/














































































































Author: Павлович С.А.
Tags: биология клетки и субклеточных частиц цитология магнетизм живые организмы
Year: 1985




Text
С. А. ПАВЛОВИЧ
МАГНИТНАЯ
ВОСПРИИМЧИВОСТЬ
ОРГАНИЗМОВ
МИНСК
«НАУКА И ТЕХНИКА:
1985
УДК 576.8.095.13
Павлович С. А. Магнитная восприимчивость организ-
мов.— Мн.: Наука и техника, 1985.— 110 с.
Обобщены данные исследований магнитной восприимчи-
вости живых объектов. Доказано, что она обусловлена коли-
чественным распределением диа-, пара- и ферромагнетиков,
в частности, магнетитом, который может определять магни-
тотаксис бактерий, а также служить своеобразным компасом
для ориентации животных (насекомых, птиц) по геомагнит-
ному полю. Отмечается, что в основе гетерогенности магнит-
ной восприимчивости микроорганизмов, органов и тканей жи-
вотных лежит различие в содержании ферритина (бактерио-,
фитоферритина), ферредоксинов, других металлопротеидных
соединений и ферментов. Намечены перспективы практиче-
ского применения магнитной восприимчивости.
Книга рассчитана на экологов, биофизиков, микробиоло-
гов, физиотерапевтов.
Табл. 14. Ил. 11. Библиогр.: с. 100—109.
Научный редактор
Ю. А. Холодов, д-р биол. наук
Рецензенты:
А. П. Дубров, д-р биол. наук,
К. О. Гранстрем, канд. мед. наук
2001040000—029
П----------------68—85
М316—85
© Издательство
«Наука и техника», 1985.
ВВЕДЕНИЕ
Магнетизм известен с древних времен. Ссылки на чудодейст-
венные свойства «рудного притягивающего камня» найдены в свя-
щенных папирусах древних индусов, датированных тысячей лет до
нашей эры. Примерно в это же время в Китае начали широко ис-
пользовать магнитную стрелку. Халдеи, евреи и египтяне смотрели
на магнит как на средство, с помощью которого можно достигнуть
бессмертия. Греки и римляне применяли магнитный железняк для
лечения болезней.
Первое объяснение природы магнетизма, основанное на атоми-.
стических представлениях о строении веществ, дал Лукреций Кар
(99—55 гг. до н. э.) в поэме «О природе вещей», а первой экспери-
ментальной работой по магнетизму, описывающей почти все основ-
ные свойства магнита, была «Натуральная магия» Джиамбаттиста.
делла Порто, вышедшая в 1589 г.
Научное изучение магнетизма начато Вильямом Гильбертом.
В своей знаменитой книге, изданной в 1600 г., «О магните, магнит-
ных телах и о большом магните — Земле» он показал, что сама
Земля представляет собой огромный шарообразный магнит, ничем
не отличающийся от любого другого магнита, «от любой намагни-
ченной терреллы» (землицы). Ученый старался слить в общее пред-
ставление магнитность Земли и ее вращение вокруг оси. В его
время уже получила известность теория Н. Коперника, а X. Ко-
лумб обнаружил западное магнитное склонение, что и позволило
В. Гильберту доказать существование на Земле магнитных полюсов,
не совпадающих с географическими. Исследователь также устано-
вил невозможность разъединения разноименных полюсов магнита,
обнаружил, что магнит теряет свои свойства при сильном нагрева-
нии, а потом опять их восстанавливает.
Крупный вклад в нарождающуюся науку внесла Петербургская
академия, в частности особое значение вопросам земного магнетиз-
ма и теоретической разработке этого учения придавал М. В. Ло-
моносов.
М. Фарадей заложил основы магнитохимии и может считаться
основоположником биомагнетизма. Основывая свои исследования
на трудах А. Ампера, X. Эрстеда и других ученых, М. Фарадей в
1845 г. показал, что все тела в природе обладают магнитными
свойствами и могут притягиваться или отталкиваться магнитным
полем. Притягивающиеся к магниту вещества он назвал парамаг-
нитными (от греч. para — вдоль, около), отталкивающиеся — диа-
магнитными (от греч. dia — поперек). Почти одновременно было
установлено, что жидкости и газы тоже диа- или парамагнетики
3
и в магнитном поле ведут себя подобно твердым телам. При этом
большинство веществ и газов являются диамагнетиками, в частно-
сти все элементы, участвующие в восстановительных процессах.
К парамагнетикам относится ограниченный круг элементов и сое-
динений. В органических субстратах, например, парамагнитные
свойства проявляют кислород, калий, многие металлсодержащие
соединения и ферменты, а в живых системах, кроме того, свободные
радикалы, образующиеся в окислительных реакциях.
Современная наука исходит из того, что носителями магнетизма
являются электроны и в меньшей мере протоны и нейтроны. Маг-
нитные свойства вещества тесно связаны с движением элементар-
ных частиц и взаимодействием их друг с другом. Диамагнитные
свойства проявляются у атомов и молекул с целиком заполненными
электронными оболочками, у ионов и элементов, обладающих чет-
ным числом электронов, у которых магнитный момент равен нулю,
парамагнитные — у атомов и молекул, имеющих нечетное число
электронов и магнитный момент больше нуля.
Живые системы в основном состоят из диамагнетиков, и вслед-
ствие этого магнитная восприимчивость биомассы органических
веществ оказывается отрицательной, однако степень выраженности
диамагнетизма у разных клеток, органов и тканей организмов в
сравнительном плане не изучалась. Имеющиеся на сегодняшний
день сведения по магнитовосприимчивости клеток ограничиваются
общими указаниями на то, что ее величина определяется процент-
ным соотношением в них диа- и парамагнитных соединений, осо-
быми магнитосомами (Blakemore, 1975), электромагнитным полем,
генерируемым клеткой (Сент-Дьёрдьи, 1960), характером и интен-
сивностью энергообмена (Жолкевич и др., 1971). При этом отме-
чается, что у живых и мертвых объектов она различна (Bauer.
Raskin, 1936; Mulay, Mulay, 1964), отражает степень жизнеспособ-
ности клеток (Новицкий, 1974) и является своеобразным барометром
диссоциативных сдвигов у бактерий при изменении геомагнитного
поля (Червинец и др., 1979). Из этого вытекает, что магнитная
восприимчивость связана со многими жизненно важными функци-
ями организмов (Холодов, 1975), возможно, она даже обеспечива-
ет их способность ориентироваться в пространстве и мигрировать
по силовым линиям геомагнитного поля (Walcott, Green, 1974;
Blakemore, 1975). Рассмотрению всех этих вопросов и некоторым
аспектам практического использования магнитной восприимчивости
организмов посвящена наша книга.
1----------------------
ГЛАВА
ФИЗИКО-ХИМИЧЕСКИЕ ОСНОВЫ
ДИА- И ПАРАМАГНЕТИЗМА
Все элементы, соединения и вещества проявляют
магнитную активность, которая определяется магнитны-
ми моментами элементарных частиц, входящих в состав
каждого атома. Ввиду того что магнитные моменты про-
тона и нейтрона (ядра) на три порядка ниже наимень-
шего магнитного момента электрона, то, следовательно,
магнитные свойства атома целиком определяются элект-
ронами. Полный магнитный момент атома представляет
собой геометрическую сумму орбитальных и спиновых
магнитных моментов электронов. При этом орбитальный
магнитный момент электрона создается в результате его
движения по некоторой замкнутой орбите внутри атома,
а спиновый магнитный момент связан с собственным
моментом импульса частицы. Следовательно, атомы и
состоящие из них тела — это своеобразные магниты.
В зависимости от поведения во внешнем магнитном
поле неорганические и органические вещества условно
подразделяют на две группы — диамагнетики и пара-
магнетики.
В диамагнетике возникающая намагниченность на-
правлена навстречу намагничивающему полю, т. е. внеш-
нее магнитное поле экранируется встречным внутренним
магнитным полем вещества. Намагничиваясь против
поля, диамагнетики выталкиваются из него.
В основе диамагнетизма лежит действие внешнего
магнитного поля на орбитальное движение электронов,
вследствие чего он является неотъемлемым свойством
практически всех молекул и веществ, но в некоторых из
них может перекрываться парамагнетизмом. Отличаясь
рядом особенностей в различных видах веществ, в изоли-
рованных атомах или ионах диамагнетизм обусловлен
ларморовой прецессией электронных оболочек. ' Диамаг-
5
нитные свойства проявляются у тех веществ, у которых
магнитный момент атома или молекулы равен нулю.
Для этого нужно, чтобы все орбитальные и спиновые
магнитные моменты были полностью скомпенсированы,
ято возможно лишь в том случае, если они у двух элект-
ронов одинаковы по величине и противоположны по на-
правлению. Указанная компенсация имеёт место в цели-
ком заполненных электронных оболочках гелия, неона,
аргона, криптона, ксенона, радона. Помимо благородных
газов диамагнитными являются многие ионы, ряд эле-
ментов, обладающих четным числом электронов и очень
малым парамагнитным эффектом, который не может
перекрыть диамагнитный (Си29, Ag47, Sb51, Bi83). Диамаг-
нитными свойствами обладают также золото, свинец,
цинк, ртуть, углерод, сера, натрий, хлор, многие газы,
например, водород и углекислый газ, вода, бензол, стек-
ло, кварц, каменная соль, алмаз, большинство органиче-
ских соединений.
Высокий диамагнетизм органических веществ обу-
словлен размерами их молекул и большими радиусами
орбит наиболее удаленных электронов, которые опреде-
ляют диамагнитный момент молекулы.
Диамагнитная восприимчивость органических ве-
ществ имеет отрицательную величину и во многих моле-
кулах, как, например, у ароматических соединений, об-
наруживает заметную анизотропию. Так как твердое
тело в среднем содержит 1025—1026 атомов в 1 м3, то объ-
емная восприимчивость Хт должна иметь порядок 10~5—
10~6, т. е. равна величине, действительно измеряемой
экспериментально для диамагнитных твердых тел. Диа-
магнитная восприимчивость не зависит от температуры.
Парамагнетики — это вещества, намагничивающиеся
по направлению магнитного поля и втягивающиеся в
него. Парамагнитными свойствами обладают все атомы
и молекулы, имеющие нечетное число электронов, пол-
ный магнитный момент которых отличен от нуля.
Парамагнетиками являются калий, другие щелочные
металлы, алюминий, окись азота, воздух, элементы пере-
ходной группы периодической системы (хром, марганец,
платина, железо при температуре выше точки Кюри),
многие редкоземельные элементы, смешанные соедине-
ния. Парамагнитна также молекула кислорода, так как
в ней имеются два неспаренных электрона. Магнитные
6
моменты атомов парамагнитных элементов составляют
несколько магнетонов Бора. При этом величина, назы-
ваемая магнетоном Бора и обозначаемая 0, равна
*3,27-10 24 А/м2. У молекулы кислорода магнитный мо-
мент, например, составляет 2,8 магнетона Бора.
В отсутствие магнитного поля магнитные моменты у
парамагнетиков обычно ориентированы хаотически по
всем направлениям, и макроскопическая намагниченность
отсутствует. Однако в магнитном поле эти моменты ори-
ентируются преимущественно в направлении поля, что
приводит к суммарной намагниченности вещества, т. е.
внутри парамагнитного тела к действию внешнего поля
прибавляется действие возникающего в образце магнит-
ного момента (намагниченности).
Ввиду того что магнитные моменты ориентируются в
направлении поля и тем самым увеличивают его, пара-
магнитная восприимчивость — существенная положи-
тельная величина, т. е. больше нуля. К этому нужно до-
бавить, что парамагнетизм характеризуется не только
природой самих магнитных моментов атомов, но и темпе-
ратурной зависимостью суммарной намагниченности об-
разца.
В отличие от ферро- и антиферромагнетизма для па-
рамагнетизма характерно отсутствие остаточной намаг-
ниченности, выраженного ориентирующего взаимодейст-
вия между магнитными моментами атомов или молекул
вещества; парамагнетики очень слабо намагничиваются
даже в очень сильных магнитных полях.
Ферромагнетики (от лат. ferrum — железо) — сильно
магнитные элементы и вещества с постоянной спонтан-
ной намагниченностью, степень которой резко возрастает
даже в очень слабых магнитных полях. Одновременно
внешнее магнитное поле обеспечивает высокую степень
ориентации элементарных моментов атомов.
Ферромагнитными свойствами в обычных условиях
обладают железо, никель, кобальт, уран, гадолиний, а
при низких температурах — эрбий, диспрозий, тулий,
гольмий, тербий. Ферромагнетизм может возникать так-
же в сплавах ферромагнитных элементов друг с другом
и с неферромагнитными, в гейслеровых сплавах из не-
ферромагнитных элементов (Си2МпА1), в некоторых
парамагнитных веществах при понижении температуры.
Помимо постоянного магнетизма (состояния намаг-
7
ниченности в отсутствие внешнего поля) ферромагнети-
ки отличаются своеобразной зависимостью магнитной
проницаемости от напряженности внешнего магнитного
поля и способностью утрачивать ферромагнитные свой-
ства при некоторой температуре, называемой точкой
Кюри (Тс). Так, например, магнитная проницаемость
(р) железа резко возрастает по мере увеличения напря-
женности (Н) намагничивающего поля, достигая макси-
мума при //«2,5 Э, дальнейший рост Н вызывает умень-
шение ц, значение которой приближается к 1 в маг-
нитных полях порядка 30—50 Э. Ферромагнитная точка
Кюри для железа равняется 770 °C, т. е. железо, нагре-
тое выше этой температуры, становится парамагнитным
и к магниту практически не притягивается. Таким обра-
зом, при температуре Кюри намагниченность ферромаг-
нетика обращается в нуль. Объясняется это полным на-
рушением спинового упорядочения из-за теплового коле-
бания атомов. Полная потеря ферромагнетизма происхо-
дит, правда, не сразу, а поэтапно. При повышении тем-
пературы от абсолютного нуля, при котором отмечается
полное упорядочение всех спинов, намагниченность фер-
ромагнетика убывает сначала очень медленно. Затем
при более высоких температурах спад усиливается, пока,
наконец, резко не исчезнет в ферромагнитной точке Кю-
ри. Выше ее ферромагнитные твердые тела имеют боль-
шую восприимчивость.
По Ланжевену, твердое тело становится полностью
упорядоченным в магнитном отношении под действием
внешнего поля при абсолютном нуле температуры; у
ферромагнетика же такое упорядочение имеет место уже
при Т=ТС. Следовательно, термические флюктуации при
температуре выше Тс в ферромагнетике должны были бы
играть почти такую же роль, какую они играют в обыч-
ном парамагнетике при температуре выше 0 °C.
Ферромагнетизм проявляется только в кристалличе-
ческих телах. Он обусловлен прежде всего наличием в
.атомах ферромагнетиков нескомпенсированных спиновых
магнитных моментов электронов. У железа, например, в
недостроенной оболочке имеется 4 нескомпенсированных
спина, у кобальта — 3, у никеля — 2. Но это условие
еще не является достаточным для возникновения ферро-
магнетизма. Определяющим фактором оказывается стро-
ение кристалла, а точнее, отношение параметра кристал-
8
лической решетки к диаметру электронной орбиты, на
которой находится электрон с нескомпенсированным
спином. Это отношение должно быть больше 1,5, т. е.
где d — параметр решетки, R — радиус орбиты элект-
рона с нескомпенсированным спином.
Согласно современным представлениям, ферромаг-
нитный кристалл при температуре ниже точки Кюри со-
стоит из большого, но предельно определенного числа
очень малых доменов (участков) размером 10~2—
IO-5 см. Домены спонтанно намагничены до насыщения.
При этом магнитные моменты определенных доменов
направлены различно, вследствие чего при отсутствии
внешнего магнитного поля полный магнитный момент
ферромагнетика равен нулю. Иными словами, магнитное
поле замыкается внутри кристалла ферромагнетика.
Таким образом, доменная структура ферромагнитного
кристалла обеспечивает устойчивое состояние системы,,
которой соответствует минимум свободной энергии.
Решающую роль в создании состояния спонтанного
намагничивания ферромагнетиков играют силы обменно-
го взаимодействия между электронами (обмен атомов-
электронами), которые имеют чисто квантовую природу.
Наличие обменного взаимодействия приводит к измене-
нию энергии системы, причем энергия взаимодействия
электронов будет различной при параллельной и анти-
параллельной ориентации спинов. Ферромагнитному со-
стоянию соответствует параллельное расположение спи-
нов, а антиферромагнитному — антипараллельная ори-
ентация спинов взаимодействующих электронов.
В основе намагничивания ферромагнетика лежит воз-
действие внешнего магнитного поля на магнитные мо-
менты доменов, что в свою очередь приводит к смеще-
нию границ между доменами и повороту направления
спинов внутри их. Оба эти эффекта вызывают так назы-
ваемое техническое намагничивание, сопровождающееся
изменением формы и, главное, размера тела, что приня-
то называть магнитострикцией.
Количественно магнитострикция определяется отно-
шением удлинения образца при намагничивании к его
длине (константа магнитострикции). Изменение разме-
&
ров образца по направлению поля может быть положи-
тельным и отрицательным. Положительной продольной
магнитострикции соответствует отрицательная попереч-
ная магнитострикция.
Свойством магнитострикции обладают все тела, но у
ферромагнетиков оно проявляется наиболее отчетливо.
У классических ферромагнетиков и сплавов из них
константа продольной магнитострикции составляет со-
тые доли процента, причем у железа в зависимости от
напряженности внешнего магнитного поля она положи-
тельна или отрицательна, а у никеля и кобальта —
только отрицательна. У редкоземельных элементов поло-
жительная продольная магнитострикция может быть
необычайно большой и у поликристаллов тербия и дис-
прозия достигает 0,3%, а в монокристаллах — до 2%.
Необычайными магнитострикционными свойствами
редкоземельные элементы обязаны особенностями строе-
ния их атомов, образующих кристаллическую решетку.
Электронные облака этих элементов имеют сильно вы-
тянутую, несферическую форму и к тому же ведут себя
как «жесткие», недеформируемые. Под действием внеш-
него магнитного поля электронное облако каждого атома
поворачивается и раздвигает соседние атомы, сильно
деформируя, растягивая всю кристаллическую решетку.
В других ферромагнетиках деформация значительно
меньше — их атомы имеют сферической формы «мяг-
кую», легко деформируемую электронную оболочку, ко-
торая не вызывает большого растяжения или сжатия ре-
шетки.
Эффект магнитострикции обратим и сопровождается
изменением намагниченности магнитострикционного ма-
териала.
Представленный здесь механизм технического намаг-
ничивания ферромагнетиков без учета анизотропии маг-
нитных свойств ферромагнитных кристаллов на самом
деле является значительно более сложным. По разным
осям кристалла для его намагничивания до насыщения
требуется затратить неодинаковую энергию — меньшую
вдоль направления легкого намагничивания и большую
по оси трудного намагничивания.
Антиферромагнетики — спонтанно намагниченные
вещества со свойствами и структурой ферромагнетиков,
но в отличие от последних нескомпенсированные спины
10
электронов на недостроенных оболочках их атомов
(ионов) внутри домена имеют противоположную упоря-
доченность, т. е. антипараллельную ориентацию спинов.
Антиферромагнетиками являются, например, MnO, MnS,
NiCr, Сг2О3, VO2 и многие другие соединения.
Для антиферромагнетиков, так же как и для ферро-
магнетиков, существует определенная температура, на-
званная точкой Нееля, при которой вещество теряет ан-
тиферромагнитные свойства и ведет себя как обычный
ферромагнетик.
Антиферромагнитные кристаллы обнаруживают по-
ложительную магнитную восприимчивость при всех тем-
пературах: при низких она имеет ничтожно малую вели-
чину, с повышением температуры, когда нарушается
строгая попарная антипараллельность спинов, воспри-
имчивость увеличивается и достигает максимума в точ-
ке Нееля. Дальнейшее повышение температуры выше
критической точки Нееля сопровождается уменьшением
магнитной восприимчивости, как у всякого парамагнети-
ка. Лишь при абсолютном нуле спиновая восприимчи-
вость равна нулю.
Особые магнитные свойства имеют соединения, на-
званные ферритами (ферримагнетиками), которые пред-
ставляют собой ионные кристаллы химического состава
MxFeyOz (М — металл). Их рассматривают как неском-
пенсированные антиферромагнетики с одинаковой орга-
низацией подрешеток кристалла, связанных антиферро-
магнитно, но обладающих разным по величине магнит-
ным моментом. Пояснение сказанного представлено на
рис. 1.
В состав ферритов входят оксиды лития, цинка, ни-
келя, кадмия, меди, магния, марганца и других метал-
лов. Кроме простых ферритов бывают двойные, в состав
которых входят два окисла металла.
Так как ферриты обладают ферромагнитными свой-
ствами, то часто говорят о ферромагнетизме ферритов.
Максимальная магнитная индукция ферритов значитель-
но ниже, чем у металлических ферромагнетиков, и со-
ставляет в среднем 0,3—0,4 Т (против 0,8—2,5 Т для
металлов).
Наиболее известным соединением из группы ферри-
тов является магнетит — Fe3O4. Ввиду того что этот ми-
нерал вносит большой вклад в остаточный магнетизм
И
земных пород и, по мнению ряда авторов, обусловливает-
высокую магниточувствительность некоторых живых су-
ществ, следует кратко описать его кристаллографию.
По данным Ч. Уэрта и Р. Томсона (1969), в химиче-
ской структуре магнетита, исходя из условия электрон-
ного равновесия, содержатся двух- и трехзарядные ионы
железа (Fe2+O2-, Fe3+Of~). Рентгенографические иссле-
дования выявили, что в элементарной ячейке содержатся:
Магнитные моменты
Рис. 1. Модели подрешеток: на вершинах куба вместо атомов и
ионов величина магнитных моментов и направления намагниченно-
сти в ферромагнетике (а), антнферромагнетике (б) н ферримагне-
тике (в)
56 атомов: 32 атома кислорода и 24 атома железа (8
ионов Fe2+ и 16 — Fe3+), расположение которых и опре-
деляет постоянный магнитный момент магнетита. Нуж-
но отметить, что ионы железа в элементарной ячейке
занимают позиции двух типов. В одной из них ион желе-
за имеет в окружении 4 иона кислорода с тетраэдриче-
ской симметрией, в другой — 6 ионов кислорода с окта-
эдрической симметрией (16 таких позиций). 16 ионов
Fe3+ располагаются в позициях обоих типов (по 8 тетра-
эдрических и октаэдрических), остальные 8 ионов Fe2+
занимают октаэдрические позиции.
Спиновые моменты 8 ионов Fe3+, расположенных в
тетраэдрических позициях, направлены противоположно
спиновым моментам 8 ионов Fe3+, занимающих октаэд-
рические позиции, и упорядочены антиферромагнитно.
Эти моменты компенсируют друг друга, и ионы Fe3+ не
вносят своего вклада в суммарную намагниченность все-
го твердого тела. Она определяется магнитными момен-
тами 8 ионов Fe2+, ориентированных в одном направле-
12
иии. Таким образом, суммарная намагниченность тела
обусловлена неполной компенсацией спиновых систем
антиферромагнитного упорядочения.
Намагниченность насыщения для Fe3O4 подсчитыва-
ется следующим образом. Каждый ион железа Fe2+ об-
ладает суммарным моментом, равным 4 магнетонам Бо-
ра. Ребро элементарного куба Fe3O4, по данным рентге-
ноструктурного анализа, равно 8,37 А. Следовательно,
магнетит имеет в целом 4X8/(8,37• 10~10)3 магнетона
Бора на 1 м3, а М = 0,5-106 Ампер-виток/м. В двойном
феррите ионы металла занимают в решетке те же пози-
ции, что ионы Fe2+, и все их моменты параллельны друг
другу. Отсюда легко вычислить суммарную намагничен-
ность любого феррита. Например, намагниченность фер-
рита NiFe2O4 составляет ~0,25-166 Ампер-виток/м.
Следовательно, все разнообразие магнитных свойств
элементов и соединений объясняется особенностями
взаимодействия орбитальных и спиновых моментов
электронов. В зависимости от ориентации спинов, их упо-
рядоченности, скомпенсированности спиновых моментов
и свободной энергии системы вещества проявляют не-
одинаковую спонтанную намагниченность, по-разному
ведут себя в магнитном поле и при изменении энергети-
ческого состояния под действием внешней температуры.
Так, диамагнетизм, присущий почти всем веществам,
особенно органическим, составляющим основу биологи-
ческих объектов, не зависит от температуры, диамагне-
тики при намагничивании выталкиваются из внешнего
магнитного поля, в то время как парамагнетики под
действием магнитного поля втягиваются в него, увеличи-
вая его своей намагниченностью, которая зависит от
температуры, т. е. свободной энергии системы. Особое
состояние парамагнетизма обнаруживается у ферромаг-
нетиков и антиферромагнетиков, которые представляют
собой своеобразные кристаллы, имеющие доменную
структуру, характеризующиеся спонтанной намагничен-
ностью, очень легкой намагничиваемостью со смещением
доменов и спинов внутри них, что в свою очередь сопро-
вождается изменением их размера, формы и магнитной
восприимчивости. Можно допустить, что живая система
содержит ионные соединения металлов и металлоидов,
аналогичных ферримагнетикам, которые тоже обладают
ферромагнитными свойствами.
2 ---------------------
ГЛАВА
ПРИРОДА БИОМАГНЕТИЗМА
Со времен М. Фарадея известно, что диамагнетизм
клеток, органов и тканей — величина очень непостоян-
ная, значительно варьирующая у разных видов организ-
мов. Логично было предположить, что степень намагни-
чиваемости биологических объектов должна определять-
ся соотношением содержащихся в них диа- и пара-
магнитных веществ или элементов, а применительно к
живой системе — типом метаболизма и характером био-
химических реакций (Дубров, 1974; Холодов, 1975).
В общих чертах такую концепцию биомагнетизма при-
нимал в расчет П. В. Савостин (1937), объясняя маг-
ниторостовые реакции у растений.
Прямое измерение магнитной восприимчивости био-
массы истинных дрожжей, кишечной палочки и вуль-
гарного протея, проведенное Е. Bauer, A. Raskin (1936),
выявило возрастание диамагнетизма у погибших микро-
бов и непосредственную связь биомагнетизма с обменом
веществ. Правда, изложенные в резюме результаты этих
исследований не дают какого-либо представления об
истинной величине диамагнетизма у бактерий и говорят
лишь о том, что магнитная восприимчивость живых и
мертвых микробов различна и не зависит от способа
разрушения клеток (разбивание, повторное заморажи-
вание и оттаивание).
В 1960 г. А. Сент-Дьёрдьи, объясняя эффект Пасте-
ра, т. е. механизм перехода дрожжей от ферментации
глюкозы к окислению при смене анаэробиоза на аэро-
биоз, предположил, что сдвиг баланса производства
энергии дрожжевых клеток зависит от возмущения
электромагнитного поля и парамагнитную составляющую
надо искать среди продуктов биологического окисления.
С этой целью предлагалось измерить магнитную воспри-
имчивость аэробов, живых размножающихся клеток и
14
сравнить ее с восприимчивостью анаэробов и погибших
клеток.
Поставленные в этом аспекте исследования В. Н.
Жолкевича и др. (1971) полностью подтвердили пред-
сказания Сент-Дьёрдьи. Измеренная методом М. Фара-
дея магнитная восприимчивость тионовых бактерий
Thiobacillus thiooxidans, окисляющих тиосульфат кисло-
родом, корней, листьев и семян высших растений оказа-
лась большей, чем у факультативных и облигатных ана-
эробов Saccharomycodes ludvigi и Clostridium butyricum.
При убивании аэробных бактерий и растительных кле-
ток их диамагнетизм возрастал. Так, например, у от-
мерших высших растений он увеличивался более чем в
два раза. Магнитная восприимчивость убитых факуль-
тативно анаэробных дрожжей снижалась незначитель-
но, а у погибших клостридий с облигатно анаэробным
типом обмена и у вируса табачной мозаики, лишенного
энергообмена, не изменялась. Кроме того, авторы пока-
зали, что парамагнитная составляющая магнитной вос-
приимчивости растущей клетки положительно коррели-
рует с энергетической эффективностью окисления, а не с
интенсивностью обмена вообще.
По данным Ю. И. Новицкого (1971, 1974), магнитная
восприимчивость живых и мертвых клеток растений рав-
нозначна. Более того, у зерен кресс-салата, ржи Гибрид-
ная-2 она практически неразличима и при измерении по
методу Гун соответствует —0,656-10~6 и —0,650-10-6.
Примерно такой же величиной диамагнетизма характе-
ризовался первичный корень 6-дневных темновых про-
ростков бобов сорта Кузьминские, выросших в геомаг-
нитном и искусственном магнитном поле напряженностью
3,6-105 А/м и 1,5-106 А/м. Относительным парамагнетиз-
мом обладала лишь верхушка корня. Из этого следует,
что степень магнитной восприимчивости растений опре-
деляется также активностью биохимических процессов.
Дж. Ингрэм (1961) полагает, что парамагнетизм
биологических объектов обусловлен свободными радика-
лами, играющими главную роль в энергообмене аэробов.
Известный вклад в парамагнитную составляющую,
очевидно, вносят также триплетные возбужденные элект-
ронные состояния (Evans, 1965).
Хорошо подкрепляет такие взгляды исследование
О. П. Самойловой и Л. А. Блюменфельда (1961), пока-
15
завшее изменение «псевдоферромагнетизма» в динамике
роста Saccharomyces cerevisiae. Выращивая дрожжи воз-
душно-приточным способом по 10-часовой схеме, авторы
через 30 мин после начала опыта регистрировали чрез-
вычайно сильный сигнал магнитного резонанса. По ме-
ре развития культуры его интенсивность нарастала, но
после 7 ч резко падала и через 8—10 ч, когда в основном
заканчивался процесс деления и роста дрожжей, возвра-
щалась к исходному уровню.
Относительно того, как сказывается на магнитной
восприимчивости химический состав организмов, можно
лишь предполагать. Между тем магнитным материалом
клетки может оказаться ферритин, хромопротеиды (ге-
моглобин, цитохромы), ферредоксины, другие металло-
протеиды, в том числе металлоферменты, химические
соединения и элементы, обладающие высоким уровнем
парамагнетизма.
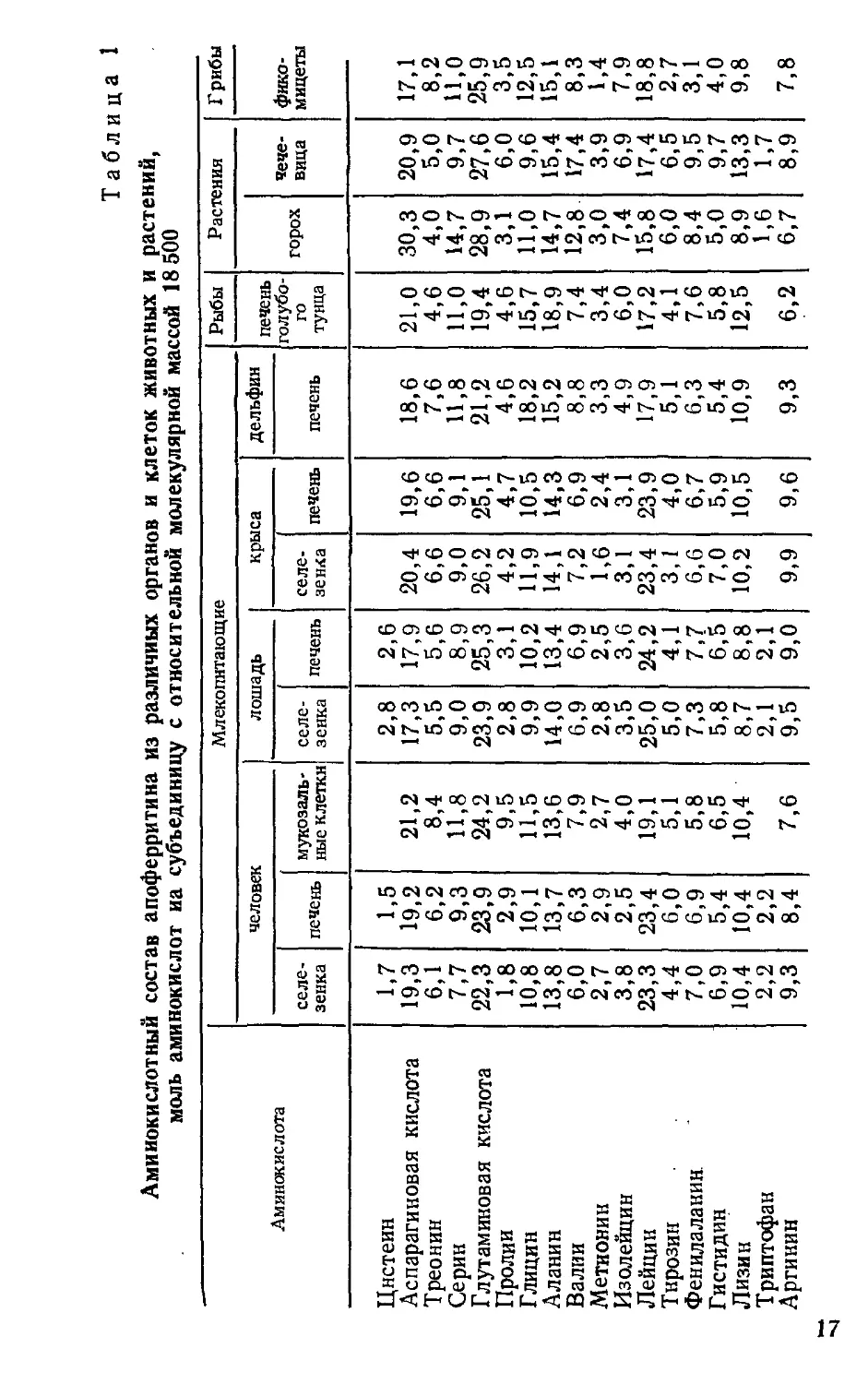
2.1. МАГНЕТИЗМ КЛЕТОЧНОГО ФЕРРИТИНА
Ферритин представляет собой железосодержащий
белок. Он был открыт в 1894 г. N. Schmideberg, а в кри-
сталлическом виде получен в 1934 г. V. Laufberger. В со-
став ферритина входят 18 аминокислот, количество ко-
торых незначительно варьирует в различных клетках,
органах и тканях человека, животных и растений
(табл. 1). Так, например, у ферритина гороха количест-
во аспарагина намного больше, чем у чечевицы, печени
и селезенки человека, лошади, крысы, дельфина и других
организмов. По содержанию пролина наиболее богаты
мукозальные клетки. Наименьшее количество тирозина,
фенилаланина и гистидина имеется у фикомицетов. Со-
держание железа в ферритине колеблется от 12 до 231%',
вследствие чего он может иметь разнообразные цветовые
оттенки — от красноватого до темно-коричневого. Элект-
ронномикроскопические исследования показывают, что
белковая часть ферритина, известная как апоферритин,
состоит из 24 субъединиц. Химическая структура связей
железа точно не известна, но, по-видимому, это фосфор-
ные соединения (FeOOH)g-(FeO-ОРОзНг), возможно,
азотные и другие композиции, которые в виде малень-
ких мицелл (55—60 А) заключены внутри апоферритина
16
Таблица 1
Аминокислотный состав апоферритина из различных органов и клеток животных и растений,
моль аминокислот на субъединицу с относительной молекулярной массой 18 500
Аминокислота Млекопитающие Рыбы Растения Грибы
человек лошадь крыса дельфин печень голубо- го тунца горох чече- вица фико- мицеты
селе- зенка печень мукозаль- ные клетки селе- зенка печень селе- зенка печень печень
Цистеин Аспарагиновая кислота 1,7 19,3 1,5 19,2 21,2 2,8 17,3 2,6 17,9 20,4 19,6 18,6 21,0 30,3 20,9 17,1
Треонин 6,1 6,2 8,4 5,5 5,6 6,6 6,6 7,6 4,6 4,0 5,0 8,2
Серин 7,7 9,3 11,8 9,0 8,9 9,0 9,1 11,8 11,0 14,7 9,7 11,0
Глутаминовая кислота 22,3 23,9 24,2 23,9 25,3 26,2 25,1 21,2 19,4 28,9 27,6 25,9
Пролии 1,8 2,9 9,5 2,8 3,1 4,2 4,7 4,6 4,6 3,1 6,0 3,5
Глицин 10,8 10,1 11,5 9,9 10,2 11,9 10,5 18,2 15,7 11,0 9,6 12,5
Аланин 13,8 13,7 13,6 14,0 13,4 14,1 14,3 15,2 18,9 14,7 15,4 15,1
Валии 6,0 6,3 7,9 6,9 6,9 7,2 6,9 8,8 7,4 12,8 17,4 8,3
Метионин 2,7 2,9 2,7 2,8 2,5 1,6 2,4 3,3 3,4 3,0 3,9 1,4
Изолейцин 3,8 2,5 4,0 3,5 3,6 3,1 3,1 4,9 6,0 7,4 6,9 7,9
Лейцин 23,3 23,4 19,1 25,0 24,2 23,4 23,9 17,9 17,2 15,8 17,4 18,8
Тирозин 4,4 6,0 5,1 5,0 4,1 3,1 4,0 5,1 4,1 6,0 6,5, 2,7
Фенилаланин 7,0 6,9 5,8 7,3 7,7 6,6 6,7 6,3 7,6 8,4 9,5 3,1
Гистидин 6,9 5,4 6,5 5,8 6,5 7,0 5,9 5,4 5,8 5,0 9,7 4,0
Лизин 10,4 10,4 10,4 8,7 8,8 10,2 10,5 10,9 12,5 8,9 13,3 9,8
Триптофан Аргинин 2,2 9,3 2,2 8,4 7,6 2,1 9,5 2,1 9,0 9,9 9,6 9,3 6,2 1,6 6,7 1,7 8,9 7,8
(Farrant, 1954; Bjork, Fish, 1971; Crichton, 1975; Treffry,
Banyard, 1977; Adelman, Drysdale, 1977).
Ферритин — один из самых древних белков, который
постоянно обнаруживается в брюхоногих моллюсках,
хитонах и у них, очевидно, является предшественником
магнетита (Lowenstam, 1962; Towe, Lowenstam,
1963). У животных этот белок находят преи-
мущественно в селезенке, печени и костном мозге
(Granick, 1943), а у человека — во многих органах
и тканях, включая мукозальные клетки тонкого кишеч-
ника, клетки яичек, почек, сердца, скелетных мышц, лег-
ких, поджелудочной железы, щитовидной железы, пла-
центы, надпочечников (Granick, 1946; Allfrey et al.,
1967; Arora et al., 1970). Недавно ферритин нашли в сы-
воротке крови. Помимо млекопитающих ферритин обна-
ружили в организме голубого тунца (Shinjo, Shimada,
1968; Kato, Shimada, 1970), в яйцах и эмбрионах лягу-
шек (Brown, Caston, 1962), в моллюсках (Towe et al.,
.1963), в печеночно-поджелудочной и половой системе
улиток (Heneine et al., 1969), в спруте и осминоге (Nardi
et al., 1971), в хлорогенных клетках кольчатых червей
Arenicola marina (Roche et al., 1961), в червях (Lumbri-
cuc terrestris), в растениях и грибах (Hyde et al., 1963;
Seckbach, 1968; Robards, Robinson, 1968; Robards,
Humpherson, 1969; Laulhere et al., 1973; Crichton, 1973;
Crichton, 1975). В наибольшем количестве фитоферри-
тин встречается в клетках, которые не ассоциированы с
активным фотосинтезом, как, например, в плодах яблок,
Таблица 2
Относительная молекулярная масса и коэффициент осаждения
апоферритинов
Органы Коэффициент седиментации Относительная молекулярная масса
олигомера субъединиц
Селезенка лошади Печень лошади Селезенка человека Печень человека Печень крысы Ткань морской свинки 17,12—17,6 16,9 16,8 17,3 17,0 17,7—18,0 430000—500000 18300—25000 18400—18800 18300—18500 18000—18700 19000 17000
18
Таблица 3
Содержание железа в различных видах ферритина
Объект исследования Содержание Fe (вмг) на 1 мг белка Количество атомов Fe на 1 молекулу, тыс.
Крыса печень 0,38 3,000
почки 0,30 2,350
селезенка 0,41 3,240
сердце 0,40 3,170
бедренная мышца 0,38 3,000
гепатома 0,10—0,44 0,800—3,500
Селезенка лошади 0,32 2,550
0,35 2,730
0,30 2,370
0,52 4,120
0,53 4,200
Селезенка дельфина 0,21 1,660
Печень кролика 0,19 1,520
Печень голубого тунца 0,16 1,160
Яйца лягушки 0,13 1,030
корнях гороха и бобов, стрелке меристемы, в тканях
листьев, имеющих низкую фотосинтетическую актив-
ность, хлоропластах. У грибов ферритин находится глав-
ным образом в спорах, спорангиях, спорангиофорах
(David, Easterbrook, 1971; Spartalian et al., 1975).
В зависимости от видовой принадлежности относи-
тельная молекулярная масса ферритина колеблется от
443 000 до 500 000, а молекулярная масса субъединиц
апоферритина—от 18 300 до 25 000 (Bryce, Crichton,
1971; Crichton, 1973). К этому нужно добавить, что у
различных людей, животных и растений белковые субъ-
единицы ферритина могут быть гибридными (табл. 2).
Столь же значительно отличаются размеры молекул
ферритина и содержание в них атомов железа (табл. 3).
Структура ферритина подобна небольшим икоса-
эдральным вирусам, но по размеру он в 3—4 раза мень-
ше любого из них (рис. 2). В частности, диаметр моле-
кулы ферритина селезенки лошади равен 12—13 нм, а
внутренняя полость, содержащая 4500 атомов железа,
приблизительно 8 нм. A. Treffry et al. (1977) внутри
ферритина нашли соли ртути, иОг+, Tb3+, CH3Hg+,
(Nb6Cli2)2+, Zn. По сравнению с вирусами молекула фер-
ритина отличается очень высокой устойчивостью, и даже
19
грубые химические и физические воздействия не вызы-
вают ее деструкции или деполимеризации. Это объясня-
ется наличием прочных связей между протеиновыми
субъединицами ферритина, с одной стороны, апоферри-
тином и железом — с другой (Massover, 1977).
Основываясь на результатах исследований физиче-
ских свойств ферритина лошади, гороха, чечевицы,
R. R. Crichton (1973), R. R. Crichton et al. (1978) пола-
гают, что молекулярная масса фитоферритина и его бел-
ковых субъединиц, размеры молекул и количество в них
атомов железа приблизительно такие же, как у ферри-
тина всех млекопитающих (табл. 4). Ярче контрасти-
руют размеры железосодержащих сердцевин микоферри-
Рнс. 2. Структура молекулы ферритина и апоферритиновых субъ-
единиц
20
Таблица 4
Физические свойства ферритина
Показатель Горох Чечевица Селезенка лошади
Содержание железа (атомов на молекулу) 2140 2130 1980
Относительная молеку- лярная масса моле- кулы ’463 700 +5000 498 500+5000 443 000
Относительная молеку- лярная масса субъ- единиц 20 300+200 21 400+400’ 18 500
Радиус круга, нм 5,27 5,48 4,90
Максимальный радиус круга, нм 6,80 7,07 5,17
тина и ферритина лошади. Как показали К. Spartalian
et al. (1975), величина железистых пучков Phycomyces
blakesleeanns на два порядка меньше, чем в ферритине
лошади. Интересно, что в мицелии этого вида гриба,
растущего на агаровой среде с наличием в ней изото-
па 57Fe, значительное количество железа находилось
вне ферритина и было вкраплено в оболочки и стенки
гиф. Ферритиновое железо находилось в спорангиях.
В культуре гриба, растущей при низкой концентрации
кислорода, во время спорообразования 80% ферритина
переходит в споры. Селективное накопление ферритина
в спорах может иметь важное значение при их прора-
стании, так как освобождающееся при этом железо не-
посредственно включается в биосинтез микроба (David,
1974).
Третичная и четвертичная структура ферритинов
одинакова.
Ферритин — антиферромагнетик с точкой Нееля
20±3° К (Schoffa, 1965). Магнитный момент ферритина
клеток живого и растительного происхождения состав-
ляет 3,81—3,85 магнетона Бора. Низкое значение маг-
нитного момента ферритина может зависеть от того,
что атомы железа находятся в довольно необычном со-
стоянии, имея только три иеспаренных электрона и кис-
лород в качестве лиганда в квадратно-плоскостной кон-
фигурации. Недавно A. Blaise et al. (1965) обнаружили
заметную зависимость магнитной восприимчивости фер-
21
ритина от величины поля и получили магнитный момент
5,08 магнетона Бора при высокой напряженности поля.
Они также показали, что ядра ферритина проявляют
супермагнетизм.
Особого внимания заслуживает бактериоферритин.
.Он обнаружен W. A. Bulen et al. (1973), выделившими из
,Azotobacter vinelandii особый белок, в составе которого
был найден уникальный цитохром 0-типа, содержащий
20% негеминового железа. Одновременно авторы отмети-
ли, что азотобактер имеет цитохром 3557,5- Выделив его
в кристаллическом виде, Е. I. Stiefel, G. D. Watt (1979)
показали, что цитохром 0557,5 содержит большое количе-
ство негеминового железа и является ферритиноподоб-
ной субстанцией. В очищенном бактериоферритине-ци-
тохроме железа несколько меньше, чем в ферритине се-
лезенки лошадей (13—20% от общей массы белка). Глав-
ное отличие бактериоферритина от других ферритинов,
пожалуй, только в низкой молекулярной массе субъеди-
ниц апоферритина. По другим свойствам он идентичен с
ферритином млекопитающих. В молекуле бактериофер-
ритина может содержаться то же количество атомов
железа Fe3+, что и в ферритине селезенки лошадей. Маг-
нитный момент бактериоферритина 3,7 магнетона Бора.
Авторы допускают, что бактериоферритин-цитохром
азотобактера участвует в регуляции дыхания, биосинтезе
и фиксации азота. В связи с этим было выдвинуто пред-
положение, что бактериоферритины вырабатываются
также другими видами бактерий, содержащих цитохромы
типа 0, и в частности отдельными разновидностями эше-
рихий, сенной и чудесной палочками, псевдомонадами
(Pseudomonas putida), многими бактериями актиноми-
цетной линии, как, например, стрептомицетами (Str.
griseus, Str. fradiae), некоторыми микрококками (Af. ly-
sodeikticus, M. denitrifleans), коринеформными бакте-
риями, микобактериями, аспергилловыми плесенями,
истинными дрожжами, фотосинтетическими бактериями
(Hadihara et al., 1975; Habernkorn et al., 1979). •*
В заключение необходимо указать, что недавно
Е. R. Bauminger et al., (1979), D. P. E. Dickson, S. Rot-
tern (1979) выделили из эшерихий и Proteus mirabilis
железосодержащие белки с молекулярной массой около
500 000. Их синтез происходил на минимальной среде с
глицеролом, обогащенной изотопом железа 57Fe. Боль-
22
шая часть железосодержащих белков находилась во
внешней мембране бактерий. Строение протеинов было
таким же, как и у ферритина, но по физико-химическим,
магнитным и электрическим свойствам они отличались
от всех известных белков.
Образование ферритина можно представить следу-
ющим образом: Ре2+-|-О2+апоферритин-> ферритин.
Рис. 3. Пространственная модель ферритина с сайтами окисления
Fe2+ в Fe3+ на субъединицах апоферритина
Предложено несколько моделей ферритина. Прост-
ранственная модель W. Niederer (1970) допускает на-
личие в протеиновой оболочке апоферритина пор, через
которые внутрь ее молекулы могут проникать маленькие
молекулы и ионы. Предполагают, что на внутренней по-
верхности апоферритина находятся сайты активации (ка-
тализа) окисления Fe2+ в Fe3+ (рис. 3).
Альтернативную модель ферритина предложили
L. Pape, J. S. Multani, С. Stitt, R. Saltman (1968). По их
мнению, его формирование начинается со сборки апо-
ферритиновых олигомеров вокруг предварительно синте-
зированных мицелл с железом.
Ферритин — внутриклеточное депо железа Fe3+ в
нетоксичной форме. У растений этот протеин обеспечи-
вает синтез ферментов, содержащих геминовое и неге-
миновое железо и участвующих в фотосинтезе (Crichton
23
et al, 1978), а у млекопитающих и, возможно, кольчатых
червей (Roche et al., 1961)—синтез гемоглобина.
Тканевый ферритин находится в динамическом рав-
новесии с железом плазмы, которое транспортируется к
трансферрину и далее на рецепторы клеточных мембран
зрелых ретикулоцитов для синтеза гемоглобина. Железо
может освобождаться из интактных молекул или из
ядер ферритина после разрушения белковой оболочки.
Мобилизация железа осуществляется путем восстанов-
ления до более растворимых ионов Fe2+. В этом процес-
се участвует цистеин, аскорбиновая кислота, глутатион,
НАД-Н, а в отсутствие восстановителей — специфиче-
ские низкомолекулярные хелаты. Восстановители и хе-
латирующие соединения мобилизуют, однако, низкий
процент железа (около 15% в течение многих часов и
дней), поэтому трудно представить, что в организме
этот механизм является основным. В печени за восста-
новление и мобилизацию железа ответственна ксантино-
ксидаза (Mazur et al., 1958; Green, Mazur, 1967). При
этом восстановление железа ферментом ведет к измене-
нию магнитной восприимчивости ферритина (Tanaka,
1956). Физиологическим стимулом для освобождения
железа из ферритина может быть тканевая гипоксия,
сопровождающаяся возрастанием количества красных
кровяных клеток и уровня ксантиноксидазы в печени
(Mazur et al., 1958).
В последнее время найден специфический фермент
мобилизации железа с ферритина, названный феррире-
дуктазой. Представленные на рис. 4 и 5 схемы мобили-
зации железа с трансферрина на ферритин показывают,
что включение и удаление железа с ферритина проис-
ходит при его окислении и восстановлении.
Столь детальное описание ферритина и обмена желе-
за не случайно. Если ферритин действительно магнито-
рецепторное вещество, то всякое количественное и каче-
ственное его изменение в организме должно отразиться
на магнитовосприимчивости клеток и тканей. Ввиду это-
го самого пристального внимания заслуживают нару-
шения синтеза ферритина.
У здоровых взрослых людей и животных, как ука-
зывалось выше, ферритин депонируется в ретикулоэндо-
телиальных клетках паренхиматозных органов, но мо-
жет находиться также в сыворотке. При этом молекулы
24
ферритина одной и той же ткани почти однотипны, но в
различных органах отличаются неодинаковой молеку-
лярной массой отдельных субъединиц. Имеются сооб-
щения, что различные ферритины имеют неодинаковое
количество железа.
М. S Linder et al. (1975) из различных органов и тка-
ней выделили три вида ферритина, отличающиеся ско-
. восстановители
г- __ р- 9^*
ге -трансферам+хелаты —: ..... ге •хелаты
3 + 9 +
Fe -ферритин ~ Fe —ферритин
Рис. 4. Модель переноса железа из трансферрина на ферритин с
участием восстановителей и хелатов
Ге2++ окислители апоферритин НАД+ + Fe^
ферридуктазп
восстановители ферритин НАДН + Н+
Рис. 5. Схема включения железа в апоферритин с участием фер-
риредуктазы
ростью миграции в геле, аминокислотным и пептидным
составом, содержанием сульфгидрильных групп, соот-
ношением белковых субъединиц в апоферритине. Так,
ферритин крыс в отличие от ферритина лошади содержит
большие олигомеры с молекулярной массой 18 000—
19 000 и малые — с молекулярной массой 13 000—
14 000.
J. W. Drysdale et al. (1975) выделили 15 типов фер-
ритина, в апоферритине которых белковые олигомеры
имели молекулярную массу от 7000 до 24 000. Однотип-
ным оказался ферритин селезенки и печени, почек и
сердца. В еще большей мере отличается ферритин у боль-
ных людей. В частности, изменение структуры ферритина
происходит при циррозах печени (Reissman, Dietrich,
1956), при вторичном гемохроматозе (Arosio et al.,
1977), при злокачественных новообразованиях (Drysdale
25
ct al., 1975; Linder et al., 1975; Munro et al., 1975; Huzard,
Drysdale, 1977). Тщательно изучив это явление при опу-
холях почек, поджелудочной железы, толстого кишечни-
ка, L. W. Powell et al. (1977) предполагают, что вариа-
ции строения ферритина обусловлены формированием
белка на разных полирибосомах.
Издавна известно, что воспаление блокирует осво-
бождение из тканей железа, а это в свою очередь про-
является снижением насыщения им трансферрина и раз-
витием гипохромной анемии с возрастанием эритроци-
тов, содержащих порфирин (Freireich et al., 1957; Noyes
et al., 1960; Cartwright et aL, 1971; Fillet et aL, 1974;
Hershko et aL, 1974). При экспериментальном асептиче-
ском воспалении у крыс, вызванном внутримышечными
инъекциями скипидара, в печени возрастал синтез фер-
ритина, задерживался выход железа и на фоне увеличе-
ния уровня фибриногена и других протеинов в сыворотке
снижалось его количество (Hershko, Konijn, 1977).
Интересно, что бактериостатическое действие сыво-
ротки при снижении в ней концентрации железа усили-
вается. Это, в частности, показали А. М. Ganzoni,
М. Puschmann (1977) на мышах, предварительно инфи-
цированных сублетальной дозой эшерихий, которые вы-
зывали спустя 10 ч уменьшение в крови уровня железа
с 365,0 до 38,0 мг/100 мл. Последующее заражение жи-
вотных сальмонеллами мышиного тифа вызывало лег-
кую инфекцию без летальных исходов. Наоборот, вну-
трибрюшинное инфицирование мышей теми же сальмо-
неллами с одновременным введением 100 мг железа
l[NH4Fe(SO4)2- 12Н2О] приводило к их гибели в течение
48 ч, что, возможно, обусловлено усиленным синтезом
железотранспортных систем (сидерохромов). При этом
важно подчеркнуть, что активация вирулентности саль-
монелл достигалась не всеми железосодержащими соеди-
нениями. В этом отношении ферритин, содержащийся
в селезенке лошади, оказался инертным веществом.
Увеличение синтеза ферритина в тканях происходит
при талассемии, апластической, сидеробластической,
серповидноклеточной анемиях и других гемоглобинопа-
тиях как следствие лизиса собственных или трансфузи-
онных эритроцитов (Cerami et al., 1977). Вместе с этим
при талассемии в клетках эритроидного ряда в 37 раз
увеличивается синтез апоферритина и нарушается соот-
26
ношение ферритина к гемоглобину (Eylar, Matioli,
1965).
Повышается уровень ферритина и железа в организ-
ме людей с наследственным гемосидерозом и гемохро-
матозом (Drysdale et al., 1975). Имеются и противопо-
ложные данные. Так, L. W. Powell et al. (1975) у боль-
ного гемохроматозом обнаружили резкое снижение
ферритина, а в почках и поджелудочной железе его пол-
ное исчезновение.
Избыточное отложение ферритина в тканях и органах,
как правило, коррелирует с высоким его уровнем в
плазме. Ферритинемия сопровождает многие болезнен-
ные состояния, в частности шок и гипертензию (Shorr,
1956), отек (Shorr et al., 1950; Shricanlia, Gopalan, 1959),
гепатоцеллюлярный некроз (Reissman, Dietrich, 1956;
Augunst, 1968), тяжелую гемолитическую анемию. Боль-
шое количество ферритина в плазме отмечается у боль-
ных злокачественными новообразованиями. При раке
мобилизация ферритина идет из пораженной ткани, а
при нераковых заболеваниях — из селезенки и печени
(Drysdale et al., 1975).
Подводя итоги сказанному, надо отметить, что в плаз-
ме крови здоровых людей находится 35,6 нг/мл ферри-
тина, при железодефицитных анемиях его концентрация
снижается до 5,4 нг/мл, а при болезнях с нарушением
обмена железа и избыточным образованием тканевого
ферритина увеличивается до 1700 нг/мл (Beamish et al.,
1972).
2.2. МАГНЕТИЗМ МЕТАЛЛОПРОТЕИДОВ КЛЕТОК
Хромопротеиды — это железосодержащие белки с
наличием в них окрашенных простетических групп раз-
личных классов органических соединений. К хромопро-
теидам относят гемоглобин, миоглобин, . цитохромы и
подобные им металлоферменты. Самую высокую намаг-
ничиваемость из них следует, естественно, ожидать от
гемоглобина, состоящего из белка глобина и 4 атомов
железа, входящих в состав гемов (96 и 4)% от массы
молекулы соответственно). Молекулярная масса гемо-
глобина эритроцитов человека равна 64 458; в одном
эритроците находится около 400 млн. молекул гемогло-
бина. Присоединяя кислород, гемоглобин превращается
27
в оксигемоглобин. У этих производных гемоглобина же-
лезо двухвалентное. Действие на гемоглобин окислов
азота, феррицианида калия, хинонов вызывает окисле-
ние Fe2+ до Fe3* и образование метгемоглобина. При
этом изменяются магнитные свойства гемоглобина.
В частности, М. Cerdonio et al. (1978, 1980) наблюдали
уменьшение парамагнетизма деионизированного карбо-
ксигемоглобина карпа при добавлении в слабокислый
раствор (pH 5,6—6,7) гексафосфата иннозитола. Диа-
магнетизм цианметгемоглобина оставался неизменным,
что свидетельствует против возможной трансформации
молекулы глобина.
Вклад миоглобина в парамагнетизм тканей (мышц,
сердца), по-видимому, небольшой, поскольку он содер-
жит всего лишь одну полипептидную или глобиновую
цепь и гем, в котором имеется один атом двухвалентно-
го железа.
Магнитовосприимчивость организмов лишь в очень,
небольшой мере может зависеть от цитохромов а, b и с
ввиду их ничтожно малых количеств в клетках, органах
и тканях (Shimizu et aL, 1981; Thomson et al., 1981).
В этом же аспекте нужно рассматривать вклад в пара-
магнетизм металлоферментов типа пероксидаз и ката-
лаз, идентичных по характеру простетических групп
цитохромам (Harbury, Marks, 1978) или белков-фермен-
тов, не имеющих порфириновых структур и координаци-
онно связанных с атомами разных металлов, как, напри-
мер, содержащую медь полифенолоксидазу, связанную
с цинком карбоангидразу, вырабатывающуюся ацинар-
ными клетками поджелудочной железы и некоторыми
бактериями карбоксипептидазу, выделяющуюся из почек
свиньи лейциноаминопептидазу и другие металлопротеи-
ды и пептидазы (Ludwig, Lipscomb, 1978; Malkin, 1978).
Исключение может представлять каталаза у М. lysodeik-
ticus, составляющая 1—2% сухой массы культуры микро-
ба. D. Herbert, J. Pinsent (1948) подсчитали, что это при-
близительно 10 доз каталазы, имеющейся в человечес-
ких эритроцитах. Одна особь микрококка может содер-
жать 10—20-Ю3 молекул каталазы. Около 0,3% сухой
массы каталаза составляет у Rhodopseudomonas sphe-
roides, если культура вырастает в аэробных условиях
(Clayton, 1959).
Ферредоксины и другие жолезо-серусодержащие бел-
28
зги как магнитный материал содержат от одного (рубре-
доксин клостридий) до восемнадцати (ДФН-Н деги-
дрогеназа митохондрий) атомов железа. Являясь фермен-
тами, они участвуют в окислительно-восстановительных
реакциях всех известных форм жизни и при этом
меняют магнитную восприимчивость, оптические и дру-
гие характеристики в строгом количественном соответ-
ствии с числом перенесенных электронов (Lovenberg,
1974; Thomson et al., 1981; Johnson et al., 1981).
Молекулярная масса ферредоксинов и железосодер-
жащих белков колеблется от 6000 (Chlorobium) до
760 000 (сульфитредуктаза кишечной палочки).
Бактериальные ферредоксины — прототип азотфик-
сирующих энзимов (Orme-Johnson, 1978), а у некоторых
видов микробов полностью им тождественны, как на-
пример у Klebsiella pneumoniae. Этот вид микроба со-
держит настоящую нитрогеназу (Johnson et al., 1981).
Полученная из клебсиелл нитрогеназа, как и у азот-
фиксирующих бактерий Azotobacter vinelandii, состоит
из малого протеина Кр 2, содержащего 4 иона железа и
4 кислотнолабильных атома серы и большого протеина
Кр 1, имеющего высокий уровень железа, серы, а также
молибдена (Stephens et al., 1981).
Негеминовое железо — составная часть ферментов
дыхательной цепи дрожжей, грибов, микобактерий, азот-
фиксирующих бактерий, кишечной палочки и многих
других видов бактерий (Bragg, 1974).
Уровень негеминового железа в дыхательных фермен-
тах микробов может быть во много раз большим, чем в
цитохромах и флавиновых компонентах клеток живот-
ного происхождения. Отношение негеминового железа
к геминовому в зависимости от исходного количества
железа в среде может составлять у коринебактерий диф-
терии 4,3 : 1, 33 : 1, у Micrococcus denitrificans — 7,8 : 1,
35:1, у микобактерий превышает уровень цитохромов в
200 раз. Нечто подобное наблюдается и у других видов
микроорганизмов.
Среди организмов широко распространены ферредок-
синоподобные протеины, в частности сильно проявляю-
щие парамагнетизм дегидрогеназы. Особенно богаты
ими бактерии семейств Thiorhodaceae (Thiocapsa ffori-
dona, Chromatium), Athiorhodaceae (Rhodospirillum
rubrum), Chlorobacteriaceae (Chlorobium, Pelodictyon),
29
Т аблица 5
Железосодержащие протеины и ферменты микроорганизмов
Субстанция Распространение, источник выделения Автор исследования
Кислородсвязывающие белки гемоглобиноподобиые Ограниченно распространенные J. Lascelles, 1964
гемоглобин Rhizobia N. Ellfolk, 1972
Железодепонирующие белки ферритин Гидропероксидазы каталаза Phycomyces blakeslecanus К- Spartalian et al., 1975
Широко распространена у аэробов, особенно много D. Herbert, J, Pinsent, 1948;
пероксидаза у М. lysodeikticus и Rhodopseudomonas spheroides Пивные дрожжи, эшерихии, псевдомонады, серные бактерии, некоторые грибы и актиномицеты, A. vinelandИ Bas. subtilis, Staph, pyogenes, Saccharomyces ce- R. K. Clayton, 1959 T. Vonetani, 1974
Переносящие электроны проте- ины цитохромы N. J. Jacobs, 1974
Негеминовые белки железо-серусодержащие reviziae, другие аэробы, облигатные анаэробы Desulfovibrio, Vibrio succinogenes, Bacteroides Дрожжи, грибы, коринебактерии, микобактерии, R. D. Brogg, 1974
гидрогеназы микрококки, эшерихии и многие другие виды бак- терий Широко распространены среди многих семейств L. E. Mortenson, J.-Ch. Chen,
нитрогеназа бактерий Азотфиксирующие виды, Klebsiella pneumoniae 1974 W. H. Orme-Johnson, 1978;
M. K, Johnson, et al. 1981,
глутаматсинтетаза
риборедуктаза
Железо-флавопротеины
Dihydroorotate
dehydrogase
сукцинатдегидрогеназа
нитроредуктаза
ксантин оксидаза
НАД Н-дегидрогеназа
малат-витамин К-редуктаза
аденилилсульфатредуктаза
НАДФН-сульфитредуктаза
форматдегидрогеназа
Оксигеназы
геминового типа
триптофандиоксигеназа
негеминового типа
различные субстраты
Железоактивирующие ферменты
Эшерихии
Эшерихии
Zymobacterium oroticum
Широко распространены
Micrococcus denitrificans
Micrococcus lactilyticus
Azotobacter vinelandii
Mycobacterium phlei
Desulfovibrio vulgaris
Энтеробактерии
Pseudomonas oxalaticus
Широко распространены
Широко распространены
Эшерихии, псевдомонады
О. Karlstrom, 1972
V. Alleman, P. Handler, 1967
C. Veeger et al., 1969
P. Forget, D. V. Dervarta-
rian, 1972
S. T. Smith et al., 1967
D. V. Dervartarian, 1972
С. K. R. Kurup, A. F. Bro-
die, 1967
H. D. Peck et al., 1972
L. M. Siegel et al., 1972
Th. Hopner, A. Troutwein,
1972
M. Nozaki, Y. Ishimura, 1974
Methanomonadaceae, Pseurfomonadaceae, Spirillaceae
(Vibrio succinogen.es, Desuifovibrio desulfuricans), Azo-
tobacter iaceae (Azotobacter vinelandii, A. agile, A. chroo-
coccum), Rhizobiaceae, Enterobacteriaceae (эшерихии,
протей, 5. Oranienburg, Aerobacter aerogenes, 4. cloacae),
Micrococcaceae (Micrococcus lactilybicus, M. aerogenes.
M. denitrifleans), Lactobacillaceae, Bacillaceae (Вас.
polymyxa, Clostridium), Mycobacteriaceae, Actinomyce-
taceae, Streptomycetaceae, Spirochaetaceae (Mortenson,
Chen, 1974).
Полный перечень железосодержащих белков и фер-
ментов микробов, обладающих парамагнетизмом, приве-
ден в табл. 5.
Ферредоксины высших организмов, находящиеся в
системе гидроксилирования стероидов, альдегидоксидазе
печени, ксантиноксидазе молока, а также в митохон-
дриях, по многим признакам проявляют сходство с фер-
редоксинами растительного происхождения.
В образовании металлсодержащих соединений у
бактерий большую роль играют сидерохромы, представ-
ляющие собой специальные пептиды, предназначенные
для связывания из окружающей среды ионов железа и
других металлов. Они, в частности, продуцируются ки-
шечными палочками, сальмонеллами мышиного тифа
(энтеробактины), псевдомонадами, антракоидами, ми-
кобактериями (микобактины), актиномицетами, гриба-
ми, дрожжами. Различают в основном два главных типа
сидерохромов — феноляты и гидроксаматы. Трифено-
ляты обнаруживаются только в истинных бактериях
(энтеробактин). Гидроксаматы встречаются в грибах,
дрожжах и бактериях (феррихром). Микобактины —
гибриды гидроксаматной и трифенолятной систем. Си-
дерохромы являются антагонистами некоторых родствен-
ных им сидерохромных антибиотиков, например альбо-
мицина, ферримицина. Как показал J. В. Nejlands
(1977), особенно большое количество железотранспорт-
ных соединений вырабатывается в культурах фагорези-
стентных, антибиотикоустойчивых и не чувствительных к
бактериоцинам штаммов эшерихий и сальмонелл мыши-
ного тифа. К тому же сидерохромы ускоряют рост энте-
ро- и микобактерий повышают вирулентность эшерихий и
способствуют развитию экспериментальных микобакте-
риозов. Интересно, что сидерохромы, выделяясь во внеш-
32
нюю среду, могут утилизироваться другими видами бак-
терий и затем использоваться при транспорте железа
(Byers et al., 1977).
У высших организмов внутриклеточных металлосвя-
зывающих белков типа сидерохромов нет. Вместо них
в сыворотке крови, межклеточной жидкости, секретах
циркулируют трансферрины (сидерофилины). Сыворо-
точный трансферрин — это 0-глобулин плазмы, состоя-
щий из одной полипептидной цепи с молекулярной мас-
сой 75 000—80 000. Обычно он связывает два атома же-
леза Fe3+, но способен к мобилизации ионов цинка, хро-
ма, кобальта, марганца, кадмия, никеля, галлия, индия,
меди. В сыворотке и плазме крови железо обнаружива-
ется в микрограммах, а другие переходные металлы — в
еще меньших количествах. Больше всего в крови медь-
содержащего гликопротеина церулоплазмина. Молеку-
лярная масса церулоплазмина 150 000—160 000; в его
молекуле имеется 6—7 атомов меди. У человека церуло-
плазмин (а2-глобулин) содержит 0,3% меди и около 8%
углеводов. В 100 мл плазмы человека обычно содержит-
ся 20—40 мг церулоплазмина. Он проявляет оксидазное
действие по отношению к ряду полиаминов, полифенолов
и иону Fe2+. При дефиците меди захват железа транс-
феррином ухудшается.
2.3. МАГНЕТИЗМ МИНЕРАЛЬНО-
ОРГАНИЧЕСКИХ СОЕДИНЕНИЙ КЛЕТОК
Элементарный состав микроорганизмов, клеток жи-
вотных и растений зависит от вида, органа, ткани, пита-
ния, возраста, стадии развития и окружающей среды.
В органах и тканях различных животных и человека
больше всего жизненно важных органогенов. Так, в про-
центах к массе тела кислород составляет 62,43, углерод—
21,15, водород — 9,86, азот — 3,10. Много в организме
зольных элементов: кальция—1,90;%, фосфора — 0,95,
калия — 0,23, серы — 0,16, хлора и натрия.— по 0,08,
магния — 0,027%. Среди микроэлементов больше всего
йода и фтора (около 0,01%), железа (0,05|%), цинка и
брома (а; 0,02 — 0,03%), алюминия и кремния (по
0,001%.).
Приведенные данные показывают, что в клетках, тка-
нях, биологических жидкостях содержится несколько
33
парамагнетиков (кислород, калий, алюминий) и отно-
сительно большое количество железа (3—5 г в организ-
ме человека), обладающего ферромагнитными свойст-
вами.
Установлено, что количество микроэлементов в ана-
логичных органах и тканях различных видов животных
неодинаково. Порядок распределения их в организме
одного и того же вида неоднотипный. Процентное содер-
жание микроэлементов зависит от физиологического
состояния, возраста, сезона года и некоторых других
факторов. Как правило, микроэлементы накапливаются
в активно функционирующих клетках и тканях с высо-
ким типом метаболизма. Может быть, поэтому наиболее
богата ими печень. Возрастная динамика количествен-
ных изменений элементов часто имеет разнонаправлен-
ный и непредсказуемый характер, но содержание кобаль-
та и никеля, которые, как и железо, наделены ферро-
магнитными свойствами, в зрелом возрасте увеличивает-
ся. Так, по данным Г. А. Бабенко (1965), уровень ко-
бальта в крови здоровых людей среднего возраста до-
стигает 8% на свежее вещество, тогда как у юношей и
пожилых может составлять 4%. Содержание никеля в-
крови детей и юношей находится в пределах 6—28(%:, а у
здоровых взрослых— 11,6—37,7% (Шостко, 1959).
Динамика содержания меди может идти, наоборот,,
по убывающей. В печени эмбриона, например, имеется
70 мг меди на 100 г свежего вещества, у маленьких де-
тей— 2,4 мг/100 г, а в печени взрослых людей не пре-
вышает 0,4—0,8 мг/100 г.
Из сказанного следует, что найти определенные за-
кономерности в качественном и количественном распре-
делении микроэлементов в клетках не удалось и до на-
стоящего времени. Исключение составляют лишь некото-
рые органы с преимущественным депонированием одного*
или нескольких микроэлементов. В частности, цинк акку-
мулируется в островковой части поджелудочной железы,
молибден — в почках, стронций — в костях, йод — в щито-
видной железе, барий — в сетчатой оболочке глаза, мар-
ганец, бром и хром — в гипофизе. Лучше всего изучено
распределение железа (табл. 6). В организме человека и
животных приблизительно 65(% его находится в гемогло-
бине ( в крови 48 мг%) и миоглобине, остальная часть
локализована в ферритине, гемосидерине и только 1%
34
Распределение железосодержащих протеинов у взрослого здорового человека
Таблица 6
Белок Относи* тельная молеку- лярная масса Содержа- ние белка, г Содержание железа Природа же- леза (Я—ге- миновое, N— негеминовое) Число ато- мов желе- за в мо- лекуле Валент- ность же- леза Функция
Г % от об- щего коли- чества Fe
Гемоглобин 68000 750 2,60 65 н 4 Fe2+ Транспорт кисло-
Миоглобин 17000 40 0,13 6 н 1 Fe2+ рода Аккумулирует кис- лород в мышцах
Трансферрин 76000 20 0,007 0,2 N 2 Fe3+ Аккумулирует же- лезо в плазме
Ферритин 444000 2,4 0,52 13 N 0—4700 Fe3+ Аккумулирует же- лезо в клетках
Гемосидерин 1,6 0,48 12 N 5000 Fe3+ Аккумулирует же- лезо в клетках
Каталаза 280000 5,0 0,004 0,1 Н Fe2+ Метаболизм Н2О»
Цитохром с Пероксидаза Цитохромы и окси- дазы Флавопротеин де- гидрогеназ, ок- сидаз и окси- геназ 12500 44100 0,8 0,004 0,02 0,1 0,5 н н н N 1 Fe2+ /Fe3+ Fe2+/Fe?+ Fe2+ Завершает окисле- ние Метаболизм Н2О2 Завершает окисле- ние Окислительные реакции, присоеди- нение молекуляр- ного кислорода
железа приходится на ферменты и транспортные системы
(Highly, 1971; Crichton, 1973).
Введенное с пищей железо и, очевидно, другие метал-
лы в основном адсорбируются в тонком кишечнике муко-
зальными клетками. При этом первичным акцептором
железа является ферритин. Активно, с помощью низко-
молекулярных переносчиков неферритиновой природы
поглощает железо плацента. В крови человека и живот-
ных железо ассоциировано с трансферрином, из которо-
го, как указывалось выше, оно транспортируется для
синтеза гемоглобина эритроцитов.
В эритроцитах человека содержится 30—41>% гемо-
глобина (в цельной крови 13—16%) и в среднем 105 мг%
железа. Помимо этого, в эритроцитах имеется около
470 мг|% парамагнитного калия. Остальные компоненты
эритроцитов — типичные диамагнетики: вода (57—
68 мц%), натрий (80 мП%), остаточный азот н азот ами-
нокислот (40—50 мг%), глютатион (75—120 мг|%), ли-
пиды (410—780 мг'%), нейтральные жиры и холестерин
(около 325 мг'%), лецитины (350 мг]%), глюкоза (60—
70 мг|%), мочевина, мочевая кислота, креатин, креатинин
(29—45 мп%), хлор и угольная кислота (по 200 мр'%),
фосфор (100—120 мг%), кальций, магний, медь и цинк
.(всего 7 мг>%).
Органно-тканевой уровень железа более низкий, чем
эритроцитарный. По данным Р. Д. Габовича и др.
(1977), применивших для определения содержания пе-
реходных металлов у крыс спектрографический метод,
больше всего его в печени — 33,1 мг|% на сырую массу.
В два-три раза меньший процент железа отмечался в се-
лезенке (14,6 мр%), головном (14,2 мг%), костном мозге
(11,4 мг%) и почках (9,4 мг%). Миокард содержал
2,8 мг|% железа, кожа — 2,2, а скелетные мышцы — всего
лишь 0,3 мг%.
Большая часть железа находится в белково-связан-
ной форме преимущественно в цитоплазме клеток. В мем-
бранных структурах его содержится не более 40 мкг/г
влажной ткани (Коваленко и др., 1981). Свободного же-
леза в клетках очень мало, не более 5—6 мкг на 1 г
влажной печени мышей и крыс независимо от возраста
животных. При этом А. Ф. Ванин и др. (1981) указыва-
ют, что в быстрорастущей гепатоме 22 А, асцитном раке
Эрлиха и раке молочной железы концентрация свобод-
36
кого железа на всех стадиях развития опухолей умень-
шается по сравнению с нормой в два раза. Ввиду того
что в эмбриональной печеночной ткани, отличающейся
интенсивной пролиферацией клеток, уровень свободного
железа был таким же, как в норме, то всякое его сниже-
ние авторы расценивают в аспекте злокачественного пе-
рерождения клеток.
Уровень других микроэлементов в клетках очень низ-
кий и, как правило, измеряется в мкг%; на сырую массу.
Депонирует их в основном печень. Так, например, Р. Д.
Габович и др. (1977) в печени крыс нашли 131,9 мк/%
марганца, 721,3 мкг% меди и 41,2 мкг%' молибдена.
В других органах и тканях концентрация марганца коле-
балась от 2,3 до 21,7 мкр'%1, меди — от 21,2 до 203,2, а
молибдена — от 2,5 до 23,4 мкг%.
Подобное явление наблюдается при рассмотрении ко-
личественного содержания микроэлементов в органах и
тканях самых разнообразных видов животных. Кратко
обобщая результаты этих исследований, остановимся на
распределении самых важных из них. В частности, мар-
ганец в печени млекопитающих содержится в пределах
0,12—0,17 (мартышка, кролик, человек)—0,30—0,34 мг
на 100 г свежего вещества (собака, кошка); в почках его
количество было . меньшим в 2—5, а в легких — в 5—
30 раз. Такое же соотношение марганца наблюдалось в
печени, почках и легких птиц. У рыб он обнаруживался
только в печени (0,40—0,89 мг/100 г вещества).
Кобальт и никель обычно сопутствуют друг другу.
Больше всего кобальта содержит поджелудочная желе-
за, тимус и печень (около 50 мкц%).
Медь в различных органах и тканях человека обнару-
живается тоже в минимальных концентрациях — от
0,64—0,89 мг/100 г сухого вещества (мышцы, селезенка,
легкие, поджелудочная железа) до 4,02 мг/100 г (пе-
чень). Несколько богаче клетки цинком. В печени чело-
века его содержится 21,9—28,8 мг/100 г сухого вещества,
в печени лягушки — 7,2—10,09, птицы — 12, в остальных
органах и тканях — от 3,6 (мозг, почки) до 22,6 мг/100 г
(мышцы). Приблизительно столько же цинка имеется у
растений и, в частности, шляпочных грибов.
В заключение нужно сказать, что в клетке содержит-
ся большое количество белка. Самый высокий процент
протеидов находится в селезенке, легких и поперечнопо-
37
лосатых мышцах (80—84;% от сухой ткани). В почках
находится 72% белка, в сердце, печени, коже и пищева-
рительном тракте— 57—63, в костях — 28%. Процент
белка от массы свежей ткани в органах животных в сред-
нем составляет 14—18. Исключением является мозг
(7,9%). Семена растений имеют 10—13%, стебли и ли-
стья— 1,5—3, корни — 0,5—3, фрукты — 0,3—1% белка
от массы свежей ткани.
Белки составляют существенный компонент биологи-
ческих жидкостей. В норме плазма крови содержит 90—
91% воды и около 6,5—8,51% белка, из которого на долю
альбумина приходится 4—5%, глобулина — 2,3—3, фиб-
риногена — 0,2—0,4%'. Остальная часть плазмы (0,5—
3,5%)—это липиды, нейтральные жиры, холестерин,
лецитин, глюкоза, глюкозамин, мочевина, молочная кис-
лота, следы других органических кислот, креатинина,
креатина и биологически активных веществ. Микроэле-
менты плазмы находятся в белково-связанной форме:
медь в купропротеинах и церулоплазмине, цинк — в
угольной ангидразе и других комплексах, кобальт как
компонент витамина Вг2 и в соединении с белками-пере-
носчиками. Кроме того, в ней содержится около 1,5 мг/л
связанного с трансферрином железа (сыворотка содер-
жит 12,5—30,4 мкмоль/л, или 70—170 мкг/100 мл). Су-
хой остаток плазмы составляет 9—10%.
Химический состав лимфы менее сложен. В ней со-
держится 94% воды, 2—4% белка, 410 мг%‘ хлора, 100—
ПО мг% глюкозы, 330 мг% натрия и 38 мг% калия, каль-
ция и магния. Сухая масса лимфы — 6%.
Минерально-органический состав микроорганизмов
тоже разнообразен. В них содержится от 75 до 85% воды,
около 50% белка, 12—28% углеводов, 0,2—41% липидов
(микобактерии туберкулеза и другие кислотоустойчивые
бактерии). Общее содержание минеральных веществ у
микробов колеблется от 2 до 14% от массы сухой клетки.
Основными элементами являются фосфор (у микобак-
терий туберкулеза 19—45%), натрий, калий, кальций,
магний, медь, кремний, сера, хлор и железо. Кроме ука-
занного в бактериальной клетке имеются микроэлемен-
ты: фтор, марганец, цинк, кобальт, никель и многие дру-
гие. Несмотря на внешнюю простоту химизма, даже у
одного и того же вида микроба состав отдельных компо-
нентов резко меняется в зависимости от физиологическо-
38
го состояния культуры, питательного субстрата, наличия
металлотранспортных систем, условий существования.
В этом плане лучше всего изучен обмен железа. Боль-
шинство бактерий, как, например, эшерихии, сальмо-
неллы, шигеллы, протей, псевдомонады и другие, хорошо
растут при наличии всего лишь 0,02—0,03 мг железа на
1 л питательной среды. Считается, что если в 1 г сухой
культуры содержится 0,1 мг железа, то она нуждается в
2 мг железа на 1 л среды. По-видимому, 10 мг железа/л
может удовлетворить потребности любого вида бак-
терий.
По данным М. Stephenson (1949), выросшие на обыч-
ном мясопептонном агаре высушенные культуры бакте-
рий содержат от 0,0036 до 0,0175% железа, в частности
в сухом остатке эшерихий его находилось 0,013—0,014%.
В перерасчете на одну клетку это в 430 раз меньше, чем у
магнитотаксического кокка (Neilands, 1974). Близкие
результаты получил Н. J. М. Bowen (1966). В планктоне
было найдено 3,5 мг железа на 1 г сухой массы, у бакте-
рий — 0,25, у грибов — 0,13 мг/г, а меди на один порядок
меньше (0,2; 0,042 и 0,015 мг/г соответственно).
Приблизительно такие же показатели содержания
железа у микробов были отмечены в самой ранней ра-
боте О. Warburg (1927). Использовав в качестве тест-
микроба Torula utilis, в 1 г сухой культуры он обнару-
жил приблизительно 0,1 мг железа. Однако в популяции
его количество у большинства клеток не было одинако-
вым и находилось в пределах 0,01—0,1 мг. Позже С. А.
Elvehjem (1931) установил, что дрожжи могут иметь око-
ло 24 мг железа на 1 г сухой культуры.
Большой процент железа ассимилируют анаэробные
бактерии родов Desulfovibrio и Desulfotomaculum. Как
показали Н. Е. Jones et al. (1976), в культурах различных
видов этих сульфатредуцирующих микробов, вырастав-
ших на глюкозо-пептонной дрожжевой среде с большим
количеством солей, через 48 ч после момента посева об-
наруживалось от 407 до 2670 мкг железа/г клеток, 37—
392 мкг меди, 180—5520 мкг цинка, около 345 мкг мар-
ганца, 337 мкг свинца, 200 мкг серебра, т. е. по железу
в 2—10 раз, а по другим металлам в несколько десятков
раз больше, чем у псевдомонад, эшерихий, клостридий и
микрококков. Электронномикроскопический анализ по-
казал, что цитоплазма сульфатредуцирующих бактерий
39
насыщена многочисленными аморфными электронноплот-
ными частицами диаметром от 0,01 до 0,05 мкм. У энте-
робактерий подобные частицы в небольшом числе опре-
делялись между цитоплазматической мембраной и кле-
точной стенкой.
Различие в количестве железа или других металлов,
по мнению G. Anderegg et al. (1963), F. T. Last, R. S.
Warren (1972), J. B. Neilands (1977), J. B. Neilands,
R. R. Wayne (1977), зависит от наличия у них сидерохро-
мов и их способности извлекать из окружающей среды
ионы.железа. Последнее, как показано Н. J. М. Bowen
(1966), обусловлено температурой. В частности, образо-
вание энтеробактина у сальмонелл мышиного тифа
прогрессивно снижается, начиная с 31 до 40,3 °C. Одно-
временно указывается, что потеря продукции сидерохро-
мов ведет к понижению вирулентности микробов. В свя-
зи с этим в заключение важно подчеркнуть, что у мо-
лочнокислых бактерий нет сидерохромов и в их культу-
ре не обнаруживается железа (Nejlands, 1974).
Обмен других элементов у микробов изучен недоста-
точно. Между тем даже минимальные количества мар-
ганца, кобальта, никеля, подобно железу, обладают фер-
ромагнетизмом, что может в значительной мере отразить-
ся на магнитной восприимчивости микробной популяции.
Это очень ярко продемонстрировано в опытах К- Н.
Evans, W. Р. Tew (1981), которые с целью повышения
магниточувствительности биологических объектов ис-
пользовали ЕгС1з. Благодаря высокому значению маг-
нитного момента катиона эрбия (9,5 магнетона Бора)
авторы намагничивали высокополимерные органические
материалы, дрожжи, бактерии, вирус Висна (Tew, 1979)
и с помощью магнита сумели извлечь их из растворов.
Способность намагничивать клетки и ткани имеют так-
же другие элементы, как, скажем, гольмий, значение маг-
нитного момента у которого выше, чем у эрбия, и равно
10, 3 магнетона Бора.
Таким образом, природа биомагнетизма имеет очень
сложный характер и магнитовосприимчивость живого
(органического) вещества, очевидно, обусловлена энер-
гетической эффективностью окисления, активностью био-
химических реакций, веществами и элементами клеток,
обладающих высоким уровнем парамагнетизма. К таким
веществам прежде всего можно отнести клеточный фер-
40
ритин, находящийся в организме животных, растениях,
спорах, спорангиях грибов и некоторых бактериях. Прав-
да, количество этого железосодержащего протеина в
клетках млекопитающих колеблется в зависимости от
их функциональных особенностей, потребностей орга-
низма в железе и общего состояния здоровья, а у расте-
ний и микробов — от видовой принадлежности, типа,
характера питания, наличия железотранспортных си-
стем. При этом допускается, что магнитовосприимчивость
молекул ферритинов неодинакова из-за различия в ко-
личестве атомов железа у них. К изменению магнитной
восприимчивости ферритина также ведет ферментатив-
ное восстановление железа или его освобождение при
разрушении апоферритина.
Большой вклад в парамагнетизм крови вносит гемо-
глобин, в котором находится 2/3 всего железа организ-
ма, т. е. эритроцитарный уровень железа намного выше,
чем органно-тканевой. При этом основная его часть со-
держится в белково-связанной форме в цитоплазме кле-
ток, но некоторое количество железа может находиться
в свободном состоянии и при этом проявляет ферро-
магнетизм.
Вклад других металлсодержащих протеинов и фер-
ментов в парамагнетизм клеток животного происхожде-
ния, по-видимому, очень небольшой вследствие малого
их количества.
У растений и микроорганизмов магнитным материа-
лом могут являться ферредоксины и другие железо-се-
русодержащие белки. В частности, магнитная восприим-
чивость грибов и бактерий должна определяться в ос-
новном сильно проявляющими парамагнетизм ферредок-
синоподобными протеинами и многочисленными металл-
содержащими ферментами.
Кроме этого, уровень биомагнетизма предопределяет
минерально-солевой состав и в первую очередь такие
парамагнетики, как кислород и калий. Величина сум-
марной магнитовосприимчивости клеток в значительной
мере может зависеть от соотношения и общего содержа-
ния в них пара- и ферромагнитных микроэлементов, в
частности марганца, алюминия, кобальта и никеля. При-
стального внимания заслуживают сообщения о легком
намагничивании редкоземельных элементов, которые
имеют очень высокое значение магнитного момента.
--------------------з
ГЛАВА
ГЕОМАГНИТОТРОПИЗМ ОРГАНИЗМОВ
Начало экспериментальному исследованию геомаг-
нитотропных реакций было положено в серии работ, про-
веденных F. A. Brown et al. (1960) на улитке Nassarius
obsolete, обитающей в морском иле. Перемещаясь в
геомагнитном поле по определенным координатам, эти
улитки всякий раз изменяли направление своего передви-
жения при воздействии на них искусственного магнит-
ного поля, напряженность которого превышала природ-
ное в 7—10 раз. Улитки реагировали также на естест-
венные флюктуации магнитного поля Земли и
периодически меняли направление движения в связи с
синодическим вращением Солнца и сменой фаз Луны.
В такой же геофизической среде подобным образом вели
себя филогенетически родственные с улитками плоские
черви Dugesia dorotocephala и простейшие Paramecium
caudatum (Brown, 1962).
Планарии, например, резко изменяли направление
движения при переходе из земного или близкого к нему
поля (14 А/м) в поле более высокой напряженности
(8-102 А/м ) и наоборот. Они не только реагировали на
естественные пульсации геомагнитного поля, но в то же
время могли дифференцировать поля с различным по
отношению к длинной оси их тела направлением сило-
вых линий или отличать северный полюс магнита от
южного.
Эксперименты с парамециями были тоже удачными.
В магнитном поле напряженностью 1,04-102 А/м про-
стейшие изменяли свойственный им характер миграции
в зависимости от направления силовых линий.
В дни новолунья и полнолунья двигательные реакции
планарий и простейших изменялись так, как во время
возмущения магнитного поля.
42
По мнению F. A. Brown (1960), ориентация по сило-
вым линиям геомагнитного поля — естественное свойст-
во организмов, имеющих «магнитный компас».
Для объяснения магнитной чувствительности организ-
мов в разное время были предложены различные гипо-
тезы: 1) оптическая модель чувствительности, связываю-
щая магниторецепцию с латеральными глазами живот-
ных (Leask, 1977); 2) электрическая модель, согласно ко-
торой у животных имеется электропроводник в виде пет-
ли, где возникает электродвижущая сила при быстром
движении в магнитном поле Земли (Kalmijn, 1966);
3) химическая модель, которая предполагает, что детек-
тор магнитного поля обладает парамагнитными свойст-
вами, т. е. в магнитном поле молекулы детектора созда-
ют собственное поле, воспринимающееся нервной систе-
мой или изменяющее обменные реакции организма
(Schulten, Schulten, 1977); 4) глубже других изучена
компасная модель, объясняющая магниточувствитель-
ность наличием в организме (клетке) органических фер-
ромагнетиков (постоянных магнитов), обладающих спо-
собностью поворачиваться и устанавливаться вдоль си-
ловых линий геомагнитного поля, как компасная стрелка.
3.1. МАГНИТОСОМЫ
И МАГНИТОТАКСИС БАКТЕРИЙ
Магнитный материал впервые был найден J. Т. Tom-
linson (1959), Н. A. Lowenstam (1962, 1967); К. М. To-
we, Н. A. Lowenstam (1967), К. М. Towe, W. F. Bradley
(1967) в зубах брюхоногих морских моллюсков хитон
(Polyplacophora), обитающих в прибрежной зоне и пи-
тающихся мелкими скальными водорослями. Было пока-
зано, что тонко измельченные минералы зубов Chiton
tuberculatus, Criptochiton steltere, Acantopleura echina-
tum, Acantopleura spiniger влияния на стрелку компаса
не оказывают, но в магнитном поле ведут себя подобно
опилкам железа, располагаясь вдоль силовых линий.
Рентгеноструктурный анализ позволил обнаружить в
составе зубов хитона магнетит (Ре2Оз-РеО), апатит и
.лепидокрокит. Кроме железа в зубах хитонов обнару-
живались медь, марганец, алюминий, барий, стронций,
силиций, магний, кальций, а у некоторых из них — мик-
родозы никеля, свинец, олово.
43
Исследования Н. A. Lowenstam долгое время оста-
вались незамеченными. Внимание к ним было привлече-
но в связи с открытием в 1975 г. R. Р. Blakemore не-
скольких видов микроаэрофильных водных бактерий,
обладающих природным магнитотаксисом, т. е. мигриру-
ющих по силовым линиям геомагнитного поля и меня-
ющих направление движения в момент приближения и
удаления искусственного магнита (рис. 6). Убитые пара-
ми четырехокиси осмия бактерии соединялись в корот-
кие цепочки и, подобно железным опилкам, выстраива-
лись в направлении силовых линий искусственного маг-
нитного поля.
В природе эти бактерии обитают во многих пресных,
и морских водоемах. Являясь микроаэрофилами, они
живут придонно на границе ил—вода. Если взмутить
воду, вследствие чего эти микробы попадают в верхние
ее слои, богатые губительным для них кислородом, то,
находясь в северном полушарии, они быстро мигрируют
в направлении магнитного севера, погружаясь при этом
вниз в бескислородную зону («север-ищущие» штаммы).
В южном полушарии магнитотаксические бактерии,
стремясь к месту своего обитания в иле, плывут также
вниз и в том же направлении, которое соответствует
здесь магнитному югу («юг-ищущие» варианты). Иными
словами, строго ориентированная миграция бактерий,
обладающих магнитотаксисом, происходит по вертикаль-
ной составляющей геомагнитного поля (Blakemore et al.,
1980; Kirschvink, 1980; Frankel et al., 1981). На экваторе,
где такой механизм магнитотаксиса не срабатывает, как
показали исследования R. В. Frankel et al. (1981), маг-
нитотаксические бактерии мигрируют вдоль горизон-
тальной составляющей геомагнитного поля, что для вы-
живания штаммов любой полярности создает более бла-
гоприятные условия, чем беспорядочное движение или
миграция вверх к поверхности воды, где существовать
они не могут. При этом оказалось, что популяции маг-
нитотаксических бактерий, выделенных из пресной воды
и морских осадков на геомагнитном экваторе, состоят из
равных количеств особей обеих полярностей, которые в
однородном магнитном поле 240 А/м мигрируют в раз-
ных направлениях.
Полагают, что в периоды геомагнитной реверсии,
когда поле в каждой точке Земли становится горизон-
44
тальным, магнитотаксические бактерии закрытых пре-
сноводных водоемов определенной полярности, как пра-
вило, вымирают. И действительно, в далеко отстоящих
кратерных озерах такие бактерии отсутствуют. Выжи-
вают лишь морские виды магнитотаксических микробов,
меняющих полярность. В последующем они могут при-
спосабливаться к существованию в пресных водах. Это
Рис. 6. Кинематографическая последовательность проявлений маг-
нитотаксиса бактерий под микроскопом: а — скопление магнито-
таксических бактерий на северном магнитном полюсе водяной капли,
бив — миграция магнитотаксических бактерий в противополож-
ном направлении под действием постоянного магнита через 0,5 с
(б) и 1 с (в). Отрезок=1 мкм
45
подтверждают эксперименты R. Р. Blakemore et aL
(1979), показавших, что северная разновидность магни-
тотаксических бактерий, пассируемая в южном полуша-
рии, приобретает способность мигрировать к южному
полюсу Земли. То же достигалось при искусственной
имитации условий южного полушария. Правда, для пре-
вращения северной расы в южную в этих случаях потре-
бовался длительный период культивирования, который
в некоторых опытах продолжался 5—7 недель.
Для доказательства того, что вертикальная составля-
ющая геомагнитного поля определяет полярность маг-
нитотаксических бактерий, R. В. Frankel et al. (1981)
«север-ищущие» штаммы бактерий помещали в экраны
из мю-металла, снижая естественную напряженность поля
приблизительно в 100 раз. В этих условиях в культуре
магнитотаксических бактерий постепенно возникали
особи с обратной полярностью, и их количество нара-
стало до тех пор, пока популяция не стала содержать
одинаковое количество «север-» и «юг-ищущих» бакте-
рий. Микробные особи контрольного штамма, находя-
щиеся вне экрана, первоначальной полярности не изме-
няли.
Выделить магнитотаксические бактерии в чистой
культуре столь же трудно, как и многие другие анаэроб-
ные литотрофы. С этой целью используют методику
Т. Т. Moench, W. A. Konetzka (1978), основанную на при-
тягательной способности магнита. При поднесении его
к сосуду через 15 мин у полюсов скопляется большое
количество магнитотаксических бактерий, откуда их
извлекают пастеровской пипеткой. Позже эти бактерии
в точках наибольшей напряженности поля агглютини-
руют и выпадают в осадок. Из жидкой среды магнито-
таксические бактерии выделяют вместе с сильными
маленькими магнитами, предварительно погруженными
в нее.
Выращивают магнитотаксические бактерии при тем-
пературе 22—30 °C в анаэробных условиях с понижен-
ным содержанием кислорода (6%) или в сосудах, за-
полненных азотом, на полусинтетических жидких средах,
основу которых в зависимости от их происхождения со-
ставляет пресноводная или морская вода с примесью
ила и добавлением янтарной кислоты, натриевой соли
азотной кислоты и сульфата железа.
46
Для культивирования бактерий, обладающих магни-
тотаксисом, D. L. Balkwill et al. (1980) предложили ис-
пользовать среду, в которую на 100 мл воды добавляют
0,25 мл 0,01 М окиси пятивалентного железа (железис-
тая соль хинной кислоты), 1,0 мл витаминного эликси-
ра, 1,0 мл минерального эликсира, 0,5 мл 1,0 М КН2РО4,
0,2 мл 0,01% резазурина, 0,75 г винной кислоты, 0,01 г
нитрата натрия, 0,005 г тиогликолата натрия. Чистую
культуру получают на полужидкой и плотной средах с
добавлением 130 мг и 0,85 г ионагара на 100 мл соот-
ветственно.
Лучше растут магнитотаксические бактерии на спе-
циально подобранных солевых средах при pH 6,7, реду-
цированных в атмосфере азота с использованием в ка-
честве восстановителя цитрата титана. Жидкая среда,
по, прописи R. Р. Blakemore et al. (1979), содержит
7Q мл дистиллированной воды, 10 мл фильтрата иловой
воды, 1 мл витаминного эликсира, 0,5 мл буферного ка-
лий фосфата, 5 мг витамина В]2, 25 мг NH4CI, 10 мг
натрий ангидридного ацетата, 0,2 мг резазурина, 90 мг
ионагара № 2.
Количество магнитотаксических бактерий в воде ко-
леблется от 200 до 1000 клеток на 1 мл. На жидких ис-
кусственных питательных средах через 2—3 месяца их
число возрастает от нескольких десятков тысяч до 106—
107/мл. Полностью выросшие культуры образуют черно-
го, цвета осадок, который прикрывается ржавым слоем
окисленного железа. В полужидкой среде колонии име-
ют средние размеры и дисковидный характер, на плот-
ной— круглые, выпуклые, с ровными краями.
Выделено несколько неидентифицированных видов
морских и пресноводных бактерий, обладающих магни-
тотаксисом. Палочковидные формы, полученные R. Р.
Blakemore (1975) из морских осадков, как и эшерихии,
обладают подвижностью, имеют размеры 3,2 мкм в
длину и 0,5 мкм в ширину, по Граму окрашиваются от-
рицательно. Пресноводные кокки, изолированные Т. Т.
Moench, W. A. Konetzka (1978), Т. Т. Moench (1979), по
диаметру (1,6±0,2 мкм) напоминают макрококки, но в
отличие от последних грамотрицательны, подвижны, со-
держат много поли-р-гидрооксибутиратных гранул и вы-
сокое молярное содержание Г + Ц в нуклеоиде (61,7±
±0,7%). У всех вариантов магнитотаксических бактерий
47
имеются два пучка асимметрично расположенных жгу-
тиков, которые у кокков берут начало от дископодоб-
ных структур, находящихся по 10—15 штук на упло-
щенной стороне (рис. 7).
Лучше других изучены магнитотаксические пресно-
водные микроаэрофильные спириллы штамма MS-1 с
биполярными жгутиками, которые отнесены к роду Aqu-
aspirillum (Blakemore et al., 1979; Balkwill et al., 1980).
Рис. 7. Магнитотаксические кокки с цепочкой гранул магнетита
между двумя пучками жгутиков. Отрезок =1 мкм
Они грамотрицательны, имеют 1—8 закрученных слева
направо завитков (в среднем 3). Размеры спирилл MS-1
колеблются от 2,3 до 11,1 мкм в длину и 0,28—0,36 мкм
в ширину (в среднем 4,3X0,32 мкм). Длина волны за-
витка варьирует в пределах 0,9—2,4 мкм, амплитудные
расстояния 0,14 — 0,44 мкм, жгутиков—1,5 — 2,5 мкм
диаметром 22 нм. Содержание Г+Ц в нуклеоиде спи-
рилл равнялось 64,5—64,9. Время генерации составляло
6—15 ч, длительность культивирования 4—6 дней (0,2—
0,5 г влажной культуры на 1л). Рост спирилл прекра-
щался при высоком содержании окислительно-восстано-
вительного потенциала среды (гН + 40 тпВ).
48
Под электронным микроскопом в цитоплазме магни-
тотаксических бактерий и спирилл штамма MS-1 обна-
руживается цепочка или две электронноплотных бога-
тых железом включений (рис. 8), каждое из которых
имеет форму куба, а по уточненным данным К. М. To-
we, Т. Т. Moench (1981), параллелепипеда со средней
длиной 99,3±8,7 и шириной 62,3±6,1 нм и отношением
ширины к длине, равным 0,63. В одной цепочке в сред-
Рис. 8. Магнитотаксические спириллы штамма MS-1 с цепочкой
гранул магнетита в цитоплазме
нем насчитывается 10—15 (изредка больше) частиц.
Каждое зерно находится в электроннопрозрачном слое
мембраны и в цепочке расположено конец в конец (не-
которые могут лежать поперек). После разрушения
(озвучивания) бактерий интрацеллюлярные частицы
конфигурации не меняют и вне клеток тоже остаются в
цепочках (рис. 9). С помощью рентгеноструктурного
анализа, метода дифракции нейтронов и мёссбауэров-
ской спектроскопии R. В. Frankel et al. (1978), R. В.
Frankel et al. (1979), D. L. Balkwill et al. (1980), К. M.
Towe, T. T. Moench (1981) установили, что внутрикле-
точные электронноплотные зерна магнитотаксических
бактерий представляют собой кристаллы магнетита с
небольшой примесью у-РегОз и минералов глины (ТЮг,
А12О3, SiO2). Отмечено, что в кристаллах магнетита
имеются пустоты, некоторые из них заполнены органи-
ческим материалом, возможно, железосодержащим про-
теином (ферритином). Допускается, что в них могут на-
ходиться и другие ферромагнетики, как например пир-
ротит, связанный с серой (0,534%).
49
Изолированные кристаллы магнетита имеют домен-
ную структуру (Butler, Banerjee, 1975; Bean, 1979), об-
ладают сильным естественным остаточным магнетизмом,
приблизительно равным 9-10-4 электромагнитной едини-
цы на 1 г сухой массы (Blakemore et al., 1979; Denham
et al., 1980), и являются ферромагнетиками. Цепочка
магнетита в цитоплазме бактерий представляет собой
единый магнитный диполь с моментом, равным сумме
Рис. 9. Цепь кристаллов магнетита вне бактерий; некоторые с по-
ристым центром, содержащим органическое вещество
моментов отдельных кристаллов. По подсчетам R. В.
Frankel, R. Р. Blakemore (1980), для клетки, содержа-
щей 22 частицы магнетита (среднее у спирилл), общий
магнитный момент М=1,3-10-12 электромагнитной еди-
ницы. В геомагнитном поле 40 А/м общая магнитная
энергия клетки МН = 6,6-10~13 эрг, т. е. на порядок боль-
ше, чем термальная энергия кТ (4,1-10“14 эрг при
300 °К). Очевидно, цепь кристаллов магнетита функцио-
нирует как очень чувствительный биомагнитный компас,
с помощью которого магнитотаксические бактерии ми-
грируют по силовым линиям геомагнитного поля. В экс-
перименте клетки, содержащие магнетит, ориентируются
50
в однородном поле напряженностью 8 А/м (Frankel,
Blakemore, 1980).
Реагирует в магнитном поле также магнетит, выде-
ленный из бактерий, правда, в дисперсном состоянии
агрегирует очень медленно из-за большой вязкости сре-
ды (Towe, Moench, 1981). Попытка размагнитить эти
бактерии не удалась. После обработки в постоянном
магнитном поле напряженностью 4,8-103 и 8-103А/м
при частоте 50 Гц R. В. Frankel, R. Р. Blakemore (1980),
R. В. Frankel et al. (1981) наблюдали, что однородная
популяция магнитотаксических бактерий стала содер-
жать приблизительно равное количество особей с проти-
воположными направлениями движения. Так как изменя-
лась только полярность магнитотаксической культуры,
то можно полагать, что магнитный диполь бактерий яв-
ляется по существу одним магнитным доменом и в силу
этого не размагничивается.
Еще ранее A. J. Kalmijn, R. Р. Blakemore (1978) по-
казали, что изменение полярности и ориентации у прес-
новодных магнитотаксических бактерий могло быть до-
стигнуто под воздействием импульсного магнитного по-
ля от 37,5 до 40 мТ, а у морских видов в диапазоне от
52,5 до 55 мТ в импульсе. Возможно, что в основе
этого лежат различия в анизотропии доменов у морских
и пресноводных видов магнитотаксических бактерий. По
расчетам авторов, один домен магнитотаксических бак-
терий в зависимости от его величины содержит от 2 до
6-10“15 г магнетита. У бактерии, имеющей 10—20 таких
частиц, количество магнетита колеблется от 2 до
12-10-14 г. Содержание магнетита в популяции прежде
всего определяется биомассой. При наличии в 1 мл сре-
ды 100 000 особей в рыхлом осадке будет содержаться
от 2 до 12-10-9 г бактериального магнетита, что при-
близительно соответствует намагниченности порядка
2—12-10-7 А/м. В уплотненных осадках намагничен-
ность может достигать 4 —24-10~9 А/м. Общее количе-
ство железа у магнитотаксических бактерий колеблется
от 2 до 3,5% на сухую массу культуры (Moench, Ко-
netzka, 1978; Blakemore et aL, 1979).
Следует отметить, что до сих пор исчерпывающих
сведений о регионах распространения магнитотаксиче-
ских бактерий нет, но с большой вероятностью можно
сказать, что они явились основным поставщиком магне-
51
тита в коре Земли, которая формировалась одновремен-
но с геомагнитным полем.
Теперь установлено (Frankel, 1979; Blakemore et al.,
1979), что популяция бактерий, обладающих магнито-
таксисом, содержит небольшое количество особей, у ко-
торых отсутствует магнетит, а по данным D. L. Balkwill
et al. (1980), цепей магнетита не имеют 19% клеток.
Более того, при выращивании магнитотаксических бак-
терий в синтетических средах с ограниченным содержа-
нием железа (3,6 мкМ) они утрачивают магнетит или
содержат его в таком ничтожно малом количестве
(1/10—1/20 часть), что не реагируют на изменение на-
пряженности магнитного поля (утрачивают магнито-
таксис).
В целом магнитные клетки популяции содержат в
10 раз больше железа, чем немагнитные, и в 100 раз
больше, чем другие виды бактерий, культивированные
«а солевой среде с наличием железистой соли хинной
кислоты.
В последнее время показано, что образование маг-
нетита не является простым процессом минерализации,
как это происходит в природе при вулканических извер-
жениях или формировании горных пород, а осуществля-
ется биогенным путем. В частности, в зубах хитонов он
появляется не сразу (Towe, Lowenstam, 1967; Kirsch-
vink, Lowenstam, 1979). Вначале зубы моллюсков
состоят из бесцветного органического остова, сформиро-
ванного одонтобластами. На второй стадии зубы хито-
нов покрываются феррогидритом (5Ее2Оз-9Н2О) и при-
обретают коричневый или красно-коричневый цвет.
Магнетит появляется на третьей стадии и в количест-
венном отношении увеличивается вплоть до завершаю-
щего этапа развития зубов. Накопление магнетита ви-
зуально проявляется сменой коричнево-красного цвета
на черный.
Полностью минерализованные зубы покрыты эпите-
лиальными клетками и тонкой пленкой феррогидрита,
разделены тонкими прослойками (<0,05 мкм) органи-
ческого матрикса и выглядят, как медовые соты с плот-
но упакованными относительно друг друга кристаллами
(доменами) магнетита размером 0,1 мкм.
Ввиду такого строения зубов намагниченность маг-
нетита характеризуется слабой устойчивостью к размаг-
52
ничиванию переменным магнитным полем (12 мТ). На-
оборот, изолированные зерна магнетита довольно устой-
чивы к размагничиванию. Так, для размагничивания
породы, содержащей магнетит, недостаточно даже
40 мТ.
По данным Н. A. Lowenstam (1967), предшественни-
ком магнетита является феррогидрит. Трансформация
последнего в магнетит происходит в клетках наружного
слоя эпителия радулы, плотно соприкасаемой с поверх-
ностью зуба, и контролируется несколькими фермента-,
ми. То же можно сказать о Формировании магнетита у
магнитотаксических бактерий. Как справедливо отмеча-
ют Т. Т. Moench, W. A. Konetzka (1978), трудно пред-
ставить, что железосодержащие интр ацеллюлярные
включения являются продуктом простого окисления Fe2+.
Да и возможно ли образование такого количества же-
лезосодержащих соединений в процессе энергообмена?.
Ведь для синтеза 1 г магнетита, как считают эти авто-
ры, потребовалось бы окислить 1300 г железа.
Другую гипотезу высказывают К. М. Towe, Н. A. Lo-
wenstam (1967), J. L. Kirschvink, Н. A. Lowenstam
(1979), допуская, что магнетит образуется вследствие
биохимического превращения в тканях радулы ферри-
тина и гемосидерина. Кстати говоря, ферритин может
обнаруживаться даже в зубах млекопитающих. В част-
ности, М. Р. Frank et al. (1960), Е. J. Roith (1961),
E. J. Runham (1963) обнаружили его в амелобластах
резцов крыс, прилегающих к зоне полного созревания
эмали. Как и в радуле хитона, глыбки ферритина име-
ли диаметр 60 А и были окружены мембранами. Прав-
да, роль железа в эмалевом органе остается неизвест-
ной. По мнению Н. A. Lowenstam (1967), у хитона, как
и у магнитотаксических бактерий, магнетит является
биологическим компасом.
Зубы различных видов хитонов состоят из 65% маг-
нетита и 35% гидроферрофосфата, в среднем содержат
4,7-10~5 г магнетита с уровнем природной намагничен-
ности, примерно равной 0,1 Ам2/кг, что намного ниже
значения насыщения (93 Ам2/кг). Представляется, что
высокую намагниченность морских осадков в основной
обусловливают хитоны и магнитотаксические бактерии.
53
3.2. БИОФИЗИЧЕСКИЕ ОСНОВЫ
НАВИГАЦИИ ОРГАНИЗМОВ
Навигация включает прежде всего ориентацию, т. е.
определение направления необходимого движения, а
также установление местоположения исходного и конеч-
ного пунктов движения.
Первое предположение о возможности использова-
ния геомагнитного поля как источника навигационной
информации птиц было сделано известным русским ака-
демиком А. Миддендорфом в 1855 г.
Почти 100 лет эта гипотеза оставалась вне поля зре-
ния экспериментаторов. Внимание к ней привлек Н. L.
Yeagley (1947), который отметил резко выраженное на-
рушение ориентации в пространстве у домашних голу-
бей, окольцованных маленькими магнитиками. В срав-
нении с контрольной группой птиц, к которым прикреп-
лялись пластинки меди, примерно треть «магнитных»
голубей, выпущенных вдали от места их обитания, до-
стигали гнездовий с большим опозданием, а некоторые
из них в голубятню так и не возвращались. В последу-
ющем воспроизвести эти опыты не удалось (Yeagley,
1951) из-за несовпадения, как теперь думается, погод-
ных условий экспериментов, что надолго задержало ис-
следование навигации птиц по геомагнитному полю.
Начатые Н. L. Yeagley работы нашли успешное про-
должение в исследованиях А. Л. Эльдарова и Ю. А. Хо-
лодова (1964), обнаруживших резкое нарушение локо-
моторной активности перелетных птиц в магнитном по-
ле слабой напряженности порядка 52—135 А/м. Не-
сколько позже W. Т. Keeton (1969, 1971, 1972) показал,
что небольшие магниты массой 3 г, прикрепленные на
спине у основания шеи голубей общей напряженностью
2-104 и 36 А/м вокруг головы, действительно нарушают
ориентационный полет птиц, выпущенных на больших
расстояниях от голубятни. Особенно резко сказывается
влияние магнита на ориентации домашних голубей в об-
лачные дни. При безоблачном небе выпущенные на рас-
стоянии 27—50 км птицы, несущие магниты, в контроль-
ные сроки времени безошибочно находили места своего
гнездовья. В этих условиях навигация нарушалась
лишь у некоторых голубей, выпущенных на расстоянии,
превышающем 50 км. Исходя из этого, автор полагает,
что у птиц есть по крайней мере два источника навига-
54
ционной ориентации — солнечно-звездный и магнитный.
Совершая перелеты в безоблачные дни и ночи, опреде-
ляя свое местоположение, птицы, по-видимому, ориенти-
руются по астрономической карте, а мигрируя в пасмур-
ную погоду, используют другой механизм информации,
которым является геомагнитное поле. Естественно, что
искусственное магнитное поле, искажающее напряжен-
ность и векторные параметры земного магнитного поля,
вызывает изменения направления полета птиц. То же
происходит во время флюктуаций геомагнитного поля
порядка 40—70 нТ, вызываемых повышенной солнцедея-
тельностью (Keeton, Cobert, 1970; Keeton et al., 1974).
При этом ориентационная способность резче наруша-
лась при увеличении индекса К. Во время сильной маг-
нитной бури полет птиц прекращается, и они проявляют
такое же беспокойство, как во время полного солнечно-
го затмения.
По данным F. R. Moore (1977, 1980), птицы, совер-
шающие дальние полеты, реагируют на любые отклоне-
ния естественных флюктуаций геомагнитного поля. Из-
менения в направлении их полета коррелируют с повы-
шением геомагнитного возмущения как по индексу К,
рассчитываемому за 3-часовые интервалы времени, так
и по суммарной оценке за день различных составляю-
щих геомагнитного поля. Автор допускает также, что
магнитная возмущенность может влиять на полет птиц
непрямым образом, через посредство изменения метео-
рологических условий во время повышения солнечной
активности.
Остроумные опыты для доказательства гипотезы
ориентации по силовым линиям геомагнитного поля
провели С. Walcott, R. Р. Green (1974), использовав
Кольца Гельмгольца из никелированной проволоки, одно
из которых укреплялось как ожерелье вокруг шеи, а
другое — над головой в виде шляпки. Питался элект-
ромагнит от миниатюрной батарейки, создавая напря-
женность от 40 до 112 А/м. С таким электромагнитом,
южным полюсом направленным кверху, птицы хорошо
ориентировались в пространстве и находили дорогу до-
мой независимо от того, на каком расстоянии, где и в
какие дни выпускались. Правда, в пасмурную погоду
возвращение голубей несколько задерживалось во вре-
мени. Птицы с северной полярностью магнита дом на-
55
ходили лишь в солнечные погожие дни. При облачной
погоде стая голубей разлеталась в разные стороны, за-
частую в противоположную от голубятни. Такой же
дезориентирующий эффект на голубей, как недавно вы-
яснилось (Walcott, 1978), оказывают аномальные маг-
нитные поля напряженностью 80 нТ, часто встречающи-
еся во многих районах земного шара на путях сезонных
миграций птиц. Как полагают, перелетные птицы вос-
принимают не только региональные, но и локальные
магнитные аномалии площадью 10 км2, обусловленные
залежами магнитных пород у поверхности земли. Об
этом, в частности, говорится в работе М. Е. Шумакова
(1967), который изучал поведенческие реакции 5 видов
перелетных птиц вначале на Балтике, а далее в г. Губ-
кине, находящемся в районе Курской магнитной анома-
лии, где их локомоторная активность при облачном небе
возрастала в 2—3 раза. Изменение вертикальной со-
ставляющей земного поля в специально сконструирован-
ном тоннеле выявляло такие же реакции у голубей
(Bookman, 1977).
Результаты исследований R. Р. Larkin, Р. J. Suther-
land (1977) указывают, что мигрирующие птицы ощу-
щают самой низкой интенсивности переменные электро-
магнитные поля уже в течение нескольких секунд. Авто-
ры доказали это, используя низкосиловые следящие ра-
дары во время ночных перелетов многих птиц. В част-
ности, над работающими антеннами мгновенно меня-
лись и высота, и направление их полета.
По-видимому, геомагнитное поле служит источником
навигационной информации не только для голубей, но
и для других птиц, совершающих сезонные миграции.
В этом аспекте очень большой интерес представляют
наблюдения F. W. Merkel, Н. G. Fromme (1958), Н. G.
Fromme (1961), F. W. Merkel et al. (1964), F. W. Mer-
kel, W. Wiltschko (1965), W. Wiltschko (1965), F. W.
Merkel (1971), W. Wiltschko (1968), W. Wiltschko,
R. Wiltschko (1972), которые показали, что европей-
ская малиновка и подобные ей певчие птицы при мигра-
ции ориентируются по звездам. В естественной геомаг-
нитной обстановке с имитацией ночного неба в мигра-
ционные периоды года они занимают в вольерах всегда
одно и то же предпочтительное положение на север, ко-
торое, однако, быстро меняется при искусственных ва-
56
риациях напряженности магнитного поля от 36 до 12 А/м
или от 27 до 67 А/м. То же происходило при изменении
направления векторных составляющих магнитного поля
Земли. В частности, медленная перемена вызывала из-
менение ориентационного положения малиновки в пред-
сказуемом направлении. В искусственном поле с силь-
ной горизонтальной и нулевой вертикальной составляю-
щими активность малиновки становилась беспорядочной.
Интересно, что если птицам давалась возможность в-
течение 3—5 дней приспособиться к необычной магнит-
ной обстановке, то они вновь занимали в клетках есте-
ственное для них положение. По мнению W. Wiltschko
(1968), система восприятия магнитного поля у малинов-
ки настроена на естественные флюктуации геомагнитно-
го поля, что автор подтвердил в специальных экспери-
ментах, где с помощью колец Гельмгольца имитирова-
лись условия колебаний интенсивности земного магне-
тизма вокруг клеток.
Приблизительно так же вели себя овсянки (Emlen,
1970, 1975; Emlen et al., 1976). Осенью эти птицы в клет-
ках ориентировались к югу, а весной — к северу. Когда
горизонтальную составляющую геомагнитного поля от-
клоняли по часовой стрелке на 120°, северная ориента-
ция птиц постепенно изменялась на восточно-южную.
Эти результаты также говорят, что птицы не только
воспринимают поле, но и используют его в миграцион-
ный период при перелетах. В межмиграционные перио-
ды способность к восприятию поля может резко пони-
жаться.
Значение геомагнитного поля в ориентации чаек убе-
дительно продемонстрировано в серии опытов W. Е. So-
uthern (1969, 1971, 1972, 1974). Поместив их в цирку-
лярные клетки диаметром несколько метров, он отме-
тил, что молодые (2—20-дневные) и взрослые птицы при
самой различной погоде занимают предпочтительно юго-
восточное направление, которое в осеннее время опре-
делит для птенцов путь миграции. Другие опыты выяс-
нили, что слабая возмущенность геомагнитного поля
стабилизировала юго-восточную ориентацию чаек, а
сильные магнитные бури нарушали ее и вызывали пере-
ориентацию на север. Автор даже определил степень
флюктуации магнитного поля, вызывающую нарушение
ориентации чаек. Она оказалась равной 20—70 нТ, что
57
приблизительно эквивалентно 1/1000 напряженности гео-
магнитного поля. В дни таких магнитных бурь изменя-
лись маршрут полета, ритмичность взлетов и повадки
птиц.
Интересные данные в последнее время получили
R. Wiltschko, W. Wiltschko (1982). Они нашли, что го-
луби во время первых вылетов и перелетов ориентиру-
ются по геомагнитному полю и только позже, приобре-
тая опыт наблюдения за дневным движением Солнца,
используют «солнечный компас» навигации. Для дока-
зательства своей теории авторы выращивали одних
птенцов в птичниках, затененных от солнца в первой
половине дня, а других — в открытых для солнца волье-
рах. Голуби, незнакомые с полным ходом солнца по не-
босклону, летели к месту обитания прямолинейно, не
огибая преград и препятствий, встречаемых на пути,
т. е. по силовым линиям геомагнитного поля. Прикреп-
ленные к спине таких голубей магнитики, как и ожида-
лось, нарушали способность находить родной птичник.
Полет голубей, выросших в птичниках, освещаемых
солнцем круглый день, в безоблачные дни происходил
нормально и не зависел от искусственного поля магнита.
Высказывается предположение, что раннее восприятие
магнитного поля Земли может играть ключевую роль в
последующем обучении птиц ориентированию по Солн-
цу. Подтверждением этому явились опыты с европей-
ской малиновкой. Выращивание в искусственном маг-
нитном поле привело к утрате природной ориентации
птиц по звездам. Показав хорошую ориентацию птиц
при низких значениях напряженности магнитного поля
в эксперименте (25 А/м, 36 А/м, 84 А/м), W. Wiltschko
(1974) полагает, что магнитный компас у них адапти-
рован к естественным флюктуациям геомагнитного по-
ля, которые могут встречаться на пути перелета. При
этом птицы-мигранты, особенно трансэкваториальные,
воспринимают обе составляющие геомагнитного поля.
Использовать полярность магнитного поля они не могут,
очевидно, потому, что в филогенезе происходила неод-
нократная реверсия с полной сменой магнитных полю-
сов Земли.
Влияние искусственных магнитных полей на ориен-
тацию и поведенческие реакции птиц априорно с самого
начала объяснялось нарушением функционирования
58
компасной системы или каких-то детекторов восприятия
геомагнетизма.
Ввиду того что дезориентация голубей резче всего
проявлялась в экспериментах с креплением маленьких
магнитов на голове или шее, высказывались предполо-
жения, что магнитный материал у них может находить-
ся в мозговой ткани, глазах или слуховом аппарате.
Наличие супермагнитного вещества в голове и шее
голубей было подтверждено с помощью сверхчувстви-
тельного магнитометра SQUID (сверхпроводящий кван-
тово-интерферрационный датчик), а вскоре С. Walcott
et al. (1979) описали его структуру, локализацию и
свойства. Оказалось, что магнитный материал распола-
гается латерально в маленьком кусочке ткани размером
1—2 мм между твердой мозговой оболочкой и черепной
коробкой и настолько с нею тесно связан, что выделить
его в обособленном состоянии оказалось невозможно.
Природный остаточный магнетизм исследуемой ткани
(поле, обусловленное суммой отдельных доменов) рав-
нялся 10~7—1СН электромагнитной единицы, а индуци-
рованный остаточный магнетизм как мера общего коли-
чества магнитного материала—от 10~6 до 10~5 электро-
магнитной единицы.
При электронной микроскопии магнитодетекторной
ткани выявлялось необычно густое сплетение соедини-
тельных и нервных волокон, обильно нашпигованных
гроздьями электронноплотных структур длиной 0,08—
0,15 мкм и шириной 0,02—0,04 мкм. С помощью элект-
ронографии было установлено, что супермагнитный ма-
териал ткани составлен из отдельных кристалликов
магнетита, обладающих свойством доменов. Магнитный
момент каждого домена приблизительно равнялся 3-10~14
электромагнитной единицы. По данным Е. D. Yorke
(1981), в кусочке ткани размером 2X1 мм содержится
от 107 до 108 доменов магнетита, что, несмотря на их
относительно большую рассеянность, может обеспечить
восприятие вариаций напряженности геомагнитного по-
ля порядка 400 нТ. Подчеркивая это, автор считает, что
птицы, обладая таким огромным количеством детектор-
ного материала, должны несравненно быстрее и лучше
ориентироваться по геомагнитному полю, чем магнито-
таксические бактерии, имеющие всего лишь два десятка
доменов магнетита.
59
Магнитная ткань содержала также желтые неста-
бильные кристаллы железосодержащего белка феррити-
на диаметром 6 мкм. Как полагают К. М. Towe, Н. А.
Lowenstam, М. Н. Nesson (1963), ферритин принимает
участие в синтезе магнетита. Кристаллы ферритина име-
ют такое количество железа, которое необходимо для
синтеза приблизительно 2-10-5 частиц магнетита. Из
этого, однако, не следует делать вывод, что остаточный
магнетизм детекторномагнитной ткани голубей обуслов-
лен ферритином. В количественном отношении этот же-
лезосодержащий протеин, как и никель, цинк, свинец,
обнаруженные в магнитной ткани птиц, составлял всего
лишь крошечную ее часть.
Исследования D. Presti, J. D. Pettigrew (1980) под-
твердили наличие индуцированного остаточного магне-
тизма в отсеченных и тщательно отмытых фосфатным
буфером головах голубей Columba livia. По их данным,
он составлял от 1,0-10-6 до 1,5-10—5 электромагнитной
единицы. Авторы нашли, что магнитный материал диф-
фузно рассеян по всей поверхности черепа и очень тесно
ассоциирован с ним. Кроме этого, магнитный материал
обнаруживался в шейных мышцах голубей. Показатель
индуцированного остаточного магнетизма в них равнял-
ся 1—1,5-10_6 электромагнитной единицы. Далее ока-
залось, что выраженный остаточный магнетизм присущ
многим перелетным птицам. В частности, у белых вен-
ценосных воробьев Zonotrichia leucophrys он составлял
2-10~6— 6-10-7 электромагнитной единицы, у древес-
ных ласточек Iridoprocne bicolor—5-10~6, у западной
поганки Aechmorphorus occidentalis — 9-10-6, у уток ши-
лохвость Anas acuta — 6-10-6, у мигрирующих венценос-
ных белых воробьев — от 2—4-Ю-4 до 6-10~4 (в голо-
ве) и 1—4-10~4 (в шее), а у остальных 38 видов птиц
колебался от 10-4 до 1СН5 электромагнитной единицы.
В препаратах из мускулатуры шеи голубей и белых
венценосных воробьев был выявлен диффузно рассеян-
ный магнетит, реже в виде черных кристаллов с оран-
жевыми глыбками ферритина.
По мнению Р. Presti и J. Pettigrew, детектор магнит-
ного поля у птиц может представлять собой сцепление
магнетита с нервными волокнами мышц.
Неожиданное и очень интересное сообщение сделал
В. Schreiber (1981). Изучая костные ниши губчатого
60
вещества черепа, в которых был обнаружен магнетит,
он установил, что эти локусы представляют собой актив-
ную эритропоэтическую зону, содержащую огромное ко-
личество ядерных эритроцитов, окруженных черной суб-
станцией. По-видимому, они представляют собой эрит-
робласты в сидероклеточной фазе или, другими слова-
ми, незрелые кровяные клетки, содержащие свободные
ионы железа или ферромагнитный протеин, который и
обусловливает чувствительность голубей к вариациям
магнитного поля.
Некоторые исследователи считают, что у птиц и жи-
вотных могут существовать и другие магниторецепторы,
как, например, латеральные глаза (Leask, 1977) или
мембраны полукружных каналов в лабиринте внутрен-
него уха (Jungerman, Rosenblum, 1980).
Магнитовосприимчивой структурой может оказаться
также эпифиз, участвующий, как известно, в регуляции
циркадианного ритма и являющийся важным звеном
компасно-солнце-часовой системы, активирующей локо-
моторно-миграционную деятельность птиц. Об этом го-
ворят исследования Р. Semm, Т. Schneider, L. Vollarth
(1980). С помощью микроэлектродной техники они уста-
новили, что магнитное поле напряженностью 40 А/м за
25 мин вызывает резкое угнетение (на 30—50%) им-
пульсной активности 21,3% нейронов эпифиза, которое
сохраняете^ в течение 30 мин. Повторные магнитные
стимулы эффекта не вызывали, а поле обратной поляр-
ности восстанавливало активность нервных клеток до
исходного уровня. Важно подчеркнуть, что для этого
потребовалось магнитное поле напряженностью всего
лишь 8 А/м.
Оценивая эпифиз как магнитодетекторный орган,
авторы отмечают, что у высших животных он тесно свя-
зан через супрахиазматическое ядро и симпатическую
нервную систему с латеральными глазами.
Геомагнитотропизм насекомых начали изучать в свя-
зи с обнаружением предпочтительного положения их
тела в состоянии покоя по направлению север — юг или
запад — восток. Так как при компенсации геомагнитно-
го поля ориентация становилась вполне определенной,
то G. Becker (1964) выдвинул гипотезу взаимодействия
геомагнитного поля с магнитным полем тела насекомых.
Н. D. Picton (1966), используя разработанную F. А,
61
Brown (1960) схему исследования, показал изменение-
курса дрозофилы, ползущей вверх по наклонной плос-
кости под влиянием очень слабого постоянного магнит-
ного поля. В это же время М. Lindauer, Н. Martin
(1968) высказали предположение, что полет пчел проис-
ходит по силовым линиям земного поля.
Одновременно В. Б. Чернышев (1966, 1969) сообщил,
что интенсивность лёта многих насекомых на свет в
большой степени связана с возмущениями геомагнитно-
го поля. В дни магнитных бурь искажался также, су-
точный ритм жука Trogoderma.
Далее было выявлено несколько магнитотропных ре-
акций у медоносных пчел, в частности ориентированный
по магнитному полю Земли танец, который сейчас рас-
сматривается как своеобразный вектор полета за некта-
ром (Martin, Lindauer, 1977), строительство ими сот в
северо-южном направлении (Lindauer, Martin, 1972),
зависимость циркадианного ритма жизнедеятельности
пчел от земного магнетизма и выраженное его измене-
ние в искусственном магнитном поле (Lindauer, 1976).
Изучив эти явления, J. L. Gould et al. (1978), J. L.
Gould (1979) показали, что ошибка вектора танца пчел
за взяткой составляет 5—10°. Однако в возмущенном
поле она резко возрастает и столь же быстро нивелиру-
ется при компенсации постоянной составляющей гео-
магнитного поля. Поворот улья и установление сот в
горизонтальном положении приостанавливает танец или
делает его крайне неупорядоченным. В этих случаях
векторный характер танца пчел восстанавливается лишь
по истечении длительного времени, необходимого для
геомагнитной адаптации роя пчел, а возможно, лишь
после обновления всей популяции. Ориентационный ха-
рактер танца медоносных пчел нарушается также при
перемещении ульев по географической широте. Правда,
упорядочение танца в этих случаях, как показал М. Lin-
dauer (1976), происходит в ближайшие 7 дней.
По данным J. L. Gould et al. (1980), геомагнитотроп-
ные реакции пчел, как у бактерий и голубей, обуслов-
лены магнетитом. Однако у пчел этот ферромагнетик
локализован не в голове, а в ткани передней трети
брюшка. Общее количество магнетита у пчел неодина-
ково и у различных особей колеблется от 1,5-107 до
2-108 доменов размером 300—350 А.
62
Естественный остаточный магнетизм у живых пчел в
среднем составлял 1,2-10~6 электромагнитной единицыС
(что эквивалентно полю, которое создает 1,5 млн. доме-
нов магнетита), а индуцированный предваритель-
ной обработкой в искусственном магнитном по-
ле напряженностью 5,6-104 А/м — 1,7-10—6 электромаг-
нитной единицы. Магнитные поля при этом были ори-
ентированы поперек оси тела пчел. У взрослых куколок
природный и индуцированный магнетизм был идентичным
и практически не отличался от показателей магнетизма
взрослых особей (1,5-10-6 электромагнитной единицы).
Юные куколки, личинки и яйца остаточного магнетиз-
ма обычно не имели, реже он составлял лишь 0,2—।
0,4-10-6 электромагнитной единицы.
Предположив, что кристаллы магнетита подвержены
действию внешнего поля, авторы с помощью специаль-
ного электромагнита переменного тока напряженностью
1,68•105 А/м сумели добиться полного размагничивания
вылетающих из улья пчел. Среднее значение остаточно-
го магнетизма у демагнетизированных пчел составляло
0,01 • 10~6 электромагнитной единицы.
По данным авторов, пчелы воспринимают колебания
напряженности геомагнитного поля порядка 8-10~3 А/м,
и ввиду этого намагничивание, размагничивание, при-
клеивание магнитных пылинок, прибавляемых к пище
пчел, должны нарушать не только их танец, но и ори-
ентацию в пространстве.
В описании J. L. Kirschvink (1981) размеры и фор-
ма кристаллов магнетита у медоносных пчел такие же,
как и у магнитотаксических бактерий с отношением-ши-
рины к длине а? 0,8. По его мнению, гипотетический
магниторецептор состоит из кристаллов магнетита, окру-
женных миелином, и свободно вращается в плотной
мембране. Предполагается, что в брюшке пчел содержит-
ся три такие взаимноперпендикулярно расположенные
структуры. Поскольку магнетит имеет самую низкую
электрическую устойчивость (5-10~3 Ом-см) из всех из-
вестных биологических материалов, то это может вы-
зывать пассивную деполяризацию нервной мембраны и
обеспечивать восприятие магнитного поля лишь по на-
правлению оси.
Обнаружить магнетит у пчелы не удается даже
сверхчувствительными магнитометрами, что говорит о
63-
его крайне малом содержании. С помощью СКВИД,
разрешающая сила которого 18-8 электромагнитных еди-
ниц, выявление магнетита было бы возможным при на-
личии 20 000 кристаллов, каждый из которых имеет
магнитный момент, равный 5,14-10-13 электромагнитной
единицы. Для выбора направления полета, по расчетам
Е. D. Yorke (1979), J. L. Kirschvink, J. L. Gould (1981),
у пчелы должно быть несколько сотен магнетитовых
органелл.
О геомагнитотропных реакциях других насекомых
можно догадываться, приняв во внимание их относи-
тельно высокий остаточный магнетизм. Так, у домашней
мухи Musca domestica, темного жука, жука с развет-
вленными мандибулами, таракана Periplaneia americana
и еще у 14 видов насекомых естественный остаточный
магнетизм был выше 0,5-10~6 электромагнитной едини-
цы, что в два-три десятка раз превышает фоновые зна-
чения магнетизма «пустых» органических оболочек
(Gould et aL, 1980).
Косвенные доказательства в пользу наличия магни-
торецепторов у насекомых приводит G. Becker (1980).
Изучив динамику суточной активности термитов Hetero-
termes indicola, больших личинок домашних жуков дре-
воточцев Hylotrupes bajulus, личинок Cerambyciden, он
установил четкую корреляцию ритма их питания с уров-
нем геомагнитной возмущенности и тесную взаимосвязь
этого процесса с синодическим вращением Солнца.
Появилось сообщение В. Mac Fadden и J. Douglas
(1981) о находке магнетита в королевской бабочке, со-
вершающей ежегодные миграции с востока Северной
Америки в центральую Мексику.
Присутствие магнитовосприимчивого вещества мож-
но ожидать в организме термитов. О регуляции их ак-
тивности геомагнитным полем говорят многие факты
(Frisch, 1974) и прежде всего то, что гигантские тер-
митники Австралии и Южной Африки имеют настолько
отчетливую северо-южную ориентацию, что некоторые
из них, как например Amitermes meridionalis Froggat,
издавна называют магнитными или меридианальными
холмами. Кстати сказать, такую же ориентацию имеют
не только австрало-африканские термитники (Becker,
1979), но и их обитатели, располагающиеся внутри тер-
митника параллельно силовым линиям магнитного поля
64
Земли. Интересно, что и матка термитов занимает поло-
жение, параллельное силовым линиям север—юг (De-
oras, 1949). Термиты утрачивают способность ориенти-
роваться, если их помещают в железную коробку; под
действием магнита, помещенного под нее, термиты вы-
страиваются вдоль силовых линий внешнего поля.
Будучи однотипными по магнитной ориентации и
структуре, термитники, однако, неоднородны по намаг-
ниченности. Измеряя ее в термитниках Индии, встреча-
ющихся под кварцево-магнетитными скалами, Е. А. V.
Prasad, А. С. Narayana (1981) показали, что интенсив-
ность намагничивания (намагниченности) в разных на-
правлениях варьирует в очень больших пределах, а это
косвенно говорит о значительных различиях в магнит-
ной восприимчивости составляющих структур.
Миграция рыб по геомагнитному полю была пред-
угадана W. N. Thorton (1931), а затем подтверждена
многими учеными. Лучше других изучены рыбы, имею-
щие электроцепторы, в частности подкласс пластиножа-
берных (акулы, скаты), хрящекостные осетрообразные
рыбы, семейства электрических сомов и гимнарховых,
населяющие Нил и реки Западной Африки, семейство
гимнотовых, клюворылые рыбы, живущие в Африке в
медленно текущих илистых мутных водах рек, озер и
болот. Они генерируют электрические поля двух катего-
рий: а) биоэлектрические поля электрических органов и
б) биоэлектрические поля других органов, свойственные
не только всем животным, но и растениям. С помощью
электрических полей все эти рыбы могут при различных
обстоятельствах устанавливать свое местоположение и
ориентироваться в пространстве, отличать живое от не-
живого, обнаруживать особь своего вида, хищника или
добычу.
Электрическое поле может возникать всякий раз при
пересечении геомагнитного поля потоком воды или дви-
жущейся рыбой. Так как в открытом океане поток воды
пересекает главным образом вертикальную составляю-
щую поля Земли, то восприятие изменения электричес-
кого поля для электрочувствительных животных может
служить ориентиром восходящего и нисходящего на-
правлений. Пересечение геомагнитного поля движущей-
ся рыбой создает в соответствий с законом Фарадея
внутреннюю дорзо-вентральную электромагнитную ин-
65
дукцию. Как; считает A. J. Kalmijn (1966, 1971), скаты
и акулы реагируют на самые незначительные колебания
магнитного и электрического поля Земли через так на-
зываемые ампулярные каналы Лоренцини (Murray,
1962), которые связывают кожу с глубжележащими
электрочувствительными клетками, способными отвечать
на поле в 1 мкВ/м (Murray, 1962). По подсчетам A. J.
Kalmijn (1978), акула, движущаяся со скоростью всего'
лишь несколько см/с, генерирует такую электродвижу-
щую силу, которая более чем достаточна для восприя-
тия естественных изменений в напряженности и направ-
лении магнитного поля. В другом эксперименте было
отмечено, что скаты (Urolophus halleri) реагируют на
однородные электрические поля океанической силы
(0,01—0,5 мкВ/см).
Н. W. Lissmann (1958), Н. W. Lissmann, К. Е. Ma-
chin (1958) показали, что подобной реакцией обладает
пещерная саламандра (Gymnarchus niloticus), отвечаю-
щая внезапным двигательным рефлексом на поднесение
к затемненному резервуару небольшого магнита, сооб-
щающего на расстоянии 50 см от него магнитное поле
напряженностью 0,8 А/м, при градиенте потенциала,
равном 3 электромагнитным единицам, или 0,03 мкВ/см.
Авторы отмечают, что магниточувствительность пещер-
ной саламандры выше, чем у неэлектрических рыб, в
10 000 раз. Интересно, что у семейства гимнотовых
(Gymnotus сагаро) был выработан пищевой условный
рефлекс на магнитное поле напряженностью 8-102 А/м,
При снятии этого поля питание рыбы сразу же прекра-
щалось. Передвигаясь со скоростью 10 см/с, она гене-
рировала электроток плотностью 10~4 мА/см2. Дальней-
шие исследования J. В. Pnillips (1977) подтвердили на-
личие у саламандры механизма реагирования на изме-
нения напряженности естественного поля Земли и ори-
ентации рыбы по его силовым линиям.
Скаты, клюворылые и гимнотовые рыбы, проявляю-
щие высокую чувствительность к электрическому и маг-
нитному полю, передвигаются весьма своеобразно, без-
помощи движений хвоста, поэтому их тело во время
плавания вытянуто по прямой линии и почти неподвиж-
но. По мнению Н. W. Lissmann, при этом не искажается
картина действующего поля и облегчается анализ при-
ходящей в мозг информации.
66
По данным J. A. Berge (1979), наиболее чувства*
тельны к флюктуациям геомагнитного поля и связан-
ным с ним пульсациям земных электрических потоков
угри. При этом американский угорь реагирует на элект-
роток, направленный перпендикулярно к оси тела, а
европейский — на электроток любого направления. Важ-
но подчеркнуть, что передняя часть угря в 4—5 раз чув-
ствительнее задней. Полагают, что угри мигрируют па-
раллельно электрическим токам Земли.
J. Zoeger et al. (1981) обнаружили магнетит в голове
тихоокеанских дельфинов (Delphinus delfts), обладаю-
щих, как известно, уникальными навигационными спо-
собностями. Местом локализации магнитного материала
оказалась серповидная складка мозга, прилегающая к
своду черепа и располагающаяся на 2 см сзади от мес-
та соединения затылочной, теменной и лобной костей.
Магнитный момент ткани, содержащей магнетит, был
равен 2-10—8 А-м2. Интересно, что полное размагничива-
ние мозговой ткани дельфинов происходило в очень сла-
бом переменном магнитном поле напряженностью при-
близительно 102 А/м, что превышает земное всего лишь
в 2 раза. Такой силы поле существенно не сказывалось
даже на величине естественного остаточного магнетизма
палеонтологических пород.
Электронномикроскопическое исследование и химиче-
ский анализ выявили в ткани серповидной складки дис-
ковидные, непрозрачные, анизотропные железосодержа-
щие частицы магнетита, отличающиеся су пер магнетиз-
мом. Глыбки магнетита были оплетены густой сетью
нервных окончаний, которые, по-видимому, являются со-
ставными элементами магниторецепторного аппарата.
По предположению авторов, магнитодетекторная ткань
дельфинов настолько хрупка, нежна и пластична, что
способна деформироваться в земном магнитном поле и,
значит, воспринимать самые незначительные изменения
в величине и направлении геомагнитного поля в целом
и его составляющих в отдельности.
Приведенный краткий обзор данных об ориентацион-
ных эффектах магнитного поля на самых разных пред-
ставителей животного мира подкрепляет компасную мо-
дель магнитной чувствительности. У птиц и дельфинов,
обладающих высокими навигационными способностями,
у магнитотаксических бактерий обнаружен и достаточ-
67
но хорошо изучен субстрат магнитной чувствительности,
содержащий кристаллы магнетита. У птиц эти кристал-
лы включают огромное количество доменов и образуют
цепь, которая может служить биомагнитным компасом.
Ткань, содержащая магнетит, у птиц и дельфинов тесно
связана с головным мозгом и, по-видимому, составляет
основу органа восприятия магнитного поля. Геомагни-
тотропные реакции пчел тоже обусловлены наличием у
них магнетита, причем размеры и форма его кристаллов
такие же, как и у магнитотаксических бактерий, хотя
содержание этого материала у тех и других относитель-
но невелико. Можно ожидать, что магнетит будет обна-
ружен и у термитов, проявляющих чувствительность к
магнитному полю. Поиски магнетита у высших живот-
ных и человека пока не увенчались убедительным успе-
хом. Органно-тканевой магнетизм у них может, как нам
кажется, определяться и другими субстратами, в част-
ности ферритином — наиболее возможным предшествен-
ником магнетита, который в особенно больших количе-
ствах обнаруживается в селезенке и печени, отличаю-
щихся, по нашим данным, относительно низким уров-
нем диамагнетизма.
--------------- 4 -----------------
ГЛАВА
КЛЕТОЧНАЯ И ОРГАННО-ТКАНЕВАЯ
МАГНИТОВОСПРИИМЧИВОСТЬ
4 ноября 1845 г. М. Фарадей открыл диамагнетизм,
а 18 декабря на Королевском обществе он сделал до-
клад «Новое магнитное действие», в котором отметил,
что магнит отталкивает «множество биологических ве-
ществ, таких, как дерево, кость, сухую телятину, свежее
и сушеное мясо, свежую и сухую кровь, кожу и яблоко».
С этих исследований М. Фарадея началось изучение
биомагнетизма и магнитной восприимчивости.
4.1. МЕТОДЫ ИЗМЕРЕНИЯ
МАГНИТНОЙ ВОСПРИИМЧИВОСТИ ОРГАНИЗМОВ
Измерение магнитной восприимчивости (х) слабо-
магнитных органических веществ можно проводить, ис-
пользуя два основных метода (Чечерников, 1969). Пер-
вый из них (метод Фарадея — Секстима и его модифи-
кации) позволяет получить абсолютные показатели маг-
нитной восприимчивости и основан на измерении меха-
нической силы, действующей на образец, помещенный в
неоднородное магнитное поле; второй (метод Гуи и его
аналоги) дает относительные величины магнитной вос-
приимчивости, регистрируя механическую силу, воздей-
ствующую на объект в однородном поле.
В неоднородном поле с высоким градиентом напря-
женности на тело с магнитным моментом М действует
сила, составляющая которой по оси a (a=xyz) равна:
где М — магнитный момент, Н — напряженность магнит-
dH
ного поля, — градиент поля.
69
Так как магнитный момент М определяется форму-
лой
М = <jtn = tny^H,
где т — масса образца, о — удельная намагниченность,
X — магнитная восприимчивость, Н—напряженность
магнитного поля, то первоначальное соотношение можно
отразить в виде
„ L/ dH
Frj. =
deft
а если учесть восприимчивость воздуха —
= (Xi — Х2)тН
da
где Xi и Х2 — соответственно магнитная восприимчивость
* rjdH
образца и воздуха, п-г —величина максимального
(I ОС
градиента поля. Измерив силу F и зная величину мак-
симального градиента поля, можно найти значение маг-
нитной восприимчивости исследуемого вещества.
Определение величины магнитной восприимчивости в
неоднородном поле представляет большие трудности
. и dH .
ввиду того, что область, где значение И приблизи-
тельно постоянно, имеет очень ограниченные размеры.
Помимо подбора минимального диаметра исследуемых
объектов при измерении абсолютных показателей маг-
нитной восприимчивости в неоднородном магнитном по-
ле требуется специальное устройство, передвигающее и
фиксирующее их в точке с максимальным градиентом
поля. Все эти сложности Делают метод Фарадея — Сек-
стима и его модификации неприемлемыми для быстрого
й широкого использования.
Легче измерить относительную величину магнитной
восприимчивости в сильном однородном магнитном поле,
где величина Н практически не имеет никакого зна-
чения и определять которую нет надобности. В этих
случаях используют эталонное вещество с известной
70
магнитной восприимчивостью. Тогда определение маг-
нитной восприимчивости производится по формуле
у — 221 ,Уп Fa-
Л — Ло /г, х >
т (Лх)о
где х и хо — удельная магнитная восприимчивость, а т
и /п0—массы исследуемого образца и эталона соответ-
ственно.
Применяя относительный метод Гуи, порошком тка-
ни, крови, высушенной массой микроорганизмов или
растений заполняют стеклянную пробирку длиной 100 мм
и диаметром 8 мм. Магнитную восприимчивость жидко-
стей измеряют в пробирке 150,0X20,0—30,0 мм. Если
замеры магнитной восприимчивости проводятся при раз-
личных температурах, то нижняя часть пробирки долж-
на иметь небольшой балластный объем, вследствие чего
ее вакуумируют. При измерениях магнитной восприим-
чивости верхняя часть пробирки с порошком помещает-
ся в область максимального поля, а нижняя, вакууми-
рованная, должна находиться вне поля, а точнее, в точ-
ке, где его напряженность приблизительно равна нулю.
В этом случае сила, действующая на образец, направле-
на вдоль его длины:
Fa = ~^S,
где S — площадь поперечного сечения образца.
Для определения силы F, действующей на образец в
магнитном поле, использовали аналитические весы
АДВ-20, к левой чаше которых прикреплялась тонкая
медная проволока для подвешивания закрытой притер-
той пробкой вакуумированной пробирки с биомассой.
После уравновешивания рабочей пробирки и ее стаби-
лизации включали электромагнит с напряженностью
поля в средней точке зазора 4-Ю5 А/м. Отсчет отклоне-
ний стрелки весов по делениям оптической шкалы до и
после включения электромагнита производился не менее
5 раз.
Магнитная восприимчивость биомассы определялась
в нескольких измерениях с учетом магнитовосприимчи-
вости пустой пробирки и биологического объекта. В ка-
честве абсолютного эталона при проверке вебов упот-
реблялась дважды вымороженная бидистиллированная
71
вода (/=—0,721-10-6). Эталоном для относительных из-
мерений служил химически чистый глицерин (х='
= —0,563-10~6). Во всех случаях навеска опытного ве-
щества составляла 0,5 г тонко измельченного в фарфо-
ровой ступке порошка. Материал, подлежащий измере-
нию магнитной восприимчивости, извлекался и при
надобности измельчался неферромагнитными стеклянны-
ми или пластмассовыми инструментами. Кровь, молоко,
органы и ткани животных и человека, биомасса расте-
ний и культуры микроорганизмов высушивались до по-
стоянного веса в химически чистых стеклянных чашках
Петри или фарфоровых ступках в автоматически регу-
лируемых сухожаровых камерах при одном и том же
температурном режиме, начиная с 50 и заканчивая 80 °C.
Точность измерения магнитной восприимчивости в на-
ших опытах находилась в пределах ±0,006-10-6.
4.2. МАГНИТОВОСПРИИМЧИВОСТЬ МИКРООРГАНИЗМОВ
Для получения наиболее полных представлений о
магнитной восприимчивости клеток изучались самые
разнообразные тест-микробы, относящиеся к различным
таксономическим группам и вследствие этого отличаю-
щиеся по многим биологическим и физико-химическим
свойствам: пекарские, винные и пивные дрожжи, три
вида почвенных актиномицетов — Act. griseus, Act. сое-
licolor, Act. globisporus, эталонная культура Mycobacte-
rium tuberculosis H37Rv, изониазидоустойчивый штамм
микобактерий туберкулеза, два возбудителя микобакте-
риозов — М. intracellularе, М. avium, микобактерии-сап-
рофиты— М. phlei, М. smegmatis, М. штамм 607,
М. штамм МГУ, Corynebacterium volutans, эшерихии
штамма К-12, сальмонеллы мышиного тифа, золотистый
стафилококк 209 Р, его ультрафиолетовый мутант с де
фектом дыхания стафилококк УФ-3, Micrococcus lyso-
deikticus, четыре вида спороносных почвенных бацилл —
Вас. cereus, Вас. subtilis, Вас. megaterium, Вас. mycoi‘
des. Таким образом, кроме исследованных нами ранее
14 видов и штаммов энтеробактерий, кокков и микрокок-
ков в опыт отбирали бактерии актиномицетной линии и
бациллы, которые, имея очень активные железотранс-
портные системы типа сидерохромов, могут в больших
количествах аккумулировать из питательной среды фер-
ромагнетики.
72
Стафилококк УФ-3 и Micrococcus lysodeikticus слу-
жили индикаторами возможного намагничивания ме-
таллсодержащих протеинов, которые могут вносить оп-
ределенный вклад в парамагнетизм микробной клетки.
При этом у биохимического мутанта стафилококка УФ-3,
как и у раковых клеток, отмечался дефект дегидрогеназ
(Гаузе и др., 1957), а микрококки в отличие от других
бактерий, по данным D. Herbert, J. Pinsent (1948), со-
держали приблизительно в 3 раза больше каталазы, чем
Rhodopseudomonas spheroides, или 10 доз фермента, име-
ющегося в человеческих эритроцитах (1—2%' сухого
веса).
Для получения 0,5 г сухой биомассы свежевыросшие
культуры эшерихий, сальмонелл, кокков, коринебакте-
рий газоном засевались на простые или сахарные пита-
тельные агары, залитые в чашки Петри; микобактерии
выращивались на пластинчатой среде Левенштейна—
Йенсена, антракоиды вследствие трудности получения
культуры на агаре из-за глубокого врастания в среду —
на питательном бульоне в матрацах, актиномицеты — на:
жидкой солевой среде Гаузе № 1 тоже в матрацах. Вы-
росшие культуры взвешивали в бидистиллированной во-
де и тщательно в ней эмульгировали. Гомогенную взвесь
микроорганизмов путем повторных центрифугирований
дважды отмывали в бидистиллированной воде и затем
высушивали. Статистическую обработку материала про-
изводили методом непрямых разностей по Монцевичю-
те — Эрингене.
Как и ожидалось, суммарно-популяционная магнит-
ная восприимчивость биомассы всех видов бактерий
оказалась отрицательной, т. е. органические вещества
микробного происхождения проявляли диамагнитные
свойства. Однако величина диамагнетизма у разных ви-
дов микроорганизмов варьировала. В частности, иссле-
дования В. М. Червинца и др. (1979), С. А. Павловича
(1981) выявили (табл. 7), что магнитная восприимчи-
вость эшерихий однозначна и колеблется от —0,416-10~6
до —0,454-Ю”6. Исключение составлял штамм Р 678 с
низкой степенью диамагнетизма (—0,329-10~6). Близкие
величины диамагнетизма обнаруживались у родствен-
ных эшерихиям сальмонелл, серратий, фекального ще-
лочеобразователя, вульгарного протея (от —0,453-10~е
до —0,496-10~б). Статистически достоверное снижение
73
Таблица ?
Суммарно-популяциониая магнитная восприимчивость бактерий, М+т
Тес т-объект Магнитная восприимчивость, xio-' Тест-объект Магнитная восприимчи- > вость, ХЮ“'
Энтеробактерии Микобактерии
Escherichia Salmonella Pseudomonas aeruginosa Serratia marcescens Alcaligenes faecalis Proteus vulgaris от —0,329+0,008 шт. (Р678) до —0,454+0,008 шт. (К-12) от —0,391+0,004 до —0,483+0,004 —0,388+0,017 —0,453+0,008 -0,464+0,003 —0,496+0,008 М. tuberculosis H37RV М. tuberculosis Is' М. avium- м. intracellulare М. smegmatis М. штамм 607 М. штамм МГУ —0,410+0,006 —0,508+0,007 —0,422+0,012 —0,241+0,008 —0,381+0,001 —0,402+0,007 —0,419+0,015
Кокки Актиномицеты
Staph, aureus 209P Staph, aureus UV-3 Micrococcus luteus Micrococcus lysodeikticus —0,466+0,007 —0,515+0,003 —0,470+0,078 —0,339—0,003 Act- griseus Act. coelicolor Act. globisporus —0,447+0,002 —0,484+0,008 —0,504+0,001
Бациллы
Вас. subtilis Вас. mycoides Вас. megaterium —0,327+0,007 —0,167+0,008 —0,499+0,006
диамагнетизма регистрировалось только у псевдомонад
и сальмонелл чумы свиней.
Магнитная восприимчивость золотистого стафилокок-
ка и желтого микрококка была такой же, как у энтеро-
бактерий. Необычно высоким парамагнетизмом отли-
чался Micrococcus lysodeikticus (—0,339-10-6).
Результаты первых измерений суммарно-популяцион-
ной магнитной восприимчивости бактерий, таким обра-
зом, оказались мало информативными. В конечном ито-
ге они свидетельствовали, что показатели диамагнетиз-
ма у энтеробактерий, золотистого стафилококка и жел-
того микрококка при выращивании этих бактерий на
обычных питательных средах одинаковы, как, между
прочим, сходен их минерально-органический состав и
клеточный биохимизм. Salm, cholerae-suis u Е. coli
штамм Р678 с низким диамагнетизмом могли оказаться
случайными парамагнитными клонами неоднородной по
магниточувствительности популяции. Пристального вни-
мания заслуживал лишь М. lysodeikticus, высокий уро-
вень парамагнетизма которого, очевидно, обусловлен
большим количеством в его клетках каталазы как же-
лезосодержащего протеина. Все это дало повод предпо-
ложить, что любая бактериальная популяция по магнит-
ной восприимчивости гетерогенна, и нацелило на даль-
нейшее изучение магнитной восприимчивости у микро-
организмов, отличающихся по типам метаболизма от
энтеробактерий и кокков.
Углубленные исследования в этих направлениях по-
казали следующее.
Видовое различие в величине диамагнетизма наибо-
лее ярко выявлялось у почвенных сибиреязвенноподоб-
ных бацилл. В частности, самый низкий диамагнетизм
определялся у Вас. mycoides (—0,167-10~6). В сравне-
нии с этим показатели диамагнетизма у Вас. subtilis
были в два, а у Вас. megaterium — в три раза боль-
шими.
Магнитная восприимчивость микобактерий была то-
же очень разнообразной. Самый низкий диамагнетизм
отмечался у М. intracellulare (—0,241-IO-6). Большие
показатели диамагнетизма регистрировались у сапрофи-
та М. smegmatis. Магнитная восприимчивость у других
микобактерий составляла от —0,402-Ю-6 (М. 607) до
—0,422-10-6 (М. avium). Специально выделенный от
75
больных изониазидоустойчивый авирулентный штамм
микобактерий туберкулеза имел существенно более вы-
сокий диамагнетизм, чем эталонная культура М. tuber-
culosis H37Rv, вызывающая при заражении морских сви-
нок диссеминированный туберкулез.
Сравнительно небольшим, но статистически достовер-
ным было различие в величине магнитной восприимчи-
вости у актиномицетов: —0,447-10-6 у Act. griseus,
—0,484-10~6 у Act. coelicolor и —0,504-10~6 у Act. globi-
sporus. У биохимического мутанта стафилококка УФ-3-
с дефектом дегидрогеназ по сравнению с эталонным
штаммом золотистого стафилококка она заметно снижа-
лась (—0,515-Ю-6 против —0,466-10~6).
Исследование случайно отобранных клонов популя-
ции показало, что каждый вид микроорганизмов имеет
определенный процент особей, магнитная восприимчи-
вость которых отличается от среднесуммарной. В опы-
тах А. И. Жмакина (1982), например, популяционная
магнитная восприимчивость сальмонеллы мышиного ти-
фа, быстро вовлекающейся в диссоциацию при пассиро-
вании в измененном геомагнитном поле, составляла
—0,495-10-6, у 4 клонов из 16 изученных она колебалась
в пределах от —0,406-10-6 до —0,326-10-6, а у одного
имела больший показатель диамагнетизма, чем у хими-
чески чистого глицерина (—0,650-10-6).
Наши исследования выявили, что популяции микро-
бов содержат разный процент особей, магнитная вос-
приимчивость которых отличается от суммарно-видового
показателя. Больше всего их обнаруживалось в культу-
рах пекарских дрожжей (33%) и С. volutans (20%).
В популяциях Е. coli, Вас. cereus, М. phlei и пивных
дрожжей таких клеток было около 10%. При этом кло-
нов с низким диамагнетизмом (обладающих относитель-
ным парамагнетизмом) встречалось в 2—5 раз больше,
чем клонов с повышенным уровнем диамагнетизма.
Здесь следует также отметить, что отклонение показа-
телей магнитовосприимчивости от среднесуммарной ве-
личины по парамагнетизму было в 2—3 раза большим,
чем по диамагнетизму (табл. 8). Так, например, у от-
дельных клонов Вас. cereus, пивных и пекарских дрож-
жей диамагнетизм снижался (увеличивался парамагне-
тизм) в 1,5—3 раза, а повышался всего лишь на 10%.
Из очень чувствительной к магнитным полям популяции
76
Таблица 8
Сравнительные показатели суммарно-популяционной и
клональной магнитовосприимчивости микроорганизмов, М+т
Тест-объект Магнитная восприимчивость, хЮ’1
суммарная клональная
Е. coli К-12 Вас. cereus М. phlei С. volutans Дрожжи пивные пекарские —0,472+0,008 —0,451+0,019 —0,513+0,009 —0,430+0,009 —0,478+0,008 —0,455+0,018 от —0,397 до —0,492 от —0,298 до —0,490 от —0,441 до —0,534 от —0,024 до —0,467 от —0,324 до —0,511 от —0,160 до —0,480
С. volutans могут выделяться клоны с исключительно
высокой степенью парамагнетизма (—0,024 -10~6). Наи-
меньшие отклонения от среднесуммарной величины диа-
магнетизма отмечались у клонов Е. coll и М. phlei, ко-
торые в противоположность другим бактериям с трудом
вовлекаются в магнитную диссоциацию.
Гетерогенность микробной популяции по магнитной
восприимчивости резко возрастала при вовлечении
культур в магнитную диссоциацию, что закономерно до-
стигалось при длительном их пассировании в ослаблен-
ном (//=0,8 Д/м), сверхслабом (7/=8-10-4 А/м) и
постоянном (Я=80 А/м) магнитных полях. При этом каж-
дая третья-четвертая колония, вовлекающаяся в магнит-
ную диссоциацию, обладала выраженным парамагне-
тизмом (табл. 9). Так, на 17-м пассаже из пяти отоб-
ранных диссоциантов Вас. subtilis только один имел
такую же степень диамагнетизма, как у контрольной
культуры. Ослизненные М-колонии отличались сравни-
тельно высоким уровнем диамагнетизма, у /?-диссоциан-
тов он составлял —0,205-10-6, что по диамагнетизму в
1,5 раза меньше, чем в контроле.
Приблизительно в такой же пропорции по частоте и
величине диа- и парамагнетизма расщеплялись магнит-
ные диссоцианты Вас. megaterium.
Исключение составляли эшерихии, диссоциация у
которых не наступала к 20-му пассажу в ослабленном,
сверхслабом и постоянном поле.
Интересные данные получали при исследовании маг-
нитных диссоциантов стафилококка УФ-3 (Павлович,
77
Арцукевич, 1983). Уже на 6-м пассаже в ослабленном
магнитном поле напряженностью 0,8 А/м около 5—7%|
особей (колоний) его популяции утрачивали присущую
биохимическому мутанту оранжевую окраску, а вместе
с ней и ауксотрофность. Показатель диамагнетизма у
таких ревертантов снижался с —0,515-10~6 до
—0,480-Ю-6, т. е. становился почти таким же, как у ро-
дительского штамма золотистого стафилококка 209Р.
Исследование высушенной микробной биомассы на
радиоспектрометре ERS-220 (v~9,3 ГГц) выявило, что
в ЭПР-спектре изученных тест-бактерий присутствует
сигнал в виде синглета, g-фактор которого почти пол-
ностью совпадает с g-фактором свободного электрона
(рис. 10). У магнитных диссоциантов Вас. subtllis и Вас.
megaterium, показатели магнитовосприимчивости кото-
рых сдвигались к парамагнетизму, ЭПР-спектр пред-
ставлял собой суперпозицию двух сигналов — синглета
Таблица 9
Магнитная восприимчивость магнитных
диссоциантов бактерий, М+т
Тест-объект Пассаж Форма колоний (R-, S-, М-) и магнитная восприимчивость их биомассы, х10”в
контрольных <77 40 А/м) пассированных в магнитных полях
77=0,8 А/м /7=8-10~* А/м 77=80 А/м
Вас. subtilis 17 R-типичная —0,327+ +0,002 Ослизненная (М-) —0,388+ +0,002 R-изменен- ная —0,205+ +0,002 R-изменен- ная —0,363+ +0,004 Ослизненная (М-) —0,398+ +0,003
Вас. megate- rium 17 R-типичная —0,499+ +0,003 R-типичная —0,405+ +0,002 Ослизненная (М-) —0,421+ +0,001 Ослизненная (М-) —0,499+ +0,002
Е. coli 20 S-типичная —0,432+ +0,002 S-типичная —0,438+ +0,001 S-типичная —0,432+ +0,002 S-типичная —0,441 + +0,002
Staph, aureus UV-3 6 S-оранжевая, влажная —0,515+ +0,001 S-беспиг- ментная, влажная —0,480+ +0,001 RS-оранже- вая —0,522+ ±0,001 RS-оранже- вая —0,500+ +0,002
78
и секстета, что, по мнению G. Н. Reed, М. Kohn (1970),
характерно для иона марганца, находящегося в составе
белка. По-видимому, у пассированных в ослабленном и
сверхслабом магнитных полях субштаммов бацилл, от-
личающихся низким уровнем диамагнетизма, происхо-
дит глубокая перестройка обмена, в результате чего-
।----------1-----------1----------1-----------f
2,36 2,52 2,68 2,8k 3,00
------* Магнитное поле>х1О5 А/м
Рис. 10. ЭПР-спектр высушенной биомассы Вас. megaterium: 1 —
контрольной культуры (МВ=—0,499-Ю-6), 2 — пассированной в
сверхслабом МП (МВ = — 0,421 • 10~6)
индуцируются или появляются парамагнитные марган-
цезависимые ферменты.
Исследование ревертантов стафилококка УФ-3 с от-
клонением магнитной восприимчивости в сторону пара-
магнетизма показало, что их ЭПР-спектр приблизитель-
но такой же, как у золотистого стафилококка 209Р
(рис. И), что может говорить о нормализации биохими-
ческих процессов, а точнее, об обратной мутации к ис-
ходному родительскому виду.
Таким образом, проведенные нами исследования еще
раз показали, что биомасса бактерий, как и многих
других организмов, диамагнитна. При этом показатель
79
суммарной магнитной восприимчивости у родственных
видов, например представителей семейства энтеробак-
терий, варьирует в очень узком диапазоне (от
—0,423-Ю*6 до —0,452-10-6), и вследствие этого его
трудно или невозможно использовать при дифферен-
циации эшерихии, сальмонелл и, по-видимому, других
—*- Магнитное поле }*105А/м
Рис. 11. ЭПР-спектр высушенной биомассы стафилококка: 1 —
контрольный штамм УФ-3, 2, 3, 4 — штаммы УФ-3, пассированные
соответственно в сверхслабом, постоянном и ослабленном МП, 5 —
эталонный штамм 209 Р
кишечных бактерий. То же самое можно сказать о кок-
ках и актиномицетах.
В противоположность этому суммарная магнитная
восприимчивость почвенных спорообразующих бацилл
отличалась большим разнообразием. Наиболее низкий
уровень диамагнетизма имела культура Вас. mycoid.es
(—0,167-10~6), а самый высокий — Вас. cereus.
Некоторый интерес представляют видовые различия
в суммарной величине магнитной восприимчивости сре-
ди микобактерий, в частности относительный парамаг-
нетизм М. intracellularе (—0,243 -10~6) и высокий по
сравнению с эталонным штаммом Л1. tuberculosis H37RV
уровень диамагнетизма у изониазидоустойчивого штам-
ма микобактерий туберкулеза. Нарастание величины
80
диамагнетизма у мутанта стафилококка УФ-3 с дефек-
том дегидрогеназ,, утратившего вирулентность, и, наобо-
рот, сдвиг в сторону парамагнетизма у его ревертанта
дают серьезное основание считать, что активность био-
химических процессов одновременно определяет патоген-
ность и уровень магнитной восприимчивости. О корреля-
тивной связи между магнитной восприимчивостью и
интенсивностью обмена свидетельствует появление у
магнитных диссоциантов антракоидов с относительным
парамагнетизмом марганцезависимых ферментов и, воз-
можно, других парамагнитных соединений. С определен-
ными оговорками подобную точку зрения можно экстра-
полировать на другие магнитные диссоцианты и гетеро-
генные по магнитной восприимчивости особи микробных
популяций. Отдельно необходимо подчеркнуть, что не-
однородность по магнитовосприимчивости в сравнении с
гетерогенностью по другим признакам и свойствам про-
является у микробов с очень высокой частотой. Это объ-
ясняет, почему бактерии отличаются высокой магнито-
чувствительностью и быстро вовлекаются в магнитную
диссоциацию, которая, надо думать, в свою очередь об-
условлена процентным содержанием в популяции особей
с относительно высоким уровнем парамагнетизма. По-
следний, как это было сказано выше, у микробов за-
висит главным образом от типа обмена, наличия у них
металлотранспортных систем и, следовательно, количе-
ства парамагнитных элементов (Neilands, 1974). При-
менительно к пекарским дрожжам большой процент ге-
терогенности по магнитной восприимчивости, отмечен-
ный в наших опытах, мог быть обусловлен необычно
большой электропроводностью и высоким электрокине-
тическим потенциалом их клеток (Дерканосов, Громков-
ская, 1979).
4.3. МАГНИТОВОСПРИИМЧИВОСТЬ ГРИБОВ
И РАСТЕНИЙ
В качестве тест-объектов изучались низшие и выс-
шие растения: шляпочные грибы, зерновые и бобовые,
плоды и овощи, кора древесины и листья, трава, лишай-
ники и мох, а также магнитная восприимчивость почвы.
Результаты исследований суммированы в табл. 10 и 11.
Данные показывают, что магнитная восприимчивость
шляпочных грибов находится в таких же пределах, как
81
Таблица 10
Суммарная магнитная восприимчивость грибов, М+т
Тест-объект Магнитная восприимчивость, Х1О“«
шляпок ножек
Мухомор —0,420+0,002 —0,460 +0,006
Рядовка краснеющая —0,499+0,006 —0,485+0,001
Опенок ложный —0,450+0,003 —0,477+0,003
Волнушка розовая —0,450+0,003 —0,450+0,003
Сыроежка пищевая Сыроежка зеленовато- —0,386+0,002 —0,483+0,002
бурая —0,468-0,003 —0,501+0,002
Рыжики —0,468 -0,007 —0,479—0,002
Лисички —0,430-0,005 —0,466+0,008
Свинушка тонкая —0,483-0,002 —0,477+0,002
Моховик желто-бурый —0,440—0,002 —0,486—0,002
Подберезовик обыкно- венный —0,436+0,001 —0,491+0,006
Белый гриб —0,412+0,003 —0,440+0,003
у бактерий, с той лишь особенностью, что биомасса
шляпок имела более низкий уровень диамагнетизма, чем
биомасса их ножек. Исключение составляла рядовка
краснеющая, у которой различные структуры обладали
одинаковой магнитовосприимчивостью.
Пытаясь глубже уяснить, в какой мере уровень диа-
магнетизма зависит от аморфности материала, мы ис-
следовали тщательно очищенную от верхнего слоя дре-
весину коры липы. При этом было установлено, что ее
магнитная восприимчивость равна —0,657 • 10~6, т. е.
близка к показателю абсолютного эталона дважды вы-
мороженной воды. Верхний слой коры, покрытый зеле-
ной массой биообрастаний, имел очень высокий уровень
парамагнетизма (+11,70-10-6). Это вызвало потреб-
ность в замерах магнитной восприимчивости лишайни-
ков и мхов — основных симбионтов зеленой раститель-
ности стволов деревьев. Уже первые опыты показали,
что величина парамагнетизма у них такая же, как у об-
росшей ими коры. Тот же показатель намагниченности
отмечался у мхов и лишайников, собранных на почве.
Естественно было предположить, что парамагнетизм
биообрастаний связан с наличием в них примесей земли
или ферромагнитных элементов. Проведенные в этом
направлении исследования выявили, что магнитная вос-
приимчивость образцов почвы, взятых из-под лишайни-
82
Таблица 11
Суммарная магнитная восприимчивость растений, М+т
Тест-объект Магнитная восприим- чивость» хЮ“*
Кора липы
Внутренний слой древесины
Верхний слой с биообрастаниями
—0,657+0,004
+ 11,70+0,002
Почвенный покров
Лишайники
Почва под лишайниками
Мох, выросший на почве и отмытый
Почва под мхом
Мох, выросший на пне и отмытый
Трава осенняя
Листья
Кленовые осенние
Кленовые весенние
Липовые осенние
Липовые весенние
+ 13,07+0,004
+6,60+0,002
+24,89+0,004
4-10,97+0,002
+2,66+0,001
—0,192+о,004
—0,242+0,005
—0,422+0,004
—0,310+0,004
—0,473+0,003
Зерновые
Пшеничная мука высшего сорта
Пшеничная мука 1-го сорта
Ячмень промытый
Ячмень нелромытый
Гречка промытая
Гречка непромытая
Манная крупа из бумажного пакета
Манная крупа из железной банки
Рис
Пшено
Бобовые
Горох
Фасоль
Овощи и фрукты
Картофель
Яблоки
—0,462+0,008
—0,121+0,006
—0,503+0,005
—0,187+0,006
—0,520+0,007
—0,507+0,005
—0,547+0,004
—0,417+0,005
—0,547+0,004
—0,534+0,003
—0,499+0,005
—0,576+0,005
—0,537+0,005
—0,542+0,008
83-
ков и мхов, колебалась от +6,60-10~6 до +10,97-10~6,
т. е. была меньшей (иди равной) магнитной восприим-
чивости их биомассы, но не превышала ее, как нами
ожидалось.
Далее было отмечено, что показатель парамагнетиз-
ма биомассы мхов снижается в несколько раз после
длительного промывания в проточной воде. И наконец,
оказалось, что уровень парамагнетизма мхов и мохооб-
разных коррелирует с величиной парамагнетизма пита-
ющей их основы. Так, у биомассы тщательно отмытых
мхов, выросших на гниющих остатках пня, показатель
парамагнетизма составлял + 2,660-10~6, а у мхов, соб-
ранных вокруг строений,---1-24,890-10-6, т. е. в два раза
превышал показатель парамагнетизма навесок почвы.
Для составления объективной оценки полученных ре-
зультатов в качестве контроля определяли магнитную
восприимчивость травянистой растительности. Ее вос-
приимчивость оказалась диамагнитной, правда, в два ра-
за меньшей, чем у шляпочных грибов (—0,192-10~6).
В последней серии опытов изучалась магнитная вос-
приимчивость биомассы листьев по сезонам. Снятая с
липы и клена в октябре зелено-желтая, желтая, багря-
но-красная осенняя листва оказалась в 1,5 раза менее
диамагнитна, чем ранневесенняя, майская.
Показатели намагничиваемости хорошо промытых и
высококачественных зерновых, бобовых, картофеля и
яблок существенно не различались, и биомасса всех изу-
ченных нами образцов была высоко диамагнитной, по
величине приближаясь к показателю магнитной воспри-
имчивости химически чистого эталонного глицерина.
Диамагнетизм биомассы зерна и круп может, одна-
ко, резко снижаться, как, например, в случаях измере-
ния магнитной восприимчивости непромытого фуражно-
го ячменя, манной крупы, находившейся в железной та-
ре, или первого сорта пшеничной муки, по-видимому, с
какими-то примесями.
Вышеотмеченное, таким образом, показывает, что
растительные организмы по магнитной восприимчивос-
ти очень разнообразны. Так, биомасса шляпочных гри-
бов диамагнитна в такой же мере, как большинство
энтеробактерий, кокков и актиномицетов. Правда, их
плодовые тела более парамагнитны, чем ножки. В боль-
шей степени диамагнитны зерновые и бобовые. Необыч-
84
но большой диамагнетизм проявляет вещество коры,
что, по-видимому, можно использовать при оценке со-
стояния древесины. Динамика сезонного изменения диа-
магнитных свойств листвы может говорить о значитель-
ной роли геомагнетизма в биохимических процессах
растительных клеток. Полученные нами факты с боль-
шой убедительностью свидетельствуют, что парамагне-
тизмом обладают не только магнитотаксические бакте-
рии, описанные R. Р. Blakemore, но и симбиоз грибов и
водорослей, составляющих лишайники, а также некото-
рые мхи и мохообразные как представители примитив-
ных высших растений. Высокий парамагнетизм лишай-
ников, очевидно, связан с наличием в симбиотических
грибах особых сидерохромов, способных извлекать из
субстратов (почвы) трехвалентные ионы железа и дру-
гие ферромагнетики (Anderegy et al., 1963). Механизм
насыщения и образования металлсодержащих соедине-
ний у мхов остается неизвестным. Еще более загадочна
сущность парамагнетизма у этих растений.
4.4. МАГНИТОВОСПРИИМЧИВОСТЬ
ХОЛОДНОКРОВНЫХ И НАСЕКОМЫХ
Среди холоднокровных магнитную восприимчивость
изучали у трех видов морских рыб (ставрида, минтай,
зубан), пресноводной щуки, прудовой лягушки и ужа.
Для получения представления о магнитной восприимчи-
вости насекомых в качестве тест-объектов использова-
ли медоносных пчел и муравьев, которые обладают вы-
сокой степенью геомагнитотропизма и способны синте-
зировать магнетит. У рыб и лягушек определяли
органно-тканевую магнитовосприимчивость. Ужа расчле-
няли на три части по длине с последующими измерения-
ми магнитной восприимчивости различных сегментов его
тела. При исследовании пчел и муравьев вначале раз-
дельно измерялась магнитная восприимчивость брюшек
и остальных частей их тела, а вслед за этим — магнит-
ная восприимчивость всей биомассы насекомых.
Исследования выявили (табл. 12), что магнитная
восприимчивость органов и тканей рыб и амфибий от-
личалась большим разнообразием. Это прежде всего
выразилось в существенных различиях показателей диа-
магнетизма сердца, мышечной ткани и особенно мозга.
85
Органно-тканевая магнитовосприимчивость
Магнитная
Вид крови] | печени | селезенки | легких почек |
Рыбы
—0,478+
±0,008
—0,272+
+0,028
Ставрида
Зубан
Минтай
Щука
—0,480+
±0,003
—0,455±
±0,004
—0,548±
±0,002
Амфибии
Прудовая
лягушка
—0,348±
±0,013
—0,237±
±0,025
—0,454±
±0,001
—0,526±
±0,005
В общем, у морских рыб отмечалось четко выраженное
снижение уровня диамагнетизма этих тканей в сравне-
нии с пресноводными (щукой и лягушкой). Очень силь-
,но изменялась магнитная восприимчивость кишечника.
У щуки, например, она составляла —0,494-10-6, а у ля-
гушки— минус 0,115-10-6. Несколько большим парамаг-
нетизмом обладали печень и жабры зубана, жабры и
мышцы ставриды, печень лягушки.
Магнитная восприимчивость разных сегментов ужа
оказалась более или менее одинаковой и от головы к
хвосту изменялась от —0,343-10-6 до —0,397-Ю-6. Боль-
шим диамагнетизмом обладала только кожа
(-0,426-10-6).
Магнитная восприимчивость общей биомассы медо-
носных пчел в среднем составляла —0,405-10-6, а му-
равьев—минус 0,436-10-8. Четкие различия определялись
лишь при сравнении магнитовосприимчивости раз-
ных сегментов тела насекомых. Точнее говоря, пока-
затель диамагнетизма брюшек муравьев был в 1,5 раза
меньшим, чем остальной части тела (—0,402-Ю-6 про-
тив —0,602-Ю-6), а у пчел — такой же, как всей био-
массы насекомых.
.86
Таблица 12
холоднокровных, М+т
восприимчивость, х ю-«
| сердца | кишечника | мышц мозга глаз жабр
Рыбы
—0,422+ —0,315± —0,530± —0,475± —0,336±
+0,003 ±0,025 +0,029 ±0,007 +0,023
—0,412+ —0,461± —0,439± —0,406+ —0,283+
+0,003 ±0,015 +0,008 ±0, 0 +0,035
—0,388+ —0,450+ —0,557± —0,440± —0,488+ —0,455+
+0,009 +0,004 +0,018 +0,003 ±0,029 ±0,004
—0,518+ —0,494+ —0,622+ —0,602+ —0,445+ —0,463+
±0,004 ±0,001 +0,017 +0,005 ±0,001 +0,001
Амфибии
—0,516+ —0,115+ —0,530+ —0,565+ —0,476±
+0,002 ±0,001 ±0,014 +0,004 +0,002
4.5. МАГНИТОВОСПРИИМЧИВОСТЬ животных,
ПТИЦ И ЧЕЛОВЕКА
Определяя магнитную восприимчивость клеток, жи-
вотного происхождения, в опыт отбирали органы и ткани
мелких лабораторных животных, кошек, собак,
свиней, коров, кур и умерших людей. При этом
большей частью подход к исследованию был эмпириче-
ским, однако для выявления степени парамагнетизма
металлсодержащих протеинов целенаправленно изуча-
ли кровь, содержащую гемоглобин, органы, богатые
ферритином (селезенка, печень) или особенно чувстви-
тельные к воздействию магнитных полей (надпочечники,
семенники), черепные кости домашних голубей, где, воз-
можно, синтезируется магнетит как органический фер-
ромагнетик.
Полученные данные свидетельствуют, что органы и
ткани теплокровных животных и птиц диамагнитны.
•Степень их диамагнетизма, как видно из табл. 13, нахо-
дится в таком же диапазоне, как у микробов и растений.
Самый большой уровень диамагнетизма имела моз-
говая ткань, незначительно варьируя у большинства
видов от —0,513-10-6 до —0,531-Ю-6. Биомасса мозга
87
Таблица 13
Органно-тканевая магнитовосприимчивость животных, птиц и человека, М+т
Вид Магнитная восприимчивость, xlO_f
кровн печени селезенки легких । почек сердца МЫШЦ мозга глаз кишечника
Мышь -0,349+ —0,409+ —0,318+ —0,495+ —0,502+ —0,494+ -0,509+ —0,531 + —0,403+
+0,001 +0,002 +0,003 +0,003 +0,005 +0,005 +0,002 +0,002 +0,005
Крыса -0,329+ —0,422+ —0,456+ —0,485+ —0,476+ —0,450+ —0,539+ —0,608+
+ 0,002 +0,005 +0,003 +0,004 +0,004 +0,003 +0,005 +0,002
Морская —0,359+ -0,512+ —0,285+ —0,555+ —0,550+ —0,510+ —0,547+ —0,602+ —0,498+
свинка +0,001 +0,002 +0,035 +0,014 +0,004 +0,004 +0,005 +0,005 +0,003
Кролик —0,348+ —0,486+ —0,546+ —0,532+ —0,513+ —0,473+ —0,477+ —0,526+ —0,519+ —0,539+
+0,018 +0,008 +0,001 + 0,017 +0,004 +0,002 +0,003 +0,016 +0,005 +0,011
Кошка —0,318+ —0,470+ —0,350+ —0,414+ —0,454+ —0,519+ -0,493+ —0,565+ —0,527+ —0,445+
+0,002 +0,002 +0,012 +0,016 +0,004 +0,018 +0,003 +0,003 +0,004 +0,005
Собака —0,339+ —0,408+ —0,297+ —0,404+ —0,475+ —0,492+ —0,473+ —0,513+ —0,451 + —0,496+
+0,003 +0,003 +0,003 +0,002 +0,002 +0,003 +0,002 +0,005 +0,002 +0,001
Свинья —0,377+ —0,423+ —0,265+ —0,497+ —0,474+ —0,490+ —0,529+ —0,516+
+0,001 +0,002 +0,001 +0,003 +0,002 +0,006 +0,002 +0,001
Корова —0,324+ —0,479+ —0,164+ —0,454+ -0,488+ -0,491+ —0,493+ —0,527+
+0,001 +0,002 +0,003 +0,003 +0,003 +0,003 +0,002 +0,001
Курица —0,437+ —0,494+ —0,482+ —0,486+ —0,476+ —0,547+ —0,525+ —0,550+ —0,506+
+0,002 +0,006 +0,004 +0,004 +0,004 +0,003 +0,001 +0,015 +0,003
Голубь —0,358+ —0,206+ —0,188+ —0,376+ —0,420+ —0,502+ —0,537+ —0,521+
+0,002 +0,007 +0,011 +0,002 +0,001 +0,001 +0,009 +0,008
Человек -0,336+ —0,274+ —0,281 + —0,432+ —0,475+ —0,427+ —0,508+ —0,676+
+0,002 +0,028 ±0,028 +0,011 +0,008 +0,003 +0,015 +0,005
кошек по диамагнетизму не отличалась от чистого гли-
церина, а у крыс и морских свинок была еще большей
(минус (0,602—0,608) • 10-6).
Очень высокие показатели диамагнетизма получали
при измерении магнитной восприимчивости мышечной
ткани бедра мышей, крыс, морских свинок, свиней, птиц
и некоторых органов морской свинки, кролика и кошки.
Суммарная магнитная восприимчивость легких (кроме
голубя), почек, сердца, кишечника, глаз, надпочечников
у одного и того же вида животных и птиц почти не от-
личалась, а у разных видов колебалась от —(0,403—<
—0,414) -10~6 (кишечник мыши, легкие кошек) до
— (0,550—0,555) • 10-6 (почки и легкие морской свинки).
Средние показатели диамагнетизма печеночной тка-
ни кролика, кошки, коровы, морской свинки, курицы
были такими же, как у других органов, и составляли
— (0,470—0,512)-Ю-6. По сравнению с этим диамагне-
тизм печени мышей, собак снижался, падая до уровня
—0,408-10-6, а у голубя — до —0,206-10-6.
Наименьший диамагнетизм обнаруживался в ткани
селезенки: у кошки —0,350-10~6, у мыши — 0,318-Ю-6,
у свиньи, собаки и морской свинки от —0,297-10-6 до
—0,265-Ю-6, а в селезенке голубя и коровы —0,188-10"*
и —0,164-10~6 соответственно, т. е., сдвигаясь в сторону
парамагнетизма, увеличивался по отношению к показа-
телям других внутренних органов и мозговой ткани жи-
вотных в 1,5—3 раза.
Магнитная восприимчивость крови у разных живот-
ных варьировала от —0,318-Ю-6 до —0,358-Ю-6 и толь-
ко у курицы и свиньи была заметно ниже.
Неоднократные замеры магнитной восприимчивости
надпочечников свиньи и коровы ничего определенного
не дали. Их биомасса имела такие же показатели диа-
магнетизма, как легкие, сердце и почки разных живот-
ных. Некоторый сдвиг к парамагнетизму обнаруживали,
исследуя плоские кости черепа (—0,412-Ю-6) и семен-
ники (—0,361-10~6) домашних голубей, взятых для ин-
дикации возможного наличия у них парамагнитного ма-
териала.
Отдельного рассмотрения заслуживает органно-тка-
невая магнитовосприимчивость человека. Эти данные
были получены при исследовании секционного материа-
ла, взятого от трупа средних лет мужчины, погибшего
89
от несчастного случая. Как видно из таблицы, его диа-
магнитная восприимчивость в среднем была меньшей,
чем у животных, совпадая лишь по крови, почкам, мыш-
цам и в меньшей мере по уровню намагничиваемости
селезенки. Легкие, сердце, кишечник и надпочечники
отличались сравнительно низким уровнем диамагнетиз-
ма. Наоборот, ткань мозга человека характеризовалась
необычно высоким диамагнетизмом, в частности, магнит-
ная восприимчивость мозжечка составляла —0,575-10~6,
а мозговой ткани больших полушарий---------0,676-10~6,
что всего лишь на 0,045 меньше, чем у дважды дистил-
лированной воды.
Сопоставляя результаты исследований, видим, что са-
мый высокий диамагнетизм присущ органам и тканям
кролика — в среднем —0,488-10-6 (без учета магнитной
восприимчивости кишечника, глаз и прочих органов).
К тому же все органы этого животного имеют равно-
значные показатели магнитной восприимчивости. То же
можно сказать об органно-тканевой магнитной воспри-
имчивости крысы (—0,471 • 10~6) и морской свинки
(—0,490-Ю-6) , правда, если не считать показателя ткани
селезенки. В органах остальных животных высокий диа-
магнетизм одних тканей перекрывался относительным
парамагнетизмом других и наоборот. Величины диамаг-
нетизма при этом составляли от —0,451 -10—6 (мышь) до
—0,425-Ю-6 (корова) и —0,382-Ю-6 (голубь).
Есть основание полагать, что органно-тканевое соот-
ношение диа- и парамагнетизма в процессе жизни меня-
ется. Прежде всего это касается органов, богатых ме-
таллсодержащими белками. У молодых животных, на-
пример, диамагнитная восприимчивость печени больше,
чем у взрослых. Неодинакова она также у разных полов.
Нами, в частности, установлено, что у самок крыс маг-
нитная восприимчивость ткани печени составляет
—0,422- IO-6, а у самцов — около —0,442-10-6.
Резкие сдвиги в диамагнетизме тканей отмечаются в
организме человека при заболеваниях. Так, диамагнит-
ная восприимчивость печени умершего больного, стра-
давшего атеросклерозом и сахарным диабетом, повыша-
лась в 1,5 раза, сердца—более чем на 20%, легких —
на 10%. Наоборот, показатели диамагнетизма ткани се-
лезенки, почек и мозга снижались (на 20—30%). Неиз-
менной оставалась лишь магнитная восприимчивость
90
мышечной ткани. Несколько иные, но тоже существенные
изменения диамагнетизма наблюдались в органах и тка-
нях умерших от подострого септического эндокардита и
перитонита. В частности, диамагнетизм их печени, где
наблюдались выраженные явления цирроза, возрастал
на 30—60%, а селезенки — чуть ли не в два раза. Ха-
рактерно, что и у этих умерших людей резко снижался
диамагнетизм мозговой ткани (до —0,424-Ю-6 против
—0,676- Ю-6 у погибшего от несчастного случая).
Выраженные изменения уровня диамагнетизма у за-
болевших могут происходить со стороны крови. Ее сум-
марная магнитная восприимчивость в норме — величина
постоянная и у разных доноров изменяется от
—0,322-10-6 до —0,339-10-8 с редкими отклонениями.
По нашим подсчетам, магнитная восприимчивость крови
здорового человека равна (—0,333 ± 0,004) • 10-®. При
эритремиях показатель диамагнетизма крови возрастал
до—(0,355—0,415) -IO—6 и в среднем составлял
(—0,374±0,011) IO*6 (р<0,01). Наоборот, при лейкозах
и лимфогрануломатозах диамагнетизм крови у всех 10
больных становился ниже — 0,293-Ю-6, а у одного из
них снижался даже до —0,160-10~®. В среднем магнит-
ная восприимчивость крови больных этой группы была
равна (—0,276±0,013) • IO-® (Р<0,01).
Отклонения в показателях магнитной восприимчиво-
сти крови у ряда больных вирусным гепатитом после
статистической обработки оказались недостоверными
(Р<0,1).
В одном случае была изучена кровь больного гемо-
сидерозом, при котором происходят избыточное образо-
вание гемосидерина (нерастворимый железосодержащий
белок-пигмент) и отложение его в тканях. Было обнару-
жено, что суммарная магнитная восприимчивость крови
при этом заболевании меньше, чем в норме
(—0,361-10-8), магнитовосприимчивость сыворотки име-
ла нижний показатель нормы, а сгустка крови — в 1,8
раза меньше ее.
Заметно изменялась магнитовосприимчивость крови
у некоторых умерших людей, например у длительно бо-
левшего атеросклерозом и сахарным диабетом и погиб-
шего от инфаркта мозга (—0,370-10~6) или страдавшего
раком почки, портальным циррозом печени и язвенной
болезнью (—0,266-10-8).
91
В последней серии опытов сопоставляли показатели
магнитной восприимчивости крови, органов и тканей
теплокровных, холоднокровных и птиц с количеством:
ионов железа в них. С этой целью после измерения маг-
нитовосприимчивости биомасса сжигалась в муфельной
печи с постепенным повышением температуры до 800 °C.
Количество двухвалентного железа определяли на спек-
трофотометре «Perkin-EJmer-402», используя раствор»
1,10 фенантролина. Измерение оптической плотности
проводилось в кювете толщиной 1 см при 510 нм (Пар-
кер, 1972). После восстановления Fe3+ до Fe2+ избытком
аскорбиновой кислоты определяли общее железо. Далее,
вычитая из последнего двухвалентное железо, находили
количество трехвалентного.
Сравнительные исследования (табл. 14) показали,
что количественное содержание ионов железа в органах
и тканях животных было обратно пропорциональным
величине диамагнетизма, т. е. уменьшалось с ее ростом
и, наоборот, увеличивалось при сдвиге диамагнетизма к
парамагнетизму. Эта зависимость контрастнее всего
проявилась при сравнении образцов биомасс с крайне
большими различиями в содержании железа. Так, при
низких показателях диамагнетизма (от —0,164-10“® до
—0,408-10“6), свойственных биомассе крови, печени и
селезенки, находили от 9,5 мкг до 22,6 мкг же-
леза на 1 г золы, а при высоких (от —0,479 -10 6 до
—0,565-10 6), характерных для мышц, сердца, мозга,—
от 0,5 мкг до 4,8 мкг/г золы.
В общем, та же тенденция просматривалась при со-
поставлении уровня магнитовосприимчивости и количе-
ственного содержания железа в биомассе различных
органов отдельно взятых животных, будь то корова, го-
лубь или лягушка.
Таким образом, органы и ткани животных, человека,
птиц, холоднокровных и насекомых имеют разную вели-
чину диамагнетизма, что, очевидно, связано с условиями
обитания, специфическими особенностями обмена орга-
низмов, функциональной активностью клеток и тканей,
наличием в них парамагнитных соединений и элементов.
Это, в частности, подтверждают высокие показатели
парамагнетизма биомассы селезенки и печени, богатых,,
как указывалось выше, ферритином, гемосидерином и
многими другими металлсодержащими протеинами, фер-
92
Таблица 14
Показатели органно-тканевой магнитовосприимчивости
и наличие железосодержащих веществ в биомассе, М+т
Тест- система Животное Магнитная вос- приимчивость, Х10-« Содержание железа мкг/г ЭОЛЫ
Fe2++Fe»+ I Fe2+ | Fe’+
Кровь Корова —0,324+0,001 13,9 1,1 13,8
Собака —0,339+0,003 20,0 4,0 16,0
Г олубь —0,358+0,002 10,6 0,5 10,1
Лягушка —0,348+0,013 14,5 3,0 11,5
Печень Корова —0,479+0,002 1,6 0,4 1,2
Собака —0,408+0,003 19,0 2,5 16,5
Г олубь —0,206+0,007 9,5 0,2 9,3
Лягушка —0,237—0,025 12,6 1,0 11,6
Селезенка Корова —0,164—0,003 18,5 1,4 17,1
Собака —0,297—0,003 22,6 6,0 16,6
Г олубь —0,188-0,011 11,3 3,0 8,3
Легкие Корова —0,454+0,003 8,8 0,8 8,0
Собака —0,404+0,002 9,2 2,0 7,2
Г олубь —0,376 +0,002 4,7 0,4 4,3
Почки Корова —0,488+0,003 0,7 0,4 0,3
Собака —0,475—0,002 10,0 2,0 8,0
Г олубь —0,420--0,001 5,1 0,3 4,8
Лягушка —0,526—0,005 10,2 0,5 9,7
Сердце Корова —0,491+0,003 2,5 0,8 1,7
Собака —0,492+0,003 4,8 1,2 3,6
Г олубь —0,502+0,001 2,4 0,6 1,8
Лягушка —0,516+0,002 4,8 0,2 4,6
Мышцы Корова —0,493+0,002 2,7 0,8 1,9
Собака —0,473+0,002 5,0 1,8 3,2
Голубь —0,537+0,009 2,5 0,8 1,7
Лягушка —0,530+0,014 5,0 0,4 4,6
Мозг Корова —0,527+0,001 0,5 0,4 0,1
Собака —0,513+0,005 3,0 0,6 2,4
Г олубь —0,521+0,008 1,4 0,5 0,9
Лягушка —0,565+0,004 0,5 0,4 0,1
ментами, ионами и солями, обладающими парамагнит-
ными свойствами. В таком же аспекте следует рассмат-
ривать парамагнетизм крови, несомненно, обусловленный
гемоглобином. Если судить по железу, то показатели
магнитовосприимчивости клеток не очень строго корре-
лируют с общим количеством в них парамагнетиков. Так,
например, в крови его содержится в два-три раза боль-
ше, чем во внутренних органах, но уровень парамагне-
тизма эритроцитов нередко оказывается таким же или
даже чуть меньшим, как у ферритиносодержащих клеток
93
селезенки и печени. Нужно думать, что это связано с
различиями в количестве ионов-парамагнетиков и сво-
бодных радикалов, образующихся при окислительно-вос-
становительных процессах. И наконец, относительный
парамагнетизм некоторых клеток, органов и тканей, по-
видимому, обусловлен ферромагнитным материалом.
Пытаясь выявить его, мы тщательно изучалд магнитную
восприимчивость черепа голубей, брюшек медоносных
пчел и муравьев, в которых С. Walcott et al. (1979) и
J. L. Gould (1980) нашли магнетит. При этом оказа-
лось, что их биомасса имела сравнительно низкие уров-
ни диамагнетизма (от —0,402-10~6 до —0,412-10-6).
Правда, различий в магнитовосприимчивости брюшек и
остальных сегментов тела у медоносных пчел не выяв-
лялось. Относительный парамагнетизм брюшек обнару'
живался лишь у муравьев. Далее, взяв во внимание-
данные J. L. Kirschvink (1981), обнаружившего в ин-
тактной ткани надпочечников человека, в среднем мозге
и мозолистом теле обезьян-резус высоко коэрцитивный
ферромагнитный материал (1—10 млн. доменов/г), мы
определили, что показатель диамагнетизма биомассы
надпочечников погибшего от несчастного случая челове-
ка был таким же, как у брюшек насекомых. Исследуя
больных и умерших людей, нами было установлено, что
различные патологические процессы сопровождаются
большими изменениями магнитной восприимчивости кле-
ток. Так, при эритремии повышается, а при лейкозе и
лимфогрануломатозе снижается диамагнетизм крови.
В постмортальном периоде в наибольшей мере изменя-
лась магнитовосприимчивость мозга, печени и крови..
Нет сомнений, что эти изменения в органно-тканевой
магнитовосприимчивости являются следствием деструк-
ции клеток, патологических сдвигов в активности фер-
ментных систем и перераспределения микро- и макро-
элементов во время болезней и после смерти.
ЗАКЛЮЧЕНИЕ
Многочисленными исследованиями доказано, что ми-
кробы, клетки животного и растительного, происхожде-
ния, органы и ткани многоклеточных организмов харак-
теризуются отрицательной магнитной восприимчивостью,
степень которой определяется соотношением в них диа-
и парамагнетиков. Точнее говоря, уровень диамагнетиз-
ма живой системы зависит от общего содержания в ней
элементов и веществ, обладающих парамагнитными
свойствами. При этом большое значение могут иметь
качественные особенности парамагнетиков и прежде все-
го величины их магнитных моментов. Следовательно»
диамагнетизм, являющийся неотъемлемым свойством
биологических объектов, может иметь разную степень
выражения. Среди клеточных парамагнетиков, перекры-
вающих диамагнетизм, в первую очередь следует выде-
лить широко распространенное в различных клетках и
тканях организмов железо и его соединения. Так, отно-
сительный парамагнетизм крови животных, несомненно,
обусловлен гемоглобином эритроцитов, а селезенки, пе-
чени, других органов и тканей — железосодержащим:
ферритином и гемосидерином.
У бактерий и грибов магнитным материалом служат
проявляющие значительный парамагнетизм ферредокси-
ны или ферредоксиноподобные протеины, бактериофер-
ритины, фитоферритины и многочисленные железосодер-
жащие ферменты. Низкий уровень диамагнетизма Mic-
rococcus lysodeikticus, по-видимому, обусловлен боль-
шим количеством в клетках каталазы. Убедительным
подтверждением этого явились опыты, в которых четко
регистрировалась обратно пропорциональная зависи-
мость величины диамагнетизма органов и тканей живот-
ных от количественного содержания в них ионов железа
или ферритина.
95
Вклад других металлсодержащих соединений в био-
магнетизм клеток, по сути дела, не изучен, но ферро-
магнитные свойства присущи многим элементам, как,
например, кобальту, никелю, хрому, марганцу, которые
и в микродозах могут существенно изменять магнито-
восприимчивость организмов. Об этом, в частности, го-
ворят наши исследования на радиоспектрометре ERS-220
(ГДР), выявившие у магнитных диссоциантов бактерий
с относительно низким диамагнетизмом повышенное ко-
личество ионов марганца.
Благодаря высокому значению магнитного момента
еще большие сдвиги в сторону парамагнетизма биомас-
сы, по-видимому, могут вызываться редкоземельными
элементами, легко намагничивающимися и извлекаемы-
ми из растворов с помощью магнитов.
Парамагнетизм может возникать также в ряде со-
единений, не содержащих ферромагнитных элементов и
образующихся в свободнорадикальных реакциях.
Особые ферромагнитные свойства имеют соединения,
названные ферримагнетиками. Наиболее известным сре-
ди них является магнетит, найденный в цитоплазме маг-
нитотаксических бактерий, в плоских костях черепа и
шейных мышцах домашних голубей, белого венценосно-
го воробья и многих других перелетных птиц, в перед-
ней трети брюшка медоносных пчел, у королевской ба-
бочки, совершающей ежегодные миграции из Северной
Америки в Центральную ^Мексику, в серповидной склад-
ке мозга тихоокеанских дельфинов. Исследования под
электронным микроскопом показали, что кристаллы
магнетита внутри клеток заключены в мембрану, а не-
которые из них заполнены органической субстанцией,
напоминающей ферритин, что может свидетельствовать
о биогенной трансформации железосодержащих протеи-
нов в магнетит. Последний, обладая сильным остаточ-
ным магнетизмом, обусловливает высокий парамагне-
тизм клеточного вещества и, по-видимому, играет роль
очень чувствительного биомагнитного компаса, который
обеспечивает ориентацию и миграцию организмов по си-
ловым линиям геомагнитного поля. Поиски магнетита у
млекопитающих, если не считать дельфинов, пока- не
увенчались успехом J. L. Kirschvink (1981) обнаружил
ферромагнитный материал в интактной ткани надпочеч-
ника человека, в среднем мозге и мозолистом теле
96
обезьян-резус, но его значение в этих органах остается
невыясненным. Правда, некоторые авторы считают, что
большое количество ферромагнетиков определяет высо-
кую чувствительность ткани к магнитному полю.
Неоднородность суммарно-популяционной магнитной
восприимчивости клеток, связанная с магнетизмом со-
ставляющих ее веществ, в организме животных отлича-
ется одним и тем же характером проявления. В част-
ности, самый большой диамагнетизм имеют мозг, мыш-
цы, сердце, а самый низкий — селезенка, кровь и печень.
Наиболее высокий уровень диамагнетизма отмечался у
кроликов (—0,488-10-6), органы и ткани которых к то-
му же имели более или менее одинаковые показатели
магнитной восприимчивости. У других теплокровных и
птиц высокий диамагнетизм одних тканей (мозг) пере-
крывался относительным парамагнетизмом других (се-
лезенка, кровь), а величины магнитной восприимчивос-
ти организма в среднем составляли от —0,451-10~6
(мышь) до —0,382-IO-5 (голубь). Столь же резко конт-
растировала органно-тканевая магнитовосприимчивость
у холоднокровных. Показатель диамагнетизма клеток
человека, погибшего от несчастного случая, был мень-
шим, чем у животных. Необычно высоким диамагнетиз-
мом характеризовалась лишь ткань мозга (—0,676-10~fi).
У умерших после различных болезней органно-тканевая
магнитовосприимчивость заметно изменялась. Особенно
большие сдвиги происходили в уровне диамагнетизма
крови при эритремии, лейкозах и лимфогранулематозе,
что предполагает практическое использование метода
определения магнитовосприимчивости в диагностике за-
болеваний.
Магнитовосприимчивость растений очень разнообраз-
на. Так, биомасса шляпочных грибов диамагнитна в та-
кой же степени, как почки и легкие животных, правда,
парамагнетизм шляпок оказался несколько больше, чем
ножек, что, по данным К. Spartalian et al. (1975), мож-
но объяснить высоким содержанием в плодовых телах
железосодержащего ферритина.
Высоким показателем диамагнетизма отличались
пшеничная, ячменная, гречневая, рисовая и пшенная
мука, горох, фасоль, яблоки, высушенная биомасса кар-
тофеля и особенно чистая древесина коры (—0,657-10~6).
Диамагнетизм продуктов может, однако, резко снижать-
97
ся вследствие попадания в них ионов металлов и метал-
лоидов, обладающих ферромагнитными свойствами, что
в свою очередь предполагает возможность применения
метода измерения магнитной восприимчивости при оцен-
ке качества пищевых продуктов. Большой интерес пред-
ставляют лишайники и мхи. Ввиду того что их магнит-
ная восприимчивость имеет положительный знак, они с
полным правом могут называться парамагнитными ор-
ганизмами. Очевидно, лишайники и мхи, как и магнито-
таксические бактерии, содержат магнетит или другие
ферромагнетики и способны реагировать иа самые незна-
чительные флюктуации магнитного поля Земли. В связи
с этим следует указать на сезонные изменения магнетиз-
ма листьев растений, что, вероятно, является следстви-
ем влияния геомагнитного поля на биохимизм расти-
тельных клеток.
Среди микроорганизмов истинным парамагнетизмом
должны обладать магнитотаксические бактерии. По-
скольку в их составе содержится столько же железа,
как и в поверхностных слоях почвы, то показатели их
магнитовосприимчивости могут составлять приблизи-
тельно +6,60-10-6---Ь 10,97• Ю-6. Магнитная восприим-
чивость изученных нами бактерий оказалась отрицатель-
ной. У родственных видов энтеробактерий и актиноми-
цетов ее показатель варьировал в очень узком диапазо-
не, а у кокков, антракоидов и микобактерий колебался
от —0,167-10-6 до 0,515-Ю-®. Тут важно отметить нали-
чие обратной связи между величиной диамагнетизма и
вирулентностью стафилококка и микобактерий туберку-
леза, т. е. у авирулентных штаммов этих бактерий воз-
растал диамагнетизм.
Одновременно с этим было установлено, что в попу-
ляциях разных видов микробов содержится от 10 до 30%''
клонов, магнитная восприимчивость которых отличается
от среднесуммарной. Клоны с низким диамагнетизмом
в культурах встречались в 2—5 раз чаще, чем клоны с
повышенным уровнем диамагнетизма, показатели от-
клонения к парамагнетизму были в два-три раза боль-
ше показателей отклонения к диамагнетизму. Гетероген-
ность микробных популяций по магнитовосприимчивости
резко увеличивалась при диссоциации культур, пассиро-
ванных в слабом и сверхслабом магнитных полях. При
этом ЭПР-спектр магнитных антракоидов-диссоциантов
98
с высоким уровнем диамагнетизма ие менялся. У суб-
штаммов с низким диамагнетизмом ЭПР-спектр выяв-
лял наличие парамагнитных маргаицезависимых фер-
ментов, т. е. у бактерий-диссоциантов происходила глу-
бокая перестройка метаболизма, обусловившая сдвиг к
парамагнетизму.
Установленная нами связь между магнитовосприим-
чивостью и биохимической активностью клетки прямо
указывает на то, что магнитоизменчивость опосредуется
через парамагнетизм клеток. Иначе говоря, в магнитную
диссоциацию прежде всего вовлекаются клоны, отлича-
ющиеся низким уровнем диамагнетизма. Отсюда выте-
кает возможность селекции штаммов, способных реаги-
ровать на магнитную возмущенность окружающей среды.
Таким образом, магнитная восприимчивость различ-
ных организмов и клеток неодинакова. Неоднородность
магнитовосприимчивости вместе с ионными токами (био-
токами) является основным источником биомагнитных
полей, которые, как сейчас доказано, объективно отра-
жают общее состояние организма, отдельных его орга-
нов и систем (магиитокардиограмма, магнитоэнцефало-
грамма). Вместе с этим степень магнитовосприимчивос-
ти может явиться индикатором магниточувствительности
клеток, что важно брать во внимание при оценке биоло-
гического и лечебного действия внешних магнитных по-
лей, а также при объяснении более частных вопросов,
как, например, изменчивости микроорганизмов, пассиру-
емых в искусственных магнитных полях. Магнитная вос-
приимчивость как инструмент познания может найти
применение в диагностике болезней, идентификации
бактерий, в оценке их биохимической активности, при
отборе штаммов с повышенным содержанием парамаг-
нитных элементов и соединений, в санитарно-гигиениче-
ских исследованиях и судебно-медицинской экспертизе.
ОСНОВНАЯ
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Бабеико Г. А. Микроэлементы в экспериментальной и клиниче-
ской медицине.— Киев, 1965.— 183 с.
Вании А. Ф., Варич В. Я., Коваленко О. А., Тарасова Н. И. Со-
держание свободного железа в пролиферирующих тканях живот-
ных.— В кн.: Магнитный резонанс в биологии и медицине. Черного-
ловка, 1981, с. 205.
Габович Р. Д., Михалюк И. А., Козярии И. П., Шутенко О. И.
Обмен и межорганное распределение железа, меди, молибдена, мар-
ганца и никеля при воздействии на организм электромагнитных по-
лей промышленной и сверхвысоких частот.— Гигиена и санитария,
.1977, № 7, с. 26—33.
Гаузе Г. Ф., Кочеткова Г. В., Владимирова Г. Б. Биохимические
Мутанты стафилококков с поврежденным окислением как тест-объ-
екты при изысканиях противораковых антибиотиков.— Докл. АН
СССР, 1957, т. 117, № 4, с. 720—722.
Деркаиосов Н. И., Громковская Л. К. Электрические свойства
дрожжей.— Изв. высш. учеб, завед.: Пищевая технология, 1979, № 6,
с. 92—94.
Дубров А. П. Геомагнитное поле и жизнь.— Л., 1974.— 175 с.
Жмакин А. И. Клональная магнитная восприимчивость сальмо-
нелл.— В кн.: Применение лазерного излучения и магнитного поля
в биологии и медицине. Минск, 1982, с. 77—78.
Жолкевич В. Н., Волков Д. И., Прудников В. И., Жегловская
И. Л. К вопросу о природе связей между дыханием и энергопотреб-
ляющими физиологическими процессами.— Докл. АН СССР, 1971,
т. 197, № 5, с. 1210—1213.
Коваленко О. А., Тарасова Н. И., Ванин А. Ф., Прусаков В. Е.
Обмен свободного железа в тканях животных: хронические и острые
эксперименты.— В кн.: Магнитный резонанс в биологии и медицине.
Черноголовка, 1981, с. 139—140.
Новицкий Ю. И. Магнитная восприимчивость корней бобов,
выросших в постоянном магнитном поле.— В кн.: Магнитное поле в
медицине. Фрунзе, 1974, с. 136—138.
Павлович С. А. Магниточувствительность и магнитовосприимчи-
вость микроорганизмов.— Минск, 1981.— 172 с.
Павлович С. А., Арцукевич А. Н. Антимутагенное действие
ослабленных магнитных полей на стафилококк 209 УФ-3.— В кн.:
Применение лазеров и магнитов в биологии и медицине. Ростов-на-
Дону, 1983, с. 91.
Паркер С. Фотолюминесценция растворов.— М., 1972.— 510 с.
100
Савостин П. В. Магнитно-физиологические эффекты у расте-
ний.— Тр. Моск. Дома ученых, 1937, вып. 1, с. 111—119.
Самойлова О. П., Блюменфельд Л. А. Изменение магнитных
свойств культуры дрожжей в процессах роста и клеточного деле-
ния.— Биофизика, 1961, Xs 6, вып. 1, с. 15—19.
Сент-Дьёрдьи А. Биоэнергетика.— М., 1960.— 198 с.
Холодов Ю. И. Реакции нервной системы на электромагнитные
поля.— М., 1975.— 207 с.
Червинец В. М., Новицкий Ю. И., Павлович С. А. Магнитная
восприимчивость микроорганизмов.— Журн. гигиены, эпидемиологии,
микробиологии н иммунологии, 1979, т. 23, Xs 3, с. 230—233.
Чернышев В. Б. Поведение насекомых и электромагнитные по-
ля.— В кн.: Материалы И Всесоюз. совещ. по изуч. влияния магнит,
полей на биолог, объекты. М., 1969, с. 248—251.
Чечерннков В. И. Магнитные измерения.— М., 1969.— 386 с.
Шумаков М. Е. Эксперименты по определению возможности
магнитной ориентации птиц.— В кн.: Вопросы бионики. М., 1967,
с. 519.
Эльдаров А. Л., Холодов Ю. А. Влияние постоянного магнитно-
го поля на двигательную активность птиц.— Журн, общ, биол.,
1964, т. 25, X» 3, с. 224.
Adelman Т. G., Drysdale J. W. Synthesis of ferritin subunits by
free and membrane bound polysomes.— fn: Proteins of iron metabo-
lism / Ed. by E. B. Brown, P. Aisen, J. Fielding, R. R. Crichton. N. Y.,
1977, p. 49—54.
Balkwill D. L., Mara tea D., Blakemore R. P. Ultrastructure of a
magnetotactic spirillum.—J. Bacteriol., 1980, vol. 141, N 3, p. 1399—
4408.
Bauminger E. R., Cohen S. G., Dickson D. P. E. et al. Observati-
on of iron-storage protein in E. coli by mossbauer spectroscopy.— J.
J. Physique, 1979, vol. 40, N 2, p. 523—525.
Bauer E., Raskin A. Increase of diamagnetic susceptibility on the
death of living cells.— Nature, 1936, vol. 138, N 3497, p. 801.
Bean С. P. Single domain particles and magnetotactic bacteria.—
Biophys. J„ 1979, vol. 25, N 2, pt 2, p. 216.
Becker G. Reaktion von Insecten auf Magnetfelder, elektrische
Felder und Atmospherics.— Z. angew. Entomol, 1964, Bd 54, N 1—2,
S. 75—88.
Becker G. Magnetfeld-Einfluss auf Aktivitat und Richtungaver-
halten von Termiten beim Bau senkrechter Galerien —Material und
Organismen, 1979, Bd 44, N 2, S. 81—92.
Becker G. Abhangigkeit der Frassaktivitat von Hausbockkafer-
Larvenwie bei Termiten — von der geomagnetischen Aktivitat.— Na-
turwissenschaften, 1980, Bd 67, N 6, S. 313.
Becker G. Reaction der Holzbohrassel Limnoria tripunctata auf
Magnetfeld-Intensitatsgradienten —Naturwissenschaften, 1980, Bd 67,
N 1, S. 42—43.
Becker G. Untersuchungen Uber die Korrelation der Termiten-
Frassaktivitat zu den durch die Sonnenaktivitat bedingten und ande-
ren Anderungen des geomagnetischen Felds.—Z. angew. Entomol.,
1980, Bd 84, N 4, S. 401—418.
Berge J. A. The perception of weak electric A. C. currents by the
european eel Anguilla anguilla.—Compar. Biochem. and Physiol.,
1979, vol. 62 A, N 4, p. 945—919
101
Bjork I., Fich W. W. Native and subunit molecular weights of
apoferritin.— Biochemistry, 1971, vol. 10, N 15, p. 2844—2848.
Blaise A., Chappert J., Girardet J. Observation par mesures mag-
netiques et effet Mossbauer d’un antiferromagnetisme de grains fins
dans la ferritine.— C. r. hebd. seanc. Acad. Sci., 1965, vol. 261,
p. 2310—2313.
Blakemore R. Magnetotactic bacteria.— Science, 11975, vol. 190,
N 4212, p. 377—379.
Blakemore R. P., Maratea D., Wolfe R. S. Isolation and pure
culture of a freshwater magnetic spirillum in chemically defined me-
dium.— J. Bacteriol., 1979, vol. 140, N 2, p. 720—729.
Blakemore R. P., Frankel R. B., Kalmijn A. J. South-seeking mag-
netotactic bacteria in the Southern Hemisphere.— Nature, 1980,
vol. 286, N 5771, p. 384—385.
Bookman M. A. Sensitivity of homing pigeons to an earth-
strenght magnetic held.— Nature (L.), 1977, vol. 267, p. 340—342.
Bragg P. D. Nonheme iron in respiratory chains.— In: Microbial
iron metabolism: A comprehensive treatise / Ed. by J. B. Neilands,
N. Y., 1974, p. 303—348.
Brown F. A. Response to pervasive geophysical factors and the
biological clock problem.— In: Cold Spring Harbor Symposia on
Quantitative Biology. Baltimore, 1960, vol. 25, p. 57—79.
Brown F. A. Responses of the planarian Dugesia and the proto-
zoan Paramecium to veri weak horizontal magnetic fields.— Biol
Bull., 1962, vol. 1123, N 2, p. 264—281.
Brown F. A., Bennet M. F., Webb H. M. A magnetic compass
response of an organism.— Biol. Bull., 1960, vol. 119, p. 65—74.
Bryce C. F. A., Crichton R. R. The subunit structure of horse spleen
appoferritin.— J. Biol. Chem., 1971, vol. 246, p. 4198—4205.
Bulen W. A., LeComte J. R., Lough S. A hemoprotein from azo-
tobacter containing non-heme iron: isolation and crystallization.—
Biochim. Biophys. Res. Commun, 1973, vol. 54 p. 1274—1281.
Butler R. F., Banerjee S. K. Theoretical single-domain grain size
range in magnetite and titanomagnetite.— J. Geophys. Res., 1975,
vol. 80, p. 4049—4058.
Byers B. R., Arceneaux J. E. L., Haydon A. H., Aswell J. E, Si-
derophore transport in Basillus megaterium.— In: Proteins of iron
fnetabolism / Ed. by E. B. Brown, P. Aisen, J. Fielding, R. R. Crichton.
N. Y„ 1977, p. 371—378.
Cerami A., Gradi R. W., Peterson С. M. et al. The development of
new iron chelating drugs for the treatment of patients with thalas-
semia.— In: Proteins of iron metabolism / Ed. by E. B. Brown,
P. Aisen, J. Fielding, R. R. Crichton. N. Y., 1977, p. 439—442.
Cerdonio M., Congiu-Castellano A., Calabress L. et al. Room-tem-
perature magnetic properties of oxy- and carbonmonoxyhemoglobin
(iron-oxygen complex magnetic susceptibility).— Proc. Nat. Acad.
Sci. USA, 1978, vol. 75, N 10, p. 4916—4919.
Cerdonio M., Morante S., Vitaie S. et al. Variability of the mag-
netic moment of carbon monoxide hemoglobin from carp.— Proc. Nat.
Acad. Sci. USA, 1980, vol. 77, N 3, p. 1462—1465.
Crichton R. R. Structure and function of ferritin.— Angewandte
chemie, 1973, vol. 12, N 1, p. 57—65.
Crichton R. R. Ferritin.— Structure and Bonding, 1973, vol. 17,
p. 67—134.
102
Crichton R. R. Structural studies on horse spleen apoferritin.—
In: Proteins of iron storage and transport in biochemistry and medi-
cine / Ed. by R. R. Crichton. Amsterdam, 1975, p. 183—185.
Crichton R. R., Pance-Ortiz V., Koch M. H. J. et al. Isolation and
characterization of phytoferritin from pea (Pissum sativum) and len-
til (Lens esculenta). Biochem. J., 1978, vol. 171, p. 349—356.
David C. N. Ferritin and iron metabolism in phycomyces.— In:
Microbial iron metabolism: A comprehensive treatise / Ed. by J. B.
Neilands. N. Y„ 1974, p. 149—158.
Denham C. R., Blakemore R. P., Frankel R. B. Bulk magnetic
properties magnetotactic bacteria.— IEEE Trans. Magn., 1980, vol. 16,
N 5, p. 1006—1007.
Dickson D. P, E., Rottem S. Mossbauer spectroscopic studies of
iron in Proteus mirabilis.— Eur. J. Biochem, 1979, vol. 101, N 1.
p. 291—295.
Drysdale J. W., Arosio P., Adelman T. et al. Microheterogeneity
of tissue ferritins in iron storage disease.— In: Proteins of iron sto-
rage and transport in biochemistry and medicine I Ed. by R. R. Crich-
ton. Amsterdam, 1975, p. 359—366.
Emlen S. T. The influence of magnetic information on the orien-
tation of the Indigo Buuting.—• Passerina cyanea. Anim. Behav., 1970a,
vol. 18, p. 215—224.
Emlen S. T. Celestial rotation: Its importance in the development
of migratory orientation.— Science, 1970b, vol. 170, p. 1198—1201.
Emlen S. T. Migration: orientation and navigation.— In: Avian
Biology/Ed. by D. S. Earner, J. R. King. N. Y., 1975, vol. 5, p. 129—
219.
Emlen S. T., Wiltschko W., Demong N. et al. Magnetic direction
finding in migratory Indigo Buntings.— Science, 1976, vol. 193,
p. 505—508.
Evans С. H., Tew W. P. Isolation of biological materials by hse
-of erbium (111)—induced magnetic susceptibilities.—Science, 1981,
-vol. 213, N 4508, p. 653—654.
Farrant J. L. An electron microscopic study of ferritin.— Biochim.
biophys. acta, 3954, vol. 13, p. 569—576.
Frankel R. B., Blakemore R. P., Wolfe R. S. Magnetite in fresh-
water magnetotactic bacteria.— Science, 1979, vol. 203, N 4387,
p. 1355—1356.
Frankel R. B., Blakemore R. P. Navigational compass in magne-
tic bacteria.—J. Magn. Magn. Mater, 1980, vol. 15—18, p. 1562—1564.
Frankel R. B., Blakemore R. P., Araujo F. F. T. et al. Magneto-
tactic bacteria at the geomagnetic equator.—Science, 11981, vol. 212,
N 4500, p. 1269—1270.
Fromrne H. Untersuchungen uber das Orientierungsvermogen
nachtlich ziehender Kleinvogel Erithacus rubecula, Sylvia communis —
Z. Tierpsychol., 1961, Bd 18, S. 205—220.
Ganzoni A. M., Puschmann M. Iron status and host defense.—
In: Proteins of iron metabolism / Ed. by E. B. Brown, P. Aisen,
J. Fielding, R. R. Crichton. N. Y„ 1977, p. 427—432.
Gould J. L. Perceptive bees, birds and bacteria.— Natural Histo-
ry, 1979, vol. 88, N 9, p. 25—36..
Gould J. L. The case for magnetic field sensitivity in animals
{such as it is).—Am. Sci., 1980, vol. 68, p. 256—267.
103
Gould J. L., Kirschvink J. L„ Deffeyes K. S. Bees have magnetic
remanence.— Science, 1978, vol. 201, N 4360, p. 1026—1028.
Gould J. L., Kirschvink J. L., Deffeyes K. S., Brines M. L. Orien-
tation of demagnetized bees.— J. Exptl Biol., 1980, vol. 86, p. 1—8.
Habernkorn R., Michel-Beuerle M. E., Marcus R. A. On spin-
exchange and electron-transfer rates in bacterial (magnetic field ef-
fects / reaction center/bacteriochlorophyll dimer/bacteriopheophytin).—
Proc. Nat. Acad. Sci. USA, 1979, vol. 76, N 9, p. 4185—4188.
Hagihara B., Sato N., Yamanaka T. Type b cytochromes.— In:
The Enzymes / Ed. by P. D. Boyer. N. Y., 1975, vol. 11, pt A, p. 550—
593.
Hershko C., Konijn A. M. Pathogenesis of impaired iron release
in inflamation.— In: Protein of iron metabolism / Ed. by E. B. Brown,
P. Aisen, J. Fielding, R. R. Crichton. N. Y., >1977, p. 417—425.
Jacobs N. J. Biosinthesis of heme.— In: Microbial iron metabo-
lism: A comprehensive treatise/Ed. by J. B. Neilands. N. Y., 1974,
p. 125—148.
Johnson M. K., Thomson A. J., Robinson A. E. et al. Low-tempe-
rature magnetic circular dichroism spectra and magnetisation curves
of 4Fe clusters in iron-sulphur proteins from Chromatium and Clostri-
dium pasteurianum.— Biochim. biophys. acta, 1981, vol. 667, N 2,
p. 433—451.
Johnson M. K., Thomson A. J., Robinson A. E-, Smith В. E. Cha-
racterization of the paramagnetic centres of the molybdenum-iron pro-
tein of nitrogenase from Klebsiella pneumoniae using low tempera-
ture magnetic circular dichroism spectroscopy.— Biochim. biophis.
acta, 1981, vol. 671, N '.1, p. 61—70.
Jones H. E., Trudinger P. A., Chambersa L. A., Pyliotis N. A.
Metal accumulation by bacteria with particular reference to dissimila-
tory sulphate-reducing bacteria.— Z. Allg. Microbiol., 1976, vol. 16,
p. 425—435.
Jungerman R. L., Rosenblum B. Magnetic induction for the sen-
sing of magnetic fields by animals — an analisis.— J. theor. Biol.,
1980, vol. 87, p. 25—32.
Kalmijn A. J. Electro-perception in sharks and rays.— Nature
(L.), 1966, vol. 212, p. 1232—1233.
Kalmijn A. J. The electric sense of sharks and rays.— J. Exp.
Biol., 1971, vol. 55, p. 371—383.
Kalmijn A. J. Experimental evidence of magnetic orientation in
elasmobranch fishes.— In: Animal Migration, Navigation and Ho-
ming/Ed. by K. Schmidht-Koenig, W. T. Keeton. N. Y., 1978,
p. 354—355.
Kalmijn A. J., Blakemore R. P. The magnetic behavior of mud
bacteria.— Proceedings in Life Sciences / Ed. by K. Schmidt-Koenig.
N. Y„ 1978, p. 355—356.
Keeton W. T. Orientation by pigeons; is the sun necessary? —
Science, 1969, vol. 165, p. 922—928.
Keeton W. T. Magnets interfere with pigeon homing.— Proc. Nat.
Acad. Sci. USA, 1971, vol. 68, N 1, p. 102—106.
Keeton W. T. Effects of magnets on pigeon homing.— In: Animal
orientation and navigation / Ed. by S. P. Galler, K. Schmidt-Koenig,
G. J. Jacobs, R. E. Belleville. Washington, 1972, p. 579—594.
Keeton W. T., Cobert A. Orientation by untrained pigeons requi-
res the sun.— Proc. Nat. Acad. Sci. USA, 1970, vol. 65, p. 853—856.
104
Keeton W. T., Larkin T. S., Windson D. M. Normal fluctuations
in the earth’s magnetic field influence pigeon orientation.— J. Comp.
Physiol., 1974, vol. 95, N 2, p. 95—'103.
Kirschvink J. L. South-seeking magnetic bacteria.— J. Exptl.
Biol., 1980, vol. 86, p. 345—347.
Kirschvink J. L. The horizontal magnetic dance of the honeybee is
compatible with a single-domain ferromagnetic magnetoreceptor.—
Biosystems, 1981, vol. 14, N 2, p. 193—203.
Kirschvink J. L. Ferromagnetic crystals (magnetite?) in human
tissue.— J. Exptl Biol., 1981, vol. 92, p. 333—335.
Kirschvink J. L., Lowenstam H. A. Mineralization and magneti-
zation of chiton teeth: paleomagnetic, sedimentologic and biologic
implications of organic magnetite.— Earth Planet. Sci. Lett., 1979,.
vol. 44, p. 193—204.
Kirschvink J. L., Gould J. L. Biogenic magnetite as the basis of
magnetic field sensitivity in animals.— Biosistems, 1981, vol. 13,
p. 181—201.
Larkin R. P., Sutherland P. J. Migrating birds respond to project
seafarer’s electromagnetic field.— Science, 4977, vol. 195, p. 777—779.
Leask M. J. M. A physicochemical mechanism for magnetic field
detection by migratory birds and homing pigeons.— Nature, 1977,
vol. 267, p. 144—145.
Lindauer M. Orientierung der Tiere.— Verh. Dtsch. Zool. Ges.,
1976, S. 156—183.
Lindauer M. Recent edvances in the orientation and learning of
honeybees.— In: Proceedings of XV International Congress of Ento-
mology. Washington, 1976, p. 450—460.
Lindauer M., Martin H. Die schwereorientierung der bienen under
dem einfluss des erdmagnetfeldes.— Z. Vergl. Physiol., 1968, Bd 60,
S. 219—243.
Lindauer M., Martin H. Magnetic effect on dancing bees.— In:
Animal orientation and navigation / Ed. by S. R. Galler, K. Schmidt-
Koenig, G. J. Jacobs, R. E. Belleville. Washington, 1972, p. 559—567.
Linder M. C., Munro H. N. Ferritin and free iron in iron absorp-
tion.— In: Proteins of iron storage and transport in biochemistry and
medicine / Ed. by R. R. Crichton. Amsterdam, 1975, p. 395—400.
Linder M. C., Munro H. N., Vulimiri L., Catsimpoolas N. Struc-
ture of isoferritins from normal and malignant rat tissues.— In: Pro-
teins of iron storage and transport in biochemistry and medicine /Ed.
by R. R. Crichton. Amsterdam, 1975, p. 201—208.
Lissmann H. W. On the function and evolution of electric organs
in fish.—J. Exptl Biol., 1958, vol. 35, N 1, p. 156—194.
Lissmann H. W., Machin К. E. The mechanism of object location
in Gymnarchus niloticus and similar fish.—J. Exptl Biol., 1958,
vol. 35, N 2, p. 451—485.
Lovenberg W. Ferredoxin and rubredoxin.— In: Microbial iron
metabolism: A comprehensive treatise/Ed. by J. B. Neilands. N. Y.,
1974, p. 161—164.
Lovenstam H. A. Magnetite in denticle capping in recent chitons
(Polyplacophora).—Geol. Soc. Am. Bail., 1962, vol. 73, p. 435—438.
Lovenstam H. A. Lepidocrocite, an apatite mineral, and magne-
tite in teeth of chitons (Polyplacophora).— Science, 1967, vol. 156,
N 3780, p. 1373-1375.
Martin H., Lindauer M. Der einfluss der erdmagnetfelds und die
105
schwercorientierung der honigbiene.— J. Comp. Physiol., 1977, Bd 1122,
S. 145—187.
Massover W. H. Ultrastructural studies of disrupted ferritin.—
In: Proteins of iron metabolism / Ed. by E. B. Brown, P. Aisen,
J. Fielding, R. R. Crichton. N. Y., 1977, p. 23—30.
Merkel T. W. Orientation behavior of birds in Kramer cages un-
der different physical cues.— Ann. N. Y. Acad. Sci., 1971, vol. 188,
p. 283—294.
Merkel T. W., Fromme H. Untersuchungen fiber das orientierungs-
vermogen nachtlich ziehender Rotkehlchen.— Naturwissenschaften,
1958, Bd 45, S. 499—500.
Merkel T. W., Fromme H. C., Wiltschko W. Nichtvisuelles orien-
tierungsvermogen bei nahtlich zugunruhigen Rotkehlchen.— Vogel-
warte, 1964, Bd 22, S. 168—173.
Merkel F. W., Wiltschko W. Magnetismus und richturgsfinden
zugunruhiger Rotkehlchen.— Vogelwarte, 1965, Bd 23, S. 71—77.
Moench T. T. Distribution, isolation and characterization of a
magnetotactic bacterium.— Diss. Abstr. Intern., 1979, vol. 40, N 2,
p. 606.
Moench T. T., Konetzka W. A. A novel method for the isolation
and study of a magnetotactic bacterium.— Arch. Microbiol., 1978,
v. 119, p. 203—212.
Moore F. R. Geomagnetic disturbance and the orientation of
nocturnally migrating birds.— Science, 1977, vol. 196, p. 682—684.
Moore F. R. It the homing pigeon’s map geomagnetic? — Nature,
1980, vol. 285, N 5760, p. 69—70.
Mortenson L. E., Chen J.-Sh. Hydrogenase.— In: Microbial iron
metabolism: A comprehensive treatise/Ed. by J. B. Neilands. N. Y.,
1974, p. 231—282.
Munro H. N., Linder M. C., Haberman M., Catsimpoolas N. Ferri-
tins from normal and malignant human tissues.— In: Proteins of
iron storage and transport in biochemistry and medicine I Ed. by
R. R. Crichton. Amsterdam, 1975, p. 223—230.
Murray R. W. The response of the ampullae of Lorenzini of
elasmobranchs to electrical stimulation.— J. Exp. Biol., 1962, vol. 39,
p. 119—128.
Neilands J. B. Iron and its role in microbial physiology.— In:
Microbial iron metabolism: A comprehensive treatise / Ed. by J. B.
Neilands. N. Y., 1974, p. 3—34.
Neilands J. B. Siderophores: Biochemical ecology and mechanism
of iron transport in enterobacteria.— Adv. Chem., 1977, vol. 162,
p. 3—32.
Neilands J. B., Wayne R. R. Membrane receptors for microbial
iron transport compounds (siderophores).— In: Proteins of iron me-
tabolism /Ed. by E. B. Brown, P. Aisen, J. Fielding, R. R. Crichton.
N. Y., 1977, p. 365—369.
Nozaki M., Ishitnura Y. Oxigenases.— In: Microbial iron meta-
bolism: A comprehensive treatise / Ed. by J. B. Neilands. N. Y., 1974,
p. 417—453.
Phillips J. B. Earth’s magnetic field and orientation of salaman-
ders.—J. Comp. Physiol., U977, vol. 121, N 1, p. 273—288.
Picton H. D. Some responses of Drosophila to weak magnetic
and electrostatic fields.— Nature, 1966, vol. 211, N 5046, p. 303—304.
Powell L. W., McKeering L., Halliday J. W. Microheterogeneity
106
of tissue ferritins in iron storage disease.— In: Proteins of iron sto-
rage and transport in biochemistry and medicine/Ed. by R. R. Crich-
ton. Amsterdam, 1975, p. 367—370.
Powell L. W., Halliday J. W., McKeering L. B., Tweedale R. Alte-
rations in serum and tissue isoferritins in disease states: II: Hemo-
chromatosis and malignant diseases.— In: Proteins of iron metabo-
lism / Ed by E. B. Brown, P. Aisen, J. Fielding, R. R. Crichton. N. Y.,
1977, p. 61—64.
Presti D., Pettigrew J. D. Ferromagnetic coupling to muscle re-
ceptors as a basis for geomagnetic field sensitivity in animals.— Na-
ture, 1980, vol. 285, N 5760, p. 99—101.
Prasad E. A. V., Narayana A. C. Magnetic orientation in termite
mounds.— Current Sci., 1981, vol. 50, 13, p. 588—589.
Quinn T. P. Evidence for celestial and magnetic compass orien-
tation in lake migrating sockeye salmon fry.— J. Comp. Physiol.,
1980, vol. 137, p. 243—248.
Reed G. H., Cohn M. Electron paramagnetic resonance spectra
of manganese (II)-protein complexes.— J. Biol. Chem., 1970, vol. 245,
N 3, p. 662—664.
Reith E. J. The ultrastructure of ameloblasts during matrix for-
mation and the maturation of enamel.—J. Biophys. Biochem. Cytol.,
H961, vol. 9, N 4, p. 825—839.
Remsen С. C., Watson S. W., Waterbury J. B., Truper H. G. Fine
structure of Ectothiorhodospira mobilis Pelsh.— J. Bacteriol., 1968,
vol. 95, p. 2374—2392.
Roche J., Bessis M., Breton-Gorius J., Stralin H. Molecules d’he-
moglobine et de ferritine dans les cellules chloragogenes d’Arenicola
marina L.— Compt. Rend, des seances de la Societe de biologie et
de ses filiales, 1961, vol. 155, p. 1790—1803.
Semm P., Schneider T., Vollrath L. Effects of an Earth-strength
magnetic field on electrical activity of pineal cello.— Nature, 1980,
vol. 288, N 5791, p. 607—608.
Schoffa G. Der antiferromagnetismus des ferritins bei massungen
der magnetichen suszeptibilitat in temperaturbereich von 4, 2 bis.
300°.—Z. Naturforschung, 1965, Bd 32, N 2, S. 167—172.
Schreiber B. Nucleus of erythrocytes clad with magnetite in the
pigeon-skull considered as a further instrument of magnetic percep-
tion in birds.— Monit. zool. ital., 1981, vol. 15, N 4, p. 325.
Shimizu T., Lizuka T., Mitani F. et al. Magnetic and natural cir-
cular dichroism spectra of cytochromes P-450n and P-450Sec purified
from bovine adrenal cortex.— Biochim. biophys. acta, 1981, vol. 669,
N il, p. 46—59.
Southern W. E. Gull orientation by magnetic cues: A hypothesis
revisited.— Ann. N. Y., Acad. Sci., 1971, vol. 188, p. 295—311.
Southern W. E. Influence of disturbances in the earth’s magne-
tic field on ring-billed gull orientation.— Condor, 1972, vol. 74,
p. 102—105.
Spartalian K-, Oosterhuis W. T., Smarra N. Mossbauer effects
studies in the fungus Phycomyces.— Biochim. biophys. acta, 1975,
vol. 399, p. 203—212.
Stephens P. J., McKenna С. E., McKenna M. C. et al. Circular
dichroism and magnetic circular dichroism of reduced molybdenum-
iron protein of Azotobacter vinelandii nitrigenase.— Biochemistry,
1981, vol. 20, N 10, p. 2857—2864.
107
Stiefel E. I., Watt G. D. Azotobacter cytochrome bssr.s is a bac-
terioferritin.—Nature (L.), 1979, vol. 279, N 5708, p. 81—83.
Theil E. C. Red blood cell ferritin: a possible role in develop-
ment.— In: Proteins of iron storage and transport in biochemistry
and medicine/Ed. by R. R. Crichton. Amsterdam, 1975, p. 371—376.
Theil E. C., Brown J. E. Red cell ferritin and iron storage during;
animal development.— In: Proteins of iron metabolism I Ed. by E. B.
Brown, P. Aisen, J. Fielding, R. R. Crichton. N. Y., 1977, p. 99—106..
Thomson A. J., Johnson M. K., Greenwood C., Gooding P. E-
A study of the magnetic properties of haem a3 in cytochrome c oxi-
dase by using magnetic-circular-dichroism spectroscopy.— Biochem..
J., 1981, vol. 193, N 3, p. 687—697.
Thomson A. J., Robinson A. E., Johnson M. K. et al. Low-tem-
perature magnetic circular dichroism evidence for the conversion of
four-iron-sulphur clusters in a ferredoxin from Clostridium pasteuria-
num into three-iron-sulphur clusters.— Biochim. biophys. acta, 1981,.
vol. 637, N 3, p. 423—432.
Thomson A. J., Robinson A. E., Johnson M. K. et al. The three-
iron cluster in a ferredoxin from Desulphovibrio gigas: A low-tem-
perature magnetic circular dichroism study.— Biochim. biophys. acta,
1981, vol. 670, N 1, p. 93—100.
Thorton W. N. Electrical perception by deep sea fish.— Proc.
Univ. Durham Phil. Soc., 1931, vol. 8, p. 301—308.
Towe К. M., Lovenstam H. A., Nesson M. N. Invertebrate ferritin:
occurence in mollusca.— Science, 1963, vol. 142, N 3588, p. 63—64.
Towe К. M., Lovenstam H. A. Ultrastructure and development of
iron mineralization in the radular teeth of Cryptochiton stelleri (mol-
lusca).— J. Ultrastruct. Res., 1967, vol. 17, p. 1—13.
Towe К. M., Bradley W. F. Mineralogical constitution of colloidae
«hydrous ferric oxides».— J. Colloid Interface Sci., 1967, vol. 24,
p. 382—392.
Towe К. M., Moench T. T. Electron-optical characterization of
bacterial magnetite.— Earth. Planet. Sci. Lett., 1981, vol. 52, p. 213—
220.
Treffry A., Banyard S. H., Hoare R. J., Harrison P. M. Structure-
and ion-binding properties of ferritin and apoferritin.— In: Proteins
of iron metabolism./ Ed. by E. B. Brown, P. Aisen, J. Fielding, R. R.
Crichton. N. Y., 1977, p. 3—11.
Walcott C. Anomalies in the earth’s magnetic field increase the
scatter of pigeons’ Vanishing bearings.— In: Animal migration, navi-
gation, and homing/Ed. by K. Schmidt-Koenig, W. T. Keeton. N. Y.„
1978, p. 143—151.
Walcott C., Green R. P. Orientation of homing pigeon altered
by a change in the direction of an applied magnetic field.— Science,
1974, vol. 184, N 4133, p. 180—182.
Walcott C., Gould J. L., Kirschvink J. L. Pigeons have magnets.—
Science, 1979, vol. 205, N 4410, p. 1027—1029.
Witschko W. Uber den Einfluss statischer Magnetfelder auf die
Zugorientierung der Rotkchlchen.— Z. Tierpsychol, 1968, Bd 25,
S. 537—558.
Wiltschko W. Der Magnetcompass der Gartengrasmiicke (Silvia
borin).—J. Ornithol, 11974, Bd 115, N 1, S. 1—7.
Wiltschko W., Wiltschko R. Magnetic compass of European ro-
bins.— Science (N. Y.), 1972, vol. 176, p. 63—64.
108
Wiltschko W., Wiltschko R. Grasmticken Benutzen den Magnet-
kompass auch bei Sternsicht.— Naturwissenschaften, 1973, Bd 60,
N 12, S. 112—117.
Yamanaka T., Okunuki K. Cytochromes.— In: Microbial iron me-
tabolism: A comprehensive treatise/Ed. by J. B. Neilands. N. Y.,
1974, p. 349—400.
Yeagley H. L. A preliminary study of a physical basis of bird na-
vigation.— J. Appl. Phys., 4947, vol. 18, N 12, p. 1035—1063.
Yeagley H. L. A preliminary study of a physical basis of bird
navigation. Pt. II.— J. Appl. Phys., 1951, vol. 22, N 11, p. 746.
Yonetani T. Hydroperoxidases.— In: Microbial iron metabolism:
A comprehensive treatise / Ed. by J. B. Neilands. N. Y., 1974, p. 401—
-415.
Yorke E. D. A possible magnetic transducer in birds.— J. Theor.
Biol., 1979, vol. 77, p. 101—105.
Yorke E. D. Sensitivity of pigeons to small magnetic field varia-
tions.— J. Theor. Biol., 1981, vol. 89, N 3, p. 533—537.
Zoeger J., Dunn J. R., Fuller M. Magnetic material in the head
•of the common pacific dolphin.— Science, 1981, vol. 213, N 4510,
p. 892—894.
ОГЛАВЛЕНИЕ
Введение .................................................. 3
Глава 1
Физико-химические основы диа- и парамагнетизма 5
Глава 2
Природа биомагнетизма 14
2.1, Магнетизм клеточного ферритина........................16
2.2. Магнетизм металлопротеидов клеток.....................27
2.3. Магнетизм минерально-органических соединений клеток 33
Глава 3
Геомагнитотропизм организмов 42
3.1. Магннтосомы и магнитотаксис бактерий..................43
3.2. Биофизические основы навигации организмов ... 54
Г л а в а 4
Клеточная и органно-тканевая магнитовосприимчивость 69
4.1. Методы измерения магнитной восприимчивости организмов 69
4.2. Магнитовосприимчивость микроорганизмов .... 72
4.3. Магнитовоспрнимчивость грибов и растений .... 81
4.4. Магнитовосприимчивость холоднокровных и насекомых 85
4.5. Магнитовосприимчивость животных, птиц и человека . . 87
Заключение.............................................95
Основная цитируемая литература........................100
СЕРГЕЙ АЛЕКСАНДРОВИЧ ПАВЛОВИЧ
МАГНИТНАЯ
ВОСПРИИМЧИВОСТЬ
ОРГАНИЗМОВ
Заведующий редакцией Д. Ф. Санько
Редактор Л. Г. Усенкова
Художник М. И. Шел кович
Художественный редактор Л. И. Усачев
Технический редактор Т. В. Л е т ь е н
Корректор 3. М. Р а й н е с
ИБ № 2310
Сдано в набор 03.10.84. Подписано в печать 19.02.85.
АТ 08175. Формат 84ХЮ8'/з2. Бум. тип. № 2. Гар-
нитура литературная. Высокая печать. Усл. печ.
л. 5,88. Усл. кр.-отт. 6,20. Уч.-изд. л. 5,9. Тираж
1050 экз. Зак. № 1252. Цена 70 к.
Издательство «Наука и техника» Академии наук
БССР и Государственного комитета БССР по де- .
лам издательств, полиграфии и книжной торгов-
ли. 220600. Минск, Ленинский проспект, 68. Типо-
графия им. Франциска (Георгия) Скорины изда-
тельства «Наука и техника». 220600. Минск, Ле-
нинский проспект, 68.