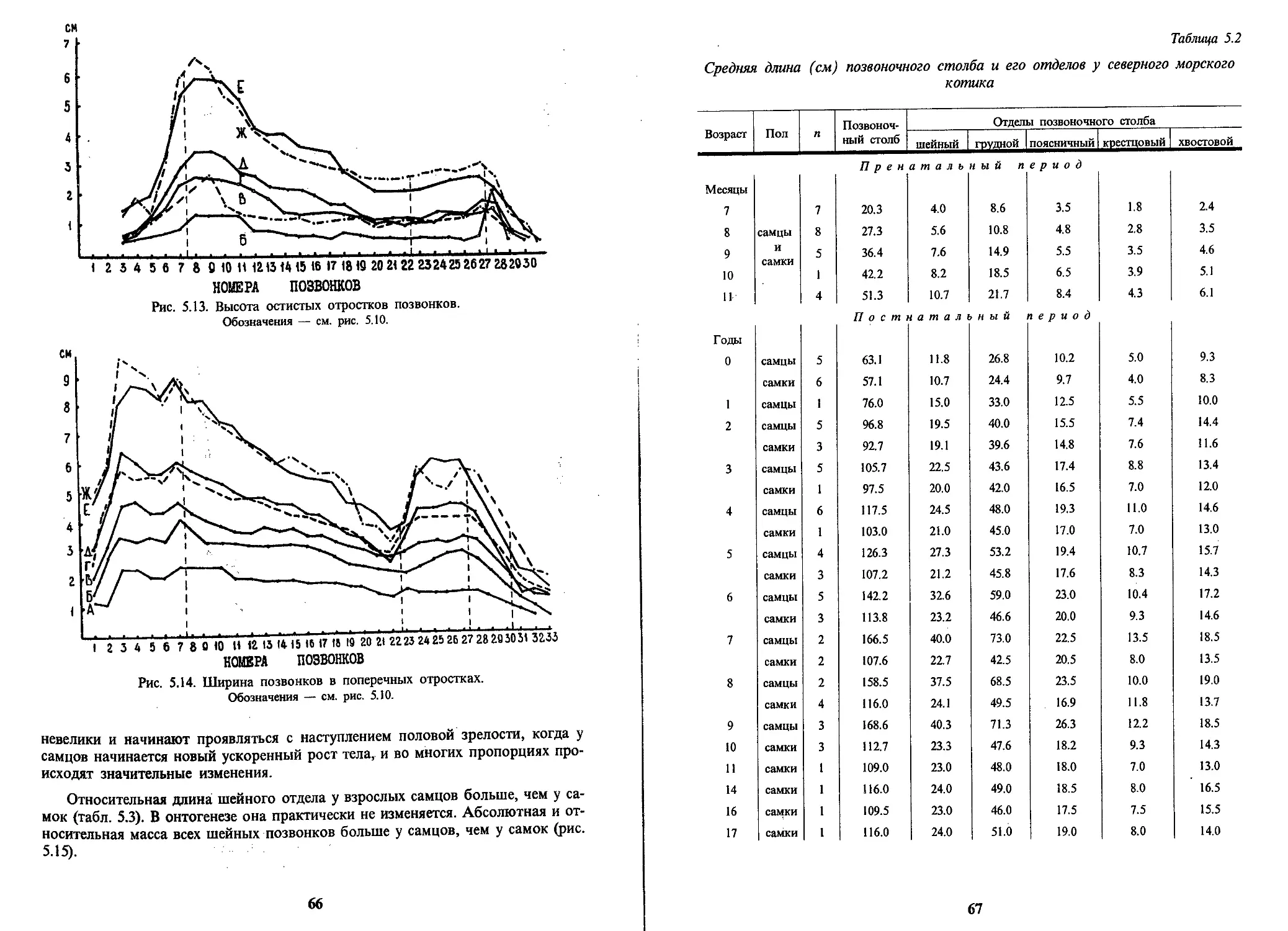
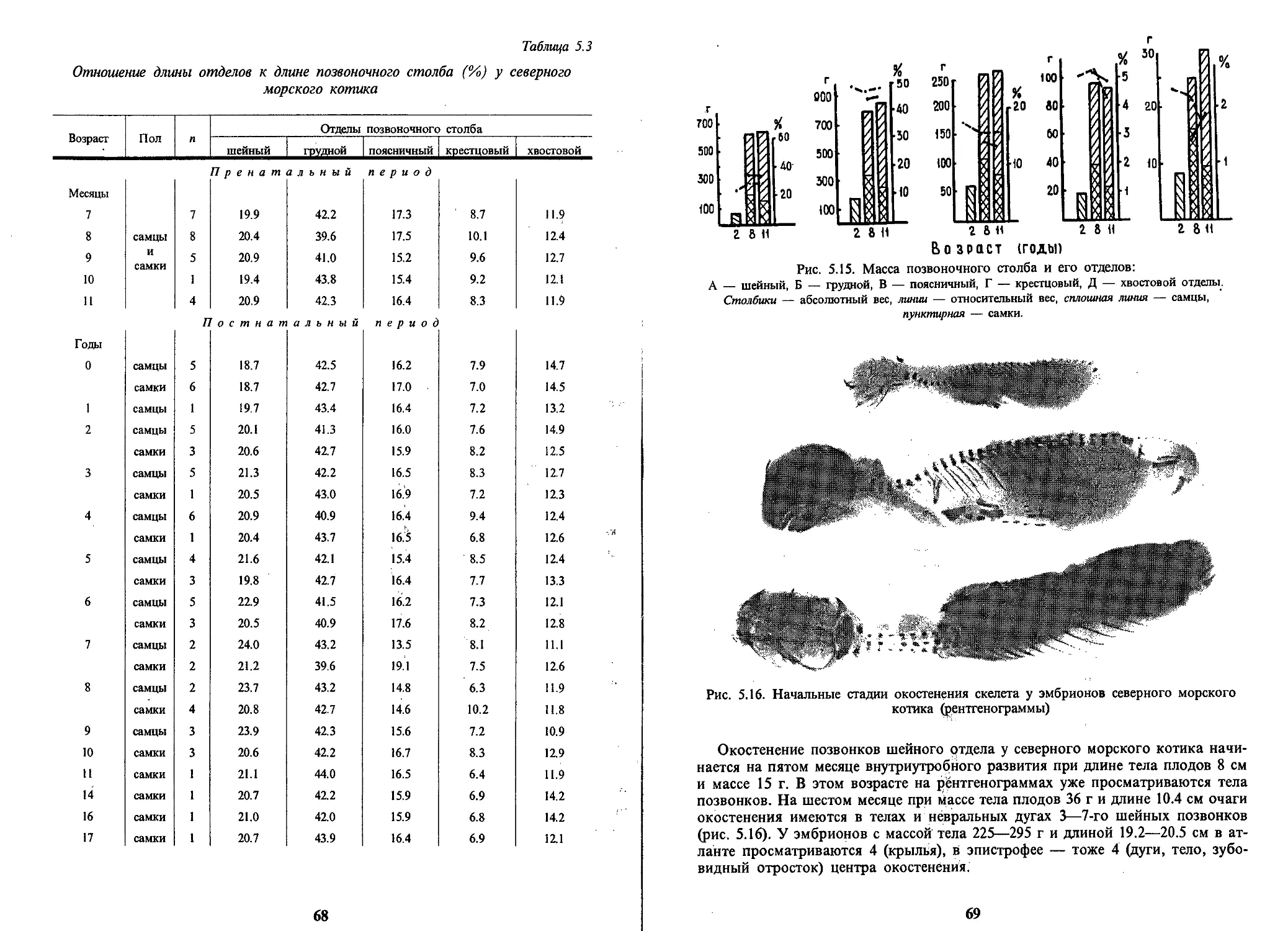
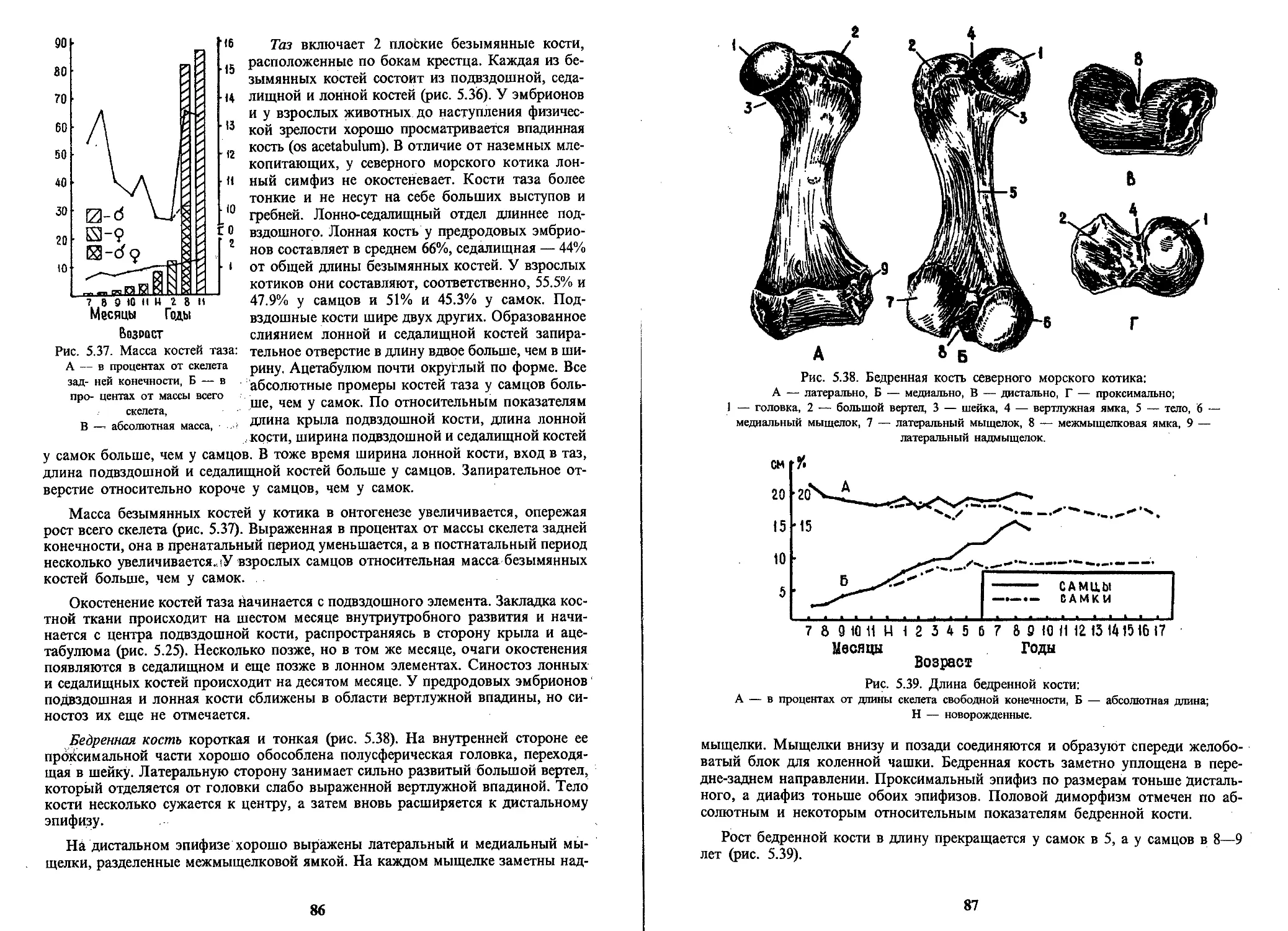
/










































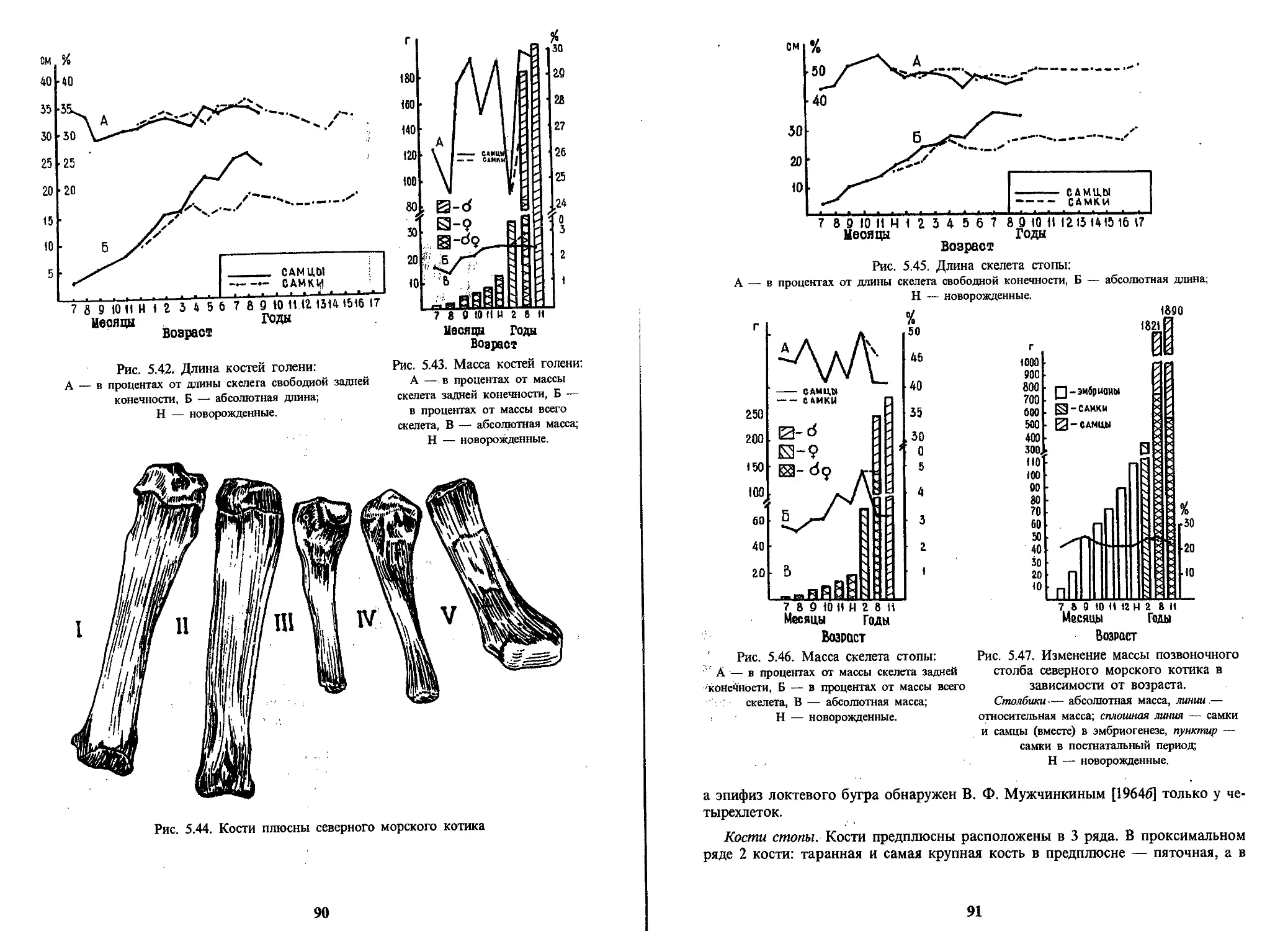







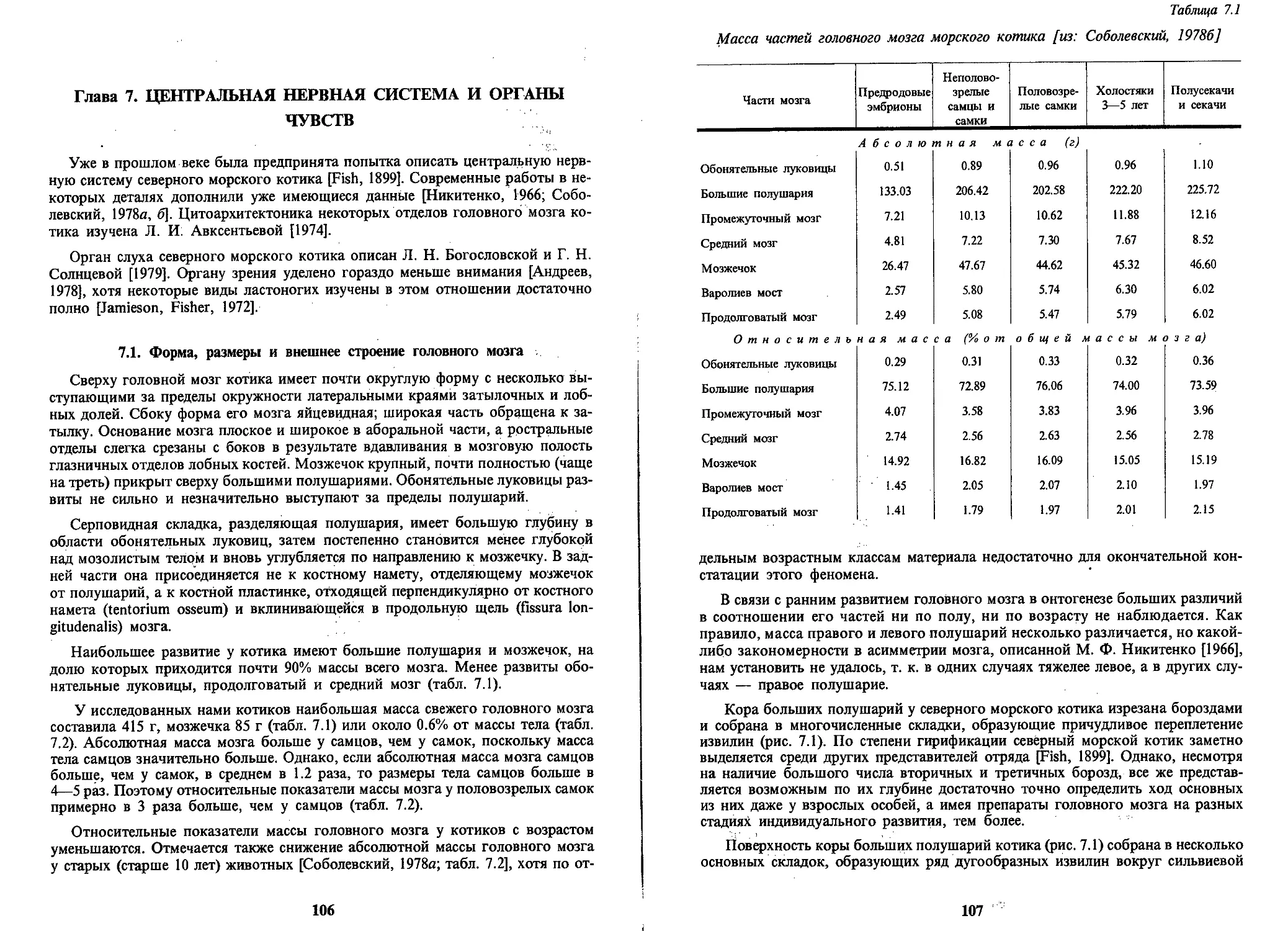























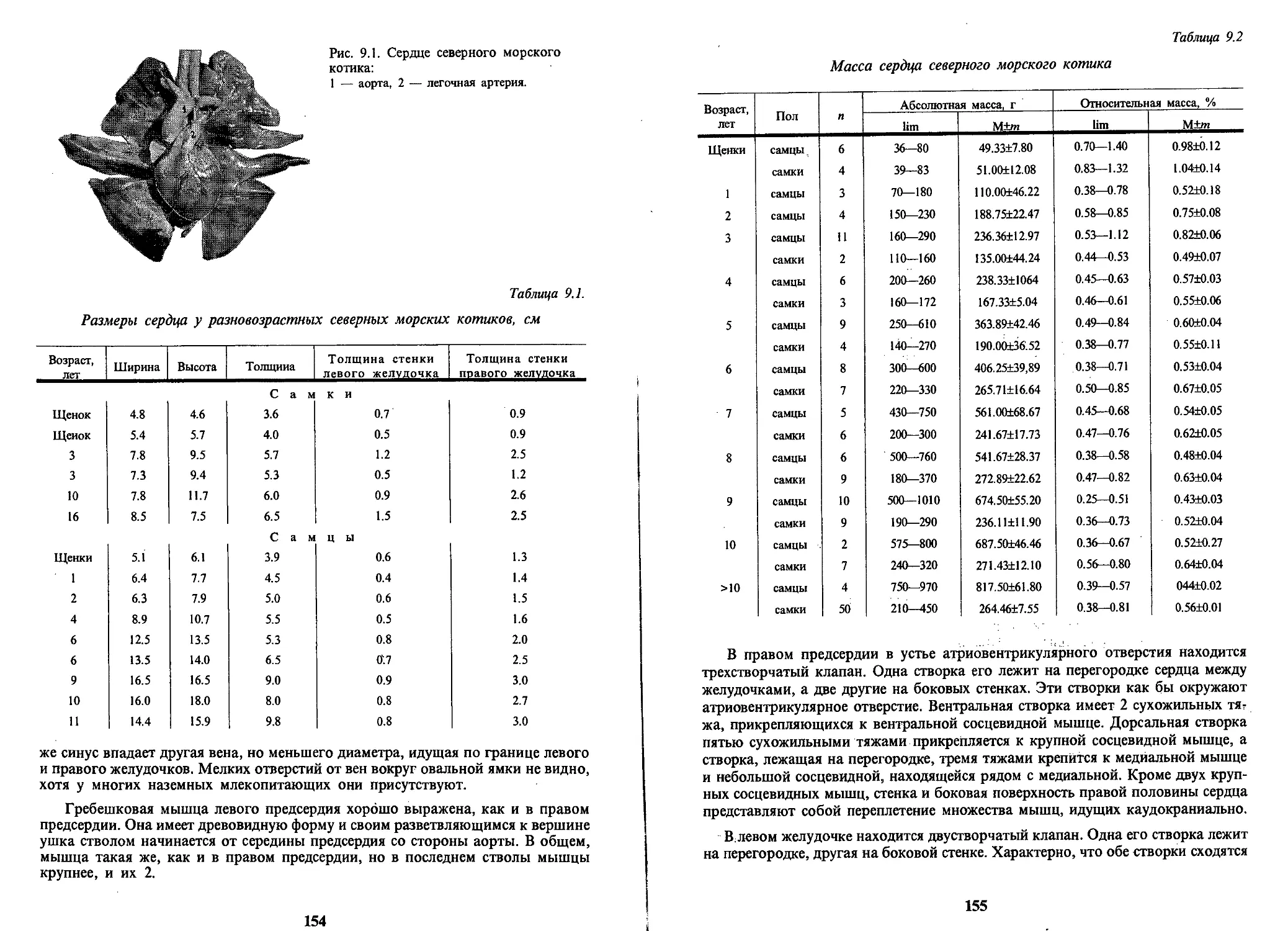




















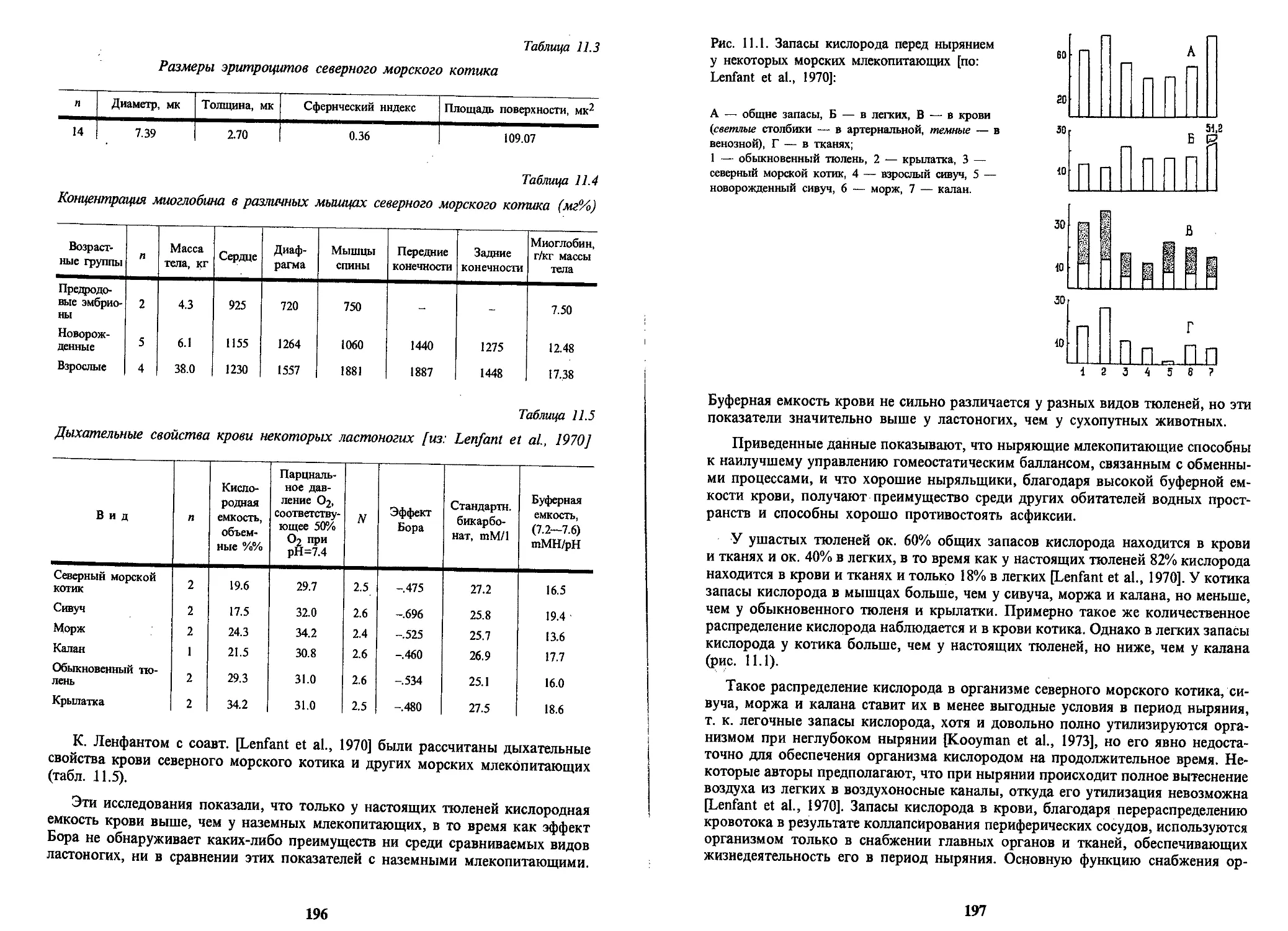





































































































Text
II
A. E. Кузин
СЕВЕРНЫЙ МОРСКОЙ КОТИК
СОВЕТ ПО МОРСКИМ МЛЕКОПИТАЮЩИМ
ТИХООКЕАНСКИЙ НАУЧНО-ИССЛЕДОВАТЕЛЬСКИЙ
РЫБОХОЗЯЙСТВЕННЫЙ ЦЕНТР (ТИНРО-центр)
A. E. Кузин
северный морской
котик
Москва
1999
УДК 599.745.1 (265)
А. Е. Кузин. Северный морской котик. — М., 1999. 000 с.
Обобщены результаты современных знаний по экологии и промыслу мор-
ского котика. Представлено описание и развитие в онтогенезе основных органов
и тканей.
Табл. 83, ил. 218, библ. ООО с.
Рецензент д. б. н. В. А. Бычков
Научный редактор к. б. н. А. А. Аристов
Издание подготовлено при финансовой поддержке
Комиссии по морским млекопитающим США
(The Marine Mammals Comission of the United States)
© Совет по морским млекопитающим
© A. E. Кузин
© А. А. Аристов, научная редактура
ПРЕДИСЛОВИЕ
Северный морской котик является одним из ведущих объектов научных ис-
следований в изучении возможности организации промысловых хозяйств. Кон-
центрация основной части поголовья в летний период на островах в значитель-
ной степени способствует положительному решению этого вопроса. Котик как
модельный объект часто привлекается учеными для теоретических разработок
динамики численности долгоживущих видов, для изучения адаптивных особен-
ностей водных млекопитающих и социальной организации сообществ полигам-
ных животных.
Можно с уверенностью сказать, что северный морской котик является наи-
более изученным из морских млекопитающих, поток научной информации по
которому не иссякает. Во многом этому способствовали межгосударственные меры
регулирования промысла и охраны, закрепленные Международными Конвенциями
1911 и 1957 гг. В рамках международных программ на протяжении многих де-
сятков лет проводились широкомасштабные круглогодичные исследования всего
жизненного цикла этого вида, а результаты этих работ находили отражение в
ежегодно представляемых отчетах.
Большая часть биологических данных, положенных в основу настоящей
книги, была собрана автором в многочисленных экспедициях, выполненных в
соответствии с Планом ежегодных мероприятий, рекомендуемых исполнитель-
ным органом Международной Конвенции — Котиковой Комиссией. Использо-
ван большой архивный статистический материал, представленный в отчетах
стран участниц Конвенции (США, Японии, Канады и России), и многочислен-
ные публикации по виду.
Северный морской котик является самым многочисленным и наиболее цен-
ным промысловым видом в отряде ластоногих, обитающих в северной части Ти-
хого океана. Основным видом продукции, получаемым при промышленном ис-
пользовании котика, являются шкуры, из которых изготовляют первоклассное
меховое сырье. Остальные части туши используются как побочные продуты про-
мысла. Мясо и кости перерабатываются на корм животным в клеточном зве-
роводстве, жир и печень применяются в фармацевтической промышленности.
Исследованиями последних лет установлено, что многие внутренние органы и
железы внутренней секреции котика по содержанию в них биологически актив-
ных веществ во много раз превосходят таковые наземных млекопитающих,
а полученные из них медицинские препараты очень эффективны.
Автор благодарит “Общественный совет по морским млекопитающим” за изыс-
кание возможности опубликования настоящей книги, оказание помощи в под-
боре научного редактора и рецензентов, а также А. В. Яблокова и В. П. Шун-
това, инициировавших ее написание. Признателен коллегам по работе, вместе
со мной участвовавшим в многочисленных экспедициях и оказывавшим помощь
в сборе научного материала, и всем сотрудникам ТИНРО, которые морально
поддерживали меня в период подготовки рукописи к изданию.
Рисунки для книги выполнены инженерно-техническим персоналом Лабора-
тории по изучению морских млекопитающих ТИНРО А. М. Николюком, О. Ю.
Тюрневой, Е. Г. Базылевой и А. А. Прохоровой. Часть фотографий была пе-
редана Ф. Г. Челноковым и Г. М. Косыгиным. Всем им большое спасибо.
3
Глава 1. СИСТЕМАТИКА И ВНУТРИВИДОВАЯ СТРУКТУРА
1.1. Систематика
Северный морской котик (Callorhinus ursinus Linnaeus, 1758) относится к от-
ряду ластоногих (Pinnipedia), надсемейству ушастых тюленей (Otarioidea), семей-
ству ушастых тюленей (Otariidae), подсемейству морских котиков (Arctocepha-
linae), роду северных морских котиков (Callorhinus).
Род СЕВЕРНЫЕ МОРСКИЕ КОТИКИ — CALLORHINVS Gray, 1859
Половой диморфизм резко выражен. Длина тела самцов до 2.2 м, масса до
320 кг, самок, соответственно, до 1.4 м и 71 кг. У самцов передняя часть тела
массивная и несет “гриву” из удлиненных волос. У самок телосложение изящное.
Ушная раковина маленькая, в виде небольшого конуса, голая, пигментирован-
ная. Голова небольшая относительно тела; морда короткая, заостренная, наруж-
ный нос не вытянут вперед и не вздернут.
Когти на передних конечностях отсутствуют; вместо них видны лишь неболь-
шие роговые бляшки или только углубления на кожном покрове ластов. На зад-
них конечностях крайние пальцы несут рудиментарные, а на трех средних паль-
цах хорошо развитые когти.
Волосяной покров состоит из ости и пуховых волос. Новорожденные окра-
шены в более темные тона. На передних конечностях волосяной покров дости-
гает только запястья, кисть обеих поверхностей ластов голая.
Кондилобазальная длина черепа у самцов не превышает 280 мм. Передний
отдел черепа короткий, процион не сильно выступает вперед.
Мозговая коробка объемистая, высокая. Гребни на черепе развиты не сильно.
Скуловые дуги незначительно выступают в стороны. Носовые отростки меж-
челюстных костей в задних отделах расширены. Задний край небных костей чаще
всего имеет форму острого угла. Щечные зубы некрупные, одновершинные и
однокоренные, кроме верхнего первого моляра, который обычно двувершинный.
Верхушка бакулюма имеет вентральное и дорсальное расширение и при взгля-
де в торец напоминает восьмерку. От родственного рода Arctocephalus отлича-
ется отсутствием меха на дорсальной стороне всех ластов, который достигает
только запястья, лицевым углом черепа (у Callorhinus менее 125°), расстоянием
контакта межчелюстных костей с носовыми, который составляет менее полови-
ны расстояния от носовых костей до резцов.
Полигамия и сезонные миграции выражены сильнее, чем у других родов под-
семейства. Обитает в умеренных водах северной части Тихого океана и сопредель-
ных морях, где встречается от 33° до 65° с. ш. В западной части Тихого океана
район обитания несколько уже (36°—60° с. ш.). В настоящее время лежбища об-
разуют на островах Командорских, Прибылова (Берингово море), Сан-Мигель
4
(Калифорния), Алеутских (о. Богослов), Курильских (Каменные Ловушки, Сред-
него), Тюленьем (Охотское море).
В роде один вид, северный морской котик, С. ursinus L., 1758.
СЕВЕРНЫЙ МОРСКОЙ КОТИК —
CALLORHINUS URSINUS Linnaeus, 1758
Вид описан К. Линнеем и назван им “ursinus” на основе подробных данных
Г. Стеллера (1751, с. 346—359), который изучил его на о. Беринга (Командорские
о-ва) летом 1742 г.
Род северных морских котиков в течение 140 лет оставался монотипичным.
Д. Джордан и Г. Кларк [Jordan, Clarck, 1898] разделили линнеевский вид на три:
Callorhinus ursinus (командорский котик), С. alascanus (прибыловский котик) и
С. curilensis (котики с о. Тюлений и Курильских о-вов). В основу нового сис-
тематического подхода ими были положены незначительные различия в окраске
меха, которая, как теперь доказано, имеет большую возрастную, половую и се-
зонную изменчивость [Wilke, 1951; Scheffer, 1962; Бычков, 1968], и присутствие
или отсутствие когтей на передних конечностях.
Более поздние исследователи [Stejneger, 1936; Hall, 1940; Allen, 1942] без осо-
бого анализа свели эти виды в 3 подвида, изменив их названия: С. и. ursinus,
С. и. mimicus, С. и. cynocephalus (соответственно, командорский, сахалинский,
прибыловский). С. И. Огнев [1935] привел описание этих подвидов. В. Шеффер
[Scheffer, 1958], основываясь на данных миграций, смешиваемости популяций (по
данным мечения), морфологии, в том числе краниологии, представленных Ф. Тей-
лором с соавт. [Taylor et al., 1955], подвидов не выделял. Ю. Кинг [King, 1964],
учитывая обособленность мест размножения котиков, допускала наличие под-
видов. К. К. Чапский [1976], несмотря на сильное сомнение в правомочности
существования трех подвидов, все-таки повторил их описание.
1.2. Внутривидовая структура
К настоящему времени накоплен значительный материал по смешиваемости
северных морских котиков разных репродуктивных группировок на всех основных
лежбищах, анализ которых может быть положен в основу внутривидового под-
разделения этих животных. Основополагающими сведениями для такого анализа в
настоящей работе послужили данные по возврату меток за период 1958—68 гг.
Как следует из данных, представленных в левой части таблицы 1.1, смеши-
ваемость меченых котиков на разных островах не пропорциональна.
Обращает внимание изолированность группировок этих животных на остро-
вах Прибылова и Тюлений, которые, в целом, не более, чем на 1—4% попол-
няются особями из других группировок. В то же время командорская группи-
ровка котиков в значительной степени “разбавлена” мигрантами из других груп-
пировок (в основном из прибыловской).
5
Таблица 1.1
Смешиваемость котиков на островах по данным возврата меток за
1958—68 гг. [из: Фрисман и др., 1985]
Остро- ва Воз- раст, лет Состав меченых котиков, % Распределение котиков по островам в зависимости от их численности, %
П К Т П К Т
п 2 99.0 1.0 0 98.2 1.8 0
к 16.0 82.0 2.0 9.4 90.5 0.1
т 0 0.3 99.7 0.1 0.3 99.6;
п 3 99.8 0.2 0 96.5 3.3 0.2
к 20.0 79.0 1.0 2.6 6.8 0.6
т 1.2 0.8 98.0 0.03 0.6 99.4
п 4 99.7 0.3 0 92.6 7.0 0.4
к 33.0 66.0 1.0 3.4 96.0 0.6
т 3.5 0.8 95.7 к- 0.1 1.3 98.6
п 5 ! 99.0 1.0 0 98.6 5.8 0.6
к 36.0 62.0 2.0 9.9 89.5 0.6
т 3.5 0.4 96.1 0.2 2.3 97.5
Примечание. П — о-ва Прибылова, К
— Командорские о-ва, Т — о. Тюлений.
Таблица 1.2
Смешиваемость котиков на отдельных лежбищах Командорских о-вов
Воз- раст, лет Лежбище Состав меченых котиков, % Доля (%) котиков, рожденных и , встреченных на данном лежбище
С С-3 Ю-В С С-3 Ю-В
2 Ю-В 6 0 94 4.7 4.5 90.8
3 С 95 2 3 90.2 . 7.4 2.4
С-3 23 62 ' 14 6.1 91.6 • 2.3
Ю-В 4 1 95 3.6 4.1 92.3
4 С 93 4 3 93.7 4.4 1.9
С-3 20 73 7 11.2 83.0 0.8
Ю-В 2 0 98 0.3 2.1 97.6
Примечание. С — Северное, С-3 — Северо-Западное, Ю-В — Юго-Восточное лежбища.
6
В среднем за 1958—68 гг. доля самцов, рожденных на Командорских о-вах
и впоследствии встреченных здесь же, уменьшается с 82% в 2-летнем возрасте
до 62% в 5-летнем.
Доля котиков с о. Тюлений на Командорских о-вах не превышает 2%, а доля
прибыловских котиков увеличивается в названных возрастных интервалах с 16
до 36%. Описанные для средних многолетних величин тенденции сохраняются
и для большинства поколений. Качественно такие же результаты по процентам
смешиваемости котиков на Тюленьем были получены В. А. Владимировым
[1978а] и Ф. Г. Челноковым [19826] по котикам Командорских о-вов и о. Тю-
лений. Однако приведенные выше наши данные и материалы названных иссле-
дователей не дают количественных оценок обменных процессов особями между
разными группировками котиков. То, что котики с о. Тюлений или с других
составляют незначительный процент на каком-нибудь из островов ничего не го-
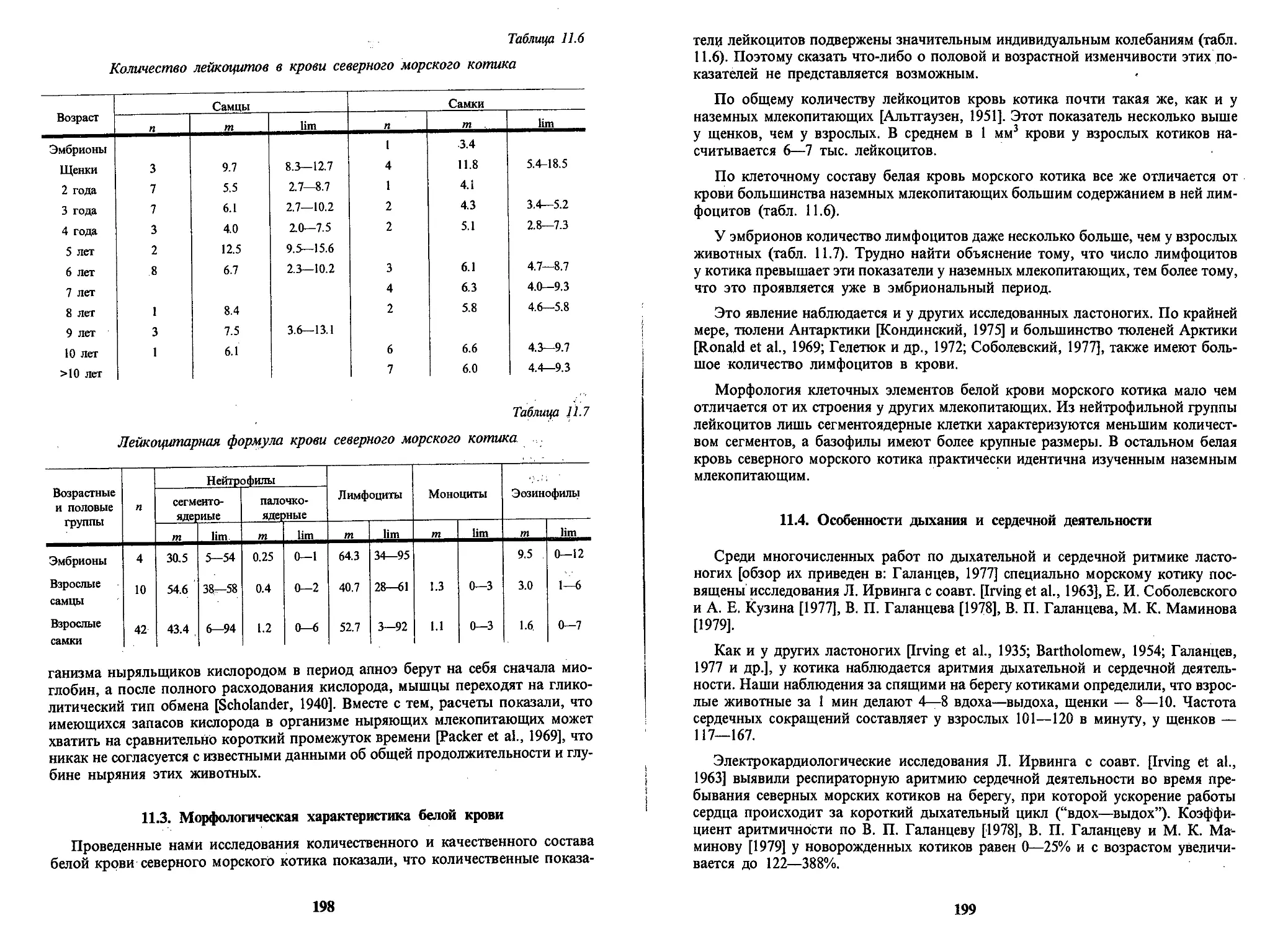
ворит. Например, даже небольшой процент котиков, мигрировавших с о-вов
Прибылова на Тюлений может составить значительную часть тюленьевской
группировки и, наоборот, большой процент мигрантов с Тюленьего на о-ва При-
былова может составить ничтожную долю в общей численности котиков, оби-
тающих на по-следних островах.
Расчетные данные с учетом численности рождающихся и помеченных на ос-
тровах котиков по поколениям, с учетом их выживаемости, промысла и потери
меток представлены в правой части таблицы 1.1. Как видно, основная часть ко-
тиков в том или ином возрасте остается на том острове, на котором они ро-
дились. Наибольшее абсолютное число котиков, приходящих с Командорских
о-вов на о-ва Прибылова (9.9%), составили пятилетки. В свою очередь, с Ко-
мандорских на о-ва Прибылова приходят до 7% четырехлеток. В других воз-
растных классах мигрантов меньше. Для ясности скажем, что, например, из по-
коления 1971 г. на Командорских о-вах находились ок. 3.5 тыс. самцов-четы-
рехлеток, рожденных на о-вах Прибылова. В среднем на каждое командорское
лежбище выходили ок. 1 тыс. прибыловских котиков в возрасте 4 лет. В то же
время максимальное число 4-летних самцов, пришедших на другие лежбища,
внутри командорской группы лежбищ составляло не более 50 особей. Из числа
родившихся в 1959 г. на о. Тюлений самцов котиков и доживших до 3-летнего
возраста в количестве 6329 особей, на о-ва Прибылова мигрировали 12 (0.2%)
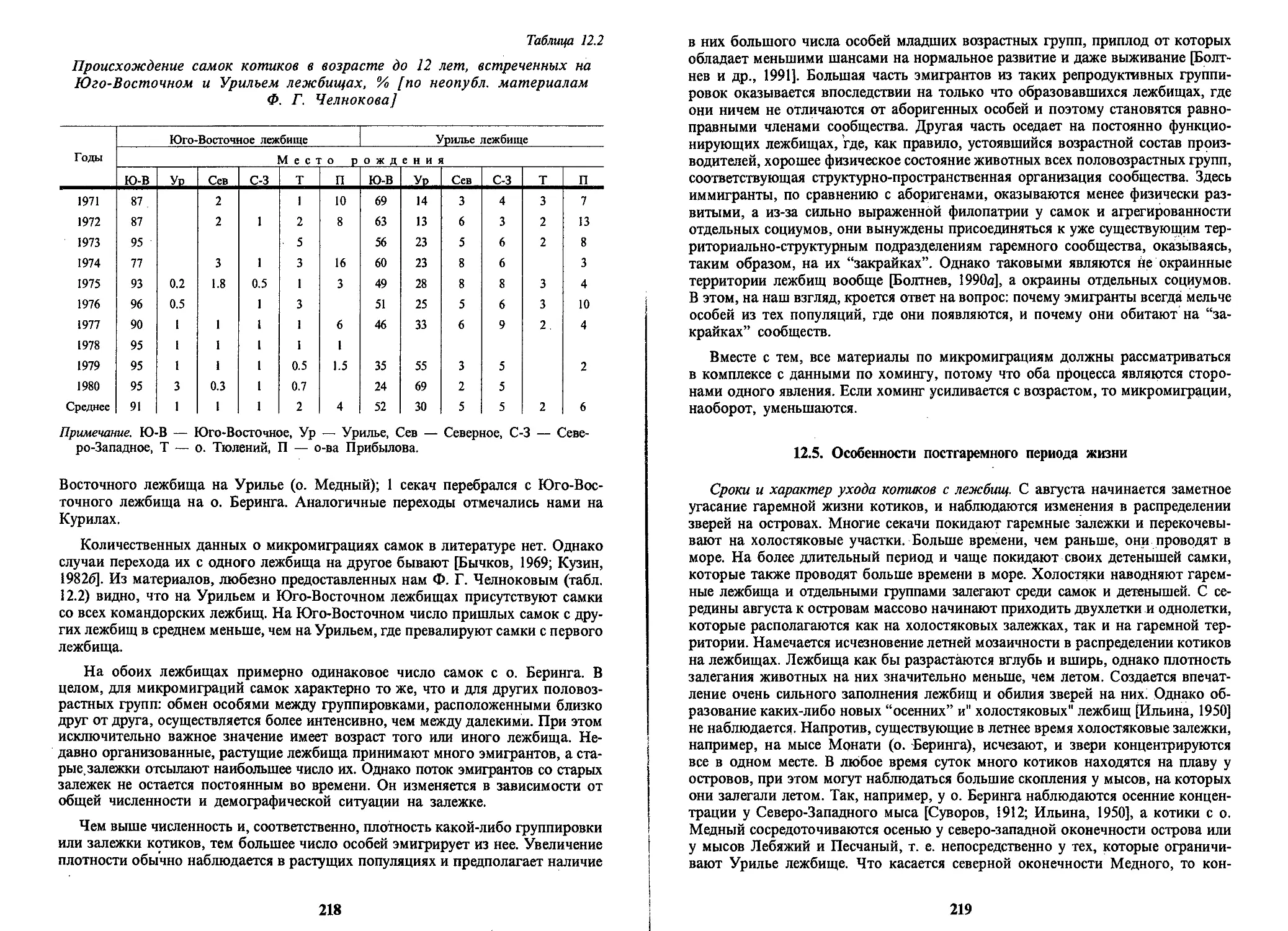
особей, на Командорские — 35 (0.5%), на о. Тюлений остались 6283 (99.3%) особи.
Таким образом, представленные данные позволяют достоверно оценить сте-
пень изолированности отдельных репродуктивных группировок котиков и на
этой основе высказать суждение о внутривидовой системе северного морского
котика. Наиболее изолированной представляется репродуктивная группировка
котиков на о. Тюлений. Средняя доля эмигрантов на этом острове до 2-летнего
возраста составляет 0.4%, до 3-летнего — 0.6%, до 4-летнего — 1.4%, до 5-летне-
го — 2.5%. Тем самым подтверждается ранее сделанный В. А. Владимировым
[1978а] вывод о правомочности выделения этой группировки северного морского
котика в отдельную популяцию. В несколько большей степени разбавлена миг-
рантами прибыловская репродуктивная группировка котиков, где они в 2-летнем
возрасте составляют 1.8%, в 3-летнем — 3.5%, в 4-летйем — 7.3%, в 5-летнем —
6.3%. Соответственно, командорская группировка принимает из других репродук-
тивных группировок 2-летних котиков 9.5%, 3-летних — 13.2%, 4-летних — 4%,
7
а 5-летних — 10.5%. Практически 95% эмигрантов с о-вов Прибылова обнару-
живаются на Командорских о-вах. В свою очередь, по интенсивности эмиграции
командорская репродуктивная группировка котиков сравнима с прибыловской,
и котики с Командор в основном уходят на о-ва Прибылова. В связи с этим,
по данным мечения, вроде бы нет основания считать группировки котиков, раз-
множающихся на островах Прибылова и Командорских, отдельными популя-
циями. Однако такой вывод представляется преждевременным, т. к. речь идет
о животных, не участвующих в размножении. Известно [Baker et al., 1995], что
с возрастом хоминг и филопатрия у северных морских котиков усиливаются,
и у взрослых животных (особенно у самок) возврат к местам своего рождения
приближается к 90%.
Интересно, что группировки котиков, залегающие на том или ином лежбище
Командорских о-вов, изолированы между собой, в общем, не меньше, а неко-
торые даже более, чем командорская и прибыловская группировки в целом
(табл. 1.2). Об этом же ранее писал Д. И. Чугунков [1966].
В недавние годы были предприняты попытки изучения популяционной
структуры морского котика с привлечением генетических [Okumoto, 1972; Бог-
данов и др., 1977] и фенетических методов [Нестеров, 1977, 1983, 1985; Нестеров,
Болтнев, 1984; Соболевский, 1984]. Н. Окумото [1972] и Л. В. Богданов с соавт.
[1977] обнаружили полиморфизм по трансферрину (Тг), щелочной фосфатазе
(Ар), эстеразе плазмы крови (EsPl), эритроцитам (EsEr). По данным последней
названной работы, “котики командорской и сахалинской популяций достоверно
отличались по частотам генов локуса Тг и EsPl, а котики из акватории тихо-
океанских берегов Японии обнаружили более выраженное сходство по Ар с ко-
тиками командорской, чем тюленьевской популяции”. На этом основании сде-
лано удачное предположение о возможном влиянии на результаты исследований
присутствие значительного количества котиков с о-вов Прибылова в данном
районе исследований.
Действительно, как показали последующие исследования [Ащепков, Кузин,
1986а, б, 1987], в тихоокеанских водах Японии численно преобладают котики
с островов Прибылова и Командорских, которые в репродуктивный период, на-
ходясь на островах, также смешиваются между собой, и в связи с этим их ге-
нетическое сходство не вызывает сомнения.
Что касается фенетического подхода к изучению структуры вида, то здесь
следует упрекнуть авторов [Нестеров, 1977, 1983, 1985; Нестеров, Болтнев, 1984;
Соболевский, 1984] в малочисленном наборе признаков для убедительного суж-
дения о внутривидовой дифференцировке северного морского котика и в отсут-
ствии достаточно полного анализа этих признаков в зависимости от пола, воз-
раста и особенно состояния численности, хотя выводы практически всех иссле-
дователей о популяционной структуре вида созвучны таковым, полученным в
результате мечения животных. Многолетний сбор данных по фенетике популя-
ции котиков о. Тюлений выявил значительную изменчивость признаков в череде
поколений и достоверные отличия их друг от друга. Не подтвердилось насле-
дование признаков и по линии “мать-плод” [Кузин, 1996], о котором писал Е. И.
Соболевский [1984]. В этой связи определенный скептицизм и неуверенность в
достоверности результатов, полученных на основе фенетики, остались.
8
Глава 2. ВНЕШНИЙ ВИД, РАЗМЕРЫ, ОКРАСКА, ПРОПОРЦИИ
И ГИДРОДИНАМИЧЕСКАЯ ХАРАКТЕРИСТИКА ТЕЛА
2.1. Внешний вид и размеры
Форма тела северного морского котика обтекаемая, сигарообразная. Относи-
тельно небольшая голова с острым, не сильно выступающим вперед носом плав-
но переходит в хорошо развитую шею, которая у самцов несет на себе выра-
женную гриву из удлиненных остевых волос.
Грудная клетка массивная. В грудной части тело котика имеет наибольший
диаметр. К хвостовой части оно постепенно становится тоньше и без заметных
перехватов в пояснице заканчивается задними конечностями. Хвост мало заме-
тен. Его длина у взрослых животных не превышает 7—10 см.
На верхней губе в несколько рядов расположены вибриссы, цвет которых
меняется в зависимости от возраста животного: от черного — у молодых, до
белого — у взрослых. У самцов-секачей длина наибольших вибрисс может дос-
тигать ?0—35 см.
Ушные раковины небольшие, но хорошо заметные. Они скручены в трубочку
в виде конуса высотой 5—6 см, неплотно прилегающего к бокам головы, вер-
шиной направленного назад и вниз.
Глаза крупные (до 5 см), почти округлые. Цвет радужной оболочки глаза
от темно-коричневого до черного.
Конечности пастообразные, причем из кожного покрова выступают только
дистальные их отделы — кисть и стопа. Кисть имеет крылообразную форму,
лишена волосяного покрова. Когти на ней или рудиментарны, или совсем от-
сутствуют, и тогда вместо когтей на коже заметны лишь небольшие углубления,
расположенные на некотором удалении от края дистальной, более тонкой кром-
ки ласта. Длина передней конечности составляет ок. 30% длины тела, а ширина
их вдвое меньше длины. Стопа пастообразной формы. Пальцы длинные и имеют
хрящевое продолжение. Между пальцами имеется тонкая эластичная перепонка.
Как и передние ласты, они лишены волосяного покрова. Когти имеются на всех
пальцах, однако на первом и пятом они недоразвиты. По общей длине задние
ласты лишь немного больше передних. Хрящевая оторочка составляет около 40%
общей длины стопы. Длина задней конечности в 2 с лишним раза больше ее
ширины. Задние конечности котика способны подгибаться под туловище и ис-
пользоваться при передвижении по твердому субстрату. Пальцы задней конеч-
ности могут сгибаться в суставах; особенно подвижны суставы дистальных фа-
ланг.
Сосков 4, они имеются у самок и у самцов. Расположены на брюшной стороне
тела и плохо просматриваются под густым меховым покровом.
Северный морской котик самый мелкий представитель семейства ушастых
тюленей, обитающих в северной части Тихого океана. Максимальная длина сам-
9
Рис. 2.1. Самец и самка северного морского котика
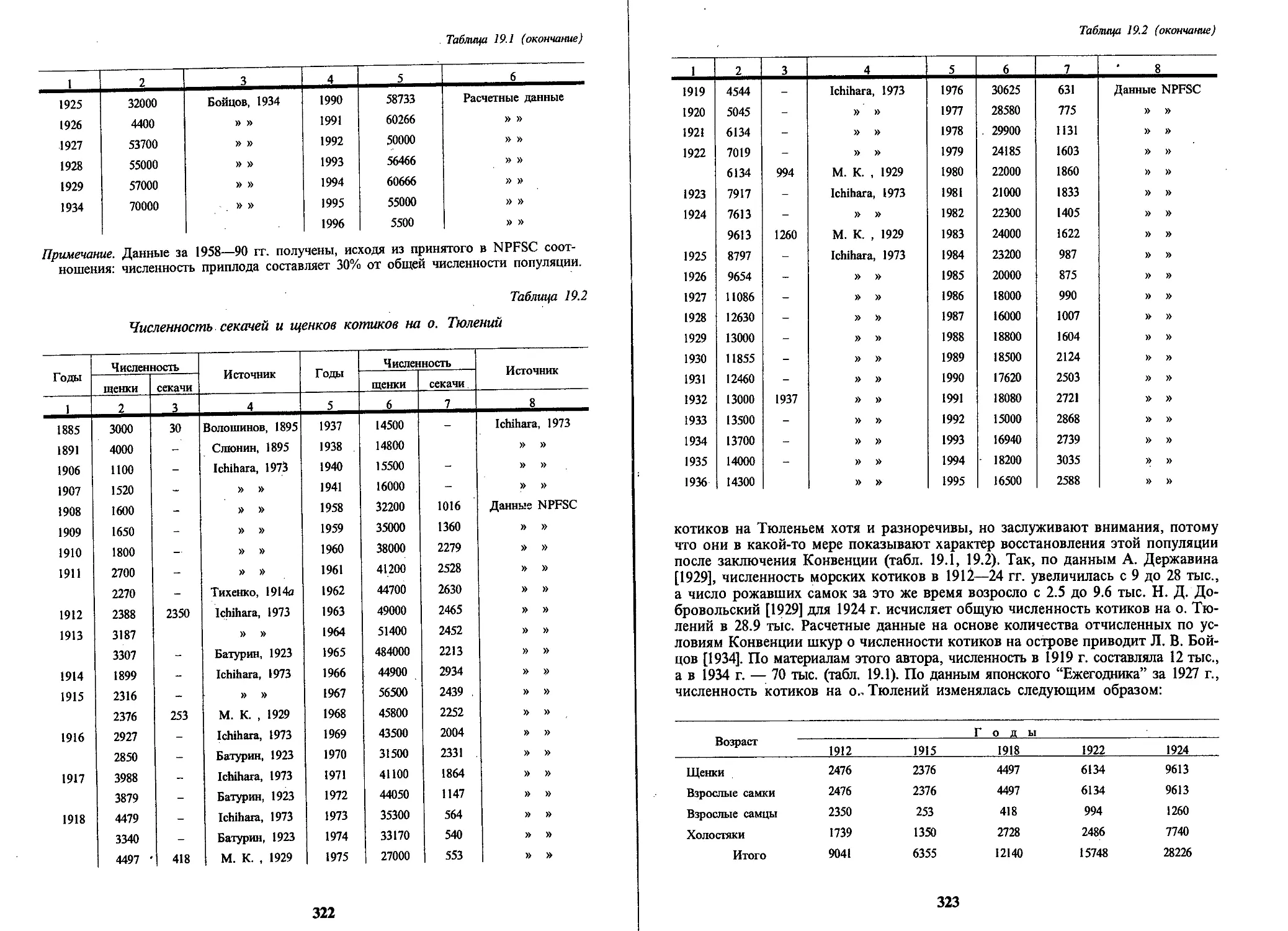
цов может достигать 2.2 м. К данным Н. Н. Сушкиной [1954] и А. Догерти [Do-
ugherty, 1965], определивших длину самцов в 2.4 и 2.5 м, следует отнестись кри-
тически.
Наибольшая масса тела самцов-секачей, по данным С. П. Крашенинникова
[1949] — 320 кг. В. Шеффер и Ф. Уилке [Scheffer, Wilke, 1953], С. В. Дорофеев
[1964], А. Догерти [Dougherty, 1965] определили массу взрослых самцов, соот-
ветственно, в 278, 285, 280 кг. По данным других авторов, масса тела секачей
составляет менее 250 кг. Масса добытых нами зимой в Японском море секачей
не превышала 320 кг. В летнее время секачи теряют более трети своей массы.
Самки котиков значительно меньше самцов (рис. 2.1). Длина их тела, по мно-
гочисленным литературным данным и собственным материалам, не превышает
1.4 м, а масса тела, также как и у самцов, сильно колеблется в зависимости
от времени взятия пробы и физиологического состояния. Видимо поэтому при-
веденные разными авторами сведения о массе и длине тела самок различаются.
Масса самки, взвешенной В. А. Бычковым [19716], составила 71 кг. С. В. До-
рофеев [1964] считает, что масса самок не превышает 63 кг, а В. Шеффер и
Ф. Уилке [Scheffer, Wilke, 1953] — 53 кг. В наших материалах масса самок не
превышала 68 кг. Осенью, с окончанием периода размножения, масса самок
уменьшается в 1.5 раза.
Таким образом, для северного морского котика характерен сильно выражен-
ный половой диморфизм в размерах тела. Самцы по длине тела больше самок
в 1.5—2 раза, а по массе тела в 4—5 раз. Подобные различия в размерах у других
млекопитающих наблюдаются чрезвычайно редко. Ярко выраженный димор-
10
физм в размерах тела является результатом сильно выраженной полигамии этих
животных.
2.2. Окраска
В. Шеффер [Scheffer, 1962] и В. А. Бычков [1968] подробно описали окраску
волосяного покрова северного морского котика с применением цветовых стан-
дартов или шкал. При написании настоящей главы нами использовались данные
этих двух авторов.
Окраска морского котика определяется цветом его мехового покрова, в част-
ности, цветом остевого волоса. Окраска подпуши резко отличается от окраски
остевых волос, но она почти не просматривается через высокую и густую ость.
Новорожденные (рис. 2.2). Окраска самок и самцов одинаковая; общий фон
ее черный.
На вершине головы и на шее разбросаны редкие беловершинные волосы. Во-
лосы в углах рта могут быть серовато-серыми с желтоватым налетом или черные.
В подмышечных впадинах волосяной покров окрашен в более светлые грязно-
серые или ржавые цвета. У некоторых щенков вокруг пениального отверстия
и реже вокруг сосков наблюдаются белые волосы [Scheffer, 1962].
Рис. 2.2. Новорожденный северный морской котик, еще не освободившийся от
плаценты
Щенки котиков в возрасте 3—4 мес. линяют и приобретают дефинитивный
волосяной нокров, который, однако, несколько отличается цветом от последу-
ющего. Следует согласиться с В. Шеффером [Scheffer, 1962], что отличить по
окраске самку от самца среди перелинявших первогодков (“серых котиков”)
очень трудно из-за сильно выраженной индивидуальной, изменчивости. Общий
11
фон окраски “серых” котиков коричневато-серый, с более светлым горлом и брю-
хом. Верхушка морды светлого серовато-коричневого цвета, иногда с примесью
желтоватых тонов. Верхняя губа, скулы серовато-коричневые. В области глаз
мех коричневато-серый. Шея, плечи, крестец коричневато-серые, но на крестце
хорошо заметны пятна более светлого, чем на спине, коричневато-серого цвета,
иногда с примесью едва заметных розовых тонов. Основание ластов с дорсаль-
ной и вентральной сторон темного серовато-желтовато-коричневого цвета. Ниж-
няя губа и подбородок желтовато-коричневые. Горло коричневато-серое у одних
и серовато-серое у других особей. Передняя часть груди от светло-серого до жел-
товато-коричневого цвета, задняя ее часть между ластами и передняя часть брю-
ха от светло-серого до желтовато-коричневого.
Годовалые и двухлетки. Общий фон окраски дорсальной части туловища ко-
ричневато-серый. Верхушка морды и лоб светлого коричневато-серого цвета, а
скулы от светло-серого до светло-коричневого цвета. Задняя часть шеи, плечи,
спина и крестец светлые, коричневато-серые. На крестце заметны пятна, окра-
шенные в более светлые тона, чем спина. Бока окрашены так же, как и спина,
но имеют менее насыщенную тональность. Основание ластов с дорсальной и
вентральной сторон серовато-коричневое. Общий фон окраски вентральной сто-
роны туловища серовато-коричневый. Нижняя губа, подбородок светлые, жел-
товато-коричневые, горло коричневато-серое. Передняя часть груди от светло-
серого до желтовато-коричневого цвета. Задняя часть груди между ластами се-
ровато-коричневая. Подмышки умеренно-коричневые. Брюхо в передней части
серовато-коричневое, в задней — от светло-серого до желтовато-коричневого цвета.
Общий фон окраски самок отличается от окраски самцов лишь в таких де-
талях как более интенсивно выраженная желтизна в местах, окрашенных у сам-
цов в рыжевато-желтые тона и присутствием более светлых оттенков серых
тонов. Двухлетние особи по окраске почти сходны с годовиками, но преобла-
дающим фоном окраски меха являются серые тона.
Трех—пятилетки. У самцов дорсально общий фон окраски меха изменяется
от свинцово-серого до буро-серого с оттенками от темно-бурого до черного. По
всему телу много серебристых волос, особенно сверху и по бокам гривы.
Кончик морды от серо-желтоватого до буровато-серого с оттенками от жел-
товато-бурого до буро-черного. У 3-леток окраска верхней губы в углах рта и
над глазами может быть беловатой. Цвет щек, лба, темени у трехлеток свет-
ло-серый или желтовато-коричневый, темнеющий с возрастом. У 5-леток они
становятся буро-серыми со слабым черно-бурым оттенком. Спина свинцово-се-
рого цвета с примесью темно-бурых тонов, к бокам эта окраска светлеет. Седина
наблюдается по всей верхней части тела. Крестец от пепельно-серого цвета с
серовато-желтоватым оттенком до буровато-серого с черновато-бурым оттен-
ком. Более светлая пятнистость в области крестца, как у 1—2-леток, заметна
только у 3-леток.
Общий фон окраски вентральной части туловища светлых серых или рыже-
ватых тонов, но она не однотонна на разных участках тела. Нижняя губа и под-
бородок от бледно-рыжего до серо-бурого цвета с различными оттенками ос-
новных тонов, а горло и передняя часть шеи серые с примесью коричневатых
тонов. Задняя часть шеи и передняя часть груди у трехлеток гораздо светлее,
чем у 4—5-леток, и имеет у первых желтовато-серый цвет, образуя хорошо за-
12
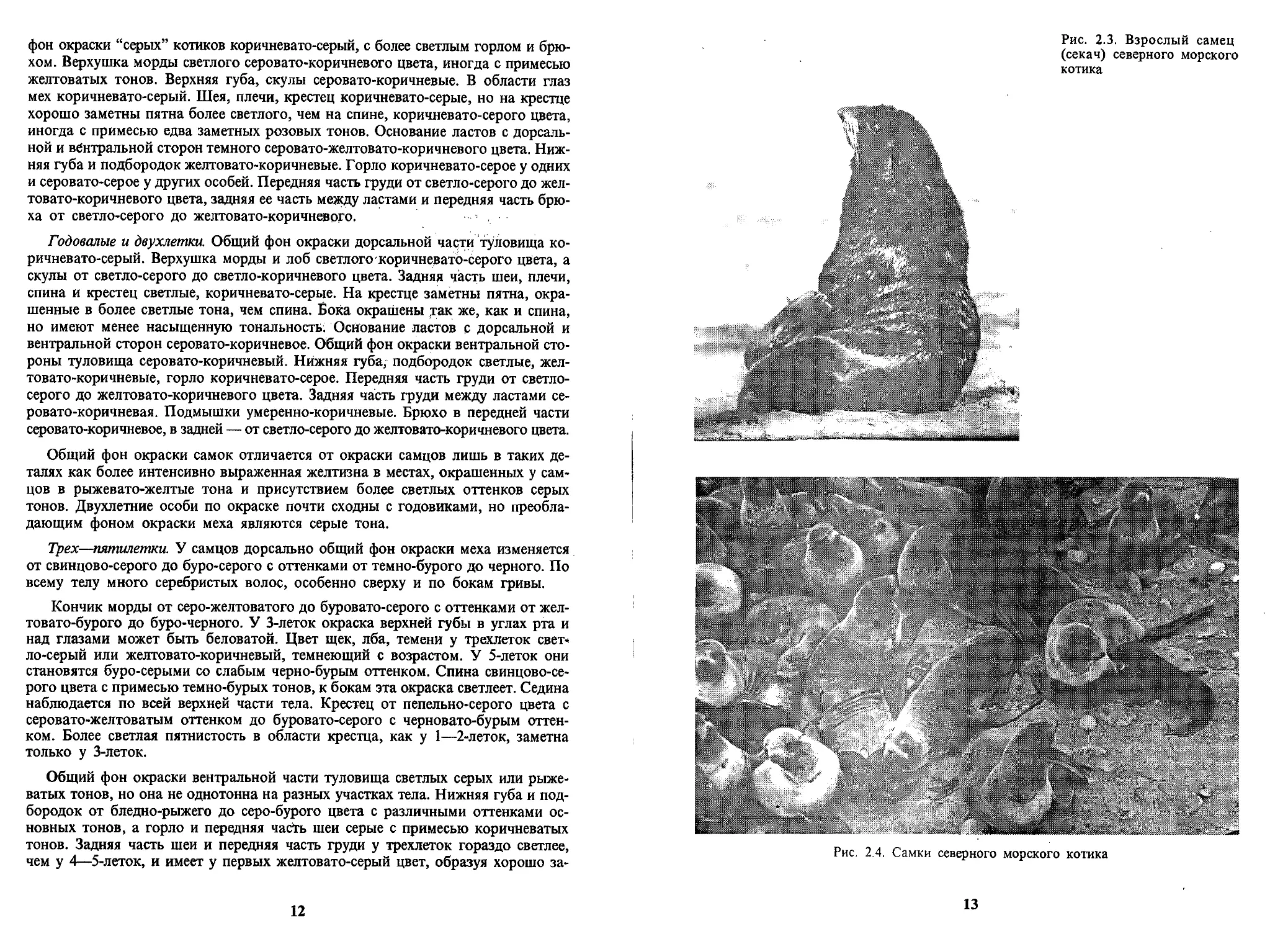
Рис. 2.3. Взрослый самец
(секач) северного морского
котика
Рис. 2.4. Самки северного морского котика
13
метное пятно, а у последних — темно-серый с оттенками от светло-бурого до
черно-бурого. Грудь от серовато-коричневого до коричнево-бурого цвета. Под-
мышки умеренно коричневые. Передняя часть живота у 3—5-леток имеет ок-
раску от темно-песчаного до темно-серого с буроватыми оттенками различной
интенсивности. Задняя часть живота от бледно-песчаной до каштановой с ры-
жеватыми, розоватыми и коричневатыми оттенками.
У 3—4-летних самок общий фон окраски дорсально темно-серый с бурова-
тыми оттенками, а брюшная поверхность имеет светло-серый с желтоватыми от-
тенками тон. В деталях среди молодых животных окраска у самок более сходна
с окраской у 3-летних самцов, но в тех местах, где у самцов преобладают бу-
ровато-рыжеватые тона, у самок они имеют розоватые оттенки.
Взрослый самец (рис. 2.3). Общий фон окраски дорсальной поверхности тела
темный, серо-коричневый. Встречаются особи с усиленной или ослабленной на-
сыщенностью общего тона окраски. Кончик морды, верхняя губа, щеки, шея
окрашены в серовато-желтовато-коричневые тона. Верх шеи светлый, желтова-
то-коричневый. Плечи светло-коричневые, и этот цвет, постепенно темнея, пе-
реходит на спину и бока, которые имеют окраску от желтовато- до темно-ко-
ричневой. Крестец серовато-коричневый. Основания ластов светло-коричневые.
С вентральной стороны тело серовато-коричневое. Нижняя губа и подбородок
серовато-желтовато-коричневые. Подмышки без присутствия серых тонов.
Рис. 2.5. Щенок-альбинос
северного морского котика
Взрослая самка (рис. 2.4). Общий фон окраски дорсальной стороны тела тем-
но-серый с рыжеватым оттенком. Кончик морды светло-коричневый. Верхняя
губа и щеки умеренно-коричневые. Лоб светло-коричневый. Макушка и верх
шеи серовато-коричневые. Низ шеи, лопатки, спина и крестец серовато-корич-
невые. Бока светло-коричневые. Вентральная сторона светло-коричневого цвета.
Ниж-няя губа, подбородок и горло серовато-коричневые. Передняя часть груди
светлая, серовато-коричневая; задняя несколько темнее. Подмышки умеренно-
коричневые; брюхо — серо-коричневое.
Окраска волосяного покрова северного морского котика имеет возрастную,
половую и индивидуальную изменчивость. В общей массе животные кажутся ок-
рашенными одинаково. Особенно резко выделяются лишь 2 типа окраски: чер-
ная — у новорожденных и в основном серая с примесью коричневатых тонов
различной интенсивности — у взрослых. У самок с возрастом окраска меха тем-
неет, а у самцов появляется больше коричневатых тонов. В летний период, когда
14
котики подолгу находятся на берегу, их окраска сильно меняется: мех пачкается
экскрементами, грунтом, а у молодых — и молоком. Кроме того, на окраску
волосяного покрова оказывает влияние солнечная радиация под влиянием, ко-
торой шерсть приобретает рыжеватые оттенки.
Северному морскому котику присущи и аберрации окраски. Среди нормально
окрашенных особей встречаются альбиносы и хромисты (рис. 2.5).
По наблюдениям В. Шеффера [Scheffer, 1958], на 100 тыс. новорожденных
приходится 1 альбинос. Иногда наблюдаются частичный альбинизм или хро-
мизм, когда часть туловища или ласты имеют отличную от обычной окраску;
соответственно меняется цвет глаз.
2.3. Пропорции тела
По данным многократных измерений тела северных морских котиков (рис.
2.6) были построены графики, на которых размеры частей тела сравнивались
либо с общей длиной зверя вне зависимости от возраста, либо определялись от-
носительные (чаще от длины тела) показатели той или иной части тела в каждом
возрастном классе.
Рис. 2.6. Схема измерений тела северного морского котика
В основе формирования дефинитивных размеров пропорций тела лежит раз-
ная интенсивность их роста. 5-месячные эмбрионы котиков лишь отдаленно на-
поминают взрослых особей. Передние и задние конечности у них по форме сход-
ны с конечностями наземных хищных. Кисть имеет типичный фронтальный изгиб.
На обеих конечностях отсутствуют хрящевые оторочки, и когти расположены
на дистальных концах пальцев. В дальнейшем в результате ускоренного роста
первого и замедленного роста остальных пальцев формируется типичная кры-
ловидная передняя конечность взрослого животного (рис. 2.7).
15
Рис. 2.7. Изменение формы
и пропорций ластов
морского котика в
онтогенезе:
А — передняя, Б — задняя
конечности;
а, б — пятый, в — шестой, г —
седьмой, д — восьмой, е — де-
вятый месяцы внутриутробного
развития, ж — новорожденный.
Рис. 2.8. Контуры тела
взрослого (пунктирная
линия) и новорожденного
морских котиков
(сплошная линия).
Контур взрослой особи —
рис. по фото из: Scheffer, 1962.
16
50 70 90 110 130 150 170 190 200 50 70 90 110 130 150 170 190 200
ДЛИНА Т В 1 A, CM
Рис. 2.9. Изменение некоторых измерений морского котика в зависимости от длины
его тела:
А — кончик носа—передний край подмышечной впадины, Б — нос—пупок, В —
пупок—анус, Г — длина передней конечности по переднему краю, Д г— длина
передней конечности по заднему краю, Е — ширина передней конечности,- Ж —
длина заднего ласта до конца плавательной перепонки, 3 — длина задних
конечностей до когтей, И — обхват головы, К — обхват шеи, Л — обхват тела за
передними конечностями, М — обхват тела на уровне пупка.
17
В задней конечности опережающим темпом растет хрящевое продолжение
пальцев и кожистая оторочка, которая у взрослых животных составляет не менее
40% от общей длины ласта. Когти при этом остаются на дорсальной стороне
ластов на значительном удалении от края.
В целом, во внутриутробный период интенсивность роста конечностей выше,
чем других частей тела, причем темп роста у задней конечности выше, чем у
передней. Несколько ниже темп роста передней части туловища (головы, шеи
и переднего отдела груди) и самый низкий он у задних отделов тела [Кузин,
1971а]. Новорожденный по многим пропорциям тела практически не отличается
от взрослого животного (рис. 2.8).
Половой диморфизм по большинству пропорций тела во внутриутробный
период не выражен, хотя и намечаются различия в абсолютных и относительных
показателях длины задней конечности, которая больше у самок, и передних ко-
нечностей, которые несколько длиннее у самцов [Scheffer, Wilke, 1953; Кузин,
Набережных, 1982а, 6].
В постнатальный период лишь уменьшаются относительные размеры голо-
вы, утолщаются шея и грудь, но остаются неизменными относительные размеры
ластов и их положение относительно туловища животного. В постнатальном он-
тогенезе эмбриональные скорости роста многих частей тела у котика сохраня-
ются. При этом характерно, что у разнополых, но одноразмерных животных
нет различий по темпам роста отдельных частей тела, тогда как у одновозраст-
ных самцов и самок в относительных показателях таковые намечаются’ (рис. 2.9,
2.10). Из сопоставления продольных измерений корпуса (рис. 2.9) видно, что на-
ибольшей скоростью роста характеризуется передняя часть тела, в которой ин-
тенсивнее растет грудной отдел. Часть тела животного от пупка до анального
отверстия с возрастом относительно уменьшается, а передняя часть тела отно-
сительно увеличивается, в то время как часть тела от кончика носа до переднего
края подмышечной впадины остается практически неизменной. Соответственно,
с возрастом уменьшаются относительные размеры тела от пупка до пениса и
от пупка до ануса.
Свойственная всем новорожденным млекопитающим “болыпеголовость” с воз-
растом у северного морского котика уменьшается (у самцов быстрее^ чем у самок).
Измерения разных частей тела по окружности характеризуют массивность
зверя. Тело котика имеет наибольшую площадь поперечного сечения в области
груди (рис. 2.9). Поэтому, естественно, с ростом организма интенсивность уве-
личения размеров туловища в этой области наибольшая. Примечательно, что
у новорожденных котиков массивность туловища в грудной части больше, чем
у неполовозрелых особей. У самок с возрастом происходит пропорциональное
изменение линейных размеров толщины тела в области груди, а у самцов, на-
против, объем грудной клетки с возрастом увеличивается быстрее, чем линейные
размеры тела. В то же время, в области брюха и таза относительные размеры
тела взрослых самцов и самок почти одинаковые (рис. 2.10). У новорожденных
и неполовозрелых животных туловище в области брюха относительно толще.
Таким образом, корпус тела новорожденных и молодых котиков характеризуется
компактностью (“сбитостью”). С возрастом животные становятся прогонистее.
В постнатальном онтогенезе сильнее изменяется характер роста конечностей.
Преобладающим становится интенсифицирующийся рост дистальных отделов,
что, впрочем, характерно уже и для второй половины внутриутробного развития.
18
ВОЗРАСТ (ГОДЫ)
Рис. 2.10. Изменение относительных показателей некоторых частей тела (сплошная
линия — самцы, пунктирная линия — самки).
Обозначения те же, что на рис. 2.7.
Рис. 2.11. Форма конечностей взрослых самца (А) и самки (Б):
слева — передняя, справа — задняя конечности (рис. по фото из: Scheffer, 1962).
19
Рост конечностей в ширину много медленнее, чем в длину. В целом, относи-
тельный рост передних конечностей хорошо скоррелирован с ростом всего тела
как у самок, так и у самцов. Достоверных половых различий по размерам пе-
редних конечностей нет, но едва заметная тенденция увеличения с возрастом
темпов их роста у самок и уменьшения у самцов намечается. Относительные
показатели ширины ластов несколько больше у самцов. Вследствие неравномер-
ности роста отдельных частей передней конечности, у самцов-секачей в дефи-
нитивном состоянии они выглядят треугольными, а у самок — ланцетовидными
(рис. 2.11).
Формирование пропорций задней конечности сходно с передней, однако в
постнатальном онтогенезе темп ее роста несколько замедляется после достиже-
ния животными длины тела 130 см. Самки в это время практически прекращают
расти, а самцы еще растут. В связи с этим относительные показатели длины
задней конечности у самок с возрастом несколько увеличиваются, а у самцов,
наоборот, уменьшаются. Примечательно, что у самок относительно длиннее, чем
у самцов, хрящевая оторочка заднего ласта, а не скелетные образования. Нет
также различий и в относительных показателях ширины задней конечности у
разнополых особей (рис. 2.10).
Таким образом, формирование большинства дефинитивных пропорций тела
северного морского котика завершается во внутриутробный период. В постна-
тальном онтогенезе большинство частей тела развиваются пропорционально.
Лишь относительные размеры головы во второй половине эмбриогенеза и в пос-
тнатальном онтогенезе растут медленнее других частей тела, да происходит до-
формирование соответствующих пропорций, связанных с объемными показате-
лями тела, которые отставали в росте от линейных.
2.4. Гидродинамическая характеристика
Строение тела и его пропорции у северного морского котика как у водного
животного характеризуются определенными параметрами, которые способству-
ют созданию наилучшей плавучести и поступательному движения прежде всего
в воде. Плавучесть обеспечивается приспособлениями как гидростатического по-
рядка (уменьшением удельного веса), так и формой тела, снижающей сопротив-
ление воды при движении. Листообразные движители способны осуществлять
максимальную эффективность локомоции при наименьших затратах энергии.
По нашим данным, удельный вес северного морского котика, как и других
тюленей [Соколов, 1955; Мордвинов, 1969], близок к 1 и составляет летом 0.97.
Зимой с накоплением жировых запасов он еще больше приближается к 1. Спе-
циальных приспособлений гидростатического порядка, таких как воздушные
мешки у моржа и крылатки [Слепцов, 1940; Fay, 1960], у котика нет. В увели-
чении плавучести котика основную роль играют густой волосяной покров, объ-
емистые легкие и “жесткая” трахея, в которой постоянно присутствует некоторое
количество воздуха [Соколов, 19696; Кузин, Соболевский, 1975], небольшой от-
носительный вес скелета [Кузин и др., 1972; Косыгин, Кузин, 1979], соответст-
вующая форма и пропорции туловища и органов локомоции. По крайней мере,
этих приспособлений, по-видимому, достаточно для того, чтобы животные мог-
ли по несколько часов находиться без видимых движений во время сна на воде.
20
Таблица 2.1
Изменение относительной наибольшей высоты (Н), относительной
наибольшей ширины (I), наибольшего диаметра (D) и показателя формы
корпуса (Y) в зависимости от размеров северного морского котика
[Мордвинов, 1972а; наши данные]
Длина тела, см п Н, % KL CV I, % Acv D, % vL Acv Y Y (наши данные)
п M Um
60-70 13 20.8 24.8 22.8 0.58 5 0.48 0.46-0.49
71-80 9 20.7 25.4 23.6 0.57 16 0.45 0.38-0.49
91-100 1 0.47
101-100 9 0.47 0.43-0.48
111-120 3 24.0 30.0 27.0 0.53 12 0.46 0.41-0.53
121-130 5 26.9 32.2 29.5 0.50 6 0.48 0.43-0.52
131-140 2 26.2 30.0 28.0 0.51 4 0.49 0.45-0.50|
141-150 5 27.1 32.7 29.0 0.49 7 0.49 0.37-0.61
151-160 3 26.7 32.0 29.3 0.48 7 0.46 0.36-0.50
161-170 4 26.2 31.5 28.8 0.47 5 0.51 0.48-0.54
171-180 1 0.48
181-190 2 27.1 33.0 30.0 0.46
191-200 3 0.49 0.46-0.53
Поступательное движение в воде котик осуществляет в основном с помощью
передних конечностей [Howell, 1930]. Они же выполняют функции рулей и тор-
мозов. Задние конечности используются животными как стабилизаторы и рули,
но в определенных условиях они также могут брать на себя функцию движите-
лей. Активная ролыв локомоции животного принадлежит корпусу [Мордвинов,
1969]. Расположение основных движителей в передней части корпуса считается
малоэффективным [Ray, 1963], но котик способен развивать скорость до 18
узлов [Scheffer, 1958; Ray, 1963; Евдокимов, 1964]. Однако, по мнению этих авто-
ров, с предельной скоростью он может плыть лишь в момент броска. По нашим
наблюдениям, с попутной волной котик может двигаться со скоростью 11 узлов
в течение 20—30 мин.
Исследования формы корпуса северного морского котика показали, что она
обладает рядом характеристик, способствующих снижению сопротивления дав-
ления и сопротивления трения [Мордвинов, 1969, 1972а, б]. Полученные харак-
теристики (табл. 2.1) свидетельствуют о том, что показатель формы корпуса (Y)
характеризуется высокими Значениями, которые, как известно из аэродинамики,
наиболее выгодны для предотвращения отрыва пограничного слоя воды на до-
вольно большом участке поверхности тела, благодаря чему сохраняется лами-
21
Таблица 2.2
Относительная прогонистость тела северного морского котика
Длина тела, СМ Самцы Самки
п М lim п М lim
60-70’ 2 9.42 8.9-10.1
71-80 2 9.50 9.2-9.6 6 9.7 9.2-10.1
91-100 6 9.90 9.2-10.6 14 9.9 8.9-12.8
101-110 9 10.30 9.1-11.7 18 10.2 8.8-11.8
111-120 11 10.0 9.1-13.1 43 9.9 8.5-11.7
121-130 7 9.90 9.2-11.0 53 9.9 8.3-11.2
131-140 4 10.50 9.6-11.0 26 10.1 8.9-11.1
141-150 7 10.1 9.6-10.5
151-160 3 9.1 9.1-9.9
161-170 5 10.2 9.1-11.2
181-190 3 9.9 8.9-10.6
192-200 2 9.5 9.3-9.7
парный характер обтекания пограничного слоя и снижается сопротивление дви-
жению животного. Определенная роль в этом принадлежит волосяному покрову,
который способствует обтеканию корпуса, уменьшая гидродинамическое сопро-
тивление [Мордвинов, 1972а].
Показатель формы корпуса (Y), по данным Ю. Е. Мордвинова [1972а], у бе-
ломорского лысуна и каспийского тюленя ниже (соответственно, 0.53—0.41 и
0.45—0.54), чем у котика (табл. 2.1).
Из данных Ю. Е. Мордвинова [1972а] следует, что с возрастом показатель
формы корпуса (Y) снижается, а относительная высота (Н), ширина (I) и диаметр
(D) увеличиваются, т. е. тело котика с возрастом приобретает компактность и
округлость, а значит с возрастом ухудшаются гидродинамические свойства тела.
Наши данные не подтверждают этого. Показатель формы корпуса, относитель-
ные показатели таких измерений как обхват шеи, тела за передними конечнос-
тями, в области пупка и бедер, по нашим данным, после 2-летнего возраста ос-
таются практически неизменными. Не подтверждается это и показателями от-
носительной прогонистости тела морского котика (табл. 2.2). Напротив, отно-
сительное удлинение корпуса с возрастом несколько увеличивается. Сказанное
позволяет утверждать, что с возрастом гидродинамические свойства тела котика
улучшаются.
Представленные данные по относительной прогонистости тела морского ко-
тика (табл. 2.2) близки к таковым белухи [Клейненберг и др., 1964].
Помимо названных показателей, о теле животного как несущей плоскости
можно судить по отношению наибольшей его высоты к наибольшей ширине.
У взрослого котика это соотношение составляет 0.57. У гренландского тюленя
и каспийской нерпы, соответственно, 0.80—0.87 и 0.82—0.91 [Мордвинов, 1969].
22
Таблица 2.3
Изменение показателей горизонтальной fZmin, Zmax? и вертикальной fzmin,
zmaJ динамической устойчивости с возрастом (Lcv, см) у северного морского
котика [из: Мордвинов, 1972а]
Длина тела, Lcv, см Zmm Zmax Zmin Zmax
70.5 -0.020 -0.018 -0.021 -0.019
114.0 -0.018 -0.017 -0.017 -0.017
129.0 -0.014 -0.012 -0.013 -0.012
143.0 -0.010 •0.011 -0.010 -0.012
162.0 -0.007 -0.009 -0.006 -0.008
166.0 -0.005 -0.007 -0.005 -0.005
Судя по представленным данным, корпус тела котика обладает лучшими гид-
родинамическими качествами, чем корпус тела настоящих тюленей.
Форма основного движителя (переднего ласта) северного морского котика
и форма крыла самолета сходны. Фронтальная часть ласта котика тоже утол-
щена, а дистальная сходит на нет. Используя формулу, применяемую в аэроди-
намике для определения относительного удлинения крыла самолета Л = 12/S, где
1 — размах крыльев, aS — площадь крыла, мы рассчитали этот показатель для
переднего ласта морского котика. Он равен 0.45—0.47, что превышает то
минимальное значение относительного удлинения аэродинамического крыла
(Л = 1/30), при котором уже возникает подъемная сила.
Из наблюдений в природе известно, что северный морской котик обладает
большой маневренностью при движении в воде. Расчет динамической устойчи-
вости и поворотливости котика (табл. 2.3) показали, что у него такие показатели
как Zmax и выше значений Zmin, zmin, что говорит об уменьшении динами-
ческой устойчивости, причем Zmin и Zmm с возрастом приближаются к нулю, т. е.
стабилизирующий момент уменьшается, а, значит, увеличивается поворотли-
вость.
Zmax И Zmax мало различаются, следовательно, вертикальная и горизонтальная
поворотливости развиты одинаково хорошо [Мордвинов, 1972а]. Морфологи-
чески это обеспечивается уменьшением с возрастом расстояния между центром
тяжести и центром динамического равновесия, уменьшением относительных ве-
личин продольных пропорций тела животных.
Показатели вертикальной и горизонтальной устойчивости у котика ниже, чем
у представителей настоящих тюленей, а значит выше маневренность [Мордви-
нов, 1972а].
Таким образом, представленные данные по пропорциям тела, органов локо-
моции и их гидродинамическим характеристикам позволяют утверждать, что се-
верный морской котик обладает рядом преимуществ, по сравнению с некото-
рыми изученными видами ластоногих, и говорит о его прекрасной приспособ-
ленности к обитанию в водной среде.
23
Глава 3. ОПРЕДЕЛЕНИЕ ВОЗРАСТА И РОСТ
3.1. Определение возраста
Продолжительный интенсивный промысел северных морских котиков позво-
лил в прошлом опытным промышленникам, а вслед за ними и натуралистам,
научиться легко распознавать отдельные возрастные группы котиков по внеш-
ним признакам, таким как длина и масса тела, размер ластов, цвет вибрисс, по-
явление “гривы” на шее и передней части туловища у самцов, размер клыков
и другим [Jordan, Clark, 1898]. Эта практика определения возраста котиков у
промысловиков существует по настоящее время. Однако еще в начале текущего
столетия стало известно, что длина тела самых крупных меченых особей из низ-
шего возрастного класса может быть много больше длины тела животного из
высшего возрастного класса [Бойцов, 1934]. Тем не менее, разработанные воз-
растные стандарты северных морских котиков, подлежащих промыслу, по длине
тела в 1913 и 1920 гг. на островах Прибылова и Командорских еще долгое время
были в ходу как у промысловиков, так и у ученых, определявших нормы выбоя
и прогнозирование промысла [Бойцов, 1934].
В. Шеффер [Scheffer, 1950с] исследовал большую серию бакулюмов северных
морских котиков с целью выяснения возможности определения по длине кости
пениса истинного возраста животных и времени наступления у них половой зре-
лости. Однако из-за большой трансгрессии показателей у одновозрастных жи-
вотных использовать этот показатель в качестве возрастного критерия оказалось
нецелесообразным.
Дальнейшие поиски в этом направлении привели В. Шеффера [Scheffer, 1950а]
к открытию на корневой части клыков хорошо заметных валиков, число ко-
торых точно соответствует возрасту меченых особей (рис. 3.1). Однако у жи-
вотных старше 10 лет точно определить возраст по валикам оказалось невоз-
можно из-за нечетко различимых наслоений последних валиков. Вместе с тем
эта работа сразу же обратила на себя внимание других исследователей и по-
высила интерес к изучению структуры зубов других морских млекопитающих
как индикатору роста. Позже [Laws, 1953] было изучено гистологическое строе-
ние клыков морского слона (Mirounga leonina), и был сделан вывод, что валики
на внешней стороне зубов есть ни что иное, как чередующиеся слои дентина
разной плотности.
При замедлении темпа роста организма зимой откладывается светлый слой
дентина, а летом — темный. Переломным моментом в отложении слоев дентина,
по мнению К. Куботы с соавт. [Kubota et al., 1963], следует считать апрель, когда
заканчивается время нагульного периода жизни котиков, и они мигрируют к
местам размножения. По мнению В. Шеффера и Б. Крауса [Scheffer, Kraus, 1964],
этот период наступает осенью и инициируется началом процесса линьки у жи-
вотных.
Таким образом, на гистологических срезах или аншлифах за годовой слой
принимают полоску темного и светлого дентина (рис. 3.2).
24
Рис. 3.1. Валики иа верхнем клыке северно-
го морского котика как критерий возраста
(фото из; Scheffer, 1962)
Рис. 3.2. Аншлиф продольного распила
верхнего клыка северного морского коти-
ка (фото из: Scheffer, 1962)
Особенностью наслоений дентина в зубах северных морских котиков, как и
в зубах других ушастых тюленей (в отличие от настоящих), является отложение
дентина по принципу “конус в конус”. Нарастание нового слоя происходит со
стороны пульпы. Как правило, полное заполнение полости происходит у самок
котиков после 10-летнего, а у самцов — после 14—15-летнего возраста. У старых
животных последние слои дентина накладываются на предыдущий слой в виде
прямой линии, а иногда даже возможно образование выпуклости в противопо-
ложную сторону от нижележащего слоя дентина.
В практике определения возраста котиков обычно пользуются подсчетом
слоев под бинокуляром на аншлифах зубов. При этом опытные исследователи
могут ошибиться до ±1.93%, а неопытные — до ±22% в ту или иную сторону
от истинного возраста [Anas, 1970].
Р. Бауэр с соавт. [Bauer et al., 1964] предприняли попытку определения воз-
раста северного морского котика по весу хрусталика глаза. Оказалось, что хотя
масса хрусталика и увеличивается всю жизнь, но имеет большой диапазон из-
менчивости, и ошибки в определении возраста этим методом могут достигать
1.5 для самцов и 1.7 раза — для самок в ту или иную сторону от истинного
возраста при 95% доверительном интервале.
3.2. Рост
Данные по массе и длине тела эмбрионов котиков представлены в работах
Ф. Тейлора с соавт. [Taylor et al., 1955] и В. Шеффера [Scheffer, 1960а, 1962]. Более
подробно рост котиков в период их внутриутробного развития изучен А. Е. Ку-
зиным [1970а], А. Е. Кузиным и И. А. Набережных [1982а]. Т. И. Чупахина [1977]
25
Таблица 3.1
Изменение массы (г) и длины (см) тела эмбрионов северного морского
котика в зависимости от возраста
Месяц добычи самок (в скобках возраст эмбрио- нов, мес.) Пол п Масса Длина
М±ли CV Mim CV
Декабрь (5) самцы 16 33.02i3.92 11.85 9.68±0.57 23.55
самки 16 28.46i4.04 14.18 9.07±0.57 25.24
Январь (6) самцы 42 157.97i20.61 84.56 16.50i0.67 26.60
самки 43 122.00il0.82 58.45 15.30±0.55 23.92
Февраль (7) самцы 13 626.53i65.26 37.61 28.26i0.31 4.31
самки 11 542.72±55.37 33.87 27.49il.09 13.27
Март (8) самцы 50 1359.84i37.65 19.56 43.71i0.49 8.00
самки 50 1194.60i31.27 18.50 42.4810.49 8.30
Апрель (9) самцы 50 2379±77.33 22.79 55.59i0.76 9.69
самки 50 2029.20i69.87 24.34 53.37Ю.58 7.79
Май (10) самцы 50 3388.40i46.78 9.76 58.3OiO.58 7.08
самки 50 3182.60i58.74 13.05 56.63i0.64 7.99
Июнь (11) самцы 50 4827.00il28.91 18.88 61.02i0.62 7.22
самки 50 4494.40+103.92 16.34 58.62±0.45 5.86
Примечание. Возраст эмбрионов считали от средней даты зачатия, условно отнесен-
ной на 15 июля.
опубликовала данные о весовой и линейной характеристике щенков котиков в
подсосный период. Рост котиков также изучали В. Шеффер и Ф. Уилке [Scheffer,
Wilke, 1953], В. А. Бычков [19716], Ф. Г. Челноков и Д. И. Чугунков [1971].
Процесс индивидуального развития млекопитающих обычно разделяют на
2 крупных периода: пренатальный и постнатальный. Последний, в свою очередь,
включает несколько фаз: ювенильную, полового созревания, стационарного (взрос-
лого) состояния организма. Каждый из названных периодов характеризуется сво-
ими особенностями роста и развития, а в целом имеет видовую специфику.
Пренатальный период. По данным В. Шеффера [Scheffer, 1960а], у самок ко-
тиков, добытых 27 октября на о-вах Прибылова, нашли бластоцисты диаметром
0.37—2.0 мм. 8 декабря были обнаружены 3 эмбриона, длина тела которых со-
ставляла 10,10 и 20 мм, а диаметр — 7.5, 8.5 и 17.0 мм, соответственно. Наиболее
ранние, добытые нами 16 декабря в западной части Тихого океана 2 самки,
имели* эмбрионы массой 2.3 и 8.5 г [Кузин, 1970а]. Средняя масса эмбрионов
за декабрь (пятый месяц внутриутробного развития) составила 30 г (п = 32) или
0.6% от средней массы детенышей при рождении и 0.07% от средней массы взрос-
лой самки. К середине января масса эмбрионов котиков увеличивается более
чем в 2 раза (табл. 3.1). В июне перед рождением эмбрионы обоего пола имели
среднюю массу тела ок. 5 кг (12.5% от средней массы взрослой самки).
26
Таблица 3.2
Отношение массы и длины тела эмбрионов северного морского котика
разного возраста к величине этих показателей на пятом месяце
внутриутробного развития, принятых за единицу
Время добычи Возраст, мес Самцы Самки .
масса, г длина, см масса, г длина, см
Январь 6 4.78 1.70 4.28 1.68
Февраль 7 18.97 2.91 19.06 3.03
Март 8 41.17 4.51 41.97 4.69
Апрель 9 72.05 5.74 71.30 5.88
Май 10 102.61 6.02 111.82 6.24
Июнь 11 146.18 6.30 157.91 6.46
Рис. 3.3. Изменение коэффициентов массы и длины тела эмбрионов котиков в
зависимости от возраста
Из данных по отношению массы и длины тела эмбрионов котиков разного
возраста к величине этих показателей в декабре, принятых за единицу, видно,
что масса эмбрионов увеличивается во много раз больше, чем длина (табл. 3.2).
Так, к периоду рождения длина тела увеличивается только в 6 раз, а масса
в 150 раз, по сравнению с пятым месяцем внутриутробного развития.. Уже на
ранних стадиях развития самцы несколько крупнее самок. Из таблицы также
видно, что, начиная с мая, интенсивность прироста массы самцов меньше, чем
самок. Для более полной характеристики интенсивности роста в период внут-
риутробного развития северных морских котиков воспользуемся коэффициента-
ми роста (рис. 3.3).
27
Таблица 3.3
Масса (г) разновозрастных эмбрионов северного морского котика
Возраст (декады) Самцы Самки .
п масса 1оК V CV cvt п масса 1ой V CV cvt
3 1 3 0.4771 2.457 8.599 1 9 0.9542 1.386 4.852
4 15 35 1.5441 0.955 ь.тп 12 36 1.5563 0.679 3.396
6 7 91 1.9590 2.050 15.376 9 71 1.8513 2.357 17.680
9 8 707 2.8494 0.277 2.636 6 750 2.8751 0.168 1.592
10 3 933 2.9699 0.091 0.958 4 887 2.9479 0.017 0.176
11 7 1261 3.1007 0.145 1.663 4 902 2.9552 0.360 4.141
12 12 1454 3.1635 0.179 2.239 16 1293 3.1116 0.220 2.754
13 14 1743 3.2413 0.191 2.580 17 1612 3.2073 0.127 1.716
14 15 2110 3.3243 0.155 2.234 15 1830 3.2625 0.254 3.686
15 17 2463 3.3915 0.277 4.297 15 2360 3.3729 0.146 2.259
16 20 3250 3.5119 0.030 0.498 19 2730 3.4362 0.043 0.707
17 12 3350 3.5250 0.069 1.209 18 2850 3.4548 0.119 2.083
18 15 3590 3.5550 0.181 3.344 18 3210 3.5065 0.092 1.704
19 10 4300 3.6335 0.014 0.269 4 3520 3.5465 0.085 1.648
20 6 4360 3.6393 0.278 5.707 3 3830 3.5832 0.248 5.093
21 10 5760 3.7604 0.054 1.163 18 4910 3.6911 0.046 0.896
22 15 6080 3.7839 15 5130 3.7101
3.596 3.467
Примечание. Возраст считали от средней даты имплантации бластоциста, условно от-
несенной на 20 ноября.
28
Таблица 3.4
Отношение массы и длины тела эмбрионов северного морского котика к
величине этих показателей у взрослых животных (%)
Время добычи Возраст, мес Самцы Самки
масса длина масса длина
Декабрь 5 0.02 0.48 0.07 0.69
Январь 6 0.08 0.83 0.29 1.17
Февраль 7 0.33 1.42 1.29 1.11
Март 8 0.73 2.20 2.84 3.26
Апрель 9 1.27 2.80 4.83 4.10
Май 10 1.82 2.93 7.57 4.35
Июнь 11 2.59 3.07 10.70 4.50
Взрослые 100 186 кг 100 1985 см 100 42 кг 100 1300 см
Как видно на рисунке, весовой прирост и увеличение линейных размеров раз-
вивающихся эмбрионов постепенно снижается, т. е. эмбрионы интенсивней рас-
тут на первых этапах своего развития. Наиболее заметное уменьшение коэффи-
циентов роста происходит до марта, после чего наблюдается последовательное
их падение, а, значит, рост эмбрионов идет равномернее, чем в предыдущий пе-
риод. Примечательно то, что периоды весового и линейного роста по интен-
сивности как бы сменяют друг друга. Так, если в возрасте 6—7 и 10—11
мес. эмбрионы более быстро увеличиваются в массе, то в эти же периоды за-
медляется линейный рост. И, наоборот, если в возрасте 8—9 мес. увеличивается
интенсивность линейного роста, то падает интенсивность весового.
Исследование весового роста котиков во внутриутробный период, выпол-
ненное нами по формулам И. И. Шмальгаузена [1935а], также показало, что про-
цесс этот неравномерен (табл. 3.3). За весь цикл внутриутробного развития на-
блюдается несколько периодов ускоренного роста. В большинстве случаев после
ускоренного роста наступает заметное падение его, что может быть связано с
процессами дифференцировки частей тела и органов эмбрионов [Шмальгаузен,
19356]. В этом исследовании весового роста наибольшая скорость отмечалась
в начале периода; в дальнейшем она неравномерно падала. Последнее увели-
чение скорости роста наблюдалось в июне, перед рождением. Характерно, что
в большинстве случаев периоды ускоренного роста наблюдаются в одних и тех
же или смежных декадах у самцов и самок, что говорит об однотипности этих
процессов у разнополых особей. Константа роста за исследованный период эмб-
рионального развития составила для самцов 3.60 и 3.48 — для самок. Тем самым
еще раз подтверждается, что интенсивность роста у самцов выше, чем у самок.
Интересен тот факт, что у исследованных нами и другими авторами эмбрио-
нов-двоен самцы также были крупнее самок, хотя условия для развития разно-
полых особей, казалось бы, должны быть одинаковыми [Niggol, Fiscus, 1960; Fis-
cus et al., 1964; Peterson, Reeder, 1966; Кузин, 1969а].
29
Для того, чтобы судить о темпе роста эмбрионов и их физической зрелости
к периоду рождения, мы массу и длину взрослых особей приняли за 100% и вы-
числили относительные показатели первых на каждом этапе их внутриутробного
развития (табл. 3.4). Представленные данные позволяют сказать, что линейные
размеры у эмбрионов-самцов несколько раньше достигают конечной величины,
чем весовые. У эмбрионов-самок наблюдается обратная картина: весовой рост
опережает линейный. Можно сказать, что развитие самок во внутриутробный
период идет скорее, чем самцов, поскольку перед рождением они имели более
приближенные к конечным размерам данные по длине и массе тела. Это вполне
закономерное явление для животных с хорошо выраженным половым димор-
физмом, у которых самцы имеют более продолжительный период роста, и у них
половая и физическая зрелость, как правило, наступает позже, чем у самок.
Сопоставление данных по росту эмбрионов котиков, добытых в северо-за-
падной и северо-восточной частях Тихого океана и принадлежащих животным
разных популяций, показало полное их совпадение [Scheffer, 1962; Кузин, 1970а].
Сходство столь высоко, что прослеживается не только в абсолютных измерениях,
но и в коэффициентах вариации.
Относительный рост котиков во внутриутробный период изучали по форму-
лам аллометрического роста y=bL. Для этого воспользовались измерениями от-
дельных частей тела, уже использованных нами при описании пропорций тела
котиков. Коэффициенты L некоторых основных измерений частей тела эмбри-
онов котиков представлены ниже в таблице.
Пол Промер (номер по рис. 2.6)
2 6 8 12 15 16 17 18 20
Самки 1.23 0.97 0.89 0.83 1.44 1.16 1.25 1.67 1.85
Самцы . 1.28 0.99 0.88 0.82 1.61 1.38 1.19 1.70 1.76
Данные аллометрии, также как и представленные выше коэффициенты роста,
показывают, что интенсивность роста разных частей тела эмбрионов котиков
не одинаковая. Темп роста конечностей выше других частей тела, причем у за-
дних выше, чем у передних. Темп роста передних отделов туловища выше, чем
у задних.
Размеры новорожденных. Сведения о размерах новорожденных котиков встре-
чаются в работах натуралистов прошлого века [Вениаминов, 1840; Elliott, 1875;
Волошинов, 1887] и у исследователей более позднего времени [Гребницкий, 1902;
Суворов, 1912; Бойцов, 1934; Ильина, 1950; Сушкина, 1954]. Однако все назван-
ные авторы не сообщают о том, как скоро после рождения были измерены и
взвешены детеныши. Возможно, поэтому длина тела новорожденных в их ма-
териалах колеблется от 24 см [Гребницкий, 1902] до 60 см [Ильина, 1950], а масса
тела от 2 кг [Сушкина, 1954] до 4.8 кг [Суворов, 1912].
По данным В. Шеффера и Ф. Уилке [Scheffer, Wilke, 1953], длина новорожденных
котиков составляет 54.6—75.4 см (в среднем 65.9 см — у самцов и 63.1 см —
у самок), а масса тела 3.3—7.1 кг (в среднем 5.4 кг — у самцов и 4.8 кг — у
самок). По данным Д. И. Чугункова [1982], на Командорских о-вах новорож-
денные самцы в возрасте до суток имеют массу тела 6.0 кг (4.3—7.0 кг), а сам-
30
ки 5.4 кг (3.8—6.7 кг). По нашим данным, полученным в 1976 г. на о. Тюлений,
длина новорожденных в возрасте 1—3 суток составляет в среднем у самок 63.1 см
(55.2—68.7 см) и 65.9 см — у самцов (55.4—68.9 см), а масса — 5.8 кг (4.1—6.3 кг)
и 6.1 кг (4.5—7.1 кг), соответственно. Некоторые различия в результатах, воз-
можно, связаны с тем, что измерения щенков производились на разных островах
и в разное время. На эти показатели возможно влияние интрапопуляционных
параметров.
В целом, представленные данные показывают, что размеры новорожденных
самцов больше размеров самок. Обращает на себя внимание и тот факт, что
масса новорожденных котиков составляет значительную долю от массы матерей,
что, в общем-то, характерно для всех ластоногих. Так, у лахтака это соотно-
шение равно 1:5 [Косыгин, 1966], у акибы — 1:8 [Федосеев, 1964], у обыкно-
венного тюленя — 1:6, у морского слона — 1:15, а, например, у льва — 1:100
и у медведя — 1:600 [Laws, 1959]. Рождение более крупного детеныша у лас-
тоногих, по сравнению с наземными хищными, является результатом более про-
должительной беременности у первых.
3.3. Фаза ювенильного роста
По данным Т. И. Чупахиной [1977], масса детенышей северного морского
котика за период с июня по первую декаду сентября увеличивается более чем
вдвое (табл. 3.5).
Скорость весового роста щенков на этом промежутке времени неодинаковая.
Они растут более интенсивно в первое время после рождения, когда самки чаще
кормят их молоком. К концу периода молочного кормления рост щенков за-
медляется. Константа роста детенышей в период молочного питания составила
0.4874 для самцов и 0.4543 — для самок. В целом, интенсивность роста щенков
много ниже, чем эмбрионов. Однако рост самцов в период молочного питания
выше, чем самок, и проявление полового диморфизма в ранний период пост-
натального развития усиливается.
Данных по росту “серых” щенков мало. Добытые 4—5 октября 1975 г. “се-
рые” самцы на о. Тюлений имели массу тела в среднем 15.5 кг, а добытые там
же 9—11 октября 1976 г. весили в среднем 16.1 кг (11—24 кг); самки, соот-
ветственно, 14.7 кг (12—17.5 кг). Длина тела этих животных составляла 87.9 см
(79—98 см). У добытых на Командорских о-вах в конце октября—начале нояб-
ря 1971—72 гг. “серых” котиков масса тела была 10—30 кг, длина 72—99 см.
Заметим, что приведенные данные по “серым” котикам относятся к разным го-
дам и разным районам добычи и, возможно, не в полной мере характеризуют
ростовые процессы, т. к. хорошо известно, что существует межгодовая измен-
чивость в размерах тела молодых животных [Scheffer, 1955].
3.4. Фазы полового созревания и стационарного состояния
Абсолютный рост. Как следует из данных предыдущих исследований [Schef-
fer, Wilke, 1953; Бычков, 1971 б', Челноков, Чугунков 1971; Мужчинкин, 1976; Ку-
зин, Набережных, 19826] и материалов, представленных на рис. 3.4, увеличение
31
Таблица 3.5
Изменение массы детенышей северного морского котика в первые месяцы
жизни (из: Чупахина, 1977, с дополнениями)
Месяц, декада Самцы Самки
п lim М п lim М
Июнь
I декада 40 4.4-8.3 6.4 27 4.3—7.1 6.0
II декада 36 5.1—9.2 7.0 12 ... 4.9—7.9 6.4
III декада 48 4.8—14.0 7.8 20 4.8—8.2 6.5
Июль
I декада 32 5.4—15.0 9.8 17 6.0—12.0 8.5
II декада 70 6.7—15.6 9.9 52 6.2—10.1 8.7
III декада 12 9.7—15.9 10.7 13 7.6—12.5 9.6
Август
I декада 11 11.0—16.0 12.0 10 8.0—11.5 9.8
II декада 16 10.5—16.0 12.9 9 8.2—14.4 10.0
III декада 20 11.0—16.2 13.1 9 9.4—14.9 11.2
Сентябрь
I декада 13 11.2—17.9 14.7 11 10.5-15.3 13.0
Октябрь
I декада 377 9.0—20.0 15.4 79 4.0—17.0 13.7
II декада 158 11.0—24.0 16.1 42 12.0—17.5 147
СМ КГ
200 • -САМЦЫ
° -САМКИ
И
150 300
100. 200
50-100
Б
инн»
А
1-f l "1-1-1-1-Г—Т-1 I I VII
3 5 7 9 II 13 15
ВОЗРАСТ /ГОДЫ/
о! | *
Н I
Рис. 3.4. Изменение массы
и длины тела северных мор-
ских котиков в постнаталь-
ном онтогенезе:
А — длина тепа, Б — масса
тела, Н — новорожденные.
32
массы и длины тела котиков в фазах полового созревания и стационарного со-
стояния происходит неравномерно.
Темп роста на первом году жизни ниже, чем на последующих, в чем наши
данные хорошо согласуются с данными В. Шеффера и Ф. Уилке [Scheffer, Wilke,
1953]. Переход с молочной диеты на самостоятельное питание в трудных усло-
виях первой самостоятельной зимовки безусловно сказывается на ростовых про-
цессах. Это закономерное явление для многих млекопитающих. Из приведенных
выше данных по массе и длине тела “серых” котиков (табл. 3.5) видно, что они
могут достигать 30 кг, в то время как масса тела годовиков не превышает 21 кг.
От 1 года до 3 лет темп роста самцов и самок одинаковый. Однако у самцов
старше 3 лет наблюдается новая волна ускоренного роста, которая продолжается
до 8—9 лет, когда практически уже прекращается увеличение массы и длины
тела (рис. 3.4). По данным вышеупомянутых авторов, акселерация роста самцов
начинается с 7—8 лет, что не согласуется с нашими данными.
Рост самок котиков начинает замедляться после 3-летнего возраста и пре-
кращается к 7—8 годам. Изменения в ростовых процессах обусловлены физио-
логическими перестройками в организме, связанными с наступлением половой
зрелости. Половина самок в 4-летнем возрасте становятся беременными. Длина
их тела в этом возрасте в среднем составляет 68% от таковой физически зрелых
животных. Среди самцов в 3-летнем возрасте половозрелыми становятся менее
половины животных [Кузин, 1978; Серебренников, 1982], тогда как в 4-летнем
возрасте — практически все. В этом возрасте длина их тела составляет 73—75%,
а масса только 35% от минимальных размеров гаремных секачей. Участвовать
в размножении самцы начинают только с 7—8 лет, когда средняя их длина со-
ставляет 85%, а масса 61% от показателей физически зрелых особей. Таким об-
разом, в размножении участвуют разнополые особи с близкими показателями
относительных размеров тела. Следует заметить, что длина тела беременных
самок, впервые вступающих в размножение, на 3—5% больше, чем небеременных
в том же возрастном классе. Аналогичная картина наблюдается и у самцов, среди
которых одновозрастные половозрелые и неполовозрелые особи различаются по
размерам тела. Таким образом, ростовые процессы заметно сказываются на сро-
ках наступления половой зрелости и, соответственно, участии в размножении
животных.
До 5—6-летнего возраста доверительные интервалы рядов массы и длины
тела самок и самцов значительно перекрываются в смежных возрастных классах,
причем с возрастом амплитуда трансгрессии увеличивается [Бычков, 19716]
(рис. 3.4). Выделенные естественные периоды замедленного и ускоренного роста
тела морского котика хорошо описываются константами роста (К) И. И. Шмаль-
гаузена [1935]. Для массы тела самцов 1—3 лет К равняется 1.05, 4—9 лет —
2.0, после 9 лет — 0.30; для массы тела самок в возрасте 2—5 лет — 1.05, 5—7
лет — 0.52, старше 7 лет — 0.07. Для длины тела самцов 1—4 лет — 0.49, 4—7
лет — 0.65, 7—9 лет — 0.45, старше 9 лет — 0.20; для самок до 5 лет— 0.34,
старше 5 лет — 0.12.
Данные по росту частей тела холостяков Ф. Г. Челнокова и Д. И. Чугункова
[1971] и наши, представленные на рис. 3.5, показали, что рост каждой из частей
имеет свой темп.
33
200 НО <00 о! ?• 0 9 ООО” О О
• • • • • 9 О
90- о ° О
О о
го М2 О
50 О О • • • • • • 9
30 9 • о О
О О
<40 ЫЗ 0 оО
<00 о О 8« • • • • • • • •
60- О •
20 О О о
<6- О о о
о о ° 0
t - о»
О о О
К >15 О О О о о
П о О
О
2 1: Мб _ О О О О О О О
а 3 90- О® О» Л о 0 $•• • • • • • • О О • О
3 20 Ы7 о О
о С-Э 50- 30; 9 8 • • • • • •
«о о
то ЙИ 9 О оО о
SO- 30: ee 98 8»• • • • • •
№ оО’°° °0 О
<0- S - о 8 • 8 8 *•• •• • • •
»: о ’° ° о О
10- Й13 о0
is- о • • • • •
<oi
<1. ||Ц о о °0 О о
<0- о оj 2 8••• • «
я . 8»
• <0' 0° О О О
SO >115 of
30- »• О $ • • • • • • • •
О 0 о
no-
ад
10
«0
40
<00
>18
0
о
о
ООО
У
35
IS
<»'
30-
JO-
IS-
о O°
У20 °
O ° • . •••• •
*
? 7..............
H 1 2 5 4 5 0 T J 9 10 <0*
о - САМЦЫ
• -СШИ
Возраст, гады
Рис. 3.5. Изменение размеров частей тела северных морских котиков в зависимости
-от возраста:
1—20 — номера промеров по рис. 2.6.
34
Таблица 3.6
Коэффициенты аллометрии промеров тела (номера — по рис. 2.6) северных
морских котиков [из: Кузин, Набережных, 19826]
Номер промера Самки Самцы Номер промера Самки Самцы
1 фаза 2 фаза 1 фаза 2 фаза
2 0.99 0.92 0.68 11 0.56 0.55 0.59
3 1.03 0.99 0.64 12 0.53 0.54 0.33
4 0.85 1.33 13 0.58 0.56 0.25
5 1.06 0.62 14 0.56 0.62 0.51
6 0.76 1.05 0.71 15 1.05 0.99 0.89
7 0.61 1.21 1.33 16 1.02 0.98 0.69
8 0.82 0.52 1.33 17 0.79 0.97 1.03
9 0.61 0.42 0.97 18 1.11 1.02 0.64
10 0.82 0.68 0.64 19 0.96 0.85 0.52
20 0.86 0.87 0.89
Темп роста объемных показателей задней части туловища у самок (промеры
9, 10 на рис. 2.6) от новорожденных до 3-летнего возраста выше, чем у самцов,
что, по всей вероятности, связано с более ранним сроком созревания самок и
подготовкой их организма к плодоношению. Проявляющаяся в период полового
созревания акселерация роста тела самцов неодинаково выражена в отдельных
частях тела. Более того, не все части тела ее испытывают. Акселерация не про-
являет себя в росте задних частей тела (промеры 4—6), головы (11—14), ширины
переднего ласта (17) и длине заднего ласта до когтя (19). Остальные части тела
растут с ускоренным темпом, но акселерация для каждой из них начинается в
разном возрасте. Передняя часть туловища (от носа до передних ластов, промеры
2—3) увеличивает темп роста с 3 лет. С этого же возраста изменяется темп роста
передних конечностей в длину (15, 16) и задних конечностей в ширину (20). Длина
задней конечности до конца плавательной перепонки (18) начинает ускоренно
расти с 5 лет. Своеобразна закономерность акселерации объемных показателей
туловища. В области бедер у самцов она начинается с 2 лет (промер 10), брюха
в области пупка — с 3 лет (9), груди (8) — с 4 лет и шеи (7) — с 5 лет. Вместе
с тем, для каждой части туловища характерен свой темп и продолжительность
ускоренного роста. Половые различия в росте у самцов особенно ярко начинают
проявлять себя в период наступления половой зрелости (3—4 года). Представ-
ляется, что все отмеченные особенности роста тела и его частей связаны с под-
держанием определенных пропорций и формы тела, обеспечивающих найлучшие
гидродинамические показатели и терморегуляцию.
Относительный рост. В связи с тем, что кривая абсолютного роста самцов
сигмовидна, аллометрические зависимости роста для этой половой группы рас-
считывали в каждом отрезке кривой отдельно. Точкой разделения общей кривой
35
роста самцов принята длина тела 146 см [Кузин, Набережных, 19826]. Для самок
условно считали, что их рост монофазный.
Как видно из таблицы 3.6, большинство коэффициентов аллометрии (L) у
самок и самцов в первой фазе роста имеют сходные величины. Это говорит о том,
что рост большинства частей тела у них идет одинаково. Однако уже в молодом
возрасте у самцов намечается тенденция к увеличению толщины шеи (промер
7 на рис. 2.6), ширины переднего ласта (17), длины поясничного и крестцового
отделов (5, 6), по сравнению с самками. В свою очередь, у самок больше, чем
у самцов, коэффициенты аллометрии таких промеров тела, как обхват брюха
(9,10), длина задних ластов до когтей (19). У самцов во второй фазе наблюдается
ускоренный рост объемных показателей тела (шеи, груди, брюха), а также ли-
нейных параметров поясничного отдела (4, 7—9).
Можно выделить 3 группы промеров участков тела: а) рост которых ниже
роста общих размеров тела, б) рост которых коррелирован с общими раз-
мерами тела и в) увеличение которых происходит быстрее, чем общих раз-
меров тела. К первой группе промеров с показателями коэффициентов ал-
лометрии L=0.53—0.82 относятся промеры головы и тела, связанные с ростом
объемных параметров последнего (обхваты). Сюда же у самок вошел промер
“пупок—анус” и ширина переднего ласта. Во вторую группу (L=0.84—1.10)
вошли промеры, связанные с ростом линейных параметров передней части те-
ла — передних и задних конечностей у самок и самцов, а также задних отделов
туловища у самцов. В третью группу (L=1.12—1.33) вошел единственный промер
у самцов, характеризующий рост толщины шеи.
Во второй фазе во многих частях тела самцов заметно снижается темп роста,
соответственно падают показатели аллометрии. Абсолютно не связан с ростом
тела (L=0.25—0.53) рост головы (промеры 12—13). Замедляется рост промежутка
от кончика носа до передних конечностей, поясничной и тазовой частей тела,
длины переднего ласта по заднему краю и длины заднего ласта. Однако рост
таких частей тела, как объем шеи и груди, усиливается, и эти промеры переходят
в третью группу.
Таким образом, как и у других млекопитающих, рост северного морского
котика в антенатальный период характеризуется наивысшими показателями на
его ранней стадии и в последующие периоды неравномерно падает. В период
внутриутробного развития интенсивность весового роста во много раз выше,
чем линейного, однако периоды интенсивного роста как весовых, так и линей-
ных показателей испытывают чередующиеся друг с другом периодичности. От-
мечено замедление прироста массы и длины тела перед рождением, причем наи-
более выраженное в линейных показателях. Важно и то, что снижение темпа
прироста весовых показателей перед рождением происходит за счет мягких тка-
ней; скелет продолжает расти с той же интенсивностью (см. главу 5). Анало-
гичные явления роста эмбрионов отмечены у овец [Третьяков, 1959а] и у свиней
[Петрищев, Бородин, 1986] и, вероятно, характерны для всех млекопитающих.
Интенсивность роста у самцов на всех стадиях их развития выше, чем у
самок, и половой диморфизм в размерах тела начинает проявляться уже на ран-
них стадиях внутриутробного развития. Безусловно, интенсификация ростовых
36
процессов у самцов детерминирована генетически и физиологически подкреп-
ляется гормональной деятельностью самих плодов. Это хорошо прослежено по
особенностям роста разнополых особей в многоплодных пометах. Однако есть
данные [Петской 1953а, 6], которые показывают, что самцы в многоплодных
пометах копытных во внутриутробный период получают больше питательных
веществ от материнского организма через систему парабиотических связей, что
не связано ни с генетическими, ни с гормональными процессами. Вместе с тем,
развитие самок в антенатальный период идет быстрее, чем самцов, и они при
рождении имеют более приближенные к конечным размерам (взрослые особи)
показатели по длине и массе тела. Большинство органов и тканей к периоду
рождения у самок более дифференцированы, чем у самцов. Представляется, что
самки должны быть более подготовлены к постнатальному существованию; и
это подтверждается тем, что на первом году жизни их выживаемость выше, чем
самцов [Lander, 1975].
Процесс постнатального роста сложный и характеризуется ярко выражен-
ными этапами, при прохождении которых организм приобретает как видоспе-
цифические черты, так и внутривидовые различия по половым признакам. Это
достигается за счет изменения темпов роста всего организма или отдельных его
частей. Примечательно, что участвовать в размножении самки и самцы начи-
нают при одинаковых относительных размерах тела, хотя рост самок идет с опе-
режающим темпом. Ростовые процессы в значительной степени связаны с про-
цессами развития систем органов и тканей и определяют время наступления по-
ловой зрелости животных. Интересно и то, что акселерации подвержены не все
части тела, а в тех, где она проявляется, ее начало и окончание не согласовано
ни с ростом других частей тела, ни с ростом общих размеров тела.
37
Глава 4. РАЗВИТИЕ И ВОЗРАСТНО-ПОЛОВЫЕ ОСОБЕННОСТИ
КОЖНО-ВОЛОСЯНОГО ПОКРОВА
Шкуры северного морского котика как отличный меховой товар стали по-
пулярны вскоре после открытия островов, на которых эти животные обитают.
Возможно поэтому лидерство в изучении кожно-волосяного покрова котика при-
надлежало не биологам, а технологам [Stevenson, 1904; Bowker, 1931; Иванов,
Уразов, 1951; Зубин, Пчелина, 1951].
Исследование кожного покрова котика как морфологической структуры, не-
посредственно контактирующей с внешней средой и обеспечивающей стабиль-
ность внутренней среды организма,было изучено позже [Scheffer, 1962; Бычков,
1971а; Соколов, 1973]. Данные по строению кожно-волосяного покрова успешно
использовались при рассмотрении вопросов как эволюционных преобразований
самой структуры, так и становления северного морского котика как водного
млекопитающего [Белкин, 1963, 1964; Scheffer, 1964а; Ling, 1970; Соколов, 1973].
Подробное описание гистогенеза кожного покрова северного морского ко-
тика в эмбриональный период, начиная с восьмого месяца внутриутробного раз-
вития, приведено А. Н. Белкиным [1964]. В. Шеффер [Scheffer, 1962] детально
рассмотрел процесс появления волосяного покрова котика в онтогенезе. Нами
исследованы стадии развития кожного покрова 5—8-месячных эмбрионов север-
ного морского котика с длиной тела 20—40 см.
Развитие кожного покрова. В коже исследованных нами декабрьских (5-ме-
сячных) эмбрионов базальный слой эпидермиса представлен кубическими клет-
ками с крупными хорошо окрашенными ядрами, расположенными в один ряд.
Шиповатый слой состоит из четырех, а роговой слой из двух рядов клеток. Во-
лосяные фолликулы только начинают зарождаться. Они представлены скопле-
нием мезенхимных клеток в эпидермальном слое, в силу чего базальная мем-
брана в этих местах прогибается, а роговой слой эпидермиса выпячивается, на-
рушая гладкость поверхности кожи.
На шестом месяце внутриутробного развития наблюдается значительное уве-
личение слоев клеток в эпидермисе (табл. 4.1). Однако в ростковом слое коли-
чество рядов клеток остается прежним, а сами клетки становятся цилиндричес-
кими. Ядра также теряют округлые формы и уплощаются с боков. Клетки ши-
поватого слоя достигают больших размеров; ядра в них округлые и очень круп-
ные. Роговой слой состоит из нескольких рядов клеток. Волосяные фолликулы
продолжают внедряться в глубину дермы, причем уровень их погружения не-
одинаков (рис. 4.1). Тело эмбриона становится серого цвета со значительным
потемнением в области головы. В отдельных глубоколежащих фолликулах на-
чинается развитие волос, представленных небольшим остроконечным конусом,
занимающим '/3 или 2/3 волосяного фолликула по длине. В дерме начинают по-
являться хорошо развитые тонкие нитевидные коллагеновые волокна; слоис-
тость ее не выражена. Зачатки кожных желез и mm. arrectores pilorum не про-
сматриваются. У эмбрионов несколько больших размеров на голове уже имеются
38
Таблица 4.1
Возрастные изменения толщины кожи и ее слоев у эмбрионов северного
морского котика
Возраст, мес. Длина тела, см Толщина мк Источник
ростковый слой эпидер- мис дерма кожа коллагено- вые пучки
6 20 11.2 37.7 313.0 386.7 14.4 Ориг. данные
7 27 14.0 49.9 926.0 976.7 17.5 » »
8 41 38.5 68.0 1071.0 1139.0 51.0 » »
8 44 58.0 80.0 1558.0 1638.0 82.0 Белкин, 1964
9 50 34.0 75.0 2550.0 2625.0 - » »
11 65 26.0 51.0 2638.0 2689.0 - » »
Рис. 4.1. Три ранних волося->
ных фолликула, внедряющих-
ся в дерму кожи эмбриона
северного морского котика.
Масса эмбриона 161 г. [из:
Scheffer, 1962].
отдельные волосы длиной 0.8 мм с луковицами шириной 66.6 мк. Волосы сильно
пигментированы.
У эмбрионов на седьмом месяце при длине тела 20—30 см продолжается уве-
личение всех слоев кожи, и начинается закладка потовых желез несколько ниже
самых глубоких волосяных фолликулов. По данным А. Н. Белкина [1964], в этом
возрасте у эмбрионов северного морского котика хорошо выражены мышцы,
поднимающие волосы (mm. arrectores pilorum). Однако ни В. Шеффер [Scheffer,
1962], ни мы в исследованной серии гистологических срезов кожи эмбрионов
котиков mtn. arrectores pillorum не нашли. Дальнейшее увеличение всех слоев
кожного покрова происходит до восьмого месяца внутриутробного развития,
когда начинается прорезывание вторичного волосяного покрова через поверх-
ность эпидермиса.
39
С восьмого месяца начинается коренная перестройка всего кожного покрова,
суть которой заключается в том, что массовое прорезывание вторичных волос
вызывает отслоение верхних слоев эпидермиса, редукцию кожных желез и пер-
вичных волос, рост зачатков, предназначенных для вторичных волос [Белкин,
1963]. Клетки нижележащих слоев эпидермиса уплощаются, в результате чего
общая толщина эпидермиса к периоду рождения уменьшается в 1.5 раза, по срав-
нению с началом восьмого месяца внутриутробного развития. Дерма на про-
тяжении всего эмбрионального развития котика продолжает утолщаться (табл.
4.1), а вязь коллагеновых волокон в ней усложняться.
У 2-летних животных толщина эпидермиса меньше, чем у предродовых эмб-
рионов, однако на последующих стадиях развития его размеры увеличиваются
в основном за счет рогового слоя, и у секача он в 4 раза толще, чем у двухлеток
[Соколов, 1959].
Толщина дермы с возрастом тоже увеличивается. У секачей она толще в 6
раз, чем у предродовых эмбрионов и “черных” котиков, и в 4.7 раза, чем у трех-
леток. Толщина сосочкового слоя дермы с возрастом почти не изменяется, и
только у секачей она резко возрастает [Соколов, 1973]. У старых самок не на-
блюдается такого явления [Scheffer, 1962]. В свою очередь, толщина сетчатого
слоя увеличивается постепенно. Основные слагающие элементы сосочкового и
сетчатого слоев кожи — коллагеновые и эластиновые пучки — с возрастом также
становятся толще, причем в сетчатом слое их диаметр несколько больше, чем
в сосочковом. С возрастом количество эластиновых волокон уменьшается, но
толщина отдельных пучков увеличивается. Кожа секача отличается от кожи ко-
тиков младших возрастных групп слабо развитой сетью эластиновых волокон
[Соколов, 1959].
Пучки гладкомышечных волокон, встречающиеся в сосочковом слое, увели-
чиваются в числе в нижележащих слоях кожи. С возрастом они становятся
толще. Сеть пучков гладкомышечных волокон примерно одинакова по густоте
у молодых и средневозрастных животных, в то время как у секачей она редкая
[Соколов, 1973].
Подкожная жировая клетчатка имеет тенденцию изменяться в толщину не
только с возрастом, но и по сезонам. Размер жировых клеток у взрослых особей
может достигать 117—126 мк [Соколов, 1959], а прослоек коллагеновых пучков
до 45 мк. Подкожно-жировой слой толще у самцов, чем у самок, а зимой у всех
возрастно-половых групп животных толще, чем летом.
Несколько иначе развивается кожный покров на ластах котика. У эмбрионов
на шестом месяце внутриутробного развития (при длине тела 20—30 см) эпи-
дермис на ластах состоит из тех же слоев, что и эпидермис туловища (табл. 4.2).
Роговой слой тонкий — 13—14 мк. На восьмом месяце внутриутробного раз-
вития (длине тела 40—50 см) в строении эпидермиса конечностей появляются
отличия от эпидермиса туловища [Белкин, 1964]. В этом возрасте в эпидермисе
ластов можно видеть начальные этапы его дифференцировки — образование
сначала зернистого, а потом и блестящего слоев.
После достижения структуры трех слоев эпидермис ластов как бы приоста-
навливается в развитии, но на самых поздних стадиях внутриутробного развития
темп роста его резко возрастает. У предродовых эмбрионов эпидермис ластов
40
Таблица 4.2
Возрастные изменения эпидермиса ластов эмбрионов северного морского
котика
Длина тела, см Толщина, мк Высота сосочков Источник
эпидермис роговой слой блестящий слой зернистый слой ростковый слой
20 45.5 14.2 - - 31.3 - Ориг. данные
27 44.4 13.0 - - 31.4 - » »
50 68.0 21.0 - - 47.0 - Белкин, 1964
55 94.2 22.0 13.2 15.0 44.0 61.6 » »
64 260.0 127.6 22.0 44.0 66.0 147.5 » »
Рис. 4.2. Эмбриональная линь-
ка северного морского котика:
1 — остаток первичного волоса,
2 — волосяной конус детского
волоса, 3 — волосяная луковица,
4 — эпидермис [из: Белкин, 1964].
в 5 раз, а его ростковый слой только в 2 раза толще, чем на туловище. Уве-
личение толщины эпидермиса на поздних стадиях внутриутробного развития
происходит, в основном, за счет рогового, блестящего и зернистого слоев. Об-
разование эпидермальных сосочков на коже ласта котика начинается у эмбри-
онов на восьмом—девятом месяцах. Сначала появляется волнистость нижнего
слоя эпидермиса, а затем типичные сосочки. При длине тела эмбриона 55 см
они имеют высоту 61.6 мк. Эпидермис толще на нижней стороне ластов, чем
на верхней. Это достигается за счет сильного развития рогового и блестящего
слоев. Напротив, зернистый и ростковый слои развиты слабо. Сосочки также
более мощные на нижней стороне ластов. В коже нижней поверхности ластов
вязь коллагеновых волокон более густая, а их толщина достигает 35 мк, в то
время как в коже верхней поверхности их толщина 13 мк. Мышечная ткань раз-
вита одинаково хорошо на обеих сторонах ластов. Отдельные мышечные про-
слойки у эмбрионов длиной 50—60 см могут достигать 1 мм в толщину. Ха-
рактерной особенностью эпидермиса ластов является наличие в базальных клет-
ках росткового слоя пигментных клеток, которые появляются у 6—7-месячных
эмбрионов.’Гранулы меланина имеют разную форму и величину, находятся как
в цитоплазме клеток, так и между ними. В коже верхней части ластов пигмент
развит мощнее, чем в нижней.
41
Сеть кровеносных сосудов в коже ластов развита чрезвычайно сильно. Уже
у 5—6-месячных эмбрионов отдельные сосуды могут достигать 50—80 мк в тол-
щину; у эмбрионов с большей длиной тела — 210 мк. У предродовых эмбрионов
в дерме дорсальной поверхности ластов обнаруживаются потовые и сальные же-
лезы. Потовые железы кубического типа с хорошо развитыми дифференциро-
ванными секреторными отделами и извитыми протоками [Scheffer, 1962; Белкин,
1964]. •
Развитие волосяного покрова. В начале февраля (седьмой месяц внутриутроб-
ного развития) глубина залегания фолликулов достигает 0.5—0.7 мм, и начи-
нается массовое прорезывание волос через эпидермис. Волосы уже хорошо за-
метны на голове и горле. К концу месяца ими покрывается все тело [Scheffer,
1962].
На девятом месяце внутриутробного развития (март) начинается эмбрио-
нальная линька, подрастание новых (детских) волос и выпадение рунных (пер-
вичных) волос [Белкин, 1963, 1964]. У эмбрионов длиной 35 см и массой 1 кг
несколько снижается глубина залегания фолликулов (0.6—0.7 мм), однако их
длина продолжает увеличиваться (0.9—1.1 мм). Первые детские волосы появ-
ляются на голове, затем на хвосте и пупке и т. д. (рис. 4.2).
Первичные (рунные) волосы эмбрионов котиков имеют форму роговых плас-
тинок с заостренной вершиной. Средняя часть стержня волоса в 2—2.6 раза шире
ее нижней части и у корня [Белкин, 1964]. В одних волосах пигмента много,
в других мало. У большинства волос вершина, основание стержня и корни ли-
шены пигмента. Сердцевинный слой отсутствует в одних и присутствует в других
волосах; там, где есть, он имеет прерывистый характер. По наличию сердцевины
волосы можно разделить на 2 категории. Волосы первой категории (с сердце-
виной) более длинные и имеют более широкую среднюю часть, чем волосы вто-
рой категории (без сердцевины) [Белкин, 1964].
Кутикула первичных рунных волос некольцевидного типа, образована чешуй-
ками зубчатого типа на кончике и со сглаженными зубчиками в гранах. Стер-
жень имеет кутикулу зубчатого типа.
Порядок появления волос на теле эмбрионов котиков показан на рис. 4.3.
У 40-сантиметровых эмбрионов детские волосы на голове достигают длины 0.5—
0.7 см, вокруг глаз и на подбородке — 0.3—0.4 см. У более крупных эмбрионов
(до 50 см) волосы на голове вырастают до 1.3 см, на загривке — до 0.45 см.
У эмбрионов до 55 см наверху головы — 1.5 см, на загривке — 0.67 см; у пред-
родовых они достигают на голове 2.1 см, на груди — 1.8 см, в подмышечной
впадине — 0.5 см.
Меховой покров новорожденного котика состоит на 75% из остевых и на
25% из пуховых волос. Волосы размещены в пучках, в которых впереди всегда
остевой, а сзади 1—3 пуховых или промежуточных волос. Глубина их залегания
в коже около 1 мм. Каждый волос в пучке имеет собственное влагалище, в свою
очередь, весь пучок также имеет общее влагалище [Scheffer, 1962].
Крупные остевые волосы новорожденного котика могут достигать длины бо-
лее 2 см. Базальная половина таких волос почти прямая и лишена пигмента.
Терминальная зона расширена в грану, почти прямая и имеет черный цвет. Волос
42
только у корня округлый, остальная часть до граны уплощена. Поперечник стержня
23><82 мк, корня — 25x42 мк, граны — 18x157 мк. Пигмент в волосах распределен
неравномерно. <В корковом слое волоса он в виде гранул; в гране пигмента боль-
ше, чем в стержне, и расположен он здесь как в виде гранул, так и “пакетами”.
Кончик волоса лишен сердцевины. Она начинается в гране, прерывается у ее
основания, а затем вновь появляется. Корень также лишен сердцевины. Кутикула
кончика волоса некольцевидного типа, зубчатая, со сглаженными вершинами.
В гране сглаженность зубчиков усиливается, и они становятся неправильной
формы [Scheffer, 1962].
Промежуточные волосы новорожденного котика имеют длину 10—15 мм. Они
тоньше остевых, грана у них короче и уже. Окраска граны коричневая, стержень
серый. Сердцевина прерывистого типа. Основание волоса вздутое и по диаметру
равно гране. Кончик волоса лишен сердцевины. В самой широкой части граны
сердцевина прерывистая, узкая; корень лишен сердцевины. Кутикула сходна с
кутикулой остевого волоса.
Пуховые волосы плоские, извитые, тонкие, без граны; их длина 6—10 мм.
Терминальная часть этих волос золотисто-коричневого цвета, базальная — поч-
ти белая, бесцветная. Поперечное сечение стержня 12—14x20—22 мк. Базальная
часть вздутая; ее диаметр 24x40 мк. Диаметр корня — 8x10 мк [Scheffer, 1962].
Сердцевина имеется. Кутикула в виде лепестков со сглаженными краями.
Осенью, после вылинивания детских волос, котик приобретает волосяной
покров взрослого типа, который более однороден по составу. Кроющие волосы
у взрослых имеют менее острую и лишенную пигмента вершину. На расстоянии
трети от вершины волосы имеют изгиб в каудальном направлении. Терминаль-
ная часть граны коричнево-черная. В среднем длина остевого волоса равна 18—
20 мм (табл. 4.3, 4.4), вершины — 1.5 мм, граны — 8—10 мм, стержня — 3—4 мм,
корня — 0.8—1.0 мм. Диаметр граны — 35x150 мк, стержня — 32—118 мк, кор-
ня — 32—55 мк. Пигмент в гранулах и “пакетах” гранул в виде полосок и рав-
номерно распределен по всей медулле, кроме терминальной и базальной зон во-
лоса. Вершина волоса без сердцевины. В гране сердцевина хорошо выражена
и составляет в среднем 118 мк. В средней части волоса сердцевина становится
прерывистой. Ниже она снова появляется. Корень лишен сердцевины. Кутикула
некольцевидного типа.
Промежуточные волосы отличаются от остевых размерами и более бледной
окраской. Они слегка волнистые; их средняя длина 14 мм, Грана узкая; ее длина
ок. 3 мм, диаметр 14x90 мк. Диаметр стержня 14—80x30—35 мк. Его базальная
часть утолщена (25x70 мк). Пигмент находится в гранулах, расположенных пол-
осами. Кончик волоса лишен пигмента, которого нет и в базальной части стер-
жня. Размеры сердцевины волоса и ее наличие зависят от размеров волоса. В
самых длинных волосах она отсутствует в вершине, в гране — прерывистая и
исчезает в средней одной третьей стержня, но вполне различима у основания
волоса. Кутикула в вершине волоса в виде неправильной волнистой Мозаики,
а в стержне имеет ромбовидные чешуйки со сглаженными краями. В основании
волоса чешуйки кутикулы такие же, как и в вершине.
Пуховые волосы плоские, несколько шире в терминальной части, чем в ба-
зальной; почти бесцветные у основания и бледно-коричневые в терминальной
43
Таблица 4.3
Длина волос (мм) самок северного морского котика, добытых в июне—июле на о. Тюлений [из: Бычков, 1968]
Возраст, лет п Загривок Спина П о я с н и ц а Бок тела Брюхо
М±т lim М±т lim М±т lim М±т lim М±т lim
С 2 18.5+1.06 17—29 17.5+0.35 О с т е в 17—18 я е в о л о 18.5±0.35 С ы 18—19 16.5+1.06 15—18 14.0+0.70 13—15
1 3 18.0+0.15 18—20 17.7+0.28 17—18 18.0+0.15 17—19 17.7+0.55 17—19 13.7+0.28 13—14
2 2 17.5+0.35 17—18 16.5+0.35 16—17 16.0+0.70 15—17 16.5+0.35 16—17 10.5+0.35 10—14
3 9 18.9+0.48 17—22 16.5+0.06 17—17 16.4+0.33 15—18 16.1+0.53 14—19 11.6+0.30 10—13
4 18 19.5+0.39 16—23 17.3+0.36 15—21 16.2+0.31 13—19 15.5+0.30 13—18 10.5+0.33 8—13
5 20 20.7+0.29 17—23 - 17.7+0.28 15—20 15.8+0.23 13—18 16.2+0.30 14—20 10.2+0.27 8—13
6 7 22.7+0.72 20—26 19.3+0.33 18—21 17.9+0.47 16—20 17.7+0.44 16—20 13.9+0.37 12—15
£ 7 4 22.7+1.51 19—27 19.0+0.62 17—20 18.0+0.36 17—18 17.3+0.54 16—19 12.7+0.36 11—13
>7 5 23.5+1.18 21—28 20.5+0.59 18—22 15.3+0.76 12—17 13.3+0.61 11—13 11.0+0.28 10—12
С 2 13.5+1.06 12—15 13.0+0.00 П у х о в ь 13—13 г е воле 12.0±1.00 С ы 11—13 12.5+0.35 12—13 9.0+0.70 8—10
1 3 13.7+0.55 13—14 13.0+0.39 12—14 12.7+1.00 11—15 11.7+0.04 11—12 9.3+0.73 8—10
2 2 15.0+0.00 14—15 14.5+0.35 14—15 13.5+0.35 13—14 13.5+0.35 13—14 7.5+0.35 7—8
3 9 14.0+0.09 13—16 14.3+0.43 12—16 12.4+0.31 11—14 12.1+0.47 11—16 7.3+0.21 6—8
4 18 14.2+0.30 11—16 13.5+0.29 11—15 11.2+0.06 10—13 12.3+0.27 10—14 6.8+0.32 5—10
5 20 12.3±0.17 11—14 11.1+0.15 10—12 10.6+0.18 9—12 10.5+0.29 8—13 7.3+0.34 4—10
6 7 11.8+0.59 10—15 12.3+0.39 11—14 10.3+0.33 9—12 11.0+0.35 10—13 7.3+0.44 5—9
7 4 14.3+0.89 12—17 13.5+0.43 12—14 11.7+0.07 11—12 11.3+0.41 10—12 5.5+0.56 4—7
>7 5 11.210.33 10—12 10.8+0.52 10—13 8.8+0.69 6—11 10.7+0.59 9—13 5.7+0.53 5—6
Примечание. С — “серые” котики.
Длина волос (мм) самцов северного морского котика, добытых в июне—июле на о.
Таблица 4.4
Тюлений [из: Бычков, 1968]
Возраст, лет 3 а г р ИВОК С п < н а Пояс н и ц а Бок тела Б р в ЭХО
п М±т lim М±т lim М±т lim М±т lim М±т lim
О с т е в я е в о л о С ы
Н 2 17.3 12.5
С 6 20.2+0.44 19—22 18.6+0.57 16—20 18.0+0.41 16—19 18.2+0.49 16—20 15.2+0.49 13—17
1 5 17.7+0.43 16—19 18.2+0.52 16—19 15.7+0.10 15—17 15.6+0.07 15—16 13.0+0.94 11—16
2 25 19.3+0.28 16—22 16.7+0.26 15—19 14.9+0.26 12—18 15.9+0.19 15—18 10.3+0.19 8—13
3 67 19.7+0.20 16—25 16.9+0.18 13—20 14.9+0.13 12—18 15.8+0.12 14—18 10.2+0.14 7—13
4 39 22.8+0.28 18—30 18.1+0.23 15—21 15.4+0.18 13—19 16.3+0.21 13—20 10.5+0.22 7—13
5 18 26.2+1.04 21—36 19.2+0.29 17—21 15.8+0.22 15—17 17.0+0.24 15—18 10.9+0.29 9—14
6 13 30.7+1.12 24—41 19.5+0.46 17—22 16.2+0.36 14—19 17.2+0.44 15—20 11.0+0.30 9—13
7 8 38.6+1.75 32—44 24.6+0.43 20—29 17.2+0.70 15—20 18.8+0.64 17—21 13.2+0.54 11—16
>7 5 58.8+4.53 42—70 27.5+2.70 20—37 17.7+0.27 17—19 18.5+0.71 17—21 13.0+0.79 10—15
П у х о в ь 1е воле С ы
Н 2 8.5 6.0
С 6 14.6+0.37 13—16 14.0+0.41 12—15 13.3+0.37 12—15 13.4+0.50 11—15 10.4+0.26 9—11
1 5 14.0+0.45 12—15 13.5+0.59 11—15 12.0+0.79 10—14 13.0+0.63 11—15 8.5+0.05 8—9
2 25 14.9+0.19 13—17 13.6+0.16 12—15 11.6+0.29 8—14 12.8+0.21 10—14 7.1+0.28 5—9
3 67 14.7+0.13 12—17 13.4+0.16 10—16 11.6+0.16 9—16 12.4+0.14 10—15 7.0+0.19 4—10
4 39 15.4+0.19 13—19 13.9+0.20 10—16 11.5+0.19 9—15 123+0.28 10—15 6.7+0.23 3—9
5 18 15.5+0.36 13—20 14.2+0.22 12—15 11.0+0.33 9—13 12.7+0.28 11—15 7.2+0.31 5—10
6 13 14.8+0.30 13—18 14.2+0.38 11—16 12.8+0.39 11—15 13.6+0.33 12—16 7.2+0.36 5—9
7 6 ’ .11.8+0.72 10—15 13.6+0.51 12—16 12.6+0.62 10—15 13.2+0.55 11—15 .7.4+0.41 6—9
>7 5 18.5+1.53 13—22 15.0+1.22 10—18 11.2+0.65 9—13 12.7+0.65 11—15 7.5+0.59 5—9
Примечание. Н — новорожденные, С — “серые” котики. Данные по новорожденным из: Scheffer, 1962.
части. Располагаются пучками по 30—50 штук позади остевого волоса. Длина
вершины этих волос ок. 1 мм, ширина менее 1 мк. Терминальная треть в по-
перечнике 6—7 мк. Пигмент распределен в терминальной трети в виде темно-
коричневых гранул и полосок. Он отсутствует в остальных двух третях стержня.
Сердцевины нет. Кутикулярный слой на вершине волоса состоит из волнистых
чешуек, края которых ровные. Вдоль стержня чешуйки гребенчатого типа. Зубец
каждой- чешуйки заворачивается наружу.
В процессе роста волосяной покров котиков претерпевает изменения. У сам-
цов, начиная с 5-летнего возраста, остевые волосы сильно удлиняются на за-
гривке и спине, образуя “гриву”, хотя длина остевых волос не одинакова и на
других участках тела животного. Как правило, они короче на пояснице, боках
и брюхе. Наиболее однородным мех бывает у “серых” котиков (4—5 мес.).
Длина пуховых волос изменяется у самцов северного морского котика не-
значительно; она всегда больше на верхней стороне тела, чем на брюхе. У самок
ость и даже пух на верхней стороне тела короче, чем у самцов. На брюхе длина
волос у самцов и самок почти одинакова (табл. 4.3, 4.4).
4.1. Линька
В онтогенезе северного морского котика наблюдаются 2 типа линьки: внут-
риутробная и сезонные (постнатальные), первая из которых отличается от вто-
рой рядом последовательных микропроцессов [Белкин, 1964].
Внутриутробная линька. По данным А. Н. Белкина [1964], внутриутробная
линька начинается с того, что клетки волосяных сосочков и производящие клет-
ки рунных волос становятся не деятельными. Одновременно в области шейки
волосяных корней появляются разрывы, и нижняя часть с луковицей отделяется
от верхней части волоса. Остаток рунного волоса удерживается в верхних слоях
кожи продолжительное время, тогда как нижняя часть и волосяная луковица
подвергаются резорбции. Установлено также, что образование новых фоллику-
лов детских волос достигается несколькими способами. Редуцированный воло-
сяной мешок первичного (рунного) волоса сначала как бы приостанавливается
в развитии, но потом начинает быстро погружаться вглубь кожи. Со временем
на его месте возникает фолликул детского волоса. Образование нового воло-
сяного фолликула может также происходить за счет скопления новых жизне-
деятельных клеток в основании прежнего волосяного фолликула рунного волоса.
Характерной особенностью развития новых фолликулов является то, что они
могут отпочковываться от первичных фолликулов. Этот процесс начинается с
того, что в области впадения протоков потовых желез клетки волосяного вла-
галища начинают размножаться, в результате чего в этом месте фолликула сна-
чала образуется небольшое, а затем отчетливо заметное выпячивание стенки ре-
дуцирующегося волосяного мешка. Новообразованная почка обнаруживает
склонность к быстрому внедрению в дерму, в ходе которого она может допол-
нительно расщепляться на 2—3 эпителиальных тяжа. Последние, по всей веро-
ятности, являются зачатком вторичных фолликулов детских волос. Зачатки фол-
ликулов быстро растут, некоторые из них обгоняют в росте фолликулы, раз-
вивающиеся на основе первичного волоса.
46
Рис. 4.3. Последовательность (1—7) эмбриональной линьки морского котика
[из: Белкин, 1964].
Вновь образованные конусы вторичных (детских) волос как бы скользят по
стенке материнского фолликула и позднее выходят на поверхность кожи рядом
с ним (рис. 4.2). Другая часть фолликулов детских волос образуется за счет пог-
ружения в дерму клеток росткового слоя эпидермиса. Уже на самых начальных
стадиях внутриутробной линьки наблюдается тенденция к сгущению волосяного
покрова. Так, если число рунных волос на 1 см2 было 800, то детских волос
на 1 см2 — 3000 [Белкин, 1964].
На ранних стадиях развития детские волосы приобретают отчетливую кег-
леобразную форму. Однако у более старших эмбрионов кеглеобразность исче-
зает. При этом диаметр шейки у расширенной части корня волоса уменьшается
в 3 раза, а диаметр волосяной луковицы в 1.5 раза.
В процессе отмирания первичного волосяного покрова котиков первичные
сальные и потовые железы разрушаются, при этом дегенерирует большая часть
протока потовой железы. Железы, образующиеся на ранних стадиях развития
новых волос, долгое время остаются недоразвитыми. Развитие потовых желез
приостанавливается на стадии образования индифферентного протока, и лишь
значительно позже (после окончательного оформления корневой части детского
волоса) он погружается в дерму ниже уровня волосяных луковиц. У предродовых
эмбрионов потовые железы были обнаружены только на верхней стороне ластов,
в губах и на верхней части головы. Недоразвитые сальные железы присутствуют
только на голове.
Необходимо заметить, что выпадение рунных и появление детских волос на
теле котиков в эмбриональный период, по данным А. Н. Белкина [1964], под-
чинено определенной топографической последовательности, которая может
быть названа стадиями эмбриональной линьки (рис. 4.3).
Линька эмбрионов начинается по достижении ими длины тела 39—40 см и
массы 0.95—1.3 кг, что соответствует возрасту 8 мес. Скорость протекания линьки
и развития детского волоса несколько выше у эмбрионов-самок. После 20 апреля
(возраст 9 мес., длина тела 53—59 см) большая часть рунных волос вылинивает.
Большинство самцов заканчивает линьку в первых числах мая (возраст 10 мес.,
длина тела 53—61 см).
Вылинявшие рунные волосы котиков находятся в околоплодной жидкости.
При заглатывании эмбрионом жидкости часть волос попадает в желудочно-ки-
шечный тракт и скапливается в прямой кишке [Белкин, 1964].
47
Таблица 4.5
Начало линьки щенков северного морского котика на о. Тюлений в 1958—62 гг.
(в % на 1000 обследованных) [из: Бычков, 19646]
Месяц, декады Г оды lim М
1958 1959 I960 1961 1962
Июль 1 — _ ' 0.8 0.9 0.8—0.9 0:9
2 - 0.2 1.7 4.1 3.5 0.2—4.1 2.4
3 3.3 0.4 2.4 5.6 3.8 0.4—5.6 3.1
Август
1 10.0 8.1 7.9 8.8 6.9 6.9—10.0 8.3
2 39.1 28,3 32.7 45.3 41.0 28.3-45.3 37.3
3 52.1 64,2 59.5 66.7 62.4 52.1—66.7 61.0
Вполне вылинявшие эмбрионы, добытые в мае (возраст 10 мес.), имели блес-
тящий, черного цвета волосяной покров
У новорожденных котиков начинается значительная перестройка составных
частей глубинных слоев кожи, связанная с заменой детского мехового покрова
на взрослый. В каждом фолликуле волосяного пучка начинается усиленное де-
ление клеток, что приводит к зарождению новых отдельных пуховых фоллику-
лов. Они образуются позади или между пуховыми фолликулами детских волос.
По мнению А. Н. Белкина [1965], в процессе этой перестройки волосяных фол-
ликулов может происходить почкование фолликулов детских волос, т. е. наблю-
дается такой же процесс дифференцировки волосяных фолликулов, как это было
при замене рунного волоса на детский во время внутриутробной линьки. Другая
часть фолликулов может образовываться вновь путем впячивания эпидермаль-
ных зачатков вглубь кожи с последующей дифференцировкой и развитием всех
сопутствующих элементов волосо-сальных пучков взрослого котика.
Время начала интенсивной линьки новорожденных котиков зависит от кли-
матических условий года. Так, по данным В. А. Бычкова [19646], в 1961—63 гг.
на о. Тюлений, несмотря на суровую ледовую обстановку в начале лета, затруд-
нившую выход и начало щенения самок, количество солнечных дней в июле—
августе было больше, и линька новорожденных котиков, соответственно, про-
текала дружнее и раньше, чем в 1958—60 гг. (табл. 4.5).
Как видно из таблицы, начало линьки щенков котиков приходится на июль
и характеризуется слабым вылиниванием детских остевых волос и интенсивным
ростом новых пуховых, численность которых увеличивается до 20—30 в каждом
волосяном пучке. Новые остевые волосы в виде черных закладок заметны только
в глубине кориума. Таким образом, рост пуховых волос предшествует росту ос-
тевых волос.
В августе, наряду со щенками, находящимися на первой стадии линьки, встре-
чаются щенки с более глубокими изменениями в волосяном покрове. К концу
48
месяца более половины детенышей находятся на второй стадии линьки, которая
характеризуется пышным развитием светло-коричневого пуха; толщина волося-
ного покрова составляет И—15 мм. Пуховые волосы извитые; в расправленном
ваде имеют длину 15—17 мм. К этому времени большинство остевых волос уже
прорезываются через эпидермис, но кончики самых длинных из них еще не до-
стигают верхнего уровня пухового слоя. Вершины остевых волос депигментиро-
ваны. Из-за неравномерного подрастания остевых одни из них еще только про-
резываются через эпидермис, другие — уже полностью обнажили грану. В этот
период на одних участках еще сохраняется прежний меховой покров, а на других
он уже почти полностью отсутствует, поэтому щенки в таком меховом наряде пегие.
В сентябре у щенков происходит массовая линька. Большинство их находится
на второй стадии линьки, однако уже имеются и вылинявшие особи. У выли-
нявших щенков на третьей стадии линьки отдельные старые остевые волосы со-
храняются на спине и огузке. На боках и в подмышечной впадине их число зна-
чительное. Новые остевые волосы имеют длину 3—19 мм; многие из них уже
достигают верхнего уровня подпуши, и на поверхности видны их светлые вер-
шины [Scheffer, 1962].
В октябре большинство щенков полностью вылинивают. Подрастает ость; ее
длина составляет 6—20 мм, а длина пуха 16—18 мм. Соотношение остевых и
пуховых волос в пучке 1:24—41 [Бычков, 19646]. В конце октября новорожденные
котики начинают покидать места рождения и уходить в открытое море на зи-
мовку. Окончательное дозревание меха, видимо, происходит в ноябре, когда жи-
вотные уже ведут пелагический образ жизни.
Вместе с тем, необходимо заметить, что хотя и считается, что “серый” котик
приобрел взрослый меховой покров, он в некоторых деталях отличается от ме-
хового покрова, который приобретают животные в процессе последующих линек
(табл. 4.3, 4.4).
О дальнейшей судьбе волосяного покрова “серых” котиков можно судить по
интересному эксперименту, проведенному в зоопарке Сиэтла (США) [Scheffer,
1962]. 24 ноября 1952 г. у “серых” самки и самца были выстрижены отдельные
участки меха. В результате эксперимента выяснили, что новые пуховые волосы
начинают расти в начале июля и почти заканчивают рост к середине августа.
Новые остевые начали расти примерно в середине августа и полностью отросли
к концу сентября.
На о. Тюлений в октябре еще также не регистрировали годовалых котиков
с вполне сформировавшимся новым волосяным покровом [Бычков, 1964а]. До-
зревание меха у котиков этой возрастной группы, видимо, происходит в нояб-
ре—декабре.
Детальное исследование линьки котиков о-вов Прибылова позволило выя-
вить сложную картину этого процесса [Scheffer, Johnson, 1963]. За начало линьки
была принята активизация корней волос, образование первичных луковиц пу-
ховых волос и подрастание их волосяных конусов до поверхности эпидермиса.
В этот период замечается образование луковиц остевых волос. Начальный пе-
риод линьки определен в 2 недели. Продолжительность следующей стадии линь-
ки, названной “ранней”, составила 6 недель. В этот период прорезываются пу-
ховые волосы. Отдельные остевые волосы возвышаются над поверхностью кожи,
49
Таблица 4.6
Число пуховых волос в волосяном пучке у северного морского котика в
зависимости от возраста, пола и сезона [из: Scheffer, Johnson, 1963]
Воз- раст, лет Месяцы Средн.
' 1 2 3 4 5 в 7 . 8 9 10 11 12
н — Сам ц ы — 16 16 17 — 16.3
1 - 14 19 21 - 22 17 - 27 31 - - 22.2
2 25 27 26 23 23 26 25 28 30 28 33 - 26.7
3 33 39 29 32 26 32 33 29 31 36 43 - 33.0
4 31 - - 29 . - 33 40 32 31 39 42 - 34.2
5 - - - - 37 - 31 - 38 44 - 37.5
>5 - - - 46 42 41 48 47 62 38 59 - 48.6
Средн. 29.7 26.7 24.7 30.2 32.0 32.5 32.3 34.0 33.7 33.1 38.8 32.4
п Н 3 3 3 5 4 6 Сам 6 к и 4 11 17 7 19 5 24 20.0
I 16 - 20 19 18 - 22 - - 21 - - 19.3
2 28 - 27 25 - - - 28 36 31 31 - 29.4
3 33 27 32 30 34 28 32 33 30 34 46 - 32.6
4 32 27 37 27- 37 35 34 34 33 37 - 37 33.6
5 ; 34 33 38 35 - 40 29 38 37 51 36 - 37.1
>5 35 41 43 43 37 40 35 36 43 45 36 42 39.8
Средн. 29.7 32.0 34.3 29.8 31.5 35.7 30.4 34.1 32.7 34.0 34.6 40.3 33.0
п 6 4 7 6 : 4 4 5 6 6 7 5 3
Примечание. Н — новорожденные.
другие еще только достигают ее. Большинство остевых волос прежней генерации
еще удерживаются в коже. Хорошо заметно потемнение мездры в местах заклад-
ки новых волос. Следующая стадия (“поздняя линька”) длится 4 недели и ха-
рактеризуется созреванием нового волосяного покрова. Кончики новой ости до-
стигали или уже выходили за уровень подпуши. Старые остевые волосы интенсивно
выпадали. Потемнение мездры незначительное. Окончательная стадия линьки длит-
ся 3 недели. Корни волос регрессируют. В основном все остевые волосы выросли.
Невооруженным глазом линька не заметна. Однако корни волос имеют незначи-
тельное количество пигмента, что говорит о продолжающемся процессе подрас-
тания волос. Хотя общая продолжительность линьки у северного морского котика
составляет не менее 8 мес., линные особи встречаются как в июле, так и в конце
января. Продолжительность линьки у отдельной особи не превышает 22 недель.
В среднем расчетное время продолжительности линьки 15 недель. Линька пуховых:
волос всегда предшествует линьке остевых примерно на 1 неделю.
50
Исследование шкур разновозрастных котиков обоего пола, добытых в разное
время года, показало, что середина линьки у них наступает не одновременно.
С увеличением возраста животных различия в наступлении середины линьки воз-
растает. Расчеты показали, что середина периода линьки у самцов наступает на
3.4, а у самок — на 5.5 дня позже с каждым годом старения животного. В то же
время разница между серединой линьки самок и самцов увеличивается примерно
на 2 дня с каждым возрастным классом животных. Кроме того, у животных стар-
ших возрастных групп линька начинается несколько позже, и она более про-
должительна, чем у молодых. Так, у исследованных годовиков этот процесс
длится 106 дней, а у животных старше 10 лет — 177 [Scheffer, Johnson, 1963].
Изучение кожно-волосяного покрова морских котиков позволило установить
феномен невыпадения волос старой генерации и накопление их в коже животных
с возрастом. Первоначально на этот факт обратили внимание при выстригании
и окраске волос несмывающимися красками в процессе изучения экологии [Schef-
fer, 1962; Бычков, 19646]. Микроскопическое изучение кожно-волосяного покро-
ва подтвердило наблюдаемые ранее факты невыпадения и накопления волос в
коже котиков. Установлено, что в среднем ок. 25% остевых волос у взрос-
лых котиков в январе являются волосами старой генерации [Scheffer, John-
son, 1963].
Количество волосяных пучков с двумя остевыми волосами больше у старых
котиков, чем у молодых. Так, если у двухлеток 19.6% волосяных луковиц имели
по 2 остевых волоса, то у четырехлеток их было 45%, а у более старых животных
40—41%. Однако не выясненной остается причина отмеченной зависимости со-
хранения волос старой генерации от возраста животных. Предполагается, что
либо у взрослых животных процесс выпадения старых волос более продолжи-
телен, либо количество волос новой генерации у них больше, чем у молодых
[Scheffer, Johnson, 1963].
Примерно такая же картина наблюдается и с пуховыми волосами (табл. 4.6).
Как видно из таблицы, количество пуховых волос в пучке у молодых животных
меньше, чем у старых; у годовалых их 14—24, у взрослых самцов и самок мак-
симально может достигать 51 и 61, соответственно.
4.2. Вибриссы
У северного морского котика на морде имеются 2 категории вибрисс — глаз-
ные и губные. Глазных вибрисс 2, из них верхняя всегда длиннее нижней.
Вибриссы у котика появляются на 4.5—5 месяце эмбрионального развития,
(декабрь) при массе тела 20—30 г. У эмбрионов с массой тела в 3—5 г (при-
близительно четвертый месяц внутриутробного развития) на верхней губе уже
бывают заметны маленькие белые точки — места будущих вибрисс. Их ровно
столько, сколько потом вырастет настоящих вибрисс. Никаких рудиментарных,
дополнительных вибрисс нет. Первыми появляются вибриссы нижнего ряда, а
в нижнем ряду — задние. Каждая пробивающаяся через эпидермис вибрисса одета
тонкой просвечивающейся оболочкой — эпидермальным выпячиванием кожно-
го покрова. Прорезывающиеся вибриссы имеют эллиптическое основание, ко-
нусовидную форму, темно-коричневый или черный цвет, верхушка белая.
51
Таблица 4.7
Число вибрисс у северного морского котика (с одной стороны морды)
п Номер ряда вибрисс от линии разреза рта Всего
1 2 3 4 5 6
62 ‘ 3 5—6 4—6 4—6 2-4 1—0 19—25
Таблица 4.8
Изменение длины (см) наиболее длинных вибрисс у северного морского
котика в онтогенезе*
Возраст Самцы Самки
М Нт п М Пт
э м б р и о н ы
5 мес. 1 0.18 1 0.13
6 мес. 4 0.24 0.18—0.25 6 0.34 0.20—0.50
7 мес. 8 1.35 0.70—1.80 7 1.28 0.80—1.70
8 мес. 4 г. 2.22 2.00—2.60 4 1.72 1.40—2.00
9 мес. 2 4.00 3.50—4.50 2 4.15 4.00—4.30
11 мес. 5 6.60 4.90—7.00 3 6.33 5.00-8.00
1 • з р о с л ы е
1 год 1 10.4 - - -
2 года 5 11.8 9.5—13.4 8 9.8 7.7—12.0
3 года 20 . . 12.4 11.1—14.4 9 9.4 8.0—11.5
4 года . 24 13.3 9.6—15.5 10 П-1 9.5—14.2
5 лет 28 14.9 11.0—22.2 10 11.3 7.9—14.1
6 лет 22 16.3 11.5—21.2 4 11.7 10.6—12.7
7 лет 6 18.5 14.4—20.4 3 15.0 12.4—16.4
8 лет 4 19.9 13.2—25.5 2 14.3 11.8—16.8
9 лег 1 30.6 1 12.0
10 лег : 1 28.6 1 12.0
>10 лег 4 25.9 21.7—33.4 5 14.5 10.4—22.0
* По эмбрионам — наши данные, по взрослым — из: Scheffer, 1962.
52
Губные вибриссы расположены в 5 или 6 рядов. Их общее число 19—25.
Количество вибрисс по рядам не одинаково (табл. 4.7). Самое постоянное их
число в первом ряду, наибольшее разнообразие отмечено во втором—пятом ря-
дах, причем у самок вариабельность выражена сильнее, чем у самцов. В шестом
ряду вибриссы чаще отсутствуют или имеется только одна. В среднем у самок
и самцов насчитывается по 21 губной вибриссе с каждой стороны морды, в т. ч.
по рядам — 3-5-5-3-1(0).
Длина вибрисс изменяется с возрастом. У старых самцов они могут достигать
30—35 см, у самок — 20—22 см (табл. 4.8). Часто наблюдается стирание или
обламывание вибрисс. Количество и топография вибрисс с возрастом остается
неизменным. Диаметр у основания вибрисс в среднем составляет у новорожден-
ных котиков 0.45x0.75 мм, у взрослых самок — 1.05x1.28 мм, у взрослых сам-
цов — 1.15x1.35 мм.
Цвет вибрисс изменяется с возрастом животных. В основном наблюдается
их посветление. Совершенно белыми или желтовато-белыми они становятся
только у очень старых животных [Abegglen et al., 1958]. У самцов посветление
вибрисс наступает несколько позже, чем у самок, что происходит, видимо, из-за
их более позднего развития. Обесцвечивание происходит в результате утраты
пигментными клетками фолликулов вибрисс своей функции, а также в результате
маскировки пигмента пузырьками газа, находящимися в корковом слое [Scheffer,
1962].
Цвет вибрисс издавна используется промысловиками для приблизительного
определения возраста животных при производстве промысловых операций. Име-
ются данные, показывающие возможность использования количественных ха-
рактеристик вибрисс в качестве популяционного признака [Косыгин, 1968].
53
Глава 5. РАЗВИТИЕ СКЕЛЕТА В ОНТОГЕНЕЗЕ И ЕГО
СТРОЕНИЕ
5.1. Череп
Череп морского котика как по внешнему виду, так и по числу костей сходен
с черепом наземных млекопитающих (рис. 5.1)
Есть указания на отсутствие слезных костей в черепе взрослых особей морского
котика [Чапский, 1963]. Они присутствуют у эмбрионов [Кузин, 1971а; King, 1971],
но в постнатальном онтогенезе бесследно срастаются с прилежащими костями.
Кондилобазальная длина черепа у взрослых самцов морских котиков в сред-
нем составляет 250 мм, а у самок 194 мм [Бычков, 1965]. Мозговой отдел у взрос-
лых самцов превышает в среднем на 23%, а у самок на 34% длину лицевого
отдела черепа. Межглазничное пространство узкое. Надглазничные отростки
развиты хорошо. Носовая часть лицевого черепа короткая и круто обрывается
при переходе к межчелюстным костям. Процион выступает за пределы резцов.
Межчелюстные кости узкие, но в местах их соприкосновения с носовыми кос-
тями сильно расширены. Носовые кости на вершине имеют вырезку по сагит-
тальной линии, куда вклиниваются лобные кости. Скуловые дуги умеренной ши-
рины, и расстояние между ними занимает лишь немного больше половины
длины черепа. Орбиты крупные. Основание лицевого черепа имеет легкую изог-
нутость, но небные кости плоские. Латеральный край небных костей длиннее
медиальных. На основной затылочной кости ярко выражены пальцевые вдав-
ления. Алисфеноидный канал имеется. Сросшиеся яремный и мастоидный от-
ростки хорошо выражены. Костные слуховые пузыри малы и уплощены. Внут-
ренний угол, образованный линией “процион—носовые кости” и большой осью
черепа, составляет более 60° [Scheffer, 1958], а лицевой угол — менее 125° [Re-
penning et al., 1971]. Сагиттальный гребень не развит, а затылочные гребни раз-
виты хорошо.
В процессе онтогенетического развития конфигурация и пропорции черепа
сильно меняются [Кузин, 1968]. У 6-месячных эмбрионов мозговой отдел округ-
лый и круто поднимается вверх от лицевого. Лобные кости выше надглазничных,
отростков выпуклые (рис. 5.2). Между лобными и теменными, также как и между
верхнезатылочной и теменной костями, имеются обширные роднички (фонти-
кулы или фонтанели). У более крупных эмбрионов того же месяца развития фон-
танели имеют форму наконечника стрелы, обращенного в сторону затылочной
кости (рис. 5.2). Вершины носовых костей овальные, а конфигурация их лате-
ральных отделов характеризуется значительной индивидуальной изменчивостью.
Сагиттальный шов погружен в мозговую полость, и на его месте образуется по-
логий желоб.
Череп 7-месячных эмбрионов внешне похож на череп 6-месячных, но лобные
кости менее выпуклые. У черепов с одинаковой основной длиной роднички мо-
гут быть разной величины. Боковые затылочные кости неплотно прилегают к
окружающим их костям. Сосцевидная часть каменистой кости не развита. Ос-
54
Рис. 5.1 Череп северного морского котика. Самец, 11 лет.
нОвная затылочная кость и базисфеноид имеют округлые контуры, а аборальный
конец основной затылочной кости островершинный. Барабанные кости полук-
руглые, как у низших позвоночных [Шмальгаузен, 1922], и они еще не срослись
с каменистыми костями. Резцовая часть межчелюстной кости с внешней стороны
округлая, процион не выступает вперед.
У 8-месячных эмбрионов лицевой отдел черепа плавно переходит в мозговой.
Лобные й теменные кости сомкнуты, венечный шов при этом имеет разнооб-
разную конфигурацию (рис. 5.3). Процион выступает вперед. Сосцевидная часть
каменистой кости почти полностью развилась и подходит вплотную к теменной
и боковой затылочным костям. На аборальном конце основной затылочной
кости намечается вырезка.
У 9-месячных эмбрионов в результате интенсивного роста мозгового отдела
черепа в ширину уменьшаются его относительные размеры в высоту, и он как
Рис. 5.2. Череп 6-месячного эмбриона морского котика с дорсальной (слева) и
вентральной (справа) сторон
Рис. 5.3. Конфигурация венечного шва у 8-месячного эмбриона северного морского котика
бы уплощается. Особенно интенсивно увеличивается в размерах затылочная об-
ласть. Основная затылочная кость за линией, соединяющей каудальные края ба-
рабанных костей, перегибается, и на ее аборальном конце образуется выемка.
Барабанные кости сливаются с каменистыми, но швы еще заметны. В результате
прогрессирующего роста верхней затылочной кости, ее чешуя становится частью
крыши черепа.
56
Таблица 5.1
Изменение пропорций черепа морского котика в онтогенезе (пренатальный
период — ориг. данные, постнаталъный период — рассчитано по данным из:
Мужчинкин, 1968)
В о з р а С Т
Показат Пренатальный период, мес. Постнатальный пе. ?иод, годы
ели 11 щенки 3 8 9
6 7 8 9 10 самцы самки самцы самки
3 проце игах от основно я длинь
1 60.3 60.2 59.0 55.9 57.6 56.7 36.6 39.1 37.5 45.7 40.3
2 100.0 90.6 94.8 87.3 83.1 83.2 82.1 64.5 65.6 57.5 60.8
3 42.6 40.6 38.2 36.4 40.9 40.1 41.1 38.5 36.9 37.1 38.1
4 65.7 62.2 63.4 61.5 63.1 62.8 58.9 53.8 53.5 57.5 52.8
5 53.5 46.3 45.5 38.2 35.9 32.8 29.5 20.1 21.0 14.9 17.0
В процент ах от М2 1СТОИДНО Й ШИрИ] 4Ы
6 188.8 199.8 194.4 191.6 187.2 188.0
7 166.1 170.7 177.4 175.6 178.1 195.5 205.5 204.8 192.9 207.5
8 125.8 131.7 130.5 133.5 126.3 125.2 128.8 105.5 104.4 89.0 113.3
9 119.3 129.8 126.0 116.0 109.4 106.2 119.7 116.5 109.9 112.6 122.6
Примечание. 1 — длина лицевого отдела, 2 — длина мозгового отдела, 3 — длина
верхнего зубного ряда, 4 — мастоидная ширина, 5 — межглазничная ширина за
надглазничными отростками, 6 — наибольшая ширина, 7 — кондилобазальная
длина, 8 — наибольшая ширина, 9 — скуловая ширина.
На 10-ом месяце у эмбрионов крыша черепной коробки еще больше упло-
щается. Задняя часть мозгового отдела становится значительно шире передней.
Процион уже сильно выступает вперед за пределы резцовой кости. На основании
затылочной кости образуются пальцевые вдавления.
У предродовых эмбрионов окончательно формируется костный череп. На-
чинается формирование отростков костных элементов. Значительно увеличива-
ется в размерах лицевой отдел.
Изменение основных пропорций черепа в процессе онтогенеза представлены
в таблице 5.1.
Как видно из таблицы, индекс лицевого отдела черепа к концу внутриут-
робного периода развития котика уменьшается. У взрослых животных его по-
казатели несколько больше, чем у щенков. Такая картина изменения индекса
длины лицевого отдела черепа объясняется усиленным ростом мозгового отдела
черепа во внутриутробный период и в первые годы постнатального развития.
На изменение этого показателя оказывает влияние также ускоренный рост про-
циона. Вместе с тем индекс мозгового отдела заметно уменьшается с возрастом
животного, хотя казалось бы, что он должен изменяться обратно пропорцио-
57
нально длине лицевого отдела черепа. Диспропорция в изменении показателей,
видимо, возникает за счет различной интенсивности роста некоторых костей,
составляющих оба отдела. Как уже отмечалось, на 9-ом месяце внутриутробного
развития начинается усиленный рост основания черепа. Затылочное отверстие
перемещается кверху по отношению к основной плоскости черепа, а верхняя за-
тылочная кость уже не нависает над затылочным отверстием. В свою очередь,
основная затылочная кость на уровне слуховых пузырей перегибается. В резуль-
тате различного темпа роста костей граница между лицевым и мозговым от-
делами топографически изменяется. Она отодвигается назад. У взрослых самцов
череп относительно длиннее в лицевой части, а у самок — в мозговой.
Увеличение длины нижнего зубного ряда происходит почти одинаково с рос-
том основания черепа. Мастоидная щирина черепа, выраженная в процентах от
основной длины, имеет обратно пропорциональную зависимость с продольными
измерениями черепа, например, кондилобазальной длиной, но изменения ее по-
казателей происходят пропорционально общим размерам черепа. Половые раз-
личия в относительных показателях мастоидной ширины черепа достигаются за
счет более сильного развития мастоидных отростков у самцов. Межглазничная
ширина черепа в абсолютном выражении почти не отличается у взрослых особей
от ее размеров у новорожденных котиков, поэтому индекс этого измерения у
взрослых животных много меньше, чем у новорожденных котиков. У взрослых
самок вследствие того, что лицевой отдел черепа относительно короче, чем у
самцов, межглазничная ширина черепа больше. В процессе онтогенетического
развития северного морского котика рост его черепа в длину происходит зна-
чительно интенсивней, чем в ширину (табл. 5.1). Относительная масса черепа
у котика с возрастом уменьшается. На 7-ом месяце внутриутробного развития
масса черепа составляет 50% массы всего скелета, а у взрослых особей всего
лишь 10%. При этом масса нижнечелюстных костей изменяется пропорциональ-
но массе всего скелета, а масса осевого черепа уменьшается.
Окостенение черепа у котика начинается на ранних стадиях внутриутробного
развития [Кузин, 19706]. У 5-месячных эмбрионов (декабрь) костные центры име-
ются почти во всех элементах лицевого отдела черепа (рис. 5.4). Больше других
по размерам они в верхне- и нижнечелюстных элементах. В лобных элементах
костная ткань формируется по их орбитальному краю и радиально распрост-
раняется на фронтальную часть черепа. В теменных элементах очаги окостенения
присутствуют в местах стыка со скуловыми отростками височных и лобных кос-
тей. Костный очаг верхнезатылочного элемента имеет форму узкой полоски, ле-
жащей у будущего основания кости. Небольшие центры окостенения присутст-
вуют в основной и боковых затылочных элементах. К концу 5-го месяца внут-
риутробного развития костные центры имелись во всех скелетных элементах че-
репа. Крыша черепа полностью формируется, но еще присутствуют небольшие
теменные роднички. Основная затылочная кость почти вплотную подходит к
затылочному отверстию. Между всеми костными элементами затылочной облас-
ти черепа еще имеются значительные неокостеневшие пространства. Это же ха-
рактерно и для основания черепа.
На шестом месяце внутриутробного развития продолжается формирование
костного черепа. У одних эмбрионов к концу месяца полностью формируется
крыша костного черепа, у других остаются роднички.
58
Рис. 5.4. Окостенение черепа эмбриона северного морского котика.
Слева — 5 месяцев, в центре — середина 6-го месяца, справа — 6 месяцев;
А — сверху, Б — снизу, В — сзади;
1 — верхняя затылочная кость, 2 — боковая затылочная кость, 3 — теменная кость, 4 —
основная затылочная кость, 5 — барабанная кость, 6 — скуловой отросток височной кости,
7 — базисфеноид, 8 — крыловидная кость, 9— скуловая кость, 10 — лобная кость, 11 —
небиая кость, 12 — верхнечелюстная кость, 13 — межчелюстная кость.
В возрасте 7 месяцев большинство костей плотно прилегают друг к другу.
Процесс окостенения несколько затягивается в скелетных элементах основания
черепа, и контуры костных очагов здесь еще имеют округлые очертания. Ба-
рабанные кости еще не срастаются с каменистыми (рис. 5.4).
59
Рис. 5.5. Дополнительные
кости (extraoccipitale) в черепе
эмбриона северного морского
котика (указано стрелкой):
Ехос — exoccipitale, Р — parietale,
Soc — supraoccipitale.
На последующих стадиях внутриутробного развития продолжается смыкание
костных элементов и уплотнение швов. К моменту рождения все кости сфор-
мированы и плотно прилежат друг к другу.
В период заполнения фонтанелей у эмбрионов котиков в затылочной области
черепа между теменными, верхней и боковой затылочными костями заклады-
ваются дополнительные или вставочные кости — extraoccipitale (рис. 5.5). Они
отмечены у 40% эмбрионов [Кузин, 1971а]. Найдены они и у других ушастых
тюленей [Dout, 1942]. Аналогичные описания были представлены Ю. Кинг [King,
1956, 1964] и А. А. Кузьминым и А. С. Перловым [1978].
Измерения черепов котиков из командорской [Мужчинкин, 1968] и тюлень-
евской [Бычков, 1965] популяций, проведенные в разное время, показали, что
промеры черепов из этих популяций различаются. Однако, сделанный В. А. Быч-
ковым анализ показал, что скорее это не популяционные различия, а хроногра-
фическая изменчивость. В. Шеффер [Scheffer, 1955] сравнил черепа прибыловских
котиков, добытых в 1945—46 и в 1916—20 гг. При этом выяснилось, что раз-
личия в краниометрических показателях были настолько велики, что они пре-
вышали различия между популяциями.
5.2. Позвоночный столб
Дж. Аллен [Allen, 1880] первым привел формулу позвоночного столба север-
ного морского котика, измерил длину отделов четырех животных (2 самки и
2 самца). Позднее уточнялось количество позвонков по отделам [Scheffer, 1958;
King, 1964; Fay, 1967; Антонюк, 1972] и исследовались изменения весовых и ли-
нейных параметров отделов позвоночного столба в онтогенезе [Кузин, 1971а,
1972а; Кузин, Маминов, 1978].
Шейный отдел. У северного морского котика 7 шейных позвонков.
Атлант (рис. 5.6) образован дорсальной и вентральной дугами. Дорсальная
дуга тоньше вентральной и имеет хорошо выраженный бугорок, соответству-
ющий остистому отростку других позвонков. Вентральный бугорок не выражен.
Передний край вентральной дуги толще заднего. Крылья атланта у самцов удли-
ненные, у самок — округлые и направлены вентролатерально. Основания крыль-
ев прободают межпоперечные отверстия, которые могут быть незамкнутыми и
выглядеть как краниодорсальные вырезки. Краниальные и каудальные сустав-
60
Рис. 5.6. Первый шейный позвонок (атлант):
А — сверху, Б — снизу, В — сзади;
1 — дорсальный бугорок, 2 — дорсальная арка, 3 — вентральная арка, 4 — каудальная
суставная площадка, 5 — крыло, 6 — межпоперечное отверстие, 8 — краниальная суставная
поверхность, 9 — крыловая ямка.
Рис. 5.7. Второй шейный позвонок (эпистрофей):
А — сбоку, Б — спереди;
1 — гребень, 2 — тело, 3 — зуб, 4 — краниальная, 5 — каудальная суставные площадки,
6 — межпоперечное отверстие, 7 — межпозвоночное отверстие, 8 — каудальный суставной
отросток, 9 :— поперечнореберный отросток, 10 — каудальная суставная поверхность.
61
3
Рис. 5.8. Третий шейный позвонок:
А — спереди, Б — сбоку, В — сзади;
1 — тело, 2 — дуга, 3 — остистый отросток, 4 — поперечнореберный отросток, 5 —
головка, 6 — ямка, 7 — краниальная, 8 — каудальная суставные площадки, 9 —
краниальный суставной отросток, 10 — межпоперечное отверстие.
ные площадки, соединяющие атлант с черепом и вторым шейным позвонком,
вынесены за пределы дуг, вследствие чего на дугах образуются выемки, за счет
которых достигается свобода движения головы в горизонтальной плоскости.
Эпистрофей короткий, но высокий (рис. 5.7). Зубовидный отросток цилин-
дрический и у взрослых особей не выступает за пределы вентральной дуги, в
то время как у новорожденных он выходит за интерцентр. Каудально по бокам
отростка расположены 2 суставные треугольные площадки с закругленными края-
ми, направленные краниолатерально. Поперечные отростки тонкие, направлены
вершиной назад; их основания пронизывают межпоперечные отверстия. Остистый
отросток мощный в виде вытянутого вдоль позвоночной дуги гребня. Его передний
край сильно нависает над зубовидным отростком, перекрывая краниальное меж-
дуговое пространство. Задний край остистого отростка расширен в виде треуголь-
ника. Каудальные суставные фасетки направлены вентролатерально. На вентраль-
ной поверхности тела позвонка имеется гребень.
Третий—шестой позвонки имеют сходное строение (рис. 5.8). Их остистые
отростки направлены дорсокаудально. Поперечнореберные отростки неодина-
ковой формы. Если у третьего позвонка хорошо выражен дорсальный бугорок,
62
4
Рис. 5.9. Седьмой шейный позвонок:
А — сзади, Б — сбоку, В — спереди;
1 — тело, 2 — головка, 3 — ямка, 4 — остистый отросток, 5 — краниальный, 6 —
каудальный суставные отростки, 7 — реберный отросток, 8 — поперечный отросток.
то у последующих позвонков постепенно обособляются дорсальный, краниаль-
ный и каудальный бугорки как составные части поперечнореберных отростков.
Краниальные суставные отростки расставлены несколько шире, чем каудальные,
и их суставные площадки обращены вентролатерально. Основания поперечно-
реберных отростков всех четырех позвонков пронизывают межпоперечные от-
верстия, образующие поперечный канал, идущий вдоль шейного отдела позво-
ночного столба.
У седьмого шейного позвонка (рис. 5.9) остистый отросток направлен до-
рсально. Тело позвонка короче, чем у четырех предыдущих. На поперечноре-
берных отростках не выражен каудальный бугорок. Межпоперечные отверстия
отсутствуют. Нет также площадок для крепления головок первых ребер, что ха-
рактерно для седьмых позвонков многих наземных млекопитающих.
В онтогенезе наибольшему изменению подвержен атлант. У эмбрионов и но-
ворожденных котиков его крылья лежат почти в вертикальной плоскости. На
более поздних стадиях развития в результате постепенного изменения позы го-
ловы и преобладающего роста позвоночника в переднем направлении они не-
сколько разворачиваются и приобретают горизонтальное положение. Другие
63
НОМЕРА ПОЗВОНКОВ
Рис. 5.10. Длина тел позвонков.
На верхнем графике длина зубовидного отростка.
А — эмбрион (8 мес.), Б — щенок, В — самка (2 года), Г — самец (4 года), Д — самка
(15 лег), Е — самец (7 лег), Ж — самец (11 лег).
позвонки шейного отдела в раннем онтогенезе представляют собой уменьшенные
копии их дефинитивных форм с менее выраженными отростками.
Как видно на рис. 5.10, самым длинным позвонком шейного отдела является
второй позвонок, но значительная часть его длины приходится на зубовидный
отросток, филогенетически принадлежащий первому позвонку [Шмальгаузен,
1947].
Длина последующих позвонков увеличивается к 6-му. Длина 7-го позвонка
меньше, чем у предыдущего. Эта закономерность изменения длины шейных поз-
вонков у северного морского котика наблюдается на протяжении всего онто-
генеза (рис. 5.10).
Высота тел позвонков увеличивается от 2-го к 7-му (рис. 5.11). Мас-са поз-
вонков изменяется иначе (рис. 5.12). У эмбрионов самый легкий четвертый поз-
вонок, у молодых животных — третий, у старых — второй, т. е. темп весового
роста второго позвонка в онтогенезе значительно ниже, чем других шейных поз-
вонков. У взрослых животных наиболее массивным становится шестой позвонок,
тогда как у эмбрионов — первый. Высота остистых отростков позвонков на всех
стадиях онтогенеза животных увеличивается от 1-го к 7-му (рис. 5.13), а ширина
в поперечных отростках увеличивается от 1-го к 3—4-му позвонкам (рис. 5.14).
Поперечные отростки 5—6-го позвонков по размерам не меньше предыдущих,
но поскольку вершины их направлены ближе к сагиттальной плоскости, то ши-
64
Рис. 5.11. Высота тел позвонков.
Обозначения — см. рис. 5.10.
Рис. 5.12. Масса сырых позвонков. Крестцовые позвонки взвешены вместе.
Обозначения — см. рис. 5.10.
рина этих позвонков в поперечнореберных отростках меньше, чем у 3—4-го. По-
перечнореберные отростки 7-го шейного позвонка у молодых особей расстав-
лены шире, Чем у старых.
Отверстие позвоночного канала уменьшается от 1-го к 3-муг позвонку, затем
расширяется к 5-му, а у 7-го позвонка вновь уменьшается в диаметре.’ Расши-
рение позвоночного канала соответствует утолщению спинного мозга.
Шейный отдел составляет в среднем 21% общей длины позвоночного столба
(табл. 5.2). У взрослых самцов шейный отдел на 30—40% длиннее, чем у самок.
В пренатальный период и в первые годы постнатальной жизни эти различия
65
НОМЕРА ПОЗВОНКОВ
Рис. 5.13. Высота остистых отростков позвонков.
Рис. 5.14. Ширина позвонков в поперечных отростках.
Обозначения — см. рис. 5.10.
невелики и начинают проявляться с наступлением половой зрелости, когда у
самцов начинается новый ускоренный рост тела, и во многих пропорциях про-
исходят значительные изменения.
Относительная длина шейного отдела у взрослых самцов больше, чем у са-
мок (табл. 5.3). В онтогенезе она практически не изменяется. Абсолютная и от-
носительная масса всех шейных позвонков больше у самцов, чем у самок (рис.
5.15).
66
Таблица 5.2
Средняя длина (см) позвоночного столба и его отделов у северного морского
котика
Возраст Пол п Позвоноч- ный столб Отделы позвоночного столба
шейный грудной поясничный крестцовый хвостовой
Месяцы П р е н а т а л ь ный п е р и о д
7 7 20.3 4.0 8.6 3.5 1.8 2.4
8 самцы 8 27.3 5.6 10.8 4.8 2.8 3.5
9 и самки 5 36.4 7.6 14.9 5.5 3.5 4.6
10 1 42.2 8.2 18.5 6.5 3.9 5.1
11 Годы 4 51.3 Пост 10.7 натал 21.7 ь н ы й 8.4 период 4.3 6.1
0 самцы 5 63.1 11.8 26.8 10.2 5.0 9.3
самки 6 57.1 10.7 24.4 9.7 4.0 8.3
1 самцы 1 76.0 15.0 33.0 12.5 5.5 10.0
2 самцы 5 96.8 19.5 40.0 15.5 7.4 14.4
самки 3 92.7 19.1 39.6 14.8 7.6 11.6
3 самцы 5 105.7 22.5 43.6 17.4 8.8 13.4
самки 1 97.5 20.0 42.0 16.5 7.0 12.0
4 самцы 6 117.5 24.5 48.0 19.3 11.0 14.6
самки 1 103.0 21.0 45.0 17.0 7.0 13.0
5 самцы 4 126.3 27.3 53.2 19.4 10.7 15.7
самки 3 107.2 21.2 45.8 17.6 8.3 14.3
6 самцы 5 142.2 32.6 59.0 23.0 10.4 17.2
самки 3 113.8 23.2 46.6 20.0 9.3 14.6
7 самцы 2 166.5 40.0 73.0 22.5 13.5 18.5
самки 2 107.6 22.7 42.5 20.5 8.0 13.5
8 самцы 2 158.5 37.5 68.5 23.5 10.0 19.0
самки 4 116.0 24.1 49.5 16.9 11.8 13.7
9 самцы 3 168.6 40.3 71.3 26.3 12.2 18.5
10 самки 3 112.7 23.3 47.6 18.2 9.3 14.3
11 самки 1 109.0 23.0 48.0 18.0 7.0 13.0
14 самки 1 116.0 24.0 49.0 18.5 8.0 16.5
16 самки 1 109.5 23.0 46.0 17.5 7.5 15.5
17 самки 1 116.0 24.0 51.0 19.0 8.0 14.0
67
Таблица 5.3
Отношение длины отделов к длине позвоночного столба (%) у северного
морского котика
Возраст Пол п Отделы позвоночного столба
шейный ГРУДНОЙ ПОЯСНИЧНЫЙ крестцовый ХВОСТОВОЙ
П р е н а т а л ь н ы й п е р и о д
Месяцы
7 1 19.9 42.2 17.3 8.7 11.9
8 самцы 8 20.4 39.6 17.5 10.1 12.4
9 и 5 20.9 41.0 15.2 9.6 12.7
самки
10 1 19.4 43.8 15.4 9.2 12.1
И 4 20.9 42.3 16.4 8.3 11.9
Г о с т н а гг а л ъ н ы й период
Годы
0 самцы 5 18.7 42.5 16.2 7.9 14.7
самки б 18.7 42.7 17.0 7.0 14.5
1 самцы 1 19.7 43.4 16.4 7.2 13.2
2 самцы 5 20.1 41.3 16.0 7.6 14.9
самки 3 20.6 42.7 15.9 8.2 12.5
3 самцы 5 21.3 42.2 16.5 8.3 12.7
самки 1 20.5 43.0 16.9 7.2 12.3
4 самцы 6 20.9 40.9 16.4 9.4 12.4
самки 1 20.4 43.7 16.5 6.8 12.6
5 самцы 4 21.6 42.1 15.4 8.5 12.4
самки 3 19.8 42.7 16.4 7.7 13.3
б самцы 5 22.9 41.5 16.2 7.3 12.1
самки 3 20.5 40.9 17.6 8.2 12.8
7 самцы 2 24.0 43.2 13.5 8.1 11.1
самки 2 21.2 39.6 19.1 7.5 12.6
8 самцы 2 23.7 43.2 14.8 6.3 11.9
самки 4 20.8 42.7 14.6 10.2 11.8
9 самцы 3 23.9 42.3 15.6 7.2 10.9
10 самки 3 20.6 42.2 16.7 8.3 12.9
11 самки 1 21.1 44.0 16.5 6.4 11.9
14 самки 1 20.7 42.2 15.9 6.9 14.2
16 самки 1 21.0 42.0 15.9 6.8 14.2
17 самки 1 20.7 43.9 16.4 6.9 12.1
68
Рис. 5.15. Масса позвоночного столба и его отделов:
А — шейный, Б — грудной, В — поясничный, Г — крестцовый, Д — хвостовой отделы.
Столбики — абсолютный вес, линии — относительный вес, сплошная линия — самцы,
пунктирная — самки.
Рис. 5.16. Начальные стадии окостенения скелета у эмбрионов северного морского
котика (рентгенограммы)
Окостенение позвонков шейного отдела у северного морского котика начи-
нается на пятом месяце внутриутробного развития при длине тела плодов 8 см
и массе 15 г. В этом возрасте на рентгенограммах уже просматриваются тела
позвонков. На шестом месяце при Массе тела плодов 36 г и длине 10.4 см очаги
окостенения имеются в телах и невральных дугах 3—7-го шейных позвонков
(рис. 5.16). У эмбрионов с массой тела 225—295 г и длиной 19.2—20.5 см в ат-
ланте просматриваются 4 (крылья), в эпистрофее — тоже 4 (дуги, тело, зубо-
видный отросток) центра окостенения.
69
Рис. 5.17. 12—15-й грудные позвонки северного морского котика:
А — сбоку, Б — сверху;
1 — тело, 2 — остистый отросток, 3 — поперечный отросток, 4 — реберные ямки, 5 —
межпозвоночная вырезка, 6 — суставные реберные фасетки, 7 — краниальный, 8 —
каудальный суставные отростки.
На седьмом месяце при массе тела эмбриона 530 г и длине 28 см очаги окос-
тенения отчетливо видны в телах и дугах всех шейных позвонков.
У 9-месячных эмбрионов при длине тела 59 см и массе 2.7 кг присутствуют
оба эпифиза одинакового размера во всех шейных позвонках. Синостозирование
эпифизов с телами позвонков происходит лишь после наступления половой зре-
лости животных.
Грудной отдел состоит из 15 (97% особей) или 16 (3%) позвонков, уменьшаю-
щихся.в размерах от 1-го к 11—12-му и вновь увеличивающихся к последнему
(рис. 5.10). По бокам позвонков находятся реберные ямки, которые на первых
двух расположены не у стыка с прилежащими позвонками, а на некотором уда-
лении от их краев. Первые 2 ребра причленяются головкой не на границе двух
смежных позвонков, а к их телам. Реберные фасетки последующих позвонков
располагаются на стыке смежных позвонков.
70
Высота остистых отростков грудных позвонков уменьшается от первого к
последнему (рис. 5.11), а ширина их у основания, наоборот, уменьшается от пер-
вого к последнему. Вершины отростков утолщены. Все они наклонены каудаль-
но. Антиклинальность не выражена, хотя более или менее прямо поставлены
остистые отростки 11—12-го позвонков. Каждый остистый отросток предыду-
щего позвонка нависает над телом последующего позвонка. Поперечные отрост-
ки хорошо развиты и наиболее выражены у первого позвонка, а у последующих
их размеры уменьшаются (рис. 5.17).
Хорошо развиты на первых грудных позвонках краниальные суставные от-
ростки, которые постепенно уменьшаются в размерах на последующих позвон-
ках и уже на 4—6-ом фактически исчезают. Соответственно, краниальные сус-
тавные площадки перемещаются на дорсальную поверхность невральных дуг.
Однако, начиная с 19—20-го позвонков, краниальные суставные площадки вновь
смещаются на медиальную поверхность суставных отростков, обособляющихся
от поперечных отростков. Каудальные суставные отростки уменьшаются в раз-
мерах от 1-го к 12—13-му грудным позвонкам, и на следующих позвонках они
выглядят как простые суставные площадки, расположенные на вентрокаудаль-
ной поверх-ности основания остистых отростков.
У эмбрионов все грудные позвонки одинаковой длины (рис. 5.10). На по-
следующих стадиях онтогенеза наблюдается более интенсивный рост 1—10-го
и 13—22-го позвонков. Иначе говоря, у северного морского котика, как и у дру-
гих исследованных млекопитающих [Бровар, 1944], наблюдается передне-задний
градиент роста с центром в 11—12-ом грудных позвонках. Высота тел грудных
позвонков у взрослых животных уменьшается от первого к последнему, а у мо-
лодых они почти одинаковой высоты (рис. 5.11).
Масса грудных позвонков у взрослых особей уменьшается от первого к по-
следнему (рис. 5.12). Однако у эмбрионов и неполовозрелых особей лишь первые
3 позвонка тяжелее остальных. Высота остистых отростков и ширина в попе-
речных отростках грудных позвонков заметно уменьшаются от первого к по-
следнему, и эта последовательность менее выражена на ранних стадиях развития
котика, чем на последующих (рис. 5.13). Диаметр позвоночного канала умень-
шается от первого к последнему позвонку отдела, при этом у разновозрастных
животных его размеры мало различаются, особенно в каудальной части грудного
отдела. По результатам измерений высоты и ширины позвоночного канала,
грудное расширение его выявляется плохо, а поясничное вовсе не выражено.
Грудной отдел составляет в среднем 42.3% (39.6—44.0%) длины позвоночного
столба (табл. 5.3). По абсолютным размерам достоверные половые различия на-
чинают проявляться с 5-летнего возраста, а по относительным показателям поло-
вых различий нет. На протяжении всего онтогенеза относительная длина груд-
ного отдела остается практически одинаковой (табл. 5.3). Абсолютная масса поз-
вонков грудного отдела у взрослых самцов вдвое больше, чем у одновозрастных
самок, однако по относительным показателям масса грудного отдела позвоноч-
ника у самок больше, чем у самцов (рис. 5.15).
Начальные стадии окостенения грудных позвонков у эмбрионов котиков про-
являются на пятом месяце. Уже у эмбрионов с массой тела 36 г и длиной 10.4 см
окостеневают тела и невральные дуги всех позвонков (рис. 5.16). Синостоз не-
вральных дуг и тел позвонков грудного отдела начинается у 8-месячных эмб-
71
Рис. 5.18. Поясничный отдел позвоночного столба {вверху) и третий поясничный
позвонок {внизу) северного морского котика:
А — сбоку, Б — сзади, В — сверху;
1 — остистый отросток, 2 — каудальный, 3 — краниальный суставные отростки, 4 —
поперечный отросток.
рионов и завершается к периоду рождения. Интенсивность оссификации нарас-
тает от грудных позвонков к шейным. У исследованного 9-месячного эмбриона
с массой тела 2.7 кг и длиной 59 см присутствовали значительные по размерам
костные эпифизы. Полное срастание эпифизов с телами позвонков происходит
в постнатальном онтогенезе.
Поясничный отдел (рис. 5.18). Поясничных позвонков у северного морского
котика 4—5, чаще 5 (97% особей). По размерам самый маленький первый, самые
крупные — 2 средние. Краниальный и каудальный суставные отростки развиты
сильно, особенно краниальные, на которых рельефно выражены сосцевидные от-
ростки. Суставные площадки краниальных суставных отростков обращены ме-
диально, каудальных — латерально, как у последних грудных позвонков. Ос-
тистые отростки направлены вертикально; они плоские и широкие. Их высота
увеличивается от первого позвонка к последнему. Поперечнореберные отростки
плоские; направлены они вентрокраниально. На первом позвонке поперечные
отростки лишь обозначены. На последующих позвонках их длина увеличивается,
и на последнем она не уступает длине остистого отростка.
72
Рис. 5.19. Крестцовый отдел позвоночника северного морского котика:
А — сбоку, Б — сверху, В — спереди;
1 — основание, 2 — вершина, 3 — остистые отростки, 4 — суставные отростки, 5 — крыло,
6 — ушковая поверхность, 7 — дорсальное крестцовое отверстие.
Поясничные позвонки эмбрионов и новорожденных практически не отли-
чаются один от другого и сходны по длине с грудными позвонками (рис. 5.10),
но с возрастом длина 1—3-го позвонков увеличивается более интенсивно, чем
других; в результате у взрослых самым длинным является 2-ой и 3-й позвонки,
а самым коротким последний. Характер весового роста поясничных позвонков
сходен с линейным (рис. 5.12). По диаметру тела поясничные позвонки прак-
тически не отличаются друг от друга (рис. 5.11). Высота остистых отростков
всех поясничных позвонков у новорожденных котиков одинаковая, а на после-
дующих этапах роста и 'развития наблюдается передне-задний градиент изме-
нения их размеров (рис. 5.13). У взрослых котиков самый большой остистый
отросток имеет последний позвонок. Аналогичным образом изменяются разме-
ры поперечных отростков (рис. 5.14). Диаметр позвоночного канала постепенно
уменьшается к хвостовому отделу. У взрослых заметно небольшое расширение
его на уровне последнего позвонка; это лучше проявляется в ширине позвоноч-
ного канала, чем в высоте.
Поясничный отдел составляет в среднем 16.2% позвоночного столба (табл. 5.3).
По абсолютным размерам он больше у самцов, а по относительным показателям
различий практически нет (табл. 5.2, 5.3).
Окостенение позвонков поясничного отдела начинается на шестом месяце внут-
риутробного развития при массе тела эмбрионов 36—37 г и длине 10.4—10.6 см
(рис. 5.16). Очаги окостенения закладываются в невральных дугах и в телах поз-
вонков. Их синостоз начинается на восьмом месяце. На девятом месяце появ-
ляются краниальные и каудальные эпифизы позвонков (рис. 5.16).
Крестцовый отдел (рис. 5.19). Крестцовых позвонков у северного морского
котика 3—4, чаще 3 (91% особей). Они слиты между собой у большинства взрос-
лых и раздельные — у эмбрионов и некоторых новорожденных.
73
Невральные дуги первого крестцового позвонка несут хорошо выраженные
краниальные суставные отростки с суставными площадками для сочленения с
последним поясничным. Образованные слившимися поперечными отростками
крылья крестцовой кости направлены латерально. Вентральная поверхность пер-
вого позвонка широкая и гладкая, а на последующих имеются овальные гребни.
В целом крестцовый отдел имеет дорсальную выпуклость. В местах стыков поз-
вонков крылья пронизывают крупные замкнутые отверстия для выхода крест-
цовых нервов, кроме отверстия для выхода 3-го крестцового нерва, которое не
замкнуто. На дорсальной поверхности крестца располагаются 3 или 4 остистых
отростка со свободными или слившимися вершинами. Все морфометрические
показатели крестцового отдела уменьшаются от первого позвонка к последнему.
Крестцовый отдел в среднем составляет 8% длины позвоночного столба (табл.
5.3). У самцов он абсолютно длиннее, чем у самок; по относительным показа-
телям половых различий нет. В пренатальный период относительная длина
крестцового отдела несколько больше, чем у взрослых особей.
Окостенение тел крестцовых позвонков начинается на шестом месяце внутри-
утробного развития при длине тела эмбрионов 13.5 см и массе 80—90 г. В этом
же месяце при длине тела 20.5 см и массе 255t очаги окостенения появляются в
невральных дугах. Эпифизарные центры окостенения появляются у 9-месячных
эмбрионов при длине тела 59 см и массе 2.7 кг.
Хвостовой отдел. Хвостовых позвонков у северного морского котика 8—10.
Морфологические структуры типичного позвонка сохраняются у первых трех—
четырех. У последующих нет поперечных отростков и гемальных дуг, и они ста-
новятся похожими на цилиндрики. Остистые отростки исчезают с 4—5-го поз-
вонка, а позвоночный канал представлен на них как желобок.
Длина хвостового отдела составляет в среднем 11.5% позвоночного столба
(табл. 5.3) По абсолютным размерам заметных отличий между самками и сам-
цами нет, также как нет их и по относительным. Масса хвостового отдела со-
ставляет в среднем 2.2% массы позвоночного столба. X самок масса хвостового
отдела относительно больше, чем у самцов (рис. 5.15).
Окостенение хвостовых позвонков начинается у 6-месячных плодов. В телах
первых четырех позвонков у эмбрионов с массой тела НО г и длиной 15.4 см
имеются очаги окостенения. Шестой позвонок окостеневает при длине тела эм-
брионов 20.5 см и массе 255 г, а окостенение 7-го позвонка начинается на седь-
мом месяце при массе 530 г и длине 28 см. Последующие хвостовые позвонки
могут не окостеневать всю жизнь. Эпифизарные центры окостенения первых
двух позвонков возникают на восьмом месяце при длине тела эмбрионов 49 см
и массе 2.7 кг, третьего — у эмбрионов с массой тела 4.5 кг и длиной 53 см,
а краниальный эпифиз 4-го позвонка появляется у эмбрионов с массой тела
4.8 кг и длиной 53 см. Новорожденные котики имеют эпифизы в 5 хвостовых
позвонках. Срастание эпифизов с телами хвостовых позвонков происходит на
поздних стадиях постнатального онтогенеза, а у последних позвонков может не
происходить вовсе.
Ребра. У северного морского котика 15 пар ребер (рис. 5.20), лишь изредка
встречается 16-я пара. Стернальных ребер чаще 9 (92.8%), но может быть и 10
(7.2%) (рис. 5.21). Астернальных, соответственно, 6 или 5. Самое короткое ребро
74
Рис. 5.20. Ребра северного морского котика.
Цифрами обозначены номера ребер.
первое, самое длинное — восьмое. У первых
ребер головка меньше бугорка, а шейка плохо
выражена. Дистальный конец утолщен и почти
округлый. Как и у наземных хищных, на реб-
рах имеется сосудистый и мышечный желобки.
Тело в проксимальной части незначительно
утолщено. У 2—12-го ребер выражены все об-
разования, свойственные типичным ребрам.
Тела 2—4-го ребер несколько сплюснуты, у
последующих пяти они в поперечнике почти
овальные, а у последних трех вновь уплоща-
ются, причем на этих же ребрах присутствует
только по одной суставной площадке на го-
ловках. Масса ребер нарастает от первого к
9—10-му, а потом убывает.
Провизорные очаги окостенения в ребрах
обнаружены уже у 5-месячных эмбрионов при
массе тела 8 г и длине 6 см. Центры окосте-
нения возникают в средней части и распрост-
раняются проксимально и дистально. К мо-
менту рождения проксимальные эпифизы еще
не окостеневают.
Рис. 5.21. Грудина с различным
числом стернальных ребер.
Стрелкой обозначен дополнительный
очаг окостенения в грудине с
избыточным числом стернальных
ребер.
Грудина (рис. 5.21) состоит из рукоятки, тела и мечевидного отростка. Ру-
коятка посередине имеет суставные площадки для ребер, а ее часть, выступа-
ющая вперед, изогнута книзу. Тело состоит из 6 сегментов призматической
формы. Мечевидный отросток плоский, сужается к середине, а каудально вновь
расширяется. Мечевидный хрящ лопатообразной формы.
Окостенение грудины начинается несколько позже, чем ребер. В каждом сег-
менте грудины возникает по одному центру окостенения. Последним окостене-
75
Рис. 5.22. Лопатка северного морского котика:
А — с медиальной стороны, Б — с латеральной стороны;
1 — ость, 2 — гребень предостной ямки, 3 — предостная ямка, 4 — заостная ямка, 5 —
акромион, 6 — краниальный угол, 7 — каудальный угол, 8 — вентральный угол, 9 —
подлопаточная ямка, 10 — подлопаточный гребень.
вает мечевидный отросток. В зависимости от количества стернальных ребер мо-
жет изменяться тип крепления к грудине (рис. 5.21), а также число очагов окос-
тенения в ней. У животных, имеющих 10 пар стернальных ребер, количество
сегментов в грудине не изменяется, но при окостенении мечевидного отростка
в нем возникают 2 очага оссификации, которые потом срастаются друг с другом
без видимых следов.
Обычно чем подвижнее животное, тем оно имеет меньше ребер [Астанин,
1958]. В свою очередь, ширина и длина грудины стоит в прямой зависимости
от степени подвижности ребер и объема органов грудной полости [Блехман, 1949].
У северного морского котика число ребер больше, чем у собаки (13), свиньи
(14), овцы (12), крупного рогатого скота (13) [Воккен и др., 1961], но позвоноч-
ный столб котика обладает гораздо большей гибкостью, чем у наземных мле-
копитающих. Нам представляется, что гибкость тела животных определяется,
кроме всего прочего, толщиной межпозвоночных дисков. Что касается количест-
ва ребер, то экологические условия существования котиков требуют, чтобы ор-
ганы грудной полости были хорошо защищены от внешних воздействий при
жизни животного на берегу. Мощное развитие ребер и грудины важно и для
общего укрепления переднего пояса и передних конечностей, являющихся ос-
новным органом локомоции котика как на берегу, так и в воде.
5.3. Периферический скелет
Подробных описаний строения периферического скелета северного морского
котика в литературе нет. Общие сведения о скелете конечностей имеются в ра-
боте В. Шеффера (Scheffer, 1958] и Ю. Кинг [King, 1964]. Окостенение скелета ко-
нечностей достаточно подробно описано В. Ф. Мужчинкиным [19646] и А. Е.
Кузиным [1969в, 1971а]. В ряде работ рассмотрен весовой и линейный рост пе-
76
Рис. 5.23. Длина лопатки
северного морского котика:
А — в процентах от длины скелета
передней конечности, Б — абсолют-
ная длина.
риферического скелета в пренатальный
[Кузин, 1971а, 1972а] и постнатальный пери-
оды [Allen, 1880; Соколов, 1971; Кузин, Ма-
минов, 1978].
Скелет передней конечности. Несмотря
на внешнюю непохожесть передних конеч-
ностей северного морского котика и назем-
ных хищных, их скелеты по числу костей и
общей топографии практически не различа-
ются. Ластообразность конечности достига-
ется разновеликостью костных и хрящевых
элементов ее дистального отдела, сильным
развитием соединительнотканной прослойки
и мощным развитием кожного покрова. В
костях передних конечностей котика нет
мозговой полости, в отличие от большинст-
ва наземных млекопитающих. Полость кос-
тей заполнена тончайшими костными плас-
тинками.
Лопатка (рис. 5.22). Краниальный край
лопатки прямоугольный, каудальный скошен.
На латеральной поверхности, кроме хорошо
выраженной ости, в передней части имеется
еще дополнительный гребень. Предостная
часть имеет не вогнутую, а выпуклую повер-
хность. Акромион выражен хорошо. Меди-
альная поверхность не плоская, а несет на се-
бе 2 пологих гребня. Вентральный угол от-
делен от остальной части хорошо выражен-
ной шейкой. Гленоидная ямка яйцевидной
формы.
Месяцы Годы
Возраст
Рис. 5.24. Масса лопатки северного
морского котика:
А — в процентах от массы скелета
передней конечности, Б — в процентах
от массы всего скелета, В — абсолютная
масса;
Н — новорожденные.
Длина лопатки относится к ее ширине как 1:1.2. Характерно, что это соот-
ношение сохраняется в течение всего онтогенеза (рис. 5.23). Поскольку ширина
предостной ямки и длина краниального края лопатки значительно превосходят
77
Рис. 5.25. Окостенение скелета передней и задней конечностей северного морского
котика.
А — начало 6-го месяца внутриутробного развития, Б — конец 6-го месяца, В — 11-й месяц.'
аналогичные размеры в заостной части, то площадь предостной ямки почти в
2 раза больше площади заостной части. Гребень предостной части короче ости
и втрое ниже его. В связи с сильно выраженным половым диморфизмом в раз-
мерах тела, абсолютные размеры лопатки у самцов больше, чем у самок. Однако
лопатка у самок относительно шире, чем у самцов, и относительные показатели
измерений, связанные с сопоставлением длины и ширины лопатки, больше у самок.
Ость лопатки у самок относительно длиннее, но по высоте уступает таковой
у самцов. Высота предостной части в среднем также относительно ниже у самок.
Рост лопатки в длину у котика прекращается одновременно с ростом тела
(рис. 5.23). Ее длина по отношению к длине скелета передней конечности из-
меняется мало в процессе онтогенеза. Относительная длина несколько больше
у самцов, но половые различия недостоверны. Масса лопатки, выраженная в
процентах от массы всего скелета, в онтогенезе постоянно увеличивается, а по
отношению к массе скелета передней конечности снижается ко времени рожде-
ния и вновь несколько увеличивается у взрослых животных (рис. 5.24).
Окостенение лопатки начинается на шестом месяце внутриутробного разви-
тия в ее средней части ближе к шейке (рис. 5.25). Рост костной ткани в даль-
нейшем идет в дорсальном и вентральном направлениях, причем в вентральном
быстрее, чем в дорсальном, и заканчивается, примерно, на месяц раньше (при-
близительно восьмой месяц внутриутробного развития). В 9 месяцев окончатель-
но формируется позвоночный край лопатки.
Плечевая кость у северного морского котика короткая, мощная, S-образной
формы, что достигается за счет сильного развития большого бугорка и его греб-
ня (рис. 5.26). По этой же причине проксимальная половина кости представ-
ляется несколько сжатой в медиолатеральном, а вентральная — в краниолате-
ральном направлении. Головка плечевой кости выпуклая и хорошо отделена от
тела шейкой. Большой бугорок располагается выше головки; его мощный гре-
бень имеет обширную дельтовидную шероховатость. Дистальный эпифиз, об-
разующий блок для соединения с костями предплечья, имеет неярко обособлен-
ный латеральный и хорошо выраженный медиальный мыщелки.
78
Рис. 5.26. Плечевая кость северного морского котика:
А — проксимальная поверхность, Б — дистальная поверхность, В — латеральная поверхность,
Г — медиальная поверхность, Д — каудальная поверхность;
1 — головка, 2 — большой бугорок, 3 — межбугорковый желоб, 4 — малый бугорок, 5 —
шейка, 6 — гребень большого бугорка, 7 — дельтовидная шероховатость, 8 — гребень
латерального надмыщелка, 9 — медиальный надмыщелок, 10 — латеральный надмыщелок,
11 — мыщелок, 12 — большое питательное отверстие.
По абсолютным и по многим относительным показателям плечевая кость у
взрослых самцов больше, чем у самок. Однако по относительным размерам эпи-
физов, длине головки и ширине блока плечевая кость у самок больше таковой
у самцов. Рост плечевой кости в длину прекращается у самок в 4—5, у сам-
цов — в 9—10 лет. Относительные показатели весового и линейного роста в
онтогенезе изменяются мало (рис. 5.27, 5.28).
: В первой половине внутриутробного развития относительная длина плечевой
кости уменьшается, т. е. темп ее роста ниже, чем у дистальных отделов конеч-
ности, однако относительно всего скелета она растет быстрее (рис. 5.27). От-
79
Рис. 5.28. Масса плечевой
кости:
А — в процентах от массы
скелета передней конечности,
Б — в процентах от массы всего
скелета, В — абсолютная масса;
Рис. 5.27. Длина плечевой кости северного морского
котика:
А — в процентах от длины скелета свободной передней
конечности, Б — абсолютная длина;
Н — новорожденные.
Н — новорожденные.
носительные показатели массы и длины плечевой кости у самцов несколько
больше, чем у самок (рис. 5.27, 5.28).
Первичные очаги окостенения в плечевой кости у северного морского котика
появляются на пятом месяце внутриутробного развития (рис. 5.25). Процесс окос-
тенения у самок идет быстрее, чем у самцов. В возрасте 8 мес. у большинства
эмбрионов-самок уже имеются проксимальный и дистальный эпифизы, а у сам-
цов только проксимальный [Кузин, 1969в]. У 4-летних самок и 8-летних самцов
еще заметны границы между эпифизами и диафизом плечевой кости [Мужчинкин,
19646].
Кости предплечья. Лучевая и локтевая кости не срастаются друг с другом
(рис. 5.29). Локтевая кость на ‘/4 длиннее лучевой. Это проявляется за счет сильно
развитого локтевого отростка. Обе кости несколько уплощены с боков. Прок-
симальная головка лучевой кости с уплощенной суставной поверхностью. Дис-
тальный конец тела значительно расширен. Суставная площадка дистального
эпифиза уже, чем проксимального. По абсолютным размерам лучевая кость у
самцов больше, чем у самок (рис. 5.30). По соотношению передне-заднего и попе-
речного диаметров лучевая кость у самок характеризуется большей уплощенностью,
чем у самцов. Эпифизы относительно шире у самцов. Рост кости в длину за-
вершается у самок в 4—5, у самцов — в 7—8 лет (рис. 5.30). Относительные размеры
лучевой кости у новорожденных не отличаются от таковых у взрослых особей.
На рентгенограммах провизорно очаги окостенения лучевой кости обнару-
живаются у эмбрионов на пятом месяце [Кузин, 1969в]. Как видно на рис. 5.25,
проксимальный эпифиз появляется на девятом месяце, а к концу этого месяца
появляется и дистальный. Процесс оссификации у самок идет быстрее, чем у
самцов.
80
Рис. 5.29. Лучевая и локтевая кости северного морского котика:
А — медиальная поверхность, Б — латеральная поверхность, В — проксимальная
поверхность; I — лучевая, II — локтевая кости;
1 — головка, 2 — шейка, 3 — тело, 4 — грифельный отросток локтевой кости, 5 —
грифельный отросток лучевой кости, 6 — локтевой отросток, 7 — локтевой бугор, 8 —
блоковая вырезка, 9 — ямка головки лучевой кости.
Рис. 5.31. Масса костей
предплечья:
А — в процентах от массы
скелета передней конечности, Б —
в процентах от массы всего
скелета, В — абсолютная масса;
Н — новорожденные.
Месяцы Года
Воарвот
Рис. 5.30. Длина лучевой кости;
А — в процентах от длины передней конечности, Б —
абсолютная длина, Н — новорожденные.
81
Локтевая кость выступает за пределы лучевой в виде сильно развитого лок-
тевого отростка (рис. 5.29), длина которого относится к ширине как 1:2. Гребень
отростка несколько утолщен и медиально изогнут. Клювовидный отросток, яв-
ляющийся верхней границей полулунной вырезки, выражен в равной степени,
как и венечный, являющийся вентральной границей полулунной вырезки. Рас-
положенная ниже полулунной вырезки сигмовидная вырезка почти равна ей по
глубине; но первая превышает вторую по площади. Уплощенное с боков тело
почти не изменяется по толщине до проксимального конца, и лишь дистальный
эпифиз незначительно расширяется. Грифельный отросток выражен. По всем аб-
солютным параметрам локтевая кость у самцов больше, чем у самок, но от-
носительные показатели площади суставной поверхности для соединения с пле-
чевой костью и глубины полулунной вырезки локтевой кости больше у самок.
Рост локтевой кости в длину прекращается у самцов к 7, а у самок к 5 годам
(рис. 5.30). Относительная длина кости уменьшается к концу внутриутробного
развития и увеличивается в постнатальном онтогенезе у самок к 3, а у самцов
к 5 годам. Масса костей предплечья, выраженная в процентах от массы скелета
передней конечности, в онтогенезе изменяется неравномерно, в то время как от-
носительно всего скелета увеличивается (рис. 5.31).
Первичные очаги окостенения в диафизах локтевой кости возникают у эмб-
рионов на пятом месяце. Закладка дистального эпифиза у самок наблюдается
в начале, у самцов в конце 11-го месяца развития. Апофиз локтевого бугра по-
является у самок лишь на четвертом году жизни [Мужчинкин, 19646].
Рис. 5.32. Строение запястья:
А — северный морской котик, Б — крылатка, В — лахтак, Г — стопоходящее наземное
млекопитающее.
Кисть северного морского котика имеет типичное для наземных хищных
строение. Radiale, centrale, intermedium срастаются между собой и образуют еди-
ную кость. Закладка этих костей у котика происходит во внутриутробный пе-
риод, а срастание лишь к концу второго или на третьем году постнатального
онтогенеза [Мужчинкин, 19646]. У собак, например, закладка их происходит в
возрасте 20—30 дней постнатальной жизни, а срастание — в 3.5—4.5 мес. [Из-
майлова, 1935]. Отсутствие эпифизов когтевых фаланг на всех стадиях онтогенеза
является специфической особенностью северного морского котика.
Кости запястья котика располагаются в 2 ряда (рис. 5.32). Проксимальный
ряд составляет комплексная кость (radiale+centrale+intermedium) и ulnare, а дис-
тальный — 1, 2, 3, 4+5 запястные кости. Комплексная кость служит опорой для
лучевой кости. Запястная локтевая, располагающаяся между лучевой и локтевой
костями, имеет на дорсальной поверхности 2 суставные площадки для соеди-
82
Рис. 5.35. Масса костей кисти:
А — в процентах от массы скелета
передней конечности, Б — в
процентах от массы всего скелета,
В — абсолютная масса; Н —
Рис. 5.34. Длина кисти северного морского котика:
А — в процентах от длины скелета свободной передней
конечности, Б — абсолютная длина;
Н— новорожденные.
новорожденные.
нения с названными костями. Первая—третья запястные кости имеют суставные
площадки для соединения с комплексной костью, а 4+5 еще и с ulnare. Прок-
симальные участки пястных костей имеют не одинаковое число суставных пло-
щадок. С волярной поверхности проксимального ряда лежит добавочная кость
(os carpi accessorium).
83
Кости пясти уменьшаются в размерах от первой к пятой, однако по массе
пятая пястная кость тяжелее четвертой, (рис. 5.33).
Кости пальцев имеют по 3 фаланги, кроме первого пальца, имеющего две.
Фаланги плоские. Они также, как и кости пясти, уменьшаются в размерах от
первого к пятому пальцам.
Рост1 скелета кисти в длину заканчивается у самок к 4, а у самцов к 7 годам
(рис. 5.34).
Относительная длина кисти, выраженная в процентах от длины свободной
конечности в пренатальный период изменяется неравномерно (рис. 5.34). Наи-
высшие показатели отмечены для 9-месячных эмбрионов (47%). В постнатальном
онтогенезе они наибольшие у 4—5-летних животных. Масса кисти в пренаталь-
ный период увеличивается очень интенсивно, и к периоду рождения она больше
массы плечевой кости, но у взрослых кости кисти легче плечевой кости (рис.
5.35). Относительная масса скелета кисти, рассчитанная от массы всего скелета,
увеличивается в пренатальный период более интенсивно, чем масса скелета про-
ксимальных костей конечности, но во взрослом состоянии они уступают им по
этому показателю. В то же время относительная масса скелета кисти, выражен-
ная в процентах от массы скелета передней конечности, увеличивается нерав-
номерно от ранних стадий внутриутробного развития к периоду рождения. У
взрослых животных эти показатели ниже, чем у предродовых эмбрионов и ново-
рожденных.
Окостенение скелета кисти начинается в начале 6-го месяца внутриутробного
развития [Кузин, 1969в; рис. 5.25]. Первыми появляются костные очаги в первой
и второй пястных костях. Одновременно с окостенением последующих метакар-
пальных элементов очаги окостенения возникают в первых фалангах 1—3-го паль-
цев. К концу 6-го месяца у эмбрионов в передней конечности на рентгенограм-
мах хорошо просматриваются все metacarpalia и первый ряд фаланг пальцев.
В основном ряду костная ткань отсутствует в фалангах 4-го и 5-го пальцев. С
этого времени процесс окостенения у самок идет быстрее, чем у самцов.
На седьмом месяце у эмбрионов-самцов костная ткань присутствует во всех
метакарпальных элементах, первых фалангах 1—5-го и вторых фалангах 2—4-го
пальцев (рис. 5.25). На стадиях закладки находится третья фаланга пальца. У
одновозрастных эмбрионов-самок процесс окостенения продвинут дальше. У них
хорошо просматриваются все metacarpalia, проксимальные фаланги 1—5-го, сред-
ние фаланги 2—4-го, дистальные фаланги 1—4-го пальцев.
На восьмом месяце развития у одних эмбрионов-самцов в стадии закладки
кост-ной ткани находятся когтевые фаланги 2—4-го пальцев, у других они еще
отсутствуют. У эмбрионов-самок в этом возрасте костная ткань отсутствует только
в дистальной фаланге 5-го пальца.
На девятом месяце эмбриогенеза начинается окостенение карпальных элементов
(рис. 5.25). У самцов в стадии закладки находятся radiale и intermedium, 1 и 4+5
carpalia distalia. У эмбрионов-самок уже хорошо заметны radiale и intermedium, все
четыре carpalia distalia, а эпифизы первой и третьей фаланг 1-го пальца и дисталь-
ные эпифизы фаланг 2—4-го пальцев — в стадии закладки или оформившиеся.
На десятом месяце эмбриогенеза, по данным В. Ф. Мужчинкина [19646], у
эмбрионов-самцов имеются radiale и intermedium, все 4 карпальные элемента,
оба эпифиза первой и дистальные эпифизы второй и третьей Метакарпальных
84
Рис. 5.36. Безымянные кости северного морского котика:
А — латерально, Б — медиально;
1 — подвздошная кость, 2 — седалищная кость, 3 — лонная кость, 4 — тело подвздошной
кости, 5 — крыло подвздошной кости, 6 — подвздошный гребень, 7 — краниальный бугорок
подвздошной кости, 8 — большая седалищная вырезка, 9 — малая седалищная вырезка, 10 —
седалищный бугорок, 11 — лонный бугор, 12 — суставная впадина, 13 — запирательное
отверстие, 14 — ушковидная шероховатость.
костей, закладка проксимальных эпифизов основных фаланг первых двух паль-
цев.
У 11-месячных эмбрионов (рис. 5.25) имеется полный набор окостеневших
карпальных элементов, проксимальный эпифиз первого метакарпального эле-
мента и третья фаланга первого пальца, дистальные эпифизы второго—пятого
метакарпальных элементов и проксимальный эпифиз основной фаланги второго
пальца. У эмбрионов-самок, кроме того, имеются очаги окостенения в прокси-
мальных эпифизах основных фаланг 3-гр и 4-го пальцев, а у некоторых и ког-
тевая фаланга мизинца. Однако, по данным В. Ф. Мужчинкина [19646], послед-
ние 2 фаланги мизинца у отдельных особей могут окостеневать лишь на третьем
году жизни. В 2-летнем возрасте у самцов окостеневает эпифиз второй фаланги
4-го пальца. Начавшееся еще у сеголеток слияние radiale и intermedium пол-
ностью осуществляется в 2 года, а в 3-летнем возрасте к ним прирастает centrale.
У 2-летних самок имеются центры окостенения во второй фаланге мизинца и
едва различимая закладка костной ткани в когтевой фаланге этого же пальца
[Мужчинкин, 19646]. По данным этого же автора, у 8-летнего самца еще четко
различима граница между эпифизами и диафизами костей кисти, кроме дисталь-
ных эпифизов метакарпальных костей.
Скелет задней конечности. Задняя конечность северного морского котика, как
и передняя, видоизменена в ласт. Пальцы имеют хрящевое продолжение, между
пальцами находится перепонка. Задние ласты способны подгибаться под туло-
вище и участвуют в передвижении по твердому субстрату.
85
Возраст
А — в процентах от скелета
зад- ней конечности, Б — в
про- центах от массы всего
скелета,
В —• абсолютная масса,
Таз включает 2 плоские безымянные кости,
расположенные по бокам крестца. Каждая из бе-
зымянных костей состоит из подвздошной, седа-
лищной и лонной костей (рис. 5.36). У эмбрионов
и у взрослых животных до наступления физичес-
кой зрелости хорошо просматривается впадинная
кость (os acetabulum). В отличие от наземных мле-
копитающих, у северного морского котика лон-
ный симфиз не окостеневает. Кости таза более
тонкие и не несут на себе больших выступов и
гребней. Лонно-седалищный отдел длиннее под-
вздошного. Лонная кость у предродовых эмбрио-
нов составляет в среднем 66%, седалищная — 44%
от общей длины безымянных костей. У взрослых
котиков они составляют, соответственно, 55.5% и
47.9% у самцов и 51% и 45.3% у самок. Под-
вздошные кости шире двух других. Образованное
слиянием лонной и седалищной костей запира-
Рис. 5.37. Масса костей таза: тельное отверстие в длину вдвое больше, чем в ши-
рину. Ацетабулюм почти округлый по форме. Все
абсолютные промеры костей таза у самцов боль-
ше, чем у самок. По относительным показателям
длина крыла подвздошной кости, длина лонной
кости, ширина подвздошной и седалищной костей
у самок больше, чем у самцов. В тоже время ширина лонной кости, вход в таз,
длина подвздошной и седалищной костей больше у самцов. Запирательное от-
верстие относительно короче у самцов, чем у самок.
Масса безымянных костей у котика в онтогенезе увеличивается, опережая
рост всего скелета (рис. 5.37). Выраженная в процентах от массы скелета задней
конечности, она в пренатальный период уменьшается, а в постнатальный период
несколько увеличивается..У взрослых самцов относительная масса безымянных
костей больше, чем у самок.
Окостенение костей таза Начинается с подвздошного элемента. Закладка кос-
тной ткани происходит на шестом месяце внутриутробного развития и начи-
нается с центра подвздошной кости, распространяясь в сторону крыла и аце-
табулюма (рис. 5.25). Несколько позже, но в том же месяце, очаги окостенения
появляются в седалищном и еще позже в лонном элементах. Синостоз лонных
и седалищных костей происходит на десятом месяце. У предродовых эмбрионов
подвздошная и лонная кости сближены в области вертлужной впадины, но си-
ностоз их еще не отмечается.
Бедренная кость короткая и тонкая (рис. 5.38). На внутренней стороне ее
проксимальной части хорошо обособлена полусферическая головка, переходя-
щая в шейку. Латеральную сторону занимает сильно развитый большой вертел,
который отделяется от головки слабо выраженной вертлужной впадиной. Тело
кости несколько сужается к центру, а затем вновь расширяется к дистальному
эпифизу.
На дистальном эпифизе хорошо выражены латеральный и медиальный мы-
щелки, разделенные межмыщелковой ямкой. На каждом мыщелке заметны над-
86
Рис. 5.38. Бедренная кость северного морского котика:
А — латерально, Б — медиально, В — дистально, Г — проксимально;
1 — головка, 2 •— большой вертел, 3 — шейка, 4 — вертлужная ямка, 5 — тело, 6 —
медиальный мыщелок, 7 — латеральный мыщелок, 8 — межмыщелковая ямка, 9 —
латеральный надмыщелок.
7 8 9 10 II Н 1 2 3 4 5 б 7 8 9 10 II 12 13 141516 17
Месяцы Годы
Возраст
Рис. 5.39. Длина бедренной кости:
А — в процентах от длины скелета свободной конечности, Б — абсолютная длина;
Н — новорожденные.
мыщелки. Мыщелки внизу и позади соединяются и образуют спереди желобо-
ватый блок для коленной чашки. Бедренная кость заметно уплощена в пере-
дне-заднем направлении. Проксимальный эпифиз по размерам тоньше Дисталь-
ного, а диафиз тоньше обоих эпифизов. Половой диморфизм отмечен по аб-
солютным и некоторым относительным показателям бедренной кости.
Рост бедренной кости в длину прекращается у самок в 5, а у самцов в 8—9
лет (рис. 5.39).
87
месяцы годы
возраст
Рис. 5.40. Масса бедренной
кости морского котика:
А — в процентах от массы ске-
лета задней конечности, Б — в
процентах от массы всего скелета,
В — абсолютная масса;
Н — новорожденные.
В пренатальный период ее относительная
длина уменьшается, а в постнатальный период
развития почти не изменяется. Относительная
длина бедренной кости у самцов несколько боль-
ше, чем у самок (рис. 5.39). У взрослых самцов
масса бедренной кости более чем вдвое тяжелее,
чем у самок. К периоду рождения масса бедрен-
ной кости составляет 13.6% от массы всего скелета
и потом почти не изменяется (рис. 5.40). Относи-
тельно скелета задней конечности масса бедрен-
ной кости в пренатальный период уменьшается,
и эта тенденция сохраняется в постнатальный пе-
риод.
Окостенение бедренной кости начинается в на-
чале января (шестой месяц внутриутробного раз-
вития). Во второй половине восьмого месяца на-
чинается окостенение эпифизов, из которых пер-
вым проявляется проксимальный, а вслед за ним
дистальный. К периоду рождения диафиз и эпи-
физы сближены, но сращение их еще не наступает.
Кости голени (рис. 5.41). Большая и малая бер-
цовые кости почти равны между собой по длине.
У многих наземных хищных это самые длинные
кости в скелете, а у котиков они или равны, или
даже немного уступают по длине локтевой кости.
Тело большой берцовой кости имеет небольшую S-образную изогнутость.
Эпифизы расширены, проксимальный больше дистального. На вершине прокси-
мального эпифиза хорошо выражены суставные площадки для соединения с мы-
щелками бедренной кости. Каждая суставная площадка завершает латеральный
и медиальный мыщелки кости. Дистальный эпифиз завершается суставным бло-
ком для сочленения с костями предплюсны. Большеберцовый гребень не сильно
развит. Медиальная лодыжка четко обозначена. Оба эпифиза кости значительно
шире в латеромедиальной, чем в краниокаудальной плоскости, а диафиз почти
одинаков в этих измерениях. ,У самок проксимальный эпифиз более вытянут в
передне-заднем направлении, чем у самцов. Относительная длина большебер-
цовой кости больше у самцов, чем у самок.
Рост костей голени в длину продолжается у самцов и у самок до 8-летнего
возраста (рис. 5.42). Во внутриутробный период интенсивность их роста выше,
чем бедренной кости. Относительная длина в первых двух третях эмбриогенеза
уменьшается, а в последствие — увеличивается. Масса костей голени, выражен-
ная в процентах от массы всего скелета, интенсивно увеличивается от ранних
стадий внутриутробного развития до рождения, а в постнатальном онтогенезе
практически не изменяется. У взрослых самок относительная масса скелета го-
лени несколько больше, чем у самцов. Наибольшую относительную массу от
скелета задней конечности кости голени имеют у предродовых эмбрионов
(рис.1 5.43).
88
3
1
2
Рис. 5.41. Кости голени северного морского котика:
А — латерально, Б — медиально, В — проксимально, Г — дистально;
I — большая берцовая кость, II — малая берцовая кость;
I — латеральный мыщелок, 2 — медиальный мыщелок, 3 — межмыщелковый бугор, 4 —
гребень большой берцовой кости, 5 — головка малой берцовой кости, 6 — межмыщелковая
впадина, 7 — медиальная лодыжка, 9 — подколенная вырезка.
Окостенение большой берцовой кости начинается у 6-месячных эмбрионов
(рис. 5.25). В конце восьмого месяца появляется проксимальный, а в начале де-
вятого — дистальный эпифиз. Эпифизы и диафиз большеберцовой кости синос-
тозируют в постнатальном онтогенезе.
Малая берцовая кость (рис. 5.41) прилежит к большеберцовой кости с ла-
теральной стороны и тоньше ее почти вдвое. Ее проксимальный конец утолщен
и имеет головку с суставной поверхностью для соединения с большеберцовой
костью. Дистальный эпифиз также утолщен и имеет 2 суставные фасетаи для
соединения с большеберцовой и таранной костями. Проксимальный эпифиз
тоньше дистального. Передне-задний и поперечный диаметры почти равны. По
относительным показателям малоберцовая кость самцов заметно превосходит
таковую самок только по размерам эпифизов в поперечнике. По другим пока-
зателям половые различия выражены слабо.
Окостенение малоберцовой кости начинается на шестом месяце эмбриональ-
ного развития. Дистальный эпифиз закладывается перед рождением (рис. 5.25),
89
Рис. 5.42. Длина костей голени:
А — в процентах от длины скелета свободной задней
конечности, Б — абсолютная длина;
Н — новорожденные.
Возраст
Рис. 5.44. Кости плюсны северного морского котика
Рис. 5.43. Масса костей голени:
А —в процентах от массы
скелета задней конечности, Б —
в процентах от массы всего
скелета, В — абсолютная масса;
Н — новорожденные.
90
Рис. 5.45. Длина скелета стопы:
А — в процентах от длины скелета свободной конечности, Б — абсолютная длина;
Н — новорожденные.
Возраст Возраст
Рис. 5.46. Масса скелета стопы:
' А — в процентах от массы скелета задней
'конечности, Б — в процентах от массы всего
скелета, В — абсолютная масса;
Н — новорожденные.
Рис. 5.47. Изменение массы позвоночного
столба северного морского котика в
зависимости от возраста.
Столбики — абсолютная масса, линии —
относительная масса; сплошная линия — самки
и самцы (вместе) в эмбриогенезе, пунктир —
самки в постнатальный период;
Н — новорожденные.
а эпифиз локтевого бугра обнаружен В. Ф. Мужчинкиным [19646] только у че-
тырехлеток.
Кости стопы. Кости предплюсны расположены в 3 ряда. В проксимальном
ряде 2 кости: таранная и самая крупная кость в предплюсне — пяточная, а в
91
среднем ряду одна центральная кость. Дистальный ряд состоит из четырех пред-
плюсневых костей. Кости предплюсны различны по форме и числу суставных
фасеток, что определяется их местоположением и числом контактов с приле-
жащими костями.
Все 5 плюсневых костей уплощены и сходны по строению с пястными кос-
тями. Самая длинная из них — первая (рис. 5.44). Самые короткие — 3 средние.
Соответствующим образом и масса этих костей изменяется от первой к пятой:
соответственно, 34, 9, 9, 10, 16 г у взрослого самца и 12, 3, 2, 2, 3 г у взрослой
самки. Фаланги пальцев на задней конечности сходны по строению с таковыми
на передней. По массе каждая последующая фаланга меньше предыдущей. Од-
нако фаланги 1-го и 5-го пальцев мощнее одноименных фаланг трех средних
пальцев.
Рост скелета стопы в длину прекращается у самок в возрасте 4—5 лет, у сам-
цов — в 7—8 лет. Интенсивность роста скелета стопы во внутриутробный период
выше, чем проксимальных скелетных элементов. К концу внутриутробного пе-
риода развития стопа составляет более половины, а во взрослом состоянии при-
мерно половину длины свободной конечности. При этом у самок, в среднем,
относительная дайна скелета стопы больше, чем у самцов (рис. 5.45).
Увеличение массы скелета стопы за период пренатального развития проис-
ходит очень интенсивно, но уступает по этому показателю скелету кисти. На
ранних стадиях онтогенеза масса костей кисти составляет 1.5% от массы всего
скелета, а стопы — 2.3%.
К концу внутриутробного периода развития масса скелета кисти достигает
3.8%, а стопы — 3.6% от массы всего скелета. Взрослые животные имеют почти
одинаковую относительную массу дистальных отделов передней и задней ко-
нечностей (рис. 5.35, 5.46).
Масса скелета стопы, выраженная в процентах от массы скелета задней ко-
нечности, несколько уменьшается от месяца к месяцу в пренатальный период.
У взрослых самок относительная масса костей стопы выше, чем у самцов (рис.
5.46).
В эмбриогенезе окостенение в стопе начинается позже, чем в кисти (рис. 5.25).
В начале 6-го месяца внутриутробного периода развития провизорные очаги
окостенений не просматриваются. У более крупных эмбрионов на том же месяце
развития закладываются метатарсальные элементы. Так же, как и в кисти, про-
цесс окостенения элементов стопы идет несколько быстрее у самок, чем у самцов.
К концу 6-го месяца в стопе у самцов имеются центры окостенения во всех ме-
татарсальных костях, проксимальных, второй—четвертой основных фалангах
пальцев. В стадии закладки находятся третья фаланга 1-го и основная фаланга
5-го пальцев, которые у самок уже хорошо заметны.
В течение 7-го месяца эмбрионального развития центры окостенения появ-
ляются во всех скелетных элементах предплюсны и фалангах, кроме когтевых
фаланг 1-го и 5-го пальцев у самцов.
На 8-ом месяце окостеневают пяточная и таранная кости (рис. 5.25). У не-
которых эмбрионов обоего пола отсутствует закладка когтевой фаланги 5-го
пальца.
92
У эмбрионов-самцов в конце 9-го месяца в дистальном ряду предплюсны
окостеневают кубовидная кость, а также третья и первая tarsalia distalia. У эмб-
рионов-самок имеются костные центры в дистальных эпифизах 2—5 metatarsalia
и в проксимальных эпифизах 1-го и 5-го пальцев (рис. 5.25).
На 10-ом месяце [Мужчинкин, 19646] в стопе эмбрионов-самцов окостенения
имеются в кубовидной, латеральной и медиальной клиновидных костях, дисталь-
ных эпифизах первых трех метакарпальных костей и проксимальных эпифизах
основных фаланг соответствующих пальцев.
У предродовых эмбрионов-самцов присутствует полный набор костей пред-
плюсны (рис. 5.25). Центры окостенения отмечаются в проксимальных эпифизах
первой плюсневой кости, в первых фалангах всех пальцев, в виде точечных оча-
гов в основных фалангах 2—5-го пальцев. Дистальные эпифизы имелись во 2—5
плюсневых костях. У всех эмбрионов-самок окостеневают предплюсневые кости.
Кроме костных элементов, указанных для самцов, у самок хорошо заметны окос-
тенения в эпифизах 2—5 основных фаланг.
У самцов на третьем месяце постнатального развития и у уже хорошо пла-
вающих детенышей кости стопы цолностью сформированы, но первая метатар-
сальная кость не слита со своим дистальным эпифизом. У самок они сливаются
[Мужчинкин, 19646].
Общая схема строения и развития скелета северного морского котика в он-
тогенезе, в принципе, не отличается от таковых у изученных наземных млеко-
питающих, в особенности хищных. Тем не менее, длительный путь эволюции
котика в специфических условиях приспособления к водному образу жизни не
мог не вызвать соответствующих модификаций определенных частей скелета и
системы в целом.
Сравнительно быстро закладываются и потом интенсивно растут те скелет-
ные элементы, которые испытывают функциональную нагрузку в тот или иной
период индивидуального развития [Лесгафт, 1881; Воккен, 1950]. Известно, что
у млекопитающих, цервой окостеневает нижняя, а несколько позже верхняя че-
люсть [Harris, 1936; Третьяков, 1959в; Воккен, и др., 1961]. Раннее окостенение
челюстей обычно связывают с началом амниотрофного питания эмбрионов, а
прилежащих к ним костей — с функционированием жевательной мускулатуры.
Это подтверждается и нашими, данными изучения окостенения черепа северного
морского котика. Характерно также, что закладка костной ткани во вторичных
элементах, составляющих, главным образом, основание черепа, сильно задер-
живается у котика, как и у других животных [Третьяков, 1959в]. Раннее окос-
тенение покровных скелетных элементов и высокий темп их оссификации ста-
вится некоторыми авторами в функциональную связь с ранним окостенением
челюстей [Третьяков 1959в], что, на наш взгляд, маловероятно. Скорее всего это
объясняется различиями в способе закладки костной ткани и, в этой связи, раз-
личиями в дальнейшем темпах ее роста. Вторичные скелетные элементы исто-
рически возникли позже [Шмальгаузен, 1922]. Возможно, здесь мы видим ис-
торический процесс становления морфологических структур. Безусловно, фун-
кции стимулируют возникновение и изменение темпа развития тех или иных мор-
фологических образований. Однако морфологическая зрелость органов и
включение их в функциональную деятельность не совпадают во времени. Сис-
93
темы начинают действовать задолго до их морфологической готовности по прин-
ципу минимального обеспечения [Анохин, 1948].
Краниометрические исследования северного морского котика показали, что
существуют значительные различия в процессе развития составляющих его кос-
тей. Одни скелетные элементы растут быстрее, другие испытывают задержку в
росте. Одни показатели пропорций черепа достигают своей окончательной ве-
личины на ранних стадиях постнатального онтогенеза, другие изменяются
вплоть до достижения животными физической зрелости. В целом видоспецифи-
ческие черты формируются в результате гетерохронного развития его элементов.
Возрастно-половые особенности скелета головы хотя и начинают проявляться
к концу внутриутробного периода развития котика, но наиболее значимые из
них формируются после становления видовых признаков.
Выявленная значительная хронографическая изменчивость пропорций черепа
в зависимости от фазы развития численности популяций при использовании ее
показателей в целях изучения популяционных различий должна сочетаться со
знаниями динамики численности.
Из наиболее ярких признаков строения позвоночника котика, сближающих
его с наземными хищными, является мощное развитие крыльев атланта, распо-
ложенных во фронтальной плоскости, концы которых простираются за каудаль-
ные суставные площадки. У тех и у других сильно развит гребень эпистрофея,
далеко выдающийся вперед и нависающий над зубовидным отростком. Попе-
речные отростки эпистрофея тонкие и направлены назад. Третий—шестой поз-
вонки имеют сложное строение, т. к. их поперечные отростки включают кра-
ниальный, каудальный и дорсальный бугорки, к которым прикрепляется хорошо
развитая шейная мускулатура. Седьмой позвонок не имеет реберной фасетки.
В грудном отделе позвоночного столба, как и у наземных хищных, наблюдается
постепенное уменьшение суставных отростков и смещение краниальных сустав-
ных площадок на дорсальную поверхность невральных дуг, а каудальных — на
вентральную поверхность основания остистых отростков. Поясничные позвонки
несут крупные уплощенные поперечные отростки, которые не выражены лишь
на первом позвонке. Крестцовый отдел представлен неподвижно сросшимися поз-
вонками, имеющими дорсальную выпуклость, с несросшимися или частично
сросшимися остистыми отростками. Первые хвостовые позвонки сохраняют
части типичного позвонка, постепенно исчезающие к концу хвоста.
Порядок закладки костной ткани в позвоночном столбе северного морского
котика и наземных хищных также сходен. В каждом позвонке, кроме атланта
и эпистрофея, закладываются по 5 очагов окостенения (2 в невральных дугах,
1 в теле и 2 в эпифизах). В атланте закладываются 4 центра окостенения (2 в
невральных дугах и 2 в интерцентре). В эпистрофее, кроме обычных для типич-
ного позвонка центров, отдельно закладываются центры окостенения зуба и его
эпифизов.
Первые костные центры раньше появляются в средних грудных позвонках,
и затем волна окостенения в позвоночнике распространяется в переднем и заднем
направлениях. Позже окостеневают первые шейные позвонки, а последними —
крестцовые и первые хвостовые. Последние хвостовые позвонки могут не окос-,
теневать всю жизнь; это является специфической особенностью северного мор-
ского котика.
94
У котика, как и у других млекопитаю-
щих, процесс оссификации позвонков про-
исходит в следующей последовательности:
срастание частей невральных дуг, форми-
рование остистых отростков, прирастание
дуги к телу, слияние эпифизов с телами
позвонков.
Основные отличия котика от типич-
ных хищных обнаруживаются уже в поз-
воночной формуле. Для него характерно
большее число грудных позвонков и
меньшее число поясничных и хвостовых.
На атланте у котика нет крыловой вырез-
ки, а его крылья гораздо дальше прости-
раются за каудальные суставные площад-
ки. На первых двух грудных позвонках
отсутствуют реберные полуфасетки, и
слабо выражены сосцевидные и дополни-
тельные отростки. Имеются различия в
соотносительных размерах тел позвонков,
их остистых и поперечных отростков по
отделам и в позвоночнике в целом. У ко-
тика длина грудного отдела в среднем со-
ставляет ок. 50% длины позвоночного
столба, в то время как у наземных млеко-
питающих она не превышает 35—40%
[Третьяков, 19596]. Лишь у крупных куньих
месяцы годы
Рис. 5.48. Относительный рост
осевого (А) и периферического (Б)
отделов скелета северного морского
котика:
Н — новорожденные.
[Соколов, Соколов, 1970] его относительные размеры близки к показателям у
котика. В то же время у котика значительно меньше длина поясничного отдела.
Если по относительной массе позвонков шейного отдела еще наблюдается не-
которое сходство с куньими [Пилипчук, 1980], то по другим отделам этого нет.
У всех наземных хищных относительная масса поясничных позвонков значитель-
но превышает таковую грудных; у котика она меньше. В целом, показатели от-
носительной массы позвоночного столба у северного морского котика больше,
чем у наземных хищных, в связи с тем, что периферический отдел скелета у него
облегчен (рис. 5.47, 5.48).
Не менее яркие отличия в позвоночном столбе у северного морского котика
можно видеть не только от экологически удаленных форм млекопитающих, ка-
кими являются наземные, но и от представителей того же отряда, например,
от настоящих тюленей. Различия выражаются, прежде всего, в соотносительных
размерах отделов позвоночника. У настоящих тюленей меньше относительная
длина шейного и грудного отделов, но больше длина поясничного [Кузин, Ма-
минов, 1978]. Остистые отростки грудных позвонков у котика значительно выше
поясничных, в то время как у настоящих тюленей наблюдается обратная кар-
тина.
Атлант настоящих тюленей, в целом, имеет форму овала, а у котика — тре-
угольника. У настоящих тюленей крылья атланта лежат ближе к горизонтальной,
95
а у котика — к вертикальной плоскости. Форма крыльев у настоящих тюленей
овальная, а у котика приближается к четырехугольнику. По ширине крылья у
котика составляют около половйны позвонка, в то время как у настоящих тю-
леней они равны позвонку. Гребень эпистрофея у котика менее развит. Тело
этого позвонка несет на себе единый поперечнореберный отросток, а у настоя-
щих тюленей он раздвоен.
Относительная масса позвонков (% от массы позвоночного столба) у котика
и настоящих тюленей различна. У котика больше масса шейных и 1—11-го груд-
ных позвонков, а у настоящих тюленей больше относительная масса 12—15-го
грудных, всех поясничных и крестцовых. В целом, ход кривых изменения от-
носительной массы шейных и 1—10(11) грудных позвонков у котика и настоящих
тюленей сходен между собой и похож на ход такой же кривой у наземных ку-
ницеобразных [Пилипчук, 1980]. Однако, если в последующих позвонках сход-
ство у настоящих тюленей и куницеобразных сохраняется, то у котика кривая
имеет совершенно иную направленность.
Не менее значительные отличия северного морского котика от наземных мле-
копитающих наблюдается и в сроках окостенения позвоночного столба. Заклад-
ка эпифизов позвонков начинается у эмбрионов котика в 7—8-месячном воз-
расте, а у собаки с 17-го дня постнатальной жизни [Жеденов, 1965]. Синосто-
зирование невральных дуг с центрами позвонков у котика завершается во всех
позвонках, кроме атланта, к периоду рождения, а у собаки оно начинается толь-
ко с 5-го месяца постнатальной жизни. В то же время синостоз эпифизов поз-
вонков с их телами у котика заканчивается в 14—15 лет (кроме последних хвос-
товых), у собаки этот процесс завершается к 7—8 годам.
В целом, изучение позвоночного столба северного морского котика пока-
зало, что становление дефинитивных форм, размеров и пропорций происходит
в течение большей части онтогенеза. Такая “открытая” на протяжении продол-
жительного периода онтогенеза система способна в случае необходимости легко
модифицироваться под воздействием изменяющихся условий существования.
Формирование видоспецифических черт строения позвоночника у морского ко-
тика в разных его частях начинается неодновременно и происходит с разной
скоростью. Это хорошо видно на графиках изменения длины и массы отделов
позвоночника, размеров поперечных и остистых отростков позвонков. Многие
из названных морфологических структур еще на средних стадиях пренатального
развития характеризуются одноразмерностью, как бы сохраняя следы первичной
метамерии. Завершающий этап формирования размеров и пропорций элементов
и частей позвоночника происходит уже после достижения животными половой-
зрелости.
Скелет передней конечности северного морского котика по общему принци-
пу строения не отличается от скелета многих наземных хищных. Однако спе-
цифика экологии, связанная с обитанием в водной среде и на суше, особенности
локомоции и ведущая роль передних конечностей в передвижении у котиков ска-
зались как на их форме, так и на особенностях роста и развития. Кости передних
конечностей короткие, но мощные. По относительным размерам, развитию диа-
физов, бугров, гребней лопатка, ;плечо и предплечье котика обнаруживают наи-
большее сходство с аналогичными костями роющего животного, в особенности
96
крота. Как у того, так и у другого на лопатке имеются 2 гребня. Плечевая кость
мощная, короткая, с сильно развитыми буграми и гребнями. Кости предплечья
расположены параллельно друг другу и одинаково хорошо развиты. Локтевая
кость несет на себе длинный лучевой отросток, значительно выступающий за
пределы сустава. Между костями предплечья сохраняется подвижность. Обна-
ружение конвергентного сходства в строении передней конечности этих живот-
ных безусловно вызвано однотипностью их функции, связанной с выполнением
гребных движений. В тоже время кисть северного морского котика не имеет ана-
логичного сходства с кистью роющего животного в связи с разной функцией,
присущей этому отделу конечности сравниваемых животных. Кисть котика пред-
назначена для осуществления гребных движений при плавании в воде и пос-
тупательного движения по земле. Она развернута на 90° по отношению к са-
гиттальной плоскости тела. В этой связи соотношение костей запястья и пясти
видо-изменено. Локтевая кость у котика опирается на пятую пястную, a car-
pale 4+5 — на комплексную кость (рис. 5.32). У наземных хищных [Манзий,
1957] и у настоящих тюленей [King, 1956; Кузин, 1971а] ulnare служит опорой
сагра- 1е 4+5, а последняя — четвертой и пятой пястным костям. Описанная пе-
рестройка скелета запястья привела к тому, что передний край ласта значительно
укрепился, первые пальцы имеют более мощный скелет, подвижность их огра-
ничена. В то же время край ласта приобретает очень большую подвижность,
а животные большую способность маневрировать при передвижении в воде.
Формирование кисти в онтогенезе проходит несколько стадий. У ранних эмб-
рионов (начало 5-го месяца) пальцы равновелики, имеют типичный фронталь-
ный изгиб и ничем не отличаются от этой же части передней конечности на-
земных хищных [Кузин, 1971а]. Однако на последующих месяцах эмбриогенеза
скорость роста от пятого к первому пальцу увеличивается, в результате чего
к моменту рождения кисть приобретает крыловидную форму. Процесс окосте-
нения начинается с проксимального переднего края (первых метакарпальных
костей), затем постепенно охватывает другие скелетные элементы и заканчива-
ется на заднем дистальном крае (мизинце). Фактически создаются условия для
соответствующего роста и развития скелета во времени, позволяющие сохранять
необходимые пропорции органов движения и тела.
У большинства наземных млекопитающих кости задней конечности — это
прежде всего важные рычаги, толкающие тело вперед. Они мощные и длинные
[Воккен и др., 1961]. У северного морского котика кости задних конечностей
короткие и тонкие и в основном служат для поддержания тела при передвижении
по земле, для стабилизации корпуса при прямолинейном движении и в качестве
рулей при маневрировании в воде. Задние конечности морского котика не ис-
пытывают значительных нагрузок, составляющие их кости тонкие, лишены вы-
раженных бугров и более просто устроены, чем кости передних конечностей.
Сравнительное исследование таза млекопитающих П. А. Осинского [1959]
показало, что ластоногие характеризуются относительно коротким тазом. Ко-
роткий таз имеют также четвероногие прыгуны (заяц, кролик) и близкие к ним
формы. У всех этих животных при движении размах конечностей увеличивается
большой подвижностью позвоночного столба. Массивность таза определяется,
главным образом, количеством и развитостью мышц заднего пояса. У северного
97
Таблица 5.4
Абсолютные и относительные показатели длины скелета конечностей
северного морского котика
Возраст Пол п Длина позвоночного столба, см Свободная передняя конечность ' Свободная задняя конечность
СМ % ОТ длины ПОЗВОНОЧНОГО столба см % от длины позвоночного столба
П р е н а т а л ь ный период
Месяцы
7 7 20.3 8.0 39.40 8.5 41.87
8 самцы н 8 27.3 10.8 35.56 12.3 45.05
9 самки 5 36.4 17.9 49.17 19.2 52.74
11 4 51.3 23.4 45.61 25.9 50.48
П с с т н а т а л ь ный период
Годы
0 самцы 5 63.1 28.4 45.00 32.2 51.03
самки 6 57.1 26.8 46.94 27.9 52.01
1 самцы 1 76.0 36.0 47.37 39.0 51.32
2 самцы 5 96.8 . 43.8 45.25 46.5 48.04
самки 3 92.7 36.5 39.37 40.3 43.47
3 самцы 5 105.7 45.9 43.42 39.6 46.93
самки 1 97.5 45.7 48.71 47.5 48.71
4 самцы 6 117.5 50.6 44.06 56.2 47.83
самки 1 103.0 46.0 44.46 51.0 49.51
5 самцы 4 126.3 58.1 46.00 60.3 : 47.74
самки 3 107.2 44.7 41.70 47.6 44.40
6 самцы 5 142.2 61.0 42.89 64.5 45.36
самки 3 113.8 44.8 38.66 46.7 41.04
7 самцы 2 166.5 71.0 42.64 74.5 44.74
самки 2 107.2 43.8 40.86 46.6 43.47
8 самцы 2 158.5 70.0 44.16 75.5 47.63
самки 4 116.0 49.5 44.67 53.1 45.78
9 самцы 3 168.6 70.6 41.87 72.4 42.94
10 самки 3 112.7 48.7 43.21 54.0 47.91
11 самки 1 109.0 48.0 44.04 52.0 47.71
14 самки 1 116.0 52.0 44.82 53.5 46.12
16 самки 1 ,109.5 49.0 44.74 . 52.0 47.49
17 самки 1 ,116.0 46.0, 39.66 56.0 48.28
98
морского котика тазовые конечности короткие и облегченные. Действительно,
при передвижении по суше у него основная тяжесть тела падает на передние
конечности. Задняя часть тела сильно изогнута вентрально и является, грубо
говоря, продолжением задних конечностей. Размах задних конечностей сильно
ограничен из-за соответствующего строения мышц тазобедренного сустава, в
особенности бицепса. При движении по суше возрастает роль позвоночного
столба и соответствующих мышц брюшного пресса. В целом, мышцы задней
конечности котика [Соколов, 1966] развиты не сильно. У настоящих тюленей
они развиты лучше, и поэтому у них тазовые кости более массивные, а под-
вздошные кости даже имеют своеобразные выступы, отсутствующие у ушастых
тюленей. Правда, у настоящих тюленей большое значение в развитии костей таза
играют мышцы туловища, обеспечивающие колебательные движения задней
части корпуса при плавании.
Бедренная кость у котика самая короткая из костей задней конечности, а
кости голени самые длинные. Темп роста задней конечности в длину и увели-
чение их массы в онтогенезе происходит от проксимального конца к дисталь-
ному. Несмотря на более позднюю закладку задней конечности, по сравнению
с передней, темп ее линейного роста выше. Уже у 7-месячных эмбрионов длина
задней конечности несколько больше передней. Ко времени рождения эти раз-
личия еще больше увеличиваются (табл. 5.4).
В то же время темп роста массы передней конечности на всех стадиях он-
тогенеза выше темпа роста задней конечности. Эти данные говорят о том, что
линейный и весовой рост конечностей не согласованы, и правильное толкование
функциональной связи частей скелета в ряде случаев лучше делать на основе
весовых показателей. Примечательно, что окостенение скелета задней конечнос-
ти, в особенности скелета стопы, у северного морского котика начинается не-
сколько позже, чем кисти, однако впоследствии темп дифференцировки костной
ткани в стопе выше, чем в кисти. Предплюсневые центры окостенения появля-
ются примерно на месяц раньше, чем запястные. Темп роста эпифизарных цент-
ров окостенений в проксимальных скелетных элементах задней конечности го-
раздо выше темпа роста гомодинамных элементов передней конечности. К мо-
менту рождения задняя конечность представлена большим числом костных эле-
ментов, чем передняя.
Ускоренное развитие задней конечности по сравнению с передней у эмбрио-
нов, по-видимому, связано с условиями дальнейшего существования животного.
Северные морские котики большую часть жизни проводят в воде, используя при
плавании в основном передние конечности. Поэтому раннее окостенение скелета
передних конечностей и, соответственно, замедление или прекращение их роста
привело бы к нарушению соотносительных размеров тела и основных органов
локомоции, что отрицательно сказалось бы на способности и результатах пе-
редвижения в воде [Мужчинкин, 19646]. Любопытно, что у настоящих тюленей
[Воккен, 1964; Кузин, 1971а], у которых ведущим локомоторным органом яв-
ляются задние конечности, темп дифференцировки костной ткани ниже в этих
скелетных элементах, чем в передней конечности, хотя на начальных стадиях
внутриутробного развития у представителей обоих семейств ластоногих закладка
костной ткани в скелетных элементах конечностей происходит в одни и те же
99
сроки [Кузин, 1971а]. В основе отмеченных особенностей дифференцировки кост-
ной ткани скелета конечностей представителей разных семейств ластоногих ле-
жат причины как экзогенной, так и эндогенной природы.
“Изучение процессов дифференцировки костного скелета млекопитающих, —
писал Г. Г. Воккен [1949], — показало, что последовательность в закладке кост-
ных очагов зависит от исторически сложившихся функций кости. Функции ор-
ганов или части могут меняться в течение онтогенеза, но изменение функции
может быть и филогенетическим. Поэтому порядок закладки очагов окостенений
зависит и от особенности смены функции, произошедшей в течение филогене-
тической эволюции вида”. Рассматривая гетерохронии в закладке органов поз-
воночных животных, С. В. Емельянов [1963] отмечал, что сроки их закладки
обладают большой устойчивостью. Справедливость сказанного ярко подтвер-
ждается примером окостенения скелета конечностей ластоногих. Нет основания
предполагать, что во внутриутробный период развития конечностям ластоногих
присущи различные функции. Скорее всего одновременность закладки костной
ткани в гомодинамных скелетных элементах представителей разных семейств в
первой половине внутриутробного развития говорит о их филетической близос-
ти. Дальнейшее дивергентное развитие скелета конечностей ластоногих в про-
цессе формирования дефинитивных органов у ушастых и настоящих тюленей
происходит в результате изменения темпа роста и развития [Кузин, 1971а] и за-
висит от экологической ниши, занимаемой каждым видом. Аналогичным об-
разом на основе гетерохроний происходит формирование и некоторых других
адаптивных признаков ластоногих [Слепцов, 1968; Кузин, 1971а].
Сравнение времени закладки костной ткани в элементах конечностей мор-
ского котика и собаки, по данным И. В. Измайловой [1935], показало сходство
8-месячного эмбриона котика с новорожденным щенком собаки. Выше было от-
мечено аналогичное сходство в сроках оссификации позвоночного столба этих
животных. Если исключить диапаузу внутриутробного развития из общей про-
должительности беременности у северного морского котика, то описанные ста-
дии оссификации скелета котика и собаки будут приходится примерно на одни
и те же периоды индивидуального развития. Однако дальнейший процесс ос-
сификации конечностей и позвоночного столба у сравниваемых животных су-
щественно различается. У ластоногих (в частности, у котика) срастание эпифизов
с диафизами сильно задерживается. По мнению Н. Н. Третьякова [1959в], на сроки
срастания эпифизов и диафизов движителей значительное влияние оказывет
опорно-двигательная функция. Частично можно согласиться с этим положением.
Однако к нему необходимо добавить особенности среды обитания организмов,
разнообразие способов их локомоции, различия в размерах тела и общей про-
должительности жизни. Уже сам процесс окостенения скелета конечностей у ко-
тика не позволяет ограничиваться только ролью опорно-двигательной функции.
Различная интенсивность роста скелетных элементов была положена в ос-
нову разделения скелета на осевой и периферический отделы, разные у позво-
ночных как по филогенетическому, так и по индивидуальному развитию, и ко-
торые различно реагируют на внешнее и внутреннее воздействие [Бровар, 1944].
Осевой и периферический отделы скелета растут с разной, обычно с противо-
положной, интенсивностью. На этом основании было выделено 3 типа постэмб-
100
рирнального роста скелета: 1) с преобладанием интенсивности роста в осевом
скелете, 2) с преобладанием интенсивности роста в периферическом скелете,
3) с одинаковой интенсивностью роста в обоих отделах скелета.
Позже С. Н. Боголюбским [1948] было установлено, что инверсия роста от-
делов скелета наступает в определенном возрасте. Сейчас уже известно, что у
копытных переломный момент в росте отделов скелета наступает перед рожде-
нием, у хищных — после рождения [Бровар, 1944; Боголюбский, 1948; Черепа-
хина, 1957; Третьяков, 1959в; Воккен и др., 1961]. У котика, как и у наземных
хищных, инверсия роста осевого и периферического отделов скелета приходится
на постнатальный период развития (рис. 5.48).
В целом, рост скелета у северного морского котика в пренатальный период
идет неравномерно. До 9-го месяца темп роста скелета и общей массы тела эм-
бриона сходен, позже в течение двух месяцев быстрее нарастает масса тела, но
перед рождением ее нарастание замедляется, а масса скелета продолжает уве-
личиваться с той же интенсивностью. Для изменения массы тела это явление
давно известно и даже используется для определения периода деторождения,
когда нет возможности исследовать непосредственно новорожденных [Федосеев,
1964]. Факт увеличения относительной массы скелета у новорожденных, по срав-
нению со взрослыми, тоже известен и рассматривается как приспособление де-
тенышей к условиям гравитации для усиления функции кроветворения, чтобы
обеспечить организм необходимым количеством кислорода в новых условиях
жизни [Коржуев, 1964, 1971].
101
Глава 6. РОЛЬ МЫШЕЧНОЙ СИСТЕМЫ В ПЕРЕДВИЖЕНИИ
Мышечная система северного морского котика почти не изучена. В. Ф. Муж-
чинкин [19716] привел схематические рисунки топографии мышц туловища и ко-
нечностей котика без описания и указания мест их крепления со ссылкой на
идентичность таковых у морского льва, описанных ранее [Howell, 1929]. Опи-
сание мышц свободных конечностей морского котика сделано А. Инглишем [En-
glish, 1976], а краткие сведения о массе мышц движителей приведены А. С. Со-
коловым [1973].
При сохранении общего плана строения мышечной системы, присущего на-
земным млекопитающим, скелетная мускулатура северного морского котика
имеет ряд отличительных черт. Это в основном касается степени развития мышц
и мест их прикрепления, что связано с особенностями изменением формы тела
и его пропорций. Наиболее заметные отличия наблюдаются в строении мышц,
связанных с плаванием. Плавание для котика стало основной формой локомо-
ции. Но, не будучи исключительно водным животным, северный морской котик
сохранил возможность передвигаться по земле с помощью всех четырех конеч-
ностей. При этом в перемещении задней части тела большое значение имеет по-
ясничный отдел. Основная опора тела при движении животного по твердому
субстрату приходится на запястье, пясть и пяточную кость. Передние ласты раз-
вернуты на 90° по отношению к сагиттальной плоскости тела. В меньшей сте-
пени, но аналогичным образом, развернуты дистальные отделы задней конеч-
ности. Все проксимальные части конечностей (кроме кисти и стопы) скрыты в
корпусе под кожно-мышечным слоем. Движение задних конечностей в сагитталь-
ной плоскости, в особенности при выносе их вперед, ограничивается широким
и хорошо развитым бицепсом и подкожной мускулатурой. В связи с этим, задняя
часть тела северного морского котика при статолокомоции и при движении рас-
положена под прямым углом к фронтальной плоскости тела, и короткие прок-
симальные костные элементы задней конечности служат в большей степени под-
порками для туловища, в то время как у большинства наземных млекопитающих
они выполняют ведущую локомоторную функцию.
Передние конечности котика, как и других ушастых тюленей [Соколов, 1973;
English, 1976], — основной локомоторный орган и при движении в воде, и при
движении по суше. Поступательное движение котика в воде осуществляется, по
данным Ю. Е. Мордвинова [1968], гребными движениями передних конечностей,
включающими несколько фаз: а) ласты отводятся от тела вперед и вверх в го-
ризонтальном положении или при некотором угле атаки (5—10°); б) ставятся
под углом, близким к прямому, по отношению к продольной оси тела; в) при-
водятся вниз и назад, сгибаясь при этом в дистальных частях; г) прижимаются
к телу. Затем этот цикл движений повторяется.
Задние ласты при этом вытянуты вдоль тела, повернуты вертикально и не
принимают участие в создании движущей силы. При выключении из работы пе-
редних конечностей функцию движителей берут на себя задние конечности и
корпус. Движения в этом случае осуществляются с помощью отведения обоих
102
Таблица 6.1
Масса мышц конечностей некоторых ластоногих [из: Соколов, 1973]
Вид п Масса мышц ластов, в % от массы тела Масса мышц туловища, в % от общей массы мышц переднего и заднего ластов
передний задний вся туловищная мускулатура мышцы спины
Сивуч 5 11.8 1.4 19.6 1.1
Северный морской котик 5 10.0 1.1 20.6 8.6
Морж 3 8.3 1.7 35.7 12.7
Лахтак 5 5.3 1.9 100.0 55.7
Крылатка 5 4.9 1.3 96.8 62.0
Кольчатая нерпа 3 3.9 1.5 64.7 39.0
ластов в сторону с одновременным изгибанием дистальных отделов туловища.
При этом тот ласт, в сторону которого совершается движение, несколько собран
и согнут в стопе, противоположный же ласт в этот момент максимально рас-
правлен. Затем ласты и задняя часть корпуса продолжают движение в проти-
воположную сторону. Иногда оба ласта отводятся в стороны, и ими произво-
дится резкий толчок. Скорость движения только с помощью задних конечностей
невысокая. По-видимому, такой способ передвижения используется котиками
крайне редко [Мордвинов, 1969].
При движении по суше используются шаг или галопирование. При движении
шагом один из передних ластов выбрасывается вперед, и вслед за ним подтя-
гиваются или вместе, или попеременно задние конечности. При галопировании
выбрасываются сразу оба передних ласта, вес тела переносится на передние ко-
нечности, и подтягиваются задние конечности.
Описанные локомоторные акции реализуются действием соответствующих
мышц конечностей и туловища.
Для наилучшего понимания особенностей двигательной активности приведем
данные по массе мышц настоящих тюленей и моржа (табл. 6.1), из которых пер-
вым присущ способ плавания с помощью задних конечностей [Gambaqan, Ка-
rapetjan, 1961], а второй использует при движении в воде обе пары конечностей,
но большее значение имеют все же задние [Kenneth, 1981].
Масса мышц конечностей северного морского котика составляет 11.1% от
массы тела, в то время как у настоящих тюленей только 6—7%. При этом масса
мышц передней конечности в значительной степени превосходит таковую зад-
ней, что в гораздо меньшей степени выражено у настоящих гюленей. Масса
мышц туловища составляет у котика лишь 20% от массы мышц передних и зад-
них конечностей. У настоящих тюленей туловищная мускулатура в 3—5 раз тя-
желее. Соответственно мышцы спины развиты у северного морского котика зна-
чительно хуже, чем у настоящих тюленей (табл. 6.1). У котика лучше развиты
103
О 100% О 400%
Рис. 6.1. Весовые соотношения групп мышц конечностей
некоторых ластоногих [по: Соколов, 1973]:
I — сивуч, II — северный морской котик, III — морж,
IV — лахтак, V — крылатка, VI — кольчатая нерпа.
А — мышцы переднего ласта: 1 — переднего пояса, 2 — плечевого сустава, 3 — локтевого
сустава, 4 — кисти и пальцев;
Б — мышцы заднего ласта: 1 — тазобедренного сустава, 2 — коленного сустава, 3 — голе-
ностопного сустава, 4 — стопы и пальцев.
мышцы живота, тогда как у настоящих тюленей — мускулатура спины (длин-
нейшая и подвздошнореберная) [Соколов, 1973]. Представленные в таблице 6.1
данные обращают внимание на почти равные показатели относительной массы
мышц заднего ласта у котика и настоящих тюленей; у первого они обслуживают
второстепенные движители, а у вторых — главные. Однако, как было показано
выше, высокий относительный показатель массы туловищной мускулатуры у на-
стоящих тюленей говорит о том, что они вместе с мышцами задней конечности
принимают активное участие в локомоции колебательными движениями всего
крестцово-поясничного отдела туловища.
Из анализа весового соотношения мышц конечностей (рис. 6.1) видно, что
у котика мышцы пояса передних конечностей менее развиты, чем у сивуча, и
значительно уступают по массе большинству мышц настоящих тюленей. Однако
мышцы плечевого сустава имеют наибольший относительный вес среди иссле-
дованных тюленей.
Что касается мышц дистальных отделов конечностей, то они имеют пример-
но одинаковые показатели у всех исследованных видов ластоногих [Соколов,
1973]. Однако, если у ушастых тюленей из группы мышц переднего пояса лучше
развиты поверхностная и глубокая грудные, то у настоящих тюленей наиболь-
шего относительного веса достигают широчайшая мышца спины и брюшная
часть грудной. Соответственно, первые из названных мышц принимают самое
активное участие в загребных движениях передней конечности, а вторая группа
мышц вместе с другими туловищными — в создании боковых колебательных
движений задней части корпуса.
Среди мышц задней конечности у ластоногих наибольшего развития дости-
гают мышцы тазобедренного сустава (рис. 6.1). Хотя масса их у представителей
разных семейств тюленей существенно не различается, но роль их в локомоции
104
у ушастых и настоящих тюленей различна. Ушастые тюлени используют задние
конечности при движении по земле. У настоящих тюленей задние конечности
не способны подгибаться под туловище и не принимают участие в передвиже-
нии по твердому субстрату. Поэтому ягодичная группа мышц сильнее развита
у ушастых (у северного морского котика 6%), чем у настоящих тюленей (3%).
У ушастых тюленей сильнее, чем у настоящих, развиты аддукторы и абдукторы.
В связи с тем, что двуглавая, полусухожильная и стройная мышцы у настоящих
тюленей короткие, у них уменьшен рычаг приложения сил.
Мышцы стопы и пальцев развиты лучше у настоящих тюленей, чем у ушас-
тых. Именно ими осуществляется регулирование площади плавательной пере-
понки и управление дистальной частью ласта. Так, например, общий сгибатель
пальцев у настоящих тюленей в 3—5 раз тяжелее, чем у Otariinae [Соколов, 1973].
Для более ясного представления о развитости мышц передней и задней ко-
нечностей морского котика воспользуемся данными В. Ф. Мужчинкина [19716].
Сгибатели и разгибатели плечевого сустава примерно равны по массе, но втрое
уступают подлопаточной мышце, приводящей плечо. Разгибатели предплечья в
5 раз слабее сгибателей. Ягодичные мышцы втрое слабее абдукторов, в то время
как сгибатели и разгибатели голени примерно одинаково развиты. Разгибатели
голеностопного сустава в 1.5 раза сильнее сгибателей, а для пальцев задней ко-
нечности характерна значительная развитость сгибателей и разгибателей.
Приведенные данные показывают, что проксимальные отделы конечностей
северного морского котика обслуживаются более мощной мускулатурой, чем дис-
тальные. При этом в передней конечности эта закономерность выражена ярче.
Сгибание передней и задней конечностей разнообразней, чем разгибание.
Так, на пальцах (от 1-го к 5-му) передней конечности при сгибании действуют,
соответственно, 4-4-5-4-3 сухожильные тяжа, а разгибают только 3-2-1-2-3.
Сгибают пальцы задней конечности 3-2-2-2-4, а разгибают 2-1-1-1-1 сухо-
жилия. В общем, сгибание и разгибание задней конечности менее разнооб-
разны, чем передней. В задней конечности более подвижны крайние пальцы,
в передней более разнообразны движения при сгибании средних, а при раз-
гибании — крайних пальцев. Описанная картина связей сухожильных голОвок
мышц с пальцами не постоянна. Могут быть широкие вариации [Мужчинкин,
19716], но закономерным остается преобладание сухожильных связей пальцев
передней конечности над задней и преобладание мышц-сгибателей над раз-
гибателями в обеих конечностях.
105
Глава 7. ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА И ОРГАНЫ
ЧУВСТВ
Уже в прошлом веке была предпринята попытка описать центральную нерв-
ную систему северного морского котика [Fish, 1899]. Современные работы в не-
которых деталях дополнили уже имеющиеся данные [Никитенко, 1966; Собо-
левский, 1978а, б]. Цитоархитектоника некоторых отделов головного мозга ко-
тика изучена Л. И: Авксентьевой [1974].
Орган слуха северного морского котика описан Л. Н. Богословской и Г. Н.
Солнцевой [1979]. Органу зрения уделено гораздо меньше внимания [Андреев,
1978], хотя некоторые виды ластоногих изучены в этом отношении достаточно
полно [Jamieson, Fisher, 1972].
7.1. Форма, размеры и внешнее строение головного мозга
Сверху головной мозг котика имеет почти округлую форму с несколько вы-
ступающими за пределы окружности латеральными краями затылочных и лоб-
ных долей. Сбоку форма его мозга яйцевидная; широкая часть обращена к за-
тылку. Основание мозга плоское и широкое в аборальной части, а ростральные
отделы слегка срезаны с боков в результате вдавливания в мозговую полость
глазничных отделов лобных костей. Мозжечок крупный, почти полностью (чаще
на треть) прикрыт сверху большими полушариями. Обонятельные луковицы раз-
виты не сильно и незначительно выступают за пределы полушарий.
Серповидная складка, разделяющая полушария, имеет большую глубину в
области обонятельных луковиц, затем постепенно становится менее глубокой
над мозолистым телом и вновь углубляется по направлению к мозжечку. В зад-
ней части она присоединяется не к костному намету, отделяющему мозжечок
от полушарий, а к костной пластинке, отходящей перпендикулярно от костного
намета (tentorium osseum) и вклинивающейся в продольную щель (fissura lon-
gitudenalis) мозга.
Наибольшее развитие у котика имеют большие полушария и мозжечок, на
долю которых приходится почти 90% массы всего мозга. Менее развиты обо-
нятельные луковицы, продолговатый и средний мозг (табл. 7.1).
У исследованных нами котиков наибольшая масса свежего головного мозга
составила 415 г, мозжечка 85 г (табл. 7.1) или около 0.6% от массы тела (табл.
7.2). Абсолютная масса мозга больше у самцов, чем у самок, поскольку масса
тела самцов значительно больше. Однако, если абсолютная масса мозга самцов
больше, чем у самок, в среднем в 1.2 раза, то размеры тела самцов больше в
4—5 раз. Поэтому относительные показатели массы мозга у половозрелых самок
примерно в 3 раза больше, чем у самцов (табл. 7.2).
Относительные показатели массы головного мозга у котиков с возрастом
уменьшаются. Отмечается также снижение абсолютной массы головного мозга
у старых (старше 10 лет) животных [Соболевский, 1978а; табл. 7.2], хотя по от-
106
Таблица 7.1
Масса частей головного мозга морского котика [из: Соболевский, 19786]
Части мозга Предродовые эмбрионы Неполово- зрелые самцы и самки Половозре- лые самки Холостяки 3—5 лет Полусекачи и секачи
А б с о л ю т н а я м с с с а (г)
Обонятельные луковицы 0.51 0.89 0.96 0.96 1.10
Большие полушария 133.03 206.42 202.58 222.20 225.72
Промежуточный мозг 7.21 10.13 10.62 11.88 12.16
Средний мозг 4.81 7.22 7.30 7.67 8.52
Мозжечок 26.47 47.67 44.62 45.32 46.60
Варолиев мост 2.57 5.80 5.74 6.30 6.02
Продолговатый мозг 2.49 5.08 5.47 5.79 6.02
Относителъ н а я мае с а (% о т общей л а с сы Mt з г а)
Обонятельные луковицы 0.29 0.31 0.33 0.32 0.36
Большие полушария 75.12 72.89 76.06 74.00 73.59
Промежуточный мозг 4.07 3.58 3.83 3.96 3.96
Средний мозг 2.74 2.56 2.63 2.56 2.78
Мозжечок 14.92 16.82 16.09 15.05 15.19
Варолиев мост ' 1.45 2.05 2.07 2.10 1.97
Продолговатый мозг 1.41 1.79 1.97 2.01 2.15
дельным возрастным классам материала недостаточно для окончательной кон-
статации этого феномена.
В связи с ранним развитием головного мозга в онтогенезе больших различий
в соотношении его частей ни по полу, ни по возрасту не наблюдается. Как
правило, масса правого и левого полушарий несколько различается, но какой-
либо закономерности в асимметрии мозга, описанной М. Ф. Никитенко [1966],
нам установить не удалось, т. к. в одних случаях тяжелее левое, а в других слу-
чаях — правое полушарие.
Кора больших полушарий у северного морского котика изрезана бороздами
и собрана в многочисленные складки, образующие причудливое переплетение
извилин (рис. 7.1). По степени гирификации северный морской котик заметно
выделяется среди других представителей отряда [Fish, 1899]. Однако, несмотря
на наличие большого числа вторичных и третичных борозд, все же представ-
ляется возможным по их глубине достаточно точно определить ход основных
из них даже у взрослых особей, а имея препараты головного мозга на разных
стадиях индивидуального развития, тем более.
Поверхность коры больших полушарий котика (рис. 7.1) собрана в несколько
основных складок, образующих ряд дугообразных извилин вокруг сильвиевой
107
Таблица 7.2
Масса мозга северного морского котика
Возраст, лет Пол п Абсолютная масса, г Относительная масса, % от массы тела
lim M±m lim Mim
Новорожденные самцы 1 120 120 1.7 1.7
самки 4 151—187 165110.20 1.80—2.20 l.S+10.04
1 самцы 1 220 220 1.10 110
3 самцы 1 290 290 1.12 1.12
самки 180—200 190 0.67—0.73 0.70
4 самцы 2 200—220 210 0.36—0.52 0.44
самки 1 160 160 0.53 0.53
5 самцы 1 190 190 0.38 0.38
самки 1 200 200 0.50 0.50
6 самки 3 200—220 21118.40 0.47—0.50 0.510.0
7 самцы 1 235 235 0.19 0.19
самки 3 170—235 205127.31 0.38—0.71 0.5210.1
8 самцы 2 260—270 265 0.19 0.19
самки 6 200—290 235115.68 0.42—0.76 0.5610.6
9 самцы 5 210—300 254119.31 0.14—0.25 0.1710.02
самки 6 190—240 210+8.86 0.33—0.60 0.46Ю.05
10 самцы 1 270 270 0.14 0.14
самки 4 185—300 232132.30 0.40-0.67 0.5310.08
>10 самки 20 105—300 216113.84 0.20-0.73 0.47Ю.03
борозды (sulcus lateralis sylvii). В отличие от многих наземных млекопитающих,
у северного морского котика сильвиева борозда образование сложное, поэтому
ее чаще называют псевдосильвиевой бороздой или ямкой [Fish, 1899]. Она за-
нимает почти вертикальное положение, начинаясь от латеральной области обо-
нятельного мозга, и фактически не выходит на дорсальную поверхность полу-
шарий. В глубине псевдосильвиевой ямки находится очень короткая сильвиева
борозда и сильно редуцированная сильвиева извилина (girus sylvius). На дне
сильвиевой борозды можно различить слабовыраженный островок (insula). S. ес-
tosylvius anterior отсутствует, a s. ectosylvius posterior скрыта в стенке псевдо-
сильвиевой ямки и с внешней стороны не видна. От эктосильвиевой извилины
сохранилась лишь s. ectosylvius posterior, занимающая довольно-таки обширное
поле височной области коры мозга.
Располагающаяся над сильвиевой извилиной супрасильвиева борозда (s. supra-
sylvius) в одних случаях образует завершенную арку, в других — образует 3 части:
ss. suprasylvius dorsalis, medialis, ventralis [Fish, 1899]. Супрасильвиева борозда на-
чинается несколько выше вентрального края плаща, направляется дорсо-каудально,
108
2
Рис. 7.1. Латеральная поверхность головного мозга северного морского котика:
1 — sulcus sylvii, 2 — s. suprasylvius, 3 — s. coronalis, 4 — s. postcruciatus, 5 — s. praesylvius,
6 — s. cruciatus, 7 — s. ansatus, 8 — s. marginalis, 9 — s. endolateralis, 10 — s. lateralis.
Рис. 7.2. Базальная сторона
головного мозга северного
морского котика:
I — nervus olfactorius, II — n.
opticus, III — n. oculomotorius,
IV — n. trochlearis, V — n. tre-
geminus, VI — n. abducens, VII —
n. fascialis, VIII — n. vestibulo-
cochlearis, IX — n. glossopharyn-
geus, X — n. vagus, XI — n. ac-
cessorius, XII — n. hypoglossus;
1 — tractus olfactorius, 2 —
chiasma opticus, 3 — hypophysis,
4 — corpus mamilare, 5 — lobus
piriformis, 6 — pons cerebri, 7 —
pyramis, 8 — medulla oblongata.
круто изгибается вокруг псевдосильвиевой ямки и заканчивается вилкой, кау-
дальный рожок которой направляется к псевдосильвиевой ямке. Однако в нее
не впадает, наблюдается лишь поверхностная связь. Другой рожок идет к пет-
листой борозде (s. ansatus). Расположенная над супрасильвиевой бороздой од-
ноименная извилина очень широкая в аборальной части и в отдельных местах
109
прорезана глубокими щелями, отходящими от супрасильвиевой борозды. В до-
рсальной части супрасильвиева извилина резко сужается. Иногда супрасильвиева
извилина имеет короткую щель, начинающуюся с вентральной ее части и про-
стирающуюся до трети ее высоты. Она условно названа нами постсупрасиль-
виевой бороздой. По всей площади супрасильвиевой извилины имеется много
коротких и длинных, глубоких и мелких борозд второго и третьего порядка,
следующих в самых различных направлениях.
Ограничивающая медиально супрасильвиеву извилину латеральная борозда
(s. lateralis) следует от основания мозга дорсокаудально, затем в дорсальной час-
ти резко поворачивает орально и на уровне вершины псевдосильвиевой ямки
принимает вентральное направление (рис. 7.1). Она заканчивается немного ниже
уровня вершины псевдосильвиевой ямки, переходя в петлистую борозду (s. ап-
satus).
Крупная латеральная извилина в затылочной области разделена глубокой ме-
диолатеральной щелью (fissura lateralis medialis). В теменной области плаща хо-
рошо выражена маргинальная борозда (s. marginalis), переходящая с мезальной
поверхности полушария. Краниальнее начинается крестовидная борозда^. cru-
ciatus), идущая краниолатерально, от вентрального конца которой к продольной
щели идет короткая предкрестовидная борозда (s. praecruciatus), прорезающая
дорсальный край полушария. Этими двумя бороздами отграничен так называе-
мый “медвежий ромб” — характерный признак представителей отряда хищных.
У северного морского котика “медвежий ромб” хорошо выражен на всех стадиях
индивидуального развития, особенно в пренатальный период. Кроме названных
борозд, на латеральной стороне мозга морского котика ниже анзатной борозды
хорошо выражены коронарная, предкрестовидная, супраорбитальная и диаго-
нальная борозды.
Не менее сложно, чем дорсальная, устроена вентральная поверхность голов-
ного мозга. Обонятельные доли представлены едва заметным расширением обо-
нятельного тракта. Сам обонятельный тракт узкий, особенно в средней части,
но постепенно расширяется аборально, где образует обонятельный треугольник
(рис. 7.2).
Каудально обонятельный тракт у котика отделен глубокой и широкой про-
дырявленной пластинкой от грушевидных долей и, таким образом, какой-либо
видимой связи между ними нет. За обонятельным треугольником между хорошо
обособленными грушевидными долями лежит зрительный перекрест (hiazma),
позади которого расположен серый бугор с воронкой и гипофизом.
В аборальной области основания головного мозга поперек продолговатого
мозга лежит мост. Это хорошо выраженное образование (рис. 7.2, 7.3)
В начальном отделе продолговатого мозга заметны пирамиды. Мост и пи-
рамиды несут на себе продольную борозду (s. medianus). Каудально пирамиды
исчезают, и это место называется перекрестом пирамид (decussatio piramidum).
Вдоль латерального края обонятельных трактов от краниального края полу-
шарий мозга и до грушевидных долей идет неглубокая базальная или обоня-
тельная борозда (s. rhinalis), которая следует к псевдосильвиевой ямке (fossa pseu-
dosylvius). За псевдосильвиевой бороздой она продолжается к щели, отделяющей
ПО
Рис. 7.3. Сагиттальный разрез головного мозга северного морского котика:
1 — спинной мозг, 2 — мозжечок, 3 — мост, 4 — грушевидная доля, 5 — гипофиз, 6 —
зрительный нерв, 7 — girus olfactorius, 8 — обонятельный тракт, 9 — обонятельная доля,
10 — sulcus olfactorius, 11 — hippocampus, 12 — s. cruciatus, 13 — s. gennalis, 14 — s.
hippocampus, 15 — gyrus cinguli, 16 — s. marginalis, 17 — s. splenialis, 18 — s. mediolateralis,
19 — s. postsplenialis.
мозжечок or большого мозга, однако, не доходя несколько миллиметров до нее,
обрывается. Латеральнее обонятельной щели хорошо заметна очень глубокая
и рано закладывающаяся в онтогенезе надорбитальная борозда (s. supraorbitalis).
Она начинается почти от переднего края полушарий, следует, несколько изги-
баясь, в латеральном направлении к грушевидным долям, но, не доходя несколь-
ко миллиметров до обонятельной борозды, обрывается. Большая часть псевдо-
сильвиевой борозды также находится на базальной поверхности мозга. Здесь же
видны начальные отделы окружающих ее других борозд.
Мозолистое тело дорсально отграничено от близлежащих тканей глубокой
бороздой мозолистого тела (s. callosi), передний отдел которой обычно назы-
вается бороздой колена, а задний — бороздой валика (рис. 7.3). Борозда колена
в отдельных случаях может сливаться с предкрестовидной бороздой [Fish, 1899]
или подходить вплотную к ней, не соединяясь. Борозда валика огибает мозо-
листое тело и круто спускается к основанию мозга. В месте перегиба от назван-
ной борозды отходят 2 борозды, следующие к затылочной области мозга. Кау-
додорсальнее сплениальной борозды, в развиже между сплениальной и верти-
кально отходящими от нее другими бороздами, лежит маргинальная борозда,
которая также вливается в перпендикулярно идущую короткую борозду, назван-
ную нами постмаргинальной. По каудальному краю затылочной области мозга
проходит медиолатеральная борозда. Борозда гиппокампа хорошо выражена, но
не видна из-за налегающих на нее других частей мозга, формирующих его ствол.
Извилины, располагающиеся между названными бороздами, несут на себе много
вторичных и третичных борозд, идущих в самых разных направлениях.
111
Рис. 7.4. Дорсальная и базальная поверхности головного мозга эмбрионов и
новорожденного котика:
А — начало 6-го месяца внутриутробного развития (масса тела 280 г, масса мозга 4 г), Б —
7-й месяц (масса тела 540 г, масса мозга 15.5 г), В — 8-й месяц (масса тела 1 кг, масса мозга
38.6 г), Г — новорожденный (масса тела 4.5 кг, масса мозга 151 г).
7.2. Развитие головного мозга в онтогенезе
У эмбрионов северного морского котика от самок, добытых в январе (6-ме-
сяцев, масса тела 280 г, масса мозга 4 г), полушария с дорсальной поверхности
фасолевидные (рис. 7.4).
Хорошо обособлены лобные и затылочные отделы; намечается височный по-
люс. На латеральной поверхности полушарий начинается формирование широ-
кой, но еще не глубокой псевдосильвиевой ямки. Варолиев мост нечетко отделен
от продолговатого мозга, но хорошо обособлен от ножек среднего. Хорошо вы-
ражен продолговатый мозг, четверохолмие, подбугорье, в особенности серый бу-
гор, обонятельные луковицы и обонятельный тракт. Конечный мозг глубокой
продольной щелью разделен на 2 полушария, еще не несущих на себе никаких
извилин и борозд.
В среднем мозге хорошо выражено четверохолмие, в котором ярче других
структур выделяется переднее двухолмие. Ножки мозга несут на себе заметную
межножковую ямку. В промежуточном мозге ясно выражен серый бугор, в ко-
тором можно различить небольшое шишковидное тело. Эпифиз в виде незна-
чительного выступа обращен в борозду между верхним двухолмием. Перекрест
зрительных нервов и зрительный тракт представлены небольшим количеством
волокон. Мозжечок хорошо выражен. Отчетливо выступает червь, правое и
левое полушария мозжечка, на поверхности которых уже заметны борозды, раз-
деляющие его на пластины.
112
Рис. 7.5. Латеральная поверхность головного мозга 7-месячного эмбриона северного
морского котика:
1 — sulcus sylvius, 2 — s. suprasylvius, 3 — s. postsuprasylvius, 4 — s. lateralis, 6 — s. margjnalis,
7 — s. ansatus, 8 — s. cruciatus, 9 — s. coronalis, 10 — “медвежий ромб”, 13 — s. supra-
orbitalis, 16 — мозжечок, 17 — спинной мозг, 18 — s. ectosylvius anterior, 19 — тройничный
нерв, 20 — зрительный нерв, 21 — гипофиз, 22 — bulbus olfactorus, 23 — островок Рейля.
Рис. 7.6. Латеральная поверхность головного мозга 8-месячного эмбриона северного
морского котика,
Обозначения те же, что на рис. 7.5.
У эмбрионов в конце 7-го месяца внутриутробного развития (рис. 7.5) при
массе тела 540 г и массе мозга 15.5 г начинается формирование борозд и извилин
на полушариях. На латеральной поверхности формируется глубокая и широкая
псевдосильвиева ямка. Она начинается от выпуклых грушевидных долей и идет
вертикально до средней общей высоты мозга. По передней стенке псевдосиль-
113
виевой ямки намечается неглубокая эктосильвиева борозда, а над ней заклады-
вается короткая, но глубокая супрасильвиева (рис. 7.5). Спереди от нее уже за-
метна неглубокая, но широкая коронарная борозда. Формирование латеральной
борозды начинается с ее переднего участка и ее продолжения — петлистой бо-
розды. Очень хорошо сформирована крестовидная борозда и предкрестовидная
щель, а образованный ими “медвежий ромб” утопает в прилежащих частях моз-
га. Формирующиеся надкрестовидная и сигмоидная извилины кажутся исклю-
чительно выпуклыми. На базальной поверхности мозга заложились пограничная
и надглазничная борозды.
У 8-месячных эмбрионов (масса тела 1 кг, масса мозга 38 г) очень рельефно
обозначены все части мозга. Хорошо развит продолговатый мозг, отчетливо вы-
деляются оливы, пирамида и задние канатики (рис. 7.6).
Варолиев мост отделен от продолговатого и среднего мозга. Хорошо развит
мозжечок, в т. ч. его полушария. Не менее ярко выражена самая древняя часть
мозжечка — клочок.
В среднем мозге хорошо обозначено четверохолмие, а в нем — переднее дву-
холмие. Ножки мозга утолщены. Межножковая ямка неглубокая. Зрительные
бугры выражены слабо. Эпифиз хорошо развит. По-видимому, коленчатое тело
слилось с передним двухолмием. Зрительный тракт и хиазма хорошо дифферен-
цированы. Серый бугор массивен, воронка соединяется с гипофизом.
В конечном мозге четко обособлены и хорошо развиты структуры обоня-
тельных долей. Относительно крупные луковицы продолжаются широким обо-
нятельным трактом, переходящим в обонятельный треугольник. Грушевидные
доли выпуклые, спереди резко отграничены пограничной бороздой, из которых
начинается псевдосильвиева ямка. В свою очередь, пограничная борозда про-
должается каудально, но не доходит до поперечной щели, отграничивающей ко-
нечный мозг от мозжечка.
Поверхность плаща покрыта глубокими и широкими бороздами, а извилины,
лежащие между ними, рельефно выступают на поверхность. На латеральной по-
верхности плаща, ближе к передней части мозга, выделяется идущая под прямым
углом от основания мозга глубокая и широкая псевдосильвиева ямка. Она не
выходит на дорсальную сторону плаща и заканчивается примерно на середине
дорсовентральной проекции мозга. Начинается она как продолжение обонятель-
ной борозды спереди и продолжается в заднем направлении как постобонятель-
ная борозда. Из-за такого положения псевдосильвиевой ямки грушевидные доли
вклиниваются своими латеральными краями в псевдосильвиеву ямку. На дне
псевдосильвиевой ямки намечается образование короткой сильвиевой борозды.
Псевдосильвиеву ямку окружает хорошо выраженная, рельефно выступающая
эктосильвиева извилина, в которой присутствуют передняя и задняя части.
Супрасильвиева борозда очень широкая и глубокая. Ее задняя часть (s. silvius
posterior) состоит как бы из трех отдельных коротких борозд: дорсальной, сред-
ней и вентральной, каждая из которых отделена невысокими перемычками, края
которых, однако, находятся ниже уровня окружающих их извилин. Все 3 бо-
розды, являющиеся продолжением одна другой, начинаются выше пограничной
борозды, следуют краниально, огибают псевдосильвиеву ямку, в одних случаях
впадают в ее дорсальный край, в других — продолжаются самостоятельной бо-
роздой почти до пограничной борозды. У некоторых эмбрионов в этом возрасте
хорошо заметна постсупрасильвиева борозда, идущая от основания мозга и про-
114
резающая супрасильвиеву извилину примерно на трети расстояния дорсовент-
ральной проекции мозга.
Латеральная борозда следует по периферии полушария, начинаясь у его ос-
нования выше пограничной борозды, огибает супрасильвиеву извилину и в лоб-
ном отделе переходит в петлистую борозду.
В лобном отделе мозга выражена коронарная борозда, начинающаяся выше
пограничной и заканчивающаяся краниальнее анзатной небольшой вилкой.
С медиальной поверхности лобного отдела мозга на его латеральную по-
верхность переходит очень глубокая крестовидная борозда, которая следует ла-
теровентрально, где ее вентральный конец упирается в предкрестовидную бо-
розду, следующую к среднемозговой щели и прорезающую глубокой щелью по-
лушария мозга у последней. Образованное этими двумя бороздами треугольное
поле несколько вдавлено в мозг. Это особенно заметно у эмбрионов на более
ранних стадиях развития и является частью “медвежьего ромба”. Между коро-
нарной и крестовидной бороздами располагаются одна над другой 2 короткие
борозды, верхняя из которых может быть названа посткрестовидной, нижняя —
постнадорбитальной. Небольшой участок маргинальной борозды переходит с
мезальной поверхности мозга на дорсальную в области темени.
В затылочной области хорошо выражена медиолатеральная борозда, идущая
параллельно центральной борозде и поднимающаяся до уровня теменных долей.
На мезальной поверхности полушарий хорошо выражена большая щель. Бо-
розда валика глубокая и широкая, огибает мозолистое тело. Начинается она
двумя рожками над серединой мозолистого тела и следует в каудальном направ-
лении, где над валиком мозолистого тела под прямым углом спускается вен-
трально и впадает в другую щель, перед этим отдавая небольшую борозду, иду-
щую к теменно-затылочному углу. Эта короткая щель гомологична постспле-
ниальной борозде, а перпендикулярная, более длинная борозда, подходящая
своим дорсальным концом почти к медиальной щели, может соответствовать
затылочной щели других млекопитающих. Гиппокомпальная щель выходит
дугой из утолщения мозолистого тела, огибает зрительные бугры и следует к
краю грушевидной доли. По каудальному краю мезальной поверхности полуша-
рий проходит глубокая и широкая медиолатеральная борозда.
На вентральной поверхности мозга хорошо выражены обонятельная и суп-
раорбитальная щели. Начальный отдел псевдосильвиевой ямки представляет
собой выемчатое углубление.
Итак, у 8-месячных эмбрионов заканчивается образование основных борозд
головного мозга. На последующих месяцах эмбрионального развития происхо-
дит доформирование вторичных и третичных борозд, а также смыкание стенок
уже заложившихся основных, поэтому мозг новорожденных и взрослых котиков
представялет собой компактное образование, испещренное множеством изящных
линий, показывающих пути прохождения борозд. Внешняя архитектоника их
мозга почти не различается по степени тарификации, наблюдается лишь про-
грессирующая оперкуляция височного края мозга. Еще глубже погружаются в
псевдосильвиеву ямку сильвиева, передняя эктосильвиева и супрасильвиева из-
вилины.
115
Общая площадь всей коры переднего мозга у взрослых котиков составляет
45708 мм2, в т. ч. новая кора — 43937 мм2, а комплекс древней, старой и ме-
жуточной коры — 1771 мм2, т. е. новая кора составляет 96.1%, а древняя, старая
и межуточная кора — 3.9% [Никитенко, 1966]. В приведенном ряду относитель-
ной величины новой коры северный морской котик стоит ниже дельфина (97.8%),
но выше человека (95.9%). По данным Л. И. Авксентьевой [1974], кора головного
мозга северного морского котика состоит из 6 слоев. Строение ее сходно со стро-
ением коры наземных хищных. Общая толщина коры составляет 1.6—2.1 мм.
Горизонтальная и вертикальная исчерченность выражены хорошо, все 6 слоев
ясно разграничены. Клетки четвертого слоя располагаются колонками. Апику-
лярные дендриты нейронов 3—5-го слоев хорошо ориентированы, в то время как
у китообразных клетки этих слоев разрежены, разные по форме и апикальные
дендриты ориентированы под углом к вышележащим слоям [Авксентьева, 1974].
Цитологическое строение коры головного мозга северного морского котика
представляется хорошо упорядоченным. Более тщательных исследований голов-
ного мозга котика не проводилось, и поэтому, по имеющимся данным, можно
говорить о степени его цефализации лишь по аналогии с уже хорошо изучен-
ными млекопитающими. Конечно, гомологизация отдельных участков неокор-
текса у разных, пусть даже близких представителей класса на основании только
сходства их внешнего строения затруднительна, однако даже в филогенетичес-
ком ряду млекопитающих сохраняется основной план расположения корковых
зон [Ладыгина, Супин, 1974, 1978]. Границами между морфофункциональными
подразделениями коры служат борозды. Борозды первого порядка, как правило,
разделяют отдельные крупные корковые области, а мелкие — зоны представи-
тельства отдельных частей тела [Welker, Campos, 1963]. Если исходить из этих
позиций, то можно провести некоторые аналогии по поводу функционального
обоснования развития отдельных областей мозга котика и некоторых хорошо
изученных представителей хищных (кошка, собака, медведь, некоторые кунеце-
вые), с одной стороны, и вторичноводйых (дельфины), с другой.
Намечающаяся у некоторых хищных оперкуляция извилин височной области
у котика представляет завершенную картину. Как было сказано выше, в псев-
досильвиеву ямку погружены сильвиева и эктосильвиева извилины. Эта область
коры мозга у хищных считается слуховой зоной. Гистологическое исследование
участка коры мозга котика, взятого из эктосильвиевой извилины, показало при-
сутствие всех цитоархитектонических формаций, в т. ч. ширину и степень стра-
тификации коры, что позволяет говорить о достаточно высоком уровне развития
в данной области коры соответствующих функций, управляющих слуховым ана-
лизатором [Ладыгина, Супин, 1974].
Латеральная извилина у котика хорошо обозначена и занимает обширную
область теменно-затылочного комплекса. У хищных и китообразных значитель-
ную часть ее представляет зрительная проекционная зона. По-видимому, не
будет большой ошибкой говорить о хорошем развитии и значительном пред-
ставительстве в коре мозга северного морского котика отделов зрительного ана-
лизатора в связи с четкой выраженностью этой области в коре мозга.
Представляет; интерес, архитектоника коры лобного отдела мозга у котика.
Коронарная извилина у взрослых особей относительно менее выражена, чем, на-
пример, у калана [Engel, Dillon, 1972], медведя [Fish, 1899] и даже у ископае-
116
могоушастого тюленя Enaliarctos mealsi [Mitchell, Tedford, 1973]. По мнению по-
следних авторов, это связано с преобразованием соматического сенсорного поля I
от тактильных манипуляторных органов предков ушастых тюленей к органам
локомоции ныне живущих ластоногих.
Сигмовидная и посткрестовидная извилины у котика развиты относительно
слабо. У енотовых [Welker, Campos, 1963] и у медведей [Smith, 1933] эти участки
коры мозга хорошо выражены, в них соматически представлены передняя и за-
дняя конечности. Манипуляторная деятельность передних конечностей ряда на-
земных хищных хорошо известна, а у котика этим органам присуща, в основном,
функция движителей.
Таким образом, имеющиеся далеко неполные сведения о строении головного
мозга северного морского котика на макро- и микроуровнях позволяет пред-
полагать довольно сложную организацию его высшей нервной деятельности. Из-
вестно, что у водных млекопитающих, живущих в трехмерном пространстве, ор-
ганизация высшей нервной деятельности всегда выше, чем у животных, живущих
в двухмерном пространстве, а у общественных животных выше, чем у ведущих
одиночный образ жизни. Образ жизни морского котика таков, что он часть
жизни проводит на земле, где исключительно хорошо проявляется его общест-
венность, другая часть годового жизненного цикла проходит в море, где жи-
вотные не контактируют с берегом, совершая самые большие по протяженности
миграции среди ластоногих. В этой связи есть все основания предполагать, что
при более тщательном исследовании головного мозга котика будут выявлены
соответствующие особенности его строения.
7.3. Черепномозговые нервы
От вентральной стороны головного мозга отходят 12 пар черепномозговых
нервов. Существенных отличий в ходе и расположении их, по сравнению с дру-
гими млекопитающими, нет (рис. 7.2).
I пара. Обонятельный нерв (nervus olfactorius) — чувствительный. Первичные
центры обонятельного анализатора — обонятельные луковицы. Их длина у
взрослых животных составляет 10—13 мм, ширина 6—7 мм, абсолютная масса
0.96—1.1 г, что составляет от массы всего мозга 0.35—0.36%.
II пара. Зрительный нерв (n. opticus) — чувствительный. Через зрительное
отверстие он выходит из мозгового отдела черепа и вступает в мозговую по-
лость, где выше серого бугра с противоположным нервом другой половины
мозга образует зрительный перекрест (chiazma opticum). Образуемый ими угол
составляет не более 15—20°.
III пара. Глазодвигательный нерв (n. oculomotorius) — двигательный. Хорошо
развит, выходит по центру ножек большого мозга и следует к гипофизу, но затем
резко поворачивает медиально, направляясь к глазному яблоку и глазничным
мышцам, которые он обслуживает.
IV пара. Блоковый нерв (n. trochlearis) — двигательный. Тонкий нерв, от-
ходящий от мозга у латерального края назального паруса; появляется на ла-
теральной поверхности в углу между мостом и продырявленной пластинкой.
117
V пара. Тройничный нерв (n. trigeminus) — смешанный (чувствительный и
двигательный). Как и у других млекопитающих, он отходит двумя корешками
от латерального края моста, из которых медиальный — чувствительный. Выходя
из черепной коробки, последний образует мощный гессеров узел, от которого
отходят три ветви. Одна из них, соединяясь с медиальным корешком, образует
нижнечелюстной нерв, а два других обслуживают нижнюю челюсть и глаз.
VI пара. Отводящий нерв (n. abducens) развит слабо. Отходит от продолго-
ватого мозга в самом его начале и следует к глазному яблоку, где иннервирует
мышцы глаза.
VII пара. Лицевой нерв (n. facialis) — смешанный. Регулирует вкусовую ре-
цепцию, двигательную функцию головы и выполняет регуляцию секреторной
функции. Хорошо развит; по диаметру из нервов мостомозжечкового угла ус-
тупает только тройничному и слуховому.
VIII пара. Слуховой, или преддверноулитковый, нерв (n. acusticus = n. ves-
tibulocochlearis) — увствительный. Корешок нерва мощный. Выходит из кранио-
латерального угла продолговатого мозга рядом с лицевым нервом. Обслуживает
внутреннее и среднее ухо.
IX пара. Языкоглоточный нерв (n. glossopharyngeus) — смешанный: вкусовой
и двигательный. Обслуживает корень языка и глотку. Очень тонкий нерв. От-
ходит от продолговатого мозга каудальнее слухового нерва.
X пара. Блуждающий нерв (n. vagus) — смешанный. Является составной
частью парасимпатической системы и обслуживает многие органы.
XI пара. Добавочный нерв (n. accessorius) — двигательный. Формируется ко-
решками из продолговатого мозга (головная часть) и корешками из шейных нер-
вов, образующих общий ствол, выходящий через рваное отверстие из полости
черепа. Иннервирует мышцы шеи и груди.
XII пара. Подъязычный нерв (n. hypoglossus) — двигательный. Иннервирует
мышцы языка и подъязычной кости. Отходит от внутренней поверхности про-
долговатого мозга.
7.4. Мозжечок
Как и у других водных млекопиающих (Клейненберг и др., 1964; Берзин, 1971
и др.), мозжечок у северного морского котика хорошо развит. У взрослых жи-
вотных он достигает 85 г (50—85 г), что составляет в среднем 0.24% (0.03—0.26%)
от массы тела или 16—17% от массы головного мозга (табл. 7.1, 7.3).
У 8-месячных эмбрионов он меньше (0.6—0.7%), а к периоду рождения уве-
личивается, соответственно, у предродовых плодов до 15%, у новорожденных
ок. 17% от массы мозга. Абсолютная масса мозжечка больше у самцов, а от-
носительная — у самок (табл. 7.1, 7.3). Длина его достигает у взрослых особей
55 мм, ширина 84 мм, высота без моста 28 мм, а с мостом 45 мм.
По данным М. Ф. Никитенко [1966], площадь поверхности мозжечка равна
17896 мм2, что составляет 39% от поверхности всего переднего мозга. Из на-
118
Таблица 7.3
Масса мозжечка северного морского котика
Возраст, лет Пол п Абсолютные данные, г По отношению к массе тела, %
lim M+m lim М±/и
Новорожденные самцы 1 41 41.0 0.40 0.40
самки 4 33—36 35.5 0.35-0.40 0.40
1 самцы 1 60 60.0 0.30 0.30
3 самцы 1 61 61.0 0.22 0.22
самки 2 60—70 65.0 0.23—0.24 0.24
4 самцы 2 60 60.0 0.11—0.14 0.13
самки 1 40 40.0 0.13 0.13
5 самцы 1 50 50.0 0.10 0.10
самки 1 65 65.0 0.16 0.16
б самки 2 50—70 60.0 0.11—0.15 0.13
7 самцы 1 65 65.0 0.05 0.05
самки 3 20—85 51.67±27.31 0.04—0.26 0.14±0.09
8 самцы 2 65—80 72.50±4.43 0.05—0.06 0.06
самки 6 60—85 73.33+4.43 0.15—0.21 0.17±0.01
9 самцы 5 60—80 66.00±4.29 0.03—0.06 0.04±0.01
самки 6 35—80 60.83+7.97 0.06—0.20 0.14±0.02
10 самцы 1 70 70.0 0.04 0.04
самки 4 50—70 60.00±5.62 0.09—0.19 0.14±0.06
>10 самки 17 50—85 65.00+2.44 0.09—0.24 0.14±0.01
званного показателя общей площади поверхности мозжечка 84.8% приходится
на поверхность самого мозжечка и 15.2% на поверхность червячка.
Являясь центральным органом рефлекторного регулирования тонуса скелет-
ных мышц, мозжечок координирует их сокращение и обеспечивает основные
двигательные функции организма. Одновременно он является подкорковым цен-
тром вегетативной нервной системы, регулирующим обмен веществ, деятель-
ность сердечно-сосудистой, пищеварительной и других систем. Высокий уровень
развития (большая поверхность, масса) мозжечка у котика предполагает широ-
кие возможности этого органа в регуляции основных процессов организма, свя-
занных с обитанием в водной среде, что сопряжено с интенсивной подвиж-
ностью, маневренностью, продолжительностью и глубиной ныряния.
7.5. Спинной мозг
Спинной мозг северного морского котика представляет собой толстый тяж
(рис. 7.7), покрытый тремя оболочками: мягкой, паутинной и твердой, из ко-
119
Рис. 7.7. Спинной мозг северного морского котика
Таблица 7.4
Возрастные изменения спинного мозга северного морского котика
[по: Соболевский, 19786]
Возрастные группы (я) Длина мозгового конуса Масса мозга
см в % к длине тела Г в % от массы тела
Предродовые плоды (1) 24.3 С а м ц ы 42.6 9.6 0.16
Новорожденные (4) 24.7—30.2 34.911.5 11.8 —14.3 0.1510.01
I—2 года (9) 28.7—34.0 33.7±0.7 16.7—22.8 0.1010.01
3—4 года (11) 35.8—45.7 32.310.8 22.8—31.0 0.0710.00
5—7 лет (9) 45.0-55.1 30.810.7 31.0—53.2 0.0510.00
8 лет (3) 53.2—60.1 29.811.7 48.8—75.4 0.0410.00
С а м к и
Предродовые плоды (5) 21.4—23.9 38.011.9 7.8—10.2 0.1610.02
Новорожденные (6) 22.8—28.6 34.610.9 9.4—13.0 0.1610.00
2—5 лет (8) 32.0—40.0 35.511.3 20.5—26.0 0.0710.00
6—10 лет (26) 33.0—53.7 33.910.9 19.7—36.7 0.0710.00
11—22 года (14) 37.6—50.0 33.610.6 22.7—38.1 0.0610.00
торых последняя очень сильно развита. Эпидуральное пространство небольшого
размера.
Форма спинного мозга конусовидная. Имеется 2 утолщения: шейное и по-
ясничное. У котика с длиной тела 127 см толщина спинного мозга на уровне
эпистрофея равна 1 см, на уровне 9—10 грудных позвонков — 0.8 см. Топо-
графически шейное утолщение начинается на третьем и заканчивается на шестом
шейных позвонках. У новорожденных котиков оно менее выражено, чем у взрос-
лых. Поясничное утолщение развито слабо, в то время как у настоящих тюленей
оно развито лучше, чем шейное [Соболевский, 19786]. У наземных млекопитаю-
щих оба утолщения развиты одинаково хорошо, у дельфинов же шейное утол-
120
щение развито очень слабо, а поясничное вовсе отсутствует. Считается, что сте-
пень развития утолщений спинного мозга находится в прямой зависимости от
функциональной значимости движителей. Приведенные сравнения по размерам
утолщений у разных представителей ластоногих [Соболевский, 19786] согласу-
ются с общепринятыми мнениями.
Длина спинного мозга тесно коррелирует с размерами тела. У северного мор-
ского котика в онтогенезе каудальная граница спинномозгового конуса смеща-
ется в краниальном направлении. В пренатальный период мозг достигает хвос-
товой части позвоночника. У новорожденных мозговой конус заканчивается в
области 9—10, у взрослых самок — на уровне 7—10, у секачей — на 5—8 грудных
позвонков. Соответственно, с возрастом уменьшается и относительная его длина
(табл. 7.4).
В среднем у взрослых котиков относительная длина спинномозгового конуса
составляет 32% от длины тела. У медведя она равна 59.6%, у дельфинов — 29.1%,
у калана — 38.7%, у ларги — 33.6% от длины тела. Относительная масса спинного
мозга у взрослого котика составляет 0.4—0.6%, у медведя — 0.07%, у ларги — 0.05%,
у дельфинов 0.05% от массы тела. С возрастом у всех млекопитающих отно-
сительная масса спинного мозга уменьшается. У морского котика хорошо вы-
ражен половой диморфизм в размерах спинного мозга. У самцов он больше в
абсолютных, тогда как у самок — в относительных показателях (табл. 7.4).
От спинного мозга морского котика отходит 32—33 пары спинномозговых
нервов, которые в начальных отделах позвоночного столба отходят почти пер-
пендикулярно, а дальше отклоняются каудально и некоторое расстояние идут
внутри позвоночного столба, образуя так называемый “конский хвост”. Его
длина у котика составляет до 80% от длины спинного мозга. Из спинномозговых
узлов наиболее выраженными выглядят IV—VII пары шейных нервов [Соболев-
ский, 19786].
Серое вещество спинного мозга развито хуже белого; их соотношение состав-
ляет у щенка на участке CI—СП 1:4.6, на участке CIV—CVI — 1.2:7, на участке
TV—ТХП — 1:2.3. У взрослой самки соответственно, 1:4.7, 1:4.0, 1:3.0 [Собо-
левский, 19786]. Передние и задние столбы спинного мозга хорошо развиты, при-
чем в области шейного утолщения лучше развиты передние рога.
7.6. Органы чувств
Орган слуха. Ушная раковина у котика представляет собой небольшой конус.
Он скручен по продольной оси, и его края соприкасаются друг с другом. На-
ружная сторона раковины пигментирована, покрыта короткими, редкими, жест-
кими волосами; внутренняя — лишена волосяного покрова. Ушные мышцы плохо
развиты. Из них наиболее выражены m. auricularis anterior и m. auricularis pos-
terior [Боголюбская, Солнцева, 1979].
Наружный слуховой проход на расстоянии 5.5—6.0 см от входного отверстия
изгибается. Он окружен хрящевыми пластинками, большая из которых подко-
вообразная, а меньшая прямая. В области входного отверстия просвет слухового
прохода щелевидный, а в проксимальной части просвет увеличивается и ста-
новится треугольным. В области перехода хрящевого отдела в костный форма
121
просвета слухового прохода становится овальной [Боголюбская, Солнцева,
1979].
По данным названных авторов, среднее ухо котика имеет хорошо выражен-
ные составные части. Молоточек имеет небольшую головку с вытянутыми сус-
тавными поверхностями полукружной формы. Рукоятка утолщена, ее основание
треугольное. К барабанной перепонке крепится ассиметрично. Длинный отрос-
ток молоточка тонкий, эластично соединенный со стенкой барабанной полости
коротким и широким мышечным пучком, который крепится к рукоятке. Нако;
вальня имеет короткое и широкое тело с двумя отростками. Стремя треугольной
формы. Дифференцировка на дужки намечается, но междуговое пространство
отсутствует. Барабанная полость шаровидной формы, выстлана толстой слизис-
той оболочкой.
Масса молоточка и наковальни составляет по 8 мг, стремечка — 4 мг. Длина
молоточка 6 мм, длина его длинного отростка (proc, longus) — 1.5 мм, а длина
рукоятки — 4 мм. Длина наковальни 2 мм, ее ширина 2.5 мм; длина ее длтнного
отростка (proc, longus) 2 мм, короткого (proc, breve) — 2 мм. Длина стремени
2 мм, ширина 1 мм. Улитка имеет 2.5 оборота [Богословская, Солнцева, 1979].
Центральные отделы слуховой системы северного морского котика изучены
недостаточно. По данным Е. М. Стародубцевой [1972], кохлеарные ядра составляют
кохлеарный комплекс, который имеет специфическое строение. Дорсальное ядро
его сращено с флоккулярной долей мозжечка, в то время как у большинства мле-
копитающих оно расположено на наружной поверхности продолговатого мозга и
пространственно отделено от флоккулярной доли. У котика не выражена слоистость
расположения клеток дорсального ядра. Оно состоит из гранулярных клеток, среди
которых рассеяны средние и крупные нейроны (18—20 мк). Клетки имеют овальную
форму и расположены вдоль волокнистых пучков. Границы между передним и зад-
ним вентральными ядрами нечеткие, но оба ядра достаточно хорошо развиты.
Общий обьем вентральных ядер превышает объем дорсальных в 10.7 раза.
Переднее вентральное ядро состоит из крупных (17—20 мк) нейронов оваль-
ной или округлой формы и мелких гранулярных элементов. Крупные клетки ори-
ентированы по ходу волокон слухового нерва и плотно расположены. В заднем
вентральном ядре клетки не так густо расположены, как в переднем. Состав-
ляющие его нейроны разнообразны по форме, а по размерам они крупнее, чем
в переднем (20—25 мк).
В целом, как замечает Е. М. Стародубцева [1972], хотя цитоархитектоника
кохлеарного комплекса у котика достаточно сложна, она характеризуется упро-
щенностью структурного ядра и достаточно высокой степенью развития струю
тур вентрального ядра. У эхолоцирующих млекопитающих оба ядра развиты
одинаково хорошо. По мнению Л. С. Богословкой и Г. Н. Солнцевой [1979],
отмеченная особенность развития дорсального и вентрального ядер характерна
для тех млекопитающих, у которых слуховая система играет существенную роль
в коммуникативных, ориентировочных реакциях и пищедобывающей деятельности.
Изучение цитоархитектоники и нейронной организации заднего двухолмия
мозга котика как рефлекторного центра, обеспечивающего связь слуховых и дви-
гательных реакций, показало, что подкорковая часть слухового анализатора не
имеет больших отличий в строении от таковой изученных наземных млекопи-
122
тающих [Суховская, 1972]. В задних холмах выделяются клеточные группы —
латеральная и центральная. Медиальная и вентральная группы, выступающие
весьма отчетливо на каудальном уровне холма, постепенно отделяются от него
и входят в состав центрального серого вещества.
В латеральной клеточной группе отмечено слоистое расположение состав-
ляющих ее элементов, по структуре напоминающих верхние этажи коры. Ней-
роны латеральной клеточной группы ориентированы перпендикулярно афферен-
тным волокнам, а их дендриты отчетливо подразделяются на апикальные и ба-
зальные. В 0.01 мм2 латеральной группы насчитывается 151 клетка, в централь-
ной — 163. Эти показатели выше у гренландского тюленя (соответственно, 164
и 273) и собаки (158 и 245). Общее количество клеток в обеих группах заднего
холма составляет у котика 902350. Этот показатель выше, чем у обезьяны, но
ниже, чем у собаки, гренландского тюленя и дельфинов [Суховская, 1972].
Таким образом, нейронная организации заднего двухолмия северного мор-
ского котика такова, что она включает мелкие афферентные нейроны (латераль-
ная группа), являющиеся чувствительными, и крупные двигательные мультипо-
лярные элементы.
Орган зрения. Наиболее полное исследование глаза ластоногих сделано
Ф. В. Андреевым [1978], по данным которого ниже и приводится описание ор-
гана зрения северного морского котика.
Как и у большинства наземных хищных, глаза котика расположены по бокам
головы, ближе к фронтальной плоскости. Общее направление “взора” переднее.
Общее поле зрения у котика в горизонтальной плоскости составляет 240°, в вер-
тикальной — 280°. Угол монокулярного поля зрения, по горизонтальной и вер-
тикальной плоскостям — 150°, а угол бинокулярного поля зрения по горизон-
тальной плоскости —60°, по вертикальной — 20°. В отличие отнастоящихтю-
леней, котик не способен видеть предметы, находящиеся сверху и. позади него.
Как и для многих других морских млекопитающих, для северного морского
котика характерно небольшое уплощение глазного яблока по оптической оси
(рис. 7.8).
У ластоногих это достигается за счет уплощения, в первую очередь, роговицы
и, в меньшей степени, склеры впереди экватора. Так, у исследованного котика,
имевшего глаз диаметром 44 мм, донно-роговой диаметр составил 39 мм, тол-
щина склеры в районе дна — 2.5 мм, в районе экватора — 1.0 мм, роговицы,
соответственно, в центре — 0.5 мм, по периферии — 1.1 мм.
Хрусталик почти округлый; индекс его уплощения составляет 0.93. Роговица
толстая. Собственно роговой слой состоит из большого числа клеток. Перифе-
рийные части роговицы, как и у всех других млекопитающих, утолщены, по срав-
нению с центром. По окружности роговица составляет 80—85%; ороговевший
эпителий занимает всю ее наружную часть.
Собственно сосудистая оболочка состоит из 5 слоев. Характерным является
ее утолщение в районе дна глазного яблока. Клетки располагающегося под слоем
сосудов рефлекторного слоя (тапетума) эллипсоидной формы, содержат крис-
таллы гуанина. Трабекулярный остов, занимающий значительное пространство
между ресничным (цилиарным) телом и склерой, рыхлый. К нему примыкает
123
7
Рис. 7.8. Строение глаза-северного морского котика.
А — горизонтальный разрез: 1 — хрусталик, ,2 — хрусталиковая связка, 3 — роговица,
4 — ресничное тело, 5 — стекловидное тело, 6 — склера, 7 — ретина, 8 — сосудистая
оболочка, 9 — сосочек зрительного нерва, 10 — передняя камера, 11 — зрительный нерв;
Б — мускулатура: 1 — латеральная прямая - мышца, 2 — медиальная прямая мышца, 3 —
вентральная прямая мышца, 4 — дорсальная пряма мышца, 5—6 — оттягиватели глаза, 7 —
косая дорсальная мышца; В — радужная оболочка.
цилиарная мышца, которая у ушастых тюленей развита слабее, чем у настоящих.
Цилиарное тело составляет четверть длины оси глаза. Цилиарные связки вы-
ражены хорошо.
В радужине хорошо развита мускулатура. Мышечные волокна образуют
сфинктор вокруг зрачкового края. Причем они имеют 2 вертикально располо-
женных мышечных тяжа, как и у хищных. Зрачок при сужении может приоб-
ретать каплевидную форму;
Толщина сетчатки составляет 250—270 мк в центре и 140—150 мк по пери-
ферии. Пигмент в клетках имеется только в зоне, лишенной рефлекторных кле-
ток по периферии. В отличие от других млекопитающих, сетчатка ластоногих
имеет волокнистую структуру. Складчатость выражена во всех слоях, кроме пиг-
ментного эпителия и пограничной мембраны; особенно складчата сетчатка в
центральных районах, к периферии складчатость исчезает. В сетчатке наиболь-
шей толщиной обладают слои фоторецепторов и наружный ядерный. В послед-
124
нем встречаются 2 типа перикарионов, характеризующих 2 типа рецепторов: па-
лочковые и колбочковые. Общее их количество различно. Число колбочек не-
велико, и расположены они только в центральном районе сетчатки. У котика
плотность распределения клеточных элементов в сетчатке глаза примерно такая
же, как и у других ластоногих. Так, в наружном ядерном слое на 1 мм2 при-
ходится от 500 тыс. (в центре) до 350 тыс. (на периферии) клеток. Во внутреннем
ядерном слое от 75 тыс. (в центре) до 50 тыс. (на периферии) клеток. Слой ганг-
лиозных клеток содержит 220 элементов на 1 мм . Число фоторецепторов на
одну ганглиозную клетку — от 2.2 тыс. (в центре) до 1.5 тыс. (на периферии).
Сетчатка глаза ластоногих не содержит специализированного участка типа
“area”. Ганглиозный слой во всех частях сетчатки состоит из одного ряда клеток,
отделенных друг от друга значительными межнейронными промежутками. Здесь
встречаются 2 типа клеточных элементов: крупные (50—60 мк) мультиполярные
клетки с четко выраженными и хорошо окрашенными начальными отделами
дендритов и мелкие (25—30 мкм) с округлыми контурами тел. Число мелких
клеток в 8—10 раз превышает число гигантских ганглиозных клеток.
Таким образом, фоторецепторный слой сетчатки глаза северного морского
котика, как и других изученных ластоногих, состоит, главным образом, из па-
лочковых рецепторов. Однако, у ластоногих сравнительно тонкий внутренний
ядерный слой и небольшая плотность распределения клеточных элементов, где
ганглиозные клетки малочисленны. Такое соотношение ядерных структур сет-
чатки позволяет предполагать присутствие высокой степени морфологической
суммации нейронных элементов [Андреев, 1978].
Оптические свойства глаза морского котика, как и других полуводных мле-
копитающих, должны рассматриваться с точки зрения способности видеть как
в воздушной, так и в водной среде. В водной среде способность видеть опре-
деляется формой хрусталика, который, как уже говорилось, у ластоногих сфе-
рической формы, имеет значительную рефракцию, небольшое фокусное рассто-
яние и большую светосилу. Определено, что у гренландского тюленя прелом-
ляющая сила хрусталика глаза составляет 80Д [Андреев, 1978]. В воздухе реф-
ракция глаза определяется не только хрусталиком, но и роговицей. Последняя
у ластоногих может быть охарактеризована как рассеивающая линза. Кроме
того, показатели преломления сфер, окружающих роговицу, меньше ее собст-
венного показателя преломления. В целом, рефракция глаза в воздушной среде
у ластоногих почти такая же, как у человека.
Аккомодация глаза осуществляется посредством сокращения цилиарной
мышцы, что ослабляет напряжение цианновых связок. При этом радужина с по-
мощью мощного сфинктера давит на хрусталик, заставляя его передвигаться к
сетчатке, фокусируя удаленные предметы. И наоборот, когда цилиарная мышца
расслабляется, увеличивается напряжение цианновых связок, и хрусталик под-
тягивается к роговице, аккомодируя глаз на видение близких предметов.
Экспериментальные данные показали, что морской котик способен воспри-
нимать световое пятно 0.007ЛК, что ниже, чем у обыкновенного дельфина, ко-
торый способен воспринимать световое пятно 0.009ЛК [Андреев, 1978].
125
Глава 8. ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Краткое анатомическое описание отдельных органов пищеварительной сис-
темы северного морского котика было сделано еще в прошлом веке [Lucas, 1899].
Исследования в этом направлении активизировались лишь в середине текущего
столетия. Были тщательно исследованы зубы [Scheffer, 1950а; Chiasson, 1957; Schef-
fer, Kraus, 1964; Kubota, Korn uro, 1961a, b‘, Kubota, Tagawa, 1964], анатомия и
гистология языка, слюнных желез [Kubota, Horiuchi, 1963; Kubota, 1968], деталь-
но описана печень [Урманов, 1970а, б; 1971]. Большое число работ посвящено
весовым и линейным параметрам отдельных органов пищеварительной системы
[Scheffer, 19606; Соколов и др., 1969а; Соколов, Перлов, 1969; Косыгин, Кузин,
1979].
8.1. Ротовая полость
Ротовая полость северного морского котика ограничена спереди и с боков
губами и щеками, сверху — твердым и мягким небом, снизу — дном полости
и корнем языка (рис. 8.1).
Губы у котика тонкие, плотно прилегающие к зубам и деснам, снаружи по-
крыты мелкими жесткими волосами. Переходная кайма слабо пигментирована.
Верхняя губа снаружи по средней линии разделена неглубокой бороздой, про-
ходящей по направлению к кончику носа.
Щеки тонкие. На верхней губе имеются расположенные в несколько рядов
вибриссы. Здесь щеки несколько утолщаются, образуя носогубную “усовую”
подушку. Задний угол ротовой щели заканчивается на пятом заклыковом зубе.
Небо покрыто слизистой оболочкой бледно-розового цвета. Передняя часть
его сравнительно гладкая. От клыков и далее слизистая твердого неба собрана
в поперечные валики, количество, направление и высота которых подвержены
индивидуальной изменчивости. Они прерываются невысоким сагиттальным
гребнем твердого неба. За последними заклыковыми зубами поверхность неба
гладкая.
Мягкое небо представляет собой утолщенную складку, состоящую из слизис-
той оболочки, железистой и соединительной ткани. Здесь также очень хорошо
развита мышечная ткань, которая образует полную перегородку между глоточ-
ной полостью и голосовой щелью. Язычок на небной занавеске отсутствует. Соб-
ственно слизистая оболочка мягкого неба продольноморщинистая и усеяна мно-
жеством мельчайших выводных протоков слизистых желез.
8.2. Язык
По размерам язык у котика небольшой. Верхушка его утончена, посередине
он имеет неглубокую бороздку, разделяющую его на 2 равные части (рис. 8.1).
Уздечка очень короткая, из-за чего язык не может высовываться из ротовой по-
126
лости. Лисса отсутствует. Вместо нее осталось
скопление жировой ткани в мускулатуре языка.
Слизистая оболочка языка покрыта многослой-
ным чешуйчатым эпителием (рис. 8.2).
На слизистой оболочке различают желобо-
видные, грибовидные и нитевидные сосочки. Об-
ласть листовидных сосочков, которая у назем-
ных хищных занимает краевые участки корня
языка, у котика также имеет образования, напо-
минающие листовидные сосочки, но они не со-
держат ни желез, ни нервов, ни вкусовых почек
[Kubota, 1968]. Желобоватых сосочков бывает
1—4. Они располагаются на границе между
телом и корнем языка, имеют округлую форму
и зернистую поверхность. В связи с отсутствием
листовидных сосочков, желобоватым приписы-
вается основная вкусовая функция. Грибовидные
сосочки малочисленны. Расположены они на
вершине, центре и по сторонам языка сверху. Их
поверхность зернистая.
Рис. 8.1. Ротовая полость
северного морского котика
Тонкий верхний слой, покрытый чешуйчатым эпителием, обильно снабжен
пучками нервных волокон, подходящих к самой его поверхности. Грибовидные
и нитевидные сосочки снабжены ветвями нервных окончаний, отходящих от этих
нервных волокон. Но грибовидные сосочки не имеют вкусовых почек (рис. 8.2).
По нейрогистологическому строению большие желобоватые сосочки морского
котика представляют собой нечто среднее между желобоватыми и грибовидными
сосочками других млекопитающих, и хотя они не имеют ни канавок, ни желе-
зистых протоков, зато они имеют много вторичных сосочков с богатой иннер-
вацией и вкусовыми почками на поверхности, у которых протоки открываются
по способу, сходному с таковым желобоватых сосочков.
8.3. Слюнные железы
По опубликованным данным [Tucker, 1958], у водных млекопитающих слюн-
ные железы по размерам меньше, чем у полуводных. У котика имеются 3 слюн-
ные железы: околоушная, подчелюстная и подъязычная (рис. 8.3).
Околоушная железа немного меньше других. Ее размеры 3.5><2.5 см. Имеет
более аморфную консистенцию, чем две другие. По форме треугольная. Лежит
у основания ушной раковины. Краниально граничит с жевательной мышцей, ка-
удально — с плечеголовной и грудиннососцевидной. Железа связана с около-
ушной системой лицевого нерва. Ее проток начинается на передней поверхности
базальной части железы и идет вдоль жевательной мышцы и далее по щеке, где
открывается в ротовую полость между третьим и четвертым премолярами [Ku-
bota, Horiuchi, 1963].
Подчелюстная железа самая крупная; ее размеры 4x3x1.8 см. Она имеет хо-
рошо очерченную треугольную форму. С дорсальной стороны бороздкой раз-
127
Рис. 8.2. Гистологическое строение языка северного морского котика:
1 — фронтальный срез через желобовидный сосочек, 2 — рудиментарная вкусовая почка во
вторичном сосочке желобовидного сосочка, 3 — нервные сплетения в желобовидном сосочке,
4 — фронтальный срез через грибовидный сосочек, 5 — грибовидный сосочек (видны
внутрисосочковые нервные сплетения, вкусовая почка отсутствует), 6 — область листовидных
сосочков, 7 — бедно иннервированный нитевидный сосочек (из: Kubota, 1968];
b — вкусовая почка, с — нервная клетка, d — проток смешанной железы, eg —
эмбриональная железа, п — нервное волокно, ns — нервный пучок, sm — серозно-мукозная
железа, smg — серозно-мукозная смешанная железа.
128
Рис. 8.3. Околоушная и
подчелюстная области северного
морского котика:
БАМ — наружный слуховой проток,
fn — лицевой нерв, М — жевательная
мышца, MD — двубрюшная мышца,
О — отверстие околоушного протока,
ОА — затылочная артерия, PD —
околоушный проток, PG — околоушная
железа, SD — проток подчелюстной
железы, SMG — подчелюстная железа,
VJE — наружная яремная вена [по:
Kubota, Horiuchi, 1963].
делена на 2 доли. Лежит в каудальном углу нижней челюсти, примыкая к дву-
брюшному мускулу. Латеральная часть прикрыта передним краем грудинносос-
цевидной мышцы. Проток подчелюстной железы отходит от ее передней поверх-
ности на верхнем конце, пересекает двубрюшный мускул, следует под челюсть
и вместе с протоком подъязычной железы идет по подъязычному пространству,
где оба протока открываются в ротовую полость на подъязычном сосочке.
Подъязычная железа состоит из двух долей; ее размеры — 4x1.2 см. Ясно
очерчена. Цвет бледно-розовый. Одна ее доля продолговатая, другая более или
менее овальная. Часть железы находится снизу под корнем языка, справа и слева
от больших язычных мышц. Другая часть доходит до второй ветви подъязычной
кости. Многочисленными протоками открывается в ротовую полость вдоль подъ-
язычной складки.
Выводной проток околоушной слюнной железы выстлан ложностолбчатым
эпителием, ядра которого выглядят двухслойными [Kubota, Horiuchi, 1963]. Сек-
реторные отделы железы имеют клетки, сходные с апокриновыми. Между сек-
реторными клетками и базальными мембранами, окружающими альвеолы, на-
ходятся хорошо развитые миоэпителиальные клетки, напоминающие гладкомы-
шечные. Ацинусы новорожденных котиков по общему гистологическому стро-
ению напоминают таковые взрослых животных, но содержат много секреторных
гранул, лишенных пигмента. У эмбрионов ацинусы околоушной слюнной же-
лезы имеют типичную трубчато-альвеолярную структуру и состоят из клеток,
напоминающих клетки серозного типа.
Подчелюстная железа по гистологическому строению ближе других к клас-
сическому типу (рис. 8.4). Она имеет протоки всех трех типов. Вставочные про-
токи короткие и выстланы кубическим эпителием. Исчерченные протоки очень
хорошо развиты. Они длинные, сильно ветвящиеся и извитые. Выстланы однос-
лойным рядом столбчатых клеток. Секреторные отделы выстланы клетками,
сходными с мукозными, и принадлежат к слизистому типу. Секреторные отделы
подчелюстной железы эмбрионов и новорожденных котиков относятся к сме-
шанному типу. Переход от серозного к слизистому типу происходит постепенно
с возрастом животных. Наибольшее развитие исчерченных протоков в этой же-
лезе компенсирует отсутствие их в околоушной железе
129
'iff J.
al
% V
- W
к »
’4”-' йссМллл
J.X' 9.
f А*
Рис. 8.4. Гистологическое строение слюн-
ных желез северного морского котика [из:
Kubota, Horiuchi, 1963]:
1 — околоушная слюнная железа новорожденно-
го: al — ацинарные полости;
2 — околоушная слюнная железа взрослого коти-
ка: ild — крупная ветвь протока железы в меж-
лобулярной соединительной ткани, аг — артери-
альный сосуд;
3 — подъязычная слюнная железа новорожденно-
го: al — просвет ацинуса, тс — слизистая клет-
ка, sc — серозная клетка;
4 — подъязычная слюнная железа взрослого ко-
тика: sa — серозный ацинус, al — проток ацинуса;
5 — подчелюстная слюнная железа взрослого ко-
тика: sd — ингралобулярный проток, sea — ацинус.
130
Подъязычные железы содержат протоки трех порядков; вставочные, исчер-
ченные и междольковые, но в целом исчерченные протоки менее развиты, чем
в подчелюстной железе. Ацинусы относятся к смешанному типу, хотя, в основ-
ном, преобладают слизистые клетки [Kubota, Horiuchi, 1963]. Гистологическое
строение подъязычной железы у эмбрионов и новорожденных близко к таковому
подчелюстной железы, но у взрослых особей железы сильно отличаются друг
от друга. В подчелюстной железе секреторные отделы и эпителий внутридолевых
протоков близки к слизистому типу.
Таким образом, в онтогенезе котика наблюдается превращение ацинарных
отделов желез из одного типа в другой, из-за чего меняется окраска некоторых
из них. Так, окраска околоушной железы меняется с возрастом от бледно-ро-
зовой до желтовато-коричневой цвета в результате появления большого коли-
чества пигментированных гранул в клетках ацинусов. Аналогичное наблюдается
и в подчелюстной железе. Ацинусы в ней образованы слизистыми клетками, у
эмбрионов — клетки серозного типа. В связи с сильным и большим, чем у других
изученных животных, развитием исчерченных протоков в подчелюстной железе
у котиков, можно предположить особую функциональную роль этой железы по
сравнению с подчелюстной, у которой количество вставочных и исчерченных
протоков значительно меньше. С возрастом в клетках ацинусов подъязычной
слюнной железы не происходит изменения их типа. Они остаются в онтогенезе
смешанными серозно-слизистыми.
8.4. Зубы
Морские котики имеют гетеродонтную зубную систему, которой, как и зубам
большинства другим млекопитающих, присущ дифиодонтизм (рис. 8.5, 8.6, 8.7).
Молочная зубная формула: i |, с |, р | = 26 (рис. 8.6)
Зубная формула постоянных зубов: I |, С |, Р 4, М | = 36 (рис. 8.7).
В зубной формуле морского котика часты отклонения, которые выражаются
либо в отсутствии некоторых зубов “нормального” ряда, либо в присутствии
большего числа зубов [Chiasson, 1957; Kubota, Komuro, 1961а, b; Scheffer, Kraus,
1964; Мужчинкин, 1964e], о чем подробно будет сказано ниже.
У всех зубов хорошо выражена коронка, шейка и корень. Основу зуба со-
ставляет дентиновый стержень, покрытый в области коронки эмалью, а в об-
ласти корня цементом. Форма коронки и корня у разных зубов не одинаковая.
Верхние медиальные резцы имеют долотообразную, уплощенную в мезио-дис-
тальном направлении коронку и корень (рис. 8.7). Режущий край этих резцов
разделен фронтальной бороздкой на лабиальный и лингвальный выступы. Ла-
теральные резцы по форме напоминают клыки, но по размерам меньше их. Ни-
жние резцы не имеют раздвоенного режущего края. Корни их уплощены в мезио-
дистальном, а коронки в лабиально-лингвальном направлениях. При смыкании
челюстей режущие края нижних резцов входят во фронтальные бороздки верх-
них. Все резцы, как правило, однокорневые, лишь в одном из 167 осмотренных
черепов был обнаружен двукорневой резец [Scheffer, Kraus, 1964].
Клыки, как и у наземных хищных, с простой острой коронкой и одним кор-
нем (рис. 8.5, 8.7). По длине верхние и нижние клыки почти одинаковые, а по
131
Рис. 8.5. Зубы северного
морского котика [из: Scheffer,
Kraus, 1964]
форме различаются. Корень верхнего клыка более или менее округлый в ди-
аметре, а дистальная поверхность коронки образует острый режущий край. Ко-
ронка и корень нижнего клыка несколько уплощенной формы, причем наиболь-
ший поперечный диаметр зуба проходит под углом к ряду заклыковых зубов,
так что мезиальный край обращен за пределы, а дистальный — внутрь аркадной
линии.
Рис. 8.6. Молочные зубы
северного морского котика [из:
Scheffer, Kraus, 1964]
132
Рис. 8.7. Постоянные зубы
северного морского котика [из:
Scheffer, Kraus, 1964]:
вверху — самец, внизу — самка.
Рис. 8.8. Нижние резцы (А)
1-месячной самки и нижние Р3, Р4,
М| (Б) взрослого самца северного
морского котика [из: Scheffer, Kraus,
1964]
Все послеклыковые или щечные зубы сходны по форме (рис. 8.5, 8.7). Однако
первые четыре в верхней и нижней челюстях считаются премолярами. Все пре-
моляры имеют молочных предшественников, за исключением первых, которые
обычно, как и моляры, не имеют молочных предшественников, они появляются
в исключительно редких случаях [Kubota, Komuro, 1961а]. Все премоляры, как
133
Рис. 8.9. Левые верхние резщя 7-летнего
самца (А) и левые верхние М1, М2 и ни-
жний М3 3-месячной самки северного мор-
ского котика (Б) [из: Scheffer, Kraus, 1964]
правило, однокорневые. У передних из них корни несколько изогнуты кзади,
а у последнего корень прямой, колышкообразный. Коронки заострены. Их вер-
шины изогнуты назад. Лингвальная поверхность нижних и небная верхних не-
сколько срезаны, вследствие чего на зубах образуются мезиальная и дистальная
острые грани, которые лучше выражены у молодых, чем у старых животных.
У основания коронки всех премоляров хорошо выражен поясок, наиболее вы-
ступающий с лингвальной стороны. Иногда премоляры могут иметь небольшие
дополнительные зубцы на мезиальной и дистальной поверхностях (рис. 8.8), и
тогда зубы выглядят трехзубчатыми [Scheffer, Kraus, 1964].
Моляры по форме сходны с премолярами. Первый из них в верхней челюсти
двукорневой, единственный в нижней челюсти однокорневой (рис. 8.9). Корни
первых трех премоляров в верхней и нижней челюстях имеют вертикальный же-
лобок, углубляющийся к их вершине, возможно, говорящий о том, что когда-то
и эти зубы у котика были двукорневыми (рис. 8.9). У взрослых животных эти
бороздки видны хуже из-за наслоения цемента. Второй верхний моляр часто от-
сутствует, а если присутствует, то имеет уплощенную форму и считается деге-
нерирующим [Chiasson, 1957; Kubota, Togawa, 1964; Scheffer, Kraus, 1964]. Его
корень чаще всего направлен вперед, что дает возможность лучше удерживаться
в челюсти.
Молочные зубы у северного морского котика очень мелкие, за исключением
клыков, которые могут достигать 1 см (рис. 8.6). У предродовых эмбрионов с
длиной тела 52 см размеры молочных зубов составили (в мм): высота — i1 — 1.5,
i2 — 2.0, i3 — 5.0, с — 9.0, р2 — 6.0, р3 — 4.5, р4 — 4.5 в верхней челюсти;
с — 8.5, Р2 — 6.0, р3 — 5.0, р4 — 6.0 в нижней, а диаметр составил i3 — 1.0,
с — 4.0, р2 — 2.0, р3 — 2.0, р4 — 2.5 в верхней челюсти; i2 — 1.0, i3 — 1.5,
с — 3.0, р2 — 2.0, рз — 3.5, р4 — 3.0 в нижней.
Молочные зубы гаплодонтные, но на мезиальной и дистальной поверхностях
премоляров можно видеть небольшие бугорки, по которым можно предполагать,
134
Рис: 8.10. Очередность
десенного прорезывания
молочных зубов у северного
морского котика [по: Kubota,
Komuro, 1961а]
что у предков северного морского котика они были трехзубчатые. Вершины ко-
ронок всех зубов сглажены, куполообразные. Наиболее изменчивы по форме
резцы. Они могут быть шаровидной формы или в виде маленьких столбиков
с куполообразными вершинами. Премоляры чаще конусовидной формы. По два
корня имеют верхние р2, р4 и нижний рд [Kubota, Komuro, 1961а; Scheffer, Kraus,
1964]. Все молочные зубы располагаются с латеральной стороны от будущих
постоянных. По данным Танако [цит. по: Scheffer, Kraus, 1964], молочные зубы
формируются не в самих челюстных костях, а на их поверхности, т. е. все мо-
лочные зубы, кроме клыков, развиваются в мелких углублениях, а не в настоя-
щих костных альвеолах. По данным этих же авторов, гистологически кальци-
фикация нижних молочных зубов начинается у эмбрионов котиков при массе
тела 50 г. У эмбрионов с массой тела 100 г дентин и эмаль присутствуют во
всех зубах.
Рентгенографически у февральских эмбрионов (масса тела ок. 0.3 кг) каль-
цификация зубов не обнаруживается, хотя у эмбрионов, добытых в то же время,
но более крупных, кальцификацией были охвачены все верхние резцы и нижний
центральный. У эмбрионов, добытых в конце февраля, кальцификация коронок
обнаружена во всех молочных зубах. Заканчивается процесс кальцификации мо-
лочных зубов к концу апреля — за 1.5—2 мес. до рождения.
Прорезывание молочных зубов начинается вскоре после кальцификации ко-
ронок у эмбрионов с массой тела 200 г. У 400-граммовых эмбрионов этот процесс
наблюдается во всех молочных зубах [Scheffer, Kraus, 1964]. По другим данным
[Kubota, Komuro, 1961], прорезывание десен молочными зубами у эмбрионов
начинается в конце апреля. При этом верхние зубы прорезываются раньше, чем
нижцие. Из-за большой индивидуальной изменчивости трудно установить оче-
редность прорезывания всех зубов. Определенно можно говорить лишь о том,
что верхние клыки, латеральные резцы (i3) и второй премоляр (р2) прорезыва-
ются первыми, а нижние латеральные резцы — последними (рис. 8.10).
135
Таблица 8.1
Альвеолярная высота (мм) зубов северного морского котика [из: Scheffer,
Kraus, 1964]
Зубы Самка, эмбрион (длина 49 см, масса 2.8 кг) Самец, эмбрион (масса 6.0 кг), 31 июля Самка, новор. (длина 67 см), 23 июля Самка, новор. (длина 67 см), 11 августа Самка, 5 лет, и=6 Самка, 10 лет, и=7 Самец, 9— 10 лет, п-1
I1 2.0 5.8 5.1 5.3 6.95 7.87 9.9
I2 2.0 5.6 5.5 6.0 7.38 7.93 10.2
I3 1.2 8.0 8.2 8.4 10.8 11.62 15.1
С 2.0 4.0 6.8 7.8 19.2 24.3 36.0
Р1 1.9 4.7 6.9 6.9 8.78 9.47 12.7
Р2 - 3.9 6.1 4.7 8.33 8.94 11.6
рЗ - 3.9 4.0 4.6 7.77 8.21 11.6
р4 - 0.8 3.5 2.4 7.48 8.09 10.4
м1 0.2 3.6 4.3 4.1 7.03 7.81 10.4
м2 - 0.8 - 2.0 6.73 7.27 9.9
Ъ 3.9 6.3 5.9 6.2 6.95 7.18 . 8.3
13 2.9 6.3 6.2 7.0 7.25 7.63 8.3
с 0Л 0.1 7.0 6.9 16.75 21.0 ‘ 32.2
Р1 3.3 6.7 6.3 7.0 ' 7.77: ' 8.56 10.3
₽2 1.0 6.6 6.0 6.6 7.9 8.4 9.7
Рз 0.2 7.0 . 6.2 6.4 8.57 9.34 .10.9
₽4 - 1.0 3.5 3.5 8.25 8.67 10.7
М[ 0.5 2.0 2.0 41 7.42 8.63 10.7
Закрытие корней резцов и премоляров начинается у 1-килограммовых эмб-
рионов и заканчивается у 1.5-килограммовых, когда первые зубы начинают про-
резываться через десну. Корни клыков закрываются незадолго до рождения, при-
чем в нижней челюсти раньше, чем в верхней [Scheffer, Kraus, 1964].
Альвеолярная высота молочных зубов у эмбрионов, добытых 28—30 марта,
по данным В. Шеффера и Б. Крауса [Scheffer, Kraus, 1964], составила (в мм):
резцов — 1.0—3.0, клыков — 3.0—-4.8, премоляров — 1.8—4.1. Первыми из мо-
лочных зубов выпадают резцы. У апрельских эмбрионов некоторые из них уже
отсутствовали. В это время масса тела эмбрионов была ок. 2 кг. Последними
выпадают клыки и второй и четвертый премоляры. Клыки могут держаться в
челюстях более двух недель постнатального развития [Lucas, 1899; Scheffer, Kra-
us, 1964]. Выпавшие молочные зубы обнаруживаются в амниотических водах,
в кишечнике или в ротовой полости эмбрионов.
Кальцификация постоянных зубов начинается с нижних клыков у эмбрионов,
добытых в январе, с массой тела 150 г [Scheffer, Kraus, 1964]. К концу февраля
процессом кальцификации охвачены нижние клык и первый премоляр и верхний
первый премоляр [Kubota, Komuro, 1961а]. В начале марта в обеих челюстях
136
Таблица 8.2
Длина корней постоянных верхних клыков северного морского котика (мм)
[из: Scheffer, Kraus, 1964]
Возраст Самцы Самки
п М lim п М lim
Эмбрион, 6 июля 4 0.5 0.3—0.7
6—-7 мес. 7 4.5 3.2—5.5
1 год 2 9.1 9.1—9.2 3 8.9
2 года 25 (19) 11.6 (11.8) 9.5—13.0 8 12.1 10.0—13.6
3 года 8 (52) 13.6 (16.2) 12.3—15.5 20 (8) 13.7 (16.7) 11.9—17.2
4 года 17 (41) 16.2 (19.4) 13.8—17.7 20 (3) 14.7 (18.3) 12.2—17.0
5 лет 2 (36) 17.1 (22.4) 17.0—17.1 20 (6) 14.9 (19.3) 13.7—18.0
6 лет 1 (29) 21.8 (25.4) 20 (11) 16.7 (21.0) 13.7—20.6
7 лет 2 (24) 23.2 (28.5) 22.3—24.1 20 (8) 18.5 (21.5) 16.5—20.2
8 лет 1 (9) 33.1 (28.7) 18 (11) 18.3 (22.5) 15.6—20.9
9 лет (Ю) (32.4) 5 (17) 18.3 (22.9) 15.7—19.8
10 лет 3 (6) 30.3 (30.2) 28.4—33.2 21 (3) 19.8 (23.3) 17.6—23.9
11 лет 13 20.3 18.9—23.4
12 лет 2 19.6 17.6—23.0
>12 лет (17) (34.6) 9(81) 21.6 (24.2) 19.1—24.3
Примечание. В скобках — ориг. данные.
присутствовали кальцифицировавшиеся клыки и первые премоляры. В середине
марта присутствовали следующие зубы с кальцифицированными кронами: в
верхней челюсти — I1—I3, С, Р1, М‘; в нижней — Ip I3, С, Рр МР В начале
апреля: в верхней челюсти — I1—I3, С, Р1—Р4, М’; в нижней челюсти — 1р 13,
С, Р|—Р4, Мр Второй верхний моляр кальцифицируется в начале мая; нижний —
тоже в мае, но несколько позже.
Из представленных данных видно, что закладка и кальцификация молочных
и постоянных зубов у котика начинается почти одновременно. При этом следует
заметить, что темп кальцификации проходит в значительной степени быстрее
в молочных зубах, в которых начало кальцификации отмечается в конце ян-
варя—начале февраля и заканчивается к началу апреля, а постоянных — на-
чинается в начале марта и продолжается до начала июня.
Убедительных литературных данных об альвеолярном прорезывании посто-
янных зубов нет. В. Шеффер и Б. Краус [Scheffer, Kraus, 1964] пишут, что у
эмбрионов с массой тела 660 г были видны верхние клыки и первые 3 премоляра,
нижние клыки и все 5 послеклыковых. Резцы прорезываются у плодов с массой
тела 2.5 кг в начале мая. У новорожденных прорезавшимися были все посто-
янные зубы. В нашей серии эмбрионов можно было видеть прорезавшимися в
верхней и нижней челюстях в марте (масса 1.1 кг, длина 39 см) только первые
137
Таблица 8.3
Время зарастания корней постоянных зубов морского котика [по: Scheffer,
Kraus, 1964]
Возраст
Самцы Самки
месяцы годы месяцы годы
9 14 14 2* 2** 3* 3** 4 5 3 7 15 23* 3**
I1 0- + + + + + + 0 + + +
12 -0- + 0 + + + + 0 + + +
13 закрывается в 7—10 лет
С закрывается после 20 лет
Р1 -------0- ----- +
Р2 ----- - - + + +
Р3 -- + - + + + о +
Р4 - - + 0 + + ---00 +
М1 - - + 0 + + + + - - 0 + 0 +
М2 -- +0-0 + + 0 +
12 - + + + + + + + + + + + + + +
h - + + + + + + 4-4- 0 0 + + + +
С закрывается после 12 лет
Р1 + 0 + 0 + + ---00 +
р2 --- + 0 + 0 + + --- + + +
Рз — — — 0 — + 0 + + - - - - 0 +
Р4 0 +
Ml — — — 0 — + 0 + + - 0 +
Примечание. * Немного менее названного возраста; ** немного старше названного
возраста;-------корневой канал открыт; 0 — корневой канал почти закрыт; + —
корневой канал закрыт.
премоляры. У апрельских эмбрионов (масса 2.2 кг, длина 41 см) в верхней че-
люсти прорезались I1,12, Р1, М1, в нижней — 12, Рь Мр У эмбрионов, добытых
12 мая (масса 3.4 кг, дайна 58 см), в верхней челюсти были I1,12, Р1, М1, в ниж-
ней — I2, Pj, Р2, Мр У эмбрионов, добытых 12 июня (масса 4.1—4.3 кг, дайна
56—58 см), прорезались все верхние и нижние зубы.
Своеобразно положение постоянных еще непрорезавшихся I3 в верхней и 12
в нижней челюстях. Эти резцы закладываются глубже других зубов и находятся
до их прорезывания через альвеолярный край челюсти во втором внутреннем
ряду зубов. Верхний I3 находится за I2, а нижний 12 за затем они занимают
соответствующее положение в зубном ряду каждой аркады.
Альвеолярное прорезывание и последующий рост — процессы не согласо-
ванные. Наибольшей интенсивностью роста в верхней челюсти характеризуются
138
клыки (табл. 8.1). В резцах интенсивность роста снижается от третьего к первому.
Из премоляров интенсивнее других растет четвертый, вдвое ниже темп роста
у первого, а два средних растут примерно с одинаковым темпом. Очень интен-
сивно растет первый коренной, второй моляр растет в несколько раз медленнее.
В нижней челюсти интенсивнее других зубов растут клыки, при этом их темп
роста много выше, чем у верхних клыков. Последовательность уменьшения темпа
роста резцов та же, что и в верхней челюсти, но резцы верхней челюсти уве-
личиваются в большее число раз в процессе постнатального онтогенеза, чем ниж-
ние резцы. Третий премоляр нижней челюсти растет интенсивнее других пре-
моляров. Ниже темп роста у первого премоляра.
Данные Ф. Лукаса [Lucas, 1899], К. Куботы и М. Комуро [Kubota, Komuro,
1961а], В. Шеффера и Б. Крауса [Scheffer, Kraus, 1964] о времени и очередности
прорезывания постоянных зубов с учетом индивидуальных вариаций сходны.
Порядок прорезывания постоянных зубов, в целом, следующий: для верхних I*—
I3, С, Р1—Р4, М', М2 последовательность прорезывания — 2, 4, 7, 16, 6, 9, 12,
13, 14, 18 дни; для нижних — I2, 13, С, ?!—Р4, М1; последовательность — 1,
3, 15, 5, 8, 10, 11, 17 дни.
В представленных данных просматривается передне-задний градиент десен-
ного прорезывания в каждом классе зубов, в то время как у большинства на-
земных млекопитающих наблюдается обратная картина этого процесса.
Рост корней и время их закрытия для постоянных зубов у северного морского
котика представлены в таблицах 8.2 и 8.3.
Как видно из таблиц, интенсивный рост, клыков у самцов продолжается до
8-летнего, а у самок до 7-летнего возраста. Позже темп роста заметно снижается,
хотя и продолжается всю жизнь после закрытия корней. Приведенные в таблице
8.2 наши данные отражают ту же тенденцию в росте, хотя и ближе к предельным
показателям размеров корней клыков в каждом возрастном классе. Причина
этого явления ясна и объясняется разным состоянием популяций. В 50-х годах
прибыловская популяция котиков находилась на начальных стадиях естествен-
ного снижения численности, и звери из этой популяции были мельче котиков
интенсивно растущей популяции о. Тюлений. Зависимость размеров животных
и их черепов от состояния популяций была доказана В. Шеффером [Scheffer,
1955]. В свою очередь, закрытие корней у разных зубов происходит не одно-
временно. Корни раньше зарастают у зубов со слабым развитием корней (на-
пример, резцы). Вслед за резцами зарастание корней происходит у премоляров
и позже всех у клыков.
Аномалии развития зубов у северного морского котика могут выражаться
в присутствии или отсутствии отдельных зубов, а также в их повреждении в
течении жизни животного. В серии из 38 черепов Р. Чиассон [Chiasson, 1957]
обнаружил 6 черепов с различными отклонениями от типичной зубной формулы.
В серии из 123 черепов аномальные зубы были в 14 (11%) случаях. В. Ф. Мужчинкин
[1964в] исследовал 135 черепов котиков и обнаружил отклонение от нормальной
зубной формулы в каждом пятом черепе самцов и в каждом деся- том — у самок.
Обобщенные сведения названных авторов по отклонениям от “нормальной”
зубной формулы выражаются в следующих характеристиках.
139
1. (Отсутствие шестой пары заклыковых зубов с одной или обеих сторон.
2. Отсутствие пятого верхнего заклыкового зуба. 3. Отсутствие четвертой пары
заклыковых зубов с одной или обеих сторон. 4. Отсутствие третьей пары за-
клыковых зубов. 5. Присутствие добавочной пары заклыковых зубов. 6. Присут-
ствие дополнительных (одного или обоих) третьих нижних резцов. 7. Присутствие
“лишних” верхних премоляров Р2 и Р2. 8. Присутствие “лишнего” премоляра
между Клыком и первым премоляром. 9. Присутствие “лишнего” верхнего резца.
Отклонение от нормы чаще встречается в верхней, чем в нижней челюсти,
и у самцов чаще, чем у самок.
Кроме названных отклонений в зубной формуле могут быть обнаружены ано-
малии развития зубов, возникающие в процессе жизни животного. К таковым
относятся травмы черепа, влияющие потом на рост и развитие зубов, а также
непосредственно травмы зубов. Такие отклонения описаны [Scheffer, Kraus,
1964].
Большинство исследователей сходятся во мнении, что наземные предки ушас-
тых тюленей могли иметь большее число зубов, чем современные северные мор-
ские котики. Это не прослеживается по материалам развития молочных зубов,
но подтверждается отсутствием первого нижнего резца, меньшими размерами
первого, чем последующих верхних резцов, отсутствием последнего нижнего мо-
ляра. У олигоценового предка северного морского котика Enaliarctos mealsi и
миоценовых предков, начиная с Thalassoleon, имелось то же число зубов, что
и у современных котиков. Лишь род Pithanotaria из раннего миоцена имел
5 послеклыковых зубов в верхней челюсти (отсутствовал второй моляр). Но и
у современных котиков встречаются особи с 5 послеклыковыми зубами. У пред-
ков, вплоть до самых поздних находок, между коренными зубами присутство-
вала диастема, которой нет у современных котиков. В этой связи уместно вер-
нуться к особенностям развития последнего моляра у современных котиков, у
которых он закладывается позже, растет медленнее, чем предыдущий, и имеет
1 корень. Большинством исследователей последний моляр верхней челюсти счи-
тается редуцирующимся, но, по А. Н. Северцову [1939], рудиментарные органы
обычно закладываются одновременно с нормально развивающимися. Поэтому
не ясно, является ли этот зуб действительно редуцирующимся, или он приобретен
сравнительно недавно в эволюции котиков. Если допустить, что он является но-
вообразованием, то легко объяснимо исчезновение диастемы между корневыми
зубами у современных котиков.
Преобладание числа двукорневых зубов в молочной зубной системе морс-
кого котика по сравнению с их числом в постоянных зубах является веским ос-
нованием в пользу точки зрения о направленности эволюции в сторону упро-
щения зубной системы, а также правомочности таксономических построений Ч. Ре-
пеннинга и Р. Тедфорда [Repenning, Tedford, 1977] линии ископаемых предков
ушастых тюленей по строению зубной системы, с чем не согласен Е. Митчелл
[Mitchell, 1975].
Среди наземных хищных нет представителей, имеющих двувершинные рез-
цы, как у северного морского котика. Возрождая теорию Розе-Кюкенталя о про-
исхождении многовершинных зубов млекопитающих из серии одновершинных,
Б. С. Матвеев [1963] исходил из самостоятельности закладок отдельных денти-
140
новых конусов у эмбрионов, впоследствии срастающихся у основания и соот-
ветствующих каждый отдельному бугорку дефинитивного зуба. Аналогичным
образом происходит закладка дентиновых конусов медиальных резцов и у ко-
тика. Более того, закладка латеральных резцов и их развитие до времени аль-
веолярного прорезывания происходит с внутренней стороны от медиальных рез-
цов. В связи с этим правомочно предположение о возможном образовании дву-
вершинных медиальных верхних резцов у котика путем срастания одновершин-
ных. Такой путь развития подтверждается находкой черепа [Scheffer, Kraus, 1964],
у которого медиальные резцы имели двувершинные корни. Если наши рассуж-
дения верны, то правомочны предположения, что у отдельных предков котиков
могло быть большее число резцов в верхней челюсти, чем у современных.
У новорожденных котиков медиальные нижние постоянные резцы трехзуб-
чатые. Все послеклыковые зубы взрослых особей имеют передние и небольшие
задние зубчики. По этим признакам можно говорить об инволюции зубов се-
верного морского котика от сложных многобугорчатых наземных предков ушас-
тых тюленей к упрощенной конической форме современных представителей се-
мейства. Тем самым снимается предостережение В. Ф. Мужчинкина [1978а] о не-
возможности обратного развития сложных маммальных зубов к предковому со-
стоянию.
8.5. Глотка
Полость глотки у котика объемистая, но под воздействием мускулов-сжима-
телей она выглядит щелевидной трубкой размером у взрослых особей 4—7 см.
Впереди граничит с отверстием хоан, на боковых стенках которых в глотку от-
крываются парные глоточные отверстия слуховых (евстахиевых) труб. Надгор-
танник и вход в гортань образуют дно фарингиальной полости. От надгортан-
ника с каждой стороны отходят черпало-надгортанные складки, которые соеди-
няют основание относительно некрупного надгортанника со складками пище-
водной части глотки. Глоточные миндалины парные. Их длина у взрослых осо-
бей 4—5 см, ширина 1.5—2 см. Слизистая оболочка вокруг и на самих мин-
далинах морщинистая. В передней нижней трети глотки слизистая образует
складку. Между этой складкой и миндалинами образуются кармашки, занимаю-
щие половину пространства каудальной части миндалин.
Стенка глотки состоит из 4 слоев. Снаружи она одета соединительнотканной
фасцией. Мышечный слой представлен рядом поперечнополосатых мышц.
Каудальный сжиматель (m. constrictor pharyngeus caudalis). В этой мышце мож-
но выделить 2 порции, хотя по средней линии волокна зтой порции перепле-
таются. Перстневидноглоточная порция (р. cricopharyngeus) начинается от зад-
него угла перстневидного хряща и идет вперед, смешиваясь со второй порцией.
Щитовидноглоточная порция (р. thyreopharyngeus) начинается с поверхности
щитовидного хряща и, дугообразно изгибаясь, встречается с волокнами проти-
воположной стороны этой же мышцы. Передние волокна этой порции частично
перекрывают волокна среднего сжимателя.
Средний сжиматель (m. constrictor pharyngeus medius). Как и предыдущая мыш-
ца, делится на порции, но в связи с многочисленностью точек прикрепления от-
141
дельных пучков мышечных волокон и их переплетением с волокнами других
мышц затруднительно гомологйзировать ее с мышцами изученных наземных
млекопитающих. Часть волокон этой мышцы отходит от щитовидноподъязыч-
ной фасции и следует к средней линии глотки. Более широкая порция начинается
от дистального членика подъязычной кости и следует к дорсальному шву глотки
и частично к основанию черепа.
Краниальный сжиматель (m. constrictor pharyngeus cranialis) развит хуже двух
предыдущих мышц. Лежит под передней частью вышеописанных мышц.
Шилоглоточная мышца (m. stylopharyngeus) очень хорошо выражена. На-
чинается на сосцевидном отростке височной кости и заканчивается на дорсаль-
ной стенке глотки и гортани.
Фиброзная оболочка глотки развита хорошо. Слизистая оболочка, являю-
щаяся продолжением слизистой оболочки ротовой полости, продольноскладчатая.
8.6. Пищевод
У предродовых эмбрионов длина пищевода 20—26 см, у щенков — 27—35 см.
У взрослых самцов он имеет длину около метра, а у самок — 50—60 см. Ширина
пищевода в средней части у взрослых самок 2 см, у самцов — 8 см. В начальных
отделах пищевода мышечная оболочка трехслойная, в средней части — дву-
слойная. Из-за этого стенка пищевода толще в начальных отделах и тоньше в
средней части. Мышечные волокна, как и у большинства других млекопитаю-
щих, проходят спиралеобразно, накладываясь слой на слой и образуя сетку в
виде мелких параллелограммов.
8.7. Желудок
Форма желудка изменяется в зависимости от степени его наполнения пищей.
В наполненном состоянии он грушевидной формы с изогнутой пилорической
частью (рис. 8.11). При отсутствии в нем пищи он крючковидный (рис. 8.12).
Желудок лежит в левом подреберье. Передняя его стенка прилежит к диаф-
рагме и печени, а задняя — к петлям кишечника. К большой кривизне желудка
прилежит селезенка. Фиксация в полости осуществляется с помощью связи его
с пищеводом и диафрагмой в области пищеводного отверстия. Кроме того, креп-
ление его осуществляется с помощью большого сальника, отходящего от боль-
шой кривизны желудка, и малого сальника, отходящего от малой кривизны. Пер-
вый из них переходит на селезенку, а в области пилорического отдела образует
желудочно-двенадцатиперстную связку. Второй посредством печеночно-желу-
дочной связки соединяет желудок с печенью. Оба сальника почти лишены жира
и сильно васкуляризированы.
Наружная серозная оболочка гладкая, плотная, телесного цвета. Внутренняя
мышечная оболочка достаточно толстая, состоит из нескольких слоев, идущих
продольно и циркулярно.
Слизистая оболочка собрана в многочисленные, различной формы и разме-
ров складки. В пилорическом отделе желудка складки испещрены множеством
идущих в разных направлениях более мелких складок (рис. 8.12). Глубина круп-
142
Рис. 8.11. Естественное положение Рис. 8.12. Внутреннее строение желудка
желудка в брюшной полости у северного морского котика:
северного морского котика: 1 — pars cardiaca, 2 — fundus ventriculi, 3 — pars
1 — желудок, 2 — тонкий кишечник, pylorica, 4 — plicae gastricae.
3 — печень, 4 — сердце.
ных складок 1.5—2 см, мелких — 0.2—0.5 см. В пилорическом отделе слизистая
оболочка по цвету бледнее, чем в кардиальном, и также собрана в многочис-
ленные складки, но их размеры и направления иные. Они неглубокие и ори-
ентированы в поперечном направлении. В местах схода желудка в двенадцати-
перстную кишку слизистая оболочка вновь формирует несколько глубоких скла-
док, окаймляющих каудальную часть пилорического отдела. Совместно с мы-
шечной оболочкой слизистая здесь формирует клапан (m. spincter pylori).
Масса желудка у новорожденных котиков 30—70 г (0.6% от массы тела). У
взрослых самцов она может достигать почти 2 кг, а у самок ок. 900 г, что со-
ставляет, соответственно, в среднем 1.5% и 1.2% от массы тела. Различия по отно-
сительным показателям у самцов и у самок незначительны [Косыгин, Кузин, 1979].
8.8. Кишечник
В кишечнике северного морского котика, как и в кишечнике сивуча [Lucas,
1899; Перлов, Урманов, 1971] по внешнему виду легко опознаваемы лишь две-
надцатиперстная, толстая и тонкая кишки. Выделить в толстом отделе кишеч-
ника ободочную, а в тонком отделе тощую и подвздошную можно лишь условно.
Двенадцатиперстная кишка своим начальным участком прилежит к пилори-
ческому отделу желудка, граничит кранйально с печенью, латерально со стенкой
брюшной полости. Остальная часть двенадцатиперстной кишки прилежит к же-
лудку, тонкому кишечнику, корню брыжейки и ее лимфоузлам, толстому кишеч-
143
1
Рис. 8.13. Начальный отдел пищеварительного
тракта северного морского котика:
1 — желудок, 2 — двенадцатиперстная кишка, 3 —
головка поджелудочной железы, 4 — сальник, 5 —
связка поджелудочной железы, 6 — лимфатический
узел корня брыжейки, 7 — артерия.
м
30 J
СМ
толстого * L180
А САМЦЫ
Д САМКИ 4 *
А * 160
• о ° о о ° о °
О тонкого
10 Л
.120
.80
• САМЦЫ
о САМКИ
fc—। j , Г"'’Т I 1
I 3 5 7 9 10+
ВОЗРАСТ
Рис. 8.14. Длина толстого (см) и
тонкого (м) кишечника северного
морского котика
нику, почкам, слепому отростку. Кишка подковой огибает поджелудочную же-
лезу, с которой она сращена в области головки, а остальной частью соединяется
с брыжейкой (рис. 8.13).
Передняя часть двенадцатиперстной кишки от пилорического отдела желуд-
ка до каудального края места прикрепления к ней головки поджелудочной же-
лезы утолщена. Остальная ее часть почти вдвое тоньше и без видимых границ
переходит в тощую кишку. На внутренней поверхности начального отдела две-
надцатиперстной кишки в нее открываются протоки поджелудочной железы и
печени. В области их выхода слизистая утолщена и образует так называемый
фатеров дивертикул.
Тощая и подвздошная кишки образуют 7—10 петель, подвешены на длинной
брыжейке, корень которой прикрепляется к позвоночнику в области окончания
двенадцатиперстной кишки. Слизистая тонкого отдела кишечника продольно
складчатая и покрыта множеством ворсинок.
Переход подвздошной кишки в толстую обозначен присутствием очень ма-
ленького слепого отростка. Со стороны слизистой заслонка не выражена, однако
слизистая в месте перехода тонкой кишки в толстую утолщена. В тонком отделе
кишечника по ходу брыжейки на расстоянии 3—5 см от места соединения под-
144
Рис. 8.15. Печень с диафрагмальной (слева) и висцеральной (справа) сторон
[из: Урманов, 19706]:
1 — правая диафрагмальная доля, 2 — правая висцеральная доля, 3 — левая диафрагмальная
доля, 4 — левая висцеральная доля, 5 — квадратная доля, 6 — желчный пузырь, 7 —
печеночный сииус, 8 — воротная веиа.
вздошной кишки с толстым кишечником тянется продольный валик высотой 2—
3 мм. Слепая кишка короткая (1—5 см) и фактически представляет собой ци-
линдрический дивертикул.
Толстый кишечник образует несколько петель в левом подреберье. От места
своего начала тощая кишка опускается к почке, затем поднимается к диафрагме
и вновь опускается косо к сагиттальной линии, откуда (уже как прямая кишка)
следует каудально. Перед анальным отверстием прямая кишка несколько рас-
ширяется. Толстый кишечник фиксируется к позвоночнику прямой и длинной
брыжейкой. Слизистая толстого кишечника имеет мелкозернистый вид.
Длина кишечника у северного морского котика (рис. 8.14) превышает длину
тела в 12—15 раз [Косыгин, Кузин, 1979], тогда как у наземных хищных только
в 4—6 раз [Жеденов, 1965]. Большая относительная длина кишечника у котика
не является исключением; у сивуча кишечник еще длиннее. Данный факт не на-
ходит пока убедительного объяснения. Особенно длинным представляется тон-
кий отдел кишечника, который превышает длину толстого в 15—20 раз. Этот
показатель у наземных хищных выражается соотношением 6:1. Рост кишечника
в длину у котика прекращается одновременно с ростом тела: у самцов в 10—12
лет, а у самок в 6—8 лет.
8.9. Печень
Печень котика, как и печень большинства хищных, многолопастной орган
(рис. 8.15).
В ней различают 3 крупные доли: правую, среднюю и левую, первая и пос-
ледняя из которых в свою очередь делятся, соответственно, на диафрагмальную
и висцеральную и на квадратную и хвостатую доли или лопасти. Каждая доля
отделена от другой междолевыми щелями, а лопасти — бороздами (рис. 8.15).
Правая и левая междолевые щели идут сагиттально от подвижного левого
края печени по направлению к фиксированной стороне печени. В свою очередь,
правая и левая латеральные междолевые щели располагаются фронтально, па-
145
раллельно диафрагмальной поверхности. Они начинаются на висцеральной по-
верхности правой и левой долей и идут в дорсальном направлении к краю пе-
чени. Эти щели более глубокие и заметно отделяют правую и левую висцераль-
ные доли от диафрагмальных, с которыми они соединяются в области дорсаль-
ного края печени дупликатурой брюшины с элементами глиссоновой ножки и
печеночными венами [Урманов, 19706]. Каждая доля печени имеет заостренную
вершину и расширенное основание. Соединяясь, они формируют треугольной
формы железу, обращенную утолщенным дорсальным краем к позвоночному
столбу, а заостренным краем к вентролатеральной стороне брюшной стенки.
Правый угол печени скошен и закруглен, а левый более острый. Диа-фрагмаль-
ная поверхность печени выпуклая, гладкая, дорсокаудальная — вогнутая. Она
соприкасается с органами брюшной полости. В целом, печень несколько смещена
вправо.
В глубине правой медиальной междолевой щели находится желчный пузырь,
а в левой — круглая и серповидная связки печени (рис. 8.15). На дорсальном
крае печени этим щелям соответствуют справа — бороздка каудальной полой
вены и печеночный синус, слева — пищеводное вдавление.
Правая диафрагмальная доля имеет треугольную форму. Верхний ее отдел
расширен и имеет отросток, прикрывающий квадратную и левую диафрагмаль-
ную доли (рис. 8.15). Из под вентрального края отростка виден желчный пузырь.
Диафрагмальная поверхность этой доли выпуклая, висцеральная — уплощенная
и прилежит к правой висцеральной доле, начальным отделам кишечника и к
поджелудочной железе. В дорсальном отделе она сращена паренхиматозно со
средней долей печени.
Правая висцеральная доля меньше предыдущей, треугольная по форме, с за-
кругленной вершиной. Со стороны диафрагмы она прикрыта правой диафраг-
мальной долей, из-под которой видны только ее правый и дорсальный края.
Висцеральная поверхность прилежит к правой почке, надпочечнику и к хвос-
товому отростку хвостатой доли, с которой она соединяется короткой сосудис-
то-желчной ножкой. В основании печени висцеральная доля соединяется с пра-
вой диафрагмальной и средней долями только париетальной ножкой, в которую
входят кровеносные сосуды, желчные протоки и нервы [Урманов, 19706].
Левая диафрагмальная доля имеет выпуклый левый край и вогнутый правый.
Диафрагмальная поверхность прилежит к диафрагме, селезенке и брюшной стен-
ке, висцеральная поверхность уплощена и вентрально прилежит к желудку, а дор-
сально — к левой висцеральной доле, отделяясь от нее латеральной междолевой
щелью; здесь же она паренхиматозно срастается со средней долей.
Левая висцеральная доля округло-треугольная. От ее правого края отходит
небольшой отросток в сторону ворот печени. Вогнутая висцеральная поверх-
ность прилежит к желудку, а диафрагмальная, соответственно, к диафрагме.
В дорсальной части соединяется с левой диафрагмальной долей перитониально.
Квадратная доля имеет форму языка. Диафрагмальной поверхностью она
прилежит к диафрагме и брюшной стенке, соприкасаясь с боков с правой и левой
диафрагмальными долями, желчным пузырем, серповидной и круглой связками.
Висцеральная поверхность прилежит к желудку, двенадцатиперстной кишке и
поджелудочной железе. В основании печени доля соединяется с диафрагмальной
и хвостатой долями.
146
Хвостатая доля располагается дорсальнее ворот печени и имеет массивный
хвостовой и менее массивный сосковидный отростки. Хвостовой отросток срас-
тается с правой висцеральной долей и на своей правой поверхности имеет пе-
ченочное вдавление; сосковидный лежит на печеночно-желудочной связке и в
основном соединяется с хвостатой долей. Дорсальные отделы хвостатой доли
срастаются с каудальной полой веной.
Каждая доля печени имеет на своей поверхности определенное количество
насечек разной формы и глубины. Больше всего их на висцеральной поверхности
квадратной, правой и левой висцеральных долей. Щели порой прорезают сво-
бодный край долей печени, которые в форме пластин или языков чаще всего
встречаются в тех местах, где печень соприкасается с желудком. На диафраг-
мальной поверхности насечки бывают реже; они здесь мельче и уже.
На висцеральной поверхности печени располагаются портальные, а на дор-
сальной — кавальные ворота печени. Портальные ворота включают воротную
вену, находящуюся в области правого края ворот, печеночную артерию, общий
желчный и пузырный протоки, а также правую и левую ветви печеночного про-
тока. Кавальные ворота печени представлены синусом, принимающим в себя
3 крупные и несколько мелких печеночных вен [Урманов, 19706].
Желчный пузырь серо-зеленого цвета, имеет каплевидную форму и вмещает
у взрослых животных до 150 мл желчи. Он расположен в глубине правой ме-
диальной междолевой щели. Дно его обращено к диафрагме. Боковые и до-
рсальные поверхности сращены с паренхимой диафрагмальной и квадратной
долей. На висцеральной поверхности печени тело желчного пузыря продолжа-
ется в узкую шейку, прилежащую к воротам печени, где она переходит в ши-
рокий пузырный проток, сливающийся с правой ветвью печеночного протока,
формирующий общий проток, впадающий в двенадцатиперстную кишку. В са-
мом начале желчного протока в него вливается левая ветвь печеночного протока
[Урманов, 1971]. Общий желчный проток располагается между листками пече-
ночно-двенадцатиперстной связки.
Образование общего желчного протока происходит в толще печеночно-две-
надцатиперстной связки, удаленной от ворот печени. Он формируется слиянием
правой ветви печеночного протока с пузырным протоком. Вскоре в него откры-
вается левая ветвь печеночного протока. Нередко общий ствол желчного про-
тока образуется слиянием пузырного и обеих ветвей печеночного протока, где
в этом случае образуется ампулообразное расширение [Урманов, 19706].
Фиксирующий аппарат печени представлен рядом связок. Серповидная связ-
ка, образованная дупликатурой брюшины, тянется от диафрагмы и вентральной
брюшной стенки в глубину медиальной междолевой щели. В толще ее вентро-
каудального края имеется круглая связка печени. Дорсальный край серповидной
связки переходит в правую и левую венечные связки, латеральные отделы
которых образуют треугольные связки. Венечные и треугольные связки имеют
незначительную длину и тянутся от дорсального края печени к позвоночнику,
пищеводу, диафрагме и каудальной полой вене. От ворот печени начинается печеноч-
но-желудочная и печеночно-двенадцатиперстная связки, образованные дупликатурой
брюшины, которые вместе формируют малый сальник.
Печень — самый крупный внутренний орган у котика. Ее абсолютный вес
достигает у взрослых самцов 8 кг, у самок — 3 кг.
147
Характерной особенностью абсолютного роста печени является снижение
массы органа у новорожденных, по сравнению с ее массой у предродовых эмб-
рионов, и увеличение абсолютной массы органа после прекращения роста тела.
Наибольшая относительная масса печени у эмбрионов котиков, наимень-
шая — на первых годах жизни. Затем у самцов к 3 годам она достигает наи-
высших показателей для постнатального периода развития животного, и в по-
следующие годы эти показатели постепенно уменьшаются. Иначе идет развитие
у самок, у которых относительные показатели с возрастом постепенно увели-
чиваются.
Наиболее крупной долей печени является правая, составляющая в среднем
45.1% от массы всего органа, а входящие в нее диафрагмальная и висцеральная
доли составляют, соответственно, 29.35 и 15.8% от массы правой доли. Масса
средней доли печени — 19.7% от массы органа, а входящие в нее квадратная
и хвостатая — 10.5% и 9.0% от массы средней доли. Левая доля по массе меньше
правой, но больше средней доли. Ее относительная масса составляет в среднем
35.2% от массы всей печени, а ее диафрагмальная и висцеральная доли, соот-
ветственно, 18.7% и 16.5% от массы всей левой доли.
8.10. Кровоснабжение печени
Вступающая в портальные ворота воротная вена, печеночная артерия и вы-
ходящие печеночные протоки, образующие глиссонову триаду, размещаются в
области правого края портальных ворот, которые представлены неглубокой
фронтальной бороздой, разделяющей хвостатую и квадратную доли. Все 3 эле-
мента (воротная вена, печеночная артерия и общий желчный проток) в области
ворот дихотомически делятся на крупную левую и меньшую правую ветви (рис.
8.16, 8.17). Иногда это разделение общих стволов триады может быть трифур-
кационным, захватывающим либо один из сосудов, либо 2, а иногда все 3.
В этом случае третья ветвь, как правило, идет в правую диафрагмальную долю
[Урманов, 1970я]. Все названные элементы сосудистой системы глиссоновой
триады по способу прохождения в печени и типу ветвления обычно повторяют
друг друга, хотя по калибру вены крупнее артерий, среди которых своей большей
величиной отличается воротная вена. Правая ветвь воротной вены короче левой.
Она вскоре делится на 2 одинаковых сосуда, один из которых является продол-
жением основной ветви и вступает в ворота правой висцеральной доли. Вторая
ветвь отходит от правой ветви под прямым углом и следует вентрально, пог-
ружаясь в паренхиму правой диафрагмальной доли (рис. 8.16). Левая ветвь во-
ротной вены идет в поперечной борозде до левого края ворот, где делится на
2 ствола. Один из них входит в левую висцеральную, другой — в левую диа-
фрагмальную доли. По ходу от левой ветви вентрально отходит крупный ствол
в квадратную долю и несколько более мелких ответвляется в хвостатую долю.
Таким образом, сосудистая система воротной вены расположена так, что каж-
дая доля печени обслуживается ветвями второго порядка, которые потом рас-
пределяются в каждой доле, отдавая ряд ветвей третьего и последующих по-
рядков, в зависимости от величины каждой доли. Вентральная ветвь воротной
вены спускается к вентральному краю правой висцеральной доли. Ветвление ее
стволов более сложно, чем левой ветви. К левому краю этой доли идет крупный
148
6
Рис. 8.16. Расположение воротной вены (слева) и желчных протоков (справа) в
печени северного морского котика [из: Урманов, 19706]:
1 — правая диафрагмальная доля, 2 — правая висцеральная доля, 3 — левая
диафрагмальная доля, 4 — левая висцеральная доля, 5 — квадратная доля, 6 — основной
ствол воротной вены (слева) и общий желчный проток (справа).
Рис. 8.17. Расположение печеночных
вен в долях печени северного
морского котика [из: Урманов, 1970а]:
1 — правая диафрагмальная доля, 2 — правая
висцеральная доля, 3 — левая диафрагмаль-
ная доля, 4 — левая висцеральная доля, 5 —
квадратная доля, 6 — печеночный синус.
ствол, отходящий от отделов вентральной ветви. Концевые отделы стволов
имеют рассыпной тип ветвления. В висцеральной доле выделяется 2 участка (до-
рсальный и вентральный), обслуживаемые ветвями портальных сосудов. В пра-
вой диафрагмальной доле проходит второй ствол правой ветви воротной вены,
идущий к центру доли и в дорсальных отделах ветвящийся по магистральному,
а с вентральной стороны — по рассыпному типу. Квадратная доля обслужи-
вается венозной ветвью, отходящей от левой ветви воротной вены. Она в самом
начале разделяется на правую и левую ветви, идущие по соответствующим краям
доли. Однако очень часто квадратная доля обслуживается единственным веноз-
ным стволом, ветвящимся по магистральному типу.
Хвостатая доля имеет несколько источников протоков портальной крови.
От левой ветви воротной вены крупный ствол следует в хвостатый отросток
доли. Сосковидный отросток и сама доля получают несколько мелких стволов
от левой ветви воротной вены. Следовательно, в хвостатой доле выделяются
2 самостояельных участка, имеющие обособленные портальные бассейны: пер-
вый — хвостатая доля и сосковидный отросток и второй — хвостатый отросток.
Часто последний разделен на дорсальный и вентральный участки, соответству-
ющие двум лепесткам отростка. Левая висцеральная доля получает один круп-
ный венозный ствол, идущий параллельно дорсальному краю печени к левому
ее углу. По ходу он отдает несколько коротких дорсальных и более крупных
149
вентральных ветвей сосудов, распределяющихся по доле печени. В левую диаф-
рагмальную долю вступает крупный венозный ствол, вскоре делящийся на дор-
сальную и вентральную вены, первая из которых распределяется в дорсальном
отделе печени по рассыпному типу, а вторая спускается вентрально, отдавая по
ходу боковые ветви, и, в целом, представляется как сосуд магистрального типа.
Параллельно вентральной ветви по правому краю доли проходит венозный
ствол, отделяющийся от вены квадратной доли. В левой диафрагмальной доле
выделяются дорсальный и вентральный участки, в которых ветви воротной вены
распределяются обособленно. Два других элемента глиссоновой триады печени
в общих чертах повторяют ход ветвления и распределения воротной вены.
Эфферентную часть кровеносного русла печени составляет система печеноч-
ных вен. Многочисленные мелкие вены этой системы печени, сливаясь, форми-
руют 3 крупные печеночные вены: правую, среднюю и левую, направляющиеся
к дорсальному краю печени и впадающие в печеночный синус и далее в кау-
дальную полую вену.
Левая печеночная вена, дренирующая левую долю печени, самая крупная.
Ее короткий ствол формируется из 2 вен, одна из которых собирает кровь из
левой диафрагмальной доли, другая — из вентральных отделов висцеральной
доли [Урманов, 197O0J. Вена левой диафрагмальной доли — крупный сосуд, ко-
торый перед впадением в печеночный синус сливается с веной левой висцераль-
ной доли, собирающей кровь из вентральных отделов левой висцеральной доли
и ветвящейся по рассыпному типу. Вена левой висцеральной доли начинается
у самой перитонеальной ножки доли, по которой вена проходит из левой вис-
церальной в левую диафрагмальную долю, где сливаясь с веной этой доли, об-
разует левую печеночную вену. Из дорсальных отделов левой висцеральной доли
кровь оттекает по нескольким венам, самостоятельно впадающим в печеночный
синус или перед этим слившимися между собой в несколько меньшее число со-
судов. Вся система этих сосудов составляет левые дорсальные вены печени.
Средняя печеночная вена мельче и короче предыдущей. Она образуется сли-
янием вен, дренирующих правую диафрагмальную и квадратную доли. В каждой
из них обычно один, но бывают и парные венозные стволы. Вершины ветвей
этих вен древовидны, а сливаясь, приобретают типичный магистральный тип вет-
вления; главный ствол направляется дорсально, по ходу принимая в себя ветви
из всех отделов соответствующих долей.
Основной ствол средней печеночной вены в дорсальной части слегка откло-
няется влево, под прямым углом пересекает ворота печени и вливается в пе-
ченочный синус. Правая висцеральная и хвостатая доли дренируются несколь-
кими венозными коллекторами. Наиболее крупным из столов является правая
печеночная вена. Она формируется из нескольких корней в вентральных отделах
правой висцеральной доли, принимает по ходу несколько боковых ветвей и вли-
вается в правую половину печеночного синуса. Она магистрального типа. Из
дорсальных отделов правой висцеральной доли кровь оттекает по нескольким
мелким и коротким венам, самостоятельно впадающим в печеночный синус.
Хвостатая доля и ее отростки дренируются несколькими мелкими венами, впа-
дающими в правую половину печеночного синуса, из которого более крупная
вена несет кровь от хвостатого отростка. Основные стволы всех печеночных вен
располагаются ближе к диафрагмальной поверхности печени, чем элементы глис-
сонойбй триады, и по диаметру они шире их.
150
Таблица 8.4
Масса поджелудочной железы северного морского котика
Возраст, лет Пол Абсолютные данные, г Относительно массы тела, %
п lim M+m lim M+m
Щенки самцы 7 10.0-21.0 13Ю.16 0.11-0.24 0.1510.01
самки 7 10.0-23.0 16.lifi.19 0.11-0.28 0.23Ю.01
1 самцы 1 90.0 90.0 0.39 0.39
2 самцы 4 37.0-103.0 56.9ifi.18 0.14-0.38 0.22+0.07
3 самцы 10 42.0-107.0 70Ю.07 0.13-0.42 0.26Ю.03
4 самцы 3 90.0-110.0 100.0Ю.84 0.20-0.26 0.2210.01
самки 10 35.65-65.0 47.0±0.32 0.11-0.92 0.49Ю.02
5 самцы 4 120.0-190.0 155.tal.96 0.21-0.25 0.2310.01
самки 12 50.0-90.0 64.310.36 0.16-0.25 0.20Ю.01
6 самцы 3 75.0-182.0 135.7Ю.44 0.12-0.26 0.1910.08
самки 5 80.0-160.0 117Ю.17 0.18-0.47 0.28+0.06
7 самцы 3 210.0-320.0 260.010.04 0.17-0.29 0.24Ю.05
самки 3 100.0-200.0 135.0Ю.42 2.27-0.48 0.3610.09
8 самцы 5 300.0-698.0 423.610.85 0.24-0.96 0.35Ю.15
самки 5 85.0-210.0 155.010.26 0.24-0.49 0.34Ю.05
9 самцы 7 200.0-1350.0 510.311.73 0.16-0.84 0.37Ю.10
самки 5 120.0-250.0 148.010.27 0.20-0.56 0.31Ю.08
10 самцы 1 360.0 360.0 0.18 0.18
самки 2 110.0-150.0 130.0 0.30-0.38 0.34
>10 самцы 1 1158.0 1158.0 0.46 0.46
самки 14 35.0-210.0 125.710.14 0.10-0.47 0.27Ю.03
Ход и ветвление печеночных вен в основном повторяют таковые других эле-
ментов глиссоновой триады, но в самом начале печеночная артерия начинает
делиться раньше, чем вены. Поэтому к воротам печени подходят уже два ее ство-
ла. Первый из них у правого края ворот делится на артерию правой висцераль-
ной доли и хвостатого отростка и артерию правой диафрагмальной доли.
Левый ствол печеночной артерии между листками гепатодуоденальной связ-
ки отдает 2—3 дорсальные ветви в сосковидный отросток, в хвостатую" и квад-
ратную доли. В области портальных ворот ветви печеночных артерий анасто-
мозируют между собой, чего обычно не наблюдается в паренхиматозной части
печени. Пузырная артерия является ветвью правого ствола или отходит непос-
редственно от печеночной артерии, реже от ее левого ствола.
151
8.11. Поджелудочная железа
Поджелудочная железа котика исследована плохо. Имеются лишь данные о
ее массе [Scheffer, 19606; Соколов и др., 19696; Косыгин, Кузин, 1979].
Поджелудочная железа представляет собой целостный орган розовато-серого
цвета (рис. 8.13). Как и у многих других млекопитающих, она состоит из от-
дельных довольно-таки крупных железистых долек, объединенных рыхлой сое-
динительной тканью. В ней различаются головка и хвост, последний имеет крюч-
ковидный отросток. Головка железы сращена с нисходящим отделом двенадца-
типерстной кишки, а хвостовая часть и крючковидный отросток присоединяются
к этой кишке с помощью короткой брыжейки.
Головка железы своей дорсолатеральной поверхностью прилежит к печени
и стенке брюшной полости. Медиовентральная поверхность железы контакти-
рует со слепой кишкой, петлями тонкого кишечника и прикорневым брыжееч-
ным лимфоузлом. Хвостовой отдел железы каудально прилежит к переднему
полюсу левой почки, слепой и толстой кишкам, вентрально — к стенке желудка
и дорсально — к стенке брюшной полости.
По краниальному краю железы проходит желчевыводящие протоки и ворот-
ная вена. Проток поджелудочной железы открывается в двенадцатиперстную
кишку на вершине сосцевидной возвышенности. При этом часть его идет в кау-
дальном направлении в стенке кишки. Он открывается позади желчевыводящего
протока и на противоположной ему стенке кишки. Толщина поджелудочной же-
лезы не одинакова в разных частях. Наиболее толстая она в головке и в хвос-
товой части.
Масса поджелудочной железы у взрослых самцов превышает 1 кг, а у са-
мок — 250 г (табл. 8.4).
Абсолютные и относительные показатели массы поджелудочной железы пре-
кращают увеличиваться одновременно со стабилизацией массы тела. У самцов
это происходит в 10—12 лет, у самок — в 8 лет.
152
Глава 9. КРОВЕНОСНАЯ СИСТЕМА
9.1 Сердце
Сердце северного морского котика сравнительно крупный мускульный орган,
в плане напоминающий равнобедренный треугольник с закругленным апикаль-
ным углом (рис. 9.1). Лежит под грудиной между 3 и 7 ребрами. По отношению
к позвоночнику верхушка сердца находится на уровне восьмого грудного поз-
вонка. Поперечные венечные борозды, отделяющие предсердия от желудочков,
хорошо заметны. Снаружи желудочки сердца отделены друг от друга правой
и левой продольными бороздами, из которых первая более короткая, прости-
рающаяся на дорсальную сторону сердца. В области межжелудочковых борозд
проходят краниальные сосуды, и всегда имеется небольшое количество жира.
Стенки левого желудочка сердца у котика развиты лучше стенок правого. Хо-
рошо прослеживается онтогенетическое преобразование их толщины. У новорож-
денных котиков стенка левого желудочка лишь вдвое толще стенки правого же-
лудочка, в то время как у взрослых животных более чем в 3 раза (табл. 9.1).
Абсолютные показатели массы сердца увеличиваются у самок до 6-летнего
возраста, а у самцов продолжают расти и после 10 лет (табл. 9.2).
Увеличение относительной массы сердца прекращается с ростом тела живот-
ного. Наибольшей относительной массой сердца характеризуются новорожден-
ные котика. С возрастом эти показатели уменьшаются. В целом относительные
показатели массы сердца у котика меньше, чем у более специализированных к
водному образу жизни настоящих тюленей [Соколов и др., 1969а; Косыгин,
Кузин, 1979].
Из левого желудочка выходит аорта, а из правого — легочная артерия. По
выходному диаметру легочная артерия больше аорты. Впадающая в правое пред-
сердие каудальная полая вена лишь немного уже краниальной, но стенки ее толще.
На внутренней поверхности предсердий расположены гребешковые мышцы,
основная из которых в правом предсердии проходит вдоль предсердия до ос-
нования краниальной полой вены, отдавая от себя в сторону вены и к ушку
сердца мощные тяжи мышц, которые, видимо, компенсируют отсутствующий
клапанный аппарат этих вен. Второй мощный гребень проходит почти у осно-
вания предсердия наискосок по основанию ушка. Мускульные тяжи от этих гре-
бешковых мышц переплетаются между собой.
Межвенозный бугорок правого предсердия, разделяющий краниальную и ка-
удальную полые вены, хорошо выражен и напоминает валик. Латерально в сто-
рону от межвенозного бугорка у взрослых животных хорошо заметна овальная
ямка, затянутая очень тонкой перепонкой. У плодов овальная ямка открыта.
Зарастание ее происходит у щенков в возрасте 7—9 недель [Slijper, 1968]. Кау-
дально от овальной ямки находится венечный синус — место выхода большой
сердечной вены, которая, в свою очередь, идет по границе соединения предсер-
дия с желудочком. Она окружена снаружи хорошо выраженной мышцей. В этот
153
Рис. 9.1. Сердце северного морского
котика:
1 — аорта, 2 — легочная артерия.
Таблица 9.1.
Размеры сердца у разновозрастных северных морских котиков, см
Возраст, лет Ширина Высота Толщина Толщина стенки левого желудочка Толщина стенки правого желудочка
Сак к и
Щенок 4.8 4.6 3.6 0.7 0.9
Щенок 5.4 5.7 4.0 0.5 0.9
3 7.8 9.5 5.7 1.2 2.5
3 7.3 9.4 5.3 0.5 1.2
10 7.8 11.7 6.0 0.9 2.6
16 8.5 7.5 6.5 1.5 2.5
Сам ц ы
Щенки 5.1 6.1 3.9 0.6 1.3
1 6.4 7.7 4.5 0.4 1.4
2 6.3 7.9 5.0 0.6 1.5
4 8.9 10.7 5.5 0.5 1.6
6 12.5 13.5 5.3 0.8 2.0
6 13.5 14.0 6.5 07 2.5
9 16.5 16.5 9.0 0.9 3.0
10 16.0 18.0 8.0 0.8 2.7
11 14.4 15.9 9.8 0.8 3.0
же синус впадает другая вена, но меньшего диаметра, идущая по границе левого
и правого желудочков. Мелких отверстий от вен вокруг овальной ямки не видно,
хотя у многих наземных млекопитающих они присутствуют.
Гребешковая мышца левого предсердия хорошо выражена, как и в правом
предсердии. Она имеет древовидную форму и своим разветвляющимся к вершине
ушка стволом начинается от середины предсердия со стороны аорты. В общем,
мышца такая же, как и в правом предсердии, но в последнем стволы мышцы
крупнее, и их 2.
154
Таблица 9.2
Масса сердца северного морского котика
Возраст, лет Пол п Абсолютная масса, г Относительная масса, %
lim M+m lim M+m
Щенки самцы 6 36—80 49.3317.80 0.70-1.40 0.9810.12
самки 4 39—83 51.00112.08 0.83—1.32 1.04Ю.14
1 самцы 3 70—180 110.00146.22 0.38—0.78 0.52Ю.18
2 самцы 4 150—230 188.75122.47 0.58—0.85 0.75Ю.08
3 самцы 11 160—290 236.36112.97 0.53—1.12 0.82Ю.06
самки 2 110—160 135.00144.24 0.44—0.53 0.4910.07
4 самцы 6 200—260 238.3311064 0.45-0.63 0.57Ю.03
самки 3 160—172 167.3315.04 0.46—0.61 0.55Ю.06
5 самцы 9 250—610 363.89+42.46 0.49—0.84 0.60Ю.04
самки 4 140—270 190.00136.52 0.38—0.77 0.55Ю.11
6 самцы 8 300—600 406.25139,89 0.38—0.71 0.53Ю.04
самки 7 220—330 265.71116.64 0.50—0.85 0.67Ю.05
7 самцы 5 430—750 561.00168.67 0.45—0.68 0.54Ю.05
самки 6 200—300 241.67117.73 0.47—0.76 0.62Ю.05
8 самцы 6 500—760 541.67128.37 0.38—0.58 0.48Ю.04
самки 9 180—370 272.89122.62 0.47—0.82 0.63Ю.04
9 самцы 10 500—1010 674.50155.20 0.25-0.51 0.4310.03
самки 9 190—290 236.1 111 1.90 0.36—0.73 0.52Ю.04
10 самцы 2 575—800 687.50146.46 0.36—0.67 0.52Ю.27
самки 7 240—320 271.43112.10 0.56—0.80 0.64Ю.04
>10 самцы 4 750—970 817.50161.80 0.39—0.57 044Ю.02
самки 50 210-450 264.4617.55 0.38—0.81 0.56Ю.01
В правом предсердии в устье атриовентрикулярного отверстия находится
трехстворчатый клапан. Одна створка его лежит на перегородке сердца между
желудочками, а две другие на боковых стенках. Эти створки как бы окружают
атриовентрикулярное отверстие. Вентральная створка имеет 2 сухожильных тя-
жа, прикрепляющихся к вентральной сосцевидной мышце. Дорсальная створка
пятью сухожильными тяжами прикрепляется к крупной сосцевидной мышце, а
створка, лежащая на перегородке, тремя тяжами крепится к медиальной мышце
и небольшой сосцевидной, находящейся рядом с медиальной. Кроме двух круп-
ных сосцевидных мышц, стенка и боковая поверхность правой половины сердца
представляют собой переплетение множества мышц, идущих каудокраниально.
В левом желудочке находится двустворчатый клапан. Одна его створка лежит
на перегородке, другая на боковой стенке. Характерно, что обе створки сходятся
155
ле nc
Рис. 9.2. Варианты строения дуги аорты северного морского котика:
А — аорта, ЛС — левая сонная, ПС — правая сонная артерии.
на середине желудочка и образуют перегородку, которая как бы разделяет левый
желудочек в верхней его части на 2 половины, формируя выходные отверстия
в аорту и в предсердие. Створки, лежащие на перегородке, образуют 6 соеди-
нительнотканных тяжей, как и створка клапана, лежащая на стенке желудочка.
Они крепятся к двум сосцевидным мышцам на боковой стенке левого желудочка.
Дорсальная мышца длиннее, но тоньше вентральной. Однако она значительно
мощнее, чем одноименная мышца в правом желудочке.
В устье легочной артерии и аорты имеются по 3 полулунных клапана. Все
клапаны равны по ширине и глубине залегания между собой. Клапаны аорты
также равны между собой, но они находятся глубже в сердце, чем клапаны ле-
гочной артерии.
По линии соединения желудочков на вершине их в основании аорты рас-
положен легочной хрящ. На краях аортальных клапанов, в центре их свободного
края, имеются мелкие узелки. В легочной артерии их нет.
9.2. Артерии малого круга кровообращения
Малый круг кровообращения начинается легочной артерией, выходящей из
правого желудочка сердца (рис. 9.1). Она направляется дорсокаудально к корню
легких и через 3—4 см от своего начала отдает первый сосуд — артериальный
проток, облитерирующийся у щенков в возрасте 2 недель [Slijper, 1968] илй даже
остающийся открытым на протяжении первого месяца постнатальной жизни. На
уровне отхождения верхушечного бронха от магистрального легочная артерия
разделяется на правую и левую, идущие к соответствующим легким. Эти артерии
проходят вдоль магистральных бронхов с дорсальной стороны. Каудальная пра-
вая и левая легочные артерии делятся на ряд мелких артерий для соответству-
ющих долей легких [Мужчинкин, 1971а; Кузин, Соболевский, 1975].
9.3. Артерии большого круга кровообращения
Большой круг кровообращения начинается самым крупным сосудом — аор-
той. При основании аорта отдает 2 мелких венечных артерии, после чего по-
156
ворачивает влево и следует каудально, образуя дугу аорты, периметр которой
в среднем в 1.6 раза у новорожденных и в 1.8 раза у взрослых котиков шире
ее основания. В нисходящую часть дуги аорты вливается артериальный проток,
который у взрослых животных облитерируется. От дуги аорты отходят левая
и правая подключичные артерии. У отдельных особей хорошо выражена пле-
чеголовная артерия, которая впоследствии отдает левую и правую подключичные.
У других все 4 ствола артерий сильно сближены между собой, а у некоторых левая
и правая сонные артерии единым стволом отходят от дуги аорты (рис. 9.2). Имеются
многочисленные переходные варианты [Slijper, 1968; Мужчинкин, 1971а].
Общие сонные артерии направляются вперед вдоль вентролатеральной по-
верхности трахеи, не отдавая на своем пути никаких сосудов. Вблизи основания
черепа общий ствол сонных артерий делится на внутреннюю и наружную сонные
артерии. Внутренняя сонная артерия входит в череп через яремные отверстия,
пронизывает слуховую кость, появляясь в черепномозговой коробке через внут-
реннее сонное отверстие, где в области гипофиза правая и левая внутренние сон-
ные артерии анастомозируют друг с другом, образуя сплетение. Здесь же к ним
присоединяется очень тонкий стволик основной мозговой артерии как продол-
жение позвоночных и спинномозговых артерий, с которыми сонные артерии
также анастомозируют. У основания базисфеноида внутренние сонные артерии
сильно ветвятся, отдавая многочисленные ветви в боковом и дорсальном на-
правлениях, оплетая весь мозг. Наружная сонная артерия после отхождения
внутренней сонной артерии отдает ряд мелких ветвей для мышц шеи, гортани
и слюнных желез и затем переходит в лицевую артерию, где последняя сильно
ветвится, питая кровью затылочную, лицевую части головы и язык.
После отхождения правой сонной от подключичной артерии краниально вы-
ходит правая позвоночная артерия. Аналогичный сосуд отходит от левой под-
ключичной артерии. Обе позвоночные артерии направляются вперед к позво-
ночному столбу, где у 6-го шейного позвонка входят в межпоперечные отверстия
позвонка и уже далее следуют по межпоперечному каналу до 2-го шейного поз-
вонка, тело которого они огибают, и входят в отверстие крыльев атланта. По
краниальной поверхности атланта артерии идут вверх по костному желобу и
затем через затылочное отверстие входят в мозговую полость черепа, где снаб-
жают кровью, в основном, мозжечок.
Таким образом, кровоснабжение головного мозга у северного морского ко-
тика происходит, в основном, за счет внутренней сонной артерии, что отрицается
Е. И. Ивановой [1975] на том основании, что ее диаметр у эмбрионов и у взрос-
лых особей одинаков. По нашим данным, диаметр внутренней сонной артерии
в области выхода ее в черепную коробку в 3—4 раза больше, чем диаметр поз-
воночной артерии.
Следующим сосудом, отходящим от подключичной артерии после позвоноч-
ной, является внутренняя грудная артерия. Оба этих сосуда (правый и левый)
идут внутрь грудной клетки и направляются назад по бокам грудины, отдавая
ряд межреберных артерий.
Далее от подключичной артерии отходит плечешейная артерия, вскоре раз-
деляющаяся на реберношейную и шейную артерии. Обе эти артерии отдают мно-
гочисленные ветви к мышцам шеи и плеча (рис. 9.3). Наибольший из этих ар-
157
Рис. 9.3. Артерии передней конечности северного морского котика [из: Мужчинкин,
1971а].
А — артерии дистальных отделов конечности:
1 — плечевая, 2 — подкожная, 3 — глубокая плеча, 4 — локтевая, 5 — межкостная, 6 —
лучевая, 7 — срединная, 8—9 — первая и вторая пястные, 10 — третья пястная;
Б — ветвление подключичной артерии:
1 — общая сонная, 2 — позвоночный ствол, 3 — позвоночная артерия, 4 — реберношейная,
5 — верхняя межреберная, 6—8 — внутренняя и наружная грудные, 7 — шейная, 9 —
плечевая, 10 — лопаточногрудная, 11 — акромиальная, 12 — подлопаточная ветвь, 13 —
дельтовидная ветвь, 14 — боковая грудная ветвь.
В — ветвление подмышечной артерии:
1 — подмышечная, 2 — плечевая, 3 — внешняя грудная, 4 — лопаточногрудная, 5 — боковая
грудная ветвь, б — дельтовидная ветвь, 7 — подлопаточная ветвь.
териальных стволов, идущий к грудным мышцам, носит название наружной
гр!удной артерии. После отхождения вышеназванных артерий подключичная ар-
терия следует как подмышечная.
Собственно подмышечная артерия — очень короткий ствол, который вскоре
делится на подлопаточную и плечевую артерии, первая из которых ветвится на
ряд более мелких артерий, пронизывающих мышечные ткани у основания под-
ключичной и на поверхности подлопаточной мышц. Более крупная дорсальная
лопаточная ветвь отходит от подлопаточной напротив шейки лопатки и отдает
несколько веточек в трехглавую и другие мышцы, окружающие плечевой сустав.
Окружные задняя и передняя лопаточногрудные артерии снабжают кровью
мышцы плечевого пояса (рис. 9.3). В свою очередь, плечевая артерия спускается
далее в конечность. Она короткая и вскоре делится на глубокую плеча и пле-
чевую артерии. Первая из них снабжает кровью медиальную и среднюю головки
трехглавой мышцы плеча. Вторая, являющаяся основным продолжением плече-
вой артерии, вскоре ветвится на 3 сосуда: первый (передняя лучевая) следует
в жировой киль передней конечности, второй (срединная ветвь) вскоре рассы-
пается на ряд пястных артерий, третий (межкостная) продолжается в четвертую
пястную кость. Задняя локтевая, отходящая вместе с межкостной, идет по зад-
нему краю запястья и всего пятого луча. В. Ф. Мужчинкин [1971а] отметил боль-
шую вариабельность ветвления сосудов передней конечности, особенно пястных
158
Рис. 9.4. Ветвление брюшной
аорты северного морского
котика [из: Мужчинкин, 1971а].
1 — чревный ствол, 2 — передняя
брыжеечная, 3 — гастродуоденальная,
4 — желудочная, 5 — селезеночная, 6 —
почечная, 7 — каудальная брыжеечная,
8 — общая подвздошная.
Рис. 9.5. Краниальная бры-
жеечная артерия северного
морского котика
артерий. Каждая из них при переходе в пальцевые артерии пересекает фаланги,
раздваивается, направляясь вперед и назад, вступая в контакт друг с другом и
с ветвями соседних пальцев. В результате чего возможно образование парал-
лельных и связанных анастомозами сосудов по краям каждого луча (рис. 9.3).
Кровоснабжение грудной клетки происходит через ветви позвоночных ар-
терий, грудных и нескольких небольших ветвей, отходящих непосредственно от
аорты. В. Ф. Мужчинкин [1971а] отмечал, что общий принцип отхождения на-
чальных крупных стволов сосудов, снабжающих грудную клетку, весьма стаби-
лен, однако ее составляющие изменчивы: 1—3 сосуда с каждой стороны полу-
чают кровь из позвоночных стволов, следующие 2—3 сосуда начинаются единым
стволом от аорты. Возможен анастомоз между обеими группами, создающий
единый сосуд с двумя начальными, от которого отходят 5—6 межреберных ар-
терии. Изменчиво число межреберных ветвей внутренних грудных артерий.
Может наблюдаться еще более дробное распределение сосудов в межреберных
159
пространствах, в которых возможно даже отхождение сразу двух анастомози-
рующих артерий в одном межреберном промежутке или слияние артерий в одну.
Однако густых переплетений и извитости сосудов в области грудной клетки с
образованием “чудесной сети”, как у китообразных [Яблоков и др., 1972], у се-
верного морского котика не наблюдается.
При выходе из грудной клетки аорта отдает диафрагмальную артерию.
Первой крупной ветвью аорты в брюшной полости является чревная артерия,
отходящая на уровне 14—15 грудных позвонков (рис. 9.4). Через 1—1.5 см ствол
чревной артерии посылает гастродуоденальную, селезеночную и печеночную ар-
терии. Характерно, что к желудку возможно отхождение ветвей от. гастродуо-
денальной артерии. Селезеночная артерия проходит с вентральной стороны се-
лезенки, отдавая персональную ветвь к верхнему ее краю. Вентральная ветвь
селезеночной артерии направляется вниз, отдавая по пути 12—13 более мелких,
переходящих на желудок артерий для разных частей селезенки.
Ниже идет краниальная брыжеечная артерия, последовательно дробящаяся
на 8—18 ветвей, снабжающих кровью кишечник (рис. 9.5).
Далее от брюшной аорты отходят почечные артерии. Их число сильно варьи-
рует. Возможны варианты, когда к правой или левой почкам отходит от 1 до
5 сосудов (рис. 9.4). Иногда рядом с почечными артериями от брюшной аорты
отходит небольшая артерия, следующая к брюшной стенке. Последняя крупная
ветвь брюшной аорты — каудальная брыжеечная артерия, впоследствии отдает
сигмоидную и геморроидальную артерии. Заканчиваясь, брюшная аорта делится
на общий ствол подвздошных артерий и следует уже далее как крестцовая ар-
терия.
Подвздошные артерии могут иметь общий короткий ствол или отходить са-
мостоятельными сосудами наружных и внутренних подвздошных артерий (рис. 9.6).
В самом начале от наружной подвздошной артерии часто отходит крупная
окружная подвздошная артерия, направляющаяся краниолатерально и распре-
деляющаяся по брюшной стенке. Выходя из брюшной полости, наружная под-
вздошная артерия становится бедренной артерией. Отдавая небольшую глубо-
кую артерию бедра, превращается в подколенную артерию, от которой, в свою
очередь, отходит множество мелких артерий, питающих переднюю часть мышц
голени.
Отдельные ветви могут спускаться до стопы и анастомозировать с первой
метатарсальной артерией [Мужчинкин, 1971а].
Внутренняя подвздошная артерия следует в заднем направлении латерально
в полость таза. Первым ответвляющимся от нее сосудом является пупочная ар-
терия, затем небольшие седалищные и тазовые ветви, после чего подвздошная
артерия направляется к голеностопному суставу, по пути отдавая 1—2 ветви к
мышцам (лодыжковую артерию), которые идут по латеральному краю ступни
и сливаются с близлежащими метатарсальными сосудами или их ветвями. Ос-
новной ствол внутренней подвздошной артерии проходит между пяточной
костью и астрагалом и раздваивается. В свою очередь, зти два сосуда снова раз-
деляются на 2, в результате образуются 4 метатарсальных артерии, которые про-
должаются в пальцевые и затем далее проникают в кожную оторочку заднего
160
Рис. 9.6. Артерии задней конечности северного
морского котика [из.- Мужчинкин, 1971л]:
1 — общая подвздошная, 2 — крестцовая, 3 —
пупочная, 4 и 5 — наружная и внутренняя
подвздошные, 6 — окружная подвздошная, 7 —
глубокая бедренная, 8 — коленная, 9 — бедренная,
10—11 — передняя и задняя лодыжковые, 12 —
подвздошная, 13—14 — метатарсальные
ласта [Мужчинкин, 1971а]. Крестцовая артерия очень тонкая, переходящая в еще
более тонкую хвостовую артерию.
Таким образом, кровоснабжение дистальных отделов задней конечности у ко-
тика осуществляется за счет сосудов внутренней подвздошной артерии, а у на-
стоящих тюленей — за счет ветвей, отходящих от наружной подвздошной ар-
терии. У наземных хищных (собака, кошка) артериальная кровь к задней ко-
нечности также притекает по сосудам, идущим от наружной подвздошной ар-
терии.
Топография артериальной системы северного морского котика в целом соот-
ветствует основному плану ее строения у типичных наземных млекопитающих.
В описании ее нами было обращено внимание на кровоснабжение головного
мозга через внутреннюю сонную артерию и приток артериальной крови к задней
конечности по внутренней подвздошной артерии. Аналогичные варианты строе-
ния артериальной системы имеются и у сухопутных животных, но у близкород-
ственных и экологически близких форм водных млекопитающих, какими явля-
ются настоящие тюлени, кровоснабжение головного мозга в основном проис-
ходит через позвоночную артерию, а к задней конечности артериальная кровь
притекает по наружной подвздошной артерии [Иванова, 1975].
В настоящее время многие исследователи придают большое значение калиб-
ру дуги аорты. Начиная с описаний А. Бурова [Burow, 1838], Дж. Мьюри [Murie,
1874], первыми обратившими внимание на значительное расширение восходящей
части дуги аорты по отношению к ее основанию и заметное сужение нисходящей
части аорты после отхождения правой подключичной артерии, этот феномен
продолжает дискутироваться [Соколов, 1959; King, Harrison, 1961; Elsner et al.,
196.6; Slijper, 1968; Drabek, 1975; King, 1977; Кузин, Маминов, 1981]. В настоящее
время не все ластоногие исследованы в этом отношении, но у большинства изу-
161
Таблица 9.3
Диаметр дуги аорты у некоторых ластоногих
Возраст, лет Пол Диаметр аорты у сердца, мм Диаметр восходящей части аорты, мм % расширения Диаметр нисходящей части аорты, мм % сужения
Север ный мор с к о й к о т и к
Щенок самка 9.6 10.8 12.5 7.6 29.6
1 самец 10.8 13.6 25.9 8.9 34.6
2 самец 14.0 19.1 34.6 10.2 46.6
4 самец 15.9 20.4 28.3 10.8 47.1
9 самец 24.8 31.2 25.8 17.2 44.9
11 самец 23.6 29.3 24.2 17.8 39.2
С и в Р ч
Щенок самка 21.6 24.2 12.0 12.7 47.5
1 самка 24.2 32.5 34.3 16.6 48.9
6 самец 38.2 54.8 43.5 24.8 54.8
8 самка 38.2 53.5 40.1 22.6 57.8
Лар ? а
1 самка 17.2 24.2 40.7 12.4 48.8
2 самка 18.4 28.9 57.1 14.9 52.6
10 самка 21.6 35.0 53.5 162 53.7
О б ы к н о веннь 1 й tn ю л е н ь
5 самка 21.6 33.8 56.5 16.8 52.1
5 самец 21.6 38.2 46.4 18.4 51.8‘ '
Крыла т к а
6 самка 19.0 32.5 71.1 17.2 47.1
ченных видов бульбообразное расширение дуги аорты обнаружено. Правда,
Е. Слайпер [Slijper, 1968] у щенков северного морского котика (длина тела 62—
112 см, масса 4.5—9.4 кг) расширения дуги аорты не нашел. И это не было ошиб-
кой исследователя. Как видно из таблицы 9.3, у исследованных нами щенков
котиков расширение дуги аорты было очень небольшим. Однако в последующих
возрастных классах оно проявляется значительно сильнее.
Аналогичную картину изменения калибра восходящей части дуги аорты с
возрастом можно видеть у сивуча, ларги, австралийского и новозеландского
морских львов [King, 1977] и у тюленя Уэдделла [Drabek, 1975]. .
Среди исследованных нами и другими авторами ластоногих самые большие
показатели увеличения диаметра дуги аорты отмечены у новозеландского мор-
ского льва (84%). У крылатки и австралийского морского льва оно несколько
меньше (71%). У северного морского котика эти показатели самые низкие и срав-
162
ними с таковыми некоторых наземных видов [Drabek, 1975; Кузин, Маминов,
1981]. Не столь сильно выражено и сужение дуги аорты в нисходящей части у
котика, как это характерно для других видов ластоногих [Turner, 1888; King,
Harrison, 1961; наши данные].
Из представленных данных видно, что расширение дуги аорты во многом
адаптивный процесс, а не “природное явление” [King, Harrison, 1961], развива-
ющийся у ластоногих в процессе онтогенеза в связи с выработкой навыков ны-
ряния. Описывая этот феномен у взрослых ластоногих, Е. Слайпер [Slijper, 1968]
заключил, что функция его не ясна. Р. Элснер с соавт. [Elsner et al., 1966; Elsner,
1969] предполагают, что роль расширения дуги аорты — в поддержании дав-
ления крови во время ныряния. Даже допускается, что бульбообразное расши-
рение аорты действует как насос в течение диастолического состояния сердца
в период брадикардии, обеспечивая перфузию мозга кровью [Strauss, 1969]. С. Дра-
бек [Drabek, 1975] считал, что расширение аорты больше у тех видов, которые
способны нырять надолго и глубоко, что согласуется со всеми теперь имеющи-
мися данными по этому вопросу. Тем не менее, также известно, что артериальное
давление в период ныряния во многих магистральных сосудах поддерживается
на достаточно высоком уровне или даже несколько снижается, а в дуге аорты
после каждого сокращения сердца может наблюдаться временный обратный ток
крови. Снижается кровоток и в сосудах, обеспечивающих головной мозг [Га-
ланцев, 1978].
Проведенное нами сравнение соотношений размеров сосудов, отходящих к
голове (сонных и позвоночных), у разных видов тюленей с размерами дуги аорты
в разных ее частях не показало каких-либо преимуществ по этому признаку у
видов, подолгу остающихся на глубине (табл. 9.3). Более того, соотношение со-
судов у сивуча и морского котика не отличалось от таковых у исследованного
нами бурого медведя.
Нам представляется, что феномен резкого увеличения диаметра восходящей
части аорты у долго- и глубоконыряющих морских млекопитающих, также как
и заметное сужение нисходящей части аорты у этих видов, связаны не с бра-
дикардией при нырянии, а с тахикардией при выныривании. Именно сильно вы-
раженная тахикардия вызывает гипертрофию начального отдела дуги аорты, со-
здавая условия для временного размещения избыточного количества артериаль-
ной крови. Если принять такое объяснение причины возникновения расширения
дуги аорты, то тогда становится понятным его увеличение в процессе онтогенеза
у ныряющих млекопитающих и большая его выраженность у долгоныряющих
млекопитающих, по сравнению с менее специализированными к продолжитель-
ному пребыванию под водой животами.
9.4. Венозная система
По данным Е. Й. Ивановой [1972, 1975] и нашим материалам, система ярем-
ных вен у северного морского котика состоит только из наружных; внутренние
вены отсутствуют.
Наружная яремная вена образуется из многочисленных ветвей, отходящих от
головной части тела, главными из которых, непосредственно формирующими
яремную вену, является наружная и внутренняя челюстные вены (рис. 9.7).
163
Рис. 9.7. Система яремных и непарной вен се-
верного морского котика:
Г — передняя полая вена, 2 — непарная вена, 3 —
плечеголовной ствол, 4 — подключичная вена, 5 —
внутренняя грудная, 6 — яремная, 7 — плечешейная,
8 — восходящая шейная, 9 — подлопаточная, 10 —
плечевая, 11 — наружная челюстная, 12 — внутрен-
няя челюстная, 13 — ушная, 14 — глазная, 15 —
межреберная; III—XIV — позвонки.
Наружная челюстная вена собирает кровь из
многочисленных вен лицевой части головы (ли-
цевая вена), уха (ушные вены), глаза (глазные
вены), языка (язычные вены) и вен слюнных же-
лез. Внутренняя челюстная вена собирает кровь
из внутричерепного пространства через систему
венозных синусов, откуда их протоки выходят
через рваные (меньшее по диаметру) и яремные
(большие по диаметру) отверстия. У основания
черепа составляющие внутреннюю челюстную
вену ветви, анастомозируя друг с другом, при-
нимают венозные ветви от основания черепа,
подъязычной кости, кондилоидную, сливаясь,
образуют внутреннюю челюстную вену, которая
вскоре объединяется с наружной челюстной
веной, и формируют яремную вену. По пути сле-
дования вдоль шеи наружная яремная вена при-
нимает несколько мелких вен-анастомозов из
восходящей шейной вены и мышечных вен. Диа-
метр наружной яремной вены на всем пути ее сле-
дования вдоль шеи остается практически одина-
ковым. До впадения в подключичную вену на-
ружная яремная вена принимает самый крупный
анастомоз от восходящей шейной вены (рис. 9.7,9.8).
Слиянием плечешейного, подмышечного и яремного стволов формируется
плечешейная вена (рис. 9.8). Плечешейный ствол сильно выражен. Он форми-
руется из подкожной вены плеча, восходящей шейной и медиальной лопаточной
вен. Из этих трех вен наиболее крупной является восходящая шейная, которая
образует множество анастомозов с другими венами. В каудальный ее отрезок
впадает крупная вена от мускулатуры последних шейных позвонков. В восхо-
дящую шейную вену впадают вены от головы, плеча и от паротидных лимфа-
тических узлов, а краниальная часть формируется из вен поверхностной мус-
кулатуры шеи и шейных лимфатических узлов [Иванова, 1972]. Подмышечная
вена состоит из многочисленных ветвей, следующих из передней конечности, где
прохождение венозных сосудов сходно с таковыми артерий. Конечными ее ство-
. лами являются плечевая, подлопаточная и грудоспинная вены (рис. 9.7).
164
Рис. 9.8. Левая яремная подключичная система вен
северного морского котика [из: Иванова, 1975]:
1 — яремная, 2 — подключичная, 3 — подмышечная, 4 —
плечешейный ствол, 5 — восходящая шейная, 6 — подкожная,
7 — плечевая, 8 — анастомоз, 9 — наружная грудная, 10 —
подлопаточная, 11 — медиальная лопаточная, 12 —
акромиальная, 13 — окружная латеральная плечевая, 14 —
вена от головки плеча, 15 — шейные лимфатические узлы.
Передняя полая вена образуется слиянием плечеголовного венозного ствола
и непарных вен. В нее впадает общий ствол позвоночных вен, а также внутренние
грудные. Возможны варианты ветвления как конечных стволов, так и отдельных
составляющих их вен.
Система непарных вен представлена единственным стволом (рис. 9.7). На-
чинается непарная вена очень тонким сосудом в области 2-го поясничного поз-
вонка. По пути следования в непарную вену впадают межреберные, располо-
женные в одних случаях симметрично, а в других — несимметрично. На уровне
4-го грудного позвонка с правой стороны межреберные вены 2—4-го позвонков
объединяются и впадают в непарную вену единым стволом. Каждая межреберная
вена получает сосуд от экстрадуральной вены. Непарная вена воссоединяется
с передней полой на уровне сочленения 3-го и 4-го грудных позвонков.
Каудальная полая вена берет свое начало от слияния общих стволов под-
вздошных вен со срединной крестцовой веной (рис. 9.9). Это происходит ка-
удальнее почек. Далее она следует краниально вдоль позвоночного столба не-
много справа, пронизывает диафрагму и продолжается в грудной полости до
впадения ее в правое предсердие. По пути следования каудальная полая вена
принимает сначала вены от половых желез. Далее в нее впадают почечные
вены, как правило, состоящие из одиночных стволов, затем вены надпочечни-
ков. В области печени сильно расширяется и вместе с печеночными венами фор-
мирует обширный печеночный синус. При прохождении через диафрагму в нее
вливается диафрагмальная вена. Здесь же за счет диафрагмальных мышц фор-
мируется сфинктер. В грудной полости вена практически не принимает сосудов.
Система подвздошных вен начинается общим парным стволом внешних и
внутренних подвздошных вен (рис. 9.9). Еще до их расхождения общий ствол
подвздошных вен принимает поясничноподвздошную, каудальную и надчревную
вены. Наружная подвздошная вена, по описанию Е. И. Ивановой [1975], справа
имеет коллатераль, формирующуюся за счет бедренной и собственно наружной
подвздошных вен.
165
Рис. 9.9. Система каудальной
полой вены северного морско-
го котика [из: Иванова, 1975]:
1 — каудальная полая веиа, 2 —
правый и левый стволы каудаль-
ной полой веиы, 3 — окружная
глубокая подвздошная, 4 — пояс-
иичноподвздошная, 5 — каудаль-
ная надчревная, 6 — наружная под-
вздошная, 7 — подчревная, 8 —
веиа от задней конечности, 9 —
внутренняя срамная, 10 — запира-
тельная;
Я — почки, Sr — надпочечник.
Дистальные протоки каудальной бедренной и подколенной вен впадают в
основное русло бедренной вены, а краниальные протоки (проксимальная вена
колена, краниальная бедренная) вливаются в коллатераль бедренной вены (рис. 9.9).
Оба русла в подколенной области анастомозируют между собой.
Внутренняя подвздошная вена формируется сосудами, идущими от дисталь-
ных отделов конечности, в области таза, принимает много вен, сильно анас-
томозирующих между собой и собирающих кровь от задних отделов туловища.
Таким образом, венозная система северного морского котика, в основном,
построена по плану, известному для многих наземных млекопитающих. Из от-
личительных особенностей топографии венозного русла можно назвать отсут-
ствие внутренней яремной вены (у некоторых наземных млекопитающих она
тоже отсутствует), наличие крупного венозного синуса и мышечного сфинктера
в области диафрагмы, краниальное расположение расхождения каудальной
полой Вены на общие стволы подвздошных вен, более тесную связь с экстра-
дуральной веной системы непарных вен, присутствие венозных сплетений в об-
ласти основания черепа, шеи и подвздошной области.
9.5. Органы кровообразования
В нашем распоряжении имеются фрагментарные данные по селезенке и тимусу.
Селезенка. Плоский, удлиненный, с закругленными краями, симметричный
орган красно-фиолетового цвета. Лежит слева от желудка. Спереди касается пе-
чени, сзади прилежит к левой почке. С поверхности селезенка покрыта серозной
оболочкой, переходящей на большую кривизну желудка и образующей желудоч-
но-селезеночную связку, с помощью которой крепится к висцеральной поверх-
ности железы. Здесь же в селезенку впадают сосуды.
166
Таблица 9.4
Масса селезенки северного морского котика
Возраст, лет Пол п Абсолютная масса, г Относительная масса, %
lim Mim lim Mim
Щенки самцы 6 7—20 13.5012.30 0.13—0.35 0.27£0.04
самки 4 9—21 16.00±3.37 0.23—0.43 0.33Ю.06
1 самцы 3 25—60 38.33114.71 0.12—0.26 0.1810.06
2 самцы 4 48—59 53.5013.09 0.18—0.26 0.21Ю.02
3 самцы 11 38—135 68.60+9.68 0.14—0.43 0.28Ю.03
самки 1 20 20 а 08 6.08
4 самцы 6 47—150 77.00118.26 0.12—0.27 0.18Ю.03
самки 3 20—42 34.00+9.24 0.06—0.15 0.11Ю.04
5 самцы 9 60—230 141.13120.24 0.12—0.56 0.25Ю.05
самки 3 70—90 78.6718.40 0.18—0.25 0.2210.03
6 самцы 8 100—350 193.75+33.24 0.13—0.50 0.2610.05
самки 7 39—160 70.14+18.34 0.09—0.35 0.1710:04
7 самцы 5 119—400 201.80160.30 0.11—0.36 0.18+0.05
самки 6 60—110 81.67+8.86 0.13—0.26 0.2110.02
8 самцы 6 180—^470 315.00151.42 0.14—0.32 0.2410.03
самки 9 20—110 78.67110.71 0.05—0.29 0.1810.03
9 самцы 8 180—370 278.75125.27 0.13—0.25 0.16Ю.02
самки 9 51—100 73.44+5.83 0.11—0.25 0.16Ю.02
10 самцы 2 260—550 405.00 0.13—0.46 0.30
самки 7 40—190 104.57122.69 0.11—0.42 0.24Ю.05
>10 самцы 4 265—600 411.25194.10 0.13-0.36 0.2310.06
самки 44 35—200 98.4315.66 0.09—0.36 0.2110.01
Длина селезенки у взрослых самцов может достигать 30 см, ширина — до
12 см, масса — до 600 г. У самок селезенка меньше как по размерам, так и
по массе (табл. 9.4).
Будучи кровяным депо, способным задерживать до 16% циркулирующей
крови в организме [Акаевский, 1984], селезенка может сильно изменяться в объ-
еме и массе, что хорошо видно из представленных в таблице 9.4 данных. По
относительной массе орган не имеет половых и возрастных различий.
Тимус, как и у других млекопитающих, состоит из 2 частей: грудной и шей-
ной. У эмбрионов хорошо развиты обе доли, особенно грудная, размеры которой
не уступают верхушечной доли легкого [Кузин, 1971а]. С возрастом тимус ре-
дуцируется. Масса шейной части тимуса у 10-месячных эмбрионов (майские) со-
ставляет 0.5%, у июньских — 0.03%, у новорожденных — 0.07%, у взрослых —
0.002% от массы тела. Окончательно редуцируется к 5 годам.
167
Глава 10. МОЧЕПОЛОВАЯ СИСТЕМА
Большая часть опубликованных работ по мочеполовой системе северного
морского котика посвящена половым органам [Scheffer, 1949, 1950с; Scheffer, Ke-
nyon, 1963; Craig, 1964; Morejohn, 1972; Yoshida, Ichihara, 1974; Yoshida et al.,
1977, 1978; Yochida, 1982].
По органам мочеотделения имеются данные о вариантах отхождения почеч-
ных артерий от брюшной аорты и числе почечек (reniculi), составляющих почку
[Щуров, 1975а, б]. Табличные материалы о массе почек приведены в работах
В. Шеффера [Scheffer, 1960ft], А. С. Соколова с соавт. [1969], Г. М. Косыгина
и А. Е. Кузина [1979].
10.1. Органы мочеотделения
Почки у северного морского котика овальной формы, слегка сплюснуты дор-
совентрально. Расположены симметрично в краниальном отделе забрюшиннного
пространства и центральной частью прилежат к последнему грудному позвонку
(рис. 10.1). Одеты внутренним листком брюшины, через который просвечивают
мелкие поверхностные сосуды. Почечные ворота не выражены. Как и у других
ластоногих [Соколов и др., 1969<5; Косыгин, Кузин, 1979], относительная масса
почек у котика составляет 0.5—2.2% от массы тела (рис. 10.2). У новорожденных
и молодых животных их относительная масса больше, чем у взрослых, а у взрос-
лых самок больше, чем у одновозрастных самцов, что связано, видимо, с боль-
шей интенсивностью метаболизма у молодых, чем у взрослых, и у самок, чем
у самцов.
Почки северного морского котика относятся к типу множественных, состо-
ящих из многочисленных отдельных почечек, составляющих, в целом, компак-
тный орган. От почек наземных млекопитающих и настоящих тюленей они от-
личаются менее развитой интерренкулярной тканью и большей компактностью.
Не все почечки, составляющие почку котика, в отличие от почек настоящих тю-
леней [Guzsal, 1959], достигают поверхности органа. Часть из них находится
внутри его. Из-за очень плотного расположения почечек сегментарность почек
внешне не выражена.
По подсчетам В. А. Щурова [1975а], число почечек, составляющих почку ко-
тика, равно 65. Однако, по нашим данным, в среднем — вдвое больше. Часть
почечек, находящихся внутри органа, имеют по 2 сосочка, и хотя с поверхности
они выглядят как единая почечка, на гистологических препаратах хорошо видна
их самостоятельность. Возможно, что причиной образования таких сложных по-
чечек является редукция интерренкулярной ткани.
Среди ластоногих наблюдается хорошая корреляция между числом почечек
и размерами тела. У лахтака, например, число почечек вдвое больше, чем у ко-
тика [Щуров, 1975а], у южного морского слона их ок. 300 [Grahame, 1959].
168
Рис. 10.1. Топография почек
северного морского котика:
1 — почки, 2 — надпочечники, 3 —
мочеточник, 4 — почечная артерия, 5 —
почечная вена, идущая поверх двух
почечных артерий.
Рис. 10.2. Абсолютная (точки)
и относительная (кружки)
масса почек:
Н — новорожденные.
Каждая почечка по макроструктуре представляет собой простую почку в ми-
ниатюре. В ней хорошо различимы корковый, мозговой слои, почечный сосочек
и чашечка. Основу коркового вещества почечки котика составляют почечные
тельца, проксимальные и дистальные извитые канальцы и дистальные отделы
собирательных трубочек (рис. 10.3).
Толщина наружного коркового слоя в среднем ок. 688 (630—790) мк. По-
чечные тельца мелкие. Их диаметр в среднем 75.0 (54.7—99) мк. Диаметр со-
судистых клубочков равен в среднем 66 (55—72) мк (рис. 10.4).
Почечные тельца в корковом слое располагаются равномерно, и их размеры
не меняются по глубине залегания. В среднем на 1 мм2 площади гистологичес-
кого среза приходится 9—10 почечных телец. Пограничный слой тонкий, но хо-
169
Рис. 10.3. Корковое вещество
почечки северного морского
котика:
в центре — клубочек.
Рис. 10.4. Сосудистый клубочек
и радиальная артерия почки
северного морского котика
Рис. 10.5. Начальный участок
пограничной зоны (вверху)
почечки северного морского
котика
рошо выражен (рис. 10.5). Его толщина 182 (180—310) мк. Наружная зона моз-
гового вещества тоньше коркового и составляет 472 (405—540) мк. Мальпигиева
пирамида высокая — 1.7 (1.65—1.83) мк. Составляющие ее основу собирательные
170
Рис. 10.6. Собирательные тру-
бочки в вершине пирамиды
почечки северного морского
котика
трубочки и самые длинные петли Генле имеют прямую направленность, создавая
палочковидную исчерченность этой структуры (рис. 10.6). В этой части почечки
диаметр собирательных трубочек в среднем равен 27.1 мк. Наиболее суженная
часть прямых канальцев в среднем равна 12 мк, а ширина — 17 мк.
Кровоснабжение почек северного морского котика осуществляется посредст-
вом почечных артерий, отходящих от брюшной аорты на уровне 14—15 грудных
позвонков. Обращает внимание большая изменчивость и несимметричность в
числе почечных артерий. Их может быть от 1 до 5, отходящих к каждой почке
в различном сочетании (рис. 10.7), а общее число доходить до 8.
В большинстве случаев каждая почечная артерия отходит от аорты самос-
тоятельно, но наблюдаются случаи, когда 2 почечные артерии отходят из одной
точки. В тех случаях, когда от аорты отходит небольшое число почечных ар-
терий, то по пути их прохождения к почкам они разветвляются и, таким образом,
количество входящих непосредственно в почку артериальных сосудов увеличи-
вается. Наблюдается линейная зависимость между суммарным диаметром по-
чечных артерий и диаметром аорты: чем больше почечных артерий подходит
к почке, тем диаметр их меньше, и наоборот.
Анализ почек 217 котиков позволил нам выделить 17 вариантов отхождения
почечных артерий от брюшной аорты. Симметричное отхождение почечных ар-
терий наблюдается в 45.1% , несимметричное — в 55.9% случаев. Наиболее часто
от почек отходит 4, 7 и 8 артерий (рис. 10.7). Сравнительно редки варианты
с отхождением более трех артерий к каждой почке. С накоплением данных, воз-
можно, удастся использовать выявленный полиморфизм по числу почечных ар-
терий в качестве популяционного признака. По представленным материалам с
использованием коэффициентов сходства и различия Животрвского и х2 досто-
верных отличий между котиками, добытыми в разных районах, не обнаружено.
Каждая почечная артерия снабжает кровью определенный участок почки, по-
этому они могут быть названы сегментарными. Таким образом, по числу по-
чечных артерий можно говорить о числе сегментов в почке северного морского
котика. На коррозионных препаратах это хорошо видно.
171
ЗЕ О 9ЯВ < Q. Тихоокеанские воды Японии Японское море Охотское море Берингово море Курильские о-ва
«а ь 0) М’ со СО ч— |0 см ю
ь- Т“ Ч» о т~ 1 1 1 1
<0 ч- О ч- 1 1 1 1
Ю Т“ м- 1 1 1 о st 1
Т“ о Т“ 1 1 1 1
СО Т“ W °. ч- о> см 1 1 см см
СМ ч- iWr СМ 1П оо об 1 о 1
Т“ Т“ W т— cn 1 со со 1 1
О Т“ 4ft о Т“ t с с t
а> 4* см об оо об 10 см Г" о об ^т
оо * <0 О) т- СО Г" О in см о об in г* ч—
* ’t см ч- СО V 10 см т“ о со СП со ч—
СО * Г" СО 1 1 о см т—
ю чЬ ч~ 1 со <6 1 1
О) см со со 10 со со 4“ СО о ’t см 1O ’tf
СО см in О> in 1 о тг ’t
см g^B чт- СО 0) см со (6 о со т— г* т— г*
т— фв 1 О> in 1 о М;
1Я1нвибвд uiooiAiaehadioa %
Рис. 10.7. Варианты отхождения почечных артерий от брюшной аорты и частота их встречаемости в зависимости от района
обитания котиков
172
Рис. 10.8. Взаимоотношение кровеносной и выделительной систем в почечке
северного морского котика:
А, Б — артерии и мочеотводящие протоки:
1 - a. renalis II порядка, 2 — аа. praerencularis, 3 — аа. rencularis, 4 — renculus, 5 — чашечка,
6 — стебелек чашечки, 7 — a. subcorticalis;
В — артерии, веиы и мочеотводящие протоки:
1 — аа. praerencularis, 2 — аа. rencularis, 3 — аа. subcorticalis, 4 — v. interrencularis, 5 — w.
subcorticalis, 6 — ветвь мочеточника, 7 — чашечка, 8 — стебелек чашечки.
Ветвление сегментарных сосудов на артерии последующих порядков проис-
ходит по магистральному типу. Их число не соответствует числу ветвей моче-
точника. Магистральные артерии по пути следования внутри почки отдают ряд
преддольковых артерий (аа. praerencularis), от которых отходят почечковые ар-
терии (аа. rencularis). Каждая почечка получает от 2 до 4 близлежащих аа. ren-
cularis, отходящих от аа. praerencularis, которые вместе с мОчеотводящими про-
токами входят в почечку со стороны ворот. Случаев отхождения аа. rencularis
от смежных аа. praerencularis или прободения артериальными сосудами корко-
вого слоя почечки сбоку или сверху, как это наблюдается у калифорнийского
морского льва [Vrobell, 1963], нами у котика не обнаружено.
Каждая аа. rencularis внутри почечки ветвится на 3—5 аа. subcorticalis I—II
порядков, которые, в свою очередь, кустообразно рассыпаются на аа. corticalis
radiate, в комплексе создавая бокаловидное сосудистое формирование, в центре
которого находится расширенная часть мочеотводящего канальца — чашечка
(рис. 10.8).
Венозная кровь из. коркового слоя почечки выносится по многочисленным
венозным капиллярам, впадающим в 2—4 субкапсулярные вены, подходящими
173
Рис. 10.9. Мочеотводящие пути почки северного .^морского котика (рисунок с
коррозионного препарата):
1 — мочеточник, 2 — сложная лоханка, 3 и 4 краниальный и каудальный отводящие
протоки, 5 — краниальный дополнительный проток, 6 — каудальный дополнительный проток,
7 — отводящие протоки, 8 — чашечка, 9 — граница дольки, 10 — стебелек.
к междольковым венам (w. interrencularis). Как правило, венозная кровь из каж-
дой почечки принимается 3—5 междольковыми венами, которые обслуживают
одновременно 2—3 почечки (рис. 10.8). Далее в области ворот междольковые
вены постепенно собираются в вены II—III порядков, которые затем формируют
v. renalis. Тип ветвления венозных сосудов на уровне отводящих — магистраль-
ный; более мелкие междольковые вены сливаются дихотомически.
Мочеотводящие пути почечки представлены почечковой чашечкой (calicis re-
nalis), от которой отходит тонкий длинный выводящий проток (стебелек). Сте-
бельковидные протоки (3—4), сливаясь, образуют отводящие протоки I порядка
(рис. 10.9). Обычно ниже в этот же проток вливается еще 1 стебельковидный
проток от чашечки, находящийся в почечке и не; выходящий на поверхность
почки. ' •
Выводящие протоки I порядка объединяются в протоки последующих по-
рядков и формируют краниальный и каудальный отводящие магистральные
пути. Перед слиянием краниального и каудального отводящих протоков их ди-
аметр увеличивается, образуя в месте слияния широкое ампулообразное расши-
рение, напоминающее почечную лоханку многососочковой почки.
Мочеточник (рис. 10.10, 10.11), выходя из почки следует в каудальном на-
правлении с обеих сторон позвоночника под внутренним листком брюшины и
входят в мочевой пузырь со стороны его верхней стенки в области шейки.
. . ’ - Мочевой пузырь имеет грушевидную форму (рис. 10.10, 10.11). В напряженном
состоянии у взрослых самцов его длина не превышает 10 см, ширина 5 см, а
у самок, соответственно, 6.5 и 3 см. Масса мочевого пузыря у новорожденных
котиков составляет в среднем 4 г, у взрослых самцов 35—55 г; у взрослых самок
и неполовозрелых самцов 20 г.
174
Рис. 10.10. Строение мочеполовых органов самца
северного морского котика:
1 — тело полового члена, 2 — пещеристое тело, 3 —
мочеполовой канал, 4 — тазовая часть мочеиспускатель-
ного канала, 5 — предстательная железа, 6 — мочевой
пузырь, 7 — мочеточник, 8 — выводное отверстие мо-
четочника, 9 — выводное отверстие семенного каналь-
ца, 10 — ножка полового члена.
Рис. 10.11. Мочеполовые органы самца северного
морского котика:
1 — мочевой пузырь, 2 — предстательная железа, 3 —
поддерживающая связка, 4 — семенной каналец, 5 —
безымянная кость, 6 — седалищнокавернозиая мышца,
7 — бульбокавернозная мышца, 8 — ретрактор пениса,
9 — мочеточник, 10 — головка полового члена, 11 —
прямая кишка.
Стенка мочевого пузыря трехслойная. Наружный слой — серозная оболоч-
ка — является частью висцерального листка брюшины; второй — мышечный;
третий — слизистая оболочка, собранная в многочисленные продольные складки.
Крепление мочевого пузыря в полости тела происходит с помощью складкок
брюшины. Непарная срединная пузырнопупочная складка проходит с вентраль-
ной стороны мочевого пузыря к лонному сращению тазовых костей и вперед
по брюшной стенке к пупку. В ней проходит облитерированный мочевой проток.
Парные боковые складки соединяют боковые поверхности мочевого с под-
вздошными костями. В средней части эти складки перехлестываются с семяиз-
вергательными канальцами и сосудами, выходящими потом через паховый
канал.
Мочеиспускательный канал. Тазовая часть мочеиспускательного канала на-
чинается отверстием шейки мочевого пузыря, где он несколько расширяется и
у самцов вступает в область предстательной железы. Здесь в нижней части пред-
стательной железы в него открываются семяизвергательные канальцы. При вы-
ходе из предстательной железы его окружает уретральный Мышечный сфинктер
(рис. 10.10, 10.11). Далее мочеполовой канал следует по вентральной стороне
175
Рис. 10.12. Мочеполовые орга-
ны самки северного морского
котика:
1 — яичник, ,2 — яйцевод, 3 —
сумка яйцевода, 4 .— брыжейка
яичника, 5 — связка яичника, 6 —
рог матки, 7 — тело матки, 8 —
шейка матки, 9 — влагалище,
10 — отверстие мочеиспускатель-
ного канала, 11 —• предверье вла-
галища, 12 :—клитор, 13— моче-
вой пузырь.
полового члена и заканчивается на его головке ниже выступающей кости поло-
вого члена.
У самок мочеиспускательный канал короткий. В области шейки, также как
и у самцов, окружен уретральным сфинктером. Канал открывается в преддверие
влагалища у геминальной складки (рис. 10.12).
10.2. Половые органы
10.2.1. Мужская половая система
Пениальное отверстие располагается на брюшной стороне тела на уровне
бедер в 25—28 см от анального отверстия у взрослых особей (рис. 10.13).
У предродовых эмбрионов и у новорожденных котиков пениальное отверстие
окружено волосами (часто белыми), а у взрослых самцов кожа вокруг него голая,
интенсивно пигментированная.
Мошонка хорошо заметна, но не сильно выражена. Наружные покровы ее
почти лишены волос, морщинисты, темно-серого цвета. Семенники опускаются
в мошонку на 3-4 году жизни. Мышечно-эластической перегородкой мошонка
разделена на правую и левую камеры, предназначенные для левого и правого
семенников. 4
Семенники в мошонке располагаются косо. Длинная ось семенника проходит
дорсолатерокаудально (рис. 10.14). Под действием мускулатуры семенники легко
втягиваются в подвздошно-ректальную ямку, которая представляет собой узкую
176
Рис. 10.13. Общий вид половых органов
самца северного морского котика:
1 — семенник, 2 — придаток семенника,
3 — семяпровод 4 — сосудисто-нервный
пучок, 5 — половой член, 6 — ретрактор
полового члена, 7 — мочеполовое
отверстие.
Рис. 10.14. Семенник и его придаток у
северного морского котика:
1 — семенник, 2 — придаток семенника, 2* —-
головка придатка, 2^ — хвостик придатка,
3 — семяпровод 4 — сосудисто-нервный
пучок, 5 — семенной канатик, 6 — связка
семенника, 7 — мошоночная связка, 8 —
брыжейка семенника, 9 — кармашек
семенника.
продолговатую полость, окруженную мышцами тазовой области. Семенники фа-
солевидной формы, слегка сплющенные с боков. Поверхность их гладкая; раз-
меры и масса с возрастом сильно изменяются (табл. 10.1).
Особенно заметное увеличение массы семенников наблюдается с 3-летнего
возраста в связи с половым созреванием. В этом возрасте примерно половина
самцов становятся половозрелыми. Однако масса семенников не коррелирует с
присутствием сперматозоидов в эпидидимисе, хотя у самцов, в эпидидимисе ко-
торых присутствовали сперматозоиды, масса семенников была несколько боль-
ше. Рост семенников прекращается одновременно с ростом тела в 10—12 лет,
хотя трансгрессия по массе уже с 7-летнего возраста настолько велика, что тен-
денция роста в последующих возрастных классах едва прослеживается. Наи-
большая масса семенников, по данным В. Шеффера [Scheffer, 1950Ь], 151 г.
В нашей серии сборов (243 экз.) 150 г весили семенники у 9-летнего самца. Четко
выраженной асимметрии в массе правого и левого семенников нет (рис. 10.15).
У исследованных нами животных 42.7% имели правый семенник немного тяжелее
левого, у 32.7% животных наблюдалась обратная картина, а у 24% массы се-
менников не различались.
Среди самцов встречаются крипторхидные особи, которые по размерам тела
в 3 раза превосходят самку, но не имеют вторичных половых признаков (раз-
растания шейной мускулатуры, гривы из удлиненных волос на плечах и шее).
Масса семенников у таких особей составляла, по данным В. Шеффера [Scheffer,
1962], 4—10 г.
177
Таблица 10.1
Масса семенников (г) котика [по: Scheffer, 1950b, с дополнениями]
Возраст, лет п \ М±ли сг CV lim
Новорожд. 7 2.6±0.13 0:34 11.07 2.3—3.2
1 5 5.6±0.73 1.63 29.10 3.2—6.6
2 24 11.7±0.59 2.89 24.70 6.0—17.0
3 111 21.9±1.16 12.22 . -55.80 8.0—70.0
4 150 55.6± 1.39 17.02 30.60 14.0—104.0
5 . 26 79.0±3.43 17.48 22.12 38.0—106.0
6 22 94.7±4.17 19.55 20.75 64.0-137.0
7 6 103.6±8.86 21.70 20.94 , 76.0—126.0
8 4 111.2±8.42 16.84 15.14 99.0—129.0
9 7 ' 119.5±8.32 22.01 18.42 86.0—141.0
10 2 100.5±34.51 48.80 48.55 81.0—120.0
Рис. 10.15. Масса правого (а) и левого (б) семенников северного морского котика:
* Н — новорожденные.
На разрезе семенник имеет типичное для млекопитающих дольчатое строение.
Каждая доля железистой ткани отделена от соседней белочной оболочкой, про-
низывающей семенник с его поверхности. По размерам доли малы, но много-
численны. Гайморово тело хорошо выражено.
Основу семенника составляют семенные канальцы (рис. 10.16, 10.17). У эм-
брионов и щенков они выглядят тяжами диаметром 40—50 мк. С возрастом их
диаметр увеличивается. В особенности заметное увеличение диаметра и прогрес-
сирующее развитие составляющих тканей семенников наблюдается с наступле-
нием половой зрелости (табл. 10.2).
178
Рис. 10.16. Гистологический
препарат семенника 2-летнего
самца северного морского котика.
Стенки семенных'канальцев построены
нз половых клеток на ранних стадиях
зрелости. Остальную часть семенника
заполняет неоформленная
соединительная ткань.
Рис. 10.17. Гистологический
препарат семенника 8-летнего
самца северного морского котика.
В центре — семенной каналец, в
котором видны все стадии
сперматогенеза.
Таблица 10.2
Изменение диаметра семенных канальцев и клеток Сертоли в зависимости
от возраста морских котиков [из: Ащепкова, Дмитриева, 1982]
Возраст, лет Средний диаметр семенных канальцев, мк Средний диаметр ядер клеток Сертоли, мк
Эмбрион 40
Щенок 45
1 75.1Ю.17 б.ООЮ.Об
2 56.5Ю.47
3 107.0Ю.61 6.53Ю.06
4 125.2Ю.89 7.38Ю.06
5 125.StO.78 7.6510.04
б 142.0Ю.89 8.06Ю.18
7 145.211.22 8.50Ю.09
8 149.111.11 8.58Ю.09
179
Таблица 10.3
Частота встречаемости (%) разных типов семенных канальцев в зависимос-
ти от возраста в семенниках северных морских котиков, добытых 8 июля—
1 августа [по: Ащепкова, Дмитриева, 1982]
Возраст, лет Типы канальцев
I II III IV V
1 100
2 100
3 12 69 18 1
4 44 52 4
5 22 68 10
6 41 52 7
7 27 67 6
8 18 66 16
У части 3-летних и большинства 4-летних самцов отдельные семенные ка-
нальцы достигают размеров, характерных для взрослых особей, что является по-
казателем наступления половой зрелости. Наибольший диаметр семенных каналь-
цев в нашем материале имели 8-летние самцы; он составлял в среднем 150 мк.
Одновременно с увеличением диаметра семенных канальцев и развитием клеток
сперматогенного эпителия происходит увеличение диаметра клеток Сертоли.
В зависимости от степени зрелости сперматогенного эпителия различают 5 ти-
пов канальцев [Ащепкова, Дмитриева, 1982].
I. Сперматогенный эпителий характеризуется наличием сустентоцитов и спер-
матогониев.
II. Эпителий представлен сустентоцитами, сперматогониями и сперматидами
первого порядка.
III. Канальцы выстланы эпителием, который сформирован недифференци-
рованными сперматидами, базальные ряды образованы сперматидами первого
порядка, сперматогониями и сустентоцитами.
IV. Канальцы выстланы эпителием, состояние которого достигло стадии фор-
мирования сперматид. Группы дифференцирующихся сперматид локализуются
около ядер клеток Сертоли. Базальнее их располагаются сперматоциты первого
порядка и сперматогонии.
V. Эпителий содержит сустентоциты, сперматогонии, сперматоциты первого
порядка и сформировавшиеся спермин, которые составляют поверхностный ряд
эпителиального пласта.
Данные о преобладающем типе канальцев у котиков разного возраста пред-
ставлены в таблице 10.3. С 3-летнего возраста одновременно с увеличением ди-
180
аметра семенных канальцев начинается интенсивное развитие сперматогенного
эпителия (рис. 10.17) Уже в этом возрасте у самцов присутствуют семенные ка-
нальца II—V типов. Фактически уже у 3-летних самцов встречаются конечные
формы клеток сперматогенного эпителия.
У 4-летних самцов преобладают канальцы IV типа, что характерно для ин-
тенсивного процесса сперматогенеза. Уменьшение диаметра канальцев у 6—7-
летннх, возможно, свидетельствует об утилизации спермиев из канальцев семен-
ников. Эти данные хорошо согласуются с экологическими материалами по раз-
множению. Действительно, с 7-летнего возраста большинство самцов принима-
ют активное участие в размножении.
Интерстициальная ткань семенников представлена клетками Лейдига, лежащи-
ми в неоформленной соединительной ткани. У предродовых эмбрионов интерсти-
циальная ткань отчетливо выражена, в то время как у 2-недельных щенков она
регрессирует. Увеличение ее клеток начинается с 3—5-летнего возраста и наибольшего
развития достигает у 8-летних самцов. У последних в цитоплазме клеток ткани от-
мечается повышенное содержание суданофильных липидов и холестерина.
Придаток семенника имеет форму вытянутого, слегка бугристого тяжа, рас-
положенного с латеральной стороны семенника (рис. 10.15). Он возвышается над
дорсальной поверхностью последнего чуть заметно утолщенной головкой. Тело
придатка, по сравнению с головкой, немного тоньше и на заднем конце пере-
ходит в петлеобразно скрученный хвостик, откуда начинается семяпровод. Ха-
рактерно, что придаток более или менее свободно соединяется с семенником при
помощи серозной оболочки, образующей между ними кармашек.
Семенник и придаток одеты общей плотной оболочкой, которая соединяет
их с дном пазухи. Крепление семенника осуществляется широкой брыжейкой,
по латеральному краю которой проходит связка, а по медиальному краю — се-
менной канатик.
Семяпроводы у котика начинаются непосредственно от хвостика придатка
выше его каудального края и, извиваясь по брыжейке, направляются вперед, где
в области пахового канала на уровне вертлужной ямки объединяются с нерв-
но-сосудистым пучком, идущим от краниального конца семенника, формирую-
щего семенной канатик. Пройдя через паховый канал в брюшную полость, се-
мяпроводы дугообразно изменяют направление каудально, ложатся на дорсаль-
ную стенку мочевого пузыря, сближаясь друг с другом, пронизывают предста-
тельную железу, открываясь отверстиями в начальном отделе мочеиспускатель-
ного канала. Конечная часть семяпровода увеличивается в диаметре, образуя
ампулу семяпровода. Место впадения семяизвергательных канальцев в предста-
тельную железу обозначено заметным холмиком.
Пенис (рис. 10.18) скрыт под кожей. Он состоит из пещеристых тел, смы-
кающихся дорсально, между которыми вентрально проходит мочеполовой канал
и кость пениса (os penis). Развитого губчатого тела нет. Общая длина пениса
может достигать у взрослых животных 25—27.5 см.
Пещеристые тела образуют спинку полового члена. Линия соединения пе-
щеристых тел представлена плотной вертикальной пластинкой белочной обо-
лочки, переходящей с их поверхности. Каудальные части пещеристых тел хо-
рошо развиты, но по мере приближения к кости пениса они несколько умень-
шаются в диаметре, а затем перед нею вновь увеличиваются.
181
Рис. 10.18. Половой член северного
морского котика:
1 — ствол полового члена, 2 — головка,
3 — вентральный листок крайней плоти,
4 — отверстие мочеиспускательного канала.
Рис. 10.19. Кость поло-
вого члена северного
морского котика
Кость полового члена хорошо развита. Каудальный конец ее округлый, с ше-
роховатой поверхностью, а краниальный конец сильно расширяется и образует
дорсальное и вентральное утолщения (рис. 10.19). Если смотреть на кость с тор-
ца, то она напоминает восьмерку. Стержень от основания к вершине несколько
уменьшается в диаметре и имеет дорсальный изгиб. Поперечное сечение у ос-
нования овальное, либо округлое, далее становится треугольным, а к вершине
углы сглаживаются, и кость в поперечнике становится овальной.
Длина кости полового члена у новорожденного самца 2.3—3.5 см, масса —
0.1—0.3 г; с возрастом эти показатели увеличиваются равномерно. У старых сам-
цов длина достигает 14.4 см [Scheffer, Kenyon, 1963], а масса — 14.5 г (табл.
Ю.4).
Масса кости пениса хорошо коррелирует с ее длиной. Для нее не отмечено
резкого увеличения в период полового созревания животных, что характерно
для масс семенников и тела.
У крипторхидных особей кость полового члена плохо развита. Она необычно
тонкая, но в длину мало уступает таковой полноценных самцов. У 5 исследо-
ванных В. Шеффером [Scheffer, 1950с] крипторхов длина кости была 7.6—13.1
см.
В связи с тем, что губчатое тело пениса не развито у северного морского
котика, головка полового члена представляет собой, по сути, покрытую снаружи
слизистой оболочкой кость пениса, переходящей с внутреннего листка крайней
плоти. Под слизистой вдоль кости проходят многочисленные сосуды и тонкие
соединительнотканные, эластические и мышечные волокна. Характерной чертой
головки пениса у котиков является то, что слизистая оболочка, не доходя 1—2 см
до конца кости пениса, венчикообразно расширяется. Из этого вялого венчи-
кообразного расширения выступает расширенная часть кости, которая плотно
обтянута слизистой.
182
Таблица 10.4
Длина бакулюма (мм) котика [из: Scheffer, 1950с, с дополнениями]
Возраст, лет п Mim СТ CV lim
Щенки' 13 27.3Ю.17 0.6 2.19 23.0—25.0
1 11 49.411.29 4.27 8.64 42.0—45.0
2 24 56.611.07 5.24 9.25 48.0—68.0
3 111 66.5Ю.50 5.26 7.90 53.0—80.0
4 150 82.210.46 5.60 6.80 69.0—99.0
5 27 97.0+1.86 9.66 9.66 77.0—115.0
6 21 108.411.54 7.05 6.50 94.0—120.0
7 6 116.112.48 6.07 5.22 112.0-126.0
8 4 130.013.65 7.30 5.61 124.0—137.0
9 2 120.015.30 7.49 6.24 117.0—123.0
10 xi;' 1 144.0
11 1 136.0
Мочеиспускательный канал открывается с вентральной стороны головки пе-
ниса (рис. 10.18). Простатическая часть канала широкая. Его слизистая собрана
в многочисленные продольные мелкие складки и покрыта мелкими бугорками,
куда открываются выводные протоки предстательной железы. Одна из складок
в простатической части мочеполового канала сильно увеличена в размерах, в
виде валика. Она проходит по дорсальной поверхности мочеполового канала
и продолжается до середины предстательной железы.
Перепончатая часть мочеполового канала несколько уже простатической и
также собрана в мелкие продольные складки. При выходе в пещеристую часть
полового члена мочеиспускательный канал расширяется.
Обслуживает пенис группа мышц (рис. 10.10). Ретракторами пениса служат
две узкие мышцы, начинающиеся от анальных мышц и заканчивающиеся в тка-
нях препуция. Мембранозная (пещеристая) часть уретры окружена тонким слоем
мышечных волокон, которые у корня соединяются с круглыми мышцами ануса.
Седалищнокавернозная мышца начинается от седалищного бугра безымянной
, кости и направляется к корню полового члена, где сильными мышечными пуч-
ками заканчивается на белочной оболочке кавернозных тел. Бульбокавернозная
мышца хорошо выражена. Она начинается от уретрального сфинктера и закан-
чивается на корневой части полового члена, прикрывая начальный отдел мо-
чеиспускательного канала. Препуциальный мешок спереди обслуживается пар-
ными мышцами (m. praeputialis), которые широким основанием начинаются от
мышц брюшной стенки и узкими пучками крепятся на препуции.
183
Предстательная железа — непарное образование, охватывающее начальную
часть мочеиспускательного канала (рис. 10.11). Ее диаметр у взрослых самцов
составляет 3.5—5.1 см, масса — 7.4—12.0 г, у холостяков, соответственно, 1.2—
3.8 см и 7.0—9.4 г.
10.2.2. Женская половая система
Наружные половые органы самок котика вместе с анальным отверстием на-
ходятся в кожной складке, лишенной волосяного покрова (рис. 10.20).
Рис. 10.20. Наружные половые органы самки северного морского котика:
1 — вход во влагалище, 2 — клитор, 3 — анальное отверстие.
Кожа вокруг половой щели в области больших половых губ темно-бурого
цвета, морщинистая. Малые половые губы не выражены. Клитор в виде неболь-
шого возвышения (до 1 см) слизистой оболочки преддверия влагалища, окружен
двумя лепестковидными складками препуция клитора. На нижней стороне кли-
тора выделяется бороздка — остаток мочеполового канала. Пещеристые тела
клитора хорошо развиты. Продолжаясь вглубь преддверия влагалища, тело кли-
тора увеличивается в диаметре и заканчивается на седалищных буграх безымян-
ных костей двумя ножками. Клитор, как правило, не содержит костных обра-
зований, однако В. Шеффер [Scheffer, 1949] обнаружил у одной самки в возрасте
4 лет кость клитора длиной 2.1 см. При этом клитор имел ненормально большие
размеры, а яичники этой самки были очень маленькие.
Влагалище представляет собой трубчатый орган. В нем хорошо различимы
собственно влагалище и преддверие (рис. 10.12). Оба образования примерно
равны'по длине и у взрослых животных составляют вместе 10—12 см. Ширина
преддверия немного больше самого влагалища и составляет у половозрелых
самок 6—8 см. Слизистая преддверия розового цвета, собрана в многочисленные
крупные и мелкие продольные складки. Оно отграничено от влагалища хорошо
выраженной геминальной складкой, имеющей ширину 0.8 см у взрослых особей.
С вентральной стороны в непосредственной близости от геминальной складки
184
Таблица 10.5
Сезонное изменение массы (г) яичников половозрелых самок северного
морского котика
Месяцы п Яичник беременного рога матки Янчник небеременного рога матки
М±т CV М±т а CV
Декабрь 16 7.17±0.55 2.19 30.67 3.81±0.40 1.60 41.99
Январь 15 6.93±0.52 2.01 29.00 3.73±0.30 1.16 31.01
Февраль ‘2 9.07±0.83 2.87 31.64 3.88±0.32 1.10 28.55
Март ‘9 7.08±0.36 1.56 22,.03 3.84±0.30 1.30 33.85
Апрель 53 7.26±0.22 1.60 23.03 3.9Н0.14 1.01 25.83
Май 10 8.95±0.б5 2.05 22.90 6.54±1.29 4.10 62.69
Июнь 9 9.10±0.73 2.21 24.52 8.30+0.69 2.07 25.06
Июль 25 Ю.18±О63 3.16 31.11 10.38±0.64 3.20 30.82
30 Т
Й 1 2 3 4 5 6 7 6 9 10 н 12 13 14 15 16 (7 16 19 20
Возраст, лет
Рис. 10.21. Изменение массы яичников в зависимости от возраста:
Н — новорожденные.
в преддверие открывается мочеиспускательный канал. Место его выхода обоз-
начено выпячиванием слизистой оболочки.
Стенки влагалища эластичны. Крупная складчатость слизистой отсутствует.
Краниально влагалище оканчивается сводом — щелевидным пространством
между выпячивающейся сюда влагалищной частью шейки матки.
Матка имеет парные рога, хорошо выраженное тело и шейку. Размеры и
положение матки в брюшной полости меняются в зависимости от возраста и
физиологического состояния. У нерожавших особей все составляющие матки
тонкие и эластичные, а у многорожавших самок они уплотнены, продольно
складчаты и бугристы.
185
Тело матки располагается в брюшной полости между прямой кишкой и мо-
чевым пузырем. Оно образовано частично слившимися рогами. Внутренняя пе-
регородка тела, разделяющая правый и левый мюллеровы протоки, простирается
довольно глубоко, и лишь на уровне начала формирования втулкообразного су-
жения мюллеровы протоки объединяются в единый канал, формируя его устье.
Описанное строение матки позволяет считать ее не двурогой, как об этом пишет
Л. В. Бойцов [1934], а двураздельной, образованной частично слившимися ма-
точными отделами с общим выводным протоком [Кузин, 1971а].
Шейка матки короткая, выпячивающаяся во влагалище. Края ее неровные,
заходящие выступами в вырезки противоположных стенок, формируя клапано-
образно закрывающийся вход в матку (рис. 10.12).
Рога матки расходятся медиально от ее тела и имеют пологое каудодорсаль-
ное скручивание, так что дистальные концы находятся несколько краниальнее
их средних частей. По длине каждый рог значительно больше тела, и лишь не-
много уступает ему по диаметру у небеременных особей.
А. Крейг [Craig, 1964] выделял 5 слоев в стенке матки, отдельно рассматривая
сосудистый и 2 мышечных слоя (слой круговых и слой гладкомышечных про-
дольных волокон). Однако другие анатомы обычно не выделяют сосудистый
слой, а два названных мышечных слоя объединяют в один [Жеденов, 1965].
Внутренняя (слизистая) оболочка матки очень тонкая, собрана в многочис-
ленные продольные складки и пронизана множеством желез с выводными про-
токами, открывающимися в полость матки. В обычном состоянии слизистая тон-
кая, железы ее не активны. В период имплантации бластоциста и плацентации
зародыша слизистая сильно пролиферирует, железы увеличиваются до крупных
размеров, образуя причудливый лабиринт, тем самым увеличивая толщину
матки [Craig, 1964].
Мышечный слой представлен внутренним слоем круглых и наружным слоем
продольных мышц, между которыми проходят многочисленные сосуды и нервы.
В период лютеальной и фолликулярной активности яичников мускульный слой
гипертрофируется и обильно васкуляризируется. Наружная серозная оболочка
покрывает матку со всех сторон и является производным висцерального листка
брюшины. Она же продолжается в виде широкой связки матки, подвешивающей
ее к дорсальной стенке брюшной полости. Широкая связка матки объединяет
брыжейку яичников, рогов и тела матки и простирается дальше на каудальную
часть почки. К широкой связке матки дорсально прикреплена круглая связка.
Яйцеводы представляют собой парные извилистые полые трубочки, начинаю-
щиеся обширной воронкой от переднего края яичника и впадающие в конечную
часть рога матки (рис. 10.12). Слизистая оболочка внутренней поверхности их
рыхлая и снабжена большим количеством сосудов. Ампула яйцевода хорошо
выражена. По направлению к матке диаметр яйцеводов уменьшается и при впа-
дении в ее рог не образует втулки. Длина яйцевода у взрослых самок 10—12 см;
диаметр в начальной части 6 мм, в конечной — 1 мм.
Яичники расположены каудальнее почек. Окружены сумкой (воронкой), по-
лость которой сообщается с брюшной полостью отверстием до 1 см, располо-
женным каудовентрально. Крепятся яичники к дистальному концу рога матки
186
размножение самок северного морского котика.
Объяснение в тексте.
мощной связкой, являющейся продолжением поддерживающей связки рога.
Кроме того, имеется брыжейка яичника, которая является краниальной частью
широкой связки матки, переходящей потом на почку. Яичники почти округлые
(2.6х2.5 см — у взрослой самки). Форма их не меняется даже у беременных,
когда в одном из яичников развивается желтое тело беременности. Поверхность
яичника гладкая, ровная.
У новорожденных самок яичники весят в среднем 2.3 г. Заметное увеличение
их массы наблюдается до 5-летнего возраста, когда большинство самок достигает
половой зрелости, и в дальнейшем их рост практически прекращается (рис. 10.21).
Яичники у самок, рожавших из обоих рогов матки (4-летние и старше), прак-
тически имеют одинаковую массу в любом возрасте.
Масса пары яичников взрослых особей в летний период (после родов до им-
плантации и без видимых следов беременности в матке) составляет ок. 15 г. Мас-
са правого и левого яичников у небеременных и неполовозрелых животных поч-
ти не различается, однако в период беременности в связи с развитием желтого
тела в одном из яичников масса последнего быстро увеличивается, и он вдвое
превосходит по массе другой яичник (табл. 10.5).
У неполовозрелых самок в связи с тем, что фолликулярная активность яич-
ников проявляется неодновременно, наибольшие показатели массы яичников не
согласованы по срокам. Как видно на рисунке 10.22, у пятилеток наибольшая
масса яичников наблюдается в июле, у трех—четырехлеток — в августе, а у двух-
леток — в сентябре. Представленные данные хорошо согласуются со сроками
прихода самок соответствующего возраста на лежбище и очередностью вступ-
ления в размножение.
К. Ешида [Yoshida, 1982], измеривший диаметр яичников молодых самок, при-
шел к заключению, что наибольшая фолликулярная активность и наибольший
диаметр яичников у годовалых самок наблюдается в ноябре, у двухлеток —
в октябре, у трех—четырехлеток — в сентябре. Наибольшая масса яичников,
187
Рис. 10.23. Гистологическое строение яичников самки северного морского котика
разного возраста и физиологического состояния [из: Yoshida, 1982]:
I — яичник щенка 3-месячной самки, 2 — яичник неполовозрелой 2-легней самки, 3 —
яичннк половозрелой 14-летней самки, 4 — некрупный фолликул в процессе дегенерации;
VF — везикулярный фолликул, ТА — tunica albuginea, О — ооцит, PH — фагоцит, ZD —
прозрачная зона, CAT — тело атретического фолликула, СА — белое тело, FC —
функционирующее желтое тело.
по нашим данным, наблюдается в январе, а наименьший диаметр, по данным
К. Ешида, — в феврале.
Яичники имеют типичное для большинства млекопитающих строение (рис.
10.23).
Одноклеточный слой зародышевого эпителия, одевающий снаружи яичник,
инвагинирует внутрь периферической части стромы яичника через белочную обо-
лочку, образуя многочисленные подповерхностные крипты [Creig, 1964]. Размер
созревших фолликулов у северного морского котика может достигать 10 мм, и
он зависит только от фолликулярных циклов и не зависит от возраста половоз-
релых самок. Зернистый слой созревших фолликулов имеет 71 мк или 10—15
клеток в толщину. Внутренняя и внешняя оболочки составляют 100 мк в толщину
[Craig, 1964]. По данным К. Ешида [Yoshida, 1982], образование желтого тела
беременности происходит за счет зернистого слоя. Желтое тело овуляции растет
очень быстро и на 15—19 сутки после родов (или на 12—15 сутки после ову-
ляции) достигает 0.5—1.0 см в диаметре (рис. 10.24).
188
Рис. 10.24. Изменение наиболь-
шего диаметра желтого тела:
светлые кружки — желтое тело бере-
менности, темные — ложное желтое
тело, крестики — белое тело.
22
МЕСЯЦЫ
Рис. 10.25. Изменение наибольшего диаметра фолликулов [из: Yoshida, 1982]:
светлые кружки — у самок, имеющих желтое тело беременности; черные — у самок, имеющих
ложное желтое тело; крестики — у самок, неовулировавших в данном сезоне размножения.
Интенсивный рост желтого тела, сопровождающийся пролиферацией и уве-
личением размеров желтых клеток, продолжается до периода имплантации
(обычно это происходит в ноябре), после чего оно становится желтым телом
беременности. Желтое тело беременности растет в несколько замедленном темпе,
чем желтое тело овуляции до апреля.
Рост желтого тела не сопровождаемого последующей беременность (ложного
желтого тела) происходит с той же интенсивностью, что и истинного желтого
тела до периода имплантации бластоциста (ноябрь), после чего оно быстро рег-
рессирует, хотя до этого времени и по размерам и гистологическому строению
истинное и ложное тела не различаются. К апрелю размеры ложного тела ста-
новятся вдвое меньше истинного желтого тела и позже эти различия в размерах
сохраняются до периода родов (июнь—июль) [Yoshida, 1982]. Лютеиновые клет-
189
ки ложного тела начинают редуцироваться с января, к июню они полностью
исчезают, и ложное тело превращается в белое тело.
Циклы у самок, впервые вступающих в размножение и яловых в данный
сезон, и у беременных различны (рис. 10.25).
У впервые вступающих в размножение самок увеличение размеров фолли-
кулов Начинается в марте—апреле и достигает наибольших размеров в июне,
перед овуляцией. У самок, содержащих в одном из яичников ложное или ис-
тинное желтое тело, фолликулы незначительно увеличиваются в диаметре до сен-
тября, усиливают рост в октябре, а после имплантации бластоцисты, когда у
одних самок формируется истинное, а у других ложное желтые тела, рост фол-
ликулов определяется присутствием того или иного из них и, соответственно,
различается. У самок, содержащих в одном из яичников ложное желтое тело,
в другом яичнике фолликулы интенсифицируют рост от января к февралю, а
у самок, в яичнике которых имеется желтое тело беременности, рост в этот пе-
риод менее интенсивен и его ускорение наблюдается от апреля к маю. Описанное
ускорение роста фолликулов у самок в разном физиологическом состоянии (от
января к февралю или от апреля к маю), по мнению К. Ешида [Yoshida, 1982],
определяется началом регрессивных процессов в желтых телах, тем не менее к
периоду гона (июнь) фолликулы у всех половозрелых самок, готовящихся к раз-
множению в предстоящий сезон, достигают почти одинакового размера (рис.
10.25). Лишь у самок, впервые вступающих в размножение в данном сезоне, наи-
большее число крупных фолликулов наблюдается позже (сентябрь—октябрь).
К периоду размножения в Граафовы пузырьки превращаются несколько фол-
ликулов, причем у впервые вступающих в размножение самок их число вдвое
больше, чем у многорожавших. Две овуляции в одном яичнике у северного мор-
ского котика не зарегистрированы, хотя двойни встречаются и составляют 0.14%
от общего числа рождающихся детенышей [Niggol, Fiscus, 1960; Creig, 1964; Pe-
terson, Reeder, 1966; Кузин, 1969й]. Двойная овуляция — случайное явление и
может наблюдаться в яичниках самок, впервые вступающих в размножение или
неоплодотворенных в предыдущем сезоне размножения, или беременность у ко-
торых была прервана на ранних стадиях [Creig, 1964]. У котиков возможна миг-
рация бластоциста из одного рога в другой.
190
Глава 11. ВОПРОСЫ ФИЗИОЛОГИИ
Исследования физиологии северного морского котика проводились, в основ-
ном, в направлении изучения адаптаций, связанных с нырянием. Изучались мор-
фофизиологические реакции организма, связанные с обеспечением его кислоро-
дом, и системы, доставляющие кислород к тканям и органам.
Сведения об объеме, составе и дыхательных свойствах красной крови север-
ного морского котика представлены в ряде работ [Кузин, Соколов, 1970; Lenfant
et al., 1970; Кузин, 19716, 19726]. Состав белой крови расматривался в статьях
А. Е. Кузина [19716] и Г. М. Косыгина и А. Е. Кузина [1979]; удельный вес и
вязкость крови — в работе Е. И. Соболевского [1977].
11.1. Общее количество крови
Методом перфузии [Глазова, Леоненко, 1966] А. С. Соколов [1969а] опреде-
лил, что у новорожденных и 4-летнего самца котика общее количество крови
в организме составляет 175—400 г или 5.2% от массы тела (табл.: 11.1). Однако
в наших исследованиях при использовании того же метода были получены по-
казатели, вдвое превышающие названные; они составили, соответственно, в сред-
нем 13% от массы тела у новорожденных и 10.7% — у взрослых котиков. С
помощью разбавления крови непосредственно в кровяном русле анастезирован-
ного животного нами были получены показатели общего количества крови в
организме котиков несколько выше названных, но они не выходили за рамки
изменчивости данных, полученных другими исследователями, использовавшими
иные методы определения объема циркулирующей в организме животного
крови. Так, методом мечения эритроцитов хромом-51 К. Ленфант с соавт. [Len-
fant et al., 1970] получили данные, близкие к нашим (табл. 11.1).
Таким образом, по нашим данным и материалам К. Ленфанта с соавт. [Len-
fant et al., 1970], общее количество крови в организме северного морского котика
составляет 10—20% от массы тела. Данные А. С. Соколова сильно занижены.
Как и у большинства других млекопитающих, включая человека [Коржуев, 1964],
относительное количество крови в организме молодых котиков больше, чем у
взрослых.
У морских глубоко- и долгоныряющих млекопитающих количество крови в
организме больше, чем у животных, экологическая специфика которых не тре-
бует выработки этих свойств. Так, например, у крылатки и обыкновенного тю-
леня на 1 кг массы тела приходится 13.2 мл крови (всего 13.2% от массы тела)
[Lenfant et al., 1970], у северного морского слона количество крови равно 21%
от массы тела [Simpson et al., 1970], у тюленя Уэдделла — 15% (Lenfant et al.,
1970]. Таким образом, намечается общая тенденция увеличения объема крови
в зависимости от специализации животных к нырянию. Как правило, у видов
с повышенным объемом крови в ее составе больше дыхательных пигментов и
она способна больше запасать кислорода, а сами звери — дольше переносить
асфиксию.
191
Таблица 11.1
Количество крови в организме северного морского котика
Возраст Пол Масса тела, кг Кол-во крови, кг % от массы тела Источник
-Щенки самка 8.4 1.196 ’ 14.23 Ориг. данные
самка 6.3 0.812 . .. 12,90 . Кузин и др., 1972
самец 5.3 0.693 13.00 » »
самец 9.9 1.145 11.56 Ориг. данные
самец 8.5 1.718 20.20 » »
? 4.3—6.3 0.175—0.400 5.2 Соколов, 1969,,
4 года самец ? ? 10.7 » » •1
8 лет самка 41.0 4.4 Ориг. данные
Годовалый и 3 взрослых ? 27—211 17.83 11.6 Lenfant et al., 1970
11.2. Морфофизиологическая характеристика красной крови
Наиболее заметные изменения в составе крови наблюдаются в пренатальный
период жизни (табл. 11.2). Количество эритроцитов в 1 мл крови эмбрионов на
ранних стадиях внутриутробного периода развития было наименьшим, однако
к периоду рождения их число увеличивается до уровня, превышающего пока-
затели взрослых животных. Количество эритроцитов у новорожденных котиков
заметно меньше, чем у предродовых эмбрионов и взрослых. Аналогичная кар-
тина наблюдается и у других млекопитающих [Предтеченский и др., 1950]. В
среднем у взрослых котиков количество эритроцитов в крови составляет 4.6
млн/ммЗ. Характерно, что возрастной и половой изменчивости в количестве
эритроцитов в постнатальном онтогенезе у котиков не наблюдается, в то время
как индивидуальные вариации в каждом возрастном классе значительны. Сред-
ний диаметр эритроцитов у котиков в пренатальный период больше, чем у взрос-
лых (соответственно, 8.27 и 7.39 мк). Другие данные о размерной характеристике
эритороцитов представлены в таблице 11.3.
Сравнение с опубликованными данными по антарктическим тюленям [Кон-
динский, 1973а] показывает, что у котиков лишь толщина и сферический индекс
эритроцитов несколько больше, в то время как диаметр в среднем на 15%, а
площадь поверхности вдвое меньше, чем у тюленя Уэддела и южного морского
слона, и на 40% меньше, чем у морского леопарда и тюленя Росса.
Объем эритроцитов у взрослых котиков меньше, чем у антарктических лас-
тоногих [Кондинский, 1973а; Kooyman, 1973]. Только у эмбрионов котиков он
приближается к объему эритроцитов настоящих тюленей. Однако половых раз-
личий-по этому признаку у котиков не обнаружено. Концентрация гемоглобина
в эритроците и во всей эритроцитарной массе в эмбриональный период у котика
выше, чем у взрослых особей, и это следует рассматривать как эмбриональное
приспособление к обеспечению кислородом растущего организма. Поскольку ве-
личина и площадь поверхности эритроцитов является существенным физиоло-
гическим показателем, по которому можно судить об интенсивности газообмена,
192
то представлется, что северный морской котик в этом отношении стоит рангом
ниже, чем настоящие тюлени.
Общее содержание гемоглобина в крови котиков на протяжении периода он-
тогенетического развития заметно меняется (табл. 11.2). Наиболее заметное уве-
личение количества гемоглобина наблюдается во внутриутробный период. Так
у 6-месячных эмбрионов его количество в крови на 30% ниже, чем у предро-
довых. Количество гемоглобина у новорожденных уменьшается по сравнению
с предродовыми эмбрионами и на последующих этапах постнатальной жизни
вновь увеличивается. По представленным данным трудно судить о половой и
возрастной изменчивости в показателях гемоглобина в связи с большой инди-
видуальной изменчивостью. Однако сгруппированные в зависимости от массы
тела животных данные в пренатальный период показали, что насыщенность
крови гемоглобином у эмбрионов больше зависит не от пола, а от массы тела
[Кузин, Соколов, 1970]. В постнатальном онтогенезе подобная зависимость не
обнаружена, но выявлено, что у самцов кровь богаче гемоглобином в среднем
на 9%, чем у самок. Количество гемоглобина в крови беременных и неберемен-
ных самок одинаково [Кузин, 19726].
Сравнение характеристик крови эмбрионов и их матерей [Кузин, 19726] по-
казало, что количество гемоглобина в крови матери выше, а число эритроцитов
меньше, чем у предродовых эмбрионов, В соответствии с этим, и объем эрит-
роцитарной массы (гематокрит) больше у плодов. Насыщенность эритроцитов
гемоглобином особенно велика у 6-месячных эмбрионов (до 50%) и постепенно
снижается (до 28.7%) к периоду рождения. У матерей на всем протяжении пе-
риода беременности этот показатель изменяется в небольших пределах (32.5—
36.3%). По мнению К. Ленфанта с соавт. [Lenfant et al., 1970], количество ге-
моглобина и гематокрит являются хорошим показателем, на основе которого
можно говорить о способности организма переносить асфиксию.
Содержание миоглобина. Обеспеченность организма ныряющих млекопита-
ющих кислородом во многом зависит от содержания миоглобина [Scholander,
1940; Коржуев, 1964; Andersen, 1966]. Миоглобин ускоряет диффузию кислорода
[Andersen, 1966], облегчает продвижение его в недра клеток [Коржуев, Глазова,
1968]. Данные по количеству миоглобина в мышцах северного морского котика,
полученные А. С. Соколовым [1969а], показали невысокую его концентрацию
по сравнению с настоящими тюленями. Однако последующие исследования внес-
ли изменения и уточнения и показали, что количество миоглобина у котика,
хотя и не достигает уровня специализированных к продолжительному и глубо-
ководному погружению морских млекопитающих, но значительно выше, чем у
наземных млекопитающих [Lenfant et al., 1970; Кузин и др., 19736]. Этими ис-
следованиями так же показано, что количество миоглобина в разных мышцах
котика не одинаково. Его высокое содержание у взрослых животных обнаружено
в мышцах основных движителей — передних конечностях. В мышцах сердца,
диафрагме и задних конечностях миоглобина меньше (табл. 11.4).
На основании результатов изменения содержания миоглобина в разных
мышцах в процессе онтогенетического развития котика можно говорить об адап-
тивном характере становления функций организма, связанных с накоплением
дыхательных пигментов.
193
Таблица 11.2
Состав красной крови у северных морских котиков [из: Кузин, 19716, с дополнениями]
Возраст Пол п Гемоглобин по Сали, г% Гематокрит, % Число эритроцитов, млн/мм3 Объем одного эриторцита, ц Кол-во гемоглобина в эритроците, и Концентрация гемоглобина в эритроцитах, %
6 мес. самцы 8 11.3 Эмбрионы 50.8 41.9
27 2.26 126.1
9.4—13.2 22—34 1.62—286 109.1—135.8 45.0—58.0 34.7—48.5
самки 4 11.9 29 2.33 113.1 48.1 43.2
10.3—13.8 28—34 2.56—2.83 109.4—129.7 45.0—48.8 34.7—44.5
7 мес. самцы 4 13.4 40 3.23 124.9 41.4 33.4
12.2—14.2 36—45 2.82—3.47 108.4—131.2 40.6—43.3 31.2—37.8
самки 2 13.8 45 3.51 127.0 39.4 31.1
13.6—14.0 42—47 3.39—3.52 119.3—134.7 38.6—40.1 29.8—32.4
8 мес. самцы 2 13.7 42 3.95 107.0 34.7 32.7
13.0—14.4 42 3.80—4.04 104.2—128.5 33.7—35.7 31.0—34.3
самки 1 13.0 42 3.65 115.1 35.6 30.9
11 мес. самцы 3 15.1 51 5.19 99.1 29.6 29.8
13.0—18.0 45—54 ' 4.08—6.07 87.3—110.3 23.7—33.3 27.2—33.3
самки 3 14.1 45 4.96 101.5 29.5 29.1
14.0—14.4 48—50 3.74-5.59 85.9—128.3 25.0—37.4 28.8—29.2
Щенки самцы 11 12.9 39 3.70 103.9 31.6 31.1
9.4—15.8 34-^17 2.89—5.48 67.0—146.8 26.3—36.6 22.3—44.0
самки 10 11.0 35 3.87 89.2 28.6 33.3
7.2—14.4 24—52 3.50—510 68.6—147.7 20.6—39.2 28.3-44.0
В з р о с л ы е
2 года самцы 12 15.6 44 4.65 87.5 33.9 35.5
13.1—18.0 37—52 . 3.83—5.96 76.0—117.4 26.7—44.1 28.0—45.9
самки 5 15.6 45 4.59 100.4 34.8 34.6
14.3—17.1 40—48 4.00—5.68 78.9—117.1 30.0—39.0 31.1—39.5
3 года самцы 11 16.2 13.8—19.8 49 43—63 5.08 4.03—6.50 99.8 69.2—141.9 31.9 26.4—37.6 33.5 24.4—37.6
самки 4 14.4 45 4.54 102.8 32.2 31.9
11.0—17.8 41—48 3.01—5.30 86.8—136.2 26.0—36.5 27.8—39.6
4 года самцы 8 14.9 52 4.66 113.0 32.2 29.5
12.6—18.4 42—63 4.04—5.57 89.3—165.9 26.3—34.8 18.9—36.8
самки 2 17.3 48 4.60 104.3 36.5 36.1
17.0—17.6 45—51 4.40—4.80 102.5—106.3 34.5—38.6 34.5—37.8
5 лет самцы 7 14.7 45 4.56 99.5 32.4 31.4
12.8—17.0 38—50 3.77—5.51 81.8—122.0 28.8—37.6 26.8—37.8
самки 3 14.0 45 4.47 100.8 31.2 31.1
11.7—15.6 36—52 4.10—4.71 100.3—110.4 28.5—33.9 28.3—32.5
6 лет самцы 7 14.2 48 4.43 11.7 32.4 29.5
10.4—15.3 39—53 2.36—5.92 88.1—148.3 27.8—39.5 26.5—34.8
и- самки 5 14.9 45 4.21 114.3 33.5 33.9
и» 11.6—17.4 40—52 2.71—5.08 83.6—159.2 28.7—40.4 28.1—40.4
7 лет самцы 1 13.9 44 4.18 105.3 33.3 31.6
самки 5 17.0 47 4.84 98.2 35.5 36.3
15.8—19.4 39—57 4.49—5.47 86.8—107.5 31.6—39.5 31.0—41.4
8 лет самцы 3 16.8 48 4.97 96.5 33.8 35.5
14.9—18.0 44—51 4.73—5.40 81.5—106.5 31.5—36.7 31.0--40.9
самки 6 14.9 43 4.52 97.6 23.2 34.9
13.6—17.2 36—51 3.49—5.58 80.5—128.9 30.2—40.7 27.8—41.0
9 лет самцы 3 14.3 43 4.28 104.4 33.9 33.2
11.0—17.2 40—46 3.15—5.73 80.7—126.9 30.2—36.8 27.5—37.4
самки 1 17.2 53 4.96 106.8 34.7 32.5
10 лет самцы 1 11.8 43 3.56 120.8 33.1 27.4
самки 9 16.1 47 4.61 105.1 35.7 34.6
14.2—19.0 40—55 3.44—5.90 78.4—144.8 31.0—48.8 30.2—42.0
>10 лет самки 16 16.3 46 4.57 100.0 36.0 36.1
13.0—18.4 32—55 3.55—5.45 87.1—120.4 30.0—44.6 32.0—41.7
Таблица 11.3
Размеры эритроцитов северного морского котика
п Диаметр, мк Толщина, мк Сферический индекс Площадь поверхности, мк2
14 7.39 2.70 0.36 109.07
Таблица 11.4
Концентрация миоглобина в различных мышцах северного морского котика (мг%)
Возраст- ные группы п Масса тела, кг Сердце Диаф- рагма Мышцы спины Передние конечности Задние конечности Миоглобин, г/кг массы тела
Предродо- вые эмбрио- ны 2 4.3 925 720 750 - 7.50
Новорож- денные 5 6.1 1155 1264 1060 1440 1275 12.48
Взрослые 4 38.0 1230 1557 1881 1887 1448 17,38
Таблица 11.5
Дыхательные
свойства крови
некоторых ластоногих [из: Lenfant et al., 1970]
Вид п Кисло- родная емкость, объем- ные %% Парциаль- ное дав- ление О2, соответству- ющее 50% Oj при рН=7.4 Эффект Бора Стандарта, бикарбо- нат, тМ/1 Буферная емкость, (7.2—7.6) шМН/рН
Северный морской котик 2 19.6 29.7 2.5 -.475 27.2 16.5
Сивуч 2 17.5 32.0 2.6 -.696 25.8 19.4
Морж 2 24.3 34.2 2.4 -.525 25.7 13.6
Калан 1 21.5 30.8 2.6 -.460 26.9 17.7
Обыкновенный тю- лень 2 29.3 31.0 2.6 -.534 25.1 16.0
Крылатка 2 34.2 31.0 2.5 -.480 27.5 18.6
К. Ленфантом с соавт. [Lenfant et al., 1970] были рассчитаны дыхательные
свойства крови северного морского котика и других морских млекопитающих
(табл. 11.5).
Эти исследования показали, что только у настоящих тюленей кислородная
емкость крови выше, чем у наземных млекопитающих, в то время как эффект
Бора не обнаруживает каких-либо преимуществ ни среди сравниваемых видов
ластоногих, ни в сравнении этих показателей с наземными млекопитающими.
196
Рис. 11.1. Запасы кислорода перед нырянием
у некоторых морских млекопитающих (по:
Lenfant et al., 1970]:
А — общие запасы, Б — в легких, В — в крови
(светлые столбики — в артериальной, темные — в
венозной), Г — в тканях;
1 — обыкновенный тюлень, 2 — крылатка, 3 —
северный морской котик, 4 — взрослый сивуч, 5 —
новорожденный сивуч, 6 — морж, 7 — калан.
Буферная емкость крови не сильно различается у разных видов тюленей, но эти
показатели значительно выше у ластоногих, чем у сухопутных животных.
Приведенные данные показывают, что ныряющие млекопитающие способны
к наилучшему управлению гомеостатическим баллансом, связанным с обменны-
ми процессами, и что хорошие ныряльщики, благодаря высокой буферной ем-
кости крови, получают преимущество среди других обитателей водных прост-
ранств и способны хорошо противостоять асфиксии.
У ушастых тюленей ок. 60% общих запасов кислорода находится в крови
и тканях и ок. 40% в легких, в то время как у настоящих тюленей 82% кислорода
находится в крови и тканях и только 18% в легких [Lenfant et al., 1970]. У котика
запасы кислорода в мышцах больше, чем у сивуча, моржа и калана, но меньше,
чем у обыкновенного тюленя и крылатки. Примерно такое же количественное
распределение кислорода наблюдается и в крови котика. Однако в легких запасы
кислорода у котика больше, чем у настоящих тюленей, но ниже, чем у калана
(рис. 11.1).
Такое распределение кислорода в организме северного морского котика, си-
вуча, моржа и калана ставит их в менее выгодные условия в период ныряния,
т. к. легочные запасы кислорода, хотя и довольно полно утилизируются орга-
низмом при неглубоком нырянии [Kooyman et al., 1973], но его явно недоста-
точно для обеспечения организма кислородом на продолжительное время. Не-
которые авторы предполагают, что при нырянии происходит полное вытеснение
воздуха из легких в воздухоносные каналы, откуда его утилизация невозможна
[Lenfant et al., 1970]. Запасы кислорода в крови, благодаря перераспределению
кровотока в результате коллапсйрования периферических сосудов, используются
организмом только в снабжении главных органов и тканей, обеспечивающих
жизнедеятельность его в период ныряния. Основную функцию снабжения ор-
197
Таблица 11.6
Количество лейкоцитов в крови северного морского котика
Возраст Самцы Самки
п т lim п т lim
Эмбрионы 1 3.4
Щенки 3 9.7 8.3—12.7 4 11.8 5.4-18.5
2 года 7 5.5 2.7—8.7 1 4.1
3 года 7 6.1 2.7—10.2 2 4.3 3.4—5.2
4 года 3 4.0 2.0—7.5 2 5.1 2.8—7.3
5 лет 2 12.5 9.5-15.6
6 лет 8 6.7 2.3—10.2 3 6.1 4.7—8.7
7 лет 4 6.3 4.0—9.3
8 лет 1 8.4 2 5.8 4.6—5.8
9 лет 3 7.5 3.6-13.1
10 лет 1 6.1 6 6.6 4.3—9.7
>10 лет 7 6.0 4.4—9.3
Таблица 11.7
Лейкоцитарная формула крови северного морского котика
Возрастные и половые группы п Нейтрофилы Лимфоциты Моноциты Эозинофилы
сегменто- ядериые палочко- ядерные
т lim т lim т lim т lim т lim
Эмбрионы 4 30.5 5—54 0.25 0—1 64.3 34—95 9.5 0—12
Взрослые самцы 10 54.6 38—58 0.4 0—2 40.7 28—61 1.3 0—3 3.0 1—6
Взрослые самки 42 43.4 6—94 1.2 0—6 52.7 3—92 1.1 0—3 1.6 0—7
ганизма ныряльщиков кислородом в период апноэ берут на себя сначала мио-
глобин, а после полного расходования кислорода, мышцы переходят на глико-
литический тип обмена [Scholander, 1940]. Вместе с тем, расчеты показали, что
имеющихся запасов кислорода в организме ныряющих млекопитающих может
хватить на сравнительно короткий промежуток времени [Packer et al., 1969], что
никак не согласуется с известными данными об общей продолжительности и глу-
бине ныряния этих животных.
11.3. Морфологическая характеристика белой крови
Проведенные нами исследования количественного и качественного состава
белой крови северного морского котика показали, что количественные показа-
198
телу лейкоцитов подвержены значительным индивидуальным колебаниям (табл.
11.6). Поэтому сказать что-либо о половой и возрастной изменчивости этих по-
казателей не представляется возможным.
По общему количеству лейкоцитов кровь котика почти такая же, как и у
наземных млекопитающих [Алытаузен, 1951]. Этот показатель несколько выше
у щенков, чем у взрослых. В среднем в 1 мм3 крови у взрослых котиков на-
считывается 6—7 тыс. лейкоцитов.
По клеточному составу белая кровь морского котика все же отличается от
крови большинства наземных млекопитающих большим содержанием в ней лим-
фоцитов (табл. 11.6).
У эмбрионов количество лимфоцитов даже несколько больше, чем у взрослых
животных (табл. 11.7). Трудно найти объяснение тому, что число лимфоцитов
у котика превышает эти показатели у наземных млекопитающих, тем более тому,
что это проявляется уже в эмбриональный период.
Это явление наблюдается и у других исследованных ластоногих. По крайней
мере, тюлени Антарктики [Кондинский, 1975] и большинство тюленей Арктики
[Ronald et al., 1969; Гелетюк и др., 1972; Соболевский, 1977], также имеют боль-
шое количество лимфоцитов в крови.
Морфология клеточных элементов белой крови морского котика мало чем
отличается от их строения у других млекопитающих. Из нейтрофильной группы
лейкоцитов лишь сегментоядерные клетки характеризуются меньшим количест-
вом сегментов, а базофилы имеют более крупные размеры. В остальном белая
кровь северного морского котика практически идентична изученным наземным
млекопитающим.
11.4. Особенности дыхания и сердечной деятельности
Среди многочисленных работ по дыхательной и сердечной ритмике ласто-
ногих [обзор их приведен в: Галанцев, 1977] специально морскому котику пос-
вящены исследования Л. Ирвинга с соавт. [Irving et al., 1963], Е. И. Соболевского
и А. Е. Кузина [1977], В. П. Галанцева [1978], В. П. Галанцева, М. К. Маминова
[1979].
Как и у других ластоногих [Irving et al., 1935; Bartholomew, 1954; Галанцев,
1977 и др.], у котика наблюдается аритмия дыхательной и сердечной деятель-
ности. Наши наблюдения за спящими на берегу котиками определили, что взрос-
лые животные за 1 мин делают 4—8 вдоха—выдоха, щенки — 8—10. Частота
сердечных сокращений составляет у взрослых 101—120 в минуту, у щенков —
117—167.
Электрокардиологические исследования Л. Ирвинга с соавт. [Irving et al.,
1963] выявили респираторную аритмию сердечной деятельности во время пре-
бывания северных морских котиков на берегу, при которой ускорение работы
сердца происходит за короткий дыхательный цикл (“вдох—выдох”). Коэффи-
циент аритмичности по В. П. Галанцеву [1978], В. П. Галанцеву и М. К. Ма-
минову [1979] у новорожденных котиков равен 0—25% и с возрастом увеличи-
вается до 122—388%.
199
В экспериментах по искусственному предотвращению поступления воздуха
в организм и при повторном вдыхании частично использованного воздуха от-
ветные реакции организма котика были сходными. Средняя частота сердцебие-
ния снижалась, а частота сердечной аритмии возрастала. При этом удлинялись
интервалы длинных циклов сердечного ритма, а короткие оставались без изме-
нений [Irving et al., 1963].
У морских котиков, находящихся в стрессовом состоянии после их поимки
и пленения, наблюдалось возрастание частоты сердечных сокращений до 150—
200 ударов в минуту с четко выраженной аритмией. Ускорение работы сердца
сопровождалось значительным укорочением интервалов длинной фазы и неболь-
шим укорочением интервалов короткой фазы. Частота дыхания не превышала
20 раз в минуту.
Общеизвестным является наличие брадикардии в период ныряния и тахи-
кардия после выныривания у всех исследованных водных млекопитающих. Ана-
логичные реакции отмечены и у наземных млекопитающих, но количественно
они выражены ярче у водных. Эксперименты по принудительному погружению
котика [Irving et al., 1963; Галанцев, Маминов, 1979] показали, что брадикардия
ныряния у котика развивается не сразу и не у всех особей одинаково. Посте-
пенное развитие брадикардии и ее интенсивность зависят от времени погружения
в воду. До погружения размах интервалов аритмии был от 0.4 до 0.6 сек при
частоте сердцебиения 120 ударов в минуту.
Брадикардия начинает проявляться через 1—15 сек. после погружения, а при
продолжительных погружениях только спустя минуту [Irving et al., 1963]. Самый
длинный интервал между ударами сердца при погружении составил 5 сек. Под
водой у северных морских котиков коэффициент аритмии увеличивается в 11 раз
[Галанцев, Маминов, 1979]. В основном увеличение продолжительности сердеч-
ных циклов под водой связано с возрастанием диастолической фазы. После про-
должительных погружений вслед за брадикарбией наступает выраженная арит-
мия, что, по мнению Л. Ирвинга с соавт. [Irving et al., 1963], вызвано энергичным
дыханием. Увеличение сердечных ритмов при тахикардии происходит за счет
сокращения времени диастолы. При аритмии длинные интервалы между ударами
сердца еще больше удлиняются, а средняя частота сердцебиения замедляется.
Аритмия проходит сразу же за восстановлением ровного дыхания, и вслед за
ней наступает тахикардия.
Разовое 5-минутное погружение не вызывало каких-либо заметных изменений
в организме котика. Трехразовое принудительное погружение по 5 мин (каждое
с перерывами по 15 мин) в экспериментах приводило к летальному исходу пос-
ле третьего погружения. Принудительные погружения котиков на глубину 150—
200 м на 5 мин 42 сек и 7 мин 45 сек с часовым интервалом между погружениями
не приводили к летальному исходу, но общее состояние животных было плохим
[Соболевский, 1974]. После подъема котиков на поверхность дыхание было тя-
желым и глубоким, голова запрокинута назад, наблюдался тремор. Частота ды-
ханий в первую минуту составляла 13, во вторую — 17, в третью — 11 и в по-
следующие 5 мин колебалась в пределах 10—13 вдохов в минуту, после чего пришло
в норму. Частота пульса, измеренная спустя 13 сек после подъема животного из
воды, в первую минуту составила 135 ударов, во вторую — 152, в третью —
147, в четвертую — 130 с последующим интенсивным снижением. Через 9 мин
дыхание и пульс не отличались от обычных.
200
Рис. 11.2. Электрокардио-
грамма северного морс-
кого котика (самка, 4.6 кг)
[из: Галанцев, Маминов,
1979]:
1 — во время погружения в
воду (стрелкой обозначен мо-
мент погружения), 2 — перед
очередным погружением, 3 —
под водой на ЗО-й секунде, 4 —
после подъема на поверхность.
2 |/л^
1с
Из приведенных выше экспериметов будто бы следует, что 5—7 мин погру-
жения являются критическими для котика. Однако, данные исследований Дж.
Куймана с соавт. [Kooyman et al., 1976] показали, что северные морские котики
добровольно ныряют на глубину до 190 м, затрачивая на это 5.4 мин, и вряд
ли при этом расходуют все запасы кислорода. Заметим, что сердечно-сосудистые
реакции животных при принудительных погружениях и при самостоятельном
нырянии отличаются [Kooyman, Campbell, 1972]. В этой связи правомочно пред-
положить, что животные могут самопроизвольно контролировать деятельность
сердца при свободном нырянии [Галанцев, 1977].
Развитие способности к нырянию и соответствующих реакций сердечно-со-
судистой системы у северного морского котика в основном происходит в течение
постнатального онтогенеза; уже у эмбрионов-выпоротков за 1.5—2 месяца до
рождения замедлялась частота сердечных сокращений после обливания головы
водой (имитация ныряния), наблюдалась брадикардия [Соболевский, Кузин,
1977]. Не исключено, конечно, что уменьшение частоты сердечных сокращений
у эмбрионов вызвано несовершенством терморегуляции, что сопровождается со-
ответствующими реакциями организма. Однако, поскольку приспособительные
реакции сердечно-сосудистой системы в ответ на погружение являются резуль-
татом множественных функций организма, в т. ч. не на последнем месте среди
них стоит терморегуляция, можно говорить о снижении частоты сердечных со-
кращений у эмбрионов при контакте с водой как о части этого общего комплекса
реакций организма на внешние воздействия.
Исследования щенков котиков [Irving et al., 1963; Галанцев, Маминов, 1979]
показали, что все ответные реакции их организма на ныряние аналогичны та-
ковым у взрослых животных (рис. 11.2), однако интервалы сердечных ритмов
при брадикардии меньше, самый продолжительный у ныряющего щенка 3.9 сек.,
что на 2 сек короче самого длинного интервала у взрослых особей. Самый длин-
ный интервал между ударами сердца при погружении составил 5 сек.
У щенков так же, как и у взрослых особей, после продолжительных ныряний
возникала аритмия, и чем продолжительнее было ныряние, тем она проявлялась
ярче.
201
Аритмия всегда осуществляется через увеличение длинных интервалов между
ударами сердца. После подъема на поверхность у щенков развивается компен-
саторная тахикардия. Частота ударов сердца увеличивается на 5% против того,
что было до погружения [Галанцев, Маминов, 1979]. Учащение сердцебиения
происходит за счет сокращения диастолы.
Нормализация сердечного ритма после выныривания зависит от времени за-
держки дыхания, а средняя частота сердцебиения тем ниже, чем продолжитель-
нее было ныряние.
Принудительное погружение щенков котиков на 2.5 мин приводит их в со-
стояние комы. Поведение щенков в этом случае было сходным с таковым при
принудительном трехразовом 5-минутном погружении взрослых особей [Irving
et al., 1963]. Вместе с тем, щенки обнаруживают сильно выраженную толеран-
тность к асфиксии, что свойственно новорожденным большинства видов мле-
копитающих. Если взрослые особи после предельного по времени принудитель-
ного погружения теряют сознание, то не всегда выживают. Щенки после потери
сознания могут продолжительное время находится без доступа воздуха и потом
снова приходить в норму. Выносливость к гипоксии в молодом возрасте, по-
видимому, объясняется менее выраженной резистентностью их тканей к недо-
статку кислорода.
Становление рефлексов, связанных с нырянием, происходит, видимо, уже во
внутриутробном периоде и затем совершенствуется в постнатальном онтогенезе.
Брадикардия и аритмия у северного морского котика не столь ярко выражены,
как у более специализированных к водному образу жизни животных, например,
настоящих тюленей [Scholandrer, 1940; Irving et al., 1942; Harrison, Tomlinson,
1960; Elsner, 1969]. Различия между настоящими тюленями и северным морским
котиком в нормах реакции на погружение существуют не только в степени вы-
раженности возбуждений и торможений блуждающего и симпатического нервов,
ответственных за процессы регуляции сердечного автоматизма, но и в самой
функции проводимости [Галанцев, Маминов, 1979]. У настоящих тюленей, как
у более специализированных ныряльщиков, скорость перестройки работы сердца
выше, чем у котика.
11.5. Терморегуляция
Большую часть жизни северные морские котики проводят в воде, темпера-
тура которой колеблется в пределах 0—20°С, обычно они предпочитают +5—
1ГС. В летний период при организации щенных залежек на островах темпе-
ратурный диапазон внешней среды мало отличается от условий обитания в пе-
риод зимовки в море, но в отдельные периоды воздух может прогреваться до
+25°С и выше. Хорошо известно, что в естественных условиях котики способны
обитать в воде при температуре ниже 0°С. Верхняя критическая температура
среды для северного морского котика, по-видимому, не превышает +40°С. По
крайней мере, известно, что на о. Сан-Мигель при температуре воздуха +42°С
и температуре песчаного грунта свыше +50°С щенки котиков погибают от пе-
регрева.
Как и у большинства млекопитающих, внутренняя температура тела у се-
верного морского котика составляет 37—39° Partholomew, Wilke, 1956; Irving
et al., 1962), при 42—43° наступает смерть. У активных котиков температура тела
повышается до 38—40°, во время сна она снижается до 36.6° [Irving et al., 1963].
202
Рис. 11.3. Позы котиков на лежбище в зависимости от температуры воздуха.
Объяснение в тексте.
Теплоизоляционные свойства котиков обеспечиваются исключительно гус-
тым и плотным меховым покровом, не промокающим даже тогда, когда котики
постоянно находятся в воде. В зимний период у котиков накапливается значи-
тельный слой подкожного жира, который в отличие от китообразных и боль-
шинства слабоопушенных ластоногих скорее всего не играет существенной роли
в сохранении тепла.
Выведение излишков тепла из организма у котиков осуществляется через хо-
рошо васкуляризированный, имеющий развитые и многочисленные потозые же-
лезы кожный покров голых ластов, суммарная площадь которых составляет 30%
всей площади тела [Irving et al., 1963].
У новорожденных котиков иные характеристики тела. Волосяной покров
редкий, и при попадании щенка в воду или при выпадении атмосферных осадков
он сильно намокает. Кожный покров туловища лишен потовых желез. Лишь
через 3 месяца сеголетки приобретают дефинитивный волосяной покров, а в те-
чение всего этого времени они часто переохлаждаются и выживают только бла-
годаря толерантности к холоду [Irving et al., 1963].
В связи с обитанием в водной или в воздушной средах проблема поддер-
жания гомойотермии и регулирования теплообмена организма морского котика
решается несколькими путями.
При пребывании на суше в период размножения в процессах терморегуляции
большое значение имеют поведенческие акты. Наблюдениями в природе уста-
новлено [Когай, 1978], что при температуре воздуха 8.2°С и относительной влаж-
ности 98% котики подгибают задние ласты под себя, а передние вытягивают
и прижимают к бокам тела (рис. 11.3).
При температуре 14°С и влажности 81% котики приподнимают переднюю
часть тела, опираясь на ласты, запрокидывают голову назад, открывают рот и
обмахиваются, как веером, задними конечностями. Другие животные лежаться
на спину, расправляя передние ласты в стороны, а задними делают обмахива-
ющие движения. В жаркие дни большинство котиков сходят в воду, а оставшиеся
на лежбище занимают участки с субстратом, который меньше нагревается [Вла-
димиров, Лыскин, 1981]. Поведение новорожденных похоже на поведение взрос-
лых, однако, в связи с тем, что у первых нет густого мехового покрова, они
203
Рис. 11.4. Подкожная температура в
разных точках тела северного морского
котика после погружения задней части
тела в воду (самка, 4 года) [из: Irving
et al., 1963].
Рис. 11.5. Подкожная температура в
разных точках тела 48-дневного
щенка котика (масса 6.5 кг) после
погружения задней части тела в
воду [из: Irving et al., 1963].
Точки см. на рис. 11.4.
легче переносят высокие температуры, и поэтому более активны в теплые дни.
В холодные дни новорожденные скучиваются, малоактивны. При намокании у
щенков отмечается компенсационная дрожь, чего у взрослых животных не от-
мечено.
Оптимальной для пребывающих на лежбище котиков, по мнению В. М.
Когая [1978], является для секачей температура воздуха +5.0—8.3°С, для самок
— +6.0—9.3°С, для щенков — +6—1ГС.
При физических нагрузках во время перегона котиков с лежбища на забой-
ную площадку внутриполостная температура тела составила 37—39.4°С [Irving
et al., 1962]. Покрытые мехом части тела имели температуру 29.2—34.7°С, а тем-
пература лишенных волосяного покрова частей ластов понижалась к их концам,
где температура была, соответственно, выше лишь на 10—15°С и 4—9°С тем-
пературы воздуха. Волосяной покров животных сверху был мокрым, прохож-
дение тепла от тела через волосяной покров не ощущалось, и в основном его
избыток удаляется через ласты [Irving et al., 1962].
При погружении задней части тела взрослого котика в холодную воду (вода —
10°С, воздух — 1ГС) происходило некоторое охлаждение как задних ластов,
так и сухих передних, температура тела при этом, практически, не падала ниже
35°С (рис. 11.4). В аналогичных опытах на щенках снижение температуры всех
конечностей было значительным, а температура задней части тела падала до
22°С (рис. 11.5).
При погружении только головной части тела взрослых котиков (имитация
затаривания) передние ласты охлаждались, а затем при окончании погружения
температура их вновь повышалась. Охлаждение и потепление было отмечено
и в не мокрых задних ластах, но в центре их этот эффект был менее выражен,
чем на периферии. Температура тела при погружении сначала падала на 1—2°,
а по окончании погружения быстро восстанавливалась. При аналогичном по-
гружении щенков восстановление температуры тела и конечностей не наблю-
далось (рис. 11.6, 11.7).
204
Рис. 11.6. Подкожная температура в Рис. 11.7. Подкожная температура у
разных точках тела у 2-летнего самца щенка северного морского котика в
северного морского котика массой 20 двух экспериментальных погружениях,
кг в течение двух экспериментальных Обозначения см. на рис. 11.4.
погружений (погружалась только
голова).
Стрелка вниз — голова в воде, стрелка вверх —
голова над водой. Точки см. на рис. 11.4.
Исследования [Ohata, Miller, 1977] по определению температурных градиен-
тов у новорожденных и подросших щенков котиков при различных условиях
(сухие, намокшие под дождем, насильно помещенные на время в воду) показали
сходные результаты с уже известными [Irving et al., 1963] как по взрослым ко-
тикам, так и по щенкам.
Таким образом, экспериментальные данные по охлаждению тела морских ко-
тиков вследствие намокания показали, что щенки способны поддерживать тем-
пературу центральной части тела, в меньшей степени в периферических его час-
тях; быстрее всего охлаждаются конечности. Погружение щенков в воду, ими-
тирующее ныряние, вызывало у них сильное охлаждение всего тела без восста-
новления температуры после выныривания. Можно придти к выводу, что щенки
не способны регулировать температуру тела в этом возрасте.
Взрослые котики при охлаждении способны поддерживать темепературу тела
на достаточно высоком уровне при значительном снижении температуры конеч-
ностей. В теплом воздухе или при движении животного, когда в организме воз-
никает излишек тепла, температура ластов может достигать температуры тела.
Нагреванием ластов, поверхность которых достигает трети поверхности тела,
и потоотделением со всего кожного покрова выводится избыток тепла из ор-
ганизма. Эксперименты, имитирующие ныряние, показали, что связанное с ним
колебание температуры ластов происходит в результате подачи крови к пери-
ферии или прекращения периферического кровоснабжения. Вероятно, у взрос-
лых котиков, как и у китообразных [Белькович, 1962], кровоснабжение пери-
ферических частей тела (в особенности ластов) пульсирующее, связанное с ак-
тами “вдоха-выдоха” и брадикардией при нырянии.
Однако у новорожденных котиков брадикардия тоже выражена, но способ-
ность к терморегуляции в воде еще не проявляется, поэтому не выявляется связь
циркуляции крови на периферии тела с брадикардией [Irving et al., 1962]. Жиз-
необеспечение новорожденных котиков в экстремальных условиях осуществля-
ется или за счет их толерантности к переохлаждению, или за счет эндотермии.
205
Глава 12. ОСОБЕННОСТИ ОБРАЗА ЖИЗНИ
Наиболее яркой и характерной особенностью биологии северного морского
котика является разделение его годового жизненного цикла на 2 периода: пе-
лагического, когда котики постоянно находятся в воде и не контактируют с су-
шей, и берегового, когда большую часть времени они проводят на берегу, где
размножаются, воспитывают потомство, линяют, а в море сходят лишь для пита-
ния, охлаждения тела, приведения в порядок мехового покрова и в случае опас-
ности. Морской период можно считать нагульным, береговой — репродуктивным.
12.1. Подход к лежбищам и выход на берег
Несмотря на то, что в море у лежбищ первыми появляются разновозрастные
самцы, на берег раньше других выходят наиболее старые самцы-секачи (рис. 12.1).
Еще Г. Стеллер, зимовавший на о. Беринга в 1742 г., определил приход первых
секачей 29 и 30 апреля. В дальнейшем данные этих наблюдений многократно
подтверждались. Многолетние (1900—33 гг.) обобщенные сведения по времени
выхода секачей на Командорские о-ва имеются у Е. И. Ильиной [1950], по данным
которой эта возрастная группа животных выходит на каждое лежбище неод-
новременно, но наиболее ранние приходы котиков отмечались в середине апреля.
Интересные обобщенные наблюдения выхода первых котиков на Командор-
ские острова имеются у Е. К. Суворова [1912], который сообщал, что в конце
прошлого и в начале нашего столетия наиболее ранний приход секачей за 40 лет
отмечен 7 апреля (о. Беринга, 1882 г.), а самый поздний — 17 мая (о. Медный,
1908 г.). За последние 170 лет не произошло каких-либо изменений во времени
привала секачей на Командорские о-ва.
За период 1958—80 гг. наблюдались аналогичные сроки выхода первых ко-
тиков на лежбища (табл. 12.1), хотя неоднократно отмечалось прибытие оди-
ночных секачей и раньше. Так, в 1967 г. на Северном лежбище уже в конце
марта залегали 4 секача; в 1970 г. первый секач вышел на берег 1 апреля. Ранние
выходы секачей на лежбище, как правило, не сопровождаются последующим
быстрым нарастанием их численности. Не исключено, что эти визиты на леж-
бище делают животные, проводящие зиму в акватории, прилежащей непосредст-
венно к островам, где они залегали прошлым летом.
Более того, имеются наблюдения, что одиночные особи и небольшие группы
секачей проводят зиму у островов, периодически выходя на лежбище [Барабаш-
Никифоров, 1936]. В среднем за многие годы наблюдений на Командорских о-вах
численность секачей начинала увеличиваться лишь после 5 мая (рис. 12.2).
На о. Тюлений первые секачи обычно приходят несколько позже, чем на Ко-
мандоры. Их приход сдерживается льдами. С 1958 по 1980 гг. наиболее ранний
выход секачей на о. Тюлений был отмечен в 1971 г., когда на лежбище уже
8 мая залегли 2 секача. В среднем за все годы наблюдений приход первых секачей
на остров отмечался в середине мая (табл. 12.1).
206
Рис: 12.1. Первыми на лежбище приходят
секачи.
На заднем плане небольшой гарем.
Рис. 12.2. Изменение численности секачей
северного морского котика на
Юго-Восточном лежбище о. Медный
(среднее за 1962, 1963, 1971, 1975, 1977,
1980 гг.)
Приход секачей на о-ва Прибылова Г. Эллиотт [Elliott, 1875] датировал 1—5
мая, Ф. Лукас [Lucas, 1899] — 13 мая. Л. В. Бойцов [1934] писал, что начало
привала секачей на о-ва Прибылова обычно приходится на начало мая, вскоре
после того, как их акватория очищается ото льда.
Таким образом, секачи раньше выходят на острова, которые не блокируются
льдами, и несколько позже на те, которые или блокируются выносными льдами,
или море вокруг которых замерзает зимой.
Время окончания привала секачей к островам установить трудно. В те годы,
когда численность котиков на Командорских о-вах была очень низкой, и имелась
возможность учета и персонального опознания каждого самца, было зарегист-
рировано последнее появление новых животных на о. Беринга в 1910 г. — 3 июня,
на лежбище Говорушечье (о. Медный) — 4 июня, а на лежбище Урилье (о. Мед-
ный) — 13 июня [Суворов, 1912]. По данным Е. И. Ильиной [1950], привал се-
качей на Командорские о-ва заканчивается в середине июня. Эти даты близки
к максимальным показателям численности секачей, подсчитываемых регулярно
на островах в 1958—80 гг. (рис. 12.2). На рис. 12.3 видно, что сроки окончания
привала секачей на о. Тюлений сжаты (20—30 июня), а на Командорские о-ва
более растянуты (25 июня—15 июля).
Холостяки начинают выходить на лежбище вслед за секачами (табл. 12.1).
По данным Е. И. Ильиной [1950], интервал между выходом секачей и холостяков
на Северное лежбище (Командорские о-ва) достигает 22 дней. Группа холостяков
объединяет животных в возрасте 2—5 лет. Известно, что первыми из них выходят
на лежбище животные 4—5 лет, затем, в конце июня—июле — 3-летние, во вто-
рой половине июля—августе — 2-летние и годовики [Дорофеев, 1964]. Это также
207
Время выхода первых котиков на лежбища
Таблица 12.1
Годы о. Тюлений Командорские о-ва
Северное лежбище Юго-Восточное лежбище
секачи самки ХОЛОСТЯКИ секачи самки холостяки секачи самки холостяки
1960 15.V 6. VI 18.V 8.V 12.V 8.V 7.V 13. VI 3.V
1961 12.V 8. VI 15.V 18.IV 15.V 9. VI 9.V
1962 4.VI 30.V 11.V 8.VI 11.V 12. VI
1963 15.V 7.VI 10.V 10.VI 20.V 9.VI 9.VI
1964 17.V 31.V 5.V 9.VI 30.V 10. VI
1965 3.VI 5.V 10.V 8.V 1.IV 12. VI 1.IV
1966 15.V 31.V 6.V 15.V 11.V 8. VI
1967 16.V 21.V 17.V 5.V 17.V 10. VI 15.V
1968 18.V 24.V 1.V 11. VI 13. VI 16.V 9. VI 17.V
1969 24.V 15.V 11. VI 7.V 11. VI
1970 20.V 6. VI 25.VI 10.IV 10. VI 10. VI 20.V 10.VI
1971 8.V 26.V 10.V 1.V 15.V 10.VI начало мая
1972 25.V 30.V 12.V 7.VI 4.V до 10. VI
1973 25.V 30.V 25.V 7.VI
1974 30. V 17.V 30. V до 15.V
1975 27.V 29.V 27.V 10.VI
1979 10. VI до 10. VI
1980 8.VI
1981 8.VI
хорошо прослеживается по данным промысла В первых отгонах присутствуют
преимущественно 4—5-летки, а в конце промыслового сезона — 2—3-летки [Ко-
гай, 1971а]. Анализ меток на о. Тюлений показал аналогичную картину (рис. 12.4).
Как видно на рис. 12.4, в первой декаде июня на лежбище присутствовали
самцы старше 2 лет, среди которых преобладали 4—6-летние; 2-летние регист-
рировались только с третьей декады июня, а превалировали в августе.
Появление первых самок на островах обычно наблюдается в начале июня
[Ильина, 1950; Kenyon, 1960; Дорофеев, 1964 и др.] и продолжается до конца
августа, о чем можно судить по рождению щенков в этом месяце и состоянию
мехового покрова животных. У вновь приходящих животных он гораздо светлее,
чем у несколько дней находившихся на лежбище. Наиболее ранний срок прихода
самок на о-ва Прибылова — 21 мая [Вениаминов, 1840], на Командорские о-ва —
27 мая [Суворов, 1912], на о. Тюлений — 21 мая [Когай, 1971а]. По нашим дан-
ным (табл. 12.1) первые самки на Тюлений приходили в конце мая—первых чис-
лах июня, на Командоры — в аналогичные сроки, но в среднем на несколько
дней позже. Существует определенный порядок появления самок на лежбище
208
Рис. 12.3. Изменение численности секачей
северного морского котика на о. Тюле-
ний (среднее за 1960—64, 1970—74, 1980—
82 гг.)
Рис. 12.4. Изменение возрастного
состава самцов северного морского
котика на о. Тюлений в 1976 г.
(читка меток на живых животных)
как показатель разновременности их
прихода на остров
Рис. 12.5. Изменение возрастного
состава самок северного
морского котика в зависимости
от времени их добычи на о-вах
Прибылова как показатель
разновременности их прихода на
острова (по материалам NPFSC)
I”
з 4 з в 7 в е ю ю'
лммг
Рис. 12.6. Изменение возрастного состава самок северного морского котика на
о. Тюлений в 1967 г. (читка меток на живых животных) как показатель
разновременности их прихода на остров
209
Рис. 12.7. Изменение возрастного состава самок северного морского котика на
о. Тюлений в течение сезона размножения [из: Бычков, 1969]
в зависимости от возраста. Так, по данным промысла самок на о-вах Прибылова
в 1956 г., когда их добывали на всех типах лежбищ, и забою подлежали все
животные с длиной тела более 41 дюймов (более 1 м), можно сказать, что в
середине июля в пробе преобладали половозрелые особи; 3-летние — отсутст-
вовали. Последние начали появляться в забое в конце июля (рис. 12.5). Ана-
логичные данные были получены при читке меток на живых самках (рис. 12.6).
По данным В. А. Бычкова [1969], в начале сезона размножения на гаремном
лежбище в основном находились самки от 4 лет и старше. Трехлетки начинали
прибывать на гаремную территорию примерно с середины июля и регулярно
встречались в августе, а появление 1—2-летних самок было зафиксировано лишь
в августе (рис. 12.7). В целом, приход самок на острова сильно растянут во вре-
мени. Он наиболее сходен по характеру с выходом на лежбища холостяков.
Порядок появления на островах северных морских котиков после их воз-
вращения из эмиграции зависит от пола, возраста, физиологического состояния
и хорошо согласуется с очередностью их весеннего передвижения по всему миг-
рационному маршруту [Кузин, Панина, 1977; Ащепков, Кузин, 1987]. На сроки
выхода первых котиков на лежбища после возвращения из эмиграции заметное
влияние оказывают гидрометеорологические условия районов зимнего и летнего
обитания, а также, по-видимому, расстояние, отделяющее места зимовок от мест
размножения.
12.2. Формирование гаремов и холостяковых залежек
Размеры участков под будущие гаремы определяются, с одной стороны, релье-
фом местности, а с другой, врожденной привычкой сохранения дистанции — тер-
риториальностью секачей. На ровных пляжах самцы ложатся не ближе 3—5 м
друг от друга, на участках со сложным рельефом гаремы, могут быть разделены
естественными преградами (камнями, расщелинами, выступами скал); в этих слу-
чаях размеры участков могут быть или больше или меньше обычных. Позже,
когда начинается формирование гаремов, размеры участков могут отличаться
от первоначальных. Это во многом зависит от активности секача. Раннее занятие
территории не обязательно предполагает лидерство в формировании гарема; это
210
Рис. 12.8. Динамика формирования гаремных залежек на о. Тюлений
[из: Кузин и др., 1996]
зависит от времени прихода самок на остров, как правило, выходящих на участ-
ки гаремного лежбища, где они были рождены и приносили потомство.
До прихода самок самцы малоактивны: большую часть времени они спят;
территориальное поведение слабо выражено. С приходом самок поведение сам-
цов резко меняется; они начинают проявлять агрессивность, двигательную ак-
тивность и территориальность. Эти поведенческие акты самцов весьма индиви-
дуальны. Некоторые очень активны, постоянно патрулируют территорию, часто
ввязываются в драки с соседями, грубо обращаются с самками. Другие, напро-
тив, спокойны, редко ссорятся с соседями, но весьма успешно удерживают тер-
риторию и поддерживают определенный порядок в гареме. Агрессивность и вы-
раженность индивидуальной территориальности имеет мало отношения к вы-
годности выбранной территории [Kenyon, 1960].
Самки выбирают территорию, на которую они собираются выйти, безотно-
сительно к находящимся там самцам [Bartholomew, Hoell, 1953]. Самцы, распо-
лагающиеся ближе к воде, быстрее собирают гаремы. Вновь приходящие на леж-
бище самки присоединяются к уже сформировавшемуся гарему.
211
E65 66 4 год
E9U15 4года
El 3 201 4 года'
Е If i 11 4 годе?
Рис. 12.9. Размещение меченых
самок на участке лежбища на
о. Тюлений в 1964 г. [из: Быч-
ков, 1969]:
1 — место регистрации самки
(точки в каждом сектрое относятся
к одной самке, номер метки кото-
рой приведен туг же), 2 — размер
участка (м): 3 — забор.
Как показали наши исследования, скорость формирования гаремов опреде-
ляется не стадностью самок. Самки из года в год выходят на одни и те же участки
гаремного лежбища, и поэтому возникновение гаремов на них зависит не от того,
как рано вернутся на лежбище самки вообще, а как рано и на сколько дружно
придут к острову самки, родившиеся на данном участке гаремной территории
из года в год залегающие и воспроизводящие здесь свое потомство. Наибольшее
влияние на скорость формирования гаремов оказывает возраст самок.
К. Кеньон [Kenyon, 1960] так описывал поведение самцов, поздно приходя-
щих к островам. Они меньше размером, чем те, которые уже обосновались на
лежбище, и менее упитаны. В течение нескольких дней они залегали на окраине
лежбища и не проявляли территориальности. Позже некоторые из них перешли
на гаремное лежбище. Некоторые обосновались на приливно-отливной полосе
и тоже обзавелись гаремами, а другие так и не смогли включиться в гаремную
жизнь. Их часто сгоняли с территории гаремного лежбища уже закрепившиеся
там самцы.
Обычно на гаремной территории образуется несколько зачаточных точек бу-
дущих гаремов, которые позже расширяются, переполняют территорию самца,
у которого они первоначально сформировались, и постепенно заполняют тер-
риторию, контролируемую другим самцом, образуя новые гаремы. Так возни-
кают так называемые коллективные гаремы (рис. 12.8).
В это время может наблюдаться некоторое изменение первоначального пол-
ожения самцов на территории лежбища. Залегающие в глубине лежбища самцы,
еще не имеющие самок, тяготеют к территориям, где уже сформировались га-
ремы, стараясь отогнать несколько самок на свою территорию и образовать
гарем. Однако это им чаще всего не удается сделать. В дальнейшем, с увели-
чением численности самок распределение самцов по лежбищу восстанавливается.
Количество самок в гареме может быть различным и во многом определяется
топографией территории. В удобных местах, где самец охраняет только часть
периметра гарема, самцы собирают до 100 самок и более, если позволяет пло-
щадь. На лежбищах неудобных, с ограниченной территорией самцы весь сезон
содержат гаремы из 3—4 самок.
Численность самок в гаремах непостоянна и изменяется как в течение сезона,
так и на протяжении дня. Родившие самки всегда возвращаются на то место,
где они оставили щенков (рис. 12.9). Но в случае ухода щенка с места его рож-
дения на несколько метров в сторону, самка в поисках детеныша может ока-
заться в гареме другого самца.
212
Холостяковые залежки формируются не сразу. До прихода самок одиночные
холостяки и даже группы могут залегать среди секачей на гаремной территории
(впрочем, отдельные секачи тоже порой залегают на холостяковых участках).
Однако основная их масса строго придерживается территории холостяковых за-
лежек. С приходом самок холостяки оттесняются с гаремных территорий и в
течение всего сезона размножения образуют отдельные холостяковые залежки.
Численность холостяков на залежках очень динамична. Их выход на берег,
как правило, групповой. В отдельные дни или часы могут залегать одновременно
несколько сот или даже тысяч животных, но потревоженные, они все быстро
сходят в воду. Территориально холостяковые залежки устойчивы. Из года в год
эта возрастная группа залегает на одних и тех же местах. Динамика численности
холостяков определяется как общей численностью, так и погодными условиями.
Плотность размещения размножающегося сообщества котиков также играет
роль в устойчивости территории холостяковых залежек и их численности на них.
При разреженных гаремных залежках холостяки проникают на гаремную тер-
риторию и образуют холостяковые залежки среди взрослых животных.
Аналогичны тенденции залегания секачей на холостяковых залежках. Чис-
ленность их здесь непостоянна. В разгар гаремной жизни их меньше, но ближе
к концу сезона размножения на холостяковых залежках скапливается довольно
много секачей. В основном это животные, не выдержавшие конкуренции в борь-
бе за места на гаремной территории или ослабевшие в результате борьбы и усту-
пившие место на гаремной территории более сильным соперникам. Поведение
таких секачей на холостяковых залежках не отличается от поведения холостяков.
12.3. Хоминг и филопатрия
Работы по изучению хоминга и филопатрии проводились практически на
всех лежбищах котиков в северной части Тихого океана. При этом было изучено
поведение возвращения на избранные места различных половозрастных групп
этих животных. К. Кеньон [Kenyon, 1960] описывает случаи возвращения 10 се-
качей на лежбище через 6 час, откуда они были согнаны во время мечения. Ана-
логичные данные приводит Ф. Г. Челноков [1970а, 1971а] по 25 меченым в 1966 г.
секачам на гаремной территории Юго-Восточного лежбища (Командорские о-ва),
когда в процессе мечения животные были оттеснены с занимаемых ими участков.
Однако уже на следующий день 22 секача оказались на своих “микроучастках”,
либо на расстоянии 2—3 м от них, а 1 секач вернулся на свое место на следующий
день. Опыт слежения за мечеными секачами был повторен в 1969 г. На лежбище
Урильем (о. Медный) из 39 секачей, помеченных резиновыми ошейниками, уже
в тот же день вернулись на свое место 36. Один секач пытался пробиться на
свою территорию, но так и не смог этого сделать; его потом видели на холос-
тяковой залежке.
Приведенные данные описывают случаи возвращения секачей на единожды
выбранные места на лежбище в течение одного сезона. Однако есть наблюдения,
которые говорят о том, что некоторые секачи из года в год занимают один и
тот же участок на лежбище. [Вениаминов, 1840; Elliott, 1875; Lucas, 1899; Су-
воров, 1912; Кузин и др., 1996; Косыгин, личн. сообщ.].
Известно, что определенное число секачей постоянно залегает на холостяковых
залежках. Для выяснения поведения этих самцов Ф. Г. Челноков [1970а, 1971а]
213
МЕСТО МЕЧЕНИЯ УШЛИ С МЕСТА МЕЧЕНИЯ
ВЕРНУЛИСЬ
Восточный Клиф 23
Северное
Восточный Риф 7
Западное
Св. Георгия
Неизвестно
• Св. Георгия
Св. Павла
* Неизвестно
,Cd. Георгия
Восточный Клиф 14 —Св. Павла
Неизвестно
-Св. Георгия 12(52%)
Рис. 12.10. Перемещение холостяков, помеченных радиометками на лежбище
Восточный Клиф (о-ва Прибылова) [из: Baba et al., 1979]
в 1968 г. пометил 79 секачей, из которых 45 потом наблюдал. Из этих секачей
35 попадали в отгоны вместе с холостяками, но были выпущены. Все они вер-
нулись на те участки, откуда были захвачены в промысловые отгоны. 12 секачей
попадали в промысловые отгоны с одного и того же участка дважды, 1 секач
в течение 8 дней — трижды, еще 1 секач за 16 дней — 4 раза. Некоторые самцы
сразу же возвращались на свои участки, хотя рядом происходил забой холос-
тяков. Остальные, выпущенные из первого загона, переходили на другие участки
залежек, хотя 2 из них через несколько дней вновь вернулись на тот участок,
откуда были отогнаны.
Что касается хоминга у самок, то отличительной чертой их поведения яв-
ляется возвращение на тот участок лежбища в течение сезона размножения, где
самка родила щенка [Bartholomew, Hoell, 1953; Kenyon, 1960; Бычков, 1969]. Мес-
то это обычно не четко обозначено, но в пределах 20—30 м гаремной территории
есть участок, который посещается чаще, чем другие (рис. 12.9). В случае нару-
шения режима лежбища (вмешательство человека, стихийное явления) самки мо-
гут оставить место рождения щенка. Так, при попытке переселения нескольких
гаремов, в состав которых входили еще не успевшие родить щенков самки, с
о. Тюлений на мыс Георгия (о. Сахалин), все они покинули щенков и ушли с мыса,
секачи же продолжали еще долгое время оставаться на месте [Когай, 1971а]. Спу-
гивание самок с мест залежки или их переселение со щенками на другое место по-
зволило выявить американским исследователям, что примерно треть самок бросают
щенков и возвращаются на прежнее место [Bigg et al., 1977, 1978].
В. А. Бычков [1969] сообщил о случае обнаружения самок на одном и том
же участке лежбища в разные годы. Так, в 1963 г. в одном из секторов гаремной
территории о. Тюлений были зарегистрированы 16 меченых самок, которые вер-
нулись сюда же в 1964 г., а 4 самки возвращались в один и тот же сектор лежбища
в течение 1963—65 гг.
В последние годы были изучены реакции холостяков на неоднократные при-
нудительные спугивания с территории лежбища и возвращения их после само-
стоятельного ухода с залежек. Так, в 1970 г. Ф. Г. Челноков [1971а] пометил
крупными бирками 100 холостяков на Урильем лежбище (о. Медный). Позже
97 из них наблюдал. Из этого числа меченых холостяков 81% животных встре-
чались на том лежбище, где они были помечены, а 19% были встречены на других
лежбищах. Несколько дней спустя на участок, где метили животных, вернулись
214
еще,5 холостяков, перед этим побывавшие на Северо-Западном лежбище (о. Бе-
ринга). Сходные данные по возвращению холостяков на место прежнего зале-
гания были получены японскими исследователями [Baba et al., -1979] на о-вах
Прибылова (рис. 12.10). Из 23 холостяков, помеченных на лежбище Восточный
Клиф (о. Св. Георгия), вернулись на прежнее место 21.
В целом, холостякам, как и другим возрастным группам котиков, свойствен
инстинкт дома, хотя, как известно [Мараков, 1974], именно им принадлежит ини-
циатива в освоении новых территорий и образовании новых лежбищ.
Интересные данные о хоминге щенков были получены К. Кеньоном [Kenyon,
1960]. При мечении их сгоняли со своих мест на расстояние 100—150 м. Через
10 дней 86% щенков вернулись на свои места, с которых они были отогнаны.
Видимо, конфигурация залежек и мозаичность распределения животных на
территории лежбища тоже является следствием хоминга или филопатрии. В под-
тверждение сказанному приведем следующие факты. Так, на Западном лежбище
(о. Св. Павла) в течение 60 лет проводили изменение рельефа лежбища с на-
деждой улучшения условий залегания зверей, однако эти места остаются неза-
нятыми котиками, хотя по субъективным оценкам они ничем не отличаются от
соседних, где котики залегают охотно [Kenyon, 1960]. На Тюленьем с юго-вос-
точной стороны острова в течение нескольких лет существовал большой “кол-
лективный гарем”, насчитывавший ок. 2 тыс. животных; в экспериментальных
целях он был выбит в 1972 г. До сих пор на этом месте котики не залегают.
В целом, хоминг и филопатрия у котиков сильно развиты. Они начинают
проявляться с рождения и усиливаются с возрастом [Kenyon, Wilke, 1953; Baker
et al., 1970]. Благодаря этим врожденным свойствам популяции северного морс-
кого котика приобретают ряд ценных качеств, направленных на стабилизацию
мест залегания, поддержание определенных внутрипопуляционных характерис-
тик, успешное размножение, воспитание потомства и выживание.
12.4. Микромиграции
Под микромиграциями мы понимаем перемещение котиков с лежбища на
лежбище, с острова на остров в пределах пространственно хорошо обособленных
репродуктивных группировок.
Раньше мечение котиков на островах не проводилось, и наблюдателям бро-
сался в глаза консерватизм в выборе места на лежбище отдельными особями,
имеющими природные отличительные черты [Вениаминов, 1840]. Однако позже
мечение даже ограниченного числа котиков показало, что перемещение их с леж-
бища на лежбище и с острова на остров существует. Уже Л. В. Бойцов [1934]
с осторожностью писал про обособленность разных стад котиков, а И. И. Ба-
рабаш-Никифоров [1936] с уверенностью сообщал, что “старое мнение о том,
что каждый отдельный котик из года в год приходит только на одно и тоже
лежбище и даже старается улечься на один и тот же камень, совершенно не под-
тверждается. Беринговские и медновские котики представляют смешанное
стадо”. Е. И. Ильина [1950] привела показатели обмена котиками между стадами
на островах Медный и Беринга. Из 22 помеченных на о. Медный зверей 18%
были забиты на о. Беринга. Из 12 клейменых на о. Беринга 17% были добыты
на Медном.
215
о. Баринга
место мечении
чмс/опонечеиных
ссиермое лежбище
75405
сеМРО-ЗСПОЗнООЛЖВИ»
лМ е Йн biu '
юго-восточное
88378
22564
число Эобштых 2108 34 74 449 279 1014
^nomuWiblx 2.87 0115 0.08 0.73 0.32 4.49
соотношение^ 0113 1.53 354 25.77 16.02 58.24
Рис. 12.11. Перемещения котиков в пределах лежбищ на Командорских о-вах
4)8 30 5228
0.15 0.13 165
2.92 0.89 емо
Дальнейшие исследования микромиграций котиков в пределах Командорских
о-вов были изучены Д. И. Чугунковым [1966]. За период 1958—62 гг. на о. Бе-
ринга были помечены 22180 щенков, на о. Медный — 28168 или 26.1% от общего
числа живых детенышей. Из числа добытых в последующие годы холостяков
(3—5 лет) на о. Беринга помеченные там же составили 929 особей, 120 сюда
пришли с Медного, что составило 0.43% от числа помеченных на последнем.
На о. Медный забили 1546 меченых котиков, из которых 47 оказались берин-
говоморскими (0.21% от числа помеченных на о. Беринга). При этом оказалось,
что миграционные способности разных поколений были одинаковыми. Из этих
данных автор сделал заключение, что котики, обитающие на островах Медный
и Беринга, образуют слабо смешивающиеся группировки.
Изучение микромиграций котиков командорской репродуктивной группиров-
ки из поколений 1963—78 гг., проведенное нами, показало (рис. 12.11), что на
Юго-Восточном лежбище (о. Медный) из общего числа добытых 2—5-летних
холостяков за период 1968—80 гг., помеченных здесь же, вернулось 3.65% жи-
вотных; с Урильего лежбища, расположенного в 16 км от Юго-Восточного —
0.85%; с Северного и Северо-Западного лежбища (о. Беринга) — 0.15% и 0.23%
меченых особей, соответственно. Сопоставление этих данных показывает, во-
первых, что основная масса животных возвращается на те острова, где они были
помечены щенками; во-вторых то, что чем ближе расположено одно лежбище
от другого, тем большее число животных предпринимают перемещения с одного
лежбища на другое. Это подтверждается и другими данными. Так, на Северо-
Западном лежбище в 1968—80 гг. добыли 4.49% котиков, помеченных в поко-
лениях, рожденных в 1963—78 гг., в то время как с Северного лежбища здесь
добыли 0.73% котиков, а с Юго-Восточного — 0.32%. В промысловых побойках
на Северном лежбище котики, помеченные здесь же, составили 2.87%, с Севе-
ро-Западного — 0.15%, с Юго-Восточного — 0.08% (рис. 12.11).
Оценивая микромиграции котиков командорской репродуктивной группи-
ровки, можно еще раз подтвердить данные Д. И. Чугункова [1966] о том, что
котики островов Беринга и Медный представляют собой мало смешивающиеся
216
Рис. 12.12. Перемещения котиков в пределах лежбищ на Курильских о-вах
группировки, и количество котиков, эмигрировавших с о. Медный на о. Беринга,
вдвое больше, чем эмигрировавших в обратном направлении. Чем ближе рас-
положены отдельные репродуктивные группировки друг от друга, тем сильнее
выражен обмен особями между ними.
Лежбища котиков на Курилах расположены на двух группах островов: Ка-
менные Ловушки, включающие скалы Котиковая, Высокая, Низкая, Долгая и
о-ва Среднего, включающие скалы Черные и Хитрая. Находятся эти группы скал
на расстоянии 70 миль друг от друга, в то время как отдельные скалы, состав-
ляющие эти группы, разделены несколькими сотнями метров. С 1963 г. на Ку-
рильских о-вах пометили 25865 котиков, в т. ч. 15359 на Каменных Ловушках
и 10470 на о-вах Среднего. 95 вернувшихся меток местного происхождения (рис.
12.12) свидетельствуют, что котики перемещаются как в пределах близлежащих
скал, так и между сравнительно далеко отстоящими друг от друга островами.
Из общего числа добытых меченых котиков на о-вах Каменные Ловушки
69.2% составили животные, помеченные на группе этих же островов, а на группе
о-вов Среднего котики, помеченные здесь же, составили 59.3%. Соответственно,
количество эмигрантов на о-вах Каменные Ловушки — 30.8%, а на о-вах Сред-
него — 40.7%. В свою очередь, из общего числа помеченных котиков на Ка-
менных Ловушках “своих” котиков добыли вдвое больше, чем котиков, поме-
ченных и добытых на о-вах Среднего [Кузин, 19826].
Обращает на себя внимание большое количество котиков местного проис-
хождения на скалах Котиковая и Хитрая (соответственно, 26% и 12%), на ко-
торых они поселились раньше, чем на других скалах. Нельзя также не заметить
того, что количество местных котиков на Черных скалах было гораздо меньше,
а доля эмигрировавших сюда животных — больше, чем на двух предыдущих.
Недавно возникшее и интенсивно развивающееся лежбище котиков на скале Дол-
гая принимает наибольшее число эмигрантов [Кузин, Перлов, 1975в; Кузин, 19826].
Приведенные1 материалы касаются только 2—5-летних зверей (в основном
самцов). Имеются немногочисленные данные и о животных старших возрастных
групп [Челноков, 1970а, 1971а]. В 1967 г. отмечались переходы 4 секачей с Юго-
217
Таблица 12.2
Происхождение самок котиков в возрасте до 12 лет, встреченных на
Юго-Восточном и Урильем лежбищах, % [по неопубл, материалам
Ф. Г. Челнокова]
Годы Юго-Восточное лежбище Урилье лежбище
Место р о ж д е н и я
Ю-В Уо Сев С-3 т П Ю-В Уо Сев С-3 Т П
1971 87 2 1 10 69 14 3 4 3 7
1972 87 2 1 2 8 63 13 6 3 2 13
1973 95 5 56 23 5 6 2 8
1974 77 3 1 3 16 60 23 8 6 3
1975 93 0.2 1.8 0.5 1 3 49 28 8 8 3 4
1976 96 0.5 1 3 51 25 5 6 3 10
1977 90 1 1 1 1 6 46 33 6 9 2. 4
1978 95 1 1 1 1 1
1979 95 1 1 1 0.5 1.5 35 55 3 5 2
1980 95 3 0.3 1 0.7 24 69 2 5
Среднее 91 1 1 1 2 4 52 30 5 5 2 6
Примечание. Ю-В — Юго-Восточное, Ур — Урилье, Сев — Северное, С-3 — Севе-
ро-Западное, Т — о. Тюлений, П — о-ва Прибылова.
Восточного лежбища на Урилье (о. Медный); 1 секач перебрался с Юго-Вос-
точного лежбища на о. Беринга. Аналогичные переходы отмечались нами на
Курилах.
Количественных данных о микромиграциях самок в литературе нет. Однако
случаи перехода их с одного лежбища на другое бывают [Бычков, 1969; Кузин,
19826]. Из материалов, любезно предоставленных нам Ф. Г. Челноковым (табл.
12.2) видно, что на Урильем и Юго-Восточном лежбищах присутствуют самки
со всех командорских лежбищ. На Юго-Восточном число пришлых самок с дру-
гих лежбищ в среднем меньше, чем на Урильем, где превалируют самки с первого
лежбища.
На обоих лежбищах примерно одинаковое число самок с о. Беринга. В
целом, для микромиграций самок характерно то же, что и для других половоз-
растных групп: обмен особями между группировками, расположенными близко
друг от друга, осуществляется более интенсивно, чем между далекими. При этом
исключительно важное значение имеет возраст того или иного лежбища. Не-
давно организованные, растущие лежбища принимают много эмигрантов, а ста-
рые, залежки отсылают наибольшее число их. Однако поток эмигрантов со старых
залежек не остается постоянным во времени. Он изменяется в зависимости от
общей численности и демографической ситуации на залежке.
Чем выше численность и, соответственно, плотность какой-либо группировки
или залежки котиков, тем большее число особей эмигрирует из нее. Увеличение
плотности обычно наблюдается в растущих популяциях и предполагает наличие
218
в них большого числа особей младших возрастных групп, приплод от которых
обладает меньшими шансами на нормальное развитие и даже выживание [Болт-
нев и др., 1991]. Большая часть эмигрантов из таких репродуктивных группи-
ровок оказывается впоследствии на только что образовавшихся лежбищах, где
они ничем не отличаются от аборигенных особей и поэтому становятся равно-
правными членами сообщества. Другая часть оседает на постоянно функцио-
нирующих лежбищах, где, как правило, устоявшийся возрастной состав произ-
водителей, хорошее физическое состояние животных всех половозрастных групп,
соответствующая структурно-пространственная организация сообщества. Здесь
иммигранты, по сравнению с аборигенами, оказываются менее физически раз-
витыми, а из-за сильно выраженной филопатрии у самок и агрегированности
отдельных социумов, они вынуждены присоединяться к уже существующим тер-
риториально-структурным подразделениям гаремного сообщества, оказываясь,
таким образом, на их “закрайках”. Однако таковыми являются йе окраинные
территории лежбищ вообще [Болтнев, 1990а], а окраины отдельных социумов.
В этом, на наш взгляд, кроется ответ на вопрос: почему эмигранты всегда мельче
особей из тех популяций, где они появляются, и почему они обитают на “за-
крайках” сообществ.
Вместе с тем, все материалы по микромиграциям должны рассматриваться
в комплексе с данными по хомингу, потому что оба процесса являются сторо-
нами одного явления. Если хоминг усиливается с возрастом, то микромиграции,
наоборот, уменьшаются.
12.5. Особенности постгаремного периода жизни
Сроки и характер ухода котиков с лежбищ. С августа начинается заметное
угасание гаремной жизни котиков, и наблюдаются изменения в распределении
зверей на островах. Многие секачи покидают гаремные залежки и перекочевы-
вают на холостяковые участки. Больше времени, чем раньше, они проводят в
море. На более длительный период и чаще покидают своих детенышей самки,
которые также проводят больше времени в море. Холостяки наводняют гарем-
ные лежбища и отдельными группами залегают среди самок и детенышей. С се-
редины августа к островам массово начинают приходить двухлетки и однолетки,
которые располагаются как на холостяковых залежках, так и на гаремной тер-
ритории. Намечается исчезновение летней мозаичности в распределении котиков
на лежбищах. Лежбища как бы разрастаются вглубь и вширь, однако плотность
залегания животных на них значительно меньше, чем летом. Создается впечат-
ление очень сильного заполнения лежбищ и обилия зверей на них. Однако об-
разование каких-либо новых “осенних” и" холостяковых" лежбищ [Ильина, 1950]
не наблюдается. Напротив, существующие в летнее время холостяковые залежки,
например, на мысе Монати (о. Беринга), исчезают, и звери концентрируются
все в одном месте. В любое время суток много котиков находятся на плаву у
островов, при этом могут наблюдаться большие скопления у мысов, на которых
они залегали летом. Так, например, у о. Беринга наблюдаются осенние концен-
трации у Северо-Западного мыса [Суворов, 1912; Ильина, 1950], а котики с о.
Медный сосредоточиваются осенью у северо-западной оконечности острова или
у мысов Лебяжий и Песчаный, т. е. непосредственно у тех, которые ограничи-
вают Урилье лежбище. Что касается северной оконечности Медного, то кон-
219
центрации котиков там вызваны нахождением в этом районе банки Китоловная,
считающейся “кормовым полем” котиков в сезон размножения. Много зверей
залегает на отдельных камнях и скалах, находящихся у лежбищ.
Рассматривая процесс ухода котиков с островов на зимовку, Е. К. Суворов
[1912] писал, что “к середине августа уже все секачи оставляют берег и больше
не возвращаются на лежбище. В это время и матки начинают бросать гаремные
лежбища и переходить на новые места или уходить с островов”. Первыми, по
его наблюдениям, уплывают секачи, за ними бездетные самки, но еще долгое
время отдельные небольшие группы котиков, среди которых больше “черных”,
могут наблюдаться на лежбищах и плавать вблизи них. Только в конце октября,
начале или середине ноября исчезают с лежбищ наиболее запоздавшие котики.
В 1900 г. последний котик был замечен на о. Беринга 11 ноября, а на о. Мед-
ный — 7 декабря, но в последующем году к началу ноября лежбища уже были
свободными от зверей. 6 ноября 1905 г. были замечены вблизи лежбищ до 1500
котиков, не вылезавших на берег. 16 декабря 1907 г. на Северном лежбище на-
блюдали до 1000 и на воде — 5000 котиков. 31 октября 1908 г. на воде и камнях
у всех Глинковских лежбищ отмечены до 1200 животных. 19 октября 1909 г.
здесь же регистрировали до 1500 котиков, а уже 31 октября на лежбищах не
оказалось ни одного котика.
Е. И. Ильина [1950] считает, что отход котиков с островов происходит вне
зависимости от пола и возраста и что все возрастные группы этих животных
одновременно и пропорционально их численности покидают острова. Дольше
других на берегу задерживаются секачи и холостяки. В начале сентября можно
наблюдать около берега группы в 200—300 голов, которые 1—2 дня держатся
близ берегов, а затем одновременно уходят. В середине ноября начинается мас-
совый отход. Отдельные экземпляры остаются и на декабрь. Так, в 1936 г. секачи
и холостяки оставались на островах до 15 декабря, а в 1937 г. — до 11 декабря.
Последние матки с двумя “серыми” котиками ушли 4 декабря. По наблюдениям
С. В. Маракова и Г. А. Нестерова [личн. сообщ.], в последующие годы первые
признаки уменьшения численности котиков на лежбищах стали наблюдаться в
первой половине октября. В это время резко уменьшилось поголовье секачей
и холостяков. Маточное стадо и сеголетки начали уменьшаться в количестве
только в октябре—начале ноября. К середине ноября на лежбищах осталось не-
большое число котиков, среди которых преобладали самки, детеныши и секачи.
К 25 ноября на Северном лежбище оставались 4 секача, на воде — 10 сеголетков
и 4 матки. 1 декабря на воде у лежбища был замечен секач, а 28 декабря —
полусекач. На Юго-Восточном лежбище уже к 5 декабря ни на лежбище, ни
на воде котиков не было.
В первой половине августа секачи на о. Тюлений начинают покидать га-
ремное лежбище и переселяться на мысы острова. На южной оконечности ос-
трова в конце августа образуется крупная залежка 1—2-летних котиков, залежка
постоянно разрастается, и звери начинают залегать уже не только на восточной
стороне острова, но и на западной. В это время на основной гаремной терри-
тории с восточной стороны острова происходит нечто похожее на возникновение
гаремной жизни. Самки группируются вокруг секачей, но драк между послед-
ними не наблюдается. Эта осенняя активность котиков получила название “лож-
ного гона” [Мараков, 1974]. С конца октября постепенно уменьшается числен-
ность различных половозрастных групп. В ноябре интенсивность ухода живот-
ных с островов резко возрастает. Однако еще в декабре можно видеть небольшое
220
50
a
e<
&
p.
40
30
20
10
• Охотское поре
•Западная часть
Берингова поря
Восточная
-о часть Берингова
ыоря
I 3 5 7 9 >10
§
OJ
ЕГ
О
р>
и
40
30
20
10
I 3 5 7 9 >10
Тихоокеанские
• воды Японии
* Японское поре
Тихоокеанские
воды Северной
Америки
Число животных в группах
Рис. 12.13. Состав группировок морских котиков в различных районах их обитания
число котиков на острове. В основном задерживаются на острове секачи и по-
лусекачи [А. М. Николаев, личн. сообщ.].
Исследования, проведенные нами на Курильских о-вах в конце ноября, также
подтвердили данные о том, что в осенний сезон на лежбищах дольше всех ос-
таются самцы старших возрастных групп (секачи и полусекачи). Отдельные
звери (в особенности секачи) могут выходить на острова всю зиму, если острова
не блокируются льдами.
Размер и состав группировок северных морских котиков в море. С окончанием
репродуктивного сезона меняется стратегия поведения котиков. Наблюдается пе-
реход животных от многоуровневого структурированного сообщества к простей-
шему типу интеграции индивидов — элементарным группировкам. По мнению
одних исследователей, эти объединения лишены прямой генетической преемст-
венности и возникают по принципу эквипотенциальности [Wada, 1969; Влади-
миров, 1980], по мнению других, “связаны между собой близким родством... с
поддержанием в них соответствующей семейной иерархии” [Соболевский, 1984].
В акваториях у репродуктивных центров в Охотском (о. Тюлений) и Берин-
говом (острова Командорские и Прибылова) морях морские котики встречаются
поодиночке и группами. В составе групп обычно насчитывается от 2 до 8 особей.
Группы более 10 голов в этих районах встречаются относительно редко. Число
зарегистрированных одиночных животных в Охотском море у о. Тюлений и в
западной части Берингова моря у Командорских о-вов примерно равно числу
встреченных разной величины групп (рис. 12.13).
В восточной части Берингова моря у о-вов Прибылова численность одиноч-
ных животных несколько превышает число учтенных групп; Группировки по 2—
3 особи во всех частях репродуктивного ареала отмечались приблизительно в
равном числе, а по 4—6 — чаще встречались в Охотском море и в западной
221
части Берингова, чем в восточной части последнего. Более крупные группировки
повсеместно регистрировались одинаково редко.
Сезонная динамика встречаемости различных по величине групп котиков в
Охотском и Беринговом морях сходна [Кузин, 1996]. Прослеживается тенденция
встречаемости наибольшего числа одиночных котиков в период подхода к ост-
ровам размножения по возвращении с мест зимовки (май—июль) и при уходе
их с островов в районы зимовки (октябрь—ноябрь). Группы по 2 особи более
многочисленны в весенне-летнее время, нежели осенью, тогда как число групп
по 3—5 котиков, напротив, увеличивается в летне-осенние месяцы. Аналогичная
тенденция просматривается и по частоте встречаемости более крупных групп
котиков.
На местах зимовки в тихоокеанских водах Японии и у берегов Сев. Америки
число одиночных котиков меньше, чем в акватории репродуктивных центров;
соответственно, число животных в группах было больше (рис. 12.13). Приме-
чательно, что в тихоокеанских водах Японии одиночные котики регистрирова-
лись вдвое реже, чем в Японском море и у берегов Сев. Америки.
Группы по 2 особи и более в Японском море и в тихоокеанских водах Сев.
Америки встречались примерно в равном числе, при этом группы по 2 особи
на местах зимовки регистрировались реже, а по 3 — чаще, чем в районах ре-
продуктивных центров. Более крупные группировки были одинаково малочис-
ленны в обеих частях ареала. Тем не менее, в обозначенных местах зимовки
чаще, чем в акватории лежбищ, отмечались группы свыше 10 особей, а мак-
симально большие группы состояли из 20—30 котиков. В тихоокеанских водах
Японии распределение животных по группам и число встреченных групп разной
величины сильно отличались от таковых в других частях ареала (рис. 12.13).
Здесь, в целом, гораздо чаще встречались объединения животных по 4 и более
особей, а число групп более 10 особей равнялось числу встреченных одиночных
особей и в несколько раз превышало таковое в Японском море и в восточной
части Тихого океана. У берегов Японии регистрировались группы по 50—70 ко-
тиков, чего практически не отмечалось в других частях ареала.
Как показали наши исследования, сезонный аспект количественного распре-
деления северных морских котиков в Японском море, в северо-западной (у бе-
регов Японии) и северо-восточной (у берегов Сев. Америки) частях Пацифики
тесно связан с этапами миграции животных к местам зимнего пребывания. При
осенней миграции на юг (ноябрь—февраль) котики осваивают большие по пло-
щади акватории и не образуют плотных концентраций. Не менее трети живот-
ных в это время встречается поодиночке. В марте—апреле, по мере продвижения
котиков в южном направлении и освоения ими традиционных районов зимовки,
таких, как банка Ямато и Восточно-Корейский зал., акватории японских пре-
фектур Санрику и Джобан и североамериканских штатов Вашингтон и Кали-
форния, численность одиночных животных заметно сокращается, а число груп-
повых объединений увеличивается. При этом, однако, просматривается неоди-
наковая частота встречаемости разной величины групп в разных районах зи-
мовки.
Так, в Японском море группы по 2—4 особи встречались чаще, чем в ти-
хоокеанских водах Японии, в течение всего периода пребывания котиков на мес-
тах зимовки, а частота встречаемости групп по 5—7 особей была выше в ти-
222
Рис. 12.14. Встречаемость
различной величины групп
котиков в зависимости от
плотности на местах зимовки
у тихоокеанских берегов
Японии [по: Wada, 1969]
Размер групп
хоокеанских водах Японии, чем в Японском море. Другой особенностью рас-
пределения котиков по районам зимовки является то, что в Японском море прак-
тически не встречались группы более 10 котиков, в то время как в Тихом океане
у берегов Японии и Сев. Америки они постоянно присутствовали. Число таких
групп увеличивалось по мере продвижения котиков на юг и уменьшалось при
возвращении в северные широты.
Среди добытых в мбре 87 групп котиков смешанных по полу оказалось 26
(29.9%), самцовых — 6 (6.9%), самочьих — 55 (63.2%). В дифференцированных
по полу группах чаще встречались особи близких возрастных классов.
Классифицированные не по возрасту, а по массе тела самочьи группы объ-
единяли особей с разницей в показателях массы до 3 кг — 27.3%, до 5 кг —
49.1%, до 8 кг — 71%, свыше 8 кг — 29% (при колебании внутрикогортной массы
тела 6—15 кг). В смешанных по полу группировках зависимости от возраста
составляющих их особей не просматривается, однако чаще объединялись близ-
кие по массе тела животные: самцы 4—5 лет и половозрелые самки всех воз-
растных классов.
В целом, из общего числа исследованных групп одновозрастных оказалось
13 (14.9%), разновозрастных 74 (85.1%); с разницей в возрасте не более года —
28 (32.2%), не более 2 лет — 41 (47.1%), не более 3 лет — 50 (57.5%) и не более
5 лет — 63 группы (72.4%).
Таким образом, ок. 30% северных морских котиков в море встречаются по-
одиночке и более 70% объединяются в различной величины группы. Частота
встречаемости групп убывает по мере увеличения их количественного состава.
Соотношение одиночных животных в группах, а также величина групп в разных
частях ареала не одинаковы. В акватории репродуктивных центров доля жи-
вотных, объединяющихся в группы, и количественный состав групп снижаются
при удалении от лежбищ, а в акватории зимовки — к окраинным частям на-
гульного ареала [Wada, 1969; Ащепков, Колотилин, 1982]. В зонах оптимума мо-
заичность распределения животных определяется различными биотическими и
абиотическими факторами, однако независимо от стимулов в любом случае ин-
теграция усиливается с увеличением плотности (рис. 12.14).
Фактически, изученная нами сезонная изменчивость интеграции животных
в процессе миграции, как и повышенные показатели встречаемости групп осо-
223
бей, по сравнению с одиночными, а также количественного состава групп в рай-
оне тихоокеанского побережья Японии, являются отражением плотностного рас-
пределения их в связи с этапностью миграции (в первом случае) и ограничен-
ностью акватории зимовки (во втором). Из экологических факторов, определя-
ющих интеграцию котиков в море, ведущая роль безусловно принадлежит пи-
щевому. Концентрации котиков в море, как правило, наблюдаются в
биологически высокопродуктивных частях ареала, такие места давно и хорошо
известны. Анализ представленных данных по половозрастному составу группи-
ровок котиков показал, что интеграция сильнее всего выражена у самок, а в
популяциях котиков около 70% составляют именно они. Кроме того, возрастной
и половой состав групп зависит от времени и места взятия проб [Wada, 1969].
В местах, где преобладают взрослые самки группировки животных состоят
из них. С преобладанием в другом месте особей иного пола и возраста группы
образуются этими особями.
Но даже с учетом обособленного распределения животных интеграция в мес-
тах обитания направлена на образование если не близких по возрасту (чаще в
однополых группировках), то близких по массе тела (в разнополых группиров-
ках) животных, что является экологически оправданным. Объединяющиеся по
признаку эквипотенциальности животные способны одинаково противостоять
давлению окружающей среды. Важно и другое следствие из анализа данных по
половозрастному составу группировок самок: они не подтверждают мнение
Е. И. Соболевского [1984] о том, что при обитании в море группообразующим
фактором являются близкородственные связи. Почти в двух третях исследован-
ных нами групп их члены по возрастным критериям не могут быть связаны близ-
ким родством. Объединению по родственной линии препятствует ряд следующих
факторов. По окончании молочного питания матери и их потомство экологи-
чески разобщены, потому что проводят зиму в разных районах океана. Повтор-
ная их встреча на лежбище возможна у самок минимум через 2—3 года, у самцов
через 7 лет (в период наступления половой зрелости).
Во время миграции и на местах зимовки совместное пребывание в группах
близкородственных животных также проблематично в связи с их обособленным
по половозрастному критерию распределением [Кузин, Панина, 1977; Ащепков,
Кузин, 1987]. Даже в тихоокеанских водах Японии, где плотность животных до-
статочно высока, а смешиваемость особей разного пола и возраста выражена
сильнее, чем в других частях ареала зимовки, эта тенденция прослеживается. Об-
щеизвестным является факт разновременного прихода северных морских коти-
ков на лежбища в зависимости от пола и возраста. Если бы в море интеграция
особей осуществлялась на родственной основе, названная закономерность не прояв-
лялась бы. Известны многочисленные достоверные факты встреч в одной группе
взрослых особей, помеченных еще щенками в разных репродуктивных центрах или
популяциях [Соболевский, 1977; наши данные]. Сказанное позволяет считать, что
в морской период жизни группообразующие взаимодействия у северных морских
котиков конъюнктивные, а интеграция индивидов осуществляется по принципу
эквипотенциальности. Эти связи могут быть инициированы знакомством или
иными стимулами, не относящимися к близкородственным, и в дальнейшем под-
держиваться на протяжении нескольких лет [Владимиров, 1990]. Группы, по-
строенные по такому принципу, легко распадаются, могут существовать в раз-
ных количественных сочетаниях и в течении разного времени. Как правило,
такие взаимоотношения наблюдаются в период использования животными ло-
кальных ресурсов, во время миграции и в иных аналогичных ситуациях.
224
Глава 13. ПОВЕДЕНИЕ И АКТИВНОСТЬ
13.1. Взаимоотношения различных половозрастных групп северных морских
котиков
Взаимоотношения взрослых самцов и самок. Формирующееся на основе по-
лигинии размножающееся сообщество котиков характеризуется различным ти-
пом персонифицированных взаимодействий особей между собой. Для взрослых
самцов они строятся на присущей им территориальности и агрессивности при
защите самок или территории. В силу этого типологически взаимодействия жи-
вотных в репродуктивных группировках могут быть отнесены к территориаль-
но-гаремному типу [Мараков, 1974; Крушинская, Лисицина, 1983].
Взаимоотношения самок в гареме, как правило, нейтральны. Только в первые
часы после родов, охраняя детеныша, самка проявляет агрессивность к соседям.
Находящиеся в течке самки располагаются в гареме ближе к самцу, хотя это
не четко проявляется и, пожалуй, зависит от индивидуальных особенностей жи-
вотных.
Поведение самцов в гареме сильно индивидуализировано. Некоторые из них
при общении с самками никогда не хватают их зубами, чтобы удержать на своей
территории, и при похищении их из другого гарема. Такие самцы патрулируют
по границе своего гарема и удерживают самок, преграждая им путь телом и
используя при этом лишь звуковые угрожающие сигналы. Они редко ввязыва-
ются в драки с соседними секачами из-за самок и не отвечают на их прово-
кационные действия.
Другие самцы часто и энергично патрулируют свою территорию, затевая ссо-
ры с соседними секачами из-за самок; грубо обращаются с самками. Во многих
случаях они хватают самок зубами, ударяют их боковой частью головы. Такие
самцы могут расширить свою территорию, однако успех в воспроизводстве в
обоих случаях может быть равнозначным. При общении с самками самец прояв-
ляет много разных реакций как двигательного, так и звукового характера [Кру-
шинская, Лисицина, 1983]. Среди них различаются: угрожающее хорканье — ис-
пользуемый секачом звук в ответ на приближающегося соперника; предупреж-
дающий рев — длинный, громкий, модулированный по амплитуде сигнал, из-
даваемый в случае приближения к территории постороннего самца; цоканье —
низкий по интенсивности сигнал, состоящий из ряда коротких импульсов, из-
даваемый самцом и при окончании драки, и при виде дерущихся пар соперников,
и при обходе гарема. Цоканье определяет особь как самца. Иногда этот сигнал
сочетается с другими сигналами, в частности с гаремным криком — низким,
сравнительно коротким тихим звуком, напоминающим рев или рявканье мед-
ведя, который самец издает при обходе гарема, обнюхивании самки, удержании
ее в гареме, что сопровождается загораживанием дороги корпусом тела, толч-
ками ее головой, оттеснением в сторону гарема. Часто этот звук сочетается с
трясением головой в горизонтальной плоскости. Каждому звуковому сигналу со-
путствуют соответствующие позы, хотя они и не ярко выражены.
225
Рис. 13.1. Комплекс поведенчес-
ких реакций самца северного
морского котика, направленных
к самке, находящейся в эструсе
Рис. 13.2. Самка, кусающая
самца в область шеи.
Фото Ф. Г. Челнокова.
У самок звуковая сигнализация ограничивается угрожающим хорканьем —
территориальным сигналом, присущим всем половозрастным группам котиков;
рычанием — предупреждающим сигналом; зовущим криком — длинным, по-
хожим на блеяние овцы сигналом, употребляемым при поиске щенка.
Большинство действий со стороны самца в гареме направлено к беременным
и находящимся в течке самкам (рис. 13.1).
Самцы наиболее активны в Начальный период гаремной жизни. Они часто
патрулируют территорию, издают предупреждающий рев, оповещая тем самым
соперников о занятости территории. При приближении к территории гарема сам-
ки секач встречает ее, производя короткие громкие звуки, напоминающие ряв-
канье медведя (гаремный крик), перемежающийся с цоканьем. Самец обнюхивает
нос самки, преграждая ей дорогу корпусом, толкает головой. Самка, в свою оче-
226
Рис. 13.3. Спаривание северных морских котиков.
Фото Ф. Г. Челнокова.
редь, старается все время располагаться к секачу грудью, часто прикусывая ему
шею, грудь, нижнюю губу (рис. 13.1, 13.2).
Если самка не пытается уйти с территории самца, то вскоре он успокаивается.
В противном случае самец хватает ее зубами за любую часть тела и бросает
в центр гарема или переносит в зубах на свою территорию (рис. 13.1). Другие
самцы загоняют самок в свой гарем с помощью оттеснения корпусом и тол-
канием головы, не хватая ее зубами. Самку, оказавшуюся на границе двух га-
ремных территорий, каждый самец может тащить к себе. При этом самка часто
бывает сильно травмирована и впоследствии погибает.
Находящихся в течке самок самец, видимо, определяет по запаху. Иногда
ему достаточно обнюхать только нос самки для того, чтобы потом сделать садку,
в других случаях он после обнюхивания носа дополнительно обнюхивает поло-
вые органы самки и только после этого делает на нее садку. Описаны случаи,
когда самец в процессе спаривания с одной самкой проявляет интерес к соседней
самке и по завершении спаривания с первой делает садку на другую.
Специфично поведение самок, находящихся в течке. Они стараются занять
положение в гареме ближе к самцу. Если самец приближается к такой самке,
то она выгибает спину, приподнимает заднюю часть тела, разводит задние ласты
в стороны. Чаще всего самцы сразу же кроют таких самок (рис. 13.3).
Если самец не делает садку на такую самку, то она не предпринимает никаких
дополнительных действий, провоцирующих самца к совокуплению. Отмечены
случаи, когда самец регулярно спаривался в течение нескольких дней. В один
из дней секач за 5 час 17 мин покрыл 6 самок [Bartholomew, Hoell, 1953].
Как правило, при общении самки и самца первые не сопровождают свои по-
веденческие акты звуковыми сигналами. Лишь только в случаях, когда самец
227
хватает самку зубами и тащит к себе в гарем, самка, пытаясь укусить самца,
сопровождает свои действия рычанием.
Взаимоотношения самок и щенков. По многочисленным нашим наблюдениям
и литературным данным [Крушинская, Дмитриева, 1975; Крушинская, Лисицина,
1983], самка не облизывает новорожденного щенка, не перегрызает пуповину и
не поедает послед, как об этом писала Е. И. Ильина [1950]. Лишь иногда самка
помогает освободиться щенку от околоплодных оболочек. В течение первых 10 мин
самка часто обнюхивает детеныша, призывно кричит. Многократно переклады-
вает заднюю часть тела к морде щенка. Уже через 3—5 мин после первого вдоха
щенок начинает подавать голос и предпринимать попытки встать на передние
ласты. Рефлекс отряхивания уже развит, и детеныш часто трясет головой первые
10 мин после проявления активных признаков жизни. Самка подтаскивает де-
теныша к своему животу. Примерно через час детеныш начинает сосать (рис. 13.4).
Продолжительность каждого кормления — 10—15 мин. В перерывах между
кормлениями детеныш спит возле матери. На приближающихся самок родившая
матка угрожающе хоркает, но, как правило, с места не уходит. В среднем самка
неотступно находится возле детеныша 4—5 дней. На 2—3 сутки после рождения
детеныш осваивается, и его бывает трудно отличить от других, рожденных ранее.
Он хорошо передвигается, пытается отойти от матери, но она всегда удерживает
его возле себя. Спустя 4—5 дней самка делает первые попытки оставить дете-
ныша одного на лежбище и уйти в море на кормежку. Иные самки делают это,
когда детеныш спит, в других случаях детеныш пытается следовать за матерью.
В таких случаях самке приходится часто возвращаться к детенышу, который по-
стоянно призывно кричит. Вернувшись она обнюхивает детеныша; они контак-
тируют носами и вибриссами; оба часто кричат. Наконец, самка покидает но-
ворожденного и уходит в море. Покинутые самками щенки продолжительное
время бродят возле места своего рождения, кричат, но потом успокаиваются и
засыпают. Игровые навыки у них в этом возрасте еще отсутствуют. На 6—7
сутки щенки вливаются в группы ранее рожденных, в которых существуют свои
суточные ритмы активности.
228
По возвращении на лежбище самка обычно выходит на тот участок, где она
родила щенка, и многократными звуковыми сигналами, напоминающими бле-
яние овцы, призывает щенка. На голос самки отзывается сразу несколько щен-
ков, многие идут к ней навстречу, но, обнюхав их, самка продолжает продви-
гаться по участку лежбища в поисках своего. При встрече со своим щенком
самка отходит на удобное место и ложится, а щенок сразу же начинает кор-
миться. В тех случаях, когда самке не удается сразу отыскать детеныша, она
ложится на участке, где родила, но через некоторое время снова возобновляет
поиск.
В целом, материнское поведение у самок котиков проявляется только до пер-
вого ухода с лежбища в море для питания. Со взрослением щенка это поведение
проявляется все меньше и меньше. Однако это компенсируется возрастающей
активностью щенков, которые сами начинают отыскивать возвращающихся на
лежбище матерей. Большинство исследователей считают, что самка кормит толь-
ко своего щенка, хотя есть указания на одновременное кормление самкой двух
детенышей [Кирчанов, 1975; Владимиров, 1976]. Учитывая слабо развитый у са-
мок северного морского котика материнский инстинкт, следует все-таки крити-
чески отнестись к данным о возможности вскармливания самкой чужих щенков.
В месячном возрасте щенки начинают спускаться к урезу воды и предпри-
нимают попытки контакта с морем. Самки не принимают участия в обучении
щенков плаванию. Более того, свалившийся случайно в воду и еще не умеющий
плавать щенок, чаще всего не получает помощи от матери. Самка только при-
зывно кричит, подзывая щенка к себе. Однако, если она имеет возможность до-
тянуться до барахтающегося в воде щенка, то может схватить его за холку и
вытащить из воды.
К осени щенки становятся вполне самостоятельными, и к молочному питанию
уже прибавляется пища, добытая ими самими в море. Перед осенней миграцией
на зимовку самки перестают кормить щенков молоком. Щенки разбредаются
по всему лежбищу и уже не придерживаются определенных территорий, как это
было в первый месяц жизни. Многолетними наблюдениями установлено, что на
зимовку самки и щенки мигрируют порознь, а не вместе, как об этом писал
С. В. Мараков [1974]. Районы зимовки у них тоже разные.
Взаимоотношения щенков и холостяков. При оптимальном состоянии попу-
ляций северных морских котиков, когда численность разных половозрастных
групп уравновешена, в особенности, когда в популяциях имеется достаточное
количество секачей, жизнь на гаремном лежбище организована таким образом,
что холостяки и щенки имеют возможность контактировать только осенью, пос-
ле распада гаремов. Однако, когда в популяциях ощущается нехватка секачей,
холостяки проникают на гаремную территорию, и тогда контакт между щенками
и холостяками возможен. “Оказавшись на территории, не охраняемой гаремны-
ми секачами, холостяки начинают обнюхивать похищенного щенка, а при по-
пытке того отползти, преграждают ему дорогу, хватая зубами и перебрасывая
через себя, т. е. ведут себя с ним как гаремный секач с самкой. Через некоторое
время холостяк предпринимает попытку спаривания с похищенным щенком. Ще-
нок пытается обороняться, но холостяк схватывает его зубами и отбрасывает
в сторону, проделывая это несколько раз подряд. Падение оглушает щенка, и
он почти перестает двигаться. Тогда холостяк снова предпринимает попытки
229
к спариванию и продолжает охранять щенка как половозрелый самец, делая вы-
пады в сторону других холостяков, если они оказываются поблизости. Случа-
лось, что в конце концов холостяк укладывался, положив голову на щенка, тем
самым ограничивая свободу его движения. Если щенков после встряски не по-
давал признаков жизни длительное время, холостяк терял интерес и уходил в
другое место или уплывал” [Челноков, 1974]. Этим же автором описаны случаи
стаскивания щенков холостяками в воду, где щенки повергались многочислен-
ным принудительным погружениям и потоплению.
В тех случаях, когда щенок оказывался в группе холостяков, доминирующий
холостяк вел себя как гаремный секач и предпринимал попытки защиты щенка
и территории. Между холостяками в этом случае возникали типичные реакции,
сопровождаемые угрожающими выпадами, укусами, толчками грудью, “фехто-
ванием” шеями и т. п. Как и взрослые самцы, несколько холостяков могут од-
новременно схватывать щенка зубами и тянуть его каждый к себе; в этом случае
действия их были стереотипны действиям взрослых самцов в борьбе за самку.
Ближе к осени, когда щенки подрастут, характер взаимоотношений холос-
тяков и щенков меняется. Их игры носят “турнирный” характер и в большинстве
случаев напоминают игры, которые предпринимают между собой одновозраст-
ные животные, в частности холостяки. Однако детеныши неохотно принимают
участие в таких играх. По отношению к холостякам они демонстрируют угро-
жающие реакции и стараются уйти от холостяков. Не исключено, что изменение
общего характера игр между холостяками и детенышами и, в частности, утрата
сексуального элемента у первых из них связана с общим ослаблением полового
инстинкта в связи с затуханием сезонной активности половых гормонов.
Взаимоотношения холостяков и самок. Контакт между самками и холостяками
на гаремной территории чаще всего возможен в случаях нарушения половоз-
растной структуры гаремного сообщества котиков, когда численность секачей
становится недостаточной. В основном при контактах самок с холостяками пер-
вые проявляются элементы агрессивного поведения, как правило, выражающи-
еся в угрожающих выпадах с вытягиванием шеи, открыванием рта и издаванием
хоркающих звуков. Холостяки до 5-летнего возраста обычно избегают встреч
с самками. Однако 5-летние самцы, когда в конце сезона размножения агрес-
сивность секачей уменьшается, или они малочисленны на лежбище, могут вести
себя как секачи, проявляя все элементы полового поведения. Есть даже указания
на то, что 5-летние самцы способны оплодотворять самок [Ильина, 1950]. В при-
бойной полосе они часто преследуют самок и пытаются организовать гарем, но
эти агрегации неустойчивы во времени.
На лежбищах можно наблюдать, как холостяки охраняют отдельные группы
самок, залегающих без секача, от других приближающихся к ним холостяков.
Самки при этом не проявляют никакого интереса к действиям доминантных хо-
лостяков. Напротив, при непосредственном контакте холостяка с самкой, по-
следняя издает угрожающие хоркающие звуки и вынуждает доминирующего хо-
лостяка отойти на недосягаемое расстояние.
Взаимоотношения секачей и холостяков. Взаимоотношения различных возрас-
тных групп самцов зависит от места встречи и фазы репродуктивного сезона.
На холостяковых залежках холостяки и секачи мирно относятся друг к другу.
Безусловно, доминирование в выборе места залегания принадлежит более круп-
230
ному самцу. Вместе с тем, разновозрастные самцы могут лежать бок о бок, не
проявляя враждебности друг к другу. Территориальность на холостяковых за-
лежках не выражена.
В начале гаремного сезона, до прихода самок на лежбище, холостяки часто
появляются на гаремной территории и могут свободно перемещаться по леж-
бищу. В это время секачи терпимо относятся к присутствию холостяков на бу-
дущем месте гарема, хотя иногда отпугивают йх, делая выпады, сопровождаемые
предупреждающим рыком и хорканьем. С приходом самок секачи активно пре-
следуют холостяков и изгоняют их с гаремной территории, при этом они про-
являют все элементы поведения, присущие им при встрече с соперником.
13.2. Взаимоотношения северных морских котиков с другими животными
На лежбищах морские котики контактируют с птицами (чистиковые, чайки),
наземными хищными (песцы, лисицы), а из морских млекопитающих с каланами,
сивучами, калифорнийскими морскими львами, настоящими тюленями (ларгой,
обыкновенным тюленем).
Птицы являются постоянными обитателями островов, на которых залегают
котики. Они постоянно патрулируют территорию лежбища и поедают плаценты,
погибших котиков — щенков и взрослых. Подросшие детеныши проявляют ин-
терес к движущимся объектам, пытаются гоняться за чайками и кайрами. Можно
часто видеть пробирающихся к воде по телам лежащих котиков кайр. Отдельные
животные при этом угрожающе хоркают на них, другие не обращают внимания.
С другой стороны, котики активно реагируют на поведение крупных птиц. Если
большие группы птиц одновременно срываются со скалы и устремляются в море,
то часть котиков панически бросается в воду, другая часть, подняв голову, пы-
тается выяснить причину тревоги.
Во взаимоотношениях котиков с лисицами и песцами тоже не наблюдается
антагонизм. Лишь самки, когда песцы пытаются похитить щенка, делают по-
пытку отогнать хищника. Другие котики не реагируют на присутствие песцов
и лисиц на лежбище.
К настоящим тюленям, иногда залегающим непосредственно вместе с коти-
ками, последние относятся с осторожностью. При непосредственных контактах
обыкновенные тюлени и ларги угрожающе рычат на котиков, вынуждая их от-
ступить. Драки между этими видами не происходят.
Больших залежек каланов на лежбищах котиков не бывает. Лишь отдельные
экземпляры или небольшие группы каланов могут выходить на края залежек
котиков. Оба вида не проявляют какой-либо настороженности друг к другу, на-
ходясь рядом на берегу или в воде.
На о. Сан-Мигель котики постоянно контактируют с калифорнийскими мор-
скими львами. Самки калифорнийских морских львов не проявляют агрессив-
ности к котикам. Адекватными были отношения котиков к самкам и щенкам
морских львов. Если территории секачей обоих видов располагались рядом, то
чаще наблюдались лишь демонстрации агрессивных отношений, при которых
морские львы отступали. При редких драках более крупные самцы морских
львов также уступали секачам котиков. Калифорнийские морские львы не пре-
231
пятствовали проходу самок морских котиков на свои территории. Нередки слу-
чаи залегания самок котиков около морских львов [Peterson et al., 1969].
Более сложные взаимоотношения на берегу складываются между котиками
и сивучами.
Сивучей часто относили к территориальным и пищевым конкурентам коти-
ков, и поэтому считали нежелательным присутствие сивучей на лежбищах ко-
тиков и даже призывали к их истреблению [Мараков, Нестеров, 1958; Белкин,
1966]. Как отмечают большинство исследователей, при залегании сивучей-холос-
тяков на гаремном лежбище котиков прямой антагонизм между ними не наблю-
дается [Мараков, Нестеров, 1958; Нестеров, 1964; Белкин, 1966], хотя такие
ситуации редки из-за того, что котики-секачи яростно защищают свои терри-
тории и вытесняют сивучей. Тем не менее, иногда залегающие у уреза воды си-
вучи-холостяки не препятствуют уходящим в море или выходящих из воды
котикам. Котики пробираются вглубь территории прямо по лежащим сивучам.
При этом следует особо отметить, что сивучи всех возрастных групп лояльно
относятся к самкам котикам, никогда не преследуют и не кусают их. К щенкам
котиков сивучи относятся не агрессивно, а молодые сивучи даже предпринимают
попытки завязать с ними игру.
При организации гаремных залежек сивучей среди залежек котиков нередки
конфликтные ситуации. Однако гаремы котиков после их разрушения сивучами
быстро восстанавливаются.
Сивучи более крупные и сильные звери, чем котики, и в соответствующих
ситуациях легко могут оказать сопротивление. Однако такие случаи редки. При
прямых контактах сивучей и котиков у обоих видов выработались соответст-
вующие поведенческие реакции как бы предупреждающие друг друга о пред-
стоящих намерениях. Так, например, при продвижении самца сивуча по лежбищу
котиков он всегда движется медленно, производя кивающие движения головой
и издавая короткие предупреждающие рявкающие звуки. Располагающиеся на
пути сивуча котики уступают ему дорогу, но вслед за ним снова занимают об-
любованные места.
Соответствующие поведенческие реакции проявляют и самцы котиков при
передвижении через залегающих сивучей. Котики-секачи стараются передвигать-
ся к намеченному месту быстро и по кратчайшему пути. Они не издают никаких
звуков и часть пути могут пройти незамеченными, хотя и идут прямо по телам
сивучей. В случае агрессивных проявлений со стороны последних котики лишь
огрызаются на их выпады. Голова котика при продвижении сквозь строй сивучей
несколько опущена, и создается впечатление, что на его морду накинута уздечка
с короткими поводьями. В редких случаях котику-секачу не удается прорваться
сквозь заслон сивучей на место облюбованной им залежки.
Отношение к человеку-наблюдателю. Оборонительный рефлекс у новорож-
денных котиков появляется спустя несколько часов после рождения, вслед за
рефлексом отряхивания головы и сосания [Соболевский, Кузин, 1977]. Он вы-
ражается угрожающим хорканьем и открыванием рта. При непосредственном
контактировании с человеком щенок кусается. При укусах щенок обычно хватает
“мертвой хваткой” и трясет головой, как это делают взрослые особи при драках.
232
Обычно щенки проявляют беспокойство при появлении среди них человека,
однако если прекратить движение, то щенки перестают обращать на него вни-
мание и подходят вплотную. Щенок при этом может спать на лежащем наблю-
дателе. Взятый на руки щенок лишь сначала обороняется и пытается укусить,
но вскоре успокаивается, и если не совершать резких движений, то он ведет себя
как ручное животное. Побывавший на руках человека щенок, отойдя на шаг-
другой, укладывается спать. От незнакомого крупного движущегося предмета
щенок уходит, а с мелким пытается играть [Крушинская, Дмитриева, 1975].
Наиболее пугливыми и осторожными являются холостяки. При появлении
человека они сразу же бросаются в бегство. Безусловно, большое значение имеет
выраженная стадность этой возрастной группы. Обычно один побежавший хо-
лостяк может увлечь за собой многочисленную группу одновозрастных живот-
ных, залегающих на значительном участке лежбища.
Были предприняты опыты по выявлению влияния регулярного появления че-
ловека на холостяковых залежках. С этой целью спугивали всех находящихся
на залежках холостяков в течение трех дней [Челноков, 1971а]. Наблюдения по-
казали, что после первого дня на залежку вышли 87% от числа согнанных с
лежбища накануне. На соседних участках численность холостяков увеличива-
лась, причем из числа помеченных наибольшее число вышло на более удаленные
участки. После вторичного спугивания количество холостяков на опытном участке
значительно сократилось, через день вернулась половина от первоначального
числа. В последующие дни спугивание не производилось, но до конца промыс-
лового сезона (август) численность холостяков на этом участке изменилась не-
значительно. Можно с уверенностью сказать, что осторожное отношение холос-
тяков к человеку не меняется в течение сезона их пребывания на лежбищах.
Иначе ведут себя по отношению к человеку взрослые котики. Секачи до при-
хода самок на лежбище легко оставляют территорию при приближении человека
[Kenyon, 1960]. С приходом самок агрессивность самцов возрастает, и секачи
яростно защищают территорию и даже преследуют человека в пределах своих
участков. В указанной работе описывается случай, когда гаремный секач бро-
сился на грузовик, заехавший на его территорию. В конце сезона размножения
секачи легко покидают места своих залежек при появлении рядом с ними че-
ловека, однако панического бегства они никогда не предпринимают. Секачи,
залегающие на холостяковых залежках, ведут себя, как холостяки.
Недавно родившие самки агрессивны и при приближении человека яростно
защищают территорию и себя. Самки, у которых щенки уже подросли, менее
агрессивны и не проявляют оборонительных реакций.
К постоянной деятельности человека, не направленной на вмешательство в
жизнь сообщества котиков, животные привыкают и безразлично относятся к по-
явлению людей вблизи них. На о-вах Прибылова наблюдатели ежедневно ходят
по открытой эстакаде, проходящей непосредственно над лежбищем, и животных
это не тревожит. Не реагируют котики и на появление человека в окнах наблю-
дательных вышек.
В прошлом представление об отношении котиков к наблюдателю было иным.
Многие действия, которые теперь считаются дозволенными, ранее запрещались
[Суворов, 1912].
233
Рис. 13.5. Активность морских котиков в
Охотском море [из: Yoshida, 1972]
Рис. 13.6. Активность северных морских
котиков в Японском море
Интересные наблюдения за котиками по отношению к человеку, находяще-
муся в воде, описаны Ф. Г. Челноковым [19706]. Котики подплывали к человеку,
плывущему с дыхательной трубкой под поверхностью воды, на расстояние до
7—10 м. С любопытством вытягивали шеи, пытаясь разглядеть незнакомое су-
щество, а потом быстро отплывали. Под водой они стремительно, на расстоянии
2—3 м от человека, резко, почти под прямым углом, отворачивали в сторону.
Первые 10—15 мин вокруг подводника кружился настоящий хоровод. Причем,
если людей было двое, и один их них находился на поверхности воды, а другой
с аквалангом на глубине, то все внимание котиков было обращено ко второму.
Никаких признаков агрессии к человеку ни самки, ни молодые самцы котиков
не проявляли.
13.3. Ритмы активности и некоторые аспекты поведении северных морских
котиков на береговых залежках и в море
В публикациях обычно указывается, что большую часть времени холостяки
и детеныши проводят в играх, самки предпочитают спать, а секачи заняты ох-
раной гаремов и ухаживанием за самками. Точных сведений по бюджету времени
никто из исследователей не приводил.
В недавнее время осуществлены работы по визуальному наблюдению за от-
дельными особями, имеющими яркие природные метки (раны, посторонние
предметы на теле, окрашенные быстросохнущими красками, или радиомеченые
особи) [Челноков, 1978; Челноков, Лыскин, 1978; Baba et al., 1979; Gentry, Joh-
234
ВРЕМЯ СУТОК В ЧАСАХ
Рис. 13.7. Активность самок и самцов морских котиков у тихоокеанских берегов
Японии [из: Ichihara, Yoshida, 1972]
Рис. 13.8. Половые различия в
активности котиков в море [из:
Ichihara, Yoshida, 1972]
ВРЕМЯ СУТОК В ЧАСАХ
nson, 1976, 1978]. Исследования
морского периода жизни котиков до сих пор
ограничиваются подсчетом числа плавающих и спящих животных в течение от-
дельных часов суток.
Суточная активность в море. Обобщенные многолетние данные по активнос-
ти морских котиков в Охотском море представлены на рис. 13.5. Наибольшее число
спящих животных встречено в утренние часы и вечером, бодрствующих — в
вечернее время. По данным А. Т. Ащепкова и Н. Е. Колотилина [1978], из 2195
встреченных котиков в Охотском море 655 (29.8%) спали и 1540 (70.2%) были
активны. Активность в ночное время в Охотском море не исследовалась. Такие
данные были получены по наблюдениям за активностью котиков у о-вов При-
былова [Yoshida et al., 1978]. Оказалось, что характер деятельности котиков в
ночное время отличается от дневного большими затратами времени на активное
поведение (табл. 13.1)
На местах зимовки распределение времени животными было несколько иным.
Так, в Японском море с 9 до 18 часов до 40% встреченных котиков спали. Сход-
ная доля спящих отмечена в предрассветные часы и с заходом солнца. Актив-
ность котиков уменьшалась с 6 до 9 и с 15 до 18 часов, (рис. 13.6).
В тихоокеанских водах Японии (рис. 13.7) активность котиков снижалась в
дневное время (с 9 до 18 часов); В предрассветные , часы и после 20 часов все
котики бодрствовали. Активность самок несколько выше, чем самцов (рис. 13.8),
но в целом ритмы активности животных разного пола сходны.
Активность северных морских котиков в районах их летнего и зимнего пре-
бывания различна. Животные более активны в районах их летнего обитания в
дневное время, а на местах зимовки и при миграциях — в ночное.
235
Таблица 13.1
Активность котиков в дневное и ночное время на о-вах Прибылова [по:
Yoshida et al., 1978]
Дневное время Ночное время
Спящие 141 (29.2%) 3 (10.7%)
Играющие 187 (38.8%) 22 (78.6%)
Плавающие 154 (32.0%) 3 (10.7%)
Таблица 13.2
Бюджет времени и некоторые формы поведенческих реакций секачей в
гаремный период жизни [из: Челноков, 1978]
Формы поведения секачей Даты наблюдений
17—18 июня 29—30 июня 14—15 июля 22—23 июля
% к длит е л ь и о с т 1 1 н а б л ю дений
Лежание 69.1 42.4 50.0 44.2
Сидение 29.5 47.1 45.6 46.1
Передвижение в пределах участка 0.5 2.9 3.7 0.2
Передвижение на мелководье 0.9 2.8 0.2 4.5
Уход за пределы участка - 4.8 0.5 5.0
Число случа е в на 1 секача за с у т к
Броски на самок, 0.23 2.58 12.8 14.5
в т. ч. с укусами - 0.66 1.4 1.9
Броски на секачей, 13.7 33.6 26.2 15.9
в т. ч. драки 2.2 3.0 3.7 1.6
Спаривания - 0.5 0.3 0.6
Длительность спаривания (мин) - 7.3 8.1 8.2
Наличие пищи в желудках котиков, добытых в разное время суток, хорошо
согласуется с представлениями об активности. В Тихом океане наибольшее число
котиков с наполненными желудками встречается днем, в то время как в Япон-
ском море частота встречаемости таковых в дневное время меньше. В Охотском
море ок. 70% котиков, у которых в желудках была пища, отмечалось до 9 час,
днем их число было меньше, а в вечерние часы вновь увеличивалось.
Сравнение графиков суточной активности котиков с данными наполнения же-
лудков пищей позволяет вполне определенно говорить о том, что активность
котиков в море полностью определяется их режимом питания. Поскольку в раз-
ных районах обитания котиков видовой состав пищевых объектов разный, ак-
тивность котиков подчинена их жизненным циклам. В Тихом океане в питании
котиков присутствует много форм морских организмов, поднимающихся к по-
236
Рис. 13.9. Активность секачей на лежбище Адамс-Ков, Калифорния
[по: Fiscus et al., 1977]
верхности воды ночью (например, миктофиды), а в Японском море котики пи-
таются в основном рыбами, которым не свойственны суточные перемещения по
глубине (например, минтай).
Активность на береговых залежках. Немаловажную роль в активности ко-
тиков на лежбище играют экзогенные факторы, такие как погодные условия,
солнечная радиация, время суток и сезон года [Fiscus et al., 1977; Челноков, 1978;
Gentry et al., 1979].
В начале гаремного периода на островах до прихода самок секачи малоак-
тивны. Более двух третей суток они лежат, а остальное время проводят сидя
(табл. 13.2). Минимальное количество времени тратится секачом на передвиже-
ние в пределах своего участка и на пребывание в мелководье. С приходом самок
на лежбище в конце июня значительно сокращается время на лежание, и пре-
обладающей становиться поза сидения. Почти в 6 раз возрастают затраты вре-
мени на передвижения, связанные с охраной гаремов, ухаживанием за самками,
на драки с соседними секачами. Вместе с тем, секачи чаще, чем в начале периода
размножения, спускаются на мелководье [Челноков, 1978].
В разгар гаремной жизни у секачей еще больше времени уходит на передви-
жение, связанное в основном с активизацией полового поведения. Они почти
не уходят со своих участков.
В третьей декаде июля, с приходом молодых самок на лежбище, усиливается
внимание к ним, но двигательная активность при этом не увеличивается. В это
время самцы чаще, чем прежде, уходят на мелководье. Количество и продол-
жительность спариваний увеличивается (табл. 13.2).
Активность секачей в течение суток исследовалась на лежбище Адамс-Ков (юг
Калифорнии). По данным американских ученых [Fiscus et al., 1977], здесь двига-
тельная активность секачей увеличивается с наступлением сумерек (рис. 13.9) и до-
стигает пика в 21 час, после чего снижается к 2 часам ночи. Затем вновь воз-
растает в ранние утренние часы и резко снижается с 7 утра до 12 дня. Изменение
ритмов активности на данном лежбище в основном сопряжено с темпера-
турными условиями этого самого южного лежбища котиков [Fiscus et al.,
1977]. С представленными данными (рис. 13.9) хорошо согласуются ритмы ре-
237
б в /о <? /4 л го гг м t о
время суток в чмкх
Рис, 13.10. Половая активность секачей в течение суток на лежбище Адамс-Ков,
Калифорния [из: Fiscus et al., 1977]
продуктивной активности (рис. 13.10). Как и в первом случае, наибольшее число
копуляций наблюдалось в ночное время с пиком в 22 часа. Повторное увеличение
спариваний было в предутренние и утренние часы. По данным В. М. Когая [1978],
оптимальной температурой на лежбище, когда наиболее ярко проявляются все по-
веденческие реакции, связанные с размножением, является +5.0—8.9°С. По мнению
американских ученых [Bartholomew, Wilke, 1953; Fiscus et al., 1977], +12°C уже яв-
ляется критической температурой, вызывающей стрессовые реакции у секачей.
Изучение ритмов активности самок на берегу было проведено отечествен-
ными и японскими учеными с помощью регистрирующей аппаратуры и радио-
меток, поставленных на животных. Было установлено, что большинство самок
возвращается на берег с 20 до 8 часов, когда температура воздуха имеет ми-
нимальные суточные показатели. Активность самок на берегу складывается из
поиска убежища от жары и периода схода в воду для охлаждения тела. Других
типов двигательной активности у самок мало. В среднем за день период актив-
ности составляет 105 мин, с пиком от 12 до 14 часов. В зто время большинство
самок сходят в воду. Средняя частота схода в воду за день составляет 2.4 раза,
а пребывание в воде — 246 мин. Однако между 2 и 5 часами, 21 и 22 часами
не было случаев схода в воду. Комфортной для самок считается температура
воздуха +6—9°С [Когай, 1978].
Активность холостяков в период пребывания их на берегу изучалась с по-
мощью радиоаппаратуры и визуальных наблюдений [Gentry et al., 1979; Baba
et al., 1979]. Данные регистрации дневной активности трех холостяков показаны
на рис. 13.11, где ломаной линией показана их двигательная активность, пунк-
тирной — отсутствие радиосвязи. Режим дня холостяков складывается из пе-
риодов активности, продолжающихся 1—2 часа, и 2—3-часовых отдыхов. Почти
все наблюдаемые животные были наиболее активны с 16 до 19, с 21 до 23, с 0
до 2 и с 5 до 7 часов (рис. 13.11, 13.12).
Среди поведенческих актов у холостяков в период активности превалируют
игры (“турнирные бои”). Передвижения по лежбищу, как правило, занимают мало
времени; чаще они сочетаются с играми. С 13 час 23 мин 14 июля до 23 час 05 мин
238
-W-V-
-y~Y____
КОТИК U2i
-----v-----
КОТИК /720
з-в; os‘ii
4 —
—w V---------w-------
--w------------------
— котики2
&fi;0Q:59 t , ,
2 4 6 d
1S‘7;i5l55
AWW—V
в-г; iO- 00
20-7;i8<07
-w------u-v
AV—W-
21'7; 20:00
i6 id 20 22
ВРЕМЯ СУТОК в KKCKX
Ю 12 Л
4WV
a* v W U
Рис. 13.11. Активность холостяков, помеченных радиометками на лежбище
[из: Baba et al., 1979].
Объяснения в тексте.
Рис. 13.12. Активность холостяков на лежбище [из: Baba et al., 1979].
Объяснения в тексте.
15 июля холостяк прошел путь равный 140 м, в среднем по нескольким наблю-
дениям путь по прямой не превысил 150 м, а общая его длина — 400 м [Baba
et al., 1979]. Все же холостяки, как и другие возрастные группы котиков, наиболее
активны ночью.
В возрасте до 2 недель новорожденные котики малоактивны. Более 50% вре-
мени они спят или передвигаются в радиусе, не превышающем 7—12 м около
места рождения [Крушинская, Дмитриева, 1975]. В этом возрасте они неохотно
вступают в контакт со сверстниками или родившимися ранее особями. В при-
сутствии матери (в перерывах между сном) через каждые 10—15 мин щенки кор-
мятся. К 2-недельному возрасту у детенышей вырабатываются характерные для
вида поведенческие реакции, и активность заметно возрастает. Все больше вре-
мени они проводят в обществе сверстников в играх и борьбе друг с другом.
В 3-недельном возрасте они могут удаляться от места рождения до 100 м и спус-
каться к урезу воды, но в воду идут неохотно. Наиболее активны в утренние
и вечерние часы. Комфортна температура воздуха +6—1ГС [Когай, 1978].
239
В холодную ветреную погоду новорожденные собираются в тесные группы
и больше проводят времени во сне, а в жаркую погоду перемещаются в защи-
щенные от солнца места, где тоже подолгу спят.
В месячном возрасте щенки начинают сходить в воду и плескаться на мел-
ководье, но после приема “охлаждающих ванн” возвращаются на лежбище и,
сгрудившись, спят [Крушинская, Дмитриева, 1975]. В более старшем возрасте
щенки все больше времени проводят в воде и играют. Они начинают разбре-
даться по лежбищу и могут быть встречены на значительном расстоянии от
места рождения.
Периодичность пребывания котиков на берегу и в море. Хорошо известно, что
в период пребывания на берегу котики периодически сходят в воду и уплывают
в море для кормежки или для охлаждения тела. В других случаях причиной схода
в воду могут быть метеоусловия. В дождь почти все взрослые котики покидают
острова. В жаркую солнечную погоду большая часть животных тоже находится
в воде. Замечено, что эти особенности поведения меняются в зависимости от
сезона.
В среднем считается, что секачи находятся на лежбище не менее месяца, с
колебаниями от 18 до 41 дня [Bartholomew, Hoell, 1953]. По данным Ф. Г. Чел-
нокова и Н. Н. Лыскина [1978], секачи в среднем остаются на берегу несколько
меньше времени — 15.4 дня, после чего уходят в море на 2—5 дней.
Взрослые самки проводят на берегу после родов до 6.6 (2.3—10) суток, а затем
уходят в море на 8—10 суток [Бычков, 1964а]. По данным радиопрослеживания
за большой промежуток времени, самки со щенками остаются на берегу в сред-
нем 2.1 дня, а в море — 4.7 дня, хотя те, у которых погибли щенки, проводят
на берегу в среднем 6.2 дней, а в море — 11.2 [Yoshida, 1982].
Холостяки остается в море от 1 до 39 дней, между днями, проведенными
на берегу [Gentry, Johnson, 1976]. Имеются близкие к приведенным данные, по
которым холостяки остаются в море от 1 до 34 дней [Baba et al., 1979]. В пределах
указанного времени — 2 цикла перемещений котиков по маршруту “суша-во-
да”. Один длится 1—2, другой — 11—34 дня, а в среднем, соответственно, 31.8%
и 68.2% от общего числа наблюдений. По другим данным, это соотношение рав-
но 20% и 65% [Gentry, Johnson, 1975]. В среднем, долгопериодные пребывания
в море составили 25.6 дня (11—34 дня). Короткопериодные пребывания в море
связывают с температурными условиями, осадками или антропогенным вмеша-
тельством, долгопериодные — с питанием.
По-видимому, показатели суточного бюджета времени отдельных возрастных
групп котиков на разных залежках будут не совпадать, т. к. их районы питания
находятся на разном удалении от лежбищ. Имеются, пусть и не очень заметные,
различия и в гидроклиматических показателях мест обитания. Поэтому время,
затрачиваемое животными на перемещения к районам питания и их реакции на
изменяющиеся условия среды будут не одинаковыми в разных районах.
13.4. Скорость плавания и глубина ныряния
Северные морские котики прекрасные пловцы. Они предпринимают самые
длинные миграции из всех ластоногих протяженностью до 10 тыс. км. Испу-
240
Таблица 13.3
Наблюдения за частотой и глубиной ныряния самок северного морского
котика [по: Kooyman et al., 1976]
Масса тела, кг Время, час Число ныряний на определенную глубину (м) Всего ныряний
0—20' 21—50 51—80 81—110 111—140 141—170 171—200
35 167 301 314 37 9 13 2 - 676
35 92 1478 1 10 9 49 7 2 1224
35 200 54 478 89 1 4 - - 626
50 149 201 200 200 14 7 6 3 431
ганный котик развивает скорость до 15 узлов [Scheffer, 1950b]. Однако при до-
лгом преследовании скорость значительно ниже, и идущий со скоростью 8 узлов
бот догоняет животное.
О глубине ныряния долгое время судили по косвенным показателям [Kenyon,
1952], и поэтому котик считался плохим ныряльщиком, способным осваивать
глубины до 55—73 м. Позже японским исследователям удалось с помощью эхо-
лота определить траекторию и глубину ныряния свободноплавающего котика
[Ichihara, Yoshida, 1972]. Оказалось, что, питаясь, он может нырять на глубину
до 100 м.
Более обширные инструментальные исследования поведения котиков в море
с определением глубины, времени и частоты ныряний были проведены амери-
канскими учеными [Kooyman et al., 1976]. С помощью регистрирующих устройств,
закрепленных на 4 самках котиков в течение 608 часов (табл. 13.3), выяснено,
что наибольшая глубина погружения составила 190 м. Средняя скорость погру-
жения 70 м/мин. Наибольшее число ныряний сделано на глубину 20 м, длившихся
не более 1 мин. Ныряние на глубину от 20 до 140 м обычно было связано, по
мнению исследователей, с питанием. Такие погружения длились 2—5 мин и
обычно происходили сериями. Например, серия ныряний одной из самок, со-
стоящая из 13 погружений на глубину НО—140 м, была выполнена за 3.6 часа.
Каждое погружение длилось 3.3—3.4 мин с интервалами между ныряниями от
6 до 30 мин.
241
Глава 14. РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Распространение северных морских котиков в Беринговом море и в северной
части Тихого океана было известно уже в конце прошлого столетия. На картах,
составленных К. Таунсендом [Townsend, 1899], показаны места судового про-
мысла и количество добытых котиков в Тихом океане от 35° до 60° с. ш. От-
сутствовавшие в материалах К. Таунсенда сведения по распространению котиков
в Японском и Охотском морях вскоре были представлены Н. Е. Слюниным [1895]
и М. Коганемару [Koganemaru, 1903а, Ь]. В эти же годы появилась гипотеза о
путях миграций котиков с мест размножения на места зимовки [Слюнин, 1895],
которая в последствии дополнилась и детализировалась [Бойцов, 1934; Огнев,
1935; Ильина 1950 и др.].
Большое значение для наиболее полного изучения распространения, распре-
деления и миграций котиков имели исследования, предшествующие подписанию
Международной Конвенции по сохранению котиков северной части Тихого океана
в 1957 г. (NPFSC), а затем работы по массовому мечению животных на местах
размножения, проводимые в рамках NPFSC.
14.1. Распространение
Охотское море. Основная масса котиков в течение летне-осеннего сезона за-
легает на о. Тюлений. “Кормовые поля” в Охотском море располагаются не да-
лее чем в 120 милях от острова. Именно в этих пределах концентрируются ко-
тики в летнее время. Предпочитаемой в обозначенных границах является ак-
ватория юго-восточнее Тюленьего (рис. 14.1). Все встречи с котиками в других
частях Охотского моря (севернее 50° с. ш.) можно отнести к случайным заходам,
не характеризующим ареал. Так, по сообщению А. Державина [1929], котики
в 1923—29 гг. неоднократно заходили в заливы Луньский, Ныйский, Байкал,
Виахту. 20 августа на побережье Сахалина близ сел. Москальво была зарегист-
рирована самка котика. По сообщению этого же автора, котиков встречали у
Советской Гавани и у мыса Литке. В 1958 г. Э. А. Тихомиров [1964] зарегист-
рировал котиков 13 сентября в Охотском море к востоку от о. Сахалин и на
севере моря.
По-видимому, опросные данные о случайных заходах котиков на север от
Сахалина и в Татарский прол, явились основанием для предположения о су-
ществовании в прошлом лежбищ котиков в этих местах [Слюнин, 1895].
В период осенней миграции на зимовку в Японское море и Тихий океан (ок-
тябрь—ноябрь) котиков можно встретить практически на всей акватории Охот-
ского моря южнее о. Тюлений. Самое позднее время обнаружения их на Тю-
леньем — 5 декабря [А. М. Николаев, личн. сообщ]
Возвращающиеся с мест зимнего пребывания первые котики появляются в
Охотском море не ранее первых чисел мая. По наблюдениям Е. И. Евдокимова
[1964], котики,10 мая были встречены у прол. Лаперуза со стороны Японского
моря. Со стороны Тихого океана, где в отдельные годы ледовые условия бывают
242
Рис., 14.1. Распространение и
плотность котиков в Охотском море
в июне—ноябре:
1 — до 10 особей, 2 — 11—20 особей, 3 —
21—50 особей, 4 — 51—100 особей, 5 — 100
особей и более.
Рис. 14.2. Распространение и плотность котиков в западной части Берингова моря
в июне—октябре.
Обозначения см. на рис. 14.1.
менее тяжелыми, чем в прол. Лаперуза, котики проникают в Охотское море рань-
ше, чем из Японского моря. Однако раньше первой декады мая они на о. Тю-
лений никогда не встречались.
Берингово море. В летний период котики здесь концентрируются вокруг ос-
тровов Командорских и Прибылова, и большинство из них не удаляются более
чем на 120—150 миль от них (рис. 14.2). Однако неполовозрелые животные могут
243
Рис. 14.3. Распространение и плотность котиков в Японском море в январе
(1958—76 гг.).
Обозначения см. на рис. 14.1.
Рис. 14.4. Распространение и плотность котиков в Японском море в феврале.
Обозначения см. на рис. 14.1.
встречаться и дальше от островов размножения, вплоть до Алеутских. В этот
же период возможен значительный обмен животными, размножающимися на ост-
ровах Прибылова и Командорских. Вместе с тем, отдельные особи были встре-
чены на значительном удалении от островов размножения. Имеются сообщения
о встречах котиков у мыса Наварин, в Анадырском зал., в Беринговом прол. [Бер-
зин, 1959]. Д. И. Чугунков и В. Г. Прохоров [1964] встречали их в ноябре—декабре
в Олюторском зал. при отрицательной температуре воды и воздуха. Северной гра-
ницей обычного распространения является 60° с. ш.; все встречи северных мор-
ских котиков в более высоких широтах следует относить к случайным.
Японское море. В январе котики концентрируются в центральной части Япон-
ского моря на банке Ямато (ок. 40° с. ш., 134—145° в. д.) и на участке, при-
лежащем к Восточно-Корейскому зал., а также в самом заливе [Панин, Панина,
1968]. М. К. Маминов [личн. сообщ.] встречал несколько мелких групп котиков
у юго-западных берегов о. Хоккайдо. В целом, в связи с непродолжительностью
244
Рис. 14.5. Распрост-
ранение и плотность
котиков в Японском
море в марте.
Обозначения см. на рис. 14.1.
131 133 135
130 132 134 136 138 140
Рис. 14.6. Распространение и плотность котиков в Японском море в апреле.
Обозначения см. на рис. 14.1.
исследований в этом месяце, картина распространения и распределения в Япон-
ском море недостаточно полна (рис. 14.3).
В феврале котики в изобилии встречаются на банке Кито-Ямато, а также
у берегов Кореи на юг до 37° с. ш. Единичные животные встречаются и в более
северных частях моря (рис. 14.4).
В марте котики в Японском море встречаются практически по всей аква-
тории от Восточно-Корейского зал. и на восток до 136° с. ш.; на юг не идут
далее 38° с. ш. На этой обширной акватории концентрации животных отмеча-
ются в районе банки Ямато и в северной части Восточно-Корейского зал. (рис.
14.5).
В апреле котики практически не встречаются южнее 39° с. ш. Скопления жи-
вотных сохраняются в районе банки Ямато. В западной части моря животные
245
<32 <34 <33 <33 <40
Рис. 14.7. Распространение и плотность котиков в Японском море в мае.
Обозначения см. на рис. 14.1.
Рис. 14.8. Распространение
и плотность котиков в тихо-
океанских водах Японии в
декабре.
Обозначения см. на рис. 14.1.
42
41
40
39
<42 U3 144
145 <43
а
В - »
EI - 2
в - 3
широко рассредоточены в полосе вдоль северных берегов п-ова Корея (рис. 14.6).
Отдельные особи отмечены вблизи Сангарского прол., что говорит о возмож-
ности миграции небольшой части котиков, зимующих в Японском море, этим
246
«1 Ш IU ш м
Рис. 14.9. Распространение и плотность
котиков в тихоокеанских водах Японии
в январе.
Обозначения см. на рис. 14.1.
Н2 Hl «4 <U
V
«О
я
М
я
Рис. 14.10. Распространение и плотность
котиков в тихоокеанских водах Японии
в феврале.
Обозначения см. на рис. 14.1.
путем в Тихий океан. Однако основным направлением передвижения котиков
весной является северное, тяготеющее к побережью Приморья.
В мае большая часть котиков покидает южные районы зимовки и встречается
вдоль всего приморского берега. В этом месяце котиков регистрировали прак-
тически на всей акватории Японского моря севернее 40° с. ш. (рис. 14.7).
В июне основная часть котиков покидает Японское море и через прол. Ла-
перуза входит в Охотское море. Небольшая часть может еще находиться на путях
миграции от берегов Приморья до о. Сахалин. С июля по ноябрь котиков в
Японском море нет.
В целом, анализ данных по распространению котиков в Японском море по-
казывает, что животные осваивают лишь его западную и центральную части.
Восточная и южная границы распространения достаточно четко обозначены по-
ложением фронтальной зоны встречных холодного Приморского и теплого Цусим-
ского течений [Панин, Панина, 1968]. Наибольшие концентрации котиков (банки
Ямато и Кито-Ямато, акватория Восточно-Корейского зал. и прилегающие к
нему воды) отмечаются в наиболее продуктивных частях моря — зонах смешения
вод разного происхождения.
Тихоокеанские воды Японии. В северо-западной части Тихого океана, восточ-
нее и юго-восточнее о. Хоккайдо, котики появляются после окончания сезона
размножения в ноябре. В этом месяце район их обитания ограничен, и котики
не образуют значительных скоплений [Kenyon, Wilke, 1953; Teylor et al., 1955;
Wada, 1971; Арсеньев, 1971a; Кузин, Панина, 1977; Ащепков, Кузин, 1986а, б, 1987].
В последующие месяцы наблюдается увеличение их численности у побережья
Японии и постепенное их продвижение к югу.
247
IA2 Ю Ш W М
Рис. 14.11. Распространение и плотность
котиков в тихоокеанских водах Японии
в марте.
Обозначения см. на рис. 14.1.
в апреле.
Обозначения см. на рис. 14.1.
В декабре скопления котиков возникают юго-восточнее о. Хоккайдо, восточ-
нее и юго-восточнее мыса Эримо. Часть животных узким клином, придерживаясь
охлажденных течений, внедряются в прибрежные воды о. Хонсю, проникая к
югу до 40° с. ш. Отдельные особи (в иные годы даже небольшие группы) могут
проникать до 38° с. ш. и встречаться на значительной акватории восточнее Япон-
ских о-вов до 147° в. д. (рис. 14.8).
В январе котики в значительном количестве встречаются южнее о. Хоккайдо;
все большее их число перемещается ближе к о. Хонсю, вдоль которого они про-
никают до 37° с. ш. (рис. 14.9).
В феврале численность котиков севернее 40° с. ш. заметно сокращается. Ос-
новная масса их сосредоточивается в координатах 36—39° с. ш. на удалении
100—120 миль от о. Хонсю. Особенно большие концентрации отмечаются на
траверзе префектур Мияги и Фукусима (рис. 14.10).
В марте котики наиболее широко рассредоточены по акватории северо-за-
падной части Тихого океана от о. Хоккайдо до 35° с. ш. широкой полосой с
удалением от о. Хонсю до 150 миль. Наибольшая встречаемость отмечается от
36° до 38° с. ш. (рис. 14.11).
В апреле, в связи с началом миграции котиков к островам для размножения,
заметно сокращается район их обитания в южной части ареала. Животные редко
248
Рис. 14.13. Распространение и плотность котиков в тихоокеанских водах Японии в мае.
Обозначения см. на рис. 14.1.
встречаются южнее 37° с. ш. Большие скопления их регистрируются у северной
части префектуры Мияги и на траверзе префектуры Ивате (рис. 14.12).
В мае наблюдается дальнейшее передвижение котиков к северу. Заметно уве-
личивается численность их восточнее и юго-восточнее мыса Эримо (о. Хоккай-
до) — в местах, где их наблюдали в декабре во время осенних миграций на юг.
В целом, район обитания котиков в мае ограничен параллелями 38—43° с. ш.,
в пределах которого наибольшие концентрации животных наблюдаются напро-
тив префектур Ивате и Аоморе (рис. 14.13).
Южной границей обитания котиков в июне остаются те же широты, что и
в мае, а северная достигает Курильских о-вов и проникает в Охотское море через
проливы Екатерины, Фриза, Уруп и Буссоль. В иные годы котики удаляются
от Японских о-вов до 164° в. д. Встречаются котики и за пределами этой ак-
ватории, но скоплений не образуют, и проследить их дальнейшее передвижение
к островам размножения не представляется возможным (рис. 14.14).
В июле котики уходят из Тихого океана, за исключением первогодков и мо-
лодых. Незначительное их число отмечено к востоку от о. Хоккайдо у порядков
сетей, выставляемых для добычи лососевых [Report..., 1963, 1964].
В августе котики очень редки в этом районе. К. Кеньон и Ф. Уилки [Kenyon,
Wilke, 1953] отмечали небольшое число животных в акватории префектуры Ивате.
В последующие месяцы, вплоть до начала осенних миграций, котики не встре-
чаются в тихоокеанских водах Японии.
249
№ JU W « <W . W
Рис. 14.14. Распространение и
плотность котиков в тихоокеан-
ских берегов Японии в июне.
Обозначения см. на рис. 14.1
Рис. 14.15. Встречаемость
котиков в открытой части
Тихого океана по литератур-
ным данным:
цифры при точках — число
встреченных особей.
Описанные границы помесячного распространения котиков у тихоокеанских
берегов Японии не четко обозначены. Существует межгодовая изменчивость как
в темпах продвижения животных из района в район, так и в границах распро-
странения на юг и восток в зависимости от гидрометеоусловий года.
Открытая часть Тихого океана. На картах К. Таунсенда [Townsend, 1898]
одиночные встречи с котиками показаны практически на всей акватории Тихого
океана севернее 40° с. ш. от берегов Японии до берегов Сев. Америки. Наиболее
частыми они были в координатах 42—44° с. ш. и 175° в. д.—175° з. д. В ап-
реле—июне 1955—59 гг. И. Б. Бирман [1966] встретил 13 котиков в открытой
части Тихого океана на траверзе Сангарского прол, примерно в тысяче миль
от Японских о-вов. В 1966 г. А. В. Ровнин [1968] наблюдал несколько котиков
в этом же районе (рис. 14.15).
Северных морских котиков постоянно регистрирую в районе дрифтерного
лова кальмаров в открытой части Тихого океана (20—46° с. ш. и 145—170° з. д.).
Здесь концентрируются в основном неполовозрелые особи.
250
Рис. 14.16. Ареал и пути миграций северных морских котиков:
1 — лежбища, 2 — основные пути миграций, 3 — возможные маршруты неосновных
миграций, 4 — субарктический фронт, 5 — границы ареала.
Таким образом, анализ литературных данных и материалов, полученных в
результате 25-летних исследований котиков NPFSC, показал, что они являются
обитателями умеренных широт северной части Тихого океана и, как правило,
не заходят в районы полярных и субтропических зон. У тихоокеанских берегов
Японии и в Японском море в зимне-весенний период котики распространяются
на юг до 35° с. ш. У берегов Сев. Америки они встречаются до 32° с. ш. В от-
крытой части Тихого океана южной границей распространения является субар-
ктический фронт. В Охотском море котики не встречаются севернее 50° с. ш.,
а в Беринговом море — выше 60° с. ш. (рис. 14.16).
14.2. Миграции
Места зимовок отдельных популяций северных морских котиков общие, од-
нако пути миграций, количественное соотношение и абсолютная численность
животных разные. Данные возврата меток с мест зимовок позволяют оценить
эти параметры.
Всего на о-вах Прибылова с 1948 по 1982 гг. и с 1957 по 1984 гг. на лежбищах
котиков, принадлежащих бывшему СССР, были помечены металлическими мет-
ками более миллиона котиков, в т. ч. на о-вах Прибылова — 595364, на о. Тюле-
ний — 210015, на Командорских о-вах — 258894 и на Курильских — 29017 осо-
бей. В среднем ежегодно возвращается 9% (3—13%) меченых котиков от числа
добытых в море с научно-исследовательскими целями и 18% (10—22%) из про-
мысловых побоек на островах. Всего с 1958 по 1982 гг. вернулись более 2 тыс.
меток из разных частей Тихого океана, в т. ч. 443 метки из районов зимовки
котиков в его северо-восточной части, 1522 метки из районов зимовки в севе-
ро-западной части и 94 метки из Японского моря.
По данным возврата меток с учетом коэффициентов мечения и потери меток
установлено, что в Японском море зимуют 64.9% котиков с о. Тюлений, 7.3% —
с Командорских о-вов и 26.7% — с о-вов Прибылова от общей численности на-
ходящихся здесь в это время.
В тихоокеанских водах Японии проводят зиму 40.1% котиков с о. Тюлений,
37.2% — с о-вов Прибылова, 21.0% — с Командорских о-вов и 1.7% котиков —
с Курильских от общей численности зимующих здесь.
251
гы с.
Рис. 14.17. Численность котиков разных популяций на совместных местах зимовки у
тихоокеанских берегов Японии (слева) и в Японском море (справа):
1 — котики с о. Тюлений, 2 — с Командорских о-вов, 3 — с о-вов Прибылова, 4 — с
Курильских о-вов.
В восточной части Тихого океана, у берегов Сев. Америки в 1958—72 гг.
из добытых 443 помеченных котиков 10 особей были помечены на Командорских
о-вах, что составляет, с учетом коэффициентов мечения и потери меток, 0.8%
от общей численности зимующих здесь котиков. Меченые звери из других по-
пуляций в акватории Сев. Америки встречены не были.
Таким образом, сравнительно обособленными представляются районы зимов-
ки котиков в Японском море и в восточной части Тихого океана, а наибольшая
смешиваемость котиков из разных популяций наблюдается в северо-западной
части Тихого океана у берегов Японии.
В абсолютных показателях, в среднем, в тихоокеанских водах Сев. Америки
проводят зиму ок. 95% (0.98—1.2 млн.) прибыловских котиков и лишь ок. 5%
(50—73 тыс.) мигрируют в северо-западную часть Тихого океана. В свою оче-
редь, в восточную часть Тихого океана мигрируют на зимовку 5—8 тыс. котиков
с Командор. Распределение котиков в северо-западной части Тихого океана по
районам неравномерное. Количество котиков, зимующих в Японском море, в
среднем, в 5—7 раз меньше, чем в тихоокеанских водах Японии и, в целом, в
первом случае составляет 20—35 тыс., а во втором — 197—220 тыс. Количест-
венное распределение животных разных популяций по районам зимовки заметно
меняется по периодам в связи с изменением общей численности и половозраст-
ного состава на местах размножения (рис. 14.17).
Так, в тихоокеанских водах Японии численность котиков с о. Тюлений по-
следовательно убывала с 93 тыс. в 1960—64 гг. до 38 тыс. в 1980—83 гг. Чис-
ленность командорских котиков за это же время, напротив, увеличилась с 41 тыс.
до 71 тыс., а прибыловских — колебалась от 63 тыс. до 69 тыс. в 1960—79 гг.
с уменьшением к 1980—83 гг. до 49 тыс. (рис. 14.17). Из представленных ма-
териалов следует, что с 1960 по 1972 гг. в тихоокеанских водах Японии пре-
обладали котики с о. Тюлений, а прибыловских котиков было больше, чем ко-
мандорских. В.последующие годы в этом районе зимовки численно преобладали
мигранты с Командорских о-вов, а прибыловских было больше, чем тюленьев-
252
Таблица 14.1
Соотношение (%) половозрастных классов котиков различного происхожде-
ния, мигрирующих в тихоокеанские воды Японии [из: Ащепков, Кузин, 1987]
Возраст, лет Острова
Тюлений Командорские Прибылова Курильские
самки самцы самки самцы самки самцы самки самцы
1 5.1 20.3 2.4 11.8 1.3 2.7 31.9 14.9
2 13.5 44.8 6.9 33.7 5.9 13.2 3.8 54.4
3 18.6 16.3 14.9 37.4 9.6 38.6 1.9 14.9
4 16.9 8.5 17.2 13.4 7.6 30.1 14.0
5 11.5 4.3 13.9 2.6 14.6 7.5 3.8 15.8
6 8.8 5.5 14.8 1.1 21.6 0.8
7 6.6 4.6 9.6 0.6 25.4
8 3.3 6.8 5.6 1.5 16.6
9 3.4 0.3 4.9 5.1 2.6
10 2.9 3.5 1.5
11 2.1 3.5 6.4 5.0
12 1.9 2.2
13 1.2 2.5 0.9
14 0.7 3.3
15 0.9 3.8
16 0.9 3.2
17 1.1 1.3
18 0.2
19 0.6
20 0.2
21 0.2
ских. Курильские котики за все годы исследований составляли небольшую часть
зимующего здесь стада (табл. 14.1).
В Японском море численность котиков с о. Тюлений в течение исследованного
периода изменялась неравномерно. Наибольшее их число отмечалось в 1965—
72 гг. (28 тыс.), наименьшая — в 1980—83 гг. (14 тыс.). Аналогичная тенденция
изменения численности котиков с о-вов Прибылова, где в 1960—64 гг. она со-
ставила 10.8 тыс., в 1965—72 гг. ок. 12 тыс., а к 1980—83 гг. численность сни-
зилась до 6.1 тыс. Численность командорских котиков в 1960—79 гг. оставалась
примерно на одном уровне (2.8—3.2 тыс.); к 1980—83 гг. она снизилась до 1.6 тыс.
(рис. 14.17). Соотношение представителей разных популяций, зимующих в Япон-
ском море, в течение исследованного периода оставалось приблизительно оди-
наковым.
Метить котиков на Курильских о-вах начали только в 1963 г., и с 1960 по
1964 гг. они в добыче не присутствовали. В последующие годы среди курильских
253
Таблица 14.2
Соотношение (%) половозрастных классов котиков разного происхождения,
мигрирующих в Японское море *
Возраст, лет Острова
Тюлений Командорские Прибылова Всего
самцы самки самцы самки самцы самки самцы самки
1 2.56 1.52
2 7.62 4.52
3 31.74 30.35 52.33 5.31 24.55 28.58
4 22.82 4.45 10.39 । 16.99 4.19
5 25.96 18.58 13.46 10.06 19.84 17.50
6 5.90 11.36 22.21 5.18 10.69
7 3.40 5.68 8.73 4.90 5.34
8 4.69 12.0 100.0 10.06 4:22 10.26
9 55.45 18.28
13 4.69 • , 4.41
14 5.05 • 1 4.76
15 4.80 4.52
16 4.69 4.41
18 5.66 5.34
самок встречались животные всех возрастных классов, а среди самцов домини-
ровали молодые особи.
Возрастной состав котиков, мигрирующих на зимовку в Японское море, силь-
но отличался от возрастного состава мигрантов в тихоокеанских водах Японии
(табл. 14.2).
Как видно из таблицы, среди котиков о. Тюлений в течение всего периода
исследований численно преобладали молодые особи. Среди самок с Командорских
о-вов доминировали 3—6-летние животные и старше 10 лет, а среди самцов —
2—3-летние. Совершенно отсутствовали секачи (самцы старше 6 лет). Доминиру-
ющей группой самок с о-вов Прибылова были особи в возрасте 5—6 лет и старше,
а среди самцов — 3—4-летки. Примечательно, что среди прибыловских котиков
в западной части Тихого океана присутствовали несколько самцов в возрасте 7 лет
и старше, чего не наблюдалось среди животных с Командорских о-вов.
Среди самок с о. Тюлений во все годы исследований в Японском море до-
минировали особи старших возрастных классов (10 лет). Совершенно отсутст-
вовали 1—2-летние. В 60-х годах здесь зимовало сравнительно много 4—8-летних
самок, и они по численности в 1.5 раза превосходили самок старше 10 лет. Од-
нако в последующие годы численность средневозрастных групп здесь уменьши-
лась, и они стали составлять всего 30—50% от их числа в возрасте старше 10
лет. Среди мигрантов с о-вов Прибылова самки отсутствовали, а с Командор-
ских о-вов присутствовала одна за все годы исследований.
254
Таблица 14.3
Сезонная динамика соотношения (%) котиков разных стад в тихоокеанских
водах Японии [из: Ащепков, Кузин, 19866]
Месяцы Пол Тюленьевское стадо Командорское стадо Прибыловское стадо Курильское стадо
Декабрь самки 100.0 - - -
самцы 100.0 - - -
Январь самки 61.7 25.9 12.4
самцы 50.0 21.2 - 26.8
Февраль самки 48.3 33.1 13.4 5.2
самцы 51.0 6.5 42.5 -
Март самки 44.1 31.6 22.3 2.0
самцы 28.7 29.4 41.6 0.3
Апрель самки 38.6 12.6 48.3 0.5
самцы 38.8 16.4 44.2 0.6
Май самки 40.7 25.3 33.8 0.2
самцы 27.4 11.6 57.9 3.1
Июнь самки 52.9 23.3 22.9 0.8
самцы 44.1 36.7 18.5 0.7
Июль самки 39.0 61.0 - -
самцы 100.0 - - -
Самцы с Тюленьего были представлены в основном холостяками старшего
возраста, тогда как с островов Прибылова и Командорских доминировала груп-
па секачей и полусекачей.
Репродуктивные центры северных морских котиков удалены от районов сов-
местного зимнего обитания у берегов Японии и в Японском море на разное
расстояние. Поэтому появление животных из разных репродуктивных группи-
ровок здесь отмечается в разное время (табл. 14.3, рис. 14.18, 14.19). О. Тюлений,
например, удален от Японских о-вов на 380 миль. Котики с этого острова появ-
ляются у о. Хоккайдо в декабре, а котики с Командорских о-вов (удаленность
от Японии составляет 860 миль) приплывают сюда в январе, котики же с о-вов
Прибылова (удаленность 2 тыс. миль) отмечаются только в феврале. Соответ-
ственно, период пребывания животных разных популяций в районе зимовки у
берегов Японии находится в обратной зависимости от длины миграционного
пути, а скорость передвижения от мест размножения до места зимовки прямо
зависит от нее. Расчеты показали [Ащепков, Кузин, 1986], что во время миграций
прибыловские котики перемещаются со скоростью 22 мили в сутки, командор-
ские — 14.3 мили, курильские — 12.6 мили, а тюленьевские — 8.6 мили.
В связи с неодновременностью прибытия зверей разных популяций в район
зимовки у берегов Японии, изменяется во времени как общее их количество,
так и соотношение по происхождению (табл. 14.3).
В начальный период зимовки и в конце его преобладают котики с о. Тюлений.
В марте котики всех трех крупных репродуктивных группировок (островов Тю-
255
43
42
41
40
31
31
37
U
)1
м
144
US
из
ui
141
о. хоккайдо
<00
30
40
30
20
inti
so
30
20
10
30
го
ю
40
20
<0
------------<
ИЮНЬ
3S
январь
по
'1111 И Л И л л
ЯНВАРЬ ПО
МАИ
142
40
30
20
Ю
а ХОНСЮ
so
40
40
30
20
10
100
ы>
30
20
<0
ФЕВРАЛЬ
S7T
JUfL.
март
on
АПРЕЛЬ
№32
ЯНВАРЬ
п
Я . в. И
№0
30
20
10
<00
40
20
10
SO
40
30
20
10
1 Л л.. л .. ,IL J Ik JLl Я fl ФЕВРАЛЬ 1 Гм Л . л . . .
1 1 - ( fe Он J rg JU л МАРТ | 1113
п г S 4 -.-Л... 0 0 Л..л.. Ю II 12 АПРЕЛЬ 24S возраст, дет 13 М 13 М 17 п я
Рис. 14.18. Возрастной состав самок северных морских котиков из разных
популяций в тихоокеанских водах Японии. Обозначения — см. рис. 14.17.
256
i44*
M)' иг' из'
U5 ’ М ’
I г J 4 5 6 7 3 9 Ю и 12 13 14 15 16 17 и 19
Рис. 14.18 (окончание). Обозначения — см. рис. 14.17.
леньего, Прибылова, Командорских) представлены примерно одинаковым ко-
личеством. Численность прибыловских котиков максимальна в апреле—мае. В по-
следующие месяцы их численность, также как и командорских, убывает в связи
с более ранним уходом из района зимовки.
Продвижение по миграционному маршруту в пределах ареала зимовки по
времени имеет свои особенности для котиков каждой репродуктивной группи-
ровки, однако порядок продвижения в зависимости от возраста и пола одинаков
для всех. Лидирующей группой, мигрирующих на юг котиков, являются самки
старших возрастных классов с о. Тюлений. Они уже в январе—феврале достига-
ют южного района зимовки, а в марте их численность здесь наибольшая (рис. 14.18,
14.19). Численность молодых самок в южном районе продолжает нарастать до
мая—июня. В апреле часть взрослых самок (в основном беременных) уже пе-
ремещается к северу, причем авангардом мигрирующих на север по-прежнему
остаются самки старших возрастных групп, в то время как продвижение к северу
животных младших возрастных групп начинается только в мае, и они задер-
живаются в тихоокеанских водах Японии до июня [Кузин, Панина, 1977; Ащеп-
ков, Кузин, 19866]. Продвижение самцов по миграционному маршруту проис-
ходит по иной схеме. Первыми на юг плывут младшие возрастные группы, а
более старшие задерживаются на севере. При возвращении из эмиграции пер-
выми район зимовки покидают секачи, последними — самые молодые самцы
(рис. 14.18, 14.19).
Котики с Командорских о-вов мигрируют на юг вслед за тюленьевскими,
частично смешиваясь с ними. Продвижение их по маршруту миграции в зави-
симости от возраста происходит по той же схеме, что и котиков с о. Тюлений,
но по времени прихода в тот или иной район они запаздывают примерно на
месяц. Весенняя миграция командорских и тюленьевских самок на север начи-
257
Рйс. 14.19. Возрастной состав самцов котиков разных популяций в тихоокеанских водах Японии. Обозначения — см. рис.
258
Условные обозначения:
О САМЦЫ А-БА1Т
• СЕКАЧИ
□ САМКИ А-Б ACT
САМКИ Б-1ОЛЕТ
О САМКИ СТАГЫК IOAKT
Л ОБА ПОЛА IГОД
A OSA ПОЛАЭДГЯДА
Рис. 14.20. Графическая модель миграции морских котиков [по: Wada, 1971]
нается практически одновременно, но ее интенсивность у первых несколько
выше (рис. 14.18, 14.19).
Особенностью миграции прибыловских котиков, как уже сообщалось, явля-
ется их поздний приход в район зимовки у берегов Японии. Взрослые самки
этой репродуктивной группировки появляются в средней части района зимовки
у восточных берегов о. Хонсю только в феврале, а неполовозрелые — в апреле.
Продвижение на юг различных возрастных групп прибыловских котиков, хотя
и происходит по той же схеме, что и зверей из других популяций, но приход
их в южный район задерживается до мая, когда взрослые животные с островов
259
129° 131° 133° 135° 137°
Приморье
5„
3;
1:П1 II 1П1 ПМ.ПД
234 557
о.Тюлений
13»
и;
9.'
?:
5'-
'з:
!rs
Икюточно-
_ Корейский
IL залив
il .lffl п п п
2 3 4 5 6
и.Болтине
«Командор-
is
I Ам* Т1Й
Прибылова
. банка
ОКито-Ямато
тетй
5, гт банка Ямато
I ТзЦДижпии Пш питай ап птияпп пп
2 3334 4 5 6 77788814151619
5:
3:
!•
42°
40°
38°
3
Рис. 14.21. Половозрастной состав котиков разных популяций в разных районах
Японского моря
Тюленьего, Командорских и Курильских уже мигрируют на север (рис. 14.18,
14.19). Маршрут большинства возвращающихся из эмиграции прибыловских ко-
тиков в пределах тихоокеанских вод Японии заканчивается в том же районе, где
они появляются при миграции в южном направлении (примерно 38—39° с. ш.), хотя
часть 3-летних самцов проникает севернее.
В целом, возвращение северных морских котиков из эмиграции в зависи-
мости от возраста происходит в той же последовательности, что и их появление
в райо.-не зимовки во время прохождения миграционного маршрута в южном
направлении. Фактически, в том или ином месте акватории зимовки присутст-
вуют котики всех популяций, но возрастной состав их не идентичен. Разумеется,
речь Идет не о полной сегрегации животных разных популяций и их половоз-
растного состава в отдельных районах зимовки, а о превалировании среди общей
массы животных особей той или иной популяции, возрастного класса или пода
в определенном месте и в определенный промежуток времени. Уместно заметить,
что при изучении миграций котиков в тихоокеанских водах Японии К. Вада
[Wada, 1971] в графическом изображении миграционной модели (рис. 14.20) и
в описательной части своей работы отобразил продвижение котиков на юг двумя
потоками, но не смог разгадать причину этого явления. Она, как нам представ-
ляется, определяется разновременностью прихода котиков в этот район и осо-
бенностью перемещений животных разных популяций в этой акватории [Ащеп-
ков, Кузин, 19866].
Достоверную картину миграции котиков разных популяций в Японском
море воспроизвести несколько труднее, чем в тихоокеанских водах Японии в
связи с малочисленностью возврата меток котиков, помеченных на островах
Прибылова и Командорских. Тем не менее, обращают на себя внимание факты
значительного, сходства процесса миграции котиков у тихоокеанских берегов
Японии и в Японском море (рис. 14.21). В обоих районах лидирующей группой
260
мигрантов при движении на юг являются самцы старшей возрастной группы,
а по возвращении из эмиграции — половозрелые самцы и беременные самки
[Кузин, Панина, 1977]. Самцы в Японском море зимуют в основном в западной
части моря, где условия обитания более суровые. Прибыловские и командорские
котики появляются в Японском море позже, чем тюленьевские, примерно на
месяц. Меченые прибыловские котики не встречались в западной части моря,
что, возможно, объясняется их более поздним прибытием в район зимовки в
Японском море.
Из анализа данных по распространению котиков, полученных в результате
многолетнего изучения пелагического периода их жизни в разных частях ареала
по сезонам и встречаемости среди них меченых животных разного происхож-
дения, вырисовывается объективная картина миграций в северной части Тихого
океана, несколько отличающаяся от ранее опубликованной [Бойцов, 1934; Огнев,
1935; Ильина, 1950; Панин, Панина, 1968; Панин, 1970]. Это в большей мере ка-
сается количественных соотношений животных разного происхождения на сов-
местных местах зимовки, чем маршрутов передвижений. Существенным дополне-
нием к имеющемуся представлению о миграции котиков разных популяций явля-
ются данные о времени и темпах миграций животных из разных популяций и поло-
возрастной сегрегации их на местах зимовок в конкретные интервалы времени.
Итак, котики с о. Тюлений после окончания сезона размножения мигрируют
осенью на зимовку двумя путями. От 27 до 43% зверей (в зависимости от фазы
развития, общей численности и половозрастного состава популяции) через прол.
Лаперуза уходят в Японское море и, в основном, вдоль приморского берега Рос-
сии следуют в юго-западном направлении к берегам п-ова Корея и к центральной
части моря. Юго-западной границей распространения котиков в период их миг-
раций их в Японском море и последующей зимовки здесь является зона субарк-
тической конвергенции, разделяющая холодное Приморское и теплое Цусимское
течения [Панин, Панина, 1968]. Другая часть тюленьевской популяции котиков
проникает через проливы Курильских о-вов в Тихий океан и проводит зиму у
берегов Японии, спускаясь на юг в отдельные годы до 35° с. ш. Нашими данными
не подтверждаются предположения Н. Е. Слюнина [1895], Л. В. Бойцова [1934],
Е. И. Ильиной [1950], О. Аустина и Ф. Уилке [Austin, Wilke, 1950], К. И. Панина
и Г. К. Паниной [1968], К. И. Панина [1970] о возможности миграции котиков
с о. Тюлений через Сангарский прол, осенью и весной — обратно. Этим путем
проходит лишь очень небольшая их часть.
В летний период основная масса котиков с Тюленьего концентрируется в
районе острова и непосредственно на нем. Ок. 21% общего числа залегающих
на Курильских о-вах котиков составляют особи, родившиеся на о. Тюлений
[Кузин, 19826]. Единичные экземпляры тюленьевских котиков были зарегистри-
рованы на островах Командорских, Прибылова, Сан-Мигель, где они состав-
ляли, соответственно, 1.4, 0.01 и 0.1% от числа местных животных.
Пути миграции котиков с Командорских о-вов до сих пор изучены недоста-
точно, несмотря на то, что доля помеченных здесь котиков рассчитана для всех
районов зимовки, где их встречаемость определена по времени и месту. На этом
основании определено, что не менее половины, а в отдельные годы треть ко-
мандорских котиков мигрирует на зимовку в тихоокеанские воды Японии. Пред-
положительно, они следуют широким фронтом вдоль Курильских о-вов. В связи
с большой его протяженностью первых мигрантов регистрируют у берегов Япо-
нии на месяц позже, чем мигрантов с о. Тюленьего. Другая часть котиков с Ко-
261
мандорских о-вов мигрирует в восточную часть Тихого океана, где смешивается
с прибыловскими [Белкин, 19656]. Третья часть следует на юг от Командорских
о-вов и проводит зиму в открытых частях Пацифики [Перлов, Кузин, 1967].
Не все командорские котики возвращаются с мест зимовки на острова своего
рождения. Среди промысловых групп животных (2—5 лет) особи с Командор со-
ставили на о. Тюлений 0.5% [Владимиров, 1978а] и 6% — на о-вах Прибылова.
На Курильских о-вах командорские котики составили 23% залегающих здесь жи-
вотных.
Пути миграции котиков с о-вов Прибылова в западную часть Тихого океана
неизвестны. Наиболее вероятным представляется перемещение части мигрантов
в западный сектор северной Пацифики через акваторию Командорских о-вов
(до 33% от числа добываемых на Командорах холостяков составляют прибы-
ловские). Не исключено также, что миграционные маршруты другой части при-
быловских котиков пролегают через открытые пространства Тихого океана.
Являясь самой большой по численности репродуктивной группировкой котиков,
прибыловская популяция “посылает” наибольшее число мигрантов на другие ост-
рова. На о. Тюлений прибыловские котики составляют в среднем 1.7%, на Ко-
мандорах — 26.3%, на Курилах — 27% общей численности, залегающих там жи-
вотных.
Представляется, что сегодняшнее распространение и формирование популя-
ций северного морского котика является отражением климатических изменений
в плейстоцене, когда происходило становление котика как вида [Davis, 1958].
В плейстоцене климатические условия были резко отличными от предшеству-
ющих периодов. Периодические похолодания в средних и высоких широтах, вы-
зывавшие возникновение крупных континентальных оледенений, сопровождав-
шихся значительными регрессиями уровня океана и увеличением площади морс-
ких льдов, занимающих большие пространства [Будыко, 1980], вносили заметные
. коррективы в распространение существующих тогда видов животных. Как раз-
вивающийся вид, северный морской котик, по-видимому, был широко распрост-
ранен по северной Пацифике в плейстоцене, имел большую численность и был до-
статочно экологически пластичен. Только обладая такой пластичностью, вид мог
выжить в условиях резкого колебания климата в плейстоцене. По-видимому, в пе-
риод похолодания ареал вида сокращался, тем самым увеличивалась концен-
трация животных на ограниченных площадях, побуждавшая к развитию поли-
гинии и миграционных способностей в связи с добыванием пищи и изменением
термических условий [Bartholomew, 1970]. Выжили лишь те колонии, которые
поселялись в зонах с высокой продуктивностью моря [Lipps, Mitchell, 1976].
Современное распространение северных морских котиков также приурочено
к зонам высокой продуктивности моря. В зимний период, в связи со снижением
биопродуктивности высокоширотных вод Пацифики, котики откочевывают на
юг, при этом и в период миграции, и на местах зимовок придерживаются зон,
способных обеспечить их достаточным количеством пищи. Любые сезонные и
межгодовые изменения гидрологического режима, вызывающие изменения про-
дуктивности традиционных районов обитания котиков, являются побуждающим
моментом к изменениям в распределении животных по занимаемой ими аква-
тории. Сохранив экологическую пластичность, вид продолжает оставаться од-
ним из самых,широко распространенных и многочисленных в северной части
Пацифики среди ластоногих.
262
Глава 15. РОЛЬ УСЛОВИЙ СРЕДЫ ОБИТАНИЯ В
РАСПРЕДЕЛЕНИИ И ВЫЖИВАЕМОСТИ СЕВЕРНЫХ
МОРСКИХ КОТИКОВ
15.1. Гидрологическая характеристика районов обитания котиков
Охотское море. Наибольшее влияние на циркуляцию вод юго-западной части
моря, где летом концентрируются котики, оказывает циклонический круговорот,
расположенный к востоку и северо-востоку от северной оконечности о. Сахалин.
Значительную роль в формировании термических условий южной части Охот-
ского моря играет внутренний обмен. Он осуществляется поступлением холод-
ных водных масс с Восточно-Сахалинский течением, теплых Цусимских вод
через прол. Лаперуза с течением Куросио, проникновением вод через проливы
Курильской гряды и их вертикальным движением. Из 5 водных масс, выде-
ленных К. В. Морошкиным [1966] в Охотском море, поверхностная водная масса
характеризуется летней и осенней модификациями, заметно отличающимися од-
на от другой температурой и соленостью [Добровольский, Залогин, 1982]. Устой-
чивый переход среднесуточных температур через 0°С у восточных берегов Са-
халина последовательно протекает с конца марта до второй половины мая. Вес-
ной повышение температуры воды на поверхности в значительной мере обус-
ловлено солнечной радиацией. Наибольших значений в июне (до +5—6°С)
температура воды достигает в юго-западной части моря. В прибрежной части
моря она на Г выше, чем в открытой. Годовой максимум температуры вод (+12—
13°С) наблюдается в августе [Веселова, 1975]. Однако водная масса, лежащая
глубже 75—100 м, и летом имеет температуру, близкую к отрицательной [Доб-
ровольский, Залогин, 1982].
Максимальная температура воды в августе—сентябре почти повсеместно пред-
шествует моменту перехода от летнего к зимнему муссону. Этот переход сопро-
вождается не только сменой направления ветра, но и резким понижением общей
облачности и влажности воздуха. Переход среднесуточной температуры воды
через 0°С в сторону отрицательных значений происходит в водах сахалинского
побережья во второй половине ноября, а в зал. Анива — в декабре [Веселова,
1975]. К этому времени большинство котиков покидают Охотское море. Поверх-
ностная вода вблизи Сахалина сильно опреснена. Промежуточная водная масса
имеет соленость 32%о и близка к средним показателям по морю. С глубиной
соленость увеличивается. Охотское море богато кислородом, которого в южной
части меньше, чем в северной, однако и здесь его содержится 6—7 мл/л в по-
верхностных слоях. С глубиной содержание кислорода уменьшается [Доброволь-
ский, Залогин, 1982].
Южная часть моря не замерзает, но весной покрывается выносными льдами,
которые скапливаются у Курильских о-вов и мешают весенним миграциям ко-
тиков на север. Лед исчезает в конце мая—начале июня.
263
Берингово море. Район летнего обитания котиков здесь характеризуется суб-
арктической структурой вод. Вдоль южной стороны Командоро-Алеутской гряды
Аляскинское течение устойчивым и почти непрерывным потоком распростра-
няется в западном направлении до прол. Ближний. В восточной части пролива
находится основное место входа в Берингово море вод, приносимых Алеутским
течением. Между Камчаткой и о. Беринга в поверхностных слоях господствует
вынос вод из моря в океан. С юго-запада в район Ближних о-вов к Берингову
морю подходит широкой полосой течение, связанное по своему происхождению
с областью смешанных вод Куросио и Оясио. Приносимые ими воды перемеши-
ваются с водами Аляскинского течения и поступают в Берингово море [Буров, 1958].
Из-за разнообразного воздействия гидрологических факторов субарктическим
структурам вод разных районов Берингова моря присущи различные характер-
ные особенности. Водные массы Берингова моря имеют слоистую структуру. Это
расслоение в основном определяется различием температуры, а не солености.
Поверхностная водная масса летней модификации представляет собой наиболее
прогретый вертикальный слой воды до глубины 50 м и характеризуется отно-
сительной однородной температурой (+7—ГС) и соленостью (33%о). К началу
осеннего охлаждения весь верхний слой вод Берингова моря прогревается мак-
симально до +9—1ГС. Начинающаяся в сентябре—начале октября выхолажи-
вание поверхностного слоя приводит к перераспределению тепла и отдаче его
верхними слоями нижележащим [Арсеньев, 1967].
Температура воды на поверхности Берингова моря понижается с юга на север,
причем в западной части моря вода несколько холоднее, чем в восточной. Летом
температура воды на поверхности в юго-западной части равна +9—1 ГС и +8—
10°С — в восточной. Уже на глубине 50 м она понижается до +2.5°С. Зимой
поверхностный слой воды охлаждается до +2°С. Соленость поверхностных вод
моря составляет 33—33.5%о. Ее увеличение происходит до горизонта 125—150 м.
Южная часть Берингова моря не замерзает круглый год благодаря теплым
водам, поступающим через алеутские проливы, которые отжимают льды к се-
веру. В апреле кромка льда достигает южной границы, в мае лед начинает раз-
рушаться, и его кромка постепенно отступает к северу. В верхних слоях воды
в Беринговом море содержание кислорода составляет 6.7—7.6 мл/л. С глубиной
его количество уменьшается [Добровольский, Залогин, 1982].
Японское море. Гидрологический режим здесь находится под непосредствен-
ным влиянием тихоокеанского течения Куросио, западная ветвь которого про-
никает через Цусимский прол. Вдоль приморских берегов на юг спускается хо-
лодное течение. Постоянное в течение всего года Цусимское течение переме-
щается вдоль западных берегов Японии на север и является восточной пери-
ферией циклонической циркуляции вод моря. Сила пульсации Цусимского
течения изменчива по сезонам и имеет непериодические межгодовые колебания.
Зимой оно несколько ослабевает, но в это время года интенсифицируется муссон,
который способствует увеличению стока вод на юг Приморским течением [По-
кудова, Тунеголовцев, 1975]. Северная ветвь теплого Цусимского течения про-
ходит над банкой Ямато. Она и является естественной южной границей рас-
пространения котиков в Японском море.
Распределение температуры воды крайне неравномерно, причем имеется 4 типа
годового хода,” характерных для разных районов Японского моря. В общих же
264
чертах поверхностная тихоокеанская водная масса характеризуется высокими
круглогодичными температурными показателями, колеблющимися в зависимос-
ти от сезона и глубины в пределах +4—25°С, и соленостью 33.6—34.5%о. По-
верхностная водная масса представлена Приморским течением и характеризуется
колебаниями температуры от 0°С до +21°С и солености от 32 до 34%о.
Собственно фронтальных вод в Японском море нет, а имеет место зона сме-
шения вод, образующейся из-за подъема вод с глубины в результате общей цик-
лонической циркуляции [Радзиховская, 1961]. Положение этой зоны имеет се-
зонную и межгодовую изменчивость.
В климатическом отношении Японское море как район зимовки котиков от-
личается сравнительной суровостью. Среднегодовые температуры воды здесь ниже,
чем в Тихом океане. В северной части моря лед появляется в декабре. В суровые
зимы лед может образовываться даже в Восточно-Корейском зал. — в одном
из районов постоянного обитания котиков.
Тихоокеанские воды Японии. Район зимовки у восточного побережья Японии
характеризуется очень сложным гидрологическим режимом. Здесь встречаются
субарктические воды, приносимые холодным течением Оясио, идущим с севера
на юг вдоль островов Хоккайдо и Хонсю, и субтропических водных масс теплого
течения Куросио, проходящего вдоль южного побережья Японии, где на 36° с. ш.
оно отклоняется на восток. Куросио на своем пути вдоль берегов Японии и далее
образует несколько крупных стационарных меандров, конфигурация которых с
течением времени непрерывно меняется. Отдельные меандры время от времени
отделяются от главного потока, образуя “инородные” крупные циклонические
вихри, в которых происходит подъем охлажденных, богатых биогенами вод. Для
Куросио характерен резкий горизонтальный градиент океанических характерис-
тик, выражающихся в образовании холодных пятен к северу от него. Такие гид-
рологические характеристики создают подобие струйчатой системы течений.
Пограничная зона, в которой ветви Куросио вступают в контакт с водами Оясио,
характеризуются как субарктическая водная зона. С усилением Куросио субар-
ктическая фронтальная зона сужается, и, наоборот, с ослаблением течения ши-
рина фронтальной зоны увеличивается. Это явление наблюдается в сезонном и
многолетнем плане, и ширина достигает 120—210 миль [Павлычев, 1975]. В связи
с этими изменениями гидрологической обстановки соответствующим образом
изменяется район концентрации котиков у берегов Японии. Кроме этой посто-
янной системы, в определенные сезоны у берегов Японии обнаруживается до-
вольно теплое течение, которое проникает сюда через Сангарский прол, из Япон-
ского моря и затем поворачивает на юг вдоль береговой полосы.
Зимой субарктические воды господствуют в районе о. Хоккайдо, но отдель-
ные струи доходят вдоль побережья до 35° с. ш. Весной с усилением деятельности
Куросио субарктический фронт смещается к северу, однако струи и вихри ос-
таются в прежних границах [Павлычев, 1967].
Как и в других районах обитания котиков, водная масса Тихого океана к
востоку от Японских о-вов имеет слоистую структуру. Толщина поверхностной
водной массы доходит до 60—200 м. На севере рассматриваемого района тем-
пература воды в январе составляет в среднем +4°С, соленость — 33%о, содер-
жание кислорода — 7 мл/л. На юге температура и соленость повышаются до
19°С и 34%о, соответственно, содержание кислорода — до 5 мл/л. К весне происходит
изменение гидрологических характеристик. На севере района температурные гра-
265
диенты мало изменяются, на юге температура воды на поверхности доходит до
20°С; соленость понижается до 33%о; содержание кислорода увеличивается до
9 мл/л. Усиливается “пятнистость”, которая характеризуется наличием холодной
водной массы среди окружающей ее теплой с большим содержанием кислорода
(9.0—9.5 мл/л) и температурой на поверхности 6°С. На глубине 100 м температура
остается более прохладной (+2°С); соленость изменяется мало (33.0—33.5%о).
Субарктические фронтальные водные массы характеризуются повышенным
содержанием кислорода и биогенных веществ, в отличие от субтропических вод-
ных масс, приносимых течением Куросио. Циклоническая циркуляция, дивер-
генция и другие проявления динамики водных масс создают условия возникно-
вения зон повышенной биологической продуктивности [Натаров, Черный, 1966],
что привлекает сюда большие косяки рыб, служащих кормом котикам.
В пределах обширного ареала котики распространены неравномерно. На местах
зимовок в Японском море, у тихоокеанских берегов Японии и у берегов Сев. Аме-
рики наибольшие их концентрации отмечаются в области смешения теплых и хо-
лодных вод, где животные придерживаются субарктического фронта. Как правило,
в таких местах отмечается разнообразие населения пелагиали и большая биомасса
нектонных организмов, служащих кормом для котиков. Такие акватории, имеющие
сходные гидрологические и физико-химические характеристики, несмотря на то, что
расположены на значительном расстоянии друг от друга, привлекают котиков. Диа-
пазон колебания температуры воды в районах пребывания котиков составляет 0—
17°С, солености — 32—35%о, а содержания кислорода — 5—7 мл/л. Однако для
основной массы котиков естественным рубежом распространения на юг является
изотерма 15°С [Wada, 1969]. Безусловно, названные термические условия не явля-
ются критическими; т: к. известно, что в неволе после соответствующей адаптации
котики в течение всего лета способны переносить температуру воды +29°С [Na-
kajima et al.,1963; Микеладзе, 1971]. Известны также многочисленные факты зимов-
ки взрослых котиков в районах репродуктивных центров, где температура воздуха
падает до -5°С, а воды — близка к 0°С. Отмечены неоднократные встречи жи-
вотных у берегов Приморья и о. Хоккайдо при температуре воды —2°С. Зафик-
сированы плавающие среди льдов в Олюторском зал. весной при отрицательных
температурах воды секачи [Перлов, 1968]. А. М. Николаев [личн. сообщ.] наблюдал
в декабре на о. Тюлений спящих на снегу щенков котиков, а Ф. Г. Челноков [1982а],
В. Е. Пинигин и В. Г. Прянишников [1982] — зимующих на Командорских о-вах
секачей и холостяков.
Существует пространственная, межгодовая, сезонная, половая и возрастная
специфика в распределении северных морских котиков в зависимости от условий
внешней среды. В Японском море на зимовке котики отмечались в акватории
с температурой воды до +14°С, солености — 32—34.5%о, при содержании кис-
лорода — 6—7 мл/л (рис. 15.1). В течение пребывания здесь котиков с января
по июнь диапазон температур, при которых регистрировались животные, пос-
тепенно расширяется (рис. 15.2).
В январе котиков отмечали при температуре воды 0—5°, в июне — 6—13°.
Показатели наибольшей встречаемости животных при тех или иных темпера-
турах воды изменяются. Так, в январе наибольшее число котиков встречено при
температуре воды 3°С, в марте — при 4°, в апреле — при 5°, в мае — при 7°
266
Рис. 15.1. Физико-химические показатели Японского моря на разрезе (врезка в
правом нижнем углу) в районе концентрации котиков (мелкие точки)
и в июне — при 10°С. Самок регистрировали при несколько повышенных (при-
близительно на 1°С) температурах водной среды. Котики младших возрастных
групп встречались при более широком диапазоне температур, чем взрослые особи.
Причем наибольшая встречаемость первых отмечалась при пониженных темпе-
ратурах воды в названных пределах ее колебаний. В качестве примера межго-
довой изменчивости распространения котиков в зависимости от температурного
фактора показательны следующие данные. В мае 1960 г. в Японском море ко-
тиков регистрировали при 2—1 ГС с пиком встречаемости при 4—5°С, в то время
как в 1964 г. их отмечали при 6—12°С, с пиком при 8—9°С. Фактически, за
18-летний период исследований котиков в Японском море было мало лет сходных
по максимальной встречаемости зверей при тех или иных температурах воды.
В западной части Тихого океана у берегов Японии распространение котиков
в зависимости от гидрологических условий несколько отличается от такового
в Японском море (рис. 15.3). Свойственные для этого района струйчатость те-
чений и образование циклонических вихрей создают довольно-таки сложную
структуру океанических вод, характеризующуюся наползанием теплых слоев
воды на холодные ядра нижележащих слоев, имеющих разнокачественные по-
казатели основных гидрологических характеристик на различных глубинах в
пределах, которые способны осваивать северные морские котики. Поэтому скоп-
ления животных, отмеченные при повышенных температурах поверхностных
слоев воды, чаще всего встречаются в тех акваториях, где нижележащие гори-
зонты воды имеют оптимальные для животных условия (рис. 15.4, 15.5).
267
Рис. 15.3. Температура воды
на поверхности (изолинии),
места концентрации котиков
(мелкие точки), гидрологичес-
кие станции (крупные точки с
цифрами)
Рис. 15.2. Встречаемость котиков в Японском море в зависимости от температуры воды
142 144 146
В тихоокеанских водах Японии котиков регистрировали при температуре
воды на поверхности 0—17°С, солености — 33—34.7%о и содержании кислоро-
да — 5.1—7 мл/л. В северных районах ареала зимовки температура воды и со-
леность ниже, а содержание кислорода несколько выше, чем в южных. Вместе
268
разрезе, обозначенном на рис. 15.3
Рис. 15.4. Гидрохимическая характеристика вод на
цифрами 1—9.
Другие обозначения см. на рис. 15.1.
Crg Ст(6 МО Сг14 Сг.<? Ы2 Crjl Ст 10
. А’ . 7»1; • • $»>; 5-5Л’: ‘ J -З*3 К543, г, 2
Ст. £
0
10
20 W
ЗО'
I
90
Т5
юо
юч_
20 X
I
ю''4.
г
П,
«о
200
’ / */
}' 'I
а /. л
6.0 Л » У
33.4 У ЗД
ия
X
г г 1
I
г
?
г
i
✓ Х? *\ Ч
/м\<
yvi
л f fX |
г
t—
:--
45 <0
г
г
И
4
г
it 1
nV
1
£
I
2
I
2
г
г
М 2*5’ в'
г
г
г
г
г
г
г
i
г
н
111
г г г
г I г
г
I \ /) ‘ulill {
346 ЗВД 341 34?
Рис. 15.5. Гидрохимическая характеристика вод на разрезе, обозначенном на рис.
15.3 цифрами 10—18.
Другие обозначения см. на рис. 15.1.
269
Рис. 15.6. Встречаемость котиков у
тихоокеанских берегов Японии в за-
висимости от температуры воды
с тем, в одни и те же месяцы, как правило, максимум встречаемости котиков
в тихоокеанских водах Японии отмечается при более высоких температурах
воды на поверхности, чем в Японском море (рис. 15.6).
В декабре наибольшая встречаемость котиков у тихоокеанских берегов Японии
была при температуре воды на поверхности 4—5°С, в январе — при 2—3°С, в фев-
рале — при 6—10°С, в марте и апреле — при 7—1 ГС, в мае — при 6—10°С, в ию-
не — при 10—12°С. Как и в Японском море, у восточных берегов Японии самки пред-
почитали более высокие температуры, чем самцы, а молодые животные встречались
чаще, чем взрослые, при пониженных температурах.
Естественно, что на распределение котиков в море существенное влияние, по-
мимо гидрологических условий, оказывают метеоусловия. Их воздействие на
взрослых животных не так ощутимо. Более чувствительны к ним сеголетки.
Предполагается, что причиной высоких показателей смертности котиков в ран-
нем возрасте являются именно погодные условия. Предпосылкой для такого суж-
дения послужило сообщение В. Шеффера [Scheffer, 19506] о большом числе тру-
пов сеголеток, обнаруживаемых на тихоокеанском побережье Сев. Америки пос-
ле суровых изнуряющих штормов. Известные различия в показателях выживае-
мости и распределении котиков по акватории зимовки в зависимости от половой
принадлежности были положены в основу логических выводов о том, что смерт-
ность самцов по сравнению с самками выше из-за того, что первые проводят
зиму в северных широтах, где число штормовых дней и средняя скорость ветра
больше, чем в южных, где зимуют самки [Ichihara, 1974]. Одновременно этот
автор показал, что наибольшее число трупов сеголеток котиков, выбрасываемых
на побережье Сев. Америки и на острова Хоккайдо и Хонсю, наблюдается в
декабре-феврале. Это хорошо согласуется с высокими среднемесячными ско-
ростями ветра в этих регионах.
Для выяснения зависимости выживаемости сеголеток от гидрометеоусловий
нами была предпринята попытка установления корреляционных связей между
коэффициентами выживаемости котиков до 2 лет, полученных по методике Р. Лен-
дера [Lander, 1975], от таких факторов, как скорость и повторяемость ветров
разной силы в течение декады, месяца, квартала или года в районах обитания
270
Рис. 15.7. Анализ основных
биологических показателей
популяции котиков о. Тюлений:
А — численность щенков, Б —
выживаемость самцов до 2 лет, В —
остаток самцов после выбоя, Г —
численность секачей.
ПОКОЛЕНИЯ
1986 66 90
ГОЛЫ
котиков у тихоокеанских берегов Японии; повторяемость циклонов на уровне
Н500; величина геопотенциала Н500; среднемесячные температуры воды на по-
верх- ности в 5° квадратах координатной сетки, по данным спутниковой ин-
формации; площадь акваторий, занятых водой с отрицательными и положитель-
ными аномалиями; среднемесячные температуры воды на поверхности, по дан-
ным японских береговых гидрометеостанций (ГМС) Судзу, Уракава, Мияко за
период 1958—86 гг. [Кузин, Шаталина, 1990].
В результате обнаружена связь между отдельными факторами и коэффици-
ентами выживаемости котиков до 2 лет. Однако в составленное нами уравнение
регрессии ювенильной выживаемости котиков в зависимости от условий внеш-
ней среды вошли только те, которые удовлетворяли показателям достоверности:
Y = 0.66207-0.05962х1-0.00272х2+0.003х3+0.0059х4+0.00423х5+0.00063х6,
где Y — выживаемость котиков, х( — температура воды в ноябре—марте, по
данным японских прибрежных ГМС, х2 — аномалии геопотенциала Н500 в январе
на о. Хоккайдо, х3 — аномалии геопотенциала Н50О в январе над о. Хонсю, х4 —
повторяемость (в днях) атмосферных циркуляций северного направления, х5 —
повторяемость (в днях) атмосферных циркуляций в январе западного направ-
ления, х6 — повторяемость (в днях) атмосферных циркуляций в январе широт-
ного направления. Сумма квадратов в приведенном уравнении равна 0.05184,
а = 0.00864, <р = 6.02387.
По полученному уравнению регрессии были рассчитаны коэффициенты вы-
живаемости северных морских котиков из поколений 1979—85 гг., которые со-
ставили: в 1979 г. — 0.30, в 1980 г. — 0.33, в 1981 г. — 0.35, в 1982 г. — 0.30,
в' 1983 г. — 0.42, в 1984 г. — 0.47, в 1985 г. — 0.37.
В качестве проверки достоверности коэффициентов выживаемости, получен-
ных на основе приведенного уравнения регрессии, был проведен анализ основ-
ных биологических показателей популяции на о. Тюлений (рис. 15.7).
Как видно на рисунке, численность секачей в 1986—89 гг. хорошо коррели-
ровала с остатком их после выбоя из поколений 1979—82 гг. (г = 0.82). При
этом естественная смертность самцов из названных поколений рассчитывалась
на основе коэффициентов выживаемости, полученных из уравнения регрессии.
271
Рис. 15.8. План о. Тюлений
//' >'/ у^а
//,\'ЛГ v 5
// '^rJ^
3"'6У3
fitf'Jn • -холостяковые
:-'у;ч /12 залежки
¥ X «в - гаремные залежки
">./13 1-13 - участки лежбища
7 котиков
/ «о - верхнее плато
/ острова
/ и хозяйственные
у “ постройки
Можно поэтому сказать, что эти коэффициенты удовлетворительно отражают
реальные процессы, происходящие в популяции. Они могут быть использованы
в расчетах динамики численности и норм добычи с заблаговременностью до 2 лет.
В целом, оценивая роль гидрометеоусловий в создании комфортности сущест-
вования морских котиков в море при миграциях и на местах зимовки можно
констатировать, что их роль существенна. Она определяется системой взаимо-
действующих течений, балансированию которых, в определенной степени, спо-
собствуют атмосферные процессы. При этом, если термические условия среды
влияют на выживаемость котиков, в основном, через пищевые связи [Кузин, Ша-
талина, 1990], то атмосферные процессы воздействуют как непосредственно, так
и опосредованно. В частности, атмосферные циркуляции из-за ветрового волне-
ния могут оказать прямое негативное воздействие на животных. Они же могут
вносить коррективы в термический режим акваторий, в особенности в районах
стыка течений, мелководий и шельфовых зон. Можно полагать, что сила и час-
тота проявления ветров северо-западного направления, являющихся сгонными,
усиливают интенсивность прибрежного апвеллинга [Седых, 1984]. В результате
этого увеличивается уровень биологической продуктивности района осенне-зим-
него пребывания сеголеток котиков, и оптимизируется гидрологический режим,
способствующий успешному размножению и выживанию массовых видов рыб,
служащих объектами питания котиков [Кузин, Шаталина, 1990].
15.2. Острова как среда обитания северных морских котиков
Котики размножаются на островах. В этот период большая часть их насе-
ления концентрируется на лежбищах. Лежбища имеются на островах Тюлений,
Каменные Ловушки и Среднего, Командорских, Прибылова и Сан-Мигель.
О. Тюлений расположен в 12 милях от мыса Георгия на Сахалине. Он пред-
ставляет собой вытянутую с северо-востока на юго-запад узкую, местами от-
весную скалу длиной 655 м и шириной ок. 110 м. Площадь острова — 0.07 км2
(рис. 15.8). Основное гаремное лежбище находится на восточном песчано-галеч-
ном пляже шириной 30—50 м, поверхность которого ровная, постепенно пони-
272
жающаяся от скалы к морю. Литоральная часть моря со стороны лежбища мел-
ководная и простирается на несколько сот метров в виде подводных каменистых
рифов, сдерживающих прохождение крупных волн к берегу во время штормов.
Холостяковые залежки располагаются на мысах, из которых площадь южного
много больше северного. Западный берег острова представляет собой узкий га-
лечный пляж, который в большие приливы или в шторм почти заливается водой.
Здесь котики не образуют залежек.
Естественное лежбище котиков на Тюленьем в 1965 г. было искусственно рас-
ширено. Для этого часть восточного склона была срыта, и сделан пологий подъ-
ем на скалу. Была создана возможность для котиков подниматься на верхнее
плато, возвышающееся на 12—17 м над ур. м. Площадь плато, занимаемого ко-
тиками, составляет 4 тыс. м2 [Когай, 1970]. Оно располагается в северной части
острова. Вся южная часть плато занята птичьим базаром, где в основном гнез-
дятся кайры.
Климат острова континентальный. Лето холодное, пасмурное, с частыми ту-
манами, в отдельные годы до 25 дней в месяц. В начале лета нередки заморозки.
Среднемесячная температура июня +4. ГС, а самого теплого месяца (август) —
+ 12°С. В отдельные дни температура может подниматься до 25°С. Температура
воды у острова летом колеблется в пределах +7—10°С; иногда вода прогревается
до 13°. Скорость ветра летом в среднем 6 м/сек, а в штормовые дни достигает
20—25 м/сек. Осадки выпадают часто и, главным образом, в виде мелкого дождя
(“буса”). В среднем за месяц бывает 15—18 дней с дождем, и выпадают 60—80 мм
осадков.
Растительность острова осоково-злакового типа, древесная — отсутствует.
Постоянными видами, размножающимися на острове, кроме северного морского
котика, из млекопитающих можно назвать только сивуча, общая численность
которого в последние годы составляет более 1000 особей. На северном мысе за-
легают ларги, численность которых сильно колеблется по дням и порой дости-
гает 700—800 голов.
О-ва Каменные Ловушки состоят из нескольких скал (Котиковая, Долгая, Вы-
сокая, Низкая). Котики залегают на всех скалах. Скалы расположены по кругу
и представляют собой останцы вершины некогда существовавшего вулкана. Бли-
жайшая от соседнего более крупного острова Курильской гряды — Шиашкотана
скала Высокая расположена в 10 милях от его южной оконечности. На скалах
Котиковая и Низкая растительность полностью отсутствует, а на скалах Долгая
и Высокая имеются осоково-злаковые сообщества. На всех скалах, кроме ко-
тиков, залегают сивучи, причем на Долгой располагается одна из самых крупных
гаремных залежек сивучей на Курилах. На Высокой существует залежка остров-
ного тюленя.
О-ва Среднего также объединяют несколько скал, но в качестве лежбища ко-
тиками используются только скалы Черные и Хитрая, последняя из которых рас-
положена в 2.6 милях от северной оконечности соседнего более крупного о. Уши-
шир. Скалы лишены растительности. Кроме котиков, на них залегают сивучи,
причем на Хитрой, как и на Долгой, располагается одно из крупнейших их га-
ремных лежбищ. Других видов млекопитающих на островах нет.
Акватория вокруг островов Каменные Ловушки и Среднего характеризуется
стремительными приливно-отливными течениями, в результате чего здесь обра-
273
Рис. 15.9. Топография лежбищ коти-
ков на о. Беринга:
А — современные, Б — в конце прошло-
го века (пунктир — по: Гребницкий,
1902; штриховка — по: Слюнин, 1895.
Рис. 15.10. Топография лежбищ ко-
тиков на о. Медный:
А — в конце XIX века [по: Волошинов,
1886], Б — в начале XX века [по: Суво-
ров, 1912], В — современность.
зуются сильные сулои. Почти вокруг всех скал имеются большие поля водоро-
слей. Летом скалы редко освобождаются от туманов, а температура воды не
превышает +5°С, воздуха — 10—15°.
Командорские о-ва составляют 2 крупных (Беринга и Медный) и 2 небольших
острова (Топорковый и Арий Камень). Лежбища котиков имеются только на
двух первых. В настоящее время на о. Беринга существуют 2 лежбища: Северное,
занимающее территорию от бухты Фонтанка до бухты Поворотная, общей дли-
ной 2.5 км; Северо-Западное, находящееся на Северо-Западном мысе и занима-
ющее полосу береговой черты от мыса до бухты Дубовая, протяженностью 2 км
(рис. 15.9).
На о. Медный в настоящее время существуют 2 лежбища: Юго-Восточное —
на одноименном мысе и Урилье — в бухте Перешеечная (рис. 15.10). Юго-Вос-
точное лежбище занимает береговую полосу протяженностью 5.3 км от мыса
Марьинский до участка “Вторая забойная площадка”. Урилье расположено в
15 км от Юго-Восточного. Котики залегают на береговой полосе длиной 4 км
от мыса Поганый до бухты Горелая.
Структура грунтового покрытия и рельеф территорий командорских лежбищ
разнообразен [Владимиров, Челноков, 1971]. На Северном лежбище имеются
участки базальтово-аденизитового происхождения, в других частях лежбища ба-
274
зальтово-аденизитовое основание покрыто слоем рыхлых отложений из песча-
но-ракушечника, гальки или состоящие из крупной гальки и мелких валунов.
Имеются песчаные участки. Значительная часть Северного лежбища (участок
“Риф”) затопляется в прилив. Рельеф лежбища разнообразен. Котики занимают
скалистые берега Сивучьего Камня, отдельные крупные камни и каменистые гря-
ды, в отлив соединяющие Сивучий Камень с основной частью острова, и не-
большие бухты, образующиеся при впадении рек Гольцовая и Кишечная, и
пляжи (“лайды”).
Акватория, прилегающая к Северному лежбищу, изобилует подводными кам-
нями, отмелями, останцами и грядами, часть из которых осыхает в отлив. Аб-
разионная терраса (“бенч”) простирается в море до 3.5 км и является хорошим
волноломом.
Территория Северо-Западного лежбища имеет в основном песчаное покры-
тие. Лишь на мысе Песчаный и напротив отдельных участков лежбища в море
имеются камни или подводные гряды, осыхающие в отлив. В отдельных местах
лежбище простирается далеко вглубь острова по склонам песчаных дюн, а в
местах, где скалистые берега подходят близко к урезу воды, котики занимают
узкую полоску песчаных пляжей. Литораль в районе Северо-Западного лежбища
образована абразионными террасами. Защищенность лежбища от наката волн
увеличивается с юго-запада на северо-восток. В сильные штормы волны могут
обрушиваться на его южную часть, в то время как северная, прикрытая бенчем,
защищена от наката.
Лежбища котиков на о. Медный по характеру грунта и рельефу сильно от-
личаются от таковых на о. Беринга. Юго-Восточное лежбище представляет
собой неширокую лайду, покрытую обломочным материалом — от многомет-
ровых до мелких, окатанных морем валунов и гальки [Владимиров, Челноков,
1971]. Подножье коренного берега представляет собой осыпи, состоящие из гро-
мадных каменных глыб. В районе лежбища часто случаются обвалы и камне-
пады. Литоральная зона представляет собой плиту с гребнем, выступающим кам-
нями и кекурами на высоту до 20 м.
Территория Урильего лежбища покрыта галькой и мелкими валунами. Ко-
ренной берег выходит ко всем участкам лежбища обрывами или крутыми скло-
нами. С моря лежбище прикрыто скальной плитой, которая постепенно сужается
с юго-востока на северо-запад.
Климатические условия во всех местах залегания котиков на Командорских
о-вах сходны. Резких температурных контрастов между сезонами года не наблю-
дается. Летом температура воздуха самого теплого месяца (август) поднимается
до +10—12°С. Относительная влажность составляет 80—90%. Часты туманы,
особенно летом (до 25 дней в месяц); осадков выпадает мало; преобладают мел-
кие моросящие дожди. Море у островов относительно теплое, незамерзающее.
Выносные льды отмечаются только в очень суровые годы. Поэтому создаются
условия для круглогодичного обитания некоторой части поголовья животных
возле островов [Вертянкин, 1986]. Растительность тундрового типа. Самый круп-
ный наземный хищник — песец. Из морских млекопитающих обитают калан,
обыкновенный тюлень, ларга и сивуч. Сивучи на некоторых лежбищах даже за-
легают вместе с котиками.
275
Таким образом, основные репродуктивные центры северного морского ко-
тика приближены к северным границам его ареала. Все лежбища расположены
на островах, лишенных древесной растительности. На островах нет крупных
хищников. Геоморфологические особенности побережий островов не играют су-
щественной роли в организации лежбищ. Лежбища могут быть размещены на
песчаных, галечных, каменистых пляжах, на почти отвесных скалах, на торфяной
подстилке, на траве, на заливаемых в прилив камнях и т. п. Они могут нахо-
диться с любой стороны островов или скал. Литоральная зона вокруг них по-
логий, значительных размеров бенч. Для островов характерен морской климат
с невысокими температурами воды и воздуха, высокой влажностью, малочис-
ленными осадками и частыми туманами в летнее время. Вместе с тем, сходные
гидроклиматические условия характерны для многих других островов Куриль-
ской и Алеутской гряд, однако ни на одном из них гаремных лежбищ котиков
нет и не было, начиная с середины XVIII в. [Владимиров, 1972]. Правда, есть со-
общения о том, что в прошлом существовали небольшие залежки котиков на не-
которых островах Курильской гряды [Хоккей, 1897; Тихенко, 1914а; Успенский, 1955],
но достоверность этих сведений сомнительна в связи с тем, что Г. Сноу [1902], хо-
рошо знавший места обитания всех видов ластоногих на Курилах, упоминает
лишь те, что известны в наши дни, и лежбище на о. Райкоке, где котиков теперь
нет в связи с тем, что оно в прошлом веке было засыпано камнями и пеплом
из-за извержения вулкана.
Исследуя комплекс факторов среды, характерных для островов, где ныне су-
ществуют лежбища котиков, В. А. Владимиров [1972] пришел к выводу, что для
организации залежек котикам необходимо сочетание высокой (90%) влажности
и сравнительно низкой (+7—9°С) температуры воздуха. Сочетание этих пока-
зателей признано зоной оптимума; эти условия особенно важны в период появ-
ления на свет молодняка. Автором также высказано предположение, что сущест-
вующие в зоне оптимума лежбища не вырождаются, и угасание некоторых из
них на Командорских о-вах в 1930-х годах произошло из-за изменения климата
под влиянием усиления деятельности течения Куросио.
Мы не отрицаем полностью важности таких факторов, как температура И
влажность, для котиков, но, тем не менее, факт существования ежегодно уве-
личивающего численность гаремного лежбища на о. Сан-Мигель, где летом тем-
пература воздуха поднимается до 40°С, а песок прогревается до 50°, говорит
о том, что котики могут успешно существовать и в иных условиях среды. То,
что лежбища северных морских котиков имелись на территории Калифорнии
в далеком прошлом, подтверждается археологическими находками в районе зал.
Гумбольдта, датируемых возрастом 1 тыс. лет назад [Hildebrandt, 1984]. При со-
держании в неволе котики легко адаптируются к температуре воды и воздуха,
близкой к 30°С [Nakajima, 1963; Микеладзе, 1971]. Показано, что новорожденные
котики более толерантны как к низким, так и высоким температурам, чем взрос-
лые особи [Irving et al., 1962]. Таким образом, если и существуют у котиков опре-
деленные температурные предпочтения, то они наиболее важны не для щенков,
а для взрослых. Это подтверждается тем, что приоритет в выборе будущих га-
ремных лежбищ всегда принадлежит холостякам и безгаремным секачам, пер-
выми осваивающими новые места обитания. Можно с уверенностью говорить,
что если условия среды удовлетворяют требованиям этой возрастной группы,
то они будут пригодны и для щенков.
276
Трудно предположить, что в северной части Тихого океана нет больше ост-
ровов, где возможна была бы организация котиками лежбищ. Возникновение
лежбищ, по-видимому, во многом определяется не только условиями среды, а,
главным образом, историей вида и сложившимися особенностями биологии се-
верных морских котиков, основными из которых являются хоминг и филопатрия.
Закрепленные в памяти и постоянно поддерживаемые из поколения в поколение
места концентраций котиков в репродуктивный период (репродуктивные цент-
ры), пути миграции, районы зимовки, полигиния являются сдерживающими фак-
торами в организации новых залежек и популяций.
По-видимому, формирование северного морского котика как морского мле-
копитающего происходило в островных условиях при отсутствии сильно выра-
женной конкуренции за пищу, за места обитания, где не было многочисленных
врагов. Такие острова должны были существовать в прошлом, а теперь только
те, которые расположены в зонах высокой биологической продуктивности, где
имеется в пределах ограниченных акваторий в достатке пища. Подтверждение
предположению об отсутствии хищников в местах исторического формирования
котика как вида могут служить такие особенности этологии, как отсутствие за-
боты о потомстве, формирование агрегаций детенышей на периферии гаремного
сообщества, где они при присутствии хищников могли быть подвергаться на-
падению первыми, периодические отлучки взрослых особей с островов в море
для питания и в случае опасности. Детеныши в таких ситуациях остаются на
берегу и не способны ни защитить себя, ни уйти в море, в связи с тем, что еще
не умеют плавать. Безусловно внешние условия среды (температура, влажность)
играют существенную роль в определении пригодности тех или иных мест для
организации залежек, но, видимо, не столь жестко регламентируют вид в выборе
мест обитания. Для поддержания гомеостаза у котиков выработался большой
комплекс морфо-физиологических адаптаций.
277
Глава 16. ВРАГИ И КОНКУРЕНТЫ
16.1. Враги
В летний период котики обитают на островах, где нет крупных хищников,
которые могли бы относиться к врагам этих животных. На островах Прибылова
и Командорских самыми крупными хищниками являются песец и лисица, ко-
торые, естественно, не могут нанести какой-либо вред взрослым котикам, однако
нападают на новорожденных [Барабаш-Никифоров, 1936; Ильина, 1950; Челно-
ков, 19706] (рис. 16.1). :
И. И. Барабаш-Никифоров, например, считает, что песцы расправляются со
слабыми и больными новорожденными и выполняют роль санитаров на леж-
бище. Ф. Г. Челноков дает количественную оценку вреда, наносимого котикам
песцами. Так, на Юго-Восточном лежбище (о. Медный) в среднем в год с 1964
по 1968 гг. погибли 50 щенков (0.2% от общего числа новорожденных). При
этом важно отметить, что наибольшее число загрызенных песцами детенышей
наблюдалось в начальный период гаремной жизни, когда численность новорож-
денных еще незначительна. С увеличением количества новорожденных и ростом
их естественной смертности песцы поедают трупы и плаценты. Расчеты Ф. Г. Чел-
нокова показали, что за летний период песцы в среднем съедают ок. 11—13%
трупов котиков, и роль песцов как санитаров не очень велика.
С. В. Мараков [1974] называет в качестве врагов котиков воронов, которые
иногда выклевывают глаза живым детенышам и могут заклевать до смерти но-
ворожденного. Однако он же отмечает, что такое случается крайне редко, т. к. пти-
цам вполне хватает последов и трупов погибших котиков.
Значительно больше врагов у котиков в море, где они проводят больше вре-
мени. Имеющиеся литературные данные позволяют назвать в качестве врагов
котиков косатку (Orchina orca) и крупных акул [Суворов, 1912; Kenyon, Wilke,
1953; King, 1964; Бычков, 1967; Baker et al., 1970; Чапский, 1976]. Однако ве-
личина вреда, наносимого тем или видом морских животных котику, количест-
венно не определена. Так, из одного литературного источника в другой пере-
ходит описание случая нападения косаток на котиков. Думается, прав С. В. Ма-
раков [1974], который писал, что, хотя косатки и представляют некоторую опас-
ность для котиков, серьезного ущерба им они нанести не могут. С. В. Мараков,
долго наблюдавший за котиками и многократно видевший косаток у берегов
Командорских о-вов, ни разу не отмечал случаи нападения косаток на котиков.
Нам в море неоднократно приходилось видеть котиков и косаток в непосред-
ственной близости друг от друга, но нападений не наблюдалось. Безусловно,
нельзя полностью отрицать хищничество косаток, т. к. имеются достоверные слу-
чаи обнаружения останков котиков в их желудках [Rice, 1968], но нужно при-
знать, что северные морские котики никак не являются основным объектом пи-
тания косаток.
Сенсационно прозвучало сообщение американских ученых на Комиссии по
котикам северной части Тихого океана о хищничестве сивучей по отношению
278
Рис. 1'6.1. Нападение песца
на детеныша котика. Леж-
бище Урилье, о. Медный.
(Фото Ф. Г. Челнокова).
к котикам. По наблюдениям на о. Св. Георгия, в 1975 г. общие потери щенков
котиков вследствие поедания их сивучами составили 3.6—6.6% живых детены-
шей, что выразилось в 2.7—5.4 тыс. голов за сезон [Gentry et al., 1981]. С других
мест совместного обитания сивучей и котиков подобных сообщений не посту-
пало, однако случаи поедания сивучами теплокровных животных известны давно
[Тихомиров, 1959].
16.2. Пищевая конкуренция
Основу питания котиков на 75% составляют рыбы, поэтому котиков можно
отнести к ихтиофагам. Рыб поедают также акулы, тунцы, скаты, крупные пал-
тусы и представители семейств Hexagrammidae, Serranidae, Pleuronectidae, Ga-
didae и некоторых других. К сожалению, количественных оценок этих конку-
рентных взаимоотношений дать невозможно из-за плохой изученности как пи-
тания, так и численности большинства рыб. Другой группой пищевых конку-
рентов котиков являются морские птицы; их не менее 30 видов, большинство
из которых являются конкурентами первого порядка.
Для расчетов биомассы корма, потребляемого птицами в районах совмест-
ного обитания с котиками, мы использовали данные по численности птиц в гнез-
довой период в Беринговом, Охотском морях и в средней части Курильской гря-
ды [Гизенко, 1955; Успенский, 1959; Карташов, 1961; Велижанин, 1978; Sowls et
al., 1978; Bertoni, Sealy, 1979; Secora et al., 1979], а также на местах зимовок ко-
тиков [Шунтов, 1972; Manuwell, Cambel, 1979]. Сведения о количестве потреб-
ляемой пищи разными видами морских птиц разноречивы [Успенский, 1959; Ogi,
Tsujita, 1977; Harrison, Hida, 1980]. Рацион птиц, по одним данным [Успенский,
1958], на 50% состоит из рыб, по другим [Harrison, Hida, 1980] — на 86% или
до 95.9% [Ogi, Tsudjita, 1977]. По некоторым расчетам, морские птицы не спо-
собны воспитать потомство, если в их пище не будет менее 70% чистого объема
рыбы. Сопоставив все имеющиеся данные по суточным нормам потребления
пищи, по процентному содержанию рыбных объектов в рационе, мы рассчитали
для каждого района общую биомассу рыб, поедаемых птицами. При этом мы
допускали, что обитающие на тех или иных островах птицы и котики кормятся
в зоне досягаемости.
279
допускали, что обитающие на тех или иных островах птицы и котики кормятся
в зоне досягаемости.
На о-вах Прибылова обитают ок. 3 млн морских птиц, которые поедают за
летне-осенний сезон (июнь—октябрь) 52 тыс. т рыбы. Всего же в пределах на-
хождения котиков в этот период в Беринговом море от Алеутских о-вов до се-
верных границ Бристольского зал. живут почти 6.7 млн. морских птиц, которые
съедают за июль—октябрь более 150 тыс. т рыбы [Sowls et al., 1978]. Это лишь
вдвое меньше, чем потребляется морскими котиками прибыловской популяции.
На Командорских о-вах, восточном побережье Камчатки до мыса Олюторский
обитают более 4 млн птиц, которые, по нашим расчетам, съедают за июль—
октябрь 110.6 тыс. т рыбы. Если говорить о Беринговом море в целом, то здесь
обитают 55.2 млн морских птиц, которые за 5 месяцев потребляют 1.5 млн т
рыбы. Котики за это же время потребляют только 389.6 тыс. т. Уместно на-
помнить, что, по имеющимся данным [Low, 1976], промышленная добыча рыбы
в Беринговом море в 1974 г. составила менее 2 млн т.
На о. Тюлений обитают ок. 150 тыс. кайр и 1.5 тыс. других видов морских
птиц (чайки, чистики), которые за июнь—октябрь поедают 3.2 тыс. т рыбы. Все-
го на восточном побережье Сахалина (в зоне доступности котиков) обитают
75 тыс. морских птиц, которые за названный период времени съедают 19.7 тыс. т
рыбы. Котики за это же время поедают 24 тыс. т.
На побережьях штатов Вашингтон, Британская Колумбия и Аляска (США)
размножаются 216.6 тыс. пар морских птиц [Manuwell, Cambel, 1979]. Однако
численность их здесь в зимний период неизвестна. Отсутствуют данные по чис-
ленности птиц, зимующих в Японском море, т. е. в тех местах, где проводят
зиму северные морские котики. Можно лишь утверждать, что их плотность в
этих районах ниже, чем в Охотском и Беринговом морях летом [Шунтов, 1972].
В этой связи можно говорить о том, что конкурентные отношения на местах
зимовок котиков и морских птиц выражены не столь сильно, как возле лежбищ.
Из млекопитающих пищевыми конкурентами котиков следует считать всех
зубатых китов; из них наиболее многочисленной группой в районах зимовки
котиков являются дельфины. Анализируя состояние промысла и численности ки-
тообразных в мировом океане, М. Митчелл [Mitchell, 1975b] приводит данные
по численности белокрылой морской свиньи и сообщает, что она превышает
в районе Японских о-вов 30—50 тыс. особей. Аналогичные цифры указывает
М. Нишиваки [Nishiwaki, 1972]. По данным зарубежных авторов [Wilke, Kenyon,
1952; Wilke, Nichollson, 1958] и нашим материалам, рацион морской свиньи схо-
ден с таковым морских котиков у берегов Японии. Из расчета суточной нормы
потребления пищи (11.3% массы тела [Sergent, 1956]) и численности 30 тыс. голов,
средней массы тела 100 кг [Кузин, Перлов, 1975а] с декабря по май белокрылые
морские свиньи поедают 20.5 тыс. т морских организмов, т. е. почти столько
же, сколько потребляется котиками в этом районе зимовки.
Из ластоногих к пищевым конкурентам котиков, в первую очередь, следует
отнести сивучей, в меньшей мере, — ларгу и островного тюленя. Потенциаль-
ными конкурентами могут быть акиба, крылатка, морж и калан, но численность
этих видов в местах обитания котиков невелика, а спектр питания некоторых
из них отличается от такового последних.
280
Рис. 16.2. Взрослые самцы
котика и сивуча демонст-
рируют угрожающее пове-
дение. Командорские о-ва.
(Фото Ф. Г. Челнокова).
В северной части Тихого океана практически нет ни одного лежбища коти-
ков, на котором не обитали бы сивучи. Часто крупные лежбища сивучей на-
ходятся в зоне досягаемости котиков, и их трофические ареалы перекрещива-
ются. Более того, спектр питания котика и сивуча во многом сходен [Панина,
1964, 1966; Spalding, 1964; Перлов, 1975]. Однако, как показали исследования
[Кузин и др., 1977], в районах совместного обитания этих видов существует чет-
кое разделение экологических ниш, направленное на снижение конкуренции за
пищу. Основу питания сивуча в районе Курильских о-вов составлял минтай (70.7%
частоты встречаемости), в то время как у котиков он встречается только в 3.4%
желудков. Общими в питании обоих видов были кальмары, однако и в этом
случае наблюдаются различия в частоте их встречаемости [Кузин и др., 1977].
Расчеты показали, что в течение времени совместного обитания с сивучами
на Курильских о-вах котики потребляют 12.9 т морских организмов, из которых
3.3 т составляют анчоусы, 9.6 т — кальмары (Berryteuthis magister), 141.6 т —
минтай и 51 т — лососевые. Сивучи, в свою очередь, потребляют 6.9 тонн пищи,
в т. ч. 5.5 т минтая, 7.3 т окуней и 7.7 т кальмаров, из которых половину со-
ставили Todorodes pacificus.
У северо-западных берегов Сев. Америки кальмары, сельдь, треска, хек, кам-
бала, терпуг, минога и корюшка были общими в питании котика и сивуча. Од-
нако 63% пищи котиков составляли мелкие косячные рыбы, в то время как у
сивуча они составляли только 11%. Крупные виды рыб (лососи, хек и др.) со-
ставляли 12% для котика и 43% — для сивуча [Spalding, 1964].
Таким образом, основными пищевыми конкурентами северного морского ко-
тика следует считать птиц, в меньшей мере китообразных и ластоногих.
16.3. Конкуренция за места залегания
Как уже отмечалось, постоянными спутниками котиков на лежбищах явля-
ются сивучи (рис. 16.2).
Взаимоотношения этих видов на совместных лежбищах многократно обсуж-
дались в печати. Выводы исследователей весьма противоречивы. Так, С. В. Ма-
раков и Г. Д. Нестеров [1958] считали, что сивучи, кроме того, что выступают
281
как физический барьер для проникновения котиков на лежбище, давят их щен-
ков. В. Ф. Мужчинкин [1964а], Д. И. Чугунков [1968] и Ф. Г. Челноков [19716],
проводившие исследования на тех же лежбищах, что и два первых автора, при-
шли к противоположным выводам. А. Н. Белкин [1965, 1966], изучавший взаи-
моотношения котиков и сивучей на совместных лежбищах на Курильских о-вах,
вслед за С. В. Мараковым и Г. А. Нестеровым отмечал, что сивучи препятствуют
залеганию котиков, занимают лучшие места и давят детенышей. Кроме того,
А. Н. Белкин описал многочисленные случаи прямого антагонизма этих видов.
В общем, территориальная конкурентоспособность котика как вида была оце-
нена им очень низко. Основываясь на своих данных, он предлагал на гаремных
лежбищах искусственно сокращать численность сивуча как более сильного тер-
риториального конкурента котиков, но менее ценного промыслового вида.
Однако всестороннее изучение конкурентных отношений котика и сивуча
[Кузин и др., 1977] показали, что территориальные взаимоотношения у этих
видов выражены не столь сильно. Хотя они и занимают одни и те же площади
залежек, у них наблюдается четкое несовпадение репродуктивных циклов. Ко
времени “расцвета” гаремной жизни у котиков (середина июля) гаремная жизнь
у сивучей полностью заканчивается. Новорожденные сивучи в это время уже
начинают сходить в воду. Позже (в августе), когда новорожденные котики под-
растают, они тоже спускаются с лежбищ к воде и учатся плавать. К этому вре-
мени сивучата вместе с родителями покидают лежбища и размещаются на близ-
лежащих камнях или уходят на соседние скалы и острова. Таким образом, в
результате несовпадения периодов спаривания, деторождения и выращивания
молодняка создаются благоприятные условия для поочередного использования
котиками и сивучами одних и тех же площадей лежбищ. Кроме того, эти виды
на лежбищах отдают предпочтение разным участкам. Сивучи, как правило, за-
нимают более низменные, равнинные участки. Конечно, если эти места остаются
свободными, то они осваиваются котиками, а при наращивании численности
сивучи могут потеснить их. Однако, наблюдениями установлено, что на леж-
бищах всегда есть свободные участки территории, где имеется возможность раз-
меститься тому и другому виду. Так, по нашим наблюдениям, на о. Тюлений
при большой плотности котиков на лежбище в 1966—68 гг. на него выходили
до 25 сивучей, а с уменьшением численности первы число последних увеличилось
в десятки раз.
В целом, оценивая конкурентные отношения северного морского котика и
сивуча, можно сказать, что они не антагонистические. По классификации Ю. Одума
[1975], это несовершенный тип конкурентных отношений.
На некоторых лежбищах котиков залегают настоящие тюлени (ларга, остров-
ной тюлень). Так, на скалах Каменные Ловушки (Курильские о-ва) ежегодно
залегает небольшая группа (10—35 особей) островных тюленей, на Тюленьем
размещаются 500—700 ларг. Безусловно, залегающие тюлени занимают часть
территории лежбищ котиков, но численность их повсюду мала, и оказать су-
щест-венного влияния на размещение котиков они не могут. То же можно ска-
зать про каланов, которые выходят на отдельные камни вблизи лежбищ котиков,
но их залежки временные, а численность животных на них незначительная.
282
Глава 17. СТРУКТУРА ПОПУЛЯЦИЙ
Знание структуры популяций интенсивно Промышляемых видов является, той
основой, на которой базируется успех ведения промысловых хозяйств. Поэтому
уже первые исследователи северного морского котика, вольно или невольно со-
прикасавшиеся с промыслом, пытались внести элементы рационализма в его по-
становку на базе изучения отдельных элементов структуры популяций. И теперь
ежегодно составляемые прогнозы добычи котиков в своей основе содержат све-
дения о половой и возрастной структуре популяций. Не менее важны сведения
о пространственном распределении животных, которые необходимо учитывать
при организации промысловых хозяйств и при изучении основ биологии вида.
В настоящее время в северной части Тихого океана нет ни одной популяции
котиков, которая бы могла служить эталоном естественной структуры. Крупные
популяции, репродуктивными центрами которых являются острова Прибылова,
Командорские и Тюлений, находятся под воздействием ежегодного селективного
промысла, направленного в основном на самцов 3—5-летнего возраста. Котики
курильской репродуктивной группировки находятся в стадии восстановления
численности, а на о. Сан-Мигель — в стадии зарождения. Каждая из этих груп-
пировок характеризуется присущими ей миграционными способностями и при-
током эмигрантов с других лежбищ, разной смертностью животных на берегу
и в море, вследствие чего имеют разный половозрастной состав.
17.1. Половая структура
Среди исследованных более чем 10 тыс. плодов котиков, взятых от самок,
добытых по всему ареалу, 48.4% составили самцы и 51.5% — самки. В Берин-
говом море первичное соотношение полов эмбрионов близко к названному, в
т. ч. в западной части моря (у Командорских о-вов) самцов было 50.6%,
самок — 49.4%, а в восточной части (у о-вов Прибылова) самцов — 49.1%,
самок — 50.9%. У самок котиков, добытых в Японском море, несколько больше
было эмбрионов-самок (53.3%), чем самцов (46.7%), а в западной и восточной
частях Тихого океана это соотношение было примерно одинаковым (соответ-
ственно, 49.7% и 50.3% и 50.0% и 50.0%).
Пол эмбрионов зависит от возраста самок (табл. 17.1). Наибольшее число
самцов рождается у самок старше 10 лет, а у самок 8—10 лет чаще рождаются
самки, что хорошо согласуется с пространственной структурой популяций, зи-
мующих в разных частях ареала.
Вторичное соотношение полов близко 1:1 (в среднем на каждого самца при-
ходится 0.94 самки).
В первые месяцы жизни смертность самцов несколько больше, чем самок,
и уже ко времени ухода котиков на зимовку среди сеголеток больше остается
в живых самок [Chapman, 1964; Ichihara, 1974; Фрисман и др., 1985].
283
Таблица 17.1
Соотношение полов эмбрионов (самцы:самки) в зависимости от возраста
беременных самок котиков
Возраст самки Тихий океан, западная часть Японское море Всего
4-7 лет 1:1.05 1:1.23 1:1.07
8-10 лет 1:1.21 1:1.28 1:1.23
>10 лет 1:0.93 1:1.14 1:1.02
Таблица 17.2
Динамика соотношения самцов и самок в модельной популяции котиков при
разных уровнях промыслового использования
Самцы Самки Соотношение самцов и самок
Воз- раст Коэфф, выживаемости Воз- раст Коэфф, выживаемости В неопромыш- ляемых популяциях При изъятии 60% холостяков При изъятии 80% холостяков
0 0 1:1 1:1 1:1
1 0.48 1 0.50 1:1.04 1:1.04 1:1.04
2 0.76 2 0.80 1:1.09 1:1.09 1:1.04
3 0.86 3 0.86 1:1.09 1:1.79 1:1.09
4 0.87 4 0.94 1:1.18 1:1.32 1:2.28
5 0.88 5 0.96 1:1.29 1:3.50 1:7.50
6 0.85 6 0.81 1:1.23 1:3.50 1:8.15
7 0.81 7 0.85 1:1.59 1:3.40 1:7.84
8 0.74 8 0.91 1:1.97 1:4.20 1:8.19
9 0.70 9 0.87 1:2.75 1:5.25 1:10.20
10 0.62 10 0.87 1:3.95 1:7.30 1:18.25
11 0.62 11 0.86 1:3.95 1:9.19 1:25.20
12 0.46 12 0.86 1:7.20 1:18.00 1:36.00
12 0.46 13 0.86 1:13.28 1:31.00 1:93.00
14 0.42 14 0.84 1:26.00 1:78.00 1:156.00
15 0.42 15 0.84 1:66.00 1:165.00 1:330.00
16 0.42 16 0.78 1:121.00
Основные коррективы в соотношение полов в дальнейшем вносит промысел.
В последнее время промыслом изымаются 60% холостяков, приравненных к 2-
летнему возрасту (25% от численности поколения). Имеются расчеты о возмож-
ности добычи до 80% холостяков без ущерба для популяции [Фрисман и др.,
1985]. Картина соотношения полов при названных нормах изъятия представлена
в таблице 17.2.
284
10
№№ ИЮНЮ ЯМУ SI Si SS St 59 61 Si 65 67 № 71 73 75 77 79
пли
Рис. 17.1. Число самок, приходящихся на одного секача в репродуктивной
группировке котиков на о-вах Прибылова:
1 — число рожавших самок, 2 — число половозрелых самок.
В среднем соотношение полов в модельной популяции составило 1:1.47, при
изъятии 60% холостяков — 1:2.35, при изъятии 80% — 1:2.94. Соотношение сам-
цов и самок в половозрелой части популяции (самцы 7 лет и старше, самки 4 года
и старше) составит при отсутствии промысла 1:7.2, при изъятии 60% холостяков
1:12.3, при добыче 80% холостяков — 1:30.
В реальных популяциях соотношение полов в половозрелой части популяции
можно определить достаточно точно, т. к., во-первых, известна численность се-
качей, ежегодно учитываемых на лежбищах, во-вторых, по численности щенков
(тоже ежегодно учитываемых) можно определить число рожавших самок, а в-треть-
их, по данным добычи самок в море определяется доля прохолоставших по тем
или иным причинам в каждом возрастном классе.
Самый большой ряд наблюдений, касающихся соотношения полов, накоплен
по репродуктивной группировке котиков о-вов Прибылова (рис. 17.1).
Наивысшие показатели числа самок, приходящихся на одного секача, на-
блюдались в 1912—14 гг., а потом в 60-х годах. В первый период нарушение
соотношения полов было вызвано нерегламентируемым береговым и морским
промыслом перед заключением Конвенции по сохранению котиков северной
части Тихого океана в 1911 г. В связи с последующим 5-летним “запуском” про-
мысла на берегу и запрещением убоя котиков в море после заключения Кон-
венции, соотношение полов резко улучшилось и к 1917 г. составило 1:17. По-
этому в 1918—20 гг. американские промысловики целенаправленно выбили не-
сколько тысяч полусекачей и секачей для создания наилучшего соотношения
полов в репродуктивной части стада. В результате половая структура вновь была
изменена в сторону увеличения количества самок, приходящихся на самца.
Новое снижение числа самцов на о-вах Прибылова наблюдалось в конце 60-х
и начале 70-х годов, что было вызвано, отчасти, чрезмерным промыслом хо-
лостяков, а также возросшей смертностью животных всех возрастных и половых
групп в море [Eberhardt, 1981; York, Hartley, 1981; Trites, 1992].
Сравнение двух кривых на рис. 17.1 показывает, что с 1969 г. многие поло-
возрелые самки не принимали участие в размножении и не приносили потомство.
Такое положение наблюдалось при соотношении секачей и рожавших самок 1:40
и выше.
285
Рис. 17.2. Число самок, приходящихся на 1 секача в популяции котиков
Командорских о-вов (А) и иа о. Тюлений (Б):
1 — число рожавших самок, 2 — число половозрелых самок.
Сведения о половой структуре популяции котиков Командорских о-вов в про-
шлом отрывочны [Stejneger, 1898; Суворов, 1912; Бойцов, 1934], но достаточно
показательны. Так, в 1909—11 гг. количество рожавших самок на 1 секача было
от 79 до ПО, в 1912—30 гг. не превышало 30. С 1958 по 1980 гг. соотношение
полов в половозрелой части популяции не превышало 1:40 (рис. 17.2).
Характерно также и то, что в командорской популяции котиков количество
рожавших и половозрелых самок, приходившихся на 1 секача, не сильно раз-
личалось, чего нельзя сказать о других популяциях. Думается, это происходит
потому, что в этой популяции соотношение половозрелых самцов и самок не
выходит за рамки 1:40.
В популяции котиков о. Тюлений за период наблюдений с 1958 по 1980 гг.
увеличение числа рожавших самок на 1 секача отмечалось в середине 70-х годов
(рис. 17.2) В этот период число прохолоставщих самок значительно увеличилось,
поэтому ход кривых на графике во многом сходен по характеру с описанным
для прибыловской популяции. Таким образом, и для тюленьевских котиков ха-
рактерен упомянутый выше показатель соотношения полов (1:40), при изменении
которого в сторону его увеличения происходит рост числа прохолоставших
самок.
286
Итак, большое количество самок, приходящихся на 1 секача, оказывает за-
метное влияние на воспроизводительный потенциал популяций северного мор-
ского котика. Этот вопрос часто обсуждается в литературе, однако определен-
ного суждения о нормах соотношения полов нет. Одни исследователи предлагали
вести промысел таким образом, чтобы на каждого секача приходилось 10—15
самок [Волошинов, 1887; Генерозов, 1916]; другие считали, что оптимальным яв-
ляется соотношение в половозрелой части популяции 1:30—40 [Суворов, 1912;
Бойцов, 1934; Огнев, 1935; Ильина 1950; Дорофеев, Бычков, 1964; Арсеньев, 1968;
Кузин, Панина, 1977а; Челноков, 1977; Земский, Кузин, 1984]. По мнению
Н. А. Гребницкого [1902] и Н. А. Смирнова [1908], возможно поддержание со-
отношения полов до Г.50—60. Допускали также, что самец способен оплодот-
ворить до 100 самок за сезон [Bartholomew, Hoell, 1953]. Не исключено, что по-
следний показатель соотношения полов возможен при определенных условиях.
Если мы обратимся к истории, то увидим, что на островах Командорских и При-
былова в 1909—11 гг. на каждого самца приходилось более 60 оплодотворенных
самок, а общее число половозрелых особей, видимо, было не менее 80—90 голов.
На отдельных лежбищах Командорских о-вов количество самок, приходящихся
на секача, вдвое—втрое превышало названное. На Тюленьем в 70-х годах при
соотношении полов в репродуктивной части популяции 1:110 каждый самец по-
крыл свыше 60 самок. В то же время на Урильем лежбище (Командорские о-ва),
где сохраняется соотношение взрослых самцов и самок в приделах 1:6—8, каж-
дый самец покрыл в среднем только 13.9—34.2 самки [Владимиров и др., 1982].
Для сохранения высоких репродуктивных показателей самок большое зна-
чение имеет возрастной состав производителей. Наблюдениями установлено, что
все самцы 11 лет и старше участвуют в размножении, в то время как 7-летние
особи покрывают только 18.6% самок [Владимиров и др., 1982]. Половая ак-
тивность секачей в течение сезона размножения не остается постоянной. В се-
редине июля самцы наиболее активны [Bartholomew, Hoell, 1953], и в это время
у них наблюдаются наивысшие показатели сперматогенеза [Земский, Кузин,
1984]. В начале августа у секачей, добытых на холостяковых залежках, сперма-
тозоиды отсутствовали, либо их количество не превышало 1.38 млн/мм3, а у га-
ремных секачей при этом оно составляло 4 млн/мм3. В августе наблюдается
выход на лежбища молодых, впервые вступающих в размножение самок, и для
оплодотворения их всех нужно, чтобы в популяции всегда было достаточно фи-
зиологически активных самцов. В связи с этим исследователи часто рассчиты-
вают норму резервных секачей, приходящихся на одного гаремного самца. По-
этому всегда подсчитывается число гаремных и резервных секачей на лежбищах
отдельно. Исходя из такого подхода, исследователи часто рассчитывают число
рожавших самок на 1 гаремного секача для установления соотношения полов
в ©промышляемых популяциях. Однако, по нашему мнению, такой подход не
оправдан. Численность гаремных секачей определяется количеством половозре-
лых самок и площадью гаремных залежек. Даже в годы, когда количество самок,
приходящихся на 1 секача, было очень большим, число гаремов не увеличива-
лось. И наоборот, когда самок было мало и они залегали более разреженно,
секачи продолжали занимать место на гаремной территории, многие из них не
имели самок в течение всего сезона размножения, а в других гаремах находилось
всего по несколько самок. Представляется, что при расчетах потребного коли-
чества взрослых самцов в ©промышляемых популяциях котиков наиболее целесо-
287
Таблица 17.3
Половозрастная структура популяции котиков на о-вах Прибылова (% от
общей численности) [из: Бойцов, 1934]
Половозрастные группы Годы
1917-21 1922-26 1927-29
С а м ц ы
Детеныши 14.7 15.4 16.2
Годовики 9.4 8.8 8.5
Двухлетки 6.5 6.2 5.8
Трехлетки 2.6 2.6 1.3
Четырехлегки 1.6 2.1 1.3
Пятилетки 1.7 1.8 1.4
Полусекачи 1.8 0.9 1.4
Секачи 3.1 1.0 1.5
С а м и
Детеныши 14.7 15.4 16.2
Молодые (1—2 года) 15.5 15.9 15.0
Взрослые (3 года и старше) 29.3 30.8 32.3
образно иметь ввиду не число гаремных секачей, приходящихся на определенное
число самок, а общее их количество. Если не будет разделения секачей на га-
ремных и резервных, это значительно облегчит расчеты и общий подход к про-
гнозированию добычи.
Однако известно, что репродуктивные показатели самок котиков находятся
в прямой зависимости от их возрастного состава [Кузин, Панина, 1977а; Кузин,
1978]. Каждая популяция котиков характеризуется собственной спецификой воз-
раста наступления половой зрелости и цикличностью плодоношений. Поэтому
репродуктивный успех популяции прямо зависит от ее качественного состава,
а значит, при разной возрастной структуре популяций соотношение полов в них
не должно иметь одинаковых показателей. Нашими исследованиями установлено
[Кузин, Панина, 1977а; Кузин, 1978; Земский, Кузин, 1984], что при преобладании
в популяциях самок младших возрастных групп соотношение половозрелых сам-
цов и самок может составлять 1:60, а при старении популяций поддерживаться
на уровне 1:30—40.
Кроме отмеченной зависимости между соотношением полов и воспроизво-
дительными способностями популяций, половая структура оказывает соответ-
ствующее влияние на пространственную структуру. При недостатке секачей в
популяциях холостяки могут проникать на гаремное лежбище, что отрицательно
сказывается на ритме гаремной жизни и даже на выживаемости щенков [Чел-
ноков, 1974]. -
288
проценты vm
Рис. 17.3. Пирамида возрастов популяции котиков на о-вах Прибылова в 50-х
годах [по: Kenyon et al., 1954]
17.2. Возрастная структура
При исследовании возрастной структуры популяций северного морского ко-
тика ранее исходили из чисто практических целей, суть которых заключалась
в том, что промыслом изымаются только молодые самцы (3—5 лет), и для того,
чтобы оставалось достаточное количество производителей, с наибольшей тща-
тельностью велась разработка соответствующих приемов определения выжива-
емости этой возрастной группы. В связи с отсутствием промысла взрослых сам-
цов их возрастной состав не исследовался, и всех взрослых самцов объединяли
в единую группу “секачей”, куда входили животные 7—17 лет и старше. Чис-
ленность взрослых самок определяли по численности рожденных детенышей, а
молодых — по данным выживаемости одновозрастных самцов (табл. 17.3).
Рассматриваемый в таблице период времени можно охарактеризовать как
восстановительный, которому предшествовал 5-летний “запуск” промысла. При-
быловская популяция находилась в стадии интенсивного подъема численности.
В 1912 г. в ней насчитывалось 215.8 тыс., а в 1929 г. — 971.5 тыс. котиков [Бой-
цов, 1934; Scheffer, 1955]. В эти годы в популяции треть поголовья составляли
детеныши, столько же было рожавших самок. Среди самок не менее половины
приходилось на неполовозрелых. Остальную часть составляли самцы, среди ко-
торых доля производителей — 1—3.1%. Оценивая состояние популяции котиков
на о-вах Прибылова в эти годы, Л. В. Бойцов [1934] писал, что “соотношение
различных половозрастных групп котиков в американском стаде является наи-
более желательным, хотя и не идеальным”.
В 1940 г. прибыловская популяция достигла предельного уровня численности
[Roppel, Davey, 1965] и несколько лет находилась в стабильном состоянии. В
опубликованной в 1954 г. “Life table” [Kenyon et al., 1954] половозрастной состав
котиков значительно отличался от описанного выше для периода интенсивного
роста популяции (рис. 17.3).
В 50-х годах в прибыловской популяции уже преобладали взрослые особи.
На долю половозрелых самок приходилось 50% особей, в то время как непо-
ловозрелые самки составляли лишь четверть. Величина пополнения явно не обес-
печивала убыли, в связи с чем в последующие годы наблюдались спад общей
289
проценты VM
численности и перераспределение возрастного состава животных, при этом уве-
личивалась численность старых самок. Преднамеренная добыча более 300 тыс.
самок в эти годы ускорила процесс снижения численности.
В опубликованной в 1980 г. “Life table” [Kajimura et al., 1980a] просматри-
ваются признаки улучшения состояния прибыловской популяции. Наметилась
тенденция к омоложению возрастного состава, что позволило надеяться на воз-
никновение новой волны подъема общей численности (рис. 17.4)
В популяции котиков о. Тюлений в 1958—82 гг., как и в прибыловской по-
пуляции, наблюдался интенсивный рост общей численности (1958—67 гг.), ко-
роткий период стабилизации ее на высшем уровне (1968—70 гг.) и последующий
спад. Этим периодам колебания численности соответствовала определенная воз-
растная структура, полностью повторяющая описанную для популяции на о-вах
Прибылова. В годы подъема численности на Тюленьем преобладали молодые
неполовозрелые и половозрелые (до 7 лет) самки, численность которых вдвое
превышала общую численность средневозрастных (8—10 лет) и старших (более
10 лет) [Кузин, Панина, 1977а; Земский, Кузин, 1984]. Такой омоложенный воз-
растной состав самок этой популяции возник потому, что в 1943—45 гг. Япония
добыла на лежбище 45 тыс. самок репродуктивного возраста. По мере увели-
чения общей численности популяции в ней накапливались животные старших
возрастных классов, и соотношение возрастных групп постепенно выравнива-
лось. Ко времени достижения популяцией наивысшей численности количество
молодых половозрелых самок (4—7 лет) в ней равнялось числу самок старше
Этих лет. В годы снижения численности в популяции преобладали самки старших
возрастных групп (рис. 17.5).
Командорская популяция котиков за 1958—82 гг. не испытывала заметных
колебаний численности. В ней наблюдались только периоды интенсивного роста
и замедленного прироста численности. В связи с этим возрастной состав котиков
290
Рис. 17.6. Возрастной состав
самок котиков, добытых у
Командорских о-вов
Рис. 17.5. Пирамида
возрастов тюленьевской
популяции котиков:
белые столбики — 1958—67 гг.,
черные — 1968—74 гг.
процент (%)
1962,65 1966-56 1910,72 1974,7? W9 tfify
U-k-lMm ;Ш-8-Ю старше Ю/юя
здесь не испытывал резкого преобладания каких-либо возрастных групп (рис.
17.6), хотя на отдельных лежбищах этот процесс проходил, видимо, по-разному
[Челноков, 1981], отчего и не наблюдалось такого спада численности, как в дру-
гих популяциях.
Таким образом, в популяциях северного морского котика наблюдается четкая
взаимосвязь фазы развития общей численности и возрастного состава населения.
В период роста численности в популяциях преобладают животные младших
возрастных групп, и наоборот, в период спада численности в них превалируют
старшие возрастные группы. Резонно поэтому представить, что в качестве эндо-
генного фактора регуляций общей численности популяций котика выступает воз-
растная структура. Ее влияние на численность осуществляется через экологи-
ческие, этологические и физиологические параметры.
17.3. Пространственная структура
Распределение котиков на всех береговых залежках, как правило, однотипно.
Участвующие в размножении особи, занимают прибрежные части лежбища, по
окраинам которых располагаются так называемые холостяковые залежки, где
291
Рис. 17.7. Гаремные и холостяко-
вые залежки северных морских
котиков на Урильем лежбище
(о. Медный)
ЦЦ глинные еллыки
У//Л ХОЛОСТЯКОЛЫ1 МЛ1ЖКИ
присутствуют самцы всех возрастных классов, неучаствующих в размножении.
Детеныши концентрируются между гаремами, образуя “детские площадки”. То-
пография гаремных и холостяковых залежек часто определяется особенностями
рельефа. Так, например, на островах Медный и Св. Георгия, где лежбища часто
располагаются на узких пляжах, огражденных с тыловой стороны отвесными
скалами, холостяковые залежки всегда находятся на периферийных, смежных
частях пляжей (рис. 17.7). В то же время на островах Беринга и Св. Павла (реже
на других) холостяковые участки простираются вглубь островов и там расте-
каются вширь или даже занимают обособленные тыловые участки за гаремными
лежбищами.
В пределах акваторий островов, на которых котики образуют лежбища, плот-
ность распределения животных не одинаковая. При этом сегрегация по полу и
возрасту не обнаруживается. На удалении 2—3 миль от берега и далее котики
распределяются неравномерно. Так, с западной стороны о. Тюлений котики очень
редки, с юга встречаются чаще, а наибольшая встречаемость отмечается с вос-
точной стороны острова в квадрате 48°20'—48°40' с. ш. и 144°40'—145°20' в. д.
Распределение морских котиков по полу и возрасту в Охотском море изменяется
во времени в связи с неодновременностью подхода животных разного возраста
к лежбищам (рис. 17.8).
В июне—июле у островов преобладают самцы разного возраста и половоз-
релые самки. В августе к островам начинают подходить молодые самки, и их
доля увеличивается до 23%, а старых снижается до 58.4%, против 7—10% и 98%,
соответственно. В сентябре возрастной состав самок выравнивается, но преоб-
ладающей группой остается старшая. Отчасти это зависит от того, что к осени
самки все больше времени проводят в воде. В октябре в районе мыса Анива
в биологической пробе присутствовали самки всех возрастных групп, но пре-
валирующей оставалась группа самок старше 10 лет. Из общего числа добытых
в октябре 27% составляли нерожавшие особи, что почти вдвое больше, чем в
августе; среди них 89% — 2—3-летние особи. Годовалые встречались единично.
Несколько увеличилась доля нерожавших самок (10.3%); количество многоро-
жавших самок заметно уменьшилось (43%). Среди самцов присутствовали звери
всех возрастных классов с небольшим преобладанием 3—5-летних (рис. 17.8).
В ноябре в небольшой биологической пробе из Охотского моря 82.5% составляли
самки старше 10 лет.
292
49
48
Ш М5 Мб 147
Рис. 17.8. Сезонная динамика половозрастного состава котиков в Охотском море
В акватории Командорских о-вов пространственное распределение котиков
отличается от такового у о. Тюлений тем, что там имеется несколько мест наи-
большей концентрации животных, фактически приуроченных к находящимся у
этих островов лежбищам. Из них 2 лежбища находятся с северо-западной сто-
роны о. Беринга и 2 — у южной оконечности о. Медный с его западной и вос-
293
Рис. 17.9. Сезонная динамика половозрастного состава котиков в западной части
Берингова моря
294
Август
10 самки
п Г
Рис. 17.9. Продолжение
Район № 3
Июль
13 самцы
25 самки
ВОЗРАСТ (ГОЛЫ)
295
Рис. 17.9. Окончание
точной сторон. Еще одно место постоянных концентраций северных морских
котиков находится на банке Китоловная в проливе между островами Медный
и Беринга (рис 17.9).
Распределение котиков по полу и возрасту в названных местах их концент-
рации различно. В период подхода котиков весной к островам образующиеся
концентрации этих животных на банке Стелмейт характеризуются присутствием
здесь в основном секачей [Перлов, 1968]. У юго-восточной оконечности о. Мед-
ный в июне преимущественно встречаются половозрелые самки, полусекачи и
холостяки. Секачи в это время уже находятся на лежбище. На банке Китоловная
94% добытых животных составили самки старше 3 лет. Самцы были малочис-
ленны. В районе Северо-Западного лежбища (о. Беринга) также в основном
встречены половозрелые самки.
В дальнейшем, как и в районе о. Тюлений, от месяца к месяцу половозрастной
состав животных меняется в сторону его омоложения в связи с подходом к ост-
ровам неполовозрелых животных. Лишь на банке Китоловная в течение всего
сезона превалируют взрослые особи.
В акватории о-вов Прибылова плотность котиков выше с западной и южной
сторон, а сезонной динамике половозрастного состава присущи те же черты,
что и в других акваториях, прилегающих к островам размножения, в частности,
к островам Тюлений и Командорским.
Представленные графики и краткое описание пространственного распреде-
ления морских котиков в акватории островов их летнего обитания характерны
только для периода размножения. С началом миграции животных с островов
на места зимовки картина резко меняется. С распадом гаремов наступает полная
дезорганизация половозрастного состава котиков на береговых залежках и даже
в море вблизи островов. На гаремные пляжи начинают выходить холостяки. На
холостяковых залежках появляются самки и детеныши. В прибрежных аквато-
риях встречаются детеныши. Часть Животных переходит на чисто пелагический
образ жизни, и они могут быть встречены на значительном удалении от островов.
В целом, пространственная структура популяций котиков в летний период
весьма специфична: ярко выраженная сегрегация животных по полу и возрасту
на береговых залежках в гаремный период жизни и отсутствие ее после гарем-
296
кого периода как на берегу, так и в море. В пределах островов размножения
имеются места, наиболее часто посещаемые животными, которые, надо полагать,
отличаются повышенной биопродуктивностью. Таковые в прошлом назывались
у американских промышленников “sealing grounds”. Эти кормовые поля у каж-
дого из островов обитания морских котиков находятся на разном удалении от
берега и, соответственно, время, затрачиваемое животными на их посещение,
разное, что отражается на общем физическом состоянии участвующих в размно-
жении взрослых животных и цикличности выкармливания детенышей.
В осенне-зимний период морские котики предпринимают дальние миграции,
в период которых находятся в постоянном движении, осваивая значительные
морские пространства северной Пацифики. Этому периоду жизни северных мор-
ских котиков посвящена обширная литература, в которой отражены особенности
пространственного распределения животных в зависимости от пола и возраста.
Годовой жизненный цикл северного морского котика складывается из двух
периодов: берегового (репродуктивного) и морского (нагульного), в течение ко-
торых пространственная структура популяций этих животных различна. В реп-
родуктивный период морские котики концентрируются на лежбищах, где исклю-
чительно высока плотность и четко выражена половая и возрастная сегрегация,
являющаяся прямым следствием полигинии вида. В акваториях лежбищ плот-
ность животных по мере удаления от лежбищ снижается, но в пределах аква-
торий, прилежащих к островам размножения, имеются участки, наиболее часто
посещаемые животными, где они кормятся. В некоторых таких местах возможно
преобладание тех или иных половых или возрастных групп животных (например,
банка Стелмейт, банка между островами Медный и Беринга). На путях миграции
к районам зимовки концентрации животных неустойчивы и непродолжительны.
Они определяются кормностью угодий. На местах зимовки котики также пос-
тоянно находятся в движении, и наибольшие их концентрации и продолжитель-
ность пребывания в каком-либо районе определяются гидрометеоусловиями и
наличием легкодоступной пищи. С изменением условий среды и исчезновением
концентраций объектов питания, плотность котков снижается, распределение в
пространстве изменяется.
Возможно, что существующая пространственная сегрегация северных мор-
ских котиков по полу и возрасту в районах зимовки позволяет им лучше ис-
пользовать пищевые ресурсы [Мараков, 1974]. Представляется, что существую-
щая, хотя и не столь четко выраженная сегрегация по полу и возрасту в море,
мозаичность распределения по акватории в период миграции и на местах зи-
мовки, кроме зависимости от пищевого фактора, может являться как следствием
неодинакового отношения разных половозрастных групп котиков к условиям
обитания, так и накопленного опыта предшествующих миграций.
297
Глава 18. РАЗМНОЖЕНИЕ
18.1. Возраст наступления половой зрелости
Возраст наступления половой зрелости самцов. Исследователи конца прошло-
го—начала нашего столетия знали, что у 3-летних самцов северных морских ко-
тиков семенники опускаются в мошонку, и в эпидидимисе присутствуют спер-
матозоиды, что говорит о достижении половой зрелости [Elliott, 1882; Jordan,
1898—99; Гребницкий, 1902; Суворов, 1912; Бойцов, 1934]. Было также известно,
что в размножении принимают участие самцы не моложе 7 лет, способные про-
тивостоять другим самцам в борьбе за самок на гаремном лежбище.
Позже было установлено [Кузин, 1978], что не все самцы в 3 года достигают
половой зрелости. Только у 45% трехлеток сперматозоиды присутствуют в ко-
личестве не менее 1.31 млн/мм3 спермы. У 25% из них количество сперматозоидов
равно таковому у секачей в гаремный период жизни. Сперматозоиды отсутствуют
у 3-леток, масса семенников которых менее 44 г. Гистологические исследования
показали [Ащепкова, Дмитриева, 1982], что с 3-летнего возраста начинается за-
метное увеличение семенных канальцев, интенсифицируется развитие спермато-
генного эпителия, в котором встречаются конечные формы клеток. Заметные
изменения происходят в интерстициальной ткани, в которой увеличиваются раз-
меры клеток Лейдига. Все 4-летние самцы половозрелы. Количество спермато-
зоидов у них колеблется от 0.36 до 4.82 млн/мм3. Соответствующие прогрессив-
ные изменения происходят и в гистологических структурах семенников.
Наилучшего физического и физиологического развития самцы достигают в
возрасте 9—11 лет, и на их долю приходится от 21.5% до почти 40% оплодот-
воренных самок, в то время как на долю 7-летних самцов лишь 2% покрытий;
14.2% самок оплодотворяются 12—15-летними самцами [Лыскин, 1983]. Безус-
ловно, приведенные данные нельзя принимать как абсолютные. Репродуктивный
успех самцов во многом определяется фазой развития популяций и в первую
очередь зависит от численности и возрастного состава производителей. При низ-
ких показателях соотношения полов в популяции все самцы принимают участие
в размножении. На Урильем лежбище при соотношении полов среди полово-
зрелых самцов и самок 1:5-7 42.8% секачей не посещали гаремного лежбища
в течение всего репродуктивного периода, а 80% — не ежегодно. 56.6% секачей
участвовали в размножении в течение одного сезона, 32.5% — в двух, 8.4% —
в трех и 2.5% — в течение четырех сезонов. В среднем репродуктивные способ-
ности самцов при названном соотношении полов сохраняются, по мнению Н. Н. Лыс-
кина [1983], в течение 1.6 сезона. При недостатке секачей в популяции практически
все принимают участие в размножении, и на каждого приходится до 50—60 (в от-
дельные годы и больше) плодоносящих самок [Кузин, Панина, 1977а].
Самцы северных морских котиков имеют сезонную половую активность.
В зимний период сперматозоиды в эпидидимисе отсутствуют. Единичные спер-
матозоиды обнаруживаются в конце мая, и постепенно их количество возрастает
к периоду размножения (июнь—июль). В августе количество сперматозоидов у
молодых половозрелых самцов несколько больше, чем у гаремных, а у секачей
на холостяковых залежках в 3 раза меньше, чем у гаремных и молодых поло-
возрелых особей. Заметное снижение сперматогенеза начинается с сентября, и
лишь отдельные особи сохраняют половую активность до конца этого месяца
[Кузин, 1978].
Возраст наступления половой зрелости самок. В прошлом наступление поло-
вой зрелости у самок определяли с помощью визуальных наблюдений за участ-
вующими в размножении животными. Так, И. Вениаминов [1840] считал, что
самки котиков достигают половой зрелости на четвертом году жизни и впервые
приносят потомство в 5 лет. Н. Слюнин [1895], впервые изучивший макрострук-
туру яичников, определил, что самки котиков могут быть оплодотворены уже
в возрасте 3 лет, но чаще приступают к размножению на четвертом году жизни.
Однако, К. Таунсенд и Ф. Лукас [Townsend, Lucas, 1898] тоже в результате ана-
лиза яичников пришли к заключению, что часть самок прибыловской популяции
могут быть половозрелыми уже в возрасте 2 лет, а командорские самки дости-
гают половой зрелости в возрасте 3 лет. Е. К. Суворов [1912], в свою очередь,
считал, что “самки котиков Командорских островов достигают половой зрелос-
ти трех лет от роду, но полного развития достигают спустя год—два”. В при-
веденных мнениях разных авторов нет противоречий, т. к. они исследовали
самок котиков в разные годы и из разных популяций. Как теперь хорошо из-
вестно, возраст наступления половой зрелости самок не остается постоянным
из года в год, а изменяется с изменением интрапопуляционных параметров, в
первую очередь, таких как общая численность и половозрастное соотношение
репродуктивной части популяций. Это хорошо подтверждается анализом мас-
сового материала по многолетней изменчивости репродуктивного состояния раз-
ных популяций северных морских котиков. Так, в 1959—64 гг. возраст самок
котиков, добытых в северо-восточной части Тихого океана, у которых в яич-
никах присутствовали вполне развитые фолликулы, колебался от 2 до 6 лет
[Craig, 1964]. Среди 4-летних 62% были половозрелыми, среди 5-летних — 71%,
6-летние — почти все. Из этих данных следует, что возраст, при котором у боль-
шинства самок наступает половая зрелость, составляет 4 года. В то же время
в 1970—71 гг., по данным NPFSC, в той же популяции котиков в возрасте 4 лет
половозрелыми были лишь 2.2% самок, а среди 5-летних — 50%. В 70-х годах,
таким образом, большинство самок прибыловской популяции достигали поло-
вой зрелости в 5 лет.
Возраст наступления половой зрелости самок котиков, добытых в северо-за-
падной части Тихого океана, где в основном проводят зиму котики с о. Тюлений,
также колеблется в значительных пределах. Как видно на рис. 18.1, число про-
овулировавших 4-леток убывало с 80% в 1964 г. до 20% в 1970 г., а к 1982 г. их
число вновь увеличилось до 95%. Среди 5-леток наблюдалось аналогичное сни-
жение числа половозрелых особей с 100% в 1964 г. до 80% в 1971 г. У самок
более старшего возраста число впервые овулировавших оставалось более посто-
янным в течение всего этого времени.
Представленные данные по возрасту наступления половой зрелости самок из
северо-западной части Тихого океана показывают, что в 1968—76 гг. они в массе
достигали половой зрелости с 4 лет, а в предшествующие и последующие го-
ды — в 3 года.
298
299
Рис. 18.1. Доля половозрелых
самок котиков, добытых у
тихоокеанских берегов Японии:
1 — среди всех самок, 2 — среди 4-
летних, 3 — среди 5-летних, 4 —
среди 6-летних.
Таким образом, возраст наступления половой зрелости самок из разных по-
пуляций изменяется в значительных пределах. Сравниваемые популяции разли-
чались как по среднему возрасту наступления половой зрелости, так и по вре-
мени проявления высших и низших пределов этого показателя. Причину этого
явления разные авторы в разное время интерпретировали по разному. В. Шеф-
фер [Scheffer, 1955] обосновал, а Ф. Нагасаки [NagasaKi, 1961] и Д. Чепман [Chap-
man, 1961, 1964] математическим моделированием подкрепили зависимости пло-
довитости, в целом, и возраста наступления половой зрелости, в частности, от
плотности популяций. Однако дальнейшие наблюдения показали, что со сни-
жением плотности в прибыловской популяции репродуктивные показатели
снизились, а возраст первой репродукции самок увеличился [Trites, Bigg, 1992;
Trites, York, 1993]. Поэтому изменения в нормах беременности и в возрасте пер-
вой репродукции среди самок котиков на о-вах Прибылова признаны несовмес-
тимыми с парадигмой зависимости от плотности [Trites, York, 1993].
Причины изменения времени наступления половой зрелости самок котиков
из западной части Тихого океана исследовал К. Ешида [Yoshida, 1982]. Он ана-
лизировал длину и массу тела впервые вступающих в размножение самок, час-
тоту встречаемости котиков, которые имели желудки с пищей и без нее, цвет
желчи как индикатор времени и продолжительности питания животных, чтобы
выявить влияние питания на ростовые процессы. В результате выяснилось, что
в одних и тех же возрастных классах длина и масса тела половозрелых особей
больше, чем неполовозрелых, и что в годы, когда наблюдались наивысшие по-
казатели числа половозрелых животных среди четырехлеток, их размеры тела
были больше, чем в годы, когда самки позже достигали половой зрелости. Не-
смотря на то, что в последние годы содержимое желудков котиков снижалось
в объеме (это может быть показателем снижения пищевых ресурсов Охотского
моря), молодые самки стали вступать в размножение раньше, и размеры их тела
увеличились. Также установлено, что наличие большого числа половозрелых
самцов в популяции не стимулирует самок к более раннему вступлению в раз-
множение.
Таким образом, ни питание, ни соотношение полов в популяции не оказы-
вают непосредственного существенного влияния на возраст наступления половой
300
Таблица 18.1
Средние длина (L) и масса (М) 4-летних беременных и небеременных самок,
добытых в море в марте—апреле, и 3-летних холостяков котиков, добытых
в северо-западной части Тихого океана и на о. Тюлений
Годы Беременные самки Небеременные самки Холостяки
п L М п L М п L М
1958—67 467 115.8 28.8 419 111.8 24.0 5545 113.4 30.5
1968—76 26 115.1 29.6 129 111.0 24.8 1484 112.2 30.2
1977—87 58 118.0 29.5 24 114.0 25.2 1310 115.9 31.8
1988—94 246 115.5 34.4
зрелости самок. Прослеживается лишь прямая связь их полового созревания с
размерами тела.
Проведенный нами дополнительный анализ зависимости возраста наступле-
ния половой зрелости самок от их размеров тела подтвердил данные К. Ешиды.
Как следует из материалов таблицы 18.1, в одних и тех же возрастных классах
половозрелые самки крупнее неполовозрелых при любом состоянии численности
популяции и изменениях интрапопуляционных параметров.
Однако важно заметить, что описанное изменение размеров тела половоз-
релых 4-летних самок характерно не только для впервые вступающих в разм-
ножение, но и для неполовозрелых самок того же возраста и для 3-летних самцов.
Хотя проведенный анализ и приводит к выводу, что непременным условием для
вступления самок в размножение является достижение ими определенных раз-
меров тела, но в то же время показывает, что размеры тела находятся под конт-
ролем факторов, воздействующих на популяцию в целом.
Рост и развитие организмов обычно ассоциируются с трофическим фактором:
во внутриутробный и подсосный периоды способностью самок обеспечить по-
томство достаточным количеством пищи, а с переходом молодняка на самосто-
ятельное питание обилием, доступностью пищи и способностью животных удов-
летворять в ней свои потребности.
Темп роста плодов у впервые забеременевших самок ниже, чем у взрослых.
Новорожденные от таких самок тоже мельче, чем от самок старшего возраста
[Calambokidis, Gentry, 1985; Trites, 1990; Болтнев и др., 1991]. Причина этого
явления заключается в том, что молодые самки, продолжая сами расти, не обес-
печивают должного развития плода. Потомство от молодых самок, особенно
впервые рожающих, отстает по физическим показателям, которые являются
внешним отражением их общей физиологической незрелости, что снижает их
жизненные функции на последующих этапах существования. Щенки от молодых
самок рождаются в конце репродуктивного периода и, по-видимому, период мо-
лочного вскармливания у них короче, чем у родившихся раньше. Пищевые
марши с островов в море у взрослых самок короче, чем у молодых, и первые
совершают их чаще, чем вторые [Goebel, 1983]. У всех найденных на Коман-
дорских о-вах в ноябре мертвых щенков линька не была завершена [Суркина,
301
Рис. 18.2. Изменение среднеко-
гортных показателей массы
тела ювенильных котиков в
процессе их индивидуального
развития, по отношению к мас-
се тела новорожденных, взвешен-
ных на о. Тюлений 29 июля—
2 августа
личн. сообщ.]. Можно полагать, что большинство из них было рождено моло-
дыми самками.
За первый год жизни в море котики теряют до половины массы тела [Scheffer,
1981]; выживают из них преимущественно особи, имевшие наибольший вес при
рождении [Болтнев и др., 1991; Baker, Fowler, 1992]. Таким образом, возрастная
структура популяций котиков является исходной предпосылкой, определяющей
их физическое состояние и физиологическое развитие на начальных стадиях пост-
натального онтогенеза. Возможность проявления компенсации в росте на юве-
нильных и постювенильных стадиях во многом зависит от влияния многих фак-
торов. Наиболее важным из них, пожалуй, является трофический фактор.
К сожалению, из-за специфики экологии котиков самый ранний и наиболее
ответственный период их индивидуального развития, когда возможно проявле-
ние положительных тенденций в компенсаторных процессах, скрыт от исследо-
вателей, и судить о нем возможно лишь косвенно по результатам линейных из-
мерений и взвешиваний щенков, и затем только животных в возрасте 2 и более
лет, когда их начинают промышлять (рис. 18.2).
На рис. 18.2 видно, что масса тела молодых котиков от возраста к возрасту
изменяется неравномерно. Можно полагать, что экологические условия продол-
жают оказывать заметное влияние на их физическое развитие не только на ран-
них, но и на последующих стадиях постнатального онтогенеза. Однако конечный
результат взаимодействий северных морских котиков с окружающей средой из-за
неполноты имеющейся информации может быть оценен лишь в период вступ-
ления их в размножение. Одним из показателей этих взаимоотношений является
масса и длина тела, а отсюда способность оказаться конкурентоспособным за
место на гаремном лежбище у самцов и за более ранний возраст вступления
в размножение у самок.
Таким образом, возрастная структура популяций является исходной предпо-
сылкой, определяющей физическое состояние потомства и его чувствительность
к воздействию внешних факторов. При этом внешние условия в меньшей степени
влияют на ювенильных котиков, если они рождаются физически крепкими и фи-
302
зиологически зрелыми (преимущественно от родителей старшего возраста) и ухо-
дят в первую самостоятельную зимовку хорошо упитанными, и те же условия
среды, элиминируя часть приплода, снижают жизненный потенциал другой, если
большая часть приплода появляется на свет физиологически незрелой и уходит
на зимовку плохо подготовленной физически и физиологически. В этом, по на-
шему мнению, заключается однотипность действия регуляторного внутрипопу-
ляционного механизма, проявляющего себя в особенностях индивидуального
развития и, соответственно, отражающегося впоследствии на плодовитости впер-
вые вступающих в размножение самок.
18.2. Темп и ритм размножения
Рис. 18.3. Доля беременных самок в каждом возрастном классе в поколениях
1951—75 гг.
Зачерченный участок нижнего плато — период, когда в тюленьевской популяции котиков
было искусственно нарушено половое соотношение.
Данные прошлых исследований репродуктивных способностей самок север-
ного морского котика разноречивы. И. Вениаминов [1840] писал: “В первые годы
молодые самки морских котиков рожают ежегодно, а под старость через год,
и за всю жизнь могут приносить до 10—15 детей и даже больше. Третья часть
не родит. Через 22 года жизни они совсем бесполезны для приплода”. Н. Слюнин
[1895] считал, что самки за всю жизнь родят 6—8 раз, т. к. число желтых тел
в яичниках бывает не больше 7. Американские исследователи [Townsend, Lucas,
1898] также на основании исследований яичников полагали, что все половозре-
лые самки, начиная с 3-летнего возраста, рожают по 1 детенышу ежегодно и
сохраняют способность к размножению всю жизнь. Е. К. Суворов [1912] писал,
что самки достигают половой зрелости в 3 года, но от 4 до 23% из них в по-
следующие годы остаются яловыми. По мнению Л. В. Бойцова [1934], они со-
храняют способность к размножению всю жизнь. Е. И. Ильина [1950] зафик-
сировала случай родов у 22-летней самки. По данным К. Эбегглена и А. Роппела
[Abegglen, Roppel, 1959], большинство самок в возрасте 4 лет рожают впервые
и, возможно, возвращаются в гарем до тех пор, пока не утратят способность
к размножению. Максимальное число беременных зарегистрировано ими среди
7-летних. С. В. Дорофеев [1964] считал, что наилучшие воспроизводительные
способности у самок сохраняются, примерно, до 20-летнего возраста, но и в
более старшем возрасте они способны к деторождению.
303
Рис. 18.4. Доля беременных самок,
добытых в северо-западной (А) и
северо-восточной (Б) частях Тихого
океана
Рис. 18.5. Процент беременных
среди половозрелых (а) и общего
числа самок северного морского
котика (б) в разных возрастных
группах, добытых в
северо-западной части Тихого
океана
Рис. 18.6. Физиологическое
состояние самок с о. Тюлений:
1 — процент половозрелых от общего
числа самок котиков в возрасте 4 лет и
старше, 2 — процент проовулировавших от
числа половозрелых самок, 3 — процент
беременных от числа овулировавших, 4 —
процент беременных от числа
половозрелых, 5 — процент беременных от
всех самок в возрасте 4 лет и старше.
(00
90
80
100
90
80
70
60
50 ,
П * » I I I I I "I I.Г I I I I I » I I
(ОМ 66 68 7Q 72 74 П 78 80
Экспериментальных данных о ритме и темпе размножения для самок север-
ного морского котика нет. Проследить этот процесс по репродуктивным орга-
нам, как это попытался сделать Н. Слюнин, нельзя, т. к. следы от желтых тел
беременности не сохраняются всю жизнь [Craig, 1964; Yoshida, 1982], а плацен-
тарные пятна отсутствуют. В природе репродуктивный цикл (отчасти по кос-
венным признакам, вследствие чего нет уверенности в достоверности данных)
304
прослежен у отдельных самок на протяжении 7 лет, в течение которых некоторые
самки приносили потомство ежегодно [Владимиров, 1983]. Однако существова-
ние в популяциях котиков малопродуктивных поколений [Владимиров, 1983] не
подтверждается нашими данными (рис. 18.3). Все изменения репродуктивных
способностей самок из 25 поколений, показанных на рисунке, объяснимы на-
рушениями отдельных параметров популяции, ответственных за размножение.
Так, намеренное нарушение половой структуры популяции котиков на о. Тю-
лений в период 1972—76 гг. сказалось на репродуктивных показателях самок
из поколений 1961—72 гг., в каком бы возрасте они не оказались в эти годы.
Исследование ритма размножения отдельных самок представляет интерес с
точки зрения оценки потенциальной плодовитости. Для исследования плодови-
тости популяций гораздо важнее показатели репродуктивных способностей от-
дельных возрастных классов. Накопление таких данных, начавшееся в 50-х годах
[Wilke, 1951; Taylor et al., 1955; Abegglen, Roppel, 1959; Craig, 1964], было про-
должено и потом [Дорофеев, 1964; Кузин, Панина, 1977а; Yoshida, 1982; Земский,
Кузин, 1984]. Уже первый анализ массового материала по плодовитости показал,
что темп вступления в размножение самок из разных популяций не одинаков.
Кривые среднемноголетних данных о плодовитости самок котиков разных по-
пуляций хотя и имеют одинаковый вид, но смещены по шкале времени (рис. 18.4).
В северо-западной части Тихого океана самки вступали в размножение и до-
стигали наивысших показателей плодовитости раньше, чем самки из северо-вос-
точной части. Соответственно, угасание репродуктивных способностей самок из
северо-западной части Тихого океана происходило со сдвигом влево на год—два.
Плодовитость самок в зависимости от возраста не остается постоянной. Она
изменяется во времени под воздействием эндо- и экзогенных факторов. При этом
для разных популяций эти изменения не согласованы во времени (рис. 18.4). Так,
в период интенсивного роста численности тюленьевской популяции котиков
(1958—63 гг.) часть самок оплодотворялась уже в возрасте 2 лет, а доля бере-
менных среди 4-леток составляла 45%. Среди 5-летних число беременных пре-
вышало 80%. В период стабилизации численности на высшем уровне плодови-
тость молодых самок резко упала (рис. 18.5).
При нарушении половой структуры популяции (1972—76 гг.) уменьшение
плодовитости отмечалось во всех возрастных классах самок. При восстановле-
нии оптимального соотношения полов в репродуктивной части популяции пло-
довитость самок всех возрастных групп начала повышаться несмотря на то, что
популяция начала испытывать депрессию численности.
Прибыловская популяция котиков достигла пика численности в 40-х годах
[Johnson, 1975]. Начавшийся после этого процесс снижения численности продол-
жается и теперь. Имеющиеся данные по плодовитости за 1958—72 гг. показы-
вают, что в этой популяции среди 3-летних самок в 1959 г. зарегистрировано
лишь 2% беременных, а среди 4—6-летних доля беременных была в несколько
раз меньше, чем в популяции котиков на о. Тюлений. Плодовитость самок из
старших возрастных классов в обеих популяциях не испытывала таких резких
изменений в зависимости от состояния численности.
Таким образом, характерной общей чертой изменения плодовитости самок
северного морского котика является то, что в период достижения популяциями
305
Рис. 18.7. Доля неовулировавших
самок от числа половозрелых в
популяции котиков на о. Тюлений
W64, «б 66 ?0 П Ц 76 76 «О 82 ГОЛЫ
верхнего уровня численности первыми от участия в размножении устраняются
молодые, впервые вступающих в размножение особи.
Поскольку плодовитость складывается из многих параметров, часто не за-
висящих друг от друга, то представляется целесообразным более детальное их
рассмотрение. Как видно на рис. 18.5 и 18.6, доля половозрелых самок в по-
пуляции на о. Тюлений колебалась в пределах 20—100% в возрастных классах,
впервые вступающих в размножение, или в пределах 83—100% от общей чис-
ленности самок старше 4 лет.
При нормальном соотношении полов доля беременных самок составляет в
среднем 80% (рис. 18.5, 18.6), в т. ч. среди 4-летних — 67% (30—86%), 5—7-
летних — 80% (69—93%), 8—10-летних — 92% (78—99%), старше 10 лет — 78%
(69—97%). При нарушении половой структуры популяции плодовитость снижа-
ется в среднем до 64%, а в отдельных возрастных группах до низших пределов
из вышеназванных показателей (рис. 18.5, 18.6).
. Определенное число половозрелых самок не овулируют. Такие особи встре-
чаются во всех возрастных классах кроме 4-леток, потому что все неовулиро-
вавшие 4-летки относятся к неполовозрелым. В среднем в популяции о. Тюлений
неовулировавшие самки составляют 6% (рис. 18.7), в т. ч. среди 5—7-летних — 4.4%
(2.2—6.1%), среди 8—10-летних — 1.8% (1.6—6,3%), среди 11—15-летних — 2.9%
(1.2—4.2%), среди 16—20-летних — 12.8% (5.8—25.0%) и старше 20 лет — 25.8%
(16.7—50.0%). Видимо, число неовулировавших половозрелых самок в популя-
циях котиков определяется возрастным составом плодоносящих особей. Так, за-
метное увеличение числа неовулировавших особей в 1970 и 1973—78 гг. (рис.
18.7) хорошо согласуется с преобладанием в эти годы в тюленьевской популяции
котиков самок старше 10 лет (рис. 18.8).
Устранение от размножения старых самок объясняется нарушением функции
их яичников. У неовулировавших особей яичники были меньше размером и име-
ли признаки нарушения клеточной структуры [Yoshida, 1982].
Не все овулировавшие самки приносят потомство (рис. 18.6). В среднем за 1964—
82 гг. доля беременных среди проовулировавших составила 80%, (68—92%).
306
Рис. 18.8. Основные биоло-
гические показатели тюлень-
евской популяции:
А — численность разных поло-
возрастных групп и число самок
на I секача, Б — возрастной со-
став самок.
Минимальной она была в 1972—76 гг. Среди 4-леток беременные составили 68%,
5—7-леток — 83%, 8—10-леток — 88%, старше 10 лет — 83%. Причина несо-
стоявшейся беременности среди проовулировавших самок может быть вызвана
как причинами эндогенного характера, так и в результате нарушения структуры
популяции. В качестве эндогенного фактора могут выступать случаи нарушения
функции половых органов, потеря бластоцисты в период диапаузы внутриут-
робного развития, аборты, резорбция эмбрионов [Craig, 1964]. Однако эти фак-
торы, по-видимому, несильно сказываются на плодовитости котиков и, как вид-
но из вышеприведенных данных, не зависят от возраста самок. Гораздо сильнее
на плодовитости проовулировавших самок сказывается сбалансированность по-
ловозрастного состава популяций. Так, при недостатке секачей на о. Тюлений
(1972—76 гг.), когда на каждого секача приходилось более 50—60 самок, часть
проовулировавших самок не забеременела (рис. 18.J, 18.6). Недостаток секачей
испытывали 40 возрастных классов из разных поколений (рис. 18.3). Это рав-
носильно тому, что 2 поколения самок не участвовали в размножении, а общие
потери в приплоде составили в среднем 50—60 тыс., из которых 30 тыс. самок.
Таким образом, количество беременных самок в популяциях северного мор-
ского котика находится под контролем многих факторов. Изначально плодо-
витость популяции определяется качественным состоянием маточного поголовья
и получаемого от него потомства. Такая программа гарантирует самкам высо-
кий темп роста и раннее вступление в размножение, что оказывает наиболее
существенное влияние на плодовитость популяций морских котиков. Значитель-
ное влияние на продуктивность популяций оказывает также сбалансированность
307
половозрастного состава. При недостатке секачей значительная часть половоз-
релых самок остается неоплодотворенной. Этот фактор может полностью конт-
ролироваться человеком, и продолжительность его воздействия на популяцию
зависит от правильного подхода к прогнозированию норм добычи молодых сам-
цов. Гораздо сложнее поддерживать оптимальной возрастную структуру попу-
ляций, полностью контролируемую трудно регулируемым фактором “смерт-
ность-выживаемость”. Выборочный выбой малопродуктивных самок старших
возрастных групп вряд ли возможен. В связи с этим фактор плодовитости и в
будущем постоянно будет играть определенную роль в динамике численности
популяций котиков. Однако в целом его роль в этом процессе в 2—2.5 раза ниже,
чем выживаемости [Гудман, 1981; Фрисман и др., 1985}.
18.3. Сроки щенки и спаривания
Сроки щенки и спаривания у северных морских котиков тесно связаны с вре-
менем появления самок на лежбище.
Результаты многолетних наблюдений показывают, что первые самки на леж-
бища Командорских о-вов выходят в первых числах июня, на о. Тюлений —
в конце мая—начале июня, на о-ва Прибылова — в середине июня, но эти сроки
могут меняться год от года в зависимости от гидрометеоусловий как в местах
зимовки, так и в районе лежбищ. Известны годы, когда из-за тяжелой ледовой
обстановки было затруднено заполнение котиками лежбищ на о. Тюлений; в
этом случае много зверей сосредоточивалось на льдах у острова [Когай, 1971а].
Д. Джордан [Jordan, 1898] описал массовую гибель самок и щенков и даже за-
пустение лежбищ после того, как весной 1834 г. о-ва Прибылова были окружены
тяжелыми льдами.
По наблюдениям В. А. Бычкова [1964а], на Тюленьем промежуток времени
между выходом самок на гаремное лежбище и родами варьирует от 2 до 99 час,
составляя в среднем 21.9 час. На о-вах Прибылова время между выходом самок
на лежбище и родами колеблется от 2 до 75 час [Bartholomew, Hoell, 1953]. На
Командорах этот период составляет не более 3 суток [Барабаш-Никифоров, 1936;
Ильина, 1950].
Существует определенная зависимость времени прибытия самок к островам,
их выхода на лежбище и щенения от возраста. Наблюдениями за мечеными сам-
ками установлено, что беременные самки заканчивают выход на лежбище в ос-
новном к концу июля, среди которых ок. 100% составляют особи старше 13 лет,
96.4% — 13-летние, 87.3% — 6-летние, 72.4% — 5-летние, 36.6% — 4-летние и
лишь 0.8% — 3-летние [Владимиров, 1983; Владимиров, Лыскин, 1984]. Эти данные
хорошо согласуются с материалами добычи самок в районе лежбищ [Кузин, Па-
нина, 1977], которых показывают примерно такое же возрастное распределение
особей. На всех ныне существующих лежбищах котиков пик щенки приходится на
середину июля, однако отдельные роды могут происходить до середины августа.
Северный морской котик характеризуется послеродовым эструсом. Об этом из-
вестно давно [см. напр., Вениаминов, 1840], но детальных исследований ранее не
проводилось. Н. Волошинов [1887] писал, что самки котиков на о. Тюлений опло-
дотворяются лишь через 1—2 недели после родов. Н. Слюнин [1895], Л. В. Бой-
цов [1934], Е. И. Ильина [1950] считали, что на Командорских о-вах самки ста-
308
ловятся вновь оплодотворенными через 2—7 суток после родов. По наблюде-
ниям В. А. Бычкова [1964а] на о. Тюлений и Г. Бертоломью и Р. Хоелла [Bar-
tholomew, Hoell, 1953] на о-вах Прибылова, интервал между родами и покрытием
самок колеблется от 2 до 7 суток, в т. ч. на о. Тюлений в среднем 4.05 суток,
а на о-вах Прибылова — 5.1 суток.
Впервые вступающие в размножение самки появляются на лежбище лишь в
первой половине августа, практически после распада гаремов Однако, несмотря
на это, половина из них бывает оплодотворена и приносит потомство на сле-
дующий год. При недостатке секачей ситуация может меняться, и большинство
из них остаются яловыми. В этом случае, по мнению некоторых исследователей
[Бойцов, 1934; Ильина, 1950], большая роль в оплодотворении молодых самок
принадлежит полусекачам и холостякам. С возможностью участия небольшой
части полусекачей в размножении, на наш взгляд, можно согласиться, но участие
холостяков нами никогда не фиксировалось. По всей вероятности, основная
масса молодых самок оплодотворяется секачами, хотя, как известно, средний
возраст их меньше, чем самцов, участвующих в размножении в разгар гаремной
жизни [Лыскин, 1983]. Кроме того, общеизвестными являются факты повторной
(осенней) вспышки фолликулярной активности у оплодотворенных самок [Craig,
1964] и возбуждения секачей, которые вновь начинают вести себя по-гаремному.
Безусловно, во вновь организуемых гаремах присутствуют самки всех возрас-
тных классов, но покрываются только молодые, находящиеся “в охоте”. Пов-
торное возбуждение самцов и самок осенью принято считать “ложным гоном”.
Он свойствен не только северным морским котикам, но отмечается и у капских
морских котиков (Arctocephalus pusillus) [Rand, 1955].
Эстральный цикл у самок северного морского котика один в году [Bartho-
lomew, Hoell, 1953; Bigg et al., 1977, 1978]. Обычно самки после оплодотворения
покидают лежбище и возвращаются только, в среднем, через 5 дней уже без яв-
ных признаков, присущих эструсу. По наблюдениям в неволе, канадскими уче-
ными [Bigg et al., 1977, 1978; Bigg, 1979] установлено, что эструс стимулируется
присутствием самца, щенка, а его продолжительность составляет 14—21 сутки.
А. Крейг [Crage, 1964] считал, что эстральный цикл длится не более 19 суток,
а по нашим наблюдениям на лежбищах, — в среднем 7 суток (5—12 суток).
Спаривание у котиков происходит на берегу, но иногда и на мелководье, обя-
зательным условием для свершения этого акта является присутствие субстрата,
хотя бы в начале спаривания. Правда, несколько раз нами фиксировалось спа-
ривание в воде, но насколько оно было успешным, сказать невозможно. При
спаривании самец фиксирует самку тяжестью своего тела сверху. Если впослед-
ствие спаривание продолжается на воде, то самец обхватывает самку передними
ластами. Продолжительность спаривания различна и зависит как от индивиду-
альных особенностей самца, так и от степени его истощения. В начале сезона
размножения самец спаривается не более 5—7 мин; однако в конце сезона раз-
множения продолжительность спаривания увеличивается в 3—5 раз. Обычно
самец спаривается с одной самкой 1 раз, но в начале сезона размножения спа-
ривание может быть многократным. В свою очередь, самка спаривается с одним
самцом, но если после спаривания самка уходит в сторону моря мимо других
секачей, то возможно повторное спаривание с иным самцом. Ежедневная норма
копуляций 20 гаремных секачей на Китовом лежбище (о-ва Прибылова) может
достигать в июне в среднем 3.71 раза в час [Bartholomew, Hoell, 1953].
309
Наибольшее число копуляций наблюдается в середине гаремного периода
жизни, когда происходит массовый выход самок на лежбище. Число копуляций
меняется в зависимости от размера гарема или числа самок, находящихся при
секаче [Bartholomew, Hoell, 1953]. В больших гаремах секачи более активны и
производят большее число копуляций, однако “коэффициент” покрытия выше
в малых гаремах. В среднем в гаремах, состоящих из 3—14 самок, он составил
1.66, в гаремах из 22—35 самок — 1.44, в гаремах из 43—82 самок — 1.6 и в гаремах
из 108—153 самок — 0.86. По признанию исследователей, эти данные не явля-
ются решающими, по ним нельзя делать далеко идущих выводов. Единственное,
что можно сказать определенно, это то, что самец не способен покрыть более
100 самок за сезон. В свою очередь, анализ данных этих авторов показал, что
в среднем на каждого секача приходилось по 39 самок и средняя норма копу-
ляций составила 1.22 на каждую самку. По наблюдениям Н. Н. Лыскина [1983],
на Командорах средняя норма покрытий на каждого секача в сутки составляет
1.6 самки.
18.4. Продолжительность беременности и развития плода
Беременность у самок северного морского котика продолжается почти год,
за вычетом времени от родов до покрытия, составляющего в среднем 5 дней.
Лишь молодые самки, впервые вступающие в размножение, покрываются на
месяц позже и имеют, по-видимому, несколько меньшую продолжительность бе-
ременности. Столь продолжительная беременность котика включает выражен-
ную эмбриональную диапаузу, свойственную всем представителям ластоногих
[Bertram, 1940; Mensfild, 1958; Craig, 1964 и др.], длительность которой в среднем
составляет 3.5—4 мес.
ПАССА ЭМБРИОНОВ, Г
Рис. 18.9. Масса эмбрионов, взятых от самок разного возраста:
темные кружочки — 27.12.1966—01.1.1967 гг., светлые — 11—13.01.1968 г.
Рассматривая вопрос продолжительности задержки имплантации бластоцис-
ты у самок котиков, В. Ф. Мужчинкин [1971в] выдвинул ряд альтернатив. Ос-
новываясь на данных встреч недоразвитых эмбрионов у котиков [Scheffer, 1962],
310
Рис. 18.10. Эмбрионы котиков от самок, добытых в один и тот же день (декабрь
1968 г.)
а также на фактах преждевременного рождения крошечных безволосых щенков
[Scheffer, 1958; Д. И. Чугунков, личн. сообщ.], у которых отсутствовали видимые
дефекты развития, он предположил, что в этом случае было позднее зачатие,
но фактическая беременность имела нормальные по времени сроки, или про-
изошло удлинение эмбриональной диапаузы. Допускались также случаи совме-
щения запоздалого оплодотворения и удлинения эмбриональной диапаузы. Все
доводы автора правомочны и требуют всестороннего обсуждения.
Основываясь на том, что оплодотворение молодых самок происходит на
месяц позже, чем старых, нами была предпринята попытка определения времени
имплантации бластоцисты у разновозрастных самок по массе тела ранних эмб-
рионов (рис. 18.9, 18.10).
311
Как видно на рисунках, масса и размеры тела ранних эмбрионов имеют боль-
шую индивидуальную изменчивость у одновозрастных самок, как в старших,
так и в младших возрастных классах. Если темп роста эмбрионов после имплан-
тации бластоцисты одинаков у всех самок, то можно сказать, что длительность
диапаузы у котика непостоянна, во многом индивидуальна и, видимо, за счет
ее продолжительности возможны изменения сроков фактической беременности.
Из данных по эмбриональному росту котиков [Scheffer, 1962; Кузин, 1970а;
Кузин, Набережных, 1982а] известно, Что в раннем периоде внутриутробного
развития удвоение массы тела у плодов происходит в течение месяца. Можно
предположить, что, по крайней мере, месячное сокращение сроков диапаузы от
средней ее продолжительности возможно. Поэтому растянутость сроков спари-
вания у разновозрастных самок может компенсироваться за счет эмбриональной
диапаузы. Последующее спаривание происходит в обычные сроки. В этом случае
рождение недоношенных физически незрелых детенышей следует рассматривать
как отклонение от нормы в период внутриутробного развития.
18.5. Количество детенышей
Обычно у северных морских котиков рождается 1 детеныш. Двойни — яв-
ление чрезвычайно редкое (рис. 18.11).
Первое упоминание о рождении двоен у морского котика находим у И. Ве-
ниаминова [1840]. К. Ниггол и С. Фискус [Niggol, Fiscus, 1960] описали 4 случая
обнаружения эмбрионов-двоен при пелагических исследованиях котиков у бе-
регов Аляски. С. Фискус с соавт. [Fiscus et al., 1964] сообщали о нахождении
еще 3 двоен у самок из этого же района. Описаны 3 случая рождения двоен
на одной из скал о-вов Прибылова [Peterson, Reeder, 1966]. Приводились сведения
о добыче 5 беременных двойнями самок, добытых в северо-западной части Тихого
океана и в Японском море. Сведения о двойнях представлены в таблице 18.2.
Как видно из таблицы, все двойни были обнаружены у многорожавших са-
мок. В двух случаях самки были 5-летними, остальные старше 10 лет, в т. ч.
дважды двойни были найдены у самок в возрасте 16 лет. Двойни отмечались
как в одном из рогов матки, так и в разных. Двойни были однополые и раз-
нополые. К сожалению, ни в одном случае не были исследованы яичники самок,
имевших двойни.
Эмбрионы-самцы в двойнях были, как правило, крупнее самок. Масса и
длина эмбрионов-двоен несколько меньше среднего размера одинцов, добытых
в тот же день, хотя вполне укладываются в пределы колебаний последних.
Только трижды удалось наблюдать за рождением двоен и последующим по-
ведением матерей. Хотя во всех случаях оба щенка не были выращены [Peterson,
Reeder, 1966], но роды у самок прошли нормально, также как и последующее
спаривание. Этим опровергается мнение И. Вениаминова [1840] о том, что рож-
дение двойни почти всегда стоит жизни матери. Сейчас трудно делать какие-
либо выводы о возможности воспитания 2 щенков самкой. Однако можно пред-
полагать, что такая возможность сомнительна, хотя и имеются наблюдения од-
новременного кормления самкой двух детенышей [Кирчанов, 1975; Владимиров,
1976]. Подтверждением нашему сомнению служат следующие факты. Котики, как
и другие ластоногие, рожают крупного детеныша (более 50% длины и ок. 10—
312
Таблица 18.2
Сведения о двойнях морского котика
Место добычи Дата Самки-матери Эмбрионы Рог матки
воз- раст состоя- ние дли- на, см мас- са, кг ПОЛ длина, см масса, кг
двой- ни ОДИН- ЦЫ двой- ни ОДИН- ЦЫ
Тихоокеанские 30.4.59 5 м 112.0 ш 46.2 46.9 - - л
воды Японии f 42.2 46.8 - - л
m 60.5 53.1 3.28 2.34 л
» » 25.4.64 10 м 129.5 50
f 58.5 51.0 2.63 2.38 л
m 43.0 47.0 1.80 2.20 л
Японское море 16.4.64 10+ м 123.0 56
f 46.0 48.0 2.10 2.36 л
tn 55.0 60.0 3.20 4.0 п
» » 10.5.63 10+ м 121.0 50
f 51.0 57.0 2.70 3.20 л
m 48.0 49.5 2.25 2.48 п
» » » » 5 9 115.0 41
f 47.0 49.5 2.20 2.48 л
2.58
Воды Аляски 10.5.25
0.85
m 54.0 3.49 л
» » 9.5.58 11
f 53.0 3.43 п
Воды Калифор- 4.2.59 16 m 23.0 0.35 л
НИИ f 21.5 0.25 п
m 28.5 0.84 л
» » 21.2.59 7
f 27.5 0.77 п
О-ва Прибыло- 11.6.62 16 m 55.0 4.60 л
ва f 53.0 4.30 л
m 5.80
» » 1952
f 5.30
Примечания. Эмбрионы: ш — самец, f — самка; М — многорожавшие самки, Л —
левый рог, П — правый рог матки. Данные из западной части Тихого океана [по:
Кузин, 1969], из восточной и о-вов Прибылова [по: Niggol, Fiscus, 1960; Fiscus et
al., 1964; Peterson, Reeder, 1966].
15% массы тела матери). Для воспитания такого крупного детеныша требуется
большое количество молока. В период воспитания щенка самка вынуждена пе-
риодически отлучаться с острова, уходя в море на кормежку.
Самка находится на берегу в среднем 2 дня, после чего уходит в море на
8—10 дней. Таким образом, за весь период молочного кормления, длящегося
ок. 16 недель, щенок бывает с матерью всего 10—12 дней [Scheffer, Peterson, 1967].
Поведенческие реакции самки на рождение второго щенка и отношение к нему
впоследствии были иными, чем к первому. Самка негативно реагировала на его
313
Таблица 18.3
Данные о лактации самок, добытых на о. Тюлений
Период исследований п Л актирующие Нелактирующие
п % п %
21—30 июля 1977 г. 97 81 83.5 16 16.5
Июль—август 1976 г. 128 98 76.6 30 23.4
17—19 сентября 1975 г. 200 101 50.5 99 49.5
Сентябрь—октябрь 1976 г 72 22 35.6 50 64.4
3—12 октября 1977 г. 55 20 36.4 35 63.6
Таблица 18.4
Анализ содержимого желудков "серых" котиков, добытых на о. Тюлений
(по неопубл, матер. СахТИНРО)
Период исследований Число исследованных желудков Анализ содержимого желудков
Пустые Молоко Рыба Беспозвоночные
п % п % п % п %
4—5.10.75 115 68 59.1 40 34.8 7 6.1 - -
9—11.10.76 146 108 74.0 33 22.6 - - 5 3.4
3—12.10.77 841 499 57.9 267 32.7 72 8.5 8 0.9
присутствие [Peterson, Reeder, 1966]. Поэтому представляется, что кормление
самкой сразу 2 щенков на лежбище, описанное В. А. Владимировым [1976] и
В. А. Кирчановым [1975], не следует относить к явлению многоплодия. Скорее
это случаи бесконтрольного со стороны самки докармливания чужих щенков.
Имеются наблюдения В. А. Кирчанова и наши за кормлении самкой сивуча од-
новременно собственного щенка и детеныша котика.
Обычно у котиков наблюдается поочередная имплантация бластоцисты в
левом или правом рогах матки. Двойная овуляция может наблюдаться у самок,
которые овулируют впервые, у прохолоставших в прошлом году или у тех, бе-
ременность которых была прервана [Craig, 1964]. Это еще один аргумент за то,
что рождение двоен у котиков явление уникальное.
18.6. Лактация
Сведения о продолжительности лактационного периода у котиков разноре-
чивы. И. Вениаминов [1840] считал, что на о-вах Прибылова самки кормят де-
тенышей 2 мес. По мнению К. Кеньона и В. Шеффера [Kenyon, Scheffer, 1955],
самки покидают щенков после 3-месячного кормления молоком. В свою очередь,
Р. Бейкер с соавт. [Baker et al., 1970] считали, что иная пища, чем молоко, в
желудках “серых” котиков, начинает появляться в возрасте 3—4 мес., и лакта-
ционный период прекращается сразу после ухода самок с островов на зимовку.
314
На Командорских о-вах, по данным Л. В. Бойцова (1934], щенки сосут мать
до ноября, из чего следует, что лактация длится 4 мес.
На о. Тюлений, по данным Н. И. Сушкиной [1954], самки л'актируют 5—6
мес., что, конечно, неверно. По мнению В. А. Бычкова [1964], период вскарм-
ливания щенков котиков на Тюленьем продолжается до конца октября—начала
ноября (4 мес.). Нами получены новые материалы о лактации самок на Тюленьем
и о питании котиков в осеннее время. Многолетние материалы анализа питания
“черных” котиков показали, что в первой декаде августа все щенки питаются
молоком. Соответственно, добытые в конце июля самки (табл. 18.3) почти все
лактировали (83.5%). Лишь небольшая часть из них (16.5%) не имела молока
в молочных железах. Этот показатель вполне согласуется с данными о числе
ежегодно не принимающих участие в размножении самок.
Из таблицы видно, что от июля к октябрю постепенно уменьшается число
лактирующих самок. В начале октября лишь треть самок продолжала лакти-
ровать. Анализ содержимого желудков “серых” котиков, добытых на о. Тюлений
в октябре, показал (табл. 18.4), что уже в начале этого месяца часть “серых”
котиков начинает приобщаться к пище, добытой в море. Обращает на себя вни-
мание большая доля желудков, в которых пища отсутствовала. Возможно, это
объясняется тем, что самки нерегулярно кормят щенков, однако не исключено
и то, что период отлучения от молочной диеты занимает у котиков, как и у
других ластоногих, определенное время. На последних этапах лактации, видимо,
самки покидают щенков, и те в результате голодания вынуждены сами добывать
пищу в море. Такой период может быть достаточно продолжительным, т. к. из-
вестно, что щенки северного морского котика могут обходится без пищи 2—3
недели.
Особенностью лактации котиков является ее цикличность, периоды корм-
ления детенышей сменяются воздержанием из-за периодического ухода самок
в море для собственного питания. По опубликованным результатам [Bartholo-
mew, Hoell, 1953], первый уход самок в море после родов на кормежку короче
по-следующих. Определенные стимулы заставляют самку возвращаться на леж-
бище через разные промежутки времени в начале и в конце лактации. Если это
наполнение молочной железы секретом, то затухание секреции к концу лактации
будет являться результатом нахождения в море и отвыкания детеныша от мо-
лочной диеты. Тогда по присутствию молока в молочной железе самки можно
говорить об индивидуальной продолжительности лактации и ее изменчивости
в зависимости от возраста и времени рождения щенка.
Всего за период молочного кормления мать кормит щенка 10—12 раз, и каж-
дое такое кормление или пребывание детеныша вместе с матерью на берегу за-
нимает несколько дней [Scheffer, Peterson, 1967]. Из этих данных можно прийти
к выводу, что на о-вах Прибылова период молочного кормления длится 3—3.5
мес. На о. Тюлений молоко в желудках “серых” самцов, имевших массу тела
13.5 кг, и “серых” же самок с массой тела 12.9 кг присутствовало. Такой средний
вес обычно имеют детеныши в первой декаде сентября [Чупахина, 1977]. Од-
новременно у других детенышей такого же веса в желудках были обнаружены
рыба, молодь осьминогов и кальмаров. Таким образом, на Тюленьем период
молочного кормления щенков продолжается 3 мес.
315
Таблица 18.5
Состав молока северного морского котика [по: Капо et al., 1981]
Основной состав Минеральный состав
Твердые вещества 61% Na 52.1
Жир 45.6% Са 56.7
Протеин 12.% ? 119.3
Карбогитраты 2.35% I C 83.8
Витамины Mg 14.1
2790 Ие/100 г Cl 119.9
В1 0.02 мг/100 г Си 0.37
В2 0.97 мг/100 г Zn 0.90
Жирнокислотный состав (масса, %) Fe 0.35
С 12:0 Аминокислотный состав
14:0 Try °-87 2.80
14:1 Lis 0.40 1.72
15:0 His 0.70 0.78
15:0 0.4 Asp 1.15 2.62
Ьг 16:1 0-2. Thr 0.66 1.68
16:0 20.0 Scr 0.52 1.22
16:1 10.3 Gen 2.17 2.12
17:0 0.2 Pro 1.14 1.06
17:1 0.9 Gly 0.20 1.00
18:0 1.9 Ala 0.52 1.88
18:1 34.5 Cys 0.13 0.40
18:2 1.9 Vai 0.85 2.18
20:0 или 18.3 0.2 Met 0.37 0.16
20:1 6.8 Hen 0.57 0.18
20:2 1.0 Len 1.33 3.10
20:4 bkb 22:0 2.4 Tyr 0.62 0.90
22:1 1.0 Pha 0.59 1.82
20:5 4.8
24:0 0.7
22:5 1.5
22:6 3.7
316
По мнению С. В. Дорофеева [1964], щенок способен высосать до 500 мл мо-
лока. Эти показатели хорошо согласуются с данными В. Шеффера [Scheffer, 1962]
о вместимости молочных цистерн самок котиков, определенное с помощью инъ-
екции жидкости в молочную железу. У исследованных на о. Тюлений щенков
обнаружено до 1900 мл молока в желудках. Однако такое количество молока
щенок может получить не за одно кормление, т. к. он сосет самку до 6—8 раз
в день. Расчетное количество молока, потребляемое щенком за 1 кормление, со-
ставляет 600—700 мл.
Небезынтересным является факт обнаружения молока в молочных железах
у нерожавших, впервые вступающих в размножение самок, хотя подобное яв-
ление отмечалось и у других млекопитающих [Грачев, Галанцев, 1973].
Как и у всех изученных ластоногих, состав молока северного морского ко-
тика характеризуется большим содержанием жира, которое достигает 45—59%
[Wilke, 1959; Ashworth et al., 1966]. По данным японцев [Kano et al., 1981], состав
молока котика приведен в таблице 18.5.
Жирность молока у северного морского котика меняется в течении лактации
[Ashworth et al., 1966] и в конце ее (в октябре) увеличивается до 59%. У южного
морского котика жирность молока составляет 18% [Rand, 1956], лактация при
этом длится почти год. Это закономерное явление, когда при низкой жирности
молока увеличивается период лактации. Жирность молока также зависит от тем-
пературного фактора. При обитании в холодных условиях жирность повышается
[Грачев, Галанцев, 1973].
317
Глава 19. ЧИСЛЕННОСТЬ И ЕЕ ДИНАМИКА
Ориентировочные сведения о численности северных морских котиков на ост-
ровах Прибылова и Командорских были получены только в 70-х годах XIX в.
О первоначальной численности котиков на о. Тюлений и Курильских о-вах мож-
но судить лишь по далеко неполным сохранившимся сведениям промысла и по
восстановленной численности. Уже более 80 лет о численности котиков на не-
которых островах и других интрапопуляционных параметрах сведения посту-
пают регулярно, и поэтому северный морской котик часто избирается в качестве
модельного объекта для теоретических разработок, связанных с динамикой чис-
ленности промышляемых долгоживущих видов [Северцов, 1941; Kenyon et al.,
1954; Chapman, 1961, 1964; Ichihara, 1972, 1973; Ricker, 1972; Кузин и др., 1978;
Скалецая и др., 1983, 1985; Eberhardt, 1981; Smith, Polacheck, 1981; Гудман, 1983;
Фрисман и др., 1985].
19.1. Методы получения исходных данных для определения численности
котиков
Пионерами в разработке методов определения численности морских котиков
были Ч. Бриан и Г. Эллиотт, которые еще в конце 19 столетия определили
общую численность котиков на о-вах Прибылова, применив при этом метод
учета по площадям. Г. Эллиотт [Elliott, 1884] считал, что при равномерном рас-
пределении на лежбище каждый котик занимает 2 кв. фута. Общая площадь,
занимаемая размножающимися животными и их потомством, составляла, по
Г. Эллиотту, на о-вах Прибылова в 1872—73 гг. 6386840 кв. футов, следова-
тельно, общая численность размножающихся котиков равнялась 3.2 млн. Чис-
ленность неразмножающейся части популяции морских котиков на тех же ост-
ровах была определена в 1.5 млн, а общая численность — в 4.5 млн. Позже
площадной метод учета применялся еще несколько раз [напр., Слюнин, 1895]
в различных модификациях, в основном касающихся определения средней пло-
щади, занимаемой каждым котиком на лежбище, и определения количества жи-
вотных, отсутствующих на лежбище. Однако последующие исследователи [Jor-
dan et al., 1898; Суворов, 1912] отрицательно отозвались об этом методе и полу-
ченных на его основе результатах. В качестве примера разноречивости данных,
полученных на основе площадного метода, можно привести исследования Г. Хан-
на [Hanna, 1918], который специально занимался его проверкой. Он установил,
что плотность размещения котиков на лежбище не везде одинаковая и каждый
котик занимает на лежбище не 2, а 6 кв. футов. Таким образом, общая чис-
ленность котиков во времена Г. Эллиотта на о-вах Прибылова была не 4.5 млн,
а всего лишь треть от этого числа. Д. Джордан, возглавивший Международную
Конвенцию по охране котиков 1911 г., назвал данные о численности котиков
на о-вах Прибылова, приведенные Г. Эллиоттом, “наивысшим выражением пыл-
кого воображения”. Итак, площадный метод учета северных морских котиков
был опровергнут и больше никогда не применялся на практике, хотя теорети-
ческие разработки еще предпринимались [Тимофеева и др., 1977].
318
В. период сокращения численности морских котиков (1892—1911 гт.) в ходу
был глазомерный метод подсчета животных на лежбищах. Е. К. Суворов [1912]
дал вполне удовлетворительную оценку этому методу при использовании его,
когда общая численность и плотность размещения котиков на залежках неве-
лики. Сам Е. К. Суворов применял метод отгонов. Для этого всех животных
отгоняли вглубь острова, потом через узкий проход между учетчиками выпус-
кали из отгона в море и подсчитывали. Сопоставление данных, полученных при
отгонах и глазомерной оценке, дало идентичные результаты. В конце прошлого
века (1893—98 гт.) для оценки численности котиков американские ученые под-
считывали число гаремов на том или ином лежбище, которое легко определить
по числу секачей. Такой подход дал возможность установить, что число самок,
находящихся на лежбищах в гаремах, в 2—2.5 раза ниже численности щенков,
а, значит, и общей численности рожавших самок. Позже (1912—17 гг.), в связи
с неполнотой получаемой информации, от метода подсчета гаремов отказались
и перешли к поголовному учету различных возрастных групп котиков, нахо-
дящихся на лежбище. Особое внимание при этом уделялось подсчету секачей
и приплода. Данные по учету щенков, секачей и промыслу легли в основу рас-
четов общей численности и качественного состава популяций. Поголовный учет
щенков на о-вах Прибылова осуществлялся с 1912 по 1916 гг. и в 1922 г., а с
1917 по 1921 гг. подсчет вели на выборочных участках. Кроме того, был при-
менен метод клеймения котиков раскаленным железом, правда, сначала с целью
порчи шкур, которые могли достаться браконьерам, но эти же данные исполь-
зовались для расчетов смертности и возрастных стандартов. В эти же годы для
определения общих размеров стад котиков был апробирован метод фотосъемки.
Предложенные методы учета численности или сбора информации, исполь-
зуемой для последующих расчетов численности в конце прошлого века и в на-
чале нашего, не потеряли своего значения и теперь. В настоящее время в со-
ответствии с программой NPFSC Россия и США ежегодно проводят на всех
лежбищах сбор следующей информации для определения численности отдельных
группировок и популяций в целом:
1) учет численности гаремных и безгаремных секачей;
2) учет численности щенков (в России на всех лежбищах, в США — на мо-
дельных, с последующей экстраполяцией на остальные);
3) учет численности полусекачей, а на отдельных лежбищах и самок (Россия);
4) учет численности мертвых детенышей;
5) мечение щенков (металлическими или пластиковыми метками, выстрига-
нием шерсти на голове, с помощью обрезания кончиков оторочки ластов, про-
биванием отверстий в перепонке ластов и т. п.);
6) читка меток на живых котиках с использованием оптических приборов;
7) фотосъемка лежбищ (очень редко в России и США);
8) аэрофотосъемка лежбищ (очень редко в России и США);
9) определение возраста всех добываемых холостяков (в России) и опреде-
ленной доли добычи (США);
10) сбор меток с добываемых животных.
319
Учет численности гаремных и безгаремных секачей производится визуально.
При этом гаремными считаются секачи, имеющие хотя бы 1 самку. Учет чис-
ленности щенков на лежбищах в России производится или методом “прогона”
или визуально двумя—тремя учетчиками с усреднением полученных данных. В
СПТА в связи с большим числом залежек и большими площадями, занимаемыми
котиками, применить метод “прогона” нельзя. Поэтому величину приплода опре-
деляют по проценту вернувшихся меток с добытых животных промыслового воз-
раста (3—5 лет) и расчетным путем с запозданием на 2—3 года получают данные
о численности щенков. На отдельных лежбищах о-вов Прибылова используется
метод Петерсона. Для этого предварительно метят определенную часть детены-
шей, выстригая шерсть на голове (“шеринг-метод”), и по соотношению числа
меченых и немеченых в смешанных группировках определяют численность при-
плода. Мертвых щенков учитывают при санитарной чистке лежбищ. Учет чис-
ленности самок, секачей и полусекачей осуществляется визуально.
Для расчетов численности самок до 50-х годов нашего столетия достаточной
информации не было. Контроль за ней осуществлялся через ежегодно рожда-
ющихся щенков. С развитием в 50-х годах пелагических исследований появилась
возможность получать данные по возрастному составу и физиологическому со-
стоянию котиков, на основе которых оценивается их выживаемость в зависи-
мости от возраста и рассчитывается общая численность. Первые полные све-
дения о численности котиков разного пола и возраста в прибыловской попу-
ляции были представлены в 50-х годах [Kenyon et al., 1954], а потом в 1980 г.
(Lander, 1980]. По другим популяциям до последнего времени таких таблиц не
составлялось. Лишь в последние годы, в связи с математическим моделирова-
нием популяций, были сделаны оценки выживаемости и численности всех воз-
растных классов котиков, и определена их общая численность [Chapman, 1961,
1964; Ichihara, 1972; Кузин и др., 1978; Гудман, 1983; Фрисман и др., 1985]. Вместе
с тем, в качестве экспресс-метода и теперь часто используют данные о числен-
ности щенков для определения общей численности котиков, исходя из того, что
приплод составляет 26—34% (в среднем 30%). Достоверность такой оценки впол-
не удовлетворительна.
19.2. Численность популяции котиков острова Тюлений
Судя по данным добычи в первые годы эксплуатации популяции северных
морских котиков после открытия на о. Тюлений их лежбища в 1852 г. [Федоров,
1964] и по восстановленной численности (1964—68 гг.), первоначальное число
зверей здесь составляло не более 120 тыс. Бесконтрольный убой животных в
прошлом не раз приводил эту популяцию на грань исчезновения. В 1885 г.
Н. Волошинов [1887] насчитал на острове 10 тыс. самок, 3 тыс. щенков н
30 секачей (табл. 19.1).
В 1891 г. Н. Е. Слюнин (1895) учел 5 тыс. самок и 4 тыс. щенков, хотя приво-
димые им же данные Брюмера показывают для этого же года цифру 15—17 тыс.
В 1892 г., по данным Н. Е. Слюнина, на острове залегали 13—16 тыс. котиков
(по Брюмеру 15 тыс.). В 1896 г., по Л. Стейнегеру [Jordan, 1898—1890], здесь
насчитывалось не более 2 тыс. котиков. Ко времени заключения первой Меж-
дународной Конвенции по сохранению котиков северной части Тихого океана
в 1911 г. на острове обитали 6 тыс. зверей. Дальнейшие сведения о численности
320
Таблица 19.1
Общая численность котиков на о. Тюлений
Годы Численность Источник Годы Численность Источник
1 2 3 4 5 6
1885 13030 Волошинов, 1887 1941 48000 Кузнецов, Ильина, 1949
1891 15000-17000 Слюнин, 1895 1942 50000 » »
1892 13000—16000 » » 1943 50000 Расчетные данные
1896 2000 Jordan et al., 1898 1958 107333 » »
1901 10288 Тихенко, 1914а 1959 116663 » »
1902 9848 » » 1960 126666 » »
1903 7000 » » 1961 136666 » »
1905 1000 » » 1962 149000 » »
1906 4050 » » 1963 163333 » »
1907 5520 » » 1964 171333 » »
1908 5797 » » 1965 161333 » »
1909 6650 » » 1966 152666 » »
1910 6600 » » 1967 145000 » »
1911 7170 » » 1968 105000 » »
1912 8988 » » 1969 105000 » »
9041 М. К., 1929 1970 137000 » »
1913 1ОО0& Тихенко; 1914а 1971 146833 » - »
11250 Батурин,. 1923 1972 117233 » »
1914 6334 » » 1973 110566 » »
1915 3983 » » 1974 90000 » »
6355 М. к:, 1929 1975 102083- » »
1916 7208 Батурин, 1923 1976 95266 » »
1917 10515 » » 1977 99666 » »
1918 8771 » » 1978 80616 » »
12140 Державин, 1929 1979 73333 » »
12140 М. К., 1929 1980 70000 » »
1919 12000 Бойцов, 1934 1981 74333 » »
1920 14000 » » 1982 80000 » »
1921 16000 » » 1983 77333 » »
1922 19000 » » 1984 66666 » »
15748 Державин, 1929 1985 60000 » »
15748 М. К., 1929 1986 53373 » »
1923 23319 Бойцов, 1934 1987 62666 » »
1924 28226 Державин, 1929 1988 61666 » »
28226 М. К., 1929 1989 58733 » »
321
Таблица 19.1 (окончание)
1 2 3 4 5 6
1925 32000 Бойцов, 1934 1990 58733 Расчетные данные
1926 4400 » » 1991 60266 » »
1927 53700 » » 1992 50000 » »
1928 55000 » » 1993 56466 » »
1929 57000 » » 1994 60666 » »
1934 70000 » » 1995 55000 » »
1996 5500 » »
Примечание. Данные за 1958—90 гг. получены, исходя из принятого в NPFSC соот-
ношения: численность приплода составляет 30% от общей численности популяции.
Таблица 19.2
Численность секачей и щенков котиков на о. Тюлений
Годы Численность Источник Годы Численность Источник
щенки секачи щенки секачи
1 2 3 4 5 6 7 8
1885 3000 30 Волошинов, 1895 1937 14500 - Ichihara, 1973
1891 4000 - Слюнин, 1895 1938 14800 » »
1906 1100 - Ichihara, 1973 1940 15500 - » »
1907 1520 - » » 1941 16000 - » »
1908 1600 - » » 1958 32200 1016 Данные NPFSC
1909 1650 - » » 1959 35000 1360 » »
1910 1800 — » » 1960 38000 2279 » »
1911 2700 - » » 1961 41200 2528 » »
2270 - Тихенко, 1914а 1962 44700 2630 » »
1912 2388 2350 Ichihara, 1973 1963 49000 2465 » »
1913 3187 » » 1964 51400 2452 » »
3307 - Батурин, 1923 1965 484000 2213 » »
1914 1899 - Ichihara, 1973 1966 44900 2934 »
1915 2316 » » 1967 56500 2439 . » »
2376 253 М. К. , 1929 1968 45800 2252 » »
1916 2927 - Ichihara, 1973 1969 43500 2004 » »
2850 - Батурин, 1923 1970 31500 2331 . » »
1917 3988 - Ichihara, 1973 1971 41100 1864 » »
3879 - Батурин, 1923 1972 44050 1147 » »
1918 4479 - Ichihara, 1973 1973 35300 564 » »
3340 - Батурин, 1923 1974 33170 540 » »
4497 ' 418 М. К. , 1929 1975 27000 553 » »
322
Таблица 19.2 (окончание)
1 2 3 4 5 6 7 8
1919 4544 - Ichihara, 1973 1976 30625 631 Данные NPFSC
1920 5045 - » » 1977 28580 775 » »
1921 6134 » » 1978 . 29900 1131 » »
1922 7019 - » » 1979 24185 1603 » »
6134 994 М. К. , 1929 1980 22000 1860 » »
1923 7917 - Ichihara, 1973 1981 21000 1833 » »
1924 7613 - » » 1982 22300 1405 » »
9613 1260 М. К. , 1929 1983 24000 1622 » »
1925 8797 - Ichihara, 1973 1984 23200 987 » »
1926 9654 » » 1985 20000 875 » »
1927 11086 - » » 1986 18000 990 » »
1928 12630 - » » 1987 16000 1007 » »
1929 13000 - » » 1988 18800 1604 » »
1930 11855 - » » 1989 18500 2124 » »
1931 12460 - » » 1990 17620 2503 » »
1932 13000 1937 » » 1991 18080 2721 » »
1933 13500 - » » 1992 15000 2868 » »
1934 13700 - » » 1993 16940 2739 » »
1935 14000 - » » 1994 • 18200 3035 » »
1936 14300 » » 1995 16500 2588 » »
котиков на Тюленьем хотя и разноречивы, но заслуживают внимания, потому
что они в какой-то мере показывают характер восстановления этой популяции
после заключения Конвенции (табл. 19.1, 19.2). Так, по данным А. Державина
[1929], численность морских котиков в 1912—24 гг. увеличилась с 9 до 28 тыс.,
а число рожавших самок за это же время возросло с 2.5 до 9.6 тыс. Н. Д. До-
бровольский [1929] для 1924 г. исчисляет общую численность котиков на о. Тю-
лений в 28.9 тыс. Расчетные данные на основе количества отчисленных по ус-
ловиям Конвенции шкур о численности котиков на острове приводит Л. В. Бой-
цов [1934]. По материалам этого автора, численность в 1919 г. составляла 12 тыс.,
а в 1934 г. — 70 тыс. (табл. 19.1). По данным японского “Ежегодника” за 1927 г.,
численность котиков на о.. Тюлений изменялась следующим образом:
возраст 1912 1915 1918 1922 1924
Щенки 2476 2376 4497 6134 9613
Взрослые самки 2476 2376 4497 6134 9613
Взрослые самцы 2350 253 418 994 1260
Холостяки 1739 1350 2728 2486 7740
Итого 9041 6355 12140 15748 28226
323
Имеются статистические данные по численности щенков за 1906—41 гт., по
которым можно определить общую численность популяции о. Тюлений за эти
годы, исходя из того, что приплод составляет примерно 30% общей численности
стада (табл. 19.2). В 1941 г. на Тюленьем насчитывалось 48 тыс., а в
1942—43 гг. — 50 тыс. взрослых особей [Кузнецов, Ильина, 1949]. Опублико-
ванных данных за 1944—57 гг. нет. Исследовательские работы в эти годы прак-
тически не проводились, хотя промысел осуществлялся.
С вступлением в силу Конвенции по сохранению котиков северной части Ти-
хого океана в 1957 г. возобновляются регулярные исследования всех популяций,
соответственно поступают сведения об их численности (табл. 19.1, 19.2). Как видно
из таблиц, численность котиков на о. Тюлений с 1957 по 1963 гг. постоянно уве-
личивалась. Средний ежегодный прирост новорожденных был примерно таким же,
как и в годы восстановления этой популяции (1906—41 гг.), и составлял 8% в
год. Ок. 5 лет (1963—68 гг.) численность держалась на высоком уровне, а позже
начала снижаться и в 1983 г. стала меньше, чем в 1958 г. С 1986 г. по 1995 гг.
популяция находилась на устойчиво низком уровне, не испытывая каких-либо из-
менений. Фактически, с 1958 по 1995 гг; популяция котиков на о. Тюлений прошла
путь своего развития, образовав замкнутую волну, включающую стадию подъема,
стабилизации на высшем уровне, спада и стабилизации на низшем уровне числен-
ности. Время, пройденное популяцией от пика (1967 г.) до минимума численности,
вполне укладывается в так называемое “популяционное время”, равное примерно
продолжительности жизни одного поколения.
19.3. Численность котиков на Курильских островах
До 1881 г. ничего не было известно о существовании лежбищ северного морского
котика на Курильских о-вах. Капитан Г. Сноу [1902 ] был первым, кто обнаружил
3 крупных лежбища котиков и определил численность животных на них. Тогда
на о-вах Среднего и о. Райкоке обитало по 12—15 тыс., а на о-вах Каменные Ло-
вушки — 2—3 тыс. котиков. Других залежек котиков на Курилах Г. Сноу не об-
наружил. Однако, по данным Хоккена [1897], морские котики залегали на 11 ост-
ровах Курильской гряды, где было 12 лежбищ. По сведениям С. А. Тихенко
[1914а] и С. М. Успенского [1955], котики обитали на 9 островах Курильской
гряды. Последний автор отмечал, что все их залежки, кроме залежек на островах
Каменные Ловушки, Среднего и Райкоке, были небольшими, насчитывающими
десятки или сотни животных.
По оценке Н. Волошинова [1889], на о-вах Среднего залегали 5—10 тыс. ко-
тиков. В конце прошлого века популяция Курильских о-вов подвергалась силь-
ному истреблению. В 1896 г. Д. Джордан [Jordan, 1898], посетивший лежбища
котиков на Курилах, животных там не нашел. Правда, он допускал, что несколь-
ко десятков их могло сохраниться и в будущем положить начало новому стаду.
Действительно, в 1911—12 гг. на о-вах Каменные Ловушки видели несколько
десятков котиков, а в 1913 г. заметили 3 щенков [Тихенко, 1914а]. В “Вестнике
рыбной промышленности” за 1915 г. в статье “О состоянии котиковых лежбищ
в Японии” говорится, что в 1913 г. на Курилах видели 48 котиков, среди них
было 9 щенков. В 1914 г. на островах зарегистрировали 11 котиков, в т. ч. 2 щенков.
Есть сведения о том, что котики долгое время держались на островах Симушир
и Кетой [Слюнин, 1895; Успенский, 1955], но Г. Сноу, хорошо знавший Куриль-
324
Таблица 19.3
Численность морских котиков на Курильских о-вах (по неопубл, данным
ТИНРО)
Годы О-ва Среднего О-ва Каменные Ловушки Итого
секачи шенки всего секачи шенки всего секачи шенки всего
1962 200 1085*
1963 500 2050* 6305
1964 1000 3500*
1965 4300 1020
1966 91 150* 1860 250 1550 5196
1967 191 1475 3770 407 2217 5879 9469
1968 1847 4695 185* 1710 3565 10181
1969 1912 3755 1810 7178 10940
1970 1741
1971 382** 1423** 525 2443 7051
1972 1927 3680 7814
1973 639 5451 8853 441 2708 8611 17464
1974 457 5802 12273 732 5802 12159 24432
1975 802 7390 16031 1054 4857 15030 1856 12247 31061
1976 971 8943 17430 1159 5783 15405 2130 14787 32826
1977 642 11788 19337 626 6508 10939 1268 18296 30276
1978 821 11455 16741 814 6940 14949 1635 18395 312960
1979 546 5640 10241 427 2099 973 7739 14500***
1980 805 8376 5522 820 5011 11727 1625 13387 24584
1981 1063 7650 14057 1074 4580 11982 2137 12230 26039
1982 688 9771 16715 1159 6322 14384 1847 16093 31099
1983 970 7222 12840 1230 5650 14430 2200 12870 27270
1984 874 6811 15743 1346 5365 15185 2200 12176 30928
1986 1286 8012 15437 1265 6784 14434 2551 14796 30871
* Только на скале Котиковая; ** только на скалах Черных; *** без учета скалы Ко-
тиковая.
ские острова, ничего не сообщал об этих лежбищах. О дальнейшей судьбе ко-
тиковых лежбищ на Курильских о-вах ничего не известно. Новые сведения об
их обнаружении здесь были получены 60 лет спустя [Клумов, 1957], по данным
которого на скале Котиковая (о-ва Каменные Ловушки) залегали 2 тыс., а на
скале Хитрая (о-ва Среднего) несколько сот зверей. Уже в 1958 г. С. Перелешин
насчитал на скале Котиковая ок. 3 тыс., а на скале Хитрая 800 котиков. С 1962 г.
контроль за численностью котиков на Курильских о-вах осуществлялся ежегодно
(табл. 19.3). В настоящее время численность северных морских котиков достигла
уровня, известного для периода их открытия, и составляет ок. 50 тыс., хотя ко-
тики отсутствуют на о. Райкоке. Появление их здесь маловероятно, т. к. известно
[Тихенко, 1914а], что это лежбище в прошлом веке было завалено камнями в
результате землетрясения.
325
Таблица 19.4
Таблица 19.4 (окончание)
Численность котиков на Командорских о-вах
Годы О. Беринга О. Медный Всего Источник
2 3 4 5
1881 200000 Гребницкий, 1902
70000 Кузин, 1982
1884 300000 100000 130000 Волошинов, 1889
1893 60000 гс >лько на Северном лежбище Слюнин, 1895
1896 30800 34200 | 6500 Stejneger, 1898
1900 25000 без щенков Суворов, 1912
1904 41000 бе з щенков » »
1906 65000 Добровольский, 1929
1909 25000 Суворов, 1912
1911 5700 2714 8414 » »
9000 Батурин, 1923
1914 14000 » »
1916 15000 » »
1917 17000 Stejneger, 1898
1918 13267 Суворов, 1931
1919 28000 Батурин, 1923
3895 11023 14918 Добровольский, 1929
1920 3371 9595 12966 » »
1921 4339 12451 16790 » »
17000 Stejneger, 1898
1922 3642 10315 13957 Добровольский, 1929
14284 Суворов, 1931
1923 3260 12000 15260 Добровольский, 1929
14908 Суворов, 1931
1924 4691 11000 15681 Добровольский, 1929
15698 Суворов, 1931
1925 5476 9795 15271 Добровольский, 1929
16482 Суворов, 1931
1926 7275 10422 17691 Добровольский, 1929
15689 Суворов, 1931
1927 5101 8261 13365 Добровольский, 1929
18067 Суворов, 1931
1928 5738 7929 13667 Добровольский, 1929
1929 25428 Суворов, 1931
1930 26687 » »
1934 15200 Taylor et al., 1955
1935 13620 » »
1941 40000 » »
1953 73000 » »
1- 2 3 4 5
1958 62666 64600 127266 Данные NFPS
1959 67833 6900 137733 » »
1960 73553 • 75826 149380 » »
1961 79666 82076 161743 » »
1962 86890 88613 175503 » »
1963 94100 95000 189100 » »
1964 97533 97410 194973 » »
1965 89466 105700 195166 » »
1966 101150 102280 203430 » »
1967 103853 77913 181766 » »
1968 114396 88466 202862 » »
1969 107563 88370 195523 » »
1970 113086 140250 207336 » »
1971 119280 128666 221646 » »
1972 104510 98350 202860 » »
1973 96546 83080 179626 » »
1974 123716 105533 229249 » »
1975 117920 128666 246586 » »
1976 128320 93300 221620 » »
1977 125233 101333 226566 » »
1978 139016 113180 252196 » »
1979 127066 106666 233732' » »
1980 117533 100333 217866 » »
1981 129833 108633 238466 »
1982 121000 108600 229600 » »
1983 131466 117933 249399 » »
1984 122233 114033 236266 » »
1985 145600 102233 247833 » »
1986 134466 106166 240632 » »
1987 138066 103466 241532 » »
1988 141633 103000 244633 » »
1989 136400 92450 228850 » »
1990 139016 93600 232616 » »
1991 130866 101266 232132 » »
1992 128833 » »
1993 144200 68700 неполные данные » »
1994 142116 59000 201116 » »
19.4. Численность котиков на Командорских островах
Первые ориентировочные цифры численности морских котиков на Коман-
дорских о-вах были получены лишь в 80-х годах прошлого столетия [Гребниц-
кий, 1902]. По этим данным, в 1881 г. на здесь залегали 2 млн. котиков (табл. 19.4).
327
326
Таблица 19.5
Численность щенков и секачей морских котиков на Командорских о-вах
Годы О. Белинга О. Медный Всего Источник
секачи шенки... - секачи шенки секачи
1910 16 2000 48 4200 64 6200 Суворов, 1912
1911 31 3335 16 1311 27 4646 » »
1934 312 312 614 Taylor et al., 1955
1935 258 3956 » »
1940 6634 6425 13059 » »
1958 640 18800 424 19380 884 38180 Данные NPFSC
1959 627 20350 497 20970 1124 41320 » »
1960 791 22066 876 22748 1667 44814 » »
1961 905 23900 1104 24623 2009 48523 » »
1962 1524 26067 1894 26584 3418 52651 » »
1963 2221 28230 2220 28500 4441 56730 » »
1964 3159 29269 2191 29223 5350 58492 » »
1965 3326 26840 2467 31710 5823 58550 » »
1966 3333 30345 2405 30684 5738 61029 » »
1967 3273 31156 2386 23374 5642 54520 » »
1968 2820 34319 2231 26540 5051 60859 » »
1969 2045 32146 1815 26511 3860 58657 » »
1970 1654 33926 1356 31275 2923 62201 » »
1971 1333 35784 1021 30710 2354 66494 » »
1972 839 31353 882 29505 1721 60858 » »
1973 658 28964 919 24924 1577 53888 » »
1974 840 37115 1184 31660 2024 68775 » »
1975 1109 35376 1509 38600 2618 73976 » »
19776 1681 38496 1755 27990 2897 66486 » »
1977 3122 37570 2897 30400 6019 67970 » »
1978 4011 41705 3332 33954 7343 75659 » »
1979 4864 38120 3889 32000 8753 70120 » »
1980 5380 35260 3505 30100 8885 65360 » »
1981 5392 38950 3858 32590 9250 71540 » »
1982 6154 36300 3500 32580 9654 68880 » »
1983 5646 39440 3981 35380 9627 74820 » »
1984 6031 36670 4219 34210 10250 70880 » »
1985 5502 43680 4719 30670 10221 74350 » »
1986 4663 40340 4402 31850 9038 72190 » »
1987 4925 41420 4324 31043 9249 72463 » »
1988 4842 42490 нет д 1 Н Н Ы X » »
1989 4959 40920 3140 27735 8099 68655 » »
1990 3824 41705 3322 28080 7146 69785 » »
1991 4923 39260 5531 30380 10453 69640 » »
1992 5294 38650 3822 — 9116 » »
1993 6973 43260 2801 20610 9774 67870 » »
1994 5274 42635 3692 17700 8996 60335 » »
328
По данным Н. Волошинова [1885], на Командорских о-вах обитали 1.3 млн
котиков. Сопоставление данных промысла котиков на островах Прибылова и
Командорских за период 1870—1911 гг. позволили нам усомниться в правиль-
ности размеров численности, приведенных Н. А. Гребницким и Н. Волошино-
вым. По нашим расчетам [Кузин, 1982а], численность котиков на Командорах
в 1881—84 гг. была не более 700 тыс. Позже и В. А. Владимиров [1985], правда,
без всяких доказательств тоже усомнился в достоверности данных названных
авторов.
В результате неумеренного промысла к 1896 г. численность котиков на Ко-
мандорских о-вах сократилась до 65 тыс. [Jordan, 1898], а к моменту заключения
Международной Конвенции 1911 г. — до 8.5—9 тыс. [Дорофеев, 1964]. Котики
на многих лежбищах были выбиты полностью. Восстановление этой популяции
после заключения Конвенции шло очень медленно в связи с браконьерством
японцев, продолжавшимся до 1945 г. [Austin, Wilke, 1950]. По отрывочным со-
хранившимся сведениям, численность котиков с 1911 по 1934 гт. увеличилась
менее чем вдвое и составила в 1934 г. 13—15 тыс. [Taylor et al., 1955]. За по-
следующие 20 лет (к 1954 г.) она увеличилась почти в 5 раз и составила 73 тыс.
[Taylor et al., 1955]. Ко времени заключения второй Международной Конвенции
по сохранению котиков северной части Тихого океана в 1957 г. на Командорских
о-вах существовали 3 лежбища (Северное, Юго-Восточное и Урилье), числен-
ность животных на которых составляла ок. 100 тыс. [Дорофеев, 1964]. В настоя-
щее время на Командорах функционируют 4 лежбища, на которых обитают ок.
210—230 тыс. котиков (табл. 19.4, 19.5).
19.5. Причины снижения численности северных морских котиков
Причины изменений численности котиков в прошлом (вплоть до 1911 г.) ясны
и определяются сильным воздействием бесконтрольного промысла, в результате
которого полностью были уничтожены отдельные репродуктивные группировки,
нарушалась интрапопуляционная структура и воспроизводительные способности
оставшихся. Однако об этом можно судить лишь по далеко неполным данным,
т. к. систематические исследования тогда не проводились.
С 1911 г. прибыловская, а с 1957 г. командорская и тюленьевская популяции
находились под контролем ученых. Интенсивное изучение биологии и осущест-
вляемая на ее основе эксплуатация ресурсов этих зверей за истекший период
вселили оптимистические надежды на возможность управления их популяциями.
Однако на деле, как справедливо заметил Д. Чепман [Chapman, 1981], это ока-
залось намного сложнее, чем мыслилось прежде, а механизмы, контролирующие
численность северных морских котиков, пока остаются непознанными. Не менее
пессимистично выглядят выводы других американских ученых [Smith, Polacheck,
1981], пересмотревших “Life table” прибыловской популяции. По их мнению, ре-
гуляция численности осуществляется посредством изменений, по крайней мере,
двух, а возможно всех трех основных компонентов таблицы. Убедительных до-
казательств о плотностно-зависимой регуляции параметров популяции, как это
представлялось ранее [Kenyon et al., 1954], нет, соответственно, понимание ме-
ханизмов регуляции численности затруднено.
В последние годы выполнен значительный объем работ по изучению основ-
ных показателей, ответственных за динамические процессы в популяциях север-
329
ных морских котиков. Проведены оценки возможного влияния на выживаемость
загрязнения окружающей среды [Fowler, 1982; Кузин, 1990], гидрометеоусловий
[Кузин, Шаталина, 1990; Fowler, 1992], состояния кормовой базы (Trites, 1992].
Получены данные о связи выживаемости котиков до 2 лет с размерами тела
новорожденных [Болтнев и др., 1991; Baker, Fowler, 1992] и зависимости послед-
них от возрастной структуры продуцирующих самок [Caiambokidis, Gentry, 1985;
Болтнев, 19906]. Переосмыслены показатели репродуктивного состояния прибы-
ловской популяции котиков за период 1958—74 гг. с учетом изменений среднего
возраста первых родов у самок в связи с возможным влиянием на него плотности
и их физического состояния, соотношения полов производителей и социальных
факторов [Trites, York, 1993].
Значительно продвинувшись в изучении отдельных вопросов биологии, мы,
тем не менее, еще далеки от понимания механизмов регуляции как вида в целом,
так и отдельных составляющих его популяций.
В настоящее время большинство исследователей северного морского котика
сходятся во мнении, что основным фактором, определяющим численность по-
пуляций этих животных является смертность [Chapman, 1964; Lander, 1979; Кузин
и др., 1981; Eberhardt, 1981; Fowler, 1982, 1990; Trites, 1989, 1992; Кузин, 1990;
Владимиров, 1991 и др.]. Роль плодовитости значима, но чувствительность по-
пуляций к этому фактору более чем 2—2.5 раза ниже, чем к смертности [Гудман,
1983; Фрисман и др., 1985]. Увеличением нормы репродуктивных показателей
невозможно не только вывести популяции из состояния депрессии, но даже при-
близить их к уровню начала роста численности [Smith, Polacheck, 1981].
Известно, что до 70% котиков из отдельных поколений гибнут в первые
2 года жизни [Lander, 1975]. Часть щенков погибает еще на берегу (при высокой
плотности в среднем 15%, при низкой — в среднем 5%), остальные — в море,
преимущественно в первую самостоятельную зимовку [Scheffer, 19506; Ichihara,
1974; Кузин и др., 1981; Кузин, Шаталина, 1990]. Основными причинами гибели
на берегу являются травмы, болезни и голод.
Многолетними наблюдениями на о. Тюлений установлено, что относитель-
ные показатели смертности щенков котиков выше в начальный период деторож-'
дения, когда рожают преимущественно старые самки (старше 12 лет), и в конце
репродуктивного периода, когда рожают первородки. Аналогичные наблюдения
имеются по командорским котикам [Болтнев, 19906].
Выявленная связь смертности щенков на берегу с массой их тела, а массы
тела — с возрастом матери [Болтнев, 1989; Болтнев и др., 1991] позволяет думать,
что смертность щенков в первые месяцы жизни во многом определяется воз-
растной структурой популяции.
В отношении выживаемости молодых котиков в море высказано несколько
гипотез. Самая ранняя из них рассматривает зависимость снижения численности
на о-вах Прибылова от плотности, в результате которой обостряются внутри-
популяционные конкурентные отношения за пищу в районе размножения жи-
вотных [Kenyon et al., 1954]. Воплощение этих гипотетических представлений
путем снижения плотности (выбоя более 300 тыс. самок) в 50—60-х годах не
дало желаемого результата [York, Hartley, 1981].
330
Рис. 19.1. Биологические показатели прибыловской популяции (о. Св. Павла):
А — численность щенков, Б — смертность щенков на острове, В — масса тела щенков, Г —
смертность самцов до 2 лет, Д — масса клыков холостяков, Е — численность секачей
[по: Fowler, 1982].
Трофическая гипотеза была отвергнута К. Фаулером (Fowler, 1982), который
проанализировал многие параметры популяции и факторы, их определяющие,
которые в той или иной степени могли быть ответственны за состояние чис-
ленности (рис. 19.1). Так, незначительные изменения в режиме эксплуатации по-
пуляции на о-вах Прибылова (изменение длины тела добываемых самцов и сро-
ков промысла) могли сказаться только на этой группе, но недостатка секачей
на протяжении всего периода снижения численности, по мнению К. Фаулера,
не было. Автор не исследовал репродуктивные способности самок этой попу-
ляции. Однако известно, что нормы беременности самок в течение 1958—74 гг.
колебались незначительно (Smith, Polacheck, 1984].
При изучении влияний условий среды на численность популяции северных
морских котиков о-вов Прибылова К. Фаулер исходил из тех предпосылок, что
если экосистема, частью которой является котик, негативно влияет на состояние
его популяций, то такое влияние должны испытывать все члены популяции, и
331
этот эффект должен проявляться в уменьшении уровня беременности, ростовых
процессов, снижении массы тела детенышей при рождении, увеличении смерт-
ности.
Было показано [Fowler, 1982], что интенсификация рыболовства в 70-х годах
в Беринговом море не отразилась на упитанности и ростовых процессах котиков.
Большинство морфометрических показателей у отдельных котиков в эти годы
были лучше, чем до развития интенсивного рыболовства. К. Фаулер отмечал,
что пищевые марши самок в период воспитания молодняка практически не из-
менились. Более того, предполагается [Schwartzmann, Harr, 1980], что изменение
возрастной структуры главного пищевого объекта северного морского котика —
минтая привело к увеличению пищевых ресурсов котиков за счет роста биомассы
мелкоразмерных рыб.
Несоответствие между улучшением показателей отдельных животных и уве-
личением смертности послужило поводом К. Фаулеру для предположения об от-
сутствии связи между смертностью котиков в море и обеспеченностью их пищей,
и о том, что смертность в море является результатом негативного воздействия
абиотических факторов. Анализ данных по уровню токсичности среды как при-
чины возможного увеличения смертности не дал положительных результатов.
Количество вредных веществ в тканях котиков осталось на том же уровне, каким
было до снижения численности этой популяции котиков, или даже уменьшилось
в последнее время.
Проявилась позитивная связь смертности котиков в море с интенсификацией
рыболовства и загрязнением окружающей среды. Особую опасность для котиков
представляют обрывки рыболовных снастей, упаковочные материалы и прочие
отходы промышленного рыболовства, в которых животные запутываются и гиб-
нут. По расчетам К. Фаулера [Fowler, 1982], из популяции котиков о-вов При-
былова в результате загрязнения среды обитания ежегодно гибнут 50 тыс. жи-
вотных. Исследователь считает, что это одна из главных причин снижения чис-
ленности прибыловских котиков в настоящее время.
А. Трайтис [Trites, 1992] критически отнесся к мнению К. Фаулера и вновь
вернулся к трофической гипотезе, несколько видоизменив ее. По А. Трайтису,
Алеутский архипелаг представляет “бутылочное горлышко” на пути миграции
ювенильных котиков, проходя которое животные испытывают недостаток пищи
в зал. Аляска, вследствие снижения продуктивности района из-за интенсифика-
ции рыболовства.
Применительно к популяциям котиков на о. Тюлений и Командорских о-вах
В. А. Владимиров [1991] предложил гипотезу, созвучную с геоклиматической тео-
рией динамики численности животных. В соответствии с этой гипотезой депрес-
сия численности котиков в 60-х—начале 70-х годов вызвана очередной сменой
векового хода солнечной активности, и, как следствие, наступлением в регионах
очередного похолодания и экосистемных изменений, негативно повлиявших на
выживаемость молодых животных. Стабильность состояния численности коман-
дорской репродуктивной группировки котиков на фоне снижения численности
других репродуктивных группировок оправдывается интенсивной иммиграцией
на Командоры прибыловских котиков.
Несостоятельность геоклиматической гипотезы очевидна. 70—80-е годы сле-
дует отнести не к холодным, а к теплым [Шунтов, 1994]. Наиболее ярким под-
332
тверждением тому служит начавшееся в это время интенсивное размножение и
развитие крупномасштабного промысла сельди-иваси в северной части Тихого
океана. Иваси проникли даже в Берингово море и служили объ'ектом питания
котиков. При обосновании стабилизации численности командорского стада ко-
тиков на достигнутом уровне в 70—80-х годах [Владимиров, 1991] за счет им-
миграции прибыловских котиков, автор вступает в противоречие с собственны-
ми исследованиями, согласно которым при депрессивных явлениях в популяциях
эмиграционные процессы снижаются до минимума [Владимиров, 19786].
Масштабность проблемы движения населения котика как вида сведена А. Трай-
тисом [Trites, 1992] к рассмотрению лишь прибыловской популяции, поэтому с
позиции его гипотетических построений невозможно обосновать аналогичные
процессы, происходящие в других популяциях. Если подходить к проблеме вы-
живаемости с точки зрения зависимости от трофического фактора, то следует
признать, что каждая репродуктивная группировка пользуется только ей зна-
комыми миграционными путями и местами зимовок, а благополучным группи-
ровкам известны районы с высокой биологической продуктивностью акватории,
в которых сохраняются из года в год благоприятные гидро-климатические и си-
ноптические условия, обеспечивающие комфортность существования и возмож-
ность беспрепятственного использования пищевых ресурсов, гарантирующих
нормальное физическое развитие и высокую выживаемость, что мало вероятно
и не подтверждается данными по смешиваемости котиков разных популяций на
местах зимовок.
Основными объектами питания северных морских котиков в Охотском море
является минтай (36% объема и 12.7% частоты встречаемости в желудках), ло-
сосевые рыбы (16.4% объема и 5.9% частоты встречаемости), треска (8.4% объема
и 5.9% частоты встречаемости). Анализ питания сеголеток в северо-западной
части Тихого океана показал, что их рацион не отличается от такового взрослых
особей [Панина, 1970, 1978]. В районе о. Хоккайдо и северной оконечности
о. Хонсю в зоне действия холодного Курильского течения и в районе смешения
вод Куросио и Оясио в осенний и ранне-зимний периоды наибольшее значение
в питании взрослых котиков играют минтай и лемонема. Анализ содержимого
желудков сеголеток и годовиков, добытых японскими исследователями в при-
брежных водах Хоккайдо, показал [Yoshida et al., 1978], что в этом районе у
этой возрастной группы преобладающим в рационе был минтай (72.7% частоты
встречаемости и 65.6% объема).
Промышленное освоение ресурсов минтая в Охотском море началось только
с 1963 г., в т. ч. у восточного побережья Сахалина — с 1975 г. В южной части
моря японский промысел минтая хотя и существовал с 30-х годов, но заметно
интенсифицировался лишь с 50-х годов. Однако стабильные уловы минтая в
водах островов Хоккайдо и Сахалин в течение долгого периода времени сви-
детельствуют об отсутствии очень резких и масштабных колебаний численности
этой рыбы в указанных районах. В Южно-Курильском районе высокие уловы
минтая пришлись на 70-е годы и держались на этом уровне до конца 80-х годов,
что говорит о хорошем состоянии этой популяции минтая.
Запасы лососей в Сахалино-Курильском районе в 50-х годах были на низком
уровне, однако с начала 60-х годов наметилась тенденция к улучшению ситуации
и, соответственно, к возрастанию береговых уловов, которые увеличивались до
333
Рис. 19.2. Упитанность морских котиков, добытых в северо-западной части Тихого
океана (самки) и на о. Тюлений (самцы):
1 — 4-летние самки, 2 — 2-летние самцы, 3 — 3-летние самцы.
Рис. 19.3. Выживаемость (W2) северных морских котиков до 2 лет:
1 — о. Тюлений, 2 — о-ва Прибылова, 3 — Командорские о-ва.
конца 70-х годов. Специализированного лова других видов рыб (трески, окуней)
в Охотском море не было. Они встречались в качестве прилова.
Анализ состояния запасов некоторых видов рыб, служащих основными объ-
ектами питания котика в Охотском море и вокруг о. Хоккайдо, не позволяет
прийти к выводу о лимитировании его пищевых ресурсов в этом районе. Со-
ответственно, отдать приоритет пищевому фактору, определившему начало деп-
рессии численности тюленьевской популяции котиков, нет оснований. Этот тезис
подтверждается материалами упитанности животных (рис. 19.2). На общем фоне
динамичности кондиционного индекса обращает внимание лишь его заметное
увеличение у котиков-холостяков в конце 80-х годов, несмотря на то, что к этому
времени ресурсы минтая были значительно подорваны.
Как и в других популяциях, ростовые процессы в период депрессии числен-
ности котиков, на о. Тюлений увеличились. Репродуктивные показатели самок
находились на высшем из известных уровней.
334
Из причин, в определенной степени снижающих потенциальную возможность
к увеличению численности тюленьевской популяции, можно назвать несбалан-
сированность возрастной структуры самок, где все еще преобладают животные
старше 10 лет, репродуктивные показатели которых в среднем на 10% ниже, чем
самок 7—10 лет, и загрязнение среды обитания. Ежегодные учеты котиков, не-
сущих на себе инородные предметы, показали, что с 1975 по 1984 гт. число трав-
мированных сетями, веревками, упаковочными материалами, леской и другими
отходами промышленного рыболовства остается стабильно высоким и состав-
ляет 1.1—1.9% в промысловых побойках. По нашим расчетам [Кузин, 1990], в
течение года из тюленьевской популяции гибнут 3.6 тыс. котиков, из командор-
ской — 8.3 тыс., а из курильской — 1.8 тыс., что составляет в 1.5—2 раза больше,
чем добывается этих животных в последние годы.
Плохо изученными остаются вопросы влияния на сеголеток котиков гидро-
климатических и синоптических условий, хотя уже первые публикации [Scheffer,
19506; Ichihara, 1974] показали их значимую роль в выживаемости этой возраст-
ной группы. В настоящее время исследования влияния этих условий на выжи-
ваемость ювенильных котиков получили развитие [Кузин, Шаталина, 1990; Ba-
ker, Fowler, 1992]. Однако перелома в восприятии биологами значения этих фак-
торов в изучении вопросов выживаемости названные публикации не произвели.
Это и понятно, потому что определение связи между различными факторами
внешней среды и выживаемостью котиков сопряжено как со значительными
трудностями в сборе большого массива данных по синоптическим и гидроме-
теорологическим условиям в конкретных районах и в конкретные сроки, так
и в связи с трудностями учета неодинакового реагирования самих животных на
изменяющиеся условия в зависимости от их персональных способностей про-
тивостоять давлению среды. В последнем случае речь идет о качественном со-
стоянии популяций, а “колебания качества популяций в зависимости от числен-
ности и возрастной структуры — явления взаимосвязанные. Динамика качества
столь же характерное их свойство, как и динамика численности” [Шварц, 1980].
Безусловно, основной качественной характеристикой популяций является воз-
растная структура, о чем можно судить по установленной зависимости выжи-
ваемости щенков котиков в первые месяцы жизни на берегу и в последующие 2 го-
да жизни в море от физических параметров их тела, а массы и длины тела от
возраста родителей [Calambokidis, Gentry, 1985; Болтнев, 1990а, 6; Baker, Fowler,
1992). Возрастной структурой, таким образом, регламентируются рамки возмож-
ностей в подготовке потомства к последующим этапам существования и чув-
ствительности к воздействию внешних факторов. Из внешних факторов наиболее
сильное негативное воздействие на выживаемость котиков оказывают ветровые
проявления циклонических циркуляций, снижающих возможности животных в
использовании пищевых ресурсов или их доступность [Кузин, Шаталина, 1990].
Январь по метеоусловиям признан нами наиболее критическим для адаптации
сеголеток котиков к пелагическому образу жизни, о чем, кстати, свидетельствуют
и данные о их физическом состоянии. Добытые в море в январе самцы-сеголетки
весили в среднем 12.7 кг, самки — 11.0 кг [Yoshida, 1978], против 19.6 кг и 14.5 кг,
соответственно, в октябре перед уходом их с островов на зимовку.
Таким образом, из трех слагаемых, определяющих динамические процессы
в популяциях млекопитающих, плодовитость и смертность для популяций се-
верного морского котика на о-вах Прибылова и о. Тюлений являются главными
335
[Фрисман и др., 1985]. В регуляции численности репродуктивных группировок
островов Командорских и Курильских на определенных этапах их развития су-
щественна роль иммиграции [Чугунков, 1966; Кузин, Челноков, 1982а, б, 1990].
Наибольшее значение в динамике плодовитости самок котиков имеют ко-
личественные изменения впервые вступающих в размножение особей. Плодови-
тость самок, переступивших порог возраста наступления половой зрелости, ве-
личина предсказуемая. Она остается стабильной при любых экологических ус-
ловиях и разном состоянии численности популяции. На показатель плодовитости
проовулировавших особей в опромышляемых популяциях большое влияние
может оказать соотношение полов среди половозрелых особей. Однако этот по-
казатель полностью контролируется величиной промыслового изъятия. Ско-
рость вступления в размножение животных коррелятивно связана с особеннос-
тями их индивидуального развития. Но ассоциативное восприятие связи физи-
ческих параметров тела котиков с обеспеченностью их пищей в процессе ин-
дивидуального развития не подтверждается. Морфометрические показатели
плодов, щенков и среднепопуляционные размеры тела размножающихся особей
в возрасте 3—4 лет были меньше в периоды, когда основу репродуктивного ядра
популяций составляли самки младших возрастных групп. Размеры тела этих ка-
тегорий животных увеличивались в периоды, когда в популяциях преобладали
самки старших возрастных групп. Возрастная структура, таким образом, явля-
ется исходной предпосылкой, определяющей физическое состояние потомства и
ставящая его в соответствующие рамки чувствительности к воздействию внеш-
них факторов. При этом внешние условия в меньшей степени влияют на юве-
нильных котиков, если они рождаются физически крепкими и физиологически
зрелыми и уходят в первую самостоятельную зимовку хорошо упитанными. Те
же условия элиминируют часть приплода, снижают жизненный потенциал дру-
гой, если большая часть приплода появляется на свет физиологически незрмГой
и уходит на зимовку плохо подготовленной физически и физиологически, с ос-
лабленными возможностями противостоять давлению среды. В этом, по нашему
мнению, заключается однотипность действия регуляторного внутрипопуляцион-
ного механизма, проявляющего себя в особенностях индивидуального развития
и, соответственно, отражающегося впоследствии на плодовитости и смертности.
В целом, представленные данные позволяют сказать, что долгопериодные
колебания численности популяций северного морского котика являются атри-
бутом внутрипопуляционных механизмов их регуляции. Другие факторы лишь
замедляют или ускоряют течение естественных процессов. Это подтверждается
тем, что показатели выживаемости котиков до 2 лет в отдельных репродуктив-
ных группировках различны (рис. 19.3).
Разные популяции и репродуктивные группировки котиков достигли наивыс-
ших показателей численности на исследованном отрезке времени в разные годы.
На о-вах Прибылова максимальная численность щенков отмечалась в начале
50-х годов [Trites, 1992], на о. Тюлений — в 1967 г., в репродуктивной груп-
пировке на Северном и Юго-Восточном лежбищах — в 1977 г., на Урильем леж-
бище — в 1983 г. [Болтнев, Нестеров, 1996]. На общем фоне последующего сни-
жения численности во всех популяциях в одних репродуктивных группировках
численность стабилизировалась на достигнутом уровне (Урилье), в других на-
336
блюдался спад и последующий интенсивный подъем (Северное). Происходило
образование новых лежбищ, где численность увеличивалась (скала Долгая, Ку-
рильские о-ва; о. Сан-Мигель, Калифорния). Фактически каждая репродуктивная
группировка характеризуется собственной экологической судьбой. Однако эти
данные обычно игнорируются исследователями при изучении динамики числен-
ности, и отдельные залежки или репродуктивные группировки включаются в со-
став той или иной популяции. Более того, как показали наши исследования
[Кузин и др., 1996], каждое крупное лежбище представляет собой сложно струк-
турированную систему со специфической биохорологической организован-
ностью.
Формирующееся в период размножения на ограниченной территории со
сходными экологическими условиями под воздействием хоминга сообщество се-
верных морских котиков лишь на первый взгляд представляется гомогенным.
На самом деле, оно экологически и, по всей вероятности, генетически четко диф-
ференцировано на отдельные составляющие, представляющие собой субпопуля-
ционные уровни организации. Так, в пределах гаремного лежбища на о. Тю-
лений четко выделяются несколько территориально обособленных группировок,
обмен особями между которыми не превышает 6%. В основе их формирования
лежит филопатрия, благодаря чему сокращается до минимума радиус репродук-
тивной активности, что способствует интеграции индивидов на ограниченной
территории, снижению уровня панмиксии в популяции и поддержанию генети-
ческой уникальности субпопуляционных ячеек. Темп их формирования, прост-
ранственная ориентация, продолжительность существования в течение репродук-
тивного сезона, хотя и строятся по единому сценарию, но индивидуализированы
по срокам, что определяется разной динамикой экологических процессов, про-
текающих внутри каждой из них, включающих в себя показатели численности,
величины приплода, его смертности, соотношения полов производителей и, со-
ответственно, репродуктивного потенциала. Экологическая уникальность каж-
дой субпопуляционной единицы сохраняется на протяжении их многолетнего су-
ществования (на о. Тюлений прослежена за 1983—97 гг.). Проведенные нами
исследования позволили прийти к выводу, что наличие пространственно устой-
чивых и репродуктивно изолированных внутрипопуляционных группировок в
пределах единого размножающегося сообщества котиков имеет следствием из-
менение во времени основных экологических параметров, ответственных за чис-
ленность как их самих, так и популяции в целом. Представляется, что при сов-
падении позитивных тенденций в динамике этих показателей в большинстве суб-
популяционных подразделений популяция может ускорить процесс нарастания
численности, при негативных тенденциях — испытывать депрессию.
Таким образом, роль социальной организации размножающегося сообщества
в динамике его численности, по-видимому, значима. Однако исследования в этом
направлении только начаты.
337
Глава 20. ПРОМЫСЕЛ
20.1. История промысла северных морских котиков на Командорских
островах
Крупномасштабный промысел котиков был начат со времени открытия экс-
педицией В. Беринга в 1741 г. Командорских о-вов, на берегах которых, по сви-
детельству участника экспедиции штурмана С. Вакселя, залегало несметное ко-
личество этих животных. Весть о пушных богатствах Командорских о-вов быстро
разлетелась по Камчатке и Сибири. Уже в 1743 г., пригласив с собой несколько
человек из состава экспедиции В. Беринга, на Командорские о-ва отправляется
шитик под началом сержанта Нижне-Камчатской команды Басова, который зази-
мовал на островах. Басов привез из первой своей экспедиции 1200 шкур морских
бобров и 4 тыс. песцовых шкур. Из второй своей экспедиции он привез большую
партию пушнины, среди которой значились 1990 шкур котиков (табл. 20.1). Басов
еще дважды в 1747 и 1749 гг. снаряжал экспедиции на Командорские о-ва и про-
мышлял там котиков [Суворов, 1912]. В 1750 г. А. Толстых добыл на о. Беринга
840 котиков [Суворов, 1912]. В 1746 г. на о. Медный зазимовал А. Всевидов, но
добыча его не известна. В 1750 г. на Командорах побывал Югов, который вер-
нулся через 3 года с большой партией шкур, среди которых значились 2222 шку-
ры котиков [Берх, 1823]. В 1752 г. Воробьев доставил 6810 шкур “черных” и
“серых” котиков [Суворов, 1912], и в дальнейшем шкуры доставлялись с Ко-
мандорских о-вов регулярно вплоть до 1758 г. [Берх, 1823]. Однако, судя по до-
шедшим до нас статистическим данным промысла, количество заготавливаемых
шкур было небольшим (табл. 20.1). Оценивая размеры промысла котиков на Ко-
мандорах в первое десятилетие после их открытия, большинство исторических
обзоров указывает, что основное внимание промышленников в те годы было
обращено на калана и песца, а котиковый мех не имел большого спроса. Процесс
выделки шкур не был еще разработан, устойчивый рынок сбыта меховых то-
варов из котика отсутствовал. Частью шкуры оседали в России и использовалось
на обшивку кучерских кафтанов и полушубков [Волошинов, 1887], частью от-
правлялись в Кяхту и оптом обменивались на чай, шелк и другие восточные
товары. Однако, по мнению Г. Эллиотта [Elliott, 1884], заготовка шкур на Ко-
мандорских о-вах в первые годы промысла достигала таких размеров, что
“между 1760 и 1768 гг. русские совсем не били котиков и даже не знали, куда
они девались”.
Начиная с 1768 г., промысел котиков здесь развивается с новой силой. Наи-
более крупные партии шкур за последующее 20-летие были доставлены в 1778 г.
штурманом Петушковым (39500 штук), мещанином Сапожниковым в 1780 г.
(33840 штук). Общая добыча котиков в 1786 г. составила 92667 голов [Берх, 1823].
В 1786 г. подштурманом Прибыловым были открыты острова, впоследствии
названные его именем. Животный мир этих островов был идентичен Командор-
ским. С этого.времени основное внимание промышленников было сконцентри-
ровано на освоении ресурсов о-вов Прибылова.
338
Таблица 20.1
Ранний период промысла котиков на Командорских о-вах
Годы Число добытых котиков Источник Годы Число добытых котиков Источник
1745 не известно Берх, 1823 1769 1845 Берх, 1823
1746 1990 » » 1772 6350 » »
1750 300 » » 1777 1620 » »
800 Берх, 1898 1778 39500 » »
1752 7010 Берх, 1823 1779 2265 » »
1754 2222 » » 1780 33840 » »
6810 Суворов, 1912 1781 31570 » »
1755 250 Берх, 1823 1784 4715 » »
1757 10 » » 175 895 » »
1758 840 » » 1786 61792 » »
1768 3370 » »
Следует, однако, сказать, что еще в 1780 г. купцы Шелихов и Голиков ор-
ганизовали Компанию по эксплуатации пушных богатств Берингова моря. Спус-
тя 8 лет они добились у правительства права на монопольное владение про-
мыслами в Беринговом море, а еще 10 годами позже фирма преобразовалась
в Соединенную Американскую Компанию, которая в 1799 г. указом царя была
переименована в Российско-Американскую Компанию. Эта Компания просущест-
вовала до 1867 г.; ее деятельность очень слабо освещена в литературе. Все дер-
жалось в строгом секрете. По-видимому, убой котиков осуществлялся в больших
масштабах, причем добывались исключительно “серые” котики обоего пола. Так
продолжалось до 1805 г., когда административным воздействием генерала Ре-
занова было приказано остановить промысел, и в 1806—07 гг. котиков не про-
мышляли. С 1808 г. промысел приказано было возобновить, но обстоятельства
не позволили осуществлять его в полном масштабе [Вениаминов, 1840].
Отсутствие спроса на шкуры котиков привело к тому, что к 1803 г. на складах
Компании скопилось до 800 тыс. шкур. Чтобы поддержать цену на них, Компания
решила сжечь и выбросить в море до 700 тыс. шкур [Гихменев, 1861]. По данным
Е. К. Суворова [1912], такие действия Компания предпринимала неоднократно, од-
нако добыча котиков в больших масштабах продолжалась. Численность животных
катастрофически сокращалась. Известно, например, что в 1797—1821 гг. были вы-
везены 1232374 шкуры [Гихменев, 1861; Elliott, 1875].
Снижение численности котиков на островах Прибылова и Командорских
вновь заставило отказаться от их промысла, и в 1826—27 гг. на о. Св. Георгия
котиков не добывали. Еще более продолжительным был “запуск” промысла на
Командорских о-вах (1830—46 гг.) [Гихменев, 1861]. Следует, однако, сказать,
что предпринимались и другие ограничения, которые способствовали росту чис-
ленности котиков.
339
Так, с 1835 по 1867 гг. добыча самцов на островах была сокращена, а добыча
самок запрещена [Roppel, Davey, 1965]. Продолжавшаяся депрессия численности
в результате перепромысла заставила Компанию вновь отказаться от промысла
котиков на Командорских о-вах в 1843—1847 гг. Даже по истечении этого срока
в 1847 г. с о. Медный были вывезены всего 900, а в следующем году с о. Беринга
только 447 шкур [Суворов, 1912]. Всего с 1842 по 1861 гг. были заготовлены
Компанией 372894 шкуры котиков [Elliott, 1875], из них на Командорских
о-вах — 24 тыс. [Дорофеев, 1964].
В 1861—67 гг. ежегодная добыча котиков на Командорских о-вах составляла
в среднем 4 тыс. (в 1863 г. — 4.3 тыс., в 1864 г. — 5 тыс.). В 1866 г. был прекращен
убой “серых” котиков, и промысел был направлен на холостяков. Это способ-
ствовало росту численности и увеличению площадей холостяковых залежек. В ре-
зультате в последние годы деятельности Российско-Американской Компании до-
быча котиков на островах начала увеличиваться.
Данные по общей численности добытых Российско-Американской Компа-
нией котиков на островах разноречивы. Так, по данным Л. Стейнегера [Stejneger,
1898], с 1746 по 1860 гг. на Командорах добыли 362181 котика, а по данным
Л. В. Бойцова [1934], за 124 года было добыто ок. 500 тыс.
В 1867 г. Аляска с прилежащими к ней островами была продана США. Ко-
мандорские о-ва в связи с окончанием договора с Российско-Американской Ком-
панией были переданы в Приморское областное Управление. Начался, хотя и
короткий, но бурный период в промысловой деятельности частных предприни-
мателей. Командорские о-ва посещали суда разных стран, которые добывали
или скупали за бесценок у местного населения большие партии шкур котиков.
Объем промысла здесь возрос с 4 тыс. в 1861—67 гг. до 27.5 тыс. в 1870 г.
В 1870 г. Конгресс США принял закон об охране пушных зверей на Аляске
и в следующем году заключил контракт с Аляскинской торговой компанией на
20-летний период исключительного права промысла котиков на о-вах Прибы-
лова. В начале 1871 г. был подписан контракт об исключительном владении про-
мыслами котиков (кроме курильских) с торговым домом “Гутчинсон, Кооль и
К°” Фактически же Аляскинская компания и Торговый дом представляли одну
и ту же фирму.
На Командорских о-вах по контракту объем добычи котиков определялся
русской администрацией, а добыча осуществлялась местными жителями русско-
го подданства. Вывоз шкур с Командорских о-вов до 1887 г. составил в среднем
26025 штук, а после 1887 г. — 41728 [Волошинов, 1887]. Всего за 20 лет (1870—
90 гг.) Американской Коммерческой Компанией на Командорских о-вах, по дан-
ным Л. В. Бойцова [1934], было добыто 740812 котиков (по данным других ав-
торов, объем добычи составил 535—770 тыс.).
В 1891 г. котиковые промыслы на Командорских о-вах были сданы в аренду
Русскому Товариществу котиковых промыслов сроком на 10 лет. О-ва Прибы-
лова были арендованы Северо-Американской Коммерческой Компанией. Про-
мысловая деятельность обоих арендаторов была менее удачной, чем предшест-
вующих. Численность котиков падала. Главной причиной уменьшения числен-
ности зверей и снижения объема добычи был широкий промысел котиков в море.
Количество котиков, добытых на Командорских о-вах в 1891—1901 гг., со-
ставляло 8375-:-33837 голов; в общей сложности добыли 200648 котиков. По не-
340
полным данным, за 1891—96 гг. у азиатских берегов в море добыли 242758 ко-
тиков [Бойцов, 1934].
С 1902 г. промысел котиков на Командорских о-вах арендовало Камчатское
торгово-промышленное товарищество, деятельность которого продолжалась до
1911 г.
За этот период добыли 258.3 тыс. котиков [Ильина, 1950]. Деятельность То-
варищества совпала с Русско-Японской войной. Кроме того, Камчатское тор-
гово-промышленное общество с 1905 по 1910 гг. добыло ок. 35 тыс. котиков,
среди которых 40% составляли самки, чем был нанесен большой ущерб попу-
ляции.
В результате неумеренного промысла на берегу и все расширяющегося про-
мысла в море численность котиков на всех островах в северной части Тихого
океана стала заметно убывать и к 1910 г. сократилась в сотни раз. В условиях
интенсивного промысла речь шла о полном исчезновении северного морского
котика. Поэтому Россия и США уже с 1900 г. начали подготовку к выработке
Международного соглашения о прекращении морского промысла котиков.
В 1911 г. была подписана Конвенция об охране котиков между Россией,
США, Японией и Великобританией. Главным ее достижением было запрещение
морского промысла. После заключения Конвенции каждая из стран-участниц
могла проводить соответствующую политику по восстановлению стад и их ис-
пользованию. США владели о-вами Прибылова, Россия — Командорскими, а
Япония (после Русско-Японской войны) — о. Тюлений. Курильские о-ва с леж-
бищами котиков на них еще в 1875 г. отошли также Японии. Численность ко-
тиков на островах в 1912 г. составляла на о-вах Прибылова — 215900 [Baker
et al., 1970], на о. Тюлений — 6562 [М. К., 1929], на Командорских о-вах —
ок. 15 тыс. [Суворов, 1931].
После подписания Конвенции США объявили “запуск” промысла на 5 лет
на о-вах Прибылова. Этому же примеру последовала Россия. Однако, если для
США этот период стал временем интенсивного изучения биологии котиков, на-
чатого еще в годы подготовки к заключению Конвенции, то на Командорских
о-вах должного изучения биологии не было организовано. По окончании “за-
пуска” США возобновили промысел в пределах 20—95 тыс. особей в год вплоть
до 1927 г., а потом к 1940 увеличили его до 65 тыс. Практически, численность
котиков на о-вах Прибылова к 1940 г. была восстановлена и позже, вплоть до
1955 г., поддерживалась на стабильном уровне.
Гораздо сложнее дело обстояло с восстановлением численности котиков на
Командорских о-вах после заключения Конвенции 1911г. Объявленный “запуск”
несколько увеличил численность, в особенности секачей [Суворов, 1927; Бойцов,
1934; Ильина, 1950], однако общее состояние популяции нельзя было признать
удовлетворительным. Несмотря на принятые Конвенцией обязательства, хищ-
нический убой котиков на Командорах продолжался [Суворов, 1912]. Оценивая
результаты “запуска” 1911—17 гг., Е. Д. Ильина [1950] писала: “В связи с уве-
личением количества секачей и холостяков появились новые холостяковые за-
лежки, отдельные от самочьих, и холостяки перестали ложиться между самок.
Только это и явилось результатом запуска. Роста поголовья маток-производи-
тельниц не было”. По данным Л. Стейнегера [Stejneger, 1925], численность ко-
тиков в 1917 г. на Командорских о-вах не превышала 18 тыс.
341
Таблица 20.2
Промысел котиков на Командорских о-вах в 1923—44 гг. [по: Добровольский,
1929; Ильина, 1950]
Годы О. Беринга О. Медный Итого
ХОЛОС- ТЯКИ матки секачи полу- секачи всего холос- тяки матки секачи полу- секачи всего
1923 257 2 90 2 351 500 3 116 72 691 1042
1924 233 - - 2 235 437 7 139 39 622 857
1925 386 3 - 46 435 461 - 31 - 492 927
1926 324 3 34 - 361 476 2 53 - 531 892
1927 336 4 1 1 432 519 2 43 5 569 911
1928 390 - 7 1 398 472 1 15 3 491 889
1929 402 1 1 1 405 400 2 - - 402 807
1930 344 2 - - 346 303 - 1 - 304 650
1931 305 . _ - 1 306 401 1 - 1 403 709
1932 354 - - - 354 395 2 - 1 398 752
1933 402 - - - 402 427 - - - 427 829
1934 398 - - - 398 451 1 - - 452 850
1935 798 - 10 - 808 646 2 1 - 649 1457
1936 1420 1 209 105 1735 673 - 134 27 834 2569
1937 652 1 168 51 872 536 - 107 69 712 1584
1938 798 - 4 2 804 721 - 45 - 766 1570
1940 1011 - 122 1 1134 нет сведений 987 2121
1941 1243 - 65 1 1309 » » » 1065 2374
1942 1526 - 68 6 1600 » » 1345 2945
1943 2007 - 69 49 2125 1883 - 95 30 2008 4133
1944 1682 - 130 27 1839 1622 - 85 56 1763 3602
В 1917—23 гг. промысел на Командорах производился в незначительном объе-
ме, однако есть указания на то, что осуществлялся тайный убой и нелегальная
торговля котиковыми шкурами, кроме того возобновились набеги браконьер-
ских шхун на острова [Бойцов, 1930; Ильина, 1950].
С 1923 г. на Командорских о-вах в основу планирования забоя были поло-
жены подсчеты численности и расчетные коэффициенты смертности, заимство-
ванные у американцев. Вследствие “запуска” промысла на островах появилось
большое количество секачей, поэтому в 1927—29 гг. ежегодно в добыче при-
сутствовало определенное их количество, а после установления соотношения
полов в половозрелой части популяции 1:40 начали добывать только холостяков.
Объем добычи в период 1923—34 гг. осуществлялся в пределах 650—1000 особей
в год.
В последующие годы в связи с увеличением темпа прироста стада появилась
возможность увеличить добычу, которая к 1944 г. составляла уже более 3 тыс.
342
Таблица 20.3
Промысел котиков на Командорских о-вах в 1945—96 гг. [по: Дорофеев,
1964; данные NPFSCJ
Годы Самцы Самки Всего Годы Самцы Самки Всего
1945 5101 1973 4535 114 4649
1946 4610 1974 1715 31 1746
1946 4710 1975 1711 19 1730
1948 4514 1976 2742 26 2768
1949 5194 1977 2758 8 2766
1950 5836 1978 3020 12 3032
1951 5146 1979 1399 1 1400
1952 2784 1980 2536 8 2544
1953 2876 1981 5093 30 5123
1954 3629 1982 5066 9 5075
1955 3300 1983 5667 52 5719
1956 4256 1984 5244 50 5294
1957 3065 1985 5179 4 5183
1958 3021 3021 1986
1959 2261 12 2273 1987 6715 (“серые” котики)
1960 4079 2 4082 1988 8177 (“серые” котики)
1961 5023 28 5051 198.9 5600 (“серые” котики)
1962 6405 22 6427 1990 6010 (“серые” котики)
1963 7317 64 7381 1991 не добывали
1964 10491 87 10578 1992 2517 (холостяки) 3 2520
1965 11290 143 11433 3687 (“серые” ко- гики)
1966 8982 152 9134 1993 1106 (холостяки) 2 1108
1967 8381 121 8502 5000 (“серые” ко- гики)
1968 9507 135 9643 1994 3113 (холостяки) 21 3134
1969 8309 199 8508 6495 (“серые” ко гики)
1970 9073 127 9200 1995 1409 (холостяки 1 1410
1971 8296 196 8492 1996 6487 (“серые” котики)
1972 7556 619 8175
голов, а к 1951 г. — более 5 тыс. В 50-х годах добыча несколько сократилась
в связи со снижением численности котиков. Есть указания, что после денонси-
рования Конвенции 1911 г. в 1941 г. Япония вновь проводила промысел котиков
в море [Austin, Wilke, 1950]. Однако, думается, промысел в море не мог сильно
сказаться на состоянии командорской популяции котиков в эти годы (табл. 20.2).
343
Добыча котиков на Командорах островах быстрыми темпами начала увели-
чиваться после 1961 г. (табл. 20.3). Если в 1958 г. был добыт 3021 котик, то
в 1965 г. — 11433. В период 1966—72 гг. промысел осуществлялся в пределах
8—9 тыс. зверей в год, но при этом численность секачей катастрофически умень-
шалась. Поэтому в 1973—74 гг. норма добычи холостяков была снижена. Про-
мысел на о. Беринга был вообще прекращен на 5 лет. С 1981 г. он возобновился,
но в ограниченных пределах.
С 1987 г. промышлялись в основном сеголетки (щенки в возрасте 3—4 мес.).
Лишь в отдельные годы осуществлялась добыча холостяков (преимущественно
на о. Медный).
20.2. История промысла котиков на острове Тюлений
О. Тюлений как место лежбищ морских котиков был открыт китобоями
в 1852 г. (Федоров, 1964]. Ранняя история промысла котиков на острове не
ясна. А. Державин [1929] и С. В. Дорофеев [1964] писали, что японские ки-
тобои в 1852 и 1853 гг. добывали на острове 50—60 тыс. котиков, в 1854 г. —
20 и в 1855 г. — 28 тыс., после чего лежбище опустело. В течение последующих
14 лет промысел не проводился, и стадо начало восстанавливаться, но в 1870 г.
было опять разгромлено. С. Г. Федоров [1964] писал, что до 1870 г. промысел
котиков на острове не велся, затем в течение одного сезона сразу были выбиты
15 тыс. зверей, и в стаде осталось 600 животных.
Н. Волошинов [1887] считал, что до 1873 г. котиков на Тюленьем не про-
мышляли и в последующие 13 лет в среднем за год добывали по 3016 голов.
Н. А. Гребницкий [1902] сообщал, что в 1853—54 гг. котики на о. Тюлений были
истреблены английскими фирмами, причем за 2 года были выбиты 35 тыс. осо-
бей, а еще через год—два всего 300. Позже, вплоть до 1869 г., не было известно
попыток промысла на острове, но в 1870 г. американцы добыли на острове
15 тыс. котиков, через 3 года — 2700, и до 1877 г. промысел держался в пределах
2—4 тыс. голов. В 1878 г. добыли всего 780 котиков. По данным Л. В. Бойцова
[1934], первое посещение острова было в 1853 г. В 1854 г. на острове добыто
ок. 20 тыс. котиков, а в следующем — 28 тыс., после чего промысел из-за мало-
численности животных не производился. За период с 1873 по 1897 гг. торговым
домом “Гутчинсон, Кооль и К0” были добыты 45514 шкуры котиков (табл. 20.4).
Много шкур (не менее 53 тыс.) было вывезено браконьерскими шхунами с
1878 по 1895 гг., добывавших котиков на острове после окончания промыслового
сезона. По Н. А. Гребницкому [1902] количество шкур, добытых американскими
и канадскими промысловиками с 1879 по 1891 гг., составило ок. 55—72 тыс.
По данным Л. Стейнегера [Stejneger, 1898], с 1872 по 1897 гг. в Охотском
море было добыто 8 тыс. котиков. Последняя шхуна, промышлявшая котиков
у о. Тюлений, зарегистрирована в 1895 г.
О количестве шкур, добытых японцами, ничего не известно. С 1896 по 1903 гг.
добыча котиков на Тюленьем исчислялась сотнями голов (табл. 20.4), а с 1904
по 1911 гг. включительно добычи не было. После окончания Русско-Японской
войны о. Тюлений отошел к Японии, и японское правительство сразу же пред-
приняло Меры по его охране. Морской промысел вокруг острова в пределах 30-
344
Таблица 20.4
Количество котиков, добытых на о. Тюлений с 1873 до настоящего времени
Годы 1 Число добытых котиков Годы Число добытых котиков
1 2 3 4
1873 2700 1950 6529
1874—77 2-4 тыс. в год 1951 3065
1978 780 1952 3973
1883 2049 1953 3705
1885 1838 1954 5175
1890 4851 1955 4058
1891 540 1956 2000
1892 1500 1957 3651
1893 1000 1958 3127
1895 1300 1959 4885 (25)
1896 149 1960 6251 (41)
1897 332 1961 6982 (35)
1898 “запуск” 1962 7123 (86)
1899 550 1963 7339 (74)
1900 585 1964 8317 (105)
1901 524 1965 8586 (154)
1902 629 1966 9380 (192)
1903 502 1967 9003 (245)
1912 139 1968 5336 (266)
1913 547 1969 7140 (919)
1914 537 1970 7106 (64)
1915 571 1971 7036 (33)
1918 550 1972 7015 (604)
1919 555 500 (“серые” котики)
1920 555 1973 4187 (54)
1921 550 1974 2447 (288)
1922 600 200 (“серые” котики)
1923 844 1975 2483 (17)
1924 942 1976 2572 (221)
1925 868 1977 4071
1926 1322 1978 3304 (720)
1927 1608 1979 2991 (23)
1928 1521 1980 3107 (8)
1929 1700 1981 3560 (53)
1930 1712 1982 2874 (50)
1931 1701 1983 2074 (50)
1932 1700 1984 2305 (17)
345
Таблица 20.4 (окончание)
1 2 3 4
1933 1700 1985 2067 (12)
1934 2000 1986 2056 (19)
1935 2004 1987 2479 (9) (“серые” котики)
1936 2140 10 (4) (холостяки)
1937 2125 1988 1777 (30)
1938 2140 1989 1364
1939 2100 1990 1556 (13)
1940 3000 1991 1532 (11)
1941 3000 1992 1950 (11) (из них 200 секачей)
1942 6000 1993 1729 (9)
1943 10000 1994 1496 (40)
1944 30000 1995 промысла не было
1945 5000 1996 промысла не было
1946 10511 Примечание. В скобках — число добытых
1947 6348 самок.
1948 6648
1949 5864
мильной зоны с 1906 г. японским подданным был запрещен, а на острове был
объявлен “запуск” до 1912 г. Предпринятые меры охраны и прекращение мор-
ского промысла благотворно сказались на состоянии тюленьевской популяции.
Численность котиков начала быстро восстанавливаться, что позволило вскоре на-
чать промысел, сначала в ограниченных масштабах, а с 1926 г. число добываемых
животных превысило 1000 голов. К началу Второй мировой войны уже добывали
до 3 тыс. голов. В период войны Япония усилила промысел. В 1942 г. были добыты
6 тыс., в 1943 г. — 10 тыс., в 1944 г. — 30 тыс. и в 1945 г. — 5 тыс. котиков
[Кузнецов, Ильина, 1949]. В 1945 г. промысел был прерван обстрелом острова с
американского миноносца, уничтожившим все постройки на нем.
В 1945 г. о. Тюлений был возвращен СССР. Благоприятное состояние по-
пуляции, несмотря на довольно-таки большое число добытых в 1943—45 гг. жи-
вотных, в т. ч. самок, позволило осуществлять в послевоенные годы добычу в
пределах 3—10 тыс. особей ежегодно. К моменту заключения Конвенции по со-
хранению котиков 1957 г. в среднем добывали 4—5 тыс. животных в год.
Объем добычи постепенно увеличивался и в 1966 г. составил 9380 голов. Сле-
дует сказать, что с 1958 г. в добыче постоянно присутствовало все возрастающее
число самцов старших возрастных классов. Общий выбой их из отдельных по-
колений составлял до 47%. К 1973 г. в популяции осталось 546 секачей, а в по-
следующие 2 года — 540 и 553. Соотношение полов резко изменилось. Упала вос-
производительная способность самок, что в сочетании со снижением выживаемости
в последующих поколениях повело к общему упадку численности. В связи с этим
была резко снижена добыча, которая в 1974 г. составила всего 2500 голов. В по-
следующие годы норма изъятия колебалась в пределах 2.8—3.5 тыс. в год.
346
20.3. История промысла котиков на Курильских островах
Многочисленные экспедиции на Курильские о-ва в XVIII в. с целью или при-
соединения их к Российской империи, или освоения пушных богатств, как пра-
вило, были малодоходными [Полонский, 1871]. Основное место среди меховых
товаров занимали наземные виды млекопитающих, а среди морских — калан.
Впервые 190 штук котиковых шкур были закуплены у айнов на Курилах в
1776 г. [Верх, 1823].
По всей вероятности, лежбища котиков на Курильских о-вах были открыты
английским капитаном Г. Сноу в 1881 г., занимавшимся нелегальной добычей
котиков, т. к. именно с этого периода количество поступавших отсюда шкур
сразу увеличилось в несколько сот раз. Г. Сноу обнаружил и впоследствии до-
бывал котиков лишь на о. Райкоке, о-вах Каменные Ловушки и Среднего. Ви-
димо, других крупных котиковых лежбищ на Курилах не было, хотя есть ука-
зания о существовании здесь не менее 9 лежбищ котиков, начиная с о. Уруп
и кончая о. Шумшу [Успенский, 1955].
Добыча морских котиков на Курильских о-вах осуществлялась с судов, ба-
зировавшихся в Японии. В Иокогаме Г. Сноу организовал Компанию по добыче
котиков, каланов и скупке других видов пушнины у местного населения. Дея-
тельность Г. Сноу была тесно связана с японской фирмой “Окура Муми”, также
имевшей в своем распоряжении несколько парусных шхун и занимавшейся пуш-
ным делом. Известно также, что закупкой пушнины у местного населения и до-
бычей животных занимались отдельные немногочисленные частные предприни-
матели [Волошинов, 1887].
Впервые японские шхуны вышли на промысел северных морских котиков
в район Курильских о-вов в 1882 г. и добыли 2 тыс. шкур. В 1883 г. добыли
всего 400 шкур. После этого в связи с запрещением японским правительством
добычи котиков японцы не предпринимали попыток выхода на промысел [Во-
лошинов, 1887]. В основном добычей котиков на Курильских о-вах занималась
фирма Г. Сноу, за короткий период разгромившая все лежбища (табл. 20.5).
По свидетельству самого Г. Сноу [1902], только на о. Райкоке в 1883 г. он добыл
15 тыс. котиков, а всего с 1881 по 1896 гг. на Курильских островах им был добыт
24491 котик. Эта цифра близка к итогу статистических данных, собранных
Л. Стейнегером [Stejneger, 1898], и, по-видимому, соответствует действительности.
Данные таблицы 20.5 показывают, что лежбища котиков фактически были
разгромлены за 3—4 года, и после 1884 г. поступления котиковых шкур с Ку-
рильских о-вов не превышали 500 штук. Разгром котиковых лежбищ на Курилах
повлек за собой банкротство фирмы Г. Сноу в 1885 г. [Волошинов, 1887].
В 1896 г. Л. Стейнегер [Stejneger, 1898] котиков в районе Курильских о-вов
не нашел. Однако в 1911 г. около двух десятков котиков вновь видели на о-вах
Каменные Ловушки, 7 из которых тут же были добыты японскими промыш-
ленниками [Успенский, 1955]. Единичные животные здесь отмечались и в после-
дующие годы [Тихенко, 1914а].
В настоящее время лежбища котиков на островах Каменные Ловушки и Сред-
него полностью восстановлены, однако промысел здесь не ведется. Лежбище на
о. Райкоке еще в прошлом веке было разрушено в результате извержения вул-
кана, и появление там этих животных вновь проблематично.
347
Таблица 20.5
Добыча котиков на Курильских о-вах
Голы Число добытых котиков Источник Примечание
1776 190 Берх, 1883 Курильские о-ва
1842-60 29 Сергеев, 1947 » » »
1873 299 Слюнин, 1895 » » »
1874 78 » » » » » »
1875 25 » » » » » »
1876 285 » » » » » »
1877 342 » » » » » »
1878 270 » » » » » »
1879 211 » » » » » »
1880 137 » » » » » »
1881 82 » » » » » »
7700 Stejneger, 1898 о-ва Среднего
1795 » » » о-ва Каменные Ловушки
7000 Слюнин, 1895 о-ва Среднего
1882 3200 Stejneger, 1898 » » »
2000 Волошинов, 1887 » » »
1883 3 Слюнин, 1895 Курильские о-ва
400 Stejneger, 1898 о-ва Среднего
15000 Сноу, 1902 о. Райкоке
400 Волошинов, 1887 Курильские о-ва
1884 54 Слюнин, 1895 » » »
700 Stejneger, 1898 о-ва Среднего
1885 31 Слюнин, 1895 Курильские о-ва
12 Stqneger, 1898 о-ва Среднего
491 » » » о-ва Каменные Ловушки
1886 99 Слюнин, 1895 Курильские о-ва
500 Stejneger, 1898 о-ва Каменные Ловушки
1887 33 Суворов, 1912 Курильские о-ва
38 Stejneger, 1898 о-ва Среднего
1890 387 Суворов, 1912 Курильские о-ва
200 Stqneger, 1898 о-ва Среднего
1891 20 » » » » » »
40 Суворов, 1912 Курильские о-ва
1892 3 » » » » » »
3 Stejneger, 1898 о-ва Среднего
1893 1 » » » » » »
3 » » » о-ва Каменные Ловушки
1894 4 » » » » » »
348
20.4. История пелагического промысла котиков
Пионерами в освоении судового промысла котиков были канадцы, добывав-
шие их у берегов о. Ванкувер уже в 1868 г. По-видимому, промысел не имел
большого значения, т. к. количество добываемых котиков определялось потреб-
ностью в шкурах местного индейского населения, из которых в основном и фор-
мировались команды судов, промышлявших потом котиков в море [Гребницкий,
1902]. В 1880 г. пелагическим промыслом были заняты 4 канадские шхуны, каж-
дая из которых добыла по 2—3 тыс. шкур в год. В последующие годы судовой
промысел быстро расширяется. К нему приобщились американцы, японцы и
предприниматели других стран, суда которых базировались в Японии. Следует
назвать несколько обстоятельств, способствовавших его развитию. В 30-х годах
прошлого века в Англии был разработан новый способ выделки шкур и изго-
товления из них легких, прочных и носких изделий. Котиковый мех стал поль-
зоваться популярностью; цены на него поднялись. Улучшилось качество постав-
ляемого сырья в связи с сокращением сроков его доставки по трансконтинен-
тальной северо-американской железной дороге, что повысило цены на сырые
шкуры и заинтересованность поставщиков. И, наконец, к промыслу котиков в
море побуждала локализация островного промысла, ущемившего интересы Анг-
лии, которая хорошо была осведомлена о доходах, т. к. шкуры продавались с
аукциона в Лондоне. (До 1869 г. шкуры из США доставлялись в Лондон морем
через мыс Горн. От долгой транспортировки, связанной с пересечением эква-
тора, большая часть шкур приходила в негодность).
В 1882 г. добычей северных морских котиков в море занимались 8 канадских
и 2 американских шхуны. В 1886 г. их число увеличилось до 34. Своего апогея
этот промысел достиг в 1891—92 гг., когда количество промысловых шхун пре-
высило сотню, а число добываемых ими котиков возросло до 100 тыс. По дан-
ным Н. А. Гребницкого [1902], в 1895 г. в море работали 192 шхуны, которые
добыли 92437 котиков.
С развитием судового промысла стал падать уровень добычи котиков на ос-
тровах. Озабоченное этим американское правительство организовало целую
флотилию военных судов для охраны котиков и начало вести энергичные пе-
реговоры с Англией об ограничении морского промысла. Результатом этих пере-
говоров явилось заключение между США и Англией в 1891 г. соглашения о пре-
кращении добычи котиков в восточной части Берингова моря на 1892 г. Россия
к этому соглашению не примкнула и, таким образом, косвенно способствовала
дальнейшему процветанию судового промысла у своих берегов. Поэтому про-
мысел быстро перенесся в район западной части Берингова и в Охотское моря.
Если до 1892 г. промысел котиков здесь был не особенно большим, то позже
весь канадский, американский и базировавшися в Японии флот других стран,
начал истреблять незащищенных никакими законодательными актами коман-
дорских, курильских и тюленьевских котиков, охрана которых совершенно не
была налажена.
Первая появившаяся у берегов Японии и в русских водах в 1890 г. шхуна
добыла 680 котиков. На следующий год было добыто втрое больше. В 1892 г.
промысловых шхун в японских водах было уже 9, которые добыли 14 тыс.
котиков, а в 1894 г. — 70 шхун, доставивших 71667 шкур [Townsend, 1898].
С 1895 г. канадский и американский промысел котиков у берегов Японии пос-
тепенно сокращается.
349
Таблица 20.6
Число шхун и общая добыча котиков Японией в 1884—1945 гг. [по:
Гребницкий, 1902; Суворов, 1912; Федоров, 1964]
Число ШХУН Добыто котиков Голы ЧИСЛО ШХУН- Добыто .КОТИКОВ—
Z 3 965 1906 29 10176
1895 6 5771 1907 35 10420
1896 9 4967 1908 31 13355
1897 14 4616 1909 35 10465
1898 16 4840 1910 37 8309
1899 12 6518 1911 50
1900 15 7533 1943 10 2616
1901 19 6945 1944 10 2222
1902 15 7462 1945 800
1903 21 11240
1904 28 15698
1905 29 10035
По данным Н. А. Гребницкого [1902], только под английским флагом здесь
плавали 37 шхун, которые добыли в 1892 г. 17222 котика. В 1893 г. общая добыча
у азиатских берегов составила 67 тыс. котиков, а в 1894 г. — 93149, что почти
в 1.5 раза больше объема годового их промысла в море у американских берегов.
Если учесть то обстоятельство, что в эти годы численность котиков на Ко-
мандорских о-вах была в 2 раза меньше, чем на о-вах Прибылова, то хищни-
чество по отношению к азиатским популяциям котиков было, как писал Л. Стей-
негер [Stejneger, 1898], в 4 раза разрушительнее, чем у американских берегов.
Обособившись от вступления в соглашение о запрете морского убоя котиков
в 1891 г., Россия предприняла энергичные меры по охране командорских котиков
с помощью военных судов, но отсутствие соответствующего законодательства
ограничивало сферу деятельности только пределами 3-мильной акватории тер-
риториальных вод. В 1893 г. был издан закон о запрещении добычи котиков
в море для подданных России, которые и так не добывали этих животных в
море. Принятие законодательства, однако, не препятствовало иностранным
судам добывать котиков за пределами 3-мильной зоны. С принятием этого за-
кона возник ряд инцидентов в 1891—92 гг., связанных с арестом иностранных
шхун за пределами территориальных вод России. Начались интенсивные пере-
говоры по поводу этих инцидентов с Англией, и одновременно вырабатывались
позиции по заключению соглашения об ограничении добычи в море. Такое со-
глашение было заключено в 1893 г. Им предусматривалось запрещение англий-
ским подданным в течение года добычи котиков в пределах 10-мильной при-
брежной зоны вдоль российских берегов и 30-мильной зоны у Командорских
о-вов и о. Тюлений. В 1894 г. аналогичное соглашение было подписано между
США и Россией. Однако промысел котиков за пределами запретных зон про-
должался. Тем временем между США и Великобританией возник ряд спорных
вопросов, которые разбирались Международным трибуналом с участием всех
350
заинтересованных стран и представителей нейтральных государств. Решением
этого кворума США удалось добиться запрета добычи котиков в море с 1 мая
по 31 июля каждого года в восточной части Тихого океана и Беринговом море
севернее 35° с. ш. и к востоку от 180° в. д. Россия вновь встретила такое решение
сочувственно, но не примкнула к нему. В 1897 г. США издали закон, запре-
тивший своим подданным добычу котиков в море во все времена года, и ор-
ганизовали охрану котиков военными судами.
Бурное развитие котикового промысла в западной части Тихого океана по-
будило Японию также заняться его освоением. В короткий срок, изучив осо-
бенности ведения судового промысла, Япония уже в 1894 г. послала 3 шхуны
для промысла котиков в море (табл. 20.6).
Через 3 года под японским флагом плавали уже 14 шхун, а количество до-
бываемых ими котиков составляло 46.7% общего судового промысла в азиатских
водах [Бойцов, 1934].
Чтобы завоевать превосходство на мировом рынке в получении прибылей
от торговли добываемыми в море котиковыми шкурами и парализовать дея-
тельность канадских и американских предпринимателей, в Японии в 1897 г. был
издан закон, который всячески поощрял развитие промысла рыбы, котиков и
каланов в открытом море. В 1903 г. 21 японская промысловая шхуна добыла
свыше 11 тыс. котиков, и в дальнейшем морской промысел их продолжал уве-
личиваться. Часть шхун уходила к берегам Америки и добывала котиков в ак-
ватории от берегов Мексики до о-вов Прибылова и Командорских о-вов, другая
часть держалась у берегов Японии и Кореи. Только в Японском море в отдель-
ные годы добывали котиков до 14 японских шхун. Столько же шхун посещали
воды у Командорских о-вов [Суворов, 1912].
С ростом японского морского промысла сокращается число канадских и аме-
риканских шхун, действующих в азиатских водах. Так, если в 1894 г. в азиатских
водах плавало 36 канадских шхун, то в 1897 г. — только 12. Число американских
шхун сократилось с 39 до 2, а европейских — с 7 до 3 [Суворов, 1912]. С 1910 г.
в азиатских водах котиков промышляли только японцы. Канадцы еще продол-
жали промысел котиков в американских водах, куда заходили 5 шхун. Япония,
не являвшаяся участницей трехстороннего соглашения об охранных зонах вокруг
островов, соблюдала утвержденные Международным трибуналом 3-мильные зо-
ны США и России и продолжала добычу котиков за ее пределами во все сезоны
года вплоть до подписания Конвенции 1911 г. Сокращение численности котиков
на островах, сделало нерентабельным морской промысел, и это побудило япон-
цев делать “набеги” на острова, пользуясь их плохой охраной. Особенно от этого
страдали российские лежбища. Только в 1911 г. были 43 попытки высадки и
добычи котиков на Командорских о-вах. В результате таких “набегов” с 1905
по 1910 гг. японцами были полностью уничтожены Корабельновские лежбища
на Командорах [Ильина, 1950].
В общей сложности на азиатской стороне Тихого океана с 1872 по 1897 гг.,
по подсчетам Л. Стейнегера [Stejneger, 1898], добыли в море ок. 295 тыс. котиков,
из которых 8 тыс. приходится на тюленьевских котиков и 287 тыс. — на ко-
мандорских. Ежегодные потери вместе с ранеными и утонувшими при морском
промысле составили с 1891 по 1897 гг. не менее 100 тыс. голов.
351
Общая добыча котиков в море с 1868 по 1909 гг. без учета утонувших и
раненых животных у восточных берегов Сев. Америки и в восточной части Бе-
рингова моря составила ок. 1000000 голов, а в западной части Берингова моря,
в Японском, Охотском морях, у берегов Курильских о-вов и у берегов Японии
ок. 500 тыс.
Крупномасштабный судовой промысел котиков прекратил свое существова-
ние после подписания Конвенции об охране котиков в 1911 г. и не возобновлялся
до 1940 г., когда Япония сообщила странам-участницам Конвенции о своем на-
мерении денонсировать ее через год по причине увеличения численности коти-
ков, которые, по мнению японцев, стали сильно вредить рыболовству. 23 октября
1941 г. Конвенция потеряла силу, и Япония возобновила промысел котиков в
море. Им стала заниматься компания “Nippon Kaiju Kaisha”. В 1943 г. добыли
2616, в 1944 г. — 2222 и в 1945 г. — 800 котиков в море, но эти цифры внушали
большое сомнение [Taylor et al., 1955]. Кроме того, Япония, по-видимому, вела
морской промысел котиков вплоть до 50-х годов.
С 1951 по 1954 гг. Япония, Канада и США провели цикл исследований се-
верных морских котиков как у берегов Америки, так и у берегов Японии, в ре-
зультате которых вновь было показано, что влияние котиков на промышленное
рыболовство ничтожно, и всем заинтересованным странам целесообразно при-
нять соответствующие меры для сохранения и увеличения численности этих жи-
вотных.
В 1957 г. была заключена Временная Конвенция о сохранении котиков се-
верной части Тихого океана между СССР, США, Японией и Канадой. С за-
ключением этой Конвенции морской промысел котиков перестал существовать.
Трудно переоценить значение международных соглашений об охране коти-
ков, исключавших возможности ведения пелагического промысла и способст-
вовавших рациональной постановке хозяйственного использования котиков.
Процесс восстановления популяций котиков проходил не одинаково и не син-
хронно. Он был сильно заторможен на Командорских о-вах. Однако, в целом,
теперь можно сказать, что восстановление популяций северных морских котиков
происходит сравнительно успешно. Популяции на островах Прибылова, Тюлень-
ем, Курильских полностью восстановлены. Командорская популяция находится
на середине пути своего полного восстановления.
Добыча котиков происходит под контролем биологов. Колебания в добыче
из популяций, достигших своего предельного уровня, могли быть менее резко
выражены, если бы наши знания биологии котиков были более глубокими.
Уменьшение объема добычи в последние годы было связано во многом с ес-
тественными процессами снижения численности в результате изменений интра-
популяционных характеристик в пределах популяционного времени, исчисляе-
мого продолжительностью жизни одного поколения. Такие колебания числен-
ности и впредь будут иметь место; это необходимо учитывать при ведении ко-
тикового хозяйства в будущем. Однако следует признать, что определенную
лепту в ускорение процесса снижения численности или ее стабилизации на от-
дельных островах внесли биологически необоснованные рекомендации по экс-
плуатации популяций, что, в конечном итоге, негативно сказалось на их половой
структуре и плодовитости.
352
20.5. Техника промысла котиков
За 250-летнюю историю промысла морских котиков на остройах техника до-
бычи существенно не изменилась. Вместе с тем, с ростом технического осна-
щения промышленных производств значительные дополнения в обслуживание
промысла вносились на протяжении всего времени его существования. Предпри-
нимались также попытки совершенствования методов добычи котиков с при-
менением различных технических средств или сильнодействующих нервно-пара-
литических препаратов. Однако они или не оправдали себя, или просто не при-
жились по тем или иным причинам. В настоящее время котиков добывают на
островах так, как их добывали еще промышленники во времена владения про-
мыслами Российско-Американской Компанией. Изъятию подлежат те же воз-
растные категории животных.
20.6. Методы добычи котиков на лежбищах и первичная обработка тушек
Наиболее распространенным методом убоя котиков на берегу с давних вре-
мен является использование “палок-дрыгалок” (как их именуют зверобои).
Метод складывается из нескольких операций.
1. Отгон котиков с мест залегания и прогон их до места убоя — загонных
площадок.
2. Непосредственный убой ударом палки по голове.
3. Докалывание животных с целью окончательного умерщвления и обескров-
ливания.
4. Съемка шкур, охлаждение их, обезжиривание, засолка, маркировка, сорти-
ровка и упаковка.
Отгон котиков производится, как правило, в ранние утренние часы, и на боль-
шинстве островов для этого выбираются пасмурные или облачные дни. Особен-
но важно было соблюсти эти условия, когда лежбища были необорудованными,
забойные площадки находились в нескольких километрах от лежбищ,, отгоны
с некоторых из них длились более суток.
Для проведения отгонов группа загонщиков скрытно пробирается к холос-
тяковой залежке и старается отрезать залегающих животных от моря. На о. Тю-
лений, например, чтобы можно было незаметно пройти к залежке холостяков
и отрезать их от моря, под землей сделаны несколько тоннелей, по которым
загонщики выходят на поверхность для загона непосредственно у воды.
Отогнанных зверей перегоняют к забойной площадке. Если путь от залежки
до площадки большой, то животным в пути дают несколько раз отдохнуть, в
противном случае многие из них перегреваются и гибнут. Мех таких животных
становится непригодным для дальнейшей обработки. На некоторых лежбищах
отогнанных котиков помещают в загонные дворики, огороженные дощатым за-
бором, на других этого сооружения нет. И с помощью нескольких специально
выделенных наблюдателей зверей останавливают на определенных местах у за-
бойных площадок. Около площадок животным дают отдохнуть и потом неболь-
шими партиями по 10—15 голов отгоняют от основной группы и забивают уда-
353
ром палки по голове. Самцов, не относящихся к забойному возрасту, и самок,
случайно попавших в загон, отпускают и дают им возможность уйти в море.
С целью повышения качества сырья и гуманизации убоя предпринимались
попытки разработки других способов умерщвления животных. В 1959—61 гг.
на Командорских о-вах и на о. Тюлений были проведены эксперименты по забою
котиков с помощью электрического тока [Мараков, 1974]. В эти же годы на Тю-
леньем был опробован метод забоя из винтовки типа “ТОЗ” (калибр 5.6 мм).
1968 г. на Тюленьем пробовали применить мелкокалиберный пистолет системы
“Марголина”. Результат оказался тот же, что и при стрельбе из винтовки. Од-
нако ни умерщвление животных с помощью электрического тока, ни оружейные
методы добычи на берегу не прижились.
В 1963—964 гг. на Командорских о-вах были проведены эксперименты по
умерщвлению котиков при помощи курареподобного препарата дитилина [Нес-
теров, 1966]. Действие дитилина заключается в полной потере у котиков дви-
гательных функций, а при летальных дозах — к остановке дыхания и сердца.
Средняя летальная доза для котиков равна 1.659±0.067 мг сухого вещества на
1 кг массы. Для быстрого убоя тюленей без утилизации крови применялась доза,
вдвое превышающая указанную. Препарат использовался в форме 5—7.5% раст-
вора в дистиллированной воде, подогретого до 40—50°. Впрыскивание раствора
препарата производилось внутри мышечно специальным автоматическим инъек-
тором, укрепленным на 2—3-метровом древке. Инъектор имел резервуар емкостью
300 мл, что исключало его перезарядку в процессе работы. При убое котиков
дитилином качество мехового сырья было выше, при этом мясо и другие по-
бочные продукты не теряли своей пищевой ценности. В целом, метод получил
хорошую оценку и может применяться в дальнейшем на промысле котиков.
Аналогичные работы по обездвижению котиков с помощью различных тран-
квилизирующих и анестезирующих препаратов были проведены американцами
[Peterson, 1965; Keyes, 1966], но они никогда не ставили своей целью так до-
бывать котиков.
Убитых животных оттаскивают в сторону и укладывают рядами, брюхом
кверху и головой всех в одну сторону. Выполнение последующих операций по
обеловке зверя происходит с четким разделением труда между промысловиками.
Съемку шкур на разных островах производят по разному. На о. Тюлений
этот процесс механизирован. За свободную часть шкуры, снятую с головы, за-
цепляют щипцами, от которых на электрическую лебедку идут металлические
тросы. Туши удерживают с помощью вилообразного держака, которым прижи-
мают ее в области шеи к бревенчатому или досщатому настилу. Аналогичным
образом с помощью щипцов, но только вручную, производят съемку шкур на
о-вах Прибылова. На Командорах шкуры снимают вручную с помощью ножей.
Все остальные операции на всех островах аналогичны.
Снятые пластом шкуры с салом (хоровины) охлаждают в бетонных чанах,
наполненных морской, периодически или постоянно сменяемой водой. На не-
оборудованных лежбищах шкуры расстилают на земле мехом вниз. Охлаждение
шкур длится от 4 до 24 час. Замечено, что после 24-часовой промывки в воде
шкуры мездрятся лучше и легче и впоследствии имеют более товарный вид. Од-
нако никаких изменений химического состава шкур и волос при разной про-
должительности промывки не отмечается [Загуляева, 1968].
354
Дальнейший процесс обработки хоровин за время существования промысла
претерпел значительные изменения. До 30-х годов прошлого века шкуры кон-
сервировали пресно-сухим способом, связывали их в тюки по 50 штук, перево-
зили морем до Санкт-Петербурга или в Аян, а оттуда по суше — в Кяхту, где
обменивали на чай, шелк и другие товары [Волошинов, 1887].
Позже обезжиренные шкуры стали засаливать и сырыми, на судах доставляя
в Англию. Переброска шкур занимала много времени, причем при переходе
через экватор в связи с резким изменением температурных условий многие
шкуры приходили в негодность и потом браковались. Непродолжительное время
в ходу был метод неполного обезжиривания шкур; их засаливали вместе с тонким
слоем сала. При такой консервации качество поставляемого сырья было якобы
лучше. В 30-х годах нашего столетия со шкур стали снимать весь жировой слой.
Этот метод подготовки шкур к консервации сохранился до наших дней.
Удаление жирового слоя со шкур или их мездрение осуществляется вручную.
На о. Тюлений была опробована мездрильная машина, применяемая при судо-
вом промысле настоящих тюленей, но в связи с тем, что жировой слой у котиков
по физическим свойствам не идентичен таковому настоящих тюленей, и в нем
присутствует значительное количество коллагеновых волокон, мездрение с по-
мощью машины не дало положительных результатов. На отмездренных таким
образом котиковых шкурах появлялось много механических повреждений в виде
выхватов, разрывов, и сама мездра в последующем требовала дальнейшей до-
работки вручную.
Мездрение котиковых шкур вручную — трудоемкий процесс. Он выполняется
на станке-навое, представляющем собой гладко обструганную деревянную плаху,
обшитую жестью или линолеумом, или плаху изготовляют из алюминиевого по-
луцилиндра, поставленного наклонно, высота верхнего края которого регули-
руется в зависимости от роста мездрильщика. Шкура с салом (хоровина) на-
брасывается на навой, затем примерно с третьей части каудальной области счи-
щают двуручным ножом жир, потом переворачивают шкуру, закрепляют кау-
дальную часть за крючки на навой и производят “строжку” остальной части.
Мездрильный нож представляет собой либо прямой двуручник типа плот-
ницкого наструга, либо его форма овальная, повторяющая изгиб навоя. У мез-
дрильщиков, как правило, 2 ножа: менее острый, с более крутой заточкой
(“тупик”) и острый, с пологой заточкой (“острик”). Иногда оба края ножа за-
тачивают — один под острик, другой под тупик. Вначале толстый слой жира
снимают острым ножом, а затем при помощи тупого ножа соскабливают и вы-
жимают из кожи остатки жировых включений. Со шкур холостяков до 3 лет
часто сало снимают только тупиком. Во время мездрения от шкуры отделяют
ушные раковины и часть шкуры носовой области до уровня глаз. На мездрение
хоровины 2-леток обычно затрачивается 3—5 мин, полусекача — 10—15 мин,
секача — 15—20 мин [Бычков, 1964в].
Отмездренные шкуры промывают в чанах с холодной морской водой, после
чего развешивают на вешалах для стекания воды. Процесс отмывки занимает
20—30 мин, стекание воды — 5—10 мин. После этого из шкур производят вы-
жимание воды с помощью деревянной планки на ровном столе, которой несколь-
ко раз проводят по шкуре вдоль волоса, а затем ту же процедуру проделывают
355
со стороны мездры. Затем шкуры развешивают для просушки на вешалах. Это
сооружение устраивают под навесом, чтобы на шкуры не попадали солнечные
лучи. Процесс сушки на Командорах длится 2—3 часа [Ильина, 1950], на о. Тю-
лений — 30 мин [Бычков, 1964в]. Затем производят засолку шкур. Здесь же про-
исходит их предварительная сортировка по площади и порокам. Рассортиро-
ванные шкуры засаливают солью помола № 2, прошедшую предварительную
проверку на фуксин. Солят шкуры в засольных закромах, пол которых засыпают
солью слоем в 2—3 см, на которую кладут шкуры мездрой кверху. Каждую
шкуру тщательно расправляют; особенно тщательно следят, чтобы были хорошо
расправлены края шкуры. На разостланный первый слой шкур насыпают ров-
ным (2—3 см) слоем соль. Следующий слой шкур кладут перпендикулярно пер-
вому, тоже мездрой кверху и т. д. Высота такого сложенного штабеля шкур
обычно не превышает 1 м. Сверху штабель засыпают толстым слоем соли. Через
8 дней шкуры извлекают из соли и аналогичным способом засаливают второй
раз, однако теперь нижние шкуры оказываются наверху. Вторичный засол шкур
длится неделю. После окончания срока засолки их извлекают из соли, отряхи-
вают от ее остатков, вторично осматривают и сортируют. Плохо отмездренные
шкуры становятся хорошо заметными из-за пожелтения жира в результате окис-
ления. Такие шкуры отправляют на доработку. Готовые просолившиеся шкуры
свертывают конвертом, скручивают в рулон и упаковывают в деревянные бочки
с полиэтиленовым вкладышем, пересыпая их небольшим объемом соли. Каждая
бочка заполняется шкурами в соответствии с их возрастом и качеством.
В 1965—68 гг. на Командорских о-вах и о. Тюлений применялись различные
химические средства для улучшения первичной обработки шкур. Для промывки
отмездренных шкур пробовали использовать порошок “Новость” и препарат
ОП-Ю. Лучшие результаты были получены при обработке шкур препаратом ОП-10.
Опытное сырье до обработки его на меховой фабрике отличалось чистым блес-
тящим мехом. Кожевенная ткань у обычных и обезжиренных таким образом
шкур не различалась [Загуляева, 1970].
В эти же годы проводились опыты по тузлучному консервированию шкур.
Продолжительность тузлучного посола в чанах без механических мешалок со-
ставляла 44—48 часов, расход соли — 70—120% от массы шкур (при штабельном
посоле — 700%). Оказалось, что при тузлучном посоле товарные качества ме-
хового покрова оценивались выше, а качество кожевенной ткани ниже. Из-за
наблюдавшейся усадки при тузлучном посоле по существующему ГОСТу нор-
мальные шкуры переводились в размер маломерок. Однако при последующих
операциях на мехообрабатывающих фабриках площадь шкур, консервирован-
ных как штабельным, так и тузлучным способами становится одинаковой.
Применение антисептических средств при транспортировке шкур, таких как
бура, борная кислота, нафталин, кремнертутный натрий, препарат “Консердур”
не улучшали качество сырья [Загуляева, 1970]. Вместе с тем, добавление борной
кислоты при консервации шкур на о-вах Прибылова считается обязательным
[Scheffer, 1962].
20.7. Добыча котиков в море
Аборигены'североамериканского континента, камчадалы, айны и японцы из-
давна добывали котиков в море. Добыча осуществлялась с гребных весельных
356
шлюпок или каноэ, копьем или из лука. С. Скаммон [Scammon, 1874] так опи-
сывает охоту индейцев на котиков в море: “В погоне за котиком 3—4 индейца
садятся в каноэ рано утром и возвращаются лишь вечером. Приспособление до-
бычи состоит из двух копий, прикрепленных к деревянной рукоятке длиной до
15 футов. К копью прикреплена веревка, плотно соединенная с рукояткой, или
эту веревку охотник придерживает рукой, когда метает копье. В каноэ имеется
два буя из шкур котиков, которые также используются в штормовую погоду
как наплава... Если охотник, загарпунивший котика, не может удержать канат
в руках, то выбрасывается буй, и животное некоторое время плывет с ним, пока
не выбивается из сил. Тогда охотник подбирает буй и подтягивает животное
к лодке, а затем убивает его ударом дубинки по голове. Некрупных животных
сразу же подтягивают к лодке и убивают аналогичным способом”. По описанию
Хоккена [1897], японцы и айны подкрадывались к котикам в море на лодках,
помахивая лисьим хвостом, отчего караульные котики обращались в бегство,
не успев разбудить охраняемых. Чтобы можно было бесшумно подкрасться к
котикам, весла обивали кожей или обматывали тряпками, уключины — верев-
ками. Камчадалы отыскивали котиков на байдарах и били небольшими копь-
ями-гарпунами, к которым привязывали длинный ремень, за который котика
подтягивали к борту [Крашенинников, 1949].
С появлением ружей котиков стали добывать с помощью гладкоствольного
оружия крупных калибров картечью. Пулевое оружие на промысле не употреб-
ляли, т. к. это усложняло попадание. К тому же убитый пулей зверь моментально
тонул и не доставался охотнику.
С развитием судового промысла на борту парусных или паровых шхун ус-
танавливались несколько ботов или шлюпок. Канадцы добывали котиков с по-
мощью индейского населения и устанавливали на шхуну до 8—18 шлюпок, ев-
ропейцы — 6—10, максимум 12. На шлюпках команда состояла из 3 человек,
на ботах — из 2. Экипаж в зависимости от размеров шхун состоял из 12—22
человек. Экипаж японских шхун был всегда многочисленным и доходил до
30 человек [Суворов, 1912].
Обнаружив скопление котиков в море, на воду спускались шлюпки, которые
расходились друг от друга на расстояние видимости и шли параллельным кур-
сом. Судно шло вместе со шлюпками с подветренной стороны. При обнаружении
котиков парус на шлюпках спускался, к котикам подходили на веслах. Добыв
животное, снова ставили парус и шли дальше. Шлюпки возвращались только
для сдачи продукции или в случае ухудшения погоды. Обычно охота продол-
жалась весь день [Дорофеев, 1964].
В настоящее время котиков в море добывают только с научно-исследователь-
скими целями. Используются суда разного типа, но способы убоя у всех стран,
занимающихся добычей котиков, идентичны. Лишь японские исследователи в
50-х годах использовали специальные суда для добычи котиков и дельфинов,
которые в носовой части имели выступающую на 4—5 м площадку, с нее и про-
изводилась стрельба по котикам. Ученые США и России для непосредственной
добычи котиков в море спускали на воду моторные шлюпки с командой из
3 человек: рулевого, моториста и стрелка. Стрельбу вели из гладкоствольного
оружия 10—12 калибра картечью № 0 или 00. При добыче котиков непосред-
ственно с борта судна убитых животных подбирали сачком, кошкой или крюч-
357
Головной мозг — — —> — ©©©©© ©©о©©©©©©©©’
< 120 220 220 290 190 210 160 190 200 211 235 205 265 235 254 | 210 270 232 216
Печень m IT» in о ’t го оо - © ем o — so © vn oo © oo © см оо оо о ^\оечгчг--т'о^'*т'пт,^сьгчфг--г--с-‘ uS сч ем
< 3g?s 3§ § §яд|£яS5i§ я § 1Rs <n ir> Z — г- — оо ем — F5 — m — — — — © — «л —
Тонкие кишки 4.1 1.9 j 1.9 ' 2.7 2.7 2.5 24 2.2 2.4 2.1 2.0 j 1.9 1 2.2 2.3 1.4 | 1.8 1.3 1.4 1.6 2.1 1.2 2.3
< 260 313 3.13 597 791 575 952 650 1384 740 1582 720 2350 940 1937 733 2370 600 2535 904 2330 1075
Толстые кишки 1£ u-j •’t Tt тт*тт,*ггчтсчгчттттспттгчсп © © © ©’ о’ ©Ъ© ©*©©©©©©©©©©©©©
32 26 60 95 94 77 153 100 263 67 284 1 80 250 НО 423 130 450 135 435 135 435 147
Поджелудоч- ная железа m os <ч \o (Nosmoo(oo'tso'r)’tr>’- oo * « r* — cm m сч ем сч 't см еч — cNcNcnmcnmcn — © © © © © ФФ©©Ф©©ФФ©©ФФ©ФФ
< 13 16 90 57 71 100 47 155 64 135 117 260 135 423 155 510 146 360 130 1150 125
I । Желудок 0.62 0.61 1.21 1.27 1.05 1.42 1.19 1.29 1.10 1.37 1.03 1.27 1.08 1.09 1.28 1.16 1.14 1.23 1.84 1.30 1.16 1.25
< Sissgasg.sgigtgsjsgsggsi
Селезенка ц: t- m оо — оо oo oo — cm \© r- oo —’toososoQ’trc- см cn — см см Ф —« — cm cm cm —. — сч n — — — cn cm cn cm © © © © ©©©©»©©’ ©©©’©’©©©Ф©Ф©
14 16 38 54 69 20 77 34 141 78 193 70 202 81 315 78 278 73 405 104 411 98
Почки t£ — r- © Tf см^ффс-г-фл — © © © оо «л — CM — *Л Г** 0\'0>*^>Ь-00>|Л^-^>,Л’Л>0'000 —’—’©’© ©’©©ф©©ф©©©©©’^©© — ф ф
61 57 100 185 263 180 290 150 452 267 617 292 509 270 675 322 896 251 1050 440 1155 402
Дафрагма ьс 0.63 0.69 0.46 0.63 0.67 0.36 0.63 0.48 0.65 0.67 ' 0.66 0.70 0.71 0.63 0.69 0.63 0.59 0.57 0.65 0.66 0.73 1 0.59
£ 8 й 2 !$ Я f а Я Я g g g; Д 2 8 | “
Легкие t£ СМ 00 Г- 00 — — OsOsOOQsOO - гмг-©еп^ооспг-т OOsO>F'-OOb-ir)V),z'>Tf^’tir)NV5Nv5~ IT) — см’ — — — — — — — — — — — — — — — — — — — —
< 1 325 407 386 490 428 641 518 847 563 1147 590 1561 614 1744 656 2111 683 2545 724 2914 713
Сердце оо см «л смог-ЛФ’ЛтГ'-'Ч’емоог^соемсМ’^^© © © *Л Г- 00^1Л‘ЛФ‘Л‘Л©'Л’©^©^,'Л‘Л©Л,’Л ©’ — ф © фф.фффффффффффффффф
-.—©оо ©<лоог-спф©<л- — — ем © г- — г- -Ч’ $ •* = 2 Я 2 Я £ 8 2 f Я Я Я Я Я 5 Я 5g Й SS 8
Пол m f tn m f m f m f m f m f m f tn f tn f i tn f tn f
Воз- раст 0 1 2 3 4 5 6 7 8 9 10 10+
Примечание, m — самцы, f —самки.
358
ком. При убое котиков с бота животных подбирали кошкой, укрепленной на
бамбуковой палке длиной 2—3 м. Процесс обработки животных в судовых ус-
ловиях ничем не отличался от такового на берегу.
20.8. Продукция промысла
Шкуры северного морского котика являются основным, а все остальные части
туши побочными продуктами промысла. Промысловыми возрастными группами
животных считаются 3—4-летние самцы, однако в промысел ежегодно вовле-
каются наиболее крупные 2-летки, подходящие под стандартные размеры 3-леток,
и мелкие 5-летки. Поэтому здесь мы, в основном, рассматриваем продукцию,
получаемую в результате забоя только 2—4-леток. Подробные данные об отно-
сительной и абсолютной массе внутренних органов приведены в таблице 20.7.
Выход побочной продукции от котиков других возрастных групп изложены
в работе Г. М. Косыгина и А. Е. Кузина [1979]. Частично данные также имеются
в статьях В. Шеффера [Scheffer, 1960b], А. С. Соколова с соавт. [19696], А. Е.
Кузина с соавт. [1973].
Шкуры как меховое сырье. Примерно в 1825 г. был разработан способ уда-
ления грубой ости и окрашивания подпуши на шкурах котиков, что сразу по-
высило спрос на меховое сырье [Scheffer, 1962]. И хотя продукция новой выделки
была очень бедной нежели та, что теперь получают из шкур котиков, она бес-
спорно имела большое преимущество перед нещипанным мехом, когда шкуры
сбывались в обмен на восточные товары.
С 1855 по 1870 гг. проводились многочисленные работы по улучшению ка-
чества котикового меха. Одновременно этим занимались фирмы “Оппенгейм и
Ко”, “Мартин и Тичман” в Лондоне и “Герг С. Тридвел” в Албании [Scheffer,
1962]. В результате были разработаны улучшенные методы выделки и крашения,
позволившие получать исключительно тонкую, мягкую шкуру с пушистой тек-
стурой меха, окрашиваемой в темно-коричневый цвет [Scheffer, 1962]. Этот ме-
ховой товар быстро вошел в моду. Из него стали делать накидки, жакеты,
муфты, использовать для отделки верхней одежды. На котиковый мех поднялись
цены. К 1870 г. Лондон стал единоличным центром выделки котиковых шкур,
и так продолжалось до 1913 г., когда на заводе братьев Фунстен (США) в г.
Сан-Луи также стали производить выделку шкур по новой технологии. В 1916 г.
эта компания продала свой первый лот котикового мехового сырья. В 1921 г.
компания “Фунстен” влилась в компанию “Фоук”, которая до сих пор зани-
мается обработкой котиковых шкур, поступающих с о-вов Прибылова.
Шкуры котиков с Командорских о-вов до 1917 г. поступали через японские
и американские порты на Лондонский завод по выделке и крашению, где и про-
давались с аукциона. После гражданской войны и возобновления торговых от-
ношений с заграницей котиковые шкуры из СССР направлялись для реализации
в США.
В настоящее время существует отечественное производство выделки шкур и
продажа их на Ленинградском международном аукционе.
Характеристика шкур морских котиков. Парные шкуры являются сырьем для
меховой промышленности. От их качества зависит качество мехового полуфаб-
359
Таблица 20.8
Количество шкур с дефектами (%) в зависимости от места их заготовки
(1966—69 гг.)
Наименование пороков О. Тюлений О. Белинга О. Медный
Закусы 70.5 57.0 69.0
Прокусы 10.5 4.5 3.0
Болячки 2.0 1.5 2.0
Дыры 3.0 4.0 8.0
Кровоподтеки 32.5 40.0 52.5
Порезы кожи 6.0 3.0 5.0
Прирези жира 8.5 8.9 10.0
Неправильный раскрой 3.5 11.0 13.0
Нормальные шкуры без дефектов 1.5 6.5 3.5
риката, из которого потом получают меховые изделия. В свою очередь, качество
парных шкур определяется присутствием пороков, которые всегда имеются в том
или ином количестве почти на каждой шкуре.
Различают 2 вида пороков: прижизненные и посмертные. Первые появляются
на шкурах еще при жизни животного и не связаны с контактом с человеком,
вторые возникают во время забоя и первичной обработке шкур и могут быть
полностью устранены или, по крайней мере, сведены минимума при правильной
организации работ при обработке.
К прижизненным порокам относятся плешины, потертости волосяного пок-
рова, прокусы, закусы и др. Если 2 первых порока комментариев не требуют,
то о 2 других необходимо сказать следующее. Закусы обнаруживаются на шкурах
в виде кровоподтеков величиной 1.5—3 см2. Структура кожевенной ткани в мес-
тах закусов нарушается, особенно эпидермис. При переработке таких шкур на
меховой фабрике в процессе подготовки к щипке эпидермис в местах закусов
окончательно разрушается и удаляется вместе с пуховым волосом. При выделке
натурального меха без щипки ости в результате того, что процесс выделки со-
кращается, плешины в местах закусов, как правило, не возникают. Если при
закусах кожа не прокусывается насквозь, то при прокусах характерно образо-
вание двух сквозных дыр на мездре размером 3—5 мм2. При обработке шкур
в этих местах возникают те же дефекты, что и при закусах.
К производственным порокам относятся следующие: теклость волоса, раз-
рывы шкуры, кровоподтеки, неправильный раскрой, окровавленность, подрезь
или выхват, прорезь, зажиренность шкур, загибы, прелины, краснота, синее ок-
рашивание шкур, солевые пятна.
Установлено, что количество пороков на шкурах котиков не одинаково на
каждом лежбище. Первое место по количеству прижизненных пороков занимают
шкуры котиков с о. Тюлений, и менее всего им подвержены шкуры с о. Беринга
(табл. 20.8).
Наиболее массовыми дефектами шкур являются закусы и кровоподтеки, ко-
личество которых на том или ином лежбище, впрочем как и других пороков,
360
Рис. 20.1. Классификация мокросо-
леных шкур самцов котиков (преиму-
щественно 3—4 лет), добытых на
о. Св. Павла 18 июня—20 июля [из:
Scheffer, 1962]
41 43 45 „ 47 49 51 М
ДЮЙМЫ
| - МЕЛКИЕ [•••] - ОСОБО КРУПНЫЕ
- СРЕДНИЕ
Щ] - КРУПНЫЕ
- ИСКЛЮЧИТЕЛЬНО
КРУПНЫЕ
40 * 42 ' 44 ’ 46
Дюймы
—i—» »
48 50 52
Рис. 20.2. Изменение массы парных шкур холостяков в зависимости от длины их
тела [из: Scheffer, 1962]
в основном зависит только от плотности размещения животных на лежбищах
и не зависит от количества забиваемых за один день животных [Загуляева, 1970].
Справедливость сказанного подтверждается также числом бездефектных шкур,
встречающихся на том или ином лежбище.
Количество закусов на 1 шкуре может быть от 1 до 20 и более, соответ-
ственно, на готовом полуфабрикате возникает от 20 до 40 плешин. Поэтому
шкуры с большим числом закусов используются в натуральном виде. Из таблицы
20.8 также видно, что количество бездефектных шкур не превышает 7%. Час-
361
Таблица 20.9
Изменение массы (кг) шкур холостяков в зависимости от способа
консервирования [из: Загуляева, 1970]
Возраст, лет Штабельный посол Тузлучный посол
.парные соленые парные соленые
2 2.0 (1.3-2.5) 1.5 (1.0-1.8) 1.9 (1.2-2.6) 1.4 (0.9-1.8)
3 2.4 (1.4-3.2) 1.8 (1.3-2.2) 2.3 (1.4-3.1) 1.7 (1.2-2.2)
4 2.7 (1.8-3.4) 2.1 (1.6-2.6) 2.8 (2.2-3.1) 2.2 (1.6-2.7)
5 3.4 (2.5-4.5) 2.8 (2.2—3.5)
тичного облагораживания сырья можно достичь при замене солевого способа
консервации на тузлучное. Часть пороков, такие как закусы, кровоподтеки при
этом способе становятся малозаметными, а розово-красные бактериальные пятна
вообще не обнаруживаются [Загуляева, 1970].
Масса и площадь шкур промысловых групп морских котиков. Данные по массе
шкур всех возрастных классов можно найти у Г. М. Косыгина и А. Е. Кузина
[1979]. Средняя масса шкур промысловых классов составила у 2-леток — 2.1 кг,
у 3-леток — 2.4 кг, у 4-леток — 3.1 кг. В. А. Бычков [1968] определил, что масса
шкур промысловых групп котиков изменяется по годам незначительно, как и
в течение промыслового сезона. К аналогичным выводам пришел В. Шеффер
[Scheffer, 1962], показавший, что в первой четверти сезона промысла масса шкур
холостяков на о-вах Прибылова составила 5.4 фунта, во второй — 5.3, в
третьей — 5.5 и в четвертой — 5.1. В среднем за сезон масса шкур холостяков
составила 5.3 фунта (рис. 20.1).
Естественно, что размеры шкуры зависят от длины тела животного, но
здесь обращает на себя внимание то, что у холостяков с длиной тела 44 дюйма
(110 см) они наиболее вариабельны [Scheffer, 1962] в связи с тем, что в эту группу
входит примерно одинаковое количество как 3-леток, так и 4-леток, среди ко-
торых часть животных уже достигла половой зрелости, и у них наступают ак-
селеративные процессы в росте тела (рис. 20.2).
Масса шкур заметно изменяется вне зависимости от способа консервирова-
ния, однако при тузлучном способе шкуры обезвоживаются сильнее (табл. 20.9).
Имеющиеся в литературе данные по площади шкур холостяков разноречивы
[Волошинов, 1889; Генерозов, 1916; Scheffer, 1962; Бычков, 1968; Загуляева, 1970].
Как писала А. И. Загуляева [1970], парные шкуры очень эластичны, вследствие
чего определить их площадь трудно. Тем не менее такие данные приведены в
печати (табл. 20.10). В среднем площадь шкур холостяков составила 45—50 дм2.
Наиболее устойчива площадь шкур у 3-леток, наиболее вариабельна у 4-леток.
Сезонной изменчивости массы и площади шкур не обнаружено [Scheffer, 1962;
Бычков, 1968], но у 2-леток отмечалось стабильное увеличение площади шкур
от июня к июлю [Бычков, 1968]. В процессе первичной обработки и переработки
шкур котиков площадь их значительно изменяется. По данным В; А. Бычкова,
362
Таблица 20.10
Изменение площади (дм2) шкур холостяков в зависимости от года убоя на
о. Тюлений [по: Бычков, 1968; Загуляева, 1970]
Год 2 года 3 года 4 года
п М п М п М
1962 25 37.0 67 44.4 39 54.0
1963 17 40.2 61 43.5 32 52.5
1964 35 38.6 56 44.4 36 51.4
1965 60 42.0 500 48.0 100 63.0
1966 76 44.6 227 49.8 27 59.2
1967 64 45.5 209 49.9 98 57.7
1968 48 45.9 553 51.9 180 58.6
Таблица 20.11
Изменение размеров шкур котиков при различных способах консервирования
[по: Загуляева, 1970]
Возраст, лет п 2 Площадь шкур, дм Усадка шкур, дм2
парных соленых
м м М
Штабелън ый посол
3 200 47.9 43.7 4.2
4 250 63.7 62.6 1.1
5-6 300 89.0 69.6 10.4
Тузлучны й посол
2 50 42.0 32.8 9.2
3 280 48.0 36.0 22.0
4 100 63.0 48.0 15.0
площадь соленых в штабелях шкур, по сравнению с парными, увеличивается:
у 2-леток на 7.1 дм2 (18.4%), у 3-леток на 8.6 дм2 (19.4%), у 4-леток на 11.3 дм2
(22.15%), у 5-леток на 17.4 дм2 (27.2%). Шкуры тузлучного посола изменяются
по площади незначительно: у 2-леток на 2.6 дм2 (6.7%), у 3-леток на 2.2 дм2
(4.7%), у 4-леток на 1.6 дм2 (3.2%), у 5-леток на 4.0 дм2 (3.3%).
По данным А. И. Загуляевой [1970], при штабельном посоле усадка шкур
холостяков составила 1.1—4.2%, а после тузлучного посола до 19.2—25.0% (табл.
20.11)
Различия в усадке шкур в зависимости от способа консервирования объяс-
няется тем, что при штабельном посоле коллагеновые волокна кожи растяги-
ваются под тяжестью соли и массы самой шкуры и фиксируются в растянутом
положении, в то время как при тузлучном способе шкуры находятся в свободном
363
Таблица 20.12
Изменение размеров шкур тузлучного посола в процессе их обработки до
полуфабриката [из: Загуляева, 1970]
Число шкур 2 Средняя площадь, дм Усадка от пло- щади парных шкур, % Увеличение площади полу- фабриката после переработки на фабрике, %
парные соленые полуфабрикат к площади парных шкур к площади со- леных ШКУР
60 46 34 49 26.1 6.5 44.0
51 47 37 52 . 21.3 10.1 40.5
55 50 38 55 24.0 9.1 44.7
30 51 37 52 27.5 2.0 40.5
196 48.5 36.5 52.0 24.7 7.2 42.4
состоянии. Минимальные размеры шкур тузлучного консервирования составили
28 дм2, в то время как по ГОСТу-2897-68 нормальной считается площадь 41 дм2.
Из экспериментальной партии шкур тузлучного посола на мехообрабатываю-
щей фабрике 41% 2-леток, 32% 3-леток и 16% 4-леток были отнесены к мало-
мерным, что в значительной степени снизило их приемную стоимость. Вместе
с тем, при дальнейшей обработке шкур тузлучного посола площадь их вновь
увеличивается (табл. 20.12).
В среднем площадь дубленого полуфабриката увеличивается на 7.2% по от-
ношению к площади парных шкур и на 42.4% — к площади соленых.
В процессе крашения и глажения шкуры вновь дают усадку на 5—6% по
отношению к площади некрашенного полуфабриката. Однако маломерные
шкуры тузлучного посола площадью 28 дм2 после всех фабричных операций
имели площадь 36.7 дм2, т. е. считались нормальными. Идентичными были из-
менения соотношения размеров шкур на фабрике Фоук при обработке прибы-
ловских котиков [Scheffer, 1962].
Выход готового полуфабриката из сырья тузлучного посола на 9.5% выше,
по сравнению со шкурами штабельного посола [Загуляева, 1970]. Это объясня-
ется тем, что на шкурах тузлучного посола в процессе подготовки и щипки ос-
тевого волоса образовывалось меньше плешин, чем на сырье штабельного по-
сола. Для экспорта из пробных партий тузлучного сырья было отобрано 33.2%
шкур, а из штабельного — только 14.4%. Лучшим сырьем для экспорта при туз-
лучном посоле признаны шкуры 2—3-леток.
Прочность, носкость и химический состав шкур морских котиков. Р. Боукер
[Bowker, 1931] исследовал прочность готовых крашеных шкур котиков-холостяков.
Толщина мездры таких шкур составила 0.0023—0.026 дюйма, прочность на разрыв
полудюймовой полоски 37.5—46.0 фунтов. Напряженность прочности на натяжение
составляет 3.15—3.68 фунтов на кв. дюйм. Процент растяжимости до разрыва —
23.5—38.1, разрыв стежка — 6.1—9.0 фунта. Кожевенная ткань морского котика,
таким образом, может сравниться с таковой телячьей и овечьей. Разрыв
364
мездры котиковой шкуры чаще происходит не вдоль, а поперек из-за специфи-
ческой направленности составляющих ее волокон. По шкале носкости волосяной
покров шкур морского котика стоит на третьем месте после выдры, рассомахи
и бобра и оценивается в 80% или вдвое выше, чем шкур лисицы.
Химический состав шкур морских котиков следующий.
Возраст Вода Жир Белок Зола
2 года 67.2 2.5 26.2 3.5
3 года 66.8 2.7 28.5 3.5
4 года 65.6 3.7 27.0 4.2
Мясо. Мышечная ткань морского котика темно-красного цвета из-за боль-
шого содержания миоглобина. Как и у других морских млекопитающих, оно
характеризуется специфическим запахом, что в значительной мере снижает его
пищевую ценность. Однако мясо молодых котиков, а также печень, язык, го-
ловной мозг, сердце взрослых животных почти не имеют запаха и вполне пригодны
в пищу. Химический состав мяса котика следующий: вода —62.8%, жир —
14.9%, белок — 21.1%, зола — 0.3%.
Сало. Толщина сала на груди у котиков с возрастом увеличивается, соот-
ветственно увеличивается выход подкожного сала (табл. 20.13).
Таблица 20.13
Толщина сала на груди у холостяков котиков [по: Scheffer 1962; наши
данные]
Возраст Толщина сала на груди, см Выход сала с 1 шкуры, кг
п М lim М
2 года 29 1.67 0.6-3.2 3.99
3 года 51 1.75 0.5-3.1 4.56
4 года 42 1.76 0.8-3.1 7.10
5 лет 24 Z03 0.8-3.7 9.80
Таблица 20.14
Выход жира (галлоны) при переработке холостяков котиков [из: Scheffer,
1962]
Годы 1949 1950 1951 1952 1953 1954 1955 1956 1957 1958
Выход жира из одной туши 0.27 0.21 0.25 0.16 0.28 0.23 0.26 0.30 0.31 0.33
Выход жира из одной хоровины 0.60 0.65 0.55 0.55 0.57 0.60 0.59 0.64 0.52 0.56
365
На перерабатывающем заводе на о-вах Прибылова жир вытапливается от-
дельно из подкожного сала и туш. Данные о выходе жира показаны в таблице
20.14.
Физические и химические показатели жира по И. В. Кизеветтеру [1966] сле-
дующие: удельный вес (dl54) — 0.9251; коэффициент рефракции — n20d 1.4742-
1.47987; кислотность — 1-1; коэффициент омыления — 186.0-189.9; йодное чис-
ло — 141.8-156.9; неомыляемые вещества — 0.2-1.4%.
В. Шеффер [Scheffer, 1962] приводит данные по физическим и химическим
характеристикам жира морского котика, полученных при варке и холодной об-
работке. Вытопленный из кусочков подкожного сала на о-вах Прибылова жир
имел следующие характеристики: свободные жирные кислоты — 1.05%; влаж-
ность — 0.34%; нерастворимые вещества — 0.05%; йодное число — 132.5; стеа-
рины при 2ГС — 1.04%; неомыляемые вещества — 0.49%; титр — 21.2°С.
При холодной обработке жир морского котика имел следующие показатели
[Scheffer, 1962]: свободные жирные кислоты — 1.58%; йодное число — 108; неомы-
ляемые вещества — 0.64; число омыления — 196.3; удельный вес при 25°С —
0.917; индекс рефракции при 25°С — 1.4743; витамин А (1894><Е328) в целом
жире без омыляемых веществ — 306 инт. ед./г.
Выделенные из подкожного жира кислоты плавятся при температуре 25.5—
28.0°С и содержат 10—11% твердых жирных кислот. Среди непредельных жир-
ных кислот преобладают кислоты с числом углеродных атомов С20 и ССС [Ки-
зеветтер, 1953].
В подкожном жире морского котика кроме витамина А присутствует неболь-
шое количество витамина Д (20—40 инт. ед./г [Кизеветтер, 1953, 1966; Бодров
и др., 1958]. В свежем жире К. Вилбер [Wilber, 1952] нашел 4.3% фосфолипидов
и 0.24% холестерола. Содержание фосфолипидов высокое как и у всех морских
млекопитающих и связано это, видимо, с высокими темпами круговорота жи-
ровых депо.
Внутренности. Масса внутренних органов морских котиков показана в таб-
лице 20.7. Наибольшей массой обладает печень, которая к тому же является наи-
более ценным продуктом из вторичных объектов сырья, получаемого из котика.
Печень содержит 1.5—4.0% жира, 1 г которого содержит 75—200 тыс. инт. ед.
витамина А.
20.9. Использование продукции
Основной вид продукции котикового хозяйства — меховое сырье. Из шкур
морского котика готовят меховой полуфабрикат, который идет в изделия как
в натуральном виде, а также в щипаном и стриженом виде. При щипке удаляется
полностью ость, при стрижке она состригается вровень с подпушью. Щипаный
и стриженый мех красится в различные цвета. Из котикового меха изготовля-
ются различные меховые товары: воротники, шапки, муфты, пелерины, горжеты,
пальто, шубы, жакеты и т. п.
Из сала в настоящее время получают жир. На о-вах Прибылова это произ-
водство налажено непосредственно на промысле котиков, а на островах Коман-
366
дорских и Тюленьем сало солят и отправляют на материк для переработки. По-
лученный из сала жир используют в технических целях. В США часть заготав-
ливаемого сала продается фирме Фоук, которая использует его при выделке
шкур морских котиков.
Из мясо-костной тушки котиков на о-вах Прибылова изготавливают мясо-
костную муку, которую продают для подкормки птиц и скота. На островах Ко-
мандорских и Тюленьем тушки холостяков котиков вместе со всеми внутренними
органами, но без печени, продают на зверофермы, где их используют на корм
клеточным пушным зверям.
Из печени морских котиков в настоящее время получают витамин А. Однако
есть полная возможность для расширения ассортимента других лекарственных
препаратов из печени.
На о. Тюлений ежегодно заготавливают половые тракты самцов котиков, ко-
торые освобождают от жира, высушивают и продают за границу, где их исполь-
зуют как препараты тибетской медицины.
Совершенно не используется кровь, кишечник, желчь, железы внутренней сек-
реции, которые могли бы быть источником сырья как для пищевой промыш-
ленности, а некоторые из них служить сырьем для фармацевтической промыш-
ленности.
В кулинарии в натуральном виде можно использовать язык, сердце, почки,
мозг, кишечник котиков.
Таким образом, ассортимент продукции, получаемой в результате промысла
северных морских котиков, в настоящее время невелик. Наиболее перспективным
направлением в его рационализации является организация производства биоло-
гически активных веществ (БАВ) из желез органов и тканей котиком, на основе
которых возможно изготовление высокоэффективных медицинских препаратов.
Соответствующие технологические разработки в этом направлении имеются в
ТИНРО и во Владивостокском медицинском институте.
367
ЛИТЕРАТУРА
Авксентьева Л. И., 1974. Цитоархитектоника слуховой и зрительной коры морских мле-
копитающих. В кн.: Морфология, физиология и акустика морских млекопитающих.
М., Наука. С. 15—19.
Альтгаузен А. Я., 1951. Лабораторные клинические исследования. М„ Медгиз. 407 с.
Андреев Ф. В., 1978. Структурно-функциональные особенности глаза дельфинов и лас-
тоногих. В кн.: Новое в изучении китообразных и ластоногих. М„ Наука. С. 116—
132.
Анохин П. К., 1948. Систематогенез, как общая закономерность эволюционного процес-
са. — Бюлл. эксп. биологии и медицины, 26, 2, 8. С. 81—99.
Антонюк А. А., 1972. Количественные изменения позвонков в позвоночнике ластоногих. —
Тр. ВНИРО, 39. С. 317—322.
Арсеньев В. А., 1968. Методика определения промыслового использования поколений
морских котиков. — Изв. ТИНРО, 62. Тр. ВНИРО, 68. С. 53—64.
Арсеньев В. А., 1971а. Возрастно-половой состав морских котиков, зимующих в западной
части Тихого океана. В кн.: Морские млекопитающие. М., ВНИРО. С. 25—43.
Арсеньев В. А., 19716. О локальном распределении морских котиков в Японском море.
— Тр. АтлантНИРО, 39. С. 138—150.
Арсеньев В. С., 1967. Течения и водные массы Берингова моря. М., Наука. 135 с.
Астанин Л. П., 1958. Органы тела млекопитающих и их работа. М., Сов. наука. 351 с.
Ащепков А. Т., Колотилин Н. Е., 1978. О группировках котиков в море. В кн.: Морские
млекопитающие. Тез. докл. 7 Всесоюзн совещ. по изучению морских млекопита-
ющих. Симферополь. С. 19—20.
Ащепков А. Т., Колотилин Н. Е., 1982. Структура группировок северного морского ко-
тика в Беринговом и Охотском морях. В кн.: Млекопитающие СССР. Тез. докл.
3 съезда Всесоюзн. териол. об-ва. Т. 2. М. С. 197.
Ащепков А. Т., Кузин А. Е., 1986а. Численность и динамика возрастно-полового состава
котиков разных популяций на местах зимовки в Японском море и тихоокеанских
берегов Японии. В кн.: Изучение, охрана и рациональное использование морских
млекопитающих. Тез. докл. 9-го Всесоюзн. совещ. Архангельск. С. 22—24.
Ащепков А. Т„ Кузин А. Е., 19866. Смешиваемость, пути и сроки миграции котиков
разных стад в тихоокеанских водах Японии. — Биология моря, 6. С. 27—32.
Ащепков А. Т., Кузин А. Е„ 1987. Возрастно-половой состав стад морских котиков миг-
рирующих в тихоокеанские воды Япониию. — Там же, 2. С. 29—38.
Ащепкова Л. Ю., Дмитриева Т. В., 1982. Возрастная динамика репродуктивных самцов
северного морского котика. 20 с. (Рук. деп. в ВИНИТИ, N 5945-82).
Барабаш-Никифоров И. И., 1936. Ластоногие Командорских островов. Морские млеко-
питающие Дальнего Востока. — Тр. ВНИРО, 3. С. 233—237.
Белкин А. Н., 1963. Об утробной линьке морских котиков. — Сообщ. ДВ ФАН СССР,
17. С. 101—105.
Белкин А. Н., 1964. Материалы по эмбриональному развитию кожного и волосяного пок-
ровов ушастых тюленей. — Изв. ТИНРО, 54. Тр. ВНИРО, 51. С. 97—130.
368
Белкин'А. Н., 1965а. Современное состояние лежбищ и численность морских котиков и
сивучей на островах Ловушки и Среднего. В кн.: Морские млекопитающие. М.,
Наука. С. 189—200.
Белкин А. Н., 19656. К вопросу о миграции морских котиков Командорских островов.
В кн.: Аннотации научных работ по исследованию сырьевой базы рыбной про-
мышленности Дальнего Востока в 1959—1962гг. Владивосток. С. 133.
Белкин А. Н„ 1966. О взаимоотношениях сивуча и котика на совместных лежбищах раз-
множения (на примере наблюдений за восстановлением лежбищ котиков на Ку-
рильских островах). — Изв. ТИНРО, 58. С. 49—68.
Белькович В. М., 1962. Приспособительные особенности строения кожного покрова во-
дных млекопитающих. Автореф. канд. дне. М. 26 с.
Берзин А. А., 1959. Котик у мыса Наварин. — Изв. ТИНРО, 46. С. 14—15.
Берзин А. А., 1971. Кашалот. М., Пищевая пром-сть. 359 с.
Берх В., 1823. Известие о меховой торговле, производимой россиянами при островах Ку-
рильских, Алеутских и северо-западном берегу Америки. — Свет отечества, 39,
43. С. 243—264, 97—106.
Бирман И. Б., 1966. Новые данные о распространении и миграции морских котиков. —
Вопросы географии Камчатки, 4. С. 112—114.
Блехман М. И., 1949. К биоморфологии грудной клетки домашних и некоторых диких
млекопитающих. — Тр. Алма-Атинского вет.-зоол. ин-та, 166. С. 18—26.
Богданов Л. В., Черноиванов В. А., Поляковский В. И., Башурова Г. И., 1977. Биохи-
мический полиморфизм северного морского котика Callorhinus ursinus из трех райо-
нов северо-западной части Тихого океана. — Генетика, 13, 6. С. 1008—1014.
Боголюбский С. Н., 1948. О некоторых общих и частных закономерностях онтогенети-
ческого развития овец. — Изв. АН СССР, сер. биол., 3. С. 307—315.
Богословская Л. С., Солнцева Г. Н., 1979. Слуховая система млекопитающих. М., Наука.
238 с.
Бодров В. А., Григорьев С. Н., Тверьянович В. А., 1958. Техника и технология обработки
морских млекопитающих (киты, дельфины, ластоногие). М., Пищпромиздат. 585 с.
Бойцов Л. В., 1934. Котиковое хозяйство. М., Внешторгиздат. 195 с.
Болтнев А. И., 1989. Эколого-морфологические особенности развития северных морских
котиков в постнатальном онтогенезе. Автореф. канд. дис. Киев. 21 с.
Болтнев А. И., 1990а. О пространственной структуре залежек северных морских котиков
о. Беринга. — Изв. ТИНРО, 112. С. 29—34.
Болтнев А. И., 19906. Причины смертности новорожденных котиков. — Там же. С. 35—38.
Болтнев А. И., Нестеров Г. А., 1996. Исторические тенденции в численности командор-
ской популяции северных морских котиков. — Изв. ТИНРО, 121. С. 62—73.
Болтнев А. И., Шпигальский С. Н„ Шпигальская Р. Ю., 1991. Связь выживаемости се-
верных морских котиков с массой и длиной тела при рождении. В кн.: НИР по
морским млекопит. сев. части Тихого океана в 1989—1990 гг. М. С. 179—182.
Бровар В. Я., 1944. Закономерности роста скелета домашних животных. — Тр. сельскохоз.
академии им. Тимирязева, 31. С. 27—38.
Брошниовский Р„ 1898. О котиковом промысле в Беринговом море. — Вести, рыбопро-
мышленности. С. 68—76.
Будыко М. И., 1980. Климат в прошлом и будущем. Л., Гидрометеоиздат. 351 с.
369
Бурков В. А., 1958. К гидрологии командоро-камчатского района Тихого океана в ве-
сеннее время. — Тр. ИОАН, 27. С. 12—22.
Бычков В. А., 1964а. Наблюдение за самками морского котика на острове Тюленьем.
В кн.: Морские котики Дальнего Востока. М., Пищевая пром-сть. С. 91—96.
Бычков В. А., 19646. Материалы по линьке морских котиков. — Изв. ТИНРО, 54. Тр.
ВНИРО, 57. С. 131—142.
Бычков В. А., 1964e. Промысел морских котиков на острове Тюленьем. В кн.: Морские
котики Дальнего Востока. М., Пищевая пром-сть. С. 155—172.
Бычков В. А., 1965. О половой и возрастной изменчивости черепа морского котика.
В кн.: Морские млекопитающие. М. С. 183—188.
Бычков В. А., 1967. О нападении косаток на морских котиков у острова Тюленьего. —
Зоол. ж., 43, 1. С. 149—150.
Бычков В. А., 1968. Характеристика размеров и окраски волосяного покрова морских
котиков. В кн.: Ластоногие северной части Тихого океана. С. 117—133.
Бычков В. А., 1969. Состав и размещение самок морских котиков на гаремном лежбище
острова Тюленьего. В кн.: Морские млекопитающие. М., Наука. С. 117—133.
Бычков В. А., 1971а. Сезонная изменчивость густоты пуха в волосяных пучках трехлетних
холостяков морских котиков. — Тр. АтлантНИРО, 39. С. 253—262.
Бычков В. А., 1971 б. Возрастная изменчивость длины тела и веса морских котиков ©.Тю-
леньего. — Экология, 1. С. 102—104.
Велижанин А. Г., 1978. Размещение и состояние численности морских птиц на Дальнем
Востоке. В кн.: Актуальные проблемы охраны окружающей среды Дальнего Вос-
тока. Владивосток. С. 154—172.
Вениаминов И., 1840. Записки об островах Уналашкинского отдела. Ч. 2. СПб. 409 с.
Вертянкин В. В., 1986. О зимовке морских котиков на лежбищах Командорских островов.
В кн.: Териол., орнитол. и охрана природы. Тез. докл. 11 Всесоюзн. симп. биол.
пробл. Севера, 3. Якутск. С. 11—12.
Веселова А. И., 1975. Особенности годового хода температуры воды юга Охотского мо-
ря. — Тр. ДВНИГМИ, 50. С. 38—56.
Владимиров В. А„ 1972. О причинах, определяющих выбор места размножения северными
морскими котиками. — Тр. ВНИРО, 90. С. 184—193.
Владимиров В. А., 1975. Опознование самкой котика своего детеныша. В кн.: Матер.
6 Всесоюзн. совещ. по морским млекопитающим. Киев, Наукова думка. С. 86—87.
Владимиров В. А., 1976. Двойня у северных котиков. — Природа, 5. С. 142.
Владимиров В. А., 1978а. Популяционный статус стада северных котиков о. Тюленье-
го. — Зоол. ж., 57, 8. С. 1248—1256.
Владимиров В. А., 19786. О формировании популяций котиков. В кн.: Морские млеко-
питающие. М. С. 67—69.
Владимиров В. А., 1980. Эмиграционные группировки у северных котиков. Там же. С. 61—66.
Владимиров В. А., 1981. Распределение и поведение котиков на лежбищах в зависимости
от территориальных и гидрометеорологических условий. — Бюлл. МОИП., отд.
биол., 86, 2. С. 19—23.
Владимиров В. А., 1983. О биологии размножения самок северного морского котика на
Урильем лежбище острова Медного. — Там же, 88, 4. С. 52—61.
Владимиров В. А., 1985. Влияние возрастного состава самок на численность приплода
северных котиков. — Там же, 90, 2. С. 23—26.
Владимиров В. А., 1990. К этологической структуре лежбищных социумов северных ко-
тиков. В кн.: Морские млекопитающие. М., ВНИРО. С. 56—71.
370
Владимиров В. А., 1991. Современное состояние популяций северных морских котиков
в СССР и проблемы их рационального использования. В кн.: НИР по морским
млекопит. сев. части Тихого океана в 1989—1990 гг. С. 130—164.
Владимиров В. А., Лыскин Н. Н., 1981. Сезонная динамика возрастного состава самцов
котиков на гаремной территории лежбищ. В кн.: Тез. Всесоюзн. конф, молодых
ученых, посвященной 60-летию ГлавморНИИа ВНИИ морского рыбного хозяй-
ства и океанографии. М. С. 5—6.
Владимиров В. А., Лыскин Н. Н., 1984. Новые данные по размножению и структуре по-
пуляций северных котиков (С. ursinus). — Зоол. ж., 63, 12. С. 1883—1889.
Владимиров В. А., Лыскин Н. Н., Садовов В. Н., 1982. Некоторые результаты иссле-
дований биологии котиков на Урильем лежбище о. Медного в 1978—81 гг. М.,
ВНИРО. 58 с.
Владимиров В. А., Челноков Ф. Г., 1971. Лежбища морских котиков на Командорских
островах. В кн.: Морские млекопитающие. М., ОНТИ. С. 128—173.
Воккен Г. Г., 1949. Закономерности дифференцировки костного скелета млекопитающих.
Автореф. докт. дис. 39 с.
Воккен Г. Г., 1950. Костный скелет туловища и конечности плодов, новорожденных и
молодняка крупного рогатого скота. В кн.: Сб. научн. трудов Ленинградск. ин-та
усовершенств. врачей, юбилейный. Л., С. 17—27.
Воккен Г. Г., 1964. Степень дифференцировки костного скелета у различных грызунов
при рождении. В кн.: Проблемы современной эмбриологии. С. 435—430.
Воккен Г. Г., Глаголев П. А., Боголюбский С. Н„ 1961. Анатомия домашних животных.
Ч. 1. М., Высш, школа. 391 с.
Волошинов Н., 1887. Отчет по командировке на Командорские острова Генерального
штаба подполковника Волошина в 1884—85 гг. — Сб. Географ, топограф, и ста-
тист. материалов по Азии. Вып. 26. С. 260—269.
Волошинов Н., 1889. Морские котики. Ч. 1, 2. СПб. 48 с.
Галанцев В. П., 1977. Эволюция адаптаций ныряющих животных: эколого- и морфофи-
зиологические аспекты. Л., Наука. 191 с.
Галанцев В. П., 1978. Приспособительные реакции сердечно-сосудистой системы морского
котика, курильского тюленя и калана. В кн.: Морские млекопитающие. Тез. докл.
7 Всесоюзн. совещ. по морским млекопитающим. М. С. 75—76.
Галанцев В. П., Маминов М. К., 1979. Различия функции сердечного автоматизма у де-
тенышей курильского тюленя (Ph. kurilensis) и морского котика (С. ursinus). —- Зоол.
ж., 58, 10. С. 1565—1573.
Гелетюк В. 3., Маминов М. К., Анбиндер Е. М., 1972. Некоторые показатели крови мор-
ской выдры, двух видов настоящих тюленей и наземных хищных. В кн.: Тез.
докл. 5 Всесоюзн. совещ. по изучению морских млекопитающих. Ч. 2. Махачкала.
С. 59—62.
Генерозов В. Я., 1916. Котиковый и песцовый промысел на островах Прибылова. —
Матер, к познанию русск. рыболовства, 5, 3. С. 1—95.
Гизенко А. И., 1955. Птицы Сахалинской области. М., Изд-во АН СССР. 328 с.
Глазова Т. Н„ 1969. Гемоглобины крови и мышц ныряющих млекопитающих и птиц.
Автореф. канд. дис. Москва. 24 с.
Глазова Т. Н., Леоненко Е. П., 1966. Применение гемометра Сали при определении об-
щего количества крови у животных. — Лаборат. дело, 9. С. 525—527.
Голубева Е.Л. Формирование центральных механизмов регуляции дыхания в онтогенезе.
М., Наука. 1971. 289 с.
371
Грачев И. И., Галанцев В. П., 1973. Физиология лактации. Л., Наука. 460 с.
Гребницкий Н. А., 1902. Новейшие данные о жизни и промысле котиков и бобров. —
Вести, рыбной пром;сти, 5. С. 269—309.
Гудман Д., 1983. Демографический подход к жестко управляемым популяциям. М., Мир.
С. 198—224.
Державин А., 1929. Морские котики у северного Сахалина. — Рыбное хоз-во ДВК, 1.
С. 37—39.
Добровольский Н. Д., 1929. Котиковое хозяйство Командорских островов. — Там же.
С. 37—39.
Добровольский Н. Д„ Залогин Б. С., 1982. Моря СССР. М. 191 с.
Дорофеев С. В., 1964. Северные морские котики. В кн.: Морские котики Дальнего Вос-
тока. М., Пищевая пром-сть. С. 25—50.
Дорофеев С. В., Бычков В. А., 1964. Биологические предпосылки для регулирования чис-
ленности секачей на острове Тюленьем. — Тр. ВНИРО, 51. Изв. ТИНРО, 54. С. 83—90.
Евдокимов Е. И., 1964. Результаты наблюдений за котиками в Охотском море. Там же.
С. 61—63.
Емельянов С. В., 1963. Гетерохронии в закладках органов позвоночных животных. —
Тр. ИЭМЭЖ АН СССР им. А. Н. Северцова, 38. С. 17—76.
Жеденов В. П., 1965. Анатомия домашних животных. Ч. 1. М., Высш, школа". 411 с.
Загуляева А. И., 1968. Уточнение технологии первичной обработки шкур морских ко-
тиков. В кн.: Ластоногие северной части Тихого океана. М., Пищевая пром-сть.
С. 145—154.
Загуляева А. И., 1970. Товароведческая оценка шкур морских котиков, добытых на
о. Тюленьем и Командорских островах. — Изв. ТИНРО, 70. С. 215—225.
Земский В. А., Кузин А. Е„ 1984. Состояние и перспективы развития популяций котиков
в северной части Тихого океана. В кн.: Морские млекопитающие. М., Наука.
С. 177—191.
Зубин А. М., Пчелина Л. П., 1951. Особенности строения кожного покрова морского
котика. — Науч.-технич. изд-во легкой пром-сти, 2. С. 100—104.
Иванов М. Ф., Уразов И. Г., 1951. Строение кожного покрова морского котика и его
изменение в процессе производственной обработки. — Легкая пром-сть, 9. С. 33—
35.
Иванова Е. И., 1971. К морфологии сосудистой системы китообразных и ластоногих.
В кн.: Исследования морских млекопитающих. Калининград. С. 179—192.
Иванова Е. И., 1972. Общая схема венозной системы морского котика. В кн.: Тез. докл.
5 Всесоюзн. совещ. по изучению морских млекопитающих. Ч. 2. Махачкала.
С. 94—96.
Иванова Е. И., 1975. Функциональная морфология сосудистой системы млекопитающих.
Экологические аспекты. М., Наука. 56 с.
Измайлова И. В., 1935. Рентгенологическое определение возраста животных по степени
окостенений скелета (рентгеноанатомическое исследование на собаках). — Вести,
рентгенологии и радиологии, 15, 1. С. 21—35.
Ильина Е. Д., 1950. Островное звероводство. М., Международная книга. 302 с.
Карташев Н. И., 1961. Птицы Командорских островов и некоторые предложения по ра-
циональному их использованию. — Зоол. ж., 10, 9. С. 1395—1410.
372
Кизеветгер И. В., 1953. Жиры морских млекопитающих. Владивосток. 103 с.
Кизеветтер И. В., 1966. Ластоногие как промышленное сырье. В кн.: Дальневосточные
ластоногие. Владивосток. С. 96—134.
Кирчанов В. А., 1975. К изучению кормления щенков в стаде котиков на о. Тюлень-
ем. — Промысловая ихтиология (реферат, инф.), сер. 1, 7. С. 1—2.
Клейненберг С. Е., Яблоков А. В., Белькович В. М., Тарасевич М. Н., 1964. Белуха. М.,
Наука. 456 с.
Клумов С. К., 1957. Береговые лежбища котиков и места обитания каланов на Курильских
островах и ориентировочное определение их численности. — Докл. АН СССР, /77,
1. С. 153—156.
Когай В. М., 1970. Результаты искусственного расширения гаремного лежбища на о. Тю-
леньем. — Изв. ТИНРО, 70. С. 235—237.
Когай В. М., 1971а. Влияние ледового режима на подход, размещение, поведение и ход
промысла морских котиков на острове Тюленьем. — Там же, 76. С. 129—132.
Когай В. М., 19716. Некоторые наблюдения за поведением меченых щенков на о. Тю-
леньем. — Там же. С. 168—169.
Когай В. М., 1978. К вопросу влияния абиотических факторов на поведение котиков.
В кн.: Морские млекопитающие. Тез. докл. 7 Всесоюзн. совещ. М. С. 156—157.
Кондинский П. А., 1973а. Размерная характеристика эритроцитов крови антарктических
ластоногих. В кн.: Исследования по биологии рыб и промысловой океанографии.
Вып. 4. Владивосток. С. 178—181.
Кондинский П. А., 19736. Вязкость, удельный вес и гематокрит крови антарктических
ластоногих. — Изв. ТИНРО, 87. С. 216—218.
Кондинский П. А., 1975. Адаптации крови антарктических ластоногих к условиям оби-
тания. Автореф. канд. дис. Владивосток. 30 с.
Коржуев П. А., 1964. Гемоглобин. М., Наука. 287 с.
Коржуев П. А., 1971. Эволюция, гравитация, невесомость. М„ Наука. 151 с.
Коржуев П. А., Глазова Т. Н., 1968. Сравнительно-физиологическая характеристика
крови и кроветворных органов рыб и водных млекопитающих. — Биология моря,
15. С. 131—147.
Косыгин Г. М., 1966. Некоторые материалы по морфологической характеристике плода
лахтака. В кн.: Тез. докл. 3 Всесоюзн. совещ. по изучению морских млекопита-
ющих. Владивосток. С. 26.
Косыгин Г. М., 1968. Некоторые материалы по морфологической характеристике плода
лахтака. — Тр. ВНИРО, 68. Изв. ТИНРО, 62. С. 26.
Косыгин Г. М., Кузин А. Е., 1979. Справочные показатели тихоокеанских ластоногих.
Владивосток. 130 с.
Крашенинников С. П., 1949. Описание земли Камчатки. М.—Л., Изд-во Главсевморпути.
841 с.
Крушинская Н. Л., Дмитриева И. Л., 1975. Два месяца из жизни “черненьких”. — При-
рода, 10. С. 101—109.
Крушинская Н. Л., Лисицына Т. Ю., 1983. Поведение морских млекопитающих. М.,
Наука. 336 с.
Кузин А. Е., 1968. Некоторые данные по эмбриогенезу черепа морского котика. — Тр.
ВНИРО, 68. Изв. ТИНРО, 62. С. 94—108.
373
Кузин А. Е., 1969а. О двойнях у морского котика. — Зоол. ж., 48, 2. С. 303—304.
Кузин А. Е., 19696. Развитие маточного комплекса беременных самок морского котика.
В кн.: Тез. докл. 4 Всесоюзн. совещ. по изучению морских млекопитающих. М.
С. 168—171.
Кузин А. Е., 1969в. Окостенение скелета конечностей морского котика. — Там же.
С. 171—174.
Кузин А. Е., 1970а. О весовой характеристике эмбрионов морского котика. — Изв. ТИНРО,
70. С. 52—59.
Кузин А. Е., 19706. Морфологическая характеристика северного морского котика (С. ur-
sinus). Особенности роста и развития в пренатальный период. Канд. дисс. Влади-
восток. 361 с.
Кузин А. Е., 1971а. Морфологическая характеристика северного морского котика (С. ur-
sinus). Особенности роста и развития в пренатальный период. Автореф. канд. дис.
Владивосток. 28 с.
Кузин А. Е., 19716. Морфологическая характеристика крови морского котика в онто-
генезе. — Изв. ТИНРО, 80. С. 102—116.
Кузин А. Е., 1972а. Весовой рост скелета морского котика в раннем онтогенезе. В кн.:
Тез. докл. 5 Всесоюзн. совещ. по изучению морских млекопитающих. Махачкала.
С. 113—114.
Кузин А. Е., 19726. Эколого-морфологические и возрастные особенности красной крови
морских котиков. В кн.: Териология. Т. 1. Новосибирск, Наука. С. 312—318.
Кузин А. Е., 1978. Половая зрелость и воспроизводительная способность популяций се-
верного морского котика. В кн.: 2 Международн. териол. конгресс. Брно. С. 400.
Кузин А. Ё., 1982а. Сколько было котиков на Командорах? В кн.: Изучение, охрана и
рац. использование морских млекопитающих. Тез. докл. 8 Всесоюзн. совещ. Аст-
рахань. С. 186—187.
Кузин А. Е., 19826. Формирование популяций котиков Курильских островов. В кн.: Эко-
лого-фаунистические исследования некоторых позвоночных Сахалина и Куриль-
ских островов. Владивосток. С. 90—95.
Кузин А. Е., 1982в. Характеристика основных биологических показателей популяций се-
верного морского котика при разных уровнях численности. В кн.: Млекопитающие
СССР. 3 съезд Всесоюзн. териол. об-ва. Тез. докл. Т. 2. М., Наука. С. 229—230.
Кузин А. Е., 1990. Оценка смертности морских котиков в результате загрязнения океана
отходами промышленного рыболовства. — Экология, 5. С. 89—92.
Кузин А. Е„ 1996. Социодемографическая характеристика адаптаций северного морского
котика в море. — Изв. ТИНРО, 62. С. 74—85.
Кузин А. Е., Маминов М. К., 1978. Линейная характеристика скелета некоторых ушастых
и настоящих тюленей. В кн.: Морские млекопитающие. Тез. докл. 7 Всесоюзн.
совещ. по изучению морских млекопитающих. М. С. 174—175.
Кузин А. Е., Маминов М. К., 1981. К морфологической характеристике крови и арте-
риальной системы бурого медведя (Ursus arctos). — Зоол. ж., 60, 7. С. 1078—1082.
Кузин А. Е., Маминов М. К., Соболевский Е. И., 1972. К проблеме морфо-физиологи-
ческих адаптаций некоторых тихоокеанских ластоногих и калана. В кн.: Тез. докл.
5 Всесоюзн. совещ. по изучению морских млекопитающих. Махачкала. С. 115—119.
Кузин А. Е„ Маминов М. К., Соколов А. С., 1973. К морфофизиологической характе-
ристике новорожденных тюленей и калана Курильских островов. В кн.: Исследо-
вания по ‘биологии рыб и промысловой океанографии. Владивосток. С. 200—208.
374
Кузин А. Е., Набережных И. А., 1982а. Изменение пропорций некоторых частей тела
Морского котика в пренатальный период. В кн.: Изучение, охрана и рац. исполь-
зование морских млекопитающих. Тез. докл. 8 Всесоюзн. сов,ещ. Астрахань.
С. 187—188.
Кузин А. Е., Набережных И. А., 19826. Относительный рост северного морского котика
в постнатальный период. Там же. С. 188—190.
Кузин А. Е., Набережных И. А., Трухин А. М., 1996. Социодемографическая структура
и статус нетрадиционных группировок северного морского котика острова Тю-
леньего. — Изв. ТИНРО, 121. С. 85—109.
Кузин А. Е., Панина Г. К., 1977а. Возрастно-половая структура и воспроизводительная
способность популяции котиков о. Тюленьего. В кн.: Исследования по биологии
рыб и промысловой океанографии. Вып. 8. Владивосток. С. 98—105.
Кузин А. Е., Панина Г. К., 19776. Миграции и особенности распределения самок морских
котиков в северо-западной части Тихого океана в зависимости от их физиологи-
ческого состояния. В кн.: Морские млекопитающие Тихого океана. Вып. 1. Вла-
дивосток. С. 3—17.
Кузин А. Е., Панина Г. К., Перлов А. С., 1977. Взаимоотношения котика и сивуча на
совместных гаремных лежбищах Курильских островов. Там же. С. 50—66.
Кузин А. Е., Перлов А. С., 1975а. Распределение и некоторые черты морфологии белок-
рылой морской свиньи (Ph. dalli True, 1885). — Изв. ТИНРО, 96. С. 259—288.
Кузин А. Е., Перлов А. С., 19756. Новое лежбище котиков на Курильских островах.
В кн.: Морские млекопитающие. 4.1. Тез. докл. 6 Всесоюзн. совещ. Киев, Наукова
думка. С. 166—167.
Кузин А. Е., Соболевский Е. И., 1975. Дыхательная система северного морского коти-
ка. — Изв. ТИНРО, 98. С. 156—183.
Кузин А. Е., Соколов А. С., 1970. Особенности красной крови эмбрионов и новорож-
денных северных морских котиков (С. ursinus). — Изв. ТИНРО, 70. С. 44—51.
Кузин А. Е., Скалецкая Е. И., Фрисман Е. Я., 1978. Определение оптимального режима
эксплуатации северного морского котика. В кн.: Исследования по биологии рыб
и промысловой океанографии. Вып. 9. С. 153—160.
Кузин А. Е., Чугунков Д. И., Фрисман Е. Я., Скалецкая Е. И., 1987. О целесообразности
добычи серебристых котиков. Владивосток. 18 с.
Кузин А. Е., Шаталина Т. А., 1990. Выживаемость северного морского котика в зави-
симости от факторов внешней среды. — Изв. ТИНРО, 112. С. 74—87.
Кузнецов Б. А., Ильина Е. Д., 1949. Остров Тюлений и его промысловые богатства.
В кн.: Пушные богатства СССР. М., Заготаздат. С. 171—179.
Кузьмин А. А., Перлов А. С., 1978. Дополнительные кости в черепе млекопитающих.
В кн.: 2 Международн. териол. конгресс. Брно. С. 359.
Ладыгина Т. Ф., Супин А. Я., 1974. Эволюция корковых зон мозга наземных и водных
млекопитающих. В кн.: Морфология, физиология и акустика морских млекопи-
тающих. М., Наука. С. 6—14.
Ладыгина Т. Ф., Супин А. Я., 1978. К вопросу о гомологии областей коры мозга ки-
тообразных и других млекопитающих. В кн.: Морские млекопитающие. М., Наука.
С. 55—65.
Лесгафт П. Ф., 1881. О причинах, влияющих на форму костей. — Тр. общества русских
врачей. С. 127—133.
375
Лисицына Т. Ю., 1983. Поведение и звуковая сигнализация северного морского котика
(С. ursinus) на лежбище. — Зоол. ж., 52, 8. С. 1220—1228.
Лыскин Н. Н., 1983. Особенности полигамной организации размножения северных мор-
ских котиков. Автореф. канд. дис. М. 23 с.
М. К., 1929. Состояние котикового стада на о. Тюленьем. — Бюлл. рыбного хоз-ва, 5.
С. 30.
Максимов А. А., 1984. Многолетние колебания численности животных, их причины и
прогноз. Новосибирск, Наука. 250 с.
Манзий С. Ф., 1957. О приспособительной перестройке запястья млекопитающих в про-
цессе эволюции. — Зоол. ж., 36, 1. С. 139—150.
Мараков С. Ф„ 1974. Северный морской котик. М., Наука. 71 с.
Мараков С. В., Нестеров Г. А., 1958. О вреде, наносимом сивучами находящимся на леж-
бище котикам. — Рыбное хоз-во, 10. С. 27—32.
Матвеев Б. С., 1963. О происхождении гетеродонтной зубной системы млекопитающих
по данным онтогенеза. — Тр. МОИП, 10. С. 62—74.
Микеладзе Д. А., 1971. О содержании морских котиков и каспийских тюленей в Батумском
аквариуме. В кн.: Исследования морских млекопитающих. Калининград. С. 273—
279.
Мордвинов Ю. Е., 1968. Наблюдения за локомоцией некоторых ластоногих. — Зоол. ж.,
42, 9. С. 1394—1402.
Мордвинов Ю. Е., 1969. Корпус как несущая плоскость и плавучесть некоторых ласто-
ногих (Pinnipedia). — Веста, зоол., 2. С. 10—14.
Мордвинов Ю. Е., 1972а. Маневренность ластоногих. — Там же, 1. С. 22—30.
Мордвинов Ю. Е., 19726. К характеристике некоторых гидродинамических показателей
формы тела ластоногих. — Гидробиол. ж., 8, 3. С. 101—105.
Морошкин К. В., 1966. Водные массы Охотского моря. М., Наука. 67 с.
Мужчинкин В. Ф., 1964а. Распределение сивучей на Юго-Восточном котиковом лежбище
острова Медного. В кн.: Морские котики Дальнего Востока. М., Пищевая пром-сть.
С. 179—186.
Мужчинкин В. Ф., 19646. Сроки появления окостенений в конечностях морских коти-
ков. — Тр. ВНИРО, 57. Изв. ТИНРО, 54. С. 151—154.
Мужчинкин В. Ф., 1964в. Некоторые отклонения от типичной зубной формулы у морского
котика (С. ursinus) и сивуча (Е. jubatus). В кн.: Морские котики Дальнего Востока.
М., Пищевая пром-сть. С. 151—164.
Мужчинкин В. Ф., 1968. Возникновение половых различий в черепе командорских мор-
ских котиков. Тр. ВНИРО, 68. Изв. ТИНРО, 62. С. 109—116.
Мужчинкин В. Ф., 1971а. Артериальная система северных морских котиков и варианты
ее строения. — Изв. ТИНРО, 80. С. 93—101.
Мужчинкин В. Ф., 19716. Мышечная система и способ передвижения северных морских
котиков. В кн.: Морские млекопитающие. М., ОНТИ. С. 79—87.
Мужчинкин В. Ф., 1971в. О продолжительности задержки имплантации у самок морского
котика. Там же. С. 93—101.
Мужчинкин В. Ф., 1976. О некоторых особенностях роста морских котиков (С. ursinus). —
Зоол. ж„ 55, 2. С. 267—275.
376
Мужчинкин В. Ф., 1978. Блочность конструкций у млекопитающих на примере семейства
ушастых тюленей (Otariidae). — Журн. общ. биол., 39, 5. С. 777—782.
Набережных И. А., 1986. Репродуктивный успех самцов морских котиков. В кн.: Изучение,
охрана и рац. использ. морских млекопит. Тез. докл. 9 Всесоюзн. совещ. по изуч.
морских млекопитающих. Архангельск. С. 287—288.
Натаров В. В., Черный Э. И., 1966. О формировании зон повышенной биологической
продуктивности в Тихом океане. — Тр. ВНИРО, 60. С. 125—134.
Нестеров Г. А., 1964. Материалы по биологии и численности сивучей Командорских ост-
ровов. — Тр. ВНИРО, 51. Изв. ТИНРО, 54. С. 173—177.
Нестеров Г. А., 1966. Опыт применения дитилина на котиковом промысле. — Изв. ТИНРО,
58. С. 41—48.
Нестеров Г. А., 1977. Индивидуальное опознание морских котиков по внешним призна-
кам. В кн.: Морские млекопитающие Тихого океана. Вып. 1. Владивосток. С. 67—71.
Нестеров Г. А., 1983. Межродовые различия в строении папилярных линий у северных
морских котиков и сивуча. В кн.: Биологические проблемы Севера. Ч. 2. Магадан.
С. 124—125.
Нестеров Г. А., 1985. Выделение неметрических признаков-маркеров в строении оторочки
передних ластов северного морского котика. В кн.: Фенетика популяций. Матер.
3 Всесоюзн. совещ. М. С. 256—257.
Нестеров Г. А., Болтнев А. И., 1984. О дифференциации популяций северного морского ко-
тика. В кн.: Морские млекопитающие Дальнего Востока. Владивосток. С. 121—126.
Никитенко М. Ф., 1966. Морфологическая характеристика головного мозга северного
морского котика (С. ursinus). В кн.: 3 Всесоюзн. совещ. по изучению морских мле-
копитающих. Тез. докл. М., Наука. С. 34—35.
Огнев С. И., 1935. Звери СССР и прилежащих стран. Т. 3. Ластоногие. М.—Л., Изд-во
АН СССР. 740 с.
Одум Ю., 1975. Основы экологии. М., Мир. 740 с.
Осинский П. А., 1959. О характере сопряженных движений таза и позвоночника у не-
которых млекопитающих. — Зоол. ж., 38, 11. С. 1711—1718.
Павлычев В. П., 1967. Некоторые особенности гидрологического режима в районе Ку-
росио в 1965 г. — Изв. ТИНРО, 61. С. 3—10.
Павлычев В. П., 1975. Режим вод и положение субарктического фронта северо-западной
части Тихого океана (1965—1979). — Изв. ТИНРО, 96. С. 3—18.
Панин К. И., 1970. Распределение и миграции морских котиков разных стад в северной
части Тихого океана по результатам мечения. — Изв. ТИНРО, 70. С. 5—37.
Панин К. И., Панина Г. К., 1968. Экология и миграции котиков в зимне-весенний период
в Японском море. В кн.: Ластоногие северной части Тихого океана. М., Пищевая
пром-сть. С. 70—80.
Панина Г. К., 1964. Питание котиков в Японском море. — Изв. ТИНРО, 54. Тр. ВНИРО,
57. С. 67—73.
Панина Г. К., 1966. О питании сивуча и тюленей на Курильских островах. — Изв.
ТИНРО, 58. С. 235—236.
Панина Г. К., 1970. Экология котиков (Callorhinus ursinus L.) в морской период их жизни.
Докт. дис. Владивосток. 236 с.
Панина Г. К., 1971. Характер распределения котиков, зимующих в Японском море. —
Тр. ВНИРО, 82. Изв. ТИНРО, 80. С. 14—24.
377
Панина Г. К., 1977. О влиянии гидрологических условий на распределение котиков в
местах зимовки у восточного побережья о. Хонсю. В кн.: Исследования по би-
ологии рыб и промысловой океанографии. Вып. 4. Владивосток. С. 182—191.
Панина Г. К., 1978. О межгодовой изменчивости питания морских котиков. В кн.: Мор-
ские млекопитающие. Тез. докл. 7 Всесоюзн. совещ. М. С. 254—255.
Перлов А. С., 1968. Весенне-летнее распределение котиков в районе Командорских ост-
ровов. — Изв. ТИНРО, 62. Тр. ВНИРО, 68. С. 89—93.
Перлов А. С., 1975. Питание сивучей в районе Курильских островов. — Экология, 4.
С. 106—108.
Перлов А. С., Кузин А. Е., 1967. О миграции котиков о. Тюленьего, Курильских и Ко-
мандорских и местах их зимовки. Тез. докл. Владивосток. С. 25.
Перлов А. С., Урманов М. И., 1971. Об анатомии пищеварительной системы сивучей.
В кн.: Морские млекопитающие. М., ОНТИ. С. 193—206.
Петрищев Б. И., Бородин А. Д., 1986. Развитие эмбрионов кабана популяции широко-
лиственных лесов Брянской области. В кн.: Морфология и генетика кабана. М.,
Наука. С. 87—104.
Петской П. Г., 1953а. Понятие и типы эмбрионального парабиоза. — Докл. АН СССР,
нов. сер., 89, 6. С. 1123—1126.
Петской П. Г., 19536. Условия внутриутробного развития Bos taurus, вызывающие явление
фримартинизма. — Там же, 90, 4. С. 693—695.
Пилипчук С. Я., 1980. Изменение массы позвонков по отделам позвоночника у хищных
млекопитающих. — Веста, зоол., 4. С. 60—63.
Пинигин В. Е., Прянишников В. Г., 1982. Сроки появления морских котиков на лежбищах
Командорских островов. — Изучение, охрана и рац. использ. морских млекопи-
тающих. Тез. докл. 8 Всесоюзн. совещ. Астрахань. С. 287—288.
Покудова В. В., Тунеголовцев В. П., 1975. Новая схема течений Японского моря. — Тр.
ДВНИГМИ, 50. С. 24—37.
Полонский А., 1871. Курилы. — Зап. русск. географ, об-ва по этнографии, 4. С. 367—576.
Предтеченский В. Е., Боровская В. М., Марголина Л. Т., 1950. Лабораторные методы
исследований. М., Медгиз. 804 с.
Ровнин А. А., 1968. Встречи котиков в Тихом океане. В кн.: Ластоногие северной часта
Тихого океана. М., ОНТИ. С. 87—88.
Северцов А. Н„ 1939. Морфологические закономерности эволюции. М.—Л., Изд-во АН
СССР. 610 с.
Северцов С. А., 1941. Динамика населения и приспособительная эволюция животных.
М,—Л., Изд-во АН СССР. 314 с.
Серебренников Е. Я., 1982. Количество сперматозоидов у самцов котиков командорской
популяции в связи с их физическим состоянием. В кн.: Изучение, охрана и рац.
использ. морских млекопитающих. Тез. докл. 8 Всесоюзн. совещ. Астрахань.
С. 319—320.
Симашко Ю., 1851. Русская фауна или описание и изображение животных, водящихся
в Империи Российской. Ч. 2. Млекопитающие. СПб. 1161 с.
Скалецкая Е. И., Фрисман Е. Я., Ашихмина Е. В., Кузин А. Е., 1983. Моделирование
динамики половозрастного состава локальной популяции северного морского ко-
тика. Оценка коэффициентов выживаемости и стационарные режимы. В кн.: Ис-
следования по математической популяционной экологии. Владивосток. С. 28—48.
378
Скалецкая Е. И., Фрисман Е. Я., Кузин А. Е., 1985. Определение оптимального режима
эксплуатации для популяции северного морского котика на основе математической
модели динамики численности. В кн.: Математическое моделирование в популя-
ционной экологии. Владивосток. С. 3—20.
Слепцов М. М., 1940. О приспособлении к плаванию ластоногих. — Зоол. ж., 19, 3.
С. 379—386.
Слепцов М. М., 1968. Изменение темпов формирования адаптивных черт в онтогенезе
и их роль в дивергентной эволюции водных млекопитающих. В кн.: Темп инди-
видуального развития животных и их изменение в ходе эволюции. М., Наука.
С. 191—214.
Слюнин Н. Е., 1895. Промысловые богатства Камчатки, Сахалина и Командорских ост-
ровов. СПб. 117 с.
Смирнов Н. А., 1908. Очерк русских ластоногих. — Зап. Импер. Академии наук, 23, 4.
С. 1—76.
Сноу Г., 1902. Курильская гряда. — Зап. об-ва изучения Амурского края, 8, 1. 119 с.
Соболевский Е. И., 1974. Морфо-функциональная характеристика органов дыхания се-
верного морского котика. Автореф. канд. дис. Владивосток. 25 с.
Соболевский Е. И., 1976. Возрастно-половая структура и смешиваемость популяций ко-
тиков в море. В кн.: Исследования по биологии рыб и промысловой океанографии.
Вып. 7. С. 156—161.
Соболевский Е. И., 1977. Морфологическая картина кровн некоторых видов тихоокеан-
ских тюленей. В кн.: Морские млекопитающие Тихого океана. С. 72—79.
Соболевский Е. И., 1978а. К вопросу о вариабельности основных борозд головного мозга
котика. В кн.: Морские млекопитающие. Тез. докл. 7 Всесоюзн. совещ. М. С. 297—298.
Соболевский Е. И., 1978(5. Сравнительная анатомия спинного мозга полуводных, водных
и наземных млекопитающих. — Арх. анат., гистол. и эмбриол., 11. С. 74—80.
Соболевский Е. И., 1978в. Изменение весовых показателей головного мозга котика в он-
тогенезе. В кн.: Морские млекопитающие. Тез. докл. 7 Всесоюзн. совещ. М.
С. 296—297.
Соболевский Е. И., 1984. Изучение популяционной структуры северного морского котика
(С. ursinus) (Pinnipedia, Otariidae) фенетическим методом. — Зоол. ж., 63, 11.
С. 1705—1712.
Соболевский Е. И., Кузин А. Е., 1977. О формировании некоторых рефлекторных актов
у морского котика в эмбриогенезе. В кн.: Управление поведением животных. 2
Всесоюзн. совещ. по поведению животных. М., Наука. С. 271—272.
Соколов А. С., 1955. Некоторые функционально-морфологические и возрастные особен-
ности ладожского тюленя в связи с водным образом жизни. Автореф. канд. дис.
Л. 27 с.
Соколов А. С., 1959. Особенности калибра и распределение некоторых кровеносных со-
судов у водных (тюлень) и наземных (собака) млекопитающих. — Арх. анат., гнс-
тол. н эмбриол., 38, 12. С. 27—34.
Соколов А. С., 1966. Особенности мускулатуры органов движения ластоногих. В кн.: Тез.
докл. 3 Всесоюзн. совещ. по изучению морских млекопитающих. М., Наука. С. 42—43.
Соколов А. С., 1969а. Количество миоглобина и крови у некоторых ушастых и настоящих
тюленей. В кн.: Тез. докл. 4 Всесоюзн. совещ. по изучению морских млекопита-
ющих. М. С. 193—196.
379
Соколов А. С., 19696. Некоторые итоги и перспективы эколого-морфологических иссле-
дований тихоокеанских ластоногих. В кн.: Морские млекопитающие. М., Наука.
С. 237—244.
Соколов А. С., 1971. Размерная характеристика скелета конечностей северного морского
котика и сивуча. В кн.: Морские млекопитающие. М„ ОНТИ. С. 88—92.
Соколов А. С., 1973. Опыт сравнительно-морфологического изучения мускулатуры ор-
ганов плавания ластоногих. В кн.: Проблемы бионики. М., Наука. С. 483—488.
Соколов А. С., Косыгин Г. М., Кузин А. Е., Перлов А. С„ Тихомиров Э. А., Шустов
А. П., 1969. Справочные показатели по характеристике внутренних органов ти-
хоокеанских ластоногих. В кн.: Морские млекопитающие. М., Наука. С. 226—233.
Соколов А. С., Перлов А. С., 1969. Особенности весовой характеристики внутренних ор-
ганов морского котика и сивуча. В кн.: Тез. докл. 4 Всесоюзн. совещ. по изучению
морских млекопитающих. М. С. 182—185.
Соколов А. С., Соколов И. И., 1970. Некоторые особенности органов движения речной
выдры и калана в связи с образом жизни. — Бюлл. МОИП, отд. биол., 75.
С. 15—17.
Соколов В. Е., 1959. Структура кожного покрова ластоногих. Сообщ. 1. — Там же, 6.
С. 5—22.
Соколов В. Е., 1960. Структура кожного покрова ластоногих. Сообщ. 2. — Там же, 4.
С. 5—18.
Соколов В. Е., 1973. Кожный покров млекопитающих. М., Наука. 487 с.
Соловьев А. И., 1947. Курильские острова. М.—Л., Изд-во Главсевморпути. 305 с.
Стародубцева Е. М„ 1972. Особенности цитоархитектоники кохлеарного комплекса не-
которых ластоногих. В кн.: Тез. докл. 5 Всесоюзн. совещ. по изучению морских
млекопитающих. Ч. 2. Махачкала. С. 239—241.
Суворов Е. К., 1912. Командорские острова и пушной промысел на них. СПб. 324 с.
Суворов Е. К., 1927. Командорские острова в 1922 г. — Изв. отд. прикладной ихтиологии,
6, 2. С. 216—232.
Суворов Е. К., 1931. Современное состояние котикового хозяйства в Америке. — Изв.
Ленинград, ихтиол, ин-та, 13, 1. С. 25—45.
Сушкина Н. И., 1954. Путешествие на о. Тюлений. М.—Л., Изд-во АН СССР. 86 с.
Суховская Л. И., 1972. Особенности цитоархнтектоники и нейронной организации заднего
двухолмия мозга некоторых морских млекопитающих. В кн.: Тез. докл. 5 Всесоюзн.
совещ. по изучению морских млекопитающих. Ч. 2. Махачкала. С. 244—247.
Тимофеева А. Т., Чупахина Т. И., Колесник Ю. А., 1977. Определение численности ко-
тиков на острове Тюленьем. В кн.: Морские млекопитающие. Вып. 1. Владивосток.
С. 35—40.
Тихенко С. А., 1914а. Об островах Тюленьем и Курильских. — Матер, к познанию русск.
рыболовства, 3, 5. С. 62—96.
Тихенко С. А., 19146. Привал котиков, время наибольшей рождаемости и уход котиков
с о. Тюленьего. — Там же, С. 66—67.
Тихенко С. А., 1914в. Морская охрана котиков и охрана их на берегу (из отчета о поездке
на Командорские острова). — Там же, С. 1—33.
Тихенко С. А., 1914г. Отчет о поездке на Командорские острова летом 1912 г. — Там
же, С. 1-^61.
380
Тихменев П., 1861—63. Историческое обозрение образования Российско-Американской
компании и действие ее до настоящего времени. Ч. 1, 2. СПб. С. 1—386.
Тихомиров Э. А., 1959. К вопросу о питании сивуча теплокровными животными. — Изв.
ТИНРО, 47. С. 185—186.
Тихомиров Э. А., 1964. Встречаемость морских котиков в Охотском море. — Изв. ТИНРО,
54. Тр. ВНИРО, 51. С. 65—66.
Третьяков Н. Н., 1959а. Рост советского мериноса в течение внутриутробного развития. —
Тр. ИЭМЭЖ, им. А. Н. Северцова, 23. С. 5—25.
Третьяков Н. Н., 19596. Линейный рост скелета овец породы советский меринос в течение
внутриутробного развития. — Там же. С. 91—145.
Третьяков Н. Н., 1959в. Окостенение скелета овец породы советский меринос в течение
внутриутробного развития. — Там же. С. 146—187.
Урманов М. И., 1970а. Внутриорганная архитектура кровеносных сосудов и желчных про-
токов печени некоторых ластоногих. — Изв. ТИНРО, 70. С. 77—86.
Урманов М. И., 19706. К вопросу об анатомии печени ушастых тюленей. — Там же.
С. 67—76.
Урманов М. И., 1971. Ангиархитектура печени калана и ластоногих в связи с желчеот-
делительной системой. — Тр. ВНИРО, 82. Изв. ТИНРО, 80. С. 247—256.
Успенский С. М., 1955. Прошлое распространение котиков и калана на островах Куриль-
ской гряды. — Бюлл. МОИП, 4. С. 9—16.
Успенский С. М., 1959. Морские колониальные гнездящиеся птицы северных и дальне-
восточных морей СССР, их размещение, численность и их роль как потребителей
планктона и бентоса. — Там же, отд. биол., 62, 2. С. 39—52.
Федоров С. Г., 1964. Конвенционные принципы регулирования промысла котиков в се-
верной части Тихого океана. В кн.: Морские млекопитающие Дальнего Востока.
М., Пищевая пром-сть. С. 9—21.
Федосеев Г. А., 1964. Об эмбриональном, постэмбриональном росте и половом созревании
кольчатой нерпы. — Зоол. ж., 43, 8. С. 1228—1235.
Фрисман Е. Я., Скалецкая Е. И., Кузин А. Е., 1985. Математическое моделирование ди-
намики численности северного морского котика и оптимальное управление коти-
ковым хозяйством. Владивосток. 156 с.
Хоккей (Митимори), 1897. Личные наблюдения водных промыслов о. Хоккайдо. — Зап.
об-ва изучения Амурского края, 6, 1. 97 с.
Чапский К. К., 1963. Ластоногие. В кн.: Млекопитающие фауны СССР. Ч. 2. М.—Л, Изд-во
АН СССР. С. 895—964.
Чапский К. К., 1976. Отряд ластоногих. В кн.: Млекопитающие Советского Союза. М.,
Высш, школа. 718 с.
Челноков Ф. Г., 1970а. О поведении морских котиков на лежбище. — Изв. ТИНРО, 70.
С. 233—239.
Челноков Ф. Г., 19706. В воде с морскими котиками. — Вопросы географии Камчатки,
6. С. 159.
Челноков Ф. Г., 1970в. О взаимоотношениях песцов и детенышей котиков. — Там же.
С. 151—158.
Челноков Ф. Г., 1971а. Перемещение морских котиков на лежбище о. Медного. — Тр.
АтлантНИРО, 39. С. 151—156.
381
Челноков Ф. Г., 19716. О размножении сивучей на Юго-Восточном лежбище острова Мед-
ный. — Изв. ТИНРО, 80. Тр. ВНИРО, 82. С. 190—192.
Челноков Ф. Г., 1971 в. Некоторые данные о переходе морских котиков с лежбища на
лежбище. — Там же. С. 44—46.
Челноков Ф. Г., 1974. О взаимоотношениях холостяков и детенышей морских котиков
на гаремной территории. В кн.: Исследования по биологии рыб и промысловой
океа-нографии. Вып. 5. С. 137—140.
Челноков Ф. Г., 1977. Прогноз промысла котиков холостяков и динамика численности
секачей на Юго-Восточном лежбище острова Медного. В кн.: Морские млекопи-
тающие Тихого океана. Вып. 1. Владивосток. С. 18—34.
Челноков Ф. Г., 1978. Поведенческие адаптации котиков секачей к длительному пребы-
ванию на лежбище. В кн.: Морские млекопитающие. Тез. докл. 7 Всесоюзн. совещ.
Симферополь. С. 347—348.
Челноков Ф. Г., 1981. Динамика возрастного состава самок котиков на лежбищах острова
Медного в 1970—1977 годах. — Изв. ТИНРО, 105. С. 146—151.
Челноков Ф. Г., 1982а. Зимнее скопление морских котиков на о. Медном (Командорские
о-ва). В кн.: Изучение, охрана и рац. использ. морских млекопитающих. Тез. докл.
8 Всесоюзн. совещ. Астрахань. С. 400—401.
Челноков Ф. Г., 19826. О смешиваемости командорских котиков с котиками других по-
пуляций. — Вопросы географии Камчатки, 8. С. 74—76.
Челноков Ф. Г., 1990. Доля котиков прибыловской популяции в добыче на Командорских
островах. — Изв. ТИНРО, 112. С. 116—117.
Челноков Ф. Г., Лыскин Н. Н., 1978. О длительности пребывания секачей морских ко-
тиков на гаремной территории. — Тр. Кировского сельхоз. ин-та, 59. С. 65—68.
Челноков Ф. Г., Чугунков Д. И., 1971. Возрастные различия некоторых морфологических
признаков тела холостяков морских котиков. В кн.: Морские млекопитающие. М.,
ОНТИ. С. 59—64.
Чугунков Д. И., 1966. О локальности стад морских котиков, обитающих на островах Бе-
ринга и Медном. — Изв. ТИНРО, 58. С. 15—22.
Чугунков Д. И., 1968. Сивучи и котиковое хозяйство Комадорских островов. — Изв. ТИНРО,
62. Тр. ВНИРО, 68. С. 159—171.
Чугунков Д. И., 1982. О весовой характеристике новорожденных детенышей северных
морских котиков. В кн.: Изучение, охрана и рац. использ. морских млекопитаю-
щих. Тез. докл. 8 Всесоюзн. совещ. Астрахань. С. 405—406.
Чугунков Д. И., Прохоров В. Г., 1964. Новые данные о зимовке котиков в Беринговом
море. — Изв. ТИНРО, 58. С. 233—234.
Чупахина Т. И., 1977. Характеристика роста массы детенышей морского котика в период
молочного питания. В кн.: Морские млекопитающие Тихого океана. Вып. 1. Вла-
дивосток. С. 46—49.
Шварц С. С., 1980. Экологические закономерности эволюции. М., Наука. 287 с.
Шмальгаузен И. И., 1922. Основы сравнительной анатомии. 425 с.
Шмальгаузен И. И., 1935а. Определение основных понятий и методика исследования
роста. В кн.: Рост животных. М.—Л., Биомедгиз. С. 8—60.
Шмальгаузен И. И., 19356. Рост и дифференцировка. Там же. С. 74—84.
382
Шмальгаузен И. И., 1947. Основы сравнительной анатомии. М., Сов. наука. 540 с.
Шунтов В. П., 1972. Морские птицы и биологическая структура океана. Владивосток.
376 с.
Шунтов В. П., 1994. Новые данные о перестройках в пелагических экосистемах морей
Дальнего Востока. — Вести. ДВО АН, 2. С. 59—67.
Щуров В. А., 1975а. Изменчивость почечных артерий северного морского котика (С. ur-
sinus). В кн.: В сб. Исследования по биологии рыб и промысловой океанографии.
Вып. 6. Владивосток. С. 107—ПО.
Щуров В. А., 19756. К морфологии почек морских млекопитающих. В кн.: Морские мле-
копитающие. Матер. 6 Всесоюзн. совещ. Ч. 2. Киев, Наукова думка. С. 182—183.
Яблоков А. В., Белькович В. М., Борисов В. И., 1972. Киты и дельфины. М., Наука.
472 с.
Abegglen С. A., Roppel A. Y., 1959. Fertility in the Northern fur seal. — J. Wildl. Manag.,
23, 1. P. 1956—1957.
Abegglen С. E., Roppel A. Y., Wilke F., 1958. Alaska fur seal investigation. — U.S. Fish
Wildl. Serv. Bur. Comm. Fisher., Wash. 193 p.
Ainly D. G., Senger C. A., 1979. Trophic relations of seabirds in North-Western Pacific Ocean
and Bering Sea. In: Conservation of marin bird of Northern Americ. U.S. Dept, interior
fish and Wildl. Serv. Report II. Wash. D.C. P. 95—122.
Allen G. M., 1942. Extinct and vanishing mammals of the western hemisphere with the marine
species of all the ocean. — Publ. Amer. Comm. Intern., Wildl. Protection, 2. P. 1—620.
Allen J. A., 1880. History of North American pinnipeds. Wash. 758 p.
Anas R. E., 1970. Accuracy in assigning age to fur seals. — J. Wildl. Manag., 34, 3.
Andersen H. T., 1966. Phisiological adaptation in diving vertebrates. — Phisiol. Rev., 46.
P. 212—243.
Ashworth U. S., Romaich G. D., Keyes M. S., 1966. Species difference in the composition
of milk mith special reference to the Northern fur seal. — Collected Rept. Div. Biol.
Res., 1. P. 1206—1211.
Austin O. L.,Wilke F., 1950. Japanese fur sealing. — U.S. Fish and Wildl. Serv. Sci., Rept.
Wildl., 6. 91 p.
Baba N., Yoshida K., Ichihara T., 1979. An application radio wave telemetry to the study
of fur seal (Callorhinus ursinus) behavior on the breeding islands. — Far Seal Fish.
Res. Lab., Bull., 17. P. 177—196.
Baker J. D., Fowler C. W„ 1992. Pup weight and survival of Northern fur seals, Callorhinus
ursinus. — J. Zool., 227. P. 231—238.
Baker R. S., Wilke F., Baltzo C. U., 1970. The Northern fur seal. Wash. 19 p.
Bartholomew G. A., 1954. Body temperature and respiratory and heart rates in Northern ele-
phant seal. — J. Mammal., 35, 2. P. 211—218.
Bartholomew G. A., 1970. A model for the evolution of pinniped polygyny. — Evolution,
24. P. 546—559.
Bartholomew G. A., Hoell P. G., 1953. Reproductive behaviour of the Alaska fur seal, Cal-
lorhinus ursinus. — J. Mammal., 36. P. 417—436.
Bartolomew G. A., Wilke F., 1956. Body temperature in the Northern fur seal, Callorhinus
ursinus. — J. Mammal., 37. P. 327—337.
383
Bauer R. D„ Johonson A. M„ Scheffer V. B., 1964. Eye lens weight and age in the fur
seal. — J. Wildl. Manag., 28, 2. P. 374—376.
Bertoni J. C., Sealy S. G., 1979. Distibution and status of marine bird breeding along the
coasts of Chukchi and Bering seas. — Conserv. of marine birds of Northern North
America. Wildl. Res. Report, 2. D.C. P. 21—31.
Bertram G. C. L., 1940. The biology of the Weddell and Crabeater seals with a study of the
comparative behaviour of the Pinnipedia. — Scietific report of the British Graham Land
expedition 1934—1937, 1, 1. P. 1—139.
Bigg M. A., MacAski J. B., Ellis G., 1977. Studies on captive fur seal. — Arctic biol. station.
Rept No 1. Quebec. 23 p.
Bigg M. A., MacAski J. B., Ellis G., 1978. Studies on captive fur seal. — Arctic biol. station.
Rept No 2. Quebec. 21 p.
Bowker R. C., 1931. Some physical properties of fur seal skin. — Techn. Assoc. Fur Industry
J„ 2. P. 34—43.
Burow A., 1838. Uber das Geftssystem der Robben. — Muller’s Arch. Anat. Physiol. P. 230—
258.
Calambokidis J., Gentry R. L., 1985. Mortality of northern fur seal pups in relation to growth
and birth weights. — J. Wildl. Dis., 21, 3. P. 327—330.
Chapman D. G., 1961. Population dynamics of the Alaska fur seal herd. In: 26th North. Amer.
Wildl. Natur. Res. Conf. Wash. D. С. P. 356—369.
Chapman D. G., 1964. Critical study of Pribilof fur seal population estimates. — Fish Bull.,
63, 3. P. 657-669.
Chapman D. G., 1981. The Northern fur seal — an exampl of complexity. In: Problems in
management of abundant wild Mammals. Acad. Press. Inc. P. 193—204.
Chiasson R. B., 1957. The dentition of Alaska fur seal. — J. Mammal., 38, 3. P. 310—319.
Clark G. A., 1912. The fur seal census. — Science, 36. P. 894—897.
Clark G. A., 1913. Conservation of fur seals. — North Amer. Rev., 197. P. 640—644.
Craig A. M., 1964. Histology of reproduction and estrus cycle in the female fur seal, Callorhinus
ursinus. — J. Fish. Res. Board Canada, 21, 4. P. 773—811.
Davies J. L., 1958. Pleistocene geography and distribution of northern pinnipeds. — Ecology,
39, 1. P. 97—113.
Drabek С. M„ 1975. Some anatomical aspects of the cardiovascular system of Antarctic seals
their possible functional significance in diving. — J. Morphol., 745. S. 85—105.
Dout K., 1942. Review of the genus Phoca. — Ann. Carneg. Mus., 29. P. 61—125.
Eberhardt L. L., 1981. Population dynamics of the Pribilof fur seal. In: Dynamic of large
mammals population. John Wiley and Sons. P. 179—220.
Elliott H. W., 1875. A report upon the condition of affairs in terrytory of Alaska. Wash.
277 p.
Elliott H. W., 1882. A monograph of seal-islands of Alaska. — U.S. Comm. Fish. Spec. Bull.,
176. 176 p.
Elliott H. W., 1884. The habits of the fur seal. — In: Fisheries and Fishery Industries of the
United States. Sect. I, 1. P. 75—113.
Elliott H. W., 1886. Our Arctic province Alaska and island. London. 473 p.
Elsner R., 1969. Cardiovascular adjusments to diving. In: The biology of marine mammals.
London, Academic Press. P. 117—145.
384
Elsner R: W., Franklin R. L., Van Critters, Kenney D. W., 1966. Cardiovascular defense against
asphyxi. — Science, 153. P. 941—949.
Engel S., 1962. Lung structure. Illionois, USA. 125 p.
Engel D. R., Dillon L. S., 1972. Cerebrum of the sea otter. — Tex. J. Sci., 24, 2. P. 221—232.
English A. W., 1976. Functional anatomy of the hands of fur seals and sea lions. — Am.
J. Anat., 147. P. 1—18.
Fay F. H., 1967. The member of fibrs and thoracic vertebrate in pinnipeds. — J. Mammal.,
48, 1.
Fiscus С. H. G., Bains G. A., Wilke F., 1964. Pelagis fur seal investigation, Alaska waters. —
U. S. Fish Wildl. Serv. Spec. Rep. Fish., 475. 48 p.
Fiscus С. H., DeLong R. L., Antonelis G. A., 1977. Fur seal investigation, 1976. — Nat.
Marin. Fish. Serv. P. 67—70.
Fish P. A., 1899. The brain of the fur seal, Callorhinus ursinus with a comparative description
of these of Zalophus californianus, Phoca vitulina, Ursus americanus and Monachus tro-
picalis. — In: The fur seas and fur seal islands of the North Pacific ocean. Pt. 3. Wash.
P. 21—41.
Fowler C. W., 1982. Interactions of Northern fur seal and commercial fisheries. — Trans.
47th North American Wildlife and Natural Resources Conf. Wash. D. С. P. 278—292.
Fowler C. W., 1990. Density dependence in Northern fur seals {Callorhinus ursinus). — Mar.
Mamm. Sci., 6, 3. P. 171—195.
Gambarjan P. P., Karapetjan W. S., 1961. Besondersheiten im Bau des Seelowen, der Bai-
kalrobbe und des Seeotters in Anpassung und die Fortbewegung im Wasser. — Zool.
Jb. Anat., 79, 9. S. 123—148.
Gentry R. L., Johnson J. H., 1976. Predation by Northern sea lions, Fur seal investigation
in 1975. — Nat. Marine Fish Serv. P. 55—64.
Gentry R. L., Johnson J. H., 1978. Fur seal investigation, 1977. Nat. Marine Fish. Serv. 96 p.
Gentry R., Holt J., Francis J., 1979. Fur seal investigation. Nat. Marine Fish. Serv., 84 p.
Gentry R. L., Johnson J. H., 1981. Predation by sea lions on northern fur seal neonatal. —
Mammalia, 45, 4. P. 423—430.
Gentry R. L., Johnson J. H., 1976. Fur seal investigation, 1975. Nat. Marin. Fish. Serv.
P. 49—64.
Goebel M. E., 1983. Duration of feeding trips and age-related reproductive success of lactating
females, St. Paul island, Alaska. — Fur seal investigations. Techn. memo. NPFSC, 146.
P. 28—33.
Grahame T., 1959. The arterial circulation of kidneys of elephant and grey seals. — Brit. Vet.
J., 115, 11. P. 397—399.
Gray J. E., 1859. On the bear of Foster, the Ursus marinus of Steller, Arctocephalus ursinus
of aurhors. — Proc. Zool. Soc., London. P. 102—103.
Guzsal E., 1959. Beitrage zur Anatomic der Robbenniere. — Acta Vet. Acad. Sci. Hung., 9,
4. S. 419—428.
Hall E. R., 1940. Pribilof fur seal on Californian coast. — Calif. Fish and Game, 26. P. 76—77.
Hanna G. D., 1918. Fur seal censuses. Alaska Fish. Fur Industries U. S. 51 p.
Hanna G. D., 1921. Genital organs of hermaphroditic fur seal. — Amer. Nat., 55. P. 473—475.
385
Hanna G. D., 1922. What becomes of the fur seals. — Science, 55. P. 505—507.
Hanna G. D., 1923. What is future of the fur seal? — Fur Trade Rev. P. 357—359.
Harris H. A., 1936. The fetal growth of the sheep. — J. Anat., 71. P. 89—112.
Harrison C. S., Hida T. S., 1980. The status of seabird research in the Northwestern Hawaiian
Island. In: Rept. Proc. Simp. “Status of resource investigation in the Northwestern Ha-
waiian Island”. P. 17—31.
Harrison R. J., Tomlinson J. D., 1960. Normal and experimental diving in the common seal
(Phoca vitulina). — Mammalia, 24. P. 386—399.
Hildebrandt W. R., 1984. Archaeological presence of the Northern fur seal Callorhinus ursinus
along the coast of Northern California. — Murrelet, 65, 1. P. 28—29.
Howell A. B., 1929. Contribution of the comparative anatomy of the eared and earless seal
(Genera Zalophus and Phoca). — Proc. U. S. Nat. Mus., 73, 15. P. 1—142.
Howell A. B., 1930. Aquatic mammals, their adaptation to life in the water. 338 p.
Ichihara T., 1972. Maximum sustainable field in robben fur seal herd. — Bull. Far Seal. Fish.
Res. Lab., 6. P. 77—93.
Ichihara T., 1973. Comments on natural mortality rate of adult female fur seal of Robben
Island origin. — Bull. Far Seal Fish. Res. Lab., 11. 21 p.
Ichihara T., 1974. Possible effect of surface force on the sexspecific mortality of younge fur
seals in the eastern Pacific. — Bull. Far Seas Fish. Res. Lab., Shimizu, 2. P. 1—8.
Ichihara T., Yoshida K., 1972. Diving depth of Northern fur seal in the feeding time. — Rept.
Sci. Whales Res. Inst., 24. 14 p.
Irving L., Peyton L. J., Bahn С. H„ Peterson R. S., 1962. Regulation of temperature in fur
seals. — Physiol. Zool., 35, 4. P. 275—284.
Irving L., Peyton L., Bahn C., Peterson K., 1963. Action of the heart and breathing during
the development of fur seals (Callorhinus ursinus). — Ibid., 36, 1. P. 1—20.
Irving L., Scholander P. F., Grinnel S. W., 1942. The regulation of arterial blood pressure
in the seal during diving. — J. Phisiol., 135. P. 557—566.
Irving L., Solandt О. M., Solandt D. Y., Fisher К. C., 1935. The respiratory metabolism of
the seal and its adjusment to diving. — J. Cell. Comp. Phisiol., 7, 1. P. 137—151.
Jamieson G., Fisher H., 1972. The pinniped eye. In: Functional Anatomy of Marine Mammals.
Vol. 1. London, N. Y., Academic Press. P. 102—108.
Johnson A. M., 1975. The status of northern fur sea! populations. — Rapp. proc, verb.-reun.
cons. int. explor. men, 169. P. 263—266.
Jordan D. S., ed., 1898—1899. The seal and fur-seal islands of the North Pacific Ocean. —
U.S. Treasury Dep., Doc. 2017. P. 1—3.
Jordan D. C., Clark G. A., 1898. The fur seals and fur seals islands of the North Pacific
Ocean. — U.S. Treasury Dep., Doc. 2017. P. 249, 251—606, 629, 384.
Kajimura H., Lander R. H., Perez M. A., York A. E., Bigg M. A., 1980a. A life table and
biomass estimate for Alaskan fur seals Furthem analis of pelagic fur seal data collected
by the U.S. and Canada during 1958—74, Pt. 1. 94 p.
Kajimura. H., Lander R. H., Perez M. A., York A. E., Bigg M. A., 19806. Further analisis
of pelagic seal data collected by the U.S. and Canada during 1958—74, Pt. 2. 172 p.
Kano K., Kogai H., Sasaki K., 1981. Chemical composition of milk of marine mammals (Pin-
nipedia). — Rept submitted on the 24th meeting Northern Pacific Fur Seal Commission.
Tokyo. 4 p.
386
Kenneth G., 1981. Locomotor behavior of the Walruses (Odobenus). — J. Zool., 195,3. P. 349—367.
Kenyon K. W., 1952. Diving depths of the Steller sea lions and Alaska fur seal. — J. Mammal.,
33. P. 345—346.
Kenyon K. W., 1960. Territorial behavior and homing in the Alaska fur seal. — Mammalia,
24, 3. P. 431—444.
Kenyon K., Scheffer V., 1955. The seals, sea-lions and otter of the Pacific coast. — U.S. Inter.
Fish and Wildl. Serv. 32 p.
Kenyon K. W., Scheffer V. B., Chapman D. G., 1954. A population study of the Alaska fur
seal herd. — U.S. Dept, inter. Fish and Wildl. Serv., Spec. Sci. Rept Wildl., 12. 77 p.
Kenyon K. W., Wilke F., 1953. Migration of the northern fur seal, Callorhinus ursinus. —
J. Mammal., 34. P. 86—98
Keyes M. C., 1966. Immobilizing and anesthetizing drugs used fur seals. In: Conf. Biol. Sonar
and Diving Mammals. Stanford Res. Inst., California. P. 74—79.
King J. E., 1956. The monk seal (Genus Monachus). — Bull. Brit. Mus. (Nat. Hist.), 13, 3.
P. 203—256.
King J. E., 1964. Seals of the World. Brit. Muz. (Nat. Hist.). London. 136 p.
King J., 1971. The lacrimal bone in the Otariidae. — Mammalia, 35, 3. P. 465—470.
King J., 1977. Comparative anatomy of the major blood wessels of the sea lions Neophoca
and Phocarctos with comments on the differences between the otariid and phocid vas-
cular systems. — J. Zool., 181. P. 69—94.
King J., Harrison R. J., 1961. Some notes on the Hawaiian monk seal. — Pacific Sci., 15.
P. 282—293.
Kooyman G. L., 1973. Respiratory adaptation in marine mammals. — Amer. J. Zool., 13,
2. P. 457^68.
Kooyman G. L., Campbell W. B., 1972. Heart rates in freely diving Weddell seals, Letonychotes
weddelli. — Compar. Biochem. Physiol., 43, 1A. P. 31—36.
Kooyman G. L., Drobek С. M., 1968. Observations on milk, blood and urine consisthens
on the Weddell seal. — Physiol. Zool., 41, 2. P. 187—194.
Kooyman G. L., Gentry R. L., Urquhart D. L., 1976. Northern fur seal diving behavior: a
new approach to its study. — Science, 193. P. 411—412.
Kooyman G. L., Kerem D. H., Campbell W. B., Wright L. L., 1973. Pulmonary gas exchange
in freedly diving Weddell seal, Leptonychotes weddelli. — Res. Physiol., 17, 3, P. 283—
290.
Kubota K., 1968. Comparative anatomical and neurogistological observation on the tongue
of the Northern fur seal (Callorhinus ursinus). — Anat. Rec., 161, 2. P. 257—265.
Kubota K., Horiuchi H., 1963. Anatomical studies on the salivary gland of fur seal. — J.
Mammal., 44, 3. P. 349—364.
Kubota K., Komuro H., 1961a. The eruption and shedding of teeth in the fur seal. — Okajimas
fol. anat., Japan, 37. P. 331—337.
Kubota K., Komuro H., 1961Z>. The calcification of teeth of fur seals (raentgenographical exa-
mination). — Ibid. P. 389—419.
Kubota K., Togawa S., 1964. Numerical variation in the dentition of some pinnipeds. — Anat.
Rec., 150, 4. P. 487—501.
387
Lander R. H., 1975. Method of determinig natural mortality in the Northern fur seal (Cal-
lorhinus ursinus) from known pups and kill by age and sex. — J. Fish. Res. Board
Canada, 31. P. 2447—2452.
Lander R. H., 1979. Role of land and ocean mortality in yield of male Alaskan fur seal, Cal-
lorhinus ursinus. — Fish. Bull., 77, 1. P. 311—314.
Lander R. H., 1980. A life table and biomass estimate for Alaskan fur seals. — Submitted
to the 23th annual meeting of the standing scientific committe, North Pacific fur seal
Commission on 7—11 April, 1980. P. 44—57.
Laws R. M., 1953. A new method of age determination in mammals with special reference
to the elephant seal (Mirounga leonind). — Folkland Islands Dep. Sur. Sci. P. 2.
Laws R. M., 1959. Accelerated growth in seals with special reference to the Phocidae. — Norsk
Hvalf. Tid., 9. p. 425-452.
Lenfant C., Johnson K., Torance J., 1970. Gas transport and oxygen sportage capacity in
some pinnipeds and sea otter. — Resp. Physiol., Holland. P. 277—286.
Ling J. K., 1970. Pelage and molting in wild mammals with special reference to aquatic
forms. — Dept. Biol. Memor. Univ. Newfoundland, Canada. P. 16—54.
Lipps J. H., Mitchell E., 1976. Trophic model for the adaptive radiations and extinction of
pelagic marine mammals. — Paleontology, 2, 2. P. 177-199.
Low Loh Lee, 1976. Status of major demerssal fishery resources of the Northwestern Pacific;
Bering See and Aleutian Islands. — NWAFC Processed Rept. P. 116.
Lucas F. A., 1899. The fur seal and fur-seal islands of the North Pacific Ocean. — U.S. Treasury
Dept., Doc. 2017, pt. 3. P. 75—98, 9—11, 45—54, 69—74, 59—68, 4—7.
Manuwell D. A., Cambel R. W., 1979. Status and distributions of breeding seabirds of so-
utheastern Alaska, British Columbia and Washington. In: Conservation of marine bird
of Northern North America. U. S. Dept. Intern. Fish Wildl. Serv., Rept. 2. P. 73—91.
Mensfild A. W., 1958. The biology of the Atlantic walrus, Odobenus rosmarus in the eastern
Canadian Arctic. — Fish. Res. Board Canada, 653. 146 p.
Mitchell E. D., 1975. Parallelism and convergence in the evolution of Otariidae and Phoci-
dae. — Rapp, et proc-verb, reun Cons. Int. Explor. Mer., 169. P. 74—75.
Mitchel E., Tedford N., 1973. The Enaliarctinae a new group of extinct aquatic Carnivora
and a consideration of the origin of the Otariidae. — Bull. Amer. Mus. Nat. Hist.,
151, 3. 284 p.
Morejohn V. G., 1972. A phylogeny of Otariid seals based on morphology of the baculum.
In: Sympos. on the Biology of the Seal. Univer. of Guelph. P. 1—2.
Murie J., 1874. Researches upon the anatomy of the Pinnipedia. Descriptive anatomy of the
sea lion, Otaria jubata. — Trans. Zool. Soc., London, 8. P. 501—582.
Nagasaki F., 1961. Population study on the fur seal herd. — Tokai Reg. Fish. Res. Lab.,
spec, publ., 7. 60 p.
Nakajima M., Sawaura K., Oda T., 1963. On the food quantaty of the Northen fur seal. —
Contrib. Enoshima Mariland, 6, 1. P. 4—10.
Niggol K., Fiscus C., 1960. Northern fur seal twins. — Mammalia, 24, 3. P. 457—458.
Nishiwaki M., 1972. General Biology. — In: Mammals of the sea. Capt. 1. Biology and Me-
dicine. C.C. Tomas, Springfild, Illionois. P. 3—201.
Ogi H., Tsujita T., 1977. Food and feeding habits of Coomon murre and thic-billed murre
in the Okhotsk Sea in summer. — Res. Inst. North. Рас. Fish Hokkaido Univ. Spec.,
8. P. 457—517.
388
Ohata C. A., Miller L„ 1977. K. Some temperature responses of Northern fur seal (Callorhinus
u'rsinus) pups. — J. Mammal., 58, 3. P. 438—440.
Okumoto Naoto, 1980. Heredity, 34, 2. P. 58—65. (На яп. языке)
Packer В. S., Altman M., Cross С. E., McRrgangh H. V., Linta J. M., Robin E. D., 1969.
Adaptation to diving in the harbor seal: exygen stores and supply. — Amer. J. Physiol.,
217, 3. P. 903—906.
Peterson R.S. Benavior of the Northern fur seal. In: Thesis J. H. Univ. Baltimore, Maryland.
P. 214.
Peterson R. S., Leboeuf B. J., Delong R. L., 1969. Fur seal in California. — Pacific Disc.,
22, 3. P. 12—15.
Peterson R. S., Reeder W. G., 1966. Multiple births in the Northern fur seal. — Z. S3ugetierk.,
31. S. 52—56.
Rand R. W., 1955. Reproduction in the female capa fur seal, Arctocephalus pusillus (Schre-
ber). — Proc. Zool. Soc., London, 124, 4. P. 717—740.
Rand R. W., 1956. Capa fur seal. — Dept. Commer. Industry. Invest. Rept, Capetoun.
21 p.
Ray C., 1963. Locomotion in pinnipeds. — Nat. Hist, 72, 3. P. 10—21.
Repenning C. A., Peterson R. S., Hubbs C. L., 1977. Contribution to the systematic of the
Southern fur seal, with particular referenca to the Juan Fernandes and Guadalup species.
In: Antarctic Pinnipedia. Wash. D. С.. P. 1—34.
Repenning C. A., Tedford R. H., 1977. Otarioid seals of Neogene. 93 p.
Report of Japanese fur seal research, 1963. — Fish. Agency. 43 p.
Report of Japanise fur seal research, 1964. — Fish. Agency. 52 p.
Rice D. W., 1968. Stomach contents and feeding behavior of Killer whales in the eastern North
Pacific. — Norsk Hvalf. Tid., 57. P. 35—38.
Ricker В. E., 1972. Notes to study fur seal. Moscow, North Pacific fur seal Commission. 4 p.
Ronald K., Foster M. E., Johnson E., 1969. The Harp seal, Pagophilus groenlandicus (Erxleben,
1777). 2. Physical blood properties. — Can. J. Zool., 47, 3. P. 461—468.
Roppel A., Davey S., 1965. Evolution of fur seal management on the Pribilof Islands. — J.
Wildl. Manag., 29, 3. P. 448^163.
Scammon S. M., 1874. Marine mammals. San-Francisco. P. 141—163.
Scheffer V. B., 1949. The clitoris bone two pinnipeds Phoca and Callorhinus ursinus. — J. Mammal.,
30, 3. P. 269—270.
Scheffer V. B., 1950a. Growth layers on the teeth of Pinnipedia as an indication of age. —
Science, 112. P. 309—311.
Scheffer V. B., 1950Z». Winter injury to young fur seals on Northwest coast. — Calif. Fish.
Game, 36. P. 378—379.
Scheffer V. B., 1950c. Growth of testes and baculum in the fur seal, Callorhinus ursinus. —
J. Mammal., 31, 4. P. 384—394.
Scheffer V. B., 1955. Body size with relation to population density in mammals. — Ibid., 37.
P. 493—515.
Scheffer V. B., 1958. Seals, sea lions and walruses. Standford Univ. Press. 177 p.
389
Scheffer V. В., 1960a. Early embryons the Northern fur seal. — Mammalia, 24, 3. P. 482—483.
Scheffer V. B., 19606. Weights of organs and glands in the Northern fur seal. — Ibid., 24,
3. P. 476-481.
Scheffer V. B., 1962. Pelage and surface topography of Northern fur seal. — North Amer.
Fauna, 64. 206 p.
Scheffer V. B., 1964. Hair pettems in seals (Pinnipedia). — J. Morphol., 115. P. 291—303.
Scheffer V. B., 1981. Nowbom size in marine mammals. — Pacific Disc., 34. P. 19—26.
Scheffer V. B., Johonson A. M., 1963. Molt in the Northern fur seal. — U.S. Fish Wildl.
Serv. Sp. Sci. Rep. Fisher., 450. 34 p.
Scheffer V. B., Kenyon K. W., 1963. Baculum size in pinnipeds. — Z. SSugetierk., 28, 1. P. 38—41.
Scheffer V. B., Kraus B. S., 1964. Dentition of the Northern fur seal. — Fish. Bull., 63, 2.
P. 293—342.
Scheffer V. B., Peterson R. S., 1967. Growth layers in teeth of suckling fur seal. — Growth,
31. P. 35—38.
Scheffer V. B., Wilke F., 1953. Relative growth in the Northern fur seal. — Ibid., 17, 3. P. 129—145.
Scholander P. F., 1940. Expiremental investigation on the respiratory function in diving mam-
mals and birds. Hval. schrift. Norse. Vidensk. Akad., Oslo. 131 p.
Schwartzman G., Harr R., 1980. Exporing interactions between fur seal populations and fis-
heries in the Bering sea. — Mar Mem. Comm. Rep., MMC-80/01. P. 1—60.
Secora P. C., Byrd G. V., Gibson D. D., 1979. Breeding distribution and status of marine
birds in the Aleutian Islands, Alaska. In: Conservation of Marine bird of North America.
U.S. Dept. Fish and Wildl. Serv. P. 33-49.
Slijper E. J., 1968. On the heart of temporary aquatic mammals with special reference to the
embryonic pathways. — Bijdr. Dierk., 38. P. 75—84.
Smith T., Polacheck T., 1981. Re-examination of the life table for Northern fur seals and im-
plication about population regulatory mechanisms Dynamics of mammal populations.
In: Dynamic of Large Mammals Population. New York, John Wiley and Sons. P. 99—
120.
Sowls A. L., Hatch S. A., Lensing C., 1978. Catalog of Alaskan seabird colonies. — Fish,
and Wildlife Serv. U.S. Dep. Int. P. 1—161.
Spalding D. J., 1964. Comparative feeding habits of the fur seal, sea lion and harbour seal
on the British Columbia coast. — Bull. Fish. Res. Board Canada, 146. 52 p.
Steller G., 1751. De Bestiis marinis. — Novi Comm. Acad. Sci. Petropolitanae, 2. P. 289—398.
Stejneger L., 1898. The asiatic fur seal islands and fur seal industry. — In: The seal and fur
seal islands of North Pacific Ocean. Pt. 4. U.S. Treasury Dept., Doc., 2017. 384 p.
Stejneger L. Georg Wilhelm Steller. The pioneer of Alaskan natural history. Harvard Univer.
Press. 623 p.
Stejneger L., 1925. Fur-seal indastry of the Commander islands, 1897 to 1922. — Bull. Burean
Fish., 41. P. 289—332.
Stevenson С. H., 1904. The skin of fur seals. Цит no: Scheffer, 1962.
Straty P., Haight R., 1979. Interactions among marine bird and commercial fish in the eastern
Bering Sea. — In: Conservation of marine bird of Northern North America. U.S. Dep.
Inter. Fish and Wildl. Serv., Rept 2, Wash. D.C. P. 207—219.
390
Strauss M. В., 1969. Mammalian adaptations to diving. U. S. Naval Submarine Medical Center.
31 p.
Taylor F. H., Fujinaga M., Wilke F., 1955. Distribution and food habits of the seals of the
North Pacific Ocean. — U.S. Dep. Int. Fish and Wildl. Serv. 86 p.
Townsend С. H., 1898. The fur seals and fur seal islands of North Pacific ocean. — U. S.
Treasury Dept. Doc., 2017, pt. 3. P. 223—274.
Townsend С. H., 1910. Fur seals and the seal fisheries. — Bull. Fish., Wash., 28. P. 315—322.
Townsend С. H„ 1911. Pribilof fur seal herd and the prospect for its increase. — Science,
34. P. 568—570.
Trites A. W., 1989. Estimating the juvenile survival rate of male Northern fur seal (Callorhinus
ursinus). — Can. J. Fish. Aquat. Sci., 46. P. 1428—1436.
Trites A. W., 1991. Fetal growth of Northern fur seals: life-history strategy and sourse of va-
riation. — Can. J. Zool., 69. P. 2608—2617.
Trites A. W., 1992. Northern fur seals: why have the declined. — Aquatic Mammals, 18, 1.
P. 3—18.
Trites A. W., Bigg M. A., 1992. Changs in body growth of Northern fur seals from 1958
to 1974: density effects or changes in the ecosystem. — Fish oceanography, 1, 2.
P. 127—136.
Trites A. W., York A. E., 1993. Unexpected changes in the reproductive rate and mean age at
furst birth during the decline of the Pribilof northern fur seal (Callorhinus ursinus). — Can.
J. Fish. Aquatic Sci., 50, 4. P. 858—864.
Tucker R., 1958. Taxonomy of the salivary glands of vertebrates. — Syst. Zool., 7, 2. P. 74—83.
Turner W., 1888. Viscera of elephant seal. — Rep. Sci. Res. Voy. H. M. S. Challenger, Zool.,
26. P. 135—138.
Vrobell К. H., 1963. Untersuchungen Uber die Blutversorgung der Nieren des Zeelowen (Za-
lophus californiensis). — Zeitsch. Anat. Entwicklungsgesch., 123, 6. S. 577—588.
Wada K., 1969. Migration of Northern fur seal along the coast of Sanriku. — Bull. Tokai
Reg. Fish. Res. Lab., 58. P. 19—82.
Wada K.. 1971. Some comments on the migration of Northern fur seal. — Ibid., 67. P. 47—80.
Welker W. J., Campos G. B., 1963. Physiological significance of sulci in somatic sensory cerebral
cortex in mammals of the famili Procionidae. — J. Comp. Neurol., 120. P. 19—36.
Wilber C. G., 1952. Fur seal blubber. — J. Mammal., 33. P. 483—485.
Wilke F., 1951. Pelagic fur-seal research off Japan in 1950. — U.S. Fish and Wildl. Serv.
Lefl. 35 p.
Wilke F., 1959. Fat content of fur-seal milk. — Murrelet, 64. P. 40.
Wilke F., Kenyon K., 1952. Notes on the food of the seal, sea-lions and harbor por-
poise. — J. Wildl. Manag., 16. P. 396—397.
Wilke F., Nicholson A., 1958. Food of porpoises in water off Japan. — J. Mammal., 39, 3.
p. 441—443.
York A. E., Hartley J. R., 1981. Pup production following harvest of female Northern
fur seals. — Can. J. Fish Aquatic Sci., 38, 1. P. 84—90.
Yoshida K., 1982. Study for reproductive mechanism of fur seal (Callorhinus ursinus) of Asian
origin. — Bull. Far Seas Fish. Res. Lab., 424. P. 1—35.
Yoshida K., Baba N., Oya M., Mizue K., 1977. On the formation and regression of corpus
luteum in the Nortnem fur seal ovaries. — Sci. Rept. Whales Res. Inst., 29. P. 121—128.
391
Yoshida К., Baba N., Oya M., Mizue K., 1978. Seasonal changes in the ovary of the Nortnem
fur seal. — Ibid., 16. P. 93—101.
Yoshida K., Ichihara T., 1974. Some comments on reproductive condition in female fur seal. —
Bull. Far Seas Fish. Res. Lab., 11. P. 9—13.
Yoshida K., Okada K., Baba N., 1978. Interum report on the visual investigation. In: The
2lst meeting of North Pacific fur seal Commission. Canada. 8 p.
Yoshida K., Okumoto N., Baba N., 1978. Japanise pelagic investigation on fur seal, 1977.
38 p.
Оглавление
Предисловие.............................. ..................... 3
Глава 1. Систематика и внутривидовая структура...................... 4
1,1. Систематика................................................ 4
1.2. Внутривидовая структура .................................. 5
Глава 2. Внешний вид, размеры, окраска, пропорции и гидродинамическая
характеристика тела ........................................... 9
2.1. Внешний вид и размеры...................................... 9
2.2. Окраска................................................... 11
2.3. Пропорции тела............................................ 15
2.4. Гидродинамическая характеристика.......................... 20
Глава 3. Определение возраста и рост............................... 24
3.1. Определение возраста...................................... 24
3.2. Рост...................................................... 25
3.3. Фаза ювенильного роста.................................... 31
3.4. Фаза полового созревания и стационарного состояния .... 31
Глава 4. Развитие и возрастно-половые особенности кожно-волосяного по-
крова ........................................................ 38
4.1. Линька................................................ 46
4.2. Вибриссы.................................................. 51
Глава 5. Развитие скелета и его строение........................... 54
5.1. Череп .................................................... 54
5.2. Позвоночный столб . ...................................... 60
5.3. Периферический скелет.................................... 76
Глава 6. Роль мышечной системы в передвижении ............ 102
Глава 7. Центральная нервная система и органы чувств.............. 106
7.1. Форма, размеры и внешнее строение головного мозга .... 106
7.2. Развитие головного мозга в онтогенезе.................... 112
7.3. Черепномозговые нервы.................................... 117
7.4. Мозжечок................................................. 118
7.5. Спинной мозг............................................. 119
7.6. Органы чувств............................................ 121
Глава 8. Пищеварительная система.................................. 126
8.1. Ротовая полость......................................... 126
8.2. Язык ............................................. 126
8.3. Слюнные железы ..................................... 127
8.4. Зубы..................................................... 131
8.5. Глотка................................................. 141
8.6. Пищевод ............................................. 142
8.7. Желудок.................................................. 142
393
8.8. Кишечник.................................................. 143
8.9. Печень.................................................... 145
8.10. Кровоснабжение печени.................................... 148
8.11. Поджелудочная железа..................................... 152
Глава 9. Кровеносная система....................................... 153
9.1. Сердце.................................................... 153
9.2. Артерии малого круга кровообращения....................... 156
9.3. Артерии большого круга кровообращения..................... 156
9.4. Венозная система.......................................... 163
9.5. Органы кровообразования................................... 166
Глава 10. Мочеполовая система...................................... 168
10.1. Органы мочеотделения................................... 168
10.2. Половые органы.......................................... 176
10.2.1. Мужская половая система................................ 176
10.2.2. Женская половая система ............................... 184
Глава 11. Вопросы физиологии....................................... 191
11.1. Общее количество крови................................... 191
11.2. Морфофизиологическая характеристика красной крови . . . 192
11.3. Морфофизиологическая характеристика белой крови.. 198
11.4. Особенности дыхания и сердечной деятельности ............ 199
11.5. Терморегуляция........................................... 202
Глава 12. Особенности образа жизни................................. 206
12.1. Подход к лежбищам и выход на берег....................... 206
12.2. Формирование гаремов и холостяковых залежек........ 210
12.3. Хоминг и филопатрия...................................... 213
12.4. Микромиграции ........................................... 215
12.5. Особенности постгаремного образа жизни................... 219
Глава 13. Поведение и активность................................... 225
13.1. Взаимоотношения различных половозрастных групп северных
морских котиков............................................. 225
13.2. Взаимоотношения северных морских котиков с другими живот-
ными...................................................... 231
13.3. Ритмы активности и некоторые аспекты поведения северных
морских котиков на береговых залежках и в море ..... 234
13.4. Скорость плавания и глубина ныряния..................... 240
Глава 14. Распространение и миграции . .......................... 242
14.1. Распространение.......................................... 242
14.2. Миграции................................................ 251
Глава 15. Роль условий среды обитания в распределении и выживаемости
северных морских котиков..................................... 263
15.1. Гидрологическая характеристика районов обитания котиков 263
; 15.2. Острова как среда обитания северных морских котиков . . 272
Глава 16. Враги и конкуренты.................................. 278
16.1. Враги........................................... 278
16.2. Пищевая конкуренция............................. 279
16.3. Конкуренция за места залегания.................. 281
394
Глава 17. Структура популяций.................................... 283
17.1 Половая структура...................................... 283
17.2. Возрастная структура................................... 289
17.3. Пространственная структура............................. 291
Глава 18. Размножение............................................ 298
18.1. Возраст наступления половой зрелости................... 298
18.2. Темп и ритм размножения................................ 303
18.3. Сроки щенки и спаривания............................... 308
18.4. Продолжительность беременности и развития плода...... 310
18.5. Количество детенышей................................... 312
18.6. Лактация............................................... 314
Глава 19. Численность и ее динамика ............................. 318
19.1. Методы получения исходных данных для определения числен-
ности котиков............................................... 318
19.2. Численность популяции котиков острова Тюлений........ 320
19.3. Численность котиков на Курильских островах............. 324
19.4. Численность котиков на Командорских островах........... 327
19.5. Причины снижения численности северных морских котиков 329
Глава 20. Промысел............................................... 338
20.1. История промысла северных морских котиков на Командорских
островах.................................................... 338
20.2. История промысла котиков на острове Тюлений............ 344
20.3. История промысла котиков на Курильских островах...... 347
20.4. История пелагического промысла котиков................. 349
20.5. Техника промысла котиков .............................. 353
20.6. Методы добычи котиков на лежбищах и первичная обработ-
ка тушек ................................................... 353
20.7. Добыча котиков в море ................................ 356
20.8. Продукция промысла.................................... 359
20.9. Использование продукции................................ 366
Литература .................................................... 368
395
Кузин Алексей Егорович
Северный морской котик
Утверждено к печати
Советом по морским млекопитающим
Подготовка оригинал-макета — А.А.Аристов
Подписано в в печать 21.04.99. Формат 70x100 1/16. Печать офсетная.
Гарнитура литер. Усл, печ. л. 24,75. Уч.-изд. л. 26. Тираж 200 экз.
Заказ № 10.
Совет по морским млекопитающим
Москва, 117851, Нахимовский пр., 36, комн. 234.
Типография ТИНРО —центра. Владивосток, 690600, ул. Западная, 10