/




































































































































































































































































































































































































Author: Энока Р.М.
Tags: медико-биологические основы физического воспитания биомеханика управление движениями нервно-мышечная адаптация принципы тренировки двигательный аппарат и концепция кинезиология
ISBN: 966-7133-08-7
Year: 1998




Text
Р.М.ЭНОКА
ОСНОВЫ КИНЕЗИОЛОГИИ
ББК 75.0
Э66
В книге ведущего зарубежного специалиста по кинезиологии Р.М.Эноки представлены современные знания в области управления движениями. Изложены основные сведения о структуре, функциях, биомеханике двигательного аппарата, концепциях управления движениями. Рассмотрены принципы тренировки и специфика нервно-мышечной адаптации, механизмы утомления, изменении гибкости, нарушений прочности кости, стадии повреждения мышц тренировкой. Предложены тесты для оценки способностей двигательных единиц сопротивляться утомлению, методики электрической и магнитной стимуляции для изучения мышечной структуры и функции, увеличения мышечной массы и силы, ускорения лечения переломов, отсрочки развития остеопороза. Рассмотрены эффекты разминки и факторы, способствующие увеличению мышечной работы за счет использования упругой энергии мышц.
Для студентов вузов физического воспитания и спорта, физиологов, специалистов по физической реабилитации, спортивной медицине, тренеров.
Научные редакторы
Борис Кифоренко, Александр Приймаков
Перевод с английского
Георгий Гончаренко
ISBN 966-7133-08-7
© Human Kinetics, 1997
© Издательство “Олимпийская литература”, 1998
ОГЛАВЛЕНИЕ
Предисловие к русскому изданию ...........5
Предисловие к английскому изданию.........8
Введение .................................9
ЧАСТЬ I
ВЗАИМОСВЯЗЬ СИЛА-ДВИЖЕНИЕ 11
Цели ....................................12
Глава 1. Движение 13
Положение, скорость и ускорение .........13
Уравнение движения ......................15
Численный анализ.........................17
Графическое представление ...............18
Поступательное и вращательное движения .... 20
Ускорение и мышечная активность .........22
Кинематика походки ......................24
Кинематика бросания и толкания ..........29
Резюме...................................36
Вопросы с альтернативным ответом.........36
Вопросы с различным выбором ответов......38
Задачи ..................................40
Г л а в а 2. Сила 43
Законы движения Ньютона .................45
Диаграмма свободного тела ...............47
Силы движения человека...................50
Вращающий момент.........................74
Резюме...................................79
Вопросы с альтернативным ответом.........79
Вопросы с различным выбором ответов......81
Задачи ..................................84
Г л а в а 3. Анализ движения 88
Подход сила—масса—ускорение..............88
Подход импульс—количество движения.......99
Подход работа—энергия...................108
Резюме................................. 115
Вопросы с альтернативным ответом........116
Вопросы с различным выбором ответов....118
Задачи ............................... 119
ЧАСТЬ II
ЕДИНАЯ СИСТЕМА СУСТАВА 123
Цели ..................................124
Г л а в а 4. Компоненты единой системы сустава 125
Жесткое звено .........................125
Синовиальный сустав....................130
Мышца .................................132
Нейрон ................................137
Чувствительное нервное окончание ......140
Резюме................................ 145
Вопросы с альтернативным ответом.......145
Вопросы с различным выбором ответов....147
Задачи ............................... 149
Г л а в а 5. Функционирование единой системы сустава 151
Двигательная единица ..................151
Возбудимые мембраны....................159
Взаимодействие возбуждения—сокращения . . 170
Обратная связь с чувствительными нервными окончаниями...................174
Резюме................................ 183
Вопросы с альтернативным ответом.......183
Вопросы с различным выбором ответов....185
Задачи ............................... 187
Г л а в а 6. Активация единой системы сустава 189
Нервные факторы .......................189
Мышечные факторы ......................195
Резюме.................................214
Вопросы с альтернативным ответом.......214
Вопросы с различным выбором ответов....215
Задачи ................................217
3
ЧАСТЬ III
ПРИСПОСОБЛЯЕМОСТЬ ДВИГАТЕЛЬНОЙ СИСТЕМЫ 219
Цели ................................. 220
Г л а в а 7. Двигательная система 221
Супрасегментная организация .......... 221
Мышечно-скелетная система ............ 231
Стратегии движений.................... 239
Резюме................................ 253
Вопросы с альтернативным ответом...... 253
Вопросы с различным выбором ответов ..... 256
Задачи ............................... 258
Г л а в а 8. Кратковременные адаптации 260
Эффекты разминки ..................... 260
Гибкость ............................. 264
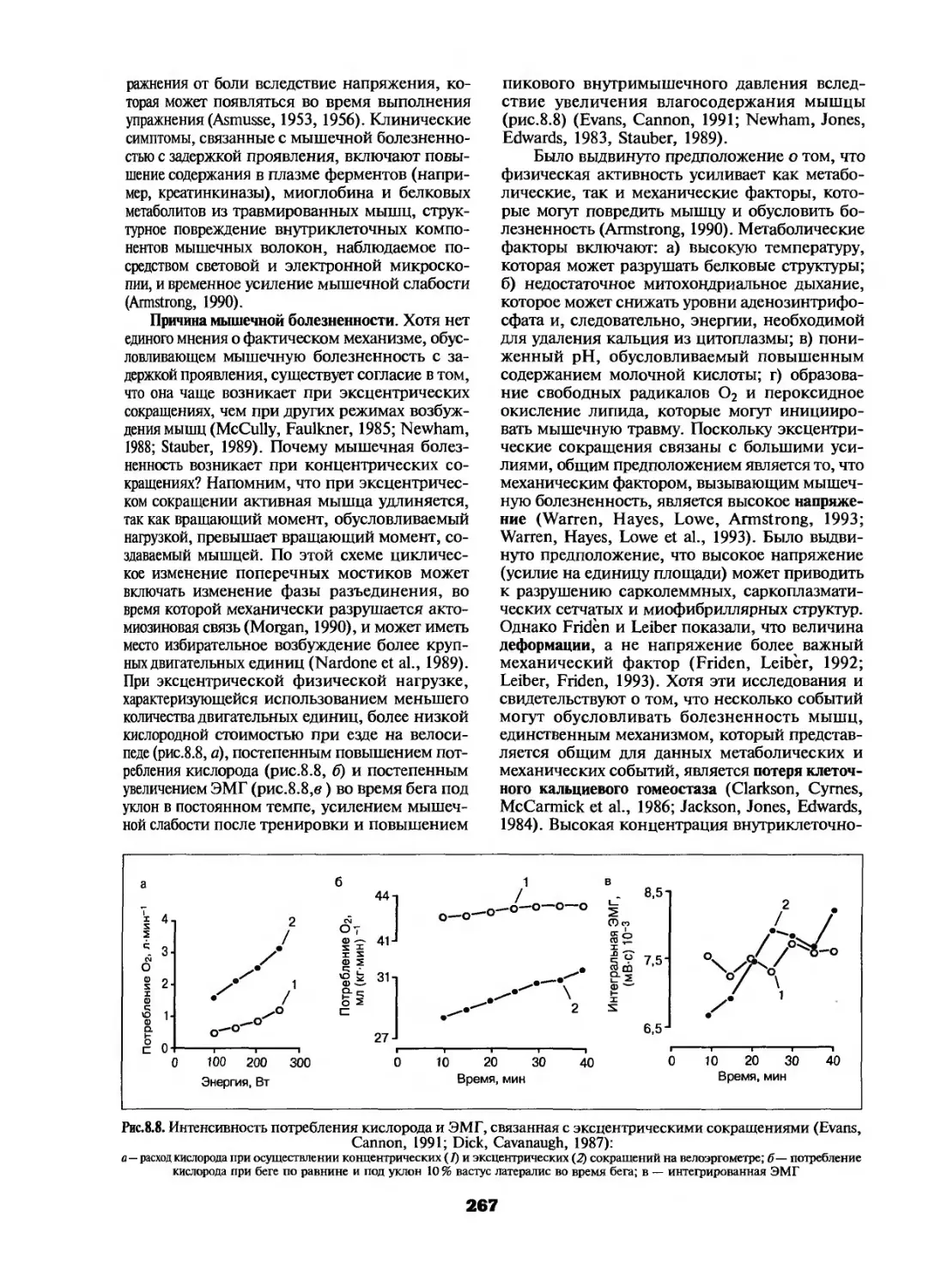
Болезненность и повреждение мышц ....... 266
Мышечное утомление ................... 270
Потенцирование мышц .................. 279
Резюме................................ 283
Вопросы с альтернативным ответом...... 283
Вопросы с различным выбором ответов ..... 286
Задачи ............................... 288
Г л а в а 9. Долговременные адаптационные реакции
290
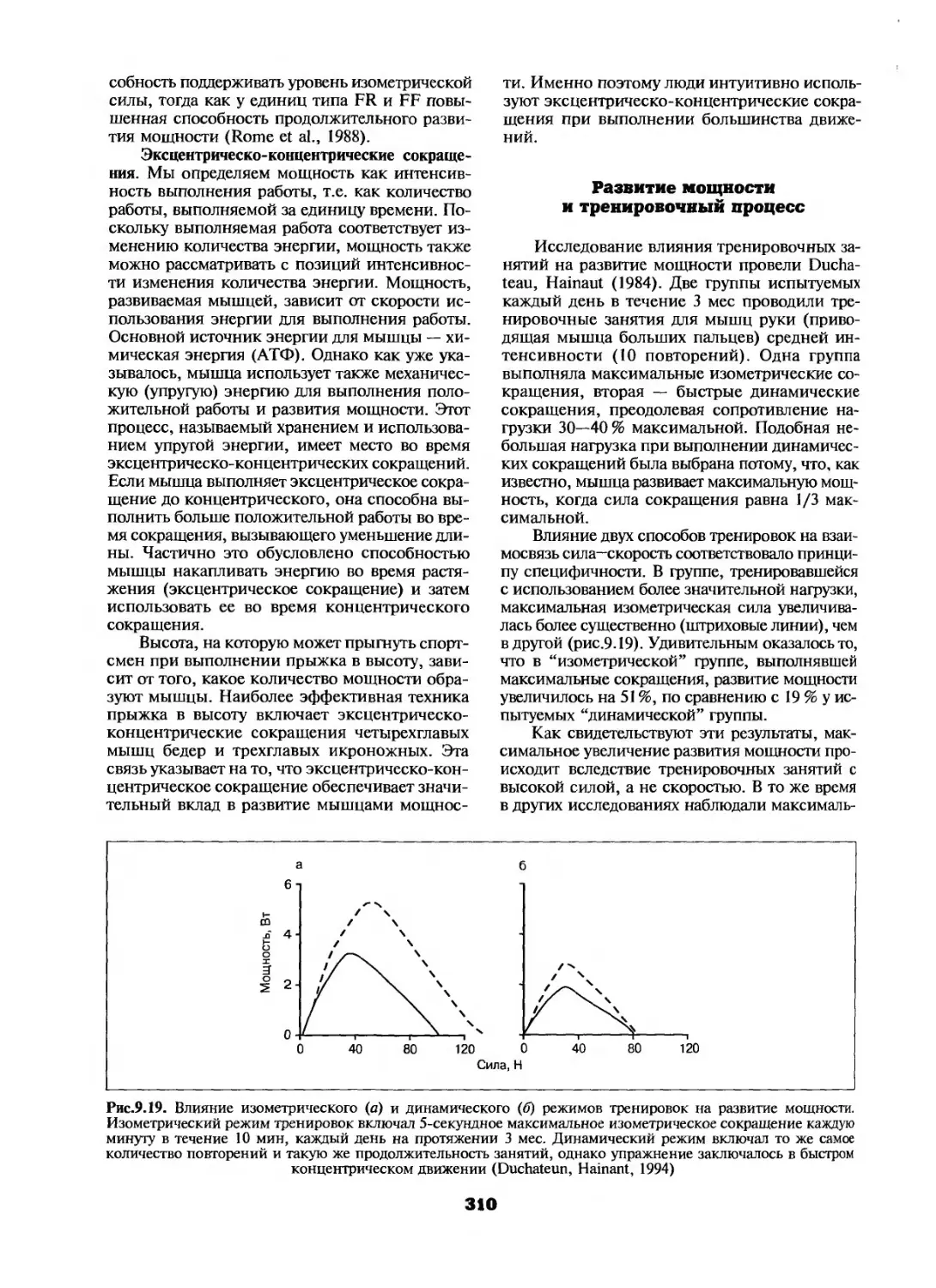
Принципы тренировки..............290
Сила ............................291
Мышечная мощность................307
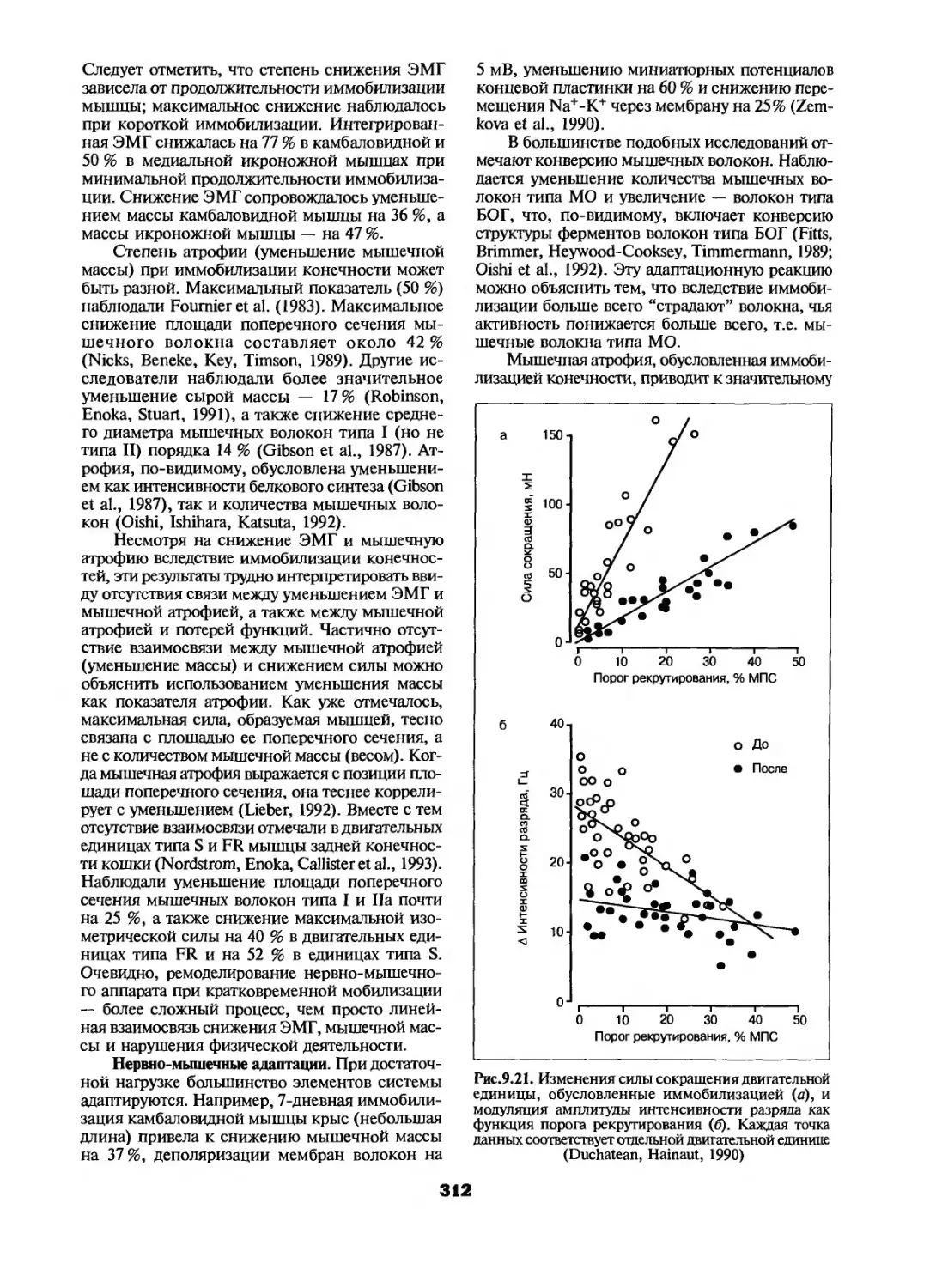
Адаптационные реакции на пониженный уровень физической активности....311
Восстановление двигательных функций после травмы.....................316
Адаптации с возрастом............319
Вопросы с альтернативным ответом.224
Вопросы с различным выбором ответов.326
Задачи .............................328
Приложение!. Система единиц СИ . . 331
Приложение 2. Векторы ..............334
Приложение 3. .Коэффициенты преобразования...................342
Приложением Параметры сегментов тела...................347
Приложение 5. Кинематические данные...........................348
Приложение 6. Сокращения............353
Словарь..........................356
Литература.......................371
Предисловие к русскому изданию
В предлагаемой вниманию читателей книге изложены основы кинезиологии — науки о движении человека и животных, основанной на понимании целенаправленного движения человека как результата сложнейшего взаимодействия элементов многозвенного мобильного объекта, каким является его тело, с окружающей средой, управляемого необычайно гибкой системой, способной в реальном масштабе времени собирать информацию о текущем состоянии всего объекта и его активных элементов, оценивать ее и формировать управляющие сигналы, исполняемые с точностью и быстродействием, необходимыми для получения совершенного конечного результата — перемещения в пространстве, столь привычно и просто называемого движением человека.
Книга адресована широкому кругу читателей. Это и студенты, впервые знакомящиеся с наукой о движении с надеждой глубже понять происходящие в человеческом организме процессы и использовать выводы и рекомендации кинезиологии в тренировочном процессе, и преподаватели, имеющие возможность обогатить свои лекции и практические занятия иллюстративным материалом и интересными задачами, и научные работники, которые найдут в книге много ценной экспериментальной информации и математических моделей, обобщающих данные зарубежных научных публикаций.
Хотя книга Р.М. Эноки написана как программированное учебное пособие, по объему и целенаправленности материала она может с полным правом быть отнесена к монографической литературе. Автор последовательно стремится придерживаться продекларированного во введении стиля изложения на базе четких основных понятий и формулировок проблем, концепций и выводов. В тех местах, где ему иногда это не удается, при редактировании внесены необходимые поправки, устраняющие стилистические, а иногда и фактические погрешности, вполне извиняемые в изложении основ столь комплексной и молодой науки, каковой является современная кинезиология, еще находящаяся в состоянии накопления и осмысления фактов и лишь дерзко замахивающаяся на основную задачу всякой естественной науки — построение теории, основанной на системе понятий постулатов и стремящейся объяснить и предсказать путем логических построений и с помощью математических методов. Одной из немногочисленных и, как нам представляется, во многом удачных попыток способствовать достижению этой цели является предлагаемая вниманию читателей монография.
5
К выигрышным особенностям книги следует отнести широкий круг вопросов, затрагиваемых автором: анализ различных концепций управления движениями, структура, функции, биомеханика двигательного аппарата, вопросы утомления, адаптивных перестроек, повреждений, диагностики и восстановления. Основной теоретический материал богато иллюстрирован удачно подобранными примерами и рисунками, что делает чтение книги весьма увлекательным занятием. Комплексный всесторонний подход к анализу движения создает у читателя целостное представление о механизмах функционирования сложнейшей интегративной структуры — “аппарата производства движения”.
Можно с уверенностью отметить, что это одна из лучших за последние годы монографий, в которой всесторонне проанализированы структура, свойства и функции двигательного аппарата. К сожалению, автор уделил мало внимания концепциям управления движениями, предложенным Н.А. Бернштейном, Р. Гранитом, В.С. Гурфинкелем, теории функциональных систем П.К. Анохина и других исследователей, многие положения которых при объяснении механизмов управления движениями остаются в настоящее время актуальными.
В I части изложены основные понятия теоретической механики и подходы в использовании ее законов и теорем при изучении движения тела человека и его элементов — перемещение в пространстве по отношению к окружающим предметам. Автор четко подчеркивает основную концепцию механики — причинно-следственную связь между силой и движением — и показывает, как на основе законов механики по результатам наблюдений движения можно определить силы, необходимые для его выполнения. Столь же подробно изложены два других подхода классической механики к изучению движения — импульсный и энергетический. Материал первых трех глав дает богатую информацию о возможностях опорно-двигательного аппарата человека и естественным образом подводит читателя к проблематике II и III частей книги, отвечающих на основной для кинезиологии вопрос, — как генерируются необходимые мышечные усилия и каким образом многообразие мышц, суставов и звеньев объединяется в совершенный аппарат управления и реализации движений.
Во II и III частях даны биомеханические, морфологические и функциональные характеристики двигательного аппарата, рассмотрена роль различных мозговых структур в управлении движениями и развитии силы, генезе утомления, освещены гипотезы управления движениями и основные стратегии, используемые нервной системой в управлении положением тела и движением, механизм быстрой передачи информации по системе, принципы тренировки и специфика нервно-мышечных адаптаций при силовой тренировке, предложены тесты для оценки способности двигательных единиц сопротивляться утомлению. Все это, несомненно, полезно для студентов, специалистов по кинезиологии, физиологии, спортивной тренировке.
Большой интерес для педагогов, специалистов в области спортивной тренировки и спортивной медицины может представить материал, в котором освещены гормональные, метаболические и механические факторы, стимулирующие мышечную гипертрофию и обусловливающие силовые возможности спортсменов, а также описаны различные методы (электростимуляция двигательных единиц, идеомоторные акты и др.) для развития силы.
Рассмотренные автором эффекты разминки, методики и механизмы, лежащие в основе изменений гибкости, факторы, способствующие накоплению дополнительной упругой энергии мышц и увеличению мышечной работы, причины, ограничивающие диапазон движения сустава, представляют как теоретический, так и практический интерес для физиологов, тренеров, медиков. К сожалению, анализируя эффект разминки, заключающийся в повышении внутренней температуры и разрушении временных связей соединительных тканей, улучшении биомеханической характеристики двигательной системы, снижении возможности деформирования мышц,
6
автор не рассматривает влияние разминки на функции центральных, управляющих, структур — силу, подвижность нервных процессов, способность к усвоению новых связей и др.
Книга представляет несомненный интерес также для специалистов по физической реабилитации, спортивной медицине в связи с тем, что рассматривая компоненты единой системы сустава, автор раскрывает механизмы нарушения прочности кости, характеризует стадии повреждения мышц тренировкой, развитие в них ригидности и болезненного ощущения, обосновывает эффективность использования электрической и магнитной стимуляции для ускорения лечения переломов, отсрочки развития остеопороза, описывает метод магнитного резонанса, открывающий большие перспективы для изучения мышечной структуры и функции.
Украинский государственный университет физического воспитания и спорта, Национальный олимпийский комитет Украины и издательство “Олимпийская литература” изыскали возможность для издания книги, отражающей современные знания в области кинезиологии, имеющей большое теоретическое и практическое значение для студентов физкультурных вузов, физиологов, специалистов по физической реабилитации, спортивной медицине, тренеров.
А.А.Приймаков, Б.Н.Кифоренко
Предисловие к английскому изданию
Обучаясь (1974 — 1981) на факультете кинезиологии Вашингтонского университета, я оказался под сильным влиянием перспективных исследований С. Хаттона, касающихся управления опорно-двигательной системой, и Д.И. Миллер по биомеханике. Одним из их новшеств была разработка для выпускников курса по кинеоэнергетике, в котором рассматривалось управление двигательной системой в том виде, в каком она развилась из нейрофизиологии, с позиций биомеханики. Хотя это был строгий курс, требовавший полного внимания от студентов, его концентрированность на движении человека вызывала их заметный энтузиазм. Одной из трудностей, с которой мы столкнулись, был подбор соответствующего учебника. Мы использовали “Понимание научных основ движения человека”А.Л. О’Коннел и Э.Б. Гарднер, но этот текст в значительной мере дополнялся материалами из исследовательской литературы и результатами, полученными д-ром Хаттоном и д-ром Миллером.
В августе 1981 г. я начал работать помощником профессора на факультете физического образования в университете Аризоны. Одной из моих обязанностей была подготовка курса для выпускников по кинезиологии. В этой работе я в значительной мере опирался на свой опыт в Вашингтонском университете, но снова столкнулся с трудностью в выборе соответствующего учебника. Я предоставил студентам несколько учебников и использовал как основной источник свои записи лекций. Каждый год объем записей лекций увеличивался. В 1988 г. издательство Хюман Кинетике Паблишерз опубликовало этот материал как первое издание “Нейромеханических основ кинезиологии”.
С 1988 по 1993 г. я использовал его как учебник в своем курсе кинезиологии для выпускников. В течение этого периода я получил много отзывов от студентов университета Аризоны, а также коллег в Северной Америке и за рубежом относительно сильных и слабых мест книги. Поскольку первое издание было написано, в основном, для удовлетворения моих собственных потребностей в учебном пособии, ряд аспектов книги могли быть улучшены. При публикации второго издания я попытался учесть эти замечания, сосредоточив основное внимание на пяти целях: а) расширение рассматриваемого в книге круга проблем, чтобы она могла удовлетворить потребности более широкой аудитории, включая лиц, занятых в реабилитации, лиц, занимающихся исследованием биомеханики, и лиц, изучающих влияние тренировки на нервно-мышечную систему; б) обновление материала с тем, чтобы он отражал текущие концепции и мнения; в) расширение перечня ссылок, чтобы заинтересованные читатели могли более подробно проследить отдельные вопросы; г) увеличение числа и типа вопросов в конце каждой главы; д) расширение иллюстративного содержания путем увеличения числа рисунков, включенных в каждую главу. Я надеюсь, что вы найдете эти особенности полезными, а само второе издание “Основ кинезиологии” ценным пособием.
Роджер М. Энока
Введение
Термин “кинезиология” образован из двух греческих слов: kinesis — движение и logos — слово, учение. В соответствии с этим определением кинезиология означает беседы о движении или, в современном понимании, изучение движения. Основателем кинезиологии был гре-~> ческий философ Аристотель (384 — 322 до н.э.), впервые описавший действие мышц и осуществивший их геометрический анализ. Хотя система, разработанная Аристотелем для объяснения движения, содержала ряд противоречий, его пионерские усилия заложили основу последующих работ Галена (131 — 201), Галилея (1564 — 1643), Ньютона (1642 — 1727) hj-Борелли (1608 — 1679). Работы этих философов и ученых привели к тому, что мы стали рассматривать движение человека как последовательность взаимодействий мышц и внешних сил, воздействующих на систему из окружающей среды. Как писал Аристотель: “Животное, которое движется, осуществляет свое изменение положения путем нажатия на то, что находится под ним” (Аристотель, 1968; Е.Форстер) (перевод, с.489). Согласно этому утверждению, изучение движения должно быть направлено на: а) характеристику физического взаимодействия между животным и его окружением и б) определение способа, которым животное организует физическое взаимодействие (нажатие, по определению Аристотеля). В этих рамках мы можем рассматривать движение как результат взаимодействия биологической системы и ее окружения. На это взаимодействие влияет ряд факторов, включая следующие:
• Структура окружающей среды — форма и стабильность.
• Поле внешних сил — ориентация относительно силы тяжести, скорость движения.
• Структура системы — расположение тела, мышечная активность, сегментная организация тела, размеры, двигательная интеграция.
• Роль психологического состояния — степень внимательности, мотивация.
• Задача, подлежащая выполнению, — ограничения на диапазон изменения параметров;_
Эти аспекты хорошо освещены Хиггинсом , (1985): “Движение неотделимо от структуры, / поддерживающей его, и от среды, определяю- ( щей его” (с. 144). """
Цель настоящего учебника — изучение ней-ромеханической основы кинезиологии, где кинезиология определяется как изучение движения. Структура книги отвечает концепции Аристотеля, согласно которой движение включает в себя взаимодействие между животным и его окружением. Соответственно текст разбит на три "части, в которых изучается механическая основа движения (часть I), разрабатывается биологическая модель для управления движением (часть II) и интегрируются механический и биологический аспекты (часть III).
Для того чтобы развивать изучение движения человека мы должны иметь в качестве основы набор четко определенных терминов и понятий. Я надеюсь, что эта книга дает именно такую основу. Движение можно рассматривать как заключительный этап последовательного включения сложных нейрофизиологических механизмов, и наша задача состоит в том, чтобы на основе строго определенных терминов и понятий механики попытаться уточнить наши представления о характере этих процессов.
Джованни Альфонсо Борелли (1608 - 1679)
Уникальные иллюстрации в начале каждой части и главы выполнены Джованни Альфонсо Борелли (1608 — 1679). итальянским физиологом и врачом, который первым объяснил мышечное движение и другие функции тела в соответствии с законами механики (статики и динамики). Преподавал математику в Мессине в 1649 г. и Пизе в 1656 г. В 1674 г. Борелли переехал в Рим, где жил под покровительством шведской королевы Кристины. Наиболее известная его работа — “De Motu Animalium” (О движении животных), опубли
кованная в 1680 г. В ней он пытался объяснить движения тела животного на основе механических принципов. Благодаря этой работе он считается основателем ятрофизической школы (ятрофизика — сочетание физики и медицины).
Из работ Борелли в области астрономии наиболее известно опубликованное в 1666 г. исследование влияния притяжения на спутники Юпитера; он также первым предположил, что кометы перемещаются по параболической траектории.
ЧАСТЬ
ВЗАИМОСВЯЗЬ СИЛА-ДВИЖЕНИЕ
Движение всегда было предметом исследований специалистов многих областей знаний. Но по мере того как границы науки расширялись, стало очевидным, что в основе движения в живой природе лежит сложный комплекс биологических и механических взаимодействий. Многие" из" тех, кто следил за этим развитием, подопипГ к йзучёДй^^ с позиций нейро-
физиологии и биомеханики,~с нейргшеханической точки зрения. Биомехани-ка — наука, использующая принципы механики при изучении биологических систем, нейромеханический подход опирается на методологию биологии и физики—
В части I дается механическая характеристика взаимодействия среды, в которой происходит движение, и движущегося тела человека или его частей (биомеханика). Вводятся необходимые термины и обсуждаются концепции, обычно используемые для описания движения. Хотя многие аспекты соотношения сила—движение проиллюстрированы числовыми примерами, важно, чтобы студенты, формально используя математические приемы, не утратили понимания физической сути явлений. Для тех студентов, которым требуется помощь в математике, в приложениях приведены необходимые сведения.
11
ЦЕЛИ
Цель монографии — описание движения как взаимодействия биологической модели с физическим миром, в котором мы живем. Задача I части — определение механических основ движения. Конкретными целями являются
• описание движения точными терминами;
• определение силы и различных ее эффектов;
• рассмотрение роли силы в движении;
• анализ движения с точки зрения различных механических аспектов.
X
ГЛАВА 1
ДВИЖЕНИЕ
Хотя оценить эстетические качества или сложность такого движения, как прыжок в полтора сальто назад с тремя винтами, нетрудно, совсем другое дело описать его точными терминами. Детальное описание движения человека связано с использованием таких понятий, как положение, скорость и ускорение. При этом удается описать лишь внешнюю картину движения, без выяснения причин, его вызывающих. Такой способ изучения движения называется кинематическим. Эти термины часто используют в повседневной речи, не заботясь, однако, об их точном значении или даже не зная его. В биомеханике, как и при любом научном подходе, наблюдения и разработанные на их основе принципы хороши постольку, поскольку хороши концепции и определения, на которых они основаны. Сложность самого процесса движения предъявляет особые требования к его анализу, который должен базироваться на глубоком понимании и правильном использовании как самих кинематических терминов, так и единиц их измерения.
ПОЛОЖЕНИЕ, СКОРОСТЬ И УСКОРЕНИЕ
Положение тела определяется его местонахождением в пространстве по отношению к определенной системе отсчета. Например, термин “трехметровая вышка для прыжков в воду” означает положение вышки над поверхностью воды. Соответственно высота планки для прыжков в высоту определяется по отношению к земле, положение финишной линии в беге описывается по отношению к стартовой линии, третья и пятая позиции в балете относятся к положению одной ноги относительно другой и т.д. Когда положение объекта меняется, он пе
ремещается, движется в пространстве. Движение не может быть обнаружено мгновенно: оно проявляется при сравнении положения объекта в один момент времени с его положением в другой момент. Следовательно, движение — явление, которое происходит в пространстве и во времени.
Когда объект описывается как испытывающий перемещение, речь идет о пространственном характере движения, т.е. об изменении положения объекта. При учете как пространственного, так и временного аспектов движения определяющим становится понятие скорости. Скорость как кинематическая характеристика двйжения показь1вает, как 'бь1стро изменяется положение тела и в каком напрдплёьли тело ‘ перемещается. Поэтому скорость является вектором, направление которого совпадает с направлением перемещения тела, а величина характеризует быстроту перемещения. Поскольку перемещение относится к изменению положения, скорость может быть описана как производная по времени от перемещения.
На рис. 1.1. представлены два наблюдения, разделенные временным промежутком 3 с, изменения вертикального положения объекта относительно некоторой базовой линии. Изменение в вертикальном направлении за период 3 с составило 2 м, поэтому скорость изменения положения составляет 2 м/3 с или 0,67 м • с-1. Это средняя скорость объекта, движущегося из положения 1 в положение 2. общем виде
скорость = А положения '.f А времени ’ v1-1/
где А — величина изменения какого-либо параметра. Поэтому графически скорость связана с углом наклона графика положение—время. Угол наклона линии характеризуется отношением разности между конечным и начальным положениями объекта (А положения) к
13
Рис. 1.1. Зависимость положение—время
промежутку времени, за которое произошло это перемещение объекта (А времени). Следовательно, наклон характеризует скорость изменения переменной. При этом чем круче наклон, тем больше скорость изменения и наоборот.
В монографии приведено много графических иллюстраций. Как правило, графики показывают зависимость, по меньшей мере, между двумя переменными. На рис. 1.1, например, показано изменение положения от времени. Вообще зависимость может быть представлена в виде линии или в виде точек, нанесенных на график. Очевидная цель графического представления данных — показать тенденцию или характер связи между переменными; более точно данные представляются в форме цифрового ряда. Зависимость между положением и временем на рис. 1.1 относительно проста, она может быть охарактеризована одним измерением — наклоном линии.
Вертикальное перемещение может изменяться не только по величине, но и по направлению (вверх или вниз). На рис. 1.2 показан соответствующий пример при регистрации положения объекта в пять разных моментов времени.
Использование соотношения (1.1) на разных участках при перемещении объекта из положения 1 в 2, из 2 в 3, из 3 в 4, из 4 в 5 дает соответственно следующие значения скорости: 0,75; 1,5; 0 и — 1,25 м-с-1. Чем более крутой наклон графика положение—время (например, при переходе из положения 2 в положение 3 — 1,50 м • с-1, при сравнении с величиной скорости при переходе из положения 1 в положение 2 — 0,75 м • с-1), тем больше скорость. Уклон вниз, например, при переходе из положения 4 к положению 5 означает, что скорость отрицательна. Отсутствие изменения положения (например, при перемещении из положения 3 в положение 4) свидетельствует, что скорость равна нулю. Этот пример иллюстрирует один
важный момент относительно скорости: знак скорости изменяется — вначале она положительна, затем уменьшается до нуля, на спуске становится отрицательной и движение меняет направление. При этом, когда направление движения изменяется, график скорость—время должен пройти через нуль. На рис. 1.2 показано, что объект вначале двигался в одном направлении (произвольно названном положительным; обратите внимание на положительный угол наклона), затем был неподвижным и, наконец, двигался в противоположном направлении (отрицательный угол наклона).
Однако не достаточно описывать движение, указывая только положение и скорость перемещения объекта. Например, шарик, удерживаемый над землей на высоте 1,23 м и затем отпущенный, достигнет земли через 0,5 с. Изменение положения составляет 1,23 м, а средняя скорость равна 2,46 м • с-1 (т.е. 1,23 м / 0,5 с). Но шарик не движется с постоянной скоростью; его скорость изменяется со временем. Начав с нулевой скорости в момент отпускания, скорость шарика возрастет до 4,91 м-с'1 перед контактом с землей. Эта величина — отношение изменения
скорости ко времени — называется ускорением. Ускорение, которое испытывает шарик во время падения, постоянно и равно 9,81 м • с-2. Если скорость объекта измеряется в метрах в секунду (м • с-1), то ускорение — изменение в метрах в
секунду каждую секунду, т.е,
А скорости ускорение =-----------
А времени
(1-2)
Например, защитник в футболе, о котором
говорят, что у него хорошее ускорение, может быстро изменять скорость (величину и направление).
Рис. 1.2. Изменения скорости, обусловленные изменениями величины и направления перемещений на графике положение—время
14
По аналогии с тем, что скорость характеризуется наклоном графика положение—время, ускорение можно оценить по наклону графика скорость—время. При этом для иллюстрации можно использовать тот же график на рис. 1.2, предположив, что по оси ординат отложена не высота в метрах, а скорость в метрах в секунду. Так, если точка 2 имела координаты 2,0 м • с-1 и 3 с, а точка 3 имела координаты 3,5 м • с-1 и 4 с, то быстрота изменения скорости при перемещении от точки 2 к точке 3, т.е. ускорение, может быть рассчитано из уравнения, аналогичного уравнению (1.2) следующим образом:
3,5-2,0 м-с-1 ускорение = ’----— =
= 1,5 м - с 2.
Подобным образом ускорение при перемещении от точки 1 к точке 2, от точки 3 к 4 и от 4 к 5 составит соответственно 0,75, 0 и -1,25 м • с-2. Как и в случае со скоростью, ускорение имеет и величину, и направление и, следовательно, является векторной величиной.
Ускорение, испытываемое шариком в предыдущем примере, вызвано гравитационным притяжением между двумя массами: массой планеты Земля и массой шарика. Сила тяжести дает постоянное ускорение, равное примерно 9,81 м-с'2 на уровне моря. В принципе, объект, на который воздействует сила, испытывает ускорение. Постоянная сила (например, сила тяжести), приложенная к не имеющему опоры объекту, дает постоянное ускорение; отсутствие действующей на объект силы означает, что объект неподвижен или перемещается с постоянной скоростью (т.е. ускорение отсутствует). Как будет выглядеть график скорость—время для постоянного и нулевого ускорений? При наличии ускорения наклон графика скорость—время отличен от нуля. Напротив, при отсутствии ускорения график скорость—время имеет нулевой наклон.
При кинематическом анализе движения особое внимание необходимо уделять единицам измерения физических величин. При этом для каждого измерения необходимо знать некоторые опорные значения, которые могут помочь при оценке его результатов. Например, если движение осуществляется со средней скоростью 2,9 м с-1, то оно быстрое или медленное? Хотя ответ зависит от деталей движения, полезно знать, что забег на 100 м, выполненный за 10 с, дает среднюю скорость 10 м • с-1. Это значение может бьпь принято за опорное для сравнения с другими значениями скорости. Подобным образом величины ускорения можно сравнивать с ускорением силы тяжести (9,81 м-с-2). Практически, однако, нет необходимости помнить больше, чем несколько основных величин, поскольку любые другие с помощью коэффициентов преобразования всегда можно получить (см. Приложение 3).
УРАВНЕНИЯ ДВИЖЕНИЯ
Из элементарных определений скорости (уравнение 1.1) и ускорения (уравнение 1.2) можно вывести алгебраические выражения, содержащие время (/), вектор, указывающий положение тела (г), векторы скорости (v) и ускорения (а), которые часто используются для описания движения. В этом разделе мы выведем три уравнения движения, которые справедливы при условии, когда ускорение постоянно (далее везде по тексту символы, представляющие векторные величины, — например, поло
жение, скорость, ускорение — выделены жирным шрифтом, а их проекции на направление движения — курсивом). В этих уравнениях буквами с индексами v, и vA обозначены соответ
ственно начальная и конечная скорости, ана
логично Г/ и гу соответствуют начальному и ко-
нечному положениям.
1. Выразить конечную скорость как функцию начальной скорости, ускорения и времени (t = Л времени).
А скорости.
А времени ’
среднее ускорение =
(1.3)
Для примера рассмотрим падение шарика. Шарик падал с высоты 1,23 м и через 0,5 с достиг земли с конечной скоростью 4,91 м • с'1, вычисленной по уравнению (1.3). Переменными, которые влияют на конечную скорость, являются начальная скорость (v,) шарика, его ускорение (а) и длительность падения (г). В этом примере v, равно нулю, а обусловлено земным притяжением (9,81 м-с~2), t составляет 0,5 с. Эти величины могут быть подставлены в уравнение (1.3) для определения конечной скорости. Допустим, мы хотим определить конечную скорость брошенного бейсбольного мяча, когда он пересекает линию. Как и раньше, мы должны знать v,-, а и t. Основной трудностью будет определение а, поскольку на мяч, кроме силы тяжести, будут действовать и другие силы. Обычно а определяют численно при видеосъемке движения. Кроме того, а можно измерить
непосредственно с помощью акселерометра.
2. Вывести формулу, определяющую положение объекта в зависимости от начальной и ко
нечной скорости, ускорения и времени.
средняя скорость =
А положения. А времени
V + vf = Гу - г,-2 t
Иначе среднюю скорость можно выразить через начальную v, и конечную Vy скорости и начальное и конечное положение г, и Гу следу
15
ющим образом. Подставив в уравнение (1.3) значение Vf, найденное из этого равенства, получим
_ у,- 4-аГ + у, f ' " 2
2v, + а/ .
гЛ - г,- = —-------Г;
f ' 2
Г/-г,- = v,r +jar2.
(1.4)
Уравнение (1.4) показывает, что изменение положения объекта или расстояние, которое объект проходит от одной точки до другой при условии, что направление не изменяется, зависит от трех переменных: его начальной скорости (vz), ускорения (а), которое он испытывает, и времени (г). Иными словами, конечное положение объекта будет другим, если любая из этих переменных изменится. Это соотношение может быть использовано для определения изменения положения объекта с изменением времени. Например, спортсмен прыгает с 10-метровой вышки (задача 119); при изменении t от О до 1,5 с с шагом 0,1 с, мы можем определить траекторию (график положение—время) прыгуна во время прыжка. Начальная скорость прыгуна равна нулю, так что уравнение (1.4) для этой задачи упрощается:
гу - г,- = jar2.
Если допустить, что влияние сопротивления воздуха настолько мало, что им можно пренебречь, ускорение будет обусловлено только силой тяготения. Набор данных положение-время (табл. 1.1) может быть определен с помощью вычислений с использованием приведенного выше уравнения при увеличении t каждый раз на 0,1 с.
3. Зависимость конечной скорости от начальной скорости, ускорения и положения.
Воспользуемся соотношениями из преды
дущего примера:
средняя скорость =
А положения. А времени
vz + У,- _ Г/ - г,-2 t
Затем найдем t из уравнения (1.3) и подставим его в предыдущее уравнение
У/+У,_ rf-fi
2 (vf-Vi)/a’
Рис. 1.3. Начальные условия в примере с жонглером
вает, что любая кинематическая переменная (в данном случае конечная скорость) может быть определена через три других параметра. Так, скорость пули на определенном известном расстоянии от пистолета (например, 10 м) зависит от скорости пули в момент выстрела (начальная скорость), ускорения, которое испытывает пуля, например, под действием сопротивления воздуха) и указанного расстояния (в данном случае 10 м).
Если начальная скорость равна нулю (т.е. объект начал движение из состояния покоя), уравнения (1.3), (1.4) и (1.5) упрощаются
V/ = ar;
г/ -г/ = 2а'2;
vf = 2а<г/ - ГД
Пример. Жонглер выступает в помещении, потолок которого находится на 2 м выше его рук. Он бросает мяч вертикально так, что тот касается потолка. На рис. 1.3 приведен эскиз траектории полета мяча:
а) С какой начальной вертикальной скоростью жонглер бросает мяч (у,)? В момент касания потолка V/ = 0:
v} = v? + 2a(rz - г, );
у2 = 0 - [2 х (-9,81) х 2];
у,- = д/Щ;
у, = 6,3 м • с-1.
б) Сколько времени требуется мячу, чтобы Достичь потолка?
2a(rz - г,) = (vz + V|)(vz - у,); <L5)
2а(гу - г,-) = (у2- - у2);
v} = v? + 2а(Г/ - г,);
Уравнение (1.5), как и (1.3) и (1.4), показы-
у f = у,- + at;
a
X’l =0’MC-
16
в) Жонглер бросает второй мяч с такой же начальной скоростью в тот момент, когда первый мяч находится у потолка. Через какое время после того, как будет брошен второй мяч, мячи пройдут один мимо другого?
Ответ на этот вопрос может быть найден путем тщательного применения приведенных выше уравнений.
Начнем с анализа начальных условий. Траектории двух мячей в основном одинаковы. Первый мяч имеет начальную скорость на пике своей траектории (в момент касания), равную нулю (0 м-с-1), и конечную скорость (в момент, когда он пойман) 6,3 м • с-1; второй мяч, напротив, имеет скорость 6,3 м • с-1 при бросании и 0 м • с-1 на пике. Поскольку траектория параболическая, она симметрична. Поэтому мячи встретятся через 0,32 с, что составляет г/2. Скорость мячей в этот момент будет около 3,1 м • с-1.
г) Когда мячи пройдут один мимо другого, на какой высоте будут они от рук жонглера?
1 7
Гу - г(- = v,/ +2 аг
Аг = (6,3 х 0,32) + 0,5[-9,81 х (0,32)2] =
= 2,02 - 0,50
Аг ® 1,5 м над руками.
ЧИСЛЕННЫЙ АНАЛИЗ
Кинематический анализ обычно основывается на данных положение—время, полученных с помощью записывающих устройств, кинокамер или видеокамер. Фильм или видеозапись движения представляет собой комплект неподвижных изображений (кадров), которые последовательно проецируются поодиночке на измерительное устройство, и определяется положение выбранных отметок по отношению к некоторому эталону. Прибор, используемый в этой процедуре и называемый диджитайзером, может определять координаты (х, у) выбранных отметок. Имея набор данных положение—время, мы можем использовать стандартную процедуру численного анализа для определения связанных с ними данных скорость—время и ускорение-время. В табл. 1.1 приведен пример таких вычислений. Дополнительно данные положение-время для ряда движений приведены в Приложении 4.
В табл. 1.1 представлены данные положение—время для абстрактного движения. 13 значений положения, записанных в различные моменты времени, представляют вертикальный путь, который объект проходит в первый период. Вначале объект поднимается над начальным положением (0,0 м) до высоты 1,0 м, затем смещается на такое же расстояние (—1,0 м) ниже исходного положения и, наконец, возвращает
Таблица 1.1. Расчет скорости и ускорения при использовании данных положение—время
Положение, м Время, с Скорость (Аположения/Авремени), м-с 1 Ускорение (Аскорости/Авремени), м-с 2
0,00 0,000 0,050 (0,59 - 0,00)/(0,100 -0,000) = 5,9 0,59 0,100 (3,6 - 5,9)/(0,150 - 0,050) = -23,0 0,150 (0,95 - 0,59)/(0,200 - 0,100) = 3,6 0,95 0,200 (1,0 - 3,6)/(0,225 - 0,150) = -34,7 0,225 (1,00 - 0,95)/(0,250 - 0,200) = 1,0 1,00 0,250 (-1,0 - 1,0)/(0,275 - 0,225) = -40,0 0,275 (0,95 - 1,00)/(0,300 - 0,250) = -1,0 0,95 0,300 (-3,6 - [—1,0])/(О,35О - 0,275) = -34,7 0,350 (0,59 - 0,95)/(0,400 - 0,300) = -3,6 0,59 0,400 (-5,9 - [—3,6])/(0,450 - 0,350) = -23,0 0,450 (0,00 - 0,59)/(0,500 - 0,400) = -5,9 0,00 0,500 (-5,9 - [—5,9])/(0,550 - 0,450) = 0,0 0,550 (-5,9 - 0,00)/(0,600 - 0,500) = -5,9 -0,59 0,600 (-3,6 - [-5,9])/(0,650 - 0,550) = 23,0 0,650 (-0,95 - [—0,59])/(0,700 - 0,600) = -3,6 -0,95 0,700 (-1,0 - |-3,6])/(0,725 - 0,650) = 34,7 0,725 (-1,00 - | —0,95])/(0,750 - 0,700) = -1,0 -1,00 0,750 (1,0 — [—3,6])/(0,775 — 0,725) = 40,0 0,775 (-0,95 - [—1,00])/(0,800 - 0,750) = 1,0 -0,95 0,800 (3,6 - 1,О)/(О,85О - 0,775) = 34,7 0,850 (-0,59 - [—0,95])/(0,900 - 0,800) = 3,6 -0,59 0,900 (5,9 - 3,6)/(0,950 - 0,850) = 23,0 0,950 (0,00 - [-0,59])/( 1,000 - 0,900) = 5,9 0,00 1,000
2 8-133
17
ся к начальному уровню 0,0 м. Скорость объекта во время этого движения рассчитывается путем применения уравнения (1.1) к выбранным интервалам времени, для которых имеется информация о положении. Например, из табл. 1.1 мы можем выбрать промежутки от 0,0 до 1,0 с, от 0,0 до 0,25 с или от 0,0 до 0,1 с. Если мы применим уравнение (1.1) к каждому из этих интервалов, средняя скорость для каждого из них будет составлять:
Л Л 1 Л — 0,0 Л “1 от 0,0 до 1,0 —----— = 0 м • с ;
1,0-0,0
Л Л Л 1,0 — 0,0 . -1
от 0,0 до 0,25 —---!— = 4 м • с ;
0,25 - 0,0
л л л 1 0,59 — 0,0 с о -1 от 0,0 до 0,1 —-----Ч- = 5,9 м • с ;
0,1-0,0
Подобным образом, если мы измерим положение только в моменты времени 0,0; 0,5 и 1,0 с, создается иллюзия того, что предмет не двигался. Действительно, чем меньше интервалы времени, в которые мы измеряем положение, тем ближе рассчитанная скорость к той, которую действительно имел объект. Однако экономя усилия целесообразно проводить измерения не слишком часто, должна быть выбрана некоторая рациональная длина интервала измерений. Параметры большинства движений человека можно адекватно измерить при частоте от 50 до 100 кадров в секунду, что соответствует промежуткам от 0,1 до 0,2 с между последовательными измерениями.
В табл. 1.1 скорость определялась для каждого интервала, на границах которого измеря-
Рис. 1.4. График кинематических данных, представленных в табл. 1 1
1 — положение; 2 — скорость; 3 — ускорение
лось положение. Например, перемещение в течение первого интервала (0,59 — 0,0 = 0,59 м) необходимо разделить на время, прошедшее в течение этого интервала (0,1 — 0,0 = 0,1 с), чтобы получить соответствующую скорость (0,59/0,1 = 5,9 м-с-1). Величина 5,9 м-с-1 представляет собой среднюю скорость в течение этого интервала и расположена в таблице против момента времени, соответствующего середине интервала (0,05 с). Аналогично первое значение ускорения (23,0 м • с-2), которое определяется с использованием уравнения (1.2), записывается в средней точке (0,10 с) первого временного интервала, на границах которого вычислялась скорость (0,05 — 0,15). С помощью этой процедуры среднее значение скорости определяется для каждого интервала положения и среднее ускорение рассчитывается для каждого интервала скорости. Таким образом, из 13 наблюдений положение—время рассчитывается 12 величин скорость—время и 11 величин ускорение—время.
Выше уже отмечалась связь между характеристиками движения (например, положение, скорость) и быстротой их изменения. Наклоны графиков положение—время и скорость-время представляют соответственно скорость и ускорение. Дальнейшее подтверждение этого соотношения приведено в табл. 1.1. Когда координата, характеризующая положение, возрастает (положения от 1 до 4 и от 10 до 13), скорость положительна, если эта координата уменьшается (позиции с 4 до 10), скорость отрицательна. Подобная зависимость существует между наклоном (увеличением или уменьшением) графика скорости и знаком ускорения (рис. 1.4).
ГРАФИЧЕСКОЕ ПРЕДСТАВЛЕНИЕ
Используя способ графического представления данных, мы можем оценить быстроту изменения кинематической величины при изменении времени. На рис. 1.5 показан этот процесс для изменения угла бедра опытного бегуна в течение одного шага, определяемого как один полный цикл (в данном примере от отталкивания до отталкивания левой ноги). Угол бедра измеряется от горизонтали. Способ замера изображен над верхней шкалой рисунка (измеренный угол относится к конечности с ботинком). Угол измеряется в радианах (1 рад = 57,3°; принятая в системе СИ единица для измерения углов; перечень единиц СИ см. в Приложении 1).
Первым шагом в построении графика скорость-время должна быть идентификация относительных минимумов и максимумов на графике положение—время. Нужно отметить
18
любые, — а их может быть несколько, — пики и впадины кривой. Эти точки определяют моменты, когда быстрота изменения равна нулю. Иными словами, наклон графика не является ни положительным, ни отрицательным, а равен нулю. На рис. 1.5 функция угол бедра—время имеет один минимум и один максимум. Из этих точек нулевого наклона и, следовательно, нулевой скорости восстанавливается перпендикулярная линия к оси времени графика скорость-время, чтобы отметить момент достижения нулевой скорости. На рис. 1.5 это моменты примерно около 0,03 и 0,30 с.
Вторым шагом является определение наклона графика положение—время между максимумами и минимумами. Наклон графика между этими точками не изменяется (остается положительным или отрицательным). Точки нулевой скорости идентифицируют моменты времени, когда кривая положение—время изменяет свой наклон (т.е. изменяет направление). В каждом интервале между минимумом и максимумом наклон может быть более или менее крутым, но направление кривой будет сохраняться. На рисунке угол бедра—время имеется один минимум и один максимум и, следовательно, три таких интервала: от начала движения до минимума, от минимума до максимума и от максимума до конца движения. Наклон графика положение—время на этих интервалах соответственно отрицателен, положителен и снова отрицателен. Таким образом, знак функции на графике скорость—время соответствует наклону функции угол бедра—время для каждого интервала. Например, для первого интервала — от начала движения до минимума — наклон графика положение—время и скорость отрицательны. Поскольку отрицательные значения скорости связываются с направленным вниз наклоном графика положение-время, отрицательные скорости на рис. 1.5 свидетельствуют об обратном вращении бедра (т.е. уменьшение измеренного угла). В целом график скорость—время на рис. 1.5 указывает на два интервала вращения бедра назад, разделенных по времени интервалом с вращением бедра вперед. Изменение скорости во времени показывает, как изменяется скорость вращения, тогда как знак (положительный или отрицательный) указывает направление вращения (вперед или назад).
Построение графика ускорение—время из графика скорость—время сопровождается такой же двухступенчатой процедурой: а) идентификацией относительных максимумов и минимумов; б) определением наклона в выделенных интервалах. Согласно рис. 1.5, кривая скорость—время имеет четыре минимума и максимума, следовательно, есть четыре случая, когда график ускорение—время должен пересекать ось абсцисс (горизонтальную коорди-
Рис. 1.5. Угловой профиль скорость—время может быть получен из графика угловое положение—время, угловой профиль ускорение—время может быть выведен из графика угловая скорость—время
натную ось). В результате на графике зависимости ускорение—время можно выделить пять интервалов с последовательным изменением знака функции. Интерпретация графика ускорение-время сложнее, чем графиков положение-время и скорость—время. На рис. 1.5 положительное ускорение означает ускорение движения бедра в направлении вперед; в течение первого интервала ускорения бедро поворачивается вначале назад, а затем вперед (это видно из графика скорость—время), но в течение всего интервала бедро испытывает ускорение в направлении вперед (вспомните пропорциональность между ускорением и силой). В принципе, объект или сегмент тела может перемещаться в одном направлении, но при этом испытывать ускорение в противоположном направлении, и не всегда можно определить направление ускорения по направлению движения.
Рассмотрим движение мяча, который жонглер подбрасывает в воздух и затем ловит. Дви-
2*
19
Рис. 1.6. Траектория мяча, брошенного жонглером, в зависимости от времени. Начало отсчета (нуль) обозначает положение рук жонглера в момент бросания
жение мяча может быть показано на графике положение—время, на котором ордината представляет высоту мяча над рукой жонглера (рис. 1.6).
Кривая изменения положения мяча имеет колоколообразную форму. Какова форма графика ускорение—время? Мы могли бы получить ответ, используя рис. 1.6, но в отличие от предыдущего случая объект в свободном падении, в данном случае мяч, брошенный в воздух, испытывает постоянное ускорение в 9,81 м-с~2. Хотя мяч движется вверх и вниз, это перемещение еще не подсказывает направление ускорения, испытываемого мячом.
С помощью процедуры построения графиков, описанной в этом разделе, мы можем определить быстроту изменения положения или скорости. Эта процедура является качественной по своей природе, при ее использовании можно получить только знак (положительный или отрицательный) для быстроты изменения и ее приближенное значение. Напротив, табл. 1.1 иллюстрирует количественный подход, при котором быстрота изменения может быть определена более точно. С помощью качественного анализа можно выяснить тип или вид изменения, тогда как количественный анализ позволяет установить, сколь велико это изменение.
ПОСТУПАТЕЛЬНОЕ И ВРАЩАТЕЛЬНОЕ ДВИЖЕНИЯ
В предыдущем разделе величина перемещения измерялась в метрах (м) или радианах (рад); различие между ними характеризует отличие соответственно между поступательным и вращательным движениями. Линейным, или поступательным, считается такое “перемещение объекта, при котором любая его часть
(и весь объект целиком) перемещается, оставаясь параллельной своему первоначальному положению в течение всего движения. Напротив, если все части объекта Испытывают неодинаковые перемещения, объект, кроме того, поворачивается. Сочетание поступательного (линейного) и вращательного (углового) движений в одноиШТОскости называется плоским движением и включает в себя вращение вокруг точки, которая сама перемещается. Если такое движение происходит более чем в одной плоскости, речь идет о перемещении в пространстве (трехмерном движении). В большинстве случаев движений человека сегменты тела испытывают как линейные, так и угловые перемещения.
Метр — единица для измерения линейного перемещения — определяется как длина, равная 1650763,76 длин волн, излучаемых крипто-ном-86 в вакууме.
Угловые перемещения измеряются в градусах или радианах. Радианное измерение угла х определяется отношением длины дуги, на которую этот угол опирается, если вершина угла совпадает с центром какой-либо окружности, к длине радиуса окружности. При этом за единицу измерения принимается радиан—угол, опирающийся на дугу, длина которой равна радиусу (1 рад 57,3°). Рассмотрим метание диска. Длина руки метателя от плеча до диска составляет 63 см. При вращении руки в плечевом суставе диск описывает круговую траекторию. Когда диск прошел по своей траектории путь 63 см (равный длине руки и тем самым равный радиусу круга), рука и диск повернулись на угол 1 рад.
При описании вращательных движений, как правило, применяют угловые единицы, а поступательных — линейные единицы. Обычно применяемые символы и связанные с ними единицы измерений для линейного и углового положения, скорости и ускорения приведены в табл. 1.2. Для обозначения линейных единиц обычно используют латинские буквы, для угловых — греческие.
Таблица 1.2. Линейные и угловые символы и единицы измерения в системе СИ
Показатель Линейные Угловые
Символ Единица Символ Единица
Скаляр Вектор
Положение Г Г М 6 рад
Перемещение д'- дг м де рад
Скорость V V мс-1 (0 рад-с-1
Ускорение а а М'С"1 а рад-с-2
20
Диаграммы угол—угол
При анализе движений обычно строится график какой-либо переменной (например, угол бедра, высота мяча) в зависимости от времени. Но поскольку движение человека осуществляется путем поворота сегментов тела друг относительно друга, часто больше информации дает изучение зависимости между различными углами во время движения. Такой график, называемый диаграммой угол—угол (Cavanagh, Grieve, 1973), обычно изображает угол (т.е. угол между двумя прилегающими сегментами тела) по отношению к абсолютному углу сегмента тела (т.е. углу относительно опорного направления в окружающей среде).
На рис. 1.7 показаны две диаграммы угол-угол для части движения при поднятии штанги (отрыв и толчок), в котором штанга поднималась из положения 1 в положение 10. Диаграмма колено—торс показывает, что движение включает в себя три отчетливо различимые фазы: а) положения 1 — 5, выпрямление коленного сустава и легкий поворот торса вперед; б) положения 5 — 8, поворот торса назад и сгибание колен; в) положения 8 — 10, распрямление коленного сустава и некоторый поворот торса
назад—вперед. Подобным образом диаграмма голеностопный сустав—бедро включает в себя три фазы: а) поворот бедра вперед—планарное выпрямление голеностопного сустава; б) постоянное дорсальное распрямление угла бедра и голеностопного сустава; в) поворот бедра вперед—планарное распрямление голеностопного сустава.
Рис. 1.7 показывает степень, до которой три фазы двух диаграмм угол—угол совпадают; движение сопровождается координированным смещением вокруг крайних нижних суставов. На изображенной на рис. 1.7 диаграмме интервал времени между точками постоянен и равен 10 мс; следовательно, чем больше расстояние между точками, тем больше скорость движения.
Кинематика углового движения
Поскольку движения человека, как правило, включают и поступательные перемещения, и вращения, необходимо знать связь между линейными и угловыми мерами положения, скорости и ускорения. Когда жесткое тело фикси
Рис. 1.7. Зависимость угол—угол во время первой фазы движения при поднятии штанги Цифры на диаграмме соответствуют положениям штангиста, показанным в верхней части рисунка. Стрелки под фигурами показывают силу, действующую на штангиста (Enoka, 1988):
F w — вес тела
21
рованной длины (г) вращается вокруг точки и переходит при этом из положения 1 в положение 2 (рис. 1,8, а), смещение (s), испытываемое концом тела, определяется уравнением
5 = г6. П-6)
Линейная скорость (у) на конце жесткого тела определяется как быстрота изменения
А? _ А(г9) At ~ At
Поскольку г имеет фиксированное числовое значение и не изменяется во времени, выражение преобразуется к виду
as __ где.
At ” At ’ (1.7)
V = Пй.
Уравнение (1.7) показывает, что линейная скорость v любой точки жесткого тела равна произведению расстояния от оси вращения до этой точки г и угловой скорости тела со. При этом с увеличением расстояния от оси вращения возрастает и линейная скорость р. Как знает каждый, кто был участником вращающейся человеческой цепи на коньках, человек, находящийся дальше всех от оси вращения, испытывает наибольшую линейную скорость. Вектор линейной скорости v всегда касателен к траектории тела (рис. 1.8, б).
Рис. 1.8. Соотношение между линейным и угловым движением твердого тела заданной длины: а — положение; б — скорость; в — ускорение
Переменные г, v и со — векторные величины, которые характеризуются величиной и направлением, как показано на рис. 1.8, б. Если направление вращения показано изогнутой стрелкой, то вектор со фактически перпендикулярен странице по направлению к читателю; уравнение (1.7) — скалярная форма зависимости между линейной и угловой скоростью. Векторное соотношение имеет вид
v = со X г. (18)
Из уравнения (1.8) следует, что линейная скорость v равна векторному произведению х угловой скорости со и радиуса-вектора точки г. Векторное произведение — это векторный оператор, используемый для умножения векторов, и результатом его является вектор, перпендикулярный плоскости векторов-сомножителей. Он равен произведению модулей этих векторов на синус угла между ними.
Для определения зависимости между линейным и угловым ускорением мы должны использовать уравнение (1.8), принимая во внимание изменения как величины, так и направления каждого вектора:
Av _ А(со х г).
At ~ At ’
а = (со х ^) + х г) = (со х у) + (а х г);
а = со х (со х г) + (а х г); (1-9)
а = а„ + ат, где ап = со х (со х г); ат = (ах г). Член ап соответствует изменению направления у, а член а1 представляет изменение величины у (рис. 1.8, в). Поскольку направление v изменяется во время углового движения, ап никогда не равно нулю, но aj может быть равно нулю, если у постоянна.
УСКОРЕНИЕ И МЫШЕЧНАЯ АКТИВНОСТЬ
Для дальнейшего рассмотрения зависимости между ускорением и силой рассмотрим следующий пример разгибания—сгибания локтя в горизонтальной плоскости, проходящей через плечевой сустав (рис. 1.9). Исходное положение: плечо и предплечье расположены в горизонтальной плоскостй и угол между ними равен 0,70 рад (40°). При непрерывном движении с умеренной скоростью плечо удерживается неподвижным, тогда как локтевой сустав разгибается в горизонтальном направлении до 3,14 рад (180°) и сгибается опять в исходное положение (0,70 рад). Как правило, сгибание сустава приводит к уменьшению угла между двумя соседними сегментами тела, которые
22
сходятся в этом суставе (в данном примере плечо и предплечье), разгибание же этот угол увеличивает. Какой вид будет иметь график положение—время, связанный с этим движением?
Плечо остается неподвижным в то время, как предплечье вращается вокруг локтя; график изменения угла в локтевом суставе во времени должен адекватно описывать этот процесс. В этом простом движении минимальный угол (0,70 рад) наблюдается в начале и конце движения, а максимальный — при полном выпрямлении (3,14 рад). График положение-время приведен на рис. 1.9.
Не используя описанные ранее соотношения между положением, скоростью и ускорением, попробуйте ответить на два вопроса (основное внимание уделите повороту локтевого сустава): Когда скорость равна нулю? Когда ускорение равно нулю? В какой момент процесса сгибания—разгибания скорость равна нулю?
Скорость равна нулю, когда нет смещения — в начале и конце движения и в момент изменения направления движения (от разгибания к сгибанию). График скорость—время на рис. 1.9 показывает, что угловая скорость обращается в ноль при максимальном смещении (3,14 рад) в момент изменения направления движения. В этом можно убедиться на основании предшествующих рассуждений о соответствии между скоростью и наклоном графика функции положение—время. Как видно из графика, скорость положительна (выше нуля), если локоть разгибается, и отрицательна при сгибании локтя. Таким образом, изменение знака скорости (например, с положительного на отрицательный) указывает на изменение направления движения.
Теперь рассмотрим ускорение. Когда ускорение равно нулю? Интуитивный ответ на этот вопрос затруднителен, так как критерии для определения нулевого ускорения не очевидны. Что касается скорости, вопрос был более ясным и заключался только в определении моментов отсутствия перемещения. Аналогично, если отсутствует изменение скорости, ускорение равно нулю.
Как видно из графика (рис.1.9), ускорение обращается в ноль в моменты, когда скорость максимальна (0,3 с) и минимальна (0,7 с). Иными словами, ускорение равно нулю, когда наклон графика скорость—время равен нулю. График ускорение—время включает в себя три этапа изменения ускорения: от начала до момента около 0,3 с движение сопровождается ускорением процесса в разгибании (ускорение положительно); затем ускорение изменяет знак и становится отрицательным, большую часть времени (от 0,3 до 0,7 с) ускорение направлено на сгибание локтя; на заключительном этапе ускоре-
Угол локтя, рад
Растяжение
Сжатие
Скорость, рад-с
Рис. 1.9. Кинематические графики и ЭМГ для простого движения сгибания—разгибания в локтевом суставе. Кинематические особенности движения в значительной степени определяются мышечной активностью
ние направлено на разгибание. Обратите внимание, что профиль ускорение—время нельзя предсказать непосредственно из графика положение-время.
МЫШЕЧНАЯ АКТИВНОСТЬ
Можно ли определить профиль ускорение-время не используя график скорость—время? Иными словами, можно ли идентифицировать моменты обращения в ноль ускорения, не зная функции скорость—время? Для ответа на этот вопрос рассмотрим график мышечной активности, сопровождающей исследуемое движение. Часто кинезиологи описывают движение в терминах мышечных групп, а не отдельных мышц. В этом примере сгибания—разгибания локтя движение включает в себя группы мышц, известные как разгибатели и сгибатели локтя. Группа
23
сгибателей включает в себя бицепс брахии, бра-хиалис и брахиорадиалис как основные мышцы, влияющие на сгибание локтя, но это не значит, что эти мышцы не могут контролировать также разгибание локтя (подробнее об этом ниже). Группа разгибателей локтя включает в себя трицепс брахии. В терминах мышечной активности вопрос о нулевом ускорении может быть переформулирован следующим образом: можно ли по динамике активности разгибателей и сгибателей локтя в процессе движения определить моменты обращения в ноль ускорения?
В части II “Единая система сустава” будет рассмотрен способ, с помощью которого нервная система активирует мышцу. В принципе, эта связь представляет собой электрохимический процесс, в котором финальная стадия — электрическая по своей природе. Поэтому чтобы определить, является ли мышца активной, мы должны только проследить ее электрическую активность. Такая техника, известная под названием электромиографии (ЭМГ), заключается в помещении электродов на кожу над мышцей для контроля электрического выхода (возбуждения) мышцы. На рис. 1.9 представлены записи ЭМГ разгибателей и сгибателей локтя в процессе его сгибания—разгибания. Хотя обе группы мышц активны в течение всего движения (явление, известное как коакги-вация), наблюдается высокая корреляция между мышечной активностью (ЭМГ) и профилем ускорение—время. Далее мы более подробно рассмотрим механическое влияние мышечной активности на сегменты тела. Один из эффектов заключается в том, что она генерирует усилие и может заставить сегменты тела поворачиваться. В движении сгибания—разгибания локтя нет других существенных горизонтальных сил, так что ускорение по отношению к локтевому суставу в значительной степени определяется активностью мышц вокруг него. Таким образом, ответ на вопрос о нулевом ускорении можно получить по картине мышечной активности.
При наличии такого соотношения между силой и ускорением график ускорение—время, показанный на рис. 1.9, должен давать представление о процессе изменения мышечной активности относительно локтевого сустава. Однако часто трудно понять, по крайней мере вначале, почему простое движение сгибания-разгибания локтевого сустава сопровождается трехфазной сменой мышечной активности. Для перемещения конечности с умеренной скоростью требуется резкое увеличение мышечной активности, чтобы ускорить перемещение конечности по направлению к цели с последующим увеличением активности противоположной мышцы, ускоряющей конечность в противоположном направлении для замедления
(торможения) ее движения. Движение сгибания-разгибания локтевого сустава включает одновременное стремление к двум различным целям, вначале — полного разгибания и затем возвращения в исходное положение. Это движение состоит из двух встречных процессов включения мышечной активности. Движение к первой цели — полное разгибание локтя — сопровождается активацией локтевого сустава, за которым следует активация его сгибателей, чтобы затормозить движение конечности. Движение ко второй цели — возврат в исходное положение — осуществляется путем активации сгибателей локтевого сустава, за которым следует активация его разгибателей для торможения движения. При взаимном наложении этих процессов результирующая активность состоит из активности разгибателя локтевого сустава, за которой следует активность его сгибателя и в заключение — активность разгибателя. Такая картина мышечной активности дает трехфазную кривую ускорение—время (рис. 1.9).
В принципе, проиллюстрированное на этом примере разгибания—сгибания локтевого сустава соотношение между активностью мышц относительно сустава и ускорением сегмента тела представляет собой особый случай. Ускорение, которое испытывает сегмент тела, — результат всех сил, действующих на сегмент. Например, если бы движение сгибания—разгибания локтя выполнялось в вертикальной плоскости, сила тяжести оказывала бы влияние на движение, и график ускорение—время отличался бы от показанного на рис. 1.9. При движении в горизонтальной плоскости, мышцы оказывают единственное существенное усилие на предплечье в этой плоскости и тем самым дают описанное выше соотношение между мышечной активностью и ускорением предплечья.
КИНЕМАТИКА ПОХОДКИ
Для того чтобы показать использование кинематических элементов (положения, скорости и ускорения) при анализе движения человека, рассмотрим некоторые кинематические характеристики походки. Походка человека включает в себя ряд чередующихся движений, при которых тело опирается вначале на одну конечность, контактирующую с землей, а затем на другую. Хотя это кажется вполне простым, управление движением достаточно сложно и до настоящего времени, несмотря на достижения современной технологии, не удалось создать техническое устройство, удовлетворительно имитирующее человеческую походку.
Походка человека имеет два режима — ходьбу и бег. Различие между ними заключается в
24
Рис. 1.10. Изменения длительности опорной и безопорной фаз по мере увеличения скорости походки (Nilsson, Thorstensson, 1987):
1 — опорная фаза; 2 — безопорная фаза
доле времени каждого цикла, в течение которого тело поддерживается при контакте ноги с землей. В процессе ходьбы (светлые символы на рис. 1.10) почти все время одна нога постоянно находится на земле, и в течение короткого периода каждого цикла обе ноги находятся на земле; ходьбу можно охарактеризовать как чередующуюся последовательность движений с опорой на одну или обе конечности. Напротив, бег (темные символы на рис. 1.10) — это чередующаяся последовательность движений с опорой и без опоры, причем доля времени опорного состояния в цикле изменяется обратно пропорционально скорости: по мере увеличения скорости длительность опоры уменьшается (рис. 1.10). Заметим, что в течение одного цик-
Рис. 1.11. События и фазы, характеризующие процессы ходьбы и бега (Voughan, 1984):
П— правая; Л— левая
ла как ходьбы, так и бега имеется последовательность наличия и отсутствия опоры. Период опоры называется опорным периодом, период отсутствия опоры — безопорным периодом. Эти интервалы разделяются двумя событиями: мгновением, в которое нога контактирует с землей — отталкиванием (FS), и мгновением, когда нога отрывается от земли — отрывом (ТО). Циклы ходьбы обычно определяются этими событиями. Например, один полный цикл от отталкивания до отталкивания одной и той же ноги определяется как скачок.
Рис. 1.11 иллюстрирует эти соотношения. Скачок состоит из двух шагов. Шаг определяется как часть цикла от отталкивания (или отрыва) одной ноги до отталкивания (или отрыва) другой. В течение скачка имеют место четыре отталкивания и отрыва, по два на каждую конечность. Это правое отталкивание (rFS), правый отрыв (гТО), левое отталкивание (1FS) и левый отрыв (1ТО). Фаза опоры существует между отталкиванием (FS) и отрывом (ТО), тогда как безопорная фаза — между ТО и FS. На рис. 1.11 показано, как длительность скачка, а также опорной и безопорной фаз изменяется со скоростью походки.
Длина и частота скачка
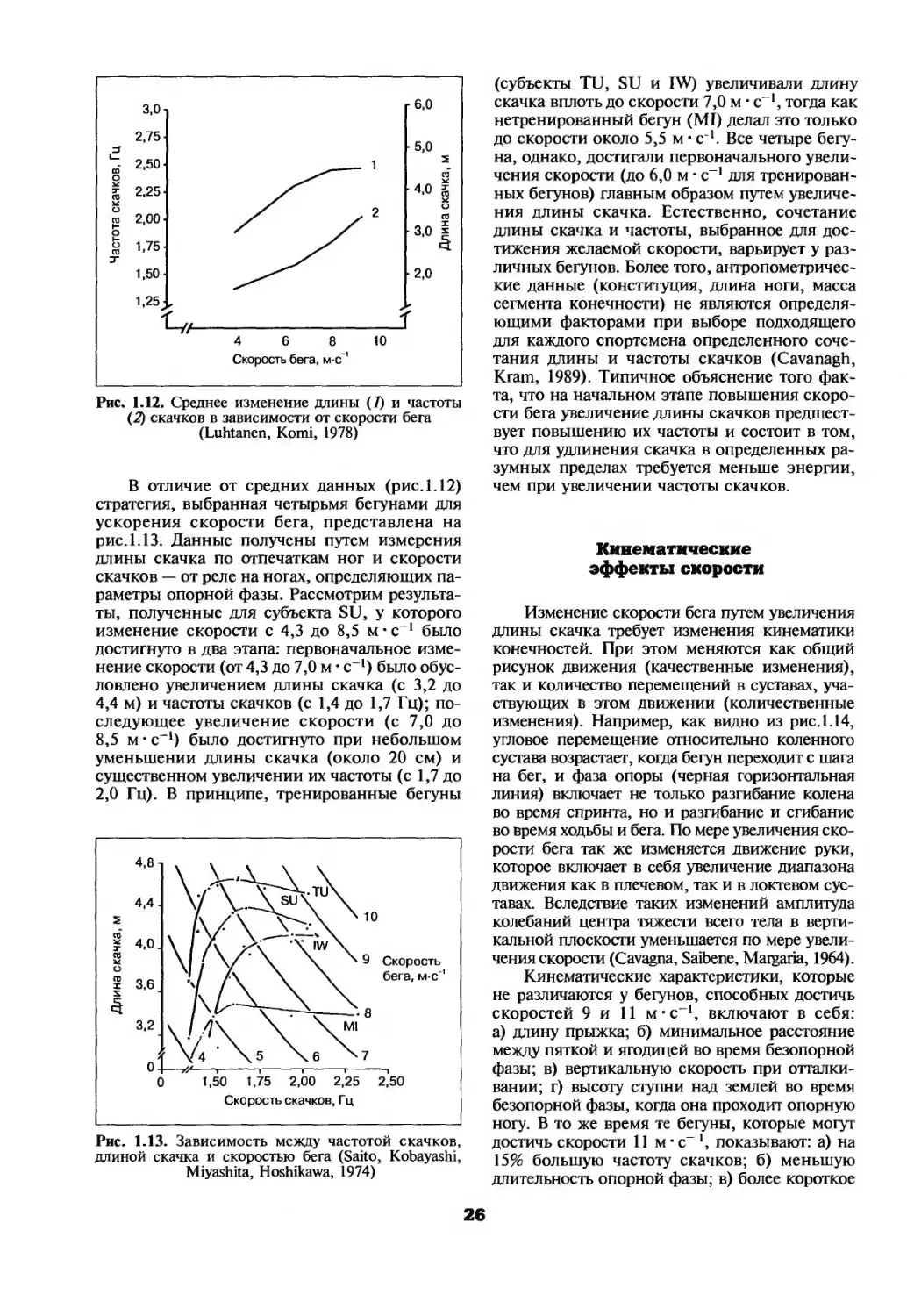
Скорость бега зависит от двух переменных — длины и частоты скачка (Voughan, 1984). Если длина скачка остается постоянной, то по мере того как длительность скачка уменьшается (т.е. возрастает частота скачков), скорость бега увеличивается. Если частота скачков остается постоянной, скорость возрастает с увеличением длины скачка. Влияние частоты и длины скачков на скорость бега показано на рис. 1.12. Желаемая скорость может быть получена при согласованном изменении длины и частоты скачков. Типичные их сочетания представлены на рис. 1.12. Например, человек, бегущий со скоростью 8 м • с-1, будет использовать частоту скачков около 1,75 Гц и длину скачка около 4,6 м. Как видно из рис. 1.12, в среднем бегун увеличивает скорость в диапазоне от 4 до 9 м с-1 путем постоянного повышения частоты скачков, хотя более медленно (наклон не столь крут) при меньших скоростях, увеличивая при этом длину скачка только при достижении скорости примерно до 8 м-с~*. Дальнейшее увеличение скорости достигается повышением частоты скачков, а их длина уже почти не изменяется. Обратите внимание, что влияние изменений длины и частоты скачков на скорость бега различно при низких и высоких скоростях; это иллюстрируется различным наклоном каждой кривой (длины и частоты скачков) при разных скоростях.
25
Рис. 1.12. Среднее изменение длины (Г) и частоты (2) скачков в зависимости от скорости бега (Luhtanen, Komi, 1978)
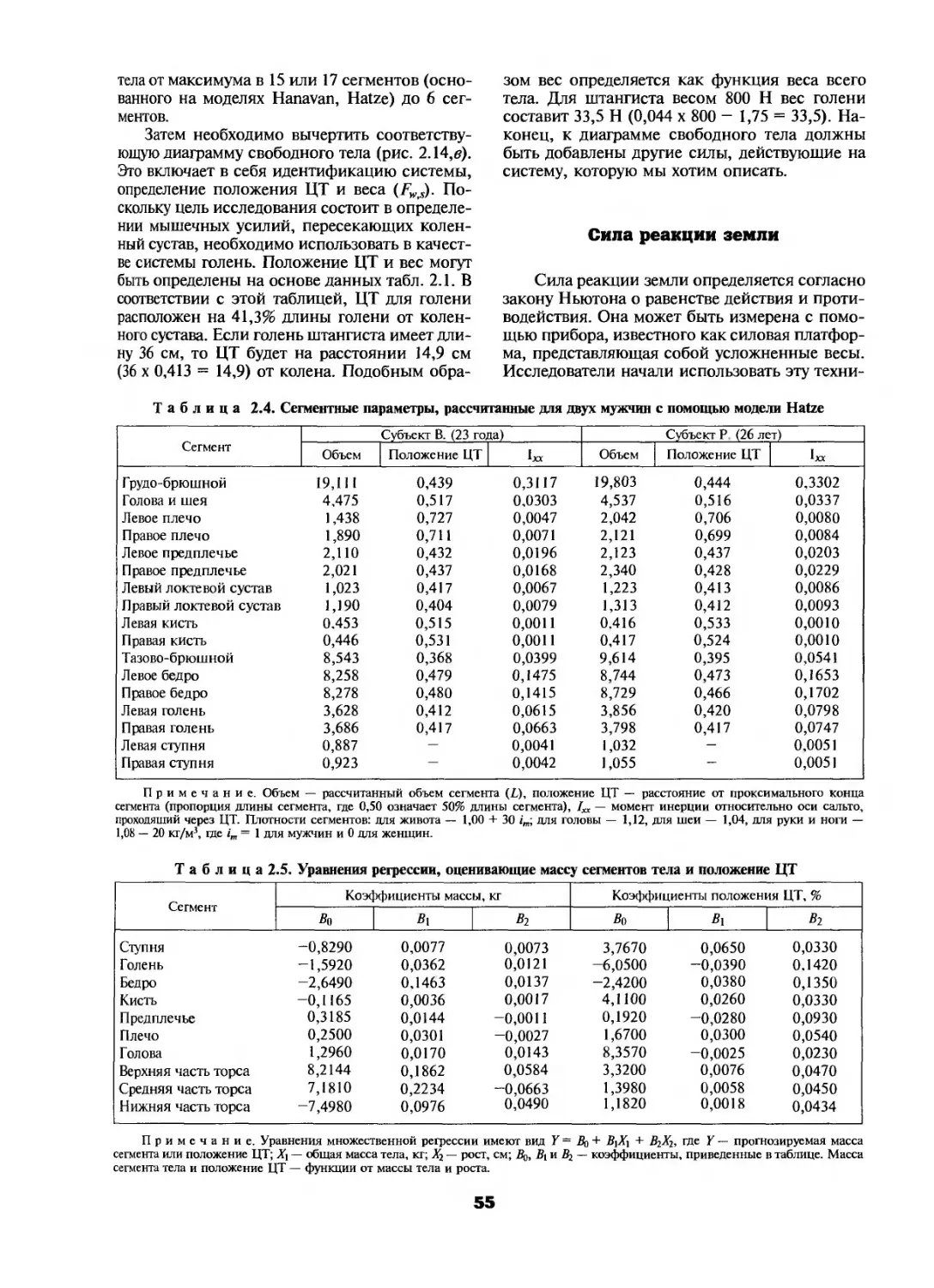
В отличие от средних данных (рис. 1.12) стратегия, выбранная четырьмя бегунами для ускорения скорости бега, представлена на рис. 1.13. Данные получены путем измерения длины скачка по отпечаткам ног и скорости скачков — от реле на ногах, определяющих параметры опорной фазы. Рассмотрим результаты, полученные для субъекта SU, у которого изменение скорости с 4,3 до 8,5 м • с-1 было достигнуто в два этапа: первоначальное изменение скорости (от 4,3 до 7,0 м • с-1) было обусловлено увеличением длины скачка (с 3,2 до 4,4 м) и частоты скачков (с 1,4 до 1,7 Гц); последующее увеличение скорости (с 7,0 до 8,5 м-с'1) было достигнуто при небольшом уменьшении длины скачка (около 20 см) и существенном увеличении их частоты (с 1,7 до 2,0 Гц). В принципе, тренированные бегуны
Рис. 1.13. Зависимость между частотой скачков, длиной скачка и скоростью бега (Saito, Kobayashi, Miyashita, Hoshikawa, 1974)
(субъекты TU, SU и IW) увеличивали длину скачка вплоть до скорости 7,0 м-с'1, тогда как нетренированный бегун (MI) делал это только до скорости около 5,5 м • с1. Все четыре бегуна, однако, достигали первоначального увеличения скорости (до 6,0 м с'1 для тренированных бегунов) главным образом путем увеличения длины скачка. Естественно, сочетание длины скачка и частоты, выбранное для достижения желаемой скорости, варьирует у различных бегунов. Более того, антропометрические данные (конституция, длина ноги, масса сегмента конечности) не являются определяющими факторами при выборе подходящего для каждого спортсмена определенного сочетания длины и частоты скачков (Cavanagh, Kram, 1989). Типичное объяснение того факта, что на начальном этапе повышения скорости бега увеличение длины скачков предшествует повышению их частоты и состоит в том, что для удлинения скачка в определенных разумных пределах требуется меньше энергии, чем при увеличении частоты скачков.
Кинематические эффекты скорости
Изменение скорости бега путем увеличения длины скачка требует изменения кинематики конечностей. При этом меняются как общий рисунок движения (качественные изменения), так и количество перемещений в суставах, участвующих в этом движении (количественные изменения). Например, как видно из рис. 1.14, угловое перемещение относительно коленного сустава возрастает, когда бегун переходит с шага на бег, и фаза опоры (черная горизонтальная линия) включает не только разгибание колена во время спринта, но и разгибание и сгибание во время ходьбы и бега. По мере увеличения скорости бега так же изменяется движение руки, которое включает в себя увеличение диапазона движения как в плечевом, так и в локтевом суставах. Вследствие таких изменений амплитуда колебаний центра тяжести всего тела в вертикальной плоскости уменьшается по мере увеличения скорости (Cavagna, Saibene, Maigaria, 1964).
Кинематические характеристики, которые не различаются у бегунов, способных достичь скоростей 9 и 11 м-с'1, включают в себя: а) длину прыжка; б) минимальное расстояние между пяткой и ягодицей во время безопорной фазы; в) вертикальную скорость при отталкивании; г) высоту ступни над землей во время безопорной фазы, когда она проходит опорную ногу. В то же время те бегуны, которые могут достичь скорости 11 м • с“ *, показывают: а) на 15% большую частоту скачков; б) меньшую длительность опорной фазы; в) более короткое
26
Рис. 1.14. Угол колена во время скачка при спринте (а), беге (б) и ходьбе (в). Угол колена 0 рад означает полное выпрямление
субъекта, у которого была ампутирована нижняя часть конечности, может быть полезным для оценки эффективности протеза в восстановлении нормальной походки. При таком анализе основное внимание уделяется сравнению формы соответствующих диаграмм угол—угол (сравните рис.1.15 и 1.16).
Для интерпретации рис. 1.15 рассмотрим кривую в направлении против часовой стрелки. Она иллюстрирует диаграмму угол—угол для одной ноги во время бега. Как указывалось выше, каждая нога во время прыжка испытывает отталкивание (FS) и отрыв (ТО). Между отталкиванием и отрывом (опорная фаза — пунктирная линия) нога находится в контакте с землей. Напротив, между отрывом и отталкиванием (безопорная фаза — сплошная линия) ступня указанной конечности не находится в контакте с землей. Кроме того, во время безо-порной фазы (от 1ТО до 1FS) другая нога вначале контактирует с землей, а затем отрывается от нее (от rFS до гТО). Соответственно рис.1.15 показывает, что: а) бедро поворачивается вперед относительно тазобедренного сустава и колено сгибается до минимального угла; б) после минимального угла колено разгибается до момента, как раз перед 1FS, тогда как бедро продолжает поворачиваться вперед, а затем начинает поворачиваться назад; в) во время опорной фазы (от 1FS до 1ТО) колено вначале сгибается, а затем разгибается, в то время как бедро поворачивается назад.
Диаграммы угол—угол имеют ряд важных особенностей, которые хорошо видны на рис. 1.15. Относительный угол (колено) изображается относительно абсолютного угла (бедро). График иллюстрирует комбинированное дейст-
горизонтальное расстояние между ногой и вертикальной проекцией центра тяжести при отталкивании; г) менее разогнутый коленный сустав при отталкивании; д) более вертикальное положение торса. Эти различия показывают, что на максимальную скорость бега, которую может развить спортсмен, влияют кинематические особенности движения.
Диаграммы угол—угол
Циклические виды деятельности, такие, как ходьба и бег, являются идеальными движениями для представления на диаграммах угол—угол, поскольку начало и конец движения находятся примерно в одной и той же точке диаграммы. Этот тип диаграмм оказался полезным при сравнении различных видов движения (Herschler, Milner, 1980а, 1980b; Miller, 1978). Например, сравнение диаграммы колено—бедро нормального субъекта во время бега с диаграммой
Рис. 1.15. Диаграмма колено—бедро левой конечности тренированного бегуна (Enoka, Miller, Bargess, 1982)
27
Угол бедра, рад
Рис. 1.16. Диаграммы колено—бедро для трех человек с ампутацией ниже колена, бегущих со скоростью от 2,7 до 3,8 м • c~l (Enoka, Miller, Burgess, 1982)
вне сгибания—разгибания колена и поворота бедра вперед—назад. На рисунке показаны три опорные оси, с помощью которых можно легче наглядно оценить диапазон перемещений при движении: а) ось 3/2 л указывает угол бедра, при котором бедро будет в вертикальном положении; б) ось л (3,14 рад) представляет угол колена при полном разгибании; в) ось л/2 показывает прямой угол (1,57 рад) коленного сустава. В соответствии с рис. 1.15 бедро проходит перед и за линией 3/2 л; коленный сустав никогда полностью не разгибается; наименьший угол колена меньше, чем прямой угол во время нормального скачка при беге со скоростью 4,2 м с-1.
В отличие от нормальной диаграммы колено-бедро опорная фаза (1FS — 1ТО), показанная на всех трех графиках рис. 1.16 для субъектов с ампутацией ноги ниже колена, существенно отличается от опорной фазы, иллюстрируемой
рис. 1.15 для здоровых лиц. Более конкретно, на диаграмме колено—бедро ампутантов можно обнаружить движение коленного сустава с постоянным углом, за которым следует сгибание, а не обычную последовательность сгибания и разгибания. Поскольку рис. 1.16 показывает диаграмму колено—бедро для протезированной конечности при ампутации ниже колена, вид графика, вероятно, не должен удивлять. Невозможность сгибания колена во время безопорной фазы, показанная на рис. 1.16 отсутствием немедленного уменьшения угла колена после FS, означает, что ампутанты использовали свои конечности как жесткие рычаги, вокруг которых осуществлялось вращение, в то время как протезированная нога была на земле. Этот тип графического изображения может быть использован в клинической практике для контроля программы реабилитации,
Рис. 1.17. Смещение вокруг правого плечевого и локтевого суставов во время бега (Li, Atwater, 1984) 1 — плечо; 2 — локоть
Рис. 1.18. Угловая диаграмма колено—плечо, основанная на данных, приведенных на рис. 1.17 (Li, Atwater, 1984)
28
направленной на корректировку походки для ее приближения к нормальной.
Диаграммы угол—угол использовались также для представления кинематики рук во время бега. Поскольку движение рук часто не ограничено сагиттальной плоскостью во время бега, необходима техника изображения, которая может учесть трехмерное движение. Если это сделать, то смещение верхней части руки относительно плеча и относительный угол между плечом и предплечьем (локтевой угол) для человека, бегущего со скоростью 11,4 м • с-1, будут иметь вид, показанный на рис. 1.17.
Положительный угол плеча означает сгибание (вперед от вертикали), отрицательный — разгибание (назад от вертикали). Конечно, амплитуда и время смещения изменяются в зависимости от скорости бега (Lusby, Atwater, 1983). Хотя рис. 1.17 дает представление о хронометрировании смещения по двум углам, то же движение целесообразнее представить в диаграмме угол—угол (рис.1.18). Смещения относительно плечевого и локтевого суставов представлены верхним правым и нижним левым квадрантами диаграммы угол—угол. Верхний правый квадрант отражает одновременное сгибание плеча и колена, тогда как нижний квадрант — одновременное разгибание плеча и колена. Опорная фаза (от rFS до гТО на рис. 1.18) в основном сопровождается одновременным сгибанием обоих суставов. Однако есть моменты, когда в обоих суставах имеют место противоположные движения. Один из примеров — фаза от 1FS до 1ТО, когда плечо разгибается, а локоть сгибается. Угловые диаграммы локоть—плечо, подобно таким же диаграммам для ноги, — качественное средство для оценки общей картины движения.
Другой полезной особенностью циклических диаграмм угол—угол, кроме качественной иллюстрации, является то, что размер диаграммы указывает диапазон перемещения в каждом суставе во время движения. Например, можно было бы ожидать, что возрастание длительности безопорной фазы по мере увеличения спринтером скорости обусловлено изменениями диапазона движения в различных суставах нижней конечности. Рис. 1.19 подтверждает эти ожидания, показывая, что при возрастании скорости (с 3,9 до 7,6 м • с-1) размах вращения как вокруг бедра, так и вокруг коленного сустава увеличивается, бульшая диаграмма угол—угол представляет большую скорость.
Хорошим упражнением для проверки усвоения способа исследования движения с помощью диаграммы угол—угол является построение диаграммы колено—бедро при движении бегуна вначале вверх по склону, а затем вниз (Milliton, Cavanagh, 1990). При этом возникает ключевой вопрос, как изменится движение по отношению к трем опорным осям. Например, было бы разумным ожидать, что по сравнению
Рис. 1.19. Диаграмма угол—угол для колена и бедра как функция от скорости бега (Miller, Enoka, McCullogh et al., 1979)
с бегуном на горизонтальной поверхности бегун, бегущий вверх по склону, будет меньше разгибать колено, меньше сгибать колено при минимальном угле и его бедро большую часть прыжка будет находиться впереди вертикали (вращение вперед). Попробуйте начертить эту диаграмму.
Диаграмма угол—угол, впервые предложенная Cavanagh и Grieve (1973), в значительной мере ограничена тем, что дает представление только о положении частей тела. Некоторые исследователи (например, Miller, 1978) экспериментировали с аналогичными диаграммами для угловой скорости и ускорения, но эти попытки были приняты неохотно, вероятно, из-за сложности интерпретации.
КИНЕМАТИКА БРОСАНИЯ И ТОЛКАНИЯ
Бросок и толчок — важные элементы движения человека. Цель обоих — придание объекту некоторой начальной скорости, обеспечивающей ему более или менее продолжительную фазу полета. Различие между ними состоит в способе ускорения объекта. В броске объект поддерживается и перемещается конечностью, обычно рукой, и увеличение его количества движения или момента (разгон) происходит в процессе проксимально-дистального движения сегментов тела (Putnam, 1993). Хотя толчок также характеризуется проксимально-дистальным
29
перемещением сегментов тела, он отличается от броска тем, что толчок — ударное движение, в котором количество движения объекта увеличивается при кратковременном взаимодействии объекта и конечности. В этом разделе мы рассмотрим некоторые основные особенности движений, связанных с бросанием и толканием, а затем изучим особенности фазы полета, которую испытывает движущийся объект. Независимо от способа, с помощью которого объект при контакте с конечностью приобретает количество движения, соответствующее бросанию или толканию, он следует по траектории, определяемой величиной и направлением полученной при этом начальной скорости и ускорением тяготения.
Бросок
Хотя бросок можно охарактеризовать как последовательное воздействие сегментов тела на количество движения бросаемого предмета (при постоянной массе изменение количества движения соответствует изменению скорости), задача может быть выполнена с помощью самых различных движений: бросок над рукой (например, бейсбол, крикет, копье, дротик), бросок под рукой (бросок в софтболе), толчок
Рис. 1.20. Последовательность движений в типичном бейсбольном броске (Feltner, Dapena, 1986)
от плеча (ядро) и бросок с натяжением (диск, молот). Кинематика типичного броска трехмерна по своей природе, особенно если бросок производится на максимальное расстояние (Feltner, 1989; Feltner, Dapena, 1986, 1989). Например, при выполнении броска над рукой (рис. 1.20) сегменты тела смещаются в вертикальном, боковом и продольном направлениях (рис. 1.21). Напротив, если задача бросания требует точности (например, метание дротика или выполнение штрафного броска в баскетболе), движение бросания, как правило, является плоскостным, и стратегия заключается в сведении к минимуму числа сегментов тела, участвующих в движении.
Последовательность движений типичного бейсбольного броска показана на рис. 1.20. Качественная оценка этой последовательности показывает, что движение включает в себя последовательный вклад в бросок различных сегментов тела, начиная от опорной площадки и далее вплоть до кисти. Бейсбольный бросок состоит из двух фаз (рис. 1.20): позиции от а до л, когда скорость мяча увеличивается в основном за счет действия ног, и позиции от м до х, когда скорость мяча возрастает за счет воздействия торса и рук. Вторая фаза, дающая большее увеличение скорости мяча, включает в себя последовательное возрастание угловой скорости сегментов тела в такой последовательности: таз, верхняя часть торса и верхняя часть руки, предплечье и кисть (Atwater, 1979). Это означает, что пик кинетического момента (момент количества движения) таза наступает раньше, чем верхней части торса и плеча; пик кинетического момента верхней части торса и плеча наступает раньше, чем у предплечья и т.д.
При этом бросок над плечом характеризуется максимальным диапазоном движений, выполняемых рукой. Такие перемещения имеют место в плечевом, локтевом и кистевом суставах, но наибольшими являются относительно плечевого сустава. Анатомия плечевого сустава допускает вращение плеча относительно трех различных осей: вокруг передне-задней оси, которое называется приведением—отведением; вокруг боковой оси, известное как сгибание—разгибание, и вокруг продольной оси, которое описывается как наружновнутреннее вращение. Рис. 1.22 показывает, что во время бейсбольного броска имеет место примерно 1,57 рад сгибания—разгибания локтя, 1,57 рад внешнего вращения и 1,0 рад внутреннего вращения. Положение р на рис. 1.20 показывает руку в положении максимального внешнего поворота. Это крайнее положение считается причиной многих травм руки, испытываемых бейсбольными питчерами.
В последовательности развития сегментной активности, которую можно видеть также в ударных движениях типа толчка, проксимальные сегменты начинают поворачиваться
30
Вид сверху
0,00
Рис. 1.21. Три вида траектории мяча во время броска над рукой. Мяч был брошен со скоростью 37,3 м • с-1 (83,5 м • ч'1), и момент выпускания обозначен как последнее зачерненное положение мяча (0,00). Зачерненные положения до выпуска означают интервалы ЮОмс. Измерения были сделаны при съемке движения (64 кадра • с-1) (Atwater, 1977)
Рис. 1.22. Диаграмма угол—угол для плеча и локтя при бейсбольном броске
Интервал между точками составляет 5 мс; чем дальше друг от друга находятся точки, тем быстрее движение. Точки л, р, ф соответствуют положениям, указанным на рис. 1.20 (Feltner, Dapena, 1986)
31
Рис. 1.23. Результирующая скорость (горизонтальная—вертикальная) мяча во время броска над рукой (Atwater, 1970)
до дистальных сегментов, и проксимальные сегменты начинают замедляться до того, как дистальные сегменты достигли пика угловой скорости. Имеется оптимальная задержка в хронометрировании поворота одного сегмента по отношению к следующему для максимизации скорости метаемого снаряда (Alexander, 1991). Результатом такой проксимально-дистальной последовательности является то, что скорость мяча не возрастает заметно до последних этапов движения. Рис. 1.23 показывает, что наибольшее увеличение скорости мяча при броске над рукой имеет место за 60 мс до момента выпуска, что соответствует интервалу, изображенному на рис. 1.20 тремя последними положениями мяча перед броском (г — t).
мени нахождения в воздухе, точности или скорости движения. Для многих видов толкания (например, ударе в футболе, волейбольной и теннисной подачи) это движение сходно с броском над плечом и включает в себя проксимально-дистальную последовательность воздействия сегментов тела на скорость до конечной точки (рука, нога, дополнение), которая нанесет удар по снаряду (рис. 1.24). Это показано на рис. 1.24: бедро достигает пика положительной (вперед) угловой скорости до того, как это делает голень, и угловая скорость бедра уменьшается в то время, как голень продолжает ускоряться до контакта с мячом. Luthanen (1988) описывает такую же последовательность для плеча, предплечья и кисти при волейбольной подаче.
Поскольку толчок передает количество движения снаряду посредством удара, важным различием между броском и толчком является степень жесткости конечности во время контакта с мячом, поэтому при выполнении ударов атлеты часто изменяют жесткость конечности. Например, изменение скорости мяча при ударе больше, если в момент контакта конечность более жесткая. Это достигается путем сжатия мышц ступни и тех, которые пересекают голеностопный сустав, так что ступня более жестко соединяется с голенью. Один из наглядных примеров изменения жесткости — пас двумя руками снизу в волейболе. Этот прием требует, чтобы игрок позволил мячу отскочить от вентральной поверхности предплечий по заданной траектории. Опытные игроки выполняют эту задачу, изменяя жесткость рук и, тем самым, определяя степень отскока мяча от предплечий. Аналогичный эффект достигается различной степенью натяжения струн ракеток, что позволяет регулировать их жесткость.
X* Д Л* A* V
Толчок
Ранее мы описывали толчок как вид удара, при котором полет является результатом кратковременного воздействия конечности (или ее дополнения) на объект. По этому критерию мы относим к толканию такие движения, как удар по футбольному или волейбольному мячу, а также удар по снаряду в видах спорта с ракеткой. Как и при броске, движение, лежащее в основе толкания, может изменяться в зависимости от целей деятельности; эти цели могут иметь отношение к горизонтальному расстоянию, вре-
0 20 40 60 80 100
Время, % длительности
Рис. 1.24. Угловая скорость голени (7) и бедра (2) бьющей ноги на последнем этапе удара (Putnam, 1991)
32
Движение снаряда
Соотношения между положением, скоростью и ускорением описывают траекторию, по которой будет перемещаться выпушенный (брошенный) предмет. Две силы действуют на траекторию снаряда: сила тяжести и сопротивление воздуха. Простейшим случаем движения снаряда является тот, в котором влияние сопротивления воздуха настолько мало, что им можно пренебречь. При наличии этого условия можно использовать следующие свойства, которые помогут изучить движение снаряда:
• под воздействием силы тяжести траектория отклоняется от прямой линии и может быть описана параболой;
• если бросок (удар, толчок) и падение происходят на одном и том же уровне, то время, требующееся снаряду для достижения пика своей траектории, равно времени движения от пика до падения;
• вертикальная составляющая скорости снаряда будет изменяться от направленной вверх (положительной) в начальный момент до нуля на пике траектории (когда меняется ее направление) до направленной вниз (отрицательной) при возвращении на землю;
• время, проводимое объектом в воздухе, зависит от величины вертикальной составляющей скорости при выпускании, (в дальнейшем в терминах “горизонтальные, вертикальные составляющие скорости, ускорения” слово “составляющие” опускается);
• единственная сила, которая действует на снаряд, — сила тяжести, и она вызывает вертикальное ускорение 9,81 м • с-2;
• поскольку нет сил, действующих в горизонтальном направлении, горизонтальное ускорение объекта равно нулю, что означает постоянство горизонтальной скорости объекта. Соответственно горизонтальное расстояние, пройденное снарядом, можно вычислить как произведение горизонтальной скорости и времени полета снаряда;
• время полета будет зависеть от вертикальной скорости в начальной точке при выпуске и от ее высоты над поверхностью приземления.
Траектория снаряда является параболической, и форма параболы зависит как от величины скорости, так и ее направления в начале движения. Три различные параболы показаны на рис. 1.25. Опираясь на законы движения снаряда, мы можем схематически представить вектор скорости в различных точках траектории, как показано, например, для копья на рис. 1.25. Если на копье в полете не действуют никакие силы (или действуют пренебрежимо малые усилия), кроме силы тяжести, его траектория определяется постоянной горизонтальной скоростью, положительной вертикальной скоростью в фазе подъема, отрицательной вертикальной скоро-
Рис. 1.25. Параболические траектории снарядов: а — прыжок в высоту; б — баскетбольный бросок;
в — метание копья
стью в фазе падения и нулевой вертикальной скоростью на пике траектории. Вертикальная скорость будет иметь наибольшее абсолютное значение в начале и в конце полета. Эти особенности относятся к полету ядра при толкании, движению гимнаста, осуществляющего прыжок, прыгуна в высоту, пролетающего над планкой, игрока в баскетбол, осуществляющего бросок в прыжке. Хотя некоторые игроки в баскетбол имеют вид зависающих в воздухе во время броска в прыжке, траектория их центра тяжести остается параболической. Иллюзия зависания создается относительным смещением рук и ног (Bushop, Неу, 1979).
Применяя определение скорости (уравнение 1.1) и ускорения (уравнение 1.2), проанализируем движение мяча при броске. Мяч брошен по углом 1,05 рад относительно горизонта и на высоте 2,5 м над землей с результирующей скоростью вдоль линии перемещения 6 м-с-1. Для решения задач о движении снаряда, удобно начать с эскиза начальных условий (рис. 1.26).
1. Сколько времени нужно мячу, чтобы достичь своей наивысшей точки? Траектория является параболической, так что мяч будет продолжать подниматься до тех пор, пока вертикальная скорость не станет равной 0:
_ ДУ vf~ vi . а = — = —-----;
Д/ t
-9,81 = (0 - 5,2)/
t = -5,2 / -9,81 t = 0,53 с,
где а среднее ускорение; v, = 6 sin 1,05 = 5,2 м-с-1
3 8-133
33
Ускорение силы тяжести ,, -9,81 мс'2
Рис. 1.26. Начальные условия для задачи о движении снаряда
2. Как высоко поднимется мяч? Максимально достигнутая высота зависит от вертикальной составляющей скорости выпуска плюс высота над уровнем земли, на которой был выпущен мяч. Мы уже знаем следующее: вертикальную скорость при выпуске, вертикальную скорость на пике траектории, вертикальное ускорение, высоту выпуска, время до достижения пика и что средняя скорость равна у:
- Дг
v = —;
д/
5,2 + 0 _ Дг .
2 “ 0,53’
Аг = 0,53(5,2.+0) =
2
Общая высота = 1,38 + 2,5 = 3,88 м.
Обратите внимание, что 2,5 — высота выпуска.
3. Какова вертикальная скорость мяча в момент перед ударом его о землю? Для ответа на этот вопрос следует рассмотреть вторую часть траектории от пика до контакта с землей. Начальная скорость равна 0 м • с-1, мяч падает на землю с максимальной высоты 3,88 м и испытывает ускорение 9,81 м • с 2:
— Ду о = —, Д/
(У г - V. )
-9 81 = 12---
t
(yf - 0)
-9,81 = —----
t
vf = -9,81/,
где a — среднее ускорение.
Зная, сколько времени потребуется мячу на путь от пика до земли, можно определить его конечную скорость перед ударом.
В качестве альтернативного варианта можно использовать одно из уравнений движения,
выведенных нами ранее. Уравнение (1.5) показывает связь между конечной скоростью, начальной скоростью, ускорением и перемещением:
у2 = vf + 2а(Дг);
v2 =2х(-9,81)хЗ,88;
v] = -76,1;
Уу = -8,73 м - с-1.
(Обратите внимание, что минус означает направление вниз.)
4. Сколько времени длится падение мяча от верхней точки траектории до поверхности земли? Мячу потребовалось 0,53 с для достижения пика, но для того, чтобы достичь земли, мяч должен дополнительно падать еще 2,5 м (высота выпуска). Общая высота от пика до земли составляет 3,88 м (—3,88 м — направление вниз):
- Дг
v = —,
Д/
Vf + Vj _ -3,88.
2 t ’
V/ + 0 -3,88
2 ~ t
Подставим в последнее равенство соотношение для Vf, полученное в задаче 3:
-9,81 х t -3,88 .
2 t ’
^2 _ —3,88 х 2
-9,81 ’
t2 = 0,79;
t = 0,89 с.
Используя полученное значение t, можно еще одним способом определить уув задаче 3: vf = -9,81/;
vf = -9,81 х 0,89;
Vf = -8,73 м • с-1.
Время, необходимое мячу для достижения земли, может быть также определено при использовании уравнения (1.3):
Vf = у,: + at = о + (-9,81/);
-8,73.
-9,81 ’
/ = 0,89 с.
5. Сколько времени мяч проводит в полете?
I = ^подъема + ^спуска = 0,53 + 0,89 — 1,42 С.
34
6. Какой горизонтальный путь пролетит мяч (т.е. как далеко он брошен)? В отличие от предыдущих вычислений, здесь нужно использовать горизонтальную, а не вертикальную составляющую скорости. Поскольку скорость при выпуске известна, мы можем определить горизонтальную скорость при выпуске. Известно также, что при отсутствии существенного сопротивления воздуха горизонтальная скорость постоянна, поскольку горизонтальное ускорение равно нулю. Отсюда
Дг т = —;
А/
vi + vf _ : Дг. 2 t ’
3 + 3 _ Дг 2 ~ 1,42 ’
Дг = 1,42 х 3 = 4,24 м.
Этот пример демонстрирует, что время полета снаряда зависит от вертикальной скорости при выпуске (или отталкивании) и высоты выпуска. Вертикальная скорость при выпуске определяет высоту, достигнутую снарядом, и время, потребовавшееся для достижения максимальной высоты. Поскольку параболические траектории симметричны, время, необходимое мячу, чтобы вернуться на высоту, с которой он был выпущен, равно тому, которое потребовалось для достижения пиковой точки. Если высота приземления отличается от высоты выпуска, время, необходимое для прохождения этого дополнительного расстояния, также необходимо учитывать при определении общего времени полета.
Если цель — максимальное увеличение дальности полета снаряда, необходимо выбрать правильное сочетание длительности полета и горизонтальной скорости, что включает выбор оптимального угла выпуска (или отталкивания). Рационально выбранный угол в большой степени зависит от относительного положения высоты выпуска и приземления. Когда высота выпуска и высота приземления одинаковы, оптимальным углом выпуска будет 0,785 рад (45°). Но если высота приземления ниже, чем высота выпуска, оптимальный угол бросания менее 0,785 рад. При высоте приземления выше высоты выпуска оптимальный угол броска больше 0,785 рад. Если скорость выпуска на данной высоте увеличивается, оптимальный угол приближается к 0,785 рад. Чем больше высота выпуска при постоянной его скорости, тем меньше (ниже 0,785 рад) оптимальный угол броска (Нау, 1985).
В этом примере мы подробно разобрали все свойства траектории полета мяча, достаточно просто используя определения для скорости и ускорения; нам не пришлось применять слож
ные формулы. Обратите внимание, что эта процедура применима только к условиям, когда влияние сопротивления воздуха настолько мало, что им можно пренебречь.
Поскольку траектория снаряда (т.е. парабола) предсказуема, это ставит под сомнение обычную практику в спорте. Часто тренеры говорят игрокам: “Следите за мячом”. Большинство атлетов интерпретируют это указание так, что нужно следить за мячом по всему его пути к бите, ракетке, ноге и т.д., для того чтобы можно было откорректировать свои действия с учетом изменений траектории снаряда. Это абсурдно. Например, допустим, что вы находитесь в защите при игре в волейбол и противник посылает на вас мяч со скоростью 100 миль в час (45 м • с-1). Кратчайшее время, за которое человек может отреагировать на стимул, — время реакции — составляет около 120 мс. За 120 мс волейбольный мяч пролетит 5,4 м. Иными словами, что бы ни случилось с траекторией мяча на последних 5,4 м, вы не сможете отреагировать достаточно быстро, чтобы изменить свои действия — вы можете с таким же успехом просто закрыть глаза во время последних 5,4 м. В этом случае рациональнее не следить за мячом весь путь до ваших рук, а скорее предсказать траекторию мяча, чтобы было достаточно времени для соответствующей реакции. Есть ли корреляция между успехами в спорте (например, волейболе, теннисе, футболе, американском футболе) и тем, как быстро игрок может предсказать траекторию снаряда? Действительно, в некоторых видах спорта, например, в волейболе, целесообразно подавать мяч без вращения, что делает его траекторию менее предсказуемой. При обтекании движущегося без вращения мяча потоком воздуха вблизи его поверхности возникают зоны отрыва потока, изменяющие величину и направление действующей на мяч аэродинамической силы, что, в свою очередь, может заметно изменять направление движения. Поскольку возникновение отрывных зон носит хаотический характер, траектория движения может отклоняться от предполагаемой на основании наблюдений в предшествующие отрыву моменты времени случайным образом.
Некоторые тренеры считают, что игрок должен следить за снарядом весь путь до контакта с ним, чтобы оценить величину своей ошибки при промахе. Например, в бейсболе есть аргумент, что баттер может “видеть”, насколько мяч разминулся с битой при отсутствии контакта. Однако нет научных свидетельств в поддержку утверждения, что можно воспринять и обработать визуальные изображения во время таких условий, так что нам не кажется правильным предлагать прослеживание снаряда на всей траектории полета, вплоть до контакта.
з*
35
РЕЗЮМЕ
Настоящая глава является основой для изучения движения человека. В ней рассмотрены строго определенные соотношения между положением, скоростью и ускорением и установлены основные понятия и определения, необходимые для точного описания движения. Кинематика — один из разделов физики, и применение ее для изучения движения челове
ка расширяет пределы понимания особенностей движения. Без четкого знания этих принципов трудно перейти к следующим уровням изучения движения человека. Поэтому в настоящей главе повторены важные концепции и приведены многочисленные графические и числовые примеры этих соотношений. Глава не является энциклопедией движения человека, но в значительной степени способствует его изучению.
iJOIIPbool С JUU ТЕГНАТИВНЫМ ОТВ1ГОМ
1. Единицей измерения смещения может быть метр. ПРАВИЛЬНО НЕПРАВИЛЬНО
2. Единица СИ для измерения массы — килограмм. ПРАВИЛЬНО НЕПРАВИЛЬНО
3. Сила — основная величина системы СИ.
ПРАВИЛЬНО НЕПРАВИЛЬНО
4. Скорость — производная величина в системе СИ. ПРАВИЛЬНО НЕПРАВИЛЬНО
5. Сокращением для секунды в системе СИ является “с”. ПРАВИЛЬНО неправильно
6. Символ “А“ означает изменение какой-либо величины.
ПРАВИЛЬНО НЕПРАВИЛЬНО
7. Километр меньше, чем миля.
ПРАВИЛЬНО НЕПРАВИЛЬНО
8. Наклон может иметь только положительный знак. ПРАВИЛЬНО НЕПРАВИЛЬНО
9. Радиан — векторная величина.
ПРАВИЛЬНО НЕПРАВИЛЬНО
10. Один градус равен 57,3 рад.
ПРАВИЛЬНО НЕПРАВИЛЬНО
11. Прямой угол равен 3,14 рад. ПРАВИЛЬНО НЕПРАВИЛЬНО
12. Перемещение описывает пространственный аспект движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
13. Скорость может быть выражена в радианах
В секунду. ПРАВИЛЬНО НЕПРАВИЛЬНО
14. Нулевая скорость означает, что объект, по крайней мере в данный момент, неподвижен.
ПРАВИЛЬНО НЕПРАВИЛЬНО
15. Скорость определяется как быстрота изменения положения по отношению ко времени. ПРАВИЛЬНО НЕПРАВИЛЬНО
16. Скалярная скорость является векторной величиной. ПРАВИЛЬНО НЕПРАВИЛЬНО
17. Направление ускорения и движения всегда совпадают. правильно неправильно
18. Ускорение всегда равно нулю, когда скорость равна нулю. ПРАВИЛЬНО НЕПРАВИЛЬНО
19. Обозначением углового ускорения является а. ПРАВИЛЬНО НЕПРАВИЛЬНО
20. Гравитационное ускорение равно 9,81 м-с-2. ПРАВИЛЬНО НЕПРАВИЛЬНО
21. Скалярная переменная характеризуется как величиной, так и направлением.
ПРАВИЛЬНО НЕПРАВИЛЬНО
22. Ускорение связано с наклоном графика скорость-время. ПРАВИЛЬНО НЕПРАВИЛЬНО
23. Объект, на который действует сила, испытывает ускорение.
ПРАВИЛЬНО НЕПРАВИЛЬНО
24. Символ 0 используется для обозначения углового положения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
25. Сочетание поступательного перемещения и вращения называется угловым движением.
ПРАВИЛЬНО НЕПРАВИЛЬНО
26. Положение может быть описано как производная от скорости.
ПРАВИЛЬНО НЕПРАВИЛЬНО
27. Единица измерения м-с-1 определяет величину ускорения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
28. Наклон графика положение—время дает информацию об ускорении.
ПРАВИЛЬНО НЕПРАВИЛЬНО
29. Когда график скорость—время имеет минимум, ускорение отрицательно.
ПРАВИЛЬНО НЕПРАВИЛЬНО
30. Символ 0 используется для обозначения углового положения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
31. Радиан определяется как отношение двух расстояний. ПРАВИЛЬНО НЕПРАВИЛЬНО
32. Нулевая скорость может свидетельствовать об изменении направления движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
33. Описание движения, которое игнорирует причины движения, известно как кинематическое описание.
ПРАВИЛЬНО НЕПРАВИЛЬНО
34. Направление ускорения совпадает с направлением движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
36
35. Одним из уравнений движения является Vx = V,2 + 2at. ПРАВИЛЬНО НЕПРАВИЛЬНО
36. Одно из уравнений движения имеет вид: rf—ri=vi+ 0,5at2.
ПРАВИЛЬНО НЕПРАВИЛЬНО
37. Одним из уравнений движения является rf = v? + 2а (rf— rj).
ПРАВИЛЬНО НЕПРАВИЛЬНО
38. Невозможно изобразить время на диаграмме угол—угол.
ПРАВИЛЬНО НЕПРАВИЛЬНО
39. Линейная скорость различных точек вдоль вращающегося тела одинакова.
ПРАВИЛЬНО НЕПРАВИЛЬНО
40. Направление вектора линейной скорости перпендикулярно траектории вращающегося жесткого тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
41. Вектор, показанный изогнутой стрелкой, имеет направление, тангенциальное к странице. ПРАВИЛЬНО НЕПРАВИЛЬНО
42. Угловая скорость является вектором.
ПРАВИЛЬНО НЕПРАВИЛЬНО
43. Векторное произведение является математическим оператором для векторов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
44. Член лэ2 показывает изменение величины со. ПРАВИЛЬНО НЕПРАВИЛЬНО
45. Направление v при угловом движении непрерывно изменяется.
ПРАВИЛЬНО НЕПРАВИЛЬНО
46. При полном разгибании угол локтевого сустава равен примерно 3,14 рад.
ПРАВИЛЬНО НЕПРАВИЛЬНО
47. Брахиорадиалис — мышца-разгибатель локтя. ПРАВИЛЬНО НЕПРАВИЛЬНО
48. Трицепс брахии — мышца-разгибатель локтя. ПРАВИЛЬНО НЕПРАВИЛЬНО
49. Единица измерения углового ускорения — рад • С 2. ПРАВИЛЬНО НЕПРАВИЛЬНО
50. Электромиография измеряет выходную электрическую активность мышцы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
51. Бицепс брахии может управлять разгибанием ЛОКТЯ. ПРАВИЛЬНО НЕПРАВИЛЬНО
52. Мышечная активность может вызывать ускорение конечности.
ПРАВИЛЬНО НЕПРАВИЛЬНО
53. При ходьбе всегда по крайней мере одна нога касается земли.
ПРАВИЛЬНО НЕПРАВИЛЬНО
54. При ускорении скорости движения опорное время сокращается.
ПРАВИЛЬНО НЕПРАВИЛЬНО
55. Прыжок состоит из двух шагов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
56. Если скорость шагов остается постоянной, то она возрастает по мере удлинения шага.
ПРАВИЛЬНО НЕПРАВИЛЬНО
57. Шаг может быть определен как интервал от отталкивания правой ногой до отталкивания правой ногой.
ПРАВИЛЬНО НЕПРАВИЛЬНО
58. Безопорная фаза длится от отталкивания до контакта ноги с землей.
ПРАВИЛЬНО НЕПРАВИЛЬНО
59. Длина прыжка для взрослого человека во время максимального ускорения имеет примерное значение 2,5 Гц.
ПРАВИЛЬНО НЕПРАВИЛЬНО
60. При беге со скоростью 8 м • с~1 частота скачков составляет примерно 20 Гц.
ПРАВИЛЬНО НЕПРАВИЛЬНО
61. Сгибание колена во время опорной фазы уменьшается со скоростью бега.
ПРАВИЛЬНО НЕПРАВИЛЬНО
62. Для увеличения длины прыжка требуется меньше энергии, чем для увеличения частоты прыжков.
ПРАВИЛЬНО НЕПРАВИЛЬНО
63. Различия в максимальной скорости бега (9 и 11 м • с-1) не зависят от длины шага, но на них влияет частота скачков.
ПРАВИЛЬНО НЕПРАВИЛЬНО
64. Угол 1,0 рад больше, чем прямой угол.
ПРАВИЛЬНО НЕПРАВИЛЬНО
65. Коленный угол никогда не достигает л рад во время нормального бега.
ПРАВИЛЬНО НЕПРАВИЛЬНО
66. Диапазон движения на диаграмме угол—угол измеряется в радианах.
ПРАВИЛЬНО НЕПРАВИЛЬНО
67. Относительный угол определяется как угол между соседними сегментами тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
68. Диаграмма угол—угол показывает диапазон движений для двух суставов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
69. Диаграммы угол—угол обычно изображают относительный угол по отношению к абсолютному углу.
ПРАВИЛЬНО НЕПРАВИЛЬНО
70. Сгибание локтя назад относится к разгибанию локтя и вытягиванию по направлению к пальцам.
ПРАВИЛЬНО НЕПРАВИЛЬНО
71. Движение руки во время бега обычно состоит из одновременного сгибания и разгибания локтя и плеча.
ПРАВИЛЬНО НЕПРАВИЛЬНО
72. Является ли толчок ударом?
ПРАВИЛЬНО НЕПРАВИЛЬНО
37
73. Кинематика броска от плеча — типичный пример дистально-проксимальной последовательности активации.
ПРАВИЛЬНО НЕПРАВИЛЬНО
74. В броске от плеча скорость мяча существенно увеличивается до последнего этапа движения. правильно неправильно
75. Во время плечевого броска смещение относительно плечевого сустава больше, чем относительно локтевого сустава.
ПРАВИЛЬНО НЕПРАВИЛЬНО
76. Во время толчка бедро достигает максимальной угловой скорости движения вперед раньше, чем голень. правильно неправильно
77. Траектория, по которой следует снаряд в воздухе, — прямая линия.
ПРАВИЛЬНО неправильно
78. Сопротивление воздуха никогда не влияет на траекторию снаряда.
ПРАВИЛЬНО НЕПРАВИЛЬНО
79. Скорость снаряда измеряется в м • с-1.
ПРАВИЛЬНО НЕПРАВИЛЬНО
80. Вертикальная скорость снаряда при выпуске определяет, как высоко поднимается снаряд. ПРАВИЛЬНО НЕПРАВИЛЬНО
81. Если пренебречь сопротивлением воздуха, горизонтальная скорость снаряда остается ПОСТОЯННОЙ. ПРАВИЛЬНО НЕПРАВИЛЬНО
82. Длительность полета зависит от горизонтальной скорости при выпуске снаряда и от высоты выпуска над поверхностью приземления. ПРАВИЛЬНО НЕПРАВИЛЬНО
83. Вертикальная скорость снаряда равна нулю на пике его траектории.
ПРАВИЛЬНО НЕПРАВИЛЬНО
84. Символом а обозначается среднее ускорение.
ПРАВИЛЬНО НЕПРАВИЛЬНО
85. Траектория снаряда является гиперболической. ПРАВИЛЬНО НЕПРАВИЛЬНО
ВОПРОСЫ С РАЗЛ1*ЧНЫМ ВЫБОРОМ ОТВЕТОВ
86. Какими будут ваш рост и масса тела в единицах СИ, если ваш рост 5 футов 5 дюймов и вы весите 150 фунтов?
а) 1,65 см, 668 Н;
б) 1,65 м, 669 кг;
в) 1,65 см, 68 кг;
г) 1,65 м, 669 Н;
д) 1,65 м, 68 кг.
87. Единицы измерения каких величин не относятся к семи основным единицам системы СИ?
а) длина;
б) сила;
в) температура;
г) время;
д) масса.
88. Что является единицей измерения для углового перемещения?
а) м;
б) рад-с-1;
в) рад;
г) м • с-1.
89. Из какой основной единицы выведен герц?
а) частоты;
б) длины;
в) времени;
г) массы.
90. Из каких единиц выведена единица для измерения скорости?
а) времени;
б) длины;
в) угла;
г) частоты.
91. Что не является единицей измерения ускорения?
а) рад • с-1 • с-1;
б) м • с' 2;
в) м/с-2;
г) рад • с-2.
92. Когда скорость имеет максимум, каким будет ускорение?
а) минимальным;
б) положительным;
в) отрицательным;
г) нулевым;
д) максимальным.
93. Какая переменная (переменные) не является векторной величиной?
а) перемещение;
б) ускорение;
в) время;
г) положение.
94. Каким символом обозначается угловая скорость?
a) v;
б) 0;
в) а;
г) со.
95. Какова будет скорость объекта, испытывающего ускорение 10 м- с-2, через 2 с? Считайте, что его начальная скорость составляла 3 м • с-1.
а) 20 м-с-1;
б) 13 м • с-1;
в) 23 м-с-1;
г) 30 м-с-1.
96. Укажите неправильные утверждения, относящиеся к рис. 1.2:
а) объект опускается от позиции 4 к позиции 5;
38
б) скорость при переходе от позиции 2 к позиции 3 меньше, чем от позиции 7 к позиции 2;
в) скорость при переходе от позиции 3 к позиции 4 равна нулю;
г) единица измерения скорости — м-с-1;
д) график дает информацию только о величине, а не о направлении вектора скорости.
97. Наклон графика какого типа указывает на скорость?
а) положение—время;
б) угол—угол;
в) сила—время;
г) ускорение—время.
98. Когда график скорость—время достигает максимума, какая переменная имеет нулевое значение?
а) смещение;
б) ускорение;
в) положение;
г) скорость.
99. Когда график скорость—время пересекает ноль, какая переменная (переменные) равна нулю?
а) ускорение;
б) скорость;
в) перемещение;
г) положение.
100. Когда график скорость—время пересекает ноль, какая переменная (переменные) имеет максимум или минимум?
а) скорость;
б) перемещение;
в) положение;
г) ускорение.
101. Какая информация не включена в диаграмму угол—угол?
а) диапазон движения;
б) скорость;
в) направление смещения;
г) время.
102. Какое уравнение отражает зависимость между линейной и угловой скоростью? a) v = га;
б) v = ли2;
в) v = гО;
г) v = га.
103. Какова единица измерения члена га2?
а) м-рад-с-1;
б) м-рад/с/с;
в) м • с-2;
г) м/с-2.
104. Безопорная фаза определяется как интервал между какими событиями?
a) FS и ТО;
б) гТО и 1ТО;
в) 1FS и гТО;
г) ТО и FS.
105. Что из указанного ниже не имеет места, когда бегун увеличивает скорость?
а) уменьшается длительность опорной фазы;
б) вначале увеличивается длина прыжка при более низких скоростях, затем возрастает частота прыжков для дальнейшего увеличения скорости;
в) уменьшается сгибание колена за толчком ноги;
г) у более быстрых бегунов частота скачков больше.
106. Какие из утверждений о кинематике походки правильны?
а) прыжок может быть определен как интервал от отталкивания правой ногой до отталкивания правой ногой;
б) первоначальное увеличение скорости бега происходит за счет одновременного увеличения длины прыжка и частоты прыжков;
в) коленный угол достигает 3,14 рад во время нормального бега (например, если смотреть по диаграммам угол-угол, колено—бедро);
г) скорость прыжков измеряется в Гц • с-1.
107. Какова типичная картина движения руки во время бега?
а) одновременное сгибание плеча и локтя;
б) разгибание плеча и сгибание локтя;
в) одновременное разгибание плеча и локтя;
г) сгибание плеча и разгибание локтя.
108. Вокруг какой оси не может вращаться плечевой сустав?
а) внешне-внутреннее вращение;
б) пронация—супинация;
в) сгибание—разгибание;
г) отведение—приведение.
109. Чем существенно отличаются бросок и толчок?
а) длительностью контакта;
б) жесткостью конечности;
в) проксимально-дистальной последовательностью активности сегментов тела;
г) целью изменения количества движения снаряда.
110. Что нехарактерно для движения снаряд, при котором сопротивлением воздуха можно пренебречь?
а) горизонтальное расстояние зависит от горизонтальной скорости при выпуске и длительности полета;
б) время полета зависит от горизонтальной скорости при выпуске и высоты места выпуска над поверхностью приземления;
39
в) вертикальная скорость снаряда во время полета остается постоянной;
г) горизонтальное ускорение, испытываемое снарядом, равно нулю.
111. Каков оптимальный угол выпуска, если высота приземления ниже высоты выпуска?
а) 0,785 рад;
б) больше 0,785 рад;
в) 1,57 рад;
г) меньше 0,785 рад.
ЗАДАЧИ
112. Определите в соответствующих единицах СИ рост человека 5 футов 8 дюймов.
113. В соответствии с “Разделом рекордов” в выпуске за март 1994 г., мировые рекорды на некоторых беговых дистанциях представлены в таблице.
Дистанция Женщины Мужчины
10,49 с 9,86 с
200 М 21,34 с 19,72 с
400 м 47,60 с 43,29 с
800 м 1 мин 53,28 с 1 мин 41,73 с
1,500 м 3 мин 50,46 с 3 мин 28,86 с
1 миля 4 мин 15,61 с 3 мин 46,39 с
3,000 м 8 мин 06,11 с 7 мин 28,96 с
5,000 м 14 мин 37,33 с 12 мин 58,39 с
10,000 м 28 мин 31,78 с 26 мин 58,38 с
Марафон 2 ч 21 мин 08,00 с 2 ч 21 мин 08,00 с
Рассчитайте среднюю скорость, с которой пробегали эти дистанции.
114. Постоянная сила (например, сила тяжести) приложена к безопорному объекту и дает постоянное ускорение, а отсутствие силы означает, что объект находится в состоянии покоя или перемещается с постоянной скоростью (т.е. ускорение равно нулю). Начертите график скорость—время для этих двух случаев.
115. Прыгун в воду выполняет прыжок с 10-метровой вышки из положения стойки на руках. Сопротивление воздуха во внимание не принимается.
А. Используя уравнение 1.4, определите время, которое потребуется прыгуну, чтобы достичь воды.
Б. Определите положение прыгуна (уравнение 1.4) с интервалами 0,1 с и постройте график положение—время.
В. Какова скорость прыгуна в момент его контакта с водой?
Г. Каково ускорение прыгуна на отметке 5 м?
116. График скорость—время показывает четыре качественно различных профиля ускорения, связанных с движением. Кратко опишите, что происходит с ускорением в каждом случае. В каких случаях ускорение равно нулю?
Время, с
117. Допустим, атлет пробегает 100 м за 9,98 с. Какова была его горизонтальная скорость в момент пересечения финишной линии? Допустить, что скорость атлета непрерывно возрастает от нуля (начало) до какой-то конечной величины. Каково было среднее ускорение атлета во время забега?
118. Какое среднее ускорение (м с-2) необходимо велосипедисту, чтобы затормозить с 22 до 7 миль в час за 51 с?
119. Постройте график ускорение—время по заданному графику скорость—время.
120. Мяч, удерживаемый на высоте 1,23 м над землей, выпускается из рук. Он достигает земли через 0,5 с.
А. Какова средняя скорость мяча (направление, величина и единица измерения) во время падения?
Б. Каково среднее ускорение, испытуемое мячом во время падения?
В. Допустим, мяч отскочил от земли до высоты 1,0 м, где был пойман. Начертите качественный график положение—время для мяча с момента его выпуска до поимки. Нанесите на оси соответствующие переменные и их единицы измерения.
40
121. Приведенные ниже данные положение-время представляют угловое движение ноги во время прыжка на лошади. Угол показывает положение бедра относительно правой горизонтали. Рассчитайте и изобразите графически соответствующие кривые угловой скорости и ускорения:
Угол, рад Время, с
2,57 0,115
2,59 0,130
2,62 0,145
2,79 0,161
3,11 0,176
3,46 0,191
3,89 0,206
4,45 0,221
5,06 0,236
5,49 0,252
5,83 0,267
5,90 0,282
5,93 0,297
5,93 0,312
6,02 0,327
122. Движения, участвующие в броске от плеча, довольно сложны. На рис. 1.22 показан профиль скорость—время для мяча во время броска.
А. Основываясь на рис. 1.22, качественно изобразите графики положение—время и ускорение—время для мяча.
Б. Какая особенность движения броска может вызвать фазу отрицательного ускорения, имеющую место примерно за 100 мс перед выпуском.
123. Приведенные ниже данные положение-время для бегуна были получены во время спринта на 50 м:
Расстояние, м 0 10 21 29 40
Время, с 0 3,1 4,3 5,5 6,6
Определите для этого бегуна
А. Среднюю скорость на 40-метровой дистанции.
Б. Ускорение на расстоянии 21 м.
В. Ускорение через 5,5 с.
124. Основываясь на связи между силой и движением, определите, какая из диаграмм правильно представляет движение объекта после его выпуска (McCloskey, 1983)? Объясните причину вашего выбора.
125. Мяч уронили с определенной высоты с нулевой начальной скоростью, он достиг земли через 0,18 с. Рассчитайте следующие переменные, относящиеся к движению мяча: А. Конечную скорость.
Б. Среднюю скорость.
В. Высоту, с которой был уронен мяч.
Г. Среднее ускорение.
126. Спортсмен выполняет бросок дротика, удерживая локтевой сустав неподвижно и поворачивая предплечье—кисть (один жесткий сегмент) вокруг локтевого сустава. Примем, что расстояние от локтевого сустава до дротика равно 41 см и предплечье-кисть имеет угловую скорость при выпуске 18 рад-с-1.
А. Рассчитайте линейную скорость дротика при выпуске.
Б. Каково ускорение, влияющее на изменение направления линейной скорости?
В. Если угол вектора линейной скорости при выпуске был равен 0,64 рад от горизонтали, какова будет горизонтальная дальность полета дротика до того, как он достигнет мишени, расположенной на той же высоте, что и точка выпуска?
127. В ходе киносъемки человека, который бежал со скоростью около 5,6 м-с-1, были получены данные, представленные в табл. Измеренные углы (рад) относятся к бедру и голени по отношению к правой горизонтали.
Интервал между кадрами фильма составлял 0,0067 с.
Нарисуйте диаграмму угол—угол для колена-бедра.
128. Копье было брошено под углом 0,78 рад к горизонтали со скоростью выпуска, направленной вдоль линии метания, 22,5 м • с-1. Каковы горизонтальная и вертикальная составляющие скорости выпуска?
129. Брошенный футбольный мяч достиг на крытом стадионе максимальной высоты 23 м. Если пренебречь эффектом сопротивления воздуха, какова будет скорость мяча в момент удара о землю?
130. Юноша ударяет мяч с подставки, находящейся на высоте 0,75 м от земли. Мяч ударен таким образом, что он смещается под углом 0,7 рад выше горизонтали. Результирующая скорость вдоль линии полета 4 м • с-1. Влиянием сопротивления воздуха пренебречь.
41
Таблица к задаче 127
Кадр Бедро Нога Колено Фаза
1 5,73 4,29 1,70 1ТО
4 5,70 4,47 1,91
7 5,65 4,75 2,24
10 5,60 4,90 2,44
13 5,51 4,95 2,58
16 5,40 4,94 2,68
19 5,30 4,81 2,65 rFS
22 5,24 4,59 2,49
25 5,20 4,40 2,34
28 5,12 4,24 2,26
31 4,94 4,08 2,28
34 4,71 3,96 2,39
37 4,49 3,89 2,54
40 4,25 3,90 2,79
43 4,12 3,91 2,93 гТО
46 4,10 3,77 2,81
49 4,12 3,53 2,55
52 4,17 3,32 2,29
55 4,27 3,08 1,95
58 4,39 2,85 1,60
61 4,49 2,72 1,37 1FS
64 4,65 2,67 1,16
67 4,88 2,66 0,92
70 5,09 2,81 0,86
73 5,30 3,10 0,94
76 5,53 3,39 1,00
79 5,67 3,74 1,21
82 5,70 4,10 1,54
85 5,71 4,27 1,70
А. Как долго мяч будет находиться в воздухе до удара о землю?
Б. Как далеко (горизонтально) пролетит мяч?
В. Почему разумно пренебречь сопротивлением воздуха?
131. В результате удара футбольный мяч получил начальную скорость 27,0 м • с-1, направленную под углом 1,15 рад к горизонтали. Мяч находился в воздухе 4,9 с, пока принимающий не принял его. Примем, что сопротивление воздуха пренебрежимо мало
и мяч достиг пиковой высоты на середине времени полета.
А. Какой наибольшей высоты достиг мяч? Б. Какое горизонтальное расстояние пролетел мяч?
132. Атлет совершает прыжок с начальной скоростью отталкивания 2,9 м-с-1.
А. Как высоко он может поднять свой центр тяжести, если прыгает вертикально вверх?
Б. Какой высоты он может достичь, если скорость отталкивания направлена под углом 0,70 рад к земле?
133. В чемпионате крокета в помещении Раль-фи Раундтри играет в финале с Минди Ма-лапроп. Минди выиграет чемпионат, если своим ударом сумеет поразить последнюю стойку. Допустим, Минди касается мяча, масса которого 1,76 кг, ударяя его так, что он приобретает начальную скорость 9,0 м-с-1. При движении по траве мяч имеет ускорение 4,0 м • с-2. Если Минди направляет мяч прямо в стойку, на каком наибольшем расстоянии она может быть от стойки, чтобы мяч попал в нее?
134. Игрок в баскетбол идет на бросок и отрывается от земли под углом 0,91 рад к горизонтали со скоростью 5,7 м-с1. Блокирующий игрок прыгает вертикально вверх со скоростью 4,8 м • с-1.
А. Какой из игроков прыгнет выше?
Б. Как долго каждый из игроков будет находиться в воздухе? Принять, что они не будут касаться друг друга.
135. Мяч для гольфа был брошен горизонтально из точки, высота которой на 3 м выше уровня дорожки. Если мяч приземлился (и остановился) на дорожке в точке, удаленной горизонтально на 150 м от исходной точки, какова была скорость мяча сразу же после удара?
ГЛАВА 2
СИЛА
Сила — это понятие, используемое для описания взаимодействия объекта с другими объектами окружающего мира. Она может быть определена как такое действие одного тела на другое, которое изменяет или стремится изменить движение: вывести тело из состояния покоя или изменить его скорость. Например, мяч, находящийся неподвижно на столе, будет оставаться в этом положении до тех пор, пока на него не подействует сила. Подобным образом человек, скользящий по льду на коньках, будет сохранять постоянную скорость, если только сила не изменит его движение, т.е. не изменит скорость или направление движения. Изучение движения, которое включает в себя понятие силы как причины изменения движения, называется динамикой.
Сила — векторная величина, характеризующаяся величиной, направлением и точкой приложения. Для того чтобы определить результирующий эффект нескольких сил (сложение) или, наоборот, разложить результирующий эффект на несколько составляющих (разложение), целесообразно применить правила векторной алгебры, используя наглядные геометрические построения. Такой метод исследования, используемый при анализе движения человека, позволяет определить суммарный эффект нескольких сил, действующих на объект или сегмент тела, и влияние каждой составляющей силы на выполнение отдельных элементов движения, например, поворота или стабилизации. Для определения суммарного (результирующего) эффекта нескольких сил строится их геометрическая сумма, называемая равнодействующей силой. Действие этой силы на объект эквивалентно сумме воздействий на объект всех ее составляющих.
На рис. 2.1 приведен пример использования сложения для определения результирующего эффекта коактивации ключичной и грудинной
частей мышцы пекторалис майор. Для проведения графического анализа действующие силы представлены стрелками (векторами), где длина стрелки указывает величину силы, а ориентация — направление действия силы. При этом вектор силы является скользящим вектором. Это значит, что его можно переносить вдоль линии действия, не изменяя при этом действие силы на объект. Техника графического построения включает в себя тщательное измерение длин и направлений стрелок и выбор масштаба, с помощью которого длина вектора соотносится с величиной силы, которую она изображает. На рис. 2.1, а показано направление и величина силы, создаваемой ключичной (Fmc) и грудинной (Fms) частями мышцы пекторалис майор. Ключичная составляющая создает силу 224 Н, направленную под углом 0,55 рад выше горизонтали, грудинная составляющая имеет величину 251 Н и действует на 0,35 рад ниже горизонтали. Графическое определение суммарного эффекта включает в себя совмещение начала одной составляющей с концом другой путем скольжения одного из векторов вдоль его линии действия (рис. 2.1,6) и затем соединения свободных концов (свободного начала со свободным концом) для получения равнодействующей (рис. 2.1,в). Равнодействующую получим, измерив длину стрелки (3 см) и преобразовав полученное значение в ньютоны (400 Н) согласно выбранному масштабу. Направление можно определить, измерив соответствующий угол транспортиром. Результирующий эффект (Fm), следовательно, имеет величину 400 Н и направление 0,1 рад над горизонталью.
Такой анализ может применяться к системам, включающим в себя несколько векторов сил. На рис. 2.2 показана нога с переломом бедренной кости, помещенная в вытяжку Рассела, состоящую из нескольких тросов и шкивов, которые поддерживают конечность. Для определения ре-
43
Рис. 2.1. Построение результирующей силы Fm, связанной с активацией как ключичной (fm>c), так и грудинной (FmJ) составляющей мышцы пекторалис майор. (Углы измерены в радианах)
зультирующей силы, воздействующей на кость, необходимо выполнить геометрическое сложение векторов сил. Для определения равнодействующей нужно вычертить вектора сил в выбранном масштабе, сохранив соотношение длин и в соответствующих направлениях. К ноге прилагаются три силы: одна через крепление на колене и две через шкив, прикрепленный к основанию вытяжки. В системе тросов и шкивов натяжение в тросе постоянное по всей длине троса, а шкивы служат для изменения направления вектора силы. Поэтому векторы сил Р, Q и 5 на рис. 2.2 имеют одинаковую величину, но^ различные направления. Равнодействующая R имеет величину, равную ее длине согласно масштабу, и направление 0,35 рад над горизонталью.
Рассмотрим процедуру графического разложения равнодействующей на составляющие.
Способ (рис. 2.3) заключается в построении параллелограмма (см. Приложение 2), где равнодействующая представлена диагональю, а стороны параллелограмма указывают величину и направление составляющих. При этом величина составляющих может быть определена при использовании масштабного коэффициента.
Во многих случаях при разложении мышечной силы составляющие направлены под прямыми углами друг к другу и параллелограмм становится прямоугольником. При этом компоненты вектора мышечной силы называются нормальной и тангенциальной составляющими. Нормальная составляющая представляет ту часть мышечной силы, которая действует перпендикулярно продольной кости сегмента и тем самым осуществляет поворот сегмента тела. Тангенциальная составляющая — часть силы, направлен
Рис. 2.2. Нога с переломом бедренной кости в вытяжке Рассела (Williams, Lissner, 1962)
Рис. 2.3. Разложение результирующей силы (Fm): а — нормальная и тангенциальная (FJ составляющие;
б — тригонометрическое соотношение между F„t„ и
44
ная вдоль кости по направлению к суставу, которая влияет на стабилизацию или сжатие сустава.
При сложении и разложении сил можно также использовать правила тригонометрии. Преимущество этого способа в том, что требуется меньшая графическая точность. Допустим, результирующая сила на рис. 2.3 равна 450 Н и направлена под углом 0,3 рад (17,2°) к плечевой кости. Рассчитаем величину составляющих. Вначале построим параллелограмм, добавим известные данные (рис. 2.3,6) и используем известные тригонометрические формулы для нахождения Fmn и Fm z:
F
а о т,п . sin 0,3 = ——;
450
Fm„ = 450 х sin 0,3 = 450 х 0,2955
Fm,„ = 133 Н;
cos 0,3 = —-т-;
450
= 450 х cos 0,3 = 450 x 0,9553;
= 430 H.
Таким образом, нормальная составляющая равна 133 Н, тангенциальная составляющая — 430 Н. При этом: а) угол натяжения большинства мышц относительно невелик, поэтому большая часть результирующей силы реализована в тангенциальном направлении; и б) результирующая представляет собой диагональ параллелограмма и поэтому должна быть наибольшей из трех величин.
Чтобы убедиться в идентичности графической и тригонометрической процедур, можно графически разложить результирующую на рис. 2.3, чтобы определить нормальную и тангенциальную составляющие. Перечертите результирующую (Fm) на миллиметровку в масштабе 1 см—25 Н и со стрелкой, направленной под углом 0,3 рад над горизонталью. Затем постройте прямоугольник так, чтобы результирующая была его диагональю. Замерьте длину двух сторон (Fm„ и Fm t) и, используя масштабный коэффициент, переведите их в силы. Насколько совпадают тригонометрический (Fmn = 133 Н и Fm t = 430 Н) и графический результаты?
ЗАКОНЫ ДВИЖЕНИЯ НЬЮТОНА
Исаак Ньютон (1642 — 1727) изучал взаимосвязь между силой и вызываемым ею движением и сформулировал три утверждения, называемые законами движения. Эти законы носят названия законов инерции, ускорения и действия-противодействия.
Закон инерции
Каждое тело продолжает сохранять состояние покоя или равномерного движения по прямой линии до тех пор, пока не будет вынуждено под действием внешних сил изменить это состояние
Иными словами, для остановки, начала или изменения движения требуется сила. Этот закон легко демонстрируется космонавтами, которые осуществляют маневры в условиях невесомости (микротяжести), например, передавая различные предметы друг другу или выполняя гимнастические упражнения. В гравитационном мире, однако, силы непрерывно действуют на тела, и изменение движения происходит там, где силы оказываются несбалансированными. При этом термин “тело” может относиться ко всему человеческому телу, части человеческого тела (например, бедру, руке, торсу) или некоторому предмету (например, ядру, бейсбольному мячу и т.п.).
Для того чтобы полностью понять этот закон, нужно знать, что такое инерция. Понятие “инерция” характеризует свойство тел сохранять неизменной скорость движения. Масса, выраженная в граммах (г), является мерой количества вещества, из которого состоит объект, и количественной мерой инерции. Рассмотрим два объекта различной массы, имеющих одинаковые скорости. Труднее изменить движение более массивного объекта, поэтому объект с большей массой обладает большей инерцией. Таким образом, инерция — это свойство тела, которое проявляется только при изменении величины или направления скорости.
В соответствии с законом инерции, движущийся объект будет продолжать равномерное прямолинейное движение (постоянная скорость), пока на него не подействует сила. Рассмотрим ситуацию в задаче 124 гл.1. Человек раскручивает над головой мяч, прикрепленный к нити. Если отпустить нить, будет ли мяч перемещаться по траектории “а” или “б”? Для ответа на этот вопрос нужно использовать закон инерции Ньютона. Если никакая сила не действует на объект, то он, благодаря своей инерции, должен перемещаться прямолинейно. Как только человек отпускает нить, на мяч не действует горизонтальная сила, и мяч будет двигаться по прямой линии (б). Но как тогда в бейсболе или софтболе можно заставить ударенный мяч отклониться от прямой линии? Очень просто. В соответствии с законом инерции на мяч с момента его выпуска начинают действовать другие силы, например, сопротивление воздуха, сила тяжести. Помните, что траектория снаряда была бы прямой линией, если бы на нее не влияли этакие силы, как тяжесть и сопротивление воздуха.
45
Поскольку при равномерном движении по прямой скорость постоянна и по величине, и по направлению, перемещение объекта по изогнутой траектории происходит под влиянием силы. Эта сила препятствует естественному стремлению объекта перемещаться прямолинейно. Это можно продемонстрировать, рассмотрев движение мяча в задаче 124 в два момента времени. Для этих двух моментов движение указывается вектором скорости (рис. 2.4).
Длина стрелки (величина вектора) одинакова в обоих положениях, что свидетельствует о том, что мяч перемещается с постоянной скоростью. Однако направление вектора различно, стрелки направлены в разные стороны. Это изменение направления может быть вызвано только действием силы. Эта сила, удерживающая мяч и направленная к центру вращения, которая изменяет направление, а не величину скорости во время вращательного движения, называется центростремительной силой. Центростремительная сила (Fc) определяется как
2
F (2.1)
с г
где т — масса; v — скорость; г — радиус кривизны траектории. Проанализируем уравнение (2.1). Как изменится Fc, если скорость мяча (v) останется постоянной, длина нити (г) такой же, а масса мяча (т) увеличится? При этих условиях центростремительная сила доля, на возрасти. Напротив, Fc уменьшится, если масса мяча и скорость останутся постоянными, а длина нити увеличится.
Рассмотрим проблему, часто решаемую специалистами по легкой атлетике. Атлеты, выполняющие метание молота, бросали молот слишком далеко и могли травмировать атлетов, бе-
Рис. 2.4. Мяч на нити, который перемещается с постоянной скоростью. Величина вектора скорости остается постоянной, но его направление изменяется
гущих по дорожке. Что нужно предпринять, чтобы уменьшить длину броска?
Поскольку длина броска зависит от скорости при выпуске, решим уравнение 2.1 относительно v и проанализируем его:
Уравнение показывает, что скорость (а отсюда и перемещение, испытываемое молотом) прямо пропорциональна Fc и г и обратно пропорциональна ш. При уменьшении Fc и /-уменьшается и v, но при уменьшении т скорость увеличивается. Поэтому для уменьшения дальности полета организаторы соревнований должны либо увеличить массу молота (т), либо уменьшить его длину (г).
Закон ускорения
Скорость изменения количества движения тела равна по величине и направлению приложенной к телу силе
Количество движения (G), которым обладает тело, определяется как произведение массы (/и) и скорости (v). Бегун с массой тела 60 кг, двигающийся с горизонтальной скоростью 8 м • с-1, имеет количество движения 480 кгм • с-1. Принимая во внимание, что
G = ту, (2.2)
скорость изменения количества движения может быть записана как
AG _ А(оту)
АГ АГ
Учитывая, что при анализе движения человека массу можно считать постоянной величиной, в соответствии с законом ускорения приложенная сила (F) пропорциональна скорости изменения количества движения:
г AG Av
F =---= т—.
АГ АГ
Далее, поскольку Av/Ar представляет собой изменение скорости во времени, оно эквивалентно ускорению и, следовательно,
F= ma. (2.3)
Уравнение (2.3) является алгебраическим выражением закона ускорения Ньютона и показывает, что сила равна произведению массы и ускорения. Вообще говоря, это связь между причиной и следствием. Левую часть выражения (F) можно рассматривать как причину, поскольку она представляет воздействие окружающей среды на движущийся объект. Правая
46
часть представляет следствие, поскольку показывает результат действия силы, ее кинематический эффект (тиа).
Закон действия и противодействия
Для каждого действия существует равное и противоположно направленное противодействие
Закон действия и противодействия утверждает, что любому действию, которое одно тело оказывает на другое, противостоит действие, оказываемое другим телом на первое. Это взаимодействие определяется силой, воздействующей на оба тела. В строгом смысле неправильно утверждать, что одно тело оказывает действие, а второе отвечает на него противодействием. Скорее два тела одновременно взаимодействуют, и это взаимодействие называется силой.
Рассмотрим игрока, осуществляющего бросок в прыжке в баскетболе. Во время прыжка человек воздействует на землю, и земля соответственно реагирует силой реакции (сила реакции земли) на прыгающего. Согласно закону действия—противодействия силы между игроком и землей (т.е. воздействие игрока на землю и земли на игрока) равны по величине, но противоположны по направлению. Следствием этого взаимодействия, как указано в законе ускорения (F = та), является то, что каждое тело (т.е. игрок в баскетбол и земля) испытывают ускорение, зависящее от их массы. Если средняя сила равна 1500 Н, и масса тела игрока 75 кг, он будет испытывать ускорение 20 м-с-2. Однако из-за большой массы планеты Земля ускорение, испытываемое Землей, будет невос-принимаемым, поскольку произведение массы и ускорения не может превысить 1500 Н. Так как масса Земли велика (по сравнению с игроком в баскетбол), ее ускорение будет ничтожно мало.
ДИАГРАММА СВОБОДНОГО ТЕЛА
Анализ целенаправленной двигательной деятельности человека представляет собой весьма сложную проблему, исследование которой требует учета большого количества переменных. Изучение механических аспектов движения с помощью диаграммы свободного тела существенно уменьшает сложность анализа. Как правило, диаграмма представляет собой схему, заменяющую исследуемый объект системой соединенных между собой отрезков. При этом все существенные для изучаемого движения влия
ния окружающей среды моделируются силами, приложенными к звеньям. Отметим, что диаграмма схематически изображает ту часть тела, движение или равновесие которой изучается. При этом влияние отброшенных частей тела заменяется силами реакций. Диаграмма свободного тела схематически представляет общий вид тела или его части как изолированного объекта в пространстве (в этом контексте телом может быть все, что занимает некоторую часть пространства и имеет инерцию).
На диаграмме все внешние силы, влияющие на систему, обозначены стрелками. Диаграмма свободного тела или его части, изображенная в виде системы взаимосвязанных элементов, схематически представляет исследуемый объект как жесткую изолированную конструкцию с действующими на нее силами. В принципе, термин “система” может применяться к объекту, характеризующемуся, по меньшей мере, одной входной и одной выходной переменной, внутренняя структура которого определяет взаимоотношения между входом и выходом (Lamarra, 1990). Например, сердце представляет собой систему, выходом которой является поток крови, а входами — механические, нервные и химические воздействия. В диаграмме свободного тела входом являются силы, действующие на систему, а выходом — движение системы. Построение соответствующих схем для различных вариантов движений в разных условиях представляет собой одну из наибольших трудностей, с которой сталкиваются начинающие студенты при анализе движения человека.
Демпстер (1961) прослеживает развитие методологии изучения движений с помощью диаграммы свободного тела и указывает, что самые ранние случаи ее использования встречаются в работах Архимеда, Леонардо да Винчи и Джиованни Борелли. Примеры работ Борелли показаны на рис. 2.5. Борелли (1680), очевидно, понял необходимость использования рычагов тела для того, чтобы сбалансировать мышечную силу и нагрузку и, тем самым, сохранить равновесие. Историческое исследование Демпстера свидетельствует, что хотя диаграмма свободного тела как технический метод была разработана в течение последних 100 лет, она основывается на более чем 350-летнем опыте классической физики.
В остальной части настоящего раздела мы рассмотрим несколько диаграмм свободного тела и обсудим способы построения системы как основы для анализа движения человека. Фактические силы, которые могут быть включены в диаграмму свободного тела, более подробно будут описаны ниже. Если диаграмма свободного тела включает в себя только две силы, они должны быть равными, противоположно направленными и параллельными.
47
Рис. 2.5. Наглядные диаграммы равновесия — одни из самых ранних примеров диаграмм свободного тела из работ Борелли
Когда диаграмма свободного тела состоит из трех или более сил, они должны быть сходящимися (т.е. их линии действия должны сходиться), так как в противном случае одна из сил будет оказывать несбалансированное воздействие на систему (Meriam, 1987).
Рассмотрим пример, в котором мы хотим определить величину и направление силы реакции земли, испытываемой бегуном (рис. 2.6,а). В соответствии с законом действия—противодействия, когда бегун отталкивается от земли, земля отталкивает его; мы хотим знать, с какой силой земля это делает. Прежде всего нужно
Рис. 2.6. Построение диаграммы свободного тела: а — реальная фигура; б — построение системы; в — система с учетом сил окружающей среды, действующих на систему
определить основные элементы системы и вычертить схему (рис. 2.6,6). Следующий шаг — идентификация всех внешних сил, действующих на систему, нанесение их на рисунок с помощью стрелок соответствующей длины и направления и введение их обозначений (рис. 2.6,в). Силы, показанные на рис.2.6,в, представляют сопротивление воздуха (Га), силу веса (Fw) и силу реакции земли (Ех). Вообще рассматриваемые силы включают в себя силу веса системы и силы, возникающие вследствие контакта с другими телами (например, силу реакции земли, сопротивление воздуха).
В диаграммы свободного тела включают только внешние силы, действующие на систему. Например, диаграмма свободного тела на рис. 2.6,в не показывает никаких мышечных сил вокруг коленного сустава, хотя мышца квадрицепс феморис, несомненно, сокращается; это не показано потому, что мышечная сила является для этой системы внутренней. Если, однако, цель анализа — изучение мышечной активности вокруг сустава во время движения, нужно формировать другую систему, учитывающую это влияние. Если мы рассматриваем конкретное взаимодействие мышц, активно участвующих в специфическом движении, мы должны определить систему так, чтобы взаимодействие происходило между системой и ее окружением. Нам нужно заново сформировать систему так, чтобы интересующая нас внутренняя сила оказалась внешней по отношению к системе. При определении мышечной активности это включает в себя условный разрез через сустав (например, коленный). Вокруг каждого сустава есть
48
ряд тканей, включая мышечную ткань, капсулу сустава, сухожилия и т.п., причем действие мышечной ткани отличается от действия на сустав других тканей. Соответственно при вычерчивании диаграммы свободного тела, которая включает в себя разрез через сустав, мы указываем результирующую мышечную силу и силу, обусловленную главным образом контактом костей (сила совместной реакции) прилегающих сегментов тела. Давайте вернемся к примеру, изображенному на рис.2.6. Для изучения взаимодействия мышц вокруг коленного сустава в момент, показанный на рис.2.6, б, соответствующая диаграмма свободного тела показана на рис.2.7.
Допустим, возникла такая ситуация: тяжелоатлет, подготовка которого включает в себя силовые подрывы (поднятие штанги от уровня колен до груди одним непрерывным движением), после нескольких месяцев тренировки начал ощущать боли в спине. Соответствующий анализ должен состоять в определении усилий, которыми нагружаются мышцы, растягивающие спину и бедро во время движения. Поскольку сила наибольшая в начале движения, предметом анализа будет начальное положение тяжелоатлета (рис.2.8, а). Цель анализа — расчет мышечной силы, поэтому система должна быть определена так, чтобы можно было идентифицировать мышечный эффект на диаграмме свободного тела. Это достигается условным разрезом системы через тазобедренный сустав так, что системой становится либо часть тела выше бедренного сустава вместе со штангой, либо часть тела ниже бедренного сустава. В первом случае фигура на схеме состоит из кружка, изображающего голову, и линии, представляющей торс и руки (рис. 2.8,6).
На следующем этапе иллюстрируются все силовые взаимодействия, схематически изображенные векторами сил (стрелками), между системой, представленной верхней частью тела штангиста вместе со штангой и ее окружением. Основные силы при этом — вес штанги Fw Z1 и вес верхней части тела штангиста (Fw „). Силы веса всегда направлены вертикально вниз. Поскольку мы условно разрезаем сустав на диаграмме свободного тела, необходимо показать как результирующую мышечную силу (Fm), так и силу реакции сустава (F,). Сила реакции сустава (Fy) представляет силы, обусловленные связками, суставными капсулами и соединениями костей между собой. Хотя вектор Fm в основном направлен так, чтобы противодействовать повороту, производимому нагрузкой (т.е. векторами Fw fc, Fw „), направление вектора F) обычно неизвестно, и он произвольно изображается в любом направлении. Но если система находится в равновесии, то силы, направленные вправо, должны быть уравновешены силами, направленными влево; силам, направ-
Рис. 2.7. Диаграмма свободного тела системы (голень и стопа), на которую воздействуют четыре внешние силы:
Fs — сила реакции земли; F,- — сила совместной реакции сус-тава; Fm — результирующая мышечная сила; Fw — вес; К— коленный сустав
ленным вверх, должны противостоять силы, направленные вниз; а силы, производящие вращение по часовой стрелке, должны уравновешиваться силами, вызывающими вращение
Рис. 2.8. Построение диаграммы свободного тела для анализа части движения по поднятию штанги: а — положение штангиста; б — система, используемая для установления сил, действующих на бедренный сустав; в — диаграмма свободного тела, показывающая взаимодействие между системой и ее окружением
4 8-133
49
против часовой стрелки. Иными словами, система сил должна быть сходящейся. Еще одна сила, связанная с этой диаграммой свободного тела, вызвана жидкостями в брюшной полости (внутрибрюшное давление F,), которое создает силу, стремящуюся вызвать растяжение относительно бедренного сустава (Н) (Andersson, Ortegren, Nachemson, 1977; Eie, Wehn, 1962; Rab, Chao, Stauffer, 1977). Это рефлекторно контролируемое усилие действует как защитный механизм и играет важную роль в поднятии тяжестей (Marras, Joynt, King, 1985). Эти пять сил (Fw/), Fw, Fy-, Fm, F() представляют основные взаимодействия между данной системой и ее окружением. Соответствующая диаграмма свободного тела показана на рис. 2.8,в.
В примере со штангистом (рис. 2.8) мышечные силы, действующие относительно бедра, были упрощенно представлены одной результирующей мышечной силой (Fm). Фактически много мышц пересекают бедренный сустав и они активны во время поднятия тяжестей. Можно начертить более подробные диаграммы свободного тела, идентифицирующие действие отдельных мышц. Рис. 2.9 дает пример такого подхода.
Диаграмма свободного тела поперечного сечения поясничного ствола на уровне позвонка L3 включает в себя 10 мышц (Fm), по 5 на каждой стороне, внутрибрюшное давление (F;), силы сжатия (Fc) и среза (Fv) в позвонке. На
Рис. 2.9. Трехмерная диаграмма свободного тела поперечного сечения поясничного ствола (Ashton-Miller, Schultz, 1988)
диаграмме представлены ректус абдоминус (Fm r), внутренняя косая (Fmj), наружная косая (Fm x), латиссимус дорси (Fm>/) и эректор спине (Fm e). Сила, оказываемая каждой мышцей, представлена одним эквивалентным вектором силы, приложенным в центре площади поперечного сечения мышцы (Nemeth, Ohlsen, 1986). Эта диаграмма свободного тела может быть использована для определения вклада мышц в результирующую силу и момент силы относительно центра межпозвоночного диска L3 (Ashton-Miller, Schulz, 1988).
Несмотря на свою элегантность, пример, показанный на рис. 2.9, не является подлинной диаграммой свободного тела. Диаграмма свободного тела должна включать в себя правильное количественное графическое решение, в котором как направление, так и величина действующих сил вычерчены в масштабе. Схема диаграммы свободного тела, приведенная на рис. 2.9, которая не включает реальные величины сил, называется трехмерной диаграммой (Dempster, 1961).
СИЛЫ ДВИЖЕНИЯ ЧЕЛОВЕКА
Определив систему с помощью диаграммы свободного тела, можем рассмотреть взаимодействия (силы), которые, вероятнее всего, существуют между системой и ее окружением. В принципе силы, включенные в диаграмму свободного тела, вызваны контактом с окружающей средой, а также гравитационными, электрическими и магнитными эффектами. При анализе движения человека в диаграмму свободного тела обычно включают восемь сил: вес, силу реакции земли, силу реакции сустава, силу мышц, внутрибрюшное давление, сопротивление жидкости, силу упругости и силу инерции.
Вес
Сила тяжести определяется законом всемирного тяготения Ньютона (законом гравитации): все тела притягиваются друг к другу с силой, пропорциональной произведению их масс и обратно пропорциональной квадрату расстояния между ними.
FocM^, (2.4)
г2
где гп\ и т2 — массы двух тел, г — расстоя- . ние между ними.
Этими силами притяжения между различными объектами обычно пренебрегают при изучении движения человека, за исключением притяжения к Земле. Вес — количественная мера
50
кавитационного взаимодействия объекта с Землей. Обычно он измеряется в ньютонах (Н). Вес изменяется пропорционально массе: чем больше масса, тем больше притяжение, но это две различные величины. Вес является силой (производной величиной), тогда как масса (основная единица в системе СИ) — мера количества вещества. Вес представляет взаимодействие между телом и Землей, и величина взаимодействия зависит от взаимодействующих масс и расстояния между ними, поэтому сила тяжести уменьшается по мере увеличения высоты над уровнем моря. Это свойство проявляется при проведении на большой высоте соревнований, в которых выступающий должен преодолевать силу тяжести (например, прыжок в длину, толкание ядра, поднятие штанги). Уменьшение тяжести, особенно в условиях микрогравитации, имеет интересные следствия для управления движением (Davis, Cavanagh, 1993).
Вес тела определяется достаточно легко путем взвешивания. Достоверность этой процедуры подтверждается простым анализом, основанным на законе ускорения Ньютона (уравнение 2.3)
LF = та,
который указывает, что сумма (Z-сигма) сил (F) вызывает в системе ускорение (а), которое зависит от массы системы. Сила и ускорение — векторные величины. Поскольку вес действует только в вертикальном направлении (z), анализ может быть ограничен теми составляющими, которые действуют вертикально. Сила веса (FK) направлена вниз и имеет отрицательное направление, а вертикальная составляющая силы реакции земли (Fgz) имеет положительное направление вверх. Решение считать FH,отрицательным, a Fgz положительным абсолютно произвольно; тем не менее важно иметь в виду эти различия в направлении. Для того чтобы учесть это, каждая диаграмма свободного тела должна сопровождаться контрольными осями, показывающими положительное направление каждой составляющей: положительные направления вертикали (z), горизонтали (х) и поворота.
Соответствующая диаграмма свободного тела показана на рис. 2.10, и анализ проводится следующим образом:
IF. = maz;
-Fw + Fgz = maz, Fw = Fgz - maz.
Поскольку во время взвешивания субъект неподвижен, az = 0. Отсюда
= F^-0;
~ Fg,z-
Направление вектора веса всегда вертикаль-
Рис. 2.10. Диаграмма свободного тела всего тела
но к центру Земли; вектор начинается в точке, называемой центром тяжести. Центр тяжести (ЦТ) представляет точку, вокруг которой равномерно распределены все частицы объекта. Это абстрактная точка, которая перемещается, когда сегменты тела сдвигаются друг относительно друга. ЦТ не ограничен физическими границами объекта; например, ЦТ бублика, несомненно, расположен внутри отверстия. Многие хорошие прыгуны в высоту могут выносить ЦТ за пределы тела или хотя бы сдвигать его к внешним границам (см. рис. 2.11, б).
Как видно на рис. 2.11, высота, которую должен преодолеть прыгун, состоит из трех компонентов: 1) высоты (Н() ЦТ над землей при отталкивании; 2) высоты (Н2), на которую прыгун может поднять ЦТ относительно Нь 3) разности между максимальной высотой, достигаемой ЦТ, и высотой планки (Нз). По отношению к Н3, хотя спортсмен может прыгнуть на 2,28 м, это не означает, что ЦТ был поднят до такой высоты, а только то, что тело прошло над планкой — оба условия не являются равнозначными. На рис. 2.11,6 показана теоретическая техника прыжка в высоту, которая делает высоту Н3 максимальной. Подобное рассуждение относится к гимнастическому прыжку (Fletcher, Lewis, 1960; Hubbord, 1980). Когда прыгун оттолкнулся от земли, путь ЦТ предопределен (вспомните детали движения снаряда в главе 1). Соответственно способность выносить ЦТ за пределы (т.е. достигать положительной Н3 служит средством для достижения большей высоты полета).
Сегментный анализ. Подобно многим силам, используемым в анализе движения человека, вес является распределенной силой. Считается, что точка приложения силы веса — центр тяжести. Например, в диаграмме свободного тела, показанной на рис. 2.9, сила, создаваемая каждой мышцей, представлена одним вектором, разме-
4*
51
a
Для прыжка 2,13 м:
Hi = 1,44 м
Н2 = 0,78 м
Из = -0,09 м 2,13 м
б
Рис. 2.11. Схематическое изображение прыжка в высоту с некоторыми значениями для прыжка на 2,13 м (а) и теоретическая техника для выноса ЦТ за пределы тела (б) (Нау, 1978)
дования на трупах для выведения простых математических выражений оценки различных антропометрических (связанных с измерением человеческого тела) сегментных размеров.
Данные Д.И.Миллера и Нельсона (Nelson, 1973) в основном состоят из измерений, выполненных на белокожих мужчинах. Результаты исследований Чандлера и его соавторов, приведенные в таблице 2.1 и 2.2, получены аутопсически.
В табл. 2.1 представлены уравнения регрессии определения веса сегмента на основе веса всего тела и местоположения ЦТ в каждом сегменте на основе данных о длине сегмента. Например, если индивидуум весил 750 Н, то вес торса = 0,532 х 750 — 6,93 = 392 Н, что дает 52 % веса тела. Подобным образом вес кисти = 0,005х х 750 + 0,75 = 4,5 Н, что составляет примерно 0,06% веса тела. Что точно имел в виду Чандлер и соавт. (1975) под обозначениями торс, кисть и т.п. указано в Приложении 4. Табл. 2.1 можно также использовать для определения местоположения ЦТ. Если длина бедра индивидуума (расстояние от бедра до колена) составляет 36 см, то, в соответствии с Чандлером и соавт., ЦТ для этого бедра будет расположен на 39,8% этого расстояния от тазобедренного сустава: местоположение ЦТ — 36 х 0,398 = 14,3 см от тазобедренного сустава.
Поскольку движение человека основывается на повороте сегментов тела относительно друг друга, распределение массы в сегменте так же важно знать, как и его массу. Параметром, пред-
щенным в центре поперечного сечения мышцы. Так же контакт между головой игрока и футбольным мячом будет изображен точкой на диаграмме свободного тела, но в действительности контакт распределен по большой площади лба. Подобным образом ЦТ представляет центр массы объекта и является точкой, вокруг которой равномерно распределена масса объекта. Соответственно, когда масса объекта перераспределяется, что обычно имеет место при движении человека, положение ЦТ также перемещается. На рис. 2.11,6 положение ЦТ смещается, когда изменяется положение прыгуна в высоту.
Для определения положения ЦТ в таких движениях, как прыжки в высоту, биомеханики разработали процедуру, известную как сегментный анализ. Она включает в себя оценку массы и положения ЦТ для каждого сегмента тела и последующее использование этой информации для определения положения ЦТ всего тела. Эта процедура рассматривается в главе 3. Здесь мы используем данные масс и ЦТ сегментов, которые представляют величину и положение вектора веса для различных сегментов тела человека.
Ряд авторов (Chandler, Clauser, McConville, Reynolds, Young, 1975; Dempster, 1955; Hanavan, 1966; Miller, Morrison, 1975) проводили иссле-
Таблица 2.1. Уравнения регрессии, оценивающие вес сегментов тела и положение ЦТ
Сегмент Вес, Н Положение ЦТ, %
Голова 0,032 ВТ + 18,70 66,3
Торс 0,532 ВТ - 6,93 52,2
Плечо 0,022 ВТ + 4,76 50,7
Предплечье 0,013 ВТ + 2,41 41,7
Кисть 0,005 ВТ + 0,75 51,5
Бедро 0,127 ВТ - 14,82 39,8
Голень 0,044 ВТ - 1,75 41,3
Стопа 0,009 ВТ + 2,48 40,0
Примечание: См. Приложение 4 для организации сегментов тела, применяемой Chandler et al. (1975). Вес сегментов тела выражен в процентах к общему весу тела (ВТ), а положение ЦТ сегментов — в процентах от длины сегмента, измеренной от проксимального конца сегмента.
Таблица 2.2. Моменты инерции всего тела относительно осей сальто, пируэта и винта, кгм2
Положение Сальто Пируэт Винт
12,55 15,09 3,83
Прогнувшись (руки в стороны) 8,38 8,98 4,79
Согнувшись (руки касаются концов ног) 8,65 6,60 3,58
В группировке 4,07 4,42 2,97
52
Таблица 2.3. Моменты инерции сегментов относительно осей сальто, пируэта и винта, кгм2
Сегмент Сальто Пируэт Винт
Голова 0,0164 0,0171 0,0201
Торс 1,0876 1,6194 0,3785
Плечо 0,0133 0,0133 0,0022
Предплечье 0,0065 0,0067 0,0009
Кисть 0,0008 0,0006 0,0002
Бедро 0,1157 0,1137 0,0224
Голень 0.0392 0,0391 0,0029
Стопа 0.0030 0.0034 0.0007
ставляющим распределение массы тела в уравнениях механики, является момент инерции. Момент инерции — это эквивалент массы как меры инерции и представляет собой меру сопротивления, которую оказывает объект изменению его движения относительно оси вращения. Момент инерции (Г) вычисляется по формуле
п
I = Ymrf’ (2.5)
1=1
где п — количество элементов (частиц) в системе; т — масса каждого элемента в системе; г — расстоянием каждого элемента от оси вращения. Момент инерции обычно определяется по отношению к трем ортогональным (перпендикулярным) осям, которые называются основными осями вращения. При анализе движения человека эти оси обычно имеют названия: “сальто” (из стороны в сторону), “пируэт” (спереди назад) и “винт” (продольная ось). В табл. 2.2 приведены моменты инерции сегментов относительно трех основных осей для тела человека в различном положении (Miller, Nelson, 1973). Обратите внимание, что момент инерции уменьшается, если масса тела приближается к оси вращения; например, для каждого положения тела момент инерции наименьший вокруг оси винта.
Кроме оценки массы и положения ЦТ, исследования на трупах позволили определить моменты инерции сегментов. Их можно получить относительно легко с помощью техники хронирования колебаний (Lephart, 1984). В отличие от уравнений регрессии в табл. 2.1 данные для моментов инерции сегментов были проведены Chandler et al. (1975) в виде средних значений (табл. 2.3). Однако Latsiorsky, Seluganov (1983, табл. 5 — 7) использовали технику сканирования излучения (описанную в этом разделе далее) для определения уравнений множественной регрессии, с помощью которых можно определить моменты инерции сегментов относительно основных осей. Как и при измерениях всего тела, момент инерции для каждого сегмента (кроме головы) одинаков относительно оси сальто и пируэта и наименьший относительно оси винта.
Чтобы дополнить данные о сегментах, полученные на трупах, некоторые исследователи использовали метод математического моделирования. При этом подходе человеческое тело представляется как набор геометрических компонентов, таких, как сферы, цилиндры и конусы (Hall, De Pauw, 1982; Hanavom, 1964, 1966; Hatze, 1980, 1981a, 1981b, Miller, 1979). Одним из первых, использовавших такой подход, был Ханаван (1964, 1966). На рис. 2.12 представлена модель Ханавана, которая разделяет тело человека на 15 простых геометрических фигур однородной плотности.
Преимуществом этой модели является то, что она требует лишь небольшого числа простых антропометрических измерений (например, длины и окружности сегментов), необходимых для ее персонализации и предсказания положения ЦТ, а также момента инерции для каждого сегмента тела. Однако три допущения, как правило, при моделировании сегментов тела, ограничивают точность оценок: сегменты принимаются жесткими, границы между сегментами принимаются четкими и считается, что сегменты имеют однородную плотность. Фактически же могут иметь место значительные смещения мягких тканей во время движения, границы между сегментами расплывчаты и плотность различна как между сегментами, так и внутри них.
Основываясь на том же подходе, Hatze (1980) разработал более детальную модель человеческого тела (рис. 2.13). Гуманоид Hatze состоит из 17 сегментов тела и для индивидуализации требует 242 антропометрических измерения.
Рис. 2.12. Модель человеческого тела Ханавана
53
Рис. 2.13. Диаграмма 17-сегментного гуманоида (Hatze, 1980):
1 — грудобрюшной сегмент; 2 — головной; 3 — левое плечо; 4 — левое предплечье; 5—левая локтевая часть; 6 — левая кисть; 7— правое предплечье; 8— правое плечо; 9 — правая локтевая часть; 10 — правая кисть; 11 — брюшно-тазовый; 12 — левое бедро; 13 — левая голень; 14 — левая ступня; 75 — правое бедро;
16 — правая голень; 17— правая стопа
Модель подразделяет сегменты на элементы небольшой массы с различной геометрической структурой, позволяя детально моделировать форму и вариации плотности сегментов. Более того, в модели не делается допущений относительно билатеральной симметрии и учитываются особенности строения мужского и женского тела, регулированием плотности некоторых частей сегментов в соответствии с содержанием подкожного жира. Модель может учесть изменения в морфологии тела, например, вызванные ожирением или беременностью, а также имитировать особенности строения тела детей. Действительно, модель использовалась для оценки массы, центра масс и момента инерции для сегментов конечностей младенцев (Schneider, Zemicke, 1992). Хотя количество вводимых в модель параметров (242 антропометрических измерения) требует много времени, выход модели дает точные оценки объема, массы, положения ЦТ и моментов инерции для определенных сегментов тела. Типичные оценки для двух мужчин показаны в табл. 2.4. Одна из заслуживающих внимания особенностей модели — подход к представлению сегментов торса и плеча (рис. 2.13).
Другой способ определения параметров сегментов тела состоит в использовании компьютерной томографии или составлении изображений с помощью магнитного резонанса (Engstrom, Loeb, Reid et al., 1991; Martin, Mungiole, Marzke,
Longhill, 1989). Примером такого подхода является использование радиоизотопов. При этом различные свойства сегментов тела определяются на основе измерений интенсивности гамма-лучей до и после прохождения через сегмент. Зацерский и Селянов (1983) выполнили эту процедуру на 100 субъектах мужского пола (возраст 23,8 — 6,2 года, рост 1,74 ± 0,06 м, вес 730 — 91 Н) и вывели уравнение регрессии (табл. 2.5), с помощью которых можно определить параметры сегментов тела для других субъектов.
Пример
Рассмотрим сегментный анализ штангиста, изображенного ранее на диаграмме свободного тела. Мы хотим определить вклад в движение мышц, пересекающих колено. Во-первых, нужно установить, сколько сегментов следует использовать для схематического представления штангиста (рис. 2.14,а). Движение, показанное сверху на рис. 1.7, указывает, что руки не сгибаются в локтях, так что руки можно рассматривать как один сегмент. Поскольку движение ограничено сагиттальной плоскостью, мы допускаем, что левая и правая стороны тела выполняют более или менее одинаковую задачу. Эти упрощения позволяют нам уменьшить количество сегментов на диаграмме свободного
Рис. 2.14. Диаграмма свободного тела для штангиста: а — изображение всего тела отрезками; б — сегментные компоненты; в — диаграмма свободного тела для ноги. Fja, FJ I: — силы реакции соответственно коленного и голеностопного суставов; Fm_a, Fnj. — результирующие мышечные силы соответственно лодыжки и колена; — вес голени
54
тела от максимума в 15 или 17 сегментов (основанного на моделях Hanavan, Hatze) до 6 сегментов.
Затем необходимо вычертить соответствующую диаграмму свободного тела (рис. 2.14,в). Это включает в себя идентификацию системы, определение положения ЦТ и веса (Fws). Поскольку цель исследования состоит в определении мышечных усилий, пересекающих коленный сустав, необходимо использовать в качестве системы голень. Положение ЦТ и вес могут быть определены на основе данных табл. 2.1. В соответствии с этой таблицей, ЦТ для голени расположен на 41,3% длины голени от коленного сустава. Если голень штангиста имеет длину 36 см, то ЦТ будет на расстоянии 14,9 см (36 х 0,413 = 14,9) от колена. Подобным обра
зом вес определяется как функция веса всего тела. Для штангиста весом 800 Н вес голени составит 33,5 Н (0,044 х 800 - 1,75 = 33,5). Наконец, к диаграмме свободного тела должны быть добавлены другие силы, действующие на систему, которую мы хотим описать.
Сила реакции земли
Сила реакции земли определяется согласно закону Ньютона о равенстве действия и противодействия. Она может быть измерена с помощью прибора, известного как силовая платформа, представляющая собой усложненные весы. Исследователи начали использовать эту техни-
Таблица 2.4. Сегментные параметры, рассчитанные для двух мужчин с помощью модели Hatze
Сегмент Субъект В. (23 года) Субъект Р (26 лет)
Объем Положение ЦТ 1хх Объем Положение ЦТ 1хх
Грудо-брюшной 19,111 0,439 0,3117 19,803 0,444 0.3302
Голова и шея 4,475 0,517 0,0303 4,537 0,516 0,0337
Левое плечо 1,438 0,727 0,0047 2,042 0,706 0,0080
Правое плечо 1,890 0,711 0,0071 2,121 0,699 0,0084
Левое предплечье 2,110 0,432 0,0196 2,123 0,437 0,0203
Правое предплечье 2,021 0,437 0,0168 2,340 0,428 0,0229
Левый локтевой сустав 1,023 0,417 0,0067 1,223 0,413 0,0086
Правый локтевой сустав 1,190 0,404 0,0079 1,313 0,412 0,0093
Левая кисть 0.453 0,515 0,0011 0.416 0,533 0,0010
Правая кисть 0,446 0,531 0,0011 0,417 0,524 0,0010
Тазово-брюшной 8,543 0,368 0,0399 9,614 0,395 0,0541
Левое бедро 8,258 0,479 0,1475 8,744 0,473 0,1653
Правое бедро 8,278 0,480 0,1415 8,729 0,466 0,1702
Левая голень 3,628 0,412 0,0615 3,856 0,420 0,0798
Правая голень 3,686 0,417 0,0663 3,798 0,417 0,0747
Левая ступня 0,887 — 0,0041 1,032 — 0,0051
Правая ступня 0,923 — 0,0042 1,055 — 0,0051
Примечание. Объем — рассчитанный объем сегмента (£), положение ЦТ — расстояние от проксимального конца сегмента (пропорция длины сегмента, где 0,50 означает 50% длины сегмента), — момент инерции относительно оси сальто, проходящий через ЦТ. Плотности сегментов: для живота — 1,00 + 30 для головы — 1,12, для шеи — 1,04, для руки и ноги — 1,08 — 20 кг/м3, где = 1 для мужчин и 0 для женщин.
Таблица 2.5. Уравнения регрессии, оценивающие массу сегментов тела и положение ЦТ
Сегмент Коэффициенты массы, кг Коэффициенты положения ЦТ, %
Во Bi в2 Во Bi в2
Ступня -0,8290 0,0077 0,0073 3,7670 0,0650 0,0330
Голень -1,5920 0,0362 0,0121 -6,0500 -0,0390 0.1420
Бедро -2,6490 0,1463 0,0137 -2,4200 0,0380 0,1350
Кисть -0,1165 0,0036 0,0017 4,1100 0,0260 0,0330
Предплечье 0,3185 0,0144 -0,0011 0,1920 -0,0280 0,0930
Плечо 0,2500 0,0301 -0,0027 1,6700 0,0300 0,0540
Голова 1,2960 0,0170 0,0143 8,3570 -0,0025 0,0230
Верхняя часть торса 8,2144 0,1862 0,0584 3,3200 0,0076 0,0470
Средняя часть торса 7,1810 0,2234 -0,0663 1,3980 0,0058 0,0450
Нижняя часть торса -7,4980 0,0976 0,0490 1,1820 0,0018 0,0434
Примечание. Уравнения множественной регрессии имеют вид Y = Вц + BtX} + В2Х2, где Y — прогнозируемая масса сегмента или положение ЦТ; А) — общая масса тела, кг; Хг — рост, см; Вд, В[ и Вд — коэффициенты, приведенные в таблице. Масса сегмента тела и положение ЦТ — функции от массы тела и роста.
55
Рис. 2.15. Вертикальные компоненты кинематики и динамики, связанные с вертикальным прыжком (Miller, 1976):
1 — сила; 2 — ускорение; 3 — скорость; 4 — положение
ку в 30-х годах XX ст. (Elfman, 1938, 1939; Fenn, 1930; Manter, 1938), хотя сама идея была предложена раньше (Amar, 1920; Магеу, 1874).
Одним из важных различий между силовой платформой и весами является то, что силовая платформа может измерять силу реакции земли в трех измерениях и делать это быстро и с минимальным искажением сигнала.
Результирующая реакция земли может быть разложена на три составляющих по трем вертикальным направлениям: вверх-вниз, вперед-назад и из стороны в сторону. Поскольку сила реакции земли, измеренная в опорной фазе, представляет собой реакцию земли на действия бегуна, которые передаются через опорную ногу, эти составляющие и представят проекцию ускорения системы на эти направления. Сила реакции земли зависит от массы выделенного сегмента и ускорения его ЦТ. Miller (1990) считает, что торс и голова составляют примерно 50% массы бегуна, тогда как каждая нога дает примерно 17% и руки около 5%.
Наглядный пример связи между силой реакции земли и соответствующей кинематикой движения приведен на рис. 2.15. Движением является вертикальный прыжок. Высота вертикального прыжка зависит от величины вер
тикальной скорости при отталкивании, которая, в свою очередь, определяется вертикальной составляющей силы реакции земли. Как видно на рис.2.15, спортсмен начинает с вертикального положения, затем опускает ЦТ примерно на 0,2 м до изменения направления движения (скорость из отрицательной становится положительной). Отталкивание происходит, когда вертикальный компонент силы реакции земли упадет до нуля (время 0,53 с). Спортсмен находился в воздухе примерно 0,41 с, в течение которых ЦТ был поднят на 0,49 м над исходным положением. Учтите, что рис. 2.15 иллюстрирует результат взаимодействия между субъектом и землей, соответственно кинематика описывает движение ЦТ всего тела.
Как видно на рис. 2.15, пиковая скорость при приседании вниз (ускорение равно 0) имеет место примерно на полпути движения вниз (время 0,19 с), а пиковая скорость при движении вверх (положительные значения; ускорение равно 0) имеет место как раз перед отталкиванием. В течение фазы полета прыжка сила реакции земли равна нулю, и вертикальная компонента ускорения составляет 9,81 мфс-2 (т.е. воздействие силы тяжести на ЦТ прыгуна). Как отмечалось ранее, сила реакции земли и графики ускорения параллельны друг другу, но смещены. Вначале ускорение системы (ЦТ всего тела) было равно нулю, а сила реакции земли была равна весу тела. По мере того как сегменты тела начали ускоряться, ускорение системы изменилось, и сила реакции земли изменилась аналогично (соответствующие кривые смещены на величину, пропорциональную весу тела). График силы реакции земли имеет четыре фазы: 1) первоначальную фазу, когда сила реакции земли меньше веса тела (отрицательное ускорение);
2) фазу, в которой сила реакции земли больше веса тела (положительное ускорение); 3) фазу полета, в которой реакция земли равна нулю (ускорение — 9,81 м • с-2); 4) фазу удара, когда прыгун возвращается на землю.
На рис. 2.16 показано изменение вертикальной составляющей (Fg^ силы реакции земли при ходьбе и беге (более подробно отличия реакции земли при ходьбе и беге см. Alexander, 1984в). Эти данные показывают, каким образом изменяется FgJ. с момента начала контакта ноги с землей (время ноль на абсциссе) до момента, когда та же нога отрывается от земли (момент, в который F&i возвращается к нулю). При анализе движения этот этап известен как опорная фаза. Fgz не равно нулю только тогда, когда нога находится в контакте с землей. Обратите внимание, что непрерывно изменяется в течение периода опоры. Вспомним из дискуссии о весе, что вес эквивалентен F&z когда система (тело) не ускоряется в вертикальном направлении. Соответственно когда Fgi отли
56
чается от веса тела, система испытывает вертикальное ускорение; если Fgi больше веса тела, вертикальное ускорение ЦТ направлено вверх, если же Fgz меньше веса тела, вертикальное ускорение ЦТ направлено вниз.
Другое различие Fgz при ходьбе и беге заключается в том, что зависимость вертикальной составляющей от времени при ходьбе имеет два пика, а при беге — один. Действительно Alexander (1984в) использовал это различие для количественного анализа изменений Fgz при изменении скорости. Можно ли по имеющейся записи изменения Fgi при ходьбе установить, когда ЦТ перемещается вниз во время опорной фазы? Как указывалось в главе 1, невозможно определить направление смещения непосредственно из графика сила—время, поскольку он дает представление лишь об ускорении (сила пропорциональна ускорению F = та), а этого недостаточно для определения смещения.
Поскольку сила Fgz — реакция земли на воздействие ноги бегуна на опорную поверхность, анализ изменения этой силы позволяет выяснить существенные особенности его движения (ускорение ЦТ) (рис. 2.17). Бегун в безопорной фазе испытывает ускорение вниз (вследствие действия тяжести) равное 9,81 м.с-2, а в момент контакта ноги с землей вертикальная составляющая (7^г) направлена вверх, для противодействия давлению ноги на землю. Более того, когда нога касается земли, она первоначально находится впереди ЦТ бегуна, что заставляет землю реагировать направленной горизонтально (назад) тормозящей составляющей Fgy. Когда ЦТ бегуна проходит над опорной ногой, горизонтальная составляющая изменяет свое направление и начинает действовать вперед (ускорение). Боковую составляющую (Fgx) объяснить труднее, но она имеет меньшую величину и более вариабельна, чем Fgл и Fgy. Однако было показано, что Fgx коррелирует (г = 0,71) с положением ноги относительно средней линии движения во время контакта с землей (Williams, 1985).
В главе 1 мы рассматривали некоторые кинематические характеристики, изменяющиеся при увеличении скорости бегуна. Эти изменения вызваны взаимодействием сегментов тела, что отражаются в силе реакции земли. Munro, Miller, Fuglevand, (1987) исследовали некоторые из этих изменений на 20 мужчинах при увеличении скорости их бега с 3,0 до 5,0 м • с_|. Они обнаружили, что в среднем длительность опорной фазы уменьшилась с 270 до 199 мс, пик Fgz увеличился с 2,51 до 2,83 F„ (где Fw — вес тела), а среднее усилие в опорной фазе возросло с 1,4 до 1,7 Fw. Эти результаты показывают, что увеличение скорости бега с 3,0 до 5,0 м • с-1, привело к большей Fgv приложенной в течение меньшего времени (рис. 2.18). Средняя сила как тормозящего, так и ускоряющего направления
Рис. 2.16. Вертикальная составляющая силы реакции земли (JLZ) во время опорного периода при ходьбе (I) и беге (2)
Fgy также возрастает по мере увеличения скорости бегуна. Подобные результаты были сообщены Nilsson, Thorstensson (1989), но они охватывали диапазон скоростей от 1,0 до 6,0 м-с-1 и включали в себя переход от ходьбы к бегу.
Центр давления. Хотя в силе реакции земли отражено действие всех сегментов тела, измерив силу реакции земли можно получить некоторое представление о действии опорной конечности. Такие измерения показали, что кинематика ноги во время опорной фазы достаточно сложна (Cavanagh, 1990; Cavanagh, Andrew, Kram et al., 1985; Cavanagh, Rodgers,
Рис. 2.17. Обобщенные кривые сила—время для трех составляющих силы реакции земли во время опорной фазы прыжка при беге. Силы выражены по отношению к весу тела (ВТ)
57
1993; Pollock, Zanda, 1977; Chao, 1986; Nigg, 1986; Vaaghan, 1984). Часть ноги, которая первоначально контактирует с землей, зависит от скорости бега, но обычно является латеральной кромкой, чаще всего между серединой ступни и пяткой, при нахождении ноги в супиниро-ванном положении (аддукция и инверсия ноги, причем инверсия относится к повороту подошвы стопы по направлению к средней линии тела). После первоначального касания нога про-нируется (абдукция и выворачивание ноги, где выворачивание относится к повороту подошвы стопы от средней линии тела), а лодыжка сгибается назад. Когда ЦТ бегуна проходит над ногой, начинают разгибаться коленный и голеностопный суставы. К моменту, когда нога отрывается от земли, точка приложения силы
Рис. 2.19. Смещение центра давления (точки приложения силы реакции земли) во время опорной фазы бега со скоростью 4,5 м • с-1: удар задней частью ноги (а); удар средней частью ноги (б) (Cavanagh, La Fortune, 1980)
реакции земли перемещается от латеральной кромки ступни при касании земли до точки на основании большого пальца или вблизи нее.
Сила реакции земли на это движение стопы лодыжки—голени может быть зарегистрирована, по меньшей мере, двумя способами: измерением центра давления и распределения давления по ступне. Центр давления представляет собой точку приложения результирующей силы реакции земли. Это просто центральная точка давления, оказываемого на ногу. Cavanagh, La Fortune (1980) разделяли бегунов на ударяющих о землю либо серединой ступни, либо ее задней частью. Это различие относится к первоначальному положению центра тяжести на стопе (рис. 2.19). Для ударяющих средней частью ноги начальным расположением центра давления была латеральная кромка в центре стопы. Это не означает, что ни передняя, ни задняя часть стопы не контактировали с землей; просто центр давления находился посреди стопы. Подобным образом у бегунов, ударявших задней частью стопы, первоначальным положением центра давления была задняя часть стопы.
Фактическое распределение давления (т.е. площадь, на которую воздействовала сила реакции земли) измерялось при использовании фотоэластичного материала или миниатюрных реле давления (Cavanagh, Ае, 1980; Hennig, Cavanagh, Albert, Macmillan, 1982). Подобно весу сила реакции земли является распределенной силой, и зона, по которой она распределена, сдвигается во время опорной фазы ходьбы или бега, а также в зависимости от типа обуви. Sanderson, Hennig (1982) сравнивали распределение давления в ботинке велосипедиста и ши-повке бегуна путем тщательного размещения в каждой обуви 12 пьезокерамических преобразователей. На рис. 2.20 показано распределение давлений, когда угол кривошипа составлял 1,57 рад после верхнего вертикального положения. Наибольшая часть нагрузки прилагалась к первому пальцу плюсны и к большому пальцу стопы. Аналогично при использовании шиповок большие давления прилагались к латеральной части переднего отдела стопы.
Конструкция обуви. Как кинематика движения, так и сила реакции земли могут быть изменены путем модификации обуви, носимой бегунами. Например, скорость сгибания колена (Frederick, 1986) и степень пронации (Zuethi, Stacoff, 1987) увеличивают последующий толчок, если подошвы беговой обуви твердые. Хорошо сконструированная беговая обувь служит двойной цели — минимизировать удар при контакте ноги с землей (т.е. амортизировать) и обеспечить устойчивость голеностопного сустава во время опорной фазы. К сожалению, увеличение амортизирующей способности ведет к снижению устойчивости и наоборот.
58
Обычно движение ноги в опорной фазе включает в себя последовательность супинации-пронации, которая распределяет силу удара по стопе и вверх по опорной конечности. Задача беговой обуви — ослабить силу удара и контролировать нормальное движение ноги. Возможность обуви обеспечить соответствующую устойчивость обычно оценивается в терминах движения задней части стопы. Такая оценка включает в себя определение угла между голенью и пяточной костью в течение опорной фазы. На рис. 2.21 показано изменение этого угла (заднего угла ноги) для трех бегунов.
У бегуна с задним углом ноги 0,14 рад более вертикальная метка пятки (пронация), а у бегуна с углом 0,07 рад более вертикальная отметка голени (супинация). Хотя диапазон изменения угла задней части ноги составляет около 0,31 рад при нормальной опорной фазе (рис. 2.21), бег в беговой обуви, очевидно, вызывает большую пронацию, чем бег босиком (Frederick, 1986). Laeshi, Stacoff (1987) показали, что плохо сконструированная обувь отрицательно влияет на устойчивость, увеличивая величину пронации во время опорной фазы примерно на 0,09 рад (5°). Этот эффект нежелателен, поскольку излишняя пронация может быть одной из наиболее важных причин беговых травм (Clement, Taunton, Smart, MacNicol, 1981).
Особенностями конструкции обуви, влияющими на устойчивость, являются твердость подошвы, высота пятки и подъема и угол расширения стопы (Frederick, 1986). Мягкая стелька допускает большую пронацию и больший диапазон движений задней части ноги. Высота пятки (расстояние от земли до пятки) и ее подъем (высота пятки над передней частью ноги), по мнению некоторых исследователей (Bates, Osternig, Mason, James, 1978; Stocoff, Kaelin, 1983), уменьшают пронацию, но другие (Clarke, Frederick, Hamie, 1983) отрицают это. Увеличение наклона пятки до 0,26 рад (15°) приводит к значительному уменьшению пронации и снижению диапазона движения задней части ноги (Frederick, 1986).
Трение. Результирующая двух горизонтальных составляющих силы реакции земли (Fgy и FgrX) представляет собой трение или силу сдвига (РЛ) между обувью и землей. Очень важно оценить силу трения движений, так как она определяет горизонтальное перемещение ЦТ. Вспомним, что три составляющих силы реакции земли представляют ускорение ЦТ всего тела, которое является суммой проекций ускорений ЦТ сегментов тела по трем направлениям. При этом Fs соответствует ускорению ЦТ всего тела в горизонтальной плоскости (т.е. в направлениях вперед-назад и из стороны в сторону).
В принципе, максимум силы трения зависит от силы, которая нормальна (перпендикулярна) к поверхности (FS z), и коэффициента (ц),
Рис. 2.20. Сравнение распределения давления в ботинке велосипедиста при педалировании (а) и шипов-ке бегуна (б)
Рис. 2.21. Изменения заднего угла ноги во время опорной фазы для трех бегунов (Edington, Frederick, Cavanagh, 1990)
1 — пронация; 2 — нейтральное положение; 3 — супинация
59
характеризующего контакт (гладкая иди шероховатая поверхность, сухая или смазанная, статический или динамический контакт) между двумя объектами
^x.max ~ (2-6)
При контакте обуви с землей коэффициент трения изменяется в зависимости от того, находится ли обувь в стационарном положении по отношению к земле (статический цх) или перемещается (динамический pd). Поскольку больше, чем ^достигает больших значений, когда обувь не движется или не скользит по земле. Это объясняет, почему труднее сделать крутой поворот, когда ботинок начал скользить по земле, р. между обувью и поверхностью определяется экспериментально путем расчета отношения Fs и Fg z в момент времени, как раз перед тем, когда обувь перемещается по отношению к земле. При таком подходе установили, что изменяется в пределах 0,3 — 2,0 со значением 0,6 для гаревой дорожки и 1,5 для травы (Nigg, Stucke, Baurdas, Baumann, 1984), а при беге на коньках этот коэффициент изменяется от 0,003 до 0,007 (de Konig, de Groot, van Ingen Schenau, 1992). Однако когда Fs меньше максимума, трение просто равно результирующей силе в горизонтальной плоскости.
По данным Miller, Nelson (1973), имеются три диапазона действия трения: 1) перед началом соскальзывания обуви по земле, когда Fs равна (максимальное трение); 2) когда обувь скользит по земле и Fs равна (меньше максимальной силы трения); 3) когда сила трения неподвижной по отношению к земле обуви возрастает еще не достигая максимума (Л,max), может быть определена как результирующая сила в горизонтальной плоскости (Fs = =Fgx2 + Fg,y2)- Эти характеристики указывают, что для данных условий обувь—земля (т.е. ц) трение возрастает пропорционально F^z и поскольку Fgz зависит, в основном, от веса тела (Fw), для более тяжелых индивидуумов трение больше. Более того, до тех пор, пока обувь ещё не скользит по земле, сила трения меньше максимально возможной (Fsmax).
Сила реакции сустава
Когда система в диаграмме свободного тела определена так, что она включает сустав, необходимо использовать понятие силы реакции сустава (Fy), чтобы учесть эффект сил, создаваемых контактом кости с костью соседних сегментов тела. Сила реакции сустава представляет суммарный эффект передачи через сустав с одного сегмента на другой усилий, вызванных мышцами, связками и контактными силами костей. При совместной активности пары мышц
агонист—антагонист сила реакции сустава — это разность активности этих мышц. Силы реакции в суставе Moiyr достигать значительных величин, например, в плечелучевом суставе при некоторых видах деятельности наблюдались значения до 3 кН. Знание этих эффектов существенно для конструирования протезов, поскольку их конструкция должна выдерживать возникающие усилия (Goodfellow, O’Connor, 1978; Rydell, 1965).
Очевидно, наименее исследован вклад, который вносят в силу реакции сустава окружающие его ткани, особенно связки. Влияние связок на Fy противоречиво; некоторые исследователи считают его относительно малым, за исключением экстремальных положений в диапазоне движений и при некоторых условиях нагружения (Amis, Dowson, Wright, 1980), тогда как другие сообщают об усилиях в связках, в три раза превышающих силу веса тела во время ходьбы (Collins, O’Connor, 1991). Напротив, определенно известно, что на Fy влияет мышечная сила (вспомните, что мышечная сила (рис. 2.3) может быть разложена на нормальную и тангенциальную составляющие и тангенциальная составляющая передается на сустав как сжимающая сила). Подобным образом, поскольку человеческое тело состоит из набора жестких сегментов, соединенных друг с другом, сила, действующая на один сегмент, может передаваться на все остальные сегменты тела. Поэтому сила реакции земли на ногу распределяется по всему телу и влияет на Fy. Любая сила, действующая на систему, может влиять на Fy. Более того, эффект, вызванный движением других сегментов тела и называемый силой инерции, может передаваться от одного сегмента к другому.
Крайне трудно измерить Fy экспериментально. Обычно Fy оценивается путем определения всех остальных сил на диаграмме свободного тела допущением, что остаточный эффект обусловлен Fy. Это можно сделать, например, если система находится в равновесии, что означает сбалансированность всех сил, действующих на систему. Такой подход будет использован в главе 3. Для определения Fy можно также использовать различные математические процедуры, например, минимизацию мышечных усилий. An, Kwak, Chao, Morrey (1984) использовали этот подход при исследовании локтевого (плечелоктевого) сустава, когда к кисти прилагалась нагрузка так, что она была перпендикулярна предплечью. Полученные значения Fy превышали нагрузку в 6 — 16 раз при изменении угла в суставе от развернутого до прямого. Если учесть нагрузки, встречающиеся в обычной повседневной деятельности, то значения Fy, равные 0,3 — 0,5 веса тела, часто встречаются в локтевом суставе. Подобным образом Ту можно определить, используя ар-
60
Рис. 2.22. Сила реакции в локтевом суставе во время выполнения изометрического сгибания (а) и разгибания (б). Угол 0 рад означает полное разгибание (Amis, Dowson, Wrait, 1980)
1 — плечелучевой; 2 — плечелопаточный; 3 — плечелоктевой; 4 — усилие на кисти
хитектуру мышцы и геометрию конечности для оценки мышечной силы. Amis, Dowson, Kait (1980) использовали эту процедуру для оценки изменений в плечелучевом, плечелопаточном и плечелоктевом суставах во время изометрического сгибания (рис.2.22, а) и растяжения (рис.22, 6) локтевого сустава. Расчетные силы реакции сустава, показанные на рис.2.22, связаны с мышечной активностью, необходимой для сопротивления усилиям, приложенным к кисти.
Опубликованы значения F7- для положения стоя, при переходе из сидячего положения в стоячее, при ходьбе, беге, поднятии тяжестей и приземлении после прыжка (Harrison, Lees, McCullogeh, Rowe, 1986). Harrison и соавт. обнаружили, что при беге со скоростью 4,5 м с-1 максимальные значения Fy- имели место в се-
Рис. 2.23. Сила реакции в коленном и голеностопном суставах во время опорной фазы бега (4,5 м • с-1) (Harrison et al., 1986):
1 — сжатие колена; 2 — сжатие голеностопного сустава;
3 — сдвиг голеностопного сустава
редине опорной фазы (рис. 2.23) и достигали пиковой силы сжатия, равной 33 значениям веса тела в коленном суставе, пиковой силы сжатия в голеностопном суставе, равной 9 значениям веса тела, и пиковой силы сдвига, равной 4 значениям веса тела в том же суставе. Даже обычная задача перейти из стоячего положения в положение сидя на корточках и затем вернуться в исходное связана с большими силами реакции в суставах. Для этого максимальная сила реакции большеберцового сустава, которая нормальна к поверхности (сила сжатия), достигала от 4,7 до 5,6 значений веса тела, тогда как тангенциальная составляющая (сила сдвига) составляла от 3,0 до 3,9 значений веса тела (Dahlkvist, Mayo, Seedhom, 1982). Тяжелоатлеты-штангисты испытывают максимальные сжимающие усилия, в 17 раз превышающие вес тела, и максимальные усилия сдвига, превышающие вес тела в 2,3 раза, в суставе между позвонками L4 — L5 во время подъема штанги (Cholewicki, McGill, Norman, 1991). Эти результаты показывают, что сила реакции в суставах существенно изменяется в зависимости от вида движений и может достигать значительных величин, особенно по сравнению с нагрузками, испытываемыми в повседневной деятельности.
Мышечная сила
Ранее мы описывали движение человека как результат механических взаимодействий между мышцей и ее окружением. При этом роль мышц заключается в создании силы, которая передается через связки, кости и вызывает поворот сегмента тела. Однако общее понятие силы (т.е. действия, изменяющего состояние или движение объекта) может быть распространено на
61
мышцу с учетом ее способности создавать только растягивающее усилие. При этом комбинация растягивающих усилий, создаваемых мышцей, может передаваться вдоль сегмента и вызывать в результате сжимающее усилие.
Вследствие способности мышцы создавать однонаправленное усилие движение относительно сустава контролируется двумя различными группами мышц со взаимнопротивоположным направлением действия. Например, при отсутствии других сил движение сгибания—разгибания локтя контролируется двумя мышечными группами: вызывающей разгибание локтя (разгибатели локтя) и управляющей сгибанием (сгибатели локтя). Однако когда на систему действуют другие силы, такие, как сила тяжести, сгибатели локтя могут управлять разгибанием, а разгибатели — сгибанием. Таким образом, функция мышцы существенно зависит от условий, в которых она работает. Тем не менее управление движением человека требует, как минимум, одной пары противоположно действующих мышц для управления каждым анатомически возможным движением (т.е. сгибание—разгибание, отведение—приведение).
Поскольку сила — это вектор, мышечное усилие схематически может быть представлено в виде стрелки, имеющей величину и направление. В этом контексте имеются четыре общепринятых допущения, которые относятся к анализу движения человека.
Во-первых, большинство мышц скелета создают усилия поперек суставов и могут вызвать поворот сегментов тела, что позволяет рассматривать многие функции тела человека с позиций теории неодушевленных машин. Crow-ninschield, Braund (1981) приписывают эю допущение Леонардо да Винчи.
Во-вторых, тело человека можно рассматривать как последовательность жестких сегментов тела; деформации мягких тканей и движение жидкостей внутри тела имеют пренебрежимо малое влияние на движение (Braune, Fischer, 1889).
В-третьих, направление вектора мышечной силы является прямой линией между проксимальным и дистальным прикреплениями, причем сила считается приложенной в точках прикрепления (Braund et al., 1982). Фактически мышца прикрепляется не в точке, прикрепление распределено по площадке конечных размеров. Однако если размеры площади пренебрежимо малы по сравнению с другими размерами системы, то усилие можно рассматривать приложенным в точке. Если площадь прикрепления мышцы значительна (например, у трапециевидной, большой грудной мышцы), то мышечное усилие может представлено несколькими линиями действия (van der Helm, Veenbaas, 1991).
Четвертое допущение состоит в том, что движение в гравитационном поле вызывается
несбалансированностью приложенных к системе сил (закон инерции). Механический анализ действия нескольких мышц, пересекающих сустав, в большинстве случаев позволяет нам определять скорее результирующее мышечное усилие, чем силу, оказываемую отдельными мышцами.
Величина мышечной силы. И величина, и направление вектора мышечной силы трудно поддаются измерению. Чтобы измерить мышечную силу, мы должны измерить силу, переданную сухожилием. В экспериментах на изолированной мышце это измерение включало в себя присоединение сухожилия к преобразователю усилия (Ralston, Inmen, Strait, Chaffrath, 1947). В экспериментах на человеке, когда сухожилие не отделено от кости, мышечное усилие может быть измерено путем помещения тензометра на сухожилие. Komi и Yarvinen (1987) использовали тензометр для измерения усилия в ахилловом сухожилии во время езды на велосипеде. Максимальные усилия порядка 700 Н они получили в правом ахилловом сухожилии во время педалирования со скоростью 90 об • мин-1; образуемая при этом мощность составляла 265 Вт.
Большинство людей, однако, не изъявляет желания добровольно участвовать в такой процедуре, поэтому для оценки мышечной силы приходится использовать косвенные измерения. Одним из широко распространенных подходов для оценки мышечной силы является измерение площади поперечного сечения мышцы в плоскости, перпендикулярной направлению мышечных волокон (Fick). Измерения могут быть выполнены на трупах или с помощью различных процедур получения изображения — ультразвук, компьютерная томография, магнитный резонанс (Narici, Roi, Landoni, 1988). В дополнение к данным, полученным для мышц, которые пересекают локтевой сустав (табл. 2.6), имеются данные для большинства других мышечных систем, таких, как бедро, голень и запястье (Brand, Pedersen, Friedrich, 1986; Clarck, Haylor, 1987; Hakkinen, Keskinen, 1989; Lieber, Faseli, Botte, 1990).
Способность мышцы генерировать силу связана с площадью поперечного сечения мышцы константой порядка 30 Н • см-2. Эта константа называется удельным натяжением, ее численное значение установлено и составляет при тщательных измерениях от 16 до 40Н • см-2 (Edgerton, Apor, Roy, 1990; Kanda, Haschisume, 1992; McDonash, Devis, 1984; Hydaard, Houston, Suzuki et al., 1983; Roy, Edgerton, 1992). Удельное натяжение, таким образом, характеризует способность мышцы генерировать усилие и не зависит от ее размера. Соотношение для оценки мышечного усилия (Fm) определяется формулой
Ли = удельное натяжение х
х площадь поперечного сечения . (2.7)
62
В соответствии с данными табл. 2.6 бицепс брахии с площадью поперечного сечения 5,8 см2 может создать максимальное усилие 174 Н. Максимальное усилие, которое может быть создано мышцами-сгибателями локтя (бицепс брахии, брахиалия, брахиорадиалия), действующими совместно, будет составлять 456 Н для поперечных сечений, указанных в табл. 2.6. Однако Edgerton et al. (1990) сообщали, что по имеющимся у них анатомическим данным наибольшее поперечное сечение составило 34,7 см2, поэтому развиваемое максимальное усилие сгибателей локтя равнялось 1297 Н.
Чем объясняется разброс константы удельного натяжения, которая изменяется в диапазоне от 16 до 40 Н • см~2? Факторами, влияющими на удельное натяжение, могут быть тип волокна, пол и строение мышцы. Некоторые данные (Schantz, Fox, Norgren, Tyde'n, 1981) позволяют предположить, что способность мышцы генерировать усилие зависит от количества быстро сокращающихся волокон, но было обнаружено, что удельное натяжение быстросо-кращаюшихся волокон (25,4 Н • см-2) ненамного отличается от соответствующего показателя мед-ленносокращаюшихся волокон (23,8 Н • см-2) (Lucas, Ruff, Binder, 1987). Данные о величинах удельного натяжения для различных мышц и разных типов волокон основываются, главным образом, на косвенных оценках при исследованиях на моторных блоках (Bodine, Roy, Eldred, Edgerton, 1987).
Одним из важных различий между измерениями на мышечном волокне и моторных блоках является то, что усилие, создаваемое изолированным мышечным волокном, может быть измерено непосредственно, тогда как усилие, создаваемое моторным блоком, измеряется на сухожилии, на которое влияют все виды соединительной ткани между стыком и сухожилием.
Поэтому, очевидно, удельное натяжение постоянно для различных типов мышечных волокон, но передача усилия от мест стыка к
сухожилию различна для разных типов моторных блоков.
Антигравитационные мышцы — те, которые участвуют в поддержании вертикального положения (например, выпрямители колена) — примерно в два раза сильнее противодействующих им мышц, но это скорее всего вызвано различиями в размерах, чем в удельном натяжении. Подобным образом тот факт, что мужчины обычно сильнее женщин (если сила определяется как способность генерировать усилие при изометрическом сокращении), обусловлен различиями в мышечной массе (рис. 2.24). Причина этих различий гормональная: тестостерон (мужской гормон) эффективнее, чем эстроген (женский гормон), стимулирует синтез протеина, что ведет к росту мышц. Для популяции, показанной на рис. 2.24, Ikai, Fiikunaga (1968) определили, что среднее значение константы удельного натяжения для сгибателей локтя равно 61,1 Н-см-2. Это значение завышено, поскольку измерялась только площадь поперечного сечения бицепс брахии. Если учесть площадь поперечного сечения всех мышц сгибателей локтя, то удельное натяжение составит от 30 до 37 Н • см-2 (Edgerton et al., 1990).
Эти наблюдения позволяют предположить, что величина мышечной силы не зависит от типа мышечных волокон или пола, а изменяется прямо пропорционально площади поперечного сечения при относительно постоянном удельном натяжении. Фактором, который может повлиять на это соотношение, является строение мышцы.
Если мы рассмотрим мышцу как простую структуру, в которой волокна расположены параллельно друг другу и продолжаются от одного конца мышцы к другому (например, бицепс брахии), то абсолютные усилия в каждой точке вдоль длины мышцы, в основном, идентичны. Соответственно усилие, создаваемое на одном конце мышцы, такое же, как и посередине мышцы, и на другом ее конце. Поскольку мышцы имеют скорее веретенообразную, чем цилиндрическую
Таблица 2.6. Сводные данные о площади поперечного сечения (ППС) и плечах сил для мышц-сгибателей и разгибателей локтя
Мышца ППС, см2 Ожидаемое усилие, Н Плечо силы, см Вращающий момент,
Нм % максимума
Бицепс брахии 5,8 174 3,8 6,6 32
Брахиалис 7,4 222 2,9 6,4 31
Брахиорадиалис 2,0 60 6,1 3,7 18
Пронатор терес 3,6 108 1,6 1,2 6
Разгибатель карпи 3,1 93 з,о 2,8 14
радиалис лонгус Трицепс брахии 23,8 536 — — —
Примечание. Ожидаемое усилие было определено путем умножения значений ППС на удельное натяжение 30 • см-2. Вращающий момент определялся как произведение ожидаемого усилия на плечо. Процент максимума определяет вклад соответствующих мышц-сгибателей локтя в общий вращающий момент сгибания локтя.
63
2,0-1
X Мужчины
О Женщины
х
*1,5-
X
5
Е 0,5-
От-------1------1-------1------1----
0 10 20
Площадь поперечного сечения, см2
30
Рис. 2.24. Зависимость между силой сгибателей локтя и площадью их поперечного сечения, полученная при исследовании 129 мужчин и 126 женщин
форму, поперечное сечение изменяется вдоль длины мышцы. Если мышечное усилие постоянно вдоль всей ее длины, а площадь поперечного сечения изменяется, то удельное натяжение также должно изменяться для сохранения постоянным произведения (уравнение 2.7); это
Рис. 2.25. Зависимость между внутримышечным давлением и вращающим моментом мышцы при изометрических сокращениях пяти мышц у одного субъекта (Sadamoto, Bonde-Peterson, Suzuki, 1983): 1 — гастрокнемиус; 2 — бицепс брахии; 3 — солеус;
4 — вастус латералис; 5 — ректус феморис
означает, что удельное натяжение должно изменяться обратно пропорционально площади поперечного сечения. Хотя еще во многом остается неясным, как передается усилие из мест присоединения на сухожилия, этот пример позволяет предположить, что архитектура мышцы влияет на мышечную силу и, возможно, обусловливает разброс данных, опубликованных для удельного натяжения.
Кроме площади поперечного сечения для оценки величины мышечной силы могут быть использованы ЭМГ и внутримышечное давление. ЭМГ позволяет измерять в мышцах электрическую активность, которая является прямой реакцией на активирующие сигналы нервной системы. При изометрических условиях величина ЭМГ в большой мере коррелирует с мышечной силой (Bigland, Lippold, 1954; Lavrens, Da Luca, 1983). Хотя это соотношение менее выражено для неизометрических условий (Calvert, Chepmen, 1977; Milner-Braun, Stein, 1975), существуют алгоритмы, с помощью которых можно определить величину мышечной силы на основе ЭМГ (Hof, Pronk, van Best, 1987; van der Berg, 1977; Marras, Sommerich, 1991a, 1991b).
Измерение внутримышечного давления с помощью катетера дает более достоверные значения мышечной силы, чем ЭМГ (Parker, Komer, Kadefors, 1984; Sadamoto, Bond-Petersen, Suzuki, 1983). Недостатком измерения внутримышечного давления является то, что это требует введения преобразователя давления в мышцу. Как и ЭМГ, внутримышечное давление возрастает линейно вместе с вращающим моментом мышцы, но наклон соответствующих графиков различен для разных мышц (рис. 2.25).
Направление мышечной силы. Если включить мышечную силу в диаграмму свободного тела, вначале нужно определить, какая группа мышц (например, сгибатели или разгибатели) создает усилие, а затем начертить вектор силы в направлении, противоположном силе реакции сустава. Рассмотрим диаграмму свободного тела, которая показывает усилие ахиллового сухожилия, измеренное Gregor et al. (1987), во время езды на велосипеде. Система (рис. 2.26) состоит из ноги (от голеностопного сустава до пальцев стопы) и взаимодействует с окружающей средой с помощью следующих сил: Fg — сила реакции земли — фактически сила, вырабатываемая педалью; Fw — вес, Fy — сила реакции сустава; Fm — мышечная сила. Обратите внимание, что Fm направлена против силы реакции голеностопного сустава, примерно в том же положении, что и ахиллово сухожилие.
Хотя, как правило, угол натяжения мышц обычно невелик, он изменяется и в ходе ряда движений может достигать значительных величин. An et al. (1984) измерили угол натяжения мышц, пересекающих локтевой сустав. Если
64
Внутрибрюшное давление
Рис. 2.26. Диаграмма свободного тела ноги, показывающая распределение сил, приложенных к стопе во время езды на велосипеде
предплечье находится в горизонтальном положении, этот угол составляет 1,4 рад для бицепс брахии, 1,2 рад для брахиалис, 0,4 рад для бра-хиорадиалис, 0,2 рад для пронатор терес и 0,05 рад для трицепс брахии. Более того, линии действия большинства мышечных сил скорее кривые, чем прямые. Рис. 2.27 показывает линии действия для шести мышц, пересекающих локтевой сустав. Линии действия были определены с помощью положения центроида в последовательных поперечных сечениях каждой мышцы (An, Hui, Моггеу et al., 1981).
Поскольку в брюшной полости много жидкости и волокнистых веществ, ее можно рассматривать как несжимаемый элемент, который способен передавать усилия от мышц, охватывающих полость, на поддерживающие структуры торса. Мышцы, окружающие брюшную полость, включают в себя брюшные мышцы спереди (ректус абдоминус, наружную и внутреннюю изогнутые и трансверсус абдоминис), диафрагму сверху и мышцы тазового дна снизу. Экспериментальное регулирование давления в брюшной полости называется процедурой Вал-салва. Внутрибрюшное давление (F,), измеряемое в паскалях (1 Па = 1 Н • м-2), может быть увеличено путем закрывания надгортанника и активизации различных мышц торса и брюшной полости (Cresswell, Grundstrom, Thorstensson, 1992). При активации мышц торса, кроме внут-рибрюшного, возрастает и внутригрудное давление. Во время поднятия тяжестей и прыжков внутригрудное и внутрибрюшное давление имеют тенденцию изменяться однонаправленно: внутрибрюшное давление при этом обычно больше (Harman, Frykman, Clagett, Kraemer, 1988).
Увеличение внутрибрюшного давления как механизма для уменьшения нагрузки на мышцы спины во время упражнений по поднятию тяжестей было описано в работах (Bartelink, 1957; Kresswell, 1993; McGill, Norman, 1993). На рис. 2.28,а показан этот эффект во время подъема массы 91 кг. Система состоит из верхней части туловища выше поясничного отдела. Она взаимодействует со своим окружением с помощью трех векторов веса — веса шеи—головы— рук (Fw>a), веса торса (FwZ) и веса 91 кг (Fw>/), которые действуют вертикально вниз, результирующего мышечного усилия мышц-разгиба-
Рис. 2.27. Центроидная линия действия для шести мышц, пересекающих локтевой сустав. Локтевой сустав показан выпрямленным и в нейтральном положении (An, Hui, Моггеу et al., 1981):
1 — бицепс брахии; 2 — брахиалис; 3 — брахиорадиалис; 4 — пронатор терес; 5 — трицепс брахии; 6 — экстенсор карпи ульнарис
5 8-133
65
Рис. 2.28. Влияние внутрибрюшного давления на силы мышц и реакции сустава, участвующие в поднятии груза 91 кг (Morris, Lucas, Bressler, 1961): а,б — этапы реакции
Рис. 2.29. Изменение углов суставов, вертикальной составляющей силы реакции земли и внутрибрюшного давления во время приседания штангиста с грузом (Lander, Betes, De-Vita, 1986):
1 — торс; 2 — колено; 3 — бедро
телей спины и верхней части бедра, силы реакции сустава и силы разгибания под действием внутрибрюшного давления. Если внутрибрюш-ное давление не учитывается, мышцы спины и бедер должны создавать усилие (Fm) 8223 Н, а сила реакции сустава должна составлять 9216 Н (F7), чтобы только поддерживать груз (рис. 2.28,а). Однако если в расчеты включается вну-трибрюшное давление 19,7 кПа (сила 810 Н), то Fm уменьшается до 6403 Н, a F7 становится 6599 Н (рис. 2.28,6).
Внутрибрюшное давление может быть измерено катетером с тензометром, введенным в брюшную полость. Оно изменяется при большинстве видов деятельности, особенно включающих в себя прыжки, подъем тяжестей и, в частности, при процедуре Валсалва (Harman et al., 1988). На рис. 2.29 показано изменение внутрибрюшного давления во время рывка — одного из упражнений при спортивном поднятии тяжестей. Спортсмен начинает его из выпрямленного положения и опускает груз, пока бедра не будут параллельны земле (углы торса, колена и бедра достигают минимальных значений), после чего возвращается в выпрямленное положение. Фигурки на рис. 2.29 схематически показывают движение, при котором штанга находится на плечах. Вследствие нагрузки (вес спортсмена и штанги), составляющий в данном примере 2200 Н, вертикальная составляющая силы реакции земли (Fg^ колеблется вокруг этой величины. Величины меньше 2200 Н показывают, что система ускоряется вниз, а больше 2200 Н соответствуют ускорению системы вверх. Изменение внутрибрюшного давления имеет общую тенденцию с изменением нагрузки на
66
мышцы торса для значений больше 2200 Н. Другие исследователи показали, что применение пояса во время поднятия тяжестей может способствовать увеличению внутрибрюшного давления (Harman, Rosenstein, Frykman, Nigro, 1989; Lander, Hundley, Simonton, 1992; McGill, Norman, 1993).
Несмотря на зависимость между различными движениями и изменениями внутрибрюшного давления, существуют определенные противоречия относительно роли этого механического эффекта (Marras, Mirka, 1992). Например, высказывалось мнение, что единственным эффектом внутрибрюшного давления является уменьшение сжимающих усилий, которые действуют на межпозвоночные диски. Однако Nachemson, Andersson, Schulz (1986) показали, что хотя маневр Валсалвы повышает внутрибрюшное давление, он может также в некоторых умеренных упражнениях увеличить давление на ядро диска L3. Тем не менее при более тяжелом упражнении, когда спортсмены наклонялись вперед до 0,53 рад, держа на вытянутых руках груз 8 кг, маневр Валсалвы увеличивал внутрибрюшное давление с 4,35 до 8,25 кПа, уменьшая давление между дисками с 1,625 до 1,488 кПа.
Как и давление в замкнутом объеме, внутрибрюшное давление действует на всю площадь поверхности брюшной полости. Усилие, которое внутрибрюшное давление оказывает на торс, обычно оценивается как произведение внутрибрюшного давления и площади поверхности диафрагмы, которая составляет, по оценке Morris et al. (1961), около 0,0465 м2 для взрослого человека. Если сопоставить это значение с пиковым внутрибрюшным давлением 25 кПа, показанным на рис. 2.29, то усилие на диафрагму вследствие внутрибрюшного давления составит во время приседания с грузом около 1,163 кН. Очевидно, что такой силой нельзя пренебречь при оценке движения человека.
Сопротивление жидкости
Как на движение человека (прыжки с трамплина, велосипедный спорт, плавание, воздушная акробатика), так и на движение снаряда (полет диска или мячика в гольфе) значительное влияние может оказывать среда (газообразная или жидкая), в которой эти явления происходят. Когда объект перемешается в жидкой или газообразной среде, он вовлекает в движение ее частицы, передавая им при этом часть своей энергии. Это приводит к возникновению сопротивления среды движению объекта. По сути, эта передача зависит от степени возмущения среды объектом. Соответственно сопротивление среды возрастает при увеличении скорости объекта.
Рис. 2.30. Прослеживание линий тока с помощью дымовых струй, показывающих перемещение воздуха вокруг обтекаемого объекта (а) и вокруг такого же объекта, разрезанного пополам (б). Воздушный поток значительно более турбулентен вокруг объекта, разрезанного пополам; линии тока прерываются
Взаимодействие между средой и объектом схематически изображено на рис. 2.30, где показан поток среды вокруг неподвижного объекта. Схематически линии потока жидкости представляют собой траектории (линии тока) частиц среды. Один слой жидкости непосредственно прилегает к объекту, второй прилегает к первому и т.д. Тонкий слой, ближайший к объекту, называется пограничным слоем. Если поместить в среду краситель, станет видно, что слои могут двигаться с различными скоростями. Как правило, у слоя, касающегося объекта, наименьшая скорость, поскольку он замедляется трением между слоем частиц и объектом; этот эффект известен под названием торможения трения.
Если форма объекта асимметрична, как, например, несимметричный кривой профиль, или когда он вращается, частицы среды, проходящие большее расстояние вокруг объекта, движутся с большей скоростью, и давление среды с этой стороны объекта меньше, чем с другой. Этот эффект известен как принцип Бернулли, согласно которому давление жидкости уменьшается при увеличении ее скорости. Когда давление на верхней поверхности крыла или другого обтекаемого объекта (например, прыгуна с трамплина в полете) меньше из-за того, что скорость потока больше, чем на нижней, сила давления толкает крыло вверх, возникающая при этом подъемная сила (Fz) представляет собой силу взаимодействия крыла и среды. Эта сила действует перпендикулярно направлению потока (рис. 2.31).
Когда линии тока плавно огибают объект, их форма может сохраняться упорядоченной.
5*
Рис. 2.31. Сила воздействия воздушного потока на прыгуна с трамплина (аэродинамическая сила). Воздух показан движущимся относительно прыгуна. Результирующий вектор аэродинамической силы (FJ разлагается на две составляющие: лобовое сопротивление (¥d) и подъемную силу (F,)
Такой поток называется ламинарным (рис. 2.30,а). Если движение частиц становится хаотическим, поток называется турбулентным (рис. 2.30,6). Основными факторами, влияющими на поток среды и его характер, являются форма и текстура объекта. Если объект (например, шлем
Рис. 2.32. Траектория и ориентация правой руки пловца во время плавания вольным стилем (Schleihauf, 1979):
а — вид спереди на траекторию, описываемую рукой; б— поток воды около руки в четырех точках во время гребка; в — вид сверху на траекторию, описываемую рукой; рука входит в воду сверху и выходит снизу
велосипедиста) имеет обтекаемую форму, это способствует сохранению ламинарности течения при больших скоростях движения. Если поток становится турбулентаым, увеличивается сопротивление среды, возрастает лобовое сопротавление (FJ. Направление вектора лобового сопротивления параллельно направлению потока среды (рис. 2.31). Лобовое сопротивление иногда называется сопротивлением давления, или сопротивлением формы, чтобы отличить его от сопротивления, вызванного трением в пограничном слое (поверхностного сопротивления).
В принципе, лобовое сопротивление препятствует движению объекта вперед, тогда как подъемная сила создает вертикальное перемещение (вверх или вниз). Однако так бывает не всегда; в плавании подъемная сила, создаваемая руками и ногами, способствует движению вперед. Фактически для успешного продвижения в плавании необходимо перемещать руки в воде таким образом, чтобы обе составляющие — лобовое сопротавление и подъемная сила — способствовали движению пловца вперед (Schleichauf, 1979). Как видно из рис. 2.32, ориентация руки пловца (показанной в виде крыла) непрерывно изменяется так, чтобы результирующая сила сопротивления жидкости (Fy) была направлена вперед в течение всего гребка. И лобовое сопротивление (F^), и подъемная сила (Fz) способствуют этому.
Полезно различать факторы, которые могут влиять на лобовое сопротавление среды. Два из них уже упоминались выше: характер течения жидкости или газа вокруг объекта (сопротавление давления) и степень трения между пограничным слоем среды и объектом (сопротивление трения). Поскольку сопротивление давления увеличивается при переходе потока от ламинарного к турбулентному, конструкторы часто делают объект обтекаемым, чтобы минимизировать тенденцию потока среды стать турбулентным.
В последние годы (после Олимпийских игр 1984 г. в Лос-Анджелесе) шлемы велосипедистов начали делать более заостренными назад, поскольку было показано, что такая конструкция уменьшает турбулентность воздушного потока. Более того, текстура поверхности объекта, например, ямки в мяче для гольфа, может влиять на ширину турбулентной спутной струи и тем самым снижать сопротивление давления, испытываемое объектом. Ямки на поверхности мяча для гольфа уменьшают ширину спутной струи и сопротивление давления путем смещения той точки на объекте, на которой струи становятся турбулентными. Результирующий эффект при этом состоит в том, что при одинаковой силе удара мячик с ямками может пролететь, например, 239 м, а гладкий мячик — только 46 м (Townend, 1984).
68
Текстура объекта влияет не только на ширину турбулентной спутной струи, но и на скорость близких к объекту слоев потока. Чем более неровная поверхность объекта, тем больше трение и, следовательно, поверхностное сопротивление. Необходимость снижения поверхностного сопротивления очевидна во многих видах спорта, например, в плавании, велосипедном спорте и гребле. Поэтому многие пловцы бреют волосы на теле и носят шапочки, а велосипедисты экспериментируют с экзотическими материалами и конструкциями в своей одежде. Поверхностное трение, однако, часто имеет значительно меньший эффект, чем сопротивление давления.
Кроме поверхностного трения и сопротивления давления, третьим фактором, влияющим на лобовое сопротивление среды, является волновое сопротивление. Этот эффект устанавливает верхний предел для скорости надводных судов и может оказаться важным при высоких скоростях, связанных с соревновательным плаванием. Эффект волнового сопротивления, вероятно, обусловлен неравномерной плотностью среды (воды или воздуха), с которой сталкивается пловец. Наличие волн, очевидно, уменьшает в среднем относительную часть тела пловца, которая находится вне воды и тем самым увеличивает эффективную плотность среды. Современная практика конструирования бассейнов заключается в сведении к минимуму волновой турбулентности путем тщательного проектирования водоотводов и разделителей дорожек.
Сопротивление среды. Сопротивление среды может быть определено из уравнения
Ff = kAv1, (2.8)
где к — константа; А — характерная площадь объекта; v — скорость среды относительно объекта. Характерная площадь — это площадь фронтальной проекции объекта. Константа к записана вместо выражения 0,5 Сд для лобового сопротивления и 0,5 р CL для подъемной силы; р — плотность среды; коэффициент сопротивления CD учитывает эффекты ламинарного и турбулентного потока, а коэффициент подъемной силы CL приблизительно пропорционален углу между потоком среды и ориентацией объекта, называемому утлом атаки.
Рассмотрим действие сопротивления воздуха на бегуна (Hill, 1928), показанное на рис. 2.6. Константа к для бегуна составляет примерно 0,55 кг • м-3. Характерная площадь бегуна, т.е. площадь фронтальной поверхности тела, на которую действует воздух, может быть оценена как 0,15, умноженное на квадрат роста в стоячем положении. Для бегуна ростом 1,55 м (рис. 2.6) А = 0,36 м2. Эта оценка, однако, несколько занижена по сравнению с данными, полученными для лыжника в позиции полуприседа (0,27 м2) и в вертикальном поло
жении (0,65 м2) (Spring, Savolainen, Erkkila et al., 1988). Тем не менее если бегун бежит со скоростью 6,5 м • с-1 при попутном ветре 0,5 м • с-1, то относительная скорость (у) будет 6,0 м • с-1. Тогда
Ff= 0,55 х (0,15 х 1,552) х 6,02 = 7,14 Н.
Согласно Shanebrook, Jaszczak (1976), при скоростях бегунов на средние дистанции (примерно 6 м-с-1) до 8 % энергии, расходуемой бегуном, тратится на преодоление сопротивления воздуха, тогда как у спринтеров эта величина может составлять до 16% общего расхода энергии. Наличие впереди бегущего лидера может уменьшить эти затраты на 80%; так, при беге со скоростью 6,0 м • с-1 следование за другим бегуном эквивалентно уменьшению скорости примерно на 0,1 м-с-1. Ward-Smith (1985) рассчитал эффект попутного и встречного ветра на время, показанное при беге на 100 м:
Скорость ветра, м-с-1 Встречный ветер, с Попутный ветер, с
1 + 0,09 -0,10
3 + 0,26 -0,34
5 + 0,38 -0,62
При встречном ветре 3 м • с-1 время при беге на 100 м будет увеличено на 0,26 с, тогда как попутный ветер 3 м • с-1 уменьшит это время на 0,34 с. Влияние попутного и встречного ветра не будет симметричным для скоростей 3 и 5 м • с-1, (т.е. потеря и выигрыш неодинаковы), поскольку суть дела заключается не в силе, связанной с ветром, а скорее в работе, выполняемой против него. Подобным образом 80% энергии, образуемой велосипедистом, едущим по ровной поверхности со скоростью 8,05 м-с-1, затрачивается на преодоление сопротивления воздуха (Gross, Kule, Malewicki, 1983). Можно уменьшить затрату энергии примерно на 30 % и все же сохранить такую же скорость при следовании за другим велосипедистом.
Лобовое сопротивление, испытываемое пловцами, определяли, измеряя усилие, необходимое для буксирования в воде пассивного субъекта. При использовании этой методики конечности фактически не выполняли плавательных движений. Лобовое сопротивление, действующее на пловца, оценивалось при этом путем измерения двигательной силы, затрачиваемой субъектом, плывущим с постоянной скоростью. При таких условиях ускорение пловца равно нулю, следовательно, силы, воздействующие на пловца, уравновешены. Таким образом, в горизонтальном направлении движущая сила равна силе сопротивления (лобовому сопротивлению). Измеренное с помощью этого метода среднее лобовое сопротивление, испытываемое пловцом, плывущим кролем,
69
Рис. 2.33. Силы, испытываемые парашютистом в свободном падении
составило 53 Н при постоянной скорости 1,48 м-с-1 (van der Vaart, 1987).
Величина сопротивления среды зависит от степени возмущения жидкости или газа движущимся объектом. Уравнение 2.8 указывает, что это возмущение пропорционально произведению характерной площади и квадрата скорости объекта относительно среды. Поскольку коэффициенты подъемной силы и лобового сопротивления зависят от ориентации объекта относительно скорости движения в среде, изменения ориентации могут использоваться для изменения подъемной силы и лобового сопротивления объекта. Например, рассмотрим спортсмена, скользящего на водных лыжах за катером. Возможность лыжника оставаться на поверхности воды зависит от усилия, которое оказывает водная среда на лыжи. Составляющая подъемной силы (FJ) должна быть при этом, по меньшей мере, такой же, как направленные вниз силы, состоящие, в основном, из веса субъекта. С увеличением скорости движения катера скорость лыж относительно воды возрастает, характерная площадь может уменьшаться, чтобы сохранять необходимую подъемную силу. Фактически такой компромисс между подъемной силой и относительной скоростью достигается во время всего заезда путем изменения ориентации лыж по отношению к направлению движения. На старте лыжник в воде направляет лыжи почти вертикально. В этом положении угол атаки и, следовательно, коэффициент подъемной силы Q лыж по отношению к воде максимизируется. Когда катер начинает двигаться и буксировать лыжника, лыжи возмущают воду и создают подъемную силу по мере того, как вода обтекает лыжи. В начале заезда угол атаки должен быть велик, поскольку он сочетается с низкой относительной скоростью для создания необходимой подъемной силы Ff и поднятия лыжника на поверхность. По мере возрастания скорости катера лыжи могут опускаться, поскольку в этом случае требуется меньший угол атаки. Когда лыжник отпускает буксировочный трос, подъемная сила
становится недостаточной из-за уменьшения скорости, и лыжник погружается в воду.
Конечная скорость. Поскольку относительная скорость объекта входит в уравнение (2.8) в квадрате, она является наиболее важным фактором, определяющим сопротивление среды. Интересно проследить роль этого фактора на примере движения парашютиста. После того как парашютист отделяется от самолета, его скорость увеличивается до некоторого конечного значения. Как определить это значение? Опять-таки, анализ следует провести на соответствующей диаграмме свободного тела (рис. 2.33).
Когда парашютист прыгает с самолета система (парашютист, парашют и связанное с ним оснащение) ускоряется под действием силы тяжести (Fw) по направлению к земле. Однако ускорение вниз системы только короткое время равно величине ускорения тяготения (9,81 м-с-2), поскольку по мере возрастания скорости падения увеличивается противодействующий эффект сопротивления воздуха. Как следует из уравнения (2.8), сопротивление воздуха возрастает пропорционально квадрату относительной скорости. Скорость системы будет увеличиваться (т.е. ускорение будет отличаться от нуля) до тех пор, пока сила сопротивления воздуха не станет равной весу системы. После того как эти две силы станут равными, скорость падения достигнет своего предельного конечного значения будет оставаться постоянной. Поскольку скорость остается постоянной (т.к. ускорение нулевое) и система находится в равновесии при конечной скорости, силы должны быть уравновешены. Таким образом,
Fw=Ff.
Поскольку, согласно уравнению (2.8),
Ff= kAv1, можно преобразовать это соотношение относительно конечной скорости
v= Ж.
V кА
Если к = 0,55 кг • м-3, А = 0,36 м2 и Fw = =750Н, то
v= I 750 у 0,55 х 0,36 и конечная скорость равняется 61,6 м-с-1 (8 миль в час).
Несмотря на то, что парашютист находится в состоянии свободного падения, обладая определенным опытом, он может довольно хорошо маневрировать, выполняя сальто, пируэты и другие движения. Каким образом он это делает? Как видно из диаграммы свободного тела (рис. 2.33), парашютист во время падения испытывает действие двух сил: веса и сопротив
70
ления воздуха. Обе эти силы являются распределенными. Это означает, что они распределены по всей поверхности системы, но вычерчиваются как действующие в одной или двух точках. Вектор веса всегда приложен в центре масс (ЦМ). Подобным образом сила, обусловленная сопротивлением воздуха, имеет центральную точку приложения, которая называется центром давления. Если линия воздействия вектора сопротивления воздуха не проходит через ЦМ, сопротивление воздуха оказывает вращающее действие (возникает вращающий момент) относительно ЦМ, что заставляет парашютиста испытывать угловое перемещение. Двигая конечностями, парашютист может изменять характерную площадь и сдвигать центр давления таким образом, чтобы вектор не проходил через ЦМ.
Аэродинамика снаряда. В главе 1 мы рассматривали случай, когда сопротивление воздуха оказывало пренебрежимое воздействие на траекторию снарядов. Но на многие движения сопротивление воздуха влияет значительно. Одно из наиболее известных влияний — воздействие сопротивления воздуха на траекторию бейсбольного, волейбольного мяча, мяча для гольфа и крикета и других снарядов. Сопротивление воздуха в дополнение к силе тяжести создает силу, которая может изменять траекторию снаряда. Сопротивление воздуха вызывает изгиб траектории бейсбольного мяча, скольжение мяча для гольфа и падение теннисного мяча. Эти воздействия на траекторию снарядов обусловлены составляющей подъемной силы сопротивления воздуха. В соответствии с принципом Бернулли существует обратная зависимость между давлением, создаваемым жидкой или газообразной средой, и ее скоростью. Давление среды уменьшается по мере возрастания скорости. Соответственно, когда среда обтекает объект, на нем возникает перепал давления, если скорости потока с разных сторон обтекаемого тела (сверху и снизу) отличаются друг от друга.
Различие в скоростях потоков, обтекающих объект, может быть вызвано формой (например, крыло) или вращением тела. Если форма несимметрична, потоки должны проходить различные расстояния вдоль поверхности объекта, и скорость потока будет большей с той стороны, где расстояние больше. Для крыла расстояние и скорость потока воздуха больше с верхней стороны, поэтому давление меньше сверху; это вызывает перепад давлений в направлении снизу вверх (т.е. подъемную силу). Подобным образом вращающийся снаряд будет создавать перепад давлений вокруг себя, поскольку вращение влияет на скорость потоков. Например, если бейсбольный мяч в полете вращается против часовой стрелки, у потоков, протекающих в направлении против часовой стрелки, будет большая скорость (вследствие уменьшенного
Рис. 2.34. Влияние составляющей подъемной силы сопротивления воздуха на траекторию бейсбольного мяча. Мяч вращается против часовой стрелки и будет поворачивать влево
поверхностного трения из-за попутного движения поверхности мяча), поэтому давление потока с левой стороны мяча будет меньше. Это создает перепад давлений поперек бейсбольного мяча справа налево и заставляет его изгибать траекторию полета влево (рис. 2.34).
Сила упругости
Растягивающее усилие может увеличить длину материала или ткани. Растяжение возможно благодаря молекулярному составу и структуре материала. Степень растяжения зависит от его природы и величины растягивающей силы. Для идеальной пружины соотношение между приложенной силой и величиной растяжения (изменением размера пружины) имеет простое выражение
F= кх, (2.9)
где F — растягивающее усилие; к — жесткость пружины; х — величина растяжения или деформации. При увеличении к жесткость пружины возрастает, и необходимо приложить большее растягивающее усилие, чтобы осуществить такую же деформацию дх. Константа к представляет собой наклон графика зависимости между F и х. Чем жестче пружина, тем круче наклон; чем более податлива пружина, тем ниже наклон (рис. 2.35, а).
Если сила, приложенная к пружине, возрастает неограниченно, пружина будет все время удлиняться и в конце концов разрушится. Этот
71
Рис. 2.35. Зависимость сила—деформация для пружины, растягиваемой в пределах ее диапазона упругости (а) и до разрушения (б) (Nordin, Frankel, 1989)
процесс показан на рис. 2.35,б. Биологические волокна (например, кости, мышцы, сухожилия, кожа) ведут себя подобным образом. Если волокно растягивается в пределах своей зоны упругости (точки от А до В на рис. 2.35, б, а затем отпускаются, оно возвращается к своей первоначальной длине благодаря упругим свойствам. Упругость волокна при этом зависит от таких факторов, как тип волокна, возраст, состояние здоровья, и определяется его молекулярной структурой (рис. 2.36). Деформация, испытываемая волокном при растяжении за пределами
Рис. 2.36. Кривая сила—деформация для сухожилия антериор круциате. Сравнивается жесткость (наклон) в здоровых коленях и коленях, которые находились в гипсе в течение 8 недель (Noyes, 1977):
1 — контроль; 2 — гипсовая повязка
упругости, постоянна в отличие от временной деформации при растяжении в пределах упругости, которая исчезает при снятии нагрузки. В области пластической деформации (точка В на рис. 2.35) будут иметь место долговременные изменения в структуре ткани. Например, если волокно растягивается до точки D (рис. 2.35,6) и затем отпускается, оно будет иметь новую длину (точка D1), которая больше первоначальной (точка А) вследствие изменений в структуре. Ткань волокна разрушается, если деформация продолжается до точки С.
Для сравнения кривых сила—деформация в различных условиях, примененные сила и деформация должны быть нормированы, т.е. выражены в виде зависимости напряжение—деформация. Напряжение (Па) — это сила, приложенная к единице площади ткани, измеренной в плоскости, перпендикулярной вектору силы. Относительная деформация (%) — отношение длины ткани к ее первоначальной длине. Верхний предел физиологических деформаций в связках и сухожилиях составляет 2 — 5%, причем сухожилия разрываются примерно при 8% (Alexander, Кег, 1990). Поскольку пиковые усилия в ахиллесовом сухожилии у человека весом 70 кг, бегущего с умеренной скоростью, составляет около 4700 Н, а поперечное сечение сухожилия примерно 89 мм2, пиковое напряжение составляет 53 МПа (Alexander, Кег, 1990), Упругая зона соотношения напряжение—деформация характеризуется модулем упругости Е, который определяется как отношение напряжения о к деформации (е)
£ = -. (2.10)
Е
Для кинезиологии важны, по меньшей мере, две особенности зависимости сила—деформация. Во-первых, степень, до которой характеристика сила—деформация ткани напоминает характеристику идеальной пружины. Эта особенность характеризует потенциальные возможности, которые имеет ткань для накопления энергии упругости (энергии деформации) во время растяжения и для последующего использования этой энергии с целью выполнения работы в окружающей среде. На пике растяжения биологическая ткань обладает определенным количеством энергии деформации благодаря силам, генерируемым молекулярной структурой ткани. Этот эффект мы называем силой упругости; она представляет собой потенциал, которым обладает ткань для возврата к своей первоначальной длине (уравнение 3.23). Например, когда прыгун подпрыгивает на трамплине, величина прогиба доски зависит от усилия отталкивания, оказываемого телом прыгуна на доску. На последней стадии прыжка сила отталкивания вниз преодолевается действующей вверх силой упругости доски, и прыгун ускоряется
72
вверх. Способность биологической ткани, особенно мышц и сухожилий, накапливать энергию деформации и впоследствии использовать ее для создания силы упругости имеет существенное влияние на движение. Это свойство учитывается при протезировании. Например, протезы ноги, конструируемые для имитации нормальной конечности, способны накапливать и возвращать упругую энергию во время опорной фазы походки (Czemiecki, Gitter, Manro, 1991).
Во-вторых, растяжение в зоне пластичности, по меньшей мере, частично изменяет структуру ткани, вызывая некоторое ослабление тонуса. Если мышцу растянуть в зону пластичности и затем отпустить, то конечная ее длина будет больше первоначальной. Это свойство используется при выполнении упражнений на гибкость, которые вносят пластичные изменения в биологические ткани, позволяя увеличивать диапазон движений относительно суставов. Ниже (глава 8) будут рассмотрены исследования для определения наиболее эффектных процедур, обеспечивающих компромисс между увеличением длины ткани и ослаблением ее тонуса, при котором происходят структурные изменения.
Сила инерции
Согласно закону Ньютона, движущийся объект продолжает двигаться прямолинейно и с постоянной скоростью, пока на него не подействует сила (закон инерции). Способность объекта сопротивляться любому изменению его движения называется инерцией. Движущийся объект благодаря своей инерции может оказывать воздействие на другой объект. Для демонстрации этого эффекта расположите одно предплечье в вертикальном положении (ладонью вверх) при горизонтальном положении плеча. Расслабьте мышцы предплечья, пересекающие сустав запястья. Медленно начните колебания вашего предплечья по небольшой дуге в направлении вперед-назад. По мере того как движение становится более энергичным, вы заметите, что ваша кисть начинает болтаться, особенно если она расслаблена. Вы подозреваете, что движение вашей кисти было вызвано движением предплечья, и вы правы. Предплечье (вследствие своего движения) оказывает инерционное усилие на кисть и вызывает ее движение. Этот простой пример подчеркивает механическую связь, существующую между сегментами тела.
Рассмотрим еще один пример силы инерции, связанной с метанием молота. Из раздела о движении снаряда (глава 1) мы узнали, что расстояние, на которое атлет может метнуть
молот, зависит от скорости молота, угла и высоты в начале броска. Для достижения мирового рекорда 86,3 м молот должен иметь скорость выпуска 29,1 м • с-1, при этом угол выпуска является оптимальным (0,78 рад, или 45°). Для достижения этой скорости выпуска атлет должен выполнить пять оборотов, перемещаясь по кругу диаметром 2,13 м. Во время этой процедуры атлет прилагает к молоту две силы (рис. 2.37): тянущую силу, которая создает угловое ускорение для увеличения скорости вращения, и центростремительную силу для сохранения вращательного движения. Согласно Brancazio (1984), для достижения скорости выпуска 29,1 м' с-1 атлет должен обеспечивать среднее тянущее усилие около 45 Н в течение всего броска и центростремительное усилие на выпуске около 2,8 кН.
Поскольку силы действуют попарно (действие-противодействие), молот должен оказывать усилие на метателя (центробежная сила), составляющее 2,8 кН при выпуске. Инерционный эффект молота можно легко представить. Допустим, атлет забыл выпустить молот и вместо этого продолжал удерживать его. Как только центростремительное усилие, оказываемое атлетом на молот, опустится ниже 2,8 кН, молот вследствие своего движения будет оказывать инерциальное усилие на атлета. В результате действия этой силы атлет будет вынужден покинуть круг для метания.
Инерционные эффекты постоянно сопровождают нашу жизнь, связанную с движением. В частности, движение одного сегмента тела, например, бедра может влиять на движение другого сегмента — голени. На рис. 2.38 показаны изменения, которые имеют место в угле колена и мышечном вращающем моменте (вращающее усилие, создаваемое мышцей) относительно тазобедренного и коленного суставов во время опорной фазы прыжка при беге (Phillips, Roberts, 1980). Мышечный вращающий момент относительно тазобедренного сустава, по сути, бифазный: он действует как сгибатель в первой половине опорной
Рис. 2.37. Силы, создаваемые метателем молота (Brancazio, 1984)
73
Рис. 2.38. Угол колена и мышечные вращающие моменты относительно тазобедренного и коленного суставов во время опорной фазы прыжка при беге со скоростью 5,2 м • с-1 (Phillips, Roberts, 1980): / — бедро; 2 — колено
фазы, а затем как разгибатель в течение второй половины. Сгибающий вращательный момент относительно тазобедренного сустава ускоряет бедро в направлении вперед, в то время как разгибающий вращающий момент ускоряет бедро в направлении назад. Подобным образом мышечный вращающий момент относительно коленного сустава (пунктирная линия на рис. 2.38), бифазен с промежуточным периодом, в течение которого вращающий момент равен нулю. Однако если вращающий момент относительно тазобедренного сустава имеет последовательность сгибания—разгибания, то результирующий мышечный вращающий момент относительно колена имеет последовательность разгибания-сгибания. На рис. 2.38 положительный вращающий момент является сгибающим моментом для бедра, но разгибающим для колена.
Роль результирующего мышечного вращательного момента относительно коленного сустава станет более ясной, если мы рассмот
рим, как изменялся угол колена во время опорной фазы. Сделаем это, сравнив два графика на рис. 2.38.
Угол колена вначале уменьшается, а затем во время опорной фазы увеличивается (рис. 2.38,7). Сгибание коленного сустава (первая половина опорной фазы) контролируется разгибающим моментом относительно коленного сустава; иными словами, сгибание управляется разгибающими мышцами. Этот процесс осуществляется в результате удлинения активных мышц разгибателей — квадрицепс феморис и рассматривается как эксцентричное сокращение. Подобным образом разгибание коленного сустава происходит в результате действия сгибающего (сухожилия, ограничивающие с боков коленную ямку) момента относительно коленного сустава.
Таким образом, смещение в коленном суставе управляется эксцентричным сокращением; сгибание колена контролируется не сгибателями, а разгибателями колена и наоборот. Причиной движения голени является сила инерции, с которой бедро воздействует на голень. Активируя мышцы, пересекающие тазобедренный сустав, можно вызывать сгибание и разгибание коленного сустава. Мышцы, которые пересекают колено, используются для контроля влияния силы инерции бедра на голень; эксцентричное сокращение служит для торможения вращения голени вперед и назад. Этот пример показывает, что сила инерции, оказываемая одним сегментом тела на другой, — важный фактор при анализе движения человека (Bizzi, Abend, 1983; Phillips, Roberts, Huang, 1983; Putnam, 1991). К этому вопросу мы вернемся в главе 3 при рассмотрении межсегментной динамики.
ВРАЩАЮЩИЙ МОМЕНТ
Все движения человека включают в себя поворот сегментов тела вокруг осей их суставов. Эти действия обусловливаются взаимодействием сил, связанных с внешней нагрузкой и мышечной активностью. В частности, движение человека — следствие несбалансированности сил, вызывающих вращение. Способность силы вызывать вращение связана с таким понятием, как вращающий момент, или момент силы относительно оси.
Рассмотрим пример тренировочных упражнений на Цибексе, связанных со сгибанием и разгибанием коленного сустава. При разгибании колена (рис. 2.39,а) коленный сустав располагается на одной линии с осью вращающегося рычага. Спортсмен толкает рычаг и вызывает его вращение. Одной из особенностей Цибекса является то, что он должен
74
измерять толкающее усилие, но все-таки возникает вопрос, что же фактически измеряет Цибекс. Для измерения силы необходимо, чтобы тензометр был расположен вдоль линии действия вектора силы.
Как видно из рис. 2.39,б, тензометр фактически расположен на некотором расстоянии от линии действия силы, оказываемой спортсменом на рычаг, в результате чего измеряется вращающий момент, а не сила.
Вращающий момент является вектором, величина которого равна произведению величины силы и расстояния между линией действия силы и осью вращения. Это расстояние измеряется длиной перпендикуляра из оси вращения к линии действия силы и называется плечом силы. Используется как скалярное, так и векторное представление плеча. В случае скалярной величины указывается расстояние между линией действия и осью вращения. В случае векторной величины учитывается и его направление, которое считается направленным от оси вращения к линии действия силы. Момент силы определяется по формуле
Т = плечо силы х F, (2.11)
где Т — вращающий момент; F — сила. Поскольку F и плечо силы в уравнении (2.11) — векторные величины, направление вектора вращающего момента перпендикулярно плоскости, в которой расположены сила и ее плечо (см. Приложение В).
Для наглядности вращающий момент обычно изображают изогнутой стрелкой в той же плоскости, что и векторы плеча силы и самой силы (см., например, рис. 2.41). На
правление изогнутой стрелки может быть определено с помощью правила правой руки, как показано на рис. 2.40. Для определения вектора вращающего момента нужно перенести векторы силы и плеча с диаграммы свободного тела, сохранив их направление (рис. 2.40, а), таким образом, чтобы их начала совпадали (рис. 2.40, б). Тогда правая рука с отставленным большим пальцем используется для удобства практического определения направления векторного произведения (рис. 2.40, в). В векторной терминологии (Приложение В) вращающий момент равен векторному произведению плеча (г) на силу (Т = г х F). Положите правую руку так, чтобы пальцы указывали направление вектора плеча, а ладонь была обращена в сторону действия силы; вытянутый большой палец покажет направление вектора вращающего момента. Применив это правило для векторов, показанных на рис. 2.40, получим, что вращающий момент направлен вверх перпендикулярно странице. При этом направление вращения против часовой стрелки и соответствующее ему направление большого пальца от страницы считаются положительными (рис. 2.40).
Вращающий момент, или момент силы, всегда определяется по отношению к конкретной оси, и эта ось должна быть четко указана. Так, для обсуждения вращательного эффекта при активации группы мышц (например, сгибателей локтя), необходимо указать ось, относительно которой будет происходить вращение, т.е. ось локтевого сустава. Поскольку вращающий момент рассчитывается как произведение силы и расстояния, единицей его измерения будут ньютонометры (Нм).
Рис. 2.39. Измерение усилий на Цибексе: а — выполнение упражнения по разгибанию колена; б — линия действия толкающей силы (контактная сила />), которую спортсмен оказывает на рычаг
Рис. 2.40. Использование правила правой руки для определения направления вектора вращающего момента
75
Пример
Человек после операции на колене, обутый в ботинок с отягощением, выполняет упражнение по выпрямлению колена в сидячем положении. Упражнение заключается в сидении на краю тренировочной скамейки и поднимании ноги (голени и стопы) из вертикального (угол колена 1,57 рад) в горизонтальное положение с последующим опусканием ноги. Какие моменты относительно коленного сустава (К) имеют место в этом упражнении? На рис. 2.41 изображена соответствующая система от коленного сустава до пальцев стопы и четыре силы — сила реакции сустава (F7), результирующая мышечная сила (Fm), вес конечности (Fw/) и вес ботинка (FwZ() (рис. 2.41, б).
Первый шаг в определении вращающих моментов заключается в вычерчивании плеч (а,Ь,с) сил, которые в данном поимере рассматриваются как скалярные величины. При этом необходимо продлить линию действия каждой силы и опустить на нее перпендикуляр из оси вращения (рис. 2.41, б). Поскольку линия действия для силы реакции сустава проходит через ось вращения, ее плечо и, следовательно, момент силы реакции сустава относительно коленного сустава равен нулю. Таким образом, для системы, показанной на рис. 2.41, имеются три силы, дающие вращающий момент относительно колен
Рис. 2.41. Диаграмма свободного тела для субъекта, выполняющего упражнения по разгибанию ноги
ного сустава. На рис. 2.41, в показан результирующий мышечный вращающий момент (Тт), вращающий момент, обусловленный весом ботинка (Tw a) и вращающий момент силы веса ноги (Т^/). Вращающий момент под действием всей нагрузки (Т/ определяется как сумма Twb и Tw/ Изогнутые стрелки на рисунке показывают направление вращений, вызываемых соответствующими моментами.
При расчете момента силы (вращающего момента) обычно используют тригонометрические функции. Допустим, что в примере, изображенном на рис. 2.41, а, вес ботинка 80 Н, а вес тела 700 Н; величина и направление вектора результирующей мышечной силы составляет соответственно 1000 Н и 0,25 рад; длина ноги равна 0,36 м, а расстояние от коленного сустава до точки приложения вектора результирующей мышечной силы — 0,05 м. Какова величина трех вращающих моментов?
Сначала рассмотрим момент мышечной силы и вычертим соответствующую диаграмму свободного тела.
sin 0,25 = ;
а = 0,05 sin 0,25 = 0,0124 м;
Тт = 1,000 х 0,0124 = 12,4 Н м.
Аналогично может быть определен момент силы, обусловленный весом конечности. Вес стопы и голени можно определить по комбинированным уравнениям регрессии в табл. 2.1 с центром тяжести (т.е. точкой приложения вектора веса) для ноги (голени и стопы), расположенным на расстоянии 43,4 % длины голени, измеряемой от коленного сустава. Отсюда следует схема:.
d= 0,36x0,434 = 0,156 м;
= (0,044 х 700 - 1,75) + + (0,009 х 700 + 2,48) = 37,8 Н;
TvA = Fwl xb = 37,8 х b-
cos 0,5 = d
b = 0,156 cos 0,5 = 0,14 m;
TwJ = 37,8 x 0,14 = 5,2 Нм.
76
Наконец, каков момент относительно коленного сустава силы веса ботинка? Ботинок расположен на расстоянии 0,36 м от коленного сустава и имеет вес 80 Н (см. схему):
Kb = F».b X С = 80 х с;
cos0’5=0J6;
с = 0,36 cos 0,5 = 0,32 м;
TWib = 80 х 0,32 = 25,3 Нм.
Обратите внимание, что вектор силы веса всегда направлен вертикально вниз. А поскольку плечо силы перпендикулярно линии ее действия, то плечо вектора веса всегда горизонтально.
Результирующий момент всех сил относительно коленного сустава в данном примере может быть определен путем суммирования величин моментов и учета направлений каждого вращающего момента. При этом момент, вызывающий вращение против часовой стрелки, считается положительным. Выше описано, как определять направление вращающего момента по правилу правой руки. Действие каждой силы на систему рассматривается отдельно путем сжатия пальцев правой руки в свободный кулак и вытягивания большого пальца. Правая рука располагается так, что согнутые пальцы указывают направление, в котором сила заставляет систему вращаться. Тогда вытянутый большой палец показывает направление вектора вращающего момента. Напомним, что вращающий момент может быть представлен вектором, направленным перпендикулярно к плоскости чертежа. Если такой вектор направлен по направлению к вам, он будет заставлять систему вращаться против часовой стрелки, что мы обычно считаем положительным направлением. В данном примере силы FWj/ и Fw Л дают вращение по часовой стрелке, и поэтому их вращающие моменты отрицательны.
Для определения результирующего эффекта этих сил на систему просуммируем (Е) моменты сил относительно коленного сустава (Мк):
™к = * а)~ (Fw,i xb)~ (Кjb хс) =
= 12,4 - 5,2 - 25,3;
YMk = -18,1 Н • м.
Таким образом, суммарный момент силы равен 18,Гг1-м и действует по часовой стрелке. Однако тот факт, что вращающий момент действует по часовой стрелке, не дает информации о направлении смещения конечности. Например, вспомним задачу о сгибании—разгибании
локтевого сустава, которую мы рассматривали в главе 1 в разделе “Ускорение и мышечная активность”. Как показано на рис. 1.9, перемещение и сила бывают направлены не только в одну, но и в разные стороны. Для получения полного описания движения нужна информация о положении, скорости, ускорении, вращающем моменте.
Поскольку вращающий момент равен произведению силы на плечо, вращающий эффект силы может быть изменен при изменении любого из этих факторов в отдельности или в сочетании. Многие анатомические структуры (пяточная кость, надколенная чашечка, локтевой отросток, вертикальные остистые отростки позвонков и др.) способны изменить длины плеч усилий соответствующих мышц. Рассмотрим присоединение надколенного сухожилия к большеберцовой кости (рис. 2.42). Если два человека могут развить одинаковое мышечное усилие в мышечной группе квадрицепс феморис, то человек с большим вертикальным расстоянием (из-за различий в надколенной чашечке) от линии действия вектора мышечной силы до оси вращения будет в состоянии генерировать больший вращающий момент разгибателя колена. Если сила рассматривается с позиций возможности создания вращающего момента, необходимо учитывать не просто величину мышечного усилия, но и плечо силы.
В принципе мышцы имеют пологий угол натяжения и размещены вблизи сустава, относительно которого они создают вращающий момент. Соответственно анатомические плечи моментов, как правило, невелики. Однако плечи моментов изменяются в процессе движения
Рис. 2.42. Механизм разгибателя колена. Вращающие моменты, развиваемые группой мышц квадрицепс феморис, зависят от трех плеч — от центра большеберцового сустава до надколенного сухожилия (d) и от надколенно-бедренного сустава до надколенного сухожилия (4/2) и до мышц квадрицепс (d|) (Grood, Suntay, Noyes, Butler, 1984):
Fjtf— сила реакции большеберцового сустава; Fjp, — сила реакции надколенно-бедренного сустава; FmJ — сила надколенного сухожилия; 7% , — сила мышц квадрицепс; Fw — вес ноги
77
Рис. 2.43. Изменение плеча мышечного усилия разгибателя колена в зависимости от угла в коленном суставе (от прямого угла до полного разгибания) (Grood, Suntay, Noyes, Butler, 1984)
Рис. 2.44. Схема упражнения по отжиманию: а — положение с прямыми руками; б— положение с согнутыми руками; в — диаграмма свободного тела; F„ — результирующая мышечная сила; Тт — вращающий момент относительно локтевого сустава равен произведению силы Fm на плечо (d)
(рис. 2.43). По данным An et al. (1981), плечи моментов для основных мышц, сгибающих локоть, удваиваются, когда локоть переходит от полностью разогнутого положения к согнутому в 1,75 рад (100°), в то время как плечо для трицепс брахии (разгибатель локтя) уменьшается примерно на одну треть при том же движении (табл. 2.7). Подобные наблюдения выполнены на мышцах, пересекающих тазобедренный сустав (Nimeth, Ohlsen, 1986), а также коленный и голеностопный суставы (Rugg, Gregor, Mandelbaum, Chin, 1990; van Zeenwen, Meskers et al., 1990).
Пример
Рассмотрим влияние изменения плеча сил для некоторых движений. Допустим, спортсмен выполняет отжимание до изнеможения. При этом основным источником энергии является мышца разгибателя локтя трицепс брахии. Допустим, что мышца максимально ак
тивна при приближении к состоянию изнеможения и, учитывая приведенные ранее данные о поперечном сечении (табл. 2.6), создает усилие 550 Н. В соответствии с данными табл. 2.7, плечо для трицепс брахии составляет примерно 2,81 см при разогнутом локте (рис. 2.44, а) и около 2,04 см, когда локоть согнут под углом 1,75 рад (рис. 2.44, б). Это плечо силы представлено отрезком d на рис. 2.44, в. При этом вращающие моменты составляют примерно 14,1 и 10,2 Нм для разогнутого и согнутого положений соответственно. Это означает, что в согнутом положении вращающий момент благодаря усилию трицепс брахии — основной действующей силе в упражнении — меньше, чем в выпрямленном положении.
Невозможность продолжать отжимания дальше скорее возникнет в согнутом положении, при котором вращающий момент минимален. В этом случае невозможность продолжать упражнение связано с неспособностью поднимать вес тела (постоянную нагрузку)
Таблица 2.7. Плечи сил сгибателей и разгибателей локтя для полностью разогнутого и согнутого под углом 1,75 рад локтя, а также для кисти в двух положениях (нейтральном и супинированном), см (An et al., 1981)
Мышца Локоть оазогнут Локоть согнут
нейтральное супинированное нейтральное супинированное
Сгибатели
Бицепс брахии 1,47 1,96 3,43 3,20
Брахиалис 0,59 0,87 2,05 1,98
Брахиорадиалис 2,47 2,57 4,16 5,19
Разгибатели
Трицепс брахии 2,81 2,56 2,04 1,87
78
вверх и вниз. Подобное рассуждение относится и к подтягиванию до изнеможения. Плечи сил для мышц сгибателей локтя минимальны при полном выпрямлении локтя, и если мышечная сила в процессе движений будет постоянной в разумных пределах, то именно в этой точке произойдет отказ от выполнения упражнения.
РЕЗЮМЕ
В этой главе описаны механические аспекта взаимодействия человека с окружающей средой, необходимые для начала движения или изменения его характера. Как указал Ньютон,
эти взаимодействия осуществляются посредством сил. Обычно при изучении движения человека рассматривается восемь сил: вес, сила реакции земли, сила реакции сустава, мышечная сила, внутрибрюшное давление, сопротивление среды, сила упругости и сила инерции. В главе охарактеризованы эти силы и их роль в движении человека. В заключение вводится понятие диаграммы свободного тела, используемой при проведении биомеханического анализа. Приведены примеры, иллюстрирующие необходимость включения каждой из перечисленных сил в диаграмму свободного тела. Поскольку движение человека состоит из поворота сегментов тела относительно друг друга, вводится понятие вращающего момента и описана соответствующая техника вычислений.
ГОПРОСК С АЛЬТЕРНАТИВНЫМ ОТВЕТОМ
1. Сила — это вектор.
ПРАВИЛЬНО НЕПРАВИЛЬНО
2. Невозможно определить равнодействующую (результирующую) нескольких сил.
ПРАВИЛЬНО НЕПРАВИЛЬНО
3. Результирующая сила может быть представлена графически как диагональ трапеции.
ПРАВИЛЬНО НЕПРАВИЛЬНО
4. Ортогональные составляющие перпендикулярны друг другу.
ПРАВИЛЬНО НЕПРАВИЛЬНО
5. Тангенциальная составляющая мышечной силы действует вдоль сегмента тела и вносит вклад в сжатие сустава.
ПРАВИЛЬНО НЕПРАВИЛЬНО
6. Нормальная составляющая всегда меньше равнодействующей.
ПРАВИЛЬНО НЕПРАВИЛЬНО
7. Закон инерции утверждает, что необходима сила для начала, конца или изменения движения. ПРАВИЛЬНО НЕПРАВИЛЬНО
8. Единица измерения инерции — грамм (г).
ПРАВИЛЬНО НЕПРАВИЛЬНО
9. Масса является мерой инерции объекта.
ПРАВИЛЬНО НЕПРАВИЛЬНО
10. Чтобы объект следовал по изогнутой траектории, на него должна действовать сила.
ПРАВИЛЬНО НЕПРАВИЛЬНО
11. Величина центростремительной силы влияет на кривизну (радиус изогнутой траектории) траектории.
ПРАВИЛЬНО НЕПРАВИЛЬНО
12. Момент определяет качество движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
13. Момент — векторная величина.
ПРАВИЛЬНО НЕПРАВИЛЬНО
14. Закон действия—противодействия утверждает, что F = та.
ПРАВИЛЬНО НЕПРАВИЛЬНО
15. В механике телом называется то, что занимает пространство и имеет инерцию.
ПРАВИЛЬНО НЕПРАВИЛЬНО
16. Входами в диаграмме свободного тела являются силы, действующие на систему.
ПРАВИЛЬНО НЕПРАВИЛЬНО
17. Все силы, действующие на диаграмму свободного тела, должны быть параллельны одна другой. ПРАВИЛЬНО НЕПРАВИЛЬНО
18. Силы инерции не могут быть показаны на диаграмме свободного тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
19. Пространственная диаграмма является схематической диаграммой свободного тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
20. Вес — это сила.
ПРАВИЛЬНО НЕПРАВИЛЬНО
21. Вектор веса всегда направлен вертикально ВНИЗ. ПРАВИЛЬНО НЕПРАВИЛЬНО
22. Центр тяжести не является фиксированной
ТОЧКОЙ. ПРАВИЛЬНО НЕПРАВИЛЬНО
23. Уравнение регрессии дает точные оценки параметров, правильно неправильно
24. Сегмент бедра имеет наибольшую массу из всех сегментов тела человека.
ПРАВИЛЬНО НЕПРАВИЛЬНО
25. Момент инерции — мера распределения массы относительно оси.
ПРАВИЛЬНО НЕПРАВИЛЬНО
79
26. Единица измерения момента инерции —
КГ • М2. ПРАВИЛЬНО НЕПРАВИЛЬНО
27. В исходном положении момент инерции наибольший относительно оси сальто.
ПРАВИЛЬНО НЕПРАВИЛЬНО
28. Fgz имеет значение, отличающееся от нуля, примерно в течение половины опорной фазы прыжка в беге.
ПРАВИЛЬНО НЕПРАВИЛЬНО
29. На составляющую Fgy силы реакции земли не влияет вес тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
30. Человек испытывает ускорение вверх только тогда, когда Fgz больше, чем вес тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
31. Средняя величина Fgz во время опорной фазы уменьшается со скоростью бега.
ПРАВИЛЬНО НЕПРАВИЛЬНО
32. Нога во время бега обычно касается земли в пронированном положении.
ПРАВИЛЬНО НЕПРАВИЛЬНО
33. Центр давления представляет собой точку приложения вектора веса.
ПРАВИЛЬНО НЕПРАВИЛЬНО
34. Центр давления измеряется в паскалях (Па).
ПРАВИЛЬНО НЕПРАВИЛЬНО
35. Беговые туфли, обеспечивающие большую амортизацию, обеспечивают и большую устойчивость. ПРАВИЛЬНО НЕПРАВИЛЬНО
36. Устойчивость ноги во время опорной фазы определяется измерением диапазона супинации-пронации движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
37. Трение между ботинком и землей — равнодействующая Fg x и Fgy
ПРАВИЛЬНО НЕПРАВИЛЬНО
38. Максимальная сила инерции пропорциональна величине силы, которая является нормальной относительно плоскости контакта. ПРАВИЛЬНО НЕПРАВИЛЬНО
39. Динамический коэффициент трения больше, чем статический коэффициент.
ПРАВИЛЬНО НЕПРАВИЛЬНО
40. Сила реакции сустава, в основном, обусловлена силами, создаваемыми связками.
ПРАВИЛЬНО НЕПРАВИЛЬНО
41. Сила реакции сустава не может быть больше веса тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
42. Трудно измерить силу реакции сустава непосредственно.
ПРАВИЛЬНО НЕПРАВИЛЬНО
43. Сила сдвига направлена тангенциально к поверхности. ПРАВИЛЬНО НЕПРАВИЛЬНО
44. Мышца не может создавать сжимающее (толкающее) усилие.
ПРАВИЛЬНО НЕПРАВИЛЬНО
45. Результирующая мышечная сила представляет собой суммарный эффект активности мышц относительно сустава.
ПРАВИЛЬНО НЕПРАВИЛЬНО
46. Мышечная сила может быть измерена непосредственно путем определения силы, передаваемой сухожилием.
ПРАВИЛЬНО НЕПРАВИЛЬНО
47. Сила, создаваемая мышцей, пропорциональна ее объему.
ПРАВИЛЬНО НЕПРАВИЛЬНО
48. Единица измерения удельного натяжения — ньютон (Н).
ПРАВИЛЬНО НЕПРАВИЛЬНО
49. Большие мышцы имеют большее удельное натяжение. правильно неправильно
50. Удельное натяжение не зависит от типа мышечного волокна.
ПРАВИЛЬНО НЕПРАВИЛЬНО
51. Удельное натяжение мышц у мужчин больше, чем у женщин.
ПРАВИЛЬНО НЕПРАВИЛЬНО
52. Архитектура мышцы может влиять на удельное натяжение.
ПРАВИЛЬНО НЕПРАВИЛЬНО
53. Мышечную силу можно определить измерением внутримышечного давления.
ПРАВИЛЬНО НЕПРАВИЛЬНО
54. Единица измерения давления — джоуль (Дж). ПРАВИЛЬНО НЕПРАВИЛЬНО
55. Мышечная сила обычно имеет малый угол натяжения, правильно неправильно
56. Внутрибрюшное давление создает усилие, которое стремится разогнуть торс.
ПРАВИЛЬНО НЕПРАВИЛЬНО
57. Внутрибрюшное давление человека может быть измерено. ПРАВИЛЬНО НЕПРАВИЛЬНО
58. Внутрибрюшное давление создает усилие, которое больше веса тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
59. Слои частиц жидкости, обтекающие объект, называются потоками.
ПРАВИЛЬНО НЕПРАВИЛЬНО
60. В соответствии с принципом Бернулли, давление жидкости возрастает с увеличением ее скорости.
ПРАВИЛЬНО НЕПРАВИЛЬНО
80
61. Подъемная сила — составляющая сопротивления жидкости — действует перпендикулярно направлению ее потока.
ПРАВИЛЬНО НЕПРАВИЛЬНО
62. Лобовое сопротивление жидкости всегда тормозит движение вперед.
ПРАВИЛЬНО НЕПРАВИЛЬНО
63. Лобовое сопротивление давления зависит от текстуры объекта, которая способствует уменьшению скорости пограничного слоя.
ПРАВИЛЬНО НЕПРАВИЛЬНО
64. Единица измерения поверхностного сопротивления — ньютон (Н).
ПРАВИЛЬНО НЕПРАВИЛЬНО
65. Волны уменьшают скорость пловца.
ПРАВИЛЬНО НЕПРАВИЛЬНО
66. Лобовое сопротивление давления — наибольшая составляющая лобового сопротивления ЖИДКОСТИ. ПРАВИЛЬНО НЕПРАВИЛЬНО
67. Наиболее существенным фактором в сопротивлении жидкости является относительная скорость взаимодействия объекта и среды.
ПРАВИЛЬНО НЕПРАВИЛЬНО
68. Среднее лобовое сопротивление, испытываемое пловцом, плывущим кролем со скоростью около 1,5 м • с-1, составляет 500 Н.
ПРАВИЛЬНО НЕПРАВИЛЬНО
69. При конечной скорости парашютист испытывает вертикальное ускорение 9,81 м • с-2.
ПРАВИЛЬНО НЕПРАВИЛЬНО
70. Составляющая силы сопротивления воздуха может изогнуть траекторию бейсбольного мяча.
ПРАВИЛЬНО НЕПРАВИЛЬНО
71. Константа к означает жесткость пружины. ПРАВИЛЬНО НЕПРАВИЛЬНО
72. Ткань возвращается к своей первоначальной длине, если она растянута в пределах пластичности. ПРАВИЛЬНО НЕПРАВИЛЬНО
73. Единица измерения относительной деформации — процент (%).
ПРАВИЛЬНО НЕПРАВИЛЬНО
74. Напряжение вызывает изменение длины ткани. ПРАВИЛЬНО НЕПРАВИЛЬНО
75. Сила упругости характеризует потенциальную способность тканей восстанавливать свою первоначальную длину.
ПРАВИЛЬНО НЕПРАВИЛЬНО
76. Единица измерения силы упругости — Н • СМ 2. ПРАВИЛЬНО НЕПРАВИЛЬНО
77. Центробежная сила делает траекторию объекта линейной. ПРАВИЛЬНО НЕПРАВИЛЬНО
78. Мышцы-разгибатели колена могут управлять сгибанием колена.
ПРАВИЛЬНО НЕПРАВИЛЬНО
79. Силой инерции называется эффект, оказываемый одним сегментом тела на другой благодаря своей массе.
ПРАВИЛЬНО НЕПРАВИЛЬНО
80. Единица измерения момента силы является — ньютонометр (Нм).
ПРАВИЛЬНО НЕПРАВИЛЬНО
81. Сила будет иметь вращающий момент относительно точки лишь в том случае, если плечо силы не равно нулю.
ПРАВИЛЬНО НЕПРАВИЛЬНО
82. Вращающий момент — скалярная величина. ПРАВИЛЬНО НЕПРАВИЛЬНО
83. Умножение используется для сложения векторных величин.
ПРАВИЛЬНО НЕПРАВИЛЬНО
84. Единица измерения плеча силы — ньютон (Н). ПРАВИЛЬНО НЕПРАВИЛЬНО
85. Изогнутая стрелка показывает вектор, перпендикулярный странице.
ПРАВИЛЬНО НЕПРАВИЛЬНО
86. Плечо силы — скалярная величина.
ПРАВИЛЬНО НЕПРАВИЛЬНО
87. Плечо силы веса всегда вертикально.
ПРАВИЛЬНО НЕПРАВИЛЬНО
88. Если мышца создает усилие — 25 Нм, то сегмент тела будет вращаться в отрицательном направлении.
ПРАВИЛЬНО НЕПРАВИЛЬНО
89. Плечо силы для мышц-сгибателей локтя наибольшее, когда локоть находится в выпрямленном положении.
ПРАВИЛЬНО НЕПРАВИЛЬНО
вопросы ггшпяыя
ВЫБОРОМ ОТВЕТОВ
90. Какое понятие используется для определения суммарного эффекта нескольких сил, действующих на систему?
а) равнодействующая;
б) состав;
в) ортогональная;
г) разрешающая способность.
91. Какой эффект оказывает нормальная составляющая мышечной силы на сегмент тела?
а) сжатие;
6 8-133
81
б) вращение;
в) коактивацию;
г) стабилизацию.
92. Какой из законов движения Ньютона утверждает, что F = та!
а) гравитации;
б) инерции;
в) действия—противодействия;
г) ускорения.
93. Какой параметр является количественной мерой инерции?
а) масса;
б) ускорение;
в) сила;
г) вес.
94. Какая переменная не влияет на центростремительную силу?
а) масса;
б) радиус;
в) ускорение;
г) скорость.
95. Момент рассчитывается как произведение каких двух переменных?
а) массы и ускорения;
б) веса и скорости;
в) веса и времени;
г) массы и скорости.
96. Когда вес описывается как распределенная сила?
а) когда система стационарна, Г&г = Fv\ б) когда он действует в центре тяжести; в) когда центр тяжести не является постоянной точкой и даже не всегда ограничен физическими пределами системы;
г) когда каждая частица системы имеет вес.
97. У какого из сегментов тела центр тяжести ближе всего к середине длины сегмента?
а) у головы;
б) у голени;
в) у предплечья;
г) у плеча.
98. Какие из утверждения относительно момента инерции правильны?
а) он представляется символом /;
б) он показывает распределение массы системы относительно ее центра тяжести;
в) он является наименьшим относительно оси кручения.
г) единица его измерения — кг • м2.
99. Какие методики применялись для получения антропометрических оценок сегментов тела?
а) радиационное сканирование;
б) изучение трупов;
в) изображение с помощью магнитного резонанса;
г) математическое моделирование.
100. Какие допущения использовал Ханаван в своей модели человеческого тела?
а) сегменты имеют одинаковую плотность;
б) левая и правая стороны симметричны;
в) сегменты являются жесткими;
г) границы между сегментами четкие.
101. Согласно какой модели разделено человеческое тело на большее число сегментов?
а) Хатце;
б) Чандлера, Клаузера, Мак-Конвиля, Рейнольдса и Юнга;
в) Зацерского и Селянова;
г) Ханавана.
102. Что происходит, если Fg z = Fw?
а) смещение центра тяжести изменяет направление;
б) центр тяжести проходит над ногой;
в) центр тяжести имеет нулевое ускорение в вертикальном направлении;
г) все сегменты тела неподвижны.
103. Какие характеристики силы реакции земли изменяются с увеличением скорости бега?
а) пик г;
б) длительность Fg>y;
в) пик
г) средняя Fgx.
104. Что является единицей измерения центра давления?
а) см;
б) Па;
в) Н;
г) Н • см"2.
105. Каково назначение хорошо сконструированных беговых ботинок?
а) максимизировать центр давления;
б) обеспечить устойчивость голеностопного сустава;
в) минимизировать удар, связанный с контактом ноги;
г) накапливать энергию упругости.
106. Какие переменные влияют на различия в трении между ботинком и землей у различных людей?
а)
-^s,max>
В) ^&z>
г) и ^.у-
107. Почему силы реакции сустава могут иметь большую величину?
82
а) сухожилия создают большие усилия между сегментами;
б) контактные усилия передаются с одного сегмента на другой;
в) мышцы имеют малый угол натяжения и поэтому большую тангенциальную составляющую мышечной силы;
г) составляющая среза обычно больше, чем составляющая сжатия.
108. Какие методики используются для определения величины мышечной силы?
а) ЭМГ;
б) площадь поперечного сечения;
в) внутримышечное давление;
г) тензометр.
109. Какое мышечное усилие будет передаваться через ахиллово сухожилие у взрослого мужчины, едущего на велосипеде с умеренной скоростью?
а)7Н;
б) 70 Н;
в) 700 Н;
г) 7000 Н.
ПО. Каково среднее значение удельного натяжения?
а) 30 Н-см-2;
б) 30 кг • см-2;
в) 30 Н • м’2;
г) 30 Па.
111. Какие факторы обусловили диапазон значений удельного натяжения, опубликованных в литературе?
а) пол;
б) техника измерения;
в) тип мышечного волокна;
г) архитектура мышцы.
112. Какова единица измерения внутрибрюшного давления?
а) Н;
б) Па;
в) Дж;
г) Нм.
113. Почему использование тяжелоатлетического пояса помогает штангисту увеличить внутрибрюшное давление?
а) активирует маневр Валсалвы;
б) вызывает параллельное повышение давления в грудной полости;
в) поддерживает активность брюшных мышц;
г) увеличивает давление между дисками.
114. Что такое принцип Бернулли?
а) жидкость вокруг объекта может быть представлена слоями частиц, обтекающими тело;
б) слой, наиболее близкий к объекту, называется пограничным слоем и имеет наименьшую скорость;
в) изменение течения жидкости с ламинарного на турбулентный вызывает увеличение сопротивления жидкости;
г) давление жидкости обратно пропорционально скорости ее частиц.
115. Какой термин используется по отношению к сопротивлению среды вследствие турбулентного потока воздуха?
а) поверхностное лобовое сопротивление;
б) подъемная сила;
в) лобовое сопротивление давления;
г) волновое сопротивление.
116. Какая переменная наиболее влияет на сопротивление среды, испытываемое объектом?
а) относительная скорость;
б) плотность среды;
в) ориентация объекта;
г) проецируемая площадь.
117. Почему брошенный бейсбольный мяч может лететь по изогнутой траектории?
а) бросающий направляет его по изогнутой траектории;
б) швы обусловливают увеличение поверхностного сопротивления;
в) вращение вызывает перепад давлений поперек мяча;
г) сила тяжести заставляет траекторию изгибаться в ту или иную сторону в зависимости от того, как брошен мяч.
118. Как нормируется зависимость сила—деформация?
а) кривой сила—длина;
б) отношением напряжение—относительная деформация;
в) жесткостью пружины;
г) соотношением энергия—работа.
119. Какая единица используется для измерения напряжения?
а) %;
б) Н;
в) мм;
г) Па.
120. Какой термин используется для описания механического эффекта, при котором движение одного сегмента тела будет влиять на соседние?
а) сила упругости;
б) сила инерции;
в) результирующий мышечный вращающий момент;
г) межсегментное давление.
6*
83
ЗАДАЧИ
121. Медиальная (М) и латеральная (Л) головки гастрокнемиус создают усилие 350 Н, которое направлено соответственно на 0,25 рад влево и вправо от осевой линии. Определите величину и направление равнодействующей.
122. Диаграмма свободного тела (рис. 2.10) показывает силы, действующие на голову для сохранения позиции согнутой шеи.
А. Используя уравнения регрессии в табл. 2.1 и 2.5, определите вес головы при условии, что человек весит 619 Н и имеет рост 162 см.
Б. Когда система находится в равновесии, силы, действующие на нее, сходятся, что показано пунктирными линиями. Сила Fm направлена под углом 0,58 рад к горизонтали. Рассчитайте 0 при условии, что эти три силы сходятся.
В. Пусть Fm равна 54 Н. Рассчитайте горизонтальную составляющую силы Fm и горизонтальную и вертикальную составляющие силы /}, которые действуют на атлантозатылочный сустав.
123. Бицепс брахии, брахиалис и брахиорадиа-лис — три мышцы-сгибатели локтя. Допустим, локтевой сустав согнут до 1,57 рад, эти мышцы создают усилия соответственно 290, 440 и 95 Н и осуществляют натяжение под углами соответственно 0,7, 0,2 и 0,3 рад. Для простоты примем, что три мышцы имеют общую точку присоединения на лучевой кости. Каковы направление и величина результирующей мышечной силы?
124. Во время разгибания колена надколенная чашечка перемещается в желобе между медиальным и латеральным мыщелками бедренной кости. Разбалансировка четырех мышц, тянущих надколенную чашечку, может привести к тому, что чашечка будет перемещаться неправильно (Grabiner, Koh, Draganich, 1994). Это может иметь место, например, если колено находилось в гипсе в течение определенного времени и вастус медиалис и латералис атрофировались в разной степени. Имея следующие данные о величине и направлениях четырех векторов мышечной силы, определите результирующий вектор:
Ректус феморис — 130 Н; 0,25 рад влево от вертикали;
Вастус латералис — 180 Н; 0,50 рад влево от вертикали;
Вастус интермедиус — 160 Н; 0,15 рад влево от вертикали;
Вастус медиалис — 210 Н; 0,62 рад вправо от вертикали.
125. Человек, выполняющий приседания при нахождении мышц-сгибателей в указанном положении, создает усилие 480 Н. Закончите диаграмму свободного тела для этой ситуации. Каковы нормальная и тангенциальная составляющие мышечной силы?
126. Сконструировано устройство для тренировки мышц-сгибателей локтя. Нарисуйте соответствующую диаграмму свободного тела для анализа усилий, создаваемых этими мышцами.
127. Тренер футбольной команды хотел бы знать, какую силу развивают разгибатели колена его игрока во время удара. Как вы определили бы систему для ответа на этот вопрос? Нарисуйте диаграмму свободного тела.
84
128. Нарисуйте диаграмму свободного тела, которая позволила бы рассчитать усилия в коленном суставе для ноги (бедро—голень—стопа) при вытяжке, показанной на рис. 2.2.
129. Человек весит 523 Н. Пользуясь уравнениями регрессии в табл. 2.1, определите вес торса этого человека и рассчитайте, какой процент это составляет от общего веса тела?
130. У человека ростом 172 см бедро и голень длиной по 36 см, а плечо и предплечье — по 25 см. Для какого из этих сегментов ЦТ расположен ближе всего к проксимальному суставу и для какого дальше всего от него?
131. Пациент с общей массой тела 78 кг, обутый в ботинок де-Лорме (4,5 кг), выполняет упражнение по разгибанию колена. В положении стоя его рост 183 см. Допустим, координаты (х, у) для различных отметок таковы:
Отметка Координаты
Коленный сустав 0,0
Голеностопный сустав -1 -44
ЦТ голени 0 -19
ЦТ стопы 8 -49
ЦТ ботинка де-Лорме 6 -52
А. Используя уравнения регрессии Zatsi-orsky, Seluyanova, определите массу голени и стопы.
Б. Используя уравнение (2.5), определите момент инерции системы (голень—стопа—ботинок де-Лорме) относительно оси сальто колена.
132. В середине фазы отрыва сальто вперед центр тяжести гимнаста (Fw = 537 Н) имеет вертикальное ускорение вверх 65,5 м • с-2. Какова Fgz в этот момент движения?
133. На ранней стадии опорной фазы во время бега составляющие силы реакции земли имеют следующие величины (рис. 2.16): Fgz = 2,6 веса тела; Fgy =1,0 веса тела; Fg^ = 0,02 веса тела. Какова результирующая сила реакции земли для человека, который весит 819 Н?
134. На основе графиков силы реакции Земли, показанных на рис. 2.15 и 2.16, перечислите переменные, которые должен измерить биомеханик, чтобы описать изменения силы реакции Земли в зависимости от скорости.
135. В один из моментов игры в ракетбол взаимодействие между ботинком и землей имело следующие характеристики: p.s = 0,85; FgtZ = 821 Н; Fgj = 594 Н; FgtX = 332 Н. Останется ли нога неподвижной или она будет скользить по земле? Для ответа на этот вопрос рассчитайте максимальную статическую силу трения (/^тах) и фактическую силу трения (Fs). Ботинок не будет скользить, если фактическая сила трения значительно меньше, чем максимальная.
136. График показывает кинематику (вертикальное положение, скорость и ускорение) ЦТ прыгуна в воду и силу реакции трамплина во время фазы отрыва для прыжка три с половиной сальто назад из положения согнувшись.
А. Когда смещение прыгуна изменяет направление? Сколько раз это имело место во время фазы отталкивания?
Б. Когда ЦТ ускоряется в направлении вверх и когда в направлении вниз?
В. Каково соотношение между кривыми ускорения и силы реакции?
Г. Каково соотношение между кривыми положения и ускорения?
Положение
Скорость
Ускорение
Сила реакции
Время, с
137. Eic (1966) измерил внутрибрюшное давление штангиста, который поднимал штангу массой 130 кг примерно на высоту колена; положение схематически показано на рис. 1.7. В этом положении внутрибрюшное
85
давление составило 225 мм рт.ст., а площадь поперечного сечения брюшной полости на уровне L5 — S1 — 608,5 см2. Какое выпрямляющее усилие оказывает внутрибрюшное давление на торс?
138. Сопротивление среды, испытываемое объектом, зависит от того, насколько объект возмущает среду. Из уравнения сопротивления среды (2.8) определите, какие две переменные оказывают существенное влияние на это возмущение.
139. Парашютист во время прыжка проходит четыре фазы: 1) начальную фазу ускорения с момента покидания самолета до достижения конечной скорости; 2) фазу полета с конечной скоростью; 3) фазу уменьшения вертикальной скорости сразу же после раскрывания парашюта; 4) окончательную фазу свободного падения с раскрытым парашютом и подготовки к приземлению. Нарисуйте график скорость-время, представляющий эти четыре фазы. Нарисуйте график сопротивление воздуха— время.
140. На рисунке показано положение, принимаемое лыжником во время скоростного спуска, при котором влияние сопротивления воздуха минимально. Определите D, F, G, L и N и объясните факторы, влияющие на каждую переменную.
141. Поведение пружины, растягиваемой в пределах ее упругости, может быть описано уравнением F = кх. Каковы единицы измерения каждой переменной в этом уравнении?
142. Удельное натяжение мышцы составляет примерно 80 Н • см-2.
А. Поперечные сечения основных сгибателей и разгибателей локтя составляют:
Мышца
Бицепс брахии Брахиадис
Брахиорадиалис Трицепс брахии
Площадь поперечного сечения, см2
4,6
7,0
1,5
18,8
Определите максимальное усилие, которое могут развить сгибатели локтя (как группа мышц).
Б. Каково удельное натяжение трицепс брахии человека, если площадь поперечного сечения 9,4 см2?
В. Когда локтевой сустав сгибается из выпрямленного положения до угла 1,75 рад, плечо силы для сгибателей локтя увеличивается вдвое. Если удельное натяжение в разогнутом положении составляет 30 Н • см 2, каково оно в согнутом положении?
143. Для каждого из следующих четырех положений начертите плечи для двух векторов веса относительно коленного сустава.
144. В каждом из следующих случаев определите момент, производимый силой относительно точки В. Жесткая балка свободно вращается вокруг точки В.
86
145. Человек на колесном кресле прилагает усилие 137 Н к ручному ободу кресла. В положении а, направление усилия касательно к ободу; т.е. перпендикулярно радиусу (26,7 см). В состоянии б сила прилагается под углом I рад к касательной обода. Каков вращающий момент относительно оси колеса в этих условиях?
146. Нарисуйте диаграмму свободного тела для каждого из показанных положений всего тела. Для каждой силы, включенной в диаграмму свободного тела, начертите плечо от силы до стопы.
'гтгпУт
ГЛАВА 3
АНАЛИЗ ДВИЖЕНИЯ
При описании движения объекта с использованием таких понятий, как положение, скорость и ускорение, движение рассматривается как результат взаимодействия между объектом и окружающей средой. Принято считать, что это взаимодействие осуществляется посредством сил, приложенных к объекту. Природу влияния сил на движение объекта определяют три закона Ньютона. Конкретная алгебраическая взаимосвязь между силой и движением зависит от цели исследования. Обычно рассматривают: а) мгновенное значение силы (подход сила — масса — ускорение); б) влияние силы, приложенной к объекту в течение некоторого времени (импульс — количество движения); в) приложение силы, вызывающей перемещение объекта на некоторое расстояние (подход работа — энергия).
ПОДХОД
СИЛА - МАССА - УСКОРЕНИЕ
При использовании указанного подхода различают два существенно разных состояния объекта. Если ускорение объекта равно нулю, он либо движется прямолинейно, равномерно и поступательно (т.е. не вращаясь), либо остается неподвижным. Условия неподвижности объекта изучает раздел механики, называемый статикой. Если же ускорение объекта отлично от нуля, его движение изучает динамика.
Статический анализ
Если в процессе исследования мы оперируем со значением силы в определенный момент времени (мгновенное значение силы), целесообразно использовать подход сила — масса
— ускорение, имеющий такое название потому, что он основан на втором законе Ньютона:
ZF = тиа.
Как мы уже знаем, левая часть уравнения (SF) отражает взаимодействие между системой и окружающей ее средой, а правая — кинематический результат этого воздействия на систему. В рассматриваемом подходе состояния системы классифицируются исходя из величины ускорения а, точнее в зависимости от того, нулевое оно или ненулевое. Статика представляет собой особый случай второго закона Ньютона, при котором ускорение нулевое и объект при этом неподвижен. В статическом анализе система находится в равновесии и сумма сил, действующих на нее, равна нулю (SF=0). Таким образом, все воздействия на систему сбалансированы, и она не испытывает никакого ускорения. Точно так же равна нулю и сумма моментов сил относительно некоторой точки 0 (SMo=O).
Условия равновесия объекта в плоской задаче статики представляют собой три независимых скалярных уравнения:
-LFX = 0; (3.1)
^Fy = 0; (3.2)
= 0; (3.3)
Уравнения (3.1) и (3.2) относятся к суммам (Z) проекций сил, действующих на систему, на два линейных направления (х и у), которые перпендикулярны друг другу. Уравнение (3.3) представляет собой сумму моментов сил относительно точки 0, которая может совпадать, а может и не совпадать с центром тяжести системы.
Ниже мы рассмотрим четыре примера статического анализа: определение величины и направления действия неизвестной силы; вычисление результирующего вращающего момента мышечных сил; разложение результирующей
88
Рис. 3.1. Распределение сил, действующих на объект
мышечной силы и силы реакции сустава на составляющие, а также нахождение центра тяжести тела с использованием сегментного анализа.
Пример 1
Рассмотрим систему, показанную на рис. 3.1, а, которая находится в равновесии и на которую действуют три известные силы и одна неизвестная. Какова величина неизвестной силы? Для начала выберем две прямых под прямым углом друг к другу и укажем положительное направление вдоль каждой прямой и положительное направление измерения углов. Выбранные направления указаны в левой нижней части рис. 3.1, а. При этом любая сила (например, R), действующая в произвольном направлении, не совпадающем с выбранными, может быть разложена на составляющие в направлениях хи у (см. рис. 2.3).
Следующий этап анализа предполагает графическое разложение силы R на горизонтальную (Rx) и вертикальную (Ry) составляющие (рис. 3.1, б). После разложения всех сил по направлениям хи у силы, действующие в каждом из этих направлений, необходимо просуммировать, чтобы из полученных соотношений определить неизвестные.
= 0;
Rx - 3 = 0;
Rx = 3 Н;
ZFy = 0;
5 + 10 - Ry = 0;
Ry = 15 H.
Определив значения Rx и Ry., можно установить величину результирующей силы (R) на основании теоремы Пифагора:
R = + R2y = л/з2 +152;
R = 15,3 Н.
Полученный результат, однако, дает лишь величину результирующей, но не ее направление. Направление можно установить различными способами, например, определить угол, который составляет сила R с горизонтальным или вертикальным направлением. Согласно правилу параллелограмма, R — диагональ прямоугольника со сторонами Rx и Ry (рис. 3.1, в). Следовательно, R„ Ry и R — стороны треугольника, из которого можно определить, используя одну из следующих формул:
R . Rv Rv
cos0 = ; sm0 = ; tan 0 = ,
R R Rx
поэтому
n RX R n Rx.
0 = arc cos —;
R
0 = arc cos -Д-;
15,3
0 = arc cos 0,1961;
0 = 1,37 рад.
Пример 2
Другой пример статического анализа приведен на рис. 2.41 (см. рис.3.2).
Цель анализа в том, чтобы определить величину и направление результирующего вращающего момента мышц относительно коленного сустава (К), обусловленного результирующей мышечной силой (Fm). На рис. 3.2 показаны три плеча сил для силы реакции сустава (Fy), веса ноги (Fw/) = 40,6 Н) и веса ботинка (F„, e = 80 Н). Допустим, человек должен удерживать конечность под углом 0,5 рад ниже горизонтальной линии. Каков будет при этом результирующий вращающий момент мышц? Сначала построим диаграмму свободного тела. Затем выберем соответствующее уравнение из системы (3.1 — 3.3) и просуммируем силы или моменты сил. Поскольку цель анализа — определение результирующего вращающего моменты мышцы, целесообразно выбрать уравнение (3.3). При суммировании моментов сил относительно оси вращения коленного сустава силой реакции сустава можно пренебречь, при этом индекс “0” в уравнении (3.3) заменим на индекс “К”,
89
Fw,b
Рис. 3.2. Диаграмма свободного тела ноги и ступни человека, выполняющего выпрямление ноги в коленном суставе (из рис. 2.41): а = 0,0124; b = 0,110; с = 0,320 м
который обозначает ось вращения. Таким образом,
ZMK = 0.
Действие трех сил вызовет вращение системы вокруг оси, проектирующейся на рисунке в точку К. Выполним следующие вычисления:
SMK = 0;
(Fm х а) - (Fwj *b)~ (FwJ) x c) = 0;
xa) = (FwJ x b) + (Fwb x c) =
= (40,6 x 0,11) + (80,0 x 0,32) = 4,47 + 25,60;
(Fm xa) = 30,07 H • m.
Итак, результирующий вращающий момент мышц есть произведение результирующей мышечной силы (Fm) на ее плечо. В нашем примере для удержания конечности в данном положении человек должен развить вращающий момент 30,07 Нм.
Пр имер 3
Студент(ка) сидит на краю скамьи и выполняет на тренажере упражнение в изометрическом режиме для укрепления четырехглавых мышц бедер. Вес ее ноги 30 Н, сила, прикладываемая к тренажеру, — 100 Н. Четырехглавая мышца прикреплена с помощью подколенного сухожилия к большеберцовой кости на расстоянии 7 см от коленного сустава; ЦТ конечности находится на расстоянии 20 см от коленного
сустава, а расстояние между коленным суставом и тренажером — 45 см. Тренажер представляет собой ремень, укрепленный на лодыжке, который с помощью каната соединен с нагруженной платформой, расположенной на спинке скамьи. Платформа может быть зафиксирована в неподвижном положении, что позволит выполнять упражнение в изометрическом режиме. Диаграмма свободного тела системы от коленного сустава (К) до пальцев ноги представлена на рис. 3.3. На рисунке показан вес ноги (FJ, нагрузка со стороны тренажера (F/), три отрезка {а, Ь, с) и три угла (6, у, Р). Также показаны нормальная (и) и тангенциальная (1) составляющие силы реакции сустава (F,). Выполним необходимые расчеты, чтобы определить
1) усилие четырехглавой мышцы бедра (Гт, <?), необходимое для сохранения заданного положения;
2) нормальную (Fy „) и тангенциальную (Fy ,) составляющие силы реакции сустава;
3) величину и направление результирующей силы реакции сустава (Fy).
Рис. 3.3. Диаграмма свободного тела ноги испытуемого, выполняющего упражнение для укрепления группы четырехглавых мышц бедра:
Fw = 30 Н; F/ = 100 Н; а = 7 см; b = 20 см; с = 45 см; 0 = 0,25 рад; р = 0,87 рад; у = 0,70 рад
90
Сначала определим два независимых направления, на которые проектируются силы, и укажем положительное направление этих осей и отсчета углов. Поскольку задача предполагает приложение сил и вращающих моментов к сегменту тела, наиболее целесообразно избрать нормальное (л) и тангенциальное (I) по отношению к сегменту направления. Они показаны в правой части рис. 3.3.
Те же три уравнения (3.1 — 3.3) можно использовать для определения Fm_ q, так как система находится в состоянии равновесия. Поскольку на систему действуют три силы неизвестной величины (Fj я; Fj Fm9), наиболее логичным представляется просуммировать моменты относительно точки К, что позволит вначале исключить силы Fy п и Fy t из уравнения 3.3 и определить Fm q. Этот процесс включает сложение моментов сил относительно точки К для силы веса ноги (Fw) и нагрузки со стороны тренажера (Fe) и приравнивание их к вращающему моменту, обусловленному активностью четырехглавой мышцы бедра (Fm> 9). Для выполнения этих расчетов нам необходимо определить плечо каждой силы, используя известные величины сил и углов с учетом их направлений (рис. 3.3):
a sin 6 = 7 sin 0,25 = 1,73 см;
fesinp = 20 sin 0,87 = 15,29 см; csinY = 45 sin 0,70 - 28,99 см.
Теперь проведем следующие расчеты:
ЕМК = 0;
(Fm? х a sin 6) - (Fw х 6sinp) - (Ft x csiny) = 0;
(Fmq x 1,73) - (30 x 15,29) - (100 x 28,99) = 0;
(Fmq x 1,73) = (30 x 15,29) + (100 x 28,99);
(F^q x 1,73) = 458,7+ 2899,0;
F = 33577 m’« 1,73 ’
Fm,g = 1941 H.
Чтобы определить Fj_ „ и FJt t, необходимо разложить каждую силу на ее нормальную и тангенциальную составляющие (рис. 3.3,6) и затем сложить проекции сил в каждом из направлений:
Fmn = Fm,g sin 0^5 = 1941 sin 0,25 = 480 Н;
Frn,t = Fm,q cos 0,25 = 1941 cos 0,25 = 1881 H;
Fw„ = Fwl sin0,87 = 30 sin 0,87 = 23 H;
= Fwj cos0,87 = 30cos0,87 = 19 H;
Fl n = F, sin 0,70 = 100 sin 0,70 = 64 H;
Fi4 = Fz cos 0,70 = 100 cos 0,70 = 77 H.
Эти значения соответствуют величинам нормальной и тангенциальной составляющих сил, приведенных на рис. 3.3,6. Тангенциальная составляющая силы реакции сустава определяется следующим образом:
EF, = 0;
Fj\t - Fmj + Av,/ - А,/ = 0;
Fyy = Fm>, - Fw>, + Fht = 1881 -19 + 77;
Fjj = 1939 H.
Аналогично находят нормальную составляющую:
SF„ = 0;
fj,„ + A„,л - Av,„ - А,л = °;
А» = -Ап,Л + Av,Л + А,„ = -480 + 23 + 64;
Fy „ = -393 Н.
Величина Fj п, согласно проведенным вычислениям, отрицательна. Это означает, что не сила отрицательна, а ее направление на рис. 3.3 выбрано неправильно. Если в результате вычисления получено отрицательное значение силы, значит, ее направление противоположно указанному на диаграмме свободного тела. Строя диаграмму, мы отмечали, что система находится в состоянии равновесия. Следовательно, сумма проекций сил на любую ось должна быть равна нулю. Что касается тангенциального направления, мы допустили, что Fj t действует в положительном направлении, это оказалось верным. Кроме того, мы предположили, что F; я также должна действовать в положительном направлении, чтобы система находилась в состоянии равновесия. Однако проведенные вычисления не подтвердили этого предположения, о чем свидетельствует отрицательное значение FJ, я. В действительности Fjt п действует в противоположном направлении, обеспечивая пребывание системы в положении равновесия. Таким образом, FJt t и Fjt п, а также результирующая Fj действуют на точку К, как показано на рис. 3.4. Зная Fj „ и
Рис. 3.4. Определение результирующей силы реакции коленного сустава для диаграммы свободного тела на рис. 3.3.
91
Fj t, можно определить Fj сустава на основании теоремы Пифагора:
FJ = + FJJ = J3932 +19392 ;
Fj = 1978 H.
Таким образом, при воздействии на лодыжку нагрузки 100 Н сила реакции коленного сустава превышает ее почти в 20 раз.
Наконец, мы можем определить направление результирующей силы реакции сустава относительно оси рассматриваемого сегмента:
tg0 = ^;
fj, t
F 0 = arc tg ;
Fj,t
393
O = arctg-^^-;
1939
0 = 0,2 рад.
Рис. 3.5. Положение ЦТ всего тела как функция положения сегментов тела:
а — границы соответствующих сегментов тела; б — положение ЦТ сегмента как процент его длины
Итак, результирующая сила реакции сустава равна 1978 Н и направлена ниже оси голени на 0,2 рад.
Пример 4
Воспользуемся описанным выше приемом для определения центра тяжести тела. ЦТ таких многосегментных объектов как тело человека, можно определить, используя статический вариант закона ускорения Ньютона (IF = = 0 и ZMo = 0). Для этого нужно найти точку, относительно которой все сегменты тела находятся в равновесии. Поскольку тело действительно находится в состоянии равновесия относительно ЦТ, сумма моментов сил веса отдельных частей тела относительно ЦТ равна нулю. Единственные силы, воздействующие в данном случае на систему, — силы веса сегментов.
Проиллюстрируем это следующим примером. Предположим, нам необходимо определить ЦТ тела гимнастки, собирающейся выполнить сальто назад (рис. 3.5). Для этого
1. Выделим соответствующие сегменты тела. Для осуществления сегментного анализа тело человека можно разделить на 14 — 17 сегментов (об этом шла речь в главе 2). Количество сегментов, используемых в анализе, определяется числом суставов, испытывающих угловое перемещение во время выполнения движения. Если, например, отсутствует вращение относительно локтевого сустава, то рука (плечо + предплечье) может быть представлена как единый сегмент. В нашем примере мы используем
14 сегментов: голову, туловище, плечи, предплечья, кисти рук, бедра, ноги (голени) и ступни. Эти сегменты показаны на рисунке 3.5, а отметками над центрами суставов, представляющими собой проксимальный и дистальный концы каждого сегмента (Chandler et. al., 1975). (Приложение Г).
2. Соединим метки центров суставов и получим фигуру (рис.3.5, б).
3. Воспользовавшись табл.2.1, определим ЦТ каждого сегмента в процентах от его длины. Длина измеряется от проксимального конца каждого сегмента (рис.3.5, б).
Сегмент Положение ЦТ, % Проксимальный конец
Голова 66,3 Верхняя часть головы
Туловище 52,2 Верхняя часть шеи
Плечо 50,7 Верхний участок грудной
клетки сбоку от шеи
Предплечье 41,7 Локоть
Кисть 51,5 Запястье
Бедро 39,8 Верхняя часть бедра
Нога (голень) 41,3 Колено
Ступня 40,0 Лодыжка
4. Определим вес сегментов в зависимости от веса тела (ВТ), использовав уравнения регрессии Chandler et al. (1975) (табл.2.1). Предположим, вес гимнаста 450 Н.
92
Сегмент Уравнение Вес
Голова 0,032 х ВТ + 18,70 = 33,10
Туловище 0,532 х ВТ- 6,93 = 232,47
Плечо 0,022 х ВТ + 4,76 = 14,66
Предплечье 0,013 х ВТ + 2,41 = 8,26
Кисть 0,005 х ВТ + 0,75 = 3,00
Бедро 0,127 х ВТ - 14,82 = 42,33
Нога (голень) 0,044х ВТ- 1,75 = 18,05
Ступня 0,009 х ВТ + 2,42 = 6,53
5. Определим ЦТ сегментов относительно осей х, у (рис.3.5,б). Расположение этих осей весьма произвольно и не влияет на ЦТ всего тела, в чем можно убедиться, произведя вычисления дважды при различном расположении осей х, у.
Сегмент Координата х, см Координата у, см
Голова 8,6 16,8
Туловище 6,9 12,8
Плечо 6,5 13,3
Предплечье 4,2 11,5
Кисть 2,8 9,6
Бедро 6,6 8,2
Нога (голень) 8,0 3,9
Ступня 8,0 0,8
6. Зная вес сегментов (Fw> s) и координаты ЦТ (х, у), складываем моменты сил веса сегментов относительно оси у (LMy = XP'w, s х xi) и оси х (LMX = XT’w, s x A’i)- Умножим на 2 вес каждого парного сегмента. Затем обратимся к следующей таблице.
Сегмент СМ К см Fws, Н м,, Нем М, Н-см
Голова 8,6 16,8 33,10 284,7 556,1 Туловище 6,9 12,8 232,47 1604,4 2975,6 Плечо 6,5 13,3 29,32 190,6 390,0 Предплечье 4,2 11,5 16,52 69,4 190,0 Кисть 2,8 9,6 6,00 16,8 57,6 Бедро 6,6 8,2 84,66 558,8 694,2 Нога (голень) 8,0 3,9 36,10 228,8 140,8 Ступня 8,0 0,8 13,06 104,5 10,5 Уму = уМх = 3018,0 5014,7
7. Найдем точку, в которой создается такой же момент силы веса всего тела относительно осей хи у, какой дает сумма моментов всех сегментов. Момент силы, обусловленный весом сегментов относительно осей у и х (соответственно 3018,0 и 5014,7 Н-см), равен моментам силы веса всего тела относительно тех же осей. Поэтому
8
7» х хцт = х xi’
i=l
8
хх/
ХЦТ ~ ~ р
‘w
Аналогично определяется координата уцт. Таким образом, координаты хи у для ЦТ всего тела будут равны:
3018 Н-см
х =------------= 6,71 см
450 Н
5014,7 Н-см 111zl у =------------= 11,14 см.
7 450 Н
На рис. 3.5,6 показана схема расположения ЦТ всего тела относительно осей х и у, выполненная в определенном масштабе.
Динамический анализ
Статический анализ — наиболее простой подход к изучению движения человека. Если же на систему действуют неуравновешенные силы, она испытывает ускорение. Использование подхода сила — масса — ускорение в ситуациях, когда ускорение не является нулевым, называется динамическим анализом. Три независимых уравнения (3.1 — 3.3), используемых в статическом анализе, применимы и в динамическом анализе с учетом того, что правая часть уравнений при этом равна произведению массы на ус-
корение. Следовательно, уравнения (3.1 — 3.3) примут вид:
Y,FX = max, (3.4)
YFy = may; (3.5)
ЕЛГ0 = /оа + mad, (3.6)
где a = ^a2 + а2 и Ef = J(EFX)2 + (Х/^)2 в плоскости (x, у). Как и в статическом анализе, эти уравнения независимы и представляют проекции сил, моментов сил и ускорений на направления двух координатных осей (х и у) и на ось вращения, проходящую через точку 0 перпендикулярно плоскости (х, у). Из уравнения (3.4) следует, что сумма (Z) проекций сил (F) на направление х равна произведению массы (т) системы на проекцию ускорения ее центра масс на направление х (ах). Из уравнения (3.5) следует то же, но только относительно проекций сил и ускорений на направление у. Согласно уравнению (3.6), сумма моментов сил относительно точки 0 состоит из двух составляющих: 1) произведения момента инерции (/„) системы на угловое ускорение системы относительно оси вращения (а); 2) произведения массы (т) системы на линейное ускорение ЦТ системы (а) и на расстояние (d) между точкой 0 и ЦТ системы.
Точка 0 может быть любой точкой, относительно которой суммируются моменты сил. Если точка 0 совпадает с ЦТ и неподвижна, ускорение ЦТ системы равно нулю, вследствие чего
93
a x, ay и mad также становятся равными нулю и уравнения (3.4 — 3.6) сводятся к Т.М0 = А>а-Если система включает отдельное жесткое тело, например, сегмент тела, вращающийся вокруг фиксированной оси, результирующий эффект сил, действующих на систему в нормально-тангенциальной системе координат, можно определить следующим образом (Meriam, Kraige, 1987):
ZFn = тгк>2; (3.7)
Y,Ft = mra.; (3.8)
SM0 = /()“, (3-9)
где га2 — центростремительное ускорение, га — угловое ускорение.
Инерцию мы определили как способность объекта сопротивляться изменению его скорости. Момент инерции (7) количественно представляет меру сопротивления объекта изменению состояния вращательного движения. Он отражает распределение массы объекта относительно оси, вокруг которой вращается объект. Как уже указывалось, единица измерения момента инерции — кг • м2. При этом чем дальше расположена масса объекта от оси вращения, тем больше момент инерции. Следовательно, момент инерции не постоянен, подобно ЦТ он может изменяться при изменении взаимного расположения звеньев системы. Например, прыгун в воду, выполняющий сальто, вращается вокруг изменяющейся (саль-товой) оси. В положении согнувшись (ноги прямые) сегменты тела находятся на большем расстоянии от оси, чем в положении сгруппировавшись, поэтому в первом случае момент инерции спортсмена относительно сальтовой оси больше (см. табл.2.2). Ось, проходящая через ЦТ спереди назад, называется осью колеса, а проходящая от головы к пальцам ног через ЦТ — осью “винта”. Как правило, момент инерции тела человека наименьший относительно оси “винта” и наибольший относительно оси “колеса” (см.табл.2.2). Момент инерции относительно сальтовой оси в значительной степени колеблется при переходе из положения сгруппировавшись в положение прогнувшись.
Если правая часть уравнений (3.4 — 3.6) не равна нулю, диаграмма свободного тела системы подобна диаграмме ускорения тела. Иными словами, согласно закону ускорения Ньютона (F= та), силы, показанные на диаграмме свободного тела, равны массам, умноженным на ускорения, показанным на диаграмме ускорения. В этом случае диаграмма свободного тела представляет левую часть уравнения, а диаграмма ускорения — правую. При этом диаграмма свободного тела определяет систему и ее взаимодействие (силы показаны стрелками) с окружающей средой. Диаграмма
ускорения показывает влияние этого взаимодействия на движение систем.
Пример 1
Тяжелоатлет поднимает штангу на грудь. Предположим, мы хотим определить вращающий момент мышц разгибателей спины и бедер в момент, когда штанга находится на уровне колена. Для этого прежде всего необходимо определить систему и изобразить вращающий момент относительно бедренного сустава как внешнее воздействие, затем учесть все другие силы, действующие на систему, и определить ее кинематику. Приняв во внимание все другие силы, действующие на систему, мы допускаем, что все то, что не сбалансировано (остаточный эффект), обусловлено вращающим моментом, вызванным усилиями мышц, проходящих через бедренный сустав. Вращающий момент, определенный таким образом, называется остаточным (результирующим) моментом силы.
В нашем примере с тяжелоатлетом соответствующая система должна включать верхнюю часть тела от пояснично-крестцового (ПК) сустава до головы (рис.3.6). При этом установлено пять сил, действующих на систему: результирующий вращающий момент мышц (Тт) относительно пояснично-крестцового сустава, сила реакции сустава (Fy), вес штанги (FK в = 1003 Н), вес системы (FK u = 525 Н), и сила, обусловленная внутрибрюшным давлением (FJ- = 1250 Н). На основании визуального анализа выполнения упражнения можно определить кинематику движения (см.рис.1.7). Задача анализа — определить Т„. Как и в статическом случае, для этого необходимо просуммировать моменты относительно ПК сустава, пренебрегая Fy. При этом получим:
Рис. 3.6. Динамический анализ (диаграмма свободного тела (ДСТ) = график ускорения массы (ГУМ)) выполнения взятия веса на грудь штангистом:
Fj = 1250 Н; F„j, = 1003 Н; FWtU = 525 Н; а = 38 см; b = 24 см; с = 9 см; d = 47 см; IL S = 7,43 кг • м2; а= 8,7 рад • с-2
94
— ^Lsa + mcicl\
(Fwb x a) + (Fwu xb)-~(Р(хс)~Тт = ILSa + mad;
Tm = (Fw,b x °) + (Av,« xfc)“
- (Л x c) - /LSa - maxdy - maydx;
Tm = (1003 x 0,38) + (525 x 0,24) -
- (1250x 0,09)-(7,43x8,7)-
- (155,8 x 0,2 x 0,40) - (155,8 x [-0,1] x 0,24);
Tm = 381 + 126-11-64-12 + 4;
Tm =424 H-m.
Главное различие между статическим и динамическим анализами, заключается в том, что при динамическом анализе в рассмотрение включают момент инерции и угловое ускорение. В данном случае эти показатели были получены без особых трудностей, хотя для определения углового ускорения, как правило, необходимо обработать много данных. В нашем примере угловое ускорение (8,7 рад • с-2) получено на основании данных положение — время из видеозаписи выполнения тяжелоатлетом движения, ускорение определено с помощью метода, описанного в табл. 1.1. Момент инерции (7,43 кг*м2) получен из уравнений регрессии (табл.2.3).
Опытный штангист выполняет упражнение приблизительно 0,4 с. Снимая выполнение упражнения со скоростью 100 кадров в секунду, мы получим 40 кадров, содержащих достаточную информацию о динамике движения. Чтобы полностью описать изменение во времени Тт, необходимо осуществить вычисления в терминах подхода сила — масса — ускорение для
Рис. 3.7. Результирующий вращающий момент мышечных усилий относительно коленного сустава при взятии веса на грудь
каждого кадра. В результате получаем мгновенные значения вращающих моментов, которые можно изобразить кривой в плоскости вращающий момент — время. На рис.3.7 приведен пример для коленного сустава, данные представляют среднее значение вращающего момента относительно коленного сустава 15 опытных штангистов при взятии штанги (1141 Н, т.е. в 1,5 раза больше веса тела) на грудь. Из графика следует, что вращающий момент мышц относительно коленного сустава достигает экстремальных значений (—100 и 50 Н-м) соответственно в направлении разгибания и сгибания. При этом положительным оказывается момент при сгибании. График вращающий момент — время соответствует графику угол—угол, приведенному на рис. 1.7.
Пример 2
При изучении движения человека часто возникает вопрос, стоит ли проводить динамический анализ, если движение настолько медленно, что его можно считать квазистатическим и, следовательно, пользоваться методами статического анализа. Rogers и Pai (1990) изучали эту проблему на простом движении, включающем перемещение человека с опоры на двух ногах на опору на одну ноту, которое наблюдается, когда человек только собирается начать ходьбу. В исходном положении вес тела переносится с двух ног на одну, в то время как другая нога сгибается в колене. Это осуществляется в результате повышения активности латеральных мышц бедра согнутой ноги, увеличения вертикальной составляющей силы реакции поверхности под согнутой ногой и смещения центра давления к опорной ноге (Rogers, Pai, 1990). Поскольку сила реакции поверхности, как отмечалось в главе 2, связана с ускорением ЦТ человека, Rogers и Pai попытались определить, насколько точно с помощью статического анализа можно определить изменение вертикальной составляющей силы реакции поверхности.
Если допустить, что начало выполнения шага можно рассматривать как квазистатичес-кое явление, то система находится в состоянии равновесия и действующие на нее силы уравновешены (рис.3.8). Теперь мы можем сложить моменты сил относительно точки приложения Fz ss (точка 0) и определить F
sm0 = 0;
(Егу х J2) - (Fw х dj) = 0;
Fvxdx
Поскольку Fv — постоянная величина, Fz f определяется величиной отношения d\ и d2. Чтобы получить данные, позволяющие проверить
95
Рис. 3.8. Квазистатическая модель, используемая для определения вертикальной составляющей силы реакции поверхности под сгибающейся ногой при переходе с опоры на двух ногах на опору на одной ноге: ср — центр давления; dt — расстояние между ср и Fw; d2 — расстояние между О и Fzj\ Fzj — вертикальная составляющая силы реакции поверхности под сгибающейся ногой; FZiSS — вертикальная составляющая силы реакции поверхности под опорой на одну ногу; Fw — вес тела; G — центр тяжести;
О — точка приложения силы Fzss
это предположение, Rogers и Pai (1990) разместили испытуемых на двух силовых платформах (одна нога на каждой платформе), что позволило измерить вертикальную составляющую силы
реакции поверхности и точку ее приложения (центр давления) на каждой ноге (рис.3.9). Обратите внимание на графики, демонстрирующие изменение Fz у со временем. Сплошной линией показано измеренное значение Fzjr прерывистой — определенное с помощью квази-статического анализа, а заштрихованный участок отражает различие между ними. Результаты показывают, что квазистатический анализ может быть использован только при медленном выполнении шага. Точно так же квазистатический анализ может быть применен при медленном поднимании штанги (Toussaint, van Buar, van Langen, de Looze, van Dieen, 1992).
Пример 3
Динамический анализ, в основном, можно осуществлять двумя способами: зная силы и вращающие моменты, определить кинематику движения и, наоборот, зная кинематику, определить силы и вращающие моменты. Первый способ связан с решением прямой задачи динамики, второй — обратной задачи. Решение обратной задачи предполагает использование данных о положении и времени для получения — скорости и ускорения во время движения. Основным недостатком этого метода является то, что погрешности в данных положение — время в значительной степени усугубляются при обработке их с целью определения ускорения. Наоборот, решение прямой задачи динамики предполагает интегрирование уравнений для сил и вращающих моментов для получения кинематической информации. Главная трудность состоит в необ-
Рис. 3.9. Изменения dj, Fz ss и Fzj со временем при начале выполнения шага — быстро (д) и медленно (б)
96
ходимости точно определить начальные условия. Альтернативой является использование обоих методов. В этом случае измерения положения, линейного ускорения и угловой скорости объединяют с целью обеспечения надежной оценки кинематики, связей и нагрузок на сустав (Ladin, Wu, 1991; Wu, Ladin, 1993).
Определение вращающего момента усилий относительно бедра в нашем примере 1 основано на решении обратной задачи динамики. В этом примере мы исходили из кинематики движения и некоторой информации о силе для определения её величины. Считается, что нервная система, планируя выполнение движений, решает обратную задачу динамики (Hollerbach, Flaih, 1982). При этом вначале она как бы задает желательную кинематику выполнения движения, а затем решает обратную задачу динамики, “вычисляет” вращающий момент мышц, необходимый для обеспечения заданной кинематики. Однако ввиду сложности подобных вычислений маловероятно, что нервная система организует выполнение движений подобным образом (Hasan, 1990).
Bobbert, Schamhardt, Nigg (1991) применили метод решения обратной задачи динамики с достаточной степенью точности для определения вертикальной составляющей силы реакции поверхности во время опорной фазы бега. Они использовали четыре видеокамеры для получения кинематической информации, необходимой для определения вертикальной составляющей ускорения ЦТ каждого сегмента тела и объединили ее с данными о массе сегментов тела (Clauser, McConvill, Young, 1969) для вычисления вертикальной составляющей силы инерции (масса х ускорение) каждого сегмента. Силы инерции каждого сегмента просуммировали, чтобы определить г, которую затем сравнили с измеренной г С помощью этого метода величину начального пика на графике F„ — время оценили с погрешностью менее 10 %, а время его появления — с погрешностью в пределах 5 мс. Вместе с тем следует отметить, что подобная точность оценки сил определяется исключительной точностью кинематических данных (Bobbert et al., 1991; Ladin, Wu, 1991).
Межсегментная динамика. Любое движение человека — это сложное действие, в котором участвует большое количество сегментов тела. В связи с этим кинематика опорно-двигательного аппарата определяется согласованным совместным перемещением всех сегментов тела, принимающих участие в конкретном движении. Рассмотрим, например, человека, собирающегося ударить по мячу ногой. Выполнение этого задания предполагает координированные движения сегментов бедра, голени и ступни. Перемещение точки касания ступни (т) зависит от относительного движения сегментов ступни, голени и бедра (рис.3.10). Перемещение точки т (Sm) можно записать как
$т = $т/Г + $Г/К + $К/Б + $Б-
Согласно этому уравнению, перемещение точки т зависит от четырех слагаемых: 5т;Г характеризует перемещение точки т относительно голеностопного сустава; 5г/к — перемещение голеностопного сустава относительно коленного; 5к/б — перемещение коленного сустава относительно бедренного; 5g — абсолютное перемещение бедренного сустава.
Точно так же, если каждый сегмент имеет свою угловую скорость (со) и ускорение, то линейная скорость (v) и ускорение (а) точки т зависят от относительной кинематики каждого сегмента в сочетании с абсолютной кинематикой бедренного сустава. Угловая скорость и ускорение каждого сегмента показаны на рис.3.10,а, где нижние индексы 1,2 и 3 соответствуют ступне, голени и бедру. Для vm можно записать уравнение, аналогичное уравнению для Sm'.
vm = Ут/Г + УГ/К + УК/Б + v£-
Поскольку линейная и угловая скорости связаны уравнением v = га,
Vm = ZjCD । +Г2(Й2 + /3Й3 +УБ,
где г! — расстояние от голени до точки т; г2 — длина голени; г3 — длина бедра. Каждая
Рис. 3.10. Кинематика нижней конечности при выполнении удара по мячу ногой:
а — ступня, голень и бедро имеют угловую скорость (<а) и ускорение (а) при перемещении конечности в точку контакта т с мячом; ступня вращается относительно голеностопного (Г) голень — относительно каленного (К), бедро — относительно тазобедренного (Т) суставов; б — кинематические факторы, влияющие на линейную скорость точки т; в — кинематические переменные, влияющие на линейное ускорение точки т
7 8-133
97
составляющая в этом соотношении направлена по касательной к траектории движения соответствующего анатомического сегмента (рис.3.10,6): г id)! по касательной к траектории точки т: г2ш2 — к траектории движения голени (конечной точке голени); г3с)3 — к траектории движения колена (конечной точке бедра).
Ускорение ат точки m также зависит от ускорения соответствующих сегментов (рис.3.10,в) и на основании соотношения между линейным и угловым ускорением (а= гео2 + га) его можно записать таким образом:
2 2 2
ят ~ г1а)1 + г1а1 + + г2а2 + Г3Ю3 + гЗаЗ + аБ ’
где гсо2 — изменение направления скорости сегмента, а га — изменение величины V. Важным моментом этого соотношения является то, что ускорение каждого сегмента в данной системе зависит от ускорения всех других сегментов. Например, преобразование этого выражения относительно ускорения голени показывает, что ускорение голени во время удара по мячу зависит от углового ускорения ступни, бедра и линейного ускорения бедра
2 2 2
г2(»2 + т2а2 - ат - Г](В] - - г3со3 - г3а3 - а£ .
Miller и Munro (1984) использовали этот метод для определения вклада каждого из сегментов тела в достижение высоты во время фазы полета при прыжке в воду с трамплина.
Кинематическая связь между сегментами обусловливает динамическое взаимодействие между ними. Исходя из известной зависимости между силой и ускорением (F=wa), можно за ключить, что отмеченное взаимодействие между ускорениями сегментов тела возникает потому, что при выполнении движения между этими сегментами действуют силы взаимодействия. В межсегментной динамике изучаются взаимодействия между сегментами в зависимости от вида движения (Hoy, Zemike, 1986; Phillips et al., 1983; Putnam, 1991). Эта область исследований связана с очевидными трудностями ввиду значительной сложности уравнений, которые необходимо четко определить, чтобы установить зависимые от вида движения взаимодействия. Методику исследований тщательно описал Putnam (1991), получивший уравнения для двухсегментной нижней конечности (бедро и голень) во время выполнения удара на основании стандартного ньютоновского анализа.
В этом анализе очень важно выяснить величины зависимых от движения воздействий. На рис.3.11,а показано изменение во времени
Рис. 3.11. Зависимые от движения эффекты по время удара по мячу ногой:
a — результирующий вращающий момент мышц относительно коленного сустава (/), зависимый от движения вращающий момент бедра, действующий на голень (2) и суммарный эффект (3) всех вращающих моментов, действующих на голень; б — результирующий вращающий момент мышц относительно тазобедренного сустава (4), зависимый от движения вращающий момент голени, действующий на бедро (5) и суммарный эффект (б) всех вращающих моментов, действующих на бедро
98
результирующего вращающего момента мышечных усилий относительно коленного сустава и вращающий момент, действующий на голень со стороны движущегося бедра, во время выполнения удара. Из графика следует, что влияние движения бедра на голень столь же значительно, как и влияние мышц, проходящих через коленный сустав. Зависимый от движения вращающий момент бедра по отношению к голени отрицателен в течение первых 50 мс выполнения удара, отрицательное направление свидетельствует о том, что бедро обеспечивает ускорение голени по направлению вращения назад. На протяжении большей части выполнения удара движение бедра обеспечивает ускорение голени по направлению вперед.
Подобным образом можно определить влияние движения голени на бедро. Оно не является зеркальным отражением влияния движения бедра на голень. Голень — дистальный сегмент в этой двухсегментной системе, а бедро — проксимальный сегмент. В этом движении нас интересует воздействие голени на бедро. На рис.3.11,5 показан вращающий момент, действующий на бедро вследствие движения голени. Величина его превышает результирующий вращающий момент мышц относительно бедренного сустава. Более того, в течение большей части выполнения удара зависимый от движения вращающий момент голени и вращающий момент мышц тазобедренного сустава действуют в разных направлениях. Если принять во внимание все вращающие моменты, действующие на бедро, окажется, что результирующий вращающий момент — небольшой и в первые 100 мс удара вызывает ускорение бедра в пе-редневращающем направлении, а затем — в зад-невращающем.
Исследование межсегментной динамики позволяет сделать вывод, что движение одного сегмента тела может вызвать значительные вращающие моменты других сегментов системы. Подобные воздействия имеют большое значение не только для всей конечности и быстрых движений (удары и метания), но даже для движений пальцев (Darling, Cole, 1990). В этом случае нервная система должна контролировать величину и характер всех воздействий, принимающих участие в движении (Smith, Zemike, 1987). Например, результирующий вращающий момент мышц относительно бедренного сустава (рис.3.11,6) должен быть достаточным, чтобы преодолеть зависимый от движения вращающий момент головы относительно бедра и обеспечить ускорение бедра сначала по направлению вперед, а затем по направлению назад. Отметим значительную сложность физиологических механизмов, с помощью которых нервная система регулирует эти зависимые от вида движения воздействия (Koshland, Smith, 1989а, 19896).
подход ИМПУЛЬС-КОЛИЧЕСТВО ДВИЖЕНИЯ
Многие движения тела человека можно рассматривать как столкновения, т.е. тело человека непосредственно или косвенно сталкивается с объектом, находящимся в окружающей среде.
Анализ столкновений обычно осуществляют с помощью такого понятия, как количество движения. Количество движения (G) движущегося'объекта определяют как произведение его массы и скорости. Единица измерения количества движения — кг м • с-1 (масса х скорость). Количество движения — векторная величина, направление которой совпадает с направлением скорости. Согласно закону ускорения Ньютона, количество движения системы изменяется при воздействии силы на систему. Действие силы на объект в течение определенного интервала времени называется импульсом силы (сила х время). Рис.3.12, иллюстрирует изменение вертикальной составляющей силы реакции поверхности (Fg во время бега в течение конечного промежутка времени.
Величину импульса можно определить графически как площадь под кривой сила—время (рис.3.12) или вычислив произведение средней силы (Н) на длительность (с). Средняя сила на рис.3.12 составляет 1,3 кН, а продолжительность действия — 0,29 с, следовательно, величина импульса — 377 Н с. Очевидно, величина импульса меняется при изменении каждой из этих величин.
Если закон ускорения Ньютона рассматривать с учетом продолжительности действия силы, его можно сформулировать так: приложение импульса силы приводит к изменению количества движения системы. Это — основа импульсного подхода к изучению движения. Количество движения определяется как масса,
Рис. 3.12. Изображение импульса как площади (заштрихованной) под кривой сила—время
7*
99
умноженная на скорость (уравнение 2.2), а момент количества движения — кинетический момент (Н) представляет собой произведение момента инерции и угловой скорости. Соотношение, связывающее импульс с количеством движения, можно получить из закона ускорения Ньютона:
EF = та;
t
lFt = m(yf -v,-); (3.10)
ZF/ = Aznv или F • t = Awv,
где ZF/ — площадь под кривой сила—время, которая равна произведению средней силы и продолжительности ее действия (F 0. Из уравнения (3.10) следует, что зная величину импульса, можно определить изменение количества движения системы. И наоборот, если известно изменение количества движения, легко определить и величину импульса.
Пример 1
Засняв выполнение удара по мячу в волейболе и измерив массу (т) мяча, можно определить величину импульса, сообщенного мячу. Для этого необходимо определить скорость мяча до (vA ) и после (va) контакта с рукой, а также продолжительность контакта руки с мячом (Z'c). Получим
vb = 3,6 м • с-1;
vc = 25,2 м с-1;
т = 0,27 кг;
tc = 18 мс.
Применим уравнение 3.10:
F‘ t = Amv = wiAv = m(ya - vft);
F • t = 0,27(25,2 - 3,6) = 5,83 H • c.
Так как нам известна продолжительность контакта (tc), можем определить среднюю силу (F), приложенную к мячу во время удара:
F-tc= 5,83 Н-с;
5,83 tc 0,018 ’
F = 324 Н.
Таким образом, несмотря на небольшую величину импульса (5,83 Н • с), кратковременная продолжительность контакта обеспечила приложение довольно значительной силы (F= 324 Н). Продолжительность контакта руки с мячом в этом примере почти идентична продолжительности контакта ноги с мячом (Asami, Nolte, 1983).
Пример 2
В большинстве контактных ситуаций, таких, как выполнение удара в волейболе, количество движения объекта изменяется вследствие приложения сил относительно большой величины в течение короткого периода времени. Наряду с этим количество движения может изменяться в результате приложения сил небольшой величины в течение более продолжительных промежутков времени (Brancazio, 1984). Каковы, например, будут последствия прыжка человека с массой тела 71 кг с 15-метрового здания на тротуар и с 15-метрового утеса в океан? В обоих случаях скорость непосредственно перед контактом составит около 17,3 м • с-1 (количество движения = т х v = 1228 кг • м • с-1). В конечном итоге скорость (а значит и количество движения) достигнет нуля. В первом случае на человека воздействуют силы большой величины в течение короткого промежутка времени (контакт с тротуаром), во втором — меньшие силы вследствие более длительного контакта с водой. В обоих случаях изменение количества движения будет одинаковым.
Пример 3
При воздействии импульса на систему количество движения системы изменяется пропорционально величине импульса и зависит от его направления. Рассмотрим изменение составляющей (Fg у) силы реакции поверхности во время опорной фазы бега (рис.2.17). Как видно из рисунка, на бегуна во время опорной фазы действуют два горизонтально направленных импульса. Сначала Fg у направлена назад, создавая тормозящий импульс, затем Fg у изменяет направление, сообщая импульс поступательному движению вперед. Ввиду противоположной направленности этих импульсов изменение количества движения бегуна (или системы) в направлении оси у зависит от разности между тормозящим и сообщающим поступательное движение импульсами. При беге с постоянной скоростью (количество движения не изменяется) два импульса равны. Чтобы увеличить скорость бега, сообщающий поступательное движение импульс должен превзойти тормозящий, и наоборот, скорость бега снижается, если преобладает тормозящий импульс. Эта взаимосвязь показана на рис.3.13. Изменение количества движения вследствие действия силы реакции поверхности равно разнице между начальным и конечным количеством движения.
Поскольку на импульсы во время движения влияет вес человека, на практике принято величины импульсов сил, действующих на тело, нормировать и рассматривать в частях от импульса, создаваемого весом тела за то же время.
100
Рис. 3.13. Изменение количества движения бегуна при тормозящем импульсе, превышающем импульс, сообщающий поступательное движение (/); при эквивалентных тормозящем и сообщающему поступательное движение импульсах (2), и при сообщающем поступательное движение импульсе, превышающем тормозящий импульс (5)
Например, если бегун весом 630 Н испытывает тормозящий импульс 15,8 Н • с продолжительностью 0,1 с, то нормированный тормозящий импульс составляет 0,25 импульса веса тела (15,8/(630 х 0,1) = 0,25). Munro et all. (1987) наблюдали тормозящий и сообщающий поступательное движение импульсы, составляющие 0,15 — 0,25 импульса веса тела, у бегунов, передвигавшихся со скоростью 3,0 - 5,0 м-с"1. Величина импульсов при этом возрастала с увеличением скорости.
Столкновения
Многие виды спорта основаны на различных видах столкновений, например, между игроками (регби, бокс), между спортсменом и снарядом (гандбол, футбол) и между снарядами (бадминтон, гольф). Если количество движения прыгуна в воду или гимнаста, выполняющего какой-либо элемент в воздухе, во время движения изменяется, то при взаимодействии двух объектов в процессе столкновения сумма их количеств движения остается неизменной. Столкновение не создает и не разрушает количество движения, происходит лишь обмен количеством движения между сталкивающимися телами. Рассмотрим, например, столкновение головами нападающего (масса 100 кг, скорость 7,2 м-с"1 вправо) и защитника (масса 85 кг, скорость 5,9 м • с'1 влево): количество движения после столкновения = количество движения до столкновения =
= (100 х 7,2) + (85 х [-5,9]) = = 720-502 = 218 кг-м-с-1.
Поскольку количество движения — вектор такого же направления, что и скорость, положительный знак перед количеством движения (218 кг-м-с-1) свидетельствует о том, что после столкновения нападающий продолжает двигаться вправо, хотя и с меньшим количеством движения, чем до столкновения. Какова скорость двух игроков после столкновения? Изменение скорости каждого игрока обратно пропорционально его массе, отсюда преимущество игроков большей массы в контактных видах спорта.
-При столкновении двух объектов они воздействуют друг на друга с одинаковой силой, однако действующей в разных направлениях. Поскольку длительность контакта при столкновении одна и та же для двух объектов и силы взаимодействия равны, то равны и импульсы, а также изменение количества движения обоих объектов. Таким образом, при столкновении имеет место сохранение количества движения. Это значит, что сумма количества движения обоих объектов (А и В) до столкновения равна сумме количества движения после столкновения:
(тАУА)д0 + {твув)до =
— А) + (^й^й)лосл<?.
Левая часть уравнения (3.11) — количество движения системы до столкновения, правая — после столкновения. Поскольку масса при движениях человека, как правило, не изменяется, можно показать, что изменение количества движения объекта А равно изменению количества движения объекта В:
(^Л^л)<)о — У после ~ (^й^й) после ~ (^B^B)doj
^л(^ЛДо — после) “ ^В^УВ,после ~ ^В,до)\
mAbvA = mBAvB;
АУл _ тв (3-12)
Ave тА
Из уравнения (3.12) следует, что отношение скоростей объекта А и В обратно пропорционально их массам. Рассмотрим, например, что происходит при ударе теннисной ракеткой по мячу. После контакта с ракеткой (столкновение) скорость мяча, как правило, намного превышает скорость ракетки. Отличие между скоростью мяча и ракетки определяется отношением их масс.
Большое значение имеет то, является ли столкновение упругим или нет, т.е. отталкиваются ли объекты друг от друга после столкновения или остаются соединенными вместе. При упругом столкновении каждый объект сохраняет определенное количество кинетической энергии (скорость после столкновения ненулевая). Упругое столкновение характеризуется коэффициентом восстановления (е). Абсолютно
101
упругое столкновение имеет коэффициент восстановления, равный 1; при этом относительные скорости до и после столкновения одинаковы. Однако для большинства столкновений при различных движениях человека коэффициент восстановления, как правило, меньше единицы:
коэффициент восстановления =
_ относительная скорость после столкновения относительная скорость до столкновения Коэффициент восстановления мяча обычно определяют, бросая мяч с известной высоты на поверхность и измеряя высоту отскока. Если мяч, брошенный с высоты 1,0 м, отскакивает вверх на 0,5 м, то коэффициент восстановления мяча равен 0,5. Коэффициенты восстановления различных мячей при скорости столкновения 24,6 м-с-1: мяч для игры в софтбол — 0,40, теннисный мяч — 0,55, мяч для игры в гольф — 0,58, баскетбольный мяч — 0,64, футбольный мяч — 0,65, “супермяч” — 0,85. Коэффициент восстановления, как правило, снижается при увеличении относительной скорости тел перед столкновением.
Коэффициент восстановления количественно зависит от степени изменения свойств материала сталкивающихся тел. После контакта ракетки с мячом скорость определяется не только их массой, но и коэффициентом восстановления. Рассмотрим контакт бейсбольного мяча с битой, который можно описать уравнениями, определяющими коэффициент восстановления и сохранения количества движения:
скорость после столкновения =
-е (скорость до столкновения)
VB,a - Vb,a = ~e(VB,b ~ *Ь,Ь ). (3-13)
где В — бита, b — мяч. Допустив, что векторы скорости мяча и биты лежат на одной прямой, как показано на рйс.3.14, преобразуем уравнение 3.13, чтобы определить скорость биты (гВя) и мяча (vbя) после контакта:
vb,« = vB,a +e<yBj> -vb,b^ (3-14)
vB,a = vb,a - e(vB,b ~ vb,b)- (3.15)
Чтобы определить скорость биты после столкновения (уВа), подставим уравнение 3.14 в выражение сохранения количества движения (уравнение 3.11), приведя в нем индексы в соответствие с рис.3.14.
mBvBJ> + mhVb,b = mBVB,a + mbvb,a '> mBv В,b + mbvb.b -
= mBvBa + /иЛ[гйя + e(yBb - гЛ /))];
vB,Amb + m в) =
= mbvb,b<l + e) + vBJ,(mB - тье); (316) mbvbJ,(l + e) + vBh(mB - mbe) ^B,u m
mb+mB
Рис. 3.14. Масса и скорость биты (В) и мяча (/>) до (yBb, vb,b> и после (vBa, vba) столкновения
Точно так же мы можем заменить выражение для vB>a уравнением (3.14) и найти vba:
msvBj> + mbvbb = mBvBa + mbvba-, mBvBJb + mbvb,b =
= mB[vbja - e(yBb - v^)] + mbvb a;
vb.a(mb+mB) =
= ^BvB,b(] + e) + vbJ,{mb + mBe); (317) _ mBVBh(\+e) + vbh{mh+mBe)
Уравнения (3.16) и (3.17) представляют скорости биты (v^a) и мяча (уЬа) после столкновения. Рассмотрим следующий пример. Зная массу мяча (ть) = 0,16 кг;
массу биты (тв) = 0,93 кг;
скорость мяча до столкновения (vbb) = —38 м-с-1 (85 миль-ч-1);
скорость биты до столкновения (увь) = 31 м-с-1 (70 миль -ч-1);
коэффициент восстановления (определенный по правилу) = 0,55,
можно определить скорость мяча и биты после столкновения:
_ mbvb,b0 + е) + vBb(mB - тье) _
VB,a ~ ~
ть + тв
-6,8(1 + 0,55) + 31(0,93 - 0,088) , с , .
0,16 + 0,93 ’ М‘С ’
mBvBb(\ + е) + vbb{mb + тве)
Vbfl т.п, “
тВ + ть
= 28,8(1+ 0,55)+ 38(0,16+ 0,51) = 64 3 м. с-1
0,16 + 0,93 “ ’ М С
Однако, как правило, скорости мяча до столкновения (vbb) и после него (гЛя) редко коллинеарны (лежат на одной прямой). Неколли-
102
неарность скорости мяча устраняется введением члена, содержащего косинус перед (уьь) в уравнениях (3.16) и (3.17).
Хотя приведенные определения скорости мяча и биты после столкновения основаны на применении законов физики, часто бывает затруднительно учесть все условия столкновения. Основные переменные, влияющие на скорость после столкновения — коэффициент восстановления, скорость тел до столкновения и их масса. В предыдущем примере мы брали массу только мяча и биты, однако эффект массы биты может быть большим, что отражено во введенном Плагенхофом понятии об эффективной ударной массе. Кроме того, весьма трудно оценить эффект увеличения массы бейсбольной биты вследствие ее связи с рукой спортсмена.
Другой аспект столкновения мяча и биты, который так или иначе связан с эффективной ударной массой, — понятие “мягкой” точки. Считается, что контакт между мячом и битой при столкновении происходит именно в “мягкой” точке, если при этом рука спортсмена не ощущает действия силы реакции. С понятием “мягкой” точки связаны три теории: центра столкновения, естественного узлового центра частоты и коэффициента столкновения (Brancazio, 1984). Наиболее популярна из них первая. При контакте мяча с битой участок биты, где не ощущается действие силы реакции (Fr) на руки, называют центром столкновения (удара) (рис.3.15). Когда сила (Ес) прикладывается в точке центра тяжести биты, она испытывает только поступательное движение (рис.3.15, а), в противном случае — поступательное и вра-
Рис. 3.15. Влияние силы, приложенной к бите, на ее движение:
а — сила соударения (Гс) приложена к ЦТ; б — приложена к другой точке, а не к ЦТ; в — приложена к центру удара
щательное (рис.3.15, б). В последнем случае в определенной точке линейный перенос и вращение взаимно компенсируется и бита не двигается. При этом точка прикосновения, обеспечивающая стационарную точку вращения (рис.3,15, в), находится на некотором расстоянии от ЦТ биты. Положение точки контакта мяча на бите относительно центра тяжести и центра столкновения определяют силу реакции, действующей на руки (рис.3.15, в).
Кинетический момент (момент количества движения)
Вращательное движение определяется вращающими моментами сил, действующими на систему. Вращающий момент представляет собой вращательное действие силы и вычисляется как произведение силы на плечо. Синоним вращающего момента — момент силы. Кинетический момент (Н) характеризует количество вращательного движения. Его вычисляют как произведение момента инерции и угловой скорости, единица измерения — кг-м2-с-1. Если на систему не действует импульс, левая часть уравнения (3.10) равна нулю, т.е. количество движения сохраняется. Это характерно для прямолинейного и вращательного движений и представляет собой закон сохранения количества движения. Вместе с тем сохранение количества движения в обоих случаях не всегда происходит в одно и то же время. В частности, это характерно для свободного падения, которое наблюдается в гимнастике (Dainis, 1981; Nissinen, Preiss, Briiggemann, 1985), прыжках в воду (Bartee, Dowell, 1982; Frohlich, 1980; Stroup, Bushnell, 1970; Wilson, 1977), которое классически демонстрирует кошка (и другие животные), брошенная с небольшой высоты и при этом всегда приземляющаяся на лапы (Капе, Scher, 1969; Laouri, Kalli-Laouri, Schwartze, 1990; Magnus, 1922, Matey, 1895).
Пример 1
После отталкивания с трамплина прыгун в воду испытывает действие двух сил — веса и сопротивления воздуха (рис.3.16). Рассмотрим, как они влияют на количество движения (G) и кинетический момент (Н) спортсмена. Используя полученное ранее соотношение импульс-количество движения, можно записать
F • t = AG;
F • t = A/nv.
Поскольку среднее значение силы включает влияние веса (Fw) и сопротивления воздуха (Ffl), F не равно нулю и имеет место изменение
103
Рис. 3.16. Силы, действующие на прыгуна в воду во время свободного падения: F„ — сопротивление воздуха; Fw — вес
количества движения, т.е. Amv^O. Следовательно, количество движения изменяется. Рассмотрим кинетический момент:
вращательный импульс = А кинетического момента
(Гх<Ц-г = дЯ;
(F х d) • t = Д/(в.
Вращательный импульс равен произведению среднего вращающего момента (/хd) на его продолжительность (/), где d — расстояние (плечо) от линии действия каждой силы до оси вращения (ЦТ). Поскольку линии действия сил FH и Fa проходят через ЦТ, плечо каждой из них равно нулю и в момент прыжка вращающий мо-
Рис. 3.17. Сохранение кинетического момента, сопровождаемое взаимными изменениями момента инерции и угловой скорости при выполнении прыжка в полтора оборота назад
мент относительно ЦТ отсутствует. Это означает, что и правая часть уравнения (Л/со) равна нулю, и, следовательно, количество движения остается постоянным. Это пример сохранения количества движения.
Поскольку кинетический момент остается постоянным и равен произведению момента инерции и угловой скорости, любое изменение одного из параметров (момента инерции или угловой скорости) влечет за собой изменение другого, сохраняя при этом величину произведения. Предположим, прыгун в воду выполняет прыжок в несколько оборотов в положении согнувшись. Если в процессе выполнения упражнения спортсмен видит, что он не может выполнить необходимое число оборотов, он может (поскольку Н остается постоянной) сгруппироваться, что позволит увеличить скорость вращений. Момент инерции (/g) прыгуна относительно оси вращения, проходящей через ЦТ, составляет около 7,5 кг • м2 в положении согнувшись и 4,5 кг • м2 в положении сгруппировавшись. Если в первом положении его угловая скорость (<в) равна 6 рад • с-1, то при переходе в положение сгруппировавшись она увеличится до 10 рад-с--1, а произведение обоих величин — кинетический момент Н (Н = Igm) остается постоянным (Н = 45 кг - м2 с-1).
В положении согнувшись
Н = 7,5 кг м2 х 6 рад • с-1;
Н= 45 кг - м2‘с_|.
В положении сгруппировавшись Я=4,5кг-м2 х 10 рад-с-1;
Н = 45 кг - m2'c-1.
Спортсмен, кроме того, может замедлить скорость вращения, увеличив момент инерции и приняв при этом более выпрямленное положение. Взаимосвязь угловой скорости и момента инерции, иллюстрирующая сохранение кинетического момента при выполнении прыжка в воду, показана на рис.3.17.
Пример 2
Соотношение импульс — количество движения можно также использовать для объяснения того, как прыгун в воду может начать выполнять винт при кажущемся отсутствии какой-либо силы, способной помочь ему в этом. На рис.3.18 показан прыгун в воду, выполняющий переворот, кинетический момент спортсмена (Hg) действует в ЦТ (рис.3.18, а). Согласно правилу правой руки, следует выяснить, в каком направлении прыгун выполняет переворот. Чтобы начать выполнять винт, спортсмен вращает руками относительно оси вращения, проходящей через грудь, правая рука идет выше головы, левая движется поперек туловища (рис.3.18,б). Поскольку руки вращаются относительно оси вращения, это не приводит к
104
Рис. 3.18. Начало выполнения винта при прыжках в воду
изменению (Нй) относительно оси переворота. Вместе с тем вращение рук создает кинетический момент относительно оси вращения (HgpyK), которому противостоит эквивалентный кинетический момент туловища, действующий в противоположном направлении относительно оси ВраЩеНИЯ (Н^туловища), Т.е. HgpyK + Н^^улсвища 0. После этого маневра руки перемешаются как показано на рисунке 3.18,6, а туловище наклонено относительно вертикали под углом 0. Поскольку спортсмен находится в состоянии свободного падения, кинетический момент остается постоянным и равным его значению в момент отталкивания от доски. Затем, вследствие изменения ориентации спортсмена, у вектора Hg возникает составляющая не только относительно оси переворота, но и относительно оси винта прыгуна, т.к. направление Hg остается постоянным, в то время как оси перемещаются вместе со спортсменом (рис.3.18,<?). Составляющая Hg относительно оси винта равна Hg sin 0, а составляющая относительно оси переворота (Hg^ определяется как Hg cos 0.
Рассмотрим угол наклона (0 — угол наклона туловища относительно вертикальной оси), который необходим для выполнения прыжка с передней позиции с полным винтом с 3-метровой вышки. Предположим, что на вышке спортсмен не начинает выполнять винт. В этом примере в момент отталкивания кинетический момент спортсмена составляет около 30 кг * м2' с-1 относительно оси переворота; Ig s — момент инерции спортсмена относительно оси переворота, проходящей через ЦТ; Igt — момент инерции пры
гуна относительно оси винта, проходящей через ЦТ; t — время, необходимое для выполнения прыжка. Подставим числовые значения:
Hg = 30 кг • м2 • с-1;
/_ t =14 кг• м2;
/g, = 1 кг • м2;
t = 1,5 с;
Hg,t =
3Osin0 = 1 х —
U
п 4,19
0 = arc sm ——;
30
0 = 0,14 рад.
При таких исходных условиях прыгуну, чтобы выполнить полный винт, необходимо наклонить туловище на 0,14 рад. Какое вращение относительно оси переворота произведет наклон на 0,14 рад? Выполним расчеты:
Нg,s ~
30 cos 0,14 = 14(о5;
29,7 = 14со5;
= 2,12 рад ’с"1;
А0 О 1 О -1
— = 2,12 рад*с ;
Д0 = 3,18 рад.
Таким образом, при угле наклона 0,14 рад прыгун выполнит прыжок винтом в полоборота (3,18 рад).
Пример 3
Анализ кинетического момента в предыдущих двух примерах имеет ограниченное применение для изучения движений человека, поскольку касается изменений по отношению к ЦТ, когда тело находится в относительно неподвижном состоянии. Вместе с тем при выполнении большинства движений сегменты тела выполняют множество движений относительно друг друга. Для подобных условий необходим анализ движений многосвязной системы, при котором определяют кинетические моменты каждого сегмента относительно его ЦТ (локальные кинетические моменты), азатем кинетические моменты ЦТ каждого сегмента относительно ЦТ системы (всего тела). Эта взаимосвязь имеет следующий вид:
HS/CS = jjBl/Cl + цВ2/С2 +
+ цИ/сз + (локальные составляющие) +
+ нс1/су+ нс2/су+ (3.18)
+ Нсз/су + ... (отдаленные составляющие),
105
Рис. 3.19. Кинетический момент гимнаста при выполнении опорного прыжка. Линии над графиком показывают длительность соприкосновения спортсмена с поверхностью или снарядом
где Н — кинетический момент; В1 — сегмент тела 1; С1 — центр тяжести сегмента 1; CS — центр тяжести системы; 5 — система; / — по отношению к.
Локальные составляющие имеют вид Igo, тогда как отдаленные состоят из векторного произведения г х mv, представляющего собой момент количества движения относительно ЦТ системы. На рис.3.19 показано изменение кинетического момента всего тела, полученное при анализе связанной системы (Hay, Wilson, Dapena, Woodworth, 1977). В этом примере гимнаст выполняет прыжок, включающий периоды его контакта с поверхностью либо с конем для выполнения опорного прыжка, а также временные промежутки отсутствия контакта. Как мы и предполагали, кинетический момент гимнаста изменялся в период контакта и оставался постоянным в фазе полета. Изменение кинетического момента обусловлено вращательным импульсом, возникающим при контакте спортсмена с опорой.
Miller и Munro (1985) анализировали вклад сегментов тела в выполнение прыжка в воду с трамплина. Они вычисляли локальные и отдаленные составляющие кинетического момента в фазе отталкивания при выполнении различных прыжков Грегом Лугонисом. Он выполнял прыжки из передней и задней стойки, с различным количеством оборотов (до 3,5). Поскольку кинетический момент во время безопорной фазы сохраняется, его величина в момент отталкивания определяет возможность выполнения задуманного прыжка. 79-93% ки
нетического момента, необходимых для выполнения каждого прыжка (—70 — 57 кг-м2'с~*), обеспечивал отдаленный кинетический момент за счет движения сегментов тела относительно ЦТ спортсмена. Вклад локального кинетического момента более значителен при выполнении прыжков из передней стойки вперед, чем из передней стойки назад. При этом вклад сегментов тела в величину кинетического момента изменяется при выполнении различных прыжков.
Пример 4
Соотношение импульс—количество движения также можно использовать для объяснения способности кошек (и других животных) приземляться на лапы после того, как их бросают вверх лапами. В связи с этим весьма интересно, каким образом кошка “приобретает” кинетический момент и выполняет вращение при отсутствии вращательного импульса, который бы мог привести к изменению кинетического момента. Специалисты в области биомеханики пока не пришли к единому мнению на этот счет. Согласно одной из теорий (Нау, 1978), вращение может осуществляться за счет последовательного изменения момента инерции различных частей тела во время падения (рис.3.20). В этом случае кошка вначале вращает верхнюю часть тела, подтягивая передние конечности ближе к туловищу, чтобы уменьшить момент инерции относительно продольной оси. В от-
106
Рис. 3.20. Процесс восстановления положения, при котором падающая с высота кошка выполняет вращение на 3,14 рад и приземляется на лапы
Рис. 3.21. Модель движения кошки во время процесса восстановления положения
вет на вращение верхней части тела нижняя часть вращается в противоположном направлении (сохранение кинетического момента). Однако ввиду того, что нижние конечности вытянуты, момент инерции больше и связанное с ним перемещение нижней части тела намного меньше, чем перемещение верхней части тела в противоположном направлении. После этого кошка выполняет вращение нижней части с небольшим моментом инерции (более значительное перемещение), вследствие чего верхняя часть ее тела “реагирует” значительным моментом инерции и небольшим перемещением. В последней фазе животное вытягивает конечности, что приводит к максимальному моменту инерции и минимальной угловой скорости. Эта теория основана на законе сохранения количества движения или кинетического момента, в основе которого — различные изменения момента инерции верхней и нижней частей тела.
Другая теория основана на законе сохранения кинетического момента верхней и нижней частей тела, обусловленном мышечным сокращением (Hopper, 1973). На рисунке 3.21, окошка изображена в виде двухсегментной системы, включающей верхнюю ((72) и нижнюю (G,) части тела, которые соединены группой мышц (PQ). Согласно этой теории, вращение возникает в результате сокращения мышц, соединяющих верхнюю и нижнюю части тела (PQ), вызывающих вращение (указанное стрелками) двух сегментов (6|И G2) относительно соответствующих продольных осей. Эти вращения можно представить в виде векторов момента количества движения для каждого сегмента (Hi и
Н2), как показано на рис.3.21, б. Согласно правилу правой руки, Н! направлен по диагонали вниз, Н2 — по диагонали вверх. Поскольку они направлены по диагонали, у них есть составляющие в горизонтальном и вертикальном направлениях. Составляющие кинетического момента в вертикальном направлении уничтожаются, а составляющие в горизонтальном направлении равняются отличному от нуля положительному числу, что свидетельствует о том, что кинетический момент, обеспечивающий вращение, направлен вправо, вследствие чего кошка имеет возможность приземлиться на лапы.
Пример 5
Рассмотрим основные двигательные навыки — ходьбу и бег. Хотя главная задача при этом состоит в перемещении ЦТ, она выполняется в результате согласованного углового смещения сегментов тела. Движение бегуна относительно оси винта можно представить в виде двухсегментной системы, включающей верхнюю и нижнюю части тела (рис.3.22, а). Когда ступня во время бега отталкивается от поверхности, на нее действует сила реакции поверхности, которая за время контакта создает вращательный импульс относительно оси винта. Бегун предотвращает угловое движение всего тела (отличный от нуля кинетический момент) относительно оси винта за счет вращательного импульса, обусловленного силой реакции поверхности, приложенной к нижней части тела в результате сокращения мышц туловища и образования про-
107
Рис. 3.22. Кинетический момент относительно оси винта при выполнении бегового шага: а — двухсегментная модель бегуна; б — кинетический момент верхней и нижней частей тела, а также всего тела
тивоположно направленного кинетического момента верхней части тела, т.е. руки и туловище вращаются в противоположном направлении относительно вращения ног. Это взаимодействие показано на рис.3.22, б: первые 40 % шага левая ступня находится в контакте с поверхностью, создавая положительный кинетический момент Нг нижней части тела, тогда как верхняя часть тела вращается с отрицательным Нг. Во второй части шага, в момент контакта правой ступни с поверхностью, все происходит наоборот. Таким образом, движение рук во время бега направлено на противодействие кинетическому моменту ног относительно оси винта (Hinrichs, 1987; Hinrichs, Cowanagh, Williams, 1987).
ПОДХОД РАБОТА-ЭНЕРГИЯ
Если объект или система передвигается на какое-то расстояние под действием силы, то при этом выполняется работа. Работа (U) — скалярная величина и определяется как произведение перемещения системы и проекции силы, действующей в направлении перемещения. Графически ее можно представить как площадь под кривой сила—перемещение (или момент — угловое перемещение). Например, человек, проходя курс реабилитации после операции на ко
лене, использует тренажер, например “Кибекс”, позволяющий измерять силу, прикладываемую к рычагу.
На рис.3.23 показана зависимость вращающего момента в коленном суставе от угла разгибания колена в диапазоне движения выше 1,57 рад. Работу (М- d), выполняемую мышцами ноги, можно определить, измерив площадь под кривой момент — угловое перемещение. Если на рычаг действует средний момент 50 Н • м, работа определяется следующим образом:
работа = момент х угловое перемещение;
М • d = 50 Н • м х 1,57 рад =
= 78,5 Дж.
Количество работы измеряется в джоулях (Дж, 1 Дж = 1 Н • м).
Суть подхода работа — энергия состоит в том, что работа, выполненная системой, равна изменению энергии в системе, т.е. выполнение работы требует затрат энергии. Если система сообщила энергию окружающей ее среде, работу выполнила система. Если перенос энергии произошел в противоположном направлении, работа выполнена окружающей средой по отношению к системе. Например, если спортсмен поднимает штангу, используя силу мышц-сгибателей локтевого сустава, они сокращаются (концентрическое сокращение), производя работу относительно штанги, физиологи называют эту работу положительной. При опуска
108
нии штанги мышцы—сгибатели локтевого сустава удлиняются (эксцентрическое сокращение), и штанга “выполняет” работу по отношению к мышцам. Такой перенос энергии называется отрицательной работой. При этом для выполнения отрицательной работы требуется намного меньше метаболической энергии, чем для положительной.
Согласно подходу работа—энергия, энергию можно представить как способность выполнять работу. При этом ее рассматривают в качестве меры имеющегося в распоряжении системы “ресурса” для выполнения работы. Действительно, многие изменяющиеся процессы в природе сопровождаются превращением энергии из одного вида в другой. При анализе движений человека особое значение имеют такие виды энергии, как потенциальная, обусловленная силой тяжести (Epg), кинетическая энергия поступательного движения (Ек1), кинетическая энергия вращения (Екг), потенциальная энергия, обусловленная деформацией (Epf), энергия, в результате использования которой образуется тепло (£А) и энергия, высвобождаемая в результате обменных процессов (Ет). Соотношение между работой и энергией можно записать в виде
работа = Д энергии;
U = ДЕ; (3.19)
U = &ЕВ р + Д£А. + &Екг + &ЕВ , + Д£А + &Ет.
Следует отметить отсутствие адекватного описания &ЕР s и Д£А при анализе движения всего тела (van Ingen Schenau, Cavanagh, 1990).
При изучении взаимосвязи между работой и энергией целесообразно в большинстве случаев использовать первый закон термодинами
ки, характеризующий взаимосвязь выполненной работы и изменения количества энергии. При этом энергетический баланс можно записать в виде
Д£т = Д£А + U. (3.20)
В биологических системах обмен энергии для выполнения работы не является абсолютно эффективным процессом, в результате чего не вся превращаемая энергия переходит в работу. Как видно из уравнения 3.20, часть энергии обменных процессов (Д£т) преобразуется в тепло (Д£А). Производительность или эффективность мышц составляет около 25 % (de Haan, van Ingen Schenau, Ettema et al., 1989; van Ingen, Schemau, Cavanagh, 1990). Это означает, что только 25 % высвобождаемой в результате обменных процессов энергии используется для выполнения работы, остальные 75 % преобразуются в тепло или используются во время восстановительных процессов. Отношение выполненной работы к изменению количества энергии характеризует эффективность (производительность) процесса. Работа, выполненная с минимальными затратами энергии, представляет собой наиболее экономное выполнение задания. Понятия эффективность и экономичность не являются синонимами. Различия между ними иллюстрируют рис.3.24: эффективность соответствует условию постоянной энергии, экономичность — постоянной работы. Максимальная эффективность (точка 2) — наблюдается при выполнении наибольшего количества работы для заданного изменения количества энергии. Максимальная экономичность (точка 5) наблюдается при выполнении работы с наименьшим изменением количества энергии.
Рис. 3.23. Кривая “вращающий момент — угловое перемещение” при выполнении разгибания ноги в коленном суставе, построенная с помощью изокинетического динамометра “Кибекс 11”
109
Эффективность
Д Энергия, Дж
Рис. 3.24. Произвольный график зависимости между работой и энергией при выполнении какого-либо задания
При изменении взаимного расположения тел, находящихся в силовом взаимодействии друг с другом, выполняется механическая работа. Если в конкретных условиях тело способно выполнить эту работу, значит, оно обладает необходимой для этого потенциальной энергией. Величина ее зависит от характера силового взаимодействия и расположения взаимодействующих тел по отношению друг к другу. При анализе движений человека, как правило, рассматривают два вида потенциальной энергии — гравитационную и упругую. Гравитационная энергия (£p>g) обусловлена положением объекта или системы в гравитационном поле по отношению к базисному уровню (например, относительно поверхности земли). Потенциальная энергия упруго деформированного тела или упругая энергия (Fps) обусловлена деформацией объекта (например, теннисной ракетки) или ткани (например, мышечной) по сравнению с начальным положением. Величину потенциальной гравитационной энергии можно определить следующим образом:
Epj,=mgh, (3.21)
где т — масса; g — ускорение, обусловленное силой тяжести (9,8 м • с-2); h — высота системы относительно базового уровня (в данном случае — поверхности земли). Поскольку g — постоянная величина, Fpg изменяется только при изменении массы системы или высоты, на которой она находится.
Потенциальная энергия упруго деформированного тела характеризует его способность возвращаться в исходное состояние. В ряде случаев при анализе двигательной деятельности человека необходимо учитывать потенциальную энергию упругой деформации объекта (теннисной ракетки, трамплина для прыжков в воду, лука) или биологической ткани (мышцы, соединительной ткани). Биологическая ткань выдерживает
небольшую деформацию без изменения свойств и может при этом выполнять работу, возвращаясь к исходной длине. Если же деформация велика и превышает способность объекта или ткани восстанавливать исходную длину, то в структуре ткани происходят пластические изменения. На рис.3.25,6 показано влияние увеличивающейся нагрузки на деформацию передней крестообразной связки; при продолжительности растяжения более 5 мин в связке происходят структурные изменения. Спортсмены, использующие упражнения на растягивание, стремятся вызвать пластические изменения в соединительной ткани для развития большей гибкости (Sapega, Quedenfeld, Moyer, Butler, 1981).
Величина деформации объекта, обусловленная приложенной силой, зависит от свойств материала, из которого он состоит. Для абсолютно упругой системы, такой, как пружина, изменение длины (х) пропорционально величине приложенной силы (F):
F = кх. (3.22)
Уравнение (3.22) представляет линейную зависимость (рис.3.25, а), при которой изменение длины (деформация) увеличивается пропорционально приложенной силе (нагрузке). Тангенс угла наклона кривой нагрузка — деформация характеризует жесткость пружины и обозначается символом к; увеличение угла соответствует более высоким значениям к, что свидетельствует о большей жесткости пружины. Количество энергии растянутой пружины (Л£д5) можно определить, вычислив работу силы £(см. уравнение (3.22) на перемещении х:
bEps=±kx2. (3.23)
Рис. 3.25. Зависимость нагрузка—деформация для пружины (а) упруговязкого элемента (6) и передней крестообразной связки (в)
110
Пружина, растянутая в допустимых для нее пределах упругости, сохраняет всю имеющуюся энергию, которая может быть использована для выполнения работы при восстановлении ее первоначальной длины, т.е. во время цикла нагрузки-разгрузки энергия не теряется (заштрихованный участок на рис.3.25, а).
В отличие от пружины мышца и соединительная ткань — упруговязкие элементы, т.е. обладают и упругими, и вязкими свойствами. Зависимость нагрузка—деформация упруговязкого элемента не является линейной, для нее характерно наличие петли гистерезиса (рис.3.25, б), свидетельствующей о том, что силы, генеризуемые элементом во время активной фазы (нагрузки) и пассивной (разгрузки), неодинаковы. Петля гистерезиса показывает, что в процессе нагрузки — разгрузки некоторое количество энергии теряется. Следовательно, не все количество энергии, содержащейся в системе во время действия нагрузки (растяжения), сохраняется для выполнения работы во время снятия нагрузки (сокращения). Эта потеря энергии показана заштрихованным участком на рис.3.25, б. Тем не менее, будучи упроговязкими элементами, мышцы и соединительная ткань могут накапливать энергию во время растягивания и использовать ее для выполнения работы при возвращении к исходной длине (Cavagna, Citterio, 1974; Schorten, 1987).
Кинетическая энергия — способность объекта выполнять работу в процессе движения. Кинетическая энергия (£}.) состоит из линейной и угловой составляющих. Ее величину можно выразить соотношением
Ек = EkJ + Екг',
F - 1 wv2 + 1 Лт2 (З-24) + у/СО ,
где т, I— соответственно масса и момент инерции системы; v и со — ее линейная и угловая скорости. Количество Ек, которым обладает система, во многом определяется ее скоростью (линейной и угловой). Об этом свидетельствуют квадратичные члены в формуле (3.24). При анализе движения человека угловая составляющая играет определяющую роль для конечностей, линейная — для головы и туловища.
Величина Ет (см. уравнение 3.19) используется для обозначения того количества энергии, которая обеспечивает реализацию биологических функций организма. Она обеспечивает энергию (АТФ), необходимую для сокращений мышц, энергию для поддержания физиологической активности сердечно-сосудистой и нервной систем, а также мышечной активности, направлен-нной на поддержание позы. Специалисты в области физиологии мышечной деятельности определяют эту энергию, измеряя количество кислорода, потребляемого во время физической деятельности (Daniels, 1985; di Prampero, 1986;
Таблица 3.1. Рекорды мира (1986) и средняя скорость в различных видах деятельности в аэробных условиях разной продолжительности (di Prampero, 1986)
Вид деятельности Дистанция, км Время, с Скорость, м-с-1 Энергозатраты кДж/км
Плавание
(вольный стиль) 1,5 895 1,68 1400
Ходьба 3,0 788 3,81 356
Бег 5,0 780 6,41 300
Скоростной бег 10,0 874 11,44 175
на коньках
Велосипедный 10,0 713 14,02 160
спорт
Joyner, 1991), когда в работе принимают участие большие группы мышц и она длится, по крайней мере, несколько минут. В табл. 3.1 сравниваются энергозатраты при выполнении упражнений аэробной направленности испытуемым с массой тела 70 кг и ростом 175 см (di Prampero, 1986). Средняя скорость, развиваемая во время выполнения того или иного вида деятельности, обратно пропорциональна энергозатратам на единицу дистанции (км).
Пример 1
Рассмотрим замкнутую систему. В этом случае энергия не поступает в систему, а переходит при выполнении системой работы из одного вида в другой. Полная механическая энергия, равная сумме кинетической и потенциальной, при этом не изменяется, оставаясь постоянной. Например, акробат с массой тела 60 кг собирается прыгнуть с 10-метровой вышки. Его потенциальная энергия — 5,88 кДж. Как только он покинет вышку, на него перестанет действовать сила реакции опоры, и под действием силы тяжести он начнет падать. Падение акробата сопровождается превращением потенциальной (Ер^ энергии в кинетическую (Ек ^. Поскольку ускорение, обусловленное силой тяжести, постоянно, скорость падения возрастает с расстоянием, которое пролетает акробат. Это согласовывается с увеличением превращения потенциальной энергии в кинетическую.
Рассмотрим энергию системы после того, как акробат пролетел 4 м. В этом положении потенциальная энергия акробата равна 3,53 кДж (уменьшилась на 2,35 кДж). Кинетическую энергию системы можно определить из уравнения
EkJ=^mv\ (3.24а)
Для определения кинетической энергии акробата после того, как он пролетел 4 м, необходимо определить его скорость (г) в этой точке. Для этого, воспользовавшись уравнением (1.4), нужно определить время (0,9045 с), потребовавшееся акробату, чтобы пролететь 4 м. Затем,
111
используя формулу для ускорения, вычислим скорость следующим образом:
Av а = —;
vf - Vi
а = —-----;
t vf - О 9,81 = -4—; 0,9045
Vf = 8,86 м • с-1,
где vit Vf — соответственно начальная (в начале падения) и конечная (на уровне 4 м) скорости. Конечную скорость (8,86 м-с-1) подставим в уравнение (3.24, а) и определим кинетическую энергию акробата на уровне 4 м.
Другая возможность определения v связана с использованием закона сохранения механической энергии [{Epg + Ekt) = {Epg + £*,,)<)], из которого следует, что сумма кинетической и потенциальной энергии в начале, в конце и на протяжении всего движения сохраняется постоянной. При этом кинетическая энергия переходит в потенциальную и наоборот, потенциальная в кинетическую. Если потенциальная энергия уменьшилась на 2,35 кДж, значит, она превратилась в кинетическую. Следовательно, кинетическая энергия на уровне 4 м известна и равна 2,35 кДж. Подставив это значение Ек) = = 2,35 кДж в формулу (3.24,а) и проведя соответствующие преобразования, получим:
Ek,t =^mv2;
х ^kj . т
v =
12x2,350.
60 ’
v = д/78,33 м2 • с 2; v = 8,85 м • с-1.
(Обратите внимание, что
Дж _Н-м_кг-м-м, кг кг кг • с
Используя первый или второй вариант, получаем значение конечной скорости = 8,85 м • с-1.
Пример 2
Первый закон термодинамики довольно трудно применить для изучения локомоции человека (Taker, 1975; Ward-Smith, 1983). На рис.3.26 предпринята попытка сделать это. Здесь схематически представлен процесс обмена энергии, который происходит во время бега на спринтерскую дистанцию (Ward-Smith, 1984). На схеме показано, что АТФ (химическая энергия) — источник энергии, обеспечи-
Рис. 3.26. Схема превращения энергии, происходящего во время бега
вающий обменные процессы в организме для поддержания жизнедеятельности и выполнения необходимой мышечной работы. При этом часть АТФ превращается в тепло (Я1). Полезным результатом при таком виде деятельности является выполнение механической работы бегуном относительно окружающей среды за счет углового движения различных сегментов его тела. За счет работы (положительной и отрицательной), выполненной мышцами, конечности получают кинетическую (энергию вращения и поступательного движения) и потенциальную энергию. Во время этого процесса некоторое количество механической энергии превращается в тепло (НТ) вследствие трения в суставах и мышцах. Механическая энергия (кинетическая и потенциальная), приобретенная конечностями, превращается в кинетическую и потенциальную энергию ЦТ бегуна и работу по преодолению сопротивления воздуха (ЙЗ). Изменение механической энергии центра тяжести в этом случае распределяется по двум направлениям. При этом кинетическая энергия Ек добавляется к горизонтальной — W1 и вертикальной — W1 составляющей, обеспечивающей выполнение работы по преодолению силы тяжести. Кинетическая и потенциальная энергия конечностей и центра тяжести, направленная на преодоление силы тяжести (И2), в конечном итоге приводит в накоплению упругой (потенциальной энергии упруго деформированного тела) энергии в мышцах (включая сухожилия), которая может
112
быть повторно использована. Это явление называется накоплением и использованием упругой энергии или циклом растяжения—сжатия. При беге вертикальное перемещение ЦТ спортсмена незначительно и, следовательно, работа, направленная на преодоление силы тяжести, не выполняется. Таким образом, АТФ, поступающий в мышцы, в конечном итоге превращается в тепло (Н\ + Н2 + + НЗ + Н4) и используется для выполнения работы, связанной с перемещением в горизонтальном направлении и для преодоления силы сопротивления воздуха (W1 + ИЗ). При этом соотношения между работой и энергией в процессе бега можно выразить с помощью уравнения (3.20).
Пример 3
Скорость, которую развивает пловец, зависит от запаса энергии спортсмена, а также от того, какой объем работы он при этом может выполнить. Большая часть выполняемой спортсменом работы идет на преодоление сопротивления воды. Затраты энергии при рекордных результатах в плавании кролем на груди колеблются от 1900 кДж-км-1 на дистанции 100 м до 1170 кДж • км-1 на дистанции 1500 м. Затраты энергии при постоянной скорости плавания оказываются наименьшими при плавании кролем на груди, более высокими при плавании кролем на спине и наибольшими при плавании брассом (di Prampero, 1986). Отношение количества выполненной механической работы к затраченной при этом энергии называется эффективностью, в отечественной литературе это отношение называется коэффициентом полезного действия. На эффективность плавания, которая колеблется от 1 до 10%, влияют такие переменные, как уровень мастерства, способ плавания, плавучесть и скорость. Эффективность можно определить следующим образом:
Эффективность =
Выполненная работа х Ю0 _ F *d х jqq Затраты энергии VO2
Обычно это соотношение выражается через мощность как величину, более доступную для измерений. Для этого числитель и знаменатель предыдущей формулы относят ко времени:
Эффективность =
(F*d)/t х 10() = х 10() VO2 /1 VO2
где VO2 — интенсивность потребления кислорода; V— скорость пловца; F— горизонтальная составляющая силы реакции воды, дейст
вующей на пловца. При постоянной скорости плавания F постоянна и равна сопротивлению воды.
Пример 4
При падении человека на землю с небольшой высоты он выполняет работу относительно поверхности (земли). При этом в момент приземления Ek>t становится равной нулю. Само приземление может быть жестким или мягким (McNitt-Gray, 1991). При жестком приземлении туловище более выпрямлено и ноги менее согнуты в коленном и бедренном суставах. DeVita и Skelly (1992) установили, что мышцы бедра и колен поглощают (отрицательная работа) больше энергии при мягком приземлении (бедро — 0,60 по сравнению с жестким — 0,39 Дж/кг; колено — 0,89 по сравнению с жестким — 0,61 Дж/кг), тогда как мышцы лодыжки поглощают больше энергии при жестком приземлении (—0,88 по сравнению с мягким — 1,00 Дж/кг). Мышцы нижних конечностей поглощают на 19 % больше кинетической энергии тела при мягком приземлении, тем самым снижая нагрузку на другие ткани тела. Большую часть энергии поглощают мышцы — сгибатели подошвы — 44%, затем следуют мышцы—разгибатели коленного сустава — 34 % и мышцы—разгибатели бедренного сустава — 22%.
Мощность
Количество работы, выполняемой за единицу времени, называют мощностью
Особенно актуально это понятие во многих видах спорта, связанных с нагрузками небольшой продолжительности (спринте, тяжелой атлетике, армрестлинге, прыжках в высоту). Продолжительность физической деятельности связана обратно пропорциональной зависимостью с необходимой для этого мощностью (рис.3.27). Мощность можно определить как работу (F-d), разделенную на количество времени (А/) или как произведение силы (F) и скорости (т):
Л/
Мощность — скалярная величина, измеряемая в ваттах (Вт); 1 кВт (1,36 лошадиных сил) — метаболическая мощность, соответ-
8 8-133
113
Рис. 3.27. Уменьшение максимального образования мощности при увеличении продолжительности физической деятельности
ствующая потреблению около 48 мл • с 'О2. Поскольку работа характеризует изменение энергии системы, мощность можно рассматривать как интенсивность использования энергии. Существует, по меньшей мере, три различных способа определения производимой мощности (DeLooze, Bussman, Kingma, Toussaint, 1992).
Прим ер 1
Вспомним пример разгибания—сгибания руки в локтевом суставе (см.рис.1.9). Учитывая, что мощность — это произведение силы и скорости, а ускорение и сила пропорциональны, профиль (график) мощность—время, характеризующий движение, можно получить, перемножив графики скорости и ускорения. В случае, когда скорость и ускорение имеют одинаковый знак (положительный или отрицательный), мощность положительна и представляет поток энергии от мышц к руке. Если же мощность отрицательна (т.е. скорость и ускорение имеют разные знаки), поток энергии идет от руки к мышцам. Это связано соответственно с выделением и поглощением энергии соответственно. Что касается разгибания—сгибания руки в локтевом суставе, положительная скорость указывает на разгибание, положительное ускорение при этом соответствует силе, направленной на разгибание, а положительная мощность свидетельствует о том, что поток энергии движется от соответствующих мышц в систему предплечье — кисть. Если график скорости или ускорения пересекает ось времени, изменяя знак, то через нуль проходит и кривая мощности. Результирующий график мощность—время для разгибания—сгибания руки в локтевом суставе характеризуется четырьмя временными
промежутками, на которых происходит последовательная смена знака функции (рис.3.28). Первые два промежутка иллюстрируют генерирование и поглощение мощности, сопровождающие процесс выпрямления руки в локтевом суставе (см. график скорость—время) и отражают периоды положительной и отрицательной работы. В первом случае мышцы выполняют работу по отношению к системе, во втором — система (вследствие инерции) выполняет работу по отношению к мышцам. Подобная последовательность (сначала образование, а затем поглощение мощности) характерна и для фазы сгибания руки в локтевом суставе.
Этот пример подчеркивает взаимосвязь понятий положительной и отрицательной работы, образования и поглощения мощности, эксцентрического и концентрического сокращения мышц.
Как видно в нижней части рис.3.28, разгибание-сгибание руки в локтевом суставе связано с четырехфазным профилем мощности—времени. Моменты образования мощности соответствуют периодам положительной работы и, следовательно, концентрическим сокращениям мышц. Поглощение мощности связано с отрицательной работой и эксцентрическими мышечными сокращениями.
Вместе с тем следует отметить, что в действительности не вся мощность во время фазы образования (особенно во второй фазе) обусловлена использованием Ет. Некоторое количество мощности, очевидно, образуется в результате упругих свойств мышц и сухожилий (Eps). Более подробно пойдет об этом речь в части II.
Рис. 3.28. Профиль мощность-время для мышц, пересекающих локтевой сустав, при сгибании-разгибании руки в локтевом суставе (см. рис. 1.9). Положительная скорость отражает разгибание; положительное ускорение — ускорение в направлении разгибания и положительная мощность — мощность, образуемую системой
114
Пример 2
Приведенную на рис.3.28 зависимость мощность-время обычно получают, вычисляя произведение угловой скорости и результирующего вращающего момента мышечных усилий относительно сустава через выбранные интервалы времени. Эта процедура, однако, включает анализ большого количества данных, поскольку скорость и вращающий момент определяют на основании измерений, производимых в процессе движения. Угловую скорость можно определить, записав на видеомагнитофон или видеокамеру выполнение движения и затем использовать метод численного анализа, приведенного в табл. 1.1. Количество измерений скорости при этом зависит от установленной частоты кадров; на практике вполне достаточна частота 50—200 кадров-с-1. Результирующий вращающий момент мышц можно определить с помощью динамического анализа, а также на основании, например, определения момента силы реакции поверхности, по отношению к интересующему суставу (см.рис.3.6). Результатом динамического анализа является ряд мгновенных значений результирующего вращающего момента мышц в процессе движения (см.рис.3.7). Последний этап определения мощности предполагает согласование полученных измерений угловой скорости с соответствующим значением результирующего вращающего момента мышц и последующее вычисление их попарного произведения.
Подобный анализ финальной фазы прыжка в высоту (рис.3.29) был проведен Huijing (1992). Главное внимание ученый обратил на голеностопный сустав и мощность, развиваемую мышцами, которые его пересекают. На рис.3.29 приведены результаты последних 300 мс прыжка, когда испытуемый перешел от положения минимального угла коленного сустава к фазе отталкивания. При минимальном угле коленного сустава (—300 мс) результирующий вращающий момент мышц был порядка 100 Н-м, а угловая скорость — 0 рад-с-1; в этом положении голеностопные суставы не генерируют мощность. Начиная с этой исходной позиции, прыжок можно рассматривать как состоящий из трех фаз: в течение следующих 150 мс (от -300 до почти —150 мс) результирующий вращающий момент мышц увеличивается до максимальной величины (240 Н-м), а угловая скорость остается небольшой (менее 2 рад • с-1), что свидетельствует о незначительной величине развиваемой мощности (положительной) — около 200 Вт. Затем, несмотря на снижение результирующего вращающего момента мышц до 75 Н • м, угловая скорость составляет 15 рад • с-1 и развиваемая при этом мощность (1700 Вт) увеличивается до макси-
Рис. 3.29. Изменение мощности голеностопного сустава во время прыжка в высоту:
а — результирующий вращающий момент мышц относительно голеностопного сустава как функция его угловой скорости (теоретическая зависимость вращающего момента—скорости напоминает зависимость Хилла, которая показана штриховой линией); б — мощность, образуемая мышцами, пересекающими голеностопный сустав
мума. Наконец, угловая скорость, результирующий вращающий момент и мощность падают до нуля в момент отталкивания спортсмена. Из этого примера следует, что при максимальном вращающем моменте мышца не развивает максимальную мощность; последняя зависит от совместного влияния вращающего момента и скорости. Более подробно этот вопрос рассматривается в главе 6.
РЕЗЮМЕ
В этой главе на основании определений, приведенных в первых двух главах, описаны биомеханические методы изучения движения. Они основаны на трех различных подходах, использующих закон ускорения Ньютона. Выбор формулировки для анализа определяется характеристиками движения, которое предстоит анализировать, а также той информацией, которую желают получить. Можно рассмат
8*
115
ривать мгновенное значение силы (подход сила—масса—ускорение), влияние силы, приложенной в течение интервала времени (подход импульс—количество движения), или приложение силы, которая вынуждает объект переместиться на какое-то расстояние (подход
работа—энергия). На основании указанных подходов можно определить взаимосвязь силы, массы и ускорения, влияние силы на количество движения системы, а также оценить, какое количество энергии необходимо для выполнения работы.
ВОПРОСЫ Г АЛЬТЕРНАТИВНЫМ ОТ lETOM
1. С помощью подхода сила—масса—ускорение можно определить мгновенные значения СИЛЫ. ПРАВИЛЬНО НЕПРАВИЛЬНО
2. Единица измерения силы в системе единиц СИ — килограмм (кг).
ПРАВИЛЬНО НЕПРАВИЛЬНО
3. Статический анализ приемлем только для стационарной (неподвижной) системы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
4. М в выражении ЕЛ/0 означает вращающий момент. ПРАВИЛЬНО НЕПРАВИЛЬНО
5. Когда система находится в состоянии равновесия, на нее не действует никакая сила.
ПРАВИЛЬНО НЕПРАВИЛЬНО
6. Понятие “момент силы” — синоним понятия “вращающий момент”.
ПРАВИЛЬНО НЕПРАВИЛЬНО
7. Величина результирующей силы всегда превышает любую из составляющих.
ПРАВИЛЬНО НЕПРАВИЛЬНО
8. Направление вектора силы можно определить с помощью угла.
ПРАВИЛЬНО НЕПРАВИЛЬНО
9. Плечо мышечных усилий относительно оси сустава обычно меньше 10 см.
ПРАВИЛЬНО НЕПРАВИЛЬНО
10. При определении ЦТ системы координату у получают из выражения ЪМУ.
ПРАВИЛЬНО НЕПРАВИЛЬНО
11. /%„ обычно меньше, чем Fm l.
ПРАВИЛЬНО НЕПРАВИЛЬНО
12. Тангенциальная составляющая силы реакции сустава направлена параллельно продольной оси сегмента тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
13. Сила реакции сустава никогда не превышает вес системы. ПРАВИЛЬНО НЕПРАВИЛЬНО
14. Центр тяжести можно определить на основании статического анализа.
ПРАВИЛЬНО НЕПРАВИЛЬНО
15. В результате внутрибрюшного давления создается сила, вызывающая сгибание туловища. ПРАВИЛЬНО НЕПРАВИЛЬНО
16. Момент силы можно измерять в Н-см. ПРАВИЛЬНО НЕПРАВИЛЬНО
17. При действии на систему не уравновешенных сил она ускоряется.
ПРАВИЛЬНО НЕПРАВИЛЬНО
18. Символ / означает инерцию. ПРАВИЛЬНО НЕПРАВИЛЬНО
19. Понятие “квазистатический” характеризует медленные движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
20. Единица измерения центра тяжести — ньютон. ПРАВИЛЬНО НЕПРАВИЛЬНО
21. Квазистатический анализ приемлем при медленном, но не быстром начале выполнения шага.
ПРАВИЛЬНО НЕПРАВИЛЬНО
22. Обратная задача динамики состоит в вычислении сил, определяющих кинематические характеристики движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
23. Единицей измерения со является м-с-1.
ПРАВИЛЬНО НЕПРАВИЛЬНО
24. га2 учитывает изменение величины v. ПРАВИЛЬНО НЕПРАВИЛЬНО
25. Межсегментная динамика играет важную роль при быстрых движениях пальцев.
ПРАВИЛЬНО НЕПРАВИЛЬНО
26. Количество движения — векторная величина. ПРАВИЛЬНО НЕПРАВИЛЬНО
27. Импульс можно представить площадью под графиком сила—время.
ПРАВИЛЬНО НЕПРАВИЛЬНО
28. Единица измерения импульса — Н • м. ПРАВИЛЬНО НЕПРАВИЛЬНО
29. Импульс вызывает изменение количества движения системы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
30. Взаимосвязь между импульсом и количеством движения вытекает из закона ускорения Ньютона.
ПРАВИЛЬНО НЕПРАВИЛЬНО
116
31. Произведение массы и скорости называется импульсом. ПРАВИЛЬНО НЕПРАВИЛЬНО
32. Н означает количество движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
33. 1 Дж = 1 Н • М. ПРАВИЛЬНО НЕПРАВИЛЬНО
34. При равных тормозящем и сообщающем поступательное движение импульсах скорость бега увеличивается.
ПРАВИЛЬНО НЕПРАВИЛЬНО
35. Вращательный импульс = А/^со.
ПРАВИЛЬНО НЕПРАВИЛЬНО
36. Количество движения объекта не изменяется до и после столкновения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
37. Продолжительность действия силы контакта при ударе мяча ногой или рукой составляет около 0,2 с.
ПРАВИЛЬНО НЕПРАВИЛЬНО
38. Коэффициент восстановления мячей всегда меньше единицы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
39. Коэффициент восстановления безразмерен.
ПРАВИЛЬНО НЕПРАВИЛЬНО
40. Масса бейсбольной биты влияет на скорость мяча и биты после их соударения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
41. Кинетический момент — момент количества движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
42. Закон сохранения количества движения имеет отношение к условиям, когда система имеет нулевое количество движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
43. Произведение угловой скорости и момента инерции постоянно во время безопорной фазы прыжка в воду.
ПРАВИЛЬНО НЕПРАВИЛЬНО
44. Наименьший момент инерции прыгуна в воду наблюдается относительно оси винта.
ПРАВИЛЬНО НЕПРАВИЛЬНО
45. При анализе многосвязной системы кинетический момент сегмента относительно его ЦТ называют отдаленным угловым моментом. ПРАВИЛЬНО НЕПРАВИЛЬНО
46. Движение рук во время бега направлено на увеличение количества движения в горизонтальном направлении.
ПРАВИЛЬНО НЕПРАВИЛЬНО
47. Работа — векторная величина.
ПРАВИЛЬНО НЕПРАВИЛЬНО
48. Единица измерения работы — Н • м.
ПРАВИЛЬНО НЕПРАВИЛЬНО
49. Выполнение работы связано с затратами энергии. ПРАВИЛЬНО НЕПРАВИЛЬНО
50. Отрицательная работа — это работа внешней среды по отношению к системе.
ПРАВИЛЬНО НЕПРАВИЛЬНО
51. Единица измерения энергии — Дж.
ПРАВИЛЬНО НЕПРАВИЛЬНО
52. Первый закон термодинамики описывает связь между работой и энергией.
ПРАВИЛЬНО НЕПРАВИЛЬНО
53. Потенциальная энергия имеет линейную и угловую составляющие.
ПРАВИЛЬНО НЕПРАВИЛЬНО
54. Объект имеет потенциальную энергию при выполнении по отношению к нему работы, направленной на изменение его количества движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
55. Мышца может накапливать упругую энергию. ПРАВИЛЬНО НЕПРАВИЛЬНО
56. Соединительная ткань теряет какое-то количество энергии во время нагрузки—разгрузки и поэтому выполняет меньший объем работы при сокращении, чем при удлинении.
ПРАВИЛЬНО НЕПРАВИЛЬНО
57. Объект может иметь как кинетическую энергию вращения, так и кинетическую энергию поступательного движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
58. VO2 отражает интенсивность поступающей в организм энергии.
ПРАВИЛЬНО НЕПРАВИЛЬНО
59. Единица измерения Дж • кг-1 — мера работы, отнесенная к единице массы тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
60. Мощность = работа х скорость.
ПРАВИЛЬНО НЕПРАВИЛЬНО
61. Мощность можно определить по интенсивности использования энергии.
ПРАВИЛЬНО НЕПРАВИЛЬНО
62. При эксцентрических сокращениях мышца генерирует мощность.
ПРАВИЛЬНО НЕПРАВИЛЬНО
63. Источником энергии для образования мощности служит только Ет.
ПРАВИЛЬНО НЕПРАВИЛЬНО
64. Максимальная мощность мышц наблюдается при максимальном вращающем моменте.
ПРАВИЛЬНО НЕПРАВИЛЬНО
65. Максимальная мощность мышцы не превышает 1 кВт.
ПРАВИЛЬНО НЕПРАВИЛЬНО
117
ВОЯРОСЬ CI зличныв-ВЬFBOPOBf ОТВЕТОВ
66. Расставьте по порядку в зависимости от степени сложности изменения следующие четыре силы:
а) силу реакции сустава;
б) вес сегмента;
в) силу реакции поверхности;
г) мышечную силу.
67. Укажите единицу измерения момента инерции:
а) кг;
б) кг - м-с"-1;
в) кг - м2;
г) Н • м • с'.
68. В каком направлении проходит ось кувыркания “колесом”?
а) с тыльной части во фронтальную;
б) от головы к пальцам ног;
в) из стороны в сторону;
г) медиально-латерально.
69. Какое утверждение(я) о принципе: сила-масса—ускорение для изучения движений человека неправильное(ые)?
а) на графике ускорения массы показаны все внешние силы;
б) согласно закону ускорения (F= та) 1Н = 1 (кг-м)с~2;
в) в динамическом анализе правая часть закона ускорения не равна нулю;
г) согласно квазистатическому анализу, ускорение системы равно нулю.
70. Какая методика используется для определения кинематики системы при заданных силах и размерах системы?
а) обратная задача динамики;
б) квазистатика;
в) прямая задача динамики.
71. Изменение скорости объекта А относительно объекта В вследствие столкновения зависит от:
а) отношения масс обоих объектов;
б) начальной скорости обоих объектов;
в) угла между направлением скоростей объектов до соударения;
г) начального количества движения объектов.
72. Выберите правильный ответ о центре удара: а) центр удара расположен дистально к центру тяжести бейсбольной биты;
б) при контакте мяча и биты в центре удара на руки не действует (не ощущается) сила реакции;
в) чем дальше от ЦТ центр удара, тем дальше полетит мяч после удара битой;
г) при контакте мяча и биты в центре удара последняя вращается относительно стационарной оси (точки) вращения.
73. Единицей измерения кинетического момента является:
а) (кг-рад)с-1;
б) (кг • м)с-2;
в) (кг - м2)с-1;
г) (кг-рад-м)с-1;
д) (кг • рад)с~2;
74. Что происходит после того как прыгун в воду оттолкнулся от доски, но еще не достиг поверхности воды?
а) сохраняется кинетический момент;
б) момент инерции постоянен;
в) на спортсмена не действует импульс;
г) сохраняется количество движения.
75. Что описывает взаимосвязь между импульсом и количеством движения?
а) влияние энергии на количество движения;
б) необходимость приложения силы, чтобы прекратить, начать или изменить движение;
в) изменение количества движения вследствие приложения силы в течение интервала времени;
г) влияние количества движения объекта на окружающую его среду;
д) изменение количества движения объекта, когда его перемещает какая-то сила.
76. Площадь под каким графиком иллюстрирует величину работы?
а) сила—время;
б) сила—скорость;
в) сила—перемещение;
г) сила—энергия.
77. При анализе движения всего тела меньше всего известно об:
В)
б) Еку,
в) Ек ь
г) EP,S-
78. Какое выражение лучше всего определяет понятие “эффективность”?
а) максимальное количество энергии, которую может образовать система;
б) выполнение работы с минимальными затратами энергии;
в) максимальное количество выполняемой работы на единицу используемой энергии;
г) максимальное количество работы, которую может выполнить система.
79. Какая из следующих форм энергии не существенна для движений человека?
118
а) кинетическая энергия поступательного движения;
б) упругая энергия;
в) теплота;
г) энергия деформации сдвига;
д) химическая энергия.
80. Что является единицей измерения деформации?
а) %;
б) мм;
в) рад;
г) Дж.
81. Укажите неправильное утверждение относительно работы и мощности.
а) выполнение работы сопряжено с затратами энергии;
б) ватт —- единица измерения мощности;
в) при эксцентрических сокращениях мышцы выполняют положительную работу;
г) при выполнении мышцей положительной работы она поглощает мощность;
д) мощность можно определить как интенсивность изменения энергии.
ЗАДАЧИ
82. Следующие системы находятся в состоянии равновесия, ускорение равно нулю, следовательно,
= 0;
^Fy = 0;
ZM() = 0.
Определить величину неизвестных сил.
83. Для каждой переменной укажите соответст-
вующую единицу измерения в системе СИ.
1. Потенциальная энергия (кг-м)с 1
2. Сила рад • с-1 Н-с
3. Импульс Н-м ватт
4. Угол кг (кг • м2)с-1
5. Вес Н Н-с'1
6. Количество движения м- с~2 Дж-с
7. Работа Дж-с 1 рад
8. Мощность градус Дж
84. Найти результирующую силу мышц—сгибателей локтевого сустава (Ли), при которой предплечье и кисть удерживались бы в горизонтальном положении. Вес предплечья и кисти — 30 Н, ЦТ системы расположен в 12 см от оси локтевого сустава (£) Результирующая мышечная сила приложена на расстоянии 5 см вдоль сегмента от Л и действует под углом 0,4 рад. Если систему дополнительно нагружают весом 100 Н, который удерживает кисть (в 25 см от оси локтевого сустава), какова должна быть результирующая мышечная сила (Ли), чтобы сохранить
85. Какой должна быть результирующая сила мышцы (Лп), действующая под углом 0,6 рад к предплечью—кисти и приложенная на расстоянии 5 см от оси локтевого (£) сустава, чтобы удержать предплечье—кисть под углом 0,2 рад над горизонтальной линией? Вес предплечья-кисти — 45 Н, точка приложения вектора веса находится на расстоянии 10 см от Е. Прежде чем приступить к вычислениям, нарисуйте диаграмму свободного тела системы. Какая результирующая мышечная сила (Fm) вызывает вращение и какая — стабилизацию?
86—87. На этом рисунке человек выполняет выпрямление ноги на тренажере “Цибекс”.
119
А. Нарисуйте диаграмму свободного тела, с помощью которой можно вычислить результирующий вращающий момент мышечных усилий относительно коленного сустава.
Б. Определите результирующий вращающий момент относительно коленного сустава, при котором испытуемый может удержать голень под углом 0,24 рад ниже горизонтальной линии, если: вес тела — 785 Н;
длина ноги
(от колена до лодыжки) — 39 см;
сила реакции (перпендикулярна голени) — 972 Н; точка приложения
силы реакции — 31 см от колена;
ЦТ конечности
(голень + стопа) — 41,3% длины голени от коленного сустава;
вес конечности
(голень + стопа) = 0,045 ВТ — 1,75.
88. Координаты х и у для нижней конечности
человека, вес которого 562 Н, таковы:
X У
Бедро 0,47 0,74
Колено 0,71 0,47
Лодыжка 0,63 0,09
Пальцы 0,73 0,00
Нанесите координаты на график и соедините их. Воспользовавшись уравнениями регрессии (табл.2.1), определите местонахождение центра тяжести нижней конечности. Затем воспользуйтесь уравнениями регрессии, приведенными в табл.2.5, и сравните полученные результаты.
89. Распространенный метод определения центра тяжести всего тела заключается в помещении испытуемого на жесткую доску спиной вниз и измерении силы реакции поверхности (7^) при известном расстоянии (х) от ступней. Воспользовавшись уравнением 3.3, получим выражение для определения положения ЦТ относительно ступней. Допустим, что вес испытуемого (Fw) 652 Н, рост — 172 см. Fgz — 311 Н, а точка ее приложения (х) находится на расстоянии 2,0 м от ступней. Определить положение ЦТ от ступней в процентах роста испытуемого.
90. Используя уравнения регрессии, приведенные в табл.2.1, и методику, приведенную в примере 4 раздела “Статический анализ”, определите положение ЦТ человека, вес которого 583 Н.
91. Человек, проходивший курс реабилитации после операции на колене, выполнял упражнение при неподвижном коленном суставе, вследствие чего ЦТ системы не испытывал линейного ускорения а. Вес голени—ступни (Fwd составил 120 Н, вес лодыжки {Fw^ — 250 Н.
А. Каков вращающий момент нагрузки в этом положении?
Б. Допустим, человек опустил ногу.
1. Какая группа мышц контролирует движение?
2. Какой максимальный вращающий момент (Тт) может произвести группа мышц, одновременно обеспечивая опускание ноги?
В. Допустим, момент инерции системы относительно коленного сустава (К) равен 0,11 кг-м. Какой вращающий момент (Тт) спортсмена обеспечит ускорение системы 9,4 рад • с-2? Подсказка-.
выполните динамический анализ, включая построение диаграммы свободного тела и ускорения.
92. На рисунке показано изменение результирующего вращающего момента относитель
но
но бедренного (сплошная линия), коленного (штриховая линия) и голеностопного (штрихпунктирная линия) суставов в опорной фазе бега со скоростью 4,5 м • с-1.
А. Почему пики вращающих моментов наступают в последовательности бедро-колено—голеностоп?
Б. Как на основании этих данных вычислить импульс вращающего момента?
Вращающий момент, Н м
93. Лухтанен (1988) измерял угловую скорость в момент контакта с мячом при выполнении подачи мяча в волейболе. Угловая скорость плеча, предплечья и кисти составила соответственно 7,18 и 38 рад-с-1. Допустим, линейная скорость плеча — 0,2 м•с-1и длина сегментов плеча — 26 см, предплечья — 25 и 7 см от запястья к центру площади контакта мяча с кистью. Определить линейную скорость кисти в момент соприкосновения с мячом.
94. Масса мяча для игры в гольф — 46 г. При ударе клюшкой его скорость изменяется от О до 60 м-с-1.
А. Как изменяется количество движения мяча?
Б. Продолжительность соприкосновения клюшки и мяча 0,5 мс.
Определить среднюю силу воздействия клюшки на мяч?
В. Если масса верхней части клюшки 200 г, а скорость клюшки перед соприкосновением с мячом 28 м • с-1, какова будет скорость верхней части клюшки после удара по мячу?
95. Передвигаясь по натянутому канату, акробат начинает падать назад. Согласно закону сохранения количества движения, в каком направлении (вперед или назад) он будет выполнять движения руками, чтобы сохранить равновесие? Объясните свой ответ.
Подсказка'.
постройте диаграмму распределения усилий акробата с вектором углового вращения для рук и отдельных частей тела).
96. Футбольный мяч весом 4,17 Н летел со скоростью 7,62 м • с-1, пока не соприкоснулся с головой игрока. После этого он полетел в противоположном направлении со скоростью 12,8 м-с-1. Если продолжительность соприкосновения мяча с головой футболиста была 22,7 мс, какая средняя сила воздействовала на мяч?
97. Согласно Стивенсону (1985), вертикальная скорость спортсмена массой 58 кг при вхождении в воду после прыжка с 10-метровой вышки уменьшается с 16,8 до 5,2 м • с-1 за 133 мс. Какая средняя сила сопротивления воды воздействует на пловца и вызывает такое изменение количества движения?
98. Вес бейсбольного мяча — 1,43 Н, а его скорость до того, как его поймает принимающий — 24,4 м/с. Какова кинетическая энергия мяча?
99. Прыгун с трамплина (вес 653 Н) стартует с верхней части трамплина, которая расположена выше площади отталкивания на 90 м. А. Определить скорость спортсмена в момент отталкивания. Трением и сопротивлением воздуха пренебречь.
Б. Дано: к = 0,55 кг/м3, А = 0,047 м2. Определить силу сопротивления воздуха сразу же после отталкивания.
100. Если допустить, что доска отталкивания равносильна линейной пружине, то количество упругой энергии, которую она может накопить, определяется уравнением 3.23. В результате отталкивания от доски (Зм) прыгуном, вес которого 575 Н, она прогибается на 0,76 м; при максимальном прогибании ее EAS равна 727 дж.
А. Какова величина “к” доски?
Б. Какова кинетическая энергия прыгуна в воду в момент отталкивания от доски?
В. Чему равна вертикальная скорость прыту-на в момент отталкивания от доски?
Г. На какую максимальную высоту переместится центр тяжести прыгуна в воду?
Д. Чему равна вертикальная скорость прыгуна в момент соприкосновения рук с поверхностью воды?
101. Ящик с оборудованием весом 89 Н находится на плоской поверхности. Допустим, что его толкают (Fp) с силой 396 Н.
121
А. Определить, какое количество работы необходимо выполнить, чтобы переместить ящик на 2 м вдоль горизонтальной поверхности в направлении приложенной силы.
Б. Допустим, |is = 1,4. Можно ли переместить ящик при Fp = 396 Н?
102. Пловец плывет кролем на груди с постоянной скоростью 1,4 м/с, сила сопротивления движения 93 Н. Интенсивность потребления кислорода пловцом 2,3 л/мин.
А. Если 1 л кислорода обеспечивает производство 20,9 Дж энергии, то какова интенсивность (Дж/с) производства метаболической энергии при данной интенсивности потребления кислорода?
Б. Какова эффективность плавания спортсмена?
В. Каковы единицы измерения эффективности?
103. Воэн (1980) получил следующие графики положения—скорости и ускорения—времени для центра тяжести акробата, выполняющего упражнение на батуте. Используйте данные графики для построения графика зависимости между мощностью и временем, использовав принцип, изложенный на рис.3.28. Построив график, объясните, что он означает. Спортсмен соприкасается с сеткой батута в промежутке между приземлением и отталкиванием.
Время, с
104. Мощность, производимая мышцей — важный фактор, характеризующий физическую деятельность.
А. Какова единица измерения мощности?
Б. Определите, что такое производство мощности. Каким образом колеблется величина производимой мышцами мощности?
В. Можно ли получить (просто!) алгебраическое выражение мощности как функции энергии. Если работа = Д энергии, какова взаимосвязь между мощностью и энергией?
ЧАСТЬ II
ЕДИНАЯ СИСТЕМА СУСТАВА
Движение человека представляет собой весьма сложное явление. Чтобы изучить нервно-механические основы движения, необходимо сделать ряд упрощенных предположений. Например, в части I “Взаимосвязь силы и движения” мы допустили, что сила, производимая мышцей, действует на неподвижные сегменты тела, проигнорировав сложность и многообразие структур и мест прикреплений. Это позволило рассмотреть некоторые основные понятия, связанные с изучением процесса движения.
Этот же подход используется и в части II, в которой тело человека описывается как система неподвижных или жестких звеньев, вращающихся с помощью мышц вокруг свободных от трения суставов. Активацию мышц контролирует и регулирует нервная система. Пять элементов (жесткое звено, синовиальный сустав, мышца, нейрон и чувствительное нервное окончание) составляют основной аппарат движения, образуя биологическую модель, которая называется единой системой сустава (рис. II. 1). Рис. II. 1 представляет собой лишь модель, включающую всего один нейрон. В действительности сотни нейронов иннервируют каждую мышцу, обеспечивая как двигательные команды (двигательные нейроны), так и сенсорную информацию (чувствительные нервные окончания). Точно так же вокруг каждого сустава находятся, как правило, группы мышц, каждая из которых осуществляет контроль движения в ограниченном количестве направлений.
В части II сначала рассмотрим упрошенное описание этих пяти элементов (глава 4 “Компоненты единой системы сустава”), затем остановимся на некоторых взаимодействиях между ними (глава 5 “Функционирование единой системы сустава”) и, наконец, рассмотрим свойства этой системы, позволяющие предельно точно контролировать мышечную силу (глава 6 “Активация единой системы сустава”).
123
ЦЕЛИ
Как уже отмечалось, цель данного учебника — описать движение в виде взаимодействия биологической модели (упрошенный вариант человека) с физическим миром. В части I мы изучили биомеханику движения, обратив внимание на взаимосвязь между силой и движением. Цель части II — определить биологическую модель и описать взаимодействия ее компонентов. К более узким целям относятся:
• описание единой системы сустава и объяснение того, почему она является адекватной моделью для изучения основных аспектов движения;
• подробное рассмотрение структурных характеристик пяти компонентов единой системы сустава;
• определение основной функциональной единицы системы;
• объяснение средств, обеспечивающих быструю передачу информации по системе;
• изложение теоретических основ методов контроля активации единой системы сустава;
• определение связи между нервным сигналом и мышечным сокращением;
• изучение нервно-анатомической основы и значения афферентной информации для функционирования системы;
• описание методов контроля мышечной силы нервной системой;
• определение механических свойств единиц, осуществляющих усилие;
• оценка влияния различного “архитектурного” строения единиц, осуществляющих усилие, на величину силы, производимой мышцей.
ГЛАВА 4
КОМПОНЕНТЫ ЕДИНОЙ СИСТЕМЫ СУСТАВА
В этой главе рассматриваются морфологические и механические характеристики компонентов единой системы сустава, а также их адаптационные способности. Поскольку все компоненты представляют собой живую ткань, то и адаптируются они в зависимости от потребностей, например, размер мышцы увеличивается в результате тренировок силовой направленности, прочность кости снижается во время пребывания в космосе, чувствительность нервных окончаний уменьшается с возрастом. В следующих главах рассматриваются взаимодействия между элементами данной системы, направленные на осуществление движения.
ЖЕСТКОЕ ЗВЕНО
Структурный каркас единой системы сустава образуют различные ткани, известные как соединительная ткань. Они включают живые (клетки) и неживые (внутриклеточный материал) вещества, которые “купаются” в тканевой жидкости. Клетки, например, фибробласты, макрофаги, жировые клетки, тучные клетки, выполняют функции, необходимые для обеспечения жизнедеятельности ткани. Внутриклеточный материал, образующий матрикс, в котором живут клетки, включает белковый коллаген, эластин и ретикулум, и определяет физические характеристики ткани. Основными соединительными элементами, образующими жесткое звено единой системы сустава, являются кость, сухожилие и связка.
Кость
Кость выполняет ряд функций, необходимых для осуществления движения:
а) обеспечивает механическую поддержку, являясь центральной структурой каждого сегмента тела;
б) образует эритроциты;
в) служит активным резервуаром ионов кальция и фосфора.
Кость — живая ткань, состоящая из белкового матрикса (в основном коллагена), на котором откладываются соли кальция (особенно фосфат). Эти минералы обеспечивают прочность кости. Вода составляет около 20 % сырой массы кости, белковый матрикс, представляющий собой в основном остеоколлагенные волокна, — около 35 %, соли — 45 %. Остеоколлагеновые волокна определяют силу и упругость кости. Основная структурная единица кости — остеон (или гаверсова система), состоящий из ряда концентрических слоев минерализованного матрикса, окружающего центральный канал, в котором находятся кровеносные сосуды и нервы. Обычный диаметр остеона — около 200 мкм. Хотя кость часто разделяют на губчатое вещество и кортикальный слой, их биомеханические свойства одинаковы и отличаются они друг от друга только степенью пористости и плотности.
Изучая биомеханику кости, ее рассматривают как материал, структуру и систему (Roesler, 1987). Механические свойства кости обычно характеризуются взаимосвязью нагрузка — деформация (глава 2). В этом случае на ткань воздействует нагрузка и измеряют имеющую место деформацию (изменение длины). Используя различную нагрузку (рис. 4.1.), можно определить такие свойства, как силу, жесткость и способность сохранять энергию. Вместо измерения абсолютной величины нагрузки и деформации, ученым следует стандартизировать эти измерения таким образом, чтобы биомеханические свойства можно было определять независимо от геометрии (например, размера) ткани.
125
Смещение Кручение Сжатие — кручение
Рис. 4.1. Виды нагрузок, которым может подвергаться такая ткань, как кость
Нагрузку стандартизируют в виде стресса (сила на единицу площади); единица измерения — максимально допустимое количество или MN/ м2. Деформацию определяют как приращение (изменение длины — как функцию начальной длины), которое является безразмерной величиной.
На рис. 4.2 приведены идеализированные кривые стресс — растяжение кортикального слоя кости, полученные в результате использования растягивающей и сжимающей нагрузки. Материальные свойства кости характеризуются измерением предела стресса (остРесс), т.е. стресса, при котором взаимосвязь изменяется от области упругих деформаций до области пластических деформаций, предельного стресса, или прочности кости (опр); критической деформации (екр); наклона области упругих деформаций (модуль упругости); наклона области пластических деформаций (модуль пластичности). На рис. 4.2 показано, как представлены эти свойства на кривой стресс-деформация максимальными показателями, точками перегиба и наклонами. Кроме того, энергию, которую может абсорбировать кость, измеряют как площадь под кривой. В табл. 4.1 приводятся типичные показатели этих параметров некоторых костей для разного возраста (Burstein, Reilly, Martens, 1976).
При изучении биомеханики кости следует рассматривать ее также как структуру. В этой связи форма и структура кости объясняются выполняемыми функциями и условиями окружающей среды, которые на них воздействуют. Например, во время бега трусцой со скоростью 2,2 м-с-1 большеберцовая кость человека больше деформируется при сжатии
(847 мкм-деформация), чем растяжении (578 мкм-деформация) (Lanyon, Hampson, Goodships, Shah, 1975). В исследованиях, проводившихся на животных, максимальная деформация возрастала при увеличении скорости локомоции. Кроме того, на бедренную кость чаще воздействовали продольно, чем перпендикулярно направленные силы. На длинные кости животных разных размеров во время локомоции, как правило, действует сжимающая сила порядка 40—80 МПа (Biewener, 1991). Именно поэтому бедренная кость характеризуется максимальным сопротивлением сжимающей нагрузки, минимальным — смещающей и средним — растягивающей. Как показывают измерения, прочность бедренной кости человека (опр) составляет 132 МПа при действии растягивающей силы вдоль длины кости (продольная) и всего 58 МПа при действии этой силы перпендикулярно кости. Точно так же прочность на сжатие бедренной кости составляет 187 и 132 МПа соответственно при продольном и перпендикуляром действии сил (Cowin, 1983). Абсолютная прочность кости колеблется в зависимости от величины стресса и деформации. Используя технические понятия, можно сказать, что кость имеет запас прочности 2—5, т.е. прочность костей в 2—5 раз превышает силы, воздействующие на них в повседневной деятельности человека (Alexander, 1984а; Biewener, 1991).
Рис. 4.2. Идеализированная взаимосвязь стресс-растяжение кортикального слоя кости человека при растяжении (а) и сжатии (6) (Carter, Spengler, 1978)
126
Таблица 4.1. Стресс-деформационные характеристики некоторых костей человека при воздействии силы растяжения (Burstein, Reilly, Martens, 1976)
Возраст, лет °сгресс> МПа о„р, МПа Модуль упругости, ГПа Модуль пластичности, ГПа ^кр Энергия, МПа
Бедренная кость
20-29 120 140 17,0 0,75 0,034 3,85
30-39 120 136 17,6 0,64 0,032 3,55
40-49 121 139 17,7 1,00 0,030 3,19
50-59 111 131 16,6 0,89 0,028 2,84
60-69 112 129 17,1 0,98 0,025 2,65
70-79 111 129 17,1 0,98 0,025 2,65
80-89 104 120 15,6 1,08 0,024 2,23
Большеберцовая кость
20-29 126 161 18,9 1,17 0,040 4,36
30-39 129 154 27,0 0,91 0,039 5,77
40-49 140 170 28,8 1,39 0,029 4,09
50-59 133 164 23,1 1,21 0,031 4,19
60-69 124 147 19,9 1,18 0,027 3,05
70-79 120 145 19,9 1,20 0,027 3,27
80-89 131 156 29,2 1,43 0,023 2,96
С позиций рассмотрения кости как структуры эти наблюдения подчеркивают, что функция оказывает основное воздействие на клеточную организацию и, следовательно, механические свойства кости. Поскольку изменяется структура зависимости стресс-растяжение, изменяются и механические свой-. ства, и клеточная организация кости. Закон Вольфа характеризует эту взаимосвязь следу-' ющим образом: “Каждое изменение... функции кости... вызывает определенные изменения... внутренней “архитектуры” и внешних параметров в соответствии с математическими законами” (Carter, 1984, с.1). В некоторых "исследованиях на основании этого закона предпринимались попытки установить соответствующие параметры, которые позволили бы описать структуру и геометрию кости. В других исследованиях внимание акцентировалось на процессах адаптации кости к условиям внешней среды, т.е. кость рассматривали как систему. Процессы, которым подвергается кость, включают развитие, укрепление и резорбцию. Они имеют собирательное название — ремоделирование, или реконструкция (Canyon, Rubin, 1984). Полный цикл ремоделирования (замены всех структур) костей конечности взрослого человека составляет около 10—20 лет (Alexander, 1984а).
Ремоделирование представляет собой равновесие между абсорбцией кости (остеокластами) и ее образованием (остеобластами). Оно постоянно изменяется и зависит от таких факторов, как физическая активность, возраст и заболевания. В научной литературе имеется сообщения о ремоделировании кости при изменении (увеличение или уменьшение) уровня
физической активности. Так, Shumski, Merten, Dzenis (1978) наблюдали более значительные отложения макроэлементов и повышенную плотность большеберцовой кости у спортсменов, чем у контрольных испытуемых. Dalen и Olsson (1974) обнаружили, что содержание макроэлементов в костяк бегунов по пересеченной местности (стаж выступлений 25 лет) было на 20 % выше в аппендикулярных участках (дистальный отдел лучевой кости, локтевая кость, пяточная кость) и менее чем на 10% выше в аксиальных участках (поясничный позвонок и головка бедренной кости), чем у контрольных испытуемых. Даже у тяжелоатлетов (опыт выступлений 6 лет) наблюдали повышенное содержание макроэлементов в основных опорных участках (поясничный отдел позвоночного столба, вертел, шейка бедренной кости) по сравнению с остальными участками тела (средняя часть лучевой кости) (Colletti, Edwards, Gordon et al., 1989). Содержание макроэлементов в костях может увеличиться уже после 6 недель физических тренировок (Beverly, Rider, Evans, Smith, 1989).
С другой стороны, главная проблема, с которой сталкиваются астронавты во время продолжительного пребывания в космосе, — потеря костной ткани (Zemicke, Vailis, Salem, 1990). Условия микроневесомости приводят к деминерализации костей, чрезмерной потере солей скелетом (Anderson, Cohn, 1985; Morey, 1979). В результате этого кости становятся менее прочными и во время значительной физической нагрузки (например, во время выполнения работ вне космического корабля) могут ломаться. Кроме того, при возвращении на Землю затрудняется процесс восстановления костей. Вслед
127
ствие этого НАСА проявляет большой интерес к разработке программ физических нагрузок, которые позволили бы свести к минимуму потери костной ткани у астронавтов. Как показывают результаты исследований, процесс ремоделирования костей более эффективно происходит при периодическом, а не постоянном воздействии физических нагрузок (Lanyon, Rubin, 1984). Кроме того, величины суставных сил в большей степени влияют на костную массу, чем количество циклов нагрузки (Whalen, Carter, Steele, 1988). Большинству из нас, естественно, не грозит потеря костной ткани вследствие пребывания в условиях невесомости, однако масса и прочность костей с возрастом снижаются (Snow-Harter, Marcus, 1991). Этот процесс называется остеопорозом, который характеризуется увеличением пористости костей, приводящей к снижению их плотности и прочности и, следовательно, повышению вероятности переломов.
Попытки определить механизмы, контролирующие ремоделирование, включали изучение электрических свойств кости (Bassett, 1968; Singh, Katz, 1989). Очевидно, если стресс достаточен, чтобы вызвать скольжение коллагеновых волокон кости друг относительно друга, в кости создаются электрические потенциалы. Образование электрических потенциалов вследствие давления получило название пьезоэлектрического эффекта кости. Предпринимались попытки установить взаимосвязь между механической нагрузкой на кость и связанными с ней электрическими эффектами. В частности, было продемонстрировано, что слабые электрические токи могут вызвать образование костной мозоли (Marino, 1984). Одним из практических последствий этого открытия явилось использование электрической и магнитной стимуляции для ускорения лечения переломов, особенно плохо срастающихся (Brighton, 1981). Некоторые ученые применяли электростимуляцию кости даже с целью отсрочить начало развития остеопороза.
Сухожилие и связка
Являясь соединительными элементами, сухожилия соединяют мышцы и кости, а связки обеспечивают соединения между костями. Главное различие между ними состоит в организации коллагеновых фибрилл, которая обусловлена их функциями. Поскольку функция сухожилия заключается в сообщении мышечной силы кости или хрящу, структура сухожилия обеспечивает его наименьшую подверженность деформации вследствие растягивающих усилий, производимых мышцей. Наоборот, очень незначительное продольносжимающее и смещающее
усилие может деформировать сухожилие. Хотя на связки действуют главным образом растягивающие силы, их главная функция — стабилизация сустава, поэтому они устроены так, чтобы обеспечивать многонаправленную стабильность сустава, и приспособлены к действию растягивающих, сжимающих и смещающих усилий (Fu, Hamer, Johnson et al., 1993).
Сухожилия и связки представляют собой плотную соединительную ткань, содержащую коллаген, эластин, протеогликаны, воду и клетки (фибробласты). Протеиновые коллаген и эластин синтезируются и выделяются фибробластами. Приблизительно 70—80% сырой массы сухожилия и связки составляет коллаген типа I — волокнистый белок, характеризующийся значительной механической устойчивостью (Buigeson, Nimni, 1992). Коллагеновая фибрилла (рис. 4.3) — основная несущая нагрузку единица как сухожилия, так и связки. Структура фибриллы от альфа-цепочек тройной спирали до скопления молекул коллагена в микрофибрилле одинаковая у сухожилия и связки. Главное различие между этими двумя соединительными элементами состоит в расположении фибрилл. В сухожилии они расположены продольно и параллельно друг Другу, что обеспечивает максимальное сопротив-
Рис. 4.3. Иерархическая структура сухожилия до уровня тройной спирали коллагена:
1 — перитендиний; 2 — сухожилие; 3 — пучок;
4, 6 — фибрилла; 5,1— микофибрилла; 8— молекула коллагена;
9 — тройная спираль
128
ление растягивающим усилиям. В связке фибриллы, как правило, располагаются параллельно, а также косо и спиралевидно, что обеспечивает сопротивление силам, действующим в различных направлениях.
Каждая молекула коллагена состоит из трех взаимопереплетенных полипептидных цепочек. Каждая цепочка включает последовательность из около 1000 аминокислот (главным образом пролин, гидроксипролин и глицин) и называется альфа-цепочкой. Молекулы коллагена располагаются конец к концу (рядами) и пять таких рядов, расположенных параллельно (бок о бок), образуют микрофибриллу. Коллагеновая фибрилла, являющаяся основной несущей нагрузку единицей сухожилия и связки, состоит из пучков микрофибрилл, соединенных биохимическими связями (поперечными сцеплениями) между молекулами коллагена; эти поперечные сцепления находятся как в пределах, так и между рядами молекул коллагена в микрофибрилле (рис. 4.3). Поскольку поперечные сцепления удерживают микрофибриллы вместе, существует мнение, что их количество и состояние значительно влияют на силу соединительной ткани (Bailey, Robins, Balian, 1974). Таким образом, функциональной основой сухожилия и связки является фибрилла — ряд единиц (микрофибрилл), соединенных друг с другом благодаря поперечным сцеплениям. От последних зависит сила фибриллы. Считают, что количество и состояние поперечных сцеплений определяются такими факторами, как возраст, пол и уровень физической активности.
Кроме коллагена и эластина, внеклеточный матрикс сухожилия и связки включает протеогликаны и воду. Вода соединяется с протеогликанами, образуя гель, вязкость которого понижается с повышением уровня физической активности. Это свойство называется тиксотропией. Сопротивление ткани удлинению при данной скорости зависит от ее вязкости: чем выше вязкость, тем больше сопротивление растяжению. При снижении вязкости ткань приспосабливается к растяжению с более высокой скоростью. Вязкость тиксотропической ткани изменяется в результате предшествующей деятельности, например, разминки или продолжительного периода бездеятельности.
Биомеханические свойства сухожилия и связки часто характеризуют взаимосвязью нагрузка-деформация в ответ на растягивающую нагрузку (рис. 4.4). Как показывают клинические наблюдения, соединительная ткань чаще разрывается, чем отрывается от кости. На рис. 4.4 показаны колебания пика силы и степени растяжения различных образцов. Например, сухожильно-костный образец срединной части надколенника растягивался на 10 мм с пиком растягивающего усилия (^стресс) порядка 3 кН до разрыва, тогда
как связочно-костный образец передней крестообразной связки растягивался на 15 мм, производя пик растягивающего усилия около 1,5 кН. Полученные результаты показывают, что сухожильно-костный образец срединной части надколенника прочнее, чем связочно-костный образец передней крестообразной связки, и оба они прочнее, чем тонкое сухожилие и широкая фасция (рис. 4.4).
При выражении нагрузки относительно единицы площади поперечного сечения, а деформации — как процента начальной длины биомеханические свойства тканей можно сравнивать как взаимосвязь стресс—растяжение. На рис. 4.5 показана идеализированная взаимосвязь стресса—растяжения коллагеновой ткани, такой, как сухожилие и связка. Данная взаимосвязь охватывает три участка: нижний, линейный и участок разрыва. Нижний участок соответствует начальному этапу взаимосвязи, когда растягиваются коллагеновые волокна,
Рис. 4.4. Взаимосвязь нагрузка—деформация образцов соединительной ткани, растянутых до разрыва (Butter, Grood, Zernicke, Hefry, 1984):
1 — тонкое сухожилие; 2 — широкая фасция; 3 —сухожилие срединной части надколенника; 4 — передняя крестообразная связка
9 8-133
129
Рис. 4.5. Идеализированная взаимосвязь стресс—растяжение коллагеновой ткани. До определенного повреждения длина ткани изменяется лишь незначительно (Butler, Grood, Noyes, Zemicke, 1978)
имеющие зигзагообразную структуру в состоянии покоя. Линейный участок характеризует эластичную способность ткани; наклон в этом участке называется модулем упругости и в более жестких тканях более крутой. За этим участком крутизна наклона уменьшается, поскольку некоторые волокна разрываются в участке повреждения. При таком растяжении в соединительной ткани происходят пластические изменения, кроме того, изменяется ее длина в состоянии покоя.
На основании взаимосвязи стресса—растяжения можно определить предельный стресс (опр), критическое растяжение (екр), модуль упругости и абсорбируемую энергию ткани (площадь под кривой стресса—растяжения). Эти свойства снижаются вследствие уменьшенного использования (например, иммобилизации), с возрастом, а также при употреблении стероидов и повышаются при постоянных физических нагрузках (Butter, Grood, Noyes, Zemicke, 1978; Noyes, 1977). Кроме того, свойства сухожилия изменяются в зависимости от функции мышцы (Shadwick, 1990).
СИНОВИАЛЬНЫЙ СУСТАВ
В теле человека насчитывается около 206 костей, которые образуют почти 200 суставов. Суставы обычно разделяют на три группы: 1) волокнистые — относительно неподвижные (например, швы черепа, межкостная мембрана между лучевой и локтевой костью или между
большеберцовой и малоберцовой); 2) хрящевые — слегка подвижные (грудино-реберные, межпозвонковые диски, лобковый симфиз); 3) синовиальные, которые свободно двигаются (локтевой, бедренный). Поскольку синовиальный сустав больше всего соответствует свободному от трения суставу согласно нашему предположению о жестком звене, именно он берется в качестве суставного компонента единой системы сустава.
Синовиальный сустав выполняет две функции: обеспечивает подвижность скелета в результате вращения одного сегмента тела относительно другого и передает усилие от одного сегмента к другому. Эти взаимодействия, включающие соприкосновение соседних костей, контролируются рядом структурных единиц, к которым относятся суставной хрящ, суставная капсула, синовиальная мембрана и геометрия костей. Поверхность костей, образующих сустав, выстлана суставным хрящем — плотной соединительной тканью белого цвета, не имеющей кровеносных сосудов, лимфатических протоков и нервов. Суставной хрящ содержит хондроциты — клетки, обеспечивающие органический компонент внеклеточного матрикса, и плотную сеть тонких коллагеновых фибрилл в концентрированном растворе протеогликанов. Основным компонентом суставного хряща является вода, большая часть которой находится вблизи поверхности. Коллагеновые фибриллы, протеогликаны и вода определяют биомеханическое поведение суставного хряща.
Суставной хрящ защищен двумя видами смазки (Моу, Proctor, Kelly, 1989): граничной, или тонкопленочной, смазкой — поглощение гликопротеид лубрицина поверхностью суставного хряща и жидкостной гидродинамической смазкой — тонкой пленкой смазочного вещества, разделяющего сочлененные поверхности. Граничная смазка, по-видимому, играет важную роль, когда контактирующие поверхности подвергаются значительным нагрузкам продолжительное время. Наоборот, жидкостная гидродинамическая смазка играет важную роль при небольших нагрузках, когда контактирующие поверхности с высокой скоростью двигаются относительно друг друга. Синовиальные суставы обладают способностью самосмазы-ваться: смазочная жидкость покрывает контактирующие поверхности, а при отсутствии нагрузки реабсорбируется (Моу et al., 1989). Смазочная жидкость, очевидно, служит для обеспечения хондроцитов питательными веществами.
Функция суставного хряща — обеспечение относительного движения поверхностей противоположных суставов с минимальным трением и изнашиванием, а также видоизменение формы кости, направленное на обеспечение лучшего контакта с соседней костью (Armstrong,
130
Таблица 4.2. Движение в основных суставах человеческого тела
Сустав Сочлененные поверхности Степень свободы Диапазон движения, рад
Позвоночный СТО Гб Атлангскйтылочная; I и II шейные позвонки СЗ-С7 Грудная Поясничная Сгибание разгибание Латеральное сгибание Сгибание—разгибание Вращение (продольное) Сгибание—разгибание Латеральное сгибание Вращение (продольное) « Сгибание—разгибание Латеральное сгибание Вращение (продольное) Сгибание —"гз'иба ие Латеральное сгибание Вращение (прод< и ьное) 0,3 0.1 0,2 0,8 0,7 сгибание, 0.4 разгибание J ' 1,5 0,06 0,2 0,06—0,16 0Д6 0,3 0,1 0,03
Плечевой Плечевая Акромиально-ключичная Грудино-ключичная Лопаточно-грудная Сгибание (сагиттальная плоскость) Разгибание (сагиттальная плоскость) Сгибание (поперечная плоскость) Выпрямление (поперечная плоскость) Отведение (передняя плоскость) Внутренне-внешнее вращение 3,14 0,9 2,4 0,9 3,14 3,14 (рука отведена и локоть при 1,57 рад)
Локтевой Плечелоктевая Плечелучевая Проксимальная лучелоктевая Плечелучевая Сги >ание—разгибание Пронация—супинация 2,4 1,2 пронация 1,5 супинация
Запястный Лучезапястная Межзапястная Сгибание—разгибание Лучелоктевое смещение 1,5 сгибание 1,4 разгибание 0,3 лучевое 0,6 локтевое
Бедренный Головка бедра-вертлужная впадина Сгибание—разгибание Отведение—приведение Внутренне—внешнее вращение 2,4 сгибание 0,3 разгибание 0,5 Отведение 0,4 приведение 1,2 внутреннее 1,5 внешнее
Коленный Тибиально-бедренная Сгибание—разгибание Внутренне-внешнее вращение Отведение—приведение 2,4 0,7 внутреннее 0,5 внешнее (колено — 1,57 рад) <0,1 (колено — 0,5 рад)
Голеностопный Большеберцовой । таранной костей Малоберцовой и таранной костей Дистальной большеберцовой и малоберцовой костей Сгибание—разгибание 0,2—0,4 сгибание назад 0,4—0,6 плантарное сгибание
Стопы Субтаранная Межплюсневая и предплюсне-плюсневая Плюснефаланговая Инверсия—эверсия Сгибание назад, плантарное сгибание Сгибание—разгибание 0,3 инверсия, 0,1 эверсия <0,1 сгибание назад 0,3 плантарное сгибание 0,5 сгибание 1,5 разгибание
Примечание. Данные диапазона движения представляют средние показатели молодых людей.
9*
131
Moy, 1980; Askew, Moy, 1978). В этой связи суставной хрящ действует подобно губке, впитывающей воду. Суставной хрящ представляет собой вязкопластичный материал. Это значит, что при постоянной нагрузке или деформации его реакция (механическое поведение) изменяется со временем. Этот процесс может включать изменение толщины хряща вследствие поступления жидкости, обусловленного нагрузкой. У более физически активных людей толщина суставного хряща больше; она увеличивается при переходе от состояния покоя к активному состоянию. Суставной хрящ может индивидуализироваться, например, в височно-нижнечелюстном и коленном суставах могут образоваться внутрисуставные диски или мениски, увеличивающие контактирующую площадь сочлененных поверхностей. Суставной хрящ выдерживает нагрузки благодаря коллаген-протеогликан-ному матриксу и сопротивлению потока жидкости через матрикс.
Сочлененные поверхности синовиального сустава заключены в суставную капсулу, прикрепленную к костям сустава, которая отделяет суставную полость от окружающих тканей. Внутренняя часть капсулы, а также участки костей, не покрытые суставным хрящем, выстланы синовиальной мембраной — васкулярной мембраной, выделяющей синовиальную жидкость в суставную полость. Эта жидкость смазывает и обеспечивает питательными веществами суставной хрящ. Суставная капсула — свободная структура, окружающая весь сустав; в некоторых местах она смыкается с околосуставными связками, которые, как считают, обеспечивают более близкое расположение сочлененных поверхностей. Как и суставной хрящ, суставная капсула и связки адаптируются к изменению структуры деятельности. Например, наибольшая опасность, обусловленная иммобилизацией суставов, заключается в адаптационных реакциях суставной капсулы и связок. Известно, что соединительная ткань имеет тенденцию адаптироваться к минимальной функциональной длине; при иммобилизации сустава суставная капсула и связки сокращаются (сжимаются), и синтезируется новая ткань, которая аккомодируется к более короткой длине. Эти изменения ограничивают подвижность сустава. При подобной иммобилизации может развиться остеоартрит (Videman, 1987).
Геометрия синовиальных суставов (т.е. архитектура сочлененных поверхностей, строение связок) во многом определяет качество движения между двумя соседними сегментами тела. Синовиальный сустав обеспечивает вращение вокруг одной — трех осей. Эти оси могут проходить через сустав с одной стороны на другую, с конца в конец и спереди назад. Каждая ось характеризуется степенью свободы. Для плечелучевого сустава характерно лишь одно
движение в плоскости сгибание — разгибание (Engin, Chen, 1987), т.е., вращение вокруг оси, проходящей через сустав с одной стороны в другую. В то же время для бедренного и плечевого суставов характерны три степени свободы (Engin, Chen, 1986, 1988; Hogfors, Sigholm, Herberts, 1987), т.е. структура сустава позволяет ему выполнять вращения вокруг каждой оси. Движения в этих суставах могут выполняться в плоскости сгибания— разгибания, отведения-приведения, а также вокруг продольной оси. В табл. 4.2. приведен краткий перечень возможных движений в некоторых суставах человеческого тела.
МЫШЦА
Благодаря протекающим в мышцах процессам происходит превращение химической энергии продуктов питания в силу. Их свойства включают: а) раздражимость — способность реагировать на стимулы; б) проводимость — способность передавать волну возбуждения; в) сократимость — способность изменять длину и г) адаптивность — ограниченный рост и регенеративная способность. Гистологически мышцы делятся на сердечную, гладкую и скелетную. Только последняя используется в анализе движения единой системы сустава. Сердечная мышца, составляющая большую часть перегородки сердца, представляет собой сеть отдельных клеток, имеющих полосы. Гладкая мышца, которая является основой стенок некоторых внутренних органов, а также кровеносных сосудов, состоит из отдельных клеток, разделенных тонкой щелью и не имеющих поперечных полос. Скелетная мышца состоит из соединительных клеток, имеющих четкие поперечные полосы (рис. 4.6). За исключением некоторых мимических мышц, скелетные мышцы, действуя поперек суставов, обеспечивают вращение сегментов тела. Следующие комментарии относятся исключительно к скелетной мышце и ее роли как производителя движения. Задача мышцы в единой системе сустава — образование силы, взаимодействующей с силами окружающей среды и обеспечивающей вращение жестких звеньев.
Мышца содержит множество элементов. После того, как вы получите достаточное представление об этой системе, обратите внимание на функции мышцы, а также на процессы, обеспечивающие выполнение этих функций. В частности, на элементы, связанные с двумя важнейшими процессами: взаимосвязью между сарколеммой и саркоплазматическим ретикулумом и компонентами саркомера. Эти процессы тесно связаны со взаимодействием между нервной системой и мышцей (сарколемма— саркоплазматический ретикулум), а также
132
Рис. 4.6. Структура скелетной мышцы от макроскопического уровня до молекулярного. (Pitman, Peterson, 1989):
а — целая мышца; б — группа мышечных волокон; в — одна миофибрилла; г — саркомер; д — один тонкий и один толстый филаменты
действуют как единая функциональная система. Иногда ее называют мышечно-сухожильной единицей; в данном учебнике понятие “мышца” включает как сократительную, так и связанную с ней соединительную ткань (в том числе и сухожилие).
Длина мышечных волокон колеблется от 1 до 4 мм, диаметр — от 10 до 60 мкм. Клеточная мембрана вокруг каждого ряда миофиламентов, включающая в себя мышечное волокно, называется сарколеммой. Подобно мембране плазмы сарколемма обеспечивает активный и пассивный избирательный транспорт — важнейшее свойство возбудимых мембран. Благодаря этому свойству сарколемма некоторым соединениям позволяет проходить сквозь нее (пассивный транспорт), а некоторым помогает проходить (активный транспорт). Толщина сарколеммы — около 7,5 нм. Жидкость, заключенная в волокне с помощью сарколеммы, называется саркоплазмой. В саркоплазме содержатся источники энергии (липидные капельки, гранулы гликогена), органеллы (ядра, митохондрии, лизосомы), ферменты (миозин, АТФаза, фосфорилаза), сократительный аппарат (пучки миофиламентов, расположенные в миофибриллах).
Кроме того, саркоплазма имеет широкую мембранную систему, которая функционально связана с поверхностью сарколеммы и помогает мышцам передавать сигналы из нервной системы. Эта мембранная система включает саркоплазматический ретикулум, латеральные мешочки (терминальные цистерны) и поперечные (Т) трубочки (рис. 4.7). Саркоплазматический ретикулум идет продольно вдоль волокна, параллельно миофибриллам, которые он окружает. В определенных участках, расположенных вдоль миофибриллы, саркоплазматический ретикулум расширяется в латеральные мешочки.
величиной усилия, производимого мышцей (саркомер). Более подробно эти процессы рассматриваются в главе 5.
Макроструктура
Мышечные волокна тесно связаны между собой трехступенчатой сетью коллагеновой соединительной ткани. Эндомизий окружает отдельные мышечные волокна, перимизий заключает пучки волокон в “гроздья”, а эпимизий “окутывает” всю мышцу (рис. 4.6 а, б). Этот матрикс соединительной ткани соединяет мышечные волокна с сухожилием и, следовательно, со скелетом (Tidball, 1991). Благодаря такой взаимосвязи мышечные волокна и соединительная ткань (в том числе и сухожилие)
Рис. 4.7. Расположение поперечных трубочек и саркоплазматического ретикулума относительно миофибрилл. На рисунке показаны 6 миофибрилл отдельного мышечного волокна (Bloom, Fawcett, 1968)
133
Перпендикулярно саркоплазматическому ретикулуму размещены поперечные трубочки, связанные с латеральными мешочками, которые представляют собой разветвленные инвагинации сарколеммы. Понятие триада относится к одной поперечной трубочке и двум терминальным цистернам саркоплазматического ретикулума, каждая из которых находится по обеим сторонам Т-трубочки. Как мы увидим дальше, эта триада способствует осуществлению быстрой связи между сарколеммой и сократительным аппаратом.
Саркомер
Волокна скелетной мышцы можно рассматривать как серию повторяющихся единиц с одинаковой характерной структурой. Гистологически такая единица определяется как саркомер и представляет зону миофибриллы от одной Z-линии до другой (рис. 4.6, г и 4.7). Миофибрилла — это серия саркомеров, соединенных концами друг с другом. Саркомер — основная сократительная единица мышцы, состоящая из набора толстых и тонких сократительных белков (рис. 4.6, г). Поскольку длина саркомера в мышце, находящейся в покое, составляет около 2,5 мкм, в 10-миллиметровой миофибрилле содержится 4000 саркомеров, соединенных концами друг с другом. Каждая миофибрилла состоит из пучков миофиламентов (тонких и толстых сократительных белков). Ее диаметр не превышает 1 мкм. Во многих мышцах довольно трудно идентифицировать миофибриллы, поэтому целесообразно говорить о миофиламентах (Hoyle, 1983).
Очевидная “полосатость” скелетной мышцы обусловлена дифференциальной рефракцией света, проходящего через сократительные белки. Зона толстых филаментов (рис. 4.6, в), включающая некоторые разобщенные тонкие филаменты, характеризуется двойной рефракцией (т.е. образованием двух преломленных лучей света из одного падающего) и имеет темную (анизотропную) линию — A-диск. В этой линии имеется зона, состоящая только из толстых филаментов. Поскольку в ней нет тонких филаментов, ее называют Н-зоной (светлая линия). Участок между A-дисками состоит, в основном, из тонких филаментов, и поскольку он характеризуется однократной рефракцией, его называют 1-диском (изотропной линией).
Каждая группа филаментов (тонких и толстых) прикрепляется к центральной поперечной полоске: толстые филаменты — к М-зоне (среднему диску — линии, расположенной в середине А-диска), тонкие филаменты — к Z-линии. На поперечном срезе A-диска видно, что каждый толстый филамент (миозин с шес
тью выступающими вперед поперечными мостиками) окружен шестью тонкими филаментами, тогда как каждый тонкий филамент — только тремя толстыми филаментами.
Миофиламевты
Миофиламент включает тонкие и толстые филаменты, которые состоят из ряда белков (рис. 4.8). В структуре такого филамента преобладает актин, а также регуляторные белки — тропомиозин и тропонин. Каждый тонкий филамент состоит из двух спиральных пучков волокнистого актина — F-актина (рис. 4.8, а). Каждый пучок F-актина представляет собой полимер (химическое соединение двух или более молекул) порядка 200 молекул глобулярного актина — G-актина (рис. 4.8, б). Молекула G-актина — белок, содержащий около 374 аминокислот.
Рис. 4.8. Структура миофиламентов (Whalen, 1985): a — тонкий филамент; б — элементы тропонина (TH); в — фрагменты молекулы миозина; г — легкие и тяжелые цепочки миозина; остальные пояснения в тексте
134
В углублении спирали F-актина находятся два переплетенных пучка тропомиозина (рис. 4.8, а, б). Структура тропомиозина представляет собой скрученную спираль двух цепочек, каждая из которых содержит около 284 аминокислот. Тропониновый комплекс (ТК) имеет шарообразную структуру, включающую три единицы (рис. 4.8, б); ТК—Т-единица связывает тропонин с тропомиозином, ТК-1 препятствует присоединению к миозину 4—7 молекул G-актина при наличии тропомиозина, ТК-С вновь присоединяет ионы Са2+ (функция концентрации кальция). ТК-С имеет четыре участка связывания — два для Са2+ и два для Са2+ или Mg2+.
Таким образом, основу тонкого филамента составляют два пучка актиновых молекул (F-актин), над которыми, окутывая и прикрепляясь, располагаются двупучковые (тропомиозин) и шарообразные (тропонин) белки. Их функции при мышечном сокращении практически одинаковы; тропомиозин и тропонин влияют на активность актина.
Набор тонких филаментов, продольно выступающих и входящих в один саркомер, соединяется в участке Z-линии с другим таким же набором, который выступает в противоположном направлении и входит в соседний саркомер. В месте этого соединения каждый тонкий филамент соединяется с четырьмя своими ближайшими “соседями”. Участок характеризуется значительной гибкостью, Z-линия изменяет свою форму в различных условиях. Ширина Z-линии различна в различных типах мышечных волокон, и, по-видимому, изменяется вследствие физических тренировок (Sjostrom, Angquist et al., 1982; Sjostrom, Kidman, Larsen, Augquist, 1982).
Из белков толстого филамента больше всеизвестно о молекуле миозина (Cooke, 1990). Она представляет собой длинный, состоящий из двух цепочек, спиралевидный хвост, который заканчивается двумя большими шарообразными головками (рис. 4.8, в). С помощью фермента (протеазы) молекулы миозина могут расщепляться на легкие меромиозиновые (ЛММ) и тяжелые меромиозиновые (ТММ) фрагменты, кроме того, последние могут разделяться на субфрагменты 1 и 2 (С1 и С2). У ЛММ-фрагмента относительно небольшая молекулярная масса (135 килодальтонов, кДа), тогда как молекулярная масса ТММ-фрагмента больше 335 кДа. Шарообразные головки молекулы миозина, одна из которых содержит АТФ-, а другая актинсвязующую часть, представляют собой С1 (115 кДа). Остальная часть ТММ-фрагмента образует С2 (60 кДа).
Поскольку ЛММ-фрагмент соединяется только с себе подобными, около 400 миозиновых молекул, объединяясь, образуют доминирующий элемент толстого филамента (Рере,
Drucker, 1979). Это объединение характеризуется четкой структурностью. Молекулы расположены парами и С1-элемент каждой молекулы ориентирован на своего “партнера” под углом 3,14 рад (180°)- Следующая пара смещается перемещением на 0,0143 мкм и вращением на 2,1 рад (120°). В результате получается упорядоченное расположение молекул миозина, при котором выступы (поперечные мостики) ТММ окружают толстый филамент (рис. 4.6, д). Каждый саркомер обычно включает два таких набора молекул миозина; вместе с тем, поскольку С1-элементы двух наборов молекул миозина направлены в противоположные стороны, ЛММ-фрагменты, соединяясь в М-по-лоске (рис. 4.6, в), образуют отдельный филамент.
В молекуле миозина есть два участка, характеризующиеся относительно большей степенью гибкости. Они находятся в местах соединения ЛММ—ТММ и Cl—С2. Вследствие этого ТММ-фрагмент может простираться от толстого филамента до тонкого (рис. 4.6, д). Благодаря способности С1 взаимодействовать с актином это выпячивание фрагмента ТММ называют поперечным мостиком. Каждый толстый филамент окружен и взаимодействует с шестью тонкими филаментами, поскольку его окружают поперечные мостики.
В четырехглавой мышце бедра человека на 1 мкм2 содержится около 1600 толстых филаментов (Claassen, Gerber, Hoppeler, et al., 1989).
Белки, образующие сократительный аппарат, можно рассматривать как производные
Таблица 4.3. Изоформы сократительных белков в скелетных мышцах взрослого человека
Семейство генов Скелетная мышца
медлен носокра-1 дающаяся быстросокра-щающаяся
Тяжелая цепоч- м Бгд, Б2Г>, Б2Х,
ка миозина t>i;o, Ьсг>
Легкая цепочка 1ма, 1мб 1 г» Зб
щелочи
Легкая цепочка 2м> 2б
дитионитробен-
зойной кислоты
Актин «sk Gsk
Тропомиозин a, ps a,ps
Тропонин С М Б
I М Б
Т М Б
Примечание. М — медленносокращающиеся; Б — быстросокращающиеся Б2д; Б2Б — два быстросокра-Шающихся волокна, определенные с помощью гистохимии; Б2Х — определены с помощью окрашивания антитела и белкового анализа; БЕо — обнаружены во внеглазной мышце взрослого человека; Бсб — супербыстрые сократительные белки мышцы челюсти.
135
различных генов. Поскольку в состав белков входят цепочки аминокислот, белок с высокой молекулярной массой (200 кДа) называют тяжелой цепочкой. Наоборот, белок с низкой молекулярной массой (меньше 30 кДа) называют легкой цепочкой. Такой белок со слегка отличающимся составом аминокислот называют изоформой; т.е. могут быть различные соформы (составы аминокислот) одного и того же белка, который является или не является производным различных генов (Babij, Booth, 1988). Изоформы иногда называют изоферментами. Существуют различные изоформы миозины тяжелой и легкой цепочки (табл. 4.3). Следует отметить изменения сократительных белков в процессе развития, а также в результате изменения уровня физической активности.
Молекула миозина состоит из двух скрученных тяжелых цепочек, легкие цепочки прикрепляются к миозиновым головкам (рис. 4.8, г). В скелетной, сердечной и гладкой мышцах изоформы отличаются. Тяжелые цепочки скелетной мышцы имеют молекулярную массу 200 кДа. Существует тесная взаимосвязь между максимальной скоростью сокращения мышечных волокон и активностью АТФазы изоформы тяжелой цепочки, содержащейся в волокне. Существует одна медленная и пять быстрых изоформ тяжелой цепочки (табл. 4.3). Четыре легкие цепочки прикреплены к шарообразным головкам, они отличаются молекулярной массой (16, 18—20 и 25—27 кДа), способностью к фосфорилированию и соединениям (щелочь или дитионитробензойная кислота), которое
отделяет их от тяжелой цепочки. Как и в случае с тяжелыми цепочками, существуют различные изоформы легких цепочек быстро- и медленносокращающихся мышечных волокон (табл. 4.3). Легкие цепочки, по-видимому, модулируют взаимодействие между актином и миозином, их функции пока недостаточно хорошо изучены.
Цитоскелет
Ряд структур, обеспечивающих физическую основу взаимодействия сократительных белков, получил название цитоскелета (Cooke, 1985). Цитоскелет состоит из двух решетчатых структур: экзосаркомерный цитоскелет обеспечивает латеральное расположение миофибрилл, ориентацию толстых и тонких филаментов в саркомере (Waterman-Storer, 1991).
Экзосаркомерный цитоскелет состоит из промежуточных филаментов, расположенных продольно и поперечно саркомерам (рис. 4.9). Промежуточные филаменты, состоящие из таких белков, как десмин, виментин и синемин, локализованы в Z-линиях и присоединяют каждую миофибриллу к соседней миофибрилле и сарколемме. Эндосаркомерный цитоскелет вместе с актином и миозином выполняет функцию третьей системы филаментов. Система включает белки тинин и небулин. Первый обеспечивает эластичность мышц в состоянии покоя, второй создает решетчатую структуру актина (Waterman-Storer, 1991).
Рис. 4.9. Структура цитоскелета скелетной мышцы. Филаменты цитоскелета соединяют соседние саркомеры (толстые и тонкие миофиламенты) друг с другом рядами и параллельно, а также с внеклеточным матриксом соединительной ткани (Thomell, Price, 1991; Waterman, Storer, 1991)
136
НЕЙРОН
Мы рассмотрели три элемента единой системы сустава: жесткое звено, образующее структурную основу системы, сустав, вокруг которого вращаются жесткие звенья, и орган — мышцу, передающий усилие жестким звеньям. Теперь мы рассмотрим нервную систему и ее клеточные компоненты, которые активируют мышцу.
Несмотря на продолжительные исследования, о функции нервной системы известно мало. Вполне естественно, что больше известно об относительно простых функциях, например, таких, как рефлексы, и значительно меньше — о более сложных процессах, таких, как изучение или контроль движения. Больше известно о свойствах отдельных нервных клеток, чем о поведении групп клеток. Наша модель (единая система сустава) включает всего несколько рудиментарных элементов нервной системы, поэтому мы можем акцентировать свое внимание на основных свойствах, имеющих отношение к контролю движения.
Существует всего два вида нервных клеток — нейроны и нейроглия. Нейрон характеризуется четкой клеточной формой, имеет внешнюю мембрану (аксолемму), способную образовывать и проводить электрический сигнал, а также уникальную структуру (синапс) для передачи информации. О нейроглиях, которых в 9 раз больше, чем нейронов, известно немного. Считают, что они обеспечивают структурный и метаболический “ремонт” после травм, а также защищают нейроны (Somjen, 1987; Varon, Somjen, 1979). Например, три основные функции, которые осуществляют или осуществлению которых содействуют глиальные клетки, — это миелинизация, фагоцитоз и метаболизм. Миелинизацию осуществляют олигодендроциты в центральной нервной системе (ЦНС) и шванновские клетки в периферической нервной системе. Во время этого процесса поверхностная мембрана глиальной клетки (олигодендроцит, или шванновская клетка) “окутывает” аксон — ответвление нейрона, принимающее участие в передаче команд (рис. 4.10). Одним из последствий этой миелинизации является то, что команда, посылаемая нейроном, идет значительно быстрее. При фагоцитозе глиальные клетки (микроглия) пролиферируют вокруг поврежденных нейронов (травмированных или дегенеративных) и превращаются в большие макрофаги, которые поглощают остатки органических веществ. Вклад нейроглии в метаболизм состоит в модуляции ионов, нейромедиаторов и метаболитов, необходимых для нормального функционирования нейронов.
Хотя морфологически нейроны представляют собой разнообразную группу клеток, их
Рис. 4.10. Миелинизация аксона шванновской клеткой:
а — поперечный разрез шванновской клетки, окружающей аксон; б — аксон, окруженный множеством шванновских клеток, что обеспечивает цельное покрытие миелином
общая функция осуществляется в три этапа: а) прием информации (ввод); б) оценка поступившей информации с целью определения, необходимо ли передавать сигнал-ответ; в) передача сигнала-ответа. Обычный нейрон состоит из четырех морфологических участков — дендритов, сомы (тела), аксона и пресинапти-ческого окончания (рис. 4.11), которые, взаимодействуя, осуществляют указанные выше задачи (Palay, Chom-Palay, 1987). Сома (соматическая клетка) включает аппарат (например, ядро, рибосомы, эндоплазматический ретикулум, аппарат Гольджи), необходимый для синтеза макромолекул. Аксон — трубчатый отросток, идущий от сомы к аксонному холмику. Одна из его функций — передача сигнала-ответа (Rail, 1987), что представляет собой электрическое явление, которое называется потенциалом действия. Аксонный холмик — наиболее возбудимый участок аксона, место инициации потенциала действия.
Как правило, аксон имеет ответвления — коллатерали. Большие аксоны окружены жировой изоляционной оболочкой — миелином. Ее образуют шванновские клетки периферических нервов. Миелин повышает интенсивность передачи потенциала действия по аксону (скорость проведения). У окончания аксон разделяется на множество тонких ответвлений, которые функционально контактируют с принимающей поверхностью клеток. Контактирующие участки называются синапсами. Окончание аксона, входящее в синапс, называется пресинаптическим окончанием. Оно обеспечивает передачу сигнала-ответа от нейрона к клетке. Во взаимодействиях между нейронами наиболее характерным “принимаю-
137
Рис. 4.11. Морфологические свойства нейрона (Ulfhake, Kellerth, 1984):
а — реконструкция двигательного нейрона спинного мозга кошки в виде большого дендритного дерева, окружающего небольшую сому и аксон; б — идеализированный нейрон с четырьмя морфологи ческими участками: дендритами, сомой, аксоном и пресинаптическим окончанием
щим” участком являются дендриты, другие отростки сомы. На них распаложено около 80% всех “принимающих участков” (синапс).
В единой системе сустава важную роль играют три функциональных класса нейронов: афферентные, промежуточные и эфферентные
Рис. 4.12. Три функциональных класса нейронов
(рис. 4.12). Эта система представляет поступление информации из окружающей среды (извне единой системы сустава) в ЦНС, также реакцию на нее. Афферентные нейроны переносят сенсорную информацию (потенциалы действия) из окружающей среды в ЦНС. Афферентные сигналы поступают в ЦНС и действуют на местном уровне; кроме того, они распространяются по ЦНС. Примерами местных, или локальных, воздействий являются простые рефлексы. Афферентный сигнал поступает в спинной мозг и вызывает ответный сигнал с этого же уровня спинного мозга. Промежуточные нейроны составляют 99 % всего количества нейронов, представляя собой компоненты ЦНС, которые модулируют взаимодействие афферентного (входящего) и эфферентного (выходящего) сигналов. Промежуточные нейроны могут вызывать возбуждающие и тормозные реакции других нейронов. Эта модуляция (возбуждение и торможение) может происходить непосредственно — промежуточный нейрон образует части цепи между афферентным и эфферентным нейронами — или косвенно — промежуточный нейрон изменяет возбудимость связи между ними (рис. 4.12). Кроме того, эфферентный нейрон может принять информацию из других структур ЦНС, а промежуточный нейрон способен модулировать это взаимодействие.
Эфферентные нейроны передают выходной сигнал (потенциалы действия) из ЦНС в орган-эффектор. В единой системе сустава таким органом является мышца. Эфферентные нейроны, иннервирующие мышцу, называются двигательными нейронами. Сомы этих нейронов находятся в головном мозгу и в сером веществе спинного мозга, а их аксоны выходят за пределы мозга и соединяются в периферические нервы, идущие к мышцам-мишеням. 43 пары нервов (12 черепных и 31 спинномозговых) в организме человека выходят из ЦНС и образуют периферическую нервную систему
138
(рис. 4.13). Спинной мозг очень часто описывают как разделенную на сегменты структуру, каждый из которых соответствует позвонкам. Между каждой парой позвонков ряд аксонов выходит, а другой ряд входит с обеих сторон (слева и справа) спинного мозга. Аксоны эфферентных нейронов выходят из спинного мозга через вентральные (передние) корешки, аксоны афферентных нейронов входят в него через дорсальные (задние) корешки (рис. 4.14). Двигательные нейроны имеют миелинизированные аксоны большого диаметра, которые идут из спинного мозга непосредственно к скелетной мышце. Тела двигательных нейронов находятся в вентральном отростке спинного мозга.
В отличие от двигательных нейронов, иннервирующих скелетную мышцу, некоторые эфферентные нейроны соединяются с сердечной и гладкой мышцей, а также с железами. Эти нейроны образуют автономную нервную систему. Они регулируют такие физиологические процессы, как артериальное давление крови, моторику и секрецию желудочно-кишечного тракта, температуру тела, потоотделение. Большинство из этих функций регулируется автономными рефлексами. Если двигательные нейроны непосредственно иннервируют скелетную мышцу, то в автономной нервной системе между ЦНС и клеткой-эффектором существует два нейрона. Тело одного из них находится в ЦНС, тело другого — за пределами ЦНС в скоплении клеток, которое называется вегетативными ганглиями. Ввиду анатомических и физиологических различий автономную нервную систему можно разделить на симпатический и парасимпатический отделы. В симпатическом отделе соматические клетки первых нейронов находятся в грудной части
Рис. 4.13. Нервная система человека (Vander, Sherman, Luciano, 1990)
спинного мозга, а вегетативные ганглии, содержащие соматические клетки вторых нейронов, обычно находятся ближе к спинному мозгу. В парасимпатическом отделе соматические клетки первых нейронов находятся в головном мозгу и сакральном участке спинного мозга, а вегетативные ганглии, содержащие
Рис. 4.14. Сегментная структура спинного мозга. На уровне каждого позвонка позвоночного столба от спинного мозга направо и налево отходит пара дорсальных и вентральных корешков
139
Рис. 4.15. Разделение мышечного нерва до уровня отдельных аксонов, иннервирующих отдельные мышечные волокна. Первичные нервные ответвления могут прервать активацию различных частей мышцы (Peters, 1989)
соматические клетки вторых нейронов, — в пределах органа-эффектора.
Аксоны двигательных нейронов, выходящие из спинного мозга через вентральный корешок, образуют периферические нервы, которые также имеют аксоны афферентных нейронов и автономную нервную систему. Дойдя до мышц, нерв вначале разделяется на первичные нервные ответвления, а затем на более мелкие ответвления до тех пор, пока отдельные аксоны не вступят в контакт с отдельными мышечными волокнами (рис. 4.15). Место соединения (синапс) аксона и мышечного волокна называется нервно-мышечным соединением, иногда его называют концевой пластинкой двигательного нерва. В нервно-мышечном соединении пресинаптическая мембрана (аксон) отделяется от постсинаптической мембраны (мышца) щелью шириной 1—2 мкм. Потенциал действия, образуемый двигательным нейроном, передается через эту щель вследствие электрохимического процесса, при котором электрическая энергия, заключенная в нервном потенциале действия, превращается в химическую в форме нейромедиатора. Возбуждение, обусловленное потенциалом действия, способствует выделению химического нейромедиатора пресинаптиче-ским окончанием (более подробно этот процесс рассматривается в главе 5).
В нервно-мышечном соединении нейромедиатором является ацетилхолин (Iversen, 1987). Нейромедиатор, в свою очередь, изменяет проницаемость и электрический статус постсинаптической мембраны, в результате чего сигнал
превращается в мышечный (сарколемный) потенциал действия. Таким образом, энергия потенциала действия двигательного нейрона превращается в химическую энергию вследствие выделения нейромедиатора и затем снова в электрическую в результате образования мышечного потенциала действия. Следовательно, двигательный нейрон представляет собой четвертый элемент единой системы сустава и позволяет контролировать величину усилия, производимого мышцей.
ЧУВСТВИТЕЛЬНОЕ НЕРВНОЕ ОКОНЧАНИЕ
Теперь мы рассмотрим пятый и последний элемент единой системы сустава — чувствительное нервное окончание. Если причина включения в единую систему сустава первых четырех элементов, в принципе, очевидна, то на каком основании в эту биологическую модель контроля движения включено чувствительное нервное окончание? Основная функция чувствительных нервных окончаний — обеспечение системы информацией о ее состоянии, а также о том, что делается в окружающей ее среде. Этот тип потока информации от чувствительных нервных окончаний в ЦНС иногда называют обратной связью. В принципе, оказывается (это характерно как для биологических систем, так и для инженерии в целом), что чем маневреннее система, тем больший объем обратной связи необходим для обеспечения ее стабильности (Hasan, Stuart, 1988). Человеческое тело, являясь биологической моделью, характеризуется высокой степенью маневренности, т.е. оно может выполнять все виды движений, для контроля которых необходим большой объем обратной связи. В действительности, количество афферентных нейронов, обеспечивающих обратную связь, значительно превышает число эфферентных нейронов, принимающих участие в активации мышц.
Чувствительные нервные окончания превращают один вид энергии в другой в результате процесса, который называется трансдукцией. Энергия может существовать во множестве различных форм, таких, как свет, давление, температура, звук, однако наиболее типичный вид энергии, образуемой чувствительными нервными окончаниями, — электрохимическая энергия в виде потенциала действия. Потенциалы действия используются ЦНС для контроля за статусом скелетно-мышечной системы. В организме человека имеется много видов чувствительных нервных окончаний, которые различаются по месту расположения (экстерорецеп-торы, проприорецепторы, интероцепторы), функции (механоцепторы, термоцепторы, фо
140
торецепторы, хеморецепторы, ноцирецепторы) и морфологии (свободные либо инкапсулированные окончания).
Единой системе сустава необходимо, по меньшей мере, два вида информации, чтобы контролировать движение. Ей необходимо знать, где она находится и, когда ее что-то “беспокоит”, что происходит в окружающей ее среде. Эту информацию обеспечивают проприо-рецепторы, выявляющие стимулы, образуемые самой системой, и экстерорецепторы, выявляющие внешние стимулы (Sanes, 1987). На основании этой информации единая система сустава способна обеспечить быструю реакцию на то, что ее “беспокоит”, установить местонахождение того, что ее “беспокоит”, а также дифференцировать самопроизводимые и внеш-невызываемые движения. К проприорецепто-рам относятся мышечные веретена, сухожильные органы и суставные рецепторы, к эксте-рорецепторам — глаза, уши и рецепторы кожи, реагирующие на температуру, прикосновение и боль.
Мышечное веретено
В мышце находится 6—1300 мышечных веретен (Hasan, Stuart, 1984; Matthews, 1972). Они имеют веретенообразную форму и располагаются параллельно волокнам скелетной мышцы (рис. 4.16). Хотя мышечное веретено — морфологически сложное нервное окончание, оно представляет собой набор миниатюрных волокон (2—12) скелетной мышцы, заключенных в капсулу соединительной ткани (рис. 4.17). Меньшие мышечные волокна называются интрафу-зальными волокнами, а те, что находятся за пределами мышечного веретена, — экстрафу-зальными. Ввиду большего содержания миофиламентов в экстрафузальном волокне, оно производит в 36 раз больше усилия, чем интрафу-зальное.
Являясь чувствительным нервным окончанием, мышечное веретено имеет афферентное снабжение, посредством которого потенциалы действия передаются в ЦНС. Афферентные аксоны разделяют на четыре группы, главным образом в зависимости от диаметра. Аксоны I группы имеют наибольший диаметр, аксоны IV группы — наименьший, чем больше диаметр аксона, тем быстрее проводится потенциал действия (табл. 4.4). Каждое мышечное веретено имеет переменное количество (8—25) афферентов групп I и П.Не все мышечные веретена имеют афференты II группы, но все имеют афференты I группы. Сомы афферентных нейронов групп I и II находятся в ганглии дорсального корешка вблизи от спинного мозга (рис. 4.12 и 4.14).
Рис. 4.16. Распределение мышечного веретена и сухожильного органа в медиальной икроножной мышце кошки (Botterman, Binder, Stuart, 1978): а — продольный разрез мышцы с окрашенными сухожильными органами (сухожильные органы соединены с апоневрозом); б — поперечный разрез мышцы, на котором видны типы мышечных волокон; в — увеличенный отдельный сухожильный орган в рядах волокон скелетной мышцы; г — продольный разрез мышцы, на котором видно распределение мышечного веретена в брюшке мышцы; д — увеличенное мышечное веретено, располагающееся параллельно волокнам скелетной мышцы
Интрафузальные волокна мышечного веретена также принимают эфферентный сигнал. Вообще волокна скелетной мышцы иннервируются тремя группами двигательных нейронов (а, р, у), отличающихся размером и видом волокон, которые они иннервируют (табл. 4.4).
Альфа(а)-мотонейроны — самые большие и иннервируют экстрафузальные мышечные во-локна; гамма(у)-мотонейроны — самые маленькие, они соединяются исключительно с ин-трафузальными мышечными волокнами, у бета(р)-мотонейронов — средние размеры, они иннервируют как экстрафузальные, так и интрафузальные мышечные волокна, р- и у-мото-нейроны контактируют с интрафузальными волокнами в их полярных участках, содержащих большое количество миофиламентов. При инициации потенциала действия в аксонном холмике р- или у-мотонейрона и его передаче в мышечное веретено сокращаются (укорачиваются) интрафузальные волокна в полярных участках и растягиваются экваториальный участок и афференты мышечного веретена (группы I и II). Растяжение вызывает активацию
141
Рис. 4.17. Схема мышечного веретена
афферентов вследствие образования потенциалов действия, передаваемых в ЦНС.
Связь в пределах афферентного элемента единой системы сустава начинается с образования генераторного потенциала в чувствительных нервных окончаниях. Так, для механорецепторов (мышечного веретена, сухожильного органа, суставного рецептора) генераторный потенциал образуется в результате механической деформации чувствительного окончания.
Таблица 4.4. Классификация нервных волокон
Тип волок на Функция* Диаметр волокна, мкм Скорость проведения, м-с“‘
Эфферентный
Скелетная мышца 15 100
АР Скелетная мышца + мышечное веретено 8 50
Ау Мышечное веретено 5 20
В Симпатическая преганглионарная 3 7
С Симпатическая пост- 1 ганглионарная (не-миелинизированная) Афферентный 1
1а Мышечное веретено 13-20 80-120
16 Сухожильный орган 13-20 80-120
II Мышечное веретено 6-12 35-75
III Глубоко расположенные в мышце рецепторы давления 1-5 5-30
IV Боль (немиелинизиро-ванные), температура 0,2-1,5 0,5-2
* Примеры функций различных классов нервных волокон.
Генераторный потенциал проводится по аксону в триггерную зону и если сигнал достаточен, возникает потенциал действия.
Мышечное веретено передает информацию об изменении длины мышцы. Поскольку мышечные веретена расположены параллельно волокнам скелетной мышцы, существует два способа активации чувствительного нервного окончания и обеспечения им афферентного сигнала относительно длины мышцы. Первый был описан выше: р- или у-мотонейроны вызывают сокращение интрафузальных мышечных волокон и растяжение экваториального участка, в котором находятся некоторые афферентные окончания I и II групп. Растяжение реверсивно деформирует афферентные окончания и приводит к образованию потенциала действия (аксонный холмик расположен вблизи чувствительного нервного окончания), который передается обратно в ЦНС. Поскольку р- и у-мото-нейроны могут активировать мышечное веретено (веретенообразной формы), их иногда называют веретенодвигательными нейронами. Второй способ активации веретена — пассивное растяжение всей мышцы.
На рис. 4.18 приведены результаты исследования активности мышечного веретена (Al-Falahe, Nagaoka, Vallbo, 1990). Измеряли разряд отдельного мышечного веретена разгибателя пальцев руки человека при синусоидальном передвижении пальца экспериментатором (рис. 4.18, а) либо самим испытуемым (рис. 4.18, б).
Как видно из представленных данных, при обоих видах движения (вынужденном или активном) разряд мышечного веретена увеличивается при растяжении мышцы—разгибателя пальцев во время фазы движения сгибания (вверх).
142
Рис. 4.18. Разряд мышечного веретена человека при пассивном (а) и активном (6) растяжении. Угол сустава мышцы предплечья направлен вверх (Al-Falahe, Nagaoka, VaUbo, 1990)
Информация о растяжении мышцы, обеспечиваемая мышечным веретеном, может служить, по меньшей мере, двум целям: во-первых, она помогает ЦНС определить положение и ориентацию единой системы сустава; во-вторых, сигнализирует о влиянии на систему условий окружающей среды. Более подробно эти процессы рассматриваются в главе 5.
активации волокон скелетной мышцы (активное растяжение) волокна коллагена сжимаются и возбуждают афферент группы 16 (рис. 4.19,6). Уровень активации сухожильного органа определяет степень проявляемой мышечной силы. Сила, необходимая для возбуждения сухожильного органа, зависит от режима активации. При пассивном растяжении требуется сила порядка 2Н,
Сухожильный орган
В отличие от мышечного веретена сухожильный орган — относительно более чувствительное нервное окончание; он состоит лишь из отдельного афферентного соединения и не содержит эфферентных соединений (Hasan, Stuart, 1984). В самом сухожилии находится всего несколько сухожильных органов. Большинство из них располагается вокруг нескольких экстрафузальных мышечных волокон, соединенных с апоневрозом (рис. 4.16). Апоневроз представляет собой сухожильные оболочки, которые простираются вдоль и вглубь брюшка мышцы. Чувствительное окончание афферентного нейрона находится в капсуле, его ответвления окружают ряд волокон коллагена, входящих в состав апоневроза (рис. 4.19, а). В капсуле обычного сухожильного органа, по данным исследований, около 10 волокон скелетной мышцы, причем каждое мышечное волокно иннервируется разными а-мотонейронами. Афферентный нейрон, связанный с сухожильным органом, называется афферентом группы 16 (табл. 4.4).
При растяжении мышцы и прикреплении соединительной ткани в результате физического (пассивного) растягивания или вследствие
Рис. 4.19. Сухожильный орган:
а — заключенный в капсулу афферент группы 16 окружает сухожилия ряда волокон скелетной мышцы; б — сокращение волокон скелетной мышцы вызывает сжатие коллагеновых волокон (сухожилия) и, следовательно, активацию афферента группы 16
143
Рис. 4.20. Разряд сухожильного органа человека при движениях пальца с нулевой (а) и небольшой (6) нагрузкой (Al-Falahe, Nagaoka, Vallbo, 1990)
тогда как при активном достаточно активности отдельного мышечного волокна — 30—90 Н (Binder, Kroin, Moore, Stuart, 1977).
Пример разряда сухожильного органа показан на рис. 4.20 (Al-Falahe et al., 1990). Испытуемый выполнял движение пальцем при нулевой (рис. 4.20, а) и небольшой (рис. 4.20, б) нагрузке. Верхняя запись на обоих рисунках соответствует углу пястно-фалангового сустава со сгибанием сустава и растяжением мышцы, показанными в виде смещения (деформации) вверх.
Следующая запись — интенсивность разряда сухожильного органа, максимальные показатели — около 40 Гц. Нижняя запись характеризует электрическую активность мышцы, осуществляющей движения пальцем. На рис. 4.20 продемонстрировано соответствие разряда сухожильного органа ЭМГ. Ввиду тесной взаимосвязи между ЭМГ и силой мышцы разряд сухожильного органа обеспечивает контроль за усилием, производимым мышцей.
Суставные рецепторы
Подобно рецепторам мышечного веретена и сухожильного органа суставные рецепторы могут функционировать как механорецепторы и обеспечивать единую систему сустава информацией, необходимой для контроля движения (Burgess, Horch, Tuckett, 1987). В то же время в отличие от рецепторов мышечного веретена и сухожильного органа суставной рецептор не является отдельной, четко определенной еди
ницей. Суставные рецепторы отличаются местонахождением (например, суставная капсула, связка, соединительная ткань), типом (окончание Руффини, окончание Гольджи, тельце Па-чини, свободные нервные окончания) и, очевидно, функцией. Эти рецепторы “обслуживают” афферентные нейроны групп II, III и IV.
Окончания Руффини, как правило, состоят из 2—6 заключенных в капсулу шарообразных телец с одним миелинизированным аксоном-родителем диаметром 5—9 мкм. Их можно разделить на статические и динамические механорецепторы, обеспечивающие информацию о положении сустава, угловой скорости и внутрисуставном давлении (Tohansson, Sjolander, Sojka, 1991). Тельца Пачини инкапсулированы с аксоном-родителем диаметром 8—12 мкм. Эти рецепторы имеют низкие пороги механической нагрузки и выявляют ускорение сустава (Bell, Bolanowski, Holmes, 1994). Окончания Гольджи представляют собой тонко инкапсулированные веретенообразные тельца, напоминающие сухожильные органы. Аксон афферентного нейрона, соединенный с окончанием Гольджи, имеет диаметр 13—17 мкм. Эти рецепторы характеризуются высокими порогами и контролируют напряжение в связках, особенно при экстремальной амплитуде движения. Свободные нервные окончания представляют собой ноцирецептор-ную систему сустава. Они имеют аксоны небольшого диаметра (0,5—5 мкм) и активны, когда сустав подвергается аномальному механическому стрессу или действию химических веществ. Рассмотренные четыре вида суставных рецепторов способны обеспечить единую систему сустава информацией о положении, перемещении, скорости и ускорении движения, а также отрицательных стимулах, воздействующих на сустав (Tohansson et al., 1991).
Значение суставных рецепторов для нормального функционирования единой системы сустава убедительно демонстрирует влияние патологии суставов на мышечную активность. Например, при экспериментальной эффузии в коленный сустав (введение жидкости в суставное пространство) у физически здоровых испытуемых значительно снижается способность активации четырехглавой мышцы бедра даже при отсутствии болевых ощущений (Stokes, Yoong, 1984; Young, Stokes, Iles, 1987). Торможение мышечной активации зависит от объема вводимой жидкости и может на 30—90% снизить максимальную произвольную активацию четырехглавой мышцы бедра. Наоборот, удаление жидкости из коленного сустава, например после менискэктомии, значительно повышает эффективность мышцы. При отсутствии болевых ощущений хроническая эффузия сустава может вызвать ослабление и атрофию мышцы. Точно так же у людей с давними разрывами крестообразной связки, как правило, отмечает
144
ся пониженная сила четырехглавой мышцы бедра и подколенных сухожилий (Grabiner, Koh, Audrish, 1992; Johansson et al., 1991).
Аксоны, иннервирующие суставные рецепторы, распределяют информацию по ЦНС от спинного до головного мозга. На уровне единой системы сустава, однако, суставные рецепторы, по-видимому, оказывают более значительное влияние на веретенообразные нейроны (у-мотонейроны), чем на а-мотонейроны. Таким образом, суставные рецепторы скорее всего косвенно воздействуют на единую систему сустава, модулируя активность мышечного веретена.
Кожные механорецепторы
В отличие от трех чувствительных нервных окончаний, о которых шла речь в этом разделе, кожные механорецепторы обеспечивают информацию исключительно о внешних стимулах, влияющих на единую систему сустава. Так, кожные механорецепторы руки и ступни дают важную информацию о взаимодействии человека с окружающей средой. Например, снижение соматосенсорной остроты, затрагивающее, очевидно, кожные механорецепторы, приводит к уменьшению контроля за положением тела у пожилых людей, а также людей с различной патологией (Horak, Shupert, Mirka, 1989). Кроме механорецепторов в коже находятся терморецепторы и болевые рецепторы, которые также могут влиять на функцию единой системы сустава и даже на функцию а-мотонейронов, активирующихся при выполнении данного задания.
Различают четыре вида кожных механорецепторов: диски Меркеля, тельца Мейсснера, окончания Руффини и тельца Пачини (Rothwell, 1987). Первые два расположены вблизи поверхности кожи, два других находятся глубже. Диск Меркеля чувствителен к локальному вертикальному давлению и не реагирует на латеральное растяжение кожи. Этот рецептор отвечает быстрым начальным разря
дом потенциалов действия, который быстро снижается до небольшой устойчивой интенсивности. Тельца Мейсснера иннервируют 2—6 аксонов, каждый аксон может иннервировать больше одного тельца. Тельца Мейсснера чувствительны к локальному, поддерживаемому давлению, однако их реакция (разряд потенциалов действия) быстро затухает. Окончания Руффини иннервируются отдельным аксоном и реагируют на растяжение кожи на большой площади. Эта чувствительность, однако, зависит от направления растяжения; окончание возбуждается при растяжении в одном направлении и ингибируется при растяжении под прямым углом в избранном направлении. Реакция окончаний Руффини медленно приспосабливается к продолжительному растяжению. Тельца Пачини — самые большие рецепторы кожи, которые иннервирует отдельный аксон. Для них характерно быстро изменяющееся давление. Эти четыре кожных механорецептора позволяют единой системе сустава выявлять стимулы, воздействующие на кожу на большой и небольшой площади в течение короткого или продолжительного периода времени.
РЕЗЮМЕ
Поскольку тело человека — весьма сложный механизм, мы разработали упрощенную модель компонентов двигательной системы, необходимых для выполнения движения. Эта упрощенная модель называется единой системой сустава. В настоящей главе рассматривались свойства пяти компонентов единой системы сустава: жесткого звена, синовиального сустава, мыщцы, нейрона и чувствительного нервного окончания. Поскольку эти пять компонентов состоят из живой ткани, они адаптируются к физическим нагрузкам повседневной жизни. Результатом является динамическая система, характеризующаяся постоянно изменяющимися свойствами.
buuPOCJ С АЛЫЕРПАТИВНЫ1. СТВЕТОЙ
1. Вода составляет 80% сырой массы кости.
ПРАВИЛЬНО НЕПРАВИЛЬНО
2. Фибробласт — основная структурная единица кости.
ПРАВИЛЬНО НЕПРАВИЛЬНО
3. Стресс — единица силы, нормализованная относительно силы (прочности) ткани.
ПРАВИЛЬНО НЕПРАВИЛЬНО
4- опред — количество энергии, которую может абсорбировать кость.
ПРАВИЛЬНО НЕПРАВИЛЬНО
5. Большеберцовая кость подвергается большей деформации при растяжении, чем при сжатии во время бега трусцой.
ПРАВИЛЬНО НЕПРАВИЛЬНО
6. Бедренная кость характеризуется максимальным сопротивлением сжимающей нагрузке.
ПРАВИЛЬНО НЕПРАВИЛЬНО
7. Кость способна выдерживать силу в 2—5 раз больше той, которой она ежедневно подвергается. ПРАВИЛЬНО НЕПРАВИЛЬНО
10 8-133
145
8. Процесс восстановления кости называется ремоделированием.
ПРАВИЛЬНО НЕПРАВИЛЬНО
9. Деминерализация представляет собой потерю костью солей.
ПРАВИЛЬНО НЕПРАВИЛЬНО
10. Ремоделирование лучше всего осуществляется в результате непрерывных нагрузок.
ПРАВИЛЬНО НЕПРАВИЛЬНО
11. Плотность костей с возрастом повышается в результате процесса, называемого остеопорозом. ПРАВИЛЬНО НЕПРАВИЛЬНО
12. При нагрузке на кость образуется электрический потенциал.
ПРАВИЛЬНО НЕПРАВИЛЬНО
13. Кость соединяется с костью с помощью связок.
ПРАВИЛЬНО НЕПРАВИЛЬНО
14. Сухожилия и связки созданы в основном для противодействия силам растягивания.
ПРАВИЛЬНО НЕПРАВИЛЬНО
15. Сухожилия и связки состоят главным образом из коллагена.
ПРАВИЛЬНО НЕПРАВИЛЬНО
16. После разминки вязкость связки изменяется. ПРАВИЛЬНО НЕПРАВИЛЬНО
17. Растягивающая деформация сухожилия возрастает линейно с увеличением силы растяжения. ПРАВИЛЬНО НЕПРАВИЛЬНО
18. Синовиальный сустав свободно двигается. ПРАВИЛЬНО НЕПРАВИЛЬНО
19. Суставной хрящ иннервируют афферентные нейроны группы III.
ПРАВИЛЬНО НЕПРАВИЛЬНО
20. Суставной хрящ представляет собой тиксотропную ткань.
ПРАВИЛЬНОНЕПРАВИЛЬНО
21. Физическая активность может изменить толщину суставного хряща.
ПРАВИЛЬНО НЕПРАВИЛЬНО
22. Суставная капсула выделяет синовиальную ЖИДКОСТЬ. ПРАВИЛЬНО НЕПРАВИЛЬНО
23. Для синовиального сустава характерны 1—3 степени свободы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
24. Для тазобедренного сустава характерна одна степень свободы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
25. Не все скелетные мышцы пересекают суставы. ПРАВИЛЬНО НЕПРАВИЛЬНО
26. Сарколемма — возбудимая мембрана.
ПРАВИЛЬНО НЕПРАВИЛЬНО
27. Волокно скелетной мышцы — отдельная клетка. правильно неправильно
28. Отдельный сократительный белок называется миофибриллой.
ПРАВИЛЬНО НЕПРАВИЛЬНО
29. Саркоплазматический ретикулум — инвагинация сарколеммы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
30. Саркомер представляет собой участок миофибриллы от одной Z-линии до другой.
ПРАВИЛЬНО НЕПРАВИЛЬНО
31. Длина саркомера около 2,5 мкм.
ПРАВИЛЬНО НЕПРАВИЛЬНО
32. Темная полоска — это А-диск.
ПРАВИЛЬНО НЕПРАВИЛЬНО
33. Н-зона находится в середине А-диска.
ПРАВИЛЬНО НЕПРАВИЛЬНО
34. Толстые филаменты соединены с М-поло-СКОЙ. ПРАВИЛЬНО НЕПРАВИЛЬНО
35. Отдельный тонкий филамент окружают шесть толстых филаментов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
36. Актин — белок.
ПРАВИЛЬНО НЕПРАВИЛЬНО
37. Тропомиозин — белок, связанный с актином. ПРАВИЛЬНО НЕПРАВИЛЬНО
38. Z-линия характеризуется высокой степенью гибкости и способностью адаптироваться.
ПРАВИЛЬНО НЕПРАВИЛЬНО
39. Легкий меромиозиновый фрагмент миозина состоит из двух субфрагментов — 1 и 2.
ПРАВИЛЬНО НЕПРАВИЛЬНО
40. Субфрагмент 1 соединяет легкие и тяжелые меромиозиновые фрагменты.
ПРАВИЛЬНО НЕПРАВИЛЬНО
41. Молекулярную массу измеряют в дальтонах. ПРАВИЛЬНО НЕПРАВИЛЬНО
42. Тяжелая цепочка имеет большую молекулярную массу.
ПРАВИЛЬНО НЕПРАВИЛЬНО
43. Каждая молекула миозина содержит тяжелую цепочку. ПРАВИЛЬНО НЕПРАВИЛЬНО
44. К шарообразной головке молекулы миозина прикрепляются четыре легкие цепочки.
ПРАВИЛЬНО НЕПРАВИЛЬНО
45. Перимизий образует цитоскелет в скелетной мышце. ПРАВИЛЬНО НЕПРАВИЛЬНО
46. Нейроглии — пассивные структуры, обеспечивающие всего лишь структурную поддержку нейронов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
146
47. Соматическая клетка нейрона называется
СОМОЙ. ПРАВИЛЬНО НЕПРАВИЛЬНО
48. Поступление сигнала в нейрон осуществляется через аксон.
ПРАВИЛЬНО НЕПРАВИЛЬНО
49. Функциональный контакт нейрона с клеткой-мишенью происходит в аксонном холмике. ПРАВИЛЬНО НЕПРАВИЛЬНО
50. Афферентные нейроны передают сенсорную информацию в ЦНС.
ПРАВИЛЬНО НЕПРАВИЛЬНО
51. Афферентные сигналы передаются непосредственно к эфферентным нейронам спинного мозга.
ПРАВИЛЬНО НЕПРАВИЛЬНО
52. Эфферентные нейроны, иннервирующие скелетную мышцу, называются двигательными нейронами.
ПРАВИЛЬНО НЕПРАВИЛЬНО
53. Аксоны эфферентных нейронов входят в спинной мозг у дорсальных корешков.
ПРАВИЛЬНО НЕПРАВИЛЬНО
54. Каждое мышечное волокно иннервирует отдельный эфферентный аксон.
ПРАВИЛЬНО НЕПРАВИЛЬНО
55. Синапс между афферентным и эфферентным нейроном называется нервно-мышечным соединением.
ПРАВИЛЬНО НЕПРАВИЛЬНО
56. Ацетилхолин — нейромедиатор.
ПРАВИЛЬНО НЕПРАВИЛЬНО
57. Мышечные толокна могут образовывать потенциал действия.
ПРАВИЛЬНО НЕПРАВИЛЬНО
58. Чувствительные нервные окончания обеспечивают обратную связь с ЦНС посредством афферетных нейронов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
59. Интрафузальные волокна меньше, чем экстрафузальные.
ПРАВИЛЬНОНЕПРАВИЛЬНО
60. Аксон группы I выполняет афферентную функцию. ПРАВИЛЬНО НЕПРАВИЛЬНО
61. Из всех эфферентных аксонов наибольший диаметр у волокна Аа.
ПРАВИЛЬНО НЕПРАВИЛЬНО
62. а-Мотонейроны могут вызвать сокращение интрафузальных волокон.
ПРАВИЛЬНО НЕПРАВИЛЬНО
63. Растяжение интрафузального волокна вызывает возникновение генераторного потенциала.
ПРАВИЛЬНО НЕПРАВИЛЬНО
64. Мышечные веретена обеспечивают информацию об изменении мышечной длины.
ПРАВИЛЬНО НЕПРАВИЛЬНО
65. а-Мотонейроны называются фузимоторны-ми нейронами.
ПРАВИЛЬНО НЕПРАВИЛЬНО
66. Сухожильный орган иннервируется р-мо-тонейронами.
ПРАВИЛЬНО НЕПРАВИЛЬНО
67. Сухожильный орган расположен последовательно с волокнами скелетной мышцы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
68. Интенсивность разряда сухожильных органов измеряется в герцах.
ПРАВИЛЬНО НЕПРАВИЛЬНО
69. Сухожильные органы не реагируют на пассивные изменения мышечной силы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
70. Окончания Руффини выявляют ускорение сустава.
ПРАВИЛЬНО НЕПРАВИЛЬНО
71. Окончания Гольджи сигнализируют о напряжении в связках.
ПРАВИЛЬНО НЕПРАВИЛЬНО
72. Накопление жидкости в суставе ингибирует мышечную деятельность.
ПРАВИЛЬНО НЕПРАВИЛЬНО
73. Кожные механорецепторы играют важную роль в контроле позы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
74. Тельца Мейсснера чувствительны к локальному вертикальному давлению.
ПРАВИЛЬНО НЕПРАВИЛЬНО
75. Тельца Пачини выявляют быстро меняющееся давление.
ПРАВИЛЬНО НЕПРАВИЛЬНО
76. Диск Меркеля реагирует быстрым начальным разрядом потенциалов действия.
ПРАВИЛЬНО НЕПРАВИЛЬНО
ВОПРОСЫ С РАЗЛИЧНЫХ ВЫБОРОМ ОТВЕТОВ
77. Что из приведенного ниже не является функцией кости?
а) обеспечение механической поддержки;
б) выделение синовиальной жидкости в суставную полость;
в) произведение эритроцитов;
г) служит резервуаром активных ионов (Са2+, Mg2+).
ю*
147
78. Какое вещество составляет большую часть сырой массы кости?
а) вода;
б) соли;
в) остеокласты;
г) остеоколлагеновые волокна.
79. Что составляет основную структурную единицу кости?
а) остеобласт;
б) остеон;
в) белковый матрикс;
г) остеокласт.
80. Каким нагрузкам лучше всего противостоит бедренная кость?
а) сжимающей;
б) вращающей;
в) растягивающей;
г) смещающей.
81. Чем объяснить понижение прочности костей астронавтов после пребывания в космосе?
а) снижением пьезоэлектрических потенциалов;
б) уменьшением фактора безопасности;
в) развитием остеопороза;
г) повышенной деминерализацией.
82. Какое свойство сухожилий и связок влияет на их зависимое от скорости сопротивление растягиванию?
а) упругость;
б) пьезоэлектрическое;
в) тиксотропия;
г) вязкость.
83. Как называется процесс смазки, при котором смазывающее вещество разделяет соприкасающиеся поверхности?
а) самосмазывание;
б) граничное смазывание;
в) жидкостное гидродинамическое смазывание;
г) вязкое смазывание.
84. Какое утверждение лучше всего объясняет изменение толщины суставного хряща вследствие чрезмерного выделения жидкости, обусловленного нагрузкой?
а) суставной хрящ ведет себя подобно впитывающей воду губке;
б) в суставном хряще отсутствуют кровеносные сосуды и нервы;
в) суставной хрящ обладает способностью самосмазываться;
г) поверхность суставного хряща может впитывать гликопротеид лубрицин.
85. Что из приведенного ниже не является свойством мышцы?
а) проводимость;
б) трансдукция;
в) возбудимость;
г) сократительная способность.
86. Какому уровню мышечной иерархии соответствует мышечная клетка?
а) пучок;
б) саркомер;
в) миофибрилла;
г) волокно.
87. Какова длина мышечного волокна человека?
а) 50 нм;
б) 50 мкм;
в) 50 мм;
г) 50 см.
88. Какая часть мышечной клетки является вытяжением сарколеммы?
а) саркоплазматический ретикулум;
б) терминальные цистерны;
в) поперечная трубочка;
г) эндомизий.
89. В какой полоске содержатся только толстые миофиламенты?
a) Z;
б) Н;
в) А;
г)1.
90. В какой полоске два набора толстых филаментов противоположной полярности (указывающих противоположные направления) соединены вместе?
а) М;
б) Н;
в) I;
r)Z.
91. Укажите регуляторные белки мышцы:
а) актин и миозин;
б) тропонин и тропомиозин;
в) легкий и тяжелый меромиозин;
г) субфрагменты 1 и 2.
92. Что означает сокращение TH?
а) титин;
б) тропонин;
в) поперечную сеть;
г) тропомиозин.
93. Какую функцию не выполняет нейроглия?
а) фагоцитоз;
б) миелинизация;
в) транспорт ионов;
г) образование потенциалов действия.
94. Укажите четыре морфологические участка нейрона:
а) аксон, пресинаптическое окончание, дендрит, сома;
б) сома, синапс, аксон, дендрит;
в) дендрит, коллатераль, пресинаптическое окончание, сома;
г) коллатераль, сома, дендрит, аксон.
148
95. В какой части нейрона образуется “выходной” сигнал?
а) аксонный холмик;
б) дендрит;
в) сома;
г) коллатераль.
96. Какой компонент не относится к одному из трех функциональных классов нейронов? а) интернейрон;
б) эфферент;
в) чувствительное нервное окончание;
г) афферент.
97. Какие нейроны называются двигательными? а) эфферентные;
б) группы II;
в) волокна типа С;
г) интернейрон.
98. Какие физиологические процессы не контролирует автономная нервная система?
а) температуру тела;
б) потоотделение;
в) сокращение скелетной мышцы;
г) давление крови.
99. Какой элемент обеспечивает эфферентную иннервацию мышечного веретена?
а) интрафузальное волокно;
б) у-мотонейрон;
в) волокно ядерной цепочки;
г) нейрон группы 1а.
100. Какие волокна иннервируют а-мотоней-роны?
а) волокна скелетной мышцы;
б) волокна ядерной цепочки;
в) коллагеновые волокна сухожильного органа;
г) волокна ядерного мешочка.
101. О чем сигнализируют сухожильные органы? а) о локальном давлении на кожу;
б) о мышечной силе;
в) о смещении сустава;
г) об изменении длины мышцы.
102 Какой афферентный аксон иннервирует сухожильный орган?
а) группы II;
б) группы 16;
в) р-мотонейрон;
г) фузимоторный нейрон.
103. Какое чувствительное нервное волокно не выполняет функцию суставного рецептора? а) диск Меркеля;
б) окончание Гольджи;
в) свободное нервное окончание;
г) тельце Пачини.
104. Какие параметры не воспринимают суставные рецепторы?
а) мышечную силу;
б) положение сустава;
в) отрицательные стимулы;
г) угловую скорость сустава.
ЗАДАЧИ
105. Рассмотрим биологическую модель, которую мы называем единой системой сустава. А. Зачем необходимо использовать эту модель?
Б. Какие 5 элементов составляют эту модель?
В. Почему модель не включает такие элементы, как легкие и сердце?
106. Допустим, вам предстоит проводить занятия с группой женщин 45 лет и старше. Какие виды физических занятий вы предложите им, чтобы свести к минимуму снижение прочности костей вследствие остеопороза?
107. Определите основную структурную единицу кости, основную несущую нагрузку, единицу сухожилий и связок, основную сократительную единицу мышцы и элемент нервной системы, активирующий скелетную мышцу.
108. Прочность сухожилий и связок зависит от состояния поперечных сцеплений. Что такое поперечные сцепления и почему прочность сухожилий и связок зависит от них?
109. Почему предварительная физическая активность влияет на величину сопротивления сухожилия и связки растягиванию?
ПО. Почему толщина суставного хряща увеличивается при переходе от состояния покоя к активному состоянию?
111. Суставной хрящ — часть элемента синовиального сустава.
А. Какую роль в функционировании суставного хряща играет смазка?
Б. Какова функция суставного хряща?
В. Что происходит с соприкасающимися поверхностями при повреждении суставного хряща?
112. Опишите виды движения для сустава, имеющего три степени свободы, и укажите оси вращения для каждого движения, а также плоскости, в которых осуществляется движение.
113. Какая разница между сарколеммой и эндо-мизием?
114. Обратимся к структуре саркомеров в мышечном волокне.
А. Какое количество саркомеров (средняя ширина 2,5 мкм) содержится в 7,2-миллиметровой миофибрилле?
149
Б. Если площадь поперечного сечения мышечного волокна 1809 мкм2, диаметр миофибриллы 0,1 мкм и они занимают 82 % площади поперечного сечения мышечного волокна, какое количество миофибрилл содержится в мышечном волокне? Если длина каждой миофибриллы 7,2 мм, какое количество саркомеров содержится в волокне?
В. Как расположено большинство саркомеров в мышечном волокне — последовательно или параллельно?
115. Какую функцию выполняет в скелетной мышце цитоскелет? Зачем он нужен?
116. Нейроны образуют сеть клеток, обеспечивающих быструю связь.
А. Укажите три фазы связи, осуществляемой нейронами.
Б. Какие морфологические участки нейрона участвуют в каждой из этих фаз?
В. Укажите три отличительных свойства афферентных и эфферентных нейронов.
117. Каким образом чувствительное нервное окончание, например, мышечное веретено, выявляет механический стимул, превращает
его в электрический сигнал и передает информацию в нервную систему?
118. Почему интрафузальные волокна не вносят значительный вклад в образование силы мышцей?
119. Число афферентных аксонов значительно превышает число эфферентных. Почему это свидетельствует о контроле единой системы сустава?
120. у-Мотонейроны обеспечивают эфферентную иннервацию мышечных веретен.
А. Как фузимоторные нейроны изменяют возбудимость мышечного веретена?
Б. Постройте график интенсивности разряда мышечного веретена в течение отрезка времени при определенном растяжении мышцы. Правильно укажите переменные и единицы измерения. Постройте реакцию мышечного веретена на два состояния: 1) обычное устойчивое состояние; 2) гипервозбудимое состояние мышечного веретена, обусловленное фузимоторными нейронами.
В. Как активируются фузимоторные нейроны?
ГЛАВА 5
ФУНКЦИОНИРОВАНИЕ ЕДИНОЙ СИСТЕМЫ СУСТАВА
Движение человека — весьма впечатляющее явление! Каждое движение — результат многочисленных и тщательных взаимодействий между нервными и мышечными клетками. В главе 5 рассматриваются четыре ключевые единицы функционирования единой системы сустава: двигательная единица — основная функциональная единица единой системы сустава; возбудимые мембраны — средства быстрой передачи информации по системе; сочетание возбуждения—сокращения — связь между нервным сигналом и сокращением мышц и обратная связь чувствительного нервного окончания — роль афферентной информации в функционировании системы.
ДВИГАТЕЛЬНАЯ ЕДИНИЦА
В части I “Взаимосвязь сила—движение” мы выяснили, что активированная мышца оказывает воздействие силой на жесткое звено и в зависимости от взаимосвязи между нагрузкой на звено и усилием, производимым мышцей, звено либо вращается, либо остается неподвижным. В этой схеме мышца характеризуется способностью производить усилие (силу), величина которого контролируется нервной системой. Таким образом, анализ движения как результата активации мышцы должен включать рассмотрение связанных с ней нервных факторов. Двигательная единица определяется как соматическая клетка и дендриты двигательного нейрона, многочисленные ответвления его аксона и мышечные волокна, которые он иннервирует. Анатомические и физиологические свойства двигательной единицы могут значительно колебаться; эти различия позволяют мышце, которая обычно включает несколько сот (100—1000) двигательных единиц (McComas, 1991; Stein, Yang,
1990), удовлетворять многочисленные потребности, а также адаптироваться к кратковременным и долговременным запросам.
Нервный компонент
Нервный компонент двигательной единицы состоит из двигательного нейрона и его дендритов (рис. 4.П). Свойства нервного компонента включают морфологию, возбудимость и распределение входящей информации. Морфологическая характеристика нервного компонента — размер двигательного нейрона, судить о котором можно по диаметру сомы, площади поверхности соматической клетки, количеству дендритов, вступающих из сомы, и диаметре аксона. Эти качества характеризуются значительной степенью корреляции: у самого большого двигательного нейрона все перечисленные показатели максимальны (Stuart, Enoka, 1983). Кроме того, двигательные нейроны, “обслуживающие” быстросокращающиеся мышечные волокна, больше, чем те, которые иннервируют медлен-носокращающиеся волокна.
Как отмечал Henneman (1957), существует тесная взаимосвязь между размером двигательного нейрона и его возбудимостью. Реобаза — непосредственная мера возбудимости, показывающая ток какой силы следует ввести в двигательный нейрон, чтобы вызвать образование потенциала действия. Реобаза небольших двигательных нейронов намного меньше реобазы больших двигательных нейронов. Скорость распространения потенциалов действия зависит от размера (диаметра) аксона. Поскольку диаметр аксонов неодинаков, у больших двигательных нейронов, иннервирующих быстросокращающиеся мышечные волокна, более высокая скорость проведения возбуждения по аксону. Небольшие двигательные нейроны более возбуди
151
мы, однако образуют и распространяют потенциалы действия с меньшей скоростью, чем большие двигательные нейроны.
Двигательные нейроны имеют обширные дендритные деревья, принимающие около 80 % входного сигнала, направленного в клетку (см. рис. 4.11). Вместе с тем входной сигнал из разных источников неодинаково влияет на образование потенциала действия двигательным нейроном. Это может быть обусловлено количеством и расположением синапсов, связанных с каждой системой ввода сигнала.
Существует три пути распределения входных сигналов (Heckman, Binder, 1990): 1) минимальный входной сигнал (наименьший эффективный синаптический ток) к наибольшим двигательным нейронам — входной сигнал из афферента группы 1а мышечного веретена; 2) равномерный входной сигнал ко всем двигательным нейронам — тормозящий входной сигнал из мышечного веретена, расположенного в мышце-антагонисте, а также из промежуточного нейрона (клетки Рен-шоу); 3) максимальный входной сигнал к наибольшим двигательным нейронам — входной сигнал из ядер ствола мозга (красных ядер) и из нерва, содержащего информацию, поступившую из рецепторов кожи. На основании изложенного можно сделать вывод, что активация двигательного нейрона зависит не только от свойственной ему возбудимости, но и от типа (распределения) принимаемого им входного сигнала. Кроме того, распределение входного сигнала влияет на интенсивность рекрутирования двигательных нейронов (Kemell, Hultbom, 1990).
Мышечный компонент
Хотя мышечное волокно иннервируется отдельным двигательным нейроном, каждый мотонейрон иннервирует более одного мышечного волокна (рис. 5.1). Количество мышечных волокон, иннервируемых отдельным двигательным нейроном, называется коэффициентом иннервации, который колеблется от 1 : 1900 (например, для икроножной мышцы, передней большеберцовой мышцы) до 1 : 15 для наружной мышцы глаза. Таким образом, один двигательный нейрон может иннервировать от 15 до 1900 мышечных волокон. Всякий раз, когда в ЦНС активируется двигательный нейрон, он образует один или больше потенциалов действия во всех своих мышечных волокнах (за исключением состояния утомления). Таким образом, чем ниже коэффициент иннервации, тем совершеннее контроль мышечной силы с точки зрения активации двигательной единицы. По коэффициенту иннервации также можно судить о количестве разветвлений, необходимых аксону, чтобы вступить в контакт со всеми своими мышечными волокнами.
На рис. 5.1 показано расположение волокон, принадлежащих отдельной двигательной единице типа FR в медиальной икроножной мышце кошки. Двигательная единица включает 500 мышечных волокон. Подобные измерения показали, что волокна отдельной двигательной единицы занимают определенный участок мышцы. Он может составлять 15% объема мышцы при плотности 2—5 мышечных волокон на 100 принадлежащих этой же двигательной единице (Burke, 1981). Это означает,что в данном участке мышцы находятся волокна 20—50 различных двигательных единиц. Следует отметить, что плотность мышечных волокон, принадлежащих одной и той же двигательной единице, увеличивается с возрастом. Это обусловлено изменением коэффициента иннервации, поскольку некоторые двигательные нейроны вырождаются и умирают, что приводит к тому, что аксоны других двигательных единиц образуют больше ответвлений и реиннервируют мышечные волокна, утратившие аксонную связь (Campbell, McComas, Petito, 1973; Hashizume, 1989; Stalberg, Fawcett, 1982). Таким образом, с возрастом количество двигательных единиц уменьшается,тогда как размер (количество мышечных волокон) тех двигательных единиц, которые остаются, — увеличивается.
Оказывается, в различных участках мышцы могут находиться определенные популяции двигательных единиц. В этой связи возникло понятие нервно-мышечного компартмента (Peters, 1989; Hamm, Stuart, 1989). Компартмент определяется как объем мышцы, который “обслуживается” основным ответвлением мышечного нерва. Компартмент содержит отдельную популяцию двигательных единиц; мышечные волокна, относящиеся к одной двигательной единице, находятся в отдельном нервно-мышечном компартменте. Количественное соотношение типов мышечных волокон в компартмен-тах одной мышцы может отличаться. Нервно-мышечные компартменты есть не во всех мышцах. Поскольку компартменты могут активироваться независимо, отдельная мышца, представляющая собой анатомическую целостность, может состоять из ряда индивидуальных участков, каждый из которых выполняет различные физиологические функции. Кроме того, существование компартментов показывает ошибочность заключения о функции мышцы, основанного на расположении ее прикреплений. Анализируя функцию мышцы следует учитывать как ее архитектуру, так и структуру иннервации.
Двигательные единицы можно сравнивать по ряду физиологических характеристик, таких, как структура разряда, скорость сокращения, величина силы и сопротивление утомлению. Сопоставления в основном осуществляют на основании свойств мышечного компонента двигательной единицы. Для оценки этих парамет-
152
Рис. 5.1. Распределение мышечных волокон на продольном (слева) и на поперечном (справа) срезах. Мышечные волокна, относящиеся к единице, ограничены субобъемом правой медиальной икроножной мышцы (Burke, Tsairis, 1973)
ров применяют два основных метода: прямой и косвенный. Прямая оценка предполагает физиологическое измерение свойств двигательной единицы. Косвенная оценка основана на гистохимических и биохимических исследованиях. Прямое определение можно осуществить, оценив характеристики разряда двигательных единиц (Gydikov, Kosarov, 1974) или механическую реакцию двигательных единиц на различные входные сигналы (Burke, Levine, Sairis, Zajac, 1973). Первый вариант мы рассмотрим в главе 6. Здесь же остановимся на механическом аспекте двигательных единиц (рис. 5.2). Результатом деятельности двигательной единицы является сокращение. Это реакция сила—время на отдельный входной сигнал (рис. 5.2, а). Выходной сигнал двигательной единицы состоит из серии суммирующихся реакций сокращения, в результате чего величина силы намного превышает таковую, связанную с отдельным сокращением. Профиль сила—время, включающий суммированные реакции сокращения, называется тетанусом (рис. 5.2, в). Одиночные сокращения представляют собой единицу реакции
мышцы на отдельный входной сигнал и характеризуется: отрезком времени до пика силы (время сокращения), величиной пика силы и отрезком времени, необходимого для того, чтобы величина силы снизилась до значения, равного половине пикового (время полурасслабления) (рис. 5.2, а). Время сокращения используется как мера скорости сократительных механизмов. При продолжительном времени сокращения двигательную единицу называют медленносо-кращающейся, при непродолжительном — бы-стросокращающейся (рис. 5.2, б).
Ученые в настоящее время не способны идентифицировать индивидуальные популяции быстро- и медленносокращающихся двигательных единиц в большинстве мышц человека. Продолжительность одиночного сокращения двигательных единиц колеблется от 20 до 72 мс (среднее значение 35 мс) в жевательной мышце, 35—85 мс (среднее 59 мс) в тенарных мышцах и 40—110 мс (среднее 76 мс) в медиальной икроножной мышце (Garnett, O’Donovan, Stephens, Taylor, 1978; Nordstrom, Miles, 1990; Thomas, Johansson, Bigland-Ritchie, 1991). Раз-
153
Рис. 5.2. Реакции сокращения и тетануса двигательных единиц мышцы задней конечности кошки: а — сокращение двигательной единицы типа FF (продолжительность сокращения СТ — 24 мс; время полурасслабления HRT — 21 мс; пик силы — 0,03 Н); б — реакции сокращения двигательных единиц типа FF, FR и S; в — прерывистый тетанус (стимулированный при 28 Гц) единицы FF (пик силы — 0,42 Н); г — сплошной тетанус (стимулированный при 200 Гц) двигательной единицы позиции в (пик силы — 1,30 Н); д — тест на утомлениедвигательных единиц типа FF и FR позиции б
личия в скорости сокращения двигательных единиц, по мнению ученых, обусловлены главным образом различиями в ферменте миозин АТФазе (привязанной к одной из SI-головок), скорости выделения и использования Са2+ саркоплазматическим ретикулумом (Kugelbeig, Thomell, 1983) и архитектуре мышцы.
При возникновении серии стимулов каждый из них вызывает отдельную реакцию сокращения и в результате образуется серия реакций. При уменьшении интервала между стимулами сокращения начинают суммироваться и возникает профиль силы, известный как тетанус. При дальнейшем уменьшении интервала между стимулами тетанус изменяется с зубчатого (прерывистого) тетануса (рис. 5.2, в) на сплошной тетанус (рис. 5.2, г). При субмаксимальной интенсивности стимулов, когда интер
вал между ними составляет 1,25 времени сокращения, прерывистый тетанус может достичь начального пика, а затем снизиться, прежде чем снова достичь пикового значения (на рис. 5.2, в показано незначительное снижение после четвертого стимула). Двигательная единица (и мышца) образуют максимальную силу во время сплошного тетануса. Разница между пиком силы одиночного сокращения и максимальной силой сплошного тетануса (Ро) называется соотношением одиночного сокращения — тетануса, которое обычно колеблется от 1 : 1,5 до 1 : 10. Следовательно, сила сплошного тетануса может превышать силу одиночного сокращения в 1,5— 10 раз. Пик тетанической силы не изменяется как функция времени одиночного сокращения; Ро колеблется в значительном диапазоне в течение данного времени одиночного сокращения и последнее может увеличиться вдвое при незначительном изменении Ро (рис. 5.3).
Сила, образующаяся в результате отдельного тетанического сокращения, со временем уменьшается, если двигательная единица производит серию тетанических сокращений. Способность двигательной единицы предотвратить такое уменьшение рассматривается как ее сопротивление утомлению. Стандартный тест утомления (Burke et al., 1973), разработанный для изучения двигательных единиц задней конечности кошек, включает стимуляцию тетанических сокращений в течение 2—6 мин со скоростью одно
Рис. 5.3. Максимальная тетаническая сила и время сокращения двигательных единиц медиальной икроножной мышцы кошки (Burke, Tsairis, 1974)
154
сокращение каждую секунду; продолжительность каждого тетанического сокращения составляет 330 мс и включает 13 стимулов. Отношение пика силы, образуемой через 2 мин после этих стимулов, к величине силы, образуемой во время начального тетанического сокращения, называется коэффициентом утомления (рис. 5.2, д). Данный тест подчеркивает связь между электрическим сигналом нервной системы и сокращением мышцы (Enoka, Stuart, 1992; Jami, Murthy, Petit, Zythicki, 1983). Это позволяет выделить невосприимчивые к утомлению типы двигательных единиц (S и FR), для которых характерен коэффициент утомления, превышающий или равный 0,75, и восприимчивые к утомлению двигательные единицы (тип FF) с коэффициентом утомления ниже 0,25. Коэффициент 0,25 указывает на то, что через 2 мин сила, образуемая единицей, составит всего 25% ее значения в начале теста. Следовательно, при продолжительных мышечных сокращениях целесообразнее активировать двигательные единицы типа S и FR, поскольку они менее восприимчивы к утомлению.
Типы двигательных единиц
Основываясь на указанных двух параметрах — снижении прерывистого тетануса и сопротивлении утомлению — двигательные единицы млекопитающих можно разделить на три группы (Burke, 1981): медленносокращающие-ся невосприимчивые к утомлению (тип S); бы-стросокращающиеся невосприимчивые к утомлению (тип FR) и быстросокращающиеся вос
приимчивые к утомлению (тип FF). Двигательные единицы типа S образуют наименьшую величину силы, тогда как единицы типа FF — наибольшую (табл. 5.1). Это различие обусловлено разным количеством мышечных волокон, входящих в состав двигательной единицы (т.е. коэффициентом иннервации), а также размером отдельных мышечных волокон (т.е. количеством сократительных белков на мышечное волокно). Несмотря на различия между мышцами, двигательные единицы типа FF, как правило, характеризуются наиболее высоким коэффициентом иннервации и наличием самых больших мышечных волокон.
Двигательные единицы можно также классифицировать косвенно, основываясь на данных гистохимических и биохимических исследований. Оба метода предполагают определение содержания ферментов в мышечных волокнах, однако один из них является качественным (гистохимический), а другой — количественным (биохимический). Поскольку ферменты — катализаторы химических реакций, измерив их количество можно определить индекс скорости или величину реакции. Таким образом, гистохимический и биохимический методы предполагают анализ механизмов, обусловливающих различные физиологические свойства (например, скорость сокращения, величину силы, сопротивляемость утомлению). Определив взаимосвязь химической и физиологической реакций, количество ферментов можно использовать как коррелят физиологической реакции. Обычно анализируют три типа ферментов: один тип показывает скорость сокращения (миозин АФТаза — показатель мак-
Таблица5.1. Характеристика двигательных единиц и мышечных волокон трех мышц задней конечности кошки (Bochine, Roy, Eldred, Edgerton, 1987; Burke, 1981; Dum, Kennedy, 1980)
Исследуемый показатель Медиальная икроножная Большой сгибатель пальцев Передняя большеберцовая
Количество мышечных волокон 170000 26000 —
Количество двигательных единиц 270 130 —
Коэффициент иннервации
S 611 180 93
FR 553 132 197
FF 674 328 255
Средняя тетаническая сила, мН
S 76 11 40
FR 287 53 101
FF 714 300 208
Площадь мышечного волокна, мкм 2
I 1980 1023 2484
Па 2370 1403 2430
Пб 4503 2628 3293
Доля мышечных волокон, %
I 25 10 11
Па 20 37 50
Пб 55 53 39
155
Рис. 5.4. Типы мышечных волокон задней большеберцовой мышцы задней конечности кошки.
Тонкий поперечный разрез мышцы окрашен для определения активности АТФазы миозина с целью выявления распределения волокон типа I (темные участки), Па (белые участки) и Пб (серые участки). Всередине находится мышечное веретено с капсулой и небольшими интрафузальными мышечными волокнами
симальной скорости сокращения) (Вагапу, 1967), тогда как два других — метаболическую основу (аэробную или анаэробную) образования энергии мышечным волокном для осуществления сокращений. К ферментам аэробного метаболизма, которые чаще всего анализируют, относят сукцинатдегидрогеназу, никотинамид-нук-леотид-дегидрогеназу, тетразолиум-редуктазу, тогда как к ферментам анаэробного метаболизма — фосфорилазу и альфа-глицерофосфат дегидрогеназу.
Основываясь на изложенном анализе ферментов мышечных волокон, их можно разде
лить на три вида. В методе, в котором волокна классифицируют, исходя из миозин АТФазы, используются обозначения тип I, тип Па и тип Пб. Различия между I и II типами мышечных волокон основаны на количестве активности АТФазы, “остающейся” в мышечных волокнах после преинкубации в ванне при pH 9,4. Тип I — медленносокращающиеся мышечные волокна, тип II — быстросокращающиеся. Мышечные волокна типа II можно разделить еще на две группы — Па и Пб после преинкубации в ванне при pH 4,3 (Па) и 4,6 (Пб) (Brooke, Kaiser, 1974). Типы волокон показаны на рис. 5.4. В другом методе, основанном на скорости сокращения (миозин АТФаза), аэробной возможности (сукцинат дегидрогеназа или никотинамид-нуклеотид-дегидрогеназа тетразолиум-редукта-за) и анаэробной возможности (фосфорилаза и альфа-глицерофосфат дегидрогеназа) используют такие понятия: медленносокращающийся окислительный (тип МО), быстросокращаю-щийся окислительно-гликолитический (БОГ) и быстросокращающийся гликолитический (БГ).
Вообще волокна типа I соответствуют волокнам типа МО, волокна типа Па — волокнам типа БОГ, а волокна типа Пб — волокнам типа БГ. Вместе с тем имеются некоторые зависящие от вида различия (Nemeth, Pette, 1981). Мышечные волокна типа МО относятся к двигательной единице типа S, волокна типа БОГ — к двигательной единице типа FR, а волокна типа БГ — к двигательной единице типа FF. Поскольку все мышечные волокна принадлежат двигательной единице одного типа, они
Таблица 5.2. Количественное соотношение типов волокон и площадь поперечного сечения избранных мышц человека
Объект исследования п Тип I Тип II
% Площадь поперечного сечения, мкм2 % Площадь поперечного сечения, мкм2
Дельтовидная мышца*
12 50 5710 50 6490
8 61 5700 49 7010
>чники 9 71 5170 29 7430
егуны 9 49 5800 51 5850
7 54 5060 46 8910
Передняя большеберцовая мышца
5 60 2216 40 3338
Мужчины*** 10 66 3463 34 4959
Трапециевидная (средняя) мышца
Женщины**** 5 63 2746 37 2270
Мужчины***** 5 67 3853 33 4647
Латеральная широкая мышца бедра******
Женщины 203 51 4044 49 3409
Мужчины 215 46 4591 54 4814
Примечание: л — количество испытуемых.
’ Tesch, Karlsson (1985); *’ Henriksson-Laison (1985); "* Henriksson-Larsen, Friden, Wretling (1985); Lindman, Eriksson, Thomell (1991); Lindman, Eriksson, Thomell (1990); Simoneau, Bouchard (1989).
156
Рис. 5.5. Физиологические и гистохимические свойства двигательных единиц медиальной икроножной мышцы кошки (Burke, Edgerton, 1975)
обладают одинаковыми свойствами и поэтому гомогенны (Edstrom, Kugelberg, 1968; Nemeth et al., 1986). Например, двигательная единица типа S содержит мышечные волокна только типа МО, которым присущи одни и те же физиологические и биохимические свойства. Это же относится и к двум другим типам двигательных единиц, и к их мышечным волокнам.
Каждая мышца человека содержит совокупность всех трех типов волокон (Edgerton et al., 1975). Состав мышцы обычно определяют с помощью пункционной биопсии, извлекая мышечную ткань, подвергая ее окраске миозин АТФазой и затем классифицируя волокна по схеме I — Па — Нб. Следует отметить различия между типами мышечных волокон в мышце человека. Эти различия, включающие численный состав типов волокон и площадь попереч
ного сечения, колеблются в зависимости от предварительного опыта физических тренировок, пола и исследуемой мышцы (табл. 5.2). Вообще мышечные волокна типа II характеризуются большей площадью поперечного сечения по сравнению с волокнами типа I; исключение составляют мышцы бегунов, а также средняя трапециевидная и латеральная широкая мышцы бедра женщин.
На рис. 5.5 суммированы физиологические и гистохимические различия между двигательными единицами типа FF, FR и S. Каждая двигательная единица показана в виде сомы, от которой отходит аксон, спускающийся вертикально, разветвляющийся и иннервирующий 4 мышечных волокна. Различия в размере (например, для сомы — диаметр аксона, площадь поперечного сечения мышечного волокна)
157
указывает на общие морфологические различия между типами двигательных единиц. Под двигательными единицами показаны соответствующие диаграммы сила—время (сокращение) и восприимчивости к утомлению. Как следует из рисунка, двигательная единица типа FF характеризуется наибольшей силой сокращения, наименьшей, его продолжительностью и наибольшей восприимчивостью к утомлению. Рис. 5.5 также иллюстрирует изменяющееся воздействие входного сигнала афферента 1а из мышечного веретена на двигательный нейрон; двигательные единицы типа FF принимают меньше афферентов 1а и, следовательно, в меньшей степени реагируют (возбудительный постсинаптический потенциал) на входной сигнал 1а. Возбудительный постсинаптический потенциал 1а представляет собой запись потенциала напряжение—время. Чем выше возбудительный постсинаптический потенциал, тем легче возбуждается двигательный нейрон в результате входного сигнала из мышечного веретена.
На рис. 5.5 приведены также гистохимические диаграммы мышечных волокон различных двигательных единиц. Каждое волокно, прикрепленное к двигательной единице, демонстрирует гистохимическую структуру окраски различных ферментов. Анализировали четыре ферментные системы: миозин АТФазу (М-АТФаза), миофибриллярную АТФазу после кислотной преинкубации (для дифференциации волокон типа Па и Пб), окислительную и гликолитическую. Интенсивность окраски была значительной (высокая плотность фермента), промежуточной и светлой. М-АТФаза коррелирует с максимальной скоростью сокращения, следовательно, у мышечных волокон с темной окраской М-АТФазы высокая скорость сокращения двигательной единицы.
Функциональное значение
Почему существуют различные типы мышечных волокон и двигательных единиц? По-видимому, их многообразие в мышце увеличивает диапазон ее возможностей. Два механизма определяют процентное соотношение белков, — наследственность и используемость. Хотя и некоторые другие молекулярные различия между мышечными волокнами влияют на определенные физиологические свойства (Groves, 1989), остальные зависят от уровня активности волокон (табл. 5.2). В табл. 5.2 приведены показатели количественного соотношения и площади поперечного сечения мышечных волокон у разных спортсменов. Как показывают данные ряда исследований, эти показатели могут изменяться в результате интенсивных тренировок в течение нескольких недель (Saltin, Gollnick, 1983), а у людей, страдающих хронической сердечной
недостаточностью, количество волокон типа Пб увеличивается в результате детренированности, а не патологических изменений (Sbllivan, Green, Cogg, 1990). Таких изменений не было бы, если бы процентное соотношение типов не зависело от их использования.
Поскольку свойства мышечных волокон и двигательных единиц зависят от вида их использования, возникает вопрос, какие переменные, связанные с видом деятельности, вызывают изменения в мышце? Проще всего ответить на этот вопрос так: любая физиологическая переменная — величина силы, скорость сокращений и утомляемость — может привести к тому, что мышца лучше выполнит поставленную задачу. Например, Monster, Chan, O’Connor (1978) исследовали степень использования 15 мышц во время обычного 8-часового рабочего дня и установили, что чаще всего “работают” мышцы, содержащие большее количество волокон типа I. Можно предположить, что у мышц с более высокой концентрацией этих волокон более высокие окислительные способности и, следовательно, они в меньшей степени подвержены утомлению вследствие метаболических факторов. Другое различие между типами мышечных волокон — величина образуемой ими мощности. Мышечные волокна типа II образуют большую величину силы, характеризуются более высокой максимальной скоростью сокращения и способны образовывать большую величину мощности. Однако хотя волокна типа I образуют меньшую величину мощности, они делают это более эффективно (Brooks, Faulkner, 1991; Rome et al., 1988).
Следует, однако, отметить, что мышечная деятельность при выполнении некоторых заданий характеризуется недостаточной степенью корреляции с количеством мышечных волокон. При изометрической нагрузке до изнеможения 20, 50 или 80% максимальной продолжительности работы характеризовалось отсутствием взаимосвязи с типами волокон в латеральной широкой мышце бедра (Maughan, Nimmo, Harmon, 1985). Не наблюдали также взаимосвязи между максимальной скоростью выпрямления ноги в коленном суставе и количеством быст-росокращающихся мышечных волокон, хотя пик ускорения характеризовался незначительной степенью корреляции с количеством быстросо-кращающихся мышечных волокон (Houston, Norman, Froese, 1988). Наряду с этим наблюдали положительную корреляцию между количеством быстросокращающихся волокон в двуглавой мышце плеча и максимальной силой сгибания руки в локтевом суставе (Nygaard et al., 1983), а также между количеством волокон типа I и МПК (Mancini et al., 1989). Эти результаты показывают, что хотя уровни мышечной деятельности человека зависят от количества волокон и двигательных единиц, на них также влияют и другие факторы.
158
ВОЗБУДИМЫЕ МЕМБРАНЫ
Двигательная единица — основная функциональная единица контроля движения. Каким образом двигательный нейрон “заставляет” мышечные волокна работать? Иными словами, посредством какого механизма осуществляется взаимосвязь между этими двумя элементами? Взаимосвязь между двигательным нейроном и его волокнами осуществляется на быстром, электрическом, уровне (потенциал действия) и на более медленном, химическом (нейротропизм). Последний довольно часто не учитывают при изучении движения.
Нейротропизм
Нейротропизм означает продолжительное воздействие одного биологического элемента, например нейрона, непосредственно на другой, например на мышечные волокна. Хотя аксон обычно изображают в виде простого цилиндра, его внутренняя структура довольно сложна и обеспечивает быстрое перемещение веществ (аксонные транспортные системы) со скоростью 250—400 мм в день. Транспорт может осуществляться от сомы до концевой пластинки (прямой транспорт) и в обратном направлении (ретроградный транспорт) (рис. 5.6). Транспортируемые вещества включают структурные элементы, белки, рибонуклеиновую кислоту и аминокислоты. Например, важный этап электрохимической передачи информации из нейрона в мышцу — выделение пузырьком нейромедиатора в синаптическую щель. Спустя несколько циклов мембране пузырька необходим “ремонт”, она транспортируется обратно в сому, где происходит “ремонт” мембраны, затем пузырек транспортируется назад в нервно-мышечное соединение.
Транспортные механизмы, лежащие в основе нейротропизма, состоят из микроканальцев, вдоль которых молекулярные “моторы” могут перемещать органеллы. Эти “моторы”, включающие ферменты кинезин и цитоплазматический динеин, прикреплены к микроканальцам и функционируют подобно поперечным мостикам в мышце (Sheetz, Stener, Schroer, 1989). Каждый двигательный нерв перемещает органеллы только в одном направлении. Кинезин осуществляет прямоходящий транспорт, а цитоплазматический динеин способствует осуществлению ретроградного транспорта. Измерения, проводившиеся с использованием луча лазера, показали, что отдельная молекула кинезина способна образовывать величину силы порядка 2 пН и перемещать органеллы со скоростью около 0,5—2,0 мкм • с-1.
Имеются достаточно веские экспериментальные доказательства нервно-мышечного и
Рис. 5.6. Диаграмма систем аксонного транспорта (Kandel, Schwartz, 1985)
мышечно-нервного трофического влияния. Например, исследования влияния денервации показали существование нервно-мышечного тро-физма. При отсечении (денервации) нерва, идущего к мышце, в нервно-мышечной системе происходят существенные изменения: снижается мышечная масса (атрофия); дегенерируют мышечные волокна вследствие перемещения ядер в центр волокна и дезинтегрируются митохондрии; уменьшается образование силы; продолжительность сокращения увеличивается; потенциал покоя деполяризуется в течение 2 ч денервации; сопротивление мембраны в покое возрастает, отражая снижение ее проницаемости; изменяется структура натриевых канальцев; сарколемма становится способной спонтанно образовывать потенциалы действия; синтезируются внесоединительные ацетилхолиновые рецепторы; снижается активность ферментной ацетилхолинэстеразы, интактные двигательные аксоны образуют ответвления. Денервация устраняет образование потенциала действия, а также аксонный транспорт, однако возникновение этих изменений зависит от того, насколько далеко от мышцы отрезан нерв. Это наблюдение показывает существенную роль аксонного транспорта в регуляции нормальных свойств нервно-мышечной системы. Реиннервация (соединение нерва с мышцей) приводит к обратимости указанных изменений.
159
Рис. 5.7. Молекулы, связанные с возбудимой мембраной (Shepherd, 1988)
Точно так же повреждение аксона указывает на существование мышечно-нервного тропизма (Lowrie, Vrbova, 1992). Об этом свидетельствуют изменения электрофизического свойства двигательного нейрона: фазы постгиперполяризации потенциала действия двигательного нейрона (Czen, Gallego, Kudo, Kuno, 1978). При искусственной стимуляции аксона в течение 14 недель на участке блокады двигательного нейрона, при которой потенциал действия не достигает мышцы, сокращается продолжительность фазы постгиперполяризации. Наряду с этим при стимуляции мышечного участка блокады и активации мышцы фаза постгиперполяризации не изменяется. Эти результаты трактуют как доказательство того, что активация мышцы, хотя и в сочетании с искусственной стимуляцией, поддерживает “здоровье” двигательного нейрона, о чем свидетельствует фаза постгиперполяризации потенциала действия. При отсечении аксона изменяются и другие электрофизиологические свойства двигательных нейронов, например, мембранный потенциал покоя, скорость проведения возбуждения аксоном (Huizar, Kuno, Kudo, Miyata, 1978).
Интенсивность аксонного транспорта с возрастом может уменьшаться (Frolkis, et al., 1985). Замедление аксонного транспорта с возрастом — основной фактор старения нейронов.
Таким образом, аксонный транспорт представляет собой механизм, посредством которого мышца оказывает нейротропное воздействие на свои двигательные нейроны, которые, в свою очередь, влияют на свойства и “здоровье” мышечных волокон. Подобно другим физиологи
ческим процессам аксонный транспорт и, следовательно, нейротропные воздействия изменяются с возрастом, а также в зависимости от физической деятельности.
Электрические потенциалы
Из двух уровней нервно-мышечного взаимодействия больше известно о быстрой, электрической, форме. Этот тип взаимодействия возможен благодаря возбудимой природе аксолем-мы и сарколеммы — мембран, окружающих соответственно аксон и мышечное волокно. Чтобы охарактеризовать образование и функционирование электрических сигналов, необходимо выяснить, почему мембраны являются возбудимыми (имеют ненулевой стабильный потенциал), а также ионную основу потенциала действия и его образование.
Стабильный потенциал. Возбудимые мембраны полупроницаемы, т.е. некоторые липидорастворимые вещества и маленькие молекулы могут проходить через них. Эти мембраны называют клеточными оболочками. Белки мембраны, составляющие 50—70 % ее структуры и выполняющие структурные, ферментные, рецепторные, канальцевые и насосные функции, определяют ее возможности накапливать, передавать и выделять энергию. На рис. 5.7 показаны некоторые молекулы, связанные с клеточной мембраной. Вокруг мембраны находится внутриклеточная (саркоплазма, аксоплазма) и внеклеточная жидкость (рис. 5.8) с различной концентрацией ионов, особенно натрия (Na+), калия (К+) и хлора (СГ). Коэффициент
160
концентрации (за пределами и внутри клетки) этих ионов неодинаков (например, в мышечной клетке теплокровного животного Na+ — 12,08; К+ — 0,03, СГ — 30.00), поэтому чистым эффектом является распределение заряда (напряжения или электрического потенциала) по обе стороны мембраны.
Определенное распределение иона представляет собой равновесие между двумя силами - электрическим и концентрационным градиентами. Электрическая сила обусловлена притяжением разноименных зарядов и отталкиванием одноименных. Сила концентрационного градиента вынуждает ионы перемещаться из участка с высокой концентрацией в участок с низкой.
Когда мембрана находится в состоянии покоя, ее потенциал остается относительно постоянным. Он называется мембранным потенциалом покоя. Мембранный потенциал покоя обычно составляет около 60—90 мВ и внутри мембраны он отрицателен по отношению к внешней части. Ионы, участвующие в установлении электрического потенциала, — это ионы, находящиеся во внеклеточной и внутриклеточной жидкостях. Распределение ионов по обе стороны мембраны представляет собой равновесие между положительными и отрицательными ионами, и обусловлено силой концентрационного градиента, воздействующей на каждый ион (рис. 5.9). В состоянии покоя натриево-калиеный насос обеспечивает транспорт Na+ из внутриклеточной жидкости во внеклеточную. Связанный с местонахождением Na+ К+ остается внутри благодаря действию натриево-калиевого насоса и электрическому притяжению к различным органическим анионам (А), представляющим собой отрицательно заряженные ионы аминокислот и белков, которые из-за своего слишком большого размера не могут проходить сквозь мембрану. Таким образом, вследствие действия натриево-калиевого насоса Na+ удерживается во внеклеточной жидкости, тогда как К+ — во внутриклеточной. Заряды, обусловленные Na+ (притяжение) и А- (отталкивание), приводят к аккумуляции С1_ во внеклеточной жидкости. Распределение этих ионов (Na+ и СГ снаружи, К+ и А" внутри) во многом определяет стабильный потенциал возбудимой мембраны.
Силы, действующие на эти ионы (Na+, К+ и СГ), а также движение ионов через мембрану в устойчивом состоянии схематически показаны на рис. 5.10. Как уже отмечалось, силы, вызывающие движение ионов через мембрану (движущие или вынужденные силы), обусловлены химическими и электрическими воздействиями. Химическая сила связана с концентрационным градиентом. Величина каждой силы, действующей на различные ионы, по-
Рис. 5.8. Распределение ионов относительно возбудимой мембраны млекопитающего
казана на рис. 5.10 длиной стрелки. Например, для Na+ характерна более высокая внешняя (за пределами) концентрация, о чем свидетельствует большой символ Na+ за пределами мембраны, следовательно, на него действует химическая сила, направляющая его вовнутрь. Заряд внутри мембраны отрицательный. На положительно заряженные ионы (Na+ и К+) действует сила, направленная вовнутрь, тогда как на отрицательно заряженные ионы (СГ) —
Рис. 5.9. Концептуальное распределение зарядов ионов внутри клетки и за ее пределами (Koester, 1985)
11 8-133
161
Возбудимая мембрана
Рис. 5.10. Результирующие движущая сила и прохождение ионов Na+, К+ и СГ’ через мембрану при устойчивом мембранном потенциале порядка —60 мВ. Размеры обозначений указывают на силу воздействия (Koester, 1985)
сила, вынуждающая их перемещаться наружу. Величину результирующей движущей силы (результирующая ДС), действующей на каждый ион, определяют, сложив химические и электрические эффекты. Например, Na+ подвержен действию направленных вовнутрь химических и электрических эффектов, что приводит к большой направленной вовнутрь результирующей ДС для Na+. На К+ действует небольшая направленная кнаружи результирующая ДС, тогда как химические и электрические компоненты, влияющие на СГ, взаимно уничтожаются, обусловливая нулевую результирующую ДС.
По результирующей движущей силе для каждого иона и его проницаемости (Р) можно определить результирующую величину, а также направление движения ионов при стабильном мембранном потенциале. Легкость, с которой ион проходит сквозь мембрану (т.е. проницаемость мембраны для данного иона), отражена размером символа; возбудимые мембраны характеризуются наибольшей степенью
проницаемости для К+ (Рк) и наименьшей -для Na+ (PNa). Вследствие этих различий в степени проницаемости количество ионов, проходящих сквозь мембрану (результирующее поступление), определяется сочетанием результирующей движущей силы, действующей на ион, и его способностью проникать сквозь мембрану. Результат (рис. 5.10, столбик результирующего притока) показывает, что в стабильных условиях через мембрану проходит приблизительно одинаковое количество Na+ и К+, тогда как ионы СГ остаются внутри. Однако после прохождения сквозь мембрану Na+ и К+ они довольно быстро возвращаются в свои исходные участки благодаря действию натриево-ка-лиевого насоса. Таким образом, электрический потенциал в стабильных условиях остается относительно постоянным.
Ионная основа потенциала действия. Несмотря на очевидную стабильность мембранного потенциала покоя, вполне возможно нарушение ионного распада распределения и изменения мембранного потенциала. Например, дви
162
жение отрицательно заряженных ионов через мембрану наружу (или положительно заряженных внутрь) приводит к снижению мембранного потенциала, т.е. деполяризации (уменьшение поляризации или приближение к нулю). И наоборот, движение отрицательно заряженных ионов вовнутрь или накопление дополнительного количества положительно заряженных ионов снаружи вызывает гиперполяризацию мембраны (увеличение поляризации).
Перемещение ионов через мембрану обусловлено изменениями в самой мембране. Они могут быть вызваны химическим (нейромедиатор) или электрическим (изменение мембранного потенциала) путем. Их описывают как изменения проницаемости мембраны, обусловленные регулирующим действием и канальцами. Например, хотя Na+ проходит через мембрану, находящуюся в состоянии покоя, со значительными трудностями, проницаемость мембраны для Na+ может измениться в 500 раз, что значительно облегчает прохождение через нее. Это изменение называют увеличением проводимости или проницаемости для Na+. После того как Na+ проходит через мембрану, его концентрационный градиент (см. рис. 5.10) смещается из внеклеточной жидкости во внутриклеточную. Как только положительно заряженные ионы попадают внутрь клетки, мембрана деполяризуется, что приводит к дополнительному увеличению проницаемости для натрия и, следовательно, к дальнейшей деполяризации и т.д. Подобные изменения проводимости для Na+ зависят от регуляции напряжения канальцами, поскольку изменения электрического потенциала (напряжения) вызывают изменения проводимости, что, в свою очередь, приводит к дальнейшим изменениям электрического потенциала и т.д. Действие канальцев, регулирующих напряжение и передачу, иллюстрирует рис. 5.11. Здесь прохождение Na+ и К+ через соответствующие канальцы, регулирующие напряжение, физически предотвращает специальная “шлюзная” (или пропускная) молекула. Ориентация этой молекулы зависит от разности потенциалов на внутренней и наружной поверхностях мембраны. В состоянии покоя (верхняя диаграмма на рис. 5.11, а) молекула расположена таким образом, чтобы блокировать канал. После деполяризации потенциала молекула изменят свое положение и канал открывается (нижняя диаграмма на рис. 5.11, а). Точно так же канал, регулирующий передачу, оказывается закрытым в стабильных условиях и открывается, когда к нему присоединяется нейромедиатор и вызывает изменение его структуры. Как Na+, так и К+ могут пройти через один и тот же канал, регулирующий передачу. Вообще эти каналы инициируют изменения мембранного потенциала, что приводит к вовлечению каналов, регулирующих напряжение.
Канал, регулирующий напряжение
№+-канал
К+-канал
Мембрана в устойчивом состоянии
Канал, регулирующий передачу
Мембрана в устойчивом состоянии
Деполяризованная мембрана
Рис. 5.11. Функции регулирующих каналов мембраны (Kandel, Siegelbaum, 1985)
Связи в пределах нервно-мышечной системы обеспечивают изменения мембранного потенциала, обусловленные действием каналов, регулирующих напряжение и передачу. Эти изменения бывают двух видов — так называемые синаптические потенциалы и потенциалы действия. Механизм возникновения обоих потенциалов одинаков — перемещение ионов через мембрану. Вместе с тем синаптический потенциал является предвестником потенциала действия. Амплитуда синаптических потенциалов снижается при прохождении вдоль мембраны, тогда как потенциал действия включает процесс самовосстановления — распространение, благодаря которому изменения амплитуды оказываются минимальными.
С физической точки зрения потенциал действия представляет собой участок поверхности мембраны, в котором происходит обращение (реверсирование) потенциала, вследствие чего ее внутренняя часть оказывается положительно заряженной относительно внешней. Этот участок обратной полярности движется вдоль возбудимой мембраны. Потенциал действия можно записать, поместив внутрь клетки датчик (электрод) и регистрируя потенциал внутри клетки до, во время и после прохождения потенциалом действия электрода. При таком вну-
11*
163
потенциала действия (а) и изменения проводимости (б)
триклеточном измерении потенциал действия регистрируется как акт напряжения—времени (рис. 5.12). Потенциал действия характеризуется быстрой деполяризацией от уровня покоя до полного обращения потенциала (внутренняя часть становится положительно заряженной относительно внешней) с последующей реполяризацией (возвращение в состояние поляризации), превышающей потенциал покоя и достигающей состояния гиперполяризации, и медленным возвращением к мембранному потенциалу покоя. Эти изменения мембранного потенциала — следствие изменения проницаемости для Na+ и К+ (gNa и gK), которая временно изменяется (рис. 5.12, б). Поскольку Na+ и К+ — положительно заряженные ионы, их одновременное прохождение через мембрану не влияет на мембранный потенциал. Однако вследствие того, что результирующий приток Na+ происходит до оттока К+, а также от того, что повышенная проводимость для Na+ длится лишь небольшой отрезок времени, мембрана подвергается сначала деполяризации, а затем реполяризации. Поэтому в любой момент времени мембранный потенциал (Ут) представляет собой дисбаланс между проводимостью Na+ и К +. Количество ионов одного вида (например, Na+ или К+), пересекающих мембрану, зависит от движущей силы и проницаемости для данного иона (например, Na+, К+, СГ). По завершении потенциала действия проводимость натрия воз
вращается к характерной для состояния покоя, а проводимость калия остается выше обычной, результирующим эффектом является гиперполяризация мембраны. Поскольку она происходит после потенциала действия, то называется постгиперполяризацией. Натриево-калиевые насосы постепенно изменяют распределение ионов, возвращая Na+ во внеклеточную жидкость, а К+ во внутриклеточную. В результате этого мембранный потенциал возвращается к стабильному уровню.
После инициации нейронного потенциала действия он распространяется вдоль аксона со скоростью до 120 м'С^1. Поскольку потенциал действия представляет собой реверсивность распределения ионов через мембрану (2 на рис. 5.13, 6), его и процесс его распространения можно представить так, как на рис. 5.13, где электрический потенциал (рис. 5.13, а) показан относительно распределения ионов (рис. 5.13, б).
Скорость распространения потенциалов действия зависит от диаметра аксона, в основном ввиду того, что большая часть поверхности больших аксонов покрыта миелином и поэтому не участвует в процессе перемещения ионов. Поэтому потенциал действия “перепрыгивает” с одного перехвата Ранвье (участок истончения оболочки миелинового нервного волокна) на другой. Это так называемое скачкообразное проведение.
Образование потенциала действия. Как уже отмечалось, физическое появление потенциала
Рис. 5.13. Распространение потенциала действия (а) и связанное с ним распределение ионов (б) (Koester, 1985)
164
действия представляет собой участок мембраны с временно обратной полярностью; изменение мембраною потенциала обусловлено изменениями проницаемости мембраны для Na+ и К+, так что данная обратимость полярности может “продвигаться” вдоль возбудимой мембраны. Однако каким образом клетки образуют потенциал действия? Чтобы ответить на этот вопрос, необходимо прежде всего рассмотреть образование потенциала действия нейроном (аксонный потенциал действия), а затем остановиться на образовании потенциала действия сарколеммой мышечного волокна (потенциал действия мышцы).
Потенциал действия представляет собой электрический сигнал по типу “все или ничего”, который клетка посылает в ответ на принимаемый сигнал. Потенциалы действия могут образовывать как нейроны, так и мышечные волокна. Сигналы, принимаемые нейроном, которые лежат в основе образования потенциала действия, имеют вид синаптических потенциалов (именно поэтому синаптический потенциал считают предвестником потенциала действия). Синаптические потенциалы бывают либо возбуждающие, либо тормозящие, они могут либо деполяризовать, либо гиперполяризовать принимающую (постсинаптическую) мембрану. Тормозящие синаптические потенциалы обусловлены повышенной проводимостью для небольших ионов, таких, как К+ и СГ. Величина синаптического потенциала зависит от ряда факторов (Burke, 1981), средняя амплитуда возбуждающих синаптических потенциалов, обусловленных отдельными афферентами группы 1а в двигательных нейронах, составляет 71 мВ для двигательных единиц типа FF, 118 мВ для единиц типа FR и 179 мВ для единиц типа S (Stuart, Enoka, 1983).
Нейрон принимает до 10 000 синапсов из различных источников, таких, как периферические чувствительные нервные окончания, интернейроны, супраспинальные центры. Эти синапсы распределяются по соме и дендритному дереву. При выделении нейромедиатора у синапса последний образует синаптический потенциал в постсинаптическом нейроне. Синаптический потенциал проходит по нейрону и затем вдоль мембраны. В последнем случае его амплитуда понижается. Чем дальше синапс от аксонного холмика, тем менее эффективно синаптический потенциал перемещает мембранный потенциал к порогу потенциала действия нейрона (уровень включения образования потенциала действия). Нейрон не образует потенциал действия всякий раз, когда принимает синаптический потенциал, образование потенциала действия основано на суммировании синаптических потенциалов на аксонном холмике. Нейрон принимает одновременно много импульсов (синаптических
Рис. 5.14. Активация регулирующего передачу (а) и напряжение каналов (б) в нервно-мышечном соединении в ответ на выделение АХ (Kandel, Siegelbaum, 1985)
потенциалов), одни из них возбуждающие, другие — тормозящие. Непрерывность поступления импульсов вызывает колебания мембранного потенциала, превышающие средний стабильный уровень. Когда мембранный потенциал в участке аксонного холмика на 10 — 15 мВ превышает стабильный уровень, достигается порог и нейрон образует потенциал действия, который распространяется вдоль аксона к клетке-эффектору. Таким образом, нейрон образует потенциал действия в ответ на непрерывное поступление импульсов (синаптических потенциалов).
Вообще синаптические потенциалы образуются и проводятся вследствие изменений, влияющих на проводимость мембраны, тогда как потенциалы действия зависят от регулирующих напряжение изменений проводимости. Это различие иллюстрирует пример образования потенциала действия сарколеммой мышечного волокна. Процесс происходит в нервно-мышечном соединении, в котором нейромедиатором выступает ацетилхолин (АХ). При выделении АХ в синаптическую щель он диффундирует через щель и прикрепляется к рецептору на регулирующем передачу Иа+-К+-канале (рис. 5.14, а). Когда канал открывается, Na+ поступает в мышечную клетку, а К+ выделяется из нее, что ведет к образованию синаптического потенци
165
ала, который называют потенциалом концевой пластинки. Поскольку потенциал концевой пластинки вызывает среднюю деполяризацию сарколеммы, это приводит к открытию регулирующих напряжение Ка+-каналов (рис. 5.14, б), что еще больше деполяризует сарколемму и смещает ее к порогу. Постепенно сарколемма в области нервно-мышечного соединения оказывается достаточно деполяризованной для достижения порога; образуется и распространяется вдоль возбудимой мембраны сарколемный потенциал действия.
Уникальной особенностью нервно-мышечного соединения является мощность потенциала концевой пластинки. В отличие от большинства синаптических потенциалов потенциал концевой пластинки крайне необходим, поскольку его возникновение обычно приводит к образованию сарколемного потенциала действия. Только в условиях утомления связь между потенциалом концевой пластинки и потенциалом действия прерывается.
Следует отметить, что нейромедиатор АХ выделяется не только в ответ на аксонный потенциал действия, в небольших количествах он выделяется постоянно. Это спонтанное выделение АХ вызывает миниатюрные потенциалы концевой пластинки с амплитудами около 0,7 мВ в быстросокращающихся мышечных волокнах и около 1,9 мВ в медленносокращающихся (Lomo, Waerhang, 1985). Интересно, что частота этих потенциалов снижается с возрастом в камбаловидной мышце, но не в длинном разгибателе пальцев (Alshuaib, Fahim, 1990).
Электромиография
Ученые разработали метод контроля электрической активности возбудимых мембран — электромиографию (ЭМГ; myo (лат.) — мышца). Поскольку ЭМГ представляет собой измерение сарколемных потенциалов действия, ее можно рассматривать как “окно” в нервную систему (Denny-Brown, 1949; Duchene, Gonbel, 1993; Hof, 1984; Loeb, Gans, 1986; Person, 1963). Чаще всего ЭМГ измеряют, помещая электрод вблизи возбудимой мембраны и регистрируя проходящие через электрод потенциалы действия. Электрод может находиться внутри мышцы, вне клетки (рис. 5.15) или же вне мышцы. Чаще всего электрод помещают на поверхность кожи.
Потенциал действия, записанный таким образом, как правило, имеет трехфазную форму (рис. 5.15, а). Эту форму объясняет график на рис. 5.16, показывающий взаимосвязь между перемещением ионов через мембрану и регистрируемым потенциалом действия. Электрод расположен вблизи возбудимой мембраны вне клетки и регистрирует потенциал в
Рис. 5.15. ЭМГ одного (а) и многих (6) потенциалов действия. Обратите внимание на временную разницу
данном участке по мере приближения, достижения и прохождения потенциалом действия электрода. На рис. 5.16 электрод на графике показан справа, это — передний фронт потенциала действия. Центральным элементом потенциала действия является участок обратимой полярности. Это обусловлено поступлением ионов Na+ и представляет отрицательную фазу потенциала действия; отрицательный потенциал соответствует поступлению положительно заряженных ионов в клетку. С каждой стороны отрицательной фазы находится отдельная положительная фаза. Начальная положительная фаза (правая часть графика) обусловлена пассивной локальной деполяризацией мембраны (т.е. мембрана менее поляризована). Это вызвано тем, что некоторые ионы Na+ направляются из участка обратной полярности во внутриклеточную жидкость вследствие электрического притяжения положительно заряженных ионов отрицательно заряженными, а некоторые отрицательно заряженные ионы перемещаются из внеклеточной жидкости в обратном направлении. Локальная активность перед участком обратной полярности воспринимается электродом как поток положительно заряженных ионов из клетки и измеряется как положительное изменение напряжения. Вторая положительная фаза потенциала (слева на графике) обусловлена оттоком К+ из клетки. Электрод, таким образом, регистрирует изменение мембранного потенциала вследствие движения положительно заряженных ионов из клетки как положительное, а обусловленное их движением в клетку — как отрицательное.
Электрод выполняет роль своеобразной антенны (Loeb, Gans, 1986) и может “работать” в однополюсном либо двухполюсном режиме. В первом случае один электрод размещается над
166
источником сигнала (например, над мышцей) и воспринимаемый им сигнал (обусловленный поступлением Na+ и оттоком К+) сопоставляется с сигналом, исходящим из отдаленного участка (земли). Во втором случае над источником сигнала устанавливают два электрода и выходной сигнал представляет собой разницу между сигналами, воспринимаемыми двумя электродами. Таким образом, при использовании однополюсного режима необходимо определять абсолютную величину сигнала, тогда как двухполюсный режим позволяет избирательно осуществить запись с локального источника сигнала.
Величина электрической активности, которую можно записать в мышце, зависит от расстояния между электродами и активными двигательными единицами, площади поверхностей, с которых осуществляется запись, и расположения электродов. При анализе движения человека ЭМГ, как правило, записывают с помощью одной пары электродов (диаметр которых около 8 мм), расположенных на расстоянии 1,5 см друг от друга на поверхности кожи над мышцей. В таком случае нельзя записать всю ЭМГ мышцы, а лишь потенциалы мышечных волокон, находящихся в 1—2 см от электродов (Fuglevaud, Winter, Patla, Stashuk, 1992). Чтобы записать ЭМГ всей мышцы, необходимо либо использовать целый ряд электродов (Chanaud, Pratt, Loeb, 1987; Thusneyapan, Zahalak, 1989), либо один электрод с большой записывающей способностью, помещенный внутрь мышцы (Stalberg, 1980). Последнее называют макроЭМГ. Этот метод позволяет записывать большинство потенциалов действия мышечных волокон активной двигательной единицы.
Естественно, анализируя движения человека мы не регистрируем потенциал действия отдельного мышечного волокна или отдельной двигательной единицы. Чаще всего ученые определяют ЭМГ многих двигательных одновременно активных единиц. Этот тип записи ЭМГ называют интерференционной структурой, поскольку имеет место наложение многих потенциалов действия (рис. 5.17; сравните шкалу времени на рис. 5.17 с такой же шкалой отдельного потенциала действия на рис. 5.15). Ученых также интересует количественное определение интерференционной ЭМГ. Эля этого существует ряд методов (Davis, Vaughn, 1993; Loeb, Gans, 1986; Perry, Bekey, 1981). Чаще всего используют метод ректификации и интеграции. Как следует из рис. 5.17, метод ректификации предполагает определение абсолютного значения ЭМГ сигнала, чтобы “перепрыгнуть” через отрицательные фазы может быть использован электронный модуль. Затем острые пики (высокие частоты), присутствующие в ректифицированной (исправленной) ЭМГ, могут
Рис. 5.16. Взаимосвязь движения ионов через сарколемму (верхний ряд) соответствующего тока (средний ряд) и формы внеклеточно записанного потенциала действия (нижний ряд) (Loeb, Gans, 1986)
быть уменьшены благодаря электронному процессу интеграции, включающему сглаживание или фильтрацию ЭМГ для снижения высокочастотного содержания сигнала. После исправления и интеграции интерференционной ЭМГ ее можно определить количественно, измерив амплитуду интегрированной ЭМГ. Целесообразность данного метода количественного определения демонстрирует сопоставление изменений интегрированной ЭМГ и силы на рис. 5.17. Для получения данных, представленных на рис. 5.17, мы постепенно увеличивали силу от нуля до определенного значения и затем измеряли интерференционную ЭМГ и силу. После исправления и интеграции запись ЭМГ соответствовала записи силы, что свидетельствует о том, что сигнал ЭМГ в этих условиях
Рис. 5.17. Интерференционная ЭМГ (7) ректифицируется и интегрируется, вследствие чего результирующий импульс (интегрированная ЭМГ) почти соответствует изменению силы (4)
167
Рис. 5.18. Интегрированная взаимосвязь ЭМГ—сила камбаловидной (/) и двуглавой (2) мышцы плеча при изометрическом сокращении. ЭМГ и сила стандартизованы относительно максимальных показателей (Bigland-Ritchie, 1981)
можно считать надежным показателем величины мышечной силы.
Между интегрированной ЭМГ и силой, образуемой мышцами, отмечается тесная взаимосвязь (Bigland-Ritchie, 1981; Fuglevand, Winter, Patla, 1993). Однако эта взаимосвязь ограничивается только изометрическими условиями, при которых мышца сокращается, не изменяя своей длины (рис. 5.18). В этих условиях между интегрированной ЭМГ и силой может наблюдаться линейная или криволинейная взаимосвязь (Lawrence, Deluca, 1983). Это означает, что
Рис. 5.19. Измерение ЭМГ и силы трехглавой мышцы плеча при различной длине. У испытуемого была ампутирована рука ниже локтя и мышца подсоединена к преобразователю силы (Heckathome, Childress, 1981)
величина ЭМГ позволяет достоверно оценить величину силы, образуемой мышцей. При изменении длины мышцы (рис. 5.19) изменяется расположение электродов относительно активных волокон, что приводит к изменению ЭМГ, не связанному с принимаемым мышцей сигналом из нервной системы (Gerilovsky, Tsvetinov, Trenkova, 1989). Именно поэтому в неизометрических условиях очень трудно количественно оценить ЭМГ (Bigland-Ritchie, 1981; Milner-Brown, Stein, 1975; Haf, van der Berg, 1981a, 19816, 1981b, 1981г).
Интерференционная ЭМГ, записанная во время физической активности, обычно изображается как функция времени (см. рис. 5.17). Ее количественное определение осуществляется путем ректификации и интеграции. Этот подход называют анализом во временной области. Альтернативный подход — анализ в частотной области — используется для определения частотного содержания сигнала ЭМГ.
Основным фактором, влияющим на частоту ЭМГ, является форма составляющих потенциалов действия (рис. 5.20). При разрядке или утомлении двигательной единицы форма потенциала действия изменяется: амплитуда снижается, а продолжительность увеличивается (рис. 5.20, а). Эти изменения проявляются в частоте сигнала в начале и в конце вызывающих утомление сокращений. Анализ в частотной области включает устранение времени между последовательными потенциалами действия с тем, чтобы они “появлялись” как периодическая функция времени (рис. 5.20, б). Каждая последовательность потенциалов действия представляется затем наиболее подходящим сочетанием синусоидально-косину-соидных функций для характеристики частоты и амплитуды сигнала; полученный результат наносится на график частоты — амплитуды. Поскольку последовательность потенциалов действия отдельной двигательной единицы на рис. 5.20 можно представить как отдельную си-нусоидально-косинусоидную функцию, ее можно охарактеризовать отдельной линией (амплитуды и частоты) в частотной области.
Поскольку интерференционная ЭМГ содержит сотни потенциалов действия различной формы и степени взаимного перекрывания, частотное содержание не ограничено отдельной частотой. Частоты образуют данный спектр, а амплитуда каждой из них колеблется. Следовательно, для представления интерференционной ЭМГ необходимо объединить множество синусоидально-косинусоидных функций различной амплитуды и частоты (интенсивность изменения). Вообще амплитуду каждой частоты измеряют как мощность, а этот вид графика называют спектральной функцией. Пример интерференционной ЭМГ и спектральной функции показан на рис. 5.21. Для характеристики спектральной функции чаще всего исполь-
168
Рис. 5.20. Анализ в частотной области потенциалов действия отдельной двигательной единицы в начале и в конце вызывающего его утомление сокращения: а — последовательность потенциалов действия в начале (верхняя запись) и в конце (нижняя запись) вызывающего утомление сокращения; б — “устранение” времени между последовательными потенциалами действия, с тем чтобы они имели вид периодических функций, которые можно охарактеризовать в частотной области на графике частоты — амплитуды
зуют среднюю частоту, т.е. центральное значение частотного спектра. С другой стороны, срединная частота — это частота, разделяющая спектр на две половины, исходя из энергетического содержания сигнала. В большинстве случаев средняя и срединная частоты дают сопоставимую информацию, хотя последняя — более чувствительная мера во время вызывающих утомление сокращений (Merletti, Knaflitz, De Luka, 1990).
Спектральный анализ обычно осуществляют, чтобы обосновать изменения ЭМГ, возникающие при утомлении (Hagg, 1992). Два основных фактора, вызывающих зависимое от активности изменение частотного содержания интерференционной ЭМГ, это — изменение формы потенциала действия (рис. 5.20) и снижение скорости разряда двигательной единицы. На форму потенциала действия больше всего влияют изменения скорости проведения возбуждения вдоль сарколеммы (De Luca, 1984; Lindstrom, Maguusson, Petersen, 1970). При снижении скорости проведения увеличивается продолжительность потенциала действия и снижается его частотное содержание, что вызывает смещение средней и срединной частоты к более низким значениям. Кроме того, скорость разряда двигательных единиц уменьшается при продолжительной активности и низкой интенсивности (8—12 Гц), что может
привести к “группированию” потенциалов действия в очевидные всплески активности (Taylor, 1962). Это не отражает синхронизацию двигательных единиц, как считают некоторые исследователи. Снижение скорости разряда вследствие этого “группирования” направлено на образование выступающего пика при данной частоте (около 10 Гц) в спектральной функции (Krogh-Lund, Jogensen, 1992). Вследствие снижения скорости проведения и разряда средняя и срединная частоты спектральной функции уменьшаются еще больше во время вызывающих утомление сокращений. Более того, это смещение влево — одно из первых явлений, происходящих при возникновении утомления мышцы.
Фономиография. Сокращение мышцы основано на ее электрической активации нервной системой. Когда мышца сокращается, возникают звуки.
Первое сообщение о “звучании” мышцы появилось в 1663 г. (цит. по Oster, 1984). Мы не слышим эти звуки ввиду их низкой частоты, невосприимчивой для уха человека. Средняя частота этих звуков составляет около 10 Гц при небольшой силе сокращения и около 22 Гц — при максимальном произвольном сокращении; для сравнения: средняя частота ЭМГ при сокращениях средней силы — около 60 Гц (Maton, Petitjean, Chockaert, 1990; Orizio, Perini, Diemont et al., 1990; Stokes, Cooper, 1992). Эти звуки, однако, можно услышать с помощью соответ-
0 1,0 2,0 3,0
Время, с
Рис. 5.21. Спектральная функция (б) интерференционной ЭМГ (а)
169
ствующего микрофона. По-видимому, они обусловлены латеральными движениями мышечных волокон при их сокращении. Поскольку звук представляет собой серию волн сжатия, его можно записать. Сигнал напоминает интерференционную ЭМГ (рис. 5.17), поэтому его можно анализировать, используя методы, применяемые для анализа ЭМГ (ректификация и интеграция). Метод записи мышечных звуков называется фономиографией (ФМГ). Другое название — звуковая, или акустическая, миография.
ФМГ, как и ЭМГ, в изометрических условиях непосредственно связана с мышечной силой; вместе с тем между ФМГ и мышечной силой наблюдается не линейная, а квадратичная взаимосвязь (Maton et al., 1990; Stokes, Moffroid, Haugh, 1988). Корреляционные анализы, однако, показывают, что ЭМГ и ФМГ изменяются независимо друг от друга при увеличении и последующем уменьшении вращающего момента и ФМГ колеблется больше, чем ЭМГ. Наблюдаются также значительные различия между ЭМГ и ФМГ при субмаксимальных сокращениях до изнеможения. При сокращениях средней интенсивности (60—80 % максимального произвольного сокращения), ЭМГ в конце сокращения больше, чем в начале, тогда как ФМГ в конце меньше, чем в начале (Orizio, Perini, Veicsteinas, 1989). Эти наблюдения свидетельствуют, что сигналы ЭМГ и ФМГ содержат различную информацию о мышечном сокращении.
ВЗАИМОДЕЙСТВИЕ ВОЗБУЖДЕНИЯ - СОКРАЩЕНИЯ
В нервно-мышечном соединении нейромедиатор ацетилхолин диффундирует через синаптическую щель и прикрепляется к рецепторам постсинаптической мембраны в течение менее 100 мс. В результате прикрепления АХ к постсинаптическим рецепторам открывается Na+-К+-канал, регулирующий передачу, ионы Na+ поступают, а ионы К+ покидают мышечное волокно. Это движение Na+ и К+ через сарколемму приводит к образованию потенциала концевой пластинки (синаптического потенциала), который “включает” возникновение потенциала действия сарколеммы. Процессы, осуществляющие превращение потенциала действия аксона в потенциал действия сарколеммы, называют нервно-мышечным распространением.
После превращения потенциала действия аксона в потенциал действия сарколеммы сигнал (потенциал действия), образуемый двигательным нейроном в результате ряда процессов, превращается в силу мышечного волокна. Эти процессы называют взаимодействием возбуждения — сокращения. Взаимодействие (рис. 5.22) вклю-
Рис. 5.22. Взаимодействия возбуждения—сокращения (Fitts, Metzger, 1988)
чает: 1) распространение потенциала действия сарколеммы; 2) распространение потенциала действия ниже поперечной трубочки; 3) взаимодействие потенциала действия с изменением Са2+-проводимости в саркоплазматическом ретикулуме; 4) выделение Са2+ из саркоплазматического ретикулума; 5) повторное поглощение Са2+ саркоплазматическим ретикулумом; 6) связывание Са2+ с тропонином и 7) взаимодействие поперечного мостика (миозинового) с актином. Большинству из этих этапов присущи явления, обеспечивающие взаимодействие актина и миозина (растормаживание Са2+), и только этап 7 действительно включает взаимодействие актина и миозина (цикл поперечного мостика).
Растормаживание Са2+
В состоянии покоя толстый и тонкий филаменты не могут взаимодействовать в результате регулирующего действия белков — тропонина и тропомиозина. Са2+ в основном содержится в саркоплазматическом ретикулуме. Этапы 1—6 на рис. 5.22 описывают явления, ведущие к его растормаживанию. Они позволяют потенциалу действия сарколеммы включить выделение Са2+ из саркоплазматического ретикулума, а также последующее торможение регуляторного действия тропонина и тропомио-
170
a
Сарколемма
Поперечная трубочка
Саркоплазматический ретикулум
Рис. 5.23. Роль Са2+ в сочетании возбуждения—сокращения (Riiegg, 1983)
тический ретикулум благодаря Са2+-насосам, прикрепленным к мембране саркоплазматического ретикулума. Интенсивность возвращения Са2+ в саркоплазматический ретикулум определяет интенсивность уменьшения силы после прекращения потенциалов действия.
Как только концентрация Са2+ в саркоплазме превысит пороговый уровень (10“7 м), Са2+ присоединяется к элементу TN—С регуляторного белка тропонина. Считают, что присоединение Са2+ к тропонину вызывает структурное изменение тонкого филамента — участок присоединения миозина на актине оказывается открытым (раскрытым) и два белка (актин и миозин) получают возможность взаимодействовать (рис. 5.24). Это “открывание” участка присоединения, очевидно, включает временную ротацию регуляторного комплекса (тропонин—тропомиозин). После прекращения потенциала действия gCa саркоплазматического ретикулума возвращается к уровню, характерному для состояния покоя, а Са2+-насосы начинают возвращать Са2+ в саркоплазматический ретикулум. Это приводит к снижению концентрации Са 2+ в саркоплазме, что, в свою очередь, вызывает торможение активности фермента (актомиозин-АТФазы), регулирующего взаимодействие актина и миозина.
зина. Конечный результат этих явлений — растормаживание.
Растормаживание Са2+ начинается с потенциала действия мышцы по мере его распространения вдоль сарколеммы со скоростью до 6 м • с-1. Интересно, что скорость распространения изменяется вследствие тренировок: у культуристов наблюдали более высокую скорость проведения возбуждения в двуглавой мышце плеча по сравнению с контрольными испытуемыми (соответственно 5,5 и 2,8 м • с-1) (Kereshi, Manzano, McComas, 1983). На рис. 5.23 продемонстрирована роль Са2+ во взаимодействии возбуждения—сокращения. В состоянии покоя потенциал действия сарколеммы отрицательный относительно внешней части и большая часть Са2+ находится в терминальных цистернах (рис. 5.23, а). Потенциал действия распространяется ниже поперечной трубочки и вызывает увеличение проницаемости для Са2+ (gCa) терминальных цистерн. При отсутствии потенциала действия gCa обычно низкая, поэтому Са2+ не так легко проникает сквозь мембрану саркоплазматического ретикулума. С увеличением gCa ионы Са получают возможность переместиться из терминальных цистерн в саркоплазму (рис. 5.23, б). Изменение gca носит временный характер и после прекращения потенциала действия (рис. 5.23, в) gCa возвращается к исходному уровню, а Са2+ возвращается в саркоплазма-
Цикл поперечного мостика
Взаимодействие актина и миозина, обусловленное растормаживанием Са2+, называют циклом поперечного мостика. Оно включает три
Рис. 5.24. Схема “раскрывания” участков присоединения (Пр) на актине после прикрепления Са2+ к С-компоненту тропонина (Тр) (Gergely, 1974): а — тонкий филамент в состоянии покоя; б —“раскрывание” участка привязывания
171
фазы: присоединение, вращение и отсоединение (рис. 5.25). В состоянии покоя актин и миозин не могут взаимодействовать (рис. 5.25, а). Как только на актине появляются участки присоединения, поперечный мостик (S1-шарообразные головки) миозина быстро присоединяется к актину (рис. 5.25, б). Каждый S1-субфрагмент содержит две шарообразные головки: одна имеет участок присоединения для взаимодействий актина и миозина, другая — ферментную миозин-АТФазу для катализа (гидролиза АТФ (АТФ -> АДФ + Рн + энергия), обеспечивающего энергию для цикла поперечного мостика (Cooke, 1990; Huxley, 1985; Lieber, 1992; Zahalak, 1990).
Мышечное сокращение представляет собой превращение химической энергии в механическую. Точный механизм превращения энергии неизвестен; считают, что после образования поперечного мостика вследствие расщепления фосфата из АТФ образуется энергия для вращения поперечного мостика (Huxley, 1985). Согласно этой схеме S1 вращается относительно S2 по направлению к фрагменту LMM (рис. 5.25, в). Поскольку вращение осуществляется при связанных актине и миозине, происходит скольжение толстого и тонкого филаментов относительно друг друга и образуется сила, действующая на Z-линии. Это так называемая теория скольжения. Движение осуществляется в каждом саркомере вдоль миофибриллы, а сила передается через окончания каждой миофибриллы и матрикс соединительной ткани вокруг каждой миофибриллы и каждого мышечного волокна. Сила, образуемая мышцей, обусловлена
цикличностью множества поперечных мостиков после растормаживания Са2+. Мы описываем эту активность как мышечное сокращение и определяем сокращение как состояние активации мышцы, при котором происходит цикличная смена поперечных мостиков в ответ на потенциал действия.
Находясь в состоянии вращения S1 присоединяет (рис. 5.25, в), другую молекулу АТФ, а миозин отсоединяется от актина (рис. 5.25, г). При отсутствии АТФ (в случаях продолжительного непотребления пиши и последующей смерти) отсоединение не происходит, и актин и миозин остаются связанными; это состояние называется трупным окоченением. После отсоединения поперечные мостики возвращаются в исходное положение и могут быть использованы для “открывания” другого участка связывания. По завершении электрических явлений в сарколемме проницаемость саркоплазматического ретикулума достигает обычного низкого уровня, Са2+-насос (расположенный в мембране саркоплазматического ретикулума) возвращает Са2+ в саркоплазматический ретикулум, а тормозящий эффект комплекса тропонин—тропомиозин восстанавливается. Последовательность этих явлений показана в табл. 5.3.
Продолжительность различных фаз, а также изменения длины поперечного мостика колеблются в зависимости от нагрузки. Если сила саркомера и нагрузка одинаковы, длина саркомера постоянна. Длина саркомера уменьшается, если его сила превышает нагрузку, и увеличивается, если нагрузка превышает силу сарко-
172
Таблица 5.3. Последовательность явлении в мышечном сокращении
1. Инициация и распространение (активное проведение) потенциала действия из аксонного холмика.
2. Выделение нейромедиатора ацетилхолина (,\Х) из нервно-мышечного соединения.
3. АХ повышает проницаемость постсинаптической мембраны (сарколеммы) для Na+ и К+, что вызывает активацию каналов, регулирующих передачу и образование потенциала концевой пластинки.
4. Потенциал концевой пластинки проводится от нервно-мышечного соединения, активируя каналы, регулирующие напряжение, и, деполяризуя сарколемму. В результате образуется потенциал действия мышцы (сарколеммы).
5. Потенциал действия мышцы распространяется вдоль сарколеммы и в систему поперечных трубочек (Т-трубочек).
6. Потенциал действия Т-трубочек вызывает увеличение проводимости саркоплазматического ретикулума для кальция (gca) и Са2+ перемещает концентрационный градиент в саркоплазму.
7. При адекватной концентрации Са2+ в саркоплазме он связывается с тропонином, растормаживая регуляторные белки.
8. Субфрагмент 1 (S1) прикрепляется к участку присоединения актина и энергия, образованная вследствие расщепления АТФ, используется для вращения поперечных мостиков.
9. Поперечный мостик отсоединяется, как только другой АТФ присоединяется к S1.
10. Цикл присоединения, вращения и отсоединения поперечных мостиков продолжается при наличии АТФ и достаточном для растормаживания количестве Са2+.
11. После прекращения потенциала действия мышцы Са2+ откачивается назад в саркоплазматический ретикулум и возвращается в латеральные мешочки.
12. Выведение Са2+ возобновляет процесс торможения регуляторными белками (тропонином и тропомиозином), и поскольку не происходит прикрепления поперечных мостиков, миофиламенты расслабляются.
мера. При увеличении длины саркомера возрастает сила каждого поперечного мостика, они могут принудительно отсоединиться, а скорость их повторного присоединения может в 200 раз превышать скорость прикрепления в обычных условиях (Lombardi, Piazzesi, 1990; Morgan, 1990).
Получение изображения с помощью магнитного резонанса
Метод получения изображения с помощью магнитного резонанса используется для регистрации мышечных сокращений. Этот метод, по
зволяющий локализовать атомные ядра, может быть использован для изучения структуры и функции единой системы сустава. Он включает три этапа: 1) экспонирование объекта в магнитном поле, которое “выстраивает” в ряд химические элементы с нечетной молекулярной массой; 2) нарушение данной структуры расположения и 3) определение скорости, так называемой скорости релаксации, с которой компоненты и ядра атомов возвращаются в исходное положение. Две основные скорости релаксации — продольная (Т1) и поперечная (Т2). Увеличение продолжительности Т1 и Т2 релаксации коррелирует с содержанием воды в ответ на некроз мышцы. Поэтому метод получения изображения с помощью магнитного резонанса используется для изучения мышечной функции.
Процедура осуществляется следующим образом. Испытуемый выполняет предписанное задание, затем часть тела, которая была активной, помещается на магнит для измерения сигнала изображения магнитного резонанса, длящегося несколько минут. Подобным образом получают информацию о кумулятивном влиянии нагрузки (активность поперечного мостика). Скорость релаксации Т2, например, характеризует обусловленное нагрузкой изменение содержания внутриклеточной воды в мышечных волокнах после мышечного сокращения (Fleckenstein, Canby, Parkey, Peshock, 1988; Fisher, Meyer, Adams et al., 1990). На продолжительность релаксации T2 могут влиять: содержание внеклеточной воды, внутриклеточный pH, уровень насыщенности крови кислородом, вязкость жидкости, температура (Yue, Alexander, Laidlaw, et al., 1994). Тем не менее этот метод открывает большие перспективы для более тщательного изучения мышечной функции.
Три фактора убедительно показывают целесообразность применение данного метода для изучения мышечной функции. Во-первых, имеет место положительная линейная взаимосвязь между интенсивностью сигнала Т2 и силой мышечного сокращения. Эта взаимосвязь характерна для концентрических и эксцентрических сокращений, а также для ряда различных мышечных групп. Например, при воздействии нагрузки на определенную группу мышц и при ее концентрических сокращениях потребление кислорода оказывается большим, и в работу вовлекается больше двигательных единиц, чем при эксцентрических мышечных сокращениях. При эксцентрических сокращениях интенсивность ЭМГ и сигнала Т2 оказывается меньшей (Adams, Duvoisin, Dudley, 1992; Shellock, Fukuhaga, Mink, Edgerton, 1991). Во-вторых, изменение изображения, полученного с помощью магнитного резонанса, характеризуется достаточной про
173
странственной разрешающей способностью, чтобы идентифицировать частичный объем мышц сгибателей пальцев (поверхностного и глубокого сгибателя пальцев), контролирующих активность отдельных пальцев (Fleckenstein, Watumull, Bertocci, et al., 1992). Это позволяет ученым определить степень функциональной компартментализации в пределах отдельной мышцы, а также в пределах группы мышц-синергистов. Кроме того, можно выяснить, вовлекает ли выполнение задания коактивацию ряда мышц агонистов-антагонистов. В-третьих, метод получения изображения с помощью магнитного резонанса позволяет неинвазивно определить состав мышечных волокон. Программа физических занятий для укрепления мышц-разгибателей коленного сустава привела к значительному увеличению содержания быстросокращающихся волокон в латеральной широкой мышце бедра. Это изменение тесно коррелировало с увеличением продолжительности релаксации Т1 и Т2 (Kuno, Katsuta, Akisada, et al., 1990). Следует отметить, однако, что последние эксперименты ставят под сомнение эффективность данного метода (Parkkola, Alanen, Kalimo, et aL, 1993).
ОБРАТНАЯ СВЯЗЬ С ЧУВСТВИТЕЛЬНЫМИ НЕРВНЫМИ ОКОНЧАНИЯМИ
Чувствительные нервные окончания обеспечивают информацию, возникающую в результате многообразия стимулов. Три типа афферентных явлений играют важную роль в контроле движения: экстерорецепция, проприоре-цепция и последовательность действий. Эту информацию обеспечивают два класса чувствительных нервных окончаний — экстероре-цепторы и проприорецепторы. Первые выявляют внешние раздражения. К ним, помимо других, относятся глаза, уши и рецепторы кожи, реагирующие на температуру, прикосновение и боль. Воспринимаемая ими информация используется для ознакомления системы с состоянием внешней среды, включая ее расположение относительно среды. Проприорецепторы воспринимают раздражители, образуемые самой системой (Sanes, Evarts, 1984), такие, как механические переменные, связанные с активацией мышцы, которые участвуют в мгновенном контроле движения. К ним относятся мышечные веретена, сухожильные органы и суставные рецепторы. Информация, которую обеспечивают экстерорецепторы и проприорецепторы, позволяет системе обеспечить быстрое реагирование на пертурбацию, определить положение конечности и провести дифферен
циацию между самопроизвольными и вынужденными движениями.
Схема рефлексов
Способность чувствительных нервных окончаний быстро реагировать на пертурбации основана на существовании быстрой связи между афферентным сигналом и двигательной реакцией (эфферентным “ответом”). Это — рефлексы, которые определяются как стереотипная двигательная реакция организма на сенсорный стимул. Простейшая нервная схема, лежащая в основе рефлекса, включает чувствительное нервное окончание, его афферентную иннервацию и группу двигательных единиц, которые принимают импульс афферента. Эта схема, однако, может быть ограничена нервными элементами, контролирующими отдельную мышцу, включать группу синергистов, взаимодействие между парой мышц агонистов-антагонистов.
Рефлексы представляют собой механизмы, способные защитить систему от непредвиденных нарушений. При возмущении системы, например, вследствие непредвиденного растяжения мышцы, рефлексы обеспечивают быструю реакцию, направленную на противодействие пертурбации. В этом смысле рефлексы рассматриваются как регуляторы, поскольку они компенсируют нарушения и поддерживают нужное состояние (Houk, 1988). Нервные схемы, обеспечивающие компенсацию расстройств со стороны связи между входным и выходным импульсом, выполняет функцию отрицательной обратной связи, при которой двигательная реакция направлена на преодоление стимула, который первоначально активировал чувствительное нервное окончание.
Рефлекс растяжения мышцы. Мышцы выполняют две основные функции — производят усилие и реагируют на нарушения. Мышца должна быть подобна пружине, чтобы реагировать соответствующим образом; этому способствует рефлекс растяжения мышцы. Когда мышца испытывает кратковременное неожиданное увеличение длины, ее реакцией является рефлекс растяжения (рис. 5.26). В данном примере испытуемый держался за ручку, которая неожиданно переместилась, что вызвало растяжение мышц-разгибателей, пересекающих запястье. Рефлекс растяжения отразился на ЭМГ мышц-разгибателей. Как следует из рис. 5.26, ЭМГ (реакция на растяжение) увеличивается сразу же после начала перемещения ручки (стимул). Рефлекс растяжения включает, по меньшей мере, два компонента (Matthews, 1991). Один из них — реакция кратковременной латентности (MJ), которую, как полагают, вызывает нервная схема, ограниченная спинным
174
мозгом (рис. 5.27). Второй компонент характеризуется долговременной латентной и более сложной структурой, которая, очевидно, включает двигательную область коры головного мозга. Иногда наблюдается и третий компонент. На рис. 5.26 показано, что различные компоненты рефлекса растяжения мышцы предшествуют начальной произвольной ЭМГ, которая подчеркивает способность рефлексов обеспечить быстрое реагирование на пертурбации. Латентность компонента Ml составляет обычно около 30 мс, М2 — около 50—60 мс, а начальная произвольная активность (ЭМГ) возникает через 170 мс.
Чувствительное нервное окончание, участвующее в рефлексе растяжения мышцы, включает, как минимум, три мышечных веретена, а также афференты группы 1а и II. Схема, включающая афферент группы 1а, приведена на рис. 5.27. Потенциалы действия образуются в афферентах мышечных веретен в ответ на растяжение мышцы и распространяются по направлению к спинному мозгу, где вызывают синаптические потенциалы в двигательных нейронах, иннервирующих мышцу, в которой находится веретено. При адекватном растяжении в двигательном нейроне образуется достаточное количество синаптических потенциалов, чтобы вызвать потенциал действия, который приведет к мышечному сокращению. Рефлекс растяжения, по-видимому, лучше всего обеспечивает аккомодацию незначительных нарушений длины мышцы. Чистым результатом схемы входного-выходного сигнала является сокращение, обусловленное растяжением (стимулом), которое сводит его к минимуму; этот тип схемы отрицательной обратной связи называют также резистентным рефлексом.
Следует отметить, что рефлекс растяжения представляет собой не только стереотипную реакцию отрицательной обратной связи, но и довольно “гибкую” реакцию, адаптирующуюся к изменяющимся условиям (Hammond, Merton, Sutton, 1956). Поскольку рефлекс растяжения возникает при любом неожиданном изменении длины мышцы, он может возникнуть и во время движения. Если во время движения имеет место несоответствие между ожидаемой и действительной длиной мышцы, этой разницы может оказаться вполне достаточно, чтобы возник рефлекс растяжения. В таких условиях, однако, могут также активизироваться сухожильные органы, а афферентные группы 16 могут посылать сигнал (тормозящий) в спинной мозг (Matthews, 1990). Таким образом, во время движения человека неожиданное растяжение мышцы может привести к передаче сигналов многими рецепторами, включая афференты групп 1а, 16 и II. В результате может возникнуть афферентная блокада, вплоть до невозможности вызвать реакцию короткой
Рис. 5.26. Рефлекс растяжения, обусловленный неожиданным растяжением (по направлению вниз) мышц-разгибателей, пересекающих запястье: (Matthews, 1991)
латентности, такой, например, когда помост, на котором стоит человек, неожиданно перемешается (Horak, Nashner, 1986; Nashner, 1977). Более того, поскольку в рефлекс растяжения мышцы вовлечена кора головного мозга, нервная система способна модулировать реакцию, распространяя ее на мышцы, которые не подвергались растяжению, включая антагонисты, если активноть является механически соответствующей (Mattews, 1991). В результате этого оба компонента рефлекса растяжения мышцы могут изменяться с тем, чтобы удовлетворить потребности, возникающие при выполнении данного задания.
Реакцией, связанной с рефлексом растяжения, является сухожильный рефлекс. Сухожильный рефлекс представляет собой разновидность рефлекса растяжения и включает активацию только афферента группы 1а и соответствующий выходной сигнал двигательного нейрона (Matthews, 1990). Поскольку он основан на моносинаптической связи афферента группы 1а, то представляет собой быструю реакцию с латентностью 25—30 мс между нанесением удара по надколенному сухожилию и регистрацией ЭМГ в четырехглавых мышцах бедра. Величину сухожильного рефлекса используют как показатель воздействия реактивности мышечного веретена, возбудимости двигательного нейрона и уровня торможения (пре-синаптического) на афференты группы 1а спинного мозга.
Реципрокный рефлекс торможения. Если неожиданное растяжение мышцы вызывает ряд “конкурирующих” (возбуждающих и тормозящих) импульсов, то импульсы ЦНС также могут быть весьма разнообразными. Примером может служить влияние импульса афферента группы 1а на двигательные нейроны, иннервирующие мышцу-антагонист. Как видно из рис. 5.27, достигая спинного мозга, афферент группы 1а разветвляется. Одно из этих ответвлений образует синапс с интернейроном — тормозящим la-интернейроном, который способен
175
Рис. 5.27. Схема, лежащая в основе рефлекса растяжения и реципрокного тормозного рефлекса
образовывать тормозные постсинаптические потенциалы в двигательных нейронах мышцы-антагониста (Day, Marsden, Obeso, Rothwell, 1984; Katz, Penicaud, Rossi, 1991). Эта связь, которая служит для торможения или уменьшения возбудимости двигательных нейронов, иннервирующих мышцу-антагонист, называется реципрокным тормозящим рефлексом. Таким образом, активация мышечных веретен в четырехглавой мышце бедра (рис. 5.27) вызывает возбуждение гомонимных (четырехглавых бедра) двигательных нейронов и одновременно тормозит двигательные нейроны мышц-антагонистов (подколенных). Поскольку результирующая активность мышцы относительно сустава обусловлена различием в активности между парой агонистов-антагонистов, реципрокный тормозной рефлекс увеличивает вероятность того, что импульс, который был воспринят одной мышцей, вызовет в ней существенную реакцию.
Рис. 5.28. Схема сгибательного и перекрестный разгибательного рефлексов
Один из способов определить силу этого рефлекса состоит в том, чтобы испытуемый выполнил сокращение мышцы в момент тестирования экспериментатором возбудимости мышцы-антагониста. Возбудимость мышцы можно определить, вызвав рефлекс Хоффманна; уменьшение рефлекса отражает снижение возбудимости двигательных нейронов, иннервирующих мышцу. При сокращении сгибателей стопы происходит снижение рефлекса Хоффманна в камбаловидной мышце вследствие реципрокного торможения (Shindo, Harayama, Kondo, et al., 1984). Однако поскольку тормозящий la-интернейрон принимает сигнал из множества источников, включая пути высших нервных центров (например, пирамидальный, вестибулоспинальный) и проприорецепторы, активация реципрокного тормозного рефлекса зависит от состояния (уровня возбудимости) интернейрона.
Сгибательный и перекрестный разгибательный рефлексы. Помимо локальных рефлексов, в которых участвуют мышечное веретено и сухожильный орган, “репертуар” взаимосвязи между поступающим и исходящим сигналом включает ряд других, обусловленных разнообразными чувствительными нервными окончаниями (например, суставные рецепторы, свободные нервные окончания) и характеризующихся различными реакциями. Примерами могут служить сгибательный и перекрестный рефлексы. Когда кожные рецепторы (афференты группы III и IV) принимают отрицательный стимул, они реагируют отдергиванием от участка стимула. Если стимул действует на тыльную часть ноги, очевидно, реакцией будет отведение ноги в сторону от действия стимула, отсюда название “сгибательный” рефлекс. Латентность реакции составляет около 100 мс. Однако поскольку сенсорные сигналы передаются через полисинаптические пути, афферентный входящий сигнал может быть изменен, вследствие чего реакция может изменяться.
Кожный афферент, участвующий в сгибательном рефлексе, разветвляется в месте вхождения в спинной мозг, распределяя поступивший сигнал по различным мишеням. Эти мишени включают двигательные рефлексы, иннервирующие мышцы-разгибатели в контрлатеральной конечности (рис. 5:28). Очень часто сгибательный рефлекс сопровождается перекрестным, который включает возбуждение двигательных нейронов, иннервирующих мышцы-разгибатели, и торможение двигательных нейронов мышц-сгибателей контралатеральной конечности. Подобная связь весьма целесообразна, поскольку если одна нога отводится (сгибается) от стимула, выпрямление контралатеральной конечности обеспечивает постуральную поддержку (опору).
Рефлекс Хоффманна. В отличие от рассмотренных рефлексов, которые могут происхо-
176
Рис. 5.29. Н-рефлекс в четырехглавой мышце бедра:
а — размещение стимулирующих электродов (С — катод; А( и А2 — два соединенных анода); б — положение испытуемого; в — Н-рефлекс, записанный как ЭМГ медиальной широкой мышцы бедра
дить в нормальных условиях движения, рефлекс Хоффманна, или Н-рефлекс, представляет собой искусственно вызываемую реакцию. Н-рефлекс представляет собой своеобразный инструмент тестирования возбудимости нервно-мышечной системы (Fisher, 1992; Hugou, 1973). Он вызывается электрошоком (стимуляцией) периферического нерва. Реакцией является сокращение мышцы, иннервированной стимулированным нервом; величину реакции можно измерить как ЭМГ или как силу сокра-
щения. Н-рефлекс обычно изучают в камбаловидной мышце, однако его можно вызвать и в других мышцах. На рис. 5.29 показана методика стимуляции Н-рефлекса в четырехглавой мышце бедра. Испытуемый ложится на спину, стимулирующий электрод помещают чуть ниже паховой связки над бедренным нервом. Поскольку Н-рефлекс зависит от избирательной активации афферентов группы 1а, большое значение имеет правильное расположение электродов относительно нерва.
Рис. 5.30. Схема Н-рефлекса. Электрический стимул воздействует на периферический нерв, и реакция измеряется в мышце, иннервированной нервом
12 8-133
177
На рис. 5.30. приведена схема Н-рефлек-са. Она включает избирательную активацию афферентов группы 1а и последующее образование потенциалов действия в двигательных нейронах, иннервирующих мышцу. Избирательная активация афферентов группы 1а достигается постепенным увеличением интенсивности стимула от невысокой до образования потенциалов действия в аксонах наибольшего диаметра; у большинства людей это афференты группы 1а. При избирательной активации афферентов группы 1а потенциалы действия распространяются по направлению к спинному мозгу (рис. 5.30, 2), где они вызывают постсинаптические потенциалы в двигательных нейронах. Вероятность того, что эти синаптические потенциалы приведут к образованию потенциалов действия, зависит от величины синаптических потенциалов и мембранного потенциала. Чем ближе мембранный потенциал к порогу образования потенциала действия, тем выше вероятность того, что стимулы (синаптические потенциалы) приведут к образованию потенциала действия. Если мембранные потенциалы большего числа двигательных нейронов оказываются ближе к порогу, стимулы вызовут образования потенциалов действия в большем количестве двигательных нейронов, и реакция (ЭМГ и сила) будет более значительной. Именно поэтому Н-рефлекс используют для тестирования уровня возбудимости пула двигательных нейронов. Чем интенсивнее реакции, тем большее количество двигательных единиц внесло свой “вклад” в нее вследствие более высокого уровня возбудимости пула двигательных нейронов.
Поскольку Н-рефлекс — мера уровня возбудимости пула двигательных нейронов, то даже отдельных стимулов (например, шума или неожиданных звуков) либо наиболее отдаленной мышечной деятельности достаточно, чтобы повлиять на реакцию. Так, Н-рефлекс, вызванный в камбаловидной мышце испытуемого, будет иметь более высокую амплитуду, если испытуемый произвольно активирует другие мышцы, например, участвующие в сжатии челюсти. Это объясняется тем, что нервный сигнал, идущий к двигательным нейронам, активирующим мышцы, сжимающие челюсть, распределяется вдоль спинного мозга и образует постсинаптические потенциалы во многих двигательных нейронах. Следовательно, мембранные потенциалы многих двигательных нейронов камбаловидной мышцы приближаются к порогу, поэтому постоянные электрические стимулы приведут к образованию потенциалов действия большим числом двигательных нейронов. Использование активности отдаленных мышц для увеличения возбудимости пула двигательных нейронов называется приемом Джендрас-сика.
Н-рефлекс изменяется при продолжительной физической активности (Nielsen, Crone, Hultbom, 1993). Например, установлено (Sale, MacDougall, Upton, McComas, 1983), что H-рефлекс, вызванный во время максимального произвольного сокращения, увеличивается после программы тренировок силовой направленности. По мнению ученых, усиление рефлекса обусловлено повышением возбудимости двигательных нейронов вследствие программы тренировок силовой направленности. В другом исследовании (Koceia, Raglin, 1992) наблюдали снижение амплитуды Н-рефлекса у группы пловцов в состоянии перетренированнос-ти и возвращения к нормальным показателям после уменьшения объема тренировочных занятий.
Когда электрический стимул, используемый для стимуляции Н-рефлекса, постепенно увеличивается, он образует потенциалы действия в аксонах меньшего диаметра (Trimble, Enoka, 1991). Следующий класс аксонов большого диаметра после афферентов группы 1а — альфа-аксоны, принадлежащие к двигательным нейронам. При образовании потенциалов действия в альфа-аксонах реакцией является сокращение кратковременной латентности (5 мс) — так называемая М-волна (рис. 5.30, 7). Стимулирование возникновения М-волны используют для определения целостности (неразрывности) схемы между участком стимула (нерв мышцы) и участком записи (как правило, ЭМГ мышцы); т.е. М-волна позволяет проверить целостность распространения (Bigland-Ritchie, Kukulka, Lippold, Woods, 1982; Enoka, Stuart, 1992).
При максимальном электрическом стимуле это приводит к синхронной активации всех мышечных волокон и представляет собой суммацию реакций всех их потенциалов действия. Если измерения проводятся при обычном (общепринятом) расположении электродов (биполярные, диаметр 8 мм, расстояние между электродами 1,5 см), запись М-волны не отражает потенциалы действия во всей мышце, а только потенциалы действия мышечных волокон, находящихся вблизи (около 1—2 см) от электродов (Fuglevand, et al., 1992). Именно этим объясняется одинаковая амплитуда М-волн в небольших и больших мышцах (Hicks, Cupido, Martin, Dent, 1992). Интересно, что амплитуда М-волны с возрастом уменьшается, очевидно, вследствие снижения возбудимости мышечных волокон (Hicks, et al., 1992).
Рефлекс тонической вибрации. Подобно Н-рефлексу рефлекс тонической вибрации представляет собой искусственно вызванную реакцию (Lance, Burke, Andrews, 1973). Схема этого рефлекса та же, что и сухожильного. Она включает мышечное веретено и гомонимные двигательные единицы, активирующиеся вследствие
178
a — активный вибратор помещен над сухожилием двуглавой мышцы плеча, который вызывает ЭМГ в мышце и образование силы запястьем; б— увеличение силы и ЭМГ, связанных с субмаксимальным сокращением двуглавой мышцы плеча, с помощью вибратора;
горизонтальные линии — период вибрации
возбуждения мышечных веретен. Вместе с тем вибрация, очевидно, активирует как моно-, так и полисинаптические пути. Мышечное веретено очень чувствительно к вибрации небольшой амплитуды с частотой 50—150 Гц. Стандартный клинический вибратор, прикладываемый к мышце, обеспечивает достаточный стимул для возбуждения многих мышечных веретен и активации достаточного количества двигательных единиц, чтобы вызвать мышечное сокращение, следствием которого будет увеличение силы и ЭМГ при субмаксимальном сокращении (рис. 5.31). Более того, вибрация приводит к снижению возбудимости двигательных нейронов, иннервирующих мышцу-антагонист, через схему реципрокного торможения. Следует отметить весьма сложное влияние вибрации. Хотя вибрация вызывает рефлекторную реакцию (ЭМГ и сила), вместе с тем она подавляет сухожильный рефлекс и Н-рефлекс.
Рефлекс тонической вибрации используют в процессе реабилитации людей, перенесших инсульт, особенно для восстановления двигательной функции верхней конечности (Bishop, 1974, 1975а, 19756).
РЕФЛЕКСЫ И ДВИЖЕНИЕ
При выполнении даже самого простого движения двигательные нейроны принимают импульсы не только из различных типов чувствительных нервных окончаний, но и из высших двигательных центров (головного мозга). Для достижения необходимой активации мышц все принятые импульсы должны быть тщательно обработаны (Fournier, Pierrot-Deselligny, 1989; Windhorst, 1988). Чтобы выяснить, как это осуществляется, необходимо обратить вни
мание по крайней мере на один из важнейших принципов, связанных с рефлексами: синтез принятых импульсов осуществляется не на уровне двигательного нейрона, а скорее на уровне интернейронов спинного мозга. Из этого следует, что рефлексы могут изменяться в процессе движения.
В процессе изучения рефлексов, по крайней мере со времен Шеррингтона (1857 — 1952), большое внимание обращали на изучение двигательного контроля. Двигательный нейрон является центральным элементом схемы, к которому поступают афферентные и супраспиналь-ные импульсы (рис. 5.32, а). Хотя двигательный нейрон по-прежнему рассматривают как заключительный общий путь, по которому импульсы передаются в мышцы, есть доказательства, что он — не главный интегрирующий элемент в контроле движения. Миллионы синаптических соединений в спинном мозгу образуют основу взаимодействий между нервными элементами. Эти взаимодействия основаны на суммировании возбуждающих (незаштрихованные кружочки) и тормозящих (заштрихованные кружочки) постсинаптических потенциалов. По-видимому, интернейрон (рис. 5.32, б), а не двигательный нейрон играет главную роль в этой интеграции (рис. 5.32) (Baldissera, Hultborn, Illert, 1981; Pearson, 1993; Windhorst, 1988). Большая часть импульсов из афферентных и супраспинальных источников “собирается” в различных интернейронах, а не идет непосредственно к двигательным нейронам. В результате общий импульс (например, импульс группы 1а, обусловленный растяжением мышцы) может измениться, прежде чем будет передан двигательным нейронам. Подобная структура обеспечивает большую гибкость взаимосвязи между входящим и исходящим импульсом (Fournier, Pierrot-Deseilligny, 1989).
12*
179
Рис. 5.32. Схемы конвергенции импульса в спинном мозгу (Burke, 1985): а — на мотонейрон; б — через интернейрон
Систематические исследования соединений спинного мозга позволили выявить ряд интересных аспектов функционирования нервной системы (Stuart, 1987а, б): схема рефлексов включает альтернативные пути, вследствие чего афференты имеют альтернативные возбуждающие и тормозящие пути к двигательным нейронам (Binder, Houk, Nichols, et al., 1982); существуют значительные различия между мышечными системами, например, между мышцами, контролирующими конечности, процесс дыхания, и мышцами головы и шеи (так, реципрокное торможение возможно только в мышцах конечностей).
Гибкость, связанную с интернейронной конвергенцией, иллюстрирует изменение рефлекса во время движения (Nashner, 1982; Stein, Capady, 1988; Yang, Stein, 1990), например, сгибательного рефлекса во время ходьбы (Сгеппа, Frigo, 1984). Болевой стимул при воздействии на кожу (в участке икроножных мышц) вызы
вает сгибательный рефлекс мышц бедра (рис. 5.33). Реакцию, которая направлена на то, чтобы отвести конечность от действия стимула, можно измерить как ЭМГ мышц бедра (латеральной широкой и двуглавой бедра). Во время ходьбы, однако, эти мышцы необходимы для удержания положения тела в определенные фазы выполнения шага. Следовательно, конечность не может быть отведена в ответ на действие стимула. Нервная система видоизменяет реакцию на стимул, чтобы обеспечить удержание положения тела, когда это необходимо; конечность отводится лишь тогда, когда это становится возможным с механической точки зрения. Такое изменение реакции двуглавой мышцы бедра показано на рис. 5.33. Реакция (величина ЭМГ) включает компоненты кратковременной и долговременной латентности. Первый (57 мс) был максимальным при переходе от стационарного положения к фазе маха, тогда как компонент долговременной латентности (132) был наибольшим в конце фазы маха. Подобные изменения присущи также Н-рефлексу во время ходьбы (Сгеппа, Frigo, 1987; Stein, Capady, 1988).
Учитывая изложенное, мы должны изменить определение рефлекса как стереотипной реакции, подчеркнув, что рефлекс — это реакция, зависящая от состояния. Это значит, что реакция, независимо от постоянного стимула, зависит от состояния активности испытуемого. Например, рефлекс растяжения самый слабый в большинстве мышц во время периодов активности (Houk, Rymer, 1989; Marsden, Merton, Morton, 1976; Vallbo, Wessbeig, 1993).
Хотя многие нервные механизмы могут изменить рефлекс, нам больше всего известно о двух из них: возвратном и пресинаптическом торможении. Возвратное торможение — схема локальной обратной связи, позволяющей изменить рефлекторные реакции благодаря интернейрону, известному как клетка Реншоу (Windhorst, 1988). Клетку Реншоу (R на рис. 5.32, б) активирует супраспинальный импульс, мышечные афференты группы III и IV, а также коллатеральное ответвление аксона а-мотонейрона. Она, в свою очередь, генерирует тормозной постсинаптический потенциал в этих же двигательных нейронах, а также других интернейронах. Схема, включающая коллатераль аксона, клетку Реншоу и двигательный нейрон, называется возвратным торможением (см. рис. 5.32). Возвратное торможение из данного пула двигательных нейронов распространяется во многие другие пулы двигательных нейронов, хотя оно является слабым или даже не достигает пулов, иннервирующих отдаленные от центра мышцы (например, кисти или стопы) (Katz, Mazzocchio, Tenicaud, Rossi, 1993). Активация возвратного торможения может вызывать снижение возбудимости двигательных нейронов. Причем оно оказывает наибольшее воздействие на возбуди
180
мость меньших двигательных нейронов. Поскольку возвратное торможение в пуле двигательных нейронов увеличивается при слабых сокращениях и уменьшается при сильных (Fournier, Pierrot-Deseilligny, 1989), более высоким оно оказывается при активности небольших двигательных нейронов, снижаясь при рекрутировании больших двигательных нейронов. Увеличение возвратного торможения может наблюдаться во время вызывающих утомление сокращений.
Помимо воздействия на двигательные нейроны, возвратное торможение вызывает тормозные постсинаптические потенциалы в у-мото-нейронах, а также в тормозном 1а интернейроне (Hultbom, Lindstnjm, Wigstrnm, 1979). Связь с у-мотонейроном означает, что возвратное торможение может изменять возбудимость мышечного веретена и, следовательно, влиять на взаимосвязь входящего-исходящего импульса рефлекса растяжения. Связь с тормозным 1а интернейроном показывает, что возвратное торможение может ингибировать (снижать возбудимость) интернейрон, являющийся посредником в реципрокном рефлексе торможение. Таким образом, возвратное торможение способно осуществлять растормаживание. Исходя из изложенного, возвратное торможение — существенный элемент адаптации рефлексов.
Пресинаптическое торможение также может изменять рефлексы при условии постоянного импульса из чувствительного нервного окончания. Многое стало известно об этом механизме при изучении способности адаптации рефлекса растяжения (Mattehews, 1990, 1991). Адаптация осуществляется наложением тормозящего воздействия на потенциалы действия, передаваемые афферентами группы I и II. Пресинаптическое торможение, снижая потенциал действия и количество выделяемого нейромедиатора у синапса афферент — двигательный нейрон, изменяет взаимосвязь между входящим и исходящим импульсами, в результате чего реакция (мышечное сокращение) на один и тот же стимул (растяжение мышцы) может быть неодинаковой (Fournier, Pierrot-Deseilligny, 1989).
Срочная и долговременная адаптация. Физическая активность может изменять взаимосвязь входящего и исходящего импульсов. Например, бег на тредбане в течение 20 мин приводит к ослаблению Н-рефлекса в трехглавых мышцах икры (Bulbulian, Bowles, 1992). Причем это ослабление более значительно при беге по наклонной вниз (-10 % наклон). При беге по наклонной вниз мышцы ног выполняют эксцентрические сокращения и появление болевых ощущений в области мышц отсрочивается. Один из лучших примеров такой “гибкости”, обусловленной продолжительной физической активностью, — изменение рефлекса растяжения и
Рис. 5.33. Реакции ЭМГ на “пагубный” стимул относительно обычного уровня ЭМГ (1 — нормальный) во время выполнения шага (Crenna, Frigo, 1984): а — реакция короткой латентности; б — реакция продолжительной латентности; в — комбинированная реакция
Н-рефлекса при соответствующей подготовке (Evatt, Wolf, Segal, 1989; Wolpaw, Carp, 1990; Wolpaw, Herchenroder, Carp, 1993). В ряде экспериментов, Wolpaw и Carp (1990) продемонстрировали возможность научить в течение 80 дней примата усиливать или ослаблять рефлекторную реакцию (рефлекс растяжения или Н-рефлекс) на постоянный стимул. Эти изменения, очевидно, обусловлены адаптацией элементов нервной схемы, воспринимающих супраспинальный импульс. Что касается Н-рефлекса и рефлекса растяжения — это синапс группы 1а на двигательном нейроне и сам двигательный нейрон. Поскольку пресинаптические импульсы из различных супраспинальных участков способны тормозить синапс группы 1а, это, по-видимому, наиболее вероятный “кандидат” долговременной адаптации рефлекса растяжения и Н-рефлекса (Wolpaw, Carp, 1990).
Значение проприорецепторов. К проприоре-цепторам относятся мышечные веретена, сухожильные органы и суставные рецепторы. Они реагируют на механические переменные, обусловленные действиями самой системы. По мнению Hasan и Stuart (1988), обратная связь с про-приорецепторами выполняет две функции в движении. Во-первых, это взаимодействие между организмом и окружающей средой, во-вторых — изменение генерируемых двигательных
181
импульсов с целью приспособления к скелетно-мышечной механике.
Одно из важных свойств единой системы сустава — способность осуществлять движение в форме, соответствующей окружающей среде. Проприорецепторы помогают в этом системе. Например, способность осуществлять движение единой системой недостаточно, она должна выполнять его в рамках ограничений со сто_ роны окружающей среды, например, непредвиденных пертурбаций, ограниченного вовлечения конечностей или мышц в выполнение задания. Проприорецепторы обеспечивают систему необходимой информацией, что позволяет ей адаптироваться к ограничениям со стороны окружающей среды. Это достигается в результате включения содействующих или резистивных рефлексов и выбора соответствующих мышечных синергий. Резистивные рефлексы представляют собой реакции, направленные на преодоление неожиданных нарушений, обусловленных условиями внешней среды; резистивным рефлексом является рефлекс растяжения. Наряду с этим проприорецепторы обеспечивают информацию, касающуюся взаимодействий единой системы сустава и условий окружающей среды, способствуя выполнению движения. Поскольку выполнению движения предшествует постуральная корректировка, проприорецепторы могут обеспечить обратную связь с тем, чтобы стабилизировать систему относительно окружающей среды. Такая корректировка называется содействующей реакцией. Кроме того, проприорецепторы обеспечивают обратную связь, благодаря которой система выбирает соответствующую группу и последовательность мышечных действий для выполнения данного задания.
Вторая роль, которую играют проприорецепторы, это — аккомодация скелетно-мышечной механики. Как мы увидим в главе 6, способность единой системы сустава осуществлять движение зависит не только от возбуждения, обеспечиваемого нервным элементом, но и от механических свойств мышцы и геометрии системы. Проприорецепторы могут обеспечить информацию, которая позволит нервной системе учесть эти особенности и послать соответствующий импульс о выполнении движения. Для достижения этой цели проприорецепторы должны обеспечивать информацию на уровнях отдельной мышцы, отдельного сустава и нескольких суставов.
Сила, образуемая отдельной мышцей, неодинакова для одного и того же импульса (команды) нервной системы. Мышечная сила зависит от таких механических характеристик, как длина мышцы и интенсивность ее изменения. Неожиданное изменение длины мышцы, таким образом, нарушает запланированный импульс. Проприорецепторы выявляют непредвиденное изменение длины мышцы и тем самым сводят к
минимуму влияние изменения длины на величину силы, позволяя нервной системе более точно контролировать образуемую мышцей силу. Подобное происходит и на уровне отдельного сустава. Поскольку плечо пары большинства мышц изменяется в диапазоне движения при пиковых значениях в середине диапазона, максимальный вращающий момент чаще всего наблюдается в промежуточном положении (см. рис. 2.42). Следовательно, чтобы данный двигательный импульс (команда) привел к образованию субмаксимальной мышечной силы, конечность должна перемещаться в одном из направлений: в любую сторону от максимального вращающего момента. Обратная связь проприорецепторов может обеспечить увеличение импульса одновременно с увеличением угла сустава, что обеспечит непрерывное увеличение двигательного импульса (команды) в диапазоне движения и отдельное определение положения каждой конечности. И, наконец, проприорецепторы обеспечивают обратную связь, способствуя осуществлению контроля за рядом суставов. Движение связанной системы, такой, как тело человека, характеризуется тем, что движение одного сегмента тела оказывает влияние на все остальные сегменты (интерсегментальная динамика). Обратная связь проприорецепторов имеет большое значение для системы, позволяя ей интерпретировать сущность этих взаимодействий и координировать деятельность всех сегментов системы.
Кинестезия
Чувствительные нервные окончания обеспечивают информацию, позволяющую системе определить положение ее конечностей и раздражитель, вызывающий движение. Эта их способность называется кинестезией. Кинестетические ощущения представляют собой сочетание импульсов различных афферентных волокон (например, мышц, кожи, сухожилий, суставных капсул, связок) и двигательных команд (импульсов) из ЦНС. В отличие от проприорецептивного импульса, являющегося более общим и включающего вестибулярные ощущения и импульсы, идущие из мышц и суставов, которые могут не восприниматься, кинестетические ощущения хорошо воспринимаются (McCloskey, 1987).
Положение и движение. Одно время полагали, что суставные рецепторы направляют основной импульс для определения положения сустава и движения, в настоящее время более важным считается импульс, идущий из мышечного веретена (Gandevia, McCloskey, Burke, 1992; Matthews, 1987; McCloskey, 1987). Исключение составляет рука, где необходимую кинестичес-кую информацию обеспечивает обратная связь со стороны сустава, кожи и мышцы. Чтобы использовать импульс мышечного веретена для
182
определения положения и движения, ЦНС должна “вычесть” фузимоторную активность из импульса мышечного веретена. Это осуществляется за счет обеспечения сенсорных центров копиями соответствующих исходящих двигательных импульсов (команд). Посредством этого механизма двигательные центры сообщают сенсорным центрам о времени и величине фу-зимоторной активности и тем самым позволяют системе интерпретировать обратную связь мышечного веретена.
Усилие и интенсивность. Когда мышца активируется и осуществляет усилие, сухожильные органы обеспечивают афферентную информацию о величине силы, образуемой мышцей. Однако мы, как правило, не используем эту информацию для получения показателя интенсивности мышечных сокращений. Например, когда человек длительное время удерживает какой-то груз (например, портфель), необходимо со временем увеличить прилагаемое усилие, чтобы удержать неизменяющийся по величине груз. Поскольку не изменяются ни величина нагрузки, ни сила, необходимая для удержания данной нагрузки, ни афферентный сигнал (импульс) из детекторов силы (сухожильных органов), почему возникает ощущение, что нагрузка стала больше? Это ощущение исходит из двигательных команд ЦНС. Двигательные центры посылают разряд в чувствительные центры и супраспинальные сенсорные центры оценивают величины двигательной команды (импуль
са). Эта оценка называется ощущением усилия. Если сравнить величину усилия при выполнении задания до и после ослабления мышцы, окажется, что несмотря на одинаковую величину силы в обоих случаях, величина усилия в ослабевшей мышце значительно больше вследствие увеличения двигательной команды из ЦНС (McCloskey, Eheling & Goodwin, 1974).
Информация, поступающая из периферических рецепторов, используется для оценки массы объекта. Ощущение тяжести (массы), по-видимому, основано на интеграции проприоцептивной обратной связи с импульсом о выполнении движения, поступающем из ЦНС. Когда человек готовится поднять объект, импульс (команда) из ЦНС постепенно усиливается. После начала перемешения объекта проприоцептивная обратная связь указывает на изменение положения, а нервная система использует это сообщение для определения величины импульса из ЦНС, направленного на поднятие объекта и, таким образом, оценку его тяжести.
РЕЗЮМЕ
В этой главе рассмотрены четыре основные аспекта, на основании которых пять элементов модели двигательной системы (единой системы сустава), взаимодействуя, осуществляют движение.
ВОПРОСЫ С ОЛЫТРНАТРШжЫМ UTBtTOM
1. Головной мозг — один из элементов единой системы сустава.
ПРАВИЛЬНО НЕПРАВИЛЬНО
2. Двигательный нейрон состоит из двигательной единицы и мышечного волокна, которые он иннервирует.
ПРАВИЛЬНО НЕПРАВИЛЬНО
3. Термин “иннервирует” относится к анатомической связи, а не к процессу активации. ПРАВИЛЬНО НЕПРАВИЛЬНО
4. Здоровая мышца человека может содержать до 10 двигательных единиц.
ПРАВИЛЬНО НЕПРАВИЛЬНО
5. Около 80 % поступающих в двигательные нейроны сигналов принимается дендритами. ПРАВИЛЬНО НЕПРАВИЛЬНО
6. Коэффициент иннервации — количество мышечных волокон, иннервируемых отдельным двигательным нейроном.
ПРАВИЛЬНО НЕПРАВИЛЬНО
7. Мышечные волокна, принадлежащие отдельной двигательной единице, как правило, заключены в отдельный нервно-мышечный компартмент. ПРАВИЛЬНО НЕПРАВИЛЬНО
8. Одиночное сокращение представляет собой реакцию двигательной единицы мышцы на отдельный потенциал действия.
ПРАВИЛЬНО НЕПРАВИЛЬНО
9. Максимальная сила, образуемая двигательной единицей, наблюдается при сплошном тетанусе. правильно неправильно
10. Двигательные единицы типа FR характеризуются сопротивляемостью утомлению.
ПРАВИЛЬНО НЕПРАВИЛЬНО
11. Различия между типами I, Па и Пб основаны на ферментной миозин АТФазе.
ПРАВИЛЬНО НЕПРАВИЛЬНО
12. В двигательной единице содержатся мышечные волокна одного типа.
ПРАВИЛЬНО НЕПРАВИЛЬНО
13. Физические тренировки не влияют на процентное соотношение типов мышечных волокон.
ПРАВИЛЬНО НЕПРАВИЛЬНО
183
14. Нейротропизм представляет собой быструю электрическую связь между нейронами.
ПРАВИЛЬНО НЕПРАВИЛЬНО
15. Отсечение нерва от мышцы называется денервацией. ПРАВИЛЬНО НЕПРАВИЛЬНО
16. Сарколемма представляет собой электрически возбудимую мембрану.
ПРАВИЛЬНО НЕПРАВИЛЬНО
17. В стабильных условиях К+, как правило, содержится в основном во внутриклеточной ЖИДКОСТИ. ПРАВИЛЬНО НЕПРАВИЛЬНО
18. Na+-K+-Hacoc представляет собой белок, расположенный в сарколемме.
ПРАВИЛЬНО НЕПРАВИЛЬНО
19. В устойчивом состоянии ионы не проходят сквозь сарколемму.
ПРАВИЛЬНО НЕПРАВИЛЬНО
20. Na+ легче проходят сквозь сарколемму, чем
К+. ПРАВИЛЬНО НЕПРАВИЛЬНО
21. Мембранный потенциал покоя отрицателен во внутренней части по сравнению с внешней. ПРАВИЛЬНО НЕПРАВИЛЬНО
22. g означает проводимость.
ПРАВИЛЬНО НЕПРАВИЛЬНО
23. Постгиперполяризация обусловлена поступлением К+.
ПРАВИЛЬНО НЕПРАВИЛЬНО
24. Потенциалы действия распространяются вдоль аксона со скоростью 120 м • с-1.
ПРАВИЛЬНО НЕПРАВИЛЬНО
25. Амплитуда потенциала действия измеряется в милливольтах (мВ).
ПРАВИЛЬНО НЕПРАВИЛЬНО
26. Амплитуда синаптических потенциалов меньше амплитуды потенциалов действия.
ПРАВИЛЬНО НЕПРАВИЛЬНО
27. Нейрон генерирует потенциал действия всякий раз принимая нейромедиатор, вызывающий синаптический потенциал.
ПРАВИЛЬНО НЕПРАВИЛЬНО
28. Нейромедиатором в нервно-мышечном соединении является калий.
ПРАВИЛЬНО НЕПРАВИЛЬНО
29. Потенциал концевой пластинки является синаптическим потенциалом.
ПРАВИЛЬНО НЕПРАВИЛЬНО
30. Амплитуду ЭМГ измеряют в милливольтах (мВ). ПРАВИЛЬНО НЕПРАВИЛЬНО
31. Интегрированная ЭМГ в изометрических условиях характеризуется наличием корреляции с мышечной силой.
ПРАВИЛЬНО НЕПРАВИЛЬНО
32. Нейромедиатором в нервно-мышечном соединении является ацетилхолин.
ПРАВИЛЬНО НЕПРАВИЛЬНО
33. Потенциал действия приводит к выделению Са2+ из Т-трубочки.
ПРАВИЛЬНО НЕПРАВИЛЬНО
34. Тропонин и тропомиозин осуществляют растормаживание, предотвращая взаимодействие между актином и миозином в состоянии ПОКОЯ. ПРАВИЛЬНО НЕПРАВИЛЬНО
35. Потенциал действия вызывает увеличение gca терминальной цистерны.
ПРАВИЛЬНО НЕПРАВИЛЬНО
36. Насосы Са2+, прикрепленные к сарколемме, осуществляют возвращение Са2+ в саркоплазматический ретикулум после прекращения потенциала действия.
ПРАВИЛЬНО НЕПРАВИЛЬНО
37. Связываясь с тропонином, Са2+ обеспечивает растормаживание и активацию циклов поперечного мостика.
ПРАВИЛЬНО НЕПРАВИЛЬНО
38. Цикл поперечного мостика обеспечивает взаимодействие актина и миозина.
ПРАВИЛЬНО НЕПРАВИЛЬНО
39. АТФ необходим для фазы отсоединения цикла поперечного мостика.
ПРАВИЛЬНО НЕПРАВИЛЬНО
40. Проприорецепторы выявляют стимулы, образуемые самой системой.
ПРАВИЛЬНО НЕПРАВИЛЬНО
41. Регуляция характеризует процесс сохранения определенного состояния или условия.
ПРАВИЛЬНО НЕПРАВИЛЬНО
42. Эффект отрицательной обратной связи характеризуется отменой или “выключением” рефлекса. правильно неправильно
43. Главная функция рефлекса растяжения — помощь мышце в образовании максимальной мощности.
ПРАВИЛЬНО НЕПРАВИЛЬНО
44. Понятие “гомонимный” означает, что элементы (например, чувствительное нервное окончание, двигательный нейрон) иннервируют разные мышцы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
45. Рефлекс растяжения представляет собой схему отрицательной обратной связи.
ПРАВИЛЬНО НЕПРАВИЛЬНО
46. Реакция, участвующая в рефлексе растяжения, характерна только для мышцы, которая была растянута.
ПРАВИЛЬНО НЕПРАВИЛЬНО
47. В сухожильном рефлексе участвуют афференты группы 16.
ПРАВИЛЬНО НЕПРАВИЛЬНО
184
48. Рефлекс реципрокного торможения представляет собой влияние афферентов группы 1а на двигательные нейроны, иннервирующие мышцу-антагонист.
ПРАВИЛЬНО НЕПРАВИЛЬНО
49. Перекрестный рефлекс вызывает тот же афферентный импульс, который вызывает сгибательный рефлекс.
ПРАВИЛЬНО НЕПРАВИЛЬНО
50. Величина Н-рефлекса зависит от уровня возбудимости пула двигательных нейронов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
51. Прием Джендрассика используется для увеличения М-волны.
ПРАВИЛЬНО НЕПРАВИЛЬНО
52. Отдельный стимул, поступающий в нерв, способен вызвать как Н-рефлекс, так и М-волну.
ПРАВИЛЬНО НЕПРАВИЛЬНО
53. Вибрация небольшой амплитуды ингибирует мышечные веретена.
ПРАВИЛЬНО НЕПРАВИЛЬНО
54. Вибрация мышцы при субмаксимальном сокращении приводит к увеличению ЭМГ и силы мышцы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
55. Вибрация мышцы может привести к реципрокному торможению мышцы-антагониста. ПРАВИЛЬНО НЕПРАВИЛЬНО
56. Понятие “заключительный общий путь” характеризует связь между двигательным нейроном и мышцей.
ПРАВИЛЬНО НЕПРАВИЛЬНО
57. Интернейрон — основной интегрирующий элемент спинного мозга.
ПРАВИЛЬНО НЕПРАВИЛЬНО
58. Поскольку рефлексы обусловлены функционированием всего нескольких нервных элементов, они стереотипны и не меняются.
ПРАВИЛЬНО НЕПРАВИЛЬНО
59. Клетка Реншоу представляет собой специальный тип двигательного нейрона.
ПРАВИЛЬНО НЕПРАВИЛЬНО
60. Клетка Реншоу принимает участие в реципрокном торможении.
ПРАВИЛЬНО НЕПРАВИЛЬНО
61. Возвратное торможение может влиять на рефлекс растяжения, действуя либо на у-мотонейрон, либо на тормозные 1а-ин-тернейроны.
ПРАВИЛЬНО НЕПРАВИЛЬНО
62. Пресинаптическое торможение предотвращает действие тормозного импульса через
афференты группы 16 на рефлекс растяжения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
63. Рефлексы могут изменяться в результате продолжительных физических тренировок.
ПРАВИЛЬНО НЕПРАВИЛЬНО
64. Рефлекс растяжения представляет собой пример содействующего рефлекса.
ПРАВИЛЬНО НЕПРАВИЛЬНО
65. Кинестетическое ощущение — результат импульсов множества различных афферентных волокон.
ПРАВИЛЬНО НЕПРАВИЛЬНО
66. Импульс суставных рецепторов позволяет определить положение сустава.
ПРАВИЛЬНО НЕПРАВИЛЬНО
67. Ощущение усилия основано на проприоцептивной обратной связи.
ПРАВИЛЬНО НЕПРАВИЛЬНО
68. Что характеризует размер двигательного нейрона?
а) диаметр аксона;
б) площадь поверхности соматической клетки;
в) реобаза;
г) степень распространения.
69. Укажите единицу измерения сопротивления импульса:
a) MW;
б) м-с-1;
в) А;
г) мс.
70. Что характерно для небольших двигательных нейронов?
а) краткий период постгиперполяризации;
б) быстрое время сокращения;
в) высокое сопротивление импульса;
г) высокая скорость аксонного проведения.
71. Чем характеризуется сокращение?
а) величиной пика силы;
б) временем полурасслабления;
в) продолжительностью сокращения;
г) степенью спада.
72. Чему равен показатель утомления для двигательных единиц типа S?
а) менее 0,25;
б) ровно 0,50;
в) более 0,75;
г) ровно 1,0.
185
73. Почему двигательные единицы типа FF образуют наибольшую силу?
а) вследствие более высокого коэффициента иннервации;
б) ввиду большей площади поперечного сечения отдельных мышечных волокон;
в) вследствие большего количества миозин АТФазы;
г) ввиду более высокого коэффициента сокращения — тетануса.
74. В каком типе мышечных волокон миозин АТФаза приобретает темную окраску?
а) БГ;
б) I;
в) Па;
г) МО.
75. Какой нервный компонент не участвует в аксонном транспорте?
а) кинезин;
б) микротрубка;
в) потенциал действия;
г) динеин.
76. Когда мембранный потенциал приближается к нулю, что более точно характеризует состояние мембраны?
а) деполяризация;
б) гиперполяризация;
в) реполяризация;
г) постгиперполяризация.
77. Что вызывает повышенную фазу потенциала действия?
а) выделение К+;
б) обусловленное регуляцией напряжения изменение дк;
в) насос Na+-K+;
г) поступление Na+.
78. Почему ЭМГ можно рассматривать как “окно” в нервную систему?
а) поскольку потенциалы действия измеряются в процессе их движения из нервно-мышечного соединения вдоль мышцы;
б) поскольку это единственный способ измерить отрицательную и положительную фазы потенциалов действия;
в) поскольку между потенциалами действия аксона и мышечного волокна существует односторонняя взаимосвязь;
г) вследствие тесной корреляции между ЭМГ и мышечной силой.
79. Откуда выделяется Са2+ во время процесса сочетания возбуждения—сокращения?
а) из сарколеммы;
б) из саркоплазматического ретикулума;
в) из Т-трубочки;
г) из терминальной цистерны.
80. Какие белки ретулируют торможение цикла поперечного мостика?
а) актин;
б) тропонин;
в) кинезин;
г) миозин;
д) тропомиозин.
81. Где не распространяются потенциалы действия?
а) сарколемма;
б) саркоплазматический ретикулум;
в) Т-трубочка;
г) аксолемма.
82. К какому белку присоединяется Са2+ в процессе сочетания возбуждения—сокращения?
а) миозин;
б) тропомиозин;
в) актин;
г) тропонин.
83. Какое выражение лучше всего объясняет понятие “сокращение”?
а) сокращение длины мышцы в ответ на потенциал действия;
б) состояние активации мышцы, при котором происходит повторяющаяся смена циклов поперечных мостиков в ответ на потенциал действия;
в) активация мышцы в ответ на зависящее от потенциала действия выделение Са2+;
г) скольжение миофиламентов (актина и миозина) вследствие растормаживания, обусловленного Са2+.
84. Какое свойство сократительной деятельности измеряется с помощью метода магнитного резонанса?
а) смена циклов поперечных мостиков;
б) аккумуляция внутриклеточной воды;
в) изменение длины мышцы;
г) потенциалы действия сарколеммы.
85. Какие три типа афферетных явлений играют важную роль в контроле движения?
а) проприоцепция;
б) последовательность действия;
в) экстерорецепция;
г) синергизм.
86. Какие рефлексы способны осуществлять отрицательную обратную взаимосвязь?
а) рефлекс растяжения;
б) реципрокный рефлекс торможения;
в) Н-рефлекс;
г) рефлекс тонической вибрации.
87. Какие чувствительные нервные окончания не участвуют в рефлексе растяжения?
а) суставные рецепторы;
б) афференты группы 1а мышечного веретена;
в) афференты группы II мышечного веретена;
186
г) афференты группы 16 сухожильного органа.
88. Какой нейрон вызывает рефлекс реципрокного торможения?
а) клетка Реншоу;
б) тормозной нейрон 1а;
в) гомономный двигательный нейрон;
г) клетка Джендрассика.
89. Возбудимость какой единицы определяют с помощью Н-рефлекса?
а) нервно-мышечного соединения;
б) пула двигательного нейрона;
в) мышечных веретен;
г) мышечных волокон.
90. Какова приблизительная латентность М-волны?
а) 100 мс;
б) 75 мс;
в) 30 мс;
г) 5 мс.
91. Какое выражение лучше всего характеризует понятие общего конечного пути?
а) интернейрон — основной интегрирующий элемент спинного мозга;
б) рефлексы — одни из основных элементов контроля нервно-мышечной системы;
в) двигательный нейрон представляет собой путь, по которому импульсы передаются мышце;
г) афференты посылают входящий импульс, а эфференты передают исходящий импульс.
92. Какие нервные элементы принимают импульс из клеток Реншоу?
а) у-мотонейроны;
б) а-мотонейроны;
в) тормозные 1а интернейроны;
г) афференты группы 1а.
93. Каково одно из известных воздействий пре-синаптического торможения?
а) торможение 1а тормозного интернейрона;
б) активация возвратного коллатераля на клетке Реншоу;
в) торможение двигательного нейрона, прежде чем он будет способен образовать потенциал действия;
г) снижение поступающего импульса в афферент группы 1а.
94. Что такое рефлекс сопротивления?
а) постуральная корректировка или движение, усиливающее эффект внешних сил;
б) реакция на неожиданное нарушение (волнение);
в) функция положительной обратной связи;
г) рефлекс, после которого происходит произвольная активация мышцы.
95. Какие восприятия основаны на кинестиче-ских ощущениях?
а) положения или движения;
б) “тяжести” (массы) предмета;
в) королларного разряда;
г) синхронизации движений.
ЗАДАЧИ
96. Почему двигательную единицу считают функциональной единицей единой системы сустава?
97. Если мышечные волокна отдельной двигательной единицы распределены по 5 волокон на 100, а их число на поперечном срезе 870 536, чему равен коэффициент иннервации двигательной единицы?
98. Что вызывает изменение скорости сокращения в двигательных единицах?
99. Почему двигательные единицы типа FF образуют наибольшую силу?
100. Чем отличаются волокна типа Т от волокон типа МО?
101. Укажите четыре морфологических и шесть физиологических различий между тремя типами двигательных единиц.
102. Волокна типа I образуют меньшую мощность, чем волокна типа П, однако делают это более эффективно. Что означает это утверждение? Что такое эффективность? Какие опыты могут подтвердить это утверждение?
103. Двигательная единица двуглавой мышцы плеча человека характеризуется продолжительностью сокращения 42 мс, пиком силы сокращения 18 мН, коэффициентом утомления 0,18, соотношением сокращение— тетанус 1 : 6 и коэффициентом иннервации 1 : 389.
А. Какой это тип двигательной единицы?
Б. Сколько мышечных волокон иннервирует двигательный нейрон?
В. Какую максимальную силу может развить двигательная единица?
Г. Какую силу может развить двигательная единица в конце теста на утомление?
104. Какова скорость транспорта материала аксонной системой; какова скорость распространения потенциалов действия? Чем объясняется столь большая разница между ними?
105. Потенциал возбудимой мембраны обусловлен неодинаковым распределением ионов во внутри- и внеклеточных пространствах.
187
А. Какие ионы вовлечены в поддержание мембранного потенциала покоя?
Б. Какие силы действуют на каждый из этих ионов?
В. Чем объясняется неравномерное распределение ионов?
Г. Постройте график потенциала действия (правильно пометьте оси) и объясните, как можно определить потенциал действия.
Д. Если один тип положительно заряженных ионов проникает в клетку, а другой покидает ее, изменение заряда равно нулю. Потенциал действия обусловлен поступлением и выходом из клетки положительно заряженных ионов, почему же изменяется мембранный потенциал во время потенциала действия?
106. Проникновение ионов через мембрану зависит от комбинированных воздействий сил, действующих на каждый тип ионов, а также степени проницаемости мембраны для различных ионов. Почему мембранный потенциал покоя обусловлен в основном распределением К+, а потенциал действия объясняют влиянием Na+?
107. Почему при образовании потенциала действия поступление Na+ в клетку характеризуют как приводящее к деполяризации, тогда как выделение К+ вызывает реполяризацию?
108. ЭМГ очень часто используют для анализа движений человека.
А. Какую информацию обеспечивает ЭМГ?
Б. В чем сущность процесса ректификации и интеграции?
В. Почему целесообразно ректифицировать и интегрировать ЭМГ?
109. Некоторые ученые используют ФМГ вместо ЭМГ.
А. Что такое ФМГ? Что она измеряет?
Б. Постройте график, показав различия между ФМГ и ЭМГ как функции мышечной силы.
В. Постройте график, показав изменения ФМГ и ЭМГ как функции времени
при продолжительном субмаксимальном сокращении.
ПО. Какие три функции, связанные с движением, выполняют экстерорецепторы и проприорецепторы?
111. Чувствительные нервные окончания способствуют образованию рефлексов?
А. Что такое рефлекс?
Б. Каким образом рефлекс выполняет функцию отрицательной обратной связи?
112. Рефлекс растяжения представляет собой “гибкую” реакцию, которая может “приспосабливаться” к изменяющимся условиям. Почему тогда этот рефлекс или сухожильный рефлекс представляют собой необходимый “инструмент” для клинических экспериментов?
113. Опишите функции следующих рефлексов: растяжения, реципрокного торможения, сгибательного и перекрестного.
114. Н-рефлекс является не естественным рефлексом, а искусственно вызываемым.
А. Чем обусловлена целесообразность использования Н-рефлекса для тестирования возбудимости единой системы сустава? Как доказать, что Н-рефлекс тестирует возбудимость системы?
Б. В чем разница между Н-рефлексом и М-волной?
115. Интернейрон рассматривается как интегрирующий элемент спинного мозга.
А. Что это означает?
Б. Как связан интернейрон с понятием заключительного общего пути?
В. Укажите три способа влияния интернейрона на функцию нервной системы.
116. Что такое пресинаптическое торможение и как оно влияет на рефлексы в процессе движения?
117. Чем отличается кинестезия от проприоцепции?
118. Что такое ощущение усилия? На чем оно основано? Какие экспериментальные данные подтверждают существование ощущения усилия?
ГЛАВА 6
АКТИВАЦИЯ ЕДИНОЙ СИСТЕМЫ СУСТАВА
Как мы уже знаем, единая система сустава состоит из пяти элементов (жесткого звена, синовиального сустава, мышцы, нейрона и чувствительного нервного окончания), представляющих собой модель двигательной системы. Поскольку движение осуществляется за счет активации двигательной системы, мы рассмотрели свойства этих элементов (глава 4) и четыре их основных взаимодействия, которые позволяют нам двигаться (глава 5). Движение человека, однако, характеризуется не случайной активацией двигательной системы (единой системы сустава), а тщательным и предельно точным контролем мышечной силы. В этой главе мы рассмотрим свойства единой системы сустава, позволяющие нам контролировать мышечную силу, в результате чего мы получаем возможность выполнять множество разнообразных движений.
НЕРВНЫЕ ФАКТОРЫ
Трансформация потенциала действия мышцы в силу мышцы называется сочетанием возбуждения — сокращения. Во время этого процесса электрические явления, сопровождающие потенциал действия, обеспечивают мышечное сокращение. В более широком смысле потенциал действия является результатом импульса нервной системы. Поскольку потенциал действия обычно связан с потенциалом действия аксона, то потенциалы действия мышцы представляют собой команду нервной системы “идти”.
В главе 5 была рассмотрена концепция, согласно которой двигательная единица представляет собой функциональную нервно-мышечную единицу, посредством которой единая система сустава контролирует мышечную силу.
Вследствие такой организации мышечная сила изменяется с изменением активности двигательной единицы (Kemell, 1992). Это осуществляется вследствие изменения либо количества активных двигательных единиц, либо интенсивности функционирования после их активации. Эти два варианта представляют собой рекрутирование двигательных единиц (процесс активации двигательных единиц) и модуляцию интенсивности разряда (частота потенциала действия).
Рекрутирование двигательных единиц
В 1938 г. Денни-Браун и Пеннибекер сделали сообщение,что определенное движение всегда выполняется в результате активации двигательных единиц в установленной последовательности. Подобную организацию активации двигательной единицы назвали упорядоченным рекрутированием. С увеличением силы, образуемой мышцей, активируются дополнительные двигательные единицы, после рекрутирования двигательной единицы она остается активной до уменьшения силы. В соответствии с этой схемой (рис. 6.1) двигательная единица I рекрутируется первой и остается активной до тех пор, пока не уменьшится сила. Увеличение силы, по крайней мере частично, осуществляется за счет продолжающегося рекрутирования двигательных единиц (еще четыре на рис. 6.1). Как видно, сила достигает плато, когда прекращается рекрутирование дополнительных двигательных единиц, а те, которые остаются активными, не изменяют интенсивности разряда потенциалов действия. С уменьшением силы двигательные единицы последовательно дезактивируются или дерекрутируются в обратной последовательности, т.е. последняя
189
Рис. 6.1. Идеализированное колебание мышечной силы вследствие упорядоченного рекрутирования и дерекрутирования двигательных единиц. Запись двигательных единиц отражает потенциалы действия, образуемые каждой двигательной единицей при изменении силы мышцы. С целью упрощения интенсивность разряда каждой д вигательной единицы показана как регулярная
Механизм. Упорядоченное рекрутирование, по всей видимости, — результат ряда физиологических процессов, а не отдельного механизма. Важным фактором, лежащим в основе упорядоченного рекрутирования, является размер двигательного нейрона, на который указывает площадь поверхности сомы и дендритов. Согласно этому так называемому принципу размера, упорядоченное рекрутирование двигательных единиц обусловлено изменениями размера двигательного нейрона: двигательная единица с наименьшим мотонейроном рекрутируется первой и наоборот (Binder, Mendell, 1990; Enoka, Stuart, 1984; Henneman, 1957, 1979). Подобная организация, как считают, характерна для двигательных нейронов, иннервирующих одну определенную мышцу. Такая группа двигательных нейронов называется пулом двигательных нейронов. Например, переднюю большеберцовую мышцу человека иннервирует приблизительно 445 ос-мотонейронов; это — размер пула двигательных нейронов мышцы. Согласно принципу размера, в пределах пула двигательные нейроны функционально организованы в соответствии с размером. Когда мышца должна произвести усилие, первым рекрутируется самый маленький двигательный нейрон пула, а самый большой рекрутируется последним. По-видимому, некоторые морфологические (количество дендритов, диаметр аксона, коэффициент иннервации), био-
рекрутированная двигательная единица дерек-рутируется первой. Однако на рис. 6.1 показан особый случай — состояние, при котором изменения силы обусловлены исключительно изменением количества активных двигательных единиц. Обычная структура активности двигательной единицы при постепенном уменьшении силы показана на рис. 6.2; при низких значениях силы такая структура включает сопутствующее рекрутирование и модуляцию интенсивности разряда.
Относительный вклад рекрутирования двигательных единиц в образование силы колеблется от мышцы к мышце. В некоторых мышцах (например, таких мышцах руки, как приводящие мышцы больших пальцев и первые тыльные межкостные) все двигательные единицы, очевидно, рекрутируются, когда сила достигает около 50 % максимальной, тогда как в других мышцах (например, двуглавой мышце плеча) рекрутирование двигательных единиц продолжается до тех пор, пока не достигнет 85% максимальной (Kukulka, Clamman, 1981). Дальнейшее увеличение силы вследствие рекрутирования двигательных единиц отражает влияние различной интенсивности разряда.
§ 60-1 S X з;
о.!40-
г? 5 20-
СО 2
з! о-1„..................................
0 2 4 6 8 10 12 14 16 18
Время, с
б
3 5------------------НИ----------------
| 4-------------MIMHI------------------
! з-------------над»-------------------
i 2 —IIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIH—
* 1 111!1111111111Ш^
Рис. 6.2. Структура рекрутирования (а) и разряда пяти (из большого числа) двигательных единиц (б) при диффренцированном сокращении до 50 % максимальной силы (Kamen, Deluca, 1989)
МПС — максимальное произвольное сокращение
190
Синаптический импульс
• Сила
Свойства мотонейронов
• Мембрана
. R , Rh, АНР
• Разряд
• Интенсивность, адаптация
Возвратное торможение
Аксон
• диаметр
• величина потенциала действия (ПД)
Рис. 6.3. Модель двух двигательных единиц, иллюстрирующая избранные морфологические и физиологические измерения, коррелирующие с размером двигательного нейрона. Морфологические колебания включают силу синаптического импульса, диаметр аксона, коэффициент иннервации, размер мышечного волокна. Функциональные последствия этих колебаний изучаются с точки зрения физиологии, путем измерения переменных, приведенных в центре рисунка. Сила означает величину постсинаптической реакции (Enoka, Stuart, 1984)
физические (сопротивление импульсу (/?N), постгиперполяризация), а также импульсные (пул клеток Реншоу, афферент группы 1а) свойства изменяются в зависимости от размера двигательного нейрона (рис. 6.3). Вместе с тем рекрутирование двигательных единиц не зависит исключительно от размера двигательных нейронов, на него влияют и другие его характеристики (внутренние факторы), а также от организации синаптического импульса (внешние факторы), поступающие на дендриты и сому пула двигательных нейронов (Burke, 1981). Внутренние факторы включают размер нейрона, чувствительность его нейромедиаторных рецепторов и электрические свойства. Последнее относится к электрическим реакциям (напряжение, ток) возбудимой мембраны вследствие изменения проводимости (например, gNa, gK). Чувствительность рецепторов и элек-тротонические характеристики двигательных нейронов могут изменяться независимо от размера двигательного нейрона. Повышение чувствительности рецепторов увеличивает возбу
димость клетки к данному импульсу, тогда как изменение электротонических характеристик повышает проводимость синаптических импульсов к аксонному холмику. Вследствие этого усиливается реакция двигательного нейрона на импульсы. Внешние факторы включают ряд синаптических окончаний на двигательном нейроне от данной системы импульсов (например, афференты группы 1а, клетки Реншоу, кожные афференты), среднее количество нейромедиаторов, высвобождаемых в каждом синапсе, а также пространственное распределение синапсов на соме и дендритах. Вполне очевидно, что чем больше число синаптических окончаний или число выделяемых нейромедиаторов, тем более возбудим двигательный нейрон. Чем ближе синапс к аксонному холмику, тем выше вероятность того, что постсинаптический потенциал вызовет потенциал действия. Эффективный синаптический ток представляет собой показатель результирующего воздействия внешних и внутренних факторов на возбудимость двигательных нейронов.
191
Рис. 6.4. Предположительная модель рекрутирования двигательных единиц, обусловленного потребностями в мощности данного задания. Рекрутирование начинается слева (при 0 % пула) и продолжается до тех пор, пока не будет рекрутировано достаточное количество единиц для образования мощности, необходимой, чтобы выполнить задание. Например, для образования достаточного количества мощности для выполнения бега необходимо рекрутировать около 48 % двигательных единиц пула
Упорядоченное рекрутирование двигательных единиц не является фиксированной структурой активации, в некоторых случаях оно изменяется.
Функциональное значение. Преимущество упорядоченного рекрутирования состоит в том, что когда мышца получает команду образовать силу, последовательность рекрутирования двигательных единиц предопределена и нет необходимости определять ее (в головном мозгу). Следовательно, головной мозг избавлен от необходимости контролировать выполнение движения на этом уровне. Вместе с тем вследствие того, что рекрутирование двигательных единиц предопределено, невозможно избирательно активировать их в другой последовательности.
Из-за предопределенного порядка количество рекрутированных для выполнения определенной задачи двигательных единиц зависит от содержания двигательных нейронов в пуле. На рис. 6.4 показана предположительная модель этой взаимосвязи, полученная в результате измерения силы икроножного сухожилия свободно передвигающихся кошек (Walmsley et al., 1978). В этой схеме количество рекрутированных двигательных единиц определяется потребностями задания. Например, бег трусцой с невысокой скоростью требует незначительных мышечных усилий. Модель показывает, что для выполнения бега трусцой необходимо рекрутирование только двигательных единиц типа S и FR. Поскольку порядок рекрутирования фиксирован, постепенное увеличение мощности
(силы) для выполнения задания основано на постепенном рекрутировании больших двигательных единиц. Однако размер двигательной единицы не увеличивается строго в зависимости от ее типа, поэтому наблюдается некоторое взаимное “перекрывание” между типом S и FR и типом FR и FF в отношении размера и порядка рекрутирования (Stuart, Enoka, 1983). Именно поэтому невозможно избирательно активировать медленно- или быстросокращающи-еся двигательные единицы. При потребности в большей силе, например, при выполнении прыжка в высоту, модель показывает, что двигательные единицы типа S рекрутируются вместе с единицами типа FR и FF в установленном порядке.
Интенсивность разряда
Сила, образуемая мышцей, частично обусловлена изменяющимся сочетанием ряда активных двигательных единиц и интенсивности образования ими потенциалов действия. При рекрутировании двигательной единицы и образовании мышцей силы интенсивность образования потенциалов действия двигательной единицей, как правило, увеличивается (см. рис. 6.2). Потенциал действия каждой двигательной единицы приводит к сокращению ее волокон; при более частом образовании потенциалов действия сокращения суммируются и образуется сила, превышающая силу одиночного сокращения. Степень суммирования сокращений зависит от интенсивности образования потенциалов действия (рис. 6.5). Взаимосвязь интенсивности (частоты) потенциалов действия и силы двигательных единиц не совсем изучена; увеличение силы вследствие повышения интенсивности потенциала действия с 5 до 10 Гц не такое же, как при увеличении интенсивности потенциала действия с 20 до 25 Гц, хотя различие в обоих случаях одинаково — 5 Гц. Этот эффект можно продемонстрировать при электростимуляции мышцы или двигательной единицы с разной частотой и измерении образующейся силы. Полученные результаты можно нанести на график (рис. 6.6). Взаимосвязь имеет S-об-разную форму, максимальное уменьшение силы наблюдается при промежуточной частоте стимуляции (7—12 Гц). Действительная взаимосвязь, показанная на рис. 6.6, зависит от длины мышцы; кривая смещается влево при большей длине мышцы (Rack, Westbury, 1969). Несмотря на то, что взаимосвязь по-прежнему характеризуется S-образной формой, максимальную силу при большой длине мышцы вызывает частота 3—7 Гц, а при короткой длине мышцы — 10-12 Гц.
Несмотря на значительные технические трудности, группе ученых удалось определить
192
Рис. 6.5. Влияние частоты раздражения на суммацию сокращений быстро- (о) и медленносокращаю-щейся (6) двигательной единицы. Медленносокра-щающаяся единица характеризуется большей продолжительностью сокращения и меньшей пиковой силой. Каждая единица произвела четыре тетанических сокращения, показывающих степень суммации реакций сокращения при различной частоте стимулирования (левая колонка). Поскольку медленносокра-щающаяся единица характеризуется более продолжительными сокращениями, наблюдалась более высокая степень суммации при низкой частоте стимуляции (Botterman, Iwamoto, Gonyea, 1986)
интенсивность разряда двигательных единиц, классифицировав их на тонические и фазные в зависимости от интенсивности разряда (Gydikov, Kosarov, 1973, 1974). Очевидным различием между ними является характер взаимосвязи мышечной силы и интенсивности потенциала действия (рис. 6.7). Для тонических двигательных единиц характерно следующее: интенсивность разряда повышается при увеличении
мышечной силы на низких уровнях, тогда как при высоких значениях силы интенсивность разряда остается постоянной. Наоборот, интенсивность разряда фазных двигательных единиц увеличивается на всем диапазоне мышечной силы (т.е. взаимосвязь линейна). Кроме того, тонические двигательные единицы образуют более низкие потенциалы действия, рекрутируются при более низких значениях силы и менее устают. Фазные двигательные единицы, по-видимому, имеют большое значение в динамических условиях и в большей мере способствуют образованию мышечной силы, чем тонические.
Степень модуляции интенсивности разряда зависит от мышцы. Если рекрутирование двигательных единиц определенной мышцы завершается при 50 % максимальной силы (например, некоторых мышц руки), то последующее увеличение силы (т.е. от 51 до 100%) должно быть обусловлено колебаниями интенсивности разряда. В то же время в мышцах, где рекрутирование продолжается до 85 % максимальной
Рис. 6.6. Взаимосвязь сила—частота: а — изменение силы камбаловидной мышцы кошки как функция частоты стимулирования при различной длине: короткой (1), промежуточной (2), длинной (J) (Rack, Westbury, 1969); б — сила двигательной единицы тенарных мышц человека как функция частоты стимулирования (Thomas, Ritchie, Johansson, 1991)
13 8-133
193
Рис. 6.7. Взаимосвязь мышечной силы и интенсивности потенциалов действия двигательных единиц мышцы, удерживаемой с постоянной длиной (Gydikov, D.Kosarov, 1974): а — снижение и плато; б — линейная
силы, только оставшиеся 15 % силы (т.е. от 86 до 100 %) обусловлены исключительно изменением интенсивности разряда. Вместе с тем начальное увеличение силы, по крайней мере до 50 % максимальной, обусловлено увеличением и рекрутирования, и интенсивности разряда (Monster, Chan, 1977; Person, Kudina, 1972). На рис. 6.8 показана активность четырех двигательных единиц в прямой мышце бедра в условиях, когда сила, образуемая мышцами-разгибателями коленного сустава, постепенно увеличивалась до 33 % максимальной. Интенсивность разряда каждой двигательной единицы увеличивалась, а затем, с изменением силы, уменьшалась. Более того, при увеличении интенсивности разряда рекрутировались другие двигательные единицы.
Структура разряда. Величина силы, образуемой мышцей, зависит не только от рекрутирования двигательных единиц и частоты разряда потенциалов действия, но и от структуры активности потенциалов действия (Windhorst, 1988). Структура разряда характе
ризует взаимосвязь во времени потенциала действия и других потенциалов действия, образуемых данными, а также другими двигательными единицами. Различают по меньшей мере три эффекта структуры разряда: мышечную “мудрость”, двойной разряд и синхронность двигательных единиц. Мышечная “мудрость” относится к изменению разряда двигательных единиц при утомлении. При вызывающих утомление сокращениях интенсивность разряда потенциалов действия двигательной единицей уменьшается (Bigland-Ritchie, Johansson, Lippold, et al., 1983; Dietz, 1978). Это обусловлено не нарушением процессов, связанных с образованием и распространением потенциалов действия, это — адаптация нервной активности к изменившимся условиям в мышце (Enoka, Stuart, 1992). Поскольку это адаптационный процесс, который, по крайней мере частично, контролируется мышцей, снижение интенсивности разряда назвали мышечной “мудростью”. Значимость этого процесса иллюстрирует рис. 6.9, где силу, образуемую мышцами-разгибателями коленного сустава в результате электростимуляции с постоянной частотой, сравнивают по мере снижения частоты электростимуляции. В те-
Рис. 6.8. Активность двигательной единицы (толстая линия) при постепенном увеличении и последующем уменьшении силы (тонкие линии). Начало каждой тонкой линии (точка слева) указывает рекрутирование, а конец линии (точка справа) — дерекрутирование. Силы рекрутирования и дерекрутирования определяются проведением вертикальной линии от конца каждой тонкой линии до ее пересечения с толстой линией. Двигательная единица I рекрутировалась с силой 18% максимальной, а дерекрутировалась с силой 10% максимальной (правая ордината); интенсивность ее разряда изменилась с 9 Гц при рекрутировании до 15 Гц при пике силы (левая ордината) (Person, Kuding, 1982):
толстая линия — мышцы-разгибатели колена; тонкие линии — изменение интенсивности разряда отдельных двигательных единиц
194
Рис. 6.9. Субмаксимальная сила, вызванная в мышцах-разгибателях коленного сустава в результате электростимулирования (Binder-Machod, Guerin, 1990):
1 — изменяющаяся частота; 2 — постоянная частота
чение 60-секундного периода времени сила при снижении частоты стимуляции уменьшилась менее значительно, чем когда частота электростимуляции оставалась постоянной.
Другая структура разряда — двойной разряд — характеризует разряд двух потенциалов
Рис. 6.10. Измерение потенциалов действия двигательной единицы. Показан прием мотонейронами двух двигательных единиц (А и Б) множества импульсов, в том числе один общий для обоих нейронов. Потенциалы действия, разряжаемые двумя двигательными единицами (изображены шкалами), записаны с помощью проволочных электродов, вставленных в мышцу. Несмотря на независимость потенциалов действия, разряжаемых двумя двигательными единицами, отмечается незначительная статистическая корреляция и, следовательно, степень синхронности, зависящие от силы общего для двигательных нейронов импульса
действия отдельной двигательной единицей в пределах 10 мс. Двигательные единицы организма человека разряжаются в диапазоне 7—35 Гц, т.е. интервал между последовательными потенциалами действия составляет 30—140 мс. Когда двигательная единица электрически стимулируется с частотой 12 Гц (интервал 82 мс) и затем двойной разряд (интервал 10 мс) вмешивается в серию стимулов, значительно возрастает сила, образуемая двигательной единицей (Burke, Rudomin, Zajac, 1970). В принципе, в двигательных единицах организма человека при произвольных движениях двойной разряд наблюдается крайне редко.
В отличие от двойного разряда синхронность двигательной единицы характеризует временную взаимосвязь потенциалов действия в двигательных единицах. Обычно эти явления изучают, записывая разряд двух двигательных единиц (рис. 6.10) и затем сравнивая статистически временное возникновение потенциалов действия. У большинства испытуемых наблюдают определенную степень синхронности разряда двигательных единиц, обусловленную единым импульсом, поступающим в двигательный нейрон из какого-либо источника (Вгетпег, Baker, Stephens, 1991а, б, в; Nordstrom, Fuglevand, Enoka, 1992). Подобная синхронность характерна для двигательных единиц, расположенных в одной мышце, а также в мышцах-синергистах.
Альтернативный метод тестирования синхронизации двигательных единиц заключается в сопоставлении разряда отдельной двигательной единицы с ЭМГ всей мышцы. На основании этого метода было установлено, что программа занятий силовой направленности повышает степень корреляции разряда двигательных единиц, относящихся к тренированной мышце (Miner, Brown, Stein, Lee, 1975). Следовательно, повышение синхронизации двигательных единиц приведет к образованию большей силы.
МЫШЕЧНЫЕ ФАКТОРЫ
Величина силы, образуемой мышцей, зависит от степени возбуждения нервной системы. Вместе с тем она обусловлена свойствами самой мышцы. К ним относятся механические свойства мышцы и структурные эффекты, обусловленные различиями мышечной архитектуры.
Мышечная механика
Потенциал действия в мышечном волокне представляет собой явление типа “все или ничего”. Однако сила, образуемая мышечным волокном после потенциала действия мышечного
13*
195
волокна, не всегда одинакова. Величина силы зависит от внутренних факторов, таких, как структура разряда, и внешних факторов — длины мышечного волокна и скорости движения. Мышечная механика представляет собой изучение внешних механических переменных (например, длины, скорости, силы, мощности) при данном внутреннем состоянии (интенсивности разряда, наличии Са2+) мышцы.
Длина мышцы. Согласно теории скольжения филаментов, образование силы мышцей сопровождается скольжением толстого и тонкого филаментов относительно друг друга. Хотя точный механизм, вызывающий это скольжение, по-прежнему неизвестен (Pollack, 1983), существует три класса теорий, пытающихся дать объяснение явлению: сокращения филаментов, электростатическая и теория независимого образования силы. Наиболее популярной является теория поперечных мостиков мышечного сокращения, которая относится к последнему классу. Согласно этой теории, поперечные мостики, идущие от толстых филаментов, способны прикрепляться к тонким филаментам, подвергаясь затем структурно-химическим преобразованиям (т.е., вращение S1 при сокращении длины мышцы), в результате которых образуется растягивающая сила. После этого поперечные мостики открепляются и могут повторить цикл сначала (Liber, 1992).
Согласно данной схеме, развитие силы зависит от этих циклов прикрепления—открепления поперечных мостиков; чем больше количество циклов происходит в одно время, тем больше образующаяся сила. Поскольку длина мышцы изменяется, а тонкий и толстый филаменты скользят относительно друг друга, изменяется и число участников прикрепления поперечных мостиков. Из этого следует, что напряжение изменяется по мере перекрывания тонкого и толстого филаментов в пределах саркомера. На рис. 6.11 показано напряжение, наблюдаемое при различной степени перекрывания миофиламентов. При длине саркомера 3,5 мкм степень взаимного перекрывания минимальна, в обычных условиях это не наблюдается. При длине саркомера 2,0 мкм наблюдается максимальное перекрывание поперечных мостиков участками прикрепления на тонком филаменте. При дальнейшем скольжении филаментов относительно друг друга длина саркомера уменьшается до 1,5 мкм и степень взаимного перекрывания миофиламентов (т.е. способность взаимодействия между поперечными мостиками и тонкими филаментами) снижается. Результатом такого изменения числа прикреплений поперечных мостиков (вследствие изменения длины мышцы) является изменение величины силы, образуемой мышцей.
Однако величина силы, образуемой мышцей, не зависит исключительно от активного
Рис. 6.11. Взаимосвязь силы сокращения, длины саркомера и взаимного перекрывания миофиламентов. Сила выражена в процентах максимальной, образуемой мышечными волокнами (Edman, 1992)
процесса циклов поперечных мостиков. В составе мышцы большое количество соединительной ткани (эндомизий, перимизий, эпимизий, сухожилие) и цитоскелетных компонентов (промежуточные филаменты, титин, небулин). При растяжении эти структуры образуют пассивную силу, которая сочетается с активной, обусловленной активностью поперечных мостиков. Вследствие этого взаимодействия сила, образуемая мышцей, обусловлена как сократительными (миофиламенты), так и структурными (соединительная ткань и цитоскелет) элементами.
Рис. 6.12 иллюстрирует вклад активных и пассивных компонентов в силу мышцы при изменении ее длины от минимальной до максимальной. Измеряли два значения силы — в
Рис. 6.12. Вклад активного и пассивного элементов в общую величину силы при изменении длины мышцы. Сила измерялась изометрически (Ralston, Inman, Sthait, Shaffrath, 1947)
196
состоянии покоя и при максимальной произвольной силе. В состоянии покоя пассивная сила (образуемая элементами соединительной ткани и цитоскелета) возрастала с увеличением длины мышцы. При выполнении максимального произвольного сокращения при каждой длине мышцы сила была обусловлена как пассивными (заштрихованные кружочки), так и активными (незаштрихованные кружочки) компонентами и изменялась, как показано на рис. 6.12. Штриховой линией показано изменение силы вследствие активного компонента как функции длины мышцы. При более короткой длине мышцы величина силы обусловлена активным компонентом (активность поперечных мостиков), тогда как при большей длине — в основном пассивным компонентом. Профиль активного компонента показывает, что максимальное взаимное перекрывание тонкого и толстого филаментов наблюдается при промежуточной длине мышцы, т.е. длине мышцы в состоянии покоя (/0).
Длина мышцы влияет также на субмаксимальную силу и даже на силу единичного сокращения (Rack, Westbury, 1969). Например, в камбаловидной мышце кошки тетаническая сила и сила одиночного сокращения достигали максимума при угле сустава порядка 1 рад (57,3°), что соответствует большей длине мышцы (рис. 6.13, а). Длина мышцы влияет не только на величину сокращения, но и на его временные параметры (рис. 6.13, б): с увеличением длины мышцы возрастает как продолжительность сокращения, так и время полурасслабления. Влияние длины на силу таково, что для образования такой же силы при уменьшении длины необходимо стимулировать мышцу при более высокой частоте (Ruck, Westbury, 1969).
Для определения функциональной значимости взаимосвязи длина—сила мышцы необходимо трансформировать его* во взаимосвязь угол—вращающий момент. Однако превращение взаимосвязи длина—сила во взаимосвязь угол—вращающий момент затрудняют, по меньшей мере, три фактора, которые могут изменять влияние длины. Во-первых, движение большинства сегментов тела (жестких звеньев в нашей единой системе сустава) контролируется группой мышц, а не отдельной мышцей. Очень часто волокна в каждой мышце такой группы располагаются неодинаково, поэтому при любом положении сустава волокна различных мышц, по-видимому, будут находиться в различных положениях кривых длина—сила. Во-вторых, поскольку некоторые мышцы (например, двуглавая мышца плеча, икроножная) проходят более чем через один сустав, на их длину и, следовательно, величину образуемой ими силы влияет положение не одного сустава. В-третьих, изменения мышечной силы не отра-
Рис. 6.13. Влияние длины мышцы на силу, образуемую камбаловидной мышцей кошки (Rael, Westbury, 1969): а — изменение тетанической силы (7), силы одиночного сокращения (2) и пассивной силы (5) как функция длины мышцы (длина мышцы увеличивалась вправо); б — изменение реакции одиночного сокращения по мере уменьшения длины мышцы от большой (угол 30") при выполнении 5-миллиметровых шагов; 30° — начальная большая длина; —5 и — 20 мм — меньшая длина
жают соответствующие изменения вращающего момента мышцы, поскольку вращающий момент — результат двух переменных (силы и плеча пары), а не влияния длины на силу; кроме того, необходимо учитывать влияние длины (угол сустава) на плечо пары. Либер (Liber, 1992) установил, что пик мышечной силы может наблюдаться при абсолютно другом угле сустава, чем пик плеча пары, вследствие этого пик вращающего момента мышцы будет наблюдаться при промежуточном угле. Вместе с тем, в принципе, мышечная сила — главный фактор, определяющий вращающий момент мышцы относительно сустава, исключение составляют камбаловидная, средняя ягодичная и прямая мышцы бедра.
Взаимосвязь угол—вращающий момент различна в разных мышцах вследствие неодинаковой структуры волокон, влияния угла сустава на плечо силы (Kulig, Andrews, Hay, 1984). Эти воздействия показаны на рис. 6.14, где изменения длины мышцы представлены изменениями
197
Угол, рад
Рис. 6.14. Три типичные формы взаимоотношения угол — вращающий момент (Kulig, Anrews, Hay, 1984): а — восходящая; б — нисходящая; в — промежуточная
угла сустава. Вообще взаимосвязь угол—вращающий момент имеет три формы — восходящую, нисходящую и промежуточную. При восходящей форме взаимодействия (сгибатели коленного сустава, приводящие мышцы бед
ра) вращающий момент повышается до плато с увеличением угла сустава. При нисходящей форме взаимодействия (отводящие мышцы бедра) вращающий момент максимален при минимальном угле сустава и снижается с увеличением угла в суставе. При промежуточном характере взаимодействия (сгибатели и разгибатели локтевого сустава, разгибатели коленного сустава) максимальный вращающий момент наблюдается при промежуточном угле сустава.
Результирующий вращающий момент относительно сустава может иметь одну из этих форм, тем не менее взаимосвязь угол—вращающий момент отдельных мышц может значительно отличаться. Это было установлено при использовании скелетно-мышечной модели ноги человека (Hoy, Zajac, Gordon, 1990). Модель, основанная на скелетно-мышечной геометрии и параметрах мышцы, определяет взаимоотношение угол—вращающий момент 18 мышц или групп мышц относительно бедренного, коленного и голеностопного суставов (рис. 6.15). Прогнозируемые показатели результирующего вращающего момента на основании модели можно сопоставить с экспериментальными измерениями; на каждом графике сравните пунктирную линию (эксперимент) со сплошной линией. Вращающие моменты измеряли
Угол бедренного сустава, рад
Угол коленного сустава, рад
Рис. 6.15. Взаимосвязь максимальный угол — вращающий момент относительно бедренного, коленного и голеностопного суставов. Графики показывают результирующую взаимосвязь (жирные линии), а также вклад отдельных мышц относительно каждого сустава (Hoy, Zajac, Gordon, 1990):
а — разгибатель бедра-подколенное сухожилие (2), большая приводящая (3), ягодичная (4) и длинная приводящая, короткая приводящая и нежная (5); б-— сгибатель бедра — прямая мышца бедра (7), подвздошно-поясничная, медиальная ягодичная и малая (2), нежная, напрягатель широкой фасции, гребешковая и портняжная (3), большая приводящая и короткая приводящая (4); в — разгибатель коленного сустава — прямая (/) и широкая мышца бедра (2); г — сгибатель коленного сустава — короткая головка двуглавой мышцы бедра (/), подколенное сухожилие (2), икроножная (3) и нежная, портняжная и напрягатель широкой фасции (штриховая линия}; о — сгибатели подошвы — камбаловидная (7), икроножная (2) и другие сгибатели подошвы (2); е — задний сгибатель голеностопного сустава
198
Рис. 6.16. Взаимосвязь чистого (результирующего) вращающего момента мышцы и вращающего момента, обусловленного нагрузкой (включая вес системы), определяющая уменьшение, увеличение или неизменность длины активных мышц
в экспериментальных условиях многие группы исследователей: разгибатели бедра — Nometh, Ekholm, Arborehus, Schuldt, 1983; сгибатели и разгибатели коленного сустава — Schuffer, 1980 и т.д. Взаимосвязь угла— вращаюшего момента в отдельных мышцах каждой группы была разной.
Изменение длины мышцы. После того как мышца активировалась и начала осуществлять вращающий момент, филаменты начинают скользить друг относительно друга, а длина активной мышцы уменьшается. Вместе с тем изменяется или не изменяется длина активной мышцы, зависит от величины вращающего момента мышцы относительно вращающего момента, обусловленного физической нагрузкой (рис. 6.16). Это соотношение (мышца — нагрузка) имеет три значения: менее 1, равное 1 и выше 1. Если соотношение равно 1, вращающие моменты одинаковы и длина всей мышцы не изменяется; это условие называется изометрическим сокращением. На рис. 6.14 и 6.15 приведены данные, касающиеся изометрических сокращений. Если же вращающие моменты разные (т.е. менее или более 1), длина мышцы изменяется. Если вращающий момент мышцы больше, соотношение превышает 1 и длина мышцы снижается, это концентрическое сокращение. Если вращающий момент, обусловленный физической нагрузкой, превышает вращающий момент мышцы, соотношение меньше 1, длина мышцы увеличивается, это эксцентрическое сокращение. Эти механические состояния — изометрическое, концентри
ческое и эксцентрическое — часто называют типами сокращений.
Взаимосвязь угол — вращающий момент, продемонстрированную на рис. 6.14, изучали при изометрических, концентрических и эксцентрических сокращениях. Вращающий момент во время эксцентрических сокращений больше, чем при концентрических, а при изометрических сокращениях его величина промежуточна между таковой при эксцентрических и концентрических сокращениях. На рис. 6.17 приведены показатели взаимосвязи угол — вращающий момент разгибателей ноги и коленного сустава при концентрических и эксцентрических сокращениях, которые характеризуют силу, образуемую ногой при выпрямлении бедренного, коленного и голеностопного суставов, когда испытуемые находились в положении сидя. Для разгибателей ноги (рис. 6.17, а) максимальный эксцентрический вращающий момент наблюдался при угле коленного сустава около 2,5 рад, тогда как концентрический вращающий момент продолжал увеличиваться до достижения максимального угла коленного
Рис. 6.17. Взаимосвязь угол — вращающий момент при концентрических и эксцентрических сокращениях. Угол колена 3,14 рад представляет полное выпрямление ноги в суставе:
а — сила, образуемая ногой при ее выпрямлении как функция угла колена (Eloranta, Komi, 1980); б — вращающий момент разгибателей коленного сустава при произвольной активации (3, 4), электростимулировании (2, 6) и сочетании активации и электростимулирования (1, 5) при эксцентрических (1—3) и концентрических (4—6) сокращениях (Westing, Seger, Thorstensson, 1990)
199
Время, с
б
О 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 1,1 Время, с
Рис. 6.18. Изменение длины мышечного волокна при изометрических (а) и эксцентрических (б) сокращениях (Griffits, 1991):
а — изометрическое сокращение в медиальной икроножной мышце; длина всей мышцы оставалась постоянной при уменьшении длины мышечного волокна и увеличении силы; длину мышечного волокна измеряли с помощью ультразвуковой техники между двумя пьезоэлектрическими кристаллами; б — эксцентрическое сокращение в медиальной икроножной мышце после касания ступней поверхности обусловлено уменьшением длины мышечного волокна; жесткое касание поверхности ступней указано вертикальной линией
сустава. На большем диапазоне движения Westimg, Seger, Tharstenson (1990) обнаружили, что вращающий момент при эксцентрических сокращениях больше, чем при концентрических. Для разгибателей коленного сустава (рис. 6.17, б) вращающий момент при эксцентрических и концентрических сокращениях был максимальным при промежуточном угле колена (2,0 рад), причем вращающий момент при эксцентрических сокращениях был больше, чем при концентрических сокращениях при всех углах колена. Кроме того, сравнивали данные разгибателей коленного сустава с взаимоотношением угол — вращающий момент, полученным при произвольной активации и электри
ческой стимуляции. Три условия концентрических и эксцентрических сокращений включали: произвольную активацию, электрическую стимуляцию и сочетание произвольной активации и электростимуляции. При концентрических сокращениях наблюдали одинаковый вращающий момент во всех трех случаях при угле коленного сустава (2,0—3,0 рад), в то же время вращающий момент при произвольной активации (кривая 4) был больше, чем при электростимуляции (кривая 6), а также при сочетании произвольной активации и электростимуляции (кривая 5). При эксцентрических сокращениях вращающий момент, обусловленный сочетанием произвольной активации и электростимуляции (кривая 7), был больше, чем вращающий момент, обусловленный электростимуляцией (кривая 2) и максимальной произвольной активацией (кривая 3). Поскольку вращающий момент, обусловленный произвольной активацией, был наименьшим при эксцентрических сокращениях, Westing (1990) предположил, что испытуемые не активировали максимально разгибатели коленного сустава при произвольных эксцентрических сокращениях.
Таким образом, направление изменения (длиннее или короче) длины мышцы зависит от соотношения между вращающим моментом мышцы и физической нагрузкой, т.е. определяется механическими условиями задания. Однако изменение длины всей мышцы, включая волокна и сухожилие, не обязательно совпадает с изменением длины мышечных волокон (Fellows, Rack, 1987; Griffiths, 1991). При электростимуляции икроножной мышцы кошки с целью осуществления изометрического сокращения, мышечные волокна могут укорачиваться на 28 % своей длины в состоянии покоя (рис. 6.18, а). Чтобы длина всей мышцы осталась постоянной (изометрическое сокращение), настолько же должна увеличиваться длина сухожилия. Такая эластичность сухожилия (mm/N) наиболее проявляется в мышцах с длинными сухожилиями. Подобное явление наблюдается не только при электростимуляции, но и во время произвольных движений, например, во время ходьбы. На рис. 6.18, б показана длина всей мышцы, длина волокон и ЭМГ медиальной икроножной мышцы кошки при выполнении цикла шага. До касания лапой поверхности земли длина мышцы и волокна увеличивается, затем уменьшается; это — стандартное концентрическое сокращение. Однако после контакта лапы с поверхностью земли длина всей мышцы увеличивается (эксцентрическое сокращение), а длина волокна уменьшается. Увеличение всей длины мышцы осуществляется в результате растяжения сухожилия. По-видимому, при изометрических и медленных эксцентрических сокращениях изменение длины мышеч-
200
кого волокна не соответствует изменению длины всей мышцы в мышцах, имеющих длинные сухожилия.
Разница между изометрическими и анизометрическими (концентрическими и эксцентрическими) сокращениями заключается в неспособности первых выполнять работу. Поскольку работа — результат силы и расстояния, неспособность переместить объект, что имеет место при изометрическом сокращении, означает, что по отношению к объекту работа не выполняется. Однако хотя по отношению к объекту механическая работа не выполняется, мышцы выполняют метаболическую работу.
Интенсивность (скорость) изменения длины мышцы. Если вращающий момент мышцы и нагрузки неодинаков, длина всей мышцы изменяется. Величина вращающего момента, который может осуществить мышца в таких условиях, зависит от величины интенсивности изменения длины — скорости мышечного сокращения. При уменьшении длины мышцы она сокращается концентрически; этот режим активности связан с меньшим вращающим моментом мышцы по сравнению с изометрическими (скорость равна 0) сокращениями; следовательно, при максимальных концентрических сокращениях вращающий момент мышцы меньше, чем при максимальных изометрических сокращениях. Кроме того, по мере увеличения скорости укорачивания мышцы, уменьшается ее вращающий момент (рис. 6.19). Хотя интенсивность циклов поперечных мостиков (прикрепление—вращение—открепление) возрастает при увеличении скорости укорачивания мышцы, средняя величина силы, образуемой каждым поперечным мостиком, уменьшается, и если длина мышцы уменьшается еще быстрее, число прикрепления поперечных мостиков значительно снижается. Поскольку количество энергии, используемой мышцей с увеличением скорости снижения длины мышцы, возрастает по мере уменьшения вращающего момента, эффективность мышцы (производительность, использование энергии) снижается и увеличивается разница между вращающим моментом мышцы и нагрузки.
Когда вращающий момент мышцы меньше вращающего момента нагрузки, длина всей мышцы увеличивается. Максимальный вращающий момент при эксцентрических сокращениях больше, чем при изометрических и концентрических. Как видно из рис. 6.19, максимальный вращающий момент изменяется следующим образом: вращающий момент при эксцентрических сокращениях больше, чем при изометрических, а при изометрических — больше, чем при концентрических. Максимальный вращающий момент при эксцентрических сокращениях в 1,5—1,8 раза больше, чем максимальная изометрическая сила. Почему наиболь-
Рис. 6.19. Взаимосвязь скорость — вращающий момент мышц-разгибателей коленного сустава. Вращающий момент измеряли при угле коленного сустава 1,0 рад при различных сокращениях с постоянной скоростью (Westing, Seger, Thorstensson, 1990);
1 — произвольная активация; 2 — электростимулирование;
3 — активация
ший вращающий момент при эксцентрических сокращениях мышцы? Очевидно, это обусловлено рядом факторов. Например, возрастание активности сократительных механизмов за счет увеличения количества выделяемого Са2+ либо растяжения менее полно активированных саркомеров в каждой миофибрилле (Hoyle, 1983).
Следует отметить, что взаимосвязь, изображенная на рис. 6.19, представляет собой упрощенный вариант влияния скорости, в частности, с точки зрения эксцентрических сокращений. Каждая точка данных на графике отражает вращающий момент при данной скорости, т.е. вращающий момент — зависимая переменная. Вращающий момент каждой точки данных измерялся при каждом отдельном сокращении (эксцентрическом или концентрическом) в диапазоне движения 1,4 рад. Например, измеряли вращающий момент при угле коленного сустава 2,62 рад от сокращения, которое “шло” от начального угла 1,74 рад до заключительного — 3,14 рад со скоростью 3,14 рад'с-1. Следующее измерение осуществляли таким же образом только при другой скорости и т.д. Поскольку сила, образуемая мышцей, изменяется в зависимости от ее длины, вращающий момент измеряли при одном и том же угле коленного сустава (2,62 рад) при каждом сокращении. Наоборот, взаимосвязь между скоростью и вращающим моментом (см. рис. 6.19) определяли при постоянной скорости.
Следует подчеркнуть это различие, поскольку известно, что сила мышцы измеряется довольно сложным образом, когда активная
201
Рис. 6.20. Изменение силы мышцы при контролируемом изменении ее длины. Длина выражена в процентах длины мышечного волокна (Lf) к длине мышц в состоянии покоя. Сила выражена в процентах максимальной изометрической силы (/%, 0). Стимулирование осуществлялось в течение времени, отмеченного горизонтальной линией внизу (McCully, Faulkner, 1985)
мышца с увеличением скорости переходит от изометрических к эксцентрическим сокращениям (Rack, Westbury, 1974). Пример этого взаимодействия иллюстрирует рис. 6.20. В этом примере длина длинного разгибателя пальцев контролировалась при стимулировании нерва, идущего к мышце, а ее сила измерялась при концентрических, изометрических и эксцентрических сокращениях (McCully, Faulkner, 1985). При каждом сокращении мышцу стимулировали с частотой 150 Гц в течение 300 мс. Через 100 мс стимуляции длина мышцы уменьшалась, была постоянной или увеличивалась. При изометрических сокращениях длина мышцы не изменялась, пока величина силы при стимулировании повышалась до плато. Вместе с тем, когда длина мышцы изменилась, изменение величины силы не соответствовало изменению ее длины. При укорачивающих сокращениях с постоянной скоростью сила уменьшалась вначале быстро, а затем намного медленнее, чем при изометрических сокращениях. В то же время при удлиняющих сокращениях с постоянной скоростью сила увеличивалась вначале быстро, а затем медленнее. При сокращениях с постоянной скоростью наклон кривой силы указывает на жесткость мышцы. При эксцентрических сокращениях жесткость вначале высокая, а затем снижается; эта начальная жесткость называется жесткостью короткого диапазона.
Пример 1
Взаимоотношение между максимальным вращающим моментом при трех видах сокращений (концентрических, эксцентрических и изометрических) часто используют при выборе упражнений. Допустим, подтягивания на перекладине входят в программу занятий по физ
культуре. Во многих группах (школьном классе, группах здоровья) некоторым не под силу выполнить упражнение, которое включает сгибание рук в локтевых суставах, чтобы подтянуть подбородок к перекладине, с последующим их выпрямлением, чтобы опустить туловище в положение виса на вытянутых руках. В этой связи рекомендуется начинать упражнение со второй части, т.е. из положения касания подбородком перекладины с последующим медленным опусканием туловища вниз. Все упражнение состоит из двух частей: концентрической активности сгибателей локтевого сустава, направленной на подтягивание туловища вверх, и эксцентрической активности, направленной на его опускание. Поскольку вращающий момент мышц при эксцентрических сокращениях больше, большинство людей могут выполнить вторую часть упражнения, т.е. опускание туловища вниз. Вместе с тем максимальный вращающий момент при эксцентрических сокращениях происходит при более низкой постоянной скорости, поэтому эксцентрические сокращения должны осуществляться медленно — это означает контролируемое опускание туловища вниз. Следует отметить, что физическая нагрузка на активную мышцу, по-видимому, вызывает практически одинаковые адаптационные реакции, независимо от того, выполняет ли мышца работу в эксцентрическом или концентрическом режиме (Komi, Buskirk, 1972). Таким образом, увеличение силовых способностей вследствие выполнения эксцентрической части упражнения положительно влияет на силовые возможности мышцы при выполнении концентрических сокращений. Поэтому исполняя вторую часть упражнения “подтягивания”, вы со временем укрепите соответствующие мышцы и сможете выполнять все упражнение.
Пример 2
Предположим, один испытуемый способен подтянуться 20 раз, а другой — только 2 раза. Предположим, они подтягиваются столько раз, сколько могут, с максимальной скоростью. Интуиция подсказывает, что первый испытуемый подтянется первый раз намного быстрее, чем второй. Почему? Объяснить это можно на основании взаимосвязи скорости—вращающего момента. Для удобства сделаем ряд предположений; предположим, что у испытуемых одинаковая масса тела, что основными движителями являются мышцы-сгибатели локтевого сустава и рассмотрим только концентрическую (собственно подтягивание) фазу. Ввиду различий в уровне силы, о чем свидетельствует вращающий момент при изометрических сокращениях (значение начального вращающего момента) (рис. 6.21), человек, способный
202
Рис. 6.21. Взаимосвязь скорость — вращающий момент мышц-сгибателей локтевого сустава у испытуемого, который подтягивается 20 раз (7) и 2 раза (2)
подтянуться 20 раз, способен образовать вращающий момент, необходимый для того, чтобы подтянуть туловище вверх (горизонтальная линия на рис. 6.21), с более высокой скоростью. В этом примере человек, способный подтянуться 20 раз, способен выполнить подтягивание туловища с любой скоростью вплоть до 4 рад • с-1, тогда как человек, способный выполнить только 2 подтягивания, может достичь максимальной скорости 2 рад - с-1. Таким образом, первое подтягивание выполняется физически сильным человеком значительно быстрее.
Пример 3
Несмотря на очевидную трудность измерения мышечной силы у испытуемых, Коми и соавт. (Greger, Komi, Browning, Tarvinen, 1991; Komi, 1990, 1992) провели серию экспериментов, в которых инвазивно прикрепляли преобразователь силы к ахилловому сухожилию добровольца и контролировали образуемую сухожилием силу при различных видах деятельности. Кроме того, исследователи засняли испытуемых во время ходьбы, бега, подпрыгиваний, прыжков и езды на велосипеде с тем, чтобы определить длину мышцы, а следовательно, и скорость. Результаты одного из экспериментов, в котором испытуемый бежал со скоростью 5,8 м-с-1, показаны на рис. 6.22. Данные характеризуют силу—скорость во время стационарной фазы; отрицательная скорость указывает на увеличение длины мышцы (включая сухожилие). При касании ступней поверхности земли длина икроножной мышцы вначале увеличивалась, о чем свидетельствовала отрицательная скорость, а сила достигала пикового значения порядка 9,500 Н. Когда длина мышцы уменьшается, сила снижается до нуля.
Эта последовательность называется эксцентри-чески-концентрической или циклом растяжения-сокращения.
Мышечная мощность. В этой главе мы описали механические свойства мышцы с точки зрения взаимосвязи длина—сила и сила—скорость. Взаимосвязь длина—сила отражает статическую способность мышцы и показывает, какова сила, образуемая мышцей при различной длине. Согласно теории поперечных мостиков, сила, образуемая мышцей, изменяется соответственно степени взаимного перекрывания толстых и тонких миофиламетов. Наряду с этим взаимосвязь сила—скорость характеризует динамическую способность мышцы, которая включает влияние интенсивности изменения степени взаимного перекрывания на мышечную силу. Другой способ характеристики динамической способности мышцы касается ее способности развивать мощность; это возможно, поскольку мощность равна произведению силы и скорости или интенсивности выполнения работы (Josephson, 1993). Когда длина мышцы уменьшается и сила, образуемая ею, вынуждает объект передвигаться, мышца выполняет положительную работу, т.е. мышца выполняет работу по отношению к объекту. С другой стороны, мышца выполняет отрицательную работу или работа выполняется по отношению к ней, когда ее длина увеличивается по мере приложения силы к объекту, который перемещается. Мышца развивает мощность, выполняя положительную работу (концентрическое сокращение), и поглощает мощность, выполняя отрицательную работу (эксцентрическое сокращение).
Скорость, % длины в покое
Рис. 6.22. Сила ахиллова сухожилия и икроножной мышцы во время стационарной фазы бега со скоростью 5,8 м • с-1. Жесткое касание ступней поверхности отмечено на графике в конечной точке справа (Komi, 1992)
203
Рис. 6.23. Приседание со штангой. На диаграмме свободного тела показана результирующая активность относительно коленного сустава. Движение осуществляется, в основном, за счет разгибателей коленного сустава (Гт)
Рассмотрим выполнение приседания со штангой (рис. 6.23). Во время выполнения этого упражнения главную роль играют мышцы-разгибатели коленного сустава. При переходе из положения стоя в положение сидя длина мышц-разгибателей колена увеличивается, поскольку они помогают опустить туловище вниз. В этот момент разгибатели сокращаются эксцентрически и выполняют отрицательную работу, т.е. работа выполняется массой системы (штанга + тяжелоатлет) по отношению к мышцам. Когда работа выполняется по отношению к мышцам (отрицательная работа), мышца поглощает мощность. Когда испытуемый подни-
Рис. 6.24. Взаимосвязь сила—скорость (сплошная линия), полученная на изолированной мышце. Кривая силы определена как результат силы и скорости
мается из положения сидя, разгибатели коленного сустава выполняют положительную работу и, следовательно, развивают мощность. В принципе, развитие мощности ограничено интенсивностью доставки энергии, необходимой для сокращения мышцы (образование АТФ), а также интенсивностью, с которой миофиламенты превращают химическую энергию в механическую работу (Fitts, McDonald, Schluter, 1991; Wess-Fogh, Alexander, 1977). Считают, что развитие мощности ограничивает работоспособность, особенно в упражнениях небольшой продолжительности (Enoka, 1988а; Ericson, 1988).
Мощность определяется как результат силы и скорости мышечного сокращения. Рис. 6.24 иллюстрирует уменьшение силы, образуемой мышцей, по мере увеличения скорости укорачивающих сокращений. Кривая силы—скорости (сплошная линия) касается осей в двух концах графика. В одном конце скорость равна 0, поэтому показатель силы отражает изометрическую силу (Fm 0). В другом конце скорость характеризуется максимальным значением (Емакс) и сила равна 0. В обеих этих точках результат силы и скорости, а следовательно и мощность, равны нулю. Поскольку между этими конечными точками сила и скорость не равны нулю, мощность изменяется в зависимости от величины произведения.
Колебание мощности как функции силы и скорости можно изучать, используя уравнение Хилла для силы—скорости мышцы. Хилл (Hill, 1938) обнаружил, что уменьшение силы как функции скорости уменьшения длины мышцы можно представить уравнением гиперболы прямоугольника, которое имеет следующий вид:
v(Fm + а) = b(Fmo - Fm);
F„v + av = 6Fmo - bFm, <61)
где v — скорость уменьшения длины;
v0 — максимальная скорость уменьшения длины при Fmt о,
Fm> о ~ максимальная изометрическая сила;
Fm — сила;
а — коэффициент тепла, образуемого при уменьшении длины 10,15 Fm 0 — 0,25
, _ Ч, рт 0; h _ постоянная (а — ~р~).
Уравнение 6.1 выражает динамическую способность мышцы как взаимосвязь работа—энергия. Левая часть уравнения (Fmv + av) соответствует интенсивности изменения энергии, правая (bFm 0 - bFm) — интенсивности выполнения работы. Ввиду определенной степени вязкости мышцы (трение жидкости) интенсивность утилизации энергии нелинейно связана с интенсивностью выполнения работы, т.е. кривая мощности на рис. 6.24 не характеризуется линей
204
ной взаимосвязью, Fmv отражает интенсивность, с которой сократительные белки выполняют работу (развивают мощность) по отношению к нагрузке, тогда как av — элемент подавления (вследствие вязкости), делающий быстрые движения расточительными. Их взаимодействие
приводит сначала к увеличению, а затем умень
шению мощности по мере увеличения скорости сокращений — кривая мощности на рис. 6.24. Уравнение Хилла (6.1) можно преобразовать, чтобы определить мощность (Fmv) более простым способом:
v(bFm<) ~ av) (v + b)
(6.2)
На основании уравнения 6.2 можно показать, что мощность (Fmv) максимальна, когда Fm v соответствуют 1/3 и 1/4 своих максимальных значений.
Поскольку взаимосвязь сила—скорость характеризует динамическую способность мышцы, эти показатели значительно влияют на выполнение движения (Fitts, et al., 1991). Вследствие этого влияния можно изменить мышечную динамику (взаимосвязь сила—скорость и мощность-скорость) и, таким образом, повлиять на способность выполнять движение.
Пример 1
Способность мышцы развивать мощность зависит от ее способности образовывать силу, а также от скорости сокращения ее длины. Поскольку площадь поперечного сечения (индекс мышечной силы) и скорость сокраще-
Рис. 6.25. Прогнозируемая взаимосвязь сила—скорость и мощность—скорость камбаловидной (/) и медиальной икроножной (2) мышц человека. Взаимосвязь сила—скорость характеризовалась пиком силы при скорости 0, затем с увеличением скорости сила снижалась. Взаимосвязь мощность — скорость характеризовалась пиком мощности при промежуточном значении скорости (1,0 см-с-1 в камбаловидной и 5,0 м • с-1 в медиальной икроножной) (Edgerton, Roy, Gregor, Rugs, 1986)
ний (отражаемая быстро- и медленносокра-щающимися мышечными волокнами) в мышцах не одинаковы, способность развивать мощность также разная в различных мышцах, что иллюстрирует рис. 6.25. Хотя камбаловидная мышца имеет большую Fm 0, а медиальная икроножная большую Имакс, пик мощности, производимой обеими мышцами-синергистами, одинаков (Wickiewich, Roy, Powell, et al., 1984).
Пример 2
Один из способов измерения взаимосвязи сила—скорость мышцы состоит в том, чтобы отсоединить один конец от кости и прикрепить к нагрузке, а затем активировать мышцу, супрамаксимально стимулируя ее нерв и измеряя силу и скорость во время сокращения. Эту процедуру называют изотонической нагрузкой. Если нагрузка между сокращениями изменяется (например, 3—75 % Fm о), получают разные значения силы и скорости; на основании полученных данных строят кривую сила—скорость (рис. 6.24). Эта процедура изотонической нагрузки позволяет нам определить взаимосвязь сила—скорость при концентрических сокращениях, а также взаимосвязь мощность—скорость при развитии мощности (рис. 6.24).
I регор и соавт. (рис. 6.26) сравнивали взаимосвязь сила—скорость и мощность—скорость, установленную с помощью данной процедуры в камбаловидной мышце, с показателями, измеренными при беге кошки со скоростью около 2,2 м • с-1 (кошки могут бегать со скоростью до 8 м • с-1) (Goslow, Reinking, Stuart, 1973). Чтобы сопоставить мышечную динамику, потребовалось измерять скорость и силу камбаловидной мышцы во время передвижения животного. Результаты приведены на рис. 6.26. Следует подчеркнуть два фактора: 1) сходство кривых передвижения у человека (рис. 6.22) и кошки (рис. 6.26, а), которое включало эксцентрическую (отрицательная скорость) и концентрическую фазы при пике силы в момент перехода от эксцентрических сокращений к концентрическим и 2) более высокие показатели силы во время бега по сравнению с условиями изотонической нагрузки, что можно объяснить использованием эксцентрическо-концентрических сокращений во время локомоции и лишь концентрическими сокращениями при изотонической нагрузке. Точно так же кривые мощность—скорость (рис. 6.26, б) включают две фазы: абсорбции (отрицательная скорость) и развитии мощности; более высокие показатели мощности камбаловидной мышцы наблюдались при беге со скоростью 2,2 м-с-1, чем во время изотонической нагрузки.
205
Рис. 6.26. Взаимосвязь сила—скорость (а) и мощность—скорость (6) камбаловидной мышцы кошки (Roy, Whitihg, Lovely et al., 1988):
1 — супрамаксимальное стимулирование и изотоническая нагрузка; 2 — скорость бега 2,2 м • с-1
Пример 3
Победителем в велогонках на спринтерскую дистанцию, как правило, становится спортсмен, развивающий наибольшую мощность, испытывая минимальное сопротивление воздуха. Как и в случае с изолированной мышцей, для ног велосипедиста характерна взаимосвязь сила-скорость, при которой максимальная сила, действующая на педали, уменьшается со снижением скорости вращения (рис. 6.27, а). При данной скорости вращения велосипедист может прилагать различную силу; максимальная сила отмечена на рис. 6.27, а линией, которая характеризует верхнюю границу. Имея эти максимальные показатели силы и скорости, можно определить взаимосвязь мощность—скорость, отражающую максимальную мощность, развиваемую при каждой скорости вращения (рис. 6.27, б). Как и в случае с изолированной мышцей, максимальная мощность развивается при промежуточном значении скорости. Оптимальная скорость у сильнейших велогонщиков составляет около 110 вращений-мин-1 при силе, действующей на педали, 500 Н.
При воздействии на спортсмена сопротивления (ветер, езда под гору) движущая сила уменьшается. Как показано на диаграмме сво
бодного тела на рис. 6.28, движущая сила (Fp) равна разнице между тангенциальными компонентами силы реакции поверхности земли (Fx ,), сопротивления воздуха (Fflj z) и векторов массы (Fw ,). В результате повышенного сопротивления и уменьшения движущей силы скорость вращения снижается, и хотя сила, действующая на педали, может увеличиваться, мощность, развиваемая велосипедистом,снижается. Чтобы воспрепятствовать этому велосипедист интуитивно меняет скорость, что ведет к увеличению скорости педалирования, скорости движения и мощности. Таким образом, наличие скоростей передач на велосипеде позволяет спортсмену изменять взаимоотношение силы—скорости и, следовательно, мощность.
МЫШЕЧНАЯ АРХИТЕКТУРА
После активации мышцы нервной системой образуемая мышцей сила зависит не только от механических свойств ее сократительных элементов, но и от архитектуры мышцы или физического расположения этих сократительных элементов. На нервно-мышечную функцию, по-видимому, значительно влияют размер и форма мышечных волокон и мышц вообще (Edgerton,
206
Рис. 6.27. Влияние скорости вращения на пик силы (а) и мощность (6), воздействующую на педали во время езды. С увеличением скорости вращения пик силы снижается; пик мощности наблюдается при промежуточной скорости вращения (Sargeant, Boreham, 1981)
Рис. 6.28. Диаграмма свободного тела системы велосипедиста:
о — система, перемещающаяся вдоль прямой, испытывает воздействие веса системы (F„), сопротивления воздуха Fo и силы реакции поверхности (Fg); движущая сила (F,) действует в направлении движения велосипедиста; б — при движении системы под гору Fw> , и F„ , противодействуют движению системы вперед, тогда как Fg , — содействует
Roy, 1991). Например, мышечная архитектура влияет как на величину максимальной силы мышцы, так и на максимальную скорость ее сокращения (укорачивания). Чтобы охарактеризовать влияние архитектуры на мышечную силу, необходимо сначала ввести модель саркомера (трехкомпонентную модель), а затем рассмотреть влияние различного строения саркомера на уровне мышечного волокна, всей мышцы и единой системы сустава.
Трехэлементная модель. Саркомер — основная функциональная единица мышцы; он содержит все элементы, необходимые для образования силы. Механическое “поведение” саркомера можно приблизительно представить в виде трехэлементной модели мышцы (рис. 6.29). Тремя элементами этой модели являются параллельный эластичный элемент (РЕ), последовательный эластичный элемент (SE) и сократительный элемент (СЭ). Эти три элемента представляют собой специфические анатомические структуры с учетом механических характеристик саркомера. SE и РЕ представляют различные типы эластичности и показаны в виде
пружин. Например, взаимосвязь длина—сила мышцы на рис. 6.12 показывает, что некоторые пассивные структуры в мышце (например, ци-тоскелетные элементы, оболочка соединительной ткани) могут образовывать пассивную силу при растяжении нестимулированной мышцы; это явление отражает РЕ. Как видно из рис. 6.12, вклад пассивных элементов (РЕ) в общую
Рис. 6.29. Трехэлементная механическая модель саркомера
207
Рис. 6.30. Упруговязкие свойства (Winter, 1990):
о — гистерезис взаимосвязи сила—длина; б — релаксация силы — величина силы снижается, тогда как длина остается постоянной;
в — увеличение длины при постоянной величине силы
величину мышечной силы возрастает с увеличением длины мышцы и максимален при самой большой длине мышцы.
Помимо эластичности РЕ некоторые структуры миофиламентов и сухожилия обладают эластичностью, которая “располагается” последовательно с единицами, образующими силу; это SE. В саркомере вклад в SE обеспечивают толстые и тонкие филаменты, поперечные мостики и Z-линии, однако наибольший вклад вносят миофиламенты (Blange, Kramer, 1972; Suzuki, Sugi, 1983). SE проявляется только во время сокращения мышцы и характеризуется меньшей степенью растяжения, чем РЕ, имея длину порядка 4—5 % /0, где /0 — длина мышцы в покое.
СЕ представляет собой способность силы-скорости саркомера и включает два компонента, один из которых обусловливает способность миофиламентов образовывать силу, а второй — связанное со скоростью влияние вязкости на силу, образуемую саркомером. О способности миофиламентов образовывать силу свидетельствуют разобщенные филаменты, характеризующие влияние взаимного перекрывания миофиламентов на величину силы. Влияние вязкости на силу можно описать с помощью уравнения F = BV, где F — сила, В — константа, V — скорость мышцы. Буфер обусловливает а Ив уравнении 6.1 и свидетельствует о том, что необходимо больше силы (и следовательно, энергии) для обеспечения скольжения миофиламентов относительно друг друга с увеличением скорости укорочения мышцы; следовательно, саркомер образует меньшую силу. Буфер — компонент модели, характеризующий пониженную эффективность быстрых сокращений.
Таким образом, механическое “поведение” активного саркомера является упруговязким, т.е. он обладает свойствами и вязкости, и упругости (эластичности). Согласно Уинтерусу (Winters,
1990), упруговязкость обусловливает: 1) гистерезис во взаимосвязи сила—длина при цикличной нагрузке, где сила, образуемая при определенной длине саркомера во время увеличения длины, больше силы, образуемой при такой же длине саркомера при сокращении длины; 2) релаксацию силы, т.е. при постоянной длине саркомера величина силы постепенно снижается; 3) потребность увеличения длины с тем, чтобы поддержать постоянную величину силы (рис. 6.30). Другим результатом последовательного характера СЕ и SE является то, что при изометрических сокращениях, даже несмотря на то, что длина всей мышцы остается постоянной, СЕ может значительно сокращаться при растяжении SE (Fellows, Rack, 1987; Griffiths, 1991).
Данные, приведенные на рис. 5.2, показывают, что пик силы, образуемой мышцей при одиночном сокращении, намного меньше, чем при тетанусе. Чем обусловлено различие в пике силы? Оно обусловлено двумя факторами: количеством кальция, выделяемого саркоплазматическим ретикулумом, и механическим поведением мышцы. При отдельном потенциале действия не выделяется достаточное количество кальция, чтобы раскрыть необходимое количество участков прикрепления для образования максимальной силы. В то же время при серии потенциалов действия (тетанусе) происходит постепенное внутриклеточное накопление кальция, которое со временем приводит к максимальному растормаживанию тропонина и тропомиозина и обеспечивает прикрепление достаточного числа поперечных мостиков для образования максимальной силы (Allen, Lee, Westerblad, 1989).
Уровень мышечного волокна.Единицы мышцы, образующие силу, по-разному расположены на уровне мышечного волокна, всей мышцы и единой системы сустава. Эти единицы на
208
AL = n(AI)
F = nf
Рис. 6.31. Архитектурные свойства мышечного волокна:
а — последовательное расположение саркомеров, каждый из которых представлен трехэлементной моделью; б — параллельное расположение саркомеров; в — влияние пеннации на вклад каждого волокна в силу и скорость укорачивания всей мышцы; р — угол пеннации; F„ — вектор мышечной силы или скорости; F„ „ — нормальная составляющая; Fra>, — тангенциальная составляющая
уровне волокна могут быть расположены последовательно (т.е. конец к концу) или параллельно (бок о бок). На рис. 6.31 показано расположение трех образующих силу единиц последовательно (рис. 6.31, а) и параллельно (рис. 6.31, б).
Миофибриллу мы описывали как ряд последовательных саркомеров. Изменение длины миофибриллы обычно обусловлено разным числом последовательно расположенных саркомеров; длина каждого саркомера в миофибрилле сильно не изменяется. Например, длина саркомера в мышцах кисти человека в среднем составляет 2,2 мкм и изменяется в данной мышце всего на 2,1 % (Lieber, et al., 1990). Когда нога занимает анатомическое положение, средняя длина саркомера в различных мышцах колеблется от 1,97 до 3,01 мкм (Cutts, 1988). Если каждый саркомер или единицу, образующую силу, представить в виде трехэлементной модели, длину миофибриллы в состоянии покоя можно изменить, изменяя число единиц в цепочке (рис. 6.31, а). Когда потенциал действия распространяется вдоль мышечного волокна, все саркомеры миофибриллы испытывают одинаковую степень возбуждения, следовательно, длина каждого саркомера изменяется в одинаковой степени (А/). В результате, чем больше число последовательно расположенных саркомеров (я), тем больше изменяется длина миофибриллы (AZ) в ответ на потенциал действия; следовательно, А/ = п (ЛГ).
Например, миофибрилла, содержащая 100 саркомеров, испытает А/ порядка 50 мкм, если каждый из них изменит свою длину на 0,5 мкм при активации потенциалом действия. Наряду с этим в таких же условиях миофибрилла, содержащая 1000 саркомеров, изменит свою длину на 500 мкм. Более того, поскольку миофибрилла может изменять свою длину на 1/3, чем больше ее длина, тем больше абсолютное изменение длины.
Другим свойством при последовательном расположении является влияние на скорость уменьшения длины. Интенсивность изменения длины (скорость) миофибриллы определяется как произведение количества саркомеров и средней скорости каждого саркомера. Чем больше число последовательно расположенных саркомеров, тем больше изменяется длина миофибриллы и тем выше интенсивность изменения длины в ответ на данный стимул; следовательно, Av = «(Av).
С другой стороны, если каждая миофибрилла образует отдельный саркомер и три таких миофибриллы расположены параллельно (рис. 6.31, б), изменение длины миофибрилл будет эквивалентно изменению длины отдельного саркомера. При таком расположении, однако, силы, образуемые каждым саркомером, объединяются, вследствие чего сила мышечного волокна (F) оказывается равной произведению числа миофибрилл (я) и средней силы, образуемой отдельной миофибриллой (/), т.е.
14 8-133
209
F = n. Следовательно, параллельное расположение единиц, образующих силу, увеличивает ее таким образом, что сила мышечного волокна оказывается прямо пропорциональной количеству параллельно расположенных миофибрилл.
Мышечное волокно состоит из параллельно расположенных миофибрилл, а миофибрилла представляет собой последовательно расположенные саркомеры; следовательно, в мышечных волокнах есть оба вида образующих силу единиц. В длинном мышечном волокне небольшого диаметра доминируют последовательно расположенные единицы, тогда как в коротком волокне большого диаметра в основном — параллельно расположенные единицы. Эти различия наблюдали при проведении измерений 25 мышц ног у пяти погибших человек (табл. 6.1). Степень различий между мышечными волокнами с точки зрения последовательной или параллельной архитектуры может быть значи
тельной. Если допустить, что длина саркомера 2,2 мкм, то среднее количество последовательно расположенных саркомеров в волокне составит от 11364 в камбаловидной (25 мм) до 203636 в портняжной (448 мм). Так же колеблется площадь поперечного сечения мышечных волокон (см. табл. 5.1 и 5.2), на которую влияют такие факторы, как возраст, пол, состояние здоровья, занятия физической культурой. Различия в площади поперечного сечения (табл. 6.1) обусловлены различиями в количестве параллельно расположенных миофибрилл.
Мышцы, обеспечивающие вертикальное положение тела, так называемые антигравитационные мышцы, как правило, вдвое сильнее, чем их антагонисты. Из этого следует, что площадь их поперечного сечения вдвое больше.
Уровень всей мышцы. Организация производящих силу единиц на уровне всей мышцы также характеризуется последовательным или параллельным воздействием на силу, образуемую
Таблица 6.1. Архитектурные характеристики мышц ноги человека
Мышца Длина волокна, мм Коэффициент длины Угол пеннации, рад Функциональная площадь поперечного сечения, см2
Мышцы бедра
короткая приводящая 103 0.70 0,00 4,7
длинная приводящая 103 0,50 0,09 6,8
большая приводящая 114 0,53 0,04 18,2
гребешковая Мышцы бедра и колена 98 0,83 0,00 2,9
двуглавая мышца бедра (длинная головка) 80 0,26 0,03 12,8
нежная мышца 264 0,83 0,04 1,8
прямая мышца бедра 64 0,20 0,13 12,7
портняжная мышца 448 0,88 0,00 1,7
полуперепончатая 63 0,27 0,27 16,9
1 п ол усухожи л ьная Мышцы колена 155 0,46 0,09 5,4
двуглавая мышца бедра (короткая головка) 130 0,52 0,37 *
широкая интермедиальная 72 0,23 0,05 22,3
широкая латеральная 72 0,23 0,12 30,6
1 широкая медиальная Мышцы колена и лодыжки 73 0,23 0,10 21,1
медиальная икроножная 37 0,16 0,25 32,4
латеральная икроножная Мышцы лодыжки 55 0,25 0,19 **
передняя большеберцовая 75 0,26 0,12 9,9
разгибатель пальцев 80 0,25 0,17 5,6
разгибатель большого пальца стопы 78 0,32 0,11 1,8
сгибатель пальцев 31 0,13 0,13 5,1
сгибатель большого пальца стопы 37 0,17 0,21 5,3
короткая малоберцовая 39 0,17 0,12 5,7
длинная большеберцовая 41 0,15 0,15 12,3
задняя большеберцовая 26 0,10 0,24 20,8
камбаловидная 25 0,08 0,48 58,0
Примечание. Коэффициент длины — отношение длины волокна к длине мышцы * Этот показатель включен в измерение длинной головки двуглавой мышцы бедра. " Этот показатель включен в измерение медиальной икроножной мышцы.
210
мышцей, т.е. изменение в расположении мышечных волокон влияет на силу, длину и скорость укорочения длины всей мышцы. В большинстве мышц отношение длины волокна к длине мышцы составляет менее 1, колеблясь от 0,08 в камбаловидной до 0,85 в некоторых мышцах ноги (портняжной, гребешковой) и запястья. Для большинства мышц отношение колеблется в пределах 0,2—0,5 (табл. 6.1). Отклонение отношения от 1 (1 означает одинаковую длину волокна и мышцы) обусловлено следующим.
Во многих мышцах образующие силу единицы ориентированы на угол к линии натяжения сухожилия, т.е. они расположены не совсем последовательно с направлением вектора силы мышцы, а прикреплены к оболочке внутримышечной соединительной ткани под углом (рис. 6.31, в). В результате длина волокна меньше, чем мышцы. Угловое отклонение волокон от линии натяжения мышцы называется углом пеннации, который обычно колеблется от 0 до 0,4 рад (табл. 6.1). Мышцы могут иметь волокна с общим углом пеннации (однопеннатные), с двумя рядами волокон с различными углами пеннации (двухпеннатные) или с несколькими рядами волокон с множеством углов (много-пеннатные). Мышечные волокна с нулевым углом пеннации характеризуют как имеющие веретеноподобное или параллельное расположение. Типы мышц показаны на рис. 6.32.
Преимущество пеннации состоит в том, что большее количество параллельно расположенных волокон может уместиться в данном объеме. Этот принцип иллюстрирует камбаловидная мышца, для которой характерно небольшое отношение длины волокна к длине мышцы, большой угол пеннации и экстенсивная площадь поперечного сечения (табл. 6.1). Количество волокон, которые могут “уместиться” в данном объеме, имеет большое значение. Например, мышцы, контролирующие движения пальцев, находятся в кисти или предплечье; расположенные в кисти — небольшие (ввиду ограниченного пространства), те же, что находятся в предплечье — большие и сильные. Если бы все мышцы, контролирующие движения пальцев, находились только в кисти, она была бы громоздкой и неуклюжей, чтобы выполнять тонкую работу.
Пеннация также снижает вклад силы волокон и скорости укорочения в показатели всей мышцы; тангенциальный компонент уступает по величине результирующему и изменяется как косинус угла пеннации (cos Р). Несмотря на то, что сила мышечного волокна (длина Fm) остается постоянной при изменении угла пеннации, тангенциальный компонент (Fm ,) уменьшается. При угле пеннации, равном 0, F,„ , равна Fm. Поскольку объем мышцы остается относительно постоянным во время сокращения, угол пеннации изменяется. Следовательно, измене-
Треугольные
Одноперистые
Двуперистые
Рис. 6.32. Традиционная классификация мышц на основании их формы и фасцикулярной организации (Gray’s Anatomy, 1980)
ние длины всей мышцы обусловлено изменением длины волокна плюс перемещение вследствие изменения угла пеннации (Gans, 1982; Huijing, 1992а; Offen, 1988). Это же характерно для скорости сокращения. При оптимальной длине мышцы изменения длины волокна и угла пеннации влияют на сокращение скорости мышцы соответственно на 84 и 6 %. Если длина мышцы меньше, влияние составляет соответственно 35 и 31 % (Zuurbier, Huijing, 1992).
Рассмотрим теперь последовательное расположение коротких зигзагообразных мышечных волокон, относящихся к отдельной двигательной единице (рис. 6.33). Onnjian и соавт. (1991) показали, что мышечные волокна отдельной двигательной единицы в передней большеберцовой мышце кошек могут распределяться вдоль всей длины мышцы и по меньшей мере один конец заканчивается в пучке.
14*
211
Рис. 6.33. Площадь поперечного сечения и относительное продольное расположение 11 волокон быстросокра-щающейся двигательной единицы передней большеберцовой мышцы кошки (Ounjian, Roy, Eldred et al., 1991) 0 — проксимальный конец мышцы
Этот конец обычно, но не всегда имеет длинное конусообразное окончание, которое, очевидно, передает силу в эндомизий (Trotter, 1990; Trotter, Purslon, 1993). Это означает, что сила, образуемая одной двигательной единицей, передается из одного конца мышцы в другой не ее волокнами, а цитоскелетными структурами и оболочками соединительной ткани. Таким образом, зигзагообразные волокна можно рассматривать как сеть генераторов силы, содержащихся в оболочках соединительной ткани. Эти же волокна повышают эластичность между сократительным элементом (СЭ) и сухожилием (рис. 6.34).
Как следует из рис. 6.33, мышечные волокна двигательной единицы, как правило, не распределяются по всей мышце. В результате последовательного или параллельного расположения мышечных волокон, относящихся к двигательной единице, активация двигательной единицы ведет к локальному сокращению в пределах мышцы.
Региональная активация мышцы выходит
Рис. 6.34. Трехэлементная модель саркомера и окружающие его эластичные слои (Huijing, 1992)
за пределы уровня двигательной единицы к уровню всей мышцы. Это явление называется компартментализацией и включает функциональное разделение мышцы на несколько единиц или компартментов, основываясь на структуре иннервации первичных нервных ответвлений (Fleckeusteru et al., 1992; Weeks, 1989; Windhorst, et al., 1989). Оказывается, некоторые мышцы не активируются как гомогенные единицы, и активация мышечных волокон может быть разной в различных частях мышцы. Эта изменяющаяся активация в пределах мышцы основана на структуре иннервации первичных нервных ответвлений. Вместе с тем, различия в иннервации не означают, что индивидуальные компартменты или синергисты обязательно активируются независимо (Grabiner, Koh, Mill, 1991).
Чем больше площадь поперечного сечения мышцы, тем большую силу может образовывать мышца. Ввиду пеннации измерение площади поперечного сечения всей мышцы затруднено. Точное измерение площади поперечного сечения предполагает измерение участка сечения, перпендикулярного к длинной оси каждого волокна. Следовательно, площадь поперечного сечения всей мышцы нельзя определить, измерив только наиболее толстый ее участок, необходимо учитывать также угол пеннации. Подобное измерение — функциональная (физиологическая) площадь поперечного сечения — позволяет более точно оценить максимальную силу, которую может образовывать мышца (Fukunaga et al., 1992; Lieber, 1992). У мышц, приведенных в табл. 6.1, функциональная площадь поперечного сечения колеблется от 1,7 до 58,0 см2.
Уровень единой системы сустава. На вращающий момент, производимый мышцей, влияют
212
еще две характеристики, включающие организацию мышц по отношению к суставу: точки прикрепления мышцы относительно сустава и пропорция длины всей мышцы, содержащей сократительную ткань. На рис. 6.35 показана геометрия мышцы отдельного сустава. Точки прикрепления определяют расстояние (с) от проксимального прикрепления мышцы к суставу, расстояние (д) от дистального прикрепления мышцы к суставу и, следовательно, угол натяжения мышцы (а), а также пропорцию мышечной силы, способствующей вращению (£с). Кроме того, на рисунке показаны плечо пары (d), угол сустава (£), сила растяжения,производимая мышцей по отношению к дистальному сегменту (Fm), а также нормальная и тангенциальная составляющие силы (F„ и Ft).
Основное воздействие различных параметров с, q и Lc на вращающий момент, производимый мышцей, включает (van Mameren, Drukker, 1979) следующие характеристики.
1. Ширину. Взаимосвязь угол—вращающий момент характеризуется более острым пиком, когда с и q равны. Если же одна из величин больше, пик более широкий, т.е. максимальные показатели вращающего момента могут наблюдаться на более широком диапазоне углов сустава (рис. 6.36, а).
2. Расположение. Пик вращающего момента достигается ближе к полному растяжению, когда сид равны, и ближе к середине диапазона (например, при угле локтевого сустава 1,57 рад), когда си q разные (рис. 6.36, 6).
3. Величину. Максимальный вращающий момент, производимый мышцей при разных с и q, значительно не возрастает при увеличении Lc. Когда же с и q равны, максимальный вращающий момент зависит от Lc. Это взаимодействие очевидно в мышцах с отношением с/д, приближающимся к 1, имеющим высокий показатель £с, тогда как в мышцах с отношением c/q меньше 1 обычно длинные сухожилия (низкие показатели £с).
Вращение одного сегмента тела относительно другого, как правило, контролируется груп-
Рис. 6.35. Геометрия односуставной мышцы (van Maneren, Drukker, 1979)
пой мышц. Довольно часто параметры с, q и Lc колеблются в мышцах одной группы, вследствие чего мышечная группа имеет широкий спектр свойств.
Изменения расстояния си q влияют на величину плеча пары мышцы относительно сустава. Изменения плеча пары, взаимодействуя с другими переменными, например, количеством последовательно расположенных саркомеров, влияют на мышечную функцию. Liber (1922) характеризует это воздействие, изучая отношение длины мышечного волокна к длине плеча пары. Величина отношения свидетельствует о степени вклада мышцы в результирующий вращающий момент мышцы на всем диапазоне движения. При высоком отношении изменение длины волокна, а следовательно и длины саркомера, небольшое относительно зависящего от угла изменения плеча пары. Согласно Либеру, к мышцам с высоким отношением относятся большая ягодичная (80), портняжная (11) и длинный лучевой разгибатель пальцев (ПО); с низким — камбаловидная (0,9), подколенное сухожилие (1,8). Подобная взаимосвязь свидетельствует, что вся архитектура мышцы — от структуры саркомеров до структуры суставов
Рис. 6.36. Влияние архитектуры на взаимосвязь угол—вращающий момент:
а — влияние на ширину S пика; б — на расположение S пика; в — влияние Ц. на величину пика вращающего момента; 1 — взаимосвязь при с = д-, 2 — с и q
213
может влиять на функциональные возможности единой системы сустава.
РЕЗЮМЕ
В этой главе мы рассмотрели факторы, влияющие на вращающий момент. Это — активация со стороны нервной системы (нервные фак
торы), а также механические и архитектурные свойства мышцы (мышечные факторы). С точки зрения механики длина мышцы и интенсивность изменения ее длины воздействуют на образуемые мышцей силу и мощность. Кроме того, длина и размер (площадь поперечного сечения) мышечных волокон, структура их расположения, а также расположение мышцы вокруг сустава влияют на вращающий момент и скорость сокращения мышцы.
ВОПРОСЫ С АЛЬТЕРНАТИВНЫМ ОТВЕТОМ
1. Всякий раз при возникновении потенциала действия двигательного нейрона образуется потенциал действия мышечного волокна. ПРАВИЛЬНО НЕПРАВИЛЬНО
2. Рекрутирование двигательных единиц и модуляция интенсивности разряда могут совпадать. ПРАВИЛЬНО НЕПРАВИЛЬНО
3. Упорядоченное рекрутирование означает, что вначале рекрутируются небольшие двигательные нейроны, а затем большие.
ПРАВИЛЬНО НЕПРАВИЛЬНО
4. Когда мышечная сила начинает спадать, первой дерекрутируется двигательная единица, которая рекрутировалась первой.
ПРАВИЛЬНО НЕПРАВИЛЬНО
5. Изменение мышечной силы никогда не обусловливается только модуляцией интенсивности разряда.
ПРАВИЛЬНО НЕПРАВИЛЬНО
6. Сила, с которой рекрутируется двигательная единица, называется порогом рекрутирования.
ПРАВИЛЬНО НЕПРАВИЛЬНО
7. Пул двигательных нейронов — группа двигательных нейронов, которые рекрутируются при выполнении определенного задания.
ПРАВИЛЬНО НЕПРАВИЛЬНО
8. Количество дендритов, коэффициент иннервации и сопротивление входящему импульсу — примеры морфологических характеристик двигательных нейронов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
9. Реобаза — показатель возбудимости двигательных нейронов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
10. Возбудимость двигательных нейронов зависит не только от размера.
ПРАВИЛЬНО НЕПРАВИЛЬНО
11. Камбаловидная мышца является медленно-сокращающейся.
ПРАВИЛЬНО НЕПРАВИЛЬНО
12. Головной мозг не определяет порядок рекрутирования двигательных единиц.
ПРАВИЛЬНО НЕПРАВИЛЬНО
13. Можно избирательно активировать либо медленно-, либо быстросокращающиеся двигательные единицы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
14. Взаимосвязь сила—частота во всей мышце линейна.
ПРАВИЛЬНО НЕПРАВИЛЬНО
15. Тонические двигательные единицы линейно увеличивают интенсивность разряда во всем диапазоне мышечной силы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
16. Единица измерения интенсивности разряда миллисекунда (мс).
ПРАВИЛЬНО НЕПРАВИЛЬНО
17. Мышечная “мудрость” характеризует снижение интенсивности разряда двигательных единиц во время вызывающих утомление сокращений.
ПРАВИЛЬНО НЕПРАВИЛЬНО
18. Двойной разряд характеризует появление двух потенциалов действия в ответ на отдельный (одиночный) потенциал действия двигательного нейрона.
ПРАВИЛЬНО НЕПРАВИЛЬНО
19 Регулярная разрядка потенциалов действия двигательной единицей называется синхронной.
ПРАВИЛЬНО НЕПРАВИЛЬНО
20. Потенциал действия мышечного волокна — это явление типа “все или ничего”.
ПРАВИЛЬНО НЕПРАВИЛЬНО
21. Теория поперечных мостиков представляет собой попытку объяснить предположение о скольжении филаментов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
22. Наибольшая мышечная сила наблюдается при самой большой длине мышцы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
23. При максимальной длине мышцы большая часть силы обусловлена активностью поперечных мостиков.
ПРАВИЛЬНО НЕПРАВИЛЬНО
214
24. Сила сокращения изменяется с изменением длины мышцы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
25. Максимальный вращающий момент мышцы наблюдается при промежуточном угле сустава (для всех мышц).
ПРАВИЛЬНО НЕПРАВИЛЬНО
26. Вращающий момент при эксцентрических сокращениях больше, чем при концентрических. ПРАВИЛЬНО НЕПРАВИЛЬНО
27. При концентрическом сокращении длина мышечного волокна может уменьшаться.
ПРАВИЛЬНО НЕПРАВИЛЬНО
28. Эффективность представляет собой количество выполненной работы за единицу использованной энергии.
ПРАВИЛЬНО НЕПРАВИЛЬНО
29. Эффективность мышечного сокращения снижается с увеличением скорости уменьшения длины.
ПРАВИЛЬНО НЕПРАВИЛЬНО
30. Ньютон (Н) — единица измерения эксцентрической силы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
31. Мышцы-сгибатели локтевого сустава — основные “двигатели” при выполнении подтягиваний на перекладине.
ПРАВИЛЬНО НЕПРАВИЛЬНО
32. Камбаловидная и икроножная мышцы “вносят свой вклад” в силу ахиллового сухожилия. ПРАВИЛЬНО НЕПРАВИЛЬНО
33. Мышца развивает мощность при выполнении положительной работы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
34. Одним из разгибателей коленного сустава является прямая мышца бедра.
ПРАВИЛЬНО НЕПРАВИЛЬНО
35. о — максимальная изометрическая сила.
ПРАВИЛЬНО НЕПРАВИЛЬНО
36. Уравнение Хилла характеризует уменьшение мышечной силы как функции скорости сокращения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
37. Вязкость характеризует жесткость мышцы. ПРАВИЛЬНО НЕПРАВИЛЬНО
38. Мощность можно определить как интенсивность изменения количества энергии.
ПРАВИЛЬНО НЕПРАВИЛЬНО
39. Единица измерения мощности — джоуль (Дж). ПРАВИЛЬНО НЕПРАВИЛЬНО
40. Разминка не влияет на скорость сокращения. ПРАВИЛЬНО НЕПРАВИЛЬНО
41. Мышечная архитектура не влияет на максимальную силу, образуемую мышцей.
ПРАВИЛЬНО НЕПРАВИЛЬНО
42. Средняя длина саркомера — около 2,2 см.
ПРАВИЛЬНО НЕПРАВИЛЬНО
43. Чем больше число последовательно расположенных саркомеров, тем выше максимальная скорость уменьшения длины мышцы. ПРАВИЛЬНО НЕПРАВИЛЬНО
44. Мышечное волокно состоит из последовательно или параллельно расположенных саркомеров.
ПРАВИЛЬНО НЕПРАВИЛЬНО
45. Антигравитационные мышцы обычно сильнее своих антагонистов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
46. Угол пеннации обычно колеблется в пределах 0—0,4 рад.
ПРАВИЛЬНО НЕПРАВИЛЬНО
47. Преимущество пеннации состоит в возможности “умещения” большего количества параллельно расположенных волокон в данном объеме.
ПРАВИЛЬНО НЕПРАВИЛЬНО
48. Зигзагообразные мышечные волокна обеспечивают последовательный эффект.
ПРАВИЛЬНО НЕПРАВИЛЬНО
49. Функциональное определение мышцы отличается от ее анатомического определения. ПРАВИЛЬНО НЕПРАВИЛЬНО
50. Мышечные компартменты различаются на основании точек прикрепления к соединительной ткани.
ПРАВИЛЬНО НЕПРАВИЛЬНО
51. При измерении функциональной площади поперечного сечения учитывается длина мышечных волокон.
ПРАВИЛЬНО НЕПРАВИЛЬНО
52. Расположение точек прикрепления мышцы к скелету может влиять на взаимоотношение угол—вращающий момент мышцы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
ВОПРОСЕ. С МЗПМЧЯЫМ ВЫБОРОМ ul JETOB
53. О чем гласит принцип размера?
а) рекрутирование двигательных единиц происходит в установленном порядке в зависимости от размера двигательного нейрона и начинается с наименьшего двигательного нейрона;
б) постепенное увеличение потребностей в мощности при выполнении задания включает постепенное рекрутирование более мощных двигательных единиц;
в) рекрутирование двигательных нейронов не зависит исключительно от размера двигательного нейрона, а от
215
других его характеристик (внутренних факторов) и организации синаптического импульса (внешние факторы) на двигательных нейронах;
г) выполнение определенного движения осуществляется вследствие активации двигательных единиц в установленном порядке.
54. Какие переменные не влияют на порядок рекрутирования двигательных единиц? а) внешние факторы;
б) биофизические характеристики;
в) время сокращения;
г) размер двигательного нейрона.
55. Почему нельзя избирательно активировать медленно- либо быстросокращающиеся двигательные единицы?
а) ввиду одинаковых внешних факторов;
б) медленносокращающиеся единицы меньше;
в) рекрутирование двигательных единиц и модуляция интенсивности разряда происходят одновременно;
г) ввиду взаимного перекрывания размера и возбудимости единиц типа S и FR.
56. Какое явление не вызывает изменения мышечной силы вследствие изменения разряда потенциала действия?
а) синхронность двигательных единиц;
б) мышечная “мудрость”;
в) взаимосвязь сила—частота;
г) двойной разряд.
57. Укажите неверное высказывание (-я) относительно активности двигательной единицы: а) рекрутирование продолжается до 85 % максимальной силы всех мышц;
б) модуляция интенсивности разряда и рекрутирование происходят одновременно;
в) все медленносокращающиеся единицы рекрутируются до рекрутирования быстросокращающихся;
г) синхронизация увеличивает силу, образуемую активными единицами.
58. О чем гласит теория поперечных мостиков? а) мышечная сила изменяется в зависимости от степени взаимного перекрывания тонких и толстых филаментов в саркомере;
б) тонкий и толстый филаменты скользят относительно друг друга при сокращении мышцы;
в) цикл поперечного мостика включает последовательность прикрепления-открепления;
г) мышечная сила прямо пропорциональна числу прикреплений поперечных мостиков.
59. Почему сила, образуемая мышцей, изменяется при изменении длины мышцы?
60.
61.
62.
63.
64.
65.
а) вследствие изменений плеча пары;
б) с увеличением длины мышцы пассивная сила также возрастает;
в) вследствие изменения степени взаимного перекрывания тонких и толстых филаментов;
г) ввиду того, что большинство мышц имеют структуру пеннации.
Чем характеризуется взаимосвязь вращающего момента мышцы и нагрузки при эксцентрических сокращениях?
а) вращающий момент мышцы меньше, чем вращающий момент нагрузки;
б) вращающие моменты одинаковы;
в) вращающий момент мышцы больше;
г) вращающий момент нагрузки меньше. Что не влияет на уменьшение мышечной силы при увеличении скорости концентрических сокращений?
а) сокращение количества прикрепленных поперечных мостиков;
б) уменьшение средней величины силы, образуемой каждым поперечным мостиком;
в) снижение эффективности сокращения;
г) повышенная интенсивность циклов поперечных мостиков.
Сила, образуемая мышцей при эксцентрических сокращениях, самая большая. Чем это обусловлено?
а) увеличением числа прикрепленных поперечных мостиков;
б) увеличением количества выделяемых Са2+;
в) изменением механизма, “ответственного” за открепление поперечных мостиков;
г) растяжением неполностью активированных саркомеров.
Когда скорость укорачивающегося сокращения равна Кикс, чему равна мощность, развиваемая мышцей?
а) нулю;
б) максимальна;
в) 1/3 максимальной;
г) определяется величиной силы.
На основании какого взаимоотношения нельзя определить мощность?
а) Дэнергия х время;
б) ^макс X Fm0 5
в) сила х скорость;
г) работа х время-1.
Сколько последовательно расположенных саркомеров (средняя длина = 2,2 мкм) находится в волокне, длина которого 11,4 см? а) 51 818;
б) 51 181;
в) 5181;
г) 518.
216
1АДАЧИ
66. Принцип размера гласит, что порядок рекрутирования двигательных единиц основан на различиях размера двигательных нейронов.
А. Что подразумевается под размером двигательного нейрона?
Б. Какие морфологические свойства двигательных единиц, по-видимому, связаны с размером двигательного нейрона?
В. Какие виды синаптического импульса, поступающего в двигательный нейрон, также изменяются в зависимости от размера?
67. Взаимосвязь интенсивности потенциала действия и силы характеризуется сигнои-дальным характером.
А. В чем качественное различие между сиг-ноидальным и линейным характером взаимосвязи?
Б. Как влияет на эту взаимосвязь длина мышцы?
В. Как отличается данная взаимосвязь в медленно- и быстросокращающихся двигательных единицах?
68. Взаимосвязь угол—вращающий момент мышц обычно имеет одну из трех форм: восходящую, нисходящую и промежуточную?
А. Что такое промежуточная взаимосвязь?
Б. Следующий график представляет наиболее типичную форму взаимоотношения угол—вращающий момент, например для сгибателей локтевого сустава. Если сгибатели локтевого сустава производят постоянную величину силы 3,5 кН в этом диапазоне движения, чему будут равны минимальные и максимальные значения плеча пары?
В. Постройте диаграмму свободного тела показав плечо пары
Г. Если график изображенный в примере Б был построен в условиях максимальных произвольных сокращений, можно ли считать, что его форма обусловлена, в основном, изменением плеча пары?
69. Понятия “изометрические”, “концентрические” и “эксцентрические” характеризуют различия механических характеристик, а не различные типы мышечных сокращений.
А. Чем отличаются эти понятия с механической точки зрения?
Б. Могут ли быть различные типы мышечных сокращений? Объясните свой ответ.
В. Понятие “изотонический” означает постоянное напряжение мышцы. Почему это не наблюдается в естественных условиях, например, при поднимании нагрузки с постоянной массой?
70. Из взаимосвязи работа—энергия мы знаем, что количество выполненной работы равно количеству затраченной энергии.
А. Какие единицы измерения работы и энергии вы знаете (системы СИ)?
Б. Каковы механические источники энергии для выполнения работы?
В. С точки зрения физиологии мы различаем положительную и отрицательную работу. С точки зрения механики мы характеризуем положительную работу как поток энергии из системы в окружающую ее среду, а отрицательную — как поток энергии в обратном направлении. Что это означает?
71. Представьте, что вы держите за ручку тяжелый портфель, ожидая автобус. Хотя вы не выполняете никакой механической работы относительно портфеля, почему вы не можете его долго удерживать?
72. Если велосипедист намерен сохранять постоянную скорость, а ему предстоит крутой подъем вверх, он меняет скорость? Если да, то почему?
73. Упруговязкость мыщцы вызывает гистерезис во взаимоотношении сила—длина мышцы (рис.6.30, а), что означает, что мышца утрачивает энергию во время повторяющихся циклов изменения своей длины. Как это показано на рисунке (работа = Д энергия)?
217
Однопеннатная мышца содержит 1167 волокон, средний угол пеннации которых 0,4 рад относительно линии натяжения сухожилия. В мышце содержится 25 % двигательных единиц типа S, 20 % типа FR и 55 % типа FF. Средняя площадь поперечного сечения первых 1370 мкм2, вторых — 2503, третьих — 4980 мкм2. Определите максимальную силу, которую может образовать мышца в направлении линии натяжения сухожилия.
75. Если каждый саркомер обеспечивает изменение длины порядка 0,5 мкм, а в мышечном волокне содержится 31,2 х 103 саркомеров, насколько изменится длина мышцы?
76. На основании данных, приведенных в табл. 6.1, и значения напряжения 30 Н/см2 определите максимальную силу, которую может образовать четырехглавая мышца бедра.
77. Почему архитектура камбаловидной мышцы сильно отличается от архитектуры большинства других мышц ноги (табл. 6.1)?
ЧАСТЬ III
ПРИСПОСОБЛЯЕМОСТЬ ДВИГАТЕЛЬНОЙ СИСТЕМЫ
В данной книге мы рассматриваем движение человека как взаимодействие биологической системы и окружающей ее среды. В части I мы определили концепции и принципы изучения движения на основе законов физики. В части II рассмотрели простую биологическую модель (односуставную систему), которая позволила нам исследовать биологические процессы, протекающие при осуществлении движения. Тем не менее организм человека более сложен, чем односуставная система. В части III модель односуставной системы расширяется для обеспечения более полного учета тех компонентов организма человека, которые непосредственно участвуют в осуществлении движения. Совместно эти компоненты называются двигательной системой и включают как двигательные, так и и чувствительные элементы. В главе 7 более подробно представлены нейронная и мышечная организации двигательной системы и описаны некоторые из стратегий, используемых двигательной системой для осуществления движения. В двух последних главах описываются кратковременная (глава 8) и постоянная (глава 9) адаптационные способности этой системы.
ЦЕЛИ
Для завершения нашего исследования нейромеханических основ кинезиологии рассмотрим адаптационные способности двигательной системы. Конкретные цели состоят в:
• описании организации супрасегментных компонентов нервной системы и потока информации, связанного с управлением движением;
• расширении описания костно-мышечной системы для рассмотрения взаимодействия множества односуставных систем;
• характеристике стратегий движения, которые были определены;
• объяснении влияния изменения внутренней температуры на работоспособность;
• указании методик, разработанных для изменения гибкости;
• изложении многофакторной основы мышечной усталости и определении процессов, которые могут быть нарушены при различных нагрузках;
• изучении сенсорных адаптаций, возникающих во время утомляющих сокращений;
• описании потенциирующих способностей мышцы;
• определении принципов выбора физических нагрузок;
• определении влияющих на работоспособность характеристик мощности и силы, а также механизмов, обусловливающих изменения этих параметров;
• обосновании адаптаций, сопровождающих периоды пониженной активности;
• оценке изменений, которые происходят со старением, и определении физиологических основ этих изменений.
ГЛАВА 7
ДВИГАТЕЛЬНАЯ СИСТЕМА
В части II мы создали модель двигательной системы, которую назвали односуставной. Используя эту модель, мы сосредоточились на некоторых основных физиологических и биомеханических особенностях организма человека, которые позволяют ему осуществлять движение. Тем не менее организм человека значительно более сложен, чем односуставная система. Например, мы не рассматривали, каким образом односуставная система инициирует движение. В главе 7 односуставная система расширяется в результате изучения роли других нервных элементов в инициировании и модуляции активности односуставной системы, рассмотрения возможностей скелетно-мышечной структуры в управлении движением и определения некоторых из стратегий, используемых двигательной системой для осуществления движения.
СУПРАСЕГМЕНТНАЯ ОРГАНИЗАЦИЯ
Ранее мы рассматривали в качестве эфферентного элемента двигательной системы только двигательный нейрон и уделяли недостаточное внимание происхождению невральных команд, управляющих мышцей. Конечно, осуществление движения не начинается и не заканчивается двигательным нейроном. Многие структуры ЦНС способствуют созданию двигательных нейронных сигналов, возбуждающих мышцу. Критическим является, конечно, головной мозг, выполняющий три основные функции, связанные с управлением движением: мотивацию, осознание и возбуждение мышцы. В инициировании и модифицировании движения важными являются мотивация и осознание. На рис. 7.1 показан поток информации через нервную систему, обусловливаю
щий возбуждение двигательных нейронов. Схема состоит из четырех компонентов: мотивации, мышления, программирования и выполнения (Cheney, 1985). В прежнем обсуждении односуставной системы мы сосредотачивались на фазе выполнения.
Лимбическая система обеспечивает мотивацию многих движений. Она управляет основными биологическими побуждениями и эмоциональным поведением, включая такие проявления, как голод, жажда, размножение, материнское поведение и установление общественных отношений. Лимбическая система может вызывать специфические эмоциональные поведенческие реакции и изменять возбудимость нейронов, непосредственно участвующих в движении (Holstege, 1992). Например, многие формы напряжения (агрессия, страх, половое возбуждение) могут индуцировать аналгезию, хотя возбудимость двигательных нейронов повышается. Лимбическая система может инициировать поведенческие реакции через сенсорно-моторную систему, которая преобразовывает мотивацию в мысль и инициирует суп-расегментные взаимодействия, завершающиеся командой на выполнение движения (в отношении дополнительных деталей этих систем см. Brooks, 1986 и Rothwell, 1987). Лимбическая система относится к функциональному комплексу, который включает ряд структур переднего мозга, связанных с гипоталамусом и частями среднего мозга. Гипоталамус — центральное звено этой системы, он регулирует многие жизненно важные функции, например, температуру тела, частоту сердечных сокращений, кровяное давление и потребление воды и пищи. Потребности, выражаемые лимбической системой, анализируются и интегрируются в мысли ассоциативной зоной коры головного мозга (префронтальной, теменной и височной долями). Мысли проецируются на сенсорно-двига
221
тельную зону коры головного мозга, мозжечок, часть базальных ядер и соответствующие подкорковые ядра для разработки программы осуществления движения.
Программирование заключается в преобразовании мысли в надлежащее усилие и схему мышечной деятельности, необходимой для желаемого движения. Основные супрасегмен-тарные центры, участвующие в программировании, включают ассоциативную зону коры головного мозга (преддвигательную), двигательную зону коры головного мозга, базальные ядра и мозжечок. Двигательная зона коры головного мозга обеспечивает связь с головным мозгом большинства двигательных нейронов. Кроме того, базальные ядра, мозжечок и кора головного мозга могут независимо влиять на двигательные нейроны через ствол мозга. Невральный выходной сигнал, который создается в процессе программирования, известен в качестве центральной команды и передается как в нижние нервные центры (ствол мозга и спинной мозг), так и назад в супрасегментарные центры, участвующие в разработке программы. Сигнал, который передается назад в супрасегментарные центры, известен в качестве результирующего разряда и является опорным сигналом, позволяющим системе интерпретировать поступающие афферентные сигналы.
Возбуждения центральной командой нижних нервных центров инициируют фазу выполнения движения. Она включает возбуждение двигательных нейронов как в мышцах, непосредственно связанных с движением, так и в тех, которые должны обеспечивать постуральную поддержку для стабилизации тела. Несмо
тря на то, что для данного движения могут быть использованы многие возможные последовательности возбуждения двигательных единиц, представляется, что усвоение обусловливает разработку конкретной последовательности возбуждения для каждой задачи. Стереотипная последовательность команд, поступающих из спинного мозга в мышцы для обеспечения конкретного поведения, известна в качестве двигательной программы. Она является результатом взаимодействий между программирующими действиями супрасегментарных центров, спинномозговых сетей и афферентной обратной связи.
В дополнение к эфферентным командам, передаваемым двигательными нейронами, фаза выполнения включает модифицирование движений посредством обратной связи от чувствительных нервных окончаний (мышечного веретена, сухожильного органа, суставного рецептора, кожного механорецептора). В результате возбуждения этих рецепторов создается афферентный входной сигнал, который действует на сегментарном уровне, а также поступает по восходящим путям в супрасегментарные центры (см. рис. 7.1). Хотя этот входной сигнал и может быть достаточным по уровню для инициирования движения (например, рефлексов), обычно он используется для обеспечения согласованности между движением и окружающей средой. Это согласование входного и выходного сигналов осуществляется сетями промежуточных нейронов, которые преобразовывают афферентный входной сигнал в соответствующие разряды двигательного нейрона, в результате чего выходной сигнал соответствует усло-
Рис. 7.1. Схема основных невральных взаимодействий, связанных с осуществлением движения (Cheney, 1985)
222
Рис. 7.2. Основные компоненты двигательной системы (Ghez, 1985)
виям, детектируемым чувствительными нервными окончаниями. Если конкретное афферентно-эфферентное преобразование происходит достаточно часто, нервная сеть может обучаться, становясь более экономичной, и возбуждаться минимальным входным сигналом. Нервные сети, способные генерировать соответствующие поведению структуры выходного сигнала в отсутствие внешних постоянных раздражителей (под действием афферентного входного сигнала), известны в качестве генераторов центральных структур.
Взяв за основу данную схему информационного потока через нервную систему, соответствующую мотивации—мышлению—программированию—выполнению, мы рассмотрим, каким образом основные компоненты двигательной системы взаимодействуют в процессе управления движением. Супрасегментар-ные центры через восходящие и нисходящие пути взаимодействуют со спинномозговыми сетями и информацией, принимаемой из окружающей среды (рис. 7.2). Структуры, которые взаимодействуют с нисходящими путями, включают сенсорно-двигательную зону коры головного мозга, ствол мозга, мозжечок и базальные ядра (Rothwell, 1987; Kandel, Schwartz, Jessell, 1991). Мы сосредоточимся на роли нисходящих путей, но поскольку людям трудно осуществлять движение как без эфферентных, так и без афферентных сигналов, кратко рассмотрим также восходящие пути и распределение афферентного входного сигнала среди супрасегментар-ных структур.
Сенсорно-двигательная зона коры головного мозга
Головной мозг состоит из левого и правого полушарий большого мозга, которые связаны аксонными пучками, называемыми комиссуара-ми. Наружная оболочка полушария большого мозга представляет собой трехмиллиметровый слой кортикальных нервных клеток, известный как кора головного мозга, которая представляет собой основной интегрирующий центр сенсорного входного сигнала и двигательного выходного сигнала. Сенсорно-двигательная зона коры головного мозга содержит участки коры головного мозга, которые располагаются непосредственно впереди и позади центральной борозды (рис. 7.3). Тремя передними компонентами являются основная двигательная зона (зона Бродмана 4), дополнительная двигательная зона (зона 6 между зонами 4 и 8 на медиальной поверхности) и преддвигательная зона (зона 6). Двумя задними компонентами являются основная соматосенсорная зона коры головного мозга (зоны 1—3) и задняя теменная зона коры головного мозга (зоны 5 и 7). Вся сенсорно-двигательная зона коры головного мозга взаимодействует с нисходящим путем, известным как кортикоспинальный тракт.
Соединения вводов и выводов сенсорнодвигательной зоны коры головного мозга являются экстенсивными, и поток информации проходит по взаимодействующим параллельным путям (Georgopoulos, Ashe, Smymis, Taira, 1992; Kalaska, Grammond, 1992; Houk, Keifer, Barto,
223
Рис. 7.3. Компоненты сенсорно-двигательной зоны коры головного мозга. На рисунке показаны левая и правая половины коры головного мозга (Ghez, 1991):
1 — предфронтальная кора головного мозга; 2 преддвигательная зона коры головного мозга; 3 — дополнительная двигательная зона; 4 — основная двигательная зона коры головного мозга; 5 — основная соматосенсорная зона коры головного мозга; 6 — задняя Теменная зона коры головного мозга
1993). В общем, выходной сигнал зоны 4 поступает в вентральную часть спинного мозга по кортикоспинальному тракту и оказывает влияние на двигательную деятельность. Выходной сигнал зон 1, 2, 3 и 5 поступает в дорсальную часть спинного мозга и модулирует периферийный сенсорный входной сигнал. Кроме того, зона 4 принимает афферентный входной сиг-
Рис. 7.4. Соединения вводов и выводов сенсорнодвигательной зоны коры головного мозга. Вводы и выводы осуществляются на обеих сторонах центральной борозды
нал от мозжечка через таламус, а зоны 1—3 принимают афферентный входной сигнал с периферии (рис. 7.4). Периферийный сенсорный входной сигнал, поступающий в основную соматосенсорную зону коры головного мозга (зоны За, 3b, 1 и 2), является дискретным. Каждая зона принимает входной сигнал от чувствительных нервных окончаний различных типов и устроена таким образом, что создается точная схема, соответствующая дерматомам всего тела. Входной и выходной сигналы сенсорно-двигательной зоны коры головного мозга имеют сомато-топическую (карта тела) структуру, в которой кортикальные нейроны, воздействующие на мышцы, и сенсорная информация от различных частей тела располагаются в упорядоченной последовательности (рис. 7.5). Заметны две особенности двигательной карты: корковые клетки воздействуют на мышцы на контралатеральной стороне тела, причем, по-видимому, больше представлены мышцы лица и кистей рук. Эти топографические карты определяются также в других областях сенсорно-двигательной зоны коры головного мозга (задняя теменная зона коры головного мозга, дополнительная двигательная зона и преддвигательная зона коры головного мозга), в мозжечке и таламусе.
Каждый участок сенсорно-двигательной зоны коры головного мозга несет основную, но не исключительную ответственность за определенные аспекты генерирования движения. Моторная зона коры головного мозга ответственна, главным образом, за правильную ориентацию
224
Рис. 7.5. Соматотопическая структура сенсорно-двигательной зоны коры головного мозга (Kandel, Jessel, 1991): а — сенсорная модель человека; 6 — двигательная модель человека; 1 — медиальная область; 2 — латеральная область; 3 — внутрибрюшная область; 4 — глотка; 5 — язык; 6 — зубы, десны и челюсть; 7 — нижняя губа; 8 — верхняя губа; 9 — лицо; 10 — нос; 11 — глаз; 12 — большой палец кисти; 13 — указательный палец; 14 — средний палец; 15 — кольцевидная структура; 16 — мизинец; 17 — кисть руки; 18 — запястье; 19 — предплечье; 20 — локоть; 21 — рука; 22 — плечо; 23 — голова; 24 — шея; 25 — туловище; 26 — бедро; 27 — нога; 28 — стопа; 29 — пальцы стопы; 30 — половые органы; 31 — лодыжка; 32 — колено; 33 — лоб;
34 — веко и глазное яблоко; 35 — губы; 36 — челюсть; 37 — глотание; 38 — жевание; 39 — слюноотделение; 40 — речь
тела и конечности перед движением и сенсорный контроль движений конечности. Эти эффекты достигаются в результате воздействия на аксиальные и проксимальные мышцы. Предмо-торная зона коры головного мозга активна также при усвоении новой задачи. Задняя теменная зона коры головного мозга интерпретирует и преобразовывает сенсорную информацию, в результате чего движение согласовывается с условиями окружающей среды. Она участвует в инициировании движений, направленных во внешнее пространство. Нейроны на участке 5 участвуют в использовании осязательной сенсорной информации для управления диагностическими движениями конечности, в то время как нейроны на участке 7 в большей степени участвуют в визуальном управлении движениями глаз и конечностей (Cheney, 1985). Повреждение преддвигательной либо задней теменной зоны коры головного мозга приводит к неспособности разрабатывать правильную стратегию движения (Jeannerod, 1988). Дополнительная двигательная зона важна для планирования движений и обеспечивает правильную последовательность активации мышц для таких движений, как игра на фортепьяно, писание и разговор. Эта дополнительная двигательная зона активна, например, во время осуществления воображаемых движений (Roland, Larsen Lassen, Skinhoj, 1980). Кроме того, дополнительная двигательная зона
оказывает влияние на соединение вводов и выводов в основной двигательной зоне коры головного мозга. Главная функция основной соматосенсорной зоны коры головного мозга — обеспечение сенсорной информацией, необходимой для конкретного планирования и инициирования движения и для модуляции непрерывного движения. Основная двигательная зона коры головного мозга синтезирует входной сигнал от других частей ЦНС, генерирует и передает центральную команду нейронам ствола мозга и спинного мозга для инициирования и модуляции движения. Кроме того, центральная команда подается на другие участки головного мозга, например, в базальные ядра и мозжечок, в виде результирующего разряда. Представляется, что центральная команда определяет мышцы, которые необходимо активировать, усилие сокращения и хронирование этого сокращения (Cheney, 1985). Пять этих субкомпонентов сенсорно-двигательной зоны коры головного мозга экстенсивно взаимодействуют для обеспечения того, чтобы произвольные движения основывались на прежних сведениях об эффекторной системе (соответствующие односуставные системы), окружающей систему среде и предполагаемых результатах движения (Ghez, 1991а; Kalaska, Grammond, 1992).
Представляется, что сенсорно-двигательная зона коры головного мозга может планировать
15 8-133
225
Рис. 7.6. Компоненты ствола мозга и положение двигательных центров (Rchmidt, 1983)
движения посредством участия отдельных мышц, групп мышц или самого движения. Анализ нейронной активности показывает, что ЦНС может смещать управление от одной нервной системы к другой, эмоциональные потребности могут изменять стратегию двигательного вывода, и двигательная зона коры головного мозга представляется наиболее необходимой для тонкой двигательной активности. Эта гибкость обеспечивается отсутствием жесткой иерархической организации между участками сенсорно-двигательной зоны коры головного мозга. Таким образом, различные участки могут становиться приоритетными в зависимости от поведенческого контекста движения (Huok et al., 1993; Kalaska, Grammond, 1992).
Ствол мозга
Ствол мозга состоит из продолговатого мозга, варолиева моста и среднего мозга (рис. 7.6). Ствол мозга в значительной степени несет ответственность за управление положением тела. Он интегрирует информацию, предоставляемую органом равновесия (вестибулярным аппаратом) и чувствительными нервными окончаниями в области шеи, вместе со входным сигналом от коры головного мозга и мозжечка. Продолговатый мозг —продолжение головного мозга, он представляет собой участок, где в головной мозг входит и выходит большинство черепных нервов. В продолговатом мозгу располагаются жизненно важные функциональнонезависимые центры в виде скопления клеток, связанные с дыханием, работой сердца и желудочно-кишечной функцией. Варолиев мост располагается между продолговатым мозгом и
средним мозгом. Большой пучок перекрещивающихся волокон, располагающийся на нижней стороне варолиева моста, связывает ствол мозга и мозжечок. Средний мозг входит спереди в таламус и гипоталамус.
В стволе мозга распределены, по меньшей мере, три двигательных центра, которые посредством эфферентных волокон воздействуют на двигательные нейроны спинного мозга (рис. 7.2 и 7.6). Эти двигательные центры включают красное ядро, латеральное и вестибулярное ядро и ретикулярную формацию. Красное ядро располагается в среднем мозгу и образует руброспи-нальный путь, который пересекает ствол мозга и воздействует н? контралатеральные сегментные центры. Этот путь имеет соматотопическую структуру (согласно карте тела), что означает, что положение аксонов в этом пути зависит от двигательных нейронов, которые они иннервируют. Например, аксоны нейронов в дорсоме-диальной части ядра располагаются в этом пути медиально и иннервируют цервикальные сегменты. При электрическом раздражении красное ядро возбуждает, главным образом, двигательные альфа- и гамма-нейроны, которые иннервируют флексорные мышцы, и ингибирует двигательные нейроны, связанные с экстензор-ными мышцами. Красное ядро принимает большую часть своего входящего сигнала от зубчатого ядра (в мозжечке) и двигательной зоны коры головного мозга. Латеральное вестибулярное ядро располагается между варолиевым мостом и продолговатым мозгом с нейронами, имеющими соматотопическую структуру в пределах ядра. Аксоны клеток, находящихся в этом ядре, образуют вестибулоспинальный тракт, который воздействует на ипсилатеральные двигательные нейроны. Латеральное вестибулярное ядро возбуждает двигательные альфа- и гамма-
226
нейроны, которые иннервируют экстензорные мышцы, и ингибируют двигательные нейроны, связанные с флексорными двигательными нейронами. Латеральное вестибулярное ядро принимает входной сигнал от мозжечка и органа равновесия (лабиринта). Ретикулярная формация образует два ретикулоспинальных тракта — компоненты, относящиеся к варолиевому мосту и продолговатому мозгу. У этих ретикулоспинальных трактов диффузная структура, заканчивающаяся между цервикальным и люмбальным сегментами. Эта ретикулярная формация принимает входной сигнал от сенсорно-двигательной зоны коры головного мозга, ядра покрышки (в мозжечке) и восходящих путей спинного мозга.
Нисходящие пути от супрасегментарных структур можно разделить на три пути: кортикоспинальный тракт и две группы (А и Б) путей ствола мозга (Kuypers, 1985). Относящийся к предцентральной борозде перекрестный компонент кортикоспинального тракта, в основном, заканчивается латерально у двигательных нейронов или вблизи двигательных нейронов, которые иннервируют дистальные мышцы (рис. 7.7). Некоторые аксоны в кортикоспинальном тракте заканчиваются вентрально у двигательных нейронов или вблизи двигательных нейронов, которые иннервируют аксиальные и проксимальные мышцы. Кортикоспинальный тракт имеет в большей степени топографическую организацию, чем любой из путей ствола мозга
(группы А и Б). Относящийся к постцентральной борозде компонент кортикоспинального тракта модулирует сенсорную информацию, поступающую от периферии, включая ту, которая необходима для управления движением.
Путь группы А располагается в вентромедиальном стволе мозга и состоит из вестибулоспинального, ретикулоспинального и покрышечноспинального путей; покрышечноспинальный путь создается нейронами в верхнем бугорке, который является участком среднего мозга, связанным с ориентацией визуального раздражителя. Покрышечноспинальный путь, вероятно, способствует реакциям ориентации головы и шеи на визуальные раздражители (Rothwell, 1987). Активация пути группы А обеспечивает постуральную поддержку направленного к цели движения с участием мышц-синергистов, в частности аксиальных и проксимальных мышц. Аксоны группы А проходят в вентральных и вентромедиальных пучках нервных клеток спинного мозга и заканчиваются билатерально у двигательных нейронов, которые иннервируют аксиальные и проксимальные мышцы (рис. 7.7). Аксоны нейронов группы А образуют многие коллатерали, которые, очевидно, необходимы для координации работы многих мышц.
Нисходящие аксоны в пути группы Б состоят из перекрестных руброспинальных и перекрестных ретикулоспинальных путей. Этот путь способствует направленной к цели двигатель-
Туловище
Рис. 7.7. Конечные зоны трех нисходящих путей. Три верхних поперечных сечения показывают положение завершений аксонов в кортикоспинальном тракте, пути группы Б и пути группы А соответственно. Нижнее поперечное сечение показывает топографическую организацию двигательных нейронов в цервикальном сегменте спинного мозга (Lawrance, Kuypers, 1968)
15*
227
Спинной мозг Спинной мозг
Рис. 7.8. Афферентный вход в мозжечок и выход через глубоко расположенные ядра мозжечка (Rothwell, 1987)
ной активности в результате воздействия на двигательные нейроны, которые иннервируют дистальные флексорные мышцы. Аксоны в этом пути имеют относительно мало коллатералей, и они заканчиваются то ли непосредственно у двигательных нейронов, то ли вблизи проприоспи-нальных аксонов спинного мозга; проприоспи-нальные нейроны являются промежуточными нейронами, которые передают информацию между спинномозговыми сегментами. Аксоны группы Б проходят в контрлатеральном дорсолатеральном пучке нервных клеток спинного мозга и заканчиваются в промежуточной зоне вблизи нейронов, воздействующих на дистальные флексорные мышцы (рис. 7.7).
Мозжечок
Мозжечок располагается ниже полушарий большого мозга и сзади ствола мозга. Эта необычная структура, содержащая более половины нейронов головного мозга, в высшей степени упорядочена и не имеет прямых эфферентных связей со спинномозговыми двигательными ядрами или двигательными ядрами ствола мозга, несмотря на свое участие в управлении движением. В мозжечке различают наружную мантию из серого вещества (кору мозжечка), внутреннее белое вещество и три пары глубоко расположенных подкорковых ядер (кровельные, промежуточные и зубчатые). Мозжечок принимает входной сигнал от периферии (соматосенсорная и визуальная информация), ствола мозга (вестибулярная информация) и коры головного мозга. Весь выходной сигнал мозжечка передается через глубоко расположенные подкорковые ядра и вестибулярное ядро в три сим
метричные пары путей, связывающих мозжечок и ствол мозга (рис. 7.8).
Мозжечок имеет три функциональных отдела, каждый из которых принимает входной сигнал и подает выходной сигнал в различные части ЦНС (Ghez, 1991): спинномозговой мозжечок, церебральный мозжечок и вестибулярный мозжечок (рис. 7.9). Спинномозговой мозжечок, который включает червь мозжечка и промежуточные полушария, принимает большую часть сенсорного входного сигнала от спинного мозга. Выходной сигнал червя мозжечка поступает к ядру покрышки, в то время как выходной сигнал промежуточных полушарий направляется к интерпонированному ядру. Выходной сигнал двух этих глубоко расположенных ядер передается в ствол мозга и спинной мозг по нисходящим путям и способствует управлению движением конечностей. Церебральный мозжечок, который относится к латеральным полушариям мозжечка, принимает входной сигнал от коры головного мозга через варолиев мост. Выходной сигнал поступает в зубчатое ядро, а затем, через таламус, — в преддвигательную и двигательную зоны коры голов-ного мозга. Благодаря этой связи церебральный мозжечок способствует планированию движений. Вестибулярный мозжечок занимает регионарно-узелковую долю и принимает входной сигнал от вестибулярных ядер в продолговатом мозгу, подавая свой выходной сигнал назад в те же ядра. Вестибулярный мозжечок способствует управлению движениями глаз и равновесию при стоянии и ходьбе.
По-видимому, мозжечок выполняет функцию компаратора; он сравнивает движения, которые хочет осуществить ЦНС, с теми, которые имеют место фактически. Эта функция выпол
228
няется мозжечком, принимающим экстенсивный афферентный входной сигнал и генерирующим выходной сигнал,который может модифицировать двигательные команды. Тем не менее отсутствует согласованность в отношении точных признаков движения, которые имеют значение для мозжечка. Rothwell (1987) выделяет три теории функционирования мозжечка: регулирующее устройство, обучающее устройство и координатор. При рассмотрении его в качестве ретулирующего устройства предполагается, что кора головного мозга инициирует движение в результате возбуждения агониста, а мозжечок прекращает это движение в требующемся положении. Мозжечок может функционировать как обучающее устройство в результате закрепления выбираемых соединений входов и выходов таким образом, чтобы со временем требовался меньший входной сигнал для генерирования надлежащего выходного сигнала. По этой схеме мозжечковые цепи модифицируют
ся опытом, в результате чего повторяющийся входной сигнал проходит через определенные синапсы, и мозжечок усваивает соответствующее движение. Мозжечок может служить также координатором работы мышц, воздейсвтвующих на различные суставы, в результате чего движения в различных суставах масштабируется для выполнения надлежащего движения конечностей. Конечно, мозжечок способен модулировать поведенческие движения; он представляется также важным для усвоения двигательных навыков.
Базальные ядра
Базальные ядра представлены пятью тесно связанными ядрами: хвостатым ядром, путаме-ном, глобус паллидусом, субталамическим ядром и черным веществом. Хвостатое ядро и пу-тамен показаны в виде одного ядра, названного
Рис. 7.9. Три функциональных подразделения мозжечка (Ghez, 1991):
с— выходные сигналы; б— входные сигналы; 1 — вестибулярный мозжечок; 2 — церебральный мозжечок (латеральная полусфера); 3 — промежуточная полусфера; 4 — червь мозжечка; 5 — спинномозговой мозжечок; 6 — ядро покрышки; 7 — к медиальным нисходящим системам; 8 — двигательное исполнение; 9 — интерпонированное ядро; 10 — к латеральным нисходящим системам; 11 — зубчатое ядро; 12 — к двигательной и преддвигательной зонам коры головного мозга; 13 — двигательное планирование; 14 — к вестибулярным ядрам; 15 — равновесие и движения глаз; 16 — входные сигналы коры головного мозга и варолиева моста; 17 — спинальные и тройничные входные сигналы; 18 — визуальные и слуховые входные сигналы; 19 — вестибулярные входные сигналы
229
Рис. 7.10. Двигательная схема базальных ядер (Cote, Crutcher, 1991):
а — полосатое тело; б — глобус паллидус; в — черная субстанция; г — субталамическое ядро; д — таламус; е — к стволу мозга и спинному мозгу; ж — преддвигательная зона коры головного мозга; з — дополнительная двигательная зона коры головного мозга; и — двигательная зона коры головного мозга; к — соматосенсорная зона коры головного мозга; л — верхняя теменная доля
на рис. 7.10 полосатым телом. Базальные ядра принимают основной входной сигнал от коры головного мозга и подают большую часть своего выходного сигнала через таламус назад в кору головного мозга (рис. 7.10). Базальные ядра представляют собой промежуточное звено между недвигательной корой головного мозга и основной двигательной зоной коры головного мозга. Базальные ядра связаны с планированием движения таким образом, что они кодируют выходные нейроны для различных аспектов движения, например, направления, амплитуды и скорости, а не уточняют возбуждение конкретных мышц и запуск двигательных программ. Хронирование нейронного разряда наводит на мысль о том, что в базальных ядрах существуют, по меньшей мере, два типа клеток: связанных с движением, относящиеся к поведенческому движению, а не к его инициированию, и активные перед началом движения и связанные с подготовкой к движению. Кроме того, базальные ядра участвуют в недвигательных поведениях, например, в познании и усвоении привычек.
Хотя базальные ядра и не имеют прямых связей со спинным мозгом, их роль в движении была хорошо изучена у пациентов с болезнью Паркинсона, болезнью Хантингтона и гемибал-
лизмом (Hallet, 1993). Согласно Cote, Cruther (1991), эти болезни вызывают недостаточность базальных ядер и характеризуются: а) непроизвольными движениями и дрожью; б) мышечной ригидностью — изменениями положения тела и мышечного тонуса; в) неподвижностью — недостаточностью и медленностью движения без паралича.
Восходящие пути
В теле человека огромное количество разнообразных чувствительных нервных окончаний. Афферентные сигналы от них поступают в спинной мозг через задние корешки и синапс к нейронам как на сегментном, так и на супра-сегментном уровнях. Группы волокон, которые распределяют информацию по супрасегментным центрам, называются восходящими путями. Соматическая сенсорная информация от рук, ног и туловища поступает через таламус в кору головного мозга двумя основными путями: по пути спинной столб — медиальный лемниск и по переднему латеральному пути. Соматическая сенсорная информация включает афферентные сигналы от механорецепторов, терморецепторов, болевых рецепторов и хеморецепторов. Два этих восходящих пути играют главную роль в ощущении, в результате чего афферентные сигналы достигают сознания и распознаются (воспринимаются).
Таламус объединяет и распределяет большую часть сенсорной и двигательной информации, поступающей в кору головного мозга. Он принимает входной сигнал о соматическом восприятии, слухе и зрении и передает информацию в мозжечок, базальные ядра и основную сенсоро-двигательную зону коры головного мозга. Афферентные аксоны в спинном столбе располагаются соматотопически со входом от крестца наиболее медиальным и входом от шеи наиболее латеральным. Соматотопическая структура афферетного входного сигнала сохраняется на всем пути к первичной (SI) и вторичной (SII) соматическим сенсорным зонам коры головного мозга и к задней теменной зоне коры головного мозга.
Передний латеральный путь передает информацию, главным образом, о боли и температуре, а также некоторую осязательную и проприоцептивную информацию. Аксоны в этом пути пересекают спинной мозг и поднимаются в латеральную часть ретикулярной формации в стволе мозга и таламус. Фактически передний латеральный путь имеет три части: спиноталамическую, спиноретикулярную и спиноцервикальную. Эти названия отражают происхождение и завершение путей. Спиноталамический и спиноретикулярный пути передают болевые и тепловые афферентные сигналы. Спиноцерви-
230
кальный путь заканчивается в зоне среднего мозга, которая содержит нейроны, участвующие в контроле боли. По этим трем путям информация передается также в первичную и вторичную соматические сенсорные зоны и заднюю теменную зону коры головного мозга.
Хотя два пути и проецируются на таламус, аксоны контактируют с отдельными популяциями нейронов. Информация от различных классов чувствительных нервных окончаний остается разделенной в спинном мозгу, стволе мозга и таламусе и не взаимодействует, пока не достигнет соматических сенсорных зон коры головного мозга. Сенсорная информация распределяется среди различных зон коры головного мозга и по подкорковым зонам, где она влияет на поток информации в нисходящих путях.
МЫШЕЧНО-СКЕЛЕТНАЯ СИСТЕМА
Мышечно-скелетная система состоит из периферийных частей двигательной системы и содержит мышцы и элементы соединительной ткани, которые образуют скелет. В дополнение к мышечно-скелетной системе двигательная система включает те компоненты нервной системы, которые участвуют в осуществлении движения. На рис. 7.12 представлены биомеханические особенности мышечно-скелетной
Ускорение
Рис. 7.12. Биомеханические следствия (электромиограмма, усилие, вращающий момент, угол, скорость, ускорение) возбуждения мышечно-скелетной системы нервной системой (Zajac, Gordon, 1989)
231
системы (Zajac, Gordon, 1989). В этой схеме сигналы, передаваемые двигательными единицами (элекгромиограмма), представляют команды, подаваемые нервной системой для возбуждения мышцы и создания усилия, которое передается скелету в виде вращающего момента относительно сустава. Усилие, создаваемое мышцей, зависит от ее механических свойств (см. главу 6), в то время как вращающий момент зависит как от вектора усилия мышцы, так и от его плеча пары относительно оси вращения сустава (см. главу 2). Тем не менее биомеханический эффект возбуждения мышцы не ограничивается суставом или суставами, пересекаемыми мышцей, а распределяется по мышечно-скелетной системе (см. “Межсегментная динамика” в главе 3). Как результат этой мышечно-скелетной организации возбуждение мышцы нервной системой может оказывать влияние на кинематические параметры (угол, скорость и ускорение) многих суставов по всему телу. Более того, при изменении кинематических параметров оно может также влиять на динамику мышцы (соотношения расстояние—усилие и усилие—скорость) и геометрию сустава (момент—длина плеча).
В схеме выделен важный вопрос. Данная команда нервной системы не всегда обеспечивает один и тот же биомеханический эффект для двигательной системы. Скорее результат невральной команды зависит от состояния мышечно-скелетной системы (например, от длины мышцы, скорости мышцы, длины плеча пары). Два этих фактора — качество и величина нервного стимула и состояние мышечно-скелетной системы — имеют, по меньшей мере, одинаковое значение при определении способности двигательной системы к осуществлению движения (Bernstein, 1984). Для выполнения желаемого движения нервная система должна знать состояние мышечно-скелетной системы. Два существенных ограничения, которые накладываются мышечно-скелетной структурой, заключаются в том, что движение обусловливается результирующей работой мышцы и что место прикрепления мышцы относительно сустава изменяется.
Результирующая работа мышцы
В главе 2 мы подчеркивали, что движение человека обусловливается, как правило, дисбалансом сил, действующих на тело, а не приложением единственного отдельного усилия. Благодаря мышечно-скелетной структуре это обобщение применимо также к влиянию работы мышцы на сустав. Обычно возбуждается не одна мышца, а группа мышц-синергистов или набор мышц-агонистов и антагонистов. В связи с таким режимом работы важно еще раз подчерк
нуть, что результирующее усилие мышцы и результирующий вращающий момент мышцы (см. главу 2) представляют собой результирующий эффект работы мышцы относительно сустава. В главе 3 рассмотрено, каким образом использовать ньютоновскую механику для расчета результирующего усилия мышцы как для статических, так и для динамических условий. С помощью этого подхода мы определили схему свободного тела для учета всех кинематических и кинетических воздействий на систему, за исключением результирующего усилия мышцы (или результирующего вращательного момента мышцы), и затем рассчитали результирующее усилие мышцы в качестве остаточного момента (см. главу 3). Это обеспечило оценку результирующей работы мышцы и вклада работы мышцы в движение.
Межкостное усилие. Ограничение в исследовании движения в качестве следствия воздействия результирующего усилия мышцы заключается в том, что мы не можем определить абсолютное усилие, создаваемое мышцей. Хотя на движение и оказывает влияние результирующее усилие мышцы, абсолютное усилие является важным фактором в таких областях, как протезирование. В главе 2 мы ввели понятие “реакция сустава” для учета результирующих усилий, которые создаются в результате костного контакта между смежными сегментами тела. Эти усилия могут создавать ткани (например, капсула, связка, мышца) и внешние агенты (например, реакция земли). В противоположность этому термин “межкостное усилие” представляет абсолютное усилие в суставе, обусловливаемое контактированием сегментов тела. Разница в величине между реакцией сустава и межкостным усилием может быть существенной.
Интересный пример этой разницы представлен Galea и Norman (1985) в исследовании межкостных усилий в голеностопном суставе во время быстрого балетного движения — прыжка с плоских стоп на носки. В этом анализе определялись как реакция сустава, так и межкостное усилие под действием сил, создаваемых мышцами, участвующими в движении. В модели, использованной для выполнения этих расчетов (рис. 7.13, а), принимались во внимание основные мышцы, пересекающие голеностопный сустав, и, следовательно, те мышцы, которые вносят вклад в зависимую от мышц составляющую межкостного усилия. Эти мышцы включали экстензор hallicus longus (EHL), tibialis anterior (ТА), флексор hallicus longus (FHL), peroneus longus (PL) и gastrocnemius/soleus (G/S). Для расчетов усилия, создаваемого каждой из этих мышц, оценивались их электромиограммы, длина и скорость изменения длины (Hof, 1984; Hof, van der Berg, 1981a, 1981b, 1981c, 198 Id).
232
Рис. 7.13. Межкостные усилия в голеностопном суставе во время полного пуанта (Galea, Norman, 1985): а — схематический вид стопы, показывающий линии действия основных мышц, пересекающих голеностопный сустав; б — разделение действий мышц на те, которые создают изгибающий назад и изгибающий плантарно вращающие моменты;
в — результирующее усилие мышц
Оценив усилия для каждой мышцы, Galea и Norman (1985) разделили усилия (рис. 7.13, б) на те, которые вносят вклад в изгибающий плантарно вращающий момент (Fm pfi, и те, которые вносят вклад в изгибающий назад вращающий момент (Fm Межкостное усилие рассчитывали как сумму касательных составляющих векторов Fm> pf и df. Однако реакцию сустава определяли как касательную составляющую результирующего усилия мышц Fm, которое представляет собой разность Fm и Fm df (рис. 7.13, в). С точки зрения векторной алгебры, которая рассматривает как величину, так и направление векторов, Fm всегда определяется как векторная сумма Fm PfH Fm df. Для одного человека Galea и Norman рассчитали среднюю реакцию сустава, обусловленную работой мышц, на уровне 732 Н во время движения и среднее межкостное усилие, составившее 6,068 Н. Это разность, соответствующая приблизительно одному порядку величины.
Работа мышцы и движение. Несмотря на разницу в абсолютном и результирующем эффектах в суставе, на движение влияет результирующее, но не абсолютное усилие мышцы. Поэтому возможность определения результирующей мышечной работы, выполняемой скелетно-мышечной системой, может оказаться полезной при тренировочной и реабилитационной деятельности. Результирующую работу мышц можно определить на основе качественного анализа движения. Например, рассмотрим человека, лежащего на скамье на
спине и выполняющего махи предплечьем (движения сгибания—разгибания локтя) с гантелью весом 120 Н в правой руке (рис. 7.14, а). Если человек выполняет медленное движение во всем диапазоне последнего, какова результирующая работа мышц при перемещении руки из позиции 1 в позицию 4? Какая группа мышц обеспечивает это движение? Действует она концен-трично или эксцентрично? Что создает нагрузку и относительно какой точки действует последняя?
Нагрузкой, которой противодействует группа мышц, является вес h, удерживаемый человеком в руке, в сумме с весом Fw а сегмента предплечья-кисти руки. Соответствующая схема свободного тела показывает, что осью вращения является локтевой сустав (рис. 7.14, б). В схему свободного тела включены также результирующее усилие мышц (Fm) и реакция сустава (7J). Анализируются вращающие моменты, действующие относительно локтевого сустава (Тт и Tt), и схема свободного тела упрощается, как показано на рис. 7.14, в. В этом упрощении вращающий момент нагрузки представляет воздействия веса системы (Fw s + FK h). Направление векторов веса определяется по правилу большого пальца правой руки; пальцы правой руки сгибаются в направлении вращения системы под действием каждого усилия (результирующее усилие мышц и результирующее усилие нагрузки), а большой палец правой руки выпрямлен. Если большой палец направлен наружу страницы, вектор вращающего момента направлен против часовой стрелки. Если же большой палеи направлен внутрь страницы, вектор вращающего момента направлен по часовой стрелке.
По этим схемам можно определить группу мышц, которая обеспечивает движение, и на-
Рис. 7.14. Определение схемы результирующей работы мышц для медленного движения:
а — выполнение движения сгибания—разгибания локтя с гантелью в правой руке; б — схема свободного тела, выделяющая сегмент предплечья-кисти руки; в — определение результирующих вращающих моментов мышц и нагрузки; г — результирующая работа мыши, определяемая сравнением направлений вращающих моментов и движения
233
правление их изменения по длине (концентричное или эксцентричное действие). Соответствующие операции включают графическое представление следующего:
1) сегмента тела в приблизительной позиции;
2) вектора вращающего момента нагрузки (7});
3) результирующего вектора вращающего момента мышц (Тт) в направлении, противоположном направлению вектора вращающего момента нагрузки;
4) направления вращения сегмента тела.
Например, рассмотрим движение сегмента предплечья-кисти руки при сгибании предплечья из позиции 3 в позицию 4. Заметим, что сегмент тела вычерчивается не в позиции 3 или позиции 4, а на полпути между двумя этими позициями (рис. 7.14, г). По этой схеме можно установить направление результирующего вектора вращающего момента мышц и определить, является результирующее действие мышц концентрическим или эксцентрическим. Для медленных движений направление результирующего вектора вращающего момента мышц противоположно направлению вектора вращающего момента нагрузки. Для определения группы мышц, представляемой результирующим вектором вращающего момента мышц, предположим, что никакие другие усилия или вращающие моменты не действуют на систему, и сегмент вращается в направлении, совпадающем с направлением результирующего вектора вращающего момента мышц. Если это направление является направлением разгибания, то этот обеспечивается экстензорами; если результирующий вектор вращающего момента мышц обусловливает сгибание, этот вектор представляет флексоры. Напомним также, что мышцы могут создавать только тяговое усилие. Следовательно, если начертить на схеме свободного тела результирующий вектор усилия мышц (Fm), можно определить, какие мышцы создают такое усилие (флексоры или экстензоры, абдукторы или аддукторы). В примере, приведенном на рис. 7.14, вращающий момент, направленный по часовой стрелке, показывает экстензоры локтя, а вращающий момент, направленный против часовой стрелки, — флексоры локтя (т.е. Тт представляет вращающий момент экстензоров локтя).
В результате сравнения направлений результирующего вектора вращающего момента мышц и вращения сегмента можно определить, является результирующее действие мышц концентрическим или эксцентрическим. Если направления результирующего вектора вращающего момента мышц и вращения сегмента одинаковы, группа мышц подвергается концентрическому сокращению. Если вращение сегмента и результирующий вращающий момент мышц имеют различные
направления (как во время движения из позиции 3 в позицию 4), результирующим действием мышц является эксцентрическое сокращение. Таким образом, движение из позиции 3 в позицию 4 осуществляется в результате эксцентричного сокращения экстензоров локтя. Вообще, если группа экстензоров осуществляет разгибание или группа флексоров — сгибание, результирующее действие мышц концентрично. В противоположность этому сгибание, осуществляемое группой экстензоров, или разгибание, осуществляемое группой флексоров, представляет эксцентрическое результирующее действие. Используя эти методики, можно определить качественно результирующее действие мышц для таких медленных движений, как сгибание предплечья, во всем диапазоне движения (табл. 7.1).
Пример сгибания предплечья человеком, лежащим на спине, подчеркивает важную особенность пути использования нами своих мышц. Допустим, человек перемещается из положения лежа на спине в вертикальное положение и выполняет подобное упражнение, поднимая и опуская вытянутую руку в пределах позиций 1 и 2 (рис.7.15). Какие схемы результирующего действия мышц необходимы для выполнения этого движения? Используя описанную методику, можно определить, что движение обеспечивается флексорами плеча, т.е. как при перемещении из позиции 1 в позицию 2, так и при перемещении из позиции 2 в позицию 1. Действие мышц концентрично при движении вверх и эксцентричо при движении вниз. Различие в схемах действия между положением лежа на спине и вертикальным положением связано с ориентацией человека (и, следовательно, движения) относительно направления силы тяготения. Гравитационная окружающая среда оказывает основное влияние на схему возбуждения мышц. Представим, что в примерах как вертикального положения, так и положения лежа на спине вертикальная линия (тяготение) проходит через ось вращения (соответственно плечо и локоть). Основное качественное различие заключается в том, что движение ни в коем случае не пересекает эту линию при выполнении упражнения в вертикальном положении, в то время как это имеет место при выполнении упражне-
Таблица 7.1. Результирующее действие мышц для упражнения сгибания предплечья, показанного на рис. 7.14
Мышцы ЛОКТЯ Движение
1—>2 2—>3 3—>4 4—>3 3—>2 2~-> 1
Флексоры К К Э Э Экстензоры Э К
Примечание: К — концентрическое; Э — эксцентрическое действие.
234
Рис. 7.15. Движение сгибания—разгибания плеча. Поскольку вращающий момент нагрузки (Fw а + Fw j) действует в одном и том же направлении в процессе всего движения, направление результирующего вращающего момента мышц (Гт) постоянно во всем диапазоне движения
ния в положении лежа на спине. Для медленных упражнений пересечение этой линии означает смену группы мышц, управляющих движением.
Заметим, что выполненный анализ применим только к условиям, которые мы называем квазистатическими, т.е. к таким, которые почти статические и в которых ускорение системы и ее различных частей довольно мало (Miller, 1980; Tousaint et al., 1992). По квазистатическим условиям мы можем аппроксимировать механическое состояние мышцы в результате выполнения статического анализа, т.е. SF = 0. Исследователи не сходятся во мнениях о границе, отделяющей квазистатическое состояние от состояния, не являющегося таковым. Так, Alexander и Vernon (1975) использовали вариант квазистатическо-го подхода для расчета результирующих враща
ющих моментов мышц относительно голеностопного и коленного суставов в вертикальной стойке для медленного движения вверх—вниз. Другие (Mann, 1981; Miller, Munro, 1985) полагают, что такие условия требуют полного динамического анализа, т.е. £F = та.
Безусловно, динамический анализ необходим для более энергичных действий, например, ударов ногой. Miller (1980) рассматривает схему действия мышц относительно коленного сустава во время выполнения футболистом удара носком (рис. 7.16). Чтобы продемонстрировать непригодность квазистатического анализа, проведем его и сравним результаты с данными динамического анализа. Четыре позиции наносящей удар ноги (с зачерненным ботинком) с рис. 7.16 переносятся на схему сначала относительно тазобедренного сустава, затем относительно коленного сустава (рис. 7.17). Заметим, что позиция голени относительно колена на рис. 7.17 такая же, как и ее первоначальная позиция на рис. 7.16. После этого определим схемы результирующего действия мышц относительно коленного сустава во время удара, как мы делали это для примера сгибания предплечья. Поскольку во время движения (позиции 1—4) ни в коем случае не пересекается вертикальная линия, проходящая через ось вращения (колено), вектор вращающего момента нагрузки и, следовательно, результирующий вектор вращающего момента мышц действуют в одном и том же направлении в процессе всего движения. Результирующее действие мышц представлено в табл. 7.2.
На основании этого квазистатического логического подхода движение относительно коленного сустава во время удара носком (рис. 7.16) обеспечивается предположительно флексорны-
Рис. 7.16. Результирующий вращающий момент мышц относительно коленного сустава наносящей удар ноги в функции коленного угла во время замаха, предшествующего контакту с мячом при выполнении футболистом сильного удара носком (Miller, 1980). Общая продолжительность замаха 170 мс с интервалом 17 мс между большими точками на графике. Интерпретацию графика начинаем с нижнего правого угла и движемся в направлении против часовой стрелки от позиции 1 к позиции 4
235
Рис. 7.17. Угловое смещение голени относительно колена во время выполнения футболистом удара носком, показанного на рис. 7.16
ми мышцами коленного сустава. Наша интуиция говорит нам, что было бы трудно убедить кого-либо в том, что флексоры коленного сустава, главным образом, управляют этим ударным действием. Очевидный недостаток такого анализа — допущение, что действие является медленным. Ясно, что оно не является медленным. В главе 2 мы рассмотрели некоторые из сил, которые обычно встречаются в движении человека. Напомним, что в результате движения объекта или сегмента создаются силы инерции. При быстрых движениях силы инерции становятся существенными и коренным образом изменяют схему действия мышц, необходимую для управления движением. Действительно, для такого движения, как сильный удар, необходимо рассмотреть, каким образом движение голени будет влиять на бедро и, наоборот, движение бедра будет влиять на голень (Putnam, 1983, 1991).
Воздействие инерции можно установить, сравнив результирующий вращающий момент мышц относительно коленного сустава на рис. 7.16 с результирующим вращающим моментом мышц, выведенным в соответствии с квази-статическим подходом. Этот график показывает несколько представляющих интерес особенностей: за исключением первой части удара, результирующий вращающий момент мышц создается экстензорами коленного сустава, а не флексорами, как предполагается в соответствии с квазистатическим анализом; большая часть движения из позиции 1 в пози-
Таблица 7.2. Результирующее действие мышц для выполняемого футболистом удара носком, показанного на рис. 7.16, на основании квазистатического анализа
Мышцы колена Движение
1 —> 2 2-» 3 3-» 4
Флексоры Экстензоры К К э
Примечание: К — концентрическое действие; Э — эксцентрическое действие.
цию 3, во время которого угол коленного сустава уменьшается, осуществляется в результате эксцентрического действия экстензоров; движение из позиции 3 до контактирования с мячом — в результате концентрического действия экстензоров и результирующий вращающий момент мышц максимален при переходе от эксцентрического действия к концентрическому. Эти наблюдения показывают, что квазистатический анализ, игнорирующий эффекты, обусловливаемые ускорением, не приемлем для определения схемы результирующего действия мышц для таких задач, как быстрый удар.
Методом, который может быть использован для качественной оценки информации, представленной на рис. 7.16, является совмещение графиков результирующего вращающего момента мышц и угловой скорости (рис. 7.18). Для иллюстрации этой методики возвратимся к примеру горизонтального разгибания-сгибания относительно локтевого сустава (см. рис. 3.28). Половина этого движения представлена разгибанием (положительная скорость), а половина — сгибанием. Изменение направления движения от разгибания к сгибанию происходит при нулевой скорости. Первая часть фазы разгибания выполняется под действием результирующего вращающего момента экстензоров; следовательно, экстензоры действуют эксцентрично. Тем не менее остальная часть фазы разгибания выполняется в результате результирующего действия флексоров, а это означает, что флексоры активно растягиваются (т.е. происходит эксцентрическое сокращение). В соответствии с подобной аргументацией фаза сгибания обеспечивается в результате концентрического (К) действия флексоров и затем в результате эксцентрического (Э) действия экстензоров (рис. 7.18). Мы получили такую же схему результирующего действия другими средствами в главе 1 (см. рис. 1.9).
В данном обсуждении результирующего действия мышц выделено важное следствие мышечно-скелетной структуры. Вследствие сегментной динамики (см. главу 3) нервная система управляет быстрым и медленным движениями различным образом, т.е. схема результирующего действия мышц изменяется со скоростью движения для согласования эффектов инерции между сегментами. Кроме того, быстрые движения обычно включают эксцен-трически-концентрическую последовательность, в которой пиковый результирующий вращающий момент мышц имеет место при переходе от эксцентрической фазы к концентрической. Тем не менее независимо от того, осуществляется движение медленно или быстро, вклад мышцы определяется результирующим действием среди мышц-синергистов и мышц-антагонистов.
236
Экстензоры
Флексоры
Время, с
Рис. 7.18. Вывод результирующего действия мышц, обеспечивающего движение разгибания—сгибания локтевого сустава в горизонтальной плоскости. Положительная скорость — разгибание локтевого сустава. Положительный результирующий вращающий момент результирующий вращающий момент экстен-зорных мышц
Прикрепление мышц
Особенностью мышечно-скелетной структуры, привлекающей значительное внимание, является распределение активности среди мышц, участвующих в движении. Термин “синергия” относится к группе мышц, вынужденных действовать в качестве одной единицы. Поскольку мышцы могут только тянуть, а не толкать, пересекать сустав для контроля каждой степени свободы (например, сгибание-разгибание, отведение-приведение) должно не менее одной пары мышц. Обычно в одном и том же дейст
вии участвуют несколько мышц. Например, основные флексоры локтя включают бицепс брахии, брахиарис и брахиорадиалис. Почему у нас так много мышц? Имеем ли мы больше мышц-чем это фактически нам необходимо?
Хотя каждая из этих трех мышц и может вносить вклад во вращающий момент флексоров локтя, они не эквивалентны генераторам усилий, так как имеют разную форму и, следовательно, различные возможности (Buchanan, Rovai, Rymer, 1989; Funk, An, Моггеу, Daube, 1987). Как следует из главы 6, различия в длине волокон мышц, площади их поперечного сечения и точках прикрепления влияют на диапазон движения, скорость сокращения, максимальное усилие и форму соотношения вращающего момента и угла. В табл. 7.3 приводятся некоторые структурные различия между тремя основными флексорными мышцами локтя; эти различия существуют во многих суставах тела (например, голеностопном, коленном, тазобедренном, позвоночном столбе, плече). Не только степень участия трех флексоров локтя может изменяться в зависимости от диапазона движения, но и вариация в точках прикрепления может позволять мышце участвовать более чем в одном механическом действии. Например, благодаря своему дистальному прикреплению бицепс брахии способен вносить вклад во вращающие моменты как флексоров локтя, так и супинаторов предплечья. Buchanan, Rowai, Rymer (1989) показали, что для множества нагрузок на запястье при сгибании-разгибании, внутреннем-внешнем вращении и супинации-пронации существует тесная связь между действием брахиалис и брахиорадиалис, но не между бицепс брахии и брахиорадиалис (рис. 7.19). Увеличение супинационных нагрузок связывалось с увеличением ЭМГ в бицепс брахии, но не в брахиорадиалис; наоборот, повышение пронационных нагрузок сопровождалось увеличением электромиограммы в брахиорадиалис, но не в бицепс брахии. Бицепс брахии обычно был наиболее активным, в то время как брахиорадиалис и брахиалис — наименее активными. При этом различии в структуре и связанной с ней функции нервная система может варьировать возбуждение мышц-синергистов в зависимости от особенностей задачи.
Таблица 7.3. Структурные особенности трех основных флексорных мышц локтевого сустава человека
Мышца Длина мышцы, см Длина волокон, см Площадь поперечного сечения мышцы, см2 Расстояние до сустава, см
проксимальное дистальное
Бицепс брахии 24,0 17,8 5,8 30 4
Брахиалис 20,1 14,4 7,4 10 3
Брахиорадиалис 23,7 14,7 2,0 6 25
Примечание: Расстояние до сустава указывает положение центра точки прикрепления (проксимальной и дистальной) относительно оси вращения локтевого сустава.
237
0,3
• Пронация
* Мягкая пронация
□ Нейтральное положение
Л Мягкая супинация
О Супинация
ЭМГ брахиорадиалис
Рис. 7.19. Сравнение изменений уровня ЭМГ для трех основных флексорных мышц локтевого сустава во время приложения различных нагрузок к запястью. Пронационные нагрузки обусловливают одновременное изменение ЭМГ в брахиалис и брахиорадиалис. В противоположность этому отсутствовало общее одновременное изменение уровней ЭМГ в бицепс брахии и брахиорадиалис при различных нагрузках (Buchanam, Rovai, Rymer, 1989)
Двусуставные мышцы. Значительное количество мышц охватывает два сустава, поэтому их называют двусуставными (например, бицепс брахии, ректус феморис). Двусуставные мышцы обеспечивают, по меньшей мере, три преимущества в управлении мышечно-скелетной системой.
Во-первых, двусуставные мышцы связывают движение двух суставов, которые они пересекают (van Ingen Schenau, Bobbert, van Soesr, 1990). Например, бицепс брахии пересекает как локтевой, так и плечевой сустав, и, таким образом, управляет сгибанием как локтя, так и плеча. Поскольку два этих движения происходят одновременно в процессе многих повседневных действий, полезно иметь мышцу, содействующую обоим действиям. К тому же эта связь может достигаться в результате сниже
ния электромиограммы односуставной мышцы и повышения активности двусуставной мышцы. Jamashita (1988) сообщал о снижении электромиограммы глютеус максимус (односуставного экстензора тазобедренного сустава) и повышении электромиограммы семимембранус (двусуставного экстензора тазобедренного сустава и флексора коленного сустава) во время одновременного разгибания тазобедренного и коленного суставов. Zajac (1993) предполагает, что односуставные мышцы создают пропульсивную энергию для прыжка в высоту, в то время как двусуставные улучшают координацию.
Во-вторых, скорость укорочения двусуставной мышцы является меньшей по сравнению с ее односуставными мышцами-синергистами (van Ingen Schenau et al., 1990). Преимущество меньшей скорости укорочения двусуставных мышц заключается в том, что у них более высокие возможности во взаимосвязи сила—скорость (см. рис. 6.19) по сравнению с односуставной мышцей и способны развивать усилие, составляющее большую долю изометрического максимума.
В-третьих, двусуставные мышцы могут перераспределять вращающий момент мышц и энергию суставов по конечности (Toussaint et al., 1992; van Ingen Schenau et al., 1990). На рис. 7.20 представлена модель ноги человека, включающая таз, бедро и голень с различными одно- и двусуставными мышцами. В этой модели мышцы 1 и 3 — односуставные экстензоры тазобедренного и коленного суставов, мышцы 2 и 4 — односуставные флексоры тазобедренного и коленного суставов, а мышцы 5 и 6 — двусуставные мышцы. Одновременное разгибание тазобедренного и коленного суставов может быть осуществлено в результате возбуждения двух односуставных экстензоров (мышцы 1 и 3). Поскольку мышца 5 (двусуставная) создает флексорный вращающий момент относительно тазобедренного сустава и экстензорный вращающий
Рис. 7.20. Модель ноги человека с шестью мышцами, располагающимися вокруг тазобедренного и коленного суставов. Мышцы 1—4 пересекают один сустав; мышцы 5 и 6 пересекают оба сустава (van Ingen Schenau, Robert, van Soest, 1990)
238
момент относительно колена, одновременное возбуждение мышцы 5 и мышц 1 и 3 обусловит уменьшение результирующего вращающего момента относительно тазобедренного сустава, но увеличение результирующего вращающего момента относительно колена. Исходя из этого взаимодействия, двусуставная мышца (мышца 5) описана в качестве перераспределяющей часть вращающего момента мышц и энергии суставов с тазобедренного сустава на колено. Наоборот, возбуждение мышцы 6 вместе с мышцами 1 и 3 обусловит перераспределение с колена на тазобедренный сустав.
Воздействия плеча пары. В части I мы обсудили представление о том, что при движении мышцы в пределах диапазона ее перемещения изменяется плечо пары от вектора усилия мышцы до оси вращения (сустава). Для флексоров локтевого сустава плечо пары максимально в среднем положении (угол локтевого сустава приблизительно 1,57 рад) и минимально при полном разгибании и полном сгибании. Исходя из этого соотношения, можно было бы предполагать, что усилие мышцы, необходимое для поддержания постоянной нагрузки, создаваемой вращающим моментом, изменяется обратно пропорционально изменениям плеча пары. Это соотношение трудно оценить непосредственно, так как трудно измерить как плечо пары, так и усилие мышцы. Вместо этого Hasan и Enoka (1985) исследовали электромиограмму и установили, что она изменяется таким образом, который можно было бы предположить для усилия мышцы при данном изменении плеча пары (рис. 7.21).
20 -
40 -
х„ х.
/Йх г х
X
40
20 -
0 J
1,57
2,62
0,52
Угол локтевого сустава, рад
Рис. 7.21. Уровень ЭМГ, необходимый для двух действующих одновременно флексорных мышц локтевого сустава с целью поддержания постоянной нагрузки, создаваемой вращающим моментом, в основном диапазоне движения локтевого сустава (Hasan, Enoka, 1985).
ЭМГ измеряли в качестве среднего уровня напряжения, необходимого для поддержания нагрузки, и выражали в процентах максимального произвольного сокращения
СТРАТЕГИИ ДВИЖЕНИЙ
Из-за сложности как взаимодействий между супрасегментными центрами, так и организации мышечно-скелетной системы для осуществления желаемого движения могут быть использованы многие различные последовательности возбуждения и комбинации мышц. Фактически существует значительно больше возможностей, чем необходимо для большинства движений (Bernstein, 1984). Какие правила использует нервная система для выбора из числа этих вариантов? Какие стратегии используем мы для осуществления различных движений? Представляется желательным наличие многих степеней свободы (скелетные, анатомические и невральные варианты) при усвоении нами нового движения. Но как только движение усвоено (например, ходьба, бросание), мы используем только подгруппу имеющихся вариантов. Тема стратегий движения касается исследования схем нервного возбуждения, связанных с различными типами движений. Общий под
ход к исследованию стратегий движения заключается в идентификации переменных, которые модулируются для управления движением (например, продолжительности и уровня электромиограммы). Такие анализы были применены к движениям в пределах от движений большого пальца кисти до взятия веса на грудь штангистом-олимпийцем. Тем не менее выбор соответствующей стратегии дополнительно усложняется необходимостью обеспечения того, чтобы система оставалась устойчивой во время движения. Следовательно, прежде чем исследовать основные стратегии движения, которые были определены, необходимо рассмотреть вопросы, связанные с положением тела.
Положение тела
Давно известно, что выполнение движения сопровождается необходимостью поддерживать устойчивое положение тела. Это сочетание описано Sherrington (1931): положение тела сопровождается движением, подобным движению тени
239
(Martin, 1977). Из-за жесткой структуры связей мышечно-скелетной системы все движения обусловливают активность, отвечающую положению тела и предназначающуюся для обеспечения устойчивости системы. Эта постуральная активность может быть локальной, связанной с возбуждением мышцы-антагониста, или более широкой, включающей возбуждение мышц туловища. Постуральная активность является автоматической, специфичной для задачи и не требует произвольного (сознательного) возбуждения мышц нервной системой для поддерживания ее устойчивости.
Устойчивость всего тела. Положение тела — нейромеханическая реакция, обеспечивающая поддержание равновесия. Система находится в механическом равновесии, если силы, действующие на систему, в сумме равны нулю (SF= 0). Эта система характеризуется устойчивостью, если после возмущения она возвращается в положение равновесия. Назначение постуральной активности заключается в поддержании устойчивости мышечно-скелетной системы. Сюда относятся поддержание положения системы относительно ее опорного основания и обеспечение требующейся ориентации сегментов тела, не участвующих в движении. Способность поддерживать вертикальное положение тела зависит от положения линии действия вектора общего веса тела относительно опорного основания. При вертикальном положении тела опорное основание определяется положением ног и включает участок под ногами и между ними. Чем шире раздвинуты ноги, тем больше опор-
Рис. 7.22. Обобщенные реакции на поворот носками вверх (а) и перемещение опорной площадки назад (б). Каждое возмущение вызывало трехфазную реакцию, которая состояла из первоначальной неустойчивой фазы (100 мс), компенсационной фазы (300 мс) и заключительной фазы коррекции (400-450 мс) (Keshner, Allum, 1990)
ное основание и больше устойчивость человека. Кроме того, устойчивость обратно пропорционально связана с высотой центра тяжести (Hayes, 1982). Человек в вертикальном положении находится в равновесии до тех пор, пока линия действия вектора веса остается в пределах границ опорного основания, и это положение устойчиво до тех пор, пока мышечно-скелетная система может воспринимать возмущения и возвращаться в равновесие.
Когда мы находимся в вертикальном положении, наше тело раскачивается назад и вперед. Мышечная активность, предохраняющая нас от потери равновесия и падения, представляет нашу автоматическую активность в отношении управления положением тела. Когда человек раскачивается назад и вперед, зрительные, соматосенсорные и вестибулярные чувствительные нервные окончания генерируют эти колебания и вызывают компенсационные реакции соответствующих мышц (Dietz, 1992). Заметим, что один из механизмов, обусловливающих увеличение количества случаев падения в преклонном возрасте, — снижение способности детектировать и контролировать раскачивание тела вперед-назад (Horak et al., 1989).
Общий метод исследования этих реакций включает наблюдение субъектов или пациентов, стоящих на площадке, которая может внезапно перемещаться в нескольких различных направлениях (Nashner, 1971, 1972). С помощью этой методики выявляют различные стратегии реагирования в зависимости от возмущения. Например, Keshner, Allum (1990) установили, что при повороте площадки передней частью вверх (рис. 7.22, а) субъекты реагировали первоначальным поворотом вперед ног и туловища и поворотом назад головы с последующим небольшим поворотом туловища назад. Эти сегментные смещения обусловливали увеличение жесткости тела с одновременным возмущением мышц на спинной поверхности при повороте носков вверх и на брюшной поверхности при повороте носков вниз. В противоположность этому при перемещении площадки назад субъекты реагировали большим поворотом ног назад и поворотом туловища вперед; т.е. два эти сегмента поворачивались в противоположных направлениях (рис. 7.22, 6). Это составило координированную многосегментную реакцию на возмущение, включающую восходящую последовательность возбуждения мышц (от дистальных до проксимальных) на брюшной поверхности тела при перемещении назад и нисходящую последовательность при перемещении вперед.
Аналогично Cordo, Nashner (1982) установили, что на корректировку положения тела влияет часть тела, обеспечивающая контактирование с окружающей средой. Когда субъект нахо-
240
Рис. 7.23. Реакция ЭМГ на возмущение положения тела. Вертикальная штриховая линия — момент движения площадки вперед. На каждом графике отражены результаты пяти опытов. Реакции в тибиалис анте-риор (д) были самыми большими для вертикального положения, в то время как реакции бицепс брахии (б) были больше, когда субъект держался за ручку (Cordo, Nashner, 1982)
дился в вертикальном положении и только ноги контактировали с окружающей средой, реакция на возмущение инициировалась в мышцах ног (рис. 7.23). Тем не менее при использовании кистей рук для обеспечения поддержки реакция на возмущение инициировалась в мышцах рук.
Возможно, сегментом, наиболее важным для устойчивости всего тела, является туловище. Повторяющиеся эргономические задачи могут обусловливать чрезмерное напряжение мышечно-скелетных элементов туловища, в частности, позвоночного столба. Для изучения этой системы туловище можно моделировать в виде двух жестких тел — грудной реберной клетки сверху и таза снизу, связанных поясничными позвонками (Bergmark, 1989). Устойчивость позвоночного столба контролируется общими мышцами, связывающими два этих жестких тела (грудную клетку и таз), и локальными мышцами и связками, прикрепленными к поясничным позвонкам. Однако общие мышцы (грудной клетки и таза) представляются наиболее важными в отношении распределения внешних нагрузок, воспринимаемых туловищем. Crisco, Panjabi показали, что позвоночные мышцы, охватывающие несколько по
звонков, и мышцы, располагающиеся наиболее латерально относительно позвоночного столба, — наиболее эффективны при его стабилизации.
Ориентация сегментов тела. В дополнение к поддержанию устойчивого положения всего тела действие, обусловливаемое его положением, связано также с обеспечением ориентации сегментов тела как в пределах конечностей, так и между ними (Hoy, Zemicke, Smith, 1985). Рассмотрим, например движение, показанное на рис. 7.24, а. Субъекта просят быстро покачать предплечьем, выполняя движение вперед-назад и удерживая в то же время плечо в горизонтальном положении. Это осуществляется в результате попеременного возбуждения флексорных и экстензорных мышц локтевого сустава. Задача требует также существенного возбуждения мышц, пересекающих плечевой сустав. Это необходимо для стабилизации плеча и минимизации инерционных эффектов движения предплечья относительно других сегментов тела. Более того, должны возбуждаться мышцы, пересекающие запястье, для управления движением кисти руки, которое могло бы изменяться вследствие неконтролируемой патологической подвижности в виде медленного колебания, с целью исключения относительного движения между запястьем и предплечьем.
Аналогично предположим, что человека, находящегося в вертикальном положении, просят возможно быстрее поднять руку в горизонтальное положение (рис. 7.24). Передняя дельтовидная мышца является основным исполнителем этой задачи. Наиболее быстро мышца может быть возбуждена (минимальное время реакции) примерно через 120 мс после подачи сигнала. Однако примерно за 50 мс до дейст-
Рис. 7.24. Распределение действия, обусловливаемого положением тела, при быстрых движениях:
а — быстрое попеременное сгибание-разгибание локтевого сустава требует обусловливаемого положением тела возбуждения мышц плеча и запястья для контроля влияния инерции, обусловливаемой движением предплечья; б— быстрому подъему руки в вытянутое горизонтальное положение предшествует действие ног, обусловленное положением тела
16 8-133
241
вия передней дельтовидной мышцы возбуждаются сухожилия, ограничивающие с боков подколенную ямку, на той же стороне тела (Belen”kii, Gurfinkel, PaF’tsev, 1967). Действие мышц ног служит, вероятно, по меньшей мере двум целям: оно обеспечивает предупредительную стабилизацию с учетом инерционных эффектов последующего движения руки и создает жесткую связь между движением конечности соответствующей реакцией земной поверхности, прилагаемой к стопам. В результате повышения жесткости сегментов тела, не участвующих в движении, предупредительное действие, обусловливаемое положением тела, очевидно, может способствовать последующему движению (Bouisset, Zattara, 1990) даже путем передачи энергии посредством межсегментной динамики.
Межконечностные реакции, обусловливаемые положением тела, заметны при использовании для поддерживания положения тела двух конечностей. Например, смещение одной ноги (возмущения) во время стойки, балансирования или ходьбы вызывает обусловливаемую положением тела двустороннюю реакцию с подобным скрытым состоянием в обеих ногах (Dietz, 1992). Двусторонняя реакция, вероятно, создает более стабильную базу компенсации возмущения. Аналогично, если воздействовать на кисть руки, сжимающую что-то в окружающей среде для поддержки, это может вызывать автоматическую реакцию, связанную с положением
Рис. 7.25. Реакции субъекта, обусловливаемые положением тела, при перемещении опорного основания (площадки) назад (а) и повороте его передней частью вверх (б). Реакцию на удлинение мышц икр голени измеряли в виде объединенной ЭМГ, влияние которой показано относительно наклона субъекта (Nashner, 1976). Для кривых как ЭМГ, так и наклона тела опыт 1 представляет верхняя линия, а опыт 4 — нижняя линия
тела (например, усиление сжатия кисти руки), в контралатеральной руке и ее кисти (Marsden, Merton, Morton, 1983). Предполагается, что такая межконечностная связь рук — один из механизмов, лежащих в основе случайных выстрелов личного огнестрельного оружия.
Приспособляемость реакций, обусловливаемых положением тела. Автоматические реакции, обусловливаемые положением тела, являются контекстно зависимыми: реакция предназначается для удовлетворения потребностей взаимодействия системы и окружающей среды. Управление положением тела не просто основывается на наборе рефлекторных реакций и не является предварительно запрограммированной реакцией, которая запускается возмущением. Скорее управление положением тела является адаптационной особенностью двигательной системы, которая основывается на интеграции афферентного входного сигнала и эфферентного выходного сигнала.
Эта приспособляемость очевидна в модуляции реакций, обусловливаемых положением тела и связанных с методикой поворота носков вверх и перемещениях назад, показанной на рис. 7.22. В эксперименте площадку в процессе выполнения четырех опытов перемещали назад (рис. 7.25, а) а затем поворачивали передней частью вверх (рис. 7.25, б). Общим эффектом обоих возмущений было удлинение, которому подвергались мышцы икр субъекта, стоящего на площадке. Хотя реакция, вызываемая удлинением, и стабилизирует положение тела субъекта, нарушенное перемещением, в результате уменьшения наклона тела вперед, она дестабилизирует положение тела субъекта после поворота носков вверх вследствие увеличения наклона тела. Следовательно, субъект учится содействовать реакции в течение четырех опытов с перемещением и приспосабливать (ослаблять) реакцию в течение четырех опытов с поворотом.
Особенностью автоматических реакций, обусловливаемых положением тела, является проявление кратковременного скрытого действия мышц, которые не удлиняются под действием возмущения. Об этом распределении автоматических реакций сообщалось для мышц ног после смещения опорной поверхности (Nashner, Woollacott, Tuma, 1979), мышц плечевых и локтевых суставов после возмущений руки (Lacquaniti, Soechting, 1986), мышц запястья после возмущения локтевого сустава (Koshland, Hasan, Gerilovsky, 1991) и билатерально после одностороннего возмущения (Dietz, Berger, 1982). Например, Koshland et al. установили, что возмущение локтевого сустава последовательно вызывало реакции в мышцах запястья, которые укорачивались в результате возмущения (рис. 7.26, а). Эта схема возбуждения, предложенная Gielen, Ramaekers, van
242
Рис. 7.26. Углы суставов и электромиограммы мышц запястья во время возмущения локтевого сустава (а) и волевого движения сгибания локтя (б). Для возмущения локтевой сустав разгибали с помощью двигателя, который создавал реакцию сгибания в запястьи. Тем не менее волевое сгибание локтя создавало небольшое первоначальное разгибание запястья с последующим его сгибанием. Однако в обоих случаях не были активными ни флексорные мышцы пальцев, ни флексорные мышцы запястья (Koshland, Hasan, Gerilovsky, 1991)
Zuylen (1988), была подобна волевому движению сгибания локтя, хотя механические воздействия на лучезапястный сустав в этих двух случаях и не были подобными (рис. 7.26, б). Приведенные наблюдения наводят на мысль, что эти автоматические реакции, обусловливаемые положением тела, не вызываются только рефлексами локального растяжения мышцы, а представляют координируемую и задаваемую двигательную схему.
Основные стратегии движения
Мы определили двигательную программу в качестве стереотипной последовательности команд, подаваемых из спинного мозга в мышцы для обеспечения конкретного поведения. Определение некоторых основных единиц, исполь
зуемых в различных стратегиях движения, предоставляют информацию о командах, шифруемых в двигательных программах.
Движение, направленное к цели. Когда человек осуществляет направленное к цели движение, например, достижение объекта, основные мышцы-агонисты и -антагонисты, управляющие движением, часто характеризуются трехимпульсной конфигурацией электромиограммы. Эта последовательность интенсивности электромиограммы очевидна в движениях со скоростью в пределах от средней до высокой. Она содержит исходный импульс электромиограммы агониста, за которым следуют импульс электромиограммы антагониста и второй импульс электромиограммы агониста (Wachholder, Altenburger, 1926). Три этих импульса обычно перекрываются, но пики располагаются в порядке агонист-антагонист-агонист (рис. 7.27).
16*
243
Рис. 7.27. Трехимпулъсная кривая интенсивности ЭМГ для движения сгибания локтя (0,5 рад) (данные электромиограммы выпрямлены и отфильтрованы) (Karst, Hasan, 1987):
а — несовпадающая последовательность; б — движение, отображающее совместное возбуждение при завершении движения;
1— антагонист, 2 — агонист
Эта последовательность была названа набором АВС, где А — импульс действия; В — импульс торможения; С — импульс фиксации (Hannaford, Stark, 1985). Импульс действия разгоняет конечность в направлении позиции цели, импульс торможения замедляет конечность по мере приближения ее к цели, а импульс фиксации фиксирует конечность в позиции цели.
Хотя наличие трехимпульсной конфигурации электромиограммы для однонаправленных движений и было известно в течение нескольких лет, не была сформулирована унифицирующая гипотеза для учета множества экспериментальных наблюдений. Несомненно, трехим-пульсная конфигурация относится к прекращению движения в позиции цели, но стратегия управления, принимаемая ЦНС, все же не была определена. Например, многие исследователи пытались определить, какие части этой последовательности предварительно запрограммированы (при управлении по разомкнутому контуру) и какие могут быть изменены посредством обратной связи (управление по замкнутому контуру). Однако два основных ограничения препятствовали полному пониманию этой последовательности электромиограммы. Во-первых, на трехимпульсную конфигурацию коренным образом влияют особенности движения, включая указания, даваемые субъекту (Gottlieb, Corcos, Agarwal, Latash, 1990a), и затруднительно систематически контролировать все переменные, влияющие на трехимпульсную конфигурацию (Karst, Hasan 1987). Во-вторых, недостаточное внимание было уделено механическим условиям, например, ориентации субъекта (плоскость, в которой осуществляется движение) и тому факту, что агонист (антагонист) для конкретного движения часто включает более одной мышцы.
Тем не менее, исходя как из экспериментальных измерений, так и моделирований на вычислительных машинах, Gottlieb et al. предложили гипотезу двойной стратегии для учета
характеристики направленных к цели движений сгибания локтя (Corcos, Gottlieb, Agarwal, 1989; Gottlieb, Corcos, Agarwal, 1989). Эта гипотеза может объяснить вариации в электромиограмме (агонист и антагонист) и результирующем вращающем моменте мышц, которые сопровождают движения, включающие различные смещения, скорости и нагрузки. Гипотеза основывается на модели возбуждения двигательного нейрона в виде прямоугольного импульса, который можно модулировать в результате варьирования его амплитуды и длительности. Модуляция амплитуды возбуждения представляет зависимую от скорости стратегию, которая включает увеличение амплитуды электромиограммы и вращающего момента и уменьшение скрытого состояния до появления электромиограммы антагониста (рис. 7.28, а). В противоположность этому модуляция длительности возбуждения представляет не зависимую от скорости стратегию, которая включает подобные исходные наклоны для электромиограммы агониста и результирующего вращающего момента мышц, увеличение продолжительности электромиограммы и вращающего момента и увеличение скрытого состояния до появления электромиограммы антагониста (рис. 7.28, б). Когда субъектам необходимо быстро выполнить задачу в пределах определенных временных ограничений, они выбирают зависимую от скорости стратегию. Однако если субъекты свободны в выборе скорости движения, они выбирают не зависимую от скорости стратегию, которая включает выбор интенсивности (амплитуды) возбуждения и последующее согласование ее с длительностью, соответствующей желаемому движению (Gottlieb et al., 1990а).
Однако движения контролируются не только преддвигательными командами ЦНС, поступающими в двигательные нейроны, но и посредством афферентной обратной связи, которая осуществляется во время движения. Поэтому Feldman (1986) предположил, что управляемым
244
параметром таких направленных к цели движений является порог (1) тонизирующего рефлекса растяжения мышцы (Latash, 1993). В этой модели, названной лямбда-моделью, управление движениями осуществляется в результате варьирования порога тонизирующего рефлекса растяжения мышцы. Latash и Gottlieb (1991а) показали возможность моделирования кривых элек-тромиограмм односуставных движений, связанных с зависящей от скорости и не зависящей от скорости стратегий, с использованием Х-модели. В этой схеме зависящая от скорости стратегия эквивалентна варьированию X при постоянной скорости для различной длительности, а не зависящая от скорости стратегия эквивалентна варьированию X при различных скоростях. Latash и Gottlieb (1991а) пришли к выводу, что некоторые характеристики кривых электромиограмм (например, появление электромиограммы антагониста) определяются исключительно центральными командами, а другие характеристики (амплитуда и длительность электромио!раммы) равным образом зависят от афферентных сигналов, генерируемых во время движения.
Схемы возбуждения семи-девяти мышц руки, участвующих в движениях (вращения плеча и локтя) в сагиттальной плоскости, навели на мысль о простых соотношениях между элек-тромиограммой, а также направлением и скоростью движения (Flanders, 1991; Flanders, Herrmann, 1992). Связанные с направлением кинематические требования к движениям удов
летворялись кривыми электромиограмм, масштабированными по амплитуде и задержанными во времени, в зависимости от направления движения. Более того, при осуществлении этих движений при различных скоростях в результате варьирования времени движения кривые электромиограмм характеризовались двумя аддитивными составляющими: тонизирующей, которая противодействовала влиянию силы тяжести, и фазовой, которая была связана с изменением скорости движения. Эти наблюдения наводят на мысль об относительно простой стратегии управления механически сложными движениями руки.
Несмотря на обоснованность данных гипотез, остается некоторая неопределенность в отношении применимости этих правил к многосуставным движениям. Движения, включающие смещения относительно более одного сустава, увеличивают возможность значительных эффектов, обусловливаемых межсегментной динамикой. Такие результаты наводят на мысль о том, что управление многосуставными движениями не может представлять собой просто суммирование односуставных стратегий (Hasan, 1991).
Совместное возбуждение. Одним из исследований, проведенных Karst и Hasan (1987), посвященных трехимпульсным конфигурациям электромиограммы, была изменчивость степени совместного возбуждения в конце движения (см. рис. 7.27). Совместное возбуждение — это одновременное действие мышц, состоящих из набора агонистов и антагонистов. Об этом процес-
Рис. 7.28. ЭМГ моделированных агонистов и антагонистов и результирующий вращающий момент мышц для быстрого движения сгибания локтя в течение четырех опытов. Для зависящей от скорости стратегии (а) амплитуда возбуждения увеличивалась в течение четырех опытов, в то время как для не зависящей от скорости стратегии (6) возрастала длительность возбуждения (Gottlieb, Corcos, Agarwal, Latash, 1990).
245
Рис. 7.29. Зависимость жесткости сустава от результирующего действия мышц (Feldman, 1986);
а — семейство соотношений угла и вращающего момента для флексоров локтевого сустава (положительные вращающие моменты) и экстензоров локтевого сустава (отрицательные вращающие моменты); штриховая линия — пассивное соотношение угла и вращающего момента; б — выбор соответствующего соотношения угла и вращающего момента для флексоров и экстензоров локтевого сустава приводит к результирующему соотношению (штриховая линия), которое характеризуется большим наклоном и, следовательно, большей жесткостью
се имеется удивительно мало информации. В частности, в значительной степени необъясне-ны условия, в которых мы используем совместное возбуждение. Совместное возбуждение оказывает механическое влияние, обусловливая увеличение жесткости сустава и затрудняя его возмущение (Baratta et al., 1988; Komecki, 1992). Жесткость проявляется в виде наклона кривой соотношения усилия и длины (или вращающего момента и угла). На рис. 7.29 показано, почему совместное возбуждение может повышать жесткость сустава. Каждая группа мышц (флексоры и экстензоры локтевого сустава) способна создавать семейство соотношений угла и вращающего момента в зависимости от уровня возбуждения (рис. 7.29, а). Из этих семейств соотношений для каждой группы мышц можно выбрать одну пару таким образом, чтобы результирующее соотношение угла и вращающего момента имело больший наклон и, следовательно, большую жесткость.
Благодаря увеличению жесткости и, следовательно, устойчивости сустава, совместное возбуждение можно было бы рассматривать как полезную особенность, когда мы усваиваем новые задачи или осуществляем движение, требующее высокой степени точности. Например, Person (1958) контролировал электромиограмму бицепс и трицепс брахии в течение двух недель обучения субъектов обработке напильником и рубке зубилом. Во время курса обучения
интенсивность электромиограммы изменялась от высокого уровня совместного возбуждения до возбуждения попеременного действия при небольшом совместном возбуждении.
Тем не менее профессиональная характеристика не отличается отсутствием совместного возбуждения. Например, Enoka (1983) установил, что профессиональные штангисты, берущие вес на грудь (см. рис. 1.7), использовали в процессе выполнения движения совместное возбуждение вастус латералис и бицепс фемо-рис (рис. 7.30). Существует, по меньшей мере, три причины использования людьми совместного возбуждения для таких задач. Во-первых, для движений, включающих изменения направления (например, смена разгибания сгибанием), представляется более экономичным модулировать уровень тонической активности мышц-агонистов и -антагонистов по сравнению с попеременным включением и выключением их. Hasan (1986) утверждает, что в определенных условиях совместное возбуждение действительно снижает напряжение, имеющее место при осуществлении движения, и некоторые эмпирические наблюдения подтверждают это предположение (Engelhom, 1983). Во-вторых, благодаря увеличению жесткости и, следовательно, устойчивости сустава, совместное возбуждение может быть желаемой стратегией, когда люди перемещают тяжелые грузы или грузы, которые могут быть возмущены. В-третьих,
246
одна из возможностей двусуставных мышц заключается в передаче энергии от одного сустава другому. Как описывалось выше, совместное возбуждение односуставного экстензора тазобедренного сустава (например, глютеус максимус) и двусуставного флексора тазобедренного сустава (например, ректус феморис) обусловливает результирующий эффект увеличения вращающего момента экстензоров коленного сустава. Следовательно, совместное возбуждение в тазобедренном суставе может увеличивать вращающий момент в коленном суставе. Эта стратегия может оказаться полезной для условий, требующих максимального усилия, или в дополнение при обусловливаемом усталостью снижении мышечной силы (Dimitrijevic et al., 1992).
Представляется, что невральная стратегия, связанная с совместным возбуждением, несколько отличается от стратегии, когда возбуждается только группа мышц-агонистов. При неожиданном изменении нагрузки, прилагаемой к флексорам или экстензорам локтевого сустава, рефлекторная реакция (разряд двигательной единицы) зависит от того, поддерживалась ли нагрузка перед возмущением в результате совместного возбуждения или нет, и от того, подана команда субъекту на освобождение от нагрузки или на противодействие ей (Gydikov, Kossel, Radicheva, Tankow, 1981). В ЭМГ рефлекса имел место максимум, когда совместное возбуждение отсутствовало и субъекту была подана команда освободиться от нагрузки. В противоположность этому при наличии совместного возбуждения имели место два максимума. Это различие было обусловлено, вероятно, ролью совместного возбуждения в повышении устойчивости сустава.
Цикл удлинения—сокращения. Общей схемой возбуждения мышц, в частности, во время выполнения задач, требующих высокой напряженности, является использование эксцентрическо-концентрической последовательности, в которой действующая мышца сначала удлиняется, а затем сокращается. Преимущество этой стратегии заключается в том, что мышца может выполнить большую положительную работу, если она активно растягивается, прежде чем сократится (Cavagna, Citterio, 1974; Fenn, 1924). В результате этого цикла удлинения-сокращения во время концетрического сокращения выполняется большее количество работы по сравнению с тем, если бы мышца сама по себе выполняла концентрическое сокращение. Экспериментальное доказательство этого вывода основывается на работе, выполняемой отдельной мышцей (рис. 7.31). Эксперимент состоял из двух частей: сначала мышца растягивалась, а затем стимулировалась, прежде чем сократиться и выполнить положительную работу (рис. 7.31, а); после этого мышца сначала стимулировалась, а затем растягивалась, прежде чем выполнить положительную работу (рис. 7.31, б). Результаты каждой части эксперимента показаны в виде графиков длина—время, усилие—время и усилие—длина. Критическое сравнение выполняется на графиках усилие-длина. Фаза с показывает изменение усилия и длины при выполнении мышцей работы. Поскольку работа определяется как произведение усилия и смещения, площадь ниже кривой усилие-длина во время фазы с представляет работу, выполняемую в течение каждой части эксперимента. Безусловно, площадь ниже этой кривой больше для второй части эксперимента, которая состояла из растягивания (удлинения)
Рис. 7.30. Результирующий вращающий момент мышц (штриховая линия) и кривые ЭМГ (сплошная линия) для коленного сустава при взятии веса на грудь при его подъеме кривые ЭМГ для бицепс феморис (BF) и вастус латералис (VL) были выпрямлены, профильтрованы и выражены в процентах соответствующих максимальных значений (Enoka, 1983):
а — профили для одного опыта; б — средние профили по данным шести опытов для одного субъекта; в — средний профиль по данным 15 опытов для троих субъектов
247
Рис. 7.31. Положительная работы, выполняемая отдельной мышцей во время последовательностей изометрическо-концентрического (а) и эксцентрическо-концентрического (6) возбуждения (Cavagna, Citterio, 1974)
действующей мышцы; это соответствует циклу удлинения—сокращения.
Соотношение между работой и энергией (см. главу 3) свидетельствует, что увеличение работы, выполняемой мышцей, требует повышения расхода энергии. Откуда могла бы поступить эта дополнительная энергия? Типичное двухэлементное логическое обоснование (например, Cavagna, 1977) таково. Во-первых, эксцентрическое сокращение нагружает последовательный упругий элемент в результате его растяжения (трехэлементная модель мышцы в главе 6), что может быть представлено в качестве передачи энергии от нагрузки последовательному упругому элементу; это представляет собой накопление упругой энергии. Например, если удерживать упругую ленту одним концом в каждой руке и затем растянуть, действие мышц кистей рук, участвующих в растягивании ленты, накапливается в ленте в виде упругой энергии. Во-вторых, после освобождения молекулярная структура упругой ленты использует эту упругую энергию для возвращения к своей первоначальной форме. Аналогичным образом, поскольку отношение мышечной силы к усилию нагрузки изменяется и мышца подвергается концентрическому сокращению, упругая энергия, накопленная в последовательном упругом элементе, может быть восстановлена и использована для содействия укорачивающему сокращению (положительная работа).
Это явление с применением первого закона термодинамики (см. главу 3) может быть выражено таким образом:
U — ДЕЬ + ДЕт + ДЕр8,
где U — работа; ДЕЬ — изменение тепловой энергии; ДЕт — изменение химической энергии; ДЕР s — изменение упругой энергии. Результатом эксцентрическо-концентрической последовательности, представленной на рис. 7.31, было увеличение работы (U), выполненной во время концентрического действия. При возбуждении мышцы в результате множества процессов обмена веществ в качестве существенного элемента химической энергии (Ет) образуется АТФ. В процессе как образования так и использования аденозинтрифосфата некоторая часть энергии расходуется в виде тепла (£/,). Согласно приведенному выше уравнению, если используемая химическая энергия и выделяющееся тепло остаются постоянными (допустим, для данного случая Eps = 0), количество выполненной работы останется таким же. Тем не менее в соответствии с рис. 7.31 выполняемая работа увеличивается при осуществлении эксцентрическо-концентрической последовательности (удлинение—сокращение). Объясняется это тем, что изменяется либо Ет, либо £/,. Однако, согласно объяснению, основывающемуся на явлении накопления и использования упругой энергии, для осуществления работы предоставляется дополнительная энергия (Ерх) наряду с той, которая предоставляется химическими средствами. В соответствии с этой точкой зрения, Ет и ЕЛ могут мало изменяться между изометрическо-концентрическим и экс-центрическо-концентрическим режимами, но большая часть дополнительной работы может быть выполнена благодаря вкладу упругой энергии (Eps).
248
На эту способность использовать накапливаемую упругую энергию оказывают влияние три переменные: время, величина удлинения и скорость удлинения. Cavagna (1977) показал, что между эксцентрическим и концентрическим сокращениями не должно быть какой-либо временной задержки; в противном случае некоторая часть накопленной упругой энергии теряется (рассеивается). Вероятно, потеря энергии обусловливается разъединением и восстановлением во время этой задержки поперечных межклеточных мостиков, вследствие чего после восстановления миофиламенты испытывают меньшее натяжение. Аналогично если удлиняющее сокращение слишком большое, после удлинения сохраняется меньше поперечных межклеточных мостиков и, следовательно, накапливается меньше упругой энергии (Fdman, Elzinda, Noble, 1978). Однако при условии, что поперечные межклеточные мостики сохраняются, чем выше скорость удлинения, тем больше накапливается упругой энергии (например, Rack, Westbuiy, 1974).
Несмотря на широко распространенное использование явления накопления и использования упругой энергии для учета увеличения положительной работы, связанной с эксцентрическо-концентрическими сокращениями, увеличение положительной работы обусловливается, вероятно, также значительным возрастанием количества предоставляемой химической энергии (Ет) (Jaric, Gavrilovic, Ivancevic, 1985). Это увеличение предоставляемой химической энергии называется преднагрузочным эффектом. Заметим, например, что в начале фазы с на графике усилие—длина, приведенном на рис. 7.31, усилие больше во время эксцентрическо-концентрического состояния, чем во время изометрическо-концентрического состояния; это соответствует крайне правому пику на графиках усилие-длина. Безусловно, в начале концентрической фазы усилие в эксцентрическо-концентрическом состоянии больше.
Относительные вклады упругой энергии и преднагрузочных эффектов можно оценить в результате рассмотрения высоты, которую субъекты могут преодолеть, используя два типа прыжков в высоту (Komi, Bosco, 1978). Прыжок согнув ноги начинается из положения на корточках (угол коленного сустава около 2 рад) и просто включает разгибание коленных и голеностопных суставов; руки удерживаются вытянутыми над головой для минимизации их вклада в прыжок. Прыжок движением в противоположном направлении начинается из вертикального положения тела и включает в течение одного непрерывного движения приседание до угла коленного сустава около 2 рад и последующее разгибание коленных и голеностопных суставов, как при выполнении прыжка согнув ноги. Основное различие между дву
мя этими способами заключается в методике использования сильных экстензоров коленных суставов, выполняющих около 50 % работы во время максимального црыжка в высоту (Hublay, Wells, 1983); а именно, прыжок согнув ноги включает только изометрическо-концентрическое сокращение экстензоров коленных суставов, в то время как прыжок движением в противоположном направлении требует эксцентрическо-концентрической последовательности.
Выполнение этих прыжков на одной или двух ногах также изменяет величину начального концентрического вращающего момента экстензоров коленного сустава. Прыжки на одной ноге создают преднагрузочный эффект, сравнимый с таковым при прыжках на двух ногах. На рис. 7.32 показано соотношение вращающего момента и угла для прыжка согнув ноги, выполняемого на одной и двух ногах. Вращающий момент представляет собой результирующий вращающий момент относительно коленного сустава для одной ноги. Во время фазы отталкивания угол коленного сустава изменяется от 2 рад до полного разгибания (3,14 рад). Основной результат, представленный на рис. 7.32, заключается в том, что начальный вращающий момент (при угле 2 рад) во время прыжка на одной ноге примерно в два раза больше, чем при прыжке на двух ногах. Во время прыжка на одной ноге эта нога должны поддерживать вес всего тела, в то время как во время прыжка на двух ногах одна нога должна поддерживать только половину этого веса. Из-за этого различия начальная нагрузка, выдерживаемая одной ногой, значительно больше при выполнении прыжка на одной ноге, и, следовательно, мы говорим, что конечность была предварительно нагружена. Кроме того, больший начальный вращающий момент, создаваемый экстензорами коленного сустава, требует больше химической энергии (£т).
Сравнивая результаты, достигаемые при выполнении прыжков в высоту (табл. 7.4), можем исследовать вклад в работоспособность
Рис. 7.32. Идеализированное соотношение между результирующим вращающим моментом мышц и углом дня одного коленного сустава во время прыжков согнув ноги на одной ноге (1) ина двух ногах (2)
249
Таблица 7.4. Высоты, достигаемые при прыжках согнув ноги и движением в противоположном направлении на одной ноге и двух ногах
Исполнение Вид прыжка
согнув ноги движением в противоположно м направлении
На одной ноге 22,1±5,9* 24,0±6,6
На двух ногах 32,4±9,1 36,4+8,5
* Эти значения являются средними величинами ± среднее квадратическое отклонение в сантиметрах для 44 субъектов.
упругой энергии (накопление и использование) и химической энергии (преднагрузочный эффект). Какая энергия (упругая или химическая) расходуется больше для достижения определенной высоты при прыжках движением в противоположном направлении? Сравнение прыжков на одной и двух ногах показывает преднагрузочный эффект (химическая энергия), в то время как различия между прыжками согнув ноги и движением в противоположном направлении включают оба эффекта (химическую и упругую энергию). Различия в высоте прыжков с четырьмя комбинациями показывают три особенности использования мышц при выполнении задачи этого типа. Во-первых, при прыжках как на одной, так и на двух ногах субъекты достигали большей высоты в прыжке движением в противоположном направлении по сравнению с прыжком согнув ноги; это наблюдение подтверждает превосходство эксцентрическо-концентрического сокращения по сравнению с изометрическо-концентрическим. Во-вторых, высоты, достигаемые при прыжках на двух ногах, были больше по сравнению с таковыми при прыжках на одной ноге из-за массы мышц (и, следовательно, химической энергии), большей при выполнении прыжков на двух ногах. В-тре-тьих, высота, достигаемая в условиях прыжков на двух ногах, не превышала в два раза высоту прыжков на одной ноге, несмотря на то, что масса мышц была в два раза больше. Иными словами, высоты, достигаемые в прыжке согнув ноги на двух ногах, составляли в среднем 147 % высот, достигаемых в прыжке согнув ноги на одной ноге. Если бы различие между двумя этими прыжками заключалось просто в количестве массы мышц, мы рассчитывали бы на высоту, достигаемую в прыжке на двух ногах, на уровне 200 % высоты, достигаемой в прыжке на одной ноге. Тем не менее поскольку высота, достигаемая в прыжке на одной ноге, превышала 50 % высоты, достигаемой в прыжке на двух ногах, повышенная работоспособность одной ноги в прыжке на одной ноге обусловливается большим количеством химической энергии (преднагрузочный эффект). Такие анализы наводят на мысль о том, что накопление и ис
пользование упругой энергии имеют меньшее значение, чем преднагрузочный эффект (увеличенная химическая энергия), при выполнении таких задач, как прыжок в высоту, но могут иметь большее значение в таких действия, как танцы (Fukashiro, Komi, 1987; van Ingen Schenau, 1984).
Многие движения человека включали использование эффекта удлинения—сокращения. Например, нормальным режимом осуществления прыжка в высоту (в баскетболе, волейболе) является сталь движения в противоположном направлении, который, как обсуждалось выше, максимизирует высоту, которая может быть достигнута. Цикл удлинения—сокращения используется также в ударах ногами и беге. В процессе бега нога сначала соприкасается с землей, и коленный сустав сгибается и разгибается, когда ЦТ всего тела проецируется на ногу (см. рис. 1.14 и 1.15). Эта схема сгибания-разгибания коленного сустава осуществляется посредством эксцентрически-концентрической последовательности действия экстензоров коленного сустава. Тем не менее этот режим возбуждения не всегда увеличивает выполняемую положительную работу; Cavagna (1977) установил, что упругая энергия, восстанавливаемая в процессе бега, обеспечивает значительные вклады в энергию, генерируемую мышцами, только при скоростях бега, превышающих примерно 6,5 м-с-1. Подобные данные получены для отталкивания при выполнении прыжков в длину и тройных прыжков (Luthanen, Komi, 1980).
Наш заключительный пример цикла удлинения-сокращения касается экспериментальной ситуации, когда штангу подвешивали к потолку таким образом, чтобы она находилась в горизонтальном положении и располагалась на высоте плеч. Субъекту предлагали стать под штангой, чтобы она касалась плеч, но без поддерживания веса штанги, и схватить концы штанги каждой рукой за конец. Находясь в этом положении, субъект получает указание повернуть штангу в горизонтальной плоскости возможно быстрее приблизительно на 1,57 рад. Каким образом он мог бы выполнить эту задачу? Crieve (1969) осуществил данный эксперимент и установил, что субъекты сначала поворачивали в желаемом направлении бедра, в то время как плечи и штанга немного поворачивались в противоположном направлении. Это действие обусловливало активное растягивание (эксцентрическое сокращение) мышц туловища, которое затем используется эксцентрически для поворота плеч и штанги. Пиковый вращающий момент, который создавался во время этого движения, приходился на эксцентрическую фазу. Результаты наблюдения подтверждают представление о работе бедрами при игре в гольф; вращающий момент, прилагаемый к клюшке для игры в гольф, будет больше, если
250
действие мышц представляет собой эксцентрическо-концентрическую последовательность, а не именно концентрическое сокращение.
Манипуляция. В нашем обсуждении стратегий движения мы сосредоточивались на таких действиях, как ходьба, бег, бросание, удары ногами, достижение и стабилизация. Тем не менее в спектре возможностей движения качественно другой тип движения включает обследование окружающей нас среды. Это отличие заключается в других функциях, выполняемых кистями рук, по сравнению с функциями рук и ног. Кисть руки и головной мозг являются близкими партнерами в способности человека обследовать физический мир и изменять его (Lemon, 1993). Как обследование, так и изменение окружающей среды зависят от точных описаний механических событий, когда объекты приводятся в тесный контакт с кистью руки. Значительная часть этой информации предоставляется механорецепторными афферентными единицами, которые возбуждают лишенную волос кожу кисти руки. Эти чувствительные нервные окончания участвуют в поведении, называемом активной пальпацией (Johansson, Cole, 1992; Phillips, 1986).
Роль кожных механорецепторов в управлении двигательным выходным сигналом изучалась в процессе исследования реакций зажимного усилия на неожиданные изменения нагрузки (Johansson, Hager, Backstrom, 1992; Johansson, Riso, Hager, Backstrom, 1992). Если объект зажат между указательным пальцем и большим пальцем щипковым захватом, неожиданное изменение тянущей нагрузки вызывает автоматическую коррекцию зажимного усилия после кратковременной задержки. Скрытое состояние реакции зажимного усилия сокращается (от 174 до 80 мс) с повышением частоты изменения тянущей нагрузки. Эта автоматическая реакция состоит из двух частей. Начальная фаза включает колоколообразную скорость изменения усилия; за ней следует вторая фаза, которая включает медленное повышение скорости изменения усилия (рис. 7.33). Начальная фаза является стандартным элементом, который характеризуется постоянной длительностью, но амплитуда которого изменяется со скоростью изменения тянущей нагрузки. Вторая фаза охватывает более продолжительные увеличения зажимного усилия и завершается при наркотизации указательного и большого пальцев. Это наводит на мысль о том, что вторая фаза зависит от афферентной обратной связи от механорецепторов в кисти руки.
Схемы возбуждения двигательных единиц. На основании исследований, которые начались в 30-е годы и продолжаются в настоящее время, установлено общее правило, касающееся последовательности возбуждения двигательных еди-
Рис. 7.33. Автоматическая реакция зажимного усилия на неожиданное изменение усилие нагрузки. Усилие нагрузки прилагалось двигателем, и субъект должен был препятствовать движению (или изменению положения). Скорость изменения зажимного усилия (верхняя кривая) характеризуется двумя фазами — начальной колоколообразной реакцией и последующим более медленным изменением (Johansson, Riso, Hager, Backstrom, 1992)
ниц: двигательные единицы используются в упорядоченной последовательности в направлении от наименьших до наибольших. Однако существуют три ограничения этого правила.
Во-первых, исследования по использованию двигательных единиц обычно ограничивались усилиями менее 30 % максимума. Вследствие этого имеет место некоторая неопределенность в отношении того, происходит ли упорядоченное использование или даже необходимо ли оно при больших усилиях (Stuart, Enoka, 1983).
Во-вторых, усилие, при котором используется двигательная единица (порог использования), отличается для изометрических условий и условий движения (концентричность и эксцентричность), и направление этого различия может изменяться в группе мышц-синергистов (Tax, Denier van der Gon, Erkelens, 1990). Эти данные наводят на мысль о том, что намерение субъекта двигаться или не двигаться может влиять на порог использования двигательной единицы.
В-третьих, порядок использования не является неизменным свойством пула двигательных единиц, имели место сообщения об изменении этого порядка. Граничные условия, которые могут изменять порядок использования, включают эксцентрические сокращения, усталость и кожную афферентную обратную связь.
Рассмотрим граничные условия, которые могут изменять порядок использования двигательных единиц. Как мы обсуждали в главе 6, усилие, создаваемое мышцей, больше во время эксцентрического сокращения, чем во время изометрического или концентрического. Несом-
251
Рис. 7.34. Действие двигательных единиц в мышце lateral gastrocnemius (LG) во время концентрического и эксцентрического сокращений. При увеличении угла голеностопного сустава (движение вверх) мышцы осуществляют концентрическое сокращение (Nardone, Romano, Schieppati, 1989):
1 — внутримышечная ЭМГ; 2 — позиция голеностопного сустава; 3 — ЭМГ soleus; 4 — ЭМГ LG
ненно, механическая характеристика мышцы различна для концентрических и эксцентрических сокращений (Morgan, 1990). Например, работоспособность мышц во время утомляющих сокращений изменяется по-разному для эксцентрического и концентрического сокращений (Enoka, Stuart, 1992). В дополнение к этим механическим различиям порядок использования
Рис. 7.35. Действие двигательных единиц в первой тыльной межкостной мышце во время изометрического сокращения, которое осуществлялось перед утомляющим сокращением и после него. Усилие, которое создается во время выполнения задачи, представляет средняя кривая. “Предусталостная” кривая показывает действие типичных двигательных единиц во время выполнения задачи. “Послеусталостные” кривые показывают три воздействия утомляющего сокращения на действие двигательных единиц: двигательная единица не была использована во время выполнения той же задачи (7); были использованы дополнительные двигательные единицы (2); разряд двигательной единицы был в большей степени переменным (3), Кривая 1 показывает, что задача была выполнена в результате возбуждения другой группы двигательных единиц (Enoka et al., 1989):
а — “предусталостная” кривая; б — усилие; в — “послеусталостные” кривые
также может отличаться во время эксцентрических сокращений, когда большие (высокопороговые) двигательные единицы могут быть использованы раньше меньших двигательных единиц (Nardone, Romano, Schieppati, 1989). На рис. 7.34 показано действие двигательных единиц в мышце lateral gastrocnemius человека во время концентрического и эксцентрического сокращений. В первом опыте (рис. 7.34, а) внутримышечный электрод не детектировал действие локальных двигательных единиц во время медленного плантарносгибающего движения (концентрическое сокращение), но двигательные единицы использовались во время эксцентрического сокращения (отрицательный наклон позиционной кривой). Во втором опыте (рис. 7.34, б) плантарносгибающее движение осуществлялось быстрее и та же двигательная единица была использована во время концентрического сокращения.
Напомним, что прежде мы различали концентрические и эксцентрические сокращения на основании отношения вращающих моментов мышцы и нагрузки. Это отношение превышало единицу для концентрических сокращений, но было меньше единицы для эксцентрических; иными словами, различие между ними было просто механическим. Однако если порядок использования для концентрических и эксцентрических сокращений отличается, то между ними имеет место также невральное различие. Вероятно, существует, по меньшей мере, две схемы использования двигательных единиц — одна для концентрических, другая для эксцентрических сокращений.
Вторым граничным условием, которое может изменять порядок использования, является утомление. Как мы обсудим в главе 8, утомление — общий термин, используемый для описания класса кратковременных эффектов, ухудшающих двигательную характеристику. Многие физиологические процессы, включая возбуждение двигательных единиц, характеризуются адаптивными способностями при осуществле
252
нии человеком утомляющего сокращения. Связанные с утомлением адаптации, касающиеся действия двигательных единиц, включают снижение скорости разряда (Bigland-Ritchie, Johansson, Lippold, et al., 1983; Dietz, 1978; Seyffarth, 1940), прекращение действия (Person, 1974) и гибкость в двигательных единицах, способствующих выполнению задачи (Enoka, Robinson, Kossev, 1989). Хотя влияние утомления на порядок использования и не было испытано непосредственно, Enoka et al. (1989) установили, что при выполнении одной и той же задачи перед утомляющим сокращением и после него двигательные единицы, участвующие в выполнении задачи, могут меняться (рис. 7.35).
Третьим граничным условием, по-видимо-му, изменяющим порядок использования двигательных единиц, являются изменения в кожной обратной связи. Garnett, Stephens (1981) установили, что усилие мышцы, при котором используется двигательная единица (порог использования), может изменяться в результате управления обратной связью от кожных афферентов (рис. 7.36). Более того, эффект определяется низким или высоким порогом использования двигательной единицы. Для низкопороговых двигательных единиц порог использования увеличивается при соответствующем кожном стимулировании (рис. 7.36, а). Для высокопороговых двигательных единиц порог использования уменьшается (рис. 7.36, б). Преимущество стратегии, посредством которой можно уменьшить порог использования высокопороговых двигательных единиц, заключается в том, что эти единицы могут быть возбуждены раньше, если условия предписывают необходимость в быстрой интенсивной реакции.
РЕЗЮМЕ
В данной главе модель односуставной системы расширяется для рассмотрения роли су-прасегментных центров, обсуждения многосуставных аспектов мышечно-скелетной структуры и идентификации стратегий движения, используемых нервной системой в управлении движением. В главе осуществлен обзор вклада супрасегментных центров, включающих сенсорно-двигательную зону коры головного мозга, ствол мозга, мозжечок, базальные ядра и вос-
Рис. 7.36. Изменения порога использования низкопороговой (а) и высокопороговой (б) двигательных единиц в мышце кисти руки при кожном стимулировании указательного пальца. Контрольные значения (предстимуляционные) порога использования приведены слева от нулевой отметки времени (Garnett, Stephens, 1981)
ходящие пути, в поток информации через двигательную систему. Рассмотрены концепция результирующего действия мышц в качестве основного определителя движения и влияние различного прикрепления мышц на выходной сигнал мышечно-скелетной системы. Идентифицированы основные стратегии, используемые нервной системой в управлении положением тела и движением. Связанные с движением стратегии включали схемы возбуждения, используемые для направленных к цели движений, применение совместного возбуждения, основу цикла удлинения—сокращения, принятие афферентной обратной связи для таких движений, как манипулирование, и общие правила, лежащие в основе возбуждения двигательных единиц.
вопросы САЛЬТЕРНл КВНЫ>1 ОТВЕТОМ
1. Мотивация важна для инициирования движения. ПРАВИЛЬНО НЕПРАВИЛЬНО
2. Ассоциативная зона коры головного мозга управляет основными биологическими
побуждениями и эмоциональным поведением. ПРАВИЛЬНО НЕПРАВИЛЬНО
3. Лимбическая система преобразовывает мотивацию в мысль.
ПРАВИЛЬНО НЕПРАВИЛЬНО
4. Ассоциативная зона коры головного мозга включает префронтальную, теменную и височные доли.
ПРАВИЛЬНО НЕПРАВИЛЬНО
253
5. Центральная команда передается в нижние нервные центры из двигательной зоны коры головного мозга.
ПРАВИЛЬНО НЕПРАВИЛЬНО
6. Результирующий разряд является двигательной командой из ствола мозга в спинной МОЗГ. ПРАВИЛЬНО НЕПРАВИЛЬНО
7. Двигательная программа представляет собой стереотипную последовательность афферентных сигналов от периферийных чувствительных нервных окончаний.
ПРАВИЛЬНО НЕПРАВИЛЬНО
8. Генераторы ЦНС генерируют команды, которые возбуждают мышцу.
ПРАВИЛЬНО НЕПРАВИЛЬНО
9. Комиссуры соединяют правое и левое полушария большого мозга.
ПРАВИЛЬНО НЕПРАВИЛЬНО
10. Основная двигательная зона — часть сенсорно-двигательной зоны коры головного мозга. ПРАВИЛЬНО НЕПРАВИЛЬНО
11. Основная соматосенсорная зона коры головного мозга находится перед центральной бороздой.
ПРАВИЛЬНО НЕПРАВИЛЬНО
12. Кортикоспинальный тракт является нисходящим путем.
ПРАВИЛЬНО НЕПРАВИЛЬНО
13. Соматотопический означает карту тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
14. Сенсорно-двигательная зона коры головного мозга содержит несколько соматотопи-ческих схем.
ПРАВИЛЬНО НЕПРАВИЛЬНО
15. Основная соматосенсорная зона коры головного мозга предоставляет сенсорную информацию для планирования и инициирования движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
16. Преддвигательная зона коры головного мозга связана, главным образом, с планированием и определением последовательности движений.
ПРАВИЛЬНО НЕПРАВИЛЬНО
17. Задняя теменная зона коры головного мозга активна во время осуществления воображаемых движений.
ПРАВИЛЬНО НЕПРАВИЛЬНО
18. Сенсорно-двигательная зона коры головного мозга может планировать движения с точки зрения участия групп мышц.
ПРАВИЛЬНО НЕПРАВИЛЬНО
19. Варолиев мост располагается в стволе мозга. ПРАВИЛЬНО НЕПРАВИЛЬНО
20. Продолговатый мозг располагается между варолиевым мостом и спинным мозгом.
ПРАВИЛЬНО НЕПРАВИЛЬНО
21. Руброспинальный путь является нисходящим путем от красного ядра.
ПРАВИЛЬНО НЕПРАВИЛЬНО
22. Ретикулярная формация является диффузной, с ядрами как в среднем, так и в продолговатом мозгу.
ПРАВИЛЬНО НЕПРАВИЛЬНО
23. Кортикоспинальный тракт заканчивается вблизи двигательных нейронов, которые иннервируют дистальные мышцы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
24. Мозжечок не имеет прямых соединений со спинным мозгом.
ПРАВИЛЬНО НЕПРАВИЛЬНО
25. Мозжечок принимает входной сигнал через кровельные, промежуточные и пильчатые ядра. ПРАВИЛЬНО НЕПРАВИЛЬНО
26. Компаратор, например, мозжечок, сравнивает желаемые движения с фактическими движениями. ПРАВИЛЬНО НЕПРАВИЛЬНО
27. Мозжечок связан исключительно с двигательным обучением.
ПРАВИЛЬНО НЕПРАВИЛЬНО
28. Базальные ядра представляют собой промежуточное звено между двигательной зоной коры головного мозга и стволом мозга. ПРАВИЛЬНО НЕПРАВИЛЬНО
29. Базальные ядра не влияют на движение.
ПРАВИЛЬНО НЕПРАВИЛЬНО
30. Восходящие пути передают афферентную информацию от периферийных рецепторов в супрасегментные центры.
ПРАВИЛЬНО НЕПРАВИЛЬНО
31. Таламус обрабатывает только сенсорную информацию.
ПРАВИЛЬНО НЕПРАВИЛЬНО
32. Передний боковой путь передает сигналы в основную соматосенсорную зону коры головного мозга.
ПРАВИЛЬНО НЕПРАВИЛЬНО
33. Биомеханический эффект возбуждения мышцы не ограничивается суставом или суставами, пересекаемыми мышцей.
ПРАВИЛЬНО НЕПРАВИЛЬНО
34. Результирующее усилие мышц представляет собой сумму усилий всех мышц относительно сустава.
ПРАВИЛЬНО НЕПРАВИЛЬНО
35. Реакция сустава учитывает результирующие усилия относительно сустава.
ПРАВИЛЬНО НЕПРАВИЛЬНО
36. Реакция сустава измеряется в Ньютонах на квадратный метр (Н • м-2).
ПРАВИЛЬНО НЕПРАВИЛЬНО
37. Межкостное усилие в суставе никогда не превышает вес тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
254
38. Для медленного движения вращающий момент мышцы противодействует вращающему моменту нагрузки.
ПРАВИЛЬНО НЕПРАВИЛЬНО
39. Если вектор усилия мышцы и смещение имеют одно и то же направление, мышца сокращается концентрически.
ПРАВИЛЬНО НЕПРАВИЛЬНО
40. Для быстрых движений вектор веса направлен перпендикулярно сегменту тела.
ПРАВИЛЬНО НЕПРАВИЛЬНО
41. Если группа флексорных мышц осуществляет разгибание, результирующее действие мышц эксцентрично.
ПРАВИЛЬНО НЕПРАВИЛЬНО
42. Если движение пересекает линию действия силы тяжести (проходящую через ось вращения) во время медленного движения, результирующий вектор мышц изменяет направление.
ПРАВИЛЬНО НЕПРАВИЛЬНО
43. Инерционные силы создают значительные эффекты во время быстрых движений.
ПРАВИЛЬНО НЕПРАВИЛЬНО
44. Нервная система управляет быстрыми и медленными движениями различным образом. ПРАВИЛЬНО НЕПРАВИЛЬНО
45. Быстрые движения обычно включают экс-центрическо-концентрическую последовательность действия мышц.
ПРАВИЛЬНО НЕПРАВИЛЬНО
46. Действие biceps brachii изменяется совместно с действием brachoradialis для многих задач. ПРАВИЛЬНО НЕПРАВИЛЬНО
47. Rectus femoris — двусуставная мышца.
ПРАВИЛЬНО НЕПРАВИЛЬНО
48. Скорость сокращения двусуставной мышцы меньше по сравнению с односуставными мышцами-синергистами.
ПРАВИЛЬНО НЕПРАВИЛЬНО
49. Результирующее воздействие возбуждения двусуставной мышцы может быть сосредоточено только на одном из двух суставов, которые она пересекает.
ПРАВИЛЬНО НЕПРАВИЛЬНО
50. Плечи пар для мышц обычно изменяются во всем диапазоне движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
51. Действие, обусловливаемое положением тела, не требует произвольного (сознательного) возбуждения мышцы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
52. Устойчивость и равновесие означают одно И ТО же. ПРАВИЛЬНО НЕПРАВИЛЬНО
53. Система в состоянии равновесия может иметь ненулевое ускорение.
ПРАВИЛЬНО НЕПРАВИЛЬНО
54. Корректировка положения тела инициируется афферентной обратной связью.
ПРАВИЛЬНО НЕПРАВИЛЬНО
55. Возмущение опорной поверхности вызывает реакцию в направлении от дистальной к проксимальной.
ПРАВИЛЬНО НЕПРАВИЛЬНО
56. Корректировка положения тела всегда начинается со стоп.
ПРАВИЛЬНО НЕПРАВИЛЬНО
57. Корректировка положения тела состоит из набора рефлекторных реакций.
ПРАВИЛЬНО НЕПРАВИЛЬНО
58. Трехимпульсная конфигурация ЭМГ имеет место в процессе медленных двунаправленных движений.
ПРАВИЛЬНО НЕПРАВИЛЬНО
59. Движения, которые осуществляются под управлением по разомкнутому контуру, изменяются посредством афферентной обратной СВЯЗИ. ПРАВИЛЬНО НЕПРАВИЛЬНО
60. Субъекты выбирают зависящую от скорости стратегию при выполнении быстрой задачи с определенными временными ограничениями.
ПРАВИЛЬНО НЕПРАВИЛЬНО
61. Не зависящая от скорости стратегия включает изменение длительности электромиограммы при постоянной амплитуде электромиограммы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
62. Совместное возбуждение повышает жесткость относительно сустава.
ПРАВИЛЬНО НЕПРАВИЛЬНО
63. Совместное возбуждение происходит лишь при усвоении человеком нового навыка.
ПРАВИЛЬНО НЕПРАВИЛЬНО
64. Мышца может выполнить большую отрицательную работу, если она активно растягивается перед сокращением.
ПРАВИЛЬНО НЕПРАВИЛЬНО
65. Работа измеряется площадью ниже графика усилие—смещение.
ПРАВИЛЬНО НЕПРАВИЛЬНО
66. Повышенная работа, выполняемая во время концентрической части эксцентричес-ки-концентрического сокращения, обусловливается наличием увеличенного количества энергии.
ПРАВИЛЬНО НЕПРАВИЛЬНО
67. Упругая энергия не может накапливаться в последовательных упругих элементах мышцы. ПРАВИЛЬНО НЕПРАВИЛЬНО
68. Соотношение работы и энергии может быть выражено в качестве первого закона термодинамики.
ПРАВИЛЬНО НЕПРАВИЛЬНО
255
69. Количество упругой энергии, которое может накапливаться в мышце, возрастает с увеличением времени между эксцентрическим и концентрическим сокращениями.
ПРАВИЛЬНО НЕПРАВИЛЬНО
70. Увеличение работоспособности в прыжке движением в противоположном направлении по сравнению с прыжком согнув ноги в большей степени обусловливается накапливанием и использованием упругой энергии, чем преднагрузочным эффектом.
ПРАВИЛЬНО НЕПРАВИЛЬНО
71. Порядок использования двигательных единиц может изменяться во время эксцентрических сокращений.
ПРАВИЛЬНО НЕПРАВИЛЬНО
72. Скорость разряда двигательных единиц повышается в период утомления.
ПРАВИЛЬНО НЕПРАВИЛЬНО
«ОПРОСЫ С PA3JПГ]1гчНЬ 'М ВЫБПГ ЭМ ОТГГТО I
73. Что вносит лимбическая система в управление движением?
а) мышление;
б) программирование;
в) мотивацию;
г) осознание.
74. Какой супрасегментарный центр подает центральную команду?
а) базальные ядра;
б) двигательная зона коры головного мозга;
в) ствол мозга;
г) мозжечок.
75. Какие компоненты ЦНС принимают результирующий разряд?
а) двигательная зона коры головного мозга;
б) мозжечок;
в) ствол мозга;
г) спинной мозг.
76. Что является двигательной программой?
а) невральная сеть, которая может генерировать соответствующую поведению схему выходного сигнала в отсутствии внешних хронирующих стимулов;
б) копия центральной команды, подаваемая из двигательной зоны коры головного мозга назад в супрасегментные центры;
в) группа мышц, которая вынуждена действовать в качестве одной единицы;
г) стереотипная последовательность команд, подаваемая из спинного мозга
в мышцы, чтобы вызвать конкретное поведение.
77. Какие участки являются частью сенсорнодвигательной зоны коры головного мозга? а) основная сенсорно-двигательная зона коры головного мозга;
б) дополнительная двигательная зона;
в) таламус;
г) средний мозг.
78. Какая зона или зоны Брод манна определяют основную двигательную зону?
а) 1, 2 и 3;
б) 4;
в) 5 и 7;
г) 8.
79. Что является соматотопией?
а) карта тела ЦНС;
б) область тела, иннервируемая одним нервом;
в) разделение двигательной и сенсорной функций в сенсорно-двигательной зоне коры головного мозга;
г) иерархическая организация супрасег-ментных центров.
80. Какая часть сенсорно-двигательной зоны коры головного мозга активна во время осуществления воображаемых движений?
а) преддвигательная зона;
б) основная соматосенсорная зона;
в) дополнительная двигательная зона;
г) задняя теменная зона.
81. Что является функцией базальных ядер?
а) возбуждение конкретных мышц и двигательных программ;
б) хронирование действия агонистов-антагонистов;
в) двигательное планирование, например, направления и амплитуды;
г) предоставление мотивации в сенсорнодвигательную зону коры головного мозга.
82. Что является осознанием?
а) интерпретация результирующего разряда;
б) афферентные сигналы, которые достигают сознания и способствуют восприятию;
в) оценка восприятия усилия;
г) любая афферентная информация, достигающая сенсорно-двигательной зоны коры головного мозга.
83. Какой параметр не влияет на состояние мышечно-скелетной системы?
а) центральная команда;
б) длина мышцы;
в) длина плеча мышцы;
г) скорость мышцы.
84. Что является самым лучшим синонимом термина “результирующее усилие мышц”?
256
а) результирующий вращающий момент мышц;
б) результирующее действие мышц;
в) общее усилие мышц;
г) равнодействующее усилие мышц.
85. Каков критически признак медленного движения, который позволяет нам определить результирующее действие мышц, связанное с движением?
а) заданное направление смещения;
б) неспособность мышцы осуществить эксцентрическое сокращение во время медленного движения;
в) направление вращающего момента нагрузки;
г) мышца может создавать только тяговое усилие.
86. Для каких условий подходит квазистатиче-ский анализ?
а) медленного бега;
б) удара ногой по мячу в футболе;
в) броска в бейсболе;
г) изометрического сокращения.
87. Какие аспекты функции мышц не подвержены влиянию вариации мест их прикрепления?
а) диапазон движения;
б) соотношение вращающий момент — угол;
в) максимальное усилие;
г) скорость сокращения.
88. В чем преимущество двусуставных мышц?
а) скорость сокращения меньше по сравнению с односуставной мышцей;
б) длина мышечных волокон больше по сравнению с односуставной мышцей;
в) вращающий момент мышцы и энергия сустава перераспределяются по всей конечности;
г) связываются общие движения.
89. Что является наилучшим определением устойчивости?
а) механическое равновесие;
б) восстановление равновесия после возмущения;
в) максимальное опорное основание;
г) неподвижная система, которая не перемещается.
90. Какая сенсорная информация важна в управлении вертикальным положением тела? а) звуковая;
б) соматосенсорная;
в) вестибулярная;
г) зрительная.
91. Какие мышцы субъект активизирует в первую очередь для выполнения задачи на время реакции, включающей подъем выпрямленной руки из бокового положения в горизонтальное положение?
а) переднюю дельтовидную мышцу;
б) туловища;
в) Quadriceps femoris;
г) сухожилия, ограничивающие с боков подколенную ямку.
92. Для каких типов движений используется трехимпульсная конфигурация ЭМГ?
а) медленных и однонаправленных;
б) быстрых и двунаправленных;
в) изометрических сокращений;
г) со скоростью от средней до высокой и однонаправленных.
93. Что пытается объяснить гипотеза двойной стратегии?
а) конфигурации ЭМГ для движений с различными смешениями, скоростями и нагрузками;
б) результирующие вращающие моменты мышц для движений, ограничиваемых горизонтальной плоскостью;
в) управление движением в результате варьирования порога тонизирующего рефлекса растяжения мышцы;
г) роль афферентной обратной связи в формировании конфигураций ЭМГ.
94. Почему труднее осуществлять управление многосуставными движениями по сравнению с односуставными?
а) свойства двусуставных мышц заметно отличаются от свойств односуставных;
б) наличие более одной оси вращения;
в) управление усложняется межсегментной динамикой;
г) смещения конечных точек сегментов значительно больше.
95. В чем преимущество совместного возбуждения, когда человек поднимает тяжелые грузы?
а) увеличение жесткости и, следовательно, устойчивости сустава;
б) уменьшение специфичности невральной команды;
в) способность передавать энергию от одного сустава к другому;
г) увеличение экономичности движений, включающих изменения направления.
96. Какие переменные влияют на способность мышцы использовать накапливаемую упругую энергию?
а) имеющаяся химическая энергия;
б) время между эксцентрическим и концентрическим сокращениями;
в) скорость растяжения;
г) величина растяжения.
97. В каких видах спорта не используются движения, включающие цикл удлинения—сокращения?
а) плавание;
б) бейсбол;
в) бег;
г) поднятие тяжестей.
17 8-133
257
98. Какие действия приводят к изменению порядка использования двигательных единиц? а) кожная стимуляция;
б) эксцентрические сокращения;
в) усталость мышц;
г) быстрые движения.
99. Какие части нервной системы могут обеспечивать мотивацию и инициировать команды, которые затем приводят к осуществлению движения?
100. Почему преобразование мысли в надлежащие интенсивность и схему действия мышц, необходимую для желаемого движения, называется программированием?
101. Почему результирующий разряд — необходимая часть схемы управления, используемой ЦНС?
102. Двигательная команда определяется как стереотипная последовательность команд, подаваемых из спинного мозга в мышцы, чтобы вызвать конкретное поведение. Какие соображения наводят на мысль о том, что данная двигательная единица может участвовать во множестве двигательных программ?
103. В чем различие между двигательной программой и центральным схемным генератором?
104. Какие супрасегментные структуры и пути (восходящие и нисходящие) имеют сома-тотопичную организацию?
105. Опишите роль следующих участков сенсорно-двигательной зоны коры головного мозга в генерировании движения:
а) преддвигательной зоны коры головного мозга;
б) основной двигательной зоны коры головного мозга;
в) задней теменной зоны коры головного мозга;
г) основной соматосенсорной зоны коры головного мозга;
д) дополнительной двигательной зоны.
106. Хотя в мозжечке находится более половины нейронов мозга, он не имеет прямых эфферентных соединений со спинномозговыми ядрами или ядрами ствола мозга.
А. Посредством каких путей мозжечок влияет на управление движением?
Б. Что является компаратором?
В. Опишите три теории, на основании которых мозжечок мог бы функционировать как компаратор.
107. Приведите какое-либо основание того, что базальные ядра участвуют в управлении движением.
108. Почему команда, поданная нервной системой, не всегда вызывает один и тот же эффект мышечно-скелетной системы?
109. Межкостное усилие представляет собой абсолютную силу в суставе и может подвергаться влиянию как усилия мышц, так и реакции земли.
А. Почему межкостное усилие значительно превышает реакцию сустава?
Б. Начертите график, способствующий объяснению, каким образом два этих усилия влияют на межкостное усилие в голеностопном суставе.
В. Один из путей определения межкостного усилия заключается в определении усилия, создаваемого каждой мышцей, пересекающей сустав. Каким образом мы можем измерить усилие, создаваемое мышцей человека?
110. Невральное управление движением зависит от скорости, с которой оно осуществляется.
А. Начертите графики временной зависимости скорости и вращающего момента для быстрого выполнения движения, показанного на рис. 7.15. Допустите, что человек использовал небольшой груз и выполнял движение возможно быстрее.
Б. Опишите результирующее действие мышц, связанное с таким режимом.
111. Двусуставные мышцы пересекают многие суставы в теле человека. Они характеризуются рядом преимуществ в отношении управления движением.
А. Почему скорость сокращения двусуставной мышцы меньше по сравнению с ее односуставными синергистами? Каково функциональное следствие этого различия?
Б. Каким образом rectus femoris (двусуставная мышца, пересекающая тазобедренный и коленный суставы) могла бы увеличить вращающий момент, создаваемый экстензорами коленного сустава?
112. Режим движения включает тесную связь между мышцами, обеспечивающими поддержание положения тела, и мышцами, обеспечивающими движение.
А. Почему корректировка положения тела рассматривается как автоматическая?
Б. Опишите условия, когда действие, обусловливаемое положением тела, связано с поддержанием устойчивости системы, и условия, когда необходимо поддерживать систему в равновесии.
258
113. Опишите движение, которое характеризовалось бы трехимпульсной конфигурацией ЭМГ. Начертите графики временной зависимости скорости и ЭМГ для этого движения.
114. Большинство исследований стратегий движения было сосредоточено на направленных к цели движениях.
А. Почему ученые анализируют конфигурации ЭМГ, пытаясь понять управление движением нервной системой?
Б. Какие параметры предлагает гипотеза двойной стратегии в качестве важных для управления движением? Почему выбираются эти параметры?
115. Общей формой возбуждения мышцы является использование эксцентрически-кон-центрической последовательности.
А. Опишите концепции накапливания и использования упругой энергии и преднагрузочного эффекта.
Б. Каким образом рефлекс растяжения мышцы мог бы, вероятно, повлиять на увеличение положительной работы, выполняемой во время цикла удлинения-сокращения? Почему рефлекс растяжения мышцы не является важным фактором, способствующим этому эффекту?
116. Типовая экспериментальная стратегия исследования цикла удлинения—сокращения заключается в сравнении характеристик прыжка согнув ноги и прыжка движением в противоположном направлении.
А. Начертите графики временной зависимости усилия для вертикальной составляющей реакции земли как при прыжке согнув ноги, так и при прыжке
движением в противоположном направлении.
Б. Почему импульс движущей силы должен превышать тормозную составляющую для фактического отделения человека от земли при выполнении прыжка движением в противоположном направлении?
В. Допустим, импульсы во время прыжка движением в противоположном направлении имели следующие значения: движущий — 320 Н • с;
тормозной — 82 Н • с.
Используйте соотношение импульса и количества движения (см. главу 3) для определения изменения скорости прыгуна (с массой тела 64 кг) во время фазы отталкивания.
117. Порядок использования двигательных единиц является довольно согласованным и в условиях соответствующего напряжения стабилен.
А. В чем преимущества использования двигательных единиц в порядке от минимальных до максимальных?
Б. Почему управление усилием мышцы может быть менее точным во время эксцентрических сокращений?
В. Какие особенности организации двигательных единиц позволяют изменять порядок их использования в результате таких событий, как утомление и кожная стимуляция?
Г. Приведите преимущество мышечно-скелетной системы для каждого из трех граничных условий, которые представляются изменяющими порядок использования.
17*
._ ИЖ ГЛАВА 8
Г С А КРАТКОВРЕМЕННЫЕ
1 АДАПТАЦИИ
Бросающейся в глаза характеристикой двигательной системы является ее адаптивность. Подвергаясь кратковременному или постоянному напряжению, двигательная система может приспосабливаться к изменяющимся потребностям использования. Эти адаптации могут быть экстенсивными и оказывать влияние на большинство как морфологических, так и функциональных характеристик системы.
В этой главе мы рассмотрим непосредственную (кратковременную) реакцию двигательной системы на напряжение, связанное с одним видом физической активности. Исследуем эффекты разминки, методик и механизмов, лежащих в основе изменений гибкости, повреждения мышц и болезненного ощущения в них, механизмом, обусловливающих утомление мышц, и явления потенцирования мышц.
ЭФФЕКТЫ РАЗМИНКИ
Часто, когда человек занимается физической деятельностью, она начинается с легких упражнений для подготовки тела к восприятию напряжения. Цель их — обеспечение эффекта разминки, включающего повышение внутренней температуры и разрушение временных связей соединительных тканей. Повышение внутренней температуры улучшает биомеханическую характеристику двигательной системы, а растягивание может уменьшить возможность деформирования мышц (Garrett, 1990; Stanish, Hubley-Kozey, 1984). Эффекты разминки следует отличать от эффектов упражнений, предназначающихся для повышения гибкости, т.е. увеличения диапазона движения относительно сустава.
Температура
Разминка оказывает значительное влияние на зависящие от температуры физиологические процессы. Повышение внутренней температуры в результате разминки может увеличивать диссоциацию кислорода из гемоглобина и миоглобина, мышечный кровоток, интенсифицировать метаболические реакции, снижать вязкость мышц, увеличивать растяжимость соединительной ткани и повышать скорость передачи потенциалов действия (Shellock, Prentice, 1985). Мы наиболее заинтересованы во влиянии разминки на механическую отдачу мышцы.
Как правило, высота, которая может быть достигнута человеком в прыжке в высоту, после разминки увеличивается. Это происходит потому, что разминка увеличивает максимальную энергию, которая может быть создана мышцей (рис.8.1), а высота прыжка зависит от количества создаваемой энергии. Улучшение показателя прыжка после разминки обусловливается влиянием температуры мышц на скорость сокращения. Так, Davies, Young (1983) установили, что повышение температуры мышц на 3,ГС уменьшает продолжительность сокращения и время полурелаксации соответственно на 7 и 22 %, но не влияет на одиночное или тетаническое напряжение. В противоположность этому снижение температуры мышц на 8,4°С (в результате погружения ноги в ледяную ванну) увеличивало как продолжительность сокращения (38 %), так и время полурелаксации (93 %). Наблюдения, подобные этим, показывают, что релаксация в большей степени зависит от температуры мышц, чем создание усилий. С точки зрения соотношения усилия и скорости изменение температуры в пределах физиологического диапазона, влияет на максимальную скорость сокращения (12 % • °C'1, но не влияет на
260
Рис.8.1. Идеализированное влияние изменения температуры в результате разминки на соотношения вращающего момента и энергии в зависимости от скорости. Соотношения энергии и скорости характеризуются колоколообразными кривыми. Повышение температуры сдвигает величину vMaKC и пиковую энергию вправо
максимальное изометрическое усилие (Binkhorst, Hoofd, Vissers, 1977). Для группы мышц с данным соотношением усилия и скорости изменение температуры мышц не влияет на величину Fm o, но сдвигает величину vmax вправо на 12 % на каждый градус (°C) повышения температуры (рис.8.1). Результирующим эффектом изменения скорости сокращения является повышение пиковой энергии.
Если изменение температуоы существенно, она может повлиять на максимальное изометрическое усилие. Например, Ranatunga, Sharpe, Tumbuel (1987) установили, что максимальное изометрическое усилие мышцы кисти руки остается относительно постоянным при охлаждении до 25°С, но уменьшается примерно на 30 % при охлаждении до 12—15°С. Bergh, Ekblom (1979) также сообщили об изменении максимального изометрического вращающего момента экстензоров коленного сустава от 262 Н • м при 30,4°С до 312 Н • м при 38,5°С (2,4 % • °C-') при измерении температуры в vastus lateralis. Эти изменения оказали влияние на показатели; спортсмены увеличили прыжок в высоту на 44 % (17 см) и максимальное создание энергии в велоспорте на 32 % (316 Вт) при повышении температуры от 30,4 до 38,5°С.
Ingjer, Stromme (1979) полагают, что наилучшей стратегией обеспечения изменений температуры мышц при измерении максимального усилия в серии опытов на “бегущей дорожке” в течение 4 мин является связанная с видом деятельности разминка (см.также Shellock, 1986; Shellock, Prentice, 1985). Активной является разминка, в процессе которой изменения температуры обусловливаются работой мышц, а не пассивным внешним источником тепла, например, теплой ванной. Ingjer, Stromme могли поднять внутримышечную температуру боковой части quadriceps femoris от 35,9°С до пример
но 38,4°С с использованием как активной, так и пассивной методик, но они привели доводы в пользу активной разминки, подтвержденные превосходными результатами во время максимального аэробного пробега по “бегущей дорожке” в течение 4 мин. Короче говоря, большая часть общей затраты энергии обеспечивается аэробными процессами, если работе предшествует активная разминка. Кроме того, эти авторы полагают, что продолжительность разминки должна превышать 5 мин и разминка должна проводиться с интенсивностью, эквивалентной темпу 7,5 мин • миля-1 для тренированного спортсмена или достаточной для потоотделения и повышения частоты сердечных сокращений у нетренированного человека. Повысившаяся в результате таких действий температура мышц снижается примерно через 15 мин после разминки; следовательно, время между разминкой и событием не должно превышать 15 мин. К тому же разминка, выполняемая для повышения внутренней температуры, необходима, вероятно, лишь в тех случаях, когда этого требует скорость выполнения задачи, и температура окружающей среды низкая.
Жесткость
Структурные элементы мышцы сопротивляются увеличению длины (растягиванию). Чем больше растягивание, тем выше пассивное сопротивление. Как отмечено в главе 6 (см.рис.6.12), между усилием мышцы и ее длиной имеет место криволинейное соотношение, если растягивается пассивная мышца. Поскольку жесткость (В • м-1 или Н мм-1) определяется наклоном кривой соотношения усилие—длина, криволинейное соотношение указывает, что жесткость растягиваемой пассивной мышцы увеличивается в функции длины. На рис.8.2 показано соотношение усилие—длина для трех мышц задней конечности кошки; тремя компонентами каждого графика являются общее, пассивное и активное усилия. На каждом графике компонент зависимости пассивного усилия от длины показан линией, которая начинает повышаться при нормированной длине 1,0, компонент зависимости активного усилия от длины представлен колоколообразной кривой, соотношение общего усилия и длины показано верхней линией (сумма пассивного и активного компонентов). Форма компонента зависимости общего усилия от длины отличается для каждой мышцы из-за различий в активном и пассивном компонентах. Мышцы, которые имеют узкую кривую зависимости активного усилия от длины и крутую кривую зависимости пассивного усилия от длины (например, soleus, lateral gastrochemius), стремятся к большей площади поперечного сечения, меньшей скорости
261
Рис.8.2. Зависимость усилия от длины для трех мышц задней конечности кошки (Gareis, Solomonow, Baratta, 1992):
а — lateral gastrochemius, б — soleus, в — экстензор digitorum longus. Усилие нормировали по пиковому усилию (Fo) при длине в состоянии покоя (Zo). Длину мышцы нормировали по длине в состоянии покоя (Zo), причем Lo — длина мышцы, при которой начинает проявляться пассивное усилие
Рис.8.3. Соотношение вращающего момента и угла, связанное с пассивным растягиванием и освобождением плантарнофлексорных мышц (Hufschmidt, Mauritz, 1985):
а — увеличение (растягивание) и уменьшение (освобождение) угла; б — пассивный вращающий момент, создаваемый мышцами; в — результирующее соотношение вращающего угла и угла, которое не перекрывается во время фаз растягивания и освобождения
сокращения и большим углам пеннации (Gareis, Solomonow, Baratta, 1992).
У людей сопротивление пассивных мышц может характеризоваться соотношением вращающего момента и угла (рис.8.3, в). Площадь, охватываемая кривой вращающий момент—угол отражает энергию, потребляемую плантарнофлексорными мышцами во время цикла пассивного растяжения—сокращения. Поскольку мышечная спастичность включает изменения в мышце, обусловливающие повышенное пассивное сопротивление угловому смещению относительно сустава, пациенты, характеризовавшиеся симптомами мышечной спастичности более одного года, имели более жесткие мышцы по сравнению со здоровыми людьми (Hufschmidt, Mauritz, 1985). Этот эффект больше для экстензорных (антигравитационных) мышц (Dietz, 1992).
Интересное свойство пассивной мышцы заключается в том, что ее жесткость (наклон
кривой вращающий момент—угол увеличивается с увеличением времени между растягиваниями (рис.8.4). Это происходит в течение 30-ми-нутного интервала покоя после возбуждения мышц (Kilgore, Mobley, 1991; Lakie, Robson, 1988). Жесткость резко увеличивается сразу же после действия, затем этот процесс становится более постепенным. Повышение жесткости можно исключить с помощью активных или пассивных движений, но не посредством изометрических сокращений (Lakie, Robson, 1988).
Этот эффект был отнесен к тиксотропному свойству мышцы. Тиксотропия — свойство, которым характеризуются различные гели, например, мышца. Гели становятся жидкими при встряхивании, перемешивании или ином возмущении и снова застывают при отстаивании. Представляется, что повышение жесткости мышцы в состоянии покоя обусловлено качественно подобным механизмом. Молекулярная перегруп-
262
Рис.8.4. Влияние переменных интервалов покоя на соотношение вращающего момента и угла, связанное с пассивным растяжением и освобождением плантарнофлексорных мышц (Hufschmidt, Mauritz, 1985)
пировка в мышце может включать образование устойчивых связей между волокнами актина и миозина. При отсутствии действия количество связей возрастает и, следовательно, увеличивается жесткость мышцы. Однако при кратковременном растягивании или в период физической активности многие, если не все связи разрушаются и жесткость мышцы снижается (Proske, Morgan, Gregory, 1993; Wiegner, 1987).
Одна из целей разминки заключается, вероятно, в минимизации жесткости мышц посредством приведения в действие большинства основных групп мышц во всем диапазоне движения (Wiktorsson-Moller, Oberg, Ekstrand, Gillquist, 1983). Разминка разрушает актин-миозиновые связи, которые образовались, и в результате этого снижает пассивную жесткость мышцы. Поскольку мы предпочитаем начинать действие или движение с разминки, это означает, что нервная система предпочитает управлять мышцей, когда она находится в состоянии минимальной пассивной жесткости. В противоположность этому повышение жесткости вследствие бездействия снижает чувствительность мышцы к возмущениям и может облегчить управление положением тела.
Мышечный тонус. Тиксотропное свойство мышцы обусловливает ее пассивную жесткость, которая противодействует изменениям ее длины. Это сопротивление релаксированной мышцы растягиванию клиницисты называют мышечным тонусом. Сопротивление растягиванию, конечно, может, дополняться рефлексом растяжения мышцы, но это не происходит у ослабленного субъекта, когда скорость растягивания характеризуется амплитудой в пределах от малой до средней.
Изменения мышечного тонуса могут быть использованы клиницистами для идентификации патологических состояний. Понижения
мышечного тонуса, известные как гипотонус, появляются у пациентов с патологическими изменениями в полушариях мозжечка и у пациентов со спинномозговыми рассечениями. Rothwell (1987) наводит на мысль о том, что гипотонус, вероятно, обусловливается пониженным уровнем возбудимости рефлекса растяжения мышцы.
Повышение мышечного тонуса, называемое гипертонусом, по-видимому, обусловливается устойчивым состоянием активности двигательных нейронов. Двумя наиболее общими формами гипертонуса являются мышечная спастичность и ригидность. Мышечная спастичность — патологически индицируемое состояние повышенной возбудимости рефлекса растяжения мышцы. Рефлекс растяжения спастической мышцы более сильный, чем рефлекс нормальной мышцы. Более того, этот повышенный рефлекс растяжения мышцы увеличивается со скоростью растягивания. В основе мышечной спастичности лежат многие механизмы, включая изменения возбудимости двигательных нейронов, постсинаптическую повышенную чувствительность к нейромедиатору, увеличение участков двигательных единиц и повышение пассивных тиксотропных свойств мышцы. Симптомы, связанные с мышечной спастичностью, включают повышенное пассивное сопротивление движению в одном направлении, повышенную активность сухожильного рефлекса, вызываемого легким ударом, принятие характерного положения соответствующей конечностью, явную неспособность релаксировать соответствующую мышцу и неспособность приводить в движение соответствующий сустав быстро или в чередующихся направлениях. Неверное представление, связанное с мышечной спастичностью, заключается в том, что изменения мышечного тонуса ухудшают способность осуществлять движение. Это неправильно. Спа
263
стичность мышцы-антагониста не является основным фактором, ухудшающим ее способность осуществлять движение. Это ухудшение обусловливается неспособностью мышцы-агониста использовать достаточное количество двигательных единиц (Sahrmann, Norton, 1977; Tang, Rymer, 1981).
Другой формой гипертонуса является ригидность. Симптомы ригидности и мышечной спастичности заметно отличаются. Симптомы, связанные с ригидностью, включают двунаправленное сопротивление движению, не зависящее от его скорости, которое наблюдается при отсутствии повышенного сухожильного рефлекса, вызываемого легким ударом. Наиболее общий пример ригидности наблюдается при болезни Паркинсона и включает постоянное сокращение мышц, которое может появляться при пассивной манипуляции в виде ряда прерывистых резких движений (шестеренная ригидность).
ГИБКОСТЬ
Часто нельзя различить разминочные упражнения и упражнения, предназначенные для повышения гибкости. Одна из функций разминочных упражнений — снижение связанной с тиксотропностью жесткости мышц, которое отличается от увеличения диапазона движения или гибкости относительно сустава. Основное отличие заключается в продолжительности эффекта. Преимущества разминки должны сохраняться во время последующей физической деятельности, в то время как задача упражнений для повышения гибкости — индуцирование более продолжительного изменения диапазона движения.
Вообще, цель упражнений для повышения
Рис.8.5. Пример фиксационно-релаксационной методики с сокращением агониста, в соответствии с которой партнер максимально растягивает сухожилия, ограничивающие с боков подколенную ямку
гибкости — увеличение диапазона движения путем пассивного (статического) растягивания мышц конечностей либо путем содействия растягиванию (динамическому) одной или нескольких мышц. Типичным примером статического растягивания является попытка увеличить диапазон сгибания тазобедренных суставов и выпрямления коленных суставов в результате наклона вперед из вертикального прямого положения тела, удерживая колени выпрямленными и пытаясь коснуться пальцами пальцев стопы. Человеку дается указание сохранять мышцы ног пассивными и удерживать позицию растяжения в течение 15—30 с. Один из вариантов этого упражнения заключается в разгибании и наклонах, с попыткой коснуться пальцев стопы, а не в выдерживании непрерывного растяжения; вариант разгибания и нагибания называется баллистическим растяжением. Аналогичное упражнение касания пальцев стопы может выполняться человеком, сидящим на полу. При выполнении данного упражнения в этой позиции сила тяжести не содействует растягиванию мышц сухожилий, ограничивающих с боков подколенные ямки.
Хотя статическое растяжение и является наиболее общеиспользуемым упражнением, улучшающим гибкость, клиницисты склонны отдавать предпочтение упражнениям, в которых растягивание сочетается с возбуждением мышц-агонистов или мышц-антагонистов. Три этих упражнения были заимствованы из реабилитационной методики, известной в качестве улучшения нервно-мышечной передачи про-приорецептивных импульсов (Knott, Voss, 1968). Фиксационно-релаксационное растяжение включает первоначальное максимальное изометрическое сокращение мышцы, подлежащей растягиванию (антагонист), с последующими релаксацией и растяжением мышцы до предела диапазона движения. Упражнение этого типа может выполняться при содействии партнера или терапевта (рис.8.5). Растягивание с сокращением агониста требует содействия партнера. Партнер перемещает конечность участника таким образом, чтобы сустав находился на пределе вращения. Участник сокоашает мышцу-агонист (например, квадрицепс феморис), в то время как партнер прилагает усилие к конечности для растягивания мышцы-антагониста (например, сухожилий, ограничивающих с боков подколенную ямку). Фиксационно-релаксационная методика с сокращением агониста является комбинацией методик фиксационно-релаксационного растяжения и растягивания с сокращением агониста.
Фиксационно-релаксационная методика предназначается для растягивания мышцы, когда двигательные альфа-нейроны наименее возбудимы, в результате чего афферентный входной сигнал от детекторов длины (мышеч
264
ных веретен), вероятно, в наименьшей степени вызывает обусловливаемое растяжением возбуждение мышцы. Для использования этой возможности непосредственно после изомет-ричных сокращений измерялись как рефлексы Хоффманна (Н), так и сухожильные рефлексы на легкий удар (Т) (Enoka, Hutton, Eldred, 1980; Moore, Kukulka, 1991). Эти данные наводят на мысль о том, что возбудимость как мышечных веретен, так и двигательных нейронов уменьшается непосредственно после изометрического сокращения, и это уменьшение продолжается в течение около 10 с (Crone, Neilsen, 1989). Более того, ослабление рефлексов аналогично для сокращений, продолжительность которых изменяется в пределах 1—30 с.
Логическим обоснованием методики сокращения агониста является возбуждение рефлекса реципрокного торможения (см. главу 5) в отношении антагониста (мышца, которая должна быть растянута) в результате сокращения агониста. В этой схеме произвольное возбуждение агониста включает возбуждение двигательных альфа- и гамма-нейронов и интернейрона (I), который является промежуточным звеном для рефлекса реципрокного торможения (рис.8.6). (Nielsen, Kagamihara, Crone, Hultbom, 1992). Этот интернейрон известен в качестве тормозящего интернейрона 1а (см.гла-ву 5). Его возбуждение обусловливает действие потенциалов, передаваемых двигательным нейроном, которые иннервируют мышцу-ан-тагонист, с последующим снижением возбудимости этих двигательных нейронов. Следовательно, механизм сокращения агониста предполагает включение возбуждения агониста и посредством реципрокного торможения релаксацию антагониста.
В процессе различных исследований сравнивалась эффективность методик растягивания (Condon, Hutton, 1987; Enture, Abraham, 1986; Guissard et al., 1988; Wallin, Ekblom, Grahn, Nordenborg, 1985). Результаты показывают, что улучшение гибкости при статическом растягивании подобно тому, которое достигается при баллистическом растягивании, но баллистические растягивания, скорее всего, обусловливают болезненность мышц (Shellock, Prentice, 1985).
Исследования методик растягивания, улучшающих нервно-мышечную передачу проприоцептивных импульсов, не определили относительного значения невральных механизмов для улучшения гибкости. Было выдвинуто предположение, что диапазон движения относительно большинства суставов ограничивается одной или несколькими структурами соединительной ткани, включая те, которые находятся в мышцах (Sapega, Quedenfeld, Moyer, Butter, 1981). Вклад различных структур суставов в жесткость сустава неодинаков: для пястно-фалангового сустава было установлено 47 % для суставной кап-
Рис.8.6. Невральные пути, возбуждаемые во время растягивания сухожилий, ограничивающих с боков подколенную ямку, в соответствии с методикой сокращения агониста. Возбуждение квадрицепс фемо-рис супрасегментарными центрами приводит к одновременному возбуждению двигательных нейронов, иннервирующих квадрицепс феморис, и тормозящего интернейрона 1а, который является промежуточным звеном для рефлекса реципрокного торможения. Это, очевидно, обусловливает релаксацию антагониста (сухожилия, ограничивающие с боков подколенную ямку) во время возбуждения агониста (квадрицепс феморис)
суды, 41 % для мышцы, 10 % для сухожилий и 2 % для кожи (Johns, Wright, 1962). Это значит, что диапазон движения пястно-фалангового сустава определяется пассивными свойствами этих тканей в данном соотношении. Неясно, каким образом на эти соотношения влияет релаксация мышц. Хотя методики улучшения нервно-мышечной передачи проприоцептивных импульсов и могут минимизировать индуцируемое растягиванием возбуждение мышц, функциональное значение этого снижения для улучшения гибкости не установлено. Альтернативно улучшения диапазона движения, обусловливаемые этими методиками, могут быть связаны с температурными эффектами.
В отсутствие неврального входного сигнала последовательные растягивания могут увеличивать длину мышечно-сухожильной единицы (Taylor, Dalton, Seaber, Garrett, 1990). В этих экспериментах мышцу задней конечности (экс-тензор-дигиторум лонгус) растягивали под исходным усилием около 2 Н до длины, которая обусловливала пассивное усилие 78 Н, и вы-
265
Рис.8.7. Индуцируемые растягиванием изменения длины (а) и усилия мышечно-сухожильной единицы (б) (Taylor, Dalton, Seaber, Garrett, 1990)
держивали при этой длине в течение 30 с. Этот цикл растяжения повторяли 10 раз. На рис.8.7, а показано, что при последующих растяжениях мышцы необходимо было растягивать до больших длин для создания усилия 78 Н; общее увеличение длины после 10 растяжений составило примерно 3,5 %. Была использована также обратная методика, в соответствии с которой мышцу растягивали на 10 % и при последующих растяжениях измеряли создаваемое усилие (рис.8.7,б). Эти измерения показывают, что при отсутствии неврального входного сигнала нагрузка на мышечно-сухожильную единицу, обусловливаемая ее пассивными свойствами (соединительная ткань), будет уменьшаться при растягиваниях до предписываемой длины.
Несмотря на неопределенность вклада невральной бессимптомности в изменения гибкости, соединительная ткань, очевидно, играет важную роль в ограничении диапазона движения (Gebhard et al., 1993; Shellock, Prentice, 1985). Поэтому упражнения на гибкость должны быть направлены на изменение длины структур со
единительной ткани. Для достижения этой цели упражнения на гибкость должны обусловливать пластичные, а не упругие изменения соединительной ткани, так как пластичные изменения обусловливают более постоянные изменения длины тканей. Относительные соотношения упругости и пластичности в пределах растягивания определяются прилагаемым усилием и продолжительностью его приложения, а также температурой тканей. Продолжительное растягивание при низком усилии оптимизирует пластичные изменения. Существует компромисс между удлинением и ослаблением: пластичные изменения, индуцируемые в ткани, обусловливают молекулярную перестройку и ослабление ткани. Тем не менее это напряжение стимулирует адаптацию ткани, и ослабление является лишь кратковременным при условии, что напряжение не слишком большое. Ткань наиболее растяжима при более высокой температуре, например, в конце тренировки, а долговременное удлинение наибольшее, если растягивание осуществляется после охлаждения ткани (Sapega et al., 1981). В некоторых случаях предлагается во время растягивания даже применение пузырей со льдом для содействия эффекту охлаждения. Следовательно, эффективное растягивание для улучшения гибкости должно предусматривать продолжительные растяжения в условиях низкого усилия и выполняться в конце тренировки.
БОЛЕЗНЕННОСТЬ И ПОВРЕЖДЕНИЕ МЫШЦ
Энергичная физическая активность может оказывать на мышцу различное влияние в пределах от внутриклеточного повреждения мышечных волокон до индуцируемых растяжением мышечных травм (деформаций). Внутриклеточное повреждение, которому подвергались наиболее активные субъекты, часто обусловливает воспитательную реакцию и связано с болезненностью мышц, которая начинается через несколько часов после выполнения упражнения. В противоположность этому деформационная травма обычно проявляется в виде острой болезненной травмы во время выполнения задач, требующих высокого напряжения, и обусловливает необходимость клинического вмешательства.
Болезненность мышц
Поскольку болезненность, связанная с внутриклеточным повреждением, не проявляется до истечения 24—48 ч после выполнения упражнения, она была описана как мышечная болезненность с задержкой проявления. Этот термин отличает болезненность после выполнения уп
266
ражнения от боли вследствие напряжения, которая может появляться во время выполнения упражнения (Asmusse, 1953, 1956). Клинические симптомы, связанные с мышечной болезненностью с задержкой проявления, включают повышение содержания в плазме ферментов (например, креатинкиназы), миоглобина и белковых метаболитов из травмированных мышц, структурное повреждение внутриклеточных компонентов мышечных волокон, наблюдаемое посредством световой и электронной микроскопии, и временное усиление мышечной слабости (Armstrong, 1990).
Причина мышечной болезненности. Хотя нет единого мнения о фактическом механизме, обусловливающем мышечную болезненность с задержкой проявления, существует согласие в том, что она чаще возникает при эксцентрических сокращениях, чем при других режимах возбуждения мышц (McCully, Faulkner, 1985; Newham, 1988; Stauber, 1989). Почему мышечная болезненность возникает при концентрических сокращениях? Напомним, что при эксцентрическом сокращении активная мышца удлиняется, так как вращающий момент, обусловливаемый нагрузкой, превышает вращающий момент, создаваемый мышцей. По этой схеме циклическое изменение поперечных мостиков может включать изменение фазы разъединения, во время которой механически разрушается актомиозиновая связь (Morgan, 1990), и может иметь место избирательное возбуждение более крупных двигательных единиц (Nardone et al., 1989). При эксцентрической физической нагрузке, характеризующейся использованием меньшего количества двигательных единиц, более низкой кислородной стоимостью при езде на велосипеде (рис.8.8, а), постепенным повышением потребления кислорода (рис.8.8, б) и постепенным увеличением ЭМГ (рис.8.8,в) во время бега под уклон в постоянном темпе, усилением мышечной слабости после тренировки и повышением
пикового внутримышечного давления вследствие увеличения влагосодержания мышцы (рис.8.8) (Evans, Cannon, 1991; Newham, Jones, Edwards, 1983, Stauber, 1989).
Было выдвинуто предположение о том, что физическая активность усиливает как метаболические, так и механические факторы, которые могут повредить мышцу и обусловить болезненность (Armstrong, 1990). Метаболические факторы включают: а) высокую температуру, которая может разрушать белковые структуры; б) недостаточное митохондриальное дыхание, которое может снижать уровни аденозинтрифосфата и, следовательно, энергии, необходимой для удаления кальция из цитоплазмы; в) пониженный pH, обусловливаемый повышенным содержанием молочной кислоты; г) образование свободных радикалов О2 и пероксидное окисление липида, которые могут инициировать мышечную травму. Поскольку эксцентрические сокращения связаны с большими усилиями, общим предположением является то, что механическим фактором, вызывающим мышечную болезненность, является высокое напряжение (Warren, Hayes, Lowe, Armstrong, 1993; Warren, Hayes, Lowe et al., 1993). Было выдвинуто предположение, что высокое напряжение (усилие на единицу площади) может приводить к разрушению сарколеммных, саркоплазматических сетчатых и миофибриллярных структур. Однако Friden и Leiber показали, что величина деформации, а не напряжение более важный механический фактор (Friden, Leiber, 1992; Leiber, Friden, 1993). Хотя эти исследования и свидетельствуют о том, что несколько событий могут обусловливать болезненность мышц, единственным механизмом, который представляется общим для данных метаболических и механических событий, является потеря клеточного кальциевого гомеостаза (Clarkson, Cymes, McCarmick et al., 1986; Jackson, Jones, Edwards, 1984). Высокая концентрация внутриклеточно
Рис.8.8. Интенсивность потребления кислорода и ЭМГ, связанная с эксцентрическими сокращениями (Evans, Cannon, 1991; Dick, Cavanaugh, 1987):
a—расход кислорода при осуществлении концентрических ( Т) и эксцентрических (2) сокращений на велоэргометре; б — потребление кислорода при беге по равнине и под уклон 10 % вастус латералис во время бега; в — интегрированная ЭМГ
267
го кальция возбуждает протеолитические и липолитические структуры, которые могут запускать автогенез — процесс разрушения клеточных структур (Armstrong, 1990).
Временное течение реакции. Структурные нарушения после тренировки включают сарко-леммное разрушение, расширение поперечной трубочной системы, деформацию миофибрил-лярных компонентов, фрагментацию саркоплазматической ретикулярной ткани, патологические изменения клеточной оболочки, повреждение цитоскелета, изменение внеклеточной миофибриллярной матрицы и набухание митохондрий (Friden, Lieber, 1992; Stauber, 1989). Многие структурные эффекты концентрических сокращений включают Z-линию. Микрофотографии показали четкую связь характера Z-линии и возможного повреждения цитоскелетных промежуточных волокон при очень сильных сокращениях (Friden, Kjorell, Thomell, 1984).
Хотя это повреждение и проявляется непосредственно после тренировки, изменения продолжаются в течение послетренировочного периода; они включают повышенное выделение специфичных для мышц ферментов и увеличение проницаемости клеток. Общий маркер повреждения мышц — фермент креатин-киназа, отражающий обмен белков, становящихся дисфункциональными под действием физического или окислительного напряжения (Evans, Cannon, 1991). Отток ферментов происходит вследствие диффузии через индуцируемые тренировкой отверстия в плазменной
мембране мышечной клетки. Послетренировоч-ное увеличение содержания ферментов ске летных мышц в крови связано с типом и интенсивностью тренировки. Оно больше при эксцентрических сокращениях, чем при концентрических, и вырастает с интенсивностью тренировки. Интересно, что эстроген может непосредственно влиять на клеточную оболочку мышцы и снижать отток ферментов скелетных мышц. Поэтому у женщин послетренировоч-ные уровни циркулирующей креатинкиназы ниже (Evans, Cannon, 1991).
Мышечная болезненность с задержкой проявления наиболее резко выражена в течение 24—48 ч после тренировки (рис.8.9). Хотя и имеют место структурные повреждения непосредственно после напряженной тренировки, значительные изменения происходят в течение нескольких дней после нее (Howell, Chleboun, Conatser, 1993). В период от 4 ч до 4 сут повышается фагоцитарная активность, что указывает на наличие воспалительной реакции. В связи с таким развитием событий во времени было выдвинуто предположение, что мышечная болезненность с задержкой проявления — следствие воспалительной реакции (Stauber, 1989). Тем не менее последствия напряженной тренировки, в частности, эксцентрических сокращений, не ограничиваются несколькими днями после нее. Исследования магнитнорезонансных изображений показали заметное опухание травмированных мышц в течение до 10 сут и увеличение интенсивности сигналов в течение до 60 сут
Рис.8.9. Изменение воспринимаемой мышечной болезненности в течение 72 ч после 40-ми яутного бега под уклон (Dick, Cavanaugh, 1987)
268
после нее (Fleckenstein, Shellock, 1991; Shellock, Fukunaga, Mink, Edgerton, 1991b).
Влияние типа волокон. Мышечные волокна типа Пб (FG) наиболее повреждаются вследствие тренировки (Friden, Seger, Ekblom, 1988). Анализ ультраструктуры мышц показал, что через 2 ч после повторяющихся забегов на короткую дистанцию 36 % мышечных волокон, исследованных в вастус латералис, характеризовались значительными изменениями, которые оказывали влияние, в основном, на Z-линию. 80% этих отклонений происходили в волокнах типа Пб. Напомним, что волокна типа Пб имеют узкие Z-линии и характеризуются низкой окислительной способностью. Основываясь на этих наблюдениях Friden, Lieber (1992) предложили следующую схему повреждения мышц: утомление волокон типа Пб (FG) обусловливает неспособность восстановления аденозинтрифосфата и развитие состояния мышечной ригидности (высокой жесткости), вследствие чего последующее растягивание приводит к механическому повреждению миофибриллярных и цитоскелетных структур.
Повреждение мышечных волокон не ограничивается сократительными белками и внутримышечным цитоскелетом. Одной из наиболее пластичных является соединительная ткань, которая также может подвергаться индуцируемому тренировкой повреждению. Влияние тренировки на соединительную ткань часто оценивается по результатам измерения уровня выделяемого мочой оксипролина — компонента созревшего коллагена. Выделение оксипролина увеличивается после тренировки, включающей эксцентрические сокращения (Stauber, 1989). К тому же мышца, подвергающаяся принудительному удлинению, характеризуется гистологическими изменениями в матрице внеклеточной соединительной ткани. Поскольку в мышечных волокнах типа II слабо развита внеклеточная матрица, они более восприимчивы к индуцируемому растяжением повреждению.
Восстановление. Armstrong (1990) идентифицировал четыре стадии, связанные с индуцируемым тренировкой повреждением мышц: начальную, автогенетическую, фагоцитарную и восстановительную. Начальная стадия — это тренировочный прием, обусловливавший повреждение. Автогенетическая стадия включает возбуждение протеолитической и липолитической систем для инициирования разрушения клеточных структур. Фагоцитарная стадия длится примерно от 4 ч до 4 сут после тренировочного приема и включает воспалительную реакцию. Заключительная стадия, во время которой происходит восстановление мышечных волокон, видимо, начинается примерно через 4 сут.
Функциональными последствиями этих реакций являются первоначальное снижение мышечной силы и уменьшение диапазона движе
ния вследствие опухания и повышения давления на болевые рецепторы (Stauber, 1989). Однако эти эффекты кратковременны, и мышечные волокна и соединительная ткань в поврежденных мышцах восстанавливаются, для чего необходимы здоровый базальный тонкий слой, непрерывное васкулярное питание, функциональная иннервация, а также, вероятно, клетки-сателлиты и процессы, подобные тем, которые протекают во время развития. Мышечные волокна могут восстанавливаться и возобновлять нормальное функционирование после повреждения, обусловливаемого тренировкой. Хотя процесс восстановления и представляется изменяющим ультраструктуру саркомеров (Friden, 1984), волокна не отличаются физиологически. Тем не менее, согласно имеющимся данным, отдельный цикл тренировки предотвращает повреждение мышц при последующих тренировках в течение около 6 недель (Byrnes et al., 1985; Newham, Jones, Clarkson, 1987). Адаптация может включать укрепление соединительной ткани (Clarkson, Tremblay, 1988). Однако ни растягивание, ни разминка перед тренировкой не предотвращают мышечную болезненность с задержкой проявления после цикла интенсивной тренировки (High, Howley, Franks, 1989).
Деформация мышц
Кроме микротравм, которые часто сопровождают эксцентрические сокращения, мышца может подвергаться кратковременным и болезненным воздействиям, например, судорогам и деформациям. Мышечная судорога — болезненное непроизвольное сокращение мышцы, запускаемое, очевидно, периферийными стимулами (Bertolasi, De-Grandis, Bongiovanni et al., 1993). Мышечная деформация — это существенное повреждение, которое сразу же распознается в качестве травмы. Мышечные деформации называют также растяжениями и разрывами. Согласно клиническим данным, эти деформации постоянно происходят в соединении мышцы и сухожилия; деформации мышц сообщались для медиал гастрокнемиус, ректус феморис, трицепс брахии, аддуктор лонгус, пекторалис майор и семимембраносус (Garrett, 1990). Наиболее подвержены такой травме двусуставные мышцы (так как они могут больше растягиваться), мышцы, которые ограничивают диапазон движения относительно сустава, и мышцы, характеризующиеся большим относительным содержанием мышечных волокон типа II (Garrett, Califf, Bassett, 1984). Более того, травмирование наиболее часто происходит во время энергичных эксцентрических сокращений, когда усилие в несколько раз превышает максимальное изометрическое усилие. Травма час-
269
Рис.8.10. Энергия деформации, поглощаемая мышечно-сухожильной единицей во время растягивания до повреждения (Garrett, Safran, Seaber et al., 1987): 7 — активная мышца; 2 пассивная мышца
то включает кровотечение и последующее накапливание крови на подкожных участках. Наиболее подходящее лечение заключается в отдыхе мышцы, приложении льда и наложении компресса. Реабилитация должна включать физиотерапию для увеличения диапазона движения и функциональные упрочняющие упражнения.
Экспериментальные исследования характеристик деформационной травмы мышц показали, что разрыв происходит вблизи соединения мышцы и сухожилия, несмотря на различия в архитектурных особенностях испытанных мышц и направлении деформации (Garrett, 1990). При пассивном растягивании различных мышечно-сухожильных единиц (мышца не сокращается) деформационная травма не имела места после постоянной деформации волокон. Тем не менее не наблюдалось какое-либо отличие в общей деформации до повреждения при растягивании данной мышечно-сухожильной единицы, когда оно было пассивным, по сравнению с возбуждением (сокращение мышцы стимулировалось электрически). Однако сокращение мышцы может удвоить энергию деформации, которая может быть поглощена мышечно-сухожильной единицей во время растягивания (эксцентрическое сокращение) (Garrett, Safran, Seaberet al, 1987). На рис.8.10,а энергия деформации показана в виде площади ниже графика зависимости усилия от длины с индуци
руемым растягиванием повреждением при пиковом усилии. На рис.8.10,б энергия деформации (площадь ниже кривой), поглощаемая активной мышцей, примерно в два раза больше энергии, поглощаемой пассивной мышцей, хотя пиковое усилие при повреждении лишь примерно на 15% больше в активной мышце. Из-за воздействия сокращения мышцы любой фактор, например, усталость или ослабление, который стремится снизить сократительную способность мышцы, может предрасположить мышечно-сухожильную единицу к деформации мышцы.
МЫШЕЧНОЕ УТОМЛЕНИЕ
Мышечным утомлением называется класс кратковременных эффектов, ухудшающих работоспособность. Они включают как двигательные, так и сенсорные процессы. Поскольку эти процессы влияют на рабочие характеристики, значительное внимание было сосредоточено на идентификации звена, которое обусловливает утомление. Тем не менее исследование, проведенное в течение 50 последних лет, четко продемонстрировало, что утомление обусловливается не ухудшением отдельного процесса, а механизмами, которые вызывают утомление и изменяются от одного сос-тояния к другому. Этот эффект был назван зависимостью утомления мышц от задачи (Enoka, Stuart, 1992). Чтобы понять утомление мышц и его причины, необходимо идентифицировать условия, которые могут ухудшать различные процессы и способствовать утомлению мышц. В данном разделе в общих чертах рассматривается то, что известно об этих условиях, и обсуждаются некоторые из сенсорных адаптаций, которые происходят во время утомляющих сокращений.
Зависимость от задачи
Когда человек выполняет задачу, требования задачи (например, величина усилия, участвующие мышцы, продолжительность действия) усиливают различные физиологические процессы, связанные с двигательной характеристикой. При изменении этих требований имеют место процессы, которые наиболее усиливаются. Переменные задачи, которые представляются оказывающими влияние на увеличивающиеся процессы, включают уровень мотивации субъекта, невральную стратегию (схему возбуждения мышц и двигательной команды), интенсивность и продолжительность действия, скорость сокращения и степень, в которой поддерживается непрерывность действия. Физиологические процессы, которые могут ухудшаться этими пере-
270
100
25 -
0 4-о
75
50-----
• 50 Гц
О Tws
О МПС
□
°П □
д
5
Рис.8.11. Уменьшение усилий при максимальном произвольном сокращении и вызываемых искусственно усилии во время утомляющего сокращения мышцы квадрицепс феморис (а) и изменение интегральной ЭМГ для квадрицепс феморис во время максимального произвольного сокращения и прерывистых субмаксимальных сокращений в процессе выполнения задачи (б) (Bigland-Ritchie, Furbush, Woods, 1986):
МПС — максимальное произвольное сокращение
Рис.8.12. Метод наложения сокращений: уменьшение амплитуды усилия судороги, вызываемой электрическим ударом, с увеличением произвольного усилия, создаваемого квадрицепс феморис (а), и уменьшение усилия судороги в функции произвольного усилия (6). Стимулирующее сокращение вызывалось при различных уровнях усилия во время произвольных сокращений. Усилие максимального произвольного сокращения составляло около 470 Н (Bigland-Ritchie, Cafarelli, V01Iestad, 1986)
а
5 ф
1
2
□ □
3
4
менными, включают возбуждение двигательных нейронов ЦНС, мышцы и двигательные единицы, которые возбуждаются, нервно-мышечное преобразование, сочетание возбуждения и сокращения, наличие метаболических субстратов, внутриклеточную среду, сократительный аппарат и мышечный кровоток.
Центральное возбуждение. Обычно максимальный уровень возбуждения, обеспечиваемого нервной системой, исследуется сравнением усилия, создаваемого во время максимального произвольного сокращения, с усилием, которое может быть вызвано искусственно при электростимуляции. Когда субъекты осуществляли непрерывное 60-секундное максимальное произвольное сокращение мышцы большого пальца кисти (аддуктор поллицис), усилие уменьшалось на 30—50 %, но это уменьшение не могло быть компенсировано электрическим ударом (Bigland-Ritchie et al., 1982). Аналогично при осуществлении субъектами прерывистого (сокра
щение в течение 6 с, отдых в течение 4 с) субмаксимального сокращения (заданное усилие составляло 50 % максимального) квадрицепс феморис, максимальное произвольное и вызываемое электрически усилия уменьшались параллельно (рис.8.11) (Bigland-Ritchie, Furbush, Woods, 1986). В этом эксперименте максимальное произвольное и вызываемое электрическим током усилия создавались периодически во время субмаксимального сокращения. Параллельное уменьшение произвольного и искусственного усилия наводит на мысль о том, что центральное возбуждение оставалось максимальным во время выполнения этих задач.
Результаты экспериментов в двух тестах искусственного стимулирования мышцы представлены на рис.8.11. В одном тесте использовалась последовательность из восьми стимулов, которые подавались с частотой 50 Гц. Часть мышцы, возбуждаемая этим стимулом, зависела от количества электрического тока, которое пропуска
271
лось через мышцу стимулирующим устройством. Для мышечных волокон, которые возбуждались, частота стимулирования 50 Гц вызывала близкое к максимальному усилие. Во втором тесте изучали наложение мышечных сокращений Tws. Он включает приложение к нерву одного-трех сверхмаксимальных электрических ударов во время максимального произвольного сокращения (МПС) и наблюдение, увеличивается ли усилие. Если усилие МПС возрастает, центральное возбуждение во время сокращения не является максимальным. В эксперименте, представленном на рис.8.12, с увеличением произвольного усилия усилие судороги, вызываемой накладываемым электрическим ударом, уменьшается. Во время прерывистого утомляющего сокращения (рис.8.11) амплитуда усилия накладываемого сокращения уменьшалась до нуля, в то время как усилие МПС достигало заданного уровня (50 % МПС); следовательно, центральное возбуждение было максимальным в конце исследования утомления.
Рис.8.13. Изменения вращающего момента (а) и ЭМГ (6) во время концентрических (незачерненные обозначения) и эксцентрических (зачерненные обозначения) утомляющих сокращений мышцы квадрицепс феморис (Tesch, Dudley, Duvoisin et al., 1990)
Исследования показывают, что невральное возбуждение мышцы, обеспечиваемое ЦНС, при утомлении не всегда максимально, и уменьшение центрального возбуждения может быть фактором, способствующим уменьшению усилия. В литературе существует, по меньшей мере, три примера неспособности людей генерировать надлежащее центральное возбуждение во время утомляющей деятельности. Во-первых, субъекты, которые не мотивируются максимально, не характеризуются параллельным уменьшением произвольного и искусственного (электрический стимул) усилий (Bigland-Ritchie et al., 1986; Gandevia, McKenzie, 1985). Эта неспособность представляется в виде отклонения линий двух усилий (МПС и 50 Гц) на рис.8.11, а. Отсутствие мотивации, вероятно, обусловливает неадекватность центрального возбуждения соответствующих двигательных нейронов. Во-вторых, представляется, что некоторые мышцы возбудить гораздо труднее, чем другие (Milner-Brown, Miller, 1989; Thomas, Woods, Bigland-Ritchie, 1989). В-третьих, утомляемость мышцы различна для концентрических и эксцентрических сокращений (рис.8.13). Субъекты осуществляли максимальные сокращения квадрицепс феморис на изокинетичес-ком устройстве, которое было настроено на диапазон движения в пределах 1,2 рад и скорость 3,14 рад'с-1. Субъекты выполняли три цикла по 32 сокращения как в концентрических, так и в эксцентрических условиях. Вращающий момент, который создавался во время концентрических сокращений, уменьшался, а крутящий момент для эксцентрических сокращений несколько увеличивался (рис.8.13,с). Более того, записи интегральной ЭМГ как для вастус латералис, так и для ректус феморис были меньше во время эксцентрических сокращений (рис.8.13, б). Диссоциация между вращающим моментом и ЭМГ для двух видов сокращений наводит на мысль о существенном различии в возбуждении мышц.
Другой подход, в соответствии с которым электрические стимулы используют для оценки роли центрального возбуждения в ограничении работоспособности в течение непрерывных действий, предусматривает воздействие гуморальных факторов в процессе циркуляции в цереброспинальной жидкости. Эти факторы, например, адреналин и нейромедиатор 5-гид-рокситриптамин, могут играть важную роль в поддержании центрального возбуждения во время выполнения утомляющих задач (Blomstrand, Celsing, Newsholme, 1988; Heyes, Garnett, Coates, 1988). Следовательно, уровни гормонов, циркулирующих в цереброспинальной жидкости, являются важным фактором поддержания центрального возбуждения.
Невральная стратегия. Результирующее мышечное усилие относительно сустава может
272
достигаться посредством множества схем возбуждения мышц. Эта гибкость имеет место в группе мышц-синергистов и, возможно, среди двигательных единиц в отдельных мышцах. Благодаря такой возможности один из путей, посредством которого двигательная система может задерживать начало уменьшения усилия (утомление), заключается в изменении вклада мышц-синергистов в результирующее мышечное усилие. Эта возможность предоставляется лишь в том случае, когда задача требует субмаксимальных усилий, но большинство видов деятельности в повседневной жизни относится к этой категории. Хороший пример такого распределения активности среди мышц-синергис-тов получен Sj^gaard et al., когда субъекты поддерживают изометрическое усилие экстензора колена на уровне 5 % усилия МПС в течение 1 ч (Sj^gaard, Kiens, Jorgensen, Saltin, 1986; Sjogaard, Savard, Juel, 1988). Несмотря на низкую интенсивность задачи, субъекты испытывали усталость, о чем свидетельствует 12%-е уменьшение усилия МПС и увеличение связанного с этим напряжения в конце часа. Во время утомляющего сокращения субъекты были способны сохранять постоянное усилие (5 % МПС), в то время как активность переключалась с ректус феморис на вастус латералис (рис.8.14). Наблюдение показывает, что невральное возбуждение мышц изменялось, в то время как результирующая производительность оставалась постоянной. Аналогичным образом, когда экстензоры колена осуществляют прерывистые сокращения низкой интенсивности до предела выносливости, имеет место постепенное увеличение совместной активности флексоров колена (Psek, Cafarelli, 1993).
Хотя двигательная активность одной конечности и может оставаться постоянной, скорость уменьшения усилия зависит от того, одна или две конечности действуют во время выполнения задачи. Например, Rube, Secher (1990) исследовали способность субъектов выполнять задачу изометрического разгибания ноги (одновременное разгибание коленного и тазобедренного суставов) одной или двумя ногами. Субъекты должны были выполнить 150 МПС для обоих условий (одна и две ноги) перед силовой тренировкой и после пяти недель тренировки. Они были разделены на три группы: контрольную группу, группу тренировки на одной ноге и группу тренировки на двух ногах. Группы тренировки как на одной ноге, так и на двух ногах увеличили силу после пяти недель тренировки, в то время как контрольная группа осталась на прежнем уровне. Более того, скорость уменьшения усилия во время 150 сокращений была меньше после тренировки (рис.8.15). Однако эффект был специфичен для режима тренировки. Группа тренировки на одной ноге подвергалась меньшему утомлению лишь во
Рис.8.14. Переменные вклады вастус латералис и ректус феморис в поддерживаемое усилие экстензоров колена. Внутримышечное давление измерялось в ректус феморис (Sjdgaard, Kiens, Jorgensen, Saltin, 1986)
время выполнения задачи для одной ноги, а группа тренировки на двух ногах подвергалась меньшему утомлению лишь во время выполнения задачи для двух ног. Эти данные наводят на мысль о том, что невральная стратегия, используемая во время выполнения задачи для одной ноги, отличалась от невральной стратегии при выполнении задачи для двух ног.
На уровне двигательных единиц два наблюдения свидетельствуют о том, что возбуждение двигательных единиц (невральная стратегия) может изменяться во время утомляющих сокращений. Во-первых, двигательные единицы, используемые во время субмаксимального изометрического сокращения, могут изменяться перед утомляющим сокращением и после него. Когда субъекты выполняли задачу сжатия и удерживания (увеличение усилия сжатия до достижения цели с последующим ослаблением сжатия до исходного уровня) с использованием мышцы кисти руки, некоторые двигательные единицы после утомляющего сокращения не использовались снова, хотя задача и была идентичной той, которая выполнялась перед задачей на утомление (см.рис.7.35) (Enoka, Robinson, Kossev, 1989). Эти данные наводят на мысль о наличии некоторой гибкости в использовании двигательных единиц, которая основывается на истории возбуждения мышцы (Person, 1974). Во-вторых, порядок использования двигательных единиц может отличаться для концентрических и эксцентрических сокращений (см.рис.7.34). (Nardone, Romano, Schieppati, 1989). Неизвестны ни диапазон усилий, в пределах которого может иметь место это различие, ни его значение для утомления.
В дополнение к этим различиям в использовании двигательных единиц их способность поддерживать активность зависит также от того, является ли сокращение изометрическим. Например, медленносокращающиеся двигательные единицы могут поддерживать изометрическое усилие дольше, чем быстросокращающиеся
18 8-133
273
(Burke et al., 1973), а медленносокращающаяся мышца (soleus) может выдерживать большее усилие во время сокращений с постоянной скоростью укорочения, чем бастросокращающаяся (экстензор дигиторум лонгус) (Brooks, Faulkner, 1991). Тем не менее быстросокращаюшаяся мышца может выдерживать большее генерирование энергии в течение 5 мин, чем медленно-сокрашающаяся (Brooks, Faulkner, 1991). Это различие обусловливается биохимическими различиями по оптимальной скорости укорочения для медленно- и быстросокращающейся мышцы. Быстросокращаюшаяся мышца характеризуется более высокой оптимальной скоростью укорочения, которая сочетается с меньшим поддерживаемым усилием для генерирования большего количества энергии. Несомненно, это различие по способности создавать усилие и генерировать энергию имеет функциональное значение, так как многие движения основываются на способности мышцы генерировать энергию, а не просто создавать усилие.
Нервно-мышечное преобразование. В преобразовании аксонного потенциала действия в сар-колеммный потенциал действия участвуют различные процессы. Их совокупность называется нервно-мышечным преобразованием. Поддерживаемая активность может ослаблять некоторые из процессов, участвующих в нервно-мышечном преобразовании, что может обусловливать уменьшение усилия, связанного с утомлением.
Возможные ослабления включают неспособность аксонного потенциала действия оказывать влияние на все ветви аксона (недостаточность в точках ветвей), истощение нейромедиатора, ослабление экзоцитоза и снижение чувствительности постсинаптической мембраны (Kugelberg, Lindegren, 1979; Spira, Yarom, Pamas, 1976).
Наиболее общий путь проверки ухудшения нервно-мышечного преобразования у людей заключается в исследовании М-волн перед утомляющим сокращением, во время него и после него. Напомним, что М-волна измеряется путем приложения электрического удара к нерву для генерирования потенциалов действия в аксонах альфа-мотонейронов и измерения по ЭМГ реакции мышцы (см.рис.5.28). Уменьшение амплитуды М-волны свидетельствует об ослаблении одного или нескольких процессов, участвующих в преобразовании аксонного потенциала действия (инициируемого электрическим ударом) в мышечный (сарколеммный) потенциал действия. На рис.8.16 показано уменьшение М-волны непосредственно после утомляющего сокращения и последующего восстановления формы волны после отдыха в течение 10 мин. Уменьшение амплитуды М-волны этого типа происходит в соответствии с тенденцией в процессе продолжительных сокращений низкой интенсивности и реже — в процессе кратковременных высокоинтенсивных сокраще-
Задача для одной ноги Задача для двух ног
Тренировка на одной ноге
А»
ЧМА /2:2
о До тренировки
• После тренировки
I---------------------------1--------------------------1----------------------------1
Тренировка на двух ногах
ф X R S .О
50 J
I----------1----------1--------п । 1 1 ।
0 50 100 150 0 50 100 150
Количество сокращений
Рис.8.15. Влияние тренировки на уменьшение усилия во время выполнения задачи разгибания одной и двух ног (Rube, Secher, 1990)
274
Рис.8.16. M-волны, полученные в мышце кисти руки (первая тыльная межкостная) в результате стимуляции локтевого нерва. М-волны получены перед утомляющим сокращением и после него, а также после восстановления в течение 10 мин. Выполнялось утомляющее сокращение, поддерживающее изометрическое усилие на уровне 35% усилия МПС, в течение возможно большего времени. Амплитуда M-волны уменьшалась непосредственно после выполнения задачи на усталость, но быстро восстанавливалась (Fuglevand, Zackowski, Huey, Enoka, 1993)
ний (Bellemare, Garzaniti, 1988; Fuglevand, Zackowski, Huey, Enoka, 1993; Milner-Brown, Miller, 1986). Согласно модельным исследованиям, ухудшение нервно-мышечного преобразования — один из механизмов, которые могут обусловливать уменьшение усилия во время выполнения задач этих типов (Fuglevand et al., 1993).
Связь возбуждения с сокращением. В нормальных условиях возбуждение нервной системой приводит к активации мышцы и соответствующему циклическому изменению поперечных мостиков. Семь процессов (Fitts, Metzger, 1988) обусловливают преобразование возбуждения (потенциал действия) в усилие мышечных волокон (см.рис.5.22): 1) распространение потенциала действия вдоль сарколеммы; 2) распространение потенциала действия по поперечному канальцу; 3) изменение обусловливаемой ионами Са2+ проводимости саркоплазматической сети; 4) перемещение ионов Са2+ в соответствии с их градиентом концентрации в саркоплазму; 5) повторное поглощение ионов Са2+ саркоплазматической сетью; 6) связывание ионов Са2+ с тропонином; 7) взаимодействие миозина и актина и выполнение работы поперечным мостиком.
Некоторые из этих процессов, как известно, подвержены влиянию внутриклеточных изменений, связанных с поддерживаемой активностью. Например, изменения во внутриклеточной среде уменьшают количество выделяемых ионов Са2т (процесс 4) и количество ионов Са2т, возвращаемых (процесс 5) в саркоплазматическую сеть (Allen et al., 1989; Westerblad, Lannergren, 1990). Уменьшение процесса 4 (количество выделяемых ионов Са2+), вероятнее всего, обусловливается ухудшением процесса выделения, возможно, вследствие инактивации каналов выделения Са2+ и истощения запасов Са2+ в саркоплазматической сети (Webb, Hibberd, Golman, Trentham, 1986). Уменьшение процесса 5 может быть обусловлено увеличе
нием внутриклеточного связывания ионов Са2+, которое понизило бы градиент концентрации и, следовательно, поток ионов Са2+ через саркоплазматическую сеть. Несмотря на эти возможности, относительное значение данных механизмов для уменьшения усилия в естественных условиях остается определить.
Один из путей иллюстрации роли ухудшения связи возбуждения с сокращением при утомлении заключается в подтверждении того, что уменьшение усилия не может быть обусловлено невральными или метаболическими факторами. Это было показано (Bigland-Ritchie, Cafarelli, V^llestad, 1986) для задачи, в соответствии с которой субъектов просили выполнить ряд 6-секундных сокращений мышцы квадрицепс феморис до усилия, уровень которого составлял 30% максимума. После каждого 6-се-кундного сокращения предусматривался 4-секундный отдых. Субъекты выполняли эту последовательность в течение 30 мин. Усилие, создаваемое во время МПС, уменьшалось параллельно возбуждаемому электрически усилию (см.рис.8.11, а), что свидетельствует о том, что субъекты произвольно создавали такое усилие, которое была способна создать мышца. Не имели места какие-либо значительные изменения количества мышечного лактата, аденозинтрифосфата или креатинфосфата, а снижение содержания гликогена было минимальным и ограничивалось мышечными волокнами типов I и Па. Следовательно, уменьшение усилия МПС не могло быть объяснено ненадлежащим невральным побуждением (М-волны не уменьшались), ацидозом или недостаточностью метаболических субстратов. Более того, судорожная реакция уменьшалась в большей степени, чем усилие тетануса или МПС, что рассматривается как доказательство ухудшения связи возбуждения с сокращением. Поэтому (Bigland-Ritchie et al., 1986) пришли к выводу, что во время выполнения данной задачи ухудшался процесс
18*
275
100-1
75-
О-о°°О
25-
ооо°° doO°d
*S88 888888888 8
п DOO ПО □ □ О □ □ о □ П
• Усилие МПС о М-волна
□ Судорожное усилие
04--------.---------1--------.--------1--------.
0 2 4 6 8 10
Время восстановления, мин
Рис.8.17. Восстановление усилия мышцы после утомляющего сокращения. Утомляющее сокращение завершается в нулевой момент времени
связывания возбуждения с сокращением, и это ухудшение обусловливало уменьшение усилия.
Другой экспериментальный подход, использованный для иллюстрации роли ухудшения связи возбуждения с сокращением при мышечном утомлении, заключался в контроле восстановления после утомления (рис.8.17). Эксперименты включали сравнение восстановления тетанического усилия, М-волн и судорожных реакций: тетаническое усилие представляет меру возбуждения, которое может быть приложено к мышце, независимо от ЦНС, амплитуды М-вол-ны — показателя стабильности нервно-мышечного преобразования, а судорожное усилие — реакции возбуждения и сокращения на отдельный импульс возбуждения. Для утомляющих сокращений, которые поддерживаются с большой продолжительностью при малом усилии, тетаническое усилие восстанавливается быстро (2 мин), восстановление после ухудшения нервно-мышечного преобразования длится несколько больше (4—6 мин), а ухудшение связи возбуждения с сокращением может продолжаться часами (Edwards, Hill, Jones, Merton, 1977; Fuglevand et al., 1993; Miller et al., 1987). Представляется, что связь возбуждения с сокращением у людей ухудшается вследствие продолжительных сокращений, но после стимулирования двигательных единиц типа FF — в течение по меньшей мере 2 мин (Burke et al., 1973; Enoka et al., 1992).
Метаболические субстраты. Двумя критическими факторами сокращения мышцы являются возбуждение мышцы нервной системой и подвод к мышце метаболической энергии. Хотя способность поддерживать сокращение мышцы не всегда зависит от наличия метаболической энергии, для некоторых задач истощение метаболического субстрата связывается со снижением производительности. Например, когда субъекты упражнялись на велоэргометре в тем
пе, соответствующем 70—80% максимальной аэробной производительности, утомление и неспособность поддерживать усилия, необходимые для выполнения задачи, совпадали с истощением гликогена в мышечных волокнах вастус латералис (Hermansen, Hultman, Saltin, 1967). Аналогично, когда тренирующиеся субъекты потребляют глюкозу или получают ее внутривенно, они способны тренироваться дольше (Coggan, Coyle, 1987; Coyle, Coggan, Hemmer, Ivy, 1986). Следовательно, наличие углеводов определяет, как долго побуждаемые субъекты могут тренироваться на велоэргометре на уровне 65—85 % максимальной аэробной производительности (Costill, Hargreaves, 1992; Hultman, Bergstrom, Spriet, Soderlund, 1990).
Как известно, на мышечное усилие влияют также продукты энергетического метаболизма. Известный фактор — концентрация ионов Н+. Хотя ионы Н+ и могут ингибировать гликолиз, это взаимодействие не представляется основным механизмом, обусловливающим уменьшение усилия. Например, снижение внутриклеточного pH (от 7,0 до 6,6) в волокнах отдельной интактной мышцы из-за повышения содержания СО2 во внеклеточной среде обусловливает лишь умеренное уменьшение количества закрепленных поперечных мостиков, усилия, создаваемого каждым поперечным мостиком, и снижение скорости циклического изменения поперечных мостиков во время укорачивающих сокращений (Edman, Lou, 1990; Lannergren, Westerblad, 1989). Вероятно, и другие продукты гидролиза АТФ (например, Mg-АДФ, Рн) могут способствовать уменьшению усилия. Например, повышение концентрации неорганического фосфата Рн может уменьшать максимальное изометричное усилие, но не оказывают влияния на максимальную скорость укорочения. Наоборот, повышение концентрации аденозиндифосфата магния (Mg-АДФ) может обусловить небольшое увеличение максимального изомет-ричного усилия и умеренное уменьшение максимальной скорости укорочения (Chase, Kushmerick, 1988; Cooke, Franks, Luciani, Pate, 1988). Условия выполнения задачи, в которых эти вещества могли бы иметь значение, пока неизвестны.
Кровоток. Ухудшение кровотока в активной мышце было одним из первых механизмов, идентифицированных в качестве таких, которые могли бы обусловливать ее утомление. Увеличение мышечного кровотока с двигательной активностью необходимо для доставки субстрата, удаления метаболитов и рассеяния тепла. Однако когда мышца активна, повышается внутримышечное давление, которое сжимает кровеносные сосуды и, в конечном счете, может перекрыть кровоток. Например, когда экстензоры колена поддерживают изометричное сокращение в течение возможно большего време
276
ни при субмаксимальном усилии (5—50 % МПС), кровоток уменьшается с повышением уровня поддерживаемого усилия (Sj^gaard et al., 1988). Тем не менее невозможно идентифицировать конкретный уровень усилия, при котором кровоток перекрывается, так как внутримышечное давление может заметно изменяться в пределах группы мышц-синергистов и даже в пределах отдельной мышцы для данной интенсивности сокращения. Однако кровоток ухудшается, вероятно, незначительно при выполнении задач, включающих менее 15 % усилия МПС (Gaffney, Sjyfgaard, Saltin, 1990; Sj^gaard et al., 1988). Когда артериальное давление искусственно поднимается выше внутримышечного, значительно увеличивается продолжительность сопротивляемости сокращениям, при которых усилие меньше 60% максимума (Petrofsky, Hendershot, 1984). При более высоких усилиях какой-либо эффект отсутствует.
Кроме этого очевидного влияния внутримышечного давления на мышечный кровоток повышенный кровоток (гиперперфузия) может снижать утомление посредством механизма, который не зависит от доставки кислорода или субстрата. Уменьшение кровотока (ишемия) влияет на механическую производительность, которая не зависит от пониженного содержания кислорода в крови (гипоксия). Увеличение кровотока, связанное с гиперперфузией, может улучшать удаление метаболитов и в результате этого снижать подавляющее влияние накапливания метаболитов (Barclay, 1986; Stainsby, Brechue, O’Drobinak, Barclay, 1990).
Сенсорные адаптации
Об изменениях сенсорных процессов во время утомляющих сокращений известно значительно меньше, чем об изменениях двигательных процессов. Проведены исследования, хотя и с противоречивыми результатами, о влиянии утомления на чувствительность проприорецеп-торных афферентов большого диаметра. Кроме того, определенное внимание было уделено основывающимся на чувствительности явлением, называемым мышечной мудростью и чувством напряжения.
Проприорецепторные афференты и рефлексы. Эксперименты, выполненные на анестезированных животных, показали, что утомление повышает чувствительность афферентов мышечного веретена (группы Та и II) в ответ на отдельные сокращения двигательных единиц (Zytnicki, Lafleur, Horcholle-Bossavit, Lamy, Jami, 1990). Однако результаты, полученные на находящихся в сознании людях, показывают, что утомление обусловливает снижение фузимотор-ного возбуждения мышечных веретен (Воп-giovanni, Hagbarth, 1990; Duchateau, Hainaut,
1993) и уменьшение веретенного разряда во время поддерживаемых изометричных сокращений (Macefield, Hagbarth, Gorman et al., 1991). Влияние утомления на афферент группы Тб сухожильного органа также не определено. У экспериментальных животных чувствительность сухожильных органов к сокращениям двигательных единиц не изменяется во время утомляющих сокращений; разряд сухожильного органа изменяется линейно с развитием усилия двигательной единицы (Gregory, 1990). В противоположность этому, как сообщалось, реакция сухожильных органов на растягивание всей мышцы с утомлением снижается (Hutton, Nelson, 1986), уменьшается также подавляющее влияние обратной связи группы Тб на двигательные нейроны (Zytnicki et al., 1990).
Несмотря на неопределенное действие утомления на проприорецепторные афференты, некоторые данные свидетельствуют, что утомление улучшает ЭМГ рефлексов и усиливает реакции двигательных нейронов на кратковременные возмущения (Darling, Hayes, 1983; Windhorst et al., 1986). Например, в эксперименте исследовали вращающий момент и реакцию в соответствии с ЭМГ флексоров локтя на растягивание, когда субъекты создавали один и тот же вращающий момент перед утомляющим сокращением и после него (Kirsch, Rymer, 1987). Задача на утомление состояла из 20 повторений 25-секундного изометрического сокращения до усилия, составляющего 50 % максимума. Хотя утомляющее сокращение и снижало существенно усилие, которое могли бы создать флексоры локтя, реакция вращающего момента на растягивание мышц была в достаточной степени подобна той, которую измеряли перед выполнением задачи на утомление. Способность субъектов реагировать сопоставимым вращающим моментом обусловливалась увеличением реакции согласно ЭМГ после упражнения (рис.8.18). Поскольку растягивание осуществлялось через 10 мин после завершения утомляющего сокращения, увеличение ЭМГ интерпретировалось как усиление неврального возбуждения, обусловленного растягиванием после утомляющего сокращения (см. также Kirsch, Rymer, 1992). Кроме того, в этом эксперименте сустав быстро смещался на 0,5 рад перед выполнением задачи на утомление и после этого. Реакция смещения была подобной перед опытами и после них.
Мышечная мудрость. Термин “мышечная мудрость” используется для описания эффекта, который наблюдался в экспериментах трех типов: мышца улучшает способность поддерживать усилие, обусловливаемое электрической стимуляцией, если частота стимулов снижается, а не остается постоянной (см. рис.6.9) (Binder-Macleod, Guerin, 1990; Marsden, Meadows, Merton, 1983); частота разрядов дви-
277
Рис.8.18. Увеличение реакции согласно ЭМГ на растягивание мышцы после утомляющего сокращения (сплошные линии) (Kirsch, Rymer, 1987).
гательных нейронов снижается во время утомляющего сокращения (Dietz, 1978; Garland, Enoka, Serrano, Robinson, 1994); утомление связывается с постепенным снижением скорости релаксации усилия (Gordon, Enoka, Karst, Stuart, 1990; Lannergren, Westerblad, 1991). Поскольку скорость релаксации уменьшается, продолжительность сокращения увеличивается, и, следовательно, та же степень слияния для тетанического сокращения (усилие) может быть достигнута при меньшей частоте возбуждения (рис.8.19). Изменение скорости релаксации обусловливается биохимическими изменениями вследствие связи возбуждения и сокращения. Следовательно, мышечная мудрость — это способность мышцы снижать разряд своих двигательных нейронов в соответствии с биохимически обусловливаемым снижением скорости релаксации.
Существует, по меньшей мере, три механизма, которые могут быть использованы сис
темой для управления снижением частоты разрядов во время утомляющего сокращения (Enoka, Stuart, 1992). Во-первых, некоторая комбинация афферентной обратной связи от периферийных источников, вероятно, ингибирует разряд промежуточных и двигательных нейронов (Bigland-Ritchie, Dawson, Johansson, Lippold, 1986; Garland, 1991). Эта обратная связь может обусловливать подавление рефлексов афферентами групп III и IV и отсутствие содействия афферентов группы 1а (т.е. ослабление управляемой фузимоторно обратной связи от мышечных веретен). Во-вторых, вероятно, имеет место адаптация возбудимости двигательных нейронов, в результате чего один и тот же синаптический входной сигнал создает меньше потенциалов действия (Kernell, Monster, 1982а, 19826). Адаптация происходит благодаря биофизическому свойству мембраны двигательных нейронов. В-третьих, разряд двигательного нейрона может уменьшаться во время утомляющего сокращения из-за снижения центрального возбуждения (Maton, 1991). Это наводит на мысль о том, что разряд двигательных нейронов может модулироваться нисходящими сигналами во время утомляющих сокращений. Вероятно, все три механизма (сенсорная обратная связь, адаптации спинномозговых нейронов и нисходящая модуляция) могут способствовать связанному с утомлением снижению частоты разрядов двигательных нейронов, но относительный вклад каждого зависит от деталей задачи, в частности, от продолжительности (Enoka, Sruart, 1992).
Чувство напряжения. Процесс двигательного программирования, осуществляемый супра-сегментными центрами, приводит к нисходящей двигательной команде, которая известна как центральная команда (см.главу 7). При осуществлении движения этот сигнал подается в нижние нервные центры (ствол мозга и спинной
Рис.8.19. Утомляющее сокращение обычно связывается с увеличением продолжительности релаксации одиночного сокращения и повышением степени слияния несливающегося тетанического сокращения
278
мозг), а также назад в супрасегментные центры в качестве результирующего разряда для содействия в интерпретировании поступающей афферентной информации. Результирующий разряд, который поступает в основную соматосенсорную область коры головного мозга, образует основу для чувства напряжения (см.главу 5). Чувство напряжения — это ощущение, которое указывает напряжение, требующееся для генерирования конкретного мышечного усилия. Оно не зависит от механизмов, уменьшающих способность мышцы создавать усилие. Для поддерживаемых субмаксимальных сокращений, осуществляемых побуждаемым субъектом, напряжение всегда будет увеличиваться, прежде чем усилие начнет уменьшаться.
Типичным путем исследования напряжения во время утомляющего сокращения является выполнение субъектом эксперимента по контралатеральному согласованию. В данном случае субъект создает поддерживаемое усилие одной (контрольной) конечностью и периодически указывает другой (согласующейся) конечностью напряжение, связанное с активацией контрольной конечности. Согласующаяся конечность обычно создает усилие до тех пор, пока ощущения, или напряжение, не будут равны для обеих конечностей. Результаты такого эксперимента показаны на рис. 8.20. Зачерненные кружки представляют нагрузку, поддерживаемую контрольной рукой. Линия, проведенная через незачерненные кружки, — нагрузка, оцениваемая согласующейся рукой, когда контрольная рука не поддерживала непрерывно нагрузку; нагрузка оценивалась через каждые 15—45 с в течение 10 мин. Линия, проведенная через незачерненные кружки, показывает нагрузку, оцениваемую согласующейся рукой, когда контрольная рука непрерывно поддерживала нагрузку и утомлялась. Увеличение со-
Рис.8.20. Увеличение напряжения во время утомляющего сокращения (McCloskey, Ebeling, Goodwin, 1974)
гласующегося усилия, когда контрольная рука поддерживала нагрузку, интерпретируется как повышение напряжения во время утомляющего сокращения.
Большинство данных указывает, что чувство напряжения во время утомляющего сокращения основывается преимущественно на результирующем разряде (Cafarelli, 1988; Jones, Hunter, 1983), причем обратная связь от периферийных рецепторов обусловливает другие ощущения (например, чувство тяжести или напряженного состояния).
ПОТЕНЦИРОВАНИЕ МЫШЦ
В противоположность снижающим производительность воздействиям утомления различные кратковременные механизмы могут повышать выходной сигнал нервно-мышечной системы (Hutton, 1984; McComas, 1977). После короткого периода активности эти механизмы могут повышать как электрическую, так и механическую выходную величину с превышением значений для состояния покоя. Примеры этих возможностей включают потенцирование моносинаптических реакций, минипотенциалы концевых пластинок, М-волны, судорожное усилие и разряд рецепторов мышечных веретен.
Мояосинаптические реакции
Представляется приемлемым принять, что большинство процессов в двигательной системе может усиливаться в течение коротких периодов активности. Это допущение оправдывается для соотношений входных и выходных сигналов спинного мозга. Lloyd (1949) подавал отдельные электрические удары на мышечный нерв экспериментального животного и измерял выходной сигнал в переднем корешке. Поскольку передний корешок был обрезан, электрический удар генерировал потенциалы действия, которые передавались по афферентным аксонам и в спинной мозг. Синаптический входной сигнал, который создавался афферентными аксонами, возбуждал двигательные нейроны, и измерялась моносинаптическая реакция (рис.8.21,а). Соотношение входного и выходного сигналов охватывало популяции афферентных и эфферентных аксонов. Моно-синаптические реакции измеряли перед тетанической стимуляцией нерва (12-секундная продолжительность при частоте 555 Гц) и после нее, причем они показаны на рис.8.21,б в нулевой момент времени. Тетаническая стимуляция увеличивала амплитуду моносинаптиче-ской реакции в семь раз по сравнению с контрольным значением (перед тетаническим
279
Рис.8.21. Потенцирование моносинаптических реакций (Lloyd, 1949): а — экспериментальная модель; б амплитуда моносинаптической реакции
сокращением). Потенцирование моносинаптической реакции прекращалось в течение 3-ми-нутного интервала (рис.8.21,6). Сообщалось о подобном эффекте в отношении потенцирования Н-рефлексов и сухожильных рефлексов после короткого периода высокочастотной электрической стимуляции (Hagbarth, 1962).
В эксперименте Lloyd (1949) при активировании различных афферентных путей после тетанической стимуляции потенцирование ограничивалось афферентным путем, который принимал тетаническую стимуляцию. Следовательно, механизм, поддерживающий это потенцирование, был предсинаптическим, т.е. он действовал до синаптического контакта с двигательными нейронами. Для данного типа стимулов, вызывающих потенцирование моносинаптической реакции, предполагается, что этот механизм охватывает афференты группы 1а (Hutton, 1984). Воздействия могут включать увеличение количества выделяемых нейромедиаторов, повышение их эффективности или уменьшение недостаточности точек аксонных ветвей по афферентам группы la (Kuno, 1964; Liischer, Ruenzel, Henneman, 1979).
Минипотенциалы концевых пластинок
В нервно-мышечном соединении спонтанное выделение нейромедиатора (ацетилхолина) обусловливает создание минипотенциалов концевых пластинок в постсинаптической мембране (см.главу 4). Минипотенциалы концевых пластинок — не постоянные события; амплитуда их больше для быстродействующих мышечных волокон по сравнению с медленнодействующими, а частота может снижаться с возрас
том (Alshuaib, Fahim, 1990; L^mo, Waerhaug, 1985). Аналогично короткий период высокочастотной стимуляции может увеличивать амплитуду и частоту спонтанно выделяющихся минипотенциалов концевых пластинок (Pawson, Grinnell, 1990; Vrbova, Wareham, 1976). Повышение частоты длится несколько минут, в то время как увеличение амплитуды, как сообщалось, продолжается несколько часов. Более того, этот эффект больше в более сильных нервно-мышечных соединениях (в данном контексте сила определяется как количество нейромедиаторов, высвобождающихся на единице длины соединения).
Представляется, что эти механизмы включают зависимое от активности повышение чувствительности постсинаптической мембраны и поступления ионов Са2+ в пресинаптическое окончание. Повышение чувствительности пост-синаптической мембраны означает, что квант нейромедиатора обусловит бо'лыпую реакцию (амплитуда синаптического потенциала) в постсинаптической мембране. Поскольку ион Са2+ необходим для слияния везикул в пресинапти-ческой мембране и последующего выделения нейромедиатора, большее поступление ионов Са2+ приведет к повышению частоты его спонтанного выделения.
Амплитуда М-волны
Процессы, участвующие в преобразовании аксонного потенциала действия в сарколем-мный потенциал действия, называются нервно-мышечным преобразованием. Один из путей проверки стабильности нервно-мышечного преобразования — генерирование М-волн (см.гла-ву 5). Проверка включает стимулирование мы-
280
Рис.8.22. Изменение ЭМГ во время 360-секундного утомляющего сокращения, включающего электрическую стимуляцию мышцы задней конечности кошки. Режим стимуляции предусматривал 330-милли-секундные последовательности 13 стимулов, которые подавались в течение каждой секунды. ЭМГ представляет суммарные потенциалы действия мышцы (подобно М-волнам), которые вызывались электрическими ударами. Амплитуда ЭМГ увеличивалась в течение первых 60 с испытания и затем уменьшалась (Enoka et al., 1992)
щечного нерва и измерение последующего общего потенциала действия мышцы. Хотя амплитуда М-волны и может уменьшаться под действием пресинаптических факторов (например, недостаточности точек ветвей, истощение нейромедиаторов), только постсинаптические факторы могут увеличивать амплитуды М-волны. К ним относятся временное рассеяние потенциалов действия мышечных волокон и увеличение амплитуды потенциалов действия отдельных мышечных волокон.
Когда человек осуществляет произвольное сокращение или мышца возбуждается электрической стимуляцией, часто происходит первоначальное временное увеличение амплитуды М-волны (рис. 8.22). Исследования отдельных мышечных волокон, которые подвергались повторяющейся стимуляции, наводят на мысль о
том, что увеличение амплитуды М-волны, по меньшей мере частично, обусловливалось зависящим от активности увеличением амплитуды потенциалов действия мышечных волокон (Hicks, McComas, 1989). Представляется, что возбуждение повышает активность насоса Na+-К+, которая снижает (гиперполяризует) мембранный потенциал покоя и увеличивает изменение напряжения (амплитуды) на мембране во время создания потенциала действия. Следовательно, некоторое первоначальное увеличение амплитуды М-волны в начале активности вызывается повышением активности насосов Na+-К+, а это, очевидно, обусловливает небольшое увеличение усилия (Enoka et al., 1992).
Посттетаническое потенцирование
Возможно, наиболее известной из реакций кратковременного потенцирования является влияние на судорожное усилие. Величина судорожного усилия является чрезвычайно изменяющейся и зависит от истории возбуждения мышцы. Судорога, вызываемая в мышце, находящейся в состоянии покоя, не представляет максимального одиночного сокращения. Скорее, судорожное усилие максимально после кратковременного тетанического сокращения; этот эффект известен как посттетаническое потенцирование одиночного усилия (Belanger, McComas, Elder, 1983; Schiff, 1858). Посттетаническое потенцирование судорожного усилия может быть существенным и вызываться как произвольными сокращениями, так и электрической стимуляцией. Более того, судорожное усилие может потенцироваться как максимальными, так и субмаксимальными произвольными сокращениями (Vandervoort, Quinlan, McComas, 1983). Особый случай потенцирования, достигаемого при субмаксимальном возбуждении, включает постепенное увеличение одиночного усилия, когда ряд судорог вызывается в плотной последовательности; это известно в качестве эффекта лестницы (Bowditch, 1871; Krarup, 1981).
По меньше мере два процесса могут участвовать в посттетаническом потенцировании судороги (Grange, Houston, 1991; Vandervoort et al., 1983). Раннее потенцирование происходит после кратковременных сокращений и относительно быстро исчезает. После интервала около 60 с протекает процесс позднего потенцирования, которое достигает максимума примерно через 200 с и затем снижается до контрольных уровней после 8—12 мин восстановления. Механизмы, лежащие в основе этих процессов, вероятно, обусловлены изменением движения кальция (Duchateau, Hainaut, 1986) и фосфорилирования легких цепей миозина (Sweeney, Bowman, Stull, 1993; Sweeney, Stull, 1990).
281
Рис.8.23. Совместное протекание потенцирования и утомления и результирующее влияние на судорожное усилие. При электрической стимуляции мышцы сокращение вызывает процессы, которые как уменьшают (утомление), так и увеличивают (потенцирование) ее усилие. Результатом является нелинейное изменение амплитуды судорожного усилия (одиночные сокращения) (Gamer, Hicks, McComas, 1989)
Потенцирование субмаксимального усилия происходит в двигательных единицах всех трех типов (S, FR и FF). Когда двигательные единицы возбуждались стимулом, который вызывал субмаксимальное тетаническое усилие, среднее потенцирование (увеличение пикового усилия) было больше среди быстродействующих двигательных единиц (50—60 % контрольного уровня для типов FR и FF) по сравнению со средним увеличением для действующих медленно двигательных единиц (20% контрольного уровня для типа S). Тем не менее, инцидентность потенцирования среди двигательных единиц была выше для устойчивых к утомлению двигательных единиц (60—75 % для типов S и FR), чем для утомляющихся двигательных единиц (40 % для типа FF). Поскольку протекание потенцирования распределялось среди всех трех типов двигательных единиц, механизмы, лежащие в основе потенцирования, отличаются от механизмов, определяющих тип двигательных единиц (Gordon, Enoka, Stuart, 1990).
Исследование посттетанического потенцирования судороги придало особое значение тому, что процессы потенцирования и утомления протекают одновременно, начиная с запуска возбуждения.
Схема взаимодействия двух этих процессов и результирующего влияния на судорожное усилие для конкретного случая показана на рис. 8.23. В эксперименте, на основании которого была создана эта схема, дорсифлексорные мышцы
голеностопного сустава добровольцев стимулировались электрически, чтобы вызвать трехсекундное субмаксимальное тетанические сокращение в течение каждых 5 с при общей продолжительности 180 с. Между тетаническими сокращениями одиночное сокращение вызывалось посредством единичного электрического удара. При этом конкретном режиме судорожное усилие увеличивалось и затем уменьшалось в течение периода стимуляции и после этого увеличивалось и уменьшалось в течение периода восстановления. Гарнер и др. (1989) предполагали, что временной ход изменения судорожного усилия обусловливался взаимодействием процессов потенцирования и утомления.
Сенсорный разряд после сокращения
В дополнение к воздействиям на двигательные процессы короткий период интенсивной активности может оказывать влияние на характеристику сенсорных процессов. Одним из примеров этого влияния является повышенная невральная активность, которая регистрировалась в задних корешках экспериментальных животных после сокращения; это явление было названо сенсорным разрядом после сокращения (Hutton, Smith, Eldred, 1973). Повышение активности задних корешков обусловливается, главным образом, увеличением разряда мышечных веретен,
282
преимущественно афферентов группы 1а. Более того, сенсорный разряд после сокращения исчезает, если мышца растягивается сразу же после сокращения. Механизмом, обусловливающим это явление, является развитие стабильных поперечных мостиков в интрафузальных мышечных волокнах. Эти поперечные мостики развиваются во время сокращения мышцы (совместное а-у-возбуждение) и сохраняются после релаксации экстрафузальных мышечных волокон, в результате чего мышечное веретено находится в состоянии повышенного напряжения и разряд в состоянии покоя является большим, чем перед сокращением (Gregory, Morgan, Proske, 1986; Hutton el al., 1973). Следовательно, при растягивании мышцы связи этих поперечных мостиков разрушаются и сенсорный разряд после сокращения исчезает.
Поскольку сенсорный разряд после сокращения увеличивает возбуждающий входной сигнал, поступающий в гомогенную группу двигательных нейронов, он влияет на последующую активность в течение периода до 15 мин с пиковым эффектом по истечении 5—20 с после сокращения. Например, он позволяет системе реагировать быстрее и сильнее на последующие изменения длины мышцы, оказывает влияние на кинестетические ощущения и оказывает сопротивление релаксации мышцы, что может оказаться желательным для растягивающих вмешательств. Уровень сенсорного разряда после сокращения может оказаться достаточным для повышения разряда двигательных нейронов в состоянии покоя (Suzuki, Hutton, 1976).
РЕЗЮМЕ
Это первая из двух глав, посвященных исследованию влияния физической активности на нервно-мышечную систему. В данной главе
внимание сосредоточено на срочную реакцию системы на одноразовую физическую нагрузку, и определены пять факторов, характеризующих адаптационные возможности системы; влияние разминки, гибкость, болезненность и повреждение мышц, мышечная усталость и потенцирование мышц. Во-первых, мы обсудили влияние разминки и описали влияние изменений температуры на механическую отдачу системы и на пассивную жесткость мышцы и соединительной ткани. Во-вторых, мы отделили разминочную активность от активности, связанной с гибкостью, и описали методы, используемые для изменения гибкости и факторов, ограничивающих диапазон движения сустава. В-третьих, мы исследовали последствия напряженной активности в отношении целостности мышцы и соединительной ткани. Это включало причину и временной ход мышечной болезненности, влияние на мышечные волокна различных типов, восстановление ткани после повреждения мышцы и следствие чрезмерных напряжений. В-четвертых, мы рассмотрели отрицательное влияние поддерживаемой активности на способность создавать усилие (мышечное утомление) и ощущаемое напряжение, связанное с активностью. Поскольку механизмы, обусловливающие мышечную усталость, изменяются с деталями выполняемой задачи, мы попытались идентифицировать условия, в которых различные механизмы могут ослабляться и, следовательно, усугублять мышечную усталость. В-пятых, мы охарактеризовали потенцирующее влияние кратковременной активности на моносинаптические реакции, минипотенциалы концевых пластинок, амплитуду М-волны, посттетаническое потенцирование судороги и сенсорный разряд после сокращения. Эти примеры предназначены не для представления полного анализа кратковременных адаптационных реакций, а для иллюстрации возможностей системы.
ВОПРОСЫ С АЛЬТЕРНАТИВНЫМ ОТВЕТОМ
1. Единственной целью разминки является повышение внутренней температуры.
ПРАВИЛЬНО НЕПРАВИЛЬНО
2. Разминка увеличивает максимальное уси- 6. лие, которое может быть создано мышцей, но не энергию, которую она может гене- 7. рировать.
ПРАВИЛЬНО НЕПРАВИЛЬНО
3. Повышение температуры мышцы, обуслов- 8. ливаемое разминкой, уменьшает как время сокращения, так и время полурелаксации.
ПРАВИЛЬНО НЕПРАВИЛЬНО
На время полурелаксации оказывает большее влияние температура, а не время сокращения. ПРАВИЛЬНО НЕПРАВИЛЬНО В результате использования пассивного внешнего источника тепла невозможно повысить температуру мышцы больше, чем на ГС. ПРАВИЛЬНО НЕПРАВИЛЬНО
Единица измерения жесткости — ньютонометр (Нм). ПРАВИЛЬНО НЕПРАВИЛЬНО Жесткость можно исследовать графически в виде наклона графика усилие—длина.
ПРАВИЛЬНО НЕПРАВИЛЬНО
Для растягивания жесткой мышцы требуется больше энергии по сравнению с менее жесткой мышцей.
ПРАВИЛЬНО НЕПРАВИЛЬНО
283
9. Мышечным тонусом является сопротивление, оказываемое растягиванию расслабленной мышцей.
ПРАВИЛЬНО НЕПРАВИЛЬНО
10. Увеличение мышечного тонуса известно в качестве гипертонуса.
ПРАВИЛЬНО НЕПРАВИЛЬНО
И. Баллистическое растягивание включает прыжковые движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
12. В соответствии с методикой растягивания с сокращением агониста растягивается мышца-антагонист.
ПРАВИЛЬНО НЕПРАВИЛЬНО
13. Мышца-антагонист релаксирует во время растягивания с сокращением агониста.
ПРАВИЛЬНО НЕПРАВИЛЬНО
14. Улучшение гибкости подобно для статических и баллистических растягиваний.
ПРАВИЛЬНО НЕПРАВИЛЬНО
15. Фиксационно-релаксационная методика с сокращением агониста обеспечивают самое большое улучшение гибкости из всех методик растягивания.
ПРАВИЛЬНО НЕПРАВИЛЬНО
16. Упражнения на гибкость включают изменение длины соединительной ткани.
ПРАВИЛЬНО НЕПРАВИЛЬНО
17. Упражнения на гибкость следует выполнять в начале тренировки.
ПРАВИЛЬНО НЕПРАВИЛЬНО
18. Соединительная ткань наиболее растяжима при высокой температуре.
ПРАВИЛЬНО НЕПРАВИЛЬНО
19. Мышечная болезненность с задержкой проявления обусловливает структурное повреждение мышечных волокон, но не потерю прочности. ПРАВИЛЬНО НЕПРАВИЛЬНО
20. Более вероятно, что мышечную болезненность с задержкой проявления обусловливает эксцентрические сокращения, а не другие типы сокращений мышц.
ПРАВИЛЬНО НЕПРАВИЛЬНО
21. Эксцентрическое упражнение создает более высокое пиковое внутримышечное давление, но снижает расход кислорода.
ПРАВИЛЬНО НЕПРАВИЛЬНО
22. Величина нагрузки, а не деформация является более важным фактором мышечной болезненности.
ПРАВИЛЬНО НЕПРАВИЛЬНО
23. Потеря внеклеточного кальция может запускать процессы, разрушающие клеточные структуры. ПРАВИЛЬНО НЕПРАВИЛЬНО
24. Мышечную болезненность часто сопровождает нарушение линии.
ПРАВИЛЬНО НЕПРАВИЛЬНО
25. Один из путей оценки степени повреждения мышцы заключается в определении количества ферментов скелетных мышц в кровообращении.
ПРАВИЛЬНО НЕПРАВИЛЬНО
26. В результате тренировки повреждаются клеточные мембраны.
ПРАВИЛЬНО НЕПРАВИЛЬНО
Т1. Мышечная болезненность с задержкой проявления может быть следствием воспалительной реакции.
ПРАВИЛЬНО НЕПРАВИЛЬНО
28. Вследствие тренировки повреждаются преимущественно мышечные волокна типа Пб, которые относятся к двигательным единицам типа S. ПРАВИЛЬНО НЕПРАВИЛЬНО
29. Вследствие тренировки могут повреждаться как внутримышечный цитоскелет, так и внеклеточная матрица соединительной ткани. ПРАВИЛЬНО НЕПРАВИЛЬНО
30. Восстановление мышечных волокон начинается примерно через четверо суток после тренировки. ПРАВИЛЬНО НЕПРАВИЛЬНО
31. Упражнения на растягивание и разминочные упражнения могут предотвратить мышечную болезненность с задержкой проявления. ПРАВИЛЬНО НЕПРАВИЛЬНО
32. Мышечные деформации обычно происходят в соединениях мышц с сухожилиями.
ПРАВИЛЬНО НЕПРАВИЛЬНО
33. Мышцы с большой долей мышечных волокон типа II более восприимчивы к мышечным деформациям.
ПРАВИЛЬНО НЕПРАВИЛЬНО
34. Наиболее часто деформации происходят во время концентрических сокращений.
ПРАВИЛЬНО НЕПРАВИЛЬНО
35. Мышечное утомление — класс постоянных эффектов, ухудшающих работоспособность.
ПРАВИЛЬНО НЕПРАВИЛЬНО
36. Центральное возбуждение считается максимальным при равенстве усилий, вызываемых произвольно и электрически.
ПРАВИЛЬНО НЕПРАВИЛЬНО
37. Методика наложения судороги включает приложение к мышце единичного удара во время произвольного сокращения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
38. Единица измерения судорожного усилия в системе СИ — килограмм (кг).
ПРАВИЛЬНО НЕПРАВИЛЬНО
284
39. Концентрические сокращения более утомительны, чем эксцентрические.
ПРАВИЛЬНО НЕПРАВИЛЬНО
40. Амфетамин может снижать утомляемость.
ПРАВИЛЬНО НЕПРАВИЛЬНО
41. Гормоны в цереброспинальной жидкости оказывают влияние на способность поддерживать центральное возбуждение.
ПРАВИЛЬНО НЕПРАВИЛЬНО
42. Мышцы-синергисты всегда возбуждаются в равной степени во время поддерживаемой активности.
ПРАВИЛЬНО НЕПРАВИЛЬНО
43. Мышцы не утомляются, если усилие меньше 10% максимума.
ПРАВИЛЬНО НЕПРАВИЛЬНО
44. Порядок использования двигательных единиц может отличаться для концентрических и эксцентрических сокращений.
ПРАВИЛЬНО НЕПРАВИЛЬНО
45. Утомление не влияет на порядок использования. ПРАВИЛЬНО НЕПРАВИЛЬНО
46. Термин нервно-мышечное преобразование относится к нескольким процессам.
ПРАВИЛЬНО НЕПРАВИЛЬНО
47. В нервно-мышечном соединении сарколемма является постсинаптической мембраной.
ПРАВИЛЬНО НЕПРАВИЛЬНО
48. М-волны вызываются в результате приложения электрического удара к нерву мышцы. ПРАВИЛЬНО НЕПРАВИЛЬНО
49. М-волна измеряется, как правило, в качестве реакции ЭМГ в мышце.
ПРАВИЛЬНО НЕПРАВИЛЬНО
50. Нервно-мышечное преобразование обычно ухудшается при кратковременных максимальных произвольных сокращениях.
ПРАВИЛЬНО НЕПРАВИЛЬНО
51. Проводимость, обусловливаемая ионами Са2+, в саркоплазматической сети в условиях отдыха обычно низка.
ПРАВИЛЬНО НЕПРАВИЛЬНО
52. Количество ионов Са2+, выделяемых из саркоплазматической сети, может уменьшаться во время утомления.
ПРАВИЛЬНО НЕПРАВИЛЬНО
53. Восстановление после нарушения связи возбуждения с сокращением длится несколько часов. ПРАВИЛЬНО НЕПРАВИЛЬНО
54. Связь возбуждения с сокращением и нервно-мышечное преобразование ухудшаются вследствие продолжительных сокращений.
ПРАВИЛЬНО НЕПРАВИЛЬНО
55. Крайняя усталость может совпадать с истощением гликогена в мышечных волокнах. ПРАВИЛЬНО НЕПРАВИЛЬНО
56. Снижение внутриклеточного pH не влияет на усилие, создаваемое отдельными поперечными мостиками.
ПРАВИЛЬНО НЕПРАВИЛЬНО
57. Мышечный кровоток не перекрывается, если только усилие мышцы не превышает 64 % максимума.
ПРАВИЛЬНО НЕПРАВИЛЬНО
58. Единственное воздействие кровотока на утомление — подвод кислорода и субстратов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
59. У людей разряд мышечных веретен снижается во время утомляющих сокращений.
ПРАВИЛЬНО НЕПРАВИЛЬНО
60. Реакция сухожильных органов на растягивание всей мышцы уменьшается с утомлением. ПРАВИЛЬНО НЕПРАВИЛЬНО
61. ЭМГ рефлекса растяжения мышцы увеличивается с утомлением.
ПРАВИЛЬНО НЕПРАВИЛЬНО
62. Частота разрядов двигательных нейронов во время утомления снижается.
ПРАВИЛЬНО НЕПРАВИЛЬНО
63. Время релаксации увеличивается с утомлением. ПРАВИЛЬНО НЕПРАВИЛЬНО
64. В соответствии с мышечной мудростью изменение скорости релаксации обусловливает изменение частоты разрядов двигательных нейронов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
65. Адаптация двигательных нейронов объясняется снижением частоты разрядов посредством ингибирующей сенсорной обратной связи. ПРАВИЛЬНО НЕПРАВИЛЬНО
66. Минипотенциалы концевых пластинок создаются спонтанно.
ПРАВИЛЬНО НЕПРАВИЛЬНО
67. После кратковременной активности изменяется частота, а не амплитуда минипотенциалов концевых пластинок.
ПРАВИЛЬНО НЕПРАВИЛЬНО
68. Увеличение амплитуды М-волны может обусловливаться возрастанием количества выделяемого нейромедиатора.
ПРАВИЛЬНО НЕПРАВИЛЬНО
69. Судорожное усилие не является максимальным в мышце в состоянии покоя.
ПРАВИЛЬНО НЕПРАВИЛЬНО
70. Судорожное усилие не может быть потенцировано стимулируемыми электрически сокращениями.
ПРАВИЛЬНО НЕПРАВИЛЬНО
71. Феномен лестницы объясняется нерегулярностями усилия во время несливающегося тетанического сокращения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
285
72. Потенцирование судорожного усилия — одна из характеристик, которые могут быть использованы для распознавания трех типов двигательных единиц.
ПРАВИЛЬНО НЕПРАВИЛЬНО
73. Сенсорный разряд после сокращения включает, главным образом, повышение разряда мышечных веретен в состоянии покоя.
ПРАВИЛЬНО НЕПРАВИЛЬНО
74. Повышение разряда мышечных веретен происходит в результате увеличения напряжения интрафузальных мышечных волокон.
ПРАВИЛЬНО НЕПРАВИЛЬНО
75. Сенсорный разряд после сокращения может обусловливать разряд потенциалов действия двигательными нейронами.
ПРАВИЛЬНО НЕПРАВИЛЬНО
Д вопросы
?J IJi ГчИЫМ ВЫГЭРОМ ОТВЕТОВ
76. Какое воздействие не обеспечивается в результате повышения внутренней температуры, обусловливаемого разминкой?
а) снижение мышечной вязкости;
б) увеличение тиксотропности;
в) увеличение растяжимости соединительной ткани;
г) увеличение мышечного кровотока.
77. Разминка показала повышение мышечной температуры и, следовательно, увеличение образования энергии. Почему?
а) упражнения на растягивание повышают гибкость:
б) возрастает vMax;
в) увеличивается время полурелаксации;
г) уменьшается время сокращения.
78. Какие единицы измерения можно было бы использовать для измерения механических последствий тиксотропности?
а) Н • м;
б) Н • мм-2;
в) Н • м-1;
г) (Н-м) рад-1.
79. Что такое мышечный тонус?
а) реакция двигательных нейронов на растягивание мышцы;
б) сопротивление растягиванию, оказываемое релаксированной мышцей;
в) отношение объема мышцы к количеству сократительных белков;
г) уровень активности мышцы в состоянии покоя согласно ЭМГ.
80. Какие рефлексы были использованы для выявления снижения возбудимости группы двигательных нейронов посредством
методики фиксационно-релаксационного растягивания?
а) рефлекс Н;
б) рефлекс рецидивного торможения;
в) рефлекс реципрокного торможения;
г) рефлекс Т.
81. Посредством каких механизмов сокращение агониста во время растягивания антагониста (методика растягивания с сокращением агониста) обусловливает релаксацию антагониста?
а) постактивационное подавление возбудимости двигательных нейронов;
б) рефлекс реципрокного торможения;
в) увеличение тиксотропности из-за повышения температуры;
г) рефлекс растяжения мышцы.
82. Что из приводимого ниже не является клиническим симптомом, связанным с мышечной болезненностью с задержкой проявления?
а) увеличение количества белковых метаболитов в травмированных мышцах;
б) структурное повреждение субклеточных компонентов;
в) непосредственное усиление боли, связанной с травмированными мышцами;
г) увеличение количества плазменных ферментов.
83. Чем эксцентричное упражнение отличается от концентрического упражнения?
а) большей болезненностью и слабостью мышц после выполнения упражнения;
б) меньшим расходом кислорода;
в) более низким внутримышечным давлением;
г) более высоким уровнем ЭМГ для одного и того же усилия.
84. Какие метаболические факторы могут приводит к мышечной болезненности?
а) неспособность удалить внутриклеточные ионы Са2+;
б) разрушение белковых структур, обусловленное снижением температуры;
в) высокое напряжение, которому подвергаются внутриклеточные структуры;
г) образование свободного кислорода.
85. Что является наиболее заметным структурным нарушением после тренировки?
а) характер Z-линии;
б) патологические изменения цитоплазматической мембраны;
в) изменение внеклеточной миофибрил-лярной матрицы;
г) распухшие митохондрии.
86. Какие типы мышечных волокон наиболее подвержены влиянию упражнения, индуцирующего мышечную болезненность с задержкой проявления?
286
а) 116;
6)1;
в) la;
г) Па.
87. Какое вещество служит показателем участия повреждения соединительных тканей в мышечной болезненности с задержкой проявления?
а) креатинкиназа;
б) ион Са2+;
в) оксипролин;
г) титин.
88. Какая стадия индуцируемого тренировкой травмирования мышцы связывается с воспалительной реакцией?
а) восстановление;
б) начальная стадия;
в) фагоцитарная стадия;
г) автогенетическая стадия.
89. Что из приводимого ниже не является переменной величиной задачи, влияющей на механизмы, связанные с утомлением мышц?
а) интенсивность сокращения;
6) мышечный кровоток;
в) возбуждение субъекта;
г) центральное побуждение.
90. Какие примеры иллюстрируют недостаточное центральное возбуждение во время утомляющей активности?
а) уменьшение амплитуды М-волны;
б) различия в утомляемости в связи с концентрическими и эксцентрическими сокращениями;
в) отсутствие возбуждения субъектов;
г) уменьшение отношения судорожных и тетанических сокращений.
91. Что происходит с электромиограммой во время утомляющего эксцентрического сокращения?
а) ЭМГ снижается параллельно усилию;
б) ЭМГ повышается и становится больше по сравнению с ЭМГ при концентрических сокращениях;
в) ЭМГ остается постоянной;
г) ЭМГ повышается, но остается ниже, чем ЭМГ для концентрических сокращений.
92. Почему быстродействующая мышца может обеспечить образование большего количества энергии по сравнению с медленнодействующей мышцей?
а) медленнодействующая мышца создает меньшее усилие;
б) быстродействующая мышца характеризуется более высокой оптимальной скоростью укорочения;
в) быстродействующая мышца быстрее утомляется;
г) медленнодействующая мышца имеет меньшую площадь поперечного сечения.
93. Что из приводимого ниже не является процессом связи возбуждения с сокращением? а) повторное поглощение ионов Са2+ саркоплазматической сетью;
б) распространение потенциала действия по сарколемме;
в) выделение нейромедиатора в нервно-мышечном соединении;
г) растормаживание тропонина.
94. Какие процессы возбуждения—сокращения ухудшаются во время утомляющих сокращений?
а) повторное поглощение ионов Са2+;
б) амплитуда М-волны;
в) выделение ионов Са2+;
г) распространение потенциала действия вдоль поперечного канальца.
95. При какой интенсивности максимальной аэробной энергии истощение гликогена совпадает по времени с крайней усталостью? а) менее 35%;
б) 100%;
в) 50-60%;
г) 65-85%.
96. Какое влияние на активность поперечных мостиков может оказывать снижение внутриклеточного pH?
а) снижение скорости циклического изменения во время укорачивающих сокращений;
б) увеличение количества закрепленных поперечных мостиков;
в) подавление нераскрытая мест связывания;
г) уменьшение усилия, создаваемого отдельными поперечными мостиками.
97. Почему перекрытие кровотока обусловливает уменьшение усилия мышцы?
а) уменьшается количество кислорода;
б) замедляется удаление метаболитов;
в) снижается мышечная температура;
г) ухудшается распространение потенциалов действия.
98. Почему разряд веретена мог бы снижаться во время поддерживаемого сокращения? а) повышение чувствительности афферентов мышечных веретен;
б) уменьшение фузимоторного возбуждения;
в) снижение возбудимости двигательных нейронов;
г) увеличение времени релаксации интра-фузальных мышечных волокон.
99. Почему способность мышцы поддерживать усилие улучшается, если частота стимулов со временем снижается?
287
а) уменьшается разряд двигательных нейронов;
б) снижается скорость релаксации мышцы;
в) сенсорная обратная связь подавляет активность двигательных нейронов;
г) адаптация возбудимости двигательных нейронов.
100. Какой сигнал используется для обеспечения ощущения напряжения?
а) двигательной программы;
б) проприорецепторной обратной связи;
в) результирующего разряда;
г) нисходящей команды.
101. Почему увеличение притока ионов Са2+ в пресинаптическое окончание нервно-мышечного соединения обусловливает повышение частоты минипотенциалов концевых пластинок?
а) увеличивается количество нейромедиатора, выделяемого в каждом кванте;
б) усиливается образование нейтромедиа-тора;
в) повышается возбудимость мембраны под действием потенциала действия;
г) повышается скорость слияния везикул с пресинаптической мембраной.
102. Какие механизмы обусловливают посттетаническое потенцирование судорожного усилия?
а) снижение жесткости мышцы;
б) повышение сарколеммной возбудимости;
в) улучшение динамики ионов Са2+;
г) улучшение биохимических реакций, включающих аденозинтрифосфат.
ЗАДАЧИ
103. Допустим, во время разминки мышца подверглась изменению температуры на 7 °C относительно первоначального значения 31 °C. Максимальный изометричный вращающий момент (Fm о) составил 238 Н • м, а максимальная угловая скорость (сотах) — 6,8 рад • с-1 перед изменением температуры. (Fm o) изменялся со скоростью 2,5 % °C, a “max — со скоростью 12,3 % °C.
А. Каковы Fmo и comQX после разминки?
Б. Постройте на миллиметровой бумаге график соотношений усилие—скорость перед разминкой и после нее. Покажите в виде прямых линий соотношения между величинами Fm o и сотах.
В. По графику линейных соотношений усилие—скорость оцените максимальную энергию, образуемую мышцей перед разминкой и после нее. Напомним, что энергию можно рассчитать в качестве произведения усилия и скорости.
104. Методика растягивания с сокращением агониста предположительно допускает растягивание мышцы, так как возбуждение рефлекса реципрокного торможения снижает возбудимость двигательных нейронов мышцы.
А. Начертите невральную схему для рефлекса реципрокного торможения.
Б. Почему рефлекс реципрокного торможения возбуждается, когда агонист не растягивается?
В. Каким образом можно отключить рефлекс реципрокного торможения?
105. Мышечная болезненность с задержкой проявления чаще связывается с эксцентрическими сокращениями.
А. Почему эксцентрические сокращения более вероятно индуцируют болезненность?
Б. Каково различие между напряжением и деформацией? Почему деформация индуцирует мышечную болезненность?
В. Почему в соответствии со схемой Фри-дена и Либера мышечные волокна типа II более восприимчивы к повреждению, индуцируемому тренировкой?
106. Элитный спринтер нередко подвергается деформации мышц сухожилий, ограничивающих с боков подколенную ямку, во время соревнования, например, спринта на 100 м. А. Постройте график вращающий момент-угол для коленного сустава во время одного махового шага для такого соревнования. Вращающий момент должен быть результирующим вращающим моментом мышц относительно коленного сустава (см.рис. 7.16 и 7.18).
Б. Укажите на графике участок наиболее вероятной мышечной деформации и объясните причину выбора этой точки.
107. Рассмотрите эксперимент на утомление, показанный на рис.8.11.
А. Что означает параллельное уменьшение усилия максимального произвольного сокращения и усилия при частоте 50 Гц?
Б. Почему ЭМГ для субмаксимальных сокращений снижается во время утомляющего сокращения?
В. Каково усилие при наложении судороги, достигающее нулевого уровня в течение примерно 5 мин?
108. Ряд процессов, связанных с мышечным утомлением, назван невральной стратегией. А. Что такое невральная стратегия?
Б. Каким образом невральная стратегия может оказывать влияние на мышечное утомление? Приведите один пример на уровне всей мышцы и другой пример на уровне двигательной единицы.
288
В. Предложите эксперимент, который исследует роль невральной стратегии во время утомляющего сокращения.
Ж М-волна является элементом, который может быть использован для контроля эффективности нервно-мышечного преобразования.
А. Что такое М-волна и каким образом она измеряется?
Б. Какие процессы могут увеличивать амплитуду М-волны?
В. Какие процессы могут уменьшать амплитуду М-волны?
Г. Когда амплитуда М-волны увеличивается и уменьшается во время поддерживающего сокращения?
ПО. Некоторые данные наводят на мысль о том, что утомление повышает ЭМГ рефлексов и реакции двигательных нейронов на кратковременные возмущения. Почему это является приемлемой кратковременной адаптацией, обеспечиваемой системой?
111. Посттетаническое потенцирование судорожного усилия показывает, что судорож
ное усилие максимально после кратковременного тетанического сокращения.
А. Поскольку потенцирование этого типа Может вызываться как произвольными сокращениями, так и электрической стимуляцией, где должен действовать этот механизм?
Б. Какие факторы определяют амплитуду судорожного усилия?
В. Каким образом изменение динамики ионов Са2+ может влиять на судорожное усилие?
Г. Объясните амплитуду и инцидентность посттетанического потенцирования судороги среди двигательных единиц трех типов, исходя из способности выделять ионы Са2+ под действием единичного потенциала действия.
Д. Что обеспечивает утомленной мышце возможность посттетанического потенцирования судороги?
112. Почему кратковременное максимальное произвольное сокращение повышает разряд покоя афферентов группы 1а?
19 8-133
ГЛАВА 9
ДОЛГОВРЕМЕННЫЕ АДАПТАЦИОННЫЕ РЕАКЦИИ
В главе 8 мы изучили адаптационные реакции двигательной системы в ответ на отдельные циклы физической нагрузки. В этой главе мы рассмотрим кумулятивную (продолжительную) реакцию двигательной системы на физические нагрузки, обусловленные продолжительной физической деятельностью.
ПРИНЦИПЫ ТРЕНИРОВКИ
В результате исследований нервно-механических основ мышечной силы был разработан ряд рекомендаций, которые называются принципами тренировки. Один из таких принципов — принцип перегрузки (Delorme, 1946), согласно которому, возникновение адаптационной реакции возможно только при преодолении определенной пороговой точки. Обычно эта точка выражается как процент максимального значения. Например, порог при физических нагрузках изометрического характера составляет около 40% максимума; т.е. адаптационная реакция будет иметь место только в том случае, если сила превысит 40 % максимальной. Поскольку максимальный вращающий момент, образуемый мышцей, со временем изменяется в результате колебаний уровня активности (тренировки и детренировки), точно так же изменяется абсолютная нагрузка, превышающая пороговые изменения. Это наблюдается у регулярно тренировавшихся испытуемых, которые затем прерывали тренировки (вследствие травмы, болезни) и после восстановления уже не могли возобновить процесс тренировок на прерванном уровне активности.
Кроме того, физические тренировки для обеспечения необходимого эффекта должны соответствовать режиму тренировочных занятий. Это так называемый принцип специфичности
(McCafferty, Horvath, 1977): адаптационные реакции специфичны для клеток и их структурных и функциональных элементов, которые подвергаются физическим нагрузкам.
Согласно этому принципу, изменение специфично величине нагрузки. Если, например, спортсмен занимается по программе занятий силовой направленности, то только для этого качества (сила), а не какого-то другого (например, выносливость) характерны адаптационные реакции (Boulay et al., 1985; Sale, MacDougall, Jacobs, Gamer, 1990). Вместе с тем, некоторые ученые установили, что занятия силовой направленности вызывают адаптационные реакции, специфичные для задания, длины и скорости мышцы, характерных для тренировочного занятия (Jones, Rutherford, Parker, 1989; Sale, MacDougall, 1981). В то же время другие исследователи отмечали отсутствие специфичности для концентрических и эксцентрических сокращений (Peterson et al., 1990), а также быстрых и изометрических сокращений (Behm, Sale, 1993).
Важным моментом принципа специфичности является то, что эффект тренировочных занятий зависит также от начального уровня подготовленности испытуемого. На рис.9.1 показано изменение результатов (поднятие веса из положения сидя) у двух групп испытуемых (спортсменов, занимающихся силовыми видами спорта и неспортсменов) в процессе программы тренировочных занятий. Группа спортсменов была представлена культуристами и спортсменами, занимающимися пауэрлифтингом. Группа неспортсменов включала тренированных испытуемых, которые использовали упражнения силовой направленности в своей программе занятий, но не участвовали в соревнованиях по силовым видам спорта. Перед началом программы тренировочных занятий спортсмены могли поднять вес (максимальный) 1400 Н, неспорт-смены — 446 Н. Как видно из рисунка у спорт-
290
Рис.9.1. Изменение результата выполнения поднятия веса из положения сидя после программы занятий силовой направленности (концентрическое выпрямление ноги в коленном суставе при нагрузке 80—100 % максимальной концентрической) (K.Hakkinen, 1985)
сменов наблюдалось более низкое абсолютное изменение, а также более низкая интенсивность изменения (наклон на графике) по сравнению с неспортсменами. Это различие было обусловлено более высоким начальным уровнем силы, а также “знакомством” с выполняемым движением у спортсменов, занимавшихся силовыми видами спорта. Начальное быстрое увеличение силы у неспортсменов отражает нервные адаптационные реакции, обусловленные программой физических занятий.
Наконец, отметим принцип обратимости, согласно которому прекращение тренировочных занятий приводит к адаптации системы к новым (пониженным) требованиям. Это так называемая детренированность (Chi et al., 1983). Один из примеров детренированности — адаптационные реакции организма астронавтов во время пребывания в условиях невесомости.
СИЛА
Динамические способности единой системы сустава можно охарактеризовать на основании взаимосвязи сила—скорость (см.рис.6.19). Данная взаимосвязь включает четыре аспекта: а) скорость равна 0 (изометрическая или Ро); б) скорость > 0 (концентрическая); в) скорость < 0 (эксцентрическая); г) максимальная скорость (гмакс). Мышечная сила определяется как величина вращающего момента, образуемого мышцей или мышцами при отдельном изометрическом сокращении неограниченной продолжительности (Atha, 1981; Enoka, 1988b). Согласно этому определению, существует только один тип силы — измеряемой в изометрических условиях. Следовательно, неправомерно использовать определения динамическая, изотоническая, изо-
кинетическая или статическая сила. Традиционно, однако, используется понятие “статическая сила” — Ро и “динамическая сила” — для измерения вращающего момента при ненулевой скорости. Мы используем только узкое определение силы, ограниченное измерением пика вращающего момента в изометрических условиях.
Виды тренировочных занятий
Согласно теории поперечных мостиков мышечного сокращения, после того как Са2+ ингибировал тормозной эффект комплекса тро-понин-тропомиозин, миозиновые поперечные мостики прикрепляются к участкам на актине и подвергаются изменению, вследствие которого образуется сила. Несмотря на то, что мышечная активация обусловлена всего одним механизмом, в литературе, посвященной тренировочным занятиям силовой направленности, рассматриваются различные типы мышечных сокращений. По нашему мнению, правильнее обратить внимание на изменения взаимосвязи вращающего момента мышцы и вращающего момента, обусловленного нагрузкой, рассмотрев изометрические, концентрические и эксцентрические условия.
Изометрический метод тренировки. Изометрическое сокращение (изо — постоянная, метрик — длина всей мышцы) определяли как условие, при котором вращающий момент, обусловленный нагрузкой, соответствует вращающему моменту, образуемому мышцей, такой же величины, но действующему в другом направлении. Хотя длина всей мышцы при изометрическом сокращении не изменяется, длина волокон уменьшается. В 50-е годы XX ст. Hettinger, Muller (1961) популяризировали физические упражнения изометрического характера как наиболее подходящие для поддержания высокого уровня физической подготовленности. Эти упражнения, как считалось, вызывали благоприятные гипертрофические реакции, занимали немного времени, были удобными и не требовали никаких материальных затрат. Схема, предложенная Hettinger и Muller характеризовалась учетом пороговых точек, уровней интенсивности и систематизацией программы физических упражнений. Например, отмечалось, что ежедневные упражнения для сгибателей локтевого сустава порядка 40—50 % максимальной интенсивности обеспечивают увеличение силы, 20—30 % — сохранение ее уровня, до 20 % — приводят к снижению силовых возможностей. Кроме того, указывалось, что при продолжительности физической нагрузки 1—2 ч ее интенсивность должна составлять 100 % максимальной, чтобы обеспечить достаточную нагрузку, тогда как
19»
291
при продолжительности 4—6 ч интенсивность должна быть меньше — 66 % максимальной. Hettinger, Muller отмечали, что одно 4—6-секундное сокращение в день с интенсивностью 40—50 % максимальной обеспечивает увеличение силы (например, на 2 % в неделю для мышц—сгибателей локтевого сустава). Подчеркивалась необходимость периодически изменять пороговые показатели.
Следует отдать должное Hettinger, Muller за то, что они подчеркнули важность количественного подхода к выбору физических упражнений. Популярность изометрических упражнений в последнее время уменьшилась, тем не менее их и теперь включают в программы физических занятий силовой направленности. Например, рекомендуют следующий режим изометрических упражнений: а) для нетренированных (тренированных) — ежедневные тренировки по 5—8 повторений для каждой мышечной группы, в которой активные мышцы образуют 40—50 % максимальной силы; б) для спортсменов высокого уровня — выполнение упражнений при различных углах сустава при силе 80—100 % максимальной, продолжительность каждого упражнения 5—10 с, а также 2—5 повторений для мышечной группы с интервалом между повторениями 30 с — несколько минут (Напе, 1982).
С упражнениями изометрической направленности связаны, как минимум, две проблемы. Во-первых, при выполнении движений преобладают концентрические и эксцентрические сокращения, при которых длина мышцы изменяется, следовательно, изометрические упражнения не специфичны целям тренировочного процесса, т.е. принцип специфичности нарушается. Например Kitai, Sale (1989) обнаружили, что увеличение силы в результате программы тренировок с использованием изометрических упражнений было характерно только для диапазона движения 0,17 рад (10°) относительно угла голеностопного сустава, при котором выполнялись упражнения. Во-вторых, когда сила, образуемая мышцей, превышает приблизительно 15 % максимальной, происходит окклюзия кровеносных сосудов и значительно увеличивается периферическое сопротивление кровотоку. В результате окклюзии повышаются частота сердечных сокращений и артериальное давление (Seals, Washburu, Hauson et al., 1983).
Пример адаптационных реакций, которые могут быть достигнуты вследствие изометрических тренировок, показан в исследовании Kitai, Sale (1989). 6 испытуемых в течение 6 недель выполняли изометрические упражнения. Они тренировались три раза в неделю, выполняя два цикла по 10 повторений. Каждое повторение представляло собой 5-секундное максимальное (100%) изометрическое сокращение мышц—сги
бателей подошвы, угол голеностопного сустава между голенью и стопой был прямым. Каждую минуту выполнялось два сокращения, интервал отдыха между повторениями составлял 2 мин. В результате занятий сила увеличилась на 18 % (25,5 Н • м) (максимальный изометрический вращающий момент) при “тренировочном” угле и на 17 и 14 % в прилегающих (0,17 рад) углах сгибания соответственно подошвы и тыльной части. Эти результаты соответствуют наблюдавшимся в других исследованиях (Davies, Parker, Rutherford, Jones, 1988; Ctarfinkel, Cafarelli, 1992). Использовав такую же методику, Alway, MacDongall, Sale (1989) обнаружили увеличение на 44 % максимального вращающего момента сгибателей подошвы в течение 16 недель тренировочных занятий.
Динамический метод тренировки. Когда вращающий момент мышцы не равен вращающему моменту, обусловленному нагрузкой, сокращения мышцы называют динамическими, поскольку ее длина изменяется. Это понятие используют, как правило, для характеристики концентрических и эксцентрических условий. Наряду с этим изометрическое сокращение можно назвать статическим, поскольку система находится в равновесии, т.е. вращающий момент мышцы и вращающий момент, обусловленный нагрузкой, одинаковы. Понятие изокинетический характеризует сокращения, обеспечивающие постоянную угловую скорость конечности; обычно это достигается с помощью какого-либо приспособления (например, изокинетического тренажера).
В современной эре силовой тренировки, DeLorme (1945), пожалуй, первым стал систематически применять динамические упражнения. Его метод предполагает выполнение циклов по 10 повторений с постепенным увеличением нагрузки в каждом цикле. Максимальную нагрузку определяют методом проб и ошибок, т.е. спортсмен, используя различный вес, подбирает такую нагрузку, которую он может поднять точно 10 раз. Ее называют нагрузкой 10 МП (МП — максимальное повторение). Первый цикл 10 повторений выполняют с нагрузкой равной 1/2 10 ПМ, второй — с нагрузкой 3/4 МП и заключительный — с нагрузкой 10 МП. Первые два цикла служат в этом случае разминкой перед максимальным усилием в третьем цикле. Существует также Оксфордский метод (Zinovieff, 1951), в котором второй цикл выполняют с 75 % 10 МП, а третий — 50 % МП. Эти два метода часто называют восходящей и нисходящей пирамидальной нагрузкой (McDonagh, Davies, 1984). Существует множество систем, характеризующихся различной величиной нагрузок, используемых в программе занятий силовой направленности (Fleck, Kraemer, 1987); это — отдельный цикл, “от небольшой к значительной”, “от значительной к
292
небольшой”, “суперцикл”, программа “блиц” и многие другие.
Поскольку динамические упражнения включают как концентрические, так и эксцентрические сокращения, было бы неправильно предположить, что нагрузка 10 МП является максимальной для всех фаз каждого повторения. На основании взаимосвязи вращающий момент-скорость (см.рис.6.19) мы знаем, что вращающий момент мышцы при эксцентрическом сокращении больше, чем при концентрическом.
Поскольку при эксцентрических сокращениях мышца образует большую силу, считается, что упражнения эксцентрического характера должны обеспечивать более значительный тренировочный импульс и, следовательно, приводить к большему увеличению силовых возможностей. Принцип увеличения нагрузки гласит, что количество стимула определяется не абсолютной величиной, а величиной силы относительно максимальной, поэтому, очевидно, эксцентрические сокращения должны вызывать более значительные адаптационные реакции, чем концентрические. Однако, как показывают результаты исследований, при использовании только концентрических или эксцентрических упражнений увеличение силы идентично. В то же время при сочетании обоих видов упражнений отмечают более значительное увеличение силовых способностей (Colliander, Tesch, 1990; Judley, Tesch, Miller, Buchanan, 1991; Hakkinen, 1985). Например, 12-недельная программа тренировок для мышц—разгибателей коленного сустава (3 раза в неделю), привела к увеличению пика вращающего момента на 18 % у испытуемых, выполнявших концентрические упражнения, и на 36 % у испытуемых, выполнявших и концентрические, и эксцентрические упражнения (Colliander, Tesch, 1990).
Как уже отмечалось, между концентрическими и эксцентрическими сокращениями существуют большие различия:
1. Субмаксимальные концентрические и эксцентрические сокращения характеризуются активацией различных двигательных единиц (Nardone et al., 1989).
2. Значительные нагрузки вследствие эксцентрических сокращений, по-видимому, влияют на поведение отдельных поперечных мостиков и саркомеров. Открепление поперечного мостика при эксцентрическом сокращении, очевидно, вызывает механический разрыв химической связи в отличие от более упорядоченного присоединения АТФ и отсоединения поперечного мостика, которое наблюдается при нормальном цикле поперечного мостика. Ввиду различий в количестве сократительных белков и степени взаимного перекрывания саркомеров, максимальная сила, образуемая каждым саркомером, значительно колеблется вдоль длины мышцы.
3. Хотя при эксцентрическом сокращении сила больше, ЭМГ значительно меньше, чем во время концентрического сокращения. Это свидетельствует о том, что индивидуумы не способны максимально активировать мышцы при эксцентрическом сокращении (Nakazawa, Kawakami, Fukunaga et al., 1993; Tesch et al., 1990; Westing, Cresswell, Thorstensson, 1991).
4. Отсроченное возникновение болезненных ощущений в области мышц чаще всего отмечается при эксцентрических сокращениях (Friden, 1984; Newham, 1988; Stauber, 1989).
5. Стационарный бег с субмаксимальной интенсивностью на тредбане связан с постоянным потреблением кислорода. Бег же по наклонной вниз (10 % наклон) с такой же интенсивностью осуществляется в результате постепенного увеличения потребления кислорода и ЭМГ в мышцах ног (Dick, Cavanagh, 1987). При беге по наклонной вниз происходит значительное использование эксцентрических сокращений мышц.
6. При беге по ровной поверхности с субмаксимальной интенсивностью на тредбане в течение 20 мин на 9 % снижается амплитуда Н-рефлекса в камбаловидной мышце. В то же время бег по наклонной вниз (под углом 10°) при такой же интенсивности приводит к уменьшению Н-рефлекса на 25 % (Bulbulain, Bowles, 1992).
7. Эксцентрические упражнения обеспечивают более эффективное стимулирование гипертрофии мышечной массы (Dudley et al., 1991).
Тренажеры. Тренажеры, в которых нагрузка контролируется системой механизмов (например, Cybex, Biodex), гидравлических цилиндров (Kincom, Lido, Omnitron) или пневматических систем (Keiser), обеспечивают аккомодирующее сопротивление, образующееся вследствие нагрузки и равное по величине, но противоположное по направлению силе, образуемой испытуемым. Одним из последствий действия систем механизмов и некоторых гидравлических тренажеров, является движение, при котором угловая скорость перемещаемого сегмента тела постоянна и, следовательно, изо-кинетична. Изокинетическое сокращение представляет собой динамическое условие (вследствие изменения длины мышцы), при котором отношение вращающего момента мышцы к вращающему моменту, обусловленному нагрузкой, равно 1. Если на систему не воздействует результирующий вращающий момент, ускорение системы равно нулю, а скорость системы постоянна. Очевидно, что оба вращающих момента (мышцы и нагрузки) не одинаковы в начале и в конце изокинетического сокращения; в противном случае движение никогда бы не началось или не закончилось, поскольку как для его начала, так и для прекращения необходимо ненулевое ускорение. Изокинетические тренажеры
293
Рис.9.2. Выполнение испытуемым изокинетического упражнения (выпрямление ноги в коленном суставе): а — упражнение на изокинетическом тренажере “Цибекс”; б — диаграмма свободного тела сил, действующих на голень; Fj — сила реакции сустава; Fv — вес конечности; F, — нагрузка тренажера; Тт — результирующий вращающий момент мышцы
предполагают выполнение концентрических и эксцентрических сокращений. При выполнении концентрического теста спортсмен преодолевает сопротивление тренажера, выполняя положительную работу по отношению к окружающей среде. При выполнении эксцентрического теста спортсмен “сопротивляется” нагрузке тренажера, поэтому имеет место выполнение отрицательной работы.
Многие исследователи используют изоки-нетические тренажеры (рис.9.2, а) для определения вращающего момента, работы и мощности, образуемой мышцами. Обычно для этого измеряют сопротивление со стороны тренажера (F/ на рисунке 9.2, б), выражая усилие в условных единицах. Соответствующая диаграмма свободного тела системы (рис.9.2, б), показывает, однако, что нагрузка трежанера — не единственный фактор, влияющий на вращающий момент, обусловленный ею; следует также учитывать вес конечности. Winter, Wells, Огг (1981) показали, что если не учитываются компоненты ускорения (ускорение, обусловленное силой тяжести, и ускорение конечности, направленное на достижение скорости, заданной тренажером), степень ошибки может составить до 500 % при максимальных скоростях при определении работы, выполненной мышцами. Вместе с тем учет компонентов ускорения позволяет провести более точные измерения (Gransberg, Knutsson, 1983). В противном случае вращающий момент, обусловленный тренажером (F/), не является мерой результирующего вращающего момента мышцы.
Недостатком изокинетических тренажеров является то, что они нарушают принцип специфичности. Естественные движения не включают постоянную угловую скорость конечности и, следовательно, не являются изокинетиче-скими. Более того, максимальная скорость многих изокинетических тренажеров (5 рад с-1) намного меньше максимальной угловой скорости, развиваемой при таких движениях, как бег, прыжки, метания. Несмотря на это, метод изо-
кинетической тренировки обеспечивает адаптационные реакции, подобные наблюдаемым при применении других тренировочных методов. Например, когда испытуемые тренировали мышцы—разгибатели коленного сустава в течение 7 недель на изокинетическом тренажере со скоростью 3,14 рад • с-1, значительно увеличивался пик вращающего момента (около 14 %) при сокращениях со скоростью 0 — 3, 14 рад • с“ *, но не при скорости 4,2 и 5,2 рад • с-1 (Lesmes, Costill, Coyle, Fink, 1978). У испытуемых, тренировавшихся в течение 10 недель со скоростью 1,1 рад-с-1, также наблюдали увеличение пика вращающего момента на 9 % (25 Н • м), тогда как у тренировавшихся со скоростью 4,2 рад с-1 увеличение пика вращающего момента составило 20 % (30 Н • м); каждое увеличение соответствовало тренировочной скорости (Ewing, Wolfe, Rogers et al., 1990). Кроме того, у спортсменов, занимающихся силовыми видами спорта, была больше площадь поперечного сечения мышцы и более значительный изокине-тический вращающий момент, чем у контрольных испытуемых (Alway, Stray-Gundersen, Gumbt, Gonyca, 1990).
Следует отметить, что специфика изоки-нетической тренировки, по-видимому, зависит скорее от намеренной, чем действительной скорости движения. Behm, Sale (1993) провели исследование, в котором испытуемые в течение 16 недель выполняли быстрые (5,23 рад-с-1) изокинетические сокращения одной ногой в одно время. Хотя цель состояла в том, чтобы выполнять быстрые изокинетические сокращения каждой ногой, одна нога не двигалась и по-этому мышцы сокращались изометрически. В результате тренировок пик вращающего момента обеих конечностей увеличился при скорости изокинетических сокращений 5,23 рад • с-1. Увеличение пика вращающего момента было более значительным, чем при более низких скоростях.
Изокинетические тренажеры имеют ряд преимуществ. Во-первых, следует отметить воз
294
можность различной нагрузки на мышечную группу в диапазоне движения. Это особенно важно в процессе реабилитации после травм. Когда в определенном участке движения возникает болевое ощущение, травмированный спортсмен может уменьшить усилие, продолжая выполнять упражнение. Более того, спортсмен может прекратить выполнение упражнения. Во-вторых, изокинетические тренажеры позволяют регулировать величину нагрузки в зависимости от индивидуальных возможностей каждого спортсмена,
В отличие от изокинетических тренажеров гидравлические и пневматические системы, обеспечивая регулирование сопротивления, не позволяют контролировать скорость движения. Гидравлические тренажеры обеспечивают сопротивление вследствие заполнения камеры жидкостью. Мышцы выполняют концентрические сокращения, противодействуя этому сопротивлению. Сопротивление изменяется в зависимости от диаметра клапана, через который вода из одной камеры переходит в другую; чем меньше диаметр клапана, тем выше сопротивление. Гидравлическая система обеспечивает аккомодирующее сопротивление. Чем больше образуемая спортсменом сила, тем выше сопротивление тренажера. Скорость движения также возрастает с увеличением прилагаемой силы. Следует отметить, что использование гидравлических тренажеров обеспечивает такой же прирост силы, как и отягощений. Например, Hoztobagyi, Katch (1990) провели исследование, в котором в течение 12 недель одна группа испытуемых тренировались с отягощениями,. а другая — с гидравлическим тренажером. Среднее улучшение результатов при выполнении жима лежа на скамье и приседаний со штангой составило 23 % в первой группе и 20% — во второй. Кроме того, изменение силы, скорости и мощности в обеих группах было практически одинаковым (~ 8 %). Пневматические системы обеспечивают изменение сопротивления с помощью специального компрессора, изменяющего давление, а с ним и сопротивление, которое преодолевает спортсмен. Изменение давления осуществляется быстро, простым нажатием кнопки. Преимущество пневматических систем заключается в возможности достаточного увеличения давления, обеспечивающего выполнение эксцентрических сокращений.
Плиометрический метод тренировки. Плио-метрические упражнения используются для тренировки определенной структуры движения — эксцентрическо-концентрической последовательности мышечной деятельности. Большинству движений человека присуща именно эта последовательность мышечной деятельности. Преимущество метода в том, что первоначальное эксцентрическое сокращение позволяет
мышце выполнить большее количество положительной работы при последующем концентрическом сокращении. Взаимосвязь работа-энергия (см.главу 3) показывает, что для выполнения мышцей большей работы, ей требуется дополнительное количество энергии. В главе 7 мы рассмотрели, как накопление и использование упругой энергии, а также предварительная нагрузка обеспечивают дополнительное количество энергии. Упругая энергия во время эксцентрического сокращения содержится в эластичных компонентах и используется во время концентрического сокращения по мере сокращения сократительного и эластичного компонентов. При предварительной нагрузке эксцентрическое сокращение вызывает увеличение мышечной силы в начале концентрического сокращения, поэтому область под графиком сила—расстояние (работа) больше.
Таким образом, несомненно, что плиоме-трические упражнения оказывают тренировочный эффект. Например, в исследовании Blattner, Noble (1979) две группы испытуемых тренировались в течение 8 недель; первая использовала изокинетические тренажеры, вторая выполняла плиометрические упражнения. В результате тренировок у спортсменов обеих групп результаты в прыжке в высоту улучшились на 5 см. Hakkinen, Komi, Alen (1985) обнаружили, что прыжковые тренировки приводят к незначительному увеличению максимальной изометрической силы мышц—разгибателей коленного сустава, однако значительно повышают интенсивность развития силы, а также площадь быстросокращающихся волокон.
Нервно-мышечное электростимулирование.
Еще в начале 70-х годов в бывшем СССР (Elson, 1974) использовали электрошок для увеличения силы мышц. Этот метод, получивший название нервно-мышечного электростимулирования, предполагает стимуляцию мышцы при минимальных болевых ощущениях (Moreno-Aranda, Siereg, 1981а, 1981b, 1981с). Нервно-мышечное электростимулирование применяют с XVIII ст. как средство реабилитации (Hainaut, Duchateau, 1992).
Основная причина эффективности нервно-мышечного электростимулирования заключается в том, что люди не в состоянии максимально активировать мышцы, а электрошок вызывает разницу между максимальной силой произвольного сокращения и максимальной способностью мышцы. Два фактора свидетельствуют о неспособности человека произвести максимальную мышечную силу при произвольном сокращении (Enoka, Fuglevand, 1993):
1. Нервная недостаточность — силу, образующуюся во время максимального произвольного сокращения, можно увеличить с помощью отдельных электрических импульсов (Hales, Gandevia, 1988) или их серии (Duchatean, Hainant,
295
Рис.9.3. Избранные режимы стимулов, используемые в процессе нервно-мышечного стимулирования (Enoka, 1988в):
а — условный ряд низкочастотных (100Гц) стимулов с шириной импульса 0,1 мс; б — структура высокочастотной стимуляции (10 кГц, синусоидальная волна), модулированной при низкой частоте (100Гц) с интервалом между последовательными стимулами 0,01 с; в — формы волн, используемые в процессе нервно-мышечной стимуляции; двуфазные формы волн колеблются относительно нулевой линии
1987, 1988; Young, McDonagh, Davies, 1985). Это свидетельствует о том, что нервного побуждения недостаточно для образования мышцей максимальной силы.
2. Нервное дополнение — сила максимального произвольного сокращения может быть дополнена с помощью изменения афферентной обратной связи. Например, Howard, Enoka (1991) изучали влияние нервно-мышечного электростимулирования правой ноги на максимальную силу произвольного сокращения мышц-разгибателей коленного сустава левой ноги. Нервно-мышечное электростимулирование четырехглавой мышцы бедра левой ноги приводило к сильным кожным ощущениям, а также образованию силы мышцами правой ноги, соответствующей 40 % максимальной. Кроме того, электростимулирование правой ноги приводило к увеличению максимальной силы произвольного сокращения левой ноги на 11 %.
Нервно-мышечное электростимулирование может применяться различным образом. Изменяемые параметры включают частоту стимулирования, интенсивность, тип и размер электродов. Простейший метод предполагает серию прямоугольных импульсов (рис.9.3, а). Недостаток данного метода в том, что для стимулирования максимальной мышечной силы необходима частота около 100 Гц, которая, однако, вызывает значительную болевую реакцию (Ban, Nielsen, Soderberg, 1986). Решение проблемы заключается в использовании высокочастотного стимулирования (10 кГц), модулированного более низкими частотами (50—100 Гц); способ был предложен Kots (1971) (рис.9.3, б). Moreno-Aranda, Seires (1981а, 19816, 1981с) проанализировали этот способ и установили, что оптимальный режим предполагает стимулирование в течение 1,5 с каждые 6 с в течение 60-секундного периода с последующим отдыхом в течение 60 с. Таким образом, значительно снижаются болевые ощущения и стимулируется образование силы, эквивалентной максимальной силе произвольного сокращения (Delitto, Brown, Strube et.al., 1989).
Кроме этого можно применять стимулирование с различной формой волны (рис.9.3,в). Изменение формы волны объясняется следующими двумя факторами. Во-первых, форма волны стимула влияет на комфорт при нервно-мышечном электростимулировании. Коммерческие электростимуляторы вырабатывают множество различных форм волны (прямоугольную, треугольную, синусоидальная), обеспечивающих электрический ток положительного (мо-нофазного) или положительно-отрицательного (бифазного) импульса. Во-вторых, стимулы общепринятой формы волны (например, бифазные прямоугольные импульсы), как известно, преимущественно активируют двигательные единицы большого диаметра, в отличие от упорядоченного (от небольшого к большому) рекрутирования, которое имеет место при произвольной активации (Fang, Mortimer, 1991; Trimble, Enoka, 1991). Хотя упорядоченное рекрутирование благоприятно в процессе силовых тренировок, а также во время восстановления после травмы, оно в то же время создает трудности в процессе функционального электростимулирования парализованной мышцы. Поскольку двигательные единицы большого диаметра характеризуются повышенной утомляемостью, стимулы общепринятой формы волны приводят к быстрому утомлению парализованной мышцы, что ограничивает возможности использования этого метода для восстановления функций (Fang, Mortimer, 1991а). Используя ква-зитрапециевидную форму волны и триполяр-ный стимулирующий электрод, Fang, Mortimer (1991в, 1991с) разработали методику преимущественного рекрутирования двигательных единиц
296
типа FR. Она обеспечивает дифференцированное блокирование (потенциалы действия не могут распространяться) вследствие гиперполяризации мембраны аксонов большого диаметра и позволяет аксонам небольшого диаметра избирательно активироваться. Методика может применяться для функционального электростимулирования парализованной мышцы.
Поскольку аксолемма легче возбуждается, чем сарколемма, электрический ток, проходящий через мышцу и вызывающий ее сокращение, обусловливает образование потенциалов действия скорее в ответвлениях внутримышечного нерва, чем непосредственно возбуждая мышечные волокна (Hultman, Sjoholm, Jadarholm-Ek, Krynicki, 1983). Следовательно, электроды с большей площадью обеспечивают более широкое распределение тока по всей мышце. Кроме того, электрод должен иметь низкий импеданс, т.е. низкое сопротивление электрическому току. Если электрод большой, плотность тока меньше, поэтому через ткань может пройти большее количество тока, не повреждая ее. В этой связи наиболее эффективны угольные прорезиненные электроды (Leiber, Kelly, 1991).
Нервно-мышечное электростимулирование включает искусственное образование потенциалов действия в периферической двигательной системе. Потенциалы действия могут также образовываться при магнитном стимулировании (Lotz, Duhne, Daube, 1989). С этой целью применяют магнитный стимулятор, образующий магнитное поле, которое изменяется во времени и создает электрическое поле и электрический ток. При значительном токе возбудимые мембраны деполяризуются и образуются потенциалы действия. Подобно нервно-мышечному электростимулированию магнитное стимулирование активирует ответвления внутримышечного нерва. В то же время порог двигательной реакции более низкий при магнитном стимулировании, поэтому этот метод менее болезненный, чем метод электростимулирования. Другое преимущество магнитного стимулирования — меньшее снижение стимула на расстояние, а также более широкое его распространение.
Метод нервно-мышечного электростимулирования, несомненно, позволяет увеличить силовые возможности здоровой и травмированной мышцы (Enoka, 1988в; Miller, Thepau-Mathien, 1993). Лучше всего использовать методику Kots (частота стимулирования 2500 Гц, модулированная “всплесками” 50 Гц) включающую 10 повторений в каждом из 15—25 тренировочных занятий. Интенсивность стимулирования устанавливается на максимально переносимом уровне. Увеличение силы (изометрическое сокращение) колеблется от 0,6 до 3,6 % за тренировочное занятие, среднее увели
чение за занятие составляет 1,6%. Подобный прирост силы наблюдается при использовании других тренировочных методов (Hainant, Duchateau, 1992).
Три аспекта, связанные с применением нервно-мышечного электростимулирования здоровой мышцы, представляют несомненный интерес. Во-первых, сопоставимое увеличение силы можно достичь при более низкой силе стимуляции, уступающей силе произвольной активации мышцы. Например, Langhman, Youdas, Garrett, Chao (1983) провели исследование, в котором две группы испытуемых участвовали в тренировочных занятиях, направленных на увеличение силы четырехглавых мышц бедра. Первая группа выполняла изометрические упражнения, во второй группе применялось нервно-мышечное электростимулирование. Несмотря на одинаковый прирост силы в обеих группах (соответственно 18 и 22 %) после 5 недель занятий, следует отметить, что средняя интенсивность занятия в первой группе была 78%, во второй — 33% максимальной. Это различие, скорее всего, обусловлено предпочтительной активацией вследствие электростимулирования двигательных единиц большого диаметра (тип II). Активация мышечных волокон абсолютно разная при произвольном сокращении и нервно-мышечном стимулировании (Adams, Harris, Woodard, Dudley, 1993).
Во-вторых, активация мышц одной конечности с помощью нервно-мышечного электростимулирования может вызвать среднее увеличение силы в противоположной конечности. Например, Laughman et al. (1983) наблюдали увеличение почти на 15 % максимальной силы произвольного сокращения четырехглавой мышцы бедра левой ноги после 5 недель тренировок (нервно-мышечное стимулирование) четырехглавой мышцы бедра правой ноги. Подобная адаптационная реакция, по-видимому, обусловлена изменениями в нервной цепи вследствие потенциалов действия, посылаемых назад в спинной мозг из искусственно активируемого нерва мышцы.
В-третьих, несмотря на сопоставимый прирост силы вследствие произвольной активации и нервно-мышечного электростимулирования Hainant, Duchateau (1992), наблюдали значительное увеличение произвольной деятельности при электростимулировании (Delitto et al., 1989). В исследовании на протяжении 140 дней (разделенных на четыре периода продолжительностью около 1 мес) участвовал опытный (стаж занятий 11 лет) тяжелоатлет. Зависимой переменной был максимальный вес, который спортсмен мог поднять при выполнении трех упражнений — толчка, рывка и упора присев. Во время второго и четвертого периода исследований обычный режим тренировочных занятий (3 ч в день) был дополнен нервно-мышечным электрости-
297
Рис.9.4. Изменение максимального веса, поднимаемого опытным тяжелоатлетом при выполнении трех упражнений. В периоды б обычные тренировочные занятия дополнялись нервно-мышечным электростимулированием четырехглавых мышц бедер (Delitto, Brown, Strube et al., 1989)
мулированием четырехглавых мышц бедра с высокой интенсивностью (200 мА). В результате электростимулирования значительно улучшились результаты выполнения трех упражнений (рис.9.4). Кроме того, установлено, что нервно-мышечное электростимулирование позволяет повысить максимальный эксцентрический вращающий момент (Wasting et al., 1990).
Нервно-мышечное электростимулирование широко применяется в процессе реабилитации после травм. По мнению ряда ученых, с помощью этого метода значительно легче укрепить
гипотрофированную мышцу, чем с помощью произвольной активации (Godfrey, Jayawardena, Welsh, 1986; Williams, Morrissey, Brewster, 1986).
Техника нагрузки
При выборе способа изменения нагрузки для обеспечения прироста силы следует учитывать ряд факторов.
Метод постепенного увеличения нагрузки. DeLorme (1945) одним из первых заинтересовался проблемой изменения величины физической нагрузки. Он предложил изменять ее от одного цикла повторений к другому. Этот метод получил название метода постепенного увеличения нагрузки упражнения. Как уже отмечалось, методика DeLorme предполагает выполнение трех циклов повторений упражнения с постепенным увеличением нагрузки с каждым циклом. В общепринятой программе занятий силовой направленности это означает увеличение веса штанги в каждом новом цикле. Однако вращающий момент мышцы зависит от плеча пары (сил) и скорости движения вдобавок к величине внешней нагрузки (вес штанги). Такие упражнения можно выполнять, не изменяя нагрузку (вес штанги), просто изменяя длину плеча пары или скорость движения. Например, на рис. 9.5, а спортсмен находится на промежуточном этапе выполнения упражнения поднятия туловища из положения лежа с согнутыми в коленном суставе ногами. В четырех положениях, указанных на рис.9.5, б, руки постепенно перемещаются из сторон за голову. Изменение положения рук не изменяет вес верхней части тела, однако приводит к перемещению ЦТ к
Рис.9.5. Выполнение поднятия туловища из положения лежа с согнутыми в коленном суставе ногами: а — диаграмма всего тела; б — изменение положения рук, влияющее на плечо пары (d) вектора веса (FK) верхней части тела относительно бедренного сустава, приводит к увеличению результирующей силы мышц бедра (fm), необходимой для медленного выполнения упражнения; (fj), (/у) — соответственно внутрибрюшное давление и сила реакции суставов
298
голове, тем самым увеличивая плечо пары веса системы относительно бедренного сустава (пунктирная линия на рис.9.5, б). В результате увеличивается вращающий момент, образуемый мышцами сгибателями бедренного сустава.
Величина нагрузки. В большинстве программ физических занятий силовой направленности цикл включает 1—8 повторений. Когда спортсмен поднимает более тяжелую штангу, результирующий вращающий момент возрастает прямо пропорционально увеличению веса и только в том случае, если кинематика движения остается постоянной. Во время тренировочного занятия тяжелоатлетов, а также в процессе реабилитации необходимо обращать внимание на форму движения, следя, чтобы нагрузка на мышцы увеличивалась прямо пропорционально и нагрузке подвергались нужные мышцы. Поскольку максимальные нагрузки, как правило, изменяют кинематику движения, в занятиях силовой направленности используют нагрузки, составляющие около 4—6 максимальных повторений (МП), выполняемые за 4 цикла (3-6) для каждого упражнения (Atha, 1981; Fleck, Kraemer, 1987; McDonagh, Davies, 1984; Sale, MacDougall, 1981). Согласно данным Sale, MacDoudall (1981), нагрузка 5—6 МП соответствует 85-90 % максимальной нагрузки (рис.9.6).
McDonagh, Davies (1984) на основании исследований с участием нетренированных испытуемых пришли к следующему заключению. Нагрузки менее 66 % максимальной не приводят к увеличению силы даже при выполнении 150 сокращений в день; нагрузки выше 66 % максимальной обеспечивают прирост силы порядка 0,5-1,0 % после каждого тренировочного занятия, а нагрузки выше 66 % максимальной при выполнении 10 повторений на каждом занятии приводят к увеличению силовых возможностей. По мнению Нагге (1982), начинающим следует использовать нагрузки 60—80 % максимальной, выполняя 8—10 повторений в каждом цикле, а сильнейшим спортсменам следует использовать нагрузки 80—100 % максимальной, выполняя за цикл 2—5 повторений.
Другой способ заключается в изменении скорости движения. Некоторые исследователи сравнивали показатели прироста силы при выполнении обычных упражнений с отягощениями, а также при выполнении “взрывных” упражнений (т.е. с высокой скоростью). Поскольку максимальная сила, образуемая мышцей, уменьшается с увеличением скорости укорачивания (взаимосвязь сила-скорость), вряд ли можно ожидать, что быстрые движения обеспечат более эффективные тренировочные стимулы. Так, Hakkinen, Komi (1986) установили, что программа занятий силовой направленности (20 недель) привела к увеличению силы у испытуемых, тогда как у испытуемых другой группы, тренировавшихся в течение такого же времени и выпол-
Рис.9.6. Зависимость количества повторений в цикле от величины нагрузки как процента максимальной (Sale, MacDongall, 1981)
нявших быстрые прыжки в высоту, увеличение силы не наблюдалось. В то же время в результате тренировки мышц—сгибателей локтевого сустава с помощью значительных нагрузок (90 % 1 МП), а также быстрых движений (10 % 1 МП) получено сопоставимое увеличение силы мышц и площади поперечного сечения мышечных волокон (Dahl, Aasemd, Jensen, 1992).
Постоянная и переменная нагрузки. Физическая деятельность силовой направленности, за исключением изометрических упражнений, включает изменение длины активной мышцы в диапазоне движения. Поскольку вращающий момент мышцы изменяется при изменении угла сустава, необходимо обращать внимание не только на величину нагрузки, но и на то, как она изменяется в диапазоне движения.
Рассмотрим упражнение сгибание предплечья, включающее чередование сгибания—разгибания руки в локтевом суставе с нагрузкой. При использовании отягощений (рис.9.7, а) нагрузка, т.е. отягощение, которое удерживает спортсмен, остается постоянной в течение всего упражнения и действует вертикально вниз. Нагрузка (/*}) при выполнении упражнения со штангой — это вес штанги. Длина вектора нагрузки остается постоянной на всем диапазоне движения (рис.9.7, а); вспомним, что длина вектора веса указывает на величину нагрузки. Однако при использовании некоторых тренажеров нагрузка, которую перемещает спортсмен, изменяется в диапазоне движения (рис.9.7, б). Нагрузка (Fz) со стороны тренажера представляет собой вращающий момент, обусловленный набором отягощений тренажера и плечом пары (d) диска. Хотя вес остается постоянным, длина плеча пары (d) от оси вращения (0) до точки, в которой цепь (подсоединенная к набору отягощений) “покидает” диск, изменяется в диапазоне движения (рис.9.7, 6). Это отражено на диаграмме свободного тела тренажера изменением длины вектора нагрузки в диапазоне движения.
299
Рис.9.7. Выполнение сгибания предплечья с постоянной (а) и переменной (б) нагрузками
Другое различие между постоянной и переменной нагрузками состоит в том, что при использовании отягощений система действует на силу (вес штанги), тогда как тренажер — против вращающего момента (набор отягощений х плечо пары).
Таким образом, при использовании обычных отягощений, нагрузка (вес штанги) остается постоянной на всем диапазоне движения, тогда как при использовании тренажеров вращающий момент (нагрузка) изменяется. Если пренебречь весом конечности, то величина нагрузки на мышцы — результат внешней нагрузки (вес штанги или вращающий момент тренажера) и расстояния плеча пары (s на рисунке 9.8). 5 значительно больше изменяется в диапазоне движения при нагрузке со стороны тренажера (рис.9.8, 6), чем при нагрузке со стороны штанги (рис.9,8, а). Понятие переменной нагрузки возникло из взаимосвязи вращающий момент—угол мышцы.
Наиболее типичная взаимосвязь (промежуточная) показывает, что максимальный враща
ющий момент имеет место на середине диапазона движения. Следовательно, тренажеры с переменной нагрузкой обеспечивают изменение нагрузки в диапазоне движения в зависимости от взаимосвязи вращающий момент-угол мышцы.
Различия в степени воздействия (стресса) на мышцу при постоянной и переменной нагрузках показаны на рис.9.9. Кривая 1 отражает взаимосвязь максимальный изометрический вращающий момент—угол для группы мышц-сгибателей локтевого сустава. Две другие кривые показывают вращающий момент сгибателей локтя, необходимый для перемещения средней нагрузки (веса) в указанном диапазоне движения с постоянной (штанга — 2) и переменной (тренажер “Наутилус” — 5) нагрузкой. Величина нагрузки (веса) в обоих случаях была одинаковой 60 % 4 МП. По мнению некоторых специалистов, использование тренажеров предпочтительнее, поскольку на мышцу оказывается более равномерное воздействие на всем диапазоне движения.
Рис.9.8. Нагрузка на мышцы относительно локтевого сустава при выполнении упражнения с отягощениями при постоянной нагрузке (а) и на тренажере (б — переменная нагрузка). Для упрощения будем считать, что переменная нагрузка направлена перпендикулярно к предплечью на всем диапазоне движения
300
Рис.9.9. Результирующий вращающий момент мышцы относительно локтевого сустава при выполнении сгибания предплечья со штангой и на тренажере “Наутилус” в сравнении с максимальным вращающим моментом, который может образоваться в диапазоне движения (Smith, 1982)
Пояснения в тексте
Адаптационные реакции нервной системы
Общим методом изучения механизмов, лежащих в основе увеличения силовых возможностей вследствие тренировочных занятий, является сопоставление временных изменений ЭМГ и размера мышц (например, площади поперечного сечения целой мышцы или мышечного волокна). Результаты проведенных исследований показывают, что изменение ЭМГ происходит до изменения размера мышцы, и оно более значительно; кроме того, программы тренировочных занятий, слишком непродолжительные, чтобы вызвать морфологические изменения, могут тем не менее изменять ЭМГ и силовые возможности (Hakkinen, Alen, Komi, 1985; Rutherford, Jones, 1986). Например, на рис.9.10 показаны временные изменения площади поперечного сечения четырехглавой мышцы бедра (определенные с помощью метода магнитного резонанса), интегрированной ЭМГ, а также максимальной изометрической силы после 60 дней тренировочных занятий и 40 дней без занятий (Narici, Roi, Landoni et.al., 1989). Увеличение ЭМГ было непропорционально больше увеличения размера мышцы. Эти результаты подтверждают вывод, что прирост силы возможен без адаптационных реакций в мышце, и не возможен без адаптационных реакций нервной системы.
Адаптационные реакции, по-видимому, имеют место на всех уровнях нервной системы. Некоторые типы адаптационных реакций, происходящих в нервной системе, приводятся ниже.
1. Воображаемые сокращения. Пожалуй, наиболее убедительное доказательство существования адаптационных реакций в нервной системе было получено при исследовании прироста силы вследствие изометрических сокращений. Одна группа испытуемых выполняла сокращения, тогда как испытуемые другой груп-
Рис.9.10. Изменение интегрированной ЭМГ широкой латеральной мышцы бедра (7) площади поперечного сечения четырехглавой мышцы бедра (2) при максимальном сокращении и сила МПС (J) во время изокинетических тренировочных занятий и в процессе детренировки (Narici, Roi, Landani et al., 1989)
301
Рис.9.11. Среднее увеличение максимальной изометрической силы вследствие тренировок тренированного (а) и нетренированного (6) 5-го пальца у трех групп испытуемых (Yne, Cole, 1992):
1 — группа, выполнявшая воображаемые сокращения; 2 — произвольные сокращения; 3 — контрольная группа
пы только представляли, что выполняют сокращения (Ync, Cole, 1992). Исследование предполагало выполнение упражнений для мышц кисти 5 раз в неделю в течение 4 недель. У испытуемых, мысленно представлявших, что они выполняют максимальные изометрические сокращения, мышца не активировалась (отсутствовала ЭМГ). Результаты исследований приведены на рис.9.11. Максимальная изометрическая сила мышц тренируемой руки увеличилась у испытуемых обеих групп (30% — в группе, выполнявшей сокращения, и 22 % — в группе мысленно представлявших выполнение сокращений), и не увеличилась в контрольной группе испытуемых, которые не выполняли упражнения. Кроме того, у испытуемых обеих групп увеличилась максимальная изометрическая сила мышц нетренируемой руки (14% — в группе выполнявших сокращение и 10% — в группе представлявших выполнение), хотя и в меньшей степени. Поскольку мышца не активировалась у испытуемых, мысленно представлявших, что выполняют сокращения, прирост силы у них, по-видимому, был обусловлен адаптационными реакциями нервной системы.
2. Координация. В исследованиях Rutherford, Jones (1986) группа испытуемых в течение 12 недель выполняла двустороннее выпрямление ноги в коленном суставе. Нагрузка составляла 6 МП, т.е. около 80% максимальной. Максимальную силу изометрического произвольного сокращения и тренировочную нагрузку регистрировали в течение всего периода тренировок. Рис.9.12 иллюстрирует изменение взаимосвязи сила—тренировочная нагрузка, имевшее место после начала тренировочных занятий. Прирост силы, показанный смешением вдоль оси х, составил 20 % у мужчин и 4 % у женщин. В то же время тренировочная нагрузка (смещение вдоль
оси у) увеличилась на 200% у мужчин и на 240 % у женщин. Поскольку величину силы тестировали при изометрических сокращениях, в то время как упражнение включало и динамические сокращения, более существенное увеличение тренировочной нагрузки, было, по-видимому, обусловлено улучшением координации при выполнении упражнения разгибания ноги в коленном суставе. Подобная адаптационная реакция, очевидно, вызвана влиянием нервной системы.
3. Коактивация. Когда испытуемые выполняют максимальное произвольное сокращение, в мышце-антагонисте, как правило, отмечается значительная активность ЭМГ (Dimitrijevic et al., 1992). Коактивация затрудняет измерение силы, поскольку сила, измеряемая при проведении силового теста, представляет собой результирующую производительность (разницу), обусловленную активацией группы мышц агонистов-антагонистов. Следовательно, очевидный прирост силы (результирующая сила) может быть обусловлен просто пониженной степенью коактивации.
4. Поперечное развитие. В процессе реабилитационных мероприятий, а также тренировочных занятий силовой направленности часто наблюдают, что физические нагрузки одной половины тела приводят к изменениям во второй половине. Эту адаптационную реакцию иллюстрирует рис.9.12, показывающий прирост силы мышц правой (нетренированной) руки в результате выполнения упражнений левой рукой. Подобная адаптационная реакция наблюдается при использовании различных программ тренировочных занятий (рис.9.13), включая мысленное представление выполнения сокращений и активацию мышцы при произвольных сокращениях и нервно-мышечном электрости-
302
Рис.9.12. Взаимосвязь силы и тренировочной нагрузки до (7) и после (2) 12-недельной программы физических занятий у женщин и мужчин (Rutherford, Jones, 1986)
мулировании. Поскольку эта реакция происходит без активации (ЭМГ отсутствует) мышц противоположной конечности (Devine, LeVean, Yack, 1981; Panin, Lindenaner, Weiss, Ebel (1961), а также без изменения площади мышечных волокон или активности ферментов (Honston, Froese, Valeriote et al., 1983), прирост силы в нетренированной конечности, обусловлен, по-видимому, адаптационными реакциями нервной системы. Более того, следует отметить увеличение амплитуды ЭМГ в нетренированной конечности, связанной с максимальным произвольным сокращением (Butler, Darling, 1990).
5. Двусторонний дефицит. Одновременная активация обеих конечностей обычно приводит к снижению силы. Ее уменьшение во время максимального произвольного сокращения в этих
условиях называют двусторонним дефицитом (Howard, Enoka, 1991; Secher, Rube, Elers, 1988), однако он ограничен гомологичными мышцами. Двусторонний дефицит (рис.9.14) — изометрическое сокращение мышц—разгибателей локтевого сустава (трехглавых мышц плеча); локтевой сустав образует прямой угол. Каждая колонка на рис.9.14 соответствует одной попытке. В первой попытке (левая колонка) испытуемый выполнял максимальное изометрическое сокращение правой рукой. В третьей попытке (правая колонка) испытуемый выполнял максимальное изометрическое сокращение левой рукой. Во второй попытке (средняя колонка) испытуемый одновременно максимально активировал разгибатели локтевого сустава обеих рук. На рис.9.14 показана ЭМГ трехглавой мышцы плеча (агони-
• Произвольные сокращения
О Электростимулирование
Сила тренированной конечности, % увеличения
Рис.9.13. Результаты 28 исследований поперечного развития демонстрируют степень взаимосвязи между приростом силы в тренированных и нетренированных конечностях. Сила, измеряемая как максимальная изометрическая сила, увеличивалась в результате произвольных сокращений или нервно-мышечного электростимулирования в нескольких различных мышцах. Точки ниже линии идентичности указывают на то, что прирост силы в тренированной конечности был выше, чем в нетренированной (Enoka, 1988b).
303
Рис.9.14. Максимальная сила и ЭМГ мышц—разгибателей локтя при максимальных изометрических сокращениях правой руки, обеих рук и левой руки (Ohtsuki, 1983):
I — сила; 2 — трехглавая мышца плеча; 3 — двуглавая мышца плеча
ста) и двуглавой мышцы бедра (антагониста). Максимальная сила, образуемая мышцами каждой руки, и количество ЭМГ были меньше во время двусторонних сокращений (т.е. двумя руками). Снижение силы вследствие двустороннего дефицита в среднем составляет — 5...—10 % максимальной. Вследствие уменьшения ЭМГ агониста и повышения степени коактивации во время двустороннего сокращения двусторонний дефицит должен быть обусловлен влиянием со стороны нервной системы (Enoka, 1988; Howard, Enoka, 1991; Koh, Grabiner, Clongh, 1993).
Хотя в большинстве исследований двусторонних взаимодействий отмечали двусторонний дефицит, они (двусторонние взаимодействия), по-видимому, изменяются вследствие тренировок. Так, Howard, Enoka (1991) сравнивали максимальную изометрическую силу при сокращениях мышц—разгибателей коленного сустава одной и двумя ногами и обнаружили двусторонний дефицит у нетренированных испытуемых (—9,5 %) и велосипедистов высокого уровня (—6,6 %), а у тяжелоатлетов — двусторонний прирост (+6,2 %). Двусторонний прирост означает, что максимальная изометрическая сила наблюдается при сокращениях в обеих конечностях. Кроме того, Howard (1987) выяснил, что 3-недельные “двусторонние” тренировочные занятия оказались достаточными, чтобы у нетренированных испытуемых двусторонний дефицит (—3,7 %) перешел в двусторонний прирост (+4,2 %).
6. Потенциация рефлексов. При возникновении максимальной М-волны в мышце, находящейся в состоянии покоя, как правило, не наблюдается Н-рефлекс. В то же время при произвольном сокращении мышцы наблюдаются две рефлекторные реакции (Vt и У2) имеет такую же латентность, как и Н-рефлекс, тогда как V2 характеризуется более продолжительной латентностью (Upton, McComas, Sica, 1971). Влияние тренировочных занятий силовой направленнос
ти на эти реакции определяли, вычисляя коэффициент потенциации, т.е. отношение амплитуд реакций И, и к амплитуде М-волны в мышце, находящейся в покое. Потенциация рефлексов наблюдается не во всех мышцах и более очевидна у тяжелоатлетов и сильнейших спринтеров; потенциация рефлексов усиливается вследствие занятий силовой направленности (Sale, 1988). Величина потенциации рефлексов, очевидно, служит показателем активации двигательных единиц при произвольном сокращении.
7. Синхронизация. Одним из наиболее часто цитируемых примеров адаптационных реакций нервной системы, обусловленных тренировочными занятиями силовой направленности, является изменение степени синхронизации различных двигательных единиц. Это свойство — мера временного совпадения потенциалов их действия. Высокая степень синхронизации означает, что двигательные единицы разряжают потенциалы действия почти в одно и то же время.
Адаптационные реакции нервной системы, описанные в этом разделе играют важную роль в начальной фазе тренировочной программы (Sale, 1988), в частности, начинающих тяжелоатлетов.
Адаптационные реакции мышц
В отличие от адаптационных реакций нервной системы, связанных с изменениями силы, вполне очевидно, что изменения силовых возможностей могут быть обусловлены механизмами, обусловливающими увеличение размера мышц (у культуристов, тяжелоатлетов, спортсменов, занимающихся силовыми видами спорта). Увеличение мышечного размера (площади поперечного сечения) может быть обусловлено гипертрофией — увеличением площади поперечного сечения отдельных мышечных волокон — или гиперплазией — увеличением количества
304
мышечных волокон. Согласно результатам исследований, наиболее типичная реакция испытуемых на занятия силовой направленности — гипертрофия; в то же время в определенных условиях может наблюдаться гиперплазия (Antonio, Gonyea, 1993; Tamaki, Uchiyama, Nakano, 1992).
Степень увеличения площади поперечного сечения зависит от ряда факторов, включая начальный уровень силы испытуемого, продолжительность и метод тренировочных занятий (изометрический, динамический, эксцентрический). У начинающих спортсменов в результате 6 недель тренировочных занятий изометрической направленности площадь поперечного сечения мышц—сгибателей локтевого сустава увеличилась на 5 % (Davies et. al., 1988), тогда как через 8 недель занятий площадь поперечного сечения четырехглавых мышц бедра увеличилась на 15 % (Garfinkel, Cafarelli, 1992). 60 дней занятий (изо-кинетических; 2,1 рад "с-1) привели к увеличению площади поперечного сечения четырехглавых мышц бедра на 9 % (Narici et al., 1989). Более того, в результате эксцентрически-концен-трических упражнений для мышц—разгибателей коленного сустава (в течение 19 недель) площадь поперечного сечения увеличилась более значительно, чем после упражнений, включавших только концентрические сокращения (Hather, Tesch, Buchanan, Jndley, 1991). В то же время 24 недели занятий в динамическом режиме не привели к увеличению площади поперечного сечения мышечных волокон двухглавой мышцы у опытных культуристов (Alway, Grumbt, Stray-Gundersen, Gonyea, 1992).
Занятия силовой направленности могут по-разному влиять на различные волокна (рис.9.15).
Например, 16 недельные тренировки (изометрические) трехглавой мышцы икры не оказали никакого влияния на состав волокон камбаловидной и латеральной икроножной мышц. В то же время после 19 недель выполнения упражнений для мышц—разгибателей коленного сустава увеличилась доля волокон типа Па и уменьшилась — волокон типа Пб в латеральной широкой мышце бедра. 16-недельная программа изометрических тренировок привела к увеличению площади поперечного сечения волокон типа I (20 %) и типа II (27 %) в камбаловидной, а также типа II (50 %), но не типа I в латеральной икроножной мышце. Эти результаты свидетельствуют, что не все мышечные волокна в активной группе мышц-синергистов подвергаются одинаковому тренировочному воздействию. Более того, увеличение площади поперечного сечения мышечных волокон оказывается большим при эксцентрически-концентрических сокращениях (тип I, увеличение 14 %; тип II, увеличение 32 %) по сравнению с концентрическими (тип II, увеличение 27 %).
Хотя тренировки силовой направленности могут вызывать гипертрофию, различия в размере мышц обусловливают не более 50 % различий в силе между испытуемыми. Например, на рис.9.16 показан разброс данных относительно линии регрессии между максимальной изометрической силой и площадью поперечного сечения четырехглавых мышц бедра. В этой связи возникает вопрос, имеют ли место другие (кроме адаптационных реакций нервной системы) изменения в мышце, обусловливающие различия в силовых показателях? Одна из возможностей касается различий в специфическом напря-
Рис.9.15. Изменение силы и площади мышечного волокна при непрерывном концентрическо-эксцентрическом режиме тренировок (а) и прерывистом концентрическом и концентрическо-эксцентрическом режиме (6). Силу измеряли как максимальную изометрическую силу разгибателей колена. Площадь мышечных волокон определяли для волокон типа I и II (Hakkinen, 1985)
20 8-1зз
305
Рис.9.16. Взаимосвязь между площадью поперечного сечения четырехглавой мышцы бедра и максимальной изометрической силой (Jones, Rutherford, Parker, 1989)
женин. Вспомним, что специфическое напряжение характеризует врожденную силу мышцы и измеряется как сила, образуемая мышцей относительно единицы площади поперечного сечения (Н • см-2). Поскольку усилие, которое может развивать мышца, равно площади поперечного сечения и специфическому напряжению, то различия в последнем могут обусловливать различия в силе между индивидуумами.
По мнению некоторых ученых, специфическое напряжение в мышечных волокнах колеблется: более высокое напряжение характерно для волокон типа II (Jones et. al., 1989). Действительно, наиболее высокое специфичное напряжение отмечали в двигательных единицах типа FF, а самое низкое — в двигательных единицах типа S (Bodine et. al., 1987; Kanda, Hashizume, 1992; McDonagh et. al., 1980). Из этого следует, что сила, развиваемая сократительным аппаратом, наибольшая в двигательных единицах типа FF. Специфическое напряжение в изолированных медленно- и быстро-сокращающихся мышечных волокнах не отличается — соответственно 24,5 и 24,3 Н • см-2 (Lucas et al., 1987). Следовательно, различие в специфическом напряжении в различных типах двигательных единиц, по-видимому, связано со способом передачи силы из мышечного волокна в мышечное сухожилие.
Считается, что для осуществления мышечной гипертрофии необходимо изменение коэффициента белкового синтеза. Механизм, регулирующий этот коэффициент, в настоящее время еще не выяснен. Потенциальными стимулами мышечной гипертрофии могут быть гормональные, метаболические и механические факторы (Jones et al., 1989). Гормональные стимулы (инсулин, гормон роста, тестостерон), вряд
ли являются ключевыми для гипертрофии. В частности, введение гормона роста не стимулировало увеличение силы и размера мышц, а также содержание белков вследствие тренировок силовой направленности (Yarasheski el al., 1992). Тем не менее гормоны могут влиять на другие факторы. Так, тиреоидный гормон (ТЗ) оказывает значительное влияние на экспрессию тяжелых цепей миозина типа I и типа Па (Caiozzo, Herrick, Baldwin, 1991, 1992).
Физическая деятельность, влияющая на метаболические факторы, как правило, приводит к повышению выносливости, а не силы. Кроме того, затраты энергии при эксцентрических сокращениях меньше, чем при концентрических, в то же время эксцентрические сокращения в большей мере стимулируют мышечную гипертрофию.
Наряду с этим, как показывают наблюдения, механические стимулы играют более важную роль в гипертрофии мышц (Vandenburgh, 1992). Механизм действия включает приложение механического стимула (например, растяжение, сокращение), который вызывает выделение второго, “посыльного”, и последующую модуляцию интенсивности синтеза и расщепления белков. Например, прерывистое растяжение клеток скелетной мышцы в культуре приводит к повышению синтеза различных простагландинов, модулирующих синтез и расщепление белков (Vandendurgh, Hatfaludy, Sohar, Shansky, 1990). Существуют два класса вторых “посыльных”: 1) молекулы внеклеточного матрикса (протеогликаны, коллаген, ламинин, фибронектин), которые окружают клетки, передают механический стимул к поверхности клетки и влияют на ядерные процессы и рост клеток; 2) вызванные растяжением изменения в плазме мембраносвязанных молекул (Na+ = К + АТ-Фаза, ионные каналы, фосфолипазы, G-белки), и связанные цитоплазматические вторые “посыльные” простагландины, ЦАМФ, внутриклеточный Са2+, протеинкиназа С.
Механические стимулы влияют не только на количество, но и на качество мышечной ткани (белок), которая синтезируется. Это осуществляется за счет регуляции выражения генов, определяющих фенотип мышечных волокон, который зависит от транскрибирующихся протеин-изоформных генов. Данный эффект изучали, определяя последствия механических стимулов в генах миозина тяжелой цепочки (Goldspink el. at., 1992). Эти гены кодируют миозиновый поперечный мостик и различные в медленно- и бысгро-сокращающихся мышцах (см.табл.4.3). Ученые пришли к заключению, что ген быстрого миозина тяжелой цепочки является дефектным и экспрессия медленного миозина тяжелой цепочки зависит от внешних механических воздействий.
Адаптационные реакции мышц определяются также эволюционным фактором. Клетки,
зов
подвергающиеся миогенезу при отсутствии нервного импульса, развивают субпопуляции миотрубочек (которые впоследствии становятся зрелыми мышечными волокнами) с ограниченной способностью изменять фенотип (Hoh, 1991; Hoh, Hughes, 1988). Кроме того, такая дифференциация типов волокон образует мышечные волокна, реагирующие на определенные физиологические пертурбации (физические занятия силовой направленности, денервацию) и не реагирующие. Таким образом, данный тренировочный стимул вряд ли вызывает одинаковую гипертрофическую реакцию во всех мышцах.
МЫШЕЧНАЯ МОЩНОСТЬ
В начале данной главы отмечалось, что сила и мощность — меры производительности двигательной системы. Чтобы различить эти два параметра, рассмотрим взаимоотношение сила-скорость мышцы (см.рис.6.19). Как известно, эта взаимосвязь включает четыре момента: скорость равна 0; сила равна 0; скорость > 0 и скорость < 0. Мы определили силу как точку на кривой сила—скорость, где скорость равна 0, тогда как мощность, развиваемая двигательной системой, соответствует участку скорости, не равному 0.
Многие исследователи традиционно определяли количественные способности физической деятельности на основании статической и динамической силы. Эти оба понятия отражают меру результирующего вращающего момента, а различие между ними — изменение или не изменение мышечной длины. Согласно взаимозависимости вращающий момент—скорость, максимальный вращающий момент, образуемый мышцей, снижается с увеличением скорости укорачивания мышцы. Это означает, что изменения динамической силы в большой степени зависят от интенсивности, с которой изменяется длина мышцы. Кроме того, тщательное определение динамической силы требует, чтобы максимальный вращающий момент измеряли при различной скорости. Более простой метод предполагает измерение пика мощности, развиваемой системой, представляющей собой сочетание силы и скорости и оказывающей максимальный механический эффект.
Какие свойства двигательной системы влияют на ее способность развивать мощность? В главе 3 мы выяснили, что мощность — результат силы и скорости. Следовательно, факторы, влияющие на мышечную силу либо на скорость сокращения мышечной длины, определяют развиваемую мощность. При условии адекватного нервного импульса основными детерминантами образования мощности являются количество параллельно активированных мышечных во
локон и интенсивность, с которой миофиламенты превращают энергию в механическую работу. Величина силы, которую может образовать мышца, прямо пропорциональна количеству образующих силу расположенных параллельно единиц; мышечная сила возрастает с увеличением площади поперечного сечения. Мощность максимальна, когда мышечная сила равна приблизительно 1/3 максимальной (см.рис.6.24); с увеличением силы мышцы (увеличение площади поперечного сечения) мощность повышается и, следовательно, показатель 1/3 возрастает. Это же относится к влиянию скорости мышцы на образование мощности. Максимальная скорость, с которой уменьшается длина мышцы (vMaKC), определяется ферментом миозин АТФазой, контролирующим скорость взаимодействия между актином и миозином и, следовательно, интенсивность циклов поперечного мостика. Активность миозин АТФазы изменяется при изменении уровня физической активности. Максимальная мощность наблюдается, когда скорость сокращения длины мышцы равна 1/4 vMaKC.
Мощность и движение
Успех во многих спортивных соревнованиях во многом зависит от способности продолжительного развития максимальной мощности. Несмотря на столь значительную роль мощности, это — наименее изученный биомеханический параметр в анализе движения человека.
Мощность и выполнение заданий для всего тела. Мощность, развиваемую двигательной системой, можно определить заданием (прыжок в высоту, поднятие штанги), используя эргометр или проведя исследование на изолированной мышце. Оценка задания дает представление о мощности всего тела. Для этой цели чаще всего используют выполнение прыжка в высоту. Испытуемый выполняет прыжок в высоту на силовой платформе, позволяющей измерить силу реакции поверхности (рис.9.17). Зная силу реакции поверхности, мы можем определить среднюю силу (F), действующую на поверхность, и среднюю скорость (v) центра тяжести и, таким образом, — среднюю мощность, развиваемую во время прыжка (Bosco, Komi, 1980). Чтобы рассмотреть только способность образования силы испытуемым, ограничим задание выполнением прыжка согнув ноги, который включает только концентрическое сокращение, а не ^эксцентри-чески-концентрическое. Получим (F), поделив величину импульса (штриховая линия (на рис.9.17) на его продолжительность. Из рис.9.17 получим следующие значения:
импульс = 179 Н • с-1 время = 0,29 с F = 617 Н.
20»
307
Рис.9.17. Вертикальная составляющая силы реакции поверхности во время выполнении прыжка из положения присев. Испытуемый выполнял прыжок в высоту на максимальную высоту
Для определения приблизительного значения средней скорости (у) во время фазы отталкивания, можем воспользоваться уравнениями из главы 1. Если допустить, что спортсмен оттолкнулся от поверхности и приземлился в одном и том же месте (положении), то половину времени “полета” он провел, поднимаясь вверх, а вторую половину — опускаясь вниз. Рассмотрим только вторую фазу “полета”. Из уравнения 1.3 мы можем определить скорость приземления (vy), поскольку нам известны ускорение, обусловленное силой тяжести (—9,81 м • с-2), начальная скорость (0) и из рис.9.17 мы можем рассчитать, что продолжительность половины полета равна 0,20 с:
vz= v, + а/ = 0 + (9,81 х 0,20);
V/= 1,96 м-с-1.
Среднюю скорость (V) во время фазы отталкивания определяют как среднее начальной и завершающей скорости:
= L96 + 0 =0>98 м.с.,
Наконец, среднюю мощность, развиваемую прыгуном во время фазы отталкивания, определяют как результат средней скорости и средней силы:
средняя мощность = F х v = = 617 х 0,98 = 605 Вт.
(1 Вт = 1 Дж *с-1 = Н-м-с-1).
Таким образом, мы установили, что средняя мощность, развиваемая прыгуном при выполнении прыжка согнув ноги (рис.9.17), была 605 Вт. Поскольку при выполнении этого прыжка руки не выполняли работу, полученный показатель характеризует только влияние мышц нижних конечностей.
Другие ученые (van Ingen Schenan, Bobbert, Huijing, Woittier, 1985) определяли взаимосвязь вращающий момент—скорость и максимальную мощность, развиваемую мышцами—сгибателями подошвы во время выполнения прыжка в высоту — 2 499. Josephson (1993) определил максимальную мощность мышцы 7—500 Вт кг-1, поддерживаемая мощность составляла 5—150 Вт - кг-1.
Мощность, развиваемую тяжелоатлетами, можно определить, установив интенсивность изменения количества механической энергии (Garhammer, 1980). Поскольку количество выполняемой работы можно определить на основании изменения количества энергии, а мощность равна интенсивности выполнения работы, то мощность можно установить по интенсивности изменения количества энергии:
. , Изменение количества энергии
Мощность =--------------т;-------------=
Время
_ Ek,t + EP,g _ 0,5/иу2 + mgh Время Время
где Ektt — максимальная кинетическая энергия; Ep>g — потенциальная энергия; время — отрезок времени до развития максимальной кинетической энергии.
Пример данного подхода приведен в таблице 9.1. В ней представлена мощность, сообщаемая тяжелоатлетом штанге, при выполнении трех упражнений: а) чистом взятии веса на грудь, т.е. движение по перемещению штанги с пола на грудь спортсмена, выполняемое за счет одного непрерывного и быстрого движения; б) упор присев—движение, при котором спортсмен со штангой у плечей перемещается из положения стоя в положение сидя (бедра параллельны полу) и затем встает со штангой; в) жим лежа на скамье, при котором спортсмен, лежа на спине, опускает штангу, которую он удерживал на вытянутых руках, на грудь и затем быстро возвращает ее в исходное положение.
Приведенные результаты свидетельствуют, что максимальный пик мощности, сообщаемый штанге, наблюдается при выполнении чистого взятия веса на грудь, минимальный — при вы-
Таблица 9.1. Мощность, сообщаемая тяжелоатлетом штанге, при выполнении трех упражнений
Показатель Чистое взятие на грудь Упор присев Жим лежа на скамье
Вес штанги, Н 1226 3694 1815
Пик скорости 2 0,30 1,54
штанги, м-с 1
Высота, м 0,30 0,30 0,06
Время до Ek t, с 0,35 1,30 0,70
Ек,ь Дж 250 17 219
Ep,g, Дж 368 1108 109
Мощность, Вт 1766 865 469
308
полпенни жима, лежа на скамье. Кроме того, данные показывают, что успешное выполнение этих двух упражнений не определяется исключительно развитием мощности. Можно предположить, что успех в большей степени связан с силовыми возможностями.
Развитие мощности относительно отдельного сустава
С помощью эргометра можно определить мощность отдельной мышечной группы (Grabiner, Jeziorowski, 1992). Эргометр позволяет определить взаимосвязь вращающий момент— угловая скорость относительно сустава, а также вычислить мощность.
Мощность, развиваемую относительно отдельного сустава, можно определить, проведя биомеханический анализ и определив вращающий момент и угловую скорость относительно сустава (табл.9.2) (Enoka, 1988а, Winter, 1983). Метод требует данных о кинематике сустава и
Таблица 9.2. Сила сгибателей локтя (Ро), максимальная угловая скорость (<откс) и пик мощности у тренированных мужчин и нетренированных мужчин и женщин (М ± т)
Показатель Тренированные мужчины Нетренированные мужчины Нетренированные женщины
Ро Н-м'1 90,0 ± 15,6 68,5 ± 11,0 42,7 ± 6,9
Омаке 17,0 ± 1,6 16,6 ± 1,5 14,9 ± 1,3
Мощность, Вт 253,0 ± 58,0 195,0 ± 46,0 111,0 ± 24,0
силе соприкосновения (например, силе реакции поверхности, силе педалирования). Например, мощность, сообщаемую испытуемым педалям велоэргометра, можно определить, вычислив силу педалирования и скорость вращения педалей или суммировав мощность бедренного, коленного и голеностопного суставов (вращающий момент х угловую скорость) (van Ingen Schenau, van Woensel, Boots, Snackers, de Groot, 1990) (рис. 9.18).
Рис.9.18. Развитие мощности во время отдельного цикла педалирования на велоэргометре (van Ingen
Schenan, van Woensel, Boots et al., 1990):
a — мощность как функция угла поворота рычага; б — мощность сустава (произведение вращающего момента и угловой скорости) бедра, колена и лодыжки и угол как функция угла поворота рычага; 1 — вычисленная мощность; 2 — суммированная мощность
Развитие мощности в изолированной мышце
Способность двигательной системы развивать мощность можно определить на уровне отдельной мышцы или мышечного волокна. Brooks, Fanlkner (1991) сравнивали способность медленносокращающейся (камбаловидной) и быстросокрашаюшейся (длинного разгибателя пальцев) мышц поддерживать образование изометрической силы и мощности. Пик изометрической силы быстросокращающейся мышцы оказался выше пика медленносокращающейся — соответственно 363 и 273 Н • м. Вместе с тем при выражении силы относительно размера мышцы (площади поперечного сечения) пика силы были одинаковыми 24,5 и 23,7 Н • м-2 соответственно в быстро- и медленносокращающейся мышце. Тем не менее быстросокращаю-щаяся мышца характеризовалась большей утомляемостью и более быстрым снижением изометрической силы при продолжительной активации; максимальная поддерживаемая сила составляла 1,38 Н • см"2 в быстросокращающейся и 4,48 Н • см-2 в медленносокращающейся. Наряду с этим при выражении развития мощности относительно мышечной массы пик поддерживаемой мощности был выше в быстросокращающейся (9,1 Вт • кг'1) по сравнению с медленносокращающейся (7,4 Вт • кг-1) мышцей. Хотя в первой мышце снижение силы было более очевидным при продолжительной активации, большая скорость сокращения длины мышцы обеспечивала более высокую мощность. Эти результаты свидетельствуют, что у двигательных единиц типа S и FR более высокая спо-
309
собность поддерживать уровень изометрической силы, тогда как у единиц типа FR и FF повышенная способность продолжительного развития мощности (Rome et al., 1988).
Эксцентрическо-концентрические сокращения. Мы определяем мощность как интенсивность выполнения работы, т.е. как количество работы, выполняемой за единицу времени. Поскольку выполняемая работа соответствует изменению количества энергии, мощность также можно рассматривать с позиций интенсивности изменения количества энергии. Мощность, развиваемая мышцей, зависит от скорости использования энергии для выполнения работы. Основной источник энергии для мышцы — химическая энергия (АТФ). Однако как уже указывалось, мышца использует также механическую (упругую) энергию для выполнения положительной работы и развития мощности. Этот процесс, называемый хранением и использованием упругой энергии, имеет место во время эксцентрическо-концентрических сокращений. Если мышца выполняет эксцентрическое сокращение до концентрического, она способна выполнить больше положительной работы во время сокращения, вызывающего уменьшение длины. Частично это обусловлено способностью мышцы накапливать энергию во время растяжения (эксцентрическое сокращение) и затем использовать ее во время концентрического сокращения.
Высота, на которую может прыгнуть спортсмен при выполнении прыжка в высоту, зависит от того, какое количество мощности образуют мышцы. Наиболее эффективная техника прыжка в высоту включает эксцентрическо-концентрические сокращения четырехглавых мышц бедер и трехглавых икроножных. Эта связь указывает на то, что эксцентрическо-концентрическое сокращение обеспечивает значительный вклад в развитие мышцами мощнос
ти. Именно поэтому люди интуитивно используют эксцентрическо-концентрические сокращения при выполнении большинства движений.
Развитие мощности и тренировочный процесс
Исследование влияния тренировочных занятий на развитие мощности провели Duchateau, Hainaut (1984). Две группы испытуемых каждый день в течение 3 мес проводили тренировочные занятия для мышц руки (приводящая мышца больших пальцев) средней интенсивности (10 повторений). Одна группа выполняла максимальные изометрические сокращения, вторая — быстрые динамические сокращения, преодолевая сопротивление нагрузки 30—40 % максимальной. Подобная небольшая нагрузка при выполнении динамических сокращений была выбрана потому, что, как известно, мышца развивает максимальную мощность, когда сила сокращения равна 1/3 максимальной.
Влияние двух способов тренировок на взаимосвязь сила—скорость соответствовало принципу специфичности. В группе, тренировавшейся с использованием более значительной нагрузки, максимальная изометрическая сила увеличивалась более существенно (штриховые линии), чем в другой (рис.9.19). Удивительным оказалось то, что в “изометрической” группе, выполнявшей максимальные сокращения, развитие мощности увеличилось на 51 %, по сравнению с 19 % у испытуемых “динамической” группы.
Как свидетельствуют эти результаты, максимальное увеличение развития мощности происходит вследствие тренировочных занятий с высокой силой, а не скоростью. В то же время в других исследованиях наблюдали максималь-
Рис.9.19. Влияние изометрического (а) и динамического (б) режимов тренировок на развитие мощности. Изометрический режим тренировок включал 5-секундное максимальное изометрическое сокращение каждую минуту в течение 10 мин, каждый день на протяжении 3 мес. Динамический режим включал то же самое количество повторений и такую же продолжительность занятий, однако упражнение заключалось в быстром концентрическом движении (Duchateun, Hainant, 1994)
310
Таблица 9.3. Экспериментальные модели, используемые для изучения влияния изменения активности
Изменение активности Тип модели
Нервные Мышечные
Повышенный уровень активности Электростимулирование Растяжение Физические нагрузки Физические упражнения Компенсаторная гипертрофия
Пониженный Денервация Тенотомия
уровень активности Тетродотоксин Кураре Колхицин Ризотомия Рассечение спинного мозга Изоляция спинного мозга Иммобилизация Подвешивание за заднюю конечность Сон вследствие барбитурата Постельный режим Погружение в воду
ное увеличение развития мощности при тренировочных нагрузках порядка 30 % максимальной по сравнению с нагрузками 0 или 100 % (Moritani, 1992).
АДАПТАЦИОННЫЕ РЕАКЦИИ НА ПОНИЖЕННЫЙ УРОВЕНЬ ФИЗИЧЕСКОЙ АКТИВНОСТИ
Наиболее популярный метод определения основных свойств двигательной системы — возмутить систему и измерить ее реакцию. Типичным возмущением системы является изменение количества ее активности. В научной литературе часто встречается предположение, что возбуждение мышцы нервной системой — главный фактор, определяющий свойства мышцы. В этой связи было разработано несколько экспериментальных моделей, направленных на изменение связи между нервной системой и мышцей, с тем, чтобы определить, какую роль играет нервная система в определении свойств мышцы (табл.9.3). Мы рассмотрим три из наиболее часто используемых.
Иммобилизация конечности
При иммобилизации конечности (наложение гипса) на несколько недель наблюдается уменьшение мышечной массы и снижение функции.
Снижение уровня физической активности. В ряде исследований, проведенных на людях и животных, изучали адаптационные реакции, обусловленные иммобилизацией конечности. При иммобилизации задней конечности крысы в течение 4 недель наблюдали значительное уменьшение активности (ЭМГ) в камбаловидной и медиальной икроножной мышцах (рис.9.20, a) (Fournier, Roy, Perhan et al., 1983).
Рис.9.20. Уменьшение активности и мышечной массы после 4 недель иммобилизации задней конечности крысы (Fournier, Roy, Perham et al., 1982):
a — снижение интегрированной ЭМГ в камбаловидной (1) и медиальной икроножной (2) мышцах при короткой иммобилизации; б — взаимосвязь между снижением ЭМГ и мышечной массой; S-S — камбаловидная, короткая длина; S-N — камбаловидная нейтральная д лина; S-L — камбаловидная, большая длина; M-S — медиальная икроножная, короткая длина; M-N — медиальная икроножная, нейтральная длина; M-L — медиальная икроножная, большая длина
311
Следует отметить, что степень снижения ЭМГ зависела от продолжительности иммобилизации мышцы; максимальное снижение наблюдалось при короткой иммобилизации. Интегрированная ЭМГ снижалась на 77 % в камбаловидной и 50 % в медиальной икроножной мышцах при минимальной продолжительности иммобилизации. Снижение ЭМГ сопровождалось уменьшением массы камбаловидной мышцы на 36 %, а массы икроножной мышцы — на 47 %.
Степень атрофии (уменьшение мышечной массы) при иммобилизации конечности может быть разной. Максимальный показатель (50 %) наблюдали Fournier et al. (1983). Максимальное снижение площади поперечного сечения мышечного волокна составляет около 42 % (Nicks, Beneke, Key, Timson, 1989). Другие исследователи наблюдали более значительное уменьшение сырой массы — 17 % (Robinson, Enoka, Stuart, 1991), а также снижение среднего диаметра мышечных волокон типа I (но не типа II) порядка 14 % (Gibson et al., 1987). Атрофия, по-видимому, обусловлена уменьшением как интенсивности белкового синтеза (Gibson et al., 1987), так и количества мышечных волокон (Oishi, Ishihara, Katsuta, 1992).
Несмотря на снижение ЭМГ и мышечную атрофию вследствие иммобилизации конечностей, эти результаты трудно интерпретировать ввиду отсутствия связи между уменьшением ЭМГ и мышечной атрофией, а также между мышечной атрофией и потерей функций. Частично отсутствие взаимосвязи между мышечной атрофией (уменьшение массы) и снижением силы можно объяснить использованием уменьшения массы как показателя атрофии. Как уже отмечалось, максимальная сила, образуемая мышцей, тесно связана с площадью ее поперечного сечения, а не с количеством мышечной массы (весом). Когда мышечная атрофия выражается с позиции площади поперечного сечения, она теснее коррелирует с уменьшением (Lieber, 1992). Вместе с тем отсутствие взаимосвязи отмечали в двигательных единицах типа S и FR мышцы задней конечности кошки (Nordstrom, Enoka, Callister et al., 1993). Наблюдали уменьшение площади поперечного сечения мышечных волокон типа I и Па почти на 25 %, а также снижение максимальной изометрической силы на 40 % в двигательных единицах типа FR и на 52 % в единицах типа S. Очевидно, ремоделирование нервно-мышечного аппарата при кратковременной мобилизации — более сложный процесс, чем просто линейная взаимосвязь снижения ЭМГ, мышечной массы и нарушения физической деятельности.
Нервно-мышечные адаптации. При достаточной нагрузке большинство элементов системы адаптируются. Например, 7-дневная иммобилизация камбаловидной мышцы крыс (небольшая длина) привела к снижению мышечной массы на 37 %, деполяризации мембран волокон на
5 мВ, уменьшению миниатюрных потенциалов концевой пластинки на 60 % и снижению перемещения Na+-K+ через мембрану на 25% (Zem-kova et al., 1990).
В большинстве подобных исследований отмечают конверсию мышечных волокон. Наблюдается уменьшение количества мышечных волокон типа МО и увеличение — волокон типа БОГ, что, по-видимому, включает конверсию структуры ферментов волокон типа БОГ (Fitts, Brimmer, Heywood-Cooksey, Timmermann, 1989; Oishi et al., 1992). Эту адаптационную реакцию можно объяснить тем, что вследствие иммобилизации больше всего “страдают” волокна, чья активность понижается больше всего, т.е. мышечные волокна типа МО.
Мышечная атрофия, обусловленная иммобилизацией конечности, приводит к значительному
Рис.9.21. Изменения силы сокращения двигательной единицы, обусловленные иммобилизацией (а), и модуляция амплитуды интенсивности разряда как функция порога рекрутирования (б). Каждая точка данных соответствует отдельной двигательной ед инице (Duchatean, Hainaut, 1990)
312
снижению силы и нарушению основных функций. Например, иммобилизация в течение 6 недель (перелом конечности) привела к снижению силы максимального произвольного сокращения на 55% и уменьшению максимального произвольного сокращения ЭМГ в мышце руки на 45% (Duchateau, Hainaut, 1991). При иммобилизации такой продолжительности ингибируется способность человека осуществлять в ЦНС побуждение, достаточное для максимальной активации мышцы руки. Тем не менее подобные изменения не нарушают способность испытуемых поддерживать максимальную силу в течение 60 с.
Последствия иммобилизации очевидны также на уровне двигательной единицы. Иммобилизация в течение 6—8 недель изменяет свойства и поведение двигательных единиц мышц руки человека (Duchatean, Hainant, 1990). При выражении порога рекрутирования относительно максимальной силы отмечали увеличение количества двигательных единиц высокого порога в иммобилизованной мышце. Однако средняя сила, образуемая этими единицами, была меньше (рис.9.21, а); кроме того, снижался пик амплитуды потенциалов действия двигательных единиц. Наблюдалось также увеличение диа-па-зона рекрутирования (рис.9.21, б) и уменьшение диапазона модуляции интенсивности разряда.
Подвешивание задней конечности
Когда астронавты впервые отправлялись в космос, тревогу специалистов вызывали две проблемы: смогут ли астронавты передвигаться
за пределами космического корабля и какие физиологические адаптации произойдут в их организме вследствие пребывания в условиях невесомости. Обе проблемы тщательно изучались. В частности, проводились биомеханические исследования динамики движений в условиях невесомости (см. главы 2 и 3). Для изучения физиологических адаптаций ученые использовали модель, согласно которой животное (обычно крыса) подвешивалось за задние конечности на несколько недель. Животное могло свободно двигать конечностями, не касаясь опоры (рис.9.22, а), и осуществлять многие из своих повседневных функций, испытывая минимальный стресс (Thomason, Booth, 1990).
Снижение уровня активности
Данная модель позволяет имитировать условия пребывания в космическом пространстве, обеспечивая краниальное смещение жидкости, деминерализацию костей, уменьшение роста и устранение необходимости постуральной активности мышц ног. Несмотря на отсутствие необходимости постурального контроля, активность ЭМГ в мышцах голени снижается незначительно (рис.9.23, а). Активность камбаловидной и медиальной икроножной мышц в первые 3 дня уменьшилась, но затем возвращалась к контрольным уровням (Alford, Roy, Hogson, Edgerton, 1987; Riley et al., 1990). Парадоксально, но наблюдалась атрофия мышц в период, когда уровни ЭМГ были нормальными (рис.9.23,б), что свидетельствует об отсутствии взаимосвязи между уровнями активности и снижением мышечной массы (рис.9.24).
Рис.9.22. Модель подвешенной конечности (Riley,Slocum, Bain et al., 1990):
a—крыса с подвешенными задними конечностями; животное может передвигаться с помощью передних конечностей; б — изменение диапазона движения камбаловидной мышцы при подвешенных задних конечностях; 1 — минимальный угол голеностопного сустава в нормальном положении стоя, 2 — нейтральное положение при подвешенных конечностях; 3 — максимальный угол при обоих
условиях
313
Отсутствие взаимосвязи между ЭМГ и атрофией мышц, очевидно, можно объяснить поведением животного (см.рис.9.22). В обычных условиях диапазон движения голеностопного сустава колеблется от 0,5 до 3,14 рад. Однако через несколько дней пребывания в подвешенном состоянии голеностопный сустав “принимает” нейтральный угол — 1,57 рад, что значительно уменьшает диапазон движения. Кроме того, вспомним, что на ЭМГ влияет длина мышц, и при данной силе ЭМГ выше при более короткой длине мышцы. Можно предположить, что при подвешивании конечности сила, образуемая мышцами, значительно снижается, несмотря на сохранение высоких уровней ЭМГ.
Другая модель для проведения исследований на людях была разработана совсем недавно (Dudley et al., 1992; Hather, Adams, Tesch, Dudley, 1992). На правую ногу испытуемые надевают ботинок с утолщенной подошвой (высота 10 см) и используют костыли. Таким образом, испытуемый не использует левую ногу. Через 6 недель такого эксперимента у испытуемых наблюдали значительное снижение площади поперечного сечения всей мышцы и мышечных волокон. Так, площадь поперечного сечения широкой мышцы бедра уменьшалась на 16 %, камбаловидной — на 17 %, икроножной — на 26 %. Биопсический анализ показывал уменьшение площади поперечного сечения волокон типа I
Рис.9.23. Результаты пребывания животного в подвешенном состоянии в течение 28 дней (Thomason, Booth, 1990);
а — интегрированная ЭМГ-активность передней большеберцовой (7), камбаловидной (2) и медиальной икроножной (J) мышц задней конечности крысы; б — уменьшение массы подошвенной (4) и камбаловидной (5) мышц
Рис.9.24. Расщепление (7), синтез (2), потери белка (4) и активность интегрированной ЭМГ (3) в камбаловидной мышце крысы в течение 28 дней пребывания в подвешенном состоянии. Чистая потеря белка во многом обусловлена его интенсивным расщеплением, несмотря на повышенную ЭМГ (Thomason, Booth, 1990)
314
на 12%, волокон типа II — на 15% в широкой мышце бедра. Эти результаты свидетельствуют о различиях в реакциях человека и крысы на условия “подвешенного состояния”.
Нервно-мышечные адаптации. Во многих исследованиях, проводившихся на животных, отмечали преимущественное влияние “подвешивания” на медленносокращающуюся мышцу (Fitts, Metzger, Riley, Unsworth, 1986; Roy et al., 1991). Поэтому в большинстве исследований основное внимание уделяли камбаловидной мышце. Несколько недель пребывания “в подвешенном” состоянии приводило к снижению пропорции волокон типа I в камбаловидной мышце, практически не влияя на волокна синергиста (медиальная икроножная) или антагониста (передняя большеберцовая). Снижалась также концентрация миозиновых и миофибрилльных белков и увеличивалось образование быстрых миозиновых изоформ. Однако это наблюдается в некоторых, но не всех мышечных волокнах типа I камбаловидной мышцы (Gardetto, Schluter, Fitts, 1989).
Указанные изменения сопровождаются определенными изменениями метаболических способностей мышечных волокон (Roy et al., 1991). Так, в камбаловидной мышце при подвешивании конечности повышаются концентрации сукцинат дегидрогеназы и цитрат синтазы (ферментов, участвующих в окислительном метаболизме). Однако через несколько недель пребывания в “подвешенном” состоянии преимущественно в быстросокращающихся мышцах (медиальной икроножной, передней большеберцовой) содержание некоторых ферментов (сукцинат дегидрогеназы), характеризующих окислительный метаболизм, понижается. В то же время в волокнах типа I и II мышц разгибателей и сгибателей количество ферментов либо не изменяются, либо повышаются (альфа-гли-церофосфат дегидрогеназа).
Эти количественные и качественные изменения содержания белков и ферментов приводят к целому ряду адаптационных реакций механических свойств мышцы. Способность образования максимальной силы камбаловидной мышцы снижается, причем намного больше, чем можно было ожидать, исходя из уменьшения мышечной массы. Это свидетельствует об изменении специфического напряжения (Herbert, Roy, Edgerton, 1988; Thomason, Booth, 1990). В то же время специфическое напряжение не изменяется в быстросокращающихся мышцах, на которые пребывание в “подвешенном” состоянии влияет в меньшей степени. На уровне мышцы—волокна наблюдается значительное снижение диаметра и пика силы волокон типа I как в камбаловидной, так и в медиальной икроножной мышцах, тогда как волокна типа Па медиальной икроножной мышцы сокращаются в диаметре, не изменяя пик силы (Gardetto et al., 1989). В то же время Dudley et
al. наблюдали снижение силы мышц—разгибателей коленного сустава у испытуемых на 21 % после 6 недель нахождения в “подвешенном” состоянии. Это снижение, однако, не включает преимущественное воздействие на мышечные волокна типа I.
Следует упомянуть также об изменении скорости сокращений (Roy et al., 1991; Thomason, Booth, 1990). Камбаловидная мышца становится быстрее, что по-видимому, обусловлено смещением миозиновых изоформ, а также уменьшением количества мышечных волокон типа I. Об этом свидетельствует увеличение максимальной скорости сокращения (гмакс) камбаловидной мышцы и некоторых ее волокон типа I; увеличение vMaKC характерно не для всех мышечных волокон. При увеличении гмакс взаимосвязь сила-скорость смещается вправо (см.рис.6.24). Кроме того, наблюдается уменьшение продолжительности сокращения и времени полурасслабления, что, очевидно, связано с изменениями кинетики Са2+. Пребывание в “подвешенном” состоянии приводит к снижению концентрации Са2+, необходимой д ля активации сократительного аппарата (Gardetto et al., 1989).
Установлено также, что пребывание в “подвешенном” состоянии не влияет на степень утомляемости камбаловидной мышцы. В то же время для медиальной икроножной мышцы крысы характерны более сложные адаптационные реакции, включая отсутствие влияния на степень утомляемости через 7 дней после пребывания в подвешенном состоянии, однако через 28 дней степень утомляемости значительно повысилась (Winiarski, Roy, Alford et al., 1987).
Рассечение спинного мозга
Рассечение спинного мозга приводит к снижению активности вследствие отделения частей нервной системы от мышцы. Его выполняют у экспериментальных животных на уровне спинного мозга, обычно это Т12-Т13.
Снижение уровня активности. Как отмечал Liber (1992), рассечение спинного мозга (спи-нализация) приводит к немедленному параличу. Через 3—4 недели после спинализации развивается мышечная спастичность, ведущая со временем к продолжительной активности разгибателей без очевидной произвольной активации мышц. В этом “парализованном” состоянии, однако, нервно-мышечная система по-прежнему может активироваться электрическим током и при соответствующей помощи, а также афферентной обратной связи животные могут выполнять локомоцию задней конечности на тредбане, причем сила, образуемая камбаловидной мышцей, практически такая же, как и у “нормальных” животных (Gregor et al., 1988; Lovely, Gregor, Roy, Edgerton, 1990).
315
Рис.9.25. Сократительные свойства камбаловидной мышцы и длинного разгибателя пальцев в нормальной (7) и спинализированной (2) задней конечности крысы (Liber, 1992):
а — отрезок времени до достижения пика силы сокращения; б— частота стимулирования, при которой тетанус становится сплошным; в — площадь поперечного сечения; г — максимальная тетаническая сила; ДРП — длинный разгибатель пальцев; КМ — камбаловидная мышца. Более существенные изменения наблюдались в камбаловидной мышце
Нервно-мышечные адаптации. Как и в случае иммобилизации или подвешивания конечности, рассечение спинного мозга приводит к мышечной атрофии с преимущественным воздействием на медленносокращающиеся мышцы, а также конверсией мышечных волокон (West, Roy, Edgerton, 1986). Наиболее очевидная конверсия мышечных волокон, а также уменьшение мышечной массы наблюдаются в мышцах одного сустава, обеспечивающих удержание веса. Камбаловидная и медиальная икроножная мышцы атрофируются соответственно на 45 и 30 % через 2 недели после рассечения спинного мозга. Снижается также доля мышечных волокон типа I и увеличивается число волокон типа П, что приводит к увеличению доли двигательных единиц типа FR в камбаловидной мышце и возрастанию количества двигательных единиц типа FF и F1 (промежуточные быстросокращающиеся) в медиальной икроножной мышце (Munson, Fochring, Lofton, Zengel, Sypert, 1986).
Конверсия типов мышечных волокон влияет также на активность ферментов, участвующих в окислительном и гликолитическом метаболизме (Roy et al., 1991). В то же время спинализация, по-видимому, не влияет на взаимосвязь между гликолитическим потенциалом мышечного волокна и его типом, определяемым миозин АТФазой.
Физиологические адаптации, наблюдаемые в спинализированных мышцах, соответствуют изменениям сократительных и метаболических белков (рис.9.25).
Спинализация не влияет на утомляемость камбаловидной мышцы (Gordon, Pattullo, 1993). Что касается медиальной икроножной мышцы, степень утомляемости может изменяться в зависимости от возраста, в котором было осуществлено рассечение спинного мозга. У молодых животных не наблюдается влияние на степень утомляемости медиальной икроножной мышцы, в то же время, если рассечение спинного мозга выполняют взрослым животным, утомляемость медиальной икроножной мышцы повышается.
В результате рассечения спинного мозга изменяются свойства и соединения нейронов спинного мозга. Так, двигательные нейроны, иннервирующие камбаловидную мышцу, становятся менее возбудимыми и начинают напоминать нейроны, иннервирующие быстросокращающу-юся мышцу. Изменения включают снижение постгиперполяризации и увеличение реобазы (Соре, Bodine, Fournier, Edgerton, 1986).
ВОССТАНОВЛЕНИЕ ДВИГАТЕЛЬНЫХ ФУНКЦИИ ПОСЛЕ ТРАВМЫ
Мы выяснили, что нервно-мышечная система обладает уникальными способностями адаптироваться. Например, может измениться структура сократительных и метаболических белков мышцы вследствие снижения активнос
316
ти, и мышца может восстановиться после повреждения, обусловленного эксцентрическими сокращениями.
Периферическая нервная система
Известно, что нервно-мышечная система способна в определенной степени восстановить функцию после повреждения периферического нерва. Нейроны в зрелой ЦНС неспособны к клеточному делению, следовательно, это восстановление зависит от способности нейронов реиннервировать соответствующие мишени. При повреждении аксона (аксотомия) дегенеративные изменения наблюдаются в аксоне дистально к месту повреждения, а также в нейроне (рис.9.26). Через 2-3 дня после аксотомии сома начинает разбухать и может увеличиться вдвое; эндоплазматический ретикулум (окрашенный как субстанция Ниссля) распадается и перемещается к соме (рис.9.26, б, в). Распад субстанции Ниссля называется хроматолизмом. Продолжительность этого процесса от 1 до 3 недель; он включает обширный ресинтез белков, необходимых д ля регенерации аксона и образования отростков. Если отросток способен “захватить” оставшиеся миелиновые фрагменты (рис.9.20, в), то, скорее всего, исходная мишень реиннервируется. “Захват” соответствующего миелинового фрагмента, по-видимому, — критическая фаза процесса восстановления функции.
Поскольку восстановление функции зависит от способности отростков реиинервиро-вать соот-ветствующие мишени, сущность травмы периферического нерва — важный показатель степени восстановления функции. Вообще при полном рассечении нерва прогноз наихудший, частичная денервация — менее серьезная травма, а ушиб имеет минимальные по продолжительности последствия. Травмы, ведущие к отсечению ствола периферических нервов, обычно сопровождаются продолжительным расстройством двигательной и сенсорной функций, характеризующихся плохой восстанавливаемостью.
Существуют специальные хирургические методы, направленные на то, чтобы помочь ре
генерирующим аксонам выпустить отростки и реиннервировать исходные мишени. Эти методы приводят к значительному числу неправильно направленных реиннерваций (табл.9.4) (Thomas, Stein, Gordon et al., 1987). Последствием этих иннерваций является то, что активация пула двигательных нейронов не приводит к избирательной активации одной мышцы. Как видно из табл.9.4, активирование пула двигательных нейронов тыльной межкостной мышцы указательного пальца приводит к активации, по меньшей мере, этой мышцы (отведение указательного пальца) и отводящей мышцы мизинца. Подобная реорганизация, несомненно, нарушает двигательную координацию при выполнении точных и тонкий действий. Подобное характерно и для сенсорных аксонов, что затрудняет локализацию сенсорного стимула. Кроме того, нарушается взаимосвязь между силой рекрутирования двигательных единиц (порог рекрутирования) и амплитудой силы сокращения. Взаимосвязь часто используют как показатель упорядоченного рекрутирования, основанного на принципе размера, согласно которому небольшие единицы рекрутируются при небольшой силе. Отсутствие взаимосвязи между порогом рекрутирования и амплитудой сокращения свидетельствует о потере способности рекрутировать двигательные единицы в соответствии с размером и, следовательно, точно дифференцировать силу, образуемую мышцей. Все изложенное выше указывает на плохую координацию тонких и точных движений.
Принципиальный механизм восстановления после частичного рассечения в некоторой степени отличается от механизма, связанного с восстановлением после полного рассечения; он включает рост аксонов и реиннервацию, однако аксоны оставшихся целыми двигательных единиц выпускают отростки, реиннервирующие мышечные волокна, которые были денервированы. Процесс называется коллатерналь-ным выпусканием отростков, поскольку отростки образуются в дистальном участке двигательного аксона вблизи мишени. Все двигательные единицы пула компенсируют такие травмы нерва коллатеральным выпусканием отростков (Rafuse, Gordon, Orozco, 1992), степень кото-
Таблица 9.4. Количество восстановившихся двигательных нейронов в реиннервированных мышцах после полного отсечения локтевого нерва на уровне запястья (Thomas et al., 1987)
Реиннервированная мышца Тыльная межкостная мышца указательного пальца Отводящая мышца мизинца Приводящая мышца большого пальца Другие
Отводящая мышца мизинца 15 (31 %) 16(34 %) 9 (18%) 8(17%)
Тыльная межкостная мышца указательного пальца 22 (39 %) 6(11 %) 12 (22 %) 16 (28 %)
317
Рис.9.26. Изменения в дистальном аксоне и нейроне после аксотомии (Kelly, 1985);
а — нормальная сома и аксон нейрона; б — через 2—3 дня после аксотомии; в — через 1—3 недели после аксотомии
рого зависит от количества волокон, которые были денервированы (рис.9.27). Чем больше степень частичного поражения и, следовательно, денервировано больше мышечных волокон, тем выше степень коллатерального выпускания отростков и больше размер оставшихся неповрежденными двигательных единиц. Коллатеральное выпускание отростков способно компенсировать “потерю” двигательных нейронов почти на 80% (рис.9.28), причем количество
двигательных единиц может увеличиваться в пять раз (Gordon, Yang, Ayer, Stein, Tyreman, 1993).
Приведенные данные свидетельствуют, что при поражении периферического нерва на 80 % нервно-мышечная система способна восстановить способность образования максимальной силы. Это восстановление обусловлено тремя механизмами: увеличением коэффициента иннервации, увеличением средней площади по-
Рис.9.27. Тетаническая сила двигательных единиц как функция степени частичной денервации (Rapuse, Gordon, Orozco, 1992):
а — двигательные единицы медиальной (Z) и латеральной (2) икроножной мышцы; б — двигательные единицы камбаловидной мышцы
318
0,0-11-------1---------1--------1---------1----—"
0 20 40 60 80 100
Денервация, % отрезанных аксонов
Рис.9.28. Максимальная тетаническая сила передней большеберцовой мышцы крысы после различной степени денервации, обусловленной частичным отсечением мышечного нерва. Величина силы выражена отношением силы, образованной денервированной мышцей, к силе, образованной контралатеральной мышцей. Показатели силы приведены для мышц контрольных животных (I), животных с частично денервированной мышцей (2), животных с рассечением спинного мозга (3) и животных с рассечением спинного мозга и частичным рассечением нерва (4) (Gordon, Yang, Ayer et al., 1993)
перечного сечения мышечных волокон или повышением специфического напряжения. Наиболее важным механизмом, по-видимому, является увеличение коэффициента иннервации, обусловленное коллатеральным выпусканием отростков (TotOsy de Zepetnek, Zung, Erdebil, Gordon, 1992).
Центральная нервная система
У ученых нет единого мнения относительно способности ЦНС восстанавливаться. Один из примеров способности реорганизации ЦНС — синаптогенез (образование новых синапсов), который наблюдается при повышенной активности (Keller, Arissian, Asanuma, 1992). Эта реорганизация, как считают, лежит в основе процессов, связанных с двигательным обучением и памятью.
Другие попытки изучения процесса выпускания отростков ЦНС касались рассмотрения способности спинного мозга компенсировать удаление избранных путей и импульсов. Существуют доказательства коллатерального выпускания отростков аксонами спинного мозга в препарата с сохраненным корешком. Кроме того, имеются очевидные примеры, восстановление функции в которых можно объяснить активацией находящегося в состоянии покоя нервного пути (или ряда синапсов), который не используется до тех пор, пока не будет прерван (нарушен) другой (основной) путь. По-видимо
му, выпускание отростков в нормальной ЦНС происходит непрерывно.
Одним из критических факторов в регенерации ЦНС является наличие или отсутствие рубцов, поскольку отростки не могут пройти через такие физические барьеры, как рубцы (шрамы), что значительно ограничивает процесс регенерации.
АДАПТАЦИИ С ВОЗРАСТОМ
Процесс старения обычно сопровождается заметным снижением способностей двигательной системы. Мы рассмотрим некоторые изменения, происходящие в двигательной системе с возрастом.
Двигательные способности пожилых испытуемых
Двигательная единица — это функциональная единица двигательной системы, обеспечивающая взаимосвязь нервной системы и мышцы. Поскольку процесс старения вызывает значительные изменения свойств двигательной единицы, он сопровождается снижением большинства аспектов движения. Эти изменения включают уменьшение силовых способностей, ослабление рефлекторных реакций, замедление быстрых реакций и т.д.
Сила. Одно из значительных воздействий процесса старения на двигательную систему — неизбежное уменьшение мышечной массы и снижение силы (Doherty et al., 1993а; Schultz, 1992). Снижение силы наблюдали в множестве различных мышц (Narici, Bordini, Ceretelli, 1991). Представляют интерес, по меньшей мере, два аспекта этой адаптационной реакции. Во многих исследованиях обнаружено, что снижение силовых возможностей человека начинается в возрасте около 60 лет и степень снижения значительно колеблется у разных испытуемых (рис.9.29). Колебание силы свидетельствует о том, что механизм или механизмы, регулирующие снижение силы у разных людей активируются в различной степени и, вероятно, могут быть приостановлены. Пожилые люди могут увеличить силовые способности посредством соответствующих тренировочных занятий, причем это увеличение частично обусловлено гипертрофией мышечных волокон типа II (Doherty et al., 1993 б; Keen, Yue, Enoka, 1994).
Утомляемость. В ряде исследований изучали влияние процесса старения на утомляемость мышц. В исследованиях, где использовали произвольную активацию мышц, не обнаружили изменений степени утомляемости с возрастом (Hicks et al., 1992). В этих исследованиях
319
выполняли субмаксимальные и максимальные произвольные сокращения, изометрические и динамические сокращения, непрерывные и прерывистые сокращения мышц ног и рук. В исследованиях с применением электростимулирования результаты были противоречивыми. Например, стимулирование в течение 30 с локтевого нерва с частотой 20 Гц (Lennmarken, Bergman, J.Larsson, L.-E.Larsson, 1985) привело к повышению утомляемости пожилых мужчин (67 лет), но не пожилых женщин (63 года), тогда как при стимуляции 30 Гц (Narici et al., 1991) наблюдали снижение степени утомляемости с возрастом (70 мужчин в возрасте 20— 91 года).
Рефлекторные реакции и быстрые реакции. С возрастом пик амплитуды Н-рефлекса ЭМГ снижается. Vandervoort, Hayes (1989) обнаружили, что амплитуда Н-рефлекса в камбаловидной мышце у группы пожилых женщин (82 года) составляет всего 43 % показателя молодых женщин (26 лет); пик амплитуды составлял соответственно 2,4 и 5,6 мВ. Это различие амплитуды Н-рефлекса можно объяснить снижением возбудимости мышечных волокон с возрастом. Точно так же отличались значения М-волны у двух групп испытуемых: 4,4 и 9,4 мВ. Вспомним, что М-волна отражает непосредственную активацию мышцы, являясь показателем возбудимости мышечных волокон (Hicks et al., 1992), тогда как Н-рефлекс характеризует активацию рефлексов двигательных нейронов в спинном мозгу. У группы пожилых испытуемых стимулирование Н-рефлекса приводило к активации 55 % мышечных волокон, тогда как в группе молодых показатель
был немного выше — 60 %. Следовательно, количество активированных стимулированием Н-рефлекса двигательных единиц пула с возрастом не изменяется.
В отличие от искусственно вызываемого Н-рефлекса наблюдали различия в сухожильном рефлексе четырехглавых мышц бедра у испытуемых двух возрастных групп — 27 и 75 лет (Burke, Kamen, Koceja, 1989). Амплитуда реакции рефлекса была больше у молодых испытуемых (30 по сравнению с 18Н) и характеризовалась менее продолжительной латентностью (60 по сравнению с 79 мс). Различия в амплитуде реакции, возможно, связаны с разницей в уровне силы у испытуемых двух групп, в то время как различия в латентности свидетельствуют об обусловленных возрастом различиях в выявлении, передаче и обработке импульсов (стимулов). Вспомним, что сухожильный рефлекс включает возбуждение мышечных веретен, распространение афферентных импульсов по направлению к спинному мозгу, активацию альфа-двигательных нейронов и возбуждение мышечных волокон. Пока не выяснено, какие из этих процессов изменяются, способствуя изменению латентности с возрастом.
В исследовании Warabi, Noda, Kato (1986) испытуемые должны были быстро реагировать на перемешения мишени. При этом измеряли продолжительность реакции, т.е. время от начала смещения мишени до начала реакции. Мишень смещалась на 0,17; 0,34 или 0,70 рад; испытуемые должны были реагировать либо глазами либо рукой. Реакцию глаз измеряли с помощью электроокулограммы (ЭОГ), реак-
Рис.9.29. Сила максимального произвольного сокращения (МПС) мышцы кисти (приводящая мышца больших пальцев руки) как функция возраста (Narici, Bordini, Ceretelli, 1991)
320
Рис.9.30. Изменения времени реакции движений глаз (а) и руки (б) как функция возраста. Амплитуда смещения цели составляла 0,17; 0,35 и 0,70 рад (Warabi, Noda, Kato, 1986)
циюруки — как движение рычага. Из рис.9.30 видно, что продолжительность реакции с возрастом увеличивалась и была большей при более значительном смещении мишени; продолжительность реакции двух различных двигательных систем (глаза и руки) была одинаковой. Поскольку двигательные компоненты этих движений (пик скорости, продолжительность, амплитуда) с возрастом не изменялись, ученые сделали вывод, что основным влиянием процесса старения на быструю реакцию было нарушение сенсорной обработки данных. Этот вывод соответствует результатам ряда других исследований, в которых отмечали снижение сенсорных способностей с возрастом (Dyck, Karnes, O’Brien, Zimmerman, 1984; Verillo, 1979).
Сохранение позы. Процесс старения обычно сопровождается пониженной способностью контролировать позу и походку (Schultz, 1992; Winter, Patta, Frank, 1990). Чаще всего это проявляется в уменьшении количества ходьбы и снижении уровня повседневной деятельности у пожилых людей. Адаптационные реакции, лежащие в основе этих уменьшений, сложны и включают как двигательные, так и сенсорные процессы. Ослабление контроля позы играет большую роль, поскольку влияет на устойчивость равновесия человека.
Сохранение вертикальной позы требует, чтобы направление вектора веса находилось в пределах основания опоры. В этом участвует несколько процессов: а) сенсорная информация для выявления ориентации и движения индивида; б) выбор соответствующей стратегии реагирования для сохранения устойчивости; в) активация мышц, способствующих преодолению постурального дисбаланса. Сенсорную информацию обеспечивают визуальные, соматосенсорные и вестибулярные источники. Требуется не вся информация, поэтому индивид может выбирать нужный сенсорный сигнал. Кроме того, при нарушении одной из сенсорных систем
индивид может легко научиться использовать две другие. Тем не менее с возрастом у человека снижается способность выбирать нужную сенсорную информацию (Horak et al., 1989). Этому в определенной мере способствует уменьшение способности контролировать постуральные движения (перемещения центра тяжести вперед—назад). По мнению Horak et al., возрастное снижение способности контролировать позу может быть обусловлено рядом различных факторов, включая аномальный выбор сенсорной информации, продолжительную латентность быстрых реакций, слабость мышц, нарушенную способность координировать активность мышц-синергистов.
Контроль субмаксимальной силы. Недостаточно хорошо изучены влияния процесса старения на более простые движения и поведение двигательных единиц, связанное с дифференциацией силы (Kamen, DuLuca, 1989). Вместе с тем исследованы контроль изометрической силы, кинематика движений рук, контроль силы хвата (Bennett, Castiello, 1994). Например, когда испытуемым двух групп (средний возраст 28 и 67 лет) предлагали поддерживать постоянный субмаксимальный уровень силы в течение 20 с (рис.9.31), наблюдали существенные колебания показателей (Galganski et al., 1993). Степень колебаний (стандартное отклонение) увеличивалась при повышении заданного уровня силы (5, 20, 35 и 50 % максимальной), а также с возрастом. При выражении колебаний силы относительно заданного показателя силы (коэффициента вариации) их степень снижалась с увеличением заданной силы. Колебания показателей силы не связаны с изменениями размера двигательных единиц и значительно уменьшаются в результате тренировочных занятий (Keen et al., неопубликованные данные). Приведенные результаты свидетельствуют, что пожилые испытуемые менее способны поддерживать постоянные субмаксимальные уровни силы.
21 8-133
321
Рис.9.31. Сила, образуемая молодым (о) и пожилым (6) испытуемым при задании поддерживать постоянный уровень силы в течение 20 с при каждом из четырех показателей “целевой” силы. Изометрическую силу образовывал указательный палец (Galganski, Fuglevand, Enoka, 1993)
Физиологические адаптации с возрастом
Как мы выяснили, движение — весьма сложное явление. Контролировать процесс движения довольно трудно, поскольку необходимо
интегрировать множество сенсорных и двигательных процессов. Многие из этих процессов изменяются с возрастом. Например, двигательные нейроны отмирают, а те, что остаются, могут реиннервировать дополнительные мышечные волокна. Другие возрастные изменения
Время, с
i ю _____________________________________I Н Ш11Л.1Ш.НПИШ11111И11НШИ
| з ______________________Ljjjjiiuiiijuiiiiiiiiimijuiiiiiiiniiiiiiiiiiiiiiiiii
I 2 _________________Illi I 111IllllllllllllllllllllllUUlllllllllllllllllllllllllllll
Д 1 ________L.l l I I I Ш11111111111111111111111И1111111иШШЛ1Ц||111НШЦЦ11Ш.Ци
Рис.9.32. Вклад двигательных единиц в силу, образуемую мышцей. Гипотетическая схема показывает увеличение мышечной силы в течение времени (внешняя линия графика сила—время), а также силу, обусловленную пятью двигательными единицами (из многих активных). На нижнем графике показаны потенциалы действия этих пяти двигательных единиц. При рекрутировании каждой двигательной единицы интенсивность ее разряда небольшая, но затем повышается
322
5 3 О \D § °
701
60-
50-
40-
30-
20-
i----------1---------1----------1----------1----------1----------1----------1
1,2 -
1,0-
0,8-
Q,6-
0,4-
---1----1 i----1----1----1----1
20 40 60 80
Возраст, лет
Рис.9.33. Доля мышечных волокон типа II (а) и относительная площадь поперечного сечения (б) широкой латеральной мышцы бедра человека как функция возраста. Данные по 100 испытуемым в возрасте 22—89 лет. Наблюдается существенное снижение доли мышечных волокон типа II, а также отношения площади поперечного сечения волокон типа II к волокнам типа I (Larsson, 1983)
включают снижение интенсивности аксонного транспорта, изменение биофизических свойств двигательных нейронов, уменьшение амплитуды М-волн и т.д.
Из этих изменений представляет интерес отмирание двигательных нейронов и его последствия. Отмирание двигательных нейронов вызывает снижение мышечной массы с возрастом, а также прогрессирующую атрофию и дезинтеграцию денервированных мышечных волокон; оставшиеся двигательные единицы вовлекаются денервированными мышечными волокнами в выпускание коллатеральных от-та внутримышечного аксона и р-н-нервацию мышечных волокон (Doherty, Vandervoort, Taylor, Brown, 1993; Rosenheimer, Smith, 1990). Вследствие этих процессов с возрастом сокращается количество двигательных единиц и увеличивается размер (коэффициент иннервации) оставшихся двигательных единиц. Уменьшение с возрастом числа двигательных единиц соответствует снижению мышечной силы (см.рис.9.29).
Реиннервация денервированных мышечных волокон оставшимися двигательными единицами означает, что коэффициент иннервации этих единиц повышается. Вспомним из главы 5, что сила, образуемая двигательной единицей, зависит от коэффициента иннервации, средней площади поперечного сечения мышечных волокон И специфического напряжения. Вследствие увеличения коэффициента иннервации пик силы, образуемой отдельными двигательными единицами, увеличивается как функция возраста (Galganski et al., 1993). Это влияет на способность человека плавно дифференцировать мышечную силу двумя способами. Во-первых, у пожилых людей по сравнению с молодыми испытуемыми значительно сокращается количество двигательных единиц небольшой силы, которые играют важную роль в тонко контролируемых движениях субмаксимальной силы. Во-вторых, плавность (отсутствие колебаний) образования силы мышцей зависит от силы, образуемой последними рекрутированными двигательными единицами (Christakos, 1982). Поскольку двигательные единицы рекрутируются при низкой интенсивности разряда, ведущего к образованию прерывистого тетануса, рекрутирование двигательной единицы способствует колебаниям чистой силы (рис.9.32). Увеличение силы двигательной единицы с возрастом приводит к более значительным колебаниям при рекрутировании каждой двигательной единицы.
По-видимому, отмирание двигательных нейронов с возрастом — неслучайный процесс, в большей мере это относится к большим двигательным нейронам. Такой вывод сделан на основании изменений распределения и доли типов мышечных волокон и двигательных единиц (рис.9.33) (Lexell, Taylor, 1991). Исходя из процесса денервации-реиннервации, с возрастом, очевидно, повышается плотность мышечных волокон двигательных единиц типа S (Kanda, Hashizume, 1989). Эти наблюдения позволяют предположить, что процесс старения связан с преимущественной дегенерацией двигательных единиц типа FF и FR и реиннервацией соседних мышечных волокон двигательными единицами типа S.
Программы силовых занятий показали прирост силы (четырехглавых мышц бедер) у 90-летних добровольцев (Fiatarone et al., 1990). Щда этом также набШОДали значительное увеличение общей площади мышц и площади поперечного сечения волокон типа I и II мышц рук (двуглавая мышца плеча) и ног (широкая латеральная мышца бедра) (Brown, McCartney, Sale 1990; Keen et al., 1994). Эти исследования подтверждают возможность гипертрофии мышц, увеличения оборота миофибриллярного белка и активации клеток-сателлитов для “ремонта” поврежденных мышечных волокон.
21*
323
вопросы
САЛЫ РЯАШВНЫМП ГВЕГОМ
1. Согласно принципу нагрузки, для возникновения адаптационной реакции необходимо превзойти пороговую точку.
ПРАВИЛЬНО НЕПРАВИЛЬНО
2. Согласно принципу обратимости, тип изменений специфичен физической нагрузке. ПРАВИЛЬНО НЕПРАВИЛЬНО
3. Мышечная сила измеряется, когда скорость равна нулю.
ПРАВИЛЬНО НЕПРАВИЛЬНО
4. Длина мышцы не влияет на изменение СИЛЫ. ПРАВИЛЬНО НЕПРАВИЛЬНО
5. Длина мышечного волокна не изменяется при изометрическом сокращении.
ПРАВИЛЬНО НЕПРАВИЛЬНО
6. Изометрические нагрузки приводят к приросту СИЛЫ. ПРАВИЛЬНО НЕПРАВИЛЬНО
7. При изокинетическом сокращении длина мышцы сокращается с постоянной скоростью. ПРАВИЛЬНО НЕПРАВИЛЬНО
8. Упражнения силовой направленности с увеличением нагрузки предполагают увеличение числа повторений в циклах.
ПРАВИЛЬНО НЕПРАВИЛЬНО
9. Эксцентрические упражнения обеспечивают больший прирост силы, чем концентрические. ПРАВИЛЬНО НЕПРАВИЛЬНО
10. Эксцентрические упражнения наиболее эффективны для стимулирования гипертрофии. ПРАВИЛЬНО НЕПРАВИЛЬНО
11. Аккомодирующийся тренажер — это тренажер, который позволяют изменять степень сопротивления в зависимости от силы, образуемой испытуемым.
ПРАВИЛЬНО НЕПРАВИЛЬНО
12. Кроме периодов начала и завершения движения скорость ускорения аккомодирующегося тренажера при изокинетическом сокращении равна нулю.
ПРАВИЛЬНО НЕПРАВИЛЬНО
13. Гидравлические тренажеры не позволяют контролировать скорость движения.
ПРАВИЛЬНО НЕПРАВИЛЬНО
14. В основе плиометрических упражнений лежат эксцентрическо-концентрические сокращения. ПРАВИЛЬНО НЕПРАВИЛЬНО
15. Плиометрические тренировки позволяют увеличить результат в прыжках в высоту.
ПРАВИЛЬНО НЕПРАВИЛЬНО
16. Согласно гипотезе нервной недостаточности, силу максимального произвольного сокращения можно увеличить, манипулируя афферентной обратной связью.
ПРАВИЛЬНО НЕПРАВИЛЬНО
17. Бифазные прямоугольные электрошоки преимущественно активируют двигательные единицы большого диаметра.
ПРАВИЛЬНО НЕПРАВИЛЬНО
18. При нервно-мышечном электростимулировании потенциалы действия образуются непосредственно в мышечных волокнах.
ПРАВИЛЬНО НЕПРАВИЛЬНО
19. Импеданс — мера сопротивления электрическому току.
ПРАВИЛЬНО НЕПРАВИЛЬНО
20. Единица измерения импеданса — mho.
ПРАВИЛЬНО НЕПРАВИЛЬНО
21. Магнитное стимулирование может вызвать потенциалы действия во внутримышечных нервных ответвлениях.
ПРАВИЛЬНО НЕПРАВИЛЬНО
22. Нервно-мышечное электростимулирование не приводит к увеличению силы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
23. Нервно-мышечное электростимулирование может сводить к минимуму атрофию четырехглавых мышц бедра у пациентов с травмами спинного мозга.
ПРАВИЛЬНО НЕПРАВИЛЬНО
24. Упражнения силовой направленности с увеличением сопротивления можно выполнять только со штангой.
ПРАВИЛЬНО НЕПРАВИЛЬНО
25. Наиболее оптимальный режим нагрузок во время занятий силовой направленности — нагрузка 8—12 МП.
ПРАВИЛЬНО НЕПРАВИЛЬНО
26. Во время тренировочных занятий силовой направленности изменение ЭМГ происходит до изменения размера мышц.
ПРАВИЛЬНО НЕПРАВИЛЬНО
27. Мысленное представление выполнения сокращений способствует приросту силы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
28. Коактивация повышает силу МПС относительно сустава.
ПРАВИЛЬНО НЕПРАВИЛЬНО
29. Степень коактивации снижается в результате тренировочных занятий силовой направленности. ПРАВИЛЬНО НЕПРАВИЛЬНО
30. Степень синхронизации измеряется сравнением амплитуды М-волны и Н-реакции.
ПРАВИЛЬНО НЕПРАВИЛЬНО
324
31. Увеличение размера мышц может быть вызвано гипертрофией или гиперплазией.
ПРАВИЛЬНО НЕПРАВИЛЬНО
32. Гипертрофия наблюдается значительно чаще вследствие тренировочных занятий силовой направленности.
ПРАВИЛЬНО НЕПРАВИЛЬНО
33. Проведение тренировочных занятий силовой направленности в течение 6 недель не приводит к увеличению площади поперечного сечения мышечных волокон у опытных культуристов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
34. Тренировочные занятия силовой направленности не приводят к гипертрофии мышечных волокон типа I.
ПРАВИЛЬНО НЕПРАВИЛЬНО
35. Специфическое напряжение максимально в двигательных единицах типа S.
ПРАВИЛЬНО НЕПРАВИЛЬНО
36. Колебания специфического напряжения двигательных единиц обусловлены различиями в плотности миофиламентов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
37. Механические стимулы могут модулировать интенсивность белкового синтеза и расщепления в мышце.
ПРАВИЛЬНО НЕПРАВИЛЬНО
38. Развитие мощности представляет собой производительность мышцы при изометрических сокращениях.
ПРАВИЛЬНО НЕПРАВИЛЬНО
39. Максимальное развитие мощности наблюдается при мышечной силе, равной 1/3 максимальной. ПРАВИЛЬНО НЕПРАВИЛЬНО
40. Мощность можно измерить как результат СИЛЫ И работы.ПРАВИЛЬНО НЕПРАВИЛЬНО
41. Средняя мощность, развиваемая при выполнении прыжка в высоту, составляет 600 Вт.
ПРАВИЛЬНО НЕПРАВИЛЬНО
42. Тяжелоатлет развивает большую мощность при выполнении жима, лежа на скамье.
ПРАВИЛЬНО НЕПРАВИЛЬНО
43. Ро = изометрический вращающий момент. ПРАВИЛЬНО НЕПРАВИЛЬНО
44. Мощность, развиваемую мышцами относительно сустава, можно вычислить как результат вращающего момента и угловой скорости.
ПРАВИЛЬНО НЕПРАВИЛЬНО
45. Медленносокращающаяся мышца способна развивать большую мощность по сравнению с быстросокращающейся.
ПРАВИЛЬНО НЕПРАВИЛЬНО
46. Наиболее оптимальная тренировочная нагрузка для увеличения мощности — 100 % МПС.
ПРАВИЛЬНО НЕПРАВИЛЬНО
47. Существует прямо пропорциональная взаимосвязь между снижением ЭМГ и уменьшением мышечной массы при иммобилизации конечности.
ПРАВИЛЬНО НЕПРАВИЛЬНО
48. Иммобилизация конечности связана с превращением быстросокращающегося мышечного волокна в медленносокращаю-щееся.
ПРАВИЛЬНО НЕПРАВИЛЬНО
49. При иммобилизации конечности утомляемость мышц повышается.
ПРАВИЛЬНО НЕПРАВИЛЬНО
50. В иммобилизационной мышце увеличивается количество двигательных единиц высокого порога.
ПРАВИЛЬНО НЕПРАВИЛЬНО
51. Поскольку при подвешивании конечности активность ЭМГ не снижается, мышца не атрофируется.
ПРАВИЛЬНО НЕПРАВИЛЬНО
52. При подвешивании задней конечности наблюдается снижение доли волокон типа I в камбаловидной мышце.
ПРАВИЛЬНО НЕПРАВИЛЬНО
53. Подвешивание задней конечности приводит к снижению активности всех ферментов, участвующих в окислительном метаболизме. ПРАВИЛЬНО НЕПРАВИЛЬНО
54. При подвешивании задней конечности специфическое напряжение в медленносокра-щающейся мышце снижается.
ПРАВИЛЬНО НЕПРАВИЛЬНО
55. Нейроны спинного мозга не имеют афферентной обратной связи при рассечении спинного мозга.
ПРАВИЛЬНО НЕПРАВИЛЬНО
56. При рассечении спинного мозга наблюдается атрофия мышц и превращение быст-росокрашаюшихся мышечных волокон в медленносокращающиеся.
ПРАВИЛЬНО НЕПРАВИЛЬНО
57. Временные изменения сокращения обусловлены изменениями кинетики Са2+.
ПРАВИЛЬНО НЕПРАВИЛЬНО
58. Спинализация вызывает изменения свойств двигательных нейронов.
ПРАВИЛЬНО НЕПРАВИЛЬНО
59. При пересечении аксона он может отпустить отростки.
ПРАВИЛЬНО НЕПРАВИЛЬНО
325
60. Перерезанные аксоны могут пустить отростки и реиннервировать мышечные волокна.
ПРАВИЛЬНО НЕПРАВИЛЬНО
61. Регенерирующие аксоны способны реиннервировать исходные мышечные волокна, которые были иннервированы до аксотомии.
ПРАВИЛЬНО НЕПРАВИЛЬНО
62. Все двигательные единицы могут выпускать коллатеральные отростки.
ПРАВИЛЬНО НЕПРАВИЛЬНО
63. Коэффициент иннервации двигательных единиц увеличивается при выпускании коллатеральных отростков.
ПРАВИЛЬНО НЕПРАВИЛЬНО
64. Коллатеральным отросткам требуется соединение с супраспинальными центрами.
ПРАВИЛЬНО НЕПРАВИЛЬНО
65. ЦНС способна к синаптогенезу.
ПРАВИЛЬНО НЕПРАВИЛЬНО
66. Деафферентация имеет место при пересечении спинного мозга.
ПРАВИЛЬНО НЕПРАВИЛЬНО
67. С возрастом количество двигательных единиц в мышце снижается.
ПРАВИЛЬНО НЕПРАВИЛЬНО
68. С возрастом средний размер двигательных единиц уменьшается.
ПРАВИЛЬНО НЕПРАВИЛЬНО
69. У пожилых испытуемых наблюдается повышенная степень утомляемости мышцы при произвольных сокращениях.
ПРАВИЛЬНО НЕПРАВИЛЬНО
70. Снижение пика амплитуды Н-рефлекса с возрастом обусловлено уменьшением с возрастом мышечной массы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
71. Латентность рефлекторных реакций с возрастом не изменяется.
ПРАВИЛЬНО НЕПРАВИЛЬНО
72. Пожилые испытуемые испытывают трудности в выборе соответствующей стратегии сохранения вертикальной позы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
73. Пожилые испытуемые испытывают трудность в сохранении постоянной субмаксимальной изометрической силы.
ПРАВИЛЬНО НЕПРАВИЛЬНО
74. Снижение мышечной массы с возрастом обусловлено, главным образом, атрофией и дезинтеграцией поврежденных мышечных волокон.
ПРАВИЛЬНО НЕПРАВИЛЬНО
75. С возрастом коэффициент иннервации двигательных единиц снижается.
ПРАВИЛЬНО НЕПРАВИЛЬНО
76. Волокна типа II, по-видимому, испытывают наибольшие изменения с возрастом.
ПРАВИЛЬНО НЕПРАВИЛЬНО
ВОН С РАЗЛИ ЧНЫМ ВЫБОР ОТ ЕХОБ
77. Что из приведенного ниже не относится к принципам тренировки?
а) обратимость;
б) способность передаваться;
в) постепенное увеличение нагрузки;
г) специфичность.
78. Какой принцип тренировки включает понятие точки порога?
а) обратимость;
б) способность передаваться;
в) постепенное увеличение нагрузки;
г) специфичность.
79. Какой параметр не влияет на мышечную силу?
а) интенсивность изменения длины мышцы;
б) плечо пары;
в) площадь поперечного сечения;
г) рекрутирование двигательных единиц.
80. В чем преимущество изометрических упражнений?
а) удобство;
б) прирост силы на всем диапазоне движения;
в) хорошая реакция гипертрофии;
г) отсутствие факторов риска.
81. При каком проценте максимальной мышечной силы происходит окклюзия кровтока? а) 5%;
б) 15%;
в) 35 %;
г) 55 %.
82. Насколько увеличивается мышечная сила после 6-недельной программы занятий изометрического характера?
а) 10%;
б) 20%;
в) 40%;
г) 80%.
83. В каких условиях вращающий момент мышцы не равен вращающему моменту, обусловленному физической нагрузкой?
а) при изометрических сокращениях;
б) при изокинетических сокращениях;
в) при динамических сокращениях;
г) при статических сокращениях.
326
84. Укажите неправильное утверждение.
а) нагрузка 1 МП ограничена силой эксцентрических сокращений;
б) эксцентрические упражнения обеспечивают больший прирост силы, чем концентрические;
в) величина нагрузки зависит от нагрузки относительно максимальных возможностей;
г) эксцентрические упражнения обеспечивают наиболее эффективное стимулирование гипертрофии.
85. Почему при эксцентрическом максимальном произвольном сокращении ЭМГ меньше, чем при концентрическом?
а) при субмаксимальных концентрических и эксцентрических сокращениях активируются различные двигательные единицы;
б) при беге по наклонной вниз амплитуда Н-рефлекса снижается в большей степени;
в) побуждение из ЦНС менее максимального;
г) ЭМГ в мышцах ног постепенно увеличивается при беге по наклонной вниз с постоянной скоростью;
86. Какие системы не обеспечивают аккомодирующее сопротивление?
а) система переключения скоростей;
б) свободные отягощения;
в) пневматические системы;
г) гидравлические цилиндры.
87. В чем преимущества изокинетического тренажера?
а) обеспечение принципа специфичности;
б) обеспечение значительно большего прироста силы, чем при использовании отягощений;
в) обеспечение опоры для испытуемого;
г) позволяет выполнять работу в ограниченном диапазоне движения.
88. Какое явление лучшего всего характеризует физиологическую основу плиометричес-ких упражнений?
а) взаимосвязь работа—энергия;
б) рефлекс растяжения;
в) взаимосвязь импульс—количество движения;
г) последовательность эксцентрических и концентрических сокращений.
89. Что обусловливает неспособность индивида проявлять максимальную силу при произвольном сокращении?
а) нервное обеспечение;
б) поперечное развитие;
в) двусторонний прирост;
г) нервная недостаточность.
90. Какова наиболее оптимальная нагрузка для тренировочного занятия силовой направленности?
а) 1 МП;
б) 2-3 МП;
в) 4-6 МП;
г) 8-12 МП.
91. Какую часть максимального значения составляет нагрузка 5—6 МП?
а) 50-60%;
б) 70-80%;
в) 80-90%;
г) 90-100%.
92. Почему отдают предпочтение тренажерам с переменным сопротивлением, а не обычным отягощениям?
а) нагрузка остается постоянной на всем диапазоне движения;
б) изменение нагрузки максимально соответствует взаимосвязи максимальный вращающий момент—угол;
в) вращающий момент, обусловленный нагрузкой относительно сустава, изменяется больше, чем вращающий момент, обусловленный отягощениями;
г) максимальный вращающий момент, обусловленный физической нагрузкой, отмечается в середине диапазона движения.
93. Какое наблюдение не характеризует значение адаптационных реакций нервной системы во время физических занятий силовой направленности?
а) прирост силы возможен вследствие тренировочных занятий небольшой продолжительности, вызывающих морфологические изменения;
б) изменение ЭМГ происходит до изменения размера мышц;
в) ЭМГ изменяется значительно больше, чем размер мышцы;
г) при выполнении мысленных сокращений прирост силы не наблюдается.
94. При каком режиме тренировок наблюдает поперечное развитие?
а) произвольные сокращения;
б) тренировки, направленные на повышение выносливости;
в) нервно-мышечное электростимулирование;
г) мысленное выполнение сокращений.
95. Что такое гиперплазия?
а) увеличение специфического напряжения;
б) увеличение количества мышечных волокон;
в) увеличение площади поперечного сечения мышечных волокон;
г) увеличение цитоскелетного матрикса.
327
96. Чем можно объяснить различие в специфическом напряжении между двигательными единицами?
а) различиями в концентрации коллагена эндомизия;
б) изменением длины мышечных волокон;
в) различиями в плотности миофиламентов;
г) разной площадью поперечного сечения.
97. Почему гормональные воздействия не являются ключевыми факторами в контроле мышечной гипертрофии?
а) ввиду их медленного действия;
б) гормоны действуют систематически и влияют на многие мышцы;
в) метаболические затраты при эксцентрических сокращениях меньше, чем при концентрических;
г) введение гормона роста усиливает изменения, происходящие вследствие тренировочных занятий силовой направленности.
98. Какие две переменные определяют количество мощности, развиваемой мышцей? а) площадь поперечного сечения мышцы; б) миозин АТФаза;
в) кинетика Са2+;
г) длина мышечного волокна.
99. Какую мощность мшут развить мышцы сгибатели стопы при выполнении прыжка в высоту?
а) 25 Вт;
б) 250 Вт;
в) 2,5 кВт;
г) 25 кВт.
100. Какая тренировочная нагрузка (от максимального произвольного сокращения), согласно принципу специфичности, обеспечивает максимальное увеличение образования силы?
а) 100%;
б) 30%;
в) 60%;
г) 90%.
101. Что не происходит при иммобилизации конечности?
а) превращение быстросокращающихся мышечных волокон в медленносокращающиеся;
б) атрофия;
в) снижение максимального произвольного сокращения ЭМГ;
г) изменения порядка рекрутирования двигательных единиц.
102. Почему мышечная масса с возрастом уменьшается?
а) двигательные нейроны отмирают и лишают мышечные волокна нервной иннервации;
б) заболевания ведут к мышечной атро-фии;в) мышца подвергается недостаточной нагрузке, чтобы поддерживать высокие уровни синтеза белков.
103. Какая адаптационная реакция, по-видимому, играет наиболее важную роль в задержке двигательных реакций у пожилых людей? а) снижение количества двигательных нейронов;
б) уменьшение количества мышечных волокон;
в) недостаточное побуждение ЦНС (из супрасегментальных центров);
г) снижение сенсорных способностей.
104. Какие факторы способствуют возрастному снижению способности контролировать позу?
а) пониженное восприятие границ устойчивого положения;
б) нарушенная способность координировать активность мышц-синергистов;
в) плохое восприятие нарушения равновесия;
г) аномальный выбор сенсорной информации.
105. Чем обусловлены большие трудности в поддержании постоянной субмаксимальной силы у пожилых людей?
а) уменьшением мышечной массы;
б) повышенной силой сокращения двигательных единиц;
в) меньшим числом двигательных нейронов;
г) потерей мышечных волокон типа II.
ЗАДАЛИ
106. Влияют ли занятия силовой направленности на выносливость и наоборот?
107. Предположим, вы измерили образование силы и мощности у 20 испытуемых. Существует ли взаимосвязь между этими двумя параметрами? Используйте для объяснения взаимосвязь сила—скорость.
108. В чем преимущества (недостатки) изометрических упражнений?
109. Сила, образуемая мышцей, больше во время эксцентрического, чем во время концентрического сокращения.
А. Почему эксцентрические упражнения не обеспечивают больший прирост силы?
Б. Почему рекомендуется включать в тренировочную программу эксцентрические упражнения для достижения гипертрофированной реакции?
В. Укажите пять различий между эксцентрическими и концентрическими сокращениями.
328
ПО. Почему аккомодирующий тренажер обеспечивает изокинетическое сокращение?
111. Почему порядок рекрутирования двигательных единиц при нервно-мышечном электростимулировании отличается от того, который имеет место при произвольной активации пула двигательных единиц?
112. Тренажеры, обеспечивающие переменную нагрузку, предназначены, очевидно, для обеспечения соответствия возможностям избранных систем суставов.
А. Нарисуйте график для локтевого сустава, показав, как должно изменяться плечо пары барабана тренажера для обеспечения соответствия результирующему вращающему моменту мышцы.
Б. Нарисуйте график, показав, как изменяется нагрузка при использовании кругового барабана в диапазоне движения.
113. Существует утверждение, что можно достичь прироста силы без адаптационных реакций в мышце, но не без адаптационных реакций нервной системы.
А. Что это означает?
Б. Какие есть доказательства этого?
В. У каких категорий испытуемых наблюдается этот эффект?
114. Некоторые адаптационные реакции нервной системы вследствие тренировок силовой направленности включает взаимодействия между конечностями. Как правило, при активности обеих конечностей, данная конечность образует меньшую максимальную силу, чем та, которая образуется при активности одной конечности. Говард (1987) изучал это явление, измеряя силу разгибателей коленного сустава до и после 3 недель тренировочных занятий силовой направленности. Силу максимального произвольного сокращения и ЭМГ (широкая латеральная мышца бедра) измеряли при выполнении сокращений одной ногой (отдельно правой и левой) и двумя ногами.
Условие эксперимента Одной ногой Двумя ногами
Правой Левой Правой Левой
До тренировок Сила, Н 782 776 762 726
После тренировок 950 897 981 966
До тренировок ЭМГ, мВ 0,55 0,51 0,52 0,51
После тренировок 0,67 0,65 0,69 0,84
Используя эти данные, а также приведенное ниже уравнение, определите двусторонний индекс (ДИ) ЭМГ и силы до и после
тренировок. “Двумя ногами” означает суммирование показателей правой и левой ноги при выполнении сокращений обеими ногами, а “правой + левой ногой” — суммирование при выполнении сокращений одной ногой. Объясните, какие результаты свидетельствуют о виде адаптационных реакций.
ДИ (%) =
_______ обе ноги_______ правая нога + левая нога
100.
115. Почему более вероятно увеличение площади поперечного сечения волокон типа ГГ, чем типа I вследствие тренировочных занятий силовой направленности?
116. В ряде исследований наблюдали тесную взаимосвязь между размером мышцы (например, площадью поперечного сечения) и максимальной силой, которую способна образовать мышца. Вместе с тем на рис. 9.16 показаны значительные колебания площади поперечного сечения четырехглавой мышцы бедра и максимальной изометрической силы. Объясните эти различия.
117. На основании линейной взаимосвязи сила-скорость определите максимальную мощность, которую могут развивать мышцы-сгибатели коленного сустава человека. Сила мышц—сгибателей коленного сустава — 237 Н • м_|, максимальная угловая скорость — 17,3 рад • с-1.
118. Брукс и Фолкнер (1991) используют для выражения мощности единицу измерения Вт-кг-1. Почему?
119. В большинстве моделей пониженной активности отмечается превращение медлен-носокращающихся мышечных волокон в быстросокращающиеся.
А. Что происходит с метаболическими ферментами? Как можно проверить эти воздействия?
Б. Что происходит с сократительными белками при пониженной активности?
В. Почему подобное превращение не происходит во всех мышечных волокнах типа I?
120. Согласно данным Тотоси де Зепетнека и др. (1992), двигательные единицы в нормальной и реиннервированной мышцах имеют следующие свойства. Определите среднюю максимальную силу, образуемую двигательной единицей каждого типа (S, FR, FF) в нормальной и реиннервированной мышцах.
121. Какие две возрастные адаптационные реакции вызывают наибольшее нарушение двигательных способностей?
329
Таблица к задаче 119
Показатель Нормальные Реиннериированные
S FR FF S FR FF
Площадь поперечного сечения мышечного волокна, мкм2 1317 2048 4134 2252 2424 3897
Коэффициент иннервации 91 91 152 ПО 97 469
Специфическое напряжение, Н ем2 27,3 24,5 23,5 24,4 25,6 25,6
122. Пик амплитуды как Н-рефлекса, так и М-волны с возрастом снижается. О каких изменениях в двигательной системе с возрастом это свидетельствует?
123. Процесс старения связан с уменьшением количества мышечных волокон типа Пб.
В то же время пониженный уровень активности приводит к превращению мед-ленносокрашаюшихся мышечных волокон в быстросокрашаюшиеся. Однако в каждом случае степень утомляемости практически не изменяется. Чем это обусловлено?
ПРИЛОЖЕНИЕ 1
СИСТЕМА
ЕДИНИЦ СИ
Система единиц СИ (Le Systems Internationale d’Unites) включает 7 основных единиц, из которых можно получить другие
ОСНОВНАЯ
ПРОИЗВОДНАЯ
1. Длина — метр (м).
Метр — длина, равная 1650763,73 длины волны изотопа криптона (8бКч).
1 дюйм = 2,54 см
1 фут = 30,48 см = 0,3048 м
1 ярд ~ 09144 м
1 миля = 1609 м = 1609 км
2. Масса — килограмм (кг).
Килограмм равен массе цилиндрической гири из платино-иридиевого сплава, хранящейся в Севре, Франция).
1 фунт = 0,454 кг
Площадь — квадратные метры (м2).
Двухмерная мера длины
1 фут 2 = 0,0929 м2
1 акр = 0,4047 гектаров (га)
Объем — кубические метры (м3).
Трехмерная мера длины. Хотя литр не является единицей системы СИ, объем часто измеряют в литрах (л)
1 мл Н2О = 1 см3
1 л = 0,001 м3
Плотность — килограмм х кубические метры-1 (кг-м3). Масса на единицу объема.
Энергия, работа — джоуль (Дж).
Энергия — способность выполнять работу, работа характеризует приложение силы на определенном расстоянии.
1 Дж = 1 Н-м
1 ккал = 4,183 кДж
1 кпм = 9,807 Дж
Сила — ньютон (Н).
Один ньютон — сила, сообщающая телу массой 1 кг ускорение 1 м * с~2 в направлении действия силы.
1 Н = 1 кг • м с"2
1 кг-силы = 9,81 Н
1 фунт-силы = 4,45 Н
Импульс — ньютон х секунда (Н • с)
Сила, приложенная в интервале времени; площадь под кривой сила—время.
Момент инерции (кг • м2).
Сопротивление объекта изменению состояния вращательного движения; мера распределения массы объекта относительно оси вращения.
Мощность — ватт (Вт).
331
3. Время — секунда (с).
Единица времени, воспроизводимая цезиевыми эталонами частоты и времени (9,192,631,770 колебаний излучения атома цезия)
4. Электрический ток — ампер (А).
Интенсивность потока заряженных частиц.
5. Температура — Кельвин (К).
Мера скорости колебаний молекул тела. От точки замерзания до точки кипения — 100 К.
0 К — абсолютный нуль
6. Количество вещества — моль.
Количество вещества, содержащего такое же количество частиц, которое находится в 12 г (1 моль) изотопа 12С.
7. Сила света — свеча (св).
Сила света 1/600 000 1 м2 абсолютно черного тела при температуре затвердевания платины (2045 К).
Скорость выполнения работы.
1 Вт = 1 Дж • с-1
1 лошадиная сила = 736 Вт
Давление — паскаль (Па).
Сила, действующая на единицу площади.
1 Па = 1 Н • м~2
1 мм рт.ст. = 133,3 Па
Вращающий момент — момент силы (Н • м).
Вращающее действие силы.
1 фут-фунт — 1,356 Н • м
Ускорение (м •с-2)
Интенсивность изменения скорости.
Ускорение, обусловленное силой тяжести — 9,807 м • с 2.
1 фут • с-2 = 0,3048 м • с'2.
Частота — Герц (Гц).
Количество колебаний в секунду.
1 Гц = 1 колебание • с-1
Количество движения (кг • м • с-1).
1 слаг • фут с-1 = 4,447 кг • м с-1.
Скорость (м • с-1).
Изменения положения во времени.
1 фут'с-1 = 0,3048 м-с-1.
1 миля • час-1 = 0,447 м • с-1 = 1,609 км • ч_|
Электрическая емкость — фарада (Ф).
Свойство проводимости и изоляции электрической системы, позволяющее накапливать электрический заряд, когда между проводниками существует разница потенциалов.
Активная проводимость — mho.
Свойство, противоположное сопротивлению, — легкость прохождения заряженных частиц через объект.
Сопротивление — ом (Ом).
Трудность прохождения потока заряженных частиц через объект.
Напряжение — вольт (В).
Разница в чистом распределении заряженных частиц между двумя точками.
Шкала Цельсия (°C)
0°С = 273,15 К
Шкала Фаренгейта (°Ф)
32°Ф = 0°С = 273,15 К °Ф = 1,8°С + 32.
Концентрация (моль • м 3)
Количество вещества в единице объема.
Люмен (Лм)
Единица светового потока.
332
ДОПОЛНИТЕЛЬНАЯ ЕДИНИЦА
8. Угол — радиан (рад).
Измерение угла в плоскости (т.е. двухмерный угол).
1 рад = 57,3°
Тэта (0) равна одному радиану, когда радиус г длина дуги а равны (рис. 1.1).
Стерадиан (стер)
Трехмерный, или пространственный, угол.
Рис.1.1. Определение радиана
Таблица 1.1. Приставки, используемые в системе единиц СИ
Приставка Сокращение Мультипликативный множитель
Тера Т 1 000 000 000 000= 1012
Гига Г 1 000 000 000= 109
Мега М 1 000 000 = 106
Кило к 1 000= 103
Гекто г 100= ю2
Деци д 0,1 = 10“'
Санти с 0,01 = 10 ~2
Милли м 0,001 = 10 ~3
Микро р 0,000 001 = 10”6
Нано п 0,000 000 001 = 10-9
Пико р 0,000 000 000 001 = 10 “12
ПРИЛОЖЕНИЕ 2
ВЕКТОРЫ
Количественные характеристики каких-либо процессов или явлений, имеющие величину и направление, называются векторами (скорость, ускорение, сила, количество движения). Те из них, которые определяются только величиной, называются скалярами (масса, длина, время, температура). Векторы можно представить графически (рис. 2.1) в виде стрелок. Длина стрелки показывает величину вектора в определенном масштабе, а его направление характеризуется линией действия и ориентацией (направление острия стрелки).
Правила параллелограмма сил
Две силы (Fp и F9), действующие на объект, могут быть заменены их равнодействующей (Fr), направленной по диагонали параллелограмма со сторонами Fp и F9 (рис.2.2). Замена системы сил
(двух и более) одной результирующей называется сложением системы сил. Понятие “разложение” означает замену данной силы двумя и более компонентами.
Предположим, например, что на бегуна, изображенного на рис. 2.6, в, действуют следующие составляющие силы реакции поверхности:
вертикальная (Fgz) 812 Н (положительная=вверх) переднезадняя (Fgy) —286 Н (положительная=вперед) боковая (FgI) 61 Н (положительная=латеральная)
1. Нарисуйте диаграмму двух составляющих и результирующей силы в каждой из сагиттальной, фронтальной и горизонтальной плоскости.
2. Определите величину и направление каждой из этих результирующих. (Ответ: сагиттальная — 861 Н; фронтальная — 814 Н; горизонтальная — 292 Н).
В. Используя теорему Пифагора, определите результирующую (FK) силу реакции поверхности (F =7/’2&г + F\y + F2gx).
(Ответ: Fg = 863 Н).
Векторная алгебра
Анализ биомеханических систем обычно включает множество векторных величин и предполагает применение различных методов. Рассмотрим некоторые из них.
Правая система координат. Механические явления по разному фиксируются в различных координатных системах. Например, изменение положения означает смещение расположения объекта относительно какого-то базиса. В простейшем случае такой базис состоит из двух
334
осей — х и у. При пространственном движении используют трехмерный базис xyz. Типичная ориентация осей показана на рис.2.3.
Если вы согнете пальцы правой руки в направлении кратчайшего поворота положительной оси х к положительной оси у, то направление вытянутого большого пальца покажет направление положительной оси z- Именно поэтому конфигурация осей xyz называется правой системой координат.
Векторные величины можно соотносить с правой системой координат. Для этой цели используют единичные векторы i, j и к. Символ i означает направление х, символ j — направление у, символ к — направление z (рис.2.4).
Рис.2.4. Единичные векторы правой системы координат
Вектор, описанный выражением 61 i — 286 j + 812 к имеет 61 единицу в направлении х, 286 — в отрицательном направлении у и 812 — в направлении z- Величину вектора можно определить на основании теоремы Пифагора: __________________
Fg = J612 +(-286)2 +8122.
Скалярное произведение. Описывая физические системы и взаимодействие их составляющих, мы очень часто манипулируем векторными величинами в алгебраических выражениях. Сложение и вычитание векторов осуществляется очень просто, путем сложения или вычитания их составляющих.
Например, рассмотрим сложение Fp и Ffl:
F, = 10 i + 28 j + 92 к и F9 = 3 i - 11 j + 46 k; Fp + F? = (10 + 3) i + (28 - 11) j + (92 + 46) k= = 13 i+ 17 j+ 138 k.
Более сложно выполняется умножение. Существует два способа умножения: скалярное и векторное.
Скалярное произведение определяется выражением
d • F- JFcosS- (2.1)
Скалярное произведение двух векторов d и F определяется как произведение d (d) на F (F) и на косинус угла (0) между двумя векторами (рис.2.5, а).
Рис.2.5. Скалярное произведение, включающее определение составляющей F (а), действующей в том же направлении, что и d (6)
Результат скалярного умножения векторов — скалярная величина. Так, например, количество работы определяется как произведение силы и перемещения. Как сила, так и перемещение являются векторами. Работа — скалярная величина — произведение составляющей силы, действующей в направлении перемещения, на величину перемещения.
Векторное произведение. Результат умножения векторов — векторная величина. При этом векторное произведение определяется по формуле
(г х F) = г Л sin 0. (2.2)
Направление вектора—произведения перпендикулярно плоскости, содержащей два других вектора. Их взаимное расположение показано на рис. 2.6.
Рис.2.6. Векторное произведение г и F является вектором Т
Рассмотрим пример, в котором сила (F) действует на систему, которая вращается отно
335
сительно оси. Расстояние от этой оси до линии действия F представлено вектором г. Произведение г и F представлено векторным произведением и показано на рис.2.6 как Т. Это произведение используется при определении вращающего момента. В этой связи важно то, что произведение (вращающий момент, Т) является вектором, направленным перпендикулярно плоскости, в которой лежат г и F. Например, если бы мышечная сила и плечо находились в плоскости данной страницы, то направление вращающего момента мышечной силы было бы перпендикулярным странице. Вектор был бы направлен либо к вам, либо от вас и зависел бы от направления г и F.
Существует несколько методов определения направления векторного произведения. Мы используем так называемое правило большого пальца правой руки. Как описывалось в главе 2, для определения направления вектора вращающего момента пальцы правой руки сгибаются в направлении вращения силы относительно оси, и вытянутый большой палец указывает направление вектора вращающего момента.
Сила — скользящий вектор. Хотя сила, действующая на жесткое тело, обычно приложена в определенной точке, вектор силы можно смещать вдоль линии ее действия, не изменяя механического воздействия силы на жесткое тело (Meriam, Kraige, 1987). В соответствии с этим принципом силу можно рассматривать как скользящий вектор. Поэтому вполне достаточно точно определить величину, направление и линию действия вектора силы, не заботясь об определении точки приложения силы.
Проведение кривой по точкам
В биомеханике очень часто исследуются функциональные зависимости одной переменной от другой. Например, мы определяем силу реакции поверхности в различные моменты времени, угол колена и соответствующий ему бедренный угол и др. В результате получается набор дискретных экспериментальных значений функций и аргумента.
При этом возникает задача аппроксимации экспериментальных данных какой-либо функциональной зависимостью. В качестве аппроксимирующих выражений часто используют конечные разности, полиноминальные функции, кубические сплайн-функции, цифровую фильтрацию и ряды Фурье (Wood, 1982). Они имеют следующий вид:
Конечные разности x(t) = > (2.3)
(2-4)
Полиноминальная функция /ч 2 п (2-5)
x(t) = aQ + att + а2г + + ant .
Цифровой фильтр
Х\пТ) = ааХ(пТ) + ахХ(пТ -Т) + а2Х(пТ - 2Т) + +Ь1Х\пТ-Т) + Ь2Х'(пТ~2Т) <2’6)
Ряды Фурье
х(0 = а0 + cos(2tu7 / T) + bx sin(2n/Y / Г)]. (2.7) 1=1
Для описания периодических зависимостей, часто встречающихся при исследовании движения, используется аппроксимация рядами Фурье.
Анализ Фурье
Переменный сигнал — это сигнал, изменяющийся во времени. Он может быть периодическим, полностью случайным или представлять собой сочетание первого и второго (рис.2.7). Кроме того, любой сигнал может включать постоянную составляющую (de), относительно которой переменный сигнал колеблется. Синусоидальная волна на рис.2.7 представляет собой низкочастотный сигнал в сравнении со случайным сигналом, который включает множество острых пиков. Кроме того, синусоидальная форма волны отражает единичную частоту, тогда как любой другой сигнал, особенно случайный сигнал на рис.2.7, содержит много частот (синусоидные и косинусоидные волны). На рис.2.8 эти сигналы показаны во временно'й (график амплитуда—время) и частотной (график амплитуда—частота) областях. В частотной области синусоидальная волна представлена единичной частотой данной амплитуды. В то же время частота случайного сигнала меняется в широком диапазоне от низкой// до высокой f2.
Анализ Фурье (преобразование Фурье), который может быть применен для периодических функций, включает представление анализируемой зависимости в виде суммы ряда членов, содержащих косинусы и синусы. Непереодический сигнал, однако, можно преобразовать в периодический и тоже подвергнуть анализу Фурье (Wood, 1982). Анализируя периодический сигнал, мы используем тригонометрические функции основной частоты (fc) и кратных ей величин. Основная частота — это частота главного члена ряда, содержащего синус и косинус и характеризующего изменения
336
ас-состовляющая
Рис.2.7. Периодический и случайный сигналы, изменяющиеся во времени (Winter, 1990):
а — периодический — синусоидная волна; б — периодический — пилообразная волна; в — случайный; г — периодический + случайный + de
сигнала на протяжении одного периода (рис.2.9). Члены ряда, представляющие колебания с кратной частотой называют гармониками высших порядков. Например, частота третьей гармоники 3 f0, девятой — 9f0. После определения основной частоты используют метод проведения кривой по точкам для определения коэффициентов гармонического многочлена, конечной части ряда Фурье, необходимого для приближенного представления сигнала
у(0 = а0 + £[а, cos(2tu7 / Т) + b, sin(2Tti7 / Г)] ,(2.8)
где у (Г) — исследуемая величина как функция времени; а0 — постоянная (средняя; обусловливает составляющую de); at, bt — коэффициенты ряда Фурье; i — номер гармоники; п — максимальный номер гармоники, учитываемый в разложении; t — время; Т — продолжительность одного периода.
Уравнение (2.8) используют для получения функциональной зависимости положения от времени. Продифференцировав его по времени получим уравнения (2.9) (функция скорость-время)
Рисунок 2.8. Преобразование сигналов Фурье из временнуй области в частотную (Winter, 1990)
22 8-133
337
Рис.2.9. Первые пять гармоник ряда Фурье
МО = sin(2n/Y / 7)(2л< /Т) + м (2.9)
+[/>, cos(2tuY / Т)(2тй / Г)]}.
Таким же образом дифференцируется по времени уравнение (2.9) для получения функции ускорение—время:
п
у(0 = ЁП-а, Sin(2nz7 / Т)(2та /Т2) +
1=1 (2.10)
+[-/>, sin(2nrt / Г)(2л/ / Г2)]}.
Определение составляющих уравнения (2.8) включает два этапа.
Первый — определение основной частоты. Понятие “частота” означает количество колебаний в секунду и измеряется в герцах (Гц). Частота 1 Гц характеризует колебание, которое совершается в течение 1 с. Один период соответствует одному полному обороту, т.е. 2 л рад (360°). Основная частота определяется сложением членов, имеющих синус и косинус этой частоты (cos 2 л t/Тк sin 2 л t/Т), где Т— продолжительность периода.
Для увеличения точности аппроксимации экспериментальных данных необходимо к фундаментальной частоте прибавить гармоники более высокой частоты.
Например, выражение 2 л it[T, где / колеблется от 1 до 5, обусловливает генерацию 5 из этих гармоник (рис.8.9). Обратите внимание, что
частота колебаний четвертой гармоники в четыре раза выше основной.
На втором этапе необходимо оценить вклад каждой из гармоник в функцию. В общем вклад снижается с увеличением номера гармоники. Вклад гармоники в выражение ряда Фурье определяется с помощью коэффициентов (й0, О/, Ь,). Эти коэффициенты определяются проведением кривой по точкам с использованием метода наименьших квадратов. На рис.2.10 показан вклад первых пяти гармоник с учетом соответствующих коэффициентов. Кумулятивный эффект суммирования первых пяти гармоник иллюстрирует рис.2.11.
Ниже приводятся коэффициенты и углы (радианы), на основании которых построены графики:
а0 = 4,8332
а{ = -0,4936 bi = -0,0478 угол! = 12,414
а2 = -0,0285 -0,0650 угол2 = 24,828
а3 = 0,0178 Z>3 — 0,0942 угол3 = 37,241
аА = 0,0195 b4 = 0,0105 угол4 = 49,655
а5 = 0,0074 bs = 0,0046 угол5 = 62,069
Цифровой фильтр
Другим методом анализа, используемым для проведения кривой по точкам, является цифровой (дискретный) фильтр (см.главу 2 — Winter, 1990). В отличие от анализа Фурье цифровой фильтр не позволяет получить уравнение сиг
338
нала. Вместе с тем подобно анализу Фурье он дает возможность сгладить сигнал и избавиться от ненужных составляющих. Как правило, эти компоненты возникают в результате ошибок в измерениях и не позволяют точно оценить данные (Winter, Sidwall, Hobson, 1974).
Выше мы рассматривали частотное содержание сигналов (анализ Фурье) и то, как каждый сигнал может быть представлен фундаментальной частотой и ее гармониками. Гармоники высшего порядка обусловливают содержание высокой частоты (острые края и пики) сигнала. Когда мы преобразуем видеозапись движения в цифровые данные, возможны ошибки вследствие таких влияний, как вибрация видеокамеры, неправильное расположение курсоров и т.п. Поскольку эти ошибки случайны, они представляют высокочастотный компонент сигнала, поэтому одна из целей методов сглаживания, на
пример, анализа Фурье, цифровой фильтрации — устранение этих ошибок—помех из сигнала. Рис.2.12 иллюстрирует методы сглаживания.
Основной этап метода цифровой фильтрации — определение частоты, разделяющей полезный сигнал и шумы; это — граничная, или пороговая, частота. К сожалению, этот этап связан со значительными трудностями, поскольку частота нужного сигнала и шумов в определенной степени “накладывается” друг на друга (рис.2.13, а). Цифровой фильтр представляет собой цифровой метод преобразования частоты спектра поступающего сигнала с целью получения выходного сигнала, включающего колебания с частотой выше/I со значительно пониженной мощностью. Влияние цифрового фильтра показано на рис.2.13, б. Граничная частота — частота, при которой мощность сигнала снижена по сравнению с первоначальным
Пятая
Четвертая
1+2+3+4+5 гармоник
1+2+3+4+5+6+7 гармоник
Рис.2.10. Влияние весовых коэффициентов на вклад первых пяти гармоник:
о — первые 5 гармоник; б — соответствие между данными и тригонометрической функцией, включающей 3, 5 либо 7 гармоник
22*
339
1 +2+3+4 гармоники
1+2+3+4+5 гармоник
Рис. 2.11. Кумулятивный эффект вклада первых пяти гармоник в ряд Фурье
значением вдвое. Возможную эффективность данного метода иллюстрирует рис.2.13, в.
Обычно используемый цифровой фильтр (фильтр Баттеруорта второго порядка) имеет следующий вид:
Х'(пТ) = а0Х(пТ) + ахХ(пТ -Т) + +а2Х(пТ - 2Т) + ЬхХ1(пТ - Т) + b2Xl(nT - 2Т),
где X— неотфильтрованные (ввод) исходные данные; X1 — отфильтрованные выходные данные; Т — временной интервал между измерениями; Х(пТ) — фильтруемая величина в момент времени пТ; а0, ах, а2, Ьх, Ь2 — коэффициенты фильтра.
Из этого выражения следует, что выходной сигнал [А1 (л 7)] — взвешенное среднее текущих
Время, с
Рис.2.12. Ускорение маркера на пальце ноги при ходьбе. Колеблющаяся линия была получена по данным положение—время. Более ровная пунктирная линия получена на основании эгих же данных после прохождения через цифровой фильтр
340
Частота
Рис.2.13. Влияние цифрового фильтра на частотное содержание сигнала:
а — исходный сигнал содержит как нужную информацию (сигнал), так и шум; шум обычно наблюдается в участке высокой частоты графика частота—амплитуда; б — влияние цифрового фильтра на амплитуду различных частот; при пороговой частоте (fc) мощность исходного сигнала снижена вдвое; в — график амплитуда—частота сигнала после прохождения через цифровой фильтр
и предыдущих неотфильтрованных данных плюс взвешенный вклад предыдущего отфильтрованного выходного сигнала.
Чтобы использовать цифровой фильтр, необходимо определить коэффициенты (а0, а2,
Ьь b2). Однако прежде всего необходимо определить fc. Для этой цели лучше всего воспользоваться методом остаточного анализа (Winter, 1990). Он предполагает сопоставление различия (остатка) между отфильтрованными и неотфиль-трованными сигналами в большом диапазоне граничных частот и выбор fc, при которой наблюдается минимальное искажение сигнала и минимальное количество шума. Для большинства движений человека наиболее оптимальный показатель fc — (> Гц. Получив его, можно определить коэффициенты, вычислив отношение выборочной частоты fxf.. Для fc = 6 Гц при использовании видеоданных, полученных при частоте 60 Гц, отношение равно 10, а коэффициенты таковы:
а0 = 0,06746;
а, = 0,13491;
а2 = 0,06746;
Ь, = 1,14298;
Ь2 = -0,41280.
Коэффициенты цифрового фильтра для различных соотношений fs,fc можно найти у Winter (1990).
Последний этап — корректировка нежелательного искажения отфильтрованных данных, обусловленного цифровым фильтром. Этот эффект представляет собой фазовое искажение, которое можно наблюдать как смещение синусоидной волны вдоль горизонтальной оси; если один цикл равен 2л рад, то искажение фазы л/2 рад равно смещению синусоидной волны на 1/4 периода. Эта величина искажения обусловлена фильтром Батгеруорта. Чтобы устранить искажение, необходимо пропустить данные через цифровой фильтр дважды — один раз в направлении вперед, второй раз — в обратном направлении.
ПРИЛОЖЕНИЕ 3
КОЭФФИЦИЕНТЫ ПРЕОБРАЗОВАНИЯ
Ускорение
1 сантиметр/секунда/секунда (см • с-2)
1 сантиметр/секунда/секунда (см • с-2)
1 фут/секунда/секунда (фут • с' 2)
1 фут/секунда/секунда (фут • с~2)
1 фут/секунда/секунда (фут • с~2)
1 километр/час/секунда (км • ч'1 • с-1)
1 километр/час/секунда (км • ч-1 • с-1)
1 метр/секунда/секунда (м • с-2)
1 миля/час/секунда
1 оборот/минута/минута
1 оборот/секунда/секунда
1 акр
1 акр
1 ар
1 цента
1 гектар (га)
1 квадратный сантиметр (см2)
1 квадратный сантиметр (см2)
1 градус в квадрате
1 квадратный дюйм (дюйм2)
1 квадратный дюйм (дюйм2)
1 квадратный фут (фут2)
1 квадратный фут (фут2)
1 квадратный фут (фут2)
1 квадратный дюйм (дюйм2)
1 квадратный километр (км2)
1 квадратный метр (м2)
1 квадратный метр (м2)
1 квадратная миля (миля2)
1 квадратный миллиметр (мм2)
1 квадратный ярд (ярд2)
1 квадратный ярд (ярд2)
= 0,036 километров/час/секунду (км ч'1 • с-1)
= 0,01 метр/секунда/секунда (м • с“2)
= 30,48 сантиметров/секунда/секунда (см • с-2)
= 1,097 километров/час/секунда (км • ч'1 • с-1)
= 0,3048 метр/секунда/секунда (м • с“2)
= 27,78 сантиметр/секунда/секунд (см • с-2)
= 0,2778 метр/секунда/секунда (м • с-2)
= 100 сантиметров/секунда/секунд (см • с-2)
= 0,447 метр/секунда/секунда (м • с~2)
= 0,001745 радиан/секунда/секунда (рад-с-2)
= 6,283 радианов/секунда/секунда (рад • с~2)
Площадь
= 4,047 квадратных метров (м2)
= 0,4047 гектар (га)
= 100 квадратных метров (м2)
= 1 квадратный метр (м2)
= 10 000 квадратных метров (м2)
= 0,0001 квадратный метр (м2)
= 100 квадратных миллиметров (мм2)
= 0,00030462 стерадиана (стер)
= 6,4516 квадратных сантиметров (см2)
= 645,16 квадратных миллиметров (мм2) = 929 квадратных сантиметра (см2)
= 0,092903 квадратных метров (м2)
= 92,900 квадратных миллиметров (мм2) = 6,452 квадратных сантиметров (см2) = 1 000 000 квадратных метров (м2)
= 0,0001 гектар (га)
= 10 000 квадратных сантиметров (см2) = 2,590 квадратных километров (км2) = 0,01 квадратный сантиметр (см2) = 8,361 квадратных сантиметров (см2) = 0,836127 квадратных метров (м2)
342
Плотность
1 фунт/кубический фут (фунт/фут3)
1 слаг/кубический фут (слаг/фут3)
1 фунт/галлон (Великобритания)
1 фунт/галлон (США)
16,01846 килограмм/кубический метр (кг • 515,3788 килограмм/кубический метр (кг-99,77633 килограмм/кубический метр (кг •
199,8264 килограмм/кубический метр (кг •
2 2 2, 2,
Электричество
1 био
1 ампер/квадратный дюйм (а • дюйм-2)
1 фарадей/секунда
1 фарадей
1 mho
= 10 ампер (А)
= 1 550 ампер/квадратный метр (А • м-2)
= 96 490 ампер (А)
= 96 490 кулонов (кулон = ампер • с)
= 1 симен (S)
Энергия и работа
1 британская тепловая единица 1 британская тепловая единица 1 британская тепловая единица 1 эрг 1 фуг = паундаль 1 фунт-силафут 1 грамм-сантиметр 1 лошадиная сила-час 1 лошадиная сила-час 1 килокалория (международная) 1 килокалория 1 килопонд-метр 1 киловатт-час = 1 055 джоуль (Дж) = 1,0548 килоджоулей (кДж) = 0,0002928 киловатт-час (кВт-ч) = 0,0001 миллиджоуль (мДж) = 0,04214 джоулей (Дж) = 1,355818 джоулей (Дж) = 0,09807 миллиджоуль (мДж) = 2 684 килоджоулей (кДж) = 0,7457 киловатт-час (кВт-ч) = 4,1868 килоджоулей (кДж) = 4 183 джоулей (Дж) = 9,807 джоулей (Дж) = 3 600 килоджоулей (кДж) Сила
1 дина 1 фут-фунт 1 фут-фунт/секунда (фут-фунт -с-1) 1 грамм (г) 1 килограмм-сила (кг-сила) 1 килопонд 1 паундель 1 фунт-сила (фунт-сила) 1 стоун (вес) 1 длинная тонна 1 метрическая тонна = 0,01 миллиньютон (мН) = 1,356 джоулей (Дж) = 0,001356 киловатт (кВт) = 9,807 миллиньютонов (мН) = 9,807 ньютонов (Н) = 9,807 ньютонов (Н) = 0,138255 ньютонов (Н) = 4,448222 ньютонов (Н) = 62,275 ньютонов (Н) = 9 964 ньютонов (Н) = 9 807 ньютонов (Н) Длина (угол)
1 ангстрем (А) 1 болт 1 сантиметр (см) 1 сантиметр (см) 1 сантиметр (см) 1 чейн 1 обхват 1 градус 1 фатом 1 фут = 0,0001 микрометр (мкм) = 36,576 метров (м) = 0,00001 километр (км) = 0,01 метр (м) = 10 миллиметров (мм) = 20,12 метров (м) = 6,283 радиана (рад) = 0,001745 радианов (рад) = 1,8288 метров (м) = 30,48 сантиметров (см)
343
1 фут = 0,3048 метров (м)
1 фут = 304,8 миллиметров (мм)
1 фарлонг = 201,17 метров (м)
1 хенд = 10,16 сантиметров (см)
1 дюйм = 2,54 сантиметра (см)
1 дюйм = 0,0254 метра (м)
1 дюйм = 25,4 миллиметра (мм)
1 световой год = 9 460 910 000 000 километров (9,46 х 1012км)
1 миля (морская) = 1,853 километра (км)
1 миля (морская) = 1 853 метра (м)
1 миля (статутная) = 1, 609 километра (км)
1 миля (статутная) = 1 609 метров (м)
1 минута (угол) = 0,0002909 радиана (рад)
1 оборот = 6,283 радиана (рад)
1 род = 5,029 метров (м)
1 сфера (пространственный угол) = 12,57 стерадианов (стер)
1 ярд = 91,44 сантиметров (см)
1 ярд = 0,9144 метров (м) Сила света
1 свеча/квадратный сантиметр = 10 000 свеча/квадратный метр (свеча • м-2)
1 свеча/квадратный фут = 10,76 свеча/квадратный метр (свеча • м~2)
1 фут = свеча = 10,764 люмен/квадратный метр (люмен • м-2)
1 фут = ламберт = 3,426 свеча/квадратный метр (свеча • м“2)
1 ламберт (L) = 3 183 свеча/квадратный метр (свеча *м-2)
1 фот = 10 000 люмен/квадратный метр (люмен • м-2) Момент инерции
1 слаг-квадратный фут = 1,35582 килограмм-квадратный метр (кг • м2)
1 фунт-квадратный фут = 0,04214 килограмм-квадратный метр (кг • м2) Масса
1 центнер = 50,8 килограмма (кг)
1 унция = 28,3495 грамма (г)
1 фунт = 0,453592 килограмма (кг)
1 слаг = 14,59 килограмма (кг)
1 тонна = 1 016 килограмма (кг)
Мощность
1 британская тепловая единица/час
1 британская тепловая единица/минута
1 калория/секунда
1 эрг/секунда
1 фут-фунт/секунда
1 фуг-фунт/секунда
1 грамм-калория
1 лошадиная сила (Великобритания)
1 лошадиная сила (метрическая)
1 килокалория/минута (ккал/мин)
1 килограмм-метр/секунда
1 килопонд-метр/минуга (кпм/мин)
= 0,2931 ватт (Вт)
= 17,57 ватт (Вт)
= 4,187 ватт (Вт)
= 10 000 000 мегаватт (МВт)
= 1,356 ватт (Вт)
= 0,04214 ватт (Вт)
= 0,001162 ватт-час (Вт • ч)
= 745,7 ватт (Вт)
= 735,5 ватт (Вт)
== 69,767 ватт (Вт)
— 9,807 ватт (Вт)
= 0,1634 ватт (Вт)
344
Давление
1 атмосфера
1 атмосфера
1 бар
1 сантиметр ртути
1 сантиметр воды
1 сантиметр воды
1 дина/квадратный сантиметр
1 фуг воды
1 дюйм ртути
1 дюйм воды (4°С)
1 килограмм-сила/квадратный метр
1 миллибар
1 миллиметр ртути (1 торр)
1 фунт-сила/квадратный фут
1 фунт-сила/квадратный дюйм
1 паундаль/квадратный фут
= 760 миллиметров ртути (мм рт.ст. при 0°С)
= 101,340 паскалей (Па)
= 100,031 паскалей (Па)
= 1 333,224 паскалей (Па)
= 0,738 миллиметров ртути (мм рт.ст.)
= 98,3919 паскалей (Па)
= 0,10 паскаля (Па)
= 2 989 паскалей (Па)
= 3 386 паскалей (Па)
= 249 паскалей (Па)
= 9,807 паскалей (Па)
= 100 паскалей (Па)
= 133,322387 паскалей (Па)
= 47,88026 паскалей (Па)
= 6,8948 килопаскалей (кПа)
= 1,488 паскалей (Па)
Температура
1 градус Цельсия (°C)
1 градус Цельсия (°C)
= (°C х 9/5) + 32 градуса по шкале Фаренгейта (°Ф) = °C + 273,18 Кельвина
Вращающий момент
1 фут-фунт
1 килопонд-метр
= 1,356 ньютон-метр (Н • м)
= 9,807 ньютон-метр (Н • м)
Скорость
1 сантиметр/секунда (см • с-1)
1 сантиметр/секунда (см • с-1)
1 фут/минута (фут • мин-1)
1 фут/минута (фут • мин-1)
1 фут/минута (фут • мин-1)
1 фут/секунда (фут • с-1)
1 фут/секунда (фут • с-1)
1 фут/секунда (фут • с-1)
1 фут/секунда (фут • с-1)
1 километр/час (км • час-1)
1 километр/час (км • ч-1)
1 узел
1 узел
1 узел
1 метр/минута (м • мин-1)
1 метр/минута (м • мин-1)
1 метр/секунда (м • с-1)
1 метр/секунда (м-с-1)
1 миля/час (миля • ч-1)
1 миля/час (миля • ч-1)
1 миля/час (миля • ч-1)
1 миля/минута (миля • мин-1)
1 миля/минута (миля • мин-1)
1 оборот/минута (об • мин-1)
1 оборот/секунда (об - с-1)
= 0,036 километр/час (км • ч-1)
= 0,6 метр/минута (м • мин-1)
= 0,508 сантиметр/секунда (см • с-1)
= 0,01829 километр/час (км-ч-1)
= 0,3048 метр/минута (м • мин-1)
= 30,48 сантиметр/секунда (см • с-1)
= 1,097 километр/час (км • ч-1)
= 18,29 метр/минута (м-мин-1)
= 0,3048 метр/секунда (м • с-1)
= 16,67 метр/минута (м • мин-1)
= 0,2778 метр/секунда (м-с-1)
= 1,8532 километр/час (км-ч-1)
= 51,48 сантиметр/секунда (см • с-1)
= 0,5155 метр/секунда (м • с-1)
= 1,667 сантиметр/секунда (см -с-1)
= 0,06 километр/час (км - ч-1)
= 3,6 километр/час (км • ч-1)
= 0,06 километр/минута (км • мин-1)
= 44,7 сантиметр/секунда (см • с-1)
= 1,6093 километр/час (км • ч-1)
= 0,447 метр/секунда (м • с-1)
= 2 682 сантиметр/секунда (см • с-1)
= 1,6093 километр/минута (км • мин-1)
= 0,1047 радиан/секунда (рад • с-1)
= 6,283 радиан/секунда (рад-с-1)
345
Объем
1 баррель (Великобритания)
1 баррель (США)
1 бушель (Великобритания)
1 бушель (США)
1 бушель (США)
1 кубический сантиметр (см3)
1 кубический сантиметр (см3)
1 кубический фут (фут3)
1 кубический фут (фут3)
1 кубический фуг/минута (фут3 • мин-1)
1 кубический фут/минуга (фут3 • мин-1)
1 кубический дюйм (дюйм3)
1 кубический дюйм (дюйм3)
1 кубический дюйм (дюйм3)
1 кубический метр (м3)
1 кубический метр (м3)
1 кубический ярд (ярд3)
1 кубический ярд (ярд3)
1 кубический ярд/минута (ярд3 • мин-1)
1 драхма
1 галлон (США)
1 галлон (США)
1 галлон
1 галлон/минута
1 джилл (Великобритания)
1 джилл (США)
I джилл (США)
1 литр (л)
1 литр (л)
1 унния (жидкая, США)
1 унция (жидкая, Великобритания)
1 унция (жидкая, США)
1 пек (Великобритания)
1 пек (США)
1 пинта
1 пинта
1 кварта
1 кварта
= 0,1637 кубических метров (м3)
= 0,11921 кубических метров (м3)
= 0,03637 кубических метров (м3)
= 0,03524 кубических метров (м3)
= 35,24 литров (л)
= 0,000001 кубических метров (м3)
= 0,001 литра (л)
= 0,02832 кубических метров (м3)
= 28,32 литра (л)
= 472 кубических сантиметров/секунда (см3 • с-1)
= 0,472 литр/секунда (л • с-1)
= 16,387 кубических сантиметров (см3)
— 0,000016 кубических метров (м3)
= 0,0164 литров (л)
= 1 000 000 кубических сантиметров (см3)
= 1000 литров (л)
= 0,7646 кубических метров (м3)
= 764,6 литров (л)
= 12,74 литров/секунда (л • с-1)
= 3,6967 кубических сантиметров (см3)
= 3785 кубических сантиметров (см3)
= 0,003785 кубических метров (м3)
= 3,785 литров (л)
= 0,06308 литров/секунду (л • с-1)
= 142,07 кубических метров (см3)
= 118,295 кубических сантиметров (см3)
= 0,1183 литров (л)
= 1000 кубических сантиметров (см3)
= 0,001 кубических метров (м3)
= 0,02957 кубических метров (м3)
= 28,413 кубических сантиметров (см3)
= 29,573 кубических сантиметров (см3)
= 9,0919 литров (л)
= 8,8096 литров (л)
= 473,2 кубических сантиметров (см3)
= 0,4732 литров (л)
= 946,4 кубических сантиметров (см3)
= 0,9463 литров (л)
ПРИЛОЖЕНИЕ 4
ПАРАМЕТРЫ СЕГМЕНТОВ ТЕЛА
Во многих биомеханических исследованиях используют организацию сегментов тела. Chandler et al. (1975) используют 14 сегментов: голова, туловище, плечи, предплечья, кисти, бедра, голени (или ноги) и ступни (рис.4.1).
Zatsiorsky и Seluynov (1983) разделяют тело на 16 сегментов: голова, верхняя часть туловища, средняя и нижняя часть туловища, плечи, предплечья, кисти, бедра, голени и ступни (4.2).
Рис.4.1 Деление туловища на сегменты по Chandler et al. (1975)
Рис.4.2. Деление туловища на сегменты по Zatsiorsky и Seluynov (1983)
347
ПРИЛОЖЕНИЕ 5
КИНЕМАТИЧЕСКИЕ ДАННЫЕ
Приложение содержит три набора кинематических данных, которые могут быть использованы учащимися при выполнении упражнений:
1. Таблица Д.1. Положение выбранных сегментов тела и штанги при взятии веса на грудь (тяжелая атлетика).
2. Таблица Д.2. Угол рук относительно горизонтальной точки при выполнении сальто вперед с разбега (гимнастика).
3. Таблица Д.З. Измерение угла коленного и локтевого суставов у бегуна во время выполнения одного шага.
Таблица 5.1. Положение избранных «ориентиров» при чистом взятии веса на грудь
Время, Плюсна Лодыжка Колено Бедро Голова Плечо Локоть Штанга
МС х* у х У х у х У х У X у х У х У
10 0,79 -0,56 0,69 -0,47 0,90 -0,13 0,45 -0,06 1,07 0,34 0,92 0,25 0,89 0,01 0,90 -0,27
20 0,79 -0,56 0,69 -0,47 0,90 -0,13 0,45 -0,05 1,07 0,34 0,92 0,25 0,89 0,01 0,90 -0,27
30 0,79 -0,56 0,69 -0,47 0,90 -0,13 0,44 -0,05 1,06 0,34 0,92 0,25 0,89 0,01 0,90 -0,27
40 0,79 -0,56 0,69 -0,47 0,89 -0,13 0,44 -0,04 1,06 0,35 0,92 0,25 0,89 0,01 0,90 -0,27
50 0,79 -0,56 0,69 -0,47 0,89 -0,13 0,44 -0,04 1,06 0,35 0,92 0,26 0,89 0,01 0,90 -0,27
60 0,79 -0,56 0,69 -0,47 0,89 -0,13 0,44 -0,03 1,06 0,35 0,92 0,26 0,89 0,02 0,90 -0,26
70 0,79 -0,56 0,69 -0,47 0,88 -0,13 0,44 -0,03 1,06 0,35 0,92 0,26 0,89 0,02 0,90 -0,26
80 0,79 -0,56 0,69 -0,47 0,88 -0,12 0,44 -0,02 1,06 0,36 0,92 0,26 0,90 0,02 0,90 -0,26
90 0,79 -0,56 0,69 -0,47 0,88 -0,12 0,44 -0,01 1,07 0,36 0,92 0,26 0,90 0,02 0,89 -0,26
100 0,79 -0,56 0,69 -0,47 0,87 -0,12 0,43 -0,00 1,07 0,36 0,92 0,27 0,90 0,02 0,89 -0,26
ПО 0,80 -0,56 0,69 -0,47 0,87 -0,12 0,43 0,00 1,07 0,37 0,92 0,27 0,90 0,03 0,89 -0,25
120 0,80 -0,56 0,69 -0,47 0,87 -0,12 0,43 0,01 1,07 0,37 0,92 0,27 0,90 0,03 0,89 -0,25
130 0,80 -0,56 0,69 -0,47 0,86 -0,12 0,43 0,02 1,07 0,37 0,92 0,28 0,90 0,03 0,89 -0,25
140 0,80 -0,56 0,69 -0,47 0,86 -0,11 0,43 0,03 1,07 0,38 0,92 0,28 0,90 0,04 0,89 -0,24
150 0,80 -0,56 0,69 -0,47 0,86 -0,11 0,43 0,04 1,07 0,38 0,92 0,28 0,90 0,04 0,89 -0,24
160 0,80 -0,56 0,69 -0,47 0.86 -0,11 0,43 0,04 1,07 0,39 0,92 0,29 0,90 0,04 0,89 -0,23
170 0,80 -0,56 0,69 -0,47 0,85 -0,11 0,43 0,05 1,07 0,39 0,92 0,29 0,90 0,05 0,89 -0,23
180 0,80 -0,56 0,69 -0,47 0,85 -0,11 0,43 0,06 1,07 0,39 0,92 0,30 0,90 0,05 0,88 -0,22
190 0,80 -0,56 0,69 -0,47 0,85 -0,11 0,43 0,07 1,07 0,40 0,93 0,30 0,89 0,06 0,88 -0,22
200 0,80 -0,56 0,69 -0,47 0,84 -0,10 0,42 0,08 1,07 0,40 0,93 0,30 0,89 0,06 0,88 -0,22
210 0,80 -0,56 0,69 -0,47 0,84 -0,10 0,42 0,09 1,08 0,41 0,93 0,31 0,89 0,07 0,88 -0,21
220 0,80 -0,56 0,69 -0,47 0,84 -0,10 0,42 0,10 1,08 0,41 0,93 0,31 0,89 0,07 0,88 -0,21
230 0,80 -0,56 0,69 -0,47 0,83 -0,10 0,42 0,11 1,08 0,42 0,93 0,32 0,89 0,08 0,88 -0,20
240 0,80 -0,56 0,69 -0,47 0,83 -0,10 0,42 0,11 1,08 0,42 0,93 0,32 0,89 0,08 0,88 -0,20
250 0,80 -0,56 0,69 -0,47 0,82 -0,09 0,42 0,12 1,08 0,43 0,93 0,33 0,89 0,09 0,88 -0,19
260 0,80 -0,56 0,69 -0,47 0,82 -0,09 0,42 0,13 1,08 0,43 0,93 0,34 0,89 0,10 0,88 -0,18
270 0,80 -0,56 0,69 -0,47 0,8) -0,09 0,42 0,14 1,08 0,44 0,93 0,34 0,89 0,10 0,87 -0,18
280 0,80 -0,56 0,69 -0,47 0,81 -0,09 0,42 0,15 1,08 0,45 0,93 0,35 0,89 0,11 0,87 -0,17
290 0,80 -0,56 0,69 -0,47 0,80 -0,09 0,42 0,16 1,08 0,45 0,93 0,35 0,89 0,11 0,87 -0,16
300 0,80 -0,56 0,69 -0,47 0,80 -0,09 0,42 0,17 1,08 0,46 0,93 0,36 0,89 0,12 0,87 -0,16
310 0,80 -0,56 0,69 -0,47 0,79 -0,09 0,42 0,17 1,08 0,47 0,93 0,37 0,89 0,13 0,87 -0,13
320 0,80 -0,56 0,69 -0,47 0,79 -0,08 0,42 0,18 1,08 0,47 0,93 0,37 0,89 0,13 0,87 -0,13
330 0,80 -0,56 0,69 -0,47 0,78 -0,08 0,42 0,19 1,08 0,48 0,93 0,38 0,89 0,14 0,87 -0,12
340 0,79 -0,56 0,69 -0,47 0,78 -0,08 0,42 0,20 1,08 0,49 0,93 0,39 0,89 0,15 0,87 -0,11
350 0,79 -0,56 0,69 -0,47 0,77 -0,08 0,42 0,20 1,08 0,49 0,93 0,39 0,89 0,16 0,87 -0,10
360 0,79 -0,56 0,69 -0,47 0,77 -0,08 0,42 0,21 1,08 0,50 0,93 0,40 0,89 0,17 0,86 -0,09
370 0,79 -0,56 0,69 -0,47 0,77 -0,08 0,42 0,21 1,08 0,51 0,93 0,41 0,89 0,17 0,86 -0,09
380 0,79 -0,56 0,69 -0,47 0,76 -0,08 0,42 0,22 1,08 0,52 0,93 0,41 0,89 0,18 0,86 -0,08
390 0,79 -0,56 0,69 -0,47 0,76 -0,08 0,42 0,22 1,08 0,53 0,93 0,42 0,89 0,19 0,86 -0,07
400 0,79 -0,56 0,69 -0,47 0,76 -0,08 0,42 0,23 1,08 0,54 0,93 0,43 0,89 0,20 0,86 -0,06
410 0,79 -0,56 0,69 -0,47 0,75 -0,08 0,43 0,23 1,08 0,55 0,93 0,44 0,88 0,2) 0,86 -0,05
420 0,79 -0,56 0,69 -0,47 0,75 -0,08 0,43 0,24 1,08 0,56 0,93 0,45 0,88 0,22 0,86 -0,04
430 0,79 -0,56 0,69 -0,47 0,75 -0,08 0,43 0,24 1,08 0,57 0,93 0,46 0,88 0,23 0,86 -0,02
440 0,79 -0,56 0,69 -0,47 0,74 -0,08 0,44 0,24 1,07 0,58 0,93 0,47 0,88 0,24 0,86 -0,01
450 0,79 -0,56 0,69 -0,47 0,74 -0,08 0,44 0,24 1,07 0,60 0,93 0,48 0,88 0,25 0,85 -0,00
460 0,79 -0,56 0,69 -0,47 0,74 -0,08 0,44 0,24 1,07 0,61 0,92 0,49 0,88 0,26 0,85 0,01
470 0,79 -0,56 0,69 -0,47 0.73 -0,07 0,45 0,24 1,07 0,62 0,92 0,50 0,87 0,27 0,85 0,02
480 0,79 -0,56 0,69 -0,47 0,73 -0,07 0,46 0,24 1,06 0,64 0,92 0,51 0,87 0,29 0,85 0,03
490 0,79 -0,56 0,69 -0,47 0,73 -0,07 0,46 0,24 ,06 0,65 0,92 0,53 0,87 0,30 0,85 0,04
500 0,79 -0,56 0,69 -0,47 0,74 -0,08 0,47 0,24 1,05 0,67 0,91 0,54 0,87 0,31 0,85 0,06
510 0,79 -0,56 0,69 -0,47 0,74 -0,08 0,47 0,23 1,04 0,68 0,91 0,55 0,87 0,32 0,85 0,07
520 0,79 -0,56 0,69 -0,47 0,74 -0,08 0,48 0,23 1,03 0,70 0,90 0,56 0,86 0,33 0,85 0,08
530 0,79 -0,56 0,69 -0,47 0,75 -0,08 0,49 0,23 1,02 0,71 0,90 0,58 0,86 0,34 0,85 0,09
349
Продолжение табл. 5.1
Время, Плюсна Лодыжка Колено Бедро Голова Плечо Локоть Штанга
МС X* У X у X у х у х У х У X у х У
540 0,79 -0,56 0,69 -0,46 0,76 -0,08 0,50 0,22 1,01 0,73 0,89 0,59 0,86 0,35 0,85 0,10
550 0,79 -0,56 0,69 -0,46 0,76 -0,08 0,50 0,22 1,00 0,75 0,89 0,60 0,86 0,37 0,85 0,11
560 0,79 -0,56 0,69 -0,46 0,78 -0,08 0,53 0,21 0,98 0,76 0,88 0,61 0,85 0,38 0,85 0,13
570 0,79 -0,56 0,69 -0,46 0,78 -0,08 0,54 0,20 0,96 0,80 0,87 0,63 0,85 0,39 0,85 0,14
580 0.79 -0,56 0,69 -0,46 0,78 -0,08 0,54 0,20 0,95 0,81 0,86 0,64 0,85 0,40 0,85 0,15
590 0,79 -0,56 0,69 -0,46 0,79 -0,08 0,56 0,19 0,93 0,83 0,85 0,66 0,84 0,41 0,85 0,16
600 0,79 -0,56 0,70 -0,46 0 80 -0,07 0,57 0,19 0,90 0,84 0,84 0,67 0,84 0,42 0,85 0,18
610 0,79 -0,56 0,70 -0,46 0,80 -0,07 0,59 0,19 0,88 0,86 0,83 0,68 0,83 0,44 0,86 0,19
620 0,79 -0,56 0,70 -0,46 0,80 -0,07 0,60 0,18 0,86 0,87 0,82 0,70 0,82 0,45 0,86 0,20
630 0,79 -0,56 0,70 -0,46 0,81 -0,07 0,62 0,18 0,84 0,88 0,81 0,71 0,82 0,46 0,86 0,22
640 0,79 -0,56 0,70 -0,46 0,81 -0,06 0,64 0,19 0,81 0,90 0,79 0,72 0,81 0,47 0,86 0,23
650 0,79 -0,56 0,70 -0,46 0,81 -0,06 0,65 0,19 0,79 0,91 0,78 0,74 0,81 0,49 0,86 0,24
660 0,79 -0,56 0,70 -0,46 0,81 -0,06 0,66 0,20 0,77 0,92 0,76 0,75 0,80 0,50 0,86 0,26
670 0,79 -0,56 0,70 -0,46 0,81 -0,06 0,67 0,21 0,74 0,93 0,75 0,76 0,79 0,51 0,86 0,27
680 0,79 -0,56 0,70 -0,46 0,80 -0,05 0,68 0,22 0,72 0,94 0,74 0,77 0,79 0,53 0,86 0,29
690 0,79 -0,56 0,70 -0,46 0,80 -0,05 0,69 0,2'3 0,71 0,94 0,73 0,78 0,78 0,54 0,87 0,29
700 0,79 -0,56 0,71 -0,45 0,79 -0,04 0,70 0,24 0,69 0,95 0,71 0,79 0,78 0,55 0,87 0,30
710 0,79 -0,56 0,71 -0,45 0,79 -0,04 0,70 0,24 0,67 0,96 0,70 0,80 0,77 0,56 0,87 0,30
720 0,79 -0,56 0,71 -0,45 0,79 -0,03 0,70 0,25 0,65 0,96 0,69 0,81 0,77 0,57 0,87 0,32
Данные соответствуют переднезаднему (х) и вертикальному (у) положению избранных «ориентиров» при поднятии штанги с пола до уровня бедер.
* Все показатели х и у указаны в метрах.
Таблица 5.2. Угол рук относительно горизонтальной точки во время выполнения сальто вперед с разбега
Время, мс Испытуемый
10 5 3 Н 2 7 1 8 4
10 0,94 1,08 1,01 0,40 0,47 -0,80 1,01 0,87 0,63 0,89
20 1,03 1,13 1,08 0,49 0,54 -0,84 1,07 0,94 0,72 0,94
30 1,12 1,20 1,15 0,56 0,61 -0,86 1,13 1,00 0,79 1,01
40 1,19 1,26 1,22 0,65 0,68 -0,89 1,19 1,07 0,86 1,07
50 1,27 1,33 1,29 0,72 0,75 -0,93 1,26 1,13 0,93 1,12
60 1,36 1,40 1,36 0,80 0,82 -0,96 1,33 1,20 1,00 1,19
70 1,43 1,45 1,43 0,87 0,89 -1,00 1,38 1,27 1,07 1,24
80 1,52 1,52 1,50 0,96 0,96 -1,01 1,45 1,33 1,13 1,39
90 1,59 1,59 1,57 1,03 1,03 -1,05 1,52 1,40 1,20 1,36
100 1,68 1,66 1,64 1,12 1,10 -1,07 1,59 1,47 1,27 1,43
ПО 1,75 1,73 1,71 1,19 1,19 -1,10 1,66 1,54 1,34 1,50
120 1,82 1,78 1,78 1,27 1,26 -1,12 1,73 1,59 1,41 1,57
130 1,89 1,85 1,85 1,36 1,33 -1,12 1,80 1,66 1,48 1,62
140 1,96 1,92 1,92 1,43 1,41 -1,13 1,87 1,73 1,55 1,69
150 2,02 1,99 1,99 1,52 1,50 -1,13 1,94 1,80 1,62 1,76
160 2,08 2,04 2,06 1,59 1,57 -1,13 2.01 1,85 1,69 1,83
170 2,15 2,11 2,13 1,68 1,66 -1.13 2,06 1,92 1,76 1,90
180 2,20 2,16 2,18 1,75 1,75 -1,13 2,13 1,97 1,82 1,97
190 2,25 2,22 2,25 1,83 1,82 -1,12 2,18 2,04 1,89 2,02
200 2,29 2,27 2,30 1,90 1,90 -1,10 2,25 2,09 1,94 2,09
210 2,34 2,32 2,36 1,97 1,97 -1,08 2,30 2,16 1,99 2,16
220 2,37 2,36 2,43 2,04 2,06 -1,07 2,36 2,22 2,06 2,23
230 2,43 2,41 2,48 2,11 2,13 -1,03 2,41 2,27 2,11 2,29
240 2,46 2,44 2,53 2,18 2,20 -1,01 2,46 2,32 2,16 2,36
250 2,50 2,48 2,58 2,25 2,27 -0,98 2,51 2,37 2,22 2,41
260 2,53 2,53 2,65 2,30 2,36 -0,94 2,57 2,43 2,27 2,48
270 2,57 2,57 2,71 2,37 2,43 -0,91 2,60 2,48 2,30 2,53
280 2,60 2,60 2,76 2,43 2,50 -0,87 2,65 2,53 2,36 2,60
290 2,64 2,64 2,83 2,48 2,55 -0,86 2,71 2,58 2,41 2,65
300 2,67 2,67 2,88 2,53 2,62 -0,82 2,74 2,64 2,44 2,71
310 2,69 2,71 2,95 2,57 2,69 -0,79 2,79 2,69 2,50 2,78
320 2,72 2,74 3,02 2,62 2,76 -0,75 2,85 2,74 2,55 2,83
330 2,76 2,78 2,67 2,81 -0,73 2,79 2,58
340 2,79 2,81 2,71 2,88 -0,70 2,85 2,64
350 2,83 2,85 2,95 2,90 2,67
360 3,02 2,95 2,71
3,00
Данные соответствуют углу плечевого сустава рук относительно правой горизонтальной линии. В каждом столбике — 2 маркера (подчерки.) в начале и в конце. Первый указывает момент контакта с силовой платформой, второй — момент отталкивания.
Таблица 5.3. Измерение угла колена и плеча у бегуна при выполнении одного шага
Время, мс Угол колена, рад Угол плеча, рад Действие
0 2,48 1,97
13 2,54 1,91 1FS
25 2,70 1,92
38 2,49 1,87
50 2,46 1,84
63 2,36 1,86
76 2,34 1,80
88 2,39 1,77
101 2,46 1,54
114 2,66 1,39
126 2,72 1,40 1ТО
139 2,57 1,40
152 2,33 1,40
164 2,02 1,51
176 1,80 1,75
189 1,50 1,96
202 1,33 2,16
214 1,17 2,36
227 1,00 2,44
239 0,78 2,54 rFS
252 0,68 2,43
265 0,61 2,40
277 0,68 2,32
290 0,75 2,21
302 0,75 2,17
315 0,82 2,20
328 0,95 2,08
340 1.U 1,97
353 1,29 1,86 гТО
365 1,55 1,82
378 1,76 1,75
391 2,04 1,62
403 2,18 1,63
416 2,38 1,79
428 2,40 1,80
441 2,47 1,92
454 2,54 1,82 1FS
466 2,53 1,97
479 2,54 1,94
Данные приведены для левой части тела бегуна 1FS — касание левой ногой; 1ТО — отталкивание левой ногой; rFS — касание правой ногой; гТО — отталкивание правой ногой.
ПРИЛОЖЕНИЕ 6
СОКРАЩЕНИЯ
а — ускорение ЕРР - потенциал концевой
а — среднее ускорение пластинки
Ох — ускорение в направлении х G - гига
ау — ускорение в направлении у G - количество движения
az — ускорение в направлении z С-актин+ — глобулярный актин
А — проектируемая площадь g — проводимость
А — ампер gCa — проводимость кальция
А-полоска — полоска анизотропная £к — проводимость калия
Альфа-ГЦФ — альфа-глицерофосфат gNa — проводимость натрия
дегидрогеназа н - тепловая энергия
АМГ — акустический миограф н - угловой момент
АНР — постгиперполяризация н+ - ион водорода
АР — потенциал действия НВ1/° — кинетический момент
АТФ — аденозин трифосфат сегментов тела I относительно
АЦХ — ацетилхолин его центра тяжести
ВПСП — возбуждающий pfCI/CS _ кинетический момент центра
постсинаптический потенциал тяжести сегмента тела I
ВТ — вес тела относительно центра тяжести
Вт — ватт системы
г — грамм HMM - тяжелый меромиозин
Группа 1а — афферент мышечного волокна HR - метод задержки—расслабления
Группа 16 — афферент сухожильного органа Н-рефлекс — рефлекс Хоффманна
Гц — герц HRT - время полурасслабления
С — химическая энергия I - момент инерции
Cl — коэффициент подъемной силы Ixx момент инерции относительно
Са2+ — ион кальция оси кувыркания
cd — свеча (кандела) Iyy момент инерции относительно
cD — коэффициент торможения оси “колеса”
Cl- — ион хлора 1ц. ~ момент инерции относительно
Да — дальтон оси кручения (винта)
Дж — джоуль k - коэффициент
е — коэффициент (упругого) пропорциональности
сопротивления К - кельвин
Е — энергия; модуль упругости K+ - ион калия
Е, — тепловая энергия кг — килограмм
Ек — кинетическая энергия л — литр
Ер — потенциальная энергия 4 пропорция длины всей
Ep,g — потенциальная энергия, мышцы, содержащей
обусловленная силой тяжести сократительный белок
Ep,s — потенциальная энергия, 1FS - жесткое касание поверхности
обусловленная деформацией левой ноги
23 8-вз
353
лм — люмен SE
4) — длины мышцы в покое
LS — пояснично-крестцовый сустав
1ТО — отталкивание левой ногой S-I
m — масса
м — метр S-II
М — мега
М — момент силы §MG _
MAD — диаграмма ускорения массы t
мкм — микрометр Т
мм — миллиметр т
М — моль Т'
МП — максимальное повторение
МПП — мембранный потенциал покоя тм
MRI — получение изображения с J m
помощью магнитного резонанса TH
MVC — максимальное произвольное ТН-Са
сокращение (обычно
изометрическое) TH-I
М-волна — двигательная реакция на
электростимуляцию нерва ТН-Т
и — нано
Н — ньютон Т-трубочка -
Na+ — ион натрия Tws —
NADH-TR — никотинамид нуклеотид Тип FF —
дегидрогеназа тетразолиум
редуктаза
ППС — площадь поперечного сечения Тип БГ —
pH — мера кислотности или
щелочности вещества
Рн — неорганический фосфат Тип БОГ —
Ро — максимальная изометрическая
сила
г — положение Тип FR —
rf — конечное положение
Г, — исходное положение
R — клетка Реншоу Тип I —
R — результирующая сила
рад — радиан Тип II -
RCP — пул клеток Реншоу
Rh — реобаза Тип S —
Rn — входное сопротивление
RT — время расслабления Тип МО —
Rx — х-составляющая
результирующей ФМГ -
Ry — у-составляющая F -
результирующей F -
Rz — ^-составляющая Fa
результирующей F-актин —
CT — время сокращения; Fc
компьютерная томография
стер — стерадиан Fd
СЭ — сократительный элемент Ff
S — смещение F.
c — секунда F,
S1 — субфрагмент 1 Fj
s2 — субфрагмент 2 F,
SD — среднеквадратическое Fm
отклонение Fm,i -
SE — стандартная ошибка
СДГ — сукцинат дегидрогеназа
последовательно расположенный упругий компонент
первичная соматосенсорная зона коры головного мозга вторичная соматосенсорная зона коры головного мозга звуковой миограф время тера вращающий момент вращающий момент, обусловленный нагрузкой тропомиозин
вращающий момент мышцы тропонин
Са2+-связывающий компонент тропонина тормозящий компонент тропонина
тропомиозин-связываюший компонент тропонина поперечная трубочка наложение сокращения быстросокращающаяся, подверженная утомлению двигательная единица быстросокращающееся гликолитическое мышечное волокно
быстросокращающееся окислительно-гликолитическое мышечное волокно быстросокращающаяся невосприимчивая к утомлению двигательная единица медленносокращающееся мышечное волокно быстросокращающееся мышечное волокно
медленносокращающаяся двигательная единица медленносокращающееся мышечное волокно фономиограф
сила
средняя сила
сила сопротивления воздуха волокнистый актин
центростремительная сила; сжимающее усилие лобовое сопротивление сила сопротивления жидкости сила реакции поверхности внутрибрюшное давление сила реакции сустава подъемная сила мышечная сила
ключичный компонент мышечной силы
354
Fm,s — грудной компонент мышечной силы V — г макс максимальная скорость сокращения длины (мышцы)
F„ — нормальная составляющая го2 - потребление кислорода
силы Vi рефлекторная реакция
FP — движущая сила короткой латентности на
Fr — сила реакции электростимулирования во
Fs — сила сдвига время произвольного
F, — тангенциальная составляющая сокращения
силы V2 рефлекторная реакция более
Fw — вес продолжительной латентности
Fx — проекция силы в направлении на электростимулирования во
X время произвольного
Fy — проекция силы в направлении сокращения
У w вес
ЦНС — центральная нервная система х — направление справа—налево
ЦТ — центр тяжести У — направление вперед—назад
ЭМГ — электромиограмма Z — вертикальное направление
эог — электроокулограмма а (альфа) — угловое ускорение;
и — работа двигательный нейрон
V — скорость Р (бета) — угол пеннации; двигательный
V — средняя скорость нейрон
VA/K — скорость точки А у (гамма) — фузимоторный нейрон
относительно точки К А (дельта) — изменение в
Vf — конечная скорость е (эпсилон)— деформация
Vi — начальная скорость 0 (тета) — угловое положение
VI — межсрединная широкая ц (мю) — коэффициент трения
мышца бедра л (пи) — 3,14 рад
VL — латеральная широкая мышца Р удельное сопротивление
бедра о (сигма) — напряжение
Kn — напряжение мембраны S сумма
VM — широкая медиальная мышца и (омега) — угловая скорость
бедра х — векторное произведение
23*
Словарь
G-актин — шарообразная молекула актина.
1-полоска — светлая полоска скелетной мышцы.
Z-полоска — интрасаркомерное соединение двух “наборов” тонких филаментов в прилегающих саркомерах.
А-полоска — анизотропная или темная полоска в скелетной мышце, обусловленная двойным преломлением лучей, идущих из одного источника.
Адаптация — снижение интенсивности разряда нейрона, несмотря на постоянный возбуждающий импульс. Снижение обусловлено биофизическими свойствами нейрона.
Адаптивный контролер — система, изменяющая элементы системы контроля, не вызывающая немедленные изменения исходящего импульса.
Аксон — трубчатый отросток на соме нейрона, который способен передавать электрические импульсы, генерируемые нейроном.
Аксонный транспорт — движение материала на внутренней части аксона. Движение может осуществляться в прямоходящем (от сомы к синапсу) и обратноходящем (от синапса к соме) направлениях.
Аксотомия — рассечение аксона.
Активное прикосновение — изучение условий окружающей среды на основании информации чувствительных нервных окончаний, связанной с движениями руки и их контролем.
Активный компонент — часть взаимосвязи длины-силы мышцы, обусловленный активностью поперечных мостиков.
Актин — основной белок тонкого филамента мышечного волокна.
Альфа(а) -мотонейрон — двигательный нейрон, иннервирующий экстрафузальные (не являющиеся мышечными веретенами) мышечные волокна.
Анализ Фурье — выведение серии членов, содержащих синус и косинус, с целью характеристики частотного содержания сигнала.
Анизометрическое — сокращение, не являющееся изометрическим.
Анион — отрицательно заряженный ион.
Антеролатеральный проводящий путь — восходящий проводящий путь, по которому передается информация, касающаяся в основном температуры и болевых ощущений, а также некоторая тактильная и проприоцептивная информация.
Антигравитационные мышцы — мышцы, производящие вращающий момент, противодействующий силе тяжести, когда человек стоит выпрямившись.
Антисипаторная (предвосхищающая) стабилизация — автоматическая постуральная адаптация, обеспечивающая производство движения и направленная на стабилизацию системы (например, конечности, тела) в ожидании механических расстройств, связанных с движением.
Антропометрия — измерения тела человека.
Аппарат Гольджи — веретенообразное тельце, напоминающее сухожильный орган, которое может воспринимать напряжение связки.
Атрофия — потеря мышечной массы.
Аутогенез — процесс разрушения клеточных структур. Наблюдается при повреждении мышцы вследствие физических нагрузок.
Афферент группы 1а — афферент мышечного веретена.
356
Афферент группы 16 — афферент сухожильного органа.
Афферент группы II — афференты с диаметром на один класс меньше, чем афференты группы I. Мышечное веретено имеет афферент труппы II.
Афферентное волокно — аксон, передающий сигналы из чувствительных нервных окончаний.
Ацетилхолин — нейромедиатор, используемый в различных участках, в частности, в нервно-мышечном соединении.
Ацетилхолинестераза — фермент в нервно-мышечном соединении, который выполняет роль терминатора активности нейромедиатора ацетилхолина.
Базальные ядра — пять тесно связанных ядер, принимающих импульс из коры головного мозга и посылающих свои импульсы через таламус назад в кору головного мозга.
Баллистическое растяжение — метод увеличения диапазона движения относительно сустава, предполагающий выполнение серии быстрых растяжений мышцы.
Бег — режим локомоции человека.
Бета((3)-мотонейрон — двигательный нейрон, иннервирующий как интрафузальные, так и экстрафузальные мышечные волокна.
Билатеральный дефицит — одновременная активация двух конечностей, которая приводит к снижению силы максимального произвольного сокращения каждой конечности.
Билатеральный прирост — одновременная активация двух конечностей, ведущая к увеличению силы максимального произвольного сокращения каждой конечности.
Биомеханика — использование инженерной механики для изучения биологических систем.
Биофизика — использование физических принципов и методов относительно биологических систем.
Брошенное тело — перемещающийся объект, не имеющий фазы опоры.
Варолиев мост — промежуточная часть ствола головного мозга.
Вектор — количество, выражающее как величину, так и направление.
Векторное произведение — метод умножения векторов.
Вентральный корешок — “выходная” зона на передней части спинного мозга. Эфферентные аксоны выходят из спинного мозга в вентральном корешке.
Вершина — точка на середине расстояния между директрисой и фокусом параболы.
Вес — выражение количества гравитационного притяжения между объектом и Землей.
Весовой коэффициент — коэффициент, используемый для модуляции амплитуды переменной и определения её вклада в выражение.
Вестибулоспинальный путь — нисходящий проводящий путь.
Взаимосвязь сила-длина — максимальная сила, производимая мышцей, изменяется как функция длины мышцы.
Взаимосвязь сила-скорость — влияние интенсивности изменения длины мышцы на величину максимальной силы, которую может произвести мышца.
Взаимосвязь сила-частота — увеличение частоты активации (например, интенсивности разряда, электростимулирование) приводит к увеличению силы.
Влияние предварительной нагрузки — увеличение силы, производимой мышцей в начале концентрической фазы во время цикла растяжения-сокращения.
Внеклеточный — за пределами клетки.
Внутрибрюшное давление — давление (Ра) внутри брюшной полости.
Внутригрудное давление — давление (Ра) внутри грудной полости.
Внутридисковое давление — давление внутри межпозвонковых дисков.
Внутримышечное давление — внутреннее давление в мышце, связанное с её сокращением.
Возбуждающий постсинаптический потенциал — деполяризующий (возбуждающий) синаптический потенциал, возникаемый в постсинаптической мембране (дистально к синапсу).
Возвратное торможение — схема локального рефлекса, при котором потенциалы действия, разряжаемые двигательным нейроном, ведут к активации через коллатерали аксона, клетки Реншоу и последующей генерации тормозного постсинаптического потенциала двигательного нейрона.
Волновое сопротивление — сопротивление волн движению пловца. Сопротивление, по всей видимости, обусловлено различиями в плотности воды и воздуха.
Волокно адерного мешочка — интрафузальное волокно, в котором ядра сгруппированы.
Волокно ядерной цепочки — интрафузальное волокно, в котором ядра расположены концами друг к другу.
Восприятие — процесс, при котором афферентные сигналы достигают сознания и способствуют возникновению ощущения.
Восходящие пути — нервные соединения, которые обеспечивают распределение информа
357
ции из чувствительных нервных окончаний в супрасегментальные центры.
Вращающий момент — вращающее действие силы; произведение силы и плеча пары (Н-м).
Вращение — движение, при котором все части системы не смещаются в одинаковой мере.
Время полурасслабления — время снижения реакции сокращения от пика наполовину.
Время реакции — минимальный отрезок времени между началом действия стимула и реакцией на него.
Время сокращения — время от начала производства силы до ее пика во время сокращения.
Вязкость — мера деформации сдвига. Изменяется в зависимости от температуры и зависит от сил сцепления между молекулами и обмена количеством движения между сталкивающимися молекулами. Единица измерения — Н с см-2. Масло характеризуется более высокой вязкостью, чем вода.
Г-актин — фиброзный актин.
Гамма (у)-мотонейрон — двигательный нейрон, иннервирующий интрафузальные (мышечное веретено) мышечные волокна.
Гармоника — кратное фундаментальной частоты.
Генераторный потенциал — электрический потенциал, который производит чувствительное нервное окончание в ответ на стимул, который оно воспринимает.
Гцдролиз АТФ — расщепление АТФ на АДФ, Р и энергию.
Гиперплазия — увеличение мышечной массы вследствие увеличения числа мышечных волокон.
Гиперполяризация — увеличение полярности мембраны, внутренняя часть становится более отрицательной, чем внешняя.
Гипертонный — увеличение мышечного тонуса, например, при спастичности и тугоподвиж-ности.
Гипертрофия — увеличение мышечной массы вследствие увеличения площади поперечного сечения мышечных волокон.
Гипотеза двойной стратегии — объяснение структур ЭМГ агонистов-антагонистов и результирующего вращающего момента мышцы, лежащее в основе движений различных смещений, скоростей и нагрузок.
Гипотонный — снижение мышечного тонуса.
Гомонимное — анатомическая связь между чувствительными нервными окончаниями и двигательными нейронами, иннервирующими одну и ту же мышцу.
Гравитационная потенциальная энергия — энергия, которую имеет система вследствие своего нахождения в гравитационном поле.
Граничная частота — частота, определенная для фильтра (цифрового или электронного), характеризующая точку, в которой частотное содержание сигнала изменяется.
Двигательная единица — соматическая клетка и дендриты двигательного нейрона, многочисленные ответвления аксона и мышечные волокна, которые он иннервирует.
Двигательная единица типа FF — быстросокра-шающаяся утомляемая двигательная единица.
Двигательная единица типа FR — быстросокра-щающаяся невосприимчивая к утомлению двигательная единица.
Двигательная единица типа S — медленносокра-щающаяся двигательная единица.
Двигательная программа — стереотипная последовательность команд, посылаемых из спинного мозга к мышцам с целью вызвать определенную структуру поведения.
Двигательный контроль — контроль движения.
Двигательный нейрон — нейрон, чей аксон непосредственно соединен с мышечными волокнами. Его называют конечным общим проводящим путем, поскольку он представляет собой заключительную стадию импульса нервной системы.
Движение — изменение положения (т) в интервале времени.
Двойной разряд — разрядка двух потенциалов действия одной двигательной единицей с интервалом около 10 мсек.
Демаскирование — восстановление функции после травмы нервной системы, может быть обусловлено способностью “спящего” проводящего пути взять на себя функцию травмированного проводящего пути. В этом случае травма демаскирует функционально “спрятанный” проводящий путь.
Деминерализация — чрезмерные потери солей костью.
Дендрит — короткий отросток, простирающийся от сомы нейрона
Денервация — состояние, наблюдаемое при пересечении нерва; мышца оказывается лишенной нервных импульсов.
Деполяризация — снижение полярности (потенциальной) через мембрану.
Дерекрутирование — инактивация двигательной единицы.
Деформация — изменение длины относительно начальной длины.
Деформация мышцы — значительная нагрузка на мышцу, сопровождающаяся болезненными ощущениями.
Диаграмма свободного тела — метод графического анализа, позволяющий определить сис
358
тему и показать её взаимодействия с окружающей средой. Диаграмма свободного тела представляет собой графический вариант левой части закона ускорения Ньютона.
Диаграмма угол—угол — график, показывающий изменения одного угла (ось у) как функции другого угла (ось х).
Диаграмма ускорения массы — графическое изображение правой части закона ускорения Ньютона; демонстрирует кинематические воздействия сил, действующих на систему.
Диапазон движения — максимальное угловое перемещение относительно сустава.
Динамический анализ — механический анализ, при котором силы, действующие на систему, не сбалансированы и, следовательно, система ускоряется. Правая часть закона Ньютона об ускорении — не равна нулю.
Директриса — фиксированная линия, от которой все точки параболы находятся на одинаковом расстоянии.
Диск Меркеля — кожный механорецептор, чувствительный к локальному вертикальному давлению.
Длина мышцы в покое — длина мышцы, при которой возникает пассивная сила.
Дополнительная двигательная зона — компонент чувствительно-двигательной зоны коры головного мозга.
Дорсальный корешок — входная зона на тыльной стороне спинного мозга. Место входа в спинной мозг афферентных аксонов.
Дробь — соотношение двух величин.
Жёсткость — крутизна (наклон) графика силы-длины; изменение силы на единицу изменения длины (Н мм). Чтобы растянуть жёсткую пружину хотя бы на несколько миллиметров, необходимо приложить значительное усилие.
Зависимая от состояния реакция — реакция, изменяющаяся в зависимости от состояния системы. Рефлексы описывают как зависимые от состояния реакции: величина определяется активностью индивида (ходьба — пребывание в неподвижном состоянии).
Зависимость от задания — например механизмы, обусловливающие возникновение утомления, зависят от деталей задания, таких, как интенсивность, продолжительность участвующих мышц и мотивации испытуемого.
Зависимые от движения взаимодействия — взаимодействующие силы между сегментами тела во время движения.
Задняя париетальная кора головного мозга — компонент чувствительно-двигательной коры головного мозга.
Закон Бернулли — давление жидкости обратно пропорционально её скорости.
Закон Вольффа — все изменения функции кости обусловлены изменениями внутренней структуры.
Закон всемирного тяготения — все тела притягивают друг друга с силой, пропорциональной произведению их массы и обратно пропорциональной квадрату расстояния между ними.
Закон действия и противодействия — для каждого действия существует равное по величине противодействие.
Закон инерции — сила, необходимая, чтобы прекратить, начать или изменить движение.
Закон ускорения — F = та.
Изокинетическое — движение, при котором угловая скорость перемещаемого сегмента тела является постоянной.
Изометрическое — механическое условие, при котором вращающий момент мышцы равен вращающему моменту, обусловленному нагрузкой, вследствие чего длина всей мышцы не изменяется.
Изотоническое — состояние, при котором мышца сокращается и выполняет работу, преодолевая постоянную нагрузку.
Изоформа — белок, синтезируемый со слегка отличительным составом аминокислот, также называется изоферментом.
Иммобилизация конечности — экспериментальный метод.
Импульс — площадь под графиком силы—времени (Н сек); интеграл силы—времени. Импульс влияет на количество движения системы.
Инерция — сопротивление объекта любому изменению своего движения.
Интеграция — математическая процедура измерения участка под кривой, например взаимосвязи напряжения—времени или силы-времени.
Интегрин — трансмембранный белок, который может служить для соединения миофибрилл с внеклеточным матриксом соединительной ткани.
Интенсивность разряда — интенсивность, с которой нейроны разряжают потенциалы действия; частота потенциалов действия. Единица измерения — Герц.
Интенсивность шагов — частота (гц), с которой выполняются шаги.
Интернейрон — нейрон, аксон которого ограничен пределами спинного мозга.
Интерсегментальная динамика — силы инерции, производимые движущимся сегментом тела по отношению к окружающей среде.
359
Интрафузальное волокно — миниатюрное волокно скелетной мышцы, образующее часть мышечного веретена.
Качественный — характеризует тип или вид.
Квадратный многочлен — многочлен второй степени.
Квазистатический — механическое состояние, при котором ускорение, испытываемое системой, достаточно небольшое, чтобы можно было его считать равным нулю.
Килограмм — единица измерения массы.
Кинезиология — наука о движении.
Кинематика — описание движения с точки зрения положения, скорости и ускорения.
Кинестезия — способность системы использовать информацию чувствительных нервных окончаний, чтобы определить положение конечностей, определить фактор, заставляющий ее двигаться, и т.п.
Кинетика — описание движения.
Кинетическая энергия — способность объекта выполнять работу вследствие своего движения.
Клетка Реншоу — интернейрон в спинном мозге, обеспечивающий тормозные постсинаптические потенциалы. Клетка Реншоу принимает импульс из коллатеральных ответвлений аксонов двигательных нейронов, а также из супраспинных источников.
Клеточная мембрана — липидный двойной слой. Коактивация — одновременная активация мышц вокруг сустава: обычно характеризует активность мышц агонистов—антагонистов.
Количественный — описывающий количество.
Количество движения — количество движения объекта; векторная величина. Единицей измерения линейного количества движения является кг м с-1; углового — кгм-с-2.
Коллатераль — ответвление аксона.
Коллатеральное ответвление — возникновение отростков (невритов) на дистальных сегментах аксона с целью реиннервации денервированной мишени. Это обычно наблюдается после повреждения или травмы клетки мишени (например, мышечного волокна, нейрона).
Компаратор — прибор или структура, сравнивающие импульсы, поступающие из различных источников.
Компартмент — субобъём мышцы, иннервированной первичным ответвлением нерва мышцы.
Конечная скорость — скорость падающего объекта, когда силы, обусловленные притяжением и сопротивлением воздуха, равны.
Конечный общий проводящий путь — выражение, характеризующее функцию двигатель
ных нейронов как путь, благодаря которому нервная система контролирует мышечную активность.
Контролер обратной связи — система, которая сравнивает выполняемую деятельность с нужной.
Контролер прямой связи — система, производящая команды без непрерывной отрицательной обратной связи.
Конусообразное ядро — одно из глубоко расположенных ядер мозжечка.
Концентрический — механический режим (состояние), при котором вращающий момент мышцы превышает вращающий момент нагрузки, вследствие чего длина активной мышцы сокращается.
Корково-спинномозговой путь — нисходящий путь с импульсом, поступающим из всей чувствительно-двигательной области коры головного мозга.
Королларная разрядка — внутренние импульсы, возникающие из двигательных команд и влияющие на восприятие (Speny, 1950).
Косеканс — тригонометрическая функция, равная гипотенузе, деленной на противоположную сторону.
Косинус — тригонометрическая функция, равная величине примыкающей стороны, деленной на гипотенузу
Котангенс — тригонометрическая функция равная примыкающей стороне, деленной на противоположную .
Коэффициент безопасности — граница безопасности до повреждения.
Коэффициент иннервации —- число мышечных волокон, иннервируемых отдельным двигательным нейроном.
Коэффициент упругого восстановления — коэффициент, характеризующий скорость объекта после столкновения по сравнению с его скоростью до столкновения.
Красное ядро — клетки, расположенные в среднем мозге.
Кратковременная ригидность — механическое свойство мышцы, высокая ригидность на первых нескольких миллиметрах растяжения и последующее значительное ее снижение.
Крутизна — интенсивность изменения одной переменной (зависимой переменной или переменной у оси) относительно другой (независимой переменной или переменной х оси).
Кубическое уравнение — многочлен третьей степени.
Ламинарное течение — однообразный поток жидкости вокруг объекта.
Латеральное вестибулярное ядро — находится между варолиевым мостом и продолговатым мозгом.
360
Латеральный мешочек — увеличение (расширение) саркоплазматического ретикулума вблизи поперечных трубочек. Другое название — терминальная цистерна.
Лёгкая цепочка — белок с низким молекулярным весом.
Лёгкий меромиозин — “хвостовой” фрагмент молекулы миозина.
Лимбическая система — ряд структур передней части головного мозга, взаимосвязанных с гипоталамусом и частями среднего мозга; иногда называют эмоциональной двигательной системой.
Линейный участок — часть кривой нагрузки-деформации с линейной взаимосвязью.
Линия обтекаемости — схематизированная линия потока жидкости вокруг объекта.
Лобовое сопротивление — составляющая вектора сопротивления жидкости, действующая параллельно потоку жидкости.
Логарифм — значение (х), к которому следует возвести число (в ), чтобы получить определенное число, например, если вх = а, то Ln а=х.
М-волна — реакция ЭМГ и силы мышцы, обусловленная активацией альфа-аксонов путём электростимулирования. Используется для тестирования целостности нервно-мышечного распространения.
М-полоска — внутрисаркомерное соединение двух рядов толстых филаментов.
Магнитное стимулирование — использование магнитного стимулирования для создания магнитного поля, вызывающего электрическое поле и потенциалы действия аксонов.
Максимумы — пики на графике.
Маневр Джендрассика — использование активности отдельной мышцы, чтобы повысить возбудимость пула двигательного нейрона и величину рефлекса Хоффманна (Н).
Мантисса — часть логарифма.
Масса — мера (кг) количества материи в объекте.
Маховая фаза — безопорная фаза во время бега или ходьбы.
Мембранный потенциал покоя — трансмембранное напряжение в стабильном положении.
Метод получения изображения с помощью магнитного резонанса — метод, предполагающий использование магнитных полей для определения расположения протонов, который может быть использован для изучения структуры и функции двигательной системы.
Метр — единица измерения длины в системе единиц СИ.
Миелин — жировая изоляционная оболочка в периферических нервах.
Микрофибрилла — удлиненный пучок, включающий пять параллельных рядов молекул коллагена, расположенных последовательно.
Миниатюрный потенциал концевой пластинки — небольшие синаптические потенциалы в нервно-мышечном соединении, которые возникают в ответ на спонтанное выделение нейромедиатора. .
Минимальные — самые низкие точки или впадины на графике.
Миозин — основной белок толстого филамента мышечного волокна; включает поперечный мостик. В отдельном толстом филаменте содержится несколько сот молекул миозина.
Миозин АТФаза — фермент, катализирующий реакцию актомиозина, связанную с циклом поперечного мостика.
Миофибрилла — единица в мышечном волокне, состоящая из последовательно расположенных саркомеров.
Миофиламент — тонкий и толстый филаменты мышечного волокна, содержащие сократительный аппарат.
Многочлен — сумма членов.
Модель лямбды (X) — понятие, согласно которому движения контролируются вследствие изменения порога тонического рефлекса растяжения.
Модуль упругости — отношение нагрузки к деформации.
Мозжечок — супрасегментная структура, участвующая в процессе контроля движения. Располагается кзади от ствола мозга.
Момент инерции — сопротивление объекта любому изменению своего углового движения. Представляет собой распределение массы объекта относительно оси вращения. Обозначается знаком I, единица измерения в системе единиц СИ — кгм2.
Момент силы — вращательный эффект силы; вращающий момент.
Мощность — интенсивность выполнения работы; интенсивность изменения количества энергии; произведение силы и скорости.
Мышечная “мудрость” — способность мышцы снижать разряд двигательных нейронов, чтобы уравновесить изменение в снижении интенсивности расслабления.
Мышечная архитектура — “дизайн” мышцы, включающей такие элементы, как длина, площадь поперечного сечения, пеннация и точки прикрепления на скелете.
Мышечная механика — изучение механических свойств, производящих силу единиц мышцы.
Мышечная сила — величина вращающего момента, производимого мышцей при отдель
361
ном максимальном изометрическом сокращении неограниченной продолжительности.
Мышечная сила — сила, производимая структурными (пассивными) и активными (поперечный мостик) элементами мышцы.
Мышечное веретено — внутримышечное чувствительное нервное окончание, контролирующее напредвиденные изменения длины мышцы.
Мышечное волокно типа I — медленносокращающееся мышечное волокно.
Мышечное волокно типа II — быстросокращающееся мышечное волокно. Волокна этого типа можно подразделить на две группы (Па и Пб) после препарации в ванной с pH 4.3 (Па) или 4.6 (Пб).
Мышечное волокно типа БГ — быстросокращающееся гликолитическое мышечное волокно в двигательных единицах типа FF.
Мышечное волокно типа БОГ — быстросокращающееся окислительное гликолитическое мышечное волокно в двигательных единицах типа FR.
Мышечное волокно типа МО — медленносокращающееся окислительное мышечное волокно в двигательных единицах типа S.
Мышечное утомление — вид острых влияний, нарушающих двигательную деятельность.
Мышечно-сухожильная единица — сочетание мышцы и структур соединительной ткани, участвующих в сообщении силы, произведенной мышечными волокнами скелету.
Мышечные судороги — болезненные, непроизвольные сокращения мышцы, обусловленные, по всей видимости, периферическими импульсами.
Мышечный тонус — пассивное сопротивление мышцы изменению ее длины.
Мышца — ткань, содержащая сократительные клетки, способные превращать химическую энергию в механическую.
Н-полоска — участок А-полоски без тонких филаментов.
Наложение сокращения — метод, используемый для тестирования максимально произвольного мышечного сокращения. Метод предполагает наложение 1—3 супрамаксимальных электрошоков мышечного нерва на произвольное сокращение, чтобы определить, увеличивается ли сила.
Насос Na+-K+ — мембранно-связанный белок, транспортирующий Na+ из внеклеточной жидкости во внутриклеточную и транспортирующий К+ в обратном направлении.
Небулин — белок в эндосаркомерном цитоскелете.
Невесомость — условия микрогравитации.
Невосприимчивые — мышечные волокна, не реагирующие на определенные виды физиологических пертурбаций (например, иммобилизация, денервация, тренировки силовой направленности).
Нейроглия — один из двух типов клеток нервной системы.
Нейромедиатор — вещество, выделяемое в незначительном количестве из окончаний нервных клеток в ответ на поступление потенциала действия.
Нейрон — один из двух видов клеток в нервной системе, способный производить и передавать электрический сигнал.
Нейротропизм — влияние одного биологического элемента на другой.
Нервная стратегия — структура двигательной команды и мышечной активации.
Нервно-мышечное распространение — процессы, участвующие в превращении потенциала действия аксона в потенциал действия сарколеммы.
Нервно-мышечное соединение — синапс между двигательным нейроном и мышечным волокном.
Нервно-мышечное электростимулирование — клинический или экспериментальный метод использования электрошока, чтобы искусственно вызвать потенциалы действия аксонов и тем самым мышечное сокращение.
Нервно-мышечный компартмент — объём мышцы, “обслуживаемый” основным ответвлением мышечного нерва.
Нисходящие проводящие пути — нервные проводящие пути, передающие информацию из супрасегментарных центров к стволу головного мозга и спинному мозгу.
Нормальная составляющая — составляющая, действующая под прямым углом к поверхности.
Ньютон (Н) — единица измерения в системе единиц СИ.
Обратная динамика — динамический анализ, используемый для определения сил и вращающих моментов, действующих на систему и основанных на кинематике движения.
Обратная связь — для у=к/х, когда х — положительная величина, которая увеличивается, у уменьшается.
Обратная связь — сигналы, поступающие из различных периферических нервных окончаний, несущие информацию в нервную систему о механических явлениях в нервно-мышечной системе.
Окончание Руффини — 2—6 шарообразных телец с одним миелинированным “отцовским” аксоном, имеющим диаметр 5—9 мм. Эти механорецепторы способны сигнализировать
362
о положении и смешении сустава, угловой скорости и внутрисуставном давлении.
Ортогональный — перпендикулярный, независимый.
Остаточный анализ — метод, используемый при подборе эмпирической кривой для сравнения различий между отфильтрованными и не-отфильтрованными сигналами.
Остаточный момент силы — оставшийся член в уравнении движения, когда все остальные члены уравнения определены. Чаще всего используется для вычисления результирующей мышечной силы.
Остеопороз — снижение костной массы.
Ось “сальто” — ось, проходящая через тело человека из стороны в сторону.
Ось колеса — ось, проходящая через тело человека спереди назад.
Ось кручения — ось, проходящая через тело человека от головы до пальцев ног.
Отведение — движение от средней линии тела или части тела.
Отрицательная обратная связь — понятие, характеризующее способность системы реагировать на расстройство таким образом, что реакция сводит на нет расстройство.
Отрицательная работа — работа окружающей среды относительно системы. Во время отрицательной работы система абсорбирует энергию окружающей среды. Мышца выполняет отрицательную работу, когда результирующий вращающий момент мышцы меньше вращающего момента, обусловленного нагрузкой.
Отсеон — основная структурная единица кости, состоящая из ряда концентрических слоев минерализованного матрикса, окружающих центральный канал.
Отсроченное возникновение болезненных ощущений в области мышц — восприятие болевых ощущений в мышцах, обусловленное субклеточным повреждением, через 24—48 часов после физической нагрузки. Болезненные ощущения могут быть обусловлены воспалительной реакцией.
Ощущение усилия — ощущение, основанное на королларном разряде, свидетельствующем о количественном изменении нисходящей команды.
Парабола — кривая, определяемая относительно пары перпендикулярных линий (Е и D) и фиксированной точки (F) на одной из линий.
Параллельный эластичный элемент — один из компонентов моделей мышцы Хилла. Обуславливает эластичность мышцы.
Пассивный компонент — часть взаимосвязи длина-сила мышцы, обусловленная структурными свойствами мышцы и сухожилия.
Пеннация — угловое отклонение между направлением мышечных волокон и линией натяжения мышцы.
Первичная двигательная зона — компонент чувствительно-двигательной коры головного мозга.
Первичная соматосенсорная зона коры головного мозга — компонент чувствительно-двигательной коры головного мозга.
Первый закон термодинамики — выполнение работы требует затрат энергии; работа= энергии.
Передняя динамика — динамический анализ для определения кинематики, проявляемой системой на основании действующих на неё сил и вращающих моментов.
Перекрёстный разгибательный рефлекс — продолжительная реакция контралатерального сустава на отрицательный стимул, вызывающий сгибательный рефлекс.
Перекрытие миофиламентов — разобщение тонкого и толстого филаментов, обеспечивающее циклы поперечных мостиков и произведение ими силы. Количество образуемых поперечных мостиков и, следовательно, производимая сила зависят от степени перекрытия.
Переменная нагрузка — тренировочная нагрузка, изменяющаяся в диапазоне движения.
Перимизий — матрикс соединительной ткани, “собирающий” мышечные волокна в пучки.
Петля гистерезиса — взаимосвязь, характеризующаяся несовпадением фаз нагрузки и разгрузки.
Плечо пары — наименьшее расстояние от линии действия вектора силы к оси вращения.
Плоский — ограниченный отдельной плоскостью.
Площадь поперечного сечения — площадь объекта (например, мышцы), рассечённой под прямыми углами к длинной оси.
Поверхностное сопротивление — сопротивление, обусловленное трением пограничного слоя жидкости и объекта.
Поглощение энергии — поток механической энергии из окружающей среды к системе. Система поглощает энергию, выполняя отрицательную работу.
Податливость — количество материала, который может быть растянут при приложении усилия. Это свойство выражается в количестве изменения длины на единицу силы, используемой для растягивания материала (мм/н).
Подвешивание задней конечности — экспериментальный метод.
363
Подъёмная сила — составляющая вектора сопротивления жидкости, действующая перпендикулярно направлению потока жидкости.
Поза — нервномеханическое состояние, касающееся сохранения равновесия.
Положение — нахождение объекта относительно какой-то определенной точки.
Положительная работа — работа, выполняемая системой по отношению к окружающей среде. Во время положительной работы система отдает окружающей среде свою энергию. Мышцы выполняют положительную работу, когда их результирующий вращающий момент превышает вращающий момент, обусловленный нагрузкой.
Поперечная трубочка, Т-трубочка — инвагинации сарколеммы, обеспечивающие быструю связь между сарколеммой (потенциалы действия) и миофиламентами, расположенными в верхней части мышечного волокна.
Поперечные соединения — биохимические связи, в результате которых молекулы коллагена содержатся вместе в соединительной ткани.
Поперечный мостик — вытяжение субфрагмента I миозиновой молекулы. Субфрагмент I представляет собой шарообразный компонент фрагмента тяжелого меромиозина.
Порог потенциала действия — мембранный потенциал, при котором клетка (нейрон или мышечное волокно) производит потенциал действия.
Порог рекрутирования — сила, при которой происходит рекрутирование (активация) двигательной единицы.
Последовательный эластичный элемент — одна из составляющих модели мышцы Хилла. Обусловливает эластичность мышцы.
Постгиперполяризация — замыкающая часть потенциала действия, когда мембрана менее возбудима. Период гиперполяризации на заключительной стадии потенциала действия.
Постоянная нагрузка — тренировочная нагрузка постоянной величины (например, штанга).
Постсинаптический — на дистальной стороне синапса.
Постсократительная сенсорная разрядка — повышение нервной активности, в основном, вследствие повышенной разрядки мышечных веретен после сокращения мышцы.
Посттетаническая потенциация — увеличение силы сокращения после кратковременного тетануса.
Поступательное движение — движение, при котором все части системы смещаются в одинаковой степени.
Потенциал действия — распространяемый импульс нейронов и мышечных волокон. Физи
чески представляет собой участок мембраны с изменяющимся распределением Na+ и К+.
Потенциал концевой пластинки — синаптический потенциал, образованный в мышечном волокне в ответ на выделение нейромедиатора.
Потенциальная энергия — энергия, которой обладает система ввиду её нахождения в стороне от более устойчивого местонахождения.
Потенциация — интенсификация реакции, несмотря на постоянный импульс.
Правило правого пальца — метод определения направления векторного произведения.
Преддвигательная зона коры головного мозга — компонент чувствительно двигательной коры головного мозга.
Препарат с одним оставшимся корешком — экспериментальная модель для изучения процесса выпускания отростков ЦНС. В этой модели отсекаются все кроме одного из дорсальных корешков, иннервирующих заднюю конечность.
Прерывистый тетанус — нерегулярная силовая реакция, обусловленная стимулированием с низкой частотой.
Пресинаптический — на проксимальной стороне синапса.
Пресинаптическое окончание — увеличение аксона, участвующего в синапсе.
Пресинаптическое торможение — гиперполяризация (торможение синаптическим потенциалом) аксона в участке непосредственно перед его синаптическим контактом с клеткой-мишенью.
Приведение — движение по направлению к средней линии тела или части тела.
Принцип обратимости — тренировочный принцип, согласно которому адаптационные реакции, обусловленные физическими тренировками, носят временный характер (являются обратимыми).
Принцип перегрузки — тренировочный принцип, согласно которому для достижения адаптационной реакции необходимо превысить пороговую точку.
Принцип размера — объяснение упорядоченного рекрутирования двигательных единиц. Согласно этому принципу, первыми рекрутируются двигательные нейроны наименьшего размера.
Принцип специфичности — тренировочный принцип, согласно которому тренировочная нагрузка должна быть специфичной (т. е, соответствовать виду вызываемого изменения).
Принцип способности передаваться — свойство, позволяющее перемещать вектор вдоль своей линии действия, не меняя механическое воздействие на жесткое тело.
364
Проба Вальсальвы — произвольное повышение давления в брюшной полости.
Проводимость — свойство мембраны, зависящее от её проницаемости для иона и от наличия иона.
Проводящий путь группы А — нисходящий проводящий путь, включающий вестибулоспинальный, ретикулоспинальный и тектоспи-нальный пути.
Проводящий путь группы Б — нисходящий проводящий путь, включающий пересеченные руброспинальный и пересеченный ретикулоспинальный пути.
Продолговатый мозг — продление головного мозга в спинном мозге.
Проектируемая площадь — член в уравнении сопротивления жидкости, обусловливающий размер (площадь поперечного сечения) движущегося в жидкости объекта.
Производство энергии — поток механической энергии из системы в окружающую среду. Система производит энергию, выполняя положительную работу.
Промежуточные филаменты — компоненты экзо-саркомерного цитоскелета; располагаются продольно вдоль и поперечно через саркомеры.
Проприорецептор — чувствительное нервное окончание (например, мышечное веретено, сухожильный орган), способное выявлять стимулы, обусловленные действиями системы.
Проприоспинальный — интернейроны, передающие информацию между спинными сегментами.
Пространственная диаграмма — схема диаграммы свободного тела, не включающая правильные величины векторов силы.
Психофизика — изучение взаимосвязи между физической переменной и соответствующим ощущением.
Пул двигательных нейронов — группа двигательных нейронов, иннервирующих отдельную мышцу.
Пьезоэлектрический — образование электрических потенциалов вследствие давления.
Работа — скалярная величина, характеризующая, насколько можно сместить объект в определенном направлении при приложении силы. Обозначается знаком U, единица измерения - джоуль (Дж).
Радиан — угол, представленный отношением расстояния окружности к её радиусу.
Разложение — процесс разложения результирующего вектора на несколько компонентов.
Распространение — проведение потенциала действия вследствие активных регенеративных процессов, сохраняющих количество потенциала.
Рассечение спинного мозга — отделение супра-спинальной нервной системы от спинной системы вследствие рассечения спинного мозга.
Растормаживание — торможение ингибитора.
Растяжение задержка—расслабление — растяжение, основанное на проприоцептивном улучшении нервно-мышечной передачи импульсов, и включающее первоначальное максимальное изометрическое сокращение мышцы, которую собираются растянуть с последующим расслаблением и растяжением до максимального диапазона движения.
Растягивающее сокращение агониста — растяжение, при котором ассистент доводит сустав до границы вращения, после чего спортсмен сокращает агонисты (например, четырехглавую мышцу бедра), способствуя дальнейшему увеличению амплитуды движения, растягивая антагонист (например, подколенное сухожилие).
Расширение — увеличение угла между двумя сегментами.
Реакция потряхивания лапы — быстрые чередующиеся сгибающе-разгибающие движения лапой с целью избавиться от раздражителя.
Регулируемые нейромедиатором — свойство некоторых канальцев в возбудимой мембране, особенно близко расположенных к синапсу. Эти каналы открывает нейромедиатор.
Регулируемый напряжением — свойство некоторых каналов возбудимой мембраны. Регулируемые напряжением канальцы открываются в результате снижения электрических потенциалов мембраны.
Регулятор — система, обеспечивающая регуляцию.
Регуляция — процесс поддержания постоянного состояния.
Резистивный импульс — электрическое сопротивление двигательного нейрона текущему импульсу, “инъекцируемому” внутриклеточным микроэлектродом.
Результирующая мышечная сила — чистая сила, производимая группой мышц относительно сустава.
Реиннервация — состояние, при котором мышца или мышечное волокно повторно присоединяются к нервной системе в результате развития функционального соединения (синапса).
Ректификация — процесс, включающий устранение или инвертирование отрицательной фазы или фаз сигнала.
Рекрутирование — процесс активации двигательной единицы.
Ремоделирование — развитие, укрепление и резорбция живой кости.
365
Реобаза — мера возбудимости нейрона, показывающая, какая величина тока может привести к образованию потенциала действия.
Реполяризация — возвращение мембранного потенциала (полярность) к устойчивому состоянию.
Ретикулоспинальный путь — нисходящий проводящий путь от ретикулярной формации, оканчивающийся вдоль длины спинного мозга.
Ретикулярная формация — диффузное скопление клеток в стволе мозга.
Рефлекс растяжения — реакция мышцы на резкое неожиданное увеличение длины мышцы. Реакция отрицательной обратной связи активирует растянутую мышцу, чтобы свести к минимуму увеличение её длины.
Рефлекс сопротивления — реакция отрицательной обратной связи.
Рефлекс тонической вибрации — реакция ЭМГ и силы вследствие высокочастотной (50— 150 гц) вибрации небольшой амплитуды. Вибрация активирует мышечные веретена, что приводит к активации рефлекса двигательных нейронов.
Рефлекс Хоффманна — реакция, искуственно вызываемая электростимулированием периферического нерва и избирательной активацией афферентов группы 1а (наибольшего диаметра). Рефлекс Хоффманна или Н-рефлекс используется для тестирования уровня возбудимости пула двигательных нейронов.
Реципрокный тормозной рефлекс — снижение возбудимости двигательных нейронов, иннервирующих мышцу-антагонист вследствие растяжения мышцы-агониста и активации афферентов группы 1а и тормозного интернейрона группы 1а.
Ригидность — состояние, включающее двухнаправленное сопротивление пассивному движению, которое не зависит от скорости движения.
Руброспинальный путь — нисходящий проводящий путь от красного ядра до двигательных нейронов спинного мозга.
Сарколемма — возбудимая клеточная мембрана мышечного волокна.
Саркомер — компонент миофибриллы, находящийся в участке от одной Z-полоски к другой. Саркомер содержит сократительный аппарат, необходимый для превращения химической энергии в механическую работу.
Саркоплазма — жидкость, заключенная в мышечном волокне сарколеммой.
Саркоплазматический ретикулум — мембраноподобная система в мышечном волокне, выступающая в латеральные мешочки вблизи поперечных трубочек.
Свободное нервное окончание — аксон небольшого диаметра, воспринимающий аномальную механическую нагрузку и химические агенты.
Связанная система — система, представленная последовательностью жестких связей. В биомеханическом анализе тело человека рассматривают в виде связанной системы.
Сгибание — уменьшение угла между двумя сегментами.
Сгибательный рефлекс — реакция сгибания ноги в ответ на отрицательный стимул, действующий на тыльную часть ноги.
Сегментный анализ — биомеханический анализ, включающий графическое разделение сегментов тела и построение графика свободного тела и уравнений движения каждого сегмента.
Секанс — тригонометрическая функция, равная гипотенузе, деленной на соседнюю сторону.
Сенсомоторная зона коры головного мозга — участки коры головного мозга.
Сжимающее усилие — сила сжатия.
Сила — механическое взаимодействие между объектом и окружающей средой. Единицей измерения в системе СИ является ньютон (н).
Сила инерции — сила, производимая объектом вследствие его движения.
Сила реакции поверхности — сила реакции, обусловленная горизонтальной поверхностью опоры.
Сила реакции сустава — чистая сила, передаваемая от одного сегмента другому вследствие контактов мышцы, связки и кости относительно.
Сила смещения — сила, действующая из стороны в сторону.
Сила тяжести — сила притяжения объекта к поверхности Земли.
Синапс — структура, посредством которой нейрон посылает сигнал клетки-мишени.
Синаптический потенциал — проводимый возбудительный или тормозящий электрический потенциал, вызываемый в нейронах и мышечных волокнах нейромедиатором. Амплитуда синаптического потенциала зависит от возбудимости клетки и количества выделяемого нейромедиатора.
Синаптогенез — образование новых синапсов.
Синергия — группа мышц, вынужденных действовать в качестве единого целого для выполнения определенного задания.
Синовиальная мембрана — сосудистая мембрана, секретирующая синовиальную жидкость в полость сустава.
Синус — тригонометрическая функция, равная величине противоположной стороны, деленной на гипотенузу.
366
Синхронность — временная связь между потенциалами действия двигательных единиц. Синхронность обусловлена синаптическим импульсом, общим для участвующих двигательных нейронов.
Система контроля — ряд субсистем и взаимосвязанных каналов, обрабатывающих информацию. Субсистема принимает поступающие сообщения и обрабатывает их, прежде чем распределить поступившую информацию по системе. Существует три вида систем автоматического контроля: контролер обратной связи, контролер прямой связи и адаптивный контролер (Houk 1988). Контрольная система, используемая главным образом для компенсации нарушений, называется регулятором.
Скаляр — переменная, которая определяется только величиной.
Скалярное произведение — метод, применяемый для перемножения векторов с целью получения скалярного количества.
Скачкообразное проведение — распространение потенциала действия вдоль миелинизированного аксона вследствие деполяризации только одной аксолеммы.
Скорость — интенсивность изменения положения относительно времени (м-сек-1); производное положения относительно времени.
Скорость проведения — скорость распространения потенциала действия вдоль возбудимой мембраны.
Скорость сокращения длины — интенсивность сокращения длины мышцы.
Смещение — изменение положения. Единицей измерения является метр (м).
Содействующие реакции — адаптационные реакции, стабилизирующие систему перед выполнением движения.
Сократительная способность — способность элемента изменять длину.
Сократительный элемент — одна из составляющих модели мышцы Хилла. Характеризует способность сократительных белков производить силу.
Сокращающее растяжение задержка-расслабление агониста — растяжение, основанное на проприоцептивном улучшении нервно-мышечной передачи импульсов. Включает сочетание задержки—расслабления и растяжение сокращений агониста.
Сокращение — состояние активации мышцы, при котором возникают поперечные мостики в ответ на потенциал действия. Длина мышцы может уменьшаться, оставаться неизменной или увеличиваться в состоянии активации.
Сома — соматическая клетка нейрона.
Сопротивление давления — сопротивление, обусловленное сущностью потока жидкости вокруг объекта.
Сопротивление жцдкости — сопротивление жидкости на двигающийся через неё объект. Величина сопротивления зависит от вида жидкости и степени её возмущения движущимся объектом.
Составление — процесс определения результирующей нескольких сил.
Сохранение количества движения — закон, согласно которому, если система не подвергается импульсу, её количество движения остается постоянным.
Сочетание возбуждения—сокращения — электрохимические процессы, участвующие в превращении потенциала действия мышцы в механическую работу, выполняемую поперечными мостиками.
Спад — прерывистый тетанус, в котором сила снижается после первых 4—8 стимулов и затем снова повышается.
Спастичность — паталогически вызванное состояние повышенной возбудимости рефлекса растяжения.
Специфическое напряжение — сила, которую может произвести мышца на единицу площади поперечного сечения (н/см2).
Сплошной тетанус — плавная реакция силы, обусловленная высокочастотной активацией.
Средний мозг — компонент ствола мозга.
Стабильность — состояние равновесия, к которому возвращается система после пертурбации.
Статический анализ — механический анализ, в котором силы, действующие на систему, сбалансированы и, следовательно, система не испытывает ускорение.
Статическое растяжение — метод увеличения диапазона движения относительно сустава, основанный на удержании мышечного растяжения в течение 15—30 секунд.
Ствол головного мозга — составляющие ЦНС между головным и спинным мозгом.
Степень свободы (сустава) — ось вращения. Сустав, имеющий три степени свободы, может вращаться относительно трёх различных осей.
Стресс (нагрузка) — сила, действующая на единицу площади (Ра).
Структура интерференции — необработанная ЭМГ, включающая взаимное перекрытие потенциалов действия сотен мышечных волокон. Иногда называют “сырой” ЭМГ.
Структура центрального генератора — нервная сеть, способная образовывать структуры двигательных импульсов при отсутствии афферентного импульса.
367
Субталамные ядра — некоторые из базальных ядер.
Субфрагмент 1 — две шарообразные головки молекулы миозина. Одна содержит АТФ-свя-зывающий участок, другая — актин-связы-вающий участок.
Субфрагмент 2 — нешарообразный участок головки тяжелого меромиозина.
Суставная капсула — ткань вокруг поверхности синовиального сустава, отделяющая полость сустава от окружающих тканей.
Суставной рецептор — класс чувствительных нервных окончаний.
Суставной хрящ — ткань, включающая упруговязкий гель, “обогащенный” коллагеном.
Сухожильный рефлекс — реакция мышцы на выполнение удара по сухожилию.
Таламус — супросегментальная структура, интегрирующая и распределяющая большую часть двигательной и сенсорной информации, идущей к коре головного мозга.
Тангенс — тригонометрическая функция, равная величине противоположной стороны, деленной на величину примыкающей стороны.
Тангенциальная составляющая — составляющая, действующая параллельно и вдоль поверхности.
Тектоспинной мозг — нисходящий проводящий путь, по всей видимости, обеспечивающий реакции ориентации головы и шеи на визуальный стимул.
Тельце Пачини — нервное окончание с родительским аксоном 8—12 мм характеризуется низким порогом механической нагрузки и, по всей видимости, выявляет ускорение сустава.
Теорема Пифагора — а2 = в2 + с2.
Теория поперечных мостиков мышечного сокращения — теория, согласно которой мышечная сила, производимая во время сокращения, обусловлена действием поперечных мостиков.
Теория скольжения филаментов — скольжение тонкого и толстого филаментов относительно друг друга во время сокращения мышцы.
Территория двигательных единиц — субобъём мышцы, где располагаются мышечные волокна отдельной двигательной единицы.
Тетанус — силовая реакция мышцы (отдельного волокна, двигательных единиц) на серию возбудительных импульсов (потенциалы действия или электрический стимул); представляет собой суммацию реакций сокращения.
Тетродотоксин — препарат, блокирующий распространение потенциалов действия, блокируя каналы Na+.
Тиксотропия — свойство, проявляемое различными гелями, такими как мышца, заключается в превращении в жидкую субстанцию при встряхивании.
Титин — белок эндосаркомерного цитоскелета. Тонический — непрерывный, поддерживаемый. Тормозной интернейрон 1а — интернейрон, возбуждаемый афферентами группы Га и суп-распинальным импульсом. Оказывает тормозное влияние на другие нейроны, особенно двигательные.
Тормозной постсинаптический потенциал — ги-перполяризированный (тормозной) синаптический потенциал, вызываемый в постсинаптической мембране (дистально к синапсу).
Трансдукция — процесс превращения одного вида энергии в другой.
Трение — сопротивление относительному движению одного тела другому, с которым оно контактирует.
Трехфазный — имеющий три фазы.
Трехэлементная модель мышцы — механическая модель мышцы, состоящая из трёх элементов: параллельно упругих, последовательно упругих и сократительных.
Триада — одна поперечная трубочка и два близлежащих латеральных мешочка саркоплазматического ретикулума.
Тригонометрия — отрасль математики.
Тройной завиток коллагена — три переплетенные полипептидные цепочки, образующие молекулу коллагена.
Тропомиозин — белок тонкого филамента, участвующий в регуляции взаимодействий между актином и миозином.
Тропонин — трёхкомпонентная молекула, образующая часть тонкого филамента и участвующая в регуляции взаимодействий между актином и миозином.
Турбулентный поток — неравномерный поток жидкости вокруг объекта.
Тяжелая цепочка — белок с высоким молекулярным весом.
Тяжелый меромиозин — фрагмент молекулы миозина, заканчивающийся головкой. Может подразделяться на субфрагменты 1 и 2 (S1 и S2).
Угловое — движение, при котором не все части объекта испытывают одинаковое смешение.
Удельная проводимость — способность распространения волны возбуждения.
Упорядоченное рекрутирование — повторяющаяся последовательность активации двигательных единиц для выполнения определенного задания.
Упражнения с постепенным увеличением нагрузки — упражнения, в которых величина на
368
грузки постепенно увеличивается от одного цикла повторений к другому.
Упругая сила — пассивное свойство растянутого материала возвращаться к исходной длине.
Упруговязкий — материал, обладающий свойством упругости и вязкости.
Уравнение линии прерывистой крутизны — у = х + b
Ускорение — интенсивность изменения скорости относительно времени (м-сек~2); производное скорости относительно времени; наклон на графике скорость—время.
Утомление — острая адаптационная реакция, нарушающая двигательную деятельность.
Участок разрушения (разрыва) — участок, в котором вследствие чрезмерной деформации материал физически повреждается.
Фаза стойки — опорная фаза при ходьбе и беге. Феномен лестницы — постепенное увеличение силы сокращения, наблюдаемое при стимулировании серии сокращений. Другое название — трепп.
Фибрилла — основная несущая нагрузку единица сухожилия и связки, состоящая из пучков микрофибрилл, удерживаемых вместе благодаря поперечным соединениям. Количество и состояние последних определяют прочность (силу) соединительной ткани.
Фильтрование — “очищение” некоторых частот сигнала.
Фокальная точка (фокус) — точка, относительно которой все точки параболы находятся на одинаковом расстоянии.
Фономиография — метод регистрации звуков, возникающих при мышечных сокращениях. Другое название — звуковая или акустическая миография.
Фузимоторный — поскольку мышечное веретено имеет веретеновидную форму, Y-мотонейроны иногда называют фузимоторными нейронами, а их аксоны — фузимоторными волокнами.
фундаментальная частота — член, содержащий синус и косинус, характеризующий изменения сигнала за один цикл.
Функциональная площадь поперечного сечения — мера площади поперечного сечения мышцы, учитывающая пеннацию мышечных волокон. Измерение выполняется перпендикулярно длинной оси мышечных волокон.
Характеристика — типичная или отличительная черта. Часть логарифма, указывающая местонахождение запятой в десятичном числе.
Хвостатое ядро — одно из базальных ядер. Ходьба — режим локомоции человека.
Хроматолиз — растворение вещества Ниссля и ресинтез белков после аксотомии.
Центр давления — точка приложения силы реакции поверхности.
Центр тяжести — точка, относительно которой равномерно распределяется масса системы.
Центр удара — участок ракетки, при попадании мяча в который на руку не действует сила реакции.
Центральная команда — нервный сигнал, посылаемый корой головного мозга к низшим двигательным центрам (стволу мозга, спинному мозгу) о выполнении движения.
Центростремительная сила — сила, направленная к оси вращения и обуславливающая изменение линейного движения на угловое.
Церебромозжечок — латеральные полушария мозжечка. Принимает импульс из коры головного мозга и посылает импульс к зубчатому ядру.
Цикл растяжения—сокращения — эксцентричес-ки-концентрическая последовательность мышечной активации. Такая последовательность позволяет мышце выполнить больший объём работы во время концентрической фазы.
Цитоскелет — ряд структур, обеспечивающих физический каркас для взаимодействий сократительных белков.
Цифровой (дискретный) фильтр — метод управления частотным спектром импульса.
Черное вещество — одно из базальных ядер.
Чувствительное тельце Мейсснера — кожный механорецептор, поддерживающий давление.
Шаг — полный цикл походки человека.
Экзосаркомеры — цитоскелетные компоненты, поддерживающие латеральное расположение саркомеров.
Экзоцитоз — процесс слияния везикулы с пре-синаптической мембраной и последующее выделение нейромедиатора.
Экономия — минимальное количество энергии, необходимой для выполнения данного объема работы.
Экспонента — степень переменной. Показывает сколько раз переменную следует умножить на её значение, например у4 имеет экспоненту 4, то есть, у следует умножить на у четыре раза.
Экспоненциальная функция — имеет общий вид у=Ьх, где х — независимая переменная.
Экстерорецептор — чувствительное нервное окончание, выявляющее избранный внешний раздражитель вследствие взаимодействий системы с окружающей средой.
Экстрафузальное волокно — волокно скелетной мышцы, не являющееся частью мышечного веретена.
Эксцентрическое — механическое условие, при котором вращающий момент, обусловленный
24 8-133
369
нагрузкой, превышает вращающий момент мышцы, и вследствие этого длина активной мышцы увеличивается.
Электрический потенциал — напряжение.
Электрод — датчик, который может быть использован для измерения физического количества, например, концентрации (ионов, метаболитов) или электрического тока.
Электромиография — измерение электрической активности мышцы (потенциалов действия), обусловленной потенциалами действия аксонов.
Эндомизий — матрикс соединительной ткани вокруг отдельных мышечных волокон.
Эндосаркомеры — цитоскелетные компоненты, поддерживающие ориентацию тонкого и толстого филаментов в саркомере.
Энергия деформации — потенциальная энергия, накапливаемая системой при её растяжении. Еденица измерения — джоуль (Дж).
Эпимизий — внешний слой соединительной ткани, покрывающий всю мышцу.
Эффективность — отношение количества выполненной работы к количеству использованной энергии. Чем большее количество работы выполняется при данных энергозатратах, тем выше эффективность системы.
Эффективный синаптический ток — чистый эффект импульса на нейрон, измеряемый при помощи внутриклеточного микроэлектрода.
Эфферентационная копия — внутренние, связанные с командой сигналы, отменяющие сенсорные разряды вследствие двигательных команд (реафферентные сигналы) и не влияющие на сенсорные сигналы (эксафферент-ные сигналы), обусловленные внешними воздействиями.
Эфферентное волокно — аксон, передающий потенциалы действия из нейронов.
Литература
Adams, G.R., Duvoisin, M.R., & Dudley, G.A. (1992). Magnetic resonance imaging and electromyography as indexes of muscle function. Journal of Applied Physiology, 73, 1578-1583.
Adams, G.R., Harris, R.T., Woodard, D., & Dudley, G.A. (1993). Mapping of electrical muscle stimulation using MRI. Journal of Applied Physiology, 74, 532-537.
Adams, G.R., Hather, B.M., Baldwin, K.M., & Dudley, G.A. (1993). Skeletal muscle myosin heavy chain composition and resistance training. Journal of Applied Physiology, 74, 911-915.
Alexander, R.M. (1981). Mechanics of skeleton and tendons. In V.B. Brooks (Ed.), Handbook of physiology: Sec. 1. The nervous system Vol. 2. Motor control. (Pt. 1, pp. 17-42). Bethesda, MD: American Physiological Society.
Alexander, R.M. (1984a). Optimal strengths for bones liable to fatigue and accidental fracture. Journal of Theoretical Biology, 109, 621-636.
Alexander, R.M. (1984b). Walking and running. American Scientist, 72, 348-354.
Alexander, R.M. (1991). Optimum timing of muscle activation for simple models of throwing. Journal of Theoretical Biology, 150, 349-372.
Alexander, R.M., & Ker, R.F. (1990). The architecture of leg muscles. In J.M. Winters & S.L.-Y. Woo (Eds.), Multiple muscle systems: Biomechanics and movement organization (pp. 568-577). New York: Springer-Verlag.
Alexander, RM., & Vernon, A. (1975). The dimensions of the knee and ankle muscles and the forces they exert. Journal of Human Movement Studies, I, 115-123.
Al-Falahe, N.A., Nagaoka, M., & Vallbo, A.B. (1990). Response profiles of human muscle afferents during active finger movements. Brain, 113, 325-346.
Alford, E.K., Roy, R.R., Hodgson, J.A., & Edgerton, V.R. (1987). Electromyography of rat soleus, medial gastrocnemius, and tibialis anterior during hind limb suspension. Experimental Neurology, 96, 635-649.
Allen, D.G., Lee, J.A., & Westerblad, H. (1989). Intracellular calcium and tension during fatigue in isolated single muscle fibres from Xenopus Laevis. Journal of Physiology (London), 415, 433-458.
Alshuaib, W.B., & Fahim, M.A. (1990). Effect of exercise on physiological age-related change at mouse neuromuscular junctions. Neurobiology of Aging, 11, 555-561.
Alway, S.E., Grumbt, W.H., Stray-Gundersen, J., & Gonyea, W.J. (1992). Effects of resistance training on
elbow flexors of highly competitive bodybuilders. Journal of Applied Physiology, 72, 1512-1521.
Alway, S.E., MacDougall, J.D., & Sale, D.G. (1989). Contractile adaptations in the human triceps surae after isometric exercise. Journal of Applied Physiology, 66, 2725-2732.
Alway, S.E., Stray-Gundersen, J., Grumbt, W.H., & Gonyea, W.J. (1990). Muscle cross-sectional area and torque in resistance-trained subjects. European Journal of Applied Physiology, 60, 86-90.
Amar, J. (1920). The human motor. London: Routledge & Sons.
Amis, A.A., Dowson, D., & Wright, V. (1980). Elbow joint force predictions for some strenuous isometric actions. Journal of Biomechanics, 13, 765-775.
An, K.N., Hui, F.C., Morrey, B.F., Linscheid, R.L., & Chao, E.Y. (1981). Muscles across the elbow joint: A biomechanical analysis. Journal of Biomechanics, 10, 659-669.
An, K.N., Kwak, B.M., Chao, E.Y., & Morrey, B.F. (1984). Determination of muscle and joint forces: A new technique to solve the indeterminate problem. Journal of Biomechanical Engineering, 106, 364-367.
Anderson, S.A., & Cohn, S.H. (1985). Bone demineralization during space flight. Physiologist, 28, 212-217.
Andersson, G.B.J., Ortengren, R., & Nachemson, A. (1977). Intradiscal pressure, intra-abdominal pressure and myoelectric back muscle activity related to posture and loading. Clinical Orthopedics Research, 129, 156-164.
Antonio, J., & Gonyea, W’J. (1993). Skeletal muscle fiber hyperplasia. Medicine and Science in Sports and Exercise, 25, 1333-1345.
Aristotle. Progression of Animals. English translation by E.S. Forster (1968). Cambridge: Harvard University Press.
Armstrong, C.G., & Mow, V.C. (1980). Friction, lubrication and wear of synovial joints. In R. Owen, J. Goodfellow, & P. Bullough (Eds.), Scientific foundations of orthopaedics and traumatology (pp. 223-232). London: William Heinemann.
Armstrong, R.B. (1990). Initial events in exercise-induced muscular injury. Medicine and Science in Sports and Exercise, 22, 429-435.
Arsenault, A.B., & Chapman, A.E. (1974). An electromuo-graphic investigation of the individual recruitment of the quadriceps muscles during isometric contraction of the knee extensors in different patterns of movement. Physiotherapy of Canada, 26, 253-261.
24*
371
Asami, T., & Nolte, V. (1983). Analysis of powerful ball kicking. In H. Matsui & K. Kobayashi (Eds.), Biomechanics VIII-B (pp. 695-700). Champaign, IL: Human Kinetics.
Ashton-Miller, J.A., & Schultz, A.B. (1988). Biomechanics of the human spine and trunk. In K.B. Pandolf (Ed.), Exercise and sport sciences reviews (Vol. 16, pp. 169-204). New York: Macmillan.
Askew, M.J., & Mow, V.C. (1978). The biomechanical function of the collagen ultrastructure of articular cartilage. Journal of Biomechanical Engineering, 100, 105-113,
Asmussen, E. (1956). Observations on experimental muscle soreness. Acta Rheumatologica Scandinavica, 2,109-116.
Asmussen, E. (1953). Positive and negative muscular work. Acta Physiologica Scandinavica, 28, 365-382.
Atha, J. (1981). Strengthening muscle. In D.I. Miller (Ed.), Exercise and sport sciences reviews (Vol. 9, pp. 1-73). Philadelphia: Franklin Institute.
Atwater, A.E. (1970, April). Overarm throwing patterns: A kinematographic analysis. Paper presented at the national convention of the American Association for Health, Physical Education and Recreation, Seattle, WA.
Atwater, A.E. (1977, June). Biomechanics of throwing: Correction of common misconceptions. Paper presented at the joint meeting of the National College Physical Education Association for Men and the National Association for Physical Education of College Women, Orlando, FL.
Atwater, A.E. (1979). Biomechanics of overarm throwing movements and of throwing injuries. In R.S. Hutton & D.I. Miller (Eds.), Exercise and sport sciences reviews (Vol. 7, pp. 43-85). Philadelphia: Franklin Institute.
Babij, P., & Booth, F.W. (1988). Sculpturing new muscle phenotypes. News in Physiological Sciences, 3, 100-102.
Bailey, A.J., Robins, S.P., & Balian, G. (1974). Biological significance of the intermolecular crosslinks of collagen. Nature, 251, 105-109.
Baker, L.L., Bowman, B.R., & McNeal, D.R. (1988). Effects of waveform on comfort during neuromuscular electrical stimulation. Clinical Orthopaedics and Related Research, 233, 75-85.
Baldissera, F., Hultborn, H., & Illert, M. (1981). Integration of spinal neuronal systems. In V.B. Brooks (Ed.), Handbook of physiology: Sec. 1. The nervous system. Vol. 2. Motor control. (Pt. I, pp. 509-595). Bethesda, MD: American Physiological Society.
Barany, M. (1967). ATPase activity of myosin correlated with speed of muscle shortening. Journal of General Physiology, 50, 197-218.
Baratta, R., Solomonow, M., Zhou, B.H., Letson, D., Chuinard, R., & D'Ambrosia, R. (1987). Muscular coactivation: The role of the antagonist musculature in maintaining knee stability. American Journal of Sports Medicine, 16, 113-122.
Barclay, J.K. ( 1986). A delivery-independent blood flow effect on skeletal muscle fatigue. Journal of Applied Physiology, 61, 1084-1090.
Barr, J.O., Nielsen, D.H., & Soderberg, G.L. (1986). Transcutaneous electrical nerve stimulation characteristics for altering pain perception. Physical Therapy, 66, 1515-1521.
Bartee, H., & Dowell, L. (1982). A cinematographical analysis of twisting about the longitudinal axis when performers are free of support. Journal of Human Movement Studies, 8, 41-54.
Bartelink, D.L. (1957). The role of abdominal pressure in relieving the pressure on the intervertebral discs. Journal of Bone and Joint Surgery, 39B, 718-725.
Basmajian, J.V., & Latif, A. (1957). Integrated actions and functions of the chief flexors of the elbow. Journal of Bone and Joint Surgery, 39A, 1106-1118.
Bassett, C.A.L. (1968). Biological significance of piezoelectricity. Calcified Tissue Research, 1, 252-272.
Bates, B.T., Ostemig, L.R., Mason, B., & James, S.L. (1978). Lower extremity function during the support phase of running. In E. Asmussen & K. Jorgensen (Eds.), Biomechanics VIB (pp. 30-39). Baltimore: University Park Press.
Bawa, P., & Calancie, B. (1983). Repetitive doublets in human flexor carpi radialis muscle. Journal of Physiology (London), 339, 123-132.
Behm, D.G., & Sale, D.G. (1993). Intended rather than actual movement velocity determines velocity-specific training response. Journal of Applied Physiology, 74, 359-368.
Belanger, A.Y., & McComas, A.J. (1981). Extent of motor unit activation during effort. Journal of Applied Physiology, 51, 1131-1135.
Belanger, A.Y., McComas, A.J., & Elder, G.B.C. (1983). Physiological properties of two antagonistic human muscle groups. European Journal of Applied Physiology, 51, 381-393.
Belen'kii, V.Y., Gurfinkel, V.S., & Pal'tsev, Y.I. (1967). Elements of control of voluntary movements. Biophysics, 12, 154-61.
Bell, J., Bolanowski, S., & Holmes, M.H. (1994), The structure and function of Pacinian corpuscles: A review. Progress in Neurobiology, 42, 79-128.
Bellemare, P., & Garzaniti, N. (1988). Failure of neuromuscular propagation during human maximal voluntary contraction. Journal of Applied Physiology, 64, 1084-1093.
Bennett, K.M.B., & Castiello, U. (1994). Reach to grasp: Changes with age. Journal of Gerontology, 49, P1-P7.
Beigh, U., & Ekblom, B. (1979). Influence of muscle temperature on maximal muscle strength and power output in human skeletal muscles. Acta Physiologica Scandinavica, 107, 33-37.
Bergmark, A. (1989). Stability of the lumbar spine: A study in mechanical engineering. Acta Orthopaedica Scandinavica, 60 (Suppl. 230), 1-54.
Bernstein, N. (1984). Biodynamics of locomotion. In H.T.A. Whiting (Ed.), Human motor actions: Bernstein reassessed (pp. 171-222). New York: Elsevier.
Bertolasi, L., De Grandis, D., Bongiovanni, L.G., Zanette, G.P., & Gasperini, M. (1993). The influence of muscular lengthening on cramps. Annals of Neurology, 33, 176-180.
Beverly, M.C., Rider, T.A., Evans, MJ., & Smith, R. (1989). Local bone mineral response to brief exercise that stresses the skeleton. British Medical Journal, 299, 233-235.
Bhagat, B., & Wheeler, N. (1973). Effect of amphetamine on the swimming endurance of rats. Neuropharmacology, 12, 711-713.
Biewener, A.A. (1991). Musculoskeletal design in relation to body size. Journal of Biomechanics, 24 (Suppl. 1), 19-29.
Bigland, B., & Lippold, O.C.J. (1954). The relation between force, velocity and integrated electrical activity in human muscles. Journal of Physiology (London), 123, 214-224.
Bigland-Ritchie, B. (1981). EMG/force relations and fatigue of human voluntary contractions. In D.I. Miller (Ed.), Exercise and sport sciences reviews (Vol. 9, pp. 75-117). Philadelphia: Franklin Institute.
372
Bigland-Ritchie, В., Cafarelli, E., & Vollestad, N.K, (1986). Fatigue of submaximal static contractions. Acta Physiologica Scandinavica, 128 (Suppl. 556), 137-148.
Bigland-Ritchie, B., Dawson, N.J., Johansson, R.S., & Lippold, O.C.J. (1986). Reflex origin for the slowing of motoneurone firing rates in fatigue of human voluntary contractions. Journal of Physiology (London), 379, 451-459.
Bigland-Ritchie, B., Furbush, F., & Woods, J.J. (1986). Fatigue of intermittent submaximal voluntary contractions: Central and peripheral factors. Journal of Applied Physiology, 61, 421-429.
Bigland-Ritchie, B., Johansson, R., Lippold, O.C.J., Smith, S., & Woods, J.J. (1983). Changes in motoneurone firing rates during sustained maximal voluntary contractions. Journal of Physiology (London), 340, 335-346.
Bigland-Ritchie, B., Johansson, R., Lippold, O.CJ., & Woods, JJ. (1983). Contractile speed and EMG changes during fatigue of sustained maximal voluntary contractions. Journal of Neurophysiology, 50, 313-324.
Bigland-Ritchie, B., Kukulka, C.G., Lippold, O.CJ., & Woods, JJ. (1982). The absence of neuromuscular transmission failure in sustained maximal voluntary contractions. Journal of Physiology (London), 330,265-278.
Binder, M.D., Houk, J.C., Nichols, T.R., Rymer, W.Z., & Stuart, D.G. (1982). Properties and segmental actions of mammalian muscle receptors: An update. Federation Proceedings, 41, 2907-2918.
Binder, M.D., Kroin, J.S., Moore, G.P., & Stuart, D.G. (1977). The response of Golgi tendon organs to single motor unit contractions. Journal of Physiology (London), 271, 337-349.
Binder, M.D., & Mendell, L.M. (Eds.) (1990). The segmental motor system. New York: Oxford University Press.
Binder-Macleod, S.A., & Guerin, T. (1990). Preservation of force output through progressive reduction of stimulation frequency in human quadriceps femoris muscle. Physical Therapy, 70, 619-625.
Binkhorst, R.A., Hoofd, L., & Vissers, A.C.A. (1977). Temperature and force-velocity relationship of human muscles. Journal of Applied Physiology, 42, 471-475.
Bishop, B. (1974). Vibratory stimulation: 1. Neurophysiology of motor responses evoked by vibratory stimulation. Physical Therapy, 54, 1273-1282.
Bishop, B. (1975a). Vibratory stimulation: 2. Vibratory stimulation as an evaluation tool. Physical Therapy, 55, 28-34.
Bishop, B. (1975b). Vibratory stimulation: 3. Possible applications of vibration in treatment of motor dysfunctions. Physical Therapy, 55, 139-143.
Bishop, R.D., & Hay, J.G. (1979). Basketball: The mechanics of hanging in the air. Medicine and Science in Sports, 11, 274-277.
Bizzi, E., & Abend, W. (1983). Posture control and trajectory formation in single- and multijoint arm movements. In J.E. Desmedt (Ed.), Motor control mechanisms in health and disease (pp. 31-45). New York: Raven Press.
Blange, T., Karemaker, J.M., & Kramer, A.EJ.L. (1972). Elasticity as an expression of crossbridge activity in rat muscle. Pflugefs Archiv, 336, 277-288.
Blattner, S.E., & Noble, L. (1979). Relative effects of isokinetic and plyometric training on vertical jumping ^rfownance. Research Quarterly, 50, 583-588.
Blinks, J.R., Rudel, R., & Taylor, S.R. (1978). Calcium transients in isolated amphibian skeletal muscle fibres:
Detection with aequorin. Journal of Physiology (London), 277, 291-323.
Blomstrand, E., Celsing, F., & Newshoeme, E.A. (1988). Changes in plasma concentrations of aromatic and branched-chain amino acids during sustained exercise in man and their possible role in fatigue. Acta Physiologica Scandinavica, 133, 115-121.
Bloom, W., & Fawcett, D.W. (1968). A textbook of histology (9th ed.). Philadelphia: Saunders.
Bobath, B. (1978). Adult hemiplegia: Evaluation and treatment. London: Heinemann Medical Books.
Bobbert, M.F., Schamhardt, H.C., & Nigg, B.J. (1991). Calculation of vertical ground reaction force estimates during running from positional data. Journal of Biomechanics, 24, 1095-1105.
Bodine, S.C., Garfinkel, A., Roy, R.R., & Edgerton, V.R. (1988). Spatial distribution of motor unit fibers in the cat soleus and tibialis anterior muscles: Local interactions. Journal of Neuroscience, 8, 2142-2152.
Bodine, S.C., Roy, R.R., Eldred, E., & Edgerton, V.R. (1987). Maximal force as a function of anatomical features of motor units in the cat tibialis anterior. Journal of Neurophysiology, 57, 1730-1745.
Bongiovanni, L.G., & Hagbarth, K.-E. (1990). Tonic vibration reflexes elicited during fatigue from maximal voluntary contractions in man. Journal of Physiology (London), 423, 1-14.
Borelli, A. (1680). De Motu Animalium. Rome: Superiorum Permissu.
Bosco, C., & Komi, P.V. (1980). Influence of aging on the mechanical behavior of leg extensor muscles. European Journal of Applied Physiology, 45, 209-219.
Botterman, B.R., Binder, M.D., & Stuart. D.G. (1978). Functional anatomy of the association between motor units and muscle receptors. American Zoologist, 18, 135-152.
Botterman, B.R., Iwamoto, G.A., & Gonyea, W.J. (1986). Gradation of isometric tension by different activation rates in motor units of cat flexor carpi radialis muscle. Journal of Neurophysiology, 56, 494-505.
Bouisset, S., & Zattara, M. (1990). Segmental movement as a perturbation to balance? Facts and concepts. In: J.M. Winters & S.L.-Y. Woo (Eds.). Multiple muscle systems: Biomechanics and movement organization (pp. 498-506). New York: Springer-Verlag.
Bowditch, H.P. (1871). Ueber die Eigenthuemlichkeiten der Reizbarkeit, welch die Muskelfasem des Herzens zeigen. Berichte Mathematische Physiologic, 23,652-689.
Boulay, M.R., Lortie, G., Simoneau, J.A., Hamel, P., Leblanc, C., & Bouchard, C. (1985). Specificity of aerobic and anaerobic work capacities and powers. International Journal of Sports Medicine, 6, 325-328.
Brancazio, P.J. (1984). Sport science: Physical laws and optimum performance. New York: Simon & Schuster.
Brand, R.A., Crowninshield, R.D., Wittstock, C.E., Pedersen, D.R., Clark, C.R., & van Krieken, F.M. (1982). A model of lower extremity muscular anatomy. Journal of Biomechanical Engineering, 104, 304-310.
Brand, R.A., Pedersen, D.R., & Friederich, J.A. (1986). The sensitivity of muscle force predictions to changes in physiologic cross-sectional area. Journal of Biomechanics, 19, 589-596.
Braune, W., & Fischer, O. (1889). Uber den Schwerpunkt des menschlichen Korpers mit Rucksicht auf die Ausrustung des deutschen Infanteristen. Abhandlungen der Mathematisch-Physischen Klasse der Saecksischen Akademie der Wissenschaften, 26, 561-672.
Bremner, F.D., Baker, J.R., & Stephens, J.A. (1991a). Correlation between the discharges of motor units
373
recorded from the same and from different finger muscles in man. Journal of Physiology (London), 432, 355-380.
Bremner, F.D., Baker, J.R., & Stephens, J.A. (1991b). Effect of task on the degree of synchronization of intrinsic hand muscle motor units in man. Journal of Neurophysiology, 66, 2072-2083.
Bremner, F.D., Baker, J.R., & Stephens. J.A (1991c). Variation in the degree of synchrony exhibited by motor units lying in different finger muscles in man. Journal of Physiology (London), 432, 381-399.
Brighton, C.T. (1981). Current concept review: The treatment of non-unions with electricity. Journal of Bone and Joint Surgery, 63A, 847-856.
Broberg, S., & Sahiin, K. (1989). Adenine nucleotide degradation in human skeletal muscle during prolonged exercise. Journal of Applied Physiology, 67, 116-122.
Brooke, M.H., & Kaiser, K.K. (1974). The use and abuse of muscle histochemistry. Annals of the New York Academy of Sciences, 228, 121-144.
Brooks, S.V., & Faulkner, J.A. (1991). Forces and powers of slow and fast skeletal muscle in mice during repeated contractions. Journal of Physiology (London), 436, 701-710.
Brooks, V.B. (1986). The neural basis of motor control. New York: Oxford University Press.
Brown, A.B., McCartney, N., & Sale, D.G. (1990). Positive adaptations to weightlifting training in the elderly. Journal of Applied Physiology, 69, 1725-1733.
Brown, G.L., & von Euler, U.S. (1938). The after effects of a tetanus on mammalian muscle. Journal of Physiology (London), 93, 39-60.
Buchanan, T.S., Rovai, G.P., & Rymer, W.Z. (1989). Strategies for muscle activation during isometric torque generation at the human elbow. Journal of Neurophysiology, 62, 1201-1212.
Bulbulian, R., & Bowles, D.K. (1992). Effect of downhill running on motoneuron pool excitability. Journal of Applied Physiology, 73, 968-973.
Burgeson, R.E., & Nimni, M.E. (1992). Collagen types. Molecular structure and tissue distribution. Clinical Orthopaedics, 282, 250-272.
Burgess, P.R., Horch, K.W., & Tuckett, R.P. (1987). Mechanoreceptors. In G. Adelman (Ed.), Encyclopedia of neuroscience (Vol. 2, pp. 620-621). Boston: Birkhauser.
Burke, D. (1988). Spasticity as an adaptation to pyramidal tract injury. Advances in Neurology, 47, 401-422.
Burke, J.E., Kamen, G., & Koceja, D.M. (1989). Long-latency enhancement of quadriceps excitability from stimulation of skin afferents in young and old adults. Journal of Gerontology, 44, M158-M163.
Burke, R.E. (1981). Motor units: Anatomy, physiology, and functional organization. In V.B. Brooks (Ed.), Handbook of Physiology: Sec. L The nervous system; Vol. 2. Motor control (Pt. I, pp. 345-422). Bethesda, MD: American Physiological Society.
Burke, R.E. (1985). Integration of sensoiy information and motor commands in the spinal cord. In P.S.G. Stein (Organizer). Motor control: From movement trajectories to neural mechanisms (pp. 44-66). Class at the Society for Neuroscience: Washington, DC.
Burke, R.E., & Edgerton, V.R. (1975). Motor unit properties and selective involvement in movement. In J.H. Wilmore & J.F. Koegh (Eds.), Exercise and sport sciences reviews (Vol. 3, pp. 31-81). New York: Academic.
Burke, R.E., Levine, D.N., Tsairis, P., & Zajac, F.E. (1973). Physiological types and histochemical profiles
in motor units of the cat gastrocnemius. Journal of Physiology (London), 234, 723-748.
Burke, R.E., Rudomin, P., & Zajac, F.E., III (1970). Catch property in single mammalian motor units. Science, 168, 122-124.
Burke, R.E., & Tsairis, P. (1973). Anatomy and innervation ratios in motor units of cat gastrocnemius. Journal of Physiology (London), 234, 749-765.
Burke, R.E., & Tsairis, P. (1974). The correlation of physiological properties with histochemical characteristics in single muscle units. Annals of the New York Academy of Sciences, 228, 145-159.
Burstein, A.H., Reilly, D.T., & Martens, M. (1976). Aging bone tissue: Mechanical properties. Journal of Bone and Joint Surgery, 58A, 82-86.
Butler, A.J., & Darling, W.G. (1990). Reflex changes accompanying isometric strength training of the contralateral limb. Society for Neuroscience Abstracts, 16, 884.
Butler, D.L., Grood, E.S., Noyes, F.R., & Zernicke, R.F. (1978). Biomechanics of figaments and tendons. In R.S. Hutton (Ed.), Exercise and sport sciences reviews (Vol. 6, pp. 125-181). Philadelphia: Franklin Institute.
Byrnes, W.C., Clarkson, P.M., White, J.S., Hsieh, S.S., Frykman, P.N., & Maughan, R.J. (1985). Delayed onset muscle soreness following repeated bouts of downhill running. Journal of Applied Physiology, 59, 710-715.
Cafarelli, E. (1988). Force sensation in fresh and fatigued human skeletal muscle. In K.B. Pandolf (Ed.), Exercise and sport sciences reviews (Vol. 16, pp. 139-168). New York: Macmillan.
Caiozzo, V.J., Herrick, R.E., & Baldwin, K.M. (1991). Influence of hyperthyroidism on maximal shortening velocity and myosin isoform distribution in skeletal muscle. American Journal of Physiology, 261, C285-C295.
Caiozzo, V.J., Herrick, R.E., & Baldwin, K.M. (1992). Response of slow and fast muscle to hypothyroidism: Maximal shortening velocity and myosin isoforms. American Journal of Physiology, 263, C86-C94.
Calancie, B. (1991). Interlimb reflexes following cervical spinal cord injury in man. Experimental Brain Research, 85, 458-469.
Calvert, T.W., & Chapman, A.E. (1977). Relationship between the surface EMG and force transients in muscle: Simulation and experimental studies. Proceedings of the IEEE, 65, 682-689.
Campbell, M.J., McComas, A.J., & Petito, F. (1973). Physiological changes in ageing muscle. Journal of Neurology, Neurosurgery, and Psychiatry, 36, 174-182.
Carolan, B., & Cafarelli, E. (1992). Adaptations in coactivation after isometric resistance training. Journal of Applied Physiology, 73, 911-917.
Carter, D.R. (1984), Mechanical loading histories and cortical bone remodeling. Calcified Tissue International, 36, S19-S24.
Carter, D.R., & Spengler, D.M. (1978). Mechanical properties and composition of cortical bone. Clinical Orthopaedics and Related Research, 135, 192-217.
Cavagna, G.A. (1977). Storage and utilization of elastic eneigy in skeletal muscle. In R.S. Hutton (Ed.), Exercise and sport sciences reviews (Vol. 5, pp. 89-129). Santa Barbara, CA Journal Publishing Affiliates.
Cavagna, G.A, & Citterio, G. (1974). Effect of stretching on the elastic characteristics of the contractile component of the frog striated muscle. Journal of Physiology (London), 239, 1-14.
374
Cavagna, G.A. Saibene, F.P., & Margaria, R. (1964). Mechanical work in running. Journal of Applied Physiology, 19, 249-256.
Cavanagh, P.R. (Ed.) (1990). Biomechanics of distance running. Champaign, IL: Human Kinetics.
Cavanagh, P.R., & Ae, M. (1980). A technique for the display of pressure distributions beneath the foot. Journal of Biomechanics, 13, 69-76.
Cavanagh, P.R., Andrew, G.C., Kram, R., Rodgers, M.M., Sanderson, D.J., & Henning, E.M. (1985). An approach to biomechanical profiling of elite distance runners. International Journal of Sport Biomechanics, 1, 36-62.
Cavanagh, P.R., & Grieve, D.W. (1973). The graphical display of angular movement of the body. British Journal of Sports Medicine, 7, 129-133.
Cavanagh, P.R., & Kram, R. (1989). Stride length in distance running: Velocity, body dimensions, and added mass effects. Medicine and Science in Sports and Exercise, 21, 476-479.
Cavanagh, P.R., & Lafortune, M.A. (1980). Ground reaction forces in distance running. Journal of Biomechanics, 13, 397-406.
Cavanagh, P.R., Pollock, M.L., & Landa, J. (1977). A biomechanical comparison of elite and good distance runners. Annals of New York Academy of Sciences, 301, 328-345.
Chandler, R.F., Clauser, C.E., McConville, J.T., Reynolds, H.M., & Young, J.W. (1975). Investigation of inertial properties of the human body (AMRL-TR-74-137 as issued by the National Highway Traffic Safety Administration). Wright-Patterson Air Force Base, OH: Aerospace Medical Research Laboratories, Aerospace Medical Division. (NTIS No. AD-A016 485)
Chanaud, C.M., Pratt, C.A., & Loeb, G.E. (1987). A multiple-contact EMG recording array for mapping single muscle unit territories. Journal of Neuroscience Methods, 21, 105-112.
Chao, E.Y.S. (1986). Biomechanics of the human gait. In G.W, Schmid Schonbein, S.L.-Y. Woo, & B.W. Zweifach (Eds.), Frontiers in biomechanics (pp. 225-244). New York: Springer.
Chaouloff, F., Kennett, G.A., Serrurier, B., Merino, D., & Curson, G. (1986). Amino acid analysis demonstrates that increased plasma free tryptophan causes the increase of brain tryptophan during exercise in the rat. Journal of Neurochemistry, 46, 1647-1650.
Chase, P.B., & Kushmerick, M.J. (1988). Effects of pH on contraction of rabbit fast muscle and slow skeletal muscle fibers. Biophysical Journal, 53, 935-946.
Cheney, P.D. (1985). Role of cerebral cortex in voluntary movements: A review. Physical Therapy, 65, 624-635.
Chi, M.M.-Y., Hintz, C.S., Coyle, E.F., Martin, W.H., Ivy, J.L., Nemeth, P.M., Holloszy, J.O., & Lowry, O.H. (1983). Effects of detraining on enzymes of energy metabolism in individual human muscle fibers. American Journal of Physiology, 244, C276-C287.
Cholewicki, J., McGill, S.M., & Norman, R.W. (1991). Lumbar spine loads during the lifting of extremely heavy weights. Medicine and Science in Sports and Exercise, 23,1179-1186.
Christakos, C.N. (1982). Astudy of the muscle force waveform using a population stochastic model of skeletal muscle. Biological Cybernetics, 44, 91-106.
Christakos, C.N., & Windhorst, V. (1986). Spindle gain increase during muscle unit fatigue. Brain Research, 365, 388-392.
Claassen, H., Gerber, C., Hoppeler, H., Lillthi, J.-M., & Vock, P. ( 1989). Muscle filament spacing and short
term heavy-resistance exercise in humans. Journal of Physiology (London), 409, 491-495.
Clark, J.M., & Haynor, D.R. (1987). Anatomy of the abductor muscles of the hip as studied by computed tomography. Journal of Bone and Joint Surgery, 69A, 1021-1031.
Clarke, T.E., Frederick, E.G., & Hamill, C.L. (1983). The effect of shoe design upon rearfoot control in running. Medicine and Science in Sports and Exercise, 15, 376-381.
Clarkson, P.M., Cymes, W.C., McCarmick, K.M., Turcotte, L.P., & White, J.S. (1986). Muscle soreness and serum creatine kinase activity following isometric, eccentric, and concentric exercise. International Journal of Sports Medicine, 7, 152-155.
Clarkson, P.M., & Tremblay, 1. (1988). Exercise-induced muscle damage, repair, and adaptation in humans. Journal of Applied Physiology, 65, 1-6.
Clauser, C.E., McConville, J.T., & Young, J.W. (1969). Weight, volume and center of mass of segments of the human body (AMRL-TR-69-70). Wright-Patterson Air Force Base, OH: Aerospace Medical Research Laboratories, Aerospace Medical Division.
Clement, D.B., Taunton, J.E., Smart, G.W., & McNicol, K.L. (1981). A survey of overuse running injuries. Physician and Sports Medicine, 9, 47-58.
Coggan, A.R., & Coyle, E.F. (1987). Reversal of fatigue during prolonged exercise by carbohydrate infusion or ingestion. Journal of Applied Physiology, 63,2388-2395.
Cole, K.J. (1991). Grasp force control in older adults. Journal of Motor Behavior, 23, 251-258.
Colleti, L.A., Edwards, J., Gordon, L., Shary, J., & Bell, N.H. (1989). The effects of muscle-building exercise on bone mineral density of the radius, spine, and hip in young men. Calcified Tissue International, 45, 12-14.
Colliander, E.B., & Tesch, F.A. (1990). Effects of eccentric and concentric muscle actions in resistance training. Acta Physiologica Scandinavica, 140, 31-39.
Collins, J.J., & O'Connor, J.J. (1991). Muscle-ligament interaction at the knee during walking. Journal of Engineering in Medicine, 205, 11-18.
Condon, S.M., & Hutton, R.S. (1987). Soleus muscle electromyographic activity and ankle dorsiflexion range of motion during four stretching procedures. Physical Therapy, 67, 24-30.
Cooke, P. (1985). A periodic cytoskeletal lattice in striated muscle. In J.W. Shay (Ed.), Cell and muscle motility (Vol. 6, pp. 287-313). New York: Plenum.
Cooke, R. (1990). Force generation in muscle. Current Opinion in Cell Biology, 2, 62-66.
Cooke, R., Franks, K., Luciani, G.B., & Pate, E. (1988). The inhibition of rabbit skeletal muscle contraction by hydrogen ions and phosphate. Journal of Physiology (London), 395, 77-97.
Cope, T.C., Bodine, S.C., Fournier, M., & Edgerton, V.R. (1986). Soleus motor units in chronic spinal transected cat: Physiological and morphological alterations. Journal of Neurophysiology, 55, 1201-1220.
Corcos, D.M., Gottlieb, G.L., & Agarwal, G.C. (1989). Organizing principles for single-joint movements: 2. A speed-sensitive strategy. Journal of Neurophysiology, 62, 358-368.
Cordo, P.J., & Nashner, L.J. (1982). Properties of postural adjustments associated with rapid arm movements. Journal of Neurophysiology, 47, 287-302.
Costill, D.L., & Hargreaves.M. (1992). Carbohydrate nutrition and fatigue. Sports Medicine, 13, 86-92.
Cote', L., & Crutcher, M.D. (1991). The basal ganglia. In E.R. Kandel, J.H. Schwartz, & T.M. Jessell (Eds.),
375
Principles of neural science (3rd ed., pp. 647-659). New York: Elsevier.
Cowin, S.C. (1983). The mechanical and stress adaptive properties of bone. Annals of Biomedical Engineering, 11, 263-295.
Coyle, E.F., Coggan, A.R., Hemmert, M.K., & Ivy, J.L. (1986). Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate, Journal of Applied Physiology, 61, 165-172.
Crago, P.E., Houk, J.C., & Hasan, Z. (1976). Regulatory actions of human stretch reflex. Journal of Neurophysiology, 39, 925-935.
Crenna, P., & Frigo, C. (1984). Evidence of phasedependent nociceptive reflexes during locomotion in man. Experimental Neurology, 85, 336-345.
Crenna, P., & Frigo, C. (1987). Excitability of the soleus H-reflex arc during walking and stepping in man. Experimental Brain Research, 66, 49-60.
Cresswell, A.G. (1993). Responses of intra-abdominal pressure and abdominal muscle activity during dynamic trunk loading in man. European Journal of Applied Physiology, 66, 315-320.
Cresswell, A.G., Grundstrom, H., & Thorstensson, A. (1992). Observations on intra-abdominal pressure and patterns of abdominal intra-muscular activity in man. Acta Physiologica Scandinavica, 144, 409-418.
Crisco, J.J., III, & Panjabi, M.M. (1990). Postural biomechanical stability and gross muscular architecture in the spine. In J.M. Winters & S.L.-Y. Woo (Eds.), Multiple muscle systems: Biomechanics and movement organization (pp. 438-450). New York: Springer-Verlag.
Crone, C., & Nielsen, J. (1989). Methodological implications of the post activation depression of the soleus H-reflex in man. Experimental Brain Research, 78, 28-32.
Crowninshield, R.D., & Brand, R.A. (1981). The prediction of forces in joint structures: Distribution of intersegmental resultants. InD.I. Miller (Ed.), Exercise and sport science reviews (Vol. 9, pp. 159-181). Philadelphia: Franklin Institute.
Cutts, A. (1988). The range of sarcomere lengths in the muscles of the human lower limb. Journal of Anatomy, 160, 79-88.
Czeh, G., Gallego, R., Kudo, N., & Kuno, M. (1978). Evidence for the maintenance of motoneurone properties by muscle activity. Journal of Physiology (London), 281, 239-252.
Czemiecki, J.M., Gitter, A., & Munro, C. (1991). Joint moment and muscle power output characteristics of below knee amputees during running: The influence of energy storing prosthetic feet. Journal of Biomechanics, 24, 63-75.
Dahl, H.A., Aaserud, R., & Jensen, J. (1992). Muscle hypertrophy after light and heavy resistance training. Medicine and Science in Sports and Exercise, 24, S55.
Dahekvist, N.J., Mayo, P., & Seedhom, B.B. (1982). Forces during squatting and rising from a deep squat. Engineering in Medicine, 11, 69-76.
Dainis, A. (1981). Amodel for gymnastics vaulting. Medicine and Science in Sports and Exercise, 13, 34-43.
Dalen, N., & Olsson, K.E. (1974). Bone mineral content and physical activity. Acta Orthopaedica Scandinavica, 45, 170-174.
Daniels, J.T. (1985). A physiologist's view of running economy. Medicine and Science in Sports and Exercise, 17, 332-338.
Darling, W.G., & Cole, K.J. (1990). Muscle activation patterns and kinetics of human index finger movements. Journal of Neurophysiology, 63, 1098-1108.
Darling, W.G., & Hayes, K.C. (1983). Human servo responses to load disturbances in fatigued muscle. Brain Research, 267, 345-351.
Datta, A.K., & Stephens, J.A. (1990). Synchronization of motor unit activity during voluntary contraction in man. Journal of Physiology (London), 422, 397-419.
Davies, C.T.M., Dooley, P., McDonagh, M.J.N., & White, M.J. (1985). Adaptation of mechanical properties of muscle to high force training in man. Journal of Physiology (London), 365, 277-284.
Davies, C.T.M., Rutherford, I.C., & Thomas, D.O. (1987). Electrically evoked contractions of the triceps surae during and following 21 days of voluntary leg immobilization. European Journal of Applied Physiology, 56, 306-312.
Davies, C.T.M., & Young, K. (1983). Effect of temperature on the contractile properties and muscle power of triceps surae in humans. Journal of Applied Physiology, 55,191 -195.
Davies, J., Parker, D.F., Rutherford, O.M., & Jones, D.A (1988). Changes in strength and cross sectional area of elbow flexors as a result of isometric strength training. European Journal of Applied Physiology, 57, 667-670.
Davis, B.L., & Cavanagh, P.R. (1993). Simulating reduced gravity: A review of biomechanical issues pertaining to human locomotion. Aviation, Space, and Environmental Medicine, 64, 557-566.
Davis, B.L., & Vaughan, C.L. (1993). Phasic behavior of EMG signals during gait: Use of multivariate statistics. Journal of Electromyography and Kinesiology, 3, 51-60.
Dawson, M.J., Gadian, D.G., & Wilkie, D.R. (1980). Mechanical relaxation rate and metabolism studied in fatiguing muscle by phosphorus nuclear magnetic resonance. Journal of Physiology (Lohdon), 299, 465-484.
Day, B.L., Marsden, C.D., Obeso, J.A., & Rothwell, J.C. (1984). Reciprocal inhibition between the muscles of the human forearm. Journal of Physiology (London), 349, 519-534.
de Haan, A., van Ingen Schenau, G.J., Ettema, G.J., Huijring, P.A., & Lodder, M.A.N. (1989). Efficiency of rat medial gastrocnemius muscle in contractions with and without an active prestretch. Journal of Experimental Biology, 141, 327-341.
de Koning, F.L., Binkhorst, R.A., Vissers, A.C.A., & van't Hof, M.A. (1985). The force-velocity relationship of arm flexion in untrained males and females and arm-trained athletes. European Journal of Applied Physiology, 54, 89-94.
de Koning, J.J., de Groot, G., & van Ingen Schenau, G.J. (1992). Ice friction during speed skating. Journal of Biomechanics, 25, 565-571.
Delitto, A., Brown, M., Strube, M.J., Rose, S.J., & Lehman, R.C. (1989). Electrical stimulation of quadriceps femoris in an elite weight lifter: A single subject experiment. International Journal of Sports Medicine, 10, 187-191.
Delitto, A., & Rose, S.J. (1986). Comparative comfort of three waveforms used in electrically eliciting quadriceps femoris muscle contractions. Physical Therapy, 66, 1704-1707.
De Looze, M.P., Busemann, J.B.J., Kingma, 1., & Toussaint, H.M. (1992). Different methods to estimate total power and its components during lifting. Journal of Biomechanics, 25, 1089-1095.
DeLorme, T.L. (1945). Restoration of muscle power by heavy-resistance exercises. Journal of Bone and Joint Surgery, 27, 645-667.
376
DeLorme, T.L. (1946). Heavy resistance exercises. Archives of Physical Medicine, 27, 607-630.
De Luca, C.J. (1984). Myoelectric manifestations of localized muscular fatigue in humans. CRC Critical Reviews in Biomedical Engineering, 11, 251-279.
Dempster, W.T. (1955). Space requirements of the seated operator (WADC-TR-55-159). Wright-Patterson Air Force Base, OH: Aerospace Medical Research Laboratory. (NTIS No. AD-87892 as issued by the United States Air Force).
Dempster, W.T. (1961). Free-body diagrams as an approach to the mechanics of human posture and motion. In G.F. Evans (Ed.), Biomechanical studies of the musculoskeletal system (pp. 81-135). Springfield, IL: Charles C Thomas.
Denny-Brown, D. (1949). Interpretation of the electromyogram. Archives of Neurology and Psychiatry, 61, 99-128.
Desmedt, J.E., & Godaux, E. (1977). Fast motor units are not preferentially activated in rapid voluntary contractions in man. Nature, 267, 717-719.
Devine, K.L., LeVeau, B.F., & Yack, H.J. (1981). Electromyographic activity recorded from an unexercised muscle during maximal isometric exercise of the contralateral agonists and antagonists. Physical Therapy, 61, 898-903.
DeVita, P., & Skelly, W.A. (1992). Effect of landing stiffness on joint kinetics and energetics in the lower extremity.
Medicine and Science in Sports and Exercise, 24, 1 OS-115.
Dick, R.W., & Cavanagh, P.R. (1987). An explanation of the upward drift in oxygen uptake during prolonged submaximal downhill running. Medicine and Science in Sports and Exercise, 19, 310-317.
Dietz, V. (1978). Analysis of the electrical muscle activity during maximal contraction and the influence of ischaemia. Journal of the Neurological Sciences, 37, 187-197.
Dietz, V. (1992). Human neuronal control of automatic functional movement: Interaction between central programs and afferent input. Physiological Reviews, 72, 33-69.
Dietz, V., & Berger, W. (1982). Spatial coordination of b.lateral leg muscle activity during balancing. Experimental Brain Research, 47, 172-176.
Dimitrijevic, M.R., McKay, W.B., Sarjanovic, 1., Sherwood, A.M., Svirtlih, L., & Vrbova, G. (1992). Coactivation of ipsi- and contralateral muscle groups during contraction of ankle dorsiflexors. Journal of the Neurological Sciences, 109, 49-55.
di Prampero, P.E. (1986). The energy cost of human locomotion on land and in water. International Journal of Sports Medicine, 7, 55-72.
Doherty, T.J., Vandervoort, A.A., & Brown, W.F. (1993). Effects of aging on the motor unit: A brief review. Canadian Journal of Applied Physiology, 18, 331-358.
Doherty, T.J., Vandervoort, A.A., Taylor, A.W., & Brown, W.F. (1993). Effects of motor unit losses on strength in older men and women. Journal of Applied Physiology, 69, 2004-2011.
Dooley, P.C., Bach, T.M., & Luff, A.R. (1990). Effect of vertical jumping on the medial gastrocnemius and soleus muscles of rats. Journal of Applied Physiology, 69,2004-2011.
Dorlochter, M., Irintchev, A., Brinkers, M., & Wemig, A (1991). Effects of enhanced activity on synaptic transmision in mouse extensor digitorum longus muscle. Journal of Physiology (London), 436, 283-292.
Duchateau, J., & Hainaut, K. (1984). Isometric or dynamic training: Differential effects on mechanical
properties of a human muscle. Journal of Applied Physiology, 56, 296-301.
Duchateau, J., & Hainaut, K. (1986). Nonlinear summation of contractions in striated muscle: 1. Twitch potentiation in human muscle. Journal of Muscle Research and Cell Motility, 7, 11-17.
Duchateau, J., & Hainaut, K. (1987). Electrical and mechanical changes in immobilized human muscle. Journal of Applied Physiology, 62, 2168-2173.
Duchateau, J., & Hainaut, K. (1988). Training effects of submaximal electrostimulation in a human muscle. Medicine and Science in Sports and Exercise, 20, 99-104.
Duchateau, J., & Hainaut, K. (1990). Effects of immobilization on contractile properties, recruitment and firing rates of human motor units. Journal of Physiology (London), 422, 55-65.
Duchateau, J., & Hainaut, K. (1991). Effects of immobilization on electromyogram power spectrum changes during fatigue. European Journal of Applied Physiology, 63, 458-462.
Duchateau, J., & Hainaut, K. (1993). Behaviour of short and long latency reflexes in fatigued human muscles. Journal of Physiology (London), 471, 787-799.
Duchene, J., & Goubel, F. (1993). Surface electromyogram during voluntary contraction: Processing tools and relation to physiological events. Critical Reviews in Biomedical Engineering, 21, 313-397.
Dudley, G.A., & Djamil, R. (1985). Incompatibility of endurance- and strength-training modes of exercise. Journal of Applied Physiology, 59, 1446-1451.
Dudley, G.A., Duvoisin, M.R., Adams, G.R., Meyer, R.A., Belew, A.H., & Buchanan, P. (1992). Adaptations to unilateral lower limb suspension in humans. Aviation, Space, and Environmental Medicine, 63, 678-683.
Dudley, G.A., Tesch, P.A., Miller, B.J., & Buchanan, P. (1991). Importance of eccentric muscle actions in performance adaptations to resistance training. Aviation, Space, and Environmental Medicine, 62, 543-550.
Dum, R.P., & Kennedy, T.T. (1980). Physiological and histochemical characteristics of motor units in cat tibialis anterior and extensor digitorum longus muscles. Journal of Neurophysiology, 43, 1615-1630.
Dyck, P.J., Kames, J., O'Brien, P.O., & Zimmerman, LR. (1984). Detection thresholds of cutaneous sensation in humans. In P.J. Dyck, P.K. Thomas, E.H. Lambert, & R. Runge (Eds,), Peripheral neuropathy (Vol 1, pp. 1103-1138). Philadelphia: Saunders.
Edgerton, V.R., Apor, P., & Roy, R.R. (1990). Specific tension of human elbow flexor muscles. Acta Physiolo-gica Hungarica, 75, 205-216.
Edgerton, V.R., & Roy, R.R. (1991). Regulation of skeletal muscle fiber size, shape and function. Journal of Biomechanics, 24 (Suppl. 1), 123-133.
Edgerton, V.R., Roy, R.R., Gregor, R.J., & Rugg, S. (1986). Morphological basis of skeletal muscle power output. In N.L. Jones, N. McCartney, & A.J. McComas (Eds.), Human muscle power (pp. 43-64). Champaign, IL: Human Kinetics.
Edgerton, V.R., Smith, J.L., & Simpson, D.R. (1975). Muscle fibre types of human leg muscles. Histochemical Journal, 7, 259-266.
Edin, B.B. (1992). Quantitative analysis of static strain sensitivity in human mechanoreceptors from hairy skin. Journal of Neurophysiology, 67, 1105-1113.
Edington, C.J., Frederick, E.G., & Cavanagh, P.R. (1990). Rearfoot motion in distance running. In P.R. Cavanagh (Ed.), Biomechanics of distance running (pp. 135-164). Champaign, IL: Human Kinetics.
377
Edman, K.A.P. (1992). Contractile performance of skeletal muscle fibres. In P.V. Komi (Ed.), Strength and power in sport, (pp. 96-114). Champaign, IL: Human Kinetics.
Edman, K.A.P., Elzinga, G., & Noble, M.I.M. (1978). Enhancement of mechanical performance by stretch during tetanic contractions of vertebral skeletal muscle fibres. Journal of Physiology (London), 281, 139-155.
Edman, K.A.P., & Lou, F. (1990). Changes in force and stiffness induced by fatigue and intracellular acidification in frog muscle fibres. Journal of Physiology (London), 424, 133-149.
Edstrom, L., & Kugelberg, E. (1968). Histochemical composition, distribution of fibres and fatiguability of single motor units. Journal of Neurology, Neurosurgery, and Psychiatry, 31, 424-433.
Edwards, R.H.T., Hill, D.K., Jones, D.A., & Merton, P.A. (1977). Fatigue of long duration in human skeletal muscle after exercise. Journal of Physiology (London), 272, 769-778.
Eie, N., & Wehn, P. (1962). Measurements of the intraabdominal pressure in relation to weight bearing of the lumbosacral spine. Journal of Oslo City Hospitals, 12, 205-217.
Elftman, H. (1938). The measurement of the external force in walking. Science, 88, 152-153.
Elftman, H. (1939). Forces and energy changes in the leg during walking. American Journal of Physiology, 125, 339-356.
Eloranta, V., & Komi, P.V. (1980). Function of the quadriceps femoris muscle under maximal concentric and eccentric contractions. Electromyography and Clinical Neurophysiology, 20, 159-174.
Elson, P. (1974, December). Strength increases by electrical stimulation. Track Technique, p. 1856.
Emerson, N.D., & Zahalak, G.L (1981). Longitudinal electrode array for electromyography. Medical & Biological Engineering & Computing, 19, 504-506.
Engelhom, R. (1983). Agonist and antagonist muscle EMG activity pattern changes with skill acquisition. Research Quarterly for Exercise and Sport, 54, 315-323.
Engin, A.E., & Chen, S.-M. (1986). Statistical database for the biomechanical properties of the human shoulder complex: 1. Kinematics of the shoulder complex. Journal of Biomechanical Engineering, 108, 215-221.
Engin, A.E., & Chen, S.-M. (1987). Kinematic and passive resistive properties of human elbow complex. Journal of Biomechanical Engineering, 109, 318-323.
Engin, A.E., & Chen, S.-M. (1988). On the biomechanics of human hip complex in vivo: I. Kinematics for determination of the maximal voluntary hip complex sinus. Journal of Biomechanics, 21, 785-795.
English, A.W. (1984). An electromyographic analysis of compartments in cat lateral gastrocnemius during unrestrained locomotion. Journal of Neurophysiology, 52, 114-125.
English, A.W., & Ledbetter, W.D. (1982). Anatomy and innervation patterns of cat lateral gastrocnemius and plantaris muscles. American Journal of Anatomy, 164, 67-77.
Engstrom, C.M., Loeb, G.E., Reid, J.G., Forrest, W.J., & Avruch, L. (1991). Morphometry of the human thigh muscles: A comparison between anatomical sections and computer tomographic and magnetic resonance images. Journal of Anatomy, 176, 139-156.
Enoka, R.M. (1979). The pull in Olympic weightlifting. Medicine and Science in Sports, 11, 131-137.
Enoka, R.M. (1981). Muscular control of a learned move
ment. Ph.D. dissertation. University of Washington, Seattle.
Enoka, R.M. (1983). Muscular control of a learned movement: The speed control system hypothesis. Experimental Brain Research, 51, 135-145.
Enoka, R.M. (1988a). Load- and skill-related changes in segmental contributions to a weightlifting movement. Medicine and Science in Sports and Exercise, 20, 178-187.
Enoka, R.M. (1988b). Muscle strength and its development: New perspectives. Sports Medicine, 6, 146-168.
Enoka, R.M., & Fuglevand, A.J. (1993). Neuromuscular basis of the maximum voluntary force capacity of muscle. In M.D. Grabiner (Ed.), Current issues in biomechanics (pp. 215-235). Champaign, IL: Human Kinetics.
Enoka, R.M., Hutton, R.S., & Eldred, E. (1980). Changes in excitability of tendon tap and Hoffmann reflexes following voluntary contractions. Electroencephalography and Clinical Neurophysiology, 48, 664-672.
Enoka, R.M., Miller, D.I., & Burgess, E.M. (1982). Below-knee amputee running gait. American Journal of Physical Medicine, 61, 66-84.
Enoka, R.M., Robinson, G.A., & Kossev, A.R. (1989). Task and fatigue effects on low-threshold motor units in human hand muscle. Journal of Neurophysiology, 62, 1344-1359.
Enoka, R.M., & Stuart, D.G. (1984). Henneman's size principle: Current issues. Trends in Neurosciences, 7, 226-228.
Enoka, R.M., & Stuart, D.G. (1992). Neurobiology of muscle fatigue. Journal of Applied Physiology, 72, 1631-1648.
Enoka, R.M., Trayanova, N., Laouris, Y., Bevan, L., Reinking, R.M., & Stuart, D.G. (1992). Fatigue-related changes in motor unit action potentials of adult cats. Muscle & Nerve, 14, 138-150.
Ericson, M.O. (1988). Mechanical muscular power output and work during ergometer cycling at different work loads and speeds. European Journal of Applied Physiology, 57, 382-387.
Etnyre, B.R., & Abraham, L.D. (1986). Gains in range of ankle dorsiflexion using three popular stretching techniques. American iournal of Physical Medicine, 65, 189-196.
Etnyre, B.R., & Abraham, L.D. (1988). Antagonist muscle activity during stretching: A paradox reassessed. Medicine and Science in Sports and Exercise, 20, 285-289.
Evans, WJ., & Cannon, J.G. (1991). The metabolic effects of exercise-induced muscle damage. In J.O. Holloszy (Ed.), Exercise and sport sciences reviews (Vol. 19, pp. 99-125). Baltimore: Williams & Wilkins.
Evatt, M.L., Wolf, S.L., & Segal, R.L. (1989). Modification of human spinal stretch reflexes: Preliminary studies. Neuroscience Letters, 105, 350-355.
Ewing, J.L., Wolfe, D.R., Rogers, M.A., Amundson, M.L., & Stull, G.A. (1990). Effects of velocity of isokinetic training on strength, power, and quadriceps muscle fibre characteristics. European Journal of Applied Physiology, 61, 159-162.
Fahrer, H., Rentsch, H.U., Gerber, N.J., Beyeler, C., Hess, C.W., & Griinig, B. (1988). Knee effusion and reflex inhibition of quadriceps: A bar to effective training. Journal of Bone and Joint Surgery, 70B, 635-638.
Fang, Z.-P., & Mortimer, J.T. (1991a). Alternate excitation of large and small axons with different stimulation waveforms: An application to muscle activation. Medical and Biological Engineering and Computing, 29, 543-547.
378
Fang, Z.-P., & Mortimer, J.T. (1991b). A method to effect physiological recruitment order in electrically activated muscle. IEEE Transactions on Biomedical Engineering, 38, 175-179.
Fang, Z.-P., & Mortimer, J.T. (1991c). Selective activation of small motor axons by quasitrapezoidal current pulses. IEEE Transactions on Biomedical Engineering, 38,168-174.
Feinstein, B., Lindegard, B., Nyman, E., & Wohifart, G. (1955). Morphologic studies of motor units in normal human muscles. Acta Anatomica, 23, 127-142.
Feldman, AG. (1986). Once more on the equilibriumpoint hypothesis (2, model) for motor control. Journal of Motor Behavior, 18, 17-54.
Fellows, S.J., & Rack, P.M.H. (1987). Changes in the length of the human biceps brachii muscle during elbow movements. Journal of Physiology (London), 383, 405-412.
Feltner, M.E. (1989). Three-dimensional interactions in a two-segment kinetic chain: 2. Application to the throwing arm in baseball pitching. International Journal of Sport Biomechanics, 5, 420-450.
Feltner, M., & Dapena, J. (1986). Dynamics of the shoulder and elbow joints of the throwing arm during a baseball pitch. International Journal of Sport Biomechanics, 2, 235-259.
Feltner, M.E., & Dapena, J. (1989). Three-dimensional interactions in a two-segment kinetic chain: I. General model. International Journal of Sport Biomechanics, 5, 403-419.
Fenn, W.O. (1924). The relation between the work performed and the energy liberated in muscular contraction. Journal of Physiology (London), 58, 373-395.
Fenn, W.O. (1930). Work against gravity and work due to velocity changes in running. American Journal of Physiology, 93, 433-462.
Fiatarone, M.A, Marks, E.G., Ryan, N.D., Meredith, C.N., Lipsitz, L.A., & Evans, W.J. (1990). High-intensity strength training in nonagenarians, lournal of the American Medical Association, 263, 3029-3034.
Fick, R. (1904). Handbuch der Anatomie des Menschen (Vol. 2). Stuttgart, Germany: Gustav Fischer Verlag.
Fisher, M.A (1992). H reflexes and F waves: Physiology and clinical indication. Muscle & Nerve, 15, 1223-1233.
Fisher, M.J., Meyer, R.A, Adams, G.R., Foley, J.M., & Potchen, E.J. (1990). Direct relationship between proton T2 and exercise intensity in skeletal muscle MR images. Investigative Radiology, 25, 480-485.
Fitts, R.H., Brimmer, C.J., Heywood-Cooksey, A. & Timmerman, R.J. (1989). Single muscle fiber enzyme shifts with hindlimb suspension and immobilization. American Journal of Physiology, 256, C1082-C1091.
Fitts, R.H., McDonald, K.S., & Schulter, J.M. (1991). The determinants of skeletal muscle force and power: Their adaptability with changes in activity pattern. Journal of Biomechanics, 24 (Suppl. 1), 111-122.
Fitts, R.H., & Metzger, J.M. (1988). Mechanisms of muscular fatigue. In J.R. Poortmans (Ed.), Principles of exercise biochemistry (pp. 212-229). Basel, Switzerland: Karger.
Fitts, R.H., Metzger, J.M., Riley, D.A., & Unsworth, B.R. (1986). Models of disuse: A comparison of hindlimb suspension and immobilization. Journal of Applied Physiology, 60, 1946-1953.
Fitzsimons, D.P., Herrick, R.E., & Baldwin, K.M. (1990). Isomyosin distribution in skeletal muscle: Effects of
altered thyroid state. Journal of Applied Physiology, 69, 321-327.
Flanders, M. (1991). Temporal patterns of muscle activation for arm movements in threedimensional space. Journal of Neuroscieitce, 11, 2680-2693.
Flanders, M., & Herrmann, U. (1992). Two components of muscle activation: Scaling with the speed of arm movement. Journal of Neurophysiology, 67, 931-943.
Fleck, S.J., & Kraemer, W.J. (1987). Designing resistance training programs. Champaign, IL: Human Kinetics.
Fleckenstein, J.L., Canby, R.C., Parkey, R.W., & Peshock, R.M. (1988). Acute effects of exercise on MR imaging of skeletal muscle in normal volunteers. American Journal of Roentgenology, 151, 231-237.
Fleckenstein, J.L., & Shellock, F.G. (1991). Exertional muscle injuries: Magnetic resonance imaging evaluation. Topics in Magnetic Resonance Imaging, 3, 50-70.
Fleckenstein, J.L., Watumull, D., Bertocci, L.A., Parkey, R.W., & Peshock, R.M. (1992). Finger-specific flexor recruitment in humans: Depiction of exercise-enhanced MRL Journal of Applied Physiology, 72, 1974-1977.
Fletcher, J.G., & Lewis, H.E. (1960). Human power output: The mechanics of pole vaulting. Ergonomics, 3, 30-34.
Foerhing, R.C., & Munson, J.B. (1990). Motoneuron and muscle-unit properties after long-term direct innervation of soleus muscle by medial gastrocnemius nerve in cat. Journal of Neurophysiology, 64, 847-861.
Foehring, R.C., Sypert, G.W., & Munson, J.B. (1987a). Motor unit properties following crossreinnervation of cat lateral gastrocnemius and soleus muscles with medial gastrocnemius nerve: 1. Influence of motoneurons on muscle. Journal of Neurophysiology, 57, 1210-1226.
Foehring, R.C., Sypert, G.W., & Munson, J.B. (1987b). Motor unit properties following crossreinnervation of cat lateral gastrocnemius and soleus muscles with medial gastrocnemius nerve: 2. Influence of muscle on motoneurons. Journal of Neurophysiology, 57, 1227-1245.
Fournier, E., & Pierrot-Deseilligny, E. (1989). Changes in transmission in some reflex pathways during movement in humans. News in Physiological Sciences, 4, 29-32.
Fournier, M., Roy, R.R., Perham, H., Simard, C.P., & Edgerton, V.R. (1983). Is limb immobilization a model of muscle disuse? Experimental Neurology, 80, 147-156.
Frederick, B.C. (1986). Kinematically mediated effects of sport shoe design: A review. Journal of Sports Sciences, 4, 169-184.
Freed, W.J., de Medinaceli, L., & Wyatt, R.J. (1985). Promoting functional plasticity in the damaged nervous system. Science, 227, 1544-1552.
Friden, J. (1984). Changes in human skeletal muscle induced by long term eccentric exercise. Cell and Tissue Research, 236, 365-372.
Friden, J., Kjorell, U., & Thomell, L.-E. (1984). Delayed muscle soreness and cytoskeletal alterations: An immunocytological study in man. International Journal of Sports Medicine, 5, 15-18.
Friden, J., & Lieber, R.L. (1992). Structural and mechanical basis of exercise-induced muscle injury. Medicine and Science in Sports and Exercise, 24, 521-530.
Friden, J., Seger, J., & Ekblom, B. (1988). Sublethal muscle fibre injuries after high-tension anaerobic exercise. European Journal of Applied Physiology, 57, 360-368.
Friden, J., Sfakianos, P.M., Hargens, AR., & Akeson, W.H. (1988). Residual muscular swelling after repetitive
379
eccentric contractions. Journal of Orthopaedic Research, 7, 142-145.
Friederich, J.A., & Brand, R.A. (1990). Muscle fiber architecture in the human lower limb. Journal of Biomechanics, 23, 91-95.
Frohlich, C. (1980, March). The physics of somersaulting and twisting. Scientific American, pp. 154-164.
Frolkis, V.V., Martynenko, O.A., & Zamostyan, V.P. (1976). Aging of the neuromuscular apparatus. Gerontology, 22, 244-279.
Frolkis, V.V., Tanin, S.A., Marcinko, V.L, Kulchitsky, O.K., & Yasechko, A.V. (1985). Axoplasmic transport of substances in motoneuronal axons of the spinal cord in old age. Mechanisms of Ageing and Development, 29, 19-28.
Frontera, W.R., Meredith, C.N., O'Reilly, K.P., Knuttgen, H.G., & Evans, W.J. (1988). Strength conditioning in older men: Skeletal muscle hypertrophy and improved function. Journal of Applied Physiology, 64, 1038-1044.
Fu, F.H., Hamer, C.D., Johnson, D.L., Miller, M.D., & Woo, S.L.-Y. (1993). Biomechanics of knee ligaments. Journal of Bone and Joint Surgery, 75-A, 1716-1727.
Fuglevand, A.J., Winter, D.A., & Patla, A.E. (1993). Models of recruitment and rate coding organization in motorunit pools. Journal of Neurophysiology, 70, 2470-2488.
Fuglevand, A.J., Winter, D.A., Patla, A.E., & Stash nk, D. (1992). Detection of motor unit action potentials with surface electrodes: Influence of electrode size and spacing. Biological Cybernetics, 67, 143-154.
Fuglevand, A.J., Zackowski, K.M., Huey, K.A., & Enoka, R.M. (1993). Impairment of neuromuscular propagation during human fatiguing contractions at submaximal forces. Journal of Physiology (London), 460, 549-572.
Fujiwara, M., & Basmajian, J.V. (1975). Electromyographic study of two-joint muscles. American Journal of Physical Medicine, 54, 234-242.
Fukashiro, S., & Komi, P.V. (1987). Joint moment and mechanical power flow of the lower limb during vertical jump. International Journal of Sports Medicine, 8, 15-21.
Fukunaga, T., Roy, R.R., Shellock, F.G., Hodgson, J.A., Day, M.K., Lee, P.L., Kwong-Fu, H., & Edgerton, V.R. (1991). Physiological cross-sectional area of human leg muscles based on magnetic resonance imaging. Journal of Orthopaedic Research, 10, 926-934.
Funk, D.A., An, K.N., Morrey, B.F., & Daube, J.R. (1987). Electromyographic analysis of muscles across the elbow joint. Journal of Orthopaedic Research, 5, 529-538.
Gaffney, F.A., Sjylgaard, G., & Saltin, B. (1990). Cardiovascular and metabolic responses to static contraction in man. Acta Physiologica Scandinavica, 138, 249-258.
Galea, V., & Norman, R.W. (1985). Bone-on-bone forces at the ankle joint during a rapid dynamic movement. In D.A. Winter, R.W. Norman, R.P. Wells, K.C. Hayes, & A.E. Patla (Eds.), Biomechanics IX-A (pp. 71 -76). Champaign, IL: Human Kinetics.
Galganski, M.E., Fuglevand, A.J., & Enoka, R.M. (1993). Reduced control of motor output in a human hand muscle of elderly subjects during submaximal contractions. Journal of Neurophysiology, 69, 2108-2115.
Gandevia, S.C., McCloskey, D.L, & Burke, D. (1992). Kinaesthetic signals and muscle contraction. Trends in Neurosciences, 15, 62-65.
Gandevia, S.C., & McKenzie, D.K. (1985). Activation of the human diaphragm during maximal static efforts. Journal of Physiology (London), 367, 45-56.
Gans, C. (1982). Fiber architecture and muscle function. In R.L, Teijung (Ed.), Exercise and sport sciences reviews (Vol. 10, pp. 160-207). Philadelphia: Franklin Institute.
Gardetto, P.R., Schluter, J.M., & Fitts, R.H. (1989). Contractile function of single muscle fibers after hindlimb suspension. Journal of Applied Physiology, 66, 2739-2749.
Gardiner, P.F., & Lapointe, M.A. (1982). Daily in vivo neuro-muscular stimulation effects on immobilized rat hindlimb muscles. Journal of Applied Physiology, 53, 960-966.
Gareis, H., Solomonow, M., Baratta, R., Best, R., & D'Ambrosia, R. (1992). The isometric length-force models of nine different skeletal muscles. Journal of Biomechanics, 25, 903-916.
Garfinkel, S., & Cafarelli, E. (1992). Relative changes in maximal force, EMG, and muscle cross-sectional area after isometric training. Medicine and Science in Sports and Exercise, 24, 1220-1227.
Garhammer, J. (1980). Power production by Olympic weight-lifters. Medicine and Science in Sports and Exercise, 12, 54-60.
Garhammer, J. (1989). Weight lifting and training. In C.L. Vaughan (Ed.), Biomechanics of sport (pp. 169-211). Boca Raton, FL: CRC Press.
Garland, S.J. (1991). Role of small diameter afferents in reflex inhibition during human muscle fatigue. Journal of Physiology (London), 435, 547-558.
Garland, S.J., Enoka, R.M., Serrano, L.P., & Robinson, G.A. (in press). Behavior of motor units in human biceps brachii during fatigue. Journal of Applied Physiology.
Gamer, S.H., Hicks, A.L., & McComas, A.J. (1989). Prolongation of twitch potentiating mechanism throughout muscle fatigue and recovery. Experimental Neurology, 103, 277-281.
Garnett, R., & Stephens, J.A. (1981). Changes in the recruitment threshold of motor units produced by cutaneous stimulation in man. Journal of Physiology (London), 311, 463-473.
Garnett, R.A.F., O'Donovan, M.J., Stephens, J.A., & Taylor, A. (1978). Motor unit organization of human medial gastrocnemius. Journal of Physiology (London), 287, 33-43.
Garrett, W.E. (1990). Muscle strain injuries: Clinical and basic aspects. Medicine and Science in Sports and Exercise, 22, 436-443.
Garrett, W.E., Califf, J.C., & Bassett, F.H. (1984). Histochemical correlates of hamstring injuries. American Journal of Sports Medicine, 12, 98-103.
Garrett, W.E., Safran, M.R , Seaber, A.V., Glisson, R.R., & Ribbeck, B.M. (1987). Biomechanical comparison of stimulated and nonstimulated skeletal muscle pulled to failure. American Journal of Sports Medicine, 15, 448-454.
Gebhard, J.S., Kabo, J.M., & Meals, R.A. (1993). Passive motion: The dose effects on joint stiffness, muscle mass, bone density, and regional swelling. Journal of Bone and Joint Surgery, 75-A, 1636-1647.
Georgopoulos, A.P., Ashe, J., Smyrnis, N., & Taira, M. (1992). The motor cortex and the coding of force. Science, 256, 1692-1695.
Gergely, J. (1974). Some aspects of the role of the sarcoplasmic reticulum and the tropomyosin-troponin system in the control of muscle contraction by calcium ions. Circulation Research, 34 (Suppl. 3), 74-82.
380
Gerilovsky, L., Tsvetinov, P., & Trenkova, G. (1989). Peripheral effects on the amplitude of monopolar and bipolar H-reflex potentials from the soleus muscle. Experimental Brain Research, 76, 173-181.
Ghez, C. (1985). Introduction to the motor systems. In E.R. Kandel and J.H. Schwartz (Eds.), Principles of Neural Science (2nd ed., pp. 429-442). New York: Elsevier.
Ghez, C. (1991 a). The cerebellum. In E.R. Kandel, J.H. Schwartz, & T.M. Jessell (Eds.), Principles of neural science (3rd ed., pp. 626-646). New York: Elsevier.
Ghez, C. (1991b). Voluntary movement. In E.R. Kandel, J.H. Schwartz, & T.M. Jessell (Eds.), Principles of neural science (3rd ed., pp. 609-625). New York: Elsevier.
Gibson, J.N.A., Halliday, D., Morrison, W.L., Steward, P.J., Hornsby, G.A., Watt, P.W., Murdoch, G., & Rennie, M.J. (1987). Decrease in human quadriceps muscle protein turnover consequent upon leg immobilization. Clinical Science, 72, 503-509.
Gielen, C.C.A.M., Ramaekers, L., & van Zuyien, E.J. (1988). Long-latency stretch reflexes as coordinated functional responses in man. Journal of Physiology (London), 407, 275-292.
Gielen, C.C.A.M., van Ingen Schenau, G.J., Tax, T., & Theeuwen, M. (1990). The activation of mono- and bi-articular muscles in multi-joint movements. In J.M. Winters & S.L.-Y, Woo (Eds.), Multiple muscle systems: Biomechanics and movement organization (pp. 302-311). New York: Springer-Verlag.
Godfrey, C.M., Jayawardena, A., & Welsh, P. (1986). Comparison of electro-stimulation and isometric exercise in strengthening the quadriceps muscle. Physiotherapy (Canada), 31, 265-267.
Goldberger, M., Murray, M., & Tessier, A. (1992). Sprouting and regeneration in the spinal cord: Their roles in recovery of function after spinal injury. In A. Gorio (Ed.), Neuroregeneration (pp. 241-264). New York: Raven.
Goldspink, G., Scutt, A., Loughna, P.T., Wells, D.J., Jaenicke, T., & Gerlach, G.F. (1992). Gene expression in skeletal muscle in response to stretch and force generation. American Journal of Physiology, 262, R356-R363.
Gonyea, W.J., Sale, D.G., Gonyea, F.B., & Mikesky, A. (1986). Exercise induced increases in muscle fiber number. European Journal of Applied Physiology, 55, 137-141.
Goodfellow, J., & O'Connor, J. (1978). The mechanics of the knee and prosthesis design. Journal of Bone and Joint Surgery, 60B, 358-369.
Gordon, D.A., Enoka, R.M., Karst, G.M., & Stuart, D.G. (1990). Force development and relaxation in single motor units of adult cats during a standard fatigue test. Journal of Physiology (London), 421, 583-594.
Gordon, D.A, Enoka, R.M., & Stuart, D.G. (1990). Motor-unit force potentiation in adult cats during a standard fatigue test. Journal of Physiology (London), 421, 569-582.
Gordon, T., & Pattullo, M.C. (1993). Plasticity of muscle fiber and motor unit types. In J.O. Holloszy (Ed.), Exercise and sport sciences reviews (Vol. 21, pp. 339-362). Baltimore: Williams & Wilkins.
Gordon, T., Yang, J.F., Ayer, K., Stein, R.B., & Tyreman, N. (1993). Recovery potential of muscle after partial denervation: A comparison between rats and humans. Brain Research Bulletin, 30, 477-482.
Goslow, G.E., Reinking, R.M., & Stuart, D.G. (1973). The cat step cycle: Hind limb joint angles and muscle lengths during unrestrained locomotion. Journal of Morphology, 141, 1-41.
Gottlieb, G.L., Corcos, D.M., & Agarwal, G.C. (1989). Organizing principles for single-joint movements; I, A speed-insensitive strategy. Journal of Neurophysiology, 62, 342-357.
Gottlieb, G.L., Corcos, D.M., Agarwal, G.C.,& Latash, M.L. (1990a). Organizing principles for single joint movements: 3. Speed-insensitive strategy as a default. Journal of Neurophysiology, 63, 625-636.
Gottlieb, G.L., Corcos, D.J., Agarwal, G.C., & Latash, M.L. (1990b). Principles underlying single-joint movement strategies. In J.M. Winters & S.L.-Y. Woo (Eds.), Multiple muscle systems: Biomechanics and movement organization (pp. 236-250). New York: Springer-Verlag.
Gowland, C., deBruin, H., Basmajian, J.V., Plews, N., & Burcea, 1. (1992). Agonist and antagonist activity during voluntary upper-limb movement in patients with stroke. Physical Therapy, 72, 624-633.
Grabiner, M.D., & Jeziorowski, J.J. (1992). Isokinetic trunk extension discriminates uninjured subjects from subjects with previous low back pain. Clinical Biomechanics, 7, 195-200.
Grabiner, M.D., Koh, T.J., & Andrish, J.T. (1992). Decreased excitation of vastus medialis oblique and vastus lateralis in patellofemoral pain. European Journal of Experimental Musculoskeletal Research, 1, 33-39.
Grabiner, M.D., Koh, T.J., & Draganich, L.F. (1994). Neuromechanics of the patellofemoral joint. Medicine and Science in Sports and Exercise, 26, 10-21.
Grange, R.W., & Houston, M.E. (1991). Simultaneous potentiation and fatigue in quadriceps after a 60-second maximal voluntary isometric contraction. Journal of Applied Physiology, 70, 726-731.
Grange, R.W., Vandenboom, R., & Houston, M.E. (1993). Physiological significance of myosin phosphorylation in skeletal muscle. Canadian Journal of Applied Sport Sciences, 18, 229-242.
Gransberg, L., & Knutsson, E. (1983). Determination of dynamic muscle strength in man with acceleration controlled isokinetic movements. Acta Physiologica Scandinavica, 119, 317-320.
Gregor, R.J., Komi, P.V., & Jarvinen, M. (1991). A comparison of the triceps surae and residual muscle moments at the ankle during cycling. Journal of Biomechanics, 24, 287-297.
Gregor, R.J., Komi, P.V., & Jarvinen, M. (1987). Achilles tendon forces during cycling. International Journal of Sports Medicine, 8, 9-14.
Gregor, R.J., Roy, R.R., Whiting, W.C., Lovely, R.G., Hodgson, J.A., & Edgerton, V.R. (1988). Mechanical output of the cat soleus during treadmill locomotion: In vivo vs in situ characteristics. Journal of Biomechanics, 21, 721-732.
Gregory, J.E. (1990). Relations between identified tendon organs and motor units in the medial gastrocnemius muscle of the cat. Experimental Brain Research, 81, 602-608.
Gregory, J.E., Morgan, D.L., & Proske, LJ. (1986). Aftereffects in the responses of cat muscle spindles. Journal of Neurophysiology, 56, 451-461.
Grieve, D.W. (1969, June). Stretching active muscles and leading with the hips. Coaching Review, pp. 3-4, 10.
Griffiths, R.I. (1991). Shortening of muscle fibres during stretch of the active cat medial gastrocnemius muscle: The role of tendon compliance. Journal of Physiology (London), 436, 219-236.
Grimby, G., & Saltin, B. (1983). The ageing muscle. Clinical Physiology, 3, 209-218.
381
Grimby, L., Hannerz, J., & Hedman, B. (1981). The fatigue and voluntary discharge properties of single motor units in man. Journal of Physiology (London), 316, 545-554.
Grood, E.S., Suntay, W.J., Noyes, F.R., & Butler, D.L. (1984). Biomechanics of the knee-extension exercise. Journal of Bone and Joint Surgery, 66A, 725-734.
Gross, A.C., Kyle, C.R., & Malewicki, D.J. (1983, December). The aerodynamics of human-powered land vehicles. Scientific American, pp. 142-145, 148-152.
Gross, T.S., & Bain, S.D. (1993). Skeletal adaptation to functional stimuli. In M.D. Grabiner (Ed.), Current issues in biomechanics (pp. 151-169). Champaign, IL: Human Kinetics.
Groves, B.K. (1989). Muscle differentiation and the origin of muscle fiber diversity. CRC Critical Reviews in Neurobiology, 4, 201-234.
Guissard, N., Duchateau, J., & Hainaut, K. (1988). Muscle stretching and motoneuron excitability. European Journal of Applied Physiology, 58, 47-52.
Gunning, P., & Hardeman, E. (1991). Multiple mechanisms regulate muscle fiber diversity. FASEB Journal, 5, 3064-3070.
Gydikov, A., & Kosarov, D. (1973). Physiological characteristics of the tonic and phasic motor units in human muscles. In A.A. Gydikov, N.T. Tankov, & D.S. Kosarov (Eds.), Motor control (pp. 75-94). New York: Plenum.
Gydikov, A., & Kosarov, D. (1974). Some features of different motor units in human biceps brachii. Pflugers Archiv, 347, 75-88.
Gydikov, A.A., Kossev, A.R., Kosarov, D.S., & Kostov, K.G. (1987). Investigations of single motor units firing during movements against elastic resistance. In B. Jonsson (Ed.), Biomechanics X-A (pp. 227-232). Champaign, IL: Human Kinetics.
Gydikov, A., Kossev, A., Radicheva, N., & Tankov, N. (1981). Interaction between reflexes and voluntary motor activity in man revealed by discharges of separate motor units. Experimental Neurology, 73, 331-344.
Hagbarth, K.-E. (1962). Post-tetanic potentiation of myotatic reflexes in man. Journal of Neurology, Neurosurgery, and Psychiatry, 25, 1-10.
Hagbarth, K.-E., Hagglund, J.V., Nordin, M., & Wallin, E.U. (1985). Thixotropic behaviour of human finger flexor muscles with accompanying changes in spindle and reflex responses to stretch. Journal of Physiology (London), 368, 323-342.
Hagg, G.M. (1992). Interpretation of EMG spectral alterations and alteration indexes at sustained contraction. Journal of Applied Physiology, 73, 1211-1217.
Hainaut, K., & Duchateau, J. (1992). Neuromuscular electrical stimulation and voluntary exercise. Sports Medicine, 14, 100-113.
Hakkinen, K. (1985). Research overview: Factors influencing trainability of muscular strength during short term and prolonged training. National Strength and Conditioning Association Journal, 7(2), 32-37.
Hakkinen, K., Alen, M., & Komi, P.V. (1985). Changes in isometric force- and relaxation-time, electromyographic and muscle fiber characteristics of human skeletal muscle during strength training and detraining. Acta Physiologica Scandinavica, 125, 573-585.
Hakkinen, K., & Keskinen, K.L. (1989). Muscle cross-sectional area and voluntary force production characteristics in elite strength-trained and endurance-trained athletes and sprinters. European Journal of Applied Physiology, 59, 215-220.
Hakkinen, K., & Komi, P.V. (1986). Training-induced changes in neuromuscular performance under voluntary and reflex conditions. European Journal of Applied Physiology, 55, 147-155.
Hakkinen, K., Komi, P.V., & Alen, M. (1985). Effect of explosive type strength training on isometric force-and relaxation-time, electromyographic and muscle fibre characteristics of leg extensor muscles. Acta Physiologica Scandinavica, 125, 587-600.
Hales, J.P., & Gandevia, S.C. (1988). Assessment of maximal voluntaiy contraction with twitch interpolation: An instrument to measure twitch responses. Journal of Neuroscience Methods, 25, 97-102.
Hall, S.J., & DePauw, K.P. (1982). A photogrammetrically based model for predicting total body mass centroid location, Research Quarterly for Exercise and Sport, 53, 37-45.
Hallett, M. (1993). Physiology of basal ganglia disorders: An overview. Canadian Journal of Neurological Sciences, 20, 177-183.
Hammond, P.H., Merton, P.A., & Sutton, G.G. (1956). Nervous gradation of muscular contraction. British Medical Bulletin, 12, 214-218.
Hanavan, E.P. (1964). A mathematical model of the human body (AMRL-TR-64-102), Wright Patterson Air Force Base, OH: Aerospace Medical Research Laboratories. (NTIS No. AD-608463 as issued by the United States Air Force)
Hanavan, E.P. (1966). A personalized mathematical model of the human body. Journal of Spacecraft and Rockets, 3, 446-448.
Hannaford, B., & Stark, L. (1985). Roles of the elements of the triphasic control signal. Experimental Neurology, 90, 619-635.
Hannan, E.A., Frykman, P.N., Clagett, E.R., & Kraemer, W.J. (1988). Intra-abdominal and intra-thoracic pressures during lifting and jumping. Medicine and Science in Sports and Exercise, 20, 195-201.
Harman, E.A., Rosenstein, R.M., Frykman, P.N., & Nigro, G.A. (1989). Effects of a belt on intra-abdominal pressure during weight lifting. Medicine and Science in Sports and Exercise, 21, 186-190.
Harre, D. (Ed.) (1982). Principles of sports training: Introduction to the theory and methods of training. East Berlin: Sportverlag.
Hamson, R.N., Lees, A., McCullagh, P.J.J., & Rowe, W.B. (1986). A bioengineering analysis of human muscle and joint forces in the lower limbs during running. Journal of Sports Sciences, 4, 201-218.
Hasan, Z. (1986). Optimized movement trajectories and joint stiffness in unperturbed, inertially loaded movements. Biological Cybernetics, 53, 373-382.
Hasan, Z. (1991). Biomechanics and the study of multijoint movements. In D.R. Humphrey & H.-J. Freund (Eds.), Motor control: Concepts and issues (pp. 75-84). Chichester, England: Wiley.
Hasan, Z., & Enoka, R.M. (1985). Isometric torqueangle relationship and movement-related activity of human elbow flexors: Implications for the equilibriumpoint hypothesis. Experimental Brain Research, 59, 441-450.
Hasan, Z., Enoka, R.M., & Stuart, D.G. (1985). The interface between biomechanics and neurophysiology in the study of movement: Some recent approaches. In R.L. Teijung (Ed.), Exercise and sport sciences reviews (Vol. 13, pp. 169-234). New York: Macmillan.
Hasan, Z., & Stuart, D.G. (1984). Mammalian muscle receptors. In R.A. Davidoff (Ed.), Handbook of the spinal cord (pp. 559-607). New York: Marcel Dekker.
382
Hasan, Z., & Stuart, D.G. (1988). Animal solutions to problems of movement control: The role of proprioceptors. Annual Review of Neurosciences, 11, 199-223.
Hasselkus, B.R., & Shambes, G.M. (1975). Aging and postural sway in women. Journal of Gerontology, 30, 661-667.
Hather, B.M., Adams, G.R., Tesch, P.A., & Dudley, G.A. (1992). Skeletal muscle responses to lower limb suspension in humans. Journal of Applied Physiology, 72, 1493-1498.
Hather, B.M., Tesch, P.A., Buchanan, P., & Dudley, G.A. (1991). Influence cf eccentric actions on skeletal muscle adaptations to resistance training. Acta Physiologica Scandinavica, 143, 177-185.
Hatze, H. (1980). A mathematical model for the computational determination of parameter values of anthropomorphic segments. Journal of Biomechanics, 13, 833-843,
Hatze, H. (1981 a). Estimation of myodynamic parameter values from observations on isometrically contracting muscle groups. European Journal of Applied Physiology, 46, 325-338.
Hatze, H. (1981b). Myocybernetic control models of skeletal muscle: Characteristics and applications. Pretoria: University of South Africa.
Hawkins, D. (1993). Ligament biomechanics. In M.D. Grabiner (Ed.), Current Issues in Biomechanics (pp. 123-150). Champaign, IL: Human Kinetics.
Hay, J.G. (1978). The Biomechanics of Sports Techniques (2nd ed.). Englewood Cliffs, NJ: Prentice Hall.
Hay, J.G. (1985). The Biomechanics of Sports Techniques (3rd ed.). Englewood Cliffs, NJ: Prentice-Hall.
Hay, J.G., Andrews, J.G., & Vaughan, C.L. (1980). The influence of external load on the joint torques exerted in a squat exercise. In J.M. Cooper & B. Haven (Eds.), Proceedings of the Biomechanics Symposium (pp. 286-293). Indiana University: Indiana State Board of Health.
Hay, J.G., Andrews, J.G., Vaughan, C.L., & Ueya, K. (1983). Load, speed and equipment effects in strengthtraining exercises. In H. Matsui& K. Kobayashi (Eds.), Biomechanics VIII-B (pp. 939-950). Champaign, IL: Human Kinetics.
Hay, J.G., Wilson, B.D., Dapena, J., & Woodworth, G.G. (1977). A computational technique to determine the angular momentum of the human body. Journal of Biomechanics, 10, 269-277.
Hayes, K.C. (1982). Biomechanics of postural control. In R.L. Terjung (Ed.), Exercise and sport sciences reviews (Vol. 10, pp. 363-391). Philadelphia: Franklin Institute.
Heckathome, C.W., & Childress, D.S. (1981). Relationships of the surface electromyogram to the force, length, velocity, and contraction rate of the cineplastic human biceps. American Journal of Physical Medicine, 60, 1-19.
Heckman, C.J., & Binder, M.D. (1990). Neural mechanisms underlying the orderly recruitment of motoneurons. In M.D. Binder & L.M. Mendell (Eds.), The segmental motor system (pp. 182-204). New York: Oxford University Press.
Heckman, C.J., & Binder, M.D. (1991). Computer simulation of the steady-state input-output function of the cat medial gastrocnemius motoneuron pool. Journal of Neurophysiology, 65, 952-967.
Henneman, E. (1957). Relation between size of neurons and their susceptibility to discharge. Science, 126,1345-1347.
Henneman, E. (1979). Functional organization of motoneuron pools: The size-principle. In H. Asanuma
& V.J. Wilson (Eds.), Integration in the nervous system (pp. 13-25). Tokyo: Igaku-Shoin.
Henning, E.M., Cavanagh, P.R., Albert, H.T., & Macmillan, N.H. (1982). A piezoelectric method of measuring the vertical contact stress beneath the human foot. Journal of Biomedical Engineering, 4, 213-222.
Henriksson-Larsen, K. (1985). Distribution, number and size of different types of fibres in whole cross-sections of female m tibialis anterior An enzyme histochemical study. Acta Physiologica Scandinavica, 123, 229-235.
Henriksson-Larsen, K. Friden, J., & Wretling, M.-L. (1985). Distribution of fibre sizes in human skeletal muscle: An enzyme histochemical study in m tibialis anterior. Acta Physiologica Scandinavica, 123, 171-177.
Herbert, M.E., Roy, R.R., & Edgerton, V.R. (1988). Influence of one week of hindlimb suspension and intermittent high load exercise on rat muscles. Experimental Neurology, 102, 190-198.
Hermansen, L., Hultman, E., & Saltin, B. (1967). Muscle glycogen during prolonged severe exercise. Acta Physiologica Scandinavica, 71, 129-139.
Hershler, C., & Milner, M. (1980a). Angle-angle diagrams in above-knee amputee and cerebral palsy gait. American Journal of Physical Medicine, 59, 165-183.
Hershler, C., & Milner, M. (1980b). Angle-angle diagrams in the assessment of locomotion. American Journal of Physical Medicine, 59, 109-125.
Hettinger, T. (1961). Physiology of strength. Springfield, IL: Charles C Thomas.
Heyes, M.P., Garnett, E.S., & Coates, G. (1985). Central dopaminergic activity influences rats' ability to exercise. Life Sciences, 36, 671-677.
Heyes, M.P., Garnett, E.S., & Coates, G. (1988). Nigost-riatal dopaminergic activity is increased during exhaustive exercise stress in rats. Life Sciences, 42, 1537-1542.
Hicks, A.L., Cupido, C.M., Martin, J., & Dent, J. (1992). Muscle excitation in elderly adults; The effects of training. Muscle & Nerve, 15, 87-93.
Hicks, A., Fenton, J., Garner, S., & McComas, A.J. (1989). M wave potentiation during and after muscle activity. Journal of Applied Physiology, 66, 2606-2610.
Hicks, A., & McComas, A.J. (1989). Increased sodium pump activity following repetitive stimulation of rat soleus muscles. Journal of Physiology (London), 414, 337-349.
Higgins, S. (1985). Movement as an emergent form: Its structural limits. Human Movement Science, 4,119-148.
High, D.M., Howley, E.T., & Franks, B.D. (1989). The effects of static stretching and warmup on prevention of delayed-onset muscle soreness. Research Quarterly for Exercise and Sport, 60, 357-361.
Hill, A.V. (1928). The air-resistance to a runner. Proceedings of the Royal Society of London, B, 102, 380-385.
Hill, A.V. (1938). The heat of shortening and the dynamic constraints of muscle. Proceedings of the Royal Society of London, B, 126, 136-195.
Hinrichs, R.N, (1987). Upper extremity function in running: 2. Angular momentum considerations. International Journal of Sport Biomechanics, 3, 242-263.
Hinrichs, R.N., Cavanagh, P.R., & Williams, K.R. (1987). Lipper extremity function in running: 1. Center of mass and propulsion considerations. International Journal of Sport Biomechanics, 3, 222-241.
Hof, A.L. (1984). EMG and muscle force: An introduction. Human Movement Sciences, 3, 119-153.
Hof, A.L., Pronk, C.N.A., & van Best, J.A. (1987). Comparison between EMG to force processing and kinetic analysis for the calf muscle moment in walking and stepping. Journal of Biomechanics, 20, 167-178.
383
Hof, A.L., & van den Berg, J.W. (1977). Linearity between the weighted sum of the EMGs of the human triceps surae and the total torque. Journal of Biomechanics, 10, 529-539.
Hof, A.L., & van den Berg, J. (1981 a). EMG to force processing: 1. An electrical analogue of the Hill muscle model. Journal of Biomechanics, 14, 747-758.
Hof, A.L., & van den Berg, J. (1981b). EMG to force processing: 2. Estimation of parameters of the Hill muscle model for the human triceps surae by means of a calfergometer. Journal of Biomechanics, 14, 759-770.
Hof, A.L., & van den Berg, J. (1981c). EMG to force processing: 3. Estimation of model parameters for the human triceps surae muscle and assessment of the accuracy by means of a torque plate. Journal of Biomechanics, 14, 771-785.
Hof, A.L., & van den Berg, J. (198Id). EMG to force processing: 4. Eccentric-concentric contractions on a spring-flywheel setup. Journal of Biomechanics, 14,787-792,
Hegfors, C., Sigholm, G., & Herberts, P. (1987). Biomechanical model of the human shoulder: I. Elements. Journal of Biomechanics, 20, 157-166.
Hoh, J.F.Y. (1991). Myogenic regulation of mammalian skeletal muscle. News in Physiological Sciences, 6, 1-6.
Hoh, J.F.Y., & Hughes, S. (1988). Myogenic and neurogenic regulation of myosin gene expression in cat jawclosing muscles regenerating in fast and slow limb muscles. Journal of Muscle Research and Cell Motility, 9, 57-72.
Hollerbach, J.M., & Flash, T. (1982). Dynamic interactions between limb segments during planar arm movement. Biological Cybernetics, 44, 67-77.
Holst, E. von. (1954). Relations between the central nervous system and the peripheral organs. British Journal of Animal Behaviour, 2, 89-94.
Holstege, G. (1992). The emotional motor system. European Journal of Morphology, 30, 67-79.
Hopper, B.J. (1973). The mechanics of human movement. New York: Elsevier.
Horak, F.B., & Nashner, L.M. (1986). Central programming of postural movements: Adaptations to altered support-surface configurations. Journal of Neurophysiology, 55, 1369-1381.
Horak, F.B., Shupert, C.L., & Mirka, A. (1989). Components of postural dyscontrol in the elderly: A review. Neurobiology of Aging, 10, 727-738.
Hortobagyi, T., & Katch, F.L (1990). Role of concentric force in limiting improvement in muscular strength. Journal of Applied Physiology, 68, 650-658.
Houk, J.C. (1988). Control strategies in physiological systems. Federation of American Societies for Experimental Biology Journal, 2, 97-107.
Houk, J.C., Keifer, J., & Barto, A.G. (1993). Distributed motor commands in the limb premotor network. Trends in Neurosciences, 16, 27-33.
Houk, J.C., & Rymer, W.Z. (1981). Neural control of muscle length and tension. In V.B. Brooks, (Ed.), Handbook: of Physiology: Sec. 1. The nervous system: Vol. II. Motor control. Part I (pp. 257-324). Bethesda, MD: American Physiological Society.
Houston, M.E., Froese, E.A., Valeriote, S.P., Green, H.J., & Ranney, D.A. (1983). Muscle performance, morphology and metabolic capacity during strength training and detraining: A one leg model. European Journal of Applied Physiology, 51, 25-35.
Houston, M.E., Norman, R.W., & Froese, E.A. (1988). Mechanical measures during maximal velocity knee extension exercise and their relation to fibre compo
sition of the human vastus lateralis muscle. European Journal of Applied Physiology, 58, 1-7.
Howard, J.D, (1987). Central and peripheral factors underlying bilateral inhibition during maximal efforts. Unpublished doctoral dissertation. University of Arizona, Tucson.
Howard, J.D., & Enoka, R.M. (1991). Maximum bilateral contractions are modified by neurally mediated interlimb effects. Journal of Applied Physiology, 70, 306-316.
Howell, J.N., Chleboun, G., & Conatser, R. (1993). Muscle stiffness, strength loss, swelling and soreness following exercise-induced injury in humans. Journal of Physiology (London), 464, 183-196.
Hoy, M.G., Zajac, F.E., & Gordon, M.E. (1990). A musculo-skeletal model of the human lower extremity: The effect of muscle, tendon, and moment arm on the moment-angle relationship of musculotendon actuators at the hip, knee, and ankle. Journal of Biomechanics, 23, 157-169.
Hoy, M.G., & Zernicke, R.F. (1986). The role of intersegmental dynamics during rapid limb oscillation. Journal of Biomechanics, 19, 867-879.
Hoy, M.G., Zernicke, R.J., & Smith, J.L. (1985). Contrasting roles of inertial and muscle moments at knee and ankle during paw-shake response. Journal of Neurophysiology, 54, 1282-1295.
Hoyle, G. (1983). Muscles and their neural control. New York: Wiley.
Hubbard, M. (1980). Dynamics of the pole vault. Journal of Biomechanics, 13, 965-976.
Hubley, C.L., & Wells, R.P. (1983). A work-energy approach to determine individual joint contributions to vertical jump performance. European Journal of Applied Physiology, 50, 247-254.
Hufschmidt, A., & Mauritz, K.-H. (1985). Chronic transformation of muscle in spasticity: A peripheral contribution to increased tone. Journal of Neurology, Neurosurgery, and Psychiatry, 48, 676-685.
Hugon, M. (1973). Methodology of the Hoffmann reflex in man. In J.E. Desmedt (Ed.), New developments in electro-myography and clinical neurophysiology (Vol. 3, pp. 277-293). Basel, Switzerland: Karger.
Huijing, P.A. (1992a). Elastic potential of muscle. In P V Komi (Ed.), Strength and power in sport (pp 151-168). Champaign, IL: Human Kinetics.
Huijing, P.A. (1992b). Mechanical muscle models. In P.V. Komi (Ed.), Strength and power in sport (pp. 130-150). Champaign, IL: Human Kinetics.
Huizar, P., Kuno, M., Judo, N., & Miyata, Y. (1978). Reaction of intact spinal motoneurones to partial denervation of the muscle. Journal of Physiology (London), 265, 175-193.
Hultbom, H., Lindstrom, S., & WigstrBm, H. (1979). On the function of recurrent inhibition in the spinal cord. Experimental Brain Research, 37, 399-403.
Hultman, E., Bergstrom, M., Spriet, L.L., & Soderlund, K. (1990). Energy metabolism and fatigue. In A.W. Taylor, P.D. Gollnick, H.J. Green, C.D. lanuzzo, E.G. Noble, G. Metivier, and J.R. Sutton (Eds.), Biochemistry of exercise VII (pp. 73-92). Champaign, IL: Human Kinetics.
Hultman, E., & Sjoholm, H. (1983). Electromyogram, force and relaxation time during and after continuous electrical stimulation of human skeletal muscle in situ. Journal of Physiology (London), 339, 33-40.
Hultman, E., Sjfiholm, H., Jaderhaolm-Ek, L, & Krynicki, J. (1983), Evaluation of methods for electrical stimulation of human skeletal muscle in situ. PflUgers Archiv, 398, 139-141.
384
Hutton, R.S. (1984). Acute plasticity in spinal segmental pathways with use: Implications for training. In M. Kumamoto (Ed.), Neural and mechanical control of movement (pp. 90-112). Kyoto: Yamaguchi Shoten.
Hutton, R.S., & Nelson, D.L. (1986). Stretch sensitivity of Golgi tendon organs in fatigued gastrocnemius muscle. Medicine and Science in Sports and Exercise, 18, 69-74.
Hutton, R.S., Smith, J.L., & Eldred, E. (1973). Postcontraction sensory discharge from muscle and its source. Journal of Neurophysiology, 36, 1090-1103.
Huxley, H.E. (1985). The crossbridge mechanism of muscular contraction and its implications. Journal of Experimental Biology, 115, 17-30.
Ikai, M., & Fukunaga, T. (1968). Calculation of muscle strength per unit cross-sectional area of human muscle by means of ultrasonic measurement. Internationale Zeitschrift fur Angewandte Physiologic Einschliesslich Arbeitsphysiologie, 26, 26-32.
hnms, F.J., Hackett, A.J., Prestidge, S.P., & Fox, R.H. (1977). Voluntary isometric muscle strength of patients undergoing rehabilitation following fractures of the lower limb. Rheumatology and Rehabilitation, 16, 162-171.
Imms, F.J., & MacDonald, I.C. (1978). Abnormalities of the gait occuring during recovery from fractures of the lower limb and their improvement during rehabilitation. Scandinavian Journal of Rehabilitation Medicine, 10, 193-199.
Ingjer, F., & Stromme, S.B. (1979). Effects of active, passive or no warm-up on the physiological response to heavy exercise. European Journal of Applied Physiology, 40, 273-282.
Inman, V.T., Ralston, H.J., Saunders, J.B. de C.M., Feinstein, B., & Wright, E.W. (1952). Relation of human electromyogram to muscular tension. Electroencephalography and Clinical Neurophysiology, 4,187-194.
Iversen, L.L. (1987). Neurotransmitters. In G. Adelman (Ed.), Encyclopedia of neuroscience (Vol. 2, pp. 856-861). Boston: Birkhauser.
Jackson, M.J., Jones, D.A., & Edwards, R.H.T. (1984). Experimental skeletal muscle damage: The nature of the calcium-activated degenerative processes. European Journal of Clinical Investigation, 14, 369-374.
Jami, L. (1992). Golgi tendon organs in mammalian skeletal muscle: Functional properties and central actions. Physiological Reviews, 72, 623-666.
Jami, L., Murthy, K.S.K., Petit, J., & Zytnicki, D. (1983). After-effects of repetitive stimulation at low frequency on fast-contracting motor units of cat muscle. Journal of Physiology (London), 340, 129-143.
Jaric, S., Gavrilovic, P., & Ivancevic, V. (1985). Effects of previous muscle contractions on cyclic movement dynamics. European Journal of Applied Physiology, 54, 216-221.
Jasmin, B.J., Lavoie, P.-A., & Gardiner, P.F. (1987). Fast axonal transport of acetylcholine in rat sciatic motoneurons is enhanced following prolonged daily running, but not following swimming. Neuroscience Letters, 78, 156-160.
Jeannerod, M. (1988). The neural and behavioural organization of goal-directed movements. Oxford: Clarendon Press.
Jiang, B., Roy, R.R., & Edgerton, V.R. (1990). Expression of a fast fiber enzyme profile in the cat soleus after spinalization. Muscle & Nerve, 13, 1037-1049.
Johansson, H., Sjfilander, P., & Sojka, P. (1991). Receptors in the knee joint ligaments and their role in the biomechanics of the joint. CRC Critical Reviews in Biomedical Engineering, 18, 341-368.
Johansson, R.S., & Cole, K.J. (1992). Sensory-motor coordination during grasping and manipulative actions. Current Opinion in Neurobiology, 2, 815-823.
Johansson, R.S., Hager, C., & Backstrom, L. (1992). Somato-sensory control of precision grip during unpredictable pulling loads: 3. Impairments during digital anesthesia. Experimental Brain Research, 89, 204-213.
Johansson, R.S., Hager, C., & Riso, R. (1992). Somato-sensoiy control of precision grip during unpredictable pulling loads: 2. Changes in load force rate. Experimental Brain Research, 89, 192-203.
Johansson, R.S., Riso, R., Hager, C., & B/jckstnxm, L. (1992). Somatosensory control of precision grip during unpredi-table pulling loads: 1. Changes in load force amplitude. Experimental Brain Research, 89, 181-191.
Johansson, R.S., & Vallbo, A.B. (1983). Tactile sensory coding in the glabrous sldn of the human hand. Trends in Neurosciences, 6, 27-32.
Johansson, R.S., & Westling, G. (1984). Roles of glabrous skin receptors and sensorimotor memory in automatic control of precision grip when lifting rougher or more slippery objects. Experimental Brain Research, 56, 550-564.
Johansson, R.S., & Westling, G. (1987). Signals in tactile afferents from the fingers eliciting adaptive motor responses during precision grip. Experimental Brain Research, 66, 141-154.
Johns, R.J., & Wright, V. (1952). Relative importance of various tissues in joint stiffness. Journal of Applied Physiology, 17, 824-828.
Jones, D.A., Rutherford, O.M., & Parker, D.F. (1989). Physiological changes in skeletal muscle as a result of strength training. Quarterly Journal of Experimental Physiology, 74, 233-256.
Jones, L.A., & Hunter, LW. (1983). Effect of fatigue on force sensation. Experimental Neurology, 81, 640-650.
Josephson, R.K. (1993). Contraction dynamics and power output of skeletal muscle. Annual Reviews of Physiology, 55, 527-546.
Joyner, M.J. (1991). Modeling: Optimal marathon performance on the basis of physiological factors. Journal of Applied Physiology, 70, 683-687.
Kalaska, J.F., & Crammond, DJ. (1992). Cerebral cortical mechanisms of reaching movements. Science, 255, 1517-1523.
Kamen, G., & DeLuca, C.J. (1989). Unusual motor unit firing behavior in older adults. Brain Research, 482, 136-140.
Kanda, K., Burke, R.E., & Walmsley, B. (1977). Differential control of fast and slow twitch motor units in the decerebrate cat. Experimental Brain Research, 29, 57-74.
Kanda, K., & Hashizume, K. (1989). Changes in properties of the medial gastrocnemius motor units in aging rats. Journal of Neurophysiology, 61, 737-746.
Kanda, K., & Hashizume, K. (1992). Factors causing difference in force output among motor units in the rat medial gastrocnemius muscle. Journal of Physiology (London), 448, 677-695.
Kandel, E.R., & Jessell, T.M. (1991). Touch. In E.R. Kandel, J.H. Schwartz, & T.M. Jessell (Eds.), Principles of neural science (3rd ed., pp. 367-384). New York: Elsevier.
Kandel, E.R., Schwartz, J.H., & Jessell, T.M. (Eds.) (1991). Principles of neural science. New York: Elsevier.
Kandel, E.R., & Siegelbaum, S. (1985). Principles underlying electrical and chemical synaptic transmission. In E.R. Kandel & J.H. Schwartz (Eds.), Principles of neural science (3rd ed. pp. 89-107). New York: Elsevier.
25 s-133
385
Kane, T.R., & Scher, M.P. (1969). Adynamical explanation of the falling cat phenomenon. International Journal of Solids and Structures, 5, 663-670.
Karst, G.M., & Hasan, Z. (1987). Antagonist muscle activity during human forearm movements under varying kinematic and loading conditions. Experimental Brain Research, 67, 391-401.
Karst, G.M., & Hasan, Z. (1991a). Initiation rules for planar, two-joint arm movements: Agonist selection for movements throughout the work space. Journal of Neurophysiology, 66, 1579-1593.
Karst, G.M., & Hasan, Z. (1991b). Timing and magnitude of electromyographic activity for two-joint arm movements in different directions. Journal of Neurophysiology, 66, 1594-1604.
Katz, R., Mazzocchio, R., Pe'nicaud, A., & Rossi, A. (1993). Distribution of recurrent inhibition in the human upper limb. Acta Physiologica Scandinavica, 149, 183-198.
Katz, R., Penicaud, A., & Rossi, A. (1991). Reciprocal la inhibition between elbow flexors and extensors in the human. Journal of Physiology (London), 437, 269-286.
Keen, D.A., Yue, G.H., & Enoka, R.M. (in press). Training-related enhancement in the control of motor output in elderly humans. Journal of Applied Physiology.
Keller, A., Arissian, K., & Asanuma, H. (1992). Synaptic proliferation in the motor cortex of adult cats after long-term thalamic stimulation. Journal of Neurophysiology, 68, 295-308.
Kelly, J.P. (1985). Reactions of neurons to injury. In E.R. Kandel & J.H. Schwartz (Eds.), Principles of neural science (2nd ed., pp. 187-195). New York: Elsevier.
Kereshi, S., Manzano, G., & McComas, A.J. (1983). Impulse conduction velocities in human biceps brachii muscles. Experimental Neurology, 80,652-662.
Kemell, D. (1992). Organized variability in the neuromuscular system: A survey of task-related adaptations. Archives Italiennes de Biologic, 130, 19-66.
Kemell, D., & Hultbom, H. (1990). Synaptic effects on recruitment gain: A mechanism of importance for the input-output relations of motoneurone pools? Brain Research, 507, 176-179.
Kernell, D., & Monster, A.W. (1982a). Motoneurone properties and motor fatigue: An intracellular study of gastrocnemius motoneurones of the cat. Experimental Brain Research, 46, 197-204.
Kemell, D., & Monster, A.W. (1982b). Time course and properties of late adaptation in spinal motoneurones of the cat. Experimental Brain Research, 46, 191-196.
Keshner, E.A., & Allum, J.H.J. (1990). Muscle activation patterns coordinating postural stability from head to foot. In J.M. Winters & S.L.-Y. Woo (Eds.), Multiple muscle systems: Biomechanics and movement organization (pp. 481-497). New York: Springer-Verlag.
Kilgore, J.B., & Mobley, B.A. (1991). Additional force during stretch of single frog muscle fibres following tetanus. Experimental Physiology, 76, 579-588.
Kirsch, R,F., & Rymer, W.Z. (1987). Neural compensation for muscular fatigue: Evidence for significant force regulation in man. Journal of Neurophysiology, 57,1893-1910.
Kirsch, R.F., & Rymer, W.Z. (1992). Neural compensation for fatigue-induced changes in muscle stiffness during perturbations of elbow angle in human. Journal of Neurophysiology, 68, 449-470.
Kitai, T.A., & Sale, D.G. (1989). Specificity of joint angle in isometric training. European Journal of Applied Physiology, 58, 744-748.
Knott, M., & Voss, D.E. (1968). Proprioceptive neuromuscular facilitation: Patterns and techniques (2nd ed.). New York: Hoeber Medical Division, Harper & Row.
Koceja, D.M., & Raglin, J. (1992). Changes in neuromuscular characteristics during periods of overtraining and tapering. Medicine and Science in Sports and Exercise, 24(Suppl.), S80.
Koester, J. (1985a). Functional consequences of passive membrane properties of the neuron. In E.R. Kandel & J.H. Schwartz (Eds.), Principles of neural science (2nd ed., pp. 66-74). New York: Elsevier.
Koester, J. (1985b). Resting membrane potential and action potential. In E.R. Kandel & J.H. Schwartz (Eds.), Principles of neural science (2nd ed., pp. 49-57). New York: Elsevier.
Koh, TJ., Grabiner, M.D., & Clough, C.A. (1993). Bilateral deficit is larger for step than for ramp isometric contractions. Journal of Applied Physiology, 74, 1200-1205.
Komi, P. V. (1990). Relevance of in vivo force measurements to human biomechanics. Journal of Biomechanics, 23(Suppl. 1), 23-34.
Komi, P.V. (1992). Stretch-shortening cycle. In P.V. Komi (Ed.), Strength and power in sport, (pp. 169-179). Champaign, IL: Human Kinetics.
Komi, P.V., & Bosco, C. (1978). Utilization of stored elastic energy in leg extensor muscles by men and women. Medicine and Science in Sports, 10, 261-265.
Komi, P.V., & Buskirk, E.R. (1972). Effect of eccentric and concentric muscle conditioning on tension and electrical activity of human muscle. Ergonomics, 15, 417-434.
Komecki, S. (1992). Mechanism of muscular stabilization process in joints. Journal of Biomechanics, 25, 235-245.
Koshland, G.F., Hasan, Z., & Gerilovsky, L. (1991). Activity of wrist muscles elicited during imposed or voluntary movements about the elbow joint. Journal of Motor Behavior, 23, 91-100.
Koshland, G.F., & Smith, J.L. (1989a). Mutable and immutable features of paw-shake responses after hindlimb deafferentation in the cat. Journal of Neurophysiology, 62, 162-173.
Koshland, G.F., & Smith, J.L. (1989b). Paw-shake responses with joint immobilization: EMG changes with atypical feedback. Experimental Brain Research, 77, 361-373.
Kots, J.M. (1971). Trenirovka mysecnoj sily metodom elektrostimuljacci. Soobscenie 1. Teoreticeskie pred-posylki. Teoriya i Praktika Fizicheskoi Kultury, 3,64-67.
Kovanen, V., Suominen, H., & Heikkinen, E. (1984a). Collagen of slow twitch and fast twitch muscle fibres in different types of rat skeletal muscle. European Journal of Applied Physiology, 52, 235-242.
Kovanen, V., Suominen, H., & Heikkinen, E. (1984b). Mechanical properties of fast and slow skeletal muscle with special reference to collagen and endurance training. European Journal of Applied Physiology, 17, 725-735.
Kranz, H., Williams, AM., Cassell, J., Caddy, D.J., & Silberstein, R.B. (1983). Factors determining the frequency content of the electromyogram. Journal of Applied Physiology, 55, 392-399.
Krarup, C. (1981). Enhancement and diminution of mechanical tension evoked by staircase and by tetanus in rat muscle. Journal of Physiology (London), 311, 355-372.
Krnevic, K., & Miledi, R. (1958). Failure of neuromuscular propagation in rats. Journal of Physiology (London), 140, 440-461.
386
Krogh-Lund, С., & Jorgensen, К. (1992). Modification of myo-electric power spectrum in fatigue from 15д maximal voluntary contraction of human elbow flexor muscles, to limit of endurance: Reflection of conduction velocity variation and/or centrally mediated mechanisms? European Journal of Applied Physiology, 64, 359-370.
Kugelbeig, E., & Lindegren, B. (1979). Transmission and contraction fatigue of rat motor units in relation to succinate dehydrogenase activity of motor unit fibres. Journal of Physiology (London), 288, 285-300.
Kugelbeig, E., & Thomell, L.-E. (1983). Contraction time, histochemical type, and terminal cistemae volume of rat motor units. Muscle & Nerve, 6, 149-153.
Kukulka, C.G., & Clamann, H.P. (1981). Comparison of the recruitment and discharge properties of motor units in human brachial biceps and adductor pollicis during isometric contractions. Brain Research, 219, 45-55.
Kulig, K., Andrews, J.G., & Hay, J.G. (1984). Human strength curves. In R.L. Terjung (Ed.), Exercise and sport sciences reviews (Vol. 12, pp. 417-466). New York: Macmillan.
Kuno, M. (1964). Mechanism of facilitation and depression of the excitatory synaptic potential in spinal motoneurons. Journal of Physiology (London), 175, 100-112.
Kuno, S.-Y., Katsuta, S., Akisada, M., Anno, L, & Matsumoto, K. (1990). Effect of strength training on the relationship between magnetic resonance relaxation time and muscle fibre composition. European Journal of Applied Physiology, 61, 33-36.
Kuypers, H.G.J.M. (1985). The anatomical and functional organization of the motor system. In M. Swash & C. Kennard (Eds.), Scientific basis of clinical neurology (pp. 3-18). London: Churchill Livingstone.
Lacquaniti, F., & Soechting, J.F. (1986). EMG responses to load perturbations of the upper limb: Effect of dynamic coupling between shoulder and elbow motion. Experimental Brain Research, 61, 482-496.
Ladin, Z., & Wu, G. (1991). Combining position and acceleration measurements for joint force estimation. Journal of Biomechanics, 24, 1173-1187.
Laforest, S., St-Pierre, D.M.M., Cyr, J., & Guyton, D. (1990). Effects of age and regular exercise on muscle strength and endurance. European Journal of Applied Physiology, 60, 104- 111.
Lagasse, P.P. (1974). Muscle strength: Ipsilateral and contralateral effects of superimposed stretch. Archives of Physical Medicine and Rehabilitation, 55, 305-310.
Lakie, M., & Robson, L.G. (1988a). Thixotropic changes in human muscle stiffness and the effects of fatigue. Quarterly Journal of Experimental Physiology, 73, 487-500.
Lakie, M., & Robson, L.G. (1988b). Thixotropy: The effect of stretch size in relaxed frog muscle. Quarterly Journal of Experimental Physiology, 73, 127-129.
Lakomy, H.K.A. (1987), Measurement of human power output in high intensity exercise. In B. Van Gheluwe & J. Atha (Eds.), Current research in sport biomechanics (pp. 46-57). Basel, Switzerland: Karger.
Lamarra, N. (1990). Variables, constants, and parameters: Clarifying the system structure. Medicine and Science in Sports and Exercise, 22, 88-95.
Lance, J.W., Burke, D., & Andrews, C.J. (1973). The reflex effects of muscle vibration. In J.E. Desmedt (Ed.), New developments in electromyography and clinical neurophysiology, (Vol. 3, pp. 444-462). Basel, Switzerland: Karger.
Lander, J.E., Bates, B.T., & De Vita, P. (1986). Biomechanics of the squat exercise using a modified center
of mass bar. Medicine and Science in Sports and Exercise, 18, 468-478.
Lander, J.E., Hundley, J.R., & Simonton, R.L. (1992). The effectiveness of weight-belts during multiple repetitions of the squat exercise. Medicine and Science in Sports and Exercise, 24, 603-609.
Lannergren, J., & Westerblad, H. (1989). Maximum tension and force-velocity properties of fatigued, single Xenopus muscle fibres studied by caffeine and high Ku. Journal of Physiology (London), 409, 473-490.
Lannergren, J., & Westerblad, H. (1991). Force decline due to fatigue and intracellular acidification in isolated fibres from mouse skeletal muscle. Journal of Physiology (London), 434, 307-322.
Lansing, R.W., & Banzett, R.B. (1993). What do fully paralyzed awake humans feel when they attempt to move? Journal of Motor Behavior, 25, 309-313.
Lanyon, L.E., Hampson, W.G.J., Goodship, A.E., & Shah, J.S. (1975). Bone deformation recorded in vivo from strain gauges attached to the human tibial shaft. Acta Orthopaedica Scandinavica, 46, 256-268.
Lanyon, L.E., & Rubin, C.T. (1984). Static vs dynamic loads as an influence on bone remodelling. Journal of Biomechanics, 17, 897-905.
Laouris, Y., Kalli-Laouri, J., & Schwartze, P. (1990). The postnatal development of the air-righting reaction in albino rats: Quantitative analysis of normal development and the effect of preventing neck-torso and torso-pelvis rotations. Behavioral Brain Research, 37, 37-44.
Larsson, L. (1983). Histochemical characteristics of human skeletal muscle during aging. Acta Physiologica Scandinavica, 117, 469-471.
Larsson, L., Sjodin, B., & Karlsson, J. (1978). Histochemical and biochemical changes in human skeletal muscle with age in sedentary males, age 22-65 years. Acta Physiologica Scandinavica, 103, 31-39.
Latash, M.L. (1993). Control of human movement. Champaign, IL: Human Kinetics.
Latash, M.L., & Gottlieb, G.L. (1991a). An equilibriumpoint model for fast, single-joint movement: I. Emergence of strategy-dependent EMG patterns. Journal of Motor Behavior, 23, 163-177.
Latash, M.L., & Gottlieb, G.L. (1991b). An equilibriumpoint model for fast, single-joint movement: 2. Similarity of single-joint isometric and isotonic descending commands. Journal of Motor Behavior, 23, 179-191.
Laughman, R.K., Youdas, J.W., Garrett, T.R., & Chao, E.Y.S. (1983). Strength changes in the normal quadriceps femoris muscle as a result of electrical stimulation. Physical Therapy, 63, 494-499.
Lawrence, D.G., & Kuypers, H.G.J.M. (1968). The functional organization of the motor system in the monkey: 2. The effects of lesions of the descending brain-stem pathways. Brain, 91, 15-41.
Lawrence, J.H., & De Luca, CJ. (1983). Myoelectric signal versus force relationship in different human muscles. Journal of Applied Physiology, 54, 1653-1659.
Lemon, R.N. (1993). Cortical control of the primate hand. Experimental Physiology, 78, 263-301.
Lennmarken, C., Bergman, T., Larsson, J., & Larsson, L.-E. (1985). Skeletal muscle function in man: Force, relaxation rate, endurance and contraction-time dependence on sex and age. Clinical Physiology, 5, 243-255.
Lephart, S.A. (1984). Measuring the inertial properties of cadaver segments. Journal of Biomechanics, 17, 537-543.
25*
387
Lesmes, G.R., Costill, D.L., Coyle, E.F., & Fink, W.J. (1978). Muscle strength and power changes during maximal isokinetic training. Medicine and Science in Sports, 10, 266-269.
Levin, M.F., & Hui-Chan, C.W.Y. (1992). Relief of hemiparetic spasticity by TENS is associated with improvement in reflex and voluntary motor functions. Electroencephalography and Clinical Neurophysiology, 85, 131-142.
Lexell, J., & Taylor, C.C. (1991). Variability in muscle fibre areas in whole human quadriceps muscle: Effects of increasing age. Journal of Anatomy, 174, 239-249.
Li, C., & Atwater, A.E. (1984, June). Temporal and kinematic analysis of arm motion in sprinters. Paper presented at the Olympic Scientific Congress, Eugene, OR.
Liberson, W.T., Holmquest, HJ., Scot, D., & Dow, M. (1961). Functional electrotherapy: Stimulation of the peroneal nerve synchronized with the swing phase of the gait of hemiplegic patients. Archives of Physical Medicine and Rehabilitation, 42, 101-105.
Lieber, R.L. (1992). Skeletal muscle structure and function. Baltimore: Williams & Wilkins.
Lieber, R.L., Fazeli, B.M., & Botte, M.J. (1990). Architecture of selected wrist flexor and extensor muscles. Journal of Hand Surgery, 15A, 244-250.
Lieber, R.L., & Fridftn, J. (1993). Muscle damage is not a function of muscle force but active muscle strain. Journal of Applied Physiology, 74, 520-526.
Lieber, R.L., Fridiin, J.O., Hargens, A.R., Danzig, L.A., & Gershuni, D.H. (1988). Differential response of the dog quadriceps muscle to external skeletal fixation of the knee. Muscle & Nerve, 11, 193-201.
Lieber, R.L., & Kelly, M.J. (1991). Factors influencing quadriceps femoris muscle torque using transcutaneous neuro-muscular electrical stimulation. Physical Therapy, 71, 715-723.
Lindman, R., Eriksson, A., & Thornell, L.-E. (1990). Fiber type composition of the human male trapezius muscle: Enzyme-histochemical characteristics. American Journal of Anatomy, 189, 236-244.
Lindman, R., Eriksson, A., & Thornell, L.-E. (1991). Fiber type composition of the human female trapezius muscle: Enzyme-histochemical characteristics. American Journal of Anatomy, 190, 385-392.
Lindstrom, L., Magnusson, R., & Petersen, 1. (1970). Muscular fatigue and action potential conduction velocity changes studied with frequency analysis of EMG signals. Electromyography, 4, 341-356.
Lloyd, A.R., Gandevia, S.C., & Hales, J.P. (1991). Muscle performance, voluntary activation, twitch properties and perceived effort in normal subjects and patients with chronic fatigue syndrome. Brain, 114, 85-98.
Lloyd, D.P.C. (1949). Post-tetanic potentiation of response in monosynaptic reflex pathways of the spinal cord. Journal of General Physiology, 33, 147-170.
Loeb, G.E., & Gans, C. (1986). Electromyography for experimentalists. Chicago: University of Chicago Press.
Lombardi, V., & Piazzesi, G. (1990). The contractile response during steady lengthening of stimulated frog muscle fibres. Journal of Physiology (London), 431,141-171.
L^mo, T., & Waerhaug, 0. (1985). Motor endplates in fast and slow muscles of the rat: What determines their differences? Journal of Physiology (Paris), 80,290-297.
Lotz, B.P., Dunne, J.W., & Daube, J.R. (1989). Preferential activation of muscle fibers with peripheral magnetic stimulation of the limb. Muscle & Nerve, 12, 636-639.
Lovely, R.G., Gregor, R.J., Roy, R.R., & Edgerton, V.R. (1990). Weight-bearing hindlimb stepping in treadmill-exercised adult spinal cats. Brain Research, 514, 206-218.
Lowrie, M.B., & Vrbova, G. (1992). Dependence of postnatal motoneurones on their targets: Review and hypothesis. Trends in Neurosciences, 15, 80-84.
Lucas, S.M., Ruff, R.L., & Binder, M.D. (1987). Specific tension measurements in single soleus and medial gastrocnemius muscle fibers of the cat. Experimental Neurology, 95, 142-154.
Luethi, S.M., & Stacoff, A. (1987). The influence of the shoe on foot mechanics in running. In B. van Gheluwe & J. Atha (Eds.), Current research in sports biomechanics (pp. 72-85). New York: Karger.
Luhtanen, P. (1988). Kinematics and kinetics of serve in volleyball at different age levels. In G. de Groot, A.P. Hollander, P.A. Huijing, & G J. van Ingen Schenau (Eds.), Biomechanics XI-В (pp. 815-819). Amsterdam: Free University Press.
Luhtanen, P., & Komi, P.V. (1978). Mechanical factors influencing running speed. In E. Asmussen & K. Jorgensen (Eds.), Biomechanics Vl-B (pp. 23-29). Baltimore: University Park Press.
Luhtanen, P., & Komi, P.V. (1980). Force-, power-, and elasticity-velocity relationships in walking, running, and jumping. European Journal of Applied Physiology, 44, 279-289.
Lusby, L.A., & Atwater, A.E. (1983). Speed-related position-time profiles of arm motion in trained women distance runners. Medicine and Science in Sports and Exercise, 15, 171.
Liischer, H.-R., Ruenzel, P., & Henneman, E. (1979). How the size of motoneurones determines their susceptibility to discharge. Nature, 282, 859-861.
MacDougall, J.D., Sale, D.G., Alway, S.E., & Sutton, J.R. (1984). Muscle fiber number in biceps brachii in body-builders and control subjects. Journal of Applied Physiology, 57, 1399-1403.
Macefield, G., Hagbarth, K.-E., Gorman, R., Gandevia, S.C., & Burke, D. (1991). Decline in spindle support to (a-motoneurones during sustained voluntary contractions. Journal of Physiology (London), 440,497-512.
Madsen, N., & McLaughlin, T. (1984). Kinematic factors influencing performance and injury risk in the bench press exercise. Medicine and Science in Sports and Exercise, 16, 376-381.
Magladery, J.W., & McDougal, D.B. (1950). Electrophysiological studies of nerve and reflex activity in normal man: I. Identification of certain reflexes in the electromyogram and the conduction velocity of peripheral nerve fibres. Bulletin of Johns Hopkins Hopsital, 86, 265-290.
Magnus, R. (1922). Wie sich die fallende Katze in de Luft umdrecht. Archives Neerlandaises de Physiologie de ГНотте et des Animaux, 7, 218-222.
Mancini, D.M., Coyle, E., Coggan, A., Beltz, J., Ferraro, N., Montain, S., & Wilson, J.R. (1989). Contribution of intrinsic skeletal muscle changes to 3IP NMR skeletal muscle metabolic abnormalities in patients with chronic heart failure. Circulation, 80, 1338-1346.
Mann, R.V, (1981). A kinetic analysis of sprinting. Medicine and Science in Sports and Exercise, 13, 325-328.
Manter, J.T. (1938). The dynamics of quadrupedal walking. Journal of Experimental Biology, 15, 522-540.
Marey, E.-J. (1874). Animal mechanism: A treatise on terrestial and aerial locomotion. New York: Appleton & Co.
388
Marey, E.-J. (1894). Des mouvemerits que certains animaux executent pour retomber sur leurs pieds, lorsqu'ils sont precipites d'un lieu eleve. Academie des Sciences, 119, 714-718.
Marino, A.A. (1984). Electrical stimulation in orthopaedics: Past, present and future. Journal of Bioelectricity, 3, 235-244.
Markhede, G., & Grimby, G. (1980). Measurements of strength of hip joint muscles. Scandinavian Journal of Rehabilitation Medicine, 12, 169-174.
Marras, W.S., & Mirka, G.A. (1992). A comprehensive evaluation of trunk response to asymmetric trunk motion. Spine, 17, 318-326.
Marras, W.S., & Sommerich, C.M. (1991a). A three-dimensional motion model of loads on the lumbar spine: I Model structure. Human Factors, 33, 123-137.
Manas, W.S., & Sommerich, C.M. (1991b). A three-dimensional motion model of loads on the lumbar spine: II, Model validation. Human Factors, 33, 139-149.
Manas, W.S., Joynt, R.L., & King, A.I. (1985). The forcevelocity relation and intra-abdominal pressure during lifting activities. Ergonomics, 28, 603-613.
Marsden, C.D., Meadows, J.C., & Merton, P.A. (1983). Muscular wisdom , that minimized fatigue during prolonged effort in man: Peak rates of motoneuron discharge and slowing of discharge during fatigue. In J.E. Desmedt (Ed.), Motor control mechanisms in health and disease (pp. 169-211). New York: Raven Press.
Marsden, C.D., Merton, P.A., & Morton, H.B. (1976). Servo action in the human thumb. Journal of Physiology (London), 257, 1-44.
Marsden, C.D., Merton, P.A., & Morton, H.B. (1983). Rapid postural reactions to mechanical displacement of the hand in man. In J.E. Desmedt (Ed.), Motor control mechanisms in health and disease (pp. 645-659). New York: Raven Press.
Marsh, E., Sale, D., McComas, A.J., & Quinlan, J. (1981). The influence of joint position on ankle dorsiflexion in humans. Journal of Applied Physiology, 51,160-167.
Martin, J.H., & Jessell, T.M. (1991). Anatomy of the somatic sensory system. In E.R. Kandel, J.H. Schwartz, & T.M. Jessell (Eds.), Principles of neural science (3rd ed., pp. 353-366). New York: Elsevier.
Martin, J.P. (1977). A short essay on posture and movement. Journal of Neurology, Neurosurgery, and Psychiatry, 40, 25-29.
Martin, P.E., Mungiole, M., Marzke, M.W., & Longhill, J.M. ( 1989). The use of magnetic resonance imaging for measuring segment inertial properties. Journal of Biomechanics, 22, 367-376.
Maton, B. (1991). Central nervous system changes in fatigue induced by local work. In G. Allan, L. Beliveau, and P. Bouissou (Eds.), Muscle fatigue: Biochemical and physiological aspects, (pp. 207-221). Paris: Masson.
Maton, B., Petitjean, M., & Cnockaert, J.C. (1990). Phonomyogram and electromyogram relationships with isometric force reinvestigated in man. European Journal of Applied Physiology, 60, 194-201.
Matthews, P.B.C. (1972). Mammalian muscle receptors and their central actions. Baltimore: Williams & Wilkins.
Matthews, P.B.C. (1987). Muscle sense. In G. Adelman (Ed.), Encyclopedia of neuroscience (Vol. 2, pp. 720-721). Boston: Birkhauser.
Matthews, P.B.C. (1990). The kneejerk: Still an enigma? Canadian Journal of Physiology and Pharmacology, 68, 347-354.
Matthews, P.B.C. (1991). The human stretch reflex and the motor cortex. Trends in Neurosciences, 14, 87-91.
Matyas, T.A., Galea, M.P., & Spicer, S.D, (1986). Facilitation of the maximum voluntary contraction in hemiplegia by concomitant cutaneous stimulation. American Journal of Physical Medicine, 65, 125-134.
Maughan, R.J., Nimmo, M.A., & Harmon, M. (1985). The relationship between myosin ATP-ase activity and isometric endurance in untrained male subjects. European Journal of Applied Physiology, 54, 291-296.
Mayer, R.F., Burke, R.E., Toop, J., Hodgson, J.A., Kanda, K., & Waimsley, B. (1981). The effect of longterm immobilization on the motor unit population of the cat medial gastrocnemius muscle. Neuroscience, 6, 725-739.
McCafferty, W.B., & Horvath, S.M. (1977). Specificity of exercise and specificity of training: A subcellular review. Research Quarterly, 48, 358-371.
McCartney, N., Heigenhauser, G.J.F., & Jones, N.L. (1983). Power output and fatigue of human muscle in maximal cycling exercise. Journal of Applied Physiology, 55, 218-224.
McCartney, N., Moroz, D., Garner, S.H., & McComas, A.J. (1988). The effects of strength training in patients with selected neuromuscul. r disorders. Medicine and Science in Sports and Exercise, 20, 362-368.
McCloskey, D.I. (1987). Kinesthesia, kinesthetic perception. In G. Adelman (Ed.), Encyclopedia of neuroscience (Vol. I, pp. 548-551). Boston: Birkhauser.
McCloskey, D.I., Ebeling, P., & Goodwin, G.M. (1974), Estimation of weights and tensions and apparent involvement of a sense of effort. Experimental Neurology, 42, 220-232.
McCloskey, M. (1983, April), Intuitive physics. Scientific American, pp, 122-130.
McComas, AJ. (1977). Neuromuscular function and disorders. Boston: Butterworths.
McComas, A.J. (1991). Motor unit estimation: Methods, results, and present status. Muscle & Nerve, 14, 585-597.
McComas, A.J., Sica, R.E., Upton, A.R.M., & Aguilera, G.C. (1973). Functional changes in motoneurons of hemiparetic patients. Journal of Neurology, Neurosurgery, and Psychiatry, 36, 183-193.
McCully, K.K., & Faulkner, J.A. (1985). Injury to skeletal muscle fibers of mice following lengthening contractions. Journal of Applied Physiology, 59, 119-126.
McDonagh, J.C., Binder, M.D., Reinking, R.M., & Stuart, D.G. (1980). A commentary on muscle unit properties in cat hindlimb muscles. Journal of Morphology, 166, 217-230.
McDonagh, MJ.N., & Davies, C.T.M. (1984). Adaptive response of mammalian skeletal muscle to exercise with high loads. European Journal of Applied Physiology, 52, 139-155.
McDonagh, MJ.N., Hayward, C.M., & Davies, C.T.M. (1983). Isometric training in human elbow flexor muscles: The effects on voluntary and electrically evoked forces. Journal of Bone and Joint Surgery, 65B, 355-358.
McGill, S.M., & Norman, R.W. (1993). Low back biomechanics in industry: The prevention of injury through safer lifting. In M.D. Grabiner (Ed.), Current issues in biomechanics (pp. 69-120). Champaign, IL: Human Kinetics.
McLaughlin, T.M., Dillman, C.J., & Lardner, T.J. (1977). A kinematic model of performance in the parallel squat by champion powerlifters. Medicine and Science in Sports, 9, 128-133.
389
McMahon, T.A. (1984). Muscles, reflexes, and locomotion. Princeton, NJ: Princeton University Press.
McNitt-Gray, J.L. (1991). Kinematics and impulse characteristics of drop landings from three heights. International Journal of Sport Biomechanics, 7, 201-224.
Melvill-Jones, G., & Watt, D.G.D. (1971). Muscular control of landing from unexpected falls in man. Journal of Physiology (London), 219, 729-737.
Meriam, J.L., & Kraige, L.G. (1987). Engineering mechanics. New York: Wiley.
Merletti, R., Knaflitz, M., & De Luca, C.J. (1990). Myoelectric manifestations of fatigue in voluntary and electrically elicited contractions. Journal of Applied Physiology, 69, 1810-1820.
Metzger, J.M., & Fitts, R.H. (1986). Fatigue from high-and low-frequency muscle stimulation: Role of sarcolemma action potentials. Experimental Neurology, 93, 320-333.
Miller, C., & The'paut-Mathieu, C. (1993). Strength training by electrostimulation conditions for efficacy. International Journal of Sports Medicine, 14, 20-28.
Miller, D.L (1976). A biomechanical analysis of the contribution of the trunk to standing vertical jump take-off. In J. Broekhoff (Ed.), Physical education, sports and the sciences (pp. 355-374). Eugene, OR: Microform.
Miller, D.L (1978). Biomechanics of running: What should the future hold? Canadian Journal of Applied Sport Sciences, 3, 229-236.
Miller, D.L (1979). Modelling in biomechanics: An overview. Medicine and Science in Sports, 11, 115-122.
Miller, D.L (1980). Body segment contributions to sport skill performance: Two contrasting approaches. Research Quarterly for Exercise and Sport, 51, 219-233.
Miller, D.L (1990). Ground reaction forces in distance running. In P.R. Cavanagh (Ed.), Biomechanics of distance running (pp. 203-224). Champaign, IL: Human Kinetics.
Miller, D.L, Enoka, R.M., McCulloch, R.G., Burgess, E.M., Hutton, R.S., & Frankel, V.H. (1979). Biomechanical analysis of lower extremity amputee extraambulatory activities (Contract No. V5244P-1540/VA). New York: Veterans Administration.
Miller, D.L, Henning, E., Pizzimenti, M.A., Jones, LC., & Nelson, R.C. (1989). Kinetic and kinematic characteristics of 10-m platform performances of elite divers: 1. Back takeoffs. International Journal of Sport Biomechanics, 5, 60-88.
Miller, D.L, & Morrison, W.E. (1975). Prediction of segmental parameters using the Hanavan human body model. Medicine and Science in Sports, 7, 207-212.
Miller, D.L, & Munro, C.F. (1984). Body segment contributions to height achieved during the flight of a springboard dive. Medicine and Science in Sports and Exercise, 16, 234-242.
Miller, D.L, & Munro, C.F. (1985). Greg Louganis' springboard takeoff: 2. Linear and angular momentum considerations. International Journal of Sport Biomechanics, 1, 288-307.
Miller, D.L, & Nelson, R.C. (1973). Biomechanics of sport. London: Kimpton.
Miller, R.G., Giannini, D., Milner-Brown, H.S., Layzer, R.B., Koretsky, A.P., Hooper, D., & Weiner, M.W. (1987). Effects of fatiguing exercise on high-energy phosphates, force, and EMG: Evidence for three phases of recovery. Muscle & Nerve, 10, 810-821.
Milliron, M.J., & Cavanagh, P.R. (1990). Sagittal plane kinematics of the lower extremity during distance
running. In P.R. Cavanagh (Ed.), Biomechanics of distance running (pp. 65-105). Champaign, IL: Human Kinetics.
Milner-Brown, H.S., & Miller, R.G. (1986). Muscle membrane excitation and impulse propagation velocity are reduced during muscle fatigue. Muscle & Nerve, 9, 367-374.
Milner-Brown, H.S., & Miller, R.G. (1989). Increased muscular fatigue in patients with neurogenic muscle weakness: Quantification and pathophysiology. Archives of Physical Medicine and Rehabilitation, 70, 361-366.
Milner-Brown, H.S., & Stein, R.B. (1975). The relation between the surface electromyogram and muscular force. Journal of Physiology (London), 246, 549-569.
Milner-Brown, H.S., Stein, R.B., & Lee, R.G. (1975). Synchronization of human motor units: Possible roles of exercise and supraspinal reflexes. Electroencephalography and Clinical Neurophysiology, 38, 245-254.
Milner-Brown, H.S., Stein, R.B., & Yemm, R. (1973). The contractile properties of human motor units during voluntary isometric contractions. Journal of Physiology (London), 228, 285-306.
Monster, A.W., & Chan, H.C. (1977). Isometric force production by motor units of extensor digitorum communs muscle in man. Journal of Neurophysiology, 40, 1432-1443.
Monster, A.W., Chan, N.C., & O'Connor, D. (1978). Activity patterns of human skeletal muscle: Relation to muscle fiber type composition. Science, 200, 314-317.
Moore, G.P., Perkel, D.H., & Segundo, J.P. (1966). Statistical analysis and functional interpretation of neuronal spike data. Annual Review of Physiology, 28, 493-522.
Moore, M.A., & Hutton, R.S. (1980). Electromyographic investigation of muscle stretching techniques. Medicine and Science in Sports and Exercise, 12, 322-329.
Moore, M.A., & Kukulka, C.G. (1991). Depression of Hoffman reflexes following voluntary contraction and implications for proprioceptive neuromuscular facilitation therapy. Physical Therapy, 71, 321-333.
Moreno-Aranda, J., & Seireg, A. (1981 a). Electrical parameters for over-the-skin muscle stimulation. Journal of Biomechanics, 14, 579-585.
Moreno-Aranda, J., & Seireg, A. (1981b). Force response to electrical stimulation of canine skeletal muscles. Journal of Biomechanics, 14, 595-599.
Moreno-Aranda, J., & Seireg, A. (1981c). Investigation of over-the-skin electrical stimulation parameters for different normal muscles and subjects. Journal of Biomechanics, 14, 587-593.
Morey, E.R. (1979). Spaceflight and bone turnover: Correlation with a new rat model of weightlessness. Bioscience, 29, 168-172.
Morgan, D.L. ( 1990). New insights into the behavior of muscle during active lengthening. Biophysical Journal, 57, 209-221.
Moritani, T. (1992). Time course of adaptations during strength and power training. In P.V. Komi (Ed.), Strength and power in sport (pp. 266-278). Champaign, IL: Human Kinetics.
Morris, A.F. (1974). Myotatic reflex effects on bilateral reciprocal leg strength. American Corrective Therapy Journal, 28, 24-29.
Morris, J.M., Lucas, D.B., & Bressler, B. (1961). Role of the trunk in stability of the spine. Journal of Bone and Joint Surgery, 43A, 327-351.
Mow, V.C., Proctor, C.S., & Kelly, M.A. (1989). Biomechanics of articular cartilage. In M. Nordin & V.H.
390
Frankel (Eds.), Basic biomechanics of the musculoskeletal system (pp. 31-58). Philadelphia: Lea & Febiger. Munro, C.F., Miller, D.I., & Fuglevand, A.J. (1987).
Ground reaction forces in running: A reexamination. Journal of Biomechanics, 20, 147-155.
Munson, J.B., Foehring, R.C., Lofton, S.A, Zengel, J.E., & Sypert, G.W. (1986). Plasticity of medial gastrocnemius motor units following cordotomy in the cat. Journal of Neurophysiology, 55, 619-633.
Nachemson, A.L., Andersson, G.B.J., & Schultz, A.B. (1986). Valsalva maneuver biomechanics. Spine, 11, 476-479.
Nakazawa, K., Kawakami, Y., Fukunaga, T., Yano, H., & Miyashita, M. (1993). Differences in activation patterns in elbow flexor muscles during isometric, concentric and eccentric contractions, European Journal of Applied Physiology, 66, 214-220.
Nardone, A., Romano, C., & Schieppati, M. (1989). Selective recruitment of high-threshold human motor units during voluntaiy isotonic lengthening of active muscles. Joumalof Physiology (London), 409, 451-471.
Narici, M.V., Bordini, M., & Ceretelli, P. (1991). Effect of aging on human adductor pollicis muscle fimction. Journal of Applied Physiology, 71, 1277-1281.
Narici, M.V., Roi, G.S., & Landoni, L. (1988). Force of knee extensor and flexor muscles and cross-sectional area determined by nuclear magnetic resonance imaging. European Journal of Applied Physiology, 57, 39-44.
Narici, M.V., Roi, G.S., Landoni, L., Minetti, A.E., & Ceretelli, P. (1989). Changes in force, cross-sectional area and neural activation during strength training and detraining of the human quadriceps. European Journal of Applied Physiology, 59, 310-319.
Nashner, L.M. ( 1971 ). A model describing vestibular detection of body sway motion. Acta Otolaryngology, 72, 429-436.
Nashner, L.M. ( 1972). Vestibular postural control model. Kybemetik. 10, 106-110.
Nashner, L.M. (1976). Adapting reflexes controlling the human posture. Experimental Brain Research, 26,59-72.
Nashner, L.M. (1977). Fixed patterns of rapid postural responses among leg muscles during stance. Experimental Brain Research, 30, 13-24.
Nashner, L.M. (1982). Adaptation of human movement to altered environments. Trends in Neurosciences, 5, 358-361.
Nashner, L.M., Woollacott, M.H., & Tuma, G. (1979). Organization of rapid responses to postural and locomotorlike perturbation of standing man. Experimental Brain Research, 36, 463-476.
Nemeth, G., Ekholm, J., Arborelius, U.P., Harms-Ringdahl, K., & Schuldt, K. (1983). Influence of knee flexion on isometric hip extensor strength. Scandinavian Journal of Rehabilitation Medicine, 15, 97-101.
Nemeth, G., & Ohlsen, H. (1986). Moment arm lengths of trunk muscles to the lumbosacral joint obtained in vivo with computed tomography. Spine, 11, 158-160.
Nemeth, P.M., & Pette, D. (1981). Succinate dehydrogenase activity in fibres classified by myosin ATPase in three hind limb muscles of the rat. Journal of Physiology (London), 320, 73-80.
Nemeth, P.M., Solanki, L., Gordon, D.A., Hamm, T.M., Reinking, R.M., & Stuart, D.G. (1986). Uniformity of metabolic enzymes within individual motor units. Journal of Neuroscience, 6, 892-898.
Newham, D.J. (1988). The consequences of eccentric contractions and their relationship to delayed onset muscle pain. European Journal of Applied Physiology, 57, 353-359.
Newham, D.J., Jones, D.A., & Clarkson, P.M. (1987). Repeated high-force eccentric exercise: Effects on muscle pain and damage. Journal of Applied Physiology, 63, 1381-1386.
Newham, D.J., Jones, D.A, & Edwards, R.H.T. (1983). Large delayed plasma creatine kinase changes after stepping exercise. Muscle & Nerve, 6, 380-385.
Nicks, D.K., Beneke, W.M., Key, R.M., & Timson, B.F. (1989). Muscle fibre size and number following immobilisation atrophy. Journal of Anatomy, 163, 1-5.
Nielsen, J., Crone, C., & Hultbom, H. (1993). H-reflexes are smaller in dancers from The Royal Danish Ballet than in well-trained athletes. European Journal of Applied Physiology, 66, 116-121.
Nielsen, J., Kagamihara, Y., Crone, C., & Hultbom, H. (1992). Central facilitation of la inhibition during tonic ankle dorsiflexion revealed after blockade of peripheral feedback. Experimental Brain Research, 88, 651-656.
Nigg, B.M. (Ed.) (1986). Biomechanics of running shoes. Champaign, IL: Human Kinetics.
Nilsson, J., & Thorstensson, A. (1987). Adaptability in frequency and amplitude of leg movements during human locomotion at different speeds. Acta Physiologica Scandinavica, 129, 107-114.
Nilsson, J., & Thorstensson, A. (1989). Ground reaction forces at different speeds of human walking and running. Acta Physiologica Scandinavica, 136, 217-228.
Nissinen, M. (1978). Kinematic and kinetic analysis of the hurdle and support phases of a running forward somersault. Unpublished master's thesis. University of Washington, Seattle.
Nissinen, M., Preiss, R., & Briiggemann, P. (1985). Simulation of human airborne movements on the horizontal bar. In D.A. Winter, R.W. Norman, R.P. Wells, K.C. Hayes, & AE. Patla (Eds.), Biomechanics IX-B (pp. 373-376). Champaign, IL: Human Kinetics.
Nordin, M., & Frankel, V.H. (1989). Biomechanics of bone. In M. Nordin & V.H, Frankel (Eds.), Basic biomechanics of the musculoskeletal system (pp. 3-29). Philadelphia: Lea & Febiger.
Nordstrom M.A, Enoka, R.M., Callister, R.J., Reinking, R.M., & Stuart, D.G. (1994). Effects of six weeks of limb immobilization on the cat tibialis posterior: 1. Motor units. Manuscript submitted for publication.
Nordstrom, M.A., Fuglevand, A.J., & Enoka, R.M. (1992). Estimating the strength of common input to motoneurons from the cross-correlogram. Journal of Physiology (London), 453, 547-574.
Nordstrom, M.A, & Miles, T.S. (1990). Fatigue of single motor units in human masseter. Journal of Applied Physiology, 68, 26-34.
Noyes, F.R. (1977). Functional properties of knee ligaments and alterations induced by immobilization. Clinical Orthopaedics and Related Research, 123, 210-242.
Noyes, F.R., Butler, D.L., Grood, E.S., Zemicke, R.F., & Hefzy, M.S. (1984). Biomechanical analysis of human ligament grafts used in knee-ligament repairs and reconstructions. Journal of Bone and Joint Surgery, 66A, 344-352.
Nygaard, E., Houston, M.E., Suzuki, Y., Jorgensen, K., & Saltin, B. (1983). Morphology of the brachial biceps muscle and elbow flexion in man. Acta Physiologica Scandinavica, 117, 287-292.
Ochs, S. (1987). Axoplasmic transport. In G. Adelman (Ed.), Encyclopedia of neuroscience (Vol. 1, pp. 105-108). Boston: Birkhauser.
O'Hara, T.E., & Goshgarian, H.G. (1991). Quantitative assessment of phrenic nerve functional recovery
391
mediated by the crossed phrenic reflex at various time intervals after spinal cord injury. Experimental Neurology, 111, 244-250.
Ohtsuki, T. (1983). Decrease in human voluntary isometric arm strength induced by simultaneous bilateral exertion. Behavioural Brain Research, 7, 165-178.
Oishi, Y., Ishihara, A., & Katsuta, S. (1992). Muscle fibre number following hindlimb immobilization. Acta Physiologica Scandinavica, 146, 281-282.
Orizio, C., Perini, R., Diemont, B., Figini, M.M., & Veicsteinas, A. (1990). Spectral analysis of muscular sound during isometric contraction of biceps brachii. Journal of Applied Physiology, 68, 508-512.
Orizio, C., Perini, R., & Veicsteinas, A. (1989). Changes in muscular sound during sustained isometric contraction up to exhaustion. Journal of Applied Physiology, 66, 1593-1598.
Oster, G. (1984, March). Muscle sounds. Scientific American, pp. 108-115.
Osternig, L.R. (1986). Isokinetic dynamometry: Implications for muscle testing and rehabilitation. In K.B. Pandolf (Ed.), Exercise and sport sciences reviews (Vol. 14, pp. 45-80). New York: Macmillan.
Otten, E. (1938). Concepts and models of functional architecture in skeletal muscle. In K.B. Pandolf (Ed.), Exercise and sport sciences reviews (Vol. 16, pp. 89-137). New York: Macmillan.
Ounjian, M., Roy, R.R., Eldred, E., Garfinkel, A., Payne, J.R., Armstrong, A., Toga, A.W., & Edgerton, V.R. (1991). Physiological and developmental implications of motor unit anatomy. Journal of Neurobiology, 11, 547-550.
Pachter, B.R., & Eberstein, A. (1986). The effect of limb immobilization and stretch on the fine structure of the neuromuscular junction in rat muscle. Experimental Neurology, 92, 13-19.
Palay, S.L., & Chan-Palay, V. (1987). Neuron. In G. Adelman (Ed.), Encyclopedia of neuroscience (Vol. 2, pp. 812-815). Boston: Birkhauser.
Panin, N., Lindenauer, H.J., Weiss, A.A., & Ebel, A. (1961). Electromyographic evaluation of the cross exercise effect. Archives of Physical Medicine and Rehabilitation, 42, 47-53.
Parker, P.A., Kbrner, L., & Kadefors, R. (1984). Estimation of muscle force from intramuscular pressure. Medical and Biological Engineering and Computing, 11, 453-457.
Parkkola, R., Alanen, A., Kalimo, H., Lillsunde, 1., Komu, M., & Kormano, M. (1993). MR relaxation times and fiber type predominance of the psoas and multifidus muscle. Acta Radiologica, 34, 16-19.
Patla, A.E. (1991). Visual control of human locomotion. In A.E. Patla (Ed.), Adaptability of Human Gait (pp. 55-97). North Holland: Elsevier Science Publishers B.V.
Pawson, P.A., & Grinnell, A.D. (1990). Physiological differences between strong and weak frog neuromuscular junctions: A study involving tetanic and postteta-nic potentiation. Journal of Neuroscience, 10, 1769-1778.
Pearson, K. (1993). Common principles of motor control in vertebrates and invertebrates. Annual Reviews in Neuroscience, 16, 265-297.
Pepe, F.A., & Drucker, B. (1979). The myosin filament IV: Myosin content. Journal of Molecular Biology, 130, 379-393.
Peiry, J., & Bekey, G.A. (1981). EMG-force relationships in skeletal muscle. CRC Critical Reviews in Biomedical Engineering, 7, 1-22.
Person, R.S. (1958). An electromyographic investigation on coordination of the activity of antagonist muscles in man during development of a motor habit. Pavlov Journal of Higher Nervous Activity, 8, 13-23.
Person, R.S. (1963). Problems in the interpretation of electromyograms: 1. Comparison of electromyograms on recording with skin and needle electrodes. Biophysics, 8, 89-97.
Person, R.S. (1974). Rhythmic activity of a group of human motoneurones during voluntary contraction of a muscle. Electroencephalography and Clinical Neurophysiology, 36, 585-595.
Person, R.S., & Kudina, L.P. (1972). Discharge frequency and discharge pattern of human motor units during voluntary contraction of muscle. Electroencephalography and Clinical Neurophysiology, 32, 471-483,
Peters, S.E. (1989). Structure and function in vertebrate skeletal muscle. American Zoologist, 19, 221-234.
Peterson, S.R., Bell, G.J., Bagnall, K.M., & Quinney, H.A. (1991). The effects of concentric resistance training on eccentric peak torque and muscle cross-sectional area. Journal of Orthopaedic and Sports Physical Therapy, 13, 132-137.
Petrofsky, J.S., & Hendershot, D.M. (1984). The interrelationship between blood pressure, intramuscular pressure, and isometric endurance in fast and slow twitch skeletal muscle in the cat. European Journal of Applied Physiology, 53, 106-111.
Petrofsky, J.S., & Stacy, R. (1992). The effect of training on endurance and the cardiovascualar responses of individuals with paraplegia during dynamic exercise induced by functional electrical stimulation. European Journal of Applied Physiology, 64, 487-492.
Pette, D., & Vrbova G. (1992). Adaptation of mammalian skeletal muscle to chronic electrical stimulation. Reviews of Physiology, Biochemistry, and Pharmacology, 120, 115-202.
Phillips, C.G. (1986). Movements of the hand. Liverpool: Liverpool University Press.
Phillips, S.J., & Roberts, E.M. (1980). Muscular and nonmuscular moments of force in the swing limb of Masters runners. In J.M. Cooper & B. Haven (Eds.), Proceedings of the Biomechanics Symposium (pp. 256-274). Bloomington, IN: Indiana State Board of Health.
Phillips, S.J , Roberts, E.M., & Huang, T.C. (1983). Quantification of intersegmental reactions during rapid swing motion. Journal of Biomechanics, 16, 411-418.
Pitman, M.I., & Peterson, L. (1989). Biomechanics of skeletal muscle. In M, Nordin & V.H. Frankel (Eds.), Basic biomechanics of the musculoskeletal system (pp. 89-111). Philadelphia: Lea& Febiger.
Pollack, G.H. (1983). The cross-bridge theory. Physiological Reviews, 63, 411-417.
Powers, R.K., Robinson, F.R., Konodi, M.A., & Binder, M.D. (1992). Effective synaptic current can be estimated from measurements of neuronal discharge. Journal of Neurophysiology, 68, 964-968.
Proske, V, Morgan, D.L., & Gregory, J.E. (1993). Thixotropy in skeletal muscle spindles: A review. Progress in Neurobiology, 41, 705-721.
Psek, J.A., & Cafarelli, E. (1993). Behavior of coactive muscles during fatigue. Journal of Applied Physiology, 74, 170-175.
Putnam, C.A. (1983). Interaction between segments during a kicking motion. In H. Matsui and K. Kobayashi (Eds.), Biomechanics VHI-B(pp. 688-694). Champaign, IL: Human Kinetics.
PUnam, C.A. (1991). A segment interaction analysis of proximal-to-distal sequential segment motion patterns.
392
Medicine and Science in Sports and Exercise, 23, 130-144.
Putnam, C.A. (1993). Sequential motions of body segments in striking and throwing skills: Descriptions and explanations. Journal of Biomechanics, 26, 125-135.
Rab, G.T., Chao, E.Y.S., & Stauffer, R.N. (1977). Muscle force analysis of the lumbar spine. Orthopedic Clinics of North America, 8, 193-199.
Rack, P.M.H., & Westbury, D.R. (1969). The effects of length and stimulus rate on tension in the isometric cat soleus muscle. Journal of Physiology (London), 204, 443-460.
Rack, P.M.H., & Westbuiy, D.R. (1974). The short range stiffness of active mammalian muscle and its effect on mechanical properties. Journal of Physiology (London), 240, 331-350.
Rafuse, V.F., Gordon, T., & Orozco, R. (1992). Proportional enlargement of motor units after partial denervation of cat triceps surae muscles. Journal of Neurophysiology, 68, 1261-1276.
Rail, W. (1987). Neuron, cable properties. In G. Adleman (Ed.), Encyclopedia of neuroscience (Vol. 2, pp. 816-820). Boston; Birkhauser.
Ralston, H.J., Inman, V.T., Strait, L.A., & Shaffrath, M.D. (1947). Mechanics of human isolated voluntaiy muscle. American Journal of Physiology, 151, 612-620.
Ranatunga, K.W., Sharpe, B., & Turnbull, B. (1987). Contractions of a human skeletal muscle at different temperatures. Journal of Physiology (London), 390, 383-395.
Rankin, L.L., Enoka, R.M., Volz, K.A., & Stuart, D G. (1988). Coexistence of twitch potentiation and tetanic force decline in rat hindlimb muscle. Journal of Applied Physiology, 65, 2687-2695.
Riley, D.A, Slocum, G.R., Bain, J.L.W., Sedlak, F.R., Sowa, T.E., & Mellender, J.W. (1990). Rat hindlimb unloading: Soleus histochemistry, ultrastructure, and electromyography. Journal of Applied Physiology, 69, 58-66.
Rios, E., Ma, J., & Gonza'lez, A. (1991). The mechanical hypothesis of excitation-contraction (EC) coupling in skeletal muscle. Journal of Muscle Research and Cell Motility, 12, 127-135.
Rios, E., & Pizaro, G. (1988). Voltage sensors and calcium channels of excitation-contraction coupling. News in Physiological Sciences, 3, 223-227.
Robinson, G.A., Enoka, R.M., & Stuart, D.G. (1991). Immobilization-induced changes in motor unit force and fatigability in the cat. Muscle & Nerve, 14, 563-573.
Rodgers, M.M. (1993). Biomechanics of the foot during locomotion. In M.D. Grabiner (Ed.), Current issues in biomechanics (pp. 33-52). Champaign, IL: Human Kinetics.
Roesler, H. (1987). The histoiy of some fundamental concepts in bone biomechanics. Journal of Biomechanics, 20, 1025-1034.
Rogers, M.W. (1991). Motor control problems in Parkinson's disease. In M.J. Lister (Ed.), Contemporary management of motor control problems (pp. 195-208). Alexandria, VA: Foundation for Physical Therapy.
Rogers, M.W., Kukulka, C.G., & Soderberg, G.L. (1992). Age-related changes in postural responses preceding rapid self-paced and reaction time arm movements. Journal of Gerontology, 47, M159-M165.
Rogers, M.W., & Pai, Y.-C. (1990). Dynamic transitions in stance support accompanying leg flexion movements in man. Experimental Brain Research, 81, 398-402.
Roland, P.E., Larsen, B., Lassen, N.A., & Skinhoj, E. (1980). Supplementary motor area and other cortical areas in organization of voluntary movements in man. Journal of Neurophysiology, 43, 118-136.
Roman, W.J., Fleckenstein, J., Stray-Gundersen, J., Alway, S.E., Peshock, R., & Gonyea, W.J. (1993). Adaptations in the elbow flexors of elderly males after heavy-resistance training. Journal of Applied Physiology, 74, 750-754.
Rome, L.C., Funke, R.P., Alexander, R.M., Lutz, G., Aldridge, H., Scott, F., & Freadman, M. (1988). Why animals have different muscle fibre types. Nature, 335, 824-827.
Rosenheimer, J.L., & Smith, D.O. (1990). Age-related increase in soluble and cell surface-associated neurite-out-growth factors from rat muscle. Brain Research, 509, 309-320.
Rossignol, S., Julien, C., Gauthier, L., & Lund, J.P. (1981). State dependent responses during movements. In A. Taylor & A. Prochazka (Eds.), Muscle receptors and movement (pp. 389-402). London: Macmillan.
Rothwell, J.C. (1987). Control of human voluntary movement. Kent, UK: Croom Helm.
Roy, R.R., Baldwin, K.M., & Edgerton, V.R. (1991). The plasticity of skeletal muscle: Effects of neuromuscular activity. In J.O. Holloszy (Ed.), Exercise and sport sciences reviews (Vol. 19, pp. 269-312). Baltimore: Williams & Wilkins.
Roy, R.R., & Edgerton, V,R. (1992). Skeletal muscle architecture and performance. In P.V. Komi (Ed.), Strength and power in sport (pp. 115-129). Champaign, IL: Human Kinetics.
Roy, R.R., Sacks, R.D., Baldwin, K.M., Short, M., & Edgerton, V.R. (1984). Interrelationships of contraction time, Vmax and myosin ATPase after spinal transection. Journal of Applied Physiology, 56, 1594-1601.
Rube, N., & Secher, N.H. (1990). Effect of training on central factors in fatigue following two- and one-leg static exercise in man. Acta Physiologica Scandinavica, 141, 87-95.
Riiegg, J.C. (1983). Muscle. In R.F. Schmidt & F. Thews (Eds.), Human physiology (pp. 32-50). Berlin: Springer-Verlag.
Rugg, S.G., Gregor, R.J., Mandelbaum, B.R., & Chiu, L. (1990). In vivo moment arm calculations at the ankle using magnetic resonance imaging (MRJ). Journal of Biomechanics, 23, 495-501.
Rutherford, O.M., & Jones, D.A. (1986). The role of learning and coordination in strength training. European Journal of Applied Physiology, 55, 100-105.
Rydell, N. (1965). Forces at the hip joint: Pt. 2. Intravital measurements. In R.M. Kenedi (Ed.), Biomechanics and related bioengineering topics (pp. 351-357). Oxford: Pergamon Press.
Sadamoto, T., Bonde-Petersen, F., & Suzuki, Y. (1983). Skeletal muscle tension, flow, pressure, and EMG during sustained isometric contractions in humans. European Journal of Applied Physiology, 51, 395-408.
Sahrmann, S.A., & Norton, B.J. (1977). The relationship of voluntary movement to spasticity in the upper motor neuron syndrome. Annals of Neurology, 2, 460-464.
Saito, M., Kobayashi, K., Miyashita, M., & Hoshikawa, T. (1974). Temporal patterns in running. In R.C. Nelson & C.A. Morehouse (Eds.), Biomechanics IV (pp. 106-1 II). Baltimore: University Park Press.
Sale, D.G., (1988). Neural adaptation to resistance training. Medicine and Science in Sports and Exercise, 20, S135-S145.
393
Sale, D.G., & MacDougall, J.D. (1981). Specificity in strength training: A review for the coach and athlete. Canadian Journal of Applied Sport Sciences, 6, 87-92.
Sale, D.G., & MacDougall, J.D. (1984). Isokinetic strength in weight-trainers. European Journal of Applied Physiology, S3, 128-132.
Sale, D.G., MacDougall, J.D., Jacobs, 1., & Bamer, S. (1990). Interaction between concurrent strength and endurance training. Journal of Applied Physiology, 68, 260-270.
Sale, D,G., MacDougall, J.D., Upton, A.R.M., & McComas, A.J. (1983). Effect of strength training upon motoneuron excitability in man. Medicine and Science in Sports and Exercise, 125, 57-62.
Sale, D.G., Quinlan, J., Marsh, E., McComas, A.J., & Belanger, A.Y. (1982). Influence of joint position on ankle plantarflexion in humans. Journal of Applied Physiology, 52, 1636-1642.
Saltin, B., & Gollnick, P.D. (1983). Skeletal muscle adaptability: Significance for metabolism and performance. In L.D. Peachey (Ed.), Handbook of physiology: Sec. 10. Skeletal muscle (pp. 555-631). Bethesda, MD: American Physiological Society.
Sanderson, D.J., & Henning, E.M. (1992, August). Inshoe pressure distribution in cycling and running shoes during steady-state cycling. Paper presented at the 16th Annual Conference of the American Society of Biomechanics, Chicago, IL.
Sanes, J.N. (1987). Proprioceptive afferent information and movement control. In G. Adelman (Ed.), Encyclopedia of neuroscience (Vol. II, pp. 982-984). Boston: Birkhauser.
Sanes, J.N., & Evarts, E.V. (1984). Motor psychophysics. Human Neurobiology, 2, 217-225.
Sapega, A.A., Quedenfeld, T.C., Moyer, R.A., & Butler, R.A. (1981). Biophysical factors in range-of-motion exercise. Physician and Sports Medicine, 9(12), 57-65.
Sargeant, A.J., & Boreham, A. (1982). Measurement of maximal short-term (anaerobic) power output during cycling. In J. Borms, M. Hebbelinck, & A. Venerando (Eds.), Women and sport (pp. 119-124). Basel, Switzerland: Karger.
Savolainen, S. (1989). Theoretical drag analysis of a skier in the downhill speed race. International Journal of Sport Biomechanics, 5, 26-39,1989.
Schantz, P., Fox, E.R., Norgren, P., & Tyde'n, A. (1981). The relationship between mean muscle fibre area and the muscle cross-sectional area of the thigh in subjects with large differences in thigh girth. Acta Physiologica Scandinavica, 113, 537-539.
Schantz, P.G., Moritani, T., Karlson, E., Johansson, E., & Lundh, A. (1989). Maximal voluntary force of bilateral and unilateral leg extension. Acta Physiologica Scandinavica, 136, 185-192.
Schiff, J.M. (1858). Lehrbuch der Physiologic des Menschen: I. Muskelund Nervenphysiologie. Lahr: Schauenburg.
Schleihauf, R.E., Jr. (1979). A hydrodynamic analysis of swimming propulsion. In J. Terauds & E.W. Bedingfield (Eds.), Swimming HI (pp. 70-109). Baltimore: University Park Press.
Schmidt, R.F., & Thews, G. (Eds). (1983). Human physiology. Berlin: Springer-Verlag.
Schneider, K., & Zemicke, R.F. (1992). Mass, center of mass, and moment of inertia estimates for infant limb segments. Journal of Biomechanics, 25, 145-148.
Schultz, A.B. (1992). Mobility impairment in the elderly: Challenges for biomechanics research. Journal of Biomechanics, 25, 519-528.
Schwartz, J.H. (1985). Synthesis and distribution of
neuronal protein. In E.R. Kandel and J.H. Schwartz (Eds.), Principles of Neural Science (2nd ed., pp. 37-48). New York: Elsevier.
Scudder, G.N. (1980). Torque curves produced at the knee during isometric and isokinetic exercises. Archives of Physical Medicine and Rehabilitation, 61, 68-72, Seals, D.R., Washbum, R.A., Hanson, P.G., Painter, P.L.,
& Nagle, F.J. (1983). Increased cardiovascular response to static contraction of larger muscle groups. Journal of Applied Physiology, 54, 434-437.
Secher, N.H., Rube, N., & Elers, J. (1988). Strength of two-and one-leg extension in man. Acta Physiologica Scandinavica, 134, 333-339.
Segal, R.L. (1992). Neuromuscular compartments in the human biceps brachii muscle. Neuroscience Letters, 140, 98-102.
Selkowitz, D.M. (1985). Improvement in isometric strength of the quadriceps femoris muscle after training with electrical stimulation. Physical Therapy, 65, 186-196.
Seyffarth, H. (1940). The behavior of motor units in voluntary contractions. Avhandlinger Utgitt Norske Videnskap-Akad Oslo. 1. Matematisk-Naturvidenskapelig Klasse, 4, 1-63.
Shadwick, R.E. (1990). Elastic energy storage in tendons: Mechanical differences related to function and age. Journal of Applied Physiology, 68, 1033-1040.
Shanebrook, J.R., & Jaszczak, R.D. (1976). Aerodynamic drag analysis of runners. Medicine and Science in Sports, 8, 43-45.
Sheetz, M.P., Steuer, E.R., & Schroer, T.A. (1989). The mechanism and regulation of fast axonal transport. Trends in Neurosciences, 12, 474-478.
Shellock, F.G. (1986). Physiological, psychological, and injury prevention aspects of warm-up. National Strength and Conditioning Association Journal, 8, 24-27.
Shellock, F.G., Fukunaga, T., Mink, J.H., & Edgerton, V.R. (1991 a). Acute effects of exercise on MR imaging of skeletal muscle: Concentric and eccentric actions. American Journal of Roentgenology, 156, 765-768.
Shellock, F.G., Fukunaga, T., Mink, J.H., & Edgerton, V.R. (1991b). Exertional muscle injury: Evaluation of concentric versus eccentric actions with serial MRI imaging. Radiology, 179, 659-664.
Shellock, F.G., & Prentice, W.E. (1985). Warming-up and stretching for improved physical performance and prevention of sports-related injuries. Sports Medicine, 2, 267-278.
Shepherd, G. (1988). Neurobiology. New York: Oxford.
Sherrington, C.S. (1931). Quantitative management of contraction in lowest level coordination. Brain, 54, 1-28.
Shindo, M., Harayama, H., Kondo, K., Yanagisawa, N., & Tanaka, R. (1984). Changes in reciprocal inhibition during voluntary contraction in man. Experimental Brain Research, 53, 400-408.
Shorten, M.R. (1987). Muscle elasticity and human performance. In B. Van Gheluwe & J. Atha (Eds.), Current research in sport biomechanics (pp. 1-18). Basel, Switzerland: Karger.
Shumskii, V.V., Merten, A.A., & Dzenis, V.V. (1978). Effect of the type of physical stress on the state of the tibial bones of highly trained athletes as measured by ultrasound techniques. Mekhanika Polimerov, 5, 884-888.
Simard, C.P., Spector, S.A., & Edgerton, V.R. (1982). Contractile properties of rat hind limb muscles immobilized at different lengths. Experimental Neurology, 77, 467-482.
394
Simoneau, J.-A., & Bouchard, C. (1989). Human variation in skeletal muscle fiber-type proportion and enzyme activities. American Journal of Physiology, 257, E567-E572.
Singh, S., & Katz, J.L. (1989). Electromechanical properties of bone: A review. Journal of Bioelectricity, 7,219-238.
Sjogaard, G., Kiens, B., Jurgensen, K., & Saltin, B. (1986). Intramuscular pressure, EMG and blood flow during low level prolonged static contraction in man. Acta Physiologica Scandinavica, 128, 475-484.
Sjogaard, G., Savard, G., & Juel, C. (1988). Muscle blood flow during isometric activity and its relation to muscle fatigue. European Journal of Applied Physiology, 57, 327-335.
Sjostrom, M., Angquist, K.-A., Bylund, A.-C., Fride, J., Gustavsson, L., & Schersten, T. (1982). Morphometric analyses of human muscle fiber types. Muscle & Nerve, 5, 538-553.
Sj&trom, M., Kidman, S., Larsen, K.H., & Angquist, K.-A, (1982). Z- and M-band appearance in different histo-chemically defined types of human skeletal muscle fibers. Journal of Histochemistry and Cytochemistry, 30, 1-11.
Sjostrom, M., Lexell, J., Eriksson, A., & Taylor, C.C. (1991). Evidence of fibre hyperplasia in human skeletal muscles from healthy young men? European Journal of Applied Physiology, 62, 301-304.
Smith, P. (1982). Dynamic variable resistance and the Universal system. National Strength & Conditioning Association Journal, 4(4), 14-19.
Smith, J.L., Betts, B., Edgerton, V.R., & Zemicke, R.F. (1980). Rapid ankle extension during paw shakes: Selective recruitment of fast ankle extensors. Journal of Neurophysiology, 43, 612-620.
Smith, J.L., & Zemicke, RF. (1987). Predictions for neural control based limb dynamics. Trends in Neurosciences, 10,123-128.
Snow-Harter, C., & Marcus, R. (1991). Exercise, bone mineral density, and osteoporosis. In J.O. Holloszy (Ed.), Exercise and sport sciences reviews (Vol. 19, pp. 351-388). Baltimore: Williams & Wilkins.
Snyder-Mackler, L., Binder-Macleod, S.A., & Williams, P.R. (1993). Fatigability of human quadriceps femoris muscle following anterior cruciate ligament reconstruction. Medicine and Science in Sports and Exercise, 25, 783-789.
Snyder-Mackler, L., Ladin, Z., Schepsis, A.A., & Young, J.C. (1991). Electrical stimulation of the thigh muscles after reconstruction of the anterior cruciate ligament. Journal of Bone and Joint Surgery, 7 ЗА, 1025-1036.
Somjen, G.G. (1987). Glial cells, functions. In G. Adelman (Ed.), Encyclopedia of neuroscience (Vol. I, pp. 465-466). Boston: Birkhauser.
Spector, S.A., Simard, C.P., Fournier, M., Stemlicht, E., & Edgerton, V.R. (1982). Architectural alterations of rat hind-limb skeletal muscles immobilized at different lengths. Experimental Neurology, 76, 94-110.
Sperry, R.W. (1950). Neural basis of the spontaneous optokinetic response produced by visual neural inversion. Journal of Comparative and Physiological Psychology, 43, 482-489.
Spira, M.E., Yarom, Y., & Pamas, 1. (1976). Modulation of spike frequency by regions of special axonal geometry and by synaptic inputs. Journal of Neurophysiology, 39, 882-899.
Spoor, C.W., van Leeuwen, J.L., Meskers, C.G.M., Titulaer, AF., & Huson, A. (1990). Estimation of instantaneous moment arms of lower-leg muscles. Journal of Biomechanics, 23, 1247-1259.
Spring, E., Savolainen, S., Erkkila, J., Hamalainen, T., & Pihkala, P. (1988). Drag area of a cross-country skier. International Journal of Sport Biomechanics, 4, 103-113.
Stacoff, A., & Kaelin, X. (1983). Pronation and sportshoe design. In B.M. Nigg & B.A. Kerr (Eds.), Biomechanical aspects of sport shoes and playing surfaces (pp. 143-151). Calgary, AB: University of Calgary.
Stainsby, W.N., Brechue, W.F., O'Drobinak, D.M., & Barclay, J.K. (1990). Effects of ischemic and hypoxic hypoxia on VO2 ; and lactic acid output during tetanic contractions. Journal of Applied Physiology, 68, 574-579.
Stalberg, E. (1980). Marcro EMG: A new recording technique. Journal of Neurology, Neurosurgery, and Psychiatry, 43, 475-482.
Stalberg, E., & Fawcett, P.R.W. (1982). Macro EMG in healthy subjects of different ages. Journal of Neurology, Neurosurgery, and Psychiatry, 45, 870-878.
Stanish, W.D., & Hubley-Kozey, C.L. (1984). Separating fact from fiction about common sports activity: Can stretching prevent athletic injuries? Journal of Musculoskeletal Medicine, 1, 25-32.
Stauber, W.T. (1989). Eccentric actions of muscles: Physiology, injury, and adaptation. In K.B. Pandolf (Ed.), Exercise and sport sciences reviews (Vol. 17, pp. 157-185). Baltimore: Williams & Wilkins.
Stein, R.B., & Capaday, C. (1988). The modulation of human reflexes during functional motor tasks. Trends in Neurosciences, 11, 328-332.
Stein, R.B., Gordon, T., Jefferson, J., Sharfenberger, A., Yang, J.F., Totosy de Zepetnek, J., & Belanger, Y. (1992). Optima] stimulation of paralzyed muscle after human spinal cord injury. Journal of Applied Physiology, 72, 1393-1400.
Stein, R.B., & Yang, J.F. (1990). Methods for estimating the number of motor units in human muscles. Annals of Neurology, 28, 487-495.
Stevenson, J.M. (1985). The impact force of entry in diving from a ten-meter tower. In D.A. Winter, R.W. Norman, R.P, Wells, K.C. Hayes, & A.E. Patla (Eds.), Biomechanics IX-B (pp. 106-111). Champaign, IL: Human Kinetics.
Stokes, I.A.F., Moffroid, M.S., Rush, S., & Haugh, L.D. (1988). Comparison of acoustic and electrical signals from erectores spinae muscles. Muscle & Nerve, 11, 331-336.
Stokes, M., & Young, A. (1984). The contribution of reflex inhibition to arthrogenous muscle weakness. Clinical Science, 67, 7-14.
Stokes, M.J., & Cooper, R.G. (1992). Muscle sounds during voluntary and stimulated contractions of the human adductor poilicis muscle. Journal of Applied Physiology, 72, 1908-1913.
Stroup, F., & Bushnell, D.L. (1970). Rotation, translation, and trajectory in diving. Research Quarterly, 40, 812-817.
Stuart, D.G. (1987a). Muscle receptors, mammalian. In G. Adelman (Ed.), Encyclopedia of neuroscience (Vol. 2, pp. 716-718). Boston: Birkhauser.
Stuart, D.G. (1987b). Muscle receptors, mammalian, spinal actions. In G. Adelman (Ed.), Encyclopedia of neuroscience (Vol. 2, pp. 718-719). Boston: Birkhauser.
Stuart, D.G., & Enoka, R.M. (1983). Motoneurons, motor units, and the size principle. In R.N. Rosenberg (Ed.), The clinical neurosciences (pp. V:471-V:517). New York: Churchill Livingstone.
Stucke, H., Baudzus, W., & Baumann, W. (1984). On friction characteristics of playing surfaces. In E.C.
395
Frederick (Ed.), Sport shoes and playing surfaces (pp. 87-97). Champaign, IL: Human Kinetics.
Sullivan, M.J., Green, H.J., & Cobb, F.R. (1990). Skeletal muscle biochemistry and histology in ambulatory patients with long term heart failure. Circulation, 81, 518-527.
Suzuki, S., & Hutton, R.S. (1976). Postcontractile moto-neuronal discharge produced by muscle afferent activation. Medicine and Science in Sports, 8, 258-264.
Suzuki, S., & Sugi, H. (1983). Extensibility of the myofilaments in vertebrate skeletal muscle as revealed by stretching rigor muscle fibers. Journal of General Physiology, 81, 531-546.
Sweeney, H.L., Bowman, B.F., & Stull, J.T. (1993). Myosin light chain phosphorylation in vertebrate striated muscle: Regulation and function. American Journal of Physiology, 264, C1085-C1095.
Sweeney, H.L., & Stull, J.T. (1990). Alteration of crossbridge kinetics by myosin light chain phosphoiylation in rabbit skeletal muscle: Implications for regulation of actin-myosin interaction. Proceedings of the National Academy of Sciences, 87, 414-418.
Tamaki, T., Uchiyama, S., & Nakano, S. (1992). A weightlifting exercise model for inducing hypertrophy in the hindlimb muscles of rats. Medicine and Science in Sports and Exercise, 24, 881-886.
Tang, A., & Rymer, W.Z. (1981). Abnormal force: EMG relations in paretic limbs of hemiparetic human subjects. Journal of Neurology, Neurosurgery, and Psychiatry, 44, 690-698.
Tax, A.A.M., Denier van der Gon, JJ., & Erkelens, C.J. (1990). Differences in coordination of elbow flexor muscles in force tasks and in movement tasks. Experimental Brain Research, 81, 567-572.
Taylor, D.C., Dalton, J., Seaber, A.V., & Garrett, W.E. (1990). The viscoelastic properties of muscle-tendon units. American Journal of Sports Medicine, 18, 300-309.
ter Haar Romeny, B.M., Denier van der Gon, JJ., & Gielen, C.C.A.M, (1982). Changes in recruitment order of motor units in the human biceps brachii muscle. Experimental Neurology, 78, 360-368.
Tesch, P.A., Dudley, G.A., Duvoisin, M.R., Hather, B.M., & Harris, R.T. (1990). Force and EMG signal patterns during repeated oouts of concentric and eccentric muscle actions. Acta Physiologica Scandinavica, 138, 263-271.
Tesch, P.A., & Karlsson, J. (1985). Muscle fiber types and size in trained and untrained muscles of elite athletes. Journal of Applied Physiology, 59, 1716-1720.
Thomas, C.K., Bigland-Ritchie, B., & Johansson, R.S. (1991). Force-frequency relationships of human thenar motor units. Journal of Neurophysiology, 65, 1509-1516.
Thomas, C.K., Johansson, R.S., & Bigland-Ritchie, B. (1991). Attempts to physiologically classify human thenar motor units. Journal of Neurophysiology, 65, 1501-1508.
Thomas, C.K., Stein, R.B., Gordon, T., Lee, R.G., & Elleker, M.G. (1987). Patterns of reinnervation and motor unit recruitment in human hand muscles after complete ulnar and median nerve section and resuture. Journal of Neurology, Neurosurgery, and Psychiatry, 50, 259-268.
Thomas, C.K., Woods, J.J., & Bigland-Ritchie, B. (1989). Impulse propagation and muscle activation in long maximal voluntary contractions. Journal of Applied Physiology, 67, 1835-1842.
Thomason, D.B., & Booth, F.W. (1990). Atrophy of the soleus muscle by hindlimb unweighting. Journal of Applied Physiology, 68, 1-12.
Thomason, D.B., Herick, R.E., & Baldwin, K.M. (1987). Activity influences on soleus muscle myosin during rodent hindlimb suspension. Journal of Applied Physiology, 63, 138-144.
Thomell, L.-E., & Price, M.G. (1991). The cytoskeleton in muscle cells in relation to function. Biochemical Society Transactions, 19, 1116-1120.
Thorstensson, A. (1977). Observations on strength training and detraining. Acta Physiologica Scandinavica, 100, 491-493.
Thusneyapan, S., & Zahalak, G.L (1989). A practical electrode-array myoprocessor for surface electromyography. IEEE Transactions on Biomedical Engineering, 36, 295-299.
Tidball, J.G. (1991). Force transmission across muscle cell membranes. Journal of Biomechanics, 24(Suppl. 1), 43-52.
Totosy de Zepetnek, J.E., Zung, H.V., Erdebil., S., & Gordon, T. (1992). Innervation ratio is an important determinant of force in normal and reinnervated rat tibialis anterior muscles. Journal of Neurophysiology, 67, 1385-1403.
Toussaint, H.M., van Baar, C.E., van Langen, P.P., de Looze, M.P., & van Dieen, J.H. (1992). Coordination of the leg muscles in backlift and leglift. Journal of Biomechanics, 25, 1279-1289.
Townend, M.S. (1984). Mathematics in sport. New York: Halstead.
Trimble, M.H., & Enoka, R.M. (1991). Mechanisms underlying the training effects associated with neuromuscular electrical stimulation. Physical Therapy, 71, 273-282.
Trotter, J.A. (1990). Interfiber tension transmission in series-fibered muscles of the cat hindlimb. Journal of Morphology, 206, 351-361.
Trotter, J.A. (1993). Functional morphology of force transmission in skeletal muscle. Acta Anatomica, 146, 205-222.
Trotter, J.A, & Purslow, P.P. (1992). Functional morphology of the endomysium in series fibered muscles. Journal of Morphology, 212, 109-122.
Tsika, R.W., Herrick, R.E., & Baldwin, K.M. (1987). Subunit composition of rodent isomyosins and their distribution in hindlimb skeletal muscles. Journal of Applied Physiology, 63, 2101-2110.
Tsujimoto, T., & Kuno, M. (1988). Calcitonin generelated peptide prevents disuse-induced sprouting of rat motor nerve terminals. Journal of Neuroscience, 8, 3951-3957.
Tucker, V.A. (1975). Aerodynamics and energetics of vertebrate fliers. In T.Y.T. Wu, C.J. Brokaw, & C. Brennan (Eds.), Swimming and flying in nature (pp. 845-867), New York: Plenum.
Ulfhake, B., & Kellerth, J.-O. (1984). Electrophysiological and morphological measurements in cat gastrocnemius and soleus a-motoneurones. Brain Research, 307,167-179.
Upton, A.R.M., McComas, A.J., & Sica, R.E.P. (1971). Potentiation of late responses evoked in muscles during effort. Journal of Neurology, Neurosurgery, and Psychiatry, 34, 699-711.
Valiant, G.A (1990). Transmission and attenuation of heelstrike acceleration. In P.R. Cavanagh, (Ed.), Biomechanics of Distance Running (pp. 225-247). Champaign, IL: Human Kinetics.
Vallbo, AB., & Wessbeig, J. (1993). Organization of motor output in slow finger movements in man. Journal of Physiology (London), 469, 673-691.
Vandenburgh, H.H. (1992). Mechanical forces and their second messengers in stimulating cell growth in vitro. American Journal of Physiology, 262, R350-R355.
396
Vandenburgh, H.H., Hatfaludy, S., Karlisch, P., & Shansky, J. (1991). Mechanically induced alterations in cultered skeletal muscle growth. Journal of Biomechanics, 24(Supplement), 91-99.
Vandenburgh, H.H., Hatfaludy, S., Sohar, I., & Shansky, J. (1990). Stretch-induced prostaglandins and protein turnover in cultured skeletal muscle. American Journal of Physiology, 259, C232-C240.
Vander, A.J., Sherman, J.H., & Luciano, D.S. (1990). Human physiology. The mechanisms of body function. New York: McGraw-Hill.
van derHelm, F.C.T., & Veenbaas, R. (1991). Modelling the mechanical effect of muscles with large attachment sites: Application to the shoulder mechanism, ioumal of Biomechanics, 24, 1151-1163.
vanderVaart, A.J.M., Savelberg, H.H.C.M., de Groot, G., Hollander, A.P., Toussaint, H.M., & van Ingen Schenau, G.J. (1987). An estimation of drag in front crawl swimming. Journal of Biomechanics, 20, 543-546.
Vandervoort, A.A., & Hayes, K.C. (1989). Plantarflexor muscle functron in young and elderly women. European Journal of Applied Physiology, 58, 389-394.
Vandervoort, A.A., Quinlan, J., & McComas, A.J. (1983). Twitch potentiation after voluntary contraction. Experimental Neurology, 81, 141-152.
Vandervoort, A.A., Sale, D.G., & Moroz, J. (1984). Com-parison of motor unit activation during unilateral and bilateral leg extension. Journal of Applied Physiology, 56, 46-51.
van Ingen Schenau, G.J. (1984). An alternative view of the concept of utilisation of elastic energy in human movement. Human Movement Science, 3, 301-336.
van Ingen Schenau, G.J., Bobbert, M.F., Huijing, P.A., & Woittiez, R.D. (1985). The instantaneous torque-angular velocity relation in plantar flexion during jumping. Medicine and Science in Sports and Exercise, 17, 422-426.
van Ingen Schenau, G.J., Bobbert, M.F., & van Soest, AJ. (1990). The unique action of biarticular muscles in leg extensions. In J.M. Winters & S.L.-Y. Woo (eds.). Multiple muscle systems: Biomechanics and movement organization (pp. 639-652). New York: Springer-Verlag.
van Ingen Schenau, G.J., & Cavanagh, P.R. (1990). Power equations in endurance sports. Journal of Biomechanics, 9, 865-881.
van Ingen Schenau, G.J., van Woensel, W.W.L.M., Boots, P.J.M., Snackers, R.W., & de Groot, G. (1990). Deter-mination and interpretation of mechanical power in human movement: Application to ergometer cycling. European Journal of Applied Physiology, 61, 11-19.
van Mameren, H., & Drukker, J. (1979). Attachment and composition of skeletal muscles in relation of their function. Journal of Biomechanics, 12, 859-867.
van Zuyien, E.J., Gielen, C.C.A.M., & Denier van der Gon, J.J. (1988). Coordination and inhomogeneous activation of human arm muscles during isometric torques. Journal of Neurophysiology, 60, 1523-1548.
Varon, S.S., & Somjen, G.G. (1979). Neuron-glia interac-tions. Neuroscience Research Progress Bulletin, 17,1-239.
Vaughan, C.L. (1980). A kinetic analysis of basic trampoline stunts. Journal of Human Movement Studies, 6, 236-251,
Vaughan, C.L. (1984). Biomechanics of running gait. CRC Critical Reviews in Biomedical Engineering, 12, 1-48.
Verillo, R.T. (1979). Change in vibrotactile thresholds as a function of age. Sensory Processes, 3, 49-59.
Videman, T. (1987). Connective tissue and immobilization: Key factors in musculoskeletal degeneration. Clinical Orthopaedics and Related Research, 221, 26-32.
Vrbova, G., & Wareham, A.C. (1976). Effects of nerve activity on the postsynaptic membrane of skeletal muscle. Brain Research, 118, 371-382.
Wachholder, K., & Altenburger, H. (1926). Beitrage zur Physiologic der willkbrlichen Bewegung: X. Mitteilung, Einzelbewegungen. Pfltingers ArchivJur die Physiologie, 214, 642-661.
Wallin, D., Ekblom, B., Grahn, R., & Nordenborg, T. (1985). Improvement of muscle flexibility: A comparison between two techniques. American Journal of Sports Medicine, 13, 263-268.
Walmsley, B., Hodgson, J.A., & Burke, R.E. (1978). Forces produced by medial gastrocnemius and soleus muscles during locomotion in freely moving cats. Journal of Neurophysiology, 41, 1203-1216.
Warabi, T., Noda, H., & Kato, T. (1986). Effect of aging on sensorimotor functions of eye and hand movement. Experimental Neurology, 92, 686-697.
Ward-Smith, A.J. (1983). The influence of aerodynamic and biomechanical factors on long jump performance. Journal of Biomechanics, 16, 263-268.
Ward-Smith, A.J. (1984). Air resistance and its influence of adverse and favourable winds on sprinting. Journal of Biomechanics, 17, 339-347.
Ward-Smith, A.J. (1985). A mathematical analysis of the influence of adverse and favourable winds on sprinting. Journal of Biomechanics, 18, 351-357.
Warren, G.L., Hayes, D.A., Lowe, D.A., & Armstrong, R.B. (1993). Mechanical factors in the initiation of eccentric contraction-induced injury in rat soleus muscle. Journal of Physiology (London), 464, 457-475.
Warren, G.L., Hayes, D.A., Lowe, D.A., Prior, B.M., & Armstrong, R.B. (1993). Materials fatigue initiates eccentric contraction-induced injury in rat soleus muscle. Journal of Physiology (London), 464, 477-489.
Waterman-Storer, C.M. (1991). The cytoskeleton of skeletal muscle: Is it affected by exercise? A brief review. Medicine and Science in Sports and Exercise, 23, 1249-1249.
Watkins, J. (1983). An introduction to the mechanics of human movement. Boston: MTP Press Ltd.
Webb, M.R., Hibberd. M.G., Golman, Y.E., & Trentham, D.R. ( 1986). Oxygen exchange between P, in the medium and water during ATP hydrolysis mediated by skinned fibers from rabbit skeletal muscle. Journal of Biological Chemistry, 261, 15557-15564.
Weeks, O.I. (1989). Vertebrate skeletal muscle: Power source for locomotion. Bioscience, 39, 791-799.
Weis-Fogh, T., & Alexander, R.M. (1977). The sustained power output from striated muscle. In T.J. Pedley (Ed.). Scale effects in animal locomotion (pp. 511-525). London: Academic.
West, S.P., Roy, R.R., & Edgerton, V.R. (1986). Fiber type and fiber size of cat ankle, knee and hip extensors and flexors following low thoracic spinal cord transection at an early age. Experimental Neurology, 91, 174-182.
Westerblad, H., & Lannergren, J. (1990). Recovery of fatigued Xenopus muscle fibres is markedly affected by the extracellular tonicity. Journal of Muscle Research and Cell Motility, 11, 147-153.
Westing, S.H., Cresswell, A.G., & Thorstensson, A. (1991). Muscle activation during maximal voluntary eccentric
397
and concentric knee extension. European Journal of Applied Physiology, 62, 104-108.
Westing, S.H., Seger, J.Y., & Thorstensson, A. (1990). Effects of electrical stimulation on eccentric and concentric torque-velocity relationships during knee extension in man. Acta Physiologica Scandinavica, 140, 17-22.
Westling, G., Johansson, R.S., Thomas, O.K., & Bigland-Ritchie, B. (1990). Measurement of contrac-tile and electrical properties of single human thenar motor units in response to intraneural motor axon stimulation. Journal of Neurophysiology, 64, 1331-1338.
Whalen, R.G. (1985). Myosin isoenzymes as molecular markers for muscle physiology. Journal of Experimental Biology, 115, 43-53.
Whalen, R.T., Carter, D.R., & Steele, C.R. (1988). Influence of physical activity on the regulation of bone density. Journal of Biomechanics, 21, 825-837.
Wickiewicz, T.L., Roy, R.R., Powell, P.L., & Edgerton, V.R. (1983). Muscle architecture of the human lower limb. Clinical Orthopaedics and Related Research, 179, 275-283.
Wickiewicz, T.L., Roy, R.R., Powell, P.L., Perrine, JJ., & Edgerton, V.R. (1984). Muscle architecture and force-velocity relationships in humans. Journal of Applied Physiology, 57, 435-443.
Wiegner, A.W. (1987). Mechanism of thixotropic behaviour at relaxed joints in the rat. Journal of Applied Physiology, 62, 1615-1621.
Wiktorsson-Moller, M., Oberg, B., Ekstrand, J., & Gillquist, J. (1983). Effects of warming up, massage, and stretching on range of motion and muscle strength in the lower extremity. American Journal of Sports Medicine, 11, 249-252.
Wilkie, D.R. (1960). Man as a source of mechanical power. Ergonomics, 3, 1-8.
Williams, K.R. (1985). Biomechanics of running. In R.L. Terjung (Ed.), Exercise and sport sciences reviews (Vol. 13, pp. 389-441). New York: Macmillan.
Williams, M., & Lissner, H.R. (1962). Biomechanics of human motion. Philadelphia: Saunders.
Williams, P.L., & Warwick, R. (Eds.) (1980). Gray's anatomy (36th ed.). Edinburgh: Churchill Livingstone.
Williams, R.A., Morrissey, M.C., & Brewster, C.E. (1986). The effect of electrical stimulation on quadriceps strength and thigh circumference in menisectomy patients. Journal of Orthopaedic and Sports Physical Therapy, 8, 143-146.
Wilson, B.D. (1977). Toppling techniques in diving. Research Quarterly, 48, 800-804.
Windhorst, U. (1988). How brain-like is the spinal cordl Berlin: Springer-Verlag.
Windhorst, U., Christakos, C.N., Koehier, W., Hamm, T.M., Enoka, R.M., & Stuart, D.G. (1986). Amplitude reduction of motor unit twitches during repetitive activation is accompanied by relative increase of hyperpolarizihg membrane potential trajectories in homonymous (a-motoneurones. Brain Research, 398, 181-184.
Windhorst. U., Hamm, T.M., & Stuard D.G. (1989). On the function of muscle and reflex partitioning. Behavioral and Brain Sciences, 12, 629-682.
Winiarski, A.M., Roy, R.R., Alford, E.K., Chiang, P.O., & Edgerton, V.R. (1987). Mechanical properties of rat skeletal muscle after hindlimb suspension. Experimental Neurology, 96, 650-660.
Winter, D.A. (1983). Moments of force and mechanical power in jogging. Journal of Biomechanics, 16, 91-97.
Winter, D.A. (1990). Biomechanics and motor control of human movement, (2nd ed.). New York: John Wiley & Sons.
Winter, D.A., Patla, A.E., & Frank, J.S. (1990). Assessment of balance control in humans. Medical Progress Through Technology, 16, 31-51.
Winter, D.A., Sidwall, H.G., & Hobson, D.A. (1974). Measurement and reduction of noise in kinematics of locomotion. Journal of Biomechanics, 7, 157-159.
Winter, D.A., Wells, R.P., & On, G.W. (1981). Errors in the use of isokinetic dynamometers. European Journal of Applied Physiology, 46, 397-408.
Winters, J.M. (1990). Hill-based muscle models: A systems engineering perspective. In J.M. Winters & S.L.-Y. Woo (Eds.), Multiple muscle systems: Biomechanics and movementorganization (pp. 69-93). New York: Springer-Verlag.
Winters, J.M., & Woo, S.L.-Y. (Eds.) (1990). Multiple muscle systems: Biomechanics and movement organization. New York: Springer-Verlag.
Woittiez, R.D., Huijing, P.A., & Rozendal, R.H. (1983). Influence of muscle architecture on the length-force diagram of mammahan muscle. Pflugers Archiv, 399, 275-279.
Wolpaw, J.R., & Carp, J.S. (1990). Memory traces in spinal cord. Trends in Neurosciences, 13, 137-142.
Wolpaw, J.R., Herchenroder, P.A., & Carp, J.S. (1993). Operant conditioning of the primate H-reflex: Factors affecting the magnitude of the change. Experimental Brain Research, 97, 31-39.
Wong, T.S., & Booth, F.W. (1990a). Protein metabolism in rat gastrocnemius muscle after stimulated chronic concentric exercise. Journal of Applied Physiology, 69, 1709-1717.
Wong, T.S., & Booth, F.W. (1990b). Protein metabolism in rat tibialis anterior muscle after stimulated chronic eccentric exercise. Journal of Applied Physiology, 69, 1718-1724.
Wood, G.A. (1982). Data smoothing and differentiation procedures in biomechanics. In R.L. Teijung (Ed.), Exercise and sport sciences reviews (Vol. 10, pp. 308-362). Philadelphia: Franklin Institute.
Wu, G., & Ladin, Z. (1993). The kinematometer: An integrated kinematic sensor for kinesiologic measurements. Journal of Biomechanical Engineering, 115, 53-62.
Yamaguchi, G.T., Sawa, A.G.U., Moran, D.W., Fessler, M.J., & Winters, J.M. (1990). A survey of human musculotendon actuator parameters. In J.M. Winters & S.L.-Y. Woo (Eds.), Multiple muscle systems: Biomechanics and movement organization (pp. 717-773). New York: Springer-Verlag.
Yamashita, N. (1988). EMG activities in mono- and bi-articular thigh muscles in combined hip and knee extension. European Journal of Applied Physiology, 58, 274-277.
Y ang, J.F., & Stein, R.B. (1990). Phase-dependent reflex reversal in human leg muscles during walking. Journal of Neurophysiology, 63, 1109-1117.
Yarasheski, K.E., Campbell, J.A., Smith, K.., Rennie, M.J., Holloszy, J.O., & Bier, D.M. (1992). Effect of growth hormone and resistance exercise on muscle growth in young men. American Journal of Physiology, 262, E261-E267.
Y oung, A., Stokes, M., & lies, J.F. (1987). Effects of joint pathology on muscle. Clinical Orthopaedics and Related Research, 219, 21-27.
Y oung, J.L., & Mayer, R.F. (1981). Physiological properties and classification of single motor units activated by intramuscular microstimulation in the first dorsal interosseus muscle in man. In J.E. Desmedt (Ed.), Motor unit types, recruitment and plasticity in health and disease (pp. 17-25). Basel, Switzerland: Karger.
398
Young, К., McDonagh, & Davies, C.T.M. (1985). The effects of two forms of isometric training on the mechanical properties of the triceps surae in man. Pfliigers Archiv, 405, 384-388.
Yue, G., Alexander, A.L., Laidlaw, D.H., Gmitro, A.F., Unger, E.C., & Enoka, R.M. (1994). Sensitivity of muscle proton spin-spin relaxation time as an index of muscle activation. Journal of Applied Physiology, 11, a84-a92..
Yue, G., & Cole, KJ. (1992). Strength increases from the motor program: A comparison of training with maximal voluntary and imagined muscle contractions. Journal of Neurophysiology, 67, 1114-1123.
Yue, G., Fuglevand, A.J., Nordstrom, M.A., & Enoka, R.M. (1992). Motor unit synchronization assessed from surface EMG increases with discharge rate of reference motor unit. Society for Neuroscience Abstracts, 18, 1406.
Zahalak, G.L (1990). Modeling muscle mechanics (and energetics). In J.M. Winters & S.L.-Y. Woo (Eds.), Multiple muscle systems: Biomechanics and movement organization (pp. 1-23). New York: Springer-Verlg.
Zajac, F.E. (1993). Muscle coordination of movement: A perspective. Journal of Biomechanics, 26, 109-124.
Zajac, F.E., & Gordon, M.E. (1989). Determining muscle's force and action in multiarticular movement. In K.B. Pandolf (Ed.), Exercise and sport sciences reviews (Vol. 17, pp. 187-230). Baltimore: Williams & Wilkins.
Zatsiorsky, V., & Seluyanov, V. (1983). The mass and inertia characteristics of the main segments of the human body. In H. Matsui & K. Kobayashi (Eds.), Biomechanics ЕШ-В(рр. 1152-1159). Champaign, IL: Human Kinetics.
Zemkova, H., Teisinger, J., Almon, R.R., Vejsada, R., Hnik, P., & Vyskocil, F. (1990). Immobilization atrophy and membrane properties in rat skeletal muscle fibres. Pfliigers Archiv, 416, 126-129.
Zernicke, R.F., Vailis, A.C., & Salem, GJ. (1990). Biomechanical response of bone to weightlessness. In K.B. Pandolf (Ed.), Exercise and sport sciences reviews (Vol. 18, pp. 167-192). Baltimore: Williams & Wilkins.
Zimny, M.L., & Wink, C.S. (1991). Neuroreceptors in the tissues of the knee joint. Journal of Electromyography and Kinesiology, 3, 148-157.
Zinovieff, A.N. (1951). Heavy-resistance exercise: The Oxford Technique. British Journal of Physical Medicine, 14, 159-162.
Zuurbier, C.J., & Huijing, P.A. (1992). Influence of muscle geometry on shortening speed of fibre, aponeurosis and muscle. Journal of Biomechanics, 25, 1017-1026.
Zytnicki, D., Lafleur, J., Horcholle-Bossavit, G., Lamy, F., & Jami, L. (1990). Reduction of lb autogenetic inhibition in motoneurons during contractions of an ankle extensor muscle in the cat. Journal of Neurophysiology, 64, 1380-1389.
Навчальне видання
ЕНОКА Роджер
Основн кin oioAorii
(росшською мовою)
Редактор — Наталия Колосок
Техшчний редактор — Тетяна Березяк Художне оформления — Олександр Придатко Комп’ютерна верстка — Володимир Томашевський Коректор — Алевтина Школаева
Пщп. до друку 30.04.98. Формат 60 х 84 1/8.
Патр офс. № 1. Гари. Таймс ЕТ. Офс. друк. Ум.-друк. арк. 46,5. Ум. фарбо-вщб 47,43.
Обл. вид. арк. 50,39. Тираж 2000 прим.
Зам. 8-133.
Видавництво “СЪпмтйська литература” при Украшському державному ушверсите-п ф!зичного виховання i спорту 252005, Ки!в-5, вул.Ф1зкультури, 1.
Кшвське акцюнерне товариство «Книга»
254655, МСП, Кшв-53, вул. Артема, 25.