


















































































































































Author: Мохан Р. Глессон М. Гринхафф П.Л.
Tags: медико-биологические основы физического воспитания физиология тренировки физкультура переводная литература издательство олимпийская литература спортивная биохимия
ISBN: 966-7133—41-9
Year: 2001




BIOCHEMISTRY
OF
EXERCISE AND TRAINING
RON MAUGHAN
University of Aberdeen
MICHAEL GLEESON
University of Birmingham
AND
PAUL L. GREENHAFF
University of Nottingham
OXFORD NEW YORK TOKYO
OXFORD UNIVERSITY PRESS
1997
Рон Мохан, Майкл Глессон,
Пауль Л. Гринхафф
БИОХИМИЯ
МЫШЕЧНОЙ
ДЕЯТЕЛЬНОСТИ
И ФИЗИЧЕСКОЙ
ТРЕНИРОВКИ
КИЕВ
ОЛИМПИЙСКАЯ ЛИТЕРАТУРА
2001
ББК 75.0я73
М86
Физиология и биохимия как учебные дисциплины неотделимы от
науки о мышечной деятельности и спорте. При написании данного
учебного пособия авторы стремились приблизиться к основам физио-
логической химии, уделив особое внимание реакциям и процессам,
протекающим в живых тканях, а также объяснить, как под влиянием
физических нагрузок изменяются метаболизм в мышечных волокнах,
физиологические адаптационные процессы при тренировке.
Для студентов вузов, специалистов по физическому воспитанию и
спорту, физической реабилитации и рекреации.
Ф1зюлопя i 6ioxiMia як навчальш дисциплши невщдшьш вщ нау-
ки про м’язову д1ялыпсть i спорт. При написанш даного навчального
поабника автори прагнули наблизитись до основ фвюлопчно! xiMii,
прид(ляючи особливу увагу реакщям i процесам, що вщбуваються в
живих тканинах, а також пояснити, як пщ впливом фгзичних наванта-
жень змшюються метаболвм у м’язових волокнах, фйюлопчш адапта-
цШш процеси пщ час тренування.
Для студенДв вуз!в, фах!вщв з фничного виховання i спорту, фь
зично! реабинтацй’ та рекреацп.
Научный редактор —
доктор педагогических наук
профессор Национального университета физического воспитания
и спорта Украины ВАЛЕРИЙ СМУЛЬСКИЙ
Перевод с английского — ВАЛЕРИЙ СМУЛЬСКИЙ
ISBN 966-7133—41-9
© Oxford University Press, 1997
© Издательство «Олимпийская литература», 2001
ПРЕДИСЛОВИЕ
Большой опыт преподавания показывает, что студенты ис-
пытывают большие затруднения при усвоении биохими-
ческой терминологии, чем физиологической. Обусловлено
это многими причинами, но не вызывает сомнений, что по-
нятийный материал в курсе физиологии человека усваива-
ется с меньшими трудностями. Вместе с тем молекулярная
основа большинства физиологических механизмов являет-
ся комплексной и поэтому тяжело воспринимается, хотя
вполне очевиден тот факт, что законы химии, применяемые
к биологическим системам, в конечном счете регулируют
все физиологические системы. Поэтому ориентировано
данное пособие, скорее, на основы физиологической хи-
мии, а не на традиционную биохимию.
В первой и второй главах изложены современные
представления о структуре, физиологии и биохимии ске-
летных мышц, а также об энергетических источниках,
обеспечивающих их сократительную активность.
Третья, четвертая и пятая главы посвящены особен-
ностям соответственно углеводного, липидного и белково-
го обменов при мышечной деятельности. Показано влия-
ние тренировки, направленной на развитие выносливости,
на метаболизм углеводов, липидов и белков во время физи-
ческих нагрузок.
Характеристика состояния энергетических систем,
обеспечивающих выполнение физических упражнений вы-
сокой интенсивности, представлена в шестой главе, а вли-
яние повышенного потребления жиров перед физическими
нагрузками и механизмы утомления — в седьмой главе.
5
В восьмой главе рассмотрены вопросы метаболичес-
кой адаптации к физическим тренировкам, направленным
на развитие выносливости, а также силовых и скоростных
возможностей. Подчеркивается, что тренировочные заня-
тия должны проводиться в течение нескольких недель или
месяцев, чтобы процессы специфической биохимической
адаптации позволили мышцам перейти на более высокий
функциональный уровень.
Авторы стремились изложить материал таким обра-
зом, чтобы он легко воспринимался, не был излишне дета-
лизирован. Вместе с тем они постарались представить ши-
рокий круг вопросов для обозрения.
Обширный иллюстративный материал позволит усва-
ивать предмет с большим интересом. Приложения и сло-
варь терминов помогут разобраться в специфической тер-
минологии, которая не очень часто встречается в научной
литературе. Студенты, проявляющие более глубокий инте-
рес к рассматриваемым темам, могут получить дополни-
тельную информацию из специальных литературных ис-
точников, приведенных в конце книги.
При написании данной книги авторы также учиты-
вали, что в результате растущего прогресса в области физи-
ологической химии, имеющей отношение к спорту, может
оказаться, что положения, которыми мы сегодня руковод-
ствуемся, назавтра могут оказаться неверными. Если фун-
даментальность, которая должна оставаться неизменной и
достаточно усвоена, то новая информация, воспринимае-
мая в рамках существующих теорий, легко может войти в
соответствие с существующими представлениями и быс-
трее поможет решению многих проблем.
Значимость изложенных в книге сведений не вызывает
сомнений. Их творческое использование спортсменами,
тренерами и врачами способно значительно повысить ка-
чество подготовки спортсменов, развить на высоком уровне
их функциональные возможности и сохранить их здоровье.
Предлагаемое читателю учебное пособие является
введением в аспекты биохимии мышечной деятельности и
предназначено для студентов-биохимиков и физиологов, а
также для специалистов в области спорта, стремящихся
глубже изучить особенности метаболизма в организме че-
ловека.
СПИСОК СОКРАЩЕНИИ
А — аденин
АДФ — аденозиндифосфат
АКРЦ -— аминокислоты с разветвленной цепью
АМФ — аденозинмонофосфат (3-,5-ц АМФ—циклическая АМФ: важный внутриклеточный посредник в действиях гор- монов)
АПБ — ацилпереносящий белок
АТФ — аденозинтрифосфат (высокоэнергетическое вещес- тво, являющееся непосредственным источником для мышечных сокращений и других энергозависимых процессов в клетке)
АТФ-аза — аденозинтрифосфатаза (энзим, обеспечивающий рас- пад АТФ до АДФ и неорганического фосфата, в резуль- тате чего освобождается энергия, которая может быть использована в биологических процессах)
АЦХ — ацетилхолин
ВЖ — внеклеточная жидкость
ВКП — вазоактивный кишечный пептид
Г — гуанин
ГАМК — гамма-аминомасляная кислота
гдф — гуанозиндифосфат
ГК — гексокиназа (энзим, катализирующий фосфорилиро- вание глюкозы)
ГЛУТ4 — переносчик глюкозы, находящийся в клеточных мем- бранах, в том числе в сарколемме мышечных волокон
ГО — газообменное отношение
Г-1-Ф — глюкозо-1-фосфат
Г-6-Ф — глюкозо-6-фосфат
ГТФ — гуанозинтрифосфат
гчл — гормончувствительная липаза
5-ГТ — 5-гидрокситриптамин (или серотонин)
ДК — дыхательный коэффициент
7
ДНК ДОФА — дезоксирибонуклеиновая кислота — дигидроксифенилаланин
1,3-ДФГ — 1,3-дифосфоглицерат
2,3-ДФГ — 2,3-дифосфогпицерат
ИМФ ИРФ кгзп кд КДО КоА — инозинмонофосфат — инсулиноподобный ростовой фактор — кальцитонингензависимый пептид — кровяное давление — коэффициент дыхательного обмена — кофермент А, который функционирует как перенос- чик ацильных групп
KoA-SH KoQ -— свободная форма коэнзима А — кофермент Q, или убихинон (обеспечивает транспорт электронов от флавопротеинов к цитохрому с в электрон-транспортной цепи)
Кр Km — креатин — константа Михаэлиса (концентрация субстрата, при которой скорость энзиматической реакции составляет половину максимальной)
КФ КФК — креатинфосфат, или фосфокреатин — креатинфосфокиназа (энзим, катализирующий пе- ренос фосфата от фосфокреатина к АДФ с образова- нием АТФ)
лдг — лактатдегидрогеназа (фермент, катализирующий об-
лнп лонп лпвп лил ратимое восстановление пирувата до лактата) — липопротеиды низкой плотности — липопротеиды очень низкой плотности — липопротеиды высокой плотности — липопротеинлипаза (фермент, катализирующий расщеп- ление триацилглицеролов в плазменных липопротеидах)
лсп ЛХАТ мдг НАД' НАДН — липопротеиды средней плотности — лецитинхолестеролацилтрансфераза — малатдегидрогеназа — никотинамидадениндинуклеотид (окисленная форма) — никотинамидадениндинуклеотид (восстановленная форма)
НАДФН — никотинамиддинуклеотидфосфат
пдг — пируватдегидрогеназа (ферментативный комплекс, ка- тализирующий превращение пирувата до ацетил-КоА
ПФ РНК — пирофосфат — рибонуклеиновая кислота (мРНК: матричная РНК; тРНК: транспортная РНК)
св сдг — сердечный выброс — сукцинатдегидрогеназа (фермент цикла трикарбоно- вых кислот)
сжк смт СР — свободные жирные кислоты — сухая масса ткани — саркоплазматический ретикулум
8
т — тиамин
ТАГ — триацилглицерол
ТКА — трикарбоксильная кислота
y(U) — урацил (также международная единица ферментатив-
ной активности: 1 U = 1 ммоль-мин-‘)
УДФ — уридиндифосфат
УТФ — уридинтрифосфат
ФАД — флавинадениндинуклеотид (окисленная форма)
ФАДН+ — флавинадениндинуклеотид (восстановленная форма)
ФДФ — фруктозо-1,6-дифосфат
ФДФаза — фруктозо-1,6-дифосфатаза
ФЕП — фосфоенолпируват
ФМН — флавинмононуклеотид (окисленная форма)
ФМНН+ — фламинмононуклеотид (восстановленная форма)
ф н — фосфат неорганический
Ф-6-Ф — фруктозо-6-фосфат
ФФК — фосфофруктокиназа (энзим, лимитирующий скорость
гликолиза)
хцк — холецистокинин
Ц — цитозин
цАМФ — циклический аденозинмонофосфат
цнс — центральная нервная система
цтк — цикл трикарбоновых кислот
чсс — частота сердечных сокращений
этц — электрон-транспортная цепь
сно — углевод
СаО2 — содержание кислорода в артериальной крови
CvO2 — содержание кислорода в венозной крови
со2 — углекислый газ
соон — карбоксильная группа
н+ — водородный ион, или протон
НСОр — бикарбонат (основной внеклеточный буфер)
nh2 — аминогруппа
NH3 — аммиак
NH.+ 4 — ион аммония
О2 — молекула кислорода
он — гидроксильная группа
Р°2 — парциальное давление кислорода
Рсо2 — парциальное давление углекислого газа
pH — показатель, отражающий кислотно-основное соотно-
шение (pH равен IglO [Н+])
к(группа) — органическая часть в молекуле аминокислоты
Углах — максимальная скорость ферментативной реакции,
когда концентрация субстратов не лимитирована
vco2 — скорость продукции углекислого газа
vo2 — скорость потребления кислорода
VO2max — максимальное потребление кислорода (МПК)
ЕДИНИЦЫ ИЗМЕРЕНИЯ
Масса
1 килограмм (кг) = 1000 грамм (г) = 106 миллиграмм (мг) = 109 мик-
рограмм (мкг)
Количество вещества
1 моль (моль) = 1000 миллимолей (ммоль) = 106 микромолей
(мкмоль)
Молярная концентрация раствора — 1 моль вещества в 1 литре
раствора (моль-л-1)
Дл ина
1 метр (м) =100 сантиметров (см) = 1000 миллиметров (мм) = 106
микрометров (мкм)
Энергия (работа, теплота)
1 килокалория (ккал) = 1000 калорий (кал) = 4184 джоулям (Дж) =
4,184 килоджоулям (кДж)
О бъ ем
1 литр (л) = 1000 миллилитров (мл) = 106 микролитров (мкл)
В р ем я
1 час (ч) = 60 минут (мин) = 3600 секунд (с)
1 секунда (с) = 1000 миллисекунд (мс) = 106 микросекунд (мкс)
Давление
Стандартное давление — 1 атмосфера (атм) = 760 мм рт.ст.
ФИЗИОЛОГИЯ И БИОХИМИЯ
СКЕЛЕТНЫХ МЫШЦ
ПРИ МЫШЕЧНОЙ ДЕЯТЕЛЬНОСТИ
1. Физиология и биохимия
физических упражнений
Физиология и биохимия как дисциплины, в сущности,
неотделимы от науки о мышечной деятельности и спор-
те. Ответы на вопросы, имеющие отношение к двига-
тельной активности и спорту, в частности такие наиболее
актуальные, как причины утомления, могут быть получе-
ны только в результате совместного изучения этих двух
дисциплин.
Безусловно, кто-то может считать, что будущее физио-
логии мышечной деятельности и спорта в наибольшей сте-
пени зависит от развития наших представлений о клеточ-
ном, субклеточном и молекулярном механизмах, объясня-
ющих, каким образом организм реагирует и адаптируется к
напряженным и постоянным физическим нагрузкам. Био-
химия обычно ориентируется на изучение процессов, про-
исходящих на субклеточном и молекулярном уровнях, что
нашло отражение и в данной книге. В частности, авторы
постарались объяснить, как под влиянием физических наг-
рузок изменяется метаболизм в мышечных волокнах. Од-
нако состояние мышц зависит также от внеклеточного
обеспечения их энергетическими источниками и кислоро-
дом, доставляемыми кровью. Поэтому всестороннее опи-
сание мышечного метаболизма должно учитывать и эти
факторы.
Тренировка изменяет метаболические реакции орга-
низма на физическую нагрузку, но эти биохимические
ответы (например, изменение активности ферментов и
способности к окислению липидов) проявляются в измене-
11
нии и физиологических показателей (например, показателей максималь-
ного сердечного выброса и максимального потребления кислорода).
По возможности мы определяли различия между физиологически-
ми и биохимическими реакциями организма на физические нагрузки, но
в настоящей главе ставилась цель представить читателю самые необхо-
димые сведения для более глубокого понимания изменений, проявляю-
щихся на субклеточном и молекулярном уровнях.
2. Скелетные мышцы
Мышцы — одна из четырех наиболее важных тканей в организме среди
таких, как нервная, соединительная и эпителиальная. Существует три ви-
да мышц: сердечная мышца; гладкая мускулатура, локализованная в стен-
ках кровеносных сосудов, дыхательных путей, кишечника и мочевого пу-
зыря; скелетная (поперечно-полосатая) мускулатура. Только скелетные
мышцы находятся под прямым волевым контролем, обеспечивающим как
перемещение частей тела, так и поддержание позы. Независимо от вида
мышечной деятельности функцией мышцы является проявление ею силы
сокращения и использование для этого необходимой энергии.
Структура, иннервация и кровоснабжение. Скелетные мышцы
отделены от соединительной ткани (перимизиума, или фасции) мембра-
нами. Соединительная ткань проникает и в полость самой мышцы,
уменьшаясь при этом по толщине (в данном случае соединительную
ткань называют эндомизиумом), в связи с чем происходит разделение
мышцы на все более и более мелкие компартменты (рис. 1.1). Наимень-
шими из них являются пучки, включающие большое количество мышеч-
ных волокон, прикрепленных друг к другу и к эндомизиуму соедини-
тельной тканью. К обоим концам мышцы эта ткань переходит в сухожи-
лия, прикрепляемые к костям скелета.
Сухожилия представляют собой плотно упакованные коллагеновые
волокна, образующие соединения между мышцами и костями. Внешняя
коллагеновая мембрана живой кости (периостеум) переходит в мембра-
ну волокон сухожилия.
Отдельные мышцы состоят из многих мышечных волокон, парал-
лельно соединенных между собой, которые могут (но не обязательно)
протягиваться по всей ее длине. Внутри мышцы соединительная ткань
также покрывает крупные кровеносные сосуды и нервы. Почти все мы-
шечные волокна иннервируются только одним нервным окончанием,
расположенным посредине волокна.
12
Концевая пластинка
Миофибрилла
Толстый
филамент
Сарколемма
<»суда
Сарколемма
Отверстия
Т-трубочек
Полоса А |
Z-линия
Z-линия *• Z-линия
Н-зона
Саркомер
Двигательный нерв
Саркоплазматический
Т-трубочки ретикулум
Т-трубочки
Тонкий
филамент
Рис. 1.1. Макро- и микроскопическая анатомия скелетной мышцы
Особый участок поверхности мышечного волокна, прилегающий к
нервному окончанию в области синапса, называется концевой пластин-
кой. Нейропередатчиком, который освобождается из нервного окон-
чания и инициирует развитие мышцей напряжения, является ацетил-
холин.
13
Кровеносные сосуды ориентированы в основном параллельно мы-
шечным волокнам, а многочисленные капилляры направлены в прос-
транство между ними. Гладкая мускулатура кровеносных сосудов сокра-
щается или расслабляется под влиянием нервных, гормональных и ло-
кальных воздействий, что обеспечивает регуляцию тока крови. Во время
выполнения физических упражнений кровоснабжение работающих
мышц может возрасти в 100 раз по сравнению с уровнем покоя.
Мышечные клетки представляют собой длинные многоядерные во-
локна (см. рис. 1.1). Их длина может варьировать от нескольких милли-
метров до 30 см, а диаметр — в пределах 10—100 мкм. Каждое мышеч-
ное волокно окружено гомогенной мембраной — сарколеммой, содер-
жащей на своей наружной поверхности коллагеновые волокна, с по-
мощью которых прикрепляется к внутримышечным элементам соедини-
тельной ткани. Внутренняя часть сарколеммы обладает свойством, бла-
годаря которому питательные вещества и шлаки могут проходить через
мембрану, в результате в мышечном волокне может возникать и распрос-
траняться потенциал действия. Инвагинированные участки сарколеммы
называються «Т-трубочки», по которым потенциал действия распрос-
траняется внутрь мышечного волокна.
Ультраструктура мышечных волокон. Внутренность мышечного
волокна заполнена саркоплазмой (цитоплазмой мышечной клетки), пред-
ставляющей собой вязкую жидкость, содержащую ядра, митохондрии,
миоглобин и около 500 нитевидных миофибрилл толщиной 1-3 мкм каж-
дая и располагающихся от одного конца мышечного волокна к другому
(см. рис. 1.1). Красный цвет саркоплазмы обусловлен присутствием в ней
миоглобина — внутриклеточного дыхательного пигмента, благодаря ко-
торому создается запас кислорода. Миофибриллы находятся в окружении
более совершенной структуры — эндоплазматического ретикулума, на-
зываемого саркоплазматическим ретикулумом (СР), который принимает
участие в процессах роста, развития и восстановления мышцы. Взаимо-
связанные мембранные трубочки находятся в узком пространстве между
миофибриллами, окружая их и располагаясь параллельно с ними.
Под микроскопом мышечные волокна обнаруживают поперечно-по-
лосатую исчерченность, связанную с уникальной организацией миофиб-
рилл. Темные полосы А сменяются светлыми дисками I по всей длине
каждой миофибриллы, которые являются сократительными элементами.
Как показано на рис. 1.1, посредине А-диска имеется более светлая по-
лоска, называемая зоной Н, которая видима только в расслабленном сос-
тоянии мышечного волокна. Сама зона Н разделяется темной линией М.
Диски I также разделяются посредине темной зоной, названной Z-
пластинками. Саркомер определяется как участок, расположенный меж-
14
ду двумя последовательными Z-пластинками и является наименьшим
сократительным элементом мышечного волокна. Каждая миофибрилла
представляет собой цепь саркомеров.
На молекулярном уровне можно обнаружить, что в паттерне мио-
фибриллярных участков внутри каждого саркомера имеется два типа
белковах филаментов (миофиламентов). Тонкие миофиламенты включа-
ют белки — актин, тропомиозин и тропонин; они простираются через
диск / и частично проникают в диск Л; толстые миофиламенты содержат
белок миозин и располагаются в Л-диске. Линия Z представляет собой
белковый слой в форме диска и служит участком, к которому прикреп-
лены тонкие миофиламенты.
Зона Н является областью, в которой толстые филаменты не пере-
крываются тонкими, поэтому под микроскопом она выглядит более
светлой по сравнению с диском А, когда миофибрилла находится в рас-
слабленном состоянии. ТИ-линия посредине зоны Н выглядит несколько
темнее из-за тонких прядей, соединяющих воедино примыкающие тол-
стые миофиламенты.
Последовательные поперечные разрезы миофибриллы обнару-
живают, что в областях, где толстые и тонкие миофиламенты перекры-
ваются, каждый толстый миофиламент окружен гексагональным распо-
ложением шести тонких миофиламентов, а каждый тонкий миофила-
мент расположен внутри треугольника, образованного тремя толстыми
миофиламентами (рис. 1.2).
Саркомер
Головка
•/полоса-----►
Рис. 1.2. Поперечные сечения (1—5) через толстые и тонкие миофиламенты по
длине саркомера
15
Молекула миозина
Участок толстого филамента
Участок тонкого филамента
Продольное сечение филаментов в пределах
одного саркомера
Рис. 1.3. Молекулярная структура толстых
и тонких филаментов
В присутствии достаточ-
ного количества кальция и
аденозинтрифосфата (АТФ)
филаменты взаимодействуют
между собой, образуя актоми-
озин, и сокращаются, скользя
относительно друг друга.
Электрическое возбуждение,
распространяясь в виде по-
тенциала действия вдоль сар-
комера и к 7-трубочкам, вы-
зывает освобождение кальция
из саркоплазматического ре-
тикулума и выход его в сар-
коплазму с последующей ак-
тивацией сокращением фила-
ментов. Возбуждение иници-
ируется достижением нервно-
го импульса мышечной мем-
браны через двигательную
концевую пластинку.
Молекулярный состав
миофиламентов. Каждый
толстый миофиламент содер-
жит около 200 молекул мио-
зина, из которых каждая, в
свою очередь, имеет стер-
жнеподобный «хвост» с дву-
мя глобулярными структура-
ми на конце, которые облада-
ют АТФ-азной активностью.
Миозиновые головки взаимо-
действуют со специфическими участками, расположенными на тонких
миофиламентах с образованием так называемых поперечных мостиков,
и генерируют развитие напряжения, приводящего к сокращению мыш-
цы. В толстом филаменте молекулы миозина связаны вместе таким об-
разом, что их «хвосты» образуют центральную часть филамента, а их
глобулярные структуры обращены наружной поверхностью в направле-
ниях, противоположных друг другу (рис. 1.3). Благодаря этому каждый
толстый филамент имеет относительно гладкую центральную секцию с
двумя концами, усеянными множеством подвижных головок миозина.
16
Тонкие миофиламенты включают в себя актин и несколько регуля-
торных белков. Глобулярные (G) мономеры актина полимеризованы
внутри длинных участков, называемых фибриллярным (F) актином. Две
актиновые нити, сплетаясь вместе, образуют основу каждого тонкого
филамента. Далее хвостоподобные молекулы тропомиозина обвивают
цепочки F-актина, чем помогают миофиламенту стать прочнее.
Другим важным белком, присутствующим в тонких филаментах, яв-
ляется тропонин, который содержит три субъединицы. Одна из них, тро-
понин I, связана с актином; другая, тропонин Т, связана с тропомиозином,
а третья, тропонин С, может быть связана с ионами кальция (см. рис. 1.3).
Механизм проявления силы мышечного сокращения. При сок-
ращении мышечного волокна его саркомеры укорачиваются, 77-зоны ис-
чезают и расстояние между ближайшими линиями Z уменьшается. Сами
же филаменты не изменяют своей длины. Поскольку тонкие филаменты
при скольжении относительно тонких частично перекрывают друг дру-
га, их длина увеличивается. Скольжение миофиламентов начинается,
когда головки миозина образуют поперечные мостики, прикрепляясь к
активным участкам актиновых субъединиц тонких филаментов. Каждое
образование и разъединение поперечных мостиков во время сокращения
происходит несколько раз храповикоподобным образом, в результате че-
го тонкие филаменты перемещаются в направлении к центру саркомера.
В результате такого процесса длина всего мышечного волокна становит-
ся короче.
Для образования миозиновых поперечных мостиков требуется при-
сутствие ионов кальция. В расслабленном мышечном волокне кальций
находится в саркоплазматическом ретикулуме и в отсутствие этих ионов
связывающие участки миозина физически блокированы тропомиозином
(рис. 1.4). Ионы кальция при освобождении из саркоплазматического ре-
тикулума (вследствие возбуждения нервным импульсом) связываются с
тропонином С и тем самым изменяют его конформацию, что приводит к
физическому перемещению тропомиозина от связывающих участков
миозина на расположенную внутри цепь актина.
Активированные, или «склеванные» головки теперь присоединяют-
ся к актину и в данном случае головная часть молекулы изменяет свою
активную конфигурацию до такого состояния своей связывающей по-
верхности, которая обеспечивает ее перемещение на тонкий филамент
путем скольжения по направлению к средине саркомера (см. рис. 1.4).
Представленная схема отражает развитие цикла поперечных мостиков и
стимуляцию его аденозиндифосфатом (АДФ) и неорганическим фосфа-
том (Фн). Как только новая молекула АТФ присоединяется к участку ми-
озиновой головки, обладающему АТФ-азной активностью, миозиновые
?
~ 0-313
17
Тонкий филамент
Проявление сократительной
активности филамента при
сгибании головки миозина
Рис. 1.4. Последовательность процессов образования поперечных мостиков
мостики отсоединяются от актина. Гидролиз АТФ до АДФ и Фн, проис-
ходящий при участии АТФ-азы, обеспечивает энергией, необходимой
для возвращения миозина в его активированное, «склевывающее» сос-
тояние, позволяющее при наличии энергетического потенциала осу-
ществлять последовательную работу поперечных мостиков. До тех пор,
пока миозин находится в активном состоянии, АДФ и Фн остаются свя-
занными с миозиновой головкой. Когда же последняя обретает способ-
ность связываться с другим участком актина, расположенного последо-
18
вательно в тонком миофиламенте, цикл связывания, развития напряже-
ния и разъединения поперечных мостиков, как и активация миозина,
повторяются. Взаимное скольжение филаментов описанным способом
может продолжаться до тех пор, пока в саркоплазме присутствуют ионы
кальция (концентрация должна быть не менее 10 ммоль). Перемещение
и обратный захват АТФ-зависимой кальциевой помпой саркоплазма-
тического ретикулума кальция восстанавливает тропомиозиновое угне-
тение образования поперечных мостиков и расслабление мышечных
волокон.
Регуляция силы мышечных сокращений. Проявление мышеч-
ным волокном силы сокращения является результатом нервной импуль-
сации от двигательного нерва, распространяющейся в виде потенциала
действия по сарколемме. Потенциал действия, достигнув двигательной
концевой пластинки, вызывает освобождение нейромедиатора ацетилхо-
лина, который перемещается специальным синапсом между нервным
окончанием и мышечным волокном (нейромышечным соединением) и
взаимодействует с ацетилхолиновыми рецепторами, расположенными
на сарколемме. Это приводит к открытию натриевых каналов, в резуль-
тате чего поток ионов натрия внутрь мышечного волокна снижает гра-
диент их концентрации. Происходит деполяризация мембраны и, как
следствие, генерация потенциала действия, который, распространяясь
по сарколемме мышечного волокна в обе стороны и внутрь Т-трубочек,
вызывает полную активацию мышечного волокна (рис. 1.5). Передача
потенциала действия к участкам, где Т-трубочки примыкают к сарко-
плазматическому ретикулуму, обусловливает последующее освобожде-
ние из него кальция (кальциевые каналы временно открываются) и его
свободная концентрация в саркоплазме становится более 10 ммоль , что
сопровождается образованием поперечных мостиков, как описано вы-
ше. Последующая активация кальциевой помпы приводит к возвраще-
нию кальция в саркоплазматический ретикулум (обычно это происходит
в пределах 30 мс), а ингибирование тропомиозина восстанавливается,
если концентрация кальция в саркоплазме становится очень низкой. Та-
кая последовательность процессов повторяется, когда другие импульсы,
поступающие от двигательного нерва, достигают двигательной конце-
вой пластинки. Если же частота импульсации высокая, то ионы кальция
продолжают высвобождаться из саркоплазматического ретикулума и
концентрация кальция в саркоплазме, окружающей миофиламенты, зна-
чительно возрастает. В таком случае мышечные волокна между последу-
ющими стимулами полностью не расслабляются и развиваемое мышцей
напряжение будет более сильным и непрерывным (вплоть до максималь-
ного) до тех пор, пока не прекратится нервная импульсация.
2*
19
, Окончание аксона
Ацетилхолин
(АЦХ)
Синаптическая щель
Т-трубочка
Сарколемма
Са
Са
Ag') Сокращение; образование миозиновых
поперечных мостиков чередуется с их
разьединением за счет использования
энергии АТФ
(2) Распространение
потенциала действия
АЦХ-рецепторы
на сарколемме
по сарколемме и
Т-тру бочкам
ЛЛ Освободившийся нейромедиатор
диффундирует через синаптическую
щель и присоединяется к
АЦХ-рецепторам на сарколемме
(3) Потенциал действия
вызывает освобождение
Са2+ из терминальных
цистерн СР через
белковые каналы
Тропомиозин блокирует возвращение
'-J к прежнему состоянию; сокращение
прекращается и мышечное волокно
расслабляется
саркоплазматическим
ретикулумом после
потенциала действия
(при участии Са -
АТФ-азы, находящейся
в мембранах СР)
Саркоплазматический
ретикулум (СР)
Терминальная
цистерна СР
/дЛ Повышение концентрации
' свободного Са2+ в среде,
окружающей миофиламенты
Присоединение ионов каль-
ция к тропонину; изменение
формы тропонина снимает
блокирующее влияние
тропомиозина, благодаря
чему.актин переходит
в'активное состояние
Рис. 1.5. Последовательность процессов возбудительно-сократительного сопряжения
Нервная импульсация. Группы мышечных волокон (одного и того
же типа) объединены в мышце их взаимосвязью с одним и тем же дви-
гательным нейроном. Каждая такая группа называется двигательной
единицей. Двигательные единицы различаются по количеству содержа-
щихся в них волокон: одни из них содержат около 50 волокон, другие —
до 1700. Мышцы, выполняющие тонкие градуальные движения (напри-
мер, мышцы глаз и рук), содержат незначительное количество двига-
20
тельных единиц; мышцы же, имеющие большую массу и выполняющие
грубые движения (например, мышцы ног), имеют большее количество
двигательных единиц.
Импульс, распространяющийся по аксону двигательного нерва, вы-
зывает деполяризацию его концевых пластинок. Все мышечные волок-
на, принадлежащие одной и той же двигательной единице, будут либо не
отвечать («), либо отвечать (б) проведением потенциала действия по мы-
шечным волокнам, что сопровождается одновременной активацией всех
волокон.
Ответы мышечных волокон на одиночные импульсы (достаточной
силы) с одиночным сокращением и расслаблением продолжаются около
30 мс. Повторяющиеся сверхпороговые стимулы при сближении их
вплотную вызывают суммацию, и когда частота стимуляции составляет
более 60 в секунду (т. е. 60 Гц), происходит слияние одиночных сокра-
щений (тетанус), при котором мышечное волокно развивает более высо-
кое напряжение.
Обычно частота стимуляции находится в пределах от 5 Гц, при ко-
торой развивается низкое напряжение, до 70 Гц. В последнем случае си-
ла сокращения мышечных волокон высокая. Так, сила сокращений це-
лостной мышцы может повышаться в результате повышения частоты
стимуляции активных двигательных единиц (а), а также вовлечением
(рекруитированием) и увеличением их количества (б).
При выполнении физических упражнений с фиксированной мощ-
ностью по мере развития утомления некоторые двигательные единицы
перестают участвовать в развиваемом мышцей напряжении, однако их
вклад в генерацию силы будет немедленно возмещаться другими двига-
тельными единицами до тех пор, пока все из них не будут рекруитиро-
ваны. При выполнении физических упражнений максимальной интен-
сивности первоначально полное (или почти полное) рекруитирование
всех двигательных единиц сопровождается в дальнейшем постепенным
снижением эффективности их участия в развитии мышцей напряжения,
т. е. проявлением утомления.
Типы мышечных волокон. Существование различных типов воло-
кон в скелетных мышцах достаточно очевидно и их изучение продолжа-
ется на протяжении длительного периода. Однако детальные физиологи-
ческие и биохимические основы этой дифференциации, а также их
функциональное значение установлены лишь в последнее время. Во
многом стимулами для этих исследований явилось понимание того, что
успех в спортивных соревнованиях, требующих способности спортсме-
на к проявлению либо максимальной силы, либо выносливости, сущес-
твенно зависит от пропорционального соотношения в мышце типов во-
21
локон. Мышечные волокна, однако, чрезвычайно пластичны, и хотя их
соотношение генетически детерминировано, реализация определенной
тренировочной программы способна во многом повлиять на метаболи-
ческий потенциал мышцы независимо от соотношения в ней типов во-
локон.
Первоначальной основой для разделения мышечных волокон на
красные, белые и промежуточные применительно к целостной мышце
послужил простой визуальный осмотр. Однако основной функциональ-
ной характеристикой дифференциации типов мышечных волокон явля-
ется скорость их укорочения и расслабления. Медленносокращающиеся
волокна характеризуются относительно продолжительным временем,
необходимым для достижения пика напряжения (около 80-100 мс для
волокон мышц человека), а также длительным полупериодом расслабле-
ния. В противоположность им для достижения пика напряжения быс-
тросокращающимися волокнами у человека требуется около 40 мс, сле-
довательно, время их расслабления соответственно короче. Эти два типа
волокон образуют отдельные группы с частичным совпадением их
свойств.
Поскольку для изучения механических свойств волокон доступ к
ткани затруднителен, а небольшие образцы мышцы относительно легко
могут быть получены с помощью пункционной биопсии, классификация
типов мышечных волокон обычно базируется на биохимическом окра-
шивании их поперечных срезов. На этой основе волокна мышц челове-
ка обычно подразделяют на три главные группы — типы I, Па и Пб, хо-
тя возможно также дальнейшее подразделение. Аналогичное деление
свойственно и мышцам животных, волокна которых классифицируются
на основе прямого определения их функциональных свойств: соответ-
ственно медленносокращающиеся, быстросокращающиеся медленноу-
томляемые и быстросокращающиеся быстроутомляемые.
Миозин различных типов мышечных волокон существует в раз-
личных молекулярных формах (изоформах), а АТФ-азная активность
миофибрилл проявляет неодинаковую чувствительность к pH. Миози-
новая АТФ-аза волокон типа II инактивируется при низких значениях
pH (менее 4,5), тогда как АТФ-азная активность миозина в волокнах
типа I остается неизменной. При значениях, превышающих pH 9, ситу-
ация изменяется: АТФ-азная активность миозина в волокнах типа II
остается стабильной, в то время как аналогичная активность в волок-
нах типа I снижается. Эти два отдельных подтипа типа II можно рас-
познать путем преинкубации волокон при значениях pH, которые нахо-
дятся в конце указанного диапазона: АТФ-азная активность миозина в
волокнах типа Па инактивируется при pH 4,6-4,8, в то время как в во-
22
Таблица 1.1. Характеристика типов мышечных волокон у человека
Тип I Тип Па Тип Пб
Характеристика Медленные, Быстрые, красные,
красные, окислительные, Быстрые, белые,
окислительные, гликолитические, гликолитические,
медленно- утомлямые медлнно- утомляемые быстроутомляемые
Размер двигательного Небольшой Большой Большой
нейрона
Частота рекруитирования Низкая Средняя Высокая
Скорость сокращения Низкая Высокая Высокая
Скорость расслабления Низкая Высокая Высокая
Максимальная сила Низкая Высокая Высокая
Выносливость Высокая Средняя Низкая
Плотность капилляров Высокая Средняя Низкая
Плотность митохондрий Высокая Средняя Низкая
Характер метаболизма Окислительный Промежуточый Гликолитический
Содержание миоглобина Высокое Среднее Низкое
Активность гликолитичео- Низкая Высокая Высокая
ких ферментов
Активность окислительных Высокая Высокая Низкая
ферментов
Содержание гликогена Низкое Высокое Высокое
Содержание триглицеридов Высокое Среднее Низкое
Содержание Низкое Высокое Высокое
креатинфосфата
Активность АТФ-азы Низкая Высокая Высокая
миозина
Активность АТФ-азы при 0 Высокая Высокая
pH 10,3
Активность АТФ-азы при 0 0 Высокая
pH 10,3 с преинкуба-
цией при pH 4,6
локнах типа Пб она не изменяется. Иногда путем преинкубации можно
выявить и волокна типа Пв, но этот тип вряд ли стоит принимать во
внимание, поскольку в мышцах человека он составляет менее 1 % всех
волокон.
Получает распространение использование гель-электрофореза для
идентификации отдельных типов мышечных волокон, основанной на
присутствии в них различных изоформ миозина. Физиологическая и
23
Таблица 7.2. Активность некоторых
гликолитических и окислительных
ферментов в различных типах волокон
скелетных мышц человека
(ммоль-мин~1т—1 сырой массы)
Фермент Тип1 Тип Па ТипПб
Фосфорилаза 2,8 5,8 8,8
Фосфофрукгоки- 7,5 13,7 17,5
наза Сукцинатде- 7,1 4,8 2,5
гидрогеназа Цитратсинтаза 10,8 8,6 6,5
биохимическая характеристики
этих основных трех типов воло-
кон представлены в табл. 1 и 2.
К типу I относятся красные
волокна, содержащие относи-
тельно медленнодействующую
миозиновую АТФ-азу и поэтому
характеризующиеся медленной
скоростью сокращения. Красный
цвет волокон обусловлен присут-
ствием миоглобина, являющего-
ся внутриклеточным дыхатель-
ным пигментом, способным свя^
зывать кислород и освобождать
его только при очень низком парциальном давлении кислорода, которое
обнаруживается вблизи митохондрий. Волокна типа I богаты митохон-
дриями, которые локализуются преимущественно ближе к периферии
волокна, к капиллярам, обеспечивающим доставку кислорода и пита-
тельных веществ. Эти волокна обладают высокими возможностями
окислительного метаболизма, устойчивостью к развитию утомления, а
также способностью поддерживать повторяющуюся напряженную дви-
гательную активность в течение длительного времени.
По сравнению с ними волокна типа II намного бледнее, так как со-
держат меньше миоглобина. В отличие от волокон типа I они обычно
крупнее в диаметре, поскольку различия в размерах зависят в основном
от особенностей двигательной активности. Волокна типа II характеризу-
ются более высокой активностью миозиновой АТФ-азы, в связи с чем их
сокращение и расслабление происходят относительно быстро, поэтому
они способны развивать приблизительно в три раза более высокую мощ-
ность по сравнению с волокнами типа I. Кроме того, они содержат мало
митохондрий и имеют недостаточное капиллярное обеспечение, однако
включают большие запасы гликогена и фосфокреатина. Высокая актив-
ность гликогенолитических и гликолитических ферментов обеспечивает
способность волокон типа II к быстрой (но относительно кратковремен-
ной) продукции АТФ, когда расход энергии превышает ее образование за
счет окислительного фосфорилирования. Другими словами, они обеспе-
чивают высокую анаэробную способность.
Следует отметить, что анаэробное дыхание (гликогенолиз и глико-
лиз) протекает без использования кислорода, но не обязательно при его
полном отсутствии (аноксии) или же в условиях низкой его доступнос-
ти (гипоксии). Волокна типа II в наибольшей степени подходят для
24
проявления быстрой и сильной активности в короткий промежуток
времени, однако утомление в них, как известно, развивается быстро
(см. главу 6).
Волокна типа Па являются красными клетками, в которых метабо-
лические и физиологические характеристики находятся между крайни-
ми значениями других двух типов волокон. Они содержат высокоактив-
ную миозиновую АТФ-азу, аналогичную волокнам типа Пб, однако ха-
рактеризуются окислительной способностью, более близкой к волокнам
типа I.
Результаты последних исследований показывают, что для большин-
ства мышечных волокон их фактическая идентификация определяется
одной и более изоформой миозина. Такая совместная экспрессия различ-
ных изоформ миозина создает предпосылки для расширения диапазона
сократительных характеристик внутри определенного типа волокон
(классифицируемых в соответствии с окраской гистологических образ-
цов) и степени их частичного совпадения между типами волокон при оп-
ределении их сократительных характеристик. В действительности мы
продолжаем оперировать сократительными и биохимическими характе-
ристиками, соответствующими двум крайним типам классификации, оп-
ределяемым как волокна типов I и Пб.
С различиями в скорости сокращения и метаболического профиля
основных типов мышечных волокон отмечается дифференциация и дви-
гательных нейронов, иннервирующих эти волокна. Так, волокна типа 1
иннервируются нейронами, имеющими малый диаметр и характеризую-
щимися низкой скоростью проведения потенциала действия, а также
низким порогом активации. Волокна типа II иннервируются нейронами
большего диаметра, которые отличаются относительно более высокой
прочностью и более высоким порогом возбудимости. Различие в поро-
гах возбудимости двигательных нейронов связано с типом мышечных
волокон, которые они иннервируют, с их рекруитированием при мышеч-
ных нагрузках, что и определяет метаболический ответ на последние.
При различных видах двигательной активности проявляется определен-
ная иерархия включения в нее двигательных единиц в зависимости от их
размера, которая, в общих чертах, соответствует волокнам типов I и II.
Это свидетельствует о том, что при легких физических нагрузках в наи-
большей степени задействованы волокна типа I, при нагрузках средней
мощности — волокна типов I и II, а при более напряженной мышечной
деятельности — все типы волокон включаются в работу (рис. 1.6).
Все мышцы организма представляют собой смесь указанных выше
трех типов волокон, хотя пропорциональное соотношение в каждой из
них и в каждом из индивидуумов обнаруживает существенное различие.
25
Интенсивность физической нагрузки
Малая —»- Средняя —»- Максимальная
Рис. 1.6. Рекрутирование мышечных волокон
типов I (медленносбкращающихся) и II (быстро-
сокращающихся) при физических нагрузках
возрастающей интенсивности
Так, мышцы, отвечающие за
поддержание позы тела (нап-
ример, m. soleus), содержат
высокий процент (обычно бо-
лее 70 %) волокон типа I,
функция которых состоит в
поддержании пролонгирован-
ного, но относительно невы-
сокого напряжения. Однако
быстросокращающиеся во-
локна типа II преобладают в
мышцах, выполняющих быс-
трые движения (например, в
мышцах руки и глаза). Другие
же мышцы, как, например, че-
тырехглавая мышца бедра,
имеют разное соотношение типов волокон. Их композиционный состав
генетически детерминирован и не поддается значительным изменениям
под влиянием тренировки. Следовательно, способность к достижению
высоких спортивных результатов является в значительной степени врож-
денной (с учетом того, что генетический потенциал индивидуума реали-
зуется при соответствующей методике тренировки и питания).
Исследование m. vastus lateralis у марафонцев высокой квалифика-
ции показало, что эта мышца содержит больший процент (около 80 %)
волокон типа I, тогда как у элитных спринтеров обнаружился более вы-
сокий процент (около 60 %) быстросокращающихся волокон типа II (см.
Коми и Карлссон (Komi and Karlsson, 1978). Безусловно, такие исследо-
вания требуют применения методики пункционной биопсии, которая
позволяет изъять около 1000 волокон в одной пробе. Относительно не-
большое количество исследований по изучению распределения волокон
в мышцах человека было проведено при вскрытии трупов. Региональная
вариативность композиционного состава мышц проявляется незначи-
тельно, хотя и отмечается тенденция к тому, что в глубоколежащих сло-
ях преобладают волокна типа I. Это подтверждается и данными, полу-
ченными с помощью достаточно надежного метода — пункционной би-
опсии. Исследования, проведенные на m. vastus lateralis у человека, сви-
детельствуют о том, что разница в показателях между пробами состав-
ляет не более 6 % (коэффициент вариации) по отношению к доминиру-
ющему типу волокон.
Типы мышечной активности. Скелетные мышцы могут проявлять
три различных типа активности: изометрический, при котором длина
26
мышцы остается постоянной; концентрический, когда мышца укорачи-
вается; эксцентрический, при котором длина мышцы, находящейся в ак-
тивном состоянии, увеличивается. Обычно подразумевается, что мыш-
цы развивают силу при сокращении. Однако тщательные исследования,
проведенные на изолированных мышцах, свидетельствуют о том, что
проявление силы не сопровождается изменением объема мышцы. Для
проявления силы мышцы стремятся сократиться (т. е. уменьшить длину
саркомеров), что в результате может иметь три основных исхода. В слу-
чае концентрической активности мышца выполняет механическую ра-
боту и развивает при этом определенную мощность. Если мышца прояв-
ляет активность в условиях изометрического или эксцентрического сок-
ращения, то мощность будет поглощаться. Такие физические упражне-
ния, выполняемые как при одном из видов активности, так и в комбина-
ции, способны вызвать нарушение гомеостаза. Наиболее важной комби-
нацией может быть цикл «удлинение-сокращение», в котором эксцен-
трической активности непосредственно предшествует концентрическая
активность. Сохранение возможности проявления высокой мощности в
начале эксцентрической фазы обеспечивается энергией, запасенной в
мышце за счет ее эластичности. Во время последующей, генерирующей
мощность, концентрической фазы вклад восстанавливающей эласти-
ческой энергии препятствует развитию утомления по сравнению со слу-
чаем чисто концентрической активности.
Повторное выполнение высокомощностных эксцентрических дей-
ствий может вызвать повреждение мышц и возникновение в них времен-
ных болезненных ощущений, которые обычно появляются через 6-12 ч
после физической нагрузки и сохраняются несколько дней. Роль различ-
ных типов волокон в эксцентрической мышечной активности еще пол-
ностью не выяснена, хотя, скорее всего, в данном случае активируется
небольшое количество волокон по сравнению с концентрическим сокра-
щением. Имеется также несколько доказательств, что волокна типа II
могут избирательно рекруитироваться во время эксцентрической актив-
ности мышц. Такая высокая нагрузка на относительно небольшое коли-
чество волокон может вызвать их локальные повреждения, связанные с
воспалительным процессом и сопровождающимися отечностью и боле-
выми ощущениями. Доказательством того, что в мышцах действительно
происходят повреждения, служит появление в крови высоких уровней
внутримышечных ферментов в дни выполнения эксцентрических уп-
ражнений, а также гистологические доказательства разрушения сарко-
меров и Z-линий в работавших мышцах.
Пластичность скелетных мышц. Скелетная мышца является чрез-
вычайно пластичной тканью: она обладает значительной способностью
27
адаптироваться к различным видам активности либо к бездействию.
Адаптация может проявляться в изменении размеров мышцы, компози-
ционного состава волокон, метаболической способности, плотности ка-
пилляризации (более подробно об адаптационных изменениях, происхо-
дящих в мышцах под влиянием тренировочных занятий, направленных
на развитие выносливости и силы, см. в главе 8).
Размер мышц и их функции с возрастом изменяются (Греймби и
Салтин (Grimby and Saltin), 1983). Максимальной силы мышцы у муж-
чин и женщин достигают в возрасте между 20 и 30 годами. К 70 годам
сила мышц снижается в среднем на 30 %. Уменьшение мышечной мас-
сы является ведущим фактором в этом процессе. Возможно также, что
происходящее с возрастом снижение мышечной силы связано с умень-
шением размеров волокон, в частности волокон типа II. Не исключено
уменьшение при этом общего количества мышечных волокон, обуслов-
ленное утратой к старости двигательных нейронов. Иннервация мышеч-
ных волокон необходима для поддержания их существования (возмож-
но, это связано с выработкой в нерве факторов роста), а денервация при-
водит к атрофии мышечных волокон и замещению их соединительной
тканью.
Связанное со старением снижение мышечной массы может проис-
ходить как за счет самих возрастных изменений, происходящих в орга-
низме, так и в результате снижения двигательной активности либо од-
новременно за счет обоих факторов. Однако ясно, что и в старом орга-
низме мышцы еще сохраняют способность адаптироваться в ответ на си-
ловую тренировку и что значительного улучшения физиологических,
структурных и физических характеристик можно достичь без реализа-
ции напряженных тренировочных программ. В молодом возрасте часто-
та, интенсивность и продолжительность упражнений являются решаю-
щими факторами, определяющими диапазон происходящих под влияни-
ем тренировки адаптационных изменений.
3. Источники энергии
для мышечных сокращений
Аденозинтрифосфат. Непосредственным источником энергии, необхо-
димой для обеспечения мышечных сокращений, является АТФ; факти-
чески, энергия, которая образуется при гидролизе АТФ, используется
для обеспечения всех форм биологических функций. В мышце энергия
образуется в результате гидролиза АТФ специфическими участками ми-
озина, обладающими АТФ-азной активностью и обеспечивающими спо-
28
собность мышцы к сокращению. Для активации обратного захвата
ионов кальция саркоплазматическим ретикулумом также требуется
АТФ. Существует четыре механизма, обеспечивающие распад и ресин-
тез АТФ:
1) энзиматический распад АТФ до аденозиндифосфата (АДФ) и не-
органического фосфата (Фп) с освобождением энергии, используемой
для обеспечения мышечной активности;
2) энзиматический распад фосфокреатина (КФ) до креатина и фос-
фата, который присоединяется к АДФ для ресинтеза АТФ;
3) превращение образующегося из мышечного гликогена или глю-
козы крови глюкозо-6-фосфата до лактата с образованием АТФ путем
анаэробного гликолиза в результате реакций субстратного фосфорили-
рования;
4) продукты метаболизма углеводов, липидов, белков и алкоголей
могут включаться в цикл трикарбоновых кислот (ЦТК, или цикл Креб-
са) в митохондриях и подвергаться окислению до углекислого газа и во-
ды. Этот процесс известен как окислительное фосфорилирование и ос-
вобождение энергии, используемой для синтеза АТФ. Некоторое коли-
чество образующегося АТФ используется для ресинтеза КФ, запасы ко-
торого истощаются при интенсивных физических нагрузках.
Использование АТФ в качестве непосредственного источника
энергии является первым из этих механизмов, тогда как остальные три
механизма, по-видимому, служат для обеспечения ресинтеза АТФ с не-
обходимой скоростью, чтобы предотвращать значительное снижение его
концентрации в мышцах. Первые три из указанных механизмов, кото-
рые являются анаэробными (т. е. протекают в отсутствие кислорода),
для образования энергии используют соответственно только один спе-
цифический субстрат — АТФ, КФ и глюкозо-6-фосфат. Хотя глицерол и
может включаться в гликолитический путь в печени и некоторых других
тканях, тем не менее, очевидно, он не является энергетическим источни-
ком для скелетных мышц, поскольку в них отсутствует фермент глице-
ролкиназа.
Аэробные процессы, протекающие в митохондриях при участии
кислорода, метаболизируют различные субстраты. Саркоплазма содер-
жит разнообразные энзимы, которые могут превращать углеводы, липи-
ды и белки в способные к использованию субстраты, преимущественно
в двууглеродную ацетильную группу, связанную с коэнзимом А (ацетил-
КоА), которая в митохондриях может полностью подвергнуться окис-
лению, в результате чего образуется АТФ. Общая информация об основ-
ных энергетических источниках и путях энергетического метаболизма
представлена на рис. 1.7.
29
Рис. 1.7. Общая схема основных путей энергетического метаболизма, использующих
углеводы, липиды и белки в качестве источников энергии
Углеводы могут участвовать как в анаэробных, так и в аэробных пу-
тях энергетического метаболизма. При гликолизе глюкозо-6-фосфат (де-
риват от гликогена или глюкозы) в анаэробных условиях распадается до
лактата, а в аэробных — до пирувата. Пируват превращается до ацетил-
КоА и полностью окисляется в ЦТК. Липиды в форме триацилглицеро-
лов гидролизуются до жирных кислот и глицерола, которые затем вклю-
чаются в гликолитический путь; жирные кислоты превращаются через
^-окисление до ацетил-КоА и последовательно подвергаются окис-
лению в ЦТК. Белковый катаболизм обеспечивается аминокислотами,
которые могут превращаться в интермедиаты ЦТК, в пируват или ацето-
ацетат и последовательно трансформироваться в ацетил-КоА.
Анаэробный метаболизм. Скелетные мышцы человека могут раз-
вивать напряжение без потребления кислорода, вследствие чего такая
способность генерировать энергию называется анаэробной. Две отдель-
30
ные системы способны обеспечить такую возможность: фосфагенная,
или высокоэнергетическая фосфатная и гликолитическая системы. Пос-
кольку гликолитическая система связана с продукцией лактата, а фосфа-
генная его не продуцирует, то эти системы иногда обозначают соответ-
ственно как лактатная и алактатная системы. Следует заметить, что тер-
мины «молочная кислота» и «лактат» взаимозаменяемы, хотя первый из
них носит в основном описательный характер, поскольку ясно указы-
вает на кислотную природу молекулы, в то время как термин «лактат»
более точный и далее именно он будет использоваться в тексте. Подроб-
нее эти энергетические системы описаны в главах 2 и 3, а в данном па-
раграфе приведен лишь их краткий обзор.
Фосфагенная система. Если мышцу отравить цианидом или йод-
ацетоуксусной кислотой, то она не сможет производить энергию ни за
счет окислительного метаболизма, ни за счет продукции лактата, однако
она еще способна выполнить сильные сокращения в течение короткого
промежутка времени до наступления утомления. Это свидетельствует о
том, что мышцы располагают еще одним источником энергии, однако
возможности этого источника лимитированы. Таким источником
являются внутримышечные запасы АТФ и фосфокреатина (КФ или из-
вестного как креатинфосфат); вместе АТФ и КФ обозначают как фос-
фагены.
Наиболее важной особенностью фосфагенов является тот факт, что
энергетический запас, который в них аккумулирован, почти немедленно
может быть использован мышцей. Реализация энергетического потенци-
ала КФ для ресинтеза АТФ в мышечных волокнах происходит с очень
высокой скоростью, что создает возможность выполнять быстрые и
мощные двигательные действия. Основной недостаток этой системы
состоит в ее ограниченной возможности обеспечивать мышечную рабо-
ту в течение продолжительного времени, и если другие энергетические
источники не поступают в мышцу, то это приводит к быстрому разви-
тию утомления. Во время спринтерского бега на дистанции 30-50 м на
последних нескольких метрах еще не обнаруживается снижение скорос-
ти движения, так как энергетические потребности обеспечиваются ис-
пользованием запасов фосфагена. На более длинных дистанциях ско-
рость бега начинает падать, так как эти запасы исчерпываются и снижа-
ется мощность мышечных сокращений. Однако скорость восстановле-
ния работоспособности после короткого спринта достаточно высокая и
повторный забег может быть выполнен с такой же скоростью, как и пре-
дыдущий, уже через 2-3 мин отдыха. После длинного спринта (100 м и
более) для проявления высокого уровня работоспособности требуется
более продолжительное время для отдыха.
31
Гликолитическая система. В обычных условиях мышца не прояв-
ляет явных признаков утомления после всего нескольких секунд напря-
жения, поскольку к ней могут поступать наряду с фосфагенами и другие
энергетические источники. Ими служат продукты, образующиеся в про-
цессе гликолиза. Этот процесс, протекающий без участия кислорода,
обеспечивает ресинтез АТФ за счет реакций субстратного фосфорилиро-
вания. Реакции, приводящие к образованию пирувата, должны быть от-
даленными; при выполнении малоинтенсивных физических упражне-
ний, для которых необходимая энергия может быть обеспечена аэроб-
ными процессами, пируват превращается до диоксида углерода и воды в
окислительном метаболизме митохондрий. В некоторых ситуациях пи-
руват превращается в лактат анаэробным путем системой, обозначаемой
как анаэробная лактатная.
Активация гликолитической системы проявляется почти сразу с на-
чалом выполнения физического упражнения несмотря на большое коли-
чество вовлекаемых при этом реакций (подробно гликолитический путь
описан в главе 3). Скорость образования лактата зависит в основном от
интенсивности физической нагрузки, однако в большей степени — от
относительной интенсивности мышечной работы (%VO2 max), чем от
абсолютной интенсивности.
Общая способность гликолитической системы к выработке энергии
в форме АТФ во многом сравнима с фосфагенной системой (табл. 1.3).
При выполнении высокоинтенсивных физических упражнений происхо-
дит быстрый распад мышечного гликогена, сопровождающийся соответ-
ственно высокой скоростью образования лактата: некоторое количество
последнего диффундирует из мышечных волокон в кровь. Бульшая
часть (но не всего) запаса мышечного гликогена может быть использова-
Таблица 1.3. Емкость и мощность анаэроб- ных энергетических систем в ресинтезе АТФ* на для обеспечения анаэ- робной энергопродукции во время высокоинтенсивной физической нагрузки и тем самым удовлетворять энерге- тическую потребность для обеспечения интенсивной мышечной работы, продол- жающейся от 20 с до 5 мин. При меньшей продолжитель- ности нагрузки главным энергетическим источником служит фосфаген, тогда как с увеличением продожитель-
Энергетическая система Емкость, ммоль АТФ х х кг“ ’ X хсмт Мощность, ммоль АТФ х х кг~1X X смт с-1
Фосфагенная Гликолитическая Комбинация 55-95 190-300 250-370 9,0 4,5 11,0
* Уровни соотнесены к I кг сухой массы мышц и основаны на определении продукции АТФ в m. vastus lateralis у человека при выполнении высокоинтенсив- ных физических упражнений
32
ности работы (или преодолеваемой дистанции) прогрессивно возраста-
ет роль окислительного метаболизма.
Хотя общая энергетическая емкость у гликолитической системы
больше, чем у фосфагенной системы, скорость, с которой она способна
продуцировать энергию (ресинтезировать АТФ), меньше (табл. 1.3). Поэ-
тому развиваемая за счет этой энергосистемы мощность мышечной рабо-
ты соответственно также оказывается ниже, в связи с чем максимальные
скорости выполнения физических упражнений не могут удерживаться бо-
лее нескольких секунд: с истощением запасов фосфагенов интенсивность
выполнения мышечной работы обязательно должна снижаться
Аэробный метаболизм: окисление углеводов, жиров и белков.
Другие пути ресинтеза АТФ связаны в основном с использованием угле-
водных или липидных субстратов из внутримышечных запасов и крови,
их последовательного распада (катаболизма) для получения энергии в
результате субстратного фосфорилирования или реакций электрон-тран-
спортной цепи и окислительного фосфорилирования. Ресинтез АТФ при
катаболизме жиров (липидов) требует использования кислорода, тогда
как катаболизм углеводов может происходить как в присутствии кисло-
рода, так и без него (гликолитическим путем, описанным выше).
Катаболизм глюкозы начинается с анаэробного гликолиза; каждая
молекула глюкозы дает две молекулы пирувата (или лактата) и в резуль-
тате гликолиза обеспечивает ресинтез двух молекул АТФ (хотя каждая
половина молекулы глюкозы дает три молекулы АТФ, если первоначаль-
ным субстратом является мышечный гликоген). При аэробном метабо-
лизме главным образом пируват (предпочтительнее, чем лактат) являет-
ся формой, которая превращается до ацетил-КоА ферментом пируватде-
гидрогеназой (см. рис. 1.7).
Катаболизм липидов в форме жирных кислот также приводит к об-
разованию ацетил-КоА (более подробно см. в главе 4). Ацетил-КоА яв-
ляется основным поставщиком энергии для ЦТК, который совершается
в митохондриях.
По сравнению с катаболизмом углеводов и липидов распад белков
представляет собой относительно незначительный источник энергии
при мышечной деятельности. В большинстве случаев вклад белкового
катаболизма составляет менее 5 % энергии, используемой мышцами при
двигательной активности. Белковый катаболизм способен обеспечить
образование как кетогенных, так и гликогенных аминокислот, которые
могут быть подвергнуты окислению в результате дезаминирования и
превращения в один из субстратных интермедиатов ЦТК, а также мета-
болизироваться до пировиноградной или ацетоуксусной кислоты и, в ко-
нечном счете, трансформироваться до ацетил-КоА (см. рис. 1.7). Однако
3 0.3,3 33
при голодании и истощении запасов гликогена катаболизм белков может
становиться все более и более важным источником энергии для мышеч-
ной деятельности.
Катаболизм углеводов, жиров и белков подробно описан в последу-
ющих главах. Однако поскольку ацетил-КоА служит общим метаболи-
том для указанных выше энергетических источников и конечные
аэробные пути ЦТК и окислительного фосфорилирования являются об-
щими для всех трех классов энергетических субстратов, в настоящей
главе целесообразно было дать общую характеристику этих важных пу-
тей энергетического метаболизма.
4. Цикл трикарбоновых кислот
Основная функция ЦТК состоит в деградации ацетил-КоА до диоксида
углерода и атомов водорода в ходе последовательных реакций, протека-
ющих в митохондриях (рис. 1.8). Атомы водорода затем подвергаются
окислению в электрон-транспортной (дыхательной) цепи, что представ-
ляет собой процесс окислительного фосфорилирования с последова-
тельным ресинтезом АТФ из АДФ.
Как показано на рис 1.9, шестиуглеродная трикарбоксильная моле-
кула цитрата образуется при конденсации четырехуглеродной молекулы
оксалоацетата с двууглеродной ацетильной группой ацетил-КоА с пос-
ледующим гидролизом. Эта реакция катализируется ферментом цитрат-
синтазой. Затем фермент аконитаза катализирует образование из цитра-
та его изомера-изоцитрата, который впоследствии подвергается окисли-
тельному декарбоксилированию до а-кетоглутарата. В процессе этой ре-
акции, катализируемой изоцитрадегидрогеназой, образуются диоксид
углерода и первая из трех молекул НАДН, продуцируемых ЦТК. Вторые
молекулы диоксида углерода и НАДН образуются в процессе последую-
щего окислительного декарбоксилирования а-кетоглутарата дегидроге-
назным ферментативным комплексом.
Энергия, освобождающаяся при распаде тиоэфирной (С-S) связи
сукцинил-КоА, используется затем для фосфорилирования гуанозинди-
фосфата (ГДФ), в результате чего образуется гуанозинтрифосфат (ГТФ)
и сукцинат. Фермент нуклеотидфосфаткиназа катализирует образование
АТФ путем переноса терминальной фосфатной группы от ГТФ к АДФ.
Следует отметить, что сукцинат представляет собой четырехуглеродное
вещество; исходный шестиуглеродный скелет молекулы цитрата подвер-
гается двум реакциям декарбоксилирования, в результате которых теря-
ется атом углерода в форме его диоксида.
34
о
(Пируват) СПз-C— СОО’
Пируватдегидрогеназа
____' ’___ О
(Ацетил-КоА) ch3-C-S -КоА
НАД
СОО'
но—А—н
СОО' НгО—S
А=о
Ан2_________
СОО Цитрат-
Оксалоацетат синтетаза
KoA-SH СОО’
СОО’
Малат
Фумараза
СОО
Малатдегидро-
геназа
НО—А—СОО
Ан
ин2
СОО’ \
Цитрат
Аконитаза
НгО
СОО
А—СОО’
НС
Аоо’
Фумарат
ФАДН2
СОО’
цис-Аконитат
НгО
ФАД
Сукцинатде-
гидрогеназа
ЦИКЛ
ЛИМОННОЙ
КИСЛОТЫ
Аконитаза
СОО’
СН2
соо
. Сукцинаттио-
киназа
Н—С—СОО’
НО—А—Н
ГТФ
СОО’
Сукцинат
СОО’
Lzti2
ГДФ + Ф,
KoA-SH
а-Кетоглутаратде-
гидрогеназа
S—КоА
Сукцинил-КоА
НАДН + И1
KoA-SH
СОО’
НАД1
сн2
Ан2
СОО’
Изоцитраг- Изоцитрат
дегидрогеназа /
СОО'
НАД4
н—с—СОО НАДН + Н
СОО’
Оксалосукцинат
СОО |со I
а-Кетоглутарат 1--
Рис. 1.8. Общая схема реакций ЦТК, демонстрирующая участки субстратного фосфо-
рилирования и восстановления НАД1 и ФАД (сх-кетоглутарат может быть обозначен и
как 2-оксоглутарат)
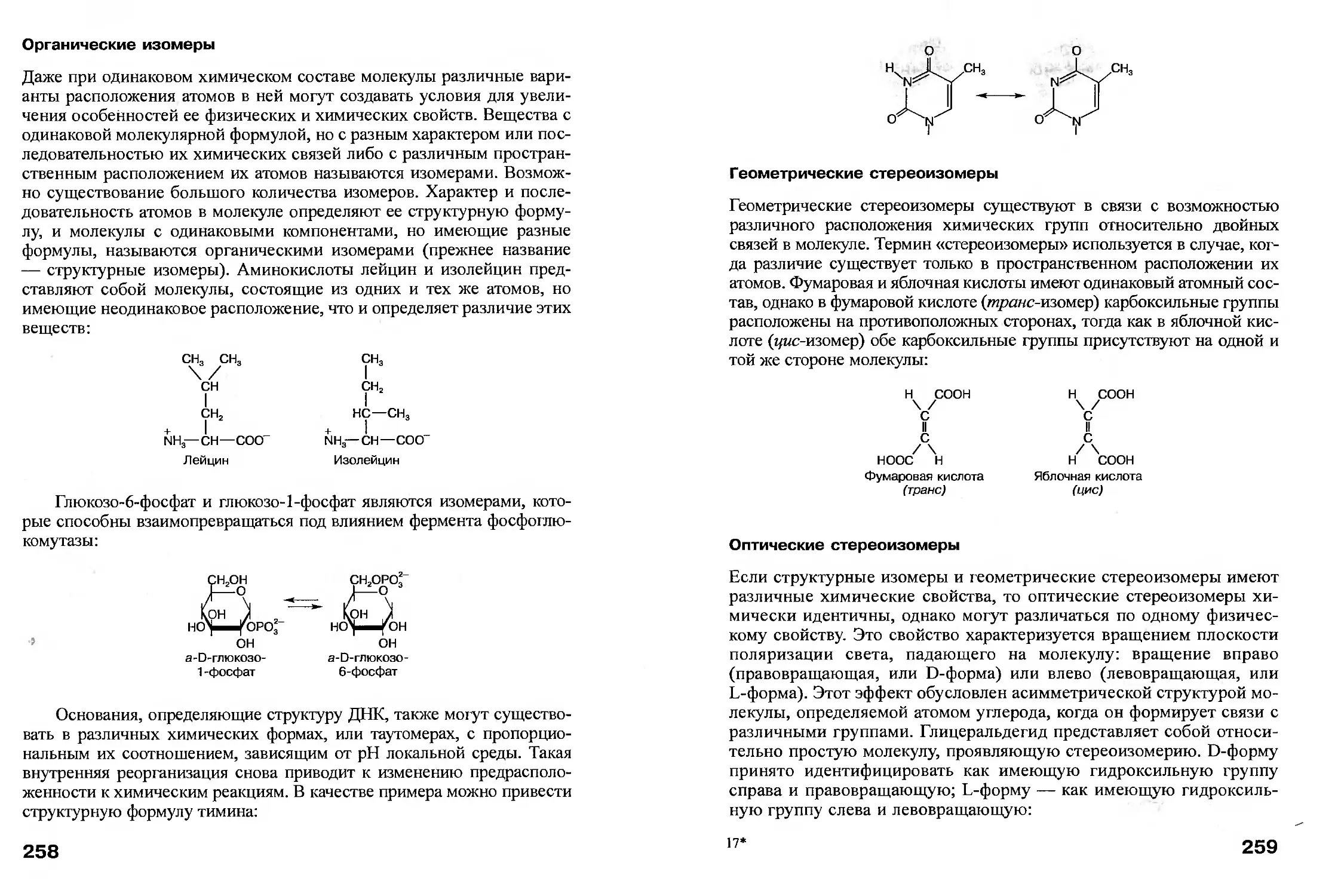
3*
35
Далее сукцинат при участии сукцинатдегидрогеназы и кофермента
— флавинадениндинуклеотида (ФАД) окисляется до фумарата с образо-
ванием ФАДН. После этого фумарат подвергается реакции гидратации,
катализируемой фумаразой, в результате чего образуется малат. Затем
фермент малатдегидрогеназа окисляет малат до оксалоацетата, что соп-
ровождается восстановлением НАД+ до НАДН. Таким образом, в дан-
ном случае происходит реформирование молекулы оксалоацетата и за-
вершается полный оборот ЦТК, в течение которого образуется одна мо-
лекула АТФ, три молекулы НАДН и одна молекула ФАДН. Следователь-
но, описанную выше реакцию для каждой молекулы ацетил-КоА можно
представить следующим образом:
Ацетил-КоА + АДФ + ЗНАД+ + ФАД —>
-> 2СО2 + АТФ + ЗНАДН + ЗН+ + ФАДН2КоА
уравнение ЦТК
Фаза
Фаза I
(ее
НАДН
ФАДН,
2Н + О
Челночный
транспорт
электронов
Пируват, образующийся
в процессе гликолиза
Электрон -тран спортн ая
цепь: восстановленные
коферментные комплексы
подвергаются окислению
Доставка водорода вос-
становленными кофер-
ментами (транспортными
НАДН
СО
НАДН
ФАДН.
НАДН
АТФ
Рис. 1.9. Схема взаимосвязи ЦТК с электрон-транспортной цепью
Ацетил-КоА
молекулами) к электрон-
транспортной цепи
36
Четырехуглеродный оксалоацетат может также образовываться из
трехуглеродного пирувата при участии фермента пируваткарбоксилазы
(включающей в качестве переносчика СО коэнзим биотин), на что затра-
чивается одна молекула АТФ. Активность этого фермента стимулирует-
ся высокой концентрацией ацетил-КоА, которая эффективно сигнализи-
рует о большей потребности в оксалоацетате. Эта реакция важна для
восполнения интермедиатов ЦТК и обеспечения субстратами глюконео-
генеза, в частности для образования глюкозы из неуглеводных пред-
шественников.
Ключевым регуляторным участком в ЦТК является реакция, ка-
тализируемая цитратсинтазой. Активность этого фермента угнетает-
ся АТФ, НАДН, сукцинил-КоА и ацетил-КоА дериватами жирных
кислот; его активность также подавляется в присутствии цитрата.
Следовательно, когда энергетические уровни в клетке высокие, то
скорость протекания ЦТК относительно невысокая, однако может
значительно возрастать, когда утилизация АТФ и НАДН увеличи-
вается, как, например, в условиях напряженной мышечной деятель-
ности.
Следует отметить, что молекулярный кислород не принимает не-
посредственного участия в реакциях ЦТК. В сущности, наиболее
важной функцией ЦТК является генерация атомов водорода для их
последующего поступления в цепь транспорта электронов с по-
мощью НАД+ и ФАД (рис. 1.9). Аэробные процессы электрон-тран-
спортного окислительного фосфорилирования ресинтезируют АТФ
из АДФ, благодаря чему определенное количество энергии, содержа-
щееся в исходных субстратах, превращается в соответсвующее коли-
чество энергии высокоэнергетических фосфатов. Пока осуществля-
ется адекватная доставка О2 и поступление субстрата, до тех пор
НАД+ и ФАД продолжают регенерироваться и возобновлять метабо-
лизм в ЦТК.
5. Электрон-транспортная цепь
Электрон-транспортная цепь (рис. 1.10) представляет собой серию
молекулярных переносчиков, которые транспортируют электроны от
водорода и, в конечном итоге, доставляют их к молекулярному кис-
лороду, который акцептирует водород с образованием воды. Большое
количество энергии, образующееся в процессе транспорта элек-
тронов, поглощается или превращается в потенциальную химическую
37
НАДН
НАДН
-дегидрогеназа
ФАДН2------*- KoQ
♦
Цитохром b
АДФ + Фв
АТФ
Цитохром с,
I
Цитохром с
♦
Цитохром а + а3
Н2О
Рис. 1.10. Электрон-транс-
портная цепь с указанием пун-
ктов, в которых происходит
синтез АТФ
энергию высокоэнергетических фосфа-
тов, но часть ее рассеивается в виде теп-
ла. В процессе описанных выше дегидро-
геназных реакций ЦТК (и гликолиза)
НАД+ восстанавливается и присоединяет
к себе водород и два электрона; осталь-
ной водород появляется во внутриклеточ-
ной жидкости в виде Н+. Флавинаденин-
динуклеотид является другим восстанав-
ливающим коэнзимом в дегидрогеназных
реакциях, который также акцептирует па-
ру электронов, при этом ФАД также при-
соединяет оба атома водорода, в резуль-
тате чего образуется ФАДН2; НАДН и
ФАДН2 представляют собой богатые
энергией молекулы, поскольку каждая из
них содержит пару электронов, облада-
ющих высоким энергопереносным потен-
циалом.
В процессе переноса этих элек-
тронов к кислороду большое количество
освобождаемой энергии может быть
использовано для фосфорилирования
АДФ, в результате чего происходит ре-
синтез АТФ. Транспорт электронов от
НАДН и ФАДН2 к молекулярному кисло-
роду осуществляется серией специфических переносчиков, локализо-
ванных на внутренней поверхности митохондриальной мембраны; эти
переносчики образуют электрон-транспортную, или дыхательную
цепь.
В первом участке электрон-транспортной цепи, катализируемом
НАДН-дегидрогеназным ферментативным комплексом, два электрона
переносятся от НАДН к флавинмононуклеотиду (ФМН), в результате
чего образуется ФМНН2, а освобождающаяся при транспорте элек-
тронов энергия используется для фосфорилирования АДФ (синтез АТФ
в пункте 1, рис 1.10). Затем электроны переносятся к коэнзиму Q (KoQ),
который также способен акцептировать электроны с образованием
ФАДН-. Таким образом, ФАДН2 включается в электрон-транспортную
цепь после первого участка синтеза АТФ. Следует отметить, что ФАДН2
находится в меньшем энергетическом состоянии по сравнению с НАДН
38
и ФМНН2, в связи с чем ФМН не может акцептировать электроны от
фадн2.
Следующими после KoQ переносчиками электронов в дыхательной
цепи служат гем- (железо-)содержащие белки, называемые цитохрома-
ми. Каждый цитохром способен переносить только один электрон (лег-
че, чем два с такими переносчиками, как НАДН, ФМНН2 и KoQH2).
Часть железа каждого цитохрома может существовать либо в окис-
ленной (Fe3+), либо в восстановленной (Fe2+) форме. При присоединении
электрона окисленная форма железа в специфическом цитохроме стано-
вится восстановленной, и наоборот, в случае отдачи электрона следую-
щему цитохрому, расположенному ниже по линии в направлении к пос-
леднему — цитохрому а, происходит отток электрона к молекулярному
кислороду.
От KoQ электроны передаются цитохрому Ъ, а затем — цитохрому
ср который является вторым пунктом синтеза молекулы АТФ (рис. 1.10,
пункт 2). Далее электроны от цитохрома с; транспортируются к цито-
хрому с и затем к цитохромам а и а3, которые существуют в виде ком-
плекса, называемого цитохромоксидазой. И наконец, цитохром а3 пере-
дает электроны молекулярному кислороду, что является третьим пун-
ктом, в котором происходит синтез АТФ (рис. 1.10, пункт 3). Следова-
тельно, каждая молекула НАДН, которая включается в цепь транспорта
электронов, обеспечивает ресинтез трех молекул АТФ, а каждая моле-
кула ФАДН2 — двух молекул АТФ. Таким образом, возвращаясь к пред-
ставленному выше уравнению ЦТК, можно прийти к заключению, что
каждая молекула ацетил-КоА подвергается полному окислению в ЦТК,
в результате чего образуется 12 молекул АТФ (включая одну молекулу
от ГТФ).
6. Окислительное фосфорилирование
Окислительное фосфорилирование является процессом, при котором
синтез АТФ происходит в результате транспорта электронов от ФАДН2 и
НАДН к молекулярному кислороду. Общепринятый механизм этого про-
цесса представлен в предложенной Митчеллом (Mitchell) химио-осмо-
тической теории, суть которой состоит в том, что перенос электронов в
электрон-транспортной цепи, локализованный во внутренней митохон-
дриальной мембране, обусловлен перекачкой водородных ионов (Н+) из
внутримитохондриального матрикса через внутреннюю митохондриаль-
ную мембрану внутрь пространства между внутренней и наружной мем-
бранами митохондрии (рис. 1.11).
39
Рис. 1.11. Схема химио-осмотического окисли-
тельного фосфорилирования
Высокая концентрация
положительно заряженных
ионов водорода в этой
внешней камере способ-
ствует тому, что ионы Н+
стремятся проникнуть об-
ратно в митохондриальный
матрикс через F^-Ft протеи-
новый комплекс, встроен-
ный во внутреннюю мем-
брану митохондрии. Поток
водородных ионов (прото-
нов) через этот комплекс
составляет протондвижу-
щую силу, которая исполь-
зуется для обеспечения син-
теза АТФ; ^-субъединица
комплекса представляет со-
бой протоковый канал, а
F'-субъединица — сопря-
женный с ней элемент, обла-
дающий АТФ-синтазной ак-
тивностью.
7. Резервы углеводов и липидов
Углеводы запасаются в организме в виде полимера глюкозы, называ-
емым гликогеном. Скелетные мышцы содержат значительный запас
гликогена в саркоплазме. Содержание гликогена в скелетной мышце
в покое составляет приблизительно 14-18 г-кг—1 сырой массы
(80-1000 ммоль-кг—1 глюкозильных единиц). Печень также содержит
гликоген; его запасы в печени взрослого человека в постабсорбтив-
ном состоянии составляет 80-110 г, что создает возможность выхода
глюкозы в кровь для поддержания ее концентрации около 5 ммоль-2
(0,9 г-л-1).
Запасы липидов представлены в виде триглицеридов (триацилгли-
церолов), содержащихся в основном в белой адипозной ткани. Первона-
чально они должны подвергаться распаду под действием липазы — рас-
щепляющему жиры ферменту, обеспечивающему выход свободных жир-
40
ных кислот (СЖК) в кровяное русло для их утилизации работающими
мышцами. Скелетные мышцы также содержат некоторое количество
триацилглицеролов, которые могут использоваться в качестве энергети-
ческого источника во время физических нагрузок, сопровождающихся
липолизом, однако этот источник может становиться относительно бо-
лее важным в послерабочем периоде. Запасов липидов в организме зна-
чительно больше по сравнению с запасами углеводов, и липиды являют-
ся более эффективной формой накопления энергии, поскольку способны
выделить 37 кДжт-1 по сравнению с 16,7 кДжт-1 — при окислении уг-
леводов.
Каждый грамм углеводов удерживает около 3 г воды, что снижает
эффективность углеводов как энергетического источника. Энергети-
ческая стоимость бега на марафонской дистанции составляет около
12000 кДж. Если бы она могла реализоваться только за счет жира, то об-
щее количество необходимых для этого липидов составило бы около
320 г, тогда как в случае использования только углеводов потребность в
них составила бы 750 г и дополнительно 2,3 кг связанной с ними воды.
Даже без учета массы, которую необходимо транспортировать, такое ко-
личество углеводов превышает общее их содержание в печени, мышцах
и крови. Общая накопительная способность для липидов чрезвычайно
большая, и для большинства практических целей количество энергии,
запасенное в форме жира, превышает требуемое для обеспечения како-
го-либо вида мышечной деятельности (табл. 1.4).
Белки не запасаются в организме, как жиры и углеводы. Они
представляют собой важные в функциональном отношении молеку-
лы (например, структурные
белки, ферменты, ионные
каналы, рецепторы, сокра-
тительные белки и т. д.).
Концентрация же большин-
ства свободных амино-
кислот во внутри- и вне-
клеточнных жидкостях ор-
ганизма довольно низкая.
Поэтому неудивительно, что
в большинстве случаев
углеводы и жиры в наи-
большей степени обеспе-
чивают потребности ресин-
теза АТФ при физических
нагрузках.
Таблица 1.4. Средние показатели энергети-
ческих запасов у человека*
Энергетический источник Масса, г Энергия, кДж
Гликоген печени 80 1280
Мышечный гликоген 350 5600
Глюкоза крови 10 160
Липиды 10500 388500
Белки 12000 204000
* Для массы тела, равной 7Q кг и содержащей 15 %
жира. Уровень глюкозы в крови включает также глю -
козу во внеклеточной жидкости. Не все белки либо
весьма незначительная их часть используются для
энергообеспечения мышечной деятельности
41
8. Методы оценки использования субстратов
Для изучения энергетического метаболизма как у человека, так и у жи-
вотных используется большое количество методов. Коэффициент дыха-
тельного обмена (КДО) в условиях устойчивого состояния позволяет
дать приемлемую оценку пропорционального соотношения участия уг-
леводов и липидов в окислении. Так, КД0, равный 1,0, указывает на то,
что в окислении участвуют только углеводы, тогда как значение этого ко-
эффициента, равное 0,7, свидетельствует о том, что единственным суб-
стратом окисления являются жиры. Величина КДО в состоянии покоя в
постабсорбтивном состоянии обычно равна примерно 0,8. Однако КДО,
который определяется путем газоанализа выдыхаемого воздуха, пред-
ставляет собой лишь непрямой показатель дыхательного коэффициента
(ДК), который является точным соотношением продукции диоксида уг-
лерода и утилизации кислорода на клеточном уровне, а поэтому точнее
отражает использование субстратной смеси. Между тем в ряде работ
подчеркивается, что КДО достаточно репрезентативен по отношению к
КДО в условиях устойчивой легочной вентиляции и кислотно-щелочно-
го баланса.
Гипервентиляция во время выполнения физических упражнений
может вызвать избыточный выход углекислого газа и тем самым повы-
сить КДО. Накопление молочной кислоты в крови будет способствовать
образованию дополнительного количества диоксида углерода из би-
карбонатов и дальнейшей стимуляции легочной вентиляции, в результа-
те чего КДО превысит 1,0. В таком случае обычно считают, что КДО ра-
вен 1,0 и что углеводы служат основным энергетическим «топливом».
Такое предположение вполне оправдано для случаев, когда накопление
лактата и гипервентиляция обычно происходят при выполнении доста-
точно высоко интенсивных динамических физических упражнений (т. е.
более 80 % VO2 max), при которых никогда нельзя достичь устойчивого
состояния.
Другие методы основаны на анализе образцов ткани и крови перед,
во время и после выполнения физических упражнений. Пробы крови
могут быть использованы для определения в ней концентрации таких
энергетических субстратов, как глюкоза, триацилглицерол, свободные
жирные кислоты, глицерол, кетоновые тела и аминокислоты. Однако оп-
ределение в крови концентрации энергетических субстратов не всегда
соответствует скорости их обмена. Так, например, полупериод сущес-
твования молекулы глюкозы составляет всего несколько минут. Тем не
менее определение скорости кровотока и одновременно с этим взятие
42
проб артериальной и венозной крови позволяет оценить поступление в
ткани либо выход из них отдельных субстратов. Введение орально или
путем инъекций таких радиоактивных или естественных изотопных ме-
ток, как глюкоза, жирные кислоты или аминокислоты, дает возможность
при анализе выдыхаемого воздуха, крови или мочи установить скорость
их поступления в ткани, скорость окисления и клиренс.
Мышечные биоптаты могут быть использованы для определения в
мышцах концентрации энергетических субстратов (гликогена, триацил-
глицеролов), а также метаболических интермедиатов. Они позволяют
получить информацию о процентном соотношении в мышце различных
типов волокон, плотности капилляров и митохондрий, информацию об
активности различных ферментов. Образцы мышечной ткани использу-
ются и в исследованиях in vitro для изучения максимальных метаболи-
ческих возможностей при окислении различных субстратов.
Последние достижения микродиализной методики применительно
к изучению мышечной и адипозной тканей позволили изучить субстрат-
ный и метаболический обмен между межклеточной жидкостью и окру-
жающими тканями. Так, например, переход глицерола в межклеточную
среду в адипозной ткани служит надежным показателем скорости липо-
лиза. Однако возможности применения этой методики ограничиваются
тем, что для взятия необходимого количества отдиализированной жид-
кости для анализа требуется, по меньшей мере, несколько минут.
Ряд методов позволяет исследовать использование субстратов в ор-
ганизме без промедления. Так, например, методика магнитно-резонан-
сной спектроскопии может быть использована для определения измене-
ния концентраций высокоэнергетических фосфатов и гликогена (а также
pH) в мышцах, а позитрон-эмиссионная томография позволяет исследо-
вать метаболизм глюкозы в головном мозге.
9. Факторы, влияющие
на утилизацию энергетических источников
при физических нагрузках
Известно много факторов, способных повлиять на выбор энергетичес-
кого «топлива» для обеспечения мышечной деятельности, в связи с чем
возможно значительное количество вариантов взаимодействия между
некоторыми из них. К таким факторам относятся доступность субстра-
та, пищевой статус, режим питания, вид, интенсивность и продолжи-
тельность физических нагрузок, композиционный состав мышц, физи-
43
Таблица 1.5. Факторы, влияющие на утилизацию субстратов
при физических нагрузках
/. Доступность субстрата
Запасы гликогена и липидов в мышцах
Выход глюкозы из печени в кровь
Липолиз в адипозной ткани, выход СЖК в кровь
Скорость кровотока в мышцах
Содержание аминокислот в мышцах
2. Доступность кислорода для транспорта электронов и окислительного
фосфорилирования
3. Активность ферментов, лимитирующих скорость метаболизма
Концентрация
Баланс между ингибиторами и активаторами
Состояние механизма обратной связи
Влияние изменений pH в мышцах
4. Уровень гормонов в плазме
На утилизацию субстратов могут также повлиять
1. Предшествующая мышечная деятельность и диета
2. Композиционный состав мышц
3. Вид, интенсивность и продолжительность мышечной работы
4. Уровень тренированности
5. Условия окружающей среды (температура, влажность, атмосферное
давление)
6. Лекарственные средства (например, кофеин)
ческое состояние либо влияние тренировки, лекарственных средств,
гормонов, предшествующей мышечной деятельности, а также таких
факторов окружающей среды, как температура и высота над уровнем
моря. В табл. 1.5 представлены основные факторы, влияющие на ис-
пользование субстратов (более подробно см. в следующем параграфе).
Интенсивность и продолжительность физической нагрузки.
Наиболее важным фактором, влияющим на выбор энергетического
«топлива» для мышечной работы, является ее интенсивность. В табл. 1.6
представлены различные соотношения энергетических запросов от ана-
эробных и аэробных источников при беге на различные дистанции с
предельной скоростью. Так, например, при спринтерском беге на 100 м
с результатом около 10 с примерно 90 % энерготрат обеспечивается за
счет анаэробных источников.
44
Для марафонцев, преодо-
левающих дистанцию 42 км
195 м быстрее 127 мин (ми-
ровой рекорд к тому време-
ни), 99 % энерготрат прихо-
дилось от аэробных источни-
ков. Таким образом, доставка
кислорода к мышцам, завися-
щая от интенсивности физи-
ческой нагрузки, и связанная
с этим продолжительность
выполнения физического уп-
ражнения являются главны-
ми факторами, определяю-
щими субстратную утили-
зацию.
Тип мышечных воло-
кон. Многочисленными гис-
Та 6 л и ца 1.6. Приблизительный вклад аэроб-
ных и анаэробных энергетических источников в
общую энергопродукцию при беге на различные
дистанции
Дистан- ция Результат, мин:с* Аэробная энергопро- дукция, % Анаэробная энергопро- дукция, %
100 м 9,84 10 90
400 м 43,29 30 70
800 м 1:41,73 60 40
1500 м 3:27,37 80 20
5000 м 12:44,39 95 5
10000 м 26:38,08 97 3
42,2 км 126:50,00 99 1
* Результаты соответствуют мировым рекордам
для мужчин на I апреля 1997 г.
тохимическими исследованиями показано, что у выдающихся спортсме-
нов, тренировки которых направлены на развитие выносливости, в
мышцах преобладает количество медленносокращающихся волокон ти-
па I, тогда как у выдающихся спринтеров большее процентное содержа-
ние волокон II типа.
Как отмечалось выше, метаболические характеристики волокон иг-
рают ведущую роль в способности каждого индивидуума к проявлению
скоростных, силовых возможностей и выносливости. Таким образом,
процентное соотношение разного типа волокон в мышце является одним
из основных детерминантов, определяющих возможность выполнения
предельных по интенсивности и продолжительности физических уп-
ражнений, а следовательно, является важнейшим фактором, определяю-
щим использование энергетических субстратов.
Питание при мышечной деятельности. Пропорциональное соот-
ношение жиров и углеводов в режиме питания, а также потребление уг-
леводов в процессе мышечной деятельности могут оказать значитель-
ное влияние на доступность субстратов и их утилизацию. Доставка суб-
страта является главным фактором, влияющим на выбор энергетичес-
кого «топлива» для обеспечения мышечной деятельности. Высокоугле-
водная диета будет способствовать поддержанию или повышению со-
держания в мышцах и печени гликогена, а также увеличению пропор-
ционального смещения энергообеспечения физических нагрузок за счет
углеводов.
45
Голодание и богатая жирами пища (с низким содержанием углево-
дов) вызывает повышение в плазме концентрации СЖК, что является
детерминирующим фактором для скорости их утилизации и будет спо-
собствовать увеличению относительного вклада липидов в окисли-
тельный метаболизм.
Большинство коферментов, принимающих участие в энергетическом
метаболизме, являются дериватами витаминов, которые не могут синте-
зироваться в организме. Дефицит таких витаминов снижает физические
возможности организма, а также влияет на утилизацию субстратов. Так,
витамин В12 и фолиевая кислота необходимы для нормальной продукции
эритроцитов в костном мозге; их дефицит может ухудшить кислород-
транспортные свойства крови, в результате чего повышается спрос на
анаэробные источники энергообеспечения мышечной деятельности.
Значимость минеральных веществ также не вызывает сомнений.
Так, например, железо — необходимый компонент гемоглобина, кисло-
род-транспортного пигмента эритроцитов и его дефицит может снизить
способность последних к транспорту кислорода.
Физическая тренировка. Адаптация организма к физической тре-
нировке также влияет на субстратный метаболизм. Так, тренировка, нап-
равленная на развитие выносливости, способствует увеличению в мыш-
цах плотности капилляров и митохондрий; увеличению обнаруживае-
мых на гистохимических образцах площади, соответствующей волок-
нам типа II; увеличению содержания в мышцах триацилглицерола, а
также повышению способности использовать липиды в качестве энерге-
тического источника для обеспечения возможности выполнения физи-
ческих упражнений субмаксимальной интенсивности.
У тренированных лиц также проявляется тенденция к повышению
возможности использования внутримышечных липидов в качестве ис-
точника энергии при мышечной деятельности. Эти влияния, а также
другие физиологические эффекты тренировки (снижение доставки кис-
лорода к работающим мышцам и изменение гормонального ответа на
физические нагрузки) снижают скорость утилизации мышечного глико-
гена и глюкозы крови, а также скорость накопления лактата при физи-
ческих нагузках субмаксимальной интенсивности. Такие адаптаци-
онные изменения способствуют значительному повышению выносли-
вости при тренировке, направленной на развитие этого двигательного
качества.
Предшествующие физические нагрузки. Указанные нагрузки вы-
сокой интенсивности и большой продолжительности приводят к исто-
щению запасов мышечного гликогена, что сопряжено с недостаточным
периодом (менее 24 часов), необходимым для восстановления его исход-
46
ного уровня, и что может привести к снижению утилизации углеводов,
повышению использования липидов при выполнении мышечной работы
на следующий день.
Фармакологические средства. Некоторые фармакологические
препараты существенно влияют на поступление и утилизацию энергети-
ческих субстратов при мышечной деятельности. Классическим приме-
ром является кофеин, который повышает мобилизацию СЖК из ади-
позной ткани, что приводит к повышению их концентрации в плазме.
После первых 30 мин выполнения физических упражнений на уровне,
соответствующем приблизительно 70 % VO2max, когда концентрация
СЖК в плазме начинает постепенно возрастать (первоначально она мо-
жет снижаться), прием кофеина в обычных дозах заметно усиливает ли-
полиз и повышает содержание в плазме СЖК, способствуя тем самым
сохранению запасов мышечного гликогена на ранних стадиях двигатель-
ной активности, содействуя большей ее продолжительности. Кофеин об-
ладает также нейротропным действием, которое может быть даже более
важным, чем его эргогенный эффект. Другие способы, с помощью кото-
рых можно повысить концентрацию в плазме СЖК (например, внутри-
венное введение гепарина), также повышают вклад окисления жиров в
энергообеспечение и тем самым способствуют сохранению запасов гли-
когена. Введение никотиновой кислоты, которая угнетает липолиз, ока-
зывает противоположный эффект. Ряд фармакологических веществ
влияет на утилизацию энергетических субстратов путем изменения
гормонального ответа на физические нагрузки либо путем воздействия
в качестве агонистов или антагонистов по отношению к действию гор-
монов.
Гормоны. Многие гормоны оказывают влияние на энергетический
метаболизм в организме (подробный обзор представлен в работе Галбо
(Galbo, 1983). Во время физических нагрузок взаимодействие между ин-
сулином, глюкагоном и катехоламинами (адреналином и норадренали-
ном) в наибольшей степени отражается на доставке и утилизации энер-
гетических субстратов. Кортизол и соматотропный гормон (гормон рос-
та) также проявляют значительный эффект. Обобщенные данные отно-
сительно гормональной и метаболической регуляции ресинтеза АТФ че-
рез анаэробные и аэробные пути энергообеспечения при мышечной де-
ятельности представлены в табл. 1.6 и 1.7. Более подробные сведения
приведены в последующих главах.
Факторы окружающей среды. Высокая температура окружающей
среды и дегидратация детерминируют физическую работоспособность и
влияют на утилизацию субстратов при мышечной деятельности. Так,
имеются данные о повышении скорости утилизации мышечного глико-
47
Т а б л и ца 1.7. Регуляция ресинтеза
Через анаэробные пути энергообеспечения
Активация распада КФ
Снижение отношения АТФ/АДФ в связи с распадом АТФ при мышечных
сокращениях
Угнетение распада КФ
Повышение отношения АТФ/АДФ в связи с увеличением аэробной продукции
АТФ или в результате снижения скорости утилизации АТФ в связи с падением
мощности мышечных сокращений
Активация гликолиза
1. Снижение отношения АТФ/(АДФ+АМФ) вследствие распада АТФ при
мышечных сокращениях
2. Снижение концентрации КФ из-за использования его для восполнения запасов
АТФ при мышечной активности
3. Увеличение концентрации кальция в связи с его выходом из саркоплазматичес-
кого ретикулума при мышечных сокращениях
4. Повышенный выход в кровь адреналина (из надпочечников) и норадреналина (в
основном из окончаний симпатических нервов), в результате чего происходит
активация фосфорилазы
5. Увеличение концентрации фруктозо-6-фосфатазы
Угнетение гликолиза
1. Повышение отношения АТФ/(АДФ+АМФ) в связи с усилением аэробной
продукции АТФ или снижением скорости утилизации АТФ в результате
падения мощности мышечных сокращений
2. Повышение концентрации цитрата в основном из-за возрастания катаболизма
СЖК и ацетил-КоА в ЦТК
3. Увеличение концентрации КФ в связи с возобновлением аэробной
продукции АТФ или из-за снижения мощности мышечных
сокращений
4. Снижение концентрации ионов кальция в саркоплазме во время расслабления
мышцы
5. Снижение pH в результате накопления лактата
6. Увеличение концентрации СЖК, угнетающее поступление глюкозы в мышцы
Через аэробные пути энергообеспечения
Активация окисления СЖК
Увеличение в плазме концентрации СЖК в связи с их повышенной мобилизацией
из адипозной ткани, что вызвано повышением в крови уровней адреналина,
норадреналина, глюкагона, гормона роста и кортизола, а также снижением в крови
уровня инсулина
48
Продолжение табл. 1.7.
Угнетение окисления СЖК
1. Повышенное накопление в крови лактата при напряженных физических
нагрузках, угнетающих липолиз
2. Повышение концентрации инсулина после приема углеводов
Активация окислительного фосфорилирования
1. Повышение в митохондриях концентрации АДФ, обусловленное распадом АТФ
при мышечных сокращениях
2. Повышение доставки кислорода к митохондриям
Угнетение окислительного фосфорилирования
1. Снижение концентрации АДФ в митохондриях из-за снижения возможности
электрон-транспортной цепи обеспечивать ресинтез АТФ
2. Снижение доставки кислорода к митохондриям
3. Снижение доставки НАДН и ФАДН2 из-за ухудшения функционирования ЦТК
гена во время мышечной работы, выполняемой на уровне 70-85 %
VO2max при температуре 41 °C, что происходило на фоне более высокой
концентрации лактата в крови. Эти эффекты могли быть обусловлены
смещением кровотока к коже и, как следствие, — снижением перфузии
работавших мышц. Снижение доставки кислорода и субстратов к мыш-
цам вызывает увеличение выхода углеводов для поддержания необходи-
мой мышечной активности за счет как анаэробных, так и аэробных про-
цессов энергообеспечения. При мышечной работе, выполняемой на
больших высотах, где парциальное давление кислорода ниже, чем над
уровнем моря, доставка кислорода к мышцам также будет занижена, в
результате чего увеличится вклад углеводов и анаэробных энергети-
ческих источников в обеспечение высокоинтенсивных физических
нагрузок. 6
10. Доставка кровью
энергетических источников и кислорода
к мышцам при физических нагрузках
Доставка экстрамускулярных энергетических источников и кислорода к
мышцам существенно влияет на метаболические реакции организма при
физических нагрузках. Поэтому физиологическому состоянию сердеч-
но-сосудистой и дыхательной систем придается большое значение при
4.з.з 49
рассмотрении мышечного метаболизма как в условиях выполнения мы-
шечной работы различной интенсивности и продолжительности, так и в
различных условиях окружающей среды. Цель дальнейшего обсужде-
ния — дать краткий обзор реакций сердечно-сосудистой и дыхательной
систем на физические нагрузки, а также дать оценку факторам, влияю-
щим на мышечный кровоток и кислородное потребление.
Распределение кровотока. В состоянии покоя скорость поступле-
ния крови к скелетным мышцам человека составляет около
20-30 мл-мин "'-кг~1 мышцы, а при открытых капиллярах — около
100 мл-мм2. Тонус симпатической нервной системы поддерживает сок-
ращение многих артериол и большинство капилляров не перфузируется.
Во время физической нагрузки, несмотря на повышение тонуса
симпатической нервной системы, кровеносные сосуды в работающих
мышцах расширяются и просвет функционирующих капилляров увели-
чивается в 5 раз по сравнению с покоем. Индуцированная физической
нагрузкой дилатация кровеносных сосудов работающих мышц вызыва-
ется локальными факторами, к которым относятся снижение рО2, повы-
шение рСО2, уменьшение pH, повышение температуры и увеличение
концентраций аденозина, оксида азота, Фн и натрия во внеклеточной
жидкости. Такая локальная вазодилатация в работающих мышцах (в не-
активных мышцах преобладает вазоконстрикция) вместе с увеличением
сердечного выброса приводит к увеличению притока крови к ним, кото-
рый может возрасти в 100 раз по сравнению с покоем. Однако в такой
степени кровоток возрастает не только в работающих мышцах: при этом
значительно увеличивается и сердечный выброс (рис. 1.12). Результаты
последних исследований показали, что увеличение мышечного кровото-
ка при физической нагрузке происходит почти мгновенно и его уровень
стабилизируется очень быстро. В связи с этим следует полагать, что ог-
раниченное потребление кислорода в начальной фазе мышечной дея-
тельности (т.е. до наступления устойчивого состояния) не связано с не-
достаточной доставкой кислорода к работающим мышцам.
Сердечный выброс и венозный возврат. Сердечный выброс в сос-
тоянии покоя у взрослого человека составляет примерно 5-6 л-мин Во
время физической нагрузки сердечный выброс должен возрастать для
обеспечения кровотока в работающих мышцах. Однако увеличение выб-
роса количества крови сердцем может быть достигнуто лишь в том слу-
чае, если ее венозный возврат от тканей к правому желудочку также бу-
дет возрастать. Более того, артериальное давление также должно увели-
чиваться: значительная вазодилатация кровеносных сосудов работаю-
щих мышц снижает периферическое сопротивление, в результате арте-
риальное давление будет резко падать, если это не будет компенсиро-
50
ваться другими эффектами.
Поддержание периферичес-
кого сопротивления и увели-
чение венозного возврата при
мышечной деятельности обу-
словлено следующими меха-
низмами:
1. Генерализованная ва-
зоконстрикция вследствие
повышения активности сим-
патических нервов ( симпати-
ческие нервы иннервируют
артериолы и вызывают сокра-
щение гладкой мускулатуры в
Предельная
физическая
Рис. 1.12. Регионарное распределение кровото-
ка в покое и во время выполнения двух физичес-
ких упражнений разной интенсивности
их стенках, следовательно,
при повышении активности
симпатических нервов диа-
метр артериол уменьшается).
2. Генерализованная кон-
стрикция вен. Последние являются емкостными сосудами системы кро-
вообращения. В состоянии, покоя около 65 % всей крови находится в ве-
нах. Некоторое количество этого объема может переходить в артериаль-
ное русло при констрикции (уменьшении диаметра и уплотнения) сте-
нок крупных вен.
3. Сокращение мышц конечностей (в частности, ног) приводит к
сдавливанию вен, расположенных параллельно мышечной ткани. Это
вызывает нагнетающий эффект, благодаря которому ускоряется возвра-
щение крови к грудной клетке. Венозные клапаны препятствуют обрат-
ному возврату крови в течение периода расслабления мышц конечности.
Этот эффект иногда называют «мышечным насосом».
4. Во время вдоха давление в грудной полости снижается, поскольку
при сокращении диафрагмы происходит ее уплощение, после чего она
опускается вниз. Это вызывает незначительное сдавливание брюшной
полости, повышение брюшного давления, сдавливание вен в органах
брюшной полости, а следовательно, перемещение крови к грудной клет-
ке (и в этом случае венозные клапаны препятствуют обратному возврату
крови). Хотя этот эффект, называемый дыхательным насосом, совершен-
но незначителен в покое, однако существенно возрастает при мышечной
работе, когда частота и глубина дыхания значительно повышаются.
Сердечный выброс во время физических нагрузок возрастает до
25-30 л-мин '1, хотя имеются данные, что у спортсменов высокой квали-
4*
51
фикации этот показатель достигает и более высокого уровня. Увели-
чение сердечного выброса зависит от интенсивности выполнения физи-
ческого упражнения (рис. 1.13) и определяется возрастанием как часто-
ты сердечных сокращений (ЧСС), так и ударного объема (объема крови,
выбрасываемого сердцем при каждом сокращении). У взрослого челове-
ка максимальная ЧСС изменяется в зависимости от возраста согласно
формуле
ЧССтах =210- (0,65 х возраст, годы) х (сокращения, мин *).
Увеличение ЧСС стимулируется повышением активности симпати-
ческих нервов, иннервирующих сино-атриальный узел (внутренний во-
дитель ритма) сердца. Эти нервы оказывают противоположное ингиби-
рующему влияние активности парасимпатических нервов, которое осу-
ществляется через п. vagus (блуждающий нерв). Наличие в циркулирую-
щей крови катехоламинов также стимулирует ЧСС. Ударный объем воз-
растает от 70 мл в покое до примерно 120 мл во время выполнения мы-
шечной работы на уровне 50 % VO2max, который далее не повышается
с увеличением мощности работы. Увеличение ударного объема достига-
ется путем большего заполнения желудочков кровью в конце диастоли-
ческой фазы сердечного цикла (за счет так называемого резидуального
объема сердца) и более мощного сокращения миокарда (систолы), что
позволяет в большей степени (по сравнению с покоем) освободить же-
лудочки от заполняющей их крови (рис. 1.14).
Во время выполнения пролонгированных физических упражнений,
в частности в условиях высокой температуры окружающей среды, отме-
чается тенденция к повышению ЧСС и снижению ударного объема по
Рис. 1.13. Изменения ЧСС и ударного объема с
увеличением интенсивности выполнения физи-
ческого упражнения
мере увеличения продолжи-
тельности физической наг-
рузки. Возможно, это связано
со снижением объема цирку-
лирующей крови и снижени-
ем давления в венах, посколь-
ку значительное количество
крови перераспределяется к
кровеносным сосудам кожи
для повышения теплоотдачи.
Таким образом, потовыделе-
ние и снижение объема плаз-
мы также могут вносить свой
вклад в этот эффект.
52
,Ударный
объем
Рис. 1.14. Объем левого желудочка сердца в состоянии покоя и при выполнении физи-
ческих упражнений различной интенсивности (Полинер и др. (Poliner et al.), 1980)
Конечный
► систолический
объем
Объем крови. В процессе выполнения физических упражнений от-
мечается тенденция к снижению объема крови в организме, что связано
в основном со снижением объема плазмы, которое в условиях очень нап-
ряженной мышечной деятельности может достигать 20 %. Такая потеря
объема плазмы происходит в результате осмотического перемещения во-
ды в сокращающиеся мышцы. Осмомолярность мышечной саркоплазмы
и межклеточной жидкости повышается в силу того, что происходит рас-
пад и метаболизм больших молекул гликогена до во много раз меньших
молекул (пирувата и лактата). К тому же усиление перфузии капилляр-
ного русла в сокращающихся мышцах повышает гидростатическое дав-
ление в капиллярах, которое способствует образованию межклеточной
жидкости из плазмы. В течение некоторого времени этот эффект компен-
сируется перемещением жидкости в кровь из тканей, в которых при мы-
шечной деятельности происходит вазоконстрикция (например, в кишеч-
53
нике, печени и почках). Степень снижения объема плазмы прямо про-
порциональна относительной (по отношению к VO2max) мощности вы-
полняемой работы. При физических упражнениях, выполняемых на
уровне 70 % VO2max, объем плазмы снижается на 10-15 % в течение
первых нескольких минут нагрузки и может частично восстанавливать-
ся при дальнейшем продолжении выполнения упражнения. Однако это
зависит от скорости поступления жидкости в организм и потовыделе-
ния, зависящих от окружающей температуры и влажности. Все опи-
санные выше факторы, обусловливающие снижение объема плазмы,
способствуют повышению концентрации эритроцитов и гемоглобина в
крови и тем самым повышают кислород-транспортные возможности,
приходящиеся на 1 л крови. Однако при этом происходит потеря обще-
го объема крови и повышается ее вязкость.
Кровяное давление. Показатели артериального давления, опреде-
ляемые во время большинства видов мышечной деятельности, проявля-
ют тенденцию к сохранению постоянства либо к некоторому повыше-
нию. Значительное увеличение систолического артериального давления
(со 120 до 180 мм рт. ст.) отмечается при выполнении динамических фи-
зических упражнений, поскольку при этом происходит выброс большо-
го объема крови в аорту за короткий промежуток времени. Однако пока-
затели диастолического артериального давления во время нагрузки про-
являют тенденцию к снижению. Диастолическое артериальное давление
отражает баланс между увеличением сердечного выброса и снижением
периферического сопротивления, обусловленного выраженной вазоди-
латацией кровеносных сосудов в работающих мышцах.
Обычно диастолическое артериальное давление в покое равно при-
мерно 80 мм рт. ст., а при выполнении динамической мышечной работы
— 60-80 мм рт. ст. При статических (изометрических) мышечных нап-
ряжениях значительно повышается как систолическое, так и диастоли-
ческое артериальное давление, хотя при этом сердечный выброс возрас-
тает несущественно. Происходит это потому, что внутримышечное дав-
ление в условиях сильного изометрического сокращения значительно
повышает систолическое артериальное давление. В данном случае при-
ток крови к мышцам незначителен или даже полностью отсутствует, од-
нако при этом заметно проявляется вызванная симпатическими влияни-
ями вазоконстрикция в других васкулярных отделах (за исключением
сердца и головного мозга), которая способствует повышению перифери-
ческого сопротивления, а следовательно, и повышению как систоличес-
кого, так и диастолического артериального давления. Из этого следует,
что для обеспечения напряженных изометрических сокращений преи-
мущественными являются анаэробные энергетические источники.
54
V021 л мин
Рис. 1.15. Соотношение легочной вентиляции и концентрации лактата в крови с пот-
реблением кислорода при возрастающей физической нагрузке до VO2max (резкий подъ-
ем линий отражает прямую зависимость между легочной вентиляцией и потреблением
кислорода при мышечной работе умеренной интенсивности)
Легочная вентиляция. Физические нагрузки значительно увели-
чивают потребление кислорода и образование углекислого газа сокра-
щающимися мышцами. Поэтому рО2 в венозной крови работающих
мышц ниже, а рСО2 — выше по сравнению с их нерабочим состояни-
ем. Газовый состав венозной крови, поступающей к правому желудоч-
ку сердца, также отражает эти изменения. Вместе с увеличением сер-
дечного выброса во время физической нагрузки это способствует уве-
личению поступления кислорода и выделению углекислого газа через
альвеолярные мембраны. При мышечной работе возрастает легочная
вентиляция, что тесно сопряжено со скоростью метаболизма и скорос-
тью образования СО2. Так, во время физической нагрузки умеренной
интенсивности (до 55 % VO2max) легочная вентиляция возрастает ли-
нейно с потреблением кислорода (рис. 1.15) и образованием углекис-
лого газа. При этом из каждых 20-25 л воздуха потребляется 1 л кис-
лорода. В таких условиях повышение легочной вентиляции достигает-
ся путем увеличения глубины дыхания, тогда как при более интесив-
ных физических нагрузках увеличение частоты дыхания приобретает
более важное значение (рис. 1.16). Такая приспособительная регуляция
55
Рис. 1.16. Изменения объема выдыхаемого при
каждом выдохе воздуха (1) и частоты дыхания
(2) в зависимости от интенсивности мышечной
работы
позволяет полностью окси-
генизировать кровь, пос-
кольку показатели альвео-
лярного (и артериального)
рО2 и рСО2 остаются очень
близкими к показателям в
состоянии покоя.
При более напряженных
физических нагрузках обра-
зующиеся в сокращающихся
мышцах ионы водорода вна-
чале буферируются присут-
ствующим в плазме бикар-
бонатом (НСО3), в результате
чего образуется большее ко-
личество СО2, но в то же вре-
мя количество водородных
ионов (и лактата) продолжает
накапливаться, а pH — снижаться. Эти изменения дополнительно сти-
мулируют легочную вентиляцию, которая ежеминутно возрастает неп-
ропорционально с повышением потребления кислорода (см. рис. 1.14).
Во время физической нагрузки с интенсивностью, близкой к VO2max, из
каждых 40 л вентилируемого легкими воздуха поглощается 1 л кислоро-
да. В связи с этим образуется большее количество углекислого газа, а
КДО превышает 1,0.
Кислородтранспортная способность крови. Кислородтранспорт-
ная способность крови определяется прежде всего концентрацией в ней
гемоглобина и рО2. При полном насыщении кислородом (когда
рО2 = 90 мм рт. ст. и более) каждый грамм гемоглобина, локализованно-
го исключительно в эритроцитах, может транспортировать 1,34 мл кис-
лорода. Таким образом, в среднем для взрослого человека, у которого
концентрация гемоглобина в крови составляет 150 г-л-1, кислородтран-
спортная способность артериальной крови на каждый литр составляет
около 200 мл О2. Известно, что концентрация гемоглобина в крови у
женщин примерно на 10 % меньше, чем у мужчин, следовательно, нас-
только же ниже и кислородтранспортная способность. Кислород высво-
бождается из гемоглобина, когда кровь протекает через капиллярное
русло тканей. Прочность связи гемоглобина с кислородом зависит от
рО2. В капиллярах, где рО2 ниже (обычно около 40 мм рт. ст.), чем в ар-
териальной крови, поскольку кислород потребляется тканями, то неко-
торое его количество (около 25 %) диссоциируется из гемоглобина и
56
таким образом становится
доступным для усвоения тка-
нями (рис. 1.17). Следует от-
метить, что рО2 в капиллярах
тканей составляет примерно
40 мм рт. ст., тогда как в ка-
пиллярах легких оно близко к
100 мм рт. ст. При повыше-
нии температуры рО2 и кон-
центрации водородных ионов
снижается способность ге-
моглобина связываться с кис-
лородом, в связи с чем кривая
диссоциации гемоглобина
Рис. 1.17. Взаимосвязь между процентным на-
сыщением гемоглобина кислородом и парциаль-
ным давлением кислорода:
• — венозная кровь, выходящая из неработающих мышц;
о — венозная кровь, выходящая из работающих мышц;
□ — артериальная кровь
смещается вправо. Это спо-
собствует более легкому ос-
вобождению гемоглобина от
кислорода в капиллярном
русле тканей. Другие факто-
ры также влияют на сродство
гемоглобина к кислороду.
Повышение рО2, температуры и концентрации водородных ионов
снижают сродство гемоглобина к кислороду, в связи с чем кислород лег-
че отдается тканям (рис. 1.17). Такие изменения происходят в метабо-
лизме мышц при физических нагрузках, и это означает, что возрастание
потребности сокращающихся мышц в кислороде автоматически удов-
летворяется увеличением освобождения кислорода из крови и создает
более благоприятные условия для поступления кислорода в митохон-
дрии, где рО2 очень низкое (возможно, менее 1 мм рт. ст.). Поддержание
высокого градиента рО2 способствует более быстрой диффузии кисло-
рода для его утилизации тканями.
11. Потребление кислорода мышцами
при физических нагрузках
Потребление кислорода мышцами в состоянии покоя или при мышечной
деятельности зависит от скорости кровотока и количества кислорода,
приходящегося на 1 л крови, которое может быть усвоено тканями. Эта
зависимость может быть выражена уравнением Фика:
57
V02 = сердечный выброс х (СаО2 - CvO2),
где VO2 — скорость потребления кислорода, мл-мин”1; сердечный выб-
рос — в мл-мин-1; СаО2. — содержание кислорода в артериальной кро-
ви, мл О2-л~' крови); CvO2, — содержание кислорода в венозной крови
(в тех же единицах, что и для СаО2). Диффузия кислорода из крови к
мышцам и скорость этого процесса зависят от величины градиента рО2
между кровью капилляров и митохондрий мышц (где кислород исполь-
зуется в окислительном фосфорилировании), а также расстояния, через
которое кислород должен диффундировать. Значимость последнего при
физических нагрузках фактически незначительна, поскольку предвари-
тельно открываются окружающие мышечные волокна неперфузируемые
капилляры. Низкое значение рО2 в мышечной саркоплазме поддержива-
ется другим кислородсвязывающим пигментом, называемым миоглоби-
ном. Этот пигмент обладает гораздо большим сродством к кислороду по
сравнению с гемоглобином и отдает кислород только при очень низких
значениях рО2 (рис. 1.18), которые несколько выше тех, которые необхо-
димы для метаболизма митохондрий. Митохондрии неравномерно рас-
пределены в мышечных волокнах и больше концентрируются на
периферии поближе к капиллярам. При максимальных физических
нагрузках мышцы не проявляют способность к утилизации кислоро-
да, который доставляется артериальной кровью. Об этом свидетельству-
ет тот факт, что венозная кровь, оттекающая от интенсивно сокращав-
шейся мышцы, еще достаточно насыщена кислородом. Процентное на-
сыщение гемоглобина кислородом в венозной крови, оттекающей от
мышцы, снижается от 50-60 % в состоянии покоя до 10 % при напря-
женных физических нагруз-
ках (рис. 1.19). Соответствен-
но венозное рО2 составляет
25-40 и 10-15 мм рт. ст., так
что работающие мышцы зна-
чительно повышают экстрак-
цию кислорода (но не всего),
который доставляется к ним.
Факторы, лимитирую-
щие потребление кислорода
во время мышечной рабо-
ты. Многие факторы, как фи-
зиологические, так и патоло-
Рис. 1.18. Кривые диссоциации миоглобина и
гемоглобина
гические, потенциально спо-
собны ограничить макси-
58
мальное потребление кисло-
рода при физических нагруз-
ках. Все эти факторы, приве-
денные в табл. 1.7, можно
разделить на две группы ме-
ханизмов — центральную
(респираторные и сердечно-
сосудистые) и периферичес-
кую (локальное кровообра-
щение и мышечные).
Как известно, спортсме-
ны, тренирующиеся в разви-
тии выносливости, имеют
высокие показатели VO2max
и все характерные функцио-
нальные и метаболические
признаки, определяющие
этот показатель. Для дости-
Кислородная
емкость
артериальной
крови
Содержание О2 в
артериальной
крови
Артерио-венозное
различие
по кислороду
Содержание О2
в смешанной
венозной крови
Содержание О2
в крови
бедренной вены
Рис. 1.19. Изменение содержания кислорода
в артериальной, смешанной венозной крови и
в крови бедренной вены во время выполнения
ногой физического упражнения возрастающей
мощности
жения высокого потребления
кислорода очень важным является эффективность функционирования
систем, обеспечивающих перенос кислорода из атмосферы к митохон-
дриям работающих мышц.
При отсутствии легочных заболеваний переход кислорода из атмос-
феры в кровь через капилляры легких обычно не является для мужчин
лимитирующим фактором, и, как показывают результаты легочного тес-
тирования элитных марафонцев, эта функция не отличается от таковой у
основной массы людей. У спортивных животных, имеющих относитель-
но большую мышечную массу, например у чистокровных лошадей, ды-
хательная функция может оказаться лимитирующим фактором, и, как
следствие, у таких животных отмечается выраженная артериальная ги-
поксемия при очень интенсивных нагрузках. Доказано, что данная осо-
бенность может встречаться и среди спортсменов высокой квалифика-
ции во время тренировок, направленных на развитие выносливости,
имеющих очень высокий показатель VO2max. У спортсменов с высоким
VO2max отмечаются и высокие показатели максимального сердечного
выброса. Потребление кислорода тканью может быть рассчитано пред-
ставленным выше уравнением Фика. Аналогично может быть определе-
но и потребление кислорода целостным организмом:
VO2организма = сердечный выброс х (СаО2 - CvO2).
59
Таблица 1.8. Факторы, лимитирующие максимальное потребление кислорода при
мышечной деятельности
Центральное ограничение
1. Респираторные факторы
Внешнее дыхание
Альвеолярная вентиляция
Диффузия кислорода
Сродство гемоглобина к кислороду
Периферическое ограничение
3. Периферическое кровообращение
Регионарное распределение крови
Кровоснабжение мышц
Плотность мышечных капилляров
Диффузия кислорода
Сродство гемоглобина к кислороду
2. Центральная циркуляция
Сердечный выброс
Артериальное давление
Концентрация гемоглобина в крови
Объем крови
4. Мышечный метаболизм
Плотность митохондрий
Активность окислительных ферментов
Доступность энергетических запасов
и субстратов
Мышечная масса
Композиционный состав мышц
Здесь Cv — концентрация кислорода в смешанной венозной крови,
так как вены получают кровь из различных тканей, содержащих не-
одинаковое количество кислорода. Разницу между содержанием кис-
лорода в артериальной и смешанной венозной крови иногда называ-
ют «показателем кислородной экстракции организма». Максималь-
ная ЧСС у выносливых спортсменов не отличается от аналогичного
показателя у лиц такого же пола и возраста, однако у таких спорт-
сменов ударный объем значительно больше, а следовательно, и боль-
ше размеры сердца. Хорошо тренированные спортсмены, специали-
зирующиеся в видах спорта, требующих проявления выносливости,
обладают и более выраженной способностью к экстракции кислоро-
да из крови. Эти и другие адаптационные изменения, происходящие
в организме под влиянием тренировки, более подробно рассмотрены
в главе 8.
Кислородтранспортная способность крови (мл О21 л-1 крови) у хо-
рошо тренированных спортсменов не отличается от физиологической
нормы. Иногда этот показатель может оказаться даже меньшим из-за
низкой концентрации гемоглобина, хотя общий объем крови больше по
сравнению с нетренированными лицами контрольной группы. При ре-
инфузировании эритроцитов хорошо тренированным спортсменам такая
процедура может временно повысить VO2max. Следовательно, для здо-
ровых и тренированных лиц доставка кислорода к работающим мыш-
цам является фактором, лимитирующим VO2max.
60
Увеличение массы работающих мышц путем дополнительного вы-
полнения работы руками к работе на велоэргометре с мощностью, соот-
ветствующей VO2max или близкой к ней, не вызывало дальнейшего уве-
личения этого показателя, что является еще одним доказательством в
пользу существования центрального сердечно-сосудистого фактора ли-
митирования VO2max
Основываясь на результатах исследований in vitro окислительной
способности мышц, Салтин (Saltin, 1985) пришел к заключению, что
окислительная способность мышц целостного организма при их полной
активации значительно превышает возможности системы кровообраще-
ния обеспечивать доставку кислорода к ним.
Во многих работах сообщалось о снижении как VO2max, так и мак-
симальной ЧСС (а следовательно, и максимального сердечного выброса)
с возрастом. Известно также, что существует тесная взаимосвязь между
изменениями ударного объема и изменениями VO2max.
Выводы
1. Скелетные мышцы представляют собой длинные многоядерные во-
локна, имеющие поперечно-полосатую исчерченность. Скелетные мыш-
цы прикреплены к костям и могут произвольно сокращаться, обеспе-
чивая движения тела и поддержание позы.
2. Миофибриллы являются сократительными элементами, состоя-
щими из цепочек саркомеров, содержащих тонкие (актиновые) и тол-
стые (миозиновые) филаменты, расположенные в правильном порядке.
Головки миозиновых молекул образуют поперечные мостики, которые
обратимо образуют связи с актиновыми филаментами и тем самым обус-
ловливают возможность филаментов скользить относительно друг друга
в направлении к средине саркомеров.
3. В регуляции сокращений скелетных мышц участвует кальций, ос-
вобождающийся из саркоплазматического ретикулума вслед за распрос-
траняющимся по сарколемме потенциалом действия. Кальций повыша-
ет активность поперечных мостиков и скольжение филаментов. Актив-
ность поперечных мостиков заканчивается, когда ионы кальция перека-
чиваются обратно в саркоплазматический ретикулум.
4. Существует три типа мышечных волокон, классифицируемых в
соответствии со скоростью их сокращения и метаболическими характе-
ристиками: тип I (медденносокращающиеся окислительные), тип Па
(быстросокращающиеся окислительные) и тип Пб (быстросокращаю-
61
щиеся гликолитические). Большинство мышц содержит разные типы
волокон.
5. Непосредственным энергетическим источником для мышечных
сокращений является АТФ, который постоянно регенерируется при мы-
шечной работе за счет фосфокреатина, анаэробного метаболизма глико-
гена или глюкозы, а также аэробного метаболизма ацетил-КоА, являю-
щегося дериватом преимущественно распавшихся углеводов или жиров.
6. Цикл трикарбоновых кислот и окислительное фосфорилирование
протекают в митохондриях. Главная роль кислорода состоит в том, что
он выполняет функцию конечного акцептора электронов в электрон-
транспортной цепи и взаимодействует с водородом с образованием
воды.
7. Существует ряд факторов, влияющих на тип субстрата, использу-
емого в качестве энергетического «топлива» для обеспечения мышечной
работы. К ним относятся доступность субстрата, диета, интенсивность
и продолжительность физической нагрузки, уровень тренированности,
гормональные влияния, предшествующая физическая нагрузка, условия
окружающей среды.
8. Во время мышечной работы сердечный выброс и легочная венти-
ляция увеличиваются в зависимости от интенсивности физической наг-
рузки. При этом увеличивающийся ток крови направляется непосредст-
венно к работающим мышцам. При максимальных физических нагруз-
ках работающие мышцы извлекают наибольшее количество кислорода,
поступающего к ним.
9. Основным фактором, ограничивающим максимальное потребле-
ние кислорода здоровым человеком, является максимальный сердечный
выброс. Следовательно, ограничение связано с возможностью системы
кровообращения доставлять кислород к работающим мышцам, а не со
способностью к извлечению кислорода из крови.
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
Galbo Н. (1983). Hormonal and metabolic adaptation to exercise. — New York: Georg
Thieme Verlag.
Grimby, Saltin B. (1983). The ageing muscle // Clin. Physiol. — 3. — P. 209-281.
Komi G.V., Karlsson J. (1978). Skeletal muscle fibre types, enzyme activities and physical
performance in young males and females //Acta Physiol. Scand. — 103. — P. 210-218.
Poliner L.R. (1980). Left ventricular performance in normal subjects: a comparison of the
responses to exercise in upright and supine positions // Circulation. — 62. — 528 p.
Saltin B. (1985). Physiological adaptation to physical conditioning // Acta Med. Scand. —
Suppl. 711. —P. 11-24.
(f ПУРИНОВЫЕ НУКЛЕОТИДЫ
Lb И ФОСФОКРЕАТИН
1. Свободная энергия
Согласно второму закону термодинамики, в результате лю-
бой биохимической реакции определенное количество
энергии должно трансформироваться в любую беспорядоч-
ную форму, недоступную для выполнения работы (энтро-
пия),при этом свободная и тепловая энергии (энтальпия) бу-
дут доступны. Автор закона, который обычно подписывался
символом G, мог использовать для работы константы темпе-
ратуры и давления. Обычно энергетическую трансформа-
цию в скелетных мышцах приводят к константе темпера-
туры и давления, поэтому изменения свободной энергии
энтальпии и энтропии могут быть описаны уравнением
AG = АЯ- ТА5,
где AG — изменение свободной энергии; АЯ — изменение
энтальпии; Т— абсолютная температура, К; AS— изменение
энтропии. Следует отметить, что здесь приведены изменения
свободной энергии, энтальпии и энтропии, которые исполь-
зуются предпочтительнее, чем их абсолютные количества.
Каждая реакция характеризуется AG. Кроме того, как
видно из приведенного выше уравнения, можно рассчитать
свободную энергию любой биохимической реакции, при-
нимая стандартные условия температуры (298 К), давления
(101 кПа) и pH (7,0).
Наиболее простым определением AG для биохимичес-
кой реакции является разность содержания свободной
энергии между реагентами и продуктами при стандартных
63
условиях. Так, в реакции А + В <-> С + D изменение AG будет меньше ну-
ля, когда сумма свободных энергий, содержащихся в С и D, меньше, чем
в А и В. В этом случае реакция будет проявлять тенденцию к спонтанно-
му смещению вправо. Если же AG имеет положительный знак, то реак-
ция не может спонтанно протекать в этом направлении, а будет происхо-
дить в случае, когда к системе добавляется энергия; без добавления
энергии реакция будет протекать в противоположном направлении. При
AD = 0 реакция не будет происходить ни в одном из направлений и, сле-
довательно, будет находиться в состоянии равновесия.
2. Аденозинтрифосфат и фосфокреатин
Свободная энергия, освобождающаяся при сгорании углеводов и липидов,
может запасаться в АТФ, поэтому термин «высокоэнергетический фосфат»,
или «фосфаген» обычно используют при описании этого вещества. Адено-
зинтрифосфат является только формой химической энергии, которая может
превращаться в другие формы энергии, используемые живыми клетками.
Так, например, при гидролизе АТФ до АДФ и Фн освобождается свободная
энергия, позволяющая генерировать силу. Реакции, связанные с распадом
фосфатных связей и освобождением Фн, катализируются ферментами, назы-
ваемыми киназами. В случае распада АТФ таким ферментом является АТФ-
аза. Кроме того, клеточные концентрации АТФ, АДФ и Фн таковы, что AG
аденилаткиназной реакции (известной также как «миокиназная реакция»,
приведенная ниже) большое по величине и отрицательное по значению, в
связи с чем освобождается достаточное количество энергии, способной
превратиться в механическую энергию для проявления силы или движения:
АТФ АДФ + Фн и АТФ АМФ + 2Фн.
Установлено, что мышцы могут выполнять до 24 кДж работы на
каждый моль распавшегося АТФ. Поэтому AG при гидролизе должно
быть больше 24 кДж-моль-'. Следует отметить, что компоненты, прини-
мающие участие в аденилаткиназной реакции, способны к взаимопрев-
ращению без каких-либо изменений AG:
АТФ АМФ о АДФ + АДФ.
Аденозинтрифосфат является нуклеотидом, вмещающим в себе пу-
риновое основание аденин, пятиуглеродный сахар рибозу и трифосфат-
ный блок (рис. 2.1).
64
Рис. 2.1. Структурная формула молекулы АТФ
Наибольшей метаболически активной формой АТФ является в ос-
новном его магниевая соль, а наиболее интересной частью ее молекулы
в плане рассмотрения метаболизма при мышечной деятельности — три-
фосфатный блок. Последовательный гидролиз двух терминальных фос-
фатных связей в аденилаткиназной реакции освобождает значительное
количество свободной энергии, которая используется для обеспечения
множества энергозависимых реакций и процессов, включая генерацию
мышечной силы, перенос ионов через мембраны и все реакции, обеспе-
чивающие синтез сложных веществ:
АТФ + Н2О АДФ + Фн + Н+: AG = -31 кДж-моль—
АДФ + Н2О АМФ + Ф„ + Н+: AG = -31 кДж-моль-1.
Следует заметить, что в представленных выше реакциях не приве-
дены частные детали, касающиеся фосфатных связей. Их важность сос-
тоит в передаче свободной энергии при гидролизе. В связи с уни-
кальной ролью в энергопродукции АТФ можно определить как энерге-
тическую валюту клетки. Однако АТФ не может накапливаться в боль-
ших количествах и внутримышечные запасы ее ограничены до
24 ммоль-кг-1 смт (рис. 2.2). Следует согласиться, что изменения в
клетке концентраций АТФ, АДФ и АМФ включаются в регуляцию мно-
гих метаболических процессов (обсуждение энергетического заряда
клетки см. далее) и было бы неразумно сводить все лишь к простому
увеличению запаса АТФ.
При максимальных физических нагрузках наличного количества
АТФ в мышце хватает приблизительно на 2 с для обеспечения их сок-
ратительной активности. Однако мышечные запасы АТФ никогда пол-
5
65
Рис. 2.2. Концентрации АТФ (А), КФ () и лактата (•) в мышцах у мужчин при
выполнении физического упражнения с разной интенсивностью (взятие мышечных би-
оптатов осуществляли последовательно через каждые 3 мин работы возрастающей
мощности)
ностью не исчерпываются, поскольку обычно он эффективно ресинте-
зируется из АДФ и АМФ с такой же скоростью, как и деградирует. При
выполнении физических упражнений субмаксимальной аэробной мощ-
ности устойчивое состояние достигается митохондриальным окисле-
нием углеводов и липидов (см. главу 7). Однако период перехода от сос-
тояния покоя до устойчивого состояния при таких же физических наг-
рузках обеспечивается, главным образом, анаэробным ресинтезом
АТФ. Показано, что в этом случае проявляется четкая взаимосвязь меж-
ду скоростью гидролиза КФ, продукцией лактата и интенсивностью
мышечной работы (см. рис. 2.2). Как указывается в главе 7, еще не яс-
но, проявляются эти ответы в результате запаздывания доставки кисло-
рода или из-за инертности активации митохондриального ресинтеза
АТФ (ЦТК и окислительного фосфорилирования) к началу сократи-
тельной активности.
При выполнении кратковременных физических упражнений макси-
мальной интенсивности величина запроса АТФ значительно превышает
способности митохондрий ресинтезировать АТФ, поэтому анаэробный
метаболизм становится доминирующим в этом процессе (см. главу 6). В
физиологической терминологии вклад анаэробного метаболизма в ре-
синтез АТФ в начальной фазе физических нагрузок субмаксимальной аэ-
робной мощности и при высокоинтенсивной мышечной работе обозна-
чается как кислородный дефицит.
Фосфокреатин находится в цитоплазме мышечных клеток, где его
концентрация составляет около 75 ммоль-кг-1 смт (см. рис. 2.2). При вы-
66
сокоинтенсивных физических нагрузках и в начале мышечной работы
субмаксимальной аэробной мощности происходит быстрый распад КФ,
поскольку потенциал его высокоэнергетической фосфатной группы
используется для ресинтеза АТФ. Это означает, что свободная энергия
гидролиза КФ (—43 кДж-моль—') больше, чем гидролиза АТФ
(-31 кДж-моль-'), в результате чего возрастает вероятность передачи
свободной энергии от КФ к АДФ и образования АТФ:
АДФ + КФ + Н+ —> АТФ + Кр.
Становится очевидным, что функция приведенной выше реакции
заключается в поддержании постоянной концентрации АТФ. Конечно,
скорость этой реакции намного ниже любой скорости реакции, связан-
ной с утилизацией кислорода в клетке, и она не является редкой для мы-
шечного запаса КФ, который может быть полностью исчерпан при мак-
симальной физической нагрузке (см. главу 6). Эта реакция называется
креатинкиназной, так как катализируется ферментом креатинкиназой.
К настоящему времени известно, что креатинкиназа имеет много
изоферментов (разновидности фермента, характеризующиеся незначи-
тельными различиями в структуре и в то же время одинаковой субстрат-
ной специфичностью), которые локализованы в различных частях клет-
ки. По крайней мере, три из них присутствуют в скелетной мышце. Так,
например, ММ-КФК локализована в участках, где образуются попереч-
ные мостики, а Mi-КФК — в митохондриальных мембранах, т. е. вблизи
участков, где образуется АТФ). Открытие существования изоферментов
КФК и их дискретной локализации в клетке позволили предположить,
что КФ может выполнять множество различных функций в скелетной
мышце. Первая и наиболее важная из них, о которой упоминалось выше,
заключается в функционировании его в роли временного буфера, обес-
печивающего поддержание концентрации АТФ и отношения АГФ/АДФ
в клетке.
Вторая функция, которая в настоящее время часто обсуждается,
состоит в том, что КФ может функционировать в качестве простран-
ственного энергетического буфера, т. е. транспортной энергетической
системой между участками, где образуется АТФ (в митохондриях), и
участками ее утилизации (например, в миофибриллах). Такая предпола-
гаемая функция привела к возникновению термина «КФ-челнок». Сто-
ронники существования такого челночного механизма считают, что
роль его как пространственного энергетического буфера для КФ может
проявляться главным образом в волокнах типа I, тогда как в волокнах
типа II ведущую роль играет энергетический буфер. В пользу такого
5*
67
предположения свидетельствует более высокая концентрация КФ в во-
локнах типа II.
Третья предполагаемая функция КФ заключается в его функцио-
нальном сопряжении с несколькими другими клеточными реакциями,
которые обеспечивают энергетический метаболизм во время мышечных
сокращений. Так, например, из приведенной выше реакции видно, что
аденилаткиназная реакция приводит к образованию Н+, а креатинкиназ-
ная — к их удалению, что представляет собой функциональное сопряже-
ние двух реакций, которое предотвращает быстрое закисление мышеч-
ных клеток с установлением сократительной активности. Аналогичным
образом быстрое освобождение Фн при гидролизе КФ во время мышеч-
ных сокращений является неотъемлемой частью процесса активации
гликогеновой фосфорилазы, благодаря чему обеспечивается поддержка
энергопродукции.
Креатинкиназная реакция (как и аденилаткиназная) является равно-
весной, а поэтому и обратимой. Обратный поцесс будет протекать вслед
мышечной активности, когда энергетический заряд клетки увеличива-
ется (энергетический заряд клетки рассмотрен далее), и при этом значи-
тельное количество свободной энергии становится доступным для ре-
фосфорилирования креатина (Кр):
АТФ + Кр -> АДФ + КФ + Н+.
Как правило, ресинтез КФ, протекающий после его полной деграда-
ции, описывается экспоненциальной зависимостью и время полуперио-
да его ресинтеза (время, необходимое для восстановления 50 % его
уровня в состоянии покоя) часто принимают за 30 с (рис. 2.3). Однако в
действительности продолжительность этого периода значительно варь-
ирует в зависимости от вида мышечной деятельности, ее продолжитель-
ности и количества повторяющихся попыток. Факторами, влияющими
на скорость ресинтеза КФ в восстановительном периоде после физи-
ческой нагрузки, являются концентрации в клетке АТФ, АДФ и КФ, ко-
торые (и это неудивительно) обусловливают равновесную природу кре-
атинкиназной реакции (влияние доступности Кр на ресинтез КФ после
приема Кр рассмотрено в главе 6). Кроме того, известно, что Н+ являет-
ся потенциальным ингибитором креатинкиназы. Поэтому низкие значе-
ния pH напряжения кислорода в мышцах и (или) снижение эффектив-
ности их кровоснабжения значительно затрудняют ресинтез КФ после
физической нагрузки. С учетом этого при проведении физиологических
и биохимических исследований часто применяют мышечную ишемию в
68
После выполнения динамического
физического упражнения при интактном
кровообращении
60
40
о
20
га
с
о
е
о
2
2
После изометрического сокращения
в условиях интактного кровообращения
Восстановление кровообращения
через 25 с
После изометрического сокращения
в условиях перекрытого кровообращения
0 1 2 4 8 12 16
Восстановительный период, мин
Физическая
нагрузка
20
Рис. 2.3. Время протекания ресинтеза фосфокреатина в мышцах в восстановительном
периоде после предельной высокоинтенсивной велоэргометрической физической наг-
рузки при интактном кровоснабжении мышц (•), после предельного изометрического
сокращения в условиях нормального мышечного кровоснабжения (о), перекрытого кро-
воснабжения () и после 25 с последующего восстановления кровотока (□) (Харрис и
др. (Harris et al.), 1976)
качестве способа, позволяющего задерживать ресинтез КФ в мышцах
после их сократительной активности (см. рис. 2.3).
К настоящему времени установлено, что ресинтез КФ в восстано-
вительном периоде после полного истощения его запасов при физи-
ческой нагрузке протекает в различных типах волокон с неодинаковой
скоростью. Более подробно этот вопрос рассмотрен в главе 6. Здесь
лишь отметим, что скорость ресинтеза АТФ в первые минуты восста-
новительного периода значительно ниже в волокнах типа II, что, веро-
ятно, связано с более выраженным в них ацидозом. Это оказывает
неблагоприятное влияние на энергопродукцию и физическую рабо-
тоспособность при повторяющемся выполнении физических упраж-
нений. По истечении нескольких первых минут отдыха ресинтез КФ в
69
волокнах типа II ускоряется, в связи с чем после 15 мин восстановле-
ния концентрация КФ становится выше по сравнению с отмечаемой в
покое. Механизм, ответственный за такое сверхвосстановление КФ,
пока не известен.
3. Свободный энергетический заряд
и аденилатный пул
Для расчета энергетического заряда клетки можно использовать показа-
тели концентраций в ней АТФ, АДФ и АМФ. Такой способ, предложен-
ный Аткинсоном (Atkinson, 1977), позволяет определить степень фосфо-
рилирования общего адениннуклеотидного пула клетки (АТФ, АДФ и
АМФ), который описывается уравнением
о „ [АТФ]+ 0,5 [АДФ]
Энергетическим заряд = [Атф] + [АДФ] + [АМФ]
Величина энергетического заряда является хорошим показателем
энергетического статуса клетки, т. е. ее способности выполнять работу.
Так, например, энергетический заряд равен единице тогда, когда весь
адениннуклеотидный пул представлен в форме АТФ, и в этом случае
клетка имеет максимальный свободноэнергетический заряд. И наоборот,
энергетический заряд будет равен нулю, когда АТФ полностью гидроли-
зуется до АМФ. Последний вариант следует рассматривать только как
теоретический пример, поскольку концентрация АТФ в скелетных мыш-
цах человека не снижается более чем на 60 % даже при выполнении уп-
ражнений максимальной мощности в условиях перекрытия мышечного
кровоснабжения.
В нормальных условиях в состоянии покоя энергетический заряд
скелетных мышц находится в пределах 0,95-0,90. Однако установлено,
что при некоторых заболеваниях (например, при сепсисе) его значения
становятся менее 0,9, а уровень ниже 0,82 указывает на необратимые
клеточные повреждения.
Установлено, что скорость ресинтеза АТФ во время физической наг-
рузки регулируется энергетическим зарядом мышечной клетки. Так,
например, снижение энергетического заряда, происходящее с началом
сократительной активности, т. е. моментальное снижение концентрации
АТФ и увеличение концентраций АДФ и АМФ, ускоряет как
анаэробный, так и аэробный ресинтез АТФ и тем самым обеспечивает
70
Аденилатный энергетический
заряд
Рис. 2.4. Скорости гидролиза и продук-
ции АТФ как функция энергетического
заряда в клетке (в точке пересечения ли-
ний х скорость образования АТФ соответ-
ствует скорости его гидролиза и клетка
находится в устойчивом состоянии энер-
гообмена)
повышение скорости энергетичес-
кого обеспечения в соответствии с
возросшей потребностью в энер-
гии. Если энергетический заряд
продолжает снижаться, то распад
АТФ будет угнетаться, т. е. в мыш-
цах будет развиваться утомление и
их работоспособность начнет па-
дать. Взаимосвязь между энергети-
ческим зарядом и скоростями ре-
синтеза и распада АТФ во время
выполнения мышечной работы по-
казана на рис. 2.4.
Скорость образования АТФ во
время мышечных сокращений бу-
дет совпадать со скоростью его
утилизации, когда две линии пере-
секутся. Можно наблюдать, что ес-
ли энергетический заряд возраста-
ет или падает по отношению к его
устойчивому состоянию, то АТФ,
АДФ и АМФ начинают функционировать как аллостерические факторы
или ингибиторы ферментативных реакций, обеспечивающих распад и
утилизацию КФ, углеводов и жиров. Так, например, креатинкиназа как
фермент, ответственный за быстрое рефосфорилирование АТФ в на-
чальной фазе сократительной активности, быстро активируется увели-
чением концентрации АДФ в цитоплазме (снижением энергетического
заряда) и угнетается повышением концентрации АТФ в клетке (увеличе-
нием энергетического заряда). Аналогично гликогеновая фосфорилаза
как фермент, катализирующий образование глюкозо-1-фосфата из глико-
гена, активируется повышением концентраций АМФ и Фп и угнетается
снижением концентрации АТФ в саркоплазме.
Однако следует быть осторожным в выводах: поскольку энергети-
ческий заряд определяется из расчета общего содержания адениннукле-
отидов в мышце, то в этом случае не учитывается та их часть, которая
может быть недоступна для участия в метаболическом контроле. Напри-
мер, метаболически активные (свободные) формы АДФ и АМФ учиты-
ваются значительно меньше, чем их соответствующие общие клеточные
концентрации (свободные + связанные). Поэтому понятно, что измене-
ния свободных форм этих веществ не всегда будут отражать изменение
энергетического заряда в целом.
71
В последнее время для определения свободных и связанных форм
фосфатов начали применять метод магнитно-резонансной спектроско-
пии, в связи с чем предпринимаются попытки решить данную проблему.
Благодаря этой методике принято условно использовать отношение кон-
центраций АТФ к АДФ и КФ к АТФ в качестве показателей энергетичес-
кого статуса клетки.
Следует учитывать различия между энергетическим зарядом и об-
щим аденилатным пулом мышцы (суммой концентраций АТФ, АДФ и
АМФ в клетке). Известно, что общий аденилатный пул может быстро
снижаться, если концентрация АМФ в клетке начинает быстро возрас-
тать в процессе развития мышечного напряжения. Это снижение проис-
ходит в основном при дезаминировании АМФ до инозинмонофосфата
(ИМФ), а также при дефосфорилировании АМФ до аденозина (см. да-
лее). Снижение уровня АМФ может первоначально противодействовать
выработке энергии по причине снижения общего аденилатного пула.
Однако следует заметить, что дезаминирование АМФ до ИМФ происхо-
дит только в условиях низкоэнергетического заряда и способности аде-
нилаткиназных реакций продолжаться, что способствует увеличению
энергетического заряда и продолжению сократительной активности
мышц. Кроме того предполагается, что свободная энергия гидролиза
АТФ будет уменьшаться при накоплении АДФ и Фн, что может впослед-
ствии отрицательно отразиться на мышечной работоспособности. Поэ-
тому существует мнение, что снижение уровня адениннуклеотидов
имеет важное значение для функционирования мышц в условиях ме-
таболического кризиса, например при выполнении физических упраж-
нений максимальной интенсивности или на последних стадиях про-
лонгированной мышечной работы, приводящей к исчерпанию запасов
гликогена.
4. Убыль адениннуклеотидов
в скелетных мышцах
Снижение содержания в скелетных мышцах адениннуклеотидов при мы-
шечной деятельности может происходить двумя путями (рис. 2.5). Пер-
вый из них, катализируемый ферментом АМФ-дезаминазой, заключается
в дезаминировании АМФ до ИМФ и аммиака. Второй путь заключается
в дефосфорилировании АМФ до аденозина при участии в этом процессе
фермента 5-нуклеотидазы. В дальнейшем ИМФ и аденозин могут распа-
даться до инозина и гипоксантина. Мышцы не содержат ферментов, не-
72
I Рис. 2.5. Убыль аденин-
Ксантин нуклеотидов в скелетных
Мочевая кислота
мышцах в результате де-
фосфорилирования и де-
заминирования
обходимых для образования инозина из гипоксантина, в связи с чем он
выводится из мышц и впоследствии подвергается в печени превращению
до мочевой кислоты, которая затем экскретируется почками.
В противоположность гипоксантину, ИМФ может подвергаться из-
менениям до аденилосукцината и, в конечном счете, — до АМФ. Деза-
минирование АМФ до ИМФ и последующее реаминирование ИМФ сос-
тавляют пуриннуклеотидный цикл, который более подробно будет рас-
смотрен ниже.
Как уже отмечалось, существует мнение, что снижение уровня аде-
ниновых нуклеотидов происходит во время выполнения интенсивных
мышечных сокращений при стремлении аденилаткиназных реакций про-
должаться, а поэтому и поддерживать высокий энергетический заряд. Та-
кая убыль адениннуклеотидов сопряжена с увеличением концентраций
ИМФ и аммиака (рис. 2.5). В связи с этим появление аммиака и гипоксан-
тина в плазме часто используют в качестве неинвазивного индикатора
убыли мышечных адениннуклеотидов при физических нагрузках.
Согласно литературным данным, доминирующие пути убыли аде-
ниннуклеотидов при мышечной деятельности у различных животных и
73
в волокнах разного типа неодинаковы. Так, например, дезаминирование
АМФ до ИМФ преобладает в быстросокращающейся мышце крыс,
содержащей преимущественно волокна типа II. В медленно сокращаю-
щейся мышце крысы, содержащей преимущественно волокна типа I,
снижение уровня адениннуклеотидов не происходит до тех пор, пока не
наступит выраженный метаболический кризис (например, мышечная
ишемия). Однако даже в таких экстремальных условиях убыль аденин-
нуклеотидов происходит преимущественно путем не дезаминирования,
а дефосфорилирования до аденозина. В мышцах человека дезаминиро-
вание АМФ до ИМФ также происходит в волокнах типа I, хотя и в мень-
шей степени, чем в волокнах типа II. Эти особенности соответствуют
приблизительно в 4 раза меньшей активности АМФ-дезаминазы в во-
локнах типа I мышц человека по сравнению с волокнами типа II с отно-
сительно более высокой активностью по сравнению с медленносокра-
щающимися волокнами в мышцах крыс. Однако не ясно, почему путь и
скорость адениннуклеотидных потерь в разных типах волокон неодина-
ковы. Одно из объяснений предполагает, что поскольку накопление
ИМФ и аммиака причастно к активации гликогеновой фосфорилазы и
фосфофруктокиназы, то их сравнительно большее содержание в быстро-
сокращающихся мышцах может инициировать и быструю активацию
гликогенолиза и гликолиза в этом типе мышц во время их интенсивной
сократительной активности. Однако активация гликогенолиза и гликоли-
за, проявляющаяся до значительного накопления ИМФ и аммиака, сви-
детельствует против такого предположения.
Альтернативным объяснением различий между снижением содер-
жания адениннуклеотидов в волокнах типов I и II является тот факт, что
образование аденозина медденносокращающимися мышечными волок-
нами ускоряется вазодилатацией, что способствует увеличению достав-
ки крови к этим волокнам. Эта гипотеза подтверждается высокой вазо-
активностью аденозина. Однако следует подчеркнуть, что для ее под-
тверждения требуются экспериментальные доказательства. Из литера-
турных данных становится ясно, что основным путем снижения содер-
жания адениннуклеотидов в скелетных мышцах человека является деза-
минирование АМФ до ИМФ и аммиака. И, наконец, следует отметить,
что присоединение АМФ-дезаминазы к миозину имеет существенное
значение для активации фермента и оно должно происходить перед де-
заминированием АМФ.
Таким образом, в покое фермент неактивен и находится в цитозоле
в свободном состоянии. Однако при развитии мышцей сильного напря-
жения значительная часть фермента связывается с миозином. Видимо,
высокая потребность в энергии для развития мышечной силы, по сущес-
74
тву, более предпочтительна, чем механизм образования поперечных
мостиков в этом связывающем процессе. Так, например, известно, что
развитие мышцей незначительного напряжения не влияет на связывание
АМФ-дезаминазы или образование ИМФ.
5. Цикл пуриновых нуклеотидов
Реакции, составляющие цикл пуриновых нуклеотидов, представлены на
рис. 2.6. Эти три реакции протекают в мышечном цитозоле и катали-
зируются АМФ-дезаминазой, аденилосукцинатсинтетазой и адени-
лосукцинатлиазой. Цикл для удобства можно подразделить на два типа
реакции: дезаминирование, катализируемое АМФ-дезаминазой, и реа-
минирование, катализируемое аденилосукцинатсинтетазой и аденило-
сукцинатлиазой. При обсуждении ранее некоторых деталей отмечалось,
что дезаминирование в цикле пуриновых нуклеотидов является важным
путем регуляции адениннуклеотидного метаболизма в мышцах. Однако
этому циклу присущи и многие другие функции, которые будут рассмот-
рены ниже.
Дезаминирование в цикле пуриновых нуклеотидов. Дезаминиро-
вание АМФ до ИМФ и аммиака предварительно уже обсуждалось в связи
с тем, что этот путь является ос-
новным механизмом снижения
содержания адениннуклеотидов
в скелетных мышцах человека
при физических нагрузках. Дав-
но известно, что сокращающиеся
мышцы продуцируют ИМФ и
аммиак в результате дезаминиро-
вания АМФ, что эти реакции об-
ратимы и что они катализируют-
ся АМФ-дезаминазой (рис. 2.6).
Исходя из изложенных выше
представлений об энергетичес-
ком заряде клетки (Аткинсон
(Atkinson)), АМФ, АДФ, АТФ и
Фп являются важными регулято-
рами АМФ-дезаминазной актив-
ности. Активность этого фер-
мента также зависит от pH, од-
нако присоединение АМФ-деза-
АТФ
Сокращение
АДФ
Фумарат
АМФ-дезаминаза Аденилосукцинатлиаза
Дценилосукцинат
АМФ
ИМФ Аденилосукцинатсинтетааа
Аспартат + гуанозинтрифосфат
Рис. 2.6. Цикл пуриновых нуклеотидов
(в прямоугольнике)
75
миназы может еще происходить и в отсутствие ацидоза. Таким образом,
метаболические условия в состоянии покоя (высокий энергетический
заряд и pH 7,0) обусловливают инактивацию фермента. Однако сниже-
ние энергетического заряда при физических нагрузках вместе с индуци-
рованным мышечной работой ацидозом, вызванным накоплением лакта-
та, приводят к активации фермента и, как следствие, — к образованию
ИМФ и аммиака. Тем не менее следует заметить, что повышение кон-
центрации АДФ и АМФ независимо от pH может активировать АМФ-
дезаминазу. Так, например, было показано, что истощение запасов мы-
шечного гликогена при пролонгированных физических нагрузках, вызы-
вающих увеличение концентрации АДФ и АМФ, но не повышающих
продукцию лактата, приводит к возрастанию активности фермента. По-
добным образом у пациентов, больных болезнью Мак-Ардля, мышцы
которых лишены гликогеновой фосфорилазы, также вырабатывают
ИМФ и аммиак несмотря на отсутствие продуцирования лактата. И на-
оборот, веским доказательством активации АМФ-дезаминазы при аци-
дозе служит наличие тесной корреляционной взаимосвязи между накоп-
лением в крови и мышцах лактата и накоплением аммиака и гипоксан-
тина у здоровых испытуемых во время возрастающей по мощности фи-
зической нагрузки. И действительно, активность АМФ-дезаминазы в
медленносокращающихся скелетных мышцах крысы повышается толь-
ко в том случае, когда pH становится ниже 6,6.
Предполагается, что для процесса дезаминирования в цикле пури-
новых нуклеотидов определенное значение могут иметь и некоторые
другие факторы. Возможно, наиболее важным из них является поддер-
жание энергетического заряда клетки при мышечной работе. Роль дан-
ного фактора частично уже обсуждалась, следовательно, нет необходи-
мости снова на этом останавливаться. Следует только добавить, что об-
разование АМФ в мышцах во время их сокращений в дополнение к под-
держанию энергетического статуса за счет снижения аденилатного пула
также обеспечивает энергетически эффективный механизм для восста-
новления аденилатного пула в восстановительном периоде после физи-
ческой нагрузки. Поскольку процесс адениннуклеотидного синтеза de
novo является энергетически неэффективным и относительно медлен-
ным, образование ИМФ представляется более предпочтительным меха-
низмом для восстановления адениннуклеотидного пула. Литературные
данные свидетельствуют о том, что обычно восстановление аденилат-
ного пула после максимальной физической нагрузки происходит в тече-
ние часа. Однако результаты последних исследований показали, что этот
процесс может протекать в течение более продолжительного периода
(более 3 дней) после 1-6 недель высокоинтенсивных тренировочных за-
76
нятий. Такая отсрочка в наибольшей степени связана с распадом ИМФ
до гипоксантина, неизбежно влекущего за собой пуриновый синтез de
novo. Интересно, что высокоинтенсивные тренировочные занятия вызы-
вают адаптационные изменения, проявляющиеся в меньшем снижении
содержания адениннуклеотидов во время мышечной работы.
Образование аммиака при дезаминировании АМФ дает основание
предполагать его важную роль в буферировании Н+, накапливающегося
в процессе выполнения интенсивных физических упражнений:
NH3 + H+ —NH4+.
Однако известно, что количество образующегося Н+ при мышечной
работе примерно в 50 раз может превышать количество образующегося
аммиака, в связи с чем такой механизм буферирования представляется
малозначительным.
Как отмечалось выше, накопление ИМФ и аммиака при физических
нагрузках связано с регуляцией утилизации углеводов. Инозинмоно-
фосфат причастен к активации гликогеновой фосфорилазы Ь, а в связи с
этим — ис гликогенолизом во время мышечной работы. Однако эта роль
ИМФ выглядит маловероятной, поскольку установлено, что накопление
ИМФ в той степени, какая отмечается при мышечной деятельности, не-
существенно для поддержания высокой скорости гликогенолиза в усло-
виях напряженных физических нагрузок. В опытах in vitro показано, что
ионы аммония (NH4+) активируют фосфофруктокиназу, поэтому их на-
копление при мышечной работе включается в регуляцию гликолиза. Од-
нако гликогенолиз и гликолиз могут ускоряться еще до накопления ИМФ
и аммиака, в связи с чем их роль в регуляции деградации углеводов нез-
начительная.
Реаминирование в цикле пуриновых нуклеотидов. Реаминиро-
вание в цикле пуриновых нуклеотидов катализируется аденилосук-
цинатсинтетазой и аденилосукцинатлиазой (см. рис. 2.6). Аденилосук-
цинатсинтетаза катализирует образование аденилосукцината из ИМФ,
аспартата и гуанозинтрифосфата. Эта реакция является звеном, лимити-
рующим скорость протекания всего цикла пуриновых нуклеотидов.
Аденилосукцинатлиаза катализирует образование АМФ и фумарата из
аденилосукцината.
Не исключено, что роль реаминирования в цикле пуриновых нукле-
отидов различна в зависимости от превалирующих метаболических ус-
ловий. Так, например, во время интесивной сократительной активности
мышц, когда преобладает дезаминирование, производительность реами-
нирования должна носить противоположный характер. В действитель-
77
ности имеется небольшое количество доказательств, свидетельствую-
щих о том, что реаминирование происходит во время проявления мыш-
цей значительной силы. Однако при выполнении пролонгированных фи-
зических упражнений малой или умеренной интенсивности следует
ожидать, что цикл пуриновых нуклеотидов будет задействован. Но, оче-
видно, при пролонгированных физических нагрузках реаминирование
может происходить в неактивных мышечных волокнах, т. е. предполага-
ется, что дезаминирование и реаминирование не являются сопутствую-
щими процессами.
На рис. 2.6 показано, что один полный оборот цикла обеспечивает
утилизацию аспартата, а также образование фумарата и аммиака без ка-
ких-либо изменений концентраций АМФ и ИМФ. Фумарат служит ин-
термедиатом ЦТК, поэтому, по мнению ряда авторов, цикл пуриновых
нуклеотидов может вносить (анаплеротически) свой вклад в снабжение
ЦТК интермедиатами, которые образуются в процессе мышечной рабо-
ты. Следовательно, цикл пуриновых нуклеотидов может способствовать
усилению митохондриального дыхания путем повышения скорости
окисления ацетил-КоА и образования НАДН, а поэтому быть, по край-
ней мере частично, ответственным за поддержание сократительной ак-
тивности мышц. Однако наличие большого количества интермедиатов
ЦТК в мышечных клетках во время выполнения высокоинтенсивных
физических упражнений, отмечаемое на фоне отсутствия реаминирова-
ния, позволяет думать, что анаплеротическая роль цикла пуриновых
нуклеотидов в митохондриальной энергопродукции незначительна. И
действительно, как свидетельствуют результаты проведенных в послед-
нее время исследований, поступление углеводов является главным де-
терминантом, определяющим поступление интермедиатов ЦТК. Более
того, нет доказательств тому, что при выполнении пролонгированных
физических упражнений происходит снижение эффективности митохон-
дриального окисления или снижение концентрации НАДН, в связи с чем
можно прийти к заключению, что доступность интермедиатов ЦТК не
лимитирует митохондриальное дыхание.
Утомление. При выполнении кратковременных интенсивных и
пролонгированных (субмаксимальной аэробной мощности) физических
упражнений отмечается тесная взаимосвязь между снижением содержа-
ния в мышцах адениннуклеотидов и развитием утомления, что указы-
вает на участие дезаминирования АМФ в механизме проявления послед-
него. Однако вероятнее всего, что эта взаимосвязь отражает снижение
поставки энергии через гидролиз КФ, гликолиз и окислительное фосфо-
рилирование, необходимой для обеспечения сократительной активности
мышц (см. главы 6 и 7). В результате такого снижения повышается кон-
78
центрация АДФ и АМФ в мышечных клетках (уменьшается энергети-
ческий заряд клетки), активизируется дезаминаза, что, в конечном счете,
приводит к развитию утомления, обусловленному локальным снижени-
ем уровня АТФ и (или) накоплением АДФ.
В пользу такого вывода свидетельствуют результаты изучения мы-
шечного метаболизма при некоторых патологических состояниях (нап-
ример, при синдроме Мак-Ардля), при которых затруднение поступле-
ния энергии происходит параллельно с повышением скорости снижения
содержания адениннуклеотидов и развитием утомления во время выпол-
нения мышечной работы. Однако результаты последних исследований
показали, что развитие утомления у пациентов с АМФ-дезаминазным
дефицитом, характеризующимся отсутствием способности к образова-
нию ИМФ в мышцах, не отличалось от испытуемых контрольной груп-
пы. Кроме того, эти пациенты продемонстрировали улучшение показа-
телей физической работоспособности в велоэргометрическом тесте пос-
ле выполнения высокоинтенсивной тренировочной программы, хотя их
адениннуклеотидный пул снизился на 20 %. На основании приведенных
данных можно сделать вывод, что развитие мышечного утомления пред-
ставляет собой многофакторный процесс, который не обязательно мо-
жет иметь отношение к энергетическому метаболизму.
Выводы
1. В ходе любой биохимической реакции освобождается свободная
энергия, которая затем может быть использована для работы. В мышцах
свободная энергия, освобождаемая при окислении углеводов и жиров,
может запасаться как химическая энергия в форме АТФ. Свободная
энергия, освобождающаяся при последующем распаде АТФ в ходе АТФ-
азных реакций, может превращаться в механическую энергию для про-
изводства работы (мышцы способны выполнять 24 кДж работы на каж-
дый моль распавшегося АТФ). Аденозинтрифосфат является един-
ственным веществом в живых клетках, которое способно превращать
энергию таким образом.
2. Фосфокреатин присутствует в цитозоле мышечных волокон, где
его концентрация примерно в три раза превышает концентрацию АТФ.
Общепризнано, что с началом сократительной активности наступает и
быстрый распад КФ в ходе креатинкиназной реакции. Освобождающая-
ся при этом свободная энергия может быть использована для ресинтеза
АТФ, благодаря чему в клетке поддерживается высокое отношение
АТФ/АДФ. Однако открытие нескольких изоферментов креатинкиназы,
79
каждый из которых характеризуется различной внутриклеточной лока-
лизацией, привело к возникновению гипотезы о том, что КФ может вы-
полнять несколько различных функций в скелетной мышце.
3. Креатинкиназная реакция обратима, и ее протекание в обратном
направлении происходит после прекращения мышечной работы, когда
достаточное количество энергии становится доступным для рефосфори-
лирования Кр. Ресинтез КФ описывается экспоненциальной кривой, од-
нако продолжительность его периода зависит от многих факторов.
4. Суммарная концентрация АТФ, АДФ и АМФ определяет общий
адениннуклеотидный пул в клетке. Степень фосфорилирования этого
пула называется энергетическим зарядом клетки, который служит хоро-
шим индикатором ее энергетического состояния. Например, снижение в
клетке концентрации АТФ при мышечных сокращениях и происходящее
параллельно увеличение концентраций АДФ и АМФ, т. е. снижение
энергетического заряда, будет сразу же стимулировать анаэробный и аэ-
робный ресинтез АТФ.
5. Общий адениннуклеотидный пул клетки будет снижаться, если
концентрация в ней АМФ при мышечной работе начнет возрастать.
Падение уровня адениннуклеотидов в клетке потенциально опасно,
поскольку при этом утрачивается возможность их фосфорилирования.
Однако этот неблагоприятный эффект превалирует в течение коротко-
го времени своим стимулирующим влиянием, которое приводит к сни-
жению концентраций АДФ и АМФ в ходе аденилатциклазных реак-
ций, в результате чего повышается энергетический заряд мышечных
клеток и становится возможным продолжение их сократительной
активности.
6. Падение уровня мышечных адениннуклеотидов происходит, во-
первых, в результате дезаминирования АМФ до ИМФ и аммиака, во-вто-
рых, при дефосфорилировании АМФ до аденозина. Как ИМФ, так и аде-
нозин в дальнейшем могут подвергнуться распаду в печени с образова-
нием инозина, а затем гипоксантина, экскретируемого почками. Основ-
ным путем для снижения уровня адениннуклеотидов у человека служит
дезаминирование АМФ до ИМФ и аммиака, однако существуют различ-
ные варианты такого снижения, зависящие от типа мышечных волокон
и вида животных.
7. Альтернативной предопределенностью для ИМФ является воз-
можность использования его для ресинтеза АМФ. Дезаминирование
АМФ до ИМФ и последующее реаминирование ИМФ формируют цикл
пуриновых нуклеотидов. Предполагается, что цикл пуриновых нуклео-
тидов может играть важную роль в мышечном энергетическом метабо-
лизме.
80
8. Существует мнение, согласно которому тесная взаимосвязь меж-
ду снижением уровня мышечных адениннуклеотидов и развитием утом-
ления при выполнении кратковременных интенсивных и пролонгиро-
ванных (субмаксимальной аэробной мощности) физических упражне-
ний указывает на участие дезаминирования АМФ в механизме проявле-
ния последнего. Однако вероятнее всего, что эта взаимосвязь отражает
снижение энергообеспечения сократительной активности мышц, обус-
ловленное многими факторами, включающими локальное исчерпание в
клетке запасов АТФ и (или) накопление АДФ.
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
Atkinson D.E. (1977). Cellular energy metabolism and its regulation. — New York:
Acad. Press.
Harris R.C., Edwards R.H.T., Hultman £. et al. (1976). The time course of phosphorylcrea-
tine resynthesis during recovery of the quadriceps muscle in man // Pflugers Arch. — 376.
— P. 137-142.
6 0-313
QJ МЕТАБОЛИЗМ УГЛЕВОДОВ
1. Роль углеводов
Нетрудно убедиться в важности роли, которую играет ме-
таболизм улеводов (СНО) в энергообеспечении мышечной
деятельности. При выполнении высокоинтенсивных физи-
ческих упражнений основная энергетическая потребность
мышц удовлетворяется благодаря расщеплению углеводов.
При более умеренных физических нагрузках, продолжаю-
щихся в течение длительного периода, уровень физической
работоспособности лимитирован в основном доставкой к
мышцам углеводов в качестве энергетического топлива.
Поэтому обеспечение углеводами и их метаболизм являют-
ся главными моментами в вопросе о способности мышц к
проявлению работоспособности.
Даже в состоянии покоя нарушение возможности под-
держания нормального уровня глюкозы в крови, которая
обеспечивает снабжение углеводным «топливом» мозг и
другие ткани, способно привести к нарушению функций
ЦНС и их прогрессированию до состояния комы и даже к
смерти.
2. Углеводное обеспечение
Углеводы поступают в организм как из пищевых продук-
тов, так и из эндогенных источников главным образом в ви-
де мышечного гликогена и гликогена печени. Организм об-
ладает также ограниченными возможностями для синтеза
глюкозы из неуглеводных предшественников. Классифика-
82
Моносахариды
Глю X Глю
Мальтоза
Глю X ФРУ /> Глю Гал Дисахариды
Сахароза Лактоза
Рис. 3.1. Структура и классификация наиболее важных углеводов, поступающих с пи-
щей, а также гликогена, представляющего собой форму запаса углеводов у животных
(Глю — глюкоза, Фр — фруктоза, Гал — галактоза)
ция поступающих с пищей углеводов более подробно описана в Прило-
жении I. К наиболее важным из них относятся моносахариды (напри-
мер, глюкоза, фруктоза, галактоза), дисахариды, состоящие из двух сое-
диненных вместе молекул сахара (например, мальтоза, лактоза) и поли-
сахариды (например, крахмал, состоящий из длинных цепочек молекул
глюкозы) (рис. 3.1). Основная часть углеводов обычно присутствует в
пище в виде сахарозы и крахмала, пропорциональное соотношение ко-
торых важно для обеспечения нормального питания и здоровья.
В какой бы форме углеводы ни поступали в организм с пищей, пе-
ред абсорбцией они должны превращаться в моносахариды. Этот про-
цесс происходит в результате гидролиза узлов, связывающих моносаха-
риды, что иллюстрировано примером деградации амилопектина (крах-
мала) на рис. 3.2.
Ограниченный гидролиз углеводов начинается уже во рту и желуд-
ке, однако в наибольшей степени он проявляется в тонкой кишке, где со-
ответствующее значение pH способствует повышению активности спе-
цифических ферментов, секретируемых в полость кишечника.
Некоторые полисахариды, как, например, целлюлоза, у которых мо-
лекулы глюкозы соединены а- 1,4-а- 1,6-связями, противостоят гидроли-
зу в верхней части кишечника у человека. Они проходят через пищева-
рительный канал большей частью не подвергнутыми пищеварению, хо-
тя некоторая бактериальная ферментация может происходить и в ниж-
ней части кишечника. Пищеварительная система жвачных животных
приспособлена для обеспечения такого ферментативного процесса, од-
нако для человека этот вид углеводов неприемлем.
6*
83
связь
Рис. 3.2. Гидролиз а-1,4- и а-1,6-связей в
крахмале в процессе пищеварения, освобож-
дающий несвязанную глюкозу, которая ста-
новится пригодной для абсорбции (сравните
с распадом гликогена в мышцах, показанном
далее на рис. 3.4).
В среднем, пища жителей
западных районов планеты, ис-
ходя из ее энергетической стои-
мости, включает менее 50 % уг-
леводов. При этом общее коли-
чество потребляемых в сутки уг-
леводов для мужчины, имею-
щего массу 70 кг и ведущего ма-
лоподвижный образ жизни, сос-
тавляет около 300 г, или около
3000 ккал. Поскольку углевод-
ные резервы организма неболь-
шие и остаются приблизительно
постоянными, это количество
углеводов должно быть близким
к показателю скорости их ути-
лизации. В среднем, в организме
мужчины, у которого масса тела
составляет 70 кг, содержится
около 300-500 г запасов углево-
дов (СНО), находящихся в ос-
новном в форме гликогена. Следовательно, количество потребляемых в
сутки углеводов приблизительно эквивалентно их нормальному запасу в
организме. Около 80-110 г СНО содержится в печени: они представля-
ют собой эффективный запас углеводов, которые могут поступать в кро-
вяное русло для транспорта в другие ткани.
Наибольшее количество углеводных запасов находится в скелетных
мышцах. Обычно женщины с небольшой мышечной массой имеют и
меньший запас СНО. Резервы мышечного гликогена недоступны для
других тканей. Общее количество глюкозы, находящейся во внеклеточ-
ной жидкости, составляет около 15 г и его нельзя рассматривать как
энергетический запас, поскольку даже незначительное снижение кон-
центрации глюкозы в крови может нарушить функцию мышечных и нер-
вных клеток. Хотя запасы мышечного гликогена и остаются почти неиз-
менными при воздержании от приема пищи, которое не сопровождается
напряженной мышечной деятельностью, однако содержание гликогена в
печени после постабсорбционного периода начинает быстро снижаться.
Молекула гликогена похожа на молекулу крахмала, за исключением
только того факта, что связи между молекулами глюкозы у них неодина-
ковые (см. рис. 3.1). У гликогена основной гликозидной связью является
«-1,4-связь с разветвленными участками в молекуле, происходящими от
84
дополнительных а-1,6-связей приблизительно через каждые 6—10 глю-
козных остатка. В молекуле крахмала присутствуют два полисахарида
— амилоза, содержащая длинные цепи молекул глюкозы, соединенных
между собой «-1,4-связями, и амилопектин, который по своей структуре
аналогичен гликогену.
3. Реакции анаэробного гликолиза
и гликогенолиза
Начальный этап распада углеводных запасов в организме происходит
без участия кислорода и поэтому является анаэробным процессом. Тер-
минология зависит от начального этапа: распад глюкозы обозначается
как гликолиз, тогда как гликогенолиз начинается с распада гликогена. За
исключением случаев, имеющих непосредственное отношение к глико-
гену, термин «гликолиз» удобнее использовать для обозначения обоих
процессов, которые протекают по общему пути после первого этапа.
Гликолиз эффективно преобразует шестиуглеродную в две трехугле-
родные молекулы глюкозы. Конечным продуктом аэробного гликолиза
является пируват, тогда как конечным продуктом анаэробного гликолиза
— лактат. Во время этого процесса некоторое количество энергии, осво-
бождаемое при разрыве связей, преобразуется в энергию АТФ. Последо-
вательность реакций, приводящих к превращению глюкозы или гликоге-
на до пирувата, представлена на рис 3.3.
Перенос молекулы глюкозы через клеточную мембрану осущест-
вляется специальным транспортным белком (ГЛУТ4). Как только моле-
кула глюкозы поступает внутрь клетки, первым этапом гликолиза стано-
вится ее необратимое фосфорилирование, предотвращающее выход
этого ценного питательного вещества из клетки: глюкоза преобразуется
до глюкозо-6-фосфата (Г-6-Ф). Этот этап протекает необратимо, по
крайней мере в мышцах. Печень имеет фосфатазный фермент, который
катализирует обратимую реакцию, позволяющую свободной глюкозе
выходить из клетки в кровяное русло. Однако в мышцах этот фермент
отсутствует.
Гексокиназа — фермент, осуществляющий фосфорилирование глю-
козы, тесно взаимосвязан с переносчиком глюкозы, благодаря чему она
легко захватывается мышечной клеткой. Гексокиназная реакция являет-
ся энергозависимой и на каждую молекулу глюкозы требуется одна мо-
лекула АТФ. Это также обеспечивает возникновение концентрационно-
го градиента глюкозы на клеточной мембране, который способствует ее
85
Глюкоза
) Гексокиназа*
Фосфоглюкомутаза
Глюкозо-6-фосфат
АТФ
АДФ
Гликоген
<фм
Фосфорилаза
" гликогена
Глюкозо-6-фосфат
Глюкозофосфатизомераза
Фруктозо-6-фосфат
АТФ---
ЛДФ » ' 6-Фосфофруктокиназа
Фруктозо-1,6-дифосфат
| Альдолаза
Дигидроацетон- —-----——. * Глицеральдегид-З-фосфат
фосфат Триозофосфат- р
изомераза
ФН’ Л Глицеральдегидфосфат-
НАДН , дегидрогеназа
1,3-Дифосфоглицерат
АДФ—х
) Фосфоглицераткиназа
АТФ-«-^|
З-Фосфоглицерат
Фосфоглицеромутаза
2-Фосфоглицерат
Енолаза
Фосфоенолпируват
АДФ--^
АТФ-*-п
Пируваткиназа
Пируват
----------1---------|z- КоА
Лактат- х—НАДН НАД+—х Пируваг-
дегидрогеназа Ч_^цдд+ НАДН-«-^ дегидрогеназа
Лактат
Ацетил-КоА + СО2
Рис. 3.3. Реакции гликолиза
транспорту. Гексокиназная реакция угнетается Г-6-Ф, накопление кото-
рого во время выполнения высокоинтенсивных физических упражнений
может лимитировать вклад глюкозы крови в углеводный метаболизм в
сокращающихся мышцах.
Если гликоген, который предпочтительнее по сравнению с глюкозой
крови, является субстратом для гликолиза, то первый этап этого процес-
86
са состоит в расщеплении
его молекулы до оди-
ночных молекул глюкозы.
Это происходит при
участии фермента глико-
генфосфорилазы. В ре-
зультате этой реакции об-
разуется глюкозо-1-фос-
фат (Г-1-Ф) и молекула
гликогена становится ко-
роче на одну глюкозную
единицу (рис. 3.4). В дан-
ном случае субстратами
служат гликоген и Фн,
причем, в отличие от гек-
сокиназной реакции, на
Рис. 3.4. Последовательность расщепления гли-
когена
этом первом этапе для
расщепления не требуется АТФ. Фосфорилаза воздействует на а-1,4-уг-
леродные связи свободных окончаний молекулы гликогена, но не может
разрушать а- 1,6-связи. Она прекращает свое действие перед каждой из
четырех глюкозных единиц перед точкой ветвления, в результате чего от
молекулы гликогена остается только сердцевина с короткими отростка-
ми (остаточный декстрин). Для дальнейшего распада молекулы гликоге-
на требуется действие другого фермента — амило-1,6-глюкозидазы, ко-
торый разрывает а-1,6-связь в точке ветвления и тем самым открывает
для действия гликогенфосфорилазы новый участок полисахаридной це-
пи. Фосфорилаза не может проявлять активность в пределах четырех
глюкозных остатков до точки ветвления, поэтому для дальнейшего рас-
щепления требуется действие фермента, разрывающего связь в точке
ветвления. Воздействие амило-1,6-глюкозидазы на точку ветвления ос-
вобождает глюкозу. Освобождающаяся глюкоза при участии гексокина-
зы быстро фосфорилируется до Г-6-Ф. Накопление свободной глюкозы
в мышечных клетках происходит только во время их высокоинтенсив-
ной сократительной активности при быстром протекании гликогеноли-
за: поскольку в таких условиях мало а-1,6-связей, то не более 10 % глю-
козных единиц появляется в виде свободной глюкозы.
Фермент фосфоглюкомутаза способствует тому, что образовавшийся
в результате воздействия фосфорилазы на гликоген Г-1-Ф быстро преоб-
разуется до Г-6-Ф, который затем включается в гликолитический процесс.
При участии фермента фосфоглюкоизомеразы из Г-6-Ф образуется
Ф-6-Ф. Этот фермент характеризуется высокой активностью и реакция
87
может протекать быстро в любом направлении. Затем следует второй
этап фосфорилирования и Ф-6-Ф преобразуется во фруктозо-1,6-дифос-
фат (ФДФ). Для данной реакции, катализируемой ферментом фосфо-
фруктокиназой (ФФК), также требуется фосфатная группа, донатором
которой служит АТФ. Активность ФФК находится под контролем мно-
гих внутриклеточных факторов, что играет важную роль в регуляции
скорости метаболического процесса, протекающего по этому пути. Фос-
фофруктокиназная реакция является первым удобным пунктом, через
который оказывается влияние на метаболизм как глюкозы, так и глико-
гена, и которую часто называют скоростьлимитирующей реакцией гли-
колиза.
При любой последовательности реакций может быть только одна из
них, которая лимитирует скорость всего процесса: он может протекать
не быстрее, чем самая медленная из них (скоростьлимитирующая реак-
ция). Поэтому ФФК обычно является участком гликолиза, определяю-
щим общую его продолжительность. Таким образом, для обеспечения
гликолиза, предназначенного для снабжения клетки энергией, требуется
две молекулы АТФ.
Фруктозо-1,6-дифосфат расщепляется ферментом альдолазой на две
трехуглеродные молекулы: глицеральдегид-3-фосфат и дигидроацетон-
фосфат. Эти две молекулы взаимопревращаются под влиянием триозо-
фосфатизомеразы и в дальнейшем их метаболизм происходит только че-
рез глицеральдегид-3-фосфат. Поэтому дигидрооксиацетонфосфат, про-
дуцируемый альдолазной реакцией, перед тем как подвергнуться даль-
нейшему метаболизму, преобразуется до глицеральдегид-3-фосфата.
Следовательно, последующие этапы гликолиза удваиваются. Ни альдо-
лаза, ни триозофосфатизомераза не лимитируют скорость гликолитичес-
кого процесса.
Глицеральдегид-З-фосфат преобразуется в 1,3-дифосфоглицерино-
вую кислоту в ходе сложной реакции, катализируемой глицеральдегид-
3-фосфатдегидрогеназой и при этом одновременно происходит переход
окисленной формы никотинам идадениннуклеотида (НАД4) в его восста-
новленную форму (НАДН) с освобождением иона водорода. При физи-
ологических значениях pH 1,3-дифосфоглицериновая кислота существу-
ет в ионизированной форме — 1,3-дифосфоглицерата (1,3-ДФГ). Присо-
единяя водородный ион, НАД4 присоединяет также два электрона. До-
полнительная фосфатная группа, входящая в 1,3-ДФГ, поступает от не-
органического фосфата и в дальнейшем АТФ не используется.
Следующим этапом гликолиза является киназная реакция, в ходе ко-
торой фосфатная группа от 1,3-ДФГ переносится к АДФ. При этом об-
88
разуется АТФ, а 1,3-ДФГ в результате реакции, катализируемой фосфо-
глицераткиназой, метаболизируется до 3-фосфоглицерата.
В ходе реакции, протекающей при участии фермента фосфоглице-
ромутазы, происходит внутренняя реорганизация молекулы 3-фосфо-
глицерата, проявляющаяся в перемещении фосфатной группы ко второй
углеродной позиции с образованием 2-фосфоглицерата. Впоследствии
эта реакция сменяется катализируемой енолазой реакцией дегидрата-
ции, в результате которой образуется фосфоенолпируват (ФЕП). На пос-
леднем этапе гликолиза происходит перенос фосфатной группы от ФЕП
к АДФ с образованием АТФ и пирувата. Эта реакция катализируется пи-
руваткиназой и является второй киназной реакцией.
Таким образом, полный эффект гликолиза заключается в превраще-
нии одной молекулы глюкозы в две молекулы пирувата с образованием
двух молекул АТФ и превращением двух молекул НАД4 в НАДН. Если
стартовым этапом этого поцесса является использование гликогена, то в
этом случае вырабатывается три молекулы АТФ, поскольку на первом
участке фосфорилирования не требуется первоначального вклада АТФ.
Хотя чистый энергетический выход и кажется незначительным, все же
относительная доступность углеводных запасов и высокая скорость про-
текания гликолиза позволяют выработать количество энергии, достаточ-
ное для обеспечения интенсивной мышечной работы. Так, при беге на
800 м общий энергетический запрос на 60 % удовлетворяется за счет
анаэробного метаболизма, при этом около 100 г углеводов (в основном
гликоген, количество которого эквивалентно 0,55 молям глюкозы) пре-
образуется до лактата менее чем за 2 мин. Количество образующегося
таким путем АТФ (три молекулы АТФ на одну молекулу распавшейся
глюкозы, всего — около 1667 ммоль) значительно превышает то его ко-
личество, которое поступает в результате гидролиза КФ. Высокая ско-
рость анаэробного метаболизма позволяет не только быстрее достигнуть
устойчивого состояния по сравнению со случаем, если бы мышечная ра-
бота обеспечивалась исключительно за счет аэробного метаболизма, но
также позволяет развить более высокий темп в начале дистанции еще до
того, как сердечно-сосудистая система приспособится к обеспечению
возможности удовлетворять доставку и утилизацию кислорода.
Реакции гликолиза, протекающие в цитоплазме, приводят к образо-
ванию пирувата, который не фосфорилируется и поэтому легко покида-
ет клетку. При высокой скорости гликолиза некоторая доля образовавше-
гося пирувата выходит из мышц, однако большая его часть впоследствии
подвергается дальнейшему метаболизму. Дальнейшая его судьба зави-
сит не только от такого фактора, как интенсивность физической нагруз-
ки, но и от метаболической способности ткани.
89
4. Регенерация НАД+
При высокой скорости гликолиза важной проблемой для клетки стано-
вится наличие в ней НАД+ — необходимого (лимитирующего) кофакто-
ра в глицеральдегид-3-фосфатдегидрогеназной реакции. Количество
НАД+ в клетке очень незначительное — всего около 0,8 ммоль-кг-1 мы-
шечной ткани (в начале развития гликолиза). Во время интенсивной мы-
шечной активности, как, например, при спринтерском беге, обращае-
мость АТФ может достигать около 125 ммоль-кг^'-мин-а при выпол-
нении кратковременных физических упражнений взрывного характера
— до 150 ммоль-кг-'-мин-’. Если образующийся в процессе гликолиза
НАДН не реокисляется до НАД+ с эквивалентной скоростью, то даль-
нейшее анаэробное энергообеспечение становится невозможным.
Существуют два механизма, благодаря которым в большинстве, но
не во всех клетках может происходить окисление НАДН и регенерация
НАД+. Это может достигаться восстановлением пирувата до лактата
(рис. 3.5). Преимущество этой реакции состоит в том, что она может
протекать в отсутствие кислорода. Лактат может накапливаться в мы-
шечных клетках до таких высоких концентраций, которых не может дос-
тигнуть ни один из гликолитических интермедиатов. Однако когда это
происходит, то присоединенные водородные ионы обусловливают сни-
жение внутриклеточного pH. Некоторое количество лактата диффунди-
рует в межклеточное пространство и постепенно начинает накапливать-
ся в крови.
Лактату, выходящему из мышечных клеток, сопутствует выход во-
дородных ионов. Несмотря на буферирование их во внеклеточном прос-
транстве, все же происходит их накопление в экстрацеллюлярной жид-
кости, вызывающее снижение pH до уровня, который может нарушить
функцию клетки. Нормальное значение pH для мышечных клеток в сос-
тоянии покоя находится около 7,1. Однако оно может достигать 6,5 и да-
же меньших значений при высокоинтенсивных физических нагрузках,
когда происходит образование большого количества лактата. Поскольку
сн3со соо
Низкое pH
Низкий уровень НАД+/
высокий уровень НАДН
+ НАДН
+ Н+
ЛДГ
Пируват
Высокое pH
Высокий уровень НАД+/
низкий уровень НАДН
СН3СН(ОН) СОО’
+ НАД+
Лактат
Рис. 3.5. Лактатдегидро-
геназная реакция, катали-
зирующая взаимопревра-
щение лактата и пирувата
и может протекать в лю-
бом направлении
90
некоторое количество водородных ионов буферируется во внутрикле-
точной и внеклеточной среде, повышение концентрации лактата и сни-
жение pH происходит в нелинейной зависимости.
При pH 6,5 сократительная способность мышечных волокон начи-
нает снижаться, что может сопровождаться угнетением активности та-
ких ключевых ферментов гликолиза, как фосфорилаза и фосфофрукто-
киназа. Снижение pH раздражает свободные нервные окончания в мыш-
цах, что вызывает болевые ощущения. Хотя негативные эффекты аци-
доза, возникающие из-за накопления лактата, часто носят затяжной ха-
рактер, все же не следует забывать, что энергия, вырабатываемая в про-
цессе анаэробного гликолиза, позволяет выполнять высокоинтенсивную
мышечную работу, которая не может быть обеспечена аэробным энерге-
тическим механизмом.
Альтернативой превращению до лактата и пирувата может служить
окислительный метаболизм до диоксида углерода (СО2) и воды. Этот
процесс протекает в митохондриях, в которые пируват переносится че-
рез митохондриальную мембрану специфическим белковым переносчи-
ком. Первым этапом в митохондриях является происходящее путем
окислительного декарбоксилирования преобразование трехуглеродного
пирувата в двухуглеродную ацетатную группу, которая присоединяется
тиоэфирной связью к коэнзиму А с образованием ацетил-КоА. Эта реак-
ция, в ходе которой НАД4 восстанавливается до НАДН, катализируется
пируватдегидрогеназным ферментативным комплексом.
Ацетил-КоА окисляется до СО2 и воды в цикле трикарбоновых кис-
лот (ЦТК) — серии последовательных реакций, известных также как
цикл Кребса, названного в честь Ганса Кребса, который впервые описал
входящие в этот цикл реакции (ЦТК еще называют циклом лимонной
кислоты, поскольку последняя является основным интермедиатом ука-
занного процесса). Эти реакции включают в себя взаимодействие
ацетил-КоА с оксалоацетатом до образования цитрата — шестиуглерод-
ной трикарбоксильной кислоты. Серии реакций приводят к последова-
тельной утрате водородных ионов и диоксида углерода, в результате че-
го происходит регенерация оксалоацетата. Ацетил-КоА служит также
интермедиатом окисления жирных кислот, поэтому конечные этапы
окислительного распада как жиров, так и углеводов одинаковые. Более
подробно ЦТК описан в главе 1.
Все приведенные выше реакции метаболизма глюкозы, приводящие
к образованию энергии, можно суммировать уравнением
Глюкоза + 6О2 + 38АДФ + 38Фн -> 6СО2 + 6Н2О + 38АТФ.
91
Общий синтез 38 молей АТФ, приходящийся на 1 моль окисленной
глюкозы при окислении восстановленных коферментов, можно предста-
вить следующим образом:
Синтезированный АТФ
Источник
2
6
24
4
2
Гликолиз
НАДН (при гликолизе)
НАДН
фадн2
ГТФ
Одной из потенциальных проблем, связанных с окислительной ре-
генерацией НАД+, является тот факт, что реакции окислительного фос-
форилирования происходят внутри митохондрий, тогда как гликолиз яв-
ляется цитоплазматическим процессом и внутренняя митохондриальная
мембрана непроницаема для НАДН и НАД+. Без регенерации НАД+ в
цитоплазме гликолиз прекращается, и поэтому должен существовать ме-
ханизм, который обеспечивал бы эффективное окисление НАДН, обра-
зующегося в процессе гликолиза. Эта сепарация преодолевается много-
численными субстратными челноками, которые транспортируют восста-
новленные эквиваленты внутрь митохондрий.
Некоторое количество пирувата может быть использовано для обра-
зования аминокислоты аланина (об этом подробно изложено в главе 5),
а часть его может также преобразовываться до четырехуглеродного ок-
салоацетата в результате взаимодействия с СО2 в реакции, катализируе-
мой пируваткарбоксилазой. Однако в мышцах эта реакция не протекает
из-за отсутствия в них указанного фермента. Происходящее таким пу-
тем образование оксалоацетата может служить первым этапом ресинте-
за глюкозы в процессе глюконеогенеза. И напротив, это может оказаться
важной анаплеротической реакцией (реакцией, обеспечивающей под-
держание внутриклеточной концентрации важнейших интермедиатов в
случае снижения их содержания).
5. Регуляция гликолиза
Скорость гликолиза должна быть регулируемой для того, чтобы обеспе-
чение снабжения аденозинтрифосфатом протекало сопряженно с его
гидролизом и с поступлением других энергетических источников. Как и
при локальной внутриклеточной регуляции, необходим согласованный
ответ различных тканей для их совместного включения в углеводный
92
метаболизм, а также для координации путей последнего с метаболизмом
жиров и белков. Поэтому эта регуляция осуществляется комплексно и
достигается интеграцией влияний многочисленных сигналов.
В углеводном метаболизме существует большое количество ключе-
вых участков. Они связаны с поступлением самой глюкозы в клетку, ко-
торое находится под контролем многих факторов. Инициирующее фос-
форилирование гексокиназой — глюкозы крови или фосфорилазой —
гликогена служит для эффективного поставления глюкозы внутрь мы-
шечной клетки. Пируватдегидрогеназная реакция также является реша-
ющей, поскольку в ходе ее образуется пируват, подвергающийся окис-
лению или дальнейшим изменениям до образования жира (ацетил-КоА
не может быть использован для ресинтеза глюкозы).
Поступление глюкозы из крови в клетки зависит от активности
транспортного белка (ГЛУТ4), локализованного в клеточной мембране и
специфичного только для скелетных мышц. Транспорт глюкозы в клет-
ку стимулируется в основном инсулином, который повышает ее внут-
риклеточные запасы после приема богатой углеводами пищи. Этот тран-
спорт стимулируется также физическими нагрузками, способствующи-
ми повышеннию доступности глюкозы как субстрата.
Регуляция скорости протекания гликолиза может осуществляться в
трех участках его пути. Первый из них является входным пунктом, свя-
занным с гексокиназной или фосфорилазной реакцией. Гексокиназная
активность стимулируется неорганическим фосфатом — одним из реак-
ционных субстратов, а ингибируется продуктом реакции — глюкозо-6-
фосфатом. Регуляция фосфорилазы носит более сложный характер
(рис. 3.6).
Фермент существует в двух формах, которые обозначаются как фос-
форилаза а и фосфорилаза Ь. Фосфорилаза а обладает намного более вы-
Адреналин
Норадреналин
Фосфорилаза b
цАМФ
Са -кальмодулин
Фосфорилаза а
Рис. 3.6. Распад гликогена,
регулируемый воздействием
катехоламинов и локальной
стимуляцией кальцием
Гликоген
Г люкозо-1 -фосфат
Гликолиз
93
сокой активностью и ее принято считать активной формой фермента, хо-
тя и форма Ъ также проявляет некоторую активность. Фосфорилаза b пе-
реходит в «-форму под влиянием фермента киназы —- фосфорилазы, ко-
торая присоединяет фосфатную группу от АТФ. Переход в ft-форму
обеспечивается фосфатазным ферментом, который удаляет фосфатную
группу от фосфорилазы а. Сама киназа фосфорилазы также существует
в активной («) и неактивной (Ь) формах; активация фермента происхо-
дит под влиянием протеинкиназы. Активность последней стимулирует-
ся адреналином: воздействие адреналина на поверхность клетки повы-
шает внутриклеточную концентрацию цАМФ, что увеличивает протеин-
киназную активность. При стрессе концентрация адреналина в крови
возрастает, что приводит к значительному повышению фосфорилазной
активности. Это проявится только в увеличении скорости гликогенолиза
при условии, если внутриклеточная концентрация кальция будет высо-
кой: этот механизм предотвращает быстрое расщепление гликогена в
состоянии покоя, когда потребность в энергопродукции осутствует,
однако обеспечивает высокую скорость гликогенолиза во время физи-
ческой нагрузки, когда активация мышц вызывает увеличение концен-
трации свободного кальция в клетке. Аналогично тому, как кальций ини-
циирует сократительный процесс, присоединяясь к кальмодулину, про-
исходит активация и протеинкиназы, благодаря чему обеспечивается
тесное сопряжение между мышечной активностью и субстратным обес-
печением.
Высокие уровни инсулина не влияют на внутриклеточную концен-
трацию цАМФ в скелетных мышцах, однако инактивируют фосфорила-
зу и снижают скорость гликогенолиза. Такие уровни гормона отмечают-
ся в основном после приема углеводной пищи, что способствует отложе-
нию углеводов в запас. Во время выполнения физических упражнений
активность фосфорилазы вначале повышается, однако впоследствии
фермент снова переходит в ft-форму. Несмотря на это, скорость гликоге-
нолиза (по непонятным пока причинам) продолжает оставаться высо-
кой. Одним из важных факторов, обусловливающих повышение гликоге-
нолиза, может быть некоторое возрастание активности фосфорилазы ft в
присутствии инозинмонофосфата (ИМФ), образующегося при распаде
АТФ.
Фосфофруктокиназа (ФФК) является ключевым регуляторным фер-
ментом, активность которого модифицируется многими различными ве-
ществами, хотя не все из них имеют какое-либо физиологическое значе-
ние. Решающе важным представляется тот факт, что ФФК-активность
ингибируется высокими концентрациями АТФ и КФ в клетке и стимули-
руется высокими уровнями АДФ и АМФ. Это означает, что активность
94
Рис. 3.7. Фосфофруктокиназа, играющая ключевую роль в регуляции скорости глико
лиза и ответственная за изменения энергетического заряда в клетке (если концентрация
АМФ возрастает, то ФФК-активность повышается; высокая концентрация АТФ оказы-
вает ингибирующий эффект, что проявляется в замедлении скорости гликолиза и прев-
ращения углеводов)
фермента понижена в том случае, когда клетка энергонасыщена, и высо-
кая — при низком энергетическом заряде (рис. 3.7). Установлено, по
крайней мере в опытах, проведенных на изолированных препаратах, что
цитрат угнетает активность ФФК, поэтому его аккумуляция в клетке
также может ингибировать активность фермента. Этот механизм функ-
ционирует в качестве связующего звена, обеспечивающего интеграцию
метаболизма жиров и углеводов. Высокая скорость окисления жирных
кислот приводит к накоплению цитрата в митохондриях и часть его бу-
дет переходить в цитоплазму, что вызовет снижение скорости гликолиза.
В результате угнетения ФФК будет происходить накопление Г-6-Ф и, как
следствие, — ингибирование активности гексокиназы, а также затрудне-
ние поступления глюкозы в мышечные клетки, что следует рассматри-
вать как неблагоприятное явление. Субстрат реакции (Ф-6-Ф), как и ее
продукт (ФДФ), могут активировать фермент.
К другим активаторам относятся аммиак и фосфат. Их концентра-
ция начинает возрастать сразу с началом мышечной работы, и такая сти-
муляция активности может быть существенной для преодоления инги-
бирующих эффектов снижения pH, которое обычно отмечается в это
время. Некоторые ключевые пункты, участвующие в регуляции распада
95
Рис. 3.8. Интеграция утилизации жиров и углеводов, достигающая координацией ско-
рости протекания многих ключевых реакций
углеводов, представлены на рис. 3.8. Интегрированный же метаболичес-
кий ответ на физическую нагрузку более подробно обсуждается в главах
6 и 7.
Пируваткиназная активность регулируется несколькими из тех же
факторов, которые воздействуют на активность ФФК: активацей высо-
кими концентрациями АДФ и угнетением АТФ и КФ.
Пируватдегидрогеназа (ПДГ) не является одиночным ферментом, а
представляет собой тройной энзиматический комплекс, который может
существовать как в активной (а) дефосфорилированной, так и неактив-
ной (Ь) фосфорилированной формах. Регуляция активности этого энзи-
матического комплекса является центральной в интеграции метаболи-
ческого ответа на физические нагрузки, однако механизм этой регуля-
ции до конца не выяснен. Как и в других подобных ферментах, взаимо-
превращение модулируется специфической киназной и фосфатазной ре-
акциями. Поэтому регуляция активности энзиматического комплекса
происходит в результате модификации активности взаимопревращаю-
щих ферментов, которая детерминирует количество активной формы
фермента. Физические нагрузки вызывают повышение в скелетных
мышцах концентрации пирувата и кальция, снижение отношения
АТФ/АДФ, увеличение отношения НАДН/НАД+, а также снижение от-
ношения ацетил-КоА/KoA-SH (KoA-SH представляет собой свободную
96
форму КоА). Все эти факторы способствуют увеличению количества ак-
тивной формы ПДГ-комплекса, что обусловлено влияниями взаимопрев-
ращающихся ферментов.
Гормональные влияния расширяют регуляцию углеводного метабо-
лизма за границы клеток, что позволяет интегрировать участие в нем
различных тканей. Подробно этот вопрос рассматривается ниже.
6. Утилизация углеводов
в различных тканях
Некоторые ткани, в наибольшей степени это касается красных кровяных
клеток (но также мозгового слоя — почки и сетчатки), не имеют мито-
хондрий, поэтому не способны к окислительному метаболизму. Следо-
вательно, эти ткани для энергообеспечения вынуждены «полагаться» ис-
ключительно на гликолиз. Большинство тканей, однако, могут использо-
вать различные виды энергетического «топлива» и имеют возможность
выбора анаэробного или аэробного метаболизма.
Скелетные мышцы служат хорошим примером ткани, которая мо-
жет удовлетворять большинство своих энергетических потребностей за
счет окислительного или анаэробного метаболизма. При этом выбор
энергетического топлива зависит от большого количества факторов,
включающих метаболические возможности ткани, поступление суб-
страта и кислорода. Хотя преобразование глюкозы до лактата и является
анаэробным процессом, тем не менее оно происходит и тогда, когда кис-
лород свободно поступает к мышцам, в связи с чем выход лактата не
обязательно предполагает неадекватное кислородное обеспечение.
Как отмечалось в главе 1, скелетные мышцы человека содержат два
основных типа волокон — I и II. Волокна типа II могут быть подразделе-
ны на два подтипа — Па и Пб. Функцию этих различных типов волокон
лучше понять, если обозначить их как медленносокращающиеся окисли-
тельные (тип I), быстросокращающиеся окислительные (подтип Па) и
быстросокращающиеся гликолитические (подтип Пб). Если мышцам не-
обходимо проявлять только низкую сократительную активность, то это
достигается рекруитированием некоторого количества волокон типа I.
Система кровообращения характеризуется инертностью и поэтому
в начале мышечной работы ее производительность возрастает медленно.
Кислородный дефицит компенсируется включением гидролиза КФ и
анаэробного гликолиза. Однако по истечении 1—2 мин физической на-
грузки поступление кислорода достигает устойчивого состояния. Высо-
7 М13 97
кая окислительная способность активных мышечных волокон способ-
ствует тому, что некоторое количество лактата, выработанного в началь-
ной фазе выполнения физического упражнения, используется этими во-
локнами для образования пирувата, который затем декарбоксилируется
до ацетил-КоА и вступает в цикл трикарбоновых кислот. Поэтому кон-
центрация лактата в крови (и мышцах) достигает пика на ранних стади-
ях выполнения физических упражнений субмаксимальной аэробной
мощности, но затем начинает снижаться даже несмотря на то, что мощ-
ность выполняемой работы остается постоянной.
Высокая способность этих волокон к окислению жиров также благоп-
риятствует тому, что потребности в энергии в наибольшей степени удов-
летворяются за счет окисления жирных кислот, а это способствует сбере-
жению ограниченных углеводных резервов. Если же данные волокна ис-
пользуют углеводы, то последние будут подвергаться полному окислению
до диоксида углерода и воды, тем самым максимизируя ресинтез АТФ.
При увеличении сократительной активности мышцы начинают рек-
руитировать некоторое количество волокон типа II: вначале подключают-
ся к сокращениям волокна подтипа Па, а затем — развивающие большую
мощность волокна подтипа Пб. При этом потребление кислорода возрас-
тает по мере увеличения мощности выполняемой работы, что свидетель-
ствует о повышении скорости окислительного метаболизма. Однако как
только к сокращениям подключаются волокна, характеризующиеся более
высокой активностью гликолитических ферментов по сравнению с ак-
тивностью окислительных энзимов, наступает момент, когда скорость
образования пирувата в процессе гликолиза в этих волокнах превышает
скорость его включения в ЦТК. Поэтому избыток пирувата должен вос-
станавливаться до лактата для обеспечения регенерации НАД+ и поддер-
жания анаэробного метаболизма. При такой высокой интенсивности фи-
зической нагрузки концентрация лактата в крови возрастает прогрессив-
но в течение всего периода выполнения упражнения, неизбежно непро-
должительного из-за развития утомления. Лактат будет продуцироваться
даже в том случае, если доставка кислорода к мышечным клеткам не ог-
раничена. Поэтому накопление лактата в крови отражает преимущест-
венно активацию мышечных волокон, в которых гликолитические воз-
можности превышают окислительный метаболизм пирувата. Неодинако-
вые способности различных типов волокон скелетных мышц человека
осуществлять распад углеводов определяются различиями в них актив-
ности фосфорилазы: ее активность в волокнах подтипа Пб выше актив-
ности в волокнах подтипа Па или типа I (Эссен и др. (Essen et al.), 1976).
Неодинаковая активность ферментов в различных типах волокон
скелетных мышц создает благоприятные условия для обмена углевод-
98
ным субстратом между волокнами в одной и той же мышце. Если в ра-
боту включаются волокна типа II и при этом происходит выход лактата
во внеклеточное пространство, то последний может быть захвачен и за-
тем подвергнут окислению присутствующими рядом волокнами типа I,
которые обладают высокой окислительной способностью. В связи с та-
ким захватом некоторое количество лактата, образующегося в процессе
физической нагрузки, не будет поступать в кровь. Миокард можно рас-
сматривать как мышцу, обладающую чрезвычайно высокой окисли-
тельной, но низкой гликолитической активностью и способной исполь-
зовать лактат в качестве энергетического топлива для окисления, если
он поступает из крови.
Печень занимает центральное место в углеводном гомеостазе, фун-
кционируя как гликогенный резервуар. Наличие в ней фермента глюко-
зо-6-фосфатазы позволяет свободной глюкозе выходить из клеток для
поддержания ее концентрации в крови. Высокая способность печеноч-
ной ткани к проявлению глюконеогенеза также позволяет печени играть
главную роль в снабжении мозга и других тканей, являющихся обяза-
тельными потребителями глюкозы, во время голодания или недостаточ-
ного поступления в организм углеводов. Даже в течение короткого пери-
ода воздержания от приема пищи, продолжающегося, например, в тече-
ние ночи, содержание гликогена в печени заметно снижается, а глюконе-
огенез ускоряется. После завершения мышечной работы, вызвавшей по-
вышение в крови концентрации лактата, значительная часть последнего
поступает в печень и используется для ресинтеза глюкозы.
7. Глюконеогенез: образование глюкозы
из неуглеводных источников
К концу физической нагрузки запасы гликогена в мышцах и печени мо-
гут истощаться. Степень этого истощения зависит от многих факторов,
главными из которых являются продолжительность и интенсивность мы-
шечной работы. При выполнении интенсивных физических упражнений,
приводящих к развитию утомления в течение нескольких минут, значи-
тельная часть запаса мышечного гликогена преобразуется в лактат, тогда
как запас гликогена в неработающих мышцах остается неизменным. При
этом происходит и некоторое снижение содержания гликогена в печени.
Более продолжительные (1-2 ч) и утомительные физические упражнения
могут привести к полному истощению запасов гликогена как в мышцах,
так и в печени. В главах 6 и 7 эти изменения описаны более подробно.
7*
99
ткань
Рис. 3.9. Глюконеогенез, происходящий прей-
мущественно в печени с использованием лакта-
та, глицерола и аминокислот, освобождающихся
из других тканей
Восполнение запасов
гликогена в мышцах и пече-
ни является существенной
частью восстановительного
процесса. При отсутствии
поступления углеводов с
пищей некоторое количес-
тво гликогена может ресин-
тезироваться за счет ис-
пользования неуглеводных
источников, однако прием
углеводсодержащих про-
дуктов существен для пол-
ного его восстановления.
Факторы питания, влияю-
щие на восстановление за-
пасов гликогена в мышцах и
печени, будут рассмотрены
в главе 7.
Глюконеогенез (образо-
вание глюкозы из неглюкоз-
ных источников) позволяет восполнить содержание некоторого коли-
чества глюкозы в организме даже в отсутствие приема пищи, что
имеет важное значение при голодании для тканей, использующих
только глюкозу в качестве энергетического топлива. Глюконеогенез
позволяет поддерживать метаболизм лактата, вырабатываемого тканя-
ми (например, эритроцитами). Обращаемость общего количества лак-
тата в организме может превышать 100 г-суг-1, и его потери с мочой
и потом чрезвычайно малы.
Процесс глюконеогенеза обеспечивается участием в нем различных
субстратов (рис. 3.9). Для этого могут быть использованы как лактат и
пируват, образующиеся в результате гликолиза, так и глицерольная осно-
ва молекул триглицеридов и углеродные каркасы некоторых амино-
кислот.
Высокоинтенсивные физические нагрузки вызывают увеличение
концентрации лактата в крови. Если после такой мышечной работы вы-
полняются физические упражнения малой интенсивности, то некоторое
количество образовавшегося лактата будет использоваться в качестве
энергетического источника для окисления в волокнах типа I, активных
преимущественно при малоинтенсивных физических нагрузках. Часть
лактата будет также поступать для окисления к сердцу.
100
Во время отдыха после мышечной работы скорость расхода энергии
снижается, при этом отпадает необходимость в энергопродукции за счет
окисления находящегося в крови лактата. В такой ситуации наибольшая
его часть будет использована для образования глюкозы.
Глюконеогенез происходит преимущественно в печени, в меньшей
степени — в почках. По мнению ряда ученых, протекание этого процесса
возможно также в скелетных мышцах. По-видимому, в какой-то мере он
все же может происходить в мышечных клетках с использованием лакта-
та в качестве исходного продукта в случае, когда концентрация последне-
го высокая, а содержание гликогена в мышцах — незначительное.
Реакции глюконеогенеза включают, где это возможно, обратимые
реакции гликолиза, однако необратимые реакции должны быть сопут-
ствующими и выдерживать определенное направление. Глюконеогенез
является энергозависимым процессом и может происходить только при
наличии в клетке АТФ.
Ниже представлены необратимые реакциии гликолиза и условия,
благодаря которым они являются сопутствующими:
Фермент
Реакция
Сопутствующий фермент
Гексокиназа
Фосфофруктокиназа
Пируваткиназа
Глюкоза —> Г-6-Ф
Ф-6-Ф -> ФДФ
ФЕП —> Пируват
Глюкозо-6-фосфатаза
ФДФ-аза
Пируваткарбоксилаза +
+ фосфоенолпируватфосфокиназа
Глицерол может подключаться к глюконеогенезу при пролонгиро-
ванных физических нагрузках, когда скорость окисления жиров, а поэ-
тому и гидролиз триглицеридов, относительно высокая. Однако в коли-
чественном отношении вклад глицерола в гомеостаз глюкозы во время
мышечной работы незначительный. В других условиях, как, например,
при длительном голодании, глицерол играет ведущую роль для поддер-
жания возможности поступления глюкозы в те ткани, для которых она
является основным энергетическим субстратом. В этом случае из жиро-
вых запасов организма освобождается около 20 г глицерола в сутки и
большая часть его используется для образования глюкозы. Глюконеоге-
нез из глицерола осуществляется в результате его поступления в гепато-
циты (молекулы глицерола имеют небольшой размер и поэтому легко
диффундируют в клетку).
Глицеролкиназа за счет использования одной молекулы АТФ
фосфорилирует поступивший в клетку глицерол, в результате чего
образуется глицерол-3-фосфат, который в дальнейшем преобразует-
ся до дигидрооксиацетонфосфата в ходе НАД+/НАДН-зависимой ре-
101
акции, катализируемой глицерол-3-фосфатдигидрогеназой. Дигид-
рооксиацетонфосфат может затем включаться в гликолитический
путь и использоваться для глюконеогенеза. Понятно, что эта реакция
может происходить только при наличии в клетке значительного ко-
личества АТФ и при допустимом значении соотношения
НАД7НАДН.
Углеродный каркас большинства аминокислот может использовать-
ся для синтеза глюкозы, в связи с чем их называют гликогенными. Эти
аминокислоты могут быть метаболизированы до пирувата или интерме-
диатов ЦТК и впоследствии использоваться для синтеза глюкозы. Мета-
болизм двух аминокислот (лейцина и лизина) приводит к образованию
ацетил-КоА, однако в данном случае отсутствует механизм, который мог
бы привести к образованию глюкозы, хотя синтез кетоновых тел и воз-
можен. По этой причине такие аминокислоты называют кетогенными.
Окисление белков во время выполнения мышечной работы способно
вносить лишь незначительный вклад в общую энергопродукцию, одна-
ко глюконеогенез позволяет использовать менее значимые для организ-
ма белки для пополнения энегретических запасов и поддержания уровня
глюкозы в крови при длительном голодании. Подробно аминокислотный
метаболизм описан в главе 5.
8. Синтез гликогена
Для восполнения запаса углеводов в печени и мышцах необходимо,
чтобы глюкоза, поступающая из пищи, так же, как и образующаяся в
результате глюконеогенеза, могла быть использована для ресинтеза
гликогена. Общим интермедиатом для глюкогенеза, происходящего в
результате использования глюкозы крови либо глюкозы, образующей-
ся в результате глюконеогенеза, служит глюкозо-6-фосфат. Первая бо-
лее предпочтительна для мышц, тогда как второй источник, очевидно,
больше подходит для печени. В дальнейшем Г-6-Ф при участии фос-
фоглюкомутазы превращается в глюкозо-1-фосфат, который затем вза-
имодействует с уридинтрифосфатом (УТФ) с образованием УДФ-глю-
козы и пирофосфата (ПФн) в ходе реакции, катализируемой Г-1-Ф-ури-
динтрансферазой (рис. 3.10). После этого глюкозный остаток прикреп-
ляется а-1,4-гликозидной связью к свободному концу ранее образован-
ной молекулы гликогена (реакция катализируется гликогенсинтетазой).
Точки ветвления вводят в структуру гликогена путем активации фер-
мента ветвления. Когда длина концевой цепочки составит около
12 глюкозных остатков, этот фермент отделяет цепочку длиной около
102
7 остатков и прикрепляет
ее к соседней цепочке
а-1,6-связью.
Активность гликоген-
синтетазы играет решаю-
щую роль для процесса
синтеза гликогена, и этот
фермент, подобно многим
другим, существует в ак-
тивной (гликогенсинтета-
заа) и неактивной (глико-
генсинтетаза б) формах,
взаимопревращение кото-
рых катализируется специ-
фическими энзимами.
Гормональный кон-
АТФ
АДФ
Глюкоза Глюкозо-6-фосфат
Г ексокиназа
Фосфоглюкомутаза
Глюкозо-1 -фосфат
УДФГ -пирофосфорилаза
УДФ-глюкоза
УДФГ-глюкозилтрансфераза
г— Гликогеннглк>кмв
Гликогенн + ,глижоэа
УДФ
Рис. 3.10. Путь начинающегося от глюкозы син-
теза гликогена (предполагается, что каркас гпико-
троль активности гликоген-
синтетазы осуществляется
адреналином, который, ак-
гена всегда доступен для присоединения дополни-
тельных глюкозных единиц)
тивируя протеинкиназу, превращает синтетазу в неактивную форму, а
также инсулином, обеспечивающим синтез гликогена. На локальном
уровне регуляция проявляется в том, что происходящее во время сокра-
щения увеличение содержания кальция в саркоплазме снижает глико-
генсинтетазную активность.
Восстановление в печени и мышцах запасов гликогена является
важной частью восстановительного процесса после пролонгирован-
ных физических нагрузок. При адекватном потреблении углеводов
для восполнения запасов обычно требуется около 24-36 часов. При
этом скорость синтеза в начале восстановительного периода в мышеч-
ных волокнах типа I выше, чем в волокнах типа II. Скорость синтеза
гликогена очень зависит от содержания углеводов в потребляемой пи-
ще, и ограничение их поступления в организм препятствует восста-
новлению нормального его запаса в тканях. Скорость гликогенового
синтеза можно повысить, если потребление высокоуглеводной пищи
начать сразу по окончании пролонгированной физической нагрузки.
Отсрочка потребления углеводов даже на 2 часа может препятствовать
достижению максимальной скорости ресинтеза. Высокая скорость
поступления глюкозы в мышцы во время выполнения мышечной ра-
боты отмечается еще некоторое время после ее окончания, что обес-
печивает доставку субстрата для ресинтеза гликогена. Потребление
фруктозы после физической нагрузки благоприятствует восстановле-
103
нию запасов гликогена в печени, но не способствует этому процессу в
мышцах, тогда как поступление в организм глюкозы более предпочти-
тельно для восполнения гликогенного резерва в мышечной ткани. Са-
хароза одинаково способствует восстановлению этого резерва как в
мышцах, так и в печени.
9. Гормональная регуляция
метаболизма углеводов
Многие гормоны играют важную роль как в регуляции скорости ути-
лизации углеводов, так и в интеграции обеспечения и утилизации уг-
леводного топлива в различных тканях в покое и во время физических
нагрузок. Концентрация глюкозы в крови должна поддерживаться в
узких пределах и недопустимо, чтобы она значительно возрастала
после приема высокоуглеводной пищи или слишком снижалась из-за
многократного увеличения скорости утилизации глюкозы работающи-
ми мышцами. В равной степени запасы мышечного гликогена являют-
ся важным энергетическим топливом для обеспечения сократитель-
ной активности мышц, однако эти запасы присутствуют в ограничен-
ном количестве и должны быть преобразованы. Интегральный мета-
болический ответ на физическую нагрузку подробно рассмотрен в
главах 6 и 7, однако гормональные факторы, влияющие на углеводный
метаболизм, будут кратко охарактеризованы в настоящем параграфе.
Поскольку влияние на углеводы нельзя рассматривать изолированно,
то ниже кратко анализируются эффекты и на другие метаболические
процессы.
Инсулин является полипептидным гормоном, состоящим из двух
пептидных цепей (A-цепи, состоящей из 21 аминокислоты, и В-цепи,
состоящей из 30 аминокислот), который играет ключевую роль в регуля-
ции углеводного метаболизма. Поскольку инсулин выполняет регулятор-
ную функцию в липидном и белковом обмене, то его можно рассматри-
вать в качестве главного гормона, обеспечивающего гомеостаз энергети-
ческого топлива в организме. Секреция инсулина 6-клетками островков
Лангерганса поджелудочной железы стимулируется повышением кон-
центрации глюкозы в крови (рис. 3.11).
Как в скелетных мышцах, так и в миокарде повышение концентра-
ции инсулина в крови вызывает увеличение количества активных пере-
носчиков глюкозы и тем самым стимулирует ее мембранный транспорт.
Гликогенсинтетаза также приходит в активное состояние в результате
104
| Глюкоза крови
Секреция инсулина
Рис. 3.11. Концентрация глюкозы в кро-
ви и секреция инсулина, регулируемые
по механизму обратной связи
воздействия инсулина на фосфа-
тазный фермент, регулирующий
взаимопереход активной и неак-
тивной форм гликогенсинтетазы. В
противоположность стимуляции в
мышцах инсулин не оказывает
влияния на транспорт глюкозы в
печени, хотя клеточная мембрана
гепатоцитов свободнопроницаема
для глюкозы. Однако при этом не
происходит увеличение активнос-
ти гексокиназы, которая фосфори-
лирует попадающую в клетку глюкозу. В некоторой степени это может
служить доказательством того, что повышение синтеза гликогена про-
исходит в результате активации гликогенсинтетазы, чем и обеспечива-
ется увеличение резерва гликогена. В такой ситуации замедляется и
скорость глюконеогенеза в печени, а также снижается гликогенолиз, что
связано с угнетением активности фосфорилазы. Поступление глюкозы
в мозг и ее поледующий метаболизм в нервной ткани нечувствительны
к инсулину.
Катехоламины (адреналин и норадреналин) проявляют много кар-
диоваскулярных и метаболических эффектов. Секрет мозгового вещес-
тва надпочечников включает в себя около 80 % адреналина и 20 % но-
радреналина, однако норадреналин, освобождающийся из нервных
окончаний, также поступает в кровь. При физических нагрузках или
других видах стресса концентрация катехоламинов в крови повышается.
В клеточной мембране адреналин активирует мембранносвязанный фер-
мент аденилциклазу, в результате чего возрастает скорость образования
цАМФ в клетках-мишенях. Повышение концентрации внутриклеточно-
го цАМФ приводит к активации фосфорилазы и возрастанию скорости
распада гликогена в работающих мышцах. При этом стимуляция глико-
генолиза в неработающих мышцах в значительной степени предотвра-
щается низкой концентрацией в клетке свободного кальция. Катехола-
мины также стимулируют гликогенолиз в печени и липолиз в адипозной
ткани, обеспечивая тем самым эффективную мобилизацию энергетичес-
кого топлива, которое может быть использовано мышцами.
Активность глюкагона — одноцепного полипептида, состоящего из
29 аминокислот, который также секретируется поджелудочной железой,
проявляется в основном в его антагонизме по отношению к эффектам
инсулина. Концентрационное отношение этих двух гормонов определя-
ет их интегральное воздействие на мобилизацию энергетических источ-
105
ников. Эффекты увеличения концентрации глюкагона проявляются в
стимуляции гликогенолиза и глюконеогенеза в печени, а также стимуля-
ции липолиза в адипозной ткани, благодаря чему повышается поступле-
ние свободных жирных кислот к работающим мышцам.
Некоторые другие гормоны и пептиды также играют важную
роль в мобилизации энергетического топлива. Гормон роста, выделя-
емый передней долей гипофиза, обеспечивает синтез белков, но так-
же стимулирует глюконеогенез в печени и снижает периферическую
утилизацию глюкозы. Этот гормон стимулирует липолиз в адипозной
ткани, однако такой эффект проявляется только в присутствии глюко-
кортикоидов.
Кортизол является глюкокортикоидным гормоном, выделяемым ко-
рой надпочечников, разнообразные влияния которого потенциально свя-
заны с эффектами других гормонов. Он повышает глюконеогенный от-
вет печени в присутствии адреналина и глюкагона, а также увеличивает
скорость липолиза в адипозной ткани в присутствии адреналина и гор-
мона роста. Тироидные гормоны — трийодтиронин и тироксин в основ-
ном стимулируют мобилизацию энергетических источников.
В связи с многочисленностью возможных регуляторных факторов и
разнообразием их взаимодействия между собой гормональный и мета-
болический ответ на какой-либо метаболический сдвиг, наступающий,
например, после приема пищи или при физической нагрузке, носит ком-
плексный характер и связан с участием в нем большого количества раз-
личных тканей, а также различных регуляторных процессов.
После приема углеводной пищи концентрация инсулина в крови
сразу же возрастает, что обеспечивает устранение глюкозы, поступаю-
щей в кровь, а концентрация глюкагона снижается.
Выделение инсулина происходит в ответ на изменение концентра-
ции глюкозы, которое контролируется 6-клетками поджелудочной желе-
зы по механизму обратной связи (см. рис. 3.11). Функционирование этой
системы более сложное, поскольку изменение в плазме аминокислот-
ного уровня также влияет на выделение инсулина. На поджелудочную
железу оказывает воздействие и изменение в плазме концентрации адре-
налина, который проявляет мощный угнетающий эффект на секрецию
инсулина, изменение концентрации в крови других гормонов, а также
симпатические и парасимпатические нервные влияния (рис. 3.12).
После приема пищи большое количество малых по размеру пептидов
выделяется кишечником, некоторые из них стимулируют инсулиновую
секрецию.
Повышение в крови уровней глюкозы, аминокислот и жирных кис-
лот (например, после приема смешанной пищи) стимулирует выделение
106
аминокислот
в плазме
| Концентрация \
глюкозы в плазме \
Симпатические
нейроны
- Поджелудочная
+ железа
Парасимпатические *—z---z—
нейроны 1
| Концентрация /
адреналина /
в плазме /
- Соматостатин
Рис. 3.12. Регуляция секреции инсулина и об-
щего метаболизма углеводов, представляющих
сложный процесс, в котором участвует много
различных гормонов
из у-клеток соматостатина,
состоящего из 14 амино-
кислот пептида. Соматоста-
тин действует локально в са-
мой поджелудочной железе,
угнетая инсулиновую секре-
цию, к тому же это вызывает
снижение перистальтики и
выделения пищеварительно-
го сока, а следовательно, за-
медляет абсорбцию пищи. В
мышцах и печени происходит
активация метаболического
пути, обеспечивающего син-
тез гликогена. Мобилизация
жирных кислот из адипозной
ткани подавляется, скорость
аминокислотного распада также снижается. Следовательно, происходит
активация анаболических и угнетение катаболических реакций.
На ранних стадиях пролонгированных физических нагрузок ути-
лизация жирных кислот из плазмы повышается. Вероятно, это проис-
ходит потому, что возрастает ток плазмы через мышечные капилляры,
в связи с чем концентрация СЖК в крови снижается. Дальнейшее
продолжение выполнения мышечной работы сопровождается возрас-
танием в плазме уровней адреналина, норадреналина, глюкагона,
кортизола и гормона роста, тогда как секреция инсулина подавляется.
Увеличение в крови концентрации катехоламинов оказывает мощное
ингибирующее влияние на секрецию инсулина, а также активирует
фосфорилазу и стимулирует липолиз. Эти изменения способствуют
выходу СЖК из адипозной ткани и повышению их концентрации в
плазме, а следовательно, и их утилизации работающими мышцами.
Снижение в крови уровня инсулина может ослабить поступление глю-
козы в мышечные клетки во время физической нагрузки, однако этому
противодействуют другие факторы, которые оказывают стимулирую-
щее влияние на переносчиков глюкозы и поддержание углеводного
обеспечения. Даже при очень низких концентрациях инсулина отмеча-
ется высокая скорость глюкозного транспорта. Происходит это, оче-
видно, потому, что увеличение внутриклеточной концентрации каль-
ция может стимулировать активность транспортеров глюкозы, скорее
всего, путем рекруитирования тех из них, которые ранее были неактив-
ными. Детальнее эти изменения описаны в главе 7.
Инсулин
107
10. Методология и достижения
в изучении углеводного метаболизма
Многое из того, что стало нам известно об углеводном метаболизме, свя-
зано с применением специфических методов исследований. В самых
ранних работах, касающихся выяснения путей перемещения интермеди-
атов в процессах распада и синтеза углеводов, использовались различ-
ные изолированные клетки или субклеточные препараты. Нарушение
структурной целостности явно предполагает и некоторую утрату инфор-
мации, важной, например, для понимания процессов, связанных с мем-
бранным транспортом. Использование очищенных препаратов фермен-
тов для изучения кинетики реакций может также привести к получению
результатов, неприемлемых в условиях живой клетки, в связи с чем к ин-
терпретации полученных таким способом данных следует подходить с
некоторой осторожностью.
Возобновление применения методики пункционной биопсии для
изучения мышечного метаболизма, которая была разработана в конце
XIX ст. (однако сразу не нашла применения), способствовало в 60-70-е
годы значительному развитию этой области биохимии. Методика прос-
та, совершенно безболезненна, лишена существенного риска и позволя-
ет производить повторные взятия образцов из крупных мышц. Именно
благодаря ей было установлено, что во время выполнения физических
упражнений происходит прогрессивная утилизация мышечного гликоге-
на; при работе только одной ногой содержание гликогена в мышцах не-
работающей конечности изменялось незначительно или оставалось не-
изменным. Скорость утилизации гликогена оказалась пропорциональ-
ной интенсивности физической нагрузки и возрастала экспоненциально
по мере возрастания мощности выполняемой работы. При выполнении
высокоинтенсивных физических упражнений развитие утомления ока-
залось не взаимосвязанным со снижением в мышцах резерва гликогена,
поскольку его содержание в утомленных мышцах продолжало оставать-
ся достаточно высоким. Было продемонстрировано, что в это же время
происходит накопление лактата, истощение запаса КФ и только незначи-
тельное снижение содержания АТФ в мышцах.
Проведенными исследованиями удалось установить, что восстанов-
ление запасов мышечного гликогена протекает относительно медленно
и что скорость этого процесса прежде всего зависит от содержания угле-
водов в потребляемой пище.
Гистохимические методы исследований применялись для изучения
изменения содержания гликогена в различных типах волокон. Позднее
108
эти методы были усовершенствованы путем применения диссекции от-
дельных мышечных волокон, которые затем подвергались биохимичес-
кому анализу, что позволяло получить количественную характеристику
утилизации гликогена в различных типах мышечных волокон при раз-
личных видах мышечной деятельности. Однако возможности примене-
ния такого метода несколько ограничены. Связано это с тем, что в слу-
чаях, когда для изучения углеводного метаболизма требуется взятие нес-
кольких образцов, наиболее приемлемым будет изъятие их на близком
расстоянии друг от друга (по крайней мере на расстоянии 2,5 см от мес-
та получения предыдущего образца). Однако сама по себе такая проце-
дура уже нарушает нормальное протекание обменных процессов в
участках мышечной ткани, где производится взятие образцов.
Успешное применение метода пункционной биопсии ограничено
при проведении исследований, связанных с изучением локальной утили-
зации жировых запасов при мышечной деятельности. Дело в том, что
определенное количество жира аккумулируется в мышечных клетках в
виде капелек, а некоторая часть его представлена в виде адипозных кле-
ток, располагающихся между мышечными волокнами. Методика пунк-
ционной биопсии, к сожалению, не позволяет определить разницу меж-
ду этими двумя жировыми депо.
В ряде работ применялась усовершенствованная методика мышеч-
ной биопсии для взятия образцов печени и определения содержания в
них гликогена у испытуемых как в состоянии покоя, так и в условиях не-
одинаковой двигательной активности и питания. Эта методика не лише-
на определенного риска, поскольку, в отличие от мышечной биопсии,
после взятия образцов печени не так легко остановить кровотечение.
В настоящее время широко применяются методы, связанные с ис-
пользованием различных изотопов, что позволяет определить вклад
энергетических источников в утилизацию субстратов при физических
нагрузках. Инфузия с постоянной скоростью изотопов глюкозы в кон-
тролируемых концентрациях позволяет определить ее количество, пос-
тупающее из печени в кровь, а обнаружение меченых атомов углерода в
выдыхаемом воздухе указывает на скорость ее окисления.
Окисление поступающих с пищей углеводов может быть изучено
аналогичным образом. Выделение меченых интермедиатов из крови и
мышц дает возможность проследить за их перемещением в процессе ме-
таболизма. Эти методы достаточно информативны для оценки скорости
окисления различных субтратов при мышечной деятельности.
Тканевой, или органный метаболизм может быть изучен путем оп-
ределения концентрационной разницы исследуемых веществ в артери-
альной и венозной крови, проходящей через ткани: если известна ско-
109
рость кровотока в ткани, то можно рассчитать и показатель субстратно-
го обмена.
Экстракция кислорода и субстратов, а также выход метаболитов из
мышц как в состоянии покоя, так и при физических нагрузках могут
быть определены путем введения канюль в бедренную артерию и бед-
ренную вену, которые дренируют переднюю группу мышц бедра. Эта
методика может быть применена в сочетании с использованием изо-
топных меток.
Современные разработки методики магнитно-резонансной спек-
троскопии расширяют возможности изучения упомянутых выше мета-
болических процессов. Эта методика позволяет неинвазивно определять
в тканях изменение концентраций гликогена, КФ, АТФ, АДФ, АМФ, а
также изменение в тканях pH.
На практике картина биохимического ответа организма на физичес-
кие нагрузки вырисовывается благодаря информации, получаемой от
всех доступных источников.
Выводы
1. Углеводы являются обычными, в наибольшей мере потребляемыми с
пищей источниками энергии для организма (около 40-60 % общего
энергопотребления). Однако запасы углеводов в организме незначитель-
ны: около 80-110 г гликогена запасено в печени и около 250-400 г — в
мышцах.
2. Углеводы используются для энергообеспечения путем анаэроб-
ного метаболизма, конечным продуктом которого является лактат, или
путем полного их окисления до диоксида углерода и воды. Чистый «вы-
ход» гликолиза заключается в преобразовании одной молекулы глюкозы
до двух молекул пирувата. Этот процесс обеспечивает ресинтез двух мо-
лекул АТФ, который приходится на одну молекулу распавшейся глюко-
зы. Если первоначальным субстратом гликолиза является гликоген, то
каждая глюкозная единица при этом обеспечивает генерацию трех моле-
кул АТФ.
3. Процесс гликолиза включает в себя восстановление НАД' до
НАДН, что приводит к снижению внутриклеточного пула НАД+. Восста-
новление пирувата до лактата способствует регенерации НАД+ из
НАДН. Альтернативным путем регенерации НАД+ служит преобразова-
ние пирувата до ацетил-КоА для субстратного окисления в ЦТК.
4. Накопление лактата в мышцах будет происходить в тех случаях,
когда скорость анаэробного гликолиза превышает скорость пируватде-
110
гидрогеназной реакции, которая преобразует пируват до ацетил-КоА.
Накопление лактата сопровождается накоплением водородных ионов,
которые могут влиять на сократительную активность мышц.
5. Ограниченные углеводные запасы организма быстро истощаются
при физических нагрузках (мышечный гликоген) или при голодании
(гликоген печени). Обычно резерв мышечного гликогена расходуется
после 1-2 часов напряженной мышечной работы. Чем выше интенсив-
ность физической нагрузки, тем быстрее снижается содержание гликоге-
на в мышцах, однако при наступлении утомления это снижение проис-
ходит не до конца.
6. Углеводы являются главным энергетическим топливом, обеспечи-
вающим способность мышц выполнять высокоинтенсивную физичес-
кую нагрузку. Время, в течение которого может поддерживаться мышеч-
ная работа определенной интенсивности, в значительной степени взаи-
мосвязано с резервом гликогена в мышцах. Это также зависит от харак-
тера предшествующей мышечной деятельности, а также особенностей
питания в предшествующие часы и дни.
7. Глюконеогенез как процесс синтеза углеводов из неуглеводных
источников происходит преимущественно в печени и может способство-
вать поддержанию обеспечения углеводами таких тканей, как мозг и
эритроциты, для которых углеводы являются основным энергетическим
топливом.
8. К интеграции и регуляции углеводного метаболизма причастны
многие гормоны, особенно инсулин, способствующий образованию уг-
леводных запасов, а также глюкагон, являющийся антагонистом инсули-
на. Адреналин и норадреналин стимулируют углеводный метаболизм
при стрессе.
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
Essen В., Jansson Е., Henriksson J, Taylor A. W., Saltin В. (1976). Metabolic characteristics
of fibre types in human skeletal muscle //Acta Physiol. Scand. — 95. — P. 153-165.
Endurance-training adaptations increase the capacity of muscle to oxidize lipid.
МЕТАБОЛИЗМ ЛИПИДОВ
1. Роль липидов
Липвды, или жиры содержат такие же структурные эле-
менты, как и углеводы, а именно — углерод, водород и кис-
лород, однако отношение количества атомов водорода к
атомам кислорода в молекулах жиров значительно выше, чем
в углеводах. Так, например, химическая формула такого рас-
пространенного липида, как трипальмитин (трипальмитоил-
глицерол), выглядит следующим образом: C5)H9gO6. Боль-
шинство липидов можно разделить на три основные группы:
простые липиды, сложные липиды и производные липидов.
Липиды присутствуют в продуктах питания как растительно-
го, так и животного происхождения и в основном малорас-
творимы в воде. Они содержат самый высокий запас хими-
ческой энергии, который может быть использован для обес-
печения разнообразных биологических процессов, включая
мышечное сокращение. Запасы липидов, депонированные в
адипозной ткани, предохраняют от охлаждения и механичес-
ких повреждений жизненно важные органы. Липиды явля-
ются важным структурным компонентом мембран. Содержа-
щиеся в пище липиды служат поставщиками в организм жи-
рорастворимых витаминов A, D, Е и К.
2. Типы липидов
Липиды могут быть классифицированы в зависимости от их
структуры. По этому признаку их подразделяют на три основ-
ных класса: простые липиды, сложные липиды и производ-
ные липидов.
112
Простые липиды. Простые липиды часто относят к «нейтральным ли-
пидам», основным составляющим компонентом которых являются триацил-
глицеролы (триглицериды). Они служат основной формой запаса жира в ор-
ганизме. Более 95 % жира в организме человека приходится на долю тригли-
церидов. Находятся они преимущественно в цитоплазме клеток белой ади-
позной ткани, хотя печень и скелетные мышцы также содержат жировые за-
пасы, имеющие важное биологическое значение. Молекула триацилптицеро-
ла (рис. 4.1) включает глицерольный трехуглеродный молекулярный остов, к
которому присоединены три молекулы жирных кислот. Все жирные кисло-
ты содержат длинную углеводородную цепь и герминальную карбоксиль-
ную кислотную группу (-СООН). Почти все они имеют четное число атомов
углерода — от 14 до 22; чаще всего встречаются жирные кислоты с 16 или
18 атомами углерода (табл. 4.1). Жирные кислоты могут быть насыщенными
или ненасыщенными. Насыщенные жирные кислоты (например, пальмити-
новая кислота) содержат углеродные атомы, соединенные между собой толь-
ко одной ковалентной связью (см. рис. 4.1). Остальные связи примыкают к
атомам водорода. Молекула считается насыщенной в связи с тем, что содер-
жит предельно возможное количество атомов водорода.
Обильными пищевыми источниками насыщенных жирных кислот
являются мясо, яичный желток и молочные продукты. К растительным
Таблица 4.1. Некоторые природные жирные кислоты
Число атомов углерода Структура Систематическое название Тривиальное название Точка плавления, °C
Насыщенные жирные кислоты
12 СН3(СН2)10СООН н-Додекановая Лауриновая 44,2
14 СН3(СН2)12СООН н-Т етрадекановая Миристиновая 53,9
16 СН3(СН2)14СООН н-Гексадекановая Пальмитиновая 63,1
18 СН3(СН2)16СООН н-Октадекановая Стеариновая 69,6
20 СН3(СН2)18СООН н-Эйкозановая Арахидоновая 76,5
24 СН3(СН2)22СООН н-Т етракозановая Лигноцериновая 86,0
Ненасыщенные жирные кислоты
16 СН3(СН2)5СН=СН(СН2)7СООН Пальмитоолеиновая -0,5
18 СН3(СН2)7СН=СН(СН2)7СООН Олеиновая 13,4
18 СН3(СН2)4СН=СНСН2СН=СН(СН2)7СООН Линолевая -5,0
18 СН3СН2СН=СНСН2СН=СНСН2СН= =СН(СН2)7СООН Линоленовая -11,0
20 СН3(СН2)4СН=СНСН2СН=СНСН2СН= =СНСН2СН=СН(СН2)3СООН Арахидоновая -49,5
113
Примеры
Ацилглицеролы
Общая структура фосфоглицеридов,
жирных кислот
НО О
НО О
Глицерол
Н
С
С
1
сн2
сн2
сн2
сн2
сн2
сн2
Н—С—он
Н—с—он
2
3
сн2
сн2
сн.
сн.
представленная в форме, отражающей
их амфипатическое свойство. Обычно
жирная кислота во втором положении
является ненасыщенной _____________
X
Н
1 -Моноацилглицерол
Н
н
н
н—с
О
О=Р-О'
Полярная
голова
о
3
ЬП,
'2
сн2
сн2
сн2
сн2
СН2
Н—С—О—С—R,
о
о
сн2
сн2
сн.
сн.
сн2
СН2
сн2
сн2
сн2
сн.
сн2
сн2
сн2
СН2
СН3
Пальмитиновая СН2
кислота I
СН3
Олеиновая
кислота
о
с=о
с=о
Н—с—он
Н
1,2-Диацилглицерол
Н
Н—С—О—С—R,
I О
Н—с—о—с—r2
о
сн2
сн2
СН2
сн.
сн.
сн2
сн2
СН2
сн.
сн2
сн2
сн2
сн.
сн.
СН2
сн.
сн
сн
Неполярные
хвосты
н
Т риацилглицерол
Н
СН2
сн2
Холестерол
Н—С—О—С—R,
I О
Н—с—о—с—r2
I О
Н—С—О—С—R3
н
о
сн2
сн.
СН2
сн2
сн.
сн2
сн3
сн2
сн2
сн2
СН2
СН2
сн.
сн3
СН3
НС —СН3
сн2
сн2
сн2
Рис. 4.1. Липидные структуры
114
источникам насыщенных липидов относятся кокосовое масло, пальмо-
вое масло и гидрогенатный маргарин. Жирные кислоты, содержащие, по
крайней мере, хотя бы одну двойную связь между атомами углерода,
классифицируются как ненасыщенные (рис. 4.1, табл. 4.1).
Если в углеродной цепи имеется только одна двойная межуглерод-
ная связь (например, в олеиновой кислоте), то такие жирные кислоты
называют мононенасыщенными. Такие кислоты содержатся в оливковом
масле. Двойные связи природных ненасыщенных жирных кислот имеют
г/пс-конфигурацию. Жирные кислоты с более чем одной двойной
связью, как, например, содержащиеся в зернах злаковых растений, сое-
вых бобах и подсолнечном масле, относятся к полиненасыщенным.
Липиды могут поступать в организм с пищей, но могут также и син-
тезироваться в организме. Исключение составляют такие незаменимые
жирные кислоты, как линолевая, линоленовая и арахидоновая, которые
поступают только с пищей и не могут синтезироваться в организме. Эти
незаменимые жирные кислоты необходимы для поддержания целост-
ности клеточных мембран, обеспечения нормального роста, репродук-
тивной функции, а также для поддержания здорового состояния кожи.
Арахидоновая кислота служит предшественником простагландинов, ко-
торые являются важными внутриклеточными переносчиками. В среднем
суточное потребление триацилглицерола жителями западных областей
планеты составляет 100-150 г. Поступающие с пищей триацилглицеро-
лы служат основным источником липидов для организма, хотя дополни-
тельное их количество может синтезироваться из углеводов, в частности
в том случае, когда содержание углеводов в пище высокое, атриацилгли-
церолов — низкое.
Сложные липиды. Сложные жиры представляют собой комбина-
цию нейтральных жиров с другими химическими веществами. К ним от-
носятся такие основные группы, как гликолипиды, сфинголипиды, фос-
фоглицериды и липопротеиды. Гликолипиды являются диацилглицеро-
лами (диглицеридами), в которых третья гидроксильная группа глице-
рольного каркаса образует гликозидную связь с сахаром. Сфинголипиды
содержат одну молекулу жирной кислоты в амидной связи с одной мо-
лекулой ненасыщенного аминоспирта сфингозина. Глицерол сфинголи-
пиды не содержит.
Самым распространенным сфинголипидом является сфингомиелин,
в головной части молекулы которого находится фосфохолин. Сфинголи-
пиды и гликолипиды входят в состав клеточных мембран; особенно бо-
гата ими нервная ткань и только незначительное их количество обнару-
живается в адипозных жировых депо.
8*
115
Фосфоглицериды (см. рис. 4.1) являются диацилглицеролами, в ко-
торых третья гидроксильная группа глицерольного каркаса эстерифи-
цирована с фосфорной кислотой, образующей вторую эфирную связь с
алкоголями или, чаще всего, с аминоалкоголями, например этанола-
мином, холином (табл. 4.2). Фосфолипиды входят в состав всех клеток
организма и являются важным компонентом мембран, формирующим
бислой (рис. 4.2), а также играют важную роль в процессе свертывания
крови.
Липопротеиды образуются в основном в печени и крови, представ-
ляя собой комбинацию триацилглицеролов, фосфолипидов, холестерола
и протеина. Липопротеиды составляют основную форму жиров, тран-
спортируемых кровью, и они необходимы, поскольку сами триацилгли-
Т а б л и ц а 4.2. Спиртовые компоненты полярных Х-групп основных фосфолипидов
Фосфоглицериды Алкогольные компоненты
Фосфатидилэтаноламин Фосфатидилхолин Фосфатидилсерин HOCH2CH2NH2 НОСН2СН2К(СН3)з HOCH2CHNH2COOH он он Н/1 Кон /н HXJ
Фосфатидилинозитол кон н/ НО н он
Фосфатидилглицерол НОСН2СНОНСН2ОН о
Фосфатидил-3-О-амино- ацилглицерол НОСН.СНОНСН.О- с I R-CH I NH2
Кардиолипин о II НОСН,СНОНСН,-О-Р-О-СН,СН—сн, I II' он о о I I о=с о=с I I R, R2
116
церолы практически не рас-
творяются в воде. Наличие
белкового слоя способствует
повышению растворимости
обрамленных липидов.
Липопротеиды плазмы
классифицируются в соот-
ветствии с их плотностью.
Липопротеиды высокой
плотности (ЛПВП) содержат
наименьшее количество хо-
лестерола и, по-видимому,
способствуют переносу хо-
лестерола из стенок артерий
к печени, где используется
для образования желчи.
Впоследствии желчь секре-
тируется в кишечник и затем
Полярная
голова
Неполярные
хвосты
Мицеллы в воде
Воздух
Монослой на водно-
воздушной границе
Мембранный бислой
Рис. 4.2. Устойчивые системы фосфолипид —
вода
экскретируется из организма
с фекалиями.
Липопротеиды низкой
плотности (ЛПНП) обычно
транспортируют 60-80 % общего холестерола, содержащегося в плазме
и проявляющего наибольшее сродство к артериальным стенкам. Отло-
жение в них холестерола в итоге приводит к пролиферации гладкой мус-
кулатуры, захвату фибробластов и содействию коагуляции. Эти измене-
ния вызывают повреждение и сужение артериальных стенок, что, если
это затрагивает коронарные артерии, снабжающие кровью сердечную
мышцу, может привести к неадекватному обеспечению ее кислородом
и развитию инфаркта (в конечном счете, к смерти или некрозу части
миокарда).
Концентрация ЛПВП и ЛПНП, а также их соотношение в плазме яв-
ляются важными факторами риска в предсказании возможности возник-
новения сердечных заболеваний. Высокое отношение ЛПНП/ЛПВП по-
вышает вероятность сокращения продолжительности болезни. Это от-
ношение можно улучшить (повысив концентрацию ЛПВП и снизив или
оставив без изменения концентрацию ЛПНП) путем потребления низко-
калорийной пищи с низким содержанием в ней насыщенных липидов и
холестерола. Физические тренировки, направленные на развитие вынос-
ливости (регулярное выполнение аэробных физических упражнений
умеренной интенсивности), также могут способствовать повышению
117
уровня ЛПВП и тем самым благоприятствовать созданию оптимального
отношения ЛПНП/ЛПВП.
Хиломикроны представляют собой самые крупные липопротеидные
частицы, находящиеся в плазме крови и содержащие самую высокую
пропорцию (приблизительно 85 %) триацилглицерола. Они происходят
только из пищевых источников триацилглицерола, продуцируются эпи-
телием кишечника и поступают в кровяное русло через лимфатическую
систему. Полупериод жизни плазменных хиломикронов у человека сос-
тавляет менее 1 часа и только несколько минут — у крыс. Однако пос-
тупление и всасывание в кишечнике пищевых липидов, а также продук-
ция и секреция хиломикронов продолжаются еще несколько часов после
еды, а следовательно, и липемия отмечается еще в течение нескольких
часов после приема жирной пищи.
Триацилглицеролы, будь они в форме хиломикронов или других ли-
попротеидов, не захватываются напрямую клетками тканей. Вначале мо-
лекулы триацилглицерола должны подвергнуться ферментативному
гидролизу до жирных кислот и глицеролу вне клетки. После этого жир-
ные кислоты могут поступать в нее; глицерол же транспортируется
кровью к печени и почкам, где и происходит его значительное усвоение.
Другие ткани, включая мышцы, обладают малой энзиматической спо-
собностью к утилизации глицерола. Внеклеточный гидролиз триацил-
глицеролов осуществляется при участии липопротеинлипазы, которая
присоединяется своей внешней поверхностью к эндотелиальным клет-
кам капилляров в таких тканях, как мышцы, адипозная ткань, сердце и
легкие. В печени этот фермент прикрепляется к внешней поверхности
гепатоцитов. Один из двух других энзимов, фосфолипаза А2, прикрепля-
ется к эндотелию капилляров, а другой, лецитинхолестеролацилтранс-
фераза (ЛХАТ), вместе с липопротеидом ЛПВП включаются в деграда-
цию хиломикронов и липопротеидов очень низкой плотности (ЛОНП).
Фосфолипаза А2 воздействует на фосфолипиды, находящиеся на по-
верхности липопротеидов. Фермент ЛХАТ присутствует в плазме и пе-
реносит жирную кислоту от лецитина (фосфатидилхолина) к неэстери-
фицированному холестеролу на ЛПВП с образованием неполярного
ацилхолестерольного эфира. Последний транспортируется от ЛПВП к
хиломикрону или к ЛОНП как триацилглицерол. Это способствует под-
держанию стабильности липопротеидных частиц, так как триацилглице-
рольный компонент начинает прогрессивно истощаться при прохожде-
нии через мышечную и адипозную ткани.
Производные липиды. Этот класс липидов включает субстанции,
являющиеся производными простых и сложных липидов. Наиболее рас-
пространенным из них является холестерол (см. рис. 4.1), который при-
118
сутствует только в тканях животных. Хотя холестерол и не содержит в
своем составе каких-либо жирных кислот, тем не менее он проявляет не-
которые физические и химические свойства, присущие липидам. Холес-
терол присутствует во всех клетках организма. Он входит в состав кле-
точных мембран, служит незаменимым предшественником для синтеза
витамина D и таких стероидных гормонов, как эстроген, тестостерон и
кортизол. Холестерол необходим для синтеза желчи, играющей важную
роль в эмульгировании жиров в пищеварительном тракте.
Самым богатым источником холестерола в пище является яичный
желток. Холестерол также содержится в мясе, рыбных и молочных про-
дуктах. Он отсутствует в растениях, в связи с чем вегетарианцы очень
мало потребляют его с пищей. Однако даже при полном отсутствии пос-
тупления с пищей холестерола синтез его в печени может составлять
0,5-2,0 г-сут—*. Возможна и более высокая его продукция, особенно ес-
ли пища содержит большое количество насыщенных липидов, которые
ускоряют синтез холестерола печенью.
3. Синтез липидов
Синтез жирных кислот. Синтез указанных кислот происходит в цито-
плазме печени, а также в клетках адипозной ткани. Из представленной
ниже обобщенной реакции видно, что в результате взаимодействия од-
ной молекулы ацетил-КоА с семью молекулами малонил-КоА образует-
ся одна молекула пальмитиновой кислоты. Восстановление в этой реак-
ции обеспечивается НАДФН:
1 ацетил-КоА + 7 малонил-КоА + 14НАДФН + 14Н+ ->
-> 1СН3(СН2)14СООН + 7СО2 + 8КоА + 14НАДФ+ + 6Н2О
Процесс синтеза сопровождается последовательным добавлением
единиц, происходящих от ацетил-КоА и содержащих два атома углеро-
да, к удлиненной углеродной цепи жирной кислоты (рис. 4.3).
Начинается он с карбоксилирования ацетил-КоА до образования ма-
лонил-КоА. Эта обратимая реакция катализируется ферментом ацетил-
КоА-карбоксилазой, который использует в качестве коэнзима биотин,
выполняющий функцию преносчика диоксида углерода. Последний
происходит из бикарбоната, а АТФ обеспечивает энергией протекание
реакции в направлении образования малонил-КоА.
На следующем этапе другая молекула ацетил-КоА и вновь образо-
вавшийся малонил-КоА соединяются посредством сульфидной связи
119
к ацилпереносящему белку (АПБ) при участии ферментов ацетил-
КоА-трансциклазы и малонил-КоА-трансциклазы, которые являются
частью энзиматического комплекса, называемого синтетазой жирных
кислот.
Ацетил-КоА
Малонил-КоА
АПБ-SH
СН3
СООН
Ацетил-КоА-карбоксилаза I
СН2
C-S-KoA
АТФ
АДФ
C-S-KoA
О
KoA-SH
Ацетил- ।
трансциклаза
СО2
Ф.
сн3
АПБ—S —С=О
О
АПБ-SH
KoA-SH
Малонеал-
трансциклаза
соон
Ацетил-S-АПБ
СН3
АПБ—S—С=О
Малонил-Б-АПБ
СН2
С=О
Конденсирующи й
фермент
СН2
СН3
АПБ—S —С=О
Ацетоацетил-8-АПБ
сн2
СН2
НАДН
НАДФ
Р-кетоацил-АПБ-
редуктаза
АПБ-8-С=О
Бутирил-Б-АПБ
СН3
СНОН
Кротонил-АПБ-
редуктаза
НАДФ
СН3
НАДФН
СН2
АПБ—S — С=О
СН
О-р-гидроксибутирил-Б-АПБ
СН
Н2О
Еноил-АПБ-
дегидролаза
Рис. 4.3. Синтез жирных кислот
АПБ-8-С=О
Кротонил-Б-АПБ
120
снг
сн2
сн2
АПБ-S — С=О
I
♦
сн.
I
(СН2)„ -
АТФ-S — С=О
Пальмитил-Э-АПБ
Г идрол итическая
деациклаза
СН3
(СН2),4
Н2О АПБ-SH СООН
Пальмитиновая
кислота
Рис. 4.4. Синтез пальмитиновой кислоты
Следующий этап катализируется другой частью энзиматического
комплекса: ацетил-малонил-АПБ-конденсирующим комплексом: аце-
тил-малонил-АПБ-конденсирующий фермент соединяет ацетил-Б-АПБ
и малонил-Б-АПБ с образованием ацетоацетил-Б-АПБ. Затем последний
восстанавливается 6-кетоацил-АПБ-редуктазой, использующей для
этого НАДФН в качестве восстанавливающего агента, в результате чего
образуется D-6-гидроксибутирил-Б-АПБ. Дальнейшее преобразование
этого продукта заключается в его дегидратации ферментом 6-гидрокси-
ацилдегидратазой (известной также под названием енол-АПБ-дегидра-
таза), приводящей к образованию кротонил-Б-АПБ. Последний при
участии кротонил-АПБ-редуктазы восстанавливается НАДФН до бути-
рил-Б-АПБ.
121
Образование бутирил-8-АПБ завершает первый из семи циклов, в
каждом из которых молекула малонил-8-АПБ присоединяется к карбок-
сильному концу растущей цепи жирной кислоты (рис. 4.4). При этом
происходит отщепление молекулы АПБ, а дистальная карбоксильная
группа малонил-8-АПБ теряется в виде СО2. Конечным продуктом явля-
ется пальмитил-8-АПБ. В конце цикла под действием гидролитической
деацилазы пальмитиновая кислота отщепляется от АПБ. Суммарное
уравнение синтеза жирных кислот можно записать следующим образом:
8 ацетил-КоА +14НАДФН +14Н+ + 7АТФ + Н2О ->
—> 1 пальмитиновая кислота + 8 КоА + 14НАДФ+ + 7АДФ + 7Фн
Несмотря на то, что синтез жирных кислот происходит в цитозоле
при участии жирнокислотного синтетазного комплекса, все же для полу-
чения ацетил-КоА требуется образование пирувата в митохондриях. Од-
нако ацетил-КоА не может непосредственно переходить через митохон-
дриальную мембрану, поэтому цитрат, образующийся из ацетил-КоА и
оксалоацетата в митохондриях и затем перешедший через митохондри-
альную мембрану, действует как переносчик двууглеродных единиц
(рис. 4.5).
Ацетил-Коа и оксалоацетат преобразуются при распаде цитрата под
влиянием АТФ-зависимого расщепляющего цитрат фермента, катализи-
рующего следующую реакцию:
Цитрат + АТФ + КоА ацетил-КоА + АДФ + Фн + оксалоацетат.
Регенерация пирувата происходит в результате превращения окса-
лоацетата до малата и от него — до пирувата. Последний может потом
возвращаться в митохондрии. Таким образом, цитрат выполняет функ-
цию переносчика ацетильных групп из митохондрий в цитоплазму (аце-
тильные группы могут также транспортироваться карнитином, но этот
путь не является главным).
Указанный челночный процесс обеспечивает также образование не-
которого количества НАДФН, необходимого для синтеза жирных кислот
из цитозольного НАДН. Остальное количество НАДФ обеспечивается
пентозофосфатным путем, в котором регенерация НАДФН от НАДФ
происходит в результате окисления глюкозо-6-фосфата до рибозо-5-фос-
фата. Активность пентозофосфатного пути очень высокая в адипозной
ткани, где значительное количество НАДФН используется для синтеза
жирных кислот. И напротив, активность этого пути в скелетных мышцах
очень низкая. Следовательно, потребность мышц в жирных кислотах в
122
большей степени удов-
летворяется утилизацией
жирных кислот, поступа-
ющих в кровь из ади-
позной ткани.
Синтез триацил-
глицерола. Триацилгли-
церолы образуются пу-
тем последовательного
присоединения трех мо-
лекул жирных кислот к
каркасу трехуглеродного
глицерола (рис. 4.6). В
цитоплазме клеток как
печени, так и адипозной
Рис. 4.5. Переход ацетил-КоА из митохондрии в ци-
тозоль при участии цитрата
ткани в процессе глико-
лиза образуется гидро-
ацетонфосфат, который
затем может быть вос-
становлен до глицерол-3-фосфата в ходе реакции, катализируемой
НАД-зависимой глицеролфосфатдегидрогеназой. Лишь в печени
присутствует глицеролкиназа, катализирующая преобразование гли-
церола до глицерол-3-фосфата. Присоединение первой молекулы
жирной кислоты, являющейся производной КоА, происходит к сво-
бодной гидроксильной группе в позиции 1 молекулы глицерол-3-фос-
фата. Вторая молекула жирной кислоты аналогичным образом присо-
единяется к позиции 2, образуя 1-фосфатидную кислоту; обе эти ре-
акции катализируются ацилтрансферазой. Фосфатидные кислоты
встречаются в клетках в следовых количествах, однако служат важ-
ными общими интермедиатами биосинтеза триацилглицеролов и
фосфоглицеридов. Для образования триацилглицеролов фосфатид-
ные кислоты подвергаются гидролизу при участии специфического
фосфатазного фермента, отщепляющего фосфатную группу от моле-
кулы диацилглицерола.
Третья жирная кислота присоединяется, как и первые две, в ходе
аналогичной, катализируемой ацилтрансферазой реакции, приводящей
к образованию триацилглицерола:
L-фосфатидная кислота + Н2О —> диацилглицерол + Фн;
КоА-производное жирной кислоты + диацилглицерол ->
-> триацилглицерол + КоА.
123
2R-C-S-K0A
Жирнокислотный ацил-КоА
2 KoA-SH
H2C-O-C-R
I И
I О
HC-O-C-R’
L-глицерол-З-фосфат
L-фосфатидиновая кислота
I О
Н2СОРО3Н2
Фосфатаза
Н2С—О—С—R
II
О
НС—О—С—R'
СН2ОН
Диацил глицерол
Рис. 4.6. Синтез триацилглицерола
(триацилглицерида)
Жирнокислотный ацил-КоА
Т'*- KoA-SH
Т риацилглицерол
В клетках кишечного эпителия, которые активно синтезируют триа-
цилглицеролы в процессе всасывания жирных кислот в кишечнике,
имеет место и другой тип реакции ацилирования. Моноацилглицеролы,
образующиеся в кишечнике в процессе переваривания пищи, могут аци-
лироваться непосредственно, минуя стадию фосфатидной кислоты:
Моноацилглицерол + КоА-производное жирной кислоты —>
—> диацилглицерол + КоА.
Триглицеролы, образующиеся в адипозной ткани, запасаются в ви-
де безводных вкраплений в цитоплазме. Они могут занимать более 95 %
124
Рис. 4.7. Основные пути образования запаса липидов, их мобилизации в адипозной
ткани человека и влияние на этот процесс гормонов
общего объема адипоцита. Распад триацилглицеролов для поставки
жирных кислот в кровь происходит при участии гормончувствительной
липазы (рис. 4.7).
Жирные кислоты плазмы крови служат важным энергетическим ис-
точником для работающих мышц, что рассмотрено далее. Триацилгли-
церолы, синтезированные в печени, могут объединяться с холестеролом,
фосфолипидами и белком с образованием липопротеидов (ЛВП и
ЛОНП), которые постепенно поступают в кровяное русло, благодаря че-
му становятся доступными для утилизации периферическими тканями,
включая и мышцы. Липопротеиды низкой плотности образуются в кро-
ви в том случае, когда ЛОНП начинают распадаться под действием ли-
попротеидлипазы, локализованной в эндотелии капилляров. Хотя содер-
жание триацилглицеролов в мышцах по сравнению с адипозной тканью
и клетками печени относительно невелико, все же они могут служить су-
щественным источником энергетического топлива для окислительного
метаболизма при пролонгированных физических нагрузках.
Триацилглицерольный запас в адипозной ткани и мышцах обеспе-
чивает наибольшую поставку СЖК, окисляющихся во время выполне-
ния мышечной работы. Обычно в нормальных условиях снабжение
плазменными триглицеролами составляет лишь незначительную про-
порцию от общего количества СЖК. Это связано с тем, что поступле-
ние триглицерола в скелетную мышцу катализируется ЛПЛ, но актив-
ность этого фермента не позволяет обеспечить потребности мышеч-
ных клеток в СЖК при физических нагрузках. Однако триглицеролы
плазмы важны для восполнения триглицеролов, утилизируемых из
адипозного и мышечного источников во время выполнения физичес-
кой работы.
125
4. Липолиз
Липиды и углеводы являются основными питательными веществами,
обеспечивающими энергетику мышечных сокращений. Липидные запа-
сы представлены преимущественно триацилглицеролом. Мышечные
клетки не способны непосредственно окислять триацилглицеролы. Вна-
чале они должны быть подвержены расщеплению до жирнокислотного
и глицерольного компонентов. Этот процесс называется липолизом и на-
чинается с гидролитического отщепления молекулы жирной кислоты от
глицерольного каркаса в положении I или 3. Этот этап катализируется
гормончувствительной триацилглицерольной липазой. Липаза, специ-
фическая для оставшегося диацилглицерола, отщепляет вторую жирную
кислоту и, наконец, следующая специфическая липаза отнимает от мо-
ноацилглицерола последнюю жирнокислотную молекулу.
Жирные кислоты и глицерол, образованные в процессе липолиза,
происходящего в адипозной ткани, постепенно поступают в кровь. Уве-
личение скорости как липолиза, так и скорости кровотока в адипозной
ткани будет влиять на скорость поступления СЖК и глицерола в кровя-
ное русло. Это не имеет существенного значения в случаях, когда про-
лонгированная мышечная работа выполняется на уровне около 50 %
VO2 max и поток крови через адипозную ткань возрастает. Однако при
нтенсивных физических нагрузках симпатическая вазоконстрикция при-
водит к снижению кровоснабжения адипозной ткани, в результате чего в
ней происходит накопление СЖК и наряду с этим — выраженное огра-
ничение поступления СЖК (и глицерола) в кровь.
Глицерол плазмы крови может захватываться печенью и затем фос-
форилироваться до глицерол-3-фосфата, который затем способен ис-
пользоваться для образования триацилглицеролов, как описано выше,
или альтернативно может быть окисленным до дигидрооксиацетонфос-
фата. Последний может включаться как в гликолитический, так и в глю-
конеогенный пути. Свободные жирные кислоты весьма слабо растворя-
ются в воде, и большинство тех, что находятся в плазме, транспортиру-
ются благодаря их непрочной связи с альбумином. В норме в состоянии
покоя концентрация СЖК в плазме составляет 0,2-0,4 ммоль-л-1. Одна-
ко во время (или сразу после окончания) пролонгированных физических
нагрузок концентрация СЖК в плазме может возрасти до 2,0 ммолыл-’.
Утилизация СЖК мышцами напрямую зависит от их концентрации в
плазме. Следовательно, липолитическая мобилизация липидных запасов
является важным способом энергообеспечения пролонгированной мы-
шечной деятельности. В плазме человека СЖК с длинной цепью пред-
126
ставлены в основном олеатом (43 %), пальмитатом (24 %), стеаратом
(13 %), линолеатом (10 %) и пальмитолеатом (5 %). Они характеризуют-
ся низкой растворимостью в воде и их транспорт обеспечивается в ос-
новном благодаря наличию в крови альбумина, являющегося наиболее
представительным белком в плазме, где его концентрация составляет
около 45 г-л~1 (приблизительно 0,7 ммоль-л-'). Каждая молекула альбу-
мина содержит три высокочувствительных (и семь низкочувствитель-
ных) для СЖК связывающих участка. Когда высокочувствительные свя-
зывающие участки заполняются (при концентрации СЖК, достигающей
2,0 ммоль-л-1), то концентрация СЖК, не связанных с альбумином, за-
метно повышается. Это сопровождается образованием жирнокислотных
мицелл, которые потенциально способны оказать повреждающее дей-
ствие на ткани из-за их детергентподобных свойств.
Гормончувствительная триацилглицероллипаза в адипозной ткани
активируется цАМФ-зависимой протеинкиназой. Присоединение адре-
налина и глюкагона к рецепторам цитоплазматических мембран в ади-
поцитах активирует аденилатциклазу и инициирует энзиматический
каскад, активирующий липазу. Высокие уровни инсулина и глюкозы уг-
нетают активность липазы, а следовательно, снижают мобилизацию
СЖК.
Скорость липолиза можно оценить по скорости освобождения жир-
ных кислот или глицерола. Однако скорость выхода СЖК фактически
отражает баланс между скоростью липолиза в адипозной ткани и ско-
ростью их обратного захвата и реэстерификации, в связи с чем она мо-
жет служить лишь приблизительным показателем скорости липолиза.
Последняя точнее определяется по скорости освобождения глицерола,
который появляется в крови только в результате липолиза. После выхо-
да из адипозной ткани глицерол уже не может метаболизироваться в
адипоцитах, поскольку последние лишены фермента глицеролкиназы.
Последние достижения методов, связанных с использованием микроди-
ализа, позволили оценить скорость липолиза in situ путем определения
поступления глицерола в межклеточную жидкость адипозной ткани.
Микродиализные исследования могут быть также использованы для ло-
кальной доставки гормонов или лекарств при изучении регуляции липо-
лиза in vivo.
Изоформы гормончувствительной липазы обнаружены и в мышцах,
где они функционируют для обеспечения распада внутримышечных
триацилглицерольных запасов. Присутствие липопротеинлипазы в эн-
дотелии капилляров тканей, в том числе и мышц, также способствует
локальному увеличению в плазме концентрации СЖК и их утилизации
мышечной тканью. Активность липопротеинлипазы повышается и под
127
влиянием цАМФ-зависимой протеинкиназы. Основным субстратом для
этого фермента служат хиломикроны или ЛОНП. Гидролиз содержаще-
гося в этих структурах триацилглицеролов снижает их липидбелковое
соотношение, приводит к образованию остаточного хиломикрона и ЛСП
(из хиломикронов и ЛОНП соответственно). Печень обладает специфи-
ческим механизмом захвата остаточного хиломикрона, а следовательно,
и наибольшей способностью метаболизировать его. Липопротеид сред-
ней плотности метаболизируется преимущественно адипозной тканью
до образования ЛИП.
Транспорт жирных кислот через сарколеммальную мембрану в мы-
шечное волокно в значительной степени обусловлен простой пассивной
диффузией. Однако результаты последних исследований свидетельству-
ют о существовании трансмембранного транспортного механизма, кото-
рый обеспечивает в плазме высокую концентрацию несвязанных жир-
ных кислот (эквивалентно около 1,5 ммоль-л-1 общей концентрации
жирных кислот).
В изолированной перфузированной скелетной мышце крысы макси-
мальная скорость поступления пальмитата возрастала при вызываемых
электрической стимуляцией сокращениях и снижалась, когда поступле-
ние углеводов строго лимитировалось. Эти данные свидетельствуют о
том, что определенные физиологические стимулы могут модифициро-
вать кинетику мембранного транспорта СЖК в мышцу. Однако СЖК в
мышечные волокна будут поступать только в том случае, когда их внут-
риклеточная концентрация будет меньше, чем во внеклеточной жидкос-
ти (< 10 ммоль-л-1). Низкая концентрация в клетке СЖК может поддер-
живаться благодаря наличию в клетке жирных кислот, связанных с бел-,
ком, возможно, аналогичных тем, что обнаруживаются в абсорбционных
клетках тонкой кишки и гепатоцитах. Из этого также следует, что ско-
рость поступления СЖК в мышечные волокна будет пропорциональна
разнице их концентраций внутри и вне клетки и будет ограничиваться в
том случае, когда мембранно-транспортный механизм становится насы-
щенным. После поступления в мышечную клетку жирные кислоты пре-
образуются до дериватов КоА под действием АТФ-связывающей жир
ацил-КоА-синтетазы (известной также как тиокиназа) и таким образом
становятся пригодными для 6-окисления — основного пути липидного
катаболизма:
RCOOH + АТФ + KoA-SH -> R-C(-O)-S-KoA + АМФ + ПФн
Следовательно, для активации каждой молекулы жирной кислоты
требуется утилизация одной молекулы АТФ.
128
5. Окисление жирных кислот
Процесс 6-окисления протекает в митохондриях и представляет собой
последовательное отщепление двууглеродных единиц от цепи жирной
кислоты и образование ацетил-КоА, который затем может включаться в
цикл трикарбоновых кислот (ЦТК). Остаток жирной кислоты молекул
ацил-КоА, находящихся в саркоплазме, транспортируются в митохон-
дрии через образование эфира жирной кислоты с карнитином
(см.рис. 4.8, а). Последний синтезируется в печени, причем обычно дос-
таточное его количество в тканях может обеспечить окисление жирных
кислот. Концентрация его в мышцах составляет примерно 1 ммоль-л-1.
Ферментом, регулирующим транспорт СЖК при участии карнитина, яв-
ляется карнитинацилтрансфераза. В мышцах существуют две формы
этого энзима: одна из них локализована на внешней поверхности мембра-
ны (для образования ацилкарнитина), а другая, расположенная на внут-
ренней поверхности митохондриальной мембраны, предназначена для
восстановления ацил-КоА и свободного карнитина (см. рис. 4.8, б). Воз-
можно, этот транспортный процесс -— основной скоростьлимитирующий
этап в утилизации жирных кислот для их использования в энергопро-
дукции в мышцах. Определение активности фермента карнитинпальми-
тоилтрансферазы обеспечивает получение достаточной информации о
скорости протекания метаболического процесса при окислении СЖК.
При выполнении высокоинтенсивных физических упражнений (на
уровне около 60 % VO2 max) скорость окисления жиров не в состоянии
обеспечить потребности в АТФ для мышечных сокращений, поэтому
для их удовлетворения энергообеспечение все в большей степени перек-
лючается на использование углеводов в анаэробном гликолизе. Энерго-
обеспечение за счет липидов не может обеспечиваться анаэробными пу-
тями: сразу же после вхождения в миохондриальный матрикс остаток
жирной кислоты молекулы ацил-КоА становится способным вклю-
чаться в 6-окисление. Карнитинацилтрансфераза ингибируется мало-
нил-КоА — предшественником жирнокислотного синтеза. В присут-
ствии достаточного количества АТФ избыток ацетил-КоА отводится от
ЦТК к малонил-КоА и, следовательно, тем самым способен уменьшать
катаболизм жирных кислот, способствовать их образованию и после-
дующему синтезу триглицеридов.
После перехода в митохондрии остаток жирной кислоты ацил-КоА
первоначально окисляется до еноил-КоА ферментом ацил-КоА-дегидро-
геназой, использующим для этого в качестве акцептора электронов фла-
винадениндинуклеотид (ФАД) (рис. 4.9). Впоследствии при участии фер-
9 оз.з 129
сн3
+1
СН3— N — сн2 — CH — сн2— соон
сн3 он Карнитин
+
R —С —S —КоА
II Жирнокислотный
О ацил-КоА
JH3
СН3 — N — сн2 — сн — сн2 — СООН
I । Жирнокислотный
3 । ацилкарнитин
С = О
I
R
+
KoA-SH
Жирная кислота
Цитозоль
Ацил-КоА
Ацил-КоА-
синтетаза
Рис. 4.8. Реакция, катализируемая ацил-КоА-карнитинацилтрансферазой (я); поступ-
ление жирных кислот в виде ацил-КоА в митохондрии через ацилкарнитин (б)
мента еноил-КоА-гидратазы происходит гидратация еноил-КоА до обра-
зования гидроксил-КоА, который затем окисляется до кетоа-
цил-КоА ферментом гидроацил-КоА-дегидрогеназой. Эта реакция явля-
ется скоростьлимитирующим участком в цикле 6-окисления жирных
кислот. Активность гидроксиацил-КоА-дегидрогеназы изменяется под
влиянием диетических манипуляций и физических нагрузок. В основе
этой регуляции лежит субстратная доступность. Конечный этап цикла ка-
тализируется ацетил-КоА-трансферазой. В ходе данной реакции проис-
ходит расщепление кетоацил-КоА, приводящее к образованию одной мо-
лекулы ацетил-КоА и КоА-эфира жирной кислоты, содержащей на два
атома углерода меньше, чем исходная жирная кислота. Образовавшийся
эфир может снова использоваться в качестве субстрата для нового ЦТК.
Выявить интермедиаты в цикле невозможно, хотя ацил-КоА и
ацетил-КоА обнаруживаются в тканях, окисляющих жирные кислоты.
Следовательно, это может быть связано с перемещением продукта каж-
дой реакции непосредственно к ферменту, катализирующему следую-
щий этап в последовательности мультиэнзимного комплекса, обеспечи-
вающего весь процесс 6-окисления. После каждого такого цикла цепь
130
Жирнокислотный R — СН2 — СН2 — СН2 — СОКоА (С,6)
ацил-КоА
Дегидрогеназа
жирнокислотного
ацил-КоА
Еноил-КоА R — СН2 — CH = СН — СОКоА
+Н2О Еноил-КоА-гидратаза
фадн2
Г идроксиацил-КоА R — СН2 — СН — СН2 — СОКоА
ОН
Г идроксиацил-КоА-
дегидрогеназа
Кетоацил-КоА
R — СН2 — С — СН2 — СОКоА
О Ацетил-КоА-
ацетилтрансфераза
Жирнокислотный ацил-КоА
(теперь укороченный
на 2 атома углерода)
R —СН2—СОКоА + ацетил-КоА
Ацетил-КоА
Ацетил-КоА
Ацетил-КоА
Ацетил-КоА
Ацетал-КоА
Ацетил-КоА
Ацетил-КоА
Рис. 4.9. Цикл Ь-окисления жирных кислот
жирной кислоты укорачивается на один двууглеродный фрагмент, от-
щепляющийся в форме ацетил-КоА, и теряет две пары атомов водорода,
переходящие к специфическим акцепторам. Пальмитиновая кислота, со-
держащая 16 атомов углерода, проходит в общей сложности семь таких
циклов, в результате чего образуется 8 молекул ацетил-КоА и 14 пар
атомов водорода. Однако активироваться для такого окисления пальми-
тиновая кислота должна только один раз, поскольку в конце каждого
цикла укороченная жирная кислота выделяется в виде соответствующе-
го КоА-эфира. Наиболее распространенные окисляемые жирные кисло-
9*
131
ты содержат 16 (например, пальмитиновая кислота) или 18 (например,
олеиновая кислота) атомов углерода в ацильной цепи.
Освобождающиеся в процессе 6-окисления пальмитиновой кисло-
ты 14 пар атомов водорода включаются в дыхательную цепь митохон-
дрий: 7 пар — в форме восстановленного флавинового кофермента
ацил-КоА-дегидрогеназы и 7 пар — в форме НАДН. Переход электронов
как от ФАДН2 к кислороду, так и от НАДН к кислороду дает ожидаемое
для каждого их этих случаев число молекул АТФ, синтезируемых в про-
цессе окислительного фосфорилирования (2 молекулы от каждого
ФАДН и 3 молекулы АТФ от каждого НАДН). Таким образом, на каж-
дую отщепленную молекулу ацетил-КоА образуется пять молекул АТФ:
1 пальмитоил-КоА + 7КоА + 7О2+ 35АДФ + 35Фн —>
—> 8 ацетил-КоА +35АТФ +42Н2О
Образовавшиеся в результате окисления пальмитиновой кислоты
8 молекул ацетил-КоА вступают в ЦТК. Баланс, включающий окисление
ацетил-КоА и сопряженные с ним реакции фосфорилирования, выража-
ется следующим уравнением:
8 ацетил-КоА + 16О2 + 96АДФ + 96Ф —>
-> 8КоА + 96АТФ + 1041^0 + 16СО2
Объединив эти два уравнения, получим суммарное уравнение
1 пальмитоил-КоА + 23 О2 + 131 АДФ + 131Ф ->
-> 1КоА + 16СО2 + 146Н,О + 131АГФ
Поскольку в самом начале на активацию свободной жирной кисло-
ты тратится одна молекула АТФ, то суммарный выход в расчете на одну
молекулу пальмитиновой кислоты составляет 130 молекул АТФ.
Жирные кислоты с нечетным числом атомов углерода в ацильной це-
пи также окисляются в ЦТК через последовательное отщепление молекул
ацетил-КоА, но до тех пор, пока не останется трехуглеродный фрагмент—
пропионил-КоА. Последний подвергается преобразованию через метилма-
лонил-КоА в сукцинил-КоА, который затем включается в цикл Кребса.
При окислении ненасыщенных жирных кислот, например олеиновой
кислоты, возникают две специфические проблемы. Во-первых, двойные
связи природных ненасыщенных кислот имеют унс-конфигурацию, тог-
да как в КоА-эфирах Д2>3-ненасыщенных кислот, являющихся промежу-
точными продуктами при окислении ненасыщенных жирных кислот,
132
двойные связи имеют упс-конфигу-
рацию. Во-вторых, двойные связи
большинства ненасыщенных жир-
ных кислот расположены в таких
положениях углеродной цепи, что
последовательное удаление дву-
углеродных фрагментов с кар-
боксильного конца до места рас-
положения первой двойной связи
дает Д3>4-ацил-КоА (рис. 4.10, а), а
не Д2’3-ацил-КоА, являющийся про-
межуточным продуктом в процессе
окисления жирных кислот.
Еноилгидратаза не может осу-
ществлять гидратацию КоА-
эфиров Д3-4-ненасыщенных жир-
ных кислот. Однако существует до-
полнительный фермент — Д3-4-г/г/с-
Д2-3-щранс-еноил-КоА-изомераза,
который катализирует перемеще-
ние двойной связи из положения
3-4 в положение 2-3 и изменение
конфигурации из цис- в транс-
еноил-КоА-изомеразу (рис. 4.10, б).
Образующийся в результате ре-
акции КоА-эфир Д2’3-щранс-ненасы-
щенной жирной кислоты представ-
ляет собой уже нормальный суб-
страт для одного из ферментов про-
цесса окисления жирных кислот, а
именно для еноилгидратазы, кото-
рая катализирует его гидратацию с
образованием гидроацил-КоА. Та-
ким образом, полное окисление оле-
иновой кислоты в цикле окисления
жирных кислот дает выход девяти
молекулам ацетил-КоА. Окисление
таких полиненасыщенных жирных
кислот требует участия в этом про-
цессе дополнительного фермента —
3-гидроксиацил-КоА-эпимеразы.
s
I
КоА
Олеил-КоА
а
О=С
Д3,4-цис-еноил-КоА Д^-транс-еноил-КоА
б
Рис. 4.10. Окислительное отщепление
трех двууглеродных фрагментов молекул
(в форме ацетил-КоА) от олеиновой кис-
лоты (а) и А3'4-лгрд//с-еноил-КоА-изоме-
разная реакция (б)
133
6. Внутримышечный триацил глицерол
Хотя основной запас липидов в организме сосредоточен в ади-
позной ткани, некоторое количество триацилглицерола также запа-
сено в скелетных мышцах. Часть последнего может находиться в
адипозных клетках, рассеянных среди мышечных волокон в ткани.
Имеются также полученные с помощью световой и электронной
микроскопии доказательства существования триацилглицерольных
вкраплений, локализованных вблизи митохондрий внутри самих во-
локон.
Результаты первых исследований обмена СЖК при мышечной дея-
тельности, проведенных с использованием меченых по углероду (14С-)
жирных кислот, показали, что при пролонгированных физических наг-
рузках СЖК плазмы крови могут только примерно на 50 % вносить
свой вклад в общее количество окисляемых жиров. Этот факт позволя-
ет предположить, что внутримышечный триацилглицерол может обес-
печить значительное количество СЖК для их окисления во время вы-
полнения пролонгированной мышечной работы. Определение измене-
ний содержания внутримышечного триацилглицерола перед началом и
после завершения физических нагрузок подтвердило эту точку зрения.
В ряде работа сообщалось о снижении на 25-30 % содержания триацил-
глицерола в мышцах после 1-2 часов мышечной работы, выполняемой
на уровне 55-77 % VO2max, хотя в более поздних исследованиях была
продемонстрирована недостоверность показателей содержания три-
ацилглицерола, определяемого в небольших биоптатах скелетных
мышц. К настоящему времени еще не установлено, то ли триацилгли-
цероловые капельки в волокнах, то ли триацилглицерол адипозных кле-
ток, находящихся между волокнами, или те и другие имеют отношение
к исчерпанию запасов триацилглицерола в мышцах при физических
нагрузках.
Скелетные мышцы человека содержат приблизительно 12 г триа-
цилглицерола на 1 кг сырой массы, однако в волокнах типа I его содер-
жится больше, чем в волокнах типа II. Установлено, что за счет сопут-
ствующего интрамускулярному липолизу окисления может быть осво-
бождено 12-25 МДж химической энергии. Внутримышечный липолиз,
по-видимому, опосредуется внутриклеточной липазой, которая анало-
гична гормончувствительной липазе адипозной ткани. Доказано, что
катехоламины регулируют мобилизацию интрамускулярных запасов
триацилглицерола.
134
7. Образование кетоновых тел
и их окисление
Образующийся в результате окисления жирных кислот ацетил-КоА вклю-
чается в ЦТК при условии наличия достаточного количества оксалоацета-
та для образования цитрата. Для этого требуется уравновешенный баланс
между расщеплением жиров и углеводов. Когда распад липидов преобла-
дает и (или) поступление оксалоацетата снижается, то ацетил-КоА пере-
ключается на образование кетоновых тел в печени (в физиологических ус-
ловиях только печень способна синтезировать кетоновые тела). Такая си-
туация возникает во время голодания, при физических нагрузках, истоще-
нии запасов гликогена и диабете. В этом случае две молекулы ацетил-КоА
соединяются между собой, образуя ацетоацетат, который впоследствии
Ацетил-КоА СН3 С — S — КоА + СН3 С — S —КоА
Ацетил-КоА-ацетилтрансфераза
СН3
Ацетон
Рис. 4.11. Синтез кетоновых тел в организме
135
восстанавливается до 3-гидроксибутирата (рис. 4.11), если отношение
НАДН/НАД в митохондриях достаточно высокое. В противном случае
ацетоацетат будет подвергаться медленному спонтанному декарбоксили-
рованию с образованием ацетона.
Образование ацетоацетата, 3-гидроксибутирата и ацетона происхо-
дит в основном в печени, и все три кетона диффундируют в кровь. Об-
щая концентрация кетоновых тел в плазме сытого человека очень низ-
кая. В отсутствие приема пищи в течение ночного сна она возрастает до
0,1 ммоль-л-1 и до 3,0 ммоль-л— 1 — после трех дней голодания.
Поступление кетоновых тел из плазмы в миокард, почки и мозг соз-
дает возможность снабжения альтернативным энергетическим топливом
эти органы в период ограниченного поступления в организм углеводов
(или голодании) и помогает сохранить уровень глюкозы в крови. Од-
нако, поскольку кетоны ацидотичны, накопление их в крови до высокого
уровня недопустимо. Низкий почечный порог для кетоновой реаб-
сорбции позволяет значительно повысить их удаление из организма с
мочой.
Кетоновые тела окисляются в митохондриях мышц, сердца, мозга и
других тканей, обладающих значительными аэробными возможностями.
Фермент 3-гидроксибутиратдегидрогеназа катализирует окисление
3-гидроксибутирата до ацетоацетата при участии НАД. Кофермент А
перемещается от сукцинил-КоА (основного источника в ЦТК) для обра-
зования ацетоацетил-КоА с помощью фермента 3-оксацид-КоА-транс-
феразы. Ацетоацетил-КоА расщепляется для образования ацетил-КоА
при участии фермента ацетил-КоА-ацетилтрансферазы (фермент, подоб-
ный тем, что используются в ацетоацетил-КоА-синтезе в печени). Обра-
зовавшийся ацетил-КоА после этого становится доступным для включе-
ния в ЦТК.
8. Регуляция метаболизма липидов
при физических нагрузках
Активация гормончувствительной липазы в адипозной ткани и липопро-
теинлипазы при мышечной деятельности обусловлены влиянием адре-
налина и глюкагона, которые выходят соответственно из мозгового ве-
щества надпочечников и поджелудочной железы. Освобожденные таким
образом жировые запасы доставляются кровью к мышцам в виде СЖК,
а также поступают из внутримышечных депо липидов. Данные СЖК яв-
ляются источником энергии, которая может быть освобождена в процес-
136
се 6-окисления и быть ис-
пользована для обеспечения
мышечной деятельности. На
протяжении коротких перио-
дов легких и умеренных фи-
зических нагрузок энергия в
организме образуется приб-
лизительно в одинаковой сте-
пени за счет окисления как
углеводов, так и липидов. Ес-
ли мышечная работа продол-
жается в течение часа и более
и при этом истощаются угле-
водные запасы, то в данном
случае для обеспечения энер-
гопродукции постепенно воз-
растает использование липи-
дов. Вероятно, это происхо-
дит из-за незначительного
снижения концентрации глю-
козы в крови и вызванного
этим увеличением выхода
глюкагона (и снижением вы-
деления инсулина) из подже-
лудочной железы.
Рис. 4.12. Потребление кислорода и энергети-
ческих продуктов питания мышцами нижней ко-
нечности во время выполнения пролонгирован-
ной физической работы умеренной интенсив-
ности (Альборд и др. (Ahlbord et al.), 1974): зате-
ненные участки — количественное соотноше-
ние вклада потребления кислорода в окисление
СЖК и глюкозы крови; незатененные — окис-
ление не поступающих из крови (внутримышеч-
ных) энергетических источников — гликогена,
триглицеридов и белков
По мере возрастания физической нагрузки повышается и концен-
трация адреналина и кортизола в плазме. Эти гормональные изменения
стимулируют мобилизацию и последующую утилизацию липидов для
образования энергии. Использование СЖК работающими мышцами
возрастает на протяжении 1-4 ч физической нагрузки умеренной ин-
тенсивности (рис. 4.12). Липолитический процесс стимулируется мы-
шечной деятельностью, однако протекает постепенно. Более того, он не
прекращается сразу же после прекращения мышечной работы.
Концентрация СЖК в крови отражает баланс между СЖК, поступа-
ющими в кровяное русло (в основном из депо адипозной ткани), и их
утилизацией различными тканями. Поскольку концентрация СЖК в
плазме крови достаточно низкая (обычно в пределах 0,2-2,0 ммоль-л-1),
их короткий полупериод пребывания в плазме (фактически, менее
2 мин) свидетельствует о быстрой скорости их утилизации тканями. На-
чало двигательной активности сопровождается раскрытием мышечных
капилляров, а следовательно, и повышением скорости утилизации СЖК.
137
Однако после прекращения работы этот процесс сразу начинает носить
противоположную направленность. В результате концентрация СЖК в
плазме обычно снижается к уровню, отмечаемому в начале физической
нагрузки, и затем постепенно возрастает. К концу мышечной работы,
когда утилизация энергетических источников резко снижается, а стиму-
ляция липолиза продолжается, концентрация в плазме СЖК обычно дос-
тигает 1-2 ммоль-л-1.
В отличие от относительно небольших запасов углеводов, запасы
липидов в организме обильны и количество их таково, что не лимити-
рует возможность выполнения пролонгированной мышечной работы.
Так, например, для преодоления марафонской дистанции за 3-5 ч тре-
буется менее 1 кг жира, запасенного в организме, при условии, если бы
для обеспечения такой физической нагрузки использовался только
жир. Последний является более эффективным энергетическим источ-
ником по сравнению с углеводами с точки зрения количества энергии,
освобождающейся при окислении 1 г вещества. При полном окис-
лении липидов в организме освобождается 37-39 кДж-г-1 , тогда как
энергетический выход при окислении углеводов составляет только
15-16 кДж-г- ’. Однако энерго продукция, приходящаяся на 1 л потреб-
ленного кислорода при окислении жиров на 8-10 % меньше, чем для
углеводов (около 19,5 кДж-л-1 О2 — для жиров и 20,9 кДж-л-1 О2 —
для углеводов).
Основная проблема относительно утилизации жира в качестве
энергетического топлива для мышечной деятельности заключается не
в физической доступности их как энергетического источника, а в ско-
рости, с которой они могут поступать и окисляться в мышцах для обес-
печения энергией. Ограниченная эффективность означает, что окис-
ление жиров может поддерживать ресинтез АТФ со скоростью, доста-
точной для поддержания возможности выполнять физическую работу
на уровне около 60 % VO2 max. Однако для того чтобы выработка АТФ
могла обеспечить возможность выполнения физических упражнений
высокой интенсивности, необходима утилизация углеводов. Их ис-
пользование как в аэробном окислении, так и в анаэробном гликолизе
позволяет генерировать АТФ значительно быстрее, чем при окислении
липидов.
Для большинства видов физических нагрузок субмаксимальной
аэробной мощности энергообеспечение мышечных сокращений осу-
ществляется за счет смешанного окисления жиров и углеводов. Оче-
видно, что чем больше жира может быть использовано в качестве
энергетического источника, тем экономнее будут расходоваться угле-
водные запасы и тем продолжительнее может выполняться мышечная
138
работа. Это было продемонстрировано результатами эксперимента, в
ходе которого мобилизацию СЖК искусственно повышали внутривен-
ным введением гепарина перед физической нагрузкой. Гепарин обла-
дает свойством антикоагулянта, однако также активирует липопроте-
инлипазу, а следовательно, стимулирует превращение плазменных
триглицеридов в СЖК. В результате концентрация в плазме СЖК до
начала мышечной работы повышается на 30 мин быстрее по сравне-
нию с отмечаемой в обычных условиях. Такая манипуляция замедля-
ет утилизацию мышечного гликогена и способствует проявлению вы-
носливости.
Потребление в течение продолжительного периода пищи с высоким
содержанием жиров и низким содержанием углеводов способствовало
повышению способности мышц окислять жиры, а следовательно, и по-
вышению выносливости у экспериментальных животных. Результаты
проведенного исследования позволяли предположить, что человек мо-
жет адаптироваться таким же образом, тренируясь на фоне потребления
пищи с высоким содержанием жира. Однако проявление такого эффекта
обнаружить не удалось.
Короткий период голодания (24-48 ч) у крыс, вызывавший до
начала физической нагрузки увеличение концентрации в плазме
СЖК в связи с влиянием возросшего уровня адреналина и кортизола
в крови, также способствовал гликогеновой защите и повышению
выносливости. Тем не менее такая стратегия для людей оказалась
неэффективной, очевидно, потому, что у них отсутствует возмож-
ность за счет окисления одних лишь жиров поддерживать скорость
ресинтеза АТФ, необходимую для выполнения мышечной работы с
интенсивностью, превышающей 60 % VO2 max. У людей кратковре-
менный период голодания снижает толерантность организма к физи-
ческим нагрузкам.
Увеличение в плазме концентрации инсулина, вызываемое прие-
мом за час до начала физической нагрузки углеводов, ослабляет воз-
растание в плазме содержания СЖК во время выполнения физическо-
го упражнения и может привести к снижению выносливости. Предпо-
ложительно, это обусловлено более высокой, по сравнению с обыч-
ными условиями, скоростью утилизации мышечного гликогена. Одна-
ко в ряде последних работ не удалось подтвердить, что прием углево-
дов за час до физической нагрузки неблагоприятен для проявления вы-
носливости. Действие инсулина обусловлено преимущественно его
стимулирующим влиянием на внутриклеточную фосфодиэстеразу, ко-
торая расщепляет цАМФ до АМФ, благодаря чему предотвращается
стимуляция цАМФ гормончувствительной липазы. В процессе мышеч-
139
ной работы концентрация инсулина в плазме обычно снижается, пос-
кольку повышение тонуса симпатической нервной системы и увели-
чение в крови концентрации катехоламинов угнетают выделение инсу-
лина. Глюкоза может непосредственно угнетать липолиз, а лактат —
снижать мобилизацию СЖК путем повышения их эстерификации в
адипозной ткани.
Некоторые препараты, ингибирующие липолитический процесс
(например, 6-адреноблокаторы и никотиновая кислота), также снижают
доступность СЖК и возможность проявления выносливости.
9. Влияние тренировки, направленной
на развитие выносливости, на метаболизм
липидов во время физических нагрузок
В главе 8 подробно рассмотрены основные физиологические адаптаци-
онные изменения, происходящие в организме под влиянием тренировки,
направленной на развитие выносливости (например, увеличение капил-
лярной плотности), а также способствующие улучшению доставки кис-
лорода и энергетических источников (глюкозы и СЖК) к работающим
мышцам, более эффективной экстракции доставляемого кислорода тре-
нированными мышцами.
Окислительная способность мышечных волокон тренированных
людей может удваиваться по сравнению с лицами, ведущими малопод-
вижный образ жизни, что обусловлено увеличением плотности мито-
хондрий. В дополнение к повышению активности ферментов электрон-
транспортной цепи возрастает активность ферментов, обеспечивающих
окисление липидов, включая тех, что связаны с 6-окислением жирных
кислот. Активность липопротеинлипазы под влиянием тренировки по-
вышается, что сопровождается широким использованием ЛПОНП для
окисления в мышцах. В процессе тренировки, направленной на развитие
выносливости, у тренирующихся внутримышечные запасы ТАГ также
более высокие.
Происходящее под влиянием тренировки увеличение плотности
6-адренорецепторов на поверхности клеток адипозной ткани повышает
чувствительность липолитического процесса к катехоламинам. Уровень
инсулина в плазме крови во время физической нагрузки у тренирован-
ных ниже и продукция лактата также ниже. Такие изменения способ-
ствуют повышению липолиза.
Доказательно также, что локальные запасы триацилглицерола в
мышцах тренированных лиц используются более эффективно. Таким
140
образом, тренировка, направленная на развитие выносливости, повы-
шает окисление липидов работающими мышцами, что способствует
сохранению углеводных запасов. Важно также отметить, что при
этом липиды могут окисляться при более интенсивных физических
нагрузках
Выводы
1. Основной формой запаса липидов в организме являются триацилгли-
церолы, большая часть которых локализована в адипозной ткани. Запа-
сы триацилглицеролов обнаруживаются также в печени, мышцах и в
крови (в составе липопротеидов).
2. Триацилглицеролы образуются в результате последовательного
присоединения трех молекул жирной кислоты к глицеролу. Для синтеза
жирных кислот из ацетил-КоА требуется АТФ и НАДФН. Он осущес-
твляется в цитоплазме клеток печени и адипозной ткани.
3. Мышцы не могут напрямую окислять триацилглицеролы. Внача-
ле молекула триацилглицерола должна быть расщеплена на жирнокис-
лотный и глицерольный компоненты в ходе процесса, называемого ли-
полизом. Последний катализируется гормончувствительной липазой,
обнаруживаемой в адипоцитах и мышечных волокнах. Липопротеинли-
паза, присутствующая в эндотелии капилляров, расщепляет триацилгли-
церолы плазмы.
4. Основным источником липидного топлива служат неходящиеся в
крови СЖК, происходящие от адипозной ткани и внутримышечного
ТАГ. Поступление СЖК в мышцы является транспортно-опосредован-
ным процессом, демонстрирующим кинетику насыщения.
5. Жирные кислоты подвергаются 6-окислению в митохондриях,
производящих ацетил-КоА, НАДН и ФАДН2. Ацетил-КоА может вклю-
чаться в ЦТК и восстанавливать коферменты, перенося свои электроны
и водород кислороду через митохондриальную дыхательную цепь.
6. Во время физической нагрузки происходит активация липолиза,
обусловленная влянием адреналина и глюкагона.
7. Вклад окисления липидов в ресинтез АТФ возрастает по мере
увеличения продолжительности выполнения мышечной работы.
8. При окислении одних лишь липидов максимальная скорость об-
разования АТФ составляет около 1 ммоль-с- 1-кг—1смт, что эквивалентно
потребности в ней при выполнении физических упражнений с интен-
сивностью, соответствующей 50-60 % VO2max. Эта потребность за счет
окисления только липидов не удовлетворяется, если упражнения выпол-
141
няются с более высокой интенсивностью. Основным лимитирующим
фактором в данном случае может быть скорость поступления СЖК в ми-
тохондрии.
9. Скорость окисления СЖК в мышцах связана с их концентрацией
в плазме, со скоростью кровотока, а также частично зависит от окисли-
тельной способности рекруитированных мышечных волокон и доступ-
ности углеводных энергетических запасов.
10. Адаптация к физическим нагрузкам, направленным на развитие
выносливости, сопровождается повышением способности мышц к окис-
лению липидов.
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
Ahlborg G., Felig Р, Hagenfeldt L., Hendler R., WahrenJ. (1974). Substrate turnover during
prolonged exercise in man И J.Clin. Invest. — 53. — P. 1080-1090.
P-FtI
МЕТАБОЛИЗМ БЕЛКОВ,
АМИНОКИСЛОТ
И РОДСТВЕННЫХ МОЛЕКУЛ
1. Роль белков
Вся информация любого вида живого организма заложе-
на в структуре ДНК, и это определяет тип и количество
белка, синтезируемого в каждой клетке. И напротив, эти
белки являются ответственными за синтез всех других
клеточных компонентов: генетический материал служит
кодом только для белков и их компонентов — амино-
кислот. Белки обеспечивают структурную основу всех
тканей и органов, и они в значительной степени опреде-
ляют присущую этим тканям форму. Однако, вероятно,
наиболее важным является то, что присутствие белков в
различных тканях наделяет каждую из них своими мета-
болическими возможностями. Присутствие или отсут-
ствие специфического фермента определяет, способна
или не способна ткань выполнить специфическую фун-
кцию, а также как быстро этот процесс может протекать.
Белки и аминокислоты также являются основой или
предшественниками многих гормонов, регуляторных
пептидов и нейропередатчиков, обеспечивают рецепцию
для этих сигнальных систем, выполняют большое много-
образие других функций.
2. Аминокислоты
Метаболизм белков и аминокислот во многих отноше-
ниях гораздо сложнее по сравнению с метаболизмом ли-
пидов или углеводов. В частности, это связано с тем,
143
Органическая
^/часгь молекулы
R а-Углерод
IZ
nh2— с—соон
/ I \
/ н \
Аминогруппа Карбоксильная
группа
Рис. 5.1. Общая структура ами-
нокислот (R — одна из возмож-
ных боковых цепей)
что в белковом и аминокислотном об-
мене участвует большое количество
различных веществ. Все 20 амино-
кислот, которые встречаются в орга-
низме, присутствуют в свободной фор-
ме, а также в виде связанных вместе
коротких цепей, образующих пептиды,
либо связанных в более длинные и бо-
лее сложные структуры, образующие
белки. Все аминокислоты, как углево-
ды и липиды, содержат углерод, водо-
род и кислород, а также азот; в состав двух аминокислот входит сера.
Базовая структура всех аминокислот включает аминогруппу (-NH2)
и карбоксильную группу (-СООН), которые присоединены к одному
атому углерода. В ней присутствует также органическая боковая цепь;
структуры этих цепей предопределяют структурные характеристики
различных аминокислот (рис. 5.1). При физиологических значениях pH
существуют они преимущественно в ионизированной форме (NH3+ и
СОО-).
Незаменимые аминокислоты не синтезируются в организме челове-
ка и поэтому должны присутствовать в пище, однако все остальные мо-
гут быть синтезированы. Отдельные аминокислоты и их символы, кото-
рыми они обычно обозначаются, представлены в табл. 5.1, а их струк-
турные формулы — на рис. 5.2.
Все аминокислоты в тканях животных, за исключением наиболее
простой и оптически не активной — глицина, встречаются в форме L-сте-
Таблица 5.1. Аминокислоты и их аббревиатуры
Название Аббревиа- тура Незамени- мость Название Аббревиа- тура Незамени- мость
Аланин Ала Нет Лейцин Лей Да
Аргинин Apr « Лизин Лиз «
Аспарагин Асн « Метионин Мет «
Аспарагинат Асп « Пролин Про Нет
Валин Вал Да Серин Сер «
Гистидин Гис Нет Тирозин Тир «
Глицин Гли « Треонин Тре Да
Глутамат Глу « Триптофан Три «
Глутамин Глн « Фенилаланин Фен «
Изолейцин Иле Да Цистеин Цис Нет
144
Глицин
некоею-
NH3
Аланин
СН2СН.СОО“
NH3
Валин
CH3^
^CH.CH.COO-
CH3
NH3
Лейцин
CH3\
^CH.CH2CH.COO~
CH3
NH3
Изолейцин
CH3CH2\
^CH.CH.COO-
CH3
NH3
Серин
HO.CH2CH.COO-
Треонин
СН3СН.СН.СОСГ
NH3
OH NH3
Лизин
H3N.CH2CH2CH2CH2CH.COO“
NH3
Аргинин
H2N
^C.NH.CH2CH2CH2CH.COO~
h2n
NH3
Г истидин
нс=с.сн2сн.соо“
I I I
HN NH NH3+
H
Аспартат
Аспарагин
оос.сн2сн.соо_
I
NH3+
H2NOC.CH2CH.COO~
I
NH3+
Цистеин
HS.CH2CH.COO~
I
NH3+
Глутамат OOC.CH2CH2CH.COO“
I
NH3+
Глутамин H2NOC.CH2CH2CH.COO_
I
NH3+
Метионин
CH3S.CH2 CH2CH.COO“
I
NH3+
Рис. 5.2. Аминокислотная структура с классификацией, соответствую-
щей структуре боковой цепи (окончание на с. 146)
*0 и зо
145
Фенилаланин
Тирозин
Н,С---СН,
I I
н,с сн.соо-
н
Пролин
Рис. 5.2 (окончание)
реоизомера, т. е. им присуща конфигурация, которая вращает плоскость по-
ляризации света влево
сосг
+ I
h3n—с—н
сн3
L-аланин
СОСГ
Н—С —H3N
СН3
D-аланин
(оптическая изомерия и другие аспекты химической структуры определе-
ны в Приложении 1).
Само по себе данное свойство не имеет особого значения, однако
оно отражает особенности формы молекулы, которая детерминирует
способность последней взаимодействовать с другими молекулами. Это
предопределяет укладку аминокислотных цепей в сложных белковых
структурах с активными участками, которые могут узнавать другие мо-
лекулы и действовать как катализаторы.
3. Структура и функция белков
Белки являются химическими структурами, представляющими собой
линейную последовательность аминокислот, сформировавшуюся в ходе
серии реакций конденсации, в которых задействованы «-карбоксильная
и «-аминная группы смежных аминокислот. Образующиеся в результате
146
RO HR' ROHR'
I II x---x II I II I I
H2N —c —C-f-OH + HTTN—c —COOH —*- h2n—c—C —N —c —COOH + H2O
I '—I III
H H H H
Пептидная связь
Рис. 5.3. Пептидные связи между аминокислотами, формирующиеся в ходе реакции
конденсации
этих реакций связи называются пептидными (рис. 5.3). Две аминокис-
лоты образуют дипептид, а более длинные цепи — полипептиды. Каж-
дая полипептидная цепь имеет одно аминное и одно карбоксильное
окончание, которые могут образовывать последующие пептидные связи
с другими аминокислотами. Многие белки состоят из более чем одной
полипептидной цепи, каждая из которых формирует субъединицу. Поря-
док, в котором аминокислоты располагаются в цепи, определяется в про-
цессе белкового синтеза последовательностью нуклеотидных оснований
в специфической ДНК, содержащей генетическую информацию, относя-
щуюся к этому белку. Последовательность аминокислот детерминирует
окончательную структуру, поскольку боковые цепи компонента амино-
кислот притягиваются, отталкиваются либо служат физическим препят-
ствием друг для друга, что «заставляет» молекулу складываться и при-
нимать окончательную, соответствующую ей форму.
Первичная структура белков определяется последовательностью в
них аминокислот. Вторичная структура формируется в виде спирали за
счет образования водородных связей между соседними группами молеку-
лы. Третичная структура характеризуется компактной формой, которая
имеет трехмерное измерение и образовалась в результате взаимодействия
боковых цепей, что предопределяет как структурные, так и каталитичес-
кие свойства молекулы. Некоторые белки состоят более чем из одной
субъединицы, что придает им четвертичную структуру. Гемоглобин, нап-
ример, содержит четыре отдельных полипептидных цепи (две — «-цепи и
две — fe-цепи). Общая структура молекулы позволяет изменять ее форму
и в зависимости от присоединения кислорода изменять чувствительность
других кислородсвязывающих участков. В результате взаимосвязь между
парциальным давлением кислорода (рО2) и связыванием кислорода носит
сигмоидный, а не линейный характер, в связи с чем значительно повыша-
ется чувствительность к небольшим изменениям локального рО2.
Активность многих ферментов регулируется аналогичным образом.
При связывании регуляторных агентов — активаторов илй ингибиторов
изменяется форма активных центров фермента и таким способом изме-
няется его чувствительность к субстрату.
10*
147
4. Нуклеиновые кислоты
и регуляция белкового синтеза
Развитие клетки детерминируется хромосомами, которые находятся в яд-
рах и содержат генетическую информацию, предопределяющую природу
клетки путем регуляции синтеза многих тысяч различных белков, кото-
рые придают присущие ей структурные и функциональные особенности.
Хромосомы состоят преимущественно из дезоксирибонуклеиновой
кислоты. Функциональная единица ДНК — дезоксирибонуклеид — сос-
тоит из молекулы пентозного сахара (дезоксирибозы), фосфатной груп-
пы и органического основания, которым может быть пурин или пирими-
дин. В ДНК присутствуют четыре различных основания: аденин (А), ти-
мин (Т), гуанин (Г) и цитозин (Ц). Каркас молекулы состоит из двух це-
пей дезоксирибозы и фосфатных групп (рис. 5.4). Типичная молекула
ДНК состоит из десятков миллионов таких единиц. Порядок расположе-
ния нуклеотидных оснований в ДНК предопределяет последователь-
ность аминокислот в синтезируемом ею белке.
0“
H2N
—О—Р—О—СН2
О
О-
Гуанин
О—Р—О—СН2
о
О"
Рис. 5.4. Короткий участок
одной из нитей молекулы
ДНК, демонстрирующий кар-
кас нуклеиновых кислот, ко-
торый состоит из чередую-
щихся остатков пентозы и
фосфорной кислоты (пурино-
вые и пиримидиновые осно-
вания составляют боковые
цепи, присоединенные к пен-
тозным единицам каркаса)
о
О-Р-О-СН, .0
Одна
дезоксинуклеотидная
единица
О
А,СН3
HN Т
Тимин
О
Цитозин
О-Р-О-СН2 О
О N'
о
О—Р —О—
О”
148
Рис. 5.5. Специфическое присоединение тимина к аденину и цитозина к гуанину, оп-
ределяющее соответствие структуры одной нити ДНК — другой
Химические свойства оснований в ДНК создают возможность обра-
зования связей между ними. Прочные водородные связи образуются
только между парами оснований: тимин — аденин и цитозин — гуанин
(рис. 5.5), которые соединяют две параллельные нити, располагающие-
ся в противоположных направлениях. Стабильность водородных связей
важна для обеспечения стабильности генетической информации, зало-
женной в этих молекулах. Однако они могут быть нарушены в процессе
транскрипции, т. е. когда заложенная информация передается белоксин-
тезирующему аппарату
В процессе транскрипции водородные связи, соединяющие основа-
ния, разрушаются и фермент РНК-полимераза формирует последова-
тельность рибонуклеотидов, комплементарную последовательности
нуклеотидов в цепи ДНК (за исключением того факта, что в РНК вмес-
то тимина присутствует урацил [У]). Последовательность оснований в
исходной молекуле ДНК (или, по крайней мере, в ее одной нити — дру-
гая нить остается неизменной). Таким способом определяется последо-
вательность оснований в молекуле РНК, обозначаемой как матричная
РНК [мРНК] (рис. 5.6). Матричная РНК перемещается от клеточных
ядер, где она образовалась, в цитоплазму, в которой находятся рибосомы
(белоксинтезирующий аппарат).
Процесс трансляции позволяет использовать информацию, зало-
женную в последовательности оснований в молекуле мРНК для установ-
ления последовательности аминокислот в синтезируемой полипептид-
ной цепи. Первой аминокислотой в этом процессе всегда является мети-
онин, а также скорость, с которой протекает этот инициирующий этап,
по-видимому, определяет скорость всего процесса белкового синтеза.
Удлинение пептидной цепи прекращается, когда неожиданно возникает
несоответствие последовательности кодонов любой из аминокислот.
149
Рис. 5.6. Процесс считывания генетической информации, содержащейся в последова-
тельности пар оснований молекулы ДНК (переписывается молекулой мРНК под кон-
тролем мРНК-полимеразы, затем на рибосомах эта информация переводится в опреде-
ленную последовательность аминокислот, которые размещаются соответствующими
антикодонами тРНК)
Каждая клетка человеческого организма содержит всю генетичес-
кую информацию, необходимую для создания всех других клеток, одна-
ко эта информация остается подавленной (частично экспрессия генети-
ческой информации мышечных клеток отличается от почечных). Адап-
тационные изменения, происходящие в мышцах под влиянием трени-
ровки, отражают изменения в экспрессии генетического материала. Так,
тренировочные занятия, направленные на развитие выносливости, при-
водят к увеличению скорости синтеза окислительных ферментов. Белко-
150
вый синтез и экспрессия генетического материала могут регулироваться
различными способами.
Транскрипторный контроль изменяет концентрацию мРНК, что,
отчасти, может быть весьма важно для печени, в которой большое коли-
чество белков имеет короткий полупериод жизни. Контроль достигается
регуляцией активности мРНК-полимеразы. Репрессорные белки, кото-
рые активируются либо угнетаются в зависимости от доступности спе-
цифических субстратов, позволяют усилить этот контроль.
Трансляционная регуляция осуществляется в пунктах сборки ами-
нокислот под контролем РНК в отсутствие каких-либо изменений ее ко-
личества. До конца не выяснено, каким образом достигается этот кон-
троль, однако возможно, что связан он с инициирующим процессом.
5. Обращаемость белков
В организме отсутствуют механизмы, которые могли бы обеспечивать
запасание в нем поступающих с пищей белков, а излишнее потребление
любых аминокислот сразу же требует их немедленного окисления и эк-
скреции азота. В общем, все структуры организма достаточно стабиль-
ны, однако многие белки тканевых компонентов имеют относительно
короткий полупериод жизни. Большинство структурных белков и фер-
ментов синтезируется и распадается с высокой скоростью, причем не
менее 20 % базального уровня энерготрат расходуется на белковый об-
мен. Вполне понятно, что этот процесс важен для восстановления пов-
режденных тканей и заживления ран, а также необходим для обеспе-
чения нормального функционирования здоровых тканей.
Полу период жизни некоторых белков чрезвычайно короткий: для
отдельных ферментов печени его продолжительность составляет менее
одного часа. Изменение количества этих ферментов является важным
фактором в регуляции их активности, в связи с чем высокая скорость их
обмена имеет существенное значение, если ткань легко реагирует на из-
менение потребностей метаболизма. В печени высокая скорость обра-
щаемости ферментов, регулирующих гомеостаз энергетических источ-
ников, позволяет быстро реагировать на доставку последних как на при-
ем пищи, так и на кратковременное голодание.
Некоторые белки проявляют более высокую стабильность и полупе-
риод их жизни определяется днями и неделями. Так, в скелетных мыш-
цах продолжительность адаптации к тренировке и деадаптации измеря-
ется днями, что может указывать на скорость обращаемости ферментов
во время протекания адаптационного процесса. Определение скорости
151
изменения активности различных ферментов показывает, что некоторые
метаболические ответы проявляются быстро. Так, при хронической
электростимуляции повышение активности гексокиназы в мышцах мо-
жет быть обнаружено в пределах 24 часов, а ее двукратное увеличение
отмечается в течение трех дней. Изменения в окислительных ферментах
происходят медленнее, а пик изменений, достигающий 10-12-кратного
возрастания активности отдельных энзимов при постоянной электро-
стимуляции мышц животных (в течение суток), наблюдался через
2-5 недель. И напротив, такие физиологические параметры, как, напри-
мер, абсолютное VO2 шах, под влиянием тренировки и детренировки из-
менялись намного медленнее. Ясно также, что изменения VO2 max в
процессе тренировки происходят в намного меньшей степени, чем изме-
нения активности этих ферментов.
Распад белков до их аминокислотных компонентов осуществляется
гидролитическими ферментами, к которым относится большое коли-
чество различных протеиназ и пептидаз. Эти ферменты образуются в
лизосомах, обладающих способностью поглощать и переваривать внут-
риклеточные структуры, но могут существовать и в растворенной фор-
ме. Механизмы, которые контролируют активность протеолитических
ферментов, достаточно не изучены, однако не вызывает сомнений, что
они находятся под влиянием инсулина, тироидного гормона и многих
других факторов. Обращаемость белков сбалансирована между скорос-
тями их синтеза и распада, регулируемых этими двумя гормонами. Но
и гормон роста, кортизол и другие гормоны также влияют на скорость
белкового синтеза. Более подробно механизмы этой регуляции рассмот-
рены далее.
Вполне очевидно, что при голодании происходит потеря белка орга-
низмом, поскольку в этом случае отсутствует поступление в организм
новых аминокислот, а распад белков и аминокислотный метаболизм
продолжаются. Уже на ранних стадиях голодания происходящий в неко-
торых тканях распад белков позволяет использовать их аминокислотный
компонент для окисления или глюконеогенеза. Установлено, что на ран-
них стадиях голодания аминокислоты обеспечивают синтез около 60 г
глюкозы в сутки. Одинаково важно и поступление незаменимых амино-
кислот, освобождаемых подвергающимися протеолизу тканями, пос-
кольку они могут быть использованы для поддержания функций других
тканей. Скелетные мышцы и ткани пищеварительного канала служат
главными источниками незаменимых аминокислот во время периодов
голодания. Эти ткани быстро распадаются при неадекватном поступле-
нии белков с пищей, но в то же время могут нормально функциониро-
вать даже при значительной потере ткани. Если голодание продолжает-
152
<
ся более нескольких дней, скорость распада белков быстро снижается.
После двух-трех недель отсутствия приема пищи глюконеогенез, проте-
кающий за счет аминокислот, может обеспечить не более 15-20 г глюко-
зы в сутки.
6. Метаболизм аминокислот
Все аминокислоты, кроме незаменимых, могут синтезироваться в орга-
низме, а затем распадаться до более простых веществ. Детальное рас-
смотрение путей синтеза незаменимых аминокислот выходит за рамки
настоящей книги, однако в ней изложены аспекты, общие для некоторых
аминокислот.
Важный первоначальный этап аминокислотного синтеза связан с
использованием интермедиата ЦТК — а-кетоглутарата: глутаматде-
гидрогеназа катализирует включение в него аммиака с одновремен-
ным окислением НАДН или НАДФН, образуя при этом глутамат
(рис. 5.7).
Глутаматдегидрогеназная реакция относится к необычным, пос-
кольку в роли водородного акцептора в ней могут выступать НАД+ или
НАДФ+, что может играть определенную роль в поддержании баланса
окислительно-восстановительного статуса этих нуклеотидов. Род-
ственные вещества — глутамин и пролин — синтезируются из глута-
мата (см. рис. 5.7). Другие метаболические интермедиаты могут учас-
твовать в синтезе аминокислот, использующих аминогруппу глутама-
та. Аланин образуется из глутамата и пирувата в реакции трансамини-
рования, и аналогичный процесс приводит к образованию аспартата из
оксалоацетата:
NH3 + а-кетоглутаровая кислота + НАДФН + Н' -» L-глутаминовая кислота + НАДФ+
NH3 + глутаминоаая кислота + АТФ -» глутамин + АДФ + Фн
НгС----СН2
О=С* CH
1 А А
СТ NH3 СОСГ
Глутамат
,СН
/ \
соон
д'-пиролин-
5-карбоксильная
Н2С----СН2
*СН2 .СН
" соон
п
Пролин
кислота
Глутаминовая кислота + пируват -> а-кетоглутаровая кислота + аланин
Рис. 5.7. Реакции биосинтеза аминокислот, связанные с использованием а-кетогпутарата
153
Аргинин
Аланин
Цистеин
Глицин
Серин
Г истидин
Глутамин
Пролин
Глутамат
Треонин
Изоцитрат
а-Кетоглутарат
Пируват
Лейцин !
Изолейцин —► Ацетил-КоАЦитрат
Триптофан у i
Ацетоацетил-КоА \
Сукцинил-КоА
Сукцинат
Оксалоацетат
Фумарат
Фенилаланин
Тирозин
Малат
Изолейцин
-*— Метионин
Валин
Тирозин
Фенилаланин
Лейцин Аспартат
Лизин Аспарагин
Триптофан
Рис. 5.8. Превращение аминокислот до пирувата, ацетата либо до медиаторов ЦТК
Незаменимые аминокислоты могут вести себя как предшественни-
ки заменимых аминокислот, образуя тирозин путем гидроксилирования
фенилаланина и цистеин — путем синтезирования из метионина.
Многие аминокислоты могут быть расщеплены в результате реак-
ции дезаминирования с получением интермедиатов ЦТК (рис. 5.8), что
может служить важной функцией в поддержании внутриклеточной кон-
центрации этих веществ.
Реакции дезаминирования приводят к образованию кетокислот из
аминокислот: акцептором аминогруппы обычно является «-кетоглута-
рат, который преобразуется до глутамата. Последний может быть реге-
нерирован под действием глутаматдегидрогеназы, высвобождающей
свободный аммиак в процесс. Дезаминирование глутамата до образова-
ния «-кетоглутарата и свободного аммиака является реакцией, противо-
положной процессу синтеза, представленного на рис. 5.7:
L-глутамат + НАД+ -> а-кетоглутарат + NH4+ + НАДН + Н+
Это весьма важно для поддержания низкой концентрации аммиака,
поскольку большая часть азота, образующегося в результате аминокис-
лотного распада, превращается в мочевину. Последняя считается отно-
сительно инертным веществом, не полярна и может экскретироваться
154
О
H2N-C~OPOf"+ 2 АДФ + Ф,
Карбамилфосфат
АТФ
л 3
(СН2)3
H-C-NHJ Орнитин
СОО" /
О
II
H2N-C-NH2
Мочевина -
O=C~NH2
NH
Митохондрия
Н2О
h2n. >н2 ар™”н
с
I
NH
(СН2)3
Н—С—NH3
СОО"
ЦИКЛ
МОЧЕВИНЫ
АМФ +
(СН2)3
HC-NHJ
Цитрулин рОО
СОО"
Аргинино-
сукцинат
СН2
I +
нс- NH3
СОО"
Аспартат
ООО"
Цитоплазма
ООО"
I
НС
II
сн
I
СОО"
Фумарат
СН2 ;
HC-NH-C''
I I
СОО NH
^NH2
(СН2)3
НС—NHJ
СОО"
Рис. 5.9. Цикл мочевины, катализирующий преобразование аминогрупп до мочевины,
благодаря чему предотвращается возможность повреждающего воздействия увели-
чения концентрации свободного аммиака
почками без какого-либо влияния на кислотно-щелочное равновесие.
Образование мочевины регулируется циклом мочевины (рис. 5.9). Этот
процесс энергозависим: при каждом обороте цикла удаляется два атома
водорода, при этом затрачивается одна молекула АТФ.
В среднем организм взрослого здорового человека в сутки выделяет
30-40 г мочевины, однако при потреблении высокобелковой пищи это
количество может возрастать в 2-3 раза. Реакции цикла мочевины про-
текают в печени. Наибольшее количество азота, доставляемого для эк-
скреции из тканей (например, из мышц), поступает в печень большей
частью в виде аминогрупп, а не аммиака. Основными аминокислотами,
освобождающимися из скелетных мышц, являются аланин и глутамин.
При пролонгированных интенсивных физических нагрузках уровень
155
аммиака в плазме повышается в результате возросшего высвобождения
его из мышц.
В качестве примера множественной роли аминокислот можно при-
вести глутамат, который способен подвергаться декарбоксилированию в
нервных клетках до образования 4-аминобутирата, известного также как
гамма-аминомасляная кислота (ГАМК), которая выполняет функцию
тормозного нейромедиатора.
Присутствие серы в молекулах некоторых аминокислот обусловли-
вает необходимость специальных механизмов для ее удаления. Одним
из конечных продуктов обмена серосодержащих аминокислот является
сульфат, который экскретируется с мочой.
7. Метаболизм белков
при мышечной деятельности
Не вызывает сомнений, что систематические занятия физическими уп-
ражнениями оказывают выраженное специфическое влияние на метабо-
лизм белков в организме. Физическая тренировка, направленная на раз-
витие силы, способствует увеличению мышечной массы и повышению
содержания в мышцах актина и миозина. В то же время тренировочные
занятия, направленные на развитие выносливости, мало влияют на мы-
шечную массу, однако повышают содержание в мышечной ткани мито-
хондриальных белков, особенно тех, которые связаны с окислительным
метаболизмом. Эти изменения носят избирательный характер и зависят
от направленности тренировочных воздействий.
Физические нагрузки способны также оказывать острые воздей-
ствия на белковый метаболизм. Проявляющиеся в ответ на напряжен-
ную мышечную деятельность реакции во многом могут быть сходны с
реакциями, характерными для острой фазы при инфекции или ранении.
Мышцы обладают ограниченной способностью к окислению ами-
нокислот. Так, скелетные мышцы млекопитающих могут окислять толь-
ко шесть из них — аланин, аспартат, глутамат, лейцин, изолейцин и ва-
лин (три последние относятся к аминокислотам с разветвленной цепью),
и их окисление мышцами приводит к возникновению проблемы устра-
нения аминогрупп, часть которых в реакции трансаминирования пере-
носится к пирувату с образованием аланина. Последний поступает в пе-
чень и затем включается в цикл мочевины (рис. 5.10).
В неактивных мышцах вклад окисления аминокислот в ресинтез
АТФ составляет не более 10 % общего количества используемых энерге-
156
Рис. 5.10. Окисление аминокислот с разветвленной цепью как важный энергетический
источник для сокращающихся мышц (аминогруппы от этих аминокислот транспорти-
руются в печень для включения в цикл мочевины)
тических источников, однако при физических нагрузках величина этого
вклада снижается. В условиях же ограничения поступления других ви-
дов «топлива» окисление аминокислот для энергообеспечения снова
приобретает более весомое значение. При этом скорость окисления от-
дельных аминокислот возрастает неодинаково (например, скорость
окисления лейцина может повыситься в пять раз). Тем не менее степень
увеличения скорости окисления лейцина требует уточнения, поскольку
использование изотопной техники в данном случае не позволяет полу-
чить достаточно надежные данные.
При пролонгированных физических нагрузках умеренной Интен-
сивности вклад белкового метаболизма в энергопродукцию составляет,
очевидно, не более 6 % общей потребности в энергии. Однако в пище
жителей западных регионов планеты в среднем около 12-15 % потреб-
ляемой энергии приходится на долю белков. Этот факт позволяет пред-
положить, что систематическая мышечная деятельность в меньшей сте-
пени повышает потребность в поступлении в организм белков по срав-
нению с потребностью поступления углеводов и жиров. При очень нап-
ряженных физических занятиях, когда для прироста мышечной массы
157
спортсмены, занимающиеся бодибилдингом, используют большое коли-
чество протеиновых добавок, все же отсутствуют доказательства, что та-
кое питание может стимулировать усвоение чрезмерно потребляемого
белка тканями организма. Однако подобного рода добавки по-прежнему
остаются популярными и применяются на фоне повышенного потребле-
ния других субстанций (включая инсулин и такие 6-агонисты, как клен-
бутерол), которые способствуют поступлению аминокислот в мышцы и
образованию из них белков.
8. Глюконеогенез и кетогенез
Углеводные каркасы аминокислот могут служить источником энергети-
ческого топлива для обеспечения окислительного метаболизма. Они мо-
гут быть использованы также для синтеза глюкозы или кетоновых тел,
которые затем расходуются для энергообеспечения другими тканями.
Аминокислоты в зависимости от того, чьими предшественниками
они являются, подразделяют на гликогенные и кетогенные. Все амино-
кислоты, за исключением лейцина и лизина, которые преобразуются до
ацетил-КоА, могут быть использованы для гликонеогенеза, протекаю-
щего преимущественно в печени и в меньшей степени — в почках. Во
время голодания образование глюкозы таким путем приобретает важное
значение, поскольку позволяет использовать большой энергетический
запас, локализованный в белке. Некоторые ткани, как, например, кишеч-
ник, достаточно лабильны и вносят основной вклад в удовлетворение
энергетических потребностей даже при относительно непродолжитель-
ном голодании.
Аланин и глутамин являются ключевыми аминокислотами в тран-
спорте азота между тканями, и наибольшее их количество, которое
мышцы теряют во время голодания, приходится на одну из этих амино-
кислот. Возникает возможность получить для ресинтеза АТФ опреде-
ленное количество энергии, аккумулированное в этих аминокислотах,
которые могут подвергаться катаболизму в скелетных мышцах с выделе-
нием потенциально токсичного аммиака. Пируват, образующийся из
глюкозы или гликогена при гликолизе, легко преобразуется до аланина в
процессе трансаминирования. Благодаря этому процессу в форме ала-
нина могут накапливаться аминогруппы из различных аминокислот. Ос-
вобождающийся аланин способен выполнять функцию важного суб-
страта для гликонеогенеза в печени во время голодания, однако в сытом
состоянии, когда гликонеогенез угнетен, его роль как субстрата весьма
незначительна. Однако даже во время голодания глюкозо-аланиновый
158
МЫШЦА
КРОВЬ
Глюкоза
Глюкоза.
\ Гликолиз
Пируват
Мочевина
ПЕЧЕНЬ
NH3
Глюконео-
генез
Дезамини-
рование
Аланин -
Т рансамини- L- Аминокислота
рование .А
- Аланин Оксокислота
Рис. 5.11. Глюкозо-аланиновый цикл, теоретически обеспечивающий возможность ис-
пользования содержащихся в мышцах белков для гликонеогенеза в печени
цикл (рис. 5.11) может иметь менее важное значение, чем это представ-
ляют, поскольку скорость гликолиза в мышцах в состоянии покоя низкая,
а освобождение лактата доминирует только в том случае, когда мышцы
находятся в активном состоянии (вероятно, он более важен для вывода
азота из мышц, когда происходит распад белков и окисление амино-
кислот).
Существует доказательство увеличения выхода аланина из сокраща-
ющихся мышц, указывающее на то, что некоторое количество образую-
щегося пирувата превращается в аланин, однако вклад этого выхода в
обеспечение поддержания уровня глюкозы в крови незначительный. По-
видимому, основная роль глюкозо-аланинового цикла заключается в
транспорте потенциально токсичного аммиака из мышц к печени для об-
разования мочевины, что более важно для обеспечения продолжения
распада белков, чем для снабжения предшественниками гликонеогенеза.
В такой ситуации цикл будет незавершенным, поскольку глюкоза пред-
почтительнее будет использоваться не мышцами, а другими тканями, в
то время как пируват, используемый для образования аланина, вероятно,
происходит скорее от окисляемых аминокислот, чем в результате глико-
лиза.
Глутамат, освобождаемый из мышц, также удаляет азот. Однако он
может служить важным энергетическим топливом для других тканей,
особенно для клеток кишечника и иммунной системы. Известно, что
глутамат является основным энергетическим источником в окисли-
тельном метаболизме для этих тканей. При прекращении приема пищи
последние могут зависеть от скелетных мышц в связи с потребностью
обеспечения энергетическим субстратом. Если потребность в глутамате
становится высокой, как, например, при травме либо инфекции, когда
возможна большая потеря мышцами глутамата, то в такой ситуации мы-
шечная масса может резко снижаться.
159
9. Регуляция обмена белков
Интегральная регуляция синтеза и распада белков представляет собой
сложный и пока недостаточно изученный процесс, однако к настоящему
времени в понимании его отдельных механизмов достигнуты опреде-
ленные успехи. Неопределенности связаны в основном с разнообразием
и сложностью реакций, в которых принимают участие аминокислоты, а
также с участием многих факторов, влияющих на азотистый баланс це-
лостного организма.
Соотношение между анаболическими реакциями, осуществляю-
щими белковый синтез, и катаболическими процессами, которые кон-
тролируют распад белков, может быть нарушено воздействиями многих
факторов, одним из которых является мышечная деятельность. Более
драматические эффекты, однако, проявляются в ответ на такие, напри-
мер, воздействия, как инфекции, при которых отмечается высокая ско-
рость распада белков и потеря азота организмом. Анаболические про-
цессы в скелетных мышцах индуцируются тяжелоатлетической трени-
ровкой, тогда как при тренировочных занятиях, направленных на разви-
тие выносливости, такой эффект не проявляется. Несмотря на то что
спортсмены, развивающие свои силовые возможности, обладают полу-
ченными опытным путем знаниями наиболее эффективных путей сти-
муляции мышечной гипертрофии, тем не менее механизмы, благодаря
которым достигаются эти эффекты, остаются невыясненными.
В организме взрослого здорового человека вырабатывается ряд гор-
монов, способных проявлять анаболическое действие. Так, гормон рос-
та, секретируемый питуитарной железой (гипофизом), находится под
контролем гипоталамуса. Значительное поступление этого гормона в
кровь наблюдается после выполнения кратковременных высокоинтен-
сивных физических упражнений, однако значение этого эффекта не ус-
тановлено. Известно, что гормон роста проявляет некоторый антагонизм
по отношению к инсулину. Воздействуя на печень, он вызывает выход в
кровь семейства пептидов, получивших название соматомедины. К ним
относятся инсулиноподобные ростовые факторы (ИРФ-I и ИРФ-П), ко-
торые обладают таким же анаболическим эффектом, как и инсулин.
Известно, что некоторые спортсмены — представители силовых ви-
дов спорта, как и специализирующиеся ь бодибилдинге, применяют
инъекции гормона роста, часто в комбинации с инсулином, стремясь тем
самым стимулировать белковый синтез, а следовательно, и повысить
свои физические кондиции. Такие попытки могут лишь отчасти способ-
ствовать достижению успеха, поскольку гормон роста более эффективен
160
при стимуляции синтеза колла-
гена, образующего соединитель-
ную ткань в мышцах, чем при
стимуляции синтеза сократи-
тельных белков — актина и мио-
зина. Поэтому улучшение фи-
зических кондиций обычно ока-
зывается менее выраженным,
чем предполагалось. Необходи-
мо также иметь в виду негатив-
ные последствия применения
гормона роста, к которым отно-
сятся рост некоторых лицевых
Таблица 5.2. Факторы, влияющие
на синтез и деградацию белков в скелетных
мышцах
Факторы Синтез Деградация
Активность — Снижение
Инсулин Повышение Снижение
Глюкагон — Повышение
Глюкоза — Снижение
Тестостерон Повышение —
Глюкокортикоиды — Повышение
Трийодтиронин Повышение —
костей черепа, костей рук, ног, а также повышение предрасположеннос-
ти к диабету.
Известно, что инсулин стимулирует белковый синтез, однако меха-
низм, с помощью которого это достигается, и специфичность проявляе-
мого эффекта до сих пор не установлены.
Тестостерон обладает выраженным анаболическим эффектом, кото-
рый важен в период ускоренного юношеского развития, но избыточное
потребление тестостерона и родственных ему анаболических стероидов
спортсменами указывает на степень, до которой рост мышц может быть
стимулирован комбинацией тренировочных занятий и применением сте-
роидов. Тем не менее одной лишь тренировкой либо только использова-
нием анаболических стероидов нельзя достичь такой степени гипертро-
фии, какую демонстрируют многие чемпионы в бодибилдинге. Однако
следует отметить, что хроническое использование стероидов не лишено
большого риска для здоровья. Смерть в молодом возрасте по причине
сердечно-сосудистых заболеваний, как и от рака печени, — относитель-
но общее явление, характерное для тех, кто пользовался подобного рода
средствами.
Исходя из известного антагонизма активностей глюкагона и инсу-
лина можно предполагать, что повышение активности глюкагона дол-
жно стимулировать распад белков (табл. 5.2). Высокие уровни тироид-
ного гормона трийодтиронина также повышают скорость белковой
деградации, хотя его низкие концентрации стимулируют синтетичес-
кий процесс. Механизм действия этих гормонов также требует выяс-
нения.
Мышечная активность оказывает значительное влияние на скорость
распада белков. Наблюдения показали, как через несколько дней иммо-
билизации мышцы происходит выраженное снижение ее массы.
11
161
10. Биологически активные аминокислоты
и их производные вещества
Все аминокислоты играют решающую роль в проявлении структурных
и функциональных свойств белков и пептидов, которые определяют не
только структуру организма человека, но и ряд других жизненно важных
функций. В частности, влияние таких веществ, как нейромедиаторы и
гормоны, позволяет интегрировать активность различных клеток, тка-
ней и органов.
Большинство нейромедиаторов, которые осуществляют взаимо-
связь между нервными клетками либо между нервными и другими
клетками, образуются из аминокислот или их метаболитов. К отдель-
ным аминокислотам, которые идентифицируются как нейромедиато-
ры, относятся глицин, глутамат, таурин и аспартат. Производными ве-
ществами, проявляющими аналогичный эффект, являются дофамин, 5-
гидрокситриптамин (5-ГТ, серотонин), гамма-аминомасляная кислота,
гистамин и многие небольшие по размеру пептиды. Кроме того, такие
наиболее широко распространенные возбуждающие агенты, как адре-
налин, норадреналин и ацетилхолин, являются дериватами амино-
кислот. Синтез и распад этих веществ представляют жизненно важный
процесс.
Ацетилхолин выполняет функцию основного нейромедиатора, свя-
зывающего активность двигательных нервов с активностью сократи-
тельного механизма в скелетных мышцах. Он синтезируется в нервных
клетках путем переноса ацетильной группы от ацетил-КоА к холину. Хо-
лин может снова образовываться из ацетилхолина, который предвари-
тельно был высвобожден в синапсе, либо может синтезироваться из фос-
фатидилэтаноламина, который является дериватом серина.
Инактивация ацетилхолина после выхода его в синаптическую щель
происходит в результате гидролиза под влиянием фермента ацетилхо-
линэстеразы. Преимущественная часть освободившегося холина захва-
тывается обратно пресинаптическим окончанием.
Во время напряженной активности количество ацетилхолина, пере-
ходящего в синаптическую щель при деполяризации пресинаптической
мембраны, снижается, однако еще не доказано, что наличие ацетилхо-
лина в пресинаптических терминалях способно лимитировать мышеч-
ную работоспособность в обычных условиях. Тем не менее известно,
что при некоторых формах проявления мышечной слабости (asthenia
gravis) снижение содержания нейромедиатора вызывает субъективное
ощущение упадка сил.
162
Т ирозин-3-монооксигеназа
Тетрагидробио- Дигидробио-
он
СНг
CHNH3+
СОО-
Дофамин
птерин, О2 птерин, Н2О
Дофаминфионооксигеназа
Аскорбат, Дегидроаскорбат,
О2 Н2О
Дегидро ксифенил-
аланин (ДОФА)
ОН
Норадреналин-N- Норадреналин
метилтрансфераза
'-у S-аденозилметионин
S-аденозилгомоцистеин
ОН
ОН
НО.СН2
CH2NH2+CH3
Адреналин
Рис. 5.12. Синтез дофамина, норадреналина и адреналина из аминокислоты тирозина
Катехоламины (адреналин и норадреналин) являются близкород-
ственными веществами. Норадреналин функционирует преимущес-
твенно как нейромедиатор, хотя некоторое его количество попадает в
кровь и проявляет соматический эффект, тогда как адреналин выделя-
ется в кровь мозговым веществом надпочечников и действует как гор-
мон. Катехоламины стимулируют гликогенолиз в печени и мышцах, а
также повышают скорость гидролиза триглицеридов в адипозной тка-
ни, тем самым способствуя повышению в плазме концентрации СЖК.
Выход инсулина также блокируется увеличением в плазме уровня ка-
техоламинов.
п* 163
Катехоламины синтезируются из аминокислоты тирозина и проме-
жуточных продуктов обмена — дигидроксифенилаланина (ДОФА) и до-
фамина, являющихся сами по себе важными веществами. Реакции, свя-
занные с синтезом, представлены на рис. 5.12.
Первый этап деградации катехоламинов катализируется аминокси-
дазой с последующей серией реакций, приводящих к образованию ко-
нечных продуктов — З-метокси-4-гидроксиманделиковой кислоты (пре-
имущественно в периферических тканях) и З-метокси-4 гидроксифенил-
гликола (преимущественно в ЦНС). Выявление этих конечных продук-
тов часто используют в качестве маркеров катехоламинового обмена.
Конечным продуктом дофаминового метаболизма является гомованили-
новая кислота.
5-Гидрокситриптамин служит важным нейромедиатором, обеспе-
чивающим широкое разнообразие установленных к настоящему време-
ни функций. Он, несомненно, играет важную роль в проявлении неко-
торых поведенческих реакций, включая пробуждение, а также поведе-
ние, связанное с приемом пищи (по крайней мере, углеводов). Получе-
ны доказательства его участия в механизме развития центрального
утомления.
Реакции, связанные с синтезом 5-ГТ из триптофана (рис. 5.13), не
являются скоростьлимитирующими. Вместо этого можно предполо-
жить, что скорость синтеза лимитируется скоростью поступления в мозг
триптофана, а это, в свою очередь, регулируется переносчиком, который
также транспортирует и другие аминокислоты, в том числе аминокис-
лоты с разветвленной цепью (АКРЦ). Поэтому отношение триптофана к
АКРЦ в плазме является немаловажным, однако представляется, что в
дальнейшем присоединение триптофана к альбумину является также ве-
сомым фактором. Только около 10 % триптофана в плазме находится в
свободной форме, и имеются доказательства, что только эта фракция
доступна для транспорта в мозг. Оставшийся триптофан связан с альбу-
мином плазмы, в то время как часть его находится в связанном состоя-
нии с жирными кислотами.
Повышение в плазме концентрации СЖК, обычно проявляющееся
во время пролонгированных физических нагрузок, будет вытеснять
триптофан из участков, связывающих его с альбумином, тем самым спо-
собствуя увеличению концентрации свободного триптофана. В то же
время поступление и окисление АКРЦ сокращающимися мышцами воз-
растает, а их концентрация в крови — снижается. Эти эффекты должны
увеличивать поступление триптофана в мозг, а следовательно, обеспе-
чение синтеза и, возможно, освобождение 5-ГТ.
164
Триптофан-5-монооксигеназа
02 Тетрагидробиоптерин
Н2О Дигидробиоптерин-*^,
Декарбоксилаза
ароматических
L-аминокислот
Рис. 5.13. Синтез нейромедиатора 5-ГТ, начинающийся с аминокислоты триптофана
Углеводная пища угнетает мобилизацию СЖК, а низкая концентра-
ция жирных кислот в плазме вызывает снижение концентрации свобод-
ного триптофана, меньшее его поступление в мозг и снижение скорости
синтеза 5-ГТ. Это обеспечивает связь между периферическими метабо-
лическими процессами и поведением. Если 5-ГТ действительно учас-
твует в механизме развития утомления, то указанные механизмы могут
обеспечивать взаимосвязь между доступностью субстратов и субъектив-
ным ощущением усталости, которое наступает при продолжительной
мышечной работе. Имеющиеся для такого утверждения доказательства,
однако, малоубедительны и дают основание предполагать, что даже зна-
чительные изменения в крови концентраций АКРЦ и триптофана могут
не вызывать изменений физической работоспособности, которые пред-
сказываются в том случае, если бы подобного рода механизм функцио-
нировал.
Родственное вещество — мелатонин — синтезируется в шишко-
видной железе в ходе дальнейшего метаболизма 5-ТГ, и это может
оказывать влияние на синхронизацию различных физиологических
процессов с изменениями продолжительности дня, а также с сезонны-
ми изменениями.
165
Другим важным нейромедиатором является ГАМК, которая син-
тезируется из глутамата и функционирует в основном как тормозной
медиатор.
К одиночным аминокислотным нейромедиаторам относятся глицин
и таурин, которые обладают ингибирующим эффектом, а также аспартат
и глутамат, оказывающие возбуждающее действие.
11. Регуляторные пептиды и белки
Очень большое количество белковых гормонов и небольших пептидных
молекул принимает участие в регуляции метаболизма, белкового синте-
за, роста тканей, а также в реакциях организма на внешние стимулы. Эти
молекулы действуют в основном как гормоны, передающие сигналы от
одних тканей к другим. Однако они могут также оказывать локальные
влияния на ткани, в которых вырабатываются. Аминокислотная структу-
ра многих таких веществ к настоящему времени установлена, а некото-
рые из них успешно синтезированы. Ряд наиболее важных гормонов
белковой природы и регуляторных пептидов кратко описаны ниже, од-
нако список их представителей нельзя считать всеобъемлющим.
Регуляторы метаболизма. Большое количество различных агентов
участвует в реакциях на изменение поступления субстратов и потреб-
ность в них. Некоторые из этих агентов и интеграция их влияний недос-
таточно изучены, другие же нуждаются в идентификации.
Инсулин (состоит из двух полипептидных цепей — A-цепи из
21 аминокислоты и В-цепи из 30 аминокислот, которые соединены меж-
ду собой дисульфидной связью; секретируется 6-клетками островков
Лангерганса поджелудочной железы) и глюкагон (состоит из 29 амино-
кислот, секретируется о-клетками) играют главную роль в интеграции
метаболизма углеводов, жиров и аминокислот. Баланс между этими дву-
мя гормонами определяет распределение питательных веществ после
приема пищи и регулирует мобилизацию субстратов после ее всасыва-
ния. Основными эффектами инсулина являются анаболическое дей-
ствие, обеспечение гликогенных запасов, снижение мобилизации липи-
дов и стимуляция белкового синтеза. Влияния глюкагона в основном
противоположны влияниям инсулина.
Важная роль инсулина отчетливо проявляется у больных сахарным
диабетом, когда прием углеводов вызывает неконтролируемое повыше-
ние в крови уровня глюкозы, что, в свою очередь, приводит к появлению
глюкозы в моче. Проявляются также другие последствия, к которым от-
166
носится связывание глюкозы с некоторыми белками, в результате чего
происходит утрата последними функциональных свойств.
Чрезмерный уровень СЖК у больных диабетом вызывает увели-
чение концентрации кетоновых тел (6-гидроксибутирата и ацетоацетата)
в крови, который, в свою очередь, приводит к метаболическому ацидозу.
Поскольку многие ткани чувствительны к влияниям инсулина и глюка-
гона — этим двум гормонам, играющим ведущую роль в регуляции ме-
таболизма, то не удивительно, что их синтез и экскреция тонко регули-
руемы.
Выделение инсулина и глюкагона происходит в ответ на метаболи-
ческие сигналы, среди которых особо важным является концентрация
глюкозы в крови, поэтому многие небольшие по размерам пептиды, вы-
деляющиеся из желудочно-кишечного тракта при приеме пищи, воздей-
ствуя на р-клетки поджелудочной железы, стимулируют выделение со-
матостатина, который, в свою очередь, угнетает выделение инсулина.
Этот эффект предотвращает чрезмерную экскрецию инсулина в ответ на
повышение концентрации глюкозы в крови.
Факторы роста. К настоящему времени идентифицировано значи-
тельное количество небольших по размерам пептидных гормонов, про-
являющих стимулирующий эффект на рост тканей, однако механизмы
их действия нуждаются в дальнейшем изучении. К ним относятся сома-
томедины, инсулиноподобные ростовые факторы (ИРФ-1 и ИРФ-2) и
другие тканево-специфические ростовые факторы, число которых пос-
тоянно увеличивается.
Пептиды кишечника. Ряд пептидов освобождается из желудка и
верхнего отдела тонкой кишки после приема мяса. К ним относятся гас-
трин, холецистокинин (ХЦК), вазоактивный кишечный пептид (ВКП),
бомбезин, секретин и другие (некоторые из них требуют идентифи-
кации). Данные пептиды играют различную роль в координации ответа
на прием пищи путем регуляции скорости опорожнения желудка, мото-
рики кишечника, а также путем стимуляции секреции пищеварительных
ферментов. Имеются доказательства роли сигналов насыщения в регуля-
ции потребления энергии. Некоторые из этих пептидов также влияют на
мобилизацию субстратов и энергетические запасы в период после прие-
ма пищи путем модулирующего воздействия на секрецию инсулина и
глюкагона поджелудочной железой.
Пептиды мозга. В мозге обнаружено большое количество пепти-
дов. Гематоэнцефалический барьер не позволяет пептидам переходить
в кровь, в связи с чем вполне возможно, что они синтезируются в ЦНС
и должны играть определенную локальную роль, функционируя то ли
как нейромедиаторы, то ли как модуляторы клеточного ответа. Некото-
167
рые из них действуют в качестве релизинг-факторов, регулирующих
секрецию гормонов, которые оказывают влияние на периферические
ткани.
Эндорфины относятся к семейству пептидов, которые в последнее
время привлекают к себе пристальное внимание, поскольку оказывают
на ЦНС такое же влияние, как и морфин, снижая ответ организма на бо-
левые стимулы. Все эндорфины являются дериватами большой белко-
вой молекулы-предшественника, состоящей из 134 аминокислот, кото-
рые разделены на активные фрагменты, соответствующие размеру энке-
фалинов (включают 5 аминокислот) и простирающиеся к 6-липотропи-
ну (включают 93 аминокислоты). Обнаруженное при физических наг-
рузках увеличение содержания энкефалинов в плазме позволяет предпо-
ложить, что эти молекулы обеспечивают способность организма повы-
шать сопротивляемость к боли или дискомфорту, а также иметь отноше-
ние к так называемому состоянию «веселости бегуна». К этим гипоте-
зам, однако, не следует относиться серьезно, поскольку гематоэнцефа-
лический барьер непроницаем для этих веществ: энкефалины, вероятно,
поступают в плазму скорее из кишечника, чем из мозга.
Выводы
1. Белки, представляющие собой последовательную цепь одной или бо-
лее аминокислот, являются основным структурным компонентом орга-
низма. Они функционируют в качестве ферментов и гормонов, а также
составляют сократительный аппарат мышц.
2. Все аминокислоты имеют общую структуру, включающую
аминную и карбоксильную группы, связанные одним атомом углерода.
Имеется также органическая боковая цепь и различные боковые группы,
обусловливающие существование 20 различных аминокислот. Некото-
рые из них не могут синтезироваться в тканях животных, хотя и синте-
зируются растениями, следовательно, должны поступать в организм с
пищей.
3. Белки состоят из линейных цепей аминокислот, соединенных
между собой в ходе реакций конденсации. Складывание этих цепей про-
исходит в результате взаимодействия между боковыми группами амино-
кислотных компонентов. Окончательная форма белковой молекулы и
способность последней изменять свою форму предопределяют свой-
ственные ей функциональные особенности.
4. Синтез белков находится под контролем генетической информа-
ции, содержащейся в ДНК. Генетическая информация детерминирует
168
установление последовательности аминокислот в белковой цепи, а так-
же инициирует и прерывает этот процесс. Изменения в структуре и
функциях организма могут происходить настолько, насколько они пре-
допределены генетически.
5. Несмотря на стабильность структуры, в организме постоянно
протекают процессы синтеза и распада белков. Полупериод существова-
ния отдельных белков варьирует от менее одного часа до нескольких не-
дель. Это определяет скорость адаптации к условиям окружающей сре-
ды, в том числе к физической тренировке.
6. Углеродный каркас аминокислот может быть использован в качес-
тве энергетического топлива для окислительного метаболизма либо для
синтеза других веществ. Белки не являются основным источником
энергии для энерго продукции при мышечной деятельности, однако сис-
тематические и напряженные физические нагрузки повышают потреб-
ность организма в пищевых белках.
7. Аминокислоты аланин и глутамин играют ключевую роль в регу-
ляции концентрации аммиака в тканях, который потенциально токсичен,
а также обеспечивают перенос азота между тканями, его транспорт в пе-
чень, где поступает в состав мочевины для экскреции почками.
8. Многие отдельные аминокислоты служат предшественниками
нейромедиаторов или сами функционируют в качестве нейромедиато-
ров. Адреналин, норадреналин, ацетилхолин, дофамин, серотонин и гис-
тамин являются дериватами аминокислот.
9. Короткие аминокислотные цепи — пептиды — играют жизненно
важную роль в регуляции метаболизма.
МЕТАБОЛИЧЕСКИЕ ОТВЕТЫ
ПРИ ВЫСОКОИНТЕНСИВНЫХ
ФИЗИЧЕСКИХ НАГРУЗКАХ
1. Ресинтез АТФ
Аденозинтрифосфат (АТФ) является единственным
энергетическим источником, который может быть не-
посредственно использован скелетными мышцами для
сокращения. Запас АТФ в скелетных мышцах человека
относительно невелик (около 24 ммоль-кг-1 смт), поэтому
постоянно должен осуществляться его ресинтез из про-
дуктов его же распада — АДФ и' АМФ (см. главу 2). Во
время выполнения мышечной работы субмаксимальной
аэробной мощности (в условиях устойчивого состояния)
адекватность ресинтеза АТФ может достигаться путем
использования запасов жиров и углеводов. Однако при
высокоинтенсивных физических нагрузках (в неустойчи-
вом состоянии) относительно медленная активация и
скорость выработки энергии за счет окислительного фос-
форилирования не могут обеспечить потребности сокра-
щающихся мышц в АТФ. В такой ситуации анаэробное
энергообеспечение работающих мышц является преиму-
щественным. Обычно для достижения устойчивого сос-
тояния требуется несколько минут, что обусловлено
большим количеством и сложностью связанных с про-
цессом окислительного процесса реакций. Иногда оно
наступает при максимальной скорости ресинтеза АТФ,
составляющей приблизительно 2,5 ммоль-кг~’смт-с—'.
Анаэробная продукция энергии, происходящая в цитозо-
ле, активируется почти мгновенно и способна обес-
печить ресинтез АТФ со скоростью, превышающей
170
11 ммоль-кг-'смт-с-Однако такая интенсивность энергопродукции
может поддерживаться лишь в течение нескольких секунд, после чего
начинает снижаться. Безусловно, окислительный и анаэробный ресин-
тезы АТФ не должны рассматриваться обособленно друг от друга. В
табл. 1.5 (глава 1) показано, как с увеличением продолжительности
выполнения физического упражнения снижается вклад энергообеспе-
чения за счет анаэробных процессов и возрастает доля аэробной энер-
гопродукции.
2. Субстраты для высокоинтенсивной
мышечной работы
Фосфокреатин. Скелетные мышцы содержат относительно высокий
резерв фосфокреатина (КФ), составляющий до 70-80 ммоль-кг—'смт в
покое. Утилизация его происходит сразу же с наступлением сократи-
тельной активности, что обеспечивает буферирование быстро накапли-
вающегося АДФ, который образуется при гидролизе АТФ в ходе мно-
жества энергозависимых процессов, связанных с мышечным сокраще-
нием и расслаблением. Моментальное увеличение концентрации АДФ
служит главным стимулом для гидролиза КФ в ходе креатинкиназной
реакции. Каждый моль распавшегося КФ обеспечивает ресинтез одно-
го моля АТФ (см. главу 2). В начале 60-х годов сложилось мнение, что
в течение первых 15 мин физической нагрузки максимальной интен-
сивности энергообеспечение осуществляется почти исключительно за
счет распада КФ. Обусловлено такое мнение тем, что КФ запасен в ци-
тозоле в непосредственной близости от пунктов, утилизирующих
энергию, а также потому, что гидролиз КФ не зависит от доставки кис-
лорода или необходимости завершения нескольких метаболических
реакций перед тем как освободится энергия, используемая для ресин-
теза АТФ.
В настоящее время принято считать, что анаэробный метаболизм
гликогена также вносит значительный вклад в ресинтез АТФ на протя-
жении первых секунд выполнения высокоинтенсивной мышечной ра-
боты. Важность гидролиза КФ заключается в чрезвычайно высокой
скорости, с которой он может осуществлять ресинтез АТФ. Это осо-
бенно характерно для кратковременных физических нагрузок макси-
мальной мощности. Так, например, на рис. 6.1 показано, как изменяет-
ся скорость ресинтеза АТФ за счет гидролиза КФ на протяжении 30 с
максимального изометрического мышечного сокращения, приводяще-
171
го к развитию утомления.
Прежде всего следует ука-
зать, что самая высокая сте-
пень утилизации КФ отме-
чается в первые 2 с сокра-
тительной активности. Од-
нако уже через 2,6 с продук-
ция АТФ за счет гидролиза
КФ снижается приблизи-
тельно на 15 %, а в последу-
ющие 10 с — более чем на
50 %. Вклад КФ в ресинтез
АТФ в течение последних
10 с тридцатисекундной мы-
шечной работы относитель-
но невелик и составляет при-
мерно 2 % первоначального
уровня. Представленные ре-
зультаты получены при оп-
ределении метаболических
Рис. 6.1. Скорость анаэробного ресинтеза АТФ
за счет гидролиза КФ, гликолиза и их совместно-
го вклада в течение 30-секундного изометричес-
кого мышечного сокращения околомакси-
мальной мощности у человека
изменений в биоптатах мышцы, подвергавшейся прерывистой элек-
тростимуляции (1,6 с — стимуляция с частотой 50 Гц и паузами про-
должительностью 1,6 с).
Механизмы, ответственные за почти немедленное снижение ско-
рости утилизации КФ во время выполнения физической работы мак-
симальной интенсивности, к настоящему времени не изучены, одна-
ко могут быть связаны с локальным снижением его доступности к
миофибриллярному аппарату. Учитывая значительный энергети-
ческий запрос при физических нагрузках максимальной интенсив-
ности, можно предположить, что очень высокая скорость утилизации
КФ в начале сократительной активности является причиной быстро-
го использования его запасов в участках срочной энергетической
транслокации (актомиозиновых поперечных мостиков). Такое пред-
положение выглядит правдоподобным, поскольку при интенсивной
мышечной работе, продолжающейся более 20 с, запасы КФ в клетке
становятся почти полностью истощенными, а скорость последующей
митохондриальной продукции АТФ не соответствует скорости гидро-
лиза КФ.
Гликогенолиз и гликолиз. Гликогенолиз представляет собой гид-
ролиз мышечного гликогена до глюкозе-1-фосфата, а гликолиз — серию
реакций, обеспечивающих распад глюкозы или глюкозо-1-фосфата до
172
пирувата или лактата (глава 3). Из предыдущих глав следует, что если
упражнения максимальной интенсивности продолжаются более нес-
кольких секунд, то должен заметно повышаться вклад гликогенолиза и
гликолиза в синтез АТФ.
Интегративная природа энергетического метаболизма обеспечивает
возможность того, что активация мышечного сокращения ионами каль-
ция и накопление продуктов гидролиза АТФ и КФ (АДФ, АМФ, ИМФ,
NH3 и Фн) действуют как стимуляторы гликогенолиза, а это гарантирует
поддержание анаэробной продукции АТФ, по крайней мере, в течение
короткого времени.
Регуляция гликогенолиза при мышечных сокращениях представ-
ляет собой очень сложный механизм, который не ограничивается
трансформацией менее активной гликогеновой фосфорилазы b в ее
более активную форму а (см. главу 3). Обнаружилось, что гликогено-
лиз может совершаться с незначительной скоростью, несмотря на поч-
ти полный переход всей фосфорилазы в форму а. И напротив, было
отмечено повышение скорости гликогенолиза во время приостановки
кровообращения при относительно низком содержании фосфорила-
зы а. На основании результатов этих и других подобных работ был
сделан вывод, что накапливающийся в процессе гидролиза АТФ и КФ
неорганический фосфат играет ключевую роль в регуляции гликогено-
литической активности фосфорилазы а, в связи с чем он служит как
бы связывающим звеном между энергетической потребностью сокра-
щающихся мышц и скоростью субстратной утилизации. Однако было
установлено, что гликогенолиз может происходить в течение первых
двух секунд мышечной работы без заметного увеличения концентра-
ции Фн и что его скорость может оказаться низкой несмотря на высо-
кие концентрации фосфорилазы а и Фн. В связи с этим есть основания
предполагать существование иных, кроме степени трансформации
фосфорилазы и доступности Фн, факторов, участвующих в регуляции
гликогенолиза.
В экспериментах, проведенных in vitro, было показано, что АМФ
способен оказывать более заметное влияние на гликогенолиз путем
увеличения гликогенолитической активности фосфорилазы а. Однако
в связи с тем, что, как предполагалось, 90 % (и даже более) общего
содержания АМФ в клетке in vivo находится в связанном с белками
состоянии, то возникал вопрос, действительно ли увеличение кон-
центрации свободного АМФ во время сокращений оказывает значи-
тельное влияние на кинетику фосфорилазы а. Тем не менее в более
поздних работах было продемонстрировано,что незначительное уве-
личение концентрации АМФ (10 ммоль-л-1) способствовало in vitro
173
существенному повышению активности этой формы фермента. Более
того, в исследованиях in vivo получены доказательства тесной взаи-
мосвязи между обращаемостью АТФ в мышцах и утилизацией глико-
гена, свидетельствующие о том, что индуцированное сократительной
активностью увеличение содержания свободной АМФ и Фн может
служить ключевым регулятором распада гликогена при мышечной
работе.
Анаэробный гликолиз включает большее количество этапов, чем
гидролиз КФ, однако по сравнению с окислительным фосфорилирова-
нием он протекает быстрее. В главе 3 отмечалось, что АТФ в процессе
гликолиза образуется путем фосфорилирования АДФ во второй поло-
вине этого процесса. На протяжении многих лет существовало мнение,
что КФ является единственным энергетическим источником, использу-
емым для инициации мышечных сокращений, а утилизация гликогена
начинается после того, как запасы КФ истощаются. Однако как ока-
залось (см. рис. 6.1), ресинтез АТФ в процессе гликолиза на протяже-
нии 30 с максимальной сократительной активности мышц, приводя-
щей к развитию утомления, наступает почти одновременно с началом
сокращений. В отличие от гидролиза КФ, продукция АТФ за счет гли-
колиза достигает своего максимума лишь после 5 с выполнения упраж-
нения, и эта высокая скорость поддерживается еще несколько секунд.
После выполнения физических упражнений в течение 30 с вклад
анаэробного гликолиза в ресинтез АТФ приблизительно в два раза пре-
вышает вклад в этот процесс гидролиза КФ. Это проявляется и в очень
высокой концентрации лактата в мышцах (более 100 ммоль-кг—1 смт),
которая достигается после 30 с и более максимальной сократительной
активности.
Относительно точных механизмов, ответственных за накопление
лактата во время интенсивных мышечных сокращений, существуют
противоречивые мнения. По мнению специалистов, высокий уровень
энергетического запроса сопряжен с неадекватным кислородным обес-
печением, т. е. когда недостаточное поступление кислорода в ткани на-
чинает ограничивать продукцию АТФ, что приводит к накоплению
НАДН в митохондриях и цитозоле, скорость протекания метаболическо-
го процесса через гликолиз и высокое отношение НАД7НАДН в цитозо-
ле поддерживается восстановлением пирувата до лактата. Однако науч-
ные данные свидетельствуют о незначительном снижении в митохон-
дриях редокс-потенциала во время мышечных сокращений. Поэтому су-
ществует мнение, что основным регулирующим механизмом продукции
лактата при интенсивной мышечной работе в условиях ишемии являет-
ся активация пируватдегидрогеназного комплекса и скорость образова-
174
ния ацетильных групп, а не доступность кислорода. Кроме того, показа-
но также, что при любой максимальной физической нагрузке накопле-
ние лактата может быть значительно изменено предрабочими вариация-
ми питания. Следовательно, субстраты и гормональный баланс являют-
ся детерминантами продукции лактата.
Как бы ни выглядели механизмы, ответственные за образование
лактата при мышечной деятельности, ясно одно: ресинтез АТФ в про-
цессе гликолиза при мышечной работе может быть значительно изменен
благодаря предшествующим диетическим манипуляциям. Подтвержда-
ется это тем, что субстратный и гормональный балансы способны детер-
минировать продукцию лактата.
И все же, какие бы механизмы ни были ответственны за образова-
ние лактата при мышечной деятельности, не вызывает сомнений, что
ресинтез АТФ в процессе гликолиза не может продолжаться неограни-
ченное время. Как видно из рис. 6.1, особенно это проявляется при вы-
полнении кратковременных физических упражненний максимальной
интенсивности. Однако механизмы, ответственные за проявление ука-
занного эффекта, не установлены, но маловероятно, чтобы они имели
отношение к истощению (локальному либо иному) запасов мышечного
гликогена, так как их уровень к концу выполнения физических упраж-
нений максимальной интенсивности остается высоким. И действитель-
но, доступность гликогена могла бы ограничивать работоспособность
до тех пор, пока его дорабочая концентрация не станет ниже
100 ммолыкг~1 смт. Опосредованное концентрацией водородных ионов
снижение активности фосфорилазы и фосфофруктокиназы (ФФК) дает
основание рассматривать его в качестве возможного механизма, ответ-
ственного за угнетение гликолиза. Фосфофруктокиназа катализирует
превращение фруктозо-6-фосфата до фруктозе-1,6-ди фосфата, что яв-
ляется «узким местом» гликолиза (см. главу 3). Тем не менее в настоя-
щее время принято считать, что возможное pH-опосредованное угнете-
ние гликолиза при максимальных физических нагрузках преодолевает-
ся в условиях in vivo накоплением активаторов ФФК (например, АМФ
и аммиаком). Как уже отмечалось, АМФ и ИМФ связаны с регуляцией
распада гликогена во время мышечной работы. Предполагалось, что как
только скорость утилизации гликогена достигает своего пика, снижение
в саркоплазме уровня свободного АМФ (вследствие снижения скорости
образования и (или) pH-индуцируемого повышения активности АМФ-
дезаминазы — фермента, катализирующего дезаминирование АМФ до
ИМФ) вызовет снижение активности фосфорилазы, что приведет к сни-
жению при работе в зоне максимальной мощности скорости гликогено-
лиза и гликолиза.
175
3. Взаимодействие фосфокреатина
и утилизации гликогена во время выполнения
кратковременных физических упражнений
максимальной интенсивности
Выше отмечалось, что высокая скорость ресинтеза АТФ за счет распада
КФ и гликогена может поддерживаться короткий промежуток времени.
Как видно из рис. 6.1, продолжительность ресинтеза превышает 30 с у
мужчин при околомаксимальных изометрических мышечных сокраще-
ниях. Скорость распада КФ становится максимальной с началом сокра-
тительной активности и начинает снижаться уже через 1,3 с. И напро-
тив, максимальную скорость гликолиз достигает только через 5 с сокра-
щения и снижается только через 20 с. Это свидетельствует о том, что
быстрая утилизация КФ может буферировать первоначальное отстава-
ние энергообеспечения, осуществляемого за счет гликолиза, и что вклад
последнего в ресинтез АТФ по мере продолжения физических упражне-
ний возрастает, а использование КФ — снижается. Приведенные данные
свидетельствуют о чрезвычайной важности КФ для начальной фазы сок-
ратительной активности мышц. Без его высокого гидролиза весьма веро-
ятно, что развиваемое мышцей напряжение будет почти сразу ослабе-
вать, что и происходит в случае, когда запасы КФ в мышцах устранялись
фармакологически или замещались креатиновым аналогом. Важно так-
же отметить, что, в конечном счете, это приводит к прогрессивному сни-
жению скорости ресинтеза АТФ за счет обоих субстратов при данном
виде мышечной нагрузки. Так, например, в течение последних 10 с вы-
полнения упражнения (см. рис. 6.1) скорость продукции АТФ за счет
гидролиза КФ снизилась примерно до 2 % максимального значения.
Аналогично соответствующая скорость ресинтеза АТФ за счет гидроли-
за гликогена снизилась примерно до 40 %.
4. Высокоинтенсивные физические нагрузки
продолжительностью более 30 с
В данной главе рассмотрены метаболические ответы на высокоинтен-
сивные физические упражнения, выполняемые в неустойчивом физио-
логическом состоянии. При выполнении таких упражнений мощность
работы после 30 с с момента ее начала снижается примерно на 40 %, по-
этому энергетическая потребность сокращающихся мышц чрезвычайно
высокая и обеспечивается в основном анаэробной утилизацией КФ и
176
гликогена. Однако доказано, что неустойчивое состояние, проявляюще-
еся хотя и менее выражение, может поддерживаться более 5 мин до нас-
тупления утомления. В таких условиях окисление субстратов может
вносить наибольший вклад в продукцию АТФ, в связи с чем эту значи-
мость нельзя недооценивать.
При выполнении утомительных физических упражнений в тече-
ние 2—3 мин окислительное фосфорилирование может обеспечивать до
80 % общей энергопродукции (см. табл. 5). Это указывает на важность
субстратного окисления при интенсивной мышечной деятельности,
что не всегда учитывают. При таких обстоятельствах мышечный глико-
ген является наиболее важным энергетическим топливом, поскольку
использование глюкозы мышцами угнетается накоплением глюкозо-6-
фосфата, а липолиз в адипозной ткани ингибируется накоплением лак-
тата. Скорость утилизации мышечного гликогена во время выполнения
высокоинтенсивного физического упражнения в течение 3-4 мин,
приводящего к развитию утомления, составляет приблизительно
45 ммоль-кг-'емт-мин-1, которая значительно ниже по сравнению со
скоростью около 160 ммоль-кг~1смт-мин~1, отмечаемой во время 30-се-
кундной мышечной работы максимальной интенсивности. Однако если
в первом примере окисляется только 50 % гликогена, то можно подсчи-
тать, что количество АТФ, ресинтезируемого при окислении гликогена,
в два раза превышает количество этого макроэрга, которое образуется
при анаэробном распаде гликогена до лактата при 30-секундной работе.
Предполагалось, что в таких условиях мышечной деятельности
внутримышечные триацилглицеролы могут принимать участие в энер-
гопродукции. Однако это маловероятно, поскольку снижение уровня сво-
бодного карнитина при высокоинтенсивной физической нагрузке будет
лимитировать поступление жирных кислот в митохондрии. Изучение
внутримышечной утилизации триацилглицеролов связано со многими
проблемами технического характера, ио вполне возможно, что снижение
содержания внутримышечных триацилглицеролов после интенсивной
мышечной работы не имеет существенного физиологического значения и
связано с быстрым повышением уровня катехоламинов в крови.
5. Повторяющиеся физические нагрузки
С началом интенсивной сократительной активности мышц происходит
быстрый и значительный гидролиз КФ и накопление лактата. Однако ес-
ли физическое упражнение выполняется в нескольких попытках через
короткие промежутки времени отдыха, то скорости гидролиза КФ и на-
копления лактата значительно снижаются. Прогрессирующее падение
утилизации КФ зависит, очевидно, от степени его ресинтеза между по-
пытками, т. е. если восстановительные процессы не в состоянии полнос-
тью обеспечить ресинтез КФ, то это будет лимитировать анаэробный ре-
синтез АТФ в последующих попытках выполнения физического упраж-
нения. Тем не менее механизмы, ответственные за снижение продукции
лактата, менее понятны. Предполагалось, что указанные особенности
метаболизма обусловлены угнетением (по механизму обратной связи)
гликолиза в результате накопления таких конечных продуктов, как водо-
род, лактат и Фн. Однако имеются данные, свидетельствующие о накоп-
лении и некоторых положительных модуляторов гликолиза во время вы-
полнения физических упражений, поэтому такое предположение не мо-
жет полностью объяснить снижение продукции лактата, проявляющееся
в случаях, когда упражнение выполняется несколько раз через короткие
промежутки отдыха. Кроме того, если интервалы отдыха между попыт-
ками продолжительны настолько, что концентрация метаболитов в
мышце существенно снижается, то это приводит к снижению накопле-
ния лактата в последующих попытках. К настоящему времени сложи-
лось мнение, что отмеченный факт обусловлен прогрессирующим воз-
растанием метаболизма через ПДГ-комплекс. Согласно этому предполо-
жению, вклад метаболизма пирувата через ПДГ-комплекс в процессе
трех 30-секундных физических упражнений максимальной интенсив-
ности, выполняемых с промежутком отдыха 4 мин, составлял 29, 33 и
63 % общей энергопродукции. Предположение о прогрессирующем воз-
растании окисления углеводов во время повторяющихся физических
нагрузок подтверждается также данными о градуальном увеличении
потребления кислорода в аналогичных условиях проведения экспери-
мента. Снижение продукции лактата в процессе выполнения повторных
нагрузок максимальной интенсивности также служит важным доказа-
тельством данного предположения.
6. Типы ответов мышечных волокон
Изложенные выводы основывались на результатах метаболических из-
менений, полученных при изучении биоптатов четырехглавой мышцы
бедра. Однако известно, что композиционный состав скелетных мышц
человека представлен, по меньшей мере, двумя различными в функцио-
нальном и метаболическом отношении типами волокон (см. главу 1). Во-
локна типа I характеризуются как медленносокращающиеся, устойчи-
вые к развитию утомления, развивающие невысокую мощность с преоб-
178
Рис. 6.2. Содержание КФ и гликоге-
на в мышечных волокнах типов I и II
в состоянии покоя и скорость распа-
да этих энергетических источников
во время бега в течение 30 с на тред-
миле с максимальной интенсивнос-
тью и при перемежающейся элек-
тростимуляции в течение 30 с (1,6 с
— стимуляция с частотой 50 Гц и
1,6 с — покой) в условиях интактно-
го и перекрытого кровотока:
1 — содержание в покое; 2 — бег в тече-
ние 30 с на тредмиле; 3 — перемежаю-
щаяся в течение 30 с электростимуляция
с перекрытым кровообращением; 4 —
электростимуляция в условиях интакт-
ного кровообращения в течение 30 с
х | 600
ф °
ладанием аэробного метаболизма для обеспечения ресинтеза АТФ. И
напротив, волокна типа II являются быстросокращающимися, быстро-
утомляемыми, развивающими высокую мощность и характеризующие-
ся преобладанием анаэробных механизмов ресинтеза АТФ. Результаты
исследований мышц животных с преобладающим составом волокон то-
го или иного типа, а также данные, полученные в одной из работ при
изучении однотипных мышечных волокон человека, свидетельствуют о
том, что быстрое и выраженное повышение с последующим снижением
способности к проявлению максимальной мощности сокращений, отме-
чаемое при интенсивной сократительной активности, может быть тесно
взаимосвязано с активацией и быстрой утомляемостью волокон типа II.
На рис. 6.2 показан распад КФ и гликогена в мышечных волокнах ти-
пов I и II при физической нагрузке максимальной интенсивности в раз-
личных условиях проведения эксперимента. Во-первых, следует отме-
тить, что концентрации КФ и гликогена в волокнах типа II выше по
сравнению с волокнами типа I; во-вторых, во время интенсивной сократи-
тельной активности мышц скорости гликогенолиза и распада КФ в волок-
нах типа II также выше, чем в волокнах типа I. Это справедливо как для
динамических физических упражнений (бег на тредмиле), так и для вызы-
ваемых электростимуляцией изометрических мышечных сокращений.
Скорость гликогенолиза в обоих типах мышечных волокон во время
бега на тредмиле и перемежающихся изометрических сокращениях в ус-
12*
179
ловиях перекрытого кровотока хорошо согласуется с Vmax фосфорила-
зы, определяемой в этих волокнах. Подтверждением может служить тот
факт, что гликогенолиз во время высокоинтенсивной сократительной ак-
тивности протекает в мышцах с околомаксимальной скоростью. Удиви-
тельным оказалось то, что при перемежающихся изометрических сокра-
щениях в условиях интактного кровотока, когда интервал отдыха между
сокращениями составлял 1,6 с, скорость гликогенолиза в волокнах типа
I была незначительной. В волокнах же типа II она достигала почти мак-
симальных значений и была такой же, как и в условиях перекрытого кро-
вотока. Изложенные факты дают основание полагать, что во время вы-
полнения высокоинтенсивных физических упражнений гликогенолиз в
волокнах типа II постоянно протекает с максимальной скоростью неза-
висимо от экспериментальных условий, тогда как скорость в волокнах
типа I, вероятно, во многом зависит от поступления кислорода в клетку
и фосфорилирующего потенциала последней.
7. Утомление
Утомление — неизменный признак высокоинтенсивных физических уп-
ражнений, и это физиологическое состояние характеризуется невозмож-
ностью продолжать мышечную работу определенной мощности.
Обычно к концу ЗО-й секунды выполнения физических упражнений
максимальной интенсивности мощность или сила мышечных сокраще-
ний снижается на 40-60 %. Однако при более продолжительной высоко-
интенсивной физической нагрузке, характеризующейся наступлением в
течение 5-минутного периода устойчивого физиологического состояния,
изменение работоспособности менее выражено, хотя и в этом случае
развивается утомление. Утомление — сложный многофакторный про-
цесс, и если физические упражнения продолжают выполнять после нес-
кольких попыток, то можно ожидать, что утомление также будет разви-
ваться до полного завершения.
Утомление, связанное с нарушением энергообеспечения. Все
представленные в настоящем параграфе данные дают основание пола-
гать, что утомление, в частности при кратковременных физических наг-
рузках максимальной интенсивности, обусловлено преимущественно
постепенным снижением анаэробной продукции АТФ или повышенным
накоплением лактата в результате снижения уровня КФ и падения ско-
рости гидролиза гликогена. Так, в одной из работ была предпринята по-
пытка установить взаимосвязь между снижением проявления целостной
мышцей максимальной силы изометрического сокращения с метаболи-
180
Рис. 6.3. Проявление силы изометрических сокращений (а) целостной мышцы (х) и
концентрации КФ (Д, А) и АТФ (И, □) в одиночных волокнах в состоянии покоя и
после 10 и 20 с перемежающей электростимуляции мышцы человека с частотой 50 Гц
(Д, □ — волокна типа I; А, — волокна типа II), а также скорость гликогенолиза (б)
в волокнах типов I и II в течение 20 и 30 с электростимуляции мышцы человека с час-
тотой 50 Гц (□ — волокна типа I; — волокна типа II)
20-30
ческими изменениями, происходящими в волокнах типов I и II. На
рис. 6.3, а показано изменение скорости утилизации КФ в этих волокнах
между 0-10 и 10-20 с максимальных мышечных сокращений, вызывае-
мых электростимуляцией. В течение первых 10 с этот показатель для во-
локон типов I и.II составил соответственно 3,3 и 5,3 ммолыкг-’смт-с~’.
Однако во втором временном промежутке стимуляции скорость утили-
зации в волокнах типа II снизилась до 2,1 ммоль-кг~'смт-с~1 (приблизи-
тельно на 60 %) и к концу сокращений.запасы КФ оказались близкими к
истощению, тогда как в волокнах типа I она оставалась относительно не-
изменной (2,8 ммоль-кг~’смт-с~’).
Как отмечалось выше, скорость гликогенолиза в целостной мышце
начинает снижаться только после 10 с интенсивной сократительной
активности (см. рис. 6.1). На рис. 6.3, б показана скорость распада
гликогена в одиночных мышечных волокнах в промежутках от 0 до 20 и
от 20 до 30 с сокращений. В течение первых 20 с скорость гликогеноли-
за в волокнах типа II была выше (6,3 ммоль-кт^'смт-с-*) по сравнению
с незначительной скоростью, отмечаемой в волокнах типа I
(0,6 ммоль-кг—’смт-с—*), и очень высокой в обоих исследуемых проме-
жутках времени, определяя максимальную скорость утилизации гликоге-
на в целостной мышце, содержащей оба типа волокон. Когда сократи-
тельная активность продолжалась до 30 с, то, по аналогии с КФ, скорость
гликогенолиза в волокнах типа II снижалась до 3,5 ммоль-кг—1смт-с—1
181
(примерно на 45 %), тогда как в волокнах типа I она оставалась на очень
низком уровне.
Как и следовало ожидать, при максимальных сокращениях проявле-
ние силы целостной мышцей также снижалось после нескольких
первых секунд сокращений и после 30 с этот показатель упал приблизи-
тельно до 40 %. Поэтому становится очевидным, что параллельно с па-
дением силы имеет место заметное снижение скорости утилизации КФ
и гликогена в волокнах типа II. После 20 с стимуляции запас КФ в этих
волокнах был почти полностью истощен, а скорость утилизации глико-
гена начинала снижаться. Отмеченные факты не указывают, вследствие
чего волокна типа II могут повышать скорость ресинтеза в них АТФ и
устранять АДФ для компенсации их энергетических путей. Поэтому
снижение скорости анаэробного ресинтеза АТФ/рефосфорилирования
АДФ не имеет существенного значения для поддержания генерации си-
лы и утомление неизбежно. Дальнейшее подтверждение высказанного
предположения получено, в частности, в ходе экспериментов, в которых
повторные физические упражнения максимальной интенсивности вы-
полнялись с короткими периодами отдыха между ними.
В результате этих исследований выявлено существование значи-
тельной взаимосвязи между степенью ресинтеза КФ в промежутке отды-
ха между попытками и последующим уровнем физической работоспо-
собности. Действительно, было показано, что если между двумя изоки-
нетическими велоэргометрическими нагрузками максимальной интен-
сивности в течение 30 с промежуток отдыха составлял 4 мин, то степень
ресинтеза КФ в восстановительном периоде проявляла тесную положи-
тельную корреляционную взаимосвязь с объемом выполненной работы
во второй попытке (г = 0,8; и = 9; р < 0,05). Кроме того, в соответствии
с предположением, что специфичное для мышечных волокон типа II ис-
тощение запасов КФ преимущественно ответственно за развитие утом-
ления, установлено, что скорость гидролиза КФ во время выполнения
первой 30-секундной попытки в волокнах типа II была на 35 % выше,
чем в волокнах типа I. Однако при повторной физической нагрузке ско-
рость гидролиза КФ в волокнах типа II снижалась на 33 %, что характер-
но для неполного ресинтеза КФ в этих волокнах в период отдыха. И на-
против, в волокнах типа I ресинтез КФ в период отдыха после первой
попытки почти полностью был завершен, а его утилизация во время вы-
полнения второй попытки оставалась неизменной (рис. 6.4).
Согласно научным данным, доступность гликогена не обязательно дол-
жна быть ответственна за проявление утомления при выполнении высоко-
интенсивных физических упражнений при условии, что дорабочий уровень
его запасов в мышцах составляет не менее 100 ммоль-кг-1 смт-с~'). Малове-
182
роятно, чтобы доступность гликогена
могла ограничивать физическую рабо-
тоспособность при перемежающихся с
отдыхом физических нагрузках, приво-
дящих к снижению гликогенолиза и на-
коплению лактата в таких условиях. К
тому же вполне возможно, что сниже-
ние скорости гликолиза, по крайней ме-
ре при кратковременных интенсивных
физических нагрузках, не имеет сущес-
твенного значения для проявления мы-
шечной силы, которое отмечается в дан-
ных условиях.
Утомление, обусловленное уг-
нетающим влиянием продуктов ре-
акций. Еще в начале XX ст. предпо-
лагалось, что накопление лактата от-
ветственно за развитие утомления во
время выполнения высокоинтенсив-
ной мышечной работы. В условиях
Первая Вторая
физическая физическая
нагрузка нагрузка
Рис. 6.4. Утилизация фосфокреатина
в мышечных волокнах типов I и II во
время выполнения с максимальной
интенсивностью двух 30-секундных
изокинетических упражнений на ве-
лоэргометре (время отдыха между по-
пытками — 4 мин) (Касей и др.
(Casey et al.), 1996)
физиологических значений pH молочная кислота почти полностью дис-
социируется на свой составной компонент — ионы лактата и ионы во-
дорода. Исследования, проведенные на препаратах скелетных мышц
животных, показали, что прямое угнетение мышечной силы может
быть достигнуто увеличением концентраций ионов лактата и водорода.
Следует также отметить, что накопление этих ионов может обусловить
развитие мышечного утомления независимо друг от друга, однако роль
водородных ионов в этом механизме просматривается чаще. В подтвер-
ждение было показано, что мышцы животных, способных проявлять
значительную мышечную силу либо переносить значительную гипок-
сию, отличаются также высокими гликолитическими и буферными воз-
можностями.
Аналогично этому у легкоатлетов-спринтеров мышцы характеризу-
ются более высокой буферной способностью по сравнению со стайера-
ми. Предполагалось, что выполнение в процессе тренировки физичес-
ких упражнений с максимальной интенсивностью может повысить
буферные свойства и тем самым противодействовать неблагоприятным
эффектам накопления водородных ионов при мышечной деятельности.
С этой же целью использовали бикарбонат натрия для снижения аккуму-
ляции ионов водорода и (или) ускорить их метаболизм во время высоко-
интенсивных физических нагрузок. Однако, хотя вполне возможно, что
183
такая аккумуляция имеет отношение к развитию утомления в мышцах,
все же маловероятно, чтобы накопление ионов лактата и водорода было
целиком ответственно за этот процесс. Так, например, исследования,
проведенные на испытуемых-добровольцах, показали, что восстановле-
ние силы мышечных сокращений после утомительных физических наг-
рузок могло происходить быстро, несмотря на то, что значение pH в
мышцах оставалось на очень низком уровне. К настоящему времени
сформировалось общее мнение, что поддержание силы мышечных сок-
ращений во время выполнения высокоинтенсивной мышечной работы
является pH-зависимым, однако первоначальное ее проявление в боль-
шей степени зависит от доступности КФ.
Роль накопления неорганического фосфата (Фн) в развитии утом-
ления впервые начали рассматривать на основании результатов исследо-
вания летательных мышц насекомых. Одним из последствий быстрого
гидролиза КФ при выполнении физических упражнений высокой интен-
сивности является накопление в мышцах Фп, который, как было показа-
но, способен оказывать прямое угнетающее влияние на сопряженность
процесса возбуждение — сокращение. Однако происходящее одновре-
менно истощение запаса КФ и накопление Фп создает трудности для раз-
деления этих эффектов in vivo. Данная проблема соприкасается с парал-
лельным повышением концентрации ионов лактата и водорода во время
интенсивной мышечной работы. Все эти метаболиты независимо при-
частны к развитию мышечного утомления.
Утомление, обусловленное факторами, предшествующими об-
разованию поперечных мостиков. Как отмечалось в главе 1, происхо-
дящий после деполяризации сарколеммы выход кальция из саркоплазма-
тического ретикулума (СР) имеет большое значение для активации в
мышце сопряженного процесса возбуждение — сокращение. Установле-
но, что во время мышечных сокращений, совершаемых на фоне утом-
ления, замедляется транспорт кальция и прогрессивно снижается каль-
циевый перенос, что является характерным признаком нарушения
обратного захвата кальция СР и (или) повышенного его связывания.
Доказательства дезинтеграции обращаемости кальция при утомлении
получены в результате исследований, продемонстрировавших, что сти-
муляция освобождения кальция из СР, вызванная введением кофеина в
изолированную мышцу, может повысить силу мышечных сокращений
даже на фоне низких значений pH. Показано также, что восстановление
после утомления, вызванного электростимуляцией, происходит значи-
тельно быстрее при применении низкочастотных раздражений по срав-
нению с высокочастотными. Замедление восстановления силы мышеч-
ных сокращений связано с нарушением сопряженности процесса воз-
184
буждение — сокращение и, поскольку активация мышц в условиях in
vivo осуществляется в низкочастотном диапазоне раздражающих стиму-
лов, особенность такой реакции может иметь определенное значение в
механизме развития утомления.
8. Питание и максимальная физическая
работоспособность
Существует мнение, что питание не имеет большого значения для про-
явления индивидуальных физических возможностей при выполнении
физических упражнений максимальной мощности. И действительно,
часто рекомендуемая необходимость высокого содержания в пище бел-
ков оказывается необязательной для проявления максимальной физичес-
кой работоспособности. Такое предположение базируется на результа-
тах исследования, показавшего, что потребность в белках у спортсме-
нов, выполняющих физические упражнения, направленные на развитие
выносливости, выше, чем у спринтеров. Кроме того, как установлено ра-
нее, маловероятно, что доступность мышечного гликогена способна ог-
раничивать работоспособность при выполнении физических упражне-
ний максимальной интенсивности до тех пор, пока его концентрация не
снизится до необычно низкого уровня. Следовательно, при адекватном
потреблении энергии и преимущественно в виде углеводов вряд ли мож-
но ожидать, что доступность последних в мышце может ухудшить физи-
ческую работоспособность.
Имеются данные, указывающие на то, что для растущего организма
поступление с пищей креатина (Кр) может быть необходимой потреб-
ностью для тех, кто желает повысить свои возможности в выполнении
высокоинтенсивных физических упражнений. Креатин, или метилгуа-
нидинуксусная кислота является веществом естественного происхожде-
ния и его общее содержание в организме взрослого человека составляет
примерно 120 г, из которых 95 % находится в мышцах. Концентрация Кр
в последних составляет около 125 ммоль-кгчсмт и в неработающих
мышцах 60 % его находится в форме КФ.
У здоровых людей Кр необратимо распадается до креатинина со
скоростью приблизительно 2 г в день, однако он постоянно восполняет-
ся синтезом эндогенного Кр и (или) его поступлением с пищей, напри-
мер с мясом. Дополнительное потребление Кр не является новым фено-
меном. Результаты исследований, выполненных еще в начале XX ст., по-
казали, что такое пищевое подкрепление восстанавливает и повышает
185
КС РХ XX ДжС 00 ЕС АЛ
ЕХ ХЛ ИС ДжВ СЛ
Испытуемые
Рис. 6.5. Общее содержание Кр у отдельных ис-
пытуемых накануне, во время и после примене-
ния креатинмоногидрата в дозе 20-30 г в день на
протяжении 3—21 дней: 21/2 — испытуемые,
принимавшие креатин через день в течение трех
недель (Харрис и др. (Harris et al.), 1992)
содержание Кр в целостном
организме, а также приводит
к незначительному увели-
чению экскреции креатинина
с мочой. Эти ранние исследо-
вания неизменно включали
периоды хронического пот-
ребления Кр. Однако в нас-
тоящее время установлено,
что потребление 20 г креати-
на в день (4 раза по 5 г) может
способствовать повышению
его общей концентрации в
мышцах, 20 % которого при-
сутствует в форме КФ (рис.
6.5). Известно также, что пре-
имущественное поступление
Кр в мышцы происходит в те-
чение первых дней подкреп-
ления, что служит подтверждением результатов ранних работ. Кроме то-
го, установлено, что если дополнительный прием Кр осуществлять в пе-
риод выполнения физических упражнений субмаксимальной мощности,
то его концентрация в мышцах возрастает на 10 %. Точный механизм,
благодаря которому осуществляется поступление и регуляция содержа-
ния Кр в мышцах, пока не установлен, однако известно, что этот процесс
является натрийзависимым. Предельная концентрация Кр в мышцах че-
ловека составляет 145-160 ммоль-кг-1 и при дополнительном его
потреблении у лиц с меньшей величиной этого показателя накопление
креатина в мышцах происходит более выражение.
Прием Кр на протяжении пяти дней (20 г в день ) выявил значитель-
ное увеличение объема работы, которую могли выполнить здоровые ис-
пытуемые-добровольцы при повторяющихся физических нагрузках мак-
симальной мощности. Такой вывод был сделан на основании результа-
тов лабораторного исследования, в котором использовались повторяю-
щиеся попытки выполнения динамической, изокинетической работы
максимальной интенсивности на велоэргометре (например, три попытки
с интервалом отдыха между ними 4 мин), а также на основании резуль-
татов «полевого» исследования, проведенного со спортсменами (напри-
мер, бег 4 х 300 м с интервалом 4 мин между попытками). Эти исследо-
вания показали, что дополнительное потребление Кр значительно повы-
шает физическую работоспособность за счет поддержания силы или
186
мощности во время выполнения физических упражнений, что проявля-
ется в увеличении общего объема выполняемой работы на 5-7 %. Точ-
ный механизм, благодаря которому увеличивается физическая работо-
способность, пока не установлен. Однако маловероятно, чтобы этот эф-
фект был обусловлен исключительно повышением в мышцах дорабоче-
го уровня КФ, поскольку степень этого увеличения (приблизительно на
8 ммоль-кг^смт) выглядит малозначительной для отмеченных улуч-
шений показателей работоспособности.
Повышение работоспособности может быть связано также со сти-
мулирующим влиянием дополнительного потребления Кр на ресинтез
КФ во время мышечной работы и в восстановительном периоде после
нее. Учитывая, что наличие КФ лимитирует физическую работоспособ-
ность при выполнении физических упражнений максимальной мощнос-
ти, все эти эффекты должны повышать сократительную способность
мышц за счет поддержания анаэробного ресинтеза АТФ. Такое предпо-
ложение подтверждается результатами исследований, которые показали,
что дополнительное потребление Кр способно снижать накопление в
плазме крови аммиака и гипоксантина, а также уменьшать степень рас-
160 -
150
140
130
120
110
______I
После
потребления
100 й '------
До
потребления
30
20
10
0
3
-J_______I_________L
20 30 40
Увеличение общей концентрации КФ
после потребления Кр
а
б
Рис. 6.6. Индивидуальные уровни общей концентрации креатина (ОКр) до и после пя-
ти дней потребления Кр (4 х 5 г в день). Здесь и на рис. 6.7 испытуемые 1—8 расположе-
ны в последовательности, соответствующей исходному содержанию в их мышцах ОКр
(а), а также индивидуальные показатели, размещенные против показателей изменения
ресинтеза КФ (рис. 6.6, 6) и показателей, отражающих увеличение объема выполненной
работы в двух попытках велоэргометрической нагрузки максимальной интенсивности
(рис. 6.7, б) (Гренхафф и др. (Greenhaff et al.), 1994)
187
пада АТФ в мышцах во время физических нагрузок максимальной ин-
тенсивности.
Известно, что лица, у которых в результате креатинового подкреп-
ления отмечается увеличение общей концентрации Кр в мышцах более
чем на 25 %, могут рассчитывать на более высокую скорость ресинтеза
АТФ в восстановительном периоде (рис. 6.6). По-видимому, такая воз-
можность существует вследствие того, что дополнительное потребле-
ние Кр поддерживает такую концентрацию его свободной фракции в
мышцах, которая выше Кт митохондриальной креатинкиназы для Кр
(около 60 ммоль-кг~1 смт) в течение всего восстановительного периода.
Благодаря этому поддерживается скорость метаболизма через креатин-
киназную реакцию, что благоприятствует ресинтезу КФ и образованию
АТФ.
При попытке использовать креатиновое подкрепление для повы-
шения физической работоспособности при выполнении физических
упражнений максимальной интенсивности часто не учитывают ряд
важных моментов. Во-первых, не для каждого человека такое под-
крепление может оказаться эффективным, поскольку у примерно 30 %
лиц поступление Кр в мышцы относительно невысокое. Во-вторых,
большинство отмеченных эффектов дополнительного приема Кр
обычно проявляется у лиц, у которых при этом достигается повыше-
ние общего его содержания в мышцах более чем на 25 % (рис. 6.7). С
Рис. 6.7. Общая концентрация креатина (ОКр) в скелетных мышцах отдельных испы-
туемых до и после пяти дней потребления КрО (4x5 г-день~’) (Касей и др. (Casey
etal.), 1996)
60
50
40 -
30 -
20 -
10 -
0 10 20 30 40
Потребление Кр, ммоль-кг-1 смт
б
188
учетом указанных моментов важно также иметь в виду, что, согласно
результатам последних исследований, потребление Кр в сочетании с
углеводами у всех лиц может способствовать повышению концентра-
ции Кр в мышцах более чем на 25 %.
Выводы
1. АТФ является единственным энергетическим топливом, которое мо-
жет непосредственно использоваться скелетной мышцей для генерации
силы. Количества этого топлива в мышце достаточно для обеспечения
максимальной сократительной активности в течение приблизительно
2 с, поэтому для продолжения возможности развивать напряжение АТФ
должен быстро ресинтезироваться из АДФ. Во время выполнения высо-
коинтенсивных физических упражнений относительно низкая скорость
ресинтеза АТФ за счет окислительного фосфорилирования приводит к
быстрой активации анаэробной энергопродукции за счет КФ и гидроли-
за гликогена.
2. Гидролиз КФ начинается сразу же с наступлением сократитель-
ной активности для буферирования быстрого накопления АДФ, которое
происходит в результате гидролиза АТФ. Однако скорость распада КФ
начинает снижаться уже через несколько секунд максимальных мышеч-
ных сокращений. Важность КФ для энергопродукции в мышцах заклю-
чается в чрезвычайно высокой скорости, с которой он способен осу-
ществлять ресинтез АТФ.
3. Гликогенолиз представляет собой гидролиз гликогена до глюко-
зе-1-фосфата. Гликолиз состоит из серии реакций, в ходе которых про-
исходит преобразование глюкозы до пирувата и лактата. Если высоко-
интенсивная мышечная работа продолжается хотя бы более нескольких
секунд, вклад гликогенолиза и гликолиза в ресинтез АТФ заметно
возрастает.
4. Наличие тесной взаимосвязи между анаэробным ресинтезом АТФ
и утилизацией гликогена дает основание полагать, что быстрое возрас-
тание свободного АМФ и неорганического фосфата может служить клю-
чевым регулятором мышечного гликогенолиза во время сократительной
активности.
5. Анаэробный гликолиз включает большее количество этапов, чем
гидролиз КФ, но по сравнению с окислительным фосфорилированием
скорость протекания этого процесса еще очень высокая. Кроме того, он
инициируется с началом мышечных сокращений, однако своей макси-
мальной скорости достигает только после 5 с, и этот уровень при рабо-
189
те максимальной интенсивности может поддерживаться еще в течение
нескольких секунд. Механизмы, ответственные за возможное снижение
интенсивности гликолиза при таких физических нагрузках, не рас-
крыты.
6. Продолжительность выполнения (в неустойчивом физиологичес-
ком состоянии) субмаксимальных высокоинтенсивных физических уп-
ражнений может достигать 5 мин до наступления явных признаков
утомления. В таких условиях мышечной деятельности окисление угле-
водов может вносить значительный вклад в продукцию АТФ, однако это
не всегда принимается во внимание.
7. При выполнении повторяющихся физических упражнений макси-
мальной интенсивности скорости гидролиза КФ и накопления лактата
снижаются. Предполагается, что этот эффект обусловлен неполным ре-
синтезом КФ в промежутках отдыха между попытками. Однако механиз-
мы, ответственные за падение скорости накопления лактата, остаются
не изученными.
8. Утомление является неизменным признаком высокоинтенсив-
ных физических упражнений, и это физиологическое состояние
характеризуется невозможностью продолжать мышечную работу оп-
ределенной мощности. Наступление мышечного утомления ассоции-
руется с нарушением энергообеспечения, угнетающим влиянием
продуктов реакций, а также с эффектами, предшествующими образо-
ванию поперечных мостиков. Предполагается, что этот процесс мно-
гофакторный.
9. Существует мнение, что питание не имеет большого значения для
проявления индивидуальных физических возможностей при выполне-
нии физических упражнений максимальной мощности. Однако имеются
данные, указывающие на то, что для растущего организма поступление
с пищей креатина может быть необходимой потребностью для тех, кто
желает повысить свои возможности в выполнении высокоинтенсивных
физических упражнений. Кроме того, пища с высоким содержанием
белков и низким содержанием углеводов способствует снижению физи-
ческой работоспособности при выполнении физических упражнений
высокой интенсивности.
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
Casey A., Howell S., Constantin-Teodosiu D, et al. (1996). The metabolic responses of type I
and II muscle fibre during repeated bouts of maximal exercise in man // Am. J. Physiol.
— 271, —E38-E43.
190
Casey A., Howell S, Constantin-Teodosiu D. et al. (1996). Effect of creatine supplementa-
tion on muscle metabolism and exercise performance // Am. J. Physiol. — 271. —
E31-E37.
Greenhaff P.L., Bodin K., Soderlund K., Halt man E. (1994). The effect of oral creatine sup-
plementation on skeletal muscle phosphocreatine resynthesis // Am. J. Physiol. — 266. —
E725-E730.
Harris R.C., Soderlund K., Hultman E. (1992). Elevation of creatine in resting and exer-
cised muscle of normal subjects by creatine supplementation // Clin. Sci. — 83. —
P. 367-374.
SoderlundK., Greenhaff P.L., Hultman E. (1992). Energy metabolism in type I and II human
muscle fibres during short term electrical stimulation at different frequenciec // Acta
Physiol. Scand. — 144. — P. 15-22.
МЕТАБОЛИЧЕСКИЕ ОТВЕТЫ
ПРИ ПРОЛОНГИРОВАННЫХ
ФИЗИЧЕСКИХ НАГРУЗКАХ
1. Энергетические источники
для пролонгированных физических
упражнений
Термин «пролонгированные физические упражнения»
обычно используют для характеристики упражнений, ин-
тенсивность выполнения которых может поддерживаться
(до ее снижения) в пределах 30-180 мин. Практически, ин-
тенсивность таких упражнений соответствует 60-85 %
максимального потребления кислорода (VO2max). Продол-
жительность упражнений, выполняемых в течение боль-
шого промежутка времени, т. е. с интенсивностью, соот-
ветствующей не более 60 % VO2max, по-видимому, не ог-
раничивается доступностью субстратов для мышц, и при
адекватном поступлении жидкости в организм такие уп-
ражнения могут выполняться в течение нескольких часов и
даже дней. В отличие от упражнений максимальной мощ-
ности, скорость продукции АТФ в мышцах, необходимой
для обеспечения пролонгированной мышечной работы, от-
носительно невысокая (менее 2,5 ммоль-кг~'смт-с—’), в
связи с чем КФ, углеводы (СНО) и жиры все вместе могут
вносить свой вклад в ресинтез АТФ. Однако СНО служат
наиболее важным энергетическим источником при таком
виде мышечной деятельности. Основное достоинство
окисления СНО, которое часто недооценивают, заключает-
ся в том, что на единицу утилизируемого кислорода они
обеспечивают самое большое количество продуцируемого
АТФ по сравнению с другими субстратами.
В начале выполнения пролонгированного физического
упражнения скорости распада КФ и накопления лактата
192
проявляют линейную зависимость от интенсивности мышечной работы.
В течение этого периода КФ и анаэробный гликолиз вносят значитель-
ный вклад в продукцию АТФ в мышцах, которой в основном свойствен-
но отставание от мышечного кровотока и поступления кислорода на фо-
не возрастающей энергетической потребности. Однако полученные в
последнее время данные дают основание предполагать, что такое запаз-
дывание может быть присуще и субстратному окислению. Без обеспе-
чения быстрой скорости доставки энергии за счет гидролиза КФ и анаэ-
робного гликолиза энергопродукция в начале выполнения мышечной ра-
боты была бы затруднительной. Однако по истечении нескольких минут
после начала выполнения физических упражнений вклад этих путей
энергообеспечения становится незначительным и устанавливается рав-
новесное состояние между энергетической потребностью и энергопро-
дукцией. В условиях устойчивого состояния окисление СНО и жиров
становится основным поставщиком энергии.
Как показывают расчеты, при VO2max, равном 3-4 л-мин-1, макси-
мальная скорость продукции АТФ за счет СНО составляет примерно
2,0-2,8 ммолыкг-1 смт, поэтому становится очевидным, что СНО в сос-
тоянии обеспечить энергопотребность при пролонгированных физичес-
ких нагрузках. Однако в обычных условиях мышечные запасы СНО (в
форме гликогена) находятся в пределах 350 ммоль-кг-1 смт, поэтому
только за их счет работа может продолжаться около 80 мин (4 л кисло-
рода окисляют 0,03 моля глюкозильных единиц и при этом образуется
1,15 моля АТФ).
Следует отметить, что СНО доставляются к мышцам в форме глю-
козы крови также из запасов, находящихся в печени, и они могут исполь-
зоваться для ресинтеза АТФ с максимальной скоростью около
1 ммоль-кг—1 смт-с—'. Если предполагаемое физическое упражнение вы-
полняется после приема пищи, а не впроголодь, то можно рассчитывать,
что запасов печеночного гликогена может хватить на обеспечение рабо-
ты в пределах 45 мин при условии, что он является единственным энер-
гетическим источником, который окисляется с максимальной скорос-
тью. Однако следует иметь в виду, что глюкоза, образующаяся в печени,
может использоваться в качестве энергетического топлива не более чем
на 45 % максимального уровня окисления углеводов, поэтому она не в
состоянии обеспечить выполнение интенсивных физических упраж-
нений.
Жиры составляют большую часть энергетического запаса в организ-
ме человека, но, к сожалению, они проявляют низкую скорость окис-
ления по отношению к скорости окисления мышечного гликогена (около
1 ммоля-кг—1 смт-с-1). Факторы, ответственные за ограничение скорос-
13„.313 193
ти окисления жиров при мышечной деятельности, к настоящему време-
ни не изучены, однако лимитирующий участок, очевидно, должен пред-
шествовать образованию ацетил-КоА, поскольку после этого этапа прев-
ращение жиров и углеводов происходит по одному и тому же пути. Вза-
имосвязь обмена жиров и углеводов более подробно рассмотрена далее.
Следует заметить, что одновременная утилизация запасов жиров и
гликогена в печени становится чрезвычайно важной для поддержания
образования АТФ в мышцах при продолжительных физических нагруз-
ках, приводящих к истощению запасов мышечного гликогена. Теорети-
чески максимальная скорость продукции АТФ при окислении жиров и
глюкозы крови должна составлять около 2 ммоль-кг^'смт-с-1, которая
примерно на 30 % ниже максимальной скорости ресинтеза АТФ при
окислении мышечного гликогена.
2. Взаимосвязь окисления
углеводов и жиров в скелетных мышцах
Механизмы, обеспечивающие интеграцию использования СНО и жиров
во время пролонгированной мышечной деятельности, довольно сложны
и недостаточно изучены. Тем не менее можно рассчитывать, что если бы
организм высококвалифицированного марафонца использовал в качес-
тве энергетического источника исключительно СНО, то он был бы из-
можден приблизительно после 90 мин бега. Мировой рекорд в марафон-
ском беге близок к 130 мин, что ясно указывает на важность «топлив-
ной» интеграции при продолжительной физической нагрузке.
По мнению ряда специалистов, участком, лимитирующим скорость
окисления СНО, является декарбоксилирование пирувата до ацетил-
КоА, которое находится под контролем пируватдегидрогеназного (ПДГ)
комплекса и служит весьма важной обратимой реакцией, обеспечива-
ющей включение пирувата в ЦТК и его окисление; ПДГ представляет
собой конгломерат трех ферментов, локализованных во внутренней ми-
тохондриальной мембране. Дополнительно к этому комплексу ПДГ
имеет также два регуляторных фермента -— фосфатазу и киназу, кото-
рые регулируют цикл активации/инактивации. Фосфатаза ответственна
за дефосфорилирование ПДГ, приводящее к трансформации ПДГ в ак-
тивную форму (ПДГа). И напротив, киназа катализирует АТФ-зависи-
мое фосфорилирование ПДК с сопутствующей этому инактивацией эн-
зиматического комплекса. Взаимопревращение активной и неактивной
форм ПДГ регулируется несколькими аллостерическими эффекторами
194
Активная форма
Фосфатаза пиру-
ватдегидрогеназы
Пируват
НАД+, KoA-SH
Ацетил-КоА
НАДН, СО2, Н
Неактивная форма
I НАД
’НАДН
t АТФ
' АДФ
t Ацетил-КоА
I KoA-SH
Рис. 7.1. Регуляция пируватдегидрогеназного комплекса, активация (дефосфорилиро-
вание) пируватдегидрогеназы фосфатазой пируватдегидрогеназы, а также активацион-
ный статус, регулируемый изменением доступности кальция и соотношений
АТФ/АДФ, ацетил-КоА/KoA-SH, НАДН/НАД+
Киназа пируват-
дегидрогеназы
АТФ
Пируват-
дегидрогеназа
и гормональным воздействием. Увеличение соотношений АТФ/АДФ,
ацетил-КоА/КоА и НАДН/НАД* активирует киназу, в результате чего
происходит угнетение активности фермента. И наоборот, снижение
приведенных выше соотношений, а также присутствие пирувата дол-
жно инактивировать киназу, пока увеличение концентрации кальция бу-
дет активировать фосфатазу, что все вместе приведет к активации ПДГ
(рис. 7.1). Таким образом, увеличение доступности кальция и пирувата
с установлением сократительной активности вызывает быструю акти-
вацию ПДГ.
Данные факторы вместе с последующим снижением соотношений
АТФ/АДФ, ацетил-КоА/КоА и НАДН/НАД*, происходящим в результа-
те продолжающихся мышечных сокращений, будут способствовать
ускорению реакции. Изменение этих соотношений в процессе регуля-
ции скорости ПДГ-реакции выглядят тесно взаимосвязанными, в связи
с чем утилизация пирувата будет всегда соответствовать потребностям
ЦТК и электрон-транспортной цепи. Так, например, в случае адекват-
ного обеспечения аденозинтрифосфатом высокое соотношение
АТФ/АДФ в митохондриях должно благоприятствовать снижению ак-
тивности ПДГа. Аналогично этому угнетающее влияние увеличения
соотношения ацетил-КоА/КоА на активность ПДГ будет способство-
вать тому, что скорость образования ацетил-КоА не ограничит скорость
утилизации ацетильных групп циклом трикарбоновых кислот. И нако-
нец, угнетающее влияние увеличения соотношения НАДН/НАД4 дол-
13*
195
будь в другом участке, например на этапе поступления глюкозы в
мышцы либо фосфорлирования гликогена. В связи с этим независимый
нейропептид — кальцитонин (КГЗП), который присутствует в мото-
нейронах и освобождается при нейромышечном выбросе в ответ на
возбуждение нерва, впоследствии угнетает инсулинстимулируемый
синтез гликогена и стимулирует фосфорилазу гликогена, распад глико-
гена, а также образование лактата в препаратах изолированных скелет-
ных мышц. Все это позволяет предположить, что КГЗП может прини-
мать участие в регуляции внутриклеточной утилизации СНО и что он
является возможным механизмом, связывающим нейромышечную ак-
тивацию с утилизацией субстратов в мышце. Следует подчеркнуть, од-
нако, что интенсивность физических нагрузок в большинстве, если не
во всех описанных выше исследованиях, имеющих отношение к ПДГ,
была достаточной для подавления активности ПДГ кальцием, поэтому
была весомее любого другого влияния ацетил-КоА и НАДН на тран-
сформацию ПДГ. Пока не установлено, действительно ли глюкозожир-
нокислотный цикл функционирует во время очень продолжительных,
малоинтенсивных физических нагрузок, когда кальциевая активация
ПДГ становится самой низкой.
Альтернативное центральное место в интеграции окисления СНО и
жиров занимает уровень малонил-КоА. Предполагалось, что увели-
чение образования ацетил-КоА во время физических нагрузок может
привести к быстрому образованию малонил-КоА. Накопление послед-
него оказывало прямое угнетающее влияние на фермент карнитинпаль-
митоилтрансферазу-1 (КПТ-1), катализирующий скоростьлимитирую-
щий участок в транспорте длинноцепочных жирных кислот ацил-КоА в
митохондрии, в связи с чем, вероятно, может происходить снижение
окисления жирных кислот. В настоящее время роли малонил-КоА в ске-
летных мышцах человека во время их сократительной активности уде-
ляется мало внимания. Однако в исследованиях, проведенных на кры-
сах, было выявлено снижение его концентрации в мышцах при сокра-
щениях. Интерпретация взаимосвязи этих данных с субстратной утили-
зацией затруднительна, поскольку отсутствуют другие, имеющие к
этому отношение, сведения. Кроме того, поскольку в мышцах крыс чув-
ствительность КПТ-1 к малонил-КоА относительно высокая, то, как
показали расчеты, даже при низких концентрациях последнего актив-
ность КПТ-1, а поэтому и окисление жиров должны полностью угне-
таться, что точно доказано. Не вызывает сомнений, что дальнейшие
исследования позволят получить больше информации о роли малонил-
КоА в интеграции утилизации энергетических субстратов при мышеч-
ной деятельности человека.
198
3. Наличие углеводов в мышцах, питание
и двигательная активность
Содержание углеводов в скелетных мышцах человека составляет около
350 ммоль кг-1 смт и в известной мере мало подвержено изменениям у
людей, ведущих малоподвижный образ жизни. Так, например, увели-
чение потребления СНО с 55 % общего энергопотребления до 80-90 %
способствовало увеличению концентрации гликогена в мышцах прибли-
зительно только на 50 ммоль-кг—1 смт. Есть основания полагать, что свя-
зано это со снижением регуляторной активности мембранного глюкозо-
транспортного белка и гликогенсинтетазы в условиях адекватного угле-
водного обеспечения. Аналогично З-Л-дневное голодание мало влияло
на запасы гликогена в мышцах. Такая особенность не присуща другим
видам млекопитающих. Так, например, у лабораторных крыс, характе-
ризующихся относительно высоким уровнем основного обмена, окис-
ление жиров происходит очень интенсивно, поэтому голодание в тече-
ние 24 часов приводит к снижению содержания гликогена в мышцах
почти до нуля. Следовательно, в случае углеводного метаболизма необ-
ходимо с осторожностью подходить к экстраполяции данных, получен-
ных в экспериментах на животных, на человека.
Представление о том, что доступность СНО должна влиять на фи-
зическую работоспособность при пролонгированных физических наг-
рузках, в наибольшей степени получило признание в XX ст. Выполнен-
ными в 30-е годы Кристенсеном и Хансеном (Cristensen and Hansen)
работами впервые было показано, что прием в течение нескольких
дней перед пролонгированной физической нагрузкой богатой углево-
дами пищи может в 2-3 раза повысить физическую работоспособность
по сравнению с приемом пищи, богатой жирами. Впоследствии Берг-
стром и Хултманом (Bergstrom and Hultman), применив в 60-е годы ме-
тодику пункционной биопсии, выявили существование тесной взаи-
мосвязи между дорабочим содержанием гликогена в мышцах и
уровнем физической работоспособности. Результаты этих работ пока-
зали, что во время выполнения физического упражнения на велоэрго-
метре с мощностью, соответствующей примерно 80 % VO2max, отме-
чается прогрессирующее снижение концентрации мышечного гликоге-
на. Примерно после 70-минутной физической нагрузки работоспособ-
ность не могла поддерживаться на прежнем уровне, что связано с ис-
тощением запасов гликогена в мышцах (рис. 7.3). Во всех случаях уп-
ражнение выполнялось до отказа, что совпадало с наступлением исто-
щения запасов мышечного гликогена.
199
Продолжительность работы, мин
Рис. 7.3. Концентрация гликогена в
m. vastus lateralis у десяти испытуе-
мых во время велоэргометрической
нагрузки с мощностью, эквивалент-
ной 80 % VO2max
Дни
Рис. 7.4. Концентрация гликогена в
m. vastus lateralis 'в предварительно
активной (•) и неактивной (о) мыш-
цах у двух испытуемых
В эти же годы было показано, что
комбинациями гликогенного истоще-
ния и высокоуглеводной диеты можно
существенно влиять на запасы мы-
шечного гликогена. Так, оказалось,
что после гликогенистощающих физи-
ческих нагрузок эти запасы можно
увеличить до супранормального
уровня (около 900 ммоль-кг-1 смт) пу-
тем потребления высокоуглеводной
пищи в дни после предшествовавших
нагрузок (рис. 7.4). Оказалось также,
что ресинтез гликогена наиболее быс-
тро протекает в первые часы после
выполненной физической работы, что,
вероятно, связано с активацией мы-
шечной гликогенсинтетазы, механиз-
мы регуляции которой полностью не
изучены. Было также четко продемон-
стрировано, что суперкомпенсация за-
пасов мышечного гликогена происхо-
дит лишь в работавших группах
мышц. Биоптаты были получены сра-
зу после выполнения на велоэргомет-
ре физического упражнения одной но-
гой, а также в последующие три дня, в
течение которых испытуемые пита-
лись высокоуглеводной пищей.
Дальнейшими исследованиями,
проведенными Бергстром и Хултма-
ном, удалось установить тесную
взаимосвязь между характером пот-
ребляемой перед физическими наг-
рузками пищей, уровнем запасов
гликогена в мышцах и физической
работоспособностью. Содержание
гликогена в мышцах вначале изменя-
ли путем применения различной
изоэнергетической диеты вслед за
гликогенистощающими физически-
ми нагрузками, после которых испы-
200
Рис. 7.5. Взаимосвязь между ис-
ходной концентрацией мышечного
гликогена и предельным временем
выполнения физического упражне-
ния у шести испытуемых при вело-
эргометрической нагрузке с интен-
сивностью, эквивалентной 75 %
VO2max, при этом каждый испытуе-
мый выполнял упражнение до отка-
за в трех случаях в течение 10 дней:
первый раз — после трех дней приема
обычной смешанной пищи (А); второй
— после трех дней приема пищи с огра-
ниченным содержанием углеводов (о);
третий — после трехдневной диеты с
высоким содержанием углеводов (•)
(Бергстом и др. (Bergstom et al.), 1967)
300
туемые выполняли на велоэргометре мышечную работу с мощнос-
тью, эквивалентной 75 % VO2max до отказа. Перед первым тестом на
выявление уровня выносливости испытуемые питались обычной сме-
шанной пищей, перед вторым — с ограниченным содержанием угле-
водов и перед последним — с повышенным содержанием СНО. В
среднем концентрация мышечного гликогена после выполнения
физических упражнений в каждом из приведенных выше случаев
составила соответственно 495, 176 и 953 ммоль-кг—*смт. Другими
словами, под влиянием диеты с ограниченным содержанием углево-
дов концентрация гликогена снизилась на 65 %, а под влиянием вы-
сокоуглеводной пищи — увеличилась на 95 %.
Параллельно с этим изменилось и предельное время выполнения
мышечной работы до отказа. Установлена тесная корреляционная взаи-
мосвязь между дорабочей концентрацией гликогена в мышцах и пре-
дельным временем физической нагрузки (рис. 7.5). Отказ от работы
всегда совпадал с истощением запасов гликогена в мышцах независимо
от особенностей предшествующего питания. Так, после приема пищи с
ограниченным содержанием углеводов предельное время выполнения
мышечной работы сократилось до 59 мин (на 55 %), а после СНО-обо-
гащенной диеты — увеличилось до 180 мин (на 50 %). По-видимому, в
последнем случае увеличение времени выполнения велоэргометричес-
кого упражнения произошло за счет удвоения в мышцах дорабочего за-
паса гликогена. Утилизация мышечного гликогена, как правило, ускоря-
201
ется тогда, когда запасы его повышаются путем манипулирования физи-
ческими нагрузками и питанием.
Результаты описанных выше работ обобщенно представляют один
из наиболее значительных исследовательских вкладов в физиологию и
биохимию мышечной деятельности. Через 30 лет, прошедших со време-
ни появления обсуждаемых публикаций, эти данные получили подтвер-
ждение во многих случаях и практика «углеводной загрузки» нашла ши-
рокое распространение среди спортсменов всего мира.
Позднее было проведено большое число фундаментальных иссле-
дований, посвященных оптимизации количества и типа потребляемых
СНО после физических нагрузок в попытке максимизировать ресинтез
мышечного гликогена. Однако основные положения, базирующиеся на
результатах работ, выполненных в 60-е годы, существенно не измени-
лись и исследования, проведенные в последние годы, преследовали цель
усовершенствовать наше понимание взаимодействия между физически-
ми нагрузками, утилизацией мышцами глюкозы и метаболизмом. Так,
например, к настоящему времени стало известно, что когда прием пищи
с высоким содержанием СНО происходит после гликогенистощающей
физической работы, то скорость ресинтеза мышечного гликогена в тече-
ние первых нескольких часов восстановительного периода примерно на
25 % выше в волокнах типа I. Затем эта скорость начинает снижаться,
однако в волокнах типа II она продолжает поддерживаться, в связи с чем
через 24 часа концентрация гликогена в обоих типах волокон становит-
ся одинаковой.
Указанные различия в скорости ресинтеза гликогена в волокнах в
начале восстановительного периода связаны с различиями в них скорос-
ти транспорта глюкозы через мембрану, которая лимитируется степенью
насыщения последней связанным с мембраной инсулинзависимым пере-
носчиком глюкозы (ГЛУТ4). Это позволило предположить существова-
ние инсулинзависимой и инсулиннезависимой фаз ресинтеза гликогена.
Включение ГЛУТ4 в мембраны мышечных волокон стимулируется фи-
зическими нагрузками и в наибольшей степени это проявляется в волок-
нах типа II. Следовательно, этот мембранный белок, очевидно, играет
жизненно важную роль в метаболизме СНО и похоже, что он является
ключевым фактором широко известного повышения инсулинчувстви-
тельности во всем организме, которое происходит под влиянием физи-
ческой тренировки. В связи с этим полученные в последнее время дан-
ные дают основание полагать, что снижение содержания в мышцах
ГЛУТ4 ответственно за снижение скорости транспорта глюкозы и ре-
синтеза гликогена, проявляющееся после вызванных физическими уп-
ражнениями (эксцентрическими) мышечных повреждений.
202
4. Наличие углеводов в печени,
питание и двигательная активность
По сравнению с метаболизмом СНО в мышцах относительно мало из-
вестно о взаимосвязи между питанием, физическими нагрузками и ме-
таболизмом СНО в печени у человека. Связано это не с недостатком ин-
тереса к этому вопросу, а с инвазивной особенностью методики пече-
ночной биопсии. Результаты ряда исследований, проведенных на здоро-
вых испытуемых-добровольцах, показали, что скорость выхода глюкозы
из печени в сытом состоянии находится в пределах 0,8—1,1 ммоль глю-
козы в минуту, что имеет существенное значение для обеспечения
потребностей только мозга. Приблизительно 70 % этого выхода извлека-
ется из запасов печеночного гликогена, а остальное ее количество синте-
зируется в процессе глюконеогенеза в печени при использовании в ка-
честве субстратов лактата, пирувата, глицерола и аминокислот. В случае
использования аминокислот это соответствует упомянутому ранее глю-
козоаланиновому циклу (см. главу 6).
Результаты проведенных в 70-е годы XX ст. экспериментальных ис-
следований показали, что аланин является не только конечным продук-
том мышечного гликолиза (образуется он при трансаминировании пиру-
вата), но вносит также значительный вклад в печеночный глюконеогенез
и используется для поддержания глюкозы в печени в условиях углевод-
ной недостаточности. Это привело к формированию представления о су-
ществовании глюкозоаланинового цикла, функционирующего между
мышцами и печенью. Цикл связан с синтезом аланина из пирувата и вы-
ходом его из мышц. Находящийся в циркулирующей крови аланин затем
экстрагируется печенью и его углеводный каркас преобразуется в глюко-
зу. Аналогично периферическое образование и печеночная экстракция
глутамина проявляются у человека, однако этот путь выглядит менее
превалирующим, чем для аланина.
Исследования, проведенные с применением метода печеночной би-
опсии, показали, что печень чрезвычайно чувствительна к изменению
содержания СНО в пище. На основании приведенных выше результатов
определения скорости освобождения глюкозы в сытом состоянии можно
рассчитать, что в течение одного дня голодания запасы гликогена в пече-
ни полностью истощаются. Этот факт был продемонстрирован Нильсо-
ном и Гултманом (1973), обнаружившим, что после однодневного голо-
дания либо ограниченного поступления СНО с пищей концентрация
гликогена в печени в среднем снижается с 270 до 30 ммоль-кг-1 сырой
массы. Дальнейшее ограничение содержания углеводов в пище поддер-
203
живает гликогеновый запас печени на таком же низком уровне и пример-
но 50 % освобождающейся из печени глюкозы образуется в процессе
глюконеогенеза. Значительно позднее с помощью метода магнитно-ре-
зонансной спектроскопии было показано, что гликогенолиз в печени
составляет около 65 % общей продукции глюкозы в течение 22 ч голода-
ния. И напротив, потребление в течение одного дня пищи с высоким со-
держанием углеводов способствовало двойному увеличению (до
500 ммоль-кг-1 сырой массы) запасов гликогена в печени и менее 10 %
глюкозы при таких условиях извлекалось из гликонеогенеза.
Скорость выхода глюкозы из печени во время мышечной работы в
сытом состоянии зависела в основном от интенсивности выполнения
физического упражнения. Однако после высокоуглеводной диеты эта
скорость проявляла тенденцию к увеличению. Захват глюконеогенных
предшественников печенью возрастал во время выполнения физических
упражнений в 2-3 раза, но наибольший (приблизительно 90 %) выход
глюкозы извлекался из печеночного глюкогенолиза, приводящего к сни-
жению и окончательному истощению запасов гликогена.
Известно, что падение в крови концентрации инсулина, а также по-
вышение уровней адреналина и глюкагона на фоне увеличения продол-
жительности выполнения физического упражнения стимулирует выход
глюкозы. Однако точные механизмы, ответственные за регуляцию этого
выхода в начале и в течение физической нагрузки, не установлены. Соз-
дается впечатление, что освобождение глюкозы начинается почти сразу
с началом мышечной работы и инициируется механизмами, чувстви-
тельными к началу и интенсивности сократительной активности мышц.
Одна из широко распространенных теорий связывает снижение локаль-
ной концентрации глюкозы в крови в начале выполнения физических
упражнений с активацией в печени фосфорилазы в результате прямой
или гормональной стимуляции. Однако результаты большинства иссле-
дований свидетельствует об увеличении в крови концентрации глюко-
зы в начале мышечной работы и поэтому опровергают эту теорию. Сог-
ласно другим предположениям, которые пока окончательно не подтвер-
ждены, регуляторные механизмы включают гормональную и автоном-
ную стимуляции. В связи с последними успехами в совершенствовании
методики магниторезонансной спектроскопии применительно к це-
лостному организму появилась надежда, что будет получена более кон-
кретная информация относительно регуляции метаболизма СНО в пе-
чени. Ясно только одно: при выполнении физических упражнений на
уровне примерно 60 % VO2max выход глюкозы из печени начинает сни-
жаться после 90 мин работы, поскольку происходит истощение пече-
ночных запасов гликогена. Показано также, что скорость ресинтеза гли-
204
когена в печени зависит от разновидности присутствующих в ней СНО.
Так, например, ресинтез гликогена при инфузии фруктозы протекает в
3—4 раза быстрее, чем при инфузии глюкозы, что связано с высокой ак-
тивностью фосфофруктокиназы в ткани печени. Это имеет важное зна-
чение для физической работоспособности, поскольку печень является
единственным источником глюкозы крови. Результаты исследований,
проведенных на животных, а также испытуемых-добровольцах, показа-
ли, что истощение запасов гликогена в печени может ограничивать фи-
зическую работоспособность при мышечной работе либо косвенно, вы-
зывая более быстрое снижение запасов мышечного гликогена, либо
напрямую за счет развития гипогликемии, которая угнетает функции
нервной системы. С учетом этого перед пролонгированными физичес-
кими нагрузками следует максимизировать запасы гликогена как в
мышцах, так и в печени.
5. Потребление углеводов
непосредственно перед физической нагрузкой
Потребление углеводов до начала мышечной работы должно способ-
ствовать оптимизации запасов гликогена в печени. Однако результаты
выполненных в 70-е годы исследований давали основание полагать, что
прием СНО перед началом физических упражнений оказывает .нежела-
тельный эффект на физическую работоспособность, в результате чего
многие спортсмены в течение многих лет стали избегать их потребление
перед тренировочными и соревновательными нагрузками. Этот эффект
был связан с преходящим увеличением в крови уровня глюкозы после
приема СНО, что вызывало быстрое освобождение инсулина и приводи-
ло к дальнейшему снижению концентрации глюкозы, угнетению осво-
бождения СЖК и преждевременному развитию утомления (инсулинри-
кошетный эффект). Основываясь на опыте прошлого, в настоящее время
можно утверждать, что такая реакция может быть вызвана большим
количеством принятых СНО перед мышечной работой (около 75 г за
30—45 мин до нагрузки). Более точные исследования показали, что при-
ем меньшего количества углеводов непосредственно перед началом уп-
ражнения не повышает в плазме концентрацию инсулина во время рабо-
ты, не вызывает гипогликемию и способствует улучшению физической
работоспособности. В результате этого прием растворов СНО прямо пе-
ред тренировочными и соревновательными нагрузками стал более рас-
пространенным.
205
Выше отмечалось, что фруктоза рекомендовалась для приема в ка-
честве энергетического источника непосредственно перед физической
нагрузкой, и это не вызывает сомнений, поскольку по сравнению с глю-
козой она проявляет менее выраженное освобождение инсулина, спо-
собствует меньшей скорости утилизации мышечного гликогена и в
3—4 раза ускоряет синтез гликогена в печени. Однако результаты изу-
чения влияния потребления фруктозы на утилизацию мышечного глико-
гена и физическую работоспособность представляются сомнительными,
что может быть связано с сообщениями о желудочно-кишечном диском-
форте, который может возникнуть после приема фруктозы. При этом
проявилась важная особенность: нецелесообразность приема пищевых
продуктов, рекомендуемых исследовательскими лабораториями, в про-
цессе соревнований. Такая практика должна предварительно апробиро-
ваться на тренировочных занятиях, поскольку она может выявить значи-
тельную индивидуальную вариативность в отношении толерантности к
углеводным напиткам.
6. Потребление углеводов
во время выполнения физических упражнений
Еще в 30-е годы XX ст. стало известно, что прием углеводных напитков
в процессе мышечной работы может способствовать повышению вы-
носливости при пролонгированных физических нагрузках. Позднее вы-
явленный эффект стали объяснять способностью потребляемых СНО
сохранять запасы мышечного гликогена, что было подтверждено резуль-
татами выполненных Бергстром и Хултманом (1976) исследований, ко-
торые в настоящее время часто недооцениваются. Авторы продемон-
стрировали, что внутривенная инфузия глюкозы нетренированным ис-
пытуемым в течение 70 мин выполнения физического упражнения на
уровне 70 % VO2max могла снижать утилизацию мышечного гликогена
на 25 %. Однако необходимо заметить, что проводимая в этом исследо-
вании инфузия вызывала увеличение концентрации глюкозы в крови от
4,6 до 21,5 ммоль-л-’, которая не является физиологичной. В более позд-
них работах было показано, что повышение в крови концентрации глю-
козы на 2-3 ммоля-л-1 в результате приема СНО во время мышечной ра-
боты может способствовать улучшению физической работоспособнос-
ти. В большинстве случаев потребление углеводов во время выполнения
физических упражнений обнаруживает возрастание их окисления при
работе, в связи с чем можно полагать, что в таком случае мобилизация и
206
окисление жиров снижается. Это выглядит вполне реально, поскольку
потребление СНО во время выполнения физических упражнений бла-
гоприятно проявляется в снижении утилизации мышечного гликогена и
улучшении физической работоспособности. Их вклад в общую энерго-
продукцию должен быть большим по сравнению с вкладом окисления
жиров.
В более поздних работах было продемонстрировано, что проявле-
ние гликогенсохраняющего эффекта потребления СНО во время пролон-
гированных физических нагрузок ограничивается исключительно мы-
шечными волокнами типа I. Как установили Тсинтзас и др. (Tsintzas et
al., 1996), прием углеводов способствовал снижению (по сравнению с
применением плацебо) утилизации углеводов на 25 % в мышечных во-
локнах типа I после 140-минутной физической нагрузки. Для утили-
зации гликогена в волокнах типа II прием СНО оказался неэффектив-
ным. Кроме того, использование углеводов способствовало увеличению
продолжительности выполнения мышечной работы на 30 мин. Эти авто-
ры пришли к заключению, что прием углеводов способствует проявле-
нию выносливости за счет увеличения вклада окислительного ресинте-
за АТФ, особенно в мышечных волокнах типа I, а также благодаря сни-
жению развития в них гликогенового истощения.
Важно также помнить, что даже если под влиянием применения
СНО утилизация гликогена оказывается неэффективной, она все же
может благоприятно отразиться на физической работоспособности
благодаря отсрочке истощения гликогена в печени либо возмещению
в ней гликогеновых запасов, когда они уже исчерпаны, в связи с чем
снижается вероятность наступления гипогликемии. И действительно,
увеличение продолжительности выполнения физического упражне-
ния до отказа в ряде случаев ассоциируется с улучшением возмож-
ности поддержания уровня глюкозы в крови. В одной из классичес-
ких работ, выполненной Коулем с соавторами (Coyle et al., 1986), бы-
ло установлено отсутствие различий в утилизации мышечного глико-
гена во время физической нагрузки в течение трех часов, когда испы-
туемые принимали соответствующего привкуса плацебо либо рас-
твор глюкозного полимера через каждые 20 мин выполнения упраж-
нения. Однако испытуемые, принимавшие раствор плацебо, смогли
проработать до отказа в среднем 182 мин и концентрация глюкозы в
крови к концу работы снизилась до 2,5 ммоль-л-1, что сопровожда-
лось снижением окисления СНО. И напротив, если испытуемые при-
нимали раствор глюкозы, то предельная продолжительность выпол-
нения ими мышечной работы в среднем составила более 240 мин, а
нормогликемия и окисление СНО поддерживались в течение всего
207
времени выполнения физического упражнения. Кроме того, во время
дополнительного часа физической нагрузки утилизация запасов мы-
шечного гликогена происходила более экономно. Предположительно,
происходило это потому, что активность гексокиназы (фермента, от-
ветственного за фосфорилирование глюкозы на мембране мышечно-
го волокна) была повышенной. Отмеченный эффект, вероятно, прояв-
лялся в связи с тем, что ингибирующее влияние гексозомонофосфата
на гексокиназную активность снижалось из-за возрастающего исто-
щения запасов гликогена в мышцах. На основании полученных ре-
зультатов авторы пришли к выводу, что прием СНО во время физи-
ческой нагрузки увеличивает предельную продолжительность выпол-
нения мышечной работы благодаря в большей мере поддержанию
окисления запасов мышечного гликогена. Следует, однако, заметить,
что указанное исследование проводилось на спортсменах высокой
квалификации при выполнении физических упражнений, направлен-
ных на развитие выносливости, и поэтому не ясно, может ли столь
высокая скорость окисления глюкозы поддерживаться за счет утили-
зации ее из крови у нетренированных лиц.
7. Влияние повышенного потребления жиров
перед физическими нагрузками
Жиры способны обеспечить все энергетические потребности мышц при
выполнении пролонгированной мышечной работы низкой интенсивнос-
ти (на уровне менее 40 % VO2max). Однако во избежание гипогликемии
при таком виде мышечной деятельности необходимо поддерживать про-
дукцию глюкозы печенью. В связи с этим особенности питания пред-
ставляют определенный интерес, в частности для представителей воен-
ных ведомств, имеющих отношение к проблеме повышения боеспособ-
ности армии.
В одном из исследований испытуемые преодолевали пешком около
40 км в день в течение четырех дней. За это время энергообеспечение за
счет углеводов составило примерно 3 % общего потребления. Несмотря
на то что утилизация жиров возросла в течение этого периода, ни у кого
из испытуемых не была отмечена гипогликемия и все они полностью
справились с поставленной двигательной задачей (рис. 7.6). Необходимо
отметить, что аналогично условиям в состоянии покоя гликогенолиз и
глюконеогенез в печени могут сочетаться в соответствии с потребнос-
тью в СНО для данного вида мышечной деятельности.
208
Рис. 7.6. Влияние диеты на изменение концентрации глюкозы в крови (М±т) при про-
должительной ходьбе в последовательные дни до и после высокоуглеводной (о) и низ-
коуглеводной (•) диет: R— пробы, взимаемые ежедневно до выполнения тестирующей
нагрузки (натощак); LI, L2, L3 — пробы, взятые после преодоления 1-го, 2-го иЗ-го
кругов дистанции соответственно (длина каждого круга составляла примерно 13 км и
преодолевалась с постоянной скоростью (Мохан и др. (Moughan et al.), 1987)
Энергетические запасы жира в организме можно считать неисчерпа-
емыми. С точки зрения энергообеспечения жиры правильнее рассматри-
вать в качестве основного энергетического источника именно при про-
лонгированной мышечной работе. В этой связи повышение способнос-
ти к выполнению физических упражнений, проявляемое после трени-
ровки, направленной на развитие выносливости, будет характеризовать-
ся увеличением вклада в энергообеспечение окисления жиров, снижаю-
щего скорость расходования гликогена. Все это привело к формирова-
нию гипотезы о том, что богатая жирами диета, предшествующая вы-
полнению физического упражнения, может способствовать повышению
физической работоспособности. Предполагалось, что этот механизм бу-
дет проявляться в ингибировании каталитической активности ФДГ и
связанного с этим снижении окисления СНО во время выполнения фи-
зических упражнений (глюкозожирнокислотный цикл). Однако, как от-
мечалось выше, нет достаточных доказательств для утверждения, что
глюкозожирнокислотный цикл функционирует в скелетных мышцах че-
ловека во время интенсивной мышечной деятельности. Кроме того, та-
кой механизм может быть характерен только для крыс, поскольку име-
14
U 315
209
ется очень мало данных, подтверждающих концепцию «жировой загруз-
ки» для человека. Вместо этого предполагается, что физические и пси-
хические возможности более вероятно улучшить стремлением повысить
доступность жиров. И действительно, исходя из эмпирических данных
становится очевидным, что окисление исключительно одного только
жира не сможет обеспечить ресинтез АТФ со скоростью, необходимой
для поддержания энергообеспечения при пролонгированной мышечной
работе на уровне, превышающем 60 % VO2max.
В большом количестве работ показано, что прием кофеина
(3-9 мг-кг-1 массы тела) перед началом выполнения физического
упражнения способствует повышению выносливости при пролонгиро-
ванных физических нагрузках. Происходящее под влиянием кофеина
увеличение мобилизации СЖК вместе с последующим снижением ути-
лизации в мышцах гликогена во время мышечной работы позволяет
предположить, что эргогенный эффект кофеина обусловлен повышени-
ем эффективности использования жиров. Однако существование такого
механизма точно пока не установлено. К альтернативным претендентам
относятся прямое влияние кофеина на ЦНС и (или) на сопряженный ме-
ханизм «возбуждение-сокращение». В последнем случае кофеин прояв-
ляет in vitro стимулирующий эффект на обратный захват кальция
саркоплазматическим ретикулумом.
8. Механизмы утомления
Несмотря на обширную информацию, свидетельствующую о том, что
доступность СНО играет очень важную роль в проявлении физической
работоспособности при выполнении интенсивных пролонгированных
физических упражнений, точные механизмы, ответственные за развитие
утомления, развивающегося на фоне истощения углеводных запасов, ос-
таются не изученными.
Результаты проведенных в последнее время исследований позволя-
ют предполагать, что истощение углеводных источников приводит к
невозможности рефосфорилирования АДФ до АТФ с требуемой скорос-
тью, в связи с чем последующее увеличение концентрации АДФ обус-
ловливает развитие утомления, возможно, в результате прямого угнета-
ющего влияния АДФ и (или) Фн на сопряженность процесса возбужде-
ние-сокращение. Подтверждением этой гипотезы являются результаты
двух различных источников экспериментальных доказательств.
Во-первых, было показано, что при истощении углеводных запасов
концентрация в мышце интермедиатов ЦТК также снижается, в связи с
210
чем возникает предположение, что это может ослабить скорость метабо-
лизма через ЦТК, а поэтому и снизить эффективность окислительного
ресинтеза АТФ. Введение во время выполнения физического упражне-
ния глюкозы замедляет снижение концентрации интермедиатов ЦТК к
концу мышечной работы, что обеспечивает поддержание отношения
АТФ/АДФ и препятствует развитию утомления. По-видимому, введение
СНО способствует тому, что доступность пирувата не лимитирует ско-
рость метаболизма через анаплеротические реакции, ответственные за
образование интермедиата цикла трикарбоновых кислот 2-оксиглутара-
та (пируват + глутамат — 2-оксиглутарат + аланин).
Вторым источником доказательства, дающего основание полагать,
что истощение запасов СНО обусловливает развитие утомления, по-
лучено в результате проведенного исследования, показавшего, что в гли-
когенистощающей мышце снижается концентрация АТФ вместе с па-
раллельным увеличением концентрации АДФ и ИМФ. Развитие мышеч-
ного утомления и возрастание концентрации ИМФ происходит из-за
неспособности процесса окислительного фосфорилирования обес-
печить рефосфорилирование АДФ до АТФ, что приводит к активации
АМФ-дезаминазы и образованию ИМФ.
Таким образом, все приведенные факты должны способствовать
представлению о том, что доступность СНО очень важна для обеспе-
чения возможности продолжать выполнение мышечной работы. Утом-
ление будет продолжать развиваться до тех пор, пока станет невозмож-
ным поддерживать скорость ресинтеза АТФ в гликогенистощенном сос-
тоянии. Это согласуется с гипотезой о том, что утомление непосред-
ственно связано с повышением внутриклеточной концентрации АДФ,
угнетающим возбудительно-сократительное сопряжение. Необходимо
также заметить, что энергетический выход в процессе гидролиза АТФ
снижается, когда концентрация продуктов этого гидролиза (АДФ и Фн)
возрастает, что также может способствовать ускорению развития утом-
ления в связи с ослаблением реакций утилизации АТФ.
Настоящая глава посвящена рассмотрению особенностей метаболи-
ческих реакций организма (преимущественно в мышцах) на пролонги-
рованные физические нагрузки, поэтому в ней мало внимания уделено
характеристике жидкостного баланса при мышечной деятельности.
Однако, по-видимому, последнее требует проявления к себе должного
внимания, поскольку дегидратация может являться наиболее важным
фактором в развитии утомления при выполнении пролонгированных
физических упражнений, особенно в условиях высокой температуры и
влажности окружающей среды. Повышение температуры ядра тела, ко-
торое может происходить во время продолжительной физической на-
14*
211
грузки, например в жаркую погоду, потенциально представляет угрозу
для жизни. Поэтому увеличение притока крови к коже, а также начало
потовыделения в таких условиях является стремлением организма уве-
личить теплоотдачу. Поддержание адекватной гидратации препятствует
повышению температуры ядра тела во время мышечной работы. Однако
при развитии дегидратации возрастающий стресс охватывает сердечно-
сосудистую и терморегуляторную системы, что необходимо для поддер-
жания температурного баланса. При этом, как показали результаты пос-
ледних исследований, снижение в таких условиях мышечного кровото-
ка может способствовать повышению скорости утилизации мышечного
гликогена, что, предположительно, будет способствовать преждевремен-
ному наступлению утомления.
Установлено, что дегидратация относительно часто проявляется у
большинства спортсменов и тем самым отрицательно сказывается на их
физической работоспособности. Однако механизм этого влияния требу-
ет своего дальнейшего изучения. Не вызывает также сомнений тот факт,
что поддержание гидратации во время выполнения физических упраж-
нений может способствовать снятию физиологического стресса, вызван-
ного дегидратацией, и тем самым повысить физическую работоспособ-
ность. В связи с этим применение низкоконцентрированных глюкозо-
электролитных растворов получило широкое распространение среди
большинства спортсменов, тренирующихся в видах спорта, требующих
проявления выносливости.
Выводы
1. Термин «пролонгированные физические упражнения» обычно ис-
пользуют для характеристики упражнений, интенсивность выполнения
которых может поддерживаться (до ее снижения) в пределах
30-180 мин. Хотя уровень потребности в АТФ при выполнении таких
упражнений относительно низкий по сравнению с упражнениями высо-
кой интенсивности, тем не менее КФ, СНО и жиры все вместе могут
вносить свой вклад в энергопродукцию.
2. В начале выполнения пролонгированного физического упражне-
ния скорости распада КФ и накопления лактата проявляют линейную за-
висимость от интенсивности мышечной работы. В течение этого перио-
да КФ и анаэробный гликолиз вносят значительный вклад в продукцию
АТФ в мышцах. Однако в условиях устойчивого состояния окисление
СНО и жиров становится основным поставщиком энергии.
212
3. В нормальных условиях только запасы СНО в мышцах при их
полном использовании могут обеспечить выполнение мышечной работы
в течение 80 мин. Однако одновременная утилизация жиров и запасов
углеводов в печени позволяет обеспечивать продукцию АТФ для выпол-
нения более продолжительной работы. В конечном счете, следует пола-
гать, что продукция АТФ может быть нарушена в результате истощения
запасов СНО в мышцах и печени, а также неспособности окисления жи-
ров возместить этот дефицит. Факторы, ответственные за ограничение
скорости окисления жиров при мышечной деятельности, к настоящему
времени не изучены, однако, очевидно, лимитирующий участок должен
предшествовать образованию ацетил-КоА, поскольку после этого этапа
превращение жиров и углеводов происходит по одному и тому же пути.
4. Принято считать, что глюкозожирнокислотный цикл регулирует
окисление СНО и жиров при пролонгированных физических нагрузках.
Однако пока это может оставаться справедливым только для неактивных
мышц. Полученные в последнее время данные свидетельствуют о том,
что глюкозокислотный цикл не функционирует в скелетных мышцах че-
ловека при пролонгированных физических нагрузках умеренной интен-
сивности и что регуляция интеграции окисления углеводов и жиров
должна находиться где-нибудь в другом участке, например на этапе пос-
тупления глюкозы в мышцы или фосфорилирования гликогена. Как сви-
детельствуют данные литературы, интеграция утилизации СНО и жиров
в скелетных мышцах при продолжительных физических нагрузках но-
сит комплексный и до настоящего времени не совсем ясный характер.
5. У людей, ведущих малоподвижный образ жизни, мышечные запа-
сы СНО мало подвержены изменениям. Однако сочетанием физических
нагрузок с диетическими манипуляциями можно существенно влиять на
их уровень. Более того, установлено существование выраженной поло-
жительной взаимосвязи между содержанием СНО в мышцах и последу-
ющей способностью к выполнению пролонгированных физических уп-
ражнений.
6. Голодание быстро приводит к снижению в печени содержания
СНО, однако мало влияет на их уровень в скелетных мышцах. Скорость
освобождения глюкозы из печени в сытом состоянии достаточна для
удовлетворения потребностей в СНО только центральной нервной
системы. Примерно 70 % этого выхода извлекается из запасов печеноч-
ного гликогена, а остальное ее количество синтезируется в процессе
глюконеогенеза.
Скорость выхода глюкозы из печени во время мышечной работы за-
висит от интенсивности выполнения физического упражнения; 90 % ос-
вобождающейся глюкозы извлекается из печеночного гликогенолиза,
213
приводящего к снижению и окончательному истощению запасов глико-
гена. Точные механизмы, ответственные за регуляцию выхода глюкозы
из печени, изучены недостаточно. Поступление глюкозы в печень после
физических нагрузок зависит, по крайней мере частично, от того, какие
углеводы присутствуют в печени.
7. Прием углеводов в течение часа до начала выполнения физичес-
кого упражнения способствует оптимальному поддержанию запасов
СНО в печени. Нет оснований полагать, что такая процедура из-за инсу-
линового эффекта может нежелательно повлиять на физическую рабо-
тоспособность. В настоящее время общепризнано, что прием углеводов
перед пролонгированной физической нагрузкой способствует улуч-
шению физической работоспособности.
8. Прием СНО во время выполнения пролонгированного физическо-
го упражнения экономит утилизацию мышечного гликогена, снижает
мобилизацию и окисление жиров, стимулирует окисление СНО и повы-
шает способность к проявлению выносливости при продолжительной
мышечной работе. Поэтому не вызывает сомнений, что вклад потребля-
емых СНО в общую энергопродукцию в этих условиях должен быть бо-
лее высоким, чем тот, что обычно извлекается при окислении жиров. Ус-
тановлено, что прием СНО во время пролонгированной физической наг-
рузки проявляет свои функциональные и метаболические эффекты пре-
имущественно в мышечных волокнах типа I. Прием углеводов может
также препятствовать развитию утомления за счет снижения скорости
истощения запасов гликогена в печени и связанного с ним развития ги-
погликемии во время мышечной работы.
9. Повышение способности к выполнению физических упражне-
ний, проявляемое после тренировочных занятий, направленных на
развитие выносливости, должно характеризоваться увеличением
вклада в энергообеспечение окисления жиров, снижающего скорость
расходования гликогена. Это привело к формированию гипотезы о
том, что богатая жирами диета, предшествующая выполнению физи-
ческого упражнения, может способствовать повышению физической
работоспособности. Однако окисление исключительно только жира
может не обеспечить ресинтез АТФ со скоростью, необходимой для
поддержания энергообеспечения при пролонгированной мышечной
работе на уровне, превышающем 60 % VO2max, и именно поэтому
следует полагать, что в таких условиях физическая работоспособ-
ность обычно начинает снижаться.
10. Точные биохимические механизмы, благодаря которым истоще-
ние мышечных запасов СНО обусловливает развитие утомления, к нас-
тоящему времени не изучены. Однако вполне возможно, что неспособ-
214
ность мышц поддерживать скорость ресинтеза АТФ в гликогенистощен-
ном состоянии приводит к накоплению АДФ и Фн и, как следствие, к
наступлению утомления.
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
Bergstrom J., Hultman Е. (1966). Muscle glycogen synthesis after exercise: an enhancing fac-
tor localised to the muscle cells in man // Nature. — 1210. — P. 309-310.
Bergstrom J., Hultman E. (1967). A study of glycogen metabolism during exercise in man
// Scand. J. Clin. Lab. Invest. — 19. — P. 218-228.
Bergstrom J., Hultman E. (1967). Synthesis of muscle glycogen in man after glucose and fruc-
tose infusion // Acta Med. Scand. — 182.— P. 93-107.
Bergstrom J., Hermansen L., Hultman E., Saltin B. (1967). Diet, muscle glycogen and physi-
cal performance //Acta Physiol. Scand. — 71. — P. 140-150.
Coyle E.F., Coggan A.R., Hemmert M. K., Ivy J.L. (1986). Muscle glycogen utilisation during
prolonged strenuous exercise when fed carbohydrate // J. Appl. Physiol. — 61. —
P. 165-172.
Maughan R.J., Greenhaff PL., Gleeson M. et al. (1987). The effect of dietary carbohydrate
intake on the metabolic response to prolonged walking on consecutive days // Eur. J. Appl.
Physiol. — 56. — P. 583-591.
Nilsson L. H, Hultman E. (1973). Liver glycogen in man — the effect of total starvation or a
carbohydrate-poor diet followed by carbohydrate feedings // Scand. J. Clin. Lab. Invest.
— 32. — P. 325-330.
Randle P. J., Garland P. B., Hales C. N., Newsholme E. A. (1963). The glucose fatty acid
cycle: its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus
// Lancet. — P. 786-790.
Tsintzas O.-K., Williams C., Boobis L., Greenhaff PL. (1996). Carbohydrate ingestion and sin-
gle muscle fibre glycogen metabolism during prolonged running in man // J. Appl.
Physiol. — 81. —P. 801-809.
11 МЕТАБОЛИЧЕСКАЯ
АДАПТАЦИЯ К ТРЕНИРОВКЕ
1. Принципы тренировки
Физиологическая и биохимическая адаптация является ре-
зультатом выполнения повторяющихся физических упраж-
нений на протяжении нескольких дней, недель или меся-
цев. Происходящие в организме адаптационные изменения
повышают способность к выполнению специфических
двигательных задач. Характер и степень этих изменений
зависят от интенсивности и продолжительности физичес-
ких упражнений, методики тренировки и частоты трениро-
вочных воздействий, а также от генетических предпосылок
и уровня предшествующей активности человека.
Для повышения эффективности адаптационных изме-
нений в организме необходимо применять повторяющиеся
специфические сверхнагрузки. Основным принципом
(табл. 8.1) является тот факт, что адаптация к тренировке
может происходить только в том случае, если физические
нагрузки по своей частоте, интенсивности и продолжи-
тельности превышают обычный уровень двигательной ак-
тивности для данного индивидуума. Соответствующая
сверхнагрузка для каждого человека может достигаться
манипулированием комбинаций метода тренировки, ее
продолжительности и интенсивности.
Другой важный принцип состоит в том, что физиоло-
гическая и метаболическая адаптация к тренировке носит
преимущественно специфический характер, соответствую-
щий применяемым сверхнагрузкам. Так, тренировка, нап-
равленная на развитие скоростных возможностей и силы,
вызывает адаптационные изменения, которые отличаются
216
Таблица 8.1. Принципы тренировки
Сверхнагрузка
Специфичность
Индивидуальность адапта-
ционных изменений
Обратимость и временность
эффектов тренировки
Вид, частота, интенсивность и продолжительность
упражнения
Задействование мышц в состоянии активности при
тренировке, направленной на развитие
выносливости или силы
Исходный уровень подготовленности
Генетическая ограниченность
Время, необходимое для восстановительных
процессов
от тех, что происходят под влиянием тренировки, направленной на раз-
витие выносливости. Основные эффекты последней в скелетных мыш-
цах проявляются в изменении их окислительной способности и капил-
ляризации. С другой стороны, силовая тренировка влияет в основном на
величину площади поперечного сечения мышцы, благодаря чему увели-
чивается сила их сокращений. Специфичность вида физических упраж-
нений также очень важна. Например, развитие способности к проявле-
нию выносливости в беге, езде на велосипеде, плавании либо гребле
наиболее эффективно достигается в случае, когда в тренировку вовлека-
ются соответствующие мышечные группы с требуемой сократительной
активностью. Это происходит потому, что систематическое выполнение
физических упражнений вызывает как центральные (например, повы-
шение функциональных возможностей сердца), так и периферические
(например, повышение локальной мышечной работоспособности) адап-
тационные изменения.
Адаптационные изменения, происходящие под влиянием трениров-
ки, носят обратимый и временный характер: уже после нескольких дней
детренировки заметно снижаются эффективность метаболизма и пока-
затели физической работоспособности, а через несколько месяцев после
прекращения регулярных тренировочных занятий эти изменения стано-
вятся выраженными. Чрезмерные тренировочные нагрузки могут при-
вести к снижению и полной утрате физической работоспособности, ус-
ловно обозначаемой как «синдром перетренировки». Для обеспечения
морфологической адаптации в процессе реализации тренировочной
программы требуется значительный восстановительный период. Мыш-
цы представляют собой чрезвычайно пластическую ткань, и хотя гене-
тические факторы являются главными детерминантами качественных и
количественных характеристик мышц каждого нетренированного чело-
века, все же под влиянием тренировки можно добиться значительных
217
изменений их морфологических, функциональных и метаболических
показателей.
В списке рекомендованной литературы читатель может найти обзо-
ры, касающиеся особенностей центральной адаптации к тренировке,
направленной на развитие выносливости. В настоящей главе основное
внимание уделено локальной адаптации, происходящей в скелетных
мышцах в ответ на скоростно-мощностно-силовой тип тренировочных
воздействий.
2. Адаптация к тренировке,
направленной на развитие выносливости
Значительные тренировочные воздействия, включающие выполнение
физических упражнений на уровне 50-80 % VO2max в течение длитель-
ного периода по несколько раз в неделю, вызывают в организме адапта-
ционные изменения, улучшающие функциональные возможности, свя-
занные с доставкой кислорода, его поступлением в ткани и утилизацией.
Возможность поддерживать выполнение таких пролонгированных
физических упражнений, как езда на велосипеде или бег, зависит от со-
ответствия скорости утилизации АТФ скорости его ресинтеза в актив-
ных мышечных волокнах. Отсутствие такого соответствия приводит к
развитию утомления: скорость утилизации АТФ начинает снижаться, а
следовательно, снижается и мощность выполняемой работы.
Во время выполнения любого физического упражнения, продолжа-
ющегося больше нескольких минут, основным путем ресинтеза АТФ
является окислительное фосфорилирование в митохондриях, утилизиру-
ющих в качестве энергетического топлива углеводы и липиды. Этот про-
Мышечная адаптация к тренировке, направленной на развитие выносливости
Избирательная гипертрофия волокон типа I
Увеличение количества капилляров, приходящихся на одно волокно
Увеличение содержания миоглобина
Повышение способности митохондрий к окислительному ресинтезу АТФ
Увеличение размеров и количества митохондрий
Повышение способности к окислению липидов и углеводов
Увеличение использования липидов как энергетического топлива
Увеличение содержания гликогена и триглицеридов
Повышение способности к проявлению выносливости
218
цесс требует адекватного обеспечения кислородом, доставляемым
кровью, и наличия адекватного количества энергетических источников.
Последние могут извлекаться из запасов, которые находятся в самих мы-
шечных волокнах (гликоген и триглицериды), а также из циркулирую-
щей крови (глюкоза и СЖК). Нарушение ресинтеза АТФ может произой-
ти в случае, когда истощаются запасы внутримышечных энергетических
источников или когда падение эффективности кровоснабжения мышц
приводит к снижению доставки к ним энергетических субстратов и кис-
лорода. Систематическое выполнение физических упражнений, направ-
ленных на развитие выносливости, вызывает мышечную и кардиоваску-
лярную адаптацию, которая влияет на эти процессы и таким образом де-
терминирует обеспечение энергетическим топливом и кислородом.
Такая адаптация, включающая как ультраструктурные, так и метаболи-
ческие (ферментативные) изменения, приводит к улучшению доставки
кислорода и его экстракции сокращающимися мышцами, а также моди-
фицирует и улучшает регуляцию метаболизма в отдельных мышечных
волокнах. Более подробно эти изменения рассмотрены ниже.
Композиционный состав мышечных волокон. У спортсменов вы-
сокой квалификации — представителей видов спорта, связанных с пре-
имущественным проявлением выносливости или силы, композицион-
ный состав мышечных волокон имеет отчетливые различия. У первых
доминируют волокна типа I, тогда как у ведущих спринтеров преоблада-
ют волокна типа II. Д.Костилл с соавторами (Costill et al., 1976) обнару-
жили, что 79 % волокон в m. gastrocnemius у бегунов на длинные дистан-
ции высокой квалификации относится к типу I (в диапазоне 50-98 %),
тогда как у неэлитных бегунов на средние дистанции этот показатель
составил 62 %, а у нетренированных мужчин — 58 %. Однако в мышцах
ведущих бегунов преобладало не только количество волокон типа I.
Площадь поперечного сечения отдельных волокон этого типа также ока-
залась на 30 % больше. Это означало, что 83 % общего поперечного се-
чения мышцы приходилось на волокна типа I. В других группах испы-
туемых размеры обоих типов волокон были одинаковы.
Результаты лонгитудинальных исследований подтвердили, что тре-
нировка, направленная на развитие выносливости, влияет на размер
отдельных мышечных волокон, вызывая избирательную гипертрофию
волокон типа I, а также может выражение изменять их окислительную
способность. Тренировка способна также сильно повлиять на функцио-
нальные и метаболические свойства мышц и компонентный состав их
волокон. В исследованиях, проведенных на людях и животных, получе-
ны доказательства, что под влиянием перекрестной иннервации или про-
лонгированной электростимуляции может происходить трансформация
219
Таблица 8.2. Изменения композиционного состава мышечных волокон под влия-
нием тренировки (перед чертой — до тренировки, после черты — после тренировки)
Тип волокон По данным П. Андерсена и Д. Хеприкссона По данным Инджейра
I 41/43 58/57
Па 37/42 (Р < 0,005) 26/32 (Р = 0,005)
Пб 19/14 (Р < 0,005) 9/3 (Р = 0,005)
типов мышечных волокон. Интенсивная интервальная тренировка, как и
тренировка, направленная на развитие выносливости, вызывает измене-
ние композиционного состава мышечных волокон у крыс. Доказано, что
реализация относительно умеренных тренировочных программ может
способствовать заметному изменению композиционного состава мышц
у человека (табл. 8.2).
Р. Андерсен и Д. Хенрикссон (1977) установили, что после 8 недель
тренировки, направленной на развитие выносливости, определенное ко-
личество волокон подтипа Пб превращалось в подтип Па. Однако эти из-
менения были весьма незначительными: в основном, по крайней мере у
части выдающихся стайеров (например, у марафонцев), мышцы кото-
рых содержат высокий процент волокон типа I (обычно 70-90 %), эта
особенность скорее предопределена генетически, чем приобретена в
процессе тренировки.
Во многих исследованиях, в которых сообщалось об изменениях
композиционного состава мышечных волокон в мышцах под влиянием
тренировки, не придавалось значения возможным изменениям в различ-
ных типах волокон. Однако Симоней с соавторами (Simoneau et al., 1985)
обнаружили, что после 15 недель высокоинтенсивной интервальной тре-
нировки увеличиваются пропорции волокон типа I и снижаются пропор-
ции волокон типа II без изменений содержатся волокна подтипа Па, при
этом площадь поперечного сечения волокон типа I и подтипа Пб возрас-
тала. Поскольку изменения в пропорции между количеством различных
типов мышечных волокон проявляться не могут, то, следовательно, при-
чина отмеченных изменений состояла только в изменении площади их
поперечного сечения.
Вполне очевидно, что существует' предел той степени, до которой
мышечные волокна могут расти в диаметре. Отчасти это может быть
связано с тем, что с увеличением диаметра увеличивается и расстояние,
через которое кислород должен диффундировать. Однако повышение ка-
пиллярной плотности может препятствовать этому ограничению, созда-
вая тем самым возможность для гипертрофии без заметного влияния на
220
среднюю величину диффузной дис-
танции для кислорода.
Плотность капилляров в мыш-
цах. Результаты изучения поперечных
срезов (рис. 8.1), а также данные, по-
лученные в течение многолетних ис-
следований (рис. 8.2), показали, что
тренировка способствует увеличению
капиллярной плотности в скелетных
мышцах, выражаемой количеством
капилляров, приходящихся на одно
волокно или на единицу площади по-
перечного сечения. Капиллярное ложе
мышцы играет важную роль в обеспе-
чении поверхности, через которую
происходит обмен между мышцей и
кровью. Увеличение количества ка-
пилляров, окружающих отдельные
мышечные волокна, будет способ-
ствовать тому, что в случае рекруити-
рования волокон во время мышечной
работы последние становятся более
доступными для снабжения кровью.
Таким образом, повышение капилляр-
ной плотности создает возможность
Рис. 8.1. Капиллярная плотность,
выражаемая количеством капилляров,
приходящихся на мышечное волокно,
и капилляров, приходящихся на 1 мм2
площади поперечного сечения m. vas-
tus lateralis у контрольных испытуе-
мых, ведущих малоподвижный образ
жизни (М), у тяжелоатлетов (Т) и
бегунов-стайеров (Б): М±гп; * —
Р < 0,01 по отношению к контроль-
ной группе испытуемых (Теч и др.
(Tesch et al.), 1984)
возрастания скорости доставки кисло-
рода, питательных веществ и удаления конечных продуктов метаболиз-
ма при высоких уровнях перфузии благодаря наличию большей площа-
ди поверхности для диффузии, а также укорочению среднего диффузи-
онного расстояния.
Более 99 % кислорода транспортируется содержащимся в эритро-
цитах гемоглобином. Повышение капилляризации будет также способ-
ствовать снижению скорости тока крови через капилляры при одной и
той же общей скорости кровотока через ткань, что увеличивает время
диффузии для кислорода, поскольку каждый эритроцит дольше находит-
ся в капилляре. Этот эффект может иметь важное значение, так как эк-
стракция кислорода осуществляется не полностью, если время тран-
спорта эритроцита через капилляр составляет менее одной секунды.
При максимальной скорости транспорта в условиях выполнения мышеч-
ной работы одной ногой время нахождения эритроцита в капилляре мо-
жет составлять менее 0,25 с. Повышение плотности капилляров позво-
221
Рис. 8.2. Изменения плотности ка-
пилляров, окружающих волокна раз-
ных типов в m. vastus lateralis до и
ляет тренированным мышцам поддер-
живать более продолжительное время
перенос кислорода, в связи с чем дос-
тигается более высокая экстракция
его даже тогда, когда скорость общего
мышечного кровотока по сравнению с
нетренированным состоянием возрас-
тает. Такая сосудистая адаптация, не-
сомненно, способна обеспечить повы-
шение кислородной экстракции, отме-
чаемое в тренированных мышцах, и
увеличение VO2max целостным орга-
низмом после реализации трениро-
вочной программы, направленной на
развитие выносливости. Результаты
ряда работ продемонстрировали про-
исходящее под влиянием тренировки
с повышением капиллярной плотное
ти параллельное увеличение VO2max.
происходящее в результате роста но-
после реализации тренировочной
программы, направленной на разви-
тие выносливости: М ± т; * •— зна-
чимость различия по сравнению с
показателями до тренировки —
Р< 0,01; + — значимость различий
вых капилляров уже в течение первых
недель тренировки. Плотность мы-
шечных капилляров проявляет тен-
денцию к прогрессивному увели-
чению в процессе тренировочных воз-
между типами волокон в одинаковых
условиях — Р < 0,01 (Инджей
(Ingier), 1979)
действий; при прекращении трениро-
вок отмечается исчезновение эффекта
замедления скорости кровотока в ка-
пиллярах (рис. 8.3).
Хотя наиболее выраженное повышение капилляризации отмечает-
ся в областях волокон подтипа Па, где она обычно самая низкая, обра-
зование новых капилляров происходит также и в области других типов
волокон.
Содержание миоглобина в мышцах. Результаты исследований,
проведенных на животных, свидетельствуют о том, что содержание ми-
оглобина в мышцах под влиянием тренировки может увеличиваться до
80 %. Следовательно, потенциальная возможность мышечного волокна,
находящегося в неактивном состоянии, к запасанию кислорода увеличи-
вается. Однако значимость возрастания этого запаса для повышения
окислительной способности мышц относительно невелика. Основной
эффект увеличения содержания миоглобина проявляется, по-видимому,
222
Рис. 8.3. Время протекания некоторых адаптационных процессов в скелетных мышцах
под влиянием тренировочных занятий, направленных на развитие выносливости, а так-
же после их прекращения (детренировки)
во время мышечной работы и связан с поддержанием низкого рО2 в
саркоплазме мышечного волокна, а также облегчением диффузии кисло-
рода в мышцы из крови. Тем не менее в скелетных мышцах человека
повышения содержания миоглобина под влиянием тренировки не наб-
людается, а может даже снизиться.
Запасы внутримышечных энергетических источников. В ряде
работ отмечается, что у хорошо тренированных лиц в состоянии покоя
обнаруживается более высокое содержание гликогена (в 2,5 раза по
сравнению с нетренированным состоянием). Увеличение запасов глико-
гена может быть обусловлено, в частности, повышением чувствитель-
ности к инсулину, что происходит под влиянием тренировки. Это спо-
собствует поступлению глюкозы (уровень транспортного белка ГЛУТ4 в
тренированных мышцах на 25 % выше по сравнению с нетренированны-
ми) и ее запасанию. В целостном организме переход неокисленной глю-
козы в мышечные клетки во время ее инфузии происходит у выносли-
вых спортсменов приблизительно на 60 % больше, чем у испытуемых
контрольной группы, ведущих малоподвижный образ жизни. Только у
тренированных лиц были обнаружены значительные запасы глюкозы и
223
гликогена в скелетных мышцах. Активность гликогенсинтетазы (общей
и инсулинзависимой) повышается под влиянием тренировки, хотя ак-
тивность ферментов, имеющих отношение к распаду гликогена (фосфо-
рилаза), также возрастает.
Тренировка крыс на тредмиле выявила 20-40 %-ное увеличение в
мышцах активности гексокиназы. Инсулин также способствует дозоза-
висимому возрастанию притока крови к инсулинчувствительной ткани,
и поскольку тренированным мышцам присуща улучшенная капилляри-
зация, этот эффект инсулина может повысить доставку кислорода к тре-
нированным мышцам. Указанные данные позволяют предположить, что
в тренированных мышцах развита повышенная способность к запаса-
нию глюкозы в виде гликогена. Однако концентрация мышечного глико-
гена будет заметно зависеть от времени, прошедшего после окончания
последней тренировочной нагрузки и последующего потребления с
пищей углеводов. Более высокое содержание мышечного гликогена у
тренированных лиц может отражать феномен гликогеновой суперком-
пенсации.
В ряде исследований также сообщалось о более высоком содержа-
нии триацилглицеролов у тренированных лиц по сравнению с нетрени-
рованными. Следовательно, под влиянием тренировки увеличивается
доступность как углеводного, так и липидного субстратов, что обеспечи-
вает достаточное время для восполнения внутримышечных запасов
энергетического топлива после предшествующей тренировочной наг-
рузки.
Гликолитические возможности. В литературе явно недостаточно
освещены вопросы о влиянии тренировочных занятий, направленных
на развитие выносливости (по сравнению с влиянием на аэробную спо-
собность) на гликолитические возможности. Хотя результаты ряда ис-
следований, проведенных на людях, и указывают на то, что активность
некоторых основных ферментов гликолиза, включая фосфофруктокина-
зу, и возрастает под влиянием такой тренировки, все же следует отме-
тить, что в большинстве других работ сообщалось либо об отсутствии
изменений, либо о незначительном снижении активности гликолити-
ческих энзимов. Содержание ферментов гликолиза в мышцах спортсме-
нов, тренирующихся в видах спорта, требующих проявления выносли-
вости, обычно низкое, однако этот факт можно объяснить преобладани-
ем в них процентного содержания волокон типа I, характеризующихся
низкой гликолитической способностью по сравнению с волокнами ти-
па II. Под влиянием хронической электростимуляции мышц кролика,
приводящей к полной трансформации волокон подтипа Пб в тип I, со-
держание гликолитических ферментов в мышце снижается приблизи-
224
тельно только на 20 % перво-
начального уровня (рис. 8.4),
что отражает различие в гли-
колитическом потенциале
между быстросокращающи-
мися и медленносокращаю-
щимися волокнами. В перио-
ды электростимуляции окис-
лительная способность мы-
шечных ферментов повыша-
лась более чем на 300 % ее
исходной величины. Ре-
зультаты этих исследований
демонстрируют максималь-
ную возможность биохими-
ческой адаптации в скелет-
ных мышцах.
Плотность митохон-
дрий в мышцах и окисли-
тельная активность фер-
Продолжительность электростимуляции, нед.
Рис. 8.4. Влияние хронической электростиму-
ляции на активность фосфофруктокиназы
(ФФК) — узлового фермента гликолиза и мито-
хондриальных энзимов ЦТК — цитратсинтазы
(ЦС), малатдегидрогеназы (МДГ) и сукцинатде-
гидрогеназы (СДГ) (Хенрикссон и др.
(Henriksson et al.), 1986)
ментов. В тренированных мышцах митохондрии характеризуются зна-
чительно более высокой способностью к окислительному ресинтезу
АТФ в процессе окислительного фосфорилирования. Окислительная
способность скелетных мышц повышена также за счет заметного увели-
чения как размера, так и количества митохондрий, приходящихся на еди-
ницу площади мышечной ткани (рис. 8.5), а также за счет площади по-
верхности митохондриальной мембраны. Взаимосвязано это с двукрат-
ным увеличением активности ферментов ЦТК и количества компонен-
тов электрон-транспортной цепи. Так, например, как показано на
рис. 8.5, в результате реализации тренировочной программы, направлен-
ной на развитие выносливости, количество митохондрий в m. vastus la-
teralis человека возросло на 120 %. Увеличение размеров митохондрий
произошло не в такой степени, как увеличение их количества. В среднем
размеры митохондрий скелетных мышц у выносливых спортсменов на
14-40 % больше по сравнению с нетренированными лицами, ведущими
малоподвижный образ жизни. Однако эта специфическая особенность
проявляется только в волокнах, задействованных в выполнении трени-
ровочного упражнения.
Под влиянием тренировки активность отдельных ферментов изме-
няется в разной степени. Так, например, под влиянием ежедневной тре-
нировки крыс в беге на тредмиле содержание цитохрома с в m. gastroc-
•5 .3.3
225
О 5 10 15 20 25 30
Недели тренировки
Рис. 8.5. Изменение количества и размеров митохондрий скелетных мышц человека
под влиянием тренировки (Кеиссинг и др. (Keissing et al.), 1971)
Размер митохондрий,
ммоль
б
nemius возросла на 120 % (Холлози и др. (Hollozy et al.), 1970), тогда как
активность ферментов ЦТК возросла на 34-101 % (рис. 8.6).
Активность изоцитратдегидрогеназы —- узлового фермента ЦТК
возросла после тренировки на 90 %. Следовательно, изменение мито-
хондриальной композиции, как и увеличение активности ферментов
ЦГК и транспорта электронов, происходят в результате тренировки, нап-
равленной на развитие выносливости. Повышение уровня активности
либо концентрации указанных выше ферментов обусловливает боль-
шую способность ЦТК и электрон-транспортной цепи генерировать
АТФ в присутствии кислорода. Ранее считали, что повышение возмож-
ностей аэробного метаболизма не используется, поскольку содержание
митохондрий считалось более чем достаточным даже в нетренирован-
ных мышцах. Однако результаты последних исследований свидетель-
ствуют о том, что увеличение содержания митохондрий под влиянием
тренировки на выносливость необходимо для повышения потенциала
окислительного ресинтеза АТФ. Эти изменения могут играть важную
роль, поскольку позволяют тренированным спортсменам поддерживать
более высокий процент аэробной производительности (VO2max) во вре-
мя выполнения пролонгированного физического упражнения.
Окислительные возможности мышечных волокон типов I и II могут
значительно повыситься под влиянием тренировки. Однако увеличение
митохондриальной плотности не специфично для волокон типа I. И дей-
226
ствительно, у бегунов высо-
кой квалификации, специали-
зирующихся на длинные дис-
танции, по сравнению с
нетренированными лицами,
способность волокон типа II
к окислению энергетических
субстратов, включая липиды,
может значительно превы-
шать окислительную способ-
ность волокон типа I.
О 20 40 60 80 100 120 140
Повышение под влиянием тренировки, %
Рис. 8.6. Активность ферментов и способность
к окислению субстратов в скелетных мышцах
под влиянием тренировочных занятий, направ-
ленных на развитие выносливости
Однако дифференциро-
ванные изменения в метабо-
лических характеристиках
различных типов мышечных
волокон могут быть обуслов-
лены различиями в направленности спортивной тренировки. Было уста-
новлено, что интервальная тренировка с использованием физических
нагрузок максимальной интенсивности либо выполнение длительных
физических упражнений субмаксимальной аэробной мощности приво-
дит к увеличению в мышцах активности сукцинатдегидрогеназы (СДГ)
на 20-30 %. Однако анализ одиночных мышечных волокон показал, что
высокоинтенсивная тренировочная программа способствовала повыше-
нию активности этого фермента в волокнах типа II на 50 %, тогда как в
волокнах типа I оставалась неизменной. В результате выполнения
тренировочной программы, в которой применялись продолжительные
беговые нагрузки, увеличение активности СДГ на 30 % обнаружилось в
волокнах типа I без изменения активности этого фермента в волокнах
типа II. Изменение окислительной активности ферментов в волокнах ти-
па II является, очевидно, наиболее важным для повышения способности
к проявлению выносливости в результате снижения возможности их пе-
реключения на анаэробный гликолиз для продукции АТФ.
3. Модификация метаболического ответа
на физическую нагрузку под влиянием тренировки,
направленной на развитие выносливости
Изменения метаболического ответа на напряженную физическую наг-
рузку после реализации тренировочной программы, направленной на
развитие выносливости, были обобщены следующим образом. Трениро-
15*
227
Изменение метаболического ответа на физическую нагрузку под влиянием
тренировочных занятий, направленных на развитие выносливости
Снижение КДО и мышечного ДК
Незначительное увеличение в плазме концентрации СЖК
Снижение скорости утилизации мышечного гликогена
Снижение утилизации глюкозы крови мышцами
Незначительное накопление в мышцах лактата
Более высокое окисление липидов по сравнению с углеводами
Повышенная утилизация внутримышечных триглицеридов
ванные мышцы проявляют более высокую способность к окислению уг-
леводов. Следовательно, большее количество пирувата может быть пре-
образовано в ацетил-КоА и пропущено через ЦТК. При этом возрастает
также способность тренированных мышц утилизировать липиды
(рис 8.7). Происходит это благодаря повышению капиллярной плотнос-
ти в мышцах, позволяющему большей площади поверхности захваты-
вать СЖК из крови, а также в связи с увеличением активности липид-
мобилизующих и липидметаболизующих ферментов. Активность ли-
попротеинлипазы (ЛПЛ) в эндотелии капилляров тренированных мышц
увеличивается так же, как и способность митохондрий к Ь-окислению
СЖК. Однако самый главный эффект энзиматических изменений, про-
исходящих в мышцах под влиянием тренировки, направленной на разви-
тие выносливости, состоит в увеличении вклада липидов и соответ-
ственно снижение вклада углеводов в окислительный энергетический
метаболизм (ресинтез АТФ) при выполнении физических упражнений
субмаксимальной аэробной мощности.
Результаты значительного количества работ свидетельствуют о том,
что под влиянием тренировки во время выполнения физических упраж-
нений происходит снижение как коэффициента дыхательного обмена
(КДО), так и определяемого в инвазивных исследованиях непосред-
ственно в работающих мышцах локального дыхательного коэффициен-
та (ДК). Отмеченное возрастание окисления липидов является, оче-
видно, следствием увеличения возможности окисления субстратов по
сравнению с гликолитической возможностью, которая проявляет менее
выраженный ответ при тренировке, направленной на развитие выносли-
вости.
Выносливые спортсмены используют больше жира и меньше угле-
водов не только при выполнении одинаковой по абсолютной мощности
мышечной работы, но и при одинаковой ее относительной мощности,
выражаемой в процентах VO2max.
228
Рис. 8.7. Влияние тренировочных занятий на аэробный потенциал скелетных мышц: а
— электрон-транспортные компоненты; б — ферменты ЦТК; в — ферменты липидно-
го метаболизма; г — способность к окислению субстратов
Под влиянием тренировки происходит снижение утилизации внут-
римышечного гликогена и глюкозы крови (рис. 8.8). В сердечной мыш-
це этот гликогензащитный эффект опосредован функционированием
глюкозожирнокислотного цикла, благодаря которому увеличение окис-
ления липидов приводит к накоплению внутриклеточного цитрата и
последующему угнетению гликолиза на уровне фосфофруктокиназы.
Однако относительно существования такого механизма в скелетных
мышцах человека мнения противоречивы.
Снижение захвата и утилизации глюкозы крови мышцами понижает
также степень гликогенолиза в печени и обеспечивает лучшее поддержание
гомеостаза глюкозы в крови во время выполнения пролонгированных физи-
229
15 30 45 60 75 90 105 120 15 30 45 60 75 90 105 120
Продолжительность физической нагрузки, мин
Рис. 8.8. Влияние тренировочных занятий в течение 12 недель, направленных на
развитие выносливости, на окисление мышечного гликогена и глюкозы плазмы крови
во время велоэргометрической нагрузки в течение 2 часов с мощностью, соответст-
вующей 60 % дотренировочного уровня VO2max; Р < 0,001 (Менденхелл и др.
(Mendenhall et al.), 1994)
ческих упражнений. Снижение скорости окисления углеводов у трениро-
ванных лиц во время выполнения физического упражнения взаимосвязано
со снижением скорости продукции лактата. При выполнении физических
упражнений субмаксимальной аэробной мощности концентрация лактата у
тренированных спортсменов ниже, чем у нетренированных лиц. Это спра-
ведливо независимо от того, выражается интенсивность выполнения физи-
ческого упражнения в абсолютных или относительных величинах. Увели-
чение лактатного клиренса, происходящего под влиянием тренировки, так-
же вносит свой вклад в снижение концентрации лактата в крови во время
выполнения физического упражнения. И хотя это продемонстрировано в
исследованиях, проведенных на крысах, все же очевидно, что отмеченный
эффект обусловлен значительным поступлением лактата и его превращени-
ем до глюкозы печенью. У человека скорость глюконеогенеза в печени во
время выполнения физического упражнения под влиянием тренировки ста-
новится ниже. Эти два важных эффекта — снижение скорости окисления
углеводов и снижение скорости продукции лактата — способствуют сохра-
нению ограниченного углеводного резерва в организме, поскольку скорость
утилизации мышечного гликогена под влиянием тренировки становится ни-
же (рис. 8.9). В связи с установлением тесной взаимосвязи между наличи-
ем мышечного гликогена как энергетического топлива и способностью к
проявлению выносливости (см. главы 3 и 7) снижение скорости утилизации
гликогена следует рассматривать в качестве главного фактора, способству-
ющего повышению физической работоспособности в видах спорта, требу-
ющих проявления этого двигательного качества.
Изменения в использовании субстратов, происходящие под влияни-
ем тренировки, могут быть также связаны с меньшим нарушением го-
меостаза АТФ во время выполнения физических упражнений: с повы-
230
шением функциональных возможностей митохондрий, происходящих
под влиянием тренировки, меньшее снижение АТФ и КФ и меньшее уве-
личение АДФ и Фн необходимы во время физической нагрузки для под-
держания баланса между скоростью ресинтеза АТФ и скоростью его
утилизации. Другими словами, с увеличением количества митохондрий
потребность в кислороде, так же как в АДФ и Фн, приходящаяся на од-
ну митохондрию, после выполнения тренировочной программы стано-
вится меньше, чем до тренировки. Меньшее возрастание концентрации
АДФ будет способствовать меньшему накоплению АМФ в процессе аде-
нилаткиназной реакции, а также меньшему образованию ИМФ и амми-
ака в ходе реакции дезаминирования. Снижение скорости гликогенолиза
и гликолиза в тренированных мышцах по сравнению с нетренированны-
ми можно объяснить менее выраженным увеличеним возрастания внут-
римышечной концентрации АМФ и Фн.
Известно, что происходящее под влиянием тренировки снижение
окисления углеводов во время выполнения мышечной работы компенси-
руется увеличением скорости окисления липидов. В течение многих лет
было принято, что это обусловлено увеличением поступления и окис-
ления плазменных СЖК, извлекаемых в процессе липолиза из адипозной
ткани. Однако результаты ряда тщательно проведенных исследований не
подтвердили эту точку зрения, а показали, что под влиянием тренировки
во время выполнения физического упражнения снижается концентрация
СЖК в плазме, а также их поступление в мышечные волокна и окис-
ление. Альтернативным источником возрастания окисления липидов,
происходящего под влиянием тренировки, могут быть внутримышечные
триацилглицерольные запасы. Доказательства такого вывода получены в
ходе исследований, в которых удалось показать, что тренировка приводит
к значительному повышению утилизации внутримышечных триацилгли-
церолов во время пролонгированной физической нагрузки (рис. 8.10). В
Рис. 8.9. Влияние тренировочных занятий в
течение 12 недель, направленных на развитие
выносливости, утилизацию мышечного гли-
когена во время велоэргометрической нагруз-
ки в течение двух часов с мощностью, соответ-
ствующей 60 % дотренировочного уровня
VO2max: * — значимость ниже по сравнению с
нетренированным состоянием; Р<0,001 (Ког-
ган и др. (Coggan et al.), 1993)
Нетрени- Трениро-
рованные ванные
231
Нетрени- Трениро-
рованные ванные
Рис. 8.10. Влияние тренировочных занятий в
течение 12 недель, направленных на развитие
выносливости, утилизацию мышечных три-
глицеридов во время велоэргометрической
нагрузки в течение двух часов мощностью, со-
ответствующей 60 % дотренировочного
уровня VO2max: * — значимость ниже по срав-
нению с нетренированным состоянием;
Р<0,001 (Харлей и др. (Hurley et al.), 1986)
одной из таких работ было установлено, что повышение внутримышеч-
ного липолиза может обеспечить все дополнительное окисление жиров
после реализации тренировочной программы, если предположить, что по
меньшей мере 8 кг мышечной ткани находилось в активном состоянии во
время выполнения физического упражнения.
Скорость гидролиза ЛОНП-триацилглицеролов также возрастает
под влиянием тренировки благодаря повышению в мышцах ЛПЛ-актив-
ности и большей площади поверхности эндотелиальных капилляров.
Однако даже в этом случае ЛОНП-триацилглицеролы не могут соста-
вить более 10 % своего вклада в общее энергообеспечение. Однако этот
эффект отчасти может иметь отношение к улучшению в крови липидно-
го профиля, проявляющегося у людей, которые после малоподвижного
образа жизни стали более активными.
Восстановление запасов внутримышечных триацилглицеролов пос-
ле пролонгированных физических нагрузок также может улучшить кли-
ренс триацилглицеролов из крови.
4. Физиологические адаптационные процессы
при тренировке, направленной на развитие
выносливости, которые влияют на метаболический
ответ во время выполнения физических упражнений
При рассмотрении влияния на метаболизм тренировочных занятий, нап-
равленных на развитие выносливости, следует учитывать центральную
кардиоваскулярную и респираторную адаптацию к тренировке, которая
влияет на доставку кислорода к работающим мышцам. Способность к
232
проявлению выносливости при физических нагрузках зависит от макси-
мально развиваемой аэробной мощности и уровня *VO2max, который
может поддерживаться. В видах физических нагрузок, когда задейство-
вана большая мышечная масса (например, бег или езда на велосипеде),
VO2max должно лимитироваться способностью мышц экстрактировать
и утилизировать кислород. Максимальный сердечный выброс проявляет
тесную взаимосвязь с VO2max и изменениями VO2max, которые проис-
ходят под влиянием тренировки и детренировки.
Периферические факторы, особенно повышение капиллярной плот-
ности в мышцах, а также увеличение окислительных возможностей (см.
выше) будет способствовать повышению утилизации липидов как мета-
болического топлива при мышечной деятельности и, возможно, прояв-
лению выносливости. Основные эффекты тренировки, направленной на
развитие выносливости, которые отражаются на состоянии сердечно-со-
судистой и дыхательной систем, кратко изложены ниже и обобщены в
табл. 8.3. Более детальное рассмотрение и обсуждение указанных изме-
нений можно найти в источниках, представленных в списке рекомендуе-
мой литературы в конце книги.
Спортсмены высокой квалификации могут достичь высоких показа-
телей VO2max, благодаря чему способны выполнять мышечную работу
в течение продожительного периода на высоком уровне потребления
кислорода (в процентном отношении к VO2max). Физиологические и
метаболические факторы, которые обеспечивают эту способность, в
настоящее время интенсивно изучаются. Для достижения высокого
уровня потребления кислорода необходимо эффективное функциониро-
вание системы его доставки из атмосферного воздуха к работающим
мышцам в сочетании с эффективной экстракцией из крови. При отсут-
ствии легочных заболеваний первая стадия этого процесса (транспорт
кислорода из атмосферного воздуха в кровь через легочные капилляры)
не является лимитирующим фактором. В сущности, при выполнении ин-
тенсивных физических упражнений достигается полное насыщение ге-
моглобина кислородом, за исключением, возможно, пожилых людей или
спортсменов с очень высоким показателем VO2max.
Потребление кислорода организмом может быть описано с по-
мощью уравнения Фика:
VO2 (мл О2-мин-') = [ЧСС (уд-мин-’) + Ударный объем (л-сокр~‘)] х
х [a-v кислородная разность (мл 02 л-1)].
Ударный (систолический) объем — это количество крови, выбрасы-
ваемое сердцем при каждом сокращении его желудочков. Ударный объ-
233
Таблица 8.3. Физиологическая адаптация к тренировочным занятиям,
направленным на развитие выносливости
Объем крови Увеличение объема плазмы и общего содержания гемоглобина
Ударный объем ЧСС Повышение, обусловленное увеличением объемов желудочков в сочетании с увеличением силы сокращения миокарда Снижение в покое и при выполнении физических упражнений субмаксимальной аэробной мощности. Максимальная ЧСС не изменяется
Сердечный выброс Кровоток и его распределение Увеличение максимального сердечного выброса, связанное с более высоким ударным объемом Повышение общего тока крови через мышцы при максимальных физических нагрузках. Снижение регионального притока крови к работающим мышцам при выполнении физических упражнений субмаксимальной аэробной мощности
Экстракция кислорода Повышение экстракции кислорода из крови, проходящей через работающие мышцы, а следовательно, увеличение артериовенозной разницы по кислороду
Артериальное кровяное давление Легочная вентиляция Снижение систолического и диастолического кровяного давления в покое и при выполнении физических упражнений субмаксимальной аэробной мощности Более высокий уровень максимальной легочной вентиляции, обусловленный увеличением как дыхательного объема, так и частотой дыхания. Снижение уровня легочной вентиляции при выполнении физических упражнений субмаксимальной аэробной мощности
ем и ЧСС определяют сердечный выброс — количество крови, выбра-
сываемое левым (правым) желудочком за 1 мин. Артериально-венозная
(a-v) разность по кислороду представляет собой различие между содер-
жанием кислорода в артериальной и смешанной венозной крови, что яв-
ляется показателем экстракции кислорода из крови при прохождении ее
через ткани.
Высокий максимальный сердечный выброс, который отмечается у
спортсменов, специализирующихся в видах спорта, требующих прояв-
ления выносливости, определяется более высоким (по сравнению с нор-
мой) этим показателем. При максимальной физической нагрузке ЧСС
обычно такая же (или несколько ниже), как и у нетренированных людей.
Высокие значения ударного объема являются результатом увеличения
объема желудочков и диастолического объема, а также повышения
сократительной способности миокарда. Сердечный выброс в состоянии
234
покоя у тренированных людей такой же, как и у нетренированных, но
поскольку у спортсменов ударный объем выше, то ЧСС у них имеет
меньшие показатели.
Во время выполнения физических упражнений субмаксимальной
аэробной мощности сердечный выброс под влиянием тренировки может
снижаться, так как в данном случае для обеспечения мышц кислородом
требуется меньший приток крови к ним в связи с повышенной экстрак-
цией кислорода. При максимальных физических нагрузках выносливые
спортсмены также проявляют более высокую способность к экстракции
кислорода из крови несмотря на более высокую скорость кровотока в
мышцах.
Кислородтранспортная способность крови (максимальное содер-
жание кислорода в артериальной крови) определяется в основном со-
держанием в ней гемоглобина. При полном насыщении каждый грамм
гемоглобина связывает 1,34 мл кислорода. Поэтому на первый взгляд
может показаться странным, что у высокотренированных, выносливых
спортсменов, как правило, отмечается низкая по сравнению с людьми,
ведущими малоподвижный образ жизни, концентрация гемоглобина в
крови. Однако увеличение его общего содержания, по крайней мере от-
части, зависит от увеличения общего объема крови с более высоким
уровнем плазмы по отношению к содержанию эритроцитов, в связи с
чем из-за большего разбавления последних их количество на 1 л крови
снижается. Тем не менее общее содержание гемоглобина в крови вы-
носливых спортсменов возрастает, а снижение его концентрации ком-
пенсируется увеличением в красных кровяных клетках содержания
2,3-дифосфоглицерата (2,3-ДФГ). Способность крови доставлять кис-
лород к мышцам под влиянием 2,3-ДФГ повышается, поскольку это ве-
щество, являющееся интермедиатом гликолиза, снижает сродство ге-
моглобина к кислороду. Меньшая концентрация эритроцитов в крови
выносливых спортсменов снижает вязкость крови, снижая тем самым
сопротивление току крови через кровеносные сосуды и облегчая рабо-
ту сердца. Таким образом, имеются убедительные доказательства, что
доставка кислорода к работающим мышцам увеличивается под влия-
нием тренировки, направленной на развитие выносливости. Возраста-
ние доставки кислорода при выполнении физических упражнений вы-
сокой интенсивности сопряжено с повышением его экстракции в тре-
нированном состоянии, а также с увеличением окислительной способ-
ности мышечной ткани, что позволяет вносить больший вклад в про-
дукцию АТФ за счет окислительного фосфорилирования и использова-
ния жирных кислот.
235
5. Продолжительность периода, необходимого
для протекания адаптационных изменений в организме
при тренировочных занятиях, направленных на развитие
выносливости, а также для детренировки
Продолжительность периода адаптации к тренировке зависит в основ-
ном от величины тренировочных нагрузок (их интенсивности, продол-
жительности и частоты) и является специфичной для мышц, задейство-
ванных в сократительной активности. Тренировочные занятия должны
проводиться на протяжении нескольких недель или месяцев, чтобы про-
исходящие под ее влиянием процессы специфической биохимической
адаптации позволили мышцам перейти на более высокий функциональ-
ный уровень (рис. 8.3). На содержание митохондрий влияет сочетание
интенсивности физических нагрузок с их продолжительностью; пик
адаптационных изменений содержания митохондрий проявляется, по-
видимому, при сокращении времени выполнения упражнений и увели-
чении их интенсивности примерно от 40 до 90 % VO2max. Доказатель-
ством благоприятного влияния реализации пролонгированной трениро-
вочной программы на повышение способности к проявлению выносли-
вости могут служить прежде всего адаптационные изменения функцио-
Проявление
выносливости
0 15 30
Дни
Рис. 8.11. Влияние 15-дневной
детренировки и последующего
возобновления шестью бегуна-
ми в течение 15 дней трениро-
вочных занятий на активность
СДГ, а также ЛДГ на проявление
выносливости и VO2max: детре-
нировка (а) вызвала значитель-
ное снижение всех параметров к
15-му дню; 15-дневный период
возобновления тренировочных
занятий (б) не восстановил изме-
ненные показатели до первона-
чального уровня (Хоустон и др.
(Houston et al.), 1979)
236
Рис. 8.12. Продолжительность
периода адаптационных измене-
ний плотности митохондрий в
скелетных мышцах в процессе
тренировки и детренировки
нального состояния сердечно-сосудистой системы, объема крови и ба-
ланса жидкости в организме, а не специфические адаптационные изме-
нения окислительной способности мышц.
Продолжительность периода изменений субстратной утилизации во
время выполнения физических упражнений, происходящих под влияни-
ем тренировки, тесно связана с периодом возрастания активности мито-
хондриальных ферментов, что отмечалось в большинстве исследований
при изучении эффектов как лонгитудинальной физической тренировки,
так и детренировки. Однако, как показали результаты работ, опублико-
ванных в последние годы, после пяти-семи дней тренировки активность
митохондриальных ферментов или их окислительная способность не из-
меняется. Вполне возможно, что на первых этапах тренировочного про-
цесса не повышение метаболической способности митохондрий, а дру-
гие механизмы ответственны за происходящие под влиянием трениров-
ки изменения в использовании субстратов во время мышечной работы.
К таким факторам может относиться раннее изменение гормонального
ответа на физическую нагрузку: имеются доказательства, что уже по ис-
течении нескольких дней тренировочных занятий катехоламиновая ре-
акция на физическую нагрузку значительно ослабевает. Так, например,
отмечалось, что только после одной недели тренировки концентрация
адреналина в плазме снизилась на 40 %, а концентрация норадреналина
во время выполнения физического упражнения с мощностью, эквива-
лентной 70 % VO2max, снизилась на 25 %.
Происходящие под влиянием тренировки на выносливость адапта-
ционные изменения могут поддерживаться только при систематическом
занятии физическими упражнениями. В течение периода прекращения
тренировочных занятий приобретенные адаптационные изменения ут-
рачиваются (рис. 8.3 и 8.11). Некоторые изменения могут возвратиться к
исходному уровню за очень короткое время. Так, например, около 50 %
237
прироста количества митохондрий под влиянием тренировки может
быть потеряно в течение 1 месяца детренировки (рис. 8.12). Возобнов-
ление тренировки позволяет восстановить адаптационные изменения,
однако времени, необходимого для возвращения прежнего уровня тре-
нированности, потребуется больше, чем потребовалось бы для достиже-
ния эффекта детренировки.
6. Гормональная адаптация
при тренировке на выносливость
Повышение окислительной способности мышц не является един-
ственным способом, благодаря которому происходит модификация суб-
стратного метаболизма под влиянием тренировки. Нейроэндокринные
ответы играют важную роль в регуляции мобилизации и утилизации
энергетических субстратов во время мышечной работы, в частности в
тех случаях, когда выполняется она, по крайней мере, в течение несколь-
ких минут. В основном, гормональные ответы на физическую нагрузку
под влиянием тренировки значительно ослабевают. Так, например, во
время выполнения физических упражнений с одинаковой абсолютной и
относительной интенсивностью в состоянии тренированности отмечает-
ся меньшее увеличение в плазме концентрации адреналина, чем до на-
чала реализации тренировочной программы. Активность симпатическо-
го отдела вегетативной нервной системы, о которой можно судить по
концентрации норадреналина в плазме, при выполнении одинаковой
мышечной работы абсолютной мощности под влиянием тренировки
снижается, оставаясь, однако, неизменной при работе одинаковой отно-
сительной мощности. Уровни адренокортикотропного гормона, кортизо-
ла, глюкагона и гормона роста во время выполнения физических упраж-
нений у тренированных лиц повышаются в меньшей степени. Концен-
трация инсулина при напряженной мышечной работе обычно падает.
Однако в состоянии тренированности это снижение менее выражено, в
связи с чем концентрация инсулина у тренированных лиц во время мы-
шечной активности проявляет тенденцию к более высоким показателям
по сравнению с нетренированными. Возможно, этот эффект тренировки
отражает, в частности, менее выраженную степень угнетения инсулино-
вой секреции под влиянием меньшей концентрации адреналина в плаз-
ме тренированных лиц во время выполнения физических упражнений.
Происходящие под влиянием тренировки указанные изменения час-
тично могут быть обусловлены увеличением VO2max, однако это не яв-
238
ляется единственной причиной, поскольку ослабление большинства гор-
мональных ответов доказано также и для тех случаев, когда тренирован-
ные испытуемые выполняли физические упражнения с одинаковой от-
носительной мощностью до возобновления тренировок (или же в срав-
нении с контрольной группой испытуемых, ведущих малоподвижный
образ жизни). Кроме того, эффекты тренировки проявлялись только тог-
да, когда в исследованиях использовались такие же физические упраж-
нения, что и в тренировочной программе, даже несмотря на то, что
VO2max могло быть одинаковым при различных условиях. У спортсме-
нов высокой квалификации в течение периода детренировки, продол-
жавшегося много недель и приведшего к снижению VO2max, катехола-
миновый ответ при выполнении физических упражнений субмаксималь-
ной аэробной мощности не изменялся. Следовательно, эти адаптаци-
онные изменения в гормональном ответе на физические нагрузки пред-
ставляют собой специфическую реакцию на тренировку и не являются
только отражением влияния сопутствующего изменения VO2max.
Физиологическая роль некоторых из этих нейроэндокринных адап-
тационных изменений достаточно изучена. Так, например, более низкие
уровни кортизола и катехоламинов должны указывать на снижение чрез-
мерного стресса, снижение ЧСС. Однако интерпретация метаболичес-
ких последствий гормональных изменений, происходящих под влияни-
ем тренировки, более затруднительна.
Проявляющееся под влиянием тренировки замедление скорости
мышечного гликогенолиза отчасти может быть обусловлено снижением
концентрации в плазме адреналина. Последнее наряду с ослаблением
снижения в плазме инсулина, по-видимому, также вносит свой вклад в
снижение скорости продукции глюкозы в печени и в менее выраженный
липолиз в адипозной ткани, которые происходят под влиянием трени-
ровки. Изложенные выводы предполагают в основном отсутствие изме-
нений в чувствительности тканей к гормональным воздействиям, одна-
ко это не всегда соответствует действительности. Например, липолити-
ческая активность адипозной ткани крыс под влиянием тренировки ста-
новится более чувствительной к влияниям адреналина.
Инсулин, хотя и является потенциальным ингибитором липолиза,
тем не менее его высокие уровни во время мышечной работы в состоя-
нии тренированности могут ослаблять освобождение СЖК из адипозной
ткани. Возможно также, что столь высокая концентрация инсулина при
физической нагрузке будет способствовать дополнительному поступле-
нию глюкозы в скелетные мышцы. Однако известно, что под влиянием
тренировки использование глюкозы плазмы во время мышечной работы
снижается. Кроме того, уровень инсулина в плазме, даже после выполне-
239
ния тренировочной программы, при физической нагрузке достаточно
низкий и, как показано выше, у человека около 85 % возрастающего пот-
ребления глюкозы во время мышечной работы обусловлено не инсу-
линопосредованными механизмами. Таким образом, отмечаемая под вли-
янием тренировки относительно более высокая концентрация инсулина
во время выполнения физического упражнения, возможно, имеет боль-
шее значение для угнетения липолиза и продукции глюкозы в печени,
чем для его предполагаемого влияния на утилизацию глюкозы мышцами.
Следует отметить, что ни одно из указанных эндокринных измене-
ний не может удовлетворительно объяснить повышение внутримышеч-
ного липолиза под влиянием тренировки. Теоретически он может возрас-
тать, если чувствительность скелетных мышц к адреналину увеличива-
ется. Однако, как свидетельствуют результаты определения плотности Ь2-
адренорецепторов в мышцах человека, этот показатель под влиянием
тренировки на выносливость не изменяется. Тот факт, что внутримышеч-
ный гликогенолиз, который стимулируется через /уадренорецепторы, в
тренированных мышцах при выполнении физических упражнений про-
текает менее активно, также является аргументом против общего увели-
чения в скелетных мышцах чувствительности к действию адреналина.
7. Адаптация к спринтерской
и силовой тренировке
Тренировочные занятия, направленные на развитие силы, мощности или
скорости, оказывает незначительное влияние (или не оказывает вообще)
на аэробные возможности и вызывает относительно небольшие адапта-
ционные изменения в сердечно-сосудистой системе. Это находится в со-
ответствии с принципом специфичности спортивной тренировки. Отно-
сительно кратковременные физические нагрузки с отягощениями либо
спринт, которые требуют проявления высокого уровня анаэробного ме-
таболизма, вызывают специфические изменения в немедленной (АТФ и
КФ) и короткоотставленной (гликолиз) системах энергообеспечения,
улучшают спринтерские способности. К последнему относится увели-
чение максимальной мощности мышечных сокращений, количества
производимой за короткий промежуток времени интенсивной работы, а
также увеличение продолжительности выполнения (выносливости) вы-
сокоинтенсивных физических упражнений.
Анаэробные возможности можно оценить непрямым способом пу-
тем определения величины кислородного долга после бега в течение
240
Адаптация мышц к силовой тренировке
Гипертрофия мышечных волокон
Увеличение площади анатомического поперечника
Повышение содержания КФ и гликогена
Повышение гликолитической способности
Повышение силы и способности к выполнению физических упражнений высокой
интенсивности
Снижение митохондриальной плотности
Улучшение буферных свойств мышц
2-3 мин бега на тредмиле до отказа. Результаты таких исследований по-
казали, что спринтерская тренировка (три занятия в неделю на протяже-
нии 6 недель) способствовала увеличению анаэробных возможностей
испытуемых на 10 %.
Кратковременная высокоинтенсивная тренировка оказывала незна-
чительное влияние (или совсем не оказывала влияния) на композицион-
ный состав мышечных волокон. Только в одной из работ сообщалось о
значительном увеличении количества волокон типа II (с 32 до 38 %) и
снижении количества волокон типа I (с 57 до 48 %) после 6 недель
спринтерской тренировки в езде на велосипеде. Изучению эффектов
спринтерской тренировки посвящено относительно небольшое число
работ, однако приведенные в них данные свидетельствуют о том, что под
ее влиянием заметной гипертрофии мышц не происходит. Возможно, это
связано с коротким периодом проведения таких исследований (обычно
6-8 недель). Известно, что гипертрофия волокон (в частности, типа II)
происходит при реализации более продолжительной тяжелоатлетичес-
кой тренировочной программы. Диаметр волокон в мышцах тяжелоатле-
тов и представителей бодибилдинга больший по сравнению с лицами,
ведущими малоподвижный образ жизни. Результаты лонгитудинальных
исследований свидетельствуют о том, что площадь поперечного сечения
волокон типа II после нескольких месяцев тяжелоатлетической трени-
ровки может увеличиться на 50 %. В экспериментах, проведенных на
животных, у которых мышечную гипертрофию вызывали путем хирур-
гического удаления мышц-синергистов, подтвердилась гипертрофия от-
дельных волокон, однако общее их количество в адаптированной мыш-
це не изменилось. Посмертные исследования мышц спортсменов, зани-
мавшихся силовыми видами спорта, также указывают на то, что мышеч-
ных волокон у них было не больше, чем у нетренированных лиц.
Анализ мышечных биоптатов, полученных до и после тренировки с
отягощениями, которая способствовала увеличению мышечной силы на
16
0-313
241
28 %, показал увеличение внутримышечной концентрации АТФ, КФ и
гликогена соответственно на 5, 10 и 10-30 %. Однако эти величины яв-
ляются средними показателями концентраций в образцах мышечных го-
могенатов и могут просто отражать относительное их увеличение в во-
локнах типа II, хотя, согласно имеющимся данным, эти волокна в состо-
янии покоя имеют более высокие концентрации фосфагена и гликогена
по сравнению с волокнами типа I. В ряде исследований не удалось обна-
ружить в состоянии покоя изменений внутримышечных концентраций
АТФ и КФ после 4-8 недель спринтерской тренировки, а также происхо-
дящей под ее влиянием существенной мышечной гипертрофии. После
такого типа тренировки запасы мышечного гликогена также повыша-
лись, что не удивительно, поскольку гликоген служит важным энергети-
ческим источником, используемым при повторных выполнениях сприн-
терских упражнений.
В случае, когда в результате спринтерской тренировки беговые
спринтерские возможности улучшались, то это сопровождалось увели-
чением обращаемости АТФ благодаря повышению вклада анаэробного
гликолиза в энергообеспечение. Количество и активность ферментов,
задействованных в гликолитическом пути (например, фосфофрукто ки-
наза), постоянно проявляют тенденцию к возрастанию под влиянием
как спринтерской, так и силовой тренировки (рис. 8.13) с наиболее вы-
раженными изменениями в волокнах типа II. Однако степень этих изме-
нений не настолько велика, как она проявляется в отношении окисли-
тельных ферментов при аэробной тренировке, направленной на разви-
тие выносливости. Так, например, в одной из работ сообщалось, что ак-
тивность ЛДГ в m. vastus lateralis у тяжелоатлетов высокой квалифика-
ции в волокнах типа II была на 62 % выше по сравнению с мужчинами,
ведущими малоподвижный образ жизни; активность этого фермента в
волокнах типа I также оказалась на 50 % более высокой по сравнению
с контролем.
Различий в миокиназной активности в волокнах типа I между тяже-
лоатлетами и испытуемыми контрольной группы отмечено не было, тог-
да как в волокнах типа II у первых она оказалась на 40 % выше. В отно-
шении изменений, касающихся аэробных (митохондриальных) фермен-
тов, то, как правило, отмечается значительная гипертрофия волокон, в
которых происходит снижение активности окислительных энзимов и
цитохромов, связаное, вероятно, с увеличением площади поперечного
сечения мышечных клеток (преимущественно волокон типа II) без адап-
тивного повышения количества митохондрий. У тренирующихся в ви-
дах спорта, требующих проявления силовых возможностей, количество
капилляров может оставаться неизменным, однако большая их поверх-
242
Г~|ффк
□ Малатдегидрогеназа
Рис. 8.13. Изменения активности ферментов в различных типах мышечных волокон
под влиянием спринтерской тренировки: * — достоверность изменений; Р < 0,05 (Таке-
кура и Йошиока (Takekura and Yoshioka), 1990)
ность между крупными мышечными волокнами обусловливает сниже-
ние капиллярной плотности, приходящейся на единицу площади сече-
ния (см. рис. 8.1).
Под влиянием тренировочных занятий анаэробной направленности
при выполнении физических упражнений максимальной интенсивности
концентрация лактата в крови может достигать более высоких значений,
что связано, очевидно, с более высоким содержанием внутримышечно-
го гликогена и ферментов гликолиза. Напряженная тренировка требует
значительной мотивации и толерантности к болевым ощущениям, воз-
никающим в результате метаболического ацидоза из-за повышения
уровня лактата в крови.
Повышение способности мышц к буферированию протонов, накап-
ливающихся в связи с накоплением лактата, также может иметь немало-
важное значение. Волокна типа II характеризуются высокими буферными
возможностями, поэтому их увеличение по сравнению с волокнами типа
I должно указывать на повышение этой способности. Несмотря на недос-
таточное количество доказательств значительного увеличения в мышцах
физико-химического буферирования, которое оценивали титрованием
после выполнения тренировочных программ, все же можно полагать, что
16*
243
если буферная способность рассчитывается на основании показателей pH
и содержания лактата, определяемых после физической нагрузки, то ее
показатель под влиянием спринтерской тренировки возрастает. Два ком-
понента буферной способности, которые относятся к бикарбонатной сис-
теме, не определяются методом титрования: это прежде всего буфер во
внутриклеточной жидкости и трансмембранный поток водородных ионов.
Последнее может служить объяснением для обнаруженного Невил с соав-
торами (Nevil et al., 1989) того факта, что после окончания мышечной ра-
боты физико-химический компонент буферирования и pH были одинако-
выми как до, так и после реализации спринтерской тренировочной прог-
раммы, вопреки тому, что послерабочая концентрация лактата в мышцах
под влиянием тренировки повышалась примерно на 20 %.
Повышение мышечной силы, по крайней мере в течение первых
нескольких недель тренировочных занятий, направленных на развитие
силовых возможностей, способствовало нейральному облегчению (рас-
торможению), приводящему, в свою очередь, к полной активации двига-
тельных единиц и мышечных групп. Первоначальный быстрый прирост
силы, отмечаемый на первых этапах тренировочного процесса, оказыва-
ется не связанным с увеличением размеров мышц и площади их физио-
логического поперечника.
Более продолжительная и напряженная тренировочная программа,
направленная на развитие силовых возможностей, приводит к гипертро-
фии мышц и дальнейшему приросту их силы. Последний эффект весьма
важен, поскольку в работающих мышцах он способствует снижению доли
проявления их максимальной сократительной активности при физической
нагрузке. Увеличение мышечной массы означает, что большее количество
мышечной ткани задействовано в выполнении работы, в результате чего
повышаются предельная мощность последней и общая энергопродукция
анаэробных систем. Физиологические и биохимические изменения, проис-
ходящие в мышцах при проявлении этих эффектов, к настоящему времени
еще недостаточно изучены. Хотя метаболические изменения, происходя-
щие в организме под влиянием аэробной тренировки, направленной на раз-
витие выносливости, достаточно широко изучены в лабораторных услови-
ях, тем не менее относительно мало работ посвящено исследованиям био-
химических изменений при анаэробной тренировке.
Следует учитывать, что эти эффекты специфичны для мышц, за-
действованных в реализации тренировочной программы, особенно для
отдельных типов мышечных волокон, рекруитированных в выполне-
ние физических упражнений. Вполне очевидно, что очень высокоин-
тенсивные тренировочные занятия требуют активации волокон типа
Пб, однако в настоящее время мы не располагаем достаточными дока-
244
зательствами, предоставляемыми спортивной наукой, чтобы давать ре-
комендации относительно интенсивности, частоты и продолжитель-
ности тренировочных воздействий, которые могли бы оптимизировать
адаптационные процессы.
8. Механизмы мышечной адаптации
к тренировке
На основании изложенных выше фактов возникают два фундаменталь-
ных вопроса относительно того, каким образом происходит адаптация
мышц к тренировочным воздействиям. Первый вопрос — что собой
представляет истинная природа стимулов (или стимула), индуцирую-
щих адаптацию? Второй вопрос — что служит молекулярным механиз-
мом, позволяющим ей совершаться? Эти вопросы начали выясняться
благодаря последним достижениям молекулярной биологии примени-
тельно к скелетным мышцам. Поскольку глубокий анализ относительно
этого не представлен в данной книге, далее проведено краткое обоб-
щение последних научных данных.
Вполне реально, что генетически обусловленные особенности мио-
зина являются ключом к пониманию мышечной пластичности. Сущес-
твует примерно семь различных версий, объясняющих наследственную
природу мышечных волокон. Теоретически мышечные клетки могут из-
менять свои сократительные свойства благодаря перестройке их мио-
фибрилл, используя для этого различные типы тяжелой цепи миозина.
Быстросокращающиеся мышечные волокна могут стать медленносокра-
щающимися в результате выключения гена, ответственного за наличие
быстромиозиновой тяжелой цепи, и включения гена, ответственного за
наличие медленной изоформы белка. Большинство генов в клетках орга-
низма включаются и выключаются непрямым воздействием таких сиг-
нальных молекул, как гормоны или факторы роста.
Протекающие под влиянием тренировки адаптационные процессы
специфичны для активных мышц; в неиспользуемых мышцах адаптация
не происходит. Таким образом, создается впечатление, что мышечные
гены регулируются преимущественно механическими и (или) метаболи-
ческими стимулами. Растяжение или сокращение мышечного волокна
воздействует на специфические гены, включая их или выключая.
Активность различных миозиновых генов может быть определена с
помощью применения ДНК-зондов. Результаты проведенных на живот-
ных исследований показали, что одиночное растяжение либо одиночное
245
сокращение, вызываемое электрическими стимулами, в быстросокраща-
ющейся мышце (например, в m. tibialis anterior) лишь незначительно
влияют на активность миозиновых генов. Однако совместное воздей-
ствие этих стимулов вызывает выраженное снижение синтеза быстроми-
озиновой тяжелой цепи и включение преимущественно медленной вер-
сии миозина.
Иммобилизация мышцы (m. soleus) у кролика медленносокращаю-
щимися волокнами возвратила быстромиозиновый ген в прежнее состо-
яние. Следовательно, вполне возможно, что быстрая форма миозина яв-
ляется «недостающим факультативом». Для медленносокращающейся
мышцы, как, например, m. soleus, с целью поддержания синтеза медлен-
номиозиновой тяжелой цепи требуются повторяющиеся растяжения.
Оба указанные проявления согласуются с нашими знаниями о влиянии
тренировок различной направленности на мышцы человека.
Растяжение мышечных волокон во время выполнения физических
упражнений является одним из возможных стимулов для адаптации. Из-
вестно, что пассивное растяжение способствует увеличению размеров
мышцы даже в отсутствие иннервации, влияний инсулина, гормона рос-
та либо адекватного питания. Передача механических сил через клеточ-
ный каркас к ядру и полирибосомам мышечных волокон может осущес-
твляться либо непосредственно, либо через мембранносвязанные, акти-
вирующиеся при растяжении ионные каналы или индуцируемые растя-
жением изменения в плазме мембранносвязанных молекул (например,
механочувствительные изоформы аденилатциклазы). Эти вызванные
растяжением сигналы могут впоследствии обусловить изменение ген-
ной экспрессии (например, мышечных факторов роста), а также изме-
нить скорость синтеза и распада белков.
Увеличение уровня цАМФ или скорости метаболизма предположи-
тельно может служить сигналом для повышения митохондриального
биогенеза под влиянием тренировки. При продолжительной электрости-
муляции скелетных мышц между повышением уровней аденилатцик-
лазной активности, концентрации цАМФ и количеством молекул мРНК
для митохондриальных белков формируется взаимосвязь. Увеличение
скорости метаболизма может сигнализироваться возрастанием соотно-
шения АДФ/АТФ либо снижением концентрации КФ.
Повреждение мышечных волокон во время выполнения физических
упражнений могут также служить стимулом для адаптации. Высокие
концентрации специфического для мышц фактора роста, освобождаю-
щегося при повреждениях, дегенерация мышечных волокон вместе с ут-
ратой контактного ингибирования между клеточными саттелитами и
жизненными процессами, протекающими внутри мышечных волокон,
246
вызывают пролиферацию клеточных саттелитов в течение первого дня
после травмирования мышцы. В связи с этим происходящие под влияни-
ем физических нагрузок изменения в миофибриллах состояния Z-линий,
клеточного каркаса или внеклеточного матрикса стимулируют пролифе-
рацию и объединение клеточных саттелитов, что приводит к гипертро-
фии. Вновь рекруитированные саттелитные клетки должны обеспечить
дополнительную потребность ядер для поддержания их плотности в ги-
пертрофированной мышце. Результаты многочисленных исследований
гибридизации ДНК свидетельствуют о депрессии регуляторного факто-
ра в ядрах мышечного волокна во время регенерации последнего и лока-
лизации мРНК тяжелых цепей миозина в фокальных плоскостях пов-
реждения, вызванного чрезмерным растяжением мышцы.
Изменения в экспрессии различных изоформ миозина, а также
изоформ других сократительных белков происходят во время явных
трансформаций типов волокон, вызываемых перекрестной иннервацией
мышц путем хирургического вмешательства, а также хронической элек-
тростимуляцией и иммибилизационной атрофией. Уровни тироидных
гормонов и гормона роста влияют на такие изменения тяжелоцепочных
форм миозина в скелетных мышцах в условиях изменения их сократи-
тельной активности. Специфическая для мышц белковая адаптация ха-
рактеризуется постепенными изменениями в группах белков. Так, нап-
ример, для специфических групп генов изменения проявляются в следу-
ющей последовательности: первым происходит изменение кальцийсвя-
зывающих белков, затем — митохондриальных; в последнюю очередь
изменяется состояние сократительных белковых структур. Такая же пос-
ледовательность изменений, очевидно, имеет место при превращении
как медленносокращающихся мышечных волокон в быстросокращаю-
щиеся под влиянием физических тренировок силовой направленности,
так и быстросокращающихся волокон в медленносокращающиеся в ре-
зультате пролонгированной электростимуляции.
9. Адаптационные изменения в крови
уровня липидов, происходящие под влиянием
тренировки
В ряде работ было показано, что систематическая тренировка, направ-
ленная на развитие выносливости, способствует снижению в крови кон-
центрации холестерола, триацилглицеролов и ЛПНП, а также увели-
чению концентрации ЛПВП (рис. 8.14). Как отмечалось в главе 4,
ЛПВП-холестерол считается протектором, предупреждающим образова-
247
Холестерин Холестерин
низкой плотности высокой плотности
| | Нетренированные
Рис. 8.14. Липидный профиль крови нетре-
нированных и тренированных (в среднем на-
бегавших 62 км в неделю) мужчин среднего
возраста (Вуд и др. (Wood et al.), 1976)
ние атеросклеротических бля-
шек и развитие коронарной бо-
лезни, тогда как высокий
уровень ЛПНП-холестерола
способствует проявлению этих
нарушений. Известно, что куре-
ние, нерациональное питание,
потребление алкоголя, наслед-
ственность, ожирение и диабет
также являются важными факто-
рами и не все исследования воз-
действия физическими нагруз-
ками не дают четкого представ-
ления о взаимосвязи мышечной
деятельности и физической под-
готовленности с уровнем липи-
дов в крови. Полученные науч-
ные данные подтверждают, что
у лиц с низкими уровнями плаз-
менного холестерола, ЛПВП и
триацилглицеролов проявляют-
ся благоприятные изменения
этих показателей под влиянием
тренировки на выносливость.
Результаты большого количес-
тва исследований указывают на
возникновение обратной взаимосвязи между уровнем функционального
состояния кардиореспираторной системы (продолжительность выпол-
нения мышечной работы или VO2 шах) и концентрацией в плазме три-
ацилглицеролов. Следует отметить, что это не взаимосвязано с прояв-
лением мышечной силы, поэтому тренировка с отягощениями может не
ассоциироваться с отмеченным выше благоприятным влиянием на
здоровье.
Спортсмены, тренирующиеся в видах спорта, требующих прояв-
ления выносливости, демонстрируют высокие показатели триацил-
глицерольного клиренса из плазмы по сравнению с нетренированными
лицами. Этот факт может быть обусловлен повышением в мышцах
ЛПЛ-активности и плотности капилляров под влиянием тренировки, ко-
торая может повысить уровень клиренса ЛПОНП-триацилглицерола из
крови. Степень триацилглицерольной деградации сопряжена со скорос-
тью синтеза ЛОНП, в связи с чем именно повышением метаболических
248
возможностей обмена триацилглицеролов можно объяснить более высо-
кое содержание в плазме ЛПВП-холестерола у тренированных лиц.
В восстановительном периоде после пролонгированной напряжен-
ной физической нагрузки (например, после марафонского бега) концен-
трация триацилглицеролов в плазме снижается, а уровень их клиренса
повышается. Результаты проведенных в последнее время исследований
свидетельствуют о том, что менее интенсивные физические нагрузки
(например, прогулка в горах) способствует ослаблению липемического
ответа на прием жирной пищи, когда физическое упражнение выполня-
лось сразу либо через несколько часов после еды. Эффект мышечной ра-
боты становился более выраженным, когда она совершалась с мощнос-
тью, эквивалентной 60 % VO2max по сравнению с мощностью, соответ-
ствующей 30 % VO2max, однако это проявление обусловлено скорее вы-
сокими энерготратами, чем интенсивностью нагрузки. Для менее выра-
женного увеличения в крови концентрации триацилглицеролов через
16 часов после приема жирной пищи требуется дополнительный расход
энергии (около 30 МДж), и этот эффект в большей мере проявляется у
тренированных лиц. Такое влияние обусловлено, по-видимому, повыше-
нием клиренса триацилглицеролов из крови для восстановления их за-
пасов, используемых во время мышечной работы.
10. Иммуносупрессия, связанная
с напряженными тренировочными нагрузками
Спортсмены, выполняющие напряженную тренировочную программу, в
частности программу, направленную не развитие выносливости, прояв-
ляют более высокую предрасположенность к инфекционным заболева-
ниям. Так, например, ангино- и гриппоподобные симптомы более прису-
щи спортсменам, чем основной популяции, и однажды инфицирован-
ный спортсмен более продолжительное время может находиться в прос-
тудном состоянии. Имеется несколько убедительных доказательств, что
такое повышение предрасположенности к инфекционным заболеваниям
обусловлено угнетением функции иммунной системы.
Основным компонентом иммунной системы являются белые кровя-
ные клетки (лейкоциты), число и функциональные способности которых
могут снижаться под влиянием повторяющихся интенсивных и пролон-
гированных физических нагрузок. Причины такого эффекта оконча-
тельно не изучены, однако предполагается, что связаны они с увеличе-
нием в крови уровня стресс-гормонов во время выполнения напряжен-
249
ной мышечной работы. Результаты ряда последних исследований дают
основание полагать, что падение в крови концентрации глутамина как
аминокислоты, необходимой для оптимального функционирования лей-
коцитов, может также вовлекаться в проявление иммуносупрессии, вы-
зываемой напряженными физическими нагрузками. Одним из таких
факторов может быть и мышечное повреждение.
Выполнение напряженной мышечной работы сопровождается
эффектами, которые во многом аналогичны эффектам, вызываемым ин-
фекцией: существенное увеличение в крови количества лейкоцитов
(преимущественно лимфоцитов и нейтрофилов) под влиянием как ин-
тенсивных кратковременных, так и продолжительных физических наг-
рузок. Наряду с этим в плазме также возрастает концентрация различ-
ных субстанций, влияющих на функции лейкоцитов, включая фактор
некроза опухоли, интерлейкины 1, 2 и 6, а также такие белки острой фа-
зы, как С-реактивный белок и комплементактивирующие фрагменты.
Под влиянием напряженной мышечной работы происходят и гормональ-
ные изменения, к которым относится увеличение в плазме концентрации
нескольких гормонов (например, адреналина, кортизола, гормона роста
и пролактина), которые, как известно, обладают иммуномодулирующи-
ми эффектами. Значительная физическая нагрузка приводит к временно-
му повышению фагоцитарной активности нейтрофилов и макрофагов, а
также к возрастанию литической активности клеток — естественных
киллеров (NK), однако при этом отмечается ослабление пролифератив-
ного ответа лимфоцитов по отношению к митогенам.
В восстановительном периоде после физической нагрузки количес-
тво NK и их активность падают ниже дорабочего уровня. Если мышеч-
ная деятельность была очень напряженной, то после ее завершения ко-
личество лейкоцитов в крови может оставаться ниже исходного показа-
теля на протяжении нескольких часов, а Т-лимфоцитное CD4+/CD8+
(хелпер/супрессорное) соотношение при этом также снижается. После
длительной и напряженной физической нагрузки выработка иммуногло-
булинов (антител) В-лимфоцитами угнетается. Эти изменения, проявля-
ющиеся в ближайшем восстановительном периоде, ослабляют потенци-
альный иммунный ответ организма на воздействие патогенных факто-
ров, в связи с чем они могут представлять собой «открытое окно» для
инфекций и обусловливать наиболее уязвимый период для спортсменов
из-за повышения предрасположенности к их приобретению.
Физическая тренировка также модифицирует иммунную систему
путем снижения ее общего функционального состояния, особенно в ус-
ловиях напряженных тренировочных воздействий. Количество содержа-
щихся в крови лейкоцитов у спортсменов в состоянии покоя обычно ни-
250
же по сравнению с людьми, ведущими малоподвижный образ жизни, а
увеличение их концентрации во время выполнения физических упраж-
нений (одинаковой абсолютной или относительной мощности) под вли-
янием тренировки происходит в меньшей степени. Как показали резуль-
таты исследования тренированных велосипедистов, фагоцитарная ак-
тивность нейтрофилов в крови оказалась заметно ниже по сравнению с
контрольной группой нетренированных мужчин такого же возраста и с
такой же массой тела. Уровень секреции иммуноглобулинов (например,
слюнного IgA) у хорошо тренированных лиц ниже, как и ниже Т-лимфо-
цитарное CD4+/CD8+ соотношение и in vitro митогенстимулируемая
лимфоцитарная пролиферация.
Известны возможные варианты ослабления иммунной функции,
обусловленные напряженными тренировками. Один из механизмов мо-
жет быть просто связан с кумулятивными эффектами повторяющихся
физических нагрузок, сопровождающихся повышенным выделением
гормонов, в частности таких, как глюкокортикоиды, которые вызывают
временную иммуносупрессию. Если физические нагрузки повторяются
часто, то время, необходимое для полного восстановления иммунной
системы, может оказаться недостаточным. Кроме того, под влиянием
повторяющихся физических упражнений, вызывающих мышечные пов-
реждения, уровень кортизола в плазме в течение нескольких дней может
оставаться завышенным.
Концентрация глутамина в плазме после физических нагрузок мо-
жет подвергаться значительным изменениям. Высокоинтенсивная ин-
тервальная тренировка существенно снижает его уровень, а повторяю-
щиеся напряженные тренировочные воздействия способствуют поддер-
жанию в плазме низкого содержания глутамина. Глутамин весьма важен
для проявления нескольких функций белых кровяных клеток, включая
их способность к делению, выработке антител и уничтожению бактерий
путем их поглощения и переваривания. Скелетные мышцы являются ос-
новным источником глутамина, выделяемого в кровяное русло, и этот
выход может играть важную роль в доставке глутамина к клеткам им-
мунной системы. Известно, что кроме физических нагрузок другие фор-
мы стресса, как, например, травмы, хирургические операции и инфек-
ция, также вызывают снижение уровня глутамина в плазме, что может
способствовать угнетению функции иммунной системы в связи с этими
стрессовыми состояниями.
Комплементарная активация также имеет место при выполнении
физических упражнений, и уменьшение комплементарной концентра-
ции в плазме при повторных физических нагрузках, сопровождающихся
мышечными повреждениями, может вносить свой вклад в ослабление
251
неспецифического иммунитета у спортсменов. У хорошо тренирован-
ных спортсменов комплементарная концентрация в плазме по сравне-
нию с контрольной группой нетренированных испытуемых имеет мень-
шие показатели.
11. Перетренировка
Спортсмены могут испытывать ущерб от перетренированное™ состоя-
ния, проявляющегося в нарушении в процессе тренировки достигнутого
уровня функциональной готовности несмотря на продолжающиеся и да-
же возрастающие тренировочные воздействия. Хотя улучшение спор-
тивной работоспособности зависит от повышения тренировочных наг-
рузок (или применения больших нагрузок), все же перетренировка —
порочный итог чрезмерных тренировочных воздействий, приводящих к
снижению работоспособности и хроническому утомлению — является
проявлением стресс-реакции на очень напряженные и частые трениров-
ки с недостаточным временем отдыха между ними, необходимого для
восстановления.
Причины, по которым некоторые спортсмены становятся перетре-
нированными, до конца не выяснены и последовательно ранжируются
от изменения функций мышц до изменения мотиваций. К патофизиоло-
гии перетренировки относятся мышечные боли и слабость, изменения
гормонального статуса и содержания гемоглобина в крови, изменчивое
настроение и психическая депрессия, такие проблемы с питанием, как
потеря аппетита и диаррея. В ряде случаев перетренированность может
быть обусловлена хронической вирусной инфекцией: известно, что не-
которые вирусы обладают способностью инфильтрировать в скелетные
и сердечную мышцы. Выраженное снижение количества лейкоцитов в
крови часто указывает на хроническую вирусную инфекцию. У спорт-
сменов с проявлениями синдрома перетренировки также часто отмеча-
ется иммуносупрессия. К этому может иметь отношение и глутамин
плазмы, поскольку по некоторым данным у перетренированных спорт-
сменов даже в состоянии покоя уровень его занижен и может оставаться
таким же в течение нескольких недель с восстановлением только при
значительном снижении объема тренировочных нагрузок. В другом слу-
чае снижение в плазме концентрации глутамина может быть вызвано ин-
фекцией, что отмечается у больных после перенесенного вирусного
стресса.
Снижение функциональной готовности может явиться результатом
происходящих под влиянием физических упражнений мышечных пов-
252
реждений. Многие спортсмены с диагнозом перетренировки жалова-
лись на ощущение боли в мышцах, которая была причиной снижения си-
лы мышечных сокращений. К последствиям вызванных физическими
нагрузками мышечных повреждений относятся болезненные ощущения
в мышцах и их закрепощенность, снижение уровня мотиваций, более
высокая по сравнению с нормой концентрация лактата в крови и его эк-
скреция при физических нагрузках, а также снижение силы мышечных
сокращений и максимальной мощности динамической работы, которое
может сохраняться 5—10 дней.
Практическим показателем мышечных повреждений у спортсме-
нов, переносящих напряженные тренировочные нагрузки, является по-
вышение в плазме крови мышечных белков (например, миоглобина, кре-
атинкиназы, лактатдегидрогеназы, фрагментов тяжелых цепей миозина).
Поврежденная мышечная ткань может инициировать активацию иммун-
ной системы, поскольку белые кровяные клетки привлекаются к пов-
режденным мышцам для устранения поврежденных волокон и иниции-
рования репаративного процесса. Однако повышение в крови уровня та-
ких стрессовых гормонов, как кортизол, может оказать угнетающее вли-
яние на лейкоциты. Эту особенность можно рассматривать в качестве
естественной реакции организма, направленной на предотвращение
чрезмерного повреждения мышц иммунной системой, тем не менее она
также может ослабить иммунный ответ на вторжение бактерий и виру-
сов, что повышает восприимчивость перетренированных спортсменов к
инфекции.
Другим детерминирующим эффектом происходящих при физичес-
ких нагрузках мышечных повреждений является замедление восстанов-
ления запасов гликогена в мышцах. После выполнения пролонгирован-
ных физических упражнений эти запасы истощаются. В поврежденных
мышцах снижается способность к экстракции глюкозы из крови для ре-
синтеза в них гликогена, в связи с чем при последующих физических
нагрузках следует ожидать меньшего проявления выносливости.
Снижение запасов мышечного гликогена, вызывемое сочетанием
тренировочных воздействий с низкоуглеводным питанием, также связа-
но с падением уровня внутримышечной и плазменной концентрации
глутамина — аминокислоты, значение которой для проявления способ-
ности лейкоцитов обезвреживать бактерии и вирусы ранее уже обсужда-
лось. Таким образом, вызываемые физическими нагрузками мышечные
повреждения могут обусловить снижение уровня спортивной работо-
способности и сделать спортсмена более восприимчивым к инфекцион-
ным заболеваниям.
253
Дополнительное потребление глутамина может способствовать
улучшению восстановления функции лейкоцитов при вызываемом
физическими нагрузками стрессе и перетренировке. Пролонгирован-
ная двигательная активность, голодание, низкоуглеводная диета, ин-
фекция и физические травмы (например, хирургические вмешатель-
ства) всегда взаимосвязаны со снижением в плазме концентрации глу-
тамина. Производимая после хирургических операций внутривенная
инфузия глутамина (0,2 г глутамина на 1 кг массы тела в течение
24 часов) сохраняла его концентрацию в мышцах и плазме, препят-
ствовала послеоперационному снижению белкового стинтеза в ске-
летных мышцах. Глутамин можно также принимать орально, его
1,5 %-ный раствор почти не имеет вкуса. Применение такого глутами-
нового напитка может значительно повысить концентрацию глутами-
на в плазме на несколько часов, поэтому он может быть рекомендован
спортсменам для предупреждения снижения в плазме концентрации
этой аминокислоты.
Адекватное поступление с пищей микронутриентов является
важным для сохранения нормального статуса иммунной системы. Из-
вестно, что дефицит витаминов А, В6 и С, а также минеральных ве-
ществ, включая цинк и железо, приводит к ослаблению иммунитета.
Однако применение их в чрезмерных дозах также таит в себе
опасность. По данным российских ученых, применение растительно-
го экстракта (Eleutherococcus senticosus, ES) снижает количество
случаев инфекционных заболеваний, а по результатам немецких спе-
циалистов, ES способствует увеличению в крови здоровых людей со-
держания лимфоцитов и NK-клеток. Механизм этого эффекта доста-
точно не изучен, однако позволяет предположить, что обусловлен он
ES-стимулируемым увеличением выработки интерферона либо повы-
шением макрофагной активности, поскольку входящие в состав ES
полисахариды могут действовать как неспецифические иммунные
стимуляторы. Для окончательного выяснения возможности примене-
ния спортсменами в качестве пищевых добавок (для поддержания
иммунной системы) ES, глутамина или цинка следует проводить
дальнейшие исследования.
Выводы
1. Существует несколько принципов, определяющих характер и степень
физиологической и метаболической адаптации к физической трениров-
ке, в частности принцип применения больших нагрузок, принцип специ-
254
фичности тренировочных воздействий и принцип обратимости эффек-
тов тренировки.
2. Адаптация к тренировке, направленной на развитие аэробной вы-
носливости, предполагает повышение капиллярной плотности, а также
увеличение количества и размеров митохондрий в тренированных мыш-
цах. Активность ферментов ЦТК и других окислительных энзимов воз-
растает на фоне сопутствующего повышения способности к окислению
липидов и углеводов.
3. Внутримышечные запасы миоглобина, гликогена и триацилглице-
рола под влиянием тренировки, направленной на развитие выносливос-
ти, возрастают.
4. Адаптация сердечно-сосудистой системы при тренировке на вы-
носливость проявляется в увеличении объема крови, ударного объема и
сердечного выброса, а также в повышении артериально-венозной разни-
цы по кислороду.
5. При выполнении физических упражнений выносливые мышцы
проявляют меньшую скорость утилизации углеводов и продукции лакта-
та, тогда как скорость окисления липидов в них более высокая по срав-
нению с нетренированными мышцами. Основным источником происхо-
дящего под влиянием тренировки увеличения окисления липидов явля-
ется внутримышечный триацилглицерол.
6. Тренировка, направленная на развитие выносливости, ос-
лабляет степень гормонального ответа на напряженные физические
нагрузки.
7. Тренировка, направленная на развитие силы, мощности или ско-
рости, оказывает незначительное влияние (или не оказывает вообще) на
аэробные возможности. Напряженные физические нагрузки с отягоще-
ниями либо спринт вызывают специфические изменения в немедленной
(АТФ и КФ) и короткоотставленной (гликолитической) системах энерго-
обеспечения и улучшают спринтерские способности.
8. Проводимая в течение нескольких месяцев напряженная трени-
ровка с отягощениями вызывает гипертрофию мышечных волокон, в
связи с чем увеличивается общая мышечная масса и максимальная мощ-
ность, которую мышца способна развить.
9. Растяжение, сокращение и повреждение мышечных волокон во
время выполнения физических упражнений обеспечивает стимуляцию
адаптационных процессов, включающих изменения экспрессии различ-
ных изоформ миозина.
10. Напряженные тренировочные нагрузки способствуют повыше-
нию восприимчивости спортсменов к инфекциям, что связано с ослаб-
лением функционирования иммунной системы.
255
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
Andersen Р, Henriksson J. (1977). Training induced changes in the subgroups of human
Type II skeletal muscle fibres// Acta Physiol. Scand. — 99. — P. 123-125.
Coggan A.R., Spiner R.J., Kohrt W.M., Holloszy J.O. (1993). Effect of prolonged exercise on
muscle citrate concentration before and after endurance training in men// Am.J.Physiol. —
275,—E215 —E220.
Costill D.L., Daniels J., Evans IV. et al. (1970). Skeletal muscle enzymes and fiber composi-
tion in male and female track athlete// J.Appl.Physiol. — 40. — P. 149-154.
Henriksson J., Chi M.M.-Y., Hintz C.S. et al. (1986). Chronic stimulation of mammalian mus-
cle: changes in enzymes of metabolic pathways// Am.J.Physiol. — 251. — C614-C632.
Holloszy J.O. (1973). Biochemical adaptations to exercise: aerobic metabolism// Exe Sports
Sci. Rey. — 1, —P. 45-71.
Holloszy J.O., Oscai L.B., Don L.J., Mole PA. (1970). Mitochondrial citric accycle and re-
lated enzymes: adaptive response to exercise// Biochem. Biophys. R Commun. — 40. —-
P. 1368-1373.
Houston M.E., Bentzen H., Larsen H. (1979). Interrelationships between skeletal muscle
adaptations and performance as studied by detraining and retraining// Acta Physiol. Scand.
— 105. —P. 163-170.
Hurley В. E, Nemeth P.M., Martin W.H. et al. (1986). Muscle triglyceride utilization during
exercise: effect of training// J.AppL Physiol. — 60. — P. 562-567.
Ingjer E (1979). Effect of endurance training on muscle fibre ATPase activity, capillary sup-
ply and mitochondrial content in man // J.Physiol. — 294. — P. 419-432.
Kiessling K., Piehl K., Lundquist C. (1971). Effect of physical training on ultrastuctural fea-
tures in human skeletal muscle// Muscle metabolism during exercise/ Ed. B.Pernow and
B-Salin. — New York: Plenum Press. — P. 97—101.
Mendenhall L.A., Swanson S.C., Habash D.L., Coggan A.R. (1994). Ten days of exercise
training reduces glucose production and utilization during moderate-intensity exercise//
Am.J.Physiol. — 266. — E136-E143.
Mole P. A.,Oscai L.B., Holloszy J.O. (1971). Adaptation of muscle to exercise. Increase in le-
vels of palmitly CoA synthetase, carnitine palmityl transferase, and palmityl CoA dehy-
drogenase, and the capacity to oxidise fatty acids// J.Clin.Invest. — 50. — P. 2323-2330.
Nevil M.E., Boobis L.H., Brooks S., Williams C. (1989). Effect of training on muscle metabo-
lism during treadmill sprinting// J.Appl. Physiol. — 67. —P. 2376-2382.
Simoneau J.-A., Lortie G., Boulay M.R. et al. (1985). Human skeletal muscle fibre alteration
with high-intensity intermittent training// Eur.J.Appl.Physiol. — 54. — P. 250-253.
Takekura H, Yoshioka T. (1990). Different metabolic responses to exercise training pro-
grammes in single rat muscle fibers// J.Muscle Res.Cell.Motil. — 11. — P. 105-113.
Tesch P.A., Thorsson A., Kaiser P. (1984). Muscle capillary supply and fiber type characteris-
tics in weight and power lifters// J.Appl.Physiol. — 56. — P. 35-38.
Wood P.D., Haskell W., Klein H. et al. (1976). The distribution of plasma lipoproteins in mid-
dle-aged male runners// Metabolism. — 25. — P. 1249-1257.
1риложение 1
ХИМИЧЕСКАЯ СТРУКТУРА
И СВЯЗИ
1. Химическая структура
Несмотря на то что электрические заряды в химических
связях удобно представлять в виде феномена «все-или-ни-
чего», тем не менее это не совсем правильно. Так, напри-
мер, кислород является более отрицательным по сравне-
нию с углеродом и электроны в углерод-кислородных
двойных связях будут находиться относительно дольше по
близости к атому кислорода, чем к атому углерода. Присут-
ствие карбоксильной группы в молекуле обусловливает по-
явление слабого отрицательного заряда на кислороде и со-
ответственно положительного заряда на углероде. Такие
частичные перераспределения зарядов не являются полны-
ми, как это происходит при ионизации. Однако они изменя-
ют форму молекулы и распределение зарядов на ней, опре-
деляя тем самым большую или меньшую ее предрасполо-
женность к взаимодействию с зарядами других молекул.
Многие молекулы могут существовать в более чем од-
ной форме с экстремумами распределения зарядов, пред-
ставляемыми в отдельных резонансной или канонической
формах, и в любом другом месте — с правильной структу-
рой между этими экстремумами:
257
Органические изомеры
Даже при одинаковом химическом составе молекулы различные вари-
анты расположения атомов в ней могут создавать условия для увели-
чения особенностей ее физических и химических свойств. Вещества с
одинаковой молекулярной формулой, но с разным характером или пос-
ледовательностью их химических связей либо с различным простран-
ственным расположением их атомов называются изомерами. Возмож-
но существование большого количества изомеров. Характер и после-
довательность атомов в молекуле определяют ее структурную форму-
лу, и молекулы с одинаковыми компонентами, но имеющие разные
формулы, называются органическими изомерами (прежнее название
— структурные изомеры). Аминокислоты лейцин и изолейцин пред-
ставляют собой молекулы, состоящие из одних и тех же атомов, но
имеющие неодинаковое расположение, что и определяет различие этих
веществ:
сн3 сн3
сн
I
+ ?Н2
NH3—СН—СОСГ
Лейцин
СН3
СН2
нс—сн3
NH — СН—СОСГ
Изолейцин
Глюкозо-6-фосфат и глюкозо-1-фосфат являются изомерами, кото-
рые способны взаимопревращаться под влиянием фермента фосфоглю-
комутазы:
он
a-D-глюкозо-
1 -фосфат
a-D-глюкозо-
6-фосфат
Основания, определяющие структуру ДНК, также могут существо-
вать в различных химических формах, или таутомерах, с пропорцио-
нальным их соотношением, зависящим от pH локальной среды. Такая
внутренняя реорганизация снова приводит к изменению предрасполо-
женности к химическим реакциям. В качестве примера можно привести
структурную формулу тимина:
258
Геометрические стереоизомеры
Геометрические стереоизомеры существуют в связи с возможностью
различного расположения химических групп относительно двойных
связей в молекуле. Термин «стереоизомеры» используется в случае, ког-
да различие существует только в пространственном расположении их
атомов. Фумаровая и яблочная кислоты имеют одинаковый атомный сос-
тав, однако в фумаровой кислоте (тирднс-изомер) карбоксильные группы
расположены на противоположных сторонах, тогда как в яблочной кис-
лоте (г/г/с-изомер) обе карбоксильные группы присутствуют на одной и
той же стороне молекулы:
н соон
\ /
с
и
с
/ \
ноос н
Н СООН
\ /
Фумаровая кислота
(транс)
/ \
Н СООН
Яблочная кислота
Щис)
Оптические стереоизомеры
Если структурные изомеры и геометрические стереоизомеры имеют
различные химические свойства, то оптические стереоизомеры хи-
мически идентичны, однако могут различаться по одному физичес-
кому свойству. Это свойство характеризуется вращением плоскости
поляризации света, падающего на молекулу: вращение вправо
(правовращающая, или D-форма) или влево (левовращающая, или
L-форма). Этот эффект обусловлен асимметрической структурой мо-
лекулы, определяемой атомом углерода, когда он формирует связи с
различными группами. Глицеральдегид представляет собой относи-
тельно простую молекулу, проявляющую стереоизомерию. D-форму
принято идентифицировать как имеющую гидроксильную группу
справа и правовращающую; L-форму — как имеющую гидроксиль-
ную группу слева и левовращающую:
17;
259
СНО 1 СНО 1
н—- с—он 1 но»—с—н 1
СН2ОН СН2ОН
D-глицер- альдегид L-глицер- альдегид
Зеркальная плоскость
Каждая из двух возможных структур называется енантиомером: ес-
ли обе формы присутствуют в равных количествах, то эта смесь называ-
ется рацемической. В данном случае эффект вращения плоскости поля-
ризации света не проявляется.
Относительно других молекул терминология может быть связана с
рядом затруднений. Так, L-аланин представляет собой енантиомер ала-
нина, который аналогичен L-глицеральдегиду и является биологически
активной формой, однако правовращающей, даже несмотря на то, что
аналогичен левовращающей форме глицеральдегида. Поэтому его сле-
дует обозначить как Ь-(+)-аланин:
соон оно
I I
н2№— с— н но»— с— н
! !
сн3 сн3
Ь-(+)-аланин Ь-(-)-глицер-
альдегид
Для преодоления этих затруднений и неопределенности в системе
терминологии Международным союзом биохимии и молекулярной биоло-
гии была принята система R/S. Эта система устанавливает для каждого
последовательного асимметрического атома углерода символ (R либо S) в
зависимости от расположения групп или атомов, с которыми он имеет
связи. Асимметрический атом углерода в D-глицеральдегиде обозна-
чается символом R.
2. Структуры молекул сахаров
Большое количество различных моносахаридов играет важную роль в
метаболизме, выполняя как структурную, так и другие функции. Моно-
сахариды классифицируются согласно количеству в их молекуле атомов
углерода. Особое внимание заслуживают моносахариды с тремя (трио-
зы), пятью (пентозы) и шестью (гексозы) атомами углерода. Альдозы яв-
260
ляются сахарами, которые имеют альдегидную группу при первом угле-
родном атоме (в связи с этим их структура аналогична глицеральдеги-
ду), а кетозы имеют при нем кетоновую группу (как дигидрооксиаце-
тон).Эти два вещества в их фосфорилированной форме следует рассмат-
ривать как продукты, образующиеся в результате воздействия альдолазы
на фруктозо-1,6-дифосфат:
СНО СН20Н
неон | С=О
СН2ОН СН2ОН
D-глицер- альдегид D-дигидро- оксиацетон
СНО СН2ОН
неон | С=О
носн НОСН
I неон неон
I неон неон I
। СН2ОН СН2ОН
D-глюкоза D-фруктоза
СНО СНО
неон неон
носн носн
I неон носн
неон неон
СН20Н СН2ОН
D-глюкоза D-галактоза
Поскольку в молекулах пентоз и гексоз содержится большое число
асимметрических атомов углерода, существует большое количество изо-
меров этих сахаров, хотя только некоторые из них представляют биоло-
гическую ценность. Глюкоза и галактоза являются диастереоизомерами,
поскольку у них не все конфигурации вокруг атомов углерода двусто-
ронние.
Пентозы и гексозы обладают альдегидной и гидроксильной группа-
ми, которые могут реагировать друг с другом, образуя при этом молеку-
261
лу с кольцевой структурой. Поэтому структурную формулу глюкозы
можно представить в виде линейной молекулы либо одного из двух воз-
можных вариантов (а и Ь) кольцевого расположения ее атомов. Шести-
членное кольцо гексоз состоит из пяти атомов углерода и одного атома
кислорода:
V0
I
н—с—он
I
но—с—н
I
н—с—он
I
н—с—он
I
СН2ОН
D-глюкоза
Н ОН
С-----
I
н—с—ОН
I
но—с—Н о
I
н—с—он
I
н—с------
I
СН2ОН
a-D-глюкоза
но /н
С---------
I
н—с—он
I
но—с—н о
I
н—с—он
I
н—с------
I
СН2ОН
b-D-глюкоза
Кольцевая структура глюкозы обычно представляется в виде, пока-
заном ниже для а-глюкозо-6-фосфата:
В глюкозофосфатизомеразной реакции, в ходе которой катализиру-
ется преобразование глюкозо-6-фосфата до фруктозо-6-фосфата, коль-
цевая структура первоначально должна быть нарушена, что могло бы
позволить реакции продолжаться, однако образующийся фруктозо-6-
фосфат возобновляет кольцевую структуру:
сно
I
нс.он
I
но.сн
I
НС.ОН ---
I
нс.он
I
СН2ОРО*
СН2ОН
с=о
I
но.сн
I
НС.ОН ---
I
нс.он
СН2ОРОз“
a-D-глюкозо-б-фосфат
a-D-фруктозо-б-фосфат
Приложение 2
ЭНЗИМАТИЧЕСКАЯ КИНЕТИКА
И РЕГУЛЯЦИЯ РЕАКЦИЙ
Энзимы повышают скорость специфических химических
реакций, благодаря чему создается возможность регуляции
в организме взаимодействия между различными метаболи-
ческими путями, обеспечивающими жизненные процессы.
Направление, в котором протекают реакции, и точка равно-
весия, которая должна быть достигнута в небиологической
системе, обусловливаются законами термодинамики. Наи-
большее количество важнейших ферментативных реакций
являются насыщаемыми: при низких концентрациях суб-
страта начальная скорость реакции возрастает линейно в
зависимости от его концентрации, однако эта скорость дос-
тигает предельных значений, приближаясь к постоянной и
не зависимой от субстратной концентрации (рис. П2.1).
Концентрация субстрата
Рис. П2.1. Влия-
ние субстратной
концентрации на
активность фер-
мента
263
1. Механизмы действия энзимов
Согласно законам термодинамики, спонтанные химические реакции
протекают только в направлении с меньшим энергетическим статусом
по сравнению с субстратами (рис. П2.2).
Ферменты функционируют в качестве повторно используемых ката-
лизаторов: они участвуют в образовании фермент-субстратного ком-
плекса как промежуточного пункта рекции. Образование этого комплек-
са снижает энергию активации. Поскольку в данном случае добавляется
меньшее количество энергии, реакция в большей степени имеет тенден-
цию к продолжению. Необходимо учитывать, что ферменты хотя и при-
нимают участие в реакции, тем не менее не расходуются и поэтому при-
сутствуют лишь в небольших количествах.
Энергетика образования фермент-субстратного комплекса изучена
недостаточно, однако установлено, что при этом образуется некоторый
вид слабой связи между субстратом и ферментом. Связь включает один
или больше активных участков фермента, которые имеют особую форму
и распределение заряда, благодаря чему создается возможность их взаи-
модействия с субстратом. Эти особенности позволяют ферментам под-
держивать скорость специфической реакции различными способами.
Там, где в реакции участвуют два или более субстратов, прикрепление к
связывающим участкам на ферменте дает возможность субстратам быть
Рис. П2.2. Взаимосвязь свобод-
ной энергии продуктов реакции
(ДО) со свободной энергией
субстратов
264
правильно ориентированными, что повышает вероятность возникнове-
ния реакции. Кроме того, присоединение к ферменту может вызвать та-
кие изменения формы молекулы субстрата, которые повысят его воспри-
имчивость к реакции.
2. Энзиматическая кинетика
Первой стадией катализируемой ферментом реакции является прикреп-
ление субстрата (S) к активному участку фермента (Е) с образованием
фермент-субстратного комплекса (ES), после чего субстрат превращает-
ся в освобождающийся продукт (Р). Выход продукта восстанавливает
фермент до его обычной свободной формы. Последовательность этой
реакции выглядит следующим образом:
E + S<^ES^E + P. (П2.1)
Первая стадия этого процесса является обратимой, вторая — необра-
тимой. Почти во всех реакциях концентрация субстрата значительно пре-
вышает концентрацию фермента. Это означает, что образование ^-ком-
плекса не вызывает заметных изменений в субстратной концентрации,
однако снижает концентрацию свободного энзима. Каждая из реакций
будет протекать со скоростью, которая определяется концентрацией про-
дукта и константой (к) скорости этой реакции. Таким образом, реакция
Е + S ES (П2.2)
будет происходить со скоростью, зависящей от концентрации Е и S, при
этом скорость изменения концентрации субстратов во времени (f) мож-
но определить следующим образом:
d[ES]/dt = £[£][£]. (П2.3)
В случае, когда концентрация реагентов равна единице (при 1 моль-1),
то скорость реакции равна ее константе (к).
Скорость образования ^-комплекса (П2.3) детерминируется кон-
стантой скорости (к^, а также концентрациями субстрата и свободно-
го фермента. Интенсивность обратной реакции зависит от константы
скорости (к2) для этой реакции и [£5]. Скорость диссоциации ком-
плекса ES для выхода свободного фермента и продукта (Л3) также за-
висит от [jESJ :
265
[Е] + [Е]<-^[ЕЕ]—^->[Е] + [Р]. (П2.4)
л2
Концентрация свободного энзима ([£]) в любое время равна общей
его концентрации ([Ео]) минус концентрация ES-комплекса. Если эта ре-
акция протекает в устойчивом состоянии, то скорость образования ES
должна быть равна скорости его распада, т. е. £2[ES] равна Л3[ЕЕ]. Из
этого также следует, что скорость образования ES (Л1[ЕСВ][5Г]) равна
сумме &2[ES] и AJES]:
^[EJEE] = /с2[ЕЕ] + А3[ЕЕ] = (к2 + Л3)[ЕЕ]. (П2.5)
Скорость общей реакции
v = £3[ЕЕ]. (П2.6)
При преобразовании
[Е5] = v/ky (П2.7)
Максимальная скорость реакции (Ктах) достигается в случае, когда
фермент полностью насыщен субстратом, т. е. когда [ЕЕ] = [Ео] и это эк-
вивалентно £3[Е0]:
[EJ = V /Е. (П2.8)
L 0J max 3 v 7
Уравнение (П2.5) можно записать следующим образом:
А1([ЕО]-[ЕЕ])[Е] = (А2 + Л3)[ЕЕ]. (П2.9)
Подставив (П2.7) и (П2.8) в уравнение (П2.9), получим
^1[^Жах -^)/Лз = (к2 + k3)v/k3. (П2.10)
Теперь обе стороны уравнения умножим на к/ку
[Е](Етах -П = ((*2 +к3Ук. +[E])v. (П2.11)
Выражение (Л2 + к3)/к}) в уравнении (П2.11) является константой
Михаэлиса (Кт). Разделив обе стороны этого уравнения на Km + [Е], по-
лучим
v = Emax[E]/(Кт+ [£])•
(П2.12)
266
Поэтому скорость реакции v определяется максимальной скоростью ре-
акции Fmax, которая является функцией концентрации фермента, суб-
стратной концентрации и константы Михаэлиса.
В процессе реакции концентрация субстрата падает и скорость ре-
акции снижается. В связи с этим v относится только к начальной скорос-
ти, когда концентрация субстрата остается высокой, а интенсивность ре-
акции распада от Р до S незначительная.
Из уравнения (П2.12) следует, что в случае, когда концентрация суб-
страта соответствует Кт, скорость реакции будет составлять половину
Ктах. Поэтому значение Кт равно такой концентрации субстрата, при ко-
торой скорость реакции равна половине ее максимального уровня.
Следовательно, высокие значения Кт указывают на низкое сродство
фермента к его субстрату, в связи с чем для достижения высокой скорос-
ти реакции, соответствующей половине ее максимального уровня, необ-
ходима высокая концентрация субстрата. Высокие скорости реакции бу-
дут достигаться только в том случае, когда субстратная концентрация бу-
дет относительно высокой. Если [S] в уравнении (А2.12) соответствует
10-кратному значению Кт, то скорость реакции составит 91 % от К ,
тогда как 99 % максимальной скорости может быть достигнуто только в
случае, когда концентрация субстрата будет в 100 раз превышать Кт.
Реакции нулевого, первого и второго порядков
Порядок реакции является выражением интенсивности, с которой концен-
трация реагента возрастает в уравнении, описывающем скорость ее проте-
кания. В случаях, когда в реакции участвуют более одного реагента, эти ин-
тенсивности суммируются. Реакция, которой сопутствует кинетика нуле-
вого порядка, является независимой от концентрации субстрата. Реакция
первого порядка проявляет линейную зависимость по отношению к увели-
чению субстратной концентрации. Если же эта зависимость является ги-
перболической, то такая реакция называется реакцией второго порядка.
3. Энзиматическая активность
Активность энзимов может быть оценена по скорости субстратной ути-
лизации или образования продуктов в стандартных условиях. Наиболее
общей единицей ферментативной активности является Международная
единица (ME или IU). Она соответствует такому количеству фермента,
которое преобразует 1 ммоль субстрата в продукт за 1 мин в специфичес-
ких для данной реакции условиях. Хотя эту единицу используют в основ-
267
ном биохимики и физиологи, оценку соответствующего показателя выра-
жают также в системе CU. Таковой является 1 катал (кат), который соот-
ветствует количеству фермента, превращающего 1 моль субстрата в про-
дукт за 1 мин при оптимальных условиях. По крайней мере, одна из при-
чин, препятствующих использованию ME, заключается в трудности оп-
ределения оптимальных условий для активности отдельных ферментов.
Факторы, влияющие на активность ферментов
Энзиматическая активность чрезвычайно чувствительна к температуре
и возрастает с ее повышением. Однако при высокой температуре актив-
ность падает резко и необратимо, поскольку в результате денатурации
белка происходят его структурные изменения. Поэтому определение ак-
тивности фермента должно осуществляться с учетом температуры, при
которой эта активность определялась. В качестве стандарта обычно ис-
пользуют температуру 25 и 37 °C. За исключением ряда заболеваний,
температура тела редко превышает 41 °C, однако это близко к уровню,
при котором некоторые ферменты и белки становятся неактивными.
Изменение ионизированного состояния фермента снижает его срод-
ство к субстрату, так как изменяет структуру или распределение заряда
его активного центра.
Локальное pH также может изменять ионизационное состояние суб-
страта. Все ферменты имеют оптимум pH (рис. П2.3), однако для раз-
личных энзимов он неодинаков и может изменяться в присутствии дру-
гих активаторов или ингибиторов. Вариативность pH в большинстве
тканей, как правило, незначительная. Наибольшие изменения этого по-
казателя отмечаются в мышцах при выполнении очень интенсивной мы-
шечной работы: pH может снижаться с 7,1 в состоянии покоя до 6,5 и да-
же ниже. Многие ферменты обычно функционируют в окружающей сре-
де, pH которой болизок к оптимуму.
Так, например, пепсин, имеющий оп-
тимум pH около 2,0, хорошо приспо-
соблен к кислой среде желудка. Одна-
ко некоторые энзимы, по крайней мере
в выделенной и очищенной форме,
имеют оптимум pH, далекий от его
значений для обычной окружающей
среды. Так, глицеролкиназа проявляет
максимум активности при pH 9,8, т. е.
Рис. П2.з. Зависимость энзимати- ПРИ условии, которое никогда не дос-
ческой активности от pH
тигается в клетке.
268
Коэнзимы, простетические группы, кофакторы и активаторы
Для многих ферментов требуется присутствие одного или более ко-
ферментов (коэнзимов) — веществ, принимающих участие в реак-
ции. Так, например, для превращения лактата до пирувата лактатде-
гидрогеназой необходимо присутствие НАД+. Коэнзимы подвержены
химическим изменениям в ходе реакции и в данном случае происхо-
дит переход НАД+ в НАДН. Поэтому коэнзимы необходимы для суб-
страта или продукта реакции, они легко регенерируются другими ре-
акциями в клетке. Некоторые коэнзимы, такие как НАД+, не прикреп-
лены связью к ферменту, однако другие (например, биотин) имеют
прочную связь и поэтому относятся к так называемым простетичес-
ким группам.
Многие ферменты в отсутствие кофакторов проявляют низкую ак-
тивность, а присутствие ионов металлов, особенно таких двувалентных
металлов, как кальций, магний, марганец и цинк, необходимо для акти-
вации целого ряда энзимов. Присоединение этих ионов изменяет рас-
пределение и форму активных центров ферментов. Выход кальция в ци-
топлазму в ответ на нервный импульс весьма важен для активации фос-
форилазы, которая обеспечивает ускорение гликолиза.
Конкурентные и неконкурентные
ингибиторы
Субстанции с химической структурой, аналогичной обычному субстра-
ту для данного энзима, могут также обладать способностью связываться
с активным центром последнего и таким образом интерферировать с
ферментативной функцией путем уменьшения количества активных
центров, доступных настоящему субстрату. Эти субстанции конкуриру-
ют с субстратом за доступ к активному центру, поэтому их относят к
конкурентным ингибиторам. Влияние последних увеличивается с воз-
растанием Кт. Повышение концентрации субстрата до определенного
уровня подавляет эффекты ингибитора и Дпах не подвергается конку-
рентному угнетению.
Неконкурентные ингибиторы связываются с энзимами другими
центрами, оставляя активные участки фермента доступными для суб-
страта, однако они изменяют конформацию белка и тем самым снижа-
ют каталитическую активность активного центра. При этом Vmax сни-
жается, но при одинаковой концентрации субстрата будет проявлять-
ся половина новой максимальной активности, т. е. Кт остается неиз-
менной.
269
Аллостерическая и ковалентная
модуляции
Под аллостерической модуляцией ферментативной активности понима-
ют обратимое присоединение небольших молекул к не активным, а дру-
гим участкам молекулы фермента с образованием конформационных из-
менений в ее структуре. Происходящие изменения формы и распределе-
ния заряда вызывают изменение (увеличение либо снижение) сродства
фермента к его субстратам или продуктам, следовательно, и его актив-
ности. Ковалентная модуляция включает фосфорилирование или дефос-
форилирование, обычно вовлекая ОН-группы серинового остатка в по-
липептидные цепи. Важность ковалентной модуляции в регуляции ак-
тивности ряда ключевых ферментов гликолиза описана в главе 3.
Изоформы энзимов
Многие ферменты существуют в более чем одной форме, и эти изо-
формы катализируют одни и те же реакции. Однако обычно они присут-
ствуют в различных тканях и могут иметь различные специфические
или каталитические особенности.
Лактатдегидрогеназа существует в двух формах, каждая из которых
состоит из четырех субъединиц. Субъединицы представлены одной из
двух форм: Н-форма обнаруживается преимущественно в сердце, тогда
как М-форма преобладает в скелетных мышцах. Возможно существова-
ние пяти различных комбинаций этих субъединиц. В мышце Н-форма
ассоциирована с тканями, обладающими высокой способностью к окис-
лительному метаболизму, и характеризуется высокими возможностями
для окисления лактата, тогда как М-форма ассоциирована с тканями,
имеющими высокую анаэробную способность относительно их окисли-
тельной активности. Многие другие энзимы также существуют в раз-
личных изоформах, однако функциональное значение этого проявления
до конца не выяснено.
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
Cornish-Bowden A., Wharton С. W. (1988). Enzyme kinetics. — Oxford: IRL Press.
Ottaway J.H. (1988). Regulation of enzyme activity. — Oxford: IRL Press.
Приложение 3
ХИМИЧЕСКИЕ БУФЕРЫ
И РЕГУЛЯЦИЯ КИСЛОТНО-
ЩЕЛОЧНОГО БАЛАНСА
1. Определения
Кислота — вещество, способное отдавать ион водорода
(Н+), например НС1, Н2СО3.
Основание — вещество, способное присоединять во-
дородный ион.
рн -— отрицательный десятичный логарифм концен-
трации свободных [Н+], т.е. десятикратное увеличение кон-
центрации свободных [Н+] на каждую единицу снижения
pH.
Более подробное рассмотрение последовательности
расчетов pH исходя из известной концентрации водород-
ных ионов может помочь пониманию последнего. Приве-
денный ниже рабочий пример демонстрирует расчет pH
раствора, содержащего 40 нмоль-л—’Н+: pH = -lg[H+],
Поскольку [Н+] = 40 нмоль-л—1 - 40 • 10—9 моль-л-1 =
= 4 • 10~8 моль-л-1, pH = -1g (4 10—8), 1g 4 = 0,60,
то pH = -1g СЮ0-6 • 10-8) = -1g IO-7-2 * 4 = - (-7,4) = 7,4.
2. Кислотно-щелочной фон
Несмотря на продукцию большого количества диоксида
углерода, что приводит к образованию водородных ионов
при взаимодействии с водой, и нелетучих кислот в орга-
низме вследствие метаболизма азота, серы и фосфора,
содержащихся в белках, внеклеточная жидкость (ВЖ) со-
держит слабые щелочи при pH 7,36 — 7,44. Органичес-
кие и неорганические компоненты вне- и внутриклеточ-
271
ной жидкости буферируют некоторое количество Н+-ионов, экскре-
цию СО2 осуществляют легкие, а оставшиеся ионы водорода удаля-
ются почками. Эти же механизмы минимизируют изменение pH во
внеклеточной жидкости при дополнительном поступлении кислот или
щелочей.
Процессы буферирования и экскреции Н+-ионов различны не
только по своему механизму, но и по скорости, с которой они проте-
кают. Следовательно, буферирование является пассивным физико-
химическим процессом, включающим почти мгновенные изменения
равновесия между Н+-ионами, щелочами, которые связывают Н+ и
Неосновным комплексом (кислотой). Это ограничивает (или буфе-
рирует) изменение свободной концентрации водородных ионов (pH)
через изменения в их связывании, однако не может предотвратить
некоторые сдвиги pH, происходящие в результате отсутствия изме-
нений общего содержания свободных водородных ионов в организ-
ме, при дополнительном поступлении кислот или оснований. Эк-
скреция СО2 или Н+ является физиологическим процессом, который
реагирует медленнее, чем буферирование (при долговременной мы-
шечной работе) для восстановления общего содержания Н+ в орга-
низме к нормальному уровню после нарушения кислотно-щелочного
равновесия.
Характер и скорость буферирования свидетельствуют о том, что это
непрерывно изменяющийся процесс, реагирующий на флуктуации Н+-
загрузки и ограничивающий колебания pH не только непосредственно
после нарушения кислотно-щелочного баланса, но и в течение периода,
когда физиологические функции направлены на восстановление нор-
мального общего содержания Н+ в организме.
3. Буферы
Существуют экстрацеллюлярные и интрацеллюлярные буферы. Ос-
новными экстрацеллюлярными буферами являются бикарбонатный
и фосфатный, а также белки плазмы. Общее количество буфера в
крови, доступное для связывания Н+, носит название «основной бу-
фер».
Внутриклеточные буферы включают бикарбонат, белки и фосфор-
ные эфиры. Важность присутствующих в клетке внутриклеточных бу-
феров для эритроцитов может быть продемонстрирована следующим
образом: если 10 ммоль Н+ добавить к одному литру плазмы, то ее pH
272
снизится с 7,4 до 5,6. Но если такое же количество Н+ добавить к I л
крови, то pH снизится только с 7,4 до 7,0.
Для каждого буфера закон действия масс формулирует, что в данном
случае должно существовать равновесие:
[Н+][В-] = Ка[ВН]. (П3.1)
Здесь [В-], [Н+] и [ВН] — соответственно концентрации основного бу-
фера, Н+ и кислоты; Ка — константа диссоциации кислоты, т. е. концен-
трация Н+, при которой концентрации В~ и ВН равны.
Приведенное уравнение может быть преобразовано в форму, назы-
ваемую «уравнение Хендерсона»:
[Н+] = К[ВН]/[В~], (П3.2)
которое выражает зависимость свободной концентрации Н+ от концен-
трации основания [В-] и кислоты. Альтернативной формой этого урав-
нения, которая используется в расчетах, является предложенное Хендер-
соном и Хассельбальчем уравнение
рН = рК + lg[B-]/[BH], (ПЗ.З)
где рКа — отрицательный логарифм для основания 10 Ка, т. е. значе-
ние pH, при котором концентрации кислоты [ВН] и основания [В ]
равны.
Если это уравнение относится к бикарбонатугольнокислотной бу-
ферной системе, то используется его модифицированная форма. Пос-
кольку угольная кислота также находится в равновесном состоянии с во-
дой и углекислым газом, то концентрация первой прямо зависит от кон-
центрации растворенного СО2 и константы равновесия этой реакции. В
результате уравнение Хендерсона-Хассельбальча для бикарбонатуголь-
нокислотной системы может быть представлено следующим образом:
pH = 6,1 + lg[HCO-]/(S CO2). (П3.4)
Здесь S — показатель растворимости СО2 в воде (1 кПа парциального
давления создает концентрацию растворенного СО2, равную
0,226 ммоль-л-1 при 37 °C, а 6,1 — константа диссоциации для реакции
между угольной кислотой и СО2 + вода.
Приблизительно средним уровнем для здоровых людей является
18 <,„3 273
7,4 = 6,1 +lg24/(0,226-5,3) (П3.5)
Иногда удобнее использовать непосредственно [Н+], чем pH:
[Н+] = крСОДНСО-]. (П3.6)
Нормальными уровнями для [Н+], рСО2 и [НСО3—] являются
40 нмоль, 5,3 kPa (40 мм Hg) и 24 ммоль-л—1 соответственно, так что
обычно
40 = к5,3/24 и к оказывается равным 181,1. (П3.7)
Приведенные расчеты показывают, что когда буферная система на-
ходится в состоянии равновесия, которое наступает очень быстро (в те-
чение нескольких секунд), то pH зависит от соотношения НСО3“ к рСО2,
которое теперь будем называть бикарбонат/СО2-отношением. Следова-
тельно, увеличение НСО3~ или падение рСО2, что повышает это отно-
шение, обусловливают увеличение pH, т. е. способствуют созданию ще-
лочной среды. Так, например, если рСО2 возрастает до 6 kPa, то для [Н+]
получаем
[Н+] = 181,1 (6,0/24) = 45 нмоль. (ПЗ. 8)
Таким образом, увеличение рСО2 приводит к возрастанию [Н+]. В
большинстве случаев нет необходимости производить расчет величины
заряда, и это может иметь определенное значение только для установле-
ния направления изменения [Н+], связанного с частичным отклонением
рСО2 или [НСО3—] от нормальных значений. В этом случае может быть
использована упрощенная формула соотношения
[Н+] рСО2/[НСО-]. (П3.9)
Из этого соотношения видно, что увеличение [Н+] способно привес-
ти к повышению рСО2 или к снижению [НСО3—] либо к тому и другому.
Бикарбонат-угольнокислотная буферная система является наиболее
важной в организме. На первый взгляд это может показаться странным,
поскольку данная система наиболее эффективно функционирует в диа-
пазоне ± 1 значения pH от рКа. Таким образом, по достижении предель-
ного значения рК, равного 6,1 , нормальное pH крови (около 7,4) выхо-
дит за пределы оптимального диапазона буферирования этой системы.
Две характерные особенности последней придают ей важное значение:
274
• существует большое количество составляющих компонентов сис-
темы, что позволяет изменять количество связанных ионов водорода при
их добавлении либо удалении;
• система является открытой, в связи с чем количество ее компонен-
тов непостоянно и может изменяться в зависимости от потребностей ор-
ганизма.
Из этих двух особенностей именно вторая представляется наиболее
важной. Значимость открытости системы можно легко представить при
сравнении свойств открытой и закрытой систем. В закрытой системе ни
один из ее компонентов не может обмениваться с окружающей средой,
как, например, если бы она находилась на плывущем корабле. Поэтому
общее количество В фиксировано, в связи с чем сумма кислот и основа-
ний, т.е. [ВН]+[В~], постоянна. При добавлении в систему Н+ последние
быстро объединяются с В-, образуя при этом ВН, в связи с чем [ВН]
быстро возрастает, а [В-] — снижается. Это продолжается до тех пор,
пока не произойдет накопление [ВН] и исчерпание [В-], что способству-
ет изменению хода реакции в обратном направлении, и достаточно для
того, чтобы противодействовать протекающему процессу. Поэтому быс-
тро устанавливается новое равновесное состояние, в котором дальней-
шее буферирование Н+, т. е. объединение с основаниями, уже не проис-
ходит, что и будет определять окончательное значение pH в системе.
В открытой системе один или более ее компонентов могут обмени-
ваться с окружающей средой, в связи с чем их концентрация может не-
зависимо варьировать от того или иного компонента. Экстрацеллюляр-
ная бикарбонатная буферная система в организме является открытой,
поскольку СО2, Н+ и НСО3— могут обмениваться с окружающей средой.
Углерод поступает в организм с пищей, метаболизируется в клетках до
СО2 и затем экскретируется легкими: Н+ входит в состав содержащихся
в пище кислот и продуктов метаболизма и экскретируется почками;
НСО3— образуется в клетках из СО2 и Н2О и может удаляться или обра-
зовываться почками. Накопление и исчерпание В и ВН, происходящее
в закрытой системе при добавлении или удалении Н+, может быть пре-
дотвращено перемещением В~ или ВН (или самих Н+) внутрь или из
системы. Это поддерживает условия, в которых буферирование Н+ мо-
жет продолжаться.
Поступление СО2 и Н+ в организм с пищей неизбежно, поэтому кис-
лотно-щелочной баланс регулируется преимущественно экскреторными
процессами. Последние могут отличаться выраженной независимостью
друг от друга физиологическими механизмами регуляции, которые спо-
собны изменять как числитель, так и знаменатель в уравнении Хендер-
сона, что обеспечивает физиологический контроль экстрацеллюлярного
18*
275
pH. В общем, регуляция экскреции СО2 в связи с ее скоростью и объе-
мом является более важной, чем изменение экскреции Н+ или концен-
трации НСО3 . Система дыхания обычно реагирует намного быстрее,
чем почки, и может дополнительно экскретировать 40 ммоль СО2-мин—1
по сравнению с 0,15 ммоль-мин~’ почками. Следует, однако, заметить,
что расположение основных хеморецепторов вне гематоэнцефалическо-
го барьера подразумевает, что начало некоторого нарушения и восста-
новление после него может задерживаться ионной разностью между
кровью и спинномозговой жидкостью. Способность почек изменять ста-
тус Н+ и бикарбонатов является жизненно важной для восстановления
экстрацеллюлярной концентрации НСО3 на длительный период и обес-
печения кислотно-щелочной регуляции уже в самом начале нарушения
дыхания, когда гиповентиляция вызывает накопление СО2.
4. Реакции в ответ на нарушение
кислотно-щелочного баланса
Наиболее просто, по крайней мере вначале, было бы предполагать, что
нарушение кислотно-щелочного баланса состоит из двух эффектов.
Первый представляет собой быстрое изменение буферирования Н+, ко-
торое препятствует изменению общей концентрации Н+ и минимизиру-
ет изменение свободной концентрации водородных ионов. Этот перво-
начальный эффект сопровождается замедлением физиологических ре-
акций — изменением экскреции СО2 легкими (респираторный ответ) и
изменением контроля статуса Н+ и НСО3— почками (ренальный ответ).
При рассмотрении влияния этих процессов учитывается, что первые
реакции вызываются простым нарушением, а именно — добавлением
кислоты к внеклеточному буферу (нарушение, аналогичное происходя-
щему при сахарном диабете).
Буферирующий (первичный) ответ
Добавление кислоты равносильно добавлению Н+ к левой части приве-
денной ниже реакции
Н+ + НСОз Н2СО3 -> Н2О + СО2
+
В“
г
вн
276
в которой В~ представляет другие (кроме НСО3—) буферные системы.
Некоторое количество добавляемого Н+ будет связываться с НСО3 с об-
разованием угольной кислоты, а другая его часть расходуется на образо-
вание СО2 и воды. Определенная доля Н+ будет буферироваться путем
связывания с В- с образованием ВН. В закрытой системе новое равно-
весное состояние будет достигнуто при снижении концентрации НСО3~
и повышении рСО2. Из приведенных в предыдущем параграфе урав-
нений видно, что изменение отношения бикарбо нат/СО2 будет соответ-
ствовать падению pH (по варианту уравнения Хендерсона-Хассельбаль-
ча). Таким образом, несмотря на присутствие буферов, не все количес-
тво добавленного Н+ становится связанным, что приводит к повышению
концентрации свободных водородных ионов в системе.
Физиологический (вторичный) ответ
Респираторная экскреция СО2. Снижение pH и повышение альвеоляр-
ного и артериального рСО2 вызывают быструю стимуляцию легочной
вентиляции, которая удаляет некоторое количество СО2 из системы, что
приводит к снижению рСО2. Последнее может происходить в связи с
тем, что функционирующая система является открытой. В результате
увеличивается соотношение бикарбонат/СО2, что способствует незамед-
лительному повышению после добавления кислоты и буферирования
pH, возвращению его к нормальному уровню. Таким образом, в данном
случае соответствующим респираторным ответом является снижение
рСО2, называемое респираторной компенсацией.
Ренальная экскреция Н+ (реабсорбция НСО3~). Аналогично тому
как дыхание может влиять на соотношение бикарбонат/СО2 путем из-
менения экстрацеллюлярной концентрации СО2, так почки могут мо-
дифицировать это соотношение за счет изменения концентрации бикар-
боната. Такой эффект является следствием имеющей важное значение
способности почек секретировать Н+-ионы. Судьба последних предоп-
ределяется по трем направлениям. Многие реакции с бикарбонатом в
жидкости почечных канальцев протекают с образованием угольной кис-
лоты, а отсюда — с образованием СО2 и воды. Последние проникают в
полость почечных канальцев, где преобразуются до Н+ и НСО3~. После
этого Н+ становится доступным для ресекреции , а НСО3— поступает в
кровь. Таким образом, наибольшее количество проникнувшего НСО3— в
нормальных условиях является «эффективно» реабсорбируемым. Сле-
дует заметить, что этот процесс не определяет Н+-экскрецию, поскольку
полностью протекает в почках, и Н+, принимающие участие в этом цик-
ле, не покидают организм.
277
В состоянии покоя определенное количество Н+ может секретиро-
ваться в виде свободных ионов (pH мочи может снижаться до 4,8), одна-
ко большинство из них экскретируется в комбинации с основаниями.
Наиболее важными из этих реакций в моче являются
НРО4“ + Н+ -> Н2РО4 и NH3 + Н+ -> NH4.
Появление NH3 во второй реакции обусловлено преимущественно
дезаминированием в почках глутамина. Необходимо обратить вни-
мание на роль угольной ангидразы: этот фермент присутствует в
цитоплазме тубулярных клеток, в проксимальных почечных канальцах
на люминальной поверхности клеток. Она катализирует превращение
воды + СО2 до угольной кислоты и эту же реакцию, протекающую в об-
ратном направлении. В отсутствие этого фермента реакция протекает
относительно медленно, что лимитирует скорость, с которой секреция
Н+ и реабсорбция НСО3— могут происходить.
Низкие значения pH стимулируют секрецию Н+ почками. Это соот-
ветствует потребности организма освободиться от избытка [Н+] и приво-
дит к реабсорбции некоторого количества бикарбоната, еще остающего-
ся в жидкости почечного канальца, а также к повышению экскреции Н+,
или в виде свободных водородных ионов, или связанными с буферами
мочи. Дополнительным следствием такого повышения потери Н+ в моче
является образование «нового» бикарбоната, и для того чтобы разоб-
раться, как это происходит, необходимо снова вернуться к механизму Н+-
секреции.
Водородные ионы, секретируемые клетками почечных канальцев,
образуются в клетках в результате диссоциации угольной кислоты, поэ-
тому данный процесс сопровождается образованием равноценного ко-
личества ионов НСО3—. Протекает он в крови несмотря на секрецию Н+-
ионов. Если последние связываются с НСО3— в жидкости почечных ка-
нальцев, то это приводит к реабсорбции воды и СО2 и в целом процесс
эквивалентен реабсорбции бикарбонатных ионов из фильтрующейся
жидкости. Однако ионы, секретируемые в жидкость почечных каналь-
цев, из которой весь бикарбонат уже реабсорбирован, экскретируются в
мочу в виде свободных ионов или связанных с буфером. В этом случае
переход бикарбоната из клеток в кровь, который происходит во время
секреции водородных ионов, не сопровождается удалением бикарбоната
из жидкости почечных канальцев, поэтому он эквивалентен добавлению
«нового» бикарбоната в кровь. С помощью такого механизма почки эф-
фективно восстанавливают запасы ранее утраченного при буферирова-
нии избытка Н+ бикарбоната.
278
Таким образом, соответствие почечного ответа, называемого ре-
нальной компенсацией, кислотной загрузке проявляется в увеличении
секреции Н+, приводящей к повышению экскреции свободных и связан-
ных ионов водорода, а также к образованию НСО3— в экстрацеллюляр-
ной жидкости и, как следствие, — к возрастанию отношения бикарбо-
нат/СО2. Следует заметить, что снижение Н+-концентрации предсказы-
вается уравнением Хендерсона. Необходимо иметь в виду, что анионы
добавляемых кислот, например С1~ или SO42-, также экскретируются в
мочу. Происходит это быстрее по сравнению с повышением экскреции
Н+ и быстрее по отношению к Na+ или К+, чем к ионам водорода.
Рассмотрим теперь другое нарушение кислотно-щелочного баланса,
проявляющееся в повышении в экстрацеллюлярной жидкости рСО2.
Последнее приводит к развитию реакции, в результате которой образу-
ются ион водорода и бикарбонат:
СО2 + Н2О -> Н2СО3 -> Н++НСО3
+
В"
г
ВН
Первичный ответ в этой ситуации проявляется в том, что другие
буферные системы быстро связывают некоторое количество Н+, мини-
мизируя таким образом происходящее изменение pH и формируя новое
равновесное состояние. Последнее характеризуется еще некоторым
увеличением рСО2, повышенными концентрациями водородных
ионов, НСО3— и ВН. Присоединение Н+ к В- фактически приводит к
увеличению количества образующегося НСО3—, поскольку при этом
происходит удаление Н+ и таким образом повышается возможность
преобразования Н2СО3 до Н+ и НСО3~. В результате должно произой-
ти увеличение отношения бикарбонат/СО2, что, в свою очередь, вызо-
вет снижение падения pH, обусловленного нарушением кислотно-ще-
лочного баланса. Следует учитывать, что в данном случае общее коли-
чество щелочного буфера не изменяется, поскольку НСО3~ служит ис-
точником для каждого используемого В-, к тому же, поскольку кон-
центрация НСО3—намного выше по сравнению с [Н+] и другими буфе-
рами, то относительное изменение концентрации НСО3— при таком ти-
пе нарушения является кратковременным и, как правило, незначитель-
ным. Поэтому при острых респираторных расстройствах можно
ожидать измений рСО2. Вновь установившееся состояние и на этот раз
279
описывается уравнением Хендерсона (или Хендерсона-Хассель-
бальча).
Вторичным эффектом в подобном случае возможен только почеч-
ный ответ, поскольку он может быть связан с ухудшением дыхательной
системы, ответственной за происхождение нарушения кислотно-щелоч-
ного баланса. Однако на протяжении длительного периода повышение
рСО2 и снижение pH во внеклеточной жидкости приводит к генерации
НСО3— почками и увеличению во внеклеточной среде концентрации
этих ионов. Если респираторная система функционирует нормально, то
легочная вентиляция будет стимулироваться повышением рСО2 и сниже-
нием pH, что приведет к увеличению экскреции СО2. В результате про-
изойдет снижение рСО2 и возрастание как соотношения бикарбо-
нат/СО2, так и pH до того, как какой-либо ренальный ответ станет оче-
видным.
Описанные механизмы функционируют подобным, но протекаю-
щим в противоположном направлении путем, когда рСО2 изменяется
под влиянием гипервентиляции. В данном случае снижение рСО2 в кро-
ви и повышение в ней pH проявляется в меньшей степени, чем это пред-
сказывается, поскольку смещение баланса реакции влево приводит к
снижению [НСО3—].
5. «Нормальные уровни» и формы проявления
нарушения кислотно-щелочного баланса
Нормальные уровни для pH и рСО2 в артериальной крови вместе с соот-
ветствующими выкладками для [НСО3 ] представлены клиническими
лабораториями и имеют следующие значения:
pH — 7,35-7,45;
рСО2 — 4,7-6,4 kPa (35—48 мм Hg);
[НСО3 ] — 23-28 ммоль-л"1.
Значения pH ниже 7,35 ([Н+], равной 10“7-35 моль-л^1 или
45 нмоль-л—') указывают на состояние, называемое ацедемией, тогда как
превышение уровня 7,45 ([Н+] менее 35 нмолыл-') указывает на алкале-
мию. Эти состояния являются выраженным нарушением нормальной
концентрации свободных ионов водорода в крови. В данном случае не
учитывается, что незначительные отклонения могут отражать сущес-
твенные изменения [Н+] во всем организме, которые компенсируются
преимущественно буферированием и физиологическими ответами. По-
280
этому весьма важно использовать показатели рСО2 и [НСО3~] для уста-
новления ведущих как внутренних, так и внешних причин нарушений
кислотно-щелочного равновесия, а также размерность соответствующе-
го ему компенсаторного ответа.
Любое повышение уровня свободных ионов водорода, даже если
оно предотвращается компенсацией, превышающей нормальный лимит,
будет указывать на проявление ацидоза, что означает увеличение обще-
го содержания Н+ в организме и формирование состояния, противопо-
ложного состоянию алкалоза. Если ацидоз сопровождается повышением
рСО2 сверх нормального уровня, то обязательно должен иметь место
респираторный ацидоз. Любое превышение нормального уровня кон-
центрации НСО3~ должно ограничивать отклонения pH и поэтому будет
частично способствовать проявлению компенсированного респиратор-
ного ацидоза. Следует напомнить, что ренальные ответы обычно замед-
лены, и если происходит компенсаторное увеличение концентрации
НСО3~ в результате явного ацидоза, то последний должен удерживаться
еще некоторое время.
Аналогично этому некоторое снижение pH, сопровождающееся
снижением [НСО3—], но не повышением рСО2, должно указывать на не-
посредственное поступление Н+, т. е. на не связанное непосредственно с
респираторной недостаточностью. Такое состояние называется метабо-
лическим ацидозом, и если сопровождается падением рСО2 ниже нор-
мального уровня, то это называется компенсированным метаболическим
ацидозом, который устраняется путем повышения легочной вентиляции.
Низкое значение pH, сопутствующее субнормальной концентрации
НСО3~, и рСО2, превышающее нормальный уровень, соответствует сос-
тоянию, обозначаемому как смешанный респираторный и метаболичес-
кий ацидоз.
0-313
19
СЛОВАРЬ ТЕРМИНОВ
Аллостерический фермент. Фермент, который изменяет свою тре-
тичную структуру в результате взаимодействия его активного цен-
тра с мелкими молекулами. Это зачастую приводит к угнетению ли-
бо активации энзима
Анаплеротическая реакция. Ферментативный механизм, обеспе-
чивающий пополнение запасов промежуточных продуктов цикла
трикарбоновых кислот (например, образование оксалоацетата из
пирувата с участием пируваткарбоксилазы)
Анаэробные процессы. Процессы, происходящие в отсутствие сво-
бодного кислорода
Аэробные процессы. Процессы, происходящие в присутствии сво-
бодного кислорода
Буфер. Вещество, препятствующее быстрому изменению в раство-
ре концентрации водородных ионов (pH)
Водородная связь. Слабое внутримолекулярное или межмолекуляр-
ное притяжение, вызванное взаимодействием атома водорода и при-
тягивающей силой парой одиночных электронов (например, кисло-
рода либо азота). Водородная связь играет важную роль в молекулах
ДНК, РНК, а также ответственна за структуру многих белков
Геометрическая изомерия. Форма стереоизомерии, при которой
различие возникает по причине противоположного вращения вок-
руг двойной связи. Ненасыщенные жирные кислоты, содержащие
одну двойную связь, имеют два изомера в зависимости от того, на-
ходятся атомы водорода на одной и той же (цис) или противополож-
ной (транс) сторонах молекулы
Гидратация. Реакция, связанная с включением воды в состав ве-
щества
282
Гидроксилирование. Реакция, в результате которой происходит присоединение
гидроксильной группы (ОН) к молекуле
Гидролиз. Реакция, в результате которой органическое вещество при взаимодей-
ствии с водой распадается до простых соединений
Гликогенолиз. Превращение гликогена до гпюкозо-1-фосфата под влиянием фос-
форилазы
Гликозидная связь. Химическая связь, в которой атом кислорода является об-
щим связующим звеном между одним и любым другим атомом углерода молеку-
лы сахара. Гликоген как полимер глюкозы представляет собой разветвленную по-
лисахаридную цепь, состоящую из молекул глюкозы, соединенных между собой
гликозидными связями
Гликолиз. Последовательная цепь реакций, превращающих глюкозу (или глюкозо-
1-фосфат) в пируват
Глюконеогенез. Образование глюкозы из таких неуглеводных предшественников,
как, например, глицерол, кетокислоты или аминокислоты
Дегидратация. Реакция, приводящая к утрате молекулы воды
Дезаминирование. Реакция, результатом которой является потеря аминогруппы
(NH2)
Декарбоксилирование. Реакция, приводящая к утрате СО2-группы
Изомер. Одно из двух или более веществ, имеющих одинаковый молекулярный
состав и относительную молекулярную массу, однако различных по структуре, что
связано с неодинаковым расположением атомов в их молекулах
Изотоп. Одно из состояний химически идентичных разновидностей атома, которые
имеют одинаковый атомный номер, но различную относительную атомную массу
(например, 12-, 13- и 14-изотопы углерода, атомные номера которых — 12, 13 и 14)
Изоформа. Химически отличимые формы ферментов, обладающие одинаковой ак-
тивностью (они еще называются изоэнзимами)
Ион. Любой атом или молекула, которые имеют электрический заряд
Ионная связь. Связь, в которой валентные электроны либо теряются, либо присо-
единяются, а атомы, имеющие противоположный заряд, удерживаются вместе
электростатически (кулоновскими силами)
Карбоксилирование. Реакция, в результате которой происходит присоединение
СО2. Она катализируется ферментом, в котором простетической группой является
биотин
Катехоламины. Биологически активные гормоны и нейропередатчики, которые
синтезируются из аминокислоты тирозина. К ним относятся, например, адреналин,
дофамин и норадреналин
Кетогенез. Синтез кетонов
19*
283
Киназа. Фермент, регулирующий реакцию фосфорилирования-дефосфорилирования
Кислота. Вещество, проявляющее тенденцию к утрате протонов (водородных
ионов)
Ковалентная связь. Химическая связь, в которой два или более атомов удержива-
ются вместе благодаря взаимодействию их внешних электронов
Ковалентная регуляция. Регуляция ферментативной активности ковалентным при-
соединением фосфатных групп к участкам, не являющимся активными центрами
Конденсация. Реакция, в результате которой соединяются две или более молекул с
отделением таких несложных радиклов, как, например, Н2О
Конформация. Форма молекулы, определяемая вращением вокруг одиночных свя-
зей, особенно в полипептидных цепях, вокруг углерод-углеродных соединений
Коэнзим. Небелковая часть некоторых ферментов, играющая важную роль в про-
явлении их активности
Липолиз. Распад триацилглицеролов до жирных кислот и глицерола
Метаболит. Продукт реакций обмена веществ
Окисление. Реакция, сопровождающаяся потерей атомом электронов. Она всегда
сопряжена с реакцией восстановления. Так, например, пируват восстанавливается до
лактата. В обратной реакции лактат окисляется НАД+ при реформировании пирувата
Основание. Вещество, обладающее свойством донатора электронной пары или ко-
ординатора электрона
Пептидная связь. Химическая связь, образующаяся в результате конденсации
аминогрупп и карбоксильных групп пары аминокислот. Пептиды состоят из боль-
шого количества аминокислот, соединенных между собой пептидными связями
Полупериод. Время, в течение которого в результате реакции половина количества
или концентрации вещества снижается или возрастает
Простетическая группа. Коэнзим, прочно связанный с энзимом
Редукция (восстановление). Реакция, в результате которой молекула присоединя-
ет электроны
Скоростьлимитирующий энзим. Энзим, который в метаболическом процессе за-
медляет протекание последнего
Стереоизомерия. Существование различных веществ, молекулы которых имеют
одинаковое сочетание атомов, но различное их расположение в пространстве
Субстрат. Молекула реактанта в реакции, катализируемой ферментом
Тиоэфирная связь. Связь, в которой кислород замещается серой (например, про-
цесс перехода КоА в ацил-КоА)
транс- Приставка, обозначающая геометрический изомер, в котором два одинаковых
радикала имеют противоположную двойную связь с ограниченным вращением
284
Трансаминирование. Реакция, обеспечивающая превращение аминогрупп (NH2)
до амино- и кетокислот
Транскрипция. Синтез рибонуклеиновой кислоты (РНК) на матрице ДНК
Трансляция. Процесс, следующий за транскрипцией и обеспечивающий синтез
белковых молекул в клетке
Флюкс. Скорость протекания метаболического процесса
Фосфаген. Термин, который относится к обоим высокоэнергетическим фосфатным
соединениям — аденозинтрифосфату и фосфокреатину
Фосфорилирование. Реакция, результатом которой является присоединение фос-
фатной (РО32-)-группы к молекуле. Так, например, в процессе окислительного фос-
форилирования АДФ ресинтезируется до АТФ
цис- Приставка, обозначающая геометрический изомер, в котором два одина-
ковых радикала имеют с одной и той же стороны двойную связь с ограниченным
вращением
Энзим (фермент). Белок, обладающий специфической каталитической активнос-
тью. Энзимы обозначают суффиксом «-аза», обычно добавляемым к типу катализи-
руемой реакции
Эпимеризация. Тип асимметрической трансформации в органических молекулах
РЕКОМЕНДУЕМАЯ
ЛИТЕРАТУРА
Глава 1
AstrandP.-O., Rodahl К. (1986). Textbook of work physiology (3rd edn). —
New York: McGraw-Hill.
Brooks G.A., Fahey T.D. (1984). Exercise physiology: human bioenergetics
and its applications. — New York: John Wiley.
Fox E.L., Bowers R. W„ Foss M.L. (1993). The physiological basis for exer-
cise and sport. W.C. Brown, Dubuque, IA.
Marieb E.N. (1993). Human anatomy and physiology (2nd edn). —
Benjamin Cummings: Redwood City, LA.
Powers S.K., Howley E.T. (1990). Exercise physiology: Theory and applica-
tion to fitness and performance/ Ed. W.C. Brown: Dubuque, IA.
Stryer L. (1988). Biochemistry (2nd edn). W.H. Freeman. — San Francisco,
CA.
Глава 2
Aragon- J.J., Tornheim K., Goodman M.N., Lowenstein J.M. (1981).
Replenishment of citric cycle intermediates by the purine nucleotide cycle in
rat skeletal muscle // Curr.Top.Cell Regul. — 18. — P. 131-149.
Broberg S., Sahlin K. (1989). Adenine nucleotide degradation in human
skeletal muscle during prolonged exercise // J. Appl.Physiol. — 67. —
P. 116-122.
Flanagan W.F., Holmes E.W., Sabina R.L., Swain J.L. (1986). Importance of
purine nucleotide cycle to energy production in skeletal muscle //
Am.J.Physiol. — 251. — C795-C802.
Hellsten Y„ Norman B„ Balsom P.D., Sjodin B. (1993). Decreased resting
levels of adenine nucleotides in human skeletal muscle after high-intensity
training // J.Appl.Physiol. — 74. — P. 2524-2528.
286
Katz A., Sahlin K, Henriksson J. (1986). Muscle ammonia metabolism during isometric con-
traction in humans // Am.J.Physiol. — 250. — C834—C840.
Lowenstein J.M. (1990). The purine nucleotide cycle revised // Int.J.Sports Med. — 11. —
P. 37-46.
McPartland A., Segal I.H. (1986). Equilibrium constants, free energy changes and coupled
reactions: concepts and misconcepts // Biochem.Ed. — 14. — P. 137-141.
Meyer R.A., Terjung R.L. (1979). Differences in ammonia and adenylate metabolism in con-
tracting fast and slow muscle // Am.J. Physiol. — 237. — Cl 11-C118.
Meyer R.A., Terjung R.L. (1980). AMP deamination and IMP reamination in working skeletal
muscle // Am.J. Physiol. — 239. — C32-C38.
Ren J.-M., Hultman E. (1989). Regulation of glycogenolysis in human skeletal muscle //
Am.J. Physiol. — 67. — P. 2243-2248.
Sahlin K, BrobergS. (1990). Adenine nucleotide depletion in human muscle during exercise:
causality and significance of AMP deamination // IntJ.Sports Med. — 11. — P. 62-67.
Sahlin K, Katz A., BrobergS. (1990). Tricarboxylic cycle intermediates in human muscle dur-
ing submaximal exercise // Am.J. Physiol. — 259. — C834—C841.
IMP accumulation in human muscle during prolonged exercise // Am.J. Physiol. — 261. —
C71-C76.
Tullson PC., Terjung RL. (1991). Adenine nucleotide mecleotide metabolism in contracting
skeletal muscle // Exerc.Sport.Sci.Rev. — 19. — P. 507—537.
Глава 3
Boobis L.H. (1987). Metabolic aspects of sprinting. — Exercise: benefits, limits and adaptions
/Ed. D.A.D. Macleod et al. — London: Spon. — P. 116-140.
Costill D.L., Miller J.M. (1980). Nutrition for endurnce sport: Carbohydrate and fluid balance
// IntJ.Sports Med. — 1. — P. 2-14.
Maughan R.J., Greenhaaff PL. (1991). High intensity exercise and acid-base balance: the
influence of diet and induced metabolic alkalosis on performance. — Advances in nutrition
and top sport /Ed. F. Brouns. — Karger: Basel. — P. 147-165.
Newsholme E.A., Leech A.R. (1983). Biochemistry for the medical sciences. — Wiley:
Chichester.
Sahlin K. (1986). Metabolic changes limiting muscle performance // Biochemistiy of exercise
vi/Ed. B. Saltin. — Human Kinetics: Champaign, IL. — P. 323-345.
Sjodin B. (1992). Anaerobic function // Sport Sci. Rev. — 1. — P. 13-27.
Глава 4
Arner P, BolinderJ, EliassonA. et al. (1988). Microdialysis of adipose tissue and blood for
in vivo lipolysis studies // Am.J.Physiol. — 225. — E737-E742.
287
TerJung R.L., Kaciuba-Uscilko H. (1986). Lipid metabolism during exercise: influence of
training. — Diabetes/Metabolism during exercise. — Exercise metabolism/ Ed.
M.Hargreaves. — Champaign, IL: Human Kinetics. — P. 99-130.
Wahrenberg H, Bolinder J, Arner P. (1991). Adrenergic regulation of lipolysis in human fat
cells during exercise // Eur.J.ClinJnverst. — 21. — P. 534-541.
Глава 5
Brooks G.A. (1987). Amino acid and protein metabolism during exercise and recovery //
Med.Sci.Sports.Exerc. — 19 (5, Suppl.). — P. 150-156.
Davis J.L. (1986). Central and peripheral factors in fatigue // J.Sports. — 13 (Special issue).
— P. 49-53.
Dohm G.L. (1986). Protein as a fuel for endurance exercise // Exerc.Sport Sci.Rev. — 1. —
P. 143-173.
Lemon P.W.R. (1991). Effect of exercise on protein requirements // J.Sports. — 9 (Special
issue). — P. 53-70.
Wagenmakers A.J.M., Beckers E.J., Brouns F. et al. (1991). Carbohydrate supplementation,
glycogen depletion, and amino acid metabolism during exercise // Am.J.Physiol. — 260. —
E883-E890.
Глава 6
Balsom P.D., Ekblom B„ Soderlund K. et al. (1993). Creatine supplementation and dynamic
high-intermittent exercise // Scand.J.Med.Sci.Sports. — 3. — P. 143-149.
Bangsbo J.. Graham T.E., Kiens B., Saltin B. (1992). Elevated muscle glycogen and anaero-
bic energy production during exercise in man // J.Physiol. — 451. — P. 205-227.
Birch R., Noble D., Greenhaff PL. (1994). The influence of dietary creatine supplementation
on performance during repeated bouts of maximal isokinetic cycling in man // Eur.J.Appl.
Physiol. — 69. — P. 268-270.
Bogdanis G.C., Nevill M.E., Boobis (1996). Contribution of phosphocreatine and aerobic
metabolism to energy supply during repeated sprint exercise // J.Appl.Physiol. — 80. —
P. 876-884.
Casey A., Short A.H., Curtis S, Greenhaff PL. (1996). The effect of glycogen availability on
power output and the metabolic response to repeated bouts of maximal isokinetic exercise in
man // Eur.J.Appl.Physiol. — 72. — P. 249-255.
Chasiotis D„ Sahlin K., Hultman E. (1983). Regulation of glycogenolysis in human muscle in
response to epinephrine infusion // J.Appl. Physiol. — 54. — P. 45—50.
Green A.L., Simpson E.J., Littlewood J. J. et.al. (1996). Carbohydrate ingestion augments cre-
atine retention during creatine feeding in humans // Acta Physiol. Scand. — 158. —
P. 195-202.
Green H.J. (1986). Muscle power: fibre type recruitment, metabolism and fatigue // Human
muscle power/ Ed. N.L. Jones, N.McCartney, A.J.McComas. — Human Kinetics: Champaign,
IL. — P. 65-79.
288
Greenhaff P.L., Ren J.-M., Soderlund K, Hultman E. (1991). Energy metabolism in single
human muscle fibres during contraction without and with epinephrime infusion //
Am.J.Physiol. — 260. — E713-E718.
Greenhaff PL., Casey A., Short A.H. et al. (1993). Influence of oral creatine supplementation
on muscle torque during repeated bouts of maximal voluntary exercise in man H Clin.Sci. —
84, —P. 565-571.
Greenhaff P.L., Soderlund K, Ren J.-M., Hultman E. (1993). Energy metabolism in single
human muscle fibres during intermittent contraction with occluded circulation // J.Physiol. —
460. — P. 443-453.
Greenhaff PL., Nevill M.E., Soderlund K. et al. (1994). The metabolic responses of human
type I and II muscle fibres during maximal treadmill sprinting П J.Physiol. — 478. —
P. 149-155.
Hultman E., Sjoholm H. (1983). Substrate availability. — Biochemistry of exercise/
Ed. H.G. Knuttgen, H.G. Vogel, J.A. Poortmans // Human Kinetics: Champaign, IL. —
P. 63-75.
Medbo J. I. Tabala I. (1989). Relative importance of aerobic and anaerobic energy release dur-
ing short-lasting exhausting bicycle exercise // J.Appl. Physiol. — 67. — P. 1881-1886.
Putman C.T., Jones N.L., Lands L.C. et al. (1995). Skeletal muscle pyruvate., dehydrogenase
activity during maximal exercise in humans // Am.J. Physiol. — 269. — E458-E468.
Ren J.M., Hultman E. (1989). Regulation of glycogenolysis in human skeletal muscle //
J.Appl. Physiol. — 67. — P. 2243-2248.
Sahlin K., Gorski J., Edstrom L. (1990). Influence of ATP turnover and metabolite changes on
IMP formation and glycolysis in rat skeletal muscle // Am. J. Physiol. — 259. — C409-C412.
Timmons J.A., Poucher S.M., Constantin-Teodosiu D. et al. (1996). Increased acetyl group
availability enhances contractile function of canine skeletal muscle during ischemia H
J.Clin.Inest. — 97. — P. 879-883.
Walker J.B. (1979). Creatine: biosynthesis, regulation and function И Adv.Enzymol. Relat.
Areas. Mol. Med. — 50. — P. 177-242.
Глава 7
BrobergS., Sahlin K. (1989). Adenine nucleotide degradation in human skeletal muscle dur-
ing prolonged exercise // J. Appl. Physiol. — 67. — P. 116-22.
Coyle E. E, HadbergJ. M., Hurley В. E et.al. (1983). Carbohydrate feeding during prolonged
strenuous exercise can delay fatigue // J. Appl. Physiol. — 55. — P. 230-235.
Dyke D. J., Putman С. T, Heigenhauser G. J. E (1993). Regulation of fat-carbohydrate inter-
action in skeletal muscle during intense aerobic cycling // Am. J. Physiol. — 265. —
E852-E859.
Putman С. T, Spriet L. L. Hultman E. et al. (1995). Skeletal muscle pyruvate dehydrogenase
activity during acetate infusion in humans // Am. J. Physiol. — 268. — E1007-E1017.
Sahlin K„ Katz A., Broberg S. (1990). Tricarboxylic cycle intermediates in human muscle dur-
ing submaximal exercise //Am. J. Physiol. — 259. — C834-C841.
289
Глава 8
Astrand P.O., Rodahl К. (1986). Textbook of work physiology/ Ed.3. — New Youk: McGraw-
Hill.
Coggan A.R., Williams B.D. (1995). Metabolic adaptations to endurance training: substrate
metabolosm during exercise. — Exercise metabolism/Ed. M.Hargreaves. — Human Kinetics:
Champaign, IL. — P. 177-210.
Fox EL, Bowers R.W., Foss M.L. (1993). The physiological basis for exercise and sport/
Ed. W.C. Brown. — Dubuque, IA.
Goldspink G. (1992). The brains behind the brawn. — New Scientist (1st August 1992). —
P. 28-33.
Power S.K., Howley E.T. (1990). Exercise physiology: Theory and application to fitness and
performance/ Ed. W.C. Brown. — Dubuque, IA.
Saltin B., Nazar K„ Costill D.L. et al. (1976). The naturt of the training response: peripheral
and central adaptations to one-legged exercise // Acta Physiol. Scand. — 96. — P. 289-305.
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ.........................................5
СПИСОК СОКРАЩЕНИЙ...................................7
ЕДИНИЦЫ ИЗМЕРЕНИЯ .................................10
Глава 1 ФИЗИОЛОГИЯ И БИОХИМИЯ СКЕЛЕТНЫХ МЫШЦ
ПРИ МЫШЕЧНОЙ ДЕЯТЕЛЬНОСТИ...................................11
1. Физиология и биохимия физических упражнений.....11
2. Скелетные мышцы.................................12
3. Источники энергии для мышечных сокращений ......28
4. Цикл трикарбоновых кислот.......................34
5. Электрон-транспортная цепь......................37
6. Окислительное фосфорилирование .................39
7. Резервы углеводов и липидов.....................40
8. Методы оценки использования субстратов..........42
9. Факторы, влияющие на утилизацию энергетических
источников при физических нагрузках.................43
10. Доставка кровью энергетических источников и кислорода
к мышцам при физических нагрузках................ 49
11. Потребление кислорода мышцами при физических
нагрузках ......................................... 57
Выводы.............................................61
Использованная литература..........................62
Глава 2 ПУРИНОВЫЕ НУКЛЕОТИДЫ И ФОСФОКРЕАТИН ...............63
1. Свободная энергия...............................63
2. Аденозинтрифосфат и фосфокреатин................64
3. Свободный энергетический заряд и аденилатный пул.70
291
4. Убыль адениннуклеотидов в скелетных мышцах........72
5. Цикл пуриновых нуклеотидов........................75
Вы воды.............................................79
Ис пользованная литература..........................81
Глава 3 МЕТАБОЛИЗМ УГЛЕВОДОВ.................................82
1. Роль углеводов....................................82
2. Углеводное обеспечение ...........................82
3. Реакции анаэробного гликолиза и гликогенолиза.....85
4. Регенерация НАД+..................................90
5. Регуляция гликолиза ..............................92
6. Утилизация углеводов в различных тканях ..........97
7. Глюконеогенез: образование глюкозы из неуглеводных
источников...........................................99
8. Синтез гликогена.................................102
9. Гормональная регуляция метаболизма углеводов.....104
10. Методология и достижения в изучении углеводного
метаболизма ........................................108
Выводы..............................................110
Использованная литература...........................111
Глава 4 МЕТАБОЛИЗМ ЛИПИДОВ ...............................112
1. Роль липидов ....................................112
2. Типы липидов.....................................112
3. Синтез липидов ..................................119
4. Липолиз .........................................126
5. Окисление жирных кислот .........................129
6. Внутримышечный триацилглицерол...................134
7. Образование кетоновых тел и их окисление.........135
8. Регуляция метаболизма липидов при физических
нагрузках ......................................... 136
9. Влияние тренировки, направленной на развитие
выносливости, на метаболизм липидов во время
физических нагрузок.................................140
Выводы..............................................141
Использованная литература...........................142
Глава 5 МЕТАБОЛИЗМ БЕЛКОВ, АМИНОКИСЛОТ И РОДСТВЕННЫХ
МОЛЕКУЛ ....................................................143
1. Роль белков......................................143
2. Аминокислоты.....................................143
3. Структура и функция белков.......................146
292
4. Нуклеиновые кислоты и регуляция белкового синтеза .148
5. Обращаемость белков ............................. 151
6. Метаболизм аминокислот ............................153
7. Метаболизм белков при мышечной деятельности........156
8. Глюконеогенез и кетогенез .........................158
9. Регуляция обмена белков............................160
10. Биологически активные аминокислоты и их производные
вещества............................................162
11. Регуляторные пептиды и белки.......................166
Выводы.................................................168
Глава 6 МЕТАБОЛИЧЕСКИЕ ОТВЕТЫ ПРИ ВЫСОКОИНТЕНСИВНЫХ
ФИЗИЧЕСКИХ НАГРУЗКАХ.....................................170
1. Ресинтез АТФ........................................170
2. Субстраты для высокоинтенсивной мышечной работы..171
3. Взаимодействие фосфокреатина и утилизации
гликогена во время выполнения кратковременных
физических упражнений максимальной
интенсивности...................................... 176
4. Высокоинтенсивные физические нагрузки
продолжительностью более 30 с.......................176
5. Повторяющиеся физические нагрузки .................177
6. Типы ответов мышечных волокон......................178
7. Утомление .........................................180
8. Питание и максимальная физическая
работоспособность...................................185
Выводы.................................................189
Использованная литература..............................190
Глава 7 МЕТАБОЛИЧЕСКИЕ ОТВЕТЫ ПРИ ПРОЛОНГИРОВАННЫХ
ФИЗИЧЕСКИХ НАГРУЗКАХ........................................192
1. Энергетические источники для пролонгированных
физических упражнений .................................192
2. Взаимосвязь окисления углеводов и жиров в скелетных
мышцах..............................................194
3. Наличие углеводов в мышцах, питание и двигательная
активность ........................................ 199
4. Наличие углеводов в печени, питание и двигательная
активность ......................................... 203
5. Потребление углеводов непосредственно перед физической
нагрузкой ..........................................205
6. Потребление углеводов во время выполнения физических
упражнений .............................. J........ 206
293
7. Влияние повышенного потребления жиров перед
физическими нагрузками..............................208
8. Механизмы утомления..............................210
Выводы..............................................212
Использованная литература...........................215
Глава 8 МЕТАБОЛИЧЕСКАЯ АДАПТАЦИЯ К ТРЕНИРОВКЕ................216
1. Принципы тренировки..............................216
2. Адаптация к тренировке, направленной на развитие
выносливости........................................218
3. Модификация метаболического ответа на физическую
нагрузку под влиянием тренировки, направленной
на развитие выносливости............................227
4. Физиологические адаптационные процессы при тренировке,
направленной на развитие выносливости, которые влияют
на метаболический ответ во время выполнения
физических упражнений ............................232
5. Продолжительность периода, необходимого для протекания
адаптационных изменений в организме при тренировочных
занятиях, направленных на развитие выносливости, а также
для детренировки....................................236
6. Гормональная адаптация при тренировке на выносливость . 238
7. Адаптация к спринтерской и силовой тренировке.....240
8. Механизмы мышечной адаптации к тренировке .......245
9. Адаптационные изменения в крови уровня липидов,
происходящие под влиянием тренировки................247
10. Иммуносупрессия, связанная с напряженными
тренировочными нагрузками...........................249
11. Перетренировка..................................252
Выводы..............................................254
Использованная литература...........................256
ПРИЛОЖЕНИЕ 1 ХИМИЧЕСКАЯ СТРУКТУРА И СВЯЗИ.......................257
1. Химическая структура.............................257
Органические изомеры .............................258
Геометрические стереоизомеры .....................259
Оптические стереоизомеры..........................259
2. Структуры молекул сахаров .......................260
ПРИЛОЖЕНИЕ? ЭНЗИМАТИЧЕСКАЯ КИНЕТИКА И РЕГУЛЯЦИЯ РЕАКЦИЙ . 263
1. Механизмы действия энзимов.......................264
2. Энзиматическая кинетика..........................265
Реакции нулевого, первого и второго порядков .....267
294
3. Энзиматическая активность ......................267
Факторы, влияющие на активность ферментов .......268
Коэнзимы, простетические группы, кофакторы
и активаторы ....................................269
Конкурентные и неконкурентные ингибиторы.........269
Аллостерическая и ковалентная модуляции..........270
Изоформы энзимов ................................270
Использованная литература..........................270
ПРИЛОЖЕНИЕ 3 ХИМИЧЕСКИЕ БУФЕРЫ И РЕГУЛЯЦИЯ
КИСЛОТНО-ЩЕЛОЧНОГО БАЛАНСА ....................................271
1. Определения.....................................271
2. Кислотно-щелочной фон...........................271
3. Буферы..........................................272
4. Реакции в ответ на нарушение кислотно-щелочного
баланса .......................................... 276
Буферирующий (первичний) ответ...................276
Физиологический (вторичный) ответ ...............277
5. «Нормальные уровни» и формы проявления нарушения
кислотно-щелочного баланса........................280
СЛОВАРЬ ТЕРМИНОВ...................................282
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА...........................286
Навчальне видання
МОХАН Рон, ГЛЕССОН Майкл, ГРИНХАФФ Пауль Л.
БЮХ1М1Я
М’ЯЗОВОТ Д1ЯЛЬНОСТ1
I Ф13ИЧНОГО ТРЕНУВАННЯ
(росййською мовою)
Редактор — Людмила Блажееич
Техшчний редактор — Тетяпа Березяк
Художне оформления — Олександр Придатке
Комп'ютерна верстка —- ОлексШ Ткач
Коректор — Галина Андреевич
Подписано до друку 23.01.2001. Формат 70x90/16. Гари. Таймс.
Друк офс. llanip офс. № 1. Ум.друк.арк. 21,65.
Ум.фарбо-вщб. 22,24. Обл.-вид.арк. 21,76.
Зам. 0-313.
Видавництво "(Ммтйська лкература"
Нацюнального ушверситету ф!зичного виховання
i спорту УкраТни
03670, Ки1в-150, вул. Ф1зкультури, 1
КиТвське АТ "Книга"
04655, КИ1В-53, вул. Артема, 25