Грызуны
11
ственное освоение этих районов не дали восстановиться числен-
ности сусликов. Крупные поселения их сохранились лишь в не-
которых местах: в предгорьях Дагестана, на Ергенях, в Сарпин-
ской низменности и Сулако-Терском междуречье. Сильно сократи-
лись поселения малого и краснощекого сусликов после распашки
целины в Казахстане. В нетронутых степях и полупустынях Ка-
захстана поселения малого суслика еще обширны. Сильно разоб-
щены они на южной границе ареала в Приаральских Каракумах,
где они занимают глинистые участки среди песков (Варшавский,
Шилов, 1957; Миронов и др., 1965; Млекопитающие Казахстана,
1969; Калязина и др., 1975; Емельянов и др., 1982).
По той же причине сельскохозяйственного освоения террито-
рии прерывистый характер носит распространение большого сус-
лика. Из некоторых мест он совсем исчез, например из южных
районов Куйбышевской области. В то же время сведение лесов на
юге Кировской области и появление на их месте луговых и остеп-
ненных участков позволили большому суслику освоить новые тер-
ритории на севере ареала (Артемьев, 1964; Некрасов, 1970; Руди,
1984).
Очаговый характер имеет распространение желтого суслика.
В Казахстане пятнистость его поселений связана с сильным ист-
реблением промысловиками; отдельные пятна, заселенные этим
видом, занимают площадь от 5 до 60 тыс. га. В Средней Азии он
живет в оазисах, занимая там пустыри, кладбища, лёссовые бугры
и другие неудобные земли. В Туркмении на возвышенности Ка-
рабиль он живет возле колодцев, площадь таких поселений, от-
стоящих друг от друга Haji—18 км, 100—150 га (Млекопитающие
Казахстана, 1969; Ермаков, Алиев, 1982).
Из-за своеобразия горных ландшафтов мозаично распростра-
нение сусликов, обитающих в горах. Размеры поселений их могут
колебаться от 0,5 га до нескольких тысяч гектаров. Для горного
суслика на Кавказе при обследовании с вертолета подсчитано
234 поселения, занимающих площадь 84 750 га из общей площади
ареала 300 тыс. га (Емельянов и др., 1982). Пятна, заселенные
реликтовым сусликом на Тянь-Шане в районе Нарынкола, не пре-
вышают 40—50 га (Афанасьев, 1960; Строганова, Чжудинь, 1961).
Размеры поселений малоазийского суслика в северо-западных
районах Армянской ССР колеблются от 5 до 1200 га (Адамян и
др., 1979).
Численность сусликов меняется по сезонам. После выхода мо-
лодняка она увеличивается в 3—4 раза. Снижение численности
наблюдается к весне, так как не все суслики благополучно перези-
мовывают. Годовые изменения численности в центральных частях
ареала обычно невелики. Сильные депрессии численности наблю-
даются только после очень засушливых лет, когда засуха начи-
нается рано и суслики не успевают нажироваться для благополуч-
ной зимовки. Значительная гибель сусликов бывает после много-
12
Rodentia
снежных зим, при сильном половодье, когда затапливаются норы
зверьков. На границах ареала наблюдаются большие изменения
численности, в Волжско-Камском крае зарегистрированы коле-
бания численности малого и большого суслика в 30—60 раз
(Артемьев, 1968).
Численность сусликов в разных частях ареала, в разных по-
селениях и биотопах различна. Она может колебаться от десятых
долей до нескольких десятков, а для некоторых видов — даже
сотен особей на гектар.
Численность длиннохвостых сусликов в разных горных масси-
вах и поясах гор зависит от распространенной там растительности.
В наиболее благоприятных лугостепных и лесостепных ландшаф-
тах их численность доходит до 20 сусликов на 1 га и более и ни-
чтожна в высокогорьях: 0,1—0,2 зверька на 1 га.
Средняя численность даурского суслика в юго-восточном За-
байкалье и приграничных районах МНР — 1—3 на га, лишь ме-
стами их численность доходит до 8—14 (Горчакова и др., 1974;
Некипелов, 1974; Дубовой и др., 1978).
Приуроченность даурских сусликов к различным раститель-
ным ассоциациям хорошо иллюстрируют учеты, проведенные в '
Восточной Монголии В. М. Липаевым (1966): в караганниках,
у юго-западной границы ареала, приходилось до 24 зверьков на
1 га, в пижмово-полынно-разнотравных степях с примесью кара-
ганы по берегу Онона— 17—21, в пижмовых степях по щебни-
стым склонам — 10—12, в дерновинно-злаковых степях с примесью
караганы — 8—10, в низкотравных ассоциациях злаковых и раз-
нотравно-злаковых степей — 5, в полынных, полынно-востряко-
вых и востряково-ковыльных степях — 2—3 и в разнотравных
ассоциациях лесостепного пояса Восточного Хэнтея — 1—2.
Численность малого суслика довольно высока на северной пе-
риферии ареала: на выгонах в Куйбышевской области бывает 30—
35, иногда 100 зверьков на 1 га (Артемьев, 1964). В степях Волго-
Донского междуречья до 30-х годов, когда поселения сусликов
были сплошными, обычна была численность в 30—40 зверьков на
1 га. После проведения здесь истребительных работ и сельскохо-
зяйственного освоения территории численность их сильно сокра-
тилась, в 60-х годах было заселено сусликами всего 9,3—18,6%
территории, со средней численностью 0,9—1,1 зверьков на 1 га.
Более высокая численность, до 20 особей на 1 га, отмечалась на
ограниченных участках: по обочинам дорог, на выгонах, по окраи-
нам посевов. Сохранялась повышенная численность и в наиболее
древних поселениях сусликов; на Ергенинской возвышенности
численность в отдельных балках доходила до 20—30 и 30—100
особей на 1 га (Миронов и др., 1965; Агафонов, 1969; Калязина
и др., 1975; Емельянов и др., 1979; Маренич, Уюмджиев, 1982).
В Волго-Уральских песках численность сусликов на «ашиках»
колеблется от 5 до 14 на 1 га (Миронов и др., 1965).
Грызуны
13
В азиатской части ареала поселения сусликов с большой плот*
ностью расположены в Уральской и Актюбинской областях, где
обычна численность 20—40 зверьков на 1 га, в целинном крае, на
участках нераспаханной степи, 8—35 зверьков (Слудский и др.,
1969; Кубанцев, 1961; Нестеров, 1980). Наиболее часто наблюдаю-
щаяся на северном побережье Аральского моря и в Южном Пред-
устюртье плотность популяции 1—3, реже 4 зверька на 1 га, на
Северном Устюрте и Зааралье —0,1—0,5 на 1 га. В Северном
Приаралье численность малого суслика выше в зоне полупустынь
(на Мугоджарах в среднем 4,4: от 2,2 до 12,3), чем в пустынной
части (в среднем от 0,5 до 4,7, максимум 7,8) (Варшавский, 1983).
Ничтожна численность сусликов на правобережье Сырдарьи, обычно
меньше 1 зверька на 1 га, только в отдельных урочищах, где ра-
стительные ассоциации включают живородящий мятлик — основ-
ное кормовое растение сусликов, численность их доходит до 5—8
и даже 10—12 на 1 га (Варшавский, Шилов, 1957; Шилов, 19666).
Численность сусликов, обитающих в горах, очень пестрая.
На Кавказе у горного суслика в отдельных поселениях она колеб-
лется от 1—5 до 20 и выше на 1 га. Максимальная численность
наблюдалась в верховьях Кубани — 57—212 на 1 га, Баксанском
ущелье — 28—68, Чегемском — 18—22 и в Черекском — 16—40
на 1 га (Акиев и др., 1972; Пилипенко и др., 1974; Петров и др.,
1976, 1978, 1979; Голубев и др., 1978; Емельянов и др., 1982).
Численность малоазийского суслика в Закавказье колеблется
от 0,5 до 32 сусликов на 1 га (Адамян и др., 1979). Для реликто-
вого суслика на Тянь-Шане обычна численность 4—5 особей на
1 га, только в наиболее благоприятных местах на 1 га приходится
10—25 зверьков (Строганова, Чжудинь, 1961).
Крапчатый и европейский суслики, ареал которых находится
в зоне интенсивного сельского хозяйства, где неосвоенных земель
практически не осталось, распространены на оставшихся неудоб-
ных для распашки местах и выпасах, приспособились они жить
и на полях многолетних трав. Площади их поселений обычно
невелики, могут быть всего 3—5 га, наиболее обширные поселения
занимают 100—150 га. Внутри таких пятен численность сусликов
может быть достаточно высока, у крапчатого суслика в Северо-
Западном Причерноморье — до 150—300 особей на 1 га, в По-
волжье — до 40—60. В Польше крупное поселение (35 га) этих
сусликов сохранилось на востоке Люблинской возвышенности,
осенью 1980 г. в нем насчитывалось 5 тыс. зверьков. В ЧССР чис-
ленность европейского суслика в некоторых местах (Южная Мо-
равия) достигает 3,7 на 1 га при 90% распаханной площади; в
Австрии в поселениях этого вида, оставшихся вдоль проселочных
дорог, на межах, сухих лугах и выпасах, плотность нор достигает
500—700 на 1 га (Попов В., 1960; Емельянов, 1983; Лобков, 1983;
Surdacki, 1980—1981).
Численность крупных сусликов подрода Colobotis невелика.
14
Rodentia
Предельная плотность обычно не выше 10 зверьков на 1 га. Чис-
ленность желтого суслика на территории Волжско-Уральских
песков 0,5—6 зверьков на 1 га, в горном Мангышлаке и на полу-
острове Бузачи 0,7—0,9, в северных Кызылкумах 2—3, Приараль-
ских Каракумах 2—10, в Муюнкумах — до 8 на 1 га. Наиболее
плотные поселения этого вида расположены в Северо-Устюртском
понижении, где численность его по краям чинков, кромкам песков
в межбугровых понижениях и по ашикам достигает 5—12 на 1 га,
на плато же его численность незначительна — 0,1—0,3 на 1 га
(Миронов и др., 1965; Шаманек и др., 1969; Млекопитающие Ка-
захстана, 1969; Гарбузов, Варшавский, 1977; Бекенов, 1983).
На большей части ареала большого суслика его численность
не превышает 1—2 на 1 га, только в местах его наибольшей кон-
центрации по западинам и озерным котловинам в Актюбинской и
Уральской областях насчитывается 12—18 особей на 1 га (Артемь-
ев, 1964; Млекопитающие Казахстана, 1969; Некрасов, 1970).
Численность краснощекого суслика также высока только в
некоторых местах: в Восточной Кулунде приходится 7 зверьков
на 1 га, в лесостепных районах Алтайского края 12—15 на 1 га,
в пределах Казахстана обычно насчитывается от 6 до 20 особей (Мле-
копитающие Казахстана, 1969; Кирьянов, 1974).
ОСОБЕННОСТИ БИОЛОГИИ
Спячка. Сезонное проявление эпизоотий и изменение воспри-
имчивости и чувствительности к инфекциям у сусликов опреде-
ляются их способностью впадать в спячку.
Продолжительность спячки у сусликов, обитающих в полупус-
тынных и пустынных районах, 8—9 месяцев. Такие сроки спячки
наблюдаются у малых, желтых, больших, краснощеких и малоазий-
ских сусликов. Эти же виды, обитающие в более умеренных ус-
ловиях, спят меньше. Например, желтые суслики в Поволжье
активны 6 месяцев, в Приаралье — 3—4. Краснощекие суслики в
пустыне Бетпак-Дала перестают выходить из нор в конце июня —
начале июля, а в Западной Сибири бодрствуют до второй половины
августа—начала сентября. У реликтового и даурского сусликов
спячка продолжается 5—6 месяцев, у кавказского горного — до 7,
крапчатые и европейские суслики спят 6—7 месяцев. Длиннохво-
стые суслики бодрствуют даже при наступлении морозов и при
наличии снегового покрова. Продолжительность спячки у С. un-
dulatus 6,5—7,0 месяцев, у С. parryi 5,5—6,5 месяцев (Громов и
ДР., 1965).
В отличие от сурков суслики зимуют поодиночке, только для
молодых горных сусликов характерна зимовка нескольких зверь-
ков в одном гнезде (Емельянов и др., 1982).
Физиологическая и биохимическая перестройка организма сус-
ликов при подготовке к спячке ведет к изменению восприимчи-
Грызуны
15
вости и чувствительности к инфекциям. Снижение температуры
тела и замедленный обмен веществ во время спячки благоприятны
для сохранения возбудителей в организме зимоспящих животных.
При пробуждении зверьков происходит активизация всех жиз-
ненных процессов, способствующая размножению в крови и тка-
нях животных микроорганизмов. Истощенные весной зверьки
восприимчивы к инфекциям. К концу лета с накоплением жиро-
вых запасов устойчивость сусликов к возбудителям болезней
повышается. При этом, поскольку взрослые особи заканчивают
нажировку раньше, чем молодые, создаются возрастные различия
в восприимчивости и чувствительности к инфекциям.
Из-за того что значительную часть года суслики проводят в
спячке, все жизненные явления активного периода проходят у
них в сжатые сроки. Сроки пробуждения сусликов от спячки оп-
ределяют сроки последующих циклов: гона, рождения детенышей,
выкармливания молодняка и его расселения. Меньше связан с
ними период залегания сусликов в спячку, больше зависящий
от погодных условий, влияющих на обеспеченность сусликов
зелеными кормами.
Суслики пробуждаются от спячки с наступлением положитель-
ных температур и таянием снега, обычно в конце марта — начале
апреля, в южных районах в феврале. Первыми пробуждаются
взрослые самцы, затем самки и последними зверьки прошлогод-
него рождения.
У горных видов в разные сроки происходит пробуждение зверь-
ков, обитающих на склонах разной экспозиции и на разной высоте,
поэтому у них гон и все последующие жизненные явления более
растянуты, чем у равнинных сусликов.
Ход весны определяет характер выхода сусликов из спячки
и последующих явлений в их жизни. При дружной весне популяция
выходит из нор в короткий отрезок времени, соответственно боль-
шинство особей одновременно размножается, а молодые выходят
из нор и расселяются в сжатые сроки.
Размножение. Гон у сусликов начинается через 3—4 дня после
пробуждения. В это время зверьки сильно истощены и подвижны.
Величина дневного пробега у даурских сусликов во время гона
1—1,5 км, они посещают за день 65, максимум 96 нор (Бибиков,
1949). В Западной Туркмении желтые суслики во время гона от-
ходят от своих нор на 800 м и больше, часто посещают чужие норы
(Никитин и др., 1962). Кроме того, весной происходит перерас-
пределение территории, так как неодновременное залегание в
спячку разных возрастных групп сусликов приводит к тому, что
на территории, занятой взрослыми особями, залегшими рано в
спячку, располагаются и норы молодых зверьков. Весной прош-
логодние особи, зимовочные норы которых оказываются располо-
женными в пределах чужих участков, вытесняются и кочуют в
поисках незанятой территории (Солдатова, 19626).
16
Rodei.lia
Размножаются суслики раз в году. Половозрелыми суслики
мелких и средних размеров становятся в возрасте одного года,
крупные (желтый, большой и краснощекий) — на второй год.
Число эмбрионов 1—17, у равнинных сусликов чаще 5—8, у гор-
ных — чаще 3. У годовалых самок процент прохолоставших выше
и помет меньше, чем у двух-трехлетних самок.
Подвижность, контакты. Во время беременности, длящейся
около месяца, и выкармливания молодняка, продолжающегося
такой же срок, зверьки живут разобщенно, придерживаясь своих
участков обитания и охраняя их. Самцы и яловые самки в это время
также малоподвижны, так как уже приступают к нажировке. Эпи-
зоотий в этот период обычно не возникает.
Величина индивидуальных участков в это время зависит от
плотности зверьков. Для малых сусликов в Западном Казахстане
и Волгоградской области она составляла при низкой плотности
(1—2 взрослых и 7—8 молодых на 1 га) у самцов 0,65—1,10 га, у
самок 0,45—0,65 га; при большей плотности (17 взрослых и 74
молодых на 1 га) участки у самцов были 0,31—0,42 га, у самок —
0,19—0,29 га (Солдатова, 1962а). На Среднем Урале при низкой
численности большого суслика (0,2 особи на 1 га) участки самцов
равнялись 3,7—6,0 га, самок 1,6 га; при численности 1—3 зверька
на 1 га участки самцов были 0,78 га, самок — 0,64 га (Некрасов,
1982). Такого же порядка размеры участка обитания сусликов,
населяющих Американский континент. Средняя площадь участка
С. leucurus за 20 дней равнялась 6 га, за 1 день — 1,6 га (Bradley,
1967). Индивидуальный участок тринадцатиполосного суслика
самца равнялся 4,7 га, самки— 1,4 га, у колумбийского сусли-
ка— в среднем 0,42 га (Murie, Harris, 1978). При повышении
плотности поселений заметно увеличивается число зверьков, сов-
местно использующих общую йлощадь (Солдатова, 1962а).
Самки после выкармливания детенышей, еще до начала их
расселения, покидают выводковые норы и поселяются отдельно.
По наблюдениям А. Н. Солдатовой (19626), за 10 самками малого
суслика они переселились за 47—225 м. Подвижность самок после
ухода от молодых возрастает. У даурского суслика величина днев-
ного пробега самки увеличивается в 4,5 раза (со ПО до 480 м
(Бибиков, 1949)).
Суслята выходят из нор обычно в середине — конце мая. Мо-
лодые зверьки в первые дни появления на поверхности держатся
вблизи материнской норы, затем постепенно расстояние, на кото-
рое они отходят, увеличивается. Наиболее подвижны суслики в
период расселения молодняка, приходящийся на июнь-июль.
Часть молодых особей оседает в пределах материнских участков,
другие бродят в поисках свободной территории.
При массовом мечении сусликов установлено, что большинство
особей встречается в пределах 100 м от места мечения, реже зверьки
уходят за 200—500 м, и лишь для единиц установлены большие
Грызуны
17
переходы: для малых сусликов — до 3—5 км, для краснощеких
сусликов — 5 км, для длиннохвостых — 20, 150 и 180 км (Зверев,
1929, 1930; Калабухов, Раевский, 1935, 1936; Медзыховский,
Маштаков, 1972).
В период расселения зверьки посещают много нор, в том числе
нежилых, где могут оставаться трупы зверьков, погибших от ин-
фекций, зараженные эктопаразиты и субстрат. Усиливаются
контакты сусликов с другими животными. На энзоотичной по чуме
территории наиболее важно увеличение контактов малых и желтых
сусликов с большими песчанками и песчанками рода Meriones,
даурских и длиннохвостых сусликов с тарбаганами, длиннохво-
стых сусликов с пищухами (см. ниже).
С окончанием расселения сокращаются контакты зверьков меж-
ду собой, молодые оседают на индивидуальных участках и присту-
пают к нажировке, а взрослые особи обычно уже залегают в
спячку.
Существует географическая и биотопическая изменчивость под-
вижности и использования территории сусликами, определяющаяся
в первую очередь кормовыми условиями. Так, участки обитания
малого суслика в северной пустыне в 2,5 раза больше, чем участки
этих сусликов в полосе южных степей. В северной пустыне участки
обитания этого вида в биотопах с хорошей обеспеченностью кор-
мами составляли 63% от размера участков в малокормных угодиях
(Варшавский, 1957).
В местах с мозаичным распределением растительности и с рас-
члененным рельефом характерны кормовые перемещения сусли-
ков. В Приаралье малые суслики в засуху перемещаются со скло-
нов долин к их днищам. В Джунгарском Алатау длиннохвостые
суслики переселяются с высоких склонов в понижения (Бекенов,
1965). В сельскохозяйственных районах суслики в конце лета кон-
центрируются по краям посевов. Обычно кормовые перемещения
невелики — 100—200 м, реже достигают 500—600 м (Афанасьев
и др., 1953; Бабенышев, Саканян, 1963; Тристан, 1965).
Годовые колебания подвижности и площади используемой тер-
ритории определяются численностью сусликов и метеорологиче-
скими условиями. Она выше при большой плотности и недостатке
влажного корма.
Возрастной состав. Суслики живут в природе до 5—6 лет.
Лишь даурских сусликов не встречали старше 3 лет (Бибиков, 1949).
Доля старых сусликов в популяции бывает незначительна. При
пробуждении от спячки большую часть (50—70%) составляют
годовалые зверьки, взрослые 2—3 лет — 20—35% и в возрасте
4 лет и старше обычно — от 2 до 10%, реже— 15—20%. К мо-
менту залегания в спячку большая часть популяции состоит из
молодых рождения текущего года (Калабухов, 1933; Наумов,
1936; Громов и др., 1965; Петровский, 1961; Залесский, 1962;
Вершинина, 1972; Емельянов и др., 1982).
18
Rodent ia
Норы. От характера убежищ зависят особенности фауны па-
разитов зверьков, видовой состав и их обилие, в субстрате нор
возможно переживание некоторых возбудителей болезней, связан-
ное с условиями температуры и влажности убежищ, определяю-
щихся их глубиной и эдафическими факторами, а также характе-
ром использования хозяином.
Все суслики живут в норах. Норы имеют вертикальные или
наклонные ходы. Соотношение таких нор у равнинных видов сус-
ликов обычно 1 : 10. Лишь у европейского суслика В. И. Пакиж
(1958) находил больше вертикальных нор. По функциональному
значению норы делятся на выводковые, зимовочные и защитные
(Кучерук, 1983). Выводковые и зимовочные норы обычно имеют
один вертикальный ход и несколько наклонных, ведущих в камеру
с гнездом из травы. Гнездовая камера у равнинных видов распо-
лагается на большой глубине (2—3 м), общая протяженность
ходов доходит до 5 м, в среднем 3 м. Летом для ночевки суслики
могут использовать норы с камерами без гнезд. Старые выводковые
и зимовочные норы могут затем служить защитными (вторичноза-
щитные норы по классификации В. В. Кучерука, 1983). Помимо _
таких глубоких нор, суслик делает массу мелких (около 30 см) —
первичнозащитных, с наклонным ходом, располагающихся у мест
кормежки.
Норы сусликов, обитающих в горах, отличаются меньшей
глубиной (до 1 м), обычно у них отсутствуют вертикальные ходы,
гнездовая камера часто расположена выше входа, что препятствует
затоплению ее талыми водами. Горные виды в качестве убежищ
используют пустоты в осыпях и скалах.
Глубина нор у длиннохвостых сусликов, обитающих в районах
с вечной мерзлотой, определяется толщиной оттаивающего слоя
(Бируля, 1936; Устьянцев, 1937; Строганова, Чжудинь, 1961;
Громов и др., 1965; Вагнер и др., 1975; Сапаров, 1982; Кучерук,
1983).
В течение лета зверьки меняют как гнездовые (вертикальные),
так и защитные норы. Самец малого суслика, за которым вели не-
прерывные наблюдения, за май сменил три таких норы. Самки
покидают выводковые норы после окончания лактации; бывает,
что и кормящие самки перетаскивают детенышей из одной норы
в другую при сильной пораженности гнезда эктопаразитами либо
потревоженные хищниками. Защитные норы меняются при смене
мест кормежки (Солдатова, 1955, 1962).
Зимовочные норы обычно используются несколько лет, суслики
их обновляют и расчищают, за счет этого растет выброс земли возле
них — курганчик. По величине его и по числу расположенных
возле него нор судят о возрасте поселений сусликов (Варшавский,
1962). Только даурский суслик в северной части ареала роет еже-
годно новые зимовочные норы, и у них при входе отсутствуют кур-
ганчики. В южной части ареала этот вид имеет обычные для сус-
Грызуны
19
ликов норы. С курганчиков суслики производят обзор местности,
горные виды используют с этой целью выступы камней.
Количество нор на 1 га зависит от численности сусликов, ха-
рактера почв, растительности, микрорельефа. В местах с повышен-
ной численностью сусликов на 1 га приходится до тысячи входов.
До проведения массовых истребительных работ на юго-западе
РСФСР такая численность наблюдалась на Черных землях,
Ергенях, в Западном Казахстане (Миронов и др., 1965).
Число нор зависит и от характера почвы, на песчаных почвах
норы скорее засыпаются и их обычно меньше, чем на плотных
грунтах, где их сохранность лучше.
Различается число нор, используемых отдельными особями.
При наблюдении за окрашенными малыми сусликами в степях
Волгоградской и Уральской областей на индивидуальном участке
взрослых самцов насчитывалось 25—45 защитных нор, у самок —
15—25. При этом за день самцы посещали 6—9 нор, самки — 3—7.
Различные норы посещались неравномерно. Самец, за которым
вели наблюдение в течение 14 суток, имевший на участке 14 защит-
ных нор, 14 — посещал 1 день, 5 — 2 дня, 3 — 3 дня, 1 — 4 дня,
2 — 6 дней. Самка, за которой вели наблюдения 12 суток, имевшая
19 защитных нор на участке, 5 использовала 1 день, 6 — 2 дня,
2 — 3 дня и по одной — 5, 7 и 9 дней. Разнится и число забегов
в норы в течение суток и время, проведенное в них. Отдельные
норы посещались самцом за время наблюдений от одного до 25 раз,
самкой — от 1 до 16 раз. Одно посещение продолжалось от 2 до
68 мин (Солдатова, 1955, 1962).
В гнездовую нору самец при низкой плотности зверьков уходил
только на ночевку, при высокой плотности посещал ее 5—7 раз
в день, самка посещала гнездовую нору от 3 до 9 раз в день, а в
период постройки гнезда — до 30 раз.
Контакты сусликов с другими позвоночными животными. Сус-
лики контактируют фактически со всеми мелкими млекопитающими,
обитающими в их же биотопах. Наиболее тесный контакт у малых
и желтых сусликов с песчанками, особенно с большими. В посе-
лениях, совместных с большими песчанками, суслики нередко
устраивают свои норы на территории их нор.
В Северном Приаралье из 14 видов мелких млекопитающих,
зарегистрированных в норах больших песчанок, малые суслики
занимают первое место, составляя 27,3% от общего числа пойман-
ных зверьков, а среди костных остатков, собранных у нор больших
песчанок, 30,7% принадлежали малым сусликам и 8,5% —жел-
тым, на долю хозяев приходилось 27,6% (Наумов, Кулик, 1955).
Наиболее велико число встреч сусликов в норах больших песчанок
в период расселения, в это время их вылавливали в 4—6 раз больше,
чем в другие периоды (Никитина, 1960).
Часто контактируют суслики и с песчанками рода Meriones.
При наблюдении за посещением нор желтого суслика уставов-
20
Rodentia
лено, что в их норы забегают грызуны шести видов, мелкие хищ-
ники (перевязки, степные хори, ласки), птицы (каменки, чеканы),
ящерицы. В Волжско-Уральских песках наиболее часто встре-
чаются гребенщиковые и полуденные песчанки и малые суслики,
составляя соответственно 34,6; 8,5 и 14,5% от всех добытых жи-
вотных, включая хозяев (Демяшев, 1964). В Центральных Кызыл-
кумах чаще контактировали с сусликами краснохвостые песчанки,
составляя 2—9% поимок в их норах (Фоттелер, 1986). В Муюнку-
мах при облове нор желтых сусликов на 100 капкано-суток прихо-
дилось 0,5—5,0% попаданий больших песчанок, 0,5—3,0% крас-
нохвостых и 0,5—1,5% полуденных и других грызунов, а при
облове нежилых нор большой песчанки 0,5—5,0% составляли
желтые суслики (Тристан, 1965). В пустынных и степных биотопах
Астраханской области межвидовые контакты были выше, чем
внутривидовые. Забеги в суслиные норы полуденных и гребенщи-
ковых песчанок были в 2—4 раза чаще, чем посещение их «чужими»
сусликами (Миронов и др., 1949).
В местах совместных поселений с сурками суслики нередко
используют их норы, поселяясь в заброшенных и забегая в жилые.
В Забайкалье в одной нежилой норе тарбагана за летний сезон
вылавливали в среднем 12,4 даурских сусликов, значительно
больше, чем других млекопитающих.
Особенно много сусликов посещает сурчины тарбагана в пе-
риод гона и расселения молодняка. При облове нежилых нор в
апреле на одну нору приходилось 4 суслика, в мае — 2, в июне —
0,4, в июле — 2, августе — 2,7, сентябре — 0,4 (Некипелов,
19596).
Суслики тесно контактируют и с пищухами. В горно-алтайском
природном очаге чумы, где монгольские пищухи служат основными
носителями возбудителя, длиннохвостые суслики часто посещают
их норы. Число забегающих зверьков особенно велико в период
расселения молодняка, в конце июля—августе. В среднем посе-
щение сусликами в период гона нежилой норы монгольской пи-
щухи 0,4, в период рождения и выкармливания молодняка 0,1,
в период расселения 1,5. Забеги 2 пищух в эти периоды соответст-
венно составляют: 3,1; 6,2; 21,6.
Контакты сусликов с песчанками, сурками и пищухами особенно
важны, так как эти виды служат основными носителями чумы в
ряде районов и способствуют распространению эпизоотий в их
совместных поселениях. Забегают в сусликовые норы тушканчики,
мыши и полевки, последние используют устья суслиных нор для
прокладывания собственных ходов и также вовлекаются в эпизоо-
тии, а домовые мыши, переселяясь в дома, при наступлении хо-
2 Забег — число зверьков, посетивших любую нежилую нору (Ешелкин,
Пуртов, 1976).
Грызуны
21
лодов могут занести туда и возбудителя, полученного в поселениях
сусликов.
Тесный контакт с сусликами отмечен и у мелких хищников,
особенно у степного хорька, в добыче которого суслики занимают
одно из первых мест. Защитные норы сусликов и устья постоянных
нор служат убежищами ежам и землеройкам. Забираются в норы
сусликов птицы-норники (чеканы, каменки-плясуньи, в норы
желтых сусликов даже огари и пеганки). Заползают в них различ-
ные рептилии: змеи, ящерицы, степные черепахи.
Суслики обмениваются эктопаразитами со всеми животными,
с которыми используют совместно норы. Особенно важное зна-
чение имеет обмен теплокровных блохами—переносчиками чумы
в очагах этой инфекции.
ЭКТОПАРАЗИТЫ СУСЛИКОВ
Блохи. Среди эктопаразитов сусликов наиболее важны в ме-
дицинском отношении блохи — основные переносчики и возможные
хранители чумы. У ряда видов блох образуется блок преджелудка,
в возникающем вследствие этого сгустке крови сохраняется и раз-
множается чумной микроб. Способность блох впадать в анабиоз
при низких температурах делает возможным сохранение чумного
микроба в них в течение спячки сусликов. При пробуждении сус-
ликов температура в гнезде увеличивается, блохи становился ак-
тивными и могут заразить животных. Возможно сохранение чум-
ного микроба и в фекалиях блох, заражение сусликов в этом случае
происходит при обновлении нор. Прослежено сохранение возбу-
дителя чумы в экскрементах блох и в мертвых блохах до 750 дней
(срок наблюдений) при температуре —10° и относительной влаж-
ности 55%. При более высоких температурах микроб погибал
(Иофф, 1941; Величко и др., 1978).
Специфические и широкораспространенные виды блох, парази-
тирующие на разных видах сусликов, представлены в табл. 1.
Таблица составлена по сведениям из определителей И. Г. Иоффа
с соавторами (Иофф, Скалой, 1954; Иофф, Тифлов, 1954; Иофф и
др., 1965), дополненные по данным сводок М. А. Микулина, 1956;
Млекопитающие Казахстана, 1969; Г. С. Летова и Г. И. Летовой,
1973; Н. Ф. Лабунец и др., 1974; Н. Ф. Лабунец и П. А. Писарской,
1977; И. В. Назаровой, 1981; 3. П. Масленниковой и др. (1982)
и др. Специфические виды составляют до 90% в сборах с сусликов.
Доминирующие виды блох могут меняться на протяжении ареала
сусликов. Так, в восточной части ареала крапчатого суслика в
Волжско-Камском крае основная блоха — С. tesquorum, в Бело-
руссии — Ct. breviatus и Ct. orientalis, а на Украине — С. simplex.
Различие в наборе видов блох, встречающихся на сусликах в
разных частях их ареалов, связано с ландшафтными и фаунистиче-
скими условиями. Наибольшее число видов — 81—обнаружено
22
Rodentia
Таблица 1. Блохи сусликов*
Вид блохи Виды сусликов
Длиннохвостый Даурский Реликтовый Малый Горный Крапчатый Малоазийский Европейский Большой Краснощекий Желтый
Специфические блохи сусликов
Neopsylla abagaitui N. setosa 0 Д Д Д Д Д Д Д Д Д Д
Cerathophyllus tesquorum Д Д — Д Д 0 — — 0 0 0
C. relicticola Д
C. gracilis c —
C. martinoi — — — — — — — 0 — — —
C. ullus c —
C. simplex — — — c — Д — Д — — —-
C. transcaucasica Д
C. trispinus — — — c — — — — c c Д
Oropsylla ilovaiskii — — 0 c 0 — 0 — Д Д 0
0. asiatica (alaskensis) 0 0 —
Ctenophthalmus pollex — — — c — — — — — — c
Frontopsylla semura — — — 0 0 0 — — — — 0
F. elatoides 0 0 — 0 — — — — — 0 —
F. kunitsky — — 0 — — — — — — — 0
Широкораспространенные блохи грызунов открытых пространств
N. pleskei 0 0 0 — — — — — — 0 о
N. bidentatiformes 0 0 0
N. teratura 0 — 0 —
N. mana 0 c
Ct. breviatus 0 0 — 0 — 0 — — 0 0 0
Ct. orientalis — — — c 0 0 — 0 — — 0
Ct. teres — 0
Ct. golovi 0
F. luculenta 0 0 — 0 —
F. hetera 0 — 0 —
F. elata c 0 — 0 —
Rhadinopsylla li 0 — — 0 0 — — — — 0 —
R. daurica 0 0 — 0 —
R. pseudodaurica 0 0 —
* Доминирующие виды блох (д) специфичны для сусликов, часто встречаются и
многочисленны; обычные виды (о) часто встречаются, но могут быть немногочисленны,
спорадические виды (с) встречаются лишь в некоторых частях ареала сусликов, но в
местах распространения могут быть многочисленными.
Грызуны
23
на длиннохвостом суслике, обладающем обширным ареалом и
заселяющем разнообразные биотопы. В отдельных местообитаниях
видов блох, паразитирующих на сусликах этого вида, обычно
около 10, реже 20—30 (Летов, Летова, 1973). Типичны всего не-
сколько видов. На малом и желтом сусликах обнаружено соот-
ветственно 41 и 44 вида, на краснощеком — 31, на крапчатом —
21, на европейском — И.
Среди блох сусликов многие виды являются переносчиками
чумы. В СССР это Cerathophyllus tesquorum, Neopsylla setosa,
C. trispinus, N. abagaitui, N. pleskey, Oropsylla asiatica, O. ilo-
vaiskyi, Ctenophthalmus breviatus, Ct. pollex, Ct. orientalis, Ct.
golovi, Frontopsylla semura, F. elatoides (Ралль, 1960; Гончаров,
1983). Наибольшее значение в эпизоотологии чумы имеют С. tes-
quorum и N. setosa, наиболее широко распространенные и много-
численные. От них выделена большая часть культур. От С. tes-
quorum в Прикаспии — 57%, в Тувинском природном очаге — 68,
в Горно-Алтайском — 70,5, в Забайкалье — 76% (Миронов и
др., 1965; Жовтый, 1975; Воронова, 1978). С. tesquorum—блоха
шерсти, массовый выплод ее происходит летом и совпадает с пе-
риодом расселения сусликов, когда развиваются наиболее мощные
чумные эпизоотии. N. setosa — блоха гнезд сусликов, обилие ее
возрастает осенью, к концу эпизоотии, и большая численность
сохраняется всю зиму и ранней весной. В связи с биологическими
особенностями этих блох им отводится различная роль в поддер-
жании эпизоотий чумы; считается, что С. tesquorum разносит
возбудителя в популяции сусликов, N. setosa участвует в сохра-
нении чумного микроба от одного эпизоотического периода до дру-
гого (Тинкер, 1940; Иофф, 1941; Лабунец, Сардар, 1961; Сазо-
нова, 1962; Шатас, 1965; и др.). В древних поселениях малого
суслика временами бывает значительна численность Ct. pollex;
при протекании чумных эпизоотий от них выделяется до 31% всех
культур, изолированных от блох (Герасимова, 1980).
Разные виды сусликов, обитающие в одних биотопах, обмени-
ваются специфическими видами блох. Так, на малых сусликах
встречается С. trispinus — специфическая блоха желтых сусликов,
О. ilovaiskii—специфическая блоха больших сусликов. Широ-
кие паразитарные контакты характерны для сусликов с другими
грызунами, что способствует вовлечению в эпизоотии разных
видов носителей. Наиболее важен обмен блохами сусликов с тар-
баганами, песчанками и пищухами. В местах совместных посе-
лений с сурками на сусликах обнаружены блохи сурков: О. si-
lantievi и Rhadinopsylla ventricosa, зарегистрированные как пере-
носчики чумы. В пустынных районах на них паразитируют блоха
тонкопалого суслика Rh. daca и блохи песчанок: Xenopsylla соп-
formis, X. gerbilli, X.hirtipes, X. nuttalli, X. skrjabini, C. aralis,
C. laeviceps, C. turkmenicus, C. tersus, Coptopsylla lamellifer, C.
bairamalensis, Ct. dolichus, Echidnophaga ochanini, Paradoxopsyl-
24
Rodentia
lus repandus, P. teretiformis, Rh. cedestis, Rh. bivirgis, Stenoponia
vlasovi, из них наиболее важное значение имеет обмен блохами
рода Xenopsylla — активными переносчиками чумы. Их обилие
бывает значительным и на сусликах.
В Алтайских очагах чумы эпизоотологическое значение имеет
обмен блохами сусликов и пищух. На длиннохвостых сусликах
встречаются блохи пищух: Amphalius runatus, С. desertus, С. sca-
loni, Ct. armatus, Ct. hirticrus, Paradoxopsyllus dashidorzhii, P.
scorodumovi, Rh. altaica, из них важными переносчиками чумы
служат A. runatus и Ct. hirticrus (54% культур выделено от этих
блох в очагах Северо-Западной Монголии) (Кошкин, Хумархан,
1974).
Обычно на сусликах в небольших количествах встречаются
блохи тушканчиков, полевок и хомячков, серых крыс и домовых
мышей. В местах соприкосновения поселений сусликов с лесными
биотопами на них единичны блохи лесных мышей, белок, бурун-
дуков и даже землероек. На пустынно-степных видах сусликов
обычны встречи блох ушастых ежей и птиц-норников. Оставляют
в норах сусликов своих паразитов и мелкие хищники, среди них -
блохи: Pullex irritans, Chaetopsylla homoeus, Ctenocephalus canis,
Ct. felis. Многие авторы придают большое значение обнаружению
на сусликах блохи Pullex irritans, нападающей и на человека.
Однако находки этой блохи всегда единичны и вряд ли могут пред-
ставлять эпидемиологическую опасность. Списки видов блох, пара-
зитирующих на американских сусликах, представлены в сводках
Ю. М. Ралля (1960) и Поллитцера (Pollitzer, 1960), они также
Включают виды, свойственные как самим сусликам, так и контак-
тирующим с ними другим животным: хомякообразным, гоферам,
белкам и бурундукам. С сусликами Неарктики связаны блохи
p.Opisocrostis (7 видов и подвидов) (Lewis, 1975). На тринадцатипо-
лосном суслике р. Ictidomys 99% сборов составляют О. bruneri
и Thrassis bacchi (Kinzel, Larson, 1973). Для сусликов Otospermo-
phillus beechey специфичны, кроме того, Diamanus montanus и
Hoplopsyllus anomalis (Ryckman, 1971). От блох сусликов в Аме-
рике также неоднократно выделяли возбудителя чумы. Наиболь-
шее значение как переносчики чумы там имеют Т. bacchi, Н. ano-
malis и D. montanus (Pollitzer, 1960).
В последние годы группа авторов, работавшая в полупустынях
Западного Казахстана, пришла к парадоксальному выводу, что
паразитарный контакт в поселениях малых сусликов не может
обеспечить развитие эпизоотий чумы в период расселения молод-
няка путем трансмиссии возбудителя через блох. Проводя мате-
матические расчеты, авторы доказывают, что трансмиссией может
быть объяснена очень небольшая часть заражений. Эта группа
ученых большую роль отводит «первичным» заражениям, имея в
виду заражение от объектов внешней среды (Солдаткин и др.,
1984; Шевченко и др., 1984).
Грызуны
25
Вши. На сусликах паразитируют три вида вшей: Neochaemato-
pinus laevisculus, Enderleinellus propinquus и E. suturalis.
Пораженность сусликов вшами достигает 90% и более. С от-
дельных особей длиннохвостых сусликов снимали по 900 экз. этих
паразитов (Вершинина и др., 1972). Значительная часть сусликов
бывает поражена вшами уже вскоре после пробуждения, причем
обычно самцы больше, чем самки. Во время лактации часть вшей
переходит на молодых (на голых детенышей вши не переходят,
только на подросших зверьков с развитым волосяным покровом).
Нередко молодые бывают больше заражены вшами, чем взрослые.
У самок без выводков зараженность вшами летом увеличивается,
у самцов такого не происходит, наблюдается ослабление поражен-
ности этими паразитами ко времени залегания в спячку. Вши
сусликов размножаются в течение всего активного периода хозяев
(Зарубина, 1963; Соснина, Артемьев, 1968; Соснина, 1982). По-
требляя большое количество крови, вши могут участвовать в цир-
куляции различных возбудителей (Бондаренко М., 1961; Олсуфьев,
1970; Бурлаченко и др., 1971; и др.). Вши тарбагана N. palearc-
ticus могут переходить на сусликов и на них питаться. Экскре-
менты зараженных вшей содержат много бактерий чумы (Жовтый,
Емельянова, 1959).
Кровососущие двукрылые. В норах желтого суслика находили
комаров Aedes caspius и Culex hortensis, мошек Simulium equinum
vlasovi и москитов (виды не указаны). Москиты встречаются в Сус-
линых норах в гораздо меньшем количестве, чем в норах больших
песчанок (Болдырев, 1959; Петрищева, 1962).
Иксодовые и аргасовые клещи. В СССР специфичен для сусли-
ков лишь один вид клещей — Rhipicephalus schulzei — гнездо-
норовый паразит, все стадии его развития проходят на сусликах.
Выплод его личинок приурочен к появлению молодых. Этот клещ
паразитирует на малом, желтом и рыжеватом сусликах в полупу-
стынях и сухих степях юго-востока РСФСР, Средней Азии, Азер-
байджана, Северного Кавказа. Для него установлено спонтанное
носительство чумы, туляремии и пироплазмоза сусликов. В экс-
перименте доказана передача чумы сусликам. В небольшом коли-
честве R. schulzei встречается на других норных грызунах. На
северо-востоке Китая обнаружен клещ Haemaphysalis verticalis,
все стадии которого проходят на даурском суслике. Значение его
не изучено.
Свойственны сусликам все гнездо-норовые клещи, распростра-
ненные в пределах их ареалов: в степных, лесостепных и полупус-
тынных районах СССР: Ixodes laguri, I. crenulatus, I. redicorcevi,
в пустынях и полупустынях — Haemaphysalis numidiana. Эти
клещи имеют широкий круг хозяев, помимо сусликов, встречаются
на всех норных теплокровных, зарегистрированы как переносчики
природноочаговых инфекций (табл. 2).
Кроме представленных в таблице видов клещей, на крапчатом
Таблица 2. Иксодовые клещи, обычно паразитирующие на сусликах в СССР
Вид клеща и паразитирующие фазы Вид суслика Частота встреч на сусликах Распространение клещей Инфекции, переносимые клещами
R. schulzei * 1, n, imago I. crenulatus ** 1, n, imago I. laguri ** 1, n, imago H. concirjna 1, n H. punctata 1, n C. pygmaeus C. fulvus C. rufescens C. pygmaeus C. suslica C. citellus C. relictus C. dauricus C. pygmaeus C. fulvus C. suslica C. citellus C. undulatus C. pygmaeus C. musicus Многочислен Обычен, но ча- ще паразитиру- ет на сурках Многочислен Местами много Обычен Сухие степи и полупустыни юга ев- ропейской части СССР, Сев. Кавка- за, Азербайджана, Казахстана, Средней Азии Предгорная и горная степь и полу- пустыня Закарпатья, Молдавии, Крыма, Закавказья, Казахстана, Таджикистана, Киргизии, Забай- калья Степи Украины, Нижнего Дона, Да- гестана, Армении, Зап. и Сев. Ка- захстана Леса и лесостепь БССР, Крыма, Закавказья, Средней Азии, Зап. и Вост. Сибири, Дальнего Востока От горных лесов до полупустынь юга европейской части СССР, Казах- стана, Средней Азии Чума, туляремия, лихорадка Ку, пироплазмоз сусликов Чума, лихорадка Ку, клещевой сыпной тиф, клещевой энцефалит, эризипелоид Чума, туляремия, риккетсиозы, геморрагическая лихорадка Туляремия, клещевой сыпной тиф, клещевой энцефалит, бру- целлез Туляремия, клещевой сыпной тиф, бруцеллез, пироплазмоз
Roden tia
Н. numidiana 1, n, imago C. pygmaeus C. fulvus Обычен Степи» полупустыни, пустыни Кры- ма, Закавказья, Казахстана, Средней Азии Чума, лихорадка Ку
D. рictus 1, n, imago C. erythro- genis Местами обы- чен Луга и лесостепь Средней Европы, Крыма, Кавказа, Закавказья, Казах- стана, Киргизии, Алтая Туляремия, лихорадка Ку, кле- щевой сыпной тиф, клещевой эн- цефалит, бруцеллез, Омская ге- моррагическая лихорадка, вирус Зап. Нила, пироплазмоз, нутта- лиоз
D. marginatus 1, n C. suslica C. pygmaeus C. fulvus C. rufescens На С. suslica много, на дру- гих редко Равнинные и горные степи и лесо- степь юга европейской части СССР, Закавказья, Казахстана, Средней Азии, южной и восточной частей Зап. Сибири Туляремия, лихорадка Ку, кле- щевой сыпной тиф, бруцеллез, листериоз, гемоспоридоз, нутта- лиоз, энцефаломиелит лошадей
D. nuttalli 1, n, imago C. undulatus Многочислен Степные участки Зап. и Вост. Си- бири Туляремия, клещевой сыпной тиф, бруцеллез, сальмонеллез, эризипелоид
H. asiaticum C. fulvus Обычен Пустыня и полупустыня Южного и Восточного Закавказья, Казахстана, Средней Азии Чума, лихорадка Ку, клещевой сыпной тиф, тейлериоз крупного рогатого скота
* Специфичный паразит сусликов.
** Обитает в норах.
Примечание. Таблица составлена по: Брегетова и др., 1955; Сердюкова, 1956; Болдырев, 1959; Пионтковская, Жмаева, 1962;
Попов, 1962; Нельзина и др., 1963; Федорова, 1968; Емельянова, Терещенко, 1971; Животягина, Иголкин, 1976; Филиппова, 1977.
Грызуны
28
Rodentia
суслике редко встречается Н. otophila, на малом — I. redicorzevi,
D. daghestanicus и R. rossicus, на малом, горном и желтом — Н.
sulcata, на желтом — R. pumilio, R. turanicus и Н. anatolicum.
Там, где длиннохвостые суслики обитают вблизи лесных биотопов,
на них зарегистрированы D. silvarum и I. persulcatus, на красно-
щеком— D. pictus и I. persulcatus, на крапчатом — I. trianguli-
ceps. Эти клещи переносят возбудителей энцефалита, риккетсиоза,
туляремии (Попов, 1962; Животягина, Иголкин, 1976; Филиппова,
1977).
В Канаде и США на сусликах паразитируют виды рода Ixodes.
Основными хозяевами суслики служат для клеща I. sculptus,
обычны на них I. marmotae, I. ochotonae, I. conperati. Кроме того,
на них встречаются: I. rugosus, I. hearlei, I. jellisoni, I. spinealis,
I. cookei, I. kingii (Колонии, 1981).
На желтом суслике в Средней Азии и Казахстане встречаются
и аргасовые клещи Ornithodorus papillipes и О. tartakovskii —
основные переносчики клещевого спирохетоза, они переносят и
возбудителя лихорадки Ку (Поспелова-Штром, 1953; Брегетова и
др., 1955; Болдырев, 1959; Филиппова» 1966; Федорова, 1968;
Шеханов, 1970). Экспериментально установлено, что в организме
О. tartakovskii чумной микроб остается жизнеспособным в течение
1138 суток (Билялов и др., 1983).
Гамазовые клещи. Наиболее свойственны сусликам Hirstio-
nyssus criceti и Haemogamasus citelli — обитатели их гнезд. Раз-
множение этих клещей связано с периодом выкармливания молод-
няка хозяев, за это время численность клещей возрастает. Зара-
женность сусликов Hir. criceti доходит до 80%. Этот клещ спосо-
бен сохранять риккетсий Бернета. Возможно участие Hg. citelli
и Eulaelaps kolpakova (клеща степных грызунов, в том числе и
сусликов) в передаче бруцелл, о чем свидетельствуют эксперимен-
тальные данные. На европейском суслике встречается мышиный
клещ Allodermanissus sanguineus — переносчик риккетсиозной оспы.
В гнездах длиннохвостого и краснощекого сусликов обнаружен
Haemogamasus mandschuricus, подозреваемый в переносе риккетсий.
Кроме того, на сусликах встречаются широкораспространенные
обитатели гнезд грызунов: Haemogamasus nidi, Hg. glasgovi, Eulae-
laps stabularis, мышиные клещи Hirstionyssus musculi, Laelaps
algericus, Myonissus decumani. В МНР на длиннохвостом суслике
широко распространен Hg. kitanoi, паразитирующий также на
пищухах и сурке (Брегетова и др., 1955; Земская, 1973; Рейтблат
и др., 1975; Шевченко и др., 1975; Животягина, Иголкин, 1976;
Иголкин, 1978; Кошкин и др., 1978; и др.).
Клещи-краснотелки. На сусликах обнаружено 15 видов: Lep-
totrombidium oblontata — на длиннохвостых и даурских сусликах,
L. schlugeri — на даурском, L. pulchrum — на малом, Neotrom-
bicula autumnalis — на длиннохвостом, малом, крапчатом и евро-
пейском сусликах; Н. japonica — на длиннохвостом, крапчатом и
Грызуны
29
европейском сусликах, N. тега — на реликтовом, N. digenula и
N. minuta — на крапчатом, N. montchadskyi — на желтом, Euschen-
gastia rotundata — на длиннохвостом и даурском, Е. taurica —
на малом, Schoutedeichia angusta — на малом, крапчатом и евро-
пейском; Chelodonia flava — на длиннохвостом и малом, С. ikaeoen-
sis — на малом и крапчатом (Кудряшова, 1979).
Наиболее распространенный клещ N. autumnalis зарегистриро-
ван как переносчик лихорадки Ку (Шлугер, 1962).
СУСЛИКИ — НОСИТЕЛИ ИНФЕКЦИЙ
Чума. Наибольшее значение имеют суслики в эпидемиологии
и эпизоотологии чумы. Суслики, обитающие в Прикаспии, на Кав-
казе, в Казахстане, Средней Азии, Забайкалье, в МНР, КНР, яв-
ляются либо основными, либо важными второстепенными носите-
лями чумного микроба. Роль их в ряде мест повысилась после ист-
ребления сурков. Ряд видов сусликов, обитающих на Американ-
ском континенте, также служат хозяевами возбудителя.
Длиннохвостый суслик участвует в поддержании чумы в За-
падной и Северо-Западной Монголии и прилегающих районах
советского Алтая и Тувы: в Центрально-Алтайском, Монголо-Ал-
тайско-Хангайском и Хэнтейском очаговых регионах, а также в
Памиро-Алайском очаге (деление на очаговые регионы дается по
В. В. Кучеруку и Д. И. Бибикову, 1980). Северная часть его ареала
лежит вне энзоотичной территории.
Значение этого суслика в разных местах неравноценно. Эпи-
зоотологическая роль его велика в местах истребления тарбагана
(Некипелов, 1962). В Центрально-Алтайском регионе длинно-
хвостый суслик участвует в эпизоотиях чумы в Монгун-Тайгинском,
Хархира-Тургенском, Верхне-Чуйском и Южно-Сайлюгемском оча-
гах. В Монгун-Тайгинском очаге сурок выбит и суслик имеет пер-
востепенное значение. Зараженность его доходит до 3%. В Тувин-
ской части очага из выделенных за 1964—1979 гг. штаммов чумы
92% получены от длиннохвостых сусликов и его эктопаразитов.
Хархира-Тургенский очаг полигостален, к основным носителям
здесь относятся тарбаган, суслик и монгольская пищуха: 70%
всех культур здесь получено от специфической блохи суслика
С. tesquorum. В Верхне-Чуйском (Горно-Алтайском) и Южно-
Сайлюгемском очагах основной носитель — монгольская пищуха,
от нее и ее блох выделено 90% штаммов. Но длиннохвостые суслики
являются важными второстепенными носителями, активизация
эпизоотий чумы происходит обычно на фоне повышенной числен-
ности пищух и сусликов в местах их совместных поселений в меж-
горных долинах. ВМонголо-Хангайском очаговом регионе по числу
выделенных культур длиннохвостый суслик занимает второе место
после сурков, от сусликов выделено 3—16% культур, от сурков —
30
Rodentia
60—90%. Эпизоотии чумы в высокогорных опустыненных степях
Западного Хангая текут на фоне высокой численности тарбаганов,
сусликов и средних и высоких индексов их блох. В Северо-Во-
сточном Xангае чума поддерживается в основном тарбаганами,
суслик вовлекается в эпизоотии в годы наивысшей их активности.
В Хэнтейском очаговом регионе значение длиннохвостого суслика
в эпизоотологии чумы мало. Возбудитель выделяется от него редко.
В Памиро-Алайском очаге длиннохвостый суслик служит носителем
чумы наравне с красным сурком (Некипелов, 1959, 1962, 1978;
Бондаренко и др., 1974, 1975; Зундуй, 1974; Варшавский и др.,
1975; Галбадрах и др., 1978; Липаев и др., 1978; Устюжина и др.,
1975; Кучерук, Бибиков, 1980).
Даурский суслик — основной носитель чумы в Даурском оча-
говом регионе (регион занимает степи Юго-Восточного Забай-
калья, крайний северо-восток Монголии и северо-западные части
Внутренней Монголии (северо-восточный район КНР)). Ареал даур-
ского суслика полностью входит в очаговую территорию.
В этом регионе первое место по числу выделенных культур
чумы до 40-х годов занимали сурки, суслики были на втором месте
(с 1911 по 1946 г. в Ононо-Аргунском очаге от сурков было выде-
лено 77 штаммов, от даурских сусликов — 21). В результате ист-
ребления сурков с 1947 по 1965 г. чумы в природе не обнаруживали.
В 1966 г. после 19-летнего перерыва были выделены культуры
чумы от даурских сусликов; эти зверьки стали основными носите-
лями. С 1966 по 1970 г. выделено 140 штаммов, из них 34 — от
сусликов и 83 — от их блох. Зараженность даурских сусликов
после истребления сурков составляла 0,3—3% (Некипелов, 19596,
1975; Солодкая, 1974; Варшавский и др., 1975; Кучерук, Бибиков,
1980).
В Маньчжурских очагах даурский суслик служит основным
носителем наравне с крысой корако (Козакевич и др., 1981).
Большая часть ареала малого суслика расположена в преде-
лах энзоотичной по чуме территории, за ее пределами находятся
только некоторые северные и западные районы распространения
вида.
Малый суслик является основным носителем в Прикаспийском
степном очаге, занимающем южные степи и полупустыни Северного
Прикаспия и Предкавказья. Этот очаг делится на несколько более
или менее автономных очагов, до недавнего времени к ним отно-
сились: Зауральский, Волго-Уральский степной, Прикаспийский
северо-западный, Дагестанский равнинно-предгорный, Терско-Сун-
женский низкогорный (Инструкция по эпиднадзору, 1979). Однако
в настоящее время в Терско-Сунженском междуречье суслик почти
полностью истреблен и очаг, возможно, будет ликвидирован, если
возбудитель не сможет укорениться на полевках, численность
которых временами бывает значительна (Сорокина и др., 1982).
Наиболее устойчивые участки энзоотии приурочены к древним
Грызуны
31
и старым поселениям сусликов: Ергеням, предгорьям Дагестана
(Лавровский и др., 1972).
Обширные эпизоотии с 1912 по 1938 г. регистрировались на
востоке Ростовской области, на севере Ставропольского края, на
юге Волгоградской области и в Калмыкии (Денисенко и др., 1981).
Работы по истреблению сусликов, проводившиеся с 1934 по 1941 г.
и после Великой Отечественной войны на больших площадях,
резко снизили численность сусликов в этих районах. С 1934 по
1941 г. только в Ростовской области было проведено истребление
сусликов на площади 10 млн га, в Волгоградской области с 1934
по 1941 г. и с 1949 по 1956 г.— на площади 11 млн га (Миронов
и др., 1965). Интенсивное сельскохозяйственное освоение терри-
тории этих областей, а также ряд лет с неблагоприятными метео-
рологическими условиями (суровые зимы с резкими колебаниями
температуры, засуха в весенне-летнее время) не давали восстано-
виться численности сусликов. В результате долгое время эпизоотии
не регистрировались. Но в 70-х годах очаги чумы активизировались
и были выделены культуры микроба от сусликов и их блох на Ер-
генинской возвышенности, в Сарпинской низменности, Сулако-
Терской равнине и в Терско-Сунженском междуречье, а также в
Зауральской степи (Лавровский и др., 1972, 1978; Кучеров и др.,
1975; Попов Н. и др., 1976; Климченко и др., 1978). В Зауральском
степном очаге увеличение эпизоотического потенциала произошло,
кроме того, за счет расселения к северу большой песчанки и об-
разованию комплексных поселений этого важного носителя чумы
с малым сусликом (Сасыкин, Поляков, 1981; Шевченко, 1982).
Однако благодаря тому, что в настоящее время на европейской
части СССР крупные поселения сусликов разобщены, приурочены
к балкам и находятся под контролем работников противочумной
системы, не ожидается таких разлитых эпизоотий, какие наблю-
дались в 30-е годы (Лавровский и др., 1974; Попов Н. и др., 1976).
В западной части Прикаспийского северо-западного природного
очага чумы с 1930 по 1984 г. под влиянием сельскохозяйственной
деятельности человека общая площадь поселений малого сус-
лика, а также размеры очага уменьшились более чем на треть.
К 1978—1982 гг. поселения сусликов практически исчезли на
сплошь распаханных массивах Волгоградской области, в Сальско-
Донских и Ставропольских степях (Варшавский и др., 1985).
В Волжско-Уральских песках и северных пустынях Среднеази-
атского равнинного очага (Северо-Восточном Прикаспии и Север-
ном Приаралье) малый суслик играет роль в поддержании очагов
чумы как второстепенный носитель (Варшавский, 1959; Бурделов
и др., 1982). В этих местах он тесно контактирует с основными
носителями чумы: в Волжско-Уральских песках — с полуденными
и гребенщиковыми песчанками, в Северо-Восточном Прикаспии и
Северном Приаралье — с большими песчанками. Здесь суслики
часто устраивают свои норы на территории жилых нор больших
32
Rodentia
песчанок, а также заселяют брошенные песчанками норы. В Се-
верном Приаралье, в южной и западной частях Каракумов неодно-
кратно выделяли культуры чумы от сусликов, но обычно в связи
с эпизоотией чумы на больших песчанках (Наумов и др., 1972).
Депрессия очагов чумы, наблюдавшаяся в Северо-Западном При-
каспии, охватила и Приаралье. Возобновление эпизоотий про-
изошло одновременно на большом пространстве. В 1971 г. в При-
аралье после 22-летнего перерыва опять регистрировалась чума
на сусликах (Варшавский, 1977). В те же годы произошла активи-
зация очагов чумы, где основным носителем служит даурский
суслик. Видимо, существуют общие закономерности проявления
очагов суслиного типа (Кузнецова и др., 1980). Некоторые авторы
связывают их с цикличностью солнечной активности (Лавровский,
1969).
Горный суслик является основным носителем чумы в Цент-
рально-Кавказском очаговом регионе (в Приэльбрусье). Регион
делится на четыре хорошо изолированных очаговых района: Верхне-
Кубанский, Кубано-Малкинский, Малко-Баксанский, Баксано-
Черекский. За этой территорией наблюдения ведутся с 1971 г.,
во всех очагах регистрировались эпизоотии на сусликах и выделя-
лись культуры возбудителя от их блох. Выделенные штаммы от-
носились к типичной сусликовой разновидности микроба чумы,
но эпизоотии в популяциях этих сусликов более растянуты, за-
хватывая весь активный период горных сусликов, который зна-
чительно длиннее, чем у равнинных малых сусликов (Акиев и
др., 1972; Абдурахманов и др., 1973; Пилипенко и др., 1974; Най-
ден, Петров, 1980; Емельянов и др., 1982).
Места стойкого укоренения чумы в Приэльбрусье характеризу-
ются вялым течением эпизоотий, зараженность зверьков в них
невелика, подсчитано, что один зараженный зверек приходится
на 400 здоровых. Чума стойко держится на блохах сусликов, от
которых выделяется более 90?4 всех штаммов (Дятлов и др., 1981).
Реликтовый суслик в Киргизии не имеет эпидемиологического
значения, хотя его поселения соприкасаются с поселениями сур-
ков (Иофф, 1949).
Желтый суслик не играет самостоятельной роли в сохранении
возбудителя чумы, так как он обычно распространен спорадично
и численность его невелика, а чувствительность к чумной инфек-
ции высокая. Но он регулярно вовлекается в эпизоотии, текущие
на других грызунах, так как поселения его и основных носителей
совпадают. В Прикаспийских очагах он обитает на одних терри-
ториях с малыми сусликами, в Волжско-Уральских песках — с
гребенщиковыми и полуденными песчанками, в Среднеазиатских
пустынях — с большой песчанкой. Выделение культур от них
зарегистрировано в Казахстане и Западной Туркмении, Северо-
Восточном Прикаспии, Северном Приаралье, на Устюрте, в Кы-
зылкумах и Муюнкумах. Там, где численность желтого суслика
Грызуны
33
высока, в его поселениях регистрируются разлитые эпизоотии.
Так, в 1964 г. на Красноводском полуострове в совместных место-
обитаниях желтого суслика и краснохвостой песчанки было вы-
делено 157 культур, из них 109 от желтого суслика и его эктопа-
разитов (46 — от суслика, 56 — от его блох, 6 — от клещей и 1 —
от вшей) (Быков, 1968; Миронов и др., 1965; Хрусцелевский, Пей-
сахис, 1969; Крылова, Шилова, 1970; Бурлаченко и др., 1971;
Наумов и др., 1972).
Основная часть ареала большого суслика лежит за пределами
распространения возбудителя чумы, только южная его часть за-
ходит на энзоотичную территорию в Западном Казахстане, где
большой суслик вовлекается в эпизоотии, текущие на малых сус-
ликах (Ралль, 1960; Миронов и др., 1965).
Краснощекий суслик играет роль второстепенного носителя
чумы в очагах Центрально-Азиатских пустынь, где основные
носители чумы — песчанки. Возбудитель чумы от этих сусликов
выделен в районе горных степей в Восточно-Гобийском аймаке
МНР во время эпизоотии среди монгольских песчанок (Некипелов,
1959; Дэмбрэл и др., 1974; Варшавский и др., 1975).
На Американском континенте возбудителя чумы выделяли от
всех представителей рода Citellus, либо от их блох, а также от
скалистых сусликов (Р. Otospermophilus), антилоповых сусликов
(р. Ammospermophilus) и иктидомисов (р. Ictidomys): от С. colum-
bianus в США (шт. Вашингтон, Орегон), С. richardsoni (Монтана,
Невада, Вайоминг) и в Канаде (Альберта, Саскачеван), С. beldingi
в США (Калифорния, Невада, Орегон), С. armatus (Монтана, Айдахо,
Юта, Вайоминг, Вашингтон) и от С. waschington (Вашингтон) и
от блох, собранных с С. townsendi и с С. brunneus (Айдахо), от
скалистых сусликов Otospermophillus variegatus (Юта, Аризона,
Колорадо, Нью-Мексико), С. beechey и О. lateralis (Калифорния)
от антилоповых сусликов Ammospermophilus leucurus (Аризона,
Калифорния), от блох иктидомисов, снятых с Ictidomys tridecem-
lineatus, I. mexicanus, I. spilosoma (Нью-Мексико и Техас) (Pol-
litzer, 1960).
Некоторые американские ученые считают, что в поддержании
очагов чумы основное значение имеют резистентные мышевидные
грызуны, а суслики вовлекаются в эпизоотию благодаря тесному
с ними контакту и способствуют ее распространению (Kartman
et al., 1958; Marchette et al., 1962).
Сезонность развития эпизоотий чумы в популяциях сусликов
определяется изменениями чувствительности зверьков, их числен-
ности и поведения. Инфекционная чувствительность сусликов к
чуме значительно изменяется по временам года и связана с физио-
логическим состоянием зверьков. Экспериментами, проведенными
с разными видами сусликов, показано, что взрослые особи наиболее
устойчивы к возбудителю в первое время после пробуждения из
зимней спячки и в осенне-зимний период. Увеличение резистент-
2 Зак. 3575
34
Rodent la
ности к осени связано с накоплением жира, витамина Е, изме-
нением состава жирных кислот и других биохимических процессов,
а также с понижением температуры тела. Высокая устойчивость
зверьков в начале весны возникает в результате несовершенной
терморегуляции и пониженного обмена, наблюдающихся вскоре
после пробуждения (Калабухов, 1958, 1969; Козакевич, 1969).
Молодые суслики обладают высокой инфекционной чувствитель-
ностью в первое время после выхода на поверхность, затем их
чувствительность снижается и даже может оказаться более низкой,
чем у взрослых зверьков. Наблюдаются большие индивидуальные
колебания чувствительности зверьков, определяющиеся разли-
чиями в упитанности, уровне обмена веществ, насыщенности крови
гемоглобином, числе лимфоцитов в крови, соотношении жирных
кислот, содержании витаминов С и Е (Козакевич, 1969).
В поселениях сусликов — основных носителей чумы: малых,
горных, длиннохвостых и даурских — эпизоотии развиваются по
общей схеме. Первые больные зверьки обнаруживаются в период
спаривания сусликов. Причиной их заболеваний может быть обо-
стрение инфекции, полученной перед залеганием в спячку, или
они могут заразиться при расчистке нор с оставшимися трупами,
инфицированными паразитами или субстратом. Распространению
эпизоотии в это время способствует большая подвижность самцов
в период гона и перемещения годовалых сусликов из зимовочных
нор, которые часто оказываются на индивидуальных участках
взрослых особей, ранее молодых ушедших в спячку. Однако ве-
сенние эпизоотии краткосрочны и редко принимают разлитой
характер, так как численность сусликов в это время невелика и
быстро устанавливается их территориальное поведение, при ко-
тором внутривидовые контакты минимальны. Популяция состоит
из взрослых маловосприимчивых особей.
Разлитые эпизоотии возникают в период расселения молодняка
(в июне-июле), так как после выхода молодых увеличивается плот-
ность населения сусликов, при расселении суслики наиболее под-
вижны, в это время наиболее часты внутри- и межвидовые кон-
такты. Расселяющиеся зверьки расчищают много заброшенных
нор, в которых могут быть трупы, голодные блохи и зараженные
экскременты. Популяция состоит в основном из молодых более
восприимчивых зверьков. К этому времени приурочено размножение
и увеличение численности блох — переносчиков чумы. Кроме
того, расселение часто совпадает по времени с засухой, которая
способствует увеличению подвижности и чувствительности зверь-
ков, ослабленных недоеданием. В засушливые годы чаще отмечали
развитие эпизоотий на сусликах (Туманский, 1935; Тинкер, 1940;
Гайский, 1949; Калабухов, 1949; Федоров, Рогозин, Фенюк, 1955;
Демяшев, 1964).
Эпизоотии затухают перед залеганием сусликов в спячку,
когда сокращаются их подвижность и контакты и увеличивается
Грызуны
35
резистентность к возбудителю одновременно с накоплением
жировых запасов.
С сезонными изменениями чувствительности сусликов к чуме
связан один из механизмов сохранения возбудителя в межэпизоо-
тический период, так называемая «латентная чума». Низкая чув-
ствительность сусликов в осенне-зимний период приводит к тому,
что зверьки, заразившиеся перед впадением в спячку, могут бла-
гополучно перезимовать, размножение микроба в их организме
начинается после пробуждения. Это доказано экспериментальными
работами по заражению сусликов малыми дозами возбудителя
или укусами блокированных блох. У сусликов, находившихся в
спячке, развивались затяжные формы инфекции, генерализация
процесса наступала после перехода зверьков в активное состояние
(Гайский, 1944, 1949; Давтян и др., 1971; Гувва и др., 1978). На-
блюдающаяся в некоторых случаях гибель сусликов от чумы в
норах в зимний период объясняется неурочным пробуждением
зверьков во время оттепелей или слишком сильных холодов, а
также ранним пробуждением плохо упитанных особей. Экспери-
ментами показано, что у ослабленных особей септицемия разви-
вается в более ранние сроки (Клец и др., 1959).
Опыты с заражением предварительно иммунизированных сус-
ликов показали, что при этом может либо произойти прорыв им-
мунитета с развитием острого генерализованного процесса и ги-
бель животного, либо гибель проникших в иммунный организм
сусликов вирулентных бактерий чумы, или длительное бессимп-
томное носительство с возможной генерализацией инфекционного
процесса в отдаленные сроки после заражения. Находки серопо-
зитивных особей в природе достаточно часты, они составляют
фонд для формирования бактерионосителей при повторных зара-
жениях (Пилипенко и др., 1980).
Существуют видовые различия чувствительности у сусликов.
Многие авторы отмечают сравнительно высокую чувствительность
к чуме желтых сусликов. Зараженные в активный период даже
малыми дозами, они погибают от септической формы чумы (Шабаев,
1946; Пейсахис и др., 1958; Гувва, 1975; Русакова и др., 1976).
Однако и у них наблюдаются сезонные изменения чувствительности
и могут быть затяжные формы чумы, заканчивающиеся выздоров-
лением (Смирнов, 1956). Высокая инфекционная чувствительность
к возбудителю чумы обнаружена и у европейского суслика (Акиев
и др., 1981). Большой суслик более устойчив к чуме, чем желтый
и малый, а горные суслики Кавказа отличаются большей рези-
стентностью по сравнению с малыми сусликами равнин Ставро-
полья (Михайлова, Ледовская, 1973). Возможно, большая рези-
стентность горных сусликов определяется их пониженным обменом
по сравнению с равнинным видом (Чугунов и др., 1956).
Существуют данные о различиях в чувствительности к возбу-
дителю зверьков одного вида сусликов, но добытых в разных
2*
36
Rodentia
местах: в эпизоотических и свободных от чумы участках. Молодые
и взрослые малые суслики, добытые на Ергенях, где протекали
интенсивные эпизоотии, оказались в 10 раз чувствительнее сус-
ликов из окрестностей Тинаки Астраханской области, где чумы
не было (Бережнов и др., 1975). В противоречии с этими данными
находятся материалы А. И. Дятлова с соавторами (1979) по изу-
чению чувствительности популяций горного суслика. Показано,
что более резистентными оказались зверьки из мест, где часто
регистрировались эпизоотии. Авторы объясняют это отбором наи-
более устойчивых особей. Возможно, противоречия в вышеприве-
денных данных объясняются тем, что суслики проявляют различия
в чувствительности при заражении их разными штаммами. Так,
эксперименты показали, что длиннохвостые суслики из Тувы
высоко восприимчивы к местным штаммам возбудителя чумы и
сравнительно резистентны к горноалтайским штаммам (Тарасова,
Иннокентьева, 1975).
Туляремия. Меньшее значение имеют суслики в очагах других
инфекций. Они могут вовлекаться в эпизоотии туляремии в очагах
степного типа, текущие на других грызунах. При массовых вспыш-
ках этой инфекции на мышевидных грызунах отмечали падеж
сусликов. Спонтанное заражение туляремией в СССР установлено
у длиннохвостого, даурского, малого,, малоазийского, большого
и краснощекого сусликов, в КНР — у даурского суслика, в МНР —
у длиннохвостого суслика. В США и Канаде возбудитель туляре-
мии обнаружен у С. beechey, С. armatus, С. townsendii, С. colum-
bianus, С. richardsoni, С. leucurus (Туманский, Колесникова, 1935;
Некипелов, 1946, 1959а; Селезнева, 1949; Олсуфьев, Дунаева,
1960; Миронов и др., 1965; Дунаева, 1970; Брикман и др., 1977;
Дубовой и др., 1978; Некипелов и др., 1981; Infectious diseases of
wild mammals, 1970).
О. А. Байтанаев и M. А. Айкимбаев (1981) считают, что желтый
суслик вовлекается в эпизоотии туляремии тугайного типа, теку-
щие в основном на домовых мышах, гребенщиковых песчанках
и зайцах толаях.
Заражение сусликов возбудителем туляремии может происхо-
дить при каннибализме и через укусы иксодовых клещей. По чув-
ствительности и восприимчивости к туляремии суслики относятся
ко второй группе животных, т. е. заражение малыми дозами (1—10
микробных клеток) вызывает их инфицирование, но не гибель.
Наименьшая смертельная доза для сусликов — 100 млн — 1 млрд
микробных клеток. Сезон года не оказывает значительного влияния
на инфекционную чувствительность сусликов к туляремии, но
одновременное их заражение другими возбудителями (стрепто-
кокком, эризипелоидом) повышало их чувствительность (Олсуфьев,
Дунаева, 1960; Дунаева, 1970).
Бруцеллез. Возможно вовлечение сусликов в эпизоотии бруцел-
леза, протекающие на сельскохозяйственных животных. От длин-
Грызуны
37
нохвостых, малых, желтых, краснощеких сусликов, живущих
на пастбищах, где выпасался бруцеллезный скот, выделены бру-
целлы, либо получены положительные серологические реакции.
Суслики оказались восприимчивыми к козье-овечьему бруцеллезу
в эксперименте. При подкожном заражении малыми дозами — 10
микробных тел (м.т.), не эффективными для мышей, у сусликов
развивалась генерализованная инфекция, часть сусликов поги-
бала. При высеве материала из лимфатических узлов, печени, се-
лезенки, костного мозга и крови наблюдался обильный рост бру-
целл. Перорально суслики заражались при введении больших
доз — 1 млрд м. т.
Для вшей сусликов N. laeviusculus установлен факт передачи
бруцелл через укус. Эксперименты, проведенные с гамазовыми
клещами и блохами — паразитами сусликов, показали, что и они
могут воспринимать бруцелл. Клещи Hg. citelli сохраняли их
24 дня, а блохи N. pleskei и С. trispinus— до 20 дней. Вдали от
неблагополучных по бруцеллезу хозяйств от сусликов и других
диких млекопитающих бруцелл не выделяли (Ременцова, 1962;
Ременцова и др., 1963; Пинигин, 1974).
Псевдотуберкулез. Суслики, видимо, широко вовлекаются в
эпизоотии этой инфекции. В Калмыкии из 51 штамма J. pseudo-
tuberculosis, выделенных от грызунов, 43 пришлось на малых
сусликов. На Чукотке псевдотуберкулез установлен у американ-
ских сусликов, от которых выделено 6 штаммов (из 8).
Кишечный иерсиниоз. Американские длиннохвостые суслики
на Чукотке оказались широко зараженными возбудителем кишеч-
ного иерсиниоза J. enterocolitica; из 29 штаммов 24 было получено
от сусликов. Обнаруженные там скопления этих зверьков на го-
родских и поселковых свалках способствуют их контакту с синант-
ропными грызунами и передаче инфекции человеку. Кроме того,
суслики могут заражать и воду, так как они заселяют и берега
городских водохранилищ (Сомов и др., 1977; Чуринов, Чуринова,
1980).
Листерии изолированы от даурских и длиннохвостых сусликов
в Юго-Восточном Забайкалье, от среднего суслика — в МНР,
на территории Южно-Гобийского аймака (Дубовой и др., 1978;
Мунхтуяа и др., 1978; Некипелов и др., 1981).
Лептоспиры выделены от длиннохвостых сусликов в Иркутской
области (L. icterochaemorragiae), от европейских сусликов — в
Румынии (L. роптопа). В СССР у краснощекого суслика обнаружены
антитела к L. hebdomadis (Карасева, Свешникова, 1971; Некипелова,
Липин, 1976). Экспериментально установлена высокая воспри-
имчивость малого и крапчатого сусликов к некоторым лептоспирам
(Земсков и др., 1950; Горшанова, 1956). Однако суслики — оби-
татели сухих биотопов — не могут иметь большого значения в пере-
носе и сохранении этих возбудителей.
Эризипелотриксы неоднократно выделяли от сусликов. Эти
38
Rode tit i a
бактерии могут вызывать падеж сусликов, хотя и в меньшей сте-
пени, чем у полевок. Широту заражения этим паразитом популяций
сусликов может иллюстрировать обследование 10 тыс. даурских
сусликов в степях Юго-Восточного Забайкалья, при котором было
получено 446 штаммов. На территории по границе с МНР, обсле-
дованной в течение 25 лет, почти ежегодно находили зараженных
сусликов, инфицированность их составляла 10%. Выделяли эри-
зипелотриксы от длиннохвостых сусликов в Бурятии и МНР (Да-
ниленко и др., 1971; Головачева и др., 1974; Дубовой и др., 1978;
Некипелов и др., 1981).
Токсоплазмоз. При исследовании на спонтанную зараженность
возбудители Т. gondii изолированы от малых и желтых сусликов;
антитела обнаружены у длиннохвостого, крапчатого, европейского,
большого и краснощекого сусликов (Калякин, 1971).
Сальмонеллы выделены от даурского суслика (Некипелов и др.,
1981).
Ку-риккетсиоз, широко распространенный у животных, обна-
ружен и у сусликов. В СССР — в Киргизии у реликтовых сусликов,
в Омской области у краснощеких, в Казахстане у малых, желтых,
больших, краснощеких и длиннохвостых, в Красноярском крае и
Бурятской АССР у длиннохвостых. В Чехословакии и Болгарии —
у европейских сусликов. Болгарские и чешские ученые считают,
что суслики имеют важное значение в эпизоотологии этой инфек-
ции (Шиндаров, 1959; Rehn, Radvan, 1959). Значительный процент
положительных результатов получен при исследовании сывороток
малых сусликов в Целинном крае и реликтовых в Киргизии (со-
ответственно 12—14 и 38%) (Карулин, Пчелкина, 1958; Прореш-
ная и др., 1960; Федорова, 1968; Вершинина и др., 1972; Рапопорт,
1982). В эксперименте показана восприимчивость сусликов при
подкожном и пероральном заражениях, а также при питании на
них зараженных имаго и нимф иксодовых клещей, при этом рик-
кетсиемия у них была продолжительной, до 6 месяцев (Федорова,
1968).
Клещевой риккетсиоз. Возбудитель R. sibirica выделен от
длиннохвостых сусликов в очагах этого заболевания в Краснояр-
ском крае и Бурятии. Суслики служат основным прокормителем
клещей — переносчиков этого заболевания Dermacentor nuttalii
и D. silvarum. Чаще выделяли возбудителя и регистрировали у
зверьков положительные серологические реакции в местах совмест-
ного распространения сусликов и D. nuttalli (Кронтовская, 1940;
Коршунова и др., 1959, 1965; Липин и др., 1983).
Американские суслики С. columbianus, С. laterails, С. leocurus
причастны к поддержанию популяции R. ricketsii —< возбудителя
лихорадки Скалистых гор (Burgdorfer et al., 1962; Vest et al., 1965,
по: Тарасевич и др., 1980).
Вирусные инфекции. Там, где длиннохвостые суслики контак-
тируют с обитателями лесных биотопов (на гарях, вырубках и
Грызуны
39
опушках), на них встречается таежный клещ I. persulcatus. В Бар-
гузинской котловине при исследовании сывороток крови у них
обнаруживали антитела к вирусу клещевого энцефалита (Верши-
нина и др., 1972).
На Чукотке от американского длиннохвостого суслика выделен
вирус, по антигенным и биологическим свойствам близкий к воз-
будителю клещевого энцефалита, так и определенный авторами
(Сомов и др., 1978). Однако, поскольку район исследования лежит
за пределами ареала клещевого энцефалита и его основных пере-
носчиков, Э. И. Коренберг и Ю. В. Ковалевский (1981) считают,
что авторы ошибочно причислили выделенный агент к клещевому
энцефалиту, на самом деле он представляет ранее неизвестный
вирус, идентификация которого вопрос будущего. Возможно того
же порядка нахождение антител к клещевому энцефалиту у даур-
ского суслика в приграничной полосе Забайкалья и МНР (Гор-
чакова и др., 1974). При экспериментальном заражении штаммом
Софьин у сусликов (большого и крапчатого) развивалась вирусе-
мия с максимальными титрами 10"3, 10“4, суслики погибали в
течение 8 дней, из мозга и крови их выделяли вирус. Эти данные
говорят о большой восприимчивости сусликов и о том, что они не
могут быть длительными носителями возбудителя (Сарманова,
Думина, 1956; Морозов, 1963).
У краснощекого суслика обнаружены антитела к омской
геморрагической лихорадке (Леонов и др., 1970).
Суслики могут заражаться бешенством, при этом заболевание
у них может проходить без видимого клинического проявления.
Возбудитель бешенства был обнаружен у сусликов в природе в
Казахстане (Коломакин и др., 1973; Титов, 1975).
У сусликов, зараженных в активный период вирусом бешенства,
время инкубации составляло 32 суток, а в период зимней спячки
147—250 суток. У павших сусликов вирус регулярно обнаружи-
вали в головном мозге, у половины животных — в буром жире и в
отдельных случаях в слюнных железах. Однако многочисленные
обследования зимоспящих сусликов в Забайкалье и Приморье
давали отрицательные результаты, поэтому нет основания счи-
тать их эпизоотологически значимыми (Черкасский, 1985).
ЭКОНОМИЧЕСКОЕ И ЭКОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ СУСЛИКОВ
Значение сусликов не ограничивается областью медицины. Они
приносят вред сельскому и лесному хозяйству, служат предметом
охотничьего промысла и играют большую средообразующую роль
в населяемых ими ландшафтах.
Суслики, обитающие вблизи посевов зерновых, бахчей и ого-
родов, поедают культурные растения и могут приносить большой
экономический урон (Колосов и др., 1961). При закладке лесных
полос суслики серьезно вредили, выедая семенной материал, по-
40
Rodentia
вреждая проростки и молодые растения (Строганова, Юдин, 1950,
1951; Башенина, 1951; Лаврова, 1955).
Мех большинства сусликов мало ценен, но принимается заго-
товительными организациями. Наилучшим мехом обладает желтый
суслик, поэтому в ряде мест он был сильно истреблен. Жир сус-
ликов употребляется в технических целях. В охотничьей про-
дукции Казахстана желтый суслик занимает 27% (Бурамбаев,
Исмагилов, 1975; Кыдырбаев, 1982).
В местах повышенной численности сусликов, где много нор
и курганчиков, наблюдается заметное их влияние на почвенный
и растительный покров, а также на микрорельеф. Наибольшее
значение в этом отношении имеет малый суслик. Влияние сусликов
не однозначно в разных условиях в зависимости от исходных почв,
глубинных слоев земли, откуда суслики выносят грунт, и перво-
начального состава растительности. В одних случаях роющая
деятельность сусликов приводит к обеднению почвы, в других она
играет положительную роль, способствуя рассолению солонцов.
В разных зонах очень различно и их значение в преобразовании
растительности (Формозов, Воронов, 1939; Ходашова, Динесман,
1961; Абатуров и др., 1969; Киселева, 1976; Варшавский, 1982;
и др.).
На энзоотичной по чуме территории проводили борьбу с сус-
ликами путем газации их нор и разбрасыванием отравленных
приманок вручную или с самолета. Применяется и обработка
их нор инсектицидами, при этом достигается оздоровление терри-
тории без уничтожения зверьков (Федоров и др., 1955).
Для уничтожения сусликов вблизи населенных пунктов пред-
лагается фостоксин (действующее вещество — фосфид алюминия,
производство фирмы «Дегеш», ФРГ), закладываемый в норы по
1 таблетке, давал гибель 85% (Шилов, 1986).
Учет сусликов проводят подсчетом курганчиков, нор, верти-
кальных и косых, и выловом зверьков на площадках капканами
(№ 0 и № 1) либо при заливании нор водой (Варшавский, 1952).
Триб2 Tamiini Black, 1963
Род Sciurotamias Miller, 1901—белкообразные бурундуки
В роде два вида, распространены в Южном и Юго-Восточном
Китае. Окраска на спине одноцветная, темная полоса проходит
только от плеча до бедра. Защечные мешки слабо развиты. Насе-
ляют широколиственные и жестколиственные леса гор и равнин.
По образу жизни сходны с бурундуками, но в спячку не впадают.
Убежища в расщелинах и пустотах под камнями. Медицинское
значение неизвестно.
Грызуны
41
Род Tamias liliger, 1811—бурундуки
18 видов (Соколов, 1977) или 24 вида (Honacki et al., 1982),
из которых один вид распространен в Евразии (Т. sibirica), а ос-
тальные — в Северной Америке. Мелкие представители наземных
беличьих, для окраски характерны темные продольные полосы на
спине. Лесные, преимущественно семеноядные зверьки, делающие
запасы корма, имеют защечные мешки, которые используют при
сборе корма. На зиму впадают в спячку, активны днем. Хорошо
лазают по деревьям, но живут в норах на земле.
У 16 видов бурундуков определяли число хромосом, у всех
оказалось 2п=38; не определено число хромосом у Т. striatus
и Т. bulleri (Орлов, 1974; Орлов, Булатова, 1983).
Из американских бурундуков наиболее широкое распростране-
ние имеют Т. striatus Linn, 1758 — восточный бурундук, занима-
ющий юго-восток Канады, северо-восток и восток США, Т. town-
sendii Bachman, 1839 — бурундук Таунсенда, обитающий на юго-
западе Канады и северо-западе США, Т. minimus Bachman, 1839—
малый бурундук, заселяющий почти всю южную часть Канады.
Более ограниченный ареал у Т. amoenus Allen, 1820 — соснового
бурундука, обитающего на юго-западе страны, и у Т. ruficaudatus
Hawell, 1920 — рыжехвостого бурундука, заселяющего несколько
штатов Канады.
В западных штатах США распространен Т. quadrivittatus Say,
1823 — колорадский бурундук, на юго-западе — Т. dorsalis Baird,
1855 — скалистый бурундук. На юго-западе США, юге Калифор-
нии, Невады и в некоторых других штатах распространены не-
сколько видов бурундуков, имеющих изолированные ареалы в
виде узких полос или небольших участков: Т. cinereicollis J. Allen,
1890 — сероворотничковый бурундук, Т. quadrimaniculatus Grey,
1867 — длинноухий бурундук, Т. speciosus Merriam, 1893 — обык-
новенный бурундук, Т. panamintinus Merriam, 1893 — панаминт-
ский бурундук, Т. umbrinus J. Allen, 1890 — унтасский бурундук,
Т. palmeri Merriam, 1897 — бурундук Пальмера, Т. alpinus Mer-
riam, 1893 — альпийский бурундук, Т. sonomae Grinnel, 1915—
сономский бурундук. Также локально распространен в Мексике
Т. bulleri J. Allen, 1889 — бурундук Буллера. В Мексике и на
двух изолированных участках в Калифорнии обитает Т. merriam
J. Allen, 1889 — бурундук Мерриама.
В СССР распространен единственный вид Т. sibiricus Laxman,
1769 — азиатский бурундук с обширным ареалом, занимающим
лесную зону северо-востока европейской части СССР, на западе
идущий до линии Архангельск — Верхняя Тойма — Вологда, на
юг — до Ветлуги, Казани, Уфы, Южного Урала; в азиатской
части занимает Северо-Восточный Казахстан, Сибирь, Дальний
Восток, кроме Камчатки, Сахалин, Шантарские острова, южные
острова Курильской гряды, Хоккайдо, северную часть МНР,
42
Rodentia
Северо-Восточный Китай, п-ов Корея; по горным лесам Китая
проникает к югу до Сычуани. Карта ареала трибы Tamiini при-
ведена И. М. Громовым с соавторами (1965), а ареал вида С. И. Ог-
невым (1940) и Г. Б. Корбетом (Corbet, 1978).
В четырех странах Западной Европы: Бельгии, Норвегии,
Швейцарии, ГДР образовались поселения бурундуков в отдельных
лесных массивах, начало которым дали зверьки, сбежавшие от
своих хозяев, державших их в качестве декоративных животных.
Бурундуки благополучно живут там уже несколько лет, в лесу
под Брюсселем с 1974 г. (Dossier, 1978). Возможно, такие поселе-
ния послужат расширению ареала этого грызуна на запад.
Границы ареала азиатского бурундука совпадают с пределами
распространения лиственницы на севере Сибири и пихты в европей-
ской части СССР. Зверек населяет хвойные и смешанные леса,
предпочитая захламленные места с обильным подростом. На юге
Тувы и в МНР по интразональным лесным угодьям в поймах рек
проникает в степь. В горах идет до верхней границы леса, селится
в гольцовом поясе, где есть заросли кедрового стланца и кустар-
ники, встречаясь по каменистым россыпям вместе с пищухами и
полевками рода Alticola. На севере заходит в криволесье, на юге —
в островные леса. Заселяет вырубки и гари, если там остаются
группы деревьев или отдельные деревья и кустарники.
Наибольшая численность бурундуков наблюдается в кедров-
никах. В горах оптимальные условия для них в среднем поясе.
В лесах Западного Саяна наибольшая численность бурундуков
наблюдалась в высокотравных кедровниках: 20 на 1 кв. км. В Се-
веро-Восточном Алтае максимальная численность бурундуков от-
мечена в кедрово-пихтовой черневой тайге, на 1 кв* км приходи-
лось до 47 зверьков до выхода молодых из нор и до 225 — после их
появления. В других типах леса их было меньше, 2—27 взрослых
и 9—71 с молодыми. Наименьшая численность наблюдается в
предгорных колках: 1—3 в июне, 2—4 в конце мая — начале
августа. В подзоне южной лесостепи в окрестностях Новосибирска
в березово-осиновых перелесках и в сосновых борах с участками
смешанного леса весной приходилось 5—19 бурундуков на 1 кв. км.
В подзоне южно-таежных лесов возле Тобольска в березово-оси-
ново-пихтовой тайге осенью было 8 зверьков на 1 кв. км, в захлам-
ленных кедровниках с подростом — 21 зверек (Штильмарк, 1963 3);
Смирнов, Равкин, 1967; Телегин, 1980 4).
Оптимальные места обитания бурундука — захламленные леса
с обильным подлеском, хорошо плодоносящие и расположенные
на легких почвах, удобных для рытья нор. В заболоченных лесах
бурундук отсутствует (Телегин, 1980). Ф. Р. Штильмарк (1966)
3 Данные Штильмарка на 1 км маршрута пересчитаны на ,1 кв. км.
4 В этой работе дано подробное описание биотопов, заселяемых бурундуком
в разных географических пунктах.
Грызуны
43
отмечает зависимость численности бурундуков от урожая корма:
при хороших урожаях кедра в лесах Западного Саяна численность
бурундуков весной была 10 на 1 га и 25—30 — после выхода мо-
лодых, при низких урожаях на 1 га приходилось только три зверька.
Основная пища бурундуков: семена хвойных и лиственных
растений. К второстепенным кормам относятся зеленые части
растений, ягоды, грибы, животные корма (в основном насекомые
и моллюски). Животные корма больше поедаются в середине лета,
когда недостает семян. Список кормовых растений превышает
60 видов (Телегин, 1980).
Бурундук делает запасы, достигающие 2—3 кг. В некоторых
местах он уничтожает половину урожая кедровых орехов (Формо-
зов, 1976). Запасы делает либо в глубоких кладовых, расположен-
ных под гнездом, либо складывает их в лесной подстилке.
У бурундука существуют зимние и летние норы. В зимних
норах обычно две шарообразные камеры, в верхней, находящейся
на глубине 48—90 см, устраивается гнездо, в котором зверек зи-
мует, а самка выводит и потомство, в нижней (на глубине 68—
130 см) — кладовая. Летние норы состоят из единственной камеры,
находящейся на глубине 54—68 см и соединяющейся с поверхно-
стью наклонным ходом. Наибольшая длина ходов бурундуков в
Амуро-Зейском междуречье 3 м 42 см (Снигиревская, 1962). Аме-
риканские восточные бурундуки в Канаде имеют норы протяжен-
ностью 6 м и больше.
Бурундук проводит в спячке более половины года: с сентября—
октября до апреля. При наступлении положительных температур
и появлении первых проталин зверьки пробуждаются. Через
2—4 дня после выхода из спячки самок начинается гон. В холодную
ветреную погоду гон задерживается. При дружной весне гон про-
ходит в короткие сроки, с середины апреля до начала мая. Бере-
менность длится 30 дней, лактация до 40 дней. Самые ранние вы-
водки появляются в половине мая. Первыми размножаются старые
самки, прошлогодние — позднее. Выход молодых из нор проис-
ходит в июне, к самостоятельной жизни зверьки переходят в воз-
расте 50—60 дней. Расселение молодых бурундуков совпадает с
периодом запасания кормов (июль — первая половина августа).
Зверьки залегают в спячку при установлении минусовых температур
еще до выпадения снега: в сентябре-октябре, реже в ноябре (Теле-
гин, 1980).
На большей части ареала бурундук размножается раз в году,
но на юге и юго-западе часть самок приносит два выводка, в При-
морье в 1977 г. 30% самок имели плацентарные пятна двух поряд-
ков, молодые первого выводка вышли в начале июня, второго —
в начале августа (Тиунов, 1981). В выводке бывает от 2 до 12 дете-
нышей, в среднем 5—6. Процент самок, участвующих в размно-
жении, зависит от урожая корма. В год, следующий за хорошим
урожаем кедра, в Западных Саянах размножалось 91—92% са-
44
Rodent la
мок, после плохих урожаев — 41%. Колебания интенсивности
размножения отражаются на возрастном составе популяции бу-
рундуков. После плохого урожая доля прошлогодних зверьков
снизилась с 65 до 38% и увеличилась доля более старших воз-
растных групп. В обычные годы к осени половину популяции
составляют молодые зверьки. При неурожае корма их доля может
уменьшиться до 5,8% (Штильмарк, 1966).
Продолжительность жизни бурундуков 3—4 года (Меженный,
1968). Для американского восточного бурундука указывается
максимальная продолжительность жизни — 7 лет (Gaishwiller,
1965). В неволе и сибирские бурундуки доживали до 8,5 лет (Те-
легин, 1980).
Бурундуки активны только в светлое время суток. Наиболее
активен бурундук в мае — в период гона и в середине августа —
при запасании корма. Перемещения бурундуков обычно невелики,
не превышают 100—200 м. Лишь отдельные зверьки уходят на зна-
чительные расстояния, установлены перемещения в период гона
до 1,5 км, при запасании кормов — на 1,0—2,5 км (Штильмарк,
1967; Телегин, 1980). При мозаичном распределении лесных ас-
социаций характерны кормовые кочевки бурундуков в зависимо-
сти от времени созревания семян разных растений, также известны
перебежки бурундуков к полям в период созревания зерновых.
Обычно эти перемещения происходят в пределах участка обитания
зверька. Наиболее подвижны бурундуки в период запасания кор-
мов (Снигиревская, 1962; Штильмарк, 1967). То же отмечено для
американских бурундуков. При мечении восточного бурундука
наблюдали смещение центров их активности к плодоносящим
кормовым растениям. Размеры участков обитания при этом не
превышали 0,4—0,8 га (Smith L., Smith D., 1971; Forsyth, Smith,
1973). Сходные размеры используемой площади — 0,6—0,9 га —
установлены и для других видов американских бурундуков: Т. town-
sendii и Т. dorsalis только при значительных снижениях численно-
сти участок Т. dorsalis увеличивался до 1,3 га (Gaschwiller, 1965;
Hart, 1976).
В период, когда запасы уже собраны, бурундуки очень привя-
заны к своим зимним убежищам и не отходят далеко от них. В это
время наблюдается изоляция индивидуальных участков и харак-
терна охрана территории возле убежищ, тогда как в другие периоды
участки бурундуков широко перекрываются (Штильмарк, 1967).
Бурундук прокармливает различные виды блох, вшей, клещей.
На нем обнаружено 30 видов блох. Из них специфичны только два
вида: Ceratophyllus tamias и Ctenophthalmus pisticus; первая
блоха составляет в сборах со зверьков 70—80%, из гнезд — до
40%, вторая соответственно — 10—12 и 20%. На них встречаются
блохи, широко распространенные среди грызунов: Neopsylla ples-
kei, N. mana, N. bidentatiformis, Frontopsylla elata, F. luculenta,
составляющие no 3—8% в сборах, и блохи, характерные для мел-
Грызуны
45
ких лесных грызунов, из них наиболее часто попадаются Neopsylla
acanthina, Pectiniceps pectiniceps, Ct. congeroides (5—9% в шерсти
зверьков). N. acantina бывает многочисленна в гнездах, до 35%
сборов. Остальные виды блох лесных мелких грызунов, белок и
птиц встречаются редко. Довольно регулярны в сборах с бурун-
дуков блохи землероек — Doratopsylla birula (до 6%), реже кро-
тов— Histrichopsylla talpae (до 1%). Там, где бурундук выходит
на опушки, на нем возможно обнаружить блох сусликов С. tesquo-
rum и пищух Ct. submarinus. Иногда на них обнаруживали блох
крыс С. fasciatus и водяных крыс С. walkeri (Попов, Иголкин,
1956; Снигиревская, 1962; Емельянова, Штильмарк, 1968; Ель-
шанская, Попов, 1968; Иголкин, 1978).
Пораженность бурундуков блохами в активный период в сред-
негорье Западного Саяна составляла 60—100%, только перед
спячкой снижаясь до 44%. Индексы были не меньше единицы, в
апреле — 5,0, в сентябре — 7,3 (Емельянова, Штильмарк, 1968)
В других местах пораженность бурундуков блохами тоже достаточ-
но высока — не менее 40%.
Видовой состав блох, обнаруженных на бурундуках, свидетель-
ствует об их широких паразитарных контактах с другими лесными
млекопитающими и обитателями опушек леса.
Специфическая вошь, паразитирующая на бурундуке, отно-
сится к виду Enderleinellus tamias Fahr. (Снигиревская, 1962).
Наибольшее значение бурундук имеет как прокормитель ик-
содовых клещей. На нем обнаружены следующие виды клещей из
подсемейства Ixodidae: Ixodes persulcatus, I. pavlovskyi, I. pome-
rantzevi, I. angustus, I. apronophorus, I. trianguliceps, из подсемей-
ства Amblyomminae: Haemaphysalis concinna, H. japonica, Der-
macentor pictus, D. silvarum (Сердюкова, 1956; Попов, 1962; Фи-
липпова, 1977).
Наиболее важна его роль как прокормителя личинок и особенно
нимф таежного клеща I. persulcatus в очагах клещевого энцефа-
лита. В некоторых районах Сибири и Дальнего Востока он про-
кармливает значительную часть популяции предимагинальных фаз
этого клеща (Попов, 1962; Тиунов, 1981). Клещи встречаются на
24—100% осмотренных бурундуков. Обычно в период максималь-
ной активности личинок и нимф их встречаемость на бурундуках
равна 100%. В хвойных лесах Приморья индексы обилия I. persul-
catus на бурундуке 29,3, с одного зверька снимали до 200 клещей
(Земская, Гроховская, 1959). В Кемеровской области встреча-
емость личинок и нимф доходит до 86%, индексы соответственно —
до 13 и 8,5 (Горчаковская и др., 1960). В северо-восточном Алтае
обычные индексы обилия клещей на бурундуке: 4,2 личинки и
5,7 нимф (Смирнов, Равкин, 1967). По подсчетам этих авторов, в
светлохвойно-мелколиственном и черневом низкогорье бурундуки
прокармливают на 1 кв. км 3—9 тыс. личинок и 4—16 тыс. нимф.
В Козульском районе Красноярского края индексы обилия личинок
46
Rodentia
этих клещей 1,5, нимф 2,0 (Никифоров, 1968). Возможно, не мень-
шую роль имеет бурундук и в прокормлении личинок и нимф
I. pavlovskyi на Алтае. На Дальнем Востоке наряду с другими мел-
кими млекопитающими бурундук прокармливает I. pomerantzevi
и I. angustus. Незначительна его роль в прокормлении I. арго-
nophorus и I. trianguliceps. Первый питается в основном на водяной
крысе, второй — на мелких грызунах и насекомоядных (Филип-
пова, 1977).
Из подсемейства Amblyomminae наибольшее значение бурундук
имеет для поддержания численности D. silvarum на юге Сибири
и в Приморье, а в Приморье еще для Н. japonica. Индексы обилия
этих клещей на бурундуке в Приморье 2,5—2,8; встречаемость
23,5—32,3; реже паразитируют на нем Н. concinna, в 6—15% слу-
чаев, с индексом обилия 1,0—3,0 (Земская, Гроховская, 1959).
D. silvarum и Н. concinna могут переносить и хранить вирус кле-
щевого и японского энцефалитов. Возможна передача бурунду-
ками через иксодовых клещей лихорадки Ку и клещевого риккет-
сиоза Северной Азии.
На бурундуках обнаружен ряд видов паразитических гамазид:
Cyrtolaelaps mucronatus, Euryparasitus emarginatus, Laelaps pav-
lovskyi, L. nuttalli, Haemogamasus mandschuricus, Hg. ambulans,
Hg. liponyssoides, Hg. serducovae, Androlaelaps glasgovi, A. casa-
lis, Eulaelaps stabularis, Hirstyonyssus pavlovskyi, Hi. isabellinus,
Hi. musculi, Hypoasois heselhausi, Pneumolaelaps marginepilosa,
Macrochelis decoloratus. Эти виды паразитируют на других мелких
млекопитающих. Кроме того, на бурундуке встречен ласточкин
клещ Dermanissus hirundinus и птичий клещ D. gallinae. Из них
наиболее часто встречаются: Hg. mandshuricus (до 50% сборов),
Hg. serdukovi (20%), Hg. ambulans (17%), Euparasitus emarginatus
(14%), Hirstyonyssus isabellinus (11%), Eulaelaps stabularis (4—
7%). Пораженность бурундуков гамазидами доходит до 50%, но
индексы обилия обычно невелики, в Приморье — 5,7 (Земская,
Гроховская, 1959; Иголкин, 1978). Возможно получение гамазо-
выми клещами ряда возбудителей природноочаговых инфекций:
туляремии, риккетсиозов, хориоменингита, клещевого энцефа-
лита. Однако передача их через укус этими паразитами проблема-
тична. Скорее возможно заражение зверьков при очесывании,
когда они скусывают клещей.
Список видов клещей-краснотелок, обнаруженных на бурун-
дуке, включает 17 видов: Miyatrombicula esoensis, Leptotrombidium
fulleri, L. orient ale, L. pallidum, L. palpale, Neotrombteula autum-
nalis, N. gardellae, N. japonica, N. microti, N. nagayoi, N. pomerant-
zevi, N. talmiensis, N. tamiya, N. zhmajevae, Ascoschoengastia
latyschevi, Euschengastia rotundata, Walchia festiva (Земская, Гро-
ховская, 1959; Кудряшова, 1979). Наиболее обильны эти клещи
в Приморье, пораженность ими бурундуков равна 71%, при сред-
нем индексе обилия 78,3, максимальном 250 (Земская, Гроховская,
Грызуны
47
1959). Возможно участие краснотелок в передаче некоторых рик-
кетсий (цуцугамуши) людям.
В природе у бурундуков обнаруживали возбудителей следую-
щих заболеваний: клещевого энцефалита, клещевого риккетсиоза,
псевдотуберкулеза, туляремии, листериоза, эризипелоида, дикро-
целиоза, гименолепидоза. Для токсоплазмоза получены положи-
тельные серологические реакции (Шеханов, 1979).
Наибольшее значение имеет бурундук в очагах клещевого эн-
цефалита как прокормитель таежного клеща, несомненно его
участие и в циркуляции вируса клещевого, и, возможно, японского
энцефалитов (Попов, Федоров, 1958; Левкович и др., 1967). Уста-
новлено, что бурундуки способны к бессимптомному носительству
вируса в центральной нервной системе до 16 дней (Карпов, Фе-
доров, 1963). В лесных очагах лихорадки Ку бурундук может
быть носителем риккетсий, от него выделяли штаммы R. sibirica
(Федорова, 1968; Шеханов, 1979). А. Д. Калмыкова с соавторами
(1962) считают, что в таежных очагах геморрагической лихорадки
в Хабаровском крае возможно заражение людей при контакте с
лесными грызунами, в том числе и с бурундуками. Предполагается,
что они могут иметь значение в очагах Омской геморрагической
лихорадки, так как распространены в пределах ареала ее возбу-
дителя (Телегин, 1980). Незначительна их роль в очагах туляремии:
по восприимчивости и чувствительности к возбудителю этого за-
болевания бурундуки относятся ко второй группе животных по
Олсуфьеву и Дунаевой (1960). Но все же известны случаи заболе-
ваний людей, связанных с промыслом бурундука (Карпов, Попов,
1956). При изучении токсоплазмоза в Васюганье, положительные
РСК получены для 17,6% бурундуков (Пестрякова и др., 1966).
Чума была обнаружена у трех видов американских бурунду-
ков: Т. minimus, Т. speciosus, Т. umbrinus. Культуры возбудителя
от бурундуков выделяли в штатах Калифорния, Невада, Юта и
Вашингтон. В 1969 г. в штате Нью-Мексико был зарегистрирован
случай заражения от бурундука Tamias sp. человека (Kartman,
1970).
Культуры возбудителя туляремии в США выделяли от Т. mi-
nimus, и Т. striatus, положительная серология была обнаружена
у Т. minimus и Т. dorsalis (Infections diseases, 1970). В Канаде от
бурундука (Tamias sp.) была выделена Pasteurella multocida (In-
fections diseases, 1970).
Шкурка бурундука не представляет большой ценности. Из меха
изготовляют жакеты, шапки, детские вещи. В СССР с 1946 по 1958 г.
заготавливали 286—1036 тыс. шкурок в год (Телегин, 1980).
Большое значение бурундук имеет как декоративное животное.
Учет численности бурундуков проводят на маршрутах по числу
встреченных зверьков, по вылову их в ловушки, расставляемые
линиями или на площадках мечения (Штильмарк, 1963; Смирнов,
Равкин, 1967). В период гона, весной, можно учитывать бурунду-
48
Rodentia
ков с манком на маршруте. Считается, что на манок откликаются
все самцы. Результат учета удваивается, поскольку самки на манок
пе отзываются (Кучерук, 1952).
ПОДСЕМЕЙСТВО XERINAE OSBORN, 1910
— ЗЕМЛЯНЫЕ БЕЛКИ
Род Spermophilopsis Blasius, 1884
— тонкопалые суслики
Единственный вид в роде S. leptodactylus Lichtenstein, 1823—
тонкопалый суслик (2п=38) — близок к африканским земляным
белкам. Распространен в песчаных пустынях Южного Казахстана
и Средней Азии, заходит на север Ирана и Афганистана. Ареал
тонкопалого суслика разорван и состоит из четырех частей: наи-
более крупных каракумской и кызылкумской, изолированных
друг от друга непригодными для него ландшафтами с глинистой
почвой и тугайной растительностью в долине Амударьи. Третья
часть ареала расположена в песках Муюнкумы, отделена от кы-
зылкумской горами. Четвертая часть находится в Семиречье, от
предыдущей также отделена горами. Долины рек в Семиречье
разделяют эту часть ареала суслика на отдельные поселения (Ко-
марова, 1980).
Распределение тонкопалого суслика мозаично, местами на се-
верной границе спорадично. Наибольшая численность его отме-
чена в крупнобугристых песках, в Каракумах и Кызылкумах, в
этих биотопах приходится от 7 до 15 зверьков на 1 га, несколько
меньше его в мелкобугристых слабозакрепленных песках 3—11,
грядовых песках 2,6—7, в лёссовой пустыне 6—8. Обычно немного
его в барханах (1 зверек на 1 га), хотя в период развития эфеме-
ровой растительности может быть до 8 зверьков на 1 га. Высока
его численность возле некоторых колодцев, в окрестностях которых
богато представлены эфемеры и эфемероиды: до 12 зверьков на
1 га. Очень низкая численность сусликов в местах перевыпаса,
не больше 1—2 зверьков на 1 га. Малочислен суслик на северной
границе ареала в Северных Кызылкумах, в районе Жанадарьи
он занимает ограниченные участки песков среди такыровидных
равнин, на площади 50 га здесь приходится 3—4 зверька. В Му-
юнкумах суслика меньше, чем в Каракумах и Кызылкумах, в
центральной части приходится 0,5—4 зверька на 1 га, в северной
0,2—0,3 на 1 га. Невысока численность тонкопалого суслика в
Прибалхашье, обычно 1—2 зверька на 1 га, лишь в наиболее бла-
гоприятных местах: крупнобугристых песках и в саксаульниках
у р. Или достигает 3—4 на 1 га (Дятлов, 1960; Карулин, 1960;
Исмагилов, 1962; Есжанов, Бекенов, 1982). Эти показатели числен-
ности характерны для отдельных биотопов, заселенных зверьками.
Грызуны
49
При пересчете их на большие площади численность сусликов
выражается обычно в более низких цифрах: для Каракумов — от
0,1 до 1,6 особей на 1 га, для Кызылкумов — 0,01—1—2 на 1 га
(Комарова, 1980).
Питается тонкопалый суслик травянистой и кустарниковой
растительностью. Список кормовых растений содержит 40 видов
(Ruzic, 1967). Большое значение в питании этого грызуна имеет
песчаная осочка и подземные части растений. Для добывания
зеленых веточек кустарников зверек нередко лазает на значитель-
ную высоту. При поисках луковиц и корневищ суслики делают
многочисленные прикопки, что вместе с норовой деятельностью
способствует развеиванию песков. Большую часть в рационе тон-
копалого суслика составляет и животная пища. Весной в 55,5%
желудков вскрытых животных встречались зеленые части расте-
ний, в 100% — подземные части, в 22,2% — зерна и в 55,5% —
животная пища (осенью соответственно — 13,3; 46,6; 60,0 и 73,3%)
(Ruzic, 1967).
Норы тонкопалого суслика уходят вглубь под углом и имеют
различную глубину (от 0,2 до 3 м) и протяженность (от 1 до 5 м)
в зависимости от характера грунта и залегания влажного слоя
песка, а также от функции норы. Наиболее просто устроены за-
щитные норы, располагающиеся у мест кормежки; более сложное
строение имеют норы, где суслики ночуют и выводят потомство
(Комарова, 1980). В Каракумах на 1 га приходилось в среднем
0,4—21 норы, максимум 27, в Кызылкумах — 0,3—30 (Карулин,
1960; Комарова, 1980). На участке одного зверька находится одна
гнездовая нора, 2—5 защитных и до 30 временно необитаемых
нор (Дубровский, Комарова, 1967). Норы очень быстро засыпаются
песком и быстро восстанавливаются (Ruzic, 1967).
Тонкопалые суслики активны круглый год. В спячку не впа-
дают. Только в экстремальных условиях, при особенно сильной
засухе, наблюдали их летнюю спячку в самую жару (июль, август).
Зимой в холодные дни с сильным ветром они не выходят из нор
по нескольку дней (Ruzic, 1967). Тонкопалые суслики активны
только в светлое время, летом они выходят из нор с рассветом и
кормятся с перерывами до захода солнца (Карулин, 1960). В хо-
лодное время года зверьки активны среди дня, обычно с 13 до 15 ч
(Ruzic, 1967).
Тонкопалые суслики очень подвижны, обладают большими
индивидуальными участками. Размеры их связаны с кормностью
угодий. В бугристых песках Туркмении, где нет выпаса скота,
кормовые участки сусликов невелики (около 500 кв. м), там же,
где растительность сильно стравлена скотом, суслики перемеща-
лись на огромной площади, в среднем 146 га, максимум 246 га (Ка-
рулин, 1960). Участок обитания одного зверька сильно изменяется
иногда в течение недели. Особенно велики передвижения суслика
весной во время гона. Г. Д. Свиденко (1963) проследил самца,
59
Rodentia
который за 5 ч пробежал 3 км. Зимой подвижность сусликов сильно
сокращается.
Размножается тонкопалый суслик весной, течка бывает в конце
февраля—марте. Детеныши родятся в конце апреля — мае.
В помете бывает 3—6 молодых. Массовый выход молодняка при-
ходится на май-июнь, расселение их — на конец июня, июль.
Зверьки становятся половозрелыми в возрасте 10—11 месяцев.
В благоприятные годы возможен 2-й и иногда даже 3-й цикл
размножения (Афанасьев и др., 19536; Петрова, 1966; Ruzic, 1967;
Свиденко и др., 1973).
Тонкопалый суслик тесно контактирует с большой песчанкой.
В местах их совместных поселений норы сусликов встречаются
на территории 74% нор песчанок, в пределах одной норы песчанок
бывает до 4 нор тонкопалого суслика (в среднем 1,3—1,7) (Дуб-
ровский, Комарова, 1967). Поселяясь в норах большой песчанки,
тонкопалый суслик чаще отрывает собственную нору, но при этом
пользуется и верхними наиболее крупными ходами хозяев. По
частоте встреч в норах большой песчанки тонкопалый суслик ус-
тупает только краснохвостой и полуденной песчанкам (Свиденко-,
1963). Большие песчанки посещают норы тонкопалых сусликов.
Из общего числа встреч разных грызунов в норах сусликов боль-
шие песчанки составляли 21% (Исмагилов, 1960). Контактируют
суслики и с другими пустынными животными, в их норах прячутся
ежи, землеройки, рептилии, забегают тушканчики, полевки, мел-
кие хищники, птицы-норники. На 100 нор тонкопалых сусликов
в Туркмении было выловлено весной 11 больших, 16 полуденных
и 4 краснохвостых песчанки и 3 тушканчика. Реже встречаются
в них желтые суслики (0,14 на 100 нор), афганские полевки (0,44),
ласки (1,2) и перевязки (Загнибородова, Свиденко, 1968). В Ка-
захстане в капканы, поставленные у нор сусликов, до 5% попа-
даний приходилось на зайцев (Млекопитающие Казахстана, 1969).
В южной части Кызылкумов 65% нор сусликов оказываются
занятыми варанами (Исмагилов, 1962). Все эти животные обмени-
ваются с тонкопалым сусликом эктопаразитами, благодаря чему
их видовой состав достаточно велик.
На тонкопалых сусликах в некоторых местах встречается до
36 видов блох. Специфичен же для них только один вид — Rhost-
ropsylla daca. В местах совместных поселений тонкопалых сус-
ликов с большими песчанками и песчанками рода Meriones на
первых встречаются в значительном количестве песчаночьи блохи
и среди них активные переносчики чумы: в западной части ареала —
Xenopsylla hirtipes, X. conformis, X. gerbilli caspica; в восточной —
X. gerbilli minax; часто встречаются (до 30%) Coptopsylla lamelli-
fer, Ctenophthalmus dolichus. Зарегистрированы на тонкопалом
•суслике и другие блохи песчанок: Ceratophyllus laeviceps, Copto-
psylla bairamalensis, Ceratophyllus turkmenicus, C. aralis, Parado-
xopsyllus teretifrons, Rhadinopsylla cedestis, Stenoponia vlasovi,
Грызуны
51
но либо их численность незначительна, либо они имеют небольшое
значение в переносе чумы. Значительно реже встречаются в норах
тонкопалого суслика блохи настоящих сусликов Neopsylla setosa
и в шерсти — Oropsylla ilovaiskii. Единичны на них блохи и других
грызунов и мелких хищников, среди последних изредка встречается
Pulex irritans, способная нападать на человека (Кондрашкин^
Дудникова, 1955; Романовский и др., 1956; Иофф и др., 1965;
Загнибородова, Свиденко, 1968; Попова, Рапопорт, 1974). По-
сравнению с норами больших песчанок в норах тонкопалых сус-
ликов блох гораздо меньше, что связано с большой подвижностью
сусликов, при которой они часто бросают свои жилища (Дудни-
кова, 1960).
На тонкопалых сусликах и в их норах встречаются иксодовые
клещи: Haemaphysalis numidiana turanica Neum, Hyalomma asia-
ticum Sch. et Sch., Ixodes redicorzevi Olenev, I. occultus Pomerant-
zev, Rhipicephalus pumilio, Dermacentor daghestanicus; из apra-
совых клещей — Ornithodorus tartacovskii Olenev (Брегетова и
др., 1955; Сердюкова, 1956; Млекопитающие Казахстана, 1969;
Филиппова, 1966; Филиппова, 1977). Эти клещи переносят и со-
храняют возбудителей чумы, туляремии, лихорадки Ку, клеще-
вого сыпного тифа и клещевого спирохетоза.
От тонкопалых сусликов выделяли возбудителей чумы, бруцел-
леза, лихорадки Ку, клещевого спирохетоза, кожного лейшмани-
оза и токсоплазмоза (Калякин, 1971; Шеханов, 1979). Кроме того,
были выявлены антитела при исследовании их сывороток на кле-
щевой риккетсиоз и крымскую геморрагическую лихорадку (Та-
расевич и др., 1977; Лебедев и др., 1980). Чуму регистрировали
у тонкопалых сусликов в Южном Прибалхашье, Кызылкумах
и Западной Туркмении (Наумов и др., 1972). Благодаря невысокой
численности и одиночному образу жизни тонкопалый суслик не
имеет существенного значения в поддержании очагов чумы, ног
передвигаясь на большие расстояния, может служить разносчиком
чумных блох. Его роль возрастает на спаде эпизоотии, когда вы-
мирают основные носители чумы и увеличивается число заражен-
ных блох (Ралль, 1960; Загнибородова, Свиденко, 1968). Экспе-
риментальными работами показано, что тонкопалые суслики вос-
приимчивы, но малочувствительны к чуме, в 9 раз устойчивее
морских свинок (Цибулевская, 1960). По чувствительности и вос-
приимчивости к туляремии тонкопалый суслик, как и суслики
р. Citellus, относится ко 2-й группе животных, восприимчивых,
но малочувствительных к возбудителю, т. е. заражение зверька
возможно небольшими дозами, меньше 10 микробных клеток (м. к.)>
но инфекция при этом не развивается, а дозы, ведущие к леталь-
ному исходу, велики—более 10 млн м. к. (Олсуфьев, Дунаева,
1960). Однако в туляремийных очагах этот вид не встречается.
На пастбищах, где пасется бруцеллезный скот, тонкопалый
суслик может получить бруцелл, но самостоятельного значения в
52
Rodentia
очагах этой инфекции он не имеет (Ременцова, 1962). В пустынных,
наиболее устойчивых очагах лихорадки Ку этот грызун может
быть разносчиком возбудителя — R. sibirica, а также членисто-
ногих — переносчиков в популяциях грызунов (Карулин, 1960).
Хотя тонкопалый суслик распространен в очагах кожного
лейшманиоза и тесно контактирует с большой песчанкой — ос-
новным носителем этого заболевания, пораженные лейшманиями
суслики встречаются редко. Ю. А. Дубровский и Л. В. Комарова
(1967) объясняют это тем, что на одного суслика в его жилых норах
приходится в 5—10 раз меньше москитов, чем на одну песчанку в
их норах. С норами больших песчанок обычно связаны лишь за-
щитные, сравнительно неглубокие норы сусликов, а москиты днем
придерживаются глубоких горизонтов, где отсутствуют колебания
температуры. В результате риск заражения сусликов лейшманио-
зом снижается в 20—40 раз по сравнению с песчанками.
Тонкопалый суслик добывается ради меха, но летняя шкурка
малоценная, а зимний промысел не развит. Местами этот суслик
вредит работам по закреплению песков, поедая посаженные ку-
старники (Афанасьев и др., 1953).
СЕМЕЙСТВО GEOMYIDAE GILL, 1872 — ГОФЕР*3'
Мелкие и средних размеров грызуны, длина тела 13—40 см,
приспособленные к подземному образу жизни (Anderson, Jones,
1967; Соколов, 1977). Тело вальковатое, с короткой шеей и корот-
кими сильными конечностями; внешне напоминает азиатских
цокоров и пластинчатозубых крыс. Для представителей семейства
характерно наличие обширных, покрытых изнутри шерстью за-
щечных мешков, которые могут выворачиваться наружу и втяги-
ваться обратно при помощи специальной мышцы. При рытье зверьки
используют резцы и лапы, снабженные широкими крепкими ког-
тями. В семействе 8 родов и 40 видов:
Число
видов
Geomys Ratinesque, 1817 восточные гоферы 8
Thomomys Wied-Neuwied, западные гоферы 6
1817
Pappogeomys Merriam, 1895 гоферы Буллера 2
Cratogeomys Merriam, 1895 желтые гоферы 12
Orthogeomys Merriam, 1895 гигантские гоферы 3
Heterogeomys Merriam, 1895 грубошерстные гоферы 2
Macrogeomys Merriam, 1895 центральноамериканские гоферы 6
Zygogeomys Merriam, 1895 мичоаканские гоферы 1
Подземный строго одиночный образ жизни при малой мигра-
ционной активности зверьков привел к существованию большого
количества подвидов, а также форм и вариаций, особенно в окраске.
Рис. 1. Распространение семейств Geomyidae (A), Heteromyidae (Б), Pedetidae (В)
Грызуны
54
Rodenlia
Только южный гофер Thomomys umbrinus имеет 214 подвидов
(Hall, Kelson, 1959).
Ареал семейства охватывает Северную и Центральную Аме-
рику от Западной Канады (54° с. ш.) до Панамы и от Атлантиче-
ского до Тихого океана с востока на запад (рис. 1А). Гоферы встре-
чаются в различных ландшафтах — осветленных лесах, дубравах,
лугах, в степях, пустынях, на полях сельскохозяйственных
культур (посевы люцерны, корнеплодов), на пастбищах и в садах.
Большинство видов требовательно к почвам, предпочитая рыхлые,
среднеувлажненные. Показано, что распространение видов рода
Geomys на юге США почти точно совпадает с распространением
песчаных почв. Гоферам свойственна аллопатрия — два вида од-
ного рода почти никогда не встречаются в одних и тех же местах
(Turner et al., 1973).
Гоферы ведут подземный образ жизни, редко выходя на поверх-
ность. Зимой часть ходов может быть проложена в толще снега.
Активны круглый год, в спячку не впадают, хотя в сильные морозы
активность значительно снижается. Нора состоит из большой
сети кормовых ходов на глубине 12—20 см и гнездовой камеры с
несколькими галереями на глубине 1—3 м (Anderson, 1980).
Основу питания составляют подземные части растений, в большом
количестве поедаются корни трав и молодых деревьев, клубни,
луковицы. Пик размножения приходится на конец зимы — вес-
ну — начало лета, хотя южные виды, вероятно, могут размно-
жаться в течение всего года. У самок половозрелость наступает
в возрасте 6—9 месяцев, детенышей в выводке от 2 до 6, хотя число
эмбрионов может достигать 13. Гоферы причиняют серьезный
ущерб сельскому хозяйству, снижая продуктивность пастбищ на
18—40% (Foster, Stubbendiek, 1980), поедая корнеплоды и корни
растений, повреждая сеянцы в садах и лесопитомниках.
Значение гоферовых для здоровья человека невелико. По мне-
нию одних авторов, они не вовлекаются в эпизоотии чумы (Stark
et al., 1966), по другим данным, в штатах Техас и Вашингтон у
Thomomys bottae выявлена чума (Meyer, 1963). Картман (Kartman,
1970) сообщает о случае заражения человека от гофера. Есть све-
дения об обнаружении туляремии у южного гофера Th. umbrinus
(Infections diseases, 1970). Однако это единичные находки, мало
меняющие картину в целом.
СЕМЕЙСТВО HETEROMYIDAE ALLEN ET CHAPMAN, 1893
— МЕШОТЧАТЫЕ ПРЫГУНЫ
Мелкие грызуны, длина тела 6—20 см, по внешнему виду на-
поминающие тушканчиков, мелких песчанок, мышей. Так же как
и гоферы, которым они систематически близко родственны, имеют
покрытые шерстью защечные мешки, способные выворачиваться
наружу. В семействе 5 родов, 65 видов, по данным некоторых
Грызуны
55
авторов — 75 (Anderson, Jones, 1967; Соколов, 1977):
Число
видов
Perognathus Wied-Neuwied, мешотчатые прыгуны 26
1839
Microdipodops Merriam, 1891 малые прыгуны 2
Dipodomys Gray, 1841 кенгуровые прыгуны 22
Liomys Merriam, 1902 колючие прыгуны 5
Heteromys Desmarset, 1817 центральноамериканские прыгуны 10
Ареал семейства охватывает юго-западную часть Канады, за-
пад США, Мексику, Центральную и Южную Америку до Эква-
дора, Колумбии, Венесуэлы (рис. 1, Б). Населяют различные
биотопы преимущественно в полуаридных и аридных областях,
но два рода — Liomys и Heteromys — заселяют влажные тропи-
ческие леса.
Прыгуны ведут преимущественно ночной образ жизни, днем
скрываясь в норах, вырытых под кустами, камнями, валежником.
У многих видов норы довольно сложные, с несколькими длинными
отнорками и камерами; они используются многочисленными ви-
дами членистоногих, а также амфибиями и рептилиями. Питаются
прыгуны семенами растений, реже другими растительными кор-
мами, поедают также насекомых и других мелких животных.
Многие виды запасают пищу, перенося ее в защечных мешках.
Некоторые прыгуны впадают в спячку. Виды родов кенгуровых
и мешотчатых прыгунов, распространенные в пустыне Аризоны,
питаются сухим зерном и не используют в пищу ни сочных кормов,
ни питьевой воды. Это объясняется способностью организма пры-
гунов использовать метаболическую воду, т. е. воду, молекулы
которой образуются в результате диссимиляции веществ пищи,
прежде всего углеводов. Сохранению воды в организме служат
дополнительные физиологические и этологические приспособле-
ния — выделение концентрированной мочи, норовый образ жизни,
ночная активность (Шмидт-Нильсен, 1972). Плодовитость сравни-
тельно невысока. Хотя южные виды могут размножаться круглый
год, за год у самки обычно бывает один, реже два выводка с 2—8,
чаще 3—5 детенышами. В неволе мешотчатый прыгун прожил более
7 лет (Жизнь животных, 1971). Прыгуны могут приносить вред
сельскому хозяйству, выкапывая и поедая посеянные семена.
Отмечено участие широко распространенных видов прыгунов
в эпизоотиях чумы (Diseases transmitted, 1963). Так, Dipodomys
or di был найден зараженным чумой, у Perognathus parvus найдены
антитела и выделена культура чумы от их специфичных блох.
Для двух видов Dipodomys ordi и D. spectabilis установлена вы-
сокая степень резистентности к чумному микробу. Считают, что
представители рода Heteromys имеют значение как звено, связы-
вающее эпизоотии в природных очагах с человеческими поселе-
ниями (Plague in the Americas, 1965; Stark et al., 1966).
56
Rodentia
В Мексике и Венесуэле от нескольких видов Heteromys выделя-
ли Leishmania mexicana (Lainson, 1982). От Perognatus parvus и
D. ordi выделен возбудитель туляремии, а у Р. formosus и D. mic-
rops обнаружены антитела (Infections diseases).
СЕМЕЙСТВО PEDETIDAE OWEN, 1847
— ДОЛГОНОГОВЫЕ
Один род с единственным видом — кафрским долгоногом Pede-
tes cafer Forster, 1778 (Соколов, 1977). Некоторые исследователи
выделяют второй вид — Р. surdaster Thomas, 1902 (Anderson, Jones,
1967). По внешнему виду зверек напоминает очень крупного, массой
до 3—4 кг тушканчика с пушистым хвостом. Распространен в Цент-
ральной и Южной Африке — от Кении и Анголы до мыса Доброй
Надежды (см. рис. 1, В). Населяет пустынные и полупустынные рав-
нины; ведет ночной образ жизни, днем скрываясь в норах. В одной
норе может жить группа семей. В других случаях норы нескольких
отдельных зверьков располагаются поблизости друг от друга. Пи-
таются сочными надземными и подземными частями растений, могут
приносить некоторый вред сельскохозяйственным культурам. Мест-
ные жители охотятся на долгоногов ради шкурки и мяса.
Долгоноги вовлекаются в эпизоотии чумы, при исследовании
12,3% оказались сероположительными (Hallet et al., 1970). Дэвис
(Davis, 1946, 1953) упоминает о случаях заражения местных жите-
лей чумой от долгоногов при охоте на них.
СЕМЕЙСТВО DIPODIDAE WATERHOUSE, 1842
— ТУШКАНЧИКИ
В семействе 29—30 видов, 11—12 родов, которые объединяются в 4 под»
семейства.
1. Euchoreutinae
Род Euchoreutes — 1 вид:
Е. naso Sclater, 1891 — Центральная Азия.
II. Cardiocraniinae *
Род Cardiocranius — 1 вид:
С. paradoxus Satunin, 1903 (2n=48) ** — Гоби, изолированные участки
в Убсунурской котловине, Бэйшане, Северном Прибалхашье. Род Salpingo-
tus — 5 видов.
1. S. crassicauda Vinogradov, 1927 (2n=46) — от Восточной Гоби до Зай-
санской котловины и пятнами в Южном Прибалхашье, Приаралье, Больших
Барсуках.
2. S. heptneri Vorontzov et Smirnov, 1969 — Северные и Северо-Западные
Кызылкумы.
3. S. kozlovi Vinogradov, 1922 — Гоби.
4. S. thomasi Vinogradov, 1928 — Южный Афганистан.
5. S. michaelis Fitzgibbon, 1966 — Западный Пакистан.
* Предлагается разделить на два самостоятельных подсемейства: Cardioc-
raniinae и Salpingotinae (Шенброт, 1985).
** Диплоидное число хромосом приведено по В. Н. Орлову (1974).
Грызуны
57
III. Allactaginae
Род Allactaga — 11 видов.
1. Al. jaculus Pallas, 1778 (2n=48) — от правобережья Днепра до Оби.
2. Al. severtzovi Vinogradov, 1925 (2n=48) — Устюрт, Северные Караку-
мы, Кызылкумы, Южное Прибалхашье.
3. Al. sibirica Forster, 1778 (2n=48) — от Восточного Прикаспии до Се-
веро-Восточного Китая.
4. Al. elater Lichtenstein, 1825 (2n=48) — от Нижнего Поволжья до
Прибалхашья и Джунгарской Гоби, Восточное Закавказье, Иран.
5. Al.williamsi Thomas, 1897 (= Al. euphratica) (2n=48) — Восточное За-
кавказье, Малая Азия, Месопотамия, Северо-Западный Иран, северо-вос-
точная часть Аравийского полуострова.
6. Al. firouzi Womochel, 1978 — Иран, Исфахан.
7. Al. hotsoni Thomas, 1920 — Иран, Керман.
8. Al. bobrinskii Kolesnikov, 1937 (2n=48) — северная часть Каракумов
и западная часть Кызылкумов.
9. Al. bullata G. М. Allen, 1925 — Гоби.
10. Al. nataliae Sokolov, 1981 — Гоби.
11. Al. tetradactyla Lichtenstein, 1823 — северо-восточная присредизем-
номорская часть Африки.
Род Alactagulus — 1 вид:
A. pygmaeus Pallas, 1778 (2n=48) — от Дона до Зайсанской котловины
и Южной Монголии, Каракумы, Кызылкумы, Прибалхашье.
Род Pygerethmus — 2 вида:
1. Р. platyurus Lichtenstein, 1823 (включает Р. vinogradovi; Громов,
Баранова, 1981) (2п=48) — Прибалхашье, Зайсанская котловина, Устюрт,
от р. Урал до Бетпакдалы.
2. Р. zhitkovi Kuznetzov, 1930 (2n=48) — Прибалхашье.
IV. Dipodinae
Род Dipus — 1 вид;
D. sagitta Pallas, 1773 (2n=48) — от Дона до северо-восточной части
КНР, включая Каракумы, Кызылкумы и Южное Прибалхашье.
Род Scirtopoda — 2 вида.
1. Sc. telum Lichtenstein, 1823 (2n=58) — от Днепра до Иртыша (не за-
ходит в песчаные пустыни Каракумов, Кызылкумов и Южного Прибалхашья).
2. Sc. andrewsi Allen, 1925 — Гоби и котловина Больших озер в МНР
(Honacki et al., 1982, считают подвидом Sc. telum).
Род Paradipus — 1 вид:
Р. ctenodactylus Vinogradov, 1929 (2n=48) — Каракумы и западная часть
Кызылкумов.
Род Eremodipus — 1 вид:
Е. 1 ichtensteini Vinogradov, 1927 (2n=48) — Каракумы, Кызылкумы,
Южное Прибалхашье.
Род Jaculus — 4 вида:
1. J. turkmenicus Vinogradov et Bondar, 1949 (2n=48) — Каракумы,
Кызылкумы.
2. J. blanfordi Murray, 1884 — Восточный Иран.
3. J. jaculus Linnaeus, 1758 (2n—48) — Северная Африка и Аравийский
полуостров.
4. J. orientalis Erxleben, 1777 — северная присредиземноморская часть
Африки.
Распространены тушканчики в Евразии и Северной Африке по
всему аридному поясу Палеарктики (рис. 2). Ареалы тушканчиков,
составленные на основе кадастровых карт распространения отдель-
ных видов, опубликованы нами ранее (Кулик, 19806). Наибольшее
58
Rode nil a
Рис. 2. Распространение семейства Dipodidae
разнообразие видов тушканчиков наблюдается в Средней Азии и
Монголии (Виноградов, 1937). Там обитает одновременно 8—9 ви-
дов этих зверьков. Наложение ареалов тушканчиков на одну карту
выявило районы их видового разнообразия, которые приурочены
к Туранской низменности, Прибалхашью, Зайсанской котловине и
Южной Монголии (Кулик, 19806). Дальнейшее уточнение система-
тики тушканчиков и новые данные об их распространении позволи-
ли И. Ф. Гачику и В. М. Неронову (1985) выделить Переднеазиат-
ский очаг видового многообразия тушканчиков, где распространено,
по новым данным, до 5 видов; большинство их — эндемики Иран-
ского нагорья. Наиболее примитивные и древние подсемейства
Euchoreutinae и Cardiocraniinae приурочены к Центральной Азии —
от Гоби до Приаралья. Более молодые подсемейства Allactaginae и
Dipodinae распространены широко по всему пустынному поясу —
от Северной Африки до северо-восточной части КНР.
Происхождение тушканчиков связывают с азиатскими пустыня-
ми — центральноазиатскими, казахстанскими, туранскими и нагор-
ными иранскими, что подтверждают и типы их ареалов. Это отли-
чает тушканчиков от другой характерной группы пустынных
грызунов — песчанок (Gerbillinae), имеющих африканское проис-
хождение (Виноградов, 1937; Кучерук, 1977; Кулик, 1980а; Гачик,
Неронов, 1985).
Тушканчики — мелкие и средних размеров зверьки, хорошо
приспособленные к обитанию в аридных условиях. Они имеют уд-
линенные задние конечности с пятью, четырьмя и тремя пальцами;
ступня удлинена, у ряда видов кости плюсны срастаются в цевку.
Зверьки передвигаются прыжками на задних ногах. У видов, оби-
Грызуны
59
тающих в песках, нижние поверхности пальцев задних лапок покры-
ты волосами той или иной длины. Хвост длинный, у большинства
видов имеет на конце «знамя» из удлиненных черных и белых волос.
Хвост служит рулем и балансиром при быстром беге. Уши обычно
длинные, у примитивных форм сросшиеся у основания в трубку.
Череп трапециевидной формы с сильно увеличенными слуховыми ба-
рабанами. Скуловые кости с двумя ветвями — горизонтальной и
вертикальной, последняя отделяет глазницу от большого предглаз-
ничного отверстия. Зубы с корнями, коренных зубов 4/3 или 3/3.
Тушканчики распространены в пустынях, полупустынях, сте-
пях, как равнинных, так и горных. Они четко разделяются на две
группы: одни виды придерживаются твердых грунтов, населяя гли-
нистые и щебнистые местообитания (роды Allactaga, Alactagulus,
Pygerethmus, Cardiocranius, Jaculus), другие обитают в песчаных
биотопах (роды Salpingotus, Dipus, Scirtopoda, Paradipus, Eremo-
dipus). Питаются семенами, плодами, луковицами, корневищами,
зелеными частями растений, поедают насекомых. Одни виды раз-
множаются раз в год (весной), молодые у них развиваются медленно
и достигают половозрелости лишь на следующий год (Al. saltator,
Al. severtzovi, Sc. telum, E. lichtensteini, C. paradoxus, S. crassi-
cauda). Другие виды могут приносить два помета в год: один вес-
ной, второй в конце лета (Al. jaculus, Al. bobrinskii, A. pygmaeus,
Р. zhitkovi, Р. platyurus, D. sagitta, P. ctenodactylus, J. turkmeni-
cus). В выводке бывает от 2 до 8 молодых, в среднем 4—5. Полово-
зрелость наступает обычно на втором году жизни. Размножение сего-
леток установлено у Р. zhitkovi и D. sagitta (Казанцева, Фенюк,
1937; Лобачев, Шенброт, 1976; Млекопитающие Казахстана, т. I,
ч. 2, 1977).
Тушканчики — ночные животные, ведут одиночный образ жизни.
Живут в норах относительно простого устройства. В их норах ни-
когда не бывает камер для запасов корма, и зверьки никогда не
затаскивают корм в норы. В качестве защитных и для дневного от-
дыха тушканчики используют неглубокие норы с одним коротким
ходом. Это обычно или бескамерные норы, или с камерами без гнез-
да, или гнездовые норы без камер для запасов корма (Кучерук,
1983). У большого тушканчика ход летних нор располагается на
глубине 20—25 см, а гнездовая камера — на глубине 50—60 см.
Летние норы обычно «временные». Например, мохноногий тушкан-
чик, меченный 60Со, почти каждую дневку проводил в новой норе.
За 6 дней наблюдений он использовал 5 нор и только в одной из
них дневал дважды. Из 30 отмеченных на опытной площадке нор в
течение двух недель наблюдений 22 норы использовались тушканчи-
ком 1 день, 6 нор — 2, 1 нора — 3, а одна нора — 5 дней (Кулик,
Карулин, 1980; Казанцева, Фенюк, 1937).
Зимовочные норы обычно более глубокие и устроены несколько
сложнее. Они часто имеют разветвленные ходы и несколько камер
на разной глубине, что позволяет зверьку выбирать оптимальные
60
Rodentia
условия в зависимости от изменений температуры почвы. У большо-
го тушканчика зимовочные норы достигают глубины 80—250 см.
Зимовочные норы земляного зайчика обычно имеют две камеры:
одну без гнезда, расположенную на глубине 30—50 см, где зверек
проводит спячку, и камеру с гнездом на глубине 80—85 см, кото-
рую он использует в периоды бодрствования во время зимних потеп-
лений (Скворцов, 1955; Кучерук, 1983).
Тушканчики, как правило, роют собственные норы. Лишь ино-
гда емуранчики устраивают свою нору на периферии норы больших
песчанок. В Северном Приаралье и на Устюрте при облове нор боль-
ших песчанок выловлены зверьки 8—14 видов, не считая большой
песчанки. Тушканчики среди них составляли от 19,3 до 47,7%. Но-
ры больших песчанок в этих районах посещают тушканчики 5 ви-
дов, но чаще других — малый и емуранчик (табл. 3). Среди костных
остатков, собранных на колониях больших песчанок, тушканчики
(тех же видов, что и выловленные в норах) составляли в Северном
Приаралье 4,8%, на Северо-Западном Устюрте 20% (Наумов, Ку-
лик, 1955; Настюков, 1975).
Таблица 3. Доля тушканчиков среди посетителей нор больших песчанок
(по Наумову, Кулик, 1955; Сабилаеву, 1970; Настюкову, 1975)
Посетители нор больших песчанок Северное Приаралье Юго-Восточный Устюрт Северо-Западный Устюрт
Все зверьки 368 228 1318
Тушканчики 71 (19,3%) 106 (46,5%) 630 (47,7%)
прыгуны — — 37
большие 3 3 19
малые 51 99 317
тарбаганчики 1 4 30
емуранчики 16 — 227
Индивидуальные участки у большинства видов занимают зна-
чительную площадь (табл. 4). Так, суточный участок меченого мо-
лодого мохноногого тушканчика в июне 1978 г. в Юго-Восточных
Каракумах занимал 1,27 га. Общая длина суточного пробега этого
зверька достигала 4678 м (Кулик, Карулин, 1980). Многодневный
участок у молодых мохноногих тушканчиков, установленный мето-
дом тропления, в середине лета имел площадь 3,75 га и 5 га, а у
взрослых зверьков (за 1,5 месяца) достигал 12—15 га (Орлов, 1957).
Гребнепалый тушканчик, по данным тропления, за ночь прошел
3300 м (Айропетьянц и др., 1978).
Тушканчик Житкова, вероятно, менее подвижен: в конце пери-
ода лактации зверьки удалялись от нор максимум на 45—50 м, а
площадь используемых участков была у самцов 750 кв. м, у самок
Грызуны
61
500 кв. м. Вылов меченых особей в течение 2 лет (15 поимок) и 3 лет
(6 поимок) выявил площадь участков этих зверьков в 0,21 га и
0,35 га (Млекопитающие Казахстана, 1977).
Таблица 4. Размеры индивидуальных участков тушканчиков
(по Лобачеву, Шенброту, 1980, с добавлениями)
Вид тушканчика Место исследования Индивидуальный участок Способ наблюде- ния *
длина, м площадь, га
Пятипалый карлико- вый Монголия 220—700 2,5—3,0 1
Жирнохвостый кар- ликовый Тушканчик Житкова Тарбаганчик Зайсан Южное Прибалхашье Калмыкия Зап. Казахстан 50—600 0,010—0,015 0,050—0,075 0,21—0,35** 0,07—2,16 1 2 2 2 1, 2
Мохноногий Каракалпакия 200—1700 3,75—5,00 12—15** 3
Юго-Восточные Кара- кумы 135—330 1,27 1
Емуранчик Приаральские Кара- кумы 20—45 0,07—0,39 2
Приаральские Кара- кумы 35—65 0,15—0,40 1
* / — визуальные наблюдения за мечеными зверьками; 2 — мечение с повторными выловами;
3 — анализ следовых меток.
** — по многосуточным наблюдениям.
Тушканчики на зиму впадают в спячку. Спячка у большинства
видов глубокая и продолжительная, до 4—5 месяцев. Однако во
время зимних оттепелей она может прерываться и зверьки выходят
на поверхность земли. У мохноногих тушканчиков в разных частях
ареала продолжительность спячки меняется от 4,5 до 1,5 месяцев.
В теплые зимы в южных районах по крайней мере часть зверьков
этого вида может не впадать в спячку. Могут проводить теплые
зимы в активном состоянии также гребнепалые тушканчики.
Численность тушканчиков относительно невелика. У крупных
видов рода Allactaga наблюдается не более 10 нор на 1 га и до 5—7
зверьков на 1 га. Мелкие тушканчики могут на отдельных участ-
ках достигать более высокой численности, например тарбаганчик
в Юго-Восточном Казахстане — 12—14 зверьков на 1 га, тушкан-
чик Житкова в Алакольской котловине — даже 30—40 зверьков на
1 га. При учетах на автомобильных маршрутах отмечали в среднем
62
Rodentia
на 10 км (Слудский и др., 1977): 1—2 больших тушканчиков (Ус-
тюрт, Южное Прибалхашье), 7 прыгунов (Зайсанская котловина),
3,7 тушканчиков Северцова (Северные Кызылкумы), а тарбаганчи-
ков до 60 зверьков (Северные Кызылкумы), малых тушканчиков —
до 12 особей (Северные Кызылкумы), мохноногих тушканчиков —
до 36 зверьков (Зайсанская котловина).
На Устюрте многочисленны малый тушканчик, тарбаганчик и
емуранчик, которые являются там фоновыми видами грызунов (На-
стюков, 1978). В этом районе тушканчики играют существенную
роль в питании хищников (Шилов, 1966а). При низкой численности
большой песчанки в добыче лисицы тушканчики составляли 63%
(в основном малый тушканчик). На стыке песчаных и глинистых
ландшафтов в рационе лисицы тушканчики составляли 16% (в ос-
новном емуранчики).
В пустынях Юго-Восточной Монголии по результатам исследова-
ния погадок хищных птиц мохноногий и карликовый пятипалый
тушканчики отнесены к многочисленным видам, местами многочис-
лен тушканчик Козлова. Однако в животном населении этих пус-
тынь тушканчики обычно занимают место лишь содоминантов, в
некоторых группировках составляя в среднем 12—29% населения
(Кучерук и др., 1980). В то же время в настоящих пустынях они
составляют основу рациона (почти половину корма) пернатых хищ-
ников. С. paradoxus, D. sagitta и S. kozlovi достигают 10—30%
каждый в добыче птиц (Кучерук, Тупикова, 1980).
ЭКТОПАРАЗИТЫ
У тушканчиков имеются свои специфические виды блох, главным
образом рода Mesopsylla: М. rothschildi, М. eucta, М. tuschkan,
М. lenis, М. hebes, М. apscheronica, а также Frontopsylla wagneri,
F. macrophtalma, Ophthalmopsylla volgensis, Oph. arnoldi, Oph.
praefecta, Oph. jettmari, Oph. kiritschenkoi (Пофф, Скалой, 1954).
Кроме того, на тушканчиках встречаются блохи песчанок — Хепор-
sylla conformis, X. gerbilli caspica, X. skrjabini, X. nuttali, X. hir-
tipes, Ceratophyllus laeviceps, Cer. tersus, Coptopsylla lamellifer,
Paradoxopsylla teretifrons и сусликов — Ceratophyllus tesquorum,
Ctenophthalmus breviatus.
Основную массу блох, найденных в шерсти тушканчиков, сос-
тавляют специфические для этих зверьков виды. Находки на туш-
канчиках блох, свойственных другим видам животных, следует рас-
сматривать как косвенное указание на посещение тушканчиками
нор этих грызунов. В пустынных, степных ландшафтах обычно до-
минируют песчанки и суслики, и именно блохи этих видов регулярно
встречаются на тушканчиках. Например, на Устюрте и в Северо-
Западных Кызылкумах осмотрено 1206 тушканчиков 8 видов и из
их шерсти собрана 1941 блоха, среди которых 13,5% (258 экз.)
были специфичные блохи большой песчанки (Сабилаев, 1970).
Грызуны
63
Среди вшей имеются специфические для тушканчиков виды рода
Eulinognathus (сем. Polyplacidae).
На тушканчиках обнаружены также клещи Hyalomma asiaticum,
Ornithodorus tartakovsky, О. alactagalis, Rhipicephalus schulzei,
Rh. pumilio, Dermacentor daghestanicus, Haemaphysalis numidiana
turanica, гамазовые клещи, краснотелки.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Несмотря на то что тушканчики — типичные пустынные живот-
ные и их происхождение и длительное развитие связано с азиатски-
ми пустынями, они не заняли ведущего положения ни в одной из
паразитарных систем природно-очаговых инфекций, не стали основ-
ными носителями возбудителей в пустынных природных очагах. Это
связано с особенностями их биологии и их местом в биоценозах пус-
тынь.
Тушканчики, хотя и широко распространены в пустынях, не
достигают нигде большой численности, а в животном населении иг-
рают в лучшем случае роль содоминантов. В пустынях Средней Азии
и Казахстана они заселяют преимущественно глинистые такыровид-
ные и щебнистые участки, а в песчаных районах — слабо закреплен-
ные пески, т. е. биотопы, где мало нор больших песчанок — основ-
ных строителей пустынных биоценозов. Тушканчики — одиночные,
ночные зверьки и живут в норах простого устройства. Главная осо-
бенность всех нор тушканчиков в том, что зверьки, находясь в норе,
единственный, как правило, вход в нору и тот забивают изнутри
земляной пробкой. Поэтому такие почти всегда изолированные от
внешней среды норы мало доступны другим животным, в том числе
теплокровным носителям возбудителей (песчанкам, сусликам, по-
левкам) и кровососущим паразитам (москитам, клещам). В норах
тушканчиков по сравнению с норами песчанок хуже условия и для
массового развития личинок блох и москитов, так как в них меньше
накапливается детрита в связи с тем, что тушканчики не запасают
корма и не затаскивают его в норы для поедания, а подстилка в гнез-
довых камерах встречается редко. В то же время сами тушканчики
посещают норы других видов, например песчанок. Там они могут
обмениваться эктопаразитами с хозяевами норы. Об этом свидетель-
ствуют регулярные находки на тушканчиках специфических блох
песчанок и сусликов. Одиночный образ жизни ограничивает внут-
ривидовые контакты тушканчиков. Они осуществляются главным
образом через последовательное использование одних и тех же нор
разными зверьками. При больших индивидуальных участках места
кормежки одного зверька почти не повторяются и не совпадают у
разных особей. Поэтому контакт зверьков на местах кормежки еще
более ограничен. Перечисленные черты экологии тушканчиков не
позволили стать им основными носителями каких-либо возбудите-
лей природно-очаговых инфекций. Однако будучи постоянными чле-
64
Rodentia
нами пустынных биоценозов, они вовлекаются в эпизоотии ряда
инфекций.
Чума. Возбудитель чумы выделен от тушканчиков 9 видов почти
во всех природных очагах этой инфекции на территории СССР, но
зараженные чумой зверьки всюду были единичными (табл. 5). Всего
исследовано в разных очагах 26 277 тушканчиков, из которых зара-
женными оказались 46 особей, или 0,17%. В Туркмении при иссле-
довании диких млекопитающих выделено 1179 культур возбудителя
чумы, из которых только 3 культуры получено от тушканчиков
(Тихомирова, 1957). В Приаральских Каракумах от тушканчиков
выделено 1,6% всех штаммов чумного микроба. Средняя заражен-
ность чумой тушканчиков в отдельные годы составляла 0,4—1,5%
(Муртазанова и др., 1964). Тушканчики высоко чувствительны к
чуме. Тарбаганчики и емуранчики погибают в короткие сроки от
дозы менее 100 микробных клеток (Шмутер и др., 1957). Мохноногие
тушканчики при заражении большими дозами от40 до 200000 микроб-
ных клеток гибли в течение 1—2 суток (Тихомирова, 1957). Тушкан-
чики Северцова, малые и тарбаганчики погибали на 2—3-и сутки от
дозы в 100 000 микробных клеток. Эти виды тушканчиков и мохно-
ногий заражались в эксперименте чумой также через укус блокиро-
ванных блох (Сабилаев, 1970).
Ю. В. Канатов с соавторами (1980) провели ретроспективную
диагностику путем регистрации специфического антигена возбуди-
теля чумы в смывах из костных остатков тушканчиков, собранных
в почве вблизи нор или извлеченных из погадок хищных птиц, и по-
казали, что тушканчики гибнут от генерализованной чумы во всех
природных очагах.
Роль тушканчиков в природных очагах чумы разные авторы оце-
нивают не одинаково, но ни в одном очаге им не отводят роль ос-
новных носителей. Б. К- Фенюк (1948) отнес тушканчиков к группе
факультативных второстепенных носителей, которые лишь вовле-
каются в эпизоотию среди основных носителей и не способны дли-
тельно сохранять возбудителя чумы в своих популяциях. Более
поздние исследователи, опираясь на фактические данные о редком
вовлечении тушканчиков в эпизоотии чумы, считают, как и
Б. К. Фенюк, этих зверьков лишь случайными носителями, не играю-
щими существенной роли в очаге (Кучерук, 1965; Ермилов и др.,
1976; Эйгелис, 1980). Высказано предположение, что тушканчики
вследствие наличия бактериемии и таким особенностям экологии, как
большие индивидуальные участки, паразитарный контакт с основ-
ными носителями, наличие зимней спячки, могут переносить возбу-
дителя чумы на большие расстояния и способствовать расширению
границ эпизоотии, а также осуществлять эпизоотическую связь
между отдельными поселениями основного носителя (Мокроусов,
1957; Муртазанова и др., 1964; Сабилаев, 1970; Настюков, 1978).
П. И. Ширанович (1972) в своем обзоре отметил, что убедительных
доказательств эти предположения не имеют и для их подкрепления
Грызуны
65
Таблица 5. Результаты исследования тушканчиков на чуму
(по Ширанович, 1972, с дополнениями)
Вид Место Годы Исследо- вано зверьков Выделено культур
Большой Волго-Уральское междуречье 1912 82 2
тушканчик То же 1950 1
Волго-Уральский песчаночий 1945—1982 3
очаг
Волго-Уральский сусликовый 1945—1982 3
очаг
Урало-Эмбинский очаг 1945—1982 1
Северо-Восточный Прикаспий 1948—1967 840 0
Северное Приаралье 1945—1956 1099 2
Тушканчик Северо-Западный Прикаспий, 1922 1
(большой)? Ергени
Малый Зауралье 1914 313 1
тушканчик » 1963—1964 2
Волго-Уральский сусликовый 1945—1982 6
очаг
Урало-Эмбинский очаг 1945—1982 4
Северо-Восточный Прикаспий 1948—1967 3081 4
Мангышлак, Приэмбинская равнина 1948—1956 4000 2
Восточное Закавказье 1953—1970 6322 17
Приараксинский очаг 77 4
Кызылкумы 1967 1
Тушканчик- Забайкалье 1926, 1927 2 2
прыгун » 1954—1981 2
Тува 1967 1
» 1969—1982 500 0
Горный Алтай 1961—1981 350 1
Приараль- Мангышлак 1948 1
ский толсто- Приэмбинская равнина 1956—1960 1
хвостый Северо-Восточный Прикаспий 1948—1967 187 1
Урало-Эмбинский очаг 1945—1982 1
3 Зак. 3575
66
Rodentia
Таблица 5 (окончание)
Вид Место Годы Исследо- вано зверьков Выделено культур
Тарбаган- Волго-Уральский сусликовый 1945—1982 4
чик очаг
Урало-Эмбинский очаг 1945—1982 4
Приэмбинская равнина 1958 1
Северо-Восточный Прикаспий 1948—1967 1908 1
Емуранчик Северо-Восточный Прикаспий 1948—1967 4160 1
Урало-Эмбинский очаг 1945—1982 8
Северное Приаралье 1945—1956 1407 1
Северо-Западный Устюрт 1965—1973 1
Северо-Западный Прикаспий 1948—1954 427 3
Западные Каракумы 1950 1
Актюбинская и Кзыл-Ордин- 1958 2
ская области Мангышлак 1948 1849 1
Мохноногий Волго-Уральские пески 1926 6 6
тушканчик То же 1962—1963 1
Волго-Уральский песчаночий 1945—1982 25
очаг
Урало-Эмбинский очаг 1945—1982 1
Центральные и Южные Кызыл- 1952—1955 167 1
кумы Западная Туркмения, Кара- 1933—1954 2
кумы Присарыкамышская равнина 1961 1
Заунгузские Каракумы 1965—1969 3
Актюбинская обл. 1959 1
Западная Туркмения, Небит- Даг 1953 2
Южное Прибалхашье 1958 1
Аксарайские пески 1928—1953 15
Тушканчик Каракалпакия, 1954 2
Лихтен- Кызылкумы
штейна Южное Прибалхашье 1958 1
Гребнепа- лый туш- канчик Западная Туркмения 1953 1
Грызуны
67
необходимы специальные целенаправленные исследования. Сообра-
жение этого автора, что тушканчики как высоко чувствительные
к чуме зверьки быстро гибнут и потому зараженные особи попада-
ются редко, опровергает А. П. Ермилов с соавторами (1976). Они
показали, что в Северо-Восточном Прикаспии среди гребенщиковых
песчанок, также высоко чувствительных к чуме грызунов, заражен-
ные зверьки встречаются в несколько раз чаще, чем среди тушкан-
чиков.
Кожный лейшманиоз. Ю. А. Дубровский (1978) относит тушкан-
чиков к случайным носителям кожного лейшманиоза в природных
очагах туранской части ареала этого заболевания. На территории
пустынной зоны Средней Азии и Казахстана исследовали на зара-
женность кожным лейшманиозом наиболее многочисленные в оча-
гах этого заболевания виды тушканчиков. Исследовано 1600 особей
четырех видов, из которых лишь один тушканчик Северцова оказал-
ся зараженным вирулентным штаммом лейшмания. У 14 зверьков
этого вида на ушах были повреждения, похожие на лейшманиоз -
ные. Такие же повреждения обнаружены у И из 740 обследованных
малых тушканчиков и у 2 из 258 мохноногих. Такая редкая вовле-
каемость тушканчиков в эпизоотии лейшманиоза объясняется, кро-
ме перечисленного выше, тем, что эти зверьки не входят в число
предпочитаемых прокормителей москитов — переносчиков лейшма-
ний. Специальные опыты показали, что тушканчик Северцова наря-
ду с домовой мышью и полуденной песчанкой входит в третью груп-
пу наименее предпочитаемых прокормителей москитов Ph. papatasi
и Ph. caucasicus — переносчиков лейшманий в Средней Азии (Стрел-
кова, Елисеев, 1983).
Другие инфекции. В Казахстане установлена (серологически) у
большого и малого тушканчиков лихорадка Ку, а в Киргизии от
большого тушканчика выделен штамм С. burneti (Жмаева др., 1956;
Евдошенко, Прорешная, 1959; Федорова, 1968).
В Туве у тушканчика прыгуна найдены антитела к клещевому
риккетсиозу, Ку-лихорадке и лептоспирам серологической группы
Icterohoemorrhagiae (Шайман и др., 1973).
На территории Калмыцкой АССР от тарбаганчика выделено 2
штамма и от малого тушканчика — 1 штамм псевдотуберкулезного
микроба (Чуринов, Чуринова, 1980).
В Забайкалье при исследовании 4477 тушканчиков-прыгунов вы-
делено 9 штаммов эризипелотрикса и 1 штамм салмонелл (Некипе-
лов и др., 1981). Сальмонеллы выделены также от емуранчика и
мохноногого тушканчика на территории Средней Азии (Алимход-
жаев, Борисов, 1979).
Исследование тушканчиков на бруцеллез дало отрицательные
результаты — в 20 пробах и серологически исследовано 90 тушкан-
чиков 4 видов: прыгун, малый, большой, мохноногий (Ременцова,
1962).
з*
68
Rodentia
Токсоплазмоз паразитологическим методом и биопробами у туш-
канчиков не установлен. В Казахстане и Средней Азии исследованы
мазки от 55 мохноногих, 13 больших и 23 малых тушканчиков с от-
рицательным результатом. Биопробы от 1 большого, 4 малых и 6
мохноногих тушканчиков также были отрицательными. Но при изу-
чении сывороток крови получены положительные данные у 2 (из
32) мохноногих и у 5 (из 8) малых тушканчиков (Левит и др., 1965).
В Туркмении токсоплазмоз серологически установлен у земляного
зайчика и малого тушканчика (Бердыев, 1976).
СЕМЕЙСТВО HYSTRICIDAE BURNETT, 1830
— ДИКОБРАЗНЫЕ
В семействе 4 рода и 11 видов (Honacki et al., 1982), распростра-
ненных в Африке, Южной Европе и Южной Азии к северу до гор
Кавказа, Средней Азии и Гималаев (рис. 3). Основные сведения по
этой группе обобщены Mop (Mohr, 1965), а также В. Е. Соколовым
(1977).
Род индонезийские дикобразы Thecurus Lyon, 1907 (3 вида)
распространен на п-ове Малакка, островах Калимантан, Суматра
и Филиппинских. Это средние по величине лесные животные (длина
тела 56—65 см). Монотипичный род длиннохвостых дикобразов
Trichys Giinther, 1976 (по другим данным в роде 5 видов — Mohr,
1965) распространен на п-ове Малакка, островах Суматра и Кали-
мантан. Это также лесные животные среднего размера (длина тела
35—45 см). Экология дикобразов обоих родов не изучена. Основные
черты образа жизни типичны для дикобразов вообще: это ночные жи-
вотные, днем скрывающиеся в разнообразных полостях (норах, дуп-
лах и т. п.), они встречаются как в девственных лесах, так и в куль-
турном ландшафте.
Кистехвостые дикобразы Atherurus F. Cuvier, 1829 .(2 вида) насе-
ляют Юго-Восточную Азию и Суматру, а также Центральную Афри-
ку — от Кении до Сьерра-Леоне. Это средние по величине лесные
животные (длина тела 40—55 см, масса 1,5—4 кг). В обоих регионах
Африки и Азии они населяют как обширные тропические леса, так
и узкие полосы галерейных лесов и небольшие участки леса до вы-
соты 3000 м над уровнем моря, по которым они проникают в зону
саванн и в горы. По данным Рама (Rahm, 1962), в Африке они ак-
тивны ночью, а днем скрываются в дуплах, расщелинах скал, тер-
митниках и других убежищах, располагающихся обычно у берега
реки или ручья. Роют неохотно. Обычно встречаются парами, а
иногда группами до 6—8 особей (иногда до 20). Участок семьи
2—5 га. Питание — всевозможные лесные плоды, а вблизи полей —
маниок, бананы, сладкий картофель.
Род дикобразов Hystrix Linnaeus, 1758 включает 5 видов, рас-
пространенных почти по всей Африке, в Италии, Передней и Сред-
ней Азии, Индии и по странам Юго-Восточной Азии, включая Боль-
Грызуны
69
Рис. 3. Распространение семейства Hystricidae
шие Зондские острова. Крупной величины животные: длина тела
60—90 см, масса до 15 кг. Все виды населяют преимущественно
горные и предгорные районы, хотя местами живут и на равнинах, в
том числе и в пустынях. Обычны рядом с сельскими населенными
пунктами, где питаются разными сорными и культурными расте-
ниями. Так, Н. indica в Пенджабе нередко живет на плантациях
(0,2—0,5 экз/га) и роет там норы с 1—5 выходами, с длиной ходов
10—20 м и глубиной до 4 м (Greaves, Khan, 1978). Все дикобразы
активны в темное время суток, днем скрываются в естественных убе-
жищах или в вырытых ими норах, часть из которых используется
многими десятками поколений, постоянно перестраивается и услож-
няется. Такие норы особенно характерны для индийского дикобра-
за (Hystrix indica=H. leucura) в аридных районах, в том числе —
нашей страны. Так, в Таджикистане общая длина подземных ходов
норы на равнине достигает 16 м, в горах — Юм (Тураев, 1971).
Индивидуальные участки обширны: дикобраз ночью может уходить
за несколько километров от своей норы и возвращаться в нее к утру.
Индийские дикобразы в норах живут парами, размножаются раз в
году, в выводке 1—2 (3) молодых, которые отделяются от взрослых
на втором году жизни. Продолжительность жизни —до 20 лет (в
неволе). В Таджикистане (Кафирниган) поедают 154 вида растений
(Тураев, 1973).
Сведения об эктопаразитах и болезнях дикобразов практически
отсутствуют. В горах Эфиопии на Hystrix galeata обнаружены клещи
Haemaphysalis tauffliebi и Rhipicephalus simus.
70
Rodentia,
В фауне СССР один вид Hystrix leucura — самый крупный пред-
ставитель нашей фауны грызунов (длина тела до 90 см, масса до
15 кг). Фауна эктопаразитов этого вида изучена слабо, известно
лишь, что в норах дикобразов в Средней Азии обычно обитают
клещи Ornithodorus papillipes — ими было заселено до 72% нор;
в них также находили О. tartacovskyi и иксодовых клещей Hyalom-
ma anatolicum, Н. asiaticum, Rhipicephalus turanicus, Dermacentor
marginatus, Haemaphysalis numidiana (Петрищева, 1951; Млекопи-
тающие Киргизии, 1972). Специфическая блоха дикобраза — Pario-
dontis riggenbachi turkestanica.
Виды рода Hystrix, как правило, бывали лишь случайными объ-
ектами эпизоотологических исследований. Они считаются естест-
венными резервуарами спирохет — возбудителей клещевого воз-
вратного тифа, которых неоднократно находили у разных видов
Hystrix в разных районах Северной Африки и Южной Азии. Цирку-
ляция возбудителя происходит в норах дикобразов, где обычно
многочисленны их специфические переносчики — клещи рода Or-
nithodorus (Петрищева, 1951; Петрищева и др., 1955). Возможно, что
дикобразы болеют вирусными зоонозами, поскольку в их норах
выплаживается и укрывается много москитов и комаров (Петрище-
ва, 1951).
СЕМ ЕЙСТВО ERETHIZONTIDAE THOMAS, 1897
— АМЕРИКАНСКИЕ ДИКОБРАЗЫ
Семейство Erethizontidae включает 5 родов с 12 видами (Honacki
et al., 1982), которые распространены по всему Новому Свету,
исключая зону тундр в Северной Америке (рис. 4). У всех предста-
вителей этого семейства в шерсти на туловище и хвосте имеются
короткие острые иглы.
Североамериканский дикобраз Erethizon dorsatum Linnaeus,
1758 — единственный представитель рода, распространен в лесных
областях Северной Америки — от Аляски до Северной Мексики.
Это сравнительно крупное животное: длина тела 60—100, масса до
18 кг. Обитает преимущественно в таежных лесах. Ночью питается
различными частями растений (зимой это только кора и хвоя, при-
чем до 90% всех погрызов приходится на хвойные деревья), днем
скрывается в различных убежищах. Так, в Нью-Брансуике (Кана-
да) на площадке в 800 га было обнаружено 69 нор (обычно под кор-
нем дерева или в прикорневом дупле) и 18 древесных гнезд (Speer,
Dilworth, 1978). Как правило, держатся поодиночке, но зимой могут
собираться небольшими группами.
При слежении за тремя мечеными зверьками установлено, что
летом каждая особь использует территорию площадью 13—14 га,
протяженность дневного хода 80 м, ночного — до 200 м; основную
часть ночи они кормятся на земле, а днем отдыхают на деревьях
на высоте 8—10 м (Marshall et al., 1961).
Грызуны
71
Плотность населения в Ми-
чигане (Канада) от 0,1 до
0,17 экз/га. Радиус суточного
участка летом от 30 до 300 м
(среднее 150 м), зимой 0—38 м
(Brander, 1973). В спячку не
впадают. Размножаются раз
в году, рождается один де-
теныш . Продолжительность
жизни до 10 лет (в неволе).
За последнее время числен-
ность во многих местах увели-
чилась, так как исчезли его
естественные враги (илька,
пума, койот) и прекратилась
его добыча из-за мяса в связи
с переселением индейцев в ре-
зервации.
Это обычное в Америке жи-
вотное не было объектом спе-
циальных эпизоотологических
исследований. При исследо-
ваниях небольшого числа осо-
бей у дикобраза обнаружены
возбудители туляремии, лихо-
радки Ку, колорадской кле-
щевой лихорадки, лихорадки
Скалистых гор (Федорова,
1968; и др.).
Специфических видов блох Е. dorsatum не имеет, других видов
блох на них бывает помалу.
Роды Coendou Lacepede, 1799 (5 видов) и Sphiggurus F. Cuvier,
1825 (4 вида) распространены в Центральной и Южной Америке —
от Мексики до Перу, Парагвая, Бразилии. Все они — мелкие и
среднего размера зверьки (длина тела 30—60 см, масса 1—4 кг).
Живут в лесах, на деревьях. Это малоподвижные (суточные переме-
щения у С. prehensillis — 50—700 м), ночные животные, питающие-
ся листьями, почками и плодами. Убежища устраивают на деревьях,
на высоте 6—10 м (Montgomery Lubin, 1978). Размер индивидуаль-
ных участков у трех особей С. prehensillis составлял 8, 10 и 38 га.
Размножаются раз в году, в помете только один детеныш.
Некоторые виды коэнду систематически исследовались в связи
с изучением природной очаговости кожных лейшманиозов в Цент-
ральной и Южной Америке. Из кожи С. rotschildi неоднократно
выделяли лейшманий вида Leishmania hertigi (Herrer et al., 1972).
Коэнду очень часто оказываются зараженными (88,5% из 104 экз.)
и выступают в качестве основного и пока единственного известного
72
Rodent i a
носителя L. hertigi на всей территории Панамы в ландшафтах всех
основных типов.
Лейшмании особого подвида L. hertigi deani не заражают чело-
века; они выделены в Бразилии в штате Пара из печени и селезен-
ки двух из трех исследованных С. prehensillis. Этого же возбудителя
выделяли не только из внутренних органов коэнду, но и из кожи
(Lainson, Snaw, 1977). Характерно, что на коэнду не паразитируют
остальные шесть видов и подвидов лейшманий, распространенных в
Южной и Центральной Америке.
Дикобразы двух монотипических родов — Echinoprocta rufes-
cens Gray, 1865 и Chaetomys subspinosus Kuhl, 1820 — сравнительно
мелкие по размеру, редкие, плохо изученные животные, обитающие
в лесах (первый) и в зарослях кустарников в саваннах (второй).
СЕМЕЙСТВО CAVIIDAE WATERHOUSE, 1839
— СВИНКОВЫЕ
Семейство включает 14 видов, относящихся к 5 родам, 4 из ко-
торых состоят из 1—3 видов.
Два вида рода мара Dolichotis Desmarest, 1820 существенно от-
личаются по экологии от остальных свинковых. Это напоминаю-
щие кроликов крупные грызуны (масса 9—16 кг), населяющие арид-
ные районы Аргентины, Боливии и Парагвая (рис. 5, А). Активны
днем, ночью скрываются в собственных норах или убежищах дру-
гих видов, где раз в год самки приносят по 2—5 детенышей. Обычно
встречаются мелкими группами (3—4 особи), иногда — более круп-
ными (до 40). Медицинское значение этих грызунов не известно.
Остальные свинковые относятся к родам:
Число
видов
Cavia Pallas, 1766 (морские свинки) 5
Kerodon F. Cuvier, 1825 (моко) 1
Galea Meyen, 1831 (куи) 3
Microcavia Gervais et Ameghino, 1880 3
(горные свинки)
Все они имеют характерный облик хорошо известной морской
свинки и распространены от Венесуэлы и Колумбии до Южной
Аргентины, исключая Чили.
В роде морских свинок Cavia имеется 5 видов, распространенных
от южных районов Колумбии и Венесуэлы до Северной Аргентины
Это мелкие зверьки, обитатели разнообразных биотопов — саванн,
мелколесья, болот, каменистых участков в горах. Питаются вегета-
тивными частями различных растений, активны в темное время
суток. Роют обычно несложные норы, в сложных иногда живут
группами по 5—10 особей. Размножение не имеет четкой сезонно-
сти. Продолжительность жизни около 8 лет.
Рис. 5. Распространение семейства Caviidae (A), Dasyproctidae (Б), Echimyidae (В)
Грызуны
74
Rodentia
Сходный с морскими свинками моко — Kerodon rupestris —
встречается в аридных районах по склонам гор и холмов, где оби-
тает в каменистых и скалистых участках; убежищем служат или
собственные норы, или расшелины между камнями (например, на
северо-востоке Бразилии моко живет только в больших нагромож-
дениях камней).
Три вида горных свинок Microcavia и три вида рода куи Galea
по экологии мало отличаются от морских свинок. М. australis в Ар-
гентине роет норы, в которых обычно живут по 3—8 особей, иногда
в той же норе обитает куи. Живут в непосредственной близости с
поселками людей, но в них не проникают. Всех свинок местное насе-
ление добывает ради мяса. Некоторые виды Cavia содержатся в не-
воле, как у нас кролики.
Наиболее подробно исследовалось значение свинок разных ви-
дов в эпизоотиях чумы (Plague in the Americas, 1965). Весьма ин-
тенсивные эпизоотии регистрировались среди Microcavia australis
в пампе и горных полупустынях Аргентины (провинции Рио-Негро,
Мендоса, Сантьяго-дель-Эстеро и др.); при них нередко гибнет около
10% зверьков. Заражение людей от этих свинок происходит при пое-
дании мяса и при контакте с трупами (дети ими играют). Опасно и
то, что эти свинки нередко живут рядом с поселками. Интенсивные
эпизоотии обеспечиваются изобилием блох на свинках: индекс этих
паразитов составляет 20 зимой и 3 летом; основные виды блох —
Craneopsylla wolffhuegeli (43%), Panallius galeanus (36%) на севере
Аргентины и Delostichus talis (45%), Hectopsylla gemina (20%) на
юге; в Боливии 97% блох этой свинки составляют Tiamastus
cavicola.
Предполагают, что С. pamparum является носителем чумы в
провинции Буэнос-Айрес. Это обитатели прерий, но весьма влаго-
любивые зверьки. Они не делают нор, а живут среди густой расти-
тельности; рядом с населенными пунктами обычно не встречаются,
мясо их едят редко, поэтому эпидемиологическое значение этого
вида меньше, чем М. australis. Эпизоотиям чумы среди С. pamparum
препятствует и низкий индекс блох — всего 0,04—0,1.
При сильных эпизоотиях среди М. australis в них нередко вовле-
каются G. musteloides. Эта свинка распространена так же, как и
М. australis, а нередко они живут совместно. Зараженные чумой
G. musteloides регистрировались в провинциях Рио-Негро, Ла-Пам-
па, Мендоса, Кордова и Сельто. Другой вид этого рода — G. spixii
нередко регистрировалась зараженной чумой в Бразилии (2,5—
9,6% от зараженных чумой грызунов всех видов). Эти зверьки не
делают нор, а скрываются в низкой и густой растительности. Смерт-
ность их от чумы бывает столь значительной, что их перестают добы-
вать охотники.
Моко Kerodon rupestris очень редко вовлекаются в эпизоотии
чумы (0,14—0,40% зараженных зверьков в Бразилии). На них очень
мало блох. Считают, что они сами по себе не могут иметь значения
Грызуны
75
в эпизоотии чумы. Однако это — потенциально опасный вид, так
как на него охотятся ради мяса и содержат в неволе, а в совместных
поселениях с хомяками на каменистых склонах гор могут существо-
вать устойчивые очаги чумы. Такие поселения местами находятся
близ населенных пунктов, что увеличивает их эпидемиологическое
значение.
Домашние свинки С. porcellus и С. арегеа нередко получают
возбудителя чумы из природных источников и затем могут передать
это человеку. Это особенно типично для очагов чумы Эквадора, где
очаги, будучи полигостальными, связаны с различными видами хо-
мяков. На хомяках и свинках, а также на крысах часто встречается
блоха Polygenis litargus, в известной мере антропофильная и играю-
щая роль основного переносчика чумы; специфическая блоха до-
машних свинок — Tiamastus cavicola, хотя нередко на них встреча-
ется много Pulex irritans.
Эпидемиологическая роль свинок в Эквадоре особенно важна
в связи с местным обычаем посылать этого зверька в качестве подар-
ка родственникам и друзьям, зачастую живущим весьма далеко.
Сходную экологическую структуру имеют очаги чумы в Перу, где
зараженными чумой были найдены дикие свинки С. tschudii (4 особи
из 16 исследованных).
Зараженность морских свинок возбудителями других инфекций
не привлекала пристального внимания. Известно, что они нередко
бывают носителями лептоспир. Так, эпизоотии лептоспироза среди
сравнительно влаголюбивых С. pamparum зарегистрированы в Ар-
гентине, от зверьков выделяли культуры лептоспир Pomona и
реже —Grippotyphosa; от С. laviaperea в Бразилии выделяли лепто-
спир Icterohaemorrhagia (Castro et al., 1961; цит. по Карасевой,
Свешниковой, 1971). Разводимые в неволе С. арегеа azarae являются
носителями лептоспир из серогруппы Javanica, а С. pamparum —
Pomona; дикая свинка С. арегеа festina зарегистрирована как носи-
тель лептоспир Hebdomadis и Pomona. От свинок неуказанного вида
и места обитания изолировали лептоспир Canicola, Sejroe, Ballum.
От домашних свинок неуказанного вида — Pomona, Hyos. Безот-
носительно к типу лептоспир указывается, что свинки могут быть
лептоспироносителями долгое время (по крайней мере до 1 года) и
постоянно выделяют микробов с мочой; последнее обеспечивает пе-
редачу возбудителя людям, в особенности ухаживающим за разво-
димыми в неволе свинками (Карасева, Свешникова, 1971).
Мелиоидоз зарегистрирован как натуральная инфекция в лабора-
торных популяциях свинок (Davis et al., 1970).
Морские свинки (очевидно, домашние) высоко восприимчивы к
возбудителю венесуэльского энцефаломиелита лошадей, патогенность
вируса для них в ряде случаев коррелирует с таковой для человека
и лошадей (Infectious diseases..., 1970).
Возбудитель болезни Чагаса обнаружен в Бразилии у трех ви-
дов семейства. С. porcellus является важнейшим источником инфек-
76
Rodentia
ции в некоторых районах Бразилии, Перу, Боливии. Куи G. spixii,
добытые в муниципии Морада-Нова (штат Сеара) и исследованные
методом ксенодиагностики, оказались зараженными в 5 случаях
из 215 (для сравнения укажем, что пораженность синантропных гры-
зунов составляет 4,6%, собак — 25, кошек — 0,7%, опоссумов —
9 зараженных из 12) (Lainson et al., 1979; Miles et al., 1979).
От содержавшихся в виварии Института технологических иссле-
дований в Куритиба (Парана, Бразилия) свинок С. porcellus из
кожных поражений, напоминавших раковые, выделены лейшмании
более крупного размера, чем все ранее известные. Они описаны как
Leishmania enriettii (Muniz, Medina, 1948). Их инокуляция другим
особям С. porcellus приводила к образованию метастаз на носу,
ушах и ногах; паразиты содержались в крови, печени, костном моз-
ге и коже. Они оказались непатогенны для мышей, собак, макак-
резусов, свинок других видов (С. арегеа) и человека; чувствительны
к ним только сирийские хомяки. В том же районе Куритиба были
обнаружены естественно зараженные лейшманиями дикие С. por-
cellus.
СЕМЕЙСТВО DASYPROCTIDAE SMITH, 1842
—АГУТИЕВЫЕ
Семейство состоит из двух родов Myorprocta Thomas, 1903
(2 вида) — акуши и Dasyprocta liliger, 1811 (12 видов) — агути.
Агутиевые распространены от Южной Мексики на юг до Южной
Бразилии и Перу, на Малых Антильских островах (рис. 5, Б). Насе-
ляют влажные места — низменные леса, берега водоемов с травянис-
той растительностью, густые кустарники; некоторые виды агути
живут в саваннах или на сухих склонах холмов. Активность пре-
имущественно дневная. Укрытием обычно служат норы, которые
эти зверьки роют самостоятельно. Питаются плодами, вегетативны-
ми частями растений. Участки обитания широко перекрываются.
Размножение не сезонно; у большинства видов два выводка в году,
по 2—4 детеныша в каждом. Продолжительность жизни —до 13—
20 лет (в неволе). Все виды семейства населением используются в
пищу.
Эпизоотологическое значение этих животных специально не
изучали; они, скорее всего, не служат основными носителями ни
при одном зоонозе. Известно, что один из видов Dasyprocta зареги-
стрирован как носитель возбудителя болезни Чагаса в Бразилии;
D. aguti отмечен как носитель псевдотуберкулеза (Davis et al.,
1970). Сравнительно внимательно агути изучали при обследовании
очагов чумы (Plague in the Americas, 1965). D. punctata обнаружена
зараженной чумой в Боливии. Однако ни в Эквадоре, ни в Венесуэле
инфицированных чумой особей не регистрировали; случаи заболе-
вания людей после контакта с агути не отмечены.
На D. variegata в Боливии из блох резко доминируют (90%)
виды рода Rhopalopsyllus, а 86% приходится на вид R. australis
Грызуны
77
СЕМЕЙСТВО ECHIMYIDAE MILLER ET GIDLEY, 1918
— КОЛЮЧИЕ ШИНШИЛЛЫ
Это наиболее богатое видами семейство дикобразоподобных.
В нем 15 родов и 55 видов (Honacki et al., 1982) или1 14 родов и
44 вида (Соколов, 1977), из которых 7 монотипичны и 6 имеют
по 2—3 вида; только в родах Proechimys и Echimys соответствен-
но 22 (12) и 10 (11) видов:
Число видов Число видов
Proechimys J. Allen, 1889 22 Echimys F. Cuvier, 1809 10
Hoplomys J. Allen, 1908 1 Dactylomys I. Geoffroy, 1838 3
Thrichomys Trouessart, 1880 (= Cercomys F. Cu- Kannabateomys Jentink, 1891 1
vier, 1829) 2 Thrinacodus Gunther, 1879 2
Mesomys Wagner, 1845 3 Clyomys Thomas, 1916 1
Lonchothrix Thomas, 1920 1 Carterodon Waterhouse, 1848 1
Isothrix Wagner, 1845 3 Euryzygomatomys Goeldi, 1
Diplomys Thomas, 1916 3 1901 Makalata Husson, 1978 1
Ареал семейства охватывает Центральную и Южную Америку —
от Никарагуа до Парагвая и Южной Бразилии, острова Порто-Рико
и Мартинику (рис. 5, В). Внешним обликом напоминают крыс. Раз-
меры обычно мелкие — длина тела до 30 см (только Diplomys круп-
нее — до 48 см). Большинство видов — ночные зверьки, обитатели
лесов, зарослей кустарников, высокотравья. Ряд видов ведет дре-
весный образ жизни, многие хорошо лазают по деревьям и кустар-
никам. Три вида (Euryzygomatomys spinosus, Clyomys laticeps,
Carterodon sulcidens) населяют открытые пространства типа саванн
и сооружают более или менее обширные и сложные норы. Экология
подавляющего большинства видов семейства не изучена. По-види-
мому, в год приносят 2 выводка по 1—3 детеныша в каждом. Некото-
рые Proechimys склонны к синантропии.
Пунарес Cercomys cunicularius (=Thrichomys apereoides), оче-
видно, играет важную роль в эпизоотиях чумы в ряде энзоотичных
районов Бразилии (Plague in the Americas, 1965). Этот зверек живет
среди камней, часто совместно с моко (Kerodon) — известным носи-
телем чумы; на пунарес обычно много блох. Эпидемиологическое
значение вида определяется тем, что пунарес добывают как источ-
ник мясной пищи. Предполагается, что в штате Гера этот вид —
основной носитель чумы. В этом штате на долю пунарес приходилось
23% зараженных грызунов, в Пернамбуко — 5, в Алагоас — 0,2%.
Зараженными чумой находили также Cercomys inermis (=Thricho-
mys apereoides).
Известно, что один из видов Echimyidae является носителем
Т. cruzi — возбудителя болезни Чагаса в Бразилии. В этой же стра-
не у Proechimys sp. обнаружены антитела к Leptospira pomona.
78
Rodentia
Более подробно изучена роль одного из самых многочисленных ви-
дов — Proechimys semispinosus в циркуляции лептоспир. В общей
сложности у них обнаружены 3 серовара из серогруппы Bataviae,
2 — из группы Cynoptery, 5 — из Hebdomadis, 1 — Icterohaemor-
rhagiae, 1—Pomona, 1—Pyrogenis, 1—Ranarum, 1—Shermani,
а всего 15 сероваров 8 серогрупп (Карасева, Свешникова, 1971).
У Proechimys sp. обнаружен III субтип вируса венесуэльского
энцефалита лошадей, который вызывает у людей субклиническую
инфекцию; он распространен вдоль северного побережья Южной
Америки — от Панамы до Амазонки; носителем этого субтипа явля-
ется также рисовая крыса Oryzomys capito (Infectious diseases,
1970).
Виды Proechimys считаются основным носителем возбудителя
лихорадки Буссуквара (флавивирус). От Proechimys guyanensis
выделяли вирус лихорадки Мукамбо (Бразилия), лихорадки груп-
пы С — Буньямвера (тоже в Бразилии), вирус Maxyno (Simpson,
1968; и др.).
На сальмонеллез в лесах Панамы исследовано 704 Proechimys
semispinosus, пораженными оказались 9 (1,2%, тогда как среди
сумчатых там же — 12—20%); в сухой период года зараженность
была в три раза выше, чем в дождливый (Taylor, 1968).
Proechimys guyanensis относится к основным хозяевам лейшма-
ний L. mexicana amazonensis в Панаме и Северо-Восточной Брази-
лии, а возможно, и в других районах Америки (Lainson, Shaw, 1968).
ОТРЯД CARNIVORA BOWDICH, 1821
— ХИЩНЫЕ
Отряд объединяет хищных (плотоядных) млекопитающих семи
семейств: Canidae — волчьи, Ursidae — медвежьи, Procyonidae —
енотовые, Mustelidae— куньи, Viverridae — виверровые1, Ну-
aenidae — гиеновые, Felidae — кошачьи. В отряде насчитывают,
согласно представлениям различных авторов, 80—99 родов и 224—
258 видов (табл. 6). Общее число хищных составляет около 8% всех 1
Таблица 6. Состав семейств в отряде хищных и их распространение *
Семейство Число Распространение
родов видов
Canidae 8—12** 5 29—35 8 Все материки, кроме Антарктиды
Ursidae 3—5 1 7—8 3 Евразия, кроме Аравийского п-ова и ост- ровов Сулавеси и Филиппинских; Северо- Западная Африка, Северная Америка на юг до центральных районов Мексики, гор- ные области северо-запада Южной Америки
Procyonidae 7—8 1 15—19 1 Северная, Центральная и Южная Амери- ка. В Азии — горные районы Гималаев, Индокитая. В СССР американский енот акклиматизирован в Закавказье, Киргизии
Mustelidae 23—29 8 63—70 Jg*»* На всех материках, за исключением Ан- тарктиды, Австралии и ряда океанических островов
Viverridae 35—36 71—75 Южная Европа, Африка, Мадагаскар, Южная и Юго-Восточная Азия, Индоне- зия, Филиппины
Hyaenidae 2—3 1 3—4 1 Африка, Передняя, Средняя и Южная Азия, Закавказье
Felidae 3—6 4 36—37 12 Евразия, Африка, Северная и Южная Америка, нет в Австралии и на Мадагас- каре
* По: Млекопитающие Советского Союза, 1967; Соколов, 1979; Каталог млекопитающих
СССР, 1981; Anderson, Jones, 1967; Honacki et al., 1982.
♦* В числителе—общее число родов и видов, в знаменателе —из них в СССР.
♦♦♦ Включая американскую норку.
1 Хонаки с соавторами (Honacki et al., 1982) разделяет на два семейства:
Viverridae — 19 родов, 34 вида и Herpestidae — 17 родов, 36 видов.
80
Carnivora
современных млекопитающих. Представители отряда обитают на
всех материках, кроме Антарктиды, встречаясь в различных ланд-
шафтных зонах — от арктических тундр до пустынь и тропических
лесов.
В отряд входят весьма разнообразные по величине (от ласки до
медведя) и строению тела звери. В основном это плотоядные формы,
активно охотящиеся за живой добычей, но почти в каждом семейст-
ве встречаются виды со смешанным характером питания или даже
преимущественно растительноядные.
Экологические типы хищников разнообразны и определяются
способами добывания пищи. Среди них есть виды, охотящиеся за
мелкими грызунами в их норах (мелкие куньи), выкапывающие или
подстерегающие ту же добычу (песец, лисица, мелкие кошки). Есть
виды, хорошо лазающие по деревьям (соболь, куницы) и хорошо
плавающие, добывающие водных или околоводных животных (нор-
ка, выдра). Крайний пример адаптации к водному образу жизни —
морская выдра, или калан, питающийся морскими беспозвоночными
и большую часть времени проводящий в воде.
Особенности питания определяют зависимость хищников от
обилия и доступности добычи, которые изменяются в различные се-
зоны и также в разные годы. У хищников наблюдаются различные
формы адаптации к переживанию неблагоприятных кормовых пе-
риодов. Зимнее прекращение активности свойственно видам со сме-
шанным характером питания — бурому и гималайскому медведям,
барсуку, енотовидной собаке, еноту-полоскуну. Пребывание в убе-
жищах может быть длительным или прерываться в теплую погоду.
Зимний сон хищников не сопровождается значительным снижением
температуры тела. Медведицы, как известно, приносят потомство
в зимние месяцы и выкармливают детенышей в берлоге.
Одним из приспособлений к переживанию неблагоприятных,
малокормных периодов служит накопление жировых запасов и спо-
собность к длительному голоданию. Звери многих видов устраивают
кратковременные запасы пищи, большинство поедает падаль. Оби-
тающие в экстремальных условиях Заполярья белый медведь и пе-
сец совершают далекие переходы в поисках пищи.
Велико значение хищников в природных биоценозах, где они
имеют тесные трофические связи со многими видами животных —
от полевых грызунов, вредящих сельскому хозяйству, до крупных
копытных и разнообразных птиц.
Многие виды хищных, имеющие густой и пушистый мех, служат
объектами промысла. Ряд видов (соболь, норка, песец, лисица)
разводятся на фермах. Некоторые виды (енотовидная собака, аме-
риканская норка, енот-полоскун и др.) акклиматизированы далеко
за пределами естественного ареала.
Участие хищников в поддержании очагов инфекций, передаю-
щихся человеку, изучено, за немногими исключениями, недостаточ-
Хищные
81
но. Имеющиеся сведения по большей части носят случайный или
отрывочный характер.
Прежде всего это связано с тем, что, по счастью, хищники не
являются основными хозяевами подавляющего большинства возбу-
дителей природно-очаговых болезней человека. По данным В. В. Ку-
черука (1977), у хищных млекопитающих обнаружена зараженность
возбудителями 18 инфекций из 30, передающихся человеку от ди-
ких животных. Но лишь для трех из них (бешенство, висцеральный
лейшманиоз и некоторые лептоспирозы) среди хищников есть ви-
ды — основные хозяева инфекции, у которых особенности патогене-
за и механизм передачи обеспечивают дальнейшее распространение
возбудителя, в том числе и к человеку. К остальным возбудителям
хищники малочувствительны и вследствие этого не могут быть ис-
точником заражения и распространителями инфекции.
Устойчивость хищников к инфекциям связана с тесными, сфор-
мировавшимися в длительном процессе эволюции, трофическими
связями их с естественными носителями возбудителей. Хищники
малочувствительны к остро протекающим болезням грызунов —
туляремии, чуме, а также к болезням копытных — сибирской язве,
бруцеллезу, ящуру. При поедании больных животных значительная
часть микробов, обсеменяющих их органы, погибает под действием
свойственной хищникам высокой кислотности желудочного сока.
В случае заражения заболевание у хищников протекает доброка-
чественно, с формированием иммунитета и слабым размножением
возбудителя в органах, не обеспечивающем дальнейшего его рас-
пространения.
В противоположность инфекциям, связанным с кормовыми объ-
ектами, хищники высокочувствительны к заболеваниям, в передаче
которых отсутствуют трофические связи. Хищники остро болеют
бешенством, заражение которым происходит через укусы, висце-
ральным лейшманиозом, передаваемым москитами, лептоспирозом
canicola, чумой плотоядных, инфекционным собачьим гепатитом,
пироплазмозом и др.
Таким образом, из обширного отряда хищных млекопитающих
лишь отдельные виды могут иметь существенное эпидемиологическое
значение. Ведущая роль некоторых хищников в поддержании эпи-
зоотий бешенства обусловила необходимость тщательного изучения
природной очаговости этой инфекции. Тесная связь природных и
антропургических очагов бешенства заставляет рассмотреть роль
домашних собак и кошек как связующего звена между природными
и антропургическими очагами. Особенное значение при этом при-
обретают сформировавшиеся во многих местах устойчивые популя-
ции одичавших собак и кошек.
82
Carnivora
СЕМЕЙСТВО CANIDAE GRAY, 1821
— ВОЛЧЬИ
В семейство входят звери собачьего облика, быстро бегающие,
активно охотящиеся за живой добычей. Размеры видов от мелких с
длиной тела 40—70 см (афганская лисица, фенек), средних размером
с лисицу до крупных с длиной тела 120—160 см (волк). В семействе
насчитывают 7—15 родов и 29—35 видов. Из них на территории СССР
обитают 8 видов, относящихся к 5 родам. Представители семейства
распространены на всех материках, кроме Антарктиды, встреча-
ясь в разнообразных ландшафтно-географических зонах — от тунд-
ры до пустынных и горных районов (табл. 7). Наиболее широко рас-
пространены и имеют важное хозяйственное значение роды Canis,
Vulpes, населяющие Евразию, Северную и Южную Америку, Аф-
рику. В Австралии обитает дикая (одичавшая) собака динго, заве-
зенная аборигенами около пяти тысяч лет тому назад, и акклимати-
зированная европейцами обыкновенная лисица.
Таблица 7. Состав и распространение семейства Canidae
Род Число видов Распространение
Nyctereutes 1 Восточная Азия — от севера Вьетнама до юга Дальнего Востока СССР. Широко акклиматизи- рована в европейской части СССР, откуда про- никла в Польшу, Румынию, ГДР, Швецию, Финляндию. Очаговое распространение в резуль- тате акклиматизации в Средней Азии, Казах- стане
Canis 6 Африка, Евразия, Северная и Центральная Аме- рика
Alopex 1 Тундровые районы и арктические острова Евразии и Северной Америки, Гренландия, Командорские, Алеутские острова
Vulpes 9 Африка, Евразия, Северная Америка
Су on 1 Азия — от гор Средней Азии. Казахстана, Южной Сибири и Дальнего Востока на юг до Индоста- на, Бирмы и Индонезии (Суматра и Ява) вклю- чительно
Urocyon 2 Северная Америка —от южных районов Канады до Центральной Америки включительно
Fennecus 1 Северная Африка, Аравия
Dusicyon 10 Южная Америка
Chrysocyon 1 То же
Lycaon 1 Африка
Speothos 1 Центральная и Южная Америка
Otocyon 1 Африка
Хищные
83
В пределах СССР распространены енотовидная собака Nycte-
reutes procyonoides Gray, 1834, волк Canis lupus Linnaeus, 1758,
шакал Canis aureus Linnaeus, 1758, песец Alopex lagopus Linnaeus,
1758; лисицы: обыкновенная Vulpesvulpes Linnaeus, 1758, афганская
V. сапа Blandford, 1877, корсак V. corsac Linnaeus, 1768, красный
волк Cuon alpinus Pallas, 1811.
По характеру питания все представители семейства плотояд-
ные звери. В их питании важное место занимают наземные позвоноч-
ные, в основном грызуны, но также и птицы, амфибии и рептилии, у
крупных видов — копытные. Нередко в рацион входят насекомые.
В летний период большинство видов поедает ягоды, плоды и другую
растительную пищу. Волк, шакал, лисица нападают на домашний
скот, птицу. Все виды охотно поедают падаль.
Билогия размножения видов сходна. Обычно это моногамы, со-
единяющиеся в пары, совместно выкармливающие потомство. Гон
происходит в зимние месяцы, рождение щенят — весной. Продол-
жительность беременности около двух месяцев. К осени щенки
переходят на самостоятельный одиночный образ жизни, но у волков
сохраняются семейные группы, состоящие из родителей, молодых
этого и прошлого (переярки) года. Половозрелыми звери становятся
на втором году жизни, енотовидные собаки — в возрасте 8—10 ме-
сяцев.
Экономическое значение имеют все широко распространенные
виды семейства. Песец, лисица, енотовидная собака служат объек-
том промысла и также разводятся на фермах. Волк, шакал и местами
енотовидная собака наносят ущерб животноводству и дичеразведе-
нию.
У представителей семейства в природных условиях установлена
зараженность (выделением культуры возбудителя) или переболева-
ние (по серологическим данным) туляремией, бруцеллезом, лепто-
спирозами, листериозом, псевдотуберкулезом, чумой грызунов,
эризипелоидом, сальмонеллезами, туберкулезом, сибирской язвой,
спирохетозом, Ку-риккетсиозом, вирусными инфекциями — бе-
шенством, ящуром, протозойными инвазиями — токсоплазмозом,
лейшманиозом.
Волчьи служат основным хозяином половозрелой стадии эхино-
кокка, многокамерного альвеококка, дифиллоботриоза и других
гельминтозов, поддерживая их циркуляцию в природных очагах
и представляя определенную опасность для заражения людей
(табл. 8).
За пределами СССР некоторые виды имеют важное значение в
распространении бешенства. В странах Европы после второй миро-
вой войны получили широкое распространение природные очаги
бешенства, поддерживаемые лисицами. Бешенство волков установ-
лено в странах Восточного Средиземноморья, шакалы служат ис-
точником бешенства в Азии и Африке. Бешенство лисиц, волков,
койотов установлено в Северной Америке.
84
Carnivora
Таблица 8. Выявление природно-очаговых инфекций и инвазий у видов
семейства Canidae в СССР (по Шеханову, 1970, с дополнениями)
Болезнь Волк Шакал Лисица Корсак Песец Енотовид- ная собака
Бруцеллез в — с — В —
Туляремия в — В — с В
Листериоз в — в В — —
Эризипелоид — — в — — —
Псевдотуберкулез — — + — — —
Чума грызунов — В в в — —
Сальмонеллезы — — с с С —
Лептоспирозы с — св — с СВ
Сибирская язва с — — — с —
Спирохетоз — в — — — —
Лейшманиоз — в — — — —
Ку-риккетсиоз — — с с — —
Бешенство в в в в в в
Токсоплазмоз с — в в в —
Трихинеллез в в в в в в
Описторхоз в — в — — в
Эхинококкоз в в в — в в
Альвеококкоз в — в — в в
Клонорхоз — — — — — в
Метагонимоз — — — — — в
Парагонимоз — — — — — в
Примечание. Обозначения: в—выделение возбудителя; с —серологическое обследо-
вание.
Там же у обыкновенной лисицы находили псевдотуберкулез, у
обыкновенной и серой лисиц были выявлены отдельные случаи
заболевания туляремией, листериозом. Туляремия обнаружена
также у койота, а листериоз — у песца (Infectious diseases...,
1970).
Из приведенного списка заболеваний первостепенное эпидемио-
логическое значение имеет, как уже говорилось, бешенство. Наибо-
лее часто источниками заражения людей служат лисицы, волки, в
меньшей степени — енотовидные собаки. Известны отдельные слу-
чаи заражения людей туляремией от укуса лисицы, при снятии
шкурки с енотовидной собаки. Енотовидные собаки служат основ-
ным хозяином трематод, вызывающих заболевания людей при упо-
треблении в пищу промежуточных хозяев паразита — рыб. Шакалы
наряду с домашними и бродячими собаками поддерживают очаги
висцерального лейшманиоза, вызывающего тяжелое заболевание
людей.
Хищные
85
Род Nyctereutes Temmink, 1832
— енотовидная собака
Енотовидная собака Nyctereutes procyonoides Gray, 1834 — един-
ственный вид в роде. Естественный ареал занимает Восточную Азию:
Северный Вьетнам, Восточный Китай, Корейский полуостров, Япон-
ские острова (Кюсю, Сикоку, Хонсю, Хоккайдо), восточную часть
МНР и советское Приамурье на запад до Аргуни. С 1927 по 1957 г.
проводился выпуск енотовидных собак более чем в 76 областях, кра-
ях и автономных республиках. В настоящее время вид имеет сплош-
ной ареал в европейской части Союза, распространившись от южной
части Кольского полуострова до Черного, Азовского и Каспийского
морей. В небольшом числе встречается в Казахстане, Томской обла-
сти, в южной части Киргизской ССР, на Алтае, в Иркутской обла-
сти, в Бурятской АССР (Громов и др., 1963; Соколов, 1979). В конце
50-х годов енотовидная собака проникла в Польшу, Румынию, ГДР,
Венгрию, Чехословакию. С 1936 г. распространилась в Финляндию,
откуда проникла в северо-восточную часть Швеции и активно рас-
селяется (Сивонен, 1979; Насимович, 1985). Карты ареала вида
приведены в сводках «Млекопитающие Советского Союза» (1967)
и Г. Корбета (Corbet, 1978).
Енотовидная собака селится в смешанных лесах, придерживаясь
участков с озерами, речными протоками. Питается преимуществен-
но мелкими грызунами, лягушками, насекомыми, рыбой, в летнее
время разоряет гнезда наземно гнездящихся птиц. Часто потребля-
ет растительную пищу, падаль, отбросы хозяйственной деятельно-
сти человека. К осени звери накапливают значительное количество
жира и на зиму (с ноября по февраль-март) впадают в зимний сон,
прерывающийся во время оттепелей. Убежища устраивает под вы-
воротами корней, в буреломах, комлевых дуплах. Использует бро-
шенные лисьи и барсучьи норы 2.
Гон наблюдается с февраля до конца апреля в зависимости от
района распространения. Беременность продолжается 59—65 дней.
Количество щенят в помете чаще 6—7, максимально 15—16. Отмеча-
ются значительные колебания величины среднего выводка в зависи-
мости от района акклиматизации и условий года (Насимович, 1985).
Распад выводков происходит в сентябре-октябре. В отличие от дру-
гих видов семейства енотовидные собаки, как правило, становятся
половозрелыми на первом году жизни, но около 12% самок остаются
яловыми. Отмечается значительная гибель молодняка, достигающая
к сентябрю-октябрю более 50% (Попов, 1956; Геллер, 1959; Юдин,
1977). Среди врагов енотовидной собаки наибольшее значение при-
надлежит волку. Легко справляются с молодыми зверями также ли-
сицы и бродячие собаки. По наблюдениям М. X. Геллера (1959),
2 Подробные сведения о размещении, экологии, хозяйственном значении
этого вида приведены в монографии «Песец, лисица, енотовидная собака».
М.: Наука, 1985.
86
Carnivora
в северо-западных районах европейской части СССР из 186 случаев
гибели енотовидных собак от хищников 64% приходилось на долю
волка, 14,3% — на лисицу и 12,8% — на бродячих собак. В Тата-
рии из 54 енотовидных собак, задавленных хищниками, в 55,6%
они были жертвами волка, в 27,8% — бродячих собак и в 11,1 % —
лисиц (Попов, 1956).
Эктопаразиты. В шерсти енотовидных собак паразитирует вла-
соед Trichodectes latus (Mallophaga), блохи Ctenocephalides canis,
Chaetopsylla trichosa, Ch. globis, Paraceras meles.
В послевоенные годы в ряде областей Украины, в Литве, в Псков-
ской и Воронежской областях наблюдалось поражение енотовидных
собак зудневым клещом Acarus siro var. vulpis (Корнеев, 1954; Пру-
сайте, 1961; Морозов, 1978). На Дальнем Востоке чесотка у еното-
видных собак не регистрируется. На Сахалине у енотовидных собак
распространено грибковое заболевание — дерматикоз (Воронов,
1972).
Гельминтозы. В пределах естественного ареала у енотовидной
собаки выявлено 29 видов гельминтов: 5 цестод, 12 трематод и 12
нематод (Козлов, 1977; Юдин, 1977). Большое количество видов
трематод, характерное для енотовидной собаки, связано с привя-
занностью хищника к влажным местообитаниям, благоприятным
для жизни моллюсков — промежуточных хозяев личиночной стадии
развития этих гельминтов. На Дальнем Востоке енотовидные соба-
ки являются распространителями опасных для человека гельминто-
зов — парагонимоза (паразит — легочная двуустка Paragonimus
westermani), клонорхоза (Clonorchis sinensis), поражающего печень,
нанофиетоза (Nanophyetus salmincola) и метагонимоза (Metagoni-
mus jocagavi), локализующихся в кишечнике. Нанофиетоз обнару-
жен у 69,6—76,4% местного населения и у 2,2—58,6% приезжих
(Мишаков, 1971). Заражение происходит при употреблении сырой
или вяленой рыбы.
В районах акклиматизации енотовидные собаки освобождаются
от свойственных дальневосточному ареалу трематод вследствие
отсутствия специфических промежуточных хозяев — моллюсков,
эндемиков Дальнего Востока. Вместе с тем у них появились новые
виды гельминтов — 3 вида цестод, 7 видов нематод и 11 видов тре-
матод, из которых 7 видов приобретены зверями в дельте Волги
(Заблоцкий, 1970). В этом регионе у енотовидных собак выявлено
заражение 21 видом гельминтов, в то время как на северо-западе —
всего 14 видов, в Татарии — 10 (Гусев, 1951; Попов, 1956). В Бело-
руссии установлена зараженность 17 видами гельминтов (Карасев,
1975). Западнее р. Оби у енотовидных собак встречается половозре-
лая стадия кошачьей двуустки (Opistorchis felineus), передающейся
человеку при поедании рыбы, пораженной метацеркариями.
Определенную опасность представляет для людей трихинеллез,
встречающийся у енотовидных собак повсеместно. Зараженность
зверей в Амурской области колеблется от 44,3 до 54,3%, в Хабаров-
Хищные
87
ском крае — от 20,9 до 60% (Юдин, 1977). В Краснодарском крае
трихинеллез обнаружен у 8,3—75% особей, в Литве — у 42,1%,
в Эстонии и Латвии — у 8,9—11,2% (Боев и др., 1978).
Практикующееся местами скармливание тушек енотовидных со-
бак свиньям обусловливает заражение последних и может привести
в дальнейшем к заражению людей. На Дальнем Востоке, а также в
некоторых районах интродукции мясо енотовидных собак употреб-
ляют в пищу и они могут явиться непосредственным источником
заражения людей трихинеллезом (Бессонов, 1972).
Бешенство. На Дальнем Востоке первое сообщение о бешенстве
енотовидных собак поступило зимой 1931/32 г., когда в парке оле-
неводческого хозяйства зарегистрирован случай заболевания оленя-
пантача, по-видимому, укушенного енотовидной собакой. Бешенст-
во в это же время наблюдали в питомнике енотовидных собак, где
они содержались для целей акклиматизации (Миролюбов, 1934).
В 1951 г. в Амурской области отмечено заболевание гидрофобией
человека, укушенного енотовидной собакой (Савицкий и др., 1980).
В 1957 г. укус енотовидной собаки вызвал заболевание человека в
Хабаровском крае (Гурьянова, Зак, 1966; Гурьянова, 1967). До
1979 г. в Хабаровском и Приморском краях зарегистрировано все-
го 5 случаев гидрофобии, источником которых были енотовидные
собаки (Савицкий и др., 1980; Ботвинкин и др., 19816). В 1976—
1980 гг. в результате целенаправленных вирусологических иссле-
дований, проведенных в ряде районов Приморья и Приамурья, было
выделено 7 штаммов вируса бешенства от енотовидных собак (а
также от лисиц, колонка и амурской кошки). Эпизоотия выявлена
в осенне-зимний период 1979/80 г. Зараженность енотовидных собак
составила 13%, лисиц — 3,5%. Учитывая высокую плотность насе-
ления енотовидных собак и их восприимчивость к вирусу бешенст-
ва, следует отнести этот вид к числу основных хозяев вируса в при-
родных очагах бешенства на Дальнем Востоке (Савицкий и др.,
1980).
Бешенство енотовидных собак в районах акклиматизации впер-
вые зарегистрировано в дельте Волги в августе 1942 г. в Астрахан-
ском заповеднике (Исаков, 1949). В декабре 1943 г. на том же участ-
ке отмечено бешенство волка, а в 1945—1946 гг. эпизоотии бешенст-
ва енотовидных собак (и лисиц) наблюдали в восточной и западной
частях дельты. Отмечалось агрессивное поведение животных, за-
болевания бешенством покусанных ими домашних собак. Падеж
енотовидных собак отмечали на большой территории дельты.
В последующие годы енотовидные собаки зарегистрированы как
носители вируса бешенства в нескольких областях РСФСР (Курс-
кой, Псковской и др.), на Украине, в Молдавии, Белоруссии, Лит-
ве, Латвии и Эстонии (Селимов, 1978). Наибольшая частота обнару-
жения бешенства у енотовидных собак наблюдается в Прибалтий-
ских республиках. Так, в Эстонской ССР с 1969 по 1974 г. выявлено
193 случая бешенства у енотовидных собак. В Литве с 1965 по
88
Carnivora
1973 г. бешенство енотовидных собак составило 36% от всех зарегист-
рированных случаев бешенства диких животных (Буракаускас,
Казакявичус, 1979). В Латвии в 1969—1976 гг. бешенство енотовид-
ных собак составляло 18,7% заболеваний диких животных.
На Украине и в Белоруссии доля участия енотовидных собак
в эпизоотиях бешенства составляла около 4%. Существенную роль
в эпизоотологии бешенства в настоящее время енотовидная собака
играет на Северном Кавказе: в Ростовской области, Краснодарском
крае, Дагестанской АССР (Черкасский, 1985).
Следует подчеркнуть, что увеличение доли участия в эпизоотии
енотовидных собак связано с резким сокращением городского бе-
шенства собак. Так, в Эстонии за 1962—1968 гг. был зарегистриро-
ван всего один случай заболевания собаки, а бешенство диких жи-
вотных, в том числе и енотовидных собак, естественно, вышло на
первое место.
Заболевания енотовидных собак бешенством тесно связано с
эпизоотией у лисиц. Отмечено, что заболевшие енотовидные собаки
появляются через 1—2 месяца после находок бешеных лисиц (Шев-
ченко, Щербак, 1980). Межвидовому распространению инфекции
способствуют контакты животных у нор. Так, при весеннем осмотре
295 нор следы лисиц и енотовидных собак были замечены у 28% нор,
а следы этих двух видов и барсуков — у 13% нор. Интенсивность
эпизоотии в популяции енотовидных собак, по-видимому, высока.
Л. С. Шевченко и Ю. Н. Щербак (1980) сообщают, что на стациона-
ре, где численность енотовидных собак и лисиц соотносилась как
1 : 10, было найдено 8 больных енотовидных собак и 11 лисиц.
В эксперименте установлена высокая восприимчивость и инфек-
ционная чувствительность енотовидных собак к вирусу бешенства
(табл. 9). Все подопытные звери погибли при заражении дозами
MLD/50 3) и выше. У погибших зверей в мозгу и слюнных железах
Таблица 9, Определение чувствительности енотовидной собаки к вирусу
бешенства (по Ботвинкину и др., 1983)
Доза MLD/50 Заражено/погиб- ло Инкубационный период, сутки Титры вируса 1g LD/5 0
Мозг Слюнные железы
5 2/2 26, 27 4,9—4,1 4,3
50 2/2 22, 32 5,1—4,9 5,1—6,1
500 2/2 17, 25 4,7—4,7 5,1—3,5
5 000 3/3 13, 15, 21 5,3—4,3 4,7—5,9
53 000 530 000 1/1 1/1 16 15 Не исследовали »
8 MLD/50 — доза, вызывающая гибель 50% белых мышей.
Хищные
89
выявлена высокая концентрация вируса. Степень восприимчивости
енотовидных собак и накопление вируса в слюнных железах у них
оказались выше, чем у лисиц, зараженных одновременно теми же
дозами (Ботвинкин и др., 1983).
Указывая на то, что опыты по заражению енотовидных собак
были проведены в ноябре-декабре и что в природных условиях не-
однократно выделяли от них вирус в зимние месяцы, авторы спра-
ведливо ставят под сомнение предположение о сохранении у этого
вида «рабической инфекции в зимний период в бессимптомном со-
стоянии» (Адамович, Бородина, 1976).
Высокая в ряде районов численность енотовидных собак, бли-
зость мест норения к населенным пунктам определяют их возмож-
ную роль как источников заражения бродячих и домашних собак и
кошек.
Енотовидные собаки были источником заражения людей в 4,2—
6,5% случаев, связанных с укусами диких животных за 1946—
1973 гг. (Лебедева, Селимов, 1975; Селимов, 1978). В целом по СССР
случаи заражения людей от енотовидных собак составили за 1976—
1980 гг. 0,8%.
В Польше в 1978—1983 гг. зарегистрировано 107 случаев бе-
шенства енотовидных собак. В ГДР в 1978 г. бешенство установлено
у двух енотовидных собак (Черкасский, 1985).
Род Canis Linnaeus, 1758 — волки
Род волков появился в верхнем плиоцене Северной Америки, в
Евразии с плейстоцена. Представители рода в пределах семейства
наиболее близки к роду лисиц и особенно к песцам.
Распространены очень широко по всей Евразии, Африке, Север-
ной Америке. Обитают во всех ландшафтах — от тундр до пустынь
и высокогорий. Представители рода составляют весьма гибкую и
жизнеспособную группу, и сокращение ареала происходит лишь в
областях с густым человеческим населением из-за активного пресле-
дования хищников, приносящих ущерб животноводству и охотни-
чьему хозяйству.
Род содержит шесть видов, один из которых — волк — встре-
чается как в диком, так и в одомашненном состоянии — домашняя
собака Canis familiaris L., 1758. Одичавшей собакой считают и
австралийского динго. В составе рода три африканских вида: эфи-
опский шакал Canis simensis Ruppel, 1835; полосатый шакал С. adus-
tus Sundevall, 1846 и чепрачный шакал С. mesomelas Schreber,
1775; затем афро-евроазиатский шакал С. aureus L., 1758; северо-
американский койот С. latrans Say, 1823; евроазиатско-североаме-
риканский волк С. lupus L., 1758.
Многие американские зоологи выделяют из С. lupus отдельный
вид рыжего волка С. niger Bartram, 1791 (юго-восточные штаты
США). Надо заметить, что репродуктивная изоляция между многи-
90
Carnivora
ми видами рода слабо выражена, при нарушении структуры попу-
ляции наблюдается гибридизация, чему способствует и одинаковый
кариотип — диплоидное число хромосом 78.
В СССР обитают два вида — волк и шакал.
Cam’s la trans Say, 1823 — койот
Койот — наиболее адаптировавшийся к человеку представитель
рода. По размерам меньше волка, но несколько крупнее шакала.
Длина тела — до 90 см, масса — до 18 кг.
Распространен по всей Северной Америке на юг до Коста-Рики,
за исключением крайнего запада Аляски и севера Канады. Расшире-
ние ареала на север с юго-запада материка проходило в историческое
время, расселение на восток до штатов Мэн и Нью-Йорк — за по-
следние 30 лет. В семействе собачьих койот, пожалуй, легче всех
приспосабливается к изменению условий и заселяет ныне различные
ландшафты — тундру, лес, степь, горы вплоть до альпийского пояса.
Расширению ареала койота способствовали истребление его конку-
рента — волка и, возможно, гибридизация с собакой.
По примерным расчетам, общая численность койотов в США око-
ло 1 млн особей. Программы контроля численности койота, иногда
нападающего на молодняк домашних животных и диких копытных,
приводят к сокращению отдельных популяций, но широкое расселе-
ние зверя компенсирует эти потери. Кроме того, сохраняющиеся
после проведения борьбы койоты увеличивают величину приплода.
Экология койота имеет много общего с волком. Однако койоты
обладают более высоким воспроизводством (размножаются с годо-
валого возраста), менее строгой территориальностью (площадь уча-
стка обитания 10—30 кв. км), более высокой подвижностью — часто
расселяются на сотни километров — и чрезвычайной способностью
адаптации к деятельности человека (использование отходов, из-
менение активности, избегание преследования).
Пища койота весьма разнообразна: мелкие позвоночные, насе-
комые и их личинки, плоды различных диких и культурных расте-
ний. Велика роль падали, в том числе и домашних животных. При
возможности успешно нападают на ягнят и домашнюю птицу, чем
вызывают повсеместную неприязнь фермеров и сохранение службы
контроля хищников (в 60-х годах в США было уничтожено за один
год 90 тыс. койотов).
Койоты высоко чувствительны к вирусу бешенства, эпизоотии
среди них отмечались в США и южных районах Канады (Sikes,
1966). При серологических исследованиях у койотов обнаруживали
антитела к возбудителю туляремии и в эксперименте установлена
малая чувствительность их к этой инфекции (Lundgren et al., 1957).
Хищные
91
Canis aureus Linnaeus, 1758 — шакал
Шакал — наименее специализированный и мелкий представи-
тель рода. Длина тела до 85 см, масса — до 13,6 кг. По общему виду
и пропорциям не только меньше волка, но легче его, короче ноги,
что является адаптацией к обитанию преимущественно в бесснеж-
ном ландшафте и питанию мелкими позвоночными, падалью, насе-
комыми и постоянно растительными кормами (плодами диких и
культурных растений).
Ареал включает Южную и Переднюю Азию, Юго-Восточную Ев-
ропу и Северную Африку. В СССР обитает в Молдавии, на Кавказе
и в Средней Азии. В горы идет до высот 1000—1100 м (Млекопитаю-
щие Советского Союза, 1967), но в последнее время есть сведения о
проникновении шакала в более высокие пояса гор и расселении его
на север в Волжско-Уральские пески. Расселение шакала связы-
вают с антропогенным изменением ландшафта, освоением пустынь
и сокращением численности волка.
В СССР обитают два подвида: туркестанский шакал—С. а.
aureus L., 1758 и С. a. moreotica Geoffroy, 1835. Оба они предпочи-
тают жить в зарослях, кустарнике, тростниках, часто по берегам
водоемов и в долинах рек. Среди открытых пространств не селится,
и этим обусловлено кружевное очертание его ареала. Тяготеют ша-
калы к оазисам и поселениям человека, где они находят корм в виде
культурных растений и падали домашних животных.
Тяготение к человеку, крайняя неразборчивость в кормах и
пристрастие к падали обусловливают медицинское значение шакала.
Шакалы сильно заражены гельминтами. В Юго-Западном Тад-
жикистане у них обнаружено 15 видов ленточных и круглых червей
и скребней. Являясь хозяевами половозрелой стадии развития лен-
точных глистов, они могут способствовать распространению у овец
и крупного рогатого скота Taenia ovis, Т. hydatigena. Шакалы за-
ражают водоемы личинками ришты Dracunculus medinensis, под-
держивая очаги дракункулеза.
По данным ВОЗ, в Азии шакал служит природным резервуаром
вируса бешенства в Индии, Пакистане, Бангладеш, Афганистане,
Иране, Ираке, странах Северной Африки и Эфиопии. В СССР бе-
шенство у шакалов отмечали на Кавказе и в Средней Азии (Черкас-
ский, 1985).
В Эфиопии за 1964—1975 гг. шакалы занимали первое место по
частоте нападения на людей, обратившихся за антирабической помо-
щью: 381 человек из 487 укушенных дикими животными. Случаи
гидрофобии, вызванной укусами шакалов, составили 12% из 66,
значительно уступая более редким, но более опасным укусам гиен,
мангуст и обезьян (Fecadu, 1981).
Шакалы наряду с домашними и бродячими собаками поддержива-
ют очаги висцерального лейшманиоза. В низовьях Сырдарьи боль-
шинство очагов висцерального лейшманиоза относится к сельской
92
Carnivora
местности и заболевания людей наблюдаются на летних пастбищах,
животноводческих фермах, на окраинах поселков.
Активность природного очага поддерживается шакалами, чис-
ленность которых заметно увеличивалась. В то же время в большин-
стве населенных пунктов в результате направленных мероприятий
снижается численность собак и среди них не регистрируются носите-
ли лейшманий (Генис, 1979).
Cam’s lupus Linnaeus, 1758 — волк
Наиболее крупный представитель рода, достигающий длины
тела 160 см, высоты в плечах 100 см и массы 80 кг. Тело мощное,
особенно шея и грудная клетка, высокие ноги. Голова тяжелая, с
сильными челюстями и широким лбом.
Восстановленный ареал волка включает практически всю Север-
ную Америку и Евразию, за исключением зоны тропических лесов.
Современный ареал сильно сокращен человеком, уничтожившим
волка на большей части Западной Европы и США (за исключением
Аляски).
По данным Рабочей группы по волку Международного союза
охраны природы и природных ресурсов, современная (начало 80-х
годов) численность волка в разных странах и регионах характери-
зуется так: Аляска — 10—15 тыс. волков, Миннесота — 1200 и
другие штаты США — 100, Канада — более 28 тыс., Мексика —
менее 50 зверей.
В Европе сохраняется менее чем по 10 волков в Норвегии, Шве-
ции и Гренландии; по 100 зверей насчитывают в Финляндии, Бол-
гарии, Италии, Португалии; 150 — в Чехословакии; по 200 — в
Польше и Испании; более 50 — в Греции; 2 тыс.— Румынии и
2—5 тыс. волков в Югославии. В СССР обитает около 7 тыс. волков,
причем около трети из них в европейской части.
Для Азии сведения неполны. На Ближнем Востоке общая чис-
ленность не превышает 1 тыс. волков, в том числе в Сирии 200—
500 и на Аравийском полуострове — менее 500 животных. В Турции
вид обычен, но численность не определена, в Иране и Афганистане
живет примерно по I тыс. волков, в Индии — менее 1 тыс., в Монго-
лии — не менее 10 тыс. зверей. В странах азиатского региона основ-
ные усилия направлены на дальнейшее сокращение численности
популяций волка, приносящего существенный ущерб животновод-
ству. В Европе же и Северной Америке наряду с программами
более или менее жесткого регулирования численности вида (СССР,
Югославия, Румыния, Канада и др.) широкое распространение полу-
чают уже осуществляемые в некоторых странах проекты Сохранения
волка от полного уничтожения на их территориях.
В XX в. в СССР дважды наблюдались большие вспышки числен-
ности волка, которые совпадали и следовали за годами войн и нару-
шений хозяйственной жизни, когда борьба с хищником ослабевала.
Хищные
93
В эти периоды общая численность, вероятно, достигала в стране
100—150 тыс. зверей и в 2—3 раза превышала минимум, который
наблюдался в конце 60-х годов. Рост численности в периоды вспышек
в основном происходил на густонаселенных сельскохозяйственных
территориях степной и лесостепной зоны европейской части СССР,
Казахстана, Западной и Южной Сибири за счет увеличения доли в
популяциях синантропной формы волка. В годы усиления борьбы
происходила элиминация волков-синантропов, причем сохранялись
преимущественно звери, питающиеся дикими копытными и обитаю-
щие в лесу и горах.
В пределах ареала волки распространены очень неравномерно.
Их размещение и численность обусловливаются обилием и доступ-
ностью корма и, конечно, интенсивностью преследования человеком.
Только благодаря усиленной борьбе к началу 70-х годов удалось
примерно на 1/3 сократить ареал волка в европейской части СССР.
Современная плотность популяций волка на огромных пространст-
вах тундр, малонарушенной тайги и пустыни обычно не превышает
1 зверя на 1 тыс. кв. км, редко где составляет 2—5 и тем более 10
зверей на 1 тыс. кв. км. В антропогенном ландшафте южной части
лесной зоны, в горах Кавказа, Средней Азии и Южной Сибири, в
районах отгонного животноводства и распространения диких копыт-
ных нередко встречается до 40 волков на 1 тыс. кв. км, а в местах
концентрации в определенные сезоны на небольших участках может
быть учтено до 100 и более зверей на I тыс. кв. км.
Основу питания волка составляют дикие копытные и домашние
животные, для синантропной формы — падаль на скотомогильни-
ках и домашние животные. Грызуны и зайцы имеют второстепенное
значение, но кое-где, например в Якутии, заяц-беляк — главная
пища волка, а в горах Средней Азии летом — сурки. Растительная
пища, плоды и ягоды,— существенная добавка к рациону преиму-
щественно на юге ареала. Географические отличия в питании волка
существенны и определяются наличием, обилием и доступностью
тех или других видов. Главные жертвы волка в тундре — домашний
и дикий северный олень; в тайге — лось, изюбрь, косуля, кабан;
в смешанных лесах и лесостепи — косуля, лось, благородный олень,
кабан; в пустыне — сайгак, джейран, кабан; в горах на юге СССР—
олени, косуля, кабан, горные козлы и бараны. Замечена специали-
зация питания отдельных семей волков определенными видами и
предпочтение добычи наиболее массовых видов. При появлении в
результате расселения новых видов потенциальных жертв проявля-
ется консерватизм, проходит несколько лет до того, как хищники
переключаются на них.
Волки — территориальные животные. Постоянство участка оби-
тания поддерживается четкой системой мечения, звуковой и визу-
альной коммуникацией. Размеры участков обитания сильно отлича-
ются в различных природных зонах и изменяются в связи с плотно-
стью зверей. Наиболее велика площадь семейных участков в тунд-
94
Carnivora
ре — нередко несколько тысяч квадратных километров. В других
природных зонах она составляет обычно 200—300 кв. км, а в антро-
погенном ландшафте может не превышать иногда 100 кв. км. При
сокращении численности популяции участки оставшихся семей
увеличиваются в 2—3 раза.
В популяции всегда имеется некоторое число более подвижных
и менее связанных с постоянной территорией обитания так называ-
емых нетерриториальных зверей. Это обычно отделившиеся от роди-
тельской семьи переярки и взрослые звери, не нашедшие террито-
рии для создания новой семьи. Такие звери более подвижны и
легко могут превратиться в волков-синантропов. Нетерриториаль-
ные звери совершают далекие перемещения вслед за мигрирующими
стадами северных оленей и сайгаков. Склонность к переключению
на питание домашними животными, протяженные миграции и спо-
собность к широким контактам из-за непостоянства их группировок
обосновывают необходимость наиболее жесткого контроля числен-
ности нетерриториальных волков, безусловно вредных в народном
хозяйстве и для здоровья человека.
Волки — моногамы, но в условиях постоянного преследования
пары постоянно разбиваются и вновь формируются. Удобных и глу-
хих мест для размножения немного, поэтому расположение логовищ
мало меняется. Даже при гибели обоих матерых хороший участок
для выводка не пустует, его занимают местные или пришлые звери.
Собственно логовище на выводковом участке обычно сменяется, но
сам участок площадью в 1—3 кв. км постоянно сохраняется. В от-
крытых ландшафтах волки устраивают логово в норе или промоине.
Нередко они расширяют норы лисиц, барсуков, сурков, дикобра-
зов. В горах и лесу логово часто располагается под камнями, в ни-
ше, под выворотнем, в старом окопе или даже открыто в густых
зарослях. Спустя 1—2 месяца после появления щенков на поверхно-
сти родители переводят их за несколько километров во временное
логово, которое обычно приближает выводок к источнику доступной
пищи.
Половозрелыми волки становятся в двухлетнем возрасте. Сроки
размножения варьируют по широте и высоте местности. Спаривание
и рождение щенков происходят на севере и в высокогорье на 1,5—
2 месяца позднее, чем на юге. Число щенков в выводке обычно 4—7,
иногда до 15. Молодые и очень старые самки имеют меньше детены-
шей. Примечательно, что в одной стае сроки течки волчиц не совпа-
дают и при гибели матерой самки выводок может принести нормаль-
но не размножающаяся подчиненная самка (субдоминант). В этом
одна из адаптаций волков к постоянному преследованию.
Эктопаразиты. Специфических эктопаразитов у волка нет. Встре-
чающиеся на нем виды свойственны собакам, шакалам, лисицам,
кошкам, барсукам, куницам, хорькам, а также его жертвам — ко-
пытным, грызунам, птицам. Широкий спектр пищевых контактов
волка с различными животными, обитание во всех ландшафтах обес-
Хищные
95
печивают разнообразие его наружных паразитов. Высокая подвиж-
ность зверя благоприятствует распространению им паразитов на
большое расстояние, что может иметь эпизоотологическое значение.
По литературным сведениям (Гребенюк, 1966; Иофф, 1947; Бром
и др., 1948; Капустин, 1955; Кондрашкина и др., 1956; Емельянова,
Жовтый, 1957; Микулин и др., 1959; Балбарын, 1962; Баваасан,
1967; Гарбузова, 1967; и др.), на волке в СССР обнаружено шесть
видов блох, три вида аргасовых и 22 вида иксодовых клещей, чесо-
точный клещ Sarcoptes scabiae (Селиванов, Машенцев, 1955) и вла-
соед Trichodectes canis (Дубинин, Дубинина, 1951).
При осмотре 72 волков, добытых летом (Бетпак-Дала), с 52 зверей
было снято 386 иксодовых клещей. Преобладали Rhipicephalus
pumilio, R. schulzei, R. rossicus (Ушакова, Кривкова, 1963). За пре-
делами СССР на волке отмечено паразитирование еще 18 видов иксо-
довых клещей. Однако следует отметить, что волк как прокормитель
этих членистоногих имеет ничтожное значение.
Широко распространена, многочисленна и связана преимущест-
венно с хищниками блоха Pulex irritans, считавшаяся прежде спе-
цифической блохой человека. Она паразитирует и на волке.
На волках и собаках (Бошко, 1956), кроме того, паразитируют
блоха хищников Chaetopsylla homoea, блоха собак Ctenocephalides
canis, блоха кошек С. felis, блоха грызунов Oropsylla silantiewi
(в местах обитания сурков), блоха копытных Vermipsylla alacurt
в сыртах Тянь-Шаня (Иофф, 1947) и в Монголии (Балбарын, 1962).
Могут встречаться на волке и другие массовые виды блох грызунов
и насекомоядных, свойственные тому или другому региону.
Гельминты. Вред, причиняемый волками народному хозяйству,
состоит не только в непосредственном уничтожении домашних и
диких животных, но и в снижении продуктивности копытных из-за
распространяемых хищниками гельминтозов (эхинококкоз, ценуроз,
альвеококкоз и др.).
У волка зарегистрировано 67 видов паразитических червей
(трематоды и скребни — 16 видов, цестоды — 24 вида, нематоды —
27 видов), в том числе на территории СССР — 50 (Морозов, 1951;
Садыхов, 1955; Беляева, 1959; Шалдыбин, 1964; Мачинский, Семов,
1968; Шлычков, 1968; Карасев, 1970; Козлов, 1977; Шималов, 1971
и др.). Данные о встречаемости гельминтов у волка вне СССР име-
ются в монографии Мича (Meeh, 1970). Заражение волков гельмин-
тами происходит преимущественно при поедании животной пищи,
содержащей инвазионных личинок биогельминтов, или путем упот-
ребления пищи и воды, загрязненных яйцами или личинками гео-
гельминтов.
Инфекции. Сведения о заболеваниях волков в природе невелики
и сводятся к регистрации отдельных случаев обнаружения того или
иного возбудителя, антител или антигена при серологических ис-
следованиях. В различных частях ареала у волка выявлено зараже-
ние бруцеллезом, туляремией, лептоспирозом, сибирской язвой,
96
Carnivora
бешенством, ящуром (Шеханов, 1970). Как видно из этого перечня,
заражение волков, за исключением бешенства, связано с поеданием
волками основных хозяев этих инфекций — копытных (бруцеллез,
сибирская язва, лептоспироз, ящур) или грызунов (туляремия, леп-
тоспироз grippotyphosa).
Малая чувствительность хищников к заболеваниям, передаю-
щимся при установившихся в процессе эволюции трофических свя-
зях, определяет несущественную их роль для популяции волков.
Гибель волков в природе установлена лишь при заболевании бешен-
ством, вызванным как классической формой возбудителя, так и его
вариететом — тундровым бешенством, или дикованием.
Имеются также сообщения о зараженности волков в Канаде и
на Аляске вирусом собачьей чумы и инфекционного собачьего гепа-
тита. В северо-западных территориях Канады сыворотки двух вол-
ков из 86 реагировали с вирусом собачьей чумы и 11 (13%) — с
вирусом инфекционного гепатита (Choquette, Kuyt, 1974). На Аляс-
ке гибель волков от этой чумы регистрировалась в 1977 и 1979 гг.
одновременно с эпизоотией чумы у собак. При серологическом ис-
следовании 57 волков из трех пунктов Аляски антитела к вирусу
собачьей чумы найдены у четырех, а к вирусу инфекционного гепа-
тита — у 54 волков (Stephenson et al., 1982). Авторы справедливо
расценивают эти результаты как свидетельство высокой чувстви-
тельности волков к собачьей чуме и широкого распространения в их
популяции вируса инфекционного гепатита. Сходные результаты
о распространении гепатита получены в Гренландии, где у 50 собак
в 100% выявлены антитела к вирусу гепатита и не было иммунных
особей к собачьей чуме (Crandell, Col, 1966).
Бруцеллез обнаружен у волков тундровой зоны, что связано
с распространением здесь бруцеллеза северных оленей — основной
пищей волков. Зараженность волков бруцеллезом составляла 9%
(Асланян и др., 1970). Культуры возбудителя бруцеллеза здесь
были выделены от 20 волков (Калиновский и др., 1971). На Аляске
при исследовании 28 волков реакция агглютинации с бруцеллезным
антигеном была положительна у 11 особей (Neiland, 1975).
Наибольшее эпизоотическое и эпидемиологическое значение волк
имеет при бешенстве. Бешеный волк чрезвычайно агрессивен, у
него притуплено чувство страха перед человеком. Перемещаясь на
большие расстояния, хищник яростно нападает на встретившихся
диких, домашних животных и человека. Широко известно нападе-
ние бешеного волка на 19 смоленских крестьян в середине 80-х
годов прошлого века, которых доставили в Париж для антирабиче-
ского лечения. Подобные происшествия изредка происходят и в
наше время. Например, в 1957 г. в Белоруссии бешеный волк за 36ч
преодолел расстояние 150 км и на пути покусал 25 человек и около
50 домашних животных (Лин, 1967; Селимов, 1978). Но В. Л. Ада-
мович (1985), проанализировав размещение 120 случаев бешенства
волков, зарегистрированных за 1954—1982 гг. в центральной части
Хищные
97
Русской равнины, пришел к выводу о незначительной подвижности
больных особей и гибели их в пределах небольшого очагового уча-
стка.
Существованию первичных природных очагов бешенства, под-
держиваемых волком, способствовали высокая инфекционная чув-
ствительность его к вирусу, продолжительный период инкубации —
9—12 дней (Грибанова и др., 1979) и агрессивность больного зверя
(Селимов, 1978). Активизация первичных очагов бешенства проис-
ходила при резком увеличении численности волков и сопровожда-
лась интенсивным вовлечением в круг циркуляции вируса домашних
животных, особенно собак. Так было после первой и второй мировых
войн (Мари, 1909; Саватеев, 1927; Канторович, 1968).
Заболевания волка бешенством зарегистрированы во всех ланд-
шафтно-географических зонах СССР на территории 90 областей и
краев.
Значение волка в эпизоотологии классического бешенства в более
южных районах СССР было различно в периоды 1945—1960 и 1961—
1980 гг., отличавшиеся состоянием его численности и распределе-
ния. В первый период бешенство волков было частым явлением,
хотя в 1945—1960 гг. заболеваемость диких животных бешенством
не подлежала обязательной регистрации в ветеринарных учрежде-
ниях (Селимов, 1972). В 40—50-х годах эпизоотии волчьего бешенст-
ва отмечались в Нижнем Поволжье (Исаков, 1949), Казахстане
(Слудский, 1954), Белоруссии (Зибицкер, Ковалев, 1968; Селимов,
1978), а также на Украине, в Молдавии, Узбекистане и во многих
областях РСФСР. Удельный вес волка как источника заболева-
ваний бешенством человека в этот период был местами велик, что
обусловливалось его высокой плотностью. Например, в Белоруссии
в 1949^-1956 гг. заражение человека от волка установлено в 28,1%
случаев (Змушко, 1967), т. е. более чем в восемь раз превышало
средний показатель по стране в те годы (Селимов, 1963).
В 1975—1978 гг. в европейской части РСФСР зарегистрировано
30 случаев бешенства волков и 36 нападений их на людей. В УССР
бешенство волков в 1964—1978 гг. составляло всего 0,8% от бешен-
ства диких животных, но наблюдающийся рост численности волков
в Полесье и Карпатах вызывает увеличение случаев бешенства (Чер-
касский, 1985).
В Казахстане в 1972—1978 гг. из 54 исследованных волков бе-
шенство установлено у 17. За это время наблюдалось 50 случаев
нападения волков на людей, 33 человека были укушены больными
зверями (Яныпин и др., 1982).
Значительное сокращение численности волка к концу 50-х го-
дов сопровождалось уменьшением его эпидемиологического значе-
ния (табл. 10).
Удельный вес волка как источника бешенства человека изменил-
ся от 3,5% в 1946—1956 гг. до 0,7—1,4% в 1958—1960 гг., а роль
лисицы возросла с 2,1 до 5,2—7,1% (Селимов и др., 1967). В после-
4 Зак. 3575
98
Carnivora
Таблица 10. Изменение соотношения источников заражения гидрофобией
в СССР (по Куче руку, 1982) (в %)
Период Собака Кошка Волк Лисица Прочие
До 1927 г. 79 2 19 0 0
1946—1956 гг. 87,1 5,3 3,5 2,1 2
1958—1963 гг. 82,2 6,7 0,9 7,0 3
1964—1975 гг. 37,5—74 2—21,5 0—7 15—38 1,1—10
дующие годы «лидерство» в распространении вируса бешенства
перешло к лисице. В 60—70-е годы бешенство распространялось
главным образом среди лисиц и они же наряду с собаками служили
основными источниками заражения людей.
В 1961—1980 гг. в Западной, Средней Сибири, Зауралье и Ка-
захстане численность волков сократилась не столь резко и за это
время бешенство было установлено у волков в Тувинской АССР
(2 случая), в Алтайском крае (6 случаев), в Красноярском крае
(3 случая), в Новосибирской и Челябинской областях (по 4 случая)
и в Омской области (3 случая) (Мальков, Грибанова, 1980; Кирья-
нов, 1962; Селимов, 1978; Джупина, Куликов, 1968; Грибанова,
Мальков, 1977; Рудаков и др., 1973; Грибанова и др., 1980; Канто-
рович и др., 1967).
В Западной и Средней Сибири после волчьих укусов бешенством
заболело три человека (1966 г.— Красноярский край, 1979 г.— Ту-
винская АССР). Нападения достоверно бешеных хищников на лю-
дей имели место в Омской, Кемеровской областях и Алтайском
крае. Сопоставление приведенных материалов с соответствующими
сведениями за предыдущие десятилетия показывают, что угроза
волчьего бешенства в Западной и Средней Сибири, хотя и снизилась
в 60—70-х годах, но все еще существует (Умов, Малокотин, 1966;
Рудаков и др., 1971, 1973).
Сокращение численности волка в степи и лесостепи Западной
Сибири привело к угасанию некоторых природных очагов бешен-
ства, в других в течение последних 20 лет регистрируются лишь
спорадические заболевания. Доля волка с лабораторно подтверж-
денным диагнозом бешенства в Челябинской, Омской, Новосибир-
ской областях в 1961—1980 гг. составила 1,6—2,6, а лисицы —
76,5—97,6% (Канторович и др., 1967; Грибанова, Мальков, 1977;
Грибанова и др., 1977). Следовательно, волк в последние десятиле-
тия стал здесь редким хозяином возбудителя.
Многолетнее изучение структуры и распределения природных
очагов бешенства в Западной Сибири показывает, что волк в пери-
од своего обилия вошел в состав вирусоносителей во вновь сформи-
ровавшихся полигостальных очагах. Центрами образования этих
Хищные
99
рецентных очагов в 40—50-х годах были, очевидно, территории со-
средоточения нескольких видов хищных зверей (волк, лисица, кор-
сак, светлый хорь), приуроченные к Тургайскому плато, Ишимской
возвышенности, Барабинской низменности и Кулундинской рав-
нине, с которыми и поныне связаны природные очаги бешенства
Западносибирско-Казахстанского очагового региона (Мальков,
1978; Мальков, Грибанова, 1980).
Сходный процесс затухания природных очагов бешенства, ранее
поддерживавшихся волком, и образования рецентных полигосталь-
ных очагов с основным хозяином — лисицей — происходит повсю-
ду, где популяции волка резко сокращены, в том числе в РСФСР,
на Украине, в Молдавии (Селимов, 1978). В этих очагах основная
роль в сохранении вируса бешенства, в передаче его человеку при-
надлежит лисице (35,6—54,1%), а значение волка ничтожно (до
0,7%). Затухания очагов пока не происходит в Казахстане, где чис-
ленность волка остается высокой на протяжении нескольких десяти-
летий. В 1969 г. в Казахстане бешенство было установлено у 32 вол-
ков. Спустя несколько лет из 47 диагностированных случаев бешен-
ства на долю волков приходилось 6%, а на лисиц — 55% (Сансыз-
баев и др., 1975).
В северных районах европейской части СССР бешенство волков,
вызванное классической формой вируса, регистрировалось в 1930^
1931 и 1955—1956 гг. в связи с распространением из более южных
районов бешенства собак. В эти годы отмечены единичные случаи
гидрофобии, вызванные укусами волков (Канторович, 1981).
О бешенстве волков, наблюдавшемся в тундровой зоне, сооб-
щают в обзорах об эпизоотиях песцов и ездовых собак Ч. Элтон
(Elton, 1931, 1942) и А. Н. Формозов (1935). Заболевания волков
отмечали в Болыпеземельской тундре, Анадырском крае, на Кам-
чатке одновременно с дикованием песцов. В Канаде упоминания о бе-
шенстве волков относятся к Северо-Западным территориям, Север-
ному Лабрадору и Баффиновой Земле. По сведениям, полученным от
эскимосов, в некоторые годы бешенство песцов и волков отмечалось
раньше, чем заболевания у собак. Фактические данные о вовлечении
волков в эпизоотии дикования немногочисленны и противоречивы.
Так, Р. А. Канторович (1963) исследовал с отрицательным резуль-
татом мозг 51 волка, добытых в Ненецком автономном округе в
1954—1960 гг. Отметим, что среди песцов здесь проходила интенсив-
ная эпизоотия в 1954/55 г., и в последующие годы вирус от них вы-
деляли регулярно. В Якутии Р. А. Колесникова (1979) диагности-
ровала бешенство у 6 волков из 10 в 1969 г. и у 6 из 22 — в 1974 г.
при сравнительно невысокой интенсивности эпизоотии у песцов.
Интересные непосредственные наблюдения за эпизоотией бешенства в
стае волков проведены в арктической части Аляски (Chapman, 1978). Автор
наблюдал 13 июля 1977 г. необычное агрессивное поведение волка, нападав-
шего в течение более чем часа на других волков, не обращая внимания на их
позы выражения покорности. На следующий день волк был убит после того,
4*
100
Carnivora
как он несколько раз подходил к палатке наблюдателя на расстояния в
10—15 и до 3 м. Бешенство у волка подтверждено лабораторными исследо-
ваниями. Между 8 и 14 августа автор нашел 6 мертвых волков из этой стаи,
в которой было 8 взрослых (старые и прошлогодние) и 2 молодых. У двух
волков диагноз бешенства установлен по реакции иммунофлуоресценции.
У одного из них и двух других, не исследованных из-за разложения трупов,
на морде обнаружены иглы дикобраза, свидетельствующие об агрессив-
ном поведении волков. Нападение на дикобраза не свойственно волкам
с нормальным поведением. Гибель этих шести волков, судя по состоянию
трупов, произошла в конце июля—начале августа, т. е. менее чем через
месяц после контакта с бешеным сочленом стаи. Все погибшие волки найдены
в пределах семейного участка, менее чем в 5 км от того места, где убит боль-
ной волк. Автор считает, что передача инфекции из одной стаи волков в дру-
гую — редкое явление вследствие низкой плотности населения волков в
тундре (1 на 180 кв. км) и территориальности волчьей стаи. Однако Рауш
(Rausch, 1958) сообщал о передвижении бешеного волка на Аляске на 24 км.
В 1942 г. в тундре Среднего Ямала бешенство двух волков наблюдали на
расстоянии примерно 40 км с промежутком в 19 дней (Т. Н. Дунаева). Пер-
вый волк, забежавший 12 мая в стадо северных оленей, стоявшее возле чума,
был найден через два дня мертвым, а покусанный им олень заболел агрессив-
ной формой болезни 28 июля и на второй день болезни его пристрелили. Вто-
рой волк, также не боявшийся людей, задавил олененка и покусал соба-
ку 30 мая. Собака заболела в начале июля, а следом за ней еще три собаки,
которые были убиты хозяином чума.
Судя по имеющимся данным, в тундровых районах среди волков
циркулируют штаммы вируса тундрового бешенства, отличающиеся
от классических отсутствием патогенности для человека. Широко
практикующийся в настоящее время ввоз в северные районы собак
из южных районов, неблагополучных по классическому бешенству,
требует чрезвычайно серьезного контроля для предотвращения по-
явления на севере опасного для людей заболевания.
Род Alopex Каир, 1829 — песцы
В роде единственный вид: песец — Alopex lagopus Linnaeus,
1758. Песец распространен циркумполярно, занимая тундру и лесо-
тундру Арктического побережья Евразии и Северной Америки,
включая арктические острова. Песцам свойственна большая по-
движность, связанная с неустойчивостью кормовых ресурсов тунд-
ры. Во время зимних миграций звери заходят в северную часть лес-
ной зоны, передвигаются на далекие расстояния по побережью
Северного Ледовитого океана, по льду достигают далеких арктиче-
ских островов. Дальние миграции—до 1000—5000 км — зареги-
стрированы не менее чем у 15% помеченных песцов, преимуществен-
но самцов (Бакеев и др., 1980). В благоприятные годы с высокой
численностью леммингов значительная часть популяции песцов
задерживается вблизи мест норения.
В материковой тундре основу питания песцов составляют тунд-
ровые грызуны, преимущественно сибирский и копытный лемминги
и в меньшей степени — серые полевки. В летнее время значительное
место в кормовом рационе составляют яйца и птенцы, ягоды. На
Хищные
ЮТ
островах и в приморской тундре песцы кормятся выбросами моря,
рыбой, беспозвоночными. Повсеместно используют отбросы промыс-
ла северных оленей, морского зверя, падаль. Несмотря на выражен-
ную полифагию, интенсивность размножения песцов находится в за-
висимости от обилия леммингов, для которых характерны цикличе-
ские изменения численности сочень широкой амплитудой. В годы
обилия леммингов размножающиеся пары песцов занимают 90—
100% нор, число щенят в выводках достигает 12—14 и более. Во
время депрессии численности леммингов, следующей за годом пика,
занятость нор сокращается до 5—10%, а в некоторые годы песцы
совсем не размножаются, и в летнее время ведут бродячий образ
жизни, передвигаясь по тундре в поисках пищи.
Промысел песца испытывает вследствие этого значительные коле-
бания. Ежегодная заготовка шкурок песцов составляет 51,9—
95,9 тыс. (Соколов, 1979).
Сообщения о болезнях песцов касаются в основном заболевания
тундровым бешенством («дикованием») и гельминтозами. Сведения
о встречаемости бактериальных инфекций носят фрагментарный
характер.
Эпизоотии дикования песцов известны на всем ареале, включая
арктические острова — Новосибирские, Врангеля, Шпицберген,
Гренландия, Медвежий (Формозов, 1935; Туревич, Тебякина, 1947;
Бернштейн и др., 1981; Elton, 1931; Wamberg, 1960; Secord et al.,
1980; 0degaard, Krogerud, 1981). Вирус дикования, впервые выде-
ленный от песца Е. И. Туревичем и А. Е. Тебякиной (1947) на о-ве
Котельном, представляет собой вариетет классического вируса
уличного бешенства, сходный с ним по антигенной структуре, но
отличающийся апатогенностью для человека. Заболевания песцов
и ездовых собак в Арктике известны более полутораста лет, но случа-
ев заболевания людей не было зарегистрировано, несмотря на тес-
ный контакт населения с заболевшими животными, в том числе и в
случае покусов (Формозов, 1935; Шерстобаев, 1949; Канторович,
1963; Строгов, 1964; Elton, 1931). Р. А. Канторович (1963) обследо-
вал в Ненецком автономном округе 5200 человек на наличие вирус-
нейтрализующих антител и получил отрицательные результаты.
Однако в 1960 г. в Гренландии зарегистрирован случай гидрофобии у
четырехлетней девочки эскимоски, покусанной собаками во время интенсив-
ной эпизоотии среди песцов и собак. Собаки, как это принято здесь, были
уничтожены. Девочка заболела 21 июля, более чем через 3 месяца после
нанесения укусов (10 апреля), и погибла от паралитической формы бешенства
9 августа в Копенгагене. Типичные тельца Негри не были обнаружены,
но в отпечатках мозга выявлены мелкие внутриклеточные включения, опре-
деленные как частицы рабического вируса. Подобная картина типична для
заболевания животных тундровым бешенством. Введение собакам, мышам и
сирийским хомякам слюны и различных участков мозга погибшей вызвало
гибель животных. У некоторых из них в аммоновом роге обнаружены ти-
пичные тельца Негри. Антирабическая сыворотка, вводимая мышам, созда-
вала 100% защиту от вируса (Wamberg, 1960; Lassen, 1962).
102
Carnivora
Выделенные в Гренландии 19 штаммов вируса (11 от песцов и
8 от собак) идентифицированы как рабические вирусы, отличающие-
ся от классического вируса бешенства отсутствием телец Негри у
животных (Crandell, 1965).
Сообщение Р. А. Канторовича и А. Г. Маркарян (1963) о пато-
генности вируса дикования для человека недостаточно убедительно,
поскольку случай гибели годовалого ребенка, покусанного на се-
верном Ямале песцом, не был исследован лабораторными методами.
Песцы высокочувствительны к вирусу тундрового бешенства.
Инкубационный период у животных, зараженных внутримышечно
10 мл суспензии мозга больных песцов, равнялся 10—18 дням. Кли-
нические проявления болезни продолжались от 2 до 5 дней и чаще
имели паралитическую форму. Водобоязни не наблюдалось (Канто-
рович, 1963; Строгов, 1964).
При исследовании диких песцов вирус в слюнных железах нахо-
дили лишь у особей с наличием вируса в мозге. Титры вируса в моз-
гу и слюнных железах варьировали от 1,5 до 6,4 1g MLD/50 (Тата-
ров, 1982).
Эпизоотии дикования широко распространяются в популяции
песцов в годы повышенной численности, повторяясь с промежутками
в 2—5 лет, чаще через 3—4 года. Так, в Якутии интенсивные эпизоо-
тии наблюдались в зимы 1943/44, 1946/47, 1950/51, 1953/54,
1957/58, 1960/61 и 1966/67 гг.; вероятно, так же протекали в 1970 г.,
отличавшемся очень высокой численностью песца, а затем были заре-
гистрированы в 1973/74 и 1976/77 гг. (Колесникова, 1973, 1979).
Во время эпизоотий в тундре находят большое количество пав-
ших песцов. Так, зимой 1955/56 г. в Ямальской тундре из 1200 пес-
цовых шкурок, сданных на факторию, более 16% было снято с пав-
ших зверей (Оленев, 1965). В Нижнеколымской тундре зимой
1950/51 г. количество больных и павших песцов составило 12—14%
от всех добытых (Соколов, Ча, 1957).
Многолетние исследования, проводившиеся в тундрах Западной
и Восточной Якутии, показали, что зараженность песцов отмечается
ежегодно, колеблясь от 0,5—2% в годы низкой численности песца
и до 8,8—11,7% в период пика численности (табл. 11).
По данным Л. М. Сюзюмовой (1965, 1967), частота выделения ви-
руса от песцов колебалась в разные годы от 0,7 до 10%. Р. А. Кан-
торович (1963) приводит очень высокие показатели зараженности
песцов, достигающие 38,5—69,0%. Вероятно, эти цифры завышены
вследствие преимущественного исследования павших песцов, достав-
ленных охотниками в период эпизоотии. В Канаде, на о-ве Банкс,
инфицированность песцов достигала 22% (Secord et al., 1980).
Дикование песцов обычно регистрируется с ноября по март.
В значительной степени это связано с тем, что в это время идет
промысел песца и от охотников поступают сведения и материал для
исследования. Высказанное Канторовичем (1963) утверждение об
исключительной приуроченности заболевания к этому периоду
Хищные
103
Таблица 11, Результаты вирусологического исследования песцов
(по Полякову, Колесниковой, 1972; Колесниковой, 1973; Татарову, 1982)
Промысловый сезон Исследовано, экз. Инфициро- вано, % Промысловый сезон Исследовано, экз. Инфици- ровано, %
1962/63 132 1,1 1967/68 149 3,0
1963/64 172 1,1 1968/69 ’ 55 1,8
1964/65 189 0,5 1974/75 273 2,2
1965/66 278 1,0 1975/76 196 2,7
1966/67 150 11,7 Декабрь 1976 440 8,8
не подтверждается наблюдениями других авторов (Дубровский А.,
1939; Рахманин, 1959; Колесникова, 1979; Бернштейн и др., 1981).
В Гренландии заболевания песцов или собак регистрировали круг-
логодично, за исключением августа-сентября (Crandell, Col, 1966).
Широкие миграции песцов определяют возможность распростра-
нения эпизоотии на обширные территории тундровой зоны, включая
и арктические острова.
Песцы, являясь основным хозяином вируса тундрового бешен-
ства, служат источником заражения и других животных тундры.
Общеизвестно широкое распространение эпизоотий «дикования»
среди собак (Elton, 1931; Формозов, 1935; и др.). В эпизоотии также
вовлекаются домашние северные олени, покусанные песцами (или
собаками). Диагностировано заболевание лисиц, волков (Канторо-
вич, 1963; Колесникова, 1979; Татаров, 1982). На одном из островов
архипелага Шпицбергена в марте—июне 1980 г. установлено за-
болевание песцов, северных оленей и тюленя (0degaard, Krogerud,
1981).
Дикование песцов обычно совпадает с пиком численности лем-
мингов, среди которых нередко отмечают падеж. Многие авторы вы-
сказывали предположение о возможности заражения песцов, а так-
же и северных оленей от этих грызунов (Дубровский А., 1939; Ме-
телева, Рубанчик, 1959; Соколов, Ча, 1958; и др.). Р. С. Колеснико-
ва (1979), основываясь на положительной реакции связывания комп-
лемента (РСК) при исследовании мозга павших и отловленных живы-
ми сибирских леммингов, предполагает, что лемминги являются не-
обходимейшим звеном в циркуляции вируса дикования 4.
Эти предположения не подтверждаются вирусологическими ис-
следованиями тундровых грызунов. Вирус тундрового бешенства не
был выделен при исследовании около 3000 зверьков в Карской тунд-
ре и 985 сибирских и копытных леммингов в Якутии (Канторович,
1963; Татаров, 1982). Об отрицательных результатах исследования
4 Применение РСК не рекомендуется для выявления рабического антигена
вследствие возможности неспецифической реакции с мозговой тканью (Ме-
тоды лабораторных исследований по бешенству. Женева: ВОЗ, 1975).
104
Carnivora
павших узкочерепных полевок на Ямале сообщает Л. М. Сюзюмова
(1965).
Добавим, что в тундровых и лесотундровых районах Западной
Сибири, где проходили эпизоотии дикования песцов, вирусологиче-
ское исследование 459 горностаев не выявило среди них инфициро-
ванных (Сюзюмова, 1965). Эти материалы еще раз подтверждают
непричастность мелких грызунов тундры — основного корма гор-
ностая — к циркуляции вируса тундрового бешенства.
Постоянное присутствие в популяции песцов хотя бы и неболь-
шого числа больных зверей, длительный инкубационный период,
высокое содержание вируса в слюнных железах, обеспечивающее
эффективность передачи через укус, определяют значение песцов
как основного хозяина в очагах тундрового бешенства.
Из бактериальных инфекций у песцов выявлено заражение бру-
целлезом (Забродин и др., 1971; Никулина, 1974), туляремией (Ал-
газин, 1980), листериозом (Nordland, 1959), сальмонеллезом, лепто-
спирозом и сибирской язвой (Шеханов, 1970). Инфицированность
бруцеллезом песцов в таймырской тундре составляла 1,7—3,6%
(Забродин, Никулина, 1976). У клеточных песцов переболевание
бруцеллезом (по серологическим данным) обнаружено в 7% из
2979 молодых зверей. Среди взрослых песцов антитела находили
у 21% нормально размножавшихся самок и у половины пропусто-
вавших (Никулина, 1974).
Песцы мало восприимчивы к штаммам бруцелл, выделенным от
северных оленей. Минимальная заражающая доза при подкожном
введении равнялась 10 млн микробных клеток, а при алиментарном
заражении — 3—6 млрд микробных клеток. У заразившихся самок
наблюдали аборты, резорбцию эмбрионов, рождение мертвых ще-
нят (Вашкевич, 1974). В годы низкой численности леммингов, когда
значительное место в питании песцов занимает падаль и отбросы
забоя оленей, возможно увеличение зараженности песцов бруцелле-
зом и проявление его патогенного действия на размножение.
Распространение туляремии среди тундровых грызунов, выяв-
ленное в недавние годы в различных регионах — на европейском
Севере, Ямале, Таймыре, в Якутии, определяет широкие возмож-
ности заражения песцов туляремией. Как и другие хищники, песцы
малочувствительны к этой инфекции, о чем можно судить по обна-
ружению у них иммунологической реакции, свидетельствующей о
переболевании их туляремией. Так, при исследовании сывороток
48 песцов на Восточном Таймыре специфические антитела были об-
наружены у 23 особей (Алгазин, 1980).
Еще более скудны сведения о других бактериальных заболева-
ниях, распространенных среди грызунов и могущих передаваться
хищникам. Описан один случай выделения культуры листерий из го-
ловного мозга и крови песца, пойманного в окрестностях форта
Черчилль. Поведение зверя было сходно с симптомами бешенства —
«дикования» (Nordland, 1959). В эксперименте листериоз у песцов
X ищные
105
развивался как бессимптомная или стертая инфекция, с накоплением
специфических антител (Жилов, 1974).
Лептоспироз у песцов может развиваться при заражении от
грызунов в южных частях тундры, где выявлены природные очаги,
поддерживаемые полевкой-экономкой и сибирскими леммингами
(Валова, 1970; Карасева и др., 1976) или при поедании органов ин-
фицированных северных оленей. На зверофермах наблюдалось за-
ражение песцов лептоспирозом, вызывавшее значительную гибель
молодых зверей. Серологические исследования выявляли антитела
к лептоспирам (ballum, pyrogenes, grippotyphosa, pomona, bata-
viae и др.) у 16—30% зверей (Солошенко, 1971; Райхлин, 1977).
У песцов обнаружена зараженность 31 видом гельминтов —
12 видов цестод, 13 видов нематод, 4 вида трематод и 2 вида скребней
(Козлов, 1977). Общая зараженность песцов гельминтами достигает
90—100% и одинаково высока в разных частях ареала (Губанов,
1964; Шиляева, 1968; Савельев, 1975; и др.). Наиболее часто песцы
заражены нематодой Toxascaris leonina. Среди ленточных глистов
наиболее обычны Tetratirotaenia polyacantha, Taenia crassiceps,
Alveococcus multilocularis, промежуточными хозяевами которых
служат лемминги и полевки. Экстенсивность заражения песцов
альвеококком повышается в годы высокой численности леммингов
(Губанов, 1964; Савельев, 1975; Овсюкова, 1974).
Освоение северных районов, расширение хозяйственной дея-
тельности оказывает воздействие на гельминтофауну песцов. Так,
на Чукотке в связи с развитием звероводства и использованием на
зверофермах мяса морского зверя повысилась зараженность диких
песцов, подбирающих остатки разделанных туш, трихинеллезом
(Овсюкова, 1972). Зараженность диких песцов вблизи фермы соста-
вила 50%, в то время как ранее равнялась 12,5%.
Из глистных инвазий песца опасность для человека может пред-
ставлять альвеококкоз. На шкурках песцов обнаруживали значи-
тельное число яиц этого паразита. При снятии и обработке шкурок
яйца глист могут попасть в организм человека.
Род Vulpes Oken, 1816 — лисицы
Род включает девять видов, из которых в СССР обитают три.
Распространение рода охватывает всю Европу, Африку, большую
часть Азии; в Северной Америке — от арктического побережья до
Калифорнии и Северной Мексики. Наиболее широко распространена
обыкновенная лисица V. vulpes Linnaeus, 1758 (см. ниже). В Афри-
ке обитают три вида лисиц: африканская V. pallida Cretz., 1826,
южноафриканская V. chama A. Smith, 1837 и песчаная V. riippeli
Schinz, 1825, ареал которой охватывает также Синайский полуост-
ров, юг Аравийского полуострова, Иран, Афганистан. В Индии,
Непале распространена бенгальская лисица V. bengalensis Schaw,
1800; в Тибете, Непале — тибетская лисица V. ferrilata Hadson,
106
Carnivora
1842. Афганская лисица V. сапа Blanford, 1872 распространена в
Восточном Иране, Афганистане, Северо-Западной Индии. В СССР
встречается в Юго-Западной Туркмении. Ареал корсака V. corsac
Linnaeus, 1768 охватывает Северо-Восточный Иран, Афганистан,
МНР, Северо-Восточный Китай. В СССР — степные и пустынные
районы — от юго-западных областей Украины (редко), Предкав-
казья, Калмыкии и Волжско-Уральского междуречья; обычен в
степях Казахстана, в равнинах Средней Азии, в степях Забайкалья.
В Северной Америке два вида — обыкновенная лисица, заселяю-
щая значительную часть континента — от Аляски до южных шта-
тов, и американский корсак V. velox Say, 1823.
Обыкновенные лисицы на протяжении своего голарктического
ареала имеют диплоидное число хромосом 2п=38, у песчаной лиси-
цы 2п=40, у бенгальской — 60, у американского корсака — 50.
Представители рода имеют важное экономическое значение как
ценные пушные звери. В СССР ежегодно добывается от 180 до 650
тыс. лисиц и от 5 до 34 тыс. корсаков.
Поедая грызунов — вредителей сельского хозяйства и распро-
странителей некоторых природно-очаговых болезней человека, ли-
сицы могут оказывать сдерживающее влияние на их численность.
Вместе с тем обыкновенная лисица, а также корсак поддерживают
существование природных очагов бешенства.
Vulpes vulpes Linnaeus, 1758 — обыкновенная лисица
Ареал вида охватывает всю Европу, Северную Африку (Алжир,
Ливия, Египет). В Азии: Малая Азия, Аравия, Ирак, Иран, Афга-
нистан, Китай, Монголия, Корейский полуостров, Япония, Индия.
В СССР — вся европейская и азиатская части страны до южной
границы тундры. В Северной Америке: от южной границы тундры
до Мексиканского залива. На этой обширной территории у лисиц
хорошо выражены географические формы, отличающиеся по разме-
рам, окраске, густоте меха.
Плотность населения лисиц в разных ландшафтах различна.
Повсеместно звери предпочитают открытые или малооблесенные
пространства. На северной границе ареала — в лесотундре и южной
тундре лисица редка и обычно селится в долинах рек с более разно-
образной и устойчивой фауной грызунов. В тайге лисица малочис-
ленна, избегает сомкнутых равнинных лесов, придерживаясь опу-
шек, вырубок, пойменных биотопов, участков леса, перемежающих-
ся полями, лугами. Наиболее благоприятны для вида богатые
грызунами открытые пространства степи и лесостепи и пустынные
биотопы. В горных районах лисица обитает во всех вертикальных
поясах, доходя до границы снегов. Особенно многочисленна в
альпийских и субальпийских лугах.
Селятся лисицы в норах, выкапывая их в оврагах, под корнями
деревьев, широко использует норы других видов — барсука, дико-
X ищные
107
браза, часто селится на колониях сурков, большой песчанки, ис-
пользует остатки блиндажей, землянок военного времени. Глуби-
на нор от 0,5 до 2,5 м, количество входов 3—5 и более. В лесной зоне
и в степях с плотными почвами норы лисиц сохраняются сотни и даже
тысячи лет (Динесман, 1968).
Гон лисиц приходится на юге ареала на декабрь—февраль, в
более северных районах на февраль—март. Детеныши рождаются
в марте-апреле. Беременность длится 48—52 дня. Количество щенят
на севере (Карелия) в среднем 3,5, максимально до 6; в южной тайге
средний выводок 4—5. В степях Предкавказья и Западной Сибири
в среднем бывает 5—6, максимально — до 13 детенышей. Лисята
рождаются слепыми, масса тела 56—ПО г, прозревают на 10—15-й
день. Лактация продолжается 1—1,5 месяца, но уже в месячном
возрасте детеныши начинают подкармливаться добычей, приносимой
обоими родителями. К 6—7-месячному возрасту молодые зверьки
приобретают облик взрослого животного и ведут самостоятельный
образ жизни, покидая место норения. Половое созревание самок
обычно к концу второго года жизни, но при благоприятных кормо-
вых условиях часть самок может спариваться уже в возрасте 9—
10 месяцев (Чиркова, 1967).
Интенсивность размножения лисиц (процент занятых нор, число
щенят) может изменяться по годам в соответствии с состоянием
численности грызунов — основного корма лисиц. Преобладающее
значение в питании лисиц имеют мелкие грызуны открытых биото-
пов — полевки, песчанки, суслики. Повсеместно важным объектом
питания лисиц стала ондатра, хатки которой лисицы разрушают
зимой, передвигаясь по замерзшим водоемам. Значение зайцев в
рационе лисиц обычно невелико, повышаясь в годы массового их
размножения и развития эпизоотий, как это наблюдалось в 1955—
1957 гг. в Якутии (Млекопитающие Якутии, 1971).
Меньше лисицы поедают птиц, преимущественно нелетных птен-
цов. В южных районах значительное место занимают насекомые,
пресмыкающиеся, плоды фруктовых деревьев, ягоды. Охотно поеде-
ют падаль.
Враги лисиц немногочисленны. Наиболее опасен волк, случаи
нападения которого на лисиц обычно отмечают возле волчьей добычи,
на приваде или падали. Изредка волки поедают лисиц, но чаще
закапывают их трупы (Гурский, 1975; Волк, 1985). Крупные хищ-
ные птицы могут нападать на лисят, реже — на взрослых зверей.
Продолжительность жизни лисиц в природе 6—7 лет.
Заготовки шкурок лисиц в СССР колеблются по годам от
180 800 до 652 100.
Эктопаразиты. Сведения об эктопаразитах лисиц скудны. На
них найдены специфические для хищников виды блох: Chaetopsylla
globiceps, Ch. trichosa, Ch.'homoeus, Echidnophaga popovi, Pulex
irritans, нередки блохи барсука — Paraceros meles, P. flabellum.
Встречаются блохи собак и кошек — Ctenocephalides canis, Ct. fe<
108
Carnivora
lis. На лисиц во время охоты переходят блохи различных грызунов:
сусликов — Ceratophyllus tesquorum, Neopsylla setosa, Frontop-
sylla semura, Ctenophthalmus polex; песчанок — Ceratophyllus
laeviceps, Rhadinopsylla sedestes, Xenopsylla conformis, X. skrjabi-
ni, X. nuttali; сурков — Oropsylla silantievi, полевок и мышей —
Ceratophyllus mokrzeckyi, C. consimilis и др. (Емельянова, Жовтый,
1957; Миронов и др., 1965).
Поскольку некоторые виды грызуньих блох известны как пере-
носчики чумного микроба, разнос их по территории лисицами может
иметь некоторое эпизоотологическое значение.
Известны случаи переноса меченых блох Р. irritans из одной
лисьей норы в другую на расстояние около 1 км (Колпакова и др.,
1968), находки блох О. silantievi, инфицированных возбудителем
чумы, на лисице, убитой в 5 км от места, где протекала эпизоотия
на сурках (Емельянова, 1957). О значении лисиц как разносчиков
инфицированных блох свидетельствуют довольно многочислен-
ные случаи выделения чумного микроба от блох, в основном грызу-
ньих, снятых с этих хищников (Варшавский и др., 1982). Следует
подчеркнуть, что блохи грызунов никогда не бывают многочислен-
ны на лисицах, в то время как специфические виды составляют до
95% блох, снятых с лисиц.
Лисицы, как и другие хищники семейства, несомненно имеют
важное значение как прокормители пастбищных иксодовых клещей,
которых они набирают с растительности во время охотничьих марш-
рутов. На лисицах обнаруживали Ixodes ricinus, I. persulcatus,
Haemaphysalis concinna, Dermacentor marginatus, D. pictus, а так-
же случайных паразитов, основными прокормителями которых
служат грызуны — Rhipicephalus rossicus, R. schulzei, Haemaphy-
salis numidiana и др. В норах лисиц проходят все стадии развития
выкармливающиеся на хищниках и сурках Ixodes crenulatus, I. kai-
seri. В Крымском заповеднике на лисицах обнаружены все фазы
развития I. ricinus и Н. concinna. С 10 осмотренных лисиц было сня-
то 35 личинок, 37 нимф и 84 имаго I. ricinus. Индекс обилия равнял-
ся 15,6. На трех из пяти осмотренных лисиц были обнаружены
158 личинок, 188 нимф и 37 имаго Н. concinna. Индекс обилия 76,6
(Мельникова, 1953).
Максимальное количество этих клещей, найденное на одной
лисице, равнялось соответственно 34 и 277.
Луговой клещ D. pictus является переносчиком губительного
для лисиц и других видов семейства заболевания — пироплазмоза
(Piroplasma canis).
На лисицах паразитирует зудневый клещ Acarus siro var. vul-
pis — полиморфный вид, паразитирующий у различных, видов жи-
вотных, а также и у человека (Дубинин, 1955). Паразитирование
чесоточного зудня вызывает воспалительный процесс, зуд, разраста-
ние эпителия, выпадение волос и может привести к гибели живот-
ных .
Хищные
109
Зудневые клещи распространены всесветно, но оптимум их оби-
тания находится в степных и лесостепных районах и совпадает с
местами наибольшей плотности населения лисиц. Стойкости очагов
зудневой чесотки способствуют благоприятные микроклиматиче-
ские условия, позволяющие клещам переживать длительное отсут-
ствие хозяина в норах хищников, где они находят мало меняющую-
ся температуру и оптимальную влажность (Чиркова, 1957). Вне
тела хозяина клещи могут здесь сохраняться до трех месяцев. В бо-
лее южных районах они погибают от недостаточной влажности, а
севернее — от низких температур.
Впервые массовое распространение зудневой чесотки у лисиц
обратило на себя внимание в окрестностях Берлина в 1899 г. В на-
шей стране первые сообщения относятся к территории Украины в
1919 г. Распространение чесотки среди диких животных могло быть
связано с большим количеством чесотки у лошадей в условиях воен-
ного времени. В 20-е годы чесотка среди лисиц распространилась
на большей части украинского правобережья Днепра, а к середине
50-х годов обнаруживалась почти на всей территории Украины, в
Белоруссии, Молдавии, Крыму, в Литве, Латвии, Эстонии и в поло-
се южных лесов и лесостепи европейской части РСФСР до Волги,
а местами и восточнее (Чиркова, 1967). В Сибири и Закавказье при
просмотре 11 тыс. лисьих шкур чесотка не была обнаружена (Ге-
расимов, 1953). С 1936 по 1954 г. поступили сведения о 8500 слу-
чаях обнаружения чесотки у лисиц. А. Ф. Чиркова (1957) указы-
вает, что заболевания и гибель лисиц от чесотки составляют в сред-
нем 55% от всех известных случаев заболеваний и смертности лисиц
в лесостепной и степной части СССР. В отдельных районах с высо-
кой плотностью населения лисиц пораженность их чесоточным зуд-
нем может быть очень велика, достигая 40—60% и более (Герасимов,
1953; Павлов, 1953; Чиркова, 1957 и др.).
Гельминтозы. В различных частях ареала у лисиц обнаружено
89 видов гельминтов: цестод 22 вида, трематод — 20, нематод —
44 и 3 вида скребней (Козлов, 1977). Количество видов гельминтов
особенно велико в южных частях ареала. В Узбекистане описано
паразитирование 51 вида (Матчанов, Муминов, 1979), в то время
как в Якутии — всего 15 видов (Губанов, 1964). Зараженность ли-
сиц большей частью поголовная, при этом обычно наблюдается мно-
жественная инвазия — от 2 до 6 видов червей и более. Сведений о
патогенном воздействии глистных инвазий на лисиц в природе мало.
Вероятна повышенная гибель молодняка при массовом заражении
щенят вблизи нор геогельминтами (Toxascaris leonina, Тохасага
canis, Uncinaria stenocephala).
Лисицы имеют определенное значение в поддержании природных
очагов альвеококкоза (Alveococcus multilocularis). Личиночную
стадию развития этого лентеца лисицы получают при поедании
промежуточных хозяев — полевок, мышей, ондатр. Инвазирован-
ность лисиц колеблется от незначительной до 80%, что, вероятно,
по
Carnivora
связано с численностью грызунов. Количество половозрелых чер-
вей, имеющих длину 3—5 мм, может достигать в кишечнике лисиц
колоссальных цифр. В Якутии среднее число паразитов равнялось
41 672 экз. В Казахстане зарегистрировано максимальное количе-
ство червей у одной лисицы — 147 000 (Губанов, 1964; Боев и др.,
1970). Загрязнение внешней среды яйцами глист может послужить
причиной попадания их в организм человека с дикорастущими тра-
вами, ягодами. Заражение может произойти также при обработке
шкурок лисиц. Развивающееся при внедрении паразитов тяжелое
поражение внутренних органов требует оперативного вмешательст-
ва. В Алтайском крае за 10 лет выявлено 411 человек, больных аль-
веококкозом (Марченко, 1971). В Башкирии альвеококкоз обнару-
жен у людей (81 случай) в 33 районах из 54 (Валиулин и др., 1979).
Немаловажное значение имеют лисицы в распространении три-
хинеллеза, известного у них повсеместно. Наиболее высока зара-
женность лисиц в Башкирии (77,7%), Воронежском заповеднике
(75%), Беловежской пуще (59,2%), в Крыму и на Кавказе (56,5—
58,5%), на Чукотке (56,4%). Несколько меньше в Горьковской, Ки-
ровской, Саратовской областях — 22—36,4% (Боев и др., 1970).
В Чехословакии трихинеллез обнаружен у 22,7% лисиц, в Поль-
ше — у 25,7, в Норвегии — у 22,8%. В Дании и Финляндии пара-
зиты обнаружены лишь у единичных зверей. В ГДР трихинеллез
не был выявлен при обследовании 337 лисиц. Высока инвазирован-
ность лисиц на Аляске — более 40% (Боев и др., 1978). Лисы зара-
жаются трихинеллезом, поедая грызунов или органы павших каба-
нов, домашних свиней, морского зверя или других плотоядных.
Скармливание тушек лисиц свиньям, практикующееся некоторыми
охотниками, может стать причиной возникновения очагов трихинел-
леза в свиноводческих хозяйствах. Одним из источников распрост-
ранения трихинеллеза среди домашних собак, кошек могут быть
лисьи шкурки, хранящиеся в местах, доступных для крыс и других
грызунов. В остатках ткани на невыделанных шкурках личинки
трихинелл сохраняют жизнеспособность до 8 лет (Куликова, 1978).
Исследования, проведенные в Казахстане, выявили различное
распространение трихинеллеза в разных ландшафтных зонах.
Исследовав в общей сложности 569 лисиц, С. Н. Боев с соавтора-
ми (1972) выявили зараженность трихинеллезом в лесостепи у
5,7%, в степи у 10,4, в полупустыне у 8,6%. Наиболее зараженными
были лисицы из пустынных — 17,1% и горных участков — 20,9%.
Авторы отмечают отсутствие синантропных очагов трихинеллеза в
обследованном районе.
Бешенство. Лисицы имеют важное значение в поддержании при-
родных очагов бешенства, получивших широкое распространение в
Европе после второй мировой войны. До середины текущего столе-
тия первое место по частоте обнаружения бешенства принадлежало
волку. Также велико было бешенство собак, с которым удалось ус-
пешно справиться благодаря вакцинации. Спорадические случаи бе-
Хищные
111
шенства лисиц регистрировались и ранее, привлекая к себе внима-
ние при нападении зверей на домашних и сельскохозяйственных
животных и редких случаях гидрофобии у людей, последовавшей
после укуса их лисицами. Проявление бешенства лисиц в виде
интенсивных эпизоотий впервые обратило на себя внимание в сере-
дине 40-х годов. В 1945—1946 гг. бешенство лисиц (волков и еното-
видных собак) наблюдалось на значительной территории дельты
Волги (Исаков, 1949). Тогда же, по сообщению А. Н. Формозова,
в Ставропольском крае отмечались многочисленные случаи заболе-
вания бешенством коров, вызванные нападением и покусами лисиц.
В 1946 г. «небывалый падеж» лисиц отмечен также в Воронежской
области, где весной было найдено 143, а в июле—сентябре еще
170 павших лисиц. Здесь же наблюдались многочисленные случаи
бешенства коров (Чиркова, 1953). В последующие годы появились
сообщения об эпизоотиях бешенства среди лисиц во многих областях
РСФСР, в Белоруссии, на Украине, в Молдавии, Латвии, Литве,
Эстонии, где в настоящее время выявлены стойкие очаги бешенства
диких животных, поддерживаемые преимущественно лисицами, а
в Прибалтийских республиках также и енотовидными собаками.
Природные очаги лисьего бешенства были выявлены в после-
военные годы во многих странах Европы, в том числе ранее сво-
бодных от этой инфекции. Предполагается, что центральноевропей-
ская эпизоотия бешенства диких животных распространилась после
второй мировой войны из очагов на территории Польши и Германии,
где бешенство регистрировалось в 30-х годах и во время войны.
В 1948 г. бешенство лисиц было зарегистрировано в Австрии, но
эпизоотия не получила распространения, и с 1959 по 1965 г. в стра-
не не было случаев бешенства. Однако в 1966 г. обнаружено девять
больных бешенством лисиц, а с 1967 по 1972 г. была зарегистриро-
вана 651 бешеная лисица (Селимов, 1978). В Венгрии после войны
отмечались небольшие вспышки бешенства городского типа, ликви-
дированные путем вакцинации собак. В 1948—1965 гг. в стране
были выявлены лишь единичные заболевания диких животных, но
уже в 1968 г. было 76 случаев бешенства лисиц и в 1967—1969 гг.
число больных диких животных достигло 1030, тогда как домаш-
них—всего 184.
Дания была свободна от бешенства на протяжении 75 лет.
В 1964 г. здесь зарегистрировано 82 случая бешенства, из них 63 —
у лисиц. Следующая эпизоотия была выявлена в 1969 г., когда из
154 случаев бешенства 122 (79,2%) приходилось на долю лисиц.
В Бельгии, Люксембурге, Швейцарии, Франции, ранее свободных
от бешенства, первые случаи зарегистрированы в 1966—1967 гг.
и в дальнейшем эпизоотии поддерживались в природных очагах
преимущественно за счет лисиц (табл. 12).
Бешенство диких животных возросло и на Американском конти-
ненте. В 1945 г. в США из 9963 зарегистрированных случаев бешен-
ства 85% приходилось на долю собак и только 4% (372 случая) —
112
Carnivora
Таблица 12. Доля бешенства лисиц от общего числа случаев бешенства
животных (по Селимову, 1978; Parker, 1975—США; Schwierz, 1979—ФРГ;
Pitzschke, 1982—ГДР; Tabel et al., 1974—Канада)
Страна Годы Число случаев бешенства Доля бешенства лисиц, % Число больных лисиц в среднем за год
всего лисиц
ФРГ 1955—1978 83 325 55 993 67,2 2333
ГДР 1953—1980 39 578 26 826 67,7 957
Франция 1968—1975 7 654 5 876 76,7 734
Австрия 1967—1972 852 651 81,0 108
Швейцария 1967—1974 4 098 3318 81,0 255
Польша 1965—1974 7 076 4 182 59,1 418
Чехословакия 1970—1974 1 961 1 403 71,5 280
Венгрия 1967—1976 4 808 4 351 90,5 435
США 1953—1972 93 419 17 820 19,0 891
Канада 1947—1972 13 261 9310* 68,8 358
* Вместе с песцами, количество которых составляет около 1%.
на долю диких животных. В 1968 г. зарегистрировано 2702 случая
бешенства диких животных, что составило 75% всех заболеваний, и
только 296 (8%) случаев бешенства собак (Parker, 1975). Бешенство
выявлено у обыкновенной (V. vulpes) и серой (U. cinereoargenteus)
лисиц. С 1953 по 1968 г. ежегодно регистрировалось от 600—800 до
1200 случаев бешенства лисиц, которые наряду со скунсами являют-
ся основными носителями инфекции.
Как видно из табл. 12, повсеместно лисицы составляют подав-
ляющее большинство животных, больных бешенством, и лишь в
США первое место по частоте регистрации случаев бешенства
они уступают скунсам.
В СССР интенсивное проявление природных очагов бешенства
совпадает с оптимальными для обитания лисиц районами, располо-
женными в европейской части Союза южнее 56° с. ш. Севернее эпи-
зоотии регистрировались лишь в Латвии и Эстонии. В 50-х годах
случаи бешенства лисиц отмечались в Ленинградской области, но
позднее они тут не повторялись (Селимов, 1978). В южной тундре
лисицы вовлекаются в эпизоотии тундрового бешенства (дикова-
ния), основным хозяином которого является песец. На Камчатке
существуют очаги сходного варианта вируса, поддерживаемые ли-
сицами (камчатская дикость лисиц) без участия песцов, изолирован-
ные колонии которых прекратили свое существование здесь в 30-х
годах (Вершинин, Лазарев, 1974). Отличительной чертой этих ва-
риантов вируса является их апатогенность для человека.
X ищные
113
В таежной зоне, по-видимому, нет самостоятельных стойких
очагов природного бешенства, что может быть связано с низкой
плотностью населения лисиц и волков.
Наибольшее значение лисицы как распространители бешенства
имеют в лесостепной и степной зонах, в окультуренных сельско-
хозяйственных ландшафтах, где звери нередко селятся вблизи на-
селенных пунктов. В Средней Азии природные очаги бешенства,
поддерживаемые лисицей (а также волком и шакалом), выявлены в
горах, полупустынях и в тугайно-камышовых зарослях пойм рек
(Маматов, 1979). Ведущее значение лисиц и других хищников се-
мейства установлено в очагах южной части Восточной Сибири (Си-
доров и др., 1979; Грибанова и др., 1980).
В СССР, в ряде республик, бешенство диких животных, в основ-
ном лисиц, занимает значительную долю среди всех регистрируемых
случаев заболевания животных. Прежде всего это связано с резким
снижением заболеваемости собак, доля которых в эпизоотологии
бешенства сократилась с 51% в 1948—1957 гг. до 16% в 1958—
1967 гг., а в последующие годы колебалась от 3,1 до 9,1 % (Селимов,
1978). Решающее значение в снижении заболеваемости собак имела
их вакцинация. На фоне уменьшения роли собак возросла доля ре-
гистрируемых случаев бешенства у диких животных. Основное зна-
чение в эпизоотологии природно-очагового бешенства приобрели
лисицы (табл. 13).
Таблица 13. Доля лисиц среди случаев бешенства диких животных в СССР
(по Селимову, 1978)
Республика Годы Бешенство лисиц, %
Украинская ССР 1968—1972 88,2
Латвийская ССР 1969—1976 70,1
Литовская ССР 1966—1973 49,5
Эстонская ССР 1969—1976 56,3
Белорусская ССР 1969—1976 82,0
В Молдавской ССР из 98 случаев бешенства диких животных
подавляющее большинство — 91 случай — относилось к бешенству
лисиц (Помирко, 1977).
Многолетние наблюдения по эпизоотологии бешенства, прове-
денные в УССР, показали, что 83% выявленных очагов бешенства
были моногостальны и поддерживались исключительно лисицами.
В тех очагах, где зарегистрированы заболевания других видов ди-
ких животных, в большинстве случаев отмечено и заболевание ли-
сиц (Щербак и др., 1978; Scherbak, 1980).
114
Carnivora
В степных и лесостепных районах Западной Сибири из 67 лабо-
раторно подтвержденных случаев бешенства диких животных 87%
составило бешенство лисиц (Грибанова, 1975). В Казахстане из
136 случаев бешенства 64% приходилось на лисиц (Сансызбаев и
др., 1975).
Приведенные материалы показывают, что повсеместно лисицы
имеют ведущее значение в эпизоотологии бешенства. Лишь в США
в ряде районов они уступают эту роль скунсам. Значение лисиц как
основных хозяев и распространителей бешенства определяется осо-
бенностями взаимоотношения с возбудителем и присущими виду
чертами биологии.
Лисицы высоковосприимчивы и чувствительны к вирусу бешен-
ства. При экспериментальном заражении половина лисиц погибала
после внутримышечного введения дозы вируса, соответствующей
10 MLD/50, в то время как для собак доза, вызывающая гибель
половины животных, была в десять раз выше. Инкубационный пе-
риод у зараженных лисиц продолжается 10—50 дней, чаще 15—25.
В отдельных случаях при малых дозах заражения инкубация про-
должалась до 109—150 дней. Длительность клинически выраженного
заболевания от одного до семи дней, реже до 10 дней, в среднем 3—
4 дня (Грибанова, 1975; Щербак и др., 1978; Ботвинкин и др., 1983;
Sikes, 1962; Winkler, 1975; Toma, Andral, 1977; Georg et al., 1980).
Развивающийся при этом заболевании энцефалит примерно в
половине случаев проявляется в виде возбуждения, агрессии, стрем-
лении нанесения укусов. У части животных наблюдается простра-
ция, параличи. Смерть наступает внезапно, в припадке бешенства
или паралича. Для лисиц характерна высокая частота и значитель-
ное содержание вируса в слюнных железах. Так, при исследо-
вании 816 лисиц, у которых вирус бешенства был найден в голов-
ном мозге, у 759 (93%) вирус обнаружен и в слюнных железах
(Wandeler et al., 1974). О несколько меньшей частоте обнаружения
вируса в слюнных железах больных лисиц сообщают Л. Я. Гриба-
нова (1975), А. Г. Татаров (1982).
Титры вируса в слюнных железах у лисиц варьируют от 1 до
6,5 1g MLD/50 0,003 мл. Преобладает высокое содержание вируса.
Так, при исследовании 56 лисиц, инфицированных в природе, у
39 из них (69,8%) концентрация вируса в слюнных железах дости-
гала 5—6 и более 1g MLD/50 и только у четырех зверей отмечены
низкие титры (Winkler, 1975). Выделение вируса со слюной проис-
ходит за несколько дней до клинических проявлений болезни. В од-
ном случае вирус был выделен из слюны за 17 дней до гибели лиси-
цы. Установлена обратная зависимость накопления вируса в слюн-
ных железах от дозы заражения: у животных, инфицированных
массивной дозой и погибающих в короткие сроки, вирус в слюнной
железе не обнаруживали или находили лишь в низких титрах. При
заражении малыми дозами проникновение вируса в слюнные железы
отмечалось у всех животных и концентрация вируса достигала вы-
Хищные
115
соких показателей (Грибанова, 1975; Грибанова и др., 1979, 1982;
Sikes, 1962; Winkler, 1975).
Эти особенности патогенеза инфекции наряду с большой плот-
ностью населения лисиц в некоторых регионах, подвижностью и
характерной для хищников склонностью к стычкам и покусам собра-
тьев, усугубляющейся при развитии заболевания, определяют роль
лисицы как основного хозяина и распространителя инфекции. В ред-
ких случаях при экспериментальном заражении лисиц отмечали
случаи переболевания их бешенством. Так, А. Д. Ботвинкин с со-
авторами (1983) сообщают о переболевании лисицы, зараженной
внутримышечно 50 MLD/50 штамма, выделенного от енотовидной
собаки (табл. 14).
Таблица 14. Результаты заражения лисиц вирусом бешенства
(по Ботвинкину и др., 1983)
Доза MLD/50 Заражено Пало Инкуба- ция, сут Титры вируса Титры антител
Мозг Слюнные железы на 30-е сут на 90-е сут
5 2 1 30 4,7 4,5 0 0
50 2 1 76 3,3 — 1:148 1:107
500 2 1 28 5,3 4,7 0
5000 2 2 23 31 4,9 3,3 4,7 4:8
У выжившей лисицы на 30-е и 90-е сутки в сыворотке крови обна-
ружены вируснейтрализующие антитела. О случаях абортивного
бешенства у экспериментально зараженных корсака, волка и собаки
сообщают Л. Я. Грибанова с соавторами (1979, 1982).
В Швейцарии при серологическом исследовании лисиц антитела
были найдены у 51% особей в энзоотичном районе и у 41% в мест-
ности, где не было эпизоотий, что свидетельствует о неспецифич-
ности реакции (Steck, 1982). Вахендорфер с соавторами (Wachen-
dorfer et al., 1978) считают, что в Центральной Европе нет случаев
латентного или абортивного бешенства у диких животных. Исследо-
вание сывороток 383 особей различных видов хищных ни в одном
случае не обнаружило антител к вирусу бешенства.
Предположения некоторых авторов (Канторович, 1971) о хро-
ническом носительстве рабической инфекции не подтверждается до-
стоверными наблюдениями.
Активизация природных очагов бешенства связана с увеличе-
нием плотности населения лисиц, вызванного снижением пресса
охоты, истреблением в ряде районов волков. Несомненное значе-
ние в военные и послевоенные годы имело ослабление профилактиче-
ских мероприятий по предупреждению бешенства у собак, рост чис-
116
Carnivora
ла бродячих собак, контактирующих с природными биотопами.
Укоренению инфекции в природных очагах не мешает усиливающее-
ся повсеместно антропогенное воздействие, поскольку лисицы легко
уживаются в сельскохозяйственном ландшафте и часто селятся в
непосредственной близости от населенных пунктов.
Инфицированность лисиц колеблется в значительных пределах,
изменяясь в зависимости от их численности, характера размеще-
ния по территории, фазы эпизоотического процесса. Так, например,
на Украине экстенсивность заражения лисиц в Полесье равнялась
10,8 на 1000, в лесостепи — 3,9, в степных районах — 1,3, а в Кар-
патах — 2,4 (Щербак и др., 1978). В Западной Сибири и Казахстане
инфицированность лисиц в период разлитой эпизоотии составляла
соответственно 11,5 и 14,7%, а в годы пониженной активности эпи-
зоотического процесса — 1,8 и 7,8 (Грибанова, 1975; Сансызбаев
и др., 1975).
В окрестностях Западного Берлина, по данным за 1953—1977 гг.,
бешенство регистрировалось примерно у 10% исследованных ли-
сиц (Nowak, 1980). Высокие показатели зараженности лисиц полу-
чены в ГДР. Из 303 лисиц, исследованных за 5 лет, бешенство диаг-
носцировано у 117, т. е. у 38,6% (Ulbrich, 1969). Следует иметь в
виду, что частота выявления бешенства зависит от характера иссле-
дуемого материала. Наибольший процент инфицированное™ на-
блюдается у лисиц, убитых при ненормальном поведении: агрессии,
потери осторожности, и среди найденных мертвыми. Так, по данным
А. Г. Татарова (1982), на территории Брянской и Тамбовской об-
ластей в 1977—1979 гг. бешенство было установлено у 106 из 476 ис-
следованных лисиц (22;2%). При этом зараженность лисиц, убитых
в населенных пунктах, была во много раз выше (58,7%), чем добы-
тых в естественных условиях (8,8%).
По наблюдениям в Украинской ССР из 302 случаев бешенства
лисиц 248 зверей были убиты на территории населенных пунктов,
животноводческих ферм и т. п. и только 54 (17,8%) — в природных
условиях (Щербак и др., 1978).
Многолетние данные, полученные при изучении эпизоотологии
бешенства лисиц в Центральной Европе, выявили неравномерность
частоты обнаружения бешенства в разных группах исследованных
лисиц (табл. 15).
Обращает на себя внимание то, что самая высокая заражен-
ность регистрируется у лисиц с ненормальным поведением, нападав-
ших на людей и животных, забегавших в населенные пункты и т. п.
Эти особенности поведения больных зверей имеют важное эпидемио-
логическое и эпизоотологическое значение, обеспечивая передачу
вируса различным видам животных. Рауш (Rausch, 1975) указыва-
ет, что из 34 больных бешенством лисиц, исследованных на Аляс-
ке, у 10 особей (29%) на морде находили много игл дикобраза.
На фотографии одной из таких лисиц можно насчитать более 100 игл,
что свидетельствует об упорном стремлении больного зверя к напа-
Хищные
117
Таблица 15. Частота обнаружения бешенства в разных группах
лисиц (по Wandeler et al., 1974)
Г руппа Число исследо- ванных Число больных лисиц
абс. %
Павшие 593 358 60,0
Задавлены на дорогах 131 15 11,4
Убиты при ненормальном поведении 2020 1729 85,5
Добыты на охоте:
отстреляны 1287 30 2,3
капканами 164 15 9,1
Всего 4195 2147
дению на дикобраза, несмотря на его активную защиту. Из 21 здо-
ровой лисицы только у одной были обнаружены иглы дикобраза.
Развитие эпизоотии в популяции лисиц сопровождается увели-
чением числа заболеваний других видов. Так, по данным, получен-
ным при исследовании бешенства в Баден-Вюртемберге (ФРГ), в на-
чале эпизоотии подавляющее число случаев бешенства приходилось
на долю лисиц (97,2% из 414). Заболевания барсуков, оленей, собак
и кошек составляли лишь доли процента. По мере развития эпизоо-
тии участие других видов повысилось до 20% (Moegle, Кпогрр,
1977). Многие авторы отмечают возникновение заболеваний сельско-
хозяйственных животных после нападения на них лисиц (Исаков,
1949; Чиркова, 1953; Гришок, 1978; Sorce et al., 1969; и др.).
Бешенство лисиц регистрируется во все сезоны, с наибольшей
частотой в январе—марте, что связано с заражением в период
гона и осенью в период повышения численности и расселения вывод-
ков (табл. 16).
Таблица 16. Сезонное распределение случаев бешенства лисиц, % *
Страна Месяцы Всего
XII —I п-ш IV-V VI-VII VIII — IX X-XI
ГДР 17,1 23,6 15,8 12,1 14,8 16,8 117
ФРГ 19,3 26,6 13,4 11,4 13,3 15,8 2609
Швейцария 19,8 23,8 14,4 10,3 14,1 17,6 1416
* По: Ulbrich, 1969; Wandeler et al., 197 4; Schwierz, 1979.
Многие авторы указывают на цикличность проявления эпизоо-
тий бешенства в популяциях лисиц. Иллюстрацией этого положения
118
Carnivora
Таблица 17. Периодичность эпизоотии бешенства лисиц в Воеводине *
Годы Всего случаев бешенства Из них лисиц Число больных лисиц в годы эпизоотии
абс. %
1953—1955 295 75 25,7 66; 5; 4
1956—1961 1962—1965 29 186 1 127 68,2 24; 95; 6; 2
1966—1971 1972—1974 9 56 56 100,0 30; 14; 12
1975—1976 1977—1978 7 338 7 309 91,4 2; 5 90; 219
* По: Dobrenov et al., 1973; Pavlovii б et al., 1976; Petrovic et al., 1980.
могут служить данные о бешенстве лисиц в Воеводине (Югославия),
где интенсивные эпизоотии лисьего бешенства впервые выявлены
в 1953 г. и в последующие годы регистрировались с промежутками
в 4—6 лет (табл. 17).
До 1953 г. в крае регистрировалось лишь бешенство домашних
животных, среди которых более 50% составляли собаки и около
40% — сельскохозяйственные животные. Во время эпизоотии 1962—
1965 гг. бешенство собак составило всего 3,7%, а бешенство сельско-
хозяйственных животных сократилось до 26,8%, что указывает на
возможность их заражения от лисиц.
Распространение бешенства в ряде стран Центральной Европы,
ранее свободных от этой инфекции, позволило установить скорость
распространения эпизоотии по территории. В течение года бешенство
распространялось на 25—50 км и даже до 80 км, как это было во
время интенсивной эпизоотии в 1953 г. в земле Гессен (ФРГ)'. Сред-
нее расстояние появления новых случаев бешенства от данных пре-
дыдущего месяца колебалось от 3,5 до 6 км. Чаще всего новые слу-
чаи бешенства регистрировались на расстоянии менее 5 км от ранее
обнаруженного очага (66,4%). Продвижение на 5—10 км отмечено
в 115 случаях (27%) и до 15 км — в 20 случаях (4,7%). Единичные
находки бешеных лисиц отмечены на расстоянии более 15—20 км.
Наиболее дальнее распространение эпизоотии наблюдалось в период
гона и в августе (Moegle et al., 1974). Авторы указывают, что ско-
рость распространения бешенства совпадает с характером подвиж-
ности здоровых лисиц, обычно (73% меченых особей) не удаляющих-
ся от места поимки далее 10 км. Привязанность лисиц к определен-
ной местности установлена путем мечения и в СССР (Бакеев и др.,
1980).
Для предотвращения развития интенсивных эпизоотий признано
необходимым сокращение плотности населения лисиц. Считается,
Хищные
119
что при численности 1 лисица на 1 кв. км инфекция получает благо-
приятные условия для распространения. При сокращении числен-
ности до 0,2 экз/кв.км возможно прекращение эпизоотии. Создание
зоны пониженной плотности населения лисиц глубиной более 60 км,
по-видимому, предотвратило распространение бешенства в Данию,
западную часть Швейцарии и часть Бельгии (Bogel, 1974).
Однако в районах со стойкими очагами бешенства инфекция
может сохраняться и при низком уровне численности лисиц. Так,
в Брянской области первые случаи бешенства лисиц были отмечены
при плотности их населения 2,3 экз. на 100 га (1969 г.), а в 1971 г.
эпизоотия регистрировалась при плотности лисиц 0,57 экз. и про-
должалась в течение шести последующих лет при плотности 0,16 экз.
на 100 га (Адамович, 1979). В Молдавии эпизоотия бешенства среди
лисиц не сокращалась при снижении численности лисиц и умень-
шении заготовок в 5,5 раз (Помирко, 1977). На Украине снижение
заготовок лисьих шкур с 215 тыс. в 1964 г. до 155 тыс. в 1973 г. не
отразилось на частоте регистрации случаев бешенства лисиц. В По-
лесье наивысший показатель зараженности лисиц — 10,8 на 1000
был отмечен при наименьшей плотности лисиц в этой зоне за 5 лет
(Щербак и др., 1978). О бешенстве лисиц при низкой плотности их
населения сообщают также авторы, изучавшие эпизоотологию бе-
шенства в Центральной Европе (Bogel, 1974; Moegle, Knorpp, 1974;
Toma, An dr al, 1977).
Подъем заболеваемости лисиц бешенством, связанный с высокой
их численностью в период 1958—1966 гг., удерживался на значитель-
ном уровне и в последующие 3 года, несмотря на резкое сокращение
поголовья, вызванное газацией нор (рис. 6). Ослабление истреби-
тельных мероприятий сопровождалось быстрым восстановлением
численности лисиц и увеличением случаев бешенства.
Предполагается, что бешенство в пик эпизоотии может привести
к гибели 40—60% популяции лисиц. Однако фактических данных
о воздействии эпизоотии на численность лисиц недостаточно. Во вся-
ком случае, многолетнее стойкое сохранение бешенства на некото-
рых территориях свидетельствует о достаточной плотности населе-
ния лисиц, обеспечивающей циркуляцию вируса.
Бешенство лисиц представляет определенную эпидемиологиче-
скую опасность. На общем фоне снижения числа случаев заражения
людей бешенством наблюдается увеличение доли лисиц как источни-
ков инфекции. В значительной степени это связано с уменьшением
роли собак в передаче бешенства, благодаря широко проводимой вак-
цинации их. Так, в 1946—1956 гг., когда основным источником зара-
жения людей были собаки (87,1% всех случаев), на долю лисиц
приходилось всего 2,1 % случаев заражения людей. В 1958—1963 гг.
лисицы были причиной заболевания людей в 7,2% случаев, а соба-
ки — в 82,8%. В 1967—1968 гг. значение собак как источников ин-
фекции снизилось до 39,2—37,5%, а лисицы были причиной зара-
жения в 31,3—37,5% случаев (Селимов, 1978). Значение лисиц в
120
Carnivora
Рис. 6. Соотношение интенсивности эпизоотии бешенства лисиц с уровнем их
добычи (по: Schwierz, 1979)
а — число больных лисиц, б — среднегодовая добыча лисиц в тысячах шкурок, в — газация
нор сплошная, г — газация нор частичная
передаче инфекции человеку связано с большой интенсивностью
эпизоотического процесса в природных очагах и особенностями кон-
такта населения с ними. Наибольшее число встреч с лисицами и слу-
чаи покусов приходятся на конец лета и связано с массовым выходом
населения в природу. Неосведомленность людей об опасности кон-
такта с близко подпускающими к себе человека зверями, причем
не всегда агрессивными, является одной из причин нанесения уку-
сов. В большинстве случаев заболевание связано с необращением
пострадавших за медицинской помощью.
Своевременная квалифицированная помощь, проведение полно-
го курса антирабической вакцинации и гаммаглобулинотерапии на-
дежно предохраняет от заболевания подвергшихся риску заражения
людей. '
Распространение бешенства в природных очагах вызывает не-
обходимость разработки мер по снижению плотности населения ли-
сиц. Координацию проведения профилактических мероприятий в
СССР осуществляет Минздрав СССР.
Для регулирования численности лисиц используется усиление
промысла. В Эстонии и Латвии введены денежные вознаграждения
за отстрел лисиц и енотовидных собак. Уничтожение бродячих собак
также входит в систему мероприятий, предупреждающих распро-
странение бешенства. Одичавшие собаки, становясь сочленами при-
Хищные
121
родных биоценозов, служат связующим звеном между природными
и антропургическими очагами.
В ряде стран (Франции, Швейцарии, Бельгии, ФРГ, ГДР, ЧССР
и др.) для уменьшения плотности населения лисиц применялась га-
зация нор. Однако эффективность этого мероприятия с годами умень-
шается, и, несмотря на все усилия, по мере восстанавления числен-
ности лисиц в этих районах снова регистрируется бешенство.
В опытах на различных лабораторных и диких животных пока-
зана возможность иммунизации живыми антирабическими вакци-
нами, при введении их через рот. В некоторых странах были
проведены полевые опыты по применению приманок, содержащих ос-
лабленный вирус бешенства. Приманки раскладывали у лисьих нор.
Возможность поедания приманок с вакциной другими видами жи-
вотных, реакция которых на живой вирус неизвестна, вызывала воз-
ражения против иммунизации лисиц в природных условиях. В экс-
перименте установлено, что вакцины ERA-Virus и Flury-Hlp-Virus
обладают остаточной вирулентностью (при введении через рот) для
горностая, ласки, хорька, водяной крысы, полевой и лесной мышей,
рыжей полевки и ежа. У некоторых видов вакцинация вызывала
клиническую форму заболевания, подтвержденную выделением ви-
руса (Bogel, 1974; Wachendorfer, 1976).
В 1972 г. ВОЗ признала ценность методики оральной иммуниза-
ции лисиц в природе и осуществляет координацию этой работы в
Европе. Успешное снижение эпизоотии путем раскладки прима-
нок — куриных голов с пластиковой капсулой, содержащей мало-
вирулентную для других видов живую вакцину, получено в Швей-
царии на площади 2700 кв. км. Планируется проведение этой работы
в земле Гессен на площади 1500 кв. км. Норма раскладки прима-
нок — 15 точек на 1 кв. км (Wachendorfer, Frost, 1983).
Важное значение для предотвращения заболевания людей имеет
система вакцинации лиц, имевших контакт с подозрительными на
бешенство животными. В целом по СССР удельный вес прививок,
проводимых людям после укусов диких животных, невелик.
В 1954—1963 гг. он составлял 0,87%, а в 1964—1976 гг.— 3%(Лебе-
дева, Селимов, 1975; Селимов, 1978).
Другие инфекции. Тесные трофические связи лисиц с грызуна-
ми определяют возможность заражения такими инфекциями, как
туляремия, лептоспирозы, токсоплазмоз. К туляремии лисицы мало-
чувствительны, и поедание больных грызунов не ведет к выраженно-
му заболеванию. В крови заразившихся зверей непродолжительное
время обнаруживают антитела к туляремийному антигену. В США
(штаты Джорджиа и Флорида) антитела были выявлены у 23,7%
обыкновенных лисиц и у 38,2% серых лисиц (McKeever et al.,
1958b).
Описаны единичные случаи заражения людей туляремией, про-
исшедшие при снятии шкурки или укусах. В этих случаях происхо-
дит контаминативное заражение при попадании возбудителя на кожу
122
Carnivora
из загрязненной остатками тканей грызунов пасти хищника. Такая
возможность передачи инфекции доказана экспериментально (Ол-
суфьев, Дунаева, 1951).
На возможность заражения лисиц лептоспирозом указывают не-
редкие случаи эпизоотий среди лисиц на фермах (Солошенко, 1971).
Заражение зверей чаще всего происходит от грызунов, загрязняю-
щих своими выделениями пищу и воду в вольерах, а также при
скармливании мяса больных сельскохозяйственных животных. Забо-
левание у хищников может протекать в различной форме — от хро-
нической до молниеносной, что зависит от дозы заражения и виру-
лентности штаммов.
В природных условиях у лисиц обнаруживали антитела к леп-
тоспирам разных серогрупп (Hebdomadis, Grippotyphosa, Icterohae-
morrhagiae) (Карасева, Свешникова, 1971). В Приморье у 57 лисиц
из 81 (70%) выявлены антитела серотипа пего (Высоцкий и др.,
1958). В США в Северной Каролине и Пенсильвании от обыкновен-
ных лисиц были выделены штаммы лептоспир grippotyphosa, icte-
rohaemorrhagiae, pomona, ballum (Infectious diseases..., 1970).
Немногочисленные случаи заражения лисиц Yersinia pestis
известны как по серологическим реакциям, так и по выделению куль-
тур возбудителя, в том числе и от погибших особей. В высокогорных
сурочьих очагах Тянь-Шаня известны всего три случая выделения
чумного микроба от лисиц. Две культуры возбудителя были выде-
лены от специфических лисьих блох Ch. homoeus и в одном случае —
от клещей I. crenulatus, снятых с лисицы. Единичные случаи выде-
ления возбудителя чумы от лисиц или их эктопаразитов отмечались
в Волго-Уральском междуречье, Приаральских Каракумах, Урало-
Эмбинском междуречье, в равнинной и горной частях п-ова Мангыш-
лак. Заражение лисиц происходит при поедании больных или пав-
ших от чумы грызунов. Эпизоотологическое значение лисиц может
выразиться в переносе блох, инфицировавшихся на грызунах, но
сами звери вряд ли служат источником заражения. Скорее следует
отметить их санитарное значение как истребителей грызунов — вре-
дителей сельского хозяйства и носителей возбудителей инфекций,
опасных для человека (Варшавский и др., 1982).
Распространение бруцеллеза среди сельскохозяйственных и
некоторых диких животных (зайцев, сайгаков, северных оленей и
др.) создает возможности для заражения лисиц, охотно поедающих
хозяйственные отбросы и падаль. У лисиц бруцеллез регистрирова-
ли при серологических исследованиях в Казахстане, Туркмении.
На фермах серебристо-черных лисиц антитела к бруцеллезному ан-
тигену обнаруживали у 9,7—11,2% особей (Ременцова и др., 1969)
Заражение лисиц токсоплазмозом происходит при поедании ор-
ганов животных (грызунов, копытных, хищных), инвазированных
цистами токсоплазм. Токсоплазмоз также может передаваться зве-
рям через яйца глист Toxascaris leonina, обычного паразита лисиц
(Калякин, 1971). Токсоплазмоз у лисиц регистрировали на зверо-
Хищные
123
фермах, в зоопарках в виде латентного носительства и в редких
случаях как острое заболевание (Новинская, 1965; Калякин, 1971).
Положительные серологические реакции выявляли у 17—52% осо-
бей. Источником заражения служили мясные продукты, на некото-
рых фермах — мясо диких животных (сайгаков, ондатр и др.).
Экспериментальное заражение лисиц массивными дозами токсо-
плазм вызывало острую форму заболевания — понос, судороги,
параличи и гибель на 6—13-е сутки после заражения (Новинская,
1965). Насколько эти данные соответствуют картине спонтанного
токсоплазмоза, судить трудно, поскольку для заражения применя-
лись массивные дозы, не встречающиеся в природных условиях.
В заключение следует отметить, что многие случаи даже массовой
гибели лисиц остаются нерасшифрованными и заключения о рас-
пространении того или иного заболевания часто не подкреплены
необходимыми лабораторными исследованиями.
Vulpes corsac Linnaeus, 1768 — корсак
Корсак — типичный обитатель сухих степей, полупустынь и
пустынь. Обычно селится в норах крупных грызунов — сурков, сус-
ликов, расширяя их, реже занимает брошенные норы лисиц, барсу-
ков. Основу питания составляют мелкие грызуны, птенцы различ-
ных птиц, пресмыкающиеся, насекомые.
На корсаках паразитируют специфические для хищников виды
блох: Pulex irrjtans, Ctenocephalides canis, Chaetopsylla homoeus.
Часто и в большом количестве встречаются блохи сурков, сусликов,
полевок и песчанок. В Забайкалье на корсаке находили 13 видов
блох, свойственных грызунам, и они составляли до 25% от общего
числа блох, снятых с корсаков (Дубинины, 1951; Бром и др., 1948).
Корсаки могут иметь значение в переносе инфицированных блох в
чумных очагах.
Болезни почти не изучены. М. В. Шеханов (1970) указывает на
зараженность корсаков бешенством, токсоплазмозом, трихинелле-
зом.
Бешенство корсаков было установлено в степи, лесостепи За-
падной Сибири, Казахстане, Узбекистане, Забайкалье (Грибанова,
Мальков, 1978; Сансызбаев и др., 1975; Маматов, 1979; Грибанова
и др., 1979). Зараженность корсаков в Западной Сибири колебалась
в разные годы от 2,6 до 5,9%, в зависимости от интенсивности эпи-
зоотии в популяции лисиц. В Казахстане зараженность корсаков
достигала 14—18% и во все годы была выше зараженности лисиц,
что, по всей вероятности, связано с большей здесь плотностью насе-
ления корсака. В Забайкалье зараженность корсаков была 3%.
За 1950—1978 гг. в Читинской области было зарегистрировано
5 случаев бешенства корсаков, что составило 13% от общего числа
зараженных бешенством диких животных (Ботвинкин и др., 1980).
Л. Я. Грибанова и др. (1979) описывают случай гибели от бешенства
124
Carnivora
кормящей самки корсака, обнаруженной в норе вместе с семью
мертвыми щенятами. Вирус бешенства был выделен из мозга и
слюнных желез самки и из мозга одного из щенков.
При экспериментальном заражении вирусом бешенства заболело
5 корсаков из 8 бывших в опыте. Инкубация длилась 9—13 дней.
Клинические проявления продолжались 1—2 дня, у одного зверя —
12 дней, а один из корсаков болел 25 дней с типичной картиной бе-
шенства — возбуждение, затем паралич. На 7-й день стал пить, на
11-й день поднялся на лапы. Исследован на 85-е сутки после зара-
жения. В мозге выявлены вируснейтрализующие антитела. За время
болезни вируснейтрализующие антитела в крови обнаружены в на-
растающих титрах — с 1 : 18 на 2-е сутки, на 10-е сутки — 1 : 625,
на 20-е сутки — 1 : 844 и на 60-е сутки — 1 : 3125 (Грибанова и др.,
1979).
Следует заметить, что заражение корсаков проводилось внутри-
брюшинно, что не соответствует естественному пути заражения.
При серологическом исследовании в Западной Сибири антитела
к вирусу бешенства в низких титрах были найдены у пяти корсаков,
но эти звери могли погибнуть впоследствии от бешенства.
СЕМЕЙСТВО URSIDAE GRAY, 1825 — МЕДВЕЖЬИ
В семействе 4 рода, 7 видов (Соколов, 1979). Род Ursus Linnaeus,
1758 объединяет бурых, черных и белых медведей и состоит из четы-
рех видов: Ursus arctos Linnaeus, 1758 — бурый медведь, U. ameri-
canus Pallas, 1780 — черный медведь (барибал), U. thibetanus
G. Cuvier, 1823 — белогрудый медведь и U. maritimus Phipps,
1774 — белый медведь. Двойное число хромосом у всех этих видов
равно 74.
Ареал рода охватывает Северную Америку, Евразию, Арктиче-
ский бассейн; южная граница распространения проходит в Север-
ной Африке, через Малую Азию, Сирию, Ирак, Южный Иран, гор-
ные области правобережья нижнего и Среднего Инда, по Гималаям и
охватывает весь Индокитай, кроме южных и средних частей Малак-
кского полуострова.
В роде Tremarctos Gervais, 1855 — очковых медведей — один
вид: Т. ornatus F. Cuvier, 1825 (2п=52), распространенный на севе-
ро-западе Южной Америки. Род Melursus Meyer, 1793 — медведей-
губачей — также состоит из одного вида — М. ursinus Shaw, 1791,
обитающего на п-ове Индостан и на о-ве Шри-Ланка. Род Helarctos
Horsfield, 1825, с одним видом (2п=74) — Н. malayanus Rafles,
1821 — распространен в Бирме, Таиланде, Индостане, Малакке,
Суматре и Калимантане, возможно, также в Южном Китае.
Крупные, однотипного медвежьего облика звери. Наименьшие
размеры у малайских медведей, масса тела которых 27—65 кг. Мед-
веди-губачи достигают массы тела 55—136 кг, очковые медведи —
до 140 кг. Масса тела бурых медведей обычно 250—300 кг, но старые
Хищные
125
особи достигают 500—640 кг (Млекопитающие Советского Союза,
1967). Белые медведи — наиболее крупные представители семей-
ства: самцы имеют массу тела 350—500 кг, наиболее крупные осо-
би — 700 и даже до 1000 кг. Самки мельче — масса тела в среднем
200—250 кг.
Распространение медведей связано с равнинными и горными ле-
сами и лишь белый медведь обитает во льдах Северного Ледовитого
океана, ведя полуводный образ жизни. Для белого медведя харак-
терно питание почти исключительно животной пищей — преиму-
щественно тюленями, в то время как другие виды всеядны и в йх
рационе значительное место занимает растительная пища — травя-
нистые растения, листья кустарников, ягоды, орехи, плоды, а так-
же различные беспозвоночные.
Потребление животной пищи бурыми медведями обычно связано
с сезонным отсутствием других кормов и наблюдается преимущест-
венно весной. В связи с особенностями питания характерно прекра-
щение активности медведей в холодное время года — зимний сон.
Залегание в берлоги на зиму наблюдается у всех особей бурых,
черных медведей и белогрудых; у белых медведей залегают беремен-
ные самки. Продолжительность пребывания в берлоге колеблется
у бурых медведей в зависимости от географического положения
местности от 2,5 до 6 месяцев. В южных районах, например в За-
кавказье, часть медведей, особенно в годы обильного урожая оре-
хов, не залегает в спячку (Млекопитающие Советского Союза,
1967). Гон у бурых медведей приходится в зависимости от места рас-
пространения на май-июнь. Продолжительность беременности 6—
8 месяцев, у черных медведей — 220 дней. Рождение детенышей
происходит в берлоге с декабря по апрель, обычно в первой полови-
не января. В помете от 1 до 5 детенышей, чаще 2—3. Масса тела
новорожденных около 500 г. На зиму медвежата залегают в берлогу
вместе с самкой. Распад семьи происходит на втором или третьем
году жизни медвежат и обычно связан с наступлением течки у самки
и спариванием ее с самцом, которого медвежата избегают и уходят
от матери. Половозрелость наступает на 3—4-м году жизни. Про-
должительность жизни бурых медведей 30—40 лет.
У белых медведей гон происходит в апреле. Беременные самки
залегают в подснежные берлоги от середины августа до октября-
ноября. Известны основные места распространения берлог: на остро-
вах архипелага Шпицберген — около 160 берлог, на Земле Франца
Иосифа — 100, на о-ве Врангеля — 180—200. Небольшое число
медведиц устраивает берлоги на Новосибирских островах, Северной
Земле, на малых островах Карского моря. В Западном полушарии —
в Гренландии, островах канадской Арктики и на юго-западном бере-
гу Гудзонова залива. Возможно часть медведиц устраивает родовые
берлоги и среди льдов (Кищинский, 1974; Успенский, 1977; Беликов,
1982).
Рождение детенышей в декабре-январе, реже — конце ноября
126
Carnivora
или в феврале. Новорожденные медвежата весят 600—800 г, прозре-
вают в возрасте около месяца. Перед выходом из берлоги в марте-
апреле масса тела медвежат колеблется от 3,8 до 17,3 кг у самцов и
от 5,7 до 16 кг у самок (Беликов, 1982). По наблюдениям, в окрест-
ностях о-ва Врангеля из 192 выводков в 70,3% было по два медве-
жонка, в 25,5% — по одному и в 4,2% — по три медвежонка. Мед-
вежата остаются с матерью на протяжении 18—22 месяцев, проводя
часть зимы с матерью в берлоге. В некоторых случаях семья дер-
жится вместе еще одну зиму, распадаясь весной (Кищинский, 1974).
Половозрелое™ самки белого медведя достигают к 4 годам,
самцы, по-видимому, позднее. Продолжительность жизни медведей
в зоопарках достигала 40 лет, но в природе обычно встречаются
звери не старше 10—12 лет, изредка добывали старых самцов 18—
24 лет и самок 17—20-летнего возраста. Обитание в экстремальных
условиях, обусловливающее значительную гибель молодых медве-
дей (12—23% до годовалого возраста), низкая репродуктивная спо-
собность — медведицы приносят всего два выводка за свою жизнь
(Беликов, 1982) и неумеренный промысел привели к уменьшению
численности белых медведей, особенно резко выявившемуся в 30-х
годах XX в. В СССР охота на белых медведей была полностью за-
прещена в 1956 г. В 1965 г. белый медведь включен в Международную
Красную книгу. Остров Врангеля — основное место концентрации
родильных берлог — объявлен заповедником.
Из заболеваний, опасных для человека, у медведей в сводке
М. В. Шеханова (1970) указаны: туляремия (бурый медведь), трихи-
неллез (бурый и белый медведи) и описторхоз (у бурого медведя).
Следует, однако, отметить, что указание на туляремию у бурого
медведя на Кавказе (Дробинский, Климухин, 1948), основанное на
случае заболевания человека после употребления в пищу медвежье-
го мяса, представляется сомнительным. Хищники малочувствитель-
ны к туляремии и пищевой путь заражения от них мало вероятен.
В полевых условиях заражение могло произойти и другим путем, и
от других источников.
Указание на бешенство у белого медведя в сводке М. В. Шеха-
нова основано, вероятно, на сообщении Н. Н. Мари (1909) о случае
гидрофобии, связанным с укусом белого медведя. Как справедливо
указывает М. А. Селимов (1978), достоверность подобных сообще-
ний проверить невозможно.
Бешенство бурых медведей зарегистрировано в Чехословакии,
Турции (Селимов, 1978) и Югославии (Petrovic et al., 1980). В СССР
единичные случаи бешенства медведей наблюдались в Краснодар-
ском крае, на Дальнем Востоке (Селимов, 1978; Ботвинкин и др.,
1981).
В Северной Америке имеются сообщение о 9 случаях бешенства
у черных медведей (U. americanus). Из них 8 бешеных медведей
отмечено в Канаде (1954—1973 гг.) и один медведь — в 1971 г. в шта-
те Аризона (Rausch, 1975). Больных медведей наблюдали в сентяб-
Хищные
127
ре—ноябре и марте—мае. Из этих девяти медведей два напали на
людей и ранили их, у четырех отмечено необычное поведение и три
были найдены мертвыми.
Заражение десяти черных медведей вирусом бешенства, выделен-
ным от естественно инфицированной лисицы на Аляске, установило
их высокую резистентность по сравнению с песцами, собаками и кой-
отами. При введении дозы в 1000 MLD/50 погиб один годовалый мед-
ведь. Клинические симптомы бешенства появились на 65-й день
после заражения, гибель наступила через 5 дней. В слюнных желе-
зах титр вируса был очень низким — всего 1,4 1g MLD/50. Три мед-
ведя, зараженные той же дозой, оставались здоровыми на протяже-
нии 6, 10,5 и 11 месяцев. Из четырех медведей, получивших дозу в
5000 MLD/50, погиб один на 20-й день. Симптомы заболевания по-
явились лишь накануне гибели. В слюнных железах вирус не обна-
ружен. Остальные звери, имевшие возраст 13, 15 и 17 месяцев, не
проявляли признаков заболевания в течение 6,5—8 месяцев. Взрос-
лый медведь, зараженный дозой в 20 000 MLD/50 (возраст 31 месяц,
масса тела 100 кг), погиб от бешенства на 30-е сутки после зараже-
ния. Клинические симптомы появились на 27-й день опыта. Титр
вируса в слюнных железах 1,2 1g MLD/50.
Три бурых медведя, зараженные дозами 5000 (один) и 10 000
MLD/50, погибли на 18—25-й день после заражения. Симптомы забо-
левания: слюнотечение, паралич — появились за 2-3 дня до гибели.
Вирус в слюнных железах не обнаружен (Rausch, 1975).
Исследования 38 бурых медведей на Аляске выявили антитела
к бруцеллезу оленьего типа у 23 особей (60,5%), что указывает на
частую возможность заражения медведей от больных бруцеллезом
северных оленей — больных или павших. При эксперименталь-
ном заражении бруцеллы обнаружены у зверей в слюнных железах,
почках (Neiland, 1975; Neiland, Miller, 1981).
Наибольшую опасность для людей представляет трихинеллез
медведей. Трихинеллез у бурых медведей распространен повсемест-
но. Экстенсивность заражения колеблется в широких пределах
(табл. 18).
В монографии А. С. Бессонова (1972) указано, что из общего
количества исследованных в СССР 280 бурых медведей заражено три-
хинеллезом оказалось 50 (18%). В более поздней сводке С. Н. Боева
с соавторами (1978) сообщается о заражении 88 медведей из 400 ис-
следованных (22%). Таким образом, каждый четвертый-пятый мед-
ведь опасен как источник заражения человека при употреблении
в пищу их мяса.
В Югославии установлена зараженность 7% медведей, в Слова-
кии — 14,3% (Боев и др., 1978). На Аляске из 43 исследованных
медведей трихинеллез установлен у 15 (37%) (Rausch, 1958).
У белых медведей трихинеллез распространен повсеместно.
Турсхауг и Рустед (Thorshaug, Rosted, 1956) сообщают, что при ис-
следовании 278 белых медведей из различных частей Ледовитого
128
Carnivora
Таблица 18. Результаты исследования бурых медведей на зараженность
трихинеллезом
Территория Количе- ство исследо- ванных Заражено Автор
абс. %
Северо-запад РСФСР 34 4 11,7 Бессонов, 1972
Северный Кавказ 33 5 15,7 »
Грузия 19 4 21,0 »
Таджикистан 73 5 6,8 »
Казахстан 20 4 20,0 »
Восточная Сибирь 23 4 17,3 »
двк 50 16 32,0 »
Хабаровский и Приморский края 123 10 8,1 Железникова, 1972
Камчатка 276 72 26,0 Синович, Соловь- ев, 1981
океана трихинеллез обнаружен у 163 особей (58,6%). На Аляске
трихинеллезом заражено было 9 медведей из 17 исследованных
(Rausch, 1958).
По данным А. С. Бессонова (1972), из 20 белых медведей, исследо-
ванных в СССР, трихинеллез установлен у четырех. С. Н. Боев
с соавторами (1972) сообщает о зараженности 11 медведей из 30
(36%) в советской Арктике и у 6 из 16 в Гренландии (37%). Общая
зараженность 231 медведя, по данным этих авторов, составляла
24%.
Фактические данные указывают на значительную пестроту экс-
тенсивности инвазии. Так, на Чукотке трихинеллез выявлен всего
у одного белого медведя из 17 исследованных, а из 19 бурых — у 11
(Овсюкова, 1966).
Источником заражения белых медведей трихинеллезом могут
быть морские млекопитающие — основной объект их питания и тру-
пы песцов, пораженность которых трихинеллезом велика, в среднем
15,2% (Бессонов, 1972). Освоение северных районов, развитие зверо-
водства и использование для кормления на зверофермах мяса мор-
ского зверя, а также часто практикующееся скармливание тушек
убитых пушных зверей или их доступность для поедания свободно
живущими хищниками в окрестностях звероферм способствуют по-
вышению зараженности последних трихинеллезом. По наблюдениям
Н. И. Овсюковой (1972), зараженность диких песцов в окрестностях
зверофермы на Чукотке повысилась с 12,5 до 50%.
Значение медведей как источников заражения людей трихинел-
лезом велико. Обычно заболевания имеют характер вспышек, по-
скольку мясо такого крупного зверя использует сразу большое коли-
чество людей. Так, в 1947 г. в Гренландии одновременно заболело
Хищные
129
более 300 человек и около 30% заболевших умерло (Успенский,
1977).
В Магаданской области в 1976 г. из 60 человек, евших мясо
убитого медведя, заболел 21 человек, в 1975 г. в Чаунском районе
заболели все 70 человек (Синович, Соловьев, 1981). В Хабаровском
крае из 65 человек, использовавших мясо трех бурых и одного бело-
грудого медведей, заболело 23 человека, в том числе 6 детей (Желез-
никова, 1972; Железникова и др., 1972).
В Якутии, на Камчатке, в районах Крайнего Севера мясо бурых
и белых медведей является единственным источником заражения
людей. В других районах значение медведей сильно колеблется в
зависимости от распространенности охоты на медведей. Всего по
СССР за 1946—1967 гг. было зарегистрировано 339 случаев зараже-
ния людей трихинеллезом от диких животных, что составило всего
2,9% от общей заболеваемости. 178 человек (52,5%) заразились от
мяса бурых медведей и 30 (8,8%) — от белых медведей (Бессонов,
1972). В последующие годы эти показатели значительно увеличи-
лись, особенно в интенсивно осваиваемых районах Крайнего Севера.
Так, в Магаданской области в 1971—1977 гг. заболело 105 человек
(Синович, Соловьев, 1981).
На Чукотке инвазированность людей, выявленная при сероло-
гическом обследовании, колебалась в отдельных населенных пунк-
тах от 13,8 до 39,3%. В среднем инвазированность составила
27,8% — 179 человек из 642 обследованных (Бессонов, 1972).
СЕМЕЙСТВО PROCYONIDAE BONOPARTE, 1850
— ЕНОТОВЫЕ
В семействе 7—8 родов с 15—20 видами (Соколов, 1979; Honacki
et al., 1982). Большая часть представителей семейства обитает в
Западном полушарии и только два вида — в Юго-Восточной Азии.
В китайских провинциях Юньнань и Сычуань, в Северной Бир-
ме, Непале, Сиккиме распространена малая панда Ailurus fulgens
F. Cuvier, 1825. Большая панда Ailuropoda melanoleuca David,
1869 имеет ареал, ограниченный провинциями Сычуань, Ганьсу
(КНР) и Тибетом. Вид занесен в Международную Красную книгу.
Хонаки с соавторами (Honacki et al., 1982) относят этот род к семей-
ству Ursidae и рассматривают его как монотипичный.
В Западном полушарии распространены виды, относящиеся к
шести родам: Bassaricyon (5 видов) — в Южной и Центральной Аме-
рике, Bassariscus (2 вида) — в Северной и Центральной Америке,
Nasuella (2 вида) — в Северной, Центральной и Южной Америке,
Potos (1 вид) — в Южной Америке, Nasua (1 вид) — от Мексики
до Бразилии и Ргосуоп (7 видов) — в Северной, Центральной и Юж-
ной Америке.
Представители семейства имеют разнообразное строение тела,
сходное с обликом медведя (большая панда), куницы, лисицы и ти-
5 Зак. 3575
130
Carnivora
пично «енотовые» (енот-полоскун). Размеры тела широко варьиру-
ют — масса тела от 800 г до 160 кг (большая панда). Населяют пре-
имущественно тропические и субтропические леса, обычно хорошо
лазают по деревьям, устраивая гнезда в дуплах. Питание разнооб-
разное, преимущественно всеядные: питаются насекомыми, ракооб-
разными, пресмыкающимися, мелкими млекопитающими и расти-
тельной пищей.
Род Procyon Storr, 1780 — еноты
В роде два вида: енот-полоскун Р. lotor Linnaeus, 1758 и енот
ракоед — Р. cancrivorus Cuvier, 1798 — ареал которого охватыва-
ет Центральную и Южную Америки.
Енот-полоскун обитает на обширной территории — от южных
районов Канады до Центральной Америки. Населяет лесные райо-
ны, заросли кустарников, обычно вблизи водоемов, но селится так-
же в болотистой местности, вдали от открытой воды. Еноты всеядны,
поедают различные плоды, ягоды, желуди, орехи, а также беспо-
звоночных, мелких млекопитающих, яйца птиц. В южных районах
активны в течение всего года, в Канаде впадают в зимний сон, про-
должительностью около четырех месяцев. Перед залеганием на зиму
звери сильно жиреют, достигая массы тела в 20 кг и более, тогда как
летом масса тела составляет 5—6 кг. Период размножения растянут,
особенно в южных районах, с февраля-марта до мая-июня. Продол-
жительность беременности 63 дня, в выводке 2—7 детенышей, в сред-
нем 4. Самки способны к размножению в возрасте 10 месяцев. Звери
ведут одиночный образ жизни на ограниченном пространстве: ин-
дивидуальный участок не превышает 4—9 кв. км. Плотность населе-
ния енотов наиболее высока в южных штатах, достигая более
100 зверей на 1000 га, а в наиболее благоприятных участках до
400 особей на 1000 га.
Еноты акклиматизированы во многих местах ФРГ и ГДР. По-
пытки акклиматизации енотов в СССР были начаты в 1936 г. Ус-
пешно прошла акклиматизация енотов в Азербайджане, где плот-
ность населения достигла 20 зверей на 1000 га и ведется промысел,
хотя и в небольших размерах (Млекопитающие Советского Союза,
1967).
В США у енотов обнаружено заражение лептоспирозами во мно-
гих частях ареала. Наиболее часто выявляли носительство лепто-
спир autumnalis и grippotyphosa (Infectious diseases, 1970). Кроме
того, у них обнаружены лептоспиры серогрупп Hebdomadis, Pomo-
na, Australis, Ballum, Icterohaemorrhagiae, Canicola.
В штате Джорджия интенсивность заражения енотов составляла
7%, а в штате Луизиана при исследовании 204 енотов носительство
лептоспир (шести серотипов) установлено у 22% особей (Gorman,
1962; Infectious diseases, 1970).
Хищные
131
В штатах Джорджия и Флорида были выявлены антитела к воз-
будителю туляремии у 181 из 618 исследованных енотов (29%).
Титры антител колебались от 1 : 20 до 1 : 2560 (McKeever et. al.,
1958b).
У енотов также обнаружены спорадические случаи заражения
листериозом, псевдотуберкулезом, пастереллезом и бруцеллезом
(Infectious diseases, 1970; Swann et al., 1980). Из вирусных инфек-
ций у енотов обнаруживали чуму плотоядных (Hoff et al., 1974)
и бешенство. Участие енотов в эпизоотиях бешенства в США при-
влекло внимание исследователей вследствие опасности этого заболе-
вания для человека и в связи с панзоотией, распространившейся
после второй мировой войны во многих регионах. Бешенство у ено-
тов впервые наблюдалось в 1936 г. в Калифорнии. С 1953 г., когда
началась регистрация заболеваний диких животных, спорадические
случаи бешенства енотов отмечались в немногих штатах, а в 12 шта-
тах вообще не было сообщений о бешенстве обитающих там енотов
(McLean, 1975). С 1953 по 1971 г. в ряде штатов — Нью-Йорк, Техас,
Миннесота, Айова, Иллинойс — регистрировались случаи бешен-
ства енотов на протяжении нескольких лет, со средней частотой
1,6—4,0 случаев в год. В двух южных штатах — Джорджии и Фло-
риде — интенсивность эпизоотии была значительно выше — около
40 случаев в год.
Распространение бешенства енотов во Флориде и Джорджии про-
слежено на протяжении 1954—1981 гг. (McLean, 1975; Sussman,
1981, 1982). Эпизоотия, первоначально наблюдавшаяся в централь-
ной части полуострова (1954—1959 гг.), в дальнейшем распространи-
лась в северные районы и в 1963—1965 гг. охватила штат Джорджия,
а в 1972 г.— Южную Каролину.
В 1954—1959 гг. на северные районы штата Флорида приходи-
лось около 25% зарегистрированных случаев бешенства енотов, а в
1960—1965 гг. здесь отмечено 48% из 247 случаев бешенства енотов.
На юге штата Джорджия в 1963 г. было 70 случаев бешенства ено-
тов, а через два года 152 больных зверя найдены на расстоянии
40 миль от прежних находок. Достоверность бурного распростране-
ния вируса в Джорджию из Флориды подтверждается исследования-
ми Вуда и Дэвиса (Wood, Davis, 1959), не обнаруживших в 1953—
1957 гг. зараженных особей среди 299 енотов, исследованных на юге
Джорджии и северной части Флориды. В 1963—1971 гг. в Джорджии
выявлено 728 больных енотов. Бешенство енотов регистрируется во
все сезоны года. Максимум заболеваемости отмечен в апреле, что
связано с повышенной активностью зверей в предшествующие ме-
сяцы — во время гона.
Участие в эпизоотии других видов хищных ограниченно. В штате
Флорида за 10 лет (1971—1980 гг.) из 721 случая бешенства на долю
енотов приходилось 66,6% (480 случаев). Бешенство лисиц состави-
ло лишь 4,4%, скунсов — 1,5%, собак и кошек — 2,9 и 3,3%. На
втором месте после енотов регистрировалось бешенство летучих
5*
132
Carnivora
мышей — 19% (Sussman, 1981). В Джорджии бешенство енотов в
годы развития эпизоотии составляло 60—90% от всех случаев бе-
шенства .
Только в этих южных штатах еноты стали основными хозяевами
вируса бешенства. На остальной территории США доля участия
енотов в эпизоотиях составляет лишь 2—6%.
Зараженность енотов вирусом бешенства в энзоотичном очаге
чрезвычайно высока и во много раз превышает зараженность их в
других частях ареала (табл. 19).
Таблица 19. Частота обнаружения бешенства у енотов в США
(по McLean, 1975)
Штат Годы Количество исследованных зверьков Зараженность
абс. %
Калифорния 1950—1969 698 9 1,3
Индиана 1961—1970 457 5 1,1
Айова 1938—1965 913 33 3,6
Нью-Йорк 1946—1963 1523 179 11,6
Флорида 1968—1969 131 81 61,8
Джорджия 1962—1970 1660 608 36,6
Причины длительного укоренения бешенства в юго-восточных
штатах США не ясны. По-видимому, это связано с благоприятными
для вида условиями обитания, высокой плотностью населения и осо-
бенностями отношения енотов к вирусу бешенства.
Еноты более резистентны к вирусу бешенства, чем лисицы.
В экспериментах регулярно наблюдалось выживание енотов при вве-
дении массивных доз вируса — от 5 до 16 тыс. 50% мышиных лета-
льных доз. Длительность инкубации колебалась от 10 до 66, в
среднем от 12 до44 дней. Клинические проявления болезни продол-
жались 2—5 и до 13 суток. В экспериментах не было выяснено, пере-
болели ли выжившие еноты бешенством, или же их порог восприим-
чивости оказался выше вводимых доз вируса.
Вместе с тем в природных условиях у енотов закономерно обна-
руживают сывороточные вируснейтрализующие антитела, не только
у особей, мозг которых заражен вирусом, но и в тех случаях, когда
исследование мозга оказывалось отрицательным. У вируспозитив-
ных енотов антитела были обнаружены у половины особей (из 16).
Титры реакции колебались от 2 до 480, в среднем — 22,3. Из 215
енотов, в мозге которых вирус не обнаружен, сывороточные антитела
найдены у 37 (17%). Титры реакции колебались от 2 до 625, но в
среднем были значительно ниже — 6,1, чем у инфицированных
особей (McLean, 1975).
Хищные
133
Наблюдения за серопозитивными особями, отловленными во
время эпизоотии во Флориде в ноябре 1970 г., установили сохране-
ние сывороточных антител у трех особей на протяжении по меньшей
мере двух лет и три енота утратили антитела после 21 месяца. В этой
же партии 2 серонегативных енота погибли от бешенства на 39-й
и 79-й день после поимки. Незадолго до гибели в крови у них были
обнаружены антитела, как это обычно наблюдается у больных
бешенством животных (Bigler et al., 1973).
Эти наблюдения подтверждают возможность несмертельного ис-
хода при заражении енотов вирусом бешенства. Маклиин (1975) ука-
зывает на возможное значение малого содержания вируса в слюне
енотов и (или) низкую вирулентность отдельных штаммов вируса.
Предполагается возможность реактивации хронической инфекции,
связанная с физиологическими изменениями организма зверей во
время периода размножения и при стрессовых состояниях. Сыворо-
точные антитела к вирусу бешенства, свидетельствующие о добро-
качественном течении заболевания, обнаруживали также у опоссу-
мов, мангустов и летучих мышей (Wood, Davis, 1959; Everard
et al., 1974).
Чума плотоядных выявлена у енотов выделением вируса и серо-
логическими реакциями в штатах Нью-Йорк, Мериленд и Флориде
(Parker et al., 1961; Hoff et al., 1974). Во Флориде в 1972—1973 гг.
заболевание чумой диагностировано у 114 енотов. Симптомы болез-
ни нередко были сходны с бешенством и возможно, что часть случаев,
зарегистрированных как бешенство, относилось к заболеванию ено-
тов чумой (Hoff et al., 1974). Инкубационный период при чуме И—
30 дней, клинические проявления 5—9 дней. Серологические реак-
ции были положительны у переболевших енотов в низких титрах.
В Европе наблюдались отдельные случаи бешенства у акклима-
тизированных енотов. За 1979—1982 гг. в ФРГ бешенство заре-
гистрировано у шести енотов, в ГДР — у двух и в Швейцарии — у
одного (Черкасский, 1985).
СЕМЕЙСТВО MUSTELIDAE SWAINSON, 1835
— КУНЬИ
В семейство входят разнообразные по величине и строению тела
звери, ведущие наземный, полудревесный, околоводный и водный
образ жизни. Система семейства окончательно не установлена. В нем
насчитывают 25—28 родов с 55 видами (Громов и др., 1963), 24 рода
и 64 вида (Соколов, 1979) или 23 рода и 63 вида (Honacki et al.,
1982). В семействе выделяют пять подсемейств (табл. 20).
Представители семейства распространены на всех материках,
кроме Австралии и ряда арктических и океанических островов,
встречаясь в разнообразных ландшафтных зонах — от тундры до
пустыни, от равнинных лесов до горной альпийской зоны.
134
Carnivora
Таблица 20. Состав и распространение семейства Mustelidae
Подсемейство Количество * Распространение
родов видов
Куньи 11 29 Евразия, Северная и Южная Амери-
Mustelinae 4 14 ** ка, Африка
Медоеды 1 1 Африка, Азия; в СССР —на юге от
Mellivorinae 1 1 Каспийского моря до Амударьи
Барсуки 4 6 Евразия, Северная Америка
Melinae 1 1
Скунсы 3 11 Северная, Центральная и Южная
Mephitinae Америка
Выдры Lutrinae 5 2 17 2 Евразия, Африка, Северная и Юж- ная Америки
* В числителе—общее число родов (видов), в знаменателе—число их в СССР.
** Включая акклиматизированную американскую норку.
Представители семейства в основном плотоядные звери, основой
питания многих видов являются мелкие грызуны — различные виды
полевок: лемминги, полевки родов Microtus, Clethrionomys, Arvicola
и др.; в южных районах песчанки — Meriones, Rhombomys, суслики.
Мелкие виды куньих добывают грызунов, проникая в ходы их нор
(ласка, горностай, хорьки, перевязка), в питании куниц, соболя
значительное место занимают обитатели древесного яруса — белки,
бурундуки, сони, яйца и птенцы наземногнездящихся и устраиваю-
щих гнезда на деревьях птиц. Ведущие околоводный образ жизни
норки и выдры в основном питаются рыбой, амфибиями и водными
беспозвоночными—моллюсками, ракообразными. Обитающий на
побережье Тихого океана калан питается преимущественно морски-
ми ежами, а также крабами, осьминогами, раками-отшельниками и
некоторыми рыбами. Росомаха, харза способны убивать крупную
добычу — зайцев, молодняк северного и пятнистого оленей, взрос-
лую кабаргу.
У многих видов значительное место в питании занимают расти-
тельные корма — ягоды, плоды диких яблонь, груш (каменная ку-
ница), кедровые орехи (соболь) и др. Хорошо роющие, имеющие
мощные когти барсуки выкапывают корневища, клубни растений,
личинок жуков, земляных червей. Медоед (Mellivora capensis) по-
едает личинок земляных ос, пчел, их мед, личинок других насеко-
мых и мелких млекопитающих, ящериц.
Большинство видов, особенно обитателей северных широт, име,
ют густой, пушистый высокоценимый мех и служат объектом промыс,
Хищные
135
ла, а некоторые виды разводятся на зверофермах. Ареалы куньих
с детальными картами подробно даны в сводке «Млекопитающие
Советского Союза» (1967).
ПОДСЕМЕЙСТВО MUSTELINAE GILL, 1872
— КУНЬИ
В подсемействе 11 родов с 29 видами (Соколов, 1979), в СССР
обитает 14 видов (включая акклиматизированную американскую
норку), относящихся к четырем родам: Martes — куницы, Gulo —
росомахи, Mustela — ласки и хорьки и Vormela — перевязки
(табл. 21).
Таблица 21. Состав и распространение основных родов подсемейства
Mustelinae
Род Число видов * Распространение
Martes 6 4 Евразия, Северная Америка. Равнинные и горные леса
Mustela 13 8 Евразия, Северная Америка, Северная Африка. Тундры, леса, степи, пустыни и тропический лес
Gulo 1 1 Евразия и Северная Америка. Северная тайга и лесотундра
Vormela 1 1 Юго-Восточная Европа, Передняя и Средняя Азия, Казахстан, северная часть Центральной Азии. Сухие степи, полупустыни и пустыни
♦ В числителе — общее число видов, в знаменателе —число видов в СССР.
В Южной и Центральной Америке распространены представители
монотипических родов: Eira — тайр, Galictis — гризонов, Lynco-
don — патагонских ласок. На Африканском континенте роды:
Ictonyx — африканские хорьки, Poecilictis — североафриканские
ласки и Poecilogale — африканские ласки, также представленные
каждый одним видом.
Род Martes Pinel, 1792 — куницы
В роде шесть видов, из них в СССР обитает четыре: соболь М. zi-
bellina Linnaeus, 1758 (2п=38), лесная куница M.martes Linnaeus,
1758 (2n=38), каменная куница M. foina Erxleben, 1777 (2n=38),
харза M. flavigula Boddaert, 1785 (2n=40). В Северной Америке рас-
пространены американская куница М. americana Turton, 1806
(2п=38) и илька М. pennanti Erxleben, 1777 (2n=38).
136
Carnivora
Восстановленный ареал соболя охватывает таежные леса Евра-
зии — от Скандинавии на западе до тихоокеанского побережья на
востоке, включая Камчатку, Сахалин, Японские острова и Корей-
ский полуостров. Южная граница выходит за пределы СССР по
лесному Алтаю, лесам Хангая и Хентэя в МНР и лесной части
Северо-Восточного Китая.
Неумеренный промысел этого ценного пушного зверя был причи-
ной сокращения численности и полного истребления его на значи-
тельной части ареала. В 1935 г. был объявлен полный запрет про-
мысла на 5 лет и организован ряд заповедников и заказников (Со-
боль, куницы, харза, 1973). С 1949 по 1970 г. было расселено около
19 тыс. соболей. Эти меры благоприятно сказались на численно-
сти зверька и восстановлении его ареала.
Ареал лесной куницы занимает всю Европу до Пиренеев на юго-
западе, исключая степную и тундровые зоны в СССР. Языком вдает-
ся в Западную Сибирь до среднего течения Оби и низовьев Иртыша,
Ишима, Тобола. По горам Малой Азии проходит в лесную часть
Кавказа, а по Эльбурсу и Загросу языками вдается в Иран.
Каменная куница распространена в Южной и Средней Европе —
от Балтийского моря на севере до побережья Средиземного моря
на юге. От Кавказа по горам Средней Азии и Южной Сибири дохо-
дит в МНР до южных склонов Хангая и восточной оконечности Го-
бийского Алтая. Через Афганистан и по Гиндукушу и Куньлуню до-
ходит до Тибета. В СССР каменная куница занимает Прибалтику,
Южную Белоруссию, Украину. Изолированные участки есть в гор-
ном Крыму, на Кавказе и языками заходят с юга в горы Средней
Азии, Казахстана и на Алтай.
Ареал харзы тянется от Индонезии (Ява, Суматра, Калимантан),
Малаккского полуострова, Индокитая до Уссурийского края и
Среднего Амура в СССР. По Гималаям узким языком доходит до Аф-
ганистана. В Южной Индии лежит изолированный участок ареала.
Два вида американских куниц населяют лесные области от се-
верной границы леса на территории Канады и Аляски до северных
штатов США на востоке, проникая на западе континента к югу по
горным лесам до севера Калифорнийского полуострова.
За исключением каменной куницы, предпочитающей открытые
горные склоны, все виды являются обитателями леса. Хорошо лаза-
ют по деревьям, устраивая убежища в дуплах или в поваленных де-
ревьях, заломах и т. п. Представители рода моногамны. Отличитель-
ной особенностью размножения является длительная диапауза в
развитии оплодотворенной яйцеклетки. Продолжительность беремен-
ности у соболя 236—297 дней, у каменной куницы 236—265 дней,
у ильки 338—358 дней, у американской куницы 220—275 дней.
Гон и спаривание зверьков происходят в летние месяцы и сильно
растянуты по срокам. У соболя гон продолжается с середины июля
до середины августа, у лесной и каменной куниц — с июня до на-
Хищные
137
чала августа. У ильки — в марте-апреле. Рождение детенышей при-
ходится на апрель-май. Число детенышей в выводках невелико: у
соболя — чаще 3—4, у куниц — 3—5, у харзы — обычно 2—3. Про-
зревают детеныши на 30—36-й день, резцы прорезываются на 38-й
день. Лактация продолжается два-три месяца. Половозрелости
самки достигают в двухлетнем возрасте. Продолжительность жизни
соболя 15—17 лет (на фермах), но в природных условиях, вероятно,
меньше. Примерно те же сроки продолжительности жизни известны
для лесной куницы и ильки (Терновский, 1977).
Болезни куниц в природе не изучены. Известны единичные
случаи выделения культур листерий, туляремии от соболей, у
лесной куницы обнаруживали эризипелоид и серологически выявля-
ли антитела к лептоспирам grippotyphosa (Олсуфьев, Дунаева,
1960; Шеханов, 1970). Наибольший интерес представляют материалы
по выяснению роли представителей рода в эпизоотиях бешенства.
Вовлечение куниц (каменной и лесной) в эпизоотии бешенства изве-
стно в Молдавии, на Украине, в Литве (Помирко и др., 1975; Щер-
бак и др., 1978; Селимов, 1978; Буракаускас, Казакявичус, 1979;
Божко и др., 1981). Доля участия куниц в эпизоотиях бешенства
в Литве составила 1,0% от 300 зарегистрированных случаев
бешенства диких животных. На Украине в 1968'—1972 гг. бешен-
ство куниц составило 2,4% от 824 случаев бешенства диких живот-
ных, а в последующие годы повысилось до 7,5%, что, по всей веро-
ятности, связано с целенаправленными поисками при изучении
природных очагов.
Бешенство куниц наблюдалось в ГДР, ФРГ, Швейцарии, Дании,
Бельгии, Австрии, Польше, Чехословакии, Франции, Люксембурге
(Селимов, 1978). Подавляющее большинство случаев бешенства от-
мечено для каменной куницы, что связано с ее большей численно-
стью и приуроченностью местообитаний к людским поселениям. Дан-
ные о вовлечении каменной куницы в эпизоотии бешенства в странах
Европы представлены в табл. 22.
Основное значение в поддержании эпизоотии повсеместно имеют
лисицы. Заболевания куниц обычно наблюдаются через 1—2 месяца
Таблица 22. Участие каменной куницы в эпизоотиях бешенства
Территория Годы Зарегистрировано бешенство Авторы
всего куниц %
ФРГ 1955—1978 59 456 2232 3,7 Schwierz, 1979
ГДР 1962—1968 354 19 5,3 Ulbrich, 1969
Швейцария 1967—1973 2316 64 2,7 Wandeler et al., 1974
Дания 1969—1970 128 6 4,6
138
Carnivora
после выявления бешенства у лисиц (Wandeler et al., 1974; Schwierz,
1979). В годы интенсивной эпизоотии лисьего бешенства увеличива-
лось число регистрируемых случаев бешенства куниц. Так, в ФРГ
(земля Рейнланд-Пфальц) в 1957—1964 гг. ежегодно регистрирова-
лось от 18 до 58 случаев бешенства лисиц (в среднем 40) и за это
время выявлено всего две куницы, больных бешенством. В 1965—
1969 гг. на фоне интенсивной эпизоотии лисьего бешенства (в сред-
нем 225 случаев в год) зарегистрировано 38 куниц, больных бешенст-
вом. В последующее пятилетие (1970—1974 гг.) при снижении
лисьего бешенства до 43—86 случаев в год сократилась и частота
обнаружения бешенства у куниц: всего выявлено 17 больных зверей.
Следующее повышение интенсивности эпизоотии лисьего бешенства,
связанное с прекращением газации их нор, снова сопровождалось
увеличением распространения заболевания среди куниц — до 10,3
случаев в год за 1975—1978 гг. (Schwierz, 1979).
Случаи бешенства куниц наблюдаются круглогодично, довольно
равномерно распределяясь по всем месяцам. Анализ распределения
2232 случаев бешенства куниц установил колебания от 6,8% (де-
кабрь-январь) до 9—10% регистрируемых случаев (апрель-май
и октябрь). Весеннее повышение заболеваемости связано с увеличе-
нием подвижности и контактов куниц в марте — в период ложного
гона.
Экстенсивность заражения куниц бешенством различна в зави-
симости от характеристики исследуемой группы зверей (табл. 23).
Таблица 23. Частота обнаружения бешенства в различных группах
каменных куниц (по Wandeler et al., 1974)
Г руппа Число исследо- ванных зверьков Случаи бешенства
абс. %
Павшие 113 11 9,7
Убиты транспортом на дорогах 36 2 5,5
Убиты при ненормальном пове- дении 139 50 35,9
Добыты на охоте 156 0 0
Наиболее высока она в группе зверьков, убитых при ненормальном
поведении: потере осторожности, агрессивности, при забегах в на-
селенные пункты и т. п. Однако следует отметить, что инфицирован-
ность в этой группе не так велика, как это установлено при анализе
частоты зараженности лисиц, достигающей 86% у зверей, убитых
при подобных обстоятельствах. Эти различия связаны, вероятно,
с выраженной синантропностью каменных куниц в европейской
части ареала. В местностях, где отсутствует бешенство, также
X ищные
139
велико число каменных куниц, убитых вблизи человеческого жилья
(Wandeler et al., 1974).
Обращает на себя внимание большое количество куниц, найден-
ных мертвыми, у которых не установлено заболевания бешенством
и не были выяснены другие причины гибели.
Больные бешенством куницы проявляют агрессивность по отно-
шению к другим животным и человеку. Из 202 каменных куниц, у
которых было установлено бешенство, 80 (40%) были убиты при на-
падении на человека и у 45 (22,5%) наблюдалась агрессивность по
отношению к другим животным (Moegle, Knorpp, 1978).
Характерной особенностью патогенеза бешенства у куниц яв-
ляется меньшая (по сравнению с лисицами и барсуками) частота
проникновения вируса в слюнные железы и меньшая его концентра-
ция. Вирус в слюнных железах обнаруживали у 50% каменных ку-
ниц, и концентрация вируса составляла всего 101’5 MLD/50 на 1 г
ткани.
Эти особенности патогенеза, наряду с невысокой плотностью
населения куниц и свойственным им одиночным образом жизни обус-
ловливают второстепенную их роль в поддержании природных оча-
гов бешенства (Bogel, 1974; Wandeler et al., 1974; Schwierz, 1979).
Род Mustela Linnaeus, 1758 — ласки и хорьки
Наиболее мелкие представители семейства куньих: длина тела
от 13—28 см (ласка) до 45—52 см (степной хорь). Зверьки имеют
однотипное строение тела — стройное гибкое вытянутое туловище,
с короткими конечностями. Мех невысокий, пушистый. За исключе-
нием ласки, все виды имеют промысловое значение и некоторые из
них разводятся на фермах, особенно с большим успехом — амери-
канская норка.
Представители рода распространены в северном полушарии—
от побережий Ледовитого океана, к югу до Северной Африки,
Северной Индии, Малайского архипелага и северной части Южной
Америки.
В роде насчитывают, по разным авторам, 13—20 видов, из них 8,
включая акклиматизированную американскую норку, обитают в
СССР (табл. 24).
Все представители рода — наземные хищники, активно добываю-
щие мелких грызунов, проникая в их норы, подснежные гнезда.
Многие виды (ласки, горностаи, хорьки и др.) в зимнее время регу-
лярно посещают стога и ометы, заселенные грызунами, и существен-
но сокращают в них численность полевок. В зависимости от размеров
хищников находится и величина основной их добычи. Мелкие виды
добывают преимущественно полевок и мышей, убивая их стереотип-
ным приемом — прокусывая тонкие кости черепа. В питании
горностая в пойменных стациях значительное место занимает водя-
ная полевка. Степной хорек добывает сусликов, пищух, хомяков.
140
Carnivora
Таблица 24. Состав и распространение рода Mustela
Вид Число хромо- сом, 2 п Распространение
Горностай Mustela erminea 42 От тундр арктического побережья Евразии и Северной Америки до Северной Африки и север- ной части Южной Америки
Ласка М. nivalis 38 От тундр арктического побережья Евразии и Северной Америки до Северной Африки и степ- ных районов Северной Америки
Колонок М. sibirica 38 Значительная часть Азии, до Северной Индии, п-ова Корея, Японии. К западу до Урала и Куйбышевской области
Солонгой М. altaica 44 Азия, от Средней Азии и Забайкалья до Север- ной Индии, Монголии, Китая, п-ова Корея. В СССР отдельными участками заходит север- ная часть ареала вида: Прибалхашье, Джун- гарское Алатау, Памир, Тянь-Шань, Алтай и др.
Черный (лесной) хорек М. putorius 40 Почти вся Европа (кроме Ирландии и северной части Фенноскандии), лесная и частично лесо- степная и степная зоны европейской части СССР на восток до Урала
Степной (светлый) хорек М. eversmanni 38 Лесостепная и степная зоны Европы и Азии, на восток до Забайкалья, МНР, Северного Ки- тая, Северной Индии
Американский хо- рек М. nigripes 38 Центральная часть Северной Америки—от юж- ных районов Канады до Техаса (включен в Крас- ную книгу)
Европейская нор- ка М. lutreola 38 Основной ареал занимает европейскую часть СССР к востоку до Иртыша и р. Урала. На север доходит до нижнего течения Северной Двины, Печоры. В Северной (Финляндия) и Во- сточной Европе
Американская норка М. vison 30 Большая часть Северной Америки, кроме арк- тического побережья и пустынных районов Те- хаса, Южной Мексики, Аризоны. В Евразии распространилась по всей Фенноскандии, многих районах европейской части СССР, Сибири и Дальнего Востока
Длиннохвостая ласка М. frenata 42 Лесные и лесостепные части Северной Америки
—
Хищные
141
Колонок, горностай и американская норка способны убивать таких
крупных грызунов, как ондатра. Норки, ведущие околоводный
образ жизни, поедают полевок, амфибий, рыб, раков. Птицы яв-
ляются случайной добычей.
Большинству видов свойственны циклические колебания чис-
ленности с 3—4-летним периодом, определяющиеся изменениями
численности грызунов. Особенно резко эта зависимость прослежива-
ется в популяциях горностая в тундровой зоне, где основным и поч-
ти единственным кормом для него являются лемминги. В пойменных
биотопах важное значение для горностая, колонка имеет водяная
полевка, состояние обилия которой определяет движение числен-
ности этих видов хищников (Формозов, 1935; Вайсфельд, 1977 и др.).
Сроки размножения у всех видов значительно изменяются в за-
висимости от широты местности и условий разных лет 5. Наиболее
лабильны сроки размножения ласок. Гон обычно бывает в марте—
апреле, но при обилии полевок ласки размножаются и в другое
время года. Выводки в гнездах находили с мая по январь. Продол-
жительность беременности ласок 34—35 дней.
Самцы горностая способны к оплодотворению с середины февра-
ля до половины июня. Рождение детенышей отмечено с марта до
июля. Продолжительность беременности у горностая колеблется от
240 до 393 дней, что обусловлено чрезвычайно длительной диапаузой
в развитии яйцеклетки, длящейся 8,5—9 месяцев. Развитие эмбрио-
нов после имплантации завершается в шесть недель.
У других видов общая продолжительность беременности колеб-
лется от 33 до 38—44 дней, у американской норки — от 30 до 92
дней. Латентный период длится 8—17 дней. Короткий период разви-
тия эмбрионов после имплантации и небольшие размеры детенышей
обеспечивают необходимую для активной охоты подвижность бере-
менных самок даже при большом количестве зародышей. Среднее
число эмбрионов у представителей рода колеблется в пределах 7—9,
а максимальное достигает 12 — 18—19 (Млекопитающие Советского
Союза, 1967; Терновский, 1977). Лактация продолжается около двух
месяцев. Половозрелости зверьки достигают в возрасте 10—11 меся-
цев.
Существенное влияние на интенсивность размножения оказы-
вает состояние кормовых ресурсов — грызунов. Существуют конку-
рентные взаимоотношения между различными видами, оказывающие
влияние на плотность населения обычно более мелких видов.
Акклиматизация американской норки была начата в СССР в
1933 г. Устойчивые популяции образовались в Белоруссии, Тата-
рии, Башкирии, Алтайском крае, Новосибирской области, Уссу-
рийском крае и Приамурье. Более крупная по размерам и экологи-
6 Подробные сведения о биологии куньих приведены в монографии Д. В. Тер-
невского (1977).
142
Carnivora
чески более пластичная, чем европейская норка, в районах совме-
стного обитания вытесняет ее, так что последней угрожает исчез-
новение (Гайдук, 1979; Терновский, 1980; Туманов, Зверев, 1986).
Описана своеобразная биологическая несовместимость европейской
и американской норок, у которых в эксперименте показана возмож-
ность скрещивания и зарегистрирована беременность, заканчиваю-
щаяся резорбцией эмбрионов. Аналогичное явление отмечено также
при скрещивании колонка и самок европейской норки (Терновский,
1977).
Сведения о распространении болезней у видов рода Mustela, за
исключением зараженности гельминтами, отрывочны. Из гельмин-
тов воздействие на состояние популяции может оказывать заражен-
ность скрябингелезом (Skrjabingylas nasicola), нематодой, локали-
зующейся в лобных пазухах, вызывая патологические изменения
костей — их истончение, вздутие, перфорацию (Контримавичюс,
1966, 1968).
Из бактериальных инфекций у представителей рода обнаруже-
ны: туляремия — у черного и степного хорьков, колонка, горностая,
ласки (Олсуфьев, Дунаева, 1970); псевдотуберкулез — у американ-
ской норки, ласки (Тимофеева и др., 1974; Адаменко и др., 1979);
лептоспирозы — у черного и степного хорьков, ласки, горностая,
колонка (Карасева, Свешникова, 1971). В северных районах Запад-
ной Сибири, в низовьях рек Пура и Таза при серологическом иссле-
довании 84 горностаев антитела к лептоспирам, преимущественно
Srippotyphosa, были выявлены у 55 зверьков (65,5%). У основных
носителей этого возбудителя — полевки-экономки, водяной полевки
и ондатры, которыми питается горностай, антитела обнаруживали
у 8, 10,5 и 16% особей (Валова, 1970).
В природных очагах чумы грызунов мелкие куньи вовлекаются
в эпизоотии, протекающие в популяциях песчанок, малых сусликов,
сурков и полевок (в закавказском высокогорном очаге). Заражен-
ность чумой установлена по бактериологическим и серологическим
исследованиям у степных хорьков в очагах Северо-Западного При-
каспия, в Волго-Уральском песчаном очаге, Тянь-Шаньском, Забай-
кальском, Тувинском, Горно-Алтайском и монгольских очагах. Чу-
ма у ласок регистрировалась в Закавказском высокогорном, Гис-
саро-Дарвазском, Приаральском очагах. У солонгоя чума установ-
лена в Забайкальском очаге, у горностая — в Горно-Алтайском и
в Монголии (Щепотьев и др., 1979; Бурделов, 1982).
Тесный контакт мелких куньих с грызунами и их норами опре-
деляет возможность заражения их чумой при поедании больных и
павших зверьков, а также переход на хищников инфицированных
блох. Так, в высокогорных сурочьих очагах чумы около 13% блох,
найденных на степном хорьке, составляют специфические блохи
сурка (Шварц, Лаврентьев, 1961). В Северо-Западных Кызылкумах
блохи больших песчанок составляли 52% блох, собранных с ласок.
Индексы обилия блох на ласке равнялись весной 6,8, а осенью —
Хищные
143
2,1, повторяя сезонные изменения численности блох на больших
песчанках (Алексеев и др., 1979).
Во многих очагах неоднократно отмечалось выделение чумно-
го микроба от блох, преимущественно свойственных грызунам, сня-
тых с мелких куньих (Варшавский и др., 1982).
А. А. Лавровский, С. Н. Варшавский и др. (1974) подчеркивают
важное значение степных хорьков в транспортировке инфицирован-
ных блох на далекие расстояния вследствие свойственной им высо-
кой подвижности и частой смены участков обитания. Авторы отме-
чают, что в природном очаге Северо-Западного Прикаспия в ряде
участков случаи выделения чумного микроба или выявление анти-
тел у степных хорьков предшествовали развитию здесь эпизоотии
в популяции малых сусликов. На территории Сарпинской низмен-
ности на участке около 30 тыс. га в 1973 г. было выявлено 15 точек,
на которых отловлены хорьки, либо зараженные чумой, либо имею-
щие антитела к чумному микробу. При исследовании 192 хорьков
было выделено 4 культуры возбудителя и у 14 зверьков обнаружены
антитела. Культуры микроба чумы были выделены также от блох,
снятых с хорька (1) и выловленных из нор, занятых хорьком (2).
От самих сусликов в это время (июнь-июль 1973 г.) культур не вы-
деляли. Индекс обилия блох на хорьках в очаге Северо-Западного
Прикаспия составлял 14,3, при этом 68,8% блох приходилось на
блох малого суслика (Топорков и др., 1979).
По многолетним материалам в чумных очагах Приаралья при ис-
следовании 3400 степных хорьков культура возбудителя чумы была
выделена в 0,3%, а при серологическом исследовании специфические
антитела обнаружены у 9,7% хорьков. Среди 1000 исследованных
ласок зараженность составила 0,2%, а антитела обнаружены в 5,6%
(Бурделов, 1982). В Северо-Западном Прикаспии при исследовании
596 степных хорьков специфические антитела были обнаружены у
30 зверьков (5,0%), с колебаниями в разные годы от нуля до 7,1%
(Топорков и др., 1979). В Муюнкумах от 783 степных хорьков за
20 лет было выделено всего 5 культур чумного микроба (Быков,
Цой, 1980). Все исследователи отмечают, что находки инфицирован-
ных или переболевших чумой мелких куньих совпадают по времени
с интенсивными эпизоотиями в популяциях основных носителей.
Эпизоотологическое значение может иметь разнос хищниками по тер-
ритории инфицированных блох, переходящих на хищников с добы-
тых грызунов или при посещении их нор (Бурделов и др., 1983).
Сами хищники не поддерживают эпизоотического процесса при чуме.
Представляет интерес выяснение роли некоторых видов этого
рода в распространении бешенства. Зараженность вирусом бешенст-
ва известна у черного и степного хорьков, колонка, горностая, нор-
ки, ласки (Рудаков, Мальков, 1970; Грибанова и др., 1977, 1981;
Wandeler et al., 1974; Schwierz, 1979).
В степных и лесостепных районах и южной тайге Западной Си-
бири с 1967 по 1976 г. бешенство было установлено у степного хорь-
144
Carnivora
ка, горностая и колонка (Грибанова и др., 1981). За этот период бы-
ло исследовано 165 степных хорьков и выделено 6 штаммов вируса
(в 107 биопробах из мозга). При исследовании мозга 215 горностаев
(в 52 биопробах) выделен один штамм вируса и при исследовании
мозга 250 колонков (в 66 биопробах) получено 4 штамма. Исследова-
ние небольшого числа ласок было отрицательно. Авторы указывают,
что сохранение возбудителя бешенства в природных очагах связано
с парцеллярными группами нескольких видов хищных млекопитаю-
щих, занимающих общую территорию. Наибольшее значение в
сохранении вируса имеют участки территории с высокой степенью
мозаичности биотопов, для которых характерно разнообразие и
стабильная численность различных видов хищных млекопитающих.
В период снижения интенсивности и распространения бешенства
вирус выделяли преимущественно (15 штаммов из 20) от животных,
обитающих в подобных участках и лишь 5 штаммов было выделено
на территории диффузных поселений хищников (Грибанова, Маль-
ков, 1978).
Зараженность степных хорьков, по данным этих авторов, мало
отличалась в годы интенсивной эпизоотии среди лисиц и при низкой
зараженности последних. Но в Казахстане инфицированность
хорьков наблюдалась лишь во время лисьих эпизоотий (Сансызба-
ев, 1981).
В эксперименте установлена малая восприимчивость хорьков
к вирусу бешенства, выделенному от лисицы. Лесные и степные хорь-
ки, зараженные внутримышечно этим штаммов в дозах 30, 300,
3000 и 30000 MLD/50, не проявили признаков заболевания, за иск-
лючением двух особей из 24 бывших в опыте. Один степной хорек
погиб от бешенства на 45-е сутки (доза 30 000 MLD/50) и один лесной
хорек погиб на 49-е сутки (доза 3000 MLD/50).
Иные результаты были получены при заражении лесных хорьков
штаммом, выделенным от горностая. Из семи хорьков, зараженных
дозами от 300 до 30 000 MLD/50, погибло от бешенства шесть. Сроки
гибели колебались от 13 до 129 суток (Грибанова и др., 1981,
1982).
Установлена также малая восприимчивость к вирусу лисьего
бешенства у хорьковМ. putoriusfuro (Blanconet al., 1982). Из сорока
хорьков погибло лишь десять, получивших наибольшие дозы
(разведения ткани мозга 10"1—10"2), причем шесть других зверьков,
получивших те же дозы, остались живы. Гибель наступала на 16—
35-е и 19—58-е сутки. Еще один хорек погиб на 87-е сутки после вве-
дения меньшей дозы. Титры вируса в мозге достигали 4,5—5,9 1g
LD/50/0,03 мл. В слюне заболевших хорьков вирус не обнаружен.
Антитела у выживших были редки и в низких титрах. Авторы счи-
тают, что хорьки являются тупиком для вируса бешенства.
О малой восприимчивости и низком содержании вируса в слюн-
ных железах у ласок и горностая сообщают Ванделер с соавторами
(Wandeler et al., 1974).
Хищные
145
Многолетние наблюдения по эпизоотологии бешенства в Цент-
ральной Европе свидетельствуют о незначительной роли мелких ку-
ньих в поддержании природных очагов бешенства. В ФРГ за 1955—
1978 гг. бешенство лесных хорьков составило 0,2% (153 больных
зверька), а бешенство ласок — всего 0,04 (36 случаев) от всех заре-
гистрированных случаев бешенства диких животных (Schwierz,
1979). В одном из округов земли Гессен через три года после пре-
кращения регистрации лисьего бешенства было исследовано 679
мелких куньих с отрицательным результатом, из чего можно заклю-
чить, что они не поддерживают очаговости бешенства в данных
условиях (Wandeler et al., 1974).
Вирусологическое исследование 459 горностаев, пров ai вшееся
в тундровых и лесотундровых районах Западной Сибири, iai же не
обнаружило среди них инфицированных вирусом тундрового бешен-
ства, которое в это время регистрировалось здесь у песцов (Сюзюмо-
ва, 1965).
Род Gulo Storr, 1775 — росомахи
В роде единственный вид — росомаха Gulo gulo Linnaeus, 1758
(2п=42). Один из самых крупных представителей семейства. Масса
тела И—16 кг, наиболее крупные особи 20—32 кг. Росомахи имеют
массивное, приземистое тело на коротких конечностях. Мех густой,
длинный, с грубой остью. Распространена в тайге, отчасти в лесотун-
дре и тундре Евразии и Северной Америки.
В питании росомах основное значение, особенно в зимний период,
имеют копытные, в основном молодняк и ослабленные, больные
животные. Часто использует остатки добычи волков, медведя, по-
едает падаль. В летнее время значительную часть рациона состав-
ляют мелкие грызуны — лесные полевки, в годы массового размно-
жения — норвежские лемминги. Росомаха поедает ягоды брусники,
голубики.
Биология размножения изучена недостаточно. Спаривание про-
исходит в конце апреля, мае и в конце июня. Рождение детенышей —
в феврале-марте. Беременность, как и у многих куньих, имеет
латентный период. Количество детенышей в помете невелико — обы-
чно 2—3 детеныша. Лактация продолжается около трех месяцев.
Половозрел ости достигают, по-видимому, в возрасте трех лет. Самки
приносят детенышей не ежегодно, и яловость взрослых самок сос-
тавляет 50—60%.
Болезни росомах не изучены. В Таймырской тундре установле-
но заражение росомах бруцеллезом (Забродин и др., 1971), который
они получают при поедании зараженных северных оленей. Зара-
женность диких северных оленей на Таймыре достигает 26,6—33,5%
(Пинигин и др., 1958). Поскольку росомахи способны добывать глав-
ным образом ослабленных, больных животных (Насимович, 1948;
Язан, 1974), санитарное их значение должно оцениваться выше, чем
146
Carnivora
небольшой ущерб, наносимый ими при разорении гнезд наземногнез-
дящихся птиц или «ограблении» охотничьих ловушек и складов.
Однако промысел росомах разрешен в течение круглого года и зверь,
ло традиции, считается вредным.
Род Vormela Blasius, 1884 — перевязки
В роде единственный вид — перевязка Vormela peregusna Giilden-
staedt, 1770, 2n=38. Небольшого размера зверек (длина тела 269—
345 мм, масса тела 370—715 г), по облику сходный с хорьками, свое-
образной яркой, пестрой, черного, белого и желтого цвета окраской.
Населяет сухие степи, полупустыни и пустыни Евразии—от Югосла-
вии до Монголии. Селится в норах грызунов —• сусликов, песчанок,
служащих основной добычей этого хищника. Численность перевяз-
ки невелика и сокращается под влиянием освоения целинных степей,
но в пустынных районах остается высокой. Промысловое значение
ничтожно. Полезна истреблением грызунов — носителей чумы.
Болезни не изучены. Известны единичные находки зараженных
чумой особей (по выделению культуры возбудителя). По серологи-
ческим данным, антитела к возбудителю чумы обнаруживали у
7,88% от 200 исследованных перевязок (Бурделов, 1982).
Описан единственный случай поражения перевязки лейшманио-
зом среди 141 исследованных особей (Файзулин, 1967).
ПОДСЕМЕЙСТВО MELLIVORINAE GILL, 1872
— МЕДОЕДЫ
В подсемействе один монотипический род Mellivora Storr, 1780.
.Mellivora capensis Schreber, 1780 — медоед, или лысый барсук.
Среднего размера зверьки, внешним видом напоминающие барсу-
ка благодаря приземистому телу и роющим передним лапам с длин-
ными когтями. Длина тела 68—75 см, масса тела самок 9 кг, самцы
,достигают 12—16 кг (Млекопитающие Советского Союза, 1967).
Ареал медоеда обширен, охватывает Африку (кроме северных час-
тей), Переднюю Азию, Индию. В СССР встречается только в Турк-
мении, между Каспийским морем и Амударьей. Всеядный хищник,
питающийся преимущественно мелкими грызунами, рептилиями,
саранчовыми, земляными пчелами, их личинками и медом. Практи-
ческого значения не имеет. Сведений о болезнях нет.
ПОДСЕМЕЙСТВО MELINAE BURMEISTER, 1762
— БАРСУКИ
В подсемействе четыре рода: Meles Brisson, 1762 (один вид, рас-
пространенный в Европе, Передней и Средней Азии, южной и средней
полосе Сибири, на юге Дальнего Востока, в Китае, Корее и Японии);
.Arctonyx F. Cuvier, 1825 (один вид, населяющий Китай, Индию,
Хищные
147
Бирму, Индокитай); Taxidea Waterhouse, 1839 (один вид, обитающий
в Северной Америке); Melogale I. Geoffroy, 1834 (три вида, населяю-
щие Непал, Индию, Индокитай, Китай).
Род Meles Brisson, 1762 — барсуки
В роде единственный вид — барсук Meles meles Linnaeus, 1758,,
2п=44. Размеры зверя средние, длина тела 60—90 см. Масса тела
10—16 кг, а при осеннем накоплении жира может достигать 20—
30 кг. Тело массивное, приземистое с короткими конечностями,
снабженными сильными слабо изогнутыми когтями. Широка
распространен на территории СССР от таежных лесов на севере
(до 65° с. ш. в европейской части и до 52° с. ш. на востоке страны)..
На юге ареал выходит за пределы СССР.
Населяет самые разнообразные биотопы во всех ландшафтно-
географических зонах, за исключением тундры. Селится в глубоких
сложных норах, вырывая их самостоятельно. Питается в основном
насекомыми и их личинками, мышевидными грызунами, лягушка-
ми, корневищами и луковицами растений. В северной части ареала
с октября-ноября по февраль—апрель впадает в зимний сон, преры-
вающийся во время оттепелей. В районах с мягкой зимой барсуки
активны в течение всего года.
Спаривание барсуков происходит в марте—июне. Беременность
протекает с длительной диапаузой и продолжается 271—374 дня
(Осмоловская, 1948). Лактация продолжается около трех месяцев.
Половозрел ость наступает у самок в двухлетнем, а у самцов —
в трехлетием возрасте.
Промысловое значение невелико. Довольно широко распростра-
нено использование барсучьего сала как целебного средства. В свя-
зи с этим необходимо упомянуть довольно высокую зараженность
барсуков трихинеллезом. Трихинеллез регистрировали во многих
участках ареала барсука. Частота выявления паразитов колеблется
в значительных пределах. Так, в Казахстане трихинеллез обнаружен
у 0,3% особей из 843, в Эстонии — у 16,6%. В Поволжье из 63 ис-
следованных барсуков трихинеллез установлен у 80%. В общей сло-
жности на всей территории страны исследовано 1118 барсуков, из
которых инвазировано 6% (Бессонов, 1972). Случаи заражения лю-
дей при употреблении мяса и сала барсуков зарегистрированы прак-
тически по всему ареалу обитания барсука в СССР.
Вовлечение барсуков в эпизоотии бешенства установлено на
значительной территории европейской части ареала. По данным
ВОЗ (Черкасский, 1985), в 1977—1982 гг. бешенство барсуков за-
регистрировано в 14 европейских странах. Всего за эти шесть лет в
них диагностировано бешенство у 2022 барсуков. Доля участия бар-
суков в эпизоотии составляла в разные годы от 1,5 до 2,3% от всех
заболевших животных. Частота выявления бешенства у барсуков в
148
Carnivora
Таблица 25. Заболевания бешенством барсуков в странах Европы за
1977—1982 гг. (по данным ВОЗ; Черкасский, 1985)
Страна Число инфицированных В среднем за год
-Австрия 565 94
ФРГ 551 90
Швейцария 360 60
Франция 118 19
.Италия 88 14
.Польша 73 12
ЧССР 62 10
ГДР 55 9
разных странах значительно различается, что, вероятно, связано с
размахом исследований (табл. 25).
В Бельгии, Венгрии, Люксембурге, Румынии, Югославии за эти
годы зарегистрированы единичные случаи бешенства барсуков (Чер-
касский, 1985). В ФРГ за 1955—1978 гг. бешенство барсуков заре-
гистрировано у 1042 особей (1,7% от всех случаев бешенства ди-
ких животных). Основную массу заболеваний среди диких живот-
ных составляли лисицы (55 993 зарегистрированных случая). Эти
.цифры в какой-то мере зависят от количественных соотношений
этих видов: добыча барсуков составляет в среднем 6,8% от числа
.добываемых лисиц, колеблясь в разные годы от 0,9 до 11,2% (Schwi-
erz, 1979). В Швейцарии на долю барсуков приходилось 2,3—4%
зарегистрированных случаев бешенства (Wandeler et al., 1974).
На территории ФРГ бешенство барсуков отмечали круглогодич-
но. Наибольшее число случаев отмечали в марте—июне (10—13%),
меньшее (5,3—6,4%) — ноябре—январе — в период наименьшей
активности зверей. Как и другие виды хищных, заболевшие бешен-
ством барсуки теряют осторожность, чаще попадаются на глаза чело-
веку, однако агрессивность их невелика: из 101 барсука с установ-
ленным заболеванием агрессивны по отношению к человеку были
всего три, а нападали на других животных — девять особей
(Moegle, Knorpp, 1978).
Вирус обнаруживали в слюнных железах у 83% больных особей
-с концентрацией 10 1g MLD/50 на 1 г ткани (Wandeler et al., 1974).
Частота обнаружения бешенства различна в разных группах ис-
следованных животных (табл. 26). Как и у других видов (см. данные
по лисице, куницам), с наибольшей частотой бешенство регистриру-
ется у животных, убитых при ненормальном поведении: потере осто-
рожности, появлении в несвойственных зверю участках и т. п. Наи-
меньшее количество больных зверей регистрируется среди животных,
.добытых на охоте.
Хищные
149
Таблица 26. Частота обнаружения бешенства в различных группах
барсуков (по Wandeler et al., 1974), Центральная Европа
Группа Число иссле- дованных Из них инфицировано
абс. %
Павшие 72 32 44,0
Убиты транспортом на дорогах 24 1 4,1
Убиты при ненормальном поведении 106 64 60,0
Добыты на охоте 119 3 2,5
Причины гибели барсуков, у которых не обнаружено заболева-
ния бешенством, обычно остаются невыясненными. По наблюдениям
в Англии, среди них отмечается довольно значительная заражен-
ность бактериями туберкулеза бычьего типа (см. ниже).
Участие барсуков в эпизоотиях бешенства находится в тесной
связи с распространением бешенства в популяции лисиц. Так, по
наблюдениям в ФРГ, на земле Рейнланд-Пфальц бешенство барсуков
регистрировалось с 1957 по 1978 г. в количестве от одного до девяти
случаев в год, что составляло от 0,5 до 6,5% от всех случаев бешен-
ства диких животных. В 1965—1969 гг. во время высокой интенсив-
ности эпизоотии среди лисиц (более 200 случаев ежегодно) было за-
регистрировано 19 больных бешенством барсуков. В период сниже-
ния бешенства лисиц до 48—86 случаев в год (1970—1974 гг.) обна-
ружен лишь один барсук, больной бешенством. Следующий подъем
заболеваемости лисиц вызвал повышение числа бешеных барсуков:
за четыре года (1975—1978) заболевание установлено у 19 зверей
(Schwierz, 1979).
Такая же зависимость частоты обнаружения бешенства у барсу-
ков от интенсивности эпизоотии в популяции лисиц прослежена на
территории Словакии. При зараженности лисиц в 27,5% (1966—
1968 гг.) бешенство было выявлено у 12 барсуков из 60 исследован-
ных (20%). В последующие семь лет зараженность лисиц колебалась
от 3,5 до 8,3%, а при исследовании 196 барсуков бешенство установ-
лено у 7 особей, т. е. у 3,5% (Matouch et al., 1976).
Исследования, проведенные главным образом в ФРГ, дают осно-
вания для вывода о значительном влиянии эпизоотии бешенства на
численность барсуков. Показано, что эпизоотия снижала числен-
ность барсуков на юго-западе ФРГ до 50% от первоначальной,
а в некоторые округах и более. Особенно губительно действует на
численность барсуков бешенство в соединении с газацией нор, широ-
ко проводившейся в 1965—1974 гг. (Wachendorfer, Schwierz, 1978).
Численность барсуков сокращается больше, чем лисиц, у которых
она быстро восстанавливается, в то время как у барсуков на про-
150
Carnivora
тяжении нескольких лет остается на низком уровне (Bogel, 1974;
Bogel et al., 1978; Schwierz, 1979).
В СССР участие барсуков в эпизоотиях бешенства зарегистриро-
вано в Эстонии — 18 случав за 1969—1976 гг., в Литве — 29 слу-
чаев за тот же период, в Латвии — 21, на Украине — 18, в Белорус-
сии — 10, в Западной Сибири — 2. На долю барсуков приходилось
1,6—3,8% от всех случаев заболевания бешенством диких животных
(Селимов, 1978). Случаи заражения людей от укусов барсуков сос-
тавляли на Украине 2,3%; в редких случаях барсуки были источни-
ком заражения сельскохозяйственных животных (Щербак и др.„
1976, 1978; Гришок, 1978).
Из других заболеваний у барсуков известны лишь немногие
случаи заражения спирохетозом, чумой грызунов, эризипелоидом
(Байрамова, 1964; Шеханов, 1970).
Представляет интерес сообщение о зараженности барсуков ту-
беркулезом (Muirhead et al., 1974). Исследования проведены в
Глостершире (Великобритания), где с апреля 1971 г. по декабрь
1973 г. возбудитель туберкулеза бычьего типа был выделен от 36-
из 165 исследованных барсуков (21,8%). Из этого числа 17 зверей
найдены мертвыми и из них заражено было 9 (53%). Среди 70 барсу-
ков, погибших от транспорта на дорогах, инфицировано было 15,7%.
Также высока была зараженность барсуков, добытых на охоте,—
16 из 78 (20,5%). Авторы связывают зараженность барсуков тубер-
кулезом с случаями заболевания крупного рогатого скота, указы-
вая на территориальную связь обнаружения инфекции.
ПОДСЕМЕЙСТВО MEPHITINAE GILL, 1872
— СКУНСЫ
В подсемействе 3 рода и 10 видов, распространенных в Северной
и Южной Америке (табл. 27, рис. 7).
Таблица 27. Состав и распространение подсемейства Mephitinae
Род Число ВИДОВ Распространение
Mephitis G. Cuvier, 1800 ч 2 М. mephitis—Северная Америка—от южных провинций Канады до Северной Мексики М. macroura — юго-западные штаты США, Мек- сика, Центральная Америка
Spilogale Gray, 1865 1 S. putorius — от Британской Колумбии в Канаде, по всей территории США до центральных райо- нов Коста-Рики
Conepatus Gray, 1837 7 Южная Америка — 6 видов. Один вид (С. meso- leucus) распространен на юго-западе США, в Мексике, Центральной Америке (до Никарагуа)
Хищные
151
Небольшие звери, имеющие
длину тела до 30—49 см, масса
тела до 1 кг (S. putorius — пят-
нистый скунс), 2,5 кг (М.
mephitis — полосатый скунс) и
до 4,5 кг (С. mesoleucus — сви-
ноносый скунс). Шерстный пок-
ров высокий, густой, пушистый,
.хвост одет длинными волосами.
Характерна яркая окраска с
светлыми полосами, пятнами, а
у свиноносого скунса — в ви-
де чепрака, идущего от лба по
спине до кончика хвоста. Очень
сильно развиты прианальные же-
лезы с резко пахнущими выде-
лениями, которые раздраженные
или испуганные зверьки могут
выбрасывать на некоторое рас-
стояние.
Населяют разнообразные био-
топы, встречаясь в лесах, кус-
тарниковых зарослях, открытых
биотопах, нередко возле челове-
ческого жилья, на фермах и т.
п. Ведут ночной или сумереч-
ный образ жизни. Всеядны: в
питании большое место зани-
мают насекомые, растительная
Рис, 7. Р аспространение подсемейст-
ва Mephitinae
пища, зимой и ранней весной чаще добывают грызунов.
Спаривание в конце зимы, продолжительность беременности у
свиноносого скунса — 42 дня, у полосатого — 63, пятнистого —
120 дней. Детеныши родятся слепыми, прозревают на 32-е сутки,
лактация продолжается около двух месяцев. Продолжительность
жизни до 8—10 лет.
У скунсов, преимущественно полосатого (М. mephitis), заре-
гистрировано заражение туляремией, листериозом, лептоспироза-
ми, лихорадкой Скалистых гор, бешенством (Infectious diseases,
1970). Серологические исследования 311 полосатых скунсов, прове-
денные в штатах Джорджия и Флорида, выявили антитела к микробу
туляремии у 79 особей (25,4%) и у двух из восьми пятнистых скун-
сов (Me Keever et al., 1958). Титры антител варьировали от 1 : 20 до
1 : 5120. Тогда же при исследовании 132 полосатых скунсов были
выделены 5 штаммов лептоспир ballum и 1 штамм pomona. В штате
Луизиана серологическое исследование 650 полосатых скунсов
выявило антитела к 8 серотипам лептоспир у 57% животных. Пре-
обладали реакции с лептоспирами pomona — 21%, для которых
152
Carnivora
полосатые скунсы являются основными хозяевами среди диких жи-
вотных: от них выделено 82% всех штаммов этой серогруппы (Свеш-
никова, Карасева, 1965). Среди других серотипов, обнаруженных у
скунсов в Луизиане, 19% отнесены к лептоспирам hyos hyos, 10%—
ballum, 6% — canicola, 2% — icterohaemorrhagiae и единичные
реакции отмечены с лептоспирами grippotyphosa u australis. В шта-
те Пенсильвания у скунсов обнаружены антитела к лептоспирам
рошопа и ballum. В Канаде — к pomona (Infectious diseases, 1970).
Важное значение скунсы имеют в эпизоотологии бешенства, из-
вестного у них с 1826г. В XIX в., в период интенсивного освоения
территории Северной Америки, неоднократно отмечались случаи
заболевания поселенцев гидрофобией, вызванные укусами скунсов.
Звери нередко кусали людей, спящих или отдыхающих на земле в
палатках, под тентами. Для защиты от нападения скунсов применя-
ли специально сконструированные тенты с бортиками (Parker, 1975).
Первый случай заражения бешенством от скунса зарегистрирован
в 1826 г. в Калифорнии. В 1858 г. погибла от гидрофобии девочка,
укушенная пятнистым скунсом в штате Техас. За 6 лет здесь заре-
гистрировано 4 случая гидрофобии, вызванной укусами этого зверя.
В 1866—1876 гг. в Канзасе было 17 случаев гидрофобии, связанной
с скунсами. Заражения от скунсов наблюдались и в тех местностях,
где не было бешенства собак (Johnson, 1971).
В начале XX в. в Аризоне зарегистрировано 10 случаев зараже-
ния людей от скунсов в течение трех лет (1907—1910 гг.).
До середины столетия наблюдались лишь спорадические случаи
бешенства скунсов. Так, например, в штате Нью-Йорк единичные
бешеные скунсы зарегистрированы в 1929, 1934, 1937 гг. С 1952 г.
наблюдалось много локальных эпизоотий бешенства среди полоса-
тых скунсов. В 1969 г. бешенство скунсов зарегистрировано в 32
штатах, причем во многих местностях они имели ведущее значение
среди всех больных животных. Так, в Калифорнии наблюдалось
бешенство 241 скунса, 8 лисиц и 3 собак; в Вайоминге — 54 скунсов,
одной кошки и одной коровы, а в некоторых участках — только у
скунсов (Johnson, 1971).
Возможной причиной распространения бешенства в популяциях
скунсов было снижение охотничьего промысла, вызванное второй
мировой войной и уменьшением стоимости шкурок. Отмечено, что к
1950 г. в ряде штатов добыча скунсов уменьшилась на 10% по срав-
нению с 1940 г., а число случаев бешенства увеличилось в 10 раз
между 1948 и 1960 г. (Parker, 1975).
С 1953 по 1972 г. в США зарегистрировано 24 021 случай бешен-
ства скунсов (преимущественно полосатого), что составило 53%
заболеваемости диких животных без летучих мышей. На долю лисиц
приходится 41% регистрированных случаев бешенства (Parker,
1975). Ежегодно регистрировалось от 319 до 2095 бешеных скунсов,
в среднем за 20 лет — 1200 случаев в год. При этом следует отметить
значительное повышение числа больных скунсов с 1961 по 1972 г.—
Хищные
153
в среднем по 1554 зарегистрированных случаев бешенства против
671 в год за 1953—1960 гг.
Заболевания бешенством пятнистых скунсов наблюдаются реже,
чем полосатых, что может быть связано с более скрытным, преиму-
щественно ночным образом жизни и меньшей вероятностью встречи
их с людьми. Значение бешенства у других видов ограничивается
малой плотностью населения и меньшей площадью ареалов.
В южных провинциях Канады, где распространены полосатые
скунсы, бешенства у них не отмечалось до 1957 г., когда был диаг-
ностирован первый случай заболевания полосатого скунса в провин-
ции Онтарио (Gunson et al., 1978). С 1961 по 1969 г. примерно 16%
случаев бешенства в Онтарио приходилось на долю скунсов (Parker,
1975). Бешенство скунсов получило широкое распространение в
Саскачеване, где в 1970—1976 гг. при исследовании 1281 скунса
бешенство было установлено у 654 особей (51,1 %). В то же время в
соседней провинции Альберта бешенство выявлено лишь у 16 из
487 (3,2%) исследованных скунсов (Gunson et al., 1978). В 1970—
1971 гг. была принята программа контроля за уменьшением плот-
ности населения скунсов для предотвращения распространения
эпизоотии.
По данным за 1968—1977 гг. в Канаде зарегистрировано бешен-
ство у 2683 полосатых скунсов, что составило 36,4% от всех случаев
бешенства у диких животных. Доля лисиц составляла 25,8%
(Bradley, 1979).
В литературе укоренилось представление о том, что скунсы менее
восприимчивы к вирусу бешенства, чем лисицы (Kaplan, 1969;
Johnson, 1971; Канторович, 1971; Черкасский, 1985). Это пред-
ставление основано на опыте Сайкса (Sikes, 1962), в котором мини-
мальная заражающая доза составляла для скунсов 1400 MLD/50,
для лисиц (обыкновенной и серой) — всего 14 MLD/50. В опыте был
использован штамм вируса, выделенный от лисицы. Следует отме-
тить, что все животные предварительно были иммунизированы про-
тив собачьей чумы и гепатита, что могло исказить их реактивность.
При использовании для заражения штамма вируса, выделенного
от скунса, доза менее чем 10 MLD/50 вызывала гибель скунсов так
же, как и лисиц (Parker, 1975). Минимальная полная смертельная
доза равнялась 1000 MLD/50 (табл. 28).
В зависимости от дозы заражения гибель скунсов наступала на
22—177-й день, в среднем на 27—84 день. Наибольшая продолжи-
тельность заболевания в 105, 137 и 177 дней наблюдалась при мень-
ших дозах заражения. Характерна для скунсов значительная про-
должительность клинических проявлений болезни — до 12—18
дней, в среднем 7—9 дней. В слюнных железах у скунсов вирус обна-
руживали за 18 дней до гибели и по меньшей мере за восемь дней до
появления клинических признаков бешенства. Титры вируса в слю-
не были высокими — 104’3—105’9, а в слюнных железах у погибших
Ю5’4—106’8.
154
Carnivora
Таблица 28. Результаты заражения скунсов вирусом бешенства
(по Parker, 1975)
Дозы MLD/50 Заражено Пало Дни гибели после заражения Длительность клиниче- ских проявлений (дней)
от —до среднее от —до среднее
10 5 2 32—137 84 1—7 4
100 6 5 30—177 56 4—12 9
1000 6 6 32—65 48 3—18 7,5
10000 6 6 22—37 28 1—12 7
100 000 6 6 25—59 27 4-11 7
Вирус в слюне находили у 15 скунсов из 20 (75%). Но из 25 по-
гибших скунсов вирус в слюнных железах найден лишь у восьми.
При естественном пути передачи во время эпизоотии в колонии
недавно пойманных скунсов, инкубационный период, прослеженный
у 10 особей, колебался от 19 до 41 дня, в среднем 29 дней. (Возмож-
но, что у двух скунсов он был короче — 11 дней, а в одном случае
вероятный контакт мог быть более отдаленным — 71 или 63 дня.)
Эпизоотия в вольерах продолжалась с конца мая до 22 октября.
Самка, положившая начало эпизоотии, была отловлена в марте,
имела гемагглютинины к вирусу бешенства, 2 мая родила, а 22 мая
погибла от бешенства (Gough, Niemeyer, 1975).
Особенности поведения в природе прослежены у 473 бешеных
и 90 здоровых особей. Установлено, что как больные, так и здоровые
звери не переходили на большие расстояния, оставаясь на одном
месте до двух недель, прежде чем были убиты. Максимальное рас-
стояние, пройденное за один день,— 4 мили (Schnurrenberger et al.,
1970). Большинство больных скунсов проявляли агрессивность или
отсутствие страха. Активное нападение на собак наблюдали у 50 бе-
шеных (10,6%) и 9 здоровых скунсов (10%). 35 бешеных скунсов
нападали на человека, 22 — на кошек и 35 — на домашний скот и
машины. 80 больных зверей и 8 здоровых не проявляли страха. При
этом 25 больных были настроены дружелюбно, а два скунса даже
играли с собаками. Отмечено много случаев нападения собак на
скунсов, в том числе в 96 случаях — на бешеных. У 76 бешеных скун-
сов (16%) наблюдалась потеря координации движений, 36 были най-
дены в состоянии летаргии, паралича или конвульсий.
Заболевание изменяет поведение скунсов. В дневные часы встре-
чено 148 бешеных и 8 здоровых зверьков. 45 бешеных и лишь один
здоровый скунс попались на глаза наблюдателям вблизи населен-
ных пунктов.
Бешенство скунсов регистрируется в течение всего года, с наи-
большей частотой — в весенние месяцы. Из 789 случаев бешенства
Хищные
155
полосатых скунсов, проанализированных в Иллинойсе, на декабрь—
февраль приходилось 3,0; 2,4; 5,5%. Наибольшая частота случаев
•отмечена в марте—июле (13,7; 19,3; 11,3; 9,9; 11,0%). В августе—
ноябре зарегистрировано 6; 8; 4,6; 5,7; 6,8% случаев заболевания
(Schnurrenberger et al., 1970).
Интенсивность эпизоотического процесса в популяциях скунсов
характеризуется следующими цифрами: в штате Иллинойс с 1958
по 1968 г. зараженность установлена у 8—9% зверьков из 577,
добытых охотниками (Parker, 1975). Но при специальных исследо-
ваниях в 1965—1968 гг. бешенство было установлено у 497 скунсов
из 808 (61,5%) (Schnurrenberger et al., 1970). В Саскачеване (Канада)
бешенство обнаруживали у 28—65% зверьков, а в провинции
Альберта — у 1,7—7,3% (Gunson et al., 1978).
Хотя скунсы составляют 60—70% от больных бешенством диких
животных, только 2% людей, обращающихся за профилактическими
прививками, имели контакт с скунсами, подозрительными на бешен-
ство. Из 250 случаев гидрофобии, зарегистрированных в США за
1946—1973 гг., скунсы были источником заражения в 9 случаях
(3,6%), но составляли 30% от числа всех диких животных (25),
ставших причиной заболевания людей.
Для сокращения численности скунсов в местах развития эпизоо-
тии бешенства используется отлов зверьков в ловушки и пере-
дача их в специальные пункты, раскладывание отравленных прима-
нок в виде куриных яиц (с эмбрионами), наполненных микстурой
из меда, стрихнина и метиленовой синьки (Parker, 1975).
ПОДСЕМЕЙСТВО LUTRINAE BAIRD, 1857
— ВЫДРЫ
В подсемействе 5 родов, с 16—17 видами. Наиболее обширен род
собственно выдр — Lutra Brisson, 1762, объединяющий И—12 ви-
дов. Представители рода обитают в Европе, Азии, Африке, Северной
и Южной Америке. Монотипичные роды: Pteronura Gray, 1837 (Юж-
ная Америка), Paraonyx Hinton, 1921 (Африка) и Enhydra Fleming,
1822 (побережья Тихого океана). В роде Aonyx Lesson, 1827, два
вида, обитающие в Африке и Юго-Восточной Азии.
В СССР обитает два вида — речная выдра Lutra lutra Linnaeus,
1758, и калан Enhydra lutris Linnaeus, 1758.
Выдра — L. lutra L. Ареал охватывает всю Евразию, за исклю-
чением тундровой зоны и пустынных междуречий Средней и Цент-
ральной Азии, Аравийского полуострова, Южного Ирана, Пакис-
тана и Индии. Изолированные участки ареала есть на самом юге
Индостана, в Шри-Ланке и в Северо-Западной Африке.
В СССР распространена почти по всей стране, кроме тундры,
безводных пустынь и полупустынь. Селится выдра по берегам рек,
пресноводных озер, водохранилищ и каналов. Плотность населения
выдр невелика. Хозяйственная деятельность, особенно вырубка ле-
156
Carnivora
сов, зарегулирование речного стока и т. п. вызывают исчезновение
выдры в ряде учстков ареала (Вайсфельд, 1977).
Сведения о болезнях выдры ограниченны. Имеются сообщения о
единичных случаях заражения выдр эризипелоидом, туляремией,
псевдотуберкулезом и сальмонеллезом на Сахалине (Тимофеева
и др., 1969,1974; Беньковскийи др., 1973). Возбудители были выде-
лены от павших или истощенных животных. В одном случае установ-
лена смешанная инфекция — туляремии и псевдотуберкулеза.
Заражение псевдотуберкулезом отмечали также в Швеции (Borg,
1967). В ГДР установлено бешенство у выдры, найденной мертвой
в феврале 1980 г. в местности, где в течение зимы наблюдали бешен-
ство 10 лисиц, двух куниц и одного хорька (Wilhelm, Vogt, 1981).
Калан — Е. lutris L. (2п=38). В СССР обитает на Командорских
островах, юге Камчатки и на Курильских островах. Вне СССР —
у берегов Японии, на островах Алеутских и Прибылова и в не-
скольких пунктах Тихоокеанского побережья до юга Калифорнии.
Адаптация к обитанию в водной среде резко выражена. Звери отлич-
но плавают, ныряют, проводя большую часть времени в воде, вклю-
чая отдых и сон. Питаются морскими беспозвоночными. Сведений
о заболеваниях каланов в доступной литературе нет.
СЕМЕЙСТВО VIVERRIDAE GRAY, 1821
— ВИВЕРРОВЫЕ
В семействе 6 подсемейств, 35 родов, включающих 71 вид.
Распространение охватывает Южную Европу, Африку, Мадагаскар,
Южную и Юго-Восточную Азию, включая Индонезию и Филиппины
(рис. 8).
Небольшие звери, длина тела от 17 до 100 см, масса от 700 г
до 15 кг. По облику сходны с куньими или кошачьими. Встречают-
ся наземные, хорошо лазающие и полуводные формы. Питаются
различными мелкими позвоночными и беспозвоночными, а также
растительной пищей.
Некоторые виды акклиматизированы (еще в XIX в.) в различных
районах, например гималайская цивета (Paguma larvata) — в Япо-
нии, малая цивета (Viverricula indica) — на Мадагаскаре, Комор-
ских островах, Филиппинах. Мангусты — род Herpestes — акклима-
тизированы на Гавайских островах и на островах Вест-Индии
(Пуэрто-Рико, Гренада, Куба и др.).
В большинстве родов (23) — по одному виду, в 10 родах — по
2—4 вида и только род генет (Genetta) и мангустов (Herpestes)
содержат соответственно 9 и 14 видов.
Биология большинства видов не изучена. Длительность бере-
менности известна у немногих видов: примерно 60 дней. В помете
от 1 до 6 детенышей. У некоторых видов бывает по 2 помета в год.
Некоторые виды живут вблизи человека и даже в его жилищах.
Известны легко приручающиеся виды. Хозяйственное значение нич-
Хищные
157
Рис. 8. Распространение семейства Viverridae
тожно. У некоторых видов секрет прианальных желез используют
в парфюмерии. Акклиматизация мангустов обычно имеет целью ист-
ребление крыс и змей, но размножившиеся в местах интродукции
зверьки нередко приносят вред истреблением мелких млекопитаю-
щих и птиц, а также распространением бешенства.
Болезни не изучены. Имеются сведения о заболевании бешен-
ством представителей родов Genetta и Cynictis в Африке (Barnard,.
1979) и рода Herpestes в Индии, Северной Нигерии и в местах интро-
дукции: на Кубе, в Доминиканской Республике, в Пуэрто-Рико и
Гренаде, в Панаме (Sikes, 1970; Everard et al., 1972, 1974; Сели-
мов, 1978).
В Южной Африке из 154 заболеваний бешенством диких живот-
ных 116 случаев (75,5%) приходилось на долю желтых мангустой
С. penicillata (Barnard, 1979). Эти небольшие зверьки, распростра-
ненные в Южно-Африканской Республике, Ботсване, Намибии и
Анголе, ведут колониальный образ жизни, устраивая сложные норы,
в которых может обитать 50 и больше особей. Несомненно, что это
способствует распространению среди них эпизоотии бешенства.
Египетский мангуст Н. ichneumon, ареал которого простирается
от Испании и Португалии до юга Африки, отмечен как хозяин виру-
са бешенства в Израиле, Зимбабве, Ботсване. В 1974 г. бешенство
мангустов в Южной Африке составило 40% от общего числа случаев»
158
Carnivora
бешенства животных (Селимов, 1978). В Эфиопии случаи гидрофобии,
вызванные укусами мангустов, составили 25,7% из 66 заболеваний,
•связанных с укусами диких животных (Fekadu, 1981).
Первая эпизоотия в Западном полушарии среди акклиматизи-
рованных индийских мангустов Н. auropunctatus была зарегистри-
рована в 1950 г. в Пуэрто-Рико (Tierkel et al., 1952). В 1969 г.
здесь установлен 21 случай бешенства у мангустов и 10 заболеваний
домашних и сельскохозяйственных животных (Селимов, 1978).
С 1952 г. известно бешенство в Гренаде, ранее здесь не выявляв-
шееся (Everard et al., 1972). С 1952 по 1967 г. здесь клинически и ла-
бораторными исследованиями подтверждено 359 случаев бешенства,
в том числе 143 (39,8%) у индийских мангустов, ввезенных на остров
еще в 1870 г. Бешенство собак и сельскохозяйственных животных в
указанные годы составляло 24,5 и 27,8%. В последующие годы
<1968—1972) из 389 зарегистрированных случаев бешенства 308
(79,2%) приходилось на долю мангустов, в то время как собачье
бешенство снизилось до 4,8%, а сельскохозяйственных живот-
ных— до 12,8% (Everard et al., 1974).
Контакты с мангустами, подозрительными на бешенство, со-
ставили более половины (56,1%) от всех случаев обращения людей
за антирабической помощью. Ежегодно от 11 до 22 человек было по-
кусано мангустами (в среднем за 5 лет — 15,6), тогда как контакты
с подозрительными на бешенство собаками в среднем равнялись
6,6 случаев в год.
В 1968—1972 гг. среди 6597 мангустов, отловленных капканами,
зараженность вирусом бешенства колебалась от 0,5 до 3,7%. Среди
мангустов с агрессивным поведением бешенство подтверждено у
'21% и по клиническим признакам — у 13,8%.
Среди отловленных мангустов случаи гибели от бешенства на-
блюдали через 2, 7, 16 и 31 день после поимки. У погибших мангустов
вирус обнаруживали в мозге, слюнных железах, в легких (у 6 осо-
бей из 7) и в одном случае — в почках. Титр вируса был значительно
выше в слюнных железах, чем в головном мозге,— 2—4,7 lg MLD/50
и 1—1,5 lg MLD/50 соответственно.
При исследовании 672 мангустов у 127 (18,9%) обнаружены сыво-
роточные вируснейтрализующие антитела. Титры антител варьиро-
вали от низких 1 : 2—1 : 20 у 48,8% зверьков до 1 : 1000—1 : 6000,
выявленных у 7,1% особей. Из 13 мангустов, не имевших клиниче-
ских признаков заболевания, но с доказанным присутствием виру-
са в головном мозге, антитела были обнаружены лишь у одного в
низком титре — 1 : 11. Возможно, что иммунизация части животных
в популяции оказывает влияние на интенсивность эпизоотического
процесса (Everard et al., 1974).
Хищные
159-
СЕМЕЙСТВО HYAENIDAE GRAY, 1869
— ГИЕНОВЫЕ
В семействе различают два подсемейства — Hyaeninae Mivart,
1882, объединяющее два рода: пятнистых — Crocuta Каир, 1828-
(1 вид) и полосатых гиен — Hyaena Brisson, 1762 (2 вида) и Proteli-
nae Mivart, 1882 — земляных волков с одним видом.
Представители семейства распространены в Африке (все 4 вида),
и в Передней, Средней и Южной Азии — полосатая гиена Н. hy-
aena Linnaeus, 1758. В СССР полосатая гиена распространена в
Азербайджане, Восточной Грузии, Копетдаге (в Бадхызе), в южной,
части Узбекистана и в полупустынях Южного Таджикистана.
Гиены имеют плотное телосложение, с массивной головой и мощ-
ными челюстями, крупными коренными зубами. Размеры тела у
более крупной пятнистой гиены колеблются от 127 до 165 см, мас-
са тела достигает 59—82 кг. Полосатые гиены меньше — длина тела
90—120 см, масса — 27—54 кг. Земляной волк — Proteles crista-
tus Sparrman, 1783 — наиболее мелкий представитель семейства:,
длина тела 55—80 см. Зверь имеет узкую морду со слабыми челюс-
тями и зубами, что связано с питанием насекомыми, их личинками
и мелкими млекопитающими, птицами и их яйцами.
Гиены в основном трупояды, поедающие падаль или остатки
добычи крупных хищников, что определяет их положительную роль
как санитаров.
Болезни не изучены. Имеются сообщения о зараженности гиен
(вид не указан) сибирской язвой во время эпизоотии среди копытных
в Южной Африке (Infectious diseases, 1970). Единичные заболевания
гиен бешенством известны в Малайзии, Кении, Танзании, Южной
Африке (Rollinson, 1956; Селимов, 1978). В Эфиопии в 1964—1975 гг.
из 487 человек, имевших контакт с дикими животными и обративших-
ся за антирабической помощью, 63 (12,9%) были укушены гиенами.
Источником заболевания людей гидрофобией гиены были в 18 слу-
чаях из 66, зарегистрированных за эти 12 лет (Fekadu, 1981).
СЕМЕЙСТВО FELIDAE GRAY, 1821
— КОШАЧЬИ
В семейство входят звери разнообразной величины (от домашней
кошки до льва), типично кошачьего облика, с гибким стройным ту-
ловищем, сильными конечностями с острыми втяжными (кроме
гепарда) когтями. В составе семейства разные авторы выделяют
от двух до 22 родов (Соколов, 1979). По современной систематике
в семействе четыре рода: Panthera Oken, 1816 — большие кошки
(4 вида), Uncia Gray, 1854 — снежные барсы (1 вид), Felis Linnaeus,
1758 — кошки (30 видов), Acinonyx Brookes, 1828 — гепарды (1 вид)
(Млекопитающие Советского Союза, 1972). До детальной ревизии
семейства это деление признано наиболее целесообразным. Однако’
поскольку между всеми кошками можно найти переходные формы,
160
Carnivora
ближе к естественной системе находится включение всех видов,
кроме гепарда, в один род — Felis (Allen, 1938; Новиков, 1956;
Соколов, 1979).
В противоположность этому взгляду высказывается предложе-
ние о выделении рысей в отдельный род Lynx с пятью видами (Ho-
nacki et al., 1982).
Диплоидное число хромосом у большинства исследованных ви-
дов равно 38, лишь у нескольких видов рода Felis, распространен-
ных преимущественно в Центральной и Южной Америке, 2п=36
(Орлов, 1974).
Представители семейства встречаются на всех материках, кроме
Антарктиды, Австралии и некоторых океанических островов, насе-
ляя разнообразные ландшафты — леса, пустыни, горы, приречные
долины, кроме тундры.
Все представители семейства — типичные хищники, активно
преследующие живую добычу — от мелких грызунов до крупных ко-
пытных, в соответствии с собственными размерами. Добычу скра-
дывают или подстерегают. Держатся звери поодиночке или семей-
ными группами. Плотность населения невелика, численность неко-
торых видов, особенно больших кошек, во многих частях ареала
резко сокращается вплоть до полного исчезновения.
Род Panthera Oken, 1816 — большие кошки
Включает четыре вида: Р. leo L., 1758 — лев, Р. tigris L., 1758 —
тигр, Р. pardus L., 1758 — леопард и Р. onca L., 1758 — ягуар.
Представители рода распространены в Африке (лев, леопард), в
южных районах Азии — от Ирана до Приморья — ив Индостане,
Индокитае (тигр, леопард). В Мексике, Центральной и Южной
Америке распространен ягуар. В Индии сохранилось небольшое
число львов (Р. leo persicus), занесенных в Красную книгу МСОП.
В Красную книгу МСОП включен обитающий в СССР амурский тигр,
численность которого на Дальнем Востоке оценивается примерно
в 150—200 особей (Животченко, 1981), в то время как в 30—40-х
годах здесь оставалось всего около 30 тигров (Капланов, 1948).
Основой питания крупных кошек служат копытные животные.
На Дальнем Востоке тигры добывают преимущественно кабана и
изюбря, реже —• пятнистого оленя, косулю (Капланов, 1948; Живот-
ченко, 1979, 1981). В течение года взрослый тигр добывает 30—36
особей крупных копытных животных.
Сведения о болезнях в природе скудны. Имеются сообщения о
заболевании сибирской язвой львов, леопардов (а также гепардов)
во время крупной эпизоотии многих видов копытных в 1959—-1960 гг.
на территории Крюгер-парка (ЮАР) и в других районах Южной
Африки (Infectious diseases, 1970). М. В. Шеханов (1970) указывает
на заболевание бешенством леопарда (по зарубежным данным).
Хищные
161
При содержании в зоопарках наблюдались заболевания псевдо-
туберкулезом, листериозом (лев, леопард), токсоплазмозом, кошачь-
им инфекционным энтеритом (Infectious diseases, 1970).
Род Felis Linnaeus, 1758 — кошки
В роде 30 видов (Млекопитающие Советского Союза, 1972).
Это типичные кошки различной величины — от мелких (масса тела
около 2 кг) до крупных, масса тела которых достигает 105 кг (пума).
Распространение соответствует ареалу семейства. Кошки населяют
различные ландшафты—от пустынь до северной границы леса, ведут
одиночный образ жизни, хорошо лазают по деревьям. Питаются
преимущественно мелкими позвоночными — грызунами, птицами.
Реже добывают зайцев, молодняк копытных животных.
Беременность у большинства видов — около 60 дней, у рыси —
67—74 дня. Лактация продолжается 2—4 месяца. Половозрелость
наступает на втором году жизни (Млекопитающие Советского Союза,
1972).
Многие виды этого рода имеют широкое распространение. В СССР
обитает семь видов: F. euptilura Elliot, 1871 — амурская кошка,
F. haus Giildenstaedt, 1776 — камышовая кошка, F. silvestris
Schreber, 1777 — дикая кошка, F. lynx Linnaeus, 1758 — рысь,
F. caracal Schreber, 1776 — каракал, F. margarita Loche, 1858 —
барханная кошка и F. manul Pallas, 1776—манул. Наиболее широко
распространена рысь, населяющая лесные районы на всей террито-
рии СССР. В настоящее время вследствие неблагоприятного воздей-
ствия антропогенных факторов на большей части ареала распростра-
нение имеет очаговый характер.
В лесных районах юга Украины, Молдавии, на Кавказе и в ту-
гайных зарослях по долинам рек Средней Азии и Казахстана распро-
странена дикая кошка 6, ареал которой вне СССР занимает Африку,
значительную часть Европы и Азию до Северо-Западного Китая.
Распространение в СССР остальных видов также представляет лишь
часть обширных ареалов, занимающих преимущественно пустыни
Африки и Азии.
Хозяйственное значение представителей рода невелико, посколь-
ку плотность их населения незначительна. Большинство видов,
обитающих в СССР, имеет малоценный мех. Ежегодный промысел
мелких видов колеблется от 5,2 до 34,0 тыс. Более высоко ценится
мех рыси, заготовки которой в 1960—1970 гг. колебались от 1,8
до 5,8 тыс. шкур (Соколов, 1979).
Болезни кошек мало изучены. Заболевание бешенством в при-
родных условиях зарегистрировано у рыси в Омской области, на
6 И. М. Громов и др. (1963), Н. А. Бобринский и др. (1965) разделяют F.
silvestris на два самостоятельных вида — дикую европейскую кошку
F. silvestris, обитающую в Европе и Малой Азии, и степную кошку F. И-
Ьуса, занимающую остальную часть ареала.
6 Зак. 3575
162
Carnivora
Украине и в Восточном Забайкалье (Рудаков, Мальков, 1970; Ре-
венко и др., 1979; Ботвинкин и др., 1980). В. А. Рудаков и
Г. Б. Мальков (1970) сообщают, что рысь, у которой вирусоло-
гически подтвержден диагноз бешенства, потеряв осторожность,
шла вслед за охотником и была им замечена на окраине дере-
вни.
Бешенство камышевого кота наблюдалось в Узбекистане (Мама-
тов, 1979). На Дальнем Востоке, в окрестностях озера Ханко,
диагностировано бешенство у амурской кошки (Ботвинкин и др.,
1981).
В Венгрии за 1967—1974 гг. зарегистрировано 29 случаев бе-
шенства у диких кошек, что составило 0,8% от заболеваний диких
животных (Koltay, 1975). В Чехословакии в 1968 г. выявлено 15
диких кошек, больных бешенством (Селимов, 1978). В Швейцарии
наблюдались единичные заболевания диких кошек (Wandeler et
al., 1974).
Бешенство диких кошек рода Felis регистрировалось в Южной и
Юго-Восточной Африке, где основными распространителями вируса
являются шакалы (Canis mesomeles) и желтые мангусты (Cynictis
penicillata). Отмечено также бешенство домашних кошек (Bar-
nard, 1979).
В США за 1950—1968 гг. было исследовано 188 рысей (L. rufus)
и у 36 из них установлено бешенство, что составило 1,7% от общего
числа (2167) бешенства диких животных (Humphrey, 1971).
Домашняя кошка имеет своим родоначальником ливийскую фор-
му дикой кошки — F. silvestris libyca Forster, 1780. Одомашнивание
произошло в Египте 3—4 тыс. лет до н. э. Отсюда кошки были выве-
зены в VI—IV в. до н. э. в Китай, Причерноморье. В XI—XII вв.
получила распространение в Европе. В Америку завезена испан-
цами.
Бешенство домашних кошек отмечается всюду, где распростра-
нена заболевание среди собак, сельскохозяйственных и диких жи-
вотных. В прошлом столетии и до середины XX в. кошки не имели
существенного эпидемиологического и эпизоотологического значе-
ния. Так, в Германии в 1887—1889 гг. было зарегистрировано
всего 13 случаев бешенства кошек, что составило 0,8% от общего
числа 1596 заболевших домашних и сельскохозяйственных живот-
ных. На долю собак приходилось 88% случаев (Герценштейн,
1891).
В России, поданным семи пастеровских станций, с 1886 по 1890 г.
число лиц, обратившихся по поводу контакта с бешеными или по-
дозрительными кошками, составило всего 6,3% от общего числа
пациентов (8023). На долю собак приходилось 88,1% (Герцен-
штейн, 1891).
В 1887 г. кошки послужили источником заражения людей
гидрофобией в 2,5% от 693 зарегистрированных заболеваний
(Мари, 1909).
Хищные
163
В настоящее время положение резко изменилось и повсеместно
увеличился не только удельный вес заболевания кошек, но и число
зарегистрированных у них случаев бешенства (табл. 29). В 1954—
1974 гг. на территории ФРГ был зарегистрирован 3801 случай бе-
шенства домашних кошек, что составило 31,4% от заболеваний до-
машних и сельскохозяйственных животных и 6,3% от общего числа
заболевших животных — домашних и диких. Заболеваемость собак
составила 4,6% от общей заболеваемости, т. е. была ниже, чем кошек
(Wachendorfer, 1975).
В ГДР доля участия кошек в заболеваемости бешенством домаш-
них и сельскохозяйственных животных составила в 1953—1956 гг.
34% (собак —49,5%), в 1967—1976 гг.—35,5% (собак —34%), в
1977—1980 гг.— 33,5%, превышая бешенство собак — 26% (Pitz-
schke, 1982).
Таблица 29. Доля кошек в заболеваемости бешенством животных
Территория Годы Число больных ко- шек Доля от всех случа- ев бешенства, % Доля от случаев бешенства домаш- них и сельскохозяй- ственных животных Авторы
ФРГ 1953—1971 923 9,8 42,2 Wandeler et al., 1974
Франция 1968—1973 127 3,3 12,4 Andral, Toma, 1973
Швейцария 1967—1973 50 1,9 40,0 Wandeler et al., 1974
Дания 1964—1970 12 5,1 31,5 Wandeler et al., 1974
Венгрия 1967—1974 147 4,8 47,7 Koltai, 1975
Польша 1971 170 12,6 45,9 Serokowa, Kreska, 1973
Чехослова- 1976—1980 94 2,9 51,3 Matouch, Polak, 1982
кия
ГДР 1977—1980 438 6,6 33,5 Pitzschke, 1982
США 1956—1957 753 7,0 11,0 Tierkel, 1959
США 1966—1968 702 5,6 19,7 Infectious diseases . . ., 1970
Удельный вес бешенства кошек в большинстве европейских стран
превышает долю бешенства собак, составив в 1977—1983 гг. от
3,6 до 5% общего числа заболевших животных. В отдельные годы
удельный вес кошек был еще выше, достигая 7,8% в ГДР, 12%
в ПНР, 13,2% в Бельгии, 15,4% в ЧССР и 23,7% в СРР (Черкас-
ский, 1985).
В США ежегодно регистрируется от 135 до 538 случаев бешен-
ства кошек. Уменьшение числа заболеваний кошек происходит мед-
леннее, чем собак. Так, если бешенство собак в 1968—1971 гг. со-
6*
164
Carnivora
кратилось по сравнению с 1953—1959 гг. в 10 раз, то бешенство
кошек только в 2,3 раза — в среднем за год с 391 до 169 случаев
(Vaughn, 1975). Вспышки бешенства кошек совпадают с акти-
визацией эпизоотии у диких животных. Во время эпизоотии лисиц
в штате Мэн в марте 1971 г. на долю кошек приходилось
27,5% от общего числа (58) зарегистрированных случаев бешен-
ства.
Тесная зависимость динамики проявления бешенства кошек и
лисиц прослежена на протяжении 1957—1961 гг. в ФРГ (Wachen-
dorfer, 1962).
В ряде стран отмечается повышение эпидемиологического зна-
чения кошек. В Швейцарии в 1967—1970 гг. контакт с бешеными
кошками был причиной обращения за антирабической помощью в 34
случаях из 50 (68%). В Польше из 16 случаев гидрофобии, зарегист-
рированных за 1961—1970 гг., в трех случаях источником заражения
были кошки (Serokova, Kreska, 1971). В ГДР кошки были причи-
ной заболевания людей в трех случаях из 34, наблюдавшихся за
1950—1974 гг. В Венгрии в 1961 г. в 15 случаях из 25 источником
заражения были кошки (Wachendorfer, 1962).
В СССР уровень заболеваемости кошек бешенством за 1966—
1975 гг. вырос в 10 раз, а удельный вес их в общем числе регистри-
руемых случаев заболеваний животных бешенством увеличился с
0,6 до 3,1%. В Украинской, Литовской, Эстонской ССР бешенство
кошек в последнее время регистрировалось чаще, чем бешенство
собак (Ведерников, Гудим, 1976). В 1946—1962 гг. случаи бешенст-
ва кошек в Латвийской ССР составили только 9,6%, а в 1963—
1979 гг. их удельный вес возрос до 29,4% от общего числа заболе-
ваний домашних и сельскохозяйственных животных (Черкас-
ский, 1985).
В Украинской ССР в 1975—1976 гг. бешенство кошек составило
21,6% от всех случаев бешенства и 36,7% среди домашних и сельско-
хозяйственных животных (Ревенко и др., 1979).
Обязательная вакцинация собак, введенная в СССР в 1956 г.>
резко снизила заболеваемость людей гидрофобией. Если в после-
военные годы (1950—1959) в среднем за год регистрировалось 385
случаев, то уже в следующем десятилетии среднегодовая забо-
леваемость сократилась до 84, в 1970—1979 гг.— до 77 случаев,
а в 1980—1983 гг. упала до 57 случаев в год (Черкасский,
1985).
На фоне уменьшения роли собак как источников гидрофобии
повысилось эпидемиологическое значение кошек. В СССР в 1946—
1956 гг. заражения от кошек составляли 5,3%, в 1958—1963 гг.—
6,6, а в 1966—1975 гг.— от 10,6 до 21,5% (Селимов, 1976; Куче-
рук, 1982).
На Украине в 1945—1955 гг. кошки были причиной заболева-
ния людей в 5,4% (собаки — в 88%), а в 1969—1977 гг. 31,8% слу-
Хищные
165
чаев гидрофобии были вызваны укусами кошек и только в 7,9%
источником заражения явились собаки (Горячева, 1959; Щербак
и др., 1978). В Молдавии в 1948—1962 гг. случаи заражения от ко-
шек составляли 3,6%, а в 1971—1979 гг.— 25%. Удельный вес
собак как источников заражения людей упал с 84,3 до 12,5% (Шля-
хов и др., 1981).
Кошки, по-видимому, менее восприимчивы к вирусу бешенства,
чем собаки. Так, из 86 кошек, которым были инокулированы разведе-
ния вируса (штаммы, выделенные от собак, лисиц, скунсов и кошек),
погибло от бешенства 26 особей. Инкубационный период 9—51 день,
в среднем 18 дней. Клинические признаки заболевания наблюдались
в течение 1—8, в среднем 5 дней. Около 70% заболевших кошек про-
являли агрессивность. В слюнных железах вирус обнаружен у 23
кошек из 26 (88%). В большинстве случаев вирус появлялся в слюне
одновременно с проявлением клинических признаков заболевания
или на вторые сутки. У пяти кошек вирус в слюне появился за сутки
и у двух кошек — через три дня после клинических признаков
бешенства. Выделение вируса со слюной продолжалось до гибели
животных. Концентрация вируса в слюне в первые дни колебалась
от 1 до 4,0 1g LD/50, а в конце заболевания достигала 6,4 1g LD/50,
в среднем 4,1 1g LD/50 (Vaughn, 1975).
В местностях, где отмечается бешенство собак и диких живот-
ных, необходимо осуществлять меры по предотвращению вовлечения
кошек в эпизоотию путем проведения вакцинации домашних кошек
и уничтожению бродячих особей. Многие авторы указывают на
важное значение кошек как звена, связывающего природные очаги
бешенства с городскими. Этому способствует большое количество
бродячих кошек, их способность к самостоятельному существова-
нию в природе вблизи населенных пунктов, где они находят доста-
точное количество пищи в виде пищевых отбросов. О количестве
бездомных кошек свидетельствуют данные об уничтожении их:
например, на Украине ежегодно уничтожалось около 30 тыс. кошек,
в Литовской ССР в 1973 г. было уничтожено 10 тыс. кошек. В Алдан-
ском районе Якутии в 1974—1978 гг. было уничтожено 14 370 ко-
шек. В Приморье в 1974 г. было вакцинировано 8700 кошек и
уничтожено 870 безнадзорных особей (Гречановская, 1966; Буракаус-
кас, Казакявичус, 1979; Омельченко и др., 1979; Егоров и др.,
1981).
Сокращение численности бродячих кошек проводится в настоя-
щее время во многих странах. В Югославии (Воеводина) в
1973—1974 гг. было уничтожено более 11 тыс. кошек (Pavlovic
et al., 1976).
Кошки имеют важное значение в распространении токсоплазмо-
за, так как они являются дефинитивными хозяевами паразита, что
было установлено лишь в начале 70-х годов (Dubey, Frenkel, 1973).
В клетках слизистой кишечника кошек происходит половое раз-
множение токсоплазм, в результате которого уже через 3—4 дня
166
Carnivora
после заражения начинается выделение ооцист (зигоцист). Выде-
ление ооцист продолжается до двух недель. За это время во внеш-
нюю среду попадает до миллиарда ооцист. Попадая с фекалиями в
почву, ооцисты при благоприятных условиях влажности и темпера-
туре около 20° через 2—5 дней спорулируют, формируя в каждой
ооцисте два спороцита с четырьмя спорозоитами в каждом. Ооцис-
ты; сохраняют инвазионность в течение многих месяцев и даже до
двух лет, что и обеспечивает возможность заражения широкого
круга животных. Отмечено, что там, где нет кошек (на некоторых
тихоокеанских островах), нет и токсоплазмоза (Лысенко,
1980).
Прохождение полового цикла токсоплазм в клетках кишечника
установлено в условиях зоопарков также у льва, тигра, гепарда,
манула, дикой кошки, оцелота, ягуарунди, пумы, бенгальской кош-
ки (Галузо, Коновалова, 1979; Поломошнов, 1979). Зараженность
в природе выявлена у рыси в БССР (Дылько, 1966), дикой кошки в
Молдавии и Казахстане (Новинская, 1965; Поломошнов, 1979).
В США у красной рыси (Infectious diseases, 1970).
В организме кошек токсоплазмы проходят также и бесполую
часть жизненного цикла, в результате которого через 1—3 недели
после заражения в различных тканях образуются цисты, сохраняю-
щиеся долгие месяцы. Развитие такой генерализованной инвазии
сопровождается формированием иммунитета и потому серопозитив-
ные кошки редко выделяют ооцисты с фекалиями.
> Пораженность кошек зависит от их контакта с грызунами, пти-
цами и интенсивностью заражения последних. Зараженность кошек
увеличивается с возрастом.
Так, по данным серологического обследования в г. Канзас-
Сити (США), из 302 котят заражено было 16,2%, а из 128 взрослых
кошек — 37,5%. Зараженность беспризорных кошек была значи-
тельно выше — 57,9% (Dubey, 1973; цит. по: Лысенко, 1980). Во
Владивостоке зараженность домашних кошек установлена в 6,5%,
а у бродячих достигала 26,6% (Яковлев, 1980). В Омской области
зараженность кошек в поселках составляла 14,8%, а в г. Омске
при исследовании фекалий от 224 домашних кошек ооцисты обнару-
жены у трех молодых особей (Рогатых, 1981). В Одессе из 209
бродячих кошек ооцисты найдены у восьми (Ковбасюк и др.,
1976).
В Вильнюсе и его окрестностях токсоплазмы были выделены от
6 кошек из 66 обследованных биологической пробой, а реакция пре-
ципитации в геле была положительна у 26,6% животных (Бута,
1980). В Казахстане (р. Или) ооцисты были найдены в фекалиях у
2 диких кошек из 75 (Поломошнов, Петешев, 1976). Наиболее север-
ное нахождение ооцист у домашней кошки, свидетельствующее о не-
давнем заражении, сделано Н. А. Рогатых в Дудинке (69° с. ш.).
При сероаллергическом обследовании коренного населения была
выявлена более высокая инфицированность жителей поселков, в
Хищные 167
которых имелись кошки, по сравнению с теми, где кошек не было.
Так, поданным реакции иммунофлуоресценции процент положитель-
ных реакций равнялся соответственно 33 и 17,8 %, а по внутрикож-
ной пробе — 44,1 и 27,6% (Рогатых и др., 1977). Зависимость часто-
ты поражения токсоплазмозом от контакта с кошками прослежена
также при обследовании беременных женщин в Москве и Баку
(Титов, 1980).
Несомненная роль кошек как источников высокоинвазивной и
устойчивой стадии паразита — ооцисты требуют усиления ветери-
нарного надзора над многочисленными популяциями домашних
кошек в населенных пунктах и особенно в пищевых объектах —
продуктовых магазинах, булочных, где нередко содержат кошек для
истребления грызунов. Бродячие кошки осуществляют связь синан-
тропных очагов токсоплазмоза с природными.
ОТРЯД CHIROPTERA BLUMENBACH, 1779
—РУКОКРЫЛЫЕ
Систематика, распространение
Отряд рукокрылых объединяет два подотряда: крыланов (Ме-
gachiroptera) и летучих мышей (Microchiroptera). В подотряде крыла-
нов одно семейство Pteropidae, насчитывающее 42 рода и 161 вид.
Все они распространены в тропических областях Старого Света.
В Восточной области обитают 25 родов и 70 видов, в Афротропиче-
ской — 13 и 39, в Австралийской — 12 и 67. В Палеарктику
незначительно заходят всего 4 вида (рис. 9, табл. 30).
В подотряде летучих мышей 16 семейств, 132 рода и 756 видов.
Этот подотряд распространен всесветно (рис. 10). Однако к Западно-
му и Восточному полушариям приурочены свои семейства и свои ро-
ды (табл. 30). Западному полушарию свойственны 6 семейств: Phyl-
lostomatidae, Desmodontidae \ Natalidae, Furipteridae, Thyrop-
teridae, Noctilionidae. Все они распространены преимущественно в
Неотропической области, и лишь 23 из 138 видов Phyllostomatidae
относятся к неарктической фауне. Восточному полушарию свойст-
венны другие 7 семейств: Rhinolophidae, распространенное по всем
материкам Старого Света (исключая Антарктиду); Megadermatidae,
обитающее в Азии, Африке и Австралии; Rhinopomatidae и Nycteri-
dae, не проникающие в Австралию; Graseonycteridae, известное из
Восточной области, и 2 монотипичные семейства—Myzapodidae,
эндемичное для Мадагаскара, и Mystacinidae, эндемичное для Новой
Зеландии. Имеется лишь три общих для обоих полушарий семейства:
Emballonuridae, Vespertilionidae, Molossidae, в двух последних
есть 5 общих родов (Myotis, Vespertilio, Nycticeius, Plecotus, Tada-
rida). Видов, общих для обоих полушарий, нет. Эю указывает на
слабые связи фаун рукокрылых Палеарктики и Неарктики, Палео-
тропики и Неотропики. В подавляющем большинстве отрядов назем-
ных млекопитающих для фаун этих областей, особенно Неарктики
и Палеарктики, есть не только общие роды и семейства, но и
общие виды.
В распределении видового разнообразия отряда рукокрылых
прослеживаются довольно четкие закономерности (рис. 11). Предста-
вители этого отряда отсутствуют в тундрах и лесотундрах. Мини-
1 Американские авторы не выделяют сем. Desmodontidae, а относят роды
Desmodus (включающий род Diaemus) и Diphylla в сем. Phyllostomidae;
в то же время роды Mormoops и Pteronotus (6 видов) выделены в самостоя-
тельное семейство Mormoopidae (Honacki, Linman, Koeppl, 1982).
Таблица 30. Распределение числа родов и видов рукокрылых по зоогеографическим областям (по: Honacki et al., 1982)
Отряд и семейства Число современных родов Зоогеографические области и число родов в них Число современных видов Зоогеографические области и число видов в них
Неотропиче- ская Неарктиче- ская Палеарктиче- ская Восточная Афротропи- ческая Австралий- ская Неотропиче- ская Неарктиче- ская Палеаркти- ческая Восточная Афротропи- ческая I Австралий- I ская
СССР Вся СССР Вся
Chiroptera 174 79 31 12 22 61 44 40 917 258 74 41 96 263 201 157
Pteropidae 42 2 25 13 12 161 4 70 38 67
Rhinopomatidae 1 1 1 1 3 3 2 2
Emballonuridae 12 8 1 1 3 4 3 48 18 1 3 10 9 15
Craseonycteridae 1 1 1 1
Nycteridae 1 1 1 1 14 1 2 12
Megadermatidae 4 1 1 2 1 5 1 2 2 2
Rhinolophidae 10 1 2 5 4 5 127 6 15 71 35 17
Noctilionidae 1 1 2 2
Mormoopidae 2 2 2 8 8 4
Phyllostomatidae 45 45 11 135 134 22
Desmodontidae 2 2 1 3 3 1
Natalidae 1 1 1 4 4 1
Furipteridae 2 2 2 2
Thyropteridae 1 1 2 2
Myzapodidae 1 1 1 1
Vespertilionidae 35 10 11 10 12 18 12 14 314 53 35 34 66 95 67 46
Mystacinidae 1 1 1 1
Molossidae 12 7 4 1 2 6 6 4 86 32 10 1 3 10 35 9
с©
Рукокрылые
Рис. 9. Распространение подотряда крыланов Megachiroptera (по: Коортап, Cockrum, 1967)
Chiroptera
Рис. 10. Распространение подотряда летучих мышей Microchiroptera (по: Коортап, Cockrum, 1967, с добавлениями)
Рукокрылые
172
Chiroptera
Рис, 11, Картосхема видового разнообразия отряда Chiroptera
мальное число видов (1—5) свойственно северо- и среднетаежным
лесам Евразии и Северной Америки, равнинным степям, пустыням
Евразии, Северной Африки, Австралии и обширным высокогорьям.
Это связано с неблагоприятным климатом (короткое лето и низкие
температуры в тайге), недостатком убежищ, регулярными сильными
ветрами и холодными ночами в равнинных степях и пустынях (Стрел-
ков, 1983). Довольно разнообразна фауна рукокрылых (11—25 ви-
Рукокрылые
173
дов) в пределах равнинных и горных широколиственно-хвойных,
широколиственных лесов, субтропических редколесий и некоторых
типов саванн. Области максимального разнообразия видов рукокры-
лых приурочены к тропическим и субтропическим лесам, а также
к саваннам с обилием деревьев. В Центральной Африке обитает
117 видов, в Малайзии — 80, на о-ве Новая Гвинея — 64, в Амазо-
нии —- более 75 видов, в Южной Африке — 57, на Сулавеси — 43
вида. Вероятно, именно тропические леса были той ареной, на кото-
рой проходили основные линии эволюции и адаптивной радиации
174
Chiroptera
Рис. 12. Кладограмма филогении от-
ряда рукокрылых (по: Smith, 1976,
с изменениями)
а — Phyllostomatidae,
б — Vespertilionidae, в — Emballonuridae,
г — Rhinolophidae; группы, распространен-
ные:
1 — в Западном полушарии,
2 — в Восточном полушарии,
3 — в обоих полушариях
рукокрылых. Напротив, тундры и лесотундры служили преградой
для распространения рукокрылых. Видимо, в связи с этим обмен
фауной рукокрылых между Палеарктикой и Неарктикой через Бе-
рингию практически отсутствовал, что подтверждается составом
паразитирующих у летучих мышей членистоногих и особенно гель-
минтов (Durette^Desset, 1982).
Так как в русской литературе практически нет публикаций по
эволюции рукокрылых, мы приводим кладограмму филогении этого
отряда (рис. 12). Из-лее видно, что основная линия развития руко-
крылых приурочена к Восточному полушарию. Emballonuridae,
Rhinolophidae, Megadermatidae, Vespertilionidae известны здесь с
эоцена, a Molossidae — с олигоцена (Громова, 1962; Smith, 1976).
В Западное полушарие рукокрылые проникали неоднократно и каж-
дое проникновение сопровождалось интенсивной дивергенцией
(Симпсон, 1983). Эндемичное для Южной Америки семейство Phyl-
lostomatidae известно там с миоцена, а в Северной Америке найдены
остатки Vespertilionidae, датируемые олигоценом и миоценом
(Smith, 1976).
На территории СССР встречаются летучие мыши 41 вида 12 ро-
дов из трех семейств: Rhinolophidae — 6 видов, Vespertilionidae —
34 вида и Molossidae — 1 вид (Каталог млекопитающих СССР, 1981):
Rhinolophus hipposideros Bechstein, 1800
Rh. blasii Peters, 1866
Rh. euryale Blasius, 1853
Rh. mehelyi Matschiae, 1901
Rh. ferrumequinum Schreber, 1775
малый подковонос (54, 56)'*
средиземноморский подково-
нос (58)
южный подковонос (58)
подковонос Метели (58)
большой подковонос (58)
* В скобках приведено диплоидное число хромосом (Орлов, 1974).
Рукокрылые
175
Rh. bocharicus Kastsch. et Akim, 1917
Myotis myotis Borkhausen, 1797
M. blythi Tomes, 1857
M. bechsteini Kuhl, 1818
M. nattereri Kuhl, 1818
M. mystacinus Kuhl, 1819
M. brandti E^ersmann, 1845
M. ikonnikovi Ognev, 1912
M. frater G. Allen, 1923
M. emarginatus E. Geoffroy, 1806
M. dasycneme Boie, 1825
M. daubentoni Kuhl, 1819
M. capaccinii Bonaparte, 1837
Plecotus auritus Linnaeus, 1758
P. austriacus Fischer, 1829
Barbastella barbastella Schreber, 1774
B. leucomelas Cretzschmar, 1830
Pipistrellus pipistrellus Schreber, 1775
P. nathusii Keyserling et Blasius, 1839
P. abramus Temminck, 1840
P. kuhlii Kuhl, 1819
P. savii Bonaparte, 1837
Nyctalus leisleri Kuhl, 1819
N. noctula Schreber, 1775
N. lasiopterus Schreber, 1780
Eptesicus serotinus Schreber, 1774
E. bottae Peters, 1869
E. nilssoni Keyserling et Blasius, 1839
E. bobrinskii Kuzyakin, 1935
Vespertilio murinus Linnaeus, 1758
V. superans Thomas, 1898
Ctonycteris hemprichi Peters, 1859
Miniopterus schreibersi Kuhl, 1819
Murina aurata Milne-Edwards, 1872
M. leucogaster Milne-Edwards, 1872
Tadarida teniotis Rafineque, 1814
бухарский подковонос
большая ночница (44)
остроухая ночница (44)
ночница Бехштейна
ночница Наттерера (44)
усатая ночница (44)
ночница Брандта
ночница Иконникова
длиннохвостая ночница
трехцветная ночница (44)
прудовая ночница (44)
водяная ночница (44)
длиннопалая ночница (44)
бурый ушан (32)
серый ушан
европейская широкоушка
азиатская широкоушка
нетопырь-карлик (42)
нетопырь Натузиуса (44)
восточный нетопырь (26)
средиземноморский нетопырь
(44)
кожановидный нетопырь (44)
малая вечерница
рыжая вечерница (42)
гигантская вечерница
поздний кожан (50)
пустынный кожан
северный кожанок
кожанок Бобринского
двуцветный кожан (38)
восточный кожан (38)
белобрюхий стрелоух (30)
обыкновенный длиннокрыл (46
малый трубконос
большой трубконос
широкоухий складчатогуб
В последней сводке по млекопитающим мира (Honacki et al.,
1982) Rh. bocharicus включен в вид Rh. clivosus Cretzschmar,
1828, a P. abramus отнесен к роду P. javanicus Grey, 1838.
Летучие мыши распространены по всей территории нашей страны,
вплоть до северного предела леса. Дальше всех на севере встречается
северный кожанок — типичный представитель таежной фауны
(табл. 31). Областям широколиственных лесов свойственны вечер-
ницы, некоторые ночницы и нетопыри. Наименее характерны лету-
чие мыши для степей (Стрелков, 1983), среди них нет типичных степ-
176
Chiroptera
Таблица 31. Число видов рукокрылых в различных ландшафтных областях
на территории СССР
Область Встречается видов
Всего типичных для области
Таежная 5 1
Широколиственнолесная европейская 26 14
Широколиственнолесная дальневосточная 20 9
Степная 6 0
Пустынная (Средняя Азия и Казахстан) 16 2
ных видов. К типичным пустынным видам можно отнести лишь два
вида — кожанов пустынного и Бобринского (Кучерук, 1959а; Ку-
лик, 1974, 1980а). В равнинных пустынях, глинистых и песчаных,
летучие мыши немногочисленны и распространены спорадически на
участках, где есть убежища. Значительно богаче фауна и выше
численность летучих мышей в долинах рек и оазисах, где зверьки
находят в достаточном количестве убежища, водопои и пищу. На-
пример, в Южной и Восточной Туркмении, занятой преимуществен-
но песчаными пустынями, отмечено лишь 4 вида, которые всюду
малочисленны или редки. Более многочисленны летучие мыши в
Северо-Западной Туркмении, где распространены останцовые воз-
вышенности, впадины, чинки Устюрта, сухие русла Узбоя; там
встречается 8 видов летучих мышей, среди которых многочислен
пустынный кожан. Значительно богаче фауна летучих мышей по
берегам рек и в оазисах, где зарегистрировано 11 видов, из которых
7 за пределами оазисов не встречаются. В оазисах обычны бухарские
подковоносы и многочисленны нетопыри-карлики и поздние кожаны.
В городах и предгорьях Копетдага и Кугитангтау, где нет недостат-
ка в убежищах, летучие мыши особенно многочисленны, здесь за-
регистрировано 19 видов (Стрелков и др., 1978).
Особенности морфологии и физиологии
Рукокрылые — единственный отряд млекопитающих, освоивший
воздушное пространство. Его представители приобрели уникальную
среди млекопитающих способность к активному полету. Согласно
новой гипотезе (Панютин, 1980), возникновение активного полета
у рукокрылых произошло через развитие и совершенствование пла-
нирования. По этой гипотезе рукокрылые произошли путем эволю-
ции мелких млекопитающих, питавшихся преимущественно насеко-
мыми и ведших первоначально малоподвижный образ жизни, под-
вешиваясь на тонких ветках; экологическая обстановка обитания
этих предков была близка к нижнему ярусу современных мангр;
Рукокрылые
177
фиксация тела с помощью задних конечностей привела к уменьше-
нию участия передних конечностей в передвижении и открыла путь
к их дальнейшему преобразованию, к смене функций. У предков
рукокрылых, обитавших над илистой поверхностью, произошло
удлинение передних конечностей и преобразование кисти; в дальней-
шем учащение махательных движений при спрыгивании вниз, уве-
личение площади кисти и развитие кожной складки от нее к бокам
тела должны были способствовать постепенному гашению скорости
при посадке. «По-видимому, на этой базе возникло крыло рукокры-
лых. Малые размеры, при которых оказывается эффективным даже
ничтожное увеличение опорной „летательной" поверхности из-за
сравнительно быстрого изменения соотношения массы и поверхно-
сти тела, и относительно высокая „энерговооруженность" мелких
животных — необходимое условие к активному полету» (Панютин,
1980, с. 286).
Активный полет современных рукокрылых обеспечивается це-
лым рядом морфологических приспособлений.
Подавляющее большинство рукокрылых — относительно неболь-
шие зверьки. Особи 80% видов имеют массу тела менее 27 г (Паню-
тин, 1980). Среди крыланов есть более крупные формы с длиной
тела до 40 см и массой тела до 900 г, а размах крыльев у некоторых
видов крыланов достигает 1,7 м.
На туловище и голове развит волосяной покров, довольно гус-
той, мягкий, средней длины. Кожная перепонка покрыта лишь ред-
кими волосами. Окраска обычно одноцветная, темных тонов. У мно-
гих видов на горле, лбу и летательной перепонке имеются специфи-
ческие кожные железы, выделяющие секрет с сильным мускусным
запахом. У самок одна пара функционирующих сосков, расположен-
ных по бокам грудной области.
Задние конечности имеют своеобразное строение. Головка бедра
опирается на поверхность суставной впадины таза не медиальной,
а латеральной частью. Вся задняя конечность оказывается как бы
вывернутой, колено обращено не вперед, а назад. Такое строение
задней конечности позволяет подтягивать тело назад. Укороченные
пальцы задней конечности снабжены когтями, которые помогают
цепко удерживаться животному в подвешенном состоянии даже на
мало шероховатых поверхностях.
Передняя конечность превращена в крыло. Эластичная кожная
летательная перепонка натянута между удлиненными пальцами пе-
редних конечностей, плечом, предплечьем, боками тела, задними ко-
нечностями и хвостом, если таковой имеется. Летательная перепонка
является на 75% производным кожи передней конечности. Первый
палец передних конечностей развит слабо, коготь имеется только
на первом пальце, иногда на втором (у крыланов).
Череп с удлиненным или укороченным лицевым отделом и отно-
сительно хорошо развитым мозговым отделом. Характерно раннее
зарастание швов между костями черепа, срастание первого грудного
178
Chiroptera
и последнего шейного позвонков; наличие на грудине небольшого
киля. Все это связано с сильным развитием мускулатуры грудного
пояса. Общее число постоянных зубов от 20 до 38; зубы дифиодонт-
ные и гетеродонтные (плоскобугорчатые у крыланов и остробугор-
чатые у летучих мышей).
Зрение и обоняние хорошо развиты только у крыланов. У лету-
чих мышей наибольшего развития достигает слух. Кроме того, все
летучие мыши обладают совершенной способностью к эхолокации.
У крыланов способность к эхолокации установлена только у пред-
ставителей рода Rousettus, живущих в пещерах. Локационные сиг-
налы у летучих мышей генерируются в гортани и испускаются в
виде коротких ультразвуковых импульсов частотой до 130 кГц
и продолжительностью 0,2—100 мс. Эхолокация позволяет летучим
мышам различать препятствия диаметром до 0,1—0,08 мм (проволо-
ка). Дальность действия эхолокации не превышает 10—15 м (Стрел-
ков, 1973).
Возникновение эхолокации у летучих мышей стало возможным
лишь после освоения животными трехмерного пространства (воз-
душной среды) в таких экологических условиях (например, пеще-
ры), где оптическими средствами было невозможно получение какой-
либо информации о наличии препятствий. «В своем развитии био-
логические сонары прошли, по-видимому, длительный путь от
непроизвольной эхолокации с использованием различных коммуни-
кационных сигналов до совершенных ультразвуковых систем с им-
пульсами, предназначенными специально для зондирования прост-
ранства» (Константинов, 1980, с. 276).
Для рукокрылых характерен короткий пищеварительный тракт,
превышающий длину тела лишь в 1,5—4,0 раза (даже у растительно-
ядных форм), и быстрый процесс пищеварения (15—35 мин) (Morri-
son, 1980). Сердце крупное, его индекс достигает 1,0—1,3% и при-
ближается к величине этого показателя у быстролетающих птиц
(Курсков, 1980). Матка простая, двурогая или переходной формы.
У многих видов эмбрион развивается всегда только в одном роге.
Семенники ингуидальные или абдоминальные, лишь в сезон размно-
жения спускающиеся во временную мошонку, образующуюся из
участка кожи, покрывающей паховую область.
У ряда видов установлен половой диморфизм размеров тела и
черепа: самки оказались в среднем крупнее самцов (Williams,
Findley, 1979). Авторы считают, что это определяется повышенным
расходом энергии во время беременности. Более крупные размеры
самок способствуют более эффективной терморегуляции (что обес-
печивает нормальное развитие эмбриона), запасанию большего
количества жиров, а также обеспечивают возможность охоты на
более крупных насекомых.
Летучие мыши — гетеротермные животные, температура их
тела вне периода активности меняется в зависимости от температуры
окружающей среды. Поэтому для них неблагоприятны как низкие
Рукокрылые
179
температуры ниже 0°, так и очень высокие, вызывающие перегрев
тела. Дневного перегрева рукокрылые избегают благодаря ночной
активности, проводя день в убежищах с благоприятным микроклима-
том. В тропиках летучие мыши активны круглый год. В умеренных
широтах они на холодный период впадают в спячку. Необходимость
спячки обусловливают не только или не столько температурные
условия, сколько отсутствие в этот период активных насекомых,
составляющих пищу летучих мышей (Saint, 1973).
Спячку летучие мыши проводят в состоянии глубокой обратимой
гипотермии в зимовочных убежищах, где поддерживается постоянно
высокая влажность и температура в пределах 0—12° (Кузякин,
1950). Например, в штольне на юге Норвегии, где зимовали летучие
мыши, температура была +5. . .+8°, относительная влажность —.
85—100% (Lauritzen et al., 1978). В Ферганской долине в шахте,
где зимовали азиатские широкоушки, температура достигала
+ 12 . . .+ 15° (Морозов, 1974).
Насекомоядные летучие мыши, обитающие в умеренных широтах,
и в летний период испытывают незакономерные колебания обилия
корма, связанные с непостоянством погоды. Благодаря гетеротер-
мии они могут при неблагоприятных погодных условиях, оставаясь
неподвижными, затормозить жизненные процессы и в течение нес-
кольких суток обходиться без пищи. Однако именно на летний ак-
тивный период у летучих мышей приходится беременность у самок,
накопление половых продуктов у самцов, линька, рост и развитие
молодых. Все эти процессы требуют от организма больших энерге-
тических затрат. В связи с этим у летучих мышей выработался ме-
ханизм поддержания в течение длительного времени высокого уров-
ня обмена и температуры тела у неподвижного зверька, не требу-
ющий больших энергетических затрат самого животного. Это — по-
веденческая терморегуляция, которая у летучих мышей развита
сильнее, чем у других млекопитающих. Например, подковоносы вы-
бирают на такое время более теплые убежища; большая и остроухая
ночницы образуют плотные скопления из нескольких десятков зверь-
ков, внутри которых температура бывает на 15—20° выше окружаю-
щего воздуха (Панютин, 1974).
Питание
Рукокрылые питаются как растительным, так и животным кор-
мом. Растительноядны крыланы (сем. Pteropidae), обитающие в
тропических областях Старого Света, и многие виды сем. Phyllo-
stomatidae, распространенного в Центральной и Южной Америке,
которые перешли на растительную пищу, вероятно, вторично (Стрел-
ков, 1973). Они поедают различные плоды или высасывают из них
сок, собирают нектар из цветков или пыльцу. Некоторые виды уз-
ко специализированы на добывании нектара из цветков и имеют
для этого морфологические приспособления, например очень длин-
180
Chiroptera
ный язык (роды Macroglossus, Syconycteris, Megaloglossus, Melo-
nycteris, Nesonycteris, Notopterus в семействе крыланов и роды
Glossophaga, Lionycteris, Lonchophylla, Platalina, Monophyllus,
Anoura, Musonycteris, Choeronycteris, Choeroniscus, Hylonycteris,
Leptonycteris, Lichonycteris в семействе американских листоносов).
Плодоядные виды, особенно крыланы, в тропических областях
Африки, Азии и Австралии временами наносят ущерб плантациям,
уничтожая созревшие фрукты. Однако положительное биоценотичес-
кое значение растительноядных рукокрылых несравненно больше
приносимого ими вреда. Поедая в большом количестве плоды, они
способствуют расселению растений, выделяя их косточки с по-
метом. Ряд растений (хироптерофилы) приспособился к опылению
с помощью рукокрылых, питающихся нектаром и пыльцой. Среди
растений наблюдается даже конкуренция за опыление рукокрылыми
(Start, Marshall, 1976; Sazima, Sazima, 1978; Kock, 1979).
Большинство видов летучих мышей подотряда Microchiroptera
питается насекомыми. Насекомые часто служат дополнительным
кормом и у растительноядных и у хищных летучих мышей. Состав
насекомых, поедаемых летучими мышами, очень разнообразен. Так,
в рационе летучих мышей Среднего Приднепровья, где среди 9 видов
доминировали рыжая вечерница, нетопыри лесной и карлик, по
числу особей преобладали кровососущие двукрылые (34,3—42,6%),
а по количеству видов первое место занимали фитофаги. Общее
же количество беспозвоночных, питающихся растениями, в добыче
летучих мышей было довольно значительно — до 41,3%. Еще
больше будет их доля по массе, так как среди них много крупных
насекомых. Таким образом, летучие мыши в этом районе имеют
немаловажное значение в уничтожении вредителей растений и
кровососущих двукрылых (Петрусенко, Сологор, 1981).
В Южной Родезии пища насекомоядных летучих мышей со-
стояла в основном из жуков, бабочек и двукрылых. Виды рода
Scotophilus питались преимущественно жуками (до 85%). У Ер-
tesicus capensis было 68% жуков и 18,5% бабочек, у Nycticeius
schlieffeni— 72% жуков и 19% двукрылых. Длиннокрылы охо-
тились в основном на околоводных двукрылых (Fenton et al., 1977).
Хищные виды, живущие за счет позвоночных животных, име-
ются только в двух семействах. В семействе Megadermatidae, рас-
пространенном в Старом Свете, 4 вида из 5 питаются в основном
мелкими летучими мышами, а также другими мелкими млекопи-
тающими, птицами, лягушками и насекомыми. В Новом Свете оби-
тают 4 вида семейства Phyllostomatidae (Phyllostomus hastatus,
Trachops cirrhosa, Chrotopterus auritus, Vampyrum spectrum), в
питании которых, кроме насекомых, значительное место занимают
мелкие позвоночные — летучие мыши, грызуны, ящерицы.
Специализированный рыбоядный вид Noctilio leporinus рас-
пространен в Центральной и Южной Америке. Поедают рыбу два
вида рода Myotis: М. vivesi, обитающая в Мексике и Калифорнии,
Рукокрылые
181
Рис. 13. Процентное соотношение видов рукокрылых, относящихся к разным
трофическим группам, по зоогеографическим областям
Процент видов, питающихся: 1 — летающими насекомыми, 2 — сидящими насекомыми
3 — плодами, 4 — нектаром, 5 — рыбами, 6 — наземными позвоночными, 7 — кровью
млекопитающих
182
Chiroptera
и, вероятно, М. macrotarsus с Филиппинских островов, а также
Megaderma lyra, распространенный в Юго-Восточной Азии.
По характеру питания выделяется семейство Desmodontidae,
к которому относятся три вида монотипичных родов: все они пита-
ются только свежей кровью позвоночных животных. Нападают
почти на всех теплокровных, главным образом на домашних и
диких копытных и птиц.
В Центральной и Южной Америке кровососущие летучие мыши
семейства Desmodontidae наносят животноводству значительный
экономический ущерб. Вампиры снижают продуктивность скота,
передают ряд возбудителей болезней домашних животных. В Ла-
тинской Америке этот ущерб оценивается в 250 млн долл, в год
(Vogt, 1973). Наиболее опасен обыкновенный вампир Desmodus
rotundus, самый многочисленный вид семейства.
Во всех зоогеографических областях среди рукокрылых пре-
обладают виды, питающиеся летающими насекомыми (рис. 13).
На эту группу приходится от 84% в фауне Палеарктики до 44%
в фауне Неотропической области. На втором месте в Австралий-
ской, Неотропической и Восточной областях стоят плодоядные
формы (от 35 до 19%), а в Афротропической, Неарктической и
Палеарктической областях — виды, собирающие насекомых с лист-
вы и земли (от 16 до 12%). На нектароядные формы приходится
не более 13% видов в Неарктической области. На каждую из всех
прочих трофических групп в любой зоогеографической об-
ласти падает менее 4% видового состава рукокрылых (Wilson,
1973).
Соответствующее место представители перечисленных выше
тропических групп рукокрылых занимают и в местных биоценозах.
С экономической точки зрения насекомоядные летучие мыши на
всех континентах весьма полезные животные, уничтожающие вре-
дителей лесов, садов, плантаций и кровососущих двукрылых,
среди последних немало переносчиков болезней человека и домаш-
них животных.
Размножение, развитие, продолжительность жизни
Размножение рукокрылых в тропических областях происходит
на протяжении всего года. У разных видов там наблюдаются три
тица размножения (Anciaux de Faveaux, 1978): 1) моноэстраль-
ный — однократный сезонный цикл с одновременным рождением
детенышей или сроки рождения молодых могут быть растянуты;
2) полиэстральный сезонный — распределение размножения на
два цикла в году, иногда один из сезонов более предпочтителен;
3) полиэстральный непрерывный — не зависит от сезона, длится
в течение всего года; в одной колонии можно обнаружить самок
на разных стадиях беременности и кормящих, самцы активны
постоянно.
Рукокрылые
183
Как у плодоядных, так и у насекомоядных видов период раз-
множения скоррелирован с пиками обилия пищевых ресурсов
(Heithaus et al., 1975; Bradbury, 1979; Fenton, Thomas, 1980).
У летучих мышей, обитающих в умеренных областях, наблю-
дается только один цикл размножения в течение года. Кроме со-
кращения числа генераций, у них произошли отклонения в сроках
образования половых продуктов. У таких видов спаривание обычно
происходит осенью, а роды весной или в начале лета. У самцов
сперматозоиды созревают к концу лета. У самок же летом и осенью
яичники не функционируют, начало овогенеза у них приурочено
к началу зимней спячки. У оплодотворенных с осени самок сперма-
тозоиды сохраняются в половых путях до полного созревания
яйцеклеток. Овуляция, оплодотворение и имплантация происходят
весной к моменту пробуждения из спячки. Часть самок спарива-
ется весной сразу после окончания спячки. В последнем случае
оплодотворение происходит сперматозоидами, перезимовавшими
в половых путях самца. Явление длительного, в течение всей
зимней спячки, сохранения зрелых сперматозоидов в половых
путях самок и самцов среди млекопитающих наблюдается только
у летучих мышей. В результате между копуляцией и родами у
разных видов проходит 150—240 дней (Кузякин, 1950; Merve,
1979; Gopalakrishna, Rao, 1977; Gopalakrishna, Madhavan, 1977;
Madhavan et al., 1978). Собственно беременность продолжается
меньший срок. В умеренных областях длительность беременности
зависит от погодных условий. У нетопыря-карлика в Великобри-
тании в 1978 г. при теплой весне беременность продолжалась 41 день,
тогда как в 1979 г. при холодной весне ее срок увеличился до 51 дня.
Различный темп развития эмбрионов был вызван разной темпера-
турой тела самки. При достаточном количестве корма в хорошую
погоду беременные самки поддерживают температуру тела на
уровне +34. . .+38°, а при голодании в холодную погоду темпе-
ратура их тела следует за изменениями температуры среды, пре-
вышая ее на 2° (Racey, Swift, 1981).
В выводке обычно бывает один детеныш, у некоторых видов —
иногда 2—3. Развитие молодых происходит очень быстро. Они
родятся без шерсти, слепыми. Первые дни детеныш висит на теле
матери, прикрепившись к соску. На кормежку самка летает пер-
вое время вместе с детенышем. Молодые начинают самостоятельно
летать и добывать корм у некоторых видов уже через 20—40 суток,
у других через 3 месяца. Половой зрелости обычно достигают на
следующий год; у ряда видов часть самок может спариваться и в
год своего рождения.
Малая плодовитость рукокрылых компенсируется большой
продолжительностью жизни. Особи даже мелких видов доживают
до 20 лет и более. Крыланы жили в неволе 15—17 лет (Кузякин,
1950). Представление о продолжительности жизни летучих мышей
в природе получены при кольцевании (Панютин, 1980). Наиболь-
184
Chiroptera
ший возраст окольцованных особей установлен у подковоносов
23,5 года), ночниц (18,5 лет) и европейской широкоушки (17 лет).
Несколько меньшая продолжительность жизни отмечена у длин-
нокрыла (12 лет), кожанов (12 лет) и еще меньшая — у нетопырей
(10 лет). После массового кольцевания американских ночниц
Myotis lucifugus в природе (Онтарио, США) два самца были пой-
маны через 29 и 30 лет (Keen, Hitghcock, 1980).
Убежища, миграции
Все рукокрылые — ночные животные. День они, как правило,
проводят в убежищах, которыми служат чаще всего пещеры и ис-
кусственные подземелья. Многие тропические виды днюют, подве-
шиваясь к веткам деревьев, где они защищены от солнечных лучей
густой листвой. Некоторые виды используют для дневки и откры-
то стоящие деревья и кустарники, не избегая прямых солнечных
лучей. При этом они заворачиваются в крылья, что защищает их
от перегрева и излишней потери влаги. Часто для дневки исполь-
зуются одни и те же деревья в течение длительного времени.
Летучие мыши сами убежищ не строят. Крыланы рода Cynopterus
иногда выкусывают центральные плоды в кисти плодов пальм и в
образовавшейся полости подвешиваются. Американские листо-
носы родов Uroderma, Artibeus могут, обкусывая листья пальм,
бананов, сооружать зонтик, защищающий их от солнца, ветра,
дождя (Foster, Timm, 1976). Некоторые виды используют для
дневки норы грызунов. В Америке в норах кроликов находили
летучих мышей родов Tonatia и Tadarida. В Африке и Азии в норах
дикобразов встречаются представители семейства Nycteridae и
рода Hipposideros. На юге Украины во время осенней миграции
находили летучих мышей (рыжую вечерницу, нетопырей лесного
и карлика) в норах емуранчика, обыкновенного слепыша и других
млекопитающих (Абеленцев, 1980). В умеренном поясе летучие
мыши в качестве убежищ используют дупла старых деревьев,
пустоты за корой, погреба, подвалы, чердаки зданий, селятся под
крышами жилых и хозяйственных построек человека, под купо-
лами церквей и мечетей, а также в пещерах и искусственных под-
земельях.
По адаптации к различным убежищам среди летучих мышей
можно выделить группу «пещерных» видов, которые в качестве
летних и зимних убежищ используют пещеры или их аналоги.
В умеренной области Евразии к ним относятся, например, длин-
нокрылы, подковоносы, некоторые ночницы. Другая группа видов
предпочитает дупла, пустоты за корой, щели в скалах, а также
постройки человека. Эту группу составляют вечерницы, нетопыри,
двуцветные кожаны (Стрелков, 1972).
Большинству видов умеренных и северных широт свойственны
сезонные миграции, связанные со сменой летних убежищ на зим-
Рукокрылые
185
ние, и наоборот. Это виды «оседлые», которые совершают миграции
в пределах одной климатической области (Стрелков, 1970). К осед-
лым видам относятся преимущественно «пещерные» виды, исполь-
зующие в качестве убежищ естественные пещеры и различные
искусственные подземелья.
В северных районах лесной зоны (Эстония) летучие мыши зи-
муют в основном в искусственных подземельях. Выявленные там
зимовочные убежища отнесены к трем типам (Мазинг, 1980): мик-
роубежища — подземные помещения с объемом до 50 куб. м (по-
греба, мелкие пещеры), их на территории Эстонии несколько ты-
сяч; макроубежища объемом свыше 1000 куб. м (большие пещеры,
обширные подземные лабиринты и штольни), их в Эстонии 8; ме-
зоубежища с объемом подземелья от 50 до 1000 куб. м — наиболее
важные для рукокрылых зимние укрытия (подвалы старых пусту-
ющих зданий, погреба с несколькими помещениями, бастионы в
старых крепостях, подземные фортификационные сооружения раз-
ных видов и некоторые пещеры), их число доходит до нескольких
сотен. В этих убежищах зимуют в основном водяная ночница,
бурый ушан, северный кожанок.
В Южной Африке (Трансвааль) длиннокрылы (М. schreibersi
natalensis) и подковоносы (Rh. clivosus) мигрируют из пещер,
где летом были материнские колонии, в другие пещеры с более
низкой температурой (Van der Merwe, 1973). В Средней Азии Ба-
харденская пещера служит убежищем для многих тысяч летучих
мышей пяти видов только летом, а на зиму все летучие мыши ее
покидают и перемещаются в другие убежища.
Летние и зимние убежища могут быть расположены на больших
расстояниях, и тогда сезонные перемещения достигают значитель-
ных масштабов. В Северной Европе, по данным кольцевания, рас-
стояние между летними и зимними убежищами водяных ночниц
достигает 330 км, в среднем — около 200 км. Большинство особей
ночниц трехцветных, усатых, больших, Наттерера и больших
подковоносов в Европе совершает сезонные миграции в пределах
100 км. Лишь отдельные особи улетают за 150 и 250 км. Европей-
ские широкоушки и поздние кожаны отлавливались летом на рас-
стоянии 40—80 км от зимних убежищ. Наиболее оседлые виды в
Европе — малые подковоносы и ушаны, которые летом удаляются
от своих зимних убежищ не более 10—20 км (Стрелков, 1970).
В Северной Америке наибольшие сезонные перемещения со-
вершают ночницы. У Myotis lucifugus максимальная зарегистриро-
ванная дальность миграции равна 270 км (Davis, Hitchcook, 1965;
Стрелков, 1970). Кольцевание Myotis grisescens показало, что эти
ночницы разлетаются от мест зимовки на расстояния до 640 км,
обычно до 200 км. Члены одной зимовочной колонии летом обитали
на площади 126 000 кв. км, более чем в 14 летних колониях (Elder,
Gunier, 1978).
Виды, использующие в качестве убежищ дупла, трещины в коре
186
Chiropter a
и скалах, в холодное время года плохо защищены от низких тем-
ператур и не могут зимовать в тех районах, где обычны сильные
морозы. Поэтому они совершают дальние перелеты в другие кли-
матические области с более мягкой зимой. Вечерницы и нетопыри
Восточной Европы на зиму улетают в Среднюю и Южную Европу,
совершая перелеты на расстояния до 1600 км, в среднем 800—900 км.
Дальние перелеты совершают североамериканские виды родов
Lasionycteris и Lasiurus (Стрелков, 1971, 1972; Панютин, 1980).
Колониальность
Летучие мыши, как правило, колониальные животные, лишь
немногие виды держатся поодиночке. Группируются рукокрылые
не только на дневках и в зимовочных убежищах, но и на кормежку
многие виды вылетают стаей.
Многотысячные колонии образуют быстролетные виды, широко-
крылые медленно летающие формы держатся более мелкими груп-
пами (Кузякин, 1950). Зимние скопления летучих мышей обычно
бывают более крупными, чем летние. Это объясняется тем, что зим-
них убежищ меньше, чем летних. Кроме того, перед залеганием
в спячку происходит спаривание и предзимние скопления летучих
мышей бывают особенно большими (Schowalter, 1980).
Самые крупные колонии в Старом Свете известны у длиннокры-
ла: 4000—40 000 особей в Бахарденской пещере в Туркмении (Ку-
зякин, 1950), 6000 зверьков — в пещере в Трансваале в Южной
Африке (Van der Merwe, 1973). Остроухая ночница образует зи-
мовочные колонии до 6000 особей. В Северной Америке Tadarida
brasiliensis mexicana образует огромные выводковые колонии в
пещерах, где скапливается до 1 млн особей (Humphrey, 1971).
У большой ночницы (Западная Украина) и бухарского подковоноса
(Средняя Азия) в колониях насчитывается до 1200—1500 особей.
Меньшие, но достаточно многочисленные колонии у малого под-
ковоноса в Западной Украине — до 150—300, большого подко-
воноса в Предкавказье и Азербайджане — до 200—300, позднего
кожана в Молдавии—до 300—500 особей. Колония нетопыря-
карлика в Великобритании содержала около 300 особей (Racey,
Swift, 1981). Колонии Hipposideros ater ater в Индии насчитывают
до 200—300 зверьков (Gopalakrishna, Madhavan, 1977). Большие
скопления на зимовках образуют американские ночницы М. 1и-
cifugus, М. volans, М. grisescens (Elder, Gunier, 1978; Schowalter,
1980).
Виды, заселяющие постройки человека, дупла и т. п., не об-
разуют крупных скоплений. Eptesicus fuscus в Канаде летом за-
селяет постройки человека, где его колонии насчитывают от 7
до 700 особей (Mills et al., 1975). Европейская широкоушка встре-
чается небольшими группами — от 2—5 до 30—40, а ушан — до
10—12 зверьков. Рыжие вечерницы летом в дуплах деревьев об-
Рукокрылые
187
разуют группы по 3—13 особей, а на зимовке встречаются по 100
и более зверьков (Материалы I Всесоюз. совещ. по рукокрылым,
1974).
Тропические виды, живущие открыто, на дневку устраиваются
на деревьях группами в несколько десятков особей. Нектароядные
летучие мыши в Америке не только отдыхают, но и питаются стаей
(каруселью). Способ питания определяет размер стаи — не более
120, обычно около 25 зверьков (Howell, 1979).
На зимовках оба пола обычно находятся вместе. Летом же в
период размножения самцы и самки многих видов ведут различный
образ жизни. У рыжей и малой вечерниц, нетопырей карлика и
лесного, американских Eptesicus fuscus, Myotis grisescens, Lasiurus
cinereus самки с детенышами образуют отдельные колонии. Самцы
же держатся поодиночке или небольшими группами и часто от
мест зимовки откочевывают лишь на небольшое расстояние. В ре-
зультате летом самцы и самки этих видов встречаются на разных
территориях. Например, взрослые самцы малой вечерницы и не-
топыря-карлика в средней полосе европейской части СССР не
встречаются. У североамериканского вида L. cinereus самцы летом
встречаются вдоль юго-западного побережья США, а самки — в
восточных и центральных частях континента (Стрелков, 1971;
Mills et al., 1975; Elder, Gunier, 1978; Findley, Jones, 1964).
В одном убежище, особенно на зимовках, могут собираться
особи разных видов. При этом один вид доминирует по числен-
ности, а другие встречаются в меньшем числе.
Несмотря на огромные скопления летучих мышей в отдельных
убежищах, особенно на зимовках, они распределены по терри-
тории неравномерно и общая численность зверьков относительно
невелика. По подсчетам, в Чехословакии плотность населения
летучих мышей в 10 раз меньше, чем мелких наземных зверьков
(грызуны, насекомоядные). В Брно общая плотность трех видов
летучих мышей, из которых наиболее многочислен нетопырь-
карлик, составляет в среднем 0,57 особи на 1 га (Gaisler, 1975, 1979).
Распределение по территории видов, живущих небольшими коло-
ниями, более равномерно, а виды, образующие большие скопле-
ния, распределены неравномерно. Например, на всей территории
обитания длиннокрыла в нашей стране известны лишь одна коло-
ния в Средней Азии, 5—б — на Кавказе и 1—2 колонии — в Крыму
(Кузякин, 1950).
Эктопаразиты
В связи с обилием современных родов (174) и видов (около 900),
обитанием на всех материках, за исключением Антарктиды, спе-
цификой жизненных форм, наличием постоянных убежищ, иногда
населяемых тысячами зверьков, рукокрылые имеют достаточно
разнообразную и высокоспецифическую фауну эктопаразитов. Спе-
188
Chiroptera
цифические паразиты рукокрылых насчитывают более 70 родов и
600 видов насекомых и более 130 родов и 800 видов клещей. Из
насекомых только на летучих мышах встречаются одно семейство
клопов (Polyctenidae), два семейства кровососущих мух (Nycteri-
biidae, Sterblidae) и одно семейство блох (Ischnopsyllidae). Из
18 семейств клещей (132 рода и 833 вида), найденных на рукокры-
лых, 4 семейства (Spinturnicidae, Spelaeorhynchidae, Chirorhyncho-
biidae, Teinocoptidae) целиком строго специфичны для этого отряда
млекопитающих, а в таких семействах, как Macronyssidae, Chiro-
discidae, Gastronyssidae, Myobiidae, подавляющее большинство
видов относится к специфическим паразитам рукокрылых (Радов-
ский, 1968; Dusbabek, 1969; Fain, 1976, 1982). Сочетание фауни-
стического многообразия с высокой специфичностью паразитов,
по всей видимости, связано не только с многочисленностью видов
рукокрылых, но также с их большой древностью и значительной
экологической изоляцией от других групп млекопитающих (Ба-
лашов, 1982).
Несмотря на большую зависимость от факторов окружающей
среды, эволюция некоторых групп убежищных кровососов проте-
кала параллельно таковой их хозяев. Наиболее часто эта тенденция
проявляется среди паразитов рукокрылых (клопы сем. Cimicidae,
гамазовые клещи сем. Macronyssidae и др.) и, вероятно, связана
с высокой степенью изоляции как самих летучих мышей, так и их
убежищ от представителей других отрядов млекопитающих.
Перейдем к детальному рассмотрению отдельных групп члени-
стоногих, паразитирующих на Chiroptera.
КЛАСС НАСЕКОМЫЕ — INSECT А
Отряд хоботные — Rhynchota
Подотряд клопы — Не terop tera
К специфическим паразитам рукокрылых относятся клопы
семейств Polyctenidae и Cimicidae.
Сем. Polyctenidae. Облигатные безотрывные эктопаразиты ру-
кокрылых, встречающиеся в тропических и субтропических рай-
онах Индии, Африки и Америки. Безглазые, мелкие насекомые
со сжатым телом и укороченными крыльями. Постоянно живут в
шерсти хозяев. Живородящие. Описано 14 родов. Биологические
особенности почти не изучены (Балашов, 1982). Сведения о пере-
даче ими возбудителей отсутствуют.
Сем. Cimicidae — постельные клопы. Относительно молодая
группа гематофагов, включающая 23 рода с 83 видами, связанными
преимущественно с рукокрылыми и в меньшей степени с птицами.
Из 6 подсемейств 4 — исключительно паразиты рукокрылых,
одно паразитирует у птиц, и только виды подсемейства Cimicinae
Рукокрылые
189
питаются на летучих мышах, птицах и человеке. Представители
подсемейства Afrocimicinae распространены в Африке и являются
специфическими паразитами крыланов. Atrocimicinae — паразиты
эндемичного южноамериканского семейства рыбоядных летучих
мышей Noctilionidae. На бульдоговых летучих мышах (сем. Molos-
sidae) паразитируют клопы рода Primacimex, считающегося наи-
более примитивным, и родов Loxaspis, Crassicimex и Striticimex.
С семейством обыкновенных летучих мышей в Голарктике связаны
клопы рода Cimex. Из 22 родов сем. Cimicidae 12 — специфические
паразиты рукокрылых и только представители рода Cimex регу-
лярно нападают, кроме летучих мышей, на птиц и других млеко-
питающих, включая человека. Случаев видовой специфичности
клопов как паразитов определенных видов летучих мышей не из-
вестно, что, вероятно, отчасти определяется характером парази-
тизма этих членистоногих, находящихся в контакте с хозяином
лишь в период кратковременного кровососания (Балашов, 1982).
Существенное значение имеет и регулярное обитание в одном убе-
жище многовидовых скоплений летучих мышей, относящихся к
разным родам.
Первичные паразиты летучих мышей Cimex lectularius и С.
hemipterus перешли к синантропному образу жизни и стали весьма
назойливыми паразитами человека, который развез их по всем
материкам. С. lectularius по происхождению — палеарктический
вид, широко расселившийся как в странах с умеренным климатом,
так и в тропиках, а С. hemipterus — выходец из Южной Азии —
стал сожителем человека в тропических странах. В тропиках,
где ареалы этих видов перекрываются, С. lectularius обитает в
городах, а С. hemipterus — в сельской местности. Паразит летучих
мышей Западной Африки Leptocimex boueti местами нападает на
человека (Usinger, 1966; Балашов, 1982). В Ираке на Pipistrellus.
kuhli kuhli, отловленных в домах и пещерах, найдены Cimex 1ес-;
tularis, С. hemipterus, С. pipistrelli, а на Asellia tridens murraiama
самка Leptocimex vespertilionis (Abul-hab, 1978).
Несмотря на теснейший контакт с человеком и длительное вы-
живание в организме постельного клопа возбудителей разных
болезней, он не имеет существенного эпидемиологического зна-
чения (Балашов, Дайтер, 1973).
Отряд двукрылые — Diptera
Кровососущие мухи семейств Nycteribiidae и Streblidae пред-
ставляют собой временных эктопаразитов исключительно руко-
крылых и неизвестны с других видов животных. Паразито-хозяин-
ная специфичность определяется возможностью контакта с опреде-
ленными видами рукокрылых, наличием подходящих мест для
развития пупариев и степенью приспособляемости мух к окружа-
ющим условиям. Большинство видов этих эктопаразитов ветре-
190
Chiroptera
чается на колониальных видах, живущих в пещерах, или же на
видах, образующих скопления в строениях человека, трещинах
скал, дуплах. На видах, живущих одиночно или использующих
для отдыха ветви деревьев, эти мухи паразитируют значительно
реже (Балашов, 1982).
Сем. Nycteribiidae — мелкие, бескрылые мухи, формой тела
напоминающие пауков. Постоянные высокоспециализированные
паразиты летучих мышей. Известно свыше 200 видов 8 родов, из
которых в СССР встречается более 17 видов 3 родов. Обитатели
пещер с летучими мышами. Большинство видов свойственно тро-
пикам и субтропикам. Биологические особенности достаточно
полно изучены только для Basilia hispida — паразита косолапого
кожана (Vespertilio pachypus) в Малайзии. Мухи постоянно живут
в шерсти хозяина. Для откладки предкуколки насекомые покидают
хозяина и помещают пупарий на стены пещер в местах дневок
кожанов. От откладки предкуколки до вылупления мух проходит
в среднем около 25 суток, причем само вылупление стимулируется
присутствием в пещере хозяев (выделение СО2 и механические
вибрации). Половая зрелость наступает через 5—6 суток после
вылупления, далее мухи откладывают пупарии с 9-дневным ин-
тервалом. Продолжительность жизни самок в среднем 5—6 недель.
Мухи приступают к кровососанию уже в первые 20 минут после
вылупления и не способны жить вне хозяина дольше 5—25 ч (Ба-
лашов, 1982).
Сем. Streblidae. Высоко специализированные временные эк-
топаразиты рукокрылых. Известно свыше 200 видов 18 родов.
Streblidae почти целиком тропическая группа, в Палеарктике встре-
чаются всего 3 рода, а в СССР 1 вид (Nycteribosca kollari). Общие
таксоны для Старого и Нового Света не известны. С первым це-
ликом связаны виды подсемейств Nycteriboscinae и Ascodipterinae,
а со вторым — Nycterophiliinae, Trichobiinae, Streblinae.
Мухи постоянно живут на теле хозяина, где обитают на голове,
туловище и перепонках крыльев. Виды, живущие на крыльях (роды
Strebla, Trichobius и др.), отличаются очень высокой подвижно-
стью.
Для видов этого семейства характерны частые приемы не-
больших порций крови. Для откладки предкуколок насекомые
ненадолго оставляют тело хозяина. Отличаются большим морфо-
логическим разнообразием. Крылья имеются или редуцированы.
Наблюдаются все степени редукции глаз до полного их отсутствия.
Есть виды с почти нормальным мушиным обликом, формы, напо-
минающие клопов поликтенид, вшей, блох, клещей, пауков, а
также уникальный род Ascodipteron с имагинальным вкожным
паразитизмом. Его вылупившиеся самки имеют нормальные крылья
и ноги. Попав на хозяина, самка отбрасывает крылья и ноги, а
иногда и жужжальца, внедряется в кожу рукокрылого, пока
снаружи не остаются лишь три последних брюшных сегмента.
Рукокрылые
191
Питающиеся мухи уже не способны покинуть хозяина и пупарии
отпадают с его тела (Балашов, 1982).
Среди Streblidae наименее специализированным считают род
Megastrebla, специфичных паразитов крыланов,— наиболее при-
митивных представителей рукокрылых (Маа, 19716). В Южной Аме-
рике у Streblidae наблюдается высокая степень привязанности к
отдельным видам летучих мышей. Среди обитающих там 87 видов
этих мух 55% приходится на моноксенные виды. Виды одного рода
редко встречаются вместе на одном и том же хозяине, а один вид
хозяина редко связан с разными видами этих паразитов в разных
частях своего ареала (Wenzel et al., 1966). На основании изучения
встречаемости 33 видов и подвидов Streblidae и Nycteribiidae на
рукокрылых Австралии установлено преобладание олигоксенных
форм, нормально паразитирующих на двух и более видах хозяев
одного рода, и в меньшей степени моноксенных форм, связанных
только с одним видом летучих мышей. Поликсенные виды отно-
сительно редки, им обычно свойственны довольно широкие ареалы
(Маа, 1971а; Балашов, 1982).
Отряд блохи — Aphaniptera
Специфическими паразитами рукокрылых являются все блохи
сем. Ischnopsyllidae, представленные 93 видами 17 родов и 3 вида
2 родов из сем. Pulicidae, что составляет примерно 5% мировой
фауны этих насекомых.
Использовав обзор Р. Е. Люиса (Lewis, 1974), мы произвели
подсчет родов и видов специфических блох, паразитирующих на
различных семействах рукокрылых. Оказалось, что наиболее
разнообразен набор специфических блох у двух семейств летучих
мышей Vespertilionidae и Molossidae, распространенных на всех
материках. На их долю приходится более 4/5 видового состава
блох рукокрылых. На обыкновенных летучих мышах паразити-
руют наиболее крупные роды: свойственный Старому Свету Ischnop-
syllus (17 видов), Nycteridopsylla (13 палеарктических и 2 неаркти-
ческих вида) и распространенный преимущественно в Западном
полушарии Myodopsylla (из 10 видов 1 палеарктический). Три
вида последнего рода обычные паразиты бульдоговых летучих
мышей. К числу их специфических паразитов относястя блохи
рода Lagaropsylla (18 видов), распространенные в Тропической
Африке, Южной Азии и Австралии, рода Araeopsylla (8 видов),
распространенного в Евразии и Африке, и свойственного Южной
Америке рода Hormopsylla (5 видов). Обращает на себя внимание
отсутствие специфических блох на видах обширного семейства
американских листоносов (Phyllostomatidae) и крайне малое ко-
личество специфических паразитов у богатых видами семейств
подковоносов, мешкокрылов и крыланов (табл. 32).
192
Chiroptera
Таблица 32, Распределение специфических родов и видов блох по
семействам рукокрылых
Отряд и семейство Рукокрылые Специфические блохи
Число современных родов Число современных видов Число родов Число видов
Chiroptera 174 917 18 96
Pteropidae 42 161 1 3
Rhinopomatidae 1 3 1 1
Emballonuridae 12 48 2 3
Craseonycteridae 1 1 — —
Noctilionidae 1 2 2 2
Nycteridae 1 14 1 1
Megadermatidae 4 5 1 1
Rhinolophidae 10 127 1 3
Phyllostomatidae 47 143 — —
Desmodontidae 2 3 1 1
Natalidae 1 4 — —
Furipteridae 2 2 — —
Thyropteridae 1 2 — —
Myzapodidae 1 1 — —
Vespertilionidae 35 314 11 51
Mystacinidae 1 1 — —
Molossidae 12 86 10 34
Анализ распределения специфических блох рукокрылых по
зоогеографическим областям мира показал, что наибольшее ви-
довое разнообразие их зарегистрировано в Палеарктике при от-
сутствии в ней эндемичных родов. По-видимому, это объясняется
как лучшей изученностью фауны блох Палеарктики, так и нали-
чием в умеренном поясе Северного полушария богатого видами
рода Nycteridopsylla — зимних блох, паразитирующих на находя-
щихся в спячке летучих мышах (Лабунец, Богданов, 1959). Видимо,
наиболее богата фауна блох рукокрылых Афротропической области,
где зарегистрировано наибольшее число родов (8), в том числе два
монотипичных эндемичных (Dampfian Oxyparius — паразиты обык-
новенных летучих мышей и щелемордов), два рода, общих с Во-
сточной и Австралийской областями (Thaumapsylla — паразиты
крыланов и Lagaropsylla — паразиты бульдоговых летучих мышей),
три рода, общих с Восточной и Палеарктической областями (Агае-
opsylla, Ischnopsyllus, Rhinolophopsylla — паразиты обыкновен-
ных, бульдоговых летучих мышей и подковоносов) и один род
общий с Палеарктикой (Chiropteropsylla — паразиты мешкокрылов,
мышехвостов и ложных вампиров). В Восточной области имеется
один монотипичный эндемичный род (Mitchella — паразит обык-
Рукокрылые
193
новенных летучих мышей), а остальные пять родов — общие с
Палеарктической, Австралийской и Афротропической областями.
В Австралийской области имеются два эндемичных рода (Porribius
с четырьмя видами, паразитирующими на эндемичных формах
обыкновенных и бульдоговых летучих мышей Австралии, о-ва Новая
Гвинея и Новой Зеландии, и монотипичный род Coorilla— паразит
бульдоговых летучих мышей). В Неарктической области нет энде-
мичных родов блох рукокрылых, два рода (Myodopsylla и Nycte-
ridopsylla) общие с Палеарктической и два (Myodopsylla и Ster-
nopsylla) с Неотропической областями. В Неотропической области
три эндемичных рода: монотипичные Ptilopsylla и Rothschildop-
sylla — паразиты бульдоговых и рыбоядных летучих мышей и род
Hopmopsylla с пятью видами, паразитирующими на бульдоговых,
обыкновенных летучих мышах и вампирах (табл. 33).
Таблица 33. Распределение родов и видов специфических блох рукокрылых
по зоогеографическим областям
Область Всего Эндемичных
родов видов родов видов
Неотропическая 6 16 4 13
Неарктическая 3 8 0 5
Палеарктическая 6 34 0 28
Афротропическая 9 24 2 22
Восточная 6 17 1 10
Австралийская 4 7 2 5
В СССР 5 21 0 0
Кроме представителей сем. Ischnopsyllidae, три вида блох,
специфических паразитов рукокрылых, есть в сем. Pulicidae.
В Неотропической области распространен эндемичный род Rhin-
chopsyllus с двумя видами (R. megastigmata и R. pulex), парази-
тирующими на бульдоговых и обыкновенных летучих мышах.
В Афротропической области обитает специфическая для рукокрылых
блоха Echidnophaga aethiops. Род Echidnophaga (21 вид) широко
распространен в Восточном полушарии. Его представители пара-
зитируют на однопроходных, сумчатых, ежах, хищных, грызунах,
зайцеобразных и бородавочниках.
В СССР известен 21 вид специфических для летучих мышей
блох, относящихся к 5 родам: Araeopsylla, Ischnopsyllus, Myodop-
sylla, Nycteridopsylla, Rhinolophopsylla,— паразитов обыкновен-
ных, бульдоговых летучих мышей и подковоносов (Емельянова,
Высоковский, 1962; Жовтый и др., 1962; Иофф, Микулин, Скалой,
1965; Волянский, Соловьев, 1967; Дубовченко, 1969; Гаджиев,
Дубовченко, 1972).
7 Зак. 3575
194
Chiroptera
КЛАСС ПАУКООБРАЗНЫЕ —ARACHNOIDEA
Отряд клещи — Acarina
Подотряд Parasitiformes
Сем. Spinturnicidae — специфические постоянные эктопаразиты
рукокрылых (10 родов с 80 видами). Распространены по всему
свету. В рассматриваемом семействе прослеживается четкая тен-
денция параллельной эволюции эктопаразитов с их хозяевами —
рукокрылыми. Одно из двух подсемейств этих клещей — Spintur-
nicinae (более древнее) возникло в Старом Свете, где распростра-
нено пять из шести входящих в него родов — Ancystropus (9 видов),
Meristaspis (9 видов), Oncoscelus (1 вид), Eyndhovenia (1 вид),
Spinturnix. Первые три рода — паразиты крыланов, четвертый —
паразит подковоносов, свойственны только Восточному полушарию.
Лишь род Spinturnix (27 видов), паразитирующий на обыкновен-
ных летучих мышах, распространен как в Старом, так и в Новом
Свете, а род Paraspinturnix (1 вид) специфичен для обыкновенных
летучих мышей, обитающих в Новом Свете.
В подсемействе Periglischrinae один род клещей Paraperigli-
schrus (7 Видов) связан с распространенным в пределах Старого
Света семейством подковоносов и евразиатско-африканским семей-
ством Nycteridae. Остальные три рода — Mesoperiglischrus (1 вид),
Periglischrus (22 вида) и Cameronieta (1 вид) — специфические эк-
топаразиты лишь незначительно заходящих в Северную Америку
южноамериканских семейств летучих мышей Natalidae и Phyllo-
stomatidae (Dusbabek, 1969; Fain, 1976; Балашов, 1982).
В СССР! установлено паразитирование на летучих мышах трех
родов (Spinturnix, Eyndhovenia, Paraperiglischrus) и девяти видов
этого семейства (Вшивков, 1963; Арзамасов, 1968; Гаджиев, Ду-
бовченко, 1967; Дубовченко, 1969).
Сем. Macronyssidae содержит паразитов рептилий, птиц и мле-
копитающих, однако большая часть видов этого семейства —
специфические паразиты летучих мышей. Подсем. Macronyssinae
состоит из 9 родов с 42 видами. В их числе монотипичные роды:
Liponysella — паразит лемуров и Acanthonyssus — паразит аме-
риканских хомяков. Все остальные 7 родов, включающие 40 видов,—
специфические постоянные эктопаразиты летучих мышей. Род Bew-
siella с двумя видами распространен в Афротропической и Авст-
ралийской областях и паразитирует на летучих мышах Nycteridae
и Hipposiderinae. Род Ichoronyssus с двумя видами распространен
в Восточном полушарии. Это паразиты ночниц и длиннокрылов.
Монотипичный род Megistonyssus свойствен афротропическим под-
ковоносам. Распространенный в обоих полушариях обширный род
Macronyssus содержит 26 видов, паразитирующих на разнообраз-
ных летучих мышах преимущественно сем. Vespertilionidae. Три
рода Parichoronyssus, Radfordiella, Macronyssoides, к которым
относятся девять видов, представляют собой южноамериканскую
Рукокрылые
195
группу, лишь незначительно проникающую в Северную Америку.
Хозяевами их являются американские листоносы и вампиры. Не-
смотря на то что только два вида Macronyssinae паразитируют не
на летучих мышах, а на других хозяевах, это подсемейство в целом
характеризуется большим морфологическим разнообразием: от
Parichoronyssus (самка в длину немногим больше 0,3 мм, с ком-
пактным телом и короткими крепкими ногами) до Megistonyssus,
который сходен с пауком, гигант 2,5 мм в длину, с ногами, дости-
гающими 1,5 мм (Радовский, 1968).
Подсем. Ornithonyssinae—11 родов с 85 видами — паразиты
млекопитающих (8 родов), птиц (2 рода), рептилий (2 рода). 6 родов
с 49 видами — преимущественные или специфические паразиты
летучих мышей. Три рода — паразиты бульдоговых летучих мышей:
Chiroptonyssus (3 вида) распространены в Западном полушарии,
a Parasteatonyssus (4 вида) и Chelanyssus (3 вида) — в Восточном.
Род Trichonyssus с двумя видами паразитирует на обыкновенных
летучих мышах в Австралийской области. Род Cryptonyssus с
пятью видами свойствен обыкновенным летучим мышам и грызунам
Голарктики. Наконец, обширный космополитный род Steatonyssus
с 32 видами паразитирует в основном на Vespertilionidae, но также
и на Rhinolophidae, Emballonuridae, Nycteridae. В его составе
есть специфический паразит птиц. Ornithonyssinae представляют
собою наиболее продвинувшуюся ветвь Macronyssidae. Они отно-
сительно однообразны, все гематофаги со значительной способно-
стью к насыщению кровью. Это подсемейство освоило огромное
разнообразие хозяев, его политипические роды паразитируют на
пресмыкающихся, птицах и различных млекопитающих, в том
числе и летучих мышах (Радовский, 1968). В связи с хорошо раз-
витой гематофагией клещи рассматриваемого семейства должны
активно участвовать в переносе возбудителей болезней.
По'Ф. Радовскому (1968), роды Bewsiella и Ichoronyssus близки
к леляптидам Notolaelaps и Neolaelaps, паразитирующим на кры-
ланах. По-видимому, от Laelaptidae, сходных с этими родами, и
произошли Macronyssidae, а дальнейшая их эволюция была свя-
зана с подотрядом летучих мышей, с которых они уже вторично
перешли на другие группы позвоночных животных. Этот переход,
вероятно, произошел сравнительно недавно, и поэтому хозяинная
специфичность паразитов других групп млекопитающих, птиц и
рептилий выражена значительно слабее, чем у их предков с лету-
чих мышей (Radovsky, 1967; Балашов, 1982).
В СССР обнаружены: Ichoronyssus scutatus, не менее 10 видов
Macronyssus, 9 видов Steatonyssus (Рыбин, 1983), Ornithonyssus
hoogstraali (Тагильцев, 1970). Сведения по биологии Macronyssus
flavus приведены А. А. Земской (1965) и В. Н. Сенотрусовой (1968).
Сем. Spelaeorhynchidae (1 род с 3 видами) — постоянные экто-
паразиты летучих мышей сем. Phyllostomatidae в Южной Америке
(Fain, 1976).
7*
196
Chiroptera
В сем. аргасовых клещей Argasidae насчитывается от 100 до
150 видов (Поспелова-Штром, 1953, 1959; Филиппова, 1966; Ба-
лашов, 1982), из них более 20 видов относятся к специфическим
паразитам рукокрылых. Из пяти родов аргасовых клещей в трех
есть виды, паразитирующие преимущественно или исключительно
на летучих мышах. К таким формам относятся оба вида американ-
ского рода Antricola, не менее десяти видов рода Ornithodoros
и около десяти видов рода Argas (все виды подродов Carios, Chi-
ropterargas, Secretoargas). По мнению Д. Константина (Constantine,
1971), все эти клещи могут нападать на людей при посещении по-
следними убежищ рукокрылых.
Виды аргасовых клещей — специфических паразитов руко-
крылых и их распространение:
Argas (Carios) vespertilionis
А. (С.) pusillus
А. (С.) australiensis
Argas (Chiropterargas) boueti
A. (Ch.) confuscus
A. (Ch.) afganistaniensis
A. (Ch.) cor diformes
Argas (Secretoargas) transgariepenus
Ornithodoros azteci
O. vignernsi
O. batuensis
O. piriformis
Ornithodoros (Reticulinasus) salahi
O. (R.) faini
O. (R.) madagascariensis
Ornithodoros (Alectorobius) concanensis
O. (A.) kelleyi
O. (A.) stageri
Antricola coprophilus
A. marginatus
Все материки Старого Света
Филиппины, Малайзия
Австралия
Евразия и Африка
Африка
Южная Палеарктика
Юго-Западная Африка
Африка
Южная Америка
» »
Индонезия, Филиппины
Индия
Юго-Западная Палеарктика
Заир
Мадагаскар
Северная Америка
» »
Северная и Южная Америка
То же
Южная Америка
Некоторые полизоидные клещи, например, Ornithodoros papil-
lipes (Al. cholodkovskyi) в массе паразитируют на летучих мышах,
обитающих в пещерах Средней Азии (Поспелова-Штром, 1979).
В СССР обнаружено два вида аргасовых клещей — паразитов
летучих мышей: Argas (Carios) vespertilionis, Argas (Chiropterargas)
boueti (Филиппова, Чунихин, 1969). Первый из них широко рас-
пространен в южных частях страны — от Карпат на западе до
Прибалхашья на востоке. В качестве его прокормителей зареги-
стрировано около 40 видов рукокрылых. Связь этих клещей с
хозяевами носит сезонный характер, паразитирование приуро-
чено к весенне-летнему периоду. Исключением является фаза ли-
чинки, которая встречается на летучих мышах круглый год (Бе-
лашова, 1966). A. vespertilionis обитает в пещерах, дуплах деревь-
Рукокрылые
197
ев, в массе заселяет чердаки построек человека. Многократно
описаны случаи заползания клещей в жилые помещения и приса-
сывание их к людям (Корелов, 1953; Галузо, 1957; Филиппова,
1966; Рыбин, 1980). От этих клещей выделены следующие возбу-
дители болезней: риккетсии Бернета (Жмаева, Пчелкина, Бела-
шова, 1966), вирусы клещевого энцефалита, Иссык-Куль, Сокулук
(Карась и др., 1976). Показана возможность передачи вируса
Иссык-Куль новорожденным белым мышам при укусе имаго и
нимф A. vespertilionis, собранных в естественных условиях (Вар-
гина и др., 1982).
Сем. иксодовые клещи Ixodidae насчитывает около 650 видов
13 родов. В фауне СССР более 80 видов 8 родов. Всего 3 вида рода
Ixodes, насчитывающего более 200 видов, специфические паразиты
рукокрылых. I. vespertilionis распространен в западной и южной
Палеарктике, Восточной и Афротропической областях, паразити-
рует на обыкновенных летучих мышах и подковоносах. I. simplex
распространен пятнами в Палеарктической, Афротропической, Во-
сточной и Австралийской зоогеографических областях, паразит
обыкновенных летучих мышей, преимущественно обыкновенного
длиннокрыла. I. kopsteini распространен в Афротропической, Во-
сточной и Австралийской зоогеографических областях, в основном
паразитирует на бульдоговых летучих мышах (Филиппова, 1972;
Колонии, 1981). В южных районах СССР распространены: I. ves-
pertilionis— от Западной Украины до Казахстана; I. simplex —
от Западной Украины до Восточного Закавказья (Белоконь и др.,
1963; Ушакова, Шаймарданов, 1979). Оба вида в СССР немного-
численны. Видимо, все иксодовые клещи, специфические паразиты
рукокрылых, при контакте могут нападать на человека, но фак-
тические сведения по этому вопросу отсутствуют. При изучении
эктопаразитов летучих мышей в СССР на них обнаружено парази-
тирование единичных I. ricinus, I. eldaricus, I. redikorzevi, Haema-
physalis concinna, H. sulcata, H. numidiana, Dermacentor margi-
natus, Rhipicephalus turanicus, Rh. rossicus, Hyalomma plumbeum
(Гаджиев, Дубовченко, 1970, 1976; Дубовченко, Гаджиев, 1976;
Филиппова и др., 1976). Д. Константин (Constantine, 1971) сооб-
щает о паразитировании на летучих мышах D. andersoni, D. reti-
culatus, Hyalomma aegyptium.
Сем. Chirodiscidae. Из 100 описанных видов 80 — специфические
паразиты рукокрылых. Питаются выделениями жировых желез
и частицами эпидермиса (Балашов, 1982). Их происхождение свя-
зывают с подковоносами или примитивными группами Vespertilio-
nidae. На крыланов они перешли уже вторично, а основная эво-
люционная линия их развития проходила на Microchiroptera.
Большинство родов этого семейства связано с несколькими семей-
ствами их хозяев и многие представлены в Старом и Новом Свете.
Из 15 специфических для рукокрылых родов сем. Chirodiscidae
9 свойственны Старому Свету. Это Asiolabidocarpus (2 вида), La-
198
Chiroptera
bidocarpoides (3 вида), Afrolabidocarpus (3 вида) — паразиты
подковоносов и Megadermatidae; Labidocarpus (3 вида) — паразиты
Megadermatidae и обыкновенных летучих мышей; монотипичные
роды — Adentocarpus (паразит обыкновенных летучих мышей),
Afrokosa и Trilabidocarpus (паразиты бульдоговых летучих мышей),
Pteropiella и Eulabidocarpus (паразиты крыланов).
Политипические роды Paralabidocarpus (10 видов), Olabidocar-
pus (11 видов), Dentocarpus (10 видов), Alabidocarpus (25 видов)
паразитируют на представителях 3; 4; 4 и 9 семейств рукокрылых
и распространены как в Старом, так и в Новом Свете. Два рода
свойственны Западному полушарию: Parakosa (3 вида) — пара-
зиты бульдоговых летучих мышей и Lawrenceocarpus (6 видов) —
паразиты сем. Phyllostomatidae (Fain, 1976).
Одним из интересных проявлений хозяинно-паразитарной спе-
цифичности у Chirodiscidae служит присутствие одного и того же
вида клещей на систематически близких, но географически разоб-
щенных хозяевах. Например, Labidocarpus rollinati — паразит
подковоносов встречается в Европе на Rh. hipposideros, Rh. fer-
rumequinum и в Южной Африке на Rh. clivosus и Rh. capensis,
a Alabidocarpus calcaratus с ночниц — в Южной Африке на Myotis
tricolar, в Южной Америке — на М. nigricans, в Северной Аме-
рике — на М. yumanensis и в СССР на М. blythi.
В СССР известен один вид Alabidocarpus calcaratus, паразити-
рующий на Myotis oxygnathus и Nyctalus noctula в Молдавии и
Азербайджане (Андрейко и др., 1968; Дубовченко, 1969).
Сем. Chirorhynchobiidae — специфические постоянные паразиты
летучих мышей, обитающие на мембранах крыльев. Питаются
выделениями жировых желез и слущивающимися частицами эпи-
дермиса. Один род с двумя видами паразитирует на обитающих в
Южной Америке летучих мышах сем. Phyllostomatidae.
Сем. Gastronyssidae — полостные эктопаразиты рукокрылых и
грызунов (в желудке, кишечнике, носовых полостях, глазах).
В этом семействе шесть родов — паразиты рукокрылых и два рода
(Seivracarus, Yunkeracarus) —• паразиты грызунов (беличьих и
мышеобразных). Среди паразитов рукокрылых три монотипичных
рода: Gastronyssus, Eidolonyssus, Mycteronyssus — специфические
паразиты крыланов. В роде Opsonyssus из 7 видов 4 паразитируют
на крыланах й 3 — на подковоносах. Из И видов рода Rodhainys-
sus 1 паразитирует на Nycteridae, 1 — на Megadermatidae, 5 — на
Vespertilionidae (4 в Восточном и 1 в Западном полушарии), 3—
на Emballonuridae (2 в Западном и 1 в Восточном полушарии)
и 1 — на американских видах Molossidae. Монотипичный род Phyl-
lostomonyssus паразитирует на южноамериканских летучих мышах
сем. Phyllostomatidae.
Сем. Sarcoptidae— чесоточные зудни. Объединяют накожных
или вкожных паразитов сумчатых, рукокрылых, насекомоядных,
грызунов, зайцеобразных, парнопалых и непарнопалых. Из 7 ро-
Рукокрылые
199
дов и 47 видов этого семейства 4 рода (Nycteridocoptes, Chirnyssoi-
des, Chirnyssus, Chirophagoides) — специфические паразиты ру-
кокрылых. Виды рода Notoedres паразитируют на летучих мышах
(18 видов), на грызунах (5 видов), на лемурах и хищных (по 1 ви-
ду) •
Предполагают, что первоначально становление этой группы
клещей происходило на рукокрылых. Наиболее примитивный род
Nycteridocoptes (13 видов) встречается только в Старом Свете.
Девять видов этого рода — паразиты крыланов, два — подково-
носов и два — обыкновенных летучих мышей. Два вида рода Chir-
nyssus паразитируют на представителях Emballonuridae и Vesperti-
lionidae Старого Света. Монотипичный род Chirophagoides — пара-
зит Mystacina tuberculata, единственного представителя эндемичного
для Новой Зеландии семейства Mystacinidae. Восемь видов рода
Chirnyssoides — специфические паразиты южноамериканских ле-
тучих мышей сем. Phyllostomatidae. В наиболее эволюционно
продвинутом роде Notoedres среди 18 видов, паразитирующих на
летучих мышах, 2 вида — специфические паразиты Emballonuridae
Восточного полушария, 8 видов — паразиты Vespertilionidae (5 в
Восточном и 3 в Западном полушарии), 8 видов — паразиты Molos-
sidae (6 в Восточном и 2 в Западном полушарии). Характерно, что
представители этого рода не встречаются на примитивных группах
рукокрылых (Pteropidae, Rhinolophidae), но зато перешли к пара-
зитированию на грызунах, приматах и хищных. Род Chirophagoides
с летучих мышей морфологически очень сходен с родом Sarcoptes,
хотя и более примитивен (Fain, 1976).
Только на крыланах встречается второе семейство чесоточных
зудней Teinocoptidae, объединяющее 3 рода с 16 видами.
Подотряд Trombidiformes
Сем. Myobiidae (волосяные клещи) — содержат около 50 родов
с 336 видами. Из них 21 род с 179 видами специфичен для руко-
крылых, 15 родов с 56 видами свойственны насекомоядным, 9 ро-
дов с 89 видами специфичны для грызунов и 4 рода с 13 видами
характерны для сумчатых (Fain, 1982). Они постоянные безотрывные
эктопаразиты. Мелкие клещи, живущие в шерсти и закрепляю-
щиеся на волосах сильно измененными ногами. Ротовой аппарат
колющесосущего типа. Питаются все активные фазы развития.
Клещи прикрепляются у основания волосяных фолликулов, про-
калывают хелицерами роговой слой эпителия и сосут лимфу. Не-
которые виды, возможно, гематофаги (Балашов, 1982).
На абсолютном большинстве родов рукокрылых паразитируют
специфические виды Myobiidae. На представителях семейства
Megadermatidae волосяных клещей не обнаружено. Также не упо-
мянуты специфические паразиты для видов семейств Rhinopoma-
tidae, Noctilionidae, Natalidae и Myzopodidae. На представителях
200
Chiroptera
всех остальных семейств рукокрылых обнаружены специфические
виды Myobiidae. На крыланах паразитирует 13 видов 3 родов Myo-
biidae. Роды Binuncus (9 видов) и Pteromyobia (3 вида) — целиком
специфичны для крыланов. Среди клещей рода Ugandobia на кры-
ланах паразитирует 1 вид, остальные 9 видов этого рода — паразиты
обитающих в Восточном полушарии летучих мышей сем. Embal-
lonuridae.
Продолжая рассмотрение Myobiidae — специфических парази-
тов подотряда Microchiroptera, отметим, что наибольшим разно-
образием клещей отличается семейство подковоносов, на которых
паразитируют 7 родов и 30 видов, в том числе: Neomyobia — 11 ви-
дов, Hipposiderobia — 8 и Metabinuncus — 7 видов. Только в
Старом Свете распространены 3 вида Calcaromyobia — паразиты
длиннокрылов и подковоносов. На видах Vespertilionidae парази-
тируют роды Acanthophthirus (46 видов) и Pteracarus (18 видов).
Оба рода встречаются как в Старом, так и в Новом Свете. На ази-
атско-африканских летучих мышах сем. Nycteridae паразитируют
клещи монотипичного рода Nycteromyobia. Монотипичный же
род Mystacobia — специфичный паразит эндемичного для Новой
Зеландии и состоящего из 1 вида семейства рукокрылых Му st а-
cinidae.
На бульдоговых летучих мышах паразитируют 17 видов из
18 видов клещей рода Ewingana и 1 вид монотопичного афро-тро-
пического рода Sohizomyobia. 9 видов рода Ewingana — специфи-
ческие паразиты Tadarida и Cheiromeles Старого Света, 8 видов
паразитируют на американских видах Tadarida и Molossus, а 1
вид — на Furipteridae Южной Америки.
Эндемичны в Западном полушарии, преимущественно в Южной
Америке, роды Eudusbabekia (25 видов — паразитов Phyllostoma-
tidae и Desmodontidae), Phyllos+omyobia (3 вида — паразитов Phyl-
lostomatidae), Expltobia (2 вида — паразиты южноамериканских
Emballonuridae) и Thyromyobia (1 вид, паразитирующий на Thy-
ropt eridae).
Таким образом, четко прослеживается разделение эволюцион-
ных ветвей Myobiidae, паразитирующих на рукокрылых. Одна,
наиболее древняя, связана с Восточным полушарием (12 родов с
59 видами), другая практически ограничена в своем развитии
Южной Америкой (4 рода с 31 видом) и только паразитирующие
на Vespertilionidae и Molossidae 3 рода (Acanthophthirus, Pteraca-
rus, Ewingana) с 83 видами имеют представителей и в Старом и в
Новом Свете.
Видимо, Myobiidae, паразитирующие на рукокрылых, про-
изошли от паразитов насекохмоядных. Высказано предположение,
что специфические для крыланов роды Binuncus и Pteromyobia
имели предками паразитов Microchiroptera. Также предположи-
тельно считается, что Myobiidae грызунов произошли от более
примитивных форм — паразитов летучих мышей, близких к совре-
Рукокрылые
201
менному роду Pteracarus, свойственному Vespertilionidae (Dusbabek,
1969; Fain, 1982).
Сем. Demodicidae — клещи-железницы относятся к постоянным
вкожным паразитам сумчатых, насекомоядных, рукокрылых, при-
матов, зайцеобразных, грызунов, хищных, непарнопалых и пар-
нопалых. Описано 134 вида, относимых к одному роду Demodex
(Балашов, 1982). Вероятно, они являются строго моноксенными
паразитами. Клещи-железницы представляют собой уникальную
группу паразитов, развивающуюся на млекопитающих с самого
начала возникновения последних и при адаптивной радиации
хозяев распространившуюся на большинство их крупных таксонов
(Балашов, 1982).
По мнению Фэна (Fain, 1976), на рукокрылых паразитируют
железницы двух родов: 2 вида рода Stomatodex и 10—11 видов
рода Demodex.
Сем. Psorergatidae — постоянные эктопаразиты или вкожные
паразиты насекомоядных, летучих мышей, грызунов, хищных и
парнопалых, обитающие на поверхности кожи, в волосяных фол-
ликулах и внутри эпидермиса. На летучих мышах паразитирует
род Psorergatoides с 11 видами (Fain, 1976; Балашов, 1982).
Сем. Ereynetidae — паразиты наземных моллюсков, амфибий,
птиц и млекопитающих. Обитают в легочной полости моллюсков
и на слизистой носовой полости позвоночных. Питаются кровью.
На рукокрылых паразитируют 3 рода: Neospeleognathopsis с 3 ви-
дами — паразитами крыланов (1 вид), обыкновенных летучих
мышей (1 вид) и бульдоговых летучих мышей (1 вид); Hipposide-
roptes с 2 видами — паразитами подковоносов (1 вид) и бульдого-
вых летучих мышей (1 вид); Speleochir с 6 видами, 5 из которых
паразитируют на Phyllostomatidae и 1 — на Nycteridae. Предста-
вители всех родов встречаются как в Старом, так и в Новом Свете.
Сем. Trombiculidae — клещи краснотелки являются времен-
ными эктопаразитами с длительным питанием (от 2—3 до 5—12
суток). Паразитирует только личиночная фаза. Известно более
100 родов и подродов и свыше 2000 видов краснотелок (Nadchat-
ram, Dohany, 1974).
По работам многих авторов (Vercammen-Grandjean, 1967, 1968,
1976; Anciaux de Faveaux, 1971; Brennan, Reed, 1975) проанализи-
рованы паразито-хозяинные отношения 68 родов и 1470 видов
краснотелок и выделены среди них виды, паразитирующие на
рукокрылых1. На представителях этого отряда найдены красно-
телки 229 видов, относящихся к 38 родам. Краснотелки 11 родов
(112 видов) встречены только на рукокрылых: род Tecomatlana,
содержащий 8 видов, Myotrombicula — 20, Chiroptella — 14, Rud-
1 За этот кропотливый анализ, предоставленные материалы и литературу мы
благодарим Н. И. Кудряшову, любезно согласившуюся помочь осветить
связи краснотелок и летучих мышей.
202
Chiroptera
nicula — 4, Riedlinia — 7, Trombigastia — 19, Perissopalla — 5,
Trisetica — 2, Whartonia — 31, Albeckia — 1, Wagenaaria — 1.
В четырех родах преобладающее число видов (27 из 36) собраны с
рукокрылых. В роде Trombicula из 16 видов 12 — паразиты руко-
крылых, остальные 4 вида паразитируют на других млекопитаю-
щих и рептилиях, в роде Boshkerria из двух видов один — паразит
летучих мышей, а второй паразитирует на других млекопитающих,
в роде Euschoengastoides из 6 видов 4 — паразиты летучих мышей,
а 2 — грызунов, в роде Sasatrombicula 10 из 12 видов — специфи-
ческие паразиты рукокрылых, а 2 найдены на грызунах и члени-
стоногих.
В 23 родах краснотелок на рукокрылых обнаружены немногие
виды: 87 из 875 видов. Большинство этих видов принадлежит
родам, характеризующимся видовым многообразием. В роде Lep-
totrombidium из 251 вида лишь 17 свойственны рукокрылым,
остальные паразитируют на широком круге хозяев: млекопитаю-
щих, птицах, рептилиях, членистоногих. В роде Microtrombicula
из 62 видов паразитируют на рукокрылых 15, а в роде Fonsecia
из 21 вида — 9, в роде Eutrombicula из 43—8. В остальных 19 ро-
дах, содержащих 498 видов, рукокрылым свойственно всего 38
и на один род приходится до 4 видов краснотелок, паразитирующих
на рукокрылых.
В СССР на летучих мышах зарегистрировано 16 видов красно-
телок, относящихся к следующим родам: Myotrombicula — 2 вида,
Chiroptellа — 4, Riedlinia — 1, Trombigastia — 2, Trombicula —
2, Leptotrombidium—1, Neotrombicula — 2, Schoutedenichia — 1
(Шлугер, 1948; Шлугер, Вшивков, 1967; Шлугер, Кудряшова,
1969; Мулярская, 1970; 1971; Мулярская, Дубовченко, 1969, 1970;
Кудряшова, Рыбин, 1974; Кудряшова, 1975, 1979).
Виды краснотелок, найденные на рукокрылых, вероятно, можно
считать если не специфичными, то характерными для этой группы
млекопитающих. На грызунах и других позвоночных эти виды
краснотелок не обнаружены. На летучих мышах в исключительно
редких случаях находили единичные экземпляры краснотелок с
грызунов, но в определенных условиях, например в каменоломнях
Молдавии, где летучие мыши прятались в щелях между камнями.
В крупных пещерах таких случаев не отмечено. Поэтому «специ-
фичность» краснотелок летучих мышей может зависеть не только
или не столько от приспособленности к паразитированию на оп-
ределенном хозяине, но и от условий обитания летучих мышей в
таких местах, где исключена возможность контакта с другими ви-
дами позвоночных.
В заключение следует отметить, что разнообразие и высокая
специфичность паразитофауны рукокрылых объясняется как древ-
ностью, всесветным распространением, обилием видов и особей
этих животных, так и образом их жизни. Способность к длитель-
ному полету вызвала необходимость специальных приспособлений
Рукокрылые
203
у паразитов, а специфические условия мест массового отдыха
рукокрылых — дупла, ветви деревьев, стены и потолки пещер,
строго определенные части построек человека резко ограничили
или полностью исключили возможность обмена паразитами с
другими группами позвоночных.
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Наибольшее число видов возбудителей болезней человека вы-
делено от рукокрылых семейств Pteropidae (6), Rhinolophidae (6),
Phyllostomatidae (8), Vespertilionidae (14), которые отличаются
наибольшим видовым разнообразием (табл. 34). Пять видов воз-
будителей установлено у кровососущих летучих мышей семейства
Desmodontidae, в которое входит лишь три вида. В шести семей-
ствах рукокрылых, бедных видами, возбудители болезней человека
не обнаружены. Из всех инфекций широкое распространение среди
рукокрылых имеют только две — бешенство, которое выявлено у
81 вида 10 семейств, и гистоплазмоз, возбудитель которого найден
у 28 видов 5 семейств.
Возбудители других болезней человека вирусной, риккетсиоз-
ной, бактериальной, протозойной этиологии обнаружены у отдель-
ных видов в разных семействах, и, по всей вероятности, летучие
мыши в их циркуляцию вовлекаются только случайно.
Кроме возбудителей, вызывающих заболевания людей, от
летучих мышей выделены вирусы, бактерии, грибки, свойствен-
ные только этим животным и циркулирующие, вероятно, лишь
в их популяциях, но все они недостаточно изучены.
Вирусные инфекции
Бешенство
Наибольшее значение летучие мыши имеют в эпидемиологии
бешенства. Кровососущие виды сем. Desmodontidae, распростра-
ненные в Центральной и Южной Америке, являются основными
носителями возбудителя в природе и переносчиками заболевания
среди домашнего скота. Они могут кусать также людей и переда-
вать им вирус. На о-ве Тринидад за 1921—1935 гг. в результате
укусов вампиров погибло от бешенства 89 человек (Greenhall,
1968). В Мексике с 1951 по 1961 г. в результате укусов летучих
мышей заболел 31 человек. Всего в Южной Америке известно
более 150 таких случаев (Atanasiu, Acha, 1975; Jones, 1976).
Кровососущие вампиры передают особую форму паралитиче-
ского бешенства, от которой страдает в большой мере скот. По
степени чувствительности к вирусу бешенства, передаваемого вам-
пирами, домашние животные составляют следующий ряд: крупный
рогатый скот, свиньи, лошади, овцы и менее всего чувствительны
собаки (Селимов, 1978). Эпизоотии бешенства скота в Латинской
204
Chiroptera
Таблица 34. Выделение возбудителей болезней человека от различных семейств
рукокрылых
Число совре-
менных
Бешенство
Число видов,
Семейства и отряд
Полушарие
Pteropidae
Rhinopomatidae
Emballonuridae
Noctilionidae
Nycteridae
Megaderma tidae
Rhinolophidae
Phyllostomatidae
Desmodontidae
NataPdae
Furipteridae
Thyrop teridae
Myzapodidae
Vespertilionidae
Mystacinidae
Molossidae
Chiroptera
42
1
12
1
1
4
10
47
2
1
2
1
1
35
1
12
174
161
3
48
2
14
5
127
143
3
4
2
2
1
314
1
86
917
30
14
72
2
1
1
1
Америке временами приобретают значительные масштабы и ох-
ватывают большие территории. Там ежегодно погибает около 500
тыс. голов крупного рогатого скота. Например, в Мексике во время
эпизоотии в 1966—1968 гг. погибло 9,6% невакцинированных и 4%
вакцинированных животных. В этой стране ежегодно погибает от
бешенства 90—100 тыс. голов крупного рогатого скота (Prieto,
Baer, 1972).
Область распространения паралитического бешенства совпа-
дает с ареалом вампиров сем. Desmodontidae (рис. 14) и охватывает
территорию от северной части Мексики на юг до центральной ча-
сти Аргентины. Наибольшее значение как переносчик бешенства
из трех видов вампиров имеет Desmodus rotundus.
Роль обыкновенного вампира как переносчика возбудителей
болезней определяется особенностями его биологии. Desmodus
rotundus распространен по всему ареалу семейства — от Северной
Мексики до Северной Аргентины и встречается как на низменно-
стях, так и в горах на высоте до 2100 м над уровнем моря. Это
Рукокрылые
205
небольшой зверек, длина тела 65—90 мм, размах крыльев 330 мм.
Для кровососания имеет специальные морфологические приспо-
собления: пара верхних резцов с очень острыми концами предна-
значена для резания; между нижними резцами имеется выемка,
по которой язык двигается при кормлении. Язык при поглощении
крови заворачивается вниз и вместе с бороздкой, имеющейся на
нижней губе, образует трубку, по которой кровь из раны поступает
в ротовую полость. Язык при этом выполняет роль поршня. Пи-
щеварительный канал (включая желудок) короткий и представ-
ляет простую трубку, приспособленную к быстрому переварива-
нию крови. В слюне содержится антикоагулянт, препятствующий
свертыванию крови.
Наблюдения за вампирами в лаборатории и в естественных ус-
ловиях, проведенные с применением инфракрасных лучей, позво-
лили проследить весь процесс питания вампиров (Vogt, 1973). На
охоту зверьки вылетают ночью. На поиски и выбор жертвы вампир
затрачивает до 1,5 ч. На. выбранное животное он садится сразу
206
Chiroptera
Рис. 14. Распространение паралити-
ческого бешенства скота и ареал ва-
мпиров сем. Desmodontidae (по: Ata-
nasiu, A ch а, 1975)
1 — область распространения бешенства,
2 — точки зарегистрированных эпизоотий
бешенства у скота с 1965 по 1974 г.,
3 — граница ареала вампиров
или заползает на него с земли. Для укуса выбирает участки тела
с поверхностной и богато разветвленной сетью капилляров, глав-
ным образом область ушей. Собственно укусу предшествует дли-
тельное (10—40 мин) смачивание волос слюной, что имеет огромное
эпизоотологическое значение. Затем зверек сгрызает волосы и
участок кожи шириной 4—5 мм и до 5 мм толщиной. Сочащуюся
из ранки кровь вампир слизывает или лакает. Сосание продол-
жается без перерыва около 30 мин, иногда более часа. За одно
кормление вампир поглощает 15—50 мл крови. Обычно он не при-
чиняет беспокойства своей жертве во время кормления. Действие
антикоагулянтов продолжается несколько часов после того, как
вампир насытится и улетит, и животное теряет еще много крови из
ранки, оставленной зверьком. После кровососания вампир отды-
хает некоторое время на дереве среди листвы, а под утро улетает
в убежище (Vogt, 1973). В качестве убежищ они используют пе-
щеры, где собираются многими сотнями особей. Как правило, вам-
пиры держатся одних и тех же мест и дальних миграций не совер-
шают (Forment et al., 1971). Увеличение в местности поголовья
скота благоприятствует увеличению численности вампиров (Green-
hall, 1972).
У вампиров заболевание бешенством протекает в виде парали-
тического энцефалита со смертельным исходом, но иногда некото-
рые особи выживают и остаются носителями вируса. Вирус в ор-
ганизме вампиров обнаруживается в центральной нервной системе,
в слюнных железах, почках, легких, сердце, в теле плода, что го-
ворит о трансплацентарной передаче возбудителя. Вирус обнаружен
в слюне и моче этих животных (Diego, Valotta, 1981).
Рукокрылые
207
Рис, 15. Распространение бешенства летучих мышей в США (по: Davis et al.t
1970; Fischman, 1976)
Распространение вируса бешенства среди вампиров может
осуществляться на дневках в пещерах через укус, при копуляции,
через молоко самок, трансплацентарно, а также путем аэрозоль-
ной передачи (Селимов, 1978; Diego, Valotta, 1981).
Для борьбы с вампирами как переносчиками бешенства приме-
няются различные методы отлова зверьков — отстрел, вылов сетя-
ми, ловушками, выкуривание из дупел и т. п. Массовое уничто-
жение вампиров проводят в пещерах с помощью динамита, отрав-
ляющих газов, электричества, но при этом гибнет много других
летучих мышей. Для отпугивания вампиров от животных исполь-
зуют искусственное освещение в ночное время загонов и помещений
для скота, но слабые источники света при этом малоэффективны.
Чтобы защитить жилые и животноводческие помещения от проник-
новения в них вампиров и других летучих мышей, ставят защит-
ные экраны металлические и пластиковые с отверстиями не более
22 мм. Для защиты скота и людей применяют также паутинные
сети, которые расставляют вокруг охраняемого лагеря или загона
со скотом. Из современных методов Комитет экспертов ВОЗ по
бешенству рекомендует метод антикоагулянтов. Антикоагулянт
(дифенадион) в небольших дозах вводят скоту, и кровь обработан-
ного животного становится смертельной для вампиров. Однако
этот метод требует дальнейшего изучения и определения степени
его безвредности. Антикоагулянт может быть применен иначе.
В смеси с вазелином он наносится на спинку отловленных вампи-
ров, которые затем снова выпускаются. В колонии, очищая друг
208
Chiroptera
друга, антикоагулянт слизывают многие зверьки и гибнут. Для
биологического контроля популяции вампиров могут быть исполь-
зованы хемостерилизанты. Все методы борьбы с вампирами тре-
буют совершенствования, дальнейшей разработки и изучения (Gre-
enhall, 1974).
Кроме кровососущих летучих мышей, вирус бешенства выделен
от многих других видов, питающихся насекомыми, плодами, нек-
таром. В Мексике бешенство установлено у 31 вида, а в Центральной
и Южной Америке — 23 видов летучих мышей из семейств Molos-
sidae, Vespertilionidae, Phyllostomatidae, Emballonuridae, Nata-
lidae, Noctilionidae (Acha, 1967a, b; Baer, 1975; Stouraitis, Salva-
tierra, 1978). Эти некровососущие летучие мыши отловлены на
территории, где распространены и вампиры. Вирус бешенства
они, вероятно, могут получить при контакте с вампирами в одних
убежищах (пещерах) или сталкиваясь с ними случайно в листве
деревьев, где последние укрываются для кратковременного отдыха.
Однако и за пределами ареала вампиров в Северной Америке
на территории США и южной части Канады также установлено
бешенство у насекомоядных летучих мышей.
В США бешенство насекомоядных летучих мышей отмечено в
47 штатах (рис. 15). Ежегодно в стране регистрируется несколько
сотен случаев бешенства у этих животных (табл. 35). Всего за 20 лет
в США установлено 5028 случаев бешенства летучих мышей.
Таблица 35. Число случаев бешенства у насекомоядных летучих мышей
в США (по Parker, 1975; Fischman, 1976)
Год Число случаев Год Число случаев
1953 8 1964 352
1954 4 1965 484
1955 14 1966 377
1956 41 1967 414
1957 31 1968 291
1958 68 1969 321
1959 80 1970 296
1960 88 1971 465
1961 186 1972 504
1962 157 1974 544
1963 303 Всего 5028
Частота выделения вируса бешенства от летучих мышей в боль-
шой мере определяется возросшим интересом исследователей к
этой проблеме. На общем фоне распространения бешенства среди
Рукокрылые
209
диких животных бешенство летучих мышей составляет относи-
тельно небольшой процент. В Канаде в 1975—1976 гг. было за-
регистрировано 4377 случаев бешенства животных (Gowan, 1976),
из них на долю диких животных пришлось 2370 случаев, в том
числе у летучих мышей — 105, что составляет 4,4% случаев бе-
шенства среди диких животных. В США в штате Канзас за 1946—
1966 гг. отмечено бешенство у 587 млекопитающих, из них было
лишь 14 летучих мышей — 2,4% (Birney, Rising, 1967). За 1953—
1972 гг. в США зарегистрировано 93 723 случая бешенства у жи
вотных (собак, кошек, лисиц, скунсов, животных на фермах),
из них на долю летучих мышей приходится 4484 случая — 4,8%
(Parker, 1975).
При специальных многолетних исследованиях летучих мышей
на бешенство во Флориде вирус был выделен в первые годы в 0,85%
случаев, а в последующие, когда применялись более совершенные
методики,— в 10,3%. В среднем вирусом бешенства поражено
3,2% летучих мышей (табл. 36).
Таблица 36. Частота выделения вируса бешенства от летучих мышей
в Северной Америке
Страна, штат Годы Всего исследо- вано зверьков Выделен вирус бешенства Авторы
число %
США Флорида 1954—1961 6 447 55 0,85 Bigler et al., 1975
Флорида 1962—1973 2 293 236 10,3 Bigler et al., 1975
Теннесси 1962 250 16 6,4 Fredrickson, Thomas,
Нью-Йорк 1974—1975 611 9 1,6 1965 Trimarchi, Debbis,
Нью-Мексико Оклахома 3 000 1 071 89 2 3,0 0,2 1977 Tierkel, 1959 пз Селимову, 1978)
Канада 1963—1967 628 44 7,0 Beauregard, 1969
Всего 14 300 451 3,2
Бешенству в Северной Америке подвержены самые различные
виды летучих мышей. Из 40 видов, встречающихся здесь, бешен-
ство зафиксировано у 38 видов, относящихся к семействам Vesper-
tilionidae, Phyllostomatidae, Molossidae. Всего у этих видов ус-
тановлено 1344 случая бешенства. Наибольшее число их отмечено
у Tadarida brasiliensis mexicana (213), Eptesicus fuscus (244) и
трех видов рода Lasiurus: L. borealis (224), L. cinereus — (173),
8 Зак. 3575
210
Chiroptera
Рис. 16. Изменение зараженности летучих мышей в США вирусом бешенства
по месяцам (по: Baer, 1975)
а — исследовано зверьков, б — выделен вирус; 1 — Eptesicus fuscus (оседлый) в Колорадо,
2 — он же в Аризоне, 3 — Lasiurus cinereus (мигрант) в Колорадо, 4 — он же в Аризоне и
Нью-Мексико, 5 — Tadarida brasiliensis mexicana (мигрант) в Нью-Мексико
L. intermedius (184) (Baer, Adams, 1970; Davis et al., 1970; Bigler
et al., 1975; Constantine, 1971, 1979; Constantine et al., 1979).
Экологически эти виды очень различны. Т. b. mexicana — коло-
ниальный вид, живущий в пещерах, совершающий сезонные миг-
рации на значительные расстояния, зимующий в Мексике и других
странах Центральной Америки. Е. fuscus тоже колониальный вид,
убежищами ему служат большей частью постройки человека, вид
оседлый, дальних миграций не совершает. Виды Lasiurus населяют
только южную часть США, живут свободно, укрываются в листве
деревьев, но на зиму улетают на юг в Центральную и Южную
Америку.
Таким образом, на зимовках многие виды имеют возможность
контактировать с вампирами и получить от них вирус. Поскольку
инкубационный период длительный, то у заразившихся на зимовке
в очагах бешенства летучих мышей болезнь проявляется, когда
они возвращаются к местам летнего обитания.
У оседлого Е. fuscus положительные находки бешенства реги-
стрировались с апреля по октябрь, у перелетного L. cinereus в
северной части ареала — с апреля по октябрь, а в южной — с
февраля по ноябрь, у совершающей дальние миграции Т. b. mexi-
cana — с февраля по декабрь с двумя максимумами в апреле и в
сентябре (рис. 16).
Американские исследователи предполагают, что насекомоядные
летучие мыши могут передавать бешенство друг другу также на
дневках в пещерах. Вирус бешенства был выделен из воздуха
пещеры (Техас, США), где в летнее время находилась колония Т. Ь.
mexicana, в которой было около миллиона самок и еще не умеющих
летать детенышей (Winkler, 1968). Описан случай, когда больная
Рукокрылые
211
бешенством Lasiurus cinereus (у нее впоследствии были найдены
тельца Негри), нападала поочередно на трех других летучих мы-
шей разных видов (Lasionycteris noctivagans, Т. brasiliensis, Ер-
tesicus fuscus), прилетавших охотиться на тот же внутренний двор
(Bell, 1980). Не исключается также вероятность передачи вируса
бешенства членистоногими, так как у летучих мышей возможна
стабильная вирусемия. В эксперименте на зараженных бешенством
летучих мышах удалось заразить иксодовых клещей разных видов
(Aubert, Andra, 1975).
Отмечено, что бешенство среди лисиц чаще встречается в тех
районах, где много пещер. Поскольку в пещерах обычно обитают
летучие мыши, было высказано предположение, что лисицы за-
ражаются от летучих мышей. Подробное изучение этого вопроса
показало, что есть положительная корреляция только между
числом пещер и частотой бешенства у лисиц, а связь заболеваемо-
сти лисиц с заболеваемостью летучих мышей оказалась слабой.
Решение этого вопроса требует убедительных доказательств как
биологической возможности передачи вируса от одного вида к
другому, так и эпидемиологического подтверждения такой пере-
Таблица 37. Выделение вируса бешенства от насекомоядных и плодоядных
рукокрылых в Европе, Азии и Африке
Вид Страна Автор
Сем. Vespertilionidae
Nyctalus noctula Югославия Nicolic, Jlesic, 1956
То же ФРГ Mohr, 1957 (по Baer, 1975)
» СССР, Казахстан Смирнова, 1969
» СССР, Узбекистан Маматов, 1976
Eptesicus serotinus ГДР Pitzscke, 1965
То же СССР, Узбекистан Маматов, 1976
Vespertilio pipistrellus То же Маматов, 1976
Sp.,? ПНР Komorowski et al., 1974
Myotis myotis Западный Берлин Hentschke et al., 1975
Vespertilio brunneus Гвинея Петручук, 1980
Сем. Rhinolophidae
Rhinolophus ferrumequinum Турция Tuncman, 1958
Сем. Megadermatidae
Lavia frons Гвинея Петручук, 1980
Сем. Pteropidae
Су nop terus brachyotis Таиланд Smith et al., 1967
Pteropus poliocephalus Индия Pal et al., 1980
8*
212
Chiroptera
Таблица 38. Частота выделения вируса бешенства от рукокрылых
в Европе, Азии и Африке
Страна Исследо- вано зверьков Выделен вирус Автор
Югославия 27 1 Nicolic, Jlesic, 1956
Румыния 304 0 Sturbu et al., 1973
Италия 504 0 Morganti, Prosper!, 1972
Швейцария 21 0 Steck, 1982
СССР 527 3 Маматов, 1976
517 0 Анина-Радченко и др., 1970
300 0 Клюева, Панютин (по Ботвинкину, 1983)
452 0 Сансызбаев, 1981
22 0 Грибанова, Мальков, 1977
151 0 Рыбин, 1980
243 0 Ботвинкин, 1983
67 0 Канторович и др., 1967
Филиппины,
о-в Лусон 133 0 Westerlund, 1966
360 0 Arambulo, 1963
о-в Негрос 562 0 Tacal, Boado, 1966
АРЕ 436 0 Sabban et al., 1967
Всего 4626 4
дачи в природе (Fredrickson, Thomas, 1965; Fischman, 1976; Fi-
schman, Young, 1976; Lineback, 1980).
Если в Центральной и Южной Америке существуют настоящие
природные очаги бешенства, в которых основным носителем и
переносчиком являются кровососущие летучие мыши, а в Северной
Америке достаточно часто выделяют вирус бешенства от насекомо-
ядных летучих мышей, то в Восточном полушарии случаи бешенства
у летучих мышей редки. Единичные находки вируса бешенства у
летучих мышей известны в Европе, Азии, Африке (табл. 37).
Обследование летучих мышей на бешенство в Восточном полу-
шарии проводили менее интенсивно и охвачено им меньшее число
особей рукокрылых, чем в Северной Америке. Тем не менее процент
зараженных среди 3483 исследованных зверьков оказался в 20 раз
ниже, чем у летучих мышей в Северной Америке, соответственно
0,14 и 3,2% (табл. 38).
Вирусологическое и серологическое исследования 436 летучих
мышей 10 видов, отловленных в различных районах АРЕ, не вы-
явило у них вируса бешенства (Sabban et al., 1967). В то же время
экспериментальное изучение восприимчивости насекомоядной ле-
тучей мыши Rhinopoma hardwickei к вирусу бешенства показало,
Рукокрылые
213
что вид полностью восприимчив к этому вирусу (Sabban et al.,
1967).
Авторы предполагают, что если эти летучие мыши столкнутся
с вирусом бешенства, то могут стать его переносчиками.
Эпидемиологическое значение бешенства насекомоядных лету-
чих мышей по сравнению с вампирами, очевидно, не столь велико,
но оно не может недооцениваться (Baer, 1975). Описаны неоднократ-
ные случаи заражения людей бешенством от насекомоядных лету-
чих мышей, когда люди брали в руки раненых или ослабевших
животных и последние их кусали (табл. 39). За 1960—1979 гг. в
Таблица 39. Случаи гидрофобии в результате контакта человека
с насекомоядными летучими мышами (Baer, 1975, с дополнениями)
Вид летучих мышей Год Страна, местность
Sp.? 1951 США Техас
Dasypterus floridanus 1953 Флорида
Tadarida brasiliensis 1956 Техас
Tadarida brasiliensis 1959 Техас
Sp.? 1959 Висконсин
Lasionycteris noctivagans 1961 Калифорния
Eptesicus fuscus 1970 Огайо
Sp.? 1971 Нью-Джерси
Sp.? 1973 Кентукки
Sp.? 1970 Канада Саскачеван
Sp.? 1985 Альберта
Miniop terns sp.? 1970 ЮАР, Трансвааль
Nyctalus noctula 1957 ФРГ, Гамбург
Vespertilio serotinus 1964 СССР Киев
Sp.? 1977 Ворошиловград
Sp.? 1985 Белгород
Sp.? 1954 Индия
США заболело бешенством 38 человек, из которых 24 были укушены
животными, в том числе четверо — летучими мышами (Anderson
et al., 1984). До 1961 г. в США было известно 6 случаев заражения
человека бешенством от летучих мышей, из них 4 от укуса. В двух
других случаях заболевшие бешенством люди не имели непосред-
ственного контакта с летучими мышами, но перед болезнью посе-
щали пещеры, в которых обитало множество летучих мышей. Ве-
роятно, в последних случаях заражение произошло аэрогенным
214
Chiroptera
путем (Rubin et al., 1969). Большинство заражений людей бешен-
ством от летучих мышей известны в Северной Америке, но такие
случаи отмечены и в Европе, и в Азии, и в Африке.
Арбовирусы
От рукокрылых на разных континентах выделены 22 арбови-
руса. Из них лишь половина патогенна для человека. Значительное
число вирусов, вероятно, видоспецифично, так как они ни разу
не были выделены от других животных (Львов, Лебедев, 1974).
Вирусы, выделенные из слюнных желез летучих мышей, еще не-
достаточно изучены, и пути их циркуляции окончательно не ус-
тановлены. В экспериментах показана восприимчивость летучих
мышей к ряду африканских арбовирусов, но требуется дальнейшее
изучение роли летучих мышей как основных хозяев этих вирусов
(Simpson, 1969).
Более подробно изучено значение летучих мышей в циркуляции
вируса венесуэльского энцефалита лошадей. Этот арбовирус пе-
редается комарами и поражает копытных, особенно губителен
для лошадей, но могут заболевать и люди. Летучие мыши также
могут быть хозяевами этого вируса. Штаммы возбудителя получены
от пяти видов летучих мышей: Uroderma bilobatum, Artibeus tur-
pis, Carollia subrufa, C. perspicillata, Desmodus rotundus. Анти-
тела к вирусу венесуэльского энцефалита найдены в Мексике и
Гватемале у десяти видов летучих мышей. В эндемичном по этому
заболеванию районе Юго-Восточной Мексики положительная РТГА
отмечена у 0,17% летучих мышей, тогда как у наземных млеко-
питающих этот процент выше — 0,43% (Scherer et al., 1971).
Инфицирование летучих мышей вирусом венесуэльского энце-
фалина может происходить как при непосредственном питании
кровью больных животных (кровососущие виды), так и через
укусы комаров — виды, питающиеся насекомыми и плодами (Cor-
rea-Giron et al., 1972; Seymour et al., 1978a).
Экспериментальное изучение венесуэльского энцефалита у пяти
видов летучих мышей (Artibeus lituratus, A. jamaicensis, Phyllo-
stomus discolor, Sturnira lilium, Carollia subrufa) показало, что у
92,5% зараженных зверьков отмечается вирусемия, но клиниче-
ское проявление болезни не наблюдается. Высокие титры виру-
семии регистрируются в первые три дня после заражения и затем
снижаются. Средняя продолжительность вирусемии от 2,3 до 5,2
дней (Seymour et al., 19786). У экспериментально зараженных жи-
вотных вирус выделяется из носоглотки (в 56%) в среднем через
1,2—2,2 дня после заражения, обычно на следующий день после
появления вирусемии. В моче и кале вирус обнаруживается реже:
он выделен только в 2 из 123 проб мочи (от 50 зверьков) !и в 2 из
86 проб кала (от 46 зверьков). Во всех случаях вирус изолирован
в первые 6 дней после заражения. Заразить здоровых летучих
Рукокрылые
215
мышей контактным или воздушно-капельным путем от зараженных
зверьков не удалось (Seymour, Dickerman, 1978).
Экспериментальное изучение японского энцефалита показало,
что летучие мыши (Eptesicus fuscus, Myotis lucifugus, Pipistrellus
subflavus) могут заражаться этим вирусом при подкожном вве-
дении, через укус комарами и при скармливании субстрата с воз-
будителем. У зараженных зверьков наблюдается вирусемия в
течение шести дней без клинических проявлений. В обстановке,
имитирующей условия спячки, летучие мыши сохраняли вирус
в течение 107 суток. При повышении окружающей температуры
у летучих мышей происходила реактивация вируса и концентрация
возбудителя в крови превышала минимальную заражающую дозу
для комаров. Зараженные в эксперименте комары передавали вирус
летучим мышам (La Motte, 1958). Эти данные говорят о возможности
вовлечения летучих мышей в эпизоотии японского энцефалита.
В Японии вирус японского энцефалита выделен от Miniopterus
schreibersi' и Rhinolophus cornutus (Мошкин, Лебедев, 1974).
Арбовирусы в СССР
Клещевой энцефалит. В Киргизии штаммы вируса клещевого
энцефалита неоднократно выделялись от летучих мышей различных
видов по всей территории республики. Из 10 пещер в южных рай-
онах Киргизии отловлено 170 экз. рукокрылых, от которых вы-
делено 25 штаммов вирусов, среди последних 7 были идентичны
вирусу КЭ. У 10,9% исследованных зверьков зарегистрирована
вирусемия. Наиболее важная роль как резервуара вируса клеще-
вого энцефалита, по-видимому, принадлежит нетопырю-карлику
и остроухой ночнице — наиболее массовым видам в южных рай-
онах Киргизии (Варгина, 1976).
В г. Фрунзе и населенных пунктах Чуйской долины и Иссык-
Кульской котловины отловлено 738 экз. летучих мышей, из которых
выделено 24 штамма арбовирусов, в том числе клещевого энце-
фалита. Наиболее пораженным видом был поздний кожан, зара-
женность которого достигала 5,53% (Варгина и др., 1976).
В Алма-Ате зимой 1975/76 г. ^исследовано 90 рыжих вечерниц
и выделено 4 штамма вирусов, из которых один оказался вирусом
клещевого энцефалита (Дробищенко и др., 1976). Штамм этого
вируса выделен также из клещей Argas vespertilionis в Киргизии
(Карась и др., 1976).
Авторы предполагают, что летучие мыши могут играть опре-
деленную роль как в рассеивании вируса по территории, чему
способствуют сезонные миграции летучих мышей, так и в сохра-
нении вируса в зимний период благодаря способности вируса дли-
тельно сохраняться при ‘низких температурах тела зимующих
летучих мышей. . '
216
Chiroptera
Вирус Иссык-Куль. На северо-западе Киргизии вирус Иссык-
Куль выделен от всех видов летучих мышей, обитающих в Чуй-
ской долине, в том числе от таких массовых видов, как остроухая
ночница, поздний кожан, рыжая вечерница, нетопырь-карлик.
За 1970—1975 гг. исследовано 917 летучих мышей и выделено
15 штаммов вируса Иссык-Куль (Карась и др., 1973; Кучук, 1977).
На юге Таджикистана исследовано 413 летучих мышей четырех
видов и от двух нетопырей-карликов (из 386) изолировано 2 штамма
вируса (Костюков и др., 1975). Вирус Иссык-Куль патогенен для
летучих мышей (нетопыря-карлика, рыжей вечерницы) и может
приводить их к гибели (Кучук, 1977). При экспериментальном
заражении у нетопырей-карликов вирусемия наблюдалась в те-
чение 19 дней, вирус обнаруживался также в буром жире, мозге
и внутренних органах (Костюков, Булычев, 1975).
Передача вируса в природе, очевидно, осуществляется крово-
сосущими членистоногими — клещами и комарами. Штаммы ви-
руса получены от Argas vespertilionis, собранных с летучих мышей
и из субстрата в их убежищах, а также от комаров (Карась и др.,
1973; Кучук, 1977). При исследовании A. vespertilionis изолиро-
вано в 25,6% биопроб 11 штаммов вируса Иссык-Куль (Карась,
и др., 1976). От 652 клещей этого вида, исследованных в 17 био-
пробах, получено 3 штамма вируса. В лабораторных условиях
показана передача вируса естественно зараженными клещами
A. vespertilionis (имаго и нимфы) через укус сосункам белых мышей
(Варгина и др., 1982). Комары также могут передавать этот вирус.
В эксперименте комары Aedes с. caspius воспринимали вирус Ис-
сык-Куль при питании на искусственно зараженных этим вирусом
нетопырях-карликах и .затем через 10 и 15 суток после заражаю-
щего кормления передавали вирус укусом сосункам белых мышей.
Коэффициент инфицирования комаров на 10-й день был достаточно
высок и составлял 55—79%. Процент передачи вируса через укус
достигал 58—63% (Костюков и др., 1982). Клещи и комары могут
как осуществлять циркуляцию вируса среди летучих мышей, так
и передавать его другим животным и человеку. Известен случай
заболевания человека в результате заражения вирусом Иссык-
Куль при сборе летучих мышей (Пак и др., 1978).
Вирус Солукук. Выделен впервые от нетопырей-карликов в Кир-
гизии (Карась и др., 1973). Он изолирован также от клещей Argas
vespertilionis (Варгина и др., 19766; Карась и др., 1976).
Вирус Тягиня. Штамм этого вируса получен от нетопыря-кар-
лика, отловленного в Таджикистане в пос. Нижний Пяндж. При
экспериментальном заражении этих летучих мышей вирусом Тя-
гиня у них наблюдается вирусемия и титр вируса в крови дости-
гает 2,7 1g LD 50,0/0,01 мл (Костюков, 1976). Вирус Тягиня пере-
дается комарами, от которых в Таджикистане выделено 4 штамма
этого вируса. Теплокровным хозяином на юге Таджикистана,
по-видимому, является нутрия. Приведенные выше эксперимен-
Рукокрылые
217
тальные данные показывают, что летучие мыши могут играть здесь
определенную роль в сохранении вируса в зимний период, особенно
нетопырь-карлик, широко распространенный на юге Таджики-
стана (Костюков и др., 1976).
Бактериальные и риккетсиозные инфекции
Лептоспироз. Роль летучих мышей в лептоспирозе еще не ясна,
но, по всей вероятности, она невелика. Есть мнение, что лепто-
спироз человека и животных в Эфиопии распространяют летучие
мыши через общие водные источники. Лептоспироносители среди
летучих мышей выявлены лишь в пяти родах двух семейств: Pte-
ropidae и Vespertilionidae. В Дании, Индонезии, Малайзии, Авст-
ралии у них найдены лептоспиры, относящиеся к трем серогруп-
пам — Cynopteri, Australis и Hebdomadis (Карасева, Свешникова,
1971; Constantine, 1971).
Кишечные инфекции. От рукокрылых в различных странах
Азии, Африки и Америки получены копрокультуры сальмонелл
43 серотипов и шигелл различных видов, среди которых часто на-
блюдаются патогенные для человека. Крылан Pteropus rufus на
Мадагаскаре является носителем Salmonella typhimurium. В ряде
колоний этих крыланов инфицированность наблюдается постоянно.
Пути передачи возбудителя среди них не ясны. Возможно, мухи
(дрозофилы) загрязняют бананы, которыми питаются зверьки (Вгу-
goo, 1974).
Клещевой боррелиоз. Возбудитель спирохетоза Borrelia recur-
rentis выделен от летучих мышей в СССР, в Европе, Передней
Азии, Африке, Северной и Южной Америке. У ряда видов вы-
звана экспериментальная инфекция, в то же время не удалось
осуществить передачу В. recurrentis свиньям с помощью Desmodus
rotundus (Constantine, 1971).
Лихорадка Ку. Известны отдельные факты выделения возбу-
дителя этого риккетсиоза от летучих мышей в различных странах.
Зараженность лихорадкой Ку установлена (получен штамм) у
Eptesicus isabellinus в Марокко (Blac, Bruneau, 1957). В СССР
С. burneti выделены от нетопыря-карлика и усатой ночницы при
исследовании 63 летучих мышей, отловленных в Ташкенте, а се-
рологически Ку-риккетсиоз установлен у позднего кожана в Алма-
Атинской области (Федорова, 1968). В зоне Панамского канала из
клещей Intercutestrix modofii, собранных с летучих мышей, также
выделен возбудитель лихорадки Ку (Yunker et al., 1975). Значение
летучих мышей в распространении этой инфекции практически
не изучено.
218
Chiroptera
Протозойные и грибковые заболевания
Трипаносомы. В тропических и субтропических странах обоих
полушарий у 52 видов летучих мышей (из 10 семейств) найдены
трипаносомы 19 видов (Constantine, 1971).
Вампиры, в основном Desmodus rotundus,— переносчики три-
паносомов копытных: муррины (возбудитель Т. hippicum) и маль-
декадерас (возбудитель Т. equinum), широко распространенных
в тропической и субтропической зонах Америки (Гоар, 1962).
Обычными переносчиками этих возбудителей служат слепни и
жигалки, которые являются типичными механическими инокуля-
торами и у которых способность передачи инфекции незараженным
особям теплокровного хозяина связана с прерванным кровососа-
нием. Роль вампиров в распространении этих инфекций среди
лошадей и рогатого скота более сложная. Трипаносомы, попав с
кровью больного животного в организм вампира, размножаются
там и выделяются со слюной. Поэтому вампиры, выполняя роль
переносчика возбудителя, в то же время являются его хозяевами.
От трипаносомозов страдают главным образом лошади, с кото-
рыми в свое время возбудитель был завезен в Америку. Истори-
чески вампиры сделались хозяевами этих трипаносом сравни-
тельно недавно. Они включились в круговорот возбудителя как
параллельный и независимый хозяин, который одновременно
служит и его переносчиком. Эпизоотологическое значение вам-
пиров определяется также тем, что в основном от них зависит
перенос возбудителя в межэпизоотический период, когда инфекция
среди лошадей затихает, но сохраняется в рогатом скоте, который
болеет бессимптомно и служит резервуаром возбудителя в это время.
Заражение вампиров от скота обеспечивается большим количест-
вом крови, которую они поглощают (Гоар, 1962).
Тридцать видов летучих мышей шести семейств (Emballonuridae,
Noctilionidae, Phyllostomatidae, Desmodontidae, Vespertilionidae,
Molossidae) инфицированы трипаносомами, морфологически сход-
ными с Т. cruzi — возбудителем болезни Чагоса (Constantine,
1971). Эти трипаносомы имеют отличия в патогенности для лабо-
раторных грызунов, но проявляют иммунологическое и биологи-
ческое сходство с Т. cruzi. Американские трипаносомы подрода
Schizotrypanum из летучих мышей по биологическим показателям
оказались близки к Т. cruzi и выделены в отдельный подвид Т.
cruzi marinkelei ssp. nov. (Baker et al., 1978). Вопрос означении
трипаносом летучих мышей для здоровья человека требует даль-
нейшего всестороннего изучения (Constantine, 1971).
Токсоплазмоз. Антитела к Toxoplasma gondii обнаружены у
Artibeus lituratus в Колумбии, у Eptesicus serotinus, Nyctalus noc-
tula в СССР (Галузо и др., 1965), у Myotis bechsteini во Франции
(Akinchina, Doby, 1968).
Другие простейшие. Летучие мыши известны как носители кок-
Рукокрылые
219
цидий (Eimeria, Isospora, Kossia) и гемоспоридий (Babesia, Plas-
modium, Nycteria, Polychromophilus, Hepatocystis), но все эти
простейшие — паразиты летучих мышей и непатогенны для че-
ловека (Constantine, 1971).
В Ираке в годы высокой заболеваемости кала-азаром проводили
исследование животных, в том числе летучих мышей, на заражен-
ность возбудителем висцерального лейшманиоза и получили отри-
цательные результаты (Sukkar, 1978).
Микозы. У летучих мышей установлены различные грибки,
которые выделяют из внутренних органов, из содержимого кишеч-
ника, из волос этих зверьков. Большинство таких грибков либо
сапрофиты, либо развиваются в живых и мертвых тканях растений
и в почве. Случаи выделения их у летучих мышей относительно
редки. Например, в Колумбии обнаружены грибки, вызывающие
иногда стригущий лишай у человека: Trichophyton mentagrophytes
(1 из 126 обследованных летучих мышей), Microsporum canis (у 1
из 1276 зверьков) (Constantine, 1971).
Дерматофиты широко распространены, и их постоянно находят
в гуано и почве в местах обитания летучих мышей.
Наибольшее значение в эпидемиологии человека имеет Histo-
plasma capsulatum, который высоко патогенен для человека. Люди
заражаются гистоплазмозом главным образом при посещении
пещер, разработке гуано или при раскапывании субстрата в убе-
жищах летучих мышей. Заболевания известны в Северной, Цент-
ральной и Южной Америке, а также в Африке (табл. 40).
Таблица 40. Случаи заболевания людей гистоплазмозом после посещения пещер *
Страна Число случаев
США, Флорида Мексика Белиз Панама Ямайка Колумбия Бразилия Африка Южная Африка 26 236 10 49 25 15 8 46 17
♦ По: Constantine, 1971; Lottenberg et al., 1979; Quinonens et al., 1978; Larrabee et. al.,
1978; Fincham, 1978.
Среди летучих мышей этот грибок широко распространен. Он
обнаружен у 9 видов в Северной Америке и у 18 видов в Централь-
ной и Южной Америке. Из 2707 летучих мышей, исследованных
220 Chiroptera
в США, гистоплазмоз выявлен у 438 экз. (16%), а из 317 зверьков,
исследованных в Центральной и Южной Америке, возбудитель
выделен от 14 особей (4,4%). Летучие мыши, собранные в пеще-
рах, поражены грибком почти поголовно (99% случаев), а собран-
ные на чердаках — лишь в 31 %. Наиболее высокий процент за-
ражения отмечен в пещерах Флориды у Myotis austroriparius
(76 %) и в пещерах Теннесси у Myotis grisescens (73%). Резервуаром
возбудителя являются не только летучие мыши, но и почва в на-
селенных ими пещерах. В штате Флорида — одном из эндемичных
по этому заболеванию районов — исследовано 148 образцов почвы
из 9 пещер и в 10 образцах обнаружены гистоплазмы. При обсле-
довании 165 экз. летучих мышей Myotis austroriparius из этих
пещер гистоплазмы отмечены у 80, что составляет 48% (Disalvo
et al., 1970).
ЛИТЕРАТУРА
Абатуров В. Д., Девятых В. Л., Зубкова Л, В. Роль роющей деятельности
сусликов (Citellus pygmaeus Pall), в перемещении минеральных веществ
в полупустынных почвах Заволжья // Почвоведение. 1969. № 12. С. 93—
99.
Абдурахманов Г. Л., Земельман Б. М., Казаков В. П. и др. О выявлении
эпизоотии чумы среди горных сусликов в Баксанском ущелье Кабардино-
Балкарской АССР // Проблемы особо опасных инфекций. Саратов, 1973.
Вып. 2(30). С. 175.
Абеленцев В. И. Об осеннем пролете рукокрылых на юге Украины // Руко-
крылые (Chiroptera). М.: Наука, 1980. С. 195—196. (Вопросы териоло-
гии).
Агафонов А. В. Сокращение поселений и численности малого суслика на
юге Волгоградской обл. под влиянием развития земледелия // Проб-
лемы особо опасных инфекций. Саратов, 1969. Вып. 3(7). С. 191 —194.
Адаменко В. И., Русакова Л. В., Бурлаченко Т. А. и др. Результаты эпи-
зоотологического обследования холмогорья Бадхыз в 1976 году // Там
же. 1979. Вып. 6(70). С. 28—30.
Адамович В. Л. Ландшафтно-экологические предпосылки к существованию
очагов рабической инфекции // Зоол. журн. 1978. Т. 57, вып. 6. С. 260—
270.
Адамович В. Л. К проблеме разрежения популяции лисицы на энзоотичной
по бешенству территории // Экологические основы охраны и рациональ-
ного использования хищных млекопитающих. М.: Наука, 1979. С. 378—
380.
Адамович В, Л. Бешенство волка (Canis lupus) в центральной части Русской
равнины // Зоол. журн. 1985. Т. 64, вып. 4. С. 590—599.
Адамович В. Л., Бородина 3. С. О роли енотовидной собаки в эпизоотологии
бешенства // Природноочаговые антропозоонозы: Тез. докл. IX Всесоюз.
конф, по природ, очаговости болезней человека и животных. Омск,
1976. С. 66—68.
Адамян А. О., Петров П. А,, Аветисян В. В. и др. Результаты карто-
графирования поселений малоазийского суслика в Армянской ССР П
Проблемы особо опасных инфекций. Саратов, 1979. Вып. 6(70). С. 37—
41.
Айрапетъянц А. Э., Никольская Н. С., Фокин И, М, Питание гребнепало-
го тушканчика (Paradipus ctenodactylus Vinogradov, 1930) в Заунгузских
Каракумах // Функциональная морфология и систематика млекопитаю-
щих. Л., 1978. С. 72—78. (Тр. Зоол. ин-та АН СССР; Т. 79).
Акиев А. КБейер А. П., Кузнецова К. А. и др. Инфекционная чувстви-
тельность к чуме европейского суслика // Эпидемиология и профилактика
природно-очаговых инфекций. Саратов: «Микроб», 1981. С. 69—73.
Акиев А. К., Голубев П. Д., Юдин Е. В. и др. О природной очаговости
чумы в районе Эльбруса // Проблемы особо опасных инфекций. Саратов,
1972. Вып. 5(27). С. 38—45.
Алгазин И. П. Туляремия в Советской Субарктике (по наблюдениям в Тай-
мырском автономном округе): Автореф. дис. . . . канд. мед. наук. М
1980. 16 с.
Алексеев А. Ф., Сержанов О. С., Соколова Т. Ю. Особенности межвидового
контакта млекопитающих в поселениях большой песчанки Северо-За-
падных Кызылкумов // 10-я Всесоюзн. конф, по природ, очаговости бо-
лезней. Душанбе, 1979. Вып. 2. С. 8—10.
222
Литература
Алимходжаев А. А., Борисов И. В. Распространение салмонелл среди
диких грызунов в Среднеазиатском очаге чумы / Тез. X науч. конф,
противочум. учреждений Средней Азии и Казахстана. Алма-Ата, 1979.
Вып. 2. С. 154—156.
Андрейко О. Ф., Скворцов В. Г., Пинчук Л. М. К вопросу о фауне парази-
тических членистоногих рукокрылых Молдавии // Паразиты животных
и растений. Кишинев, 1968. Вып. 3. С. 8—23.
Анина-Радченко Н. Д. Бешенство и борьба с ним. Кишинев: Штиинца, 1979.
97 с.
Анина-Радченко Н. Д., Корчмарь Н. Д., Глушко Л. В. и др. Бешенство
среди диких животных // Ветеринария. 1970. № 10. С. 64—65.
Арзамасов И. Т. Гамазовые клещи фауны Белоруссии. Минск: Наука и тех-
ника, 1968. 100 с.
Артемьев Ю. Т. К вопросу о размещении вида внутри ареала на примере
рыжеватого и малого сусликов (Citellus major Pall., С. pygmaeus Pall.) //
Природные ресурсы Волжско-Камского края. Животный мир. М.: Наука.
1964. С. 53-59.
Артемьев Ю. Т. Материалы по экологии и динамике численности малого и
рыжеватого сусликов И Природные ресурсы Волжско-Камского края.
Казань: Изд-во Казан, ун-та, 1968. Вып. 2. С. 35—51.
Асланян Р. Г., Кайтмазова Е. И., Курдина Д. С., Забродин В. А, К вопро-
су о природной очаговости бруцеллеза // Материлы XV Всесоюзн. съезда
эпидемиологов, микробиологов и инфекционистов: Тез. докл. М., 1970.
Ч. 1. С. 112—113.
Афанасьев А. В. Зоогеография Казахстана. Алма-Ата: АН КазССР, 1960.
259 с.
Афанасьев А. В., Бажанов В. С., Корелов М. Н. и др. Род Суслики —
Citellus Oken// Звери Казахстана. Алма-Ата: Изд-во АН КазССР, 1953а.
С. 139—167.
Афанасьев А. В., Бажанов В. С., Корелов М. Н. и др. Род тонкопалые
суслики // Там же. 19536. С. 180—181.
Бабенышев В, П., Саканян А. Б. Некоторые материалы по распростране-
нию и экологии европейского суслика (Citellus citellus L.) в Армянской
ССР // Тр. Арм. противочум. станции. Ереван, 1963. Вып. 2. С. 327—334.
Баваасан А. Монгол Алтайн тарваган тахлын голомтын судалгаанаас //
Гоц аюулт халдварт овчнцийг эсэрчууцэн судлах гасрын мэдээлээл.
Улаанбаатар, 1967. №2. С. 106—116.
Байрамова Р. А, Сравнительная роль диких и домашних животных в цир-
куляции спирохет клещевого раккуренса в Азербайджанской ССР //
Мед. паразитология. 1964. Т. 33, № 2. С. 182—183.
Байтанаев О. А., Айкимбаев М. А, Природные очаги туляремии тугайного
типа // Биологические проблемы природной очаговости болезней. Ново-
сибирск: Наука, 1981. С. 50—64.
Бакеев Н. Н., Черникин Е. М., Шиляева Л. М. Хищные // Итоги мечения
млекопитающих. М.: Наука, 1980. С. 77—94.
Балашов Ю. С. Паразито-хозяинные отношения членистоногих с наземными
позвоночными. Л.: Наука, 1982. 320 с. (Тр. Зоол. ин-та АН СССР; Т. 97).
Балашов Ю. С., Дайтер А. Б. Кровососущие членистоногие и риккетсии.
Л.: Наука, 1973. 250 с.
Балбарын Цэдэв. Вермипсиллы и вермипсиллез животных. Улан-Батор:
Наука, 1962, № 4.
Башенина Н. В. К вопросу о характере вредной деятельности малого сус-
лика и некоторых других грызунов в посадках дуба Сталинградской
области // Зоол. журн. 1951. Т. 30, вып. 3. С. 194—206.
Бекенов А. Места обитания, численность и норовая деятельность длинно-
хвостого суслика в высокогорьях Джунгарского Алатау // Изв. АН
КазССР. Сер. биол. 1965. Вып. 4. С. 49—56.
Литература
223
Бекенов А. Желтый суслик в Северном Кызыл куме // Грызуны: Материалы
VI Всесоюз. совещ. Л.: Наука, 1983. С. 288—290.
Белашова В. С. Клещи Argas vespertilionis в Казахстане // Тез. докл. I ака-
ролог. совещ. М.; Л.: Наука, 1966. С. 25—26.
Беликов С. Е. Белый медведь (Ursus maritimus Phipps) в районе острова Вран-
геля: Экология, поведение, охрана: Автореф. дис. ... канд. биол. наук.
М., 1982. 21 с.
Белоконь Е. М., Кушнирук В. А., Климишин В. С., Белинкевич Т. Д.
К фауне эктопаразитов рукокрылых западных областей Украины II
Проблемы паразитологии: Тр. IV науч. конф, паразитологов УССР.
Киев, 1963. С. 308—309.
Беляева М. Д. К изучению гельминтофауны млекопитающих Беловежской
пущи // Тр. Всесоюз. ин-та гельминтологии им. акад. К. И. Скрябина.
М.: Минсельхоз СССР, 1959. Вып. 6. С. ПО—114.
Беньковский Л. М., Головина Т. И., Щербина Р. Д. К вопросу изучения
болезней и паразитов выдры острова Сахалина// Вести, зоологии. 1973.
№ 6. С. 21—24.
Бердыев А. С. Носительство токсоплазм и обнаружение некоторых пара-
зитических простейших у диких животных Туркмении // Материалы
II Всесоюзн. съезда протозоологов. Киев, 1976. Ч. 1. С. 24—25.
Бережное А. 3., Козакевич В. П., Величко Л. Н. и др. Характер инфекцион-
ной чувствительности к чуме малых сусликов, населяющих эпизооти-
ческие и свободные от чумы участки Северо-Западного Прикаспия //
Проблемы особо опасных инфекций. Саратов, 1975. Вып. 2(42). С. 24—28.
Бернштейн А. Д., Клюева Е. В., Овсяников Н. Г., Селимов М. А. Природ-
ный очаг арктического бешенства на острове Врангеля // Вирусы и вирус-
ные инфекции человека. М., 1981. С. 151—152.
Бессонов А. С. Эпизоотология, эпидемиология и профилактика трихинелле-
за. Вильнюс: Минтис, 1972. 304 с.
Бибиков Д, И. К экологии даурского суслика (Citellus dauricus), сообщ. 1 //
Изв. Иркут, науч.-исслед. противочум. ин-та Сибири и Дальнего Восто-
ка. 1949. Т. 7. С. 147—159.
Билялов 3. А., Свиридов Г. Г., Ершова Л. С. и др. Влияние длительного
пребывания в организме клещей Ornithodorus tartakovskyi (Argasidae)
на свойства возбудителя чумы // Паразитология. 1983. Т. 17, вып. 1.
С. 75—76.
Бируля Н. Б, Экологические закономерности распределения малого суслика
в пространстве // Сб. Науч.-исслед. ин-та зоологии МГУ. 1936. Вып. 3.
С. 117—143.
Бобринский Н. А., Кузнецов Б. А,, Кузякин А. П. Определитель млекопи-
тающих СССР. М.: Просвещение, 1965. 380 с.
Боев С. Н., Бондарева В. И., Тазиева 3, X., Соколова И. Б. Значение
различных животных в распространении альвеококкоза в Казахстане I/
Вопросы природной очаговости болезней. Алма-Ата: Наука, 1970.
Вып. 3. С. 71—78.
Боев С. Н., Бондарева В. И., Соколова И. Б., Тазиева 3. X. Пути цир-
куляции трихинелл в биоценозах Казахстана // Там же. 1972. Вып. 5.
С. 169—179.
Боев С. Н., Бондарева В. И., Соколова И. Б., Тазиева 3. X. Трихинеллы
и трихинеллез. Алма-Ата: Наука, 1978. 242 с.
Божко Г. К., Гришок Л. П., Синицын А. Ю. Бешенство диких животных.
Киев: Наук, думка, 1981. 82 с.
Болдырев С. Т. Эктопаразиты желтого суслика (Citellus fulvus L.) // Зоол.
журн. 1959. Т. 38, вып. 11. С. 1757—1761.
Бондаренко А. А., Тарасова В. Е., Деревщиков А. Г. и др. Сайлюгемский
очаг чумы // Докл. Иркут, науч.-исслед. противочум. ин-та Сибири и
Дальнего Востока.. Чита, 1974. Вып. 10. С. 57—60.
Бондаренко М. Ф. Случай выделения чумного микроба от вши в Зааралье И
224
Л итература
Материалы расшир. науч, конф., посвящ. 40-летию КазССР. Алма-Ата,
1961. С. 32.
Ботвинкин А. Д. Эколого-вирусологическая характеристика природных
очагов бешенства в Забайкалье, Приамурье и Приморье: Автореф. дис.
. . . канд. биол. наук. М., 1983. 17 с.
Ботвинкин А. Д., Грибанова Л. Д., Никифорова Т. А. Бешенство у еното-
видной собаки в эксперименте // ЖМЖ 1983. № 12. С. 37—40.
Ботвинкин А. Д., Савицкий В. П., Сидоров Г. Я. и др. О природной очаго-
вости бешенства в Забайкалье // Современные методы изучения природно-
очаговых инфекций. Л.: Наука, 1980. С. 52—61.
Ботвинкин А. Д., Савицкий В. П., Сидоров Г. Н.у Юдин В. Г. К изуче-
нию природной очаговости бешенства в Приморье и Приамурье И При-
родно-очаговые болезни человека (Вопр. эпидемиологии и профилакти-
ки). Омск, 1981а. С. 122—130.
Ботвинкин А. Д., Савицкий В. П., Сидоров Г. Н., Юдин В. Г. Значение
енотовидной собаки в эпидемиологии и эпизоотологии бешенства на Даль-
нем Востоке // ЖМЭИ. 19816. № 12. С. 79—82.
Бошко Г, В. К вопросу о переносе собаками блох грызунов // Зоол. журн.
1956. Т. 35, вып. 1. С. 74—76.
Брегетова Н. Г., Буланова-3ахваткина Е. М., Волгин В. И., Дубинин В. Б.
Клещи грызунов фауны СССР. М.; Л.: Изд-во АН СССР, 1955. 457 с.
Б рикман Д. И., Мизитова Л, А., Колупаева Н. А. и др. Изучение инфек-
ционной чувствительности даурского суслика (Citellus dauricus Brandt)//
Проблемы особо опасных инфекций. Саратов, 1977. Вып. 2(54). С. 76—77.
Бром И. П., Вовчинская 3. М., Федорова Л. В. О роли хищных млекопитаю-
щих в распространении блох грызунов // Зоол. журн. 1948. Т. 27, вып. 2.
С. 167—174.
Буракаускас А., Казакявичюс П. Эпизоотологическая ситуация бешенства
диких животных в Литовской ССР // Природно-очаговые инфекции, инва-
зии: Материалы науч. конф. Вильнюс, 1979. С. 105—106.
Бурамбаев К., Исмагилов М. И. Образ жизни и межвидовые отношения
островной популяции суслика-песчаника// Биол. науки. Алма-Ата, 1975.
Вып. 2. С. 85—100.
Бурделов Л, А. Опыт сравнительной оценки популяционной чувствитель-
ности млекопитающих к чумному микробу по результатам эпизоотоло-
гического обследования // ЖМЭИ. 1982. № 6. С. 26—29.
Бурделов Л. А., Жубаназаров И. Ж., Кочкина Л. И. и др. К проблеме
межвидового паразитарного контакта в природных очагах чумы при-
аральских Каракумов // Мед. паразитология. 1983. Т. 60, № 3.
С. 46—50.
БурбеловЛ, А., Кошеное У. А., Жубаназаров И. Ж. и др. Анализ заражен-
ности млекопитающих в связи с их эпизоотологическим значением в оча-
гах чумы Приаралья // Зоол. журн. 1982. Т. 61, вып. 5. С. 755—763.
Бурлаченко Т. А., Гувва Л. А., Ефимов В. И, и др. Эпизоотия чумы среди
желтых сусликов и краснохвостых песчанок на Красноводском плато в
1964 г. // Проблемы особо опасных инфекций. Саратов, 1971. Вып. 2(18).
С. 88—97.
Бута Р. Токсоплазмоз кошек в городе Вильнюсе и прилегающих к нему
районах //IX конф. Укр. паразитол. о-ва. Киев, 1980. Ч. 1. С. 104.
Бутовский П. М., Тагильцев А. А. Возможная роль насекомоядных летучих
мышей в природной очаговости болезней // Вопросы природной очагово-
сти болезней. Алма-Ата: Наука, 1968. С. 47—58.
Быков Л. Т. Об участии второстепенных носителей чумы в эпизоотиях в
Муюнкумах // Проблемы особо опасных инфекций. Саратов, 1968. Вып. 2.
С. 85—90.
Быков Л. Т., Цой Д. Г. Об участии мелких куньих хищных в эпизоотии чумы
в Муюнкумах и восточных Кызылкумах // Проблемы изучения механизма
энзоотии чумы: Тез. докл. Всесоюз. конф. Саратов, 1980. С. 154—157.
Литература
225
Вагнер И. К., Титов Э. К., Лобачев В. С. Особенности строения нор кавказ-
ского горного суслика // Науч. докл. высш. шк. Биол. науки. 1975.
№ 3. С. 7—12.
Вайсфельд М. А. Выдра // Колонок, горностай, выдра. М.: Наука, 1977.
С. 175—204.
Валиулин С. М., Аюпов X. В., Мухаметов Р. Ю., Самигуллин Р. Н.
Распространение и основные резервенты альвеококкоза в Башкирии //
X Всесоюзн. конф, по природ, очаговости болезней. Алма-Ата, 1979.
Т. 1. С. 166—168.
Валова Г. П. Географическое распространение природных очагов лепто-
спирозов на крайнем севере Западной Сибири // Учен. зап. Свердл. и
Тюмен. пед. ин-тов. Тюмень, 1970. Сб. 145, вып. 2. С. 31—36.
Варгина С. Г. К вопросу о роли летучих мышей в экологии вируса клещевого
энцефалита на юге Киргизии II Экология вирусов: Материалы X сим-
поз. Баку, 1976. С. 81—82.
Варгина С. Г., Карась Ф. Р., Гонтарь И. А., Кучук Л. А. К вопросу о
распространении арбовирусов летучими мышами // Сб. науч. тр. Кирг.
мед. ин-та. 1976. Т. 3. С. 76—80.
Варгина С. Г., Кучук Л. А., Герштейн В. И., Карась Ф. Р. Передача вируса
Иссык-Куль клещами Argas vespertilionis в эксперименте // Экология
вирусов. М.: Ин-т вирусологии, 1982. С. 123—127.
Варгина С. Г., Стеблянко С. Н., Серополко А. А. и др. Циркуляция арбо-
вирусов в Чуйской долине // Сб. науч. тр. Киргиз, мед. ин-та. 1976. Т. 3.
С. 94—97.
Варшавский Б, С, Материалы по эпизоотологии чумы среди малых сусликов в
некоторых поселениях Северного Приаралья // Проблемы особо опасных
инфекций. Саратов, 1977. Вып. 5(57). С. 13—16.
Варшавский Б. С. Динамика численности малого суслика в некоторых по-
селениях Северного Приаралья // Экология. 1983. № 3. С. 88—90.
Варшавский С. Н. Современные методы учета численности сусликов и боль-
ших песчанок // Методы учета численности и географического распреде-
ления наземных позвоночных. М.: Изд-во АН -СССР, 1952. С. 47—67.
Варшавский С. Н. Некоторые особенности внутрипопуляционных отношений
у сусликов и их экологическое значение // Вопросы экологии: По мате-
риалам III экол. конф. Киев, 1957. Т. 2. С. 12—25.
Варшавский С. Н. Некоторые экологические причины особенностей эпизоото-
логического значения малого суслика в пустынной зоне // X совещ. по
паразитол. пробл. и природноочаговым болезням. М.; Л.: Изд-во АН
СССР, 1959. Вып. 1. С. 195—197.
Варшавский С. Н. Возрастные типы поселений и история расселения малого
суслика // Исследования географии природных ресурсов животного и
растительного мира. М.: Изд-во АН СССР, 1962. С. 59—79.
Варшавский С, Н. Зонально-географические особенности воздействия рою-
щей деятельности малого суслика на ландшафт в связи с возрастом посе-
лений// Теоретические и прикладные аспекты биогеографии. М.: Наука,
1982. С. 99—112.
Варшавский С. Н., Козакевич В. П., Лавровский А. А. и др. География и
структура природных очагов чумы в Центральной и Восточной Азии //
Международные и национальные аспекты эпиднадзора при чуме. Ир-
кутск: Науч.-исслед. противочум. ин-т, 1975. С. 80—82.
Варшавский С. Н., Попов Н. В., Шилов М. Н. и др. Численность и пло-
щадь поселений малого суслика в западной части Прикаспийского Се-
веро-Западного природного очага чумы // Эпизоотология природно-оча-
говых инфекций. Саратов: «Микроб», 1985, с. 45—49.
Варшавский С. Н., Шилов М. Н. Южная граница распространения малого
суслика в Приаралье // Материалы совещ, по зоогеографии суши. Львов:
Изд-во Львов, ун-та, 1957. С. 36—43.
Варшавский С. Н., Щепотьев Н. В., Конева 3. А. Хищные млекопитающие
226
Литература
как носители в природных очагах чумы СССР: Аннот. библиогр. указ.,
1912—1980. Ставрополь: «Микроб», 1982. 96 с.
Вашкевич Р. Б. Бруцеллез пушных зверей (норки, лисицы, песца) // Биоло-
гия и патология пушных зверей. Петрозаводск, 1974. С. 172—174.
Ведерников В. А., Гудим В. Е. К вопросу об эпидемиологическом и эпизоото-
логическом значении бешенства кошек // Бюл. ВОЛИЭВ, 1976. Вып. 26.
С. 42—46.
Величко Л. Н., Кондрашкина К. И., Ермилов А. П. и др. Экскременты
блох — естественная среда долговременного хранения микроба чумы //
Проблемы особо опасных инфекций. Саратов, 1978. Вып. 6(64). С. 51—53.
Вершинин А. А., Лазарев А. А. Биология и промысел камчатской лисицы //
Охотоведение М.: Лесн. пром-сть, 1974. С. 5—26.
Вершинина Т. А., Елтышев Ю. А., Лямкин В. Ф., Рященко С. В. Длинно-
хвостый суслик (Citellus undulatus Pall.) Баргузинской котловины
(биология и значение в формировании природных предпосылок болез-
ней человека)// Териология. Новосибирск: Наука, 1972. Т. 1. С. 290—302.
Виноградов Б. С., Тушканчики. М.; Л.: Изд-во АН СССР, 1937. 196 с. (Фауна
СССР; Т. 3, вып. 4: Млекопитающие).
Волк: Происхождение, систематика, морфология, экология / Отв. ред.
Д. И. Бибиков. М.: Наука, 1985. 606 с.
Волянский Ю. Е., Соловьев В, И. Материалы к распространению летучих
мышей и их эктопаразитов в юго-восточной части Украины // IV Межвуз.
зоогеогр. конф.: Тез. докл. Одесса, 1966. С. 52.
Воронов Г. А. Некоторые итоги опыта акклиматизации енотовидной собаки
на Сахалине: Зоолог, пробл. Сибири. Новосибирск: Наука, 1972. С. 368—
369.
Воронов Г. A. Ceratophyllus tesquorum altaicus loff, 1936 как основной
переносчик и хранитель чумного микроба в Тувинском природном очаге //
Эпидемиология и профилактика особо опасных инфекций в МНР и СССР.
Улан-Батор: Госиздат, 1978. С. 152—155.
Воронцов Н. Н., Жолнеровская Е. И., Баранов О. К. Иммуносистематика
сусликов Палеарктики (Citellus, Sciuridae, Rodentia) // Зоол. журн.
1981. Т. 60, вып. 7. С. 1070—1077.
Вшивков Ф. Н. Гамазовые клещи рукокрылых Крыма // Проблемы паразито-
логии: Тр. IV науч. конф, паразитологов УССР. Киев, 1963. С. 324—326.
Высоцкий Б. В., Малых Ф. С.у Кузнецов А. П. Промысловые животные —
дополнительные резервуары лептоспир в природе // Тр. Владивост.
НИИЭМГ. Владивосток, 1958. Сб. 1. С. 99—100.
Гаджиев А. Т., Дубовченко Т. А. Гамазовые клещи (Parasitiformes, Ga-
masoidea) рукокрылых Азербайджана // Зоол. журн. 1967. Т. 46, вып. 11.
С. 1716—1719.
Гаджиев А. Г., Дубовченко Т. А. Летучие мыши — прокормители пре-
имагинальных фаз иксодовых клещей // Тез. докл. II акаролог. совещ.
Киев, 1970. Ч. 1. С. 122—124.
Гаджиев А. Г., Дубовченко Т. А. Эктопаразиты синантропных летучих мы-
шей в Азербайджане // Проблемы паразитологии: Тр. VII науч. конф.
УРНОП. Киев, 1972. С. 174—176.
Гаджиев А. Г., Дубовченко Т. А. Летучие мыши — прокормители парази-
тических членистоногих — переносчиков вирусных инфекций в СССР И
Экология вирусов: Материалы X симпоз. Баку, 1976. С. 15—16.
Гайдук В. Е. К изучению экологии европейской и американской норок //
Экологические основы охраны и рационального использования хищных
млекопитающих. М.: Наука, 1979. С. 311—312.
Гайский Н. А. Инфекция и иммунитет у животных, залегающих в спячку //
Изв. Иркут, науч.-исслед. противочум. ин-та Сибири и Дальнего Восто-
ка. 1944. Т. 5. С. 82—123.
Гайский Н. А. Особенности чумной эпизоотии на сусликах в 1929 г. в Фур-
мановском районе Западного Казахстана // Там же. 1949. Т. 7. С. 16—29.
Литература
227
Галбадрах Д., Чинболд Л., Жамъянсурэн П., Пурэвжал Ч. Современное
состояние природного очага чумы Монгольского Алтая // Эпидемиология
профилактика особо опасных инфекций в МНР и СССР. Улан-Батор:
Госиздат, 1978. С. 42—44.
Галузо И. Г. Аргасовые клещи. Алма-Ата: Наука, 1957. 130 с.
Галузо И. Г., Высокова Л, А., Кривкова А. М. Токсоплазмоз летучих
мышей // Токсоплазмоз животных. Алма-Ата: Наука, 1965. С. 218—223.
Галузо И. Г., Коновалова С. И. О природной очаговости токсоплазмоза //
Вопросы природной очаговости болезней. Алма-Ата: Наука, 1979.
Вып. 10. С. 49—57.
Гарбузов В. К., Варшавский Б. С. Некоторые особенности экологии жел-
того суслика в Северном Приаралье // Проблемы особо опасных инфек-
ций. Саратов, 1977. Вып. 2(54). С. 18—23.
Гарбузова Л. Ф. Материалы по фауне блох южной и центральной части
Актюбинской области // Грызуны и их эктопаразиты. Саратов: Изд-во
Сарат. ун-та, 1968. С. 414—421.
Гачик И. Ф., Неронов В. М. О своеобразии фауны тушканчиков Передней
Азии И Тушканчики фауны СССР. М., 1985. С. 3—6.
Геллер М. X, Биология уссурийского енота (Nyctereutes procyonoides Gray),
акклиматизированного на северо-западе европейской части СССР //
Тр. Науч.-исслед. ин-та сел. хоз-ва Крайнего Севера. Норильск, 1959.
Т. 9. С. 55—142.
Герасимов Ю. А. Зудневая чесотка диких лисиц // Тр. ВНИИОЗ. 1953.
Вып. 13. С. 116—134.
Герасимова Н. Г. Продолжительность жизни Ctenophthalmus pollex в ла-
бораторных условиях // Эпидемиология и эпизоотология чумы. Саратов:
«Микроб», 1980. С. 40—44.
Герценштейн Г. Г, Бешенство // Энцикл. словарь Брокгауз и Ефрон. 1891.
Т. 5. С. 263—266.
Гоар С. А. Вампиры как переносчики трипанозомозов // Вопросы общей
зоологии и медицинской паразитологии. М.: Медгиз, 1962. С. 303—314.
Головачева В. Я., Жовтый И. Ф. Дальнейшие наблюдения за естественной
зараженностью эктопаразитов мелких млекопитающих возбудителями
бактериальных инфекций в природных очагах чумы Сибири и МНР //
Эпидемиология и профилактика особо опасных инфекций в МНР и СССР.
Улан-Батор: Госиздат, 1978. С. 99—100.
Голубев П, Д., Петров П, А., Дятлов А. И. Ландшафтная приурочен-
ность поселений горного малого суслика // Состояние и перспективы
профилактики чумы: Тез. докл. Всесоюз. конф. Саратов, 1978. С. 107—
ПО.
Гончаров А. И. Дополнения к списку (Ралль, 1960) видов блох, обнаружен-
ных инфицированными в природных условиях // Профилактика при-
родно-очаговых инфекций: Тез. докл. Всесоюзн. науч.-практ. конф.
Ставрополь, 1983. С. 229—231.
Горчакова Т. Ф., Ган Н. В., Шакуров Д. В. и др. Результаты эпизоотологи-
ческого обследования приграничной с Забайкальем полосы МНР //
Докл. Иркут, науч.-исслед. противочумн. ин-та Сибири и Дальнего Во-
стока. Чита, 1974. Вып. 10. С. 76—78.
Горчаковская Н. Н., Преображенская Н. К., Штильмарк Ф. Р., Тощиеин Ю. В,
Зоопаразитологическая характеристика природного очага клещевого эн-
цефалита в Кемеровской обл. // Вопросы медицинской вирусологии.
М., 1960. Вып. 4: Нейроинфекции. С. 145—151.
Горманова Е, Н. К вопросу об экспериментальном лептоспирозе сусликов
(Аннотация) // ЖМЭИ. 1956. № 2. С. 107.
Горячева О. А. К эпидемиологии бешенства на Украине // Зоонозные ин-
фекции. Киев, 1959. С. 19—27.
Гребенюк Р. В. Иксодовые клещи Киргизии. Фрунзе: Илим, 1966. 328 с.
228
Литература
Гречановская В. П. Профилактика и борьба с бешенством в УССР // Зооноз-
ные инфекции. Киев, 1966. С. 178—182.
Грибанова Л. Я. Характеристика природных очагов бешенства в районе
Западной Сибири: Автореф. дис. . . . канд. биол. наук. М., 1975. 20 с.
Грибанова Л. Я-, Мальков Г. Б. Вирусологическая характеристика при-
родных очагов бешенства в Западной Сибири // Симпоз. по бешенству.
Потсдам: (ГДР). 1977. С. 24—25.
Грибанова Л. Я-, Мальков Г, Б. Изменчивость эпизоотологической напря-
женности в природных очагах бешенства в степи и лесостепи Западной
Сибири // Вопросы природной очаговости болезней. Алма-Ата: Наука,
1978. Вып. 9. С. 107—110.
Грибанова Л. Я-, Мальков Г. Б., Ботвинкин А. Д. и др. О значении хищ-
ных млекопитающих семейства куньих в формировании очагов бешенства
в Сибири // Природно-очаговые болезни человека: Вопр. эпидемиологии,
профилактики. Омск, 1981. С. 131 —139.
Грибанова Л. Я-, Мальков Г. Б., Ботвинкин А. Д., Никифорова Т. А.
Основные результаты изучения чувствительности диких, домашних и
лабораторных животных к вирусу бешенства // Природно-очаговые бо-
лезни человека: Вопр. лаб. диагностики,эпидемиологии и профилактики.
Омск, 1982. С. 116—122.
Грибанова Л. Я-, Мальков Г. Б., Грибенча С. В., Селимов М. А. Опре-
деление чувствительности к вирусу бешенства некоторых диких, домаш-
них и лабораторных животных // Современные методы в изучении при-
родно-очаговых инфекций. Л., 1979. С. 28—33.
Грибанова Л. Н., Мальков Г. Б., Савицкий В. П. и др. Результаты комп-
лексного изучения природных очагов бешенства и оценка риска заболе-
ваний гидрофобией в районах новостроек Восточной Сибири и Дальнего
Востока // ЖМЭИ. 1980. № 7. С. 86—90.
Грибанова Л. Я-, Мальков Г. Б., Тофанюк Е. В., Поспелов Е. С., Шать-
ко П. Д. Анализ эпизоотологической обстановки по бешенству в Но-
восибирской области И Природно-очаговые болезни в Сибири: (Эпидемио-
логия и профилактика). Л., 1977. С. 27—32.
Грибанова Л. Я-, Мальков Г. Б., Чиж Г. С. и др. К анализу заболеваемости
бешенством в Омской области в 1974—1979 гг. // Современные методы
изучения природно-очаговых инфекций. Л., 1980. С. 70—75.
Гришок Л. П. Эпизоотологическая характеристика бешенства в Украин-
ской ССР: Автореф. дис. ... канд. биол. наук. Минск, 1978. 20 с.
Громов И. Л4., Бибиков Д. И., Калабухов Н. И., Мейер М. Н. Наземные
беличьи (Marmotinae). М.; Л.: Наука, 1965, 477 с. (Фауна СССР; Мле-
копитающие. Т. 3, вып. 2).
Громов И. М., Гуреев А, А., Новиков Г. А. и др. Млекопитающие фауны
СССР. М.; Л., 1963. Ч. 2. С. 740—895.
Громова В. И. Отряд Chiroptera. Рукокрылые или летучие мыши // Основы
палеонтологии: Млекопитающие. М.: Гостехиздат, 1962. С. 86—90.
Губанов Н. М. Гельминтофауна промысловых млекопитающих Якутии.
М.: Наука, 1964. 154 с.
Гувва Л. А. О чувствительности желтого суслика из Западной Туркмении к
возбудителю чумы // Проблемы особо опасных инфекций. Саратов, 1975.
Вып. 3/4(43/44). С. 41—47.
Гувва Л. А., Пейсахис Л. А., Забегалова М. Н. О переживании чумного
микроба в организме желтых сусликов, зараженных естественным пу-
тем, в период зимней спячки // Проблемы особо опасных инфекций. Сара-
тов, 1978. Вып. 6(64). С. 38—40.
Гурский И. Г. Гибридизация волка с собакой в природе // Бюл. МОИП. Отд.
биол., 1975. Т. 80, вып. 1. С. 131—136.
Гурьянова Л. И. Материалы по эпидемиологии бешенства в Хабаровском
крае // Вопросы инфекционных заболеваний и производства вакционно-
сывороточных препаратов. Хабаровск, 1967. С. 166—173.
Литература
229
Гурьянова Л. И., Зак А. Д. Случай гидрофобии человека, связанный с уку-
сом енотовидной собаки // Учен. зап. Хабар, науч.-исслед. ин-та эпиде-
миологии и микробиологии. 1966. Вып. 8/2. С. 96—97.
Гусев А. В. К паразитофауне уссурийского енота // Паразитол. сб. ЗИН
АН СССР. 1951. Т. 13. С. 96—104.
Давтян Г. Г., Матевосян Л. Ш., Навасардян Т.Ф. и др. Течение экспери-
ментальной чумы у малоазийских сусликов в зимнем сезоне // Проблемы
особо опасных инфекций. Саратов, 1971. Вып. 1(17). С. 45—50.
Даниленко А. Ф., Живоляпина Р. Р., Краминская Н. Н. Совмещенные
очаги чумы и клещевого энцефалита на юго-западе Тувы // Докл. Иркут,
науч.-исслед. противочум. ин-та Сибири и Дальнего Востока. 1971.
Вып. 9. С. 24—25.
Демяшев М. П. Видовой состав и распространение диких млекопитающих
в Уральской обл.// Материалы юбил. конф. Урал, противочум. станции.
Уральск, 1964. С. 111—122.
Денисенко И. И., Сурвилло А. В., Подсвиров А. В. К эпизоотологическому
районированию северной и центральной части Прикаспийского северо-
западного природного очага чумы // Эпидемиология и профилактика
природно-очаговых инфекций. Саратов, 1981. С. 23—29.
Джонсон X, Бешенство // Вирусные и риккетсиозные инфекции человека.
М.: Медицина, 1955. С. 303—308.
Джу пина С. И., Куликов В. Т. Эпизоотология бешенства сельскохозяйст-
венных животных в Новосибирской области // Сб. науч, работ Новосиб.
науч.-исслед. вет. станции. Новосибирск, 1968. Вып. 3. С. 163—166.
Динесман Л. Г. Изучение истории биогеоценозов по норам животных. М.:
Наука, 1968. 100 с.
Дробинский И. Р., Климухин В. К. Туляремия в горных районах // ЖМЭИ.
1948. № 1. С. 29—31.
Дробищенко П. И., Кирющенко Т. В,, Каримов С. К. и др. Циркуляция
вируса клещевого энцефалита среди летучих мышей (Nyctalus noctula
Schreber) города Алма-Ата // Экология вирусов: Материалы X симпоз.
Баку, 1976. С. 83—84.
Дубинин В. Б. Чесоточные клещи (Acariformes, Sarcoptoidae) и чесоточные
заболевания диких млекопитающих // Зоол. журн. 1955. Т. 34, вып. 6.
С. 1189—2202.
Дубинины В. Б. и М. Н. Паразитофауна млекопитающих Даурской степи //
Фауна и экология грызунов. М.: МОИП, 1951. Вып. 4. С. 98—156.
Дубовой А. А., Хамаганов С. А., Саран М. и др. Итоги многолетнего эпизоо-
тологического обследования на чуму приграничных территорий Забай-
калья (СССР), Восточного, Хэнтэйского и Хубсугульского аймаков
(МНР) // Эпидемиология и профилактика особо опасных инфекций в
МНР и СССР. Улан-Батор: Госиздат, 1978. С. 27—29.
Дубовченко Т. А. Эколого-фаунистический обзор клещей рукокрылых Азер-
байджана // Тез. докл. I акаролог. совещ. М.; Л.: Наука, 1966. С. 85—86.
Дубовченко Т. А. Материалы к изучению блох рукокрылых в Азербайджан
не // Вопросы паразитологии. Баку: Элм, 1969. С. 236—240.
Дубовченко Т. А., Гаджиев А. Т. Клещи — паразиты обыкновенного длин-
нокрыла (Miniopterus schreibersi Kuhl) в СССР // III Всесоюз. совещ. по
теорет. и прикл. акарологии: Тез. докл. Ташкент, 1976. С. 106—108.
Дубровский А. Н. Наземные млекопитающие Новой Земли // Изв. Гос. геогр.
о-ва. 1939. №3 С. 402—414.
Дубровский Ю. А. Песчанки и природная очаговость кожного лейшманиоза.
М.: Наука, 1978. 183 с.
Дубровский Ю. А., Комарова Л. В. Место тонкопалого суслика среди есте-
ственных носителей кожного лейшманиоза // Экология млекопитающих и
птиц. М.: Наука, 1967. С. 57—59.
Дудникова А. Ф. Фауна и динамика численности блох грызунов Западных
Кара-Кумов. Сообщ. 2. Блохи полуденных и краснохвостых песчанок,
230
Литература
тонкопалых сусликов и тарбаганчиков // Вопросы природной очагово-
сти и эпизоотологии чумы в Туркмении. Ашхабад, 1960. С. 335—341.
Дунаева Т, Н. Позвоночные животные — носители и источники туляремий-
ной инфекции И Природная очаговость, эпидемиология и профилактика
туляремии. М.: Медицина, 1970. 271 с.
Дылько Н. И. Токсоплазмы позвоночных Белоруссии // Проблемы токсоплаз-
моза животных: Материалы Всесоюз. конф, по токсоплазмозу животных.
Алма-Ата, 1966. С. 77—79.
Дэмбрел Ж., Краминский В. А., Некипелов Н. В. и др. Районирование чум-
ных очагов Центральной Азии // Докл. Иркут, науч.-исслед. противо-
чум. ин-та Сибири и Дальнего Востока. Чита, 1974. Вып. 10. С. 45—47.
Дятлов А. И. Материалы к биологии тонкопалого суслика (Spermophilop-
sis leptodactylus Licht.) в Каракалпакской части Кызылкумов // Тр.
Ин-та зоологии АН КазССР. 1960. Т. 13. С. 37—44.
Дятлов А. Я., Елкин Ю. М., Лунина Е. А. и др. Популяционные раз-
личия в чувствительности к чуме у малых сусликов на Кавказе И Проб-
лемы особо опасных инфекций. Саратов, 1979. Вып. 6(70). С. 55—59.
Дятлов А. Я., Петров П. А., Улуханов Ч. Б., Шпилько И. В. Количест-
венная характеристика эпизоотического процесса на участке стойкой
очаговости чумы в Приэльбрусье // Эпидемиология и профилактика при-
родно-очаговых инфекций. Саратов: «Микроб», 1981. С. 30—33.
Евдокимов А. В., Крюков И, Л., Очиров Ю.Д. и др. Ландшафтно-эпизоотоло-
гическая дифференциация территории Тувинского природного очага чу-
мы // Проблема изучения механизма энзоотии чумы: Тез. докл. Саратов:
«Микроб», 1980. С. 158—163.
Евдошенко В. ГПрорешная Т. Л. О зараженности диких животных севера
Киргизии возбудителем лихорадки Ку // X совещ. по паразитол. пробл. и
природно-очаговым болезням. М.; Л.: Изд-во АН СССР, 1959. Вып. 1.
С. 109—111.
Егоров И. Д., Сергеев А. Ф., Чернявский В. Ф. Организация и итоги анти-
рабических мероприятий в Якутии (по материалам 1974—1978 гг.) //
Природно-очаговые болезни человека: Вопр. эпидемиологии и профи-
лактики. Омск, 1981. С. 122—130.
Ельшанская Н. И., Попов М. В. Зоолого-паразитологическая характери-
стика долины р. Кенкеме (Центр. Якутия) // Изв. Иркут.' науч.-исслед.
противочум. ин-та Сибири и Дальнего Востока. Кызыл, 1968, Т. 27.
С. 316—322.
Емельянов П. Ф. Современное распространение европейского суслика //
Грызуны: Материалы IV Всесоюзн. совещ. Л.: Наука, 1983. С. 9—11.
Емельянов П. Ф., Брюханова Л, В., Глушко Н. В. и др. Некоторые стороны
экологии малого суслика в изолированных поселениях в Ставропольском
крае // Экология. 1979. № 3. С. 69—74.
Емельянов П. Ф., Вагнер И. К., Кармов А. М. и др. Биология горного
суслика Citellus musicus (Rodentia, Sciuridae) на Центральном Кавказе//
Зоол. журн. 1982. Т. 61, вып. 3. С. 419—427.
Емельянова Н. Д. К вопросу об эпидемиологической роли хищных млеко-
питающих в Монголии // Изв. Иркут, науч.-исслед. противочум. ин-та
Сибири и Дальнего Востока. Иркутск, 1957. Т. 15. С. 285—291.
Емельянова Н. Д., Высоковский Н. Н. К вопросу о зимовках летучих мышей
и их эктопаразитах в окрестностях Красноярска // Докл. Иркут, науч.-
исслед. противочум. ин-та Сибири и Дальнего Востока. Хабаровск,
1962. Вып. 3. С. 146—148.
Емельянова Н. Д., Жовтый И. Ф. Краткий обзор эктопаразитов млекопитаю-
щих монголо-забайкальского чумного очага в связи с их эпизоотологи-
ческим значениям // Изв. Иркут, науч.-исслед. противочум. ин-та Сибири
и Дальнего Востока. Иркутск, 1957. Т. 15. С. 259—283.
Емельянова Н. Д,, Терещенко О. Н. К биологии иксодового клеща Нае-
maphysalis verticalis Itagaki et al., 1944 // Докл. Иркут, науч.-исслед.
Литература
231
противочум. ин-та Сибири и Дальнего Востока. Иркутск, 1971. Вып. 9.
С. 242.
Емельянова Н. Д.у Штильмарк Ф. Р. Блохи насекомоядных грызунов и
зайцеобразных центральной части Западного Саяна // Изв. Иркутск,
науч.-исслед. противочум. ин-та Сибири и Дальнего Востока. Кызыл,
1968. Т. 27. С. 241—246.
Емчук Е. М. Фауна й еколопя 1ксодових клад в Зах1*дного Пол1сся УРСР //
35. праць Зоол. музея. Ких’в, 1954. №26. С. 40—51.
Ермаков Н. М., Алиев Л. И. Новые находки желтого суслика на возвы-
шенности Карабиль // Млекопитающие СССР: III съезд Всесоюзн. те-
риол. о-ва: Тез. докл. М., 1982. Т. 1. С. 113—114.
Ермилов А. П., Ротшильд Е. В., Постников Г. Б. и др. Носители и пе-
реносчики чумы в Северо-Восточном Прикаспии // Профилактика особо
опасных инфекций. Саратов: Изд-во Сарат. ун-та, 1976. Вып. 1. С. 3—12.
Есжанов Б., Бекенов А. Распределение и численность тонкопалого суслика//
Животный мир Казахстана и проблемы его охраны. Алма-Ата: Наука,
1982. С. 74—76.
Ешелкин И. П., Пу ртов С. М, Подвижность и контакт между зверьками в
Горно-Алтайском природном очаге чумы // Экология. 1976. № 6.
С. 87—89.
Железникова В. В. Об исследовании на трихинеллез медведей и кабанов в
Хабаровском и Приморском краях // Гельминты Дальнего Востока.
Хабаровск, 1972. С. 40—42.
Железникова В. В., Богомолова Ф. И., Зуева Н. В. О групповых заболева-
ниях трихинеллезом в Хабаровском крае в 1969—1977 гг. // Там же.
С. 33—35.
Животченко В. И. О величине годовой добычи диких копытных тиграми
одной семейной группы // Экологические основы охраны и рационального
использования хищных млекопитающих. М.: Наука, 1979. С. 246—247.
Животченко В. И, Амурский тигр юга Приморья и его охрана: Автореф. дис.
... канд. биол. наук. М., 1981. 24 с.
Животягина Н. А., Иголкин Н. И. К изучению фауны эктопаразитов
сусликов (длиннохвостого и краснощекого) на границе их ареалов //
Вопросы биологии и агрономии. Томск: Изд-во Том. ун-та, 1976.
С. 89—92.
Жизнь животных / Ред. Наумов С. П., Кузякин А. П. М.: Просвещение, 1971.
Т. 6: Млекопитающие или звери. 627 с.
Жилое М. М. Экспериментальный листериоз у песцов // Биология и пато-
логия пушных зверей. Петрозаводск, 1974. С. 178—181.
Жмаева 3. М., Карулин Б. Е., Пчелкина А. А., Шеханов М. В. Природ-
ные очаги Ку-лихорадки в районах освоения целинных земель Северного
Казахстана I/ Тез. докл. Межинститут, науч. конф, по заболеваниям с
природ, очаговостью, посвящ. 50-летию Том. НИИВС. Томск, 1956.
С. 105—106.
Жмаева 3. М., Пчелкина А. А., Белашова В. С. О спонтанной заражен-
ности риккетсиями Бернета Argas vespertilionis на юге Казахстана II
Мед. паразитология. 1966. №5. С. 595—596.
Жовтый И. Ф. Паразитологическая ситуация в природных очагах чумы на
границе Сибири и МНР // Докл. Иркут, науч.-исслед. противочум. ин-та
Сибири и Дальнего Востока. Чита, 1974. Вып. 10. С. 197—199.
Жовтый И. Ф., Емельянова Н. Д. Переносчики чумной инфекции в Монголь-
ской Народной Республике//Изв. Иркут, науч.-исслед. противочум.
ин-та Сибири и Дальнего Востока. Улан-Удэ, 1959. Т. 22. С. 72—107.
Жовтый И. Ф., Зарубина В. Н.у Прокопьев П. Н., Шведко Л. П. Об экто-
паразитах рукокрылых Юго-Восточного Забайкалья и прилежащих
районов Монгольской Народной Республики // Там же. 1968. Т. 24.
С. 338—343.
Заблоцкий В. И. Гельминтофауна енотовидной собаки н ондатры, аккли-
232
Литература
матизированных в дельте Волги // Тр. Астрахан. гос. заповедника. 1970.
Вып. 13. С. 364—381.
Забродин В. А., Вагина Л. Н., Забродина Е. Ф. Некоторые данные по
бруцеллезу диких животных Таймыра и Эвенкии // Вопросы сельского и
промыслового хозяйства Крайнего Севера. Норильск, 1971. С. 102—109.
(Тр. НИИ сел. хоз-ва Крайнего Севера; Т. 19).
Забродин В. А., Никулина В. П. Изучение бруцеллеза песцов на Таймы-
ре // Науч. тр. НИИ сел. хоз-ва Крайнего Севера. Норильск, 1976.
Т. 22. С. 59—65.
Загнибородова Е. Н., Свиденко Г. Д. Межвидовые связи тонкопалого сус-
лика в Туркмении // Грызуны и их эктопаразиты. Саратов: Изд-во Са-
рат. ун-та, 1968. С. 192—197.
Залесский А. Н. Влияние системы земледелия на динамику численности и
поведение краснощекого суслика в районе ветровой эрозии // Тр. науч.-
исслед. ин-та защиты растений КазССР. 1962. Т. 7. С. 529—575.
Зарубина В. Н, Материалы по биологии и экологии вшей даурского суслика //
Изв. Иркут, науч.-исслед. противочумного ин-та Сибири и Дальнего Во-
стока. Иркутск, 1963. Т. 25. С. 363—368.
Зверев М. Д. Биология сусликов Эверсманна и опыты по борьбе с ними
отравленными приманками // Изв. Сиб. краев, станции защиты растений.
Новосибирск, 1929. Вып. 6. С. 113—130.
Зверев М. Д. Весенние наблюдения над краснощеким сусликом И Изв.
Сиб. краев, станции защиты растений. Новосибирск, 1930. Вып. 4(7):
Зоология. С. 13—25.
Земская А. А. Паразит летучих мышей Ichoronyssus flavus Kolenati, 1956
(Gamasoidea, Dermanyssidae) // Паразиты и паразитозы человека и жи-
вотных. Киев: Наук, думка, 1965. С. 316—323.
Земская А. А. Паразитические гамазовые клещи и их медицинское значе-
ние. М.: Медицина, 1973. 167 с.
Земская А. А., Гроховская И. М. Клещи бурундуков в Приморском крае //
Мед. паразитология. 1959. Т. 28, № 2. С. 152—157.
Земсков М. В., Кругликов В. М., Шитов К. А., Холин С. Р. Эксперименталь-
ный лептоспироз сусликов // ЖМЭИ. 1950. № 8. С. 32—35.
Зибицкер Д. Е., Ковалев Е.А. Эпизоотология и эпидемиология бешенства в
Белоруссии. Минск: Ураджай, 1968. С. 3—103.
Змушко Л. С. Некоторые вопросы эпидемиологии и эпизоотологии бешенства
в Белоруссии // Вопросы борьбы с бешенством. М.: Медицина, 1967.
С. 138—143.
Зундуй Ч. Энзоотичная по чуме территория в Северо-Восточном Хангае //
Докл. Иркут, науч.-исслед. противочум. ин-та Сибири и Дальнего Во-
стока. Чита, 1974. 10. С. 65—68.
Иголкин Н. И, Комплексы эктопаразитов мелких млекопитающих юго-
восточной части Западной Сибири. Томск: Изд-во Том. ун-та, 1973. 239 с.
Инструкция по эпидемиологическому надзору в природных очагах чумы
Советского Союза. Саратов: «Микроб», 1979. 59 с.
Иофф И. Г9 Вопросы экологии блох в связи с их эпидемиологическим зна-
чением. Пятигорск: Орджоникидз. краев, изд-во, 1941. 116 с.
Иофф И. Г. Алакурт // Тр. Биол. ин-та Кирг. фил. АН СССР. 1947. Вып. 1.
С. 111 — 135.
Иофф И. Г. Aphaniptera Киргизии. М.: АМН СССР, 1949. 211 с.
' Иофф И. Г., Микулин М. А., Скалой О. И. Определитель блох Средней
Азии и Казахстана. М.: Медицина, 1965. 370 с.
Иофф И. Г,, Скалой О. И. Определитель блох Восточней Сибири и Даль-
него Востока. М.: Медгиз, 1954. 275 с.
Иофф И, Г,, Тифлов В. Е. Определитель Афаниптера юго-востока СССР.
Ставрополь: Кн. изд-во, 1954. 198 с.
Исаков ТО. А. О бешенстве у диких животных, обитающих в районе дельты
Литература
233
Волги // Вопросы краевой, общей и экспериментальной паразитологии.
М.: Изд-во АМН СССР, 1949. Т. 6. С. 82—86.
Исмагилов М. И. Материалы к фауне и экологии грызунов Южных Кызыл-
кумов // Материалы по фауне и экологии наземных позвоночных Ка-
захстана. Алма-Ата: Изд-во АН КазССР, 1962. С. 192—200.
Казанцева Ю. М., Фенюк, Б. К. К экологии мохноногого тушканчика Dipus
sagitta Pall. (Mammalia) // Учен. зап. Сарат. ун-та. Биол. сер. 1937. Т. 1
(14), Вып. 1. С. 134—166.
Калабухов Н. И. Кольцевание сусликов (Citellus pygmaeus Pall.) при изу-
чении продолжительности их жизни в природе И Учен. зап. МГУ 1933.
Вып. 1. С. 1—6.
Калабухов Н. И. Значение грызунов как фактора очаговости некоторых
инфекций // Зоол. журн. 1949. Т. 28, вып. 5. С. 389—406.
Калабухов Н. И, Особенности терморегуляции грызунов как один из фак-
торов их чувствительности к чумной инфекции // ЖМЭИ. 1958. № 9.
С. 91—97.
Калабухов Н. И. Явление длительных перерывов эпизоотологической актив-
ности природных очагов чумы и его вероятные причины // Зоол. журн.
1959. Т. 48, вып. 2. С. 165—178.
Калабухов Н. И., Раевский В. В, Изучение передвижений сусликов в степ-
ных районах Северного Кавказа методом кольцевания // Вопросы эколо-
гии и биоценологии. Л.: Биомедгиз, 1935. Вып. 2. С. 170—195.
Калабухов Н. И., Раевский В. В. Цикл жизни малого суслика // Вести,
микробиологии, эпидемиологии и паразитологии. Саратов, 1936. Вып. 1
С. 109—130.
Калиновский А. И., Пинигин А. Ф., Петухова О. С. Характеристика бру-
целл, выделенных в Восточной Сибири и на Дальнем Востоке // Докл.
Иркут, науч.-исслед. противочумного ин-та Сибири и Дальнего Востока.
Иркутск, 1971. Вып. 9. С. 132—134.
Калмыкова А. Д., Козловская, О. Л., Гарбузов М. А., Рябов Н. И. О роли
некоторых видов грызунов в эпидемиологии геморрагического нефрозо-
нефрита и мерах профилактики II Там же. 1962. Вып. 3. С. 73—76.
Калязина И. М., Донченко Г. Е., Денисов П. С. Состояние численности
малого суслика и его блох в южной части Ергеней в 1973 г. //Проблемы
особо опасных инфекций. Саратов, 1975. Вып. 3/4. С. 37—40.
Карякин В. Н. Токсоплазмоз млекопитающих // Ежегодник БМЭ. М.: Сов.
энциклопедия, 1971. Т. 3. С. 865—906.
Канатов Ю. В., Лобачев В. С., Муафихов Г. М. и др. Ретроспетивная
диагностика чумы у различных видов тушканчиков // Тушканчики
фауны СССР. М., 1985. С. 202—204.
Качтэрэзчм Р. А. Материалы по экологии дикования животных в Заполя-
рье // Бюл. МОИП. Отд. биол. 1963. Т. 68, вып. 4. С. 3—13.
Канторович Р. А. Природные очаги дикования и бешенства в СССР: Авто-
реф. дис. . . . д-ра биол. наук. М., 1965. 30 с.
Канторович Р. А. Некоторые аспекты мирового распространения и экологии
бешенства // Итоги науки и техники: Мед. география. М.: ВИНИТИ,
1968. С. 273—321.
Канторович Р. А. Бешенство // Ежегодник БМЭ. М.: Сов. энциклопедия,
1971. Т. 12. С. 165—167.
Канторович Р. А. Роль социально-экономических факторов в эпидемиологии
зоонозов (на модели бешенства) // Современные проблемы зоонозных
инфекций: Тез. докл. М., 1981. С. 180—182.
Канторович Р. А., Горбенко М. Н., Назарова 3. Д. и др. Опыт изучения
природных очагов бешенства на территории Челябинской области //
Вопросы борьбы с бешенством. М.: Медицина, 1967. С. 181—196.
Канторович Р. А., Маркарян А. Г. О патогенности вируса «дикования» для
человека (письмо в редакцию) // Вопр. вирусологии. 1963. № 6. С. 747.
Капланов Л. Г. Тигр в Сихотэ-Алине// Тигр, изюбрь, лось. М.: МОИП, 1948.
234
Литература
С. 18—45. (Материалы к познанию фауны и флоры СССР. Н. С. Отд.
зоол.; Вып. 14).
Капустин В. Ф. Атлас паразитов крови животных и клещей иксодид. М.:
Сельхозгиз, 1955. 98 с.
Карасев И. Ф. Гельминтофауна млекопитающих Березинского заповедника//
Березинский заповедник. Минск: Ураджай, 1970. Вып. 1. С. 155—179.
Карасев Н. Ф. Цестоды хищных млекопитающих // Там же. 1975. Вып. 4.
С. 221—223.
Карасева Е. В., Свешникова Н. П. Дикие позвоночные животные — носи-
тели лептоспир в природе и характер эпизоотий в их популяциях //
Лептоспирозы людей и животных. М.: Медицина, 1971. С. 163—207.
Карасева Е. В., Чернуха Ю. Г., Телицин Ю. М. и др. К изучению биологии
обских леммингов и полевок-экономок и их роли в природных очагах
лептоспирозов на Ямале// Бюл. МОИП. Отд. биол. 1976. Т. 81, вып. 6.
С. 32—39.
Карась Ф. Р., Варгина С. Г., Осипова Н. 3. и др. Изучение очагов арбови-
русных инфекций на территории Киргизии // Экология арбовирусов. М.,
1973. Вып. 1. С. 69—73.
Карась Ф. Р., Варгина С. Г., Осипова Н. 3., Львов Д. К. Материалы об-
следования на арбовирусы иксодовых клещей Киргизии // Экология виру-
сов: Материалы X симпоз. Баку, 1976. С. 84—86.
Карпов С. П., Попов В. М. Млекопитающие Западной Сибири как резервуа-
ры и источники инфекций человека // Тр. Том. ун-та. Томск, 1956. Т. 142.
С. 345—352.
Карпов С. П., Федоров Ю. В. Эпидемиология и профилактика клещевого
энцефалита. Томск: Изд-во Том. ун-та, 1963. 228 с.
Карулин Б. Е. К биологии тонкопалого суслика // Материалы к познанию
фауны и флоры СССР (МОИП). Отд. зоол. 1960. Вып. 38(53). С. 165—182.
Карулин Б. Е., Пчелкина А. А. Теплокровные животные — носители
возбудителя Q-лихорадки // Докл. АН СССР. 1958. Т. 115, № 5. С. 1054—
1056.
Каталог млекопитающих СССР. Л.: Наука, 1981. 455 с.
Кирьянов Г. И. О заболеваниях животных бешенством в Алтайском крае //
Докл. Иркут, науч.-исслед. противочум. ин-та Сибири и Дальнего Восто-
ка. Хабаровск, 1962. Вып. 3. С. 96—97.
Кирьянов Г. И. Краснощекий суслик (Citellus erythrogenys Brandt) в Алтай-
ском крае // Териология. Новосибирск: Наука, 1974. Т. 2. С. 289—290.
Киселева Н. К. Влияние малого суслика на миграцию солей в почвах При-
каспийской низменности // Почвоведение. 1976. № 1. С. 73—86.
Кищинский А. А, Белый медведь. М.: Лесн. пром-сть, 1974. 69 с.
Клец Э. И., Колесник Р. С., Хрусцелевский В. П. и др. К вопросу об экспе-
риментальной чуме у тарбаганов и длиннохвостых сусликов // Изв. Иркут,
науч.-исслед. противочум. ин-та Сибири и Дальнего Востока. Улан-Удэ,
1959. Т. 20. С. 15—30.
Климченко И. 3., Найден П. Е., Петросян Э. А., Адамян А. О. Борьба
с грызунами — одна из активных форм профилактики чумы И Эпидеми-
ология и профилактика особо опасных инфекций в МНР и СССР. Улан-
Батор: Госиздат, 1978. С. 121—123.
Ковбасюк Р. Ф., Брацлавец Н. Ф.у Мельник М. Н. и др. Роль кошек в
поддержании синантропного очага токсоплазмоза // Вопросы природной
очаговости болезней. Алма-Ата: Наука, 1976. Вып. 8. С. 105—109.
Когтева Е. 3., Морозов В. Ф. Состояние популяции лисицы и ее промысло-
вое использование на северо-западе РСФСР//Тр. ВНИИОЗ. 1972. Вып.
24. С. 170—181.
Козакевич В. П. Зависимость чувствительности малых сусликов к чуме от их
физиологического состояния // Проблемы особо опасных инфекций.
Саратов, 1969. Вып. 5(9). С. 74—80.
Козакевич В. П., Варшавский С. Н., Некипелов Н. В. География природ-
Литература
235
ных очагов чумы в Восточной Азии // Эпидемиология и профилактика
природно-очаговых инфекций. М.: М3 СССР, 1981. С. 12—23.
Козлов Д. П. Определитель гельминтов хищных млекопитающих СССР.
М.: Наука, 1977. 275 с.
Колесникова Р. С. Природная очаговость дикования и значение животных
в его распространении // Материалы по экологии и численности живот-
ных в Якутии. Якутск, 1973. С. 159—170.
Колесникова Р. С. Значение диких млекопитающих тундровой зоны Якутии
в распространении вируса дикования // Охрана и рациональное исполь-
зование животного мира и природной среды Якутии. Якутск, 1979.
С. 40—44.
Коломакин Г. А., Тимонина М. С., Сансизбаев Б. К., Ушаков И. К. О роли
сусликов и барсуков в эпизоотологии бешенства // Вопросы природной
очаговости болезней. Алма-Ата: Наука, 1973. № 6. С. 39—44.
Колонии Г. В. Мировое распространение иксодовых клещей (род Ixodes).
М.: Наука, 1981. 115 с.
Колосов А. М., Лавров Н. П., Наумов С. П. Семейство белки — Sciuridae//
Биология промысловых зверей СССР. М.: Высш, шк., 1961. С. 163—196.
Колпакова С. А., Лапина Н. Ф., Me лещу к А. 77., Шаднева X. Г. Эколо-
гические особенности блох хищных млекопитающих в Астраханской
области // Грызуны и их эктопаразиты. Саратов, 1968. С. 65—72.
Комарова Л. В. Распространение и некоторые черты экологии тонкопалого
суслика // Фауна и экология грызунов. М.: Изд-во МГУ, 1980. Вып. 14.
С. 201—231.
Кондрашкин Г. А., Дудникова А. Ф. О млекопитающих и их блохах пусты-
ни Муюн-Кумы и низовьев р. Таласа // Грызуны и борьба с ними. Сара-
тов: Кн. изд-во, 1955. Вып. 4. С. 249—262.
Кондрашкина К. И., Кукин В. М., Козин М. М. Паразитирование иксодо-
вых клещей на некоторых диких и домашних млекопитающих Западного
Казахстана в зимний период // Тр. Рост, науч.-исслед. противочумного
ин-та. Ростов н/Д, 1956. Т. 11. С. 123—126.
Константинов А. И. Причины возникновения эхолокации у рукокрылых
// Рукокрылые (Chiroptera). М.: Наука, 1980. С. 271—276 (Вопр. терио-
логии).
Контримавичус В. Л. Гельминтофауна куньих СССР // Тр. гельминтол.
лаб. АН СССР. 1966. Т. 17. С. 54—81.
Контримавичус В. Л. Пути формирования гельминтофауны куньих // Там
же. 1968. Т. 19. С. 90—98.
Корелов М, Н. Отряд Летучие мыши — Chiroptera // Звери Казахстана.
Алма-Ата: Изд-во АН КазССР, 1953. С. 37—87.
Коренберг Э. И., Ковалевский Ю. В. Районирование ареала клещевого энце-
фалита. М.: ВИНИТИ, 1981. 147 с. (Итоги науки и техники. Мед. геогра-
фия; Т. 11).
Корнеев А. П. Енотовидная собака на Украине (Результаты работ по аккли-
мации) // Тр. Зоомузея Киев, ун-та. 1954. № 4. С. 13—72.
Коршунова О. С., Пионтковская С. П.у Никитина Н. А. О природных
очагах клещевого сыпного тифа Азии в Хакассии и в центральной части
Западного Саяна // Зоол. журн. 1959. Т. 38, Вып. 3. С. 385—393.
Коршунова О. С.у Пионтковская С. П., Флинт В. Е. О природных очагах
клещевого сыпного тифа в Бурятской АССР // Там же. 1965. Т. 44, Вып. 7,
С. 980—985.
Костюков М. А. О роли летучих мышей в экологии арбовирусов в Таджи-
кистане // Экология вирусов: Материалы X симпоз. Баку, 1976.
С. 92—93.
Костюков М. А., Булычев В. П. Экспериментальное изучение восприимчи-
вости летучих мышей Vespertilio pipistrellus к вирусу Иссык-Куль //
Экология вирусов: Материалы IX симпоз. Душанбе, 1975. С. 34—35.
Костюков М. А., Булычев В. П., Данияров О. А. и др. О выделении 2
226
Литература
штаммов вируса Иссык-Куль от летучих мышей Vespertilio pipistrellus
в Таджикистане // Там же. 1975. С. 31—32.
Костюков М. А., Булычев В. П., Данияров О. А., Пак Т. П. К экологии
вируса Тягиня в Таджикистане // Сб. тр. Ин-та вирусологии АМН СССР.
1976. Вып. 4. С. 112—118.
Костюков М. А., Булычев В. П., Лапина Т. Ф. Экспериментальное зара-
жение Aedes caspius caspius Pall, на летучих мышах нетопырь-карлик
Vespertilio pipistrellus, инфицированных вирусом Иссык-Куль, с после-
дующей передачей его восприимчивым животным// Мед. паразитология.
1982. Т. 60, № 6. С. 78—79.
Кошкин С. М., Лазарева Л. Л., Ельшанская Н, И., Хумархан К. Некото-
рые итоги изучения фауны гамазовых клещей мелких млекопитающих
и птиц в очагах чумы Горного Алтая и Западной Монголии // Эпидемио-
логия и профилактика особо опасных инфекций в МНР и СССР. Улан-
Батор: Госиздат, 1978. С. 140—141.
Кошкин С. М., Хумархан К. Видовой состав, экология и эпизоотологиче-
ское значение блох монгольской пищухи северо-западной Монголии //
Докл. Иркут, науч.-исслед. противочумного ин-та Сибири и Дальнего
Востока. Чита, 1974. Вып. 10. С. 202—204.
Красная Книга СССР. М.: Лесн. пром-сть, 1978. С. 15—22.
Кронтовская (Дцимирская) М. К. Клещевой сыпной тип // Тр. Всесоюзн.
конф, эпидемиологии, микробиологии и инфекц. болезней. М.: Медгиз,
1940. С. 234—241.
Крылова К. Т., Шилова Е. С. Эпизоотологическое значение особенностей
экологии желтого суслика в Северном Приаралье // Проблемы особо
опасных инфекций. Саратов, 1970. Вып. 4(14). С. 172—177.
Кубанцев Б. С. Динамика численности и интенсивность размножения ма-
лого суслика // Учен. зап. Волгогр. пед. ин-та. 1961. Вып. 1. С. 79—82.
Кудряшова Н. И. Новые виды клещей краснотелок (Acariformes, Trombi*
culidae) с летучих мышей из Ирана // Зоол. журн. 1975. Т. 54, вып. 10.
С. 1562—1565.
Кудряшова Н. И. Современное состояние изученности клещей краснотелок
(Acariformes, Trombieulidae) фауны СССР // Итоги науки и техники.
Зоопаразитология. М.: ВИНИТИ, 1979. Т. 5. С. 5—112.
Кудряшова Н. И., Рыбин С. Н. Новые виды клещей краснотелок (Acarifor-
mes, Trombieulidae) из Киргизии // Зоол. журн. 1974. Т. 53, вып. 4.
С. 633—639.
Кузнецова К. А., Иванов В. А., Варшавский С. Н. и др. Эпизоотическое со-
стояние природных очагов чумы СССР и-дальнейшее изучение ее энзоо-
тии // Проблемы изучения механизма энзоотии чумы: Тез. докл. Саратов,
1980. С. 19—22.
Кузякин А. 77. Летучие мыши. М.: Сов. наука, 1950. 443 с.
Кулик И. Л. Сравнительный анализ фаунистических комплексов млекопи-
тающих лесной части Северной Евразии // Териология. Новосибирск:
Наука, 1974. Т. 2. С. 151 — 162.
Кулик И. Л. Взаимопроникновение фаунистических комплексов млекопи-
тающих // Современные проблемы зоогеографии. М.: Наука, 1980а.
С. 272—284.
Кулик И. Л. Распространение и типы ареалов тушканчиков (Dipodidae,
Rodentia) // Там же. 19806. С. 152—167.
Кулик И. Л., Карулин Б. Е. Опыт использования радионуклеидной метки
для наблюдения активности и поведения мохноногого тушканчика (Di-
pus sagitta) // Зоол. журн. 1980. Т. 59, вып. 10. С. 1559—1565.
Куликова Н. А. Трихинеллез млекопитающих //II съезд Всесоюзн. териол.
о-ва: Тез. докл. М.: Наука, 1978. С. 261—262.
Курское А. Н. Эколого-морфологический анализ летательного аппарата
рукокрылых // Рукокрылые (Chiroptera). М.: Наука, 1980. С. 21—26.
(Вопр. териологии).
Литература
237
Кучеров П. Л4., Оптякова А. Ф., Рожков А. А. и др. Опыт профилактики
чумы в зоне деятельности противочумной станции // Международные и
национальные аспекты эпиднадзора при чуме. Иркутск, 1975. С. 77—78.
Кучерук В. В. Количественный учет важнейших видов вредных грызунов и
землероек // Методы учета численности и географического распределения
наземных позвоночных. М.: Изд-во АН СССР, 1952. С. 9—46.
Кучерук В. В. Степной фаунистический комплекс млекопитающих и его
место в фауне Палеарктики // География населения наземных животных
и методы его изучения. М.: Изд-во АН СССР, 1959а. С. 45—87.
Кучерук В. В. Вопросы палеогенезиса природных очагов чумы в связи с
историей фауны грызунов // Фауна и экология грызунов. М.: Изд-во
МГУ, 1965. Вып. 7. С. 5—86.
Кучерук В. В. Млекопитающие — носители болезней, опасных для чело-
века // Успехи современной териологии. М.: Наука, 1977. С. 75—90.
Кучерук В. В. Итоги работ советских ученых по изучению и профилактике
природно-очаговых болезней. Сообщ. 1 // Мед. паразитология. 1982.
Т. 60, № 6. С. 9—17.
Кучерук В. В. Норы млекопитающих — их строение, использование и
типология // Фауна и экология грызунов. М.: Изд-во МГУ, 1983. Вып. 15.
С. 5—54.
Кучерук В. В., Бибиков Д. И. Сурки как хранители чумы // Сурки: Био-
ценотическое и практическое значение. М.: Наука, 1980. С. 111—164.
Кучерук В. В., Тупикова Н. В. Роль тушканчиков в питании хищных птиц
степей и пустынь восточной половины МНР // Тушканчики фауны СССР.
М., 1985. С. 204—205.
Кучерук В. В., Тупикова Н. В., Доброхотов Б. П. и др. Группировки
населения мелких млекопитающих и их территориальное размещение в
восточной половине МНР // Современные проблемы зоогеографии. М.:
Наука, 1980. С. 115—151.
Кучук Л. А. Летучие мыши — носители арбовирусных инфекций в Кирги-
зии // Сб. тр. Кирг. НИИ эпидемиологии, микробиологии и гигиены.
Фрунзе, 1977. Т. 17. С. 87—90.
Кыдырбаев X. К. Современное состояние промысла и перспективы исполь-
зования запасов суслика-песчаника в Казахстане // Животный мир Ка-
захстана и проблемы его охраны. Алма-Ата: Наука, 1982. С. 103—104.
Лабунец Н. Ф., Акиев А. К., Гончаров А. И., Голубев П. Д. О блохах гор-
ного суслика в Приэльбрусье // Проблемы особо опасных инфекций.
Саратов, 1974. Вып. 1(35). С. 82—89.
Лабунец Н. Ф., Богданов О. П. Зимние виды блох летучих мышей из Уз-
бекистана // Зоол. журн. 1959. Т. 38, вып. 2. С. 221—227.
Лабунец Н. Ф., Писарская П. О распространении блох (Siphonaptera) евро-
пейского суслика (Citellus citellus L.) в Болгарии // VII междунар. сим-
поз. по энтомофауне Средней Европы: Тез. докл. Л.: Наука, 1977. С. 56.
Лабунец Н. Ф., Сардар Е. А. Исследование блох малого суслика в чумном
очаге Дагестана // Тр. научн.-исслед. противочум. ин-та Кавказа и За-
кавказья. Ставрополь, 1961. Вып. 5. С. 5—18.
Лаврова М. Я- Грызуны лесных полезащитных полос Ставропольских и
Сальских степей // Материалы по биогеографии СССР. М., 1955. С. 108—
149. (Тр. Ин-та географии АН СССР; Т. 66, выпс 2).
Лавровский А. А. О цикличности эпизоотий в природных очагах чумы и
причинах ее обуславливающих // Проблемы особо опасных инфекций.
Саратов, 1969. Вып. 1(5). С. 3—10.
Лавровский А. А., Абдурахманов Г. А., Сорокина 3. С. и др. Особенности
активации дагестанского равнинно-предгорного очага чумы в 1975 году //
Там же. 1978. Вып. 6(64). С. 18—22.
Лавровский А. А., Варшавский С. Н., Герасимова Н. Г. и др. Эпизоотии
чумы среди малых сусликов в природном очаге Северо-Западного При-
каспия в 1972—1973 гг. / Там же. 1974. Вып. 3(37). С. 5—17.
238
Литературе?
Лавровский А. Л., Варшавский С. Н., Шилов М. Н. и др. Современные
представления о структуре природных очагов чумы сусликового типа в
Северо-Западном Прикаспии и в Предкавказье и важнейшие принципы
эпизоотологического обследования их территорий // Там же. 1972.
Вып. 5(27). С. 15—30.
Лебедев А. Д., Пак Т. П., Мелиев Л. Экологическая география вирусов
крымской геморрагической лихорадки и Конго // Географическая среда и
распространение болезней. М.: ВИНИТИ, 1980. С. 3—148. (Итоги науки
и техники. Мед. география; Т. 10).
Лебедева И. Р., Селимов М. А. К эпидемиологии бешенства природного и
городского типов // Вопр. мед. вирусологии: Тез. конф. М., 1975. С. 493—
" 494.
Левит А. В., Вустина У. Д., Губенко Л. Н. Токсоплазмоз грызунов //
Токсоплазмоз животных. Алма-Ата: Наука, 1965. С. 238—273.
Левкович Е. Н., Погодина В. В., Засухина Г, Д., Карпович Л. Г. Вирусы
комплекса клещевого энцефалита. Л.: Медицина, 1967. 245 с.
Леонов Ю. Л., Харитонов Н. Н., Сапегина В. Ф. Краснощекий суслик и его
значение в эпизоотологии омской геморрагической лихорадки // Изв.
СО АН СССР. Сер. биол. наук. 1970. № 10, вып. 2. С. 126—129.
Летов Г. С., Летова Г. И. Эктопаразиты длиннохвостого суслика (Citellus
undulatus) в связи с эпизоотологическим значением в Туве // Зоол. журн.
1973. Т. 52, вып. 4. С. 525—531.
Лин Л. Л. Битва в Миорах. М.: Знание, 1967. С. 3—80.
Липаев В. М. Материалы по стациональному размещению грызунов в сте-
пях Северо-Восточной Монголии // Изв. Иркут, науч.-исслед. противочум.
ин-та Сибири и Дальнего Востока. Иркутск, 1966. Т. 26. С. 168—174.
Липаев В. М., Бусоедова Н. М., Деревич Ш. М. Основные итоги десяти-
летнего (1967—1976 гг.) эпизоотологического обследования северо-
западной части МНР // Эпидемиология и профилактика особо опасных
инфекций в МНР и СССР. Улан-Батор: Госиздат, 1978. С. 30—31.
Липин С. И., Литвиненко Р. П., Васенин А. Л., Горин О. 3. Материалы
к эпизоотологической характеристике различных территорий Бурятской
АССР по клещевому риккетсиозу Азии // Мед. паразитология. 1983.
№3. С. 21—28.
Лисецкий А. С. Влияние орошения на численность и размещение малого
суслика (Citellus pygmaeus Pall.) в Каменско-Днепровском районе
Запорожской области // Учен. зап. Харьк. ун-та. 1959. Т. 106. С. 123—
131. (Тр. НИИ биологии и биол. фак.; Вып. 28).
Лобачев В. С., Шенброт Г. И. Особенности размножения тушканчиков //
Бюл. МОИП. Отд. биол. 1976. Т. 81, вып. 4. С. 38—52.
Лобачев В. С., Шенброт Г. И. Тушканчики // Итоги мечения млекопитаю-
щих. М.: Наука, 1980. С. 147—153.
Лобков В. А. О размножении крапчатого суслика (Citellus suslicus Giild.) в
антропогенном ландшафте Северо-Западного Причерноморья // Эколо-
гия. 1983. № 2. С. 44—49.
Лысенко А. Д. Токсоплазмоз. М.: Медицина, 1980. 64 с.
Львов Д. К., Лебедев Л. Д. Экология арбовирусов. М.: Медицину, 1974.
184 с.
Мазине М. В. О местах зимовки рукокрылых в Эстонии // Рукокрылые (Chi-
roptera). М.: Наука, 1980. С. 196—198. (Вопр. териологии).
Маковкин Л. И. Влияние амурского тигра на популяцию изюбра на юге
Приморья // Экологические основы охраны и рационального использо-
вания хищных млекопитающих. М.: Наука, 1979. С. 248—250.
Мальков Г. Б. Структура и пространственно-временные изменения природ-
ных очагов бешенства // II съезд Всесоюзн. териол. о-ва: Тез. докл. М.:
Наука, 1978. С. 263—264.
Мальков Г. Б., Грибанова Л. Я- Основные результаты изучения природной
очаговости бешенства в Сибири и на Дальнем Востоке и направления
Литература
239
дальнейших исследований // Современные методы изучения природно-
очаговых инфекций. Омск, 1980. С. 11—22.
Маматов Н. М. Вирусологическое обследование диких, домашних плотояд-
ных животных на бешенство в различных ландшафтных зонах Узбеки-
стана // Тез. докл. Юбил. конф., посвящ. 50-летию со дня основания
УзНИВИ. Тайляк, 1976. Ч. 1. С. 174—176.
Маматов Н. М. О природных очагах бешенства животных на территории
Узбекской ССР // X Всесоюзн. конф, по природной очаговости болезней.
Душанбе, 1979. Т. 2. С. 134—135.
Мамонтов И. М. Значение агротехнических и мелиоративных мероприятий
в борьбе с малым сусликом в Котельниковском районе Сталинградской
области // Вести, микробиологии, эпидемиологии и паразитологии.
1937. Т. 16, вып. 1/2. С. 198—210.
Маренич Б. И., Улюмджиев О. Ц. Состояние численности малого суслика
на западных склонах Ергенинской возвышенности // Вопросы природ-
ной очаговости зоонозов. Саратов: «Микроб», 1982. С. 64—70.
Мари Н. Н. Основы учения о зоонозах. СПб., 1909. Вып. 2: Бешенство.
222 с.
Марченко И. Д. К вопросу о круге хозяев альвеококкоза И Материалы науч,
конф. Всесоюзн. о-ва гельминтологов, 1968. М., 1971. Вып. 2. С. 153—
155.
Масленникова 3. П., Куницкая Н. Т., Золотова С. И. и др. К изучению
блох рода Frontopsylla (Siphonaptera, Leptopsyllidae) Тянь-Шаня //
Зоол. журн. 1982. Т. 61, вып. 10. С. 1597—1599.
Материалы Первого Всесоюзного совещания по рукокрылым / Под ред.
П. П. Стрелкова, А. П. Кузякина. Л.: ЗИН АН СССР, 1974. 182 с.
Матчанов Н. М., Муминов П. Гельминты домашних и диких плотоядных
животных Узбекистана и их значение в возникновении гельминтозных
инвазий // X Всесоюзн. конф, по природной очаговости болезней. Алма-
Ата, 1979. Т. 1. С. 198—199.
Мачинский А. П., Семов В. Н. К познанию гельминтофауны хищных (сем.
Canidae) Мордовии // Учен. зап. Мордов. гос. ун-та. Сер. вет. и мед.
наук. Саранск, 1968. № 75, ч. 1. С. 133—145.
Медзыховский Г, А., Маштаков В. И. О пространственных перемещениях
малого суслика на северной окраине Волго-Уральских песков // Проблемы
особо опасных инфекций. Саратов, 1972. Вып. 2(24). С. 88—93.
Меженный А. А. Биология бурундука в Южной Якутии // Материалы по
биологии и динамике численности мелких млекопитающих Якутии.
Якутск, 1968. С. 87—119.
Мельникова Т. Г. Иксодовые клещи диких и домашних животных Крым-
ского заповедника // Зоол. журн. 1953. Т. 32, вып. 3. С. 422—434.
Метелева Р. И., Рубанчик И. С. Бешенство среди оленей в Заполярье И
Ветеринария. 1954. № 1. С. 47—48.
Микулин М. А. Блохи Центрального Казахстана // Тр. Среднеаз. науч.-
исслед. противочум. ин-та. Алма-Ата, 1956. Т. 2. С. 109—126.
Микулин М. А., Мурзахметова К., Подлесский Г. И. и др. Новые афани-
птерологические находки в Казахстане и Средней Азии // Там же 1959
Вып. 6. С. 251—256.
Миролюбов И. И. Болезни пятнистого оленя // Вопросы пантового оленевод-
ства. Владивосток, 1934. Ч. 2. С. 1—22.
Миронов Н. П.у Карпузиди К. С., Клименко И. 3. и др. Источники и
переносчики чумы и туляремии. М.: Медицина, 1965. 195 с.
Миронов Н. П.у Ширанович П. М., Путница Ф. А. Контактные связи между
грызунами в условиях песчаной полупустыни Северо-Западного Прикас
пия II Реф. науч.-исслед. работ Рост, противочум. ин-та. Ростов н/Д
1949. Т. 8. С. 5—11.
Михайлова Р. С,, Дедовская А. П. Различия в чувствительности к чумному
240
Литература
микробу горных и равнинных сусликов Предкавказья // Проблемы
особо опасных инфекций. Саратов, 1973. Вып. 2(30). С. 64—69.
Мишаков Н. Г. К гельминтофауне населения Приморского края и некоторые
вопросы эпидемиологии нанофиетоза // Материалы науч. конф. Всесоюз.
о-ва гельминтологов. 1971. Вып. 22. С. 172—175.
Млекопитающие в наземных экосистемах. М.: Наука, 1985. 289 с.
Млекопитающие Казахстана. Алма-Ата: Наука, 1969. Т. 1, ч. 1: Грызуны
(сурки и суслики) / Под ред. А. А. Слудского и др. 453 с.
Млекопитающие Казахстана. Алма-Ата: Наука, 1977. Т. 1, ч. 2: Грызуны
(кроме сурков, сусликов, земляной белки, песчанок и полевок) / Под ред.
А. А. Слудского. 535 с.
Млекопитающие Казахстана. Алма-Ата: Наука, 1981. Т. 3, ч. 1: Хищные
(собачьи, медвежьи, енотовые), ластоногие (настоящие тюлени) / Под
ред. А. А. Слудского. 231 с.
Млекопитающие Казахстана. Алма-Ата: Наука, 1982. Т. 3, ч. 2: Хищные
(куньи, кошки) / Под ред. А. А. Слудского. 244 с.
Млекопитающие Киргизии / Под ред. И. М. Громова и А. И. Янушевича.
Фрунзе: Илим, 1972. 453 с.
Млекопитающие Советского Союза / Под ред. В. Г. Гептнера и др. М.: Высш,
школа, 1967. Т. 2. Ч. 1: Морские коровы, хищные. 1003 с.
Млекопитающие Советского Союза / Под ред. В. Г. Гептнера, А. А. Слуд-
ского. М.: Высш, школа, 1972. Т. 2. Ч. 2: Хищные (гиены и кошки).
551 с.
Млекопитающие Якутии / Под ред. В. А. Тавровского и др. М.: Наука, 1971.
660 с.
Мокроусов Н. Л- Экологическое размещение и численность емуранчиков по
подзонам Северо-Западного Прикаспия // Науч. конф, по природной
очаговости и эпидемиологии особо опасных инфекцион. заболеваний:
Тез. докл. Саратов, 1957. С. 259—260.
Морозов В. Ф. Результаты акклиматизации, промысел и численность еното-
видной собаки на Северо-Западе СССР // Акклиматизация охотничьих
животных в СССР. Минск, 1978. С. 125—127.
Морозов П. Н. К биологии азиатской широкоушки (Barbastella leucomelas
Cretz.) // Материалы I Всесоюзн. совещ. по рукокрылым. Л., 1974. С. 92—
93.
Морозов Ф. Н. Гельминты волков Мордовского госзаповедника 77 Тр. лаб.
ГЕЛАН СССР. М.: Изд-во АН СССР, 1951. Т. 5. С. 146—149.
Морозов Ю, В. Новые данные о восприимчивости теплокровных животных к
вирусу клещевого энцефалита // Клещевой энцефалит и вирусные гемор-
рагические лихорадки: Материалы конф. Омск, 1963. С. 203—204.
Москвин И. А. Клещевые спирохетозы Л.: Медгиз, 1960. 163 с.
Мошкин А. В., Лебедев'А. Д. Арбовирусы мира и их географическое рас-
пространение. Ч. 2. Антигенные группировки, голотипы и синтипы арбо-
вирусов, распространение их по зоогеографическим подразделениям
суши, государствам и странам мира // Итоги науки и техники. Мед.
география. М.: ВИНИТИ, 1974. Т. 6. С. 10—304.
Мулярская Л. В. Новые виды тромбикулид (Acariformes, Trombiculidae) с
летучих мышей Азербайджана // Вести, зоологии. 1970. № 5. С. 42—47.
Мулярская Л. В. Два новых вида тромбикулид (Acariformes, Trombiculidae)
с летучих мышей из Азербайджана // Зоол. журн. 1971. Т. 50, вып. 12.
С. 1877—1880.
Мулярская Л. В., Дубовченко Т. А. Материалы к познанию тромбикулид
(Acariformes, Trombiculidae) с летучих мышей Азербайджана /7 Изв.
АН АзССР. Сер. биол. наук. 1969. № 6. С. 51—56.
Мулярская Л. В., Дубовченко Т. А. Новые виды тромбикулид Trombigastia
и Riedlinia из Азербайджана// Паразитология. 1970. Т. 4, № 4. С. 365—-
370.
Мунхтула О., Жигмэд С., Ажиндай Д. Случай выделения возбудителя
Литература
241
листериоза из трупа краснощекого суслика // Эпидемиология и профи»
лактика особо опасных инфекций в МНР и СССР. Улан-Батор: Госиздат,
С. 100—102.
Муртазанова Э. Ш., Лобачев В. С., Савченков Ю. И. Эпизоотологическое
значение тушканчиков в Приаральских Каракумах // Бюл. МОИП*
Отд. биол. 1964. Т. 69, вып. 5. С. 156—157.
Назарова И. В. Блохи Волжско-Камского края. М.: Наука, 1981. 168 с.
Найден П. Е., Петров П. А. О механизме сохранения возбудителя чумы в
горных очагах Кавказа // Проблема изучения механизма энзоотии чумы*
Саратов: «Микроб», 1980. С. 75—79.
Насимович А. А. Новые данные по экологии росомахи в Лапландском запо-
веднике // Тр. Лапланд. гос. заповедника. М., 1948. Вып. 3. С. 107—
124.
Насимович А, А. Енотовидная собака // Песец, лисица, енотовидная собака*
М.: Наука, 1985. С. 116—145.
Настюков Н. 3. Изучение костных остатков животных из колоний большой
песчанки на Западном Устюрте // Проблемы особо опасных инфекций.
Саратов, 1975. Вып. 3/4. С. 115—121.
Настюков Н. 3. Фауна грызунов и некоторые вопросы эпизоотий чумы на
Западном Устюрте: Автореф. дис. ... канд. биол. наук. Саратов, 1978.
18 с.
Наумов Н. П. Определение возраста малого суслика (Citellus pygmaeus)
// Защита растений. Л., 1936. № 11. С. 131 —134.
Наумов Н. 77., Кулик И. Л. О костях млекопитающих, собранных на коло-
ниях больших песчанок // Вопросы краевой, общей, экспериментальной
паразитологии и медицинской зоологии. М.: Медгиз, 1955. Т. 9. С. 203—
217.
Наумов Н, П., Лобачев В. С., Дмитриев П. П., Смирин В. М. Природ-
ный очаг чумы в Приаральских Каракумах. М.: Изд-во МГУ, 1972.
404 с.
Некипелов Н. В. О сравнительном значении некоторых грызунов в сохране-
нии возбудителя туляремии в природе // Изв. Иркут, науч.-исслед.
противочум. ин-та Сибири и Дальнего Востока. Иркутск, 1946. Т. 6.
С. 245—257.
Некипелов Н. В. Вспышки туляремии в СССР (с 1921 по 1942 г.) // Там же.
1959а. Т. 20. С. 133—146.
Некипелов Н. В. Эпизоотология чумы в Монгольской Народной Республике//
Там же. 19596. Т. 20. С. 188—242.
Некипелов Н. В. О значении длиннохвостого суслика как носителя чумы //
Докл. Иркут, науч.-исслед. противочум. ин-та Сибири и Дальнего Во-
стока. 1962. Вып. 4. С. 116—119.
Некипелов Н. В. Эколого-эпизоотологическая обстановка в чумных очагах на
советско-монгольской границе // Там же. Чита, 1974. Вып. 10. С. 172—
176.
Некипелов Н. В. Об очагах чумы на советско-монгольской границе и под-
ходах к их оздоровлению // Международные и национальные аспекты
эпиднадзора при чуме. Иркутск, 1975. С. 18—21.
Некипелов Н. В. Об эпизоотиях чумы на советско-монгольской границе //
Эпидемиология и профилактика особо опасных инфекций в МНР и
СССР. Улан-Батор: Госиздат, 1978. С. 24—26.
Некипелов Н. В., Головачева В. Я., Гужевников И. А. и др. Природно-
очаговые инфекции в степях Юго-Восточного Забайкалья // Биологиче-
ские проблемы природной очаговости болезней. Новосибирск: Наука,
1981. С. 22—35.
Некипелова Г. А., Липин С. И. К изучению эпидемиологии лептоспирозов в
Иркутской области// Лептоспироз: VI Всесоюзн. конф, по лептоспирозу,
22—24 окт. 1975 г. в Баку: Тез. докл. М., 1976. С. 107—108.
9 Зак. 357 5
242
Литература
Некрасов Е. С. Распределение плотности большого суслика на северной
границе ареала // Экология. 1970. №1. С. 103.
Некрасов Е. С. Плотность поселения и размеры индивидуальных участков
большого суслика на Среднем Урале // Млекопитающие СССР: III съезд
Всесоюзн. териол. о-ва: Тез. докл. М., 1982. Т. 1. С. 257—258.
Нельзина Е. Н., Данилова Г. М., Чернова Н. И, Численность и простран-
ственное распределение микропопуляций кровососущих членистоногих
в микробиотопах малого суслика И Мед. паразитология. 1963. Т. 32,
. № 1. С. 45—54.
Нестеров А. П. Численность и структура поселений малого суслика в
центральных частях посевов яровой пшеницы в Западном Казахстане И
Грызуны: Материалы V Всесоюзн. совещ. Саратов, 1980. М., 1980. С. 433.
Никитин В, П., Зуйченко Н. А., Черникин С. М. К экологии желтого
суслика (Citellus fulvus Thomas) в Западной Туркмении // Изв. АН
ТССР. Сер. биол. наук. 1962. № 6. С. 58—65.
Никитина Н. А. Некоторые особенности экологии малого суслика на южной
границе ареала // Фауна и экология грызунов. М.: Изд-во МГУ, 1960.
Вып. 6. С. 5—26.
Никифиров Л. П. Млекопитающие природного очага клещевого энцефалита
в Козульском районе Красноярского края // Вопросы эпидемиологии
клещевого энцефалита и биологические закономерности в его природном
очаге. М.: Медицина, 1968. С. 43—75.
Никулина В. И. Эпизоотологические данные по бруцеллезу песцов на Тай-
мыре // Биология и патология пушных зверей. Петрозаводск, 1974.
С. 174—176.
Новиков Г. А. Хищные млекопитающие фауны СССР. М.; Л.: Изд-во АН
СССР, 1956. 294 с.
Новинская В. Ф. Токсоплазмоз хищных млекопитающих // Токсоплазмоз
животных. Алма-Ата: Наука, 1965. С. 274—283.
Овсюкова Н. И. Гельминты и основные гельминтозы млекопитающих Чу-
котки: Автореф. дис. ... канд. биол. наук. М., 1966. 18 с.
Овсюкова Н. И. Влияние деятельности человека на гельминтофауну живот-
ных Чукотки // Охотоведение. М.: Лесн. пром-сть, 1972. С. 162—168.
Овсюкова Н. И. Гельминты песцов Чукотки // Там же. 1974. С. 102—104.
Огнев С. И. Звери СССР и прилежащих стран. Грызуны. 1940. М.; Л.: Изд-во
АН СССР. Т. 4. 615 с.
Оленев В. Г. К вопросу о прямом и косвенном влиянии дикования на чис-
ленность песцов // Экология позвоночных животных Крайнего Севера.
Свердловск, 1965. С. 29—31. (Тр. Ин-та биологии Урал. фил. АН СССР;
Вып. 38).
Олсуфьев Н. Г. Беспозвоночные животные — переносчики и хранители ту»
ляремийной инфекции // Природная очаговость, эпидемиология и про-
филактика туляремии. М.: Медицина, 1970. С. 38—59.
Олсуфьев Н. Г., Дунаева Т. Н. О чувствительности к туляремии и о воз-
можном эпидемиологическом значении ласки, хорька и лисицы // Зоол.
журн. 1951. Т. 30, вып. 1. С. 82—91.
Олсуфьев Н. Г., Дунаева Т. Н. Эпизоотология (природная очаговость)
туляремии // Туляремия. М.: Медгиз, 1960. С. 136—206.
Олсуфьев Н. Г., Дунаева Т. Н. Природная очаговость, эпидемиология и
профилактика туляремии. М.: Медицина, 1970. 270 с.
Омельченко М. П., Потапов Н. М., Клепко 3. С., Кирьянов Е. А. Вспыш-
ка заболевания бешенством животных в Приморье // Инфекционные бо-
лезни животных. Омск, 1978. Вып. 36. С. 119—124.
Орлов В. Н. Кариосистематика млекопитающих. М.: Наука, 1974. 204 с.
Орлов В. Н., Булатова Н. Ш. Сравнительная цитогенетика и кариосисте-
матика млекопитающих. М.: Наука, 1983. 405 с.
Орлов О. Ю. Материалы по биологии тушканчиков в Западных Кызылку-
мах // Биология: Сб. студ. науч, работ. М.: Изд-во МГУ, 1957. С. 21—28.
Литература
243
Павлов М. П. Массовое заболевание лисиц Крыма // Тр. Всесоюзн. науч.-
исслед. ин-та охотничьего промысла. М., 1953. Вып. 13. С. 135—145.
Пак Т. П., Костюков М. Л., Данияров О. Л. и др. Арбовирусные инфекции в
Таджикистане // Арбовирусы. М., 1978. Вып. 3. С. 35—39.
Пакиж В. И. Распространение европейского суслика в Молдавской ССР
и некоторые данные по его экологии // Зоол. журн. 1958. Т. 37, вып. 1е
С. 105—119.
Панютин К. К. Гетеротермия и терморегуляторное поведение у руко-
крылых // Материалы Первого всесоюз. совещ. по рукокрылым. Л., 1974.
С. 15—17.
Панютин К. К. Происхождение полета рукокрылых // Рукокрылые (Chirop-
tera). М.: Наука, 1980а. С. 176—186. (Вопросы териологии).
Панютин К. К. Рукокрылые//Итоги мечения млекопитающих. М.: Наука,
19806. С. 23—46.
Пейсахис Л. Л., Степанов В. М., Куница Г. М. Реактивность желтых
сусликов из зоны стойкой очаговости чумы к возбудителю инфекции //
Вопросы природной очаговости болезней. Алма-Ата: Наука, 1968.
С. 144—149.
Перри Р. Мир белого медведя. Л.: Гидрометеоиздат, 1974. 158 с.
Пестрякова Т. С., Тихонова Л. Я., Шубин Н. Г., Петрова Н. К. К вопро-
су о природной очаговости токсоплазмоза Васюганья // Природа и эко-
номика Привасюганья. Томск: Изд-во Том. ун-та, 1966. С. 221—223.
Петрищева П. А. Эктопаразиты — переносчики болезней человека из
нор дикобраза // Вопросы краевой, общей и экспериментальной парази-
тологии и медицинской зоологии. М.: Изд-во АМН СССР, 1951. Т. 7.
С. 8—23.
Петрищева П. Л. Москиты // Переносчики возбудителей природно-очаговых
болезней. М.: Медицина, 1962. С. 64—93.
Петрищева П. Л., Суворова Л. Г., Кербабаев Э. Б. О спонтанной заражен-
ности спирохетами клещей рода Ornithodorus в пустынях Туркмении //
Там же. 1955. Т. 9. С. 17—25.
Петров П. Л., Дятлов Л. И., Голубев П. Д. Результаты картирования
поселений горного подвида малого суслика в верховьях Кубани I/ Проб-
лемы особо опасных инфекций. Саратов, 1976. Вып. 5(51). С. 19—22.
Петров П. Л., Дятлов Л. Я., Голубев П. Л., Пшихачев X. X. Опыт кар-
тографирования поселений горных малых сусликов в Центрально-Кав-
казском очаге чумы // Состояние и перспективы профилактики чумы:
Тез. докл. на Всесоюзн. конф. Саратов, 1978. С. 129—131.
Петров П. Л., Дятлов А. И., Голубев П. Д., Пшихачев X. X. Современ-
ный ареал и пространственная структура поселений горного подвида
малого суслика на Центральном Кавказе // Проблемы особо опасных
инфекций. Саратов, 1979. Вып. 4(68). С. 17—21.
Петрова Л. Л. Двукратное размножение в течение года тонкопалого сус-
лика (Spermophilopsis leptodactylus Licht.) В Каршинской степи //
Зоол. журн. 1966. Т. 45, Вып. 12. С. 1887—1889.
Петровский Ю. Т. Особенности экологии крапчатого суслика в Белорус-
сии // Там же. 1961. Т. 40, Вып. 5. С. 736—748.
Петрусенко Л. Л., Сологор Е. Л. К определению роли рукокрылых в эко-
системах Среднего Приднепровья // Вести, зоологии. 1981. № 6. С. 44—
47.
Петручук Е. М. Выделение вируса бешенства природного и городского типа
в г. Киндия Гвинейской Республики // Вирусы, их резервуары и пере-
носчики в Гвинейской Республике. Киндия, 1980. Вып. 2. С. 170—176.
Пилипенко В. Г., Найден П. Е., Абдурахманов Г. А. и др. Некоторые
итоги и первоочередные задачи изучения природного очага чумы Цент-
рального Кавказа // Проблемы особо опасных инфекций. Саратов, 1974.
Вып. 3(37). С. 18—24.
Пилипенко В. Г., Тарасова В. Е., Таран И. Ф. и др. Иммунитет и бактерионо-
2*
244
Литература
сительство при чуме у сусликов и значение этих факторов в сохранении
возбудителя в природе в межэпизоотические сезоны // Проблема изучения
механизма энзоотии чумы. Саратов: «Микроб», 1980. С. 125—130.
Пинигин А. Ф. Дополнительные носители бруцеллеза в Забайкалье // Докл.
Иркут, науч.-исслед. противочум. ин-та Сибири и Дальнего Востока.
Чита, 1974. Вып. 10. С. 165—167.
Пинигин А. Ф., Петухова А. П., Кокоуров А. П. Эпидемиология бруцеллеза
северных оленей в Сибири и на Дальнем Востоке. Сообщ. 1. Северный
олень как резервуар бруцеллезной инфекции // Вопросы эпидемиологии и
эпизоотологии особо опасных инфекций. Кызыл, 1978. Вып. 1. С. 198—
204.
Пионтковская С. П., Жмаева 3. М. Иксодовые клещи (Acarina, Parasitifor-
mes. Ixodidae) И Переносчики возбудителей природноочаговых болезней.
М.: Медицина, 1962. С. 196—262.
Поломошнов А. П. Дефинитивные хозяева токсоплазм // Вопросы природной
очаговости болезней. Алма-Ата: Наука, 1979, вып. 10. С. 68—72.
Поломошнов А. П., Петешев В. М. Природный очаг токсоплазмоза в пойме
реки Или // Природно-очаговые антропозоонозы: Тез. докл. IX Всесоюзн.
конф, по природной очаговости болезней человека и животных. Омск,
1976. С. 215—216.
Поляков А. В., Колесникова Р. С. Эпизоотия «дикования» в Колымо-Инди-
гирской тундре в сезон 1966/67 г. // Сб. науч.-техн. ВНИИОЗ. 1972.
Вып. 36. С. 43—50.
Померанцев Б. И. Иксодовые клещи (Ixodidae). М.; Л.: Изд-во АН СССР,
1950. 223 с. (Фауна СССР; Т. 4, вып. 2: Паукообразные).
Помирко Г. И. Эпизоотология и профилактика бешенства в Молдавской
ССР: Автореф. дис. . . . канд. биол. наук. Минск, 1977. 24 с.
Помирко Г. И., Резниченко Н. Г., Шарбан Е. К., Ивченко В. И. Вопросы
экологии, диагностики и профилактики бешенства в Молдавской ССР И
Вопросы медицинской вирусологии: Тез. конф. М., 1975. С. 501—502.
Попов В. А. Млекопитающие Волжско-Камского края. Казань, 1960. 468 с.
Попов В. М. Иксодовые клещи Западной Сибири. Томск: Изд-во Том. ун-та,
1962. 260 с.
Попов В. М., Иголкин Н. И. Фауна блох грызунов семейства беличьих
(Sciuridae), обитающих в лесной полосе Западной Сибири // Тр. Том.
науч.-исслед. ин-та вакцин и сывороток. 1956. Т. 7. С. 47—52.
Попов В. М., Федоров Ю. В. Бурундук как прокормитель лесного клеща
и носитель вируса клещевого энцефалита в Томском очаге инфекций И
Там же. 1958. Т. 9. С. 19—22.
Попов Н, В., Маренич Б. И., Куницына Р. Г., Щепотьев И. В. К эпизо-
отологической оценке современного состояния поселений малого суслика
в северной части Ергеней // Проблемы особо опасных инфекций. Саратов,
1976. Вып. 2(48). С. 16—21.
Попов Ю,. К. Материалы по экологии енотовидной собаки (Nyctereutes
procyonoides Gray) в Татарской АССР // Изв. Казан, фил. АН СССР.
1956. Вып. 5. С. 193—229.
Попова А. С. Фауна блох пустыни Муюнкумы// Грызуны и их эктопаразиты.
Саратов: Изд-во Сарат. ун-та, 1968. С. 402—406.
Попова А. СРапопорт Л. П. Материалы по фауне блох Восточных Кы-
зылкумов (в пределах Чимкентской области) // Проблемы особо опасных
инфекций. Саратов, 1974. Вып. 5(39). С. 90—93.
Поспелова-Штром М. В. Клещи-орнитодорины и их эпидемиологическое
значение. М.: Медгиз, 1953. 235 с.
Поспелова-Штром М. В. Фауна и географическое распространение клещей-
орнитодорин в странах мира // Паразитарные болезни и борьба с ними в
зарубежных странах. М.: Медгиз, 1959. С. 260—289.
Поспелова-Штром М. В. Экология Alectorobius cholodkovskyi (Pavl., 1930)
Литература
245
(Ornithodorinae) // Развитие паразитологической науки в Туркменистане.
Ашхабад, 1979. С. 84—87.
Прорешная Т. Л., Рапопорт Л. П., Евдошенко В. Г., Кичатов Э. А. Ма-
териалы по изучению природных очагов лихорадки Ку в Киргизии //
ЖМЭИ. 1960. №9. С. 32—37.
Прусайте Л. А. Звери семейства Canidae Литовской ССР: Автореф. дис. . . .
канд. биол. наук. Вильнюс, 1961. 26 с.
Радовский Ф. Эволюция и адаптивная радиация у Gamasina (Acarina: Me-
sostigmata) — паразитов позвоночных // Паразитология. 1968. Т. 2,
№2. С. 124—136.
Райхлин М. И. Лептоспирозы в Таймырском автономном округе // Проблемы
эпидемиологии и профилактики природно-очаговых заболеваний в За*
полярье. Омск, 1977. С. 100—126.
Ралль Ю. М. Грызуны и природные очаги чумы. М.: Медгиз, 1960. 224 с.
Рапопорт Л. П. Опыт районирования Южного Казахстана и Киргизии
по диким млекопитающим — носителям возбудителей трансмиссивных
болезней человека И Млекопитающие СССР: III съезд Всесоюз. тер иол.
о-ва: Тез. докл. М., 1982. Т. 1. С. 290—291.
Рахманин Г. Е, Пушной промысел Ямало-Ненецкого округа и мероприятия
по его рационализации // Тр. Салехардск. стационара Уральск, филиала
АН СССР. Тюмень, 1959. Вып. 1. С. 101 — 176.
Ревенко И. П., Мазыгула Н. М., Змиян Ю. Н. О роли диких животных
в распространении бешенства на Украине // Науч. тр. ЦСХА. 1979.
Вып. 216. С. 119—122.
Рейтблат А. Г., Лабунец Н. Ф., Емельянов П. Ф. и др. Сравнение фаун га-
мазовых клещей равнинного и горного подвидов малого суслика // Проб-
лемы паразитологии. Киев: Наук, думка, 1975. Ч. 2. С. 132—134.
Ременцова М. М. Бруцеллез животных. Алма-Ата: Наука, 1962. 269 с.
Ременцова М. М., Безукладникова Н. А., Бусалаева Н. H.f Сенотрусо-
ва В. Н. Экспериментальное заражение гамазовых клещей, вшей и блох
бруцеллами. Сообщ. 2 // Тр. Ин-та зоологии АН КазССР. 1963. Т. 19.
С. 226—233.
Ременцова М. М., Постричева О. В., Рыбалко С. И. Бруцеллез промысло-
вых животных. Алма-Ата: Наука, 1969. 154 с.
Рогатых Н. А. Эпидемиологическая характеристика токсоплазмоза в Си-
бири и биологические свойства выделенных штаммов токсоплазм: Ав-
тореф. дис. . . . канд. биол. наук. М., 1981. 22 с.
Рогатых Н. А., Коновалова С. И., Тобольская Л. В. Токсоплазмоз на
Крайнем Севере // Проблемы эпидемиологии и профилактики природно-
очаговых заболеваний в Заполярье. Омск, 1977. С. 126—134.
Романовский И. Д., Олейник К. Т., Трифонова Е. И., Крупеник Н. К.
Фауна блох Кара-Калпакии // Тр. Среднеаз. науч.-исслед. противочум.
ин-та. Алма-Ата, 1956. Т. 2. С. 127—134.
Рудаков В. А., Грибанова Л. Д., Удина Н. П. и др. Эпидемиологическая
характеристика заболеваемости бешенством в Западной Сибири за 1970—
1972 годы // Вопр. инфекц. патологии. Омск., 1973. С. 34—38.
Рудаков В. А., Мальков Г. Б. Вирусологическое обследование диких жи-
вотных на бешенство в Омской области // Там же. 1970. Вып. 2. С. 102—
103.
Рудаков В. А., Мальков Г. Б., Грибанова Л. Д. и др. Эпидемиологическая и
эколого-вирусологическая характеристика бешенства в Омской области
в 1970—1972 гг. // Там же. 1973. С. 39—43.
Рудаков В. А., Стрыгина Н. П., Тофанюк Е. В. и др. К эпидемиологической
характеристике гидрофобии в Западной Сибири // Там же. 1971. С. 114—
116.
Руди В. Н. О распространении рыжеватого суслика на Южном Урале //
Науч. докл. высш. шк. Биол. науки. 1984. № 7. С. 52—56.
Рукокрылые (Chiroptera). М.: Наука, 1980. 316 с. (Вопр. териологии).
246
Литература
Рукокрылые: III съезд Всесоюзн. териол. о-ва: Тез. докл. М., 1982. Т. 2.
С. 304—361.
Русакова Л. В., Новокрещенова Н. С., Загнибородова Е, Н. и др. Сравни-
тельная патологоанатомическая характеристика чумы у больших, крас-
нохвостых песчанок и желтых сусликов, зараженных подкожно и блоки-
рованными блохами // Проблемы особо опасных инфекций. Саратов,
1976. Вып. 6(52). С. 6—11.
Рыбин С. Н. Рукокрылые Южной Киргизии // Рукокрылые (Chiroptera).
М.: Наука, 1980. С. 87—95. (Вопр. териологии).
Рыбин С. Н. Гамазоидные клещи рукокрылых и их убежищ в Южной Кир-
гизии (Gamasina) // Паразитология. 1983. Т. 17, № 5. С. 335—360.
Рябошапка А. П., Кролевецкая Н. М. Изучение биологических свойств ви-
руса бешенства, выделенного от насекомоядной летучей мыши // Материа-
лы ХШ-й сес. Ин-та полиомиелита и вирусных энцефалитов. М., 1967.
С. 80—81.
Сабилазв А. С. Место тушканчиков (Rodentia, Dipodidae) в природном
очаге чумы на Устюрте и в Северо-Западных Кызылкумах // Зоол. журн.
1970. Т. 49, вып. 6. С. 916—920.
Саватеев А. И. Бешенство. М.; Л.: Госиздат, 1927. С. 3—199.
Савельев В. Д. Паразитические черви промысловых млекопитающих и пути
их циркуляции в тундровых биоценозах полуострова Таймыр: Автореф.
дис. . . . канд. биол. наук. Л., 1975. 23 с.
Савицкий Б. П. [Рецензия] // Мед. паразитология. 1980. Т. 49, № 2. С. 86—
87.— Рец. на кн.: Медицинская териология Отв. ред. В. В. Кучерук.
М.: Наука, 1979. 300 с.
Савицкий В. П., Ботвинкин А. Д., Белко В. И. и др. Эпидемиологические
особенности бешенства на Дальнем Востоке // Современные методы
изучения природно-очаговых болезней. Л., 1980. С. 31—41.
Садыхов И. А. Гельминтофауна пушных зверей Азербайджана: Автореф.
дис. . . . канд. биол. наук. Баку, 1955. 21 с.
Сазонова О. Н. Блохи // Переносчики возбудителей природно-очаговых
болезней. М.: Медицина, 1962. С. 179—195.
Сансызбаев Б. К. Нозоареал и структура природного очага бешенства в Ка-
захстане // Проблемы медицинской географии Казахстана: Материалы
секции мед. географии I съезда географов Казахстана. Алма-Ата, 1981.
С. 55—60.
Сансызбаев Б. К., Шарифхакова А. Н., Махметов М. М. и др. К экологии
бешенства в Казахстане // Вопросы медицинской вирусологии: Тез.
конф. Душанбе, 1975. С. 508—509.
Сапаров А. Норы реликтового суслика в горах Кетмень и Лобас (Централь-
ный Тянь-Шань) // Изв. АН КазССР. Сер. биол. 1982. № 3. С. 41.
Сарманова Е. С., Думина А. Л. Изучение вирусемии у диких животных и
птиц при экспериментальном заражении вирусом клещевого весенне-
летнего энцефалита // Вопр. вирусологии. 1956. № 3. С. 23—25.
Сасыкин Г. А,, Поляков В. К. К вопросу об эпизоотологической роли
малых сусликов в комплексных поселениях с большой песчанкой в се-
верной части Урало-Эмбинского междуречья // Эпидемиология и профи-
лактика чумы. Саратов, 1981. С. 66—68.
Свешникова Н. П., Карасева Е. В. Лептоспирозы диких млекопитающих
Северной Америки // Зоол. журн. 1965. Т. 44, Вып. 2. С. 253—265.
Свиденко Г. Д. Материалы по экологии тонкопалого суслика в Туркмении //
Изв. АН ТССР. Сер. биол. наук. 1963. № 2. С. 78—85.
Свиденко Г. Д., Свиденко А. А., Никитин В. П. и др. Размножение тон-
копалого суслика (Spermopilopsis leptodactylus) в Туркмении // Зоол.
журн. 1973. Т. 52, вып. 12. С. 1836—1842.
Селезнева А. А. Суслики как резервуар инфекции при туляремии // Тр. Том.
ин-та эпидемиологии и микробиологии. 1949. Т. 4. С. 49—50.
Литература
247
Селиванов А. А., Машенцев А. Е. Редкий случай чесотки у волка // Вете-
ринария. 1955. № 11. С. 87.
Селимов М. А. Пути ликвидации гидрофобии. М.: Медгиз, 1963. 291 с.
Селимов М. А. Некоторые вопросы современной эпизоотологии и эпидемио-
логии бешенства // ЖМЭИ. 1972. № 11. С. 129—138.
Селимов М. А. Бешенство. М.: Медицина, 1978. 334 с.
Селимов М. А., Ширвинская А. И., Семенова Е. В. Состояние заболевае-
мости гидрофобией за 10 лет (1954—1963 гг.) // Вопросы борьбы с бешен-
ством. М.: Медицина, 1967. С. 120—138.
Сенотрусова В. Н. Биологические особенности гамазового клеща Jchorony-
ssus flavus (Kolenati, 1856) // Паразитология. 1969. Т. 2, № 4. С. 339—341.
Сердюкова Г. В. Иксодовые клещи фауны СССР. М.; Л.: Изд-во АН СССР,
1956. 122 с.
Сивонен Л. Млекопитающие Северной Европы. М.: Лесн. пром-сть, 1979.
231 с.
Сидоров Г. Н., Савицкий В. П., Ботвинкин А. Д., Мальков Г. Б. Разме-
щение и динамика численности диких хищников семейства собачьих и их
роль в распространении бешенства в южной части Восточной Сибири и
Дальнего Востока // X Всесоюз. конф, по природной очаговости болез-
ней: Тез. конф. Душанбе, 1979. Ч. 2. С. 215—216.
Симпсон Дж. Великолепная изоляция: История млекопитающих Южной
Америки. М.: Мир, 1983. 256 с.
Синелыциков В. А. К фауне гамазовых клещей (Parasitiformes, Gamasoidea)
Павлодарской области Казахской ССР // Энтомол. обозрение, 1967. Т. 46,
№ 4. С. 835—844.
Синовия Л. И., Соловьев Ю. В. К эпидемиологии трихинеллеза в районах
крайнего северо-востока СССР // Вопросы природной очаговости болез-
ней. Алма-Ата: Наука, 1981. Вып. 12. С. 101 —107.
Скворцов Г. Н. Об условиях зимней спячки земляного зайчика в Туркме
нии // Грызуны и борьба с ними. Саратов: Изд-во Сарат. ун-та, 1955
Вып. 4. С. 39—51.
Слудский А. А. Роль диких млекопитающих в распространении инфекцион-
ных заболеваний домашних животных // Природная очаговость заразных
болезней в Казахстане. Алма-Ата: Наука, 1954. Вып. 2. С. 67—98.
Смирнов В. М., Равкин Ю. С. О значении бурундука в очагах клещевого
энцефалита северо-восточного Алтая // Природа очагов клещевого энце-
фалита на Алтае. Новосибирск: Наука, 1967. С. 126—133.
Смирнов В. П. Экспериментальная чума у желтого суслика // Тр. Науч.-
исслед. противочум. ин-та Кавказа и Закавказья. Ставрополь, 1956.
Вып. 1. С. 194—198.
Смирнова М. М. О роли насекомоядных летучих мышей в сохранении
очагов рабического вируса в Казахстане // VII Всесоюз. конф, по при-
родной очаговости болезней. Секция природной очаговости болезней:
Тез. докл. Алма-Ата; Самарканд, 1969. С. 60—61.
Снигиревская Е. М. Биология бурундука Амуро-Зейского плато // Зоол.
журн. 1962. Т. 41, вып. 9. С. 1395—1401.
Соколов В. Е. Систематика млекопитающих: Отряды однопроходных, сум-
чатых, насекомоядных, шерстокрылов, рукокрылых, приматов, непол-
нозубых, ящеров. М.: Высш, шк., 1973. 430 с.
Соколов В. Е. Систематика млекопитающих: Отряды зайцеобразных, грызу-
нов. М.: Высш, шк., 1977. 494 с.
Соколов В. Е. Систематика млекопитающих: Отряды: китообразных, хищных,
ластоногих, трубкозубых, хоботных, даманов, сирен, парнокопытных,
мозоленогих, непарнокопытных. М.: Высш, шк., 1979. 528 с.
Соколов Н. Н., Ча Н. И. Проблема эпизоотии «дикования» или «тундрового
бешенства» млекопитающих Крайнего Севера // Докл. на VIII науч. сес.
Якут. фил. АН СССР. Ботаника, почвоведение, зоология, зоотехния.
Якутск, 1957. С. 148—156.
248
Литература
Солдаткин И. С., Руденчик Ю. В., Шевченко Г, В. и др. О роли трансмис-
сивного пути передачи в развитии эпизоотий чумы в природных очагах //
Природные очаги чумы и других зоонозов. Саратов: «Микроб», 1984.
С. 3—10.
Солдатова А. Н. О некоторых особенностях периодических явлений жизни
малого суслика в Южном Заволжье // Материалы по биогеографии
СССР. 1955. С. 188—207. (Тр. Ин-та географии АН СССР; Т. 66, Вып. 2).
Солдатова А. Н. Влияние плотности населения на характер использования
территории малым сусликом// Зоол. журн. 1962а. Т. 16, вып. 6. С. 913—
921.
Солдатова А. Н. Особенности использования территории малым сусликом в
разные периоды его жизнедеятельности // Зоол. журн. 19625. Т. 41,
вып. 11. С. 1706—1713.
Солодкая А. Д. Современное состояние Забайкальского природного очага
чумы // Докл. Иркут, науч.-исслед. противочумного ин-та Сибири и
Дальнего Востока. Чита, 1974. Вып. 10. С. 47—51.
Солошенко И. 3. Лептоспирозы промысловых и домашних животных //
Лептоспирозы людей и животных. М.: Медицина, 1971. С. 222—249.
Сомов Г. П., Шубин Ф. Н., Гопаченко И. М. и др. Изучениезооантропоноз-
ных инфекций на Чукотке И Природно-очаговые заболевания в зонах
освоения Сибири и Крайнего Севера. Новосибирск: Наука, 1977. С. 16—
29.
Сомов Г. П., Шубин Ф. Н., Гопаченко И. М. и др. Природно-очаговые
инфекции на севере Дальнего Востока. Сообщ. 1. Выявление природных
очагов инфекций на Чукотке И ЖМЭИ. 1978. № 10. С. 101 —104.
Сорокина 3. С., Абдурахманов Г. А., Кочетов В. А., Дятлов А. И. Исто-
рия формирования и современное состояние поселений малого суслика в
Терско-Сунженском междуречье//Болезни с природной очаговостью на
Кавказе. Ставрополь, 1982. С. 130—132.
Соснина Е. Ф. О паразитохозяинных отношениях вшей и грызунов // Пара-
зитология. 1982. Т. 16, вып. 1. С. 62—68.
Соснина Е. Ф., Артемьев Ю. Т. О зараженности вшами трех видов сусликов
у границ их ареалов в Волжско-Камском крае // Там же. 1968. Т. 2,
вып. 1. С. 18—26.
Стрелков П. П. Оседлые и перелетные виды летучих мышей (Chiroptera)
в европейской части СССР. Сообщ. 1 // Бюл. МОИП. Отд. биол. 1970.
Т. 75, вып. 2. С. 38—52.
Стрелков П. П. Оседлые и перелетные виды летучих мышей (Chiroptera)
в европейской части СССР. Сообщ. 2 // Там же. 1971. Т. 76, вып. 5. С. 5—
21.
Стрелков П. П. Оседлые и перелетные виды летучих мышей (Chiroptera)
в европейской части СССР. Сообщ. 3 // Там же. 1972. Т. 77, вып. 2.
С. 27—31.
Стрелков П. П. Летучие мыши (Microchiroptera) // БСЭ. 2-е изд. 1973. Т. 14.
С. 378.
Стрелков П. П. Анализ распространения рукокрылых (Chiroptera) Ф^уны
Казахстана//Фауна, систематика и биология млекопитающих. Л., 1983.
С. 139—150. (Тр. Зоол. ин-та АН СССР; Т. 119).
Стрелков П. П., Сосновцева В. П., Бабаев X. Б. Летучие мыши (Chirop-
tera) Туркмении // Функциональная морфология и систематика млекопи-
тающих. Л., 1978. С. 3—71. (Тр. Зоол. ин-та АН СССР; Т. 79).
Стрелкова М. В., Елисеев Л. Н. Грызуны и рептилии как прокормители
москитов (в связи с их значением в эпизоотологии зоонозного кожного
лейшманиоза) // Фауна и экология грызунов. М.: Изд-во МГУ, 198о.
Вып. 15. С. 55—82.
Строганова А. С., Чжудинъ. Систематическое положение реликтового сус-
лика и материалы по его экологии в горах Кулук-Тау (Центральный
Литература
249
Тянь-Шань) // Морфология и экология позвоночных животных. М.:
Изд-во АН СССР, 1961. С. 81 — 100. (Тр. Зоол. ин-та АН СССР; Т. 29).
Строганова А. С., Юдин К. А. Вредная деятельность малого суслика в мо-
лодых посадках дуба Сталинградской области И Зоол. журн. 1959. Т. 29,
вып. 5. С. 385—388.
Строганова А. С., Юдин К. А. Суслики и зайцы как вредители гнездовых
посевов дуба в Заволжье // Там же. 1951. Т. 30, вып. 2. С. 106—110.
Строгов А. К. Вирусный арктический энцефаломиелит // Исследования
по экологии, динамике численности и болезням млекопитающих Якутии.
М.: Наука, 1964. С. 60—174.
Сюзюмова Л. М. К вопросу эндемичности тундрового бешенства на Ямале //
Экология позвоночных животных Крайнего Севера. Свердловск: Средне-
Уральское кн. изд-во, 1965. С. 3—19.
Сюзюмова Л. М. К изучению эпизоотологии бешенства песца на Ямале //
Пробл. Севера. 1967. № 11. С. 99—106.
Тагильцев А. А. О членистоногих, собранных с ночниц в Зайсанской котло-
вине // Паразитология. 1971. Т. 5, №4. С. 382—384.
Тарасевич И. В., Панфилова С. С., Фетисова Н. Ф. Экологическая геогра-
фия риккетсиозов группы клещевой пятнистой лихорадки // Итоги науки
и техники: Мед. география. М.: ВИНИТИ, 1977. Т. 8. С. 6—121.
Тарасевич И. В., Панфилова С. С., Фетисова Н. Ф. Экологическая геогра-
фия риккетсиозов группы клещевой пятнистой лихорадки. Ч. 2. Пятни-
стая лихорадка Скалистых гор, марсельская лихорадка, везикулезный
риккетсиоз, клещевой тиф Северного Квинсленда // Там же. 1980. Т. 10.
С. 149—236.
Тарасова В. К., Иннокентьева Т. И. Восприимчивость и инфекционная
чувствительность к чуме длиннохвостого суслика из Тувинского природ-
ного очага // Проблемы особо опасных инфекций. Саратов, 1975.
Вып. 2(42). С. 19—23.
Татаров А. Г. Изучение природных очагов лесного и арктического бешен-
ства: Автореф. дис. . . . канд. биол. наук. М., 1982. 24 с.
Телегин В. И. Бурундук Западной Сибири. Новосибирск: Наука, 1980. 111 с.
Терновский В. Д. Биология куницеобразных (Mustelidae). Новосибирск:
Наука, 1977. 280 с.
Терновский В. Д. Европейская норка — исчезающий вид // Влияние хозяй-
ственной деятельности человека на популяции охотничьих животных и
среду их обитания: Материалы науч. конф. Киров, 1980. Т. 2. С. 182—
183.
Терновский В. Д, Феноменальные особенности размножения горностая //
Охота и охот, хоз-во. 1983. № 1. С. 18—20.
Тимофеева А. А., Щербина Р. Д., Евсеева Т. И. К изучению структуры
очага туляремии на Сахалине // Докл. Иркут, науч.-исслед. противочум.
ин-та Сибири и Дальнего Востока. Кызыл, 1969. Вып. 8. С. 126—128.
Тимофеева А. А., Евсеева Т. И., Щербина Р. Д., Федоров Л. И. О при-
родной очаговости и эпидемиологических особенностях псевдотуберку-
леза на Сахалине и Курильских островах // ЖМЭИ. 1974. № 12. С. 11 —17.
Тинкер И. С. Эпизоотология чумы на сусликах. Ростов н/Д.: Ростведиздат,
1940. 97 с.
Титов В. Л. Вирусоносительство бешенства у некоторых диких грызунов в
Казахстане // Вопросы природной очаговости болезней. Алма-Ата:
Наука, 1975. № 7. С. 54—58.
Титов Е. Б. Результаты изучения домашних плотоядных как источников
токсоплазмозной инфекции // Научные основы гигиены окружающей
среды и инфекционной патологии. Баку, 1980. С. 113—118.
Тиунов М. П. Размножение и эпизоотологическая роль бурундука на Даль-
нем Востоке // Размножение и численность грызунов на Дальнем Восто-
ке. Владивосток, 1981. С. 92—95.
Тихомирова М. М. Итоги работ Туркменской противочумной станции за
250
Литература
20 лет (1935—1955) // Тр. Туркм. противочумной станции. Ашхабад,
1957. Т. 1. С. 5—47.
Топорков В. П.у Марин С. Н., Кондрагин Ю. И. и др. Оценка эпизоото-
логической роли степного хоря в прикаспийском северо-западном при-
родном очаге чумы по данным серологического обследования // Проб-
лемы особо опасных инфекций. Саратов, 1979. Вып. 6(70). С. 19—22.
Тристан Д. Ф. Материалы по экологии желтого суслика в Муюнкумах.
Сообщ. 2. Питание желтого суслика // Материалы IV науч. конф, по
природ, очаговости и профилактике чумы. Алма-Ата, 1965. С. 258—259.
Туманов И, Л., Зверев Е. Л. Современное распространение и численность
европейской норки (Mustela lutreola) в СССР // Зоол. журн. 1986. Т. 65,
вып. 3. С. 426—435.
Туманский В. М. О начале спонтанных эпизоотий чумы сусликов и их зна-
чении // Вести, микробиологии, эпидемиологии и паразитологии. Сара-
тов, 1935. Т. 14, вып. 4, с. 419—424.
Туманский В. М., Колесникова 3. И. Эпизоотия туляремии среди сусликов в
Джамбейтинском районе Западно-Казахстанской области весной 1934 г. //
Там же. Вып. 3. С. 263—269.
Тураев А. К. О распространении и экологии дикобраза Hystrix leucura в
Южном Таджикистане // Зоол. журн. 1971. Т. 50, Вып. 4. С. 605—607.
Тураев А. К. Некоторые данные о питании и значении дикобраза в Таджики-
стане // Учен. зап. Душанбин. гос. пед. ин-та. 1973. Т. 87. С. 78—83.
Туревич Е. И., Тебякина А. Е. Бешенство и так называемое «дикование»
животных в Заполярье // ЖМЭИ. 1947. № 2. С. 17—25.
. Тушканчики фауны СССР. М., 1985. 246 с.
Умов А. А., Малокотин С. П. Очерки истории ветеринарии Курганской
области. Челябинск, 1966. С. 3—62.
Успенский С. М. Родина белых медведей. М.: Мысль, 1973. 176 с.
Успенский С. М. Белый медведь. М.: Наука, 1977. 80 с.
Устюжина И. М., Равдоникас И. О., УспТюжин Ю. А. Некоторые итоги
изучения Тувинского природного очага чумы (1964—1974 гг.). Сообщ. 2.
Биоценотическая структура очага // Международные и национальные
аспекты эпиднадзора при чуме. Иркутск, 1975. С. 52—57.
Устьянцев М. М. К биологии суслика Эверсманна в Восточной Сибири //
Сборник трудов по защите растений Восточной Сибири. Иркутск, 1937.
Т. 5. С. 96—127.
Ушакова Г. В., Кривкова А. М. Иксодовые клещи, паразитирующие на
волках в пустыне Бетпак-Дала // Тр. Ин-та зоологии АН КазССР. 1963.
Т. 19. С. 241—242.
Ушакова Г, В., Шаймарданов Р. Т. Находка специфического паразита
летучих мышей Ixodes vespertilionis Koch. (Ixodoidea, Ixodidae) в Казах-
стане // Фауна и экология паразитических насекомых и клещей Казах-
стана. Алма-Ата: Наука, 1979. С. ПО—117.
Файзулин Ф. Г. Кожный лейшманиоз у перевязки Varmela peregusna Giild II
Мед. паразитология. 1967. Т. 36, № 4. С. 494—495.
Федоров В. Н., Рогозин И. И., Фенюк Б. К. Профилактика чумы. М.:
Медгиз, 1955. 230 с.
Федорова Н. И. Эпидемиология и профилактика Ку-риккетсиоза. М.: Ме-
дицина, 1968. 251 с.
Фенюк Б. К. Экологические факторы очаговости и эпизоотологии чумы гры-
зунов. 2. Значение второстепенных носителей чумы // Тр. науч, конф.,
посвящ. 25-летнему юбилею ин-та «Микроб». Саратов, 1948. С. 31—50.
Филиппова Н. А. Аргасовые клещи (Argasidae). М.; Л.: Наука, 1966. 252 с.
(Фауна СССР. Паукообразные. Н. С.; Т. 4, Вып. 3).
Филиппова Н. А. Новые данные по клещам рода Ixodes Latr. (Ixodoidea,
Ixodidae) — специфическим паразитам летучих мышей // Энтомол. обо-
зрение. 1972. Т. 51, №2. С. 463—475.
Литература
251
Филиппова Н. А. Иксодовые клещи, подсем. Ixodinae. Л.: Наука, 1977.
387. с. (Фауна СССР. Паукообразные; Т. 4, вып. 4).
Филиппова Н. А., Неронов В. М., Фаранг-Азад А. Материалы по фауне
иксодовых клещей (Acarina, Ixodidae) мелких млекопитающих Ирана //
Энтомол. обозрение. 1976. Т. 55, № 2. С. 467—479.
Филиппова Н. Л., Чунихин С. П. Клещ Argas (Chiropterargas) boueti
Rouband et Colas-Belcour, 1933 — новый вид фауны СССР // Зоол. журн.
1969. Т. 48, вып. 9. С. 1407—1409.
Фокин И. М. Тушканчики. Л.: Изд-во ЛГУ, 1978. 182 с.
Формозов А. Н. Колебания численности промысловых животных. М.; Л.:
КОИЗ, 1935. 107 с.
Формозов А. Н. Звери, птицы и их взаимосвязи со средой обитания. М.:
Наука, 1976. С. 11—175.
Формозов А. Н., Воронов А. Г. Деятельность грызунов на пастбищах
и сенокосных угодьях Западного Казахстана и ее сельскохозяйственное
значение // Учен. зап. МГУ. Зоология. 1939. Вып. 20. С. 3—122.
Фоттелер Э. Р. О межвидовых контактах желтого суслика в Центральных
Кызылкумах // IV съезд Всесоюзн. териол. о-ва. М., 1986. Т. 1. С. 362—
363.
Ходашова К. С., Динесман Л. Г. Роль малых сусликов в формировании
комплексного почвенного покрова в глинистой полупустыне Заволжья //
Почвоведение. 1961. № 1. С. 68—76.
Хрусцелевский В. П., Пейсахис Л. А. О некоторых ландшафтно-экологиче-
ских закономерностях природной очаговости чумы в Муюнкумах //
Проблемы особо опасных инфекций. Саратов, 1969. Вып. 1(5). С. 40—47.
Цибулевская Ф. С. Чувствительность тонкопалого суслика к чумной инфек-
ции // Вопросы природной очаговости и эпизоотологии чумы в Туркме-
нии. Ашхабад, 1960. С. 277—279.
Черкасский Б. П. Эпидемиология и профилактика бешенства. М.: Медицина,
1985. 288 с.
Чиркова А. Ф. Бешенство среди лисиц // Ветеринария. 1952. № 5. С. 39—40.
Чиркова А. Ф. Материалы по динамике численности лисиц Воронежской об-
ласти в связи с прогнозами их «урожаев» // Тр. Всесоюз. науч.-исслед.
ин-та охотничьего промысла. М., 1953. Вып. 13. С. 20—31.
Чиркова А. Ф. Распространение зудневой чесотки среди лисиц в СССР в связи
с географическими факторами // Зоол. журн. 1957. Т. 36, вып. 5. С. 773—
786.
Чиркова А. Ф. Красная лиса // Млекопитающие Советского Союза. М.: Высш,
шк., 1967. Т. 2. С. 343—383.
Чугунов Ю. Д., Кудряшова Н. И., Чугунов Н. Д. Особенности терморе-
гуляции степных и горных малых сусликов в условиях Северного Кав-
каза// Тр. Науч.-исслед. противочум. ин-та Кавказа и Закавказья. Став-
рополь, 1956. Вып. 1. С. 265—276.
Чуринов И. А., Чуринова Л. В. К вопросу о выделении возбудителя псевдо-
туберкулеза грызунов на территории Калмыцкой АССР // Эпидемиология
и эпизоотология чумы. Саратов: «Микроб», 1980. С. 28—35.
Шабаев Н. Д. Восприимчивость желтого суслика к чумной инфекции //
Изв. Иркут, науч.-исслед. противочум. ин-та Сибири и Дальнего Восто-
ка. Иркутск, 1946. Т. 6. С. 141 —148.
Шайман М. С., Райхлин М. И., Бусыгин Ф. Ф. и др. Природноочаговые
инфекции диких мелких млекопитающих Тувинской АССР // Вопросы
инфекционной патологии. Омск, 1973. С. 25—27.
Шалдыбин Л. С. Гельминтофауна млекопитающих Мордовского государст-
венного заповедника // Тр. Мордов. гос. заповедника. 1964. Вып. 2.
С. 135—180.
Шаманек П. И., Кузнецов В, И., Медведевских В. И., Солецкий Г. К.
Энзоотии чумы на северо-западном Устюрте // Проблемы особо опасных
инфекций. Саратов, 1969. Вып. 1(5). С. И —17.
252
Литература
Шатас Я. Ф. Биология некоторых видов блох в связи с их эпизоотологиче-
ским значением // Эпидемиология и эпизоотология особо опасных ин-
фекций. М.: Медицина, 1965. С. 205—227.
Шатько П. Д., Корнилова А. Л., Ермилов Д. К. Природные очаги бешен-
ства в условиях Новосибирской области // ЖМЭИ. 1963. № 6. С. 33—38.
Шварц Е. А., Лаврентьев Л. Ф. К вопросу обмена блохами между грызунами
и наземными хищниками в сыртах Тянь-Шаня // Тр. Среднеаз. науч.-
исслед. противочум. ин-та. Алма-Ата, 1961. Вып. 7. С. 293—297.
Шевченко В. Л. Современное эпизоотийное состояние зауральского степного
очага чумы в связи с усложнением его биоценотической и пространствен-
ной структуры // Млекопитающие СССР: III съезд Всесоюз. териол. о-ва:
Тез. докл. 1982. Т. 2. С. 300—301.
Шевченко Г. В., Солдаткин И. С., Мастюков М. Л. и др. Уровень па-
разитарного контакта в поселениях малого суслика в полупустынях
Западного Казахстана // Паразитология. 1984. Т. 18, № 6. С. 459—463.
Шевченко 3, Г., Стриханова Е. В., Петрова Л. Р. и др. Материалы к изу-
чению фауны гамазовых клещей Краснодарского края // Проблемы особо
опасных инфекций. Саратов, 1975. Вып. 3/4 (43—44). С. 103—107.
Шевченко Л. С., Щербак Ю. Н. Экологические предпосылки формирования
очагов бешенства природного типа в УССР // Вести, зоологии. 1980. № 4.
С. 64—69.
Шенброт Г. И. Современное состояние и перспективы изучения филогении
и систематики тушканчиков // Тушканчики фауны СССР. М., 1985. С. 21 —
25.
Шерстобаев К. Н. О камчатской дикости животных // Тр. Иркут, науч.-исслед.
вет. опытной станции. Иркутск, 1949. Вып. 1. С. 99—116.
Шеханов М. В. Естественное носительство возбудителей болезней человека
позвоночными фауны СССР И Малая Медицинская энциклопедия. М.:
Сов. энциклопедия, 1970. Т. 12. С. 262—273.
Шеханов М. В. Естественное носительство возбудителей болезней человека
представителями отрядов зайцеобразных (Lagomorpha) и грызунов
(Rodentia) фауны СССР // Медицинская териология. М.: Наука, 1979.
С. 280—293.
Шилов М., Н. К распространению и экологии лисицы в Предустюртье и на
Устюрте // Бюл. МОИП. Отд. биол. 1966а. Т. 71, вып. 5. С. 147—148.
Шилов М. Н. К экологии малого суслика на Устюрте и в Предустюртье:
Автореф. докл., прочит, на засед. Аральск, отд-ния МОИП 24 апр.
1964 г. // Там же. 19665. Т. 71, вып. 5. С. 148—149.
ШиловМ. Н. Опыт использования фостоксина для борьбы с малым сусликом//
IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 384.
Шиляева Л. М. Гельминты песца и их воздействие на популяцию хозяина в
Ненецком национальном округе // Зоол. журн. 1968. Т. 47, вып. 3.
С. 331—336.
Шималов В. Т. Гельминтофауна псовых в Белоруссии // Материалы науч,
конф. Всесоюз. о-ва гельминтологов. М., 1971. Вып. 22. С. 289—293.
Шиндаров Л. Изучение эпидемиологии и этиологии лихорадки Ку в Болгарии
по сравнению с некоторыми странами // Тез. докл. конф, по някон. зоо-
нази и с природн. огницност. заболевания. Пловдив, 1958.
Ширачэзич П. И. К вопросу об эпизоотологическом значении тушканчиков в
природных очагах чумы // Проблемы особо опасных инфекций. Саратов,
1972. Вып. 6. С. 141—152.
Ширанович П. И., Исаев Э. В., Эйгелис Ю. К. и др. О спонтанной чуме у
тушканчиков в очагах Закавказья // Там же. 1974. Вып. 2. С. 17—23.
Шлугер Е. Г. Новые краснотелки (Acari, Trombiculidae) фауны СССР И
Энтомол. обозрение. 1948. Т. 30, № 1/2. С. 157—164.
Шлугер Е. Г. Клещи — краснотелки (Trombiculidae) // Переносчики возбу-
дителей природноочаговых болезней. М.: Медицина, 1962. С. 324—342.
Шлугер Е. Г., Вшивков Ф. Н. Клещи краснотелки (Acariformes, Trombicu-
Литература
253
lidae) Крыма // Энтомологическое обозрение. 1967. Т. 46, № 4. С. 909—
915.
Шлугер Е. Г., Кудряшова Н. И. Новые и впервые обнаруженные в СССР
виды краснотелок (Acariformes, Trombiculidae) // Паразитология. 1969.
Т. 3, № 2. С. 115—122.
Шлычков М. И. К изучению гельминтофауны собак, волков и лисиц в Куй-
бышевской области // Науч. тр. Куйбышев, науч.-исслед. вет. станции.
Куйбышев, 1968. Вып. 5. С. 251—254.
Шляхов Э. Н., Прискаръ В. И., Мельникова В. М. Бешенство в Молдавии //
Современные проблемы зоонозных инфекций. М., 1981. С. 52—53.
Шмит-Нильсен К. Животные пустынь. Физиологические проблемы тепла и
воды. Л.: Наука, 1972. 308 с.
Ш му тер М. Ф., Федорова Т. В., Столченова Н. А. Восприимчивость туш-
канчиков (емуранчиков и тарбаганчиков) к чуме в зависимости от сезо-
нов года // Науч. конф, по природ, очаговости и эпидемиологии особо
опасных инфекционных заболеваний: Тез. докл. Саратов, 1957. С. 463—
464.
Штильмарк Ф. Р. Об экологии бурундука в кедровых лесах Западного Саяна//
Зоол. журн. 1963. Т. 42, вып. 1. С. 92—128.
Штильмарк Ф. Р. Динамика населения бурундука в кедровых лесах Западно-
го Саяна // Вопросы зоологии. Томск: Изд-во Том. ун-та, 1966. С. 272—
273.
Штильмарк Ф. Р. Изучение динамики населения и подвижности бурундука
(Eutamias sibiricus Laxm.) путем длительных наблюдений за мечеными
зверьками // Экология млекопитающих и птиц. М.: Наука, 1967. С. 39—
49.
Щепотьев Н. В., Варшавский С. Н., Шилов М. Н. Хищные млекопитаю-
щие как носители чумы в природных очагах СССР // Проблемы особо
опасных инфекций. Саратов, 1979. Вып. 4(68). С. 11—16.
Щербак Ю. Н. Вирусологическое изучение бешенства в УССР // Вирусы и
вирусные болезни. Киев: Здоров’я, 1984. С. 11 —16.
Щербак Ю. Н., Рябошапка А. П., Шевченко Л. С. Экологические и эпидемио-
логические проблемы бешенства природного типа // ЖМЭИ. 1978. № 12.
С. 14—21.
Щербак Ю. Н., Рябошапка А. П., Шевченко Л. С., Березкина Г. П. К проб-
леме природной очаговости бешенства в Украинской ССР // Природно-
очаговые антропозоонозы. Омск, 1976. С. 61—62.
Эйгелис Ю. К. Грызуны Восточного Закавказья и проблема оздоровления
местных очагов чумы. Саратов: Изд-во Сарат. ун-та, 1980. 260 с.
Юдин В. Г. Енотовидная собака (Nyctereutes procyonoides) Приморья и При-
амурья. М.: Наука, 1977. 162 с.
Язан Ю. П. Росомаха. М.: Леей, пром-сть, 1974. 43 с.
Яковлев А. А. Эпидемиологические и иммуногенетические аспекты изучения
токсоплазмоза на Дальнем Востоке: Автореф. дис. . . . канд. биол. наук.
Л., 1980. 25 с.
Яныиин Ю. М., Комиссаров Л, В., Унайбаев Е. Ж. Профилактика гидрофо-
бии среди лиц, покусанных волком // Здравоохранение Казахстана.
1982. №1. С. 66—68.
Abul-hab Jalil. On the bed bugs (Hemiptera, Cimicidae) in Iraq // Bull. Endemic
Diseases. 1978. Vol. 19, N 1/4. P. 65—76.
Acha P. N. Epidemiology of paralytic bovine rabies and bat rabies // Bull.
Office intern, epizoot. 1967a. Vol. 67, N 3/4. P. 343—382.
Acha P. N. Algunas consideraciones sobre las condiciones actuales de la rabia
en las Americas // Bol. hig. у epidemiol. 1967b. Vol. 5, N 3. P. 485—497.
Akinchina G. T., Doby J. M. Premiere observation microscopique directe de
Toxoplasma gondii Nicolle et Manceaux, 1909 (Protozoaire, Toxoplasatea)
chez un Chiroptere Myotis bechsteini // C. r. Acad. sci. 1968. T. 266 N 3
P. 2431—2433.
254
Литература
Alexander R., Svrcek S., Vrtiak 0., Ondrejka R., Benisek Z., Zadova I, Patogene-
sis of experimental rabies infection of foxes and cats with viruses isolated
from hamsters (C. cricetus) // Vet. med. (CSSR). 1981. Vol. 26, N 4. P. 233—
246.
Allen G. M. Bats. Cambridge: Harvard Univ, press, 1939. 386 p.
Anciaux de Faveaux M. Les cycles annuels de reproduction chez les Chiropteres ca-
vernicoles du Shaba (S-E Zaire) et du Rwanda // Mammalia. 1978. Vol. 42,
N 4. P. 413—490.
Anderson G. G. Rodent control on agricultural land in British Columbia. 1980.
Vol. 6. P. 608—613.
Anderson S., Jones J, R. Recent mammal of the world. N. Y.: Ronald, 1967.
453 p.
Anderson L. J., Nicholson R. G., Tauxe R. K, Winkler W. G. Human rabies
in the United States, 1960 to 1979: epidemiology, diagnosis and preventi-
on // Ann. Intern. Med. 1984. Vol. 100, N 5. P. 728—735.
Andral L., Toma B. Etude de 1’epidemiologie de la rage vulpine en France,
1968—1973 // Cah. med. vet. 1973. Vol. 42, N 5. P. 203—214.
Animal desease in relation to animal conservation. L., 1982. 336 p.
Arambulo P. V. A summary of zoonoses in the Philippines. Amer. Vet. Epidemi-
ol. Soc. 1971. 29 p.
Atanasiu P,, Acha P. La rage chez les vampires// Ann. speleol. 1975. Vol. 30.
N 2. P. 379—385.
Aubert M. F. A., Andral L. Role eventuel des arthropodes dans 1’epidemiolo-
gic de la rage 11 Symbioses. 1975. Vol. 7, N 1. P. 57—74.
Baer G. M. Rabies in nonhematophagous bats// The natural history of rabies.
N. Y. etc.: Acad, press, 1975. Vol. 2. P. 79—97.
Baer G. M., Adams D. B. Rabies in insectivorous bats in the United States,
1953—1965 //Public Health Rep. 1970. Vol. 85, N 7. P. 637—645.
Baker J. R., Miles M. A., Godfrey D. G., Barrett T. V. Biochemical characte-
rization of some species of Trypanosoma (Schizotrypanum) from bats
(Microchiroptera)//Amer. J. Trop. Med. and Hyg. 1978. Vol. 27, N 3.
P. 483—491.
Barnard В. I. The role played by wildlife in the epidemiology rabies in South
Africa and South-West Africa H Quderstepoort J. Vet. Res. 1979. Vol. 46,
N 3. P. 155—163.
Beauregard M. Bat rabies in Canada 1963—1967 // Canad. J. Comp. Med. 1969.
Vol. 33, N 3. P. 220—226.
Bell G. P. A possible case of interspecific transmission of rabies in insectivo-
rous bats // J. Mammal. 1980. Vol. 61, N 3. P. 528—530.
Bigler W. J., Hoff G. L., Buff E, E, Chiropteran rabies in Florida: a twenty-
year analysis, 1954—1973 П Amer. J. Trop. Med. and Hyg. 1975. Vol. 24,
N 2. P. 347—352.
Bigler W. J., McLean R. G., Trevino H. A. Epidemiologic aspects of racoon
rabies in Florida // Amer. J. Epidemiol. 1973. Vol. 98, N 5. P. 326—335.
Biology of bats I Ed. W. A. Wimsatt. N. Y.: Acad, press. Vol. 1. 1970. 417 p.;
Vol. 2. 1971. 477 p.
Birney E. C., Rising J. D. Notes on distribution and reproduction of some bats
from Kansas, with remarks on incidence of rabies // Trans. Kans. Acad.
Sci. 1967. Vol. 70. P. 519—524.
Blac G., Bruneau I. Presence chez une chauvesouris du Maroc Eptesicus isabel-
linus dun virus de Q fever // Bull. Soc. pathol. exot. 1957. T. 50, N 5.
P. 653—656.
Blancon J., Aubert M. F., Artois M. Rage experimentale du furet (M t la,
Putorius furo) // Rev. med. vet. (France). 1982. Vol. 133, N 8/9. P. 553—
557.
Bogel R. Wildlife rabies in Europe // WHO. Interegional seminar on natural
foci of zoonoses. 1974. P. 1 —11.
Bogel R., Schaal E., Moegle H. The significance of martens as transmitters
Литература
255
of wildlife rabies in Europe// Zentr.-bl. Bakteriol., Parasitenk., Infektioons-
krankh. und Hyg. I Abt. Orig. A. 1977. Vol. 238, N 2. P. 184—190.
Borg K. Silvatic pseudotuberculosis in Scandinavia // Intern. Symp. Pseudotu-
berculosis. Paris, 1967. Ser. Immunobiol. Standard. Basel; N. Y.: Karger,
1968. Vol. 9. P. 129—132.
Bougler I. R.y Porterfield J. S. Isolation of a virus from Nigerian frut bats //
Trans. Roy. Soc. Trop. Med. and Hyg. 1958. Vol. 52, N 5. P. 421—424.
Bradbury J, W. Behavioural aspects of reproduction in Chiroptera // J. Reprod.
and Fert. 1979. Vol. 56, N 1. P. 431—438.
Bradley I. A. Laboratory diagnosis of rabies in western Canada, 1968—1977 11
Canad. Vet. J. 1979. Vol. 20, N 7. P. 186—190.
Bradley W. G. Home range, activity patterns and ecology of the antelope ground
squirrel in southern Nevada // Southwest. Natur. 1967. Vol. 12, N 3.
P. 231—252.
Brander R. В. Life-history notes on the porcupine in a harwoodhemloch forest
in upper Michigan // Mich. Acad. 1973. Vol. 5, N 4. P. 425—433.
Brennan J. M., Reed J. T. A list of Venezuela chiggers, particularly of small
mammalian hosts (Acarina: Trombiculidae). Pt 1,2// Bull. Brrgham Young
Univ. Sci. Biol. Ser. 1975. Vol. 20, N 1. P. 45—75.
Brygoo E. R. Les Chiropteres reservoirs de virus d’enterobacteries pathogenes
pour 1’homme // Arch. Inst. Pasteur Madagascar. 1974. Vol. 42, N 1.
p 239________252
Cerny V. fRecensio] // Folia parasitol. 1980. Vol. 27, N 4. P. 366.— Pec.
ad op.: Meditsinskaya teriologiya (Medical theriology) / Ed. V. V. Kuche-
ruk. Moscow: Nauka, 1979. 327 p.
Chapman R. C. Rabies: decimation of a wolf pack in arctic Alaska П Science.
1978. Vol. 201, N 4353. P. 365—367.
Choquette L. P. E., Kuyt E. Serological indication of infectious caning hepati-
tis in wolves (Canis lupus L.) in northern Canada И J. Wildlife Diseases.
1974. Vol. 10, N 4. P. 321—324.
Constantine D. G. Bats in relation to the health, welfare and economy of man //
Biology of bats. N. Y.: Acad, press, 1971. Vol. 2. P. 382—477.
Constantine D. G. An updated list of rabies-infected bats in North America П
J. Wildlife Diseases. 1979. Vol. 15, N 2. P. 347—349.
Constantine D. G., Humphrey G. L., Herbenick T. B. Rabies of Myotis thysano-
des, Lasiurus ega, Euderma maculatum and Eumops perotis in Califor-
nia// Ibid. 1979. Vol. 15, N 2. P. 343—345.
Corbet G. B. The mammals of palaearctic region a taxonomic review. L.; Itaca:
Cornell Univ, press, 1978. 314 p.
Correa-Giron P., Calisher С. H., Baer G. M. Epidemic strain of Venezuelan
equine encephalomyelitis virus from a vampire bat captured in Oaxaca,
Mexico, 1970 // Science. 1972. Vol. 175, N 4021. P. 546—547.
Crandell R. A. Laboratory investigation of arctic strains of rabies virus //
Acta pathol. et microbiol. scand. 1965. Vol. 63. P. 587—596.
Crandell R. A., Col L, T. Rabies in northern Greenland: some observation on
the epizootology and etiology // Nat. Rabies Symp. Nat. Commun. Disea-
ses Center. Atlanta (Ga), 1966. P. 37—42.
Crandell R. A., Jackson W. E., Wamberg K. Serologic study for canine distem-
per and hepatites in Northern Greenland // Nord. Vet. Med. 1966. Vol. 18
P. 162—165.
Davis D. H. S. A plague survey of Ngamiland, Bechuanaland Protectorate,
during the epidemic of 1944—1945 // S. Afr. Med. J. 1946. Vol. 20
P. 462—467, 511—515.
Davis D. H. S. Plague in Africa from 1935 to 1949 // Bull. WHO. 1953. Vol. 9
P. 665—700.
Davis W. H., Hitchcoock H. B. Biology and migration of the bat Myotis luci-
fugus in New England П J. Mammal. 1965. Vol. 46. N 2.
De Diego A. /., Valotta J. R. Una enfermedad que se este extendiendo en Ar-
256
Литература
gentina: La rabia transmitida рог murciclagos 11 Ilerica. 1981. Vol. 59,
N221. P. 133—138.
Disalvo A. F., Bigler W. J., Ajello L., Johnson L. E., Palmer J. Bat and soil
studies for sources of histoplasmosis in Florida // Public Health Rep. 1970.
Vol. 85, N 12. P. 1063—1069.
Diseases trasmitted from animales to man / Ed. T. G. Hall II Ann. N. Y. Acad.
Sci. 1963. Vol. 48. P. 527—564.
Dobrenov D., Petrovic M., Palic D. Besnilo liska u Vojvodini 11 Vet. glasn. 1973.
N 7. S. 475—480.
Dossier X. Installation de l’ecureuil de coree en foret de soignes // Homme et
oiseau. 1978. Vol. 16, N 4. P. 162—163.
Dubey J. P., Miller N. L., Frenkel J. R. Characterization of the new fecal
from of Toxoplasma gondii 11 J. Parasitol. 1970. N 56. P. 447—456.
Dubey J. P., Frenkel J. R. Cyst — induced toxoplasmosis in cats 11 J. Proto-
zool. 1972. N 19. P. 155—177.
Durette-Desset M. C. Notes et discussions sur les Chiropteres et leurs parasi-
tes// Mem. Mus. nat. hist, natur. A. 1982. T. 123. P. 213—214.
Dusbabek F. To the phylogeny and zoogeography of bats (Chiroptera) based
on the study of their parasitic mites (Acarina) // Lynx. 1960. N 10. P. 19—
24.
Elder W. H., Gunier W. J. Sex ratios and seasonal movements of gray bats
(Myotis grisescens) in southwestern Missouri and adjacent states П Amer.
Midland Natur. 1978. Vol. 99, N 2. P. 463—472.
Elton Ch. Epidemics among sledge dogs in the Canadian Arctic and their rela-
tion to disease in the arctic fox // Canad. J. Res. 1931. Vol. 5, N 6. P. 673—
692.
Elton Ch. Voles, mice and lemmings. Problems in population dynamics. Ox-
ford: Clarendon press, 1942. 496 p.
Everard С. O. R., Baer G. M.f James A. Epidemiology of mongouse rabies in
Grenada // J. Wildlife Diseases. 1974. Vol. 10. P. 190—196.
Everard C. 0. R., Murray D., Gilbert P. R. Rabies in Grenada // Trans. Roy.
Soc. Trop. Med. and Hyg. 1972. Vol. 66, N 6. P. 878—888.
Fain A. Les Acarines parasites des chauves-souris, biologie, role pathogene,
specificite, evolution parallele parasites-hotes // Ann. speleol. 1976. Vol.
31. P. 3—25.
Fain A. Specificite et evolution parallele hotes-parasites chez les Myobiidae
(Acari) // Mem. Mus. nat. hist, natur. A. 1982. T. 123. P. 77—84.
Fekadu M. Epizootology of rabies in Ethiopia: Diss. Uppsala, 1981. 122 p.
Fenton M. B., Thomas D. W. Dry-season overlap in activity patterns, habitat
use and prey selection by sympatric African insectivorous bats// Biotropica.
1980. Vol. 12, N 2. P. 81—90.
Fenton M. B., Boyle G. H., Harrison T. M., Oxley D. J. Activity patterns,
habitat use and prey selection by some African insectivorous bats // Ibid.
1977. Vol. 9, N 2. P. 73—85.
Fincham A. G. Histoplasmosis in Jamaican caves // Trans. Brit. Cave Res* As-
soc. 1978. Vol. 5, N 4. P. 225—228.
Findley J., Jones C. Seasonal distribution of the hoary bat // J. Mammal. 1964.
Vol. 45, N 3. P. 306—311.
Fischman H. R. Consideration of an association between geographic distributi-
on of caves and occurrence of rabies in foxes // Amer. J. Vet. Med. Assoc.
1976. Vol. 169, Nil. P. 1207—1213.
Fischman H. R., Young G. S. An association between the occurrence of fox ra-
bies and the presence of caves. Geographic relationships// Amer. J. Epide-
miol. 1976. Vol. 104, N 6. P. 593—601.
Forment W. L., Schmidt U., Greenhall A. M. Movement and population studies
of the vampire bat (Desmodus rotundus) in Mexico // J. Mammal. 1971.
Vol. 52, N 1. P. 227—228.
Forsyth D. J., Smith D. A. Temporal variability in home ranges of eastern
Литература
257
chipmunks (Tamias striatus) in a southeastern Ontario woodlot // Amer.
Midland Natur. 1973. Vol. 90, N 1. P. 107—117.
Foster M., Stubbendieck. Effect of the plain pocket gophers (Geomys bursarius)
on rangeland // J. Range Manag. 1980. Vol. 33. N 1. P. 74—78.
Foster M., Timm R. M. Tent-making by Artibeus jamaicensis (Chiroptera:
Phyllostomatidae) with commens on plants used by bats for tents // Bio-
tropica. 1976. Vol. 8, N 4. P. 265—269.
Fredrickson L. E., Thomas L. Relationship of fox rabies to caves// Public Health
Rep. 1965. Vol. 80, N 6. P. 495—500.
Gaisler J. A quantitative study of some populations of bats in Czechoslovakia
(Mammalia: Chiroptera) // Prirodoved. pr. Ustavii CSAV Brne. 1975. Vol. 9,
N 5. P. 1—46.
Gaisler J. Results of bat census in a town (Mammalia: Chiroptera) И Vestn. Cs.
spolec. Zool. 1979. Vol. 43, N 1. P. 7—21.
Gashwiler J. S. Longevity and home range of a townsend chipmunk // J. Mam-
mal. 1965. Vol. 46, N. 4. P. 693—697.
George J. P.f George J., Blancon J. Description clinique de la rage du renard.
Etude experimental//Rev. med. vet. (France). 1980. Vol. 131, N 2. P. 153—
160.
Gopalakrishna A., Madhavan A. Breeding habits and associated phenomena in
some Indian bats. Pt 3. Hipposideros ater ater (Templeton) — Hipposide-
ridae // J. Bombay Natur. Hist. Soc. 1977. Vol. 74, N 3. P. 511—517.
Gopalakrishna A., Rao К. V. Breeding habits and associated phenomena in
some Indian bats. Pt 2. Rhinolophus rouxi (Temmingk) — Rhinolophi-
dae // Ibid. 1977. Vol. 74, N 2. P. 213—219.
Gorman G, W., McKeever «S., Grimes R. D. Leptospirosis in wild mammals
from southwest Georgia // Amer. J. Trop. Med. and Hyg. 1962. Nil.
P. 518—524.
Gounh P. M., Niemeyer C. A rabies epidemic in recently captured skunks //
J. Wildlife Diseases. 1975. Vol. 10, N 2. P. 170—176.
McGowan J. E. Bovine rabies in Canada // Bull. Office intern, epizoot. 1976.
Vol. 85, N 5/6. P. 725—729.
Greaves J. H., Khan A. A. The status and control of porcupines genus Hystrix
as forest pests // Commonw. Forest. Rev. 1978. Vol. 57, N 171. P. 25—32.
Greenhall A. M. Bats, rabies and control problems // Oryx. 1968. Vol. 9, N 4.
P. 263—266.
Greenhall A. M, The problem of bat rabies, migrator bats, Livestock and wild-
life // Trans. XXXVII N. Amer. Wildlife and Natur. Res. Conf., Mexico
City, 1972. Wash. (D. C.), 1972. P. 287—292, 292—293.
Greenhall A, M. Vampire bat control in the Americas: a review and proposed
program for action // Bull. Pan Amer. Health Organiz. 1974. Vol. 8, N 1.
P. 30—36.
Grulich I. Sysel obecny Citellus citellus L. v CSSR // Prace Brnenske zaklad.
CSAV. 1960. Sv. 32, N 11. S. 473—561.
Gunson J. R., Dorward W. J., Schowalter D. B. An evolution of rabies control
is skunks in Alberta // Canad. Vet. J. 1978. Vol. 19, N 8. P. 214—220.
Hall E. R., Kelson K. R. The mammals of North America. N. Y., 1959. Vol. 1,
2. 1082 p.
Hallet A. F., McNeill D., Meyer K. F. A serological survey of the small mam-
mals for plague in Southern Africa // S. Afr. Med. J. 1970. Vol. 44, N 18.
P. 831—837.
Hart E. В. Seasonal movements of the cliff chipmunk in northern Utah // Proc.
Utah Acad. Sci. Arts and Lett. 1976. Vol. 53, pt 1. P. 13—16.
Hattwick M. A. W., Gregg M. В. The Disease in Man // Natural History of
Rabies, New York; London, 1975. Vol. 2. P. 281—304.
Heithaus E. R., Fleming T, H., Opler P. A. Foraging patterns and resource
utilization in seven species of bats in a seasonal tropical forest // Ecology.
1975. Vol. 56, N 4. P. 841—854.
258
Л итература
Hentschke J., Hellmann E. Nachweis der Tollwut bei einer Fledermaus in Ber-
lin // Tierartzl. Wochklin. Berlin; Munchen, 1975. Bd. 88, N 5. S. 92—94.
Hemer A., Christensen H. A., Beumer R. J. Reservoir hosts of cutaneous leish-
maniasis among panamian forest mammals // Amer. J. Trop. Med. and
Hyg. 1973. Vol. 22, N 5. P. 585—591.
Hoff H. L., Bigler W. JProctor S. J., Stallings L. P. Epizootic of canine dis-
temper virus infection among urban raccoons and gray foxes 11 J. Wildlife
Diseases. 1974. Vol. 10, N 4. P. 423—428.
Honacki J. Я., Kinman К. E., Koeppl J. W. Mammal species of the world.
A taxonomic and geographic reference. Lawrence (Kans.), 1982. 694 p.
Humphrey G. L. Field control of animal rabies H Rabies / Ed. Y. Nagano,
F. M. Davenport. Tokyo, 1971a. P. 277—327.
Humphrey S. R. Photographic estimation of population size of the Mexican
free-tailed bat, Tadarida brasiliensis // Amer. Midland. Natur. 1971b.
Vol. 86, N 1. P. 220—223.
Howell D. J. Flock fraging in nectarfeeding bats: advantages to the bats and to
the host plants// Amer. Natur. 1979. Vol. 114, N 1. P. 23—49.
Infectious diseases of wild mammals I Ed. J. W. Davis, L. H. Karstad,
D. O. Trainer // Ames Jowa State Univ. Press, 1970. 421 p.
Jones C. Economics and conservation. Biology of bats of the New World Family
Phyllostomatidae. Pt 1 // Spec. Publ. Mus. Texas Tech. Univ. 1976. N 10.
P. 133—144.
Johnson H, N. General epizootology of rabies // Rabies / Ed. Y. Nagano,
F. M. Davenport. Tokyo, 1971. P. 237—251.
Kaplan M. M. Epidem ology of rabies // Nature. 1969. Vol. 221, N 5179»
P. 421—425.
Kartman L. Historical and oecological observations on plague in the United
States // Trop. and Geogr. Med. 1970. Vol. 22. P. 257.
Kartman L., Prince F. M., Quan S. F., Stark H. E. New knowledge on the
ecology of sylvatic plague // Ann. N. Y. Acad. Sci. 1958. Vol. 70. P. 668—
711.
Keen R.f Hitghcock H. B. Survival and longevity of the little brown bat (Myo-
tis lucifugus) in southeastern Ontario П J. Mammal. 1980. Vol. 61, N 1.
P. 1—7.
McKeever S., Gorman G. W., Chapman J. E., Galton M. M., Powers D. K.
Incidence of leptospirosis in wild mammals from southwest Georgia with
report of new hosts for 6 serotypes of leptospirosis // Amer. J. Trop. Med.
and Hyg. 1958. Vol. 7. P. 646—655.
McKeever S., Schubert J. H., Moody M.D., Gorman G. W., Chapman J. F.
Natural occurence of tularemia in Marsupials, Carnivores, Lagomorfs and
large Rodents in southwestern Georgia and northwestern Florida//J. In-
fect. Diseases. 1958. Vol. 103, N 2. P. 120—126.
Kinzel R. F., Larson 0. R. North Dakota fleas. V. Siphonaptera from Citellus
tridecemlineatus (Mitchill) and their East-West distribution across North
Dakota H Amer. Midland Natur. 1973. Vol. 90, N 2. P. 456—460.
Klein J. M., Alonso J. M., Baranton G., Poulet A. R., Mollaret H. H, La
peste en Mauritanic // Med. et malad. infec. 1975. Vol. 5, N 4Ь P. 198—207.
Kock D. Epauletten — Flughunde, Glieder eines tropischen Okosystems 11
Vivar. Darmstadt. 1979. N 1. S. 8—13.
Koltai L. A. Veszettseg alsulasa 1967 et 1974 kozott // Magar allatorvos. Lapja,
1975. Evf. 30, N 7/8. Old. 541—545.
Komorowski A., Szankowska Z., Ramisz A. E. Pierwzy przypadek wsciekliny
u nietoperza w Polsce // Przegl. epidemiol. 1974. T. 28, N 1. P. 41—43.
Lainson R. Leishmanial parasites of mammals in relation to human disease //
Symp. Zool. Soc. L., 1982. N 50. P. 137—179.
Lainson R., Shaw J. Leishmaniasis in Brasil // Trans. Roy. Soc. Trop.
Med. Hyg. 1968. Vol. 62, N 3. P. 385—395.
Lainson R.f Shaw J. Some problems in studies on parasites of the Leishmania
Литература
259
brasiliensis complex // Colloq. Intern. Cent. Nat. Rech. Sci. 1977. P. 83—
85.
Lainson R.t Shaw J. J., Fraiha H.f Miles M. A., Draper С. C. Chagas’s disease
in the Amason Brazil. I. Tripanosoma cruzi infections in silvatic mam-
mals, triatomine bugs and man in the stste of Para, North Brazil // Trans.
Roy. Soc. Trop. Med. and Hyg. 1979. Vol. 73, N 2. P. 193—204.
La Motte L. C. Japanese В encephalitis in bats during simulated hibernation //
Amer. J. Hyg. 1958. Vol. 67. P. 101.
Larrabee W. F., Ajello L., Kaufman L. An epidemic of histoplasmosis on the
isthmus of Panama // Amer. J. Trop. Med. and Hyg. 1978. Vol. 27, N 2,
pt 1. P. 281—285.
Lassen H. C. A. Paralytic human rabies in Greenland // Lancet. 1962. Vol. 1,
N 7233. P. 247—249.
Lauritzen S.-E., Fremming 0. R., Hardeng G. Overvintring av flaggermus i
noensydnorskegruveroggrotter//Fauna (Norge). 1978. Bd31,N 4. S. 243—
249.
McLean R. Racoon rabies // The natural history of rabies. N. Y.; L., 1975.
Vol. 2. P. 53—77.
Lewis R. E. Notes on the geographical distribution and host preferences in the
order Siphonaptera. Pt 5. Ancistropsyllidae, Chimaeropsyllidae, Ischnop-
syllidae, Leptopsyllidae and Macrosyllidae// J. Med. Entomol. 1974. Vol.
11, N 5. P. 525—540.
Lewis R. E. Notes on the geographical distribution and host preferences in the
order Siphonaptera. Pt 6. Ceratophyllidae // Ibid. 1975. Vol. 11, N 6.
P. 658—676.
Lineback N. G. A model of rabies diffusion // Southeast. Geogr. 1980. Vol. 20,
N 1. P. 1—15.
Lottenberg R., Waldman R. H., Ajello L., Hoff G. L., Bigler W., Zellner S. R.
Pulmonary histoplasmosis associated with exploration of a bat care /I
Amer. J. Epidemiol. 1979. Vol. 110, N 2. P. 156—161.
Lundgren D., Marchette H., Smart K. Tularemia in the coyote, Canis latrans
lestes Merr// J. Infect. Diseases. 1957. Vol. 101, N 2. P. 154—157.
Maa T. C, Revision of the Australian batflies (Diptera: Streblidae and Nycteri-
biidae) // Pacif. Insects Monograph. 1971a. Vol. 28. P. 1 —118.
Maa T. C. Review of the Streblidae (Diptera) parasitic on Megachiropteran
bats // Ibid. 1971b. Vol. 28. P. 213—243.
Madhavan A., Patil D. R., Gopalakrishna A. Breeding habits and associated
phenomena in some Indian bats. Pt 4. Hipposideros flavus flavus (Gray) —
Hipposideridae // J. Bombay Natur. Hist. Soc. 1978. Vol. 75, N 1. P. 96—
103.
Marchette N. J., Bushman I. B., Parcer D, D., Johnson E. E. Studies on infec-
tions diseases in wild animals in Utah. IV. A wild rodent (Peromyscus
spp.) plague focus in Utah // Zoonoses Res. 1962. Vol. 1. P. 341—361.
Marshall W. H., Gallon G. W., Schwab R. G. Early summer activites of purcu-
pines as determined by radiopositioning techniques // J. Wildlife Manag
1962. Vol. 26, N 1. P. 75—79.
Matouch O., Polak L. Rabies epizootology and control methods in Czechoslova-
kia // Comp. Immunol., Microbiol., Infect. Diseases. 1982. Vol. 5, N 1/3
P. 303—307.
Matouch O., Navensnik S., Hruska JHoryna B. Vzteklina na uzenic CSR v
prubehu preiovacho odstrelu lisek v letech 1969—1975 // Veterinarstvi
1976. Vol. 26, N 12. P. 544—545.
Meeh D. The wolf: the ecology and behavior of an endangered species. N. Y.:
Natur. Hist. Soc. press. Garden City, 1970. 834 p.
Merve M. Foetal growth curves and seasonal breeding in the Natal clinging bat,
Miniopterus schreibersi natalensis // S. Afr. J. Zool. 1979. Vol 14 N Г
P. 17—21.
260
Литература
Meyer К. F. The prevention of plague in the light of newer knowledge // Ann.
N. Y. Acad. Sci. 1963. Vol. 48. P. 429—467.
Mills R, S., Barrett G. W., Farrell M. Population dynamics of the big brown
bat (Eptesicus fuscus) in southwestern Ohio // J. Mammal. 1975. Vol. 56,
N 3. P. 591—604.
Miles M. A., Barret T. Y., Hoff R. H., Mott R. Epidemiological studies of
Chagas’s disease in Brazil // Trans. Roy. Soc. Trop. Med. and Hyg. 1979.
Vol. 73, N 2. P. 135—136.
Moegle H., Rnorpp F., Bogel R., Arata A., Dietz Д’., Diethelm F. Zur Epide-
miologie der Wildtiertollwut untersuchungen im siidlichsten Teil der
Bundesrepublik Deutschland // Zentr.-bl. Vet. Med. 1974. В 21. S. 647—
659.
Moegle H., Rnorpp F. Zur Epidemiologie der Wildtollwut, 2 Mitteilung: Beo-
bachtungen uber den Dachs// Zbl. Veterinarmed. B. 1978. Bd. 25. S. 406—
415.
Mohr E. Altweltliche Stachelschweine. Wittenberg; Lutherstadt, 1965. 164 S.
(Neue Brehmbucherei; N 350).
Montgomeri G. G., Lubin Y. D. Movements of Coendou prehensill is in the Vene-
zuelan Llanos // J. Mammals. 1978. Vol. 59, N 4. P. 886—887.
Morganti L., Prosperi S. Ricerche su focolai naturali di alcune malattie infetti-
ve// Atti Conv. ita'lo-sovietico, 13—15 genn. 1972. Bologna, 1972. P. 532—
541.
Morrison D. W. Effeciency of food utilization by fruit bats // Oecologia. 1980.
Vol. 45, N 2. P. 270—273.
Muirhead R. H., Gallagher J., Burn R. J. Tuberculosis in wild badgers in
Gloucestershire: epidemiology // Vet. Rec. 1974. Vol. 95, N 24. P. 552—
555.
Muniz J., Medina H. Leishmaniose tegumentar do cobaio (Leishmania enriet
tii n. sp.) // Histopal. Rio de J. 1948. Vol. 33 (I). P. 7—25.
Murie J. O.f Harris M. A. Territoriality and dominance in male Columbian
ground squirrels (Spermophilus columbianus) // Canad. J. Zool. 1978.
Vol. 56. Nil. P. 2402—2412.
Nadchatram M., Dohany A. L. A pictorial key to the subfamilies, genera and
subgenera of Southeast Asian chiggers (Acari, Prositgmata, Trombiculi-
dae). Lumpar (Malaysia), 1974. 67 p. (Bull. Inst. Penyelidekan Perubatan
Kuala; N 14).
Neiland R. Further observation on rangiferine brucellosis in Alaskan carnivo-
res// J. Wildlife Diseases. 1975. Vol. 11, N 1. P. 45—53.
Neiland R. A., Miller L. G. Experimental Brucella suis type 4 infections in
domestic and wild Alasca carnivores // Ibid. 1981. Vol. 17, N 2. P. 183—
189.
Nicolic M., Jlesic Z. Isolation of rabies virus from insectivorous bats in Yugo-
slavia // Bull. WHO. 1956. Vol. 14, N 4. P. 801.
Nordland O. S. Host-parasite relations in initiation of infection. 1. Occurence
of listeriosis in arctic mammals with a note on its possible pathogenesis //
Canad. J. Comp. Med. 1959. Vol. 23, N 12. P. 393—400.
Novak D. Die Bekampfung der Wildtollwut in Berlin (West) und Munchen //
Tierarztl. Wochenschr. 1980. Bd. 93, N 5. S. 93—95.
Odegaard A., Rrogerud J. Rabies in Svalbard: infection diagnosed in arctic
fox, veindeet and seal // Vet. Res. 1981. Vol. 109, N 7. P. 141 — 142.
Pal S. R., Arora B. M., Chhuttani P. N., Broor S., Choudhury S., Josh R, M.,
Ray S, D. Rabies virus infection of a flying fox bat, Pteropus poliocepha-
lus in Chandigarh, Northern India // Trop. and Geogr. Med. 1980. Vol. 32,
N 3. P. 265—267.
Parker R. L. Rabies in Skunks // The natural history of rabies. N. Y.: Acad,
press, 1975. Vol. 2. P. 41— 51.
Parker R. L., Cabasso V. JDean D. J., Cheatum E. L. Serologic evidence of
Литература
261
certain virus infections in wild animals // J. Amer. Vet. Med. Assoc. 1961
Vol. 138, N 8. P. 437—440.
Pavlovic R., Petrovic M., Ercegan M.t Dujin T., Corda A., Olah M. Besnilo
lisica u Vojovdini sa narocitim osvtrom na nacine tamanjenja lisica // Vet
glasn. 1976. N 5. S. 471—479.
Petrovic M., Panjevic D., Paukovuic C., Zeleznik Z. Besnilo u Jugoslaviyi epi-
zootioloska situacija i suzbijanie // Vet. glasn. 1980. N 34, N 7. S. 621______
632.
Pitzschke H, Tollwut bei einer Breitflugel — Fledermaus (Eptesicus serotinus)
in Thiiringen // Zentr.-bl. BakterioL, Parasitenk., Infektionskrankh. und
Hyg. 1965. Bd. 196, N 4. S. 411—415.
Pitzschke H. Stand unserer Kentnisse fiber Epizootiologie und Bekampfung der
Tollwut // Monatsh. Veterinarmed. 1982. Jg. 37, H. 1. S. 12—15.
Plague in the Americas. Wash. (D. C.); 1965. 156 p. (Pan Amer. Health
Org. Sci. Publ.; N 115).
Pollitzer R. Plague. Geneva: WHO, 1954. 698 p.
Pollitzer R, A review of recent literature on plague // Bull. WHO. 1960. Vol. 23,
N 2/3. P. 313—400.
Pool G. E., Hacker C. S. Geographic and seasonal distribution of rabies in
skunks, foxes, and bats in Texas // J. Wildlife Diseases. 1982. Vol 18
N 4. p. 405—418.
Prieto J. F., Baer G. M. An outbreak of bovine paralytic rabic in Tuxtepec,
Oaxaca, Mexico // Amer. J. Trop. Med. and Hyg. 1972. Vol. 21, N 2
P. 219—225.
Quinones F., Roplan J. P., Pike L., Staine F., A jello J. Histoplasmosis in
Belize, Central America // Amer. J. Trop. Med. and Hyg. 1978. Vol. 27,
N 3. P. 538—561.
Racey P. A., Swift S. M. Variations in gestation length in a colony of pipistrell
bats (Pipistrellus pipistrellus) from year to year // J. Reprod. and Fert
1981. Vol. 61, N 1. P. 123—129.
Radovsky F. I. The Macronyssidae and Laelaptidae (Acarina: Mesostigmata)
parasitic on bats // Univ. Cal. Publ. Entomol. 1967. Vol. 46. 288 p.
Rahm U. Biologie und Verbreitung des afrikanischen Quastenstachlers Atheru-
rus africanus Gray (Hystricomorpha) // Rev. suisse zool. 1962. Bd. 69
N 2. S. 344—359.
Rausch R. Some observations on rabies in Alaska with special reference to wild
canides // J. Wildlife Manag. 1958. Vol. 22, N 4. P. 246—260.
Rausch R. L. Rabies in experimentally infected bears, Ursus sp. with epizooto-
logie notes// Zentr.-bl. Veterinarmed. B. 1975. Bd. 22, H. 5. S. 420—437.
Rehn F., Radvan R. К некоторым вопросам природной очаговости лихорадки
Ку // Журн. гигиены, эпидемиологии, микробиологии и иммунологии
Прага, 1967. Т. 11, №2. С. 178—185.
Rollinson D. Н. L. Problems of rabies control in Africa // East Afr. Med. J
1956. Vol. 33, N 11. P. 425—433.
Rubin R. H., Gregg M. B., Sikes R. K. Rabies in citizens of the United Sta-
tes, 1963—1968: epidemiology, treatment and complications of treatment
// J. Infect. Diseases. 1969. Vol. 120, N 2. P. 268—273.
Ruzic A. Contribution a Tecologie de Spermophilopsis leptodactylus Licht,
dans le desert au Sud de Buchara // Mammalia. 1967. Vol. 31, N 2. P. 173—
185.
Ryckman R. E. Plague vector studies. Pt III. The rate deparasitized ground
squirrels are reinfested with fleas under field conditions// J. Med. Entomol
1971. Vol. 8, N 6. P. 668—670.
Sabban El M. S., Fouad M. «$., Hussein N., Sadek B. Bats and their relation
to rabies in the United Arab Republic // Bull. Office intern, epizoot. 1967.
Vol. 67, N 3/4. P. 543—557.
Saint G. Particularites du cycle annuel des chiropteres en regions tem-
perees froides // Period, biologorum. 1973. Vol. 75, N 1. P. 207—213.
262
Литература
Sazima M.f Sazima I. Bat pollination of the passion flower, Passiflora mucrona-
ta, in Southeastern Brazil // Biotropica. 1978. Vol. 10, N 2. P. 100—
109.
Schaller G. B., Crawshaw Ir. P. G. Social organisation in a capybara popula-
tion // Saugetierk. Mitt. 1981. Vol. 29, N 1. P. 3—16.
Scherer W. F., Dickerman R. IF., La Fiandra R. P., Chia C. W., Terrian I,
Ecologic studies of Venezuelan encephalitis virus in Southeastern Mexico.
IV. Infection of wild mammals // Amer. J. Trop. Med. and Hyg. 1971.
Vol. 20, N 6, pt 1. P. 980—988.
Schnurrenberger P. R., Martin R. J., Koch J. M. Rabies in Illinois skunks //
J. Amer. Vet. Med. Assoc. 1970. Vol. 157 (10). P. 1336—1342.
Schowalter D. B. Swarming, reproduction and early hibernation of Myotis
lucifugus and M. volans in Alberta, Canada // J. Mammal. 1980. Vol. 61 >
N 2. P. 350—354.
Schwierz G. Analyse des Tollwutgeschehens bei den Musteliden in der Bundes-
republik Deutschland und in dem Bundeslandern Hessen und Rheinland-
Pfalz im Zeitraum 1952 bis 1978 zugleich eine Studie fiber mogliche Ursa-
chen der Dezimierung der Dachspopulation//Tierarzt aus Lugendorf ober
Schlesien. Giessen, 1979. 160 S.
Shcherbak Yu. Epidemiology of Hydrophobia: United Nations environment
programme USSR commision for UNEP: Intern. Project on Zoonoses Ma-
nage 1980: Training Course. Moscow; Kiev; Bacu; Samarkand; 1980.
20 p.
Secord D. S., Bradley J. A., Eaton R. D., Mitchell D. Prevalence of rabies vi-
rus in foxes, trapped in the Canadian Arctic // Canad. Vet. J. 1980. Vol. 21,
Nil. P. 297—300.
SerocovaDKreska B. Wscieklina (Lyssa, Rabies) // Choroby zakazne w Polske.
1971. N 18. S. 273—286.
Serean P., Lermain M., Herve I. P. et al. Isolement du virus Lagos bat en Em-
pire Centrafricain // Bull. Soc. pathol. exot. et filiales. 1970. N 70. P. 467—
470.
Seymour C., Dickerman R. W. Venezuelan encephalitis virus infection in neot-
ropical bats. III. Experimental studies on virus excretion and non-arthropod
transmission // Amer. J. Trop. Med. and Hyg. 1978. Vol. 27, N 2, pt. 1.
P. 307—312.
Seymour C., Dickerman R. W., Martin M.S. Venezuelan encephalitis virus
infection in neotropical bats. I. Natural infection in a guatemalan enzootic
focus // Ibid. 1978a. Vol. 27, N 2, pt 1. P. 290—296.
Seymour C., Dickerman R. W., Martin M. S. Venezuelan encephalitis virus
infection in neotropical bats. II. Experimental infections// Ibid. 1978b.
Vol. 27, N 2, pt 1. P. 297—306.
Sikes R. K. Pathogenesis of rabies in wildlife. I. Comparative effect of varying
doses of rabies virus inoculated into foxes and skunks // Amer. J. Vet.
Res. 1962. Vol. 23, N 96. P. 1041 — 1047.
Sikes R. K. Wolf, fox and coyote rabies // Intern. Nat. Rabies Symp. U. S.
Dep. Health, Educ. and Welfare Nat. Communicable, Diseases Center.
Atlanta (Ga), 1966. P. 31—33.
Sikes R. K. Rabies// Infectious diseases of wild mammals. Ames (Iowa), State
Univ. Press, 1970. P. 3—19.
Simpson D. I. Arboviruses and free-living wild animals. Diseases in free-living
wild animals// Symp. Zool. Soc. L., 1968 (1969). N 24. P. 13—28.
Smith J. D. Chiropteran evolution. Biology of bats of the New World Family
Phyllostomatidae. Pt 1 // Spec. Publ. Mus. Texas Tech Univ. 1976. N 10.
P. 49—69.
Smith P. S., Lawhaswasdi K., Vick W. S., Stanton J. S. Isolation of rabies
virus from frut bats in Thailand // Nature. 1967. Vol. 216, N 5113. P. 384.
Smith L. C., Smith D. A. Chipmunk neighbours // Trail and Landscape. 1971.
Vol. 5, N 5. P. 124—129.
Л итература
263
Sorce G., Guercio V., Colcnna V. Indagni su un nuova focalio pi rabbia mei
bovin transmesso dalle volpi. Veterinar ital, 1969. Vol. 20, N 11—12.
P. 721—729.
Speer R. JDilworth T. G. Porcupine winter foods and utilization in central
New Brunswick // Canad. Field-Natur. 1978. Vol. 92, N 3. P. 271 —
274.
Stark H. S., Hudson B. IF., Pittman B. Plague epidemiology. Atlanta (Ga),
1966. 117 p.
Start A. N., Marshall A. G. Nectarivorous bats as pollinators of trees in West
Malaysia // Trop. Trees Var. Breed, and Conserv. N. Y., 1976. N 2342.
P. 141—150.
Steck F. Rabies in wildlife // Symp. Zool. Soc. London. 1972. N 50. P. 57—75.
Stephenson R. 0., Ritter D. G., Nielsen C. A. Serologic survey for canine dis-
temper and infections canine hepatitis in wolves in Alaska // J. Wildlife
Diseases. 1982. Vol. 18, N 4. P. 419—424.
Stirbu A., Filotti A., Toacsen E. The rabies epidemo-epizootic in Rumania
and the role of bats in the transmission of this disease // Stud, si cere, viru-
sol. 1973. Vol. 24. P. 95—104.
Stouraitis P., Salvatierra J. Isolation of rabies virus from bats in Bolivia //
Trop. Anim. Health, and Prod. 1978. Vol. 10, N 2. P. 101—102.
Sukkar F. Kala azar in Iraq in 1975 // Bull. Endem. Diseases. 1978. Vol. 19,
N 1/4. P. 29—38.
Surdacki S. Stanowisko susla perelkowanego Citellus suslicus (Gueldenstaedt,
1770) w Hubalach Cprzysiolek wsi Mokre k. Zamoscia // Ann. UMCS. B.
1980—1981. Vol. 35/36. P. 255—265.
Sussman 0. Animal rabies in Florida // Florida Vet. J. 1981. Vol. 10, N 2.
P. 19—20.
Sussman 0. Facts about rabies in Florida in 1981 // Ibid. 1982. Vol. 11, N 2.
P. 30—31.
Swann A. L., Schnurrenberger R. B., Brown R. R. Brucella abortus isolations
from wild animals // Vet. Rec. 1980. Vol. 108, N 3. P. 57.
Tabel H., Corner A. H., Webster W. A., Casey C. A. History and epizootology
of rabies in Canada // Canad. Vet. J. 1974. Vol. 15, N 10. P. 271 —
281.
Tacal I. V. (Jr.), Boado P. B. Examination of negri bodies of a species of
Philippine bat (Scotophilus temmincki) // Acta med. philipp. 1966. N 3.
P. 103—106.
Taylor J. Salmonella in wild animals // Symp. Zool. Soc. London. 1968. N 24.
P. 51—73.
Thorshaug K., Rosted A. Researches into the prevalence of Trichinosis in ani-
mals in Arctic and Antarctic waters // Nord. Veterinarmed. 1956. Vol. 8,
N 2. P. 115—129.
Tierkel E. S. Rabies// Adv. Vet. Sci. 1959. Vol. 5. P. 183, 226.
Tierkel E. S. Canine rabies // The natural history of rabies. N. Y.; L., 1975.
Vol. 2. P. 123—137.
Tierkel E. S., Ar bona G., Rivera A., Juan A. Mongoose rabies in Puerto-Rico //
Public Health Rep. 1952. Vol. 67. P. 274—278.
Toma B., Andral L. Epidemiology of fox rabies // Adv. Virus Res. 1977. Vol. 21.
P. 1—36.
Trimarchi С. K, Debbie J. G., Naturally occurring rabies virus and neutralizing
antibody in two species of insectivorous bats of New York State // J. Wild-
life Diseases. 1977. Vol. 13, N 4. P. 366—369.
Tuncman Z. Las recherches du virus rabique chez les chauvessouris en Turquil.
Istambul, 1958.
Turner G. T., Hansen V. H., Reid H. P., Tietjen H. P., Ward A. L. Pocket go-
phers and Colorado Mauntain Rangeland // Col. State Univ. Agr. Exp.
St. Bull. 1973. 554.
264
Литература
Ulbrich F. Uber die jahreszeit lichen Schwankungen beim Auftreten der Toll-
wutseuche und uber Zusammenhange zwischen Alter, Geschlecht und Toll-
wutinfektion beim Fuchs// Arch. Exp. Vet. 1969. Bd. 23, H. 5. S.
Ulbrich F. Tollwut bei Mustelidae (Marderartige) // Monatsh. Veterinarmed.
1969. Bd. 24. S. 780.
Usinger R. L. Monograph of Cimicidae (Hemiptera — Heteroptera). Baltimore,
1966. 585 p.
Van der Merwe. Aspects of temperature and humidity in preferred hibernation
sites of the Natal clinging bat Miniopterus schreibersi natalensis (A. Smith,
1834) // Zool. Afr. 1973. Vol. 8, N 1. P. 121—134.
Vaughn J. B. Cat rabies// The natural history of rabies. N. Y.; L., 1975. Vol. 2.
P. 139—154.
Vercammen-Grandjean P. H. Notes on the Trombiculidae // Acarologia. 1967.
T. 9, fasc. 1. P. 127—134.
Vercammen-Grandjean P. H, The chigger mite of the Far East (Acarina: Trom-
biculidae and Leeuwenhoekiidae). San-Francisco, 1968. 135 p.
Vercammen-Grandjean P, H., Langston R. The chigger mite of the world. VIII.
Leptotrombidium complex. San Francisco, 1976. 1043 p. Illustr.
298 p.
Vogt H.-H. Wie leben die Vampire? // Kosmos (BRD). 1973. Bd. 69. Nil.
S. 376, 378.
Wachendorfer G. Die Katze als epizootologischer Faktor und als Gefahr fur den
Menschen im augenblicklichen Tollwutseuchenzug// Dt. tierarztl. Wochen-
schr. 1962. Bd. 69, N 19. S. 555—559.
Wachendorfer G. Grundsatzliches zur Epidemiologie und Bekampfung der sil-
vatischen Tollwut in der Bundesrepublik Deutschland // Bundesgesund-
heitsblatt. 1975. N 18. S. 285—289.
Wachendorfer G. Gegenwartige Stand der Vakzination von Fuchsen gegen Toll-
wut // Prakt. Tierarzt. 1976. N 12. S. 801—808.
Wachendorfer G., Forster U., Menzel W., Krekel H., Scharfen E., Frost Y. W.„
Osthaff F. Serologische Untersuchungen bei wildlebenden Spezies Mitteleu-
ropas zum Nachweis neutraliseren der Antikorper gegeniiber dem Tollwut
Virus// Berl., Munch. Tierarztl. Wochenschr. 1978. Bd. 91, N 18. S. 357—
360.
Wachendorfer G., Frost J. W. Orale immuniesierung von Fuchsen gegen Toll-
wut // Wild und Hund. 1983. Bd. 85, N 26. S. 12—13.
Wachendorfer G., Schwierz G, Zur Epidemiologie und Bekampfung der Wildtoll-
wut — Studie uber mogliche Ursachen des starken Riickganges der Popula-
tion des Dachses (Meles meles) in Hessen, 1952—1977 // Dt. Tierarztl.
Wochrnschr. 1980. Bd. 87, N 4. S. 255—260.
Wamberg K. Rabies in Gronland // Nord. Vet. Med. 1960. Vol. 12, N 12.
P. 769—796.
Wandeler A., Wachendorfer G., Forster U., Krekel H., Schall W., Muller J.,
Steck F. Rabies in wild Carnivores in Central Europe // Zentr.-bl. Veteri-
narmed. B. 1974. Bd. 21. S. 735—773.
Wells K. F. Control of rabies // Canad. J. Comp. Med. 1954. Vol. 18, N 8.
P. 302—309.
Wenzel R. L., Tipton V. J., Kiewlicz A. The Streblid batflies of Panama //
Ectoparasites of Panama. Chicago, 1966. P. 405—675.
Westerlund N. C. Survey of rats and bat brains for the presence of rabies antigen
using fluorescent antibody techniques // V Epid. Flight USAF. 1966.
SEL-TR-66-5.
Wilhelm A., Vogt D. Tollwut beim Fischotter (Lutra lutra) // Monatsh. Veteri-
- narmed. 1981. Jg. 36, H. 10. S. 361.
Williams R. B. Epizootic of rabies in interior Alaska, 1945—1947 // Canad.
J. Comp. Med. 1949. Vol. 13, N 6. P. 136—143.
Williams D. F., Findley J. S. Sexual size dimorphism in Vespertilionid bats //
Amer. Midland Natur. 1979. Vol. 102, N 2. P. 113—126.
Литература
265
Wilson D. E. Reproduction in neotropical bats // Period, biologorum. 1973.
Vol. 75, N 1. P. 215—217.
Winkler W. G. Airborne rabies virus isolation // Bull. Wildlife Disease Assoc.
1968. Vol. 4, N 2. P. 37—40.
Winkler W. G. Rabies in the United States 1951—1970 // J. Infect. Diseas.
1972. Vol. 125, N 6. P. 674—675.
Winkler W. G. Fox rabies // The natural history of rabies. N. Y.; L., 1975,
Vol. 2. P. 3—22.
Wood J. E., Davis D, E. The prevalence of rabies of foxes in the southern
States// J. Amer. Vet. Med. Assoc. 1959. Vol. 135. P. 121.
Yunker С. E., Brennan J. Л4., Hughes L. E., Philip С. B., Clifford C. M.t
Peralta P. H., Vogel J. Isolation of viral and rickettsial agents from Pana-
manian Acarina // J. Med. Entomol. 1975. Vol. 12, N 2. P. 250—255.
СОДЕРЖАНИЕ
Предисловие (В. В. Кучерук)................................... 3
Отряд Rodentia — грызуны (Н. А. Никитина)..................... 7
Триба Citellini (Н. А. Никитина).......................... 7
Род Citellus — настоящие суслики........................ 8
Триба Tamiini (Н. А. Никитина)........................... 40
Род Sciurotamias — белкообразные бурундуки............. 40
Род Tamias — бурундуки................................. 41
Подсемейство Xerinae — земляные белки (Н. А. Никитина) 48
Род Spermophilopsis — тонкопалые суслики............... 48
Семейство Geomyidae — гоферы (Г. А. Сидорова) .... 52
Семейство Heteromyidae — мешотчатые прыгуны (Г. А. Си-
дорова) ................................................. 54
Семейство Pedetidae — долгоноговые (Г. А. Сидорова) . 56
Семейство Dipodidae — тушканчики (И. Л. Кулик) ... 56
Семейство Hystricidae — дикобразные (Ю. А. Дубровский) 68
Семейство Erethizontidae — американские дикобразы
(Ю. А. Дубровский)....................................... 70
Семейство Caviidae — свинковые (Ю. А, Дубровский) ... 72
Семейство Dasyproctidae — агутиевые (Ю. А. Дубровский) 76
Семейство Echimyidae — колючие шиншиллы (Ю. А. Дуб-
ровский )................................................ 77
Отряд Carnivora — хищные (Т. Н. Дунаева)..................... 79
Семейство Canidae — волчьи (Т. Н. Дунаева)............... 82
Род Nyctereutes — енотовидная собака (Т. Н. Дунаева) . . 85
Род Canis — волки (Д. И. Бибиков, Т. Н. Дунаева) . . 89
Род Alopex — песцы (Т. Н. Дунаева).................... 100
Род Vulpes — лисицы (Т. Н. Дунаева)................... 105
Семейство Ursidae — медвежьи (Т. Н. Дунаева)............ 124
Семейство Procyonidae — енотовые (Т. Н. Дунаева) . . . 129
Род Ргосуоп — еноты................................... 130
Семейство Mustelidae — куньи (Т. Н. Дунаева)............ 133
Подсемейство Mustelinae................................. 135
Род Martes — куницы................................... 135
Род Mustela — ласки и хорьки.......................... 138
Род Gulo — росомахи................................... 145
Род Vormela — перевязки.............................. 146
Подсемейство Mellivorinae — медоеды..................... 146
Подсемейство Melinae — барсуки.......................... 146
Род Meles — барсуки................................... 147
Подсемейство Mephitinae — скунсы........................ 150
Содержание 267
Подсемейство Lutrinae — выдры........................... 155
Семейство Viverridae — виверровые (Т. Н. Дунаева) ... 156
Семейство Hyaenidae — гиеновые (Т. Н. Дунаева) .... 159
Семейство Felidae — кошачьи (Т. Н. Дунаева)........... 159
Род Panthera — большие кошки.......................... 160
Род Felis — кошки..................................... 161
Отряд Chiroptera — рукокрылые (И. Л. Кулик, В. В. Кучерук) 168
Систематика, распространение ......................... 168
Эктопаразиты.......................................... 187
Медицинское значение.................................. 203
Литература................................................... 221
CONTENTS
Foreword (V. V. Kucheruk)................................. 3
Order Rodentia (N. A. Nikitina)........................... 7
Tribe Citellini (N. A. Nikitina)...................... 7
Genus Citellus...................................
Tribe Tamiini (N. A. Nikitina)....................... 40
Genus Sciurotamias................................. 40
Genus Tamias....................................... 41
Subfamily Xerinae (N. A. Nikitina)................... 48
Genus Spermophilopsis.............................. 48
Family Geomyidae (G. A. Sidorova).................... 52
Family Heteromyidae (G. A. Sidorova)............... 54
Family Pedetidae (G. A. Sidorova).................... 56
Family Dipodidae (I. L. Kulik)..................... 56
Family Hystricidae (Yu. A. Dubrovsky).............. 68
Family Erethizontidae (Yu. A. Dubrovsky)............. 70
Family Caviidae (Yu. A. Dubrovsky)................... 12
Family Dasyproctidae (Yu. A. Dubrovsky).............. 76
Family Echimyidae (Yu. A. Dubrovsky) ,............... 77
Order Carnivora (T. N. Dunaeva).......................... 79
Family Canidae (T. N. Dunaeva)....................... 82
Genus Nyctereutes (T. N. Dunaeva).................. 85
Genus Canis (D. I. Bibikov, T. N. Dunaeva) ... 89
Genus Alopex (T. N. Dunaeva)...................... 100
Genus Vulpes (T. N. Dunaeva)...................... 105
Family Ursidae (T. N. Dunaeva)...................... 124
Family Procyonidae (T. N. Dunaeva).................. 129
Genus Procyon..................................... 130
Family Mustelidae (T. N. Dunaeva)................... 133
Subfamily Mustelinae................................ 135
Genus Martes.................................... 135
Genus Mustela................................... 138
Genus Gulo...................................... 145
Genus Vormela................................... 146
Subfamily Mellivorinae.............................. 146
Subfamily Melinae................................... 146
Genus Meles......................................Д47
Subfamily Mephitinae................................ 150
Subfamily Lutrinae.................................. 155
Family Viverridae (T. N. Dunaeva)................... 156
Family Hyaenidae (T. N. Dunaeva).................... 159
Contents 269
Family Felidae (T. N. Dunaeva)....................... 159
Genus Panthera...................................... 160
Genus Felis......................................... 161
Order Chiroptera (I. L. Kulik, V. V. Kucheruk)......... 168
Sistematics, distribution...................... . 168
Ectoparasites....................................... 187
Medical importance.................................. 203
References . . *....................................... 221
НАУЧНОЕ ИЗДАНИЕ
Серия «Вопросы териологии»
МЕДИЦИНСКАЯ ТЕРИОЛОГИЯ
ГРЫЗУНЫ, ХИЩНЫЕ,
РУКОКРЫЛЫЕ
Утверждено к печати
Всесоюзным теориологическим обществом
Редактор издательства Г. М. Орлова
Художественный редактор Н. Н. Михайлова
Технический редактор Т. С. Жарикова
Корректор Е. Н. Белоусова
ИБ № 37235
Сдано в набор 14.03.89.
Подписано к печати 20.06.89
Т-09888. Формат бОХЭО1/^
Бумага типографская № 2
Гарнитура литературная
Печать высокая
Усл. печ. л. 17. Усл.-кр. отт. 17,0. Уч.-изд. л. 20,4
Тираж 2 000 экз. Заказ № 979
Цена 4 р. 20 к.
Ордена Трудового Красного Знамени
издательство «Наука»
1 17864 ГСП-7, Москва В-485
Профсоюзнал ул., 90
2-я типография издательства «Наука»
121099, Москва, Г-99, Шубинский пер., 6
Заказ № 3575
Ордена Октябрьской Революции и ордена Трудо-
вого Красного Знамени МПО «Первая Образцовая
типография» Государственного комитета СССР по
делам издательств, полиграфии и книжной тор-
говли. 1 13054, Москва, Валовая, 28
В ИЗДАТЕЛЬСТВЕ «НАУКА»
готовятся к печати:
Павлинов И. Я., Дубровский Ю. А., Россолимо О. Л.,
Потапова Е. Г.
ПЕСЧАНКИ МИРОВОЙ ФАУНЫ
—27 л.
Песчанки — важнейшие грызуны аридных регионов Старо-
го Света — имеют большое значение в природных экосистемах
и являются хранителями многих опасных для человека природ-
но-очаговых болезней. В книге обобщены данные по биологии и
распространению песчанок в объеме мировой фауны. Рассмотре-
ны особенности формирования их региональных фаунистических
комплексов. Проанализированы закономерности морфологиче.
ских адаптаций и направления специализации в основных груп-
пах семейства.
Для специалистов в области эволюции, систематики, морфо-
логии и биологии млекопитающих, медицинских зоологов.
СЕРАЯ КРЫСА:
Систематика, экология, регуляция численности
—40 л.
В книге описано и проанализировано систематическое поло-
жение серой крысы (пасюка) — вредителя полеводства, животно-
водства и складского хозяйства, носителя возбудителей многих
инфекций. В сравнительном аспекте характеризуются особен-
ности размножения, возрастного состава, питания, норострое-
ния и пр. Описано поведение крыс в искусственных и естествен-
ных условиях. Особое внимание уделено описанию вредоносной
деятельности пасюков и их медицинскому значению. Большой
раздел посвящен методам ограничения численности этого вред-
нейшего из грызунов.
Для зоологов, экологов, сотрудников медико-санитарной
службы.