/









































































































































































































Text
М. Ю. ПУНИН
КИШЕЧНАЯ
РЕГУЛЯТОРНАЯ
СИСТЕМА
БЕСПОЗВОНО ЧНЫХ
ЖИВОТНЫХ
И ЕЕ ПРЕДПОЛАГАЕМАЯ
ЭВОЛЮЦИЯ
У МНОГОКЛЕТОЧНЫХ
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА
Том 290
Выпускаются с 1932 г.
RUSSIAN ACADEMY OF SCIENCES
PROCEEDINGS OF ZOOLOGICAL INSTITUTE
Vol. 290
Publishing since 1932
1832
RUSSIAN ACADEMY OF SCIENCES
ZOOLOGICAL INSTITUTE
M. Yu. PUNIN
INTESTINAL REGULATORY SYSTEM
OF INVERTEBRATES
AND ITS POSSIBLE EVOLUTION
IN METAZOA
St. Petersburg
2001
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
М. Ю. ПУНИН
КИШЕЧНАЯ РЕГУЛЯТОРНАЯ СИСТЕМА
БЕСПОЗВОНОЧНЫХ ЖИВОТНЫХ
И ЕЕ ПРЕДПОЛАГАЕМАЯ ЭВОЛЮЦИЯ
У МНОГОКЛЕТОЧНЫХ
Санкт-Петербург
2001
УДК: 591.43:591:87:591:88:591:147
Пунин М. Ю. Кишечная регуляторная система беспозвоночных
животных и ее предполагаемая эволюция у многоклеточных. - СПб.,
2001.- 166 с.
Настоящая монография представляет собой оригинальную сводку данных по эндокрин-
ным и нервным клеткам в составе пищеварительной системы многоклеточных животных. Она
базируется на обширном литературном материале и результатах многолетних оригинальных
исследований и охватывает большинство современных таксонов Metazoa— от книдарий до
млекопитающих. Эндокринные и нервные клетки рассматриваются как образующие единую
регуляторную систему. На основании сравнительно-гистологического анализа высказывается
и обосновывается новая гипотеза об источниках возникновения и возможных путях становле-
ния и развития этой системы в филогенезе. Книга предназначена для специалистов, работаю-
щих в области сравнительной и эволюционной гистологии, морфологии, эндокринологии и
нейробиологии, а также для студентов, специализирующихся в этих направлениях.
Библиогр. 481 назв. Ил. 300. Табл. 5.
Главный редактор —
директор Зоологического института РАН
академик РАН А. Ф.Алимов
Редакционная коллегия:
В. В. Хлебович (зам. редактора), ТА. Асанович, Ю.С. Балашов,
В.Я. Бергер, С.Д. Гребельный, И. С. Даревский, В.Р. Дольник, С.Ю. Кузнецов,
В. А. Тряпицын, Я. И. Старобогатов
Рецензенты:
А.Д.Харазова, Ю.П.Лагутенко
Издание осуществлено при поддержке
Российского фонда фундаментальных исследований
по проекту № 01-04-62035
Р
И
© М.Ю.Пунин, 2001
© Зоологический институт РАН, 2001
Моему учителю,
Алексею Алексеевичу Заварзину,
и моему другу и коллеге,
Василию Константиновичу Казакову
ПРЕДИСЛОВИЕ
Эта книга представляет собой попытку подвести промежуточный итог исследованиям
по одной из тем, над которой я работаю уже более 25 лет, практически с момента окончания
в 1975 г. кафедры цитологии и гистологии Ленинградского (теперь снова Санкт-Петер-
бургского) государственного университета.
Изучая строение эпителия пищеварительного тракта некоторых морских беспозвоноч-
ных животных (этой теме посвящена специальная монография — «Гистологическая органи-
зация кишечных эпителиев приапулид, брахиопод, двустворчатых моллюсков и полихет»,
вышедшая в свет в издательстве «Наука» в 1991 г.) я буквально «наткнулся» на необычные
по морфологии клетки, сочетавшие в себе черты эндокринных и нервных. Они меня крайне
заинтересовали, я стал искать подобные элементы в пищеварительном тракте у представи-
телей различных типов Invertebrata и, как правило, находил что-либо подобное. Выясни-
лось, что присутствие таких структур, — скорее, правило, чем исключение, и беспозвоноч-
ные, очевидно, обладают собственной кишечной регуляторной системой, аналогом кишеч-
ной диффузной (энтериновой) системы, хорошо известной у позвоночных.
По мере накопления оригинальных данных и анализа литературы меня, естественно,
заинтересовал следующий вопрос — как рано в филогенезе такие клетки, названные мной
регуляторными, появляются; какие общие черты и особенности наблюдаются в организа-
ции образованных этими элементами систем в различных таксонах многоклеточных, распо-
ложенных на разных уровнях эволюционного древа; можно ли нащупать некие общие зако-
номерности в их эволюции? Существовавшие в литературе точки зрения на эти вопросы
полностью меня удовлетворить не могли, поэтому постепенно я начал строить свою кон-
цепцию (Пунин, 1986, 1993, 1996, 2000, 2001), последний вариант которой изложен в за-
ключительной главе этой книги.
Я не могу не сказать несколько слов о моих учителях, а также коллегах, без общения и
работы с которыми вряд ли я смог бы проводить мои исследования и, соответственно, под-
готовить эту книгу. Прежде всего, это— заведующий кафедрой цитологии и гистологии
Ленинградского университета Алексей Алексеевич Заварзин (младший), которого я считаю
своим главным учителем. Он во многом определил мой интерес к сравнительной и эволю-
ционной гистологии, первым обратил внимание на актуальность и значительный интерес
темы этой работы, всегда был одновременно и суровым, и доброжелательным критиком
моих статей.
С Василием Константиновичем Казаковым мы много и плодотворно работали вместе
вплоть до его безвременной кончины в 1999 г. Он был моим первым учителем в иммуноги-
5
стохимии — методе, благодаря использованию которого удалось получить многие важные
фактические данные.
Майе Сергеевне Константиновой я обязан освоению метода флуоресцентной гисто-
химии. Увы, ее трагическая гибель прервала наше короткое, но плодотворное сотрудниче-
ство.
Я хочу поблагодарить своих коллег по институту и соавторов — Светлану Александ-
ровну Филимонову и Тамару Григорьевну Маркосову, моих бывших студентов с кафедры
цитологии и гистологии СПбГУ — Светлану Будаеву, Станиславу Панкратову и Левона
Мкртчяна, чьими руками были получены многие данные, вошедшие в мои статьи и эту
книгу.
Большой объем иммунопатологических исследований был сделан во время моей го-
дичной стажировки на отделении гистологии и электронной микроскопии в Vrije
Universiteit van Amsterdam (Свободный университет Амстердама) в Нидерландах по гранту
NWO № 07-13-028. Я не могу не выразить искренней благодарности заведующему этим
отделением, профессору Harry Boer, и его сотрудникам, а также д-ру Jan van Minnen, без
активной помощи которого эта поездка не была бы возможной.
Ряд экспериментов на гидрах был проведен во время моего короткого пребывания в
центре молекулярной нейробиологии университета г. Гамбурга (Германия), и я очень бла-
годарен д-ру S. Hoffmeister и проф. Н. С. Schaller, любезно пригласившим меня для озна-
комления с исследованиями их лаборатории.
Я должен также сказать большое спасибо за постоянное внимание и поддержку всем
моим коллегам, с кем я работал на кафедре цитологии и гистологии СПбГУ (которой в на-
стоящее время заведует проф. А.Д.Харазова), в лаборатории физиологии питания Институ-
та физиологии им. И. П. Павлова РАН (у академика А.М.Уголева), в Зоологическом инсти-
туте РАН на Беломорской биологической станции (зав. проф. В.Я.Бергер) и в лаборатории
эволюционной морфологии (зав. академик РАЕН проф. Ю.В.Мамкаев), в которой и была
написана эта книга.
Ряд исследований, результаты которых вошли в эту монографию, проводился при фи-
нансовой поддержке РФФИ (проекты №№ 95-04-11838, 97-04-48175, 99-04-49789, 01-04-
63129). Настоящее издание осуществлено благодаря финансовой поддержке РФФИ по про-
екту № 01-04-62035.
«Многие из биологических наук, а в частности
гистология и анатомия, сделали такой громадный
шаг вперед со времени Биша, что пожалуй начинаю-
щему может показаться, как справедливо замечает
Раубер, будто все главное сделано, и теперь остались
лишь незначительные детали».
(В.П.Гаряев. Строение пищеварительного тракта
некоторых Cephalopoda. Харьков, 1915. С. 1.)
Глава 1
ВВЕДЕНИЕ
(ПРОБЛЕМА, ТЕРМИНОЛОГИЯ, МЕТОДЫ)
Постановка проблемы. Г истологические, гистохимические, электронно-микро-
скопические и иммуногистохимические исследования двух последних десятилетий убеди-
тельно продемонстрировали присутствие в эпителии пищеварительной системы беспозво-
ночных животных различных уровней организации и относящихся к различным системати-
ческим группам своеобразных клеток. Различаясь между собой как по многим деталям сво-
его строения, так и по содержащимся в них биологически активным веществам (моноами-
ны, регуляторные пептиды, некоторые другие соединения), все эти клетки имеют одну об-
щую черту. Она заключается в том, что данные клетки отличаются от остальных эпители-
альных тем, что не принимают непосредственного участия в процессах пищеварения (мем-
бранное пищеварение, всасывание, внутриклеточное пищеварение, секреция пищевари-
тельных ферментов, слизи и компонентов перитрофических мембран, перемешивание со-
держимого кишки и его перемещение по пищеварительному тракту и т.д.), а, очевидно, иг-
рают какую-то регуляторную, интегрирующую роль.
Некоторые из них по своему строению в высшей степени соответствуют кишечным эн-
докринным клеткам позвоночных животных, другие очень похожи на нервные элементы.
Наконец, как я покажу ниже, многие из них на современном уровне наших знаний вообще
не могут быть однозначно отнесены к тем или иным, занимая некое «промежуточное» по-
ложение. Как правило, эти клетки достаточно многочисленны, часто связаны друг с другом
отростками. По аналогии с кишечной гормональной (энтериновой) системой млекопитаю-
щих они, очевидно, так же образуют (в составе эпителиального пласта) собственную сис-
тему.
Для сравнительной и эволюционной гистологии представляет определенный интерес
провести сравнительный анализ кишечных регуляторных систем в разных систематических
группах беспозвоночных животных, сопоставить их с функционально аналогичными систе-
мами позвоночных, а также попробовать проследить вероятные пути эволюции этих систем
у многоклеточных. И хотя беспозвоночные животные (как я покажу ниже) в сравнении с
позвоночными (главным образом млекопитающими) в этом отношении изучены еще крайне
недостаточно (впрочем очевидно, что этот разрыв в степени изученности будет и далее воз-
7
растать), тем не менее, как мне кажется, уже представляется возможным и интересным вы-
сказать некоторые предположения относительно развития кишечных регуляторных систем
в филогенезе.
Некоторые замечания о терминологии. Вопросы терминологии в отношении рас-
сматриваемых кишечных клеточных элементов в действительности крайне запутанны и к
настоящему времени еще не решены. Во многом это определяется отсутствием строгих
морфологических и биохимических критериев для определения их нервной или эндокрин-
ной природы: критериев, которые были бы действенны на всех уровнях эволюционного
древа Metazoa. Попробую пояснить это замечание.
Такие морфологические критерии для выявления нервных элементов, как синапсы, не
могут быть в полной мере применены ко многим беспозвоночным, где даже в центральной
нервной системе типичные синаптические контакты часто крайне редки (например, в ганг-
лиях двустворчатых моллюсков— см. в частности: Кулаковский и др., 1990). Центральная
нервная система брюхоногих моллюсков (например, у Lymnaea stagnalis — см. работы: Boer
et al., 1992, 1994; Boer, Montagne-Wajer, 1994 и др.), состоит из значительного числа круп-
ных нейроэндокринных клеток, посылающих свои отростки с секреторными гранулами ли-
бо к тканям (органам)-мишеням, либо к соединительнотканной оболочке ганглиев, где и
осуществляется вывод секреторного продукта.
В тех случаях, когда известны конкретные регуляторные соединения, присутствующие
в тех или иных клетках (впрочем, значительно чаще в иммуногистохимических работах
речь идет о ^-подобных иммунореактивностях), эти данные все равно не могут быть ис-
пользованы в интересующих нас целях. К настоящему времени хорошо известно, что у по-
звоночных одни и те же вещества (и моноамины, и различные пептиды) могут присутство-
вать в элементах как нервной, так и эндокринной систем, играя роль и нейротрансмиттеров,
и нейрогормонов, и гормонов.
В то же время один из очень важных, интересных и даже принципиальных вопросов,
который встает при описании элементов кишечной регуляторной системы беспозвоночных
животных, это — относить их к эндокринной или нервной составляющим? [Иногда эту
проблему «обходят», рассматривая и нервную, и эндокринную системы как единую «нейро-
эндокринную» (см., в частности: Falkmer et al., 1985). Однако, если в функциональном от-
ношении противопоставление нервной и эндокринной регуляции, очевидно, часто не имеет
смысла (см.: Лейбсон, 1981, 1983), то в отношении гистогенеза составляющих ее элементов
это может оказаться принципиальным (что я постараюсь показать далее)].
Как я только что отметил, строгих, однозначных критериев для разделения нервных и
эндокринных клеток в кишечном эпителии к настоящему времени не существует. Интерес-
но, что эта проблема во многих работах решается весьма своеобразно. Если авторы изучают
нервную систему объекта и обнаруживают в кишечном эпителии какие-то сходные (напри-
мер, по демонстрируемой иммунореактивности) структуры, они автоматически зачисляют
их в нервные. Если же анализируется гистология эпителия «сама по себе», то по аналогии с
млекопитающими такие клетки, скорее всего, будут названы эндокринными. Поэтому при
дальнейшем описании (как возможные) я буду использовать чисто морфологические и дос-
таточно простые критерии: в качестве эндокринных (наверное, правильнее — эндокринопо-
добных) я буду рассматривать клеточные элементы, лишенные базальных отростков либо
имеющие короткие отростки, не образующие заметных базиэпителиальных сплетений. На-
против, клетки с длинными отростками, часто ветвящимися, имеющими характерные для
нервных варикозные расширения, иногда уходящими за пределы эпителиального пласта и
вступающими в состав субэпителиальных плексусов, я буду рассматривать как элементы
8
периферической нервной системы. В тех случаях, когда имеются данные электронно-
микроскопических исследований, будут приниматься во внимание черты сходства в тонком
строении внутриэпителиальных и субэпителиальных отростков, присутствующих в стенке
кишечной трубки и, в частности, в мышечной оболочке, и определенно являющихся нерв-
ными. Такие критерии (при всех возможных ошибках в их использовании) представляются
мне в настоящее время единственно реально работающими.
Все же такие клетки, независимо от их конкретной принадлежности к той или иной
(нервной, эндокринной) системе, я буду называть регуляторными, а совокупность таких
элементов в пищеварительном тракте — кишечной регуляторной системой.
О методах выявления кишечных регуляторных клеток. К таким методам можно
отнести ряд гистологических окрасок и гистохимических реакций, позволяющих выявлять
определенные тинкториальные и гистохимические свойства кандидатов в регуляторные
элементы, характерные для ряда эндокринных клеток позвоночных. Это могут быть арги-
рофильная и аргентаффинная реакции, окраски пседоизицианином и паральдегидфуксином
(широко применяемые для выявления Р-клеток поджелудочной железы млекопитающих),
методы импрегнации серебром для выявления нервных клеток, прижизненное окрашивание
метиленовым синим (метод, широко применявшийся в первой половине XX века для изуче-
ния нервной системы беспозвоночных животных — подробное его описание см.: Догель,
1902). Часто весьма информативными оказываются флуоресцентно-гистохимические мето-
ды — Фалька-Хилларпа (Falck et al., 1962; Falck, Owman, 1965), FAGLU (Furness et al., 1977,
1987), метод с глиоксиловой кислотой, — выявляющие катехоламин- и индоламин (серото-
нин)-содержащие структуры на основании их различного свечения. Электронная микроско-
пия позволяет выявить клетки с характерными деталями тонкого строения: в первую оче-
редь это — наличие гранул, по размерам, форме и локализации в клетках, отличных от сек-
реторных экзокринных, или особой организации апикальных частей клеток, отличающихся
от обычных эпителиальных и тем самым дающих основание рассматривать их в качестве
рецепторного аппарата. Наконец, это разнообразные иммуногистохимические методы [см.,
в частности, ставшее классическим руководство Стернбергера (Stemberger, 1979)], позво-
ляющие выявлять определенные клетки по демонстрируемой ими (нейро)гормон-подобной
иммунореактивности, а на электронно-микроскопическом уровне— определять, в каких
гранулах или иных клеточных структурах локализованы анализируемые вещества.
Естественно, что каждый из этих методов имеет свои определенные недостатки и огра-
ничения. Так, окрашивание паральдегид-фукс ином позволяет выявлять группы соединений,
богатые S-S-связями, среди которых инсулин — лишь один из таких пептидов. Аргиро-
фильная и аргентаффинная реакции, как показали некоторые работы, лишь в единичных
случаях дают положительные результаты на беспозвоночных животных.
Прижизненное окрашивание метиленовым синим в методическом плане весьма «ка-
призно», а химические механизмы выявления при его использовании именно нервных эле-
ментов до сих пор неизвестны. Детали тонкого строения клеток в действительности часто
не дают бесспорных доказательств их нервной или эндокринной природы, поскольку и
микротрубочки, и микрофиламенты являются компонентами клеток, принадлежащих к раз-
личным тканям, а относительно наличия типичных синаптических контактов я уже упоми-
нал. Такие параметры, как размеры и тонкое строение секреторных гранул, могут быть с
более или менее серьезными основаниями применены лишь при сравнении клеток между
достаточно близкими в систематическом отношении группами животных, но чем дальше
анализируемые ряды отстоят друг от друга в филогенетическом плане, тем менее достовер-
ными они будут являться.
9
Наконец, и иммуногистохимические методы, совсем недавно казавшиеся универсаль-
ными, имеют целый ряд существенных ограничений. Прежде всего, позволяя охарактеризо-
вать клетки на предмет наличия в них тех или иных регуляторных соединений, они не дают
возможности определить их нервную или эндокринную природу. Так, для позвоночных жи-
вотных убедительно показано, что одни и те же регуляторные пептиды и амины могут при-
сутствовать и в нервных, и в типичных эндокринных клетках, играя роль как медиаторов,
так и гормонов. С другой стороны, на основании только иммуногистохимических реакций,
даже проведенных со всеми необходимыми строгими контролями иммуноспецифичности,
можно говорить не более чем о подобных иммунореактивностях.
Иными словами, одну и ту же положительную реакцию могут давать как вещества, к
которым получены используемые антитела, так и отличные соединения, если они имеют
сходные антигенные детерминанты (т.е. короткие идентичные аминокислотные последова-
тельности). А поскольку очень часто в экспериментах на одних животных используются
антисыворотки, полученные к биологически активным соединениям других видов (гетеро-
логичные антисыворотки), вероятность такой ошибки всегда существует, и она тем больше,
чем дальше в филогенетическом плане отстоят между собой эти виды. Последнее ограниче-
ние метода хорошо известно работающим с ним специалистам, хотя, быть может, и не все-
гда достаточно акцентируется в публикуемых статьях.
И напротив, иногда бывают случаи, когда из-за каких-то присущих данному виду осо-
бенных тканевых свойств, несмотря на все методические ухищрения (деблокирование анти-
генных детерминант, повышение чувствительности метода и т.д.), все равно не удается ло-
кализовать анализируемое вещество, хотя биохимические данные однозначно показывают
его присутствие. Иными словами, отрицательные результаты не обязательно говорят об
отсутствии того или иного соединения.
Отмечу, что в последнее время в арсенал исследований вошел еще один мощный ме-
тод — in situ гибридизация. Однако постепенно накапливаются данные, что и он имеет свои
ограничения. Впрочем, к настоящему моменту я еще не встречал работ, где бы этот метод
был использован для изучения кишечных регуляторных клеток беспозвоночных.
И все же, при всех своих ограничениях, перечисленные выше методы на настоящем
уровне исследований дают достаточно информации если не для полной характеристики
кишечной регуляторной системы, то, по крайней мере, для выявления клеток-«кандидатов»
в ее элементы и для демонстрации сложности ее организации.
Структура книги. Данные, касающиеся кишечных регуляторных систем беспозвоноч-
ных животных, как я уже отмечал, в сравнении с общим количеством видов крайне немно-
гочисленны. Более того, в этом плане различные таксоны изучены очень неравномерно.
Наряду с такими относительно хорошо исследованными группами, как насекомые (в не-
сколько меньшей степени — моллюски и иглокожие), есть классы и даже типы, данные по
которым исчерпываются буквально несколькими замечаниями в одной-двух статьях. Есте-
ственно, что такая неравномерность изученности обусловила и разные объемы глав, и опре-
деленные отличия в их построении.
Располагая главы, в которых приводится фактический материал, я выбрал несколько
нетрадиционную схему (от высших таксонов к низшим) и завершил рассмотрение темы, во-
первых, главой о представителях плоских червей (турбелляриях) как группы, наиболее
близкой из трехслойных к гипотетическому предку многоклеточных, и, во-вторых, главой о
книдариях. Хотя я не считаю двуслойных книдарий предками других Metazoa, полагая, что
они представляют собой самостоятельную эволюционную ветвь, представлялось, тем не
10
менее, интересным дать хотя бы краткую характеристику их гастродермальной регулятор-
ной системы.
Изучая любую проблему можно идти как «вширь», так и «вглубь». Применительно к
рассматриваемой теме можно дать общее (неизбежно довольно поверхностное) описание
кишечной регуляторной системы у максимального количества групп и объектов, а можно
детально изучать различные физиологические и биохимические аспекты, особенности син-
теза тех или иных биологически активных веществ в определенных клетках у конкретного
вида и т.д. Конечно, в идеале хотелось бы и того, и другого, но, увы, физически это сделать
невозможно. Я буду (по отдаленной аналогии с выражением Л.Н.Гумилева*) рассматривать
заявленную тему «с высоты птичьего полета».
Основные условные обозначения, принятые на рисунках и микрофотографиях:
аг — аппарат Гольджи
ап — апикальные пузырьки
ац — амебоцит
бк — базальные клетки (у полихет, олигохет)
бм — базальная мембрана
бс — базиэпителиальное сплетение
вак — вакуоль
гд — гастродермис (у немертин, турбеллярий, гидр)
гэс — гладкая эндоплазматическая сеть
д — десмосома
кн — корневая нить реснички
кол — коллаген
кс — кровеносный сосуд, кишечный синус
л — липидная капля
м — мышечная оболочка, мышечная клетка
мв — микроворсинки
мет — мультивезикулярное тело
мит — митохондрия
мкс — мешок кристаллического стебелька (у дву-
створчатых моллюсков)
мт — микротрубочки
мф — микрофиламенты
нк — нервная клетка
но — нервный отросток
п — пузырек
пар — паренхима (у немертин, турбеллярий)
пр — просвет пищеварительного тракта (желудка,
кишечника и т.д.)
пщд — пищевод
р — ресничка
рг — регенерационное гнездо (у насекомых)
риб — свободные рибосомы
рк — ресничная клетка
с — внеклеточный кристаллический стебелек (у дву-
створчатых моллюсков)
сг — секреторная гранула
ск — секреторная клетка
срк — средняя кишка
цэ — целомический эпителий
шэс — шероховатая эндоплазматическая сеть
эд — эпидермис (у немертин, турбеллярий, гидр)
экз — экзоцитоз
эп — эпителий
я — ядро
ядр — ядрышко
Л.Н.Гумилев. В поисках вымышленного царства. - М., 1992, с. 14.
Глава 2
КИШЕЧНАЯ ЭНДОКРИННАЯ СИСТЕМА
ПОЗВОНОЧНЫХ ЖИВОТНЫХ
Я не претендую на то, чтобы данном разделе дать исчерпывающую характеристику же-
лудочно-кишечной регуляторной системы Vertebrata. Различным аспектам этой темы посвя-
щены многие сотни (если не тысячи) работ; особенно интенсивно разносторонние исследо-
вания проводились и проводятся на лабораторных млекопитающих и человеке. Эти работы,
естественно, имеют в первую очередь практическую, медицинскую направленность. Регу-
лярно выходят подробные обзоры, посвященные различным аспектам организации и функ-
ционирования кишечной эндокринной системы (см., в частности: Fujita, Kobayashi, 1977;
Уголев, 1978; Solcia et al., 1981a; Алешин, 1984; Яглов, Ломоносова, 1985; Шахламов, Ма-
карь, 1985; Пузырев, Иванова, 1992; Иванова, Пузырев, 1998; и многие другие).
Подробное рассмотрение данной темы выходит за рамки этой работы, и далее я приве-
ду лишь основные сведения (преимущественно морфологического плана) и лишь в том объ-
еме, насколько это необходимо для сопоставления с кишечными регуляторными системами
беспозвоночных. К сожалению, я не имею возможности рассмотреть здесь вопросы, ка-
сающиеся физиологии и биохимии этих систем.
2.1. MAMMALIA (МЛЕКОПИТАЮЩИЕ)
Впервые эндокринные клетки в составе эпителия пищеварительного тракта были опи-
саны еще в конце XIX века (Heidenhein, 1870), затем данные об этих клеточных элементах
нарастали лавинообразно.
Согласно лишь одной из классификаций эндокринных клеток, принятой в 1980 г, в га-
стро-энтеро-панкреатической системе человека (на основании в первую очередь размеров и
тонкого строения секреторных гранул и содержащихся в них биологически активных ве-
ществ) исследователи описывают, по крайней мере, 18 типов таких клеток (Solcia et al.,
1981b). Из них а- и Р-клетки (содержащие соответственно глюкагон и инсулин) присутст-
вуют только в островках поджелудочной железы, D- (соматостатин), РР- (панкреатический
полипептид) и Dr клетки — как в островках, так и эпителии желудочно-кишечного тракта.
Кроме уже отмеченных пептидных гормонов, в эндокринных клетках выявлены также гаст-
рин, холецистокинин, гастроингибирующий полипептид, энтероглюкагон, мотилин, нейро-
тензин (рис. 1, вклейка), секретин, вазоактивный интестинальный полипептид (VIP), суб-
станция Р, энкефалин, бомбезин, пептид тирозин тирозин (PYY) и ряд других, а также
серотонин (рис. 2-4, вклейка).
12
Некоторые эндокринные клетки могут содержать одновременно несколько различных
биологически активных веществ (например, Есгклетки— серотонин, субстанция Р; Ес2-
клетки — серотонин, мотилин; G-клетки — гастрин, энкефалин; Ес-клетки — серотонин,
мелатонин), что существенно затрудняет общую классификацию. У некоторых млекопи-
тающих описаны дополнительные типы клеток (например, EcL-клетки у грызунов, содер-
жащие гистамин). Клетки одного типа могут встречаться на всем протяжении желудочно-
кишечного тракта (например, Ес-клетки), другие в своем распространении ограничены его
определенными отделами (подробнее см. обзоры: Шахламов, Макарь, 1985; Яглов, Ломоно-
сова, 1985; Пузырев, Иванова, 1992; Иванова, Пузырев, 1998; Костюкевич и др., 1998).
О той большой роли, которую играет в общеорганизменной регуляции кишечная регу-
ляторная система, свидетельствует хотя бы тот факт, что суммарное количество эндокрин-
ных клеток в желудке и кишечнике намного превышает количество клеток любой из эндок-
ринных желез организма млекопитающего.
Кишечные эндокринные клетки могут выделять биологически активные вещества как
непосредственно ъ кровеносную систему (эндокринная секреция), так и в межклеточные
пространства (паракриния). В последнем случае воздействие осуществляется на соседние
клетки и/или чувствительные нервные окончания, расположенные в непосредственной бли-
зости от эпителиального пласта. В литературе обсуждается также возможность выделения
секрета непосредственно в полость кишечника, поскольку ряд гормонов выявляется радио-
иммунологическими методами в содержимом гастроинтестинального тракта.
Кишечные эндокринные клетки (для них часто используются такие термины, как эн-
докриноциты, энтериноциты) обычно располагаются поодиночке в эпителии железистых
крипт желудка, ворсинок и крипт различных отделов кишечника. Апикальные части боль-
шинства клеток подходят к просвету пищеварительного тракта (клетки «открытого» типа,
рис. 1, 2, вклейка), некоторые клетки расположены в основании эпителиального пласта и
контакта с просветом не имеют (клетки «закрытого» типа, рис. 3, вклейка).
Клетки открытого типа (рис. 5) несут на апикальной поверхности микроворсинки, по
плотности расположения и размерам обычно несколько отличающиеся от микроворсинок
соседних эпителиальных клеток и, очевидно, в данном случае представляющие собой ре-
цепторный аппарат. Клеточные органоиды, как правило, развиты умеренно. Аппарат Голь-
джи может занимать положение и над ядром, и под ним. В цитоплазме (в большинстве слу-
чаев электронносветлой) присутствуют микротрубочки и микрофиламенты. Эндокринным
клеткам свойственны обычные для эпителиального пласта клеточные контакты.
Секреторные гранулы сосредоточены главным образом в нижней половине клеток. Эк-
зоцитоз их содержимого наблюдается на базальной поверхности клеток, а также на лате-
ральных поверхностях недалеко от базальной мембраны. На апикальной поверхности экзо-
цитоза не отмечалось.
Помимо гормонов, в секреторных гранулах могут присутствовать прогормоны, дейст-
вующие на молекулы прогормонов протеолитические ферменты, не относящиеся к гормо-
нам кислые белки типа хромогранинов, некоторые металлы. Иногда внутри секреторной
гранулы различные компоненты имеют отличное расположение (Fujita et al., 1988).
Клетки закрытого типа в целом имеют сходное строение (за исключением, естественно,
отсутствия подходящей к просвету апикальной части), однако не демонстрируют заметной
полярности в своей организации.
У ряда эндокринных клеток отмечены отходящие от перикарионов базальные отростки
(рис. 2, вклейка). Иногда эти отростки могут оканчиваться на других эндокринных клетках
(см. в частности: Larsson et al., 1979).
13
Рис. 5. Общая схема строения эндокринной клетки открытого типа
в желудочно-кишечном тракте млекопитающих.
кк - кровеносный капилляр.
Fig. 5. General scheme of organization of an open-type endocrine cell
in the digestive tract of mammals.
Важно отметить, что в эпителии желудочно-кишечного тракта (как в норме, так и при
патологии) описаны так называемые «смешанные» клетки, сочетающие черты строения как
слизистых и эндокринных (рис. 4, вклейка), так и панетовских и эндокринных (Nebeyama,
1975; Пузырев, Иванова, 1986).
Помимо интраэпителиальных, эндокриноподобные клетки присутствуют также в со-
ставе lamina propria, где они могут контактировать с нервными окончаниями и клетками,
напоминающими по строению шванновские. Подобные клетки более обычны для плодов,
их количество также резко возрастает при некоторых заболеваниях желудочно-кишечного
тракта (Solcia et al., 1981а).
Первые кишечные эндокринные клетки появляются у эмбрионов на довольно ранних
стадиях развития (рис. 6). У человека в эпителии будущей двенадцатиперстной кишки они
отмечены на 3-м месяце эмбрионального развития, еще до формирования системы крипт и
ворсинок. Их количество заметно возрастает по мере развития плода; на 6-м месяце на вер-
шине ворсинок возникают скопления эндокринных клеток — «шапочки Сежи» («Segi’s
caps»), которые, однако, исчезают вскоре после рождения.
Один из важнейших и интереснейших вопросов, касающихся кишечной эндокринной
системы — это вопрос об источниках ее происхождения. Согласно концепции Пирса
(Pearse, 1968, 1969, 1973), все эндокринные клетки, диффузно расположенные в различных
тканях и органах, развиваются из мигрирующих в них элементов нейроэктодермы. Соответ-
ственно нервное происхождение должны иметь и эндокринные клетки желудочно-
кишечной системы. В основе этой концепции лежит фундаментальное сходство биохимиче-
14
вор
А Б В Г ДЕ
Рис. 6. Онтогения эндокринных клеток в эпителии двенадцатиперстной кишки человека
(по: Fujita et al., 1988):
А - 3, Б- 4, В - 5, Г- 6-8 месяцев беременности, Д- новорожденный, Е - взрослый организм.
вор - ворсинка; кр - крипта; ш - шапочка Сежи. Черным выделены эндокринные клетки.
Fig. 6. Ontogeny of endocrine cells in the duodenal epithelium of man (according to: Fujita et al., 1988).
ских процессов в пептидергических нейронах и эндокринных клетках, что и нашло отраже-
ние в ее названии: APUD ^Amine and amine Precursors Uptake and Decarboxylation, т.е. спо-
собность клеток поглощать и декарбоксилировать амины и их предшественников).
Однако в дальнейшем работы других исследователей показали, что, по крайней мере,
часть известных типов эндокринных клеток пищеварительной системы бесспорно имеет
эктодермальное происхождение (Cheng, Leblond, 1974а; Pictet et al., 1976; Le Douarin, 1978;
Andrews et al., 1982 и др.). В пользу энтодерм ал ьно го происхождения кишечных эндокрин-
ных клеток свидетельствует и существование отмеченных выше смешанных экзо-
эндокринных клеток (Nebeyama, 1975; Пузырев, Иванова, 1986). Что же касается клеток
нервного гребня, мигрирующих в стенку развивающейся кишечной трубки, то, как предпо-
лагают критики концепции Пирса, они дифференцируются в нейроны интрамуральных
ганглиев, оставаясь за пределами эпителиального пласта.
В то же время вопрос о конкретных источниках возникновения и обновления эндок-
ринных клеток в эмбриональном и постэмбриональном периодах остается неясным. Обсуж-
даются три возможных варианта: 1) образование из общих для всего эпителия камбиальных
клеток— так называемая унитарная теория (рис. 7) (Cheng, Leblond, 1974b), 2) деление
непосредственно эндокринных клеток и 3) передифференцировка экзокринных секреторных
клеток в эндокринные.
15
Рис. 7. Схема организации процессов репродукции и дифференцировки в эпителии
тонкой кишки мыши
(по: Cheng, Leblond, 1974b, из: Заварзин, 1985):
/ - стволовая мультиполярная клетка; 2-5 - полустволовые клетки; 6-9 - дифференцированные клетки;
2,6 - клетки Панета; 3,7 - всасывающие клетки; 4,8- бокаловидные клетки; 5, 9 - энтерохромафинные клетки.
Пунктирной линией обведены клетки, способные к делению. Стволовые и дифференцирующиеся клетки
располагаются в составе эпителия крипт, дифференцированные
(за исключением клеток Панета, остающихся в основании крипт)
составляют эпителиальную выстилку ворсинок.
Fig. 7. Scheme of organization of cell reproduction and differentiation processes in the mouse small intestine
(according to: Cheng, Leblond, 1974b, from: Заварзин, 1985).
16
2.2. AVES (ПТИЦЫ)
Энтериновая система у представителей этого класса позвоночных в целом принципи-
ально сходна с таковой млекопитающих. Гомологи большинства кишечных эндокринных
клеток Mammalia выявлены в различных отделах пищеварительной системы у многих видов
изученных птиц (см., в частности: Monesi, 1960; Toner, 1964; Ferreira, 1966; Pentilla, 1968;
Andrew, 1976a, b; Rawdon, Andrew, 1981; Rawdon, 1984; Salvi, Renda, 1986; Мутовкина,
Иванова, 1987; Иванова, 1995 и др.). Эндокринные клетки располагаются по всей высоте
ворсинок, а также в криптах и, как правило, встречаются поодиночке. Среди них наблюда-
ются клетки как открытого, так и закрытого типов.
Иммуногистохимическими методами показано присутствие клеток, иммунореактивных
к секретину, энтероглюкагону (Polak et al., 1974), панкреатическому полипептиду (РР)
(Alumets et al., 1978а), /ем-энкефалину (Alumets et al., 1978b), серотонину (Watanabe et al.,
1987), бомбезину (D’Este et al., 1984), субстанции P (Brodin et al., 1981; Salvi, Renda, 1989)
гастрин/холецистокинину, соматостатину, нейротензину (рис. 8-9, вклейка), мотилину
(Salvi, Renda, 1989).
Достаточно подробно изучен и онтогенез кишечных эндокринных клеток (главным об-
разом у кур). Подобно млекопитающим первые эндокринные элементы появляются в эм-
бриогенезе очень рано, еще до формирования системы крипт и ворсинок. Эти клетки лежат
в основании эпителиального слоя непосредственно на базальной мембране и обычно имеют
округлую форму; апикальные и (часто) базальные отростки появляются позднее. В разных
отделах пищеварительного тракта первые эндокринные клетки появляются в различные
сроки; при этом время появления клеток, демонстрирующих различную иммунореактив-
ность, также различается (Salvi, Renda, 1989).
В некоторых участках эпителия желудка эмбрионов цыплят присутствуют клетки, им-
мунореактивные к лютеинизирующему гормону. Своеобразие этих клеточных элементов
заключается в том, что иммунореактивный материал наблюдается не в базальных, а в рас-
ширенных апикальных частях клеток (Shirasawa et al., 1996).
2.3. REPTILIA (РЕПТИЛИИ ИЛИ ПРЕСМЫКАЮЩИЕСЯ)
В составе эпителия разных отделов желудка и кишечника у представителей различных
отрядов пресмыкающихся (чешуйчатых рептилий, крокодилов и черепах) описаны клетки,
дающие положительные аргирофильную и/или аргентаффинную реакции (Luppa, 1977).
Аргирофильные клетки обычно имеют веретеновидную форму, а клетки с одновременно
аргирофильным и аргентафинным секреторными продуктами — грушевидную форму
(Gabe, 1973). Присутствие в эпителии клеток с характерными для эндокринных мелкими
секреторными гранулами в базальных частях продемонстрировано и в электронно-микро-
скопических исследованиях (Luppa, 1977).
Иммуногистохимические исследования показали наличие эндокринных клеток, реаги-
рующих с антисыворотками к разнообразным регуляторным пептидам. Соматостатин-
иммунореактивные клетки описаны у чешуйчатых рептилий (ящерицы и змеи) (Falkmer et
al., 1978; Seino et al., 1979; Masini, 1986; D’Este et al., 1993), представителей отряда крокоди-
лов (Buchan et al., 1983; Yamada et al., 1987) и черепах (Gapp et al., 1985), бомбезин-
иммунореактивные— у ящериц (El-Salhy, Grimelius, 1981) и аллигаторов (Buchan et al.,
1982), нейротензин-иммунореактивные— у ящериц (Reinecke et al., 1980; El-Salhy,
17
Grimelius, 1981; D’Este et al., 1993). Клетки, реагирующие с антисывороткой к панкреатиче-
скому полипептиду (РР), выявлены в эпителии пищеварительного тракта ящериц (El-Salhy,
Grimelius, 1981; D’Este et al., 1993) пептиду тирозин тирозину (PYY) — у ящериц (El-Salhy
et al., 1982; D’Este et al., 1993) и черепах (Gapp et al., 1985), гастрину— у черепах (Gapp et
al., 1985) и крокодилов (Yamada et al., 1987), холецистокинину (Larsson, Rehfeld, 1977) и
инсулину (Gapp, Polak, 1985) — у черепах.
У черепах Testudo graeca и Maureniys caspica показано присутствие в эпителии желуд-
ка и кишечника секретин-, бомбезин-, нейротензин-, гастрин-, глюкагон-, соматостатин-,
пептид тирозин тирозин- и инсулин-иммунореактивных клеток, причем у второго вида —
также клеток, иммунореактивных к РР. У ящерицы Lacerta lepida, кроме всех вышеотме-
ченных, обнаружены и клетки, реагирующие с антителами к субстанции Р (Perez-Tomas et
al., 1989). У другого вида ящериц, Podarcis sicula, выявлены также серотонин- и гистамин-
иммунореактивные клетки (D’Este et al., 1993).
Если бомбезин-иммунореактивные клетки относятся к элементам закрытого типа, то
инсулин-, пептйд тирозин тирозин- и РР-иммунореактивные принадлежат к открытому ти-
пу. Клетки, реагирующие с антисывороткой к соматостатину, различны по своей морфоло-
гии. Если у клеток в составе эпителия тонкой кишки апикальные части подходят к просвету
кишечника, то в желудке аналогичные по демонстрируемой иммунореактивности клеточ-
ные элементы относятся, по-видимому, к закрытому типу (Perez-Tomas et al., 1989).
2.4. AMPHIBIA (ЗЕМНОВОДНЫЕ)
В желудке ряда представителей отряда Anura (бесхвостые амфибии) выявлены бомбе-
зин-иммунореактивные клетки закрытого типа (Lechago et al., 1978; Diaz de Rada et al.,
1987), а также нейротензин- (Reinecke et al., 1980) и холецистокинин- (Rajjo et al., 1988) им-
мунореактивные. В желудке и кишечнике Xenopus laevis описаны нейротензин/ксенопсин- и
гастрин/каерулин-иммунореактивные клетки. Интересно, что в некоторых участках пище-
варительного тракта все иммунореактивности демонстрируют одни и те же клетки, тогда
как в других это, очевидно, — самостоятельные клеточные типы (Flucher et al., 1988).
Серотонин-содержащие клетки выявлены в эпителии желудка и/или кишечника как у
взрослых бесхвостых амфибий (Geuze, 1971; Gabe, 1972; Giraud, Yeomans, 1981; El-Salhy et
al., 1982), так и головастиков Alytes obstetricans (L’Hermite et al., 1988).
Количество некоторых типов эндоринных клеток, очевидно, может меняться в течение
года. Так, в толстой кишке Rana temporaria число аргирофильных клеток в период зимней
спячки достоверно снижается (Мутовкина, Иванова, 1987). Интересно, что, как и у млеко-
питающих, у лягушек отмечено присутствие смешанных экзо-эндокринных клеток (Пузы-
рев, Иванова, 1986).
По данным ультраструктурного анализа в эпителии тонкого кишечника Rana
temporaria присутствуют 7 различных типов эндокринных клеток. Этим клеткам свойствен-
на определенная полярность в строении: базальные части заполнены главным образом сек-
реторными гранулами, тогда как клеточные органоиды смещены в апикальную часть. В
эпителии наиболее часты клетки, аналогичные Ес-клеткам млекопитающих. Интересно, что
они имеют заполненные секреторными гранулами тонкие отростки, тянущиеся вдоль ба-
зальной мембраны (Матвеева, 1998).
Имеются также данные о присутствии эндокринных клеток в желудочно-кишечном
тракте и у некоторых представителей отряда хвостатых амфибий (Conlon et al., 1985).
18
2.5. PISCES (РЫБЫ)
Данные об эндокринных клетках в пищеварительном тракте рыб особенно многочис-
ленны и разнообразны.
Такие клетки выявляются на всем протяжении пищеварительного тракта при проведе-
нии классических аргирофильной и аргентаффинной реакций (Tagliafierro, Faraldi, 1976;
Tagliafierro et al. 1976; Honma et al., 1986). При этом, как и у млекопитающих, у некоторых
хрящевых рыб (Chondrichthyes) описаны клетки, совмещающие характерное для бокало-
видных клеток строение апикальных частей и интенсивную аргентаффинную реакцию ба-
зальной цитоплазмы (Honma et al., 1986). В желудке и кишечнике различных представите-
лей хрящевых рыб выявлены интраэпителиальные клетки, имунореактивные к нейропепти-
ду Y (Cimini et al., 1992; Chiba et al., 1995), субстанции P (Holmgren, 1985; Cimini et al.,
1989), вазоактивному интестинальному полипептиду (Falkmer et al., 1980), инсулину (El-
Salhy, 1984), серотонину, гастрину/холецистокинину, соматостатину, нейротензину
(Holmgren, Nilsson, 1983), бомбезину (Holmgren, Nilsson, 1983; Cimini et al., 1989) и пептиду
гистидин-изолейцину (Cimini et al., 1989). Наличие серотонин-содержащих клеток было
показано также методом флуоресцентной гистохимии (Holmgren, Nilsson, 1983). В кишеч-
нике Scyliorhinus stellaris иммунореактивности к субстанции Р, бомбезину и пептиду гисти-
дин-изолейцину наблюдались в одних и тех же клетках (Cimini et al., 1989). У Squalus
acanthias клетки, реагировавшие с антиинсулиновой сывороткой, описаны как у взрослых
рыб, так и у личинок (El-Salhy, 1984).
Подобно млекопитающим и птицам, первые кишечные эндокринные клетки появляют-
ся в онтогенезе очень рано. В основании желез слизистой желудка ската Dasyatis cunericana
эндокринные клетки закрытого типа отмечены уже у эмбрионов (Hamlett et al., 1996). У
Scyliorhinus stellaris количество эндокринных клеток в желудке и кишечнике возрастает по
мере эмбрионального развития, причем на последней стадии этих клеток оказывается даже
больше, чем у взрослых рыб.
NPY (нейропептид У)-иммунореактивные клетки у Scyliorhinus stellaris характеризу-
ются присутствием в цитоплазме большого количества гранул, округлых (диаметром 97-230
нм) или овальных (от 72x240 до 145x436 нм) (Cimini et al., 1992).
Клетки, реагирующие с антисыворотками к серотонину (L’Hermite et al., 1985; Abad et
al., 1987; Burkhardt-Holm, Holmgren, 1989; Beorlegui et al., 1992a; Kiliaan et al., 1996) и раз-
личным регуляторным пептидам также описаны в эпителии желудка и/или кишечника у
представителей многих семейств костных рыб (Osteichthyes). Из клеточных элементов, со-
держащих пептид-иммунореактивные вещества, отмечу /пег-энкефалин-иммунореактивные
(Langer et al., 1979; Rombout, Reinecke, 1984; L’Hermite et al., 1985; Rombout et al., 1986a, b;
Elbal et al., 1988; Burkhardt-Holm, Holmgren, 1989; Beorlegui et al., 1992b), глюкагон-
иммунореактивные (Langer et al., 1979; Rombout, Taveme-Thiele, 1982; Holmgren et al., 1982;
Yoshida et al., 1983; Rombout, Reinecke, 1984; L’Hermite et al., 1985; Rombout et al., 1986a, b;
Abad et al., 1987; Elbal et al., 1988; Nozaki et al., 1988; Beorlegui et al., 1982a, b), иммунореак-
тивные к пакреатическому полипептиду (Langer et al., 1979; Noaillac-Depeyre, Hollande,
1981; Rombout, Taverne-Thiele, 1982; Rombout et al., 1986a, b; Abad et al., 1987; Elbal et al.,
1988; Burkhardt-Holm, Holmgren, 1989), пептиду тирозин тирозину (PYY) (Ali-Rochedi et al.,
1984; Beorlegui et al., 1992a, b), нейропептиду Y (NPY) (Burkhardt-Holm, Holmgren, 1989;
Beorlegui et al., 1992a, b), холецистокинину (Rajjo et al., 1988), гастрину/холецистокинину
(Noaillac-Depeyre, Hollande, 1981; Holmgren et al., 1982; Rombout, Taverne-Thiele, 1982;
19
Reifel et al, 1983; Vigna et al., 1985; Rombout et al., 1986a, b; Abad et al., 1987; Jonsson et al.,
1987; Burkhardt-Holm, Holmgren, 1989; Beorlegui et al., 1992a, b).
В пищеварительном тракте многих костистых рыб описаны также соматостатин-
иммунореактивные клетки (Langer et al., 1979; Noaillac-Depeyre, Hollande, 1981; Holmgren et
al., 1982; Yoshida et al., 1983; Noaillac-Depeyre, 1985; L’Hermite et al., 1985; Abad et al., 1987;
Elbal et al., 1988, 1991; Beorlegui et al., 1992a, b). В кишечнике Gadus morhua и Oncorhynchus
mykiss присутствуют клетки, иммунореактивные к аденилат циклаза активирующему поли-
пептиду (РАСАР) и хелоспектину (соединения, родственные вазоактивному интестиналь-
ному полипептиду— VIP). Сходные по иммунореактивности нервные отростки лежат в
стенке кишечника вне эпителия (Olsson, Holmgren, 1994). Клетки, реагирующие с антисы-
вороткой к вазоактивному интестинальному полипептиду, обнаружены в эпителии кишеч-
ника Oreochromis mossambicus (Kiliaan et al., 1996).
В целом для эндокринных клеток рыб характерно хорошее развитие шероховатой эн-
доплазматической сети, наличие в цитоплазме свободных рибосом, лизосом, микротрубо-
чек. Оболочка ядер часто имеет заметные инвагинации. Типы клеток выделяют в первую
очередь на основании размеров и тонкого строения секреторных гранул. Так, в различных
участках эпителия желудка, пилорических выростов и кишечника при электронно-
микроскопическом исследовании у Mugil saliens выявлены 9 типов эндокринных клеток
(Elbal et al., 1988). Всего у костистых рыб, имеющих желудок, выделяют (включая остров-
ковую ткань поджелудочной железы) до 10 основных типов эндокринных клеток, а у рыб,
не имеющих морфологически оформленного желудка, — до 6 (Ширкина, 1995).
2.6. CYCLOSTOMATA (КРУГЛОРОТЫЕ)
Класс круглоротых, образованный современными подклассами Myxini (миксины) и
Petromyzones (миноги), относится к надклассу Agnatha (бесчелюстные), — первой по вре-
мени появления группе позвоночных животных.
Пищеварительный тракт круглоротых характеризуется рядом особенностей по сравне-
нию с другими позвоночными, а у миног в определенной степени различается у личинок
(пескороек) и взрослых животных. Он состоит из глотки, пищевода, средней кишки, под-
разделяемой на передний и задний отделы, и задней кишки. У личинок миног в начальный
отдел кишечника открывается проток печени, у взрослых форм печень утрачивает связь с
кишечной трубкой (Barrington, 1972; Youson, 1981а, b). У круглоротых присутствует также
расположенная в подслизистой оболочке эндокринная поджелудочная железа, обычно
имеющая фолликулярное строение. Она возникает за счет обособления и специфической
дифференцировки части эпителиальных клеток дистального отдела пищевода и, в меньшей
степени, кишечника и протока печени (Barrington, 1972; Ostberg et al., 1976b). Большую
часть ее клеточных элементов составляют клетки — гомологи [3-клеток позвоночных; это
заключение было сделано на основании их положительной реакции с антисывороткой к
инсулину (Van Noorden et al., 1977). Интересно отметить, что у миксин инсулин-
иммунореактивные клетки располагаются также в эпителии желчного пузыря (Ostberg et al.,
1975, 1976а; Фалкмер, 1977). У Myxine glutinosa в фолликулах поджелудочной железы об-
наружены и редкие клетки с более мелкими (диаметром около 213 нм) гранулами. Однако
последние рассматриваются не как самостоятельный клеточный тип, а как промежуточная
стадия дифференцировки [3-клеток (Thomas et al., 1973).
20
Эндокринные клетки присутствуют у круглоротых и в кишечнике. Впервые энтеро-
хромаффинные клетки были описаны у Petromyzon planeri еще в 1938 г. (Uggeri, 1938). По
всей длине средней кишки пескороек встречаются вытянутые клетки открытого типа с рас-
положенными в базальной цитоплазме электронноплотными гранулами диаметром порядка
100 нм. Основания клеток, лежащие на базальной мембране, несколько расширены; узкая
апикальная часть подходит к просвету и несет микроворсинки, а часто и ресничку (Van
Noorden et al., 1972). Однако использование антисывороток к гастрину, глюкагону и секре-
тину у пескоройки не выявило окрашивающихся клеток (Van Noorden et al., 1972).
Эндокринные клетки в кишечнике Petromyzon marinus — открытого типа, по сравне-
нию с окружающими всасывающими имеют мало митохондрий и слаборазвитую шерохова-
тую эндоплазматическую сеть, но хорошо заметный аппарат Гольджи. Апикальная поверх-
ность несет микроворинки и ресничку (Youson, 1981а). Сходные клетки в кишечнике мик-
сины Myxine glutinosa имеют, по-видимому, только микроворсинки (Ostberg, Boquist, 1976;
Hardisty, Baker, 1982). Диаметр секреторных гранул составляет 163-178 нм (Ostberg et al,
1976а).
В центральном отделе средней кишки Lampetra fluviatilis описаны 5 типов эндокрин-
ных клеток, различающихся как по диаметру гранул (от 124 до 187 нм), так и по их тонкому
строению. Четыре типа клеток — открытого типа, а один, возможно, не контактирует с про-
светом кишечника. Всем эндокринным клеткам открытого типа свойственны некоторые
общие черты строения: аппарат Гольджи и другие органеллы расположены главным обра-
Рис. 10. Локализация инсулин- и глюкагон/гастрин-иммунореактивных клеток в пищеварительном
тракте личинки миноги (по: Ван Ноорден, Пирс, 1977, с небольшими изменениями).
ггк- глюкагон/гастрин иммунореактивные клетки; зк - зимогенные клетки; ик- инсулин-иммунореактивные
клетки. Следует отметить, что инсулин-иммунореактивные клетки могут занимать как суб-,
так и интраэпителиальное положение.
Fig. 10. Localization of insulin- and glucagon/gastrin-immunoreactive cells in the digestive tract of larval
lamprey (according to: Ван Ноорден, Пирс, 1977, with some changes).
21
зом над ядром, а гранулы сконцентрированы в нижней половине клетки (Пузырев, Иванова,
1991).
Иммуногистохимическими методами в кишечнике у взрослых особей Lampetra
fluviatilis удалось выявить клетки, цитоплазма которых реагировала с антисыворотками к
гастрину, глюкагону (рис. 10), С-концевому фрагменту пентагастрина и каерулину амфибий
(Van Noorden, Pearse, 1974; Hardisty, Baker, 1982). Однако существует мнение, что всеми
антисыворотками окрашиваются одни и те же клеточные элементы (Hardisty, Baker, 1982).
Наличие гастриноподобного вещества в кишечнике миноги было подтверждено также био-
химическими методами, показано присутствие и холецистокинин-подобного пептида
(Holmquist et al., 1979). В кишечнике Myxine glutinosa описаны клетки, иммунореактивные к
глюкагону, гастрину, пентагастрину, каерулину, панкреатическому полипептиду, нейропеп-
тиду Y, пептиду тирозин тирозину (Ostberg et al., 1976а; Cheung et al., 1991).
Завершая эту главу, можно отметить, что уже представители низших Vertebrata обла-
дают достаточно сложно организованной кишечной регуляторной системой, состоящей из
полиморфных тслеток, синтезирующих, очевидно, разнообразные регуляторные соединения,
и сопоставимой по своей организации с энтериновой системой млекопитающих.
Глава 3
КИШЕЧНАЯ РЕГУЛЯТОРНАЯ СИСТЕМА ДРУГИХ ХОРДОВЫХ
3.1. AMPHIOXI (ЛАНЦЕТНИКИ)
Класс Amphioxi (ланцетники) — единственный, образующий подтип бесчерепных
(Acrania), — насчитывает всего несколько десятков видов. Эти животные представляют
особый интерес тем, что, очевидно, очень близки к древнейшему предку позвоночных.
Пищеварительный тракт ланцетников начинается ротовым отверстием и следующей за
ним объемистой глоткой с многочисленными жаберными щелями (этот отдел среди прочих
выполняет дыхательную функцию) и двумя продольными желобками, один из которых (эн-
достиль) считается аналогом щитовидной железы позвоночных. За глоткой следует кишеч-
ник, имеющий слепой пищеварительный вырост (другие названия: печеночный вырост,
caecum), который рассматривается как предшественник печени, а возможно, и поджелудоч-
ной железы Vertebrata.
В начале 60-х годов XX века в составе эпителия так называемого «латерального» и
«дорсального» ресничных трактов кишечника Branchiostoma lanceolatum были описаны
редкие клетки, окрашивающиеся методом серебряной импрегнации (Gerzeli, 1961). Позднее
в этих же участках эпителиальной выстилки кишечника ланцетника с помощью флуорес-
центно-гистохимического метода Фалька-Хилларпа были выявлены клетки с типичной для
серотонина интенсивной желтой флуоресценцией (Hulting, 1973). Клетки и волокна в ки-
шечнике, демонстрирующие как желтое, так и зеленое (характерное для катехолами катехо-
ламинов) свечение отмечены и в другом исследовании (Салимова, 1978), однако каково их
положение (интра- или субэпителиальное), к сожалению, не указывается.
По данным ультраструктурного исследования (Hulting, 1973) в отмеченных выше
«сильно ресничных» участках эпителиальной выстилки кишечника среди реснично-
слизистых клеток располагаются клетки характерного строения, которые, по-видимому,
соответствуют отмеченным выше энтерохромафинным.
Это— вытянутые цилиндрические клетки открытого типа (рис. И), с несколько рас-
ширенным основанием и хоаноцито-подобным апикальным концом, несущим ресничку.
Шероховатая эндоплазматическая сеть развита довольно слабо. В апикальной цитоплазме
многочисленны мелкие пузырьки, аутофагические вакуоли и мультивезикулярные тела.
Аппарат Гольджи располагается над ядром, в цитоплазме много микротрубочек. Многочис-
ленные гранулы диаметром 250-300 нм имеют гало, их содержимое варьирует по электрон-
ной плотности. Гранулы встречаются по всей клетке, однако главным образом сконцентри-
рованы в базальной части, где во флуоресцентно-гистохимическом методе отмечается наи-
более интенсивное свечение.
23
Рис. 11. Схема строения эндокринной клетки эпителия
кишечника ланцетника Branchiostoma lanceolatum
(по: Hulting, 1973).
ав - автофагическая вакуоль.
Fig. 11. Scheme of an endocrine cell organization
in Branchiostoma lanceolatum intestinal epithelium
(after: Hulting, 1973).
В пищеварительном выросте Branchistoma lanceolatum также отмечено присутствие
единичных клеток с большим количеством мелких электронноплотных гранул, свободными
рибосомами и хорошо развитым аппаратом Гольджи, в зоне которого гранулы и формиру-
ются. Помимо клеток, описаны также различные по диаметру отростки. Крупные, содер-
жащие (помимо гранул) митохондрии, цистерны аппарата Гольджи и рибосомы, по-
видимому, принадлежат эндокринным клеткам. Принадлежность мелких отростков к эн-
докринной или нервной системам требует дальнейших исследований (Welsch, 1975). Два
типа эндокринных клеток выявлены в задней части средней кишки Branchiostoma japonicum
(Kataoka, Fujita, 1974). Ван Ноорден и Пирс (1977) описывают в кишечнике 3 или 4 типа
«базально гранулярных» клеток. Наиболее обычны клетки с гранулами диаметром 100-200
нм (средний диаметр около 140 нм). Клетки второго типа содержат менее электронноплот-
ные гранулы со средним диаметром 170 нм. Еще один клеточный тип характеризуется бо-
лее мелкими (до 115 нм в диаметре) гранулами. Наконец, гранулы с эксцентрично располо-
24
женным содержимым, отмеченные в некоторых клетках, возможно, представляют собой
стадию созревания секреторного продукта.
В эпителии задней части средней кишки Branchiostoma lanceolatuni показано присутст-
вие специализированных мелкогранулярных клеток, выделяющих продукт белковой приро-
ды в полость кишечника. Гистохимические данные позволили предположить, что их секрет
сходен с инсулином или проинсулином (Winbladh Biuw, Hulting, 1971). В экспериментах с
нагрузкой глюкозой, проведенных на животных этого же вида, продемонстрировано, что
клетки, сходные по морфологии с описанными выше мелкогранулярными, отвечают на это
выводом своего альдегидфуксин-положительного секрета в просвет, что подтверждает воз-
можность их сходства с инсулин-продуцирующими клетками (Полякова, Плисецкая, 1976).
В иммуногистохимических исследованиях у ланцетника выявлены интраэпителиаль-
ные клетки открытого типа, реагирующие с антисыворотками к инсулину (Van Noorden,
Pearse, 1976; Emdin, Falkmer, 1977, Ван Ноорден; Пирс, 1977; Reinecke, 1981; Reinecke et al.,
1993), а также к панкреатическому полипептиду, соматостатину, секретину, вазоактивному
интестинальному полипептиду, пентагастрину и нейротензину (Reinecke, 1981). Кроме них,
описаны также гастрин- и глюкагон-иммунореактивные клетки, причем, по-видимому, с
обеими антисыворотками реагируют одни и те же клеточные элементы (рис. 12) (Ван Ноор-
ден, Пирс, 1977).
Рис. 12. Локализация инсулин- и глюкагон/гастрин-иммунореативных клеток в пищеварительном
тракте ланцетника (по: Ван Ноорден, Пирс, 1977, с небольшими изменениями).
ггк - глюкагон/гастрин-иммунореактивные клетки; зк - зимогенные клетки;
ик - инсулин-иммунореактивные клетки; св - слепой вырост средней кишки.
Fig. 12. Localization of insulin- and glucagon/gastrin-immunoreactive cells in the digestive tract
of a lancelet (according to: Ван Ноорден, Пирс, 1977, with some changes).
Таким образом, даже имеющиеся немногочисленные данные однозначно свидетельст-
вуют в пользу наличия в кишечнике у представителей этой группы животных достаточно
сложной регуляторной системы, состоящей из нескольких типов эндокринных клеток.
25
3.2. TUNICATA (ОБОЛОЧНИКИ)
Этот тип, относящийся к группе хордовых (Chordata) и насчитывающий в своем соста-
ве более тысячи видов, состоит из 3 классов: Appendiculariacea (аппендикулярии),
Ascidiacea (асцидии) и Salpariacea (сальпы).
К сожалению, все известные мне данные по регуляторным клеткам в пищеварительном
тракте оболочников касаются только асцидий, зато последние в этом отношении изучены
достаточно хорошо.
3.2.1. Ascidiaceae (асцидии)
Пищеварительная система асцидий состоит из рта, массивной глотки (выполняющей в
том числе дыхательную функцию), короткого пищевода, желудка и кишечника, образующе-
го в теле животного двойную петлю. У некоторых видов асцидий желудок имеет вырос-
ты — так называемую «печень». Далее я рассмотрю главным образом данные, касающиеся
регуляторных клеток в пищеводе, желудке и кишечнике, хотя они присутствуют также и в
эпителии глотки, а расположенный на ее брюшной стороне желобок (эндостиль) в функ-
циональном отношении, возможно, сопоставим со щитовидной железой позвоночных.
В 1946 г. из желудка и кишечника Ciona intestinalis было экстрагировано «энтерамино-
подобное» вещество (Erspamer, 1946), позднее идентифицированное как серотонин
(Erspamer, Asero, 1952; Welsh, Loveland, 1968). Гистохимическими методами энтерохрома-
финные клетки были выявлены в составе эпителиальной выстилки пищеварительного трак-
та у целого ряда видов асцидий (Gerzelli, 1964; Fritsch, 1976; Relini Orsi, Pestarino, 1977;
Pestarino, 1979, 1982a, 1982b). Подобные клетки имеют вытянутую форму; у некоторых из
них несколько расширена базальная часть, где расположено ядро; апикальная часть подхо-
дит к просвету тракта. Следует отметить, что энтерохромафинные клетки есть, по-
видимому, только у взрослых животных: у личиночных форм их выявить не удалось
(Gerzelli, 1964).
Электронно-микроскопические исследования, проведенные на ряде видов, также вы-
явили интраэпителиальные регуляторные клетки, рассматриваемые авторами как эндокрин-
ные (Burighel, Milanesi, 1975а, 1975b, 1977; Thomdyke, 1977; Fritsch, Sprang, 1977; Tunas,
1977; Thomdyke, Bevis, 1978). Характерная черта всех таких клеток— наличие в цитоплаз-
ме электронноплотных гранул, как правило, небольшого размера. Чаще всего эти гранулы
округлые, реже — палочковидные; их размеры у некоторых из изученных видов приведены
в табл. 1. Варьирование размеров гранул в клетках разных отделов пищеварительной систе-
мы у таких видов, как Styela rustica, Styela clava, Polycarpa pomaria, Dendrodora glossularia,
указывает на то, что популяция эндокринных клеток образована несколькими клеточными
типами.
Тонкое строение эндокринных клеток у разных видов также варьирует в определенных
пределах. В эпителии желудка колониальной асцидии Botryllus schlosseri эндокринные эле-
менты имеют хорошо развитые шероховатую и гладкую эндоплазматические сети; в цито-
плазме многочисленны свободные рибосомы и полисомы. Апикальные поверхности несут
микроворсинки; ядра обычно неправильной формы. Секреторные гранулы, формирующиеся
в цистернах аппарата Гольджи, двух типов— округлые и палочковидные. Такие клетки
особенно часто расположены в тех участках выстилки, где под эпителием лежат кровенос-
ные лакуны. Это наблюдение предполагает, что секреторные продукты могут выводиться
непосредственно в циркуляторную систему (Burighel, Milanesi, 1975b).
26
Таблица 1
Размеры секреторных гранул в интраэпителиальных эндокринных клетках
пищеварительного тракта асцидий
Вид Размеры гранул, нм Литературный источник
пищевод желудок кишечник
Styela rustica 160-250 200-220* Tunas, 1977
Styela clava 100-175 50-155 50-155 Thomdyke, Bevis, 1978
Ciona intestinalis 100-200 100-200 100-200 Fritsch, Sprang, 1977
Botryllus schlosseri 80-120 Burighel. Milanesi, 1977
Polycarpa pornaria 200 170 Tunas, 1977
Dendrodora glossularia 100-150 170 100-120 Tunas, 1977
Molgula citrina 100-110 100-120 Tunas, 1977
* Клетки выявлены только в пилорической области желудка
В эндокринных клетках желудка и кишечника Styela clava много пузырьков гладкой
эндоплазматической сети и свободных рибосом (Thorndyke, Bevis, 1978). В эндокринных
клетках Ciona intestinalis, в отличие от отмеченной выше Botryllus schlosseri, шероховатая
эндоплазматическая сеть развита слабо; в цитоплазме присутствуют микротрубочки
(Fritsch, Sprang, 1977).
Хочу особо отметить, что в известной мне литературе, посвященной ультраструктур-
ному анализу эпителия пищеварительного тракта асцидий, я не встречал указаний на нали-
чие каких-либо интраэпителиальных структур, которые можно было бы рассматривать как
отростки нервных клеток.
Иммуногистохимические методы позволили выявить в эпителии пищеварительного
тракта достаточно многочисленные клетки, демонстрирующие иммунореактивности к раз-
личным регуляторным пептидам и серотонину. Как пример, интраэпителиальная нейротен-
зин-иммунореактивная клетка в желудке Styela rustica представлена на рис. 13 (вклейка). В
пищеводе, желудке и кишечнике Ciona intestinalis описаны клетки, реагирующие с антисы-
воротками к серотонину, нейротензину, субстанции Р, соматостатину, гастрину, панкреати-
ческому полипептиду, секретину и кальцитонину (в глотке также присутствуют и бомбезин-
иммунореактивные элементы) (Fritsch et al., 1978, 1980, 1982), а также инсулин-подобным
веществам (Reinecke et al., 1993). Подавляющее большинство таких клеток— открытого
типа; однако, например, кальцитонин-иммунореактивные клетки, лежащие в основании
эпителиального пласта, не контактируют с просветом (Fritsch et al., 1980). Наиболее много-
численны клетки в желудке; в кишечнике их количество убывает в дистальном направле-
нии. Распределение различных иммунореактивных клеток по отделам пищеварительной
системы у Ciona intestinalis показано на рис. 14.
Очевидно, асцидиям свойственны межвидовые различия, касающиеся клеток, демонст-
рирующих те или иные иммунореактивности. Так, секретин-подобная активность отмечена
в желудочно-кишечном тракте Dendrodora grossularia, но отсутствует у Ascidiella aspersa
(Bevis, Thorndyke, 1979).
В эпителии пищевода Styela clava описаны клетки, реагирующие с антителами к бычь-
ему инсулину. Одновременно здесь выявляются и альдегидфуксин-положительные клетки,
однако они заметно более многочисленны (Bevis, Thomdyke, 1978). Отмечу, что окраши-
вающиеся альдегидфуксином клетки ранее были описаны в эпителии желудка и кишечника
Ciona intestinalis (Davidson et al., 1971), однако, скорее всего, в данном случае выявлялись
обычные секреторные элементы. В то же время из тканей пищеварительного тракта Руига
27
Рис. 14. Распределение эндокринных клеток, демонстрирующих различные иммунореактивности,
в пищеварительном тракте Ciona intestinalis (по: Fritsch et al., 1982, с некоторыми изменениями).
б - бомбезин-, г - гастрин-, к - кальцитонин-, н - нейротензин-, пп - панкреатический полипептид-,
с - серотонин-, ск - секретин-, сп - субстанция Р-иммунореактивные клетки.
Fig. 14. Distribution of endocrine cells demonstrating different immunoreactivities in the digestive tract of
Ciona intestinalis (after: Fritsch et al., 1982, with some changes).
pachyderm at in а выделены вещества, вступающие в реакцию с антителами к инсулину сви-
ньи и стимулирующие липогенез в изолированных жировых клетках крыс. Эти инсулино-
подобные вещества представлены двумя иммунологически активными формами, одна из
которых в значительной степени близка инсулинам млекопитающих (Galloway, Cutfield,
1988), однако конкретная тканевая локализация этих соединений пока неизвестна.
28
В пищеводе Styela plicata выявлены соматостатин-иммунореактивные клетки (Pestarino,
1983), а также клетки, одновременно содержащие секретин-подобное вещество и биогенные
амины. В то же время в желудке локализованы клетки, содержащие только секретин-
подобное вещество (Pestarino, 1982с). Сходная картина отмечена и для эндокринных клеток
пищеварительного тракта Halocynthia papillosa (Pestarino et al., 1982; Pestarino, Tagliafierro,
1983).
У Styela plicata клетки, демонстрирующие различные иммунореактивности, располо-
жены в разных участках эпителиальных складок желудка. Если уже отмеченные секретин-
иммунореактивные элементы локализованы на вершине эпителиальных складок, то гаст-
рин-, глюкагон-, соматостатин- и, возможно, инсулин-иммунореактивные клетки распола-
гаются главным образом в эпителии их боковых поверхностей (Pestarino, 1979, 1982b).
В эпителии пищевода и мелких складок желудка Styela plicata выявлены клетки, реаги-
рующие с антисывороткой к свиному пептиду гистидин-изолейцину (PHI) и серотонину
(Pestarino, 1986, 1989). В то же время использование антисыворотки к вазоактивному инте-
стинальному полипептиду дало отрицательные результаты (Pestarino, 1986). В желудке у
этого вида показано наличие клеток, иммунореактивных к гастрину 2-17. Здесь следует
отметить, что иммунореактивное вещество локализовано главным образом в апикальной
цитоплазме и что такие клетки не окрашиваются антисыворотками к синтетическому гаст-
рину человека, а также гастрину 1-34 (Pestarino, Garaud, 1986; Pestarino et al., 1988), поэтому
вопрос о том, действительно ли эти клеточные элементы связаны с ситезом гастрин-
подобного вещества, требует дальнейшего изучения.
Что касается функционального значения кишечных эндокринных клеток, предполага-
ется, что они могут быть связаны с контролем как всасывания, так и секреции кишечным
эпителием пищеварительных ферментов (Burighel, Milanesi, 1975b; Tunas, 1977; Bevis,
Thomdyke, 1981). Серотонин предположительно участвует в контроле выделения слизи
(Burighel, Milanesi, 1977). Считается, что соматостатин-иммунореактивные клетки Styela
plicata паракриновым путем могут регулировать деятельность секретин-иммунореактивных
элементов (Pestarino, 1983). Кальцитонин-иммунореактивные клетки, возможно, выполняют
общеорганизменную функцию, будучи связаны с регуляцией Са++ в тунике (Fritsch et al.,
1980). Показано также, что ряд гормонов гастро-энтеро-панкреатической системы млекопи-
тающих стимулируют у Styela clava высвобождение пищеварительных ферментов из зимо-
генных клеток желудка (Thomdyke, Bevis, 1984).
Таким образом, независимо от конкретного функционального значения тех или иных
эндокринных клеток, приведенные литературные данные показывают наличие в эпителии
пищеварительной системы асцидий сложной гетерогенной регуляторной системы, элемен-
ты которой имеют значительное сходство с кишечными эндокринными клетками позвоноч-
ных.
Глава 4
ENTEROPNEUSTA (КИШЕЧНОДЫШАЩИЕ)
Кишечнодышащие вместе с крыложаберными (Pterobranchia) составляют тип полухор-
довых, или Hemichordata.
Эта своеобразная, эволюционно тупиковая группа представлена всего несколькими де-
сятками видов. Кишечнодышащие — морские свободноживущие червеобразные животные.
Их тело четко дифференцировано на три отдела — хоботок, воротничок и длинное тулови-
ще. Пищеварительная система начинается расположенным у основания хоботка ртом. От
следующей за ним короткой глотки со спинной стороны внутрь хоботка отходит слепое
выпячивание, или нотохорд, который рассматривается как зачаток хорды. Глотка переходит
в пищевод, представляющий собой в первую очередь дыхательный отдел пищеварительной
системы. Его боковые стенки пронизаны жаберными щелями, через которые полость пище-
вода сообщается с внешней средой. За глоткой следует кишечник, передняя часть которого
(так называемый «печеночный отдел») образует на спинной стороне многочисленные вы-
росты («печеночные придатки»). Вторая половина кишечника трубчатая, придатков не име-
ет. Она переходит в заднюю кишку, открывающуюся наружу анальным отверстием.
Элементы нервной системы (преимущественно нервные отростки) в пищеварительном
тракте описаны на светооптическом уровне у ряда видов кишечнодышащих (Bullock, 1945;
Silen, 1950). Однако эти наблюдения касаются главным образом эпителия глотки, пищевода
и задней кишки. В то же время у Glossobalanus marginatus в кишечнике на границе с пище-
водом обнаружены чувствительные клетки, имеющие вытянутую форму (Silen, 1950).
В ходе ультраструктурного исследования печеночного отдела кишечника Saccoglossus
mereschkovskii Атаманова (1980) выявила в основании эпителия группы отростков с элек-
тронносветлой цитоплазмой, содержащие пузырьки и электронноплотные гранулы диамет-
ром 100-300 нм. По-видимому, эти отростки относятся к интраэпителиальному нервному
слою (рис. 15).
Проведенный мной предварительный иммуногистохимический анализ кишечника
Saccoglossus mereschkovskii показал присутствие в печеночном отделе элементов базиэпите-
лиального нервного сплетения, реагирующих с антисыворотками к РМИРамиду и нейротен-
зину (рис. 16-18, вклейка), в составе которого отмечены и единичные клетки закрытого ти-
па. По своей морфологии эти сплетения, по-видимому, представляют собой часть интраэпи-
телиальной нервной системы; в некоторых участках эпителиального пласта они очень хо-
рошо выражены.
30
Рис. 15. Схема строения базальной части эпителия «печеночной области» средней кишки
Saccoglossus mereschkowskii [по электроннограммам Атамановой (1980)].
бмк — базальные части мерцательных клеток.
Fig. 15. Scheme of epithelial basal part organization of midgut «pancreatic region»
in Saccoglossus mereschkowskii [according to electronnograms from Атаманова (1980)].
Характерные клетки удалось наблюдать в эпителии центрального отдела кишечника
этого вида и после импрегнации азотнокислым серебром по Колонье (что указывает на их
нервную природу). Эти клетки имеют вытянутую форму, и их большая часть может быть
отнесена к элементам открытого типа. От базальных частей клеток отходят отростки, тяну-
щиеся в основании эпителиального пласта.
Таким образом, несмотря на отрывочные и неполные данные, можно заключить, что у
кишечнодышащих кишечная регуляторная система образована, по крайней мере, элемента-
ми нервной системы.
Глава 5
ECHINODERMATA (ИГЛОКОЖИЕ)
Иглокожие представляют собой разнородную и весьма своеобразную группу живот-
ных. Этот тип беспозвоночных располагается в основании ветви Deuterostomia и имеет в
своей организации много относительно примитивных черт.
Степень изученности кишечных регуляторных систем представителей различных клас-
сов иглокожих неодинакова. Больше всего данных к настоящему времени получено на мор-
ских звездах (Asteroidea). Морские ежи (Echinoidea), офиуры Ophiuroidea) и голотурии
(Holoturoidea) изучены в этом отношении гораздо хуже. Наконец, практически ничего не
известно об организации этой системы у морских лилий (Crinoidea), за исключением того,
что у представителей этого класса (впрочем, как и у других иглокожих) в основании эпите-
лия (по крайней мере, некоторых отделов пищеварительной системы) расположено хорошо
развитое нервное сплетение (Филимонова, 1979).
5.1. ASTEROIDEA (МОРСКИЕ ЗВЕЗДЫ)
Пищеварительная система у представителей этого класса начинается окруженным мяг-
кой кольцевой губой ртом, расположенным на оральной стороне диска. При помощи корот-
кого пищевода рот сообщается с массивным складчатым кардиальным (другое название —
оральный) желудком, способным выворачиваться наружу. Кардиальный желудок переходит
в пилорический (аборальный), от которого в стороны расходятся слепые выпячивания —
пилорические выросты. Кишечник обычно короткий, часто с небольшими ректальными
выростами, и открывается в центре аборальной стороны диска. У некоторых видов анальное
отверстие отсутствует, и пищеварительная система оказывается слепозамкнутой.
5.1.1. Данные электронно-микроскопических исследований
Эпителий пищеварительного тракта морских звезд, как и других иглокожих, характе-
ризуется присутствием многочисленных нервных отростков, тянущихся между узкими ос-
нованиями эпителиальных клеток (Bargmann, Behrens, 1968; Jangoux, Perpeet, 1972; Фили-
монова, 1979; Van der Plats et al, 1984; Martinez et al., 1989). Эти отростки обычно содержат
микротрубочки и мелкие гранулы. У Asterias rubens диаметр последних составляет 100—
160 нм (Bargmann, Behrens, 1968). Отростки образуют в основании кишечного эпителия
сплетение и мощные стволы, иногда насчитывающие в своем составе сотни отдельных от-
ростков. Такие стволы особенно хорошо заметны в кардиальном и пилорическом желудках,
32
причем иногда видны даже под световым микроскопом. Отростки, расположенные в таких
стволах по периферии, тесно контактируют с базальными частями обычных эпителиальных
клеток, однако типичных синаптических структур при этом не выявлено. Интересно, что
отмечены также контакты с эпителиальными клетками пищеварительной системы и отрост-
ков целомического (субэпителиального) сплетения (Jangoux, 1982).
Ультраструктурные данные о наличии у морских звезд клеток с признаками нервных
или эндокринных отрывочны и неполны. Так, Филимонова (1979) обнаружила в эпителии
кардиального желудка Asterias rubens клетки с мелкими гранулами диаметром 160-300 нм,
напоминающими по строению гранулы (3-клеток позвоночных. У этого же вида описаны
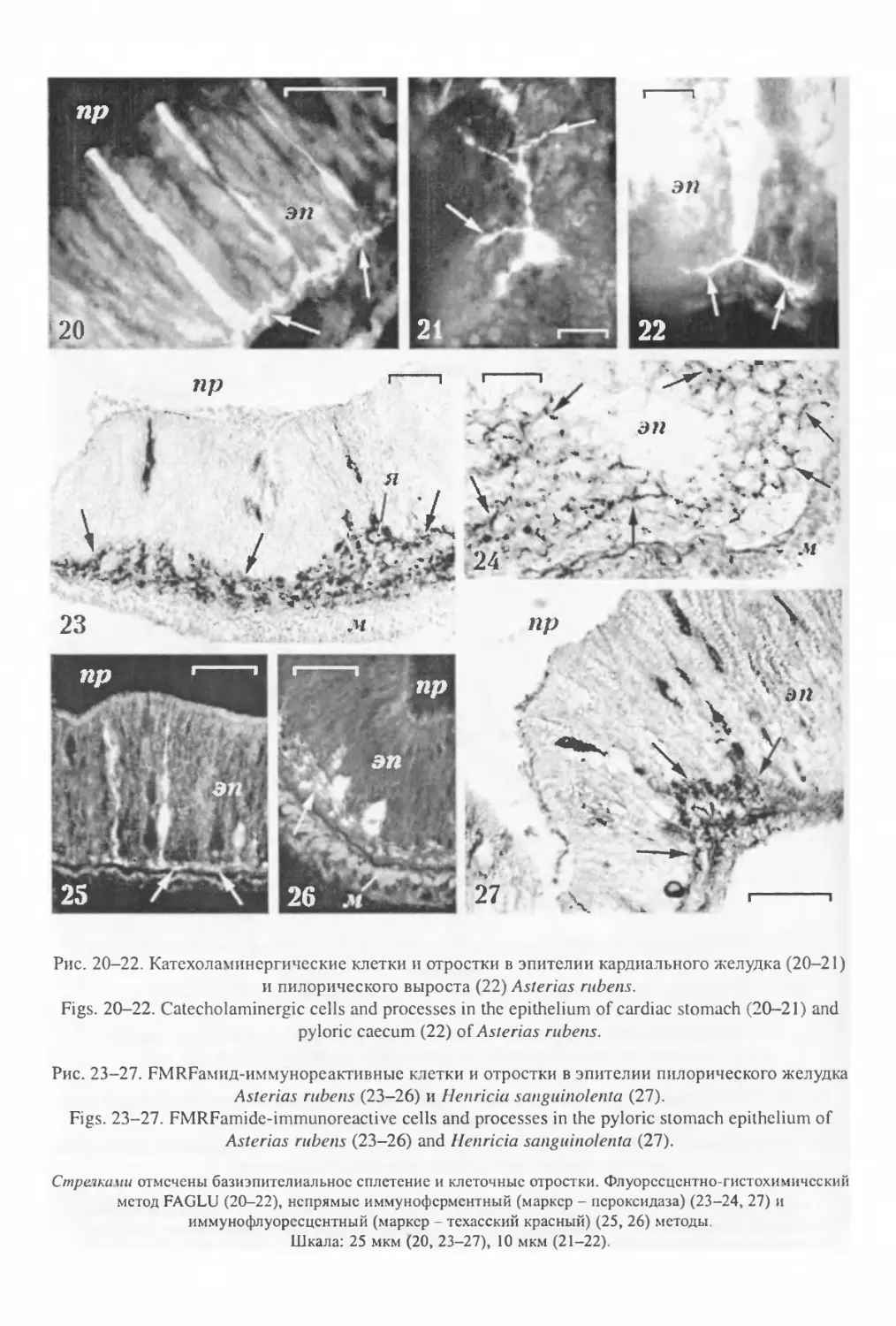
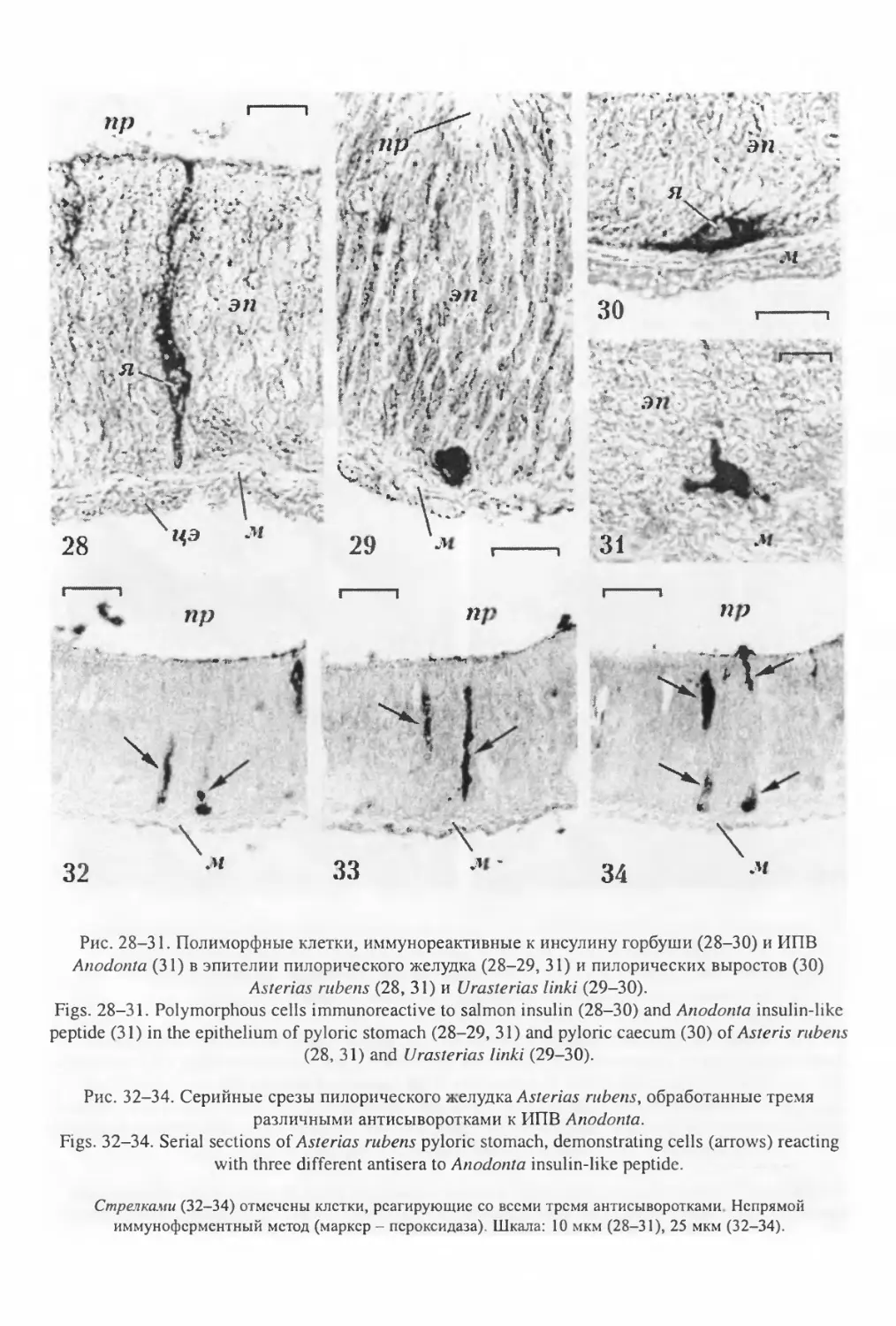
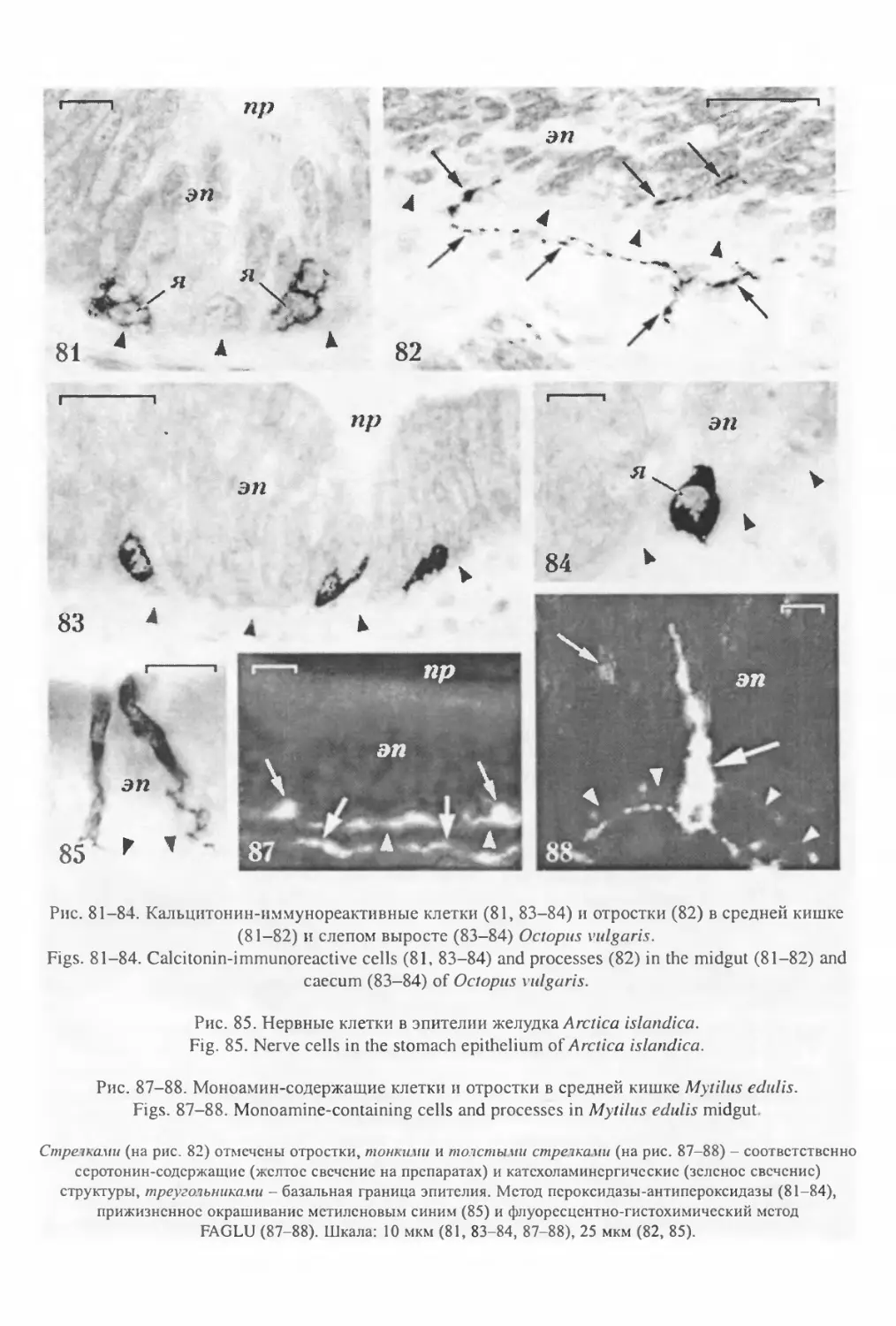
«крупные аксоны» (Bargmann, Behrens, 1968), в которых, помимо микротрубочек и поли-
морфных гранул диаметром от 100 до 160 нм, присутствуют цистерны эндоплазматической
сети и аппарата Гольджи. По-видимому, в действительности эти «аксоны» представляют
собой фрагменты интраэпителиальных эндокринных или нервных клеток, ядросодержащие
части которых не попали на срез.
В эпителии пилорических выростов Marthasterias glacialis на основании тонкого
строения описаны, по крайней мере, два типа клеток, по мнению авторов (Martinez et al.,
1989), эндокринных. Они относятся к клеткам открытого типа (тянутся от базальной мем-
браны до просвета выростов) и различаются главным образом по размерам и содержимому
секреторных гранул. Первый тип характеризуется электронноплотными гранулами диамет-
ром 135-170 нм, второй— гранулами с содержимым различной электронной плотности
Рис. 19. Взаимоотношения эндокринной клетки и нервных отростков
в эпителии пилорических выростов Martasterias glacialis
(по: Martinez et al., 1989).
Fig. 19. Interrelationships of endocrine cell and nerve processes
in the epithelium of pyloric caecum of Martasterias glacialis
(after: Martinez et al., 1989).
33
диаметром 70-100 мкм. Количество гранул может меняться от клетки к клетке, однако, как
правило, гранулы весьма многочисленны. Некоторые из этих клеток находятся в тесном
контакте с нервными отростками базального интраэпителиального сплетения (рис. 19).
5.1.2. Клетки, содержащие моноамины
Флуоресцентно-гистохимический метод FAGLU позволил выявить в составе эпители-
ального пласта пищеварительной системы Asterias rubens хорошо развитую систему кате-
холаминергических клеток и отростков (Пунин, 1996а, 1997а). Катехоламинергические
клетки, характеризующиеся специфической зеленой флуоресценцией, особенно часто
встречаются в кардиальном и пилорическом желудках, причем в некоторых участках эпите-
лиальной выстилки они исключительно многочисленны (рис. 20, вклейка), тогда как в дру-
гих могут полностью отсутствовать; закономерностей в их таком неравномерном распреде-
лении обнаружить не удалось. В протоках пилорических выростов катехоламинергические
клетки, как правило; немногочисленны, а в самих выростах — единичны.
Почти все катехоламинергические клетки относятся к клеткам открытого типа (имеют-
ся апикальные отростки, которые подходят к просвету пищеварительного тракта). Они
имеют веретеновидную форму с несколько расширенной ядросодержащей частью (узкое
вытянутое ядро обычно располагается в нижней половине эпителиального пласта) либо,
если ядро лежит у базальной мембраны, — расширенную базальную часть. Практически у
всех клеток есть базальные отростки с варикозными расширениями, отходящие от перика-
рионов в разные стороны, часто ветвящиеся и тянущиеся на значительные расстояния (рис.
21-22, вклейка). Эти отростки формируют базальное катехоламинергическое сплетение,
особенно мощно развитое в тех участках, где клетки более многочисленны.
Характерное для серотонин-содержащих структур золотисто-желтое свечение у изу-
ченного вида морских звезд отсутствовало.
В стенке пищеварительного тракта за пределами эпителия специфически флуоресци-
рующие клетки и отростки не наблюдались.
5.1.3. Клетки, содержащие ЕМИЕамид-подобное вещество
У Martasterias glacialis РМЯРамид-иммунореактивные клетки встречаются в эпителии
всех отделов пищеварительного тракта. Эти клетки — открытого типа, вытянутой формы.
Кроме них, в составе базиэпителиального нервного сплетения антисывороткой окрашива-
ются многочисленные отростки. По данным электронно-микроскопической иммуноцито-
химии иммунореактивность связана с мелкими округлыми (диаметром около 95 нм) грану-
лами с электронноплотным содержимым (Martinez et al., 1991, 1993).
У Asterias rubens клетки, реагирующие с антисывороткой, присутствуют в составе эпи-
телия кардиального и пилорического желудков (рис. 23-26, вклейка) (Пунин, 1996а). Боль-
шая часть из них относится к клеткам открытого типа и имеет различную форму: веретено-
видную, узкую цилиндрическую, с расширенной ядросодержащей базальной частью и тон-
ким апикальным отростком (рис. 23, 25, вклейка). Клетки закрытого типа единичны (рис. 26,
вклейка). Практически всем клеткам свойственно наличие тонких базальных отростков с
варикозными утолщениями, формирующих в основании эпителиального пласта хорошо
выраженное сплетение (рис. 23-24, вклейка). Как и в случае с катехоламинергическими
клетками, в некоторых участках эпителиальной выстилки желудка РМЯРамид-
иммунореактивные структуры многочисленны, в других — единичны или совсем отсутст-
вуют. В эпителии пилорических выростов иммунореактивные клетки обнаружены не были.
34
Иммунореактивные структуры не были выявлены и под эпителием во всех изученных
отделах пищеварительного тракта, хотя ранее у Asterias rubens было показано присутствие
ЕМИЕамид-иммунореактивных клеток и отростков в составе нервной системы в других
тканях и органах (Elphick et al., 1989).
РМИРамид-иммунореактивные клетки открытого и закрытого типов и базиэпителиаль-
ное сплетение выявлены мной также в эпителии кардиального и пилорического желудков
Henricia sanguinolenta (рис. 27, вклейка). К некоторым отличиям по сравнению с Asterias
rubens следует отнести присутствие единичных иммунореактивных клеток и в эпителии
пилорических выростов.
5.1.4. Клетки, иммунореактивные к вАЬМЕамиду
ЗАЬМРамиды (S1 и S2)— первые нейропептиды, выделенные из иглокожих и полно-
стью охарактеризованные (Elphick et al., 1991а, b). По данным RJA (радиоиммунологиче-
ского анализа) и иммуногистохимии, присутствие S1 типично для нервной системы Asterias
rubens (Moore et al., 1991). SI представляет собой октапептид co следующей аминокислот-
ной последовательностью: СЕ^АЬМЕамид.
Показано, что S1-иммунореактивные структуры типичны для пищеварительной систе-
мы у Asterias rubens (Moore, Thomdyke, 1993). На всем протяжении пищеварительного трак-
та в эпителии присутствуют узкие вытянутые иммунореактивные клетки и многочисленные
иммунореактивные отростки, образующие базиэпителиальное сплетение. Реагирующие с
антисывороткой к S1 отростки располагаются также в субцеломическом нервном сплете-
нии.
Практически идентичные результаты, касающиеся распределения S1-иммуно-
реактивности, были получены при изучении пищеварительной системы Marthasterias
glacialis (Martinez et al., 1993). Авторы этого исследования на основании наблюдаемой ими
перекрестной реакции между антителами к S1 и РМИЕамиду полагают, что эти антисыво-
ротки в действительности реагируют с одной и той же молекулой, предположительно S1. В
то же время они отмечают, что, несмотря на значительное сходство в характере окрашива-
ния и распределении выявляемых обеими антисыворотками структур, имеют место и неко-
торые отличия (в частности, в отсутствии иммунореактивности во многих участках субце-
ломического плексуса) (Martinez et al., 1993).
Очевидно, что вопрос о том, распознают ли антисыворотки к ЕМКГамиду и SALMF 1
амиду одни и те же молекулы, или нет, требует дальнейшего изучения. Пока лишь следует
отметить, что преабсорбция S1-антисыворотки ЕМИЕамидом не давала существенных из-
менений в характере окрашивания (Moore, Thomdyke, 1993). Это наблюдение, скорее, сви-
детельствует в пользу наличия у морских звезд обоих пептидов.
5.1.5. NO-синтетаза- и GABA-иммунореактивные клетки
В составе эпителия базиэпителиального плексуса Marthasterias glacialis было показано
наличие интраэпителиальных клеток и отростков, иммунореактивных к изоформам NO-
синтетазы млекопитающих. Иммунореактивность наблюдалась в тех же структурах, кото-
рые давали реакцию на NADPH-диафоразу (ко-фактор для NO-синтетазы), что подтвержда-
ет присутствие в них анализируемого фермента. Предполагается, что NO играет у иглоко-
жих роль нейронального мессенджера (Martinez et al., 1994).
Клетки и базиэпителиальные отростки, демонстрирующие иммунореактивность к
GABA (нейротрансмиттер, оказывающий в центральной нервной системе млекопитающих
35
ингибирующее действие), выявлены в эпителии пищеварительного тракта Asterias rubens
(Newman, Thomdyke, 1994).
5.1.6. Клетки, содержащие инсулиноподобные вещества
В начале 60-х годов прошлого века из пилорических выростов морской звезды Pisaster
ochraceous был экстрагирован гипогликемический фактор, по своим биохимическим и им-
мунологическим свойствам напоминавший инсулин млекопитающих (Wilson, Falkmer,
1965). В составе эпителия этого отдела пищеварительного тракта с помощью окраски псев-
доизоцианином и альдегидфуксином были выявлены продолговатые клетки с гранулярной
цитоплазмой, сходные по гистохимическим и тинкториальным свойствам с (3-клетками по-
звоночных (Wilson, Falkmer, 1965). В эпителии пилорических выростов Henricia
sanguinolenta также наблюдались альдегидфуксин-положительные клетки, являвшиеся, по
мнению исследователя, выявившего их (Chia, 1969), инсулинпродуцирующими. Однако
позднее было показано, что подобные клетки, по всей видимости, представляют собой же-
лезистые (слизистые) элементы (Chan, Fontaine, 1971).
Иммуногистохимический анализ с использованием антисыворотки к инсулину млеко-
питающих, проведенный на срезах пилорических выростов Marthasterias glacialis, не вы-
явил в эпителии этого отдела пищеварительного тракта иммунореактивных структур
(Martinez et al., 1989). В то же время исследования, проведенные мной на 2 видах морских
звезд (Asterias rubens и Urasterias linki), дали положительные результаты.
В этих исследованиях были использованы 3 антисыворотки к инсулиноподобному ве-
ществу (ИПВ), выделенному из двустворчатого моллюска Anodonta cygnea (подробнее об
этом веществе см.: Русаков и др., 1991), антисыворотка к С-концевому фрагменту ИПВ
брюхоногого моллюска Lymnaea stagnalis и антисыворотки к инсулинам позвоночных —
горбуши и свиньи. При использовании всех антисывороток в составе эпителия различных
Отделов пищеварительного тракта (желудок, пилорические выросты и их протоки) были
выявлены иммунореактивные клетки (рис. 28-31, вклейка). Подавляющее большинство этих
клеток имело вытянутую форму (узкие цилиндрические, веретеновидные, с расширенной
базальной частью) и контактировало с просветом пищеварительного тракта (рис. 28, вклей-
ка). В то же время были зарегистрированы редкие клетки закрытого типа, лежащие в осно-
вании эпителиального пласта (рис. 29, вклейка), иногда распластанные по базальной мем-
бране (рис. 30, вклейка). У некоторых клеток присутствовали короткие базальные отростки,
не образующие, однако, интраэпителиального сплетения (рис. 31, вклейка).
Окрашивание последовательных серийных срезов обоих видов морских звезд разными
антисыворотками продемонстрировало, что основная часть выявляемых клеток одновре-
менно реагирует с различными антителами (рис. 32-36, вклейка).
Доступность антител к инсулинам и ИПВ, выработанных у различных животных-
доноров (кролик и морская свинка), дала возможность провести как последовательную, так
и одновременную обработку двумя разными антисыворотками одних и тех же срезов. В
большинстве случаев при окрашивании обеими антисыворотками одних и тех же клеток
удается наблюдать конкуренцию между различными антителами. Так, при одновременной
обработке свечение клеток, обусловленное реакцией с антисыворотками к инсулинам или
ИПВ, выработанными у морской свинки, значительно более интенсивное (использовался
непрямой метод окрашивания с вторыми антителами, маркированными разными флуоро-
хромами), чем обусловленное реакцией с антителами, выработанными у кролика. В случае
последовательного окрашивания, при использовании в качестве первой специфической ан-
36
тисыворотки кроличьей, интенсивность свечения оказывается примерно одинаковой (рис.
37-38, вклейка), тогда как при обратной последовательности инкубаций свечение, обуслов-
ленное реакцией с антителами морской свинки, значительно интенсивнее, чем обусловлен-
ное реакцией с кроличьими антителами (рис. 39-40, вклейка).
Конкуренция между антителами к различным инсулинам и ИПВ может быть объяснена
тем, что все они реагируют главным образом с одними и теми же антигенными детерминан-
тами. Это позволяет с достаточно большой вероятностью говорить о наличии в эпителии
пищеварительного тракта морских звезд клеток, связанных с синтезом инсулиноподобных
веществ.
5.1.7. Клетки, иммунореактивные к другим регуляторным пептидам
Пищеварительная система Marthasterias glacialis была подробно изучена с использова-
нием свыше 20 антисывороток к различным регуляторным пептидам (Martinez et al., 1991,
1993). В результате удалось выявить клеточные структуры, иммунореактивные к пептиду
тирозин-тирозину- (PYY), панкретатическому пептиду (РР), а-меланоцит стимулирующему
гормону ( JMCH) и пептидилглицин монооксигеназе (РАМ).
Практически все иммунореактивные клетки принадлежат к открытому типу, и только
клетки, реагирующие с антисывороткой к PYY, большей частью расположены в основании
эпителиального пласта и имеют округлую форму. Помимо интраэпителиальных клеток, с
отмеченными выше антисыворотками реагирует также базиэпителиальное сплетение и, в
меньшей степени, субцеломическое. Лишь антисыворотка к GMCH окрашивает главным
образом последнее сплетение, хотя также выявляет клетки и отростки в эпителии. Все анти-
сыворотки, хотя и с разной частотой, окрашивают интраэпителиальные иммунореактивные
структуры на всем протяжении пищеварительной системы (кардиальный и пилорический
желудки, пилорические и ректальные выросты, кишечник).
В эпителии пилорических выростов было продемонстрировано также присутствие кле-
ток, иммунореактивных к соматостатину и глюкагону (Martinez et al., 1989). Важно отме-
тить, что базиэпителиальное нервное сплетение с этими антителами не реагировало.
С помощью антисывороток к панкреатическому полипептиду, соматостатину и глюка-
гону был изучен пищеварительный тракт 3 видов морских звезд: Asterias rubens, Henricia
sanguinolenta и Urasterias linki (Панкратова, 1993; Панкратова и др., 1995; Пунин и др.,
1996). В составе эпителия пилорического желудка (кардиальный желудок из-за обилия сек-
реторных экзокринных клеток, не специфически реагирующих со всеми антисыворотками,
оказался, к сожалению, мало удобным для детального анализа) у этих видов отмечены клет-
ки, реагирующие с использованными антителами (рис. 41-45, вклейка). За редким исключе-
нием, все они имеют сильно вытянутую форму и могут быть отнесены к открытому типу.
Клетки сравнительно редки и располагаются в эпителии поодиночке или небольшими груп-
пами. У некоторых соматостатин-иммунореактивных клеток Asterias rubens и Urasterias
linki и глюкагон-иммунореактивных клеток Asterias rubens отмечены короткие базальные
отростки. Базиэпителиальный нервный плексус, как правило, не реагирует с использован-
ными антителами; рыхлое соматостатин-иммунореактивное сплетение в некоторых участ-
ках эпителиальной выстилки наблюдалось у Asterias rubens, а у Henricia sanguinolenta сход-
ная картина была отмечена в отношении РР-иммунореактивных отростков. Следует отме-
тить, что в стенке желудка за пределами эпителия никаких иммунореактивных структур
нет.
37
В пилорических выростах иммунореактивные клетки выявлены только у Henricia
sanguinolenta (Пунин и др., 1996). В эпителии карманов Тидемана (участки протоков, со-
единяющие конечные отделы выростов — латеральные дивертикулы — с пилорическим
желудком) присутствуют редкие РР-иммунореактивные клетки без базальных отростков как
открытого (большинство), так и закрытого типа, а также глюкагон-иммунореактивные клет-
ки открытого типа (рис. 46, вклейка). Соматостатин-иммунореактивные отростки и мелкие
клетки здесь располагаются субэпителиально. В эпителии латеральных дивертикулов при-
сутствуют клетки, реагирующие со всеми тремя антисыворотками. Эти клетки не имеют
базальных отростков; многие из них, по-видимому, не граничат с просветом пилорических
выростов.
В пилорическом и кардиальном желудках, а также в пилорических выростах
Martasterias glacialis выявлено базиэпителиальное сплетение, реагирующее с антисыворот-
кой к адреномедуллину — пептиду, состоящему из 52 аминокислот и имеющему отдален-
ное сходство с CGRP (calcitonin gene related peptide). Интересно, что иммунореактивные
интраэпителиальные перикарионы отсутствовали. По-видимому, это может быть объяснено
либо тем, что клеточные тела находятся в каких-либо удаленных органах, либо тем, что
синтезированное вещество быстро перемещается в отростки (Martinez et al., 1996).
В заключение следует отметить, что далеко не ко всем распространенным регулятор-
ным пептидам позвоночных, присутствующих в том числе и у многих Invertebrata, антисы-
вортки выявляют регуляторные клетки и отростки в пищеварительном тракте морских
звезд. Так, отрицательные результаты были получены на Asterias rubens с антителами к
нейротензину (Reinecke, 1985) и на Martasterias glacialis с антителами к кальцитонину и
CGRP (Martinez et al., 1993).
5.1.8. Колокализация различных регуляторных веществ
Значительное сходство в морфологии регуляторных клеток и характере их положения в
эпителиальном пласте дало основание предполагать, что различную иммунореактивность
могут демонстрировать одни и те же клеточные элементы. Хотя этот вопрос нуждается в
дальнейших исследованиях, в отношении некоторых регуляторных соединений такие дан-
ные уже получены. Так, при изучении серийных срезов, обработанных разными антисыво-
ротками, было установлено, что в эпителии пилорических выростов Marthasterias glacialis
иммунореактивности к РР и глюкагону свойственны разным клеткам (Martinez et al., 1989).
Как уже было отмечено выше, по крайней мере, для некоторых клеток это справедливо и в
отношении проявляемых иммунореактивностей к FMRF-амиду и БАЬМРамиду 1.
В моих экспериментах на Asterias rubens и Urasterias linki были проведены как окра-
шивания последовательных серийных срезов антисыворотками к ИПВ моллюсков и FMRF-
амиду, так и одновременное окрашивание среза обоим антителами (рис. 47-48, вклейка).
Было обнаружено, что в эпителии пилорического желудка у этих видов присутствуют три
группы (разновидности) клеток и отростков: первая реагирует только с антисывороткой к
FMRFaMH/iy, вторая — только с антисывороткой к ИПВ, тогда как третья — с обеими анти-
сыворотками. При одновременном окрашивании (с использованием вторых антител, соот-
ветственно маркированных разными флуорохромами) свечение клеток и базальных отрост-
ков, обусловленное реакцией с антителами к FMRFaмидy, было значительно более интен-
сивным, если эти клетки одновременно не реагировали с антисывороткой к ИПВ.
Поразительное сходство во внешнем виде катехоламинергической и РМРБамид-
иммунореактивной систем в пилорическом желудке Asterias rubens дало основание предпо-
38
дожить, что флуоресцентно-гистохимический и иммуногистохимический методы выявляют
одни и те же структуры. Однако комбинирование обоих методов на одних и тех же срезах
показало, что РМЯРамид-иммунореактивные клетки, как правило, отсутствуют в тех участ-
ках эпителия, где наблюдаются катехоламинергические, и наоборот (Пунин, 1996а). В зо-
нах, где одновременно присутствуют и те, и другие клетки, зеленое и красное свечения ха-
рактерны для различных структур (рис. 49-51, вклейка).
5.2. ECHINOIDEA (МОРСКИЕ ЕЖИ)
Пищеварительная система морских ежей имеет вид длинной тонкостенной трубки и
описывает в теле животного сложную петлю. Обычно в ней выделяют начальную часть
(пищевод), центральную часть (среднюю кишку, сопровождаемую так называемым сифо-
ном — тонким трубчатым образованием, в своем начале и конце соединяющимся с кишеч-
ником) и дистальную часть.
Относительно кишечных регуляторных систем представители Echinoidea изучены
крайне фрагментарно. Микроскопические исследования показали присутствие в основании
эпителиального пласта мощной (особенно в пищеводе) системы отростков, по своему тон-
кому строению (наличие микротрубочек, микрофиламентов, мелких пузырьков) соответст-
вующих нервным. При этом типичные синаптические контакты между отростками и эпите-
лиальными клетками отсутствуют; только у Heliocidaris erythrogramma отмечены немного-
численные синаптические контакты между самими отростками, рассматриваемые как аксо-
аксональные (Cobb, 1969а).
5.2.1. Моноамин-содержащие клетки
С помощью флуоресцентно-гистохимического метода в составе эпителия пищевода
Heliocidaris erythrogramma были выявлены базиэпителиальные отростки и перикарионы,
содержащие моноамины (Cobb, 1969b) (по-видимому, речь идет о катехоламинах, хотя кон-
кретно об этом в работе не говорится).
Исследование эпителия пищеварительной системы другого морского ежа —
Strongylocentrotus droebachiensis, проведенное с использованием метода FAGLU (Пунин,
1996а, 1997а) показало наличие мощной базиэпителиальной катехоламинергической систе-
мы в пищеводе (рис. 52, вклейка). В этом отделе выявляются демонстрирующие зеленую
флуоресценцию стволы отростков, преимущественно тянущиеся вдоль тракта; их диаметр
на поперечных срезах может достигать 7-8 мкм. В то же время катехоламин-содержащие
перикарионы единичны, расположены в основании эпителиального пласта и, очевидно, не
граничат с просветом пищевода.
В первой половине средней кишки катехоламинергические структуры отсутствуют, а
во второй появляются единичные клетки как закрытого, так и открытого типов и редкие
отростки в основании эпителия (рис. 53, вклейка), не образующие, однако, того мощного
сплетения, которое характерно для пищевода.
В стенке пищеварительной системы за пределами эпителия катехоламинергические
структуры не наблюдались. Не было отмечено также и специфической золотисто-желтой
флуоресценции, типичной для серотонина.
39
5.2.2. Клетки, иммунореактивные к регуляторным пептидам
Иммуногистохимический анализ показал присутствие в эпителии пищеварительного
тракта Strongylocentrotus droebachiensis клеток, реагирующих с антисыворотками к FMRF-
амиду, панкреатическому полипептиду, соматостатину и глюкагону.
РМИРамид-иммунореактивные клетки выявлены на всем протяжении средней кишки,
но не в пищеводе (Пунин, 1996а). Они единичны и расположены в эпителиальном пласте
либо недалеко от базальной мембраны, либо в непосредственном контакте с последней (рис.
54, вклейка). Клетки имеют овальную или округлую форму, мелкие (около 10 мкм в диамет-
ре) и могут быть отнесены к элементам закрытого типа. Часто РМИРамид-иммуно-
реактивные клетки контактируют с интраэпителиальными нервными пучками, в составе
которых отростки не реагируют с антисывороткой. От перикарионов могут отходить тонкие
отростки, иногда проникающие в нервные пучки. За пределами эпителия иммунореактив-
ных структур не отмечено.
Среди соматостатинчиммунореактивных клеток отмечены клетки как открытого, так и
закрытого типов (рис. 55, вклейка). Редкие РР-иммунореактивные клетки могут быть отне-
сены к открытому типу (рис. 5.6, вклейка). Наконец, глюкагон-иммунореактивные клетки
(рис. 57, вклейка) — самые малочисленные, расположены у базальной мембраны. Все опи-
санные выше клетки лишены базальных отростков. Ни в эпителии пищевода и задней киш-
ки, ни в стенке пищеварительного тракта за пределами эпителия иммунореактивные струк-
туры не наблюдались (Панкратова и др., 1995; Пунин и др., 1996).
5.3. OPHIUROIDEA (ОФИУРЫ)
Отличительной чертой организации пищеварительного тракта офиур является относи-
тельная простота его анатомического строения. Пищеварительная система слепо замкнута и
состоит из обширного складчатого желудка без каких-либо выростов. Отсутствует и собст-
венно кишечник.
Кишечная регуляторная система у представителей этого класса изучена крайне слабо.
Ультраструктурные исследования показали в основании эпителиального пласта желудка
наличие системы отростков, на основании их тонкого строения рассматриваемых как нерв-
ные. У Ophiuroidenna panatnensis в них присутствуют овальные гранулы размером 100-150
х 50-100 нм (Schechter, Lucero, 1968). У Ophiopholis aculeata помимо электронноплотных
гранул диаметром до 100 нм, отмечены пузырьки с электронносветлым содержимым (Фи-
лимонова, 1979, 1980).
Мною было проведено флуоресцентно-гистохимическое исследование желудка
Ophiopholis aculeata и Ophiocanta bidentata. У обоих видов выявлены клетки, демонстри-
рующие зеленое свечение, типичное для катехоламинов (рис. 58-59, вклейка). У Ophiocanta
bidentata отмечены клетки как закрытого, так и открытого типов, тогда как у Ophiopholis
aculeata — только закрытого. Катехоламинергические клетки у обоих видов относительно
редки. Как правило, от перикарионов отходят базальные отростки, часто с варикозными
расширениями, не образующие, однако, заметного интраэпителиального плексуса. Лишь в
отдельных участках у базальной мембраны было отмечено очень рыхлое катехоламинерги-
ческое сплетение. За пределами эпителия специфически флуоресцирующих структур не
обнаружено.
Иммуногистохимический анализ желудка Ophiopholis aculeata и Ophiura robusta пока-
зал наличие в эпителии мелких клеток, реагирующих с антисыворотками к глюкагону, со-
40
матостатину (рис. 60-62, вклейка) и РР (рис. 63, вклейка) (Пунин и др., 1996). В некоторых
участках эпителиальной выстилки иммунореактивные клетки единичны, в других — отно-
сительно многочисленны. Только единичные соматостатин-иммунореактивные клетки
Ophiura robusta ориентированы перпендикулярно базальной мембране, однако не достига-
ют просвета пищеварительного тракта. Все остальные клетки обычно вытянуты вдоль ба-
зальной мембраны. Клетки имеют отростки, местами образующие хорошо заметное базаль-
ное сплетение. Под эпителием иммунореактивные структуры отмечены не были.
5.4. HOLOTHUROIDEA (ГОЛОТУРИИ)
Пищеварительная система голотурий представляет собой длинную трубку, обычно об-
разующую внутри животного петлю и лишенную каких-либо выростов. На основании мик-
роанатом ического и гистологического строения пищеварительный тракт подразделяют на
несколько отделов (Глотка, пищевод, желудок, средняя и задняя кишка); в некоторых отде-
лах, в свою очередь, выделяют дополнительные участки.
Как и у других иглокожих, в основании кишечного эпителия голотурий располагаются
отростки нервных клеток. У Cucwnaria japonica в так называемом «бульбовидном» отделе
(участок между мускулистым желудком и кишечником) такие отростки берут начало от
нервных клеток, расположенных в базальной части целомического эпителия. Они могут
быть собраны в окруженные мембраной пучки, проникающие в эпителиальный пласт и тес-
но соприкасающиеся с базальными частями эпителиальных клеток (Елисейкина, 1993; Ели-
сейкина, Лейбсон, 1996). В отростках присутствуют гранулы с электронноплотным содер-
жимым и светлые пузырьки диаметром 60-100 нм (Елисейкина, Лейбсон, 1996).
В кишечнике Holothuria mexicana, Holothuria glaberrima и Stichopus badionotus выявле-
ны клетки, демонстрирующие холецистокинин-подобную иммунореактивность (Garcia-
Arraras et al., 1991a), однако все они располагаются не в эпителии, а под ним, в соедини-
тельнотканной оболочке кишки. Такое же положение характерно и для описанных у
Holothuria glaberrima РМКРамид-иммунореактивных нейронов (Garcia-Arraras et al., 1991b).
Нам удалось продемонстрировать в кишечнике Chiridota laevis наличие интраэпители-
альных клеток, реагирующих с антисыворотками к соматостатину и глюкагону (рис. 64-65,
вклейка). Эти клетки единичны, по форме напоминают обычные эпителиальные и могут
быть отнесены к открытому типу. Они не имеют базальных отростков. Кроме интраэпите-
лиальных, клетки, реагирующие с этими антисыворотками, а также с антисывороткой к РР,
присутствуют под эпителием. Они характеризуются наличием длинных, иногда ветвящихся
отростков, которые могут вплотную подходить к эпителиальному пласту, однако не прони-
кают в него (Пунин и др., 1996).
Мне удалось также наблюдать в эпителии Chiridota laevis клетки, реагирующие с одной
из антисывороток к ИПВ Anodonta cygnea, выработанной у кролика. Эти клетки (рис. 66-67,
вклейка) на срезах имеют треугольную форму, реже распластаны вдоль базальной мембра-
ны. У них нет базальных отростков, и они не контактируют с просветом кишечника. Имму-
ногистохимический анализ с использованием антисыворотки к РМИГамиду не выявил в
эпителии иммунореактивных клеток. Отрицательные результаты были получены и в отно-
шении моноамин-содержащих структур при использовании флуоресцентно-гисто-
химического метода FAGLU.
41
5.5. ЗАКЛЮЧЕНИЕ
Приведенные выше данные убедительно демонстрируют присутствие в составе эпите-
лия пищеварительного тракта иглокожих сложной регуляторной системы. Безусловно,
представляло бы большой интерес проанализировать имеющиеся данные по разным клас-
сам этого типа беспозвоночных с точки зрения наличия или отсутствия в эпителии клеток с
теми или иными регуляторными веществами. Однако, к сожалению, корректно сделать это
не представляется возможным (и не только из-за различной степени изученности предста-
вителей разных классов) — ограничения иммуногистохимического метода позволяют дос-
таточно корректно говорить (как я уже отмечал ранее в гл. /) лишь о присутствии подоб-
ных иммунореактивностей, так как наблюдаемые положительные реакции могут быть обу-
словлены сходными антигенными детерминантами в отличных молекулах. Это тем более
важно принимать во внимание, что в большинстве описанных выше работ были использо-
ваны гетерологичные антисыворотки, т.е. полученные к веществам, выделенным из живот-
ных не просто других Ьидов, а относящихся к иным типам Metazoa.
Несколько более определенно в этом отношении можно говорить о данных, получен-
ных методом флуоресцентной гистохимии. По-видимому, с большой долей уверенности
можно утверждать о наличии у морских звезд, морских ежей и офиур катехоламинергиче-
ской системы, состоящей из клеток и отростков. При этом следует отметить, что морфоло-
гическая организация этой системы у Asteroidea и Ophiuroidea имеет ряд общих черт. В то
же время катехоламинергическая система, по-видимому, не обязательно свойственна пред-
ставителям всех классов Echinodermata (еще раз отмечу отрицательные результаты, полу-
ченные на кишечнике голотурии Chiridota laevis).
Вопрос о присутствии в кишечном эпителии серотонин-содержащих клеток остается
открытым. Весьма возможно, что катехоламины и серотонин могут находиться в одних и
тех же клеточных структурах. В этом случае зеленое свечение будет маскировать золоти-
сто-желтое, характерное для серотонина, препятствуя его обнаружению. В то же время из
опубликованных работ следует, что серотонин, являющийся важным компонентом нервной
системы личинок иглокожих (Bisgrove, Burke, 1986; Nakajima, 1988), у взрослых животных,
за исключением единичного сообщения (Dolder, 1975), не выявлен.
Возвращаясь к данным, полученным в иммуногистохимических экспериментах, и имея
в виду определенные ограничения этого метода, нельзя не отметить, что, тем не менее, эти
данные демонстрируют существенное разнообразие кишечных регуляторных систем как в
плане демонстрируемых иммунореактивностей у одного вида, так и вариаций между раз-
личными видами, а, тем более, классами типа иглокожих.
К сожалению, мало известно о конкретной физиологической роли тех или иных эле-
ментов кишечной регуляторной системы. Так, предполагается, что базиэпителиальное спле-
тение в целом может быть связано с регуляцией функционирования ресничного эпителия
(биения ресничек) (Cobb, Raymond, 1979). У морских звезд клетки, демонстрирующие им-
мунореактивность к РМИГамиду и/или БАЬМРамиду, предположительно обеспечивают
сокращение филаментов в эпителиальных клетках (Martinez et al., 1993). В то же время
вполне возможно, что, по крайней мере, часть регуляторных клеток в эпителии связана и с
регуляцией общеорганизменных функций.
Попробую охарактеризовать описанные элементы в кишечнике иглокожих с точки зре-
ния их принадлежности к нервной или эндокринной системе. Как я уже отмечал ранее, кри-
терии для этого в действительности крайней недостаточны и базируются в первую очередь
42
на чисто морфологических характеристиках (наличие базальных отростков у клеток, обра-
зование ими базального сплетения, и т.д.).
Катехоламинергическую, NO-синтетаза-, GABA-, PYY-, РАМ-, МСН-, FMRF-амид-
и/или ЗАЬМРамид S1-иммунореактивные системы морских звезд, очевидно, с большой
долей уверенности можно отнести к компонентам интраэпителиальной нервной системы.
Инсулин- и глюкагон-иммунореактивные клетки, скорее всего, представляют собой эндок-
риноподобные элементы. РР-иммунореактивные структуры у одних видов морских звезд
или в некоторых отделах их пищеварительной системы, по-видимому, являются нервными
элементами, тогда как в других случаях, скорее, напоминают эндокринные. Соматостатин-
иммунореактивную систему Asterias rubens трудно однозначно отнести к нервной или эн-
докринной составляющим: хотя клетки и имеют базальные отростки, образующие сплете-
ние, последнее развито крайне слабо.
В любом случае кажется логичным сделать следующий вывод— кишечная регулятор-
ная система морских звезд представляет собой гетерогенное образование, состоящее как из
нервных (причем-различной ергичности), так и из эндокринных (также связанных с образо-
ванием различных регуляторных веществ) элементов. Интересно при этом подчеркнуть, что
и нервные, и эндокринные клеточные элементы представлены клетками как открытого, так
и закрытого типов. Предполагаемая схема морфологической организации такой системы
представлена на рис. 68.
Рис. 68. Схема организации регуляторной системы в стенке
пищеварительного тракта морских звезд.
ст - соединительная ткань; цнп - целомический нервный плексус;
эк - эндокринные клетки; энп - кишечный базиэпителиальный нервный плексус.
Fig. 68. The scheme of regulatory system organization
in the digestive tract wall of sea stars.
43
Хотя данные по кишечным регуляторным системам в других классах иглокожих весь-
ма неполны, здесь также можно говорить р некоторых чертах их организации, общих с мор-
скими звездами. По крайней мере, у изученных морских ежей и голотурий в их состав вхо-
дят как нервные, так и эндокринные элементы. Лишь у офиур все выявленные кишечные
регуляторные клетки, скорее, относятся к нервной системе. Впрочем, возможно, это просто
связано с недостаточной степенью изученности этой группы иглокожих, и в действительно-
сти следует ожидать, что отмеченные выше черты организации кишечных регуляторных
систем являются общими для всего типа Echinodermata.
Глава 6
ARTHROPOD А (ЧЛЕНИСТОНОГИЕ)
Членистоногие — самый мнфгочисленный тип животного царства, включающий в себя
высших представителей первичноротых (насекомых), и уже только по этой причине пред-
ставляющий особый интерес для сравнительного анализа кишечных регуляторных систем.
В то же время степень изученности кишечных регуляторных элементов у представителей
различных систематических групп Arthropoda неодинакова. Если насекомые в этом отно-
шении исследованы достаточно полно, данные, известные мне по другим классам, крайне
скудны.
6.1. TRACHEATA (ТРАХЕЙНЫЕ)
Все известные мне данные о кишечных регуляторных элементах у представителей под-
типа Трахейные касаются класса насекомых (Insecta). Учитывая достаточно обширную ли-
тературу по этому вопросу, я рассмотрю имеющиеся сведения по отдельным таксонам этого
класса. i
Эктодермальная часть пищеварительной системы Insecta представлена трубчатой сред-
ней кишкой. У многих видов насекомых в ее начальную часть открываются слепые выпячи-
вания — пилорические выросты или придатки, эпителий которых по своему строению не-
сколько отличается от эпителия средней кишки. Гистологическая организация пищевари-
тельной системы Insecta подробно рассмотрена в ряде обзоров, в которых, в частности, от-
мечается и наличие в составе эпителиального пласта эндокринных клеток (см., например:
Chapman, 1985; Kramer, 1985; Dow, 1986).
6.1.1. Entognatha (скрыточелюстные)
В эпителии средней кишки Folsomia Candida (отр. Collembola — ногохвостки) и пред-
ставителей рода Japyx (отр. Diplura — двухвостки) присутствуют редкие клетки, по своему
строению напоминающие эндокринные. Если у Folsomia Candida они представлены одним
типом, то у Japyx описаны два типа клеток, один из которых имеет длинные отростки, тя-
нущиеся вдоль базальной мембраны. Все эти клетки относятся к открытому типу (Cassier,
Fain-Maurel, 1977). У Sinentomon erytanum (отр. Protura— бессяжковые) клетки с характер-
ными мелкими гранулами выявлены в выростах на границе между средней и задней кишкой
(Dallai et al., 1989).
45
6.1.2. Ectognatha (открыточелюстные)
Отр. Thysanura (щетинкохвостки). В средней кишке Petrobius maritinius эндокрин-
ные клетки присутствуют в составе эпителия регенерационных гнезд, в их центральной час-
ти, располагаясь обычно по 2 на 1 гнездо. Все они — закрытого типа. В клетках отмечены
многочисленные профили аппарата Гольджи, эндоплазматическая сеть преимущественно
гладкая (Cassier et al., 1972; Fain-Maurel et al., 1973). На основании строения и диаметра сек-
реторных гранул у этого вида выделены 3 типа эндокринных клеток (Cassier, Fain-Maurel,
1977). Эндокринные клетки описаны также в составе кишечного эпителия представителей
рода Machilis (Cassier, Fain-Maurel, 1977).
Отр. Blattoidea (тараканы). Кишечная регуляторная система представителей этого от-
ряда изучена, наверное, наиболее полно, причем главным объектом многих исследований
послужил американский таракан Periplaneta americana. На основании ультраструктур но го
анализа в эпителии кишки этого вида выделяют до 7 типов эндокринных клеток (табл. 2).
Таблица 2
Типы эндокринных клеток в кишечнике Periplaneta americana
(по: Nishiitsutsuji-Uwo, Endo, 1981; Endo, Nishiitsutsuji-Uwo, 1982a)
Тип клетки Структура и размер секреторных гранул Частота встречаемости клеток Примечание
1а Округлые электронноплотные, 200- 300 нм Редко Клетки закрытого типа
1b Округлые электронноплотные, 300- 400 нм Около 40% от общего коли- чества эндокринных клеток Клетки открытого и закрыто- го типов
1с Округлые гомогенные, низкой элект- ронной плотности, 200-400 нм Очень редко Клетки треугольной формы либо распластанные по ба- зальной мембране
1а+с Округлые электронноплотные (300- 400 нм) и электронносветлые (200- 400 нм), иногда промежуточной ве- личины (200-300 нм) До 15% от общего количе- ства эндокринных клеток Предположительно зрелые клетки 1b типа
Па Электронноплотные неправильной формы, 200-300 нм Около 25% от общего количества эндокринных клеток
ПЬ То же, 700-800 нм Предположительно зрелые клетки Па типа
Пс Неправильной формы и различной электронной плотности, 700-800 нм
Иммуногистохимическими методами у Periplaneta americana показано наличие клеток,
иммунореактивных к панкреатическому полипептиду (РР), энтероглюкагону, соматостати-
ну (Iwanaga et al., 1981; Nishiitsutsuji-Uwo et al., 1982), а также предположительно к серото-
нину (Nishiitsutsuji-Uwo et al., 1984). РР-подобную реактивность демонстрируют клетки с
большими электронноплотными гранулами неправильной формы, величиной 500-800 нм,
соответствующие клеткам типа Па (Endo et al., 1982). Клеток, иммунореактивных к таким
кишечным гормонам млекопитающих, как инсулин, гастрин, секретин, мотилин, нейротен-
зин и вазоактивный интестинальный полипептид (V1P), не обнаружено.
Другие исследователи выявили у этого вида клетки, реагирующие с антисыворотками к
уротензину I (Iwanaga et al., 1986), пептиду гистидин-изолейцину (PHI) (Yui et al., 1985),
аденокортикотропному гормону (ACTH), p-эндорфину, а-меланоцитстимулирующему гор-
мону (a-MSH), Zew-энкефалину, кортикотропин релизинг гормону (CRF) (Schols et al., 1987)
и холецистокинину (ССК) (Andries, Beauvillaiu, 1988). Относительно последних показано,
46
Рис. 69. Развитие и дифференцировка эндокринных клеток в эпителии средней кишки таракана
Periplaneta americana (по: Endo, Nishiitsutsuji-Uwo, 1982b, с изменениями):
/-направление дифференцировки пищеварительных клеток; 11 - направление дифференцировки эндокринных
клеток; мп - межклеточное пространство с развивающимися микроворсинками молодой эндокринной клетки;
ндк - недифференцированные (камбиальные) клетки регенерационного гнезда; эк! - дифференцированная
эндокринная клетка закрытого типа; эк2 - дифференцированная эндокринная клетка открытого типа.
Fig. 69. Development and differentiation of endocrine cells in the midgut epithelium of a cockroach
Periplaneta americana (after: Endo, Nishiitsutsuji-Uwo, 1982b, with some changes).
что ССК-подобную иммунореактивность в таких клетках демонстрируют гранулы со сред-
ним диаметром 312 нм.
Эндокринные клетки первоначально возникают как клетки закрытого типа и имеют ок-
руглую форму (рис. 69). Молодые клетки с немногочисленными гранулами лежат в составе
регенерационных гнезд. Более зрелые клетки располагаются недалеко от гнезд, где они по-
степенно приобретают треугольную (на срезах) форму; апикальная часть тянется к просве-
ту, на ее поверхности в межклеточном пространстве формируются микроворсинки. Затем
отростки достигают просвета; тем не менее, у зрелых эндокринных клеток открытого типа
микроворсинки более редкие и короткие, чем у составляющих основную часть эпителиаль-
ного пласта цилиндрических (пищеварительных). Интересно, что между микроворсинками
наблюдаются картины пиноцитоза. Следует отметить, что и на значительном удалении от
регенерационных гнезд встречаются клетки закрытого типа с большим количеством гранул
в цитоплазме и отростками, тянущимися вдоль базальной мембраны. Очевидно, это — так-
же зрелые эндокринные клетки. Экзоцитоз содержимого секреторных гранул отмечен и у
некоторых эндокринных клеток в составе регенерационных гнезд; это наблюдение предпо-
лагает, что зрелые клетки могут располагаться и здесь (Endo, Nishiitsutsuji-Uwo, 1982а, b, с).
В опытах с тимидиновой авторадиографией, проведенных на Periplaneta americana,
было показано, что РР-иммунореактивные клетки обновляются из источника, общего с пи-
щеварительными (по своей организации — всасывающе-секреторными) клетками (Endo et
al., 1983). Эти эндокринные клетки, очевидно, могут иметь различный по длительности
47
жизненный цикл, причем более продолжительный, чем у пищеварительных. Так, первые
меченые РР-иммунореактивные клетки закрытого типа на краю регенерационных гнезд бы-
ли зарегистрированы лишь на 14-е сутки после введения 3Н-тимидина, а первые меченые
клетки открытого типа — на 28-е сутки опыта. При исследовании ранних личиночных ста-
дий Periplaneta americana было обнаружено, что клетки, реагирующие с антисывороткой к
РР, появляются в кишечнике раньше, чем аналогичные по иммунореактивности элементы в
центральной нервной системе (Endo, 1984).
Проведенное мной окрашивание эпителия средней кишки Nauphoeta cinerea позволило
выявить достаточно многочисленные РМИЕамид-иммунореактивные клетки (рис. 70-71,
вклейка) преимущественно открытого типа. Для некоторых из них характерно наличие ко-
ротких базальных отростков.
Электронно-микррскопическое исследование средней кишки Nauphoeta cinerea дало
основание авторам (Cassier, Fain-Maurel, 1977) выделить 4 типа эндокринных клеток, кото-
рые обычно лежат по периферии регенерационных гнезд. Среди них есть клетки открытого
типа веретеновидной формы, открытого типа с базальными отростками и закрытого типа,
распластанные по базальной мембране.
В эпителии кишечника личинок и имаго Blaberus craniifer на основании различий в
структуре и размерах секреторных гранул описаны 10 типов эндокринных клеток (Andries,
Tramu, 1985b). Гранулы расположены главным образом в базальных частях клеток, где и
наблюдается экзоцитоз их содержимого. Следует отметить, что форма клеток одного и того
же типа может быть различной; некоторые клетки имеют базальные отростки. Иммуноги-
стохимический анализ показал наличие клеток, реагирующих с антисыворотками к met-
энкефалину, соматостатину, холецистокинину (ССК), соматолиберину, гонадолиберину,
субстанции Р и VIP. Среди последних пяти иммунореактивных элементов есть клетки от-
крытого типа. Показано, что клетки, демонстрирующие идентичную иммунореактивность,
могут иметь различную форму (Andries, Tramu, 1985b). В последнем исследовании-было
также продемонстрировано наличие в стенке кишечника под эпителием отростков (по-
видимому, нервных), реагирующих с антисыворотками к ССК, лпег-энкефалину и серото-
нину.
Наличие ССК-иммунореактивных клеток также показано у Eublaberus distanti (Andries,
Beauvillain, 1988).
Отр. Orthoptera (прямокрылые). Впервые у представителя этого отряда (саранчи
Locusta migratoria) в ходе электронно-микроскопического исследования эндокринные клет-
ки были описаны в эпителии дистальной части средней кишки (Aubry, 1975). Позднее на
основании тонкого строения и диаметра секреторных гранул у этого вида были выделены 3
типа эндокринных клеток (Cassier, Fain-Maurel, 1977).
Иммуногистохимическими методами у Locusta migratoria показано присутствие кле-
ток, реагирующих с антисывороткой к ССК. Продукт реакции был локализован над элек-
тронноплотными гранулами диаметром 100-280 нм (Andries, Beauvillain, 1988). Выявлены
также многочисленные клетки, иммунореактивные к нейропептиду Y (NPY — регулятор-
ному пептиду, состоящему из 36 аминокислотных остатков и родственному панкреатиче-
скому полипептиду). Аналогичные по иммунореактивности отростки (очевидно, нервные)
присутствуют в наружной мышечной оболочке кишечника, однако прямых контактов по-
следних с интраэпителиальными клетками не отмечено (Schoofs et al., 1988). В составе эпи-
телия средней кишки описаны клетки, иммунореактивные к эндотелину (Montuenga et al.,
1994).
48
По моим данным, в эпителии средней кишки и пилорических выростов (это — отходя-
щие от передней части кишечника и тянущиеся вдоль него трубчатые выросты) Locusta
migratoria многочисленны также клетки, реагирующие с антисывороткой к ЕМИРамиду.
Они могут быть как открытого, так и закрытого типов; у некоторых из них отмечены корот-
кие базальные отростки (рис. 72-73, вклейка). Не исключено присутствие в кишечном эпи-
телии этого вида и клеток, связанных с синтезом инсулиноподобного вещества. Так, с по-
мощью биохимических методов из средней кишки Locusta migratoria выделен инсулинопо-
добный белок (Benkhay et al., 1986). Исследования, проведенные мной совместно с
В.К.Казаковым и С.Ю.Будаевой, показали наличие у этого вида саранчи клеток открытого
(рис. 76, вклейка) и закрытого типов, иммунореактивных к инсулину горбуши.
В составе эпителия кишечника другого вида саранчи (Schistocerca gregaria) на основа-
нии строения секреторных гранул описаны 7 типов эндокринных клеток (Montuenga et al.,
1989). Их количество заметно возрастает по направлению к задней кишке (Montuenga,
Sesma, 1985). Все эндокринные клетки у этого вида— открытого типа, характеризуются
электронносветлой цитоплазмой. Иммуногистохимически показано присутствие клеток,
реагирующих с антисыворотками к бомбезину, ССК, гастрину 17, инсулину и РР; послед-
ние клетки наиболее многочисленны (Montuenga et al, 1989). Интересно, что эндокринные
клетки у этого вида присутствуют также в эпителии мальпигиевых трубочек (Alba et al.,
1983; Montuenga et al., 1989).
В эпителии кишечника сверчка Gryllus bimaculatus по данным ультраструктурного ана-
лиза присутствуют 5 типов эндокринных клеток (Cassier, Fain-Maurel, 1977). Среди них до-
вольно многочисленны клетки, иммунореактивные к уротензину 1 (Iwanaga et al., 1986), а
также к РМИРамиду (74-75, вклейка) и инсулину горбуши (рис. 77-78, вклейка) (собствен-
ные данные). Некоторым РМИРамид-иммунореактивным клеткам свойственно наличие не
только коротких, но и длинных базальных отростков. Среди инсулин-иммунореактивных
встречаются клетки как закрытого, так и открытого типов; ряд клеток располагается в со-
ставе регенерационных гнезд. Наконец, выявлены также единичные субэпителиальные ин-
сулин-иммунореактивные клетки округлой формы, без отростков.
Отр. Phasmoptera (палочники). В эпителии средней кишки Carausius morosus выяв-
лены клетки, иммунореактивные к холецистокинину (ССК). ССК-подобное вещество пред-
положительно локализовано в гранулах со средним диаметром 159 нм (Andries, Beauvillain,
1988).
Отр. Odonata (стрекозы). У личинки Aeshna суапеа на электронно-микроскопическом
уровне в эпителии средней кишки описаны 2 типа эндокринных клеток. На срезах они име-
ют треугольную форму; тонкий апикальный отросток тянется по направлению к просвету
пищеварительного тракта, однако, по-видимому, не достигает его. На базальной и лате-
ральной клеточных мембранах отмечены картины экзоцитоза содержимого гранул
(Komnick, Kukulies, 1987). В отличие от пищеварительных в цитоплазме эндокринных кле-
ток практически нет пероксисом (Faber, Komnick, 1989).
С помощью иммуногистохимических методов в эпителии кишечника личинки этого
вида выявлены клетки, иммунореактивные к целому ряду регуляторных пептидов: met-
энкефалину, ССК, субстанции Р, гонадолиберину, VIP, соматостатину, Zeu-энкефалину и
соматолиберину. Клетки, реагирующие с тремя последними антисыворотками, на всем про-
тяжении кишечника крайне редки; количество остальных возрастает по направлению к гра-
нице с задней кишкой. Форма эндокринных клеток различна; по крайней мере, часть из
них — открытого типа. Предполагается, что одна и та же клетка может реагировать с двумя
и более антисыворотками (Andries, Tramu, 1984, 1985а).
49
В ультраструктурном иммуноцитохимическом исследовании было показано, что ССК-
подобная иммунореактивность в клетках кишечника Aeshna суапеа связана с электронноп-
лотными гранулами диаметром 100-260 нм; гранулы другого размера и строения с антисы-
вороткой не реагировали (Andries, Beauvillain, 1988). Всего, согласно данным последней
работы, в эпителии средней кишки этого вида стрекоз присутствует до 7 типов эндокрин-
ных клеток.
Отр. Lepidoptera (чешуекрылые). Ультраструктурный анализ выявил эндокринные
клетки в кишечнике 7 видов гусениц и взрослых бабочек (Endo, Nishiitsutsuji-Uwo, 1981,
1982с). В целом строение этих клеток сходно у разных видов. Большая их часть — открыто-
го типа, несет на апикальной поверхности пучок микроворсинок. Аппарат Гольджи пред-
ставлен несколькими стопками цистерн, шероховатая эндоплазматическая сеть развита
сравнительно слабо. Секреторные гранулы имеют различные размеры. Большинство из них
диаметром около 100 нм (такие гранулы присутствуют в цитоплазме порядка 70% эндок-
ринных клеток), .но встречаются и более крупные: 150-250 и 300-500 нм. Как правило, они
электронноплотные, с гало. В клетках (главным образом у взрослых бабочек) могут присут-
ствовать также электронносветлые гранулы и гранулы, не полностью заполненные секре-
торным продуктом, диаметром 100-200 нм. Все эти наблюдения указывают на наличие не-
скольких типов эндокринных клеток. Интересно, что у взрослых бабочек количество таких
клеток обычно меньше, чем у гусениц (Endo, Nishiitsutsuji-Uwo, 1981).
В средней кишке гусениц Helions zea присутствуют клетки, иммунореактивные к
ЕМИЕамиду. Они, как правило, открытого типа, встречаются по всей длине кишечника,
однако более многочисленны в его начальной части, где реагируют также с антисывороткой
к панкреатическому полипептиду. ЕМИЕамид-иммунореактивные аксоны отмечены в мы-
шечной оболочке заднего отдела средней кишки (Jenkins et al., 1989).
У сытых гусениц количество ЕМЯЕамид-иммунореактивных клеток примерно в 2 раза
больше по сравнению с голодными. Радиоиммунологический анализ показал, что и количе-
ство ЕМЯЕамид-подобного вещества в экстрактах кишечника сытых животных достоверно
выше, чем у голодных, тогда как характер изменения его содержания в гемолимфе проти-
воположный. Предполагается, что имеет место эндокринный ответ на поступление пищи,
причем ЕМЯЕамид-подобное вещество, присутствующее в гемолимфе, поступает туда из
эндокринных клеток кишечника (Jenkins et al., 1989).
В кишечнике гусениц тутового шелкопряда Botnbyx mori выявлены клетки, иммуноре-
активные к пептиду гистидин-изолейцину (PHI) (Yui et al., 1985), а у яблонной плодожорки
Cydia pomonella обнаружены клетки, содержащие /ем-каллатостатин. Интересно, что 1еи-
каллатостатин присутствует также в нервах, локализованных вне эпителиальной выстилки
(Duve et al., 1997).
Отр. Coleoptera (жесткокрылые). Эндокринные клетки, расположенные в тесном
контакте с регенерационными гнездами, выявлены в кишечнике мучного хрущака Tenebrio
molitor (Cassier, Fain-Maurel, 1977). Единичные клетки, реагирующие с антисывороткой к
холецистокинину, отмечены в средней кишке жука-плавунца Dytiscus marginalis (Andries,
Beauvillain, 1988).
Отр. Siphonaptera (блохи). В средней кишке Xenopsylla chaeopis, Echidnophaga
gallinaceae и Tunga penetrans описаны редкие клетки со светлой цитоплазмой и мелкими
гранулами. Это — клетки открытого типа, со слабо развитым (в сравнении с другими эпи-
телиальными) базальным лабиринтом, многочисленными стопками цистерн аппарата Голь-
джи; они не имеют контакта с регенерационными гнездами (Reinhardt, 1976). Филимонова
(1985, 1986, 1990) отмечает, что у Xenopsylla chaeopis секреторные гранулы имеют диаметр
50
около 100 нм и характеризуются наличием электронноплотного центра и гало. Клеточные
отростки с такими же гранулами обнаружены в основании эпителиального пласта у базаль-
ной мембраны.
Отр. Hymenoptera (перепончатокрылые). Выявленные у пчелы Apis mellifera эндок-
ринные клетки встречаются на всем протяжении средней кишки, за исключением ее первой
четверти. Они возникают из регенерационных гнезд, характеризуются электронноплотной
цитоплазмой, широким основанием и узким апикальным отростком с короткими микровор-
синками на свободной поверхности. Секреторные гранулы различного строения (что дало
основание выделить 2 типа клеток) и мелкие пузырьки сконцентрированы в базальной ци-
топлазме, предполагая возможность выведения секреторных продуктов в гемолимфу (Raes,
1990). Иммуногистохимическими методами продемонстрировано наличие клеток, реаги-
рующих с соматостатиновой антисывороткой (Bounias, Dubouis, 1982).
Отр. Diptera (двукрылые). Эндокринные клетки описаны у ряда представителей этого
отряда. У личинки Chironomus streuzkei единичные мелкие клетки присутствуют в эпителии
передней части средней кишки, в той области пищеварительного тракта, где формируется
перитрофическая мембрана (Platzer-Schultz, Welsch, 1969). Эти клетки — открытого типа, с
короткими микроворсинками на апикальной поверхности и электронноплотными гранула-
ми диаметром около 100 нм.
В кишечнике мухи Calliphora erytrocephala выявленные эндокринные клетки характе-
ризуются электронноплотной цитоплазмой, обилием цистерн шероховатой эндоплазмати-
ческой сети и электронноплотными гранулами с гало диаметром около 100 нм. Клетки
имеют неправильные очертания; по крайней мере, часть из них — открытого типа. Предпо-
лагается, что эти клеточные элементы берут начало от общих для всего эпителия генера-
тивных клеток (Priester, 1971). У близкого вида (Calliphora vomitoria) в эпителии выявлены
мелкие клетки, иммунореактивные к панкреатическому полипептиду (Duve, Thorpe, 1982),
РМИБамиду (Cantera, Nassel, 1991), а также клетки, реагирующие с антисывороткой к 1еи-
каллатостатину (Duve, Thorpe, 1994). Каллатостатин-иммунореактивные клетки присутст-
вуют также в кишечнике Drosophila melanogaster (Yoon, Stay, 1995).
Ультраструктурные данные указывают на наличие эндокринных клеток в кишечнике
некоторых комаров (Hecker, 1977; Hecker et al., 1971a, b).
У взрослых особей комара Aedes aegypti описаны 2 типа эндокринных клеток с грану-
лами диаметром 60-120 нм. Основное различие между этими клетками заключается в нали-
чии или отсутствии гало у секреторных гранул. Клетки характеризуются сильным развити-
ем белок-синтезирующего аппарата, цитоплазма может иметь различную электронную
плотность. Базальный лабиринт, типичный для обычных пищеварительных клеток, отсутст-
вует. По-видимому, как и у описанных выше эндокринных клеток таракана Periplaneta
americana, микроворсинки клеток открытого типа развиваются в межклеточном простран-
стве до достижения апикальными выростами просвета кишечника. У зрелых клеток откры-
того типа микроворсинки заключены в гликокаликс, более электронноплотный, чем у пи-
щеварительных клеток. Некоторые клетки имеют базальные отростки. Всего в средней
кишке насчитывается около 500 эндокринных клеток, причем большая их часть сосредото-
чена в дистальной части кишечника, где преобладают клетки с гранулами без гало (Brown et
al, 1985).
Иммуногистохимический анализ показал наличие у Aedes aegypti клеток (общим чис-
лом около 250), реагирующих с антисыворотками к бычьему панкреатическому полипепти-
ду и БМЯРамиду, причем часть из них выявляется и теми, и другими антителами. Спустя 6
ч после питания как количество иммунореактивных клеток, так и интенсивность иммуноги-
51
стохимической реакции уменьшаются примерно в 2 раза. Это дает основание полагать, что
секреторные продукты выводятся в ответ на поступление в кишечник пищи и, возможно,
действуют на соседние пищеварительные клетки (паракриния), стимулируя выведение фер-
ментов (Brown et al., 1986). Есть данные, что все гранулы кишечных эндокринных клеток
Aedes aegypti выводятся в течение первых 24 ч после приема пищи, а очередная порция сек-
реторных продуктов синтезируется перед следующим питанием (VonWeymam, 1977; цит.
по Glattli et al., 1987).
Поскольку, как известно, поступление пищи в кишечник у кровососущих насекомых
стимулирует созревание ооцитов, можно ожидать, что этот процесс происходит при уча-
стии кишечных эндокринных клеток. Можно также предположить, что средняя кишка у
Aedes aegypti является самым большим эндокринным органом (Brown et al., 1985), так как
общее количество нейросекреторных клеток в центральной и периферической нервной сис-
теме у двукрылых составляет около 200 (Rowell, 1976).
Эндокринные клетки, иммунореактивные к бычьему панкреатическому полипептиду,
выявлены также у самок комара Anopheles stephensi; при этом, как и у Aedes aegypti, не все
эндокринные клетки кишечника реагируют с антисывороткой. Интересно, что различий в
тонком строении между клетками, реагирующими и не реагирующими с антисывороткой,
не обнаружено (Glattli et al., 1987).
6.2. BRANCHIATA (ЖАБРОДЫШАЩИЕ)
Пищеварительная система у представителей класса ракообразных (Crustacea) — един-
ственного класса подтипа Branchiata, как и у других артропод, состоит из трех отделов: пе-
редней и задней кишок, имеющих эктодермальное происхождение и обычно покрытых ку-
тикулой, и энтодермальной средней. В своей начальной части средняя кишка обычно имеет
железистые выросты. Данные, говорящие о присутствии регуляторных клеток в эпителии
средней кишки ракообразных, немногочисленны.
Наличие инсулиноподобной активности в экстрактах пищеварительной железы (желе-
зистых выростов средней кишки) Carcinus maenas и Homarus gammarus дало основание
предполагать присутствие в эпителии этого отдела клеток — аналогов [3-клеток позвоноч-
ных (Davidson et al., 1971). Вещество с глюкагоноподобным действием было выделено им-
мунохимическими методами из железы средней кишки речного рака Astacus fluviatilis
(Maier et al., 1975). С помощью радиоиммунного анализа и иммунохимии продемонстриро-
вано присутствие гастрин/холецистокинин-подобного вещества в средней кишке краба
Cancer magister (Larson, Vigna, 1983) и креветки Palaenion serratus (Favrel, Van Wormhoudt,
1986), однако конкретная тканевая локализация этих соединений пока неизвестна.
Ультраструктурные исследования показали, что у ряда видов ракообразных в основа-
нии эпителиального пласта средней кишки присутствуют отростки, по своему строению
напоминающие аксоны. Такие отростки описаны у балянусов (род Balanus) (Koulish, 1971)
и десятиногих раков: Penaeus aztecus (Talbot et al., 1972) и Pachygrapsus crassipes (Mykles,
1977).
В то же время у Decapoda (двух представителей рода Homarus, а также Cancer magister)
выявлены клетки, по строению напоминающие эндокринные (Mykles, 1979). В цитоплазме
этих клеток находятся многочисленные мелкие гранулы с материалом различной электрон-
ной плотности. Апикальные части клеток обычно расширены и содержат много пузырьков
и микротрубочек, базальные контактируют с базальной мембраной. Помимо клеток, в осно-
52
вании эпителия расположена система клеточных отростков (предположительно аксонов), в
которых, помимо мелких электронноплотных гранул, присутствуют микротрубочки и еди-
ничные митохондрии.
Интересные по своему строению (предположительно эндокринные) клетки описаны в
эпителии задних слепых выростов средней кишки бокоплава Orchestia cavimana (Graf,
1982). Они расположены в верхней половине эпителиального пласта, имеют грушевидную
форму и не контактируют с базальной мембраной (рис. 79). Апикальная поверхность несет
короткие прямые микроворсинки, между которыми отмечены многочисленные картины
пиноцитоза. Клетки характеризуются хорошим развитием белок-синтезирующего аппарата.
Рис. 79. Схема строения эндокринной клетки в эпителии задних слепых выростов
кишечника бокоплава Orchestia cavimana (из: Graf, 1982, с изменениями).
ок - основные клетки эпителия; пп - пиноцитозный пузырек; пц - пиноцитоз; ц - центриоль.
Fig. 78. Scheme of endocrine cell organization from posterior intestinal diverticuli epithelium of
Orchestia cavimana (from: Graf, 1982, with some changes).
53
Гранулы диаметром 40-120 нм с электронноплотным центром и гало, формирующиеся в
зоне аппарата Гольджи, располагаются главным образом по периферии клетки. Здесь отме-
чены картины экзоцитоза их содержимого в межклеточное пространство. Наличие в цито-
плазме значительного количества лизосом, аутофагосом, миелиноподобных и мультивези-
кулярных тел предполагает существование механизма кринофагии. Отмеченное выше от-
сутствие контакта клеток с базальной мембраной дало основание рассматривать их как па-
ракринные (Graf, 1982).
6.3. CHELICERATA (ХЕЛИЦЕРОВЫЕ)
Единственные известные мне данные относительно кишечных регуляторных элементов
у представителей подтипа хелицеровых касаются одного из видов отряда клещей (Acari) —
иксодового клеща Rhipicephalus appendiculatus. При ультраструктурном анализе в основа-
нии эпителиального пласта кишечника нимф этого вида описаны тонкие отростки с мелки-
ми гранулами, предположительно представляющие собой аксоны (Walker, Fletcher, 1987).
Подробно изучавший гистологию краснотелковых клещей (сем. Trombiculidae) Шатров
(2000) особенно подчеркивает, что не обнаружил эндокриноподобных элементов в составе
эпителия пищеварительной системы проанализированных им видов.
6.4. ЗАКЛЮЧЕНИЕ
Резюмируя приведенные выше весьма немногочисленные данные по ракообразным,
можно предположить, что, по крайней мере, у некоторых представителей этого класса мы
имеем дело с кишечной регуляторной системой, состоящей как из нервных, так и эндок-
ринных элементов. В отношении хелицеровых известно лишь о присутствии в кишечном
эпителии отростков нервных клеток.
В плане своего состава кишечная регуляторная система насекомых, в отличие от дру-
гих членистоногих, имеет ряд особенностей. Имеющиеся сведения морфологического ха-
рактера говорят о большом сходстве в строении ее элементов с кишечными эндокринными
клетками млекопитающих, а отсутствие как контакта с элементами периферической нерв-
ной системы, так и базального интраэпителиального сплетения, характерного для многих
других групп беспозвоночных (что будет показано в следующих главах), позволяют с
большой долей вероятности считать эту систему чисто эндокринной.
Как видно из приведенных выше данных, такая система присутствует как у насекомых
с различным типом питания, так и на разных стадиях их онтогенеза (личинки, imago).
Если попытаться дать общую характеристику строения эндокринных клеток Insecta,
получается следующая картина. Это— клетки как открытого, так и закрытого типов,
имеющие разнообразную форму (рис. 80), иногда с базальными отростками. На ультра-
структурном уровне клетки обычно характеризуются (за некоторыми исключениями) элек-
тронносветлой цитоплазмой, слабо (в сравнении с обычными эпителиальными клетками)
выраженным базальным лабиринтом (складчатостью базальной цитомембраны) и наличием
гранул, преимущественно локализованных в основании клеток. Клетки открытого типа не-
сут на свободной поверхности микроворсинки, но они более короткие, чем у обычных пи-
щеварительных клеток. Эти микроворсинки, по-видимому, представляют собой рецептор-
ный аппарат, и такие клеточные элементы следует считать рецепторно-секреторными. Сте-
пень развития белок-синтезирующего аппарата может быть различной, однако весьма веро-
54
А Б В Г Д E Ж 3
Рис. 80. Морфологическое разнообразие эндокринных клеток в кишечном эпителии насекомых.
А-Г - клетки открытого типа: А - цилиндрической формы; Б - веретеновидной формы; В - с расширенным основа-
нием; Г - с базальным отростком. Д-З - клетки закрытого типа: Д- распластанная по базальной мембране; Е -
треугольной формы; Ж - округлой формы; 3 - с базальным отростком.
Fig. 80. Morphological diversity of endocrine cells in the intestinal epithelium of insects.
ятно, что эта черта строения, как и количество секреторных гранул (а, может быть, в неко-
торых случаях и наличие или отсутствие контакта клеток с просветом пищеварительного
тракта) связана с определенными стадиями синтеза, накопления и выведения биологически
активных веществ, т.е. с секреторным циклом.
Большое разнообразие секреторных гранул как по их тонкому строению, так и диамет-
ру говорит о гетерогенности эндокринных клеток насекомых, причем, очевидно, количество
клеточных типов может быть весьма большим, сопоставимым с разнообразием кишечных
эндокринных элементов у млекопитающих. О последнем свидетельствуют и данные имму-
ногистохимических исследований. Если объединить все имеющиеся к настоящему времени
сведения, окажется, что в кишечном эпителии насекомых присутствуют клетки, иммуноре-
активные к панкреатическому полипептиду, соматостатину, холецистокинину, субстанции
Р, вазоактивному интестинальному полипептиду, соматолиберину, гонадолиберину, met-
энкефалину, /ем-энкефалину, пептиду гистидин-изолейцину, нейропептиду Y (NPY), инсу-
линам млекопитающих и рыб, бомбезину, эндотелину, гастрину, уротензину I, 1еи-
каллатостатину, ЕМКБамиду, энтероглюкагону и серотонину. Безусловно, в большинстве
случаев можно говорить лишь о подобных реактивностях. В действительности наблюдае-
мые реакции могут быть объяснены присутствием сходных антигенных детерминант в от-
личных соединениях. И, тем не менее, даже принимая во внимание возможность колокали-
зации разных биологически активных веществ в одних и тех же клеточных элементах, мож-
но однозначно утверждать о гетерогенности кишечной регуляторной системы насекомых в
отношении содержащихся в ней биологически активных веществ.
При наличии общих черт очевидно определенное разнообразие в организации кишеч-
ной регуляторной системы у представителей различных систематических групп насекомых.
Это касается наличия или отсутствия клеток, демонстрирующих ту или иную иммунореак-
тивность, соотношения клеток открытого и закрытого типов, распределения эндокринных
клеток по длине пищеварительного тракта. Обращают на себя внимание и определенные
различия в организации кишечной регуляторной системы у личинок и взрослых форм насе-
комых с полным превращением (например, Lepidoptera). Последнее, очевидно, в первую
очередь связано с изменением типа питания.
55
Насекомые — это практически единственная группа беспозвоночных, для которой
имеются убедительные данные об источниках развития эндокринных клеток. По крайней
мере, относительно части этих клеточных элементов можно утверждать, что они обновля-
ются за счет общих для эпителиального пласта генеративных клеток и, следовательно,
имеют энтодермальное происхождение.
Как обычно, один из самых трудных для решения вопросов — это вопрос о конкретных
функциях тех или иных эндокринных элементов. В настоящий момент, наверное, имеет
смысл говорить лишь о том, что они связаны не только с регуляцией собственно эпители-
ального пласта (а это особенно важно, поскольку данных о проникновении в эпителий
нервных отростков, как уже отмечалось, пока нет), но и играют определенную (и, наверное,
весьма большую) роль в регуляции общеорганизменных функций. В любом случае, очевид-
но, что в процентном отношении кишечные эндокринные клетки составляют значительную
часть от общего количества регуляторных элементов в организме насекомых.
Глава 7
MOLLUSCA (МОЛЛЮСКИ)
Моллюски, коТорых насчитывается свыше 100 тысяч видов, составляют второй по чис-
ленности (после членистоногих) тип многоклеточных. Этот тип подразделяют на 2 подтипа:
1) Amphineura (боконервные), в который входят 2 класса— Polyplacophora (панцирные или
хитоны) и Solenogastres (бороздчатобрюхие), и 2) Conchifera (раковинные) с классами
Monoplacophora (моноплакофоры), Scaphopoda (лопатоногие), Gastropoda (брюхоногие),
Bivalvia (двустворчатые) и Cephalopoda (головоногие). Мне известны сведения о кишечных
регуляторных клетках у представителей хитонов, двустворчатых, брюхоногих и головоно-
гих моллюсков, которые и будут приведены далее.
7.1. CEPHALOPODA (ГОЛОВОНОГИЕ)
Класс головоногих (порядка 700-800 видов) представляет собой наиболее высокоорга-
низованную группу моллюсков. Пищеварительная система (в сравнении с другими класса-
ми) в микроанатомическом плане значительно более сложна (подробнее см. Bidder, 1966).
Эктодермальное происхождение имеют задняя часть пищевода (у Decapoda — десятиногих)
и зоб (у Octopoda — восьминогих), желудок, слепой вырост (caecum), пищеварительная же-
леза с придатками протоков [так называемые «печень» («liver») и «поджелудочная железа»
(«pancreas»)], кишечник, чернильный мешок и железа чернильного мешка (Boletzky, 1978).
К сожалению, сведения о кишечных регуляторных элементах в этой интереснейшей группе
моллюсков единичны и отрывочны.
В экстрактах слепого выроста и кишечника осьминога Eledone cirrata была продемон-
стрирована инсулиноподобная иммунореактивность и описаны клетки, окрашивающиеся
альдегид-фуксином (Mehrotra, Falkmer, 1968; Davidson et al., 1971).
В эпителии средней кишки и слепого выроста другого осьминога {Octopus vulgaris) нам
удалось выявить кальцитонин-иммунореактивные клетки (рис. 81, 83-84, вклейка). В сред-
ней кишке они составляют до 1% от общего количества эпителиальных клеток, имеют на
срезах округлую или треугольную форму и довольно слабо реагируют с антисывороткой. В
основании эпителия слепого выроста лежат единичные интенсивно иммунореактивные
клетки с широким основанием и тянущейся к просвету узкой апикальной частью. У некото-
рых клеток отмечены короткие базальные отростки, однако интраэпителиального сплетения
нет. Иммунореактивные перикарионы и отростки, очевидно, принадлежащие перифериче-
ской нервной системе, хорошо заметны в стенке пищеварительного тракта. В средней киш-
ке отдельные отростки могут проникать из него в основание эпителиальной выстилки (рис.
57
82, вклейка), однако мы не наблюдали их контакта с интраэпителиальными иммунореактив-
ными клетками (Пунин, Казаков, 1992).
Интраэпителиальные ЕМИЕамид-иммунореактивные структуры у Octopus vulgaris, по-
видимому, отсутствуют. Лишь в «печени» удалось наблюдать субэпителиальные иммуноре-
активные отростки, которые, однако, не приближались к эпителию (Пунин, 1997в).
Что касается ультраструктурных данных, то только в эпителии выростов протоков пи-
щеварительной железы («поджелудочная железа») у некоторых каракатиц описаны отрост-
ки, напоминающие нервные (Schipp, Boletzky, 1976). Интересно, что в достаточно подроб-
ном электронно-микроскопическом исследовании пищеварительного тракта наутилуса
Nautilus pompilius (Westermann, Schipp, 1998) каких-либо подобных структур в эпителии не
отмечено.
7.2. BIVALVIA (ДВУСТВОРЧАТЫЕ)
Пищеварительная система двустворчатых моллюсков состоит из короткого пищевода,
желудка, в который открываются протоки массивной пищеварительной железы, и собст-
венно кишечника, обычно образующего в теле моллюска ряд петель, а в своей дистальной
части — прободающего перикард. Участок кишечника после перикарда обычно рассматри-
вается как задняя кишка, хотя по гистологическому строению он принципиально не отлича-
ется от средней кишки. В разных группах двустворчатых моллюсков наблюдаются вариан-
ты микроанатомического строения желудка и проксимального отдела кишечника: наличие
обособленного мешка кристаллического стебелька, степени развития и присутствия специа-
лизированных отделов (например, у корабельных червей) в пищеварительной железе. У
некоторых Bivalvia весь пищеварительный тракт может подвергаться значительной редук-
ции, вплоть до полного исчезновения (см. подробнее: Owen, 1966а, b; Purchon, 1978; Morton,
1983).
Относительно элементов кишечной регуляторной системы двустворчатые моллюски
изучены относительно хорошо. Я последовательно рассмотрю данные гистологических,
флуоресцентно-гистохимических и иммуногистохимических, а также электронно-
микроскопических исследований.
7.2.1. Данные гистологичеких исследований
В составе эпителия средней кишки Anodonta с использованием метода прижизненного
окрашивания метиленовым синим Гилев (1952) описал единичные клетки, которые рас-
сматривал как нервные рецепторные. Апикальные части этих клеток подходили к просвету
кишечника, а базальные истончались и в виде тонкого отростка покидали эпителиальный
пласт, вступая в состав субэпителиального нервного сплетения.
Позднее у Arctica islandica, Mytilus edulis и My a arenaria с помощью этого же метода
также были выявлены окрашивающиеся метиленовым синим клетки (рис. 85, вклейка),
весьма многочисленные и разнообразные по морфологии (Пунин, 1981 д, 1985; Пунин, Кон-
стантинова, 1988; Punin et al., 1991). В средней кишке Mytilus edulis можно выделить сле-
дующие типы клеток (рис. 86): 1) интраэпителиальные клетки открытого типа с одним или
несколькими (до 7) отростками, имеющими характерные варикозные расширения и не по-
кидающими эпителий; 2) аналогичные клетки, один из отростков которых уходит из эпите-
лия и вступает в состав субэпителиального нервного сплетения; 3) интраэпителиальные
58
Рис. 86. Регуляторные (нервные) клетки, окрашивающиеся метиленовым синим,
в пищеварительном тракте Mytilus edulis (А-Г), Arctica islandica (Д, Е) и Муа arenaria (Ж).
Сплошной линией отмечена апикальная, точками - базальная границы эпителия.
Рисунки (Д- вид на клетку сверху) с препаратов (прижизненное окрашивание метиленовым синим).
Шкала. 10 мкм.
Fig. 86. Regulatoty (nerve) cells in the digestive tract of Mytilus edulis (А-Г),
Arctica islandica (Д, E) and Mya arenaria (Ж) revealed by vital staining with methylene blue.
59
клетки с погруженной в подстилающие эпителий ткани базальной частью; 4) клетки, пери-
карионы которых расположены субэпителиально, а один из отростков проникает в эпителий
и тянется к просвету кишечника. Отростки клеток формируют базиэпителиальное сплете-
ние, местами очень хорошо развитое. «Капризность» использованного метода не позволяет
провести точный подсчет количества таких клеток, однако в удачно прокрасившихся участ-
ках эпителия (по ориентировочным оценкам) они могут составлять до 3% от общего коли-
чества клеток. Что касается их распространения по пищеварительной системе, то у Arctica
islandica окрашивающиеся метиленовым синим клетки наблюдались во всех ее отделах за
исключением пищеварительной железы (Пунин, 1981 д).
Альдегид-фуксин, широко применяемый для окрашивания 0-клеток в островках подже-
лудочной железы млекопитающих, выявляет в средней кишке двустворчатых моллюсков
фуксин-положительные клетки, которые обычно рассматривались как синтезирующие инсу-
лин-подобное вещество (Fritsch et al., 1976; Казаков, 1977). Важно отметить, что введение
Unio pictorum и Anodonta cygnea глюкозы вызывает значительные дегенеративные изменения
в морфологии фуксинофильных клеток: дегрануляцию цитоплазмы, пикноз ядер, а иногда и
полное их разрушение. Достоверно уменьшается также и общее количество таких клеток.
Предполагается, что эти изменения могут указывать на участие фуксинофильных клеток в
регуляции метаболизма углеводов (Казаков, 1977; Плисецкая, 1977; Plisetskaya et al., 1978).
В эпителии кишечника Mytilus galloprovincialis описаны скопления («островки») ок-
руглых клеток (Crasium et al., 1981; Madar et al., 1981). Авторы выделяют в их составе две
разновидности клеточных элементов, связанные с продуцированием соответственно инсу-
линоподобного и глюкагоноподобного веществ. Однако приводимые в этих работах доказа-
тельства не представляются мне убедительными. Судя по опубликованным микрофотогра-
фиям, это, в действительности, — скопления амебоцитов, которые, как было показано ранее
(см.: Narain, 1975), часто мигрируют в эпителиальный пласт.
7.2.2. Данные флуоресцентной гистохимии и иммуногистохимии
Клетки, содержащие моноамины. В начальном отделе средней кишки Mytilus edulis
использование флуоресцентно-гистохимического метода FAGLU позволило выявить клетки
и отростки, содержащие катехоламины и серотонин (Пунин, Константинова, 1988, 1989;
Макаренкова и др., 1988; Konstantinova, Punin, 1990; Punin et aL, 1990). Катехоламинсодер-
жащие структуры представлены главным образом клетками и отростками, расположенными
в составе субэпителиального сплетения (рис. 87, вклейка). Лишь единичные клетки, демон-
стрирующие типичную для катехоламинов зеленую флуоресценцию, лежат интраэпители-
ально. Базальные части этих клеток могут покидать эпителий и образовывать отростки в
субэпителиальном сплетении (рис. 88, вклейка). Клетки с серотонин-подобным веществом,
демонстрирующие золотисто-желтое свечение, лежат интраэпителиально. Они более мно-
гочисленны, могут быть отнесены к открытому типу. Их базальные отростки образуют ин-
траэпителиальное сплетение. Последнее и субэпителиальное сплетение, содержащее кате-
холамины, часто располагаются параллельно и в непосредственной близости друг от друга
(рис. 87, вклейка).
Иммуногистохимический анализ показал присутствие в дистальной части кишечника
(в районе перикарда) Mytilus edulis двух компактных групп интраэпителиальных серотонин-
иммунореактивных клеток открытого типа, лежащих по краям от тифлозоля, а также субэ-
пителиальных нервных стволов и отдельных отростков (Пунин и др., 1988; Punin et al.,
1991).
60
Инсулин-йммунореактивные клетки. Инсулиноподобная активность экстрактов пи-
щеварительного тракта Муа arenaria была обнаружена еще в 1923 г. (Collip, 1923). Иммуно-
гистохимическими методами присутствие в эпителии пищеварительного тракта инсулин-
иммунореактивных клеток (использовались антисыворотки к инсулину млекопитающих)
было впервые показано у Mytilus edulis (Fritsch et al., 1976), а также Unio pictorum и
Anodonta cygnea (Казаков, 1977). Эти клетки могут быть отнесены к открытому типу; они
лежат в эпителии поодиночке; в своем распространении ограничены участком средней
кишки, проходящей через пищеварительную железу (т.е. начальным отделом кишечника).
Цитоплазма выявляемых клеток дает положительную реакцию с ШИК-альциановым синим
и альдегид-фуксином {см. предыдущий раздел), однако фуксинофильных клеток в этом слу-
чае заметно больше, чем иммунореактивных. Как было показано, инсулин-иммуно-
реактивные клетки не реагируют с антисыворотками к гастрину, пентагастрину, глюкагону
и каерулину (Fritsch et al., 1976).
Позднее иммунореактивные клетки были выявлены и при использовании антисыворо-
ток, выработанных к инсулиноподобному веществу, выделенному из двустворчатого мол-
люска Anodonta (Русаков, Казаков, 1979; Казаков, 1996). У Arctica islandica некоторые из
таких клеток имеют причудливую форму и образуют интраэпителиальные базальные отро-
стки, которые удается проследить на значительном расстоянии (рис. 89). Многие иммуно-
реактивные клетки Муа arenaria и Mytilus edulis соответствуют типичным элементам от-
крытого типа (рис. 90, вклейка), хотя встречаются клетки и закрытого типа.
Кальцитонин-иммунореактивные клетки. Кальцитонин-иммунореактивные клетки
и отростки были выявлены с помощью антисыворотки к С-концевому фрагменту кальцито-
нина лосося в кишечнике Mytilus edulis, Муа arenaria, Arctica islandica и Unio pictorum (рис.
91-94, вклейка) (Пунин, Казаков, 19896, 19906; Пунин и др., 1990а). У разных видов эти
клетки различаются как по морфологии, так и по частоте встречаемости и распространению
по пищеварительному тракту. У Mytilus edulis кальцитонин-иммунореактивные клетки ред-
ки и присутствуют главным образом в эпителии большого тифлозоля начального отдела
средней кишки. Они относятся к открытому типу, часто на базальном конце имеют неболь-
шие терминальные расширения, иногда образуют варикозные отростки, отходящие от око-
лоядерной области (рис. 91, вклейка). Редкие иммунореактивные отростки присутствуют
также под эпителием.
Рис. 89. Клетка, иммунореактивная
к инсулиноподобному веществу Anodonta,
в эпителии средней кишки Arctica islandica.
Рисунок с препарата (реконструкция по серии срезов).
Шкала: 10 мкм.
Fig. 89. Cell immunoreactive \.о Anodonta insulin-like
peptide in the midgut epithelium of Arctica islandica.
61
Рис. 95. Полиморфизм кальцитонин-иммунореактивных клеток в эпителии кишечника
Arctica islandica.
Сплошной линией отмечена апикальная, точками - базальная границы эпителия, стрелкой - отросток,
покидающий эпителиальный пласт. Рисунки с препаратов. Шкала: 10 мкм.
Fig. 95. Polymorphous calcitonin-immunoreactive cells in the intestinal epithelium
of Arctica islandica.
У Arctica islandica интраэпителиальные кальцитонин-иммунореактивные клетки в на-
чальной части средней кишки значительно более многочисленны и разнообразны по форме
(рис. 95). Некоторые из таких клеток встречаются в эпителии изолированного мешка кри-
сталлического стебелька (рис. 93, вклейка'). Отмечены клетки открытого типа как без отро-
стков, так и с базальными отростками (в том числе уходящими из эпителия) и клетки за-
крытого типа. Оба типа клеток присутствуют в эпителии средней кишки Муа arenaria,
клетки открытого типа — у Uniopictorum (Пунин и др., 1990а).
Нейротензин-иммунореактивные клетки и отростки. Интраэпителиальные нейро-
тензин-иммунореактивные клетки выявлены в пищеварительном тракте Mytilus edulis и
Unio pictorum (рис. 96-97, вклейка) (Punin, Kazakov, 1991а, b, с; Пунин и др., 19926). У ми-
дии редкие клетки располагаются в начальном отделе средней кишки в составе эпителия
большого тифлозоля и примыкающего к нему участка эпителия мешка кристаллического
стебелька. Все клетки — открытого типа, веретеновидной формы либо с несколько расши-
ренной базальной частью. У клеток в эпителии мешка кристаллического стебелька с анти-
сывороткой реагирует главным образом надъядерная и апикальная цитоплазма (рис. 96,
вклейка). Сходные по морфологии с последними иммунореактивные клетки есть и в эпите-
лии задней кишки.
62
У Unio pictorum нейротензин-иммунореактивные клетки встречаются на всем протяже-
нии пищеварительного тракта за исключением пищеварительной железы. В желудке они
расположены в эпителии его вентральной поверхности; в начальном отделе кишечника — в
эпителии кишечного желобка, главным образом со стороны тифлозоля (рис. 98).
Рис. 98. Локализация нейротензин-иммунореактивных клеток
в различных участках кишечника Unio pictorum.
Поперечные срезы: А - начального отдела средней кишки, Б - собственно средней кишки,
В - задней кишки, окж- основная часть кишечного желобка, скж - складчатая часть кишечного желобка,
т - тифлозоль начального отдела средней кишки, тз - тифлозоль задней кишки.
Черным отмечены иммунореактивные клетки.
Fig. 98. Localization of neurotensin-immunorective cells in different parts of Unio pictorum intestine.
Большинство клеток имеет расширенную, интенсивно окрашивающуюся базальную
часть и узкий, подходящий к просвету апикальный отросток (рис. 97, вклейка)', в то же вре-
мя некоторые из этих клеток могут быть, по-видимому, и закрытого типа. Цитоплазма,
главным образом апикальная, у части иммунореактивных клеток в средней кишке слабо
окрашивается альциановым синим. Ни интраэпителиальных отростков, ни субэпителиаль-
ных иммунореактивных структур не обнаружено.
НАР [пептид-активатор (роста) головы гидры]-иммунореактивные структуры.
Пищеварительная система Mytilus edulis демонстрирует очень своеобразный характер рас-
пределения клеток, иммунореактивных к пептиду-активатору роста головы гидры (рис. 99).
(Подробнее о данном пептиде см. главу 14). Такие клетки отсутствуют в составе эпителия
желудка и большей части проксимального отдела средней кишки. Только ближе к концу
этого отдела первые (единичные) клетки появляются в зоне эпителия кишечного желобка
рядом с большим тифлозолем (Пунин и др., 19926).
В начале собственно средней кишки с НАР-антисывороткой реагируют практически
все эпителиальные клетки (важно подчеркнуть, что эта реакция специфична), неокрашен-
63
кж
Рис. 99. Локализация клеток, иммунореактивных к пептиду-активатору роста (головы) гидры,
в кишечнике Mytilus edulis.
Поперечные срезы: А - начального отдела средней кишки, Б - начальную, В - центральную и
Г - дистальную части собственно средней кишки, и Д- заднюю кишку, бт - большой тифлозоль;
кж - кишечный желобок; мт - малый тифлозоль; тс - тифлозоль средней кишки.
Черным отмечены иммунореактивные клетки.
Fig. 99. Localization of Hydra head activator peptide-immunoreactive cells in the intestine of
Mytilus edulis.
ной остается лишь небольшая группа клеток в желобке тифлозоля. Более того, с антисыво-
роткой часто реагирует и содержимое просвета кишечника. Очевидно, окрашивание (по
крайней мере, большей части эпителия) может быть объяснено наличием в клетках каких-то
иных пептидов, имеющих, однако, сходные с НАР антигенные детерминанты.
Окрашивание антисывороткой обычных эпителиальных клеток, очевидно, маскирует
специализированные клеточные элементы. Последние становятся заметными в дистальной
части кишечника, где цитоплазма эпителиальных клеток уже не вступает в реакцию с анти-
сывороткой. Эти клетки— открытого типа, достаточно многочисленны, имеют различную
форму (веретеновидную, цилиндрическую, грушевидную). Цитоплазма реагирует с антисы-
вороткой интенсивно; на апикальной поверхности эпителия часто видны картины, напоми-
64
нающие выведение иммунореактивного секрета в просвет кишечника. Характерной чертой,
отличающей данные клеточные элементы от остальных эпителиальных, является наличие
отростков, формирующих хорошо заметное базиэпителиальное сплетение (рис. 100,
вклейка).
В задней кишке снова появляются слабо реагирующие с антисывороткой клетки, рас-
положенные группами. Кроме них, в основании эпителия присутствуют иммунореактивные
продольно ориентированные пучки отростков, а также интенсивно окрашивающиеся клетки
открытого типа. Они имеют расширенное основание, обычно лишены базальных отростков;
центральная и особенно апикальная цитоплазма часто окрашивается также альциановым
синим.
Редкие НАР-иммунореактивные биполярные клетки (рис. 101, вклейка) и единичные
тонкие отростки, расположенные субэпителиально, встречаются на всем протяжении пище-
варительного тракта мидии. Не исключено, что субэпителиальные отростки могут контак-
тировать с интраэпителиальным сплетением.
У Unio pictorutn НАР-иммунореактивные клетки (рис. 102, вклейка) встречаются в на-
чальной части средней кишки и в задней кишке. В средней кишке все клетки расположены в
эпителии основной части кишечного желобка со стороны, противоположной тифлозолю
(рис. 103). Они довольно многочисленны, имеют расширенную базальную часть и узкий
апикальный отросток. Базальных отростков нет. В задней кишке единичные иммунореак-
Рис. 103. Локализация клеток, иммунореактивных к пептиду-активатору (роста) головы гидры,
в кишечнике Unio pictorum.
Поперечные срезы: А - начального отдела средней кишки, Б - задней кишки.
окж - основная часть кишечного желобка; скж - складчатая часть кишечного желобка;
т - тифлозоль начального отдела средней кишки, тз - тифлозоль задней кишки.
Черным отмечены иммунореактивные клетки.
Fig. 103. Localization of Hydra head activator peptide-immunoreactive cells in the intestine
of Unio pictorum.
65
тивные клетки лежат в складках по краям от тифлозоля. Их базальные части слабо окраши-
ваются антисывороткой, а апикальные— альциановым синим. Субэпителиальных НАР-
иммунореактивных структур не обнаружено (Пунин и др., 19926).
ЕМИГамид-иммунореактивные клетки. Из 6 видов {Mytilus edulis, Musculus
laevigator, Unio pictorum, Hiatella arctica, My a arenaria, Macoma baltica), лишь у первых
трех были обнаружены интраэпителиальные иммунореактивные структуры. У Масота
baltica единичные окрашивающиеся субэпителиальные отростки были отмечены в некото-
рых участках пищеварительной железы и желудка.
Редкие интенсивно окрашивающиеся интраэпителиальные клетки наблюдались у
Mytilus edulis в желудке: в районах отхождения протоков пищеварительной железы (рис.
104, вклейка) и в зоне заходящего из кишечника большого тифлозоля. По размерам они та-
кие же, как обычные эпителиальные, веретеновидной формы, иногда имеют базальные от-
ростки. В начальном отделе кишечника единичные иммунореактивные клетки лежат в эпи-
телии мешка кристаллического стебелька, рядом с большим тифлозолем, а многочисленные
отростки, ориентировайные преимущественно вдоль кишечной трубки, пронизывают осно-
вание эпителиальной стенки тифлозоля. Дистальнее в средней кишке редкие клетки и отро-
стки встречаются по всему периметру кишечной трубки. Субэпителиальных иммунореак-
тивных элементов в кишечнике нет.
У Unio puctorum редкие иммунореактивные клетки — без базальных отростков, в ос-
новном открытого типа (рис. 105, вклейка), расположены в эпителии желудка и кишечном
желобке начального отдела средней кишки. Интраэпителиальные отростки, реагировавшие
с антисывороткой, наблюдались только в одном из участков стенки желудка, где был отме-
чен их контакт с рыхлым субэпителиальным сплетением (рис. 106, вклейка). В средней
кишке субэпителиальные иммунореактивные отростки отсутствовали. Единичные клетки,
по-видимому, закрытого типа, иногда имеющие короткие базальные отростки, обнаружены
в эпителии желудка Musculus laevigator (рис. 107, вклейка). Иммунореактивное сплетение в
этой части пищеварительного тракта расположено только субэпителиально. В эпителии
кишечника отмечены лишь единичные отростки (Пунин, 1997в).
Добавлю, что в пищеварительном тракте еще у одного вида двустворчатых моллюсков
(Placopecten magellanicus) отмечены субэпителиальные отростки, реагировавшие с этой
антисывороткой, однако интраэпителиальных иммунореактивных структур нет (Too, Croll,
1995).
Клетки, демонстрирующие другие иммунореактивности. В эпителии трубочек пи-
щеварительной железы Pecten maximus описаны клетки, иммунореактивные к соматостати-
ну (LeRoux et al., 1987), однако маловероятно, что в данном случае удалось выявить дейст-
вительно соматостатин-подобное вещество. Эпителий трубочек пищеварительной железы
двустворчатых моллюсков, помимо недифференцированных клеток, состоит из высокоспе-
циализированных секреторных и пищеварительных (см. обзор: Пунин, 1991). В данном
случае окрашивание наблюдалось в цитоплазме последних и, очевидно, может быть объяс-
нено присутствием в пищеварительных клетках иного вещества, случайно имеющего сход-
ные антигенные детерминанты.
Это же замечание справедливо и для описанных в пищеварительной железе Mi-
zuchopecten yessoensis /еи-энкефалин-иммунореактивных клеток (Вараксин и др., 1990).
Иммунореактивность в этом случае наблюдалась только в надъядерной цитоплазме секре-
торных клеток, где по данным электронно-микроскопических работ расположен аппарат
Гольджи и молодые секреторные гранулы (см.: Sumner, 1966; Pal, 1971 и др.).
66
7.2.3. Данные электронно-микроскопических исследований
В разных отделах пищеварительного тракта ряда видов двустворчатых моллюсков опи-
саны расположенные в основании эпителиального пласта отростки, содержащие мелкие
электронноплотные гранулы. Эти отростки обычно рассматриваются как нервные. Они от-
мечены в кишечнике Mytilus galloprovincialis (Giusti, 1970), Tapes watlingi (Dougan, McLean,
1970) и ряда других видов (Jegla, Greenberg, 1968; Judd, 1979). У Arctica islandica отростки c
гранулами диаметром около 100 нм наблюдались во всех отделах пищеварительного тракта,
за исключением эпителия дистальных протоков и собственно трубочек пищеварительной
железы (Пунин. 1981а, д; Пунин, Филимонова, 1986). Обычно такие отростки группируются
по несколько вместе; иногда встречаются пучки, насчитывающие многие десятки таких от-
ростков (в качестве примера можно привести тифлозоль средней кишки Mytilus edulis — см.
рис. 108, вклейка). Часто отростки с мелкими гранулами и их пучки сопровождаются отро-
стками с крупными (400-550 нм в диаметре) зернистыми гранулами, по-видимому, принад-
лежащими глиальным клеткам, расположенным под эпителием. Наблюдались картины пе-
рехода небольших групп отростков из эпителия в субэпителиальные ткани, или наоборот
(Пунин, 1981д).
В эпителии кишечника Mytilus edulis отростки характеризуются значительным поли-
морфизмом гранул (рис. 109-111, вклейка). Можно выделить три основных типа гранул: 1)
электронноплотные с гало, 2) зернистые умеренной электронной плотности и 3) электрон-
носветлые; их размеры приведены в табл. 3. Соотношение гранул разных типов, как и их
общее количество, могут существенно меняться в разных отростках; в некоторых присутст-
вуют только гранулы одного типа. Кроме гранул, в отростках встречаются мелкие пузырь-
ки, продольно ориентированные микротрубочки и микрофиламенты, иногда небольшие
митохондрии, вакуоли с различным содержимым, узкие каналы с периодическими расши-
рениями, миелиноподобные структуры. Синаптических контактов ни с эпителиальными
клетками, ни с другими отростками не отмечено. Хотя в участках соприкосновения отрост-
ков друг с другом часто встречались скопления гранул, изменения толщины цели между
мембранами не наблюдалось. Зарегистрированы картины, которые можно интерпретиро-
вать как экзоцитоз электронносветлых гранул или пузырьков (рис. 112, вклейка) (Пунин,
1989; Konstantinova, Punin, 1990).
Таблица 3
Размеры гранул в интра- и субэпителиальиых отростках
и интраэпителиальных перикарионах в средней кишке Mytilus edulis
Структура Диаметр гранул, нм
электронноплотные умеренной электрон- ной плотности электронносветлые
Интраэпителиальные отростки 126±2 147±2 142±2
Субэпителиальные отростки 163±3 209±4 202±2
Интраэпителиальные перика- рионы 140±2 159±4 149±3
Под эпителием также присутствуют отростки с полиморфными гранулами, имеющими,
однако, несколько другие размеры (табл. 3). Их пучки иногда сопровождают отростки с
крупными (диаметром около 500 нм) зернистыми гранулами.
67
Кроме отростков, в эпителии выявлены также клетки характерного строения. В средней
кишке Mytilus edulis описаны клетки, содержащие в цитоплазме (главным образом в базаль-
ных частях) гранулы диаметром от 200 до 400 нм с электронноплотным центром и узким
либо широким гало. Белок-синтезирующий аппарат представлен главным образом свобод-
ными рибосомами, цистерн шероховатой эндоплазматической сети мало. Подходящие к
просвету кишечника узкие апикальные части клеток несут микроворсинки (Fritsch, Sprang,
1977). Авторы полагали, что, возможно, именно эти клетки реагировали с антисывороткой к
инсулину млекопитающих (см. предыдущий раздел этой главы).
У этого же вида я наблюдал, по крайней мере, два типа клеток. Первый (мелкограну-
лярные) характеризуется наличием небольших полиморфных гранул, близких по размерам
в уже описанных выше отростках (см. табл. 3) и многочисленными стопками цистерн аппа-
рата Гольджи (рис. 113-114, вклейка). Клетки второго типа отличаются заметно менее раз-
витым белок-синтезирующим аппаратом и также полиморфными, но несколько более круп-
ными (200-300 нм.в диаметре) гранулами (рис. 115-116, вклейка). Интересно отметить, что
отростков с гранулами такой величины я не наблюдал.
У Unio pictorum в качестве возможных элементов, синтезирующих инсулиноподобное
вещество, были описаны клетки также с более крупными (250-350 нм в диаметре) гранула-
ми без гало (Plisetskaya et al., 1978).
В задней кишке Mizuchopecten yessoensis отмечены интраэпителиальные клетки с элек-
тронноплотными гранулами диаметром 170-220 нм (Ушева, 1983). Есть указание на нали-
чие двух типов «эндокриноподобных» (мелкогранулярных и базальных клеток) у этого вида
в эпителии желудка и средней кишки (Ушева, 1988), однако подробно их тонкое строение и
отличия между собой автором, к сожалению, не описываются.
В эпителии средней кишки Arctica islandica удалось наблюдать клетки с большим ко-
личеством (главным образом в околоядерной цитоплазме) многочисленных мелких (диа-
метром 89± 10 нм) гранул с электронноплотным содержимым и пузырьков диаметром около
50 нм (рис. 117, вклейка). Ядра имеют глубокие инвагинации ядерной оболочки. Хорошее
развитие получает шероховатая эндоплазматическая сеть, многочисленны и свободные ри-
босомы. Аппарат Гольджи диктиосомного типа: на некоторых срезах вокруг ядра удается
насчитать до 11 небольших стопок цистерн. Многочисленны микротрубочки и микрофила-
менты. В качестве вариантов наблюдались клетки с расширенными цистернами эндоплаз-
матической сети и/или более электронноплотной цитоплазмой. Возможно, такие модифика-
ции связаны с существованием у этого типа клеток определенного секреторного цикла, ли-
бо могут быть объяснены старением клеток. В отходящих от клеток отростках присутству-
ют гранулы (часто даже более многочисленные, чем в перикарионах), микротрубочки и
микрофиламенты, иногда цистерны шероховатой эндоплазматической сети и митохондрии.
В целом тонкое строение отростков этих клеток в многом соответствует таковому отрост-
ков базиэпителиального сплетения (Пунин, 1981д, 1991). Предполагаемая схема строения
интраэпителиальной мелкогранулярной клетки представлена на рис. 118.
Рис. 118. Схема строения мелкогранулярной клетки в эпителии кишечника Arctica islandica.
ао — апикальный отросток погруженной мелкогранулярной клетки; бк - бокаловидная (слизистая) клетка;
кгк- крупногранулярная (глиальная?) клетка; пмк- погруженная мелкогранулярная клетка;
окгк - отростки крупногранулярных глиальных клеток; омк - отростки мелкогранулярных клеток.
Fig. 118. The scheme of organization of small granular cell in the Arctica islandica intestinal epithelium.
68
69
7.3. GASTROPODA (БРЮХОНОГИЕ)
Брюхоногие представляют собой самый многочисленный класс среди моллюсков
(свыше 85 тысяч видов). Пищеварительная система у представителей этого класса принци-
пиально сходна с таковой Bivalvia; основные отличия касаются, в первую очередь, услож-
нения строения передней, эктодермальной части пищеварительной трубки.
Сведения, касающиеся регуляторных элементов в пищеварительном тракте Gastropoda,
в сравнении с двустворчатыми не столь многочисленны.
7.3.1. Данные гистологических исследований
Обнаружение инсулиноподобной активности в экстрактах различных отделов пищева-
рительного тракта переднежаберного (Prosobranchia) моллюска Buccinum undatum дало ос-
нование искать в составе эпителия клетки— возможные продуценты инсулиноподобных
веществ. Действительно, при использовании альдегид-фуксина и некоторых других краси-
телей удалось выявить клеточные элементы, по некоторым тинкториальным и гистохими-
ческим свойствам соответствовавшие Р-клеткам позвоночных животных (Mehrotra, Falkmer,
1968; Davidson etal., 1971; Falkmer et al., 1973).
В пищеводе у представителей родов Umax, Arion, Helix и Limnaea (все— Pulmonata
или легочные) описаны довольно многочисленные, погруженные в соединительную ткань
клетки, рассматриваемые в качестве нервных. Они имеют отростки, которые проникают в
эпителий и подходят к просвету; на апикальной поверхности отростки несут реснички
(Лавров, 1941).
В эпителии пищеварительной железы голожаберного моллюска Dendronotus
arborescence (Opistobranchia или заднежаберные) обнаружены очень редко встречающиеся
своеобразные клетки. На полутонких срезах у них наблюдается отросток, тянущийся вдоль
базальной мембраны и, возможно, покидающий эпителий (Обухова, 1982).
7.3.2. Данные иммуногистохимических исследований
Основные результаты на брюхоногих моллюсках получены с использованием антисы-
вороток к FMRFaMH/iy и кальцитонину. Впервые интраэпителиальные РМЯРамид-имму-
нореактивные клетки были выявлены в желудке и кишечнике легочного моллюска Lymnaea
stagnalis. Часть из них по форме и размерам соответствовала обычным эпителиальным, дру-
гие (предположительно нейроны) имели округлую форму и были заметно крупнее (Schot,
Boer, 1982).
С помощью этой антисыворотки мной был изучен пищеварительный тракт 8 видов
гастропод (Пунин, 1997в). У всех исследованных моллюсков обнаружены иммунореактив-
ные клетки, однако проанализированные виды заметно различались как по морфологии
этих клеток, так и по частоте их встречаемости и распространению по пищеварительной
системе.
У Testudinalis tesselata (переднежаберные моллюски) интраэпителиальные иммуноре-
активные клетки и отростки присутствовали в желудке и по всей длине средней кишки.
Клетки (как открытого, так и закрытого типов) по размерам не отличаются от обычных эпи-
телиальных. В желудке их отростки образуют базиэпителиальное сплетение, а в средней
кишке тянутся преимущественно вдоль кишечной трубки. Под эпителием иммунореактив-
ных элементов не обнаружено. У другого переднежаберного моллюска (Littorina sax at dis')
70
все выявленные FMRFaM ид-иммунореактивные элементы локализованы в эпителии и стен-
ке желудка в районе отхождения протоков пищеварительной железы. Интраэпителиальные
клетки, как открытого, так и закрытого (рис. 119-120, вклейка) типов, здесь довольно мно-
гочисленны, а по размерам примерно такие же, как и другие эпителиальные.
Отростки клеток в местах их локализации образуют хорошо выраженное интраэпите-
лиальное сплетение (рис. 119, вклейка). Под эпителием также встречаются единичные им-
мунореактивные отростки. Не исключено, что они могут контактировать с интраэпители-
альным сплетением.
У изученных голожаберных моллюсков (Nudibranchia, Opistobranchia - заднежаберные)
интраэпителиальные РМЯРамид-иммунореактивные элементы редки и отмечены не у всех
видов, хотя и у Onchidoris muricata, и у Palio dubia, и у представителей рода Coryphella в
желудке присутствуют субэпителиальные отростки и перикарионы. У Onchidoris muricata
единичные иммунореактивные отростки отмечены в пищеварительной железе, где они тя-
нутся вдоль базальной границы эпителия, однако, по-видимому, в него не проникают. У
Palio dubia в этом отделе пищеварительной системы субэпителиальные отростки многочис-
ленны, некоторые из них проникают в эпителий, оканчиваясь между базальными частями
пищеварительных и секреторных клеток (рис. 121, вклейка). У этого же вида было зарегист-
рировано несколько иммунореактивных перикарионов, частично расположенных в основа-
нии эпителиального пласта, частично — субэпителиально (рис. 122, вклейка).
У другого заднежаберного моллюска (Clione limacina) интраэпителиальные FMRF-
амид-иммунореактивные клетки и отростки, напротив, многочисленны, встречаются в же-
лудке, пищеварительных выростах и средней кишке (рис. 123-125, вклейка). Все они — за-
крытого типа, округлой формы, крупнее обычных эпителиальных (в диметре около 20 мкм),
с большими ядрами. В желудке такие клетки иногда лежат небольшими группами, по 2-3. В
пищеварительных выростах клетки локализованы только в их проксимальных частях (т.е.
рядом с желудком); здесь также присутствуют и более мелкие (диаметром порядка 10 мкм)
иммунореактивные перикарионы. Отростки клеток образуют мощное единое интраэпители-
альное сплетение (рис. 124-125, вклейка), не имеющее, однако, контакта с более рыхлым
субэпителиальным.
Самое большое разнообразие РМИГамид-иммунореактивных клеток наблюдается у 2
видов, относящихся к легочным моллюскам (Pulmonata) — Planorbarius corneus и Lymnaea
stagnalis.
В желудке Planorbarius corneus присутствуют как интра-, так и субэпителиальные
РМИРамид-иммунореактивные клетки. Последние обычно крупные (диаметр перикарионов
порядка 30-40 мкм), их отростки могут проникать в эпителий. Интраэпителиальные клетки
могут быть как открытого типа, с расширенной базальной частью и узким апикальным от-
ростком (составляют основную часть иммунореактивных элементов), так и округлой формы
закрытого типа (диаметр перикарионов от 10 до 25 мкм). В некоторых участках эпители-
альной выстилки они очень многочисленны (рис. 126, вклейка) и своими базальными отро-
стками формируют мощное сплетение, в других — единичны.
Единичные мелкие клетки открытого и закрытого типов присутствуют и в эпителии
пищеварительной железы (рис. 127-128, вклейка). Хотя в железе у этих клеток наблюдают-
ся тонкие короткие отростки, базального сплетения здесь нет. Некоторые клетки частично
погружены в подстилающие эпителий ткани. В эпителии средней кишки ЕМИГамид-
иммунореактивных клеток, по сравнению с желудком, заметно меньше; слабее выражено и
базиэпителиальное сплетение. Помимо довольно крупных реагирующих с антисывороткой
71
округлых клеток закрытого типа, наблюдались и сходные по размерам неокрашивающиеся
перикарионы с подходящими к ним иммунореактивными отростками (рис. 129, вклейка).
У Lymnaea stagnalis все клетки были обнаружены только в средней кишке (рис. 130—
132, вклейка). Интраэпителиальные иммунореактивные клетки можно разделить по разме-
рам на крупные и мелкие, и (дополнительно) на клетки открытого и закрытого типов. Ос-
новную массу составляют крупные клетки открытого типа, с расширенным основанием и
узким апикальным отростком. У многих клеток имеются базальные отростки, формирую-
щие интраэпителиальное сплетение, связанное с несколько более слабо развитым субэпите-
лиальным (рис. 131, вклейка). Некоторые субэпителиальные клетки также посылают свои
отростки в эпителий (рис. 132, вклейка) (Пунин, 1997в).
Интересно, что еще у одного легочного моллюска {Helix aspersa) интраэпителиальных
иммунореактивных элементов в пищеварительном тракте не обнаружено, хотя субэпители-
альные отростки присутствуют (Lehman, Price, 1987; Marchand, Colard, 1991).
Отмечу, что попытки выявить интраэпителиальные регуляторные элементы у Helix
aspersa с использованием антисывороток к инсулину, гастрину, холецистокинину, глюкаго-
ну, соматостатину, вазоактивному интестинальному полипептиду (VIP), бомбезину, met-
энкефалину, субстанщж Р, секретину, панкреатическому полипептиду и серотонину не да-
ли положительных результатов (Abaurrea, Vazquez, 1979; Alba et al., 1988).
В эпителии средней кишки Lymnaea stagnalis удалось продемонстрировать наличие
кальцитонин-иммунореактивных клеток. В начальной части кишечника это— единичные
клетки открытого типа, такие же по размерам, как и обычные эпителиальные (рис. 133,
вклейка). По направлении к задней кишке появляются клетки закрытого типа, более круп-
ные, округлой или неправильной формы (рис. 134-135, вклейка). Их цитоплазма реагируете
антисывороткой с различной интенсивностью. Здесь также удается наблюдать редкие ин-
траэпителиальные иммунореактивные отростки (Пунин, Казаков, 1992).
Наши (совместно с В.К.Казаковым) эксперименты позволили выявить в желудке и ки-
шечнике Lymnaea stagnalis клетки, реагирующие с антисывороткой к инсулиноподобному
веществу Anodonta (рис. 136, вклейка). Эти клетки могут занимать как интра-, так и субэпи-
телиальное положение и характеризуются различными размерами. В то же время использо-
вание антисыворотки С-фрагменту инсулиноподобного вещества (MIP С), выделенного из
этого же брюхоногого моллюска (подробнее об этом пептиде см.: Smit et aL, 1988; Meester
et al., 1992), при окрашивании пищеварительного тракта не дало положительных резуль-
татов.
В кишечнике Aplisia californica отмечены субэпителиально расположенные нервные
отростки, иммунореактивные к панкреатическому полипептиду, уротензину I и пептиду
гистидин-изолейцину (PHI) (Yui et al., 1985) В то же время аналогичные по иммунореактив-
ности отростки выявлены и в основании эпителия (Fujita et al., 1988). Проникающие из су-
бэпителиального плексуса в основание эпителия отростки, реагирующие с антисывороткой
к нейропептиду F (NPF), описаны в кишечнике Helix aspersa (Leung et al., 1994).
В заключение этого раздела отмечу, что есть все основания ожидать наличие в пище-
варительном тракте брюхоногих моллюсков интраэпителиальных структур, демонстри-
рующих и иные (помимо описанных выше) иммунореактивности. Так, биохимическими и
иммунохимическими методами в тканях пищеварительного тракта Aplisia californica и Otala
lactea показано присутствие трех молекулярных форм гастрина (Straus et al., 1975), гаст-
рин/холецистокинин-подобного вещества в кишечнике у первого вида (Larson, Vigna, 1983)
и соматостатин-подобного вещества в пищеварительной железе Helix aspersa (Asaka et al.,
1987). Однако конкретная тканевая локализация этих соединений пока неясна.
72
7.3.3. Данные электронно-микроскопических исследований
В средней кишке Buccinum undatum среди всасывающих, зимогенных и бокаловидных
клеток встречаются клетки с электронноплотными гранулами диаметром 150-200 нм. В
таких клеточных элементах хорошо развиты шероховатая эндоплазматическая сеть и аппа-
рат Гольджи. Апикальные части подходят к просвету, но не имеют микроворсинок (Boquist
et al., 1971).
У Patella vulgata в дистальной части кишечника описаны специализированные базаль-
ные клетки, предположительно эндокринные. Они характеризуются присутствием в цито-
плазме крупных гранул диаметром 400-600 нм. Белок-синтезирующий аппарат развит сла-
бо, митохондрий немного. Своими основаниями такие клетки часто лежат на стволиках из
отростков (рассматриваемых как нервные) с мелкими пузырьками диаметром 40-50 нм и
электронноплотными гранулами диаметром 75-100 нм. Сходные по строению отростки
встречаются и субэпителиально. Плазматические мембраны клеток и отростков тесно при-
лежат друг к другу и разделены пространством порядка 10-12 нм (Bush, 1988).
У Helix aspersa интраэпителиальные эндокриноподобные клетки, выявленные в ходе
электронно-микроскопического исследования, редки и локализованы главным образом в
начальной и центральной частях средней кишки. Клетки характеризуются в целом слабым
развитием органоидов. Секреторные гранулы со средним диаметром 112 нм имеют содер-
жимое различной электронной плотности. Количество таких гранул в разных клетках варь-
ирует. По мнению исследователей (Alba et al., 1988), все обнаруженные клетки могут быть
отнесены к одному типу.
Как и у Patella vulgata, многие клетки своими базальными отростками контактируют с
интраэпителиальными пучками отростков (рассматриваемых как нервные), в которых при-
сутствуют электронносветлые пузырьки диаметром 50-70 нм и два типа гранул: с содержи-
мым умеренной электронной плотности диаметром 100-125 нм и более крупные (170-225
нм) электронноплотные.
Кроме чисто интраэпителиальных эндокриноподобных клеток в стенке кишечника
Helix aspersa присутствуют так называемые «нейроэндокринные комплексы» (рис. 137)
(Alba et al., 1989). Это морфологические образования из нескольких эндокриноподобных
клеток и окружающих их отростков двух типов: уже описанных нервных и предположи-
тельно глиальных, с электронноплотными гранулами диаметром порядка 500 нм. Снаружи
такое скопление клеток и отростков ограничено мембраной. Синаптических контактов в
нейроэндокринных комплексах не обнаружено, однако отмечался экзоцитоз гранул на по-
верхности отростков, обращенных к клеткам.
Некоторые эндокриноподобные клетки в составе таких образований, в свою очередь,
имеют отростки. Наблюдаемое в цитоплазме большое количество аутофагических вакуолей,
находящихся рядом с гранулами, предполагает существование в этих клетках механизма
кринофагии. Часть нейроэндокринных комплексов расположена в стенке кишечника на
значительном удалении от эпителия и полностью изолирована от него. Другие связаны с
эпителиальным пластом отростками эндокринных клеток, а окружающая эту структуру
мембрана переходит в базальную мембрану эпителия (рис. 137) (Alba et al., 1989).
Наконец, в эпителии протоков и трубочек пищеварительной железы Biomphalaria
pfeifferi описаны отростки двух типов— с электронноплотными гранулами диаметром 80-
140 нм и с гранулами диаметром 300-600 нм (Meuleman, 1972).
73
Рис. 137. Эндокринная клетка и «нейроэндокринный комплекс» в кишечнике Helix aspersa
(по: Alba et al., 1988, 1989).
огк - отростки глиальных клеток; онк - отростки нервных клеток.
Fig. 137. Endocrine cell and «neuroendocrine complex» in the intestine of Helix aspersa
(according to: Alba et al., 1988, 1989).
74
7.4. POLYPLACOPHORA (ХИТОНЫ)
Этот класс (другое название — Loricata, панцирные) насчитывает около тысячи видов.
Пищеварительная система у хитонов начинается эктодермальными по происхождению
глоткой и пищеводом; последний переходит в эктодермальную среднюю кишку. Начальный
отдел средней кишки обычно расширен, образуя мешковидный желудок, в который откры-
ваются протоки массивной двулопастной пищеварительной железы. Собственно средняя
кишка длинная, образует в теле животного многочисленные изгибы. Пищеварительный
тракт завершается короткой задней кишкой.
Иммуногистохимическим методом с использованием антисыворотки к ЕМЯГамиду
мне удалось продемонстрировать наличие иммунореактивных клеток в пищеварительном
тракте у 2 видов хитонов — Ishnochiton albus и Tonicella marmorea (Пунин, 1997в).
У первого вида довольно многочисленные ГМИГамид-иммунореактивные клетки при-
сутствуют в эпителии желудка и передней части кишечника (рис. 138, вклейка). Они распо-
лагаются в основании эпителиального пласта поодиночке и могут быть отнесены к закры-
тому типу. Обычно клетки имеют округлую форму (диаметром около 7-8 мкм) и базальные
отростки. В основании эпителия присутствует иммунореактивное сплетение, в одних участ-
ках — едва заметное, в других — получающее хорошее развитие. В эпителии выростов пи-
щеварительной железы также встречаются редкие иммунореактивные клетки как закрытого,
так и открытого типов (рис. 139-140, вклейка), меньших размеров, чем обычные эпители-
альные, часто имеющие тонкие базальные отростки.
У Tonicella marmorea интраэпителиальные ЕМЯГамид-иммунореактивные клетки еди-
ничны и отмечены только в желудке (рис. 141, вклейка) и пищеварительной железе.
Иммунореактивные клетки и отростки у обоих видов присутствуют также под эпители-
ем в стенке пищеварительного тракта. У Ishnochiton albus отростки могут располагаться
очень близко к базиэпителиальному сплетению, что не исключает возможности контакта
между ними, а у Tonicella marmorea отмечено проникновение отдельных отростков в осно-
вание эпителиального пласта (Пунин, 1997в).
Еще у одного вида хитонов (Lepidopleurus asellus) интраэпителиальные ЕМЯГамид-
иммунореактивные клетки и отростки в большом количестве отмечены в пищеводе и задней
кишке (т.е. эктодермальной части пищеварительного тракта), однако в желудке, пищевари-
тельной железе или средней кишке выявлены не были (Moroz et al., 1994).
7.5. ЗАКЛЮЧЕНИЕ
Как уже отмечалось, степень изученности регуляторных клеток в разных группах мол-
люсков различна, что затрудняет проведение серьезных сравнений между классами. Так,
относительно хитонов и головоногих моллюсков имеющиеся сведения позволяют говорить
лишь о существовании в эпителии тех или иных отделов пищеварительного тракта элемен-
тов регуляторной системы. На основании используемых мной критериев оценки интраэпи-
телиальные кальцитонин-иммунореактивные клетки Octopus vulgaris, скорее всего, — эн-
докринные (эндокриноподобные), а различные ЕМКЕамид-иммунореактивные элементы у
хитонов могут быть как эндокринными, так и нервными.
Кишечные регуляторные системы двустворчатых и брюхоногих моллюсков демонст-
рируют значительную сложность своей организации. Если взять за основу данные флуорес-
центной гистохимии и иммуногистохимии, то оказывается, что у двустворчатых моллюсков
в эпителии присутствуют два типа клеток, содержащих моноамины (серотонин и катехола-
75
мины), а также клетки, иммунореактивные к инсулину млекопитающих, инсулиноподобно-
му веществу Anodonta, кальцитонину, нейротензину, FMRF-амиду, пептиду-активатору
(роста) головы гидры (НАР) и, возможно, некоторым другим биологически активным со-
единениям. Данные электронно-микроскопических работ подтверждают полиморфизм ин-
траэпителиальных регуляторных клеток. Можно говорить, по крайней мере, о двух клеточ-
ных типах, один из которых по своему тонкому строению очень похож на нервные (или
нейросекреторные, учитывая наличие в цитоплазме секреторных гранул), а второй более
соответствует эндокринным клеткам. Подобная картина сложной организации системы на-
блюдается и у гастропод (с некоторой поправкой на то, что данные иммуногистохимии в
этом классе моллюсков не столь многочисленны).
Одной из важных черт организации регуляторной системы брюхоногих и двустворча-
тых моллюсков является наличие хорошо развитого базиэпителиального сплетения. Осно-
вываясь на тонком строении образующих его отростков, можно считать, что это — главным
образом интраэпителиальный нервный плексус. Очевидно, что у разных видов и в разных
отделах пищеварительного тракта он может быть образован отростками, демонстрирующи-
ми различную иммунореактивность и, следовательно, содержащими различные биологиче-
ски активные вещества. Важно отметить, что в состав этого плексуса, очевидно, входят и
отростки эндокринных клеток, а эндокринные клетки, не имеющие отростков, практически
всегда находятся в контакте с этим плексусом. Таким образом, и нервные, и эндокринные
элементы связаны (по крайней мере, структурно) в единую систему.
Имеющиеся к настоящему времени ультраструктурные данные, отрицающие (или, по
крайней мере, не показывающие) присутствие в интраэпителиальном плексусе синаптиче-
ских контактов, предполагают тем самым, что биологически активные вещества, содержа-
щиеся в гранулах и пузырьках, выводятся непосредственно в межклеточные пространства и
оказывают локальное действие (т.е. имеет место паракриния). В то же время вполне воз-
можно, что продуцируемые регуляторными клетками соединения могут диффундировать из
эпителия, проникать в лакуны незамкнутой кровеносной системы и проявлять свое дейст-
вие дистантно.
Обращает внимание и характерная для двустворчатых и брюхоногих моллюсков хоро-
шо развитая связь между интраэпителиальным сплетением и субэпителиальным нервным
плексусом, а также наличие клеток, частично или целиком (имеются в виду перикарионы)
погруженных в подстилающие эпителий ткани.
При общем сходстве строения регуляторной системы очевидны частные различия меж-
ду видами не только из разных классов моллюсков, но и принадлежащими к одному классу.
Это проявляется и в различном распределении однотипных по проявляемой иммунореак-
тивности клеток по отделам пищеварительной системы, и в их общем количестве, и в мор-
фологии самих клеток (открытый или закрытый типы, размеры, наличие или отсутствие
базальных отростков). В качестве иллюстрации последнего замечания на рис. 142 представ-
лено разнообразие морфологических типов регуляторных клеток на примере двустворчатых
моллюсков, а на рис. 143 — разнообразие клеток у представителей разных классов моллю-
сков, сходных по проявляемой ими иммунореактивности (на примере РМЯРамида).
Еще одной интересной чертой является то, что аналогичные по демонстрируемой им-
мунореактивности клетки у некоторых брюхоногих и двустворчатых моллюсков по морфо-
логическим критериям в одних отделах пищеварительного тракта соответствуют нервным
элементам, а в других - эндокринным. Я уже отмечал, что в реакцию с антисывороткой мо-
гут вступать различные вещества, если только они имеют отдельные сходные антигенные
детерминанты. Поэтому такие наблюдения можно объяснить двумя способами: либо при-
76
А Б ВГДЕЖЗ И
Рис. 142. Морфологическое разнообразие регуляторных клеток
в кишечном эпителии двустворчатых моллюсков:
А - погруженная в субэпитслиальные ткани клетка; Б - клетка с погруженной в субэпителиальные ткани
базальной частью; В-Д - интраэпителиальные клетки открытого типа с отростками;
Е,Ж- интраэпителиальные клетки открытого типа без отростков; 3 - клетка закрытого типа с отростками;
И - клетка закрытого типа без отростков.
Fig. 142. Morphological diversity of regulatory cells in the intestinal epithelium of bivalve molluscs.
сутствием одного и того же биологически активного вещества в клетках разных (нервной,
эндокринной) систем, либо тем, что антисыворотка в действительности выявляет различные
соединения.
Интересной особенностью, описанной пока только у одного вида брюхоногих моллю-
сков {Helix aspersa) является существование своеобразных «нейроэндокринных комплек-
сов». С определенными допущениями эти образования можно рассматривать как процесс
перехода от диффузной эндокринной системы к образованию обособленных эндокринных
(нейроэндокринных) желез и их ухода из состава эпителиального пласта.
Еще одной характерной чертой, свойственной легочным брюхоногим моллюскам, яв-
ляется присутствие в эпителиальной выстилке и стенке пищеварительного тракта крупных
клеток: по-видимому, полиплоидных нейронов или нейросекреторных клеток. Интересно,
что и ганглии у этой группы Gastropoda в значительной степени состоят из очень крупных
нейросекреторных клеток.
К сожалению, говорить о конкретных функциях тех или иных клеток кишечной регу-
ляторной системы на основании имеющихся данных очень трудно, однако об одной из воз-
можных функций у двустворчатых моллюсков я хотел бы упомянуть. Пищеварительный
тракт у этих животных не имеет самостоятельной мышечной оболочки, поэтому перемеще-
ние содержимого тракта происходит за счет занимающего большую поверхность эпители-
альной выстилки ресничного эпителия. Соответственно работа ресничного аппарата сосед-
них клеток требует исключительной согласованности, что, возможно, и обеспечивает осо-
бенно хорошо развитая в таких участках эпителия (сортировочные зоны желудка, кишеч-
ник) система клеток и отростков. В связи с этим интересно отметить, что и у брюхоногого
Clione limacina практически все ЕМИЕамид-иммунореактивные клетки и сплетение отрост-
ков располагаются в участках ресничного эпителия (желудок и средняя кишка).
77
Рис. 143. Разнообразие интра- и субэпителиальных РМИРамид-иммунореактивных структур
и их взаимоотношений в пищеварительном тракте панцирных (А - Ishnochiton albus),
двустворчатых (Б - Mytilus edulis, В - Unio pictorum, Г- Musculus laevigator)
и брюхоногих (Д - Testudinalis tesselata, Е - Palio dubia,
Ж - Clione limacina, 3 - Lymnaea stagnalis) моллюсков.
Fig. 143. Diversity of FMRFamide-immunoreactive sub- and intraepithelial structures
and their relationships in the digestive tract of Loricata (A - Ishnochiton albus),
Bivalvia (Б - Mytilus edulis, В - Unio pictorum, Г - Musculus laevigator)
and Gastropoda (Д - Testudinalis tesselata, E - Palio dubia,
Ж - Clione limacina, 3 - Lymnaea stagnalis).
И, наоборот, лишь у некоторых видов брюхоногих и панцирных моллюсков единичные
регуляторные клетки обнаружены в трубочках пищеварительной железы, функция которой
связана главным образом с внутриклеточным пищеварением и, частично, секрецией фер-
ментов для полостного пищеварения в желудке.
В целом можно заключить, что, несмотря на неполные данные и возможные ошибки
относительно природы отдельных выявляемых типов клеток, кишечная регуляторная сис-
тема (по крайней мере, двустворчатых и брюхоногих моллюсков) образована и эндокрин-
ными, и нервными элементами.
Глава 8
ANNELIDA (КОЛЬЧАТЫЕ ЧЕРВИ)
Кольчатые черви занимают одно из ключевых положений в системе животного царст-
ва, так как по современным представлениям они располагаются в основании двух основных
ветвей первичноротых — членистоногих и моллюсков. К этому типу беспозвоночных отно-
сят 3 класса— полихеты (Polychaeta), олигохеты (Oligochaeta) и пиявки (Hirudinea). Иногда
из класса полихет в качестве самостоятельного класса выделяют архианнелид
(Archiannelida).
8.1. POLYCHAETA (МНОГОЩЕТИНКОВЫЕ ЧЕРВИ, ИЛИ ПОЛИХЕТЫ)
Полихеты представляют собой самый многочисленный класс аннелид. Пищеваритель-
ный тракт у этой группы кольчатых червей в микроанатомическом плане организован отно-
сительно просто. В его составе выделяют передний, средний и задний отделы, из которых
первый и последний имеют эктодермальное происхождение. У большинства бродячих по-
лихет (п/кл. Errantia) центральная (энтодермальная) часть пищеварительной системы пред-
ставлена тонкостенной трубкой. Только у представителей сем. Aphroditidae и Myzostomidae
от кишечника отходят парные боковые выросты. У большинства видов сидячих полихет
(п/кл. Sedentaria) в начале энтодермальной части кишечной трубки выделяют желудок (бо-
лее широкий, с сильно развитой мышечной стенкой, иногда с внеклеточной структурой на
поверхности эпителия) и собственно среднюю кишку (Jenniaux,1969; Michel, Devillez, 1978).
Иногда на основании деталей гистологического строения желудок и кишечник в свою оче-
редь подразделяют на отделы (Ullman, Bookhout, 1949; Dales, 1955; Pilgrim, 1965 и др.).
Сведения по гистологической организации кишечного эпителия полихет можно найти в
ряде обзоров (Michel, DeVillez, 1978; Michel, 1988; Кагановская, 1989; Пунин, 1991 и др.).
В отношении кишечных регуляторных элементов эта группа кольчатых червей изучена
сравнительно хорошо. Я последовательно рассмотрю данные гистологических исследова-
ний на светооптическом уровне, а также результаты иммуногистохимических и электронно-
микроскопических работ.
8.1.1. Данные гистологических исследований
Еще при описании общей гистологии кишечного эпителия некоторые исследователи
отмечали присутствие в основании эпителиального пласта клеток округлой формы, не кон-
тактировавших с просветом кишечника (Ullman, Bookhout, 1949; Dales, 1955), однако рас-
сматривали их как «замещающие», т.е. обеспечивающие обновление дифференцированных
79
клеток (Dales, 1955). Наши исследования показали, что такие клетки, очевидно, разнородны
(вопрос об их соответствии камбиальным элементам я рассмотрю позднее). Так, у
Harmathoe imbricata, Lepidonotus squamatus и Nereis virens при окраске метиловым зеленым-
пиронином в цитоплазме некоторых клеток выделяются участки повышенной концентрации
РНК (рис. 144, вклейка), обычно свойственные зонам расположения шероховатой эндоплаз-
матической сети; иногда интенсивно окрашивается вся цитоплазма (Пунин, 1981 в; Пунин,
Лукьянова, 19846). Подобный полиморфизм базальных (названных так по положению в
эпителиальном пласте) клеток наблюдается и на полутонких срезах. У части клеток заметна
сходная картина окрашивания цитоплазмы толуидиновым или метиленовым синим, тогда
как у других цитоплазма остается практически неокрашенной, однако в ней видны мелкие
гранулы (Пунин, 1981 в; Punin, Kazakov, 1991а) (рис. 145, вклейка). В процентном отноше-
нии у афродитид базальные клетки в собственно средней кишке составляют порядка 3,5%
от общего количества эпителиальных клеток, а соотношение клеток с повышенной концен-
трацией РНК и обычных примерно 1:2 (Пунин, 1981 в). У Nereis virens количество базаль-
ных клеток возрастает от‘7-10% в проксимальной части средней кишки до 19-22% в дис-
тальной (Пунин, Лукьянова, 19846). Интересно, что некоторые базальные клетки Lepi-
donotus squamatus избирательно включают 3Н-глюкозу (рис. 146, вклейка), что, очевидно,
указывает на некоторые особенности их синтетической деятельности в сравнении с осталь-
ными базальными клетками и основным эпителием (Заварзин, Пунин, 1981).
Отростки базальных клеток тянутся вдоль базальной мембраны (не покидая эпители-
ального пласта) на значительные расстояния и часто объединяются в пучки. У Nereis virens
отмечена заметная концентрация отростков и базальных клеток в тех участках эпителиаль-
ной выстилки, где под эпителием лежат кровеносные синусы кишечного сплетения (Пунин,
Лукьянова, 19846) (рис. 147-148, вклейка). Сходная концентрация «замещающих» клеток у
синусов ранее была отмечена у Amphitrite johnstoni (сем. Terebellidae) (Dales, 1955).
У Hermodice carunculata (сем. Amphinomidae) описаны интраэпителиальные клетки,
рассматриваемые в качестве нервных. Они имеют вытянутую форму, их апикальные части
подходят к просвету, а базальные суживаются в одиночный отросток, вступающий в состав
расположенного в основании эпителия сплетения волокон. Это базальное сплетение автор
(Marsden, 1963) считает нервным плексусом.
Наличие в основании эпителия средней кишки нервного сплетения с редкими нервны-
ми клетками отмечал в своей сводке по полихетам еще Ливанов (1940), однако, к сожале-
нию, не приводил подробного описания и не указывал конкретных работ.
Использование метода прижизненного окрашивания метиленовым синим (в прошлом
классического метода для изучения нервных клеток) позволило выявить в эпителии кишеч-
ника Nereis virens единичные клетки (рис. 149, вклейка). В зависимости от высоты эпители-
ального пласта их форма меняется от цилиндрической до веретеновидной (рис. 150), однако
все такие клетки — открытого типа. У одних клеток от базальной части отходят отростки, у
других вся базальная часть сильно истончается. Однако ни базального сплетения, ни картин
ухода отростков за пределы эпителия не было отмечено. Интересно, что у афродитид и си-
дячей полихеты Arenicola marina (сем. Arenicolidae) результаты окрашивания метиленовым
синим были отрицательными, однако это может быть связано не с отсутствием в эпителии
подобных клеток, а с определенными трудностями в применении этого метода (Пунин,
1981д, 1985).
80
Рис. 150. Нервные клетки в эпителии средней кишки Nereis virens.
Прижизненное окрашивание метиленовым синим. Рисунки с препаратов. Шкала: 10 мкм.
Fig. 150. Nerve cells in the midgut epithelium of Nereis virens.
8.1.2. Данные флуоресцентной гистохимии и иммуногистохимии
Наличие в эпителии кишечника клеток и отростков, содержащих серотонин, впервые
было продемонстрировано у Nephtys caeca (сем. Nephtyidae) (Clark, 1966). Эти клеточные
элементы были локализованы в основании эпителиального пласта и рассматривались авто-
ром как нервные.
С использованием флуоресцентно-гистохимического метода FAGLU нами была прове-
дена попытка выявления моноамин-содержащих клеток у ряда полихет, относящихся к раз-
личным семействам: Harmathoe imbricata, Lepidonotus squamatus (сем. Aphroditidae),
Arenicola marina (сем. Arenicolidae), Nereis virens (сем. Nereidae), Scoloplos armiger (сем.
Aricidae), Amphitrite cirrata, Pista maculata (сем. Terebellidae). (Punin et al., 1990; Пунин,
1999). Лишь у первых 3 видов удалось выявить интраэпителиальные клетки, содержащие
биогенные амины. На основании демонстрируемой ими специфической зеленой флуорес-
ценции можно предположить, что подобные клетки у изученных афродитид содержат кате-
холамины. Они очень редки (в среднем 1 на 4-5 поперечных срезов кишечника, т.е. на не-
сколько тысяч эпителиальных клеток), лежат в основании эпителиального пласта и могут
быть отнесены к закрытому типу. Клетки имеют округлую или овальную форму, иногда
вытянуты вдоль базальной мембраны (рис. 151-152, вклейка). От перикарионов могут отхо-
дить тонкие, сравнительно короткие отростки, однако какое-либо заметное сплетение в ос-
новании эпителия отсутствует. Эти катехоламин-содержащие структуры наблюдались лишь
в собственно средней кишке и отсутствовали в эпителии латеральных выростов и связы-
вающих их с кишечником ресничных протоках.
81
У Arenicola marina все выявленные клетки демонстрировали золотисто-желтое свече-
ние, характерное для серотонина (рис. 153, вклейка). Большинство подобных клеток было
обнаружено в эпителии средней кишки, тогда как в желудке (расширенная начальная часть
кишечника) они крайне редки. [Подробно общая гистологическая организация пищевари-
тельного тракта у этого вида описана в серии работ Кагановской (1976, 1978а, б, 1979)]. Все
серотонин-содержащие клетки располагались в составе эпителия железисто-всасывающей
(желудок) и всасывающей (кишечник) зон, но не в ресничном эпителии продольных желоб-
ков. Подавляющее большинство таких клеток может быть отнесено к элементам закрытого
типа, хотя отдельные (найденные в собственно кишечнике) имеют вытянутую форму и ори-
ентированы перпендикулярно базальной мембране. Однако контакта их апикальных частей
с просветом наблюдать не удалось. Местами в основании эпителия отмечены рыхлые ло-
кальные сплетения золотисто-желтых отростков (Пунин, 1999).
Исследование пищеварительного тракта Nereis pelagica, Nereis virens и Lepidonotus
squamatus с использованием антисыворотки к серотонину показало следующее.
У Lepidonotus squamatus в эпителии собственно средней кишки (но не латеральных вы-
ростов) выявлены иммунореактивные клетки закрытого типа (рис. 154-156, вклейка) и от-
ходящие от них отростки (Пунин, Казаков, 1988, 1989а; Punin, Kazakov, 1991а). Нетрудно
заметить, что относительно результатов флуоресцентно-гистохимического и иммуногисто-
химического методов, примененных на этом виде полихет, имеются определенные расхож-
дения. Возможно, их можно объяснить различной чувствительностью использованных ме-
тодов.
У нереисов интраэпителиальных клеток не обнаружено, однако в мышечной стенке
средней кишки выявлена система волокон и единичных отростков с варикозными расшире-
ниями (рис. 157, вклейка). Ряд отростков из этого сплетения направляется к эпителию, но
заканчивается вблизи его, не прободая базальной мембраны. И лишь единичные отростки
проникают в эпителий (рис. 158-159, вклейка), оканчиваясь между основаниями секретор-
ных и всасывающих клеток (Пунин, Казаков, 1989а; Punin, Kazakov, 1991а).
Кишечник Nereis virens и Arenicola marina был изучен нами с использованием антисы-
воротки к нейротензину (Пунин, Казаков, 1990а; Пунин и др., 19906). Если у первого вида
интраэпителиальных иммунореактивных структур обнаружить не удалось, то у Arenicola
marina были зарегистрированы редкие клетки (менее 1% от общего количества эпителиаль-
ных клеток). Все нейротензин-иммунореактивные клетки расположены в основании эпите-
лиального пласта, ниже слоя ядер основных эпителиальных клеток (рис. 160-161, вклейка),
преимущественно в участках всасывающего эпителия на границе с ресничными зонами. В
этих участках клетки могут располагаться небольшими группами (рис. 162, вклейка). В же-
лудке подобные клетки единичны и отмечены только в составе эпителия вентрального Со-
образного ресничного канала. Все нейротензин-иммунореактивные клетки относятся к за-
крытому типу, лежат на базальной мембране и часто имеют неправильную, лопастную
форму. У многих клеток видны отростки, тянущиеся на значительные расстояния и местами
образующие локальные сплетения (рис. 160-161, вклейка).
Выявление РМИРамид-иммунореактивных клеток было проведено на 7 видах полихет
(уже упоминавшиеся Harmathoe imbricata, Lepidonotus squamatus, Nereis pelagica, Arenicola
marina, Amphitrite cirrata, а также Eteone longa (сем. Phyllodocidae) и Polydora quadrilobata
(сем. Spionidae) (Пунин, 19976). Лишь у Nereis pelagica не были обнаружены интраэпители-
альные иммунореактивные структуры. У других изученных полихет в эпителии кишечника
были выявлены клетки, содержащие РМИРамид-подобное вещество (рис. 163-164, 166,
вклейка). Независимо от вида червей, практически все клетки относятся к элементам закры-
82
того типа, и только у Harmathoe imbricata была зарегистрирована клетка с апикальным от-
ростком, тянущимся к просвету (рис. 163, вклейка). Перикарионы небольшие, округлой или
овальной формы; в последнем случае они вытянуты вдоль базальной мембраны (рис. 166,
вклейка). Клетки располагаются в эпителии по одиночке и лишь у Harmathoe imbricata и
Arenicola marina иногда по 2-3 вместе в небольших скоплениях базальных клеток. Особо
подчеркну, что большинство базальных клеток не реагирует с антителами к РМИГамиду
(рис. 164-165, вклейка).
От перикарионов обычно отходят один или несколько отростков с варикозными рас-
ширениями. Одни отростки тянутся по направлению к соседним базальным клеткам и окан-
чиваются на них, причем, как правило, не реагирующих с антисывороткой (рис. 167-169,
вклейка)-, другие заканчиваются между основаниями обычных эпителиальных клеток. Ино-
гда отростки образуют рыхлые локальные базиэпителиальные сплетения (рис. 170, вклейка),
не имеющие между собой видимой связи. У некоторых видов изученных полихет, помимо
интраэпителиальных иммунореактивных отростков, встречаются и субэпителиальные (рис.
171-172, вклейка), однако их связи с друг с другом я не наблюдал (Пунин, 19976). Предпо-
лагаемая схема организации кишечной РМИРамид-иммунореактивной системы на примере
Harmathoe imbricata показана на рис. 173.
Распределение РМИРамид-иммунореактивных интраэпителиальных клеток по пищева-
рительному тракту у исследованных полихет несколько различается. У Amphitrite cirrata
Рис. 173. РМКРамид-иммунореактивная система базальных клеток в эпителии средней кишки
Harmathoe imbricata.
обк- отростки базальных клеток. Иммунореактивные клетки отмечены черным.
Высота эпителия уменьшена, соеднительнотканная и мышечная оболочки не показаны.
Fig. 173. FMRFamide-immunoreactive system of basal cells in the midgut epithelium
of Harmathoe imbricata.
83
такие клетки встречаются по всей длине средней кишки, с некоторым преобладанием в ее
центральной части. У Eteone longa все клетки были выявлены в первой половине кишечни-
ка, тогда как у Polidora quadrilobata — главным образом в задней части средней кишки. У
афродитид РМИРамид-иммунореактивные клетки встречаются только в эпителии собствен-
но средней кишки (но не железистых выростов и протоков), а у Arenicola marina - главным
образом в желудке (Пунин, 19976).
Наконец, в кишечнике Nereis diversicolor отмечено присутствие интраэпителиальных
клеток (с тонкими отростками, тянущимися к просвету тракта), демонстрирующих иммуно-
реактивность к эндотелину (биологически активный пептид, состоящий из 21 аминокислот-
ного остатка) (Kasuya et al., 1990).
8.1.3. Данные электронно-микроскопических исследований
Проведенные электронно-микроскопические исследования подтвердили как наличие у
полихет кишечных регуляторных клеток, так и их разнообразие. У Harmathoe imbricata,
Lepidonotus squamatus и Nereis virens на основании тонкого строения можно выделить, по
крайней мере, 2 типа базальных клеток. Клетки первого типа, названные нами мелкограну-
лярными, в первую очередь характеризуются наличием в цитоплазме электронноплотных
гранул диаметром 90-110 нм у Harmathoe imbricata и 80-90 нм у Nereis virens (максималь-
ный диаметр до 130-150 нм, минимальный — 60 нм). Клетки второго типа— крупнограну-
лярные— имеют (как следует из названия) более крупные гранулы: округлые диаметром
300-400 нм (редко до 500 нм) у Harmathoe imbricata, и примерно такой же величины, но чуть
вытянутой формы — у Nereis virens (Пунин, 1981 в; Пунин, Лукьянова, 19846).
Мелкогранулярные клетки (рис. 174-175, 181, вклейка) имеют ядра сложных очерта-
ний, часто с глубокими инвагинациями ядерной оболочки. Белок-синтезирующий аппарат
развит, как правило, хорошо, цистерны шероховатой эндоплазматической сети сконцентри-
рованы в одном или нескольких участках цитоплазмы. Очевидно, эти клетки соответствуют
выявляемым на светооптическом уровне клеткам с пиронинофильными участками цито-
плазмы. Аппарат Гольджи расположен около ядра, окружен мелкими гранулами и много-
численными пузырьками. У Nereis virens он представлен несколькими стопками цистерн.
Следует отметить, что по степени развития белок-синтезирующего аппарата и количеству
гранул клетки этого типа могут различаться. Скорее всего, такие различия связаны с суще-
ствованием определенного секреторного цикла.
Крупногранулярные клетки (рис. 176-178, 182, вклейка) (скорее всего, именно их гра-
нулы видны на полутонких срезах) имеют заметно более слабо развитый белок-
синтезирующий аппарат. Содержимое гранул зернистое, умеренной электронной плотно-
сти. В цитоплазме много микрофибрилл, объединенных в пучки. У Nereis virens эти клетки
связаны с обычными эпителиальными десмосомами, типичными для эпителия этого вида, а
с базальной мембраной— полудесмосомами (рис. 182, вклейка)', у афродитид подобных
клеточных контактов не отмечено.
Рис. 179. Схема строения эпителия средней кишки Harmathoe imbricata.
кгк - крупногранулярная базальная клетка; мгк- мелкогранулярная базальная клетка;
о - отростки базальных клеток, сс - слоистые структуры в апикальных частях ресничных клеток.
Fig. 179. The scheme of midgut epithelium organization in Harmathoe imbricata.
84
85
Рис. 180. Схема строения эпителия выростов средней кишки Harmathoe imbricata.
азпк- апикальная зона пузырьков и канальцев в пищеварительных клетках; вк- вакуолизированные
(пищеварительные) клетки; кгк - крупногранулярная клетка;
ндк - недифференцированные (камбиальные) клетки.
Fig. 180. The scheme of midgut caecum epithelium organization in Harmathoe imbricata.
В эпителии собственно средней кишки афродитид присутствуют клетки обоих типов,
тогда как в боковых выростах — только крупногранулярные (общая схема организации эпи-
телия этих отделов пищеварительного тракта у Harmathoe imbricata представлена на рис.
179 и 180). Интересно, что в ресничных протоках ни один из клеточных типов не был
обнаружен (Заварзин и др., 1984).
86
И крупногранулярные, и мелкогранулярные клетки имеют отростки, которые тянутся
между узкими основаниями обычных эпителиальных клеток вдоль базальной мембраны на
большие расстояния. Отростки клеток обоих типов часто сливаются в пучки и формируют
хорошо развитое базиэпителиальное сплетение (рис. 183, вклейка). Количество гранул в
отростках, как и диаметр последних, варьируют в большом диапазоне. При этом в самых
тонких (диаметром порядка 50-100 нм) можно наблюдать лишь микротрубочки и микрофи-
ламенты. Картин ухода отростков из эпителия не отмечалось.
Следует отметить еще одну важную деталь. У Nereis virens в участках расположения
под эпителием кровеносных синусов, помимо высокой концентрации в эпителиальном пла-
сте базальных клеток и их отростков, в разделяющей эпителий и полость синуса мембране
присутствуют узкие каналы шириной около 50 нм (рис. 182, вкпейка). Таких образований в
базальной мембране вне зон расположения синусов нет. Это наблюдение позволяет предпо-
лагать возможность выведения содержимого гранул в подобных участках непосредственно
в кишечный синус (Пунин, Лукьянова, 19846). Схема организации системы базальных кле-
ток в эпителии средней кишки Nereis virens приведена на рис. 185.
Рис. 185. Схема организации базальной части эпителиального пласта средней кишки Nereis virens.
век - всасывающие клетки; г! - гранулы мелкогранулярных клеток; г2 - гранулы крупногранулярных клеток;
гЗ - гранулы в цитоплазме амебоцита; кгк - крупногранулярные клетки; мгк - мелкогранулярная клетка;
мс - мембрана кишечного синуса; пд - полудесмосомы; пор - пора в мембране кишечного синуса.
Fig. 185. The scheme of basal part epithelium organization in Nereis virens midgut.
87
Интересно, что у Arenicola marina отмечены некоторые особенности в тонком строении
базальных клеток. Кагановская (1985, 1,988) выделяет три типа таких клеток, либо три их
различных функциональных состояния. Как правило, базальные клетки у этого вида харак-
теризуются хорошо развитым белок-синтезирующим аппаратом. Ядра имеют лопастную
форму. Диаметр гранул колеблется в пределах от 100 до 800 нм, причем (что важно отме-
тить) гранулы различного диаметра могут присутствовать в одной и той же клетке. Как у
афродитид и нереиса клетки имеют базальные отростки, тянущиеся в основании эпители-
ального пласта. Подобные клеточные элементы отмечены главным образом в кишечнике.
Два типа клеточных отростков — с крупными (диаметром около 500 нм) и мелкими
(порядка 150 нм) гранулами— отмечены в основании эпителиального пласта кишечника
Nephtys ciliata (сем. Nephtyidae) (Welsch, Storch, 1970). На электроннограмме эпителия
Aphrodite aculeata, приведенной в работе Далеса и Пелла (Dales, Pell, 1971), также можно
наблюдать два типа базальных отростков.
У Nereis virens обнаружен еще один тип клеток, заметно отличающийся от базальных.
Это очень редкие клетки открытого типа, которые несут на апикальной поверхности (кроме
микроворсинок) реснички (рис. 184, вклейка). В апикальной цитоплазме многочисленны
продольно ориентированные микротрубочки, каких-либо гранул нет. Строение базальных
частей таких клеток наблюдать, к сожалению, не удалось. Поскольку обычные эпителиаль-
ные (всасывающие и секреторные) клетки у этого вида ресничного аппарата не имеют (Пу-
нин, Лукьянова, 1984а), можно рассматривать реснички этих клеток как рецепторный аппа-
рат, а сами клетки — как рецепторные. Скорее всего, эти клеточные элементы соответству-
ют уже описанным выше клеткам, окрашивающимся метиленовым синим (Пунин, 1981 д).
8.1.4. Пролиферация и обновление регуляторных клеток
У Harmathoe imbricata и Nereis virens, помимо картин деления клеток, наблюдаемых в
апикальной части эпителиального пласта и связанных с физиологической регенерацией ос-
новного эпителия, единичные метафазные пластинки (рис. 186-187, вклейка) присутствуют
и у базальной мембраны (Пунин, 1981 в; Пунин, Лукьянова, 1984а, б). По-видимому, они
принадлежат базальным клеткам одного или обоих типов, хотя не исключено, что в эпите-
лиальном пласте могут делиться и амебоциты, часто мигрирующие у полихет в кишечный
эпителий. В опытах с введением меченого предшественника синтеза ДНК 3Н-тимидина у
обоих видов также были отмечены редкие первично меченные ядра (рис. 188, вклейка),
причем принадлежащие базальным клеткам обоих типов. Индекс меченых ядер (ИМЯ) для
базальных клеток заметно ниже, чем для основных эпителиальных, а в характере изменений
индекса меченых ядер (ИМЯ) для обеих клеточных популяций нет прямой корреляции (Пу-
нин, 1991).
Эти наблюдения позволяют предполагать, что базальные клетки имеют собственные
источники физиологической регенерации, и процесс их обновления не связан напрямую с
обновлением основного эпителия. Отсутствие морфологически выраженных камбиальных
элементов в основании эпителиального пласта говорит о том, что к делению способна, по
крайней мере, часть дифференцированных функционирующих клеток.
8.2. OLIGOCHAETA (МАЛОЩЕТИНКОВЫЕ ЧЕРВИ, ИЛИ ОЛИГОХЕТЫ)
Пищеварительный тракт этой группы кольчатых червей, насчитывающей около 3500
видов, состоит из 1) эктодермальной передней части, включающей глотку, пищевод (иногда
88
зоб), и мускульный желудок; 2) энтодермальной средней кишки (трубчатой и не имеющей
каких-либо выростов) и 3) эктодермальной задней. У многих видов этих высших червей
стенка средней кишки по всей своей длине или на некотором ее протяжении с дорзальной
стороны вдается в полость, образуя тифлозоль.
Сведения о кишечных регуляторных элементах у олигохет немногочисленны и ограни-
чиваются главным образом земляными червями (сем. Lumbricidae).
8.2.1. Данные гистологических исследований
Еще в конце XIX в. у представителей этого семейства в эпителии средней кишки были
описаны свободные нервные окончания (Smirnov, 1894). С использованием метода прижиз-
ненного окрашивания метиленовым синим Невмывака (1947, 1956, 1961, 1966) выявил в
средней кишке Allolobophora calliginosa интраэпителиальные клетки цилиндрической фор-
мы с двумя базальными отростками, уходящими в подстилающие ткани. Эти клетки рас-
сматривались им как нервные рецепторные.
В средней кишке Octolasium complanatum показано наличие интраэпителиальных кле-
ток, цитоплазма которых дает положительную аргирофильную реакцию. (Отмечу, что
это — редкий, если не единственный, случай положительной реакции у беспозвоночных).
На этом основании данные клетки рассматриваются как эндокринные (Andreozzi et al.,
1990). Немного забегая вперед, замечу, что имеются данные о присутствии в тканях кишеч-
ника Lumbricus terrestris гастрин-подобного пептида (Rzasa et al., 1982) и вещества, имму-
нореактивного к адренокортикотропному гормону (Kaloustian, 1986), однако точная ткане-
вая локализация этих соединений пока неизвестна.
8.2.2. Данные флуоресцентной гистохимии и иммуногистохимии
Иммуногистохимический анализ с использованием 10 антисывороток к различным
биологически активным пептидам (в том числе панкреатическому полипептиду, глюкагону,
вазоактивному интестинальному пептиду, нейротензину) не выявил каких-либо иммуноре-
активных клеток в эпителии кишечника у Lumbricus terrestris. Лишь антисыворотки к пан-
креатическому полипептиду и PYY (пептид тирозин-тирозин) позволили обнаружить во-
локна и отростки, предположительно иннервирующие основание эпителиальной выстилки
(Curry et al., 1989). В средней кишке Eisenia foetida описано базиэпителиальное РМИРамид-
иммунореактивное сплетение, однако реагирующие с этой антисывороткой клетки отмече-
ны лишь в глотке (Fujii et al., 1989). Все эти элементы рассматриваются авторами как часть
периферической нервной системы.
Мы провели флуоресцентно-гистохимический и иммуногистохимический анализ эпи-
телия уже упомянутых люмбрицид Lumbricus terrestris и Eisenia foetida, а также Tubifex
tubifex (сем. Tubificidae). Сразу отмечу, что у последнего вида интраэпителиальных струк-
тур, иммунореактивных к серотонину, нейротензину, кальцитонину или РМИРамиду, обна-
ружить не удалось (Пунин и др., 1998). Моноамин-содержащие и нейротензин-иммуно-
реактивные клетки в составе эпителия отсутствовали и у изученных земляных червей.
В то же время и у Eisenia foetida, и у Lumbricus terrestris выявлены РМИРамид-
иммунореактивные клетки, которые имеют у обоих видов много общего как в своей струк-
туре, так и положении в эпителиальном пласте (Пунин и др., 1998, 1999). Они присутствуют
на всем протяжении кишечника, редки (в среднем 1-2 клетки на поперечный срез, насчиты-
вающий несколько сотен эпителиальных клеток) и встречаются только в эпителии вен-
тральной и латеральных стенок средней кишки, но не в тифлозоле. Клетки расположены в
89
основании эпителиальной выстилки, ниже слоя ядер обычных эпителиальных клеток (рис.
189-191, вклейка), и могут быть отнесены к закрытому типу. Перикарионы небольшие (по-
рядка 10-15 мкм), округлые, овальные, иногда вытянутые вдоль базальной мембраны. У
некоторых клеток имеются базальные отростки, но они короткие и не образуют заметных
сплетений. Единичные иммунореактивные отростки были зарегистрированы также в мы-
шечной стенке кишечника (рис. 189, вклейка), однако их контактов с интраэпителиальными
структурами мы не наблюдали. Подчеркну, что результаты наших иммуногистохимических
реакций с антисывороткой к РМИЕамиду, полученные на Eisenia foetida, заметно отличают-
ся от данных работы, цитированной выше (Fujii et al., 1989). Объяснить причины этих рас-
хождений я пока не могу.
Кальцитонин-иммунореактивные элементы, выявленные в средней кишке, напротив, у
исследованных видов различаются и по морфологии, и по положению в эпителиальном
пласте. У Lumbricus terrestris такие клетки вместе с отростками образуют сеть на границе
эпителия и подлежащих тканей (рис. 192, вклейка). Большая их часть занимает субэпители-
альное положение, однако' некоторые клетки лежат интраэпителиально и не имеют контакта
с просветом кишечника. Единичные иммунореактивные отростки присутствуют в мышеч-
ной оболочке.
Напротив, в средней кишке Eisenia foetida кальцитонин-иммунореактивные клетки за-
нимают исключительно интраэпителиальное положение. Как и РМИГамид-иммунореак-
тивные, они отсутствуют в эпителии тифлозоля, однако в целом встречаются несколько
чаще (5-7 перикарионов на поперечные срез кишечника). Большинство клеток имеет вытя-
нутую форму (рис. 193, вклейка)', их апикальные части прослеживаются почти до просвета
кишечника, что с некоторым допущением позволяет отнести данные клетки к открытому
типу. Заметно меньше клеток имеет округлую форму (в диаметре около 10-12 мкм). Они
расположены в основании эпителия, иногда по 2-3 рядом (рис. 194, вклейка). Лишь у не-
скольких клеток были отмечены короткие базальные отростки (рис. 193, вклейка).
Рис. 195. Регуляторные клетки в средней кишке земляных червей.
Их локализация (отмечена точками) (А), форма и положение в эпителии
РМИРамид- (сплошная заливка) и кальцитонин- (точки) иммунореактивных элементов
у Lumbricus terrestris (Б) и Eisenia foetida (В).
т - тифлозоль.
Fig. 195. Regulatory cells in the midgut of lumbricids. Their localization (marked by dots) (A),
form and position relative to epithelium of FMRFamide- (black) and calcitonin- (dots)
immunoreactive elements in Lumbricus terrestris (Б) and Eisenia foetida (B).
90
Положение в эпителиальном пласте и варианты морфологии БМЯРамид- и кальцито-
нин-иммунореактивных клеток у Lumbricus terrestris и Eisenia foetida показаны на рис. 195.
8.2.3. Данные электронно-микроскипических исследований
Ультраструктурный анализ показал присутствие в эпителии кишечника Eisenia foetida
мощного базиэпителиального сплетения отростков. Это сплетение часто хорошо заметно и
на полутонких срезах (рис. 196, вклейка). Отростки тянутся между тонкими основаниями
ресничных и секреторных клеток в разных направлениях, образуя стволы, насчитывающие
десятки таких структур. Диаметр отростков варьирует от 0,15 до 1,5 мкм; в самых мелких
отростках удается наблюдать лишь продольно ориентированные микротрубочки, в более
крупных присутствуют гранулы и пузырьки (рис. 197, вклейка).
На основании диаметра и тонкого строения гранул отростки можно разделить на два
основных типа. Отростки первого типа характеризуются наличием довольно крупных гра-
нул с электронноплотным зернистым содержимым диаметром 220-260 нм.
Составляющие основную часть сплетения отростки второго типа имеют мелкие пу-
зырьки диаметром 40-65 нм и полиморфные гранулы. Большинство гранул — округлые
электронноплотные с гало, с диаметром 120-170 нм; иногда сходные гранулы имеют слегка
овальную форму. Реже встречаются гранулы такого же диаметра с гало и содержимым уме-
ренной электронной плотности, еще реже — несколько более крупные (диаметром до 200-
230 нм) с электронносветлым зернистым материалом. Некоторые гранулы по своему виду,
скорее, напоминают пузырьки с электронноплотным центром.
Как уже отмечалось, в одном и том же отростке могут присутствовать гранулы различ-
ной структуры; их соотношение, как и общее количество, варьирует от отростка к отростку.
Часто наблюдалась концентрация гранул и пузырьков на стороне, обращенной к базальной
мембране (рис. 197, вклейка) или к цитомембране ресничных и секреторных эпителиальных
клеток. Были отмечены и редкие картины, напоминающие экзоцитоз пузырьков. Подчерк-
ну, что подобные отростки не встречались в эпителии тифлозоля (Пунин и др., 1999).
Отростки базиэпителиального сплетения Eisenia foetida по своему строению сходны с
описанными в мышечной стенке кишечника Lumbricus terrestris, которые рассматриваются
как нервные (Csoknya et al., 1992). Отмечу, что подобные по тонкому строению отростки
есть среди мышечных клеток стенки кишечника и у Eisenia foetida.
На основании тонкого строения в эпителии кишечника Eisenia foetida мы первоначаль-
но выделяли 3 типа регуляторных клеток (Пунин и др., 1999). Два из них располагаются в
основании эпителиального пласта и различаются по размерам гранул: соответственно мы
назвали их мелкогранулярными и крупногранулярными. Однако более подробный анализ
позволил выделить 5 подтипов или разновидностей мелкогранулярных клеток.
Мелкогранулярные клетки (самые многочисленные из регуляторных элементов
в эпителии) (рис. 198, вклейка) имеют округлую или овальную форму и располагаются либо
непосредственно на базальной мембране, либо (чаще) чуть выше ее. Часто они находятся в
контакте с уже описанными отростками. Эти клетки сами имеют отростки, вступающие в
состав базального сплетения (рис. 199, вклейка).
Первая разновидность мелкогранулярных клеток характеризуется наличием в цито-
плазме большого количества округлых гранул с электронноплотным содержимым и узким
гало. Диаметр гранул колеблется в пределах 120-170 нм. Цистерн шероховатой эндоплаз-
матической сети довольно мало, зато многочисленны свободные рибосомы. Ядра (как и у
других разновидностей этого «типа» клеток) овальные, с небольшим, но хорошо заметным
91
ядрышком и распыленным гетерохроматином. В цитоплазме довольно много вакуолей с
ламеллярным содержимым, их диаметр достигает 1 мкм.
В цитоплазме клеток второй разновидности присутствуют 2 типа гранул. Наиболее
обычны гранулы диаметром 145-190 нм с мелкозернистым содержимым умеренной элек-
тронной плотности. Более редки электронноплотные гранулы с узким гало или без него
диаметром 110-150 нм. В таких клетках хорошо развита шероховатая эндоплазматическая
сеть, многочисленны и свободные рибосомы. В области аппарата Гольджи этих клеток мно-
го пузырьков диаметром до 140 нм. По всей цитоплазме встречаются вакуоли с разнообраз-
ным содержимым и микротрубочки.
Клетки третьей разновидности очень редки. Они содержат большое количество округ-
лых (диаметром 140-200 нм) или слегка овальных гранул. Гранулы имеют широкое гало, а
их электронноплотное содержимое часто располагается эксцентрично. Встречаются также
более редкие, мелкие (диаметром 80-90 нм) гранулы с узким гало и содержимым умерен-
ной электронной плотности. Шероховатая эндоплазматическая сеть развита слабо, в цито-
плазме довольно много Вакуолей.
Клетки четвертой разновидности содержат умеренное количество электронно-плотных
гранул без гало диаметром 175-220 нм и мелкие пузырьки (до 95 нм в диаметре). Белок-
синтезирующий аппарат представлен главным образом свободными рибосомами. В цито-
плазме присутствуют микротрубочки и единичные большие вакуоли с ламеллярным содер-
жимым.
Наконец, в цитоплазме мел ко гранулярных клеток пятой разновидности присутствуют
гранулы с зернистым содержимым без гало, диаметр которых колеблется в пределах 180—
280 нм. В электронносветлой цитоплазме много вакуолей с разнообразным содержимым и,
как и у большинства других мелкогранулярных клеток, свободных рибосом.
Второй тип клеток (крупногранулярные) (рис. 200, вклейка) характеризуется
наличием зернистых электронноплотных гранул без гало, диаметром около 250 нм. Ядра
этих клеток часто имеют глубокие инвагинации ядерной оболочки и неправильные очерта-
ния. Отростки крупногранулярных клеток также вступают в состав базиэпителиального
сплетения. Клетки со сходными по строению гранулами были отмечены и под базальной
мембраной.
Наконец, было зарегистрировано также несколько клеток вытянутой формj>i,
ориентированных длинной осью перпендикулярно базальной мембране и, по-видимому,
контактирующих с просветом кишечника. Одну из таких клеток удалось проследить вплоть
до зоны апикальных клеточных контактов. В центральной цитоплазме довольно многочис-
ленны округлые электронноплотные гранулы диаметром 125-175 нм (иногда чуть вытяну-
тые или слегка неправильной формы) и электронносветлые пузырьки диаметром около 100
нм. Цистерн шероховатой эндоплазматической сети немного, преобладают свободные ри-
босомы. В апикальной цитолазме заметны продольно ориентированные микротрубочки,
гранулы и пузырьки единичны (Пунин и др., 1999).
Общая схема строения эпителия средней кишки Eisenia foetida представлена на рис.
201. На ней не выделены разновидности мелкогранулярных клеток и не показаны регуля-
торные клетки открытого типа.
Рис. 201. Схема организации эпителия средней кишки Eisenia foetida.
кгк - крупногранулярная клетка; мгк - мелкогранулярная клетка; со - интраэпителиальный ствол отростков.
Мелкогранулярные клетки на этой схеме представлены как единый тип.
Fig. 201. The scheme of the epithelium organization in Eisenia foetida midgut.
92
93
В настоящее время на основании имеющихся наблюдений трудно сопоставлять данные
ультраструктурного и иммуногистохимического анализа. Можно лишь предположить, что
последний описанный клеточный тип соответствует кальцитонин-иммунореактивным клет-
кам открытого типа, либо нервным элементам, соответствующим описанным Невмывакой
(1947, 1956, 1961, 1966) с помощью метиленового синего у Allolobophora calliginosa. Обра-
щает внимание тот факт, что количество клеток, выявленных в ходе ультраструктурного
анализа, заметно превосходит количество РМЯРамид- и кальцитонин-иммунореактивных
элементов. Подчеркну еще раз, что и при использовании этих антисывороток многие клетки
у базальной мембраны не окрашивались, а базиэпителиальное сплетение практически не
выявлялось. Эти наблюдения предполагают, что кишечная регуляторная система у Eisenia
foetida весьма гетерогенна в плане содержащихся в ее элементах биологически активных
соединений, выявить и охарактеризовать которые еще предстоит. Отмечу также, что отме-
ченный полиморфизм гранул в некоторых клетках (и особенно отростках) не исключает
также колокализации различных регуляторных веществ.
8.3. HIRUDINEA (ПИЯВКИ)
Пиявки представляют собой специализированную группу кольчатых червей, сильно
измененных потомков Oligochaeta. Среди них встречаются как эктопаразиты, так и свобод-
но живущие хищники. Класс Hirudinea подразделяют на 2 подкласса: Archihirudinea (древ-
ние пиявки) и Euhirudinea (настоящие пиявки). К сожалению, в известной мне литературе
какие-либо сведения об элементах кишечной регуляторной системы пиявок отсутствуют.
Совместно с В.К.Казаковым, С.Ф.Филимоновой и Л.Г.Мкртчяном мной были изучены
с использованием методов иммуногистохимии и электронной микроскопии 4 вида настоя-
щих пиявок: относящаяся к хоботным пиявкам Glossiphonia complanata (сем.
Glossiphonidae — плоские пиявки) и бесхоботные Hirudo medicinalis, Haemopis sanguisuga
(сем. Gnathobdellidae— челюстные пиявки) и Herpobdella {=Erpobdella) octoculata (сем.
Herpobdellidae — глоточные пиявки).
Энтодермальная часть пищеварительного тракта изученных пиявок несколько различа-
ется по микроанатомии. У эктопаразита— улитковой пиявки Glossiphonia complanata —
она представлена так называемым «желудком» с 6 парами латеральных карманов и более
короткой средней кишкой с 4 парами боковых выростов. У медицинской пиявки Hirudo
medicinalis (также эктопаразитического вида) желудок имеет 10 пар боковых карманов, то-
гда как средняя кишка лишена выростов. Принято считать, что у паразитических форм соб-
ственно процессы пищеварения идут в средней кишке, тогда как желудок выполняет глав-
ным образом роль резервуара для хранения поглощенной пищи. У двух других видов, отно-
сящихся к хищным пиявкам, боковые выросты желудка исчезают; только у большой лож-
ноконской {Haemopis sanguisuga) сохраняется одна пара боковых выростов, отходящих от
дистальной части органа.
94
8.3.1. Данные иммуногистохимического анализа
Все 4 вида пиявок были изучены нами (Пунин и др., 2000) с использованием антисыво-
роток к нейротензину, кальцитонину, ЕМКЕамиду и серотонину (табл. 4).
Таблица 4
Иммунореактивные клетки и отростки в пищеварительной системе пиявок
Виды Иммунореактивность к
серотонину РМКРамиду нейротензину кальцитонину
Glossiphonia complanata +/- -/+ -/- +/+
Hirudo medicinalis +/- -/+ +/+ -/+
Haemopis sanguisuga -/+ -/+ -/+ +/-
Herpobdella octoculata -/- +/+ +/+ +/-
Примечание: +— наличие иммунореактивных элементов;-отсутствие иммунореактивных элемен-
, знак над чертой — интра-; знак под чертой — субэпителиальное положение иммунореактивных элементов.
В составе эпителия пищеварительного тракта Glossiphonia complanata выявлены серо-
тонин- и кальцитонин-иммунореактивные клетки (рис. 202-203, вклейка). Первые крайне
редки, имеют небольшие размеры (порядка 8-12 мкм) и расположены в основании эпители-
ального пласта средней кишки. По-видимому, они могут быть отнесены к клеткам закрыто-
го типа. У некоторых клеток отмечены короткие базальные отростки, не образующие, одна-
ко, какого-либо заметного сплетения.
Кальцитонин-иммунореактивные элементы несколько более многочисленны, встреча-
ются как в желудке, так и в средней кишке, располагаются и интра-, и субэпителиально.
Интраэпителиальные клетки полиморфны, представлены элементами открытого и закрыто-
го типов. Первые могут быть цилиндрические, трапециевидные, веретеновидные; некото-
рые имеют расширенную базальную часть и тонкий апикальный отросток. Базальных отро-
стков у этих клеток не отмечено. Субэпителиальные клетки встречаются заметно чаще,
контактов между ними и интраэпителиальными элементами мы не наблюдали.
ЕМИЕамид-иммунореактивные клетки (рис. 204, вклейка) расположены только субэпи-
телиально. Они достаточно многочисленны, округлые или овальные, сравнительно крупные
(до 20 мкм) и имеют отростки. Отростки образуют сплетения, которые тянутся вдоль ба-
зальной границы эпителия, однако, по-видимому, в него не проникают.
В составе пищеварительной системы Hirudo medicinalis интраэпителиальные иммуно-
реактивные клетки крайне редки. Нам удалось наблюдать лишь единичные клетки в вырос-
тах желудка, реагировавшие с антисывороткой к нейротензину, и в средней кишке — к се-
ротонину. Последние — закрытого типа, располагаются как по одиночке, так и группами по
2-3 (рис. 205, вклейка). Под эпителием серотонин-иммунореактивных структур не отме-
чено.
Нейротензин-иммунореактивные клетки у медицинской пиявки также можно отнести к
элементам закрытого типа (рис. 206, вклейка). Они небольшие (около 10 нм), без отростков,
имеют на срезах округлую или овальную форму. Под эпителием также расположены клетки
и отростки, сходные по демонстрируемой ими иммунореактивности. Некоторые из клеток
лежат практически у базальной поверхности эпителия (рис. 207, вклейка). Иммунореактив-
ные отростки также могут подходить вплотную к эпителиальному пласту, но, по-видимому,
не проникают в него.
95
Под эпителием отмечены также FMRF-амид- и кальцитонин-иммунореактивные клетки I
и отростки, некоторые отростки тянутся вдоль базальной поверхности эпителиальной вы-
стилки.
У Haemopis sanguisuga мы наблюдали только кальцитонин-иммунореактивные интра-
эпителиальные клетки (рис. 208, вклейка). Они очень редки и встречаются в выростах же-
лудка. Эти клетки располагаются в основании эпителиального пласта обычно поодиночке,
реже — по 2, и, по-видимому, не имеют контакта с просветом выростов. Отростков у них
нет. Под эпителием кальцитонин-иммунореактивных структур не отмечено.
Субэпителиальное положение у этого вида имеют клетки и отростки, реагирующие с
антисыворотками к нейротензину, серотонину и РМКГамиду. Нейротензин-иммунореак-
тивные клетки редки, обычно округлой формы, иногда относительно крупные (до 20 мкм) и
лежат на некотором удалении от эпителиальной выстилки. Серотонин- и РМВГамид-
иммунореактивные элементы (последние заметно более многочисленны), обычно вытяну-
той формы, с отростками, могут располагаться и непосредственно под эпителием. Размеры
этих клеток колеблются в пределах от 10 до 30 мкм. Отростки образуют субэпителиальное
сплетение.
У Herpobdella octoculata обнаруженные интраэпителиальные РМКГамид-иммунореак-
тивные клетки небольшие (10-12 мкм), имеют округлую или овальную форму и встречают-
ся редко. По своему строению они могут быть отнесены к элементам закрытого типа (рис.
209, вклейка). Заметно чаще сходные клетки располагаются субэпителиально.
Интраэпителиальные нейротензин-иммунореактивные клетки у этого вида пиявок по-
лиморфны. Большая их часть представлена элементами веретеновидной формы (рис. 210,
вклейка) либо имеет расширенное основание и узкий апикальный отросток. Более редки
небольшие клетки закрытого типа округлой или овальной формы. Хотя на изученных пре-
паратах отхождения базальных отростков от перикарионов не отмечалось, в основании эпи-
телия были зарегистрированы единичные иммунореактивные отростки. Субэпителиально
расположены сходные по иммунореактивности клетки и отростки с варикозными расшире-
ниями, образующие местами локальные сплетения.
Редкие кальцитонин-иммунореактивные клетки располагаются исключительно интра-
эпителиально (рис. 211, вклейка) и представлены элементами как открытого, так и закрыто-
го типов. Базальных отростков у них не отмечено.
8.3.2. Данные электронно-микроскопических исследований
Совместно с С.А.Филимоновой мной было проведено предварительное электронно-
микроскопическое исследование эпителия кишечника у 2 видов пиявок— Glossiphonia
complanata и Herpobdella octoculata.
В эпителии желудка и боковых выростов улитковой пиявки удалось выявить редкие
клетки с характерными гранулами (рис. 212, вклейка). Эти клетки располагаются в основа-
нии эпителия поодиночке, реже — по 2, имеют электронносветлую цитоплазму. Ядра слож-
ной неправильной формы. По сравнению с составляющими основу эпителиальной выстилки
пищеварительными (всасывающе-секреторными по своей дифференцировке) клетками, они
характеризуются заметно более слабым развитием органоидов. Цистерны шероховатой эн-
доплазматической сети и свободные рибосомы единичны, митохондрии небольшие и не-
многочисленны. Аппарат Гольджи развит умеренно. Если в обычных эпителиальных клет-
ках много гликогена, то в этих клетках его заметно меньше. В цитоплазме присутствуют
микротрубочки и микрофиламенты. У ряда клеток отмечены короткие отростки, тянущиеся
96
вдоль базальной мембраны. Апикальных отростков мы не наблюдали, что позволяет отне-
сти описываемые клетки к закрытому типу.
Секреторные гранулы электронноплотные, без гало, для них характерен определенный
полиморфизм. В одной и той же клетке могут присутствовать как округлые гранулы диа-
метром от 150 до 220 нм, так и овальные (в длину достигающие 270 нм). Многие гранулы
имеют подкововидную форму или края неправильных очертаний.
В основании эпителия, помимо отростков (рис. 213, вклейка), очевидно принадлежа-
щих описанным выше клеткам, отмечены также редкие отростки с гранулами несколько
иного строения. Они также электронноплотные, без гало, однако на срезах всегда имеют
округлую форму и заметно меньше размерами: их диаметр колеблется в пределах 70-115
нм (рис. 214, вклейка). Среди таких гранул встречаются и более мелкие (70-75 нм в диамет-
ре) с содержимым умеренной электронной плотности. Поскольку в таких отростках (на по-
перечных срезах их диаметр составляет обычно порядка 1-1,5 мкм) иногда могут присутст-
вовать небольшие вакуоли, фрагменты цистерн гладкой эндоплазматической сети и мито-
хондрии, можно предположить, что срез прошел недалеко от перикариона и, соответствен-
но, в эпителии могут присутствовать и гранулярные клетки другого типа, отличные от опи-
санных выше.
Если в основании эпителия отростки с гранулами встречались поодиночке или не-
большими группами, то под эпителием отмечены их скопления, насчитывающие в своем
составе до нескольких десятков отростков и напоминающие по строению нервные стволы.
Часть отростков содержит электронноплотные гранулы диметром около 70 нм, другие —
около 200 нм. Схема организации эпителия желудка Glossiphonia complanata с элементами
кишечной регуляторной системы представлена на рис. 215.
В основании эпителиального пласта желудка Herpobdella octoculata обнаружены ред-
кие клетки с электронноплотными гранулами без гало различной формы — округлыми и
овальными (рис. 216-218, вклейка). Диаметр первых составляет 180-220 нм, вторые могут
достигать в длину 300-500 нм. Эти клетки имеют умеренно развитую шероховатую эндо-
плазматическую сеть и ядра сложной конфигурации, у некоторых отмечены длинные вет-
вящиеся базальные отростки (рис. 216-217, вклейка). Иногда базальные отростки распола-
гались достаточно близко к кишечным кровеносным синусам (рис. 219, вклейка). Контакта
клеток с просветом пищеварительного тракта не отмечалось. Кроме подобных клеточных
элементов, в эпителии обнаружены клетки со значительно более крупными гранулами (до 1
мкм), часто имеющими менее электронноплотную зернистую сердцевину (рис. 218, вклей-
ка). Учитывая весьма большие для эндокринных размеры гранул, а также наличие подоб-
ных клеток также вне эпителия, можно предположить, что эти элементы, скорее всего, яв-
ляются проникшими в эпителиальный пласт амебоцитами.
Клетки и отростки с мелкими электронноплотными гранулами диаметром 70-90 нм
(очевидно, нервные) присутствуют в мышечной оболочке кишечника малой ложноконской
пиявки, однако в самом эпителии подобных структур не отмечено.
97
98
8.4. ЗАКЛЮЧЕНИЕ
Приведенные выше гистохимические, иммуногистохимические и ультраструктурные
данные, на мой взгляд, убедительно показывают присутствие в эпителии пищеварительного
тракта представителей всех трех классов кольчатых червей регуляторных клеток. Несмотря
на то, что степень изученности этих элементов в разных классах неодинакова, кажется воз-
можным, тем не менее, сделать некоторые предварительные заключения.
Прежде всего обращает на себя внимание определенное сходство в организации регу-
ляторной системы у полихет и изученных представителей олигохет— земляных червей.
Во-первых, это наличие по крайней мере двух основных морфологических типов клеток —
крупногранулярных и мелкогранулярных, и мощного базиэпителиального сплетения отро-
стков. Во-вторых (по крайней мере у нереиса и земляных червей) в эпителиальном пласте
присутствуют клетки открытого типа, очевидно рецепторные (нервные). Большее (по срав-
нению с морфологическим) разнообразие регуляторных клеток по демонстрируемым ими
иммунореактивностям не'исключает возможности того, что при внешнем ультраструктур-
ном сходстве базальные регуляторные клетки могут содержать различные биологически
активные вещества.
В организации кишечной регуляторной системы пиявки, по-видимому, несколько отли-
чаются от олигохет и полихет. Это, в первую очередь, касается общего количества регулятор-
ных элементов. Несмотря на достаточное разнообразие как по морфологии, так и по демонст-
рируемым иммунореактивностям (рис. 220), в сравнении с другими кольчатыми червями ре-
гуляторные клетки единичны. Это, соответственно, существенно затрудняет и их анализ на
ультраструктурном уровне (вполне возможно и даже вероятно, что в действительности они
представлены более чем одним-двумя выявленными клеточными типами). Однако очевидно,
что (по крайней мере у Glossiphonia complanata и Herpobdella octoculata) по сравнению с по-
лихетами и олигохетами, в эпителии пищеварительного тракта нет столь хорошо развитого
базального сплетения отростков. [Интересно, что в подробном исследовании тонкого строе-
ния эпителия желудка и кишечника, проведенного на Hirudo medicinalis (Hammersen, Pokahr,
1972a, b), нет никаких упоминаний о специфических клетках или отростках. На мой взгляд,
это может быть объяснено тем, что подобные структуры крайне редки.]
Возможно, отмеченные отличия в организации кишечной регуляторной системы у пия-
вок связаны с особым типом питания, свойственным исходно эктопаразитическим формам
этой группы кольчатых червей.
В нашем понимании организации кишечных регуляторных систем аннелид остается
много вопросов. Так, пока неясно, в какой степени эта система у дождевых червей соответ-
ствует другим олигохетам (и имеется ли у них вообще). Предварительные данные, получен-
ные нами на представителе сем. Tubificidae (Tubifex tubifex) позволяют предполагать суще-
ственные отличия между разными семействами. Требует дальнейшего изучения и такой ас-
Рис. 215. Схема строения эпителия боковых карманов желудка Glossiphonia complanata
с элементами регуляторной системы.
гг- гранулы гликогена; гк- гранулярная клетка; глк- гликокаликс; нс - субэпителиальный нервный ствол;
омг - отростки с мелкими гранулами: пк - пищеварительные (основные) клетки; пп - пиноцитозные пузырьки.
Fig. 215. The scheme of stomach lateral diverticuli epithelium organization in Glossiphonia complanata with
elements of regulatory system.
99
пект, как характер контакта (связи) интра- и субэпителиальной нервной систем. Практиче-
ски нет данных (за исключением афродитид и Nereis virens) о физиологической регенерации
элементов этой системы. Наконец, учитывая циклический характер питания и пищеварения
у паразитических пиявок не исключена и существенная перестройка системы регуляторных
клеток в зависимости от стадии пищеварения.
Г
I 1 В 2 ш з I I 4
Рис. 220. Морфологическое разнообразие регуляторных элементов различной иммунореактивности
и варианты их положения относительно эпителиального пласта в кишечнике пиявок
Glossiphonia complanata (A), Hirudo medicinalis (Б), Haemopis sanguisuga (В)
и Herpobdella octoculata (Г).
1 - нейротензин-, 2 - РМИРамид-, 3 - кальцитонин- и 4 - серотонин-иммунореактивные клетки и отростки.
Представленная схема не отражает частоты встречаемости этих регуляторных элементов.
Fig. 220. Morphological diversity of regulatory elements demonstrating different immunoreactivities and
variety of their localization relative to epithelium in the intestine of leeches Glossiphonia complanata (A),
Hirudo medicinalis (B), Haemopis sanguisuga (В) и Herpobdella octoculata (Г).
И, тем не менее, несмотря на недостаточную изученность олигохет и пиявок, можно,
наверное, заключить, что кишечная регуляторная система кольчатых червей гетерогенна и
состоит как из эндокринных (эндокриноподобных), так и из нервных клеток.
Глава 9
SIPUNCULIDA (СИПУНКУЛИДЫ)
Сипункулиды (небольшая группа, насчитывающая около 250 видов) обычно рассмат-
риваются как дополнение к классу кольчатых червей (Annelida). Пищеварительный тракт у
этих животных представляет собой трубку, практически лишенную каких-либо заметных
выростов; эктодермальная средняя кишка образует нисходящее и восходящее колена, за-
крученные друг вокруг друга.
Рис. 221. Эндокринная клетка в эпителии
ресничной бороздки восходящей петли
кишечника Phascolopsis gouldii.
гк — гранулярная (эндокринная) клетка;
жк - железистые клетки бороздки.
По электронным фотографиям из: Michei,
DeVillez, 1984.
Fig. 221. An endocrine cell in the glandular
ciliary gutter of Phascolopsis gouldii ascend-
ing intestine.
101
В известной мне литературе имеется лишь единичное упоминание о наличии регуля-
торных клеток в кишечнике представителя сипункулид — Phascolopsis gouldii. Эти клетки
располагаются в составе вентрального ресничного желобка, тянущегося вдоль восходящей
петли средней кишки. От остальных клеток они отличаются тем, что лежат в основании
эпителиального пласта и содержат в цитоплазме мелкие электронноплотные гранулы
(Michel, DeVillez, 1984). Судя по опубликованной в цитируемой работе электроннограме,
эти клетки— закрытого типа (рис. 221). Подробно детали их тонкого строения в статье не
приводятся.
Предпринятое мной иммуногистохимическое выявление регуляторных элементов в
кишечнике беломорской сипункулиды Golfingia vulgaris с использованием антисывороток к
нейротензину и РМИРамиду пока не дало положительных результатов.
Глава 10
TENTACULATA (ЩУПАЛЬЦЕВЫЕ)
К этой группе животных, занимающих промежуточное положение между первичноро-
тыми и вторичноротыми, относятся 3 типа — мшанки (Bryozoa), форониды (Phoronidea) и
брахиоподы (Brachiopoda). Иногда эти типы рассматриваются как классы внутри типа щу-
пальцевых (Tentaculata). Я придерживаюсь первой точки зрения, хотя в данном случае это
не имеет принципиального значения.
Сведения, касающиеся кишечных регуляторных систем (как, впрочем, и гистологиче-
ской организации пищеварительного тракта) у представителей форонид и брахиопод, как я
покажу далее, очень скудны. Что касается мшанок, в известной мне литературе такие дан-
ные отсутствуют совсем.
10.1. PHORONIDEA (ФОРОНИДЫ)
Пищеварительный тракт у немногочисленных представителей этого типа имеет U-
образную форму, ротовое и анальное отверстия расположены недалеко друг от друга. Ка-
кие-либо железистые выросты по ходу пищеварительной трубки отсутствуют.
Имеются электронно-микроскопические данные о присутствии в составе эпителия ки-
шечника Phoronis vancouverensis особых клеток, характеризующихся наличием в цитоплаз-
ме достаточно мелких (диаметром 180-200 нм) гранул с электронноплотным содержимым
(Vandermeulen, 1970). К сожалению, детального описания этих клеток не приводится; на
основании косвенных данных и приведенных электроннограмм можно предположить, что
эти клетки относятся к закрытому типу. Их функция, возможно, связана с нейросекретор-
ным контролем процессов пищеварения (тем самым автор исследования, очевидно, относит
их к нейросекреторным).
В основании эпителиального пласта желудка и средней кишки Phoronis psatntnophila
описаны тонкие отростки, предположительно принадлежащие нервным клеткам (Emig,
1967, 1968).
Пытаясь выявить регуляторные элементы в кишечнике Phonoropsis harmeri и Phoronis
ovalis, я использовал антисыворотки к серотонину, нейротензину и ЕМВБамиду, однако
результаты иммуногистохимического окрашивания с этими антителами не дали положи-
тельного результата.
103
10.2. BRACHIOPODA (ПЛЕЧЕНОГИЕ)
Пищеварительная система у представителей этого типа беспозвоночных представлена
пищеводом (по крайней мере, в своей проксимальной части имеющим эктодермальное про-
исхождение), переходящим в расширенную эктодермальную часть тракта— желудок. В
него открываются массивные парные пищеварительные выросты (так называемая пищева-
рительная железа). Желудок переходит в среднюю кишку, у представителей замковых бра-
хиопод (Articulata) заканчивающуюся слепо.
У Terebratulina retusa в основании эпителия желудка и средней кишки описаны отрост-
ки, объединенные в пучки. В них присутствуют гранулы диаметром 100-130 нм с электрон-
ноплотным центром и гало. Авторы (d’Hondt, Boucaud-Camou, 1982) рассматривают эти
отростки как аксоны.
Подобные структуры я наблюдал у Heniithyris psittacea, за исключением трубочек пи-
щеварительной железы, в основании эпителиальной выстилки во всех отделах пищевари-
тельного тракта (пищевод, желудок, средняя кишка и ее терминальное слепое расширение)
(схема организации эпителия слепого расширения представлена на рис. 222) (Пунин,
1981г). Диаметр отростков колеблется от 0,1-0,2 до 1-2 мкм. В мелких отростках удается
наблюдать лишь микротрубочки, в крупных присутствуют гранулы диаметром 0,10-0,15
мкм: электронноплотные с гало или без него, умеренной электронной плотности и светлые,
а также мелкие пузырьки, иногда митохондрии. Как правило, отростки объединены в пучки
до нескольких десятков в каждом (рис. 223, вклейка), которые расположены в тесном кон-
такте с базальной мембраной либо чуть выше последней. В средней кишке эти пучки тянут-
ся преимущественно вдоль нее. Синаптических контактов ни между отростками, ни между
отростками и эпителиальными клетками я не наблюдал. Иногда отмечалась концентрация
мелких пузырьков в районе контакта отростка с базальной мембраной; при этом непосред-
ственно под эпителием какие-либо клеточные структуры отсутствовали (рис. 224-225,
вклейка). Все наблюдавшиеся отростки имели исключительно интраэпителиальное поло-
жение.
В эпителии средней кишки и ее слепого расширения удалось наблюдать редкие клетки,
названные мной мелкогранулярными, отличающиеся по своему строению от остальных эпи-
телиальных. В этих клетках отмечено несколько более сильное развитие белок-синтези-
рующего аппарата (шероховатая эндоплазматическая сеть и свободные рибосомы) и при-
сутствие немногочисленных электронноплотных гранул такого же диаметра, как и в отро-
стках. Отмечены также картины отхождения отростков от тел клеток и их вступления в со-
став пучков (рис. 226, вклейка).
Иммуногистохимический анализ показал присутствие в эпителии пищеварительного
тракта Hemithyris psittacea клеток и отростков, реагирующих с антисыворотками к нейро-
тензину, пептиду-активатору (роста) головы гидры (НАР) и ЕМИРамиду.
Нейротензин-иммунореактивные клетки выявлены в эпителии пищевода, желудка и
средней кишки (рис. 227-228, вклейка). Эти клетки очень редки, обычно имеют цилиндри-
ческую или веретеновидную форму и могут быть отнесены к открытому типу. Если в пище-
воде у этих клеток наблюдаются базальные отростки, образующие рыхлое сплетение, то в
других отделах пищеварительного тракта такие отростки практически не встречаются
(Kazakov, Punin, 1991; Пунин и др., 1992а).
104
Рис. 222. Схема организации эпителия
слепого расширения средней кишки
Hemithyris psittacea с элементами
регуляторной системы.
мгк - мелкогранулярная клетка; о - отрост-
ки с гранулями; фб - фибробласт.
Fig. 222. Epithelium organization of
midgut blind widening in Hemithyris
psittacea with regulatory system
elements.
105
Клетки, реагирующие с НАР, имеют как ряд особенностей в своем строении, так и
иную локализацию в пищеварительном тракте. Они присутствуют в желудке (главным об-
разом в участках эпителия, прилегающих к отверстиям долей пищеварительной железы)
(рис. 229, вклейка) и в средней кишке. В эпителии желудка НАР-иммунореактивные клетки
имеют преимущественно веретеновидную форму. Апикальные части подходят к просвету
тракта; базальные, сильно истончаясь, вступают в состав базального сплетения, мощно раз-
витого в зонах отхождения выростов пищеварительной железы (рис. 230, вклейка) и до-
вольно рыхлого — в других участках желудка. Образующие сплетение отростки имеют хо-
рошо заметные варикозные расширения.
В средней кишке НАР-иммунореактивные клетки очень редки (2-5 на несколько тысяч
обычных эпителиальных). Как и в желудке, они имеют веретеновидную или цилиндриче-
скую (в участках с невысоким эпителием) форму. Основное отличие их строения заключа-
ется в отсутствии у этих клеток базальных отростков (рис. 231, вклейка).
Вопрос о присутствии нейротензин- и НАР-иммунореактивных регуляторных клеток в
эпителии слепого расширения остается открытым. Весь эпителиальный пласт интенсивно
окрашивается обеими антисыворотками, причем эти реакции специфичны, так как при про-
ведении контрольных реакций с использованием антисывороток, преадсорбированных ан-
тигеном, окрашивания нет. Тем не менее, скорее всего, эти реакции связаны с каким-то ве-
ществом, синтезируемым в эпителиальных клетках слепого расширения, имеющим имму-
нологическое сродство с обеими использованными антисыворотками. Отмеченное сплош-
ное окрашивание эпителиального пласта полностью маскирует возможное присутствие в
нем специализированных клеток.
Важно отметить, что в стенке пищеварительного тракта, за пределами эпителия, струк-
туры, иммунореактивные к НАР и нейротензину, не наблюдались (Пунин и др., 1992а).
ЕМИЕамид-иммунореактивные клетки в пищеварительном тракте Hemithyris psittacea
также располагаются исключительно интраэпителиально. Подавляющее большинство этих
клеток закрытого типа, локализовано у базальной мембраны (рис. 232, вклейка) и характе-
ризуется очень небольшими размерами (около 3—4 мкм в диаметре). В некоторых участках
эпителиального пласта иммунореактивные клетки закрытого типа столь многочисленны,
что образуют практически сплошной, интенсивно окрашенный базальный слой. Клетки от-
крытого типа, напротив, весьма редки, имеют расширенную базальную часть и тонкий (не
более 0,5 мкм шириной) апикальный отросток, направленный к просвету тракта (рис. 233,
вклейка). И у тех, и у других клеток отмечены отходящие от перикарионов базальные отро-
стки, формирующие интраэпителиальное сплетение (Пунин, 1998).
Все ЕМКЕамид-иммунореактивные клетки были обнаружены в желудке, средней киш-
ке и ее слепом расширении, но не наблюдались в пищеводе и выростах пищеварительной
железы.
В желудке клетки, реагирующие с антисывороткой к ЕМИЕамиду, расположены в со-
ставе эпителия его дорзальной поверхности, в районе отхождения выростов пищеваритель-
ной железы, а также в эпителии продольного желобка на вентральной поверхности. В по-
следнем ЕМКЕамид-иммунореактивные клетки наиболее многочисленны в центральной
части и убывают (в количестве) по мере удаления от нее. Прямого контакта между этими
двумя зонами расположения иммунореактивных клеток в желудке нет.
В проксимальной части средней кишки клетки, реагирующие с антисывороткой к
ЕМКЕамиду, лежат исключительно в эпителии глубокого фронтального желобка, являюще-
гося непосредственным продолжением вентрального желобка желудка. Однако дистальнее
этот четкий порядок постепенно нарушается: все больше клеток располагается на заметном
106
удалении от желобка. Наконец, в районе перехода средней кишки в слепое расширение
ЕМИРамид-иммунореактивные клетки довольно равномерно распределены по всему пери-
метру этого отдела пищеварительного тракта.
В слепом расширении средней кишки иммунореактивные клетки также равномерно
распределены по эпителию, причем их количество уменьшается по направлению к слепому
концу, где клетки единичны (Пунин, 1998). Схема распределения РМКЕамид-иммуно-
реактивных клеток в пищеварительном тракте Hemithyris psittacea представлена на рис. 234.
Рис. 234. Распределение ЕМКЕамид-иммунореактивных клеток
в пищеварительном тракте Hemithyris psittacea:
А - схематизированный рисунок пищеварительной системы (трубочки пищеварительной железы не показаны);
Б-В - поперечные срезы через соответственно проксимальную (сечение X - X’)
и дистальную (сечение Y- У’) части средней кишки, жлд - желудок; мез - мезентерий;
опж - отверстия выростов пищеварительной железы; пщд - пищевод; ср - слепое расширение средней кишки;
фж- фронтальный желобок. Точками (А) отмечены зоны расположения иммунореактивных клеток,
черным (Б-В) - иммунореактивные клетки в составе эпителия.
Fig. 234. Distribution of FMRFamide-immunoreactive cells in the digestive tract of
Hemithyris psittacea.
У другого вида замковых брахиопод (Coptothyris grayi) пищеварительный тракт кото-
рого был изучен с помощью антисывороток к нейротензину и НАР (Пунин, Казаков, 1993),
характер распределения иммунореактивных структур и некоторые детали их строения за-
метно отличаются от Hemithyris psittacea. Клетки и отростки, реагирующие с использован-
ными антисыворотками, присутствуют только в эпителии пищевода и проксимальной части
желудка (до отхождения выростов пищеварительной железы). Сходное строение НАР- и
107
нейротензин-иммунореактивных структур и характер их локализации (рис. 235-236, вклей-
ка) дают основания предположить, что с обеими антисыворотками реагируют одни и те же
клетки и отростки.
В эпителии пищевода иммунореактивные клетки тонкие (толщиной не более 1-2 мкм),
открытого типа, довольно многочисленны (рис. 237, вклейка). Их базальные отростки обра-
зуют хорошо развитое сплетение, состоящее из многочисленных анастомозирующих пуч-
ков, преимущественно тянущихся вдоль пищевода. Интересно отметить, что подобная кар-
тина морфологии и расположения НАР- и нейротензин-иммунореактивных клеток и отро-
стков наблюдалась также в эпителии ротового отверстия и основания лофофора.
В эпителии желудка иммунореактивные клетки не столь многочисленны и представле-
ны главным образом лежащими в основании эпителиального пласта мелкими отростчатыми
клетками закрытого типа (рис. 238, вклейка). Клетки открытого типа (рис. 239, вклейка)
редки и по своему строению напоминают таковые пищевода. Базальное сплетение в этом
отделе тракта непосредственно связано со сплетением в пищеводе, однако развито не столь
сильно; пучки тоньше; хорошо заметны отдельные отростки с варикозными расширениями.
Вне эпителия на всем протяжении пищеварительного тракта иммунореактивные струк-
туры отсутствовали. Схема организации выявленной иммунореактивной системы у
Coptothyris grayi представлена на рис. 240.
Рис. 240. Нейротензин- и НАР-иммунореактивная система клеток на границе {пунктирная линия)
пищевода и желудка Coptothyris grayi.
жк - железистые клетки пищевода; зк - клетки закрытого типа; ок - клетки открытого типа;
пэ - поверхность эпителия; эпж - эпителий желудка; эпп - эпителий пищевода.
Fig. 240. Neurotensin- and HAP-immunoreactive system on the border {dotted line) of oesophagus
and stomach in Coptothyris grayi.
108
10.3. ЗАКЛЮЧЕНИЕ
К сожалению, единичные данные по форонидам позволяют лишь констатировать нали-
чие в эпителии пищеварительного тракта клеток и отростков, возможно, представляющих
собой элементы периферической нервной системы.
Что касается брахиопод, имеющиеся немногочисленные данные по регуляторным
клеткам позволяют придти к следующему заключению. Изученным видам, с одной сторо-
ны, свойственно присутствие в эпителии клеток и отростков, представляющих собой, по-
видимому, элементы периферической нервной системы. К таким элементам можно отнести
НАР- и нейротензин-иммунореактивные клетки и отростки в эпителии пищевода и желудка
Coptothyris grayi, РМИЕамид-иммунореактивные клетки в желудке, средней кишке и ее сле-
пом расширении у Hemithyris psittacea, а также НАР-иммунореактивные клетки в желудке
последнего вида. В то же время все или большая часть нейротензин-иммунореактивных
клеток и НАР-иммунореактивные клетки в средней кишке Hemithyris psittacea, если при-
держиваться используемых морфологических критериев, скорее всего, являются
эндокринными.
При сравнении организации выявленных регуляторных систем у изученных видов об-
ращают на себя внимание два факта. Первый заключается в том, что клетки, реагирующие с
одной и той же антисывороткой, по своим морфологическим признакам часто могут быть
отнесены как к нервным, так и к эндокринным элементам (НАР-иммунореактивные клетки
Hemithyris psittacea). Второй указывает на то, что у разных видов брахиопод регуляторные
вещества, обладающие сродством с одними и теми же антисыворотками, могут присутство-
вать как в разных (Hemihyris psittacea), так и, по-видимому, в одних и тех же (Coptothyris
grayi) структурах (нервных, эндокринных), а элементы, демонстрирующие одну и ту же
иммунореактивность, у этих 2 видов локализованы в разных отделах пищеварительной сис-
темы, либо участках эпителиальной выстилки.
К сожалению, имеющиеся данные недостаточны, чтобы обсуждать конкретные функции
выявленных регуляторных элементов. Лишь в отношении ЕМИРамид-иммунореактивных
клеток Hemithyris psittacea, учитывая их расположение в районах отхождения выростов пи-
щеварительной железы и вентрального желобка желудка [фронтального желобка средней
кишки, т.е. в тех участках, где эпителий имеет особо мощно развитый ресничный аппарат
(Пунин, 1981г)], позволяет предположить, что эти клетки связаны с регуляцией биения ресни-
чек и контролируют перемещение жидкости и частиц по тракту. В то же время отмеченная в
ходе ультраструктурного исследования концентрация мелких пузырьков в отростках у мем-
браны, обращенной к базальной мембране, предполагает выведение биологически активных
веществ за пределы эпителия, т.е. допуская подобие эндокринной регуляции.
Таким образом, несмотря на крайне недостаточную изученность кишечной регулятор-
ной системы брахиопод, можно, тем не менее, говорить о ее гетерогенности как в отноше-
нии клеточного состава, так и спектра продуцируемых биологически активных веществ.
Глава 11
CEPHALORHYNCHA (ГОЛОВОХОБОТНЫЕ)
В этой главе я рассмотрю имеющиеся сведения, касающиеся кишечных регуляторных
элементов у двух своеобразных групп животных с не ясным до конца систематическим по-
ложением — киноринх и приапулид. Часто их рассматривают в качестве дополнения к типу
круглых червей (Nemathelminthes). Некоторые исследователи полагают, что приапулид сле-
дует отнести к примитивным Coelomata, родственным вторичноротым и щетинкочелюст-
ным (Chaetognatha). Недавно предложено эти группы на правах классов [вместе с лорици-
ферами (Loricifera) и волосатиками (Nematomorpha)] объединить в новый тип — головохо-
ботных (Cephalorhyncha) (Малахов, 1980; Малахов, Адрианов, 1995). Здесь я буду придер-
живаться этой точки зрения.
11.1. KINORHYNCHA (КИНОРИНХИ)
Пищеварительная система киноринх (всего в этой группе беспозвоночных насчитыва-
ют около 100 видов) состоит из выстланной кутикулой эктодермальной передней кишки,
энтодермальной средней кишки и задней кишки, как и передней, образованной кутикуляр-
ным эпителием (Neuhaus, 1994; Малахов, Адрианов, 1995).
Сведения о кишечных регуляторных клетках, существующие к настоящему времени,
исчерпываются очень краткими данными по 3 видам: Zelinkaderes floridensis, Pycnophyes
kielensis и Pycnophyes dentatus (Neuhaus, 1994). В составе эпителия средней кишки
Zelinkaderes floridensis выявлено всего 5 клеток, имеющих характерное строение. Три из
них расположены примерно на границе первой и второй четверти кишечной трубки, две
остальные — рядом с началом задней кишки. У видов рода Pycnophyes подобных клеток
только 2: одна— в первой трети кишечника, другая (кстати, единственная из всех, имею-
щая базальный отросток), как и у Zelinkaderes floridensis, — на границе с задней кишкой.
Все отмеченные выше клетки имеют ряд общих черт строения (рис. 241): расширенная яд-
росодержащая часть расположена в основании эпителия, а тонкий апикальный отросток
подходит к просвету кишечника и несет ресничку. В околоядерной цитоплазме располага-
ются многочисленные гранулы с гало. Следует отметить, что нервные отростки в эпителий
не проникают и, соответственно, с этими клетками не контактируют.
Поскольку все остальные клетки эпителиальной выстилки ресничек не имеют, данные
клетки рассматриваются как рецепторно-эффекторные, действующие (по-видимому, путем
выведения содержимого гранул на базальной клеточной поверхности и через базальную
мембрану) на клетки мышечной оболочки кишечника (Neuhaus, 1994).
ПО
Рис. 241. Рецепторная клетка в эпителии средней кишки Pycnophyes
(по: Neuhaus, 1994).
рцк - рецепторная клетка.
Fig. 241. Receptory cell in the midgut epithelium of Pycnophyes
(according to: Neuhaus, 1994).
11.2. PRIAPULIDA (ПРИАПУЛИДЫ)
Всего в настоящее время насчитывают около 10 видов приапулид. В микроанатомиче-
ском плане пищеварительная система у этих животных достаточно проста. За эктодермаль-
ными глоткой и пищеводом следует эктодермальная средняя кишка, относительно широкая
в передней части (этот отдел часто называется желудком) и сужающаяся по направлению к
заднему концу. Пищеварительный тракт завершается короткой задней кишкой.
Не только данные, касающиеся регуляторных клеточных элементов, но и сведения о
гистологической организации собственно эпителия у представителей этой интересной
группы беспозвоночных единичны и касаются 4 видов: Priapulus caudatus, Halicryptus
spinulosus (Заварзин, Пунин, 1979; Пунин, Заварзин, 1979), Tubilichus pliilippinensis и
Tubilichus corallicola (Storch, Alberti, 1985; Higgins, Storch, 1989). Что касается отрывочных
данных по интраэпителиальным регуляторным клеткам, все они получены мной на
Priapulus caudatus и Halicryptus spinulosus.
Электронно-микроскопический анализ выявил в эпителии средней кишки Priapulus
caudatus очень редко встречающиеся клетки характерного строения (рис. 242, вклейка), на-
званные мной мелкогранулярными (Пунин, 19816). Их основная черта— присутствие в ци-
топлазме многочисленных небольших гранул (рис. 243, вклейка) с мелкозернистым элек-
тронноплотным содержимым. На срезах эти структуры имеют форму овала длиной 0,10—
0,20 и шириной 0,06-0,12 мкм. Второй отличительной чертой этих клеток является строе-
ние их ядер. Располагаясь среди ядер других эпителиальных клеток, они имеют многочис-
ленные инвагинации ядерной оболочки. Третьей особенностью является положение в клет-
111
ке аппарата Гольджи. Как и у других клеток, он представлен несколькими стопками цис-
терн, однако последние не рассредоточены по всей цитоплазме (Пунин, Заварзин, 1979), а
собраны в одной зоне, обычно под ядром. Наконец, только в мелкогранулярных клетках
отмечены единичные микротрубочки.
Мелкогранулярные клетки располагаются либо в непосредственном контакте с базаль-
ной мембраной, либо чуть выше ее; в последнем случае к базальной мембране тянутся 1-2
тонких отростка. Цистерны шероховатой эндоплазматической сети и митохондрии немно-
гочисленны (последние могут присутствовать и в отростках), встречаются единичные ва-
куоли с зернистым и ламеллярным содержимым, липидные капли. Клетки связаны с сосед-
ними типичными для эпителиального пласта клеточными контактами. Подхода апикальных
частей клеток к просвету не отмечено.
В основании эпителиального пласта, в контакте с базальной мембраной или чуть выше
ее, отмечены редкие отростки с такими же овальными гранулами, по-видимому, принадле-
жащие мелкогранулярным клеткам. Иногда отростки объединены в маленькие пучки. Схема
строения мелкогранулярной клетки Priapulus caudatus представлена на рис. 244.
Наши совместные с В.К.Казаковым попытки выявить кишечные регуляторные элемен-
ты иммуногистохимическими методами с использованием антисывороток к серотонину,
ЕМИРамиду, нейротензину, кальцитонину, пептиду-активатору (роста) головы гидры (НАР)
и некоторым другим регуляторным соединениям не дали положительных результатов, при-
чем (что важно отметить) окрашивание не наблюдалось и в других тканях и органах. Лишь
использование флуоресцентно-гистохимического метода FAGLU позволило предположить
Рис. 244. Схема строения мелкогранулярной клетки
в эпителии средней кишки Priapulus caudatus.
кк - клеточные контакты; о - отросток мелкогранулярной клетки.
Fig. 244. Organization of a small granular cell
in Priapulus caudatus midgut epithelium.
112
наличие в эпителии средней кишки Halicryptus spinulosus и Priapulus caudatus клеток, со-
держащих катехоламины и серотонин (Пунин, 1999).
У Halicryptus spinulosus в эпителии присутствуют клеточные элементы как демонстри-
рующие типичную для катехоламинов зеленую флуоресценцию, так и характеризующиеся
золотисто-желтым свечением, свойственным серотонин-содержащим структурам. Послед-
ние исключительно редки и представлены клетками округлой или слегка неправильной
формы, лежащими в основании эпителия у базальной мембраны (рис. 245, вклейка). В стен-
ке пищеварительного тракта за пределами эпителиального пласта каких-либо структур с
золотисто-желтым свечением не отмечено.
Катехоламин-содержащие клетки и отростки располагаются, главным образом, в мы-
шечной стенке средней кишки, причем в отдельных участках отростки образуют довольно
заметное сплетение. В то же время единичные (но более многочисленные, чем содержащие
серотонин) клетки зарегистрированы и в составе эпителиального пласта (рис. 246-248,
вклейка). Интересно, что, несмотря на уже отмеченную редкую встречаемость, местами в
поле зрения микроскопа удается наблюдать до 5-6 таких клеток (рис. 246, вклейка).
Большинство катехоламинергических клеток имеет вытянутую форму и ориентировано
перпендикулярно базальной мембране (рис. 247, вклейка), однако контакта их апикальных
частей с просветом не отмечено (возможно, это связано с отсутствием в апикальной цито-
плазме катехоламинов). Некоторые клетки имеют короткие тонкие базальные отростки.
Местами в основании эпителия выявляются очень рыхлые, небольшие по занимаемой пло-
щади, локальные сплетения тончайших катехоламинергических отростков.
В средней кишке Priapulus caudatus были зарегистрированы лишь катехоламинергиче-
ские клетки, напоминающие аналогичные структуры у Halicryptus spinulosus, однако бы-
строе падение интенсивности свечения не дало возможности наблюдать детали их строения.
К сожалению, крайняя скудость данных не позволяет серьезно обсуждать организацию
кишечной регуляторной системы у представителей этой своеобразной группы Invertebrata.
Пока можно лишь утверждать, что такая система у приапулид имеется.
Строение интраэпителиальных катехоламинергических клеток Halicryptus spinulosus и
Priapulus caudatus (в первую очередь наличие базальных отростков и базиэпителиального
сплетения, правда, развитого очень слабо) позволяет предположить их принадлежность к
периферической нервной системе. На основании этих же критериев серотонин-содержащие
клетки Halicryptus spinulosus, скорее всего, являются эндокринными. По уже отмеченному
наличию базальных отростков и положению в эпителиальном пласте катехоламинергиче-
ские клетки обеих изученных приапулид имеют определенное сходство с мелкогрануляр-
ными клетками средней кишки Priapulus caudatus. Однако гранулы последних слишком
крупны для содержащих моноамины и напоминают, скорее, пептидные гранулы.
Можно ожидать, что дальнейшие исследования продемонстрируют существенную ге-
терогенность клеточного состава кишечной регуляторной системы приапулид. В связи с
этим мне кажется, что отрицательные результаты иммуногистохимического анализа, ско-
рее, связаны с какими-то биохимическими особенностями тканей этих животных, требую-
щими (для получения нормальной реакции) подбора специальных модификаций стандарт-
ных методик. Трудно предполагать, что Priapulida являются исключением, и у них нет ней-
ротензин- или ЕМИЕамид-подобных веществ, широко распространенных у представителей
разных систематических групп беспозвоночных. В пользу этого предположения говорит и
факт выявления серотонин-содержащих клеток у Halicryptus spinulosus флуоресцентно-
гистохимическим методом при отсутствии положительного результата в экспериментах с
использованием соответствующей антисыворотки.
Глава 12
NEMERTINI (НЕМЕРТИНЫ)
Положение этого типа беспозвоночных, насчитывающего около тысячи видов, в систе-
ме животного царства до конца не ясно. Отсутствие целома, строение покровов и некоторые
другие черты сближают этих животных с ресничными червями (Turbellaria, Plathelminthes),
тогда как наличие замкнутой кровеносной системы и сквозного пищеварительного тракта
указывают на прогрессивную эволюцию этой группы. Более того, существует точка зрения,
что кровеносные сосуды, полость хоботка (ринхоцелом), мешки гонад представляют собой
модифицированный целом, и, следовательно, немертин все же следует относить к
Coelomata (Remane et al., 1980; Turbeville, Ruppert, 1985). Этот тип по наличию или отсутст-
вию стилетов на выворачивающемся хоботке разделяют на 2 класса (или подкласса): Enopla
(вооруженные немертины) и Anopla (невооруженные немертины).
Пищеварительная система немертин состоит из эктодермального пищевода, длинной
эктодермальной средней кишки [как правило, с повторяющимися парными боковыми вы-
ростами (карманами), между которыми у половозрелых особей расположены гонады] и эк-
тодермальной задней кишки. Эпителий кишечника (или, как его часто называют у этой
группы, гастродермис) образован столбчатыми ресничными клетками и, в меньшей степе-
ни, железистыми (Jennings, 1960, 1969; Jennings, Gibson, 1969, Gibson, Jennings, 1969 и др.).
Сведения о присутствии в пищеварительном тракте немертин регуляторных элементов, за
исключением упоминания о наличии нервных и так называемых «глиальных» отростков
(Turbeville, Ruppert, 1985), в известной мне литературе отсутствуют.
Использование метода импрегнации азотнокислым серебром по Колонье дало возмож-
ность мне (совместно с О. В.Зайцевой) выявить в средней кишке целого ряда видов —
Lineus viridis (=gesserensis), Lineus ruber, Cephalotrix linearis, Cerebratulus sp. (все — Anopla)
и Amphiporus lactifloreus (Enopla) — большое количество интраэпителиальных нервных
клеток и мощного сплетения отростков, расположенного на границе эпителия и подсти-
лающей его мышечной оболочки (рис. 249-253, вклейка). Более подробно к настоящему
времени изучен первый вид — Lineus viridis, у которого удалось выделить несколько мор-
фологических типов рецепторных клеток (Зайцева, Пунин, 2001): 1) малодифференциро-
ванные рецепторные клетки, по форме и размерам сходные с обычными эпителиальными;
2) рецепторные клетки с дифференцированным чувствительным периферическим отрост-
ком; 3) рецепторные клетки с одним или несколькими периферическими отростками, вет-
вящимися в эпителии (рис. 254). Как правило, эти клетки несут на апикальной поверхности
микроворсинки, однако некоторые, возможно, имеют пучки длинных цилий (ресничек).
Помимо рецепторных клеток, в стенке кишечника выявляется большое количество
свободных нервных окончаний, а также отдельные нейроны, расположенные в эпителии и
114
Рис. 254. Нервные элементы в эпителии боковых карманов средней кишки Linens viridis.
нк- интраэпителиальный нейрон; рк! - малодифференцированные рецепторные клетки;
рк2 - рецепторная клетка с дифференцированным чувствительным отростком;
ркЗ - рецепторная клетка с периферическим отростком, ветвящимся в эпителии;
цо - центральные отростки рецепторных клеток.
Импрегнация азотнокислым серебром по Колонье, рисовальный аппарат. Шкала: 10 мкм.
Fig. 254. Nerve elements in the midgut lateral pouches epithelium of Lineus viridis.
не имеющие контакта с просветом кишечника; отростки последних преимущественно лежат
в базальной части эпителиальной выстилки.
Центральные отростки рецепторных клеток (отростки, отходящие от базальных частей)
образуют на границе эпителия и подстилающих структур уже отмеченное мощное сплете-
ние. Оно представляет собой кольца вокруг кишечника, связанные между собой более тон-
кими пучками и/или отдельными отростками (рис. 253, вклейка). На толстых целлоидино-
вых срезах его локализацию — интра- или субэпителиальную — точно определить, к сожа-
лению, невозможно, однако предварительный электронно-микроскопический анализ сред-
ней кишки Lineus viridis показал, что, по крайней мере, частично сплетение занимает интра-
эпителиальное положение (рис. 255, вклейка).
От базального сплетения группы отростков вместе с центральными отростками рецеп-
торных клеток, расположенных в эктодерме, заходят в боковые (латеральные) нервные
стволы, тянущиеся вдоль всего тела животного. При этом часть отростков оканчивается в
нейропиле стволов, а часть проходит нейропиль «транзитом», отдавая в него только корот-
кие колатерали.
Пищеварительный тракт Lineus viridis также был изучен нами иммуногистохимическим
методом с использованием антисывороток к ЕМИЕамиду и нейротензину.
Антитела к ЕМИЕамиду не выявили в эпителии кишечника каких-либо иммунореак-
тивных структур, хотя отдельные нейроны и отростки были отмечены в боковых нервных
стволах. (Отмечу, что сходная картина окрашивания была получена и на Amphiporus
lactifloreus.)
115
В то же время нейротензин-иммунореактивные клетки присутствовали в эпителии ки-
шечника (рис. 256-257, вклейка), причем заметно чаще — в его второй половине. Эти, как
правило, единичные клетки располагаются поодиночке, имеют вытянутую форму с апи-
кальным концом, направленным к просвету кишки и, по-видимому, достигающим его. Ядра
расположены в нижней трети эпителиального пласта, иногда довольно близко к базальной
мембране. Базальные отростки у этих клеток отсутствуют.
Хотя нейротензин-иммунореактивные отростки были также отмечены в нейропиле бо-
ковых нервных стволов (рис. 256, вклейка), их контактов с клетками кишечника не наблю-
далось.
Таким образом, наши наблюдения позволяют предполагать наличие в кишечнике не-
мертин достаточно сложно организованной регуляторной системы. Ее основную часть со-
ставляют нервные (в первую очередь — рецепторные) элементы. В то же время нельзя ис-
ключить присутствие в эпителии и эндокриноподобных элементов: выявленные у Lineus
viridis нейротензин-иммунореактивные клетки по своей морфологии более соответствуют
таковым.
Глава 13
TURBELLARIA (ТУРБЕЛЛЯРИИ, ИЛИ РЕСНИЧНЫЕ ЧЕРВИ)
Ресничные, черви на правах класса входят в тип плоских червей (Plathelminthes) — наи-
более просто устроенных двусторонне симметричных животных (Bilateria). К настоящему
времени данные по элементам кишечной регуляторной системы у Plathelminthes этим клас-
сом и исчерпываются.
Ресничные черви представляют особый интерес в плане изучения кишечных регуля-
торных систем. Связано это с тем, что турбеллярии среди современных форм, очевидно,
наиболее близки к общему предку многоклеточных. Немаловажно и то, что у представите-
лей этого класса мы наблюдаем различные варианты морфофункциональной организации
пищеварительной системы — от пищеварительной паренхимы у бескишечных турбеллярий
(Acoela) до вполне микроанатомически и гистологически дифференцированного кишечника
(см. подробнее: Иванов, Мамкаев, 1973; Мамкаев, 1979). Таким образом, можно ожидать
обнаружения у представителей этой группы животных стадии возникновения кишечной
регуляторной системы, наиболее близкие к исходным. Однако к настоящему времени в
этом аспекте пищеварительная система турбеллярий изучена крайне недостаточно.
13.1. ДАННЫЕ ГИСТОЛОГИЧЕСКИХ
И ИММУНОГИСТОХИМИЧЕСКИХ ИССЛЕДОВАНИЙ
Субэпителиальный нервный плексус в кишечнике, образованный только нервными от-
ростками, описан с использованием гистологических и гистохимических методов у некото-
рых видов рода Dugesia (отр. Tricladida) (Baguna, 1974; Baguna, Ballester, 1978) и у Acerotisa
sp. (отр. Polycladida) (Котикова, 1983). Кишечный плексус, состоящий как из отростков, так
и из небольших нейронов, отмечен у представителей отрядов Catenulida, Seriata,
Lecithoepitheliatha и Rhabdocoela (Reisinger, 1976), однако, к сожалению, из описания его
точное (интра- или субэпителиальное) положение остается неясным.
Присутствие кишечного плексуса, реагировавшего с антисывороткой к нейропептиду F
(NPF), было показано у Dugesia tigrina (Reuter et al., 1995a) и Microstomuni lineare (отр.
Macrostomida) (Reuter et al., 1995b). В кишечнике Polycelis nigra описаны отростки, имму-
нореактивные к FMRFaмидy (Wikgren et al., 1986). Однако в этих исследованиях, как и в
отмеченных выше работах, нет указаний на их точное положение относительно эпителия.
Теперь остановлюсь на имеющихся литературных данных, касающихся интраэпители-
альных клеток — возможных кандидатов в регуляторные.
117
Интраэпителиальные ЕМИРамид-иммунореактивные клетки впервые были продемон-
стрированы у Microstomum lineare (Reuter et aL, 1984; Wikgren et al., 1986) и у Geratrix
hermaphroditicus (отр. Neorhabdocoela, п/отр. Kalyptorhynchia) (Reuter et al., 1988). У первого
вида описаны также клетки, реагировавшие с антисыворотками к бычьему панкреатическо-
му полипептиду (РР) и вазотоцину (Reuter, Palmberg, 1987). Однако здесь следует отметить
два момента: первый заключается в том, что с антителами к FMRFамиду и РР, очевидно,
реагируют одни и те же клетки; второй, более важный для нас, касается природы этих кле-
ток. Было отмечено, что иммунореактивные клетки содержат в своих апикальных частях
неокрашивающиеся гранулы, тогда как продукт пероксидазной реакции (в качестве маркера
использовалась пероксидаза) присутствует над базальными частями. На основании этих
наблюдений авторами исследования был сделан вывод, что в данном случае выявлялись не
какие-то специализированные клетки, а типичные железистые (так называемые «granular-
club cells»), с характерным расположением шероховатой эндоплазматической сети в осно-
вании клетки и околоядерной зоне и секреторных гранул - в апикальной цитоплазме. Толь-
ко вазотоцин-иммунореактивные клетки отличались своей формой от секреторных (причем
были отмечены клетки как закрытого, так и открытого типов: оба без базальных отростков),
однако насколько наблюдавшаяся реакция была специфичной, осталось неясным, посколь-
ку контрольные окрашивания не проводились (Reuter, Palmberg, 1987).
Совместно с Т.Г.Маркосовой и В.К.Казаковым мы провели иммуногистохимическое
выявление возможных регуляторных элементов у 13 видов турбеллярий: Notoplana atomata,
Notoplana humilis (отр. Polycladida), Procerodes littoralis, Uteriporus vulgaris, Planaria torva,
Dendrocoelum lacteum (отр. Tricladida), Macrostomum sp. (отр. Macrostomida), Monocelis fusca
(отр. Proceriata), Plagiostoniuni vitatum (отр. Prolecithophora), Macrorhynchus crocea,
Provortex karlingi, Astrothorynchus bifudus (отр. Neorhabdocoela) и Diceratocephala boschma
(отр. Temnocephalida). Для анализа были использованы антисыворотки к серотонину, FMR-
Рамиду и нейротензину. Полученные результаты частично опубликованы (см.: Маркосова,
Пунин, 1997; Markosova et al., 1997; Punin, Markosova, 2000; Пунин, Маркосова, 2001).
Несколько замечаний о строении кишечника турбеллярий: в микроанатомическом пла-
не у представителей этих отрядов он заметно различается. Наиболее сложную дифференци-
ровку кишечник имеет у Polycladida и Tricladida. У первых от глотки берет начало главная
кишка с передним и двумя боковыми выростами. Эти выросты, в свою очередь, образуют
многочисленные ответвления, пронизывающие все тело червя. Три ветви кишечника, разде-
ляющиеся на многочисленные выросты, отходят от глотки и у Tricladida. У представителей
других отрядов кишечник организован значительно проще и не имеет такой системы вы-
ростов. Общим для всех турбеллярий является то, что пищеварительная система слепо
замкнута.
Результаты нашего иммуногистохимического исследования представлены в табл. 5.
Далее я опишу полученные данные по конкретным видам несколько подробнее.
Отр. Polycladida. У Notoplana atomata, при обилии РМРРамид-иммунореактивных
клеток и отростков (расположенных главным образом субэпителиально) в глотке, в самом
кишечнике эти элементы не столь многочисленны и встречаются преимущественно в глав-
ной кишке. В конечных выростах иммунореактивные клетки и отростки единичны.
Реагирующие с антисывороткой клетки и тонкие отростки с характерными варикозны-
ми расширениями встречаются как в эпителии (у турбеллярий его часто называют гастро-
дермисом’, в дальнейшем я буду использовать оба термина как синонимы), так и под ним,
среди мышечных клеток и в паренхиме (рис. 258-263, вклейка), где они формируют сплете-
ние, окружающее кишку. Из этого сплетения отростки могут проникать в эпителий и под-
118
ходить к просвету кишечника (рис. 262, вклейка). Большинство иммунореактивных элемен-
тов— это уни-, би- и мультиполярные субэпителиальные клетки, расположенные как на
некотором удалении от эпителия, так и очень близко к нему. От некоторых из них отростки
идут непосредственно в гастродермис (рис. 259, вклейка). Часть клеток (как правило, уни- и
биполярных) лежит на границе между эпителиальным слоем и мышцами (насколько это
можно точно определить под световым микроскопом) (рис. 258, 263, вклейка). Интраэпите-
лиальные клетки довольно редки, и могут быть как открытого (с базально расположенным
ядром и узким апикальным отростком) (рис. 261, вклейка), так и закрытого (биполярные или
овальной формы) (рис. 260, вклейка) типов. Их перикарионы небольшие (порядка 10 мкм в
длину), высота клеток открытого типа соответствует высоте эпителия (т.е. колеблется в
пределах от 20 до 100 мкм).
Таблица 5
Интра- и субэпителиальные элементы, демонстрирующие различные иммунореактивности,
______.________________ в кишечнике ресничных червей____________________________
Виды Иммунореактивность к
РМИРамиду нейротензину серотонину
Notoplana atoniata +/+ +/- +/+
Notoplana humilis +/+ +/-
Procerodes littoralis +/+ +/+ +/+
Plana ria torva +/+ -/+ -/+
Dendrocoelum lacteum +/+ ?/+
Uteriporus vulgaris +/+
Macrorhynchus crocea +/+ -/- -А
Provortex karlingi +/+ -/- -А
Astrothorynchus bifundus +/- -А -А
Macrostomum sp. +/+ -А
Monocelis fusca +/+ -А -А
Plagiostomum vitatum +/+ -А -А
Diceratocephala boschma -/- -А -А
Примечание. + — наличие имунореактивных клеток и отростков;---------отсутствие иммунореактивных
клеток и отростков; ? — окрашивание предположительно железистых клеток; знак над чертой — гастродермаль-
ное (интраэпителиальное) положение; знак под чертой — субгастродермальное и пограничное положения.
Нейротензин-иммунореактивные элементы кишечника Notoplana atoniata представле-
ны главным образом интраэпителиальными клетками, отростки единичны. Как и FMRF-
амид-иммунореактивные элементы, они встречаются в основном в главной кишке и началь-
ных участках ее выростов. При этом интересно отметить, что в глотке подобные элементы
очень редки.
По своей морфологии нейротензин-иммунореактивные клетки могут быть разделены
на несколько типов, или разновидностей (рис. 264-267, вклейка). Две из них представляют
собой элементы открытого типа: 1) продолговатые клетки шириной 2-3 мкм, с вытянутым
ядром, расположенным в центральной части, без отростков; 2) более широкие (порядка 8
мкм) цилиндрические клетки с ядром, расположенным в нижней половине, с 2-3 короткими
базальными отростками. У этих клеток с антисывороткой реагирует преимущественно ба-
зальная цитоплазма. Клетки третьей разновидности — закрытого типа. Они имеют
продолговатую форму, ориентированы параллельно базальной границе эпителия; отростков
у них не обнаружено.
Мы наблюдали лишь единичные интраэпителиальные нейротензин-иммунореактивные
отростки вне прямой связи с перикарионами. В отличие от ГМИРамид-иммунореактивных
119
они толще и не имеют варикозных расширений. По-видимому, эти отростки принадлежат
описанным выше клеткам, не попавшим на срез.
Нужно отметить, что в непосредственной близости от эпителия нейротензин-
иммунореактивные структуры отсутствовали.
Интраэпителиальные серотонин-иммунореактивные клетки редки, однако встречаются
и в главной кишке, и в боковых выростах. Они сравнительно крупные; ядра располагаются
в центральной части гастродермиса, апикальный отросток подходит к просвету (рис. 268,
вклейка). Субэпителиальные клетки — биполярные и мультиполярные — многочисленны,
имеют короткие отростки, которые, однако, не доходят до основания гастродермиса.
В кишечнике Notoplana huniilis ЕМИЕамид-иммунореативные структуры, по сравнению
с предыдущим видом, встречаются значительно реже. В главной кишке отмечен лишь до-
вольно рыхлый субэпителиальный плексус с небольшими редкими перикарионами, а в бо-
ковых выростах — только единичные отростки. В гастродермисе удалось наблюдать крайне
редкие мелкие мультиполярные клетки закрытого типа.
Характер распределения нейротензин-иммунореактивных элементов также в целом
сходен с наблюдавшимся у Notoplana atomata. Что касается клеточного состава, нам уда-
лось наблюдать лишь две разновидности интраэпителиальных клеток открытого типа; эле-
менты закрытого типа отсутствовали.
Отр. Tricladida. У Procerodes littoralis ЕМИЕамид-иммунореактивные элементы в ки-
шечнике очень редки и представлены только отростками. Эти отростки лежат преимущест-
венно субэпителиально, образуя рыхлое сплетение. Некоторые из них тянутся между осно-
ваниями эпителиальных клеток или вдоль базальной границы эпителия. Наконец, единич-
ные интраэпителиальные отростки с характерным варикозными расширениями направлены
к просвету кишечника, однако, по-видимому, его не достигают (рис. 269, вклейка).
Нейротензин-иммунореактивные структуры более многочисленны и разнообразны по
форме; они встречаются как в эпителии, так и под ним. (Интересно, что в глотке подобные
структуры отсутствуют.) Субэпителиальные клетки небольшие (перикарионы не превыша-
ют в длину 10 мкм), обычно биполярные, лежат на границе между эпителием и мышцами.
Клеточные отростки длинные и образуют субэпителиальное сплетение. Некоторые из них
проникают в эпителий и походят к просвету кишки, другие уходят глубже в мышцы.
Интраэпителиальные клетки представлены элементами как закрытого, так и открытого
типов (рис. 270-271, 273, вклейка). Среди первого отмечены 2 разновидности: 1) неболь-
шие, имеющие на срезах овальную или треугольную форму, и 2) более крупные, округлые
(диаметром 15-20 мкм), с ядрами неправильных очертаний. Иногда у этих клеток отходят
короткие отростки, которые, по-видимому, оканчиваются в основании эпителиального пла-
ста. Клетки открытого типа имеют веретеновидную или цилиндрическую форму, только у
некоторых из них отмечены 1-2 базальных отростка.
Некоторые из этих отростков тянутся в основании эпителия, другие покидают его и
вступают в состав субэпителиального плексуса; точное положение многих отростков под
световым микроскопом определить невозможно.
Наконец, в кишечнике выявлена и еще одна разновидность нейротензин-иммуно-
реактивных клеток. Их небольшие перикарионы занимают неясное (интра-, субэпителиаль-
ное) положение; один из отростков тянется вдоль базальной границы эпителия, а другой
направлен к просвету кишечника (рис. 272, вклейка).
Серотонин-иммунореактивные элементы у Procerodes littoralis весьма многочисленны,
однако расположены главным образом под эпителием (рис. 274-275, вклейка). Это — бипо-
лярные и мультиполярные клетки с длинными ветвящимися отростками, образующими
120
сплетение вокруг кишечника. Некоторые отростки проникают в эпителий, где либо оканчи-
ваются между основаниями гастродермальных клеток, либо достигают просвета кишки
(рис. 275, вклейка). Сплетение наиболее хорошо заметно недалеко от глотки, в дистальных
ответвлениях кишки выражено слабее. Интраэпителиальные клетки единичны, присутству-
ют только в начале кишечника; они представлены элементами закрытого типа.
У Planaria torva ЕМИЕамид-иммунореактивные клетки отмечены по всей длине кишки.
Все они — закрытого типа (рис. 276, вклейка), некоторые имеют отростки. Кишечник окру-
жает субэпителиальное сплетение, образованное биполярными и мультиполярными клетка-
ми; некоторые из их отростков проникают в гастродермис, оканчиваясь в его основании.
Нейротензин-иммунореактивные элементы представлены единичными субэпителиаль-
ными клетками и отростками, тянущимися вдоль границы с гастродермисом. Возможно,
некоторые из отростков могут проникать в его основание. Сходная картина наблюдается и в
отношении серотонин-иммунореактивных структур.
ЕМИЕамид-иммунореактивные элементы в кишечнике Dendrocoelum lacteuni встреча-
ются как в гастродермисе, так и за его пределами. Интраэпителиальные клетки очень редки.
Они могут быть отнесены к закрытому типу и отростков, как правило, не имеют. Окру-
жающие кишечник клетки и образующие сплетение отростки, напротив, достаточно много-
численны. Они встречаются по всему кишечнику и могут вплотную прилегать к гастродер-
мису. Часть отростков из этого сплетения проникает в гастродермис, некоторые из них
оканчиваются рядом с основаниями железистых эпителиальных клеток.
Под эпителием присутствуют нейротензин-иммунореактивные клетки (рис. 278, вклейка)
с длинными отростками, формирующими сплетение, окружающее кишечник. В самом гаст-
родермисе (главным образом в начале кишки) отмечено значительное количество иммуноре-
активных клеток, в том числе с отростками. Однако их размеры, форма и характер окрашива-
ния цитоплазмы, скорее, соответствуют железистым гастродерм ал ьным элементам.
Нейротензин-иммунореактивные элементы Uteriporus vulgaris представлены, с одной
стороны, субгастродермальным сплетением вокруг кишечника, с другой — интраэпители-
альными клетками (рис. 279-280, вклейка). Последние — вытянутой формы, открытого ти-
па, встречаются преимущественно в главной кишке. Отростки из субгастродермального
сплетения могут проникать в эпителий, некоторые из них доходят до просвета кишечника.
Отр. Neorhabdocoela. У всех изученных 3 видов этого отряда (Macrorhynchus сгосеа,
Provortex karlingi и Asthrothorynchus bifudus) в гастродермисе и вблизи кишечника нейро-
тензин- и серотонин-иммунореактивные клетки и отростки отсутствовали, хотя в глотке
отмечалась положительное окрашивание.
Что касается ЕМЯГамид-иммунореактивных структур, у Asthrothorynchus bifudus они
представлены единичными отростками в основании гастродермиса. У других 2 видов отме-
чено состоящее из клеток и отростков сплетение, расположенное вдоль базальной поверх-
ности эпителия и на некотором удалении от него. У Provortex karlingi некоторые отростки
из этого сплетения проникают в основание гастродермиса. Отмечены также единичные
мелкие интраэпителиальные клетки закрытого типа.
Другие отряды. У представителей других исследованных отрядов мы не выявили в
кишечнике и непосредственной близости от него серотонин- (кроме Macrostomum sp., кото-
рый не анализировался) и нейротензин-иммунореактивных элементов (а у Diceratocephala
boschma — также и ЕМИЕамид-иммунореактивных структур), хотя в глотке наблюдались
отростки и перикарионы, интенсивно реагировавшие с этими антителами.
У Macrostomum sp. (отр. Macrostomida), Monocelis fusca (отр. Proceriata) и Plagiostomum
vitatum (отр. Prolecithophora) выявлены элементы ЕМЯЕамид-иммунореактивного сплетения
121
вокруг кишечника. Отличия между видами заключаются в степени его развития и соотно-
шении би- и мультиполярных клеток в его составе. У Monocelis fusca наблюдались также
единичные отростки, проникающие в гастродермис и достигающие просвета кишки. В гаст-
родермисе отмечены также единичные мелкие клетки: закрытого типа — у Macrostomum sp.
и Plagiostomum vitatum, открытого — у Monocelis fusca. _
В завершение этой части главы отмечу, что при исследовании Notoplana atomata и
Procerodes littoralis мы использовали также антисыворотку к кальцитонину. В кишечнике и
вблизи него реакция была отрицательной, хотя в паренхиме и (особенно) половой системе
этих червей удалось наблюдать довольно многочисленные окрашивающиеся клетки и отро-
стки.
13.2. ДАННЫЕ ЭЛЕКТРОННО-МИКРОСКОПИЧЕСКИХ ИССЛЕДОВАНИЙ
В известной мне литературе данные электронной микроскопии имеются лишь для од-
ного вида — Microstomum lineare (отр. Macrostomida). В составе эпителия кишечника этого
червя описаны 4 типа клеток (рис. 281): фагоцитирующие, гранулярные (секреторные), не-
областы (недифференцированные клетки) и так называемые гастродермалъные нейроны
(Reuter, Palmberg, 1987). Последние характеризуются электронносветлой цитоплазмой;
большинство из этих клеток имеет форму, близкую к цилиндрической и ориентировано
перпендикулярно базальной мембране. Однако апикальные части нейронов, по-видимому,
Рис. 281. Эпителий кишечника Microstomum lineare с гастродермальным нейроном
(по: Reuter, Palmberg, 1987, с некоторыми изменениями).
гн - гастродермальный нейрон; жк - железистая клетка; нб - необласт;
фк - фагоцитирующие (пищеварительные) клетки.
Fig. 281. Intestinal epithelium of Microstomum lineare with a gastrodermal neuron
(from: Reuter, Palmberg, 1987, with some changes).
122
не достигают просвета кишечника. Интересно, что не все клетки, согласно наблюдениям
авторов, контактируют и с базальной мембраной.
В цитоплазме гастродермальных нейронов отмечены свободные рибосомы, полисомы
и разбросанные цистерны шероховатой эндоплазматической сети, а также многочисленные
стопки цистерн аппарата Гольджи; гранулы с электронноплотным центром имеют диаметр
от 70 до 100 нм.
Отростки с аналогичными по размерам и тонкому строению гранулами встречаются
как у базальной мембраны, так и между фагоцитирующими клетками. Предполагается, что
они принадлежат гастродермальным нейронам. Отмечены как обычные (односторонние)
синапсы на фагоцитирующих (пищеварительных) клетках, так и двусторонние синапсы ме-
жду такими отростками (Reuter, Palmberg, 1987).
Наши электронно-микроскопические исследования кишечника Procerodes littoralis,
Planaria torva, Dendrocoelum lacteum, Macrostomum sp. и Provortex karlingi выявили значи-
тельное количество нервных отростков и тела нервных клеток. Большая их часть располага-
лась под гастродермисом, иногда в прямом контакте с базальной мембраной. Единичные
отростки и перикарионы были зарегистрированы и в гастродермисе (рис. 282-283, вклейка).
В цитоплазме клеток присутствовали в большом количестве мелкие гранулы с электронно-
плотным центром диаметром около 100 нм и пузырьки. Аппарат Гольджи в этих клетках
представлен несколькими стопками цистерн, расположенных вокруг ядра; цистерны шеро-
ховатой эндоплазматической сети обычно собраны в компактные группы. Митохондрии, в
отличие от обычных гастродермальных клеток, мелкие. В целом строение этих клеток соот-
ветствовало строению нейронов, лежащих в паренхиме и в составе нервных стволов.
В то же время в гастродермисе были отмечены отростки, отличные как по морфологии,
так и форме и размерам присутствующих в них гранул (рис. 284, 286-288, вклейка). Интерес-
но, что, помимо интраэпителиальных, отличные от нервных отростки встречались и непо-
средственно под гастродермисом (рис. 289, вклейка). У Dendrocoelutn lacteum и Planaria torva
в основании гастродермиса несколько раз были отмечены фрагменты клеток с гранулами,
принципиально иные, чем в железистых клетках и нейронах. Некоторым из этих клеток свой-
ственно хорошее развитие белок-синтезирующего аппарата (рис. 285, вклейка). К сожалению,
крайне редкая встречаемость таких элементов не позволила дать их подробное описание.
13.3. ЗАКЛЮЧЕНИЕ
Анализируя имеющиеся данные по кишечным регуляторным элементам ресничных
червей, можно придти к заключению, что основу рассматриваемой системы в этой группе
животных составляют нервные клетки и отростки. Очевидно, что суб- и интраэпителиаль-
ные элементы морфологически тесно связаны друг с другом и образуют в кишечнике еди-
ную сеть. Степень развития этой системы, по-видимому, в значительной степени коррели-
рует со степенью микроанатомической дифференцировки кишечника. Так, наибольшее
морфологическое разнообразие интра- и связанных с ними субгастродермальных нервных
элементов мы наблюдаем у исследованных представителей отрядов Polycladida и Tricladida
(рис. 290). Интересно, что и общее количество интрагастродермальных нейронов в этих
отрядах, по-видимому, больше, чем в других.
Несмотря на то, что набор использованных для выявления регуляторных элементов ан-
тисывороток к настоящему времени весьма невелик, нетрудно заметить, что кишечная
нервная система турбеллярий достаточно сложна и в отношении состава биологически ак-
123
Рис. 290. Морфологическое разнообразие РМИГамид- (A-В), нейротензин- (Г-Д) и серотонин- (Е-Ж)
иммунореактивных элементов и их различное положение относительно гастродермиса в кишечнике
Notoplana atomata (А, Г, Ж), Procerodes littoralis (Б, Д-Е) и Planaria torva (В).
Fig. 290. Morphological diversity of FMRFamide- (A-В), neurotensin- (Г-Д) and serotonin-
(Е-Ж) immunoreactive elements and their different position relatively to gastrodermis
in the intestine of Notoplana atomata (А, Г, Ж), Procerodes littoralis (Б,Д-Е)
and Planaria torva (B).
тивных соединений. Подчеркну, что и в этом плане наибольшее разнообразие демонстри-
руют представители отрядов с более дифференцированным пищеварительным трактом. Тот
факт, что (за небольшим исключением) элементы, демонстрирующие одинаковые иммуно-
реактивности, имеют и интра-, и субэпителиальное положение, также свидетельствует о
том, что они образуют в кишечнике единую систему.
В то же время есть основания полагать, что, помимо нервных элементов в составе гаст-
родермиса (по крайней мере некоторых изученных нами червей) присутствуют клетки по
своей общей морфологии и тонкому строению отличающиеся от типичных нервных и более
соответствующие эндокриноподобным. К таким клеткам можно предположительно отнести
в первую очередь нейротензин-иммунореактивные клетки Notoplana atomata и некоторые
клетки, реагирующие с этой антисывороткой, у Procerodes littoralis. Важно отметить, что
такие эндокриноподобные клетки опять наблюдаются у представителей отрядов с хорошо
микроанатомически (и гистологически) дифференцированным кишечником.
Таким образом, мы видим, что достаточно хорошо сложившаяся кишечная регулятор-
ная система может быть обнаружена уже у таких сравнительно низкоорганизованных мно-
гоклеточных, как ресничные черви.
Глава 14
CNIDARIA (КНИДАРИИ)
Книдарии в известной степени оказываются «исключением» среди всех многоклеточ-
ных, поскольку вместе с Ctenophora (гребневиками) представляют двухслойных животных,
образованных двумя эпителиальными пластами: эктодермальным и энтодермальным, раз-
деленными мезоглеей. Соответственно все регуляторные элементы (нервные клетки, обра-
зующие рыхлую сеть в основании эпителия) могут располагаться либо в одном из этих сло-
ев, либо в обоих (Davis, 1972; Josephson, 1974).
Хотя я считаю книдарий самостоятельной, рано обособившейся ветвью многоклеточ-
ных, тем не менее мне кажется интересным привести некоторые данные по организации их
энтодермальной нервной системы.
Принято считать, что нервная система книдарий состоит из нескольких основных кле-
точных элементов (рис. 291): 1) чувствительные (сенсорные) клетки открытого типа с рес-
ничками, среди которых выделяют также нейросекреторные клетки, дополнительно харак-
теризующиеся присутствием в базальных частях секреторных гранул; 2) ганглиозные клет-
Рис. 291. Нервная система Hydra (по: Lentz, 1968).
гк - ганглионарные клетки; гп - гастральная полость; жк - железистая (секреторная) клетка;
мг - мезоглея; рнк - рецепторные нервные клетки.
Fig. 291. The nerve system of Hydra (according to: Lentz, 1968).
125
ки закрытого типа, биполярные и мультиполярные, расположенные в основании эпители-
альных слоев; среди них также дополнительно выделяют нейросекреторные элементы
(Lentz, 1966, 1968; Lentz, Barmett, 1965; обзор: Chapman, 1974). Однако у Hydra, согласно
данным более поздних электронно-микроскопических исследований, нет принципиальной
разницы между рецепторными и ганглиозными клетками, так как практически все они со-
держат секреторные гранулы, образуют синаптические контакты и являются одновременно
мото- и интернейронами (Westfall, 1973; Westfall, Kinnamon, 1978).
Нервные элементы развиваются из интерстициальных клеток (Davis, 1971; Chapman,
1974). Контакты в нервной сети между отростками или отростками и клетками преимуще-
ственно синаптические; синапсы могут быть как поляризованными (с синаптическими пу-
зырьками только в одной из составляющих контакт), так и неполяризованными двусторон-
ними (когда пузырьки присутствуют по обе стороны синапса) (Josephson, 1974). В нервных
элементах гидры (класс Hydrozoa - гидрозои) выявлены многочисленные биологически ак-
тивные пептиды: гастрин/холецистокинин, субстанция Р, нейротензин, бомбезин, оксит-
цин/вазопрессин, FMRF-амид и другие (см.: Spencer, 1989). Моноамины не выявлены, и
считается, что нервная система кишечнополостных исключительно пептидергическая
(Grimmelikhuijzen et al., 1996). Отмечу, однако, что в большинстве случаев присутствие от-
меченных выше соединений показано в нервных клетках эктодермы.
У морской анемоны Calliactis parasitica (кл. Anthozoa — коралловые полипы) выявлено
наличие в энтодерме «рецепторных» клеток и базиэпителиального сплетения отростков,
иммунореактивных к анто-КБамиду (Grimmelikhuijzen et al., 1989, 1992). У другого пред-
ставителя этого класса (актинии Hydractinia echinatd) в области гипостома присутствуют
клетки открытого типа с короткими базальными отростками, имунореактивные к
CLWaMHfly— одному из новых нейропептидов, обнаруженных у кишечнополостных
(Schmich et al., 1998).
Особый интерес представляет локализация у гидры четырех соединений-морфогенов:
активатора и ингибитора (роста) головы и активатора и ингибитора (роста) подошвы, под
контролем которых находятся рост и дифференцировка всего животного (Schaller et al.,
1979; Schmidt et al., 1980). Активатор головы (результаты использования антисыворотки к
которому на многих беспозвоночных приводились в предыдущих главах) представляет со-
бой пептид из 11 аминокислотных остатков со следующей последовательностью: pGlu-Pro-
Pro-Gly-Gly-Ser-Lys-Val-Ile-Leu-Phe. Этот пептид впервые был выделен из гидры и дру-
гих кишечнополостных (Schaller, 1973), а позднее, что интересно, выявлен также в мозге и
кишечнике млекопитающих (Bodenmuller, Schaller, 1981; Schaller et al., 1989). Больше всего
активатора головы обнаруживается в головном конце животного, причем биохимическими
и цитологическими методами показано, что он содержится в секреторных гранулах нервных
клеток (Schaller, Gierer, 1973).
У гидры активатор выводится в межклеточное пространство и действует, с одной сто-
роны, как митоген на все типы делящихся клеток, а с другой — определяет специфическую
дифференцировку нервных клеток (Hoffmeister, Schaller, 1987; Schaller et al., 1989).
Используя антисыворотку к синтетическому пептиду-активатору головы, нам удалось
продемонстрировать конкретную локализацию клеток, содержащих этот пептид (Punin,
Kazakov, 199Id). Все иммунореактивные элементы располагались в энтодерме, но больше
всего их наблюдалось в области гипостома (рис. 292, вклейка), где клетки формировали во-
круг него своеобразное кольцо (рис. 294, вклейка). Сходная картина отмечалась и у моло-
дых, еще не отпочковавшихся гидр (рис. 293, вклейка). По направлению к подошве количе-
ство клеток резко падало, и уже в центральной части животного реакции не отмечалось.
126
Небольшие скопления иммунореактивных клеток также встречались в участках формиро-
вания почек (вегетативное размножение гидр). Таким образом, характер локализации эле-
ментов, содержащих вещество, иммунореактивное к головному активатору, полностью со-
ответствовал данным биохимических исследований.
Клетки, реагирующие с антисывороткой к активатору головы, имеют (как и другие
нервные элементы) очень небольшие размеры. Среди них встречаются клетки как открыто-
го, так и закрытого типов (рис. 295-296, вклейка); в районе гипостома наблюдается хорошо
выраженное базиэпителиальное сплетение отростков.
Интересно отметить еще один факт. У лишенных нервных клеток линии гидр, полу-
чаемых в результате специальной обработки колхицином, активатор головы (как и три дру-
гих морфогена) тем не менее присутствует, причем иногда даже в больших, чем у интакт-
ных животных, концентрациях, и обеспечивает нормальный рост и регенерацию животного.
В этом случае синтез морфогенов осуществляют обычные гастродермальные клетки. У
«нормальных» животных этот процесс, очевидно, подавляется присутствием нервных эле-
ментов (Schaller et al., 1980).
Таким образом, у кишечнополостных в энтодерме расположена система нервных эле-
ментов, содержащих разнообразные биологически активные соединения как пептидной, так
и непептидной природы (отмеченные выше ингибиторы роста). В морфологическом отно-
шении она состоит из клеток открытого и закрытого типов, а регуляторные вещества могут
как выступать в роли медиаторов и модуляторов, так и оказывать паракринное действие
(т.е. действовать подобно биологически активным веществам эндокринных клеток) на со-
седние клетки.
Глава 15
РАСПРОСТРАНЕНИЕ КИШЕЧНЫХ РЕГУЛЯТОРНЫХ
СИСТЕМ У МНОГОКЛЕТОЧНЫХ
И ИХ ПРЕДПОЛАГАЕМАЯ ЭВОЛЮЦИЯ
В предыдущих главах были приведены известные мне литературные данные и резуль-
таты собственных наблюдений, касающиеся кишечных регуляторных систем и их элемен-
тов у представителей различных таксонов животного царства. В этой небольшой заключи-
тельной главе я хочу провести некоторые сравнения организации этих систем в разных
классах и типах многоклеточных, а также (в пределах имеющихся данных), предложить
гипотезу о происхождении компонентов кишечной регуляторной системы и ее предпола-
гаемой эволюции (последовательные этапы ее формулировки отражены в ряде обзоров —
см: Пунин, 1986, 1993, 19966, 2000, 2001).
Еще раз повторю замечание, уже сделанное в первой главе. Все приведенные выше ос-
новные данные касались главным образом строения этой системы и ее элементов. В опре-
деленной степени это не случайно. Дело в том, что, если энтериновая (кишечная гормо-
нальная) система позвоночных достаточно хорошо изучена как с точки зрения физиологии,
так и биохимии, относительно других групп многоклеточных такие сведения фрагментарны
или отсутствуют совсем, поэтому я буду сопоставлять организацию кишечных регулятор-
ных систем главным образом с точки зрения их морфологического строения.
Фрагментарность наших знаний о кишечных регуляторных системах во многих таксо-
нах (в первую очередь это касается беспозвоночных животных) ограничивает меня возмож-
ностью высказывать лишь предположения. Очевидно, что дальнейшие исследования смогут
многое изменить в трактовке организации этих систем и, соответственно, существенно
скорректировать, а, возможно, даже опровергнуть некоторые из изложенных далее
предположений.
15.1. РАСПРОСТРАНЕНИЕ И НЕКОТОРЫЕ ОБЩИЕ ЧЕРТЫ ОРГАНИЗАЦИИ
КИШЕЧНЫХ РЕГУЛЯТОРНЫХ СИСТЕМ
На рис. 297 показано распространение элементов рассматриваемой системы в ряду
многоклеточных животных.
Как видно из этой схемы, интраэпителиальные кишечные системы и их элементы
встречаются у представителей большинства систематических групп многоклеточных, рас-
полагающихся на разных ярусах эволюционного древа. Группы животных, у которых такие
элементы отсутствуют, единичны. Впрочем, я полагаю, что в действительности это связано
с недостаточно изученной в этом отношении пищеварительной системой, и дальнейшие
128
VERTEBRATA
Рис. 297. Распространение кишечных регуляторных систем у многоклеточных.
Жирным выделены названия типов и классов животных, у которых в составе эпителия
эндодермальной части пищеварительного тракта выявлены нервные и/или эндокринные клетки.
Взрослые Pogonophora не имеют пищеварительного тракта, лишь у одного представителя
Chelicerata отмечены интраэпителиальные нервные (?) отростки; Cnidaria в схему не включены.
Fig, 297. Distribution of intestinal regulatory systems in Metazoa,
исследования покажут присутствие эндокринных и/или нервных клеток у многих предста-
вителей и этих классов и типов. В этой связи особенно интересными для такого углублен-
ного анализа представляются такие группы, как Chaetognatha (щетинкочелюстные),
Chelicerata (хелицеровые) и Nemathelminthes (круглые черви). Однако я не исключаю, что в
каких-то группах беспозвоночных элементы кишечной регуляторной системы могут полно-
стью отсутствовать.
Важно отметить, что кишечные регуляторные элементы присутствуют у животных не-
зависимо от преобладающего типа пищеварения (внутриклеточное, полостное) и могут
встречаться в разных отделах пищеварительной системы, вне зависимости от их конкретной
функциональной роли в процессе пищеварения. При этом нетрудно заметить определенные
различия как в характере их распределения по разным отделам пищеварительного тракта,
так и морфологии составляющих их клеток и набору регуляторных веществ, ими продуци-
руемых (к последнему замечанию следует подходить достаточно осторожно, в пределах
129
достоверности гистохимических и иммуногистохимических методов) не только у предста-
вителей достаточно далеко отстоящих друг от друга таксонов, но даже у близкородствен-
ных видов.
Кишечные регуляторные системы, очевидно, гетерогенны не только в отношении ти-
пов клеток, синтезирующих различные регуляторные вещества, но и в отношении того, что
в большинстве систематических групп они образованы и нервными, и эндокринными (эн-
докриноподобными) элементами. (Исключение в этом плане составляют вторичноротые,
начиная с асцидий, и насекомые, у которых нервные клетки и отростки расположены вне
эпителиального пласта.) В то же время кишечные регуляторные системы разных групп бес-
позвоночных могут существенно различаться как по соотношению этих элементов, так и по
их связи с субэпителиальной нервной системой (субэпителиальным плексусом).
У представителей практически всех таксонов встречаются регуляторные клетки и от-
крытого, и закрытого типов. При этом часто оба типа клеточных элементов, по-видимому,
могут содержать одни и те же регуляторные вещества (или, по крайней мере, демонстриру-
ют сходные иммунореактивное™).
Еще одной важной общей чертой кишечных регуляторных систем является то, что их
элементы (как правило, разнородные по химизму) часто структурно объединены в единую
сеть за счет базальных отростков (что особенно хорошо прослеживается у животных сред-
них ярусов эволюционного древа— двустворчатых и брюхоногих моллюсков, полихет,
олигохет, морских звезд). Важно подчеркнуть, что такая картина характерна не только для
нервных элементов: ряд эндокринных клеток также имеет базальные отростки, связываю-
щие их друг с другом.
Обращается на себя внимание и тот факт, что общее количество интраэпителиальных
регуляторных элементов у большинства представителей различных систематических групп
весьма велико. Как я уже отмечал, хорошо известно, что у позвоночных животных кишеч-
ная диффузная эндокринная система играет существенную роль в общеорганизменной ре-
гуляции, поэтому можно предположить, что и у многих беспозвоночных животных ее
функция не ограничивается регуляцией деятельности только пищеварительного тракта (на
последнее указывают и отмеченные ранее специфические взаимоотношения регуляторных
клеток и их отростков с синусами кишечного кровеносного сплетения у кольчатых червей).
15.2. ПРЕДПОЛАГАЕМАЯ ЭВОЛЮЦИЯ
КИШЕЧНЫХ РЕГУЛЯТОРНЫХ СИСТЕМ
Проблема эволюции кишечных регуляторных систем тесно связана с вопросом о гисто-
генетических источниках ее элементов.
Как уже отмечалось ранее (см. гл. 2), Пирс (Pearse, 1968, 1969, 1973) в своей концепции
APUD-системы рассматривал все эндокринные клетки пищеварительной системы как про-
изводные нейроэктодермы. Однако позднее в свете новых данных он пересмотрел эту часть
своей теории, признавая энтодермальное происхождение эндокринных элементов гастро-
энтеро-панкреатической системы (Pearse, 1987). После этого концепция в известной степени
потеряла свою стройность и законченность; впрочем, и изначально в ней мало внимания
было уделено эволюционным аспектам.
Японский исследователь Т.Фудзита (Т.Fujita), первоначально придерживавшийся кон-
цепции APUD-системы, в дальнейшем стал ее критиком. Принимая во внимание, что не все
нейроны и пептид-продуцирующие эндокринные клетки обладают способностью погло-
щать и декарбоксилировать предшественники аминов, что эндокринные клетки могут иметь
130
различные эмбриональные источники, что четкие критерии для нейронов и эндокринных
клеток в действительности отсутствуют, им была выдвинута концепция паранейрона
(paraneuron) (Fujita et al., 1980, 1988).
Согласно этой концепции к паранейронам следует относить клетки, которые: 1) проду-
цируют вещества, сходные или идентичные нейротрансмиттерам или нейрогормонам;
2) содержат в цитоплазме гранулы и/или пузырьки, по строению напоминающие нейросек-
реторные или синаптические, и 3) являются по своей природе рецепторно-секреторными.
Парнейроны могут иметь различные эмбриональные источники своего происхождения,
большинство из них способно митотически делиться. Будучи различными по происхожде-
нию, паранейроны структурно и функционально сходны между собой и нейронами вследст-
вие конвергентной дифференцировки.
В данном случае для нас в этой концепции представляет интерес та ее часть, которая
касается эволюции кишечных интраэпителиальных нейронов и паранейронов.
Фудзита обращает внимание на то, что уже у гидры в гастродермисе и эпидермисе при-
сутствуют клетки как закрытого («ганглионарные»), так и открытого («чувствительные»)
типов (рис. 298). Последние Фудзита рассматривает как «архипаранейроны» (archipara-
neurons), которые, являясь типичными рецепторно-секреторными элементами, по своему
тонкому строению в высшей степени соответствуют базально-гранулярным (эндокринным)
клеткам в эпителии кишечника позвоночных, включая человека, а также пищеварительного
тракта многих беспозвоночных. «Ганглионарные» клетки эпидермиса и гастродермиса гид-
ры являются «архинейронами» (achineurons). И те, и другие клетки мультифункциональны,
Рис. 298. Стенка тела гидры (А) и эволюция кишечных паранейронов (Б) (по: Fujita et al., 1988).
апн - «архипаранейрон»; гк - ганглионарные клетки («архинейроны»); рек - рецепторно-секреторные клетки
(«архинейроны»); эк1-эк2 - кишечные эндокринные клетки (паранейроны) у первичноротых
и вторичноротых соответственно.
Fig. 298. Body wall of Hydra (A) and evolution of intestinal paraneurons (Б)
(according to: Fujita et al., 1988).
131
демонстрируют одновременно как паракринное (нейросекреторное) выведение содержимо-
го секреторных гранул, так и контакты типа синаптических, оканчивающиеся на соседних
клетках.
В ходе эволюции эти два типа мультифункциональных клеток дают большое разнооб-
разие нейронов и паранейронов, причем в результате специализация приводит к превраще-
нию мультифункциональных клеток в унифункциональные. Что касается «архинейронов»,
то, возможно, именно они впоследствии образуют элементы интрамуральной автономной
системы кишечника позвоночных животных (Fujita et al., 1988).
На мой взгляд, если принимать концепцию Фудзиты целиком, очевидны ее определен-
ные неточности и даже противоречия. По крайней мере, это касается нейрональных и пара-
нейрональных элементов в кишечных эпителиях. Одно из принципиальных возражений
связано с тем, что в основу филогенетической части концепции в качестве исходного звена
положены кишечнополостные (гидра). Я придерживаюсь, как уже отмечал, точки зрения,
что двуслойные кишечнополостные представляют собой самостоятельную ветвь эволюции
Metazoa, и их нельзя рассматривать как прямых предков других многоклеточных. Другое
возражение связано с тем, что уже исходный (открытый или закрытый) тип клетки опреде-
ляет в дальнейшем ее принадлежность к тому или иному ряду (нейрональному или пара-
нейрональному). Такой подход требует дополнительных пояснений и поправок, поскольку,
как было показано в предыдущих главах, среди эндокринных клеток (паранейронов) много-
численны элементы закрытого типа, и, наоборот, интраэпителиальные нейроны открытого
типа не представляют собой исключения. Кроме того, если современные эндокринные
клетки являются прямыми потомками «архипаранейронов», мы должны наблюдать в эво-
люционном ряду непрерывные, плавные переходы от одних интраэпителиальных параней-
ронов к другим, что в действительности далеко не всегда имеет место.
На основании приведенных выше данных я хочу предложить другой вариант возник-
новения и эволюции кишечных регуляторных систем. Прежде всего, отмечу, что в отличие
от концепции паранейрона в основу филогенетических построений как животных, наиболее
близких к примитивным многоклеточным, я помещаю плоских червей (турбеллярий).
Именно в этой группе собственно и наблюдаются различные этапы появления и дифферен-
цировки пищеварительной системы [этот вопрос подробно рассмотрен в работе Мамкаева
(1979)].
Очень важно, что уже у турбеллярий в составе эпителия пищеварительной системы
(часто еще весьма слабо дифференцированного, способного терять многие признаки эпите-
лиального пласта во время процесса пищеварения) мы наблюдаем интраэпителиальные (га-
стродермальные) нервные элементы, а также отдельные клетки, имеющие признаки эндок-
ринных (эндокриноподобных). Еще раз отмечу, что положение нервных элементов относи-
тельно эпителиального пласта у турбеллярий оказывается исключительно разнообразным
(см. рис. 290). (Среди других многоклеточных сходную картину можно наблюдать, навер-
ное, только у двустворчатых и брюхоногих моллюсков.) Я полагаю, что это разнообразие
может отражать различные этапы возникающих контактов между нейрональными элемен-
тами и дифференцирующимся эпителием пищеварительного тракта, в том числе миграцию
нервных клеток в эпителиальный пласт. (Тогда как у моллюсков мы наблюдаем, возможно,
обратный процесс — выселение нервных элементов из эпителия).
Факт присутствия нервных клеток в эпителии кишечника олигохет (Невмывака, 1947) и
двустворчатых моллюсков (Гилев, 1952) (см. подробнее гл. 7 и 8) дал основание Невмываке
(1956, 1961) и Беклемишеву (1964) предположить возможность энтодерм ал ьно го происхож-
дения у беспозвоночных части нервных элементов. Отмечу, что концепция полигенеза
132
нервной системы, предложенная Сахаровым (1974), также предполагает такой путь, однако
прямых доказательств (по крайней мере относительно кишечных регуляторных элементов)
этого пока нет. Мне представляется более вероятным предположить миграцию нервных
элементов в состав энтодермы в ходе эмбрионального развития организма.
Теперь остановлюсь на вопросе о возможных гистогенетических источниках кишечных
эндокринных клеток. Теоретически можно допустить, что некоторые интраэпителиальные
клетки {архинейроны по Фудзита) могли в определенной степени трансформироваться (ли-
шиться отростков, синаптических контактов, потерять прямую морфологическую связь с
нервной системой), приобретая тем самым «внешний облик» эндокринных. Однако мне
представляется более вероятным происхождение кишечных эндокринных элементов непо-
средственно из эпителиальных.
Академиком А.М.У голевым была предложена экскреторная теория происхождения
внешней и внутренней секреции (Уголев, 1979, 1985). Согласно этой теории секреция воз-
никает из неспецифической экскреции (рис. 299). Последний процесс не идеален, и вместе с
продуктами распада во внешнюю (по отношению к клетке) среду поступают не только про-
дукты катаболизма, но и некоторое количество биологически полезных и физиологически
активных молекул. При соответствующих условиях такая потеря может стать полезным
нэ
Рис. 299. Формирование различных типов внешней и внутренней секреции
на основе неспецифической экскреции (по: Уголев, 1979, 1985).
нэ - неспецифическая эксреция; ир1-ир2 - формирование внутренней секреции регуляторов;
иф1-иф2 - формирование внутренней секреции ферментов; ср1-ср2 - формирование внешней
секреции регуляторов; сф1-сф2 - формирование внешней секреции ферментов.
Черные кружки - молекулы ферментов, светлые кружки - молекулы регуляторов,
треугольники - экскретируемые вещества.
Fig. 299. Formation of different types of internal and external secretion based
on unspecific excretion (according to Уголев, 1979, 1985).
133
признаком, и на основе усиленного выделения во внешнюю или внутреннюю среду опреде-
ленного продукта происходит формирование секреторного процесса. Синтез и экспорт тех
или иных молекул становятся доминирующей функцией данного типа клеток (Уголев,
1985).
Из этой концепции следует принципиально важный вывод, что внешняя и внутренняя
секреция могут формироваться в процессе филогенеза на основе неспецифической экскре-
ции многократно и в различных по своему происхождению структурах (тканях).
Применительно к кишечным регуляторным системам из этого логично вывести не
только возможность энтодермального происхождения кишечных эндокринных клеток (что,
как было ранее показано, имеет и экспериментальные подтверждения), но и то, что опреде-
ленные типы (по синтезируемым ими биологически активным веществам) эндокринных
клеток могут возникать (и, очевидно, также и исчезать) многократно и независимо в разных
группах многоклеточных.
Возможность синтеза биологически активных веществ, типичных для регуляторных
элементов, обычными эпителиальными можно наблюдать (как это уже отмечалось в гл.
14) у гидры (продукция пептида-активатора роста головы у гидр, лишенных нервных кле-
ток). Имеются данные, что в задней кишке гусениц бабочки Ostrinia nubilis все эпителиаль-
ные клетки бифункциональны: обеспечивают как транспорт ионов, так и секрецию гормона
проктодона (Alexander, Fahrenbach, 1969). В эпителиальных клетках желудка рака Cancer
magister при демонстрируемой ими иммунореактивности к гастрину и холецистокинину в
цитоплазме отсутствуют какие-либо секреторные гранулы (Scalise et al., 1984). Такое со-
стояние, наверное, можно рассматривать как начало дифференциации одного из типов эн-
докринных клеток.
Потенциальную способность «не нервных» тканей генерировать эндокринные элемен-
ты можно проиллюстрировать таким примером. Группы кардиомиоцитов в предсердиях
млекопитающих, помимо миофиламентов, содержат в цитоплазме характерные для эндок-
ринных клеток гранулы и связаны с синтезом пептидного гормона — так называемого пред-
сердного натрийуретического фактора (Кантен, Жене, 1986). (Интересно, что этот гормон
действует на самые различные ткани и органы, включая регуляторные образования голов-
ного мозга). Таким образом, в гистологическом плане эти клетки представляют собой сме-
шанные мышечно-эндокринные.
Снова вернемся к особенностям организации кишечных регуляторных систем на раз-
ных уровнях эволюционного древа. У наиболее близких к примитивным многоклеточным,
расположенных в основании филогенетического древа современных турбеллярий (по край-
ней мере, некоторых из них) мы уже наблюдаем кишечную регуляторную систему, образо-
ванную как разнообразными по строению и положению относительно эпителия нервными
клетками, так и единичными эндокринными (эндокриноподобными). (рис. 300). У живот-
ных более высоких ярусов эволюционного древа (иглокожие, моллюски, брахиоподы, коль-
чатые черви) происходит усложнение этой системы, главным образом за счет большего
разнообразия эндокринных элементов (как относительно спектра синтезируемых биологи-
чески активных веществ, так и морфологии — появление клеток и открытого, и закрытого
типов). Важно подчеркнуть, что на этом этапе и нервные, и эндокринные клетки, очевидно,
непосредственно контактируют друг с другом, а также могут быть связаны и с субэпители-
альными нервными элементами (моллюски, кольчатые черви).
Наконец, следующий и высший этап — пространственное разделение эндокринных
клеток (эпителий) и нервных элементов (субэпителиальные ткани). Это состояние мы на-
блюдаем в ряду вторичноротых (начиная с оболочников — асцидий) и первичноротых (на
134
CHORDATA
(Vertebrata - Timicata)
ARTHROPODA
(Insecta)
ECHINODERMATA
(Asteroidea, Echinoidea)
ANNELIDA
(Polychaeta, Oligochaeta)
II
PLATHELMINTHES (Turbellaria)
Рис. 300. Предполагаемая эволюция кишечной регуляторной системы многоклеточных:
1 - возникновение системы; 7/- система у многоклеточных средних ярусов эволюционного древа;
777- система у высших многоклеточных. Черным отмечены нервные клетки и отростки,
белым - эндокринные (эндокриноподобные) клетки.
Fig. 300. Proposed evolution of intestinal regulatory' system in Metazoa.
135
секомые). Каким образом происходит этот процесс, — в результате исчезновения нервных
клеток в эпителиальном пласте или их миграции (погружения) в подстилающие эпителий
ткани (этапы подобной миграции, возможно, демонстрируют двустворчатые и брюхоногие
моллюски) — пока неясно. Добавлю, что тенденция к пространственному разделению
нервных и эндокринных элементов, возможно, наблюдается и в третьей линии, ведущей к
головоногим моллюскам (поскольку фактических данных по этим моллюскам крайне мало,
это предположение нуждается в дальнейшей проверке). И, наконец, только позвоночные
демонстрируют дальнейшее усложнение системы — появление (помимо диффузно распре-
деленных эндокринных элементов) компактных эндокринных желез.
Таким образом, я полагаю, что разделение исходно гетерогенной регуляторной систе-
мы, появляющейся практически одновременно с эпителиализированным кишечником, на
эндокринную и нервную составляющие, и вынесение последней за пределы эпителиального
пласта шли независимо и параллельно в ряду первичноротых и вторичноротых животных.
В свете имеющихся к настоящему времени данных предложенная здесь гипотеза пред-
ставляется мне наименее противоречивой.
ЗАКЛЮЧЕНИЕ
Завершая эту книгу попробую сформулировать ее основные положения.
Под кишечной регуляторной системой подразумевается совокупность клеток и отрост-
ков, расположенных в составе эпителия энтодермальной части пищеварительного тракта и
имеющих морфологические, гистохимические и иммуногистохимические признаки эндок-
ринных (эндокриноподобных), нервных и нейросекреторных элементов.
Такие кишечные регуляторные системы (или, по крайней мере, их элементы) известны
у представителей многих групп беспозвоночных животных, занимающих различные ярусы
эволюционного древа. К настоящему времени они выявлены у членистоногих (насекомые,
ракообразные), сипункулид, моллюсков (хитоны, двустворчатые, брюхоногие, головоно-
гие), кольчатых червей (полихеты, олигохеты, пиявки), мшанок, брахиопод, приапулид,
киноринх, иглокожих (морские звезды, морские ежи, офиуры, голотурии), оболочников
(асцидии), полухордовых (кишечнодышащие), немертин и плоских червей (турбеллярий).
Как у представителей разных систематических групп, так и у одного вида кишечные
регуляторные клетки могут существенно различаться по своей общей морфологии (форма,
размеры, наличие или отсутствие базальных отростков, контакт с просветом пищеваритель-
ного тракта), тонкому строению (в первую очередь по степени развития белок-
синтез ирующе го аппарата и аппарата Гольджи, по строению и размерам секреторных гра-
нул и пузырьков), а также по характеру синтезируемых биологически активных соединений
(различные пептиды, моноамины).
Кишечная регуляторная система отмечена у животных с различными преобладающими
типами пищеварения (полостное, внутриклеточное, смешанное). Элементы этой системы
могут присутствовать в отделах пищеварительного тракта, выполняющих самые разные
функции (проводящую, сортирующую, секреторную, функции внутриклеточного перевари-
вания, пристеночного пищеварения, всасывания и т.д.).
Кишечная регуляторная система наблюдается уже у таких представителей низших
многоклеточных, как турбеллярий (ресничные черви). Это означает, что она возникает
практически одновременно с появлением микроанатомически дифференцированного и эпи-
телизированного кишечника. У турбеллярий данная система образована главным образом
нервными элементами, хотя в эпителии, по-видимому, присутствуют и эндокринные (эн-
докриноподобные) клетки. Интраэпителиальная система связана отростками с субэпители-
альными нервными элементами.
У представителей многих таксономических групп беспозвоночных средних ярусов эво-
люционного древа (моллюски, кольчатые черви, брахиоподы, иглокожие) наблюдается ус-
ложнение системы за счет большего разнообразия клеточных элементов. Кишечная регуля-
торная система образована здесь как нервными, так и гетерогенными (по морфологии и хи-
137
мизму) эндокринными клетками. Общим в этих группах беспозвоночных является то, что
элементы системы объединены интраэпителиальными отростками в единую сеть. У дву-
створчатых и брюхоногих моллюсков, полихет и олигохет кишечная регуляторная система
в той или иной степени непосредственно связана с субэпителиальным нервным сплетением,
а у иглокожих и брахиопод является, по-видимому, исключительно интраэпителиальной.
У высших первичноротых (насекомых) и в ряду вторичноротых, начиная с оболочни-
ков (асцидии), при увеличении общего разнообразия интраэпителиальных регуляторных
клеток наблюдается пространственное разделение эндокринных и нервных элементов. Вто-
рые занимают исключительно субэпителиальное положение и теряют прямой морфологиче-
ский контакт с интраэпителиальными.
Общее количество кишечных регуляторных клеток у большинства представителей бес-
позвоночных весьма велико и сопоставимо с относительной массой клеток кишечной гор-
мональной системы позвоночных. Это, очевидно, указывает на то, что, как и у Vertebrata,
кишечные регуляторные системы Invertebrata играют не только местную (внутриэпители-
альную), но и общеорганизменную интегрирующую роль.
Таким образом, возникая первоначально на основе нервных элементов, кишечная регу-
ляторная система развивается и усложняется в результате появления эндокринных клеток,
очевидно дифференцирующихся непосредственно из эпителиальных. Возникновение ин-
траэпителиальных эндокринных клеток происходит, по-видимому, многократно и незави-
симо в разных систематических группа беспозвоночных. Пространственное разделение ис-
ходно гетерогенной кишечной регуляторной системы на интраэпителиальную эндокринную
и субэпителиальную нервную составляющие, очевидно, идет независимо в ряду первично-
ротых и вторичноротых животных.
SUMMARY
Histological, histochemical, electron microscopic and immunohistochemical studies for the
recent decades have revealed the presence of peculiar cells in the epithelium of the alimentary
system (in its entodermal part) of representatives of many invertebrate taxons located at different
levels of the evolutionary tree. These cells differ from each other by both many structural details
and biologically active substances they contain (monoamines, peptides, some other substances).
Nevertheless, they have a common feature. As judged from their morphology, they seem to take
no direct assistance in digestive processes (membrane and intracellular digestion, absorption of
nutrients, secretion of digestive enzymes, mucus, components of peritrophic membranes,
intermixing of intestinal contents and its translocation along the intestinal tract, etc.); they rather
perform some integrative, regulatory functions.
According to their structure some of these cells correspond significantly to intestinal
endocrine cells of vertebrates, others, to neurons. At last, the third type of cells cannot be
unanimously considered as belonging to the former or the latter, so they occupy some
“intermediate” position. I will call all these cells regulatory cells. By analogy with the intestinal
hormonal (enterinic) system of mammals, they also are likely to form their own regulatory
system(s) in the digestive tract.
This monograph presents a review on data obtained from studies of many authors as well as
from original investigations concerning morphology and distribution of intestinal endocrine
(endocrine-like), nerve and related elements. The review is completed by a new hypothesis, which
explains possible appearance and subsequent evolution of intestinal regulatory systems in
Metazoa.
VERTEBRATA. The enterinic system of Mammalia is well investigated. It is formed by
endocrine cells of many different types. Thus, in the human stomach, intestine and pancreas
according to one of classifications at least 18 cellular types are identified. This identification is
based predominantly on the size and fine structure of secretory granules and on their biologically
active substances. In the endocrine cells such peptide hormones as somatostatin, pancreatic peptide
(PP), gastrin, cholecystokinin (CCK), gastroinhibiting polypeptide, enteroglucagon, motilin,
neurotensin, secretin, vasoactive intestinal polypeptide (VIP), substance P, enkephalin, bombesin,
peptide tyrosine-tyrosine (PYY) and several others as well as serotonin are found.
The intestinal endocrine cells usually are located individually in the epithelium. In most cells
the apical parts face the digestive tract lumen (so-called “open-type” cells) while others are located
in the base of epithelial layer and have no contact with the lumen (cells of “closed” type).
On the apical surface of open-type cells there are microvilli which seem to be a receptor
apparatus. Numerous secretory granules are mainly concentrated in the lower half of the cells.
Exocytosis of their content occurs on the basal surface as well on the lateral cell surfaces near
139
basal membrane. The closed-type cells have basically a similar structure, however there is no
marked polarity in their organization. Also the “combined” cells with structural features of the
mucous and endocrine as well as of the Paneth and endocrine cells are described. The endocrine
cells secrete their substances both into the blood capillaries located under the epithelium
(endocrinia) and into intracellular space (paracrinia). In the latter case their action is directed
towards the neighboring epithelial cells and/or sensory nerve terminals located closely to the
epithelium.
The gastrointestinal system in representatives of other vertebrate classes (Aves, Reptilia,
Amphibia, Pisces) basically does not principally differ from that of mammals both in the
morphologic diversity of endocrine cells and by the variety of biologically active substances
present in them. The organization of the intestinal endocrine system in Cyclostomata is also rather
complex. Several types of endocrine cells are known in the digestive tract of some lancelets
(subtype Acrania). These cells also produce different hormones.
TUNIC AT A. Representatives of the class Ascidiaceae demonstrate a well developed
intestinal endocrine system. In the digestive tract of these animals intraepithelial cells both of the
open and of the closed types are described, some of them can have basal processes which,
however, do not leave the epithelial layer. The fine structure of the regulatory cells is somewhat
different not only in different species, but also within one species, which suggests the existence of
several cell types. This is confirmed by immunohistochemical data.
Before considering the other animal groups one remark is worthy to make. In all known cases
the intestinal regulatory systems of CHORDATA are purely intraepithelial systems formed, as a
rule, by diffusely spread cells which only occasionally contact with each other. The nerve cell
processes can come close to the epithelium but do not penetrate it. In all cases the nerve ending
and the endocrine cell are separated by the basal membrane.
HEMICHORDATA. For some representatives of Enteropneusta, on the contrast with
Chordata, only intraepithelial nerve processes and cells are known.
ECHINODERM AT A. One of the peculiar features of organization of alimentary tract
epithelium in representatives of all classes of this type - Asteroidea, Echinoidea, Ophiuroidea,
Holoturoidea and Crinoidea - is the presence of well developed basiepithelial nerve plexus. In
starfishes this plexus is formed by the processes of intraepithelial cells mainly belonged to the
open-type. It was demonstrated that the intestinal nerve system can contain different substances
such as FMRFamide, SALMFamide, PYY, PP and some others. Nevertheless, cells of the open
and closed types sharing morphological features of the endocrine ones are revealed in the
epithelium too. They also contain different regulatory substances: somatostatin, PP, insulin-like
peptides. Electron-microscopic data confirm the existence of several types of the endocrine cells.
In some investigated sea urchins the basiepithelial nerve plexus consists of monoamine-
containing cells and processes. At the same time cells of the open and closed types more
resembling endocrine ones and demonstrating immunoreactivities to different regulatory peptides
are also revealed.
Data on the digestive system of brittle stars deal mainly with intraepithelial nerve elements.
The neurons and their processes seemingly contain PP-, glucagon-, somatostatin-like substances as
well as catecholamines. In sea cucumbers the intraepithelial plexus is partly formed by the
processes of neurons located in the coelomic epithelium. Nevertheless, the endocrine-like cells
demonstrating immunoreactivities to somatostatin, glucagon and insulin-like peptide of molluscs
were also revealed.
140
Summarizing data mentioned above it may be concluded that at least in Asteroidea,
Echinoidea and Holoturoidea the intestinal regulatory system is a heterogeneous formation
consisting of both the nerve and polymorphous endocrine cells.
ARTHROPODA. The intestinal regulatory system elements are described in many
representatives of class Insecta (subtype TRACHEATA), belonging to both suborders -
Entognatha and Ectognatha. The regulatory cells in insects in many aspects are similar to the
endocrine elements of vertebrates. Among them cells of the open and closed types are seen.
Usually the cells have no basal processes, however in some of them the processes are present.
Nevertheless, the basiepithelial plexus cannot be revealed. The secretory granules are concentrated
in the basal cytoplasm, their content is released by exocytosis across both the basal and lateral
cytomembranes. Based on their morphology several (up to 10 in one of cockroaches) types of
endocrine cells can be identified. It have been shown that at least some of types of endocrine cells
develop from undifferentiated cells of regenerative nests, i.e they have common sources of
phisiological regeneration and, hence, like other epithelial cells, the entodermal origin. The
diversity of the insect endocrine cells has been confirmed by immunohistochemical data. In
different species cells immunoreactive to PP, enteroglucagon, somatostatin, serotonin, urotensin I,
peptide histidine isoleucin (PHI), CCK, somatoliberin, substance P, VIP and some others were
revealed. While nerve processes sometimes come close to the epithelium there is no evidence for
the presence in it of any elements considered as the nerve ones. Thus, it should be emphasized
again that the insect gut regulatory system turns out to be rather morphologically similar to the
enterinic system of vertebrates.
Unfortunately the data on the gut regulatory elements of other arthropods are fragmentary.
Nevertheless, they allow to suggest substantial differences in organization of their systems as
compared to insects. Among some species from subtype CHELICERATA in the base of intestinal
epithelium thin processes, presumably axons, have been revealed. Similar processes were found in
the midgut of some Crustacea (subtype BRANCHIATA). At the same time in a few species of the
former subtype the cells structurally reminding the endocrine ones were described.
MOLLUSCA. In many representatives of class Bivalvia histochemical, immunohisto-
chemical and electron-microscopic investigations indicate a complicated intestinal regulatory
system formed by both the nerve and endocrine cells. The nerve component is well developed, it
consists of different cells variable in their morphology. Cell processes form a prominent
basiepithelial plexus connected with the subepithelial one. Ultrastructural data also indicate the
existence of intraepithelial processes and cell bodies resembling in their fine structure the nerve
ones. Nevertheless, some cells with different morphology more typical for endocrine elements
were also revealed. In different bivalves cells immunoreactive to insulin, insulin-like peptides,
calcitonin, hydra head activating peptide (HAP), neurotensin, FMRFamide and serotonin have
been described. Some of them form by their processes an intraepithelial plexus while other lack
the processes.
Gastropoda species are studied less completely; however the data available show that their
intestinal regulatory system is basically similar to that of bivalves. Unfortunately only few remarks
can be made about Cephalopoda and Polyplacophora, where some cells more resembling
endocrine ones were revealed by means of immunohistochemistry.
ANNELIDA. In some representatives of class Polychaeta at least two different types of the
regulatory cells were observed. They belong to the closed-cell type. Processes running from
perikarya form a well developed basiepithelial plexus. Nevertheless, this plexus in some detail
differs from a typical nerve one, and many intraepithelial cells participating in its formation more
resemble the endocrine-like elements. Rare nerve processes from a subepithelial plexus can
141
penetrate the epithelial layer; they end somewhere between the bases of common epithelial cells.
Single typical receptor (open-type) cells were found in some species.
Two similar closed-type cells and a receptor one were revealed during electron-microscope
investigations of Oligochaeta species. Immunohistochemical investigations demonstrated in earth
worms the presence of cells immunoreactive to FMRFamide, calcitonin and processes (obviously
nerve) immunoreactive to PP and PHY were detected.
Immunihistochemical and electron-microscopic investigations of some Hirudinea species
have shown significant differences in the intestinal regulatory system organization compared with
Polychaeta and Oligochaeta. The regulatory cells in leeches seem to be much more rare; they more
resemble the endocrine cells. While some typical processes with small granules were described
during electron-microscopic investigations, a developed basiepithelial plexus was not revealed.
There is evidence for existence of endocrine-like cells in SIPUNCULIDA, and intraepithelial
nerve processes as well as seemingly regulatory cells in some species of PHORONIDAE.
BRACHIOPODA. The intestinal regulatory system in this animal group is mainly formed by
nerve elements. The processes of intraepithelial nerve cells form an easily detectable plexus but
never leave the epithelial layer. HAP-, FMRFamide and neurotensin-immunoreactive cells were
revealed; most of them were presented by the open- and closed-type elements. Nevertheless, some
of these cells without processes share more similarities with the endocrine-like cells.
CEPHALORHYNCHA. Among the classes of this type extremely rare regulatory (more
likely - endocrine) cells were revealed in Kinorhyncha. In some investigated species of
Priapulida intraepithelial monoamine-containing cells with short processes were detected.
Electron microscopic analysis showed the presence of specific cells with small processes and oval
granules. In the structure these cells more resemble the endocrine ones.
NEMERTINI. These worms possess a well developed basiepithelial nerve plexus formed by
the processes of numerous polymorphous intraepithelial cells. The plexus is associated with nerve
structures outside the intestine. Immunohistochemical data also demonstrate the presence of
neurotensin-immunoreactive cells of the open type. These cells lack processes and more
correspond in their structure to the endocrine ones.
PLATHELMINTHES. Representatives from different orders of class Turbellaria
demonstrate significant variety in the organization of the intestinal regulatory system. The most
complicated conditions are revealed in orders Polyclada and Triclada, where in comparison with
other turbellarian groups one can see more differentiated in microanatomical and histological
aspects intestine. The regulatory system of these flatworms composes mainly of the nerve
elements. Polymorphous intra- and subepithelial neurons are combined into a common network
with the aid of the processes. Nevertheless, some cells more corresponding to the endocrine-like
were also detected. Only rare nerve elements are known for orders with more primitive
organization of intestine.
CNIDARIA represent two-layered animals with the nerve system located in the epidermis
and gastrodermis (epithelium of the gastral cavity). The gastrodermal part of Hydra nerve system
consists of neurons and neurosecretory cells of the open as well as of the closed types. There is no
evidence for the presence of gastrodermal endocrine cells, nevertheless it was demonstrated that in
experiments common gastrodermal cells can start producing biologically active substances (HAP)
typical for some neurons.
As it was shown above, intraepithelial regulatory systems (or at least their elements) can be
found in representatives of most large taxons located on different levels of the evolutionary tree.
There are only few taxonomic groups for which these elements are unknown. It should be noted
that the regulatory cells are present in animals with different dominant type of digestion (luminal,
142
intracellular, mixed). They can be observed in quite different regions of the alimentary tract
regardless of their specific functional role in digestive processes.
Attention has also been paid to the fact that the relative number of intestinal regulatory cells
in most invertebrate representatives is rather high and comparable with the relative mass of cells of
the vertebrate enterinic system. This indirectly indicates that the intestinal regulatory systems in
invertebrates, like in vertebrates, perform not only the local (intraepithelial) but also the whole-
organism regulatory function.
Speculating about possible roots of the origin and following evolution of the intestinal
regulatory systems I would like to place in the base of phylogenetic construction turbellarians (fig.
300), as they are the closest to the primitive three-layered animals. (My viewpoint is that two-
layered Cnidaria are an independent brunch of evolution of Metazoa and therefore this animal
group cannot be considered as direct ancestor of other multicellular organisms.) Already in this
group the intraepithelial element with morphological features of both the nerve and endocrine
(endocrine-like) ones* are observed. The diversity of position of the nerve elements towards the
epithelial layer probably reflects migration of neurons from parenchima to gastrodermis.
In animals from higher levels of the evolutionary tree (brachiopods, annelids, molluscs,
echinoderms) the intestinal regulatory system is submitted to a complication: on the one hand due
to a more pronounced development of the “neuronal” component, while on the other hand due to a
greater diversity of the endocrine elements (with respect to both their morphology and the
spectrum of synthesized biologically active substances). Many endocrine cells have a direct
contact with the nerve processes (although the interregulation is most likely occurs by the principle
of paracrinia).
Finally, in the higher Protostomata (insects) and in a branch of Deuterostomata, beginning
from tunicates (ascidians), with increase of diversity of the intraepithelial regulatory cells, the
spatial separation of endocrine and nerve elements takes place. The latter acquire the exclusively
subepithelial location and lack the direct morphological contact with intraepithelial cells. So far it
is yet unclear whether this occurs due to extrusion of the nerve elements from the epithelium or
due to their direct disappearance and what provides this process. The histogenetical source of
intestinal endocrine cells (or at least of some of them) most likely is entoderm itself.
Thus, the intestinal regulatory systems, having initially appeared based upon predominantly
nerve elements, developed and complicated as a result of appearance of endocrine cells that
originated directly from the epithelial ones. The appearance (and probably disappearance) of the
intraepithelial endocrine cells probably occurred repeatedly and independently in different taxons.
The spatial separation of the initially heterogeneous system into the intraepithelial endocrine and
subepithelial nerve components came in parallel in the branches of Protostomata and
Deuterostomata.
ЛИТЕРАТУРА
Алешин Б.В. Проблема нейроэндокринных клеток и гипотезы «диффузной эндокринной системы» И Успехи
соврем, биол., 1984. Т. 98. Вып. 1. С. 116-133.
Атаманова М.В. Морфофункциональные особенности кожного покрова и центральной паренхимы бески-
шечных турбелярий, кожного и кишечного эпителиев немертин и кишечнодышащих: Автореф. канд. дисс. - Л.,
1980. 20 с.
Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. Т. 2. - М.: Наука, 1964. 446 с.
Ван Ноорден С., Пирс А.Г.Е. Локализация иммунореактивности к инсулину, глюкагону и гастрину в кишеч-
нике ланцетника (Branchiostoma lanceolatum). И Эволюц. эндокринология поджелудочной железы. - Л.: Наука,
1977. С. 97-104.
Вараксин А.А., Ушева Л.Н., Зверков И.В. Локализация иммунореактивного лейэнкефалина в пищеваритель-
ной железе приморского гребешка Mizuchopecten yessoensis (Jay) И Арх. анат. гистол. эмбриол., 1990. Т. 98. № 1.
С. 71-74.
Гилев Ф.Д. О чувствительной иннервации кишечника беззубки И Докл. АН СССР, 1952. Т. 87. С. 1059-1061.
Догель А.С. Техника окрашивания нервной системы метиленовой синью. - СПб., 1902. 48 с.
Елисейкина М.Г Морфофункциональные состояния и физиологическая регенерация выстилающего эпителия
пищеварительного тракта у японской кукумарии: Автореф. канд. дисс. - Владивосток, 1993. 26 с.
Елисейкина М.Г., Лейбсон Н.Л. Ультраструктурная характеристика кишечного эпителия японской кукумарии
Cucumaria japonica И Биол. моря, 1996. Т. 22. № 2. С. 102-109.
Заварзин А.А. Основы сравнительной гистологии. - Л.: Изд-во ЛГУ, 1985. 400 с.
Заварзин А.А., Пунин М.Ю. Исследование организации кишечного эпителия приапулид. I. Гистохимический и
авторадиографический анализ И Цитология, 1979. Т. 21. № 6. С. 717-721.
Заварзин А. А., Пунин М.Ю. Кинетика репродукции и дифференцировки клеток в эпителии кишки и ее произ-
водных у афродитид. I. Эпителий средней кишки И Цитология, 1981. Т. 23. С. 283-291.
Заварзин А.А., Пунин М.Ю., Цветков А.Г Кинетика репродукции и дифференцировки клеток в эпителии
кишки и ее производных у афродитид. III. Эпителий железистых выростов И Цитология, 1984. Т. 26. С. 52-59.
Зайцева О.В., Пунин М.Ю. Система регуляторных элементов кишечника немертин И Механизмы функциони-
рования висцеральных систем: Тез. докл. - СПб, 2001. С. 133.
Иванов А.В., Мамкаев Ю.В. Ресничные черви (Turbellaria). Их происхождение и эволюция. - Л.: Наука, 1973.
221 с.
Иванова О.В. Гистотопография эндокриноцитов в эпителии прямой кишки кур в онтогенезе И Морфология,
1995. Т. 108. Вып. 1.С. 76-78.
Иванова В.Ф., Пузырев А.А. Дифференцировка эндокриноцитов толстой кишки некоторых позвоночных жи-
вотных и человека И Морфология, 1998. Т. 114. Вып. 4. С. 107-111.
Кагановская Э.А. Кинетика репродукции и дифференцировки клеток в кишечном эпителии полихеты
Arenicola marina И Цитология, 1976. Т. 18. С. 171-177.
Кагановская Э.А. Тонкое строение железисто-всасывающей зоны эпителия желудка полихеты Arenicola
marina// Проблемы эволюц. морфол. - Л.: Изд-во ЛГУ, 1978а. С. 86-103.
Кагановская Э.А. Электронно-микроскопическое исследование ресничных и всасывающих клеток и их взаи-
моотношений в эпителии желудка полихеты Arenicola marina// Цитология, 19786. Т. 20. С. 882-890.
Кагановская Э.А. Ультраструктур ное исследование эпителия средней кишки полихеты Arenicola marina //
Цитология, 1979. Т.21. С. 650-656.
Кагановская Э.А. Организация репродукции и функциональной дифференцировки клеток в кишечном эпите-
лии полихеты Arenicola marina: Автореф. канд. дисс. - Л., 1985. 21 с.
144
Кагановская Э.А. Ультраструктура базальных клеток эпителия желудка и средней кишки полихеты Arenicola
marina// Цитология, 1988. Т. 30. С. 891-894.
Кагановская Э.А. Некоторые морфофункциональные особенности пищеварительной системы полихет// Арх.
анат. гистол. эмбриол., 1989. Т. 96. Вып. 4. С. 84-92.
Казаков В.К. О содержащих инсулиноподобные вещества клетках в кишечном эпителии моллюска Unio
pictorum//Журн. эвол. биохим. физиол., 1977. Т. 13. С. 439-442.
Казаков В.К. Иммуногистохимическое исследование регуляторных клеток в эпителии пищеварительного
тракта двустворчатых моллюсков: Автореф. канд. дисс. - СПб., 1996. 16 с.
Кантен М, Жене Ж. Сердце как эндокринная железа // В мире науки, 1986. № 4. С. 40-43.
Костюкевич С.В., Пузырев А.А., Иванова В.Ф. Эндокринный аппарат эпителия слизистой оболочки червеоб-
разного отростка человека// Морфология, 1998. Т. 113. № 2. С. 21-35.
Котикова Е.А. Нервная система Acerotisa sp. (Policladida, Cotylea, Turbellaria) 11 Зоол. журн., 1983. T. 62. № 8.
С. 1148-1154.
Кулаковский Э.Е., Макаренкова Е.П., Прочуханова А.Р. Ультраструктура цереброплеврального ганглия ми-
дии Mytilus edulis И Цитология, 1990. Т. 32. С. 599-604.
Лавров А.К. Концевые отделы периферической нервной системы. - Ростов-на-Дону: Ростведиздат, 1941.
192 с.
Лейбсон Л.Г Успехи и проблемы эволюционной эндокринологии //Журн. эвол. биохим. физиол., 1981. Т. 17.
С. 116-127.
Лейбсон Л.Г Происхождение и эволюция эндокринной системы. И Эволюционная физиология. Ч. 2. - Л.:
Наука, 1983. С. 3-52.
Ливанов Н.А. Класс полихет (Polychaeta). И Руководство по зоологии. Т. 2. - М.-Л.: Изд-во АН СССР, 1940.
С. 10-136.
Макаренкова Е.П., Кулаковский Э.Е., Пунин М.Ю. Электронно-микроскопическое и флуоресцентно-
гистохимическое изучение регуляторных клеточных элементов в цереброплевральном ганглии и кишечнике дву-
створчатого моллюска Mytilus edulis И Простые нервные системы. - Л., 1988. С. 176-180.
Малахов В.В. Cephalorhyncha— новый тип животного царства, объединяющий Priapulida, Kinorhyncha,
Gordiacea, и система первичнополостных червей И Зоол. журн., 1980. Т. 59. N 1. С. 485-499.
Малахов В.В., Адрианов А.В. Головохоботные (Cephalorhyncha) — новый тип животного царства. - М.: КМК
Scientific Press Ltd., 1995. 200 с.
Мамкаев Ю.В. О гистологической организации пищеварительной системы турбеллярий. И Эволюц. морфоло-
гия беспозвоночных. - Л.: Зоол. ин-тАН СССР, 1979. С. 13-24.
Маркосова Т.Г., Пунин М.Ю. РМИРамид-подобная иммунореактивность в кишечнике турбеллярии Notoplana
atomata И Докл. РАН, 1997. Т. 353. N 5. С. 697-699.
Матвеева О.Н. Эндокринный аппарат эпителия слизистой оболочки тонкой кишки летней травяной лягушки
И Морфология, 1998. Т. 113. № 2. С. 88-94.
Мутовкина Т.Н., Иванова В.Ф. Содержание эндокринных клеток в эпителии толстой кишки у различных
представителей позвоночных И Архив анат. гистол. эмбриол., 1987. Т. 42. N 4. С. 93-95.
Невмывака Г.А. Иннервация кишки у дождевого червя (Allolobophora calliginosa) И Докл. АН СССР, 1947.
Т. 56. С. 533-536.
Невмывака Г.А. О возможных источниках происхождения нервных элементов кишки беспозвоночных И Реф.
научн. работ Молотовск. (Пермского) отд. Всесоюзн. о-ва анатомов, гистологов и эмбриологов. - Молотов
(Пермь), 1956. С. 97-98.
Невмывака Г.А. К вопросу об источниках происхождения нервной системы И Тр. IV Всесоюзн. съезда анато-
мов, гистологов и эмбриологов. - Харьков, 1961. С. 846-848.
Невмывака Г.А. Нервная система дождевого червя. - М.-Л.: Наука, 1966. 133 с.
Обухова Е.В. Изучение кинетики клеточных популяций эпителия пищеварительной железы голожаберного
моллюска Dendronotus arborescence И Цитология, 1982. Т. 24. С. 1177-1184.
Панкратова С.Ю. Иммуногистохимическое выявление эндокриноподобных клеток в пищеварительном трак-
те некоторых иглокожих И Тез. XXXVI конфер. студентов и аспирантов морфол. кафедр и лабораторий Санкт-
Петербургских ВУЗов и НИИ. - СПб., 1993. С. 21.
Панкратова С.Ю., Казаков В.К, Пунин М.Ю. Регуляторные клетки в пищеварительном тракте иглокожих:
иммуногистохимический анализ И Нейроэндокринология-95: Тез. докл. IV Всероссийск. конфер. - СПб., 1995.
С. 96.
Полякова Т.И., Плисецкая Э.М. Влияние глюкозы на клетки кишечного эпителия ланцетника Branchiostoma
lanceolatum//Журн. эвол. биохим. физиол., 1976. Т. 12. С. 184-187.
Плисецкая Э.М. Инсулин у моллюсков и его роль в регуляции углеводного обмена И Журн. эволюц. биохим.
физиол., 1977. Т. 13. С. 600-606.
145
Пузырев А.А., Иванова В.Ф. «Смешанные» гландулоциты эпителия двенадцатиперстной кишки некоторых
позвоночных животных и человека// Архив анат. гистол. эмбриол., 1986. Т. 90. Вып. 4. С. 48-54.
Пузырев А.А., Иванова В.Ф. Микроскопическая организация эндокринных клеток эпителия слизистой желу-
дочно-кишечного тракта низших позвоночных. 111. Эндокринные клетки эпителия средней кишки миноги Lampetra
fluviatilis И Цитология, 1991. Т. 33. N 3. С. 30-34.
Пузырев А.А., Иванова В.Ф. Гастроэнтеропанкреатическая система (развитие, строение, регенерация) И Мор-
фология, 1992. Т. 102. № 1. С. 5-28.
Пунин М.Ю. Эпителий протоков пищеварительной железы Cyprina islandica. Авторадиографический и элек-
тронно-микроскопический анализ И Цитология, 1981а. Т. 23. С. 134-144.
Пунин М.Ю. Мелкогранулярные клетки в составе эпителия средней кишки Priapulus caudatus И Вестник ЛГУ,
19816. №9. С. 110-113.
Пунин М.Ю. Кинетика репродукции и дифференцировки клеток в эпителии средней кишки и ее производных
у афродитид. II. Система базальных клеток в эпителии средней кишки // Цитология, 1981 в. Т. 23. С. 378-385.
Пунин М.Ю. Исследование организации эпителия пищеварительного тракта замковой брахиоподы Hemithyris
psittacea. II. Электронномикроскопический анализ//Цитология, 1981г. Т. 23. С. 1227-1234.
Пунин М.Ю. Нервные клетки в кишечном эпителии полихеты Nereis virens и двустворчатого моллюска
Arctica islandica И Биол. моря, 1981 д. № 4. С. 48-54.
Пунин М.Ю. Нервные клетки в кишечном эпителии двустворчатых моллюсков и полихет// Простые нервные
системы. Ч. 2. - Казань, 1985. С. 73-75.
Пунин М.Ю. Эндокриноподобные элементы в кишечном эпителии беспозвоночных животных И Цитология,
1986. Т. 28. № 10. С. 1035-1055.
Пунин М.Ю. Нейроподобные отростки в стенке кишечника двустворчатого моллюска Mytilus edulis И Цито-
логия, 1989. Т. 31. С. 154-159.
Пунин М.Ю. Гистологическая организация кишечных эпителиев приапулид, брахиопод, двустворчатых мол-
люсков и полихет. - СПб.: Наука, 1991. 248 с.
Пунин М.Ю. Система регуляторных элементов в эпителии пищеварительного тракта беспозвоночных живот-
ных И Цитология, 1993. Т. 35. № 2. С. 3-26.
Пунин М.Ю. Клетки, содержащие катехоламины и FMRFaM ид-подобное вещество, в эпителии пищевари-
тельного тракта морской звезды и морского ежа// Цитология, 1996а. Т. 38. № 10. С. 1048-1054.
Пунин М.Ю. Система регуляторных (нервных и эндокринных) клеток в кишечном эпителии беспозвоночных
животных И 1(Х1) Международное совещ. по эволюц. физиол. - СПб., 19966. С. 198-199.
Пунин М.Ю. Катехоламины в эпителии пищеварительного тракта иглокожих И Докл. РАН, 1997а. Т. 353.
№ 1.С. 131-133.
Пунин М.Ю. РМИРамид-иммунореактивные клетки в эпителии средней кишки полихет И Журн. эвол. био-
хим. физиол., 19976. Т. 33. С. 212-217.
Пунин М.Ю. РМИРамид-иммунореактивные клетки и отростки в пищеварительном тракте моллюсков //
Журн. эвол. биохим. физиол, 1997в. Т. 33. С. 648-656.
Пунин М.Ю. РМИРамид-подобная иммунореактивность в эпителии пищеварительного тракта брахиоподы
Hemithyris psittacea// Цитология, 1998. Т. 40. С. 347-352.
Пунин М.Ю. Моноамин-содержащие структуры в кишечном эпителии приапулид и полихет И Биол. моря,
1999. Т. 25. №4. С. 306-310.
Пунин М.Ю. Кишечные регуляторные системы беспозвоночных животных и их вероятная эволюция у много-
клеточных //Журн. эволюц. биохим. физиол., 2000. Т. 36. № 6. С. 586-599.
Пунин М.Ю. Кишечные регуляторные системы и их вероятная эволюция И Механизмы функционирования
висцеральных систем: Тез. докл. - СПб., 2001. С. 299 - 300.
Пунин М.Ю., Заварзин А.А. Исследование организации кишечного эпителия приапулид. II. Ультраструктура
основных клеток эпителия желудка и средней кишки и организация процессов репродукции И Цитология, 1979.
Т. 21. №8. С. 887-894.
Пунин М.Ю., Казаков В.К. Серотонин-иммунореактивные клетки и отростки в стенке кишечника полихет
Nereis virens, N.pelagica и Lepidonotus squamatus 11 Простые нервные системы. - Л.. Наука, 1988. С. 246-249.
Пунин М.Ю., Казаков В.К. Содержащие серотонин-подобное вещество клетки и отростки в стенке кишечника
полихет// Цитология, 1989а. Т. 31. С. 277-281.
Пунин М.Ю., Казаков В.К. Кальцитонин-иммунореактивные эндокриноподобные клетки в кишечнике дву-
створчатых моллюсков И Тез. докл. Всесоюзн. конфер. - Апатиты, 19896. С. 51.
Пунин М.Ю., Казаков В.К. Нейротензин-подобная иммунореактивность в кишечнике Arenicola marina И Про-
блемы изучения, рационального использования и охраны природных ресурсов Белого моря: Тез. докл. IV регион,
конфер. - Архангельск, 1990а. С. 109-110.
Пунин М.Ю., Казаков В.К. Кальцитонин-иммунореактивные клетки в кишечном эпителии двустворчатых
моллюсков И Докл. АН СССР, 19906. Т. 311. С. 736-738.
146
Пунин М.Ю., Казаков В.К. Кальцитонин-иммунореактивные структуры в пищеварительном тракте брюхоно-
гих и головоногих моллюсков И Цитология, 1992. Т. 34. № I. С. 54-57.
Пунин М.Ю., Казаков В.К. Клетки и клеточные отростки, иммунореактивные к пептидам нейротензину и ак-
тиватору головы гидры, в эпителии пищеварительной системы брахиоподы Coptothyris grayi И Цитология, 1993.
Т. 35. № 8. С. 24-29.
Пунин М.Ю., Константинова М.С. Эндокриноподобные клетки в кишечном эпителии двустворчатого мол-
люска Mytilus edulis И Цитология, 1988. Т. 30. С. 795-801.
Пунин М.Ю., Константинова М.С. Использование флуоресцентно-гистохимического метода Фернесса для
выявления моноамин-содержащих элементов у беспозвоночных животных И Цитология, 1989. Т. 31. С. 861-863.
Пунин М.Ю., Лукьянова Е.Л. Организация эпителия средней кишки бродячей полихеты Nereis virens. I. Вса-
сывающие и секреторные клетки И Цитология, 1984а. Т. 26. С. 60-68.
Пунин М.Ю., Лукьянова Е.Л. Организация эпителия средней кишки бродячей полихеты Nereis virens. 11. Ба-
зальные клетки И Цитология, 19846. Т. 26. С. 188-195.
Пунин М.Ю., Маркосова Т.Г Кишечная регуляторная система ресничных червей. И XII Международное со-
вет. по эвол. физиол. - СПб., 2001. С. 124.
Пунин М.Ю., Филимонова С.А. Организация эпителия пищевода, желудка и кишечника двустворчатого мол-
люска Arctica islandica// Цитология, 1986. Т. 28. С. 165-172.
Пунин М.Ю., Константинова М.С., Казаков В.К. Эндокриноподобные клетки в стенке пищеварительного
тракта двустворчатых моллюсков: Тез. докл. III Всесоюзн. конфер. по нейроэндокринологии. - Л., 1988. С. 197.
Пунин М.Ю., Казаков В.К, Кулаковский Э.Е. Кальцитонин-иммунореактивные структуры в кишечнике и в
центральной нервной системе двустворчатых моллюсков И Цитология, 1990а. Т. 32. С. 611-619.
Пунин М.Ю., Казаков В.К, Кочергинский Е.Б., Калихевич В.Н., Ардемасова З.А. Нейротензин-
иммунореактивные клетки в кишечном эпителии полихеты Arenicola marina И Цитология, 19906. Т.32. С. 1002-
1005.
Пунин М.Ю., Казаков В.К., Кочергинский Е.Б. Иммуногистохимическое выявление регуляторных клеточных
элементов в пищеварительном тракте брахиоподы // Цитология, 1992а. Т 34. № 1. С. 50-53.
Пунин М.Ю., Казаков В.К, Кочергинский Е.Б. Клетки, иммунореактивные к нейротензину и пептиду-
активатору роста головы гидры, в пищеварительном тракте и центральной нервной системе двустворчатых моллю-
сков И Цитология, 19926. Т. 34. № 6. С. 45-54.
Пунин М.Ю., Казаков В.К, Панкратова С.Ю. Иммуногистохимическое выявление регуляторных (эндокрин-
ных и нервных) клеток в пищеварительном тракте иглокожих // Цитология, 1996. Т.38. № 10. С. 1055-1061.
Пунин М.Ю., Казаков В.К, Мкртчян Л.Г Регуляторные клетки в эпителии средней кишки олигохет// Докл.
РАН, 1998. Т. 362. С. 718-720.
Пунин М.Ю., Филимонова С.А., Казаков В.К, Мкртчян Л.Г Регуляторные клетки в эпителии кишечника зем-
ляных червей И Цитология, 1999. Т 41, № 11. С. 932-938.
Пунин М.Ю., Казаков В.К, Мкртчян Л.Г Иммуногистохимическое выявление регуляторных клеток в пище-
варительной системе пиявок И Журн. эвол. биохим. физиол., 2000. Т 36. № 2. С. 141-146.
Русаков Ю.И., Казаков В.К. Получение инсулиноподобного вещества моллюсков и антисыворотки к нему И
Журн. эволюц. биохим. физиол., 1979. Т. 15. С. 617-619.
Русаков Ю.И., Бондарева В.М., Карасев В.С., Лейбуш Б.Н., Баркан Р.С., Перцева М.Н. Некоторые биохими-
ческие характеристики инсулиноподобного вещества двустворчатого моллюска Anodonta cygnea И Биохимия, 1991.
Т. 56. Вып. 4. С. 718-726.
Салимова Н.Б. Локализация биогенных аминов у ланцетника Branchiostoma lanceolatum И Докл. АН СССР,
1978. Т. 242. С. 939-941.
Сахаров Д.А. Генеалогия нейронов. -М.: Наука, 1974. 183 с.
Уголев А.М. Энтериновая (кишечная гормональная) система. -Л.: Наука, 1978. 314 с.
Уголев А.М. Эволюция пищеварения и некоторые принципы эволюции функций И Журн. эвол. биохим. физи-
ол., 1979. Т. 15. С. 239-248.
Уголев А.М. Эволюция пищеварения и принципы эволюции функций: элементы современного функциона-
лизма. -Л.: Наука, 1985. 544 с.
Ушева Л.Н. Гистоморфология и пролиферация клеток эпителия задней кишки у приморского гребешка И
Биол. моря, 1983. № 3. С. 17-24.
Ушева Л.Н. Морфология и пролиферация эпителиев органов пищеварительной системы приморского гре-
бешка: Автореф. канд. дисс. - Владивосток, 1988. 19 с.
Фалкмер С. Выработка островковых гормонов у беспозвоночных, круглоротых и примитивных челюстноро-
тых. И Эволюц. эндокринология поджелудочной железы. - Л.. Наука, 1977. С. 90-96.
Филимонова Г.Ф. Функциональная морфология пищеварительной системы иглокожих. - Л.: Наука, 1979.
120 с.
147
Филимонова Г.Ф. Адаптивные черты в строении пищеварительного тракта офиуры Ophiopholis aculeate
(Echinodermata: Ophiuroidea)//Журн. общ. биол., 1980. Т. 41. №1. С. 118-124.
Филимонова С.А. Изменение ультраструктуры эпителия средней кишки Xenopsylla chaeopis (Siphonaptera,
Pulicidae) в процессе пищеварения И Паразитол. сб., 1985. Т. 33. С. 149-158.
Филимонова С.А. Ультраструктурные изменения кишечного эпителия имаго Xenopsylla chaeopis
(Siphonaptera) после выхода из кокона и начала питания И Паразитология, 1986. Т. 20. № 2. С. 99-105.
Филимонова С.А. Структура и функциональные особенности эпителия средней кишки блох (Siphonaptera):
Автореф. канд. дисс. - Л., 1990. 16 с.
Шатров А.Б. Краснотелковые клещи и их паразитизм на позвоночных животных. - СПб: Изд-во СПбГУ,
2000. 282 с.
Шахламов В.А., Макаръ В.И. Энтероэндокринные клетки, их структура и функция И Архив анат. гистол. эм-
бриол., 1985. Т. 89. Вып. 9. С. 7-17.
Ширкина Н.И. Строение и гистотопография гастроэнтеропанкреатической системы костистых рыб И Морфо-
логия. 1995. Т. 108. Вып. 3. С. 78-83.
Яглов В.В., Ломоносова Г.А. Диффузная эндокринная система. Итоги и перспективы исследования И Успехи
соврем, биол., 1985. Т. 99. Вып. 2. С. 264-276.
Abad М.Е., Peeze Binkhorst M.M.t Elbal M.T., Ronibout J.H.W.M. A comparative immunocytochemical study of the
gastro-entero-pancreatic (GEP) endocrine system in a stomachless and stomach-containing teleost // Gen. Comp.
Endocrinol. 1987. Vol. 66. P. 123-136.
Abaurrea A., Vazques J. J. Estudio estructural у ultraestructural del intestino de Cryptophallus aspersa 11 Morphol.
Norm. Pathol. Sect. A. 1979. Vol. 3. P. 403^17.
Alba Y., Villaro A.C., Ortiz A., Sesma P., Lopez J. Estudio histologico de les tubulos malpigio de Schistocerca
gregaria (Forscal). I. Microscopia de luz// Morphol. Norm. Patol. Sect. A. 1983. Vol. 7. P. 146-162.
Alba Y., Villaro A.C., Sesma P., Vazquez J.J., Abaurrea A. Gut endocrine cells in the snail Helix aspersa // Gen.
Comp. Endocrinol. 1988. Vol. 70. P. 363-373.
Alba Y. Villaro A.C., Sesma P., Vazquez J J. Neuroendocrine complexes in the gut of the snail Helix aspersa // Gen.
Comp. Endocrinol. 1989. Vol. 199. P. 313-325.
Alexander N.J., Fahrenbach W.H. Fine structure of endocrine hindgut cells of a lepidopteran, Ostrinia nubilalis
(Hubn) // Z. Zellforsch. 1969. Bd 94. S. 337-345.
Ali-Rochedi A., Varndell I.M., Adrian T.E., Gapp D.A., Van Noorden S., Bloom S.R., Polak J.M. Peptide YY (PYY)
immunoreactivity is co-stored with glucagon-related immunoreactants in endocrine cells of the gut and pancreas //
Histochemistry. 1984. Vol. 80. P. 487^91.
Alumets J., Hakanson R., Sundler F. Distribution, ontogeny and ultrastructure of pancreatic polypeptide (PP) cells in
the pancreas and gut of the chicks // Cell Tissue Res. 1978a. Vol. 194. P. 377-386.
Alumets J., Hakanson R., Sundler F, Chang K. Leu-enkephalin-like material in nerves and enterochromaffin cells in
the gut. An immunohistochemical study// Histochemistry. 1978b. Vol. 56. P. 187-196.
Andreozzi G., Antonucci R., Battaglibi P., Gargiulo G., Russo P. Sulla presenza e destribuzione de cellule SNED nell
intestino di OctoIasium complanatum (Annelide, Oligochete). Nota preliminare // Boll. Soc. Natur. Napoli. 1990. Vol. 97.
P. 37-51.
Andrew A. Intestinal endocrine cells of chicks around the time of hatching // Cell Tissue Res. 1976a. Vol. 172.
P. 541-551.
Andrew A. Endocrine cells of the stomach of chiks around the time of hatching // Cell Tissue Res. 1976b. Vol. 172.
P. 553-561.
Andrews A., Kramer B., Rowdon B.B. The embryonic origin of endocrine cells in the gastrointestinal tract // Gen.
Comp. Endocrinol. 1982. Vol. 47. P. 249-265.
Andries J.C, Beauvillain. Ultrastructural study of cholecystokinin-like immunoreactivity in endocrine cells of the
insect midgut // Cell Tissue Res. 1988. Vol. 254. P. 75-81.
Andries J.C., Tramu G. Detection immunohistochemique de substances apparantees a des hormones peptidiques de
mammiferes dans le mesenteron d’Aeshna cyanea (Insecte, Odonate) // C. R. Acad. Sci. Ser. 3. 1984. T. 299. P. 181-184.
Andries J.C., Tramu G. Distribution patterns of mammalian-like peptide immunoreactive cells in the midgut of
Aeshna cyanea (Insecta, Odonata) // Experientia. 1985a. Vol. 41. P. 500-503.
Andries J.C., Tramu G. Ultrastructural and immunohistochemical study of endocrine cells in the midgut of the
cockroach Blaberus craniifer (Insecta, Dictyoptera) П Cell Tissue Res. 1985b. Vol. 240. P. 323-332.
Asaka L, Marchand C.R., Strosser M.T., VanDorsselaer A. Mise evidence biochimique d un peptide
immunologiquement apparente a la somatostatine-14 dans Г hepatopancreas de 1’escargat Helix aspersa // C. r. Soc. Biol.
1987. N. 181. P. 187-193.
Aubry R. Sur 1’existence d’un type cellulaire mal connu, d’allure endocrine dans le mesenteron de Locusta
migratoria // C. R. Acad. Sci. Ser. D. 1975. T. 281. P. 1405-1407.
148
Baguna J. A demonstration of a peripheral and a gastrodermal nervous plexus in planarians // Zool. Anz. 1974.
Bd 193. S. 240-244.
Baguna J., Ballester R. The nervous system in planarians: peripheral and gastrodermal plexuses, pharynx
innervation, and the relationship between central nervous system structure and the acoelomata organization // J. Morphol.
1978. Vol. 155. P. 237-252.
Barginann W., Behrens B. Uber die Pylorusanhange des Seestems (Asterias rubens L.) insbesondere ihre Innerva-
tion // Z. Zellforsch. 1968. Bd 84. S. 563-584.
Barrington E.J.W. The pancreas and intestine. // The biology of lampreys. - London; New-York. Vol. 2. 1972.
P. 135-169.
Benkhay M., Moreuau R., Gourdoux L, Dutriieu J. Effects of intestinal insulin-like substance on glucose catabolism
in locust// Bull. Soc. Zool. Fr. 1986. T. 111. P. 33.
Beorlegui C., Martinez A., Sesina P. Endocrine cells and nerves in the pyloric caeca and the intestine of
Onchorhynchus mykiss (Teleostei): an immunocytochemical study // Gen. Comp. Endocrinol. 1992a. Vol. 86. P. 483-495.
Beorlegui C, Martinez A., Sesma P. Some peptide-like colocalizations in endocrine cells of the pyloric caeca and the
intestine of Onchorhynchus mykiss (Teleostei) // Cell Tissue Res. 1992b. Vol. 269. P. 353-357.
Bevis P.J.R., Thomdyke M.C. Endocrine cells in the oesophagus of the ascidian Styela clava, a cytochemical and
immunofluorescence study//Cell Tissue Res. 1978. Vol. 187. P. 153-158.
Bevis P.J.R., Thorndyke M.C. A cytochemical and immunofluorescence study of endocrine cells in the gut of the
ascidian Styela clava // Cell Tissue Res. 1979. Vol. 199. P. 139-144.
Bevis P.J.R., Thorndyke M.C. Stimulation of gastric enzyme secretion by porcine cholecystokinin in the ascidian
Styela clava // Gen. Comp. Endocrinol. 1981. Vol. 45. P. 458-464.
Bidder A.M. Feeding and digestion in Cephalopoda. // Physiology of mollusca. Vol. 2. - New-York; London. 1966.
P. 97-124.
Bisgrove B.W., Burke R.D. Development of serotoninergic neurons in embryos of the sea urchin Strongylocentrotus
purpuratus // Dev. Growth Differ. 1986. Vol. 28. P. 569-574.
Bodenmuller H., Schaller H.C. Conserved amino acid sequence of a neuropeptide, the head activator, from
coelenterates to humans // Nature. 1981. Vol. 293. P. 579-580.
Boer H.H., Montagne-Wajer C. Functional morphology of the neuropeptidergic light-yellow-cell system in
pulmonate snails // Cell Tissue Res. 1994. Vol. 277. P. 531-538.
Boer H.H., Montagne-Wajer C, van Minnen J., Ramkema M., de Boer P. Functional morphology of the
neuroendocrine sodium influx-stimulating peptide system of the pond snail, Lymnaea stagnalis // Cell Tissue Res. 1992.
Vol. 268. P. 559-566.
Boer H.H., Montagne-Wajer C, Smith F.G., Parish D.C., Ramkema M., Hoek R.M., van Minnen J., Benjamin P.R.
Functional morphology of the light yellow cell and yellow cell (sodium influx-stimulating peptide) neuroendocrine system
of the pond snail Lymnaea stagnalis И Cell Tissue Res. 1994. Vol. 275. P 361-368.
Boletzky S. von. Gut development in cephalopods: a correction // Rev. Suisse Zool. 1978. Vol.85. P. 379-380.
Boquist L., Falkmer S., Mehrotra B.K. Ultrastructural search for homologues of pancreatic В-cells in the intestinal
mucosa of the mollusc Buccinum undatum // Gen. Comp. Endocrinol. 1971. Vol. 17. P.236-239.
Bouinas M., Dubois M. Inhibition par la somatostatine de faction hypoglycemiante de 1’insuline chez 1’Abeille in
vivo et localisation cytochemique d’un antigene apparante a la somatostatine dans 1’epithelium digestif ceinsi que les tubes
de Malphighi de 1’Abeille // C. R. Acad. Sci. Ser. 3. 1982. T. 294. P. 1029-1034.
Brodin E., Alumets J., Hakanson R., Leander S., Sundler F. Immunoreactive substance P in the chicken gut:
distribution, development and possible functional significance // Cell Tissue Res. 1981. Vol. 216. P. 455^469.
Brown M.R., Raikhel A.S., Lea A.O. Ultrastructure of midgut endocrine cells in the adult mosquito Aedes aegypti //
Tissue Cell. 1985. Vol. 17. P. 709-721.
Brown M.R., Crim J.W., Lea A.O. FMRFamide- and pancreatic polypeptide-like immunoreactivity of endocrine cells
in the midgut of a mosquito // Cell Tissue Res. 1986. Vol. 18. P. 419-428.
Buchan A.M.J., Lance V., Polak J.M. The endocrine pancreas of Alligator missisipiensis: an immunocytochemical
investigation // Cell Tissue Res. 1982. Vol. 224. P. 117-128.
Buchan A.M.J., Lance V., Polak J.M. Regulatory peptides in the gastrointestinal tract of Alligator missisipiensis: an
immunocytochemical study // Cell Tissue Res. 1983. Vol. 231. P. 439-449.
Bullock Т.Н. The anatomical organization of the nervous system of Enteropneusta // Quart. J. Mier. Sci. 1945.
Vol. 86. P. 55-111.
Burighel P., Milanesi C. The cytology of the gut epithelium in the colonial ascidian Botryllus schlosseri // Boll. Zool.
1975a. Vol.42. P. 439.
Burighel P., Milanesi C. Fine structure of the gastric epithelium of the ascidian Botryllus schlosseri. Mucous,
endocrine and plicated cells // Cell Tissue Res. 1975b. Vol. 158. P. 481^496.
Burighel P., Milanesi C. Fine structure of the intestinal epithelium of the colonial ascidian Botryllus schlosseri // Cell
Tissue Res. 1977. Vol. 182. P. 357-369.
149
Burkhardt-Hohn P., Holmgren S. A comparative study of neuropeptides in the intestine of two stomachless teleosts
(Poecilia reticulata, Leuciscus idis melanotus) under conditions of feeding and starvation // Cell Tissue Res. 1989. Vol.
255. P. 245-254.
Bush M.S. The ultrastructure and function of the intestine of Patella vulgata // J. Zool. 1988. Vol. 215. P. 695-702.
Cantera R., Nassel D.R. Dual peptidergic innervation of the blowfly hindgut: a light- and electron microscopic study
of FMRFamide and proctolin immunoreactive fibres // Comp. Biochem. Physiol. 1991. Vol. C99. P. 517-525.
Cassier P., Fain-Maurel M.A. Sur la presence d’un systeme endocrine diffus dans la mesenteron de queiques
insectes // Arch. Zool. Exp. Gen. 1977. T. 118. P. 197-209.
Cassier P., Alibert J., Fain-Maurel M.A. Sur la presencede cellules de type endocrine dans I’intestine moyen de
Petrobius maritimus Leach (Insecte apterygote, Thysanoure) // C. R. Acad. Sci. Ser D. 1972. N. 275. P. 2691-2693.
Chan V.G., Fontaine A.R. Is there a В-cell homolog in starfish? // Gen. Comp. Endocrinol. 1971. Vol. 16. P. 183—
191.
Chapman D.M. Cnidarian histology. // Coelenterate biology. - New York; etc.: Academic Press, 1974. P. 1-92.
Chapman R.H. Structure of the digestive system // Comp. Insect Physiol. Biochem. Pharmacol. 1985. Vol. 4. P. 165—
211.
Cheng H., Leblond C.P. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small
intestine. III. Enteroendocrine cells // Amer. J. Anat. 1974a. Vol. 141. P. 503-520.
Cheng H, Leblond C.P. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small
intestine. V Unitarian theory of the origin of the four epithelial cell types // Amer. J. Anat. 1974b. Vol. 141. P. 537-562.
Cheung R., Andrews P.C., Plisetskaya E.M., Younson J.H. Immunoreactivity to peptides belonging to the pancreatic
polypeptide family (NPY, aPY, PP, PYY) and to glucagon-like peptide in the endocrine pancreas and anterior intestine of
adult lampreys, Petromyzon marinus: an immunohistochemical study // Gen. Comp. Endocrinol. 1991. Vol. 81. P. 51-63.
Chia F.S. Histology of the pyloric caeca and its changes during brooding and starvation in a starfish, Leptasterias
hexatilis//Biol. Bull. 1969. Vol. 136. P. 185-192.
Chiba A., Honma Y., Oka S. Ontogenetic development of neuropeptide Y-like immunoreactive cells in the
gastroenteropancreatic endocrine system of the dogfish // Cell Tissue Res. 1995. Vol. 282. P. 33-40.
Cimini V., Van Noorden S., Polak J.M. Co-localization of substance P-, bombesin- and peptide histidine isoleucin
PHI-like peptides in gut endocrine cells of the dogfish Scyliorhinus stellaris // Anat. Embriol. 1989. Vol. 179. P. 605-614.
Cimini V., Van Noorden S., Sansone M. Neuropeptide Y-like immunoreactivity in the dogfish gastroenteropancreatic
tract: light and electron microscopical study // Gen. Comp. Endocrinol. 1992. Vol. 86. P. 413-423.
Clark M.E. Histochemical localization of monoamines in the nervous system of the polychaete Nephtys // Proc. Roy.
Soc. London, 1966. Vol. B165. P. 308-325.
Cobb J.L.C. The innervation of the oesophagus of the sea urchin Heliocidaris erythrogramma // Z. Zellforsch. 1969a.
Bd 98. S. 323-332.
Cobb J.L.C. The distribution of monoamines in the nervous system of echinoderms // Comp. Biochem. Physiol.
1969b. Vol. 28. P. 967-971.
Cobb J.LC, Raymond A.M. The basiepithelial nerve plexus of the viscera and coelom of eleutherozoan
Echinodermata//Cell Tissue Res. 1979. Vol. 202. P. 155-163.
Collip J.B. The demonstration of insulin-like substance in the tissues of the clam (Mya arenaria) // J. Biol. Chem.
1923. Vol. 55. P. XXXIX.
Conlon J.M., Bal 11 nan n M., Lamberets R. Regulatory peptides (glucagon, somatostatin, substance P and VIP) in the
brain and gastrointestinal tract of Amblistoma mexicanus // Gen. Comp. Endocrinol. 1985. Vol. 58. P. 150-158.
Crasium C, Molnar B., Madar I., Sildan N., Рога E. Histological evidence of the action of some alfa- and beta-
cytotoxic compounds upon the islet-like formations in the intestinal mucosa of Mytilus galloprovincialis (L.)// Rev. Roum.
Biol. Ser. Biol. Anim. 1981. T. 26. P. 149-153.
Csoknya M., Lengravi I., Benedeczky I., Hamori J. Immunohistochemical and ultrastructural study of the enteric
nervous system of earthworm, Lumbricus terrestris // Acta Biol. Hung. 1992. Vol. 43. P. 241-251.
Curry W.J., Fairweather I., Lohnston C.F., Halton D.W., Buchanon K.D. Immunochemical demonstration of
vertebrate neuropeptides in the earthworm Lumbricus terrestis (Annelida, Oligochaeta) // Cell Tissue Res. 1989. Vol. 257.
P. 577-586.
Dales R.P. Feeding and digestion in terebellid polychaetes // J. Mar. Biol. Ass. U.K. 1955. Vol. 34. P. 55-79.
Dales R.P., Pell J.S. The origin and nature of the brown substance in the gut caeca of the polychaetes Aphrodite
aculeata and Gattyana cirrosa//J. Zool. 1971. Vol. 163. P. 413-419.
Dallai R., Yin W.-Y., Xue L, Francois J. The gut structure of Sinentomon erythanum Yin (Protura: Sinentomidae)//
Int. J. Insect. Morphol. Embryol. 1989. Vol. 18. P. 173-184.
Davidson J.K., Falkmer S., Mehrotra B.K., Wilson S. Insulin assay and light microscopical studies of digestive
organs in Protostomian and Deuterostomian species and in Coelenterates // Gen. Comp. Endocrinol. 1971. Vol. 17. P. 388—
401.
Davis L.E. Differentiation of ganglionic cells in Hydra// J. Exp. Zool. 1971. Vol. 176. P. 107-128.
150
Davis L.E. Ultrastnictural evidence for the presence of nerve cells in the gastrodermis of Hydra // Z. Zellforsch.
1972. Bd 123. S. 1-17.
D'Este L, Campo S., Salvi E., Renda T. Ontogenesis of bombesin like immunoreactive cells in the chicken
proventriculus//Bas. Appl. Histochem. 1984. Vol. 28. P. 143-150.
D'Este L., Buffa L, Casu C, Carboni N., Pelagi M., Siccardi A.G., Renda T. Immunohistochemical localization of
chromogranin A and В in endocrine cells of the alimentary tract of the adult lizard Podarcis sicula // Cell Tissue Res. 1993.
Vol. 273. P. 335-344.
d'Hondt J-L., Boucaud-Camou E. Etude de 1’intestin et de la glande digestive de Terebratulina retusa (L.)
(Brachiopode). Ultrastructure et recherche d’activites amilasiques et protealisiques // Bull. Soc. Zool. France. 1982. Vol.
107. P. 207-216.
Diaz de Rada O., Sesma P., Vazquez J.J. Endocrine cells of the gastric mucosa of Rana temporaria L. // Histol.
Histopathol. 1987. Vol. 2. P. 119-128.
Dolder H. An ultrastnictural and cytochemical study of neuromuscular junctions in echinoderms // Histochemistry.
1975. Vol. 44. P. 313-322.
Dougan D.F.H., McLean J.R. Evidence for the presence of dopaminergic nerves and receptors in the intestine of a
mollusc, Tapes watlingi // Comp. Gen. Pharmacol. 1970. Vol. 1. P. 33-46.
Dow J.A.T. Insect midgut function//Adv. Insect Physiol. 1986. Vol. 19. P. 187-328.
Duve H., Thorpe A. The distribution of pancreatic polypeptide in the nervous system and gut of the blow-fly
Calliphora vomitoria (Dyptera) // Cell Tissue Res. 1982. Vol. 227. P. 67-77.
Duve H., Thorpe A. Distribution and functional significans of Leu-callatostatins in the blowfly Calliphora vomito-
ria // Cell Tissue Res. 1994. Vol. 276. P. 367-379.
Duve H., Johnsen A.H., Maestro J.-L, Scott A.G., Crook N., Winstanley D., Thorpe A. Identification? Tissue
localization and physiological effect in vitro of a neuroendocrine peptide identical to a dipteran Leu-callatostatin in the
codling moth Cydia pomonella (Torticidae: Lepidoptera) // Cell Tissue Res. 1997. Vol. 289. P. 73-83.
El-Salhy M. Immunocytochemical investigation of the gastro-entero-pancreatic (GEP) neuro-hormonal polypeptides
in the pancreas and gastrointestinal tract of the dogfish Squalus acanthias // Histochemistry. 1984. Vol. 80. P. 193-205.
El-Salhy M., Grimelius L. The endocrine cells of the gastrointestinal mucosa of a squamate reptile, the grass lizard
(Mabuya quinquetaeniata): a histological and immunohistochemical study // Biomed. Res. 1981. Vol. 2. P. 639-658.
El-Salhy M., Grimelius L, Lundberg J.M., Tatemoto K., Terenius L Immunocytochemical evidence for the
occurrence of PYY, a newly isolated gut polypeptide in endocrine cells in the gut of amphibians and reptiles // Biomed.
Res. 1982. Vol. 3. P. 303-306.
Elbal M.T., Lozano M.T., Aguielleiro B. The endocrine cells in the gut of Mugul saliens Risso, 1810 (Teleostei): an
immunocytochemical and ultrastnictural study // Gen. Comp. Endocrinol. 1988. Vol. 70. P. 231-246.
Elbal M.T., Garcia Ayala A., Aguielleiro B. Light and electron microscopic immunocytochemical demonstration of
the coexistence of somatostatin 14- and somatostatin 25-like peptides in the endocrine cells of the stomach of Sparus aurata
(Teleostei)//Gen. Comp. Endocrinol. 1991. Vol. 84. P. 36-43.
Elphick M.R., Emson R.H., Thorndyke M.C. FMRFamide-like immunoreactivity in the nervous system of the starfish
Asterias rubens//Biol. Bull. 1989. Vol.177. P. 141-145.
Elphick M.R., Price D.A., Lee T.D., Thorndyke M.C. The SALMFamides: a new family of neuropeptides isolated
from an echinoderm// Proc. Roy. Soc. Lond. 1991a. Vol. B243. P. 121-127.
Elphick M.R., Reeve J.R., Burke R.D., Thorndyke M.C. Isolation of the neuropeptide SALMFamide-1 from starfish
using a new antiserum// Peptides. 1991b. Vol. 12. P. 455-459.
Emdin S.O., Falkmer S. Phylogeny of insulin // Acta paediat. 1977. Vol. 270. Suppl. P. 15-25.
Emig C.C. Ultrastructure de tube digestif de Phoronis psammophila Cori. I. Estomac (Cellules ciliees) // Arch. Zool.
Exp. Gen. 1967. Vol. 108. P.617-632.
Emig C.C. Ultrastructure de tube digestif de Phoronis psammophila Cori. II. Pre-estomac et intestin // Arch. Zool.
Exp. Gen. 1968. Vol. 109. P.145-154.
Endo Y. Ontogeny of endocrine cells in the gut of the insect Periplaneta americana // Cell Tissue Res. 1984. Vol. 238.
P. 421-423.
Endo Y., Nishiitsutsuji-Uwo J. Gut endocrine cells in insects: the ultrastructure of the gut endocrine cells of the
lepidopterous species// Biomed. Res. 1981. Vol. 2. P. 270-280.
Endo Y., Nishiitsutsuji-Uwo J. Exocytotic release of secretory granules from endocrine cells in the midgut of insects
//Cell Tissue Res. 1982a. Vol. 222. P. 515-522.
Endo Y., Nishiitsutsuji-Uwo J. Fine structure of developing endocrine cells and columnar cells in the cockroach
midgut// Biomed. Res. 1982b. Vol. 3. P. 637-644.
Endo Y., Nishiitsutsuji-Uwo J. Gut endocrine cells i insects. // The ultrastructure and functioning of insect cells. -
Tokyo, 1982c. P. 105-106.
Endo Y., Nishiitsutsuji-Uwo J., Iwanaga N., Fujita T. Ultrastructural and immunohistochemical identification of
pancreatic polypeptide-immunoreactive endocrine cells in the cockroach midgut // Biomed. Res. 1982. Vol. 3. P. 454-456.
151
Endo Y., Sugihdra H, Fujita T, Nishiitsutsuji-Uwo J. Kinetics of columnar and endocrine cells in the cockroach
midgut// Biomed. Res. 1983. Vol. 4. P. 51-60.
Erspamer V. Presenza di enteramina о di una sostanza enteraminosimile neddli estratti gastrointestinali e spleneci dei
pesci e negli estratti gastroenterici delle Ascidie// Esperientia. 1946. Vol.2. P. 369-371.
Erspamer V., Asero B. Identification of enteramine, the specific hormone of the enterochromaffin cell system, as 5-
hydroxytryptamine//Nature. 1952. Vol. 169. P. 800-801.
Faber V., Komnick H. Peroxisomes in the midgut epithelium, malpigian tubules and fat body of larvae of the
dragonfly, Aeshna cyanea //Tissue Cell. 1989. Vol. 21. P. 917-923.
Fain-Maurel M.A., Cassier P., Alibert J. Etude infrastructurale et cytochemique de Г intestin moyen de Petrobius
maritimus Leach en rapport avec ses fonctions excretrices et digestives // Tissue Cell. 1973. Vol. 5. P. 603-631.
Falck B., Hillarp N.-A., Thieme C., Thorp A. Fluorescence of catecholamines and related compounds condensed with
formaldehyde//J. Histochem. Cytochem. 1962. Vol. 10. P. 348-354.
Falck B., Owman C. A detailed description of the fluorescence method for cellular demonstration of biogenic
monoamines // Acta Univ. Lund. 1965. Sect. 2. N 2. P. 1-23.
Falkmer S., Emdin S., Havn N., Lundgren G., Marques M., Ostberg Y., Steiner D.F., Thomas H.W. Insuli
invertebrates and cyclostomes // Amer. Zool. 1973. Vol. 13. P. 625-638.
Falkmer S., Eide R.P., Hellerstrom C, Peterson B. Phylogenetic aspects of somatostatin in the
gastroenteropancreatic (QEP) endocrine system// Metabolism. 1978. Vol. 27. P. 1193-1197.
Falkmer S., Fahrenkrug J., Alumets J., Hakanson R., Sundler F. Vasoactive intestinal polypeptide (VIP) in epithelial
cells of the gut mucosa of an elasmobranch cartilaginous fish, the ray // Endocrinol. Japan. S.R. 1980. Vol. 27. Suppl.
Vol. l.P. 31-35.
Falkmer S., Gustafsson M.K.S., Sundler S. Phylogenetic aspects on the neuroendocrine system // Nord. Psykiatr.
Tidsskr. 1985. Vol. 39. Suppl. 11. P. 21-30.
Favrel P., Van Wormhoudt A. Hormones gastrointestinales et enzymes digestives chez les crustaces decapodes //
Cah. Biol. Mar. 1986. Vol. 27. P.377.
Ferreira N. On the existence of two ultrastructural types of enterochromaffin cells in the duodenum of the chick
embryo// Arch. Port. Sci. Biol. 1966. Vol. 15. P. 75-86.
Flucher B.E., Lenglachner-Bachinger C, Feurle G.E. Immunocytochemical evidence for the co-localization of
neurotensin/xenopsin- and gastrin/caerulin-immunoreactive substances in Xenopus laevis gastro-intestinal tract // Gen.
Comp. Endocrinol. 1988. Vol. 72. P. 54-62.
Fritsch H.A.R. The occurrence of argirophilic and argentaffin cells in the gut of Ciona intestinalis L. // Cell Tissue
Res. 1976. Vol. 175. P. 131-135.
Fritsch H.A.R., Sprang R. On the ultrastructure of polypeptide hormone-producing cells in the gut of the ascidian,
Ciona intestinalis L. and the bivalve, Mytilus edulis L. // Cell Tissue Res. 1977. Vol. 177. P. 407-413.
Fritsch H.A.R., VanNoorden S., Pearse A.G.E. Cytochemical and immunofluorescence investigations on insulin-like
producing cells in the intestine of Mytilus edulis L. (Bivalvia) // Cell Tissue Res. 1976. Vol. 165. P. 365-369.
Fritsch H.A.R., VanNoorden S., Pearse A.G.E. Localization of somatostatin- and gastrin-like immunoreactivity in the
gastrointestinal tract of Ciona intestinalis L. // Cell Tissue Res. 1978. Vol. 186. P. 181-185.
Fritsch H.A.R., VanNoorden S., Pearse A.G.E. Calcitonin-like immunoreactivity in the alimentary tract of Ciona
intestinalis L. // Cell Tissue Res. 1980. Vol. 205. P. 439-444.
Fritsch H.A.R., VanNoorden S., Pearse A.G.E. Gastrointestinal and neurohormonal peptides in the alimentary tract
and cerebral complex of Ciona intestinalis (Ascidiaceae). Their relevance to the evolution of the diffuse neuroendocrine
system//Cell Tissue Res. 1982. Vol. 223. P. 369-402.
Fujii K., Ohta N., Sasaki T, Sekizava J., Yamada C, Kobayashi H. Immunoreactive FMRFamide in the nervous
system of the earthworm, Eisenia foetida // Zool. Sci. 1989. Vol. 6. P. 951-961.
Fujita T, Kobayashi S. Structure and function of gut endocrine cells // Int. Rev. Cytol. 1977. Suppl. 6. P. 187-233.
Fujita T, Kobayashi S., Yui R., Iwanaga T. Evolution of neurons and paraneurons. // Hormones, adaptation and
evolution. - Tokyo; Berlin: Jap. Sci. Press, Springer Verlag, 1980. P. 35-43.
Fujita T, Kanno T, Kobayashi S. The paraneuron. - Tokyo; etc., 1988. 367 p.
Furness J.B., Costa M., Wilson A.J. Water-stable fluorophores, produced by reaction with aldehyde solutions, for
histochemical localization of catechol- and indolamines // Histochemistry. 1977. Vol. 52. P. 159-170.
Furness J.B., Costa M., Llewellyn-Smith I. J. Localization of monoamines by aldehyde-induced fluorescence in the
peripheral nervous system. // Monoaminergic neurons. - J.Willey and sons; Chichester etc., 1987. P. 1-27.
Gabe M. Donnees histologiques sur les cellules endocrines gastrointestinales des amphibiens // Arch. Histol. Japan.
1972. Vol. 35. P. 51-81.
Gabe M. Contribution a 1’histologie des cellules endocrines duodenales des sauropsides // Acta Anat. 1973. Vol. 85.
p. 434-449.
Galloway S.M., Cutfield J.F. Insulin-like matherial from the digestive tract of the tunicate Pyura pachydermatina (sea
tulip)//Gen. Comp. Endocrinol. 1988. Vol. 69. P. 103-113.
152
Gapp D.A., Polak J.M. Localization of insulin to gastro-entero-pancreatic cells in the turtle gastrointestinal tract //
Gen. Comp. Endocrinol. 1985. Vol. 78. P. 48-55.
Gapp D.A., Kenny M.P., Polak J.M. The gastro-entero-pancreatic system of the turtle, Chrysemiys picta // Peptides.
Vol. 6. 1985. Suppl. 3. P. 347-352.
Garcia-Arraras J.E., Torres-Avillan I., Ortiz-Miranda S. Cells in the intestinal system of Holoturians
(Echinodermata) express cholecystokinin-like immunoreactivity//Gen. Comp. Endocrinol. 1991a. Vol.83. P. 233-242.
Garcia-Arraras J.E., Enamorado-Ayala I., Torres-Avillan I., Rivera V. FMRFamide-like immunoreactivity in cells
and fibers of the holoturian nervous system// Neurosci. Lett. 1991b. Vol. 132. P. 199-202.
Gerzelli G. Presence of enterochromaffin cells in the gut of amphioxus // Nature. 1961. Vol. 189. P. 237-238.
Gerzelli G. The enterochromaffin cells in the oozooids and blastozooids of tunicates // Folia Histochem. Cytochem.
1964. Vol. 3. P. 225-231.
Geuze J. J. Light and electron microscopic observations on the gastric mucosa of the frog Rana esculenta. 1. Normal
structure//Z. Zellforsch. 1971. Bd 117. S. 87-102.
Gibson R., Jennings J.B. Observations on the diet, feeding mechanisms, digestion and food reserves of the entocom-
mensal rhynchocoelan Malacobdella grossa // J. Mar. Biol. Ass. U. K., 1969. Vol. 49. P. 17-32.
Giusti F. The fine structure of the style sac and intestine in Mytilus galloprovincialis Lam. // Proc, malacol. Soc.
London. 1971. Vol. 39. P. 95-104.
Giraud A., Yeomans N.D. Fine structure of the gastric mucous and endocrine cells of the toad, Bufo marinus // Cell
Tissue Res. 1981. Vol. 218. P. 663-668.
Glattli E., Rudin W., Hecker H. Immunoelectron microscopic demonstration of pancreatic polypeptide in midgut
epithelium of hematophagous dipterans//J. Histochem. Cytochem. 1987. Vol. 35. P. 891-896.
Graf F. Les cellules paracrines des caecums posterior de crustace: un nouveau type cellulaire de systeme endocrine
diffus du tube digestif// C. R. Acad. Sci. Ser. 3. 1982. T. 294. P. 319-324.
Grimmelikhuijzen C.J.P., Graff D., McFarlane I.D. Neurons and neuropeptides in coelenterates // Arch. Histol.
Cytol. 1989. Vol. 52. Suppl. P. 265-276.
Grimmelikhuijzen C.J.P., Carstensen K, Danner D., Moosler A., Nothacker H.-Р., Reischeid R.K., Schmutzler C,
Vollert H., McFarlane I., Rinehart K.L. Coelenterate neuropeptides: structure, action and biosynthesis // Amer. Zool. 1992.
Vol. 32. P. 1-12.
Grimmelikhuijzen C.J.P., Leviev /., Carstensen K. Peptides in the nervous systems of cnidarians: structure, function,
and biosysnthesis // Int. Rev. Cytol. 1996. Vol. 167. P. 37-89.
Hamlett W.C, Musik J.A., EulittA.M., Jarrell R.L, Kelly M.A. Ultrastructure of fetal alimentary organs: stomach and
spiral intestine in the southern stingray, Dasyatis americana // Canad. J. Zool. 1996. Vol. 74. P. 1431-1442.
Hammersen E, Pokahr A. Elektronenmikroskopische Untersuchungen zur Epithelstruktur im Magen-Darmkanal von
Hirudo medicinalis L. I. Mitteilung: Das Epithel der Divertikel. //Z. Zellforsch. 1972a. Bd. 125. P. 378-403.
Hammersen F., Pokahr A. Elektronenmikroskopische Untersuchungen zur Epithelstruktur im Magen-Darmkanal von
Hirudo medicinalis L. II. Mitteilung: Das Epithel der Enddarmes. // Z. Zellforsch. 1972b. Bd 125. P. 532-552.
Hardisty M.W., Baker B.I. Endocrynology of lampreys. // The biology of Lampreys. Vol. 4b. - London; etc., 1982.
P. 1-115.
Hecker H. Structure and function of midgut epithelial cells in Culicidae mosquitoes (Insecta, Diptera) // Cell Tissue
Res. 1977. Vol. 184. P. 321-341.
Hecker H, Freyvogel T.A., Briegel H, Steiger R. Ultrastructural differentiation of the midgut epithelium in female
Aedes aegypti L. (Insecta, Diptera) imagines // Acta trop. 1971a. Vol. 28. P. 80-104.
Hecker H., Freyvogel T.A., Briegel H, Steiger R. Ultrastructural differentiation of the midgut epithelium in Aedes
aegypti L. (Insecta, Diptera) males // Acta trop. 1971b. Vol. 28. P. 275-290.
Heidenhein H. Untersuchungen uber den Bau der Lobdrusen // Arch. mikr. Anat. 1870. Bd 6. S. 368-396.
Higgins R.P., Storch V. Ultrastructural observations on the larva of Tubilichus corallicola (Priapulida) // Helgoland.
Wiss. Meeresuntersuch. 1989. Bd 43. S. 1-11.
Hojfmeister S.A.H., Schaller H.C. Head activator and head inhibitor are signals for nerve cell differentiation in hydra
// Developm. Biol. 1987. Vol. 122. P. 72-77.
Holmgren S. Substance P in gastrointestinal tract of Squalus acanthias // Moll. Physiol. 1985. Vol. 8. P. 119-130.
Holmgren S., Nilsson S. Bombesin-, gastrin/ССК-, 5-hydroxytryptamine-, neurotensin-, somatostatin-, and VIP-like
immunoreactivity and catecholamine fluorescence in the gut of elasmobranch, Squalus acanthias // Cell Tissue Res. 1983.
Vol. 234. P. 595-618.
Hordgren S., Vaillant C. Dimaline R. VIP-, gastrin/ССК-, bombesin-, somatostatin- and glucagon-like
immunoreactivities in the gut of the rainbow trout, Salmo gairdneri // Cell Tissue Res. 1982. Vol. 223. P. 141-153.
Holdquist A.L., Dockray G.J., Rosenquist C.L., Walsh J.H. Immunocytological characterization of cholecystoki
like pepties in lamprey gut and brain // Gen. Comp. Endocrinol. 1979. Vol. 37. P. 474—481.
Honma Y., Nakazawa K, Isobe H. Endocrine cells in the gut of a surfperch and some elasmobranchs I I Rep. Sado
Mar. Biol. Stat., Niigata Univ. 1986. N 16. P. 9-19.
153
Hutting G. Intestinal enterochromaffin cells in Branchiostoma lanceolatum// Acta Zool. Vol. 54. 1973. P. 173-178.
IwanagaT., Fujita T., Nishiitsutsuji-Uwo J., Endo Y. Immunochemical demonstration of PP-, somatostatin-,
enteroglucagon- and VIP-like imihunoreactivites in the cockroach midgut// Biomed. Res. 1981. Vol. 2. P. 202-207.
Iwanaga T., Fujita T., Takeda N., Endo Y. Urotensin l-like immunoreactivity in the midgut endocrine cells of the
insect Grillus bimaculatus and Periplaneta americana // Cell Tissue Res. 1986. Vol. 244. P. 565-568.
Jangoux M. Digestive system: general considerations. // Echinoderm nutrition. - Rotterdam: Balkema, 1982. P? 185-
187.
Jangoux M., Perpeet A.C. Etude comparative de la structure fine des caecums pyloriques de troix especes
d’Asteriidae (Echinodermata, Asteroidea)//Cah. Biol. Mar. 1972. Vol. 13. P. 401-420.
Jegla T.C., Greenberg M.J. Structure of bivalve rectum. II. Notes on cell types and innervation // Veliger. 1968.
Vol. 10. P. 314-319.
Jenkins A.C., Brown M.R., Crim J.W. FMRF-amide immunoreactivity and the midgut of the com worm (Heliotys
zea)//J. Exp. Zool. 1989. Vol. 252. P.71-78.
Jenniaux C. Nutrition and digestion. // Chemical Zoology. Vol. 4. - N-Y; London, 1969. P.69-91.
Jennings J.B. Observations on the nutrition of the rhynchocoelan Lineus rubber (O.F. Muller) // Biol. Bull. 1960.
Vol. 119. P. 189-196.
Jennings J.B. Ultrastnictural observations on the phagocytic uptake of food materials by the ciliated cells of the
rhynchocoelan intestine// Biol. Bull. 1969. Vol. 175. P. 176-485.
Jennings J.B., Gibson R. Observations on the nutrition of seven species of rhynchocoelan worms // Biol. Bull. 1960.
Vol. 136. P. 405-433.
Jonsson A.C., Hormgren S., Holstein B. Gastrin/CCK-like immunoreactivity in endocrine cells and nerves in the
gastrointestinal tract of the cod, Gadus morhua, and the effect of peptides of gastrin/ССК family on cod gastrointestinal
smooth muscle//Gen. Comp. Endocrinol. 1987. Vol. 66. P. 190-202.
Josephson R.K. Cnidarian neurobiology. // Coelenterate biology. - New York; etc.: Academic Press, 1974. P. 245-
280.
Judd W. The secretion and fine structure of bivalve crystalline sacs // Ophelia. 1979. Vol. 18. P. 205-233.
Kaloustian K.V. Immunochemical evidencs for AKTH-like immunoreactivity in tissues of the earthworm Lumbricus
terrestris//Comp. Biochem. Physiol. 1986. Vol. A85. P. 351-353.
Kasuya Y., Kobayashi H, Uemura H. The distribution of an immunoreactive endothelin-like substance in the nervous
system of the nereid, Neanthes di versicolor // Cell Tissue Res. 1990. Vol. 261. P. 485-492.
Kataoka K, Fujita H. The occurrence of endocrine cells in the intestine of the lancelet, Branchiostoma japonicum.
An electronmicroscopy study // Arch, histol. Jap. 1974. Vol. 36. P. 401-406.
Kazakov V.K., Punin M.Yu. Hydra head activating peptide- and neurotensin-immunoreactive structures in the
digestive tract of brachiopod Hemithyris psittacea. // Sympler nervous systems (Abstracts of Regional Meeting of the
International Society for Invertebrate Neurobiology, Minsk, 1991). - Moskow: Nauka, 1991. P. 41.
Kiliaan A.J., Scholten G., Groot J.A. Ultrastructural study of the presence of vasoactive intestinal polypeptide and
serotonin in mucosal nerve fibres and endocrine cells of the intestine of goldfish (Carassius auratus) and tilapia
(Oreochromis mossambicus)//Cell Tissue Res. 1996. Vol. 283. P. 143-150.
Komnick, H, Kukulies J. Cytology of the mid-gut epithelium of Aeshna larvae (Insecta, Odonata) // Zoomorphology.
1987. Vol. 107. P. 241-253.
Konstantinova M.S., Punin M.Yu. Endocrine-like cells in the digestive tract of bivalve molluscs // Anat. Anz. 1990.
Bd 166. Suppl. S. 439-440.
Koulish S. Fine structure of the basal surface of intestinal epithelium in the mid-gut region of the Balanidae, with
special reference to “neural-like” processes // J. Morphol. 1971. Vol. 135. P. 1-12.
Kramer KJ. Vertebrate hormones in insects // Comp. Insect Physiol. Biochem. Pharmacol. 1985. Vol. 7. P. 511-536.
Langer M.S., Van Noorden S., Polak J.M., Pearse A.G.E. Peptide hormone-like immunoreactivity in the
gastrointestinal tract and endocrine pancreas of eleven teleost species // Cell Tissue Res. 1979. Vol. 199. P. 493-508.
Larsson L.-L, Rehfeld J.F. Evidence for a common origin of gastrin and cholecystokinin // Nature. 1977. Vol. 269.
P 335-338.
Larsson B.A., Vigna S.R. Species and tissue distribution of cholecystokinin/gastrin-like substances in some
invertebrates//Gen. Comp. Endocrinol. 1983. Vol. 50. P. 469-475.
Larsson L.-I., Goltermann N., De Magistris L., Rehfeld, J.F., Schwartz T.W. Somatostatin cell processes as pathways
for paracrine secretion // Science. 1979. Vol. 205. P. 1393-1395.
Le Douarin N.M. The embryological origin of the endocrine cells associated with the digestive tract: experimental
analysis based on the use of a stable cell marking technique. // Gut hormones. - Edinburg; etc.: Churchill Livingstone,
1978. P. 49-56.
Lechago J., Holmquist A.L., Rosenquist G.L, Walsh J.H. Localization of bombesin-like peptides in frog gastric
mucosa//Gen. Comp. Endocrinol. 1978. Vol. 36. P. 553-558.
154
Lehman H.K., Price D.A. Localization of FMRFamide-like peptides in the snail Helix aspersa // J. Exp. Biol. 1987.
Vol. 131. P. 37-53.
Lentz T.L The cell biology of hydra. - Amsterdam: North-Holland Publishing Company, 1966. 199 p.
Lentz T.L. Primitive nervous systems. - New Haven, London: Yale Univ. Press, 1968. 148 p.
Lentz T.L., Barrnett R.J. Fine structure of the nervous system of Hydra // Amer. Zool. 1965. Vol. 5. P. 341-356.
LeRoux S., Bellon-Humbert C., Lucas A. Mise en evidence par iftimunocytochemie d’un substance apparentee la
somatostatine dans le systeme nerveux, le rein et la glande digestive de juveniles de Pecten maximus (Mollusque
Bivalve)//C. R. Acad. Sci. Ser. 3. 1987. N. 304. P. 115-118.
Leung P.S., Shaw C, Johnston C.F., Irvine G.B. Immunocytochemical distribution of neuropeptide F (NPF) in
gastropod mollusc, Helix aspersa, and in several other invertebrates // Cell Tissue Res. 1994. Vol. 275. P. 383-393.
L*Hermite A., Ferrand R., Dubois M.P., Anderson A.C. Detection of endocrine cells by immunofluorescence method
in gastro-entero-pancreatic system of the adult eel, glass-eel, and leptocephalic larva (Anguilla anguila L.) // Gen. Comp.
Endocrinol. 1985. Vol. 58. P. 347-359.
L'Hermite A., Hourdry J., Ferrand R. Investigations of endocrine cells in the gastrointestinal tract and pancreas
during metamorphosis of an anuran (Alytes obstetricans L.,): histochemical detection of APUD cells // Gen. Comp. Endo-
crinol. 1988. Vol. 381-390.
Luppa H. Histology of the digestive tract. // Biology of the Reptilia. Vol. 6. - London; New York: Acad. Press, 1977.
P. 225-313.
Madar J., Sildan N., Рога E.A., Crasiuni C, Molnar E. Quelques aspects du reglage endocrinien du metabolisme
glicidique chez Mytilus galloprovincialis L. de la Mer Noire // Rapp. Comm. Int. Mer. Medit. 1981. N. 27. P. 153-161.
Maier V., Kroder A., Groner E., Keller E., Pfeiffer E.F. Glucagon-like activity (GL1) in the intestine of Porcus
domesticus and Astacus fluviatilis // Acta Endocrinol. 1975. Vol. 78. Suppl. 193. P. 41.
Marchand C.R., Colard C. Presence of cells and fibres immunoreactive towards antibodies to different peptides or
amines in the digestive tract of the snail Helix aspersa // J. Morphol. 1991. Vol. 207. P. 185-190.
Markosova T.G., Punin M.Yu., Kazakov V.K. Regulatory cells in the intestine of turbellarians. // Simpler Nervous
Systems. Abstr. of the 5lh East Europ. Conf of the Int. Soc. for Invertebr. Neurobiol. - Moskow, 1997. P. 49.
Marsden J.R. The digestive tract of Hermodice carunculata (Pallas) (Polychaeta: Amphinomidae) // Canad. J. Zool.
1963. Vol. 41. P. 165-184.
Martinez A., Villaro A.C., Sesma P. Microscopic study of the pyloric caeca of the starfish Marthasterias gracialis
(Echinodermata): finding of endocrine cells //J. Morphol. 1989. Vol. 202. P. 151-164.
Martinez A., Lopez J., Montuenga L, Villaro A.C. Regulatory peptides in gut endocrine cells and nerves in a starfish
//Gen. Comp. Endocrinol. 1991. Vol. 82. P. 280.
Martinez A., Lopez J., Montuenga L., Sesma P. Regulatory peptides in gut endocrine cells and nerves in the starfish
Marthasterias glacialis // Cell Tissue Res. 1993. Vol. 271. P. 375-380.
Martinez A., Riveros-Moreno V., Polak J.M., Moncada S., Sesma P. Nitric oxide (NO) synthase immunoreactivity in
the starfish Marthasterias glacialis // Cell Tissue Res. 1994. Vol. 275. P. 599-603.
Martinez A., Unsworth E.J., Cuttitta F. Adrenomedullin-like immunoreactivity in the nervous system of the starfish
Marthasterias glacialis // Cell Tissue Res. 1996. Vol. 283. P. 169-172.
Masini M.A. Immunohistochemical localization of gut peptides in the small intestine of snakes // Bas. Appl.
Histochem. 1986. Vol. 30. P. 317-324.
Meester I., Ramkema M.D., Van Minnen J., Boer H.H. Differential expression of four genes encoding molluscan
insulin-related pepetides in the central nervous system of the pond snail Lymnaea stagnalis // Cell Tissue Res. 1992. Vol.
269. P. 183-188.
Mehrotra B.K., Falkmer S. Occurrence of insulin-producing cells in the gastrointestinal tract of invertebrates //
Diabetologia. 1968. Vol. 4. P. 244.
Meuleman E.A. Host-parasite interrelationships between the fresh-water pulmonate Biomphalaria pfeifferi and the
trematode Schistosoma mansoni // Neth. J. Zool. 1972. Vol.22. P. 355-427.
Michel C. Intestine and digestive glands // Microfauna marina. 1988. Vol. 4. P. 157-175.
Michel C, DeVillez E.J. Digestion. // Physiology of Annelids. - N-Y., 1978. P.509-554.
Michel C, DeVillez E.J. Cytological and histochemical observations on the secrertion in the digestive tract of the
sipunculan, Phascolopsis gouldii (Portales, 1951)//Cell. Mol. Biol. 1984. Vol. 30. P. 257-267.
Monesi V. The appearence of enterochromaffin cells in the intestine of the chick embryo // Acta Anat. 1960. Vol. 41.
P. 97-114.
Montuenga L.M., Sesma P. Sistema endocrino difuso de artropodos // Acta microscop. 1985. Vol. 8. P. 337-343.
Montuenga LM., Barrenechea M.A., Sesma P., Lopez J., Vazquez J.J. Ultrastructure and immunocytochemistry of
endocrine cells in the midgut of the desert locust Schistocerca gregaria (Forscal) // Cell Tissue Res. 1989. Vol. 258. P. 577-
583.
Montuenga L.M., Prado M.A., Springall D.R., Polak J.M., Sesma P. Endothelin-like immunoreactivity in midgut
endocrine cells of the desert locust Locusta migratoria // Gen. Comp. Endocrinol. 1994. Vol. 81. P. 433-441.
155
Moore S.J., Thomdyke М.С. Immunocytochemical mapping of the novel echinoderm neuropeptide SALMFamide I
(SI) in the starfish Asterias rubens // Cell Tissue Res. 1993. Vol. 274. P. 605-618.
Moore S.J., Elphick M.R., Thorndyke M.C. Mapping SALMFamide neuropeptides in echinoderms // Regul. Pept.
1991. Vol.35. P.250.
Moroz L, Nezlin L, Elofsson R., Sakharov D. Serotonin- and FMRFamide immunoreactive nerve elements in the
chiton Lepidopleurus asellus (Mollusca, Polyplacophora) // Cell Tissue Res. 1994. Vol. 275. P. 277-282.
Morton B. Feeding and digestion in Bivalvia. // The Mollusca. Physiology. Vol. 5. Pt. 2. - New-York, etc., 1983.
P. 65-147.
Mykles D.L. The ultrastructure of the posterior midgut caecum of Pachygrapsus crassipes (Decapoda, Brachyura)
adapted to low salinity//Tissue Cell. 1977. Vol. 9. P. 681-691.
Mykles D.L. Ultrastructure of alimentary epithelia of lobsters, Homarus americanus and H.gammarus, and crab
Cancer magister//Zoomorphology. 1979. Vol. 92. P. 201-215.
Nakajima Y. Serotonergic nerve cells of starfish larvae. // Echinoderm biology. - Balkema: Rotterdam, 1988. P. 235-
239.
Narain A.S. Anatomical studies of the trans-pericardial alimentary tract of a pelecypod // Arch. Biol. 1975. Vol. 86.
P. 377-389.
Nebeyama A. Presence of cell combining features of two different cell types in the colonic crypts and pyloric glands
of the mouse // Amer. J. Anat. 1975. Vol. 142. P. 471-483.
Neuhaus B. Ultrastructure of alimentary canal and body cavity, ground pattern, and phylogenetic relationships of the
Kinorhyncha // Microfauna marina. 1994. Vol. 9. P. 61-156.
Newman S.J., Thorndyke M.C. Localization of gamma aminobutyric acid (GABA)-like immunoreactivity in the
echinoderm Asterias rubens // Cell Tissue Res. 1994. Vol. 278. P. 177-185.
Nishiitsutsuji-Uwo J., Endo Y. Gut endocrine cells in insects: the ultrastructure of the endocrine cells in the
cockroach midgut// Biomed. Res. 1981. Vol. 2. P. 30-44.
Nishiitsutsuji-Uwo J., Endo Y., Iwanaga T, Fujita T. Immunohistochemical demonstration of brain-gut endocrine
system in insects. // The ultrastructure and functioning of insect cells. - Tokyo, 1982. P. 107-108.
Nishiitsutsuji-Uwo J., Takeda M., Saito H. The production of an antiserum to serotonin and serotonin-like
immunoreactivity in the cockroach brain-midgut system// Biomed. Res. 1984. Vol. 5. P. 211-224.
Noaillac-Depeyre J. Localization by immuno-electron microscopy of somatostatin-containing cells in the gastric
mucosa of a teleost fish, Perea fluviatilis // Biol. Cell. 1985. Vol.53. P. 263-268.
Noaillac-Depeyre J., Hollande E. Evidence for somatostatin-, gastrin- and pancreatic polypeptide-like substances in
the mucosa cells of the gut in fishes with and without stomach // Cell Tissue Res. 1981. Vol.216. P. 193-203.
Nozaki M., Migata K., Oota Y., Gorbman A., Plisetskaya E.M. Colocalization of glucagon-like peptide and glucagon
immunoreactivities in pancreatic islets and intestine of salmonids // Cell Tissue Res. 1988. Vol. 253. P. 371-375.
Olsson C, Holmgren S. Distribution of PACAP (putuitary adenylate cyclase-activating polypeptide)-like and
helospectin-like peptides in the teleost gut // Cell Tissue Res. 1994. Vol. 277. P. 539-547.
Ostberg Y., Boquist L. Ultrastructural and fluorescence microscopical characterization of the intestinal endocrine
cells in a cyclostome, Myxine glutinosa // Acta Zool. 1976. Vol. 57. P. 41-57.
Ostberg Y., Van Noorden S., Pearse A.G.E. Cytochemical, immunofluorescence and ultrastructural investigations on
polypeptide hormone localization in the islet parenchyma and bile duct mucosa of a cyclostome, Myxine glutinosa // Gen.
Comp. Endocrinol. 1975. Vol. 25. P. 274-291.
Ostberg Y., Van Noorden S., Pearse A.G.E., Thomas N.W. Cytochemical, immunofluorescence and ultrastructural
investigations on polypeptide hormone containing cells in the intestinal mucosa of a cyclostome, Myxine glutinosa // Gen.
Comp. Endocrinol. 1976a. Vol. 28. P. 213-227.
Ostberg Y. Boquist L, Van Noorden S., Pearse A.G.E. On the origin of islet parenchymal cells in a cyclostome,
Myxine glutinosa. A fluorescence microscopical and ultrastructural study with particular reference to endocrine cells in bile
mucosa//Gen. Comp. Endocrinol. 1976b. Vol. 28. P. 228-246.
Owen G. Feeding. // Physiology of Mollusca. Vol. 2. - New-York, London, 1966a.. P. 1-52.
Owen G. Digestion. // Physiology of Mollusca. Vol. 2. - New-York, London, 1966b. P. 53-96.
Pal S. The fine structure of digestive tubules of Mya arenaria L. I. Basiphilic cell // Proc, malacol. Soc. London.
1971. Vol. 39. P. 303-308.
Pearse A.G.E. Common cytochemical and ultrastructural characteristics of cells producing polypeptide hormones
(the APUD series) and ultimobranchial C-cells and calcitonin // Proc. Roy. Soc. Biol. 1968. Vol. 170. P. 71-80.
Pearse A.G.E. The cytochemistry and ultrastructure of polypeptide hormone-producing cells of the APUD series and
emryologic, physiologic and pathophysiologic implications of the concept // J. Histochem. Cytochem. 1969. Vol. 17.
P. 303-313.
Pearse A.G.E. Cell migration and the alimentary system: endocrine contribution of the neural crest to the gut and its
derivates//Digestion. 1973. Vol. 8. P. 372-385.
156
Pearse A.G.E. Endocrine-autonomic relationships: neural crest and placodal contributions to the enteric nervous
system. // Histochem. and cell biol. autonom. neurons and paraganglia. - Berlin, 1987. P: 201-207.
Pentilla A. Enterochromaffin cells in the chicken duodenum during prenatal development // Z. Zellforsch. 1968.
Bd 91. S. 380-390.
Perez-Tomas R., Ballesta J., Pastor L.M., Madrid J.F., Polak J.M. Comparative immunohistochemical study of the
gastroenteropancreatic system of three reptiles // Gen. Comp. Endocrinol. 1989. Vol. 76. P. 171-191.
Pestarino M. Osservazioni istochimiche preliminari selle cellule endocrine gastroesophagee di Styela plicata Les.
(Urochordata, Ascidaceae) // Bas. Appl. Histochem. 1979. Vol.23. Suppl. P. XXXVI.
Pestarino M. Histomorphology and histochemistry of the digestive apparatus of the ascidian Microcosmus
polymorphus. FV Intestine// Boll. Zool. 1982a. Vol. 49. P. 125-133.
Pestarino M. Cytochemical investigation on endocrine-like cells of the digestive tract of the ascidian Styela plicata //
Bas. Appl. Histochem. 1982b. Vol. 26. P. 107-116.
Pestarino M. Occurrence of different secretin-like cells in the digestive tract of the ascidian Styela plicata
(Urochordata, Ascidiaceae)//Cell Tissue Res. 1982c. Vol. 226. P. 231-235.
Pestarino M. Immunofluorescent localization of somatostatin-like cells in the oesophagus of a protochordate // Arch,
anat. microsc. morphol. exp. 1983. Vol. 72. P. 273-278.
Pestarino M. Localization and distribution of immunoreactive glucagon-like, peptide histidine isoleucine-like, and
vasoactive intestinal petide-Jike cells in a protochordate// Canad. J. Physiol. Pharmacol. 1986. Vol. 64. Suppl. P. 27-28.
Pestarino M. Localization of serotonin immunoreactivity in the ascidian Styela plicata // Bas. Appl. Histochem.
1989. Vol. 33. P. 84.
Pestarino M., Garaud J.-C. Gastrin-like peptides in the alimentary tract of a protochordate // Bull. Soc. Zool. Fr.
1986. Vol. 111. P. 56.
Pestarino M., Tagliafierro G. Immunocytochemical localization of secretin-like cells in the digestive tract of the
ascidian Halocynthia papillosa// Bas. Appl. Histochem. 1983. Vol. 27. P. 141-148.
Pestarino M., Tagliafierro G., Lamoretti M. Gastric secretin-like cells in the ascidian Halocynthia papillosa // Bas.
Appl. Histochem. 1982. Vol. 26. Suppl. P. CXI.
Pestarino M., Garaud J.-C., Ravera F. Localization of immunoreactive gastrin-like cells in the alimentary tract of the
ascidian Styela plicata // Gen. Comp. Endocrinol. 1988. Vol. 69. P. 13-18.
Pictet R.L., Rail L.B., Phelps P., Rutter W.J. The neural crest and the origin of insulin-producing and other
gastrointestinal hormone producing cells//Science. 1976. Vol. 191. P. 191-192.
Pilgrim M. The functional anatomy and histology of the alimentary canal of the maldanid polychaetes Clymenella
torquata and Euclymene oerstedy // J. Zool. 1965. Vol. 147. P. 387-405.
Platzer-Schultz L, Welsch U. Zur Entstehung und Feinstructur der peritrophischen Membran der Larven von
Hironomus strenzkei Httkau (Diptera) //Z. Zellforsch. 1969. Bd 100. S.594-605.
Plisetskaya E., Kazakov V.K., Soltitskaya L, Leibson L.G. Insulin-producing cells in the gut of fresh-water bivalve
molluscs Anodonta cygnea and Unio pictorum and the role of insuli in the regulation of their carbohydrate metabolism //
Gen. Comp. Endocrinol. 1978. Vol. 35. P. 133-145.
Polak J.M., Pearse A.G.E., Adams C. Garaud J.C. Immunohistochemical and ultrastructural studies on the
endocrine polypeptide (APUD) cells in the avian gastrointestinal tract// Experientia. 1974. Vol. 30. P. 564-567.
Priester W. de. Ultrastructure of the midgut epithelial cells in the fly Calliphora erytrocephala // J. Ultrastr. Res.
1971. Vol. 36. P. 783-805.
Punin M.Yu., Kazakov V.K. The system of nerve and endocrine-like elements in the gut epithelium of Polychaeta. //
Sympler nervous systems. - Manchester Univ. Press, 1991a. P. 51-59.
Punin M.Yu., Kazakov V.K. Cells, immunoreactive to neurotensin and hydra head activating peptide, are present in
the central nervous system and the intestine of bivalve molluscs. // Abstr. reg. meet. Int. Soc. for invertebr. neurobiolgy. -
Moskow: Nauka, 1991b. P. 78.
Punin M.Yu., Kazakov V.K. Immunohistolchemical localization of neurotensin and hydra head activating peptide in
the central nervous system and the intestine of bivalve molluscs. // The peptidergic neuron. Иm Int. Symp. on
Neurosecretion. - Amsterdam, 1991c. P. 39.
Punin M.Yu., Kazakov V.K. Immunohistochemical localization of hydra head activator peptide in hydra. // Abstr. of
4lh Int. Workshop on Hydroid Development. - Schloss Reisenburg, 199Id. P. 68.
Punin M.Yu., Markosova T.G. Regulatory cells in the intestine of turbellarian worms and possible evolution of
intestinal regulatory system in Metazoa //Trudy Zool. Inst. Ross. Acad. Nauk.Vol. 286. 2000. P. 121-128.
Punin M.Yu., Konstantinova M.S., Korolkova E.D. Monoamine containing structures in the intestines of bivalve
molluscs and a polychaete annelid revealed by fluorescence histochemistry // Cell Tissue Res. 1990. Vol. 259. P. 389-391.
Punin M.Yu., Konstantinova M.S., Kazakov V.K. A regulatory system in the digestive tract epithelium of bivalve
molluscs. // Sympler nervous systems. - Manchester Univ. Press. 1991. P. 60-73.
Purchon R.D. The biology of Mollusca. - Oxford; etc., 1978. 594 p.
Raes H. Two types of of endocrine cells in the midgut of the honeybee // Belg. J. Zool. 1990. Vol. 120. P. 101.
157
Rajjo I.M., Vigna S.R., Crim J.W. Cholecystokinin immunoreactivity in the digestive tract of bowfin (Amia calva),
bluegill (Lepomis macrochirus), and a bullfrog (Rana catesbeiana) // Gen. Comp. Endocrinol. 1988. Vol. 70. P. 133-144.
Rawdon B.B. Gastrointestinal hormones in birds: morphological, chemical and developmental aspects // J. Exp. Zool.
1984. Vol. 232. P. 659-670.
Rawdon B.B., Andrew A. An immunocytochemical survey of endocrine cells in the gastrointestinal tract of chicks at
hatching // Cell Tissue Res. 1981. Vol. 220. P. 279-292.
Reifel C.W., Marin-Soresen M., Samloff I.M. Gastrin-immunoreactive cells in the gastro-intestinal tracts from four
species of fish // Canad. J. Zool. 1983. Vol. 61. P. 1464-1468.
Reinecke M. Immunohistochemical localization of polypeptide hormones in endocrine cells in the digestive tract of
Branchiostoma lanceolatum//Cell Tissue Res. 1981. Vol. 210. P. 445-456.
Reinecke M. Neurotensin: immunohistochemical localization in the central and peripheral nervous system and in
endocrine cells and its functional role as neurotransmitter and endocrine hormone // Progr. Histochem. Cytochem. 1985.
Vol.16. P. 1-175.
Reinecke M., Almasan K., Carraway R., Halmstaedter V., Forssmann W.G. Distrubution patterns of neurotensin-like
immunoreactive cells in the gastrointestinal tract of higher vertebrates // Cell Tissue Res. 1980. Vol. 205. P. 383-395.
Reinecke M., Betzler D., Drakenberg K., Falkmer S., Sara V.R. Occurrence of members of the insulin superfamily in
central nervous system and digestive tract of protochordates // Histochemistry. 1993. Vol. 99. P. 277-285.
Reinhardt C.A. UJtrastructural comparison of the midgut epithelia of fleas with different feeding behavior patterns
(Xenopsylla chaeopis, Echidnophage gallinaceae, Tunga penetrans, Siphonaptera, Pulicidae) // Acta trop. 1976. Vol. 33.
P. 105-132.
Reisenger E. Zur Evolution des stomatogastrischen Nervensystems bei den Plathelminthen // Z. Zool. Syst.
Evolutionsforsch. 1976. Bd. 14. S. 241-253.
Relini Orsi L., Pestarino M. Aspects of specialization of the alimentary tract of Microcosmus polymorpha
(Ascidiacea, Pyuridae) // Boll. Zool. 1977. Vol. 44. P. 189-193.
Remane A., Storch V., Welsch U. Systematische Zoologie - Stuttgart: Gustav Fischer Verlag, 1980. 682 p.
Reuter M., Palmberg I. An ultrastructural and immunocytochemical study of gastrodermal cell types in
Microstomum lineare (Turbellaria, Macrostomida) // Acta Zool. 1987. Vol. 68. P. 153-163.
Reuter M., Karhi T, Schot L.P.C. Immunocytochemical demonstration of peptidergic neurons in the central and
peripheral nervous system of the flatworm Microstomum lineare with antiserum to FMRF-amide // Cell Tissue Res. 1984.
Vol. 238. P. 431-436.
Reuter M., Lehtonen M., Wikgren M. Immunocytochemical evidence of neuroactive substances in flatworms of
different taxa - a comparison // Acta Zool. 1988. Vol. 69. P. 29-37.
Reuter M., Gustafsson M., Sheiman I.M., Terenina N., Halton D.W., Maule A., Shaw C. The nervous system of
Tricladida. II. Neuroanatomy of Dugesia tigrina (Paludicola, Dugesiidae): an immunocytochemical study // Invertebr.
Neurosci. 1995a. Vol. 1. P. 133-143.
Reuter M., Maule A., Halton D., Gustafsson M., Shaw C. The organization of the nervous system in Plathelminthes.
The neuropeptide F-immunoreactive pattern in Catenulida, Macrostomida, Proseriata // Zoomorphology. 1995b. Vol. 115.
P. 83-97.
Rombout J.H.W.M., Reinecke M. Immunohistochemical localization of (neuro)peptide hormones in endocrine cells
and nerves of the gut of a stomachless teleost fish, Barbus conchonius (Cyprinidae) // Cell Tissue Res. 1984. Vol. 237.
P. 57-65.
Rombout J.H.W.M., Taverne-Thiele J. J. Immunocytochemical and electron microscopical study of endocrine cells in
the gut and pancreas of a stomachless teleost fish, Barbus conchonius (Cyprinidae) // Cell Tissue Res. 1982. Vol. 227.
P. 577-593.
Rombout J.H.W.M., van der Grinten C.P.M., Peeze-Brinkhorst F.M., Taverne-Thiele J.J., Schooneveld H.
Immunocytochemical identification and localization of peptide hormones in the gastro-entero-pancreatic (GEP) endocrine
system of the mouse and stomachless fish, Barbus conchonius // Histochemistry. 1986a. Vol. 84. P. 471-483.
Rombout J. H.W.M., Bol J., Taverne-Thiele J. J. Ultrastructural characterization of 6 immunoreactive entero-endocrine
cells in Barbus conchonius (Teleostei, Cyprinidae) // Histochemistry. 1986b. Vol. 85. P. 467-473.
Rowell H.F. The cells of the insect neurosecretory system: constancy, variability, and the concept of the unique
identifiable neuron // Adv. Insect Physiol. 1976. Vol. 12. P. 63-213.
Rzasa P., Kaloustian K.V., Prokop E.K. Immunochemical evidence for a gastrin-like peptide in the intestinal tissues
of the earthworm Lumbricus terrestris // Comp. Biochem. Physiol. 1982. Vol. A71. P. 631-634.
Salvi E., Renda T. Immunohistochemical studies on the ontogenesis of some endocrine cells in the chicken antrum
and duodenum// Bas. Appl. Histochem. 1986. Vol. 30. P. 307-316.
Salvi E., Renda T. Ontogenesis of endocrine cells in the chicken intestine: an immunohistochemical study // Cell.
Mol. Biol. 1989. Vol. 35. P. 17-26.
Scalise F.W., Larson B.A., Wigna S.R. Localization of a peptide identified by antibodies to gastrin/CCK in the gut of
Cancer magister// Cell Tissue Res. 1984. Vol. 238. P. 113-119.
158
Schaller H.C. Isolation and characterization of a low molecular weigh susbstance activating head and bud formation
in hydra // J. Embr. Exp. Morphol. 1973. Vol. 29. P. 27-38.
Schaller H.C., Gierer A. Distribution of the head activating substance in hydra and its localization in membranous
particles in nerve cells // J. Embr. Exp. Morphol. 1973. Vol. 29. P. 39-52.
Schaller H.C., Schmidt T, Grimmelikhuijzen C.J.P. Separation and specificity of action of four morphogens from
Hydra// Wilhelm Roux’s Arch. 1979. Vol. 186. P. 139-149.
Schaller H.C., Schmidt T, Grimmelikhuijzen C.J.P., Rau T, Bode H. Morphogenetic substances in nerve-free hydra.
// Developmental and cellular biology of coelenterates. - Amsterdam; etc.: Elsevier, 1980. P.389-394.
Schaller H.C., Hoffmeister S.A.H., Dubel S. Role of the neuropeptide head activator for growth and development in
hydra and mammals // Development. 1989. Suppl. P. 99-107.
Schechter J., Lucero J. A light and electronmicroscopic investigation on the digestive system of Ophiuroiderma
panamensis (Brittle star) // J. Morphol. 1968. Vol. 124. P. 451-482.
Schipp R., Boletzky S. von. Pancreatic appendages of dibranchiate cephalopods. I. The fine structure of the
«pancreas» in Sepioidea // Zoomorphology. 1976. Bd 86. S. 81-98.
Schmich J., Rudolf R., Trepel S., Leitz T. Immunohistochemical studies of CLWamides in Cnidaria // Cell Tissue
Res. 1998. Vol. 294. P. 169-177.
Schmidt T, Grimmelikhuijzen C.J.P., Schaller H.C. Morphogenetic substances in hydra. // Developmental and
cellular biology of coelenterates. - Amsterdam; etc.: Elsevier, 1980. P. 395-399.
Schols D., Verhaert P., Huybrechts R., Vaudry H., Jegon S., De Loof A. Immunocytochemical demonstration of
proopiomelanocortin- and other opioid-related substances and CRF-like peptide in the gut of the american cockroach,
Periplaneta americana L. // Histochemistry. 1987. Vol. 86. P. 345-351.
Schoofs L, Danger J.M., Jegon S., Pelletier G., Huybrechts R., Vaudry H, DeLoof A. NPY-like peptides occure in
the nervous system and midgut of the migratory locust, Locusta migratoria and in the brain of the gery fleshfly, Sarcophaga
bullata// Peptides. 1988. Vol. 9. P. 1027-1036.
Schot LP.C, Boer H.H. Immunocytochemical demonstration of peptidergic cells in the pond snail Lymnaea
stagnalis with antiserum to the molluscan cardioactive tetrapeptide, FMRFamide // Cell Tissue Res. 1982. Vol. 225.
P. 347-354.
Seino Y., Porte D., Smith P.H. Immunohistochemical localization of somatostatin-containing cells in the intestinal
tract: a comparative study// Gen. Comp. Endocrinol. 1979. Vol. 38. P. 229-233.
Shirasawa N., Shiino M., Shimizu Y., Nogami H, Ishii S. Immunoreactive luteinizing hormone (ir-LH) cells in the
lung and stomach of chick embryons // Cell Tissue Res. 1996. Vol. 283. P. 19-27.
Silen L On the nervous system of Glossobalanus marginatus Meek (Enteropneusta) // Acta Zool. 1950. Bd 31.
S. 149-175.
Smirnov A. Uber freie Nervendigungen im Epithel des Regenwurms // Anat. Anz. 1894. Bd 9. S. 570-578.
Smit A.B., Vrengden E., Ebberink R.H.M., Geraerts W.P.M., Klootwijk J., Joose J. Growth-controlling molluscan
neurons produce the precusor of an insulin-related peptide // Nature. 1988. Vol. 331. P. 535-538.
Solcia E., Capella C, Bujfa R., Usellini L., Fiocca R., Sessa F. Endocrine cells of the digestive system. // Physiology
of gastrointestinal tract. Vol. 1. - New York: Raven Press, 1981a. P. 39-58.
Solcia E., Creutzfeld W., Falkmer S., Fujita T, Creider M.H., Grossman M., Grube D., Yakanson R., Larsson LI.,
Lechago J., Lewin K., Polak J., Rubin W. Human gastroenteropancreatic endocrine-paracrine cells: Santa Monica 1980
classification. // Cellular basis of chemical messengers in the digestive system. - London; etc.: Acad. Press, 1981b. P. 159-
165.
Spencer A.N. Neuropeptides in the Cnidaria // Amer. Zool. 1989. Vol. 29. P. 1213-1225.
Stemberger LA. Immunocytochemistry. - N.-Y.: Wiley, 1979. 354 p.
Storch V., Alberti G. Zur Ultrastruktur des Darmtraktes von Tubilichus philippiensis (Tubiluchidae, Priapulidae) //
Zool. Anz. 1985. Bd 214. S. 262-272.
Straus F. Yalow R.S., Gainer H. Molluscan gastrin: concentration and molecular forms // Science. 1975. Vol. 190.
P. 687-689.
Sumner A.T. The fine structure of digestive gland cells of Anodonta // J. Roy. microsc. Soc. 1966. Vol. 85. P. 417—
423.
Tagliafierro G., Faraldi G. Differentiation of the endocrine cells of the gastric epithelium of Sciliorhinus stellaris //
Boll. Zool. 1976. Vol. 43. P. 75-85.
Tagliafierro G., Faraldi G., Raineri M. Aspetti morphologici ed istochimici delle cellule argentaffini ed argentofile
nell’epiteliogastrico di Sciliorhinus stellaris and Sciliorhinus canicula // Riv. Istochim. Norm. Patol. 1976. Vol. 20. P. 217-
230.
Talbot P., Clark W.H., Lawrence A.L. Fine structure of the mid-gut epithelium in the developing brown shrimp,
Penaeus aztecus//J. Morphol. 1972. Vol. 138. P. 467-486.
Thomas N.W., Ostberg Y., Falkmer S. A second granular cell in the endocrine pancreas of the hagfish Mixine
glutinosa//Acta Zool. 1973. Vol. 54. P. 201-207.
159
Thorndyke M.C. Observations on the gastric epithelium of ascidians with special reference to Styela clava // Cell
Tissue Res. 1977. Vol. 184. P. 539-550.,
Thorndyke M.C., Bevis P.J.R. Endocrine cells in the gut of the ascidian Styela clava // Cell Tissue Res. 1978.
Vol. 187. P. 159-165.
Thorndyke M.C., Bevis P.J.R. Comparative studies on the effects of cholecystokinin, caerulin, bombesin 6-14
nonapeptide, and physalaemin on gastric secretion in the ascidian Styela clava // Gen. Comp. Endocrinol. 1984. Vol. 55.
P. 251-259.
Toner P.G. The fine structure of argyrophil and argentaffin cells in the gastrio-intestinal tract of the fowl //
Z. Zellforsch. 1964. Bd 63. S. 830-839.
Too C.K.L., Croll R.P. Detection of FMRFamide-like immunoreactivities in the sea scallop Placopecten
magellanicus by immunohistochemistry and Western blot analysis // Cell Tissue Res. 1995. Vol. 281. P. 295-304.
Tunas T.T. Histologische, histochemische und elektronenmikroskopische Utersuchungen am Verdaungstrakt der
Ascidien // Zool. Jahrb. Anat. 1977. Bd 98. S. 505-543.
Turbeville J.M., Ruppert E.E. Comparative ultructucture and evolution of nemertines // Amer. Zool. 1985. Vol. 25. P.
53-71.
Uggeri B. Ricerche sulle cellule enterochromaffini e cellule argentofile dei pesci // Z. Zellforsch. 1938. Bd 28. S.
648-673.
Ullman A., Bookhout C.G. The histology of the digestive tract of Clymenella torquata (Leidy) // J. Morphol. 1949.
Vol. 84. P. 31-48.
Vandenneulen J.H. Functional morphology of the digestive tract epithelium in Phoronis vancouverensis (Bixell): an
ultrastnictural and histochemical study // J. Morphol. 1970. Vol. 130. P. 271-286.
Van der Plas A.J., Jangoux M., Voogt P.A. A light and electron microscopical study of the pyloric caeca of Asterias
rubens (Echinodermata: Asteroidea) 11 Neth. J. Zool. 1984. Vol. 34. P. 144-158.
Van Noorden S., Pearse A.G.E. Immunorective polypeptide hormones in the pancreas and gut of the lamprey // Gen.
Comp. Endocrinol. 1974. Vol. 23. P.311-324.
Van Noorden S., Pearse A.G.E. The localization of immunoreactivity to insulin, glucagon and gastrin in the gut of
Amphioxus (Branchiostoma lanceolatum). // Evolution of the pancreatic islets. - Oxford, 1976. P. 163-177.
Van Noorden S., Greenberg J., Pearse A.G.E. Cytochemical and immunofluorescence investigations on polypeptide
hormone localization in the pancreas and gut of the larval lamprey // Gen. Comp. Endocrinol. 1972. Vol. 19. P. 192-199.
Van Noorden S., Ostberg Y., Pearse A.G.E. Localization of somatostatine-like immunoreactivity in the pancreatic
islets of the hugfish, Myxine glutinosa and the lamprey, Lampetra fluviatilis // Cell Tissue Res. 1977. Vol. 177. P. 281—
285.
Vigna S.R., Fischer B.L, Morgan J.L.M., Rosenquist G.L. Distribution and molecular geterogenety of
cholecystokinin-like immunoreactive peptides in the brain and gut of the rainbow trout, Salmo gairdneri // Comp. Biochem.
Physiol. 1985. Vol. 82. P. 143-146.
Walker A.R., Fletcher J.D. Histology of digestion in nymphs of Rhipicephalus appendiculatus fed on rabbit and
cattle naive and resistant to the ticks // Int. J. Parasitol. 1987. Vol. 17. P. 1393-1411.
Watanabe T, Chikazawa H., Chungsamarnyart N., Fujioka T, Yamada J. Serotonin-storing cells of the chicken
duodenum: light, fluorescence and electron microscopy, and immunohistochemistry 11 Cell Tissue Res. 1987. Vol. 247.
P. 25-32.
Welsch U. The fine structure of the pharinx, cyrtopodocytes and digestive caecum of Amphioxus (Branchiostoma
lanceolatum) // Symp. zool. Soc. London. 1975. N 36. P. 17-41.
Welsch U., Storch V. Histochemical and fine structural observations on the alimentary tract of Aphroditidae and
Nephtyidae (Polychaeta, Errantia) // Mar. Biol. 1970. Vol. 6. P. 143-147.
Welsh J.H., Loveland R.E. 5-hydroxytryptamine in the ascidian Ciona intestinalis // Comp. Biochem. Physiol. 1968.
Vol. 27. P. 719-722.
Westermann B., Schipp R. Cytological and enzyme-histochemical investigations on the digestive organs of Nautilus
pompilius (Cephalopoda, Tetrabranchiata) // Cell Tissue Res. 1998. Vol.293. P. 327-336.
Westfall J.A. Ultrastnictural evidence for a granule containing sensory-motor-interneuron in Hydra littoralis //
J. Ultrastr. Res. 1973. Vol. 42. P. 268-282.
Westfall J.A., Kinnamon J.C. A second sensory-motor-interneuron with secretory granules in Hydra 11 J. Neurocytol.
1978. Vol. 7. P. 365-379.
Wikgren h^C., Reuter M., Gustafsson M. Neuropeptides in free-living and parasitic flatworms (Platyhelminthes). An
immunocytochemical study // Hidrobiologia. 1986. Vol. 132. P. 93-99.
Wilson S., Falkmer S. Starfish insulin // Canad. J. Biochem. 1965. Vol. 43. P. 1615-1624.
Winbladh Biuw L, Hutting G. Fine-grained secretory cells in the intestine of the lancelet, Branchiostoma
(Amphioxus) lanceolatum, studied by light microscopy // Z. Zellforsch. 1971. Bd 120. S. 546-554.
160
Yamada J., Campos V.J.M., Kitamura N., Pachecco A.C., Yamashita T., Yanaihara N. An immunohistochemical
study of the endocrine cells in the gastrointestinal mucosa of the Caiman latirostris // Arch. Histol. Japan. 1987. Vol. 50.
P. 229-241.
Yoon. J.B., Stay B. Immunocytochemical localization of Diploptera punctata allatostatin-like peptide in Drosophila
melanogaster// J. Comp. Neurol. 1995. Vol. 363. P. 475-488.
Yoshida K., Iwanaga T., Fujita T. Gastroenteropancreatic (GEP) system of the flatfish, Paralichtys olivaceus: an
immunocytochemical study// Arch. Histol. Japan. 1983. Vol. 46. P. 259-266.
Youson J.H. The alimentary canal. // The biology of lampreys. Vol. 3. - London; etc., 1981a. P. 95-189.
Youson J.H. The liver. 11 The biology of lampreys. Vol. 3. - London; etc., 1981b. P. 263-332.
Yui R., Iwanaga T., Kuramoto H., Fujita T. Neuropeptide immunocytochemistry in protostomian invertebrates with
special reference to insects and molluscs // Peptides. 1985. Vol. 6. Suppl. 3. P. 411-415.
ОГЛАВЛЕНИЕ
Предисловие..................................................................................... 5
Глава 1. Введение (проблема, терминология, методы).............................................. 7
Глава 2. Кишечная эндокринная система позвоночных животных....................................... 12
2.1. Mammalia (млекопитающие)........................................................... 12
2.2. Aves (птицы)....................................................................... 17
2.3. Reptilia (рептилии или пресмыкающиеся)............................................. 17
2.4. Amphibia (земноводные)............................................................. 18
2.5. Pisces (рыбы)...................................................................... 19
2.6. Cyclostomata (круглоротые)..........................................................20
Глава 3. Кишечная регуляторная система других хордовых..........................................23
3.1. Amphioxi (ланцетники)...............................................................23
3.2. Tunicata (оболочники)...............................................................26
3.2.1. Ascidiaceae (асцидии).........................................................26
Глава 4. Enteropneusta (кишечнодышащие).........................................................30
Глава 5. Echmodermata (иглокожие)...............................................................32
5.1. Asteroidea (морские звезды).........................................................32
5.1.1. Данные электронно-микроскопических исследований...............................32
5.1.2. Клетки, содержащие моноамины..................................................34
5.1.3. Клетки, содержащие ЕМИЕамид-подобное вещество.................................34
5.1.4. Клетки, иммунореактивные к 8АЬМЕамиду 35
5.1.5. NO-синтетаза- и GABA-иммунореактивные клетки..................................35
5.1.6. Клетки, содержащие инсулиноподобные вещества..................................36
5.1.7. Клетки, иммунореактивные к другим регуляторным пептидам.......................37
5.1.8. Колокализация различных регуляторных веществ..................................38
5.2. Echinoidea (морские ежи)............................................................38
5.2.1. Моноамин-содержащие клетки....................................................38
5.2.2. Клетки, иммунореактивные к регуляторным пептидам..............................40
5.3. Ophiuroidea (офиуры)................................................................40
5.4. Holoturoidea (голотурии) 41
5.5. Заключение..........................................................................42
Глава 6. Arthropoda (членистоногие).............................................................45
6.1. Tracheata (трахейные)...............................................................45
6.1.1. Entognatha (скрыточелюстные)..................................................45
6.1.2. Ectognatha (открыточелюстные)................................................ 46
6.2. Branchiata (жабродышащие)...........................................................52
6.3. Chelicerata (хелицеровые)...........................................................54
6.4. Заключение..........................................................................54
Глава 7. Mollusca (моллюски)....................................................................57
7.1. Cephalopoda (головоногие)...........................................................57
7.2. Bivalvia (двустворчатые)............................................................58
7.2.1. Данные гистологических исследований...........................................58
7.2.2. Данные флуоресцентной гистохимии и иммуногистохимии...........................60
7.2.3. Данные электронно-микроскопических исследований...............................67
162
7.3. Gastropoda (брюхоногие)............................................................ 70
7.3.1. Данные гистологических исследований.......................................... 70
7.3.2. Данные иммуногистохимических исследований.................................... 70
7.3.3 Данные электронно-микроскопических исследований............................... 73
7.4. Polyplacophora (хитоны)............................................................ 75
7.5. Заключение......................................................................... 75
Глава 8. Annelida (кольчатые черви)............................................................ 79
8.1. Polychaeta (многощетинковые черви, или полихеты) 79
8.1.1. Данные гистологических исследований.......................................... 79
8.1.2. Данные флуоресцентной гистохимии и иммуногистохимии.......................... 81
8.1.3. Данные электронно-микроскопических исследований.............................. 84
8.1.4. Пролиферация и обновление регуляторных клеток................................ 88
8.2. Oligochaeta (малощетинковые черви, или олигохеты).................................. 88
8.2.1. Данные гистологических исследований.......................................... 89
8.2.2. Данные флуоресцентной гистохимии и иммуногистохимии.......................... 89
8.2.3. Данные электронно-микроскопических исследований.............................. 91
8.3. Hirudinea (пиявки)................................................................. 94
8.3.1. Данные иммуногистохимических исследований.................................... 95
8.3.2. Данные электронно-микроскопических исследований.............................. 96
8.4. Заключение......................................................................... 99
Глава 9. Sipunculida (сипункулиды)............................................................ 101
Глава 10. Tentaculata (щупальцевые)........................................................... 103
10.1. Phoronidea (форониды)............................................................ 103
10.2. Brachiopoda (брахиоподы)......................................................... 104
10.3. Заключение....................................................................... 109
Глава 11. Cephalorhyncha (головохоботные)....................................................... 110
11.1. Kinorhyncha (киноринхи)............................................................ 110
11.2. Priapulida (приапулиды)............................................................ 111
Глава 12. Nemertini (немертины) 114
Глава 13. Turbellaria (турбеллярии, или ресничные черви)........................................ 117
13.1. Данные гистологических и иммуногистохимических исследований........................ 117
13.2. Данные электронно-микроскопических исследований.................................. 122
13.3. Заключение....................................................................... 123
Глава 14. Cnidaria (книдарии)................................................................. 125
Глава 15. Распространение кишечных регуляторных систем у многоклеточных и их предполагаемая
эволюция...................................................................................... 128
15.1. Распространение и некоторые общие черты организации кишечных регуляторных систем... 128
15.2. Предполагаемая эволюция кишечных регуляторных систем............................. 130
Заключение.................................................................................... 137
Summary 139
Литература.................................................................................... 144
CONTENTS
Preface.................................................................................................... 5
Chapter 1. Introduction (problem, terminology, methods).................................................. 7
Chapter 2. Intestinal endocrine system of vertebrates.................................................... 12
2.1. Mammalia..................................................................................... 12
2.2. Aves......................................................................................... 17
2.3. Reptilia..................................................................................... 17
2.4. Amphibia?.................................................................................... 18
2.5. Pisces....................................................................................... 19
2.6. Cyclostomata.................................................................................20
Chapter 3. Intestinal regulatory system of other Chordata................................................23
3.1. Amphioxi.....................................................................................23
3.2. Tunicata.....................................................................................26
3.2.1. Ascidiacea.............................................................................26
Chapter 4. Enteropneusta.................................................................................30
Chapter 5. Echinodermata.................................................................................32
5.1. Asteroidea...................................................................................32
5.1.1. Data of electron-microscopic investigations............................................32
5.1.2. Monoamine containing cells.............................................................34
5.1.3. Cells containing FMRFamide-like substance..............................................34
5.1.4. Cells immunoreactive to SALMFamide.....................................................35
5.1.5. NO-synthase and GABA-immunoreactive cells..............................................35
5.1.6. Cells containing insulin-like substances...............................................36
5.1.7. Cells immunoreactive to other regulatory peptides......................................37
5.1.8. Co-localization of different regulatory substances.....................................38
5.2. Echinoidea...................................................................................39
5.2.1. Monoamine-containing cells..............................................................39
5.2.2 Cells immunoreactive to regulatory peptides..............................................40
5.3. Ophiuroidea..................................................................................40
5.4. Holoturoidea.................................................................................41
5.5. Conclusion...................................................................................42
Chapter 6. Arthropoda....................................................................................45
6.1. Tracheata....................................................................................45
6.1.1. Entognatha.............................................................................45
6.1.2. Ectognatha.............................................................................46
6.2. Branchiata...................................................................................52
6.3. Chelicerata..................................................................................54
6.4. Conclusion...................................................................................54
Chapter 7. Mollusca......................................................................................57
7.1. Cephalopoda..................................................................................57
7.2. Bivalvia.....................................................................................58
7.2.1. Data of histological investigations................................................... 58
7.2.2. Fluorescent histochemical and immunohistochemical data.................................60
7.2.3. Data of electron microscopic investigations............................................67
164
13. Gastropoda..................................................................................... 70
7.3.1. Data of histological investigations...................................................... 70
7.3.2. Data of immunohistochemical investigations............................................... 70
7.3.3. Data of electron microscopic investigations.............................................. 73
7.4. Polyplacophora................................................................................. 75
7.5. Conclusion..................................................................................... 75
Chapter 8. Annelida........................................................................................ 79
8.1. Poly chaeta.................................................................................... 79
8.1.1. Data of histolodical investigations...................................................... 79
8.1.2. Fluorescent histochemical and immunohistochemical data.................................. 81
8.1.3. Data of electron microscopic investigations.............................................. 84
8.1.4. Proliferation and physiological renewal of regulatory cells.............................. 88
8.2. Oligochaeta.................................................................................... 88
8.2.1. Data of histolodical investigations...................................................... 89
8.2.2. Fluorescent histochemical and immunohistochemical data................................... 89
8.2.3. Data of electron microscopic investigations.............................................. 91
8.3. Hirudinea........*............................................................................. 94
8.3.1. Immunohistochemical data................................................................. 95
8.3.2. Data of electron microscopic investigations.............................................. 96
8.4. Conclusion..................................................................................... 99
Chapter 9. Sipunculida.................................................................................... 101
Chapter 10. Tentaculata................................................................................... 103
10.1. Phoronidea................................................................................... 103
10.2. Brachiopoda.................................................................................. 104
10.3. Conclusion................................................................................... 109
Chapter 11. Cephalorhyncha................................................................................ 110
11.1. Kinorhyncha.................................................................................. 110
11.2. Priapulida................................................................................... 111
Chapter 12. Nemertini..................................................................................... 114
Chapter 13. Tur bell aria................................................................................. 117
13.1. Histological and immunohistochemical data.................................................... 122
13.2. Data of electron microscopic investigations.................................................. 123
13.3. Conclusion
Chapter 14. Cnidaria...................................................................................... 125
Chapter 15. Distribution of intestinal regulatory systems in Metazoa and their proposed evolution......... 128
15.1. Distribution and some common features of organization of intestinal regulatory systems....... 128
15.2. Proposed evolution of intestinal regulatory systems.......................................... 130
Conclusion.................................................................................................. 137
Summary 139
Literature.................................................................................................. 144
Научное издание
МИХАИЛ ЮРЬЕВИЧ ПУНИН
КИШЕЧНАЯ РЕГУЛЯТОРНАЯ СИСТЕМА БЕСПОЗВОНОЧНЫХ
И ЕЕ ПРЕДПОЛАГАЕМАЯ ЭВОЛЮЦИЯ У МНОГОКЛЕТОЧНЫХ
Труды Зоологического института РАН
Том 290
Утверждено к печати
редакционно-издательским советом
Зоологического института РАН
План 2001 г.
Редактор Т.А. Асанович
Компьютерная верстка ИМ. Беловой
Подписано в печать 20.09.2001. Формат 70х 100‘/i6-
Бумага офсетная. Печать офсетная.
Печ. л. 10,5 + 2 вклейки
Тираж 400 экз. Заказ 33 •
Типография издательства СПбГУ, 199061, СПб., Средний пр., 41
ВКЛЕЙКА
Рис. 1-4. Клетки, иммунореактивные к нейротензину (1) и серотонину (2-4) в эпителии тонко?
кишки ежа (1), золотистого хомячка (2-3) и крысы (4).
Figs. 1-4. Cells immunoreactive to neurotensin (1) and serotonin (2-4) in the small intestine
epithelium of hedgehog (1), golden hamster (2-3) and rat (4).
Рис. 8-9. Нейротензин-иммунореактивные клетки в эпителии двенадцатиперстной кишки
цыпленка.
Figs. 8-9. Neurotensin-immunoreactive cells in the duodenal epithelium of chicken.
Рис. 13. Нейротензин-иммунореактивная клетка в эпителии желудка Styela rustica.
Fig. 13. Neurotensin-immunoreactive cell in the stomach epithelium of Styela rustica.
Рис. 16-18. FMRFaMim- (16) и нейротензин- (17-18) иммунореактивные сплетения в эпителии
печеночного отдела кишечника Saccoglossus mereschkovskii.
Figs. 16-18. Intraepithelial FMRFamide- (16) and neurotensin- (17-18) immunoreactive plexus in th
pancreatic part of Saccoglossus mereschkovskii intestine.
Стрелками (8, 16-17) отмечены имунореактивныс структуры, треугольниками - базальная граница
эпителия, звездочкой (4) - апикальная цитоплазма, окрашивающаяся альциановым синим. Непрямой
иммуноферментный метод (маркер - пероксидаза). Шкала: 10 мкм (1-5, 9, 13, 16, 18), 25 мкм (8, 17).
Рис. 20-22. Катехоламинергические клетки и отростки в эпителии кардиального желудка (20-21)
и пилорического выроста (22) Asterias rubens.
Figs. 20-22. Catecholaminergic cells and processes in the epithelium of cardiac stomach (20-21) and
pyloric caecum (22) of Asterias rubens.
Рис. 23-27. РМИРамид-иммунореактивные клетки и отростки в эпителии пилорического желудка
Asterias rubens (23-26) и Henricia sanguinolenta (27).
Figs. 23-27. FMRFamide-immunoreactive cells and processes in the pyloric stomach epithelium of
Asterias rubens (23-26) and Henricia sanguinolenta (27).
Стрелками отмечены базиэпителиальное сплетение и клеточные отростки. Флуоресцентно-гистохимический
метод FAGLU (20-22), непрямые иммуноферментный (маркер - пероксидаза) (23-24, 27) и
иммунофлуорссцснтный (маркер - техасский красный) (25, 26) методы.
Шкала: 25 мкм (20, 23-27), 10 мкм (21-22).
Рис. 28-31. Полиморфные клетки, иммунореактивные к инсулину горбуши (28-30) и ИПВ
Anodonta (31) в эпителии пилорического желудка (28-29, 31) и пилорических выростов (30)
Asterias rubens (28, 31) и Urasterias linki (29-30).
Figs. 28-31. Polymorphous cells immunoreactive to salmon insulin (28-30) and Anodonta insulin-like
peptide (31) in the epithelium of pyloric stomach (28-29, 31) and pyloric caecum (30) of Asteris rubens
(28, 31) and Urasterias linki (29-30).
Рис. 32-34. Серийные срезы пилорического желудка Asterias rubens, обработанные тремя
различными антисыворотками к ИПВ Anodonta.
Figs. 32-34. Serial sections of Asterias rubens pyloric stomach, demonstrating cells (arrows) reacting
with three different antisera to Anodonta insulin-like peptide.
Стрелками (32-34) отмечены клетки, реагирующие со всеми тремя антисыворотками Непрямой
иммуноферментный метод (маркер - пероксидаза). Шкала: 10 мкм (28-31), 25 мкм (32-34).
Рис. 35-36. Два серийных среза пилорического желудка Urasterias linki, обработанные
антисыворотками к инсулину горбуши (35) и ИПВ Anodonta (36).
Figs. 35-36. Two serial sections of Urasterias linki pyloric stomach, demonstrating cells (arrows),
reacting with salmon insulin (35) and Anodonta insulin-like peptide (36).
Рис. 37—40. Срезы пилорического выроста Asterias rubens, последовательно окрашенные
антисыворотками к С-фрагменту ИПВ Lymnaea stagnalis (37) и инсулину горбуши (38), инсулин
горбуши (39) и С-фрагменту ИПВ Lymnaea stagnalis (40).
Figs. 37-40. Sections of Asterias rubens pyloric ceacum stained step by step with antisera to C-fragme
of Lymnaea stagnalis insulin-like peptide (37) and salmon insulin (38), salmon insulin (39) and C-fragn
of Lymnaea stagnalis insulin-like peptide (40).
Стрелками отмечены клетки, реагирующие с обеими антисыворотками. Непрямые иммуноферментный
(маркер - пероксидаза) (35-36) и иммунофлуорссцснтный [маркеры - техасский красный, красное свечею
(37, 40) и FITC, зеленое свечение (38-39)] методы. Шкала: 25 мкм.
Рис. 41-46. Клетки, иммунореактивные к панкреатическому полипептиду (41), глюкагону (42 43,
45^46) и соматостатину (44) в эпителии пилорического желудка (41^45) и кармана Тидемана (46)
Henricia sanguinolenta (41, 46), Urasterias linki (42-43) и Asterias rubens (44-45).
Figs. 41-46. Cells, immunoreactive to PP (41), glucagon (42-43, 45-46) and somatostatin (44) in the
epithelium of pyloric stomach (41-45) and Tiedemann’s pouches (46) of Henricia sanguinolenta (41, 46),
Urasterias linki (42-43) and Asterias rubens (44-45).
Рис. 47-48. Одновременное выявление ИПВ Anodonta- (47) и FMRFaMWj- (48) иммунореативных
элементов в эпителии пилорического желудка Asterias rubens.
Figs. 47^4-8. Simulteneous detection of Anodonta insulin-like peptide- (47) and FMRFamide- (48)
immunoreactive elements in the pyloric stomach epithelium of Asterias rubens.
Стрелкой (47-48) отмечена клетка, реагирующая с обеими антисыворотками. Метод меченого авидин-биотина
(маркер - пероксидаза) (41—46), непрямой иммунофлуоресцснтный метод [маркеры - FITC, зеленое свечение
(47), техасский красный, красное свечение (48)]. Шкала: 25 мкм.
Рис. 49-51. Эпителий пилорического желудка Asterias rubens (49). Одновременное выявление
катехоламинов (50) и РМКРамид-подобного вещества (51).
Figs. 49-51. Pyloric stomach epithelium of Asterias rubens (49). Simulteneous detection of
catecholamines (50) and FMRFamide-like substance (51).
Рис. 52-57. Катехоламинергические отростки (52-53), РМИРамид- (54), соматостатин- (55),
панкреатический полипептид- (56) и глюкагон- (57) иммунореативные клетки в эпителии
пищевода (52) и кишечника (53-57) Strongylocentrotus droebachiensis.
Figs. 52-57. Catecholaminergic processes (52-53), FMRFamide- (54), somatostatin- (55), PP- (56) and
glucagon- (57) immunoreactive cells in the epithelium of oesophagus (52) and intestine (53-57) of
Strongylocentrotus droebachiensis.
Стрелками отмечены катехоламинергические (50, 52-53) и РМИРамид-иммунорсактивныс (51) структуры.
Флуоресцентно-гистохимический метод FAGLU (50, 52-53), непрямые иммунофлуорссцснтный (маркер -
техасский красный) (51) и иммуноферментный (маркер - пероксидаза) (54) методы, метод меченого авидин-
биотина (маркер - пероксидаза) (55-57). Шкала: 25 мкм (49-51, 55-57), 10 мкм (52-54).
1
Рис. 58-63. Катехоламинергические клетки (58-59) и клетки, иммунореактивные к соматостатину
(60-62) и панкреатическому полипептиду (63) в эпителии желудка Ophiocantha bidentata (58),
Ophiopholis acideata (59, 61) и Ophiura robusta (60, 62-63).
Figs. 58-63. Catecholaminergic cells (58-59) and cells, immunoreactive to somatostatin (60-62) and PP
(63) in the stomach epithelium of Ophiocantha bidentata (58), Ophiopholis acideata (59, 61) and
Ophiura robusta (60, 62-63).
Рис. 64-67. Клетки, иммунореактивные к соматостатину (64), глюкагону (65) и ИПВ Anodonta
(66-67) в эпителии средней кишки Chiridota laevis.
Figs. 64-67. Cells, immunoreactive to somatostatin (64), glucagon (65) and Anodonta insulin-like
peptide (66-67) in Chiridota laevis midgut epithelium.
Стрелками (57-58) отмечены катехоламинергические клетки, звездочкой - автофлуорссцснция в апикальных
частях эпителиальных клеток. Флуоресцентно-гистохимический метод FAGLU (58-59), метод меченого авидин-
биотина (маркер - пероксидаза) (60-63) и непрямой иммуноферментный метод (маркер - пероксидаза) (64-67).
Шкала: 25 мкм (58-60, 64-65), 10 мкм (61-63, 66-67).
Рис. 70-78. Клетки, иммунореактивные к РМИРамиду (70-75) и инсулину горбуши (76-78) в
эпителии средней кишки (70-71, 74-75, 77) и пилорических выростов (72-73, 76, 78) Nauphoeta
cinerea (70-71), Locusta migratoria (72-73, 76) и Gryllus bimaculatus (74-75, 77-78).
Figs. 70-78. Cells, immunoreactive to FMRFamide (70-75) and salmon insulin (76-78) in the epithelium
of midgut (70-71, 74-75, 77) and pyloric caeca (72-73, 76, 78) of Nauphoeta cinerea (70-71), Locusta
migratoria (72-73, 76) and Gryllus bimaculatus (74-75, 77-78).
щк - щеточная каемка (микроворсинки). Стрелками отмечены клеточные отростки. Непрямой
иммуноферментный метод (70-75) и метод меченого авидин-биотина (76-78) (маркер - пероксидаза).
Шкала: 10 мкм.
Рис. 81-84. Кальцитонин-иммунореактивные клетки (81, 83-84) и отростки (82) в средней кишке
(81-82) и слепом выросте (83-84) Octopus vulgaris.
Figs. 81-84. Calcitonin-immunoreactive cells (81, 83-84) and processes (82) in the midgut (81-82) and
caecum (83-84) of Octopus vulgaris.
Рис. 85. Нервные клетки в эпителии желудка Arctica islandica.
Fig. 85. Nerve cells in the stomach epithelium of Arctica islandica.
Рис. 87-88. Моноамин-содержащие клетки и отростки в средней кишке Mytilus edulis.
Figs. 87-88. Monoamine-containing cells and processes in Mytilus edulis midgut.
Стрелками (на рис. 82) отмечены отростки, тонкими и толстыми строчками (на рис. 87-88) - соответственно
серотонин-содержащие (желтое свечение на препаратах) и катехоламинергические (зеленое свечение)
структуры, треугольниками - базальная граница эпителия. Метод псроксидазы-антипероксидазы (81-84),
прижизненное окрашивание метиленовым синим (85) и флуоресцентно-гистохимический метод
FAGLU (87-88). Шкала: 10 мкм (81, 83-84, 87-88), 25 мкм (82, 85).
Рис. 90-94. Клетки, иммунореактивные к ИПВ Anodonta (90) и кальцитонину (91-94) в эпителии
средней кишки (90-92, 94) и мешка кристаллического стебелька (93) Mytilus edulis (90-91), Муа
arenaria (92), Arctica islandica (93) и Unio pictorum (94).
Figs. 90-94. Cells, immunoreactive to Anodonta insulin-like peptide (90) and calcitonin (91-94) in the
epithelium of midgut (90-92, 94) and style sac (93) of Mytilus edulis (90-91), Mya arenaria (92),
Arctica islandica (93) and Unio pictorum (94).
Рис. 96-97. Нейротензин-иммунореактивные клетки в эпителии мешка кристаллического стебелька
Mytilus edulis (96) и желудка Unio pictorum (97).
Figs. 96-97. Neurotensin-immunoreactive cells in the epithelium of Mytilus edulis style sac (96) and
Unio pictorum stomach (97).
Рис. 100-102. НАР-иммунореактивные клетки в средней кишке Mytilus edulis (100-101)
и Unio pictorum (102).
Figs. 100-102. HAP-immunoreactive cells in the midgut of Mytilus edulis (100-101)
and Unio pictorum (102).
Стрелками отмечены клеточные отростки (91, 93) и базиэпителиальное сплетение (100), треугольниками -
базальная граница эпителия, звездочкой (93) - нсспецифическос окрашивание. Метод пероксидазы-
антипсроксидазы. Шкала: 10 мкм (90-94, 97, 100-102), 25 мкм (96).
Рис. 104-107. РМИРамид-иммунореактивные клетки и отростки в желудке (104, 106-107) и
средней кишке (105) Mytilus edulis (104), Unio pictorum (105-106) и Musculus laevigator (107).
Figs. 104-107. FMRFamide-immunoreactive cells and processes in the stomach (104, 106-107) and
midgut (105) of Mytilus edulis (104), Unio pictorum (105-106) and Musculus laevigator (107).
Рис. 108. Ствол отростков в основании эпителия тифлозоля средней кишки Mytilus edulis.
Fig. 108. A trunk of processes in the basal part epithelium of Mytilus edulis midgut typhlosole.
отр - отросток, вступающий в состав ствола; эк - фрагменты эпителиальных клеток. Треугольниками (104-107)
отмечена базальная граница эпителия. Непрямой иммуноферментный метод (маркер - пероксидаза) (104-107) и
элсктроннограмма (108). Шкала: 25 мкм (104-106), 10 мкм (107), 1 мкм (108).
Рис. 109-112. Большой тифлозоль средней кишки Mytilus edulis. Отростки базиэпителиального
сплетения (109), полиморфизм гранул в отростках (110-111) и предполагаемый экзоцитоз
содержимого пузырька (112).
Fig. 109-112. Midgut major typhlosole of Mytilus edulis. Processes of basiepithelial plexus (109),
polymorphous granules in the processes (110-111) and exocytose (112).
эк — фрагменты эпителиальных клеток тифлозоля. Стрелками отмечены тонкие отростки без гранул (109) и
экзоцитоз (112), треугольниками - базальная граница эпителия. Шкала: 1 мкм (109), 0,25 мкм (110-112).
Рис. 113-117. Фрагменты мелкогранулярных клеток (113-114, 117) и регуляторной клетки с
крупными гранулами (115-116) в эпителии большого тифлозоля средней кишки Mytilus edulis
(113-116) и Arctica islandica (117).
Figs. 113-117. Small granular cells (113-114, 117) and regulatory cell with large granulers (115-116) in
the epithelium of midgut major typhlosole of Mytilus edulis (113-116) and Arctica islandica (117).
Треугольниками (115) отмечена базальная граница эпителия.
Шкала: 1 мкм (113), 0,5 мкм (117), 0,25 мкм (114-116). •
Рис. 119-129. РМКРамид-иммунореактивные клетки и отростки в желудке (119-120, 123-124, 126),
выростах пищеварительной железы (121-122, 127-128) и средней кишке (125, 129) Littorina saxatilis
(119-120), Palio dubia (121-122), Clione limacina (123-125)
и Planorbarius corneus (126-129).
Figs. 119-129. FMRFamide-immunoreactive cells and processes in stomach (119-120, 123-124, 126),
digestive diverticula (121-122, 127-128) and midgut (125, 129) of Littorina saxatilis (119-120), Palio
dubia (121-122), Clione limacina (123-125) and Planorbarius corneus (126-129).
эж - эпителий желудка; эпж - эпителий выростов пищеварительной железы. Стрелками отмечены
иммунореактивные отростки, треугольниками - базальная граница эпителия. Непрямой иммуноферментный
метод (маркер - пероксидаза). Шкала: 25 мкм (119, 124, 126), 10 мкм (120-122, 127-129), 50 мкм (123, 125).
Рис. 130-136. Клетки и отростки, Ихммунореактивные к РМКРамиду (130-132), кальцитонину (133—
135) и ИПВ Anodonta (136) в средней кишке Lymnaea stagnalis.
Figs. 130-136. Cells and processes, immunoreactive to FMRFamide (130-132), calcitonin (133-136) and
Anodonta insulin-like peptide (136) in the midgut of Lymnaea stagnalis.
Рис. 138-141. РМКРамид-иммунореактивные клетки в эпителии средней кишки (138), выростов
пищеварительной железы (139-140) и желудка (141) Ishnochyton albus (138-140)
и Tonicella marmorea (141).
Figs. 138-141. FMRFamide-immunoreactive cells in the epithelium of midgut (138), digestive diverticu-
lum (139-140) and stomach (141) of Ishnochyton albus (138-140)
and Tonicella marmorea (141).
Стрелками отмечены иммунореактивные отростки, треугольниками - базальная граница эпителия, звездочкой -
пигментный слой в стенке кишечника (133-134). Непрямой иммуноферментный метод (маркер - пероксидаза)
(130-132, 138-141) и метод псроксидазы-антипсроксидазы (133-136). Шкала: 25 мкм (130-132), 10 мкм (133-
136, 138-141).
Рис. 144-149. Базальные клетки и отростки (144-148) и нервные клетки (149) в эпителии средней
кишки Harmathoe imbricata (144-145), Lepidonotus squamatus (146) и Nereis virens (147-149).
Figs. 144-149. Basal cells and processes (144-148) and nerve cells (149) in the midgut epithelium of
Harmathoe imbricata (144-145), Lepidonotus squamatus (146) and Nereis virens (147-149).
Рис. 151-153. Катехоламин- (151-152) и серотонин- (153) содержащие клетки в эпителии средней
кишки Harmathoe imbricata (151—152) и Arenicola marina (153).
Figs. 151-153. Cathecholamine- (151-152) and serotonin- (153) containing cells in the midgut epithelium
of Harmathoe imbricata (151—152) and Arenicola marina (153).
обк - отростки базальных клеток. Стрелками отмечены пиронинофильныс шапочки (144), два типа базальных
клеток (145) и клеточные отростки (151-153), треугольниками - базальная граница эпителия. Парафиновый
срез, окраска метиленовым синим (144), полутонкис срезы, окраска толуидиновым синим (145, 147-148),
автограф после введения 3Н-глюкозы (146), целлоидиновый срез после прижизненного окрашивания
метиленовым синим (149) и флуоресцентно-гистохимический метод FAGLU (151-153). Шкала: 10 мкм (144-148,
151-153), 50 мкм (149).
1
г
1
Рис. 154-162. Серотонин- (154-169) и нейротензин- (160-162) иммунореактивные клетки и
отростки в эпителии и стенке средней кишки Lepidonotus squamatus (154-156),
Nereis virens (157—159) и Arenicola marina (160—162).
Figs. 154-162. Serotonin- (154-169) and neurotensin- (160-162) immunoreactive cells and processes in
the midgut epithelium and wall of Lepidonotus squamatus (154—156), Nereis virens (157—159)
and Arenicola marina (160—162).
Стрелками отмечены суб- и интраэпителиальные иммунореактивные отростки (на 158-159 - проникающие в
основание эпителия), треугольниками - базальная граница эпителия, звездочкой (154) - нсспсцифичсскос
окрашивание. Метод псроксидазы-антипсроксидазы (154, 157-162) и меченого авидин-биотина (маркер -
пероксидаза) (155-156). Шкала: 25 мкм (154, 157), 10 мкм (155-156, 158-162).
Рис. 163-172. РМКРамид-иммунореактивные клетки и отростки в эпителии и стенке средней кишки
Harmathoe imbricata (163-168), Lepidonotus squamatus (169), Amphitrite cirrata (170)
и Eteone longa (171-172).
Figs. 163-172. FMRFamide-irnmunoreactive cells in the epithelium and midgut wall of Harmathoe
imbricata (163-168), Lepidonotus squamatus (169), Amphitrite cirrata (170)
and Eteone longa (171-172).
Тонкими стрелками отмечены иммунореактивные структуры, толстыми стрелками - базальные клетки, не
реагирующие с антисывороткой, треугольниками - базальная граница эпителия. Непрямой иммуноферментный
метод (маркер - пероксидаза) (163, 166, 169-172), непрямой иммунофлуорссцентный метод (маркер - техасский
красный) (164, 167) и те же срезы в проходящем свете (165, 168).
Шкала: 10 мкм (163, 169-172), 25 мкм (164-168).
Рис. 174-178. Тонкое строение базальных регуляторных клеток в эпителии средней кишки
Harmathoe imbricata. Мелкогранулярные клетки (174) и их гранулы (175), крупногранулярная
клетка (176), ее аппарат Гольджи (177) и секреторная гранула (178).
Figs. 174-178. Fine structure of basal regulatory cells in the midgut epithelium of Harmathoe imbricata
Small granular cells (174) and their granules (175), large granular cell (176), its Golgi apparatus (177)
and secretory granule (178).
окг - отростки с крупными гранулами; окэ - основные клетки эпителия.
Шкала: 1 мкм (174, 176), 0,5 мкм (175, 177-178).
Рис. 181-184. Тонкое строение регуляторных элементов в эпителии средней кишки Nereis virens.
Фрагменты мелкогранулярной (181) и крупногранулярной (182) клеток, базиэпителиальное
сплетение (183), апикальная часть нервной рецепторной клетки (184).
Figs. 181-184. Fine structure of regulatory elements in Nereis virens midgut epithelium. Fragments of
small granular (181) and large granular (182) cells,’basiepithelial plexus (183), apical part of nerve receptor
cell (184).
кгк- крупногранулярная клетка; окг- отростки с крупными гранулами; омг - отростки с мелкими гранулами;
пд- полудесмосо.мы, пор - пора в мембране кишечного синуса. Шкала: 0,5 мкм (181-182), 1 мкм (183-184).
Рис. 186-188. Митозы (186-187) и первично меченная клетка (6 ч после введения 3Н-тимидина)
(188) в основании эпителиального пласта средней кишки Harmathoe imbricata (186, 188) и
Lepidonotus squamatus (187).
Figs. 186-188. Mitosis (186-187) and primary labelled cell (6 h after injection of 3H-thymidine) (188) in
the basal part of Harmathoe imbricata (186, 188) and Lepidonotus squamatus (187) midgut epitelium.
Рис. 189-194. РМКРамид- (189-191) и кальцитонин- (192-195) иммунореактивные клетки в
эпителии средней кишки Lumbricus terrestris (189, 190, 192) и Eisenia foetida (191, 193-195).
Figs. 189-194. FMRFamide- (189-191) and calcitonin- (192-194) immunoreactive cells in the midgut
epithelium of Lumbricus terrestris (189, 190, 192) and Eisenia foetida (191, 193, 194).
Рис. 196. Базиэпителиальный тяж отростков в средней кишке Eisenia foetida.
Fig. 196. Basiepithelial trunk of processes in the midgut of Eisenia foetida.
хэ - хлорагогенный эпителий. Стрелками отмечены митозы (186-187), первично меченная клетка (188),
субэпитслиальный отросток (189), базальный клеточный отросток (193) и базиэпителиальный тяж (196),
треугольниками - базальная граница эпителия. Парафиновые срезы, докраска гсмалауном (186-188), непрямой
иммуноферментный метод (маркер - пероксидаза) (189-194) и полутонкий срез, окраска толуидиновым синим
(196). Шкала: 10 мкм (186-189, 193-194, 196), 25 мкм (190-192).
Рис. 197-200. Элементы регуляторной системы в эпителии средней кишки Eisenia foetida. Отростки
у базальной мембраны (197), мелкогранулярные (198-199)
и крупногранулярная (200) клетки.
Figs. 197-200. Elements of regulatory system in Eisenia foetida midgut epithelium. Processes near basal
membrane (197), small granular (198-199) and large granular (200) cells.
Стрелками (199) отмечен клеточный отросток, вступающий в состав базального сплетения.
Шкала: 0,5 мкм (197), 1 мкм (198-200).
Рис. 202-211. Интра- и субэпителиальные серотонин- (202, 205), кальцитонин- (203, 208, 211),
FMRFaMna- (204, 209) и нейротензин- (206-207, 210) иммунореактивные клетки и отростки в
желудке (204, 206-208) и средней кишке (202-203, 205, 209-211)
Glossiphonia complanata (202-204), Hirudo medicinalis (205-207), Haetnopis sanguisuga (208) и
Herpobdella octoculata (209-211).
Figs. 202-211. Intra- and subepithelial serotonin- (202, 205), calcitonin- (203, 208, 211), FMRFamide-
(204, 209) and neurotensin- (206-207, 210) immunoreactive cells and processes in the stomach (204, 206-
208) and midgut (202-203, 205, 209-211) of Glossiphonia complanata (202-204), Hirudo medicinalis
(205-207), Haemopis sanguisuga (208) and Herpobdella octoculata (209-211).
Стрелками отмечен отросток, отходящий от перикариона (202), и субэпитслиальные отростки (204 и 210),
треугольниками - базальная граница эпителия. Непрямой иммуноферментный метод (маркер - пероксидаза).
Шкала: 25 мкм (202, 211), 10 мкм (203, 205-210), 50 мкм (204).
Рис. 212-214. Гранулярная клетка (212) и отростки двух типов (213-214) в эпителии боковых
выростов желудка (212) и кишечника (213-214) Glossiphonia complanata.
Figs. 212-214. Granular cell (212) and two types of cell processes (213-214) in the stomach lateral
diverticuli (212) and intestinal (213-214) epithelium of Glossiphonia complanata.
Рис. 216-219. Гранулярные клетки (216, 218) и отростки (217, 219) в эпителии кишечника
Herpobdella octoculata.
Figs. 216-219. Granular cells (216, 218) and cell processes (217, 219) in the intestinal epithelium of
Herpobdella octoculata.
бг - большие гранулы (предположительно амебоцита).
Шкала: 1 мкм (212, 218), 0,5 мкм (213-214, 217-218), 5 мкм (216).
Рис. 223-229. Отростки (223-225), мелкогранулярная клетка (226), нейротензин- (227-228) и НАР-
(229) иммунореактивные клетки и отростки в эпителии пищевода (227),
желудка (223-225, 229), средней кишки (228) и ее слепого расширения (226) Hemithyris psittacea.
Figs. 223-229. Cell processes (223-225), small granular cell (226), neurotensin- (227-228) and HAP-
(229) immunoreactive cells and processes in the oesophageal (227), stomach (223-225, 229), midgut (228)
and midgut blind widening (226) epithelium of Hemithyris psittacea.
Стрелками отмечена концентрация пузырьков у базальной мембраны (224—225), отросток, отходящий от
псрикариона (226), базальные отростки и сплетение (227, 229). Элсктроннограммы (223-226) и метод меченого
авидин-биотина (маркер - пероксидаза) (227-229).
Шкала: 0,5 мкм (223-225), 1 мкм (226), 10 мкм (227-228), 25 мкм (229).
Рис. 230-233. НАР- (230-231) и РМКРамид- (232-233) иммунореактивные клетки и отростки в
эпителии желудка (230, 232-233) и средней кишки (231) Hemithyris psittacea.
Figs. 230-233. НАР- (230-231) and FMRFamide- (232-233) immunoreactive cells and processes in the
stomach (230, 232-233) and midgut (231) epithelium of Hemithyris psittacea.
Рис. 235-239. Нейротензин- (235, 237) и HAP- (236, 238-239) иммунореактивные клетки и отростки
в эпителии пищевода (237) и желудка (235-236, 238-239) Coptothyris grayi.
Figs. 235-239. Neurotensin- (235, 237) and HAP- (236, 238-239) immunoreactive cells and processes in
the oesophageal (237) and stomach (235-236, 238-239) epithelium of Coptothyris grayi.
Стрелками отмечены иммунореактивные клетки и отростки, треугольниками - базальная граница эпителия,
звездочкой (235-236) - нсспсцифичсское окрашивание. Метод меченого авидин-биотина (маркер - пероксидаза)
(230-231, 235-239) и непрямой иммуноферментный метод (маркер - пероксидаза) (232-233).
Шкала: 10 мкм (230-233, 237-239), 25 мкм (235-236).
Рис. 242-243. Базальная часть мелкогранулярной клетки (242) и ее гранулы (243) в эпителии средне
кишки Priapulus caudatus.
Figs. 242-243. Basal part of a small granular cell (242) and its granules (243) in the midgut epithelium
of Priapulus caudatus.
Рис. 245-248. Серотонин-содержащая (золотисто-желтая флуоресценция на препаратах) (245) и
катехоламинергические клетки и отростки (зеленая флуоресценция) (246-248) в средней кишке
Halicryptus spinulosus.
Figs. 245-248. Serotonin-containing (yellow fluorescence) (245) and catecholaminergic cells and process
(green fluorescence) (246-248) in the midgut of Halicryptus spinulosus.
о - клеточные отростки. Стрелками отмечены катехоламинергические клетки (246, 248) и отростки (247),
треугольниками - базальная граница эпителия, звездочкой - нсспсцифичсская флуоресценция.
Элсктроннограммы (242-243) и флуоресцентно-гистохимический метод FAGLU (245-248).
Шкала: 1 мкм (242), 0,25 мкм (243), 10 мкм (245), 25 мкм (246), 12,5 мкм (247-248).
Рис. 249-253. Нервные клетки в кишечнике Cerebratulus sp. (249), Amphiporus lactiflroreus (250),
Cephalotrix linearis (251), Linear ruber (252) и Linens viridis (253).
Figs. 249-253. Nerve cells in the intestine of Cerebratulus sp. (249), Amphiporus lactiflroreus (250),
Cephalotrix linearis (251), Lineur ruber (252) and Linens viridis (253).
Рис. 255-257. Базиэпителиальное кишечное сплетение (255) и нейротензин-иммунореактивные
клетки и отростки (256-257) Linens viridis.
Figs. 255-257. Intestinal basiepithelial plexus (255) and neurotensin-immunoreactive cells and processes
(256-257) of Lineus viridis.
бнс - боковой нервный ствол; бс - базиэпителиальное сплетение; км - кольцевой мышечный слой;
кне - кольцевое нервное сплетение; пм - продольный мышечный слой; ерк - средняя кишка; хо - хоботок;
эпк - эпителиальные клетки кишечника. Стрелками отмечены нервные тяжи и отростки (249-251, 253),
иммунореактивные отростки в нейропиле нервного ствола и клетка в гастродермисе (256), треугольниками -
базальная граница гастродермиса. Целлоидиновые срезы, импрегнация азотнокислым серебром по Колонье
(249-253), элсктроннограмма (255) и непрямой иммуноферментный метод (маркер - пероксидаза) (256-257).
Шкала: 200 мкм (249, 252, 256), 50 мкм (250-251), 25 мкм (253, 257), 5 мкм (255).
Рис. 258-268. FMRFaMim- (258-263), нейротензин- (264-267) и серотонин- (268) иммунореактивные
клетки и отростки в кишечнике Notoplana atoniata.
Figs. 269-280. FMRFamide- (258-263), neurotensin- (264-267) and serotonin- (268) immunoreactive
cells and processes in the intestine of Notoplana atomata.
жк - железистые (секреторные) клетки гастродермиса. Стрелками отмечены иммунореактивные отростки,
треугольниками - базальная граница гастродермиса.
Непрямой иммуноферментный метод (маркер - пероксидаза). Шкала: 10 мкм.
Рис. 282-289. Элементы регуляторной системы в кишечнике Provortex karlingi (282), Procerodes
littoralis (283, 286-287), Macrostomum sp. (284), Dendrocoehun lacteum (285)
и Planaria torva (288-289).
Figs. 282-289. Elements of regulatory system in the intestine of Provortex karlingi (282), Procerodes
littoralis (283, 286-287), Macrostomum sp. (284), Dendrocoehun lacteum (285)
and Planaria torva (288-289).
жк - жсзсзистая (секреторная) клетка гастродермиса. Стрелками отмечены отросток в основании железистой
клетки (282) и отростки у базальной мембраны (284). Шкала: 1 мкм (282-287, 289), 0,5 мкм (288).
Рис. 292-296. Клетки, иммунореактивные к пептиду-активатору (роста) головы гидры (НАР),
у Hydra fusca (292-293) и Hydra oligactis (294-296).
Figs. 292-296. Cells, immunoreactive to hydra head activator peptide (HAP), in Hydra fusca (292-293)
and Hydra oligactis (294-296).
гс - гипостом, мг - мезоглея; щ - щупальца. Непрямой иммунофлуорссцснтный метод (маркер - флуоресцеин)
(292-293) и непрямой иммуноферментный метод (маркер - пероксидаза) (294-296).
Стрелками отмечены иммунореактивные структуры. Шкала: 200 мкм (292-293), 10 мкм (294-296).
from Biological Faculty (Chair of Cytology
and Histology) of Leningrad State University
in 1975. Doctor of Philosophy (1983).
Senior Research Fellow in the Laboratory
of Evolutionary Morphology, Zoological
Institute of Russian Academv of Sciences.
ПУНИН Михаил Юрьевич родился
в 19531 в Ленинграде В 1975 г. окончил
Биолого-почвенный факультет Ленинградского
государственного университета. Работал
на кафедре цитологии и гистологии ЛГУ.
затем в лаборатории физиологии питания
Института физиологии им. И. П. Пав лова
АН СССР. С 1986 г в штате Беломорской
биологической станции Зоологического
института АН СССР. В настоящее время
старший научный сотрудник лаборатории
эволюционной морфологии этого института.
Кандидат биологических наук (1983г.).
Автор более 90 научных работ, в том числе
монографии «Гистологическая организация
кишечных эпителиев приапулид, брахиопод,
двустворчатых моллюсков и полихет» (СПб..
«Наука», 1991) и соавтор коллективной
монографии «Мембранный гидролиз и
транспорт. Новые данные и гипотезы» (Л.,
«Наука», 1986). Область научных интересов
сравнительная и эволюционная гистология,
м< >рфология, эндокринология и нейробиология
беспозвоночных животных.
Present Address: Zoological Institute of
Russian Academy of Sciences, University skay a
nab., 1, 199034, St. Petersburg, Russia
Phone: (812) 3280612
E-mail: morphol@zin.ru
Рабочий адрес: 199034, Санкт-Петербург,
Университетская наб., д. 1. Зоологический
институт РАН.
Телефон: (812) 3280612
E-mail: morphol@zin.ru