/










































































































































































































Author: Федотов М.Д.
Tags: биология клетки и субклеточных частиц цитология биология эволюционный процесс беспозвоночные филогенез
Year: 1966




Text
Ц.М.Федотов
ЭВОЛЮЦИЯ И ФИЛОГЕНИЯ БЕСПОЗВОНОЧНЫХ животных
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ
МОРФОЛОГИИ животных
им. А. Н. СЕВЕРЦОВА
8
Д. М. ФЕДОТОВ
Эволюция
и филогения
беспозвоночных
животных
ИЗДАТЕЛЬСТВО «НАУКА»
МОСКВА-1966
УДК 576.12 : 592
Дмитрий Михайлович Федотов
Эволюция и филогения
беспозвоночных животных
Утверждено к печати
Институтом морфологии, животных им. А. И. Северцова
Академии наук СССР
Редактор О. М. Мартынова
Редактор издательства Э. А. Фролова
Художник А. Д. Смеляков.
Технический редактор А. П. Ефимова
Сдано в набор 21/VII 1966 г. Т-14865.
Подп. к пе.ч. 2/XI 1966 г. Формат 70 X 108716-
Печ. л. 25,25+1 вкл. (0,25 п. л.). Усл. печ. л. 35,7.
Уч-изд. л. 36,9. Тираж 1700 экз. Изд. № 1060/66.
Тип. зак. 63Г1. Цена 2 р. 44 к.
Издательство «Наука»,
Москва, К-62, ' Подсосенский пер., 21
2-я типография издательства «Наука»,
Москва, Г-99, Шубинский пер.,' 10
2-10-6
59-БЗ-25-66
Содержание
От автора............................................... 6
Современные подходы к разработке проблемы эволюции
и филогении животных............................... 7
Губки (Porifera)...................................... 16
Кишечнополостные (Coelenterata).........................26
Гребневики (Ctenophora).................................57
Плоские черви (Plathelminthes)..........................66
Первичнополостные черви (Askelminthes)................. 89
Немертины (Nemertini)..................................115
Моллюски (Mollusca) .... ; : 124
Общая часть.....................................' 124
Брюхоногие моллюски..............................139
Двустворчатые моллюски . ...................148
Лопатоногие моллюски.............................155
Головоногие моллюски . . . . , 156
Заключительная часть............................ 169
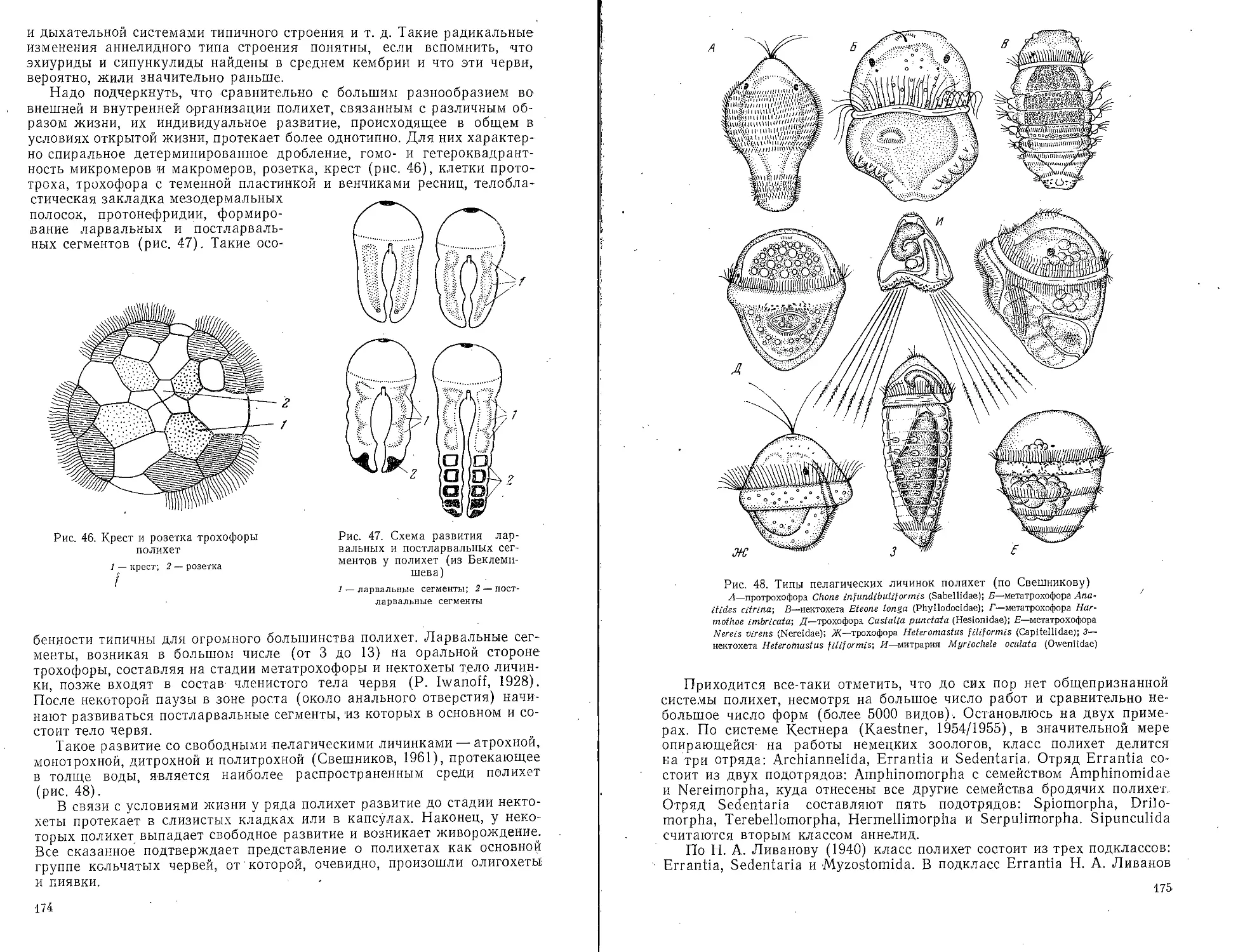
Аннелиды (Annelida)................................ 171
Тардиграды (Tardigrade)................................190
Пентастомнды (Pentastomida).......................... 194
Онихофоры (Onychophora)................................199
Членистоногие (Arthropoda)........................* . 208
Форониды (Phoronida)...................................223
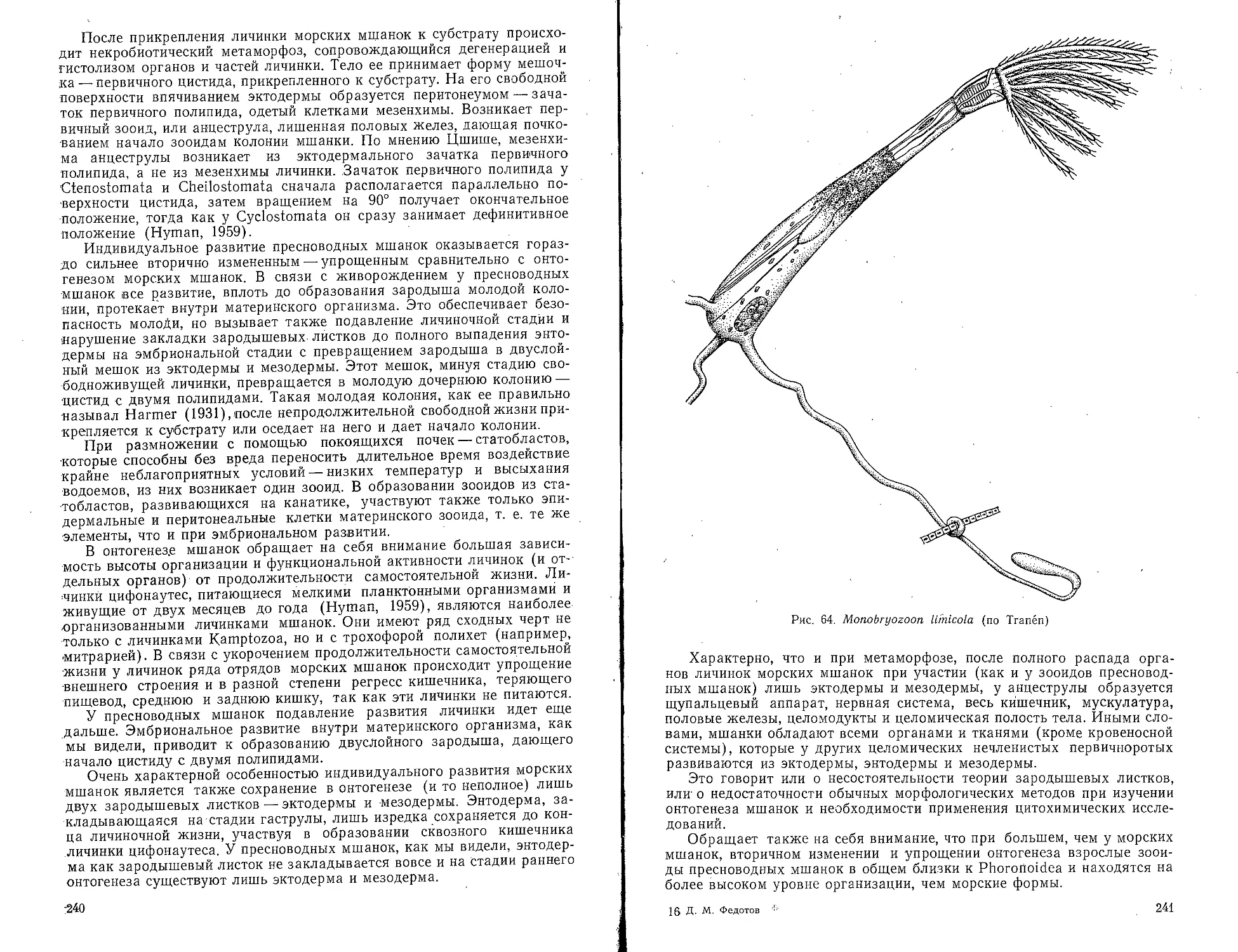
Мшанки (Bryozoa).......................................235
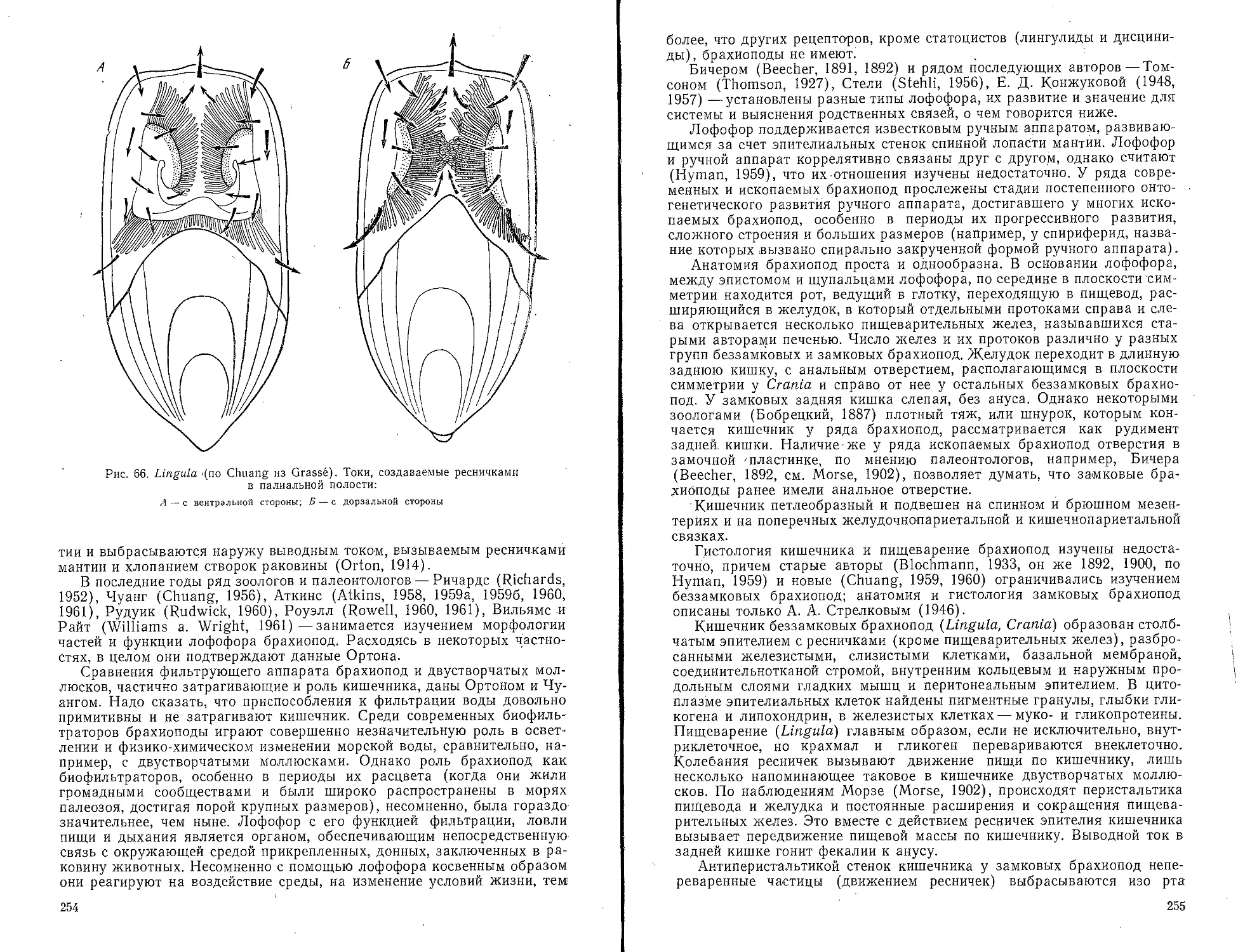
Брахиоподы (Brachiopoda)...............................250
Кишечнодышащие.. (Enteropneusta).......................267
Птеробранхии (Pterobranchia)...........................281
Граптолиты (Graptolitha) ..............................288
Иглокожие (Echinodermata)..............................291
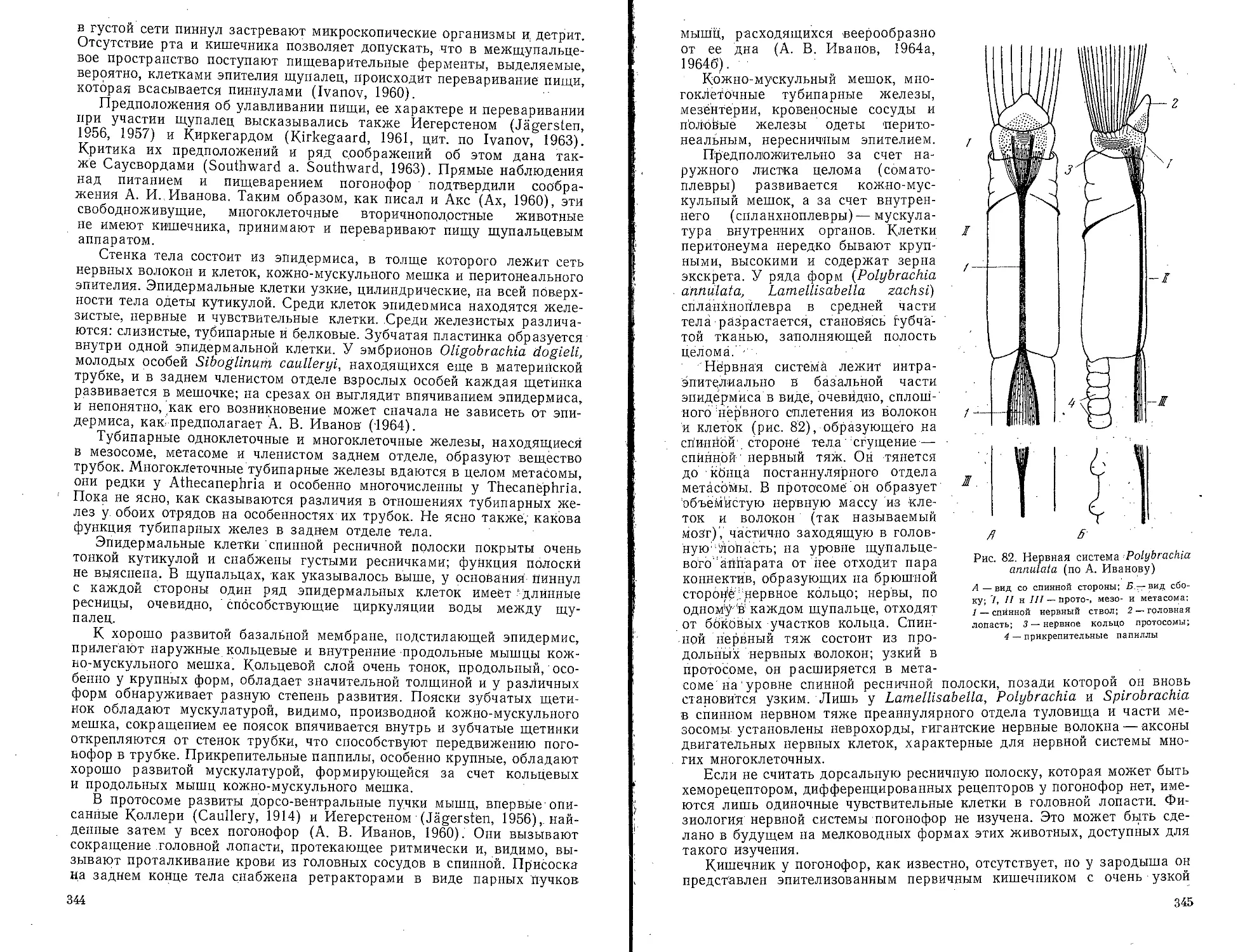
Погонофоры (Pogonophora)...............................340
Филогенетические отношения типов животных . . . 354
Литература.............................................362
От автора
Необходимость написать труд об эволюции и о
филогении беспозвоночных ощущалась давно, но
возможность к этому появилась для меня лишь в
последние годы. Интерес к эволюции и филогении
возник у меня тогда, когда я еще учился и работал
в Петербургском университете, где эта проблема ин-
тересовала многих зоологов, особенно профессора
В. М. Шимкевича. Теперь, однако, эволюция и фи-
логения животных не проблема зоологии и палеон-
тологии (метод тройного параллелизма), а пробле-
ма комплексная, и не только большинства биологи-
ческих дисциплин, но и геохимических, геологиче-
ских и других наук о Земле. В разработке ее как
комплексной проблемы предстоит еще большая ра-
бота ученых и особенно с позиций диалектико-мате-
риалистической концепции развития.
Я много обязан дружеской помощи академика
Ю. А. Орлова, облегчившего трудности подготовки
рукописи к выходу в свет, и очень признателен про-
фессорам А. В. Иванову, А. А. Парамонову и
Г. А. Шмидту за критический просмотр всей рукопи-
си, а также палеонтологам Е. А. Ивановой,
Г. Г. Астровой, В. Н. Шиманскому, Ю. А. Арендту
и А. Н. Соловьеву за критические замечания по от-
дельным главам. За помощь я признателен и кол-
лективу лаборатории морфологии беспозвоночных
Института морфологии животных им. А. Н. Север-
цова АН СССР. Приношу глубокую благодарность
Э. Д. Давыдовой и Н. А. Румянцевой, помогавшим
мне в работе по эволюции и филогении беспоз-
воночных, и О. М. Месснер, оказавшей мне очень
ценную помощь в подготовке рукописи к печати.
Проф. Д. Федотов
«...не забывать' основной исто-
рической связи, смотреть на каждый
вопрос с точки зрения того, как из-
вестное явление в истории возникло,
какие главные этапы в своем разви-
тии это явление проходило, и с точ-
ки зрения этого его развития смот-
реть, чем данная вещь стала теперь».
(Ленин, т. 29, стр. 436, изд. 4).
СОВРЕМЕННЫЕ ПОДХОДЫ К РАЗРАБОТКЕ
ПРОБЛЕМЫ ЭВОЛЮЦИИ И ФИЛОГЕНИИ
ЖИВОТНЫХ
Филогения животных в настоящее время представляет собой пробле-
му не только зоологии, но и диалектико-материалистической концепции
развития, так как мир организмов есть часть объективного мира, раз-
вивающегося по законам движения материи, одной из высших форм
которой является живой белок. Вся история животного мира говорит о
его развитии от низшего к высшему, от простого к сложному, идущему
на противоречивом единстве внутреннего и внешнего, под направляю-
щим воздействием рреды и при взаимодействии между организмом и
условиями его жизни.
Гению Дарвина и его бессмертным трудам мы обязаны, как изве-
стно, научным, естественноисторическим объяснением эволюции и про-
исхождения животного мира. Родство между всеми животными, о чем
говорил уже Ламарк, Дарвин- показал на громадном фактическом ма-
териале.
С последней трети XIX века, благодаря теории Дарвина, анатомия
и эмбриология стали разрабатываться в историческом направлении,
сравнительный метод в них стал преобладать над описательным, все
шире начал применяться экспериментальный метод. Направляющим те-
чением в анатомии и эмбриологии делалось филогенетическое: выясне-
ние родственных отношений, происхождения, филогении и эволюции
животных стало задачей работ прогрессивных зоологов. Описательный
характер палеонтологических работ начал уступать место эволюцион-
ному, а направленность работ зоологов й палеонтологов стала сходиться.
Разработка филогении животных проводилась по методу тройного па-
раллелизма— сравнительной анатомией, эмбриологией и палеонтологи-
ей. предложенному Геккелем (Haeckel, 1874), автором и самого термина
филогения.
Дарвин ясно показал значение этих трех дисциплин для выяснения
происхождения животных и в ранних своих работах и тем более в «Про-
исхождении видов», но данные этих дисциплин тогда были бедны
7
и часто недостоверны, а методы весьма примитивны. Теория эволюции
Дарвина,- вызвавшая подъем морфологических дисциплин, и сама полу-
чила опору и подкрепление благодаря этому подъему.
Ведущее значение в разработке филогении занимает эмбриология,
так как реальные родственные связи между особями животных, потом-
ками и предками осуществляются размножением и развитием в цепи
поколений, связывающих огромное число современных и вымерших осо-
бей вида. Факты онтогенеза животных, имеющие филогенетическое зна-
чение, были известны эмбриологам и раньше —вспомним эмбриологию
позвоночных, созданную русскими академиками Вольфом, Пандэром и
Бэром; но лишь после трудов Дарвина, с созданием сравнительной
эмбриологии, чем наука обязана тоже русским ученым—А. О. Ковалев-
скому и И. И. Мечникову — были установлены закономерности онтоге-
неза, общие для беспозвоночных и позвоночных: дробление яйца, за-
кладка зародышевых листков, источники происхождения органов и
систем органов (органогенез), развитие эмбрионов и личинок, превра-
щение и формирование из них взрослых организмов. Был установлен
исторический характер индивидуального развития животных, г. е. про-
явление черт предков в развитии потомков (F. Muller, 1864), что отме-
чал еще Дарвин и что получило оформление в биогенетическом законе
Э. Геккеля (Haeckel, 1874) как повторение филогении онтогенией.
В позднейших работах некоторых зарубежных ученых, а главным обра-
зом работами А. Н. Северцова (1939) и ряда его учеников односторон-
ность биогенетического закона была преодолена и было показано, что
не только филогения творит онтогению, как говорил Геккель, но и об-
ратно— онтогения творит филогению, т. е. они взаимно связаны и вза-
имно обусловлены. В нашей литературе в зоологии это показано
И. И. Ежиковым (1939, 1940), Б. С. Матвеевым (1937, 1947, 1957), в
палеонтологии — В. Е. Руженцовым (1939, 1940), А. В. Ивановым (1939)
и др. (см. Федотов, 1959).
В работах А. Н. Северцова, как известно, показана связь между ин-
дивидуальным и историческим развитием (между онтогенезом и фило-
генезом) и значение эмбриологии для эволюции; последняя идея и рань-
ше развивалась рядом ведущих русских зоологов — В. М. Шимкевичем,
В. В. Заленским и др. К сожалению, в вопросе о взаимоотношении
эмбриологии и филогении А. Н. Северцов недооценивал значения воз-
действий условий жизни на взрослые организмы в процессе изменения
филогении и эволюции животных, которое признавал еще Дарвин. Влия-
ние на ход эмбрионального развития потомков воздействий условий
жизни на взрослых особей предков показано в работах Д. М. Федотова
о развитии, регрессивных форм высших насекомых (1945, 1946, 1954,
1955; Fedotov, 1956, 1957) и А. В. Иванова о развитии паразитических
моллюсков (1937, 1949; см. гл. «Немертины»).
Значение эмбриологии для понимания родственных, филогенетиче-
ских отношений животных общеизвестно. Оно особенно ясно видно при
изучении развития свободноживущих форм, со своеобразным, загадоч-
ным характером строения, и паразитических, сильно измененных. Не
приводя массы данных, особенно многочисленных по позвоночным, огра-
ничусь некоторыми примерами из позднейших советских работ о бес-
позвоночных. Так, у погонофор, червеобразных организмов, живущих в
трубках, преимущественно на абиссальных глубинах океана, во взрос-
лом состоянии внешне похожих на кольчатых червей, но лишенных ки-
шечника и имеющих необычайно своеобразное строение (А. В. Иванов,
1957, и др.; см. гл. «Погонофоры»), во время развития возникает стадия
с трехчленистым телом и тремя сегментами вторичной полости тела.
Наличие такой стадии развития сближает погонофор с группой типов,
кишечножаберных и хордовых.
8
Еще более яркий пример значения эмбриологии для понимания род-
ства животных дает изучение развития эндопаразитов. Среди брюхоно-
гих моллюсков (Грузов, 1957, 1963, 1965а, 19656; см. гл. «Моллюски»)-
со спирально закрученной, тяжелой известковой раковиной и сложной
анатомией имеются эндопаразиты морских огурцов (голотурий) и звезд,
крайне измененные паразитизмом. Такие взрослые моллюски, как, на-
пример, Entocolax, Parenteroxenos, Asterophila и др. (Шванвич, 1917;
А. В. Иванов, 1937, 1940, 1945—1949, 1952, 1953; см. гл. «Моллюски») —
червеобразные мягкие организмы, лишенные почти всех органов моллюс-
ка: раковины, нервной системы, органов чувств, дыхания, выделения,
кровообращения и т. д„ они обладают лишь сильно развитым половым,
гермафродитным или женским, аппаратом; если имеются самцы, то они
упрощаются до состояния семенников, теряя.почти все черты организма,
и живут в самках. При изучении развития таких форм (Шванвич, 1917;
А. В. Иванов, 1937, 1949) обнаружено, что у них образуются, личинки с
раковинкой и со всеми упомянутыми внутренними органами, присущими
типичным брюхоногим моллюскам. Когда же личинка от свободного об-
раза жизни переходит к паразитизму и забирается в органы хозяина,
вся организация такой типичной личинки брюхоногого моллюска рег-
рессирует, почти .нацело исчезают внешние и внутренние черты моллюс-
ка. У такого организма, утратившего почти все признаки и класса брю-
хоногих моллюсков и всего типа моллюсков, его родственные связи и
происхождение выясняются эмбриологией при изучении развития даже
одной формы, в то время как анатомии требуется для этого сравнитель-
ное изучение ряда представителей.
Примеров, может быть, не таких ярких, значения эмбриологии для
выяснения родства и происхождения беспозвоночных достаточно как в
зарубежной, так и советской литературе. Упомяну еще лишь работы о
паразитических червях школы К- И. Скрябина (1949—1954, 1949—1964),
работы В. Л. Вагина (1938—1957; см. Федотов, 1960) по паразитическим
ракообразным, живущим в морских звездах.
Важное значение для филогении имеют работы по эксперименталь-
ной эмбриологии (хотя они проводятся в гораздо’меньшем масштабе).
Еще в 1907 г. П. П. Иванов начал исследования кольчатых червей, при
этом он учитывал данные эмбриологии и регенерации. В советское время
он широко развернул эту методику исследований, применив ее и на дру-
гих типах животного мира. Он установил закономерности формирова-
ния членистого тела животного из сегментов двух родов, различных по-
развитию, строению и регенерации: ларвальных и постларвальных. По.
различию числа ларвальных сегментов и ряду черт их развития он на-
шел новые критерии родства разных классов кольчатых червей, мол-
люсков, членистоногих (мечехвостов, насекомых, многоножек), хор-
довых и др.
Работы по механике развития (экспериментальной эмбриологии) на-
чались за рубежом в 80-х годах XIX века (В. Ру, продолжили Гербст,
Шпеман и др., а у нас Д. П. Филатов). Филатов отмечал, что механика
развития близко подходит к основам исторического развития (1937).
Эволюционно-филогенетическая же направленность экспериментальных
исследований характеризует ленинградскую (петербургскую) школу
(В. М. Шимкевич, П. П. Иванов, К. Н. Давыдов); она выражена в ра-
ботах и некоторых зарубежных ученых (Runnstrom, 1925).
Говоря о роли эмбриологии в решении вопросов филогении, напом-
ню, что Гроббен (Grobben, 1908) и Гатчек (Hatschek, 1888, 1911) с по-
мощью эмбриологии установили две главные филогенетические ветви
двубокосимметричных животных: первичноротых и вторичноротых.
У первых (черви, моллюски, членистоногие) первичный рот взрослого-
животного, у вторых (иглокожие, кишечножаберные, хордовые, щетинко-
челюстные) — рот животного образуется независимо от первичного
рта. Полость тела у первых (целом) образуется расхождением клеток
в сегментах мезодермальных пластинок, у вторых — выпячиванием сте-
нок первичного кишечника.
Позже характеристика первичноротых и вторичноротых была допол-
нена данными эмбриологии и анатомии Гейдером (Heider, 1914). В на-
стоящее время эти две филогенетические ветви высших типов животных
охарактеризованы не только еще рядом новых особенностей эмбриоло-
гии и анатомии, но и биохимии, физиологии, экологии (Федотов, 1933,
1935, 1936, 1938; Fedotov, 1956, 1959); теперь эти ветви признаются боль-
шинством зоологов, за немногими исключениями (например, Н. А. Ли-
ванов).
Анатомия современных животных как область, развившаяся ранее
эмбриологии, вносит немалый вклад в выяснение филогении животных.
Сравнения строения, основанные на общности происхождения,— а мы
знаем, что и организация и развитие животных зависят не только от
условий современной жизни, но обусловлены и исторически — дают
критерии для понимания филогении и эволюции животных. Рас-
смотрение всех вопросов сравнительной анатомии позвоночных на
основе эволюционных идей Дарвина позволило В. М. Шимкевичу
(1905, 1923), а затем И. И. Шмальгаузену (1947) создать курсы сравни-
тельной анатомии на эволюционном, филогенетически более высоком
уровне, чем это сделали немецкие анатомы Гегенбауэр и Видерсгейм в
начале XX столетия.
Несколько иное положение наблюдается в анатомии беспозвоночных.
Беспозвоночные животные, как известно, представлены преобладающим
большинством современных видов (более 1 500 000 против 70 тысяч),
чрезвычайно разнообразных по строению сравнительно с позвоночными.
Продолжающиеся находки не только новых отрядов, но новых подтипов
или типов (погонофоры) и даже живых представителей примитивного
класса моллюсков, считавшегося вымершим в древнем палеозое (Neopi-
lina—Lemche, 1957), позволяют в известной мере выяснять вопросы
родства беспозвоночных даже описательно-анатомическим методом.
Однако преобладающая роль в этом сейчас принадлежит сравнительной
анатомии; таково положение и за рубежом, и у нас.
Приведу некоторые примеры из нашей литературы. Рядом анатоми-
ческих работ (нередко с учетом экологии), частью начатых до револю-
ции и широко проведенных в наше время, сделано немало для выяснения
родства, филогении и эволюции многих классов и типов беспозвоночных.
Таковы работы по низшим ресничным червям — турбелляриям (Н. В. На-
сонов и В. Н. Беклемишев), эктопаразитическим сосальщикам (Б. Е. Бы-
ховский), немертинам, полихетам, олигохетам и пиявкам (Н. А. Ливанов
с учениками и Г. А. Шмидт), по мизостомидам — паразитическим
кольчатым червям, иглокожим и паразитическим моллюскам (Д. М. Фе-
дотов, Б. Н. Шванвич и А. В. Иванов); многоножкам и насекомым
(Э. Г. Беккер) и т. д. В. Н. Беклемишевым впервые созданы основы
сравнительной анатомии беспозвоночных (1944, 1952, 1964). Разработке
филогенетических отношений беспозвоночных, в том числе и однокле-
точных, посвящены работы В. А. Догеля (1930, 1951), А. П. Маркевича
(1964), и др.
Данным анатомии и эмбриологии современных животных мы придаем
историческое, филогенетическое значение, остатки же ископаемых жи-
вотных являются фактическими историческими документами и притом
хронологического значения. Поэтому палеонтология, даже примитивного
уровня времен Дарвина и создания метода тройного параллелизма Гек-
келя, была важной частью этого метода. Эволюционная теория Дарвина,
как известно, помогла перевести палеонтологию на более высокий уро-
10
Фень. Основы эволюционной палеонтологии, заложенные В., О. Ковалев-
ским, разрабатывались прогрессивными зарубежными и особенно рус-
скими палеонтологами. Это А. П. Карпинский, С. Н. Никитин, А. П. Пав-
лов, Н. Н. Яковлев и особенно А. А. Бори'сяк и их-прямые и косвенные
ученики (см. Давиташвили, 1940, 1948). Требованиям эволюционной
теории Дарвина курс палеонтологии А. А. Борисяка (1905) отвечает в
.большей степени, чем позже изданные курсы других наших палеонтоло-
тов, за исключением курса исторической геологии Н. М. Страхова (1948),
в котором находим глубокое понимание эволюции животных и растений.
Им же (см. 1954; см. Федотов, 1959) показана причинная связь между
осадкообразованием и жизнедеятельностью животных и растений,
между изменением характера и природы осадкообразования и возраст-
тающим воздействием живого вещества на химию моря (данные
литологии свидетельствуют о поступательной эволюции жи-
вотных) .
Как в сравнительной анатомии современных позвоночных животных,
так и по ископаемым сделано много для выяснения их филогении и эво-
люции. Вспомним давно известные работы скандинавских палеонтоло-
гов Киера и Стеншио (продолжающего работать и сейчас) по вымершим
низшим водным позвоночным; их данные находятся в согласии с рабо-
тами А. Н. Северцова по современным формам; работы Д. В. Обручева
важны для понимания происхождения низших бесчелюстных; работы
шведского палеонтолога Ярвика вскрыли пути филогении основных
классов четвероногих позвоночных из разных групп кроссоптеригий.
Эволюция и филогения низших групп земноводных и рептилий во мно-
гом выяснены И. А. Ефремовым и его сотрудниками. Работы А. Г. Во-
логдина по археоциатам, Б. С. Соколова, Е. Д. Сошкиной и Т. А. Добро-
любовой по палеозойским кораллам, М. И. Шульги-Нестеренко,
В. П. Нехорошева и Г. Г. Астровой по мшанкам, Т. Г. Сарычевой,
Е. А. Ивановой по брахиоподам, В. Е. Руженцева и В. Н. Шиманского
.по головоногим, Н. Н. Яковлева, Р. Ф. Геккера и Д. М. Федотова по древ-
нейшим прикрепленным и свободноподвижным иглокожим, А. В. Мар-
тынова, Б. Б. Родендорфа, О. М. Мартыновой, Е. Э. Беккер-Мегдисовой,
А. Г. Шарова по насекомым (см. «Основы палеонтологии» под ред.
Ю. А. Орлова 1960—1964), и’ряд работ других советских палеонтологов
и многих зарубежных, на которых нет возможности останавливаться,
.дали немало для понимания родства, филогении и эволюции разных ти-
пов и классов беспозвоночных и позвоночных животных. Картины фило-
гении даны также в статье Д. М. Федотова (1935) и в коллективном
труде сотрудников Палеонтологического института (под ред. Геккера,
1947).
Однако сейчас метод тройного параллелизма — сравнительная ана-
томия, эмбриология и палеонтология — уже недостаточен для разработ-
ки проблемы филогении. Еще в начале XX столетия чешский биолог
Ружичка, а затем и другие, выдвинули теорию химической наследствен-
ности, химической специфичности вида; началось изучение родства жи-
вотных методикой преципитиновых реакций и серологических исследо-
ваний— Нуталл и Фридентал,' в 30-х годах и позже Байер, Байер и
Бойден, Вольф, Леоне и др. (см. Федотов, 1954). К сожалению, у нас эти
методы изучения родства применялись редко. Можно назвать работы
А. С. Щепотьева (Schepotieff, 1913) по выяснению родства между клас-
сами низших червей и Д. Н. Талиева и А. Я. Базикаловой (1934) между
представителями губок, многощетинковых червей и ракообразных Бай-
кала и других наших водоемов. В настоящее время этой методикой
удалось выяснить разные степени филогенетического родства многих
групп и классов животных в тех случаях, когда морфология была
бессильной.
11
В 30-х годах XX века сначала за рубежом, а затем и у нас (работы
Крепса, 1933, 1963, и его сотрудников) начинают развиваться новые ме-
тоды выяснения филогении животных. Так оказалось, что у первичноро-
тых (черви, моллюски, членистоногие) источником мышечного сокраще-
ния служит аргининфосфат, а у вторичноротых (иглокожие, кишечно-
жаберные, щетинкочелюстные и хордовые) —креатинфосфат.
Роль биохимии и физиологии в решении вопросов родства и филоге-
нии животных продолжает нарастать. Важное значение в этом отноше-
нии имеют исследования дыхательных пигментов крови (см. сводку
Опарина, 1957). По данным П. А. Коржуева (1949, 1964), у представи-
телей первичнороТых дыхательными пигментами крови являются гемо-
глобин, гемэритрин, гемоцианин и хлорокруорин, а у вторичноротых —
гемоглобин.
По исследованиям П. В. Макарова (1938), представители первично-
ротых, кроме высших классов членистоногих, обладают большой стой-
костью к наркозу; напротив, вторичноротые животные, даже представи-
тели низших типов, обнаруживают слабую стойкость к нему.
Работы по эволюционной физиологии, даже если ограничиться ссыл-
кой на работы школы Л. А. Орбели, его учеников — А. Г. Гинецинского
и Е. М. Крепса с сотрудниками (см. Гинецинский, 1945; Крепе, 1933,
1963, Kreps, 1958), вносят ряд новых критериев для познания филогении
и эволюции животных. Они уже привели важные факты по индивидуаль-
ному и историческому развитию (онто- и филогенезу) многих функций
животных, существенные для филогении. Физиологи не только пользуют-
ся филогенетическими построениями зоологов, но и подкрепляют их»
например, Е. М. Крепе (1933) и П. А. Коржуев (1949, 1964) —филогению
Д. М. Федотова (1935), а Проссер (Prosser, 1952) —филогению Гаймен
(Hymen, 1940) и т. д. Физиологи считают проблему филогении своей
проблемой (Prosser, 1952).
В связи с этим неслучайно и в работах биохимиков, особенно рус-
ских, например, школы А. И. Опарина (1957, 1965, Oparin, 1964), зоологи
находят данные об эволюционном развитии типов обмена веществ, ко-
торые позволяют признать более древним возникновение амебоидных
организмов с гетеротрофным питанием сравнительно с происхождением
жгутиковых форм с автотрофным питанием, т. е. считать класс сарко-
довых более древним и примитивным, чем класс флагеллят.
Выше я говорил о теории химической наследственности и химической
специфичности вида, возникшей в начале XX столетия. Многолетние об-
ширные исследования А. П. Виноградова и его школы (1933, 1937, 1944,
1953, Vinogradov, 1953) об элементарном химическом составе морских
организмов позволили ему принять химическую характеристику вида,
вскрыть химические черты, характеризующие разные классы животных
и дополняющие морфологическую характеристику. Им установлены так-
же направления в характере обмена веществ, например, в образовании
скелета и опорной ткани в разных группах животного мира. Кремневый
скелет развит у наиболее древних классов простейших и губок, карбо-
натный тип скелета преобладает среди классов первичноротых, а карбо-
натно-магниево-фосфатный скелет — у вторичноротых, к которым отно-
сятся и позвоночные животные.
После этих кратких данных о роли биохимии, физиологии и химии,
в разработке вопросов филогении, отмечу нарастающее значение для
этой проблемы экологии в широком смысле. Так, в работе М. С. Гиляро-
ва, разработавшего основы учения о почве как среде обитания животных
(1949а, 19496 и позже), данными экологии подкрепляются филогенети-
ческие построения в типе членистоногих.
Дополнительные данные для филогении дают также исследования
паразитизма. А. Г. Кнорре (1937а, 19376) отмечает,, что явление парази-
12
тизма широко распространено среди классов первичноротых (у двух
третей классов)., среди же вторичноротых оно отсутствует. Он связы-
вает это со спецификой химизма внутренней ' среды организмов,
которая благоприятствует или препятствует переходу организмов к па-
разитизму. • . -
В установлении родства и начальных этапов филогении мелких
таксономическйх единиц важную роль начинают играть работы генети-
:ков, которые уже не противопоставляют наследственность организму и
не видят в ней нечто, контролирующее обмен веществ .в организме и от
него не зависящее — как справедливо иронизирует "индийский биолог
Мани (Mani, 1953, 1954).
Напомню, что современный мир животных, как и его геологическая
история, говорит о неразрывной связи животных с развитием мира рас-
тений.
Примерно со второй четверти XX столетия в разработке проблемы
филогении сильно возросла роль палеонтологии. Улучшение методики
изучения ископаемых животных—'Применение срезов, слепков, рекон-
-струкций — позволило восстанавливать состояние внутренних органов
вплоть до деталей строения нервной системы, головного мозга и т. д.,
что привело к созданию сравнительной анатомии в палеонтологии. Воз-
растающее число находок прекрасной сохранности (амфибии, насеко-
мые и т. д.) позволило изучать гистологическое строение, например,
покровного эпителия, мускулатуры, желез, трахей и т. д.; в палеонтоло-
гии возникает гистология ископаемых организмов. Химический анализ
привел к появлению палеофизиологии (Орлов, 1949) и установлению
геохимической деятельности организмов. Возрастающие возможности
изучения индивидуал'ьного развития ископаемых (археоциат, полипов,
аммонитов, трилобитов, строения личинок последних, сегментации их
головы и туловища, возрастных стадий развития палеозойских ракоскор-
пионов и некоторых четвероногих позвоночных) привели к созданию
палеоэмбриологии. Благодаря применению методов вариационной ста-
тистики и биометрии значительно улучшились результаты систематики
ископаемых. Нарастающая разработка экологического направления, за-
.ложенного еще В. О. Ковалевским, привела к созданию палеоэкологии,
позволяющей восстанавливать не только условия и образ жизни иско-
паемых животных, но и режим целых ископаемых морей (Геккер и сотр.,
1962).
Таким образом, современная палеонтология, как и зоология, рас-
полагает почти всеми методами ряда биологических дисциплин: систе-
матики, анатомии, эмбриологии, экологии, биохимии, отчасти физиоло-
гии, но, имея дело с вымершими организмами, она ограничена в этих
возможностях, зато это подлинно историческая наука. Все это сильно
смягчает «неполноту» палеонтологической летописи, ведь примерно из
1 500 000 описанных видов на долю известных ископаемых приходится
всего 80 000 видов (Мур, 1957).
И все же приходится признать, что в проблеме филогении мы не мо-
жем ограничиться даже комплексом методов биологических наук, не
говоря уже о тройном параллелизме, которым еще до сих пор нередко
ограничиваются даже у нас.
Положение о взаимодействии между организмом и условиями жизни
является важным в разработке проблемы филогении. Выше были приве-
дены некоторые примеры формирующего воздействия среды на организм.
Еще в 80-х годах В. В. Докучаев, основоположник учения о почве как
особом естественноисторическом теле, являющемся результатом слож-
ных взаимосвязей между организмами и средой, указывал на роль орга-
низмов в почвообразовании. Работами русских почвоведов, агрономов,
микробиологов, а теперь и зоологов показано, что почвенные организмы,
13
в том числе и животные, не только изменяют своей жизнедеятельностью.-
свойства почвы, но и создают почвенные разности.
В -меныпей мере жизнедеятельность животных и других организ-
мов меняет условия среды в Мировом океане. Напомню -роль животных-
биофильтраторов (губки, моллюски, оболочники и др.), раскрытую в
работах французских, а также и советских (Зенкевич, 1944, 1951, 1963
и др.) исследователей. Так, небольшие двустворчатые моллюски — ми-
дии (Воскресенский, 1948) —образуют у побережий барьеры, меняющие
рельеф дна и направление токов воды моря; они участвуют в образова-
нии илистых отложений, изменяют прозрачность воды и т. д. Давно из-
вестна роль коралловых полипов в образовании атоллов и рифов, но их.
распространение ныне ограничено теплыми морями, тогда как деятель-
ность биофильтраторов охватывает весь Мировой океан.
В. И. Вернадский в ряде работ, начиная с 1916 г., и особенно в совет-
ское время, показал, что благодаря дыханию, питанию и размножению-
животных и растений существует непрерывная материальная связь меж-
ду живым и косным (атмосфера, гидросфера и литосфера) веществами-
биосферы, происходит миграция атомов химических элементов из косных
тел биосферы в живые организмы и обратно. В результате жизнедеятель-
ности организмов меняются физические, химические и механические-
свойства геологической оболочки нашей планеты. По В. И. Вернадскому,
живое вещество является геологической силой планетного значения.
А. П. Виноградов, о значении работ которого для филогении было от-
части сказано выше, раскрыл геохимическую роль жизнедеятельности
современных морских организмов и показал зависимость биохимических
провинций от жизнедеятельности растительных организмов.
Очевидно, что при существовании тесных взаимозависимости и вза-'
имодействия между животными и окружающей средой, в частности с
земной корой, понять эволюцию и филогению животного мира нельзя,
ограничиваясь лишь методами биологических дисциплин и игнорируя,
развитие нашей планеты. И к ней, как ко всему объективно существую-
щему, относится положение диалектического материализма о движущей-
ся материи, при этом понимается изменение и развитие. В ходе истории
Земли, изменения материков и океанов нашей планеты, идет увеличение
континентов и нарастающее изменение их рельефа, уменьшение Миро-
вого океана и увеличение его глубин (Амалицкий, 1896). Эти изменения
на нашей планете, сильно усложняющие физико-географические условия
жизни, не могут не вызывать глубоких изменений животного мира, а сле-
довательно, и его филогении.
В настоящее время явления геотектоники — поднятия и опускания
земной коры, с которыми связаны давно известные громадные изменения
в животном мире в прошлом, не сводятся к физическим, химическим и
механическим изменениям земной коры эндогенного происхождения.
В. И. Вернадский (1935, 1939) подчеркивал неразрывную связь между
геологическими процессами и солнечной радиацией. Некоторые ученые
(Личков, 1945, 1954, 1956; Стовас, 1951; Эйгенсон, 1954), используя
идею Канта о замедлении скорости вращения Земли трением приливов
Мирового океана, развивают положение об экзогенных причинах геотек-
тоники и связанных с ней изменений климата, прежде всего солнечной
активности, с ритмом которой связаны и ритмы размножения животных.
Отмечу, что Н. С. Щербиновский (1952) связывает периодические,
вспышки массовых размножений пустынной саранчи с активностью сол-
нечной радиации.
Огромным изменениям в животном мире, происходившим в периоды
резких тектонических явлений, тектонических революций по Б. Л. Лич-
кову, этот автор дает объяснение. Они зависят от того, что на известной
фазе устанавливается засушливый климат, сильно уменьшается количе-
14
ство осадков, повышается содержание в почве и растениях солей, идет
засолонение почв, прекращается пйтание илов морей и океанов. Такие
изменения условий жизни после благоприятных и относительно устойчи-
вых условий, продолжавшихся в длительный геотектонический покойный
период, усиливали борьбу за существование, приводили к гибели групп
животных, приспособившихся к прежним условиям, и, напротив, стиму-
лировали эволюцию групп менее специализированных. Таким образом,
геолог дает биологическое объяснение зависимости между изменениями
животного мира и средой в эпохи поднятия и опускания земной коры,
упрекая палеонтологов (например, Давиташвили), не. разгадавших ее
(Дичков, 1956).
Изменения среды и условий жизни несомненно вызвали изменения в
отправлениях, потребностях, в природе и организации, т. е. наследствен-
ности организмов. Мало того, в отличие от своего прежнего мнения (Фе-
дотов, 1959), нахожу, что нельзя отрицать и роли космических воздей-
ствий на возникновение мутаций, приводящих, по Шиндевольфу (Schin-
dewolf, 1950, 1954, 1956, 1962), к гибели одних групп и эволюции других.
Изучение современных животных дает нам ясное представление о
равных уровнях прогрессивной эволюции: низком — губки, кишечнопо-
лостные, высшем — млекопитающие, головоногие моллюски, высшие от-
ряды насекомых. Палеонтология дает картины последовательного хода
и нарастания темпов прогрессивной эволюции. Геология и комплекс род-
ственных ей наук позволяют причинно связывать крупные изменения в
ходе эволюции и филогении животных с изменениями нашей планеты.
В решении проблемы филогении, как и других крупных проблем науки,
возрастает необходимость комплексирования методов ряда дисциплин
и роль пограничных областей между ними.
Губки (Porifera)
Без симметрии или с неопределенным числом ее элементов прикреп-
ленные к субстрату морские, редко пресноводные, животные, не имеющие
органов и тканей, обладающие лишь поклеточной дифференцировкой.
Тело состоит,-главным образом, из мезенхимы или мезоглеи с разнооб-
разными клетками, включая, по-видимому, и нервные, с неорганическим
кремневым, известковым и роговым скелетом и пронизано ирригационной
системой полостей с вводными микропорами и выводным отверстием.
Тело покрыто пинакоцитами, реже вся внутренняя поверхность, чаще
лишь камеры, одеты жгутиковыми, воротничковыми клетками, несущими
пищеварительную функцию. Размножение бесполое (почками или гем-
мулами) и половое. Онтогенез примитивный, зародышевые листки не
выражены. Современные формы распространены по всему Мировому
океану, ископаемые многочисленны, известны с докембрия.
Тело губок представляет собой бокаловидное или мешковидное обра-
зование, без рта, с выводным отверстием наверху (оскулум), слепым кон-
цом прикрепленное к субстрату, с гетерополярной главной вертикальной
осью, с многочисленными мельчайшими порами или поровыми каналами
с мезоглеей с многочисленными разнообразными клетками. Чрезвычайно
простая организация губок выражается в том, что у них (как нетканевых
организмов) нет еще стабильной тканевой дифференцировки и органов,
а лишь поклеточная дифференцировка (П. П. Иванов, 1937; Ливанов,
1945, 1955), но тем не менее им свойственна определенная видовая спе-
шифичность. Клетки тела губок обладают большой самостоятельностью
й легко дедифференцируются до амебоидного состояния (типа археоци-
та) и вновь претерпевают специализацию. Всем этим обусловлен и ха-
рактер онтогенеза губок, в котором первичные органы, или зародышевые
листки, эктодерма и энтодерма, еще не выражены. По мнению А. А. За-
варзина (1945), во время развития губок процессы эмбриогенеза ограни-
чиваются дроблением и дифференцировкой бластомеров.
Губки, будучи донными, прикрепленными и тем не менее широко .
распространенными обитателями Мирового океана, входят в состав мно-
гих его биоценозов и находятся в определенных взаимоотношениях с
представителями растительного и особенно животного мира (Резвой,,
1937; Нутпап, 1940—1959). Эти взаимоотношения выражаются в форме
мутуализма, комменсализма и паразитизма; сожительство губок с дру-
гими организмами, в частности, представлены синойкией, эпиойкией и
энтоойкией (Догель, 1962). Среди губок распространено прирастание не
только к подводным скалам, но и к различным водным растениям и
особенно широко к подвижным и малоподвижным беспозвоночным.
Реже встречается прикрепление других животных к губкам, например,
кишечнополостных, мшанок, циррипедий. Особенно разнообразны
и часты взаимоотношения, включая и паразитизм, между губками всех
классов и ракообразными, от низших и кончая высшими раками (Arndt,
1933, 1937; см. также Hyman, 1940—1959). Кроме ракообразных, в теле
•16
губок живут также разные полихеты (например, Spinther') и офиуры: го-
ловоногий моллюск Possia откладывает яйца в ткань. :губки Mycale.
Сверлящие губки (сем. Clionidae) являются паразитами двустворчатых
моллюсков (например, мидий, устриц и других) и ряда коралловых по-
липов. Они сверлят ходы в раковинах своих хозяев, обусловливая ги-
бель последних.
Будучи аэробами, губки живут в чистых, хорошо аэрируемых водах;
они не выносят недостатка кислорода в воде и гниющих водах и в таких
условиях погибают. Их существование зависит также от степени соле-
ности и химизма воды, температуры, характера грунта, в частности его
заиленности.
Форма тела губок разнообразна; они бывают в виде бокальчика, ци-
линдра, бочонка, воронки, чаши, шара, кустисторазветвленными и, обра-
стая субстрат, корковидными.
Окраска губок тоже разнообразна: тускло-серая, ярко-красная, жел-
тая, оранжевая, изумрудная, голубая, фиолетовая, черная. Она обуслов-
лена наличием в мезоглее пигментных клеток. Зеленая окраска губок
пресноводного семейства Spongillidae обязана зоохлореллам, находя-
щимся в археоцитах, известковые губки пигмента не имеют.
Размеры губок различны: от 10—15 мм до 1,5 м в высоту и до 1—2 м
в диаметре, например, у Poterion neptuni, Sphecio spongio j. У Monorha-
phis chuni тело губки, достигающее 1 м в длину, сидит на огромной спи-
куле до 3 м в длину.при диаметре 1 см.
Губки, кроме пресноводных Spongillidae и Lubomirskiidae, обитатели
моря; они населяют (преимущественно известковые) главным образом
мелководье, от зоны отлива до 50 м глубины; среди демоспонгий и гек-
сактинеллид, распространенных на больших глубинах, имеются ультра-
абиссальные виды. Наибольший диапазон в вертикальном распростране-
нии характерен для стеклянных губок (в общепринятом объеме этого
класса), обитающих от полосы прибоя и отлива до ультраабиссали —
8610—8660 м (см. Беляев и др., 1960), при максимальной численности на
глубинах от 200 до 500 м. Стеклянные губки живут во всех морях от
Арктики до Антарктики, но они наиболее многочисленны в тропиках.
Очень широко распространены географически и вертикально кремневые
демоспонгии, встречающиеся на глубинах и более 4000 м. Из них убикви-
стами являются пресноводные спонгиллиды. Роговые губки также рас-
пространены почти по всему земному шару, среди них немало убиквистов
й биполярных видов. Наиболее многочисленны они в теплых морях,
в частности в морях индо-австралийского архипелага. Известковые губ-
ки обитатели, как было сказано, мелководья, особенно полосы приливов
и отливов, лишь немногие живут на глубине 4—18 м.
По особенностям строения (отсутствию тканей и органов, черт харак-
терных для многоклеточных животных, и отсутствию первичных орга-
нов— зародышевых листков в онтогенезе) наиболее обосновано противо-
поставление губок как подцарства Parazoa с одним типом Porifera
( = Spongia) всем остальным многоклеточным животным, объединяемым
в подцарство Eumetazoa (Sollas, 1884, 1888; Biitschli, 1884). Я придержи-
ваюсь классификации, по которой тип губок делится на три класса —
Demospongia, Hexactinellida и Calcispongia (Яковлев, 1925,1937; Lauben-
fels, 1955; А. Н. Muller, 1958, 1961),—и считаю, что эту систему надо на-
чинать, учитывая особенно их хронологию, с демоспонгий и заканчивать
известковыми губками, как это принимает Н. Н. Яковлев (1925).
Согласованного лгнения о числе видов современных и ископаемых
губок нет. Современных видов чаще насчитывают до 5000 (Kaestner,
1954/1955), реже даже до 10 000 (Laubenfels, 1955), иногда же считается,
что их всего две с небольшим тысячи (Easton, 1960). Мало определенных
данных и о числе ископаемых видов губок.. Как увидим ниже, губки в
мезозое (юра, мел) были массовыми формами, широко распространен-
17
2 Д. М. Федотов
ними в морях того времени; в нынешнюю эпоху губки, несмотря на при-
митивный уровень организации, прикрепленный образ жизни, на рассе-
ление лишь с помощью ресничных стадий (если не считать геммул),.
очень широко расселились по Мировому океану и являются,, как было'
сказано, сочленами многих биоценозов.
У губок с их основными типами организации — аскона, сикона и
лейкона (известковые губки), рагона и лейкона (остальные губки)
:(рис. 1) -—главную массу тела составляет мезоглея, преимущественно с
минеральным скелетом и весьма разнообразными клетками очень мелких
Рис. 1. Основные типы строения гуоок (из Grasse)
А — аскон; Б — сикон; В—лейкон; /— атриум; 2 —оскулум; 3 — хоаноциты
размеров (Farkas, 1929; Заварзин, 1945, 1947; Tuzet, 1948, 1963; Levi,.
1956, 1957 и др.). Из них колленциты (соединительнотканые клетки по-
Заварзину, 1945, 1947) звездчатой формы, связанные друг с другом свои-
ми отростками, выделяют студенистое вещество — коллагеновую субстан-
цию мезоглеи. В мезоглее находятся археоциты—амебоциты зернистые
и незернистые" (эозинофильные, базофильные, гиалиновые, лофоциты)
(Ankel u. Wintermann-Kilian, 1952), пигментные клетки, миоциты, скле-
робласты, спонгиобласты, половые клетки и, по данным французской
школы Тюзе (Tuzet, 1963; Tuzet et cet., 1952, 1953; Pavans de Ceccatty,
1955, 1959), нервные клетки. В мезоглее находятся также мигрировавшие
в нее и дедифференцировавшиеся покровные клетки — пинакоциты и
пищеварительные клетки — хоаноциты. Исходной формой клеток являет-
ся амебоидный археоцит; все клетки, как известно, легко упрощаются до
амебоидного состояния и могут возвращаться к специализированному со-
стоянию (редифференцироваться).
Склеробласты образуют минеральный (кремневый или известковый),
а спонгиобласты роговой скелет. Каждая кремневая и известковая
игла образуется одним спикулобластом, а многолучевая — ассоциацией
нескольких спикулобластов. Известковый скелет губок, как и элементы
скелета иглокожих, представлен кристаллами кальцита со свойственны-
ми последним оптическими осями, фигурами вытравливания и двойным
лучепреломлением (Догель, 1938). Вместе с тем иглы губок — это био-
кристаллы с основой из органического вещества. Известковые и кремне-
вые спикулы бывают одно-трех- и четырехлучевыми, разной величины
(макро- и микросклеры) и очень разнообразной формы и степени
развития лучей. Среди микросклер нередки многолучевые звездообраз-
ные формы. Спикулы известковых губок лежат свободно, как исключе-
нйе они спаиваются друг с другом известковым цементом. У ряда кремне-
вых губок, например, у стеклянных губок Hexactinellida, иглы срастаясь,
18
образуют правильной формы решетчатый скелет..У кремнероговых губок
кремневые спикулы склеиваются органическим веществом — спонгином
(протеином), который образуется спонгиобластами; у этих форм наблю-
дается эволюция частей скелета: от -кремневого, спаянного небольшим
количеством спонгина, до чисто органического, рогового. Лишь немногие
губки лишены скелета.
Спикулярный характер скелета губок, как подчеркивает А. А. Завар-
зин (1945, 1947), является очень примитивным и наблюдается у наиболее
древних низкоорганизованных многоклеточных. Из двух типов минераль-
ного скелета кремневый — более древний; его возникновение связано с
физико-химическими условиями докембрийского моря (Виноградов,
1933, 1944, 1953). Позже, судя по развитию соотношений между этими
элементами скелета, у разных групп демоспонгий появился органический
роговой скелет. Известковый скелет появился еще позже.
Сократимые элементы мезоглеи—-миоциты находятся на очень низ-
ком уровне дифференцировки. Они образуют род сфинктеров, вызываю-
щих сужение устьев, приводящих-и выводящих каналов, причем сокра-
щения сфинктеров происходят очень медленно. Открывание и замыкание
пор пороцитов обусловлено, видимо, лишь раздражимостью и сократи-
мостью цитоплазмы.
Судя по работам Тюзе (Tuzet et cet., 1952, 1953, 1963) и особенно
Паван. де Секкатти (Pavans de Ceccatty, 1955, 1957, 1959), в мезоглее
имеются биполярные и мультиполярные паукообразные и пузыревидные
нервные клетки, одно-двух- и тринейронные нервные дуги, синаптиче-
ские связи нервных клеток с другими клетками, например, с пинакоцита-
ми и хоаноцитами, и асйнаптические связи нервных клеток друг с другом.
Паван де Секкатти предполагает, что нервные клетки несут функцию
трофической регуляции. Однако эти утверждения требуют доказательств,
и вопрос о нервной системе губок окончательно еще не решен, так как
пока нет для этого, как указывает Г. И. Роскин (1957), физиологической
аргументации. Небоходимы поиски физиологически активных веществ —
медиаторов, ацетилхолина и адреналиноподобных веществ, с которыми
связаны процессы возникновения и распространения возбуждения. Это
тем более необходимо, поскольку возникновение и распространение воз-
буждения установлены и у одноклеточных, например, инфузорий, и на
донервной стадии онтогенеза у голожаберных моллюсков (Коштоянц,
1957, 1963). Ничего не известно и о гормонах у губок.
Нужно подчеркнуть, что клетки, описанные французскими авторами
как нервные (Tuzet, Loubatieres, Pavans de Ceccatti, 1952), по их же дан-
ным, способны претерпевать дедифференцировку, т. е. они так же ла-
бильны, как и все другие клеточные элементы губок, что совершенно-
чуждо клеткам нервной системы. Можно еще отметить, что ряд физиоло-
гов (Prosser a. Brown, 1961; Jones, 1962) вообще не признают у губок
нервной системы.
Мезоглея с ее клетками и скелетом составляет главную массу тела
большинства губок; она является (Ливанов, 1945, 1955) ведущей особен-
ностью губок и выполняет функции механической и химической связей,
опоры, защиты (токсинами, запахом, противным вкусом) и, видимо,
трофических нервных связей (Tuzet, 1963). Эволюция губок, как жи-
вотных мезенхиматозного типа, тесно связана с развитием и дифферен-
цировкой мезоглеи, ее клеток и скелета. Для губок характерно отсут-
ствие границ между пограничными элементами и мезоглеей, которую
условно называют тканью внутренней среды.
Поверхность тела губок, приводящих и отводящих каналов и полостей
ирригационной системы покрыта пинакоцитами, которые не образуют на-
стоящего покровного эпителии, у многих губок они способны уходить в
глубь тела и превращаться в амебоидные клетки; среди пинакоцитов на-
ходятся еще пороциты с внутриклеточными порами, миоциты и желези-
2* 1»
захватывают и переваривают
Рис. 2. Схема циркуляции воды
у Sycon (из Grasse)
Стрелки указывают направление
циркуляции
стые клетки. По сравнению с пинакоцитами морфологически более обо-
соблены хоаноциты, жгутиковые, воротничковые клетки общей полости
тела у губок типа аскона и жгутиковых камер большинства губок. Одна-
ко и хоаноциты не образуют кишечника как органа пищеварения. Они
пищу каждый независимо друг от друга,
после чего утрачивают воротнички и
жгуты и превращаются в амебоциты'
(зернистые амебоциты) и уходят в ме-
зоглею, где переваривают пищу. Как
известно, при неблагоприятных усло-
виях и длительном раздражении хоа-
ноциты также теряют жгуты, ворот-
нички и эпителиальное расположение.
Электронномикроскопические ис-
следования хоаноцитой (Rasmont,
1959; Atzelius, 1961) показали, что во-
ротничок хоаноцитов не сплошной, а
состоит из четырех десятков цитоплаз-
матических ворсинок, в то время как
жгутик несет двустороннее оперение и
состоит из 9 периферических волоко-
нец и двойного осевого. Такое строение
характерно только для жгутиков хоа-
нофлагеллят и губок, что может гово-
рить об их родстве.
Очень характерной системой губок
как биофильтраторов является иррига-
ционная система (рис. 2) аскоидного,
сикоидного и лейкоидного типов, при-
чем последний наблюдается примерно
у 98% всех губок (Laubenfels, 1955).
Ирригационную систему образуют кле-
точные поры пороцитов и канальцы,
приводящие каналы, жгутиковые ка-
меры (рис. 3), отводящие каналы и
устье или многие устья.
Поступление воды в тело губки и
выведение ее наружу вызывается движением жгутиков хоаноцитов
(рис. 4). Ирригационная система обеспечивает не только пассивное пи-
тание губок, но, кроме того, удаление непереваренных частиц, экскретов
и дыхание. Она является фильтрующим аппаратом губок, пищей кото-
рых служат преимущественно бактерии, а также диатомовые, флагелля-
ты, инфузории, частицы детрита. Например, у Leucandra aspera (7 см в
длину и 1 см в поперечнике); имеется 2,25 миллионов жгутиковых камер
с общей поверхностью 52,5 см2-, 81 000 приводящих и 5 200 отводящих
каналов; такая губочка за 24 часа фильтрует 22,5 л воды (Kaestner,
1954/1955).
.. Сказанное выше относится к строению почти всех губок, кроме пред-
ставителей класса Hexactinellida, которые по ряду черт строения стоят
особняком. На наружной и внутренней поверхностях тела у последних
нет пинакоцитов. Тело состоит не из мезоглеи, а из сети трабекул, обра-
зованных разветвленными колленцитами с археоцитами и удлиненными,
наперсткообразными жгутиковыми камерами, расположенными, в ряд,
параллельно наружной и внутренней поверхностям тела губки. Жгутико-
вые воротничковые клетки синцитиально слиты базально, кремневый
скелет состоит из трехосных, обычно шестилучевых, игл, спаянных друг
€ другом кремневым цементом, чего нет у других губок.
.720
Питание и пищеварение губок и-вообще их физиология изучены не-
достаточно. В их экстрактах обнаружены разнообразные протеолитиче-
ские ферменты, сходные с трипсином, пепсином, эрепсином, а также ли-
паза, инвертаза, амилаза и др;. (Hyman, 1940). По поводу экскреции
можно сказать только, что у них не найдены мочевина, мочевая кислота
и другие продукты азотистого распада, обычные для высших животных.
Дыхание губок—аэробное.
Рис. 3. Жгутиковая камера пресноводной губки Ephydatia
(из Kaestner)
1 — отводящий канал; 2 — отверстие, соединяющее жгутиковую
камеру с каналом; 3 — археоцит; 4— хоаноциты
Рис. 4. Хоаноциты стек-
лянных губок (из Gras-
se). Основания клеток
слиты
Размножение происходит почкованием, наружным и внутренним, с
помощью геммул у пресноводных и некоторых морских демоспонгий или
при помощи соритов у ряда гексактинеллид и некоторых демоспонгий.
Образование геммул происходит в конце жизни пресноводных губок,
и наличие их является приспособлением к переживанию холодного перио-
да года в умеренном поясе и периода засухи в тропиках, тело при этом
дегенерирует.
Ряд экспериментов (протирание губок через мельничий газ, разделе-
ние их тела на мелкие кусочки 1,5—2 мм в поперечнике и др.) выявил
высокую регенеративную способность губок (Galtsoff, 1932; Korotkova,
.1962, 1963). Это вполне согласуется с поклеточной, лабильной диффе-
ренцировкой тела губок, хотя видовая специфичность сохраняется после
диссоциации тела нескольких видов губок — восстанавливается их видо-
вая специфичность.
Губки чаще гермафродиты, иногда раздельнополы; как исключение —
они яйцекладущи (например, сверлящие СНопа), обычно живородящи;
развитие до возникновения ресничного зародыша протекает в теле мате-
ри. Оогонии и сперматогонии — амебоидные, подвижные клетки, возни-
кающие из незернистых амебоцитов, а по некоторым авторам (Tuzet,
1963), из дедифференцированных хоаноцитов. Питание и рост полбвых
клеток сопровождается фагоцитозом. Несмотря на такой характер поло-
вых клеток считают, что оогенез и сперматогенез протекает путем, типич-
ным для половых клеток многоклеточных (см. Duboscq et Tuzet, 1937;
Tuzet, 1948; Levi, 1956). Некоторые авторы (Burton, 1949) допускают,
что глубоководные губки размножаются почти исключительно бесполым
путем. ।
21
Основными данными по эмбриологии демоспонгий и известковых гу-
бок мы обязаны работам конца прошлого столетия (Metschnikoff 1874,
1876, 1879; Schulze, 1875—1881; Sollas, 1884, 1888; Heider, 1886; Maas,
1892, 1893, 1898, 1900, 1909; Minchin, 1896, и др.; см. также Давыдов,
1914, Dawydoff, 1928; П. П. Иванов, 1937, 1944; Захваткин, 1949; Шмидт,
1951, 1953; Иофф, 1962).
Наши знания об эмбриологии губок существенно обогатились за счет
недавних работ французских зоологов (Duboscq et Tuzet, 1937; Tuzet,
1947, 1948, 1963; Levi, 1956). Небольшое исследование Окада (Okada,
Рис. 5. Развитие Sycon raphanus
Л-г-.плавающая'личинка; Б— прикрепившаяся личинка (из Kaestner); / — покровные
клетки; 2 — воротничковые клетки
1928) посвящено развитию стеклянных губок, которые изучены крайне
слабо. По'данным Варбуртона (Warburton, 1961), у губок в образовании
зародыша, помимо бластомеров яйца, участвуют амебоидные-клетки ма-
тери, т, е. образуется химера.
Дробление яиц полное, неравномерное или равномерное в зависимо-
сти от количества желтка в яйце, радиального типа. У многих известко-
вых губок оно правильное, причем первые три:плоскости дробления- —
меридиональные, четвертая — экваториальная. Восемь бластомеров
верхнего полушария — мелкие (микромеры), они делятся быстрее вось-
ми гораздо более крупных, зернистых бластомеров (макромеров) ниж-
него полушария. У части известковых и у большинства остальных губок,
в том числе таких, как Halisarca и Oscarella, которые считаются самыми
примитивными губками (Levi, 1956, 1957), дробление вскоре становится
неправильным.
В результате дробления образуется целобластула или стомабластула,
превращающаяся в амфибластулу (многие известковые губки): или па-
ренхимулу (часть известковых и большинство остальных губок — крем-
невые, роговые); личинки покидают тело матери и с помощью жгутиков
плавают (рис. 5).
Личинка прикрепившись передним концом к субстрату, сильно упло-
щается, превращаясь в куколку, тело которой по мере размножения ее
клеток приподнимается над. субстратом. На верхнем конце прорывается
выводное отверстие — оскулум, пинакоциты одевают поверхность тела,
хоаноциты выстилают внутреннюю полость, возникает у известковых гу-
бок стадия аскона, которая у большинства, переходит в стадии сикона и
лейкона. У громадного большинства губок развитие начинается со стадии
рагона, установленной Солласом (Sollas, 1888), которая.переходит непо-
средственно в стадию лейкона.
22
/ Забывая о том, что губки в сформированном состоянии'не имеют тка-
ней и органов, лишены стабильной тканевой дифференцировки, обладают
лишь поклеточной дифференцировкой, многие авторы (например, Бекле-
мишев, 1964; Tuzet et cet., 1961 и др.) ошибочно признают в онтогенезе
губок эктодерму, энтодерму и мезенхиму.
Следуя взгляду Мааса (Maas, 1898) и Деляжа (Delage, 1898), ряд
авторов признает у губок извращение зародышевых листков и считает
их вывороченными наизнанку многоклеточными, что раньше признавал и
я (Федотов, 1938). Дюбоск и Тюзе показали, что в онтогенезе губок имеет
место явление инкурвации, подобное тому, какое наблюдается в онтоге-
незе Volvox — представителя не животного, а растительного мира
(Duboscq et Tuzet, 1937). Правильно подчеркнув наличие примитивных
черт в онтогенезе губок и «протозойные» черты в их организации (в отли-
чие от «метазойных» в организации кишечнополостных)-А. А. Захваткин
(1949) все же говорит о зародышевых листках у губок, экто-, мезо-и
энтобласте губок и кишечнополостных, что стоит в противоречии с низ-
ким поклеточным уровнем эволюции губок.
В действительности же в онтогенезе губок (на стадии бластулы)
имеются лишь временно эпителиально расположенные ресничные клетки
и некоторое время (на стадии личинки) — личиночный орган (Remane,
1954, 1959). Далее происходит поклеточная дифференцировка, развитие
из части погрузившихся внутрь ресничных клеток воротничковых жгути-
ковых клеток, за счет зернистых клеток, не имеющих или утративших
реснички —1 пинакоцитов, пороцитов, пигментных клеток, миоцитов, желе-
зистых клеток и, видимо, первичных склеробластов. Из части бластоме-
ров дробления, не подвергшихся дифференцировке на ресничные поверх-
ностные клетки, образуются амебоциты, точнее, археоциты, за счет кото-
рых возникают мезоглея и многочисленные категории ее клеток.
В онтогенезе губок нет образования зародышевых листков, нет ни
эктодермы, ни энтодермы, следовательно, нет и среднего зародышевого
листка. Идет лишь поклеточная дифференцировка клеток разных кате-
горий, слагающих организм губки и сохраняющих, как известно, у сфор-
мировавшихся губок способность вновь дедифференцироваться.
Имея разные взгляды на онтогенез губок, все:исследователи едино-
душно принимают их как Parazoa, стоящими в самом низу родословного
древа многоклеточных. Но ведь исторически развиваются не только сами
животные, план строения, анатомия, гистология, физиология, но и. их
онтогенез, так как в ходе эволюции любой группы происходит эволюция
и ее онтогенеза. Если, как увидим ниже, онтогенез низко организованнр-
го типа кишечнополостных в ряде характерных черт не достиг уровня, ти-
пичного для остальных групп животных, то это тем более относится к
онтогенезу губок, у которых зародышевые листки еще не возникли.
Вместе с тем губки в основном не являются колониями, как подчер-
кивает Тюзе (Tuzet, 1963). Им свойственна видовая специфичность, о чем
свидетельствуют как старые опыты, например, Вильсона (Wilson, 1907,
1911), так и современные эксперименты с протиранием тела разных ви-
дов губок через мюллеровский газ (стр. 21).
Картина исторического развития губок недостаточно ясна, хотя в раз-
ные эпохи ископаемые'губки были массовыми формами; так известны из
нижнекаменноугольных отложений Великобритании слои толщиной
105 м,. состоящие из изолированных спикул губок. Стеклянные губки
в мальме (ГДР, ФРГ, Франция) были рифообразователями.
Эволюция губок началась в протерозое; из докембрия известны де-
моспонгии, из раннего кембрия — гексактинеллидеи, некоторые палеон-
тологи (без достаточных оснований) допускают в раннем кембрии на-
личие даже известковых губок; бесспорные известковые губки найдены
однако лишь в раннем девоне (может быть, в силуре). Исходя из того,
что кремневая функция живого вещества наиболее древняя и. среди
23
многоклеточных ' она останавливается на губках (Виноградов, 1933,
1944, 1953), кажется более вероятным, что известковые губки возникли
значительно позже кремневых. К сожалению, строение скелета известко-
вых губок, состоящего из игл (у большинства не спаянных друг с дру-
гом), неблагоприятствовало сохранению их в целом виде. Наиболее ча-
стыми ископаемыми были демоспонгии (особенно литистиды) и гексак-
тинеллиды.
Как сказано выше, демоспонгии, в основном кремневые губки, по-
видимому известны из докембрия. В кембрии они еще бедно представ-
лены, но уже в ордовике и силуре они достигли известного расцвета,
претерпев в девоне упадок. В перми они вновь увеличиваются в численнб-
Рис. 6. Схема примерного количественного распределения
ископаемых губок (из A. Milller)
4—Demospongia (Litisthida—271 род); Б—Hexactinellida (279 родов);
В—Calcispongia (94 рода)
сти, но в триасе претерпевают резкий упадок, после которого расцветают
и достигают максимума развития в верхнем меле (А. Н. Miiller, 1961).
Кремневые губки — демоспонгии и из них, главным образом, литистиды,
с их прочно спаянным кремневым скелетом известны в ископаемом со-
стоянии. Кремнероговые губки, от которых произошли роговые, изве-
стны с юры и особенно из мела.
Гексактинеллиды достигают некоторого расцвета в ордовике, еще
большего в девоне и карбоне, далее переживают кризис, становясь ред-
кими в перми и триасе (рис. 6), а затем снова наступает их расцвет, до-
стигающий максимума в верхнем меле. С третичного времени число видов
этих губок уменьшается, резко сокращаясь (рис. 6) в современной фау-
не (А. Н. Miiller, 1958, 1961; Laubenfels, 1955). Следует однако отметить,
что, согласно данным Рида (Reid, 1963), Heteractinida не стеклянные
губки, a Demospongia и они, видимо, древнее Hexactinellida.
Историческое развитие известковых губок известно недостаточно. Не-
сомненно известковые губки (из фаретронных) найдены в нижнем дево-
не, может быть даже в нижнем силуре (Соколов, 1962). Однако некото-
рые палеонтологи, например, Лаубенфельс (Laubenfels, 1955) и Мюллер
(А. Н. Miiller, 1958, 1961), на основании данных конца XIX столетия, до-
пускают существование известковых губок в кембрии, что представляет-
ся однако мало достоверным. Самым важным отрядом, к которому отно-
сятся более половины (67) родов этого класса, по Мюллеру, являются
фаретронные губки, историческое развитие которых, благодаря хорошей
сохранности в ископаемом состоянии (прочный, спаянный известковым
цементом скелет), хорошо известно. Эти губки малочисленные в палео-
зое, испытывают расцвет в триасе, юре и особенно в меле и вымирают
в эоцене, за исключением немногих существующих ныне родов — Pet-
rostoma, Murrayana, Minchinella. Большинство ныне существующих от-
24
рядов со свободными, неспаянными иглами известны из поздней юры
(мальм): Homocoela Полежаева (Polejaeff, 1884), Asconosa или Solenida
Лаубенфель-еа и Heterocoela- Полежаева (Syconosa или Labetida, Лау-
бенфельса). Исторические данные о них бедны вследствие плохой их
сохранности.
Трудности восстановления геологической истории губок вообще вы-
зываются еще и тем, что у ископаемых губок происходит переотложе-
ние кремневых элементов скелета известковыми и обратно (Яковлев,
1937). Кроме того, кремневый скелет участвует в образовании таких ми-
нералов, как роговик, халцедон, кремень (Резвой, 1934).
Палеонтологическая летопись свидетельствует, что наиболее мощны-
ми, долговечными и разнообразными являются демоспонгии; стеклянные-
губки (Hyalospongea) им в этом уступают.. Демоспонгии и сейчас сохра-
няют вместе с чисто -роговыми губками господствующее положение. Бо-
лее поздняя, в прошлом мощная группа известковых фаретронных губок
резко уступала демоспонгиям, а в современной фауне известковых губок
она уступила место отрядам, у которых известковые иглы свободно ле-
жат в теле. В настоящее время максимум развития достигает отряд He-
terocoela, или Syconosa.
Встречаются ископаемые губки в очень различных отложениях — в
известняках, песчаниках, писчем меле, глубоководных глинах и т. д., что
говорит о разнообразных условиях жизни губок, причем глубина их оби-
тания изменялась от мелководья в палеозое (кембрии) до ультраабис-
сали в настоящее время (Соколов, 1962). В итоге надо считать, что в:
основе типа губок стоят кремневые—Demospongia, к ним примыкают
Hyalospongea, связанные друг с другом через Heteractinida. Непосред-
ственной связи известковых губок с двумя предыдущими классами отме-
тить пока нельзя, исторически они появились позже последних и стоят
не в начале системы губок, а в конце ее.
Родственные связи между классами губок остаются др сих пор далеко
не уточненными, хотя они составляют единый тип, т. е. имеют общего
предка, отошедшего от родоначального ствола многоклеточных до по-
явления кишечнополостных, настоящих многоклеточных, с которыми у
них нет близости и в серологическом отношении (Tuzet et cet., 1961)..
По совокупности всех данных общей организации, гистологии, онтогенеза
и биологии губки действительно являются подцарством Parazoa; они бо-
лее примитивны и отличны от всех остальных типов, составляющих под-
царство Eumetazoa и, вероятно, произошли от хоанофлагеллятных про-
стейших.
Кишечнополостные (Coelenterata)
. Кишечнополостные (или жгущиеся)—радиально-симметричные с
элементами двулучевой и двусторонней симметрии, прикрепленные
(полипы) и свободновзвешенные (медузы) водные организмы. Тело об-
разовано двумя слоями клеток — эпидермисом и гастродермисом со
стрекательными капсулами в наружном, реже и во внутреннем слое, и
мезоглеей, являющейся продуктом выделения клеточных слоев. Скелет —
наружный в виде хитиноидного перисарка, известковый или роговой.
Рот, обычно окруженный венчиком щупалец, ведет в мешковидную или
поделенную септами гастральную или гастроваскулярную полость, без
анального отверстия. Нервная система— диффузная сеть нервных кле-
ток и волокон в стенках тела и гастральной полости, рецепторы — в виде
глазков и статоцистов лишь у медуз. Мускулатура в .основном образо-
вана эпителиально-мышечными клетками. Полости тела, органов дыха-
ния, кровообращения и выделения нет. Дифференцированных половых
органов нет, половые клетки развиваются в связи с эктодермой и энто-
дермой. Размножение — бесполое (почкованием или делением) и половое
(обычно со сменой поколений). Типы дробления яиц и образования за-
родышевых листков— эктодермы и энтодермы—щеустановивщиеся,
характерной личинкой является планула. Кишечнополостные — свобод-
ноживущие, как исключение паразитические, морские, редко пресндвбд-
ные, широко распространенные в Мировом океане животные, богато
представленные и ископаемыми формами, начиная с кембрия.
Гаймен (Hyman, 1940—1959) дала почти исчерпывающий обзор ли-
тературы по кишечнополостным; известны важнейшие исследования
зарубежных авторов последних 20—30 лет и рцд имен русских и совет-
ских зоологов, изучавших кишечнополостных; это А. В. Белоусов (1960—
1963), И. А. Ветохин (1926а, б), А. П. Владимирский (1913, 1915),
к. П. Иванова (1949), Н. А. Иванцов (Iwantzow, 1928), В. М. Исаев
(1925), И. И. Канаев (1949, 1952—1954, Kanajew, 1930), А. О. Ковалев-
ский (1873), Г. Корсакова (1949), И. И. Мечников (Metschnikoff, 1874,
1882, 1885, 1886, 1950), Д. В. Наумов (1953, 1957, 1960, 1961), А. А. Ти-
хомиров (1887), В. Н. Ульянин (1876), Я- П. Щелкановцев (1905).
Существенные различия в строении и онтогенезе и отсутствие пере-
ходных форм заставляют считать кишечнополостных Coelenterata, или
Cnidaria и гребневиков Ctenophora самостоятельными типами (см. ниже).
Известно, что классификации кишечнополостных, предлагаемые зооло-
гами и палеонтологами, в общем сходны, но отличаются частностями.
Я считаю возможным придерживаться следующей классификации. Тип
Coelenterata содержит: класс Hydrozoa с подклассами Hydroidea (отря-
ды Thecaphora, Athecata, Trachylina, Hydrida) и Siphonophora (отряды
Disconantha, Siphonantha), класс Scyphozoa с подклассом Scyphomedusae
(отряды Stauromedusae, Cubomedusae, Coronotae, Semeastomeae, Rhizo-
stomeae и Conulatae f ), класс Hydroconozoa f (установленный недавно
Кордэ, 1963), класс Anthozoa с подклассами Tabulata f , Octocorallia (от-
ряды Alcyonacea, Gorgonacea, Pennatalucea и др.), Rugosa f и Hexaco-
26
Tallia (отряды Ceriantharia, Antipatharia, Scleractinia, Zoantharia, Actinia-
ria). Я не ввожу в тип кишечнополостных классы Protomedusae и Dipleu-
.rozoa, так к-ак природа ’этих докембрийских и нижнекембрийских
ископаемых крайне неясна.
Кишечнополостные широко распространены в Мировом океане, встре-
чаются от прибрежной зоны до сверхабиссали, редкое солоноватоводной
и пресноводной среде; это или прикрепленные бентосные или пелагиче-
ские плавающие, почти исключительно свободноживущие животные. Из
"9 000—9 500 современных видов кишечнополостных известно всего 25 па-
разитических видов, притом из них 20 относится к наркомедузам (Cuni-
па, Cunocantha и др.) и являются паразитами гастроваскулярной полости
других медуз. Из гидрополипов Polypodium hydriforme является эндопа-
разитом яиц стерляди, a Hydrichthus hydriforme — эктопаразитом рыбы
Cyclothone signata (Догель, 1962).
Классические примеры сожительства (эпиойкии) известны среди раз-
личных актиний и раков-отшельников. Обитание некоторых гидрозоев на
раковинах и поверхности различных животных в ряде случаев связано
со случайным оседанием их личинок, в то время как жизнь ряда поли-
лет, ракообразных и даже рыб в субумбреллярной полости корнеротых,
видимо, можно считать симбиозом. Хотя оседание личинок кишечнопо-
лостных (например, гидрозоев) на субстрат может иметь случайный ха-
рактер, однако участие их в образовании разных-биоценозов определяет-
ся исторически сложившимися причинами. Среди разных групп морских1
кишечнополостных распространен симбиоз с желтыми зооксантеллами,
а у гидр — с зелеными зооксантеллами, которые, по-вйдимому, являются-
для хозяев источниками образования витаминов и гормоноподобных ве-
ществ. При сравнительно большом количестве современных видов у
кишечнополостных, естественно, наблюдается большое разнообразие в
размерах тела. Так, Protohydra измеряется 1 — 1,5 мм, тдуоид Branchio-
cerianthus imperator — 2,23 м, актиния Stoichactis -имеет-1,5 м в
диаметре. Многие гидромедузы имеют диск меньше 1 см в диаметре,
тогда как у сцифомедузы Cyanea polaris диаметр диска 2 м,-а длина щу-
палец более 30 м. Сифонофора Apolemia свыше 3 м в длину, колониаль-
ный коралловый полип Cirripathes (Antipathatia) до 6 м в высоту и т. д.
Окраска многих кишечнополостных (некоторые гидроидные полипы,
сифонофоры, восьмилучевые коралловые полипы — Alcyonacea, Gorgo-
nacea, Pennatulacea, Шестилучевые коралловые полипы — Actiniaria и
ряд других, часть сцифомедуз — Stauromedusae, Coronatae) представле-
на всеми цветами спектра: встречаются черные, чернофиолетовые, фио-
летовые, синие, голубые, зеленые, оранжевые, красные и пурпурные
формы. Некоторые формы обладают хроматофорами, обеспечивающими
способность изменять окраску. Многие гидромедузы и сцифомедузы
прозрачны. Некоторые кишечнополостные (гидрополипы,, сифонофоры,
морские перья) люминисцируют. У ряда форм окраска связана с. нали-
чием Пигмента в склеритах.
Радиальная симметрия кишечнополостных, обычно объединяемых
.вместе с типами губок, археоциат и ктенофор в раздел Radiata,-— неуста-
повившаяся четырехлучевая с элементами двулучевой и двусторонней
-симметрии; ее развитие обусловлено у полипов прикрепленным, сидячим
образом жизни, а у медуз — пелагическим.
Неустановившаяся, четырехлучевая симметрия у большинства гидро-
идных полипов с их простой неразделенной на отделы гастральной по-
лостью определяется расположением и числом щупалец, собранных .в
виде одного венчика (например, Obelia из Thecaphora)wiH в виде двух
й трёх венчиков (Athecata), причем число щупалец в венчиках, разбро-
санных по гидранту в беспорядке (например, Clava), может быть очень
большим и нефиксированным. Реже у ряда форм четырехлучевая сим-
метрия заменяется трех- и пятилучевой. Число щупалец может умень-
27
шаться до двух (например, Proboscidactyla) и даже до одного (Mono-
brachium), что придает им двустороннюю симметрию, которая наблю-
дается и у других гидроидных полипов, как одиночных (Branchiocerian-
thus), так и у колониальных (например Sertularidae, Plumularidae). Дву-
сторонняя симметрия возникает в связи с косым положением гидранта
на ножке или на стволе колонии (В. Н. Беклемишев, 1952, 1964). У гид-
•рид (Protohydra) с утратой щупалец образуется неопределенно много-
лучевая симметрия. Также лабильна радиальная симметрия у гидроме-
дуз, причем у них происходит как полимеризация, так и олигомеризация
щупалец и радиальных каналов гастроваскулярной системы. У антоме-
дузы Turritopsis lata число краевых щупалец возрастает до 60—130, у
лептомедузы Ptychogena plactea — до 300—500, у трахимедузы Crossotc^
brunnea их может быть 640 (Kramp, 1961). Чаще происходит незначи-
тельное увеличение числа радиальных каналов гастроваскулярной систе-
мы (6, 8, 11), но в отдельных случаях число последних может возрастать
до 60—80, и даже до 160 (трахимедуза — Aequorea aequorea, Ае. macro-
dactyla), причем у некоторых форм гидроидных медуз образуются слепые
каналы, отходящие от кольцевого (Kramp, 1961). У гидромедуз Spiroco-
don и Polyorchis радиальные каналы имеют разветвленные боковые вет-
ви (Uchida, 1963). Другое направление в изменении гастроваскулярной
системы гидроидных медуз заключается в олигомеризации радиальной
симметрйи; возникают формы двулучевые с двумя щупальцами и даже,
билатеральные с одним щупальцем, с одной плоскостью симметрии, с
уменьшенным числом радиальных каналов (например, Corymorphq nu-
tans и др.).
Значительно более глубокие изменения, чем у Hydrozoa] радиальная
симметрия претерпевает у коралловых полипов, как ископаемых, так и
современных. У многих из них внутренняя организация имеет черты
двулучевой и двусторонней симметрии. Это наблюдается даже у тех:
(например, Actiniaria), которые внешне остаются радиальносимметрич-
ными. У одиночных вымерших кораллов Rugosa, у ряда современных
одиночных шестилучевых кораллов, например, Cerlantharia, у колони-
альных Antipatharia происходит изгибание тела и приобретение им дву-
лучевой симметрии, вызванной условиями жизни, например, боковым
прикреплением полипа к субстрату (Яковлев, 1928) или расположением',
полипов на ветвях колонии (Antipatharia).
У современных Anthozoa — восьмилучевых, шестилучевых и с сим-
метрией, кратной шести, а также, вероятно, и у вымерших Rugosa —
четырехлучевых и с симметрией, кратной четырем, существует двулуче-
вая и двусторонняя симметрия. У них имеются щелевидный рот, глотка,
сдавленная с боков в плоскости симметрии, с одним (для дыхания) или
с двумя (для питания и дыхания) сифоноглифами, мышечные валики на
мезентериях с различным расположением у восьми- и шестилучевых
полипов и дифференцировка мезентериев, часть которых становится на-
правленными (спинными). Наиболее совершенная, фиксированная че-
тырехлучевая симметрия, к которой добавляется неполная восьмилуче-
вая, установилась у сцифомедуз. Она захватила внешнюю форму и вну-
треннее строение, распространяясь на краевые лопасти, ротовые щупаль-
ца, гонады, радиальные каналы гастроваскулярной системы и т. д.
В онтогензе, как будет сказано ниже, становление лучистой снимет*
рии проходит стадии двулучевой и двусторонней симметрии. Большин-
ство авторов, как и Беклемишев (1944, 1952, 1964), считают лучистую
симметрию у кишечнополостных первичной, а начиная с Хаджи (Hadzi,
1944), Д. М. Федотов и другие — вторичной (см. ниже).
Кишечнополостные, как выше отмечено, образованы двумя зароды-
шевыми листками: эктодермой и энтодермой и располагающейся между
ними их производной — мезоглеей (рис. 7) . О морфологическом и функ-
циональном значении последней будет сказано ниже.
28
Эпидермис (эктодерма) состоит из однослойного ряда плоских, приз-
матических, узких или крайне длинных'цилиндрических эпителиальных
клеток со жгутиками,, а иногда эпидермис принимает синцитиальный ха-
рактер. По-видимому у-многйх кишечнополостных жгутики эктодермаль-
ных клеток в ходе онтогенеза утрачиваются. Эпителий эктодермальной
глотки восьмилучевых полипов (например, Alcyonaria) имеет реснички,
а у шестилучевых полипов (например, у актиний) — жгутики (Полян-
ский, 1958). Часть эктодермальных клеток несет по жгутику и у взрос-
лых сцифомедуз, например, Aurelia aurita (A. J. Southward, 1955).
Рис. 7. Схема продольного разреза
•стенки тела Hydra (из Borradaile)
J —эктодерма; 2~ энтодерма; 3 — мезоглея
Рис. 8. Схема строения стрекательной клетки
(по Mergner)
Л — в покое; Б — с вывернутым стрекательным
аппаратом
Наиболее типичными являются эпителиально-мышечные клетки, со-
-ставляющие главную толщу покровов, их мышечные отростки направ-
лены преимущественно вдоль главной оси тела. Между эпителиально-
мышечными клетками разбросаны железистые (слизистые и белковые)
и чувствительные клетки, причем последние связаны с нижележащим
эктодермальным нервным сплетением; у основания эпителиально-мышеч-
ных клеток находятся стрекательные клетки (рис. 8).
Стенка гастральной (гастроваскулярной) полости образована энто-
.дермальными эпителиально-мышечными клетками с мышечными отро-
стками, направленными поперек главной оси тела. Из них часть клеток —
пищеварительные и абсорбирующие пищу, часть — железистые. Среди
эпителиальных клеток находятся чувствительные клетки, связанные с
ассоциативными клетками и эктодермальным нервным сплетением. Стре-
кательные клетки находятся в гастральных нитях сцифомедуз, мезенте-
риальных нитях и аконциях коралловых полипов.
Стрекательные клетки, характернейшая особенность кишечнополо-
стных, расположены батареями и возникают из интерстициальных, ле-
жащих группами у основания эпителиально-мышечных клеток, вблизи
базальной мембраны. Стрекательные клетки разнообразны по строению
и функции. Стрекательных капсул различают до 16—17 катего-
рий -(Hyman, 1940—1959). Четыре категории их установлено у пресно-
..водных гидр (Полянский, 1958; Канаев, 1952). Стрекательные клётки
29
Рис. 9. Схема строения актинии
(из Borradaile)
/ — рот; 2—ротовой сфинктор; 3 — щу-
пальце', 4 — мышца-ретрактор; 5 — акон-
ция; 6—мезентериальные нити; 7—гонады
расположены, главным образом, на щупальцах, трубчатые нити одних,
пробивают покровы добычи, других — обвивают, третьих — прилипают
к телу добычи, при этом вещество стрекательных капсул, или книд, ток-
сичное для животных, парализует добычу или служит для защиты от
врага. Ожоги крупных сифонофор, актиний и сцифомедуз и даже неко-
торых мелких гидромедуз (например,.
Gonionemus — 4—5 см в диаметре) не
только крайне болезненны для челове-
ка, но даже и опасны для жизни. Хи-
мическая природа токсинов не выясне-
на. Клетка, выбросившая нить, поги-
бает. Выбрасывание нити — рефлек-'
торный акт, который вызывается, преж-
де всего, естественными раздражите-
лями, например, животными, являющи-
мися пищевыми объектами кишечно-
полостных. В жидкости капсул обнару-
жены ацетилхолин, гистамин, адрена-
лин, серотонин, т. е. вещества, являю-
щиеся передатчиками нервно-мышеч-
ного возбуждения и торможения
(Коштоянц, 1957; Welsh, 1957; см. Кош-
тоянц, 1963). Четкое представление,
однако, не перекрывающее определет
ний X. С. Коштоянца о строении и
функции стрекательных клеток, дано
Мергнером (Mergner, 1964).
По устаревшим данным интерстициальные клетки возникают доволь-
но поздно, отделяясь от клеток эктодермы. По позднейшим данным
Мергнера (Mergner, 1957), они образуются в результате деления одного
или нескольких бластомеров еще до сформирования зародышевых ли-
стков, Морфогенетическое значение интерстициальных клеток толкуется,
однако, по-разному. По мнению некоторых авторов, из этих клеток воз-
никают не только книдобласты, но и половые, нервные и эпителиально-
мускульные клетки (Brien, 1951), а рост, регенерация и размножение-
гидры происходит за счет интерстициальных клеток; причем все клетки
тела гидры, возникающие из последних, обновляются за 45 дней. В про-
тиворечии с этим, по позднейшим экспериментам (Haynes a. Burnett,
1963), у Hydra viridis интерстициальные клетки образуются дедиффе-
ренцировкой клеток энтодермы, которые способны дифференцироваться
в культуре в любую клетку этой гидры. Однако согласованности в дан-
ных авторов о гистогенезе при регенерации кишечнополостных пока нет.
Так, по Цвилингу (Zwilling, 1963), у полипов Cordylophora lacustrls и
у сцифистомы Aurelia aurita, наоборот, энтодерма возникает путем де-
дифференцировки эктодермы.
Гидры, гидромедузы, сифонофоры, сцифомедузы и актинии (за не-
многими исключениями) не имеют минерального скелета (рис. 9). Меду-
зы и актинии на 99% состоят из воды (Виноградов, 1953).
Скелет у гидроидных и коралловых полипов — эктодермальный. Хи-
тиноидная кутикула (тека или перисарк) покрывает всю колонию гид-
роидов, образуя чашечку, гидротеку вокруг головки гидранта Thecaphora,
отсутствующую у Athecata. Такой же характер носит и тека сифонофор.
У гидр тека отсутствует. У гидроидных полипов Milleporina и Stylaste-
rina развивается массивный известковый скелет. У альционарий склеро-
бласты уходят из эктодермы в мезоглею, где образуют скелет из араго-
нита с примесью до 17% магнезии (MgCOg), состоящий из отдельных
склеритов; у других Octocorallia (например, Gorgonaria), имеющих ро-
говой скелет с примесью СаСОз, эктодермальный эпителий подошвы,
30
впячиваясь внутрь колонии, образует плотный скелет, проходящий по-
оси ветвящейся колонии. Известковый арагонитовый скелет с примесью
магнезии у шестилучевых .колониальных и немногих одиночных корал-
ловых полипов, более сложный. Клетками эктодермы подошвы сначала
образуется пластинка, а'затем, между мезентериями или мягкими септа-
ми возникают склеросепты; кольцевая складка подошвы дает также теку..
Таким образом, скелет Anthozoa является производным эктодермы, хотя
по положению он внутренний.
В эктодерме или в энтодерме взрослых кишеч-
нополостных лежат половые клетки,'которые ско-
рее всего возникают из бластомеров дробяще-
гося яйца до формирования зародышевых лист-
ков. Интересные эксперименты Вернера (Werner,
1963) по развитию у гидроидных половых клеток:
из соматических, к сожалению, не сопровожда-
ются цитохимическим анализом.
У гидрид, псаммофильных актинулид и в го-
ловках гидрантов колониальных гидрополипов,,
у которых мешковидное тело полипа до краев рта
одето эктодермой, рот переходит непосредст-
венно в простую гастральную полость, не поде-
ленную на отделы, причем у Protohydra (AbonyiA
1929; Westblad, 1935) нет и щупалец. У коп^-
Рис. 11. Схема строения Velella (из Borradaile)
1 — пневматофор; 2 — ценосарк; 3 — дактилозоид; 4 — гонозоидг
5 — гастрозоид; 6 — медузоид
Рис. 10. Схема строения си-
фонофоры (из Kfihn)
/ — пневматофор; 2 — плаватель-
ные колокола; 3— арканчик;
4 — гастрозоид; 5 — медузоид
ниальных гидрополипов, как и у ряда сифонофор (рис. 10), гастральная
полость продолжается в ствол (ценосарк) и его ветви. Но у крупных оди-
ночных форм, например, Corymorpha, Branchiocerianthus, гастральная
полость головки гидранта продолжается в виде нескольких (до 12) кана-
лов. Некоторая дифференцировка гастральной полости состоит в том,.'
что заглоченная пища не поступает и не переваривается в щупальцах
Hydrozoa, даже у тех групп, у которых щупальца полы и не имеют оси
из столбика эктодермальных клеток.
Значительную дифференцировку претерпевает гастральная полость
гастрозоидов ряда сифонофор; у одних из них энтодерма образует про-
дольные валики, увеличивающие ее поверхность (Apolemia), у других
(Velella) (рис. 11) гастральная полость центрального гастрозоида
31
образует направляющиеся кверху переплетающиеся каналы (так назы-
ваемая печень), в которых циркулирует жидкая пища (Догель, 1938).
У Anthozoa, гидромедуз и сцифомедуз происходит разделение га-
стральной. полости на центральную и периферическую. У гидромедуз,
животных чаще мелких размеров, от небольшой центральной части, в
которую открывается ротовой хоботок, отходит, как правило, четыре.ра-
диальных канала, возникающие в онтогенезе как выросты гастральной
дюлости и впадающие в кольцевой канал.
Рис. 12. Продольная мускулатура септ (по Рах)
А — Stichopathes ceylonensis; Б — Antipathes longibrachiata
У сцифомедуз происходит значительная эволюция гастральной поло-
•сти. Четырехгранный ротовой хоботок, выстланный энтодермальным*
клетками, по углам рта вытянут в четыре ротовые лопасти и ведет й
желудок, в который вдаются четыре септы с продольными мышечными
.лентами эктодермального происхождения. Септы на свободных краях
несут гастральные филаменты — нити с железистыми клетками и нема-
тоцистами. От желудка отходит система радиальных каналов 1, 2, и
.3-го порядков — перрадии, интеррадии и адрадии, соединяющиеся коль-
цевым каналом и закладывающиеся в виде простых каналов; в ходе он-
тогенеза перрадиальные и интеррадиальные каналы разветвляются.
•Особенно сложную систему анастомоз они образуют у медуз отряда
Rhizostomeae.
У Anthozoa происходит не только дифференцировка эктодермальной
гастральной полости на центральный и периферический отделы (Бекле-
мишев, 1952, 1964), но впервые среди многоклеточных животных обра-
зуется эктодермальная глотка, т. е. дифференцируется передняя и сред-
няя кишка (Догель, 1938). Глотка сплющена с боков в плоскости сим-
метрии, снаружи одета энтодермой; она имеет мерцательный желобок
(сифоноглиф): или один, располагающийся на брюшной стороне (вось-
милучевые коралловые полипы), или два (шестилучевые коралловые
полипы). Восемь мезентериев (или саркосепт) расположены на равных
расстояниях друг от друга, мускульные валики, образованные верти-
кальными мышцами, расположены симметрично от спинной к брюшной
стороне полипа (Octocorallia). У Hexacorallia мезентерии расположены
попарно, число их в типичном случае —шесть пар или кратное щести
и может превышать 100, они полные или неполные (не прирастающие к
глотке); мускульные валики в двух парах мезентериев направительных
камер направлены друг от друга (рис. 12), у остальных четырех пар —
друг к другу. Полные мезентерии продырявлены в верхней части цирку-
моральным круговым и краевым отверстиями (Pantin, 1960).
Мезентерии подразделяют гастральную полость на центральную и
периферическую части. Последняя на уровне глотки состоит из раДиаль-
32
ных камер, выстланных .простым эктодермальным эпителием. Ниже глот-
ки мезентерии свободно вдаются в центральную часть гастральной по-
лости; их свободные края несут железистые утолщения — гастральные
нити, богатые пищеварительными и стрекательными клетками, а нижние
концы могут продолжаться в нитевидные аконции, которые выбрасыва-
ются наружу. При пищеварении свободные края мезентериев сближают-
ся и отграничивают центральную часть кишечника от периферической;
физиологически и временно анатомически возникает кишечник и вокруг
него полости тела (Беклемишев, 1952, 1964).
Добычей книдарий, которую они поражают стрекательными клетка-
ми щупалец и ротового хоботка, служат личинки и яйца различных жи-
вотных, мелкие ракообразные, двустворчатые и брюхоногие моллюски,
нематоды, кольчатые черви, рыбы. В общем, размеры добычи соразмер-
ны с величиной хищника. Эфиры и медузы аурелии питаются мелкой
добычей, поступающей в рот благодаря токам, создаваемым ресничками
на поверхности тела, однако пресноводные гидры, питающиеся также
мелкими животными, могут заглатывать, например, нематод, мало усту-
пающих по величине самой гидре (Westblad, 1947).
Пищеварение как у низших, так и у высших кишечнополостных ,в на-
чальной фазе — полостное, внеклеточное; оно ^происходит под действием
протеазы типа пепсина, эрепсина, трипсина и других ферментов в щелоч-
ном растворе, выделяемом пищеварительными эктодермальными клетка-
ми гастральной полости, у сцифомедуз септальных, у Anthozoa — мезен-
териальных нитей. Жиры при этом эмульгируются, но углеводы остаются
неизмененными. Лишь у сифонофор дело ограничивается только внекле-
точным пищеварением, у остальных кишечнополостных после предвари-
тельного полостного пищеварения измельченные частицы пищи захваты-
ваются эктодермальными клетками и перевариваются внутриклеточно в
пищеварительных вакуолях. Часть фагоцитов переходит в мезоглею,
распространяясь в последней. Непереваренные остатки пищи и продукты
обмена веществ, экскреты, выбрасываются наружу через рот. Движение
пищи при переваривании в гастральной полости в основном обеспечи-
вается движением жгутиков эктодермальных клеток.
Правильная циркуляция жидкости с частицами пищи, размельченны-
ми после предварительного внеклеточного переваривания в желудке, про-
исходит у сцифомедуз (Ветохин, 1926а; Southward, 1955; Borradaile et
cet., 1959). Из карманов желудка жидкость движением жгутиков гонится
по неразветвленным адрадиальным; каналам в кольцевой канал, откуда
по разветвленным перрадиальным и йнтеррадиальным каналам она по-
ступает в выводящие желобки ротового стебелька и выводится с непере-
варенными частицами и экскретом наружу. Продукты, обмена веществ
поступают в кольцевой канал через отверстия в адрадиальных каналах
(Thiel, 1959). Именно у медуз оправдывается название гастральной поло-
сти как гастроваскулярной системы, несущей функции пищеварения
и распределения.
В движении пищи у гидрополипов и Anthozoa принимают участие,
помимо жгутиков клеток гастральной полости, перистальтические сокра-
щения стенок тела и эктодермальной глотки (у Anthozoa).
Мускулатура при наименее дифференцированном состоянии образо-
вана продольными мышечными отростками эпителиально-мышечных
клеток эктодермы и кольцевыми — энтодермы, как в теле полипа, так и
в его щупальцах, когда они остаются полыми. При наличии такой муску-
латуры и отсутствии скелета возможно сокращение тела гидроида во всех
направлениях и максимальное его уменьшение при удалении воды из
гастральной полости. Развитие теки обусловливает обеднение мускула-
туры эктодермальных клеток и утрату ее эктодермальными клетками вет-
вей колонии (при сохранении ее гидрантами), что обеспечивает функции
3 Д. М. Федотов
33
ловли, удержания, глотания добычи и защиты (колониальные Athecata-
и Thecaphora) . По-видимому, это еще в большей степени выражено у из-
вестковых Milleporina и Stylasterina.
Эволюция мускулатуры в других группах кишечнополостных, особен-
но у способных к активному передвижению, выражается в меньшем или
большем обособлении (возможно путем деления клетки сократимых
отростков эпителиально-мышечных клеток) в самостоятельные мышеч-
ные волокна, что придает им большую подвижность, в развитии попереч-
нополосатой структуры, повышающей скорость сокращения, и в совер-
шенствовании мышечной системы — двигательного, по терминологии-
В. Н. Беклемишева (1952), вернее, нервно-мышечного аппарата. У си-
фонофор, например, обладающих большой подвижностью, мышцы пол-
ностью обособляются и приобретают поперечнополосатое строение
(Schaeppi, 1898; Iwantzow, 1928); сильное развитие мускулатуры есте-
ственно происходит в стенках пневматофоров и плавательных колоколов,
служащих для движения колоний, а также гастрозоидов и столона.
Исходной общей особенностью мускулатуры гидроидных и коралло-
вых полипов является продольное расположение мышечных волокон эпи-
телиально-мышечных эктодермальных клеток и кольцевое — эктодер-
мальных. Однако у коралловых полипов и сцифомедуз мышечные отро-
стки в эктодерме становятся уже самостоятельными и образуют слой
продольных мышц; эктодермальная мускулатура у них развита слабее
эктодермальной; последняя часто оставаясь в виде эпителиально-мышеч-
ных клеток, особенно сильно развивается в верхней части полипа.
Эктодермальные мышечные волокна в ротовом диске актинии, рас-
положенные радиально, образуют расширители рта. Эктодермальная
кольцевая мускулатура ниже края диска образует сфинктор, а эктодер-
мальные радиальные и кольцевые мышцы в подошве составляют ее-
мускулатуру. Расширители рта играют у актиний важную роль при за-
глатывании добычи, а сфинктор — в защите от врагов и от воздействия
окружающей абиотической среды, так как при сокращении его и про-
дольных мышц тело актинии становится плоской лепешкой; с помощью
мускулатуры подошвы происходит присасывание ко дну и ползание.
В коррелятивной связи с развитием эктодермальной глотки и системы
перегородок гастральной полости стоит усложнение у коралловых поли-
пов мускулатуры. В стенке эктодермальной глотки снаружи развивается
кольцевая мускулатура, возникающая из эктодермальных клеток, а из
эктодермального эпителия образуется продольная мускулатура. В пере-
городках из эктодермальной продольной мускулатуры возникают мощ-
ные ретракторы, которые в виде валиков расположены в определенном,,
но различном у восьми- и шестилучевых коралловых полипов порядке
на одной стороне перегородок. При их сокращении происходит впячива-
ние щупалец ротового диска, укорачивание и утолщение всего тела акти-
нии. На стороне полных перегородок, противоположной ретракторам,
развиваются радиальные мышцы, сокращение которых вызывает расши-
рение глотки. Ниже глотки эти мышцы прикрепляются к подошве, уча-
ствуя в образовании ее мускулатуры (Беклемишев, 1952, 1964; Pantin,
1960).
У примитивных актиний — Protanthea (Nyholm, 1958—1960) и, веро-
ятно, у восьмилучевых полипов — Protalcyonaria мускулатура в соответ-
ствии с более простой общей организацией значительно проще описанной
выше; имеются два слоя эктодермальных и эктодермальных мышц, нет
сфинкторов, базилярных мышц, слабо развиты ретракторы и т. д.
Наличие скелета и прикрепленный образ жизни, естественно, ограни-
чивают также и функциональное значение мускулатуры, сохраняя за ней
функции захватывания добычи и защиты. У одиночных, бесскелетных
шестилучевых полипов функциональное значение мускулатуры возра-
34
стает, она участвует в передвижении', ряда форм актиний по субстрату,
а некоторые из низших'их представителей, например, Protanthea (Ny-
holm, 1958—1960), используют при ползании подобно гидрам щупальца
и подошву.
Отсутствие скелета в мезоглее и небольшая 'ее плотность, характер-
ные для большинства актиний, обусловливает то, что находящаяся в га-
стральной полости морская вода приобретает опорное значение поло-
стной жидкости, подобно тому, что имеет место у первичнополостных и
вторичнополостных червей (см. ниже). При помощи токов, создаваемых
ресничками поверхности тела, щупалец и сифонослифа вода поступает
в гастральную полость актинии; при закрытом рте она находится там
под довольно высоким давлением, при этом сжимается и глотка, кото-
рая, становясь клапаном, не дает жидкости вытекать из гастральной
полости. При таком положении происходит повышение давления жидко-
сти, сокращение продольной мускулатуры тела вызывает его укорочение,
а сокращение кольцевой — его вытягивание. Наоборот, при открытых рте
и глотке сокращением мускулатуры тела вода выталкивается из акти-
нии, все тело крайне уплощается, что усиливается еще сокращением
мышц верхней части тела. Таким образом, как было сказано выше, акти-
ния защищается от врагов и внешних неблагоприятных воздействий
(Беклемишев, 1952, 1964). Все это позволяет считать актиний гидроста-
тическими животными, тем более, что гидромеханическое значение имеет
и субэпителиальная жидкость, находящаяся у актиний между эпителием
и мускулатурой (Robson, 1957).
Естественно, что в связи со свободнопрдвижным образом жизни му-
скулатура у медуз построена несколько иначе, чем у сидячих форм.
На эксумбреллярной стороне зонтика гидромедуз эпителиально-мышеч-
ных клеток почти нет; в парусе, играющем важную роль в плавании
гидромедуз, эпителиально-мышечные клетки образуют сильное мускуль-
ное кольцо, поперечно-полосатой структуры. У сцифомедуз в самой суб-
умбрелле на нижней стороне зонтика находится кольцевая мышечная
лента, восемь радиальных лент, а также мышцы краевых лопастей зон-
тика. Мышечные отростки эпителиально-мышечных клеток поперечно-
полосатые. В ротовом стебельке (или манубриуме) у гидромедуз разви-
ты мышечные отростки эктодермальных и эктодермальных эпителиально-
мышечных клеток. У сцифомедуз сильные продольные мышцы развиты
в ротовом стебельке и его лопастях.
Нервно-мышечный механизм обеспечивает ритмические сокращения
зонтика, вызывающие выбрасывание из него воды, осуществляя тем са-
мым реактивное (толчками) движение медузы. Из сцифомедуз лишь лю-
пернарии, перешедшие к сидячему образу жизни, утратили способность
к плаванию и перешли к ползанию с помощью ножки и щупалец, как
гидры; в связи с этим кольцевая мускулатура зонтика и другие ее муску-
лы развиты слабо, но сильно развиты продольные мышцы сократимой
ножки.
Распрямление зонтика, особенно у высших медуз, обладающих мощ-
но развитой мезоглеей (у ряда форм с консистенцией гиалинового хря->
ща), происходит благодаря упругим свойствам мезоглеи.
По ничтожному количеству двигательной мускулатуры, менее 10%
ко всему объему тела, кишечнополостные могут быть отнесены к олиго-
миариым животным (Зенкевич, 1944) и оказываются ниже всех осталь-
ных групп многоклеточных. Низок и уровень дифференцировки их муску-
латуры, или сократительно-двигательного аппарата (Беклемишев, 1952,
1964)-
Такому состоянию мускулатуры кишечнополостных вполне соответ-
ствовало (по исследованиям 30-х годов зарубежных и советских ученых)
установление в качестве источника энергии мышечного сокращения неор-
3*
35
панических фосфатов, например, ортофосфата у сцифомедузы Pilema
pulmo (Вержбинская, 1935 и др.). Это также оправдывало'положение
кишечнополостных в основании филогенетического ствола всех двусто-
роннесимметричных типов, у которых и в более поздних исследованиях
были установлены аргининофосфат у первичноротых и креатинофосфат
у вторичноротых. Такому распространению фосфагенов многие (в том
числе Федотов, 1936, 1.938, 1959) придают филогенетическое значение.
Некоторые авторы (Roch a. Robin, 1954; Roch, Thoai a. Robin, 1957)
утверждают, что у кишечнополостных и даже у губок обнаруживаются
аргинин и креатин, однако, по Болдуину (Baldwin, 1957), у первичноро-
тых образуется лишь аргинин, у позвоночных — креатин, а у ряда других
вторичноротых (иглокожих, кишечнодышащих) — и аргинин и креатин.
По.Болдуину, аргинин образуется у сцифомедуз и актиний, но у них
отсутствует креатин; оба эти вещества отсутствуют у губок. В тканях
позвоночных свободного аргинина нет, но предполагают, что креатин
образуется из аргинина.
По современным представлениям (Baldwin, 1957) аденизинтрифос--
фатаза играет роль не только в сокращении мышц, но и в общем обмене
веществ. Аргинин является сравнительно широко распространенной ами-
нокислотой, встречающейся, вероятно, в протеинах тканей всех типов
животных. Сокращение мышц связано с аргинином у первичноротых и
некоторой части вторичноротых. У части последних и у всех позвоноч-
ных он замещается креатином.
Нервная система среди многоклеточных сформировывается впервые
у кишечнополостных; в соответствии с низким уровнем организации и
поведения она примитивна и по строению и по отправлениям. Она состоит
из общего диффузного сплетения, чувствительных клеток — рецепторов,
би- и мультипслярных ассоциативных клеток и из двигательных или
эффекторных клеток. Рецепторные клетки находятся среди клеток по-
кровного эпителия — эктодермы и среди клеток гастральной полости —
энтодермы. Нервное сплетение, возникающее за счет эктодермы, в обла-
сти рта переходит в нервное сплетение, развивающееся за счет энто-
дермы.
Поскольку кишечнополостные, в отличие от губок, питающихся мель-
чайшими взвешенными частицами,— хищники, питающиеся крупной до-
бычей (до рыб включительно), которую они захватывают ртом, вокруг
последнего, например, у актиний (Robson, 1963), происходит значитель-
ная концентрация нервного сплетения с ганглиозными клетками. У сидя-
чих форм ’(гидрополипов и Anthozoa) сохраняется диффузный характер
нервной системы, нервная сеть объединяет особей колонии в единое це-
лое. У сифонофор она также должна бы связывать всю колонию, что в
старых работах (Schaeppi, 1898) оставалось невыясненным. Недавно
установлено у Velelta (Mackie, 1960) наличие в эктодерме двух незави-
симых систем с нейрочувствительными клетками; «закрытой», состоящей
из сети толстых волокон, и, «открытой», состоящей из тонких волокон.
У одиночных подвижных форм — гидр и актиний происходит сгущение
нервного сплетения еще и на подошве.
В отличие от преимущественно колониальных прикрепленных или
одиночных малоподвижных, обычно придонных гидрополипов и Anthozoa
у свободноподвижных, плавающих медуз происходит значительная диф-
ференцировка нервной системы и развитие органов чувств. Кроме общего
подкожного нервного сплетения, у гидроидных медуз развиваются одно
кольцевидное сгущение с ганглиозными клетками на субумбреллярной,
а другое — на эксумбреллярной стороне кольцевого канала гастроваску-
лярной системы. Два кольцевидных нервных сплетения (Horridge a. all.,
1962) имеются и у сцифомедуз; из них главное, находящееся на суб-
умбреллярной стороне, вызывает пульсацию колокола, благодаря сокра-
36
щениям кольцевых и радиальных мускулов,'и другое, более диффузное,
расположенное в субумбреллярном эпидермисе, обеспечивает пищевую
реакцию и торможение сокращения колокола.
Дифференцировка нервной системы медуз связана с развитием у
Anthomedusae светочувствительных пятен — глазков, глазного пузыря,
у Leptomedusae— статоцистов С эктодермальными статолитами, у Тга-
chylina — с эктодермальными статолитами. У сцифомедуз развиваются
комплексы органов чувств в-краевых тельцах, или ропалиях, в виде орга-
нов световой чувствительности, вплоть до высоко дифференцированного
глаза (Schewiakoff, 1889) и органов статического равновесия.
Примитивность нервной системы кишечнополостных заключается в
отсутствии дифференцировки ее на центральный и периферический отде-
лы, специализации среди ассоциативных клеток и глиальной оболочки
у аксонов (голые аксоны), В распространении раздражения по всем на-
правлениям, в медленном проведении раздражения сравнительно со
скоростью проведения у других многоклеточных. Примитивность нервной
системы выражается и в том, что кусочек тела полипа или зонтика ме-
дузы обладает самостоятельным рефлексом и отвечает мышечными со-
кращениями на раздражение.
Даже у актиний величина реакции нервной системы различных частей
тела находится в прямой зависимости от силы механического раздраже-
ния и частоты раздражений, при этом ацетилхолин и адреналин не
играют той роли в процессе нервного возбуждения, как у других выше-
стоящих групп многоклеточных животных. Все же взгляд на диффузный
характер нервной системы кишечнополостных, в которой клеточные эле-
менты связаны друг с другом анастомозами, изменился благодаря новым
данным о наличии синаптических связей между ее элементами и химиче-
ской природой раздражения (Коштоянц, 1957; Horridge a. MacKay,
1962).
Что касается функции краевых органов, то ритмические сокращения
зонтика гидромедуз и сцифомедуз, имеющие важное приспособительное
значение, вызываются химической стимуляцией со стороны краевых те-
лец, хотя сокращения зависят также и от периферической нервной сети.
Установлены также факторы, усиливающие и ускоряющие рефлектор-
ную деятельность мышц зонтика медуз.
Отмеченные общие особенности деятельности нервной системы ки-
шечнополостных были установлены работами многих физиологов (Бек-
лемишев, 1952, 1964; Коштоянц, 1957, 1963). И все же приходится отме-
тить, что у кишечнополостных остается невыясненной природа передат-
чиков нервного возбуждения и торможения, так как установлены лишь
некоторые факты физиологического значения передатчиков. Между тем,
в настоящее время (Коштоянц, 1963) показана физиологическая роль
ацетилхолин-холинэстеразы и у инфузорий, а также значительно уве-
личено число обнаруженных веществ, передатчиков нервного возбужде-
ния и торможения, у ряда групп многоклеточных.
Эмбриологически кишечнополостные являются двуслойными живот-
ными (рис. 13). так как у них существует лишь два зародышевых ли-
стка— эктодерма и энтодерма. Лежащая между ними мезоглея, а имен-
но базальная мембрана, студенистое вещество, коллагеновые волокна,
свободные амёбоциты образуются в постэмбриональный период клетками
эктодермы и энтодермы. Третий зародышевый листок (мезодерма в виде
зачатка), возникающий на ранних этапах эмбрионального периода,
раньше других многоклеточных появляется у гребневиков. На этом осно-
вании кишечнополостных надо считать двуслойными, как' предложил
Рай Ланкестер (Ray Lankester, 1873), а остальных многоклеточных —
трехслойными (Hyman, 1940; Pantin, 1960 и др.). Мезоглея кишечнопо-
лостных, вопреки мнению А. А. Заварзина (1945), не мезодерма по про-
37
исхождению, а мезенхима (Давыдов, 1914) или первичная мезенхима
(Шмидт, 1953). У большинства медуз, даже у высших сцифомедуз, мезо-
глея сохраняется в виде студенистого вещества, но у некоторых, преиму-
щественно сцифомедуз, она настолько уплотняется, что резать ее нож-
ницами приходится с напряжением, и тогда по консистенции она упо-
добляется гиалиновому хрящу, не становясь действительно таковым
(вопреки мнению Заварзина, 1945). Судя по работам последнего време-
ни на гидре, ряде сцифомедуз и Anthozoa (Chapman, 1953; Hess, Cohen
a. Robson, 1957; Robson, 1957; Grimstom, Horne, Pantin a. Robson, 1958;
Рис. 13. Стадии онтогенеза различных Hydrozoa (из Hennig)
А — молодая планула со сплошной энтодермой; Б — продольный разрез
через зрелую планулу Gonothyraea sp_; В, Г— осевшая планула Seriu-
larella polyzonias начинает образовывать полип
Pantin, 1960), взгляд А. А. Заварзина подтверждается, но лишь отчасти.
Мезоглея кишечнополостных — ткань внутренней среды, она может быть
плотной, похожей на гиалиновый хрящ (у некоторых сцифомедуз и акти-
ний), ее химическая природа — протеины (в виде аминокислот); ее во-
локна гистологически и химически сходны с коллагеном позвоночных.
В ходе эволюции, как известно, у высших классов кишечнополостных
идет дифференцировка мускулатуры тела; мускульные волокна эпите-
лиально-мускульных клеток отделяются и становятся самостоятельной
мускулатурой, порой даже поперечнополосатой.
Все это дает некоторое право говорить о кишечнополостных, не исклю-
чая и .гидроидных, как о трехслойных животных, а их мезоглею назы-
вать тканью внутренней среды, но все же не позволяет (Pantin, 1960)
проводить гомологию этой ткани кишечнополостных с таковой высших
типов многоклеточных животных и даже низших плоских червей — тур-
беллярий.
Переходя к онтогенезу кишечнополостных, следует отметить, что
низкая, вернее, невысокая, степень интеграции взрослых особей гидроид-
ных проявляется и в особенностях их онтогенеза. На ранних стадиях,
когда отдельные бластомеры сравнимы с одноклеточными особями
(Захваткин, 1949), они обладают большой независимостью друг от дру-
38
чего положение и плоскости сим-
6
Рис. 14. Развитие Sagartia troglody-
tes (по Nyholm)
А — lQ-тиклеточная стадия; Б — свободно-
плавающая личинка
га; изолированные под ударами -волн в природе или в эксперименте бла-
стомеры развиваются дальше и обычно, дают нормальную бластулу лишь
несколько уменьшенных размеров (рис,; 14). С низкой степенью инте-
грации гидроидов связано также необычайное разнообразие форм
.дробления яиц: от аморфного, анархического до строго радиального, не-
редко с признаками спирального, точнее, ложно-спирального (Nyholm,
1942—1944), от радиального до близкого к билатеральному, от дробле-
ния на самостоятельные бластомеры до поверхностного и синцитиального
(Mergner, 1957). Примитивность .онтогенеза низших кишечнополостных
проявляется и в том, что плоскости деления бластомеров располагаются
•самым различным образом, вследствие
метрии в начале дробления не .фик-
сированы. В результате дробления
•получается жгутиковая целобласту-
ла, которая при униполярной мигра-
ции клеток на заднем конце, мульти-
полярном выклинивании их на всей .
поверхности или при деляминации
становится двухслойной паренхи-
мулой.
Продольная ось зародыша отве-
чает главной или первичной оси
тела. Анимальный полюс, на кото-
ром происходит образование экто-
дермы, является аборальным, бога-
тым нервными элементами, и у сво-
бодноплавающей личинки он на-
правлен вперед; личинка прикреп-
ляется аборальным концом. Вегета-
тивный полюс, где происходит обра-
зование энтодермы, является ораль-
ным, при плавании личинки он об-
ращен назад. По данным А. А. За-
хваткина (1949) и Рейзингера (Reisinger, 1957), это относится и к греб-
невикам. Гетерополярная ось с анимальным и вегетативным полюсами
характерна и для яиц других многоклеточных (Беклемишев, 1952, 1964).
Первичный метаморфоз гидроидных состоит в переходе пелагической
или придонной целобластулы или паренхимулы к сидячему образу жиз-
ни, в перестройке ее организации, в оформлении двух пластов погранич-
ной ткани, становящихся далее эктодермой и энтодермой и оформлении
тканей внутренней среды — по мнению А. А. Захваткина (1949) —«тре-
тьего пласта», в формировании радиально симметричного полипа с ани-
мальным способом питания. Среди жгутиковых клеток эктодермы диф-
ференцируются железистые, эпителиально-мышечные, нейроэпителиаль-
ные (чувствительнее) клетки, а также клетки нервного сплетения.
За счет интерстициальных клеток, выделяющихся в виде крупных скоп-
лений, образуются стрекательные клетки. В энтодерме происходит диф-
ференцировка на пищеварительные (с двумя, иногда пятью жгутиками)
и абсорбирующие клетки (те и другие с мускульными отростками), на
ацидофильные и базофильные железистые клетки, и, наконец, на чув-
ствительные клетки. Между клетками стенки гастральной полости обна-
руживаются небольшие группы интерстициальных клеток. За счет энто-
дермы развивается также нервное сплетение с его ассоциативными
клетками.
Дальнейшее развитие протекает следующим образом. Мерцательный
покров эктодермы сбрасывается. Эктодерма выделяет теку или пери-
сарк, при этом у одних форм Thecaphora личинка ложится на субстрат
39
параллельно, его поверхности, превращаясь в зачаток стелящейся гид-
роризы, у других — прикрепляется передним полюсом, превращаясь в
диск,- центральная часть которого выпячивается кверху и становится
первым гидрантом (первичным зооидом); в первом случае первичная
ось личинки утрачивается, во втором — сохраняется. У гипогенетических
Athecata метаморфоз почти отсутствует, так как прикрепившаяся к суб-
страту передним концом планула непосредственно переходит в полипа.
Далее наступает вегетативное размножение зооида: почкованием,
продольным и поперечным делением, происходит развитие одиночного
зооида (гидранта) в колонию (морфо-физиологическое целое), состоя-
щую из ствола, системы столонов и гидрантов, возникающих при поч-
ковании. Вегетативное размножение приводит также к половому размно-
жению; почкованием на бластостиле, измененном гидранте, образуются
у различных форм гидроидных все переходы от двуслойной почки с
амебоидноподвижными половыми клетками (гонофора) до свободнопла-
вающих медуз со сменой бесполого и полового поколений. Наличие в
почке половых клеток определяет ее судьбу — превращение в медузу.
Такой ход онтогенеза наблюдается у форм, яйца которых бедны желт-
ком. У форм с большим количеством желтка (например, Eudendrium,
Tubularia и др.) происходит эмбрионизация онтогенеза — стадия гастру-
лы и даже планулы протекает внутри оболочки яйца. В последнем слу-
чае из яйца выходит своеобразная личинка — актинула с двумя венчи-
ками щупалец. Актинула ползает по дну и после прикрепления абораль-
ными щупальцами, превращается в полипа, дающего почкованием
начало колонии.
У гипогенетических медуз (трахимедуз) планула превращается в ме-
дузу, минуя стадию полипа, причем у ряда форм происходит вегетатив-
ное размножение почкованием. Из почек, возникающих на краю зонтика,
у основания щупалец, на ротовом стебельке, на радиальных каналах
медуз, образуются дочерние медузы, отрывающиеся от материнской осо-
би (Pflugfelder, 1962).
• У паразитических наркомедуз (Metschnikoff, 1882, 1886; Ульянин,
1876; Щелкановцев, 1905; см. Шмидт, 1953; Hanitzsch, 1921), часть раз-
вития которых протекает в некоторых метагенетических и гипогенетиче-
ских медузах, наблюдается смена бесполого и полового поколений, раз-
множение идет путем почкования личинок, паразитирующих в медузах.
У гидр амебоидноподвижная яйцеклетка, питающаяся другими яйце-
клетками, находящимися по соседству в стенке тела, после оплодотво-
рения претерпевает полное, почти равномерное дробление. Гаструла
образуется из бластулы (у ряда форм из целобластулы) при мультипо-
лярной иммиграции и деляминации; она становится двуслойной плану-
лой, которая превращается в гидру (Brien, см. Gfasse et cet, 1961).
Молодая гидра все же проходит постэмбриональное развитие; у одних
видов гидра выходит из яйца, не имея щупалец, у других — с одним, у
третьих с четырьмия щупальцами; у некоторых же видов щупальца на-
чинают возникать еще в скорлупе (Канаев, 1952). После выхода из яйца
происходит и гистологическая дифференцировка клеток. Половое раз-
множение протекает осенью и зимой; весной и летом происходит веге-
тативное размножение почкованием.
Мелкие псаммофильные одиночные свободные гидроидные, впервые
в 1927 г. найденные Ремане, составляют самостоятельный отряд Actinu-
lides (см. Swedmark et Teissier, 1959). У них (Halammohydra и Otohydra,
сравнимых во взрослом состоянии с актинулой) развитие, как и у боль-
шинства гидр, почти прямое. Синцитиальная деляминация образует га-
струлу, причем при развитии щупалец у одних наблюдается четырех-
радиальная, у других — билатеральная симметрия (Swedmark et Teis-
sier, 1959).
40
У ряда свободноплавающих форм' сифонофор. дробление яйца, раз-
вивающегося в гонофорах' или медузоидных особях, приводит к делями-
нации на стадии морулы, превращающейся' в жгутиковую планулу;.
внутренняя масса такой планулы состоит из крупных клеток провизор-
ной энтодермы, из которых позже образуется дефинитивная энтодерма,,
выстилающая гастральную полость. Планула удлиняется, по своей фор-
ме и возникновению почек на брюшной стороне она двусторонне симмет-
рична и двумя перетяжками разделена, на три части. На переднем ее-
полюсе образуется из эктодермы закладка пневматофора, на противо-
положном (с прорывом ротового отверстия) первичный полип — гастро-
зоид. Средняя часть, на брюшной стороне которой из почки возникает
арканчик, удлиняется, становясь у многих сифонофор длинным столо-
ном. Развитие происходит почкованием в двух зонах: в верхней (почко-
вание по типу медуз) образуются плавательные колокола, гонофоры и
медузоиды, в нижней зоне (по типу полипов) —гастрозоиды, арканчики,.
цистозоиды, бластостили, щиты и т. -д. Такой тип онтогенеза с личинкой
сифонулой нередко, правда со многими значительными особенностями,,
в связи со строением сформированных колоний, характерен для сифоно-
фор отряда Siphonantha. В отряде же Disconantha образуется личинка
(конария) типа полипа с парой личиночных щупалец, позже исчезаю-
щих. На переднем полюсе простым впячиванием покровов образуется
пневматофор, на противоположном — рот гастрозоида, остающегося
единственным первичным полипом колонии. Вокруг последнего выпочко-
вываются многочисленные бластостили, гонозоиды с ротовым отверстием,,
несущие гонофоры, а по периферии — дактилозоиды. Личинки и взрос-
лые сифонофоры отряда Disconanta радиально симметричны (Беклеми-
шев, 1952, 1964).
Разные точки зрения о природе колоний сифонофор, их компонентов,
(как органов или особей) и разные взгляды на происхождение сифоно-
фор хорошо разобраны В. Н. Беклемишевым (1944, 1952, 1958, 1964)..
Поэтому подчеркну лишь, что ход онтогенеза говорит о сифонофорах
как колониях гидроидных, возникших из планулы путем почкования по-
липоидных и медузоидных особей, достигших в животном мире едва ли-
не высшей ступени колониальной индивидуальности, с низведением осо-
бей у ряда форм до уровня частей или органов, а самой колонии до со-
стояния организма. В эволюции сифонофор несомненно огромную роль,
сыграли отсутствие прикрепления к субстрату и плавающий образ жиз-
ни в подвешенном положении ртом книзу, подобно тому, как плавает гид-
роид Pelagohydra mirabilis (Kaestner, 1954/1955). Этим усиливалось зна-
чение медузоидных особей колонии. В основе строения и развития сифо-
нофор лежит полипоидный организм.
Разнообразен онтогенез особенно в постэмбриональном периоде и,
у сцифомедуз, хотя в значительно меньшей мере, чем у гидроидных. Так,,
например, у взрослых Stauromedusae (Lucernariidae) в результате пол-
ного дробления образуется стеррогаструла, которая, вытягиваясь в дли-
ну, превращается в ползающую планулу без жгутиков, с эктодермаль-
ными клетками, расположенными в один ряд. Планула образует почки,,
становящиеся в свою очередь свободными планулами. Посце некоторого-
периода свободной жизни первичная планула и дочерние почки при-
крепляются к водорослям и превращаются в бокаловидные полипы, а за-
тем в стауромедуз.
Онтогенез остальных медуз, изученный главным образом у низших
Semaeostomeae, во многом близок к таковому гидроидных, но имеет ряд.
значительных отличий, связанных с более высокой организацией взрос-
лых форм. Наряду с превращением целобластулы в гаструлу путем-
миграции бластомеров внутрь бластоцеля, у многих сцифомедуз гастру-
ляция происходит инвагинацией (Nausithoe, по Мечникову, 1950; Met-
44
-schnikoff, 1886; Chrysaora, Aurelia, Cyanea — ряд авторов). Однако спо-
соб гаструляции не является еще строго постоянным; как известно, у
Aurelia) гаструляция происходит иммиграцией, деляминацией и инва-
гинацией, причем это (подобно раннему развитию гидроидных) зависит
от условий внешней среды и может варьировать даже у одного вида
Aurelia.
Разнообразен и ход постэмбрионального развития у сцифомедуз.
Прикрепившаяся к субстрату планула, подобно полипоидной стадии
гидроидных медуз, превращается в стадию полипа — сцифистому. У ряда
«форм развитие сцифистомы сопровождается последовательным образо-
ванием у нее сначала одной пары щупалец, затем второй пары, появляю-
щейся под прямым углом к первой паре. Далее образуются тениоли,
-септы гастральной полости, щупальца третьей и четвертой пар и т. д.
Характерным для онтогенеза ряда сцифомедуз до превращения их во
взрослых медуз и достижения 16-лучевой симметрии является прохож-
дение стадий с двух- и четырехлучевой симметрией (Захваткин, 1949),
однако в стадии сцифистомы у ряда форм сразу возникает четыре щу-
пальца. Развитие метагенетических сцифомедуз сопровождается сменой
бесполого полипоидного и полового медузоидного поколений, первое раз-
множается бесполым путем и обладает большой регенеративной способ-
ностью, взрослые половозрелые сцифомедузы обычно утрачивают спо-
собность к бесполому размножению и к регенерации.
Сцифистомы обычно размножаются многократно следующим друг
за другом поперечным делением — стробиляцией, ведущей к образова-
нию свободноплавающих медуз — эфир; однако, как исключение, у Aure-
lia наблюдается превращение сцифистомы в эфиру без стробиляции.
Сцифистомы, кроме того, могут делиться продольно, а также путем
почкования. Почки возникают на боковой стенке сцифистомы, на подош-
ве ее ножки, на боковых столонах; отпочковываясь, они превращаются
в самостоятельных сцифистом, или, покрываясь ресничками, в свободно-
плавающую планулу. У некоторых сцифомедуз на подошве ножки сци-
фистомы образуются покоящиеся почки (подоцисты), одетые капсулой
и превращающиеся в сцифистом. Старые данные о метаморфозе сцифо-
медуз и бесполом размножении (Давыдов, 1914) дополнены рядом новых
исследований (Percival, 1923; Berrill, 1949; Uchida a. Nagao, 1963).
У Nausitho'e, по данным Комаи (Komai, 1935), сцифистомы трубчатой
-формы, одетые кольчатой перидермой или текой, рядом черт отличаю-
щиеся от сцифистом других сцифомедуз, иногда у некоторых видов обра-
зуются разветвленные колонии. Однако и они типичной стробиляцией
.дают начало эфирам. В онтогенезе гипогенетических сцифомедуз, на-
пример, Pelagia, отряда Semaeostomeae, куда относятся Aurelia и Cya-
nea, полипоидное поколение может выпадать, планула превращается не-
посредственно в медузу (П. П. Иванов, 1937; Pflugfelder, 1962), что,
вероятно, связано с условиями жизни.
При однообразной экологии взрослых шестилучевых полипов, как
правило, донных одиночных или колониальных прикрепленных форм,
реже свободных и передвигающихся при помощи щупалец, биология их
онтогенеза разнообразна. Все развитие, вплоть до сформирования акти-
нии, может протекать или (реже) в гастральной полости матери или
(чаще) вне материнского организма. Яйца нередко лежат на дне моря,
все развитие от целобластулы, свободноплавающей личинки, до превра-
щения в полип протекает быстро. У некоторых форм, например, Hal-
сатра (рис. 15), Edwardsiidae, личинка, лишенная ресниц, выйдя из сту-
денистой оболочки или капсулы яйца, лежащего на дне, ползает по дну
и, прикрепившись, развивается в полипа. У ряда шестилучевых полипов
{например, Sagartia) свободноплавающая личинка уже через несколько
дней после выхода из яйца прикрепляется ко дну. Чаще свободнопла-
-42
Рис. 15. Halcatnpa duodecimcirrata. Перед-
няя часть личинки с 8 мезентериями, соот-
ветствует стадии Edwardsia (по Nyholm)
полипов в зависимости от количе-
жающие личинки живут-месяцами, обеспечивая тем самым распростране-
ние вида. У некоторых-форм (например,~P.ea.chia) существуют медузо-
фильные личинки; которые, оказываясь .в субумбрелле медуз, расселяют-
ся с помощью последних (Nyholm, 1942—1.944, 1949, 1958—1960).
Борозды дробления бластомеров у ряда цериантид и актиний (Ny-
holm, 1942—1944) являются «врезающимися», а у некоторых форм (на-
пример, Pachycerianthus, Sagartia) на стадии восьми бластомеров
дробление спиральное. Такие же особенности раннего развития были
установлены у гидроидных, как было сказано выше И. И. Мечниковым
(Metschnikoff, 1886), который дал бороздам дробления название «вре-
зающиеся» (см. также Захваткин,
1949). Характерно, что у некоторых
актиний (например, Sagartia troglo-
dytes) образуются даже квартет
макромеров и квартет микромеров.
'Однако, как правильно считает Ни-
гольм (Nyholm, 1942—1944), это —
.ложное спиральное дробление, так
как смещение бластомеров происхо-
дит без наклонов веретен дробле-
ния, что характерно для истинного
спирального дробления червей и
моллюсков.
Стадия бластулы у шестилуче-
.вых полипов лишена жгутиков или
ресничек и в известной мере отве-
чает представлению о ней А. А. За-
.хваткина (1949) как о «синзоо-
споре»;
Характер дробления коралловых
ства желтка в яйце бывает полным равномерным (Sympodium из вось-
милучевых, Metridium из шестилучевых полипов), полным неравномер-
ным (Sagartia), поверхностным (Urticina, Bolocera из Шестилучевых —
отряд Actiniaria и у некоторых восьмилучевых). У восьмилучевых
(Sympodium) в результате дробления образуется стерробластула, на-
ружный слой которой дает эктодерму, внутренний — энтодерму, зародыш
сильно вытягивается в длину, покрывается ресницами, превращаясь в
свободноплавающую планулу, прикрепляющуюся передним (абораль-
ным) концом к субстрату.
У шестилучевых полипов образуется целобластула, превращающаяся
в гаструлу инвагинацией или деляминацией; у одних еще в теле матери
гаструла развивается в молодого полипа, у других она становится планк-
тонной личинкой. У представителя примитивной группы (трибы) акти-
ний— Protanthea (Nyholm, 1958—1960) пелагическая личинка со сплош-
ным ресничным покровом, непохожая на личинок других актиний, несет
апикальный султан из длинных ресниц. Удлиненные личинки других
шестилучевых полипов — zoanthella, zoanthina из отряда Zoantharia
(Hyman, 1940; Pflugfelder, 1962) — имеют ресничные полосы или кольца,
с помощью которых они плавают.
Развитие мадрепоровых полипов изучено слабо. В результате син-
цитиальной деляминации и миграции клеток периферия личинки или
планулы одевается эктодермальным эпителием с крупными ядрами,
стенку гастральной полости выстилают эктодермальные клетки с мел-
кими ядрами — Pocillopora, Seriatopora, Goniastrea, Favia (Давыдов,
1937, см. Бляхер, 1963). Пелагическая личинка аборальным концом при-
крепляется к субстрату. Впячиванием эктодермы на месте бластопора
образуются щелевидное ротовое отверстие и сдавленная с боков
43
эктодермальная глотка с одним или двумя сифоноглифами, определяю-
щими двулучевую симметрию молодых коралловых полипов. Однако у
примитивной актинии Protanthea (Nyholm, 1958—1960) молодая бентос-
ная стадия радиальносимметрична, щелевидный рот при питании ста-
новится круглым, сифоноглифы не образуются, симметрия оказывается
переходной между 'радиальной и двулучевой. Известно, что затем у вось-
милучевых полипов из складок стенок гастральной полости сразу воз-
никает восемь септ, тогда как у шестилучевых септы образуются посте-
пенно и строго определенно для каждого семейства (Беклемишев, 1952,
1964), при этом характерно, что молодая актиния и внешне не радиаль-
носимметрична, у нее сначала возникает непарное, спинное щупальце,
а затем два боковых. Соответственно сначала в плоскости симметрии
образуется одна пара септ, делящая кишечную полость на меньшую,
брюшную и большую, спинную части, а затем образуется вторая пара
септ, далее — третья и четвертая пары; шестилучевой коралловый полил
проходит стадию Edwardsia с восемью септами: после паузы начинается
образование пятой и шестой пар септ (или промезентериев), что приводит
к развитию шестилучевой симметрии (щупалец по числу радиальных
камер образуется также шесть).
Полипчик Protanthea (триба Nynantheae) сначала так ж& имеет
шесть щупалец, но в отличие от других актиний полных септ у них обра-
зуется не двенадцать, а только восемь; развиваются также сплошной
эпидермально-мышечный слой и нервная сеть по всему стволу, но не воз-
никают сифоноглифы, базилярные мускулы, сфинктор и ряды жгутико-
вых клеток на септальных нитях, характерные для других актиний.
В онтогенезе мадрепоровые полипы (склерактинии) также проходят
стадию Edwardsia с двенадцатью полными септами и четырьмя непол-
ными, за исключением Асгорога и Porltes, которые сохраняют такое
строение и во взрослом состоянии. Затем происходит дальнейшее нара-
стание числа септ, кратное шести. Сходно идет развитие и склеросепт,
тесно связанное с развитием мягких септ (Hyman, 1940). Как и у других
групп современных шестилучевых коралловых полипов, у мадрепоровых,.
не имеющих сифоноглифов, и в развитии, и во взрослом состоянии наблю-
дается двулучевая и шестилучевая симметрии. В связи с постоянным'
ростом у большинства шестилучевых кораллов (актинарий) происходит
увеличение числа вторичных септ (метамезентериев), в соответствии .с
этим идет и нарастание числа щупалец, возникает неполная, кратная
шестилучевая симметрия.
Не останавливаясь на довольно разнообразном ходе постэмбриональ-
ного развития других отрядов шестилучевых кораллов, надо отметить,
что колониальные формы и часть одиночных теряют способность к по-
стоянному росту.
Следует напомнить, что склеросепты у ругоз тоже возникают после-
довательно, сначала образуется одна пара, затем вторая, третья и т. д.
В расположении склеросепт у ругоз вместе с четырехлучевой сохраняет-
ся и двулучевая симметрия.
Следовательно, и в онтогенезе (развитие щупалец, промезентериев
или первичных септ, склеросепт) и в организации взрослых (сдавленная
с боков глотка, сифоноглифы) у современных и ископаемых Anthozoa
элементы двулучевой или двусторонней симметрии комбинируются с ра-
диальной— четырех-, восьми- и шестилучевой.
Как уже.отмечалось выше, для кишечнополостных характерно очень
большое разнообразие особенностей онтогенеза в строении яиц, типов
дробления, характере личинок и жизненных циклов, участия в последних
полипоидных и медузоидных особей, смены поколений и т. д. Рядом
экспериментов в течение последних нескольких лет (1955—1963) у гид-
роидов (Athecata, Thecaphora, Anthomedusa, Leptomedusa) в жизненном.
44
цикле вскрыта зависимость .формообразовательных процессов от усло-
вий жизни: например, зависимость характера'"развития колокола медуз
от пищи, формы колоний от фактора пространства, почкования медуз
у метагенетических видов на полипах и выпадения его от температуры
<и даже зависимость развития соматических и первичных половых клеток
от температуры (Werner, 1963). Несомненно, что распространение та-
кого экспериментального анализа онтогенеза на все группы кишечно-
полостных позволит выяснить закономерности индивидуального разви-
тия, их эволюцию, а вместе с тем поможет выяснить родственные отно-
шения классов этого типа, включая и ряд вопросов об их строении,
например, такой группы, как сифонофоры. Высокая лабильность онто-
генеза или жизненного цикла гидроидов, вместе с лабильностью их тка-
ней у взрослых особей говорит об исходной примитивности, а не об упро-
щении этой группы, об их низком эволюционном уровне.
В связи с бесполым размножением и почкованием среди кишечнопо-
лостных широко распространена регенерация, прежде всего на ранних
•стадиях развития, например, у сифонофор, гидромедуз и сцифомедуз.
Наиболее высокой регенеративной способностью, регуляцией и морфол-
лаксисом обладают гидры и гидрополипы с их необычайно сильной те-
.кучестью и подвижностью клеточного состава (см. Белоусов, 1960а,
19606, 1961, 1963; Канаев, 1952 и ряд зарубежных авторов). У гидр из-
вестны способность восстановления целого организма из ’/гоо части,
образование химер из частей-разных видов (Исаев, 1925) и редукция под
воздействием неблагоприятных условий в круглый комочек и т. д. Коло-
нии сифонофор утратили регенеративную способность. Гидромедузы и
сцифомедузы репарируют обычно лишь удаленные или утраченные части
тела — края зонтика и ротовых щупалец, ропалии. Однако стауромеду-
зы, подобно гидрополипам (Hyman, 1940), сохраняют высокую репара-
тивную способность. Достаточно широко регенерация распространена
среди коралловых полипов, особенно актинарий.
Из основных классов кишечнополостных наиболее многочисленны
(более 6000 видов) коралловые полипы, процветающие в настоящее вре-
мя, из них примерно 2300 видов восьмилучевых и 3760 (наиболее широко
распространенных), шестилучевых. По уровню организации, гистологи-
ческой дифференцировки и характеру онтогенеза коралловые полипы
стоят выше остальных классов, хотя прикрепленный образ жизни и от-
части колониальность неблагоприятно сказались, если сравнивать их со
-сцифомедузами, на развитии их нервной системы и органов чувств. Важ-
ной особенностью коралловых полипов является сохранение у ряда оди-
ночных форм свободного подвижного придонного существования;
Другим многочисленным классом кишечнополостных являются гид-
розон, представленные бблее 2700 современными видами полипов и ме-
дуз, из которых последних, по любезно сообщенным мне данным
Д. В. Наумова, примерно 660 видов. Для гидрозоев характерны отсут-
ствие узкой специализации и простота строения полипов, в частности
необычайная текучесть их клеточных элементов, почти неограниченная
обновляемость последних и экологическая пластичность, выражающаяся
в переходе ряда атекат из морской среды в солоноватоводную и пресно-
водную. Для многих гидромедуз (часть которых, принадлежащая атека-
там, заходит в пресную воду и существует даже в условиях аквариумов)
характерно непостоянство общей четырехлучевой симметрии. Наблю-
.дается, с одной стороны, олигомеризация числа щупалец и. радиальных
каналов, с другой стороны, увеличение их числа. Гидрозои, находящиеся
во многом на низком уровне эволюции, по численности видов уступают
.лишь коралловым полипам.
Наиболее ограниченным является класс сцифомедуз, вся организа-
ция которых во взрослом состоянии носит черты крайней специализации,
45
при высокой приспособленности их строения и функций к жизни во взве-
шенном состоянии в толще воды моря. Одиночные, неспособные, к бес-
полому размножению (почкованию), с ограниченной репарацией уча-
стков тела, представленные всего 200 современными видами, появляю-
щимися, однако, в массовом количестве особей, и, по-видимому, бедные-
по числу видов в прошлые геологические эпохи сцифомедузы являются
классом кишечнополостных, гораздо менее пластичным, чем гидрозон-
и коралловые полипы.
В историческом развитии кишечнополостных возникновение гидро-
полипов и коралловых полипов наметилось в раннем кембрии, а может
быть еще в докембрии. У гидрополипов, защищенных лишь органическим
скелетом — хитиноидной кутикулой (текой или перисарком) — успех в
борьбе за существование достигался образованием колоний, возникав-
ших при бесполом размножении сохранением связей дочерных особей-
с материнскими. Возникала колония, состоящая из многочисленных осо-
бей, которая при прикрепленном образе жизни древних форм обеспечи-
вала существование, питание и сопротивляемость неблагоприятным воз-
действиям абиотических и биотических факторов. Общепризнано, что
колониальность влечет за собой ослабление индивидуальности особей
колоний, утрачивающих способность к самостоятельному существованию..
Наличие кембрийских представителей гидроидов — Thecaphora, остаю-
щихся и ныне чисто морскими формами, говорит в пользу очень древнего
возникновения колониальности.
Начиная с юры, появляются колониальные гидроиды (миллеггорины
и стиластерииы), образующие уже более надежный известковый скелет..
Значительно позже, в нижнем мелу и текафоры становятся известковыми.
Атекаты — преимущественно морские формы, часть которых в ходе эво-
люции очень недавно стала приспосабливаться к жизни в солоноватовод-
ной и пресноводной среде.
У гидроидных с образованием колониальности (столониального, мо-
ноподиального с терминальным гидрантом, моноподиального с терми-
нальной зо.ной роста и симподиального типов) возникает разделение-
труда зооидов, полиморфизм, дифференцировка на бесполых (гидран-
тов) и половых (медузоидов). Среди первых возникают гастрозоиды,
дактилозоиды, бластозоиды (бластостили), нематофоры, шипы и т. д.;
половые же зооиды остаются связанными с колонией или становятся
свободноживущими гидромедузами. Под воздействием условий жизни,
особенно в результате приспособления к жизни в солоноватоводной и
пресноводной средах, происходит исчезновение у ряда атекат крупных
колоний и колониальности вообще, вплоть до возникновения одиночных
полипов, свободно передвигающихся и даже пелагических.
Другим направлением в историческом развитии полипов является
эволюция коралловых полипов, известных пока только со среднего ордо-
вика. Для них характерно вначале появление одиночных особей и широ-
кое использование растворенных в морской воде солей для образования
защитного известкового скелета. С самостоятельным образом жизни
древнейших одиночных коралловых полипов связано усложнение их
строения, особенно гастральной полости. Позже на базе бесполого раз-
множения у коралловых полипов чаще путем продольного деления, реже
образованием почек на полипе или на ценосарке, возникла колониаль-
ность. Типы колоний, особенно у палеозойских коралловых полипов, до-
статочно многообразны. В историческом развитии от одиночных корал-
ловых полипов возникают колониальные формы и вновь одиночные от
колониальных, а у ряда высших современных шестилучевых кораллов —
бесскелетных анемон — возникают почти исключительно одиночные фор-
мы, достигающие порой громадных размеров (например, Stoichactis sp..
46
достигает 1,5 м в диаметре). Однако в отличие.-'от гидрополипов, у-
которых происходит уменьшение'’величины, упрощение организации,,
значительная утрата индивидуальности особями''колонии, полипы ко-
лониальных кораллов сохраняют высокий уровень организации и строе-
ние, типичное для Anthozoa,'.- хотя полипы и становятся мельче оди-
ночных. ’
При решении вопроса о филогенетических отношениях групп кишеч-
нополостных в основном приходится пользоваться материалами по со-
временным формам, так как историческая летопись их. неполна. Исклю-
чение представляет лишь часть коралловых полипов — табуляты, четы-
рехлучевые и шестилучевые.
Гидроидные полипы Athecata с голыми гидрантами, имеющими раз-
личное число венчиков щупалец, порой беспорядочно расположенных,
морфологически ниже гидроидных полипов Thecaphora, приобретших,
теку и претерпевших олигомеризацию щупальцевого аппарата, состоя-
щего из одного венчика щупалец. Среди атекат существует немало оди-
ночных полипов, как мелких форм примитивной организации, например,
Tricyclusa, Arum и др. (см. Rees, 1957), так и гигантов вроде Branchioce-
rianthus, претерпевших осложнение наружного и внутреннего строения.
Наряду с этим у атекат наблюдается развитие более разнообразной,
чем у текафор, формы колоний. Видимо, также, и среди медузоидного по-
коления антомедузы с глазками (особенно такие, как Sarsia) более при-
митивны, чем лептомедузы со статоцистами.
При таком, даже крайне ограниченном сравнении атекат и текафор-
первых приходится считать менее специализированными формами. Одна-
ко для окончательного решения этого вопроса не хватает хронологиче-
ских данных. При одинаковом характере наружного скелета, если не-
считать отсутствия теки у Athecata, способность к сохранению у вымер-
ших представителей обеих групп как будто одинакова. И тем не менее-
ископаемые формы текафор известны с кембрия, а атекат — с мезозоя.
По-видимому, ископаемые остатки атекат по каким-то причинам в палео-
зое не сохранялись, это более вероятно, чем предположение о более-
позднем появлении атекат сравнительно с текафорами.
В пользу примитивности атекат говорит их большая, сравнительно-
с текафорами, биологическая пластичность.
Некоторые виды гидроидов атекат становятся пелагическими форма-
ми, среди которых гидры вообще и в особенности такие, как Microhydra,.
претерпели крайнее упрощение и в организации и в жизненном цикле.
Именно в группе атекат и медузы приспосабливаются к солоноватым'
и пресноводным условиям жизни (Зенкевич, 1940; Rees, 1957).
Атекатный характер гастрозоидов сифонофор позволяет думать, как.
предполагается многими, что сифонофоры возникли от каких-то прими-
тивных атекат. Но, к сожалению, пока ископаемые атеката известны
с мезозоя, а сифонофоры — со среднего ордовика.
Положение в системе и родственные связи сифонофор с гидроидным»-
полипами установлены давно-; считать ли их отрядом или подклассом —
подкласса или класса Hydrozoa или отдельным классом — вопрос не-
принципиальный. Однако нет еще согласованного мнения в толковании
некоторых категорий особей или компонентов сифонофор (Беклемишев,
1952, 1964), причем особенно важным является вопрос о природе пнев-
матофора. У одних сифонофор его считают производным медузоида,
у других не измененной особью, а органом — гидростатическим аппара-
том колонии: пневматофор развивается как впячивание стенки абораль-
ного конца первичного полипа (Metschnikoff, .1874) и в его развитии
(Garstang, 1946; Hadzi, 1954) нет черт медузоидного узелка радиальных,
каналов, кольцевых каналов и гомолога манубриума, т. е. всего того,
что характеризует медузоид.
47
По системе, данной Геккелем (Haeckel, 1888, см. Беклемишев, 1952,
.1964), сифонофоры делятся на Siphonantha и Disconantha. Хаджи
(Hadzi, 1954) придерживается деления сифонофор на Calycophorae
Leuckart, 1854 — сифонофоры без пневматофора — и Pneumatophorae
Chun, 1897 — сифонофоры с пневматофором. Хаджи считает, что у Caly-
cophorae, первично не имевших пневматофора, гидростатическую функ-
цию несет плавательный колокол — медузоид, у которого развиваются
олеоцисты. По мнению Хаджи, Calycophorae очень рано отделились от
главной линии сифонофор, до возникновения у последних пневматофора,
и являются самой примитивной группой. По Хаджи, эволюция сифонофор
шла путем прогрессивного развития новообразования — пневматофора.
Porpita и Vetella из Chondrophorae ( = Disconantha) с очень сложным
строением пневматофора — по Хаджи, наиболее высоко стоящие сифо-
.нофоры.
С такой трактовкой сифонофор трудно согласиться. Она сугубо одно-
сторонняя, поскольку учитывает только строение пневматофора и игно-
рирует основные черты организации сифонофор; ясно, что такая класси-
-фикация оказывается неестественной, а предполагаемый путь эволюции
маловероятным.
Следует иметь в виду, что все сифонофоры типа сифонантов билате-
ральносимметричные, с длинным стеблем или стволом, реже, например,
у Physalia, ствол становится укороченным, как правило, с большим на-
бором особей, видоизмененных медузоидсв и преимущественно поли-
поидов, собранных в кормидии. Среди сифонант много очень крупных
форм со стволом до 3 л длиной и огромным пневматофором, 30 см в диа-
метре.
Другой тип строения характерен для сифонофор — дисконантов или
хондрофорид. Эти сифонофоры радиальносимметричны, с элементами
.двулучевой (Velella) и восьмилучевой (Discalia) симметрии, с диском,
.несущим центральный гастрозоид и венчики полипоидов — бластозоидов
(гонозоидов) и дактилозоидов.
Таким образом, у сифонантов и дисконантов различно не только
строение пневматофоров, различны и типы их организации (архитекто-
ника), они далеки друг от друга, будучи, очевидно, разного происхожде-
ния. Не случайно дисконанты (Velella, Porpita) рассматриваются порой
видоизмененным полипом, а не колонией. На самом деле они явля-
ются колонией с аборальным гидростатическим органом (крайне эксцес-
сивно развитым пневматофором), с единственным первичным гастро-
зоидом, с наружным венчиком дактилозоидов (Picard, 1955), видоизме-
ненных полипоидов, а не щупалец, и внутренним венчиком бластозоидов,
дающих начало медузам (Chrysomitra).
Рис (Rees, 1957), как известно, различает у атеката две ветви эво-
люции: одну для надсемейства Corynoidea, другую для надсемейства
Tubularoidea. Дактилозоиды и бластостили дисконантов обладают рядом
-сходных черт с таковыми медуз и полипов гидрозоев семейства Pterone-
matidae (Pteronetna, Pteroclava, Zanclea, Cladoryne), относящихся к пер-
вой ветви. Общими признаками у них являются тип книдома, строение
медуз, типы полипов, строение перисарка. От этой ветви гидрозоев и
надо, видимо, выводить дисконантов.
У сифонантов же наблюдаются общие черты в строении полипов и
.медуз с таковыми другой ветви гидрозоев — надсемейства Tubularoidea,
отдаленные, примитивные формы которого могли быть предками сифо-
нантов. В отличие от дисконантов, небольшой, малочисленной группы,
'сифонанты— многочисленная, разнообразная группа типичных сифоно-
'фор, колонии которых, как известно, достигают наиболее совершенной
интеграции среди колониальных животных мира. Вместе с тем, как
давно отмечалось, полипоиды, дисконанты и сифонанты построены по
48
Рис. 16. Схема примерного количественного распределения ископаемых коралловых
полипов (из A. Mflller)
А — все коралловые полипы (794 рода); Б — только Tabulata (105 родов); В — только Rugosa
(326 родов)
типу полипов атеката, что и позволяло всех сифонофор выводить от пред-
ков, взявших начало от гидрозоев атекат.
Однако в отличие от мнения Хаджи я считаю (исходя из основ строе-
ния колонии), так же как и Гарстанг (Garstang, 1964), более примитив-
ными дисконантов, хотя они и обладают крайне развитым пневматофо-
ром, а более совершенными, прогрессивными — сифонантов.
Данных об историческом развитии сифонофор мало: в ископаемом
состоянии, благодаря прочной кутикуле пневматофоров, известны только
дисконанты, остатки которых сохранились из ордовика, силура и девона
(Moor, 1956, см. Соколов, 1962); ископаемые сифонанты пока не из-
вестны.
Родственные отношения между крупными группами коралловых по-
липов все еще недостаточно выяснены, отчасти потому, что ископаемых
восьмилучевых кораллов известно мало, а бесскелетные шестилучевые
кораллы в ископаемом состоянии пока вообще не описаны. Геологиче-
ская история хорошо раскрыта для табулят, ругоз и склерактиний
(рис. 16). Эволюция табулят и ругоз протекала в палеозое и закончилась
в мезозое, эволюция восьмилучевых и шестилучевых, начавшись в мезо-
зое, развернулась в нашу эпоху. В происхождении коралловых полипов
основную роль играли табуляты. Известные ib количестве 105 родов, пред-
ставленные редкими примитивными формами в среднем и позднем кемб-
рии, табуляты становятся довольно многочисленными й разнообразными
в среднем и позднем ордовике; в силуре и девоне они переживают морфо-
биологический прогресс (число родов максимально, строение разнооб-
разно, распространены по всему Мировому океану), затем в конце палео-
зоя начинается упадок и как подкласс табуляты вымирают в меловом
4 Д. М. Федотов
49
периоде, когда появляются восьмилучевые кораллы. У табулят как древ-
нейших (со среднего кембрия) коралловых полипов (Соколов, 1955),
почкованием дававших начало мономорфным колониям с мелкими корал-
литами (0,5—4,0 мм), радиальные склеросепты были почти недиффе-
ренцированными, количество их было разнообразно и кратно 4, 6 и 8,
без определенного порядка в их закладке.
Несомненно, обосновано мнение Б. С. Соколова (1955, см. 1962)
о близком родстве табулят и ругоз. В строении септального аппарата,
в сходстве главных направлений его развития, в последовательности
усложнения днищ и связанных с ними скелетных частей, в общности на-
чальных стадий онтогенеза и в ряде других черт, как указывает Б. С. Со-
колов, обнаруживается близкое родство табулят и ругоз как ветвей,
возникших от общего ствола.
Установление родства табулят с ругозами не снимает вопроса об их
филогенетических связях с другими классами или подклассами коралло-
вых полипов — восьмилучевых и шестилучевых. Я присоединяюсь к
взгляду Мозли, хорошо обоснованному в 1924 г. Сардессоном (Соколов,
1962) и принятому очень многими исследователями, особенно палеонто-
логами (Соколов, 1955),. о происхождении и близком родстве с табуля-
тами восьмилучевых коралловых полипов (или альционарий).
Как и табуляты, восьмилучевые кораллы почти исключительно коло-
ниальные формы с полипами небольшой величины, с относительно про-
стыми мезентериями, как и у ряда табулят — в числе восьми. Историче-
ское развитие восьмилучевых коралловых полипов начинается в мезозое,
а их эволюция протекает в нашу эпоху. Вполне вероятно, что именно
в мезозое от каких-то табулят с восемью септами произошли древние
восьмилучевые коралловые полипы, а из поздних ругоз, как это доста-
точно убедительно показано рядом палеонтологов (а не из восьмилуче-
вых), возникли древнейшие шестилучевые кораллы.
Как большие самостоятельные группы (подклассы или классы) вось-
милучевые и шестилучевые кораллы стали формироваться в мезозое:
первые — в юре, вторые — в триасе; это также противоречит мнению
о происхождении шестилучевых кораллов от восьмилучевых, тем более,
что шестилучевые (склерактинии) достигли расцвета и были очень мно-
гочисленными тогда, когда восьмилучевые кораллы были еще небольшой,
малочисленной группой.
Известно, что в онтогенезе современных восьми- и шестилучевых
кораллов при общности основных черт находим ряд достаточно крупных
особенностей (способы развития мезентериев, их строение). Правда,
в онтогенезе шестилучевых кораллов обнаружены стадии развития, вос-
производящие черты строения восьмилучевых кораллов, но этот факт
говорит не о происхождении шестилучевых от восьмилучевых, а лишь
о родстве тех и других, об общих корнях их, на что указывают палеон-
тологические данные, говорящие о происхождении шестилучевых корал-
лов от ругоз.
Современных восьмилучевых кораллов известно' примерно 2300, ше-
щтилучевых — около 4000 (3950) видов. Первые наиболее многочисленны
в теплых береговых водах, вторые распространены по всему Мировому
океану и на всех глубинах. По строению, онтогенезу и экологии восьми-
лучевые кораллы однообразнее и более специализированы, чем шестилу-
чевые, хотя одиночные примитивные формы найдены и среди альциона-
рий (Grasse et cet., 1961).
Известно, что ругозы, появившись в низах среднего ордовика, т. е. не-
сколько позже табулят, процветали в палеозое; они дали большое коли-
чество родов (326 по А. Н. Miiller, 1961), громадное разнообразие строе-
ния одиночных и колониальных форм, чрезвычайно широко распростра-
нившихся в палеозое по морям всего земного шара. Ругозы являются
50
исключительно ярким примером, морф.обиОлогического прогресса. Одна-
ко, как и у других групп, в историческом развитии ругоз периоды, или
фазы, расцвета сменялись периодами упадка, несомненно связанными
с изменениями условий существования. Мощно развившись в силуре, они
испытывают одну фазу расцвета в среднем девоне, другую — в раннем
карбоне.
Знаниям о ругозах —• их строении, онтогенезе, пространственном и
хронологическом распространении и филогенетических отношениях, мы
обязаны работам Ведекинда, Шиндевольфа (Schindewolf, 1939, 1942) и
особенно советских палеонтологов — Т. А. Добролюбовой (1948, 1952,
1958, 1960), Е. Д. Сошкиной (1941, 1960), Н. В. Кабакович (см. Соколов,
1962) и Т. Г. Ильиной, (1963а, 19636). При разнообразии строения всего
скелета у ругоз наблюдается развитие нескольких типов септ — клино-
видных, пластинчатых, губчатых, шиповидных, образование септальных
конусов, развитие днищ—полных, неполных, простых, пузырчатых,
простого осевого столбика или сложной центральной зоны и т. д. На ко-
нечных стадиях онтогенеза ругоз установлено появление новых призна-
ков и переход их закладки в ходе филогенеза на начальные стадии.
Установлено наличие среди ругоз стенофациальных и эврифациальных
форм. В филогенезе ругоз возникло несколько крупных ветвей, напри-
мер, отряды Cystiphyllida, Streptelasmatida и др., с характерными осо-
бенностями формы, строения, палеоэкологии и хронологии.
Вопреки широко распространенному мнению четырехлучевые корал-
лы (Rugosa) не вымерли в конце перми, в триас перешло несколько их
семейств, в том числе плерофиллиды, которые являются настоящими
переходными формами между четырех- и шестилучевыми кораллами не
по отдельным, а по целому комплексу признаков (Ильина, 19636).
После стадии с шестью склеросептами у этих четырехлучевых корал-
лов развитие происходит билатеральносериально, около главной склеро-
септы с обеих сторон и около боковых склеросепт со стороны противо-
положных квадрантов. У шестилучевых после стадии с шестью склеро-
септами дальнейшее развитие склеросепт происходит радиальноциклично
во всех шести септах (Ильина, 1963а).
При таких четких различиях ругоз и склерактиний у позднепалеозрй-
ских и раннетриасовых ругоз, например, плерофиллид, появляются черты
шестилучевых кораллов; так увеличивается дифференциация септ, воз-
никают внешняя радиальность и цикличность, уменьшаются главные
квадранты и т. д. ‘В то же время у ряда древних, триасовых склерактиний
проявляются черты, общие с ругозами, — нет правильной цикличности,
протосепты закладываются последовательно, попарно, как у ругоз, а не
одновременно, как у Hexacorallia, два сектора около противоположной
септы по размерам и по числу септ отстают сравнительно с другими,
заметна билатеральная симметрия и т. д.
Эти данные позволяют присоединиться к взгляду Шиндевольфа
(Schindewolf, 1939, 1942), Соколова (1955, 1962) и Т. Г. Ильиной (1963а,
19636) о происхождении шестилучевых кораллов от четырехлучевых.
• Мнение Хилл (1963) об отсутствии родства между этими группами
Anthozoa кажется ошибочным, особенно, если учесть, что автор к опре-
делению этих групп кораллов подходит статически, а не динамически,
т. е. по существу отрицает их эволюцию.
Среди шестилучевых кораллов склерактинии, или мадрепоровые ко-
раллы, имеют очень важное значение, объединяя ископаемые (381 род,
5000 видов) и многие современные формы (2500 видов). Сменив в на-
чале мезозоя ругоз, склерактинии быстро эволюировали. В их историче-
ском развитии наблюдались три фазы процветания (А. Н. Muller, 1961):
первая, слабая, протекала в среднем и позднем триасе, вторая, .самая
сильная, начавшись в средней юре, проходила -в позднем меле, третья —
4*
51
в миоцене. И в настоящее время склерактиний представлены многочис-
ленными как колониальными, рифообразующими, так и одиночными
формами, обладают большой биологической пластичностью й дают
яркие примеры взаимозависимости организма и условий жизни. Рифо-
вые кораллы, живущие в мелких водах тропической и субтропической
зон (между 28° с. ш. и 28° ю. ш.), при температурном оптимуме между
25—29° могут переносить колебания суммарной солености от 27 до 40%,'
непродолжительное пребывание на воздухе (см.-Соколов, 1962). Одиноч-
ные склерактинии, родственные рифовым, хотя и живут в мелководье
(50—75 м), но наиболее распространены на глубине 180—550 м, заходят
и в сверхабиссальные глубины (8000 м).
Сравнительно с прикрепленными к субстрату специализированными
склерактинйями близко родственные им анемоны морфологически и эко-
логически более пластичны. Наряду с преобладающим большинством-
высоко дифференцированных (как и склерактиний) типичных актинарий,
принадлежащих к Nynantheae, некоторые формы анемон обнаруживают
примитивное строение и сохраняют способность передвигаться, прикреп-
ляясь к субстрату то подошвой, то ротовым диском, то щупальцами
(Protantheae). Среди Nynantheae, как известно,: имеются даже свобод-
ноплавающие формы (семейство Minyadidae), обладающие гидростати-
ческим аппаратом.
Актинарии живут во всех морях, многие формы распространены очень
широко, некоторые являются даже циркумполярными видами. Однако
крупнее и многочисленнее они в теплых водах, а некоторые являются
исключительно тропическими формами. Будучи наиболее многочйслен-
нымй-'в Мелких, прибрежных водах, часть анемон живет и размножается
в сверхабиссальных глубинах — до. 10 710 м (Беляев и др., 1960).
-'"Ряд данных говорит как бы в пользу взгляда Гаймен (1940) о про-
исхождении склерактиний от анемон, но палеонтологические доказатель-
ства свидетельствуют о непосредственном происхождении их от ругоз.
Можно допустить, что какие-то вымершие, одиночные склерактинии,
возможно в третичном периоде, утратили скелет, и их потомки, приспо-
собившись к контрастным условиям жизни, дали начало бесскелетным
шестилучевым кораллам-—Actiniaria, широко распространившимся по
Мировому океану и находящимися в расцвете и в настоящее время.
Обычно с анемонами филогенетически связывают внешне сходных
с ними Zoantharia, небольшую группу (около 300 видов) современных
Мягких одиночных и колониальных коралловых полипов. По строению
они .значительно отличаются от анемон наличием толстой кутикулы, ка-
налов в мезоглее стенки тела, сообщающихся, с гастральной полостью,
двух зон роста мезентериев в межкамерных пространствах по бокам
пары направляющих перегородок, примыкающих к сифоноглифу. Свое-
образны у них и личинки — zoanthina и zoanthella. Но все же из бесске-
летных шестилучевых кораллов зоантарии наиболее близки к анемонам
и им родственны.
Филогенетические отношения между Antipatharia и Ceriantharia
(небольшими малочисленными группами шестилучевых кораллов) почти
не выяснены. Во-первых, ископаемых остатков их не обнаружено даже
для антипатарий, колонии которых нередко бывают 2 л в длину (напри-
мер, длина Stichopath.es gracilis достигает 2,64 м) и снабжены твердым,
напоминающим рог, осевым скелетом. Во-вторых, не изучен онтогенез
этих кораллов, известны лишь отрывочные данные о формировании ме-
зентериев. Не изучена и их биология. Известно только, что церианта-
рии — одиночные, бесскелетные шестилучевые коралловые полипы с тер-
минальной порой на заднем конце тела, с большим неопределенным
числом мезентериев, не расположенных парами, с одним сифоноглифом,
с продольной мускулатурой длинного тела. В отличие от цериантарий
52
антипатарии— колониальные щестилучевые коралловые , полипы с осе-
вым скелетом из рогоподобного вещества, с , венчиком из невтягиваю-
щихся шести щупалец, расположенных билатеральносимметрично,
с поперечным ротовым отверстием, с двумя сифоноглифами, с 6, 10, 12
полными мезентериями, расположенными парами, с-мезоглеей в виде тон-
кого гомогенного слоя, со слабой мускулатурой (Hyman, 1940; Kaestner,
.1954/1955; Jagersten, 1959).
До сих пор, начиная с конца XIX столетия, на основании некоторых
общих: черт онтогенеза антипатарии считают родственными церианта-
риям и даже объединяют их вместе в Ceriantipatharia (Pax, 1918; Franz,
1924; Hyman, 1940). Однако слишком велики различия между антипа-
тариями и цериантариями, чтобы можно было их объединять, а относи-
тельно их филогенетических связей можно лишь сказать, что антипата-
рии, и цериантарии, как, и все остальные отряды, образующие группу
шестилучевых кораллов, связанных друг с другом происхождением от
каких-то общих предков, и по данным строения и онтогенеза противопо-
ставляются группе восьмилучевых кораллов.
Представления о филогенетических отношениях между Современными
и ископаемыми сцифомедузами . .предположительны. Стауромедузы, судя
по их строению и онтогенезу, ряду сходных черт с полипами, возникших,
очевидно, под воздействием донного сидячего образа жизни, являются
уклонившейся ветвью сцифомедуз, эволюция которых как бы приоста-
новилась на пути развития остальных сцифомедуз. Эта небольшая груп-
па,-преимущественно мелких сцифомедуз, обитающая, главным образом,
в холодных частях земного шара.
Четырехлучевые кубомедузы.с кубическим зонтиком, с четырьмя ро-
палиями и щупальцами, четырьмя парами гонад, мешковидным желуд-
ком с четырьмя карманами также обладают некоторыми чертами поли-
па. Это—-небольшая группа примитивных небольших размеров сцифо-
медуз, обитающих в. мелководье тропических и субтропических областей.
В ископаемом состоянии известны с поздней юры.
Не совсем ясны также родственные отношения медуз короната с
характерной формой колокола, поделенной кольцевой бороздой на верх-
ний, прочный как хрящ, отдел, и периферический, с лопастями, с широ-
ким синусом гастральной системы. Сцифистомы некоторых коронат, на-
пример, Nausithoe (стр. 42), образуют длительно существующую много-
ветвистую колонию полипоидов, считавшихся раньше самостоятельным
организмом. Это тоже небольшая группа, примерно из 20, большею
частью мелких, меньше 5 см в диаметре (обычно 0,9—4 см), сцифомедуз,
живущих в основном на больших глубинах океана.
Наиболее типичными, многочисленными, порой достигающими огром:
ных размеров (как Cyanea polaris), обычно обитателями умеренных об-
ластей океана являются. Semaeostomeae. Они населяют береговые воды
всех зон Мирового океана и нередко встречаются в громадных количе-
ствах. Эти медузы составляют большую часть из 200 современных видов
сцифомедуз. Близко к ним стоят сходные с ними Rhizostomeae сцифо-
медузы с колоколом от 4 до 80 см в диаметре, обитающие в мелководье
преимущественно тропической и субтропической зон, главным образом
индо-пацифической области. Именно эти две группы сцифомедуз по
своей организации, численности и широкому распространению в совре-
менном океане являются среди сцифомедуз наиболее эволюировавшими,
известными в ископаемом состоянии, как кубомедузы и коронаты, с верх-
ней юры.
Отсутствие скелета у сцифомедуз делает находки ископаемых форм
крайне редкими, а описания их лишь по внешней форме не вскрывают
истинной природы ископаемых. Наличие в онтогенезе сцифомедуз поли-
поидной стадии или поколения сцифистомы, дающей стробиляцией
53
начало поколению медуз, а у некоторых разветвленную колонию поли-
поидных особей (Nauslthoe), надо считать указанием на происхождение
сцифомедуз от примитивных кораллоподобных предков. Об этом говорят
также и палеонтологические данные. Четырехлучевые палеозойские за-
ходящие в триас Conulatae обладали хабитусом полипа, в строении
имели общие черты с сцифомедузами и их сцифистомами и были донны-
ми обитателями — прикрепленными или свободными во взрослом состоя-
нии (Соколов, 1962).
Допущение филогенетической связи сцифомедуз с коралловыми по-
липами подтверждается также недавним открытием нового класса
Hydroconozoa (Корде, 1963) одиночных, обладающих скелетом, радиаль-
но-симметричных, с элементами двулучевой симметрии древнепалеозой-
ских кишечнополостных, с чертами строения ругоз и сцифистом совре-
менных сцифомедуз. Поэтому не исключена возможность, что в отдален-
ном геологическом прошлом в жизненном цикле коралловые полипы
вначале были связаны со сцифомедузами так, как связаны между собой
гидрополипы и гидромедузы.
Каковы родственные отношения и вообще существуют ли они между
современными сцифомедузами и так называемыми медузами Protome-
dusae и Dipleurozoa (см. -Соколов, 1962) из докембрийских отложений
Южной Австралии, Юго-Западной Африки и АнУлии — мы не знаем. Эти
остатки, например, Cyclomedusa, Pseudochizostomites,-Beltanella,. ыряд,
ли относятся действительно к сцифомедузам, так как, кроме круглой
формы, ничего общего с медузами . у них нет. Rangea, описанная
М. Ф. Глесснером, по боковым ответвлениям несколько напоминает
морское перо. Однако до проверки природы этих остатков современными
методами исследования (применение изотопов, химического анализа
и т. д.) лучше не торопиться относить их к сцифомедузам и коралловым
полипам. Однако по наличию в этих докембрийских отложениях остат-
ков кольчатых червей и ходов существование в них кишечнополостных
и губок вполне вероятно. Удивительно лишь то, что колониальные гид-
розон с достаточно прочным скелетом так мало известны в ископаемом
состоянии. (В ходе исторического развития кишечнополостных, с одной
стороны, шло приспособление от ползания по дну к прикрепленному, с
другой стороны, к свободноплавающему образу жизни, с чем и связано
развитие двух основных жизненных форм — полипа и медузы.
Известно, что, по мнению одних зоологов [из советских — В. М. Шим-
кевича (1908, 1918), В. А. Догеля (1930), В. Н. Беклемишева (1944, 1952,
1964), Н. А. Ливанова (1945, 1955), А. А. Захваткина (1949), Д. В. На-
умова (1953, 1960) и ряда зарубежных], у гидрозоев первичным, более
древним поколением являются полипы; по мнению других зоологов
(см. Hyman, 1940, многие американские авторы),—-медузы. В последнее
время (Werner, 1963) высказывается мнение о равнозначности в жиз-
ненном цикле как тех, так и других.
Надо помнить, что половой зачаток, или первичные половые клетки
формируются из бластомеров яйца до дифференциации зародышевых
листков и, очевидно, на стадии двуслойной личинки планулы они еще
не приурочены ни к одному из последних. Стадия планулы характерна
для всех групп кишечнополостных, и едва ли можно оспаривать ее фило-
генетическое значение. С появлением рта она становится близкой к по-
липу, и, кроме обладания единой гастральной полостью, по архитекто-
нике медузы имеют мало общего с nejo. В ходе индивидуального разви-
тия, когда из планулы образуются гидрополипы, гидроидные • медузы,
сцифомедузы и коралловые полипы, происходит не возникновение, а
лишь перемещение первичных половых клеток, установление связи с на-
ружной эктодермальной стенкой организма (или гастродермисом) и
развитие из них гонад. У колониальных гидрозоев, как правило, половые
54
железы развиваются в полипах (бластостилях), причём для развития
половых клеток в гонады гастральная полость колонии вступает в тес-
ный контакт с половыми клетками и это обусловливает развитие гоно-
форов, которое часто заканчивается образованием медузоида. Превраще-
ние последнего в свободноживущую медузу для прикрепленных колоний
гидроидных полипов является важной приспособительной особенностью,
обеспечивающей размножение и распространение вида. Поэтому такая
смена поколений, естественно, получила широкое распространение у
огромного числа текафор и атекат. В настоящее время среди гидрозоев
нередки случаи, когда свободные, самостоятельные, медузы не образу-
ются, что является результатом упрощения, редукции развития, вызван-
ного воздействиями условий жизни. Во всяком случае для пресноводных
гидр вряд ли кто сомневается в том, что они утратили поколение медуз
и колониальность. Но среди типичных, исконно морских гидрозоев, не-
сомненно, первичные родоначальные формы были одиночными, а не ко-
лониальными организмами; колониальность возникла у них в ходе эво-
люции как приспособление для защиты от вредных внешних воздействий
и для обеспечения ловли добычи и лучшего питания. Вряд ли можно
сомневаться в том, что у трахилид имело место обратное явление —•
утрата поколения полипов и переход к развитию медузы непосредственно
из планулы. Все согласны, что из организации полипа легко вывести
медузу, полип по строению близок к червеобразной плануле, напоми-
нающей родоначальную форму кишечнополостных и, наоборот, медуза
крайне далека от нее. Своеобразие строения и лучистая симметрия медуз
причинно связаны со свободным плавающим образом жизни (Гиляров,
1944) и являются вторичной особенностью, как медузоидная форма ряда
плавающих голотурий. Все это подтверждает точку зрения о том, что
в жизненном цикле кишечнополостных поколение медуз появилось поз-
же полипов. Мало понятно поэтому противоположное мнение в основном
американских зоологов (Hyman, 1940) и палеонтологов (например,
Shrock a. Twenhofel, 1953; Moor, 1952).
За вторичный характер медузоидов сравнительно с полипоидами
говорят также и численные соотношения у современных форм: на 660 ви-
дов гидромедуз приходится 2000 и более видов гидрополипов, на 200 ви-
дов сцифомедуз приходится 6000 и более видов коралловых полипов.
Строение двухслойной стадии планулы, характерной для онтогенеза
всех кишечнополостных (несмотря на очень большое разнообразие типов
дробления яйца и модусов образования зародышевых листков, собствен-
но энтодермы), имеет филогенетическое значение. В преобладающем
числе случаев планула является полой, энтодерма выстилает стенку
гастральной полости, несмотря на то, что низшие кишечнополостные
даже во взрослом состоянии сохраняют необычайную текучесть, подвиж-
ность клеточного состава (см. Белоусов, 1960—1963; Канаев, 1952 и ряд
зарубежных исследователей). Очевидно, на такой примитивной стадии
существования полостное пищеварение для целентерат более выгодно,
чем внутриклеточное при сплошной массе энтодермы.
Не менее важными для филогенетических обобщений являются дву-
сторонняя симметрия планулы и то, что лишь на дальнейших этапах
онтогенеза происходит развитие лучистой симметрии, в типичном случае
после прикрепления планулы к субстрату.
В итоге я согласен с мнением Хаджи (Hadzi, 1944, 1963) и Егерстена
(Jagersten, 1955, 1959) о вторичном характере лучистой симметрии ки-
шечнополостных, первично билатеральносимметричных животных, о чем
я уже писал. И думаю, что не надо даже приводить доводы против
попыток Хаджи считать коралловые полипы произошедшими от турбел-
лярий, а сцифомедуз и гидроидных — от кораллов, так как для этого нет
никаких фактов из всей истории кишечнополостных и червей.
55
Мое представление об-исходной, примитивной, родоначальной форме-
.кишечнополостных как о свободноподвижном, придонном билатерально-
симметричном организме, в основном сходится со взглядом Егерстена.
У большинства кишечнополостных о такой родоначальной, предковой
форме говорит планула, которая обычно, и это надо подчеркнуть, возни-
кает без прохождения инвагинационной гаструлы. В последнем я рас-
хожусь с Егерстеном, теория билатерогастреи которого возникла под
-влиянием- взглядов Геккеля о гастрее. Под влиянием Геккеля, как из-
вестно, многие зоологи в толковании эмбриологии кишечнополостных
.ошибочно применяли мерки и подходы, установленные в эмбриологии
высших групп многоклеточных (Захваткин, 1949). Считаю, что Егерстен
ошибочно приписывает своей билатерогаструле кишечнополостных на-
личие гастральных карманов, сначала расположенных метамерно, а да-
лее, при перестройке всей организации, дающих начало периферическо-
му отделу гастральной полости кораллов. Проводить гомологию между
периферическими отделами гастральной полости кораллов и энтероцель-
ными карманами архентерона, дающими целом у вторичноротых, нельзя
уже по одному тому, что у кишечнополостных еще не возник зачаток
третьего зародышевого листка — мезодермы, которая появляется лишь у
следующих, вышестоящих групп многоклеточных, с чем связано. отсут-
ствие у кишечнополостных многих органов. Не прав Егерстен и в том,
что, как и ряд-зоологов (Ulrich, 1949, 1951; Remane, 1954, 1959; Marcus,
1958)-, следуя неудачному почину Хаджи (Hadzi, 1944), при рассматри-
вании филогенетического ряда: коралловые полипы — сцифомедузы — ,
гидрозон считает наиболее примитивными коралловых полипов, называя
их исходной группой. Допустим, что гидроиды стали наиболее упрощен-
ными по строению и гистологии, произойдя от морских форм, переселив-
шихся через солоноватоводную в пресноводную среду. Однако и мор-
ские, сложноколониальные гидрозои остаются простыми, необычайно
текучими в своем клеточном строении формами, способными к регене-
рации, морфоллаксису и регуляции. И коралловые полипы, и сцифомег
дузы гораздо более высокоорганизованы, гораздо менее пластичны и'
в морфологическом и экологическом отношениях. При переходе организт
мов от свободного существования к паразитическому, от подвижного
к прикрепленному образу жизни известно много случаев возникновения
развития от сложных форм к простым, упрощения строения взрослых
форм и хода их онтогенеза. Однако все это не оправдывает взглядов '
Хаджи и Егерстена на происхождение кишечнополостных от вышестоя-
щих трехслойных многоклеточных — турбеллярий и о ходе эволюции
по линии Anthozoa — Scyphozoa — Hydrozoa.
Ни в онтогенезе кишечнополостных, ни в их организации взрослых
форм нет никаких данных о вторичном упрощении этих животных, как
нет и фактов, указывающих на то, чтобы какие-либо черты турбеллярий
проявлялись как древние, исходные особенности в любом классе кишеч-
нополостных. Примитивный характер онтогенеза кишечнополостных и
возможность истолкования его, исходя из закономерностей онтогенеза
высших групп, достаточно обоснованы А. А. Захваткиным (1949). Если
у плоских червей кишечник целентерический (с ротовым и без анального
отверстия), подобно таковому кишечнополостных, то все остальные си-
стемы — кожно-мускульный мешок, паренхима, нервная, выделительная
и половая — у кишечнополостных еще не представлены (Hand, 1959;
Pantin, 1960 и др.). Это понятно, так как, как было сказано раньше
(стр. 37), в эмбриональном развитии кишечнополостных нет зачатка
среднего зародышевого листка — мезодермы, и даже у высших групп —
коралловых полипов и сцифомедуз-—ткань внутренней среды образова-
на не средним зародышевым листком, а клетками, выходящими из экто-
дермы и энтодермы.
58
Гребнейики (Ctenophora)
Гребневики — малочисленная группа чисто морских одиночных сво-
бодноподвижных, преимущественно пелагических животных, с удельным
весом, близким к удельному весу морской воды, благодаря наличию в
бесскелеФном, обычно прозрачном, богатом мезоглеей теле более 99%
воды. Форма тела сферическая, с главной осью, на верхнем абораль-
ном пблюсе находится аборальный ,орган чувств — аппарат, тонизирую-
щий и регулирующий движение, на нижнем оральном полюсе — рот.
Восемь меридиональных рядов гребных пластинок являются органами
движения. Пара щупалец, снабженных клейкими клетками, служит ор-
ганом защиты и ловли добычи этих хищных животных. Нервная систе-
ма •—диффузное нервное сплетение, сгущающееся под меридиональны-
ми рядами гребных пластин,ок и связанное с аборальным органом
чувств; Имеется гладкая мускулатура в стенках тела и в кишечнике.
Кишечник состоит из большой эктодермальной глотки и эктодермаль-
ных желудка и системы каналов. Органов дыхания и выделения нет.
Полоёые железы развиваются на стенках меридиональных каналов ки-
шечника, полы соединены. Неравномерное,, детерминированное, мо-
заичное дробление с преформир.ованными в яйце плоскостями симметрии
будущего гребневика ведет к образованию характерной цидиппидной
личинки, превращающейся во взрослое животное. В онтогенезе хорошо
выражены эктодерма, энтодерма и третий зародышевый листок' (мезен-
хима). Кроме полового размножения наблюдается размножение деле-
нием и регенерация. Гребневики — свободноживущие организмы, среди
них известна лишь одна паразитическая форма — Gastrod.es.
Гребневики — небольшая группа, всего 90. видов в современной фау-
не. Большею частью это небольшие животные (Tenerfe суапеа), длиной
всего в 2—2,5 мм, многие платиктениды крупнее, 5—10 мм в плоскости
щупалец и как исключение — 60—70 мм, реже это более крупные фор-
мы, так у Вегоё высота колокола достигает 20 см, a Cestiim вытяги-
вается до 1,5 м в длину. Гребневики — преимущественно прозрачные,
нередко светящиеся животные, глубоководные формы Вегоё обычно
красного цвета (Mortensen, 1927); многие ползающие гребневики —
платиктениды — окрашены в различные цвета: молочно-белый, серый,
желтый, оливковый, красный, нередко с различным рисунком (Komai.
1922; Dawydoff, 1938, 1946).
Благодаря прекрасному, очень полному использованию литератур-
ных данных в книге Гаймен (Hyman, 1940) можно органичиться напо-
минанием лишь русских и советских авторов, не рассмотренных ею: это
работы И. И. Мечникова (1885, 1955; Metschnikoff, 1885) и А. О. Кова-
левского (Kowalevsky, 1886) о развитии типичных гребневиков,
А. О. Ковалевского (Kowalevsky, 1880) о целоплане, А. Коротнева
(Korotneff, 1886) о ктеноплане, Д. К. Третьякова (1936, Tretjakoff,
1930) о гистологии'мезоглеи и происхождении гребневиков, Г. И. Роски-
на о строении мускульных клеток и К. Н. Давыдова (Dawydoff, 1933,
1936, 1938, 1946) о платиктенидах.
57
Тип гребневиков делится на два класса: со щупальцами — Tentacula-
ta, состоящего из четырех отрядов: Cydippida, Lobata, Cestida и Platy:
ctenida и без щупалец — Nuda, с отрядом Beroida. Гребневиков надо
считать типом, четко ограниченным по всей совокупности признаков
строения и развития от типа Coelenterata, или Cnidaria. Ископаемые
формы не известны.
Преобладающая форма гребневиков, как пелагических плавающих
животных,— шаровидная, грушевидная или цилиндрическая (взрослые
цидиппиды, лобаты, цидиппидная стадия развития остальных гребне-
виков). У цидиппид тело в поперечном разрезе круглое, у змеевиднопла-
вающих цестид оно вытягивается в глоточной плоскости в длинную уз-
кую ленту, у бероид вытягивается по главной оси и несколько сжато
в плоскости глотки, у платиктенид в связи с ползающим образом жизни
главная ось чрезвычайно укорачивается, тело сплющивается в дорсо-
вентральном направлении. При плавании с помощью гребных пласти-
нок (основного двигательного аппарата) прозрачные пелагические греб-
невики двигаются ртом вперед, в спокойную погоду держась в поверх-
ностных слоях воды, в бурную — опускаясь в глубину. Некоторые виды
встречаются на глубинах до 3000 м. Платиктениды, ползающие по суб-
страту (по водорослям, колониальным кишечнополостным), становятся
планариеобразными, тело их удлиняется в плоскости щупалец, и при
ползании одно щупальце направлено вперед, другое назад. По мне-
нию В. М. Шимкевича (1918), длинная ось платиктенид отвечает
продольной оси турбеллярий и всех двустороннесимметричных жи-
вотных.
Гребневики встречаются в разных областях океана, например, бе-
роиды характерны для холодных вод, платиктениды приурочены к тро-
пическим и субтропическим зонам.
По наличию двух щупалец, двух щупальцевых каналов, двух гло-
точных каналов, двух выделительных пор, четырех каналов в области
аборального .органа чувств, восьми ребер с мерцательными гребешка-
ми и восьми меридиональных каналов у гребневиков сочетаются дву-
лучевая, четырехлучевая и восьмилучевая симметрии (Беклемишев,
1952, 1964). Элементы симметрии, как видно из сказанного выше, в свя-
зи с характером движения претерпевают у разных групп значительные
изменения. Наибольшее уклонение от лучистой симметрии и приближе-
ние к двусторонней и в форме тела и во внутреннем строении, естествен-
но, происходит у платиктенид.
Характерной чертой организации гребневиков являются восемь ме-
ридиональных рядов гребных пластинок, служащих , двигательным ап-
паратом, связанным с аборальным органом чувств, тонизирующим и
регулирующим его работу. Они присутствуют у всех гребневиков и об-
наруживают большое разнообразие строения в связи с изменениями
формы тела (Dawydoff, 1946). У ползающих платиктенид, в связи с
переходом к донному образу жизни, гребные пластинки подвергаются
значительной деградации, а некоторые платиктениды даже полностью
утрачивают гребешки (Komai, 1922). Характерно, что некоторые пела-
гические гребневки, цидиппиды, например, молоденькие особи Pleu.ro-
brachia pileus (Remane, 1956), взрослые особи Lampetia могут ползать
на вывороченной наружу глотке, и наряду с этим Ctenoplana из платик-
тенид не только ползают, но и плавают.
Для гребневиков характерным органом является и пара щупалец,
находящихся во влагалищах в щупальцевой (трансверзальной) плоско-
сти, ближе к аборальной стороне. Щупальца — сильно растяжимые и
сократимые, с большим количеством в их эпидермисе клейких клеток
очень характерного, специфического только для гребневиков строения.
Щупальца, нередко со многими разветвлениями, являются мощным ор-
58
таном защиты и ловли добычи. Они отсутствуют или,, вернее, исчезают
лишь у бероид, ловчим аппаратом которых становится большой рот,
разросшейся до громадных размеров глотки-. У паразитичёских Gastro-
des щупальца, естественно, становятся рудиментарными.
Стенка тела гребневиков состоит из эпидермиса, к которому приле-
тают продольные и кольцевые мышцы и часть нервной системы, распо-
ложенной в мезоглее. Эпидермис синцитиальный или образован плоски-
ми, кубическими и столбчатыми клетками, несущими в определенных
местах реснички, за счет которых образуются и гребные пластинки.
У некоторых платиктенид, не имеющих гребешков (очевидно, утратив-
ших их), весь эпидермис становится ресничным (Komai, 1922). В эпи-
дермисе находятся чувствительные клетки — рецепторы, многочислен-
ные эозинофильные и цианофильные железистые, клейкие и пигментные
клетки. В глубоких слоях эпидермиса, по-видимому, имеются (Komai,
1922) и интерстициальные клетки. Мезоглея в виде бесструктурного
студенистого вещества заполняет у большинства пелагических гребневи-
ков промежутки между каналами гастроваскулярной системы; в ней
располагаются мускулатура и нервная сеть. У ползающих гребневиков
мезоглея настолько обедняется студенистым веществом, что уподоб-
ляется паренхиме плоских червей, как ее и называют, например, Комаи
(Komai, 1922). Она содержит также соединительнотканные звездчатые
и пигметные клетки, которым платиктениды обязаны своим разнообра-
зием окраски и рисунков (Komai, 1922; Dawydoff, 1938).
Мускулатура гребневиков образована гладкими волокнами (Роскин,
1923) мезенхимного происхождения, ’не проходящими, как отмечает
Н. А. Ливанов (1955), стадию эпителиально-мышечной клетки, и состо-
ит из достаточно сформированной системы мускулов, расположенных
в мезоглее. В основном в мускулатуре тела имеются продольные и коль-
цевые мышцы, пересекающиеся под прямым углом, кольцевые и про-
дольные мышцы глотки, радиальные мышечные волокна, идущие от
глотки к стенкам тела, мощные продольные мускулы щупалец, сфинкто-
ры рта и на границе глотки с желудком. У лобат в лопастях их тела из
веерообразно расходящихся и взаимопересекающихся волокон разви-
вается мускулатура, сокращением которой обеспечивается передвиже-
ние этих форм. У лент,обидных цестид развивается кожно-мускульный
мешок из поперечных, продольных и диагональных или косых мышц,
благодаря сокращению которого гребневик (например, Cestum) плавает,
змеевидно изгибаясь. Еще большую дифференцировку и развитие испы-
тывает мускулатура платиктенид, ползающих и обладающих способ-
ностью легко изменять форму тела (см. Dawydoff, 1933). К сожалению,
даже в такой капитальной работе, как работа Комаи (Komai, 1922),
гистология мускулатуры платиктенид изучена недостаточно. Однако
бесспорно, что у гребневиков вместо эпителиально-мышечных эктодер-
мальных и эктодермальных клеток кишечнополостных развились глад-
кие мышцы (Роскин, 1923) и что мускулатура гребневиков ближе к
мускулатуре плоских червей, чем к таковой кишечнополостных.
Нервная система гребневиков состоит из аборального центра (або-
ральный орган чувств) (рис. 17), связанного с субэпителиальным нерв-
ным сплетением, обладающим синаптическими связями и образован-
ным двумя типами нейронов: ассоциативцо-двигательными, располо-
женными в мезоглее, и рецепторными, расположенными в эпидермисе
и в околоротсвом кольце. Уровень дифференцировки и функции нервной
системы гребневиков обусловлены большой подвижностью этих жив от -
ных, большой сократимостью их тела и щупалец и работой их двига-
тельного аппарата — гребных пластинок. Как известно, на куполе
колокола помещается аборальный орган чувств, от основания пружи-
нок которого отходят четыре мерцательных шнура или желобка,
59
расходящихся по восьми мерцательным ребрам. Субэпителиальное,спле-
тение образует соответственно- восемь меридиональных- сгущений. И.$
него же возникают-, околоротовое кольцо и сильное сгущение — нервные
тяжи щупалец (Беклемишев, 1952, 1964; Heider, 1927; Hanstrom, 4928;
Korn, 1959; Pavans de Geccatty, Hernandes et Thiney, 1962). При этом, по
данным Корна, у Pleurobranchia. pileus нервное сплетение имеется и под
эпителием энтодермального желудка.
;-У ползающих гребневиков (рис. 18) (см. Беклемишев, 1952, 1964)
в области аборального органа происходит образование четырех ганглиоз-
ных скоплений. Этот орган считается центром, координирующим и то-
низирующим работу гребных пластинок, его удаление нарушает работу
последних.
Рис. 17. Строение аборального органа (из. Hyman)
1 — статолит; 2 — балансеры; 3 — ресничная бороздка
Рецепт.орные клетки эпидермиса тела, особенно щупалец, восприни-
мают различные раздражения, на которые щупальца отвечают рефлек-
торным путем —резким сокращением и растягиванием. У пелагических
(Вегоё) и тем более у ползающих гребневиков механическое или иное
раздражение одной стороны тела вызывает сокращение мышц другой
стороны [например, у Coeloplana bockii (Moore, 1936)]. Рецепрокной
иннервацией и функциональной особенностью чувствительно-двигатель-
ного аппарата гребневики, особенно ползающие, сближаются с турбел-
ляриями (Коштоянц, 1957). В связи с этим А. А. Заварзин (1950), и ряд
других исследователей допускают происхождение турбеллярий от пла-
тиктенид, делая черезчур далеко идущие отождествления аборального
органа чувств и меридиональных сгущений нервной сети гребневиков с
эндомом и продольными коннективами турбеллярий.
В эволюционном и филогенетическом отношениях несомненно важно,
что реципрокная иннервация, как отмечает Мур (Moor, 1936), характер-
ная для червей, моллюсков, ракообразных и игл.окожих, существует у
гребневиков и, видимо, отсутствует у кишечнополостных. Все же для
окончательных выводов приходится признать, что нервная, система у
гребневиков изучена еще недостаточно, новые работы (Korn, 1959; Pa-
vans de Ceccatty, Hernander et Thiney, 1962) отрывочны, строение и
взаимоотношения аборального органа чувств, околоротового и меридио*
нальных сгущений нервной системы, как отмечал X. С. Коштоянц (1957),
недостаточно выяснены.
Кишечник (или гастроваскулярная система) гребневиков, являющих-
ся хищниками, образован эктодермальной глоткой с широким, ротовым,
отверстием, сдавленным в сагиттальной (или медиальной) плоскости,
и энтодермальными желудком, узким сообщающимся с глоткой пище-,
водом, сдавленным в латеральной или щупальцевой плоскости, и систе-
мой эктодермальных каналов. Один канал отходит от желудка, кверху
60
и дает под аборальным орга-
ном четыре короткие ветви, с
концами, вздутыми в виде, ам-
пул, две из которых открыва-
ются выделительными порами
наружу для вывода воды и не-
переваренных остатков пищи.
Два канала (глоточные) отхо-
дят от желудка по бокам глот-
ки и направляются к ротовому
полюсу, два канала над ними
отходят в плоскости щупалец,
непарная ветвь которых, или
щупальцевый канал, идет к ос-
нованию щупалец, а парные
(трансверзальные каналы) вет-
вясь, образуют восемь мери-
диональных каналов, идущих
параллельно мерцательным
ребрам. Таким образом, в ки-
шечнике выражены одновре-
менно и дву- и восьмилучевая
симметрии.
В связи с образом жизни и
характером пищи у гребневи-
ков наблюдаются значитель-
ные видоизменения кишечника.
Строение, описанное выше, ха-
рактерно, главным образом,для
цидиппид, питающихся мелки-
ми планктонными организмами
(ракообразными, личинками,
яйцами рыб), которых они ло-
Рис. 18. Ползающий гребневик Coeloplana
mesnili (по Давыдову)
ВЯТ разветвленными щупаль- Г —папиллы; 2 —гонады; 3 — щупальце; 4-ста-
цами И богатыми клейкими же- тоцист; 5 —глотка
лезами, необычайно растяжи-
мыми: например, у гребневика высотой в 7,5 мм растягиваются до 75 см
в длину (Kaestner, 1954/1955). У бероид, питающихся крупной добычей,
другими гребневиками, в том числе и цидиппидами, но не ракообразны-
ми (Камшилов, 1955, 1960), необычайно увеличиваются в размерах рот
и особенно глотка, которая у Вегоё занимает большую часть внутренней
полости; рот превращается в ловчий аппарат, которым гребневик захва-
тывает и заглатывает свою добычу целиком. Меридиональные и глоточ-
ные каналы образуют тонкие ветвящиеся разветвления и соединяются
кольцевым каналом. У платиктенид глотка становится еще больших раз-
меров, ее дистальная часть, как и у способной ползать цидиппиды Lam-
petia (Hyman, 1940), становится ползателыюй подошвой тела; меридио-
нальные каналы их образуют густую сеть разветвлений.
В глотке, выстланцрй однослойным мерцательным эпителием, по
обеим плоским сторонам двойные валики несут железистые клетки, с
помощью ферментов которых происходит внеклеточное пищеварение
(полостная фаза пищеварения). Из желудка в каналы кишечника посту-
пает жидкая, частично переваренная пища. Их эндодермальный эпите-
лий на наружной стороне состоит из клеток с ресничками и нереснич-
ных амебоидного характера клеток с вакуолями, служащих для внутри-
клеточного пищеварения (внутриклеточная фаза), а возможно и для
экскреции. Циркуляция жидкости в кишечнике вызывается движением
61
ресничек и напоминает циркуляцию крови в кровеносных сосудах
(Komai, 1922). В каналах кишечника, очевидно, происходит и газовый
обмен, так как органов дыхания у гребневиков нет, как нет и выдели-'
тельных органов. Экскреторную функцию допускают у ресничных розе-
ток, что, однако, экспериментально не проверено.
Гребневики — гермафродиты, половые железы у них находятся в
меридиональных каналах кишечника; яичники—на одной стороне, се-
менники— на другой. Происхождение первичных половых клеток в
онтогенезе не прослежено, однако связь их с эктодермальными стенка-
ми меридиональных каналов несомненно вторичная и вызвана питани-
ем половых клеток.
Рис. 19. Начальные стадии развития гребневиков (из Heider)
/—микромеры; 2 — макромеры; 3 — мелкие эктодермальные клетки; 4~ энтодерма;
5 — эктодерма
Размножение гребневиков — полов,ое. У большинства яйца пелагиче-
ские и оплодотворяются в воде. Половое созревание и размножение у
некоторых видов цидиппид, как известно, наступает у личинок, которые,
достигнув сформированного состояния, вновь приступают к размноже-
нию (явление диссогонии).
Половые продукты выводятся обычно через рот, только у платикте-
нид семенники имеют протоки, открывающиеся наружу, а яйца,' не ста-
новясь пелагическими, развиваются на брюшной стороне матери: на-
пример, у Coeloplana bockii (Komai, 1922).
Эмбриологии типичных гребневиков, главным образом Callianira и
Вегоё, посвящено большое количество работ (Metschnikoff, 1885; Kowa-
levsky, 1880, 1886; Chun, 1880; Ziegler, 1898; Fischel, 1887—1898; Spek,
1926; Yatsu, 1911, см. Давыдов, 1914; Pflugfelder, 1962; П. П. Иванов,
1937; Шмидт, 1951, 1953; Us, 1932). Исследования по развитию пла’тикте-
нид принадлежат Комаи (Komai, 1922), а краткие наблюдения К. Н. Да-
выдову (см. Бляхер, 1963).
Дробление яиц (включая и платиктенид) двусимметричное, детер-
минированное, мрзаичное, что обусловлено строением яйца (Komai,
1922; Давыдов, см. Бляхер, 1963). Первая борозда дробления проходит
по будущей сагиттальной плоскости гребневика, вторая — по латераль-
ной плоскости. Бластомеры на 8-клеточной стадии расположены дву-
симметрично. Третья борозда дробления, проходящая несколько нз-
62
искось, дает 16-клетрчную стадию из 8 макромеров и 8 микромеров,
составляющих четыре-квадранта из 2 макрбмеров и 2. микромеров каж-
дый. Микромеры делятся (рис. 19), а. макромеры поворачиваются на
180°, отделяют каждый по микромеру и поворачиваются еще раз, но уже
на 90°. Микромеры при этом смещаются к анимальному полюсу и об-
разуют крестообразный зачаток эктодермальной мезенхимы. Микроме-
ры— клетки эктодермы, делясь, обрастают макромеры — клетки энто-
дермы. Впячиванием клеток эктодермы образуется рот и эктодермаль-
ная глотка. Клетки энтодермы образуют зачаток первичного кишечни-
ка, дающего желудок и зачаток кишечных каналов сначала в виде четы-
рех эктодермальных карманов. Впячиванием эктодермы на аборальном
полюсе зародыша возникает аборальный орган чувств с балансерами
и статоцистами. В латеральной плоскости зародыша из эктодермы об-
разуется зачаток двух щупалец (кроме бероид) с влагалищем (цидип-
пиды) или без него (лобаты). Кроме энтрмезенхимы, имеются еще клет-
ки эктомезенхимы, которые отделяются от эктодермы глотки позднее,
в течение всего постэмбрионального развития (см. Шмидт, 1953), и уча-
, ствуют в образовании мезоглеи и ее клеточных элементов. И. И. Меч-
ников >(1955, Metschnikoff, 1885) считал, цто из латеральных частей кре-
стообразного зачатка — энтрмезенхимы (по терминологии Мечникова
«мезодермы») образуется мускулатура щупалец, а из медиальных —
блуждающие клетки мезоглеи, он ошибочно признавал у ктенофор ме-
зодерму. К сожалению, и в настоящее время нет точных эмбриологиче-
ских данных о происхождении нервной системы, мускулатуры стенок
тела и кишечника гребневиков. Врзможно, что эктомезенхима дает му-
скулатуру стенок тела, а энтомезенхима — мускулатуру кишечника.
Из яиц всех гребневиков, если не считать паразитического Gastro-
des, у которого образуется личинка — планула, вылупляется цидиппид-
ная личинка у цидиппид, вырастающая с развитием половых желез во
взрослую ктенофору, а у остадьных претерпевающая сложную пере-
стройку (в сущности метаморфоз) в соответствии со строением взрослых
форм, особенно таких, как Cestida, Platyctenida и Beroida.
В онтогенезе гребневиков, не говоря об особенностях организации
детерминированных яйца и стадий дробления, проявляются черты двулу-
чевой и четырехлучевой симметрии. Эктодерма образует четыре зачат-
ка будущих ребер с гребными пластинками, бластопор имеет крестооб-
разный просвет, клетки эктодермальной мезенхимы дают крестообраз-
ную закладку, энтодема^льный кишечник образует четыре кармана,
дающих систему гаструлярных каналов.
Экспериментальные данные (Fischel, 1887—1898; Yatsu, 1912а, 1912b;
Spek, 1926, цит. по Farfaglio, 1963) говорят о детерминированности бла*
стомеров дробящегося яйца ктенофор, определяющих развитие, напри-
мер, гребных пластинок, и известной регулятивной способности после
удаления части бластомеров до восстановления цельной, но уменьшен-
ной личинки.
По более новым данным, главным образом К- Н. Давыдова (см.
Бляхер, 1963), индокитайские платиктениды обладают небольшой ре-
генерационной способностью и не могут размножаться путем деления.
Филогенетические отношения между отрядами гребневиков сравни-
тельно просты. Основной, наиболее типичной и примитивной группой
являются цидиппиды. Наиболее близки к ним — лопастные, отличаю-
щиеся от цидиппид в основном развитием лопастей в оральной части
тела. Близкородственными к ним надо считать платиктенид, хотя у по-
следних организация, вследствие перехода к ползанию, заметно изме-
нена: они приобрели планариеобразный облик. Эти три отряда по ко-
личеству видов составляют главную массу гребневиков; остальные пред-
ставлены единичными видами. Удлинение тела в виде ленты и змеевидное
63
движение привели к значительному изменению организации цестид (по
сравнению с таковой цидиппид); отсутствие или, вернее, утрата щупалец
и приобретение формы ’высокой шапки, или скуфьи, вызвало сильное из-
менение в строении бероид. Однако наличие у всех групп цидиппидной’
личинки и формирование в ходе онтогенеза из нее взрослых форм делают
ясным возникновение отрядов гребневиков от общей прародительской
цидиппидной формы.
Известно, что еще Гатчек (Hatschek, 1888) считал гребневикрв рав-
ноценными книдариям и губкам. Некоторые зоологи, из советских Лива-
нов (1945, 1955), из зарубежных Hyman (1940), Storer a. Usinger
(1957) и другие, считают гребневиков и целентерат (книдарий) само-
стоятельными типами. Если ограничиваться рассмотрением только не-
которых черт (таких, как радиальная с элементами двух-, четырех- и
восьмилучевой симметрии, единая — гастральная с ротовым отверстием
полость, простота строения стенок тела и гонад, низкий уровень строе-
ния и физиологии нервной системы, отсутствие выделительных органов,
сохранение главной оси зародыша во взрослом состоянии), книдарий и
гребневиков можно было бы считать двумя подтипами одного типа.
Однако совокупность ряда черт строения и развития ктенофор рез-
ко отличает их от кишечнополостных, или книдарий. Так, надо отме-
тить наличие двигательного аппарата из восьми меридиональных ребер
с гребными пластинками, двух пл,отных мускулистых щупалец с клей-
кими клетками и аборального органа чувств, связанного с восемью ра-
диальными сгущениями нервной сети. Чертами онтогенеза, специфич-
ными для гребневиков, являются детерминированный, мозаичный харак-
тер дробления с образованием микромеров (эктодермы), макромеров
(энтодермы) и микромеров, дающих мускулатуру, почти прямое превра-
щение личинок во взрослых особей, без резких перестроек, характерных
для метаморфоза, развитие мускулатуры стенок тела и кишечника за
счет мезенхимы и т. д. На основании всего этого надо считать гребне-
виков самостоятельным типом, стоящим значительно выше, чем тип ки-
шечцополостных.
Выработав организацию, так хорошо приспособленную к пелагиче-
скому, свободному существованию, лишь несколько изменяющуюся в
соответствии с характером плавания, гребневики сохранили достаточ-
ную пластичность для перехода от плавания к придонной жизни, к пол-
занию. Изменения организации с переходом к ползанию таковы, что
они дали основание А. О. Кавалевскому (Kowalevsky, 1880), Лангу
(Lang, 1884) и ряду последующих авторов считать платиктенид пред-
ками турбеллярий. В настоящее время однако (Komai, 1922; Kaestner,
1954/1955) достаточно ясно, чтр платиктениды, благодаря сходному
образу жизни, лишь уподобились в ряде отношений плоским червям.
У них нет настоящей двусторонней симметрии, дифференцировки нерв-
ной системы на ортогон и коннективы, паренхимы, кожно-мускульного
мешка из сложно расположенных слоев мышц, органов выделения —
протонефридиев, полового аппарата со сложной системой дополнитель-
ных желез и половых протоков. У них нет и спирального типа дробле-
ния и развития с настоящим метаморфозом.
Вопрос о происхождении и филогенетических связях гребневиков с.
кишечнополостными и плоскими червями обсуждался многими зоолога-
ми. Гаймен (Hyman, 1940) и некоторые другие авторы, признающие
основной, родоначальной формой для кишечнополостных медузу, произ-
водят гребневиков от медузоидной формы, при этом (например, Яковлев,
1964), признавая исходным образо?и жизни для этих животных планк-
тонный, считают основным стволом эволюции не книдарий, а пелагиче-
ских гребневиков. Другие авторы рассматривают гребневиков как не-
отенические формы, возникшие от личинок коралловых полипов (Kors-
64
chelt u. Heider, 1890—1893; Franz., 19^4 и др.)' или от личинок медуз
(Третьяков, 1936). Последний-автор опирался при этом на сходству в
строении мезоглеи сцифомедуз и гребневиков (Treti'akoff, 1930), но не-
доучитывал совокупности различий в характере строения и развития
этих групп. ' • '
Однако организация и тип онтогенеза гребневиков, поразительно при-
способившихся к пелагическому образу жизни, настолько отличны от
таковых кишечнополостных (см. стр. 26), что нам представляется невоз-
можным выводить ни гребневиков от кишечнополостных, ни наоборот.
Во всяком случае несомненно, что эволюционный уровень гребневикрв
выше уровня кишечнополостных и они являются более поздними фор-
мами. К сожалению, пока не найдены ископаемые гребневики, нет воз-
можности судить об их геологическом возрасте.
Взгляд на родство между гребневиками и плоскими червями — турбел-
ляриями — ведет свое начало от работы А. О. Ковалевского (Kowalev-
sky, 1880) над Coeloplana и Ланга (Lang, 1884) и был принят многими
зоологами. Однако надо считать маловероятной гипотезу о происхожде-.
нии гребневиков путем неотений от пелагической личинки поликлад
(MacBride, 1914, а также Hadzi, 1929, 1957, 1958, 1963; Beer, 1954), Ни
онтогенез, ни анатомия не дают оснований для такого взгляда. Вслед-
ствие перехода от исконного пелагического к донному, ползающему об-
разу жизни гребневики, как было сказано, лишь уподобляются плоским
червям — турбеллярием; по всей совокупности черт строения и отогене-
за они стоят на более низком эволюционном уровне, чем турбелдярии,
и не мргут быть их потомками. Составляя самостоятельный тип, стоя-
щий в ряде отношений выше кишечнополостных по уровню эволюции,
они ближе к последним.
5 Д. М. Федотов
Плоские черви (Plathelminthes)
Плоские черви составляют единый тип низших паренхиматозных
червей, основной, родоначальной группой которого являются турбелля-
рии, почему в последние годы некоторые зоологи (Remane, 1954, 1959)
называют его Turbellariomorpha.
Двустороннесимметричное, обычно плоское или удлиненночервеоб-
разное тело паренхиматозное, без полости тела, покрыто ресничным
эпителием у взрослых свободноживущих червей и у личиночных стадий
или кутикулой во взрослом состоянии у паразитических форм. Система
субэпителиальных и паренхимных мышц кожно-мускульного мешка
обеспечивает червеобразное поступательное движение тела, лишенного,
как правило, наружных органов движения. Ортогонального типа нерв-
ная система, лежащая в паренхиме, реже йнтра- или субэпителиально,
состоит из 6—2 продольных нервных стволов и церебрального ганглия.
Из слаборазвитых органов чувств наиболее обычны примитивного строе-
ния глазки на переднем конце тела. Кишечник — со ртом и средней киш-
кой, но без задней кишки и анального отверстия, с внутриклеточным и
частично полостным пищеварением, реже вторично упрощенный до син-
цитиальной массы без просвета у бескишечных турбеллярий или нацело
исчезнувший у цестод. Органы кровообращения и дыхания отсутствуют.
Функцию распределения несет кишечник. Органами осморегуляции и
выделения служат протонефридии, чаще всего в виде двух продольных
каналов, пронизывающих паренхиму своими разветвлениями и откры-
вающиеся наружными порами. Половая система, в противоположность
общей организации и примитивному строению большинства органов,
сложна, высоко дифференцирована, как правило гермафродитная, она
состоит обычно из яичников и семенников, часто желточников, ряда до-
полнительных желез, половых протоков и совокупительных органов:
оплодотворение — внутреннее. Развитие в исходном состоянии проте-
кает со спиральным детерминированным дроблением яиц и образова-
нием квартетов бластомеров (турбеллярий Polyclada, Rhabdocoela),
упрошенных до дуэтов (Acoela) с прямым и свободным личиночным раз-
витием, с типичным образованием зародышевых листков или без зак-
ладки последних и развития зародыша червя из синцитиальной массы
(сосальщики, цестоды), образованной нередко макромерами, мезомера-
ми и микромерами и с образованием свободноподвижных личиночных
временно ресничных стадий, нередко со сложной сменой поколений и
сменой временных и дефинитивных хозяев (паразитические классы).
Среди плоских червей встречаются как свободноживущие подвижные,
мелкие (до 3 мм длиной) формы, плавающие с помощью ресничек, так
и крупные формы, ползающие благодаря сокращениям кожно-му-
скульного мешка. Большинство плоских червей — наружные или вну-
тренние паразиты, питающиеся слизью, кровью, соками и тканями сво-
их хозяев; встречаются и хищники, питающиеся, соразмерно величине,
мелкой пищей или различными беспозвоночными.
66
Редко (в семействе цестод Microstomidae) наглодается бесполое раз-
множение поперечным делением, но многочисленность цестод, по суще-
ству, является не доведенным до конца поперечным делением (Бек-
лемишев, 1952, 1964). Высокая степень регенерации и регуляции — вос-
становление организации целого организма со всеми системами органов
из ничтожного куска тела характерна для многих турбеллярий, особен-
но бескишечных, рабдоцелид и триклад. Экспериментально установлено
также, что при голодании появляется упрощение и распад многих орга-
нов— полового аппарата и паренхимы (Шульц, 1908).
Свободноживущие плоские черви, как пелагические, так и бентос-
ные формы, исцользуют разнообразные биотопы Мирового океана, а
также пресные воды и влажные места суши. Паразитические формы,
связанные со свободноживущими через эктокомменсалистов, объеди-
няют экто- и эндопаразитов, паразитирующих во временных или по-
стоянных хозяевах, принадлежащих почти ко всем типам животного
мира.
Тип плоских червей, до недавнего времени подразделявшийся на три
класса — Turbellaria, Trematoda и Cestoda, теперь разбит на значительно
большее число классов, о чем говорится ниже.
Отсутствие ископаемых остатковг заставляет при восстановлении пу-
тей эволюции и филогении плоских червей ограничиваться заключения^
ми, построенными на изучении современных форм.
Несмотря на часто значительные особенности внешней формы, на гро-
мадное разнообразие деталей строения половой системы, общность основ
организации классов плоских червей позволяет считать их единым ти-
пом— Plathelminthes.
Турбеллярий как родоначальная группа, а все остальные классы
плоских червей как паразиты позвоночных (в том числе и человека)
и ряда различных классов беспозвоночных привлекают внимание мно-
гих исследователей нашегр столетия. Трудно дать достоверные сведе-
ния о количестве видов плоских червей.' Если недавно (Kaestner,
1954/1955) для всего типа насчитывалосьи%олее 5500, то в последнее
время одних моногенетических сосальщиков насчитывается около
1500 видов (по устному сообщению Б. Е. Быховского), а дигенетиче-
ских (Hyman, 1959) свыше 5000. В меньшей степени, но все же продол-
жает нарастать число описанных видов и для турбеллярий. Среди пара-
зитических классов плоских червей увеличивается число новых отрядов,
подклассов и даже классов, в основном в результате лучшего изучения
и пересмотра признаков организации; созданы, например, классы —
Udonelloidea, Monogenoidea и др. Среди турбеллярий нахождение та-
ких форм, как Xenoturbella, Gnathostomula, раздвигает рамки Класса
турбеллярий и делает, видимо, необходимым создание нового класса для
Xenoturbellida и нового отряда для Gnathostomulida, не укладывающе-
гося в рамки класса турбеллярий.
Диапазон в размерах плоских червей велик. Размеры турбеллярий
вообще незначительны, они колеблются у бескишечных между 0,4—
1,2 мм, изредка достигая 12 мм (Anaperus sulcatus из Каспия — Век-
lerniSchew, 1915). Рабдоцелиды также мелкие черви, от 0,3 до 2—3 м.м
длиной. Поликлады более крупные, их размеры колеблются от 2—3 мм
до 16 см. Триклады крупнее всех остальных турбеллярий, среди них са-
мые мелкие формы длиной 2—3 мм, а самые крупные достигают порой
относительно громадной величины, 50—60 см в длину (ряд наземных
триклад, Bipalium и др.).
Среди дигенетических сосальщиков, размеры которых колеблются
между 1—2—8 см, встречаются как редкость гиганты вроде Rhinoboth-
rium filarina, достигающие 1 —1,5 м в длину. Более мелкими формами
являются Моногенетические сосальщики, размеры которых колеблются
5*
67
между 0,03—20 мм в длину и не превышают 30 мм (Squalonocochotyle
borealis'). Среди цестод, в общем наиболее крупных среди плрских чер-
вей и, достигающих 1.2—-15 м в длину (Diphyllobothrium latum), встре-
чаются и карликовые формы вроде Davainea proglottina— 0,5 мм или
Echinococcus granulosus 2,5—6 мм длиной.
За . редкими исключениями, вроде дигенетических сосальщиков
Wedlia и Didymocystis, имеющих мешковидную форму (Hyman, 1951),
плоская червеобразная форма тела четко сохраняется в типе плоских
червей. Даже членистые цестоды, порой состоящие из 4000'—4500 прр-
глоттид, не нарушают эту характерную черту типа. Среди Cestodes из-
вестны черви, отдельные проглоттиды которых крайне подвижны и бы-
стро передвигаются, например, Railietina cesticillus (Hyman, 1951).
Более того, у цестоды Trilocularia (Догель, 1959) отделившаяся проглот-
тида в кишечнике акулы сильно увеличивается, в ней развиваются поло-
вые продукты, на одном ее конце возникают, крючья, которыми такой
червь-проглсттида прикрепляется к стенке кишечника хозяина. Отдель-
ные проглоттиды у Phyllobothrium musteli (Fuhrmann, 1928—1933) по-
парно спариваются, как два самостоятельных червя. Эти факты служат
также подтверждением взгляда В. Н. Беклемишева (1952, 1964) о том,
что членистость цестод есть результат недоведенного до конца бесполо-
го размножения поперечным делением.
В ходе регрессивной эволюции, у немертин, например, Maldcobdella,
наблюдаются случаи приобретения внешней формы ниже' стоящего
типа — сосальщиков-. У кольчатых червей мы знаем пример утраты
ряда черт наружной и отчасти внутренней организации, что делает их
сходными с низшими червями, например, Donophilus, Protomyzostomum
и др. .Среди моллюсков также известны случаи упрощения организации
и уподобления турбелляриям, например; Rhodope, Pseudoverm.es и т. д.
Занимая низшую ступень в системе Bilateria, плоские черви, есте-
ственно, в случаях регрессивного развития внешне уподобляются еще бо-
лее низкрорганизованным формам. Действительно, малые размеры,
утрата полости кишечника и превращение его в паренхиму, скопление
клеток, захватывающих микроскопически мелкую пищу (диатомовые,
мелкие животные) и внутриклеточно переваривающих ее, делают'похо-
жими Acoela на инфузорий. Это было причиной увлечения Хаджи
(Hadzi, 1944, 1958, 1963), Штейнбока (Steinbock, 1958) и Ганзона (Han-
son, 1958) неудачной идеей о происхождении турбеллярий — Acoela от
инфузорий.
Никто не сомневается, что турбеллярий по своему строению, онтоге-
незу, физиологии, экологии (хищничество, комменсализм и пара-
зитизм, различные биотопы моря, пресных вод и влажной почвы), явля-
ются основным классом типа плрских червей (рис. 20). Различны взгля-
ды на направление эволюции этой группы, на то, являются ли современ-
ные Acoela ob origine примитивными формами или вторично упростив-
шимися. Многие зоологи, из современных особенно В. Н. Беклемишев,
А. В. Иванов, Гаймен приводят как будто убедительные доводы в пользу
первичной примитивности бескишечных турбеллярий. Другие зоологи,
начиная с С. М. Переяславцевой (1892, Perejaslavceva, 1885), Мак-
Брайд (MacBride, 1914), а из современных Н. А. Ливанов (1955),
С. И. Богомолов (1960, 1964), Маркус (Marcus, 1958), Ремане (Remane,
1954, 1959), Беттгер (Boettger, 1952), Акс (Ах, 1961), на мой взгляд,-
резонно считают бескцшечных турбеллярий вторично регрессировавши-
ми формами, но обладающими и рядом примитивных черт. Несомненно,
по состоянию тканевой дифференцировки и уровню организации (кроме
строения половой системы) бескишечные турбеллярий стоят ниже всех
двустороннесимметричных форм. Установленная недавно необычайно
сильная регенеративная способность этих турбеллярий (Steinbock a. all.,
6.8
Рис. 20. Схема строения различных турбеллярий (из Hennig)
.4—Acoela; Б — Polycladida; В — Neorhabdocoela; Г — Triclada; / — кишечник; 2 — гермарий; -5-
семенник; 4 — экскреторный орган; 5 — копулятивный аппарат; 6— половое отверстие; 7 —щупальце;
8 — глаз; 9 — нервная система; 10— желточник; // — глотка; 12— фронтальный орган; 13 — перифе-
рическая паренхима; 14 — кишечная паренхима; 15 — рот и половое отверстие самца
пит. по Hyman 1959) в некоторой степени подкрепляет мнение об их
первичной примитивности.
Если принять, что необособленность покровных клеток от паренхи-
мы и отсутствие базальной перепонки, наличие нервной системы в виде
сплетения без дифференцировки на продольные стволы и отсутствие
кишечника являются исходным состоянием Acoela, тогда этих турбел-
лярий можно считать не только низшими червями, но и животными ме-
нее организоваными, чем кишечнополостные. Однако следует помнить,
что наряду с примитивными чертами организации Acoela обладают уже
сложной половой системой, характерной для типа плоских червей, нали-
чие которой заставляет думать, что многие их кажущиеся примитивны-
ми черты являются результатом вторичного упрощения.
Соблазнительно, как делает В. Н. Беклемишев (1952, 1964), считать
турбеллярий произошедшими от Acoela путем прогрессивного развития
покровного эпителия от ресничных клеток, сомкнутых лишь на поверх-
ности тела (не отграниченных от паренхимы, но непосредственно свя-
занных с наружной паренхимой), к типичному покровному эпителию,
подостланному базальной мембраной, от субэпителиального нервного
сплетения к централизованной нервной системе, лежащей в паренхиме
в виде головного мозга и нескольких пар продольных стволов, от бес-
форменной пищеварительной паренхимы лишь с ротовым отверстием к
трубчатому кишечнику с мускулистой глоткой; от разбросанных или
собранных лишь в скопления мужских и женских половых клеток в па-
ренхиме без яйцеводов к сформированным семенникам и яичникам, оде-
тым оболочкой, от спирального дробления дуэтами к образованию
квартетов с отчетливо выраженными зародышевыми листками и т. д.
69
Рис. 21. Схема строения покровов Fasciola
hepatica (no Threadgold)
/ — базальная мембрана; 2 — кольцевые мышцы;
3 — продольные мышцы; 4— клетки паренхимы
Вполне возможно счи-
тать описанное состояние
ресничных клеток покровов,
диффузный характер нерв-
ной системы, отсутствие
сформированного кишечни-
ка —примитивными черта-
ми строения, если исходить
из теории И. И. Мечникова
о происхождении многокле-
точных от гипотетической
фагоцителлы и если мино-
вать в истории развития
многоклеточных эволюцию
типов кишечнополостных и
гребневиков. Однако, счи-
тая, что родоначальные фор-
мы кишечнополостных и
плоских червей были близ-
кими, учитывая, что для
всех типов многоклеточных
животных характерным яв-
ляется наличие кишечника,
а его отсутствие всегда, а
тем более у типов Bilateri'a,
бесспорно — явление вторич-
ное, надо признать вторич-
ным отсутствие кишечника и
у Acoela. Наличие у ряда
Acoela глотки, при отсутст-
вии кишки, как справедливо
отмечает Н. А. Ливанов
(1955), говорит о редукции у них энтодермальной кишки. Об этом гово-
рит и онтогенез Acoela, у которых в прошлом столетии С. М. Переяслав-
цева (1892, Perejaslavceva, 1885) и некоторые другие, а недавно С. И. Бо-
гомолов (1960, 1964) наблюдали гаструляцию и гастральную полость,
почему Переяславцева с полным правом переименовала Acoela в Pseu-
doacoela.
В прльзу вторичного упрощения тела могут говорить и ничтожные
размеры, плавание с помощью ресничек, питание микроскопически мел-
кой добычей; все это могло привести у ряда Acoela к упрощению строе-
ния нервной системы. Это тем более вероятно, что даже у ряда члени-
стоногих, например, клещей, в связи с ничтожными размерами тела
наблюдаются упрощение строения и утрата мальпигиевых сосудов, ки-
шечника, кровеносной системы.
Примитивность в строении покровного эпителия Acoela бесспорно
заключается в том, что у них, как подчеркивает В. Н. Беклемишев (1915,
1916, 1925, 1937, 1952, 1964), имеется три типа строения покровного эпи-
телия: погруженный эпителий, немертиновый и нормальный. В отдель-
ности эти типы эпителия реализуются у разных отрядов турбеллярий.
У некоторых Acoela имеются даже эпителиально-мышечные клетки по-
кровного эпителия, которые, как известно, типичны для ряда кишечно-
полостных, но как примитивная черта встречаются и у других типов, на-
пример, иглокожих.
Благодаря жизни во влажных биотопах почвы наземные триклады
сохраняют ресничный покровный эпителий, а остальные классы (пара-
зитические плоские черви) в ходе эволюции или сохраняют ресничный
70
эпителий только у личинок (дигенетические, моногенетические сосаль-
щики, цестоды), или на всех стадиях развития приобретают кутикуляр-
ные покровы и погруженный характер эпителия (рис. 21).
В жизни турбеллярий большое приспособительное защитное значе-
ние при необходимости сохранения нежных ресничных покровов имеет
развитие белковых и слизистых кожных желез; рабдиты, например,
образуют чехол вокруг тела; прилипая к добыче или врагу, они задер-
живают их движение. Кроме того, слизистые нити кожных желез помо-
гают движению турбеллярий (Беклемишев, 1952, 1964; Hyman, 1959).
Среди также сильно развитого головного комплекса желез у личинок и
зародышей дигенетических сосальщиков и цестод выделяются железы,
секрет которых вызывает лизис тканей хозяина, позволяющий паразиту
проникнуть внутрь хозяина (Hyman, 1951, 1959).
Многие ученые, из советских Е. Д. Логачев (1957а, 19576, 1961),
Б. Е. Быховский (1937, 1957) и некоторые другие, отрицают наличие
эктодермального эпителия у паразитических плоских червей, поскольку
эмбриональная эктодерма в ходе их онтогенеза сбрасывается. Другие
исследователи, Кестнер (Kaestner, 1954/1955), А. В. Иванов (1952а,
19526), в том числе и я, считают покровы паразитических червей на-
стоящим погруженным эпителием. Опираясь на данные онтогенеза (см.
ниже), я не могу признать отсутствия у паразитических плоских червей
эктодермальных покровов и мезодермального или паренхимного харак-
тера последних. Моя точка зрения подтверждается и наличием у всех
классов плоских червей, начиная с низших турбеллярий, субэпителиаль-
ных эктодермального происхождения слоев мускулатуры, сквозь кото-
рые опускаются базальные концы клеток погруженного эпителия, по-
мимо основной части кожно-мускульного мешка паренхимного происхож-
дения (см. также Threadgold, 1963).
Как известно (Беклемишев, 1944, 1952, 1964; Зенкевич, 1944; Ливанов,
1955), у плоских червей впервые в животном мире возникает кожно-
мускульный мешок, обеспечивающий сохранение, или изменение формы
тела, растягивание последнего в ниточку или сокращение с превраще-
нием в шарик, а также червеобразное изгибание тела и прямолинейное
движение — ползание по дну и плавание (Беклемишев, 1952). Поступа-
тельное движение при помощи ресничек тела сохраняется лишь у мел-
ких турбеллярий, преимущественно Pseudoacoela, у остальных оно за-
меняется работой мускулатуры тела. У низших олигомиарных форм
турбеллярий относительная масса мускулатуры невелика (Зенкевич,
1944), у мезомиарных (поликлад и триклад) она значительнее, но все
же редко превышает 30% массы тела. Вместе с тем идет эволюция всего
мышечного аппарата, который у наземных триклад состоит, по Граффу,
из 12 групп двигательных мускулов (Зенкевич, 1944).
У паразитических классов плоских червей при общем регрессе муску-
латуры тела наблюдается высокая специфическая дифференцировка
мускульного аппарата органов прикрепления, что особенно характерно
для прикрепляющихся моногенетических сосальщиков и цестод. Органы
прикрепления (церкомеры) признаются у моногенетических сосальщиков
и цестод гомологичными друг другу, но они отличаются по положению
у зародыша тем, что прикрепительный аппарат у первых развивается
на заднем конце, а у вторых —на переднем (Быховский, 1937; Ogren,
1956, 1957).
Паренхима, которой плоские черви обязаны названием — паренхима-
тозных и которая впервые появляется в животном мире в этой группе
червей, представлена синцитием из звездчатых, многоветвистых клеток
основного вещества и несколькими типами свободных клеток, производ-
ными мелких базофильных амёбоцитов (Prenant, 1922, Заварзин, 1945,
1947; Логачев, 1957а, 19576, 1961).
71
Впервые у плоских червей вместе с выделительными и фагоцитар-
ными клетками паренхимы возникает ретикуло-эндотелиальная систе-
ма, но в отличие от мнения А. А. Заварзина и Е. Д. Логачева в ее оцен-
ке Я' исхожу не от теории фагоцителлы И. И. Мечникова, а от данных
индивидуального развития поликлад как исходного для всего типа пло-
ских червей.
Я подчеркиваю, что более поверхностная часть паренхимы по проис-
хождению— эктомезенхима, а остальная (большая) часть паренхимы —
энтомезодермальная, т. е. в строении тела плоских червей участвуют
эктодерма, энтодерма и мезодерма. Однако я считаю, что В. В. Зален-
ский (Salensky, 1907) был не прав, когда на основании наличия у плоских
червей мезенхимы и мезодермы, считал их деградированными потомками
целомических форм (см. стр. 76).
Необходимо подчеркнуть как важную особенность рабдоцелид на-
личие в паренхиме схизоцельных полостей, которые у типа первичнопо-
лостных червей (Askelminthes) являются источником происхождения
полости тела.
Кишечник плоских червей обнаруживает разные ступени эволюции и
состоит из эктодермальной глотки разных типов строения (простой,
складчатой или массивной), открывающейся ротовым ,отверстием, и
трубчатой, мешковидной дву- или многоветвистой энтодермальной киш-
ки. Задняя кишка и анальное отверстие отсутствуют.
Большинство зоологов считает тацое строение кишечника (с ротовым
отверстием как для введения пищи, так и для выведения фекалий) исход-
ным, примитивным, целентерическим по выражению Н. А. Ливанова
(1955). Некоторые же зоологи, преимущественно немецкие (например,
Boettger, 1952; Remane, 1959; Steinbock, 1958), не учитывая сравнитель-
но-анатомических и онтогенетических данных, считают, что плоские чер-
ви утратили заднюю кишку и анальное отверстие, которые вследствие
смены функции превратились в бурсу. Поры, открывающиеся наружу,
встречающиеся в кишечнике ряда турбеллярий, моногенетических и ди-
генетических червей (у некоторых из последних существует даже сооб-
щение кишечника с мочевым пузырем), не функционируют как аналь-
ное отверстие (Беклемишев, 1952, 1964) и вряд ли имеют какое-либо
отношение к отсутствующим у плоских червей задней кишке и поро-
шице.
Я считаю правильным трактовать отсутствие кишечника или поло-
сти в энтодермальной пищеварительной паренхиме у Acoela, как явле->
ние вторичное^, Не могу согласиться с точкой зрения Штейнбока или
Ремане об утрате задней кишки с анусом или о превращении части сред-
ней или задней кишки в бурсу.
Пищеварение у плоских червей с кишечником — преимущественно
внутриклеточное, реже полостное, энзиматозное, у цестод—парэнте-
ральное. Сильно разветвленный кишечник у крупных турбеллярий не-
сет, кроме пищеварительнрй, и распределительную функцию.
Турбеллярий — хищники, часть — типичные, микрофаги. Основная
масса' питается (в зависимости от размеров) гидрами, мелкими личин-
ками, насекомыми, рачками, олигохетами, моллюсками. Некоторые тур-
беллярии, например, Kalyptorhynchidae, пользуются для поражения
добычи хоботом со стилетом, другие для этого применяют стилет пениса,
открывающегося в ротовую полость (Kaestner, 1954/1955). Среди Acoela
и особенно Rhabdocoela немало форм, ведущих эндопаразитический об-
раз жизни. Дигенетические и моногенетические сосальщики питаются
через рот слизью, кровью, клетками тканей, частью пищи хозяина и об-
ладают, по-видимому, преимущественно полостным пищеварением. Це-
стоды, обладающие парэнтеральным пищеварением, абсорбируют пищу
непосредственно поверхностью тела.
72
Нервная система плоских червей, находящаяся на низком уровне
развития, представлена диффузным интра- или субэпителиальным,спле-
тением у низших турбеллярий; у большинства она уходит в паренхиму,
претерпевает централизацию и в известной степени цефализацию, при-
обретая строение ортогона с резкой степенью развития головного моз-
га, снабжающего передний конец тела с его органами чувств нескольки-
ми парами нервов, и с продольными нервными стволами в количестве от
6 до 1 пары, соединенными поперечными комиссурами (Беклемишев,
1952, 1964; Коштоянц, 1957). Ортогональный тип строения характери-
зует нервную систему и паразитических классов: у сосальщиков 3—
2 пары, у цестод 2—5 пар продольных нервов.
В целом эволюция нервной системы плоских, особенно своб.одножи-
вущих червей связана отчасти с размерами тела, а, главным образом, с
развитием и состоянием мускулатуры тела, с подвижностью и хищниче-
ством турбеллярий. Происходит дифференциация чувствительных и дви-
гательных разделов нервной системы — головного мозга, перифериче-
ской сети и продольных нервных стволов. Обращает однако на
себя внимание сильная концентрация нервной системы (до одной пары
нервов) и развитие головного мозга у Macrostomum из отряда низших
турбеллярий (Беклемишев, 1964), а также и положение ее у ряда Acoela
в паренхиме, а не поверхностно (Ах, 1961).
Головной м,озг турбеллярий является центром, регулирующим вза-
имоотношения с наружной средой: он контролирует нормальное движе-
ние, ловлю и заглатывание добычи, ориентацию и общую чувствитель-
ность, а также процессы регенерации (Коштоянц, 1957). К сожалению,
данных о физиологии нервной системы паразитических классов еще мало,
хотя в последние годы усиленно развиваются гистохимические и биохи-
мические исследования, характеризующие специфику обмена веществ
у этих классов (Hyman, 1959).
Из органов чувств, развитых у плоских червей в общем слабо, наи-
более характерны: орган равновесия, статоцист, имеющийся у Acoela,
Catenulida и Proseriata, и одна или две пары простых глазков на перед-
нем конце тела (многие турбеллярий, личиночные стадии дигенетических
сосальщиков и взрослые формы моногенетических). Глаза и статоцисты
по своему строению примитивнее, чем у кишечнополостных и гребневи-
ков, хотя, статоцисты, по мнению В. Н. Беклемишева, в пределах этих
групп являются гомологичными образованиями.
Система протонефридиев в целом однотипна, хотя в числе и располо-
жении главных каналов, их соединений, мочевых пузырей и нефропоров
в разных классах наблюдается большое разнообразие. Обычно отмеча-
ют наибольшее сходство протонефридиев у далиеллид и паразитических
классов. Во всяком случае бесспорно, что наличие двух боковых каналов
протонефридиев, как и парность для большинства органов плоских чер-
вей и всех Bilateria, надо считать исходным и примитивным состоянием.
С протонефридиями как органами выделения и осморегуляции у разных
групп плоских червей связаны особые клетки — паранефроциты и ар-
троциты, из которых последние особенно сильно развиты при метамер-
ном расположении у Udonelloida (А. В. Иванов, 19526). Отсутствие
протонефридиев у Acoela ряд авторов, например Акс (Ах, 1961), по мое-
му мнению, не без основания считает вторичным. Экскреция обеспечи-
вается не только терминальными частями протонефридиев, но и клетка-
ми кишечника; продукты распада, кроме того, откладываются в стенках
тела, что обусловливает нередко яркую окраску тела турбеллярий. Тон-
чайшее строение и функция терминальных частей протонефридиев тре-
матод, выяснены с помощью электронного микроскопа (Kiimmel, 1958).
При невысоком, сравнительно с другими типами двустороннесим-
метричных животных, уровне организации большинства органов
73
плоских червей их половая система,< как общеизвестно, очень сложна и
разнообразна. Раздельнополость является лишь исключениями в каж-
дом классе (Sabussowia dioica и Cercyra teissieri—из морских триклад,
Didymozoonidae и Schistosomatidae — среди дигенетических сосальщи-
ков, Dioecocestus — среди цестод). В основном плоские черви — герма-
фродиты, обладающие внутренним оплодотворением; и их спермин, как
отмечают, далеко не примитивного строения. Основной тип строения
половой системы проходит через все классы, детали же строения на-
столько разнообразны, что решающими видовыми признаками Служат
частности ее строения.
Для многих отрядов классов плоских червей характерно наличие
двух — одного семенников и одного, реже двух яичников, половые отвер-
стия большей частью непарные, только Macrostomum обладает раздель-
ными мужскими и Женскими половыми отверстиями, а поликлады —
парными мужскими половыми отверстиями. Характерным для плоских
червей является эволюция женских половых желез: от яичников, обра-
зующих энтолецитальные яйца через гермовителлярии, к раздельным
собственно яичникам, или гермариям, и желточникам с экзолецитальны-
ми яйцами. Разнообразные дополнительные железы, сложные выводные
протоки, копулятивные органы, общий половой атриум или отдельные
мужские и женские половые отверстия, нередко наличие генитоинтести-
нального канала, порой сообщения мужского полового протока с ротовой
полостью, например, у Prorhynchus st agnails из Lecithoepitheliata (Lu-
ther, 1960), до некоторой степени дают представление о сложности и
разнообразии строения половой системы этих классов.
Несмотря на вторичное упрощение многих черт строения бескишеч-
ных турбеллярий, все же вряд ли можно оспаривать, что наличие у них
разбросанных мужских половых элементов в паренхиме и состояние
яичникрв, не одетых оболочкой, возникающих концентрацией вначале
разбросанных в паренхиме яйцевых клеток, является примитивным со-
стоянием. Вероятно, таким путем шла эволюция половых желез у древ-
них примитивных предков плоских червей, и у Acoela примитивное
строение могло сохраниться.
В общем, в эволюции полрвой системы плоских червей наблюдается
развитие от множественности половых желез к олигомерии. У поликлад
и Acoela семенников и яичников много, у рабдоцелид и у связанных с
ними происхождением остальных групп турбеллярий (дигенетических
и моногенетических сосальщик,ов, цестод и удонеллид) имеется два или
один яичник и во многих случаях пара или один семенник. Но наряду с
этим, например, у триклад и цестод, имеется множество семенников,
а у некоторых рабдоцелид рода Multipeniata путем физиологической ре-
генерации происходит множественное образование копулятивных органов
(Nassonov, 1932); у некоторых же Aco.ela (Anaperus gardineri)—мно-
жество семенных бурс (Hyman, 1951). Более того, у ряда триклад, на-
пример, Procerodes, метамерность настолько охватывет почти все сис-
темы органов, что напоминает метамерию кольчатых червей. Согласно
принципу олигомеризации В. А. Догеля (1954), Б. Е. Быховский (1957),
в отличие от большинства авторов и, на мой взгляд, в противоречии с
данными экологии о связях паразитов и систематическом положении их
хозяев, считает в классе Monogenoidea подкласс Polyonchoinea с боль-
шим количеством крючьев церкомера более примитивным, чем подкласс
Oligonchoinea. Несомненно, олигомеризация гомологичных органов яв-
ление широко распространенное, но оно далеко не закономерно, так как
имеется много примеров перехода и от олигомеризации к полимеризации.
Особенностью плоских червей, связанной с наличием в половой си-
стеме ряда желез, является внутреннее оплодотворение и откладывание
не просто яиц, а яйцевых коконов с несколькими яйцами или с яйцом и
74
желточными клетками. Более примитивным типом онтогенеза в целом
обладают турбеллярий, имеющие яичники с энтолецитальными яйцами,
Polyclada, Acoela, низшие Rhabdocoela — Catenulidae, Macrostomidae.
Измененный тип онтогенеза характерен для турбеллярий, имеющих яич-
ники и желточники и экзолецитальные яйца, это многие Rhabdocoela,
Alloeocoela, Triclada и классы паразитических червей.
Насколькр пластична и неустойчива организация взрослых турбелля-
рий разных отрядов (что влечет за собой продолжающуюся перестройку
классификации этих червей, уничтожение рядом авторов таких отрядов,
как Rhabdocoela, Alloeocoela и даже небольшого отряда Acoela), на-
столько же пластичны и неустойчивы и типы дробления. Характерным
для Acoela является спиральное дробление с .образованием дуэтов (Бо-
гомолов, 1960, 1964). Однако у одного и того же вида возможны прц
известных условиях дробление с образованием квартетов и даже одно-
лучевое дробление. Такая же неустойчивость спирального дробления
характерна и для Rhabdocoela с простыми яичниками и с энтолециталь-
ными яйцами. У одного вида — Macrostomum lineare Ben.— С. И. Бого-
молов, проследивший достаточно полно дробление яйца, уверенно опи-
сывает образ,ование квартетов бластомеров, вплоть до 64 клеточной ста-
дии. Однако следы спирального дробления С. И. Богомолов находит в
дроблении яиц и ряда Rhabdocoela с гермовителляриями и экзолециталь-
яыми яйцами (Graffillidae) и с гонадами, дифференцированными на
яичники и желточники (Phaenocoridae).
В отличие от данных С. И. Богомолова (1949) немецкий зоолог
Зейлерн-Аспанг (Seilern-Aspang, 1957) у другого вида того же рода —
Macrostomum. appendiculatum — описывает спиральное дробление, то с
образованием дуэтов, как у Acoela, то с образованием квартетов, как у
Polyclada. После 16-клеточной стадии характер дробления становится
сходным с таковым турбеллярий с экзолецитальными яйцами. Это го-
ворит не против филогенетического значения характера онтогенеза, в
частности типа дробления, как думает Зейлерн-Аспанг, а о большой
пластичности, неустойчивости, продолжающемся в настоящее время
энергичном процессе эволюции и у взрослых турбеллярий и, естествен-
но, их. стадий онтогенеза. Это затрудняет систематиков, подходящих к
изучению турбеллярий с позиций статики, к установлению у них от-
рядов и подотрядов с фиксированными границами. Мы не знаем нена-
рушенного спирального развития без квартетов бластомеров, знаем
среди Protostomia и членистоногих немало примеров утраты харак-
терных черт этого дробления и образования вместо квартетов дуэтов
среди малощетинковых червей, некоторые особенности в виде утраты
морфогенетических способностей микромерами и макромерами — клет-
ками четвертого квартета — в образовании кишечника, кроме 4d у по-
ликлад, в остальном сохраняющих типичный характер гетероквадрант-
ного спирального дробления и т. д. Я считаю, что С. М. Переяславцева
(1892, Perejaslavceva, 1885) была совершенно права, когда, открыв
дуэтный характер образования бластомеров у Acoela, не считала эту
черту онтогенеза, как и отсутствие у них кишечника,— примитивными.
Позднейшие исследователи, главным образом Бресслау (Bresslau, 1909)
и его прямые и косвенные ученики — немецкие зоологи, оказались в пле-
ну идей Зейлерн-Аспанга и не разобрались в истинной природе ни дро-
бления, ни строения Acoela, считая их первично примитивными.
У поликлад, как известно, наблюдается типичное спиральное, детер-
минированное, гетероквадрантное дробление и хорошо выраженное раз-
витие трех зародышевых листков — эктодермы, энтодермы и среднего
листка в виде эктодермальной мезенхимы, производной второго и треть-
его квартетов микромеров и энтомезодермы, возникающей за счет
макромера 4d, дающего также и среднюю кишку. Резорбция макромеров
75
4а —4с'и 4А— 4D, используемых развивающимся : зародышем, являет-
ся особенностью развития прликлад, отличающих их от большинства
классов, принадлежащих к вышестоящим типам Spiralia. Сравнительно
с развитием многих из них становление тела поликлад идет более при-
митивно, без разрушения частей личинки; тело червя возникает йз тела
зародыша при прямом развитии, а при наличии пелагических личинок,
геттевской и мюллеровской, из тела последних путем роста в размерах
и удлинения их главной оси в продольную ось червя. Субэпителиальная
мускулатура кожно-мускульного мешка червя — эктодермального проис-
хождения, а паренхимная мускулатура и большая часть паренхимы об-
разуются за счет производных 4d. Важными примитивными чертами он-
тогенеза поликлад являются радиальносимметричная закладка эктоме-
зодермы (эктомезенхимы), головного мозга и двустороннесимметричная
закладка энтомезодермы (энтомезенхимы), временно образующей две
клеточные полоски, распадающиеся на клетки паренхимы.
Производными такого основного типа квартетного спирального дррб*
ления и органогенеза поликлад являются типы онтогенезов у Acoela и
низших Rhabdocoela с эндолецитальными яйцами. Наиболее полными и
достоверными данными об индивидуальном развитии Acoela мы обяза-
ны русским эмбриологам: С. Переяславцевой (1892, Perejaslavceva,
.1885), отчасти В. Репяхову (Repjachow, 1885) и С. И. Богомолову, (1960,
1964). Спиральное дробление, образование микромеров и макромеров
дуэтами, развитие эктодермальных покровов, эктомезенхимы (эктомезо-
дермы) за счет второго дуэта (2ab, 2cd), энтомезодермы — за счет кле-
ток 4cd четвертого дуэта, возникновение в связи с условиями развития
у одного и того же вида (например, Convoluta borealis — Богомолов,
1960) квартетного и даже однолучевого дробления говорят о вторичном)
производном, но не примитивном ходе онтогенеза Acoela. Вместе с этим у
Convoluta гаструляция и образование полости кишки, стенки которой
образованы макромерами 5АВ, 5CD и микромерами пятого дуэта, сви-
детельствуют о том, что наличие сплошной синцитиальной пищевари-
тельной паренхимы вместо кишки с полостью у Acoela — явление вто-
ричное.
Ясно выраженное типичное спиральное дробление с образованием
квартетов установлено и старыми и новыми (например, Богомолов,
1949, 1960, 1964) исследователями, но, к сожалению, без выяснения
детальной генеалогии и судьбы бластомеров у ряда низших Rhabdocoela,
Catenulida, Macrostomida, имеющих яичники и эндолецитальные яйца.
У ряда высших Rhabdocoela, обладающих гермовителляриями, или са-
мостоятельными яичниками, желточниками и экзолецитальными яйцами,
происходит исчезновение спирального дробления, от которого сохраня-
ются в большей или меньшей мере лишь следы.
Приходится отметить, что описание хода эмбрионального развития-
Acoela у Гаймен (Hyman, 1951) резко отлично от только что сказанно-
го; по Гаймен, у Acoela нет следов кишечной полости и первичным яв-
ляется стереогаструла. Это расхождение, с другими авторами объяс-
няется отчасти игнорированием работ С. Переяславцевой, отчасти не-
знанием работ С. И. Богомолова. К сожалению, Гаймен не разбирает
эмбрионального развития низших Rhabdocoela.
Дальнейшей эволюцией онтогенезов турбеллярий, удаляющихся еще
далее от исходного типа онтогенеза морских форм с эндолецитальными
яйцами, являются онтогенезы турбеллярий с экзолецитальными яйцами,
которые откладываются в кокон с большим, иногда громадным, коли-
чеством желточных клеток. При сильно измененном ходе онтогенеза
у ряда пресноводных высших Rhabdocoela и Alloeocoela все же сохра-
няются следы спирального дробления яиц. В ходе развития желточные
76
клетки образуют синцитий, по которому расползаются бластомеры. Наи-
более сильно уклоняется от исходного типа онтогенеза турбеллярий раз-
витие, идущее по инкапсулированному (Шмидт, 1953) ,личиночному
типу, пресноводных Triclada. Дробление яйца оказывается беспорядоч-
ным, анархическим. Образуются зародышевые или провизорные экто-
дерма, глотка для заглатывания желтка и эктодермальная кишка сфе-
рической личинки. Дефинитивные органы и ткани будущего червя,
позже вылупляющегося из кокона, возникают в недифференцированной
клеточной массе в виде скоплений клеток, без выраженных зародыше^
вых листков. Желточные клетки, число которых у некоторых пресновод-
ных триклад может достигать, как известно, 10 000, образуют питатель-
ную среду, в которой развитие дробящегося яйца протекает как бы в
культуре тканей, в питательной среде, созданной материнским организ-
мом, под защитой яйцевого кокона, из которого вылупляется уже вполне
сформированный молодой червь.
Несмотря на краткость обзора основных моментов онтогенеза тур-
беллярий все же видно, что наиболее примитивный ход онтогенеза на-
блюдается у поликлад, имеющих пелагические личинки; онтогенез бес-
кишечных и части прямркишечных турбеллярий, при сохранении ряда
исходных черт (основы спирального дробления, зародышевые листки),
обнаруживает вторичные изменения.
Еще более измененный ход онтогенеза наблюдается у остальных
групп турбеллярий, обладающих экзолецитальными яйцами; дробление,
сохраняя у некоторых форм следы спирального, становится беспорядоч-
ным, анархическим, а зародышевые листки оказываются почти не вы-
раженными. Все это позволяет понять основные черты организации от-
рядов турбеллярий и их положение в системе.
• . Если так сильно изменяется характер индивидуального развития у
турбеллярий с переходом их предковых форм от жизни в море к жизни
в пресных водах, понятно насколько сильно вторично изменен онтоге-
нез у остальных паразитических классов плоских червей, хотя и проис-
шедших, как общепризнано, от рабдоцелоподобных предков и состав-
ляющих с турбелляриями естественную группу — тип плоских червей.
В отличие от турбеллярий, паразитические плоские черви обладают
громадной плодовитостью. Каждая особь производит огромное коли-
чество яиц. В связи с этим их экзолецитальные яйца, по о дно.му за-
ключенные в яйцевой кокон, снабжены малым количеством желточных
клеток: от нескольких — у дигенетических сосальщиков (Trematoda) до
одной — у ряда цестод.
Дробление яиц у дигенетических и моногенетических сосальщиков
и цестод беспорядочное, неравномерное, причем образуется небольшое
число бластомеров. В отличие от дробления у части рабдоцелид, АПоео-
coela и триклад, бластомеры у сосалыцикрв не расползаются, но со-
ставляют компактную группу макро- и микромеров. Согласно старым
работам конца XIX — начала XX столетий у дигенетических и моноге-
нетических сосальщиков один или несколько макромеров окружены
снаружи микромерами. Ряд новых работ (Hyman, 1951) и особенно
исследование Риса (Rees, 1940) внесли много существенного в наши
знания об онтогенезе дигенетических сосальщиков.
Детерминированный характер дробления яиц дигенетических сосаль-
щиков выражается в том, что при первом дроблении яйца несколько
меньший бластомер — пропагативный—дает в ходе онтогенеза половые
клетки и более мелкие клетки, образующие часть тела зародыша. Дру-
гой, более крупный эктодермальный бластомер отделяет более мелкий
бластомер, делящийся на два. Несколько дериватов мелких бластоме-
ров отделяются от зародыша, образуя его оболочку; продукты деления
этого крупного бластомера и более мелких участвуют в образовании
77
тела зародыша или мирацидия. По аналогии с онтогенезом цестод
(Spatlich, 1926) развитие дигенетических сосальщик,ов (Rees, 1940}
можно истолковать так: больший бластомер можно считать макроме-
ром, а два более мелких его производных — мезомерами, которых Рис
и другие исследователи правильно считают эктодермальными бласто-
мерами. Вероятно, они дают начало покровам, нервной системе, прото-
нефридиям, субэпителиальной мускулатуре. Пропагативный бластомер
является другим макромером, энтомезодермальным, дающим половые
клетки и половые органы, а с его дериватами — микромерами — связа-
но развитие кишечника, паренхимы и паренхимной мускулатуры.
К сожалению, дробление
Рис. 22. Два типа свободноплавающих личинок мо-
ногенетических сосальщиков (по Быховскому,
1957)
и органогенез у моногенети-
ческих сосальщиков (Мопо-
genoidea) не подвергались
новым исследованиям. Надо
думать, что они окажутся
сходными с таковыми диге-
нетических сосальщиков. Су-
щественным отличием раз-
вития моногенетических со-
сальщиков, кроме биологии
и жизненных циклов, яв-
ляется гораздо большее
сходство организации их ли-
чиночных стадий с таковой
рабдоцелид-далиеллид, а
также образование на их
заднем конце органа при-
крепления с хитиноидными
крючками (рис. 22), гомо-
лога церкомера цестод
(Janicki, 1921; Быховский,
1937, 1957).
Об эмбриональном раз-
витии цестод (Cestoidea),
кроме старых, имеются новые работы (Spatlich, 1926; Silverman, 1954;
Clarke, 1954; Ogren, 1956, 1957). По Шпетлиху образуются макромеры,
мезомеры и микромеры. Два макромера с желтком и их производное —1
3 мезомера — образуют (наружную) первую и вторую оболочки заро-
дыша, 2 гранулированных макромера дают более крупные наружные,
а несколько микромеров, потомки этих макромеров с желтком, дают бо-
лее мелкие внутренние клетки зародыша, или онкосферы, с 3 парами
хитиновых крючков. В синцитиальной массе онкосферы, поверхность ко-
торой образует третью оболочку эмбриона, одни ядра содержат РНК,
другие ДНК (Silverman, 1954). В эмбрионе, или онкосфере, развива-
ются нервная система, субэпителиальные мышцы, железы, паренхимные
мышцы кожно-мускульного мешка, паренхима, протонефридии и гонады;
как и у дигенетических сосальщиков, зародышевые листки не выражены,
во всяком случае они трудно различимы.
Некоторые авторы (например, Hyman, 1951) считают, что онкосфера
образована мезоэнтодермой, по мнению других (Silverman, 1954;
Clarke, 1954; Ogren, 1956, 1957),— мезодермой. Цестоды, дигенётиче-
ские и моногенетические сосальщики, как считают последние авторы,
лишены эктодермальных покровов, поскольку в ходе онтогенеза у сво-
бодноплавающих личинок сбрасываются эмбриональные оболочки и да-
же ресничный эпителий. Так называемый погруженный эпителий этих
червей, по мнению Е. Д. Логачева (1957а, 19576, 1961) и Б. Е. Быховско-'
78
го (1937, 1957),— клетки желез паренхимы.
Думаю, что такие соображения не отвечают действительности, а
лишь говорят о примитивности или беспомощности применяемых мор-
фологических методов. Цитохимическое изучение и, может быть, мето-
дика мечения клеток изотопами, я уверен, установят тождество бласто-
меров эмбрионального развития сосальщиков и ленточных глист с бла-
стомерами поликлад, дающими эктодерму, энтодерму и мезодерму.
Кажется также маловероятным, чтобы при таком резком различии в
эмбриональном развитии между рабдоцелидами, моногенетическими и
дигенетическими сосальщиками и цестодами (утерявшими кишечник)
совершенно независимо и из других клеточных источников могли воз-
никать организация и характерные органы, типичные для их предковых
форм — рабдоцельных турбеллярий. Подтверждением родства этих
групп могут служить факты рабдоцельного строения свободноподвиж-
ных личиночных стадий дигенетических и моногенетических сосальщиков
и сходство, вернее, гомология церкомеров моногенетических сосальщи-
ков и цестод.
Провести достоверное сравнение бластомеров и приурочить появле-'
ние зачатков частей зародыша у сосальщиков и ленточных червей с та-
ковым.и. поликлад, бескишечных турбеллярий и примитивных-р-абдоце-
лид трудно, поскольку онтогенез у сосальщиков и цестод является тело-
бластическим уже по тому, что у паразитических классов число бла-
стомеров очень ограничено, дробление неправильное, а развитие органов
зародыша происходит в синцитиальной массе. Из работы Шпетлиха
(Spatlich, 1926) ясно, что макромеры с желтком образуют не толькс
первую эмбриональную оболочку, а их производные (мезомеры)—вто-
рую эмбриональную оболочку, но и мелкие клетки (микромеры), уча
ствуюЩие в образовании задорыша, а гранулированные. макромеры
образуют большие клетки тела зародыша (см. стр. 78);
На основании этого, судя по происхождению и судьбе более крупных
и более мелких клеток зародыша цестод, мы имеем право считать их
эктодермальными и энтомезодермальными бластомерами. Они дают по-
кровы (погруженный эпителий), мускулатуру, паренхиму, протонефри-
дии и половые органы взрослых цестод. Из сказанного выше о развитии
дигенетических и, видимо, моногенетических сосальщиков ясно, что и у
них имеются эктодермальные и энтомезодермальные бластомеры, при-
чем и те и другие участвуют в образовании тела зародыша. Если думать,
что сбрасывание оболочек и покровного эпителия в ходе онтогенеза дей-
ствительно ведет к утрате у этих классов эктодермы, тогда развитие у
них нервной системы и пр.отонефридиев надо считать мезодермальным,
что маловероятно. Во всяком случае выпадение ряда делений бластоме-
ров и ограничение их числа наблюдается и у турбеллярий, в направле-
нии от поликлад к бескишечным турбелляриям (рабдоцелидам) —и на-
конец к трикладам. В этом же ряду идет постепенное ослабление типич-
ного развития зародышевых листков, почти не выраженных в онтогенезе
триклад. Кроме того, даже у поликлад, как было сказано выше, 4 мак-
ромера и 3 микромера 4а—4с идут на питание личинки и лишь бласто-
мер 4d дает начало кишечнику и мезодерме, как и у цестод, у кото-
рых энтомезодерма возникает при делении всего двух бластомеров.
По положению в зародыше, или онкосфере, более крупные клетки,
производные гранулированных макромеров, казалось должны отвечать
эктомезенхиме, тем более, что, видимо, субэпителиальная мускулатура
тела, нервная система и протонефридии образуются ими, а более мелкие,
глубже расположенные клетки, потомки микромеров, должны быть эн-
томезодермой (или энтомезенхимой), поскольку, очевидно, они дают
главную массу паренхимы, паренхимную мускулатуру и основные части
половой системы. Мне кажется, что вернее отрицать не вообще наличие
79
эктодермальных клеток, а утрату ими морфогенетической способности
формировать кишечник.
Заканчивая рассмотрение онтогенеза цестод, необходимо вспомнить
не укладывающиеся в обычные рамки данные Чайльда (Child, 1906),
описавшего развитие спермы из соматических клеток у Moniezia,
В. Р. Вейцмана (1939, 1953а, 19536) о развитии у Taenia solium и Т. cras-
slcollis онкосфер из мелких соматических клеток стенки матки без уча-
стия спермы с предшествующей дегенерацией последней, а также о пол-
ном распаде у этих цестод яичников, желточников и клеток, их состав-
ляющих. Исследования Е.Д. Логачева (1957а, 19576, 1961) подтверждают
данные В. Р. Вейцмана в том, что у ряда турбеллярий, дигенетических
сосальщиков и цестод половые клетки образуются из базофильных амё-
боцитов паренхимы.
Наряду с этим у ремнецов и лентецов, в частности у Lingula,
В. Р. Вейцман (1940) наблюдал обычный ход развития яиц и желточ-
ных клеток.
Как ни странно, до сих пор еще нет классификации турбеллярий,
на которой бы сходилось большинство зоологов. Это отчасти объясняет-
ся продолжающимися находками новых форм, порой не укладываю*
щихся в существующие отряды и надотряды, и отчасти различным
толкованием природы, например, Acoela, Rhabdocela и т. д. Вместо
4—5 отрядов (Polyclada, Acoela, Rhabdocoela, Alloeocoela и Triclada)
Кестнер (Kaestner, 1954/1955) принимает 7 отрядов: Acoela, Catenuli-
da, Macrostomida, Polycladida, Alloeocoela, Neorhabdocoela, Temno-
cephalida. В. H. Беклемишев (1964) принимает 11 отрядов — Acoela,
Xenoturbellida, Proplicastomata, Polyclada, Notandropora, Macrostomi-
da, Griathostomulida, Alloeocoela, Triclada, Rhabdocoela и Terrtnocepha-
la. Гаймен (Hyman, 1951) раньше принимавшая 5 отрядов,— Acoela,
Rhabdocoela, Alloeocoela, Triclada и Polycladida, позже (1959), учиты-
вая мнения ряда специалистов, принимает 11 следующих отрядов —
Acoela, Catenulida, Macrostomida, Neorhabdocoela, Gnathostomulida,
Archeophora, Lecithoepitheliata, Holocoela, Seriata, Tricladida и Poly-
Hadida. Таким образом, два крупнейших знатока турбеллярий—
В. Н. Беклемишев и Гаймен согласны включать в отряд Rhabdocoela
3 самостоятельные отряда — Catenulida ( = Notandropora), Macrostomi-
da ( = Opisthandropora) и Rhabdocoela ( = Lecithophora, или Neorhab-
docoela), но расходятся в понимании объема Acoela. В. Н. Беклемишев
считает самостоятельными отрядами — Xenoturbellida, Nemertodermati-
da и Proplicastomata, хотя и допускает принадлежность двух последних к
Acoela, тогда как Гаймен включает их, несмотря на наличие кишечника,
в отряд Acoela.
Я считаю, что Гаймен права. Высказываясь против примитивности
явления отсутствия кишечника у Acoela, объективно она сама опровер-
гает это положение, включая Nemertodermatida и Proplicastornata в от-
ряд Acoela почти во всем сходных с последними, но обладающих кишеч-
ником. Иными словами, С. М. Переяславцева (1892) с полным основа-
нием называла Acoela Pseudoacoela.
С другой стороны, В. Н. Беклемишев сохраняет единство отряда
Alloeocoela, тогда как Гаймен, следуя некоторым авторам, делит этот от-
ряд на 4 —Archeophora, Lecithoepitheliata, Holocoela и Seriata.
В классификации, недавно разработанной отчасти Карлингом (Каг-
iing, 1940), а в основном Вестбладом (Westblad, 1950) и принятой Лю-
тером (Luther, 1960) и Аксом (Ах, 1961), Polyclada, Catenulida и Mac-
rostomida из Rhabdocoela, Acoela и некоторые небольшие группы, при-
мыкающие к Acoela, объединены в Archeophora, а остальные турбелля-
рии — в Neoophora. Первые характеризуются рядом общих признаков
строения, и прежде всего отсутствием желточников, эндолецитальными
80
яйцами и правильным, точнее, по-моему, спиральным дроблением. Neo-
ophora объединяются также рядом других общих черт строения и на-
личием гермовителляриев или яичник,ов и желточников, экзолециталь-
ными яйцами и беспорядочным дроблением. Мне представляется такая
классификация рациональной, надо лишь отказаться от названия Arche-
ophora, ранее данного Рейзингером для Proporoplana, тем более, что,
по мнению Гаймен (Hyman, 1951), Рейзингер неправильно понимал при-
роду Proporoplana.
К характеристике Archeophora надо добавить и экологические осо-
бенности: все эти турбеллярий, кроме Catenulida, морские формы, тогда
как среди Neoophora преобладают пресноводные формы, наряду с искон-
ными морскими (ряд собственно Rhabdocoela), вторичноморскими Tricla-
da, Maricola и наземными. Прежний отряд Alloeocoela представлен Leci-
thoepitheliata, Holocoela и Seriata, а отряд Triclada — Maricola, Paludi-
cola и Terricola.
Ряд особенностей строения (в основном строение половой системы)
Xenoturbella такрвы, что вместе с Егерстеном (Jagersten, 1959) и Ремане
(Remane, 1959) я не могу поместить их в класс турбеллярий. Системати-
ческое положение Xenoturbella не выяснено, но во всяком случае, во-
преки мнению Вестбляда, В. Н. Беклемишева и Гаймен, это не турбелля-
рия и, конечно, не неотеническая личинка Coelomopora (Deuterstomia),
как это предполагает Рейзингер (Reisinger, 1961). Кроме некоторых
сходств в строении покров,ов, частностей нервной системы, статоциста,
с которыми оперирует Рейзингер, по существу своей организации Xeno-
turbella— низший, плоский, паренхиматозный червь, без задней кишки
и анального отверстия, не имеющий никаких существенных черт, свой-
ственных личинкам или стадиям развития Deuterostomia. Видимо, такие
разногласия во взглядах на природу Xenoturbella связаны с недостаточ-
ностью материала; необходимо сначала выяснить природу этих организ-
мов, а затем уже говорить об их систематическом положении.
Таким образом, надо считать, что среди морских исходных групп
турбеллярий на первом месте стоят Polyclada, они обладают наиболее
полным набором черт, характерных для свободноживущих плоских чер-
вей, и являются наиболее высокоорганизованными турбелляриями, кон-
вергентно в ряде отношений они сходны с платиктенидами. Polyclada
обнаруживают отсутствие черт регресса и сохраняют в наиболее чистом
виде тип эмбрионального развития со спиральным детерминированным
характером дробления яиц, с закладкой трех зародышевых листков и с
пелагическими личинками. Acoela, как было сказано, по уровню строе-
ния стоят значительно ниже Polyclada, но вместе с тем анатомически и
эмбриологически они вторично сильно упрощены. Было бы необъективно
отрицать необычайно примитивный уровень организации бескишечных
'турбеллярий, обладающих при этом согласно недавним исследованиям
Гаймен (Hyman, 1959) необычайно высокими регенеративными способ-
ностями: целый организм может регенерировать из мельчайших кусоч-
ков эпидермиса или внутренней синцитиальной ткани. Однако они зани-
мают самое низкое место среди Bilateralia как по примитивным чертам,
так и в силу вторичного упрощения ряда органов и тканевой органи-
зации.
Основываясь на том, что многие Acoela мельче инфузорий и внешне
весьма сходны с последними (эктодермальный синцитий, захватываю-
щий пищу, напоминает эндоплазму инфузорий), Хаджи (Hadzi, 1944,
1958, 1963), а затем Штейнбока (Steinbock, 1958) и Гансон (Hanson,
1958) выводили бескишечных турбеллярий из инфузорий, о чем было
сказано уже выше (стр. 68).
Прямокишечные турбеллярий по общей организации, строению ки-
шечника, протонефридов, нервной системы, половой системы достаточно
.<5 Д. М. Федотов
81
четко отличаются от Polyclada и Acoela. По пластичности своей орга-
низации, по экологии (обитание в море, пресной воде, комменсализм,,
зктопаразитизм, эндопаразитизм), по численности видов рабдоцелиды,.
даже в- новом сокращенном объеме отряда, являются, как общепризна-
но, основной группой класса турбеллярий, давшей начало остальным
отрядам этого класса, всем другим классам паразитических плоских чер-
вей и классам первичнополостных червей.
Взгляды зоологов расходятся на трактовку Temnocephala. Одни про-
должают считать этих червей самостоятельным классом (А. В. Иванов,
1952а, 19526; Быховский, 1957), другие (Беклемишев, 1952, 1964), преи-
мущественно зарубежные зоологи,— подотрядом Rhabdocoela. По ана-
томическим данным Temnocephala очень сходны с Rhabdocoela, что осо-
бенно подчеркивается особенностями строения рода Didymorchis из
Temnocephala, который раньше относили к далиеллидам. Все это дает
мне основание присоединиться к точке зрения Беттгера (Boettger, 1952,
1957) и рассматривать Temnocephala как подотряд Rhabdocoela.
Нет достаточной ясности о природе и положении в системе Gnatosto-
mulida, небольшой, недавно установленной группы низких псаммофиль-
ных червей (Ах, 1956, 1960, 1961). Их особенности строения (перетяжкой
отделен от тела головной отдел, небольшой, но ясно выраженный хво-
стовой отдел, жгутик,рвый характер ресничного покрова, парные кути-
кулярные с зубчиками челюсти в глотке со специальной поперечно-
полосатой мускулатурой, отсутствие протонефридиев, общий тип поло-
вой системы с яичником и бурсой, далеко занесенной в передний конец
тела, и семенниками с копулятивным органом в задней половине тела)
в целом не типичны для турбеллярий. Вряд ли можно, как это делает
Ю. В. Мамкаев (1961), уподоблять или сравнивать парные челюсти в-
глотке гнатостомулид с щипчиками хоботка Kalyptorhynchia. Насто-
ящие челюсти гнатостомулид отсутствуют и в глотке плоских червей и у
немертин. На этом основании нельзя гнатостомулид помещать, как это
делает Ю. В. Мамкаев, в класс турбеллярий. По-видимому, нет также-
достаточных данных видеть в них, подобно Штейнбоку (Steinbock, 1958),
гастротрих.
Для выяснения природы и положения в системе этой группы своеоб-
разных червей необходимы поиски новых представителей и прежде все-
го изучение их индивидуального развития.
Подводя итоги по классификации турбеллярий, очевидно, нельзя не
прийти к выводу, что морфологические методы недостаточны для опре-
деления систематического положения и родственных отношений многих
групп турбеллярий; для этого не,обходимо применение методов биохи-
мии, и прежде всего серологического анализа.
Если классификация турбеллярий все же близка к естественной-и
разногласия касаются толкования, например, природы Acoela или при-'
падлежности к классу турбеллярий таких групп, как Xenoturbellida или
Gnathostomulida, то положение с классификацией паразитических клас-
сов сложнее, хотя за последние десятки лет сосальщики и цестоды, среди
которых много опасных паразитов' не только высших теплокровных жи-
вотных, но и человека, интенсивно изучаются.
Классификацию дигенетических сосальщиков одни зоологи, медики и
ветеринары строят на основе данных организации взрослых червей (Пав-
ловский, 1937; Скрябин, 1947—1962; Hyman, 1951), другие, и притом
большинство (La Rue, 1957),— на признаках строения церкарий.
Еще в начале XX столетия русский зоолог Д. Ф. Синицин (1905,
1911), изучавший строение церкарий, придавал им основное таксоно-
мическое и филогенетическое значение. Реконструируя организацию
предковой формы, по его термину — пропартениты, он приписал ей на-
личие не только полости тела (существующей у спороцист и церкарий);.
82
которую ан считает целомической, но и кишечника с анальным отверсти-
ем и ряд других черт, свойственных первичнополостньим червям, в силу
чего он и придавал главенствующее значение среди дигенетических со-
сальщиков партените-церкариям' и подчиненное— взрослым сосальщи-
кам. По этим же основаниям Д. Ф. Синицин отрицал филогенетические
связи сосальщиков с турбелляриями. Большая часть всех этих сообра-
жений Д. Ф. Оиницина имеет исторический интерес; но несомненно его
исследования морфологии и экологии церкарий и значение для класси-
фикации дигенетических сосальщиков строения церкарий повлияли на
взгляды многих зарубежных зоологов-паразитологов. Так, Ля Рю (Ба
Rue, 1957), используя работы ряда предшественников, построил класси-
фикацию дигенетических сосальщиков как подкласса Trematodes.
Мы знаем примеры классификации личинок, например, для иглоко-
жих Мортенсена (Mortensen, 1920; см. гл. «Иглокожие») и для насеко-
мых М. С. Гилярова (1964), причем они в общем подтверждают клас-
сификации, построенные для взрослых животных. Совпадение классифи-
каций по взрослым' формам и по личинкам является подтверждением
естественности классификации.
По-видимому, классификация дигенетических сосальщиков, построен-
ная Ля Рю (La Rue, 1957), по церкариям, лучше старых классификаций,
построенных по взрослым формам, но вряд ли это значит, что не следует
искать путей для рациональной классификации взрослых сосальщиков.
Общепринятое деление дигенетических сосальщиков на основании поло-
жения ротового отверстия на Gasterostomata и Prosostomata и включе-
ние в Prosostomata огромного большинства их семейств, несомненно, яв-
ляются искусственными и не оправдываются особенностями индивидуаль-
ного развития. Пока естественной классификации этих червей нет, что
отчасти связано с необычно большим и продолжающим расти количе-
ством описываемых видов (Hyman, 1959).
В настоящее время широко распространилась тенденция повышать
многие существующие классы в ранг надклассов, отряды — в подклассы
и классы и т. д.
Сосальщики Aspidogastridae, обладающие рядом признаков строе-
ния, общих с таковыми взрослых дигенетических сосальщиков, а по
развитию резко отличающихся от последних (постепенным развитием
эмбриона во взрослого червя без прохождения стадий- спороцисты, ре-
дки и церкарии), составляют, как резонно считает Ля Рю (Ба Rue, 1957),
самостоятельный класс Aspidogastrea. Однако многие, игнорируя особен-
ности онтогенеза, рассматривают их лишь как семейство Aspidogastri-
dae дигенетических сосальщиков.
Точно так же, согласно доводам А. В. Иванова (1952), несмотря
на бедность количества форм, по своей организации и прямому разви-
тию, -без личинки, семейство Udonellidae надо считать самостоятельным
классом плоских червей — Udonelloidea, хотя издавна по внешним при-
знакам этих червей помещали в маногенетические сосальщики, не имея
па то серьезных оснований.
Несмотря на почти полное отсутствие сведений (если не считать ста-
рых исследований по онтогенезу единичных видов) о ранних стадиях
индивидуального развития моногенетических сосальщиков, классифика-
ция Monogenoidea близка к естественной. Она основывается преимуще-
ственно на данных о строении взрослых червей. Классификация моноге-
нетических сосальщиков, разработанная Б. Е. Быковским (1937, 1957),
наиболее рациональна. Она основана на комплексном исследовании
строения взрослых, некоторых моментов позднего онтогенеза, строения
свободноподвижных личиночных стадий, развития прикрепительного ор-
гана (церкомера), экологии, особенно связей между паразитом и хозяи-
ном.
6;
83
Большинство авторов придерживается старой точки зрения, объеди-
няя дигенетических и моногенетических сосальщиков в класс Tremato-
da. Соглашаясь с доводами Б. Е. Быховского о том, что моногенети-
ческие (Monogenoidea) и дигенетические (Trematoda) сосальщики — два
самостоятельных класса и что по некоторым признакам первые стоят
ближе вторых к классу цестод, нельзя все же признать правильным
объединение моногенетических сосальщиков и цестод в надкласс. Осно-
вы организации, жизненный цикл, экология и характер индивидуального
развития обоих классов настолько различны, что объединять их лишь по
наличию церкомера вряд ли возможно.
Значительно лучше, чем для дигенетических сосальщиков, обстоит
дело с классификацией цестод. Мне кажется, что лучшей, хотя неполной,
является классификация, разработанная О. Фурманом (Fuhrmann,
1930—1931), построенная им на учете данных строения индивидуально-
го развития и экологии, принятая В. Н. Беклемишевым (1944, 1952,
1964). Как известно, Фурман в подкласс Cestodaria относил два отря-
да — Amphilinidea и Gyrocotylidea, а в подкласс Cestoda пять отрядов —
Tetraphyllidea, Diphyllidea, Tetrarhynchidea, Pseudophyllidea и Cyclophyl-
lidea.
Некоторые авторы, например Линч и Б. Е. Быхо.вокий (1957), созда-
ют для Gyrocotylidea самостоятельный класс, якобы близкий к Mono-
genoidea. Однако отсутствие кишечника, наличие ликосферы с 6 крючья-
ми, эндопаразитизм в кишечнике хозяина, по моему мнению, определяют
положение Gyrocotylidea как отряда класса цестод, который по некото-
рым признакам строения (Быховский, 1957) лишь немного больше схо-
ден с монотеистическими сосальщиками, чем другие Cestodaria.
Классификация цестод, приведенная Гаймен (Hyman, 1951), близка
к таковой Фурмана, особенно если вспомнить, что отряд Aporidea—
нечленистые черви, близкие к цестодам Taenioidea или Cyclophyllidea,—
не был им создан. Последующие изменения классификации заключаются
в том, что отряд Фурмана Tetraphyllidea последующими паразитологами
разбит на три отряда — Tetraphyllidea, Lecanicephaloidea и Proteocepha-
loidea — и создан новый отряд Nippotaeniidea. Все это мелкие неприн-
ципиальные изменения классификации Фурмана. Природа же Nemato-
parataenia и Gastrotaenia, паразитов кишечника лебедя, настолько
неотчетлива, что для выяснения ее нужны еще серьезные исследования.
К сожалению, в многочисленных сводках о сосальщиках и цестодах, соз-
данных К. И. Скрябиным и его учениками, классификация не перерабо-
тана. Чтобы улучшить наши представления о родственных связях как
для турбеллярий, так и для паразитических классов плоских червей, при-
ходится ждать биохимических и серологических исследований. Несмотря
на все возрастающее количество работ по биохимии и обмену веществ;-
по экспериментальному изучению паразитических классов, судя по Гай-
мен (Hyman, 1951, 1959), серологических исследований этих классов еди-
ницы, и притом они выполнены не на современном уровне (Eisenbrandt,
1938; Wilhelmi, 1940).
Поскольку данных об ископаемых турбелляриях, об их историче-
ском развитии, у нас нет, наши^филогенетичеокие построения для типа
плоских червей, как отмечал Бреослау (Bresslau, 1928, 1933), являются
в основном спекулятивными. Учитывая примитивные черты строения и
онтогенеза современных низших турбеллярий, их сохранение мы можем
приписать очень медленным темпам эволюции и допускать геологиче--
скую древность этих групп. Некоторые косвенные данные, связанные с
историческим прошлым их позвоночных хозяев, позволяют, как это уже
отмечалось выше (стр. 67), предполагать, что моногенетические сосаль-
щики и цестоды происходят из древнего палеозоя (Fuhrmann, 1928—
1933; Быховский, 1937, 1957). •
.84
Все известное нам об историческом развитии многоклеточных, начи-
ная с губок и кишечнополостных, об эволюционных изменениях кишеч-
ника в разных типах Bilateria позволяет считать, что предковые формы
всех многоклеточных обладали кишечной полостью и что отсутствие ки-
шечника у представителей разных типов Protostomia и Deuterostomia—
явление вторичное.
Из всех современных свободноживущих плоских червей Xenoturbella
почти во всех отношениях ближе к родоначальному прототипу плоских
червей. Будем ли мы помещать Xenoturbella в класс турбеллярий или
считать их самостоятельным классом, родословную типа плоских червей
надо начинать именно с Xenoturbellida.
Современные Acoela, обладающие примитивными и вторично упро-
щенными чертами и строения и онтогенеза, потерявшие просвет кишеч-
ника (т. е. по существу не Acoela, по Ульянину, a Pseudoacoela, по Пе-
реяславцевой), не являются родоначальной группой для остальных тур-
беллярий. Это боковая, слепо кончающаяся ветвь, стоящая в основании
родословного древа плоских червей недалеко от Xenoturbellida. Надо ду-
мать, что родоначальником турбеллярий и через них всех классов плос-
ких червей был мелкий червеобразный, ресничный, придонный морской
организм, питавшийся микроскопической растительной и животной пи-
щей; он обладал мешковидным кишечником без ануса, гермафродитной
половой системой, с половыми клетками, разбросанными в паренхиме,
не оформленными в виде половых желез, эндолецитальными яйцами,
спиральным типом дробления и развитием без метаморфоза.
От родоначального ствола, недалеко от ответвления Pseudoacoela,
отошла ветвь поликлад, представленная прогрессивно развившимися
крупными морскими хищными турбелляриями, сохранившими в онтоге-
незе много исходных черт, характерных для типов Spiralia, частично ут-
раченных или измененных у мелких Pseudoacoela и примитивных групп
рабдоцелид— Macrostomida и Catenulida. Эти два отряда, близко свя-
занные рядом черт с двумя предыдущими, очевидно, отошли от родо-
начального ствола, недалеко от последних. Самый факт перехода от жиз-
ни в море к существованию в пресных водах говорит о позднейшем про-
исхождении Catenulida сравнительно с Macrostomida. Объединение всех
этих отрядов в Archeophora, на мой взгляд, имеет не только классифика-
ционный, но и филогенетический смысл.
Продолжением, а не боковыми ветвями, как большинство отрядов
Archeophora, главного ствола являются рабдоцелиды. Классификацион-
но Macrostomida и Catenulida отделены от собственно Rhabdocoela, или,
как некоторые их называют, Neorhabdocoela. Они однако связаны друг с
другом филогенетически как родственные группы. Даже и в таком обед-,
ненном объеме рабдоцелиды, как общепризнано, являются по строению,
индивидуальному развитию и экологии центральной группой, давшей на-
чало остальным отрядам турбеллярий и классам паразитических плоских
червей.
Мне представляется возможным, несколько упрощая и схематизируя,
считать, что прежние Alloeocoela (Lecithoepitheliata, Cumulata, или Но-
iocoela, и Seriata) и Triclada (Maricola, Paludicola и Terricola) филогене-
тически связаны с рабдоцелидами (или Neorhabdocoela). и объединение
этих групп в Neoophora имеет и классификационные и филогенетические
основы.
Подробных обоснований для такого понимания филогенетических
связей отрядов турбеллярий я не даю. Они в значительной мере вы-
текают из всего текста, посвященного плоским червям, и, кроме того,
они даны рядом авторов (Meixner, 1926; Bresslau, 1928, 1933; Hyman,
1951; Беклемишев, 1937, 1952, 1964; Marcus, 1958; Remane, 1954, 1959;
Ax, 1960, 1961, и др.).
85
Что касается вопроса об эволюции и филогении паразитических
плоских червей, то общепринято считать далиеллид родоначальной
группой для дигенетических и моногенетических сосальщиков, удонел-
лид и цестод. С этим согласны многие авторы: В. Н. Беклемишев (1937,
1944, 1952, 1964), А. В. Иванов' (1952), Б. Е. Быховский (1957), Мейкс-
нер (Meixner, 1928), Бреслау (Bresslau, 1928, 1933), Гаймен (Hyman,
1951). Это подтверждается сходством между ними в строении всех ос-
новных систем органов, в строении свободноплавающих личинок ряда
паразитических классов червей и наличием среди далиеллид, помимо сво-
бодноживущих— комменсалистов, и внутренних паразитов разных
групп морских беспозвоночных. Степени сходства и различия между
классами дигенетических сосальщиков, удонеллид, моногенетических со-
сальщиков и цестод таковы, что их по праву рассматривают происшед-
шими если не от одной общей, то от разных рабдоцелоподобных пред-
ковых форм. В пользу взгляда о происхождении цестод от далиелли-
доподобных предков говорит также наличие среди далиеллид ряда эндо-
паразитических семейств, в частности Fecampiidae, у которого во взрос-
лом состоянии наблюдается регресс глаз, ротового отверстия, глотки
и т. д.
Большинство зоологов принимают систематическую и филогенетиче-
скую близость дигенетических и 'моногенетических сосальщиков, объеди-
няя их в класс Trematoda. Все основные черты строения взрослых чер-
вей, кроме некоторых частностей анатомии, бесспорно общие. В том
числе у них одинаковое устройство половой системы, отличное от поло-
вой системы цестод (как отмечает Беклемишев, 1952), обладающей
большой простотой сравнительно с таковой моногенетических сосаль-
щиков (Remane, 1959). Вместе с тем, наличие,двух присосок, сложный
жизненный цикл, смена партеногенетических и половых поколений и сме-
на промежуточных и дефинитивных хозяев, эндопаразитизм в кишечном
тракте дефинитивных хозяев, в основном млекопитающих и птиц, явля-
ются существенными особенностями дигенетических сосальщиков. Нали-
чие на заднем конце прикрепительного органа — церкомера с хитиноид-
ными крючьями, эктопаразитизм в основном на жабрах и коже элазмо-
бранхий, морских и пресноводных костистых рыб и в редких случаях на
четвероногих, жизненный’цикл без чередования поколений и смены хо-
зяев также достаточно четко отличают моногенетических сосальщиков
от ди-генетических и могут быть основаниями для разделения последних
на два класса, как делает Б. Е. Быховский, высказывающийся за фи-
логенетическую близость моногенетических сосальщиков и цестод,
несмотря на отсутствие у последних кишечника и на стадиях развития,
и у взрослых червей и наличие его у моногенетических и дигенетических
сосальщиков.
О возможной филогенетической близости моногенетических сосальщи-
ков и цистод говорят не столько данные строения и развития, сколько
экологии.
Моногенетические сосальщики, как и низшие цестоды, являют-
ся почти исключительно паразитами водных позвоночных, в частности,
паразитами низших рыб. Вполне допустимо, что в эволюции древних мо-
ногенетических сосальщиков, эктопаразитов на селяхиях, могли произой-
ти изменения условий жизни, когда некоторые их представители стали
сначала паразитами ротовой полости, а затем самого пищеварительного'
тракта хозяина из элазмобранхий. Далее, можно допустить, что подоб-
ные родоначальные моногенетические формы цестод, став паразитами
кишечника, подверглись регрессивным изменениям, потеряли кишечник
и в ходе эволюции превратились из нечленистых в ленточных членистых
червей и приобрели сложный жизненный цикл со сменой временного и-
дефинитивного хозяев.
86
Считая вероятным, что исторически паразитизм моногенетических со-
сальщиков на элазмобранхиях, а примитивных цестод'в элазмобранхиях,
как .думают Б. Е. Быховский (1937, 1957) и Фурман (Fuhrmann, 1928—
1933), возник в отдаленное геологическое время— в раннем палеозое,
можно понять утрату кишечника и приобретение сколекса с ботридия-
ми и членистости тела предковыми формами цестод. Можно отсюда
допустить и близость в строении древнейших моногенетических сосаль-
щиков и цестод.
'Эндопаразитизм дигенетических сосальщиков в кишечном тракте и
его железах у рыб, амфибий, рептилий и особенно у млекопитающих и
птиц говорит о том, что эволюция дигенетических сосальщиков шла в на-
правлении приспособления к жизни в этих группах животных, а следова-
тельно, исторически гораздо позже, чем начиналась эволюция моногене-
тических сосальщиков и цестод, как паразитов; с этим согласуется также
мощное видообразование у дигенетических сосальщиков, число видов ко-
торых (5000), как исторически более молодой группы, во много превы-
шает численность видов моногенетических сосальщиков (700) и цестод
(1500 видов).
Все сказанное является косвенными соображениями в пользу взгля-
да Б. Е. Быховского о филогенетических -связях между моногенетиче-
скими сосальщиками и цестодами. Однако- к этому надо добавить, что
нужно изучение раннего индивидуального развития примитивных мор-
ских моногенетических сосальщиков и необходимы биохимические иссле-
дования (включая серологические) -примитивных моногенетических со-
сальщиков и -низших цестод.
Сначала В. В. Заленский (Salensky, 1907), затем Ремане (Remane,
1954, 1958, 1959) и Маркус (Marcus, 1958), видимо, знавшие не взгляды
Заленского, а лишь мнение Боаса (Boas, 1917) и Нэфа (Naef, 1931,
цит. по-Ремане), развивают взгляд -о происхождении плоских червей от
вторичнополостных предков. По мнению Ремане, эти Archicoelomata
обладали тремя отделами целома, кишечником с ротовым и анальным
отверстиями, кровеносными сосудами, мезодермальными воронками вы-
делительных органов и др.
Гипотетическая родоначальная форма Ремане довольно близко от-
вечает нашему представлению о предковой форме иглокожих, кишечно-
дышащих, вероятно и всей ветви Deuterostomia, но она чужда организа-
ции типов группы Protostomia. Ремане воскрешает идею Седжвика
(Sedgwick, 1884) о происхождении целома из гастральных карманов
актиниеподобных предков и, не зная этого, идею В. В. Заленского
(1915) об эволюции спирального, детерминированного дробления с тело-
бластической закладкой листков и органов из недетерминированного ра-
диального дробления и энтероцельной закладки целома. Разбор этих
взглядов будет дан -в другом месте. Здесь отмечу, что все, что мы знаем
о современных низших плоских червях, не дает -оснований считать их
деградированными предками вторичнополостных организмов.
Примитивный ресничный покров, паренхимат-озность, отсутствие пер-
вичной и вторичной полости тела, -низкий уровень строения нервной си-
стемы и органов чувств, отсутствие органов дыхания и кровообращения,
сквозного кишечника и преимущественно внутриклеточное пищеварение,
примитивность органов выделения, отсутствие у свободноживущих
форм признаков регресса в ходе онтогенеза, кроме некоторых специфиче-
ских для плоских червей, и ряд других -особенностей подчеркивают
исходную примитивность этого типа. Этот тип по своей исконной приро-
де стоит на низшем этапе эволюции Spiralia, но вместе с тем некоторые
группы плоских червей Acoela или лучше Pseudoacoela, несомненно, ут-
ратили просвет кишечника и оказались вторично как -бы проще кишечно-
полостных -и гребневиков.
87
Не останавливаясь на разборе различных теорий о происхождении
турбеллярий (Граффа, Гаймен, Беклемишева и др., см. Ах, 1961), отме-
чу, что признание предками плоских и первичнополостных червей и не-
мертин двусторонне-симметричных, придонных, ползающих планулооб-
разных организмов с просветом в эктодермальном слое кажется наи-
более правдоподобным.
Рис. 23. Самец Wedlia sp., «паразити-
рующий» на самке (из Hyman)
1 — самец; 2 — самка
Для начальных этапов эволюции
турбеллярий-рабдоцелид характер-
но хищничество, питание микроско-
пическими водорослями и животны-
ми, мелкими беспозвоночными, не-
большие размеры тела, паренхима-
тозность, кожно-мускульный мешок,
кишечник без задней кишки, герма-
фродитная сложная половая систе-
ма, спиральное детерминированное
дробление и т. д. и переход к пара-
зитизму, приведший к формирова-
нию паразитических классов плос-
ких червей.
Другое направление в эволюции
исходных планулообразных предко-
вых форм, приведшее к формировав
нию первичнополостных червей, ха-
рактеризуется широким распростра-
нением перехода от свободного су-
ществования к паразитизму (рис.
23) после приобретения сквозного
кишечника с ротовым и анальным
отверстиями, развитием из щелей в
паренхиме первичной полости тела, преобладающим развитием отдель-
ных пучков мышц тела, простым строением половой системы и раздель-
нополостью, сохранением в онтогенезе лишь следов спирального дробле-
ния и т. д.
Третье направление ведет к развитию немертин — червей, питающих-
ся крупной добычей, добываемой активным хищничеством, ловлей добы-
чи хоботом, а не ртом, крупными размерами тела, сквозным кишечником,
очень большой подвижностью, благодаря сильному развитию кожно-
мускульного мешка, развитием кровеносной системы, простым строени-
ем половой системы, раздельнополостью, спиральным, детерминирован-^
ным дроблением, часто образованием пелагических личинок.
В итоге длительной дивергентной эволюции возникли три типа чер-
вей — плоские, первичнополостные и немертины.
Первичнополостные черви (Askelminthes)
Червеобразные, двустороннесимметричные, часто круглые в попереч-
ном сечении, преимущественно микроскопических и мелких размеров —
свободные обитатели моря, пресных вод, почвы, паразиты растений, жи-
вотных и человека; имеют кутикулярный покров тела, клеточный или
синцитиальный эпидермис (или гиподерму), сквозной кишечник с пе-
редней, средней и задней кишкой; передняя кишка—ю мускулистой
глоткой трехгранного просвета; нервная система—• субэпителиальная,
типа сртогона; органы чувств — в виде малодифференцированных тан-
го-, хемо- и фоторецепторов; обычно отсутствуют кожно-мускульный ме-
шок, органы дыхания и кровообращения, мезенхиматозная соединитель-
ная ткань редуцирована, полость тела первичная. Осморегуляторно-экс^
креторный аппарат — в виде слаборазвитых протонефридиев или в виде
комплекса видоизмененных кожных желез. Гонады мешковидной или
трубчатой формы, с собственными эпителиальными стенками. Неболь-
шое число клеточных делений в онтогенезе, более или менее выраженная
мозаичность билатерального со следами спирального дробления. Разви-
тие, как правило, прямое или с личинками и с признаками метамор-
фоза.
Все это составляет характерные особенности строения первиччопо-
лостных червей. Тип Askelminthes, установленный Гроббеном (Grobben,
1908), объединяет следующие классы: Gastrotricha, Nematodes, Rotatoria
и Kinorhyncha, а также Nematomorpha, или Gordiacea; к первичночоло-
стным червям более или менее близки Acanthocephala.
Некоторые гастротрихи (Macrodasyoidea и часть Chaetonotoidea)
являются донными обитателями моря; большая часть Chaetonotoidea
живет в пресных водах; киноринхи — донные обитатели моря, неболь-
шая часть коловраток живет в море, преобладающее большинство на-
селяет пресные воды, иногда— почву; нематоды живут в море, солоно-
ватоводной среде, пресной воде, в почве, многие являются паразшами
растений и животных.
По наиболее распространенной классификации первичнополостные
черви — тип. Реже, и то в последнее время, нематоды (В. G. Chitwood
and М. В. Chitwood, 1950) и даже киноринхи (В. G. Chitwood, 1959) воз-
водятся в самостоятельные типы с несколькими классами. Если прини-
мать эту точку зрения, классы следует характеризовать не совокуп-
ностью черт организации и развития, а отдельными признаками, что осо-
бенно касается такой бедной группы, как киноринхи. Я считаю более
обоснованным видеть в гастротрихах, киноринхах, коловратках и нема-
тодах классы одного типа.
Малочисленный класс гастротрих состоит из отрядов Macrodasyoidea
и Chaetonotoidea. Еще меньшего объема класс киноринх состоит из трех
отрядов — Cyclorhage, Conchorhage и Homalorhage или, по Хиггинсу
(Higgins, 1962), из двух — Cyclorhagida и Homalorhagida. Более обшир-
ный класс коловраток образован тремя отрядами: Seisonidea, Monogo-
89-
nonta и Bdelloidea. Для громадного по численности класса нематод обос-
новано деление на два подкласса — Adenophorea ( = Aphasmidia) и Se-
cernenta. ( = Phasmidia) с несколькими отрядами в каждом подклассе.
Из четырех классов первичнополостных червей наибольшее количе-
ство исследований, которые все время продолжают появляться, относит-
ся к нематодам, и особенно к паразитическим формам последних. Работ
такое количество, что мет возможности привести труды даже наиболее,
выдающихся исследователей. Приходится ограничиться ссылками на
книгу Торна (Thorne, 1961), в которой обрисовано значение основопо-
ложников нематодологии, в том числе И. Н. Филиппьева, а также на
книгу Гаймен (Hyman, 1951), в которой с большим искусством исполь-
зована литература всего мира. Выдающееся значение имеют работы
К. И. Скрябина и его школы по зоонематодам (см. «Основы немато-
дологии», т. I—IX, 1949—1963, «Определитель паразитических нематод»,
т. I—IV, 1949—1954, «Строительство Советской гельминтологии», 1946,
«Строительство гельминтологической науки и практики в СССР», т. I,
1962) и по фитонематодам работы А. А. Парамонова (1962, 1964).
Основные черты внешней морфологии первичнополостных червей сле-
дующие: тело удлиненное, несколько уплощенное на брюшной стороне
в связи с ползанием (гастротрихи и часть коловраток, киноринхи), или
длинное нитевидное, веретенообразное, цилиндрическое круглое в раз-
резе (нематоды), или короткое, мешковидное или шарообразное (плапк-
тичеокие коловратки). Тело состоит из головного, туловищного и хво-
стового отделов. Головной отдел с его танго-, хемо- и фоторецепторами,
хорошо выражен у коловраток, поскольку несет коловращательный аппа-
рат, и у киноринх, у которых он вооружен шипами и впячивается внутрь
как хоботок. Туловищный отдел, собственно тело, лишь у киноринх со-
стоит из зонитов. Их кутикула разбита на метамерные пластинки и не-
сет метамерно расположенные шипы и щетинки. У остальных классов
первичнополостных туловище не расчленено, ио.кутикула бывает поделе-
на на кольца (нематоды, часть коловраток). У гастротрих на поверхнос-
ти тела метамерно расположены клейкие трубочки (или хватательные
ножки) и чувствительные волоски (Macrodasyoidea), а у нематод — ще-
тинки. Задний отдел тела хорошо дифференцирован: в виде, хвоста у
нематод, в виде ноги у коловраток и некоторых гастротрих, у которых он
раздвоен подобно вилке или реже вытянут в длинный тонкий хвост
(Wilke, 1954). У большинства гастротрих тело на конце суживается и
несет лишь большое количество клейких трубочек. У киноринх задний
конец тела не обособлен как хвост и несет .пару длинных щетинок.
Размеры тела у гастротрих колеблются от 0,07 до 1,5 мм, у кино-
ринх— от 0,2 до 1 мм, у коловраток — от 0,04 до 2 мм (как исключе-
ние Aspianchna— 5 мм). Нематоды, также преимущественно микроско-
пические и мелкие черви, от 0,02 до 10 мм в длину, иногда, однако, до-
стигают огромных размеров (например, Anisakls skrjabini, паразит кита,
в длину более 5 м, а паразит кашалота — Placentonema gigantissima—
8,4 м длиной).
Стенка тела состоит из кутикулы, эпидермиса (гиподермы) без
сплошного мышечного слоя. Кожно-мускульный мешок развит лишь у
нематод. Кутикула возникает перерождением наружных слоев прото-
плазмы эпидермальных клеток (Беклемишев, 1944, 1952, 1964) и обра-
зована не хитином, а склерой ротондами. У нематод в нее входят аль-
бумины, глюкопротеиды, фиброид, коллаген и кератин (Chitwood and
Chitwood, 1950; Парамонов, 1962).
Кутикула чаще гладкая, порой с ребрами, орнаментированная, реже
кольчатая, состоящая из метамерно расположенных колец (нематоды от-
ряда Chromadorida и др.). Сбрасывание кутикулы в. онтогенезе связано
с ростом и наблюдается у нематод и киноринх.
90
Кутикула у гастротрих, коловраток, киноринх и свободных морских
и пресноводных нематод' и многих фитонематод очень тонкая, гибкая.
Кутикула у нематод особенно прочная, эластичная, крайне растяжимая.
У многих зоонематод она становится очень толстой, состоящей из не-
скольких (до девяти) слоев, например, аскаридаты, стронгиляты
(Chitwood and Chitwood, 1950; Богоявленский и Балагина, 1962). Мож-
но сказать, что кутикула нематод многофункциональна; она очень устой-
чива к химическим и другим воздействиям; у многих форм она в разной
степени полупроницаема («фито- и зоонематоды). Помимо того, что она
несет защитную функцию, через нее поддерживается связь с внешней
средой, она сохраняет внутренний тургор, й служит опорой для мышц;
ее эластичность играет большую роль в движении червей (Парамонов,
1962).
Производными кутикулы у первичнополостных червей являются щеч-
тинки, иглы, папиллы, у самцов нематод — спикулы, спинные и брюш-
ные пластинки или склериты зонитов киноринх.
Эпидермальный слой — реже клеточного строения, с одноклеточными
железами (примитивные нематоды —свободные морские, частью прес-
новодные), чаще в виде синцития (большинство нематод, гастротрихи,
киноринхи и коловратки), причем у ряда панцирных коловраток во
взрослом состоянии синцитий исчезает (Pejler, 1962).
Реснички обычно отсутствуют, лишь у гастротрих имеется две рес-
ничные полосы на брюшной стенке, ресницы коловращательного аппа-
рата у коловраток и в передней части брюшной стороны у наиболее
примитивных коловраток.
Гиподерма у гастротрих образует боковые продольные утолщения, у
нематод—два сильные боковые и более слабые спинной и брюшной
утолщения —валики или поля, неудачно называемые некоторыми авто-
рами (Hyman, 1951; Парамонов, 1962) «хордами».
У всех названных классов, произшедших, вероятно, от ресничных па-
ренхиматозных червей (рабдоцелид), наблюдается редукция ресничек
эпидермиса, утрата клеточного строения, усложняющееся развитие и
многофункциональность кутикулы, связанные с условиями жизни в пес-
ке, почве и с паразитизмом.
С гиподермой связаны сериально расположенные вдоль тела одно-
клеточные кожные железы, наиболее сильно развитые у свободноживу-
щих нематод, шейная железа, железы амфид, фазмиды и хвостовые же-
лезы, несущие, как будет видно дальше, разные функции. Хорошо разви-
ты кожные железы и у гастротрих — сериально расположенные по телу
железистые клетки и железы клейких трубочек, или хватательных ножек,
выделяющих слизь. Кожные железы у киноринх представлены клеевыми
железами пары клейких трубочек, выделяющих слизь. У коловраток кож-
ных желез мало; к ним относятся ретроцеребралыные органы неизвест-
ной функции и цементные или клейкие железы ноги, секрет которых
служит для закрепления на субстрате, а у ряда коловраток участвует
в образовании их трубочек и домиков.
Лишь у нематод, непосредственно под гиподермой, подостланной ба-
зальной мембраной, расположен слой продольных мышц (кожно-мус-
кульный мешок), число которых у одних (Meromyaria) очень мало и
строго постоянно, боковые валики гиподермы делят мышцы на два поля,
а у других (огромное большинство свободноживущих нематод) мышеч-
ных клеток много и они поделены на шесть-семь полей (Polymiaria).
Мышечные клетки очень характерного строения: веретеновидные с мио-
фибриллами, расположенными в виде продольного желоба, внутри кото-
рого лежит саркоплазма с ядром. Отростки саркоплазмы связаны со
спинным и брюшным продольными нервами. Попеременные сокращения
спинных и брюшных мышц (рис. 24), антагонистами которых являются
91
кутикула и тургор .полости тела, обеспечивают скользящие, змеевидные-
движения нематод взад и вперед. Отсутствие кольцевых мышц и силь-
ное развитие боковых полей гиподермы определяют движения нематод,
не на (брюшной стороне, а на боку, так что боковая сторона у них
(Филипьев, 1918—1921) функционально замещает брюшную сторону.
Нар-яду с таким, наиболее распространенным способом движения, у раз-
ных экологических групп нематод, особенно фитопематод, наблюдаются-
Рис. 24. Пристеночная мускулатура $ Proales daphnicola (по Властову)
и другие виды движения, вплоть до утраты способности к передвиже-
нию при полной редукции мускулатуры тела, как у ряда эндопаразитов;
растений. Скользящие движения на боку, коррелятивно связанные с уд-
линенной, цилиндрической формой тела без наличия каких-либо вспомо-
гательных наружных органов движения, очень характерны для нематод.
Такой способ передвижения, видимо, был причиной исчезновения у не-
матод кольцевых мышц стенок тела, так как для червей как двусторон-
несимметричных организмов типичным является наличие по меньшей
мере продольных и кольцевых слоев кожно-мускульного мешка.
Однако надо отметить, что некоторые морские свободные немато-
ды— Draconema, Rhabdogaster (хромадориды)—ползают по субстра-
ту, как гусеницы пядениц, почему И. И. Мечников по этой особенности .
даже противопоставлял их всем остальным нематодам как ползающих с
(Щепотьев, 1908; Филипьев, 1918—1921; Парамонов, 1962).
Очень многие свободные нематоды прикрепляются с помощью хво-
стовых желез к субстрату и производят энергичные движения передним
концом тела (Филипьев, 1918—1921).
У гастротрих, передвигающихся .при помощи ресничек, мускулатура
тела олигомеризована, в основном с каждой стороны тела сохраняются
2—4 продольные мышцы. Из них вентро-латеральные обеспечивают
движение передних и задних прикрепительных или хватательных трубо-
чек, а дорсальные мышцы, прикрепляющиеся к головному отделу и .
глотке, действуют как ретракторы.
У коловраток мускулатура тела более дифференцирована, чем у -
гастротрих и киноринх, что связано с расчленением тела на голову,
туловище и ногу и, соответственно, большей подвижностью. Мускулату-
ра используется для ловли добычи ртом,- для изгибания тела в случае
его расчленения, для втягивания его отделов наподобие подзорной тру-
бы (по выражению Беклемишева, 1952, 1964), для ползания. Большое
число разнообразных мышц при мягких покровах обеспечивает сильную
92
подвижность тела и верткость/ необходимую при ловле подвижной до-
бычи и для опасения от врагов.
Мускулатура тела коловраток состоит из кольцевых, лежащих под
ними цельных или прерванных на брюшной или спинной сторонах, попе-
речных и продольных мышц. Кольцевые мышцы в передней части
тела образуют сфинктор, смыкающийся над втянувшимся коловра-
щательным аппаратом, а в задней части тела — сфинктор ноги. Часть
кольцевых и продольные мышцы обеспечивают указанные выше
сокращения тела и его частей. Специальная система мышц обслужи-
вает коловращательный аппарат, парные спинные и брюшные ретрак-
торы втягивают голову. Слабые кожно-висцеральные мышцы распо-
лагаются между стенками тела и внутренними органами. Мускулатура
коловраток преимущественно поперечнополосатая. Преобладающий
способ передвижения — плавание с помощью коловращательного ап-
парата.
У киноринх мускулатура тела наиболее сложна, что связано с рас-
членением их тела на кольца (зониты) со спинными и брюшными пла-
стинками. Продольные дорсо-латеральные и вентро-латеральные мышцы
метамерно разделены на пучки, идущие от одного зонита к другому; от
спинной стенки зонита к брю-шной идут дорсо-вентральные пучки мышц.
Сокращением кольцевых мышц двух первых зонитов и дорсо-вентраль-
ных мышц других зонитов, благодаря увеличению тургора в полости
тела, голова вытягивается, сокращением же ретракторов она впячивает-
ся. Киноринхи двигаются медленно; выпятив голову и закрепившись
ее шипами в субстрате, они подтягивают к ней туловище, двигаясь по
гидравлическому способу (Л. А. Зенкевич, 1944),
У нематод нервная система лежит субэпителиально и обладает по-
стоянным и небольшим числом клеток, что, как известно, характерно
для всей организации нематод. Построена она по типу ортогона. У сво-
бодных форм (Филипьев, 1918—1921; Парамонов, 1962) она состоит из
окологлоточного фибриллярного кольца и отходящих от него вперед,
к головным органам, шести нервов и десяти продольных нервов или
тяжей, отходящих назад и соединенных анальным кольцом; из них
брюшной—наиболее мощный. Впереди и позади от окологлоточного
кольца лежат скопления ганглиозных клеток. Гистологическое строенио
нервной системы изучено лишь у аскарид Гольдшмидтом (Goldsch-
midt, 1908, 1909, 1910).и Д. И. Дейнекой (Deineka, 1908). Важные дан-
ные о нервной системе фитонематод получены Парамоновым (1962,
1964), установившим у них густую сеть сериально расположенных ганг-
лиозных клеток. К сожалению, гистологическое строение нервной систе-
мы свободных нематод мало известно. Физиология нервной системы изу-
чена недостаточно, но, по-видимому, она примитивна (Парамонов, 1962.
1964).
С большим своеобразием строения мышечных клеток, длинные сар-
коплазматические отростки которых подходят к спинному и брюшному
нервным тяжам (или к коннективам), связано то, что концевой мотор-
ный аппарат нервной системы становится очень коротким, не нервы под-
ходят у нематод к мышцам, а ' мышцы — к нервам (Hanstrom, 1928).
В нервной системе аскарид Ганстрем отмечает ряд особенностей, кото-
рые указывают на вторичные изменения в строении нервной системы
в исходном ортогональном типе: например, образование брюшного нер-
ва слиянием первоначально двух. Мне кажется, это говорит не только
о становлении нервной системы нематод из ортогонального типа, но и в
пользу возможного происхождения, как думает В. Н. Беклемишев
{1952, 1964), их от турбелляриеподобных предков.
.Нервная система гастротрих лежит субэпителиально и состоит из
надглоточного ганглия, седловидно налегающего на глотку, и отходя-
93
щих от него двух продольных брюшных нервных тяжей. Допустимо,,
учитывая разные формы нервной системы турбеллярий, что и.нервная
система гастротр/их принадлежит к ортогональному типу.
У киноринх «.мозг» — окологлоточное кольцо с ганглиозными клет-
ками — охватывает переднюю часть глотки. От него отходит брюшной
нервный тяж. с метамерно расположенными ганглиозными клетками по-
середине каждого туловищного зонита. Группы ганглиозных клеток на-
ходятся в боковых и медиодорсальных утолщениях эпидермиса (гипо-
дермы) в каждом зоните. .На этом осно-
Рис. 25. Схема строения перед-
него конца примитивно постро-
енной нематоды (из Hennig)
1 — амфида; 2 — губы; 3 — го-
ловные щетинки; 4 — наружные
и внутренние губные щетинки
вании, хотя здесь нет выраженных нерв-
ных тяжей, в литературе высказывается
мнение о наличии у киноринх четырех
продольных нервных тяжей. Нервную си-
стему киноринх в таком случае можно
отнести к ортогональному типу. Но впол-
не вероятно, что у киноринх мы имеем,
дело с тем, что найдено у ряда фитонема*
тод (Парамонов, 1962). Гистологическая
нервная система киноринх настолько сла-
бо дифференцирована, что она едва от-
личима от эпидермиса, с которым тесно
связана.
Нервная система, особенно у очень
подвижных и активных коловраток отря-
да Monogononta, высоко дифференциро-
вана. В основном, она состоит из мозга
или церебрального ганглия, лежащего
над мастаксом (глоткой), чувствительных
и двигательных нервов, идущих от него-
к органам чувств головы, к различным ее мышцам, и двух вентро-лате-
ральных нервных тяжей с ганглиозными клетками, заходящими в ногу.
С мозгом связана и вегетативная нервная система ,в виде сплетения нер-
вов и ганглиев в стенке мастакса.
В связи с низким уровнем организации первичнополостных червей-
органы чувств у них развиты слабо и представлены примитивными ре-
цепторами. У «нематод они сконцентрированы преимущественно на пе-
реднем,. головном конце тела. Это головные и губные тангорецеп-
торы в виде щетинок и папилл (рис. 25) у свободных морских и пресно-
водных нематод, папилл у почвенных и паразитических форм. Головные-
и губные тангорецепторы интересны тем, что их контакт с жертвой вы-
зывает глотательный рефлекс нематоды (Парамонов, 1962). Тангорецеп-
торы располагаются и вдоль тела, на его заднем конце, у самцов они
связаны с половой жизнью червя.
Хе,мо|рецепторы свободных аденофорей — амфиды лежат по бокам
головы; у сецернентов они сильно редуцированы, но у хищных, очень
подвижных форм они развиты хорошо (Парамонов, 1962). К типу хе-
морецепторов, как железисто-чувствительных органов, относят и фаз-
миды сецернентов.
Фоторецепторы в виде пары глазков с кутикулярной линзой и пиг-
ментом на переднем конце тела или в виде пигментных пятен распростра-
нены среди свободных морских и пресноводных нематод (Филипьев,
1918—1921; Wieser, 1953; Murphy, 1963).
У ряда гастротрих, по бокам головного конца, имеется пара реснич-
ных ямок, которые являются и хеморецепторами и гомологичны, по-
А. А. Парамонову (1962), амфидам нематод и обонятельным ямкам тур-
беллярий (Беклемишев, 1944, 1952, 1964). У немногих гастротрих име-
ются парные глазки, сходные с глазками нематод, и мало дифференци-
94
раванные тангорецепторы в виде осязательных щетинок, расположенных:
продольными рядами на туловище.
У киноринх существуют простые глазные пятна и та1нго'рецепторы и
виде осязательных щетинок, расположеиных продольными рядами на
спинной стороне и на боках туловища.
У коловраток органы чувств многочисленны и, в связи с сильным'
развитием коловращательного аппарата и укорачиванием тела, концент-
рируются на переднем конце последнего. Пара глазков с красным пиг-
ментом погружены в ткань мозга, они сравнимы с глазками нематод;
у некоторых имеется, кроме того, пара латеральных и пара апикальных
глазков на боках короны. Парные ресничные ямки коловращательного
аппарата, по-видимому, являются хеморецепторами. Папиллы, щетинки,
находящиеся в разных местах переднего конца тела, так называемая
спинная антенна, пара латеральных и некоторые другие, вероятно, функ-
ционируют как тангорецепторы.
Органы чувств гастротрих, киноринх и коловраток изучены, к сожа-
лению, далеко не достаточно.
Кишечник состоит из передней — эктодермальной кишки, средней —
эктодермальной и задней — эктодермальной. Обычно он в виде трубки
вытянут по длинной оси тела (нематоды, гастротрихи, киноринхи). Рог
терминальный (нематоды, гастротрихи, ' киноринхи) или лежит на
брюшной стороне (коловратки), у нематод анальное отверстие нахо-
дится на брюшной стороне или открывается в клоаку, у гастротрих оно1
лежит на брюшной или спинной стороне, у коловраток — открывается
в клоаку, у киноринх — терминальное.
Передняя кишка, начинающаяся ротовой полостью, состоит из глот-
ки и пищевода. Глотка обычно сохраняет строение массивной глотки
турбеллярий — трубки с трехгранньим просветом и радиальной мускула-
турой (нематоды, гастротрихи, киноринхи) или становится своеобраз-
ным жевательным аппаратом — мастаксом (коловратки), вооруженным
твердыми кутикулярными пластинками, образующими наковальню и мо-
лоточек, и снабженным системой поперечнополосатых мышц. Он слу-
жит для дробления и перетирания пищи и захватывания добычи.
Ротовая полость у нематод ограничена шестью губами, несу-
щими губные рецепторы. Строение губ очень -разнообразно, у ряда
форм губы сливаются в три губы, но в онтогенезе таких специализиро-
ванных форм имеются стадии со строением губ, типичным для взрослых
особей более примитивных видов и групп (Osche, 1957). В связи с важ-
ной ролью в захвате очень разнородной пищи ротовая полость у нема-
тод претерпевает многочисленные, необычайно разнообразные приспосо-
бительные изменения в строении, в вооружении и отправлениях, особен-
но у свободных нематод. У ряда нематод ротовая полость вооружена
зубами или копьем (одонтостилем или стоматостилем), образующимся
за счет стенок стомы (Steiner, 1956). По характеру питания и функции
ротовой полости — это формы хищные, нехищные, колющие, сосущие,
кусающие, глотающие, схватывающие или скребущие (Wieser, 1953).
У ряда зоонематод ротовая полость становится мощной присоской, с по-
мощью которой паразит прикрепляется к стенкам органов хозяина и вы-
сасывают жидкую пищу из ткани органа — Soboliphyme (Petrov, 1930),
Syngamus (Рыжиков, 1949), стронгиляты.
Нередко глотка образует одно или несколько мускулистых вздутий,
или луковиц. Вместе с ротовой полостью и стилетами с капиллярным'
просветом глотка нематод становится сосущим аппаратом необычайной
силы.
У ряда фитонематод (Мюге, 1958а, 19586), апирурат, аскаридат я
филариид задний отдел мускулистой глотки теряет мускулатуру и ста-
новится железистым органом.
95
У ряда нематод (сецерненты) в ротовую полость, порой в глотку,-
открываются одна спинная и две или пять дагеро-вентральных железис-
тых клеток (эноплида) — эктоферментативные железы (Парамонов,
1962), у гастротрих (Chaetonotoidea) в глотку открываются две лары
желез, возможно слюнных, пара или более желез открывается у кино-
ринх в пищевод и на границе пищевода и желудка. У коловраток пара
слюнных желез открывается в мастакс, на месте соединения пищевода
с желудком лежит пара так называемых пищеварительных желез, откры-
вающихся в желудок. Функциональное значение бесспорно доказано,
по-видимому, лишь для желез глотки нематод (Мюге, 1958; Пара-
монов, 1962).
Глотка открывается или непосредственню в среднюю кишку, как у
турбеллярий (нематоды), или переходит сначала в пищевод (гастротри-
хи, киноринхи, коловратки). Средняя кишка или вытянута в длинную
прямую трубку (нематоды, гастротрихи, киноринхи), или дифференциро-
вана на железистый желудок и собственно кишку.
Стенки средней кишки образованы высокими цилиндрическими клет-
ками со слоем тончайших палочек на внутренней поверхности, вероят-
но, являющимися измененными ресницами (нематоды), столбчатыми
или кубическими клетками (гастротрихи), призматическими клетками
(киноринхи), крупными многоугольными ресничными клетками (боль-
шинство коловраток) и реже синцитиальным слоем. Вследствие утраты
первичнополостными червями паренхимы тела, средняя кишка их не
имеет в своей стенке слоя мускулатуры. У киноринх и коловраток сетью
из кольцевых и продольных мышечных волокон, а у гастротрих тонкими
кольцевыми мышцами она окружена снаружи, однако, по Ремане (Re-
mane, 1935—1936), у ряда гастротрих кольцевые мышцы лежат и непо-
средственно в стенке кишки.
Задний конец средней (энтодермальной) кишки переходит в заднюю
или прямую кишку, которая возникает путем впячивания покровов тела
и открывается порошицей на заднем конце тела. Порошица открывается
или независимо от половых и выделительных отверстий самки нематод,
гастротрих и киноринх, или превращается в клоаку, в которую впадают
экскреторные и половые протоки (самки коловраток), или мужской поло-
вой проток и сумка спикул (самцы нематод) (Voltzenlogel, 1902).
Для нематод характерны разнообразие пищи и способов питания,
встречаются и свободные хищники, и сапробионты, и паразиты растений,
тканей, в том числе крови и разных органов животных, особенно позво-
ночных. Свободные морские, пресноводные и почвенные нематоды пита-
ются детритом с бактериями, простейшими и крупной добычей — тур-
белляриями, коловратками, нематодами, мелкими аннелидами и другими
водными животными. Хищные нематоды, например Seinura, высасывают
других нематод (Mulvey, 1961), Mononchus, Anatonchus высасывают
или заглатывают некоторых нематод целиком (Парамонов, 1962).
Гастротрихи и киноринхи питаются диатомовыми, бактериями, детри-
том. Некоторые коловратки являются седементаторами (Догель, 1930),
улавливающими при помощи токов воды, вызываемых коловращатель-
ным аппаратом, микроскопические организмы — жгутиконосцев, одно-
клеточных водорослей. Другие коловратки — хищники, питающиеся
крупной добычей, которую они захватывают частями коловращательного
аппарата, и мастаксом. Коловратки играют важную роль в пищевой
цепи пресных вод и сами являются пищей для мелких червей, ракооб-
разных и т. д.
Пищеварение у нематод — внекишечное и происходит под воздействи-
ем секрета эктоферментативных желез, ферменты которых расщепляют
сахар, гликоген, пептиды и белки (Мюге, 1958а, 19586). В средней киш-
ке пищеварение завершается, в ней происходит- главным образом вса-
96
сЫвание 'продуктов внекишечнаго .пищеварения слоем тончайших пало-
чек кишечного эпителия. В клетках средней кишки происходит накапли-
вание питательных веществ (см. Парамонов, 1962).
У коловраток пищеварение идет внеклеточно в полости желудка за
счет секрета пищеварительных желез и заканчивается внутриклеточно в
пищевых вакуолях синцитиальных клеток желудка. Пищеварение вооб-
ще, и особенно гастротрих и киноринх, изучено недостаточно.
У ряда зоонематод паразитический образ жизни вызывает pespeoc
частей кишечника. У многих Dracunculidae нет задней кишки и анально-
го отверстия; у Leptosoma средняя кишка дегенерирует полностью; умер-
митид средняя кишка превращается в мешок, слепо замкнутый на обоих
концах. У Buddenbrockia, эндопаразита Plumatella, исчезает весь пище-
варительный тракт. Редукция его происходит и у карликовых, рудимен-
тарных самцов коловраток, но по другим причинам.
Органов дыхания и кровообращения у первичнополостных червей нет.
Дыхание происходит через стенки тела. Оно аэробное, но у зоонематод
из сецернентов, кроме трихонеллид и диоктофимат, преобладает ана-
эробное дыхание. В связи с разными типами дыхания у разных групп
нематод наблюдается известное различие в образовании продуктов об-
мена веществ — жирные кислоты у одних, жиры — у других (Мюге,
1958а, б, 19586). У коловраток заглатываемая ртом вода поступает через
стенки желудка в полость тела, доставляя туда кислород.
Типичные органы выделения—пара протонефридиев с концевыми
клетками с мерцательным пламенем — имеются у коловраток, киноринх
и части гастротрих (макродазиодеи). У нематод они замещены кожны-
ми железами, что является очень характерной особенностью нематод.
Протонефридии у киноринх открываются в девятом зоните тела, у гаст-
ротрих—-в средней части тела, у коловраток протоки протонефридиев
впадают в пульсирующий мочевой пузырь, открывающийся в клоаку
вблизи полового отверстия и порошицы. Протонефридии наиболее разви-
ты у коловраток. Два главных канала тянутся по всей длине их тела,
образуя петли; число концевых клеток с мерцательным пламенем с каж-
дой стороны обычно от двух до восьми. Протонефридии выводят экс-
креты и воду, заглатываемую ртом и поступающую в полость тела.
Можно напомнить, что протонефридии плоских червей (Беклемишев,
1952, 1964) выполняют двоякую функцию — экскреторную и осморегуля-
торную, при этом считается, что филогенетически первичной была осмо-
регуляторная функция, т. е. удаление воды, постоянно поступающей в
организм. Двоякую функцию признают и за протонефридаями.киноринх,
гастротрих и коловраток, постоянная пульсация мочевого пузыря кото-
рых наглядно -говорит об этом.
Отношения аппаратов осморегуляции и экскреции у нематод более
сложные и несомненно вторичные, возникшие позже, чем в других клас-
сах (рис. 26).
Рациональные данные о строении, отправлении фагоцитарноэкскре-
торного аппарата и его эволюции у нематод мы находим у Н. В. Насоно-
ва (Nassonov, 1897, 1900), Е. П. Головина (1901, 1902) и И. Н. Филипь-
ева (1918—1921). Эти органы изучались многими зарубежными исследо-
вателями, но наиболее четкие морфо-эколого-физиологические данные
об осморегуляции и экскреции мы находим у А. А. Парамонова (1962,
1964). По данным последнего, важнейшую роль в этих процессах играют
кожные (гиподермальные) железы, составляющие полимеризованный
аппарат в виде многочисленных одноклеточных желез (так называемых
паралатеральных), сериально расположенных вдоль боковых валиков
гиподермы, шейной железы (ренетты)—компактной, массивной одно-
клеточной железы, лежащей вентрально у заднего конца глотки с корот-
ким протоком, открывающимся наружу, и три одноклеточные хвостовые
7 Д. М. Федотов
97
А — Rhabditis sp.; Б—Апсг/losioma sp.;
В — Oesophagoslomuni sp.
мы в полости тела лежат
железы. Таков тип осморегуляторно-экскреторногр аппарата у наиболее
примитивных, свободных морских и пресноводных, с тонкой кутикулой
легко проницаемой для растворов солей, оксифильных червей, нематод
из аденофорей. Это "связано с их жизнью в чистой воде, характеризую-
щим режимом.
У нематод-сецернентов (фазмидие-
вые) — почвенных, сапробиотических,
паразитов растений и животных с
трудно проницаемой, нередко толстой
кутикулой, живущих обычно в среде*
бедной кислородом,— вырабатывается
олигомеризованный (по А. А. Парамо-
нову) осморегуляторно-осмотический
аппарат. Он в основном состоит из од-
ной разветвленной железистой клет-
ки — шейной железы (ренетты) с
длинными внутриклеточными канала-
ми; с одним передним и одним задним
или с двумя задними, или с двумя пе-
редними и двумя задними каналами;
каналы с внутриклеточными просвета-
ми, которые проходят в боковых гипо-
дермальных полях. Шейная железа
длинным экскреторным протоком, от-
крывается наружу порой по срединной
брюшной линии. .
В эту систему входят также фаз-
миды — пара гиподермальных клеток,
с экскреторной функцией у одних и с
чувствительной у других форм (Пара-
монов, 19546, 1962). У ряда паразити-
ческих зоонематод на боковых валиках
четыре, реже шесть, крупных звездча-
тых клеток, наиболее полно описанных Н. В. Насоновым (Nassonov,
1897, 1900). Они захватывают нерастворимые продукты распада, абсор-
бируют введенные в полость тела краски и другие посторонние вещества,,
являясь (по Догелю, 1962), таким образом, почками накопления.
Наблюдения над развитием шейной железы у нематод показывают,
что исходным состоянием для обоих типов является массивная, компакт-
ная клетка (Hyman, 1951), тогда как разветвленная клетка с длинными
выделительными каналами является производной и связана с усилением
обмена веществ (Парамонов, 1962), что особенно характерно для нема-
тод-сецернентов, живущих в кишечнике (сапробиосе) и других частях
тела позвоночных, т. е. в средах, бедных кислородом. . .
В процессах осморегуляции и экскреции у нематод участвуют также
фазмиды, стенки глотки и кишки (Парамонов, 1962, 1964).
Сравнительно с протонефридиями киноринх, гастротрих и коловраток
осморегуляторно-экскреторный аппарат нематод очень своеобразен, но-
общим для всех является то, что эти аппараты или органы — произвол--
ные кожных покровов, т. е. эктодермы.
Что касается продуктов азотистого распада у свободных и паразити-
ческих нематод, то они (Rothstein, 1963) представлены в основном аммо-
нием и аминокислотами; мочевая кислота, мочевина, аллантоин, или
креатинин, образуются в незначительном количестве.
Очень характерной чертой всех Askelminthes является наличие перг
вичной полости тела, схизопеля, не имеющей эпителиальной выстилки и
являющейся по развитию полостью бластоцеля. У микроскопически мел-
98
Рис. 28. Aphelenchus sacchari (из Парамо-
нова)
А — самка, тотально; Б — головной конец;
В — хвост
ких гастротрих и киноринх она мала и представлена щелями между стен-,
ками тела и внутренними органами; она хорошо выражена у коловраток
и особенно у нематод. Впрочем у примитивных свободных морских .и
пресноводных нематод полость тела настолько бывает заполнена много-
численными свободными клетками и отростками мышечных клеток, что-,
ее почтц нет (Филипьев, 1918—1921). В полости тела у первичнополост-
ных червей развита нежная, сетчатая ткань, вроде паренхиматозной,,
являющаяся, вероятно, остатками когда-то существовавшей у предков
паренхимы. Амебоидные клетки полости тела, по-видимому, являются
фагоцитарными и экскреторными.
Полость тела (Парамонов, 1962) —не просто щели между органами,
и стенкой тела. Она, будучи наполненная полостной жидкостью, обеспе-
чивает тургор, участвует в регуляции осмотического давления, в ней,
как в органе внутренней среды, протекают процессы обмена веществ и
происходит их распределение по телу. В полостной жидкости собираются
продукты обмена веществ, в том числе такие, характерные для аскарид,
как валериановая, молочная и другие кислоты. У ряда нематод в полос-
ти ив стенках тела обнаружен гемоглобин (Hyman, 1951).
.Нематоды раздельнополы, половой диморфизм касается размеров
тела и формы заднего конца; среди рабДитид встречается гермафроди-
тизм, у некоторых из них и .мермитйд наблюдается партеногенез. В связи
с цилиндрической формой тела гонады у свободных нематод трубчатые.
7* 99..
(рис. 27, 28), парные, симметрично расположенные (аденофореи), у па-
разитических нематод (сецернентов), особенно паразитов позвоночных,
очень длинные половые трубки расположены несимметрично, чаще две
у самок, одна — у самцов.
Половые трубки самок и самцов — сложное образование, в котором
собственно яичники и семенники возникают из продуктов деления первич-
ной толовой клетки зародыша; яйцеводы, матка, семеприемник, вагина
самок, семепроводы, семенной пузырек, семеизвергательный канал и спи-
кулы самцов являются производными эктодермальных бластомеров S;
и S5 (Pai, 1928). В сформированном состоянии половой аппарат пред-
ставлен трубками без ясных границ между частями. Половое непарное
отверстие у самок открывается у переднего или заднего конца тела на се-
редине его брюшной стороны. У самцов семяизвергательный канал от-
крывается в клоаку рядом с анальным отверстием, в клоаку же откры-
вается и сумка с двумя спикулами.
Оплодотворение внутреннее, .сперматозоиды очень своеобразны, аме-
боидноподвижны. Наблюдается попарное прочное соединение дву< по-
лов (Syngamus trachea), паразитирование мелких самцов во влага-
лище самок {Trichosomoides crassicauda). Численное соотношение по-
лов у нематод, как правило, равное, хотя у многих пресноводных из-
вестны лишь самцы. Индивидуальная плодовитость свободных нематод
и многих фптонематод незначительна, что, однако, компенсируется
крайней быстротой развития (см. ниже); у зоопаразитов, наоборот, она
колоссальная. Так, Trichinella spiralis ограждает до 10 тысяч личинок,
Ascaris lumbricoides за 5—6 месяцев продуцирует 50—60 миллионов
яиц, за год жизни вес отложенных яиц превышает вес червя в 1700 раз.
У Sphaerularia bombi матка, наполненная громадным количеством яиц
и вышедшая наружу, по объему превышает тело червя в 15—20 тысяч
раз (Догель, 1962).
Яйца, особенно у зоопаразитов, защищены плотной скорлупой, у ас-
карид состоящей из пяти оболочек, крайне стойкой к воздействиям
внешней среды и сильных химических реагентов.
Гастротрихи обычно гермафродиты (Macrodasyoidea и часть Chaeto-
notoidea), у большинства последних известны лишь самцы. Парные или
непарные яичники и семенники лежат по сторонам или над средней
.кишкой, яичники в виде простого конгломерата овоцитов (Remane,
1935—1936), семенники длинные, мешковидные. Яйцевод с семяприем-
ником, совокупительной сумкой, открывается впереди или вместе с
анальным отверстием. Парные семяпроводы открываются непарным от-
верстием впереди женского отверстия или вместе с последним и поро-
шицей. Оплодотворение внутреннее, индивидуальная плодовитость са-
мок незначительная.
Киноринхи раздельнополы, половой диморфизм незначительный; го-
нады парные, мешковидные; яичники с половыми и питательными клет-
ками и семенники лежат по сторонам кишечника, короткие половые про-
токи парными отверстиями открываются наружу на заднем конце, в три-
надцатом зоните. У самцов имеется два копулятивных органа в виде
подвижных кутикулярных шиповидных стерженьков, яйцеводы с семя-
приемниками. Оплодотворение внутреннее. Индивидуальная плодови-
тость самок малая.
Коловратки раздельнополы, степень выраженности полового димор-
физма сильно варьирует. У Seisonidae, например, самцы мало отлича-
ются от самок; у Monogononta наблюдаются разные степени дегенерации
самцов вплоть до карликовых, утративших, кроме половых, почти все ор-
ганы и ставших, по выражению Б. В. Властова (1955), самостоятельно
двигающимся мужским половым аппаратом; у Bdelloidea происходит
полное исчезновение самцов и переход к партеногенезу. У самок в зад-
100
ней части тела, под кишечником, лежит чаще один (ио парный по проис-
хождению) мешковидный яичник, состоящий из гермария и вителдярия-,.
с коротким яйцеводом, Открывающимся в клоаку. Непарный семенник у
самцов с непарным семяпроводом, который снабжен у одних форм цир-
русом, у других — пенисом, открывается наружу на заднем конце тела;
Оплодотворение происходит реже введением копулятивного органа в
клоаку самки, чаще пиподермальной импрегнацией. Индивидуальная
плодовитость самок невысокая, 10—20 яиц, но развитие протекает быст-
ро. Коловратки откладывают яйца, реже — живородящи.
У морских коловраток Seisonidea сохраняются .исходные отношения,
яйца оплодотворяются, из них возникают самки и самцы. У большин-
ства коловраток (Monogononta), ставших пресноводными, в связи с рез-
ко выраженными сезонами года, возникают смены поколений, половых
и партеногенетических и разное число жизненных циклов (виды моно-
ди- и 1пол1и1ци1клические). Размножение сопровождается образованием
трех родов яиц. Крупные яйца с системой плотных оболочек (покоящие-
ся) после длительного покоя (порой до года) весной дают партеногенети-
ческих самок. Они летом размножаются партеногенетическм и произво-
дят тонкостенные яйца; из крупных яиц возникают самки, из мелких —
самцы.
Как показали экспериментальные исследования, особенно А. О. Тау?
сои (1923а, 19236; Tauson, 1924, 1927), появление самок полового
поколения обусловлено рядом природных условий и может регули-
роваться экспериментально. После спаривания самки откладывают опло-
дотворенные, покоящиеся яйца, дающие вновь поколение партеногене-
тических самок. У Bdelloidea, часто обитателей почвы, в связи ср
своеобразной экологией, происходят дальнейшие, еще большие измене?
ния в жизненном цикле —самки размножаются исключительно партено?
генетически, самцы вовсе исчезают.
Об эмбриональном развитии зоонематод мы знаем из старых клас-
сических работ Бовери (Boveri, 1892, 1899) и Цур-Штрассена (Zur-Stras-
sen, 1896) по Rhabditis и из более новых работ Мартини (Martini, 1903,
1906) по Camallanus ( — Cucullanus) и Пая (Pai, 1928) по Turbatrix
(=Anguillula), а также из последней работы Цур-Штрассена (Zur-Stras-
sen, 1959) по механизму эмбрионального развития нескольких родов
паразитических нематод. Мало изучен ранний онтогенез фитонематод,
не изучено эмбриональное развитие свободных нематод.
Дробление у зоонематод полное, детерминированное, типично мо-
заичное. Судьба отдельных бластомеров, давно получивших буквенные
обозначения, как показала прослеженная их генеалогия, фиксиро-
вана.
Продукты деления бластомера Si дают бластомеры эктодермы, деле-
нием бластомера Ро образуются бластомеры эктодермы (стомодеум),
энтодермы, мезодермы и первичная половая клетка, зародышевый путь
которой выражен с начала дробления яйца.
На стадии двух бластомеров фиксированы продольная ось тела, пе-
редний и задний концы, спинная и брюшная стороны тела будущего
червя, например, у Turbatrix aceti (Pai, 1928). Характерно также распо-
ложение бластомеров на стадии четырех в виде буквы Т и ромба.
Клетки эктодермы дают начало эпидермису с его полями, нервной си-
стеме, осморегуляторно-экскреторному аппарату, задней клинике с клоа?
кой; клетки энтодермы образуют среднюю кишку, клетки мезодермы —
продольную мускулатуру тела и соединптельнотканые клетки первичной
полости тела. Первичная половая клетка дает зачаток яичника и семен-
ника (см. стр. 100); яйцевод, матка, семяприемник самок, семепровод,
семенной пузырек, семяизвергательный канал самцов возникают за счет
продуктов деления бластомера S5, а вагина (вульва) самки и спикулы
самца — за счет деления бластомера Sj.
101
' ‘ Мезодерма возникает из двух источников (хотя некоторые авторы на-
считывают три источника). Передний зачаток мезодермы происходит от
бластомера EMSt вместе с зачатком энтодермы и стомодеума и образует
пару мезодермальных однослойных полосок, дающих мускулатуру тела.
Задний зачаток мезодермы возникает вместе с клетками эктодермы.
Можно называть первый — энтомезенхимой, второй— эктомезенхимой.
Данные относительно происхождения передней кишки, особенно глот-
ки, расходятся: по одним — она эктодермального происхождения (на-
пример, по Goette, 1881), по другим (в том числе и Pai, 1928) —эктодер-
мального, так как возникает за счет продуктов деления бластомера
EMSt, но обычно передняя кишка считается эктодермальной.
У ряда зоонематод, например, Syngamus merulas, Cuathostoma bon-
larti из семейства Syngamidae (Рыжиков, 1954, см. Скрябин, 1949—
1964), на стадии восьми бластомеров отчетливо выражен спиральный
тип дробления.
Как известно, число клеток в онтогенезе нематод, как и в органах
взрослых, ограниченное, небольшое, не превышающее 1000 и постоянное
у разных форм. Это говорит о вторичном характере онтогенеза нема-
тод. Детерминированный мозаичный характер дробления типичен и для
фитонематод, причем и у них наблюдается линейное расположение
четырех первых бластомеров — Rodolphtis sitnilis (Weerdt, 1960 — цит. по
Парамонову, 1962), но ранний эмбриональный период развития фито-
нематод изучен недостаточно. К сожалению, не изучено эмбриональное
развитие примитивных нематод — свободных морских и пресноводных,
у которых, надо думать, лучше сохранилось спиральное дробление.
В постэмбриональном развитии нематоды проходят пять личиночных
возрастов, сопровождаемых четырьмя линьками, во время которых про-
исходит постепенное формирование внутренних органов, однако корен-
ные изменения претерпевает лишь половая система (Парамонов, 1962).
У многих нематод и, видимо, свободных фитонематод индивидуальное
развитие протекает необычайно быстро, порой в течение нескольких
часов.
У многих паразитических нематод имеются рабдитовидные личинки,
что указывает на родственные отношения со свободными рабдитидами,
причем личинки ведут самостоятельный, наземный образ жизни и отли-
чаются необычайной устойчивостью против воздействия крайне небла-
гоприятных естественных и экспериментальных условий (Догель, 1962).
Из яйца вылупляется молодой червь, отличающийся от взрослого
меньшими размерами, неразвитой половой системой и некоторыми част-
ностями строения. Эту стадию развития обычно называют неправильно,
как считает Гаймен (Hyman, 1951), личинкой. Однако здесь мы имеем
то, что наблюдается у насекомых с неполным метаморфозом, у которых
мы говорим о личинках или нимфах. Откидывая последний термин,
по-моему мало необходимый, можно применительно к нематодам гово-
рить о личинках, тем более, что они проделывают четыре линьки и про-
ходят пять возрастав, лишь после чего становятся половозрелыми.
Насколько мало работ об эмбриональном развитии нематод, настоль-
ко много исследований по постэмбриональному развитию самых разно-
образных групп нематод, особенно паразитов животных и растений, при-
чем установлены примеры, когда личинки всех возрастов обладают ти-
пичной для взрослых организацией и когда они, напротив, резко отли-
чаются от взрослых. У нематод наблюдаются разные жизненные циклы:
развитие без чередования и без смены хозяев (аскариды), чередование
поколений без смены хозяев (Rhabdias bufonis), когда паразитирующее
поколение представлено партеногенетической самкой, свободноживу-
щее — раздельнополыми особями, без чередования поколений с одно-
кратной сменой хозяев, когда взрослый червь паразитирует в одном хо-
102
зяине, личинка—-в другом (многие Spirurata, Dractineulus medinensis').
У многих нематод жизненный цикл включает двух промежуточных хо-
зяев (дополнительный резервуарный хозяин); взрослый червь Gnatosto-
та spinigerum паразитирует в стенке желудка плотоядных, первым про-
межуточным хозяином личинки является циклоп, вторым — рыбы, ля-
гушки и змеи (Догель, 1962).
Онтогенез киноринх почти не изучен, что значительно затрудняет по-
нимание природы этой группы. Известно только, что яйца у них разви-
ваются после оплодотворения, партеногенеза нет, вылупившаяся малень-
. кая личинка постепенно, с несколькими линьками (Nyholm, 1947) пре-
вращается во взрослого червя.
Развитие гастротрих изучено несколько лучше (Beauchamp, 5929;
Sacks, 1955); у некоторых гастротрих, как у коловраток (Hyman, 1951),
яйца двух родов — subitaneous и dormant с толстой -вооруженной оболоч-
кой и развиваются партеногенетическш Дробление полное, почти равно-
мерное, несколько детерминированное; на стадии 8 бластомеров .спираль-
ного, далее — билатерального типа. На брюшной стороне 32-клеточной
целобластулы в ее обширную полость погружаются две клетки, зачаток
антодермальной кишки, у заднего конца которой (делением этих клеток
На четыре?) ложатся две первичные половые клетки. Передняя и задняя
кишка образуются впячиванием эктодермы; эктодерма дает начало эпи-
дермису, мускулатуре, нервной системе, протонефридиям. Зародыш в
яйце загнут на брюшную сторону. Все развитие протекает очень быст-
ро— 22—31 час (Sacks, 1955).
Развитие гастротрих, как находит А. А. Парамонов (1962), однако,
лишь в известной мере сходно с таковым нематод, что обусловлено зна-
чительным различием в строении этих классов. Но приходится подчерк^
нуть сильное отличие онтогенеза гастротрих и от такового турбеллярий —
рабдоцелид, с которыми из всех классов первичнополостных червей гаст-
ротрихи, как известно, наиболее близки. Работ по онтогенезу коловраток
довольно много (П. П. Иванов, 1937; Dawydoff, 1928; С. Переяславцева,
1885; Tannreuther, 1920; Nachtwey, 1925). Дробление полное, неравномер-
ное, с признаками спирального типа, детерминированное (детерминиро-
ваны продольная ось коловратки, передний и задний концы). К. Н. Да-
выдов находит сходство первых стадий с дроблением аннелид, что, одна-
ко, мало убедительно. Полость дробления, из которой образуется первич-
ная полость тела, небольшая. Из четырех первых бластомеров А, В, С и
D последний самый крупный, первые три и их дериваты дают эктодерму,
как и d1, d2, d3, отделившиеся от 3D. Энтобласт 3D является зачатком
средней кишки и половой железы. Эктодермальные бластомеры дают на-
чало эпидермису, нервной' аистеме, передней и задней кишке, клоаке с
анусом, экскреторным и половым отверстиями, мускулатуре, протонефри-
диям. Амебоидные клетки полости тела, видимо, являются эктомезенхи-
, мой. Зародыш в яйце загнут на брюшную сторону, развитие протекает
без метаморфоза, число клеточных делений небольшое, ограниченное, что
связано с небольшим, постоянным числом клеток взрослых коловраток.
Для их онтогенеза характерно (что отмечала С. М. Переяславцева,
- 1885) очень раннее и очень быстрое обособление отдельных зачатков
органов, что типично и для нематод. Старые авторы (Переяславпева,
1885) в онтогенезе коловраток находили зачаток мезодермы, что не под-
тверждают позднейшие исследования.
Индивидуальное развитие нематод, гастротрих и коловраток своеоб-
разно уже потому, что средний зародышевый листок, поскольку у них
нет целома, не играет в нем роли, так что вся организация этих червей
построена из эктодермы и энтодермы, с небольшим участием экю- и
эндомезенхимы. Сохранение бластоцеля в виде дефинитивной, первич-
ной, полости тела взрослого червя является также очень характерной
103
чертой этой группы. Отсюда ясно, что эмбриональное развитие первич-
нополостных червей не похоже на таковое паренхиматозных червей, и
тем более вторичнополостных или целомических червей. Однако-было
бы странным, если бы в онтогенезе первичнополостных червей не
нашлось бы ничего общего с таковым паренхиматозных червей.
Следы спирального дробления у нематод, гастротрих и коловра-
ток, полоски мезодермы у нематод и, видимо, коловраток можно счи-
тать общими чертами онтогенеза' их и турбеллярий. При невысоком
уровне организации, сравнимой в ряде отношений с организацией раб-
доцелид, можно было ожидать в онтогенезе именно гастротрих больше
сходства с онтогенезом рабдоцелвд, чем у других первичнополостных
червей. Но, очевидно, исторически более раннее формирование гаст-
ротрих как уже первичнополостных червей отложило достаточно четкий
отпечаток и на их онтогенез. К сожалению, данных экспериментальной
эмбриологии, чтобы пояснить родственные связи этих классов, мало, и
они касаются лишь нематод.
Очень характерно, что при громадном числе видов и при необычай-
ном разнообразии экологии нематоды обнаруживают1 удивительную
стойкость организации класса, подвергаясь лишь незначительным при-
способительным прогрессивным или репрессивным изменениям в -трое-
нии, и вместе с тем приобретают многообразие жизненных циклов.
И микроскопически мелкие свободные морские нематоды с малой инди-
видуальной плодовитостью и крупные, порой гигантские, нематоды с гро-
мадной плодовитостью, паразиты позвоночных сохраняют типичный план
строения первичнополостнык круглых червей.
Известно (Парамонов, 1962), что нематоды распространены по всему
земному шару, многие из них настоящие убиквисты. Они живут во всех
биоценозах бентоса и стациях океанов и морей от Северного полюса до
Антарктиды включительно, в солоноватых и пресных, стоячих и текучих
водах, в горячих источниках (Coninck. 1962). Они живут в почве, будучи
самыми многочисленными по числу особей ее обитателями (до 3 биллио-
нов в акре). Нематоды-—свободные черви, сапробионты, факультатив-
ные и облигатные паразиты, паразитирующие в большинстве беспозво-
ночных, позвоночных, включая человека, во многих культурных и диких
растениях, нередко способны к анабиозу.
Обращает на себя внимание, что при громадной численности, повсе-,
местном распространении и разнообразии экологии, врагов у нематод
немного — это некоторые бактерии, грибки и простейшие: амебы, аме-
боидные стадии миксомицетов, некоторые хищные нематоды и энхитреи-
ды, клещи, аптериготы (Парамонов, 1962).
Несомненно (Парамонов, 1962) нематоды относятся к классам живот-
ных, переживающих в настоящее время биологический прогресс в самом
широком его понимании; в сущности говоря, на глазах идет у них видо-
образование, дифференциация, и становление систематических категорий
разного таксономического значения.
Понятно, что с ними несравнимы в этом отношении не только ма-
лочисленные, микроскопически мелкие донные прибрежные обитатели
моря и пресных вод — гастротрихи и киноринхи, но и ротатории. Послед-
ние биологически достаточно пластичны, но в основном все же это оби-
татели пресных вод, среди которых не мало убиквистов, широко распро-
страненных по Земле благодаря способности к анабиозу и образованию
стойких, покоящихся, легко разносимых яиц; но они не приспособились
к паразитизму.
Переходя к краткому рассмотрению классификации классов первич-
нополостных червей, можно ограничиться лишь самым многочисленным,
теоретически и в практическом отношении самым важным классом — не-
матодами. Отмечу, что нет единого мнения о числе видов этого класса.
104
По данным одних авторов (Kaestner, 1954/1955), нематод 5000 видов,
других (Storer a. Usinger, 1957) — 12000 й третьих (Hyman, 1951; Па-
рамонов, 1962) —не менее 500000 и даже больше.
Нет единого мнения и о числе отрядов нематод и их размещении в
системе. Гаймен (Hyman, 1951) необоснованно принимает 17 отрядов —
Enoploidea, Dorylaimoidea, Mermithoidea, Chroriiadoroidea, Araeolaimoidea,
Monhysteroidea, Desmoscolecoidea, Rhabditoidea, Rhabdiasoidea, Oxyuroi-
dea, Ascaroidea, Strongyloidea, Spiruroidea, Draciinculoidea, Filarioidea,
Trichinelloidea, Dioctophymoidea. С моей точки зрения, такое разделение
на отряды неестественно, поскольку два последних отряда должны нахо-
диться перед отрядом рабдитид жак принадлежащие подклассу адено-
форей; формы Rhabdiasoidea, как известно, настолько близки к рабди-
тидам, что нет основания считать их самостоятельным отрядом, за раб-
цитидами- должны следовать стронгилиды, а не оксиураты и аска-
риды.
Другую крайность допускают, слишком укрупняя отряды, многие
крупнейшие исследователи паразитических нематод, преимущественно
так называемых зоогельминтов. Так, Читвуды (В. G. Chitwood and
М. В. Chitwood, 1950) всех аденофорей объединяют в два отряда —
Chromadorida и Enoplida, а сецернентов— в Rhabditida и Spirurida и не-
правильно начинают классификацию не со свободных примитивных не-
матод, а с групп измененных паразитизмом. К- И. Скрябин и его школа
также принимают два отряда Chromadorida и Enoplida для аденофорей
и три отряда —• Rhabditida, Ascaridida и Spirurida для сецернентов. Недо-
статком обеих классификаций является отнесение паразитических групп
Dioctophymata и Trichocephalata в отряд Enoplida.
Торн (Thorne, 1961) устраняет эти крайности, принимая 6 отрядов,
сецернентов (Tylenchida, Rhabditida, Strongylida, Ascaridida, Spirurida
и Camallanida) и 5 отрядов аденофорей (Dorylaimida, Chromadorida,
Monhysterida, Enoplida и Dioctophymatida). К сожалению, он также оши-
бочно начинает классификацию с паразитических групп.
В настоящее время гельминтологами принято деление нематод на два
подкласса — Adenophorea (=Aphasmidia) и Secernentea ( = Phasmidia),
при этом все основные черты строения, целостность организации (архи-
тектоника, по Беклемишеву, 1944, 1952, 1964) класса круглых червей со-
храняются в обоих подклассах, с рядом преимущественно вторичных из-
менений, вызванных паразитизмом, в основном во втором подклассе.
С точки зрения архитектоники, сравнительной морфологии и основ
физиологии можно обойтись и без деления нематод на два подласса, как
и обходятся до сих пор многие зоологи. Однако углубленное, тщательное
изучение нематод обнаруживает у них наличие исторически скоордини-
рованных систем морфо-физиолого-экологических признаков (Филипьев,
1918—1921; Парамонов, 1962, 1964 и более ранние его работы), которые
обосновывают деление нематод на подклассы аденофорей и сецернен-
тов, а внутри их на группы отрядов свободных нематод, фитонематод и
зоонематод; такое деление в известной степени подтверждается и фило-
генетическими отношениями. Приходится однако признать, что в целом
последний вопрос еще не разрешен, вследствие неполноты знаний по
сравнительной анатомии, микроскопической анатомии и онтогенезу
(М. В. Chitwood, 1957) и вследствие трудностей в изучении поведения и
физиологии такой важной группы, как зоонематоды — эндопаразиты жи-
вотных (М. В. Chitwood, 1957), а также из-за отсутствия палеонтологи-
ческих данных.
Айне представляется возможным принять следующую классификацию
нематод: 1-й подкласс — Adenophorea (=Aphasmidia) с отрядами Enopli-
da, Chromadorida, Monhysterida, Dorylaimida, Mermithida, Trichocephalida
и Dioctophymatida и 2-й подкласс Secernenta ( = Phasmidia) с отрядами
105
Rhabditida, Tylenchida, Strongylida, Spirurida и Ascaridida.
Отмечая ряд основных особенностей аденофорей, о них можно ска-
зать следующее: это наиболее богатая, разнообразная группа нематод
с полной и примитивной организацией, включающая морских, почвенных
и меньше пресноводных оксифильных, микроскопически мелких, мелких,
небольших, свободных (за редким исключением .паразитических) типич-
ных круглых червей. Кутикула их тонкая, легко проницаемая для рас-
творов солей и красок; гиподерма в основном сохраняет клеточное строе-
ние и богата железистыми клетками в виде многочисленных, сериально
•расположенных (паралатеральных) и трех терминальных хвостовых же-
лез. Органы чувств хорошо развиты, особенно в головном отделе, и пред-
ставлены танго-, хемо- и фоторецепторами. В связи с жизнью в водной,
подвижной среде х-еморецепторы и амфиды—• карманообразные, спи-
ральные и круглые, как исключение поровидные, с большим наружным
отверстием. Нервная система примитивна, богата ганглиозными клетка-
ми, серийно расположенными, порой в виде цепочки. Мускулатура не
специализирована, полимиарная. Осморегуляторно-экскреторный аппа-
рат полимеризованный, представлен многочисленными экскреторными
клетками гиподермального происхождения, свободными фагоцитарными
клетками полости тела и шейной железой в виде одной массивной, труб-
чатой, грушевидной, мешковидной клетки без боковых каналов. Пище-
варительный тракт (глотка и средняя кишка) без выростов и диверти-
кулов; анальных желез обычно нет.
В связи со способом питания и характером пищи возникает громадное
разнообразие в приспособительных особенностях строения рта и ротовой
полости и их вооружения. Нередко наблюдается примитивность поло-
вой системы — наличие двух яичников и семенников. Бурсальных крыль-
ев у самцов, как правило, нет. Половой диморфизм не резкий.
Среди этих, типичных свободных водных оксифильных нематод лишь
некоторые группы переходят к паразитизму: часть дорилаймид становит-
ся паразитами растений, мермитиды делаются Эндопаразитами насеко-
мых в личиночном состоянии (во взрослом — это свобо'дноживущие чер-
ви); трихоцефалиды и диоктофиматиды— паразиты внутренних органов
и тканей позвоночных. Характерно, что даже последние сохраняют фи-
зиологическую черту подкласса — оксифильность. Живя в кишечнике и
других внутренних органах млекопитающих и птиц, они головным кон-
цом внедряются в ткань органа, богатую кислородом крови (Рыжиков,
см. Парамонов, 1964).
Особое место среди аденофорей занимают плектиды (Plectidae)
мз хромадорид; будучи в основном типичными формами этого подкласса,
они проникают через пресные воды в почву и факультативно способны
к сапробиосу; некоторые из них поселяются в прикорневой системе рас-
тений. Они приобретают признаки сецернентов в строении глотки, в на-
личии анальных (ректальных) желез, в склеротизации протока шейной
железы на веем протяжении (Парамонов, 1962). Это позволяет думать,
что Plectidae играют важную роль в филогении подкласса сецернентов
(или фазмидиевых).
Из важнейших черт подкласса Secernentea, крайне богатой, разнооб-
разной, многочисленной группы нематод, можно отметить следующие:
небольшое число свободных и большинство весьма многочисленных фи-
тонематод и часть паразитов позвоночных — это мелкие и микроско-
пические мелкие формы. Многие зоопаразиты — черви средней и боль-
шой величины (до 1 м 20 см), с тенденцией к увеличению размеров (До-
гель, 1962; Парамонов, 1962). В связи с паразитизмом их организация,
сохраняя основную архитектонику класса круглых червей, подвергается
значительным вторичным, в том числе регрессивным, изменениям.
106
Кутикула, чаще кольчатая, чем гладкая, становится толстой, состоя-
щей из нескольких (до девяти) слоев и крайне стойка к вредным, даже
сильно ядовитым веществам как во внешней среде, так и в эксперимен-
тах. Гиподерма теряет клеточное строение, превращаясь в синцитий.
Характерной чертой является наличие пары одноклеточных боковых
хвостовых желез железистой и экскреторной пр ироды (фазмиды). Гипо-
дермальные железы их осморегуляторно-экскреторного полимеризован-
ного аппарата редуцируются, вследствие чего (а не путем слияния
отдельных компонентов) последний олигомеризуется. Основным элемен-
том этого аппарата делается шейная железа, становящаяся разветвлен-
ной клеткой с системой внутриклеточных каналов, проходящих в боко-
вых полях гиподермы. Происходит регресс головных органов чувств —
танго-, хемо-фоторецепторов. Амфиды специализируются, становясь
поровидными, очень мелкими или исчезают полностью. Утрачиваются
глазки. Головные щетинки замещаются папиллами, но головные и губ-
ные тангорецепторы (как органы восприятия вкуса), важные в захвате
добычи или приеме пищи, развиты хорошо. Под влиянием эндопарази-
тизма, вследствие регресса, обедняются сети ганглиозных и сериально
расположенных клеток нервной системы, сохраняющихся еще у ряда
фитонематод, уменьшается также число тангорецепторов на поверхно-
сти тела. У ряда форм полностью редуцируется мускулатура тела в ре-
зультате утраты подвижности этих червей (самки сем. Heteroderidae).
Паразитизм вместе с тем вызывает появление прИ1Спооабительных
черт' в пищеварительном тракте. Вследствие усиления сосущеколющей
функции ротовой полости (стомы) развивается их вооружение — губы,
стилет и мускулатура, особенно усиливается мускулатура глотки, в
которой дифференцируются бульбусы. У ряда эндопаразитов позвоноч-
ных, например, аскаридит, глотка дифференцируется на мускулистую и
железистую части, железистая часть образует выросты или диверти-
кулы. Возникают выросты и в передней части средней кишки. У самцов
сохраняется всегда одна гонада, у самок в ряде случаев одна гонада
редуцируется; для обеспечения спаривания у самцов развивается бур-
сальный аппарат с мускулатурой и бурсальными крыльями и тангорё-
цепторы. Для многих почвенных нематод и фитонематод характерна
необычайно высокая скорость индивидуального развития, порой закан-
чивающегося в течение нескольких часов, чем достигается быстрое на-
растание численности червей при наличии лишь нескольких синхронно
созревающих яиц в яичниках. У многих зоонематод развиваются необы-
чайно высокая продуктивность яичников (стр. 100) и сложные жизнен-
ные циклы.
В противоположность аденофореям сецернентов почти нет в море,
много в пресных водах; для них характерна жизнь в средах, бедных
кислородом,— олигосапробных водоемах, почве, сапробиотичеокой сре-
де, растительных и животных организмах. Среди почвенных форм ши-
роко распространен анабиоз.
Очень важно отметить, что некоторые плектиды из хромадо-
рид, проникая из моря через пресные воды в почву, в среду, более бед-
ную кислородом, приобретают ряд черт, общих с сецернентами (Пара-
монов, 1962), а среди рабдитид, как известно, имеются свободные фор-
мы, сапробиотические, паразиты растений и животных. Поэтому мож-
но считать, что правы авторы, считающие рабдитид за узловую группу
в эволюции сецернентов (Скрябин и др.; 1946—1962; Chitwood and Chit-
wood, 1950; Парамонов, 1962, 1964). Это кажется справедливым, хотя,
к сожалению, о ходе эволюции нематод нет палеонтологических данных.
Лишь для зоонематод, паразитов позвоночных, мы можем говорить и
геологической древности и предполагаемом ходе их филогении, хотя
бы по косвенным данным, по их связям с хозяевами-позвоночными,
107
т. е. так, как приходится поступать в отношении и паразитических плос-
ких червей.
Работами И. Н. Филипьева, К- И. Скрябина и его школы, А. А. Па-
рамонова и многих зарубежных авторов обосновано признание свобод-
ных морских групп нематод наиболее примитивными формами этого
класса, от которых взяли начало как паразитические формы подкласса
аденофорей, так и все остальные отряды сецернентов. Я придерживаюсь
мнения И. Н. Филипьева о том, что отряд эноплида с гладкой кутикулой,
с карм ан собранными амфидами и глоткой без бульбусов — наиболее
примитивный отряд, которым следует начинать естественную классифи-
кацию нематод. Отряд хромадорид с их кольчатой кутикулой, глот-
кой с бульбусом, разными, но преимущественно спиральными амфидами
уже по этим чертам организации охватывает формы измененные, эво-
люировавшие, сравнительно с простыми, примитивными представителя-
ми эноплид. И эколого-физиологически хромадориды более разнообраз-
ны. Если для свободных нематод характерно в целом отсутствие склон-
ности к сапробиозу, что полностью относится к эноплидам, то среди хро-
мадорид, например, плектид (Парамонов, 1962), имеются формы, перехо-
дящие из моря через пресные воды к жизни в почве и обладающие склон-
ностью к сапрабиотической жизни в прикорневой системе растений. Для
эноплид, типичных обитателей богатой кислородом морской среды, это
несвойственно и, очевидно, формами отряда хромадорид приобретено
впоследствии.
Для двустороннесим1метричных нематод несомненно, что кармано-
образные симметричные амфиды также являются исходным примитивным
признаком; уже вторичной позднейшей особенностью является спираль-
ная закручена ость амфид хромадорид и других отрядов.
Высокая оксифильность эноплид, живущих почти исключительно ®
чистой морской воде, и ряд особенностей их организации говорят в поль-
зу мнения об энаплидах как наиболее примитивной группе нематод.
Хромадориды во многих отношениях опециализованы, менее требова-
тельны к чистоте морской воды; а формы вроде плектид, как отмечалось
выше,.переходят к жизни в почве и сапробиосу. Используя эти разли-
чия между энаплидами и хромадоридами, можно видеть в них две фи-
логенетически расходящиеся ветви. Одна ветвь (эноплиды) дала начало
таким отрядам аденофорей, как дорилаймиды и мермитиды. Многий
авторы на основании данных морфологии относят к отряду эноплид
трихинеллид и диоктофимат. Это нарушает естественный характер клас-
сификации; снижать же эти две группы до подотрядов нет оснований;,
во многом отличаясь от эноплид, они являются самостоятельными отря-
дами. Но их оксифильность, как указывалось выше, является важной
физиологической чертой, подкрепляющей принадлежность трихинеллид.
и диоктофимат к группе высоко оксифильных эноплид, дорилаймид. и
мермитид. Высокай оксифильность характеризует и Dorylaimida, пара-
зитирующих на растениях и животных.
Другую большую ветвь свободных нематод составляют хромадори-
ды, монхистериды с плектидами, дес м оскол ециды и ареоламиды, кото-
рых можно считать, как делают Гаймен (Hyman, 1951) и Гудей (Goo-
<теу, 1963), отрядами.
Если для форм первой ветви характерна жизнь в чистых прибреж-
ных водах моря и мало типично обитание в солоноватых и пресных
водах и в почве, то во второй ветви, гораздо более пластичной во мно-
гих отношениях, наблюдается переход к сапробиотической жизни и рас-
пространение в почве (плектиды).
Как отмечалось выше, почвенные плектиды приобретают черты
сходства со свободными рабдитатами — сецернентами; и это говорит о
возможной филогенетической связи ветви хромадорид через рабдитид
108
^Парамонов, 1962; 1964; Gerlach, 1963), включающих свободные сапро-
биотические формы и пара®ито.в растений и животных, с остальными
типичными паразитическими сецернентйми. Напомню при этом, что уже
то, что И. Н. Филипьев давно (1918—1921), по неполным еще данным
того времени, относил все паразитические группы этого подкласса к от-
ряду Anguillulata (= Rhabditida), говорит о морфологической близости
и родстве этих групп, о филогенетической связи их с рабдитидами. Пра-
вы И. Н. Филипьев (1918—1921), К. И. Скрябин и его школа (1946,
1962), А. А. Парамонов (1954а, 19546, 1962, 1964) и ряд зарубежных
авторов, которые считают рабдитид этапом эволюции сецернентов, груп-
пой, родоначальной для всех сецернентов.
Так как, по моему мнению, наиболее обоснованные соображения'of.
эволюции и филогенении фито- и зоонематод намечены И. Н. Филипье
вым и развиты К- И. Скрябиным и А. А. Парамоновым, я присоеди
няюсь к .ним. Конечно, не современные рабдитиды, а их вымершие, при
митивные формы были предками паразитических групп сецернентов.
При этом очень важно, что среди современных форм Rhabditida одни
экологически и морфологически близки к зоонематодам, другие — к фи-
тонематодам.
Как известно, гельминтологами (Chitwood a. Chitwood, 1950; Thorne,
1961; Goodey, 1963) признается небольшое число отрядов зоонематод и
«фитонематод, тогда как зоологами обычно больше (до 6—7). Здесь
можно ограничиться наиболее крупными, важными группами — рабди-
тидами, стронгилидами, оксиуратами, спируридами, аскаридитами и ти-
денхидами, используя филогенетические соображения И. Н. Филипьева,
К. И. Скрябина и А. А. Парамонова.
Филогенетические отношения зоонематод и особенно фитонематод
выясняются совокупностью данных: внешняя форма, строение кишечни-
ка, особенно ротовая полость и глотка и отчасти средняя кишка, шей-
гая железа, гонады, продуктивность гонад, особенности строения и функ-
ции копулятивных органов, данные индивидуального развития, жизнен-
ных циклов, отчасти физиология, особенно же экология,'положение в си-
стеме хозяев и историческое развитие последних (преимущественно по-
звоночных) .
От примитивных паразитических кишечных рабдитид (точнее их
ближайших предков) взяли начало стронгилиды, обладающие рядом
черт строения, .сходных с рабдитидами и свободными рабдитовидными
.личинками гипотетических Protostrongylata. Стронгилята — большая
группа зоонематод, паразитов чаще разных отделов пищеварительного
тракта, органов дыхания, кровеносной системы, преимущественно мле-
копитающих (включая человека), всех сельскохозяйственных и промыс-
ловых животных, реже птиц, рептилий и амфибий. Наиболее примитив-
ные представители этого отряда Trichostrongyloidea —первичные энтеро-
теогелыминты.
Филогенетически более близкими к ним считают и оксиурат, обла-
дающих рядом общих черт с рабдитидами в отношении строения пище-
вода, шейной железы, но утративших свободные личинки. Они, как и
примитивные стронгилиды, также первичные энтерю- и геогельминты,
паразиты кишечника млекопитающих, птиц, рептилий и амфибий. От
первичных энтерогельминтов (геогельминтов) — Trichostrongyloidea —
развилась многочисленная ветвь биогельминтов, перешедших из кишеч-
ника в легкие и кровеносную систему хозяев (надсемейство Metastron-
.gyloidea). Именно в этой группе стронгилид возникла, как полагает
К. И. Скрябин, с одной стороны, ветвь вторичных геогельминтов — па-
разитов кишечника, отчасти легких и тканей (надсемейство Strongyloi-
dea), и остальная масса зоонематод: биогельминты тканевополостные
-филарииды и перешедшие вторично в кишечник хозяев вторичные энте-
109
рогельминты—биогельминты спируриды и вторичные геогельмингы—г
аскариды (Скрябин и др., 1946—1962). Опируриды, как известно, пара-
зиты всех органов и тканей хозяев—рыб, реже амфибий, рептилий,,
птиц и млекопитающих, имеющие в ряде случаев промежуточных хо-
зяев, обычно насекомых. Аскаридиды— паразиты пищеварительного
тг-акта белее двух десятков отрядов морских и пресноводных рыб, вклю-
чая акул, окатов, химер, осетровых, амфибий, многих отрядов рептилий,,
более 20 отрядов птиц, включая таких, как пингвины, страус нанду
и 13 отрядов млекопитающих, в том числе и человека.
Филогенетические связи спирурид и аскаридид все же неясны.
К. И. Скрябин считает, что эти группы имеют общих предков, вышедших
из семейства стронгилид — Pseudaliidae, которые, разошлись на две вет-
ви— спируриды и аскаридиды. Отмечают, что спируриды по ряду
признаков строения ближе к свободным рабдитидам, чем аскари
диды, которые во многом, и в том числе в филогенетическом отношении^
значительно обособившаяся от других группа зоонематод. Но о филоге-
нетических связях аскаридид, может быть, лучше, чем для других,
отрядов, судить по их организации, рекапитуляции в онтогенезе-
ряда признаков (например, строения губ), по распространению в хозяе-
вах —наличию более примитивных семейств в примитивных древних
морских рыбах (акулах, скатах, химерах), по эволюции строения (на-
пример, пищеварительного тракта) и, видимо, физиологии. Аскариды,
от форм, паразитирующих в костистых рыбах, амфибиях, рептилиях,,
идут, к паразитизму в птицах и млекопитающих (Мозговой, 1953а—в;
Hartwich, 1957; Osche, 1957). Можно думать, что предками спирурид и.
аскаридид были разные формы, так как паразитизм аскаридид—явле-
ние исторически более раннее; у спирурид таких древних морских хо-
зяев, как у аскаридид, нет, поздним явлением надо -считать и включе-
ние насекомых в качестве промежуточных хозяев, что наблюдается у
спирурид. На этом основании я затрудняюсь объединять общим пред-
ком спирурид и аскаридид и думаю, что вторые произошли исторически,
раньше спирурид, и общим в их происхождении является лишь то, что-
их корни, как и всех остальных зоопаразитов, идут от вымерших мор-
ских свободных рабдитид.
Не могу я также согласиться, что предки спирурид и аскаридид
взяли начало от Pseudaliidae из надсемейства Metastrongyloidea,.
уже потому, что хозяева Pseudaliidae — зубастые киты и наземные хищ-
ники (куницы) —появились много позже, во всяком случае позже хозяев
аскаридид — акул, скатов, химер. В известной мере это относится и к
спируридам, часть которых является исконными водными обитателями^
хотя и не столь древними, как хозяева аскаридид.
Паразитирование примитивных аскаридид в древних морских при-
тонных рыбах -неслучайно, вероятно заражение их происходило еще:
какими-то примитивными рабдитидами, исконной средой обитания ко-
торых было море, как и для всех групп аденофорей. Ясной паразитиче-
ской связи спирурид на заре истории их становления именно с древ-
ними морскими позвоночными нет. Очень вероятно, что свое проис-
хождение они -ведут от паразитических рабдитид, уже переселившихся
из морской среды в пресную воду или почву. К сожалению, для выяснен
ния филогенетических связей спирурид их отношения с хозяевами дают
мало.
Филогенетические связи фитонематод с рабдитидами гораздо непо-
средственнее, чем у зоонематод. Паразитизм среди фитойематод, надо-
думать (Парамонов, 1962, 1964. и более ранние работы, см. список ли-
тературы), ведет свое начало от приспособления почвенных рабдитид
к жизни в прикорневой системе растений, затем нематоды по-
падали в ткани растений и использовали для питания продукты распада
ПО
органического материала, который вызывался сапрофитной флорой —
грибами и бактериями. Со временем, в'связи с усилением деятельности
эктоферментативных желез, фитонематоды уще самостоятельно начали
использовать мицелий грибов сапробиоса. Это привело к эктопарази-
тизму фитонематод. Следующей ступенью эволюции их паразитизма
было проникновение в растения, ослабленные микозами и бактериоза-
ми. Далее шло освоение ими организма растения, его тканей и органов
как среды обитания, что было связано с приапособителБНЫми измене-
ниями особенно в физиологии (приобретение способности самостоятель-
но расщеплять белки и углеводы) и в строении нематод, становящихся
настоящими паразитами, нередко крайне опасными, разных групп выс-
ших цветковых растений, в усилении формообразования и расселении
на все большей территории (т. е. к биологическому прогрессу).
При этом очень правдоподобным ходом эволюции фитонематод,
является следующий: по-видимому, от потомков рабдитид произошли
группы форм надсемейства Cephaloboidea. Это'—преимущественно поч-
венные сапробионты с кутикулой повышенной непроницаемости, при-
способившиеся к среде, обедненной кислородом, и частично перешедшие
к паразитированию в органах растений. Дальнейший шаг в филогении
фитонематод наблюдается в развитии группы форм, объединенных в от-
ряд Tylenchida; одни из них (семейство Aphelenchidae и другие надсе-
мейства Aphelenchoidea) по ряду черт организации и биоэколопии еще
близки к рабдитидам, а другие (формы надсемейства Tylenchoidea)
по своей организации, физиологии и экологии становятся уже типичны-
ми фитонематодами (Парамонов, 1962).
О филогенетических отношениях разных групп коловраток прихо-
дится судить по данным отроения, экологии и лишь отчасти онтогенеза,
так как последний изучен недостаточно, а данные палеонтологии отсут-
ствуют.
Общепризнана примитивность чисто морского отряда Seisonidea,
очень небольшой группы червеобразных без пальцев коловраток, не
имеющих смены поколений и обладающих одинаково хорошо развиты-
ми как самками, так и самцами с парными половыми железами: самки
без дифференцировки яичников на гермарий и вителлярий отклады-
вают лишь одного рода яйца, развивающиеся после оплодотворения.
Очевидно, это коловратки, живущие на примитивных, древних ракооб-
разных Nebalia, ползающие наподобие пиявок с прикреплением при-
соской,—-реликты прежде более многочисленной группы примитивных
морских коловраток. Простота строения коловращательного аппарата,
органов чувств, жевательного аппарата, сравнительно с уровнем раз-
вития этих органов у большинства других коловраток, является резуль-
татом отчасти сохранения примитивного состояния, отчасти воздействия
на них образа жизни..
Основной группой класса является отряд Monogononta, вероятно,
взявший начало от каких-то примитивных морских форм, сходных или
близких к Seisonidea, но давно перешедших к жизни в пресных водах.
Эта группа коловраток в настоящее время достигает расцвета: у нее
наблюдается наибольшее разнообразие внешней формы, организации,
поведения, образа жизни, сред обитания (реки и замкнутые водоемы,
море и солоноватые воды, псаммон, почва; свободнотсдвижные фор-
мы— планктонные, ползающие донные, полусидячие, сидячие, эктопа-
раЗ'Итические, колониальные и т. д.).
Наиболее примитивными среди Monogononta являются удлиненные,
червеобразные придонные формы бентоса с низко дифференцированным
коловращательным аппаратом в виде поля ресничек на передней части
брюшной стороны тела (семейство Notommatidae, Беклемишев, 1952,
1964); таковы формы рода Proales (Властов, 1959), обладающие ясно
Ш
выраженной 'мультифункциональностью 1колюаращателБного аппарата,
который служит у них для плавания, парения, передвижения по субст-
рату, для захвата пищи. Нога у них — это орган прикрепления к суб-
страту, орган передвижения и т. д. У рода Proales в различные фазы
жизни возникают адаптации в виде следующих форм: натантной, эррант-
пой, парения, предохранительно-защитной (/благодаря координирован-
ным сокращениям и расслаблениям мускулатуры, изменениям в состоя-
нии стенок тела, в тургоре полостной жидкости). <Все это может послу-
жить для понимания /происхождения филогенетических изменений
(Властов, 1959).
Приспособлением к жизни в пресных .водоемах с неустойчивым конт-
растным режимом (смена сезонов года) является у коловраток отряда
Monogononta развитие быстро следующих друг за другом многих поко-
лений и партеногенеза, сопровождаемое подавлением самцов, становя-
щихся (карликовыми. С этим также связаны у самок дифференцировка
становящегося непарным яичника, обеспечивающая развитие полноцен-
ных яиц, возникновение покоящихся яиц, анабиоза и т. д. С переходом от
придонного к пелагическому образу жизни шло развитие и совершен-
ствование |колов(ращательйого аппарата как органа передвижения, при-
обретение /короткомешковидной (например, Asplanchnidae), дорсо-
вентрально сплющенной формы тела с панцирем (сем. Synchaetidae, Вга-
chionidae) и овально-шаровидной (как Trochosphaera) формы, напоми-
нающей трохофорных личинок кольчатых червей. При переходе ряда
Monogononta из водоемов к обитанию в почве и ползанию в ней еще
более усилилось развитие удлиненной, червеобразной формы.
Такой переход от бентосного водного к наземному, почвенному об-
разу жизни происходит и в другом, гораздо меньшем и однообразном
отряде Bdelloidea, сохранившем, однако, парные яичники, но совсем по-
терявшем самцов, перешедшем к /развитию исключительно с помощью
партеногенетических яиц, к широкому развитию анабиоза—:/пережива-
нию в обезвоженном состоянии периодов засухи (высыхания водоемов)
и низких температур (замерзания). Они приспособились также к жиз-
ни среди м<хов, лишайников, в почве и смогли распространиться по вер-
тикали на /высоту до 5500 м над уровнем моря. В этом небольшом от-
ряде более примитивными по 'ряду признаков являются семейства Habro-
f.rochidae и Philodioidae, давшие начало другим семействам отряда
(Н. Schulte, 1954).
Сл'абая дифференцировка коловращательного аппарата, нервной си-
стемы, отсутствие мочевого пузыря, парность половых желез, удлинен-
ная червеобразная форма тела сближают Bdelloidea с морскими Seiso-
noidea. Большую часть этих признаков Bdelloidea, как и ряд признаков
Notommatidae из Monogononta, можно считать примитивными, сохра-
нившимися от червеобразных предков коловраток. Переход к жизни в
пресных водах у коловраток, вероятно, произошел путем освоения через
солоноватые воды почвенных вод прибрежной полосы, затем бентоса
пресных водоемов. Это обусловило сохранение червеобразной формы
тела и передвижение путем ползания, что характерно для Bdelloidea и
части Monogononta; большая часть последних, как было сказано, широка
использовала толщу пресноводных водоемов и эволюировала как типич-
ные ее обитатели.
Филогения первичнополостных червей давно привлекает внимание
многих исследователей, однако вопрос этот очень трудный и в настоя-
щее время далек от разрешения. Вопреки мнению Ремане (Remane,
1959) и Маркуса (Marcus, 1958), я считаю, что и за последние годы нет
анатомических и эмбриологических данных, подтверждающих родство
первичнополостных червей с какими-либо вторичнополостными кольча-
тыми червями. Лишь у киноринх метамерный характер пластинок зони-
112
тов, дорео-вентральных мышц и скоплений .ганглиозных клеток в брюш-
ном нерве соответственно числу зонитов напоминает метамерию аннелид.
Однако .возникновение этих черт, .вероятно, связано со способом пере-
движения киноринх, так как у них нет ни основных компонентов сег-
мента, ни ряда других черт организации аннелид. Метамерность .кино-
ринх'относится к широко распространенному явлению среди многих
групп .беспозвоночных — к метамерии, возникающей путем упорядочения
и дифференцировки (Беклемишев, 1952, 1964). План строения киноринх
в целом .говорит о принадлежности их к первич.нополостны1м червям, но
не позволяет сближать их конкретно с какой-нибудь группой этих червей.
Для выяснения этого вопроса нужны дополнительные эмбриологические,
биохимические и серологические исследования.
Некоторые авторы (Marcus, 1958) находят общие черты и родство
коловраток с архианнелидами, например с Diurodrilus (Gerlach, 1952)
и Psammodrilus (Swedmark, 1955, см. гл. «Аннелиды», 1955). Внешнее
сходство, в частностях морфологии, между последними и некоторыми ко-
ловратками, 'безусловно, имеется, но эти черви являются метамерными,
вторичнополостными.червями с примитивными и деградированными чер-
тами аннелид, лишенными даже намека на сложный половой аппарат
коловраток. Вряд ли можно .найти что-нибудь общее в онтогенезе архи-
аннелид и коловраток, они так же резко отличны друг от друга, как и
планы строения этих двух групп. Считать, что коловратки деградирован-
ные потомки каких-то кольчатых целомических червей, нет основания.
Более рациональным является взгляд, который наиболее убедительно
обоснован А. А. Парамоновым (1957, 1962) и Беклемишевым (1944, 1952,
1964), что первичнополостные черви — потомки низших ресничных па-
ренхиматозных червей — турбеллярий — рабдоцелид и являются этапом
их исторического развития, а не деградации. В ходе эволюции предки
первичнополостных червей приобрели гибкий, прочный кутикулярный по-
кров тела вместо ресничного и движения путем змеевидного изгибания
тела, а не с помощью ресничек, сквозной кишечник вместо слепого, пер-
вичную полость тела с полостной жидкостью, возникшей за счет щелей
в паренхиме и редукции последней. Это сделало внутренние органы (ки-
шечник, пбловой аппарат) .более подвижными и хорошо обеспечило в них
обмен веществ. Мало дифференцированная сложная система кожно-мус-
кульного мешка паренхиматозных предков превратилась в систему от-
дельных оиышц, связанных с разными органами, и лишь у нематод
сохранился слой продольных мышц .в стенке тела. В связи с .быстрыми
движениями и ловлей подвижной добычи — хищничеством —-стоит и пе-
рестройка ортогональной нервной системы, богатой нервными стволами
и обильным нервным подкожным -сплетением паренхиматозных предков.
Нервная система нематод, сохраняющая основные черты ортогона, обед-
няется числом комиссур, ганглиозных сплетений и количеством клеток,
становящимся постоянным и небольшим (всего 162). У других .классов
ортогональный тип нервной системы в значительной мере утрачивается,
центральной частью нервной системы становится головной ганглий.
Сложная, гермафродитная половая система, наиболее близкая к таковой
ресничных червей, у гастротрих упрощается, теряя многие вспомогатель-
ные части; полы становятся раздельными. С утратой паренхимы, запол-
нявшей все промежутки между органами, упрощается юсморегуляторно-
экскреторная система. Разветвленность протонефридиев уменьшается,
или протонефридии функционально замещаются кожными железами
(гастротрихи—.Macrodasyoidea и нематоды). Дробление становиго®
билатеральным, сохраняющим лишь следы спирального, развитие
делается прямым, а при наличии личинок оно без настоящего метамор-
фоза и, как правило, происходит путем постепенной, без коренной лом-
ки перестройки органов личинки в таковые имаго.
8 Д. М. Федотов j 13
Коловратки, приспособившись в основной массе к планкгической
жизни, уклонились от червеобразной исходной формы, стали менее чер-
веобразными, чем гастротрихи, нематоды и киноринхи. Гастротрихи наи-
более (близки к турбелляриям (Парамонов, .1957, 1962; Беклемишев, 1944,
1952, 1964; Steinbock, 1958). Вместе с тем, по строению почти всех систем
органов, кроме половой, близкой с таковой турбеллярий, и по характеру
индивидуального развития гастротрихи несомненно близки к нематодам
(Парамонов, 1957, 1962). Общность плана строения (или архитектоника)
и характер индивидуального развития делают более обоснованным
предположение о происхождении ротаторий, гастротрих, нематод и, ви-
димо, киноринх от общих турбелляриобразных предков путем посту-
пательного развития и достижения нового этапа эволюции сравнитель-
но с историческим развитием паренхиматозных червей, а не эволюцией —
путем утраты основных черт вторичнополостных, хотя и примитивных,
кольчатых червей. Эволюция первичнополостных червей, как отмечалось
выше, сопровождалась также и утратой ряда примитивных черт их
турбелляриобразных предков. Соображения о происхождении первично-
полостных червей не имеют доказательств исторического характера;
лишь по связям паразитических нематод с хозяевами — позвоночными —
можно считать геологически древнее происхождение нематод, а отсюда
не менее древнее происхождение гастротрих, коловраток и, вероятно,,
киноринх.
.В настоящее время биологическое положение 'классов первичноиоло-
стных червей; как известно, очень различно. Главенствующего поло-
жения достигли нематоды, как говорилось выше, широко расселившиеся
по всей Земле (в морях, пресных водах, почве), ставшие паразитами,,
нередко крайне опасными, особенно позвоночных животных и человека.
В связи с необычайно разнообразной экологией нематод, класса, пере-
живающего биологический прогресс, происходит энергичное видообра-
зование, формирование новых родов, семейств и других более крупных
таксонов; нематоды уже сделались одним из наиболее многочисленных
в видовом отношении классов животных, объединяющем червей длиной
в несколько долей миллиметра и более 8 л в длину. При этом характер-
но и теоретически очень важно, что нематоды сохраняют основной план
строения, изменяясь и приспосабливаясь к условиям существования
лишь в частностях морфологии, имеющих, однако, важное функциональ-
ное значение. Описано несколько десятков тысяч видов, а допускают
существование 500 000 и более (Hyman, 1951; Парамонов, 1962).
Коловратки, почти исключительно овободноживущие, .микроско-
пически мелкие формы, довольно богаты видами (1500—2000 видов),
.имея, как и нематоды, «сходной средой обитания море, они освоили и
заселили пресные воды, распространившись очень широко, и (стали, бла-
юдаря биологическим приспособлениям (например, анабиозу, образова-
нию покоящихся стойких яиц), убиквистами (по выражению Парамо-
нова, 1962).
История эволюции гастротрих и киноринх бедна и несравнима не
только с таковой нематод, но и коловраток.
Микроскопически мелкие (0,3—1,5 мм) обитатели бентоса и псаммо-
на в основном моря, частью пресных вод, гастротрихи сохранили среди
первичнополостных червей наибольшее родственное сходство с турбел-
ляриеобразными предками и даже с современными турбелляриями
(Steinbock, 1958), благодаря условиям жизни, в ряде черт строения (Па-
рамонов, 1957, 1962); они ближе к нематодам, чем к коловраткам. Эво-
люция гастротрих идет .медленными темпами, это небольшая группа
из двух отрядов и 300 видов.
Еще беднее эволюция класса киноринх, таких же микроскопически
мелких, как и гастротрихи, свободноживущих червей, населяющих мор-
ской прибрежный бентос и представленных всего примерно 100 видами.
Немертины (Nemertini)
Немертины — плоские, паренхиматозные, длинные, ресничные, сво-
бодноживущие, дышащие |паверх|ностью тела черви, имеющие головную
лопасть, хобот — орган нападения на добычу и защиты, необычайно
мощную мускулатуру тела, центральную нервную систему из двух спин-
ных и двух брюшных головных ганглиев и двух боковых стволов, труб-
чатый кишечник с анальным отверстием, кровеносную систему, мешко-
видные гонады. Характерной чертой многих немертин является также
метамерное расположение боковых карманов (или ветвей) средней киш-
ки, мешковидных гонад, поперечных сосудов, соединяющих спинной с бо-
ковыми кровеносными сосудами, а также нередко поперечных комиссур
нервной системы ,и мешковидных выростов влагалища хобота. Онтогенез
характеризуется спиральным, детерминированным дроблением яиц, пря-
мым развитием с примитивной личинкой или развитием с пелагической
личинкой — пилидием—и некробиотичесиим метаморфозом. У немер.тин
наблюдаются бесполое размножение, необычайно сильная регенератив-
ная способность, инцистирование, иногда живорождение, комменсализм,
и отсутствие настоящего паразитизма. Немертины — морские и, как ред-
кое исключение, пресноводные и наземные животные. Это очень подвиж-
ные придонные ползающие и пелагические плавающие черви. Немерти-
ны— хищники, активно охотящиеся за добычей; питаются обычно жи-
выми и реже мертвыми животными. Размеры колеблются от нескольких
миллиметров (Carinella galatheae— 7 мм) до 30 м (Linetis longissimus)..
Представленные одним классом и 550—750 видами, они являются скорее
самостоятельным типом, чем подтипом плоских червей. Геологическая
история неизвестна, однако возможны нахождения следов жизни вьы
мерших роющих форм.
Как известно, в середине XIX столетия, когда немертины были изу-
чены недостаточно, Шульце (М. Schulze, 1851, 1852 — цит. по Burger,.
1897—1907) делил немертин на невооруженных и вооруженных — Anopla
и Enopla. Несколько позже Губрехт (Hubrecht, 1879, см. Burger, 1897—
1907) установил три их отряда (Palaeonemertini, Schizonemertini и Нор-
lonemertini) также лишь по немногим признакам. Бюргер, разносторонне
изучивший немертин, в том числе и анатомо-гистолопически, на основа-
нии комплекса признаков строения.и развития, предложил, с моей точки
зрения, близкую к естественной классификацию, разделив немертин на
четыре отряда — Protonemertini, Mesonemertini, Metanemertini и Hetero-
nemertini.
К сожалению, некоторые позднейшие авторы (Bohmig, 1929; Hyman,.
1951: Kaestner, 1954/1955; Ливанов, 1955) вновь вернулись к классифика-
ции М. Шульце и делят немертин на неестественные группы, выделяя
подкласс Anopla, включающий как наиболее примитивных Palaeonemer-
tini, так и наиболее высокоорганизованных Heteronemertini, и подласс
Enopla, включающий как вооруженных Hoplonemertini, так и Bdello-
nemertini, имеющих невооруженный хобот. Иными словами, названные
8*
115-
исследователи повторяют, ошибку М. Шульце, придавая большое таксо-
номическое значение присутствию или отсутствию стилета в хоботе, а не
совокупности признаков всей организации и онтогенеза: например, раз-
витию прямому или с метаморфозом.
На мой взгляд, от названия отряда Palaeonemertini, предложенного
Губрехтом, следует отказаться, так как никаких древних ископаемых
немертии нет, возможно лишь, что относящиеся сюда немертины древ-
нее, чем остальные группы. Однако несомненно, что эти немертины наи-
более примитивны по своей организации и название Бюргера — Proto-
nemertini отвечает их природе более, чем название Palaeonemertini.
Вопрос заключается в том, следует ли сохранить отряд Бюргера Meso-
nemertini для другой группы, также низкостоящих немертин, включаю-
щей семейства Carinomidae и Cephalothrichidae, или последних надо
включить в отряд Protonemertini? Поскольку название Mesonemertini
ясно указывает на их относительно низкое положение в системе, его
следовало бы, с моей точки зрения, сохранить.
Перечисленные авторы сходятся в том, что Hoplonemertini близки к
Bdellohemertini, но они вносят недопустимый беспорядок в систему, от-
нося Malacobdellidae с единственным родом Malacobdella — семейство не
имеющее и, судя по данным индивидуального развития (Hammarsten,
1918; Iwata, 1960), не имевшее стилета в хоботе,—к вооруженным не^
мертинам. Если эту группу немертин назвать Metanemertini, как сделал
это Бюргер, то в ней можно различать только Enopla и Anopla и делить
Enopla на Monostylifera и Polystylifera.
Продольные нервные стволы у метанемертин лежат глубже, чем у ге-
теронемертин; у первых имеется слепой вырост средней кишки, тогда
как у вторых его нет; в остальном же ('развитие спинных ганглиев, ку-
тиса, кровеносной системы, кожно-мускульного мешка) гетеронемертины
обнаруживают более высокую степень организации. Вместе с тем их
характеризует и более сложный тип онтогенеза (личинка пилидий и не-
кробиотический метаморфоз) по сравнению с метанемертинами, как, на-
пример, Protonemertini и Mesonemertini, имеющими прямое развитие и
более простой тип становления организации взрослого червя, у которого
рост в длину идет по продольной оси первоначального-короткого оваль-
ного зародыша.
Все это является для меня основанием придерживаться классифика-
ции немертин, установленной Бюргером, как наиболее близкой к есте-
ственной. 'Сравнительное однообразие внешней формы и почти полнее от-
сутствие (каких-либо наружных придатков вынуждают зоологов при си-
Стематико-фаунистических исследованиях немертин детально изучать их
анатомию и гистологию. Благодаря этому, анатомия и гистология взрос-
лых немертин давно и хорошо изучены, что сильно способствовало выяс-
нению хода эволюции организации этой группы.
У немертин более, чем у какой-либо другой группы беспозвоночных,
вся систематика и классификация, начиная от вида, основаны на анато-
мо-гистологических данных. В связи с этим и в филогенетических по-
строениях (см. Burger, 1897—1907, отчасти Brinkmann, 1917; Коротке-
вич, 1955 и др.) авторы оперируют с родами немертин, не ограничиваясь
семействами.
Известно, что если рассматривать немертин от низших форм к выс-
шим, то наблюдается усложнение строения стенок тела: от слабого слоя
гомогенной гиалиновой соединительной ткани к мощному слою волок-
нистого кутиса, в добавление к одноклеточным слизистым и белковым
железам эпидермиса развиваются многочисленные пакеты многокле-
точных желез, лежащие глубоко в стенках тела. Наблюдается прогрес-
сивное развитие центральной нервной системы, особенно спинных ганг-
лиев головного мозга, дифференцировка в них групп ассоциативных
116
клеток, формирование структур, 'близких к гломерула'м аннелид, и нали-
чие в головном мозге нейросекреторных клеток (Lechenault, 1962, 1963,
цит. по Gersch, 1964 и рукописи А. А. Панова). Прослеживается раз-
витие крайне важных хеморецепторов—фронтального и церебральных
органов—и их связей со спинными ганглиями головного мозга, проис-
ходит изменение положения мозга и нервных стволов от примитивного
положения в эпидермисе к погруженному под эпидермис, между слоями
кожно-мускульного мешка и глуб-
же в паренхиму. Наблюдается
дифференцировка кишечника от
простой трубчатой формы к раз-
витию метамерно расположенных
карманов или выступов в средней
кишке, и прогрессивное развитие
кожно-мускульного мешка от
двухслойного к трехслойному и
возникновение слоя мышц под ба-
зальной мембраной эпидермиса, а
также развитие системы диаго-
нальных и дорсо-вентральных
мышц тела. Наблюдается также
усложнение кровеносной системы
от лакун и двух боковых сосудов
без правильного кровообращения
у примитивных форм к системе,
состоящей из спинного сосуда,
соединенного с боковыми сосуда-
ми поперечными сосудами с пра-
вильной циркуляцией крови, дви-
жущейся спереди назад в боко-
вых сосудах и сзади наперед —
в спинном (рис. 29). Высокой
гистологической дифференциров-
ки достигают кровеносные сосу-
ды, имеющие эндотелиальную вы-
стилку^ соединительнотканный
слой, слой кольцевых и порой
продольных мыши, и .наружный
слой паренхимных клеток.
Наблюдается у немертин и
А
Рис. 29. Кровеносная система немертин
(нз Hyman)
А — схема строения у Carino/na- Б — разрез
через сократимый кровеносный сосуд Malaco-
bdella; / —головная лакуна; 2—боковой крове-
носный сосуд; 3 и 4 — кровеносные сосуды хо-
бота; 5 — нефридий; 6 — коннективы; 7 — эндо-
телий; 8 — кольцевые мышцы; 9— наружный
клеточный слой
эволюция онтогенезов. Для большинства низших немертин и метанемер-
тин характерно прямое или примитивное личиночное развитие, заклю-
чающееся в росте личинки по длинной оси и становлении из организации
личинки организации взрослого червя. У большинства гетеронемертин
основным типом онтогенеза является развитие с пелагической личин-
кой— пилидием, из части которого путем некробиотического метамор-
фоза формируется взрослая немертина. В связи с условиями жизни
путем эволюции этого основного типа развития возникает у гетеронемер-
тины Linens desori иеличиночный (дезоровский) тип онтогенеза, при
котором из яйца, богатого желтком, вылупляется молодая сформирован-
ная немертинка, а у другого вида гетеронемертин Linens ruber образует-
ся несвободный или инкапсулированный тип развития, при котором
формирование личинки, протекающее внутри кокона (рис. 30), проис-
ходит за счет поглощения личинкой погибающих яиц (Шмидт, 1946,
1951, 1953, 1956, 1962а, 19626; Gontscharoff, 1960; Schmidt, 1963).
Онтогенез немертин обладает рядом особенностей, многие из кото-
рых являются примитивными. Спиральное, детерминированное дробле-
117
Рис. 30. Развитие немертин (из Беклемишева)
.4 — Carinotna tremaphoros (Protonemertini), ранний зародыш с теменной пластинкой на абораль-
ном полюсе и бластопором на оральном; Б— то же, более поздний зародыш сбоку, в оптическом
разрезе, первичный рот сместился на брюшную сторону тела (турбелляриеобразная стадия); В— то
же, личинка со сформированным дефинитивным ртом и эктодермальной глоткой, но еще без аналь-
ного отверстия; Г — Oerstedia dorsalis (Metanemertini), удлиненный зародыш с теменной пластин-
кой на аборальном полюсе; Д— то же, более поздняя стадия, в оптическом разрезе, виден отшн^'-
ровавшийся кишечник и зачаток рннходеума; Е— то же, поздняя стадия с мозгом н 4 глазами,
ринходеум открылся спереди в среднюю кишку, на месте бывшего бластопора образовался задний
проход (по Сое); Ж — пилидий (личинка Heteronemertini), вид слева; 3 — образование немерти-
ны (по В. В. Заленскому); / — мозг; 2— дефинитивный рот; 3— глотка; 4 — средняя кишка;
5 — рииходеум; 6 — зародышевые диски; 7— мышца; 8— прототрох
ъие немертин отличается от такового других (паренхиматозных червей
(поликлад), а также аннелид тем, что оно равномерное, гомоквадрантное,
и бластомеры, являющиеся по своей судьбе микромерами и микромера-
ми, одинаковы по размерам, а число квартетов большое — 6. Велика м-но-
гоклстоимость стадий дробления, бластоцель—обширная полость, гаст-
руляция инвагинационная и происходит на стадии девятого или десятого
деления бластомеров (512—1024 клетки, см. Шмидт, 1953, 1962; S. Нбг-
stadius, 1937). Первые три квартета бластомеров дают эктодерму, произ-
водные второго квартета — эктомезенхиму, четвертый — энтомезенхиму,
пятый и шестой — энтодерму. Производные первого квартета дают клет-
ки креста, но последний не образует головных ганглиев. Личинка, в част-
118
пости пилидий,— с двумя лопастями, имеет сферическую форму, очень
слаборазвитую посторальную часть (типосферу) и кишечник со слепой
энтодермальной кишкой. Производные бластомера 4d образуют рассеян-
ные, отдельные клетки энтомезенхимы или две небольшие мезенхимные
полооки (П. П. Иванов, 1937). Несколько неожиданными, противореча-
щими примитивному строению Protonemertini, являются данные
К. Н. Давыдова (Dawydoff, 1928) о наличии у них лишь трех квартетов
микромеров, перехода спирального дробления 28-клеточной стадии к бес-
порядочному, об отсутствии креста и розетки и т. д. Очевидно, необходи-
мо детальное изучение раннего онтогенеза этой группы.
Как известно из работ разных авторов над развитием ряда Protone-
mertini (палеонемертин, Беклемишев, 1952, 1964), становление организа-
ции червя идет путем роста в длину овального тела личинки с теменной
пластинкой на одном и с бластопором на другом полюсе. Смещением
бластопора на брюшную сторону и впячиванием 'эктодермы образуется
рот и эктодермальная передняя кишка. На заднем конце образуется
анальное отверстие. У метанемертин бластопор, не смещаясь, замыкает-
ся, на его .месте образуется анальное отверстие; задняя кишка возникает
как вырост энтодерм ал ын ого кишечника зародыша, на месте соприкосно-
вения которого с эктодермой образуется анальное отверстие.
Некоторые авторы, например, В. В. Заленский (Salensky, 1912, 1914),
вопреки данным большинства эмбриологов, ошибочно считает переднюю
кишку энтодермальной. Существуют разногласия и о природе задней
кишки немертин, некоторые авторы приписывают ей, явно по ошибке,
эк! одермалыное происхождение.
Вследствие одинаковой величины и большого числа клеток дробления
у немертин генеалогия бластомеров,, а также их участие в развитии ряда
органов точно не прослежены. В частности, не выяснена природа крове-
носной системы, образование которой во всем животном мире связано с
развитием мезодермы. Среди низших первичноротых или нецел омических
Spiralia кровеносная система, впервые появляющаяся у немертин, обла-
дает двойственным характером. Ее основной частью являются сократи-
мые кровеносные сосуды с высокой дифференцировкой строения и от-
правления, а другой частью — лакуны, или щели в паренхиме без соб-
ственных стенок и тем более без эндотелия. Как отмечалось, сократимые
кровеносные сосуды имеют эндотелиальную выстилку, соединительно-
тканный слой, слой кольцевых (по В. Н. Беклемишеву) и продольных
мышц и неклеточный слой или слой клеток паренхимы, отделяющий со-
суды от соседних тканей (Oudemans, 1885; Burger, 1897-—1907; Bohmig,
1898; Salensky, 1912, 1914; Hyman, 1951, и др.).
Если в отношении лакунарной части кровеносной системы немертин
представление о происхождении ее как системы лакун и щелей, а крови
как жидкой ткани основного вещества мезенхимы вполне допустимо, то
этого нельзя сказать о сократимых кровеносных сосудах немертин.
В. В. Заленский, Нуссбаум и Окснер (Nussbaum u. Охпег, 1912),
К. Н. Давыдов, а их поддерживают В. А. Догель (1938), П. П. Иванов
(1937) и Маркус (Marcus, 1958), считают, что боковые кровеносные
сосуды немертин тесно связаны с протонефридиями и целомами, но с
этим не согласны В. Н. Беклемишев, Н. А. Ливанов и Г. А. Шмидт.
Я считают, что доказательств, опровергающих этот взгляд, нет. Разви-
тие боковых кровеносных сосудов интимно связано с энтомезенхимой,
представленной разрозненными клетками или двумя клеточными полос-
ками. Оно не имеет ничего общего с образованием околокишечного сину-
са у аннелид и отлично от развития лакунарной части кровеносной си-
стемы как щелей в мезенхиме. Это, вместе с характерным гистологи-
ческим строением, заставляет считать боковые сосуды негомологич.тыми
лакунарной части кровеносной системы, а ее в целом — гетерогенным
119
образованием. Однако, по Нуссбауму и Окснеру (Nussbaum и Охпег,
1913), спинной сосуд, когда он имеется, развивается из щелей в мезен-
химе, по их выражению из остатков бластоцеля; по В. В. Заленскому
(Salensky, 1914), происхождение спинного сосуда неясно, однако его
строение в сформированном состоянии такое же, как и боковых сосудов.
К сожалению, и последние работы Ивата (F. Iwata, 1957, 1958) о раз-
витии гетеронемертины Micrura dkkeshiensis не разрешают вопроса.
Автор не проследил 4d и его производных. Он говорит лишь о четырех
группах мезодермальных (очевидно, мезенхимных) клеток, видимо про-
изводных второго квартета, и о том, что они не дают соматоплевру и
спланхноплевру, типичных для аннелид. Не проливают света на развитие
мезодермальных органов и схематичные рисунки Ивата, изображающие
формирование молодого червя. Между тем и те авторы, которые отри-
цают у немертин образование целомических полостей (Ливанов, 1955;
Шмидт, 1946, 1958, 1962; Schmidt, 1937), различают у них производ-
ные и 2d и 4d.
Экология немертин довольно однообразна. За редкими исключения-
ми (стр. 115)—это свободноживущие обитатели моря (от литорали до
абиссальных глубин — 7210—7230 м) (см. Г. М. Беляев и др., 1960), в ос-
новном придонные, ползающие благодаря перистальтическим сокраще-
ниям мускулатуры тела, ресничкам и слизи, выделяемой эпидермисом.
Реже встречаются роющие формы. Мелкие виды плавают только с по-
мощью ресничек тела. Ряд семейств метанемертин ведет пелагический
образ жизни вплоть до глубин более 3000 м (батипелагические формы),
плавая также с помощью волнообразных изгибаний тела благодаря со-
кращениям кожно-мускульного мешка. Именно на пелагических немерти-
нах, как, правда, и на ряде многих форм из разных классов и типов жи-
вотных, особенно ярко сказывается формирующее воздействие условий
жизни. У пелагических немертин, по одним авторам (Brinkmann, 1917),
имеющих монофилетическое происхождение, по другим (Короткевич,
1955) —полифилетическое, возникает сплющенное, торпедо образной
формы укороченное тело, прозрачность тканей, на переднем конце пара
щупалец, туловищные и хвостовой плавники, уменьшается число' слоев
кожно-мускульного мешка, появляется дифференцировка их на спинные
и брюшные ленты, возникает в мышцах поперечная полосатость, появ-
ляется половой диморфизм и т. д. (Короткбвич, 1955; Сое, 1943).
Все немертины, как отмечалось,— хищники; мелкие виды питаются
простейшими; большинство — охотники, активно добывающие, часто
ночью, крупную добычу — турбеллярий, нематод, моллюсков, полихет,
ракообразных и даже рыб. Крупных, мало уступающих им по величине
полихет они заглатывают целиком или же добычу высасывают (Gont-
scharoff, 1948). Гетеронемертины обвивают тело добычи хоботом, подав-
ляя ее подвижность, после чего добычу проглатывают. Метанемертины
(например, Amphiporus) уколом стилета вводят в тело добычи алкалоид
из группы никотина, вызывающий у добычи паралич, а затем ее высасы-
вают (К. В. Беклемишев, 1955).
Переваривание пищи полостное и очень быстрое. Вместе с тем, не-
мертины могут жить, голодая более года. При неблагоприятных усло-
виях у них наблюдаются инцистирование (Кайзер, 1925) и разрыв
тела на мелкие куски, а затем регенерация до целых червей (Сое,
1943). Некоторыми исследователями (Nussbaum и. Охпег, 1912, 1913),
и особенно К- Н. Давыдовым (1910, 1915), экспериментально доказана
необычайная регенеративная способность немертин.
Данных о физиологии немертин очень мало. Известно, что удаление
церебральных органов вызывает полный паралич червя (Kaestner,
1954/1955). Раздражение переднего конца червя вызывает волну сокра-
щений от переднего конца к заднему и возвращение этой волны в обрат-
120
'.юм направлении. Фронтальный и церебральные, органы 'Считаются хе-
морецепторами; последние, кроме того, как думают, служат для ориен-
тировки немертин в пространстве. Плохо известна и физиология крове-
носной системы; ее основной функцией считается трофическая (Лива-
нов, 1955), однако у немертин гемоглобин имеется и в плазме,, и в клет-
ках крови (Коржуев, 1949, 1964), и даже в ткани нервной системы.
Природу немертин, несколько изменяя и дополняя положения
Н. А. Ливанова (1940, 1955; см. гл. «Моллюски»), можно охарактеризо-
вать следующим образом-. Примитивными чертами у них являются:
ресничный эпидермис, сильное развитие паренхимы, протонефридии,
простые мешкообразные гонады. Хищничество, активная ловля добычи
и большая подвижность немертин обусловили развитие ряда про-
грессивных черт организации: удлиненного червеобразного тела, муску-
латуры тела столь мощной, что по количеству двигательной мускулатуры
немертины стоят среди животных на одном из первых мест, приближаясь
к гипермиарности (более 50% к объему тела) (Зенкевич, 1944), стабили-
зации рта и образования головной лопасти, трубчатого кишечника с
анальным отверстием и с полостным пищеварением, головного мозга с
группами ассоциативных клеток в спинных ганглиях, близких к гломеру-
лам, и церебральными органами, кровеносной системы с правильным
кровообращением и, наконец, специфического органа ловли добычи и на-
падения — хобота. Несомненно (как и считает Н. А. Ливанов) с форми-
рованием этой группы достигнут новый этап в эволюции червей, возник
новый подтип или даже тип червей — немертины.
Нельзя согласиться с Н. А. Ливановым лишь в том, будто бы образо-
вание хобота является «эисцессивногадаптивной» чертой и будто в связи
с этим эволюция немертин завершена. Вряд ли справедливо это утверж-
дение, тем более, что мы знаем, как велика способность немертин регене-
рировать из мельчайших частей тела, 'разрезанного экспериментатором
(Nussbaum и. Охпег, 1912, 1915; Давыдов, 1910, 1915) или распавшегося
в неблагоприятных условиях жизни в природе. Хобот, «эксцессивно-адап-
тивный» орган, образуется заново из части тела немертины, лишенной
каких-либо следов этого органа. Вместе с тем хобот — несомненно
очень своеобразная особенность организации немертин, недаром
М. Шульце (1851, 1852—см. Burger, 1897—1907) называл их Rhyncho-
coel а.
Бюргер (Biirger, 1897—1907), у которого мы находим обзор взгля-
дов на филогению немертин, на основании сравнительно-анатомических
данных, считал отряд Protonemertini родоначальным для остальных не-
мертин. От него он выводил, с одной стороны, Mesonemertini, давших на-
чало Metanemertini, с другой стороны, Heteronemertini. К этим филоге-
нетическим построениям, которые разделяют ряд зоологов, например,
Бемиг (Bohmig, 1929), Франц (Franz, 1924), можно присоединиться, но
теперь их необходимо проверить и уточнить, хотя бы применив совре-
менную методику серологических исследований.
Как известно, в родстве немертин с турбелляриями никто не сомне-
вается, спор идет о происхождении немертин.. Большинство зоологов
считает, что немертины произошли от примитивных форм турбеллярий,
вроде надсемейства Dalyelliidea. Основные примитивные черты строения
и индивидуального развития немертин, общие с таковыми свободножи-
.вущих, паренхиматозных червей, безусловно говорят о возможном про-
исхождении немертин от турбеляриеподобных предков. Это подкрепляет-
ся исследованиями А. С. Щепотьева (Schepotieff, 1913) с преципитивной
или вассермановской реакцией. Таково мнение В. А. Догеля (1938, 1940),
П. П. Иванова (1937), В. Н. Беклемишева (1952), Н. А. Ливанова
(1955), Бюргера (Burger, 1897'—1907), Бемита (Bohmig, 1929), Бресслау
(Bresslau, 1932), Франца (Franz, 1924, 1943), Кестнера (Kaestner
1954/1955), Гаймен (Hyman, 1951) и многих др.
121
Значительно меньше сторонников взглядов о происхождении немер-
тин от аннелид: (Заленский (Salensky, 1912, 1914), Нуссбаум и Окснер
(Nussbaum и. Охпег, 1913), Давыдов, 1915, 1928, Маркус (Marcus,
1958}, Ремане (Remane, 1954, 1959), причем многие из них, не отрицая
родства между немертинами и тур-бел лярия ми, считают немертин и тур-
беллярий деградированными потомками полихет. В онтогенезе и во
взрослом состоянии ряда немертин имеются целомические полости.
В процессе регенерации у немертин образуются септы, делящие целом
на .метамерные полости. Отсутствие метамерии в онтогенезе и в строении
немертин, по мнению Ремане, не мешает считать их потомками аннелид,
.так как у Echiuroidea, нечленистых во взрослом состоянии, следы мета-
мерии проявляются в онтогенезе. Иными словами, по мнению этой груп-
пы зоологов, немертины могли утратить метамерию.
В целом уровень организации немертин и характер их онтогенеза
настолько примитивнее, чем у аннелид, что нет никаких оснований счи-
тать немертин вторично упрощенными потомками кольчатых червей.
Лишь боковые кровеносные сосуды -с их эндотелиальной выстилкой,
возникающие за счет производных бластомера 4d, подобно сегментиро-
ванным целомическим полостям аннелид и полости перикардия и гонад
моллюсков, несмотря на возражения ряда зоологов, все же можно счи-
тать производными целомических полостей. По ряду примитивных черт
организации, отмеченных выше (стр. 121), немертины, бесспорно, близки
к типичным плоским червям, турбелляриям. И вместе с тем, по комплек-
су прогрессивных черт они стоят гораздо выше последних.
В группе низших, плоских червей немертины занимают, как обще-
признано, высшее место. В. Н. Беклемишев (1952) думает, что они
являются слепой ветвью, не давшей потомков; по мнению Н. А. Лива-
нова (1955) немертины стоят между плоскими и кольчатыми червями,
и немертиноподобные формы были предками кольчатых червей. Я счи-
таю. что низшие, плоские паренхиматозные черви — немертины и ан-
нелиды — связаны друг с другом родством и общими предками. Разли-
чия в их организации выражают разные уровни эволюционного разви-
тия. За их родство говорит общий характер раннего онтогенеза —
спиральное, детерминированное дробление, источники происхождения
трех зародышевых листков и главнейших их производных. Но как в ор-
ганизации взрослых, так и в ходе онтогенезов наблюдаются разные
уровни эволюции этих групп червей. Мне представляется вполне вероят-
ным происхождение предков кольчатых червей в отдаленное геологиче-
ское время от примитивных плоских (немертинообразных) червей с
мсевдометамерией, еще без хобота, которые в ходе длительной эволю-
ции приобрели членистость, или метамерию тела. Допущение обратного
хода эволюции — от кольчатых червей к немертинам и далее к турбел-
ляриям— надо считать ошибочным. При свободном образе жизни, ак-
тивном хищничестве, низком уровне организации, большом числе при-
митивных черт строения и онтогенеза и при отсутствии признаков
регресса у немертин и турбеллярий для меня непонятно утверждение
об аннелидном происхождении этих групп.
Когда-то Губрехт (Hubrecht, 1887) находил сходство в строении и
родство между немертинами и хордовыми. Как ни странно, это необо-
снованное положение возрождается вновь Шторером и Юзингером
(Storer a. Usinger, 1957), которые с серьезным видом говорят о сходст-
ве между спинным и боковыми нервными стволами позвоночных и не-
мертин, мозгом позвоночных и головными ганглиями немертин, влага-
лищем хобота немертин и нотохордом гемихордовых. Можно лишь по-
жалеть трату времени на такие сравнения.
Однако надо отметить особенности немертин, в целом чужлые
Protostomia, а именно: отсутствие наружного, эктодермального, кути-
122
кулярного скелета, наличие соединительнотканного кутиса, который
вместе с кожно-мускульным мешком составляет своего рода внутрен-
ний скелет, образование задней кишки за счет энтодермы, а не эктодер-
мы, возникновение у ряда немертин на месте замкнувшегося бластопора
анального отверстия, а также, гаструлы путем инвагинации на поздних
многоклеточных (512—1024 бластомеров) стадиях дробления. Все эти
черты свойственны не паренхиматозным и целомическим Protostomia,
a Deuterostomia.
Учитывая все сказанное о немертинах, я склонен считать, как
И другие зоологи, немертин самостоятельным типом червей, а не под-
типом плоских, стоящим по своему строению и развитию и по положе-
нию в системе между типами плоских и кольчатых червей.
Моллюски (Mollusca)
Общая часть
Моллюски характеризуются компактным типом строения двусторон-
несимметричного тела, состоящего из головы, туловищного, или внут-
ренностного мешка, одетого на спинной стороне раковиной и мантией,
ограничивающей мантийную полость, и ноги — мускулистого выроста
на брюшной стороне. В мантийной полости на заднем конце тела на-
ходится так называемый паллиальный комплекс органов—анальное
отверстие, парные экжреторные и половые отверстия, пара ктенидиев
и пара осфрадиев. Центральная нервная система состоит из несколь-
ких пар ганглиев, соединенных друг с другом коннективами и комис-
сурами, и связана с рядом эффекторов—органами зрения, обоняния,
осязания, равновесия, слуха и др. Кишечник с радулой и парной пе-
ченью, реже это прямая короткая трубка, чаще — длинная, образующая
изгибы и петли. Органы дыхания — часто пара ктенидиев. Кровеносная
система незамкнутая, с дыхательными пигментами (гемоглобином или
гемоцианином), имеет центральный пульсаторный орган — сердце, на-
ходящийся в целомическом перикардии, с которым сообщается пара
экскреторных органов — почек или целомодуктов. Полости парных го-
над целомического происхождения, с парными выводными протоками
(целомодуктами); полы чаще разделены. Перикардий и гонады нахо-
дятся в задней части тела. Полости тела как таковой нет; промежутки
между органами заполнены паренхимой, сложной системой отдельных
мышц и большими без собственных стенок кровеносными синусами.
Индивидуальное развитие характеризуется спиральным, детерминиро-
ванным дроблением яиц и сложным метаморфозом с личинками, тро-
хофорой и велигером, а также ростом тела в течение всей жизни.
А4одлюски — это многочисленные, разнообразные, преимущественно
морские, распространенные от литорали до оверхабиосальных глубин
(10 710 м), придонные, грунтовые, реже планктичеокие, порой экто- и
эндопаразитические, пресноводные, наземные, растительноядные, реже
хищные, крайне разнообразные по величине, форме, окраске, строению
и экологии животные, дневные, сумеречные или ночные. Живя нередко
огромными сообществами и быстро размножаясь, моллюски дают яр-
кие примеры взаимозависимости организма и среды, выступая мощными
биофакторами, преобразующими среду, и одновременно дают прекрас-
ные примеры формирующего воздействия на организм условий жизни.
Экспериментально-морфологические, биометрические, физиологиче-
ские, биохимические, химические и экологические исследования по мол-
люскам вместе с плодотворным изучением ископаемых форм объединен-
ными усилиями зоологов и палеонтологов дали основные знания о типе
моллюсков.
Резко отличные по особенностям и степени совершенства организа-
ции, физиологии и экологии от кольчатых червей, но близко сходные с
последними по особенностям раннего онтогенеза моллюски являются
124
одной из наиболее изученных групп животного мира. Это хорошо ил-
люстрируется тем, что строение «живого ископаемого моллюска»
Neopilina во многом совпало' с представлениями Пельзенера (Pelseneer,
1900) и Плате (Plate, 1895) о строении гипотетичного урмоллюска, ос-
иованны'ми на данных начала 90-х годов XIX столетия о строении и раз-
витии представителей разных классов этого типа.
Среди всех типов животного мира моллюски по количеству (120 000)
видов занимают после членистоногих второе место. Обращает на себя
внимание большой диапазон колебаний размеров тела, чрезвычайная
пластичность организации, громадные различия особенностей строения,
поведения и физиологии от примитивного до уровня, почти не уступаю-
щего позвоночным, а также богатство палеонтологической летописи.
По всем этим особенностям моллюски далеко опережают почти все типы
беспозвоночных, включая и кольчатых червей.
Согласованных данных о числе современных и ископаемых видов
моллюсков нет. Соленогастров насчитывают 140 видов, хитонов — 1000,
-с рюхоногих — 85 000, двустворчатых — 25 000, лопатоногих — 300, голо-
чюногих — 600 (Kaestner, 1954/1955), из них для брюхоногих известно
11 300, а для двустворчатых— 15000 ископаемых видов. Кроме того,
.надо добавить к лопатоногим 200, к хитонам 300, а к головоногим при-
близительно 14 000 ископаемых видов: около 2500 наутилоидей, пример-
но 9000 аммоиоидей, 2500 белемиоидей.
Представление о размерах моллюсков дают следующие примеры;
среди соленогастров известны формы от 0,5 мм до 30 см в длину
(Epimenia verrucosa), среди хитонов — от 2,5мм (Lepidochiton margina-
tus) до 33 см в длину (Cryptochiton stelleri), среди лопатоногих рако-
вина самого крупного вида имеет 12 см в длину, некоторые ископаемые
формы (например, Antalis raymondi, верхний карбон) достигали 60 см
в длину и 40 мм в диаметре. Среди брюхоногих моллюсков наблюдаются
-формы от 0,5 мм до 60 см в длину, так, Punctum pygmeum имеет в дли-
ну 1,6 мм, Semifuscus proboscidiforus — 60 см, Aplysia— 40 см,
Dendronotus giganteus — 25 см, Vaginula schwelgae— 35 см, червеоб-
разный эндопаразит голотурии Par enter oxenos достигает 128 см в длину.
Значительные колебания в размерах (ют 1 мм до 120 см) наблюдаются
у двустворчатых, при этом Tridacna gigas, достигающая в длину от
100 до 120 см, весит до 220—225 ка; среди ископаемых двустворчатых
известны громадные формы, так, Haploscapha достигала 1,5 м в длину
,и до 1 .и в ширину. Наибольший размах в колебаниях размеров характе-
рен для головоногих, длина которых колеблется от 1 см (Idiosepius
pygmaeus) до 22 м (Architeuthis), считая руки и туловище; среди иско-
паемых головоногих также известны большие крайности, так, нижне-
кембрийская Volborthella была в длину меньше 1 см; ордовичский
Orthoceras из наутилоидей имел 4—4,5 м в длину, а меловой спирально
свернутый аммонит Pachydiscus seppenradensis-достигал 2,55—3 м в ди-
аметре.
В эволюционном отношении моллюски важны необычайным разно-
образием и пластичностью своей организации. Низшие представители '
их, как соленюгастры, еще не приобрели ряда характерных черт строе-
ния типа, у ряда же высших представителей (из классов Conchifera)
под воздействием условий жизни происходят очень сильные изменения
и нередко вторичная утрата многих важнейших черт компактного типа
строения, в том числе и раковины, которую некоторые зоологи напрасно
рассматривают как узкоадаптивную особенность моллюсков (Ливанов,
1955).
Среди свободноживущих брюхоногих нередко наблюдаются утрата
/компактного типа строения, асимметрия (по В. И. Беклемишеву —
диссимметрия) туловищного мешка и раковины, например, двусторон-
125
1
несимметричные Dorididae из заднежаберных моллюсков. Среди пос-
ледних известны также червеобразные, плоские, турбелляриеподюбные
формы вроде Pseudovermis, Rhodope и др., без головы, раковины, ноги,
радулы, челюстей, жабр, сердца и т. д. В ряде форм переднежаберных
(комменсалисты, эктопаразиты, эндопаразиты) наблюдаются переходы
от почти типичного брюхоногого через разные степени регресса орга-
низации к червеобразному организму, со сложным половым аппаратом,
но без остальных органов типичного моллюска.
Как некоторые примеры приспособительных изменений такого ти-
пичного для брюхоногих органа, как мускулистая, с ползательной по-
дошвой нога, можно упомянуть превращение ее у пелагических
Heteropod a (Pterolrachea) в вертикальный плавник, у некоторых перед-
нежаберных (Amalthea) в орган прикрепления к субстрату и как ред-
кое пиление редукция ноги (Vermetidae, Pteropoda) и даже полное ис-
чезновение ноги (например, Rhodope). Среди необычайно разнообраз-
ных изменений, претерпеваемых раковиной до полного ее исчезновения,
у брюхоногих обращает на себя внимание • развитие двустворчатой
раковины у таких заднежаберных, как Berthelinia (рис. 36) и Midorigai.
Это не только говорит о высокой пластичности организации брюхоногих,
ио и напоминает о родственных связях брюхоногих и двустворчатых
моллюсков.
Приведенные примеры, хотя они касаются и очень существенных
черт организации, Дают лишь некоторое представление о пластичности
строения брюхоногих, самого многочисленного по количеству видов,
самого разнообразного по биоэкологии и самого распространенного в
Мировом океане, пресных водах и на суше класса моллюсков.
Современные двустворчатые моллюски, даже с учетом таких своеоб-
разных ископаемых представителей, как рудисты, обнаруживают зна-
чительную стойкость в сохранении общего типа строения своего класса.
Изменения, раков ины, ее замка и мантии и перехода от димиарного
состояния через гетеромиарное к мономиарному отчетливо связаны с
условиями жизни. Происходящие при этом изменения внутренней орга-
низации значительны. Однако изменения двустворчатых, несравнимы
с тем, что наблюдается у брюхоногих. Это объясняется малоподвижным
образом жизни в толще грунта или на дне моря, часто прикрепленным
образом жизни, пассивным способом улавливания мелкой пищи и филь-
трацией через жабры. Однако в связи со способом питания у двуствор-
чатых происходят значительные изменения и эволюция жабр, от гре-
бенчатых ктенидиев через нитевидные жабры к жаберным пластинкам
(Yonge, 1949). Лишь среди позднейших немногих двустворчатых на-
блюдается регресс раковины, становящейся внутренней вследствие ее
обрастания мантией, регресс замка, а также' редукция и даже полная
утрата мускулов-аддукторов — Chlamydoconcha, Entovalva, Brechites и
ряд др.
Идея Н. А. Ливанова об узкоадаптивном значении раковины мол-
люсков и подтверждается и опровергается на примере эволюции голо-
воногих. В течение нескольких сот миллионов лет сохраняя наружную,
нередко тяжелую раковину как орган пассивной защиты, хищные по-
движные, преимущественно плавающие головоногие пережили несколько-
периодов расцвета и длительное время в разные геологические периоды
в фауне прошлых морей были господствующими беспозвоночными.
В отношении уровня организации, физиологии и ряда биологических
особенностей (с превращением раковины во внутреннюю и еще более
с ее редукцией) в настоящее время головоногие стоят выше всех осталь-
ных беспозвоночных.
Нейросекреторный аппарат у моллюсков изучен пока недостаточно,
нет данных для соленогастров и хитонов. Наибольшее число исследо-
126
ваний посвящено разным группам брюхоногих моллюсков, особенно
легочным. Нейросекреторные клетки найдены во многих ганглиях
центральной нервной системы, причем , у легочных и заднежаберных
моллюсков, они имеются и в ганглиях щупалец. Секрет нейросекретор-
ных клеток, по-видимому, по аксонам поступает в кровеносные сосуды.
В ряде случаев установлен годичный цикл изменений секреторной
активности нейросекреторного аппарата, связанный с годичным циклом
размножения. Показано также влияния секрета этого аппарата на об-
мен веществ (например, на гепатопанкреас, на водный обмен), на разви-
тие спермы и овоцитов. Эндокринным органом является, по-видимому,
и фолликулярная железа, находящаяся в церебральных ганглиях
(Gabe, 1951, 1953, 1955; Ferrissia-Lever, 1957; Altman und Kuhnen-Glau-
sen, 1959; Lever, 1958'; Lemche, 1957; Krause, 1960; Van Mol, 1960;
Hekstra and Lever, 1960; Lever and Josse, 1961; Jungstand, 1962; San-
chez et Sablier, 1962; Sanchez, 1962; Pelluet and Lane, 1961, и др.— цит.
no Gersch, 1964 и сводке, любезно составленной А. А. Пановым,).
Сравнительно немного работ по этому аппарату посвящено предста-
вителям разных групп двустворчатых моллюсков. У них установлены
нейросекреторные клетки в ганглиях центральной нервной системы,
сезонные изменения наполнения их секретом в течение года, паралле-
лизм с ходом полового цикла, движение секрета по аксонам и выход
через оболочки клеток и т. д. Сравнительно с брюхоногими, двустворча-
тые моллюски изучены хуже. Этими исследованиями занимались мно-
гие авторы (Gabe, 1954; Lubet, 1955, 1956; Fahrmann, 1961; Barenyi a.
Salanki, 1963; Antheunisse, 1963, цит. no Gersch, 1964 и сводке
А. А. Панова).
Работ по нейросекреторному аппарату лопатоногих почти нет, не-
достаточно он изучен и у головоногих. У последних эндокринными желе-
зами признают зрительную железу—-группу клеток на зрительных сте-
бельках, с которой связано созревание гонад, и задние слюнные
железы — выделяемый ими тирамин действует на тонус мышц, нервной
системы и на активность хроматофоров. К эндокринным органам отно-
сятся также бранхиальные железы, управляющие силой и ритмом
сердечных -сокращений и артериальным давлением, и эпистеллярные
железы, оказывающие тоническое действие.
Вещества, выделяемые спиральной кишкой, вызывают активацию
гепатопанкреаса (Wells and Wells, 1959; Joly, 1962 — цит. no Gersch,
1964 и Панову).
Приходится с сожалением признать слабую изученность нейросекре-
торного аппарата и эндокринной системы моллюсков, что не позволяет
получить представление об истинном положении и эволюции их у
моллюсков.
Ввиду того, что сравнительно-анатомические и экологические типы из-
вестны как для моллюсков в целом (Беклемишев, 1944, 1952, 1964), так
и для отдельных его классов — хитонов, двустворчатых, брюхоногих и
головоногих (Boettger, 1956), остановлюсь детальнее только на солено-
гастрах, о природе которых до сих пор нет единого мнения, и на «жи-
вом ископаемом» — Neopilina galatheae Lemche, имеющем исключитель-
ное значение для понимания всего типа. Но перед этим необходимо рас-
смотреть, что дают'для выяснения филогенетических связей моллюсков
данные онтогенеза, в том числе о становлении их организации.
Известно, что ранний онтогенез (начиная -от спирального детермини-
рованного дробления яйца с определенным положением плоскостей
симметрии будущего зародыша в самом начале дробления до образо-
вания т-рохофоры) очень сходен с таковым аннелид-полихет. Многие
эмбриологи (Давыдов, 1914; MacBride, 1914; Korschelt a. Heider,
1890—1893, 1936; и другие сторонники аннелидного происхождения
127
сколько большие, чем это обычно считают
Спиральное дробление у моллюсков
6
Рис. 31. Развитие Epimetiia verrucosa (из Kaest-
ner)
А—24-часовая трохофора; Б — 96-часовая трохофора;
/ — эписфера; 2 — гипосфера; 3 — гиподермальиые па-
пиллы; 4 — зачаток брюшной бороздки
моллюсков) считали, что дробление и (кроме судьбы клеток креста и ро-
зетки) образование зародышевых листков и трохофорной личинки у
обеих, групп проходит тождественно.
На самом деле в раннем эмбриогенезе различия между моллюсками
и кольчатыми червями, как правильно отмечает Г.А. Шмидт (1951), не-
многие эмбриологи.
или гомоквадрантное, или
далеко не столь гетеро-
квадрантное, как у анне-
лид. Бластомер D мало
отличается от бластоме-
ров А, В и С, а микромер
2d — от других клеток
второго квартета микро-
меров; закладка эктодер-
мальной мезенхимы, воз-
никающая из третьего
квартета микромеров, не
радиальная, как у анне-
лид, а билатерально-сим-
метричная, не говоря уже
о различии у моллюсков
и аннелид мезодермаль-
ных полосок, производных
бластомеров 4d. Имеются
различия в способах ин-
вагинации и степени фор-
мирования организации
трохофоры.
Можно еще отметить,
что моллюски отличаются
от аннелид и в том, что
у первых в яйцах имеется лишь одна вегетативная полярная плазма
и телобластическая закладка касается лишь мезодермальных органов,
тогда как у вторых существуют анимальная и вегетативная полярная
плазмы, а телобластический характер развития распространяется, кро-
ме того, на эктодермальные органы и целомические сомиты (Шмидт,
1951).
Характерной чертой индивидуального развития большинства мол-
люсков (Seidel, 1960), как при развитии со свободной пелагической
личинкой (большинство классов Conchifera), так и при прямом разви-
тии из яйца, богатого питательным материалом (преобладающая масса
головоногих), является раннее обособление в виде отдельных зачатков
главнейших частей тела — мантии, 'раковинной железы и раковины, ту-
ловищного мешка, ноги, зачатков перикардия и сердца, почек, гонад
Ь т. д.
Ни у трохофорной личинки соленогастров и хитонов, ни тем более у
велигера остальных моллюсков нет ни телобластической закладки за-
родышевых листков, ни зоны роста на заднем конце личинки, дающей
сегменты туловища и метамерные полости целомов, что так характерно
для аннелид-полихет.
Деление на низших (соленогастры и хитоны) и высших (брюхоно-
гие, двустворчатые, лопатоногие и головоногие) моллюсков, проводимое
на основании сравнительно-анатомических данных, подтверждается
(если не считать головоногих) и данными онтогенеза (рис. 31, 32).
У низших моллюсков, соленогастров и хитонов, имеется, как и у
.аннелид, одна трохофорная личинка, которая непосредственно превра-
128
щается в моллюска. Гипосфера трохофоры путем роста удлиняется и
становится вытянутым телом моллюска; за счет эписферы трохофоры
дифференцируется голова последнего. У хитонов далее развиваются ха-
рактерные черты внешней организации моллюска; на спинной стороне
головы и туловища личинки формируются мантия и пластинки ракови-
ны, на брюшной стороне — плоская нога с мускулистой подошвой.
В онтогенезе соленогастров (Baba, 1938, 1940а, 19406), как будет
сказано ниже более подробно, также происходит превращение тела
трохофоры в тело моллюска, но без образования зачатков мантии, ра-
ковины, ноги и туловищного мешка.
5
Рис. 32. Схема строения трохофорных личинок (из Kuhn)
Л—Annelida; Б— моллюск Teredo; 1—аборальная пластинка; 2 — рот; 3— протонефри-
дйи; 4 — сегменты тела червя; 5 — метатрох; 6 — прототрох; 7 — раковина
У всех высших моллюсков (за исключением головоногих) имеются
две личиночные стадии — трохофора и велигер (с парусом или велю-
мом, который образуется за счет прототрохальной части трохофоры с
резкой перестройкой всей организации личинки). Спинная сторона ги-
посферы личинки, на которой образуются мантия и раковинная железа,
дающая раковину (сначала эмбриональную), благодаря усиленному
росту вздувается, становится туловищным, или внутренностным меш-
ком, в который вовлекаются развивающиеся внутренние органы.
На нижней стороне гипосферы в виде мускулистого выроста'возникает
зачаток ноги и вместе с головой, образующейся из эписферы личинки,
составляет головоногу моллюска. Так формируется компактный тип
организации моллюсков Conchifera — голова, нога и внутренностный
мешок, одетый мантией и раковиной, в мантийной полости располагает-
ся паллиальный комплекс органов (анальное отверстие, ктенидии,
осфрадии, половые и выделительные отверстия).
У брюхоногих, несмотря на громадные различия в экологии взрослых
моллюсков (свободноживущих морских, пресноводных, наземных и па-
разитических форм, включая и эндопаразитов), в онтогенезе образуется
типичная личиночная стадия как при свободном личиночном, так и при
инкапсулированном развитии. Многие наземные легочные моллюски
откладывают в землю яйца, одетые известковой скорлупой, которые у
некоторых форм, например у Strophochilus spixx при величине моллюска
в 1.4 см достигают величины небольших куриных яиц до 5 см в длину.
Надо отметить, что господствующее представление об онтогенезе
головоногих, как о прямом развитии из яйца сформированного молодого
9 Д. М. Федотов
129
моллюска, оказывается неверным. У одного -из кальмаров Architeuthana
(Кондаков, 1940) эмбрион на стадии обрастания бластодермой желтка
имеет ресницы, напоминающие прототрох личинок других моллюсков, а
по данным работы Гренахера (Grenacher, 1874), изложенной Бальфуром
(Balfour, 1880), эта форма имеет пелагические личинки. Развитие дру-
гого глубоководного представителя Teuthoidea —• Vampyroteuthis infer-
nalis (Pickford, 1940, 1949, 1950) также идет с метаморфозом. Из яйца
вылупляется (по Пикфорду) личинка 5,5 мм длиной; взрослое голо-
воногое формируется, проходя четыре личиночные стадии. Для ряда
кальмаров известно, что они откладывают плавающие в воде большие
студенистые кладки с огромным количеством мелких яиц.
Особенностью раннего эмбрионального 'развития головоногих, кро-
ме того, является то, что рост зародышевого диска, находящегося на
брюшной стороне, идет не как у всех Protostomia в длину от рта к анусу,,
а перпендикулярно к поверхности яйца (Kaestner, 1954/1955).
Трохофоры кольчатых червей и низших моллюсков (соленогастров-.
и хитонов) стоят на одном уровне организации. Существенные различия
появляются только при формировании тела взрослых особей. У полихет
с начала субтерминального линейного роста гипосферы трохофоры
развиваются сначала ларвальные, затем постларвальные сегменты ме-
татрохофоры и возникает членистое тело червя. У соленогастров и хито-
нов ни ларвальных сегментов, ни зоны роста на заднем конце тела не
образуется, возникает более или менее удлиненное нечленистое тело-
моллюска.
У высших моллюсков с превращением трохофоры в велигер станов-
ление из него моллюска происходит иным, правда, более сложным пу-
тем, .чем у. аннелид и низших моллюсков. Происходит рост личинки и
ио горизонтальной, передне-задней, и по дорсовентральной оси, прохо-
дящей через зачатки мантии и ноги велигера. Благодаря такому спо-
собу морфогенеза возникает организация компактного типа высших
моллюсков.
Однако ни при (в основном) линейном росте в длину тела личинки
низших моллюсков, ни при сложном морфогенезе высших моллюсков
никаких следов метамерии или сегментации, как известно, не-
наблюдается.
Большой . интерес представляет целомическое происхождение пери-
кардиальной и половой полостей и экскреторных и половых протоков,
составляющих одну из характернейших черт строения всех моллюсков,
в чем многие и старые, и новые (например, Seidel, 1960) авторы видят
указание на родство моллюсков с аннелидами.
Однако в то время -как у кольчатых -червей мезодермальные полоски,
продукты деления бластомера 4d, дают лишь начало метамерным соми-
там целома, у моллюсков, как известно, лишь в задней части небольших
и недолговечных мезодермальных полосок образуется пара (или две)
целомических полостей, дающих начало, неточно выражаясь, урогени-
тальным органам; остальная (главная) масса мезодермальных полосок
распадается и вместе с производными клеток эктодермы участвует в
образовании соединительной ткани.
При отсутствии каких-либо зачатков сегментов тела в онтогенезе-
моллюсков, естественно, у них нет и расчленения мезодермальных
полосок на метамер-ные целомические полости. Предположение
А. О. Ковалевского о наличии целомических полостей у хитонов не
подтверждено последующими авторами. Временные щели в мезодер-
мальных полосках, найденные А. А. Захваткиным (1949) у пресновод-
ного переднежаберного -моллюска Bithynia ventricosa Gray, являются
не целомическими полостями, а лишь результатом временного расхож-
дения клеток этих полосок, распадающихся при дифференцировке на'
-130
клетки паренхимы, что вовсе не указывает на когда-то существовавший
морфогенез целома.
В стремлении отыскивать в онтогенезе моллюсков черты родства с
аннелидами некоторые зоологи пытаются без оснований рассматривать
в закладке целомических полостей перикардия и гонад наличие двух
сегментов, а сами эти целомические полости — как редуцированные
части некогда обширного целома аннелидных предков. Некоторые ав-
торы (например, Ливанов, 1955) видят в комплексе перикардий, гонады
и почки наличие олигомерии у моллюсков и сходство их с ларвальными
сегментами. Даже если бы у моллюсков было действительно когда-то
сегментированное тело и указанные органы были оставшимися частями
некогда существовавших метамер, то по положению в задней части
тела и по наличию в них половых желез эти метамеры были постлар-
вальными, а никак не ларвальными. Однако даже открытие Neopilina,
на мой взгляд, не решает вопроса в пользу признания метамерии у мол-
люсков.
Хотя преобладающее число работ говорит о целомическом, мезодер-
мальном происхождении перикардия и полости гонад у моллюсков,
отдельные авторы считают их эктодермальными. Последние соображе-
ния были использованы сторонниками взгляда о происхождении мол-
люсков от платодных предков (Plate, 1897, 1899, 1901; Thiele, 1891,
1895, 1902; Nierstrasz, 1909, 1910; Odhner, 1912, и другие). По мнению
Гофмана (Hoffmann, 1937), Гаммерстен и Рунстрема (Hammersten и.
Runnstrom, 1926а, 19266), также не признающих целомического харак-
тера органов, в онтогенезе моллюсков судьба мезодермальных полосок
сходна с таковой поликлад. Мезодермальные полоски, распадаясь, об-
разуют паренхиму, в которой возникают зачатки перикардия, сердца,
выделительных органов и гонад, причем, полость перикардия не являет-
ся целомом.
Вместе с тем Гофман, признавая сходство в раннем развитии и в
личинках аннелид и моллюсков и относя их к группе трохофорных жи-
вотных, резонно отрицает наличие метамерии у моллюсков и их про-
исхождение от сегментированных предков. С этим я вполне согласен,
что же касается толкования Гофманом судьбы мезодермальных полосок
и отрицания им у моллюсков органов и полостей целомического харак-
тера, то считаю их необоснованными. Как известно, у ряда групп, стоя-
щих выше, чем аннелиды, имеются органы и полости целомического
происхождения, но нет целома как общей полости тела. Кроме того, на-
пример, у иглокожих и кишечнодышащих, помимо энтероцельного об-
разования целома, он образуется и схизоцельным путем, т. е. расхож-
дением клеток.
Данные индивидуального развития, таким образом, говорят об отсут-
ствии у моллюсков на всех этапах онтогенеза сегментов тела, сомитов
целома, целома как общей полости тела и о -мезодермальном, целомиче-
ском происхождении полостей перикардия и гонад.
Образование в онтогенезе выделительных органов и половых прото-
ков как выпячивания стенки целома (перикардия) было основанием для
Гудрича (Goodrich, 1895, см. 1945) считать их у моллюсков целомодук-
тами и отрицать наличие метанефридиев, которые, как и протонефри-
дии, в типичном случае развиваются у аннелид из эктодермы. Как из-
вестно, одни зоологи считают выделительные органы моллюсков нефри-
диями (Ливанов, 1955, и многие зарубежные авторы), другие —
целомодуктами, вторично получившими экскреторную функцию (Бек-
лемишев, 1944, 1952, 1964, и мн. др.).
Явление меторизиса В. М. Шимкевича (1908) распространено очень
широко среди разных, групп животного мира и затрагивает не только
выделительные, но и очень многие другие органы. Протонефридии
9;
131
Я 6
Рис. 33. Cryptoplax evanescens, чер-
веобразная Placophora с рудимен-
тарными пластинками раковины (А),
и Proneomenia antarctica (Aplacopho-
ra) (5) (из Hennig)
наблюдаются у ряда личинок моллюсков, хотя и реже, чем у личинок
аннелид. В позднем онтогенезе, когда развиваются выделительные и
половые протоки, развитие мантии и раковины, охватывающих спинную
и боковые стороны тела, и мускулистой ноги на брюшной стороне, а
также сильная паренхиматизация у моллюсков сравнительно с кольча-
тыми червями (с их линейным ростом тела в одной плоскости) крайне
изменяют топографию выделительных органов. Вследствие этого у мол-
люсков зачатки целома принимают непосредственное участие в разви-
тии органов выделения, в связи с чем правильнее признавать у моллюс-
ков пару выделительных органов и пару половых протоков.
Большой интерес и много споров
вызывает толкование природы и по-
ложения соленогастров (рис. 33); по
мнению одних (Thiele, 1935; R. Hoff-
mann, 1937; Boettger, 1956; Remane,
1954, 1959, и др.), это самые примитив-
ные по строению и по развитию мол-
люски; по. мнению других, регрессив-
ные черты у соленогастров преобла-
дают над примитивными (Korschelt и.
Heider, 1936; Беклемишев, 1944, 1952,
1964; S. Hoffmann, 1949; Догель, 1959,
и др.).
К сожалению, ни старые (Pruvot,
1890, 1892), ни новые данные (Baba,
1938, 19406) о развитии Epimenia не
позволяют выяснить, насколько близко
,спиральное детерминированное дроб-
ление соленогастров к таковому
остальных моллюсков и аннелид. Судя
по наблюдениям Баба за разви-
тием Epimenia, за 16-клеточной ста-
дией следует инвагинационная гаст-
рула, превращающаяся в трохофору с
теменным султаном, покрытую мелки-
бластопором, перемещающимся вперед.
Туловище возникает в виде почки на нижнем конце гипосферы трохо-
форы, которая увеличивается и несет на заднем конце телотрох. Ни до,
ни после вылупления трохофоры у Epimenia никаких черт, свойственных
трохофоре и велигеру других моллюсков, не появляется, возникает лишь
продольный медиальный желобок, который Баба называет ногой, хотя
сам отмечает отсутствие признаков того, что это рудимент ползательной
подошвы. И, действительно, как указывает П. П. Иванов (1945), произ-
водные 2d, образующие спинную и боковую стенки тела, не дают начала
ни мантии, ни ноге; средняя часть бластопора закрывается не зачатком
ноги, как у остальных моллюсков, а как у аннелид — только производ-
ными третьего квартета; сильно удлиняющийся бластопор превращается
в продольный медиальный желобок. Трохофора, увеличиваясь в разме-
рах, становится червеобразным соленогастром. Однако в развитии Epi-
menia нет никаких намеков на метамерию, на зачатки, как у полихет,
ларвальных и постларвальных сегментов. Лишь внешне поздняя трохо-
фора Epimenia (с ее удлиненным телом с прото- и телотрохом) напоми-
нает мета- и нектотрох некоторых полихет (Свешников, 1961; Korschelt
u. Heider, 1936), но последние, в отличие от трохофоры Epimenia, явля-
ются членистыми.
Никаких закладок типичных органов моллюсков — мантии, раковин-
ной железы, раковины, внутренностного мешка, ноги и последующего их
132
ми ресницами, с прототрохом и
регресса в развитии Epimenia не происходит. Основной чертой онтоге-
неза Epimenia (Solenogastres), общей с таковой хитонов (Polyplacop-
hora), является непосредственное развитие тела моллюска из трохофоры
путем роста ее гипосферы, подобно развитию сегментированного тела
аннелид. Сходство с хитонами заключается еще в развитии в покровах
известковых спикул. Таким образом, в становлении внешней организа-
ции у Epimenia нет характерных черт, свойственных не только высшим
моллюскам Conchifera, но и типичным Amphineura — хитонам, нет и
признаков их регрессивного развития, что подчеркивает, как признавал
и П. П. Иванов (1945), примитивность соленогастров.
В связи с этим надо напомнить о большой стойкости онтогенезов
моллюсков (особенно брюхоногих с уклоняющимся строением) даже при
утрате в случаях эндопаразитизма почти всех черт наружной и внут-
ренней организации класса и типа. Признаки трохофоры, велигера и
зачатки многих внутренних органов проявляются (хотя затем и исчеза-
ют) в ходе развития.
К сожалению, не изучено развитие внутренних органов, но по строе-
нию центральной нервной системы, наличию радулы, элементов печени,
перикардия, сердца, полового аппарата и выводных протоков нельзя
сомневаться в принадлежности соленогастров к моллюскам; вопрос за-
ключается лишь в том, как рассматривать внутреннее строение солено-
гастров: как примитивное или как вторично упростившееся.
Червеобразная двусторюннесимметричная форма соленогастров, ис-
ходя из данных онтогенеза, является первичной чертой, а не результа-
том упрощения; вторичным сравнительно с типичными лишь удлиненно-
овальными хитонами является сильное вытягивание тела в длину. Среди
последних (например, Cryptoplax и другие), в связи с частичной редук-
цией восьми пластинок раковины и большей подвижностью, также на-
блюдается сужение и вытягивание тела в длину.
Известно, что, благодаря развитию ноги, в онтогенезе моллюсков
сохраняются кольцевые и продольные мышцы кожно-мускульного меш-
ка лишь на брюшной стороне. На спинной стороне личинки или зароды-
ша, в связи с развитием раковины, дифференцируются обособленные,
отдельные мускулы — мантийные, ножные и др. На этом основании нали-
чие у соленогастров кожно-мускульного мешка говорит об его первич-
ном характере, в отличие от бесспорно вторичного возникновения его
у ряда голых заднежаберных моллюсков, у которых это связано с утра-
той раковины и приобретением турбелляриеобразного двустороннесим-
метричного тела. Отсутствие в онтогенезе у соленогастров зачатков ра-
ковины и ноги заставляет считать, что кожно-мускульный мешок у них
сохранился от предков, а не возник вторично, как предполагают неко-
торые авторы. (Беклемишев, 1952). 1
Кишечник соленогастров в виде короткой прямой трубки с малодиф-
ференцированной радулой, сидящей непосредственно на кутикуле глот-
ки, а не на базальной мембране, как у всех остальных моллюсков, на-
чиная с хитонов, и слабо дифференцированная печень носят примитив-
ный характер. Можно напомнить, что у двустороннесимметричных,
примитивных, сравнительно с другими моллюсками, хитонов (рис. 34)
и у Neopilina печень хорошо дифференцирована, является парной, с
парными протоками, а средняя кишка образует несколько петель, что
нарушает их двустороннюю симметрию тела.
Физиология пищеварения соленогастров мало известна. Многие из
них, если не все, в отличие от преимущественно растительноядных хито-
нов, соскребывающих водоросли со скал, питаются животной пищей.
Соленогастры, живущие на гидроидных и восьмилучевых коралловых
полипах, поедают полипов и ценосарк. С животной пищей соленогастров
в известной мере связана короткая трубчатая форма кишечника.
133
Рис. 34. Внешний вид >(Б) и схема строения (Л, В) Placophora (Б — Chiton
comingsii) (из Hennig)
1 — рот; 2 — мантийный желобок; 3 — гонада; 4 — жабры; 5 — гонодукт; 6 — почка;
7 — перикардий; 8 — анус; 9 — кишечник; 10 — нога; 11 — тегментум; 12 — перитонеум
Центральная нервная система соленогастров дифференцирована луч-
ше, чем у хитонов; наблюдется ее ганглионизация; вместо тяжей у хито-
нов хорошо выражены головные ганглии. Это едва ли не единственная
прогрессивная черта в строении соленогастров сравнительно с хитонами.
Такая дифференцировка нервной системы, имеющей, однако, тот же
тип строения, что и у хитонов, и особенно образование головных ганг-
лиев, связдны с большей подвижностью соленогастров по сравнению с
малоподвижными хитонами, прикрытыми раковиной. Соленогастры
достаточно хорошо передвигаются и изгибаются, благодаря сокраще-
нию кожно-мускульного мешка, обладают подвижным головным концом.
Кровеносная система соленогастров несовершенна: сердце — часть
спинного кровеносного синуса, без предсердий, нередко в виде незам-
кнутого впячивания стенки перикардия, вместо кровеносных сосудов
имеются синусы, циркуляция крови очень слабая. Низкий уровень стро-
ения кровеносной системы соленогастров, несомненно, связан с отсут-
ствием у большинства их ктенидиев. Газовый обмен, очевидно, происхо-
дит через стенки тела, 'преимущественно брюшной стороны. У некоторых
форм на ней образуются кожные складки с анальной полостью (зачаток
мантийной полости?), а у Chaetodermatidae пара ктенидиев, которые
следует считать гомологичными ктенидиям прочих моллюсков и задней
паре ктенидиев хитонов.
У нас нет никаких данных о редукции органов дыхания у солено-
гастров; у большинства открытый образ жизни допускает газовый об-
134
мен через стенки тела. Ктенидии возникают у Chaetodermatidae, веду-
щих роющий образ жизни в грунте.
Целом у соленогастров — единая полость, не разделенная на два
отдела: перикардиальный и половой, как у хитонов и остальных мол-
-люсков, кроме ряда головоногих. Кишечник участвует в экскреции;
целомодукты не приобрели выделительной функции.
Отдельные анатомические и гистологические особенности соленога-
стров (S. Hoffmann, 1949) интересны, но они не являются доказатель-
ствами того, что многие черты строения соленогастров, в том числе
перикардий, сердце, гонады и их протоки, выводятся из таковых хито-
нов, стоящих якобы ниже соленогастров. С. Гофман, скорее под влияни-
ем точки зрения Пельзенера (Pelseneer, 1890, 1906) и Вирена (Wiren,
1892, 1893) находит у соленогастров в покровах брюшной стороны, в
брюшной складке и, брюшном желобке мантийную складку, мантийную
полость и ногу хитонов. В онтогенезе Epimenia, по часам прослеженно-
му Баба (Baba, 19-406), как мы видели, нет и следов зачатков этих
органов. Исходя из основного общего типа строения соленогастров,
хитонов и остальных моллюсков, мы можем лишь сказать, что покровы
тела, брюшные складки и брюшной желобок соленогастров принадле-
жат тем частям тела, которые у хитонов более элементарны, а у
остальных моллюсков более сложным путем дают начало таким орга-
нам, как мантийная складка, раковина и нога. В особенностях экологии
соленогастров мы также не находим условий, которые способствовали
бы исчезновению этих органов.
Некоторые авторы (Gegenbauer, 1898; Hatschek, 1888; Thiele, 1895,
1902, 1929) относят соленогастров (на основании их строения) к червям;
другие же зоологи относят их к Amphineura. Одни, в том числе Бёттгер
(Boettger, 1956), считают соленогастров примитивнее хитонов, дру-
гие— наоборот. Коршельт и Гейдер (Korschelt u. Heider, 1936), преиму-
щественно на основании данных онтогенеза, резонно считают, что нель-
зя выводить ни соленогастров из хитонов, ни хитонов из соленогастров.
Гофман (Н. Hoffmann, 1937) считал соленогастров нетипичными
моллюсками, помещая их в этот тип как Paramollusca и противопостав-
ляя им всех остальных моллюсков (Eumollusca — Placophora и Соп-
chifera).
Некоторые современные авторы, в основном немецкие (например,
Franz, 1943; Remane, 1954, 1959), разобщают соленогастров и хитонов,
причем Ремане делит всех моллюсков на три подтипа: Solenogastres,
Placophora и Conchifera, к такому же взгляду близок и Кестнер
(Kaestner, 1954/1955).
Я думаю, что такое деление типа моллюсков обоснованно и лучше,
чем общепринятое, выражает родственные отношения в пределах этого
типа, но считаю, что в один ряд с Solenogastres надо ставить и группу
Aplacophora (что подчеркивает безраковинность как очень характерную
черту этих моллюсков), как делает Кестнер.
Из данных по развитию и строению соленогастров вытекает, что это
самые примитивные представители типа моллюсков, еще не получившие
таких важных органов, как мантия, раковина, нога, имеющие примитив-
ное строение кишечника, кровеносной системы, кожно-мускульного меш-
ка и в большинстве еще не приобретшие ктенидиев и выделительной
функции протоков целома, выполняемой у них кишечником.
Имея ряд общих черт с хитонами, соленогастры по уровню своего
строения и онтогенезу стоят, очевидно, гораздо ближе, чем хитоны, к
червеобразному, нечленистому предку моллюсков, обладавшему цело-
мическими перикардием и гонадами, расположенными в заднем конце
тела (важной чертой строения, присущей всем моллюскам). Можно
допустить, что предки соленогастров ответвились от родоначального
135
ствола моллюсков на этом начальном периоде его эволюции, а несколь-
ко позже, уже на более высоком уровне дифференцировки, ответвились
хитоны. Можно думать, что древние соленогастры жили сначала на.ко-
лониальных полипах, позже часть их перешла к жизни в илу и песке,
вследствие чего у них сохранились основные примитивные черты стро-
ения.
Остатки ископаемых соленогастров, к сожалению, не найдены; у нас
нет никаких данных об их геологической истории. Возрастающее усовер-
шенствование методов палеонтологии допускает возможность раскрыть
геологическую историю соленогастров, и, надо думать, она подтвердит
такое понимание природы этой группы.
Преобладающее большинство хитонов живет в приливно-отливной
полосе, и лишь некоторые примитивные формы (отряд Lepidopleurida)
являются обитателями абиссали. Все они малоподвижные, преимущест-
венно растительноядные формы, присасывающиеся к скалам и твердому
грунту мускулистой ногой. В этой группе, очевидно, на протяжении
всей ее истории, начиная с ордовика и по настоящее время, сохранилось
то, с чего начинали остальные классы моллюсков — жизнь в прибреж-
ной полосе, движение с помощью ноги с плоской подошвой, двусторон-
несимметричное тело и развитие с одной личинкой — трохофорой, ра-
стущей в длину в линейном передне-заднем направлении; на спинной
стороне личинки образуется мантия с пластинками раковины, а на
брюшной — мускулистая нога. Общепринято и обосновано признавать
множественность ктенидиев хитонов результатом полимеризации, ве-
роятно, вызванной необходимостью максимально использовать кислород
воды, медленно обновляющийся при малоподвижном образе жизни хи-
тонов. Мало специализированные боковые стенки тела хитонов, несом-
ненно, сохраняют достаточные формативные способности, чтобы на воз-
действие условий жизни отвечать формообразованиями — развитием
новых ктенидиев, повышающих интенсивность дыхания. Думаю, что мне-
ние старых авторов о гомологии только одной пары ктенидиев хитонов
(расположенной непосредственно позади отвёрстий выделительных орга-
нов) паре ктенидиев остальных классов — правдоподобно. За полимери-
зацию жабр у хитонов говорит также небольшое число их (6 пар)
у низших хитонов Lepidopleurida и нарастание числа (до 88 пар) в отря-
де Chitonida.
К ряду черт организации, хорошо отграничивающих хитонов от клас-
сов Conchifera, относится раковина из 8 сочлененных пластинок. Эту
особенность надо рассматривать как приспособление, придающее зна-
чительную гибкость телу хитонов и позволяющее им свертываться.
Л1алоподвижный пассивный образ жизни хитонов, сохранившийся, судя
по строению раковины ископаемых форм, на всем протяжении их геоло-
гической истории, начиная с ордовика, не способствовал прогрессивной
эволюции этих моллюсков. Ни в одном геологическом периоде они ни-
когда не были многочисленными. Они стояли и стоят в стороне от боль-
шой дороги эволюции высших моллюсков — Conchifera.
Рассмотрение филогенетических отношений классов Conchifera надо
начать с определения природы и систематического положения Neopilina
galatheae Lemche. Подробно на организации Neopilina нет необходимости
останавливаться, поскольку она детально описана в работах Лемхе и
Вингстранда (Lemche a. Wingstrand, 1959, 1960), Лемхе (Lemche, 1957,
1959а—в), Акса (Ах, 1960), Беклемишева (1959, 1964) и других авторов,
дающих интерпретацию строения и положения в системе этого
моллюска.
Двусторонняя симметрия внешней формы и внутреннего строения,
симметричное положение по отношению к продольной оси тела на пе-
реднем конце тела рта и на заднем конце тела анального отверстия,
136
сериальное расположение 5 пар ктенидиев, 8 пар ретракторов ноги,.
6 пар целомодуктов, парных семенников и яичников, симметричное стро-
ение центральной нервной системы с церебральным тяжом и двумя
парами продольных стволов и парной печени являются, бесспорно, ос-
новными признаками примитивности Neopilina.
Наряду с этим Neopilina не лишена и признаков вторичного упро-
щения (подавленное развитие многих органов чувств на голове, тонко-
стенная раковина), в известной степени связанных с глубоководным,,
малоподвижным образом жизни. Гребневидное строение ктенидиев,
вместо типичного двоякоперистого строения их у большинства мол-
люсков, также является вторичным, а не исходным — примитивным
(Yonge, 19576).
Если бы можно было не считаться с сериальной повторностью рет-
ракторов ноги, ктенидиев и целомодуктов и отсутствием мантийной по-
лости на заднем конце, можно было бы согласиться с В. Н. Беклеми-
шевым (1959) в том, что Neopilina подтверждает прототип Prorhipi-
doglossa Пельзенера (Pelseneer, 1890, 1900), Praerhipidoglossa Плате
(Plate, 1895) и недавние представления о нем Ионга (Yonge, 1947, 1954,
19576), Грээма (Graham, 1957) и др. Но нельзя сказать, чтобы под-
тверждалось предположение Пельзенера о происхождении моллюсков
ог полихет, вроде Eunicidae, и мнение Лемхе и других исследователем
о метамерном строении Neopilina.
Вопрос о метамерном строении Neopilina решается просто, если при-
держиваться концепции Ремане о том, что все двустороннесимметрич-
ные Protostomia и Deuterostomia являются метамерными ab origine, и
признавать его дейтомеры и тритомеры. Мне кажется, что и концепция
Гудрича о сегментах, или метамерах как сериально расположенных
участках, или кольцах тела с эктодермальными придатками-конечностя-
ми, жабрами, с парой ганглиев и нервами брюшной нервной цепочки
(невросомит Ливанова), с участками кожно-мускульного мешка
(миосомит его же), парным целомом, парой нефридиев или целомодук-
тов, участком кровеносной системы (ангиосомит Тимофеева) остается
в силе (Федотов, 1960). Обосновано также и положение П. П. Иванова
о ларвальных и постларвальных сегментах.
Несколько сложнее обстоит дело с метамерией упорядочения
(рис. 35) В. Н. Беклемишева, наблюдаемой и у моллюсков. Мы имеем
бесспорные примеры ее среди потомков кольчатых червей, например
динофил и мизостомид, тело которых построено из ларвальных сег-
ментов (5) остатка тела личинки с зачатками половых желез, сегмента-
ция которого подавлена, но без которого такие олигомерные черви не
могли бы стать половозрелыми. Наряду с этими существуют многие
черви, хотя бы из немертин, турбеллярий и некоторых других, с мно-
жественным числом сериальных органов, однако не имеющие среди
своих предков сегментированных форм. Очевидно, что существо и про-
исхождение такого «метамерного» строения этих форм различны.
Характерной чертой настоящей метамерии даже в случаях подавле-
ния или выпадения некоторых компонентов сегмента является корреля-
тивная связь частей сегмента как отдела тела. У моллюска с единой
колпачковидной раковиной (вдобавок еще имеющей спирально сверну-
тую эмбриональную раковинку) 8 пар .ретракторов ноги, 5 пар ктени-
диев, 10 пар коннективов, связывающих продольные нервные стволы,
6 пар нефридиев, или целомодуктов, при одной или двух парах половых
желез не составляют метамеров. Нервная система не имеет следов ме-
тамерных парных ганглиев, они при якобы метамерной организации
Neopilina отсутствуют. Кровеносная система, хотя и недостаточно изу-
ченная, вероятно, в силу плохой сохранности материала, также’ немета-
мерна, ее центральный орган — сердце в перикардии — лежит в заднем
137
’ в
Рис. 35. Метамерия упорядочения у моллюсков (из Беклемишева)
A—Tonica fastigiata (Loricata), молодая форма с дорзальной стороны; Б—Boreochiton cinereus (Loricata)
метамерное расположение ктенидиев; B—Fissurella maxima (Prosobranchia); Г—Tritonia llneata' (Opistho-
branchia)
конце тела моллюска. Все это подчеркивает, что коррелятивные связи,
подчиненность рассмотренных органов сегменту как целому, отсутству-
ют. Даже число пар «нефридиев», или целомодуктов, не согласуется с
числом половых желез. Таким образом, надо считать вместе с Бёттге-
ром) утверждение Лемхе о наличии пяти метамер у Neopilina
ошибочным. ов)
Neopilina и хитоны, бесспорно, сходны по примитивной двубокрт
симметрии строения, в том числе и нервной системы. Однако гораздо
больше сходства наблюдается между Neopilina и брюхоногими мол-
люсками. Спиральная эмбриональная раковина, колпачковидная дефи-
нитивная раковина, длинная, сложенная несколькими петлями, несим-
метричная 'Средняя кишка, хрустальный конус — вот основные черты
Neopilina, свойственные и низшим представителям переднежаберных.
Однако наличие симметричной ab origine центральной нервной си-
стемы, в остальном по своему уровню дифференцировки близкой к
нервной системе низших Prosobranchia, говорит о том, что Neopilina
можно рассматривать как брюхоногое, не испытавшее торзиона и сто-
ящее по многим признакам на самом примитивном уровне организации.
В сущности говоря, спиральная эмбриональная раковинка и колпачко-
138
видная, к тому же несколько асимметричная, дефинитивная раковина
сразу определяют принадлежность Neopilina к классу брюхоногих,
тогда как внутреннее строение подтверждает это положение.
Видимо, необходимо, как уже считал Маркус (Marcus, 1958), соз-
дать новый подкласс гастропод — Orthoneura и противопоставлять его
подклассам Streptoneura и Euthyneura, заменив этим названием неудач-
ное Monoplacophora. Это поставило бы Neopilina на- место в системе и
подчеркнуло бы ее филогенетическую связь с остальными брюхоногими.
В построении классификации или системы животных надо исходить из
рациональных основ природы организмов, а не из несовершенных правил
международной номенклатуры. Термин Monoplacophora в его зна-
чении самостоятельного' класса был введен Однером, когда о древних
моллюсках Tryblidiacea было еще слишком мало известно, чтобы иметь
возможность сопоставить их с Polyplacophora или Placophora. Отсут-
ствие близких отношений между хитонами и неопилиной, несомненное
родство неопилины и ее ископаемой родни с гастроподами и наличие
среди переднежаберных ряда групп с простой, не завитой спиралью
дефинитивной раковиной и со спиральной эмбриональной раковинкой
делают название Monoplacophora малообоснованным.
Я считаю также нерациональным сохранение деления Gastropoda
на Amphigastropoda, Archaeogastropoda, Mesogastropoda и Neogastro-
poda, о чем скажу ниже. Считаю также ненужным сохранять и название
Amphigastropoda для низших гастропод, предлагая для них название
Orthoneura.
Брюхоногие моллюски
Брюхоногие моллюски, с которыми обычно связывается представле-
ние как об очень пассивных, медленно ползающих слизняках или улит-
ках и организация которых легла в основу учения о компактном типе
строения моллюсков (стр. 124), характеризуются высокой пластичностью
организации, разнообразием экологии и наличием в онтогенезе трохо-
форы и велигера. В литературе справедливо отмечается (Kaestner,
1954/1955), что брюхоногие по разнообразию способов передвижения,
добычи пищи, характеру пищи, по овладению разнообразными биото-
пами Мирового океана, от береговой зоны до сверхабиссальных глубин
(10415—10687 м) (Беляев и др., 1960), пресных вод и суши, от Арктики
до тропиков включительно, от равнин и до горных цепей (выше 5000 м
над уровнем моря) соперничают с членистоногими и позвоночными.
Все это, а также большая древность брюхоногих, известных, начиная с
раннего кембрия, богатство палеонтологической летописи (известно
более 15000 ископаемых видов) и громадная численность видов
(85000) дают некоторое представление об этом наиболее разнообраз-
ном и многочисленном классе современных моллюсков.
Возникновение, значение и особенности скручивания (тррсиона) и
раскручивания, столь характерные для брюхоногих, исследовались мно-
гими, как старыми, начиная со Шпенгеля (Spengel, 1881), так и новыми,
.особенно английскими, зоологами (Garstrang, 1928; Crofts, 1955; Mor-
ton, 1958), так что об этих явлениях можно много не говорить. Однако,
на мой взгляд, нельзя, как делают Гарстанг и его последователи (Yonge,
Knight, Crofts и др.), сводить происхождение скручивания к мутацион-
ному изменению морфогенеза личинки-велигера и отрывать его от ус-
ловий жизни взрослых моллюсков. Скручивание туловищного, мешка,
перемещение мантийной полости с комплексом органов, редукция пер-
воначально правых органов мантийного комплекса, хиастоневрия, свя-
занное со всем этим спиральное закручивание раковины надо рассмат-
ривать как результат длительного приспособления ряда поколений
139'
брюхоногих к ползающему образу жизни, сказавшееся и на ходе мор-
фогенеза. Благодаря скручиванию достигается большая устойчивость
и подвижность животного, большая компактность раковины и своеоб-
разие формы, облегчающие передвижение моллюска по субстрату.
При высоком, коническом внутренностном мешке, рост которого связан-
с удлинением петли кишечника, увеличением размеров его желез (пече-
ни), развитием половой железы и толстой прочной раковины для
защиты внутренних органов, скручивание, -ведущее к изменениям фор-
мы тела и строения внутренних органов, имеет важное приспособитель-
ное значение. Смещение органов паллиального комплекса вперед .у
более подвижных и активных переднежаберных, сравнительно с пред-
ковыми, двустороннесимметричными формами имеет важное функци-
ональное значение. В частности, положение мантийной полости на пе-
реднем конце обеспечивает необходимый вводный ток воды слева к
осфрадию и ктенидию, а справа — выносящий ток с экскретами и фека-
лиями (Morton, 1958).
Многое из сказанного в той или иной форме выдвигалось разными
авторами, занимавшимися изучением скручивания (или торсиона) у
брюхоногих. Формирование внутренней организации после метаморфо-
за и оседания велигера на субстрат, далее рост тела, не. ограниченный
сроком, и значительная продолжительность жизни брюхоногих заставля-
ют считать, что происхождение скручивания и всех изменений в строе-
нии мягкого, тела и в форме раковины брюхоногих является результа-
том длительных приспособлений к условиям придонного существования..
Изменение морфогенеза на зародышевых или личиночных стадиях — не-
причина скручивания и всех связанных с ним изменений -строения взрос-
лых моллюсков, а последствия воздействий на последние условий жизни.
Если раковина головоногих моллюсков, ее форма и строение изучены
достаточно подробно с позиций статики и динамики (стр. 156), то отно-
сительно закономерностей механики развития, строения и движения ра-
ковины брюхоногих мы почти ничего не знаем. В известной мере мы
учитываем особенности строения раковины брюхоногих, коррелятивно-
связанные с теми или другими их чертами мягкого тела, но в целом
раковина изучена еще недостаточно, что хорошо демонстрируется на
примере системы, предложенной Термие (Н. et G. Termier, 1952), соз-
давших только по раковине неестественные группировки ископаемых и
современных переднежаберных и заднежаберных брюхоногих.
Выше было показано (стр. 138—139), что Neopilina galatheae Lemche
и, очевидно, и Neopilina (Vema) ewingi (Clarke a-. Menzies, 1959) по ос-
новным признакам организации наиболее близки к примитивным брю-
хоногим, в чем мое мнение совпадает со взглядом Бё'ттгера (Boettger,
1952, 1954). Я лишь не согласен с ним, как и с Венцем (Wenz, 1938,
1940), в необходимости создания подкласса Amphigastropoda для
Tryblidiacea и Bellerophontacea. Tryblidiacea — брюхоногие моллюски
с мантийной полостью на заднем конце тела, Bellerophontacea — с ман-
тийной полостью на переднем конце, как у переднежаберных. Современ-
ная классификация брюхоногих моллюсков, давно построенная зооло-
гами по признакам внутренней организации, учитывает строение цент-
ральной нервной системы. Еще старые классики—Иерииг (Ihering,.
1877) и Шпенгель (Spengel, 1881) придавали важнейшее значение для
естественной классификации брюхоногих строению нервной системы:
Иерингом был даже введен термин Orthoneura для ряда брюхоногих
моллюсков (современных Opisthobranchia и Pulmonata), у которых он
ошибочно, как показал Шпенгель (Spengel, 1881), признавал, a.b origine
ортогональную нервную систему.
140
Строение Neopilina показывает, что это не представитель самостоя-
тельного класса,’а брюхоногое, не подвергшееся торсиону, т. е. настоящее
Orthoneura, ' как, вместо Monoplacophora," следовало бы называть и
всех Tryblidiacea.
Как известно, Истон (Easton, 1960) и Камерон (Cameron, 1960)
помещают мюноплакофор в подкласс Protogastropoda, установленный
Шаймером и Шроком (Shimer a. Shrock, 1944).
В этот подкласс отнесены не только собственно протогастроподы,
каковыми являются современные и ископаемые Tryblidiacea, но и при-
митивные, с рядом черт двубокой симметрии стрептонеура, что делает
подкласс сборным. На этом основании я считаю сохранение подкласса
Protogastropoda неоправданным и более рациональным установление
для Tryblidiacea и родственных форм подкласса Orthoneura.
Для меня также являются неубедительными соображения польского
.зоолога Феликсяка (Feliksiak, 1959) о филогении моллюсков, который
считает моноплакофора не подклассом брюхоногих, а группой, равноцен-
ной всем раковинным моллюскам. В связи с этим зоологическая клас-
сификация брюхоногих, в которой различали подклассы Prosobranchia,
Opisthobranchia и Pulmonata, должна быть пополнена подклассом
Orthoneura. Для большей стройности классификации следует вместо
Prosobranchia применять название подкласса Streptoneura Spengel, а
для Opisthobranchia и Pulmonata — подкласса Euthyneura Spengel.
К сожалению, в настоящее время, в меньшей мере среди зоологов,
но главным образом среди палеонтологов, применяется система, предло-
женная Тиле (Thiele, 1935) и Венцем (Wenz, 1938), по которой брюхоно-
гие подразделяются на Archaeogastropoda, Mesogastropoda и Neogast-
ropoda в соответствии с геологической историей (палеозой, мезозой и
кайнозой), вместо деления на Diotocardia, Monotocardia (Taenioglossa
и Stenoglossa). Против такой классификации гастропод возражал еще
Бёттгер (Boettger, 1952, 1955). Классификацию Тиле следовало бы
отвергнуть, так как естественная классификация животных организмов
вообще строится на основании черт, присущих им как организмам, а не
по их геологической истории. Разделяя ряд брюхоногих моллюсков на
Archaeogastropoda, Mesogastropoda и Neogastropoda, авторы точно
отказываются считать заднежаберных и легочных брюхоногими мол-
люсками.
Тиле и Венц проделали с классификацией переднежаберных то, что
сделал в свое время Ведекинд, поделивший аммоноидей на Palaeo-,
Meso- и Neoammonoidea (цит. по Соболеву, 1924), и что в свое время
палеонтологами было отвергнуто.
Если исходить из цифровых, хотя и неполных, данных И. А. Коробко-
ва (1960), то отряд Diotocardia (Archaeogastropoda) представлен при-
мерно 48 семействами, существовавшими только в палеозое, 14—
в палеозое и мезозое и 12 — только в мезозое; из 29 семейств современной
фауны небольшая часть известна в мезозое, главным образом из мела.
В целом эта группа брюхоногих существует с кембрия, в котором она
еще бедно представлена; в ордовике, силуре, карбоне и перми она
достаточно обильна, в девоне и триасе многочисленна, а в третичное
время и ныне наиболее многочисленна (например, надсемейства
Trochacea, Neritacea). Таким образом, к характеристике примитивных
брюхоногих отряда Diotocardia (Archaeogastropoda), несомненно, надо
лишь добавить, что они являются древними; преимущественно палезой-
скими формами. Monotocardia-Taenioglossa, с характерной лентовидной
радулой, с 7 зубами в каждом ряду (Mesogastropoda), представлены
7 вымершими семействами и 69 ныне существующими. Из последних
Cerithiacea и Cyclophoracea известны с палеозоя, 17 семейств — с мезо-
зоя, большая же часть — с палеогена. Зта группа очень многочисленна
141
и в третичное время и в кайнозое, а не в мезозое; она не характеризует’
мезозой и не мезозоем характеризуется; к ней относится наибольшее-
число современных «новых» переднежаберных моллюсков.
Monotocardia-Stenoglossa, с узкой радулой с 1—3 зубами в каждом
ряду (Neogastropoda), представлены 3 небольшими вымершими семей-
ствами (одно мезозойское и два палеогеновых) и 29 существующими;,
большинство этих семейств становится известным с конца мезозоя, и.
наиболее обильно распространено в современной' фауне. Следова-'
тельно, по геологическому распространению нельзя противопоставлять
Monotocardia-Stenoglossa — Neogastropoda, a Monotocardia-Taenioglos-
sa — Mesogastropocla.
Для понимания эволюции и природы организмов необходимо знание-
их геологической истории, но заменять рациональные названия больших
групп, объединенных по совокупности характерных особенностей строе-
ния, искусственными, вроде Archaeogastropoda, Mesogastropoda и Neo-
gastropoda, тем более, если они не совпадают с геологическим распро-
странением, не следует. Между тем, многие палеонтологи применяют эту
неудачную классификацию. Имея дело лишь с раковинами брюхоногих
и мало зная строение вымерших форм, они обычно следуют по пути
Тиле и Венца. Между тем, когда нет преемственности между вымершими
в основном палеозойскими и современными семействами, для выяснения
природы брюхоногих моллюсков недостаточно знания лишь морфологии
раковины. Уже этим подчеркивается необходимость объединения иссле-
дований зоологов, строящих свою классификацию в основном на сово-
купности данных анатомии и эмбриологии (правда, сравнительно огра-
ниченного числа современных форм), и палеонтологов, имеющих дело
в основном с наружным скелетом —• раковиной, но большим числом форм
и притом на разных этапах исторического развития.
В целом классификация брюхоногих, разработанная зоологами, яв-
ляется наиболее естественной, отражающей их филогенетические отно-
шения, чем мы многим обязаны ряду крупных исследований, преимуще-
ственно английской школы и Бёттгера (Boettger, 1952, 1955). Палеонто-
логи неправы, разделяя и противопоставляя часть отряда Monotocardia-
Taenioglossa как Mesogastropoda другой части его, именно Stenoglossa
как Neogastropoda, так как обе группы этих моллюсков неразрывно свя-
заны друг с другом. Хотя многие палеонтологи, в том числе И. А. Короб-
ков (1960), продолжают придерживаться классификации Тиле, некото-
рые палеонтологи начинают уже отказываться от этой системы (Сох,.
1960а — в; Сох a. Knight, 1960). В упрек зоологам надо поставить только
отсутствие порой стройности классификации. Так, некоторые употреб-
ляют таксоны «триба» или «надсемейство» наряду с подотрядом (напри-
мер, Иванов, 1940а; Kaestner, 1954/1955). Палеонтологи более строги в
отношении формы, в их системе семейства объединяются в надсемейства
(Коробков, 1960; Moore a. all., 1952; Knight a. all., 1954), а последние —
в подотряды (Сох, 1960; Сох a. Knight, 1960).
Значительные разногласия у палеонтологов наблюдаются в толкова-
нии природы и положения в системе некоторых надсемейств, что имеет
существенное значение и для филогении. Я приведу лишь два примера
из работ последнего времени: И. А. Коробкова (1960) и Кокса и Найта-
(Сох a. Khight, 1960).
К Archaeogastropoda (Diotocardia) И. А: Коробков относит 16 над-
семейств: Bellerophontacea, Pleurotomariacea, Raphistomacea, Euompha-
lacea, Patellacea, Trochonematacea, Amberleyacea, Neritacea, Cocculina-
cea, Trochacea, Murchisoniacea, Nerineacea, Loxonematacea, Subulitacea,.
Pseudomelaniacea, Pyramidellacea.
В новой классификации Кокса и Найта (Сох a. Knight, 1960) этот
отряд делится на 7 подотрядов (Bellerophontina, Macluritina, Pleuroto-
142
mariina, Trochina, Patellina, Neritojjsin^ Murchisonina), причем эта клас-
сификация свободна от ряда погрешностей, допущенных И. А. Коробко-
вым и другими палеонтологами.
Например, существенным недочетом, у И. А. Коробкова, как и ряда
других (палеонтологов, является то, что ошибочно, только на основании
признаков строения раковины, игнорируя данные зоологов по анатомии,
он относит надсемейство Pyramidellacea (девон?, мел — ныне) к Archaeo-
gastropoda, т. е. к Diotocardia. Это надсемейство, созданное лишь по
особенностям строения раковины, подлежит ликвидации, так как в него
палеонтологами включены представители и семейства Melanellidae, и се-
мейства.Stiliferidae (из типичных переднежаберных, или Streptoneura), и
семейство Pyramidellidae (из заднежаберных или Euthyneura) (Boettger,
1955; Fretter a. Graham, 1954; Knight'a. all., 1954).
Насколько относительна таксономическая ценность раковины брю-
хоногих, говорит различие в понимании природы и положения в системе
ряда палеозойских надсемейств этих моллюсков. Так, по мнению
И. А. Коробкова (1960) и ряда зарубежных палеонтологов, надсемей-
ства Loxonematacea и Subulitacea относятся к Archaeogastropoda, т. е.
к Diotocardia. Кокс (Сох, 1960а) же находит у Loxonematacea
сходство с Cerithiacea и относит, это надсемейство к Chaenogastropoda
(по Коксу Monotocardia-Taenioglossa), а надсемейство Subulitacea
(Knight a. all., 1954, 1958) по признакам раковины считает Neogastro-
poda, т. е. высшими переднежаберными (Stenoglossa).
Уже это немногое говорит о необходимости для понимания родства -
и филогении брюхоногих серьезной ревизии существующих классифика-
ций ископаемых брюхоногих, их дополнительных исследований и согла-
сования ‘в этой области работ зоологов и палеонтологов.
Эволюция брюхоногих, как соответственно и других классов моллюс-
ков, начинается двустороннесиммётричными по внешней форме и по
внутреннему строению примитивными моллюсками, близкими к Trybli-
diacea (нижний кембрий — триас, ныне). Выше были отмечены неко-
торые черты вторичного характера у Neopilina сравнительно со строе-
нием гипотетической исходной формы и с состоянием ряда органов у не-
которых примитивных современных форм брюхоногих. Это делает
вероятным существование в протерозое форм более примитивных, чем
Tryblidiacea, и подчеркивает необходимость поисков протерозойских
представителей брюхоногих и изучения форм вроде Уолктовской Chuaria
circularis (Walcott, 1899).
В кембрии и ордовике протекает эволюция примитивных брюхоногих,
наряду с Tryblidiacea; возникают двусторонне-симметричные формы с
симметричной плоскоспиральной раковиной в виде надсемейств Helcio-
nellacea (нижний — средний кембрий) и Bellerophontacea (нижний кем-
брий— начало триаса), представленного семью семействами, вымерши-
ми к триасу, а.в верхнем кембрии — надсемейства Pleurotomariacea
(поздний кембрий — ныне), с асимметричной, свернутой по винтовой
спирали раковиной. Судя по строению раковины у Bellerophontacea, их
мантийная полость с паллиальным комплексом органов, как и у Pleuro-
tomariacea, 'была расположена впереди, а нервная система обладала
хиастоневрией, т. е. Bellerophontacea уже 'претерпели скручивание или
торсион (Yonge, 1947; Knight a. all., 1954; Сох, 1960а, б), а их пред-
ковыми формами были Helcionellacea (Knight a. all., 1958; Сох. 1960).
Однако природа Bellerophontacea окончательно не выяснена. Некоторые
авторы [Зиморт (Simroth), Венц (Wenz), Бёттгер (цит. по Сох, 1960).
В. Н. Беклемишев и другие] отрицают торсион у этого надсемейства.
Кроме того, обращает на себя внимание сходство плоскоспиральных
раковин ряда Bellerophontacea с таковыми древнепалеозойских на-
утилид.
143
Судя по данным палеонтологии, Bellerophontacea не играли сущест-
венной роли в эволюции брюхоногих, в отличие от Pleurotomariacea, ко-
торые заняли центральное положение в родословном древе Streptoneura
и Euthyneura.
На основании морфологии раковины, палеонтологи (Коробков, 1960)
с известным правом считают, что от Pleurotomariacea произошли Trocho-
nematacea (силур — карбон), Murchisoniacea (ордовик—-триас), Loxo-
nematacea (ордовик—лора) и Subulitacea (ордовик — триас), причем
промежуточной группой между Pleurotomariacea и Trochacea, Neritacea
и Cocculinacea были Trochonematacea.
Низшие переднежаберные, или Streptoneura — Pleurotomariacea, Ра-
tellacea (палеозой — ныне), Neritacea (пермь — ныне), Trochacea (три-
ас— ныне), Cocculinacea (миоцен — ныне), входящие вместе с Pleuroto-
mariacea в отряд Diotocardia, в процессе эволюции в разной степени
отклоняются от двустороннесимметричного строения, в наибольшей- сте-
пени сохраняющегося у Pleurotomariacea (два двоякоперистых ктенидия,
сердце с двумя предсердиями, две почки). У наиболее примитивных пред-
ставителей Diotocardia имеются еще церебральные тяжи, а не ганглии,
появляющиеся позже, длинные педальные тяжи с правильно отходящими
нервами, кроме того, у них (Hanstrom, 1928) имеются лабиальная, суб-
радулярная и субцеребральная комиссуры. Слабая ганглионизация и
диффузность частей центральной нервной системы сочетается с мало-
подвижным образом жизни (ползание по дну моря), с питанием расти-
тельной пищей (микроскопическими водорослями), с наличием у ряда
-форм внутриклеточного пищеварения, сортирующего отдела желудка
(Morton, 1953), хрустального столбика и ряда других примитивных черт.
В основном это обитатели прибрежной полосы моря. Благодаря жизни
в приливно-отливной зоне, среди Neritacea начинают формироваться оби-
татели солоноватых и пресных вод и даже суши [семейства Helicinidae
(верхний мел — ныне) и Hydrocenidae (плейстоцен—-ныне)], что в более
значительной степени происходит уже среди Monotocardia.
Этот отряд примитивных переднежаберных является и наиболее
древним. Из 10 вымерших надсемейств только в палеозое существова-
ло 6, в палеозое и частью мезозое—4, из существующих ныне надсе-
мейств 3 известны с палеозоя, 1 — с юры. Если ряд надсемейств дости-
гал расцвета в ордовике и силуре, то, как было сказано выше, ряд сов-
ременных надсемейств (Trochacea, Neritacea) богато представлен в тре-
тичное время и ныне.
Отряд Monotocardia (или Pectinibranchia), включающий Taenioglossa
и Stenoglossa, своими низшими представителями непосредственно свя-
зан с Diotocardia. Эти брюхоногие испытывают дальнейшую прогрессив-
ную эволюцию в строении, развитии, экологии и становятся наиболее
обширной группой современных Streptoneura.
Большинство (12 из 19 современных надсемейств) Monotocardia-
Taenioglossa становится известным с конца мезозоя (преимущественно
с верхнего мела), два — Littorinacea и Naticacea — с триаса и пять Vivi-
paracea, Rissoacea, Cerithiacea, Scalacea и Xenophoracea — с юры (Ко-
робков, 1960). Эволюцию этих так называемых Mesogastropoda харак-
теризует не мезозой, а кайнозой. В третичное и четвертичное время они
были наиболее многочисленными и разнообразными, особенно надсемей-
ства Cerithiacea, Naticacea и Strombacea.
Низшие Taenioglossa (Cyclophoracea, Rissoacea) и некоторые дру-
гие, подобно Diotocardia, являются растительноядными микрофагами,
соскабливающими радулой водоросли со скал, или ' планктоноядными
прибрежными формами. Они сохраняют сортировочный отдел желудка,
полость которого не участвует в переваривании пищи, внутриклеточное
пищеварение, хрустальный столбик, например, семейства Turritellidae,
144
Calyptraeidae, Vermetidae, Capulidae, Struthiolariidae, (Morton, 1951),
ресничнослизистые механизмы передвижения пищи по" кишечнику (Mor-
ton, 1953). У них. наблюдаются длинные, педальные с правильно распо-
ложенными комиссурами и плевровисцеральные тяжи, сравнительно
слаборазвитые ганглии центральной нервной системы.
Как было сказано выше, это в значительной мере прибрежные и мел-
ководные моллюски. С распространением в Мировом океане, с освое-
нием новых, разнообразных биотопов связана эволюция таких надсе-
мейств, как Strombacea, Naticacea, Cypraeacea, Топпасеа. Увеличиваются
подвижность и активность, происходит переход к плотоядному питанию,
к активному хищничеству, с чем связана дифференциация пищевари-
тельного тракта. Совершенствуется строение, увеличиваются размеры
гребенчатых ктенидиев и осфрадиев, усиливаются их функции (Yonge,
1947; Morton, 1958). Со всем этим связана эволюция нервной системы,
происходит развитие ганглиев, особенно церебральных, их сближение,
совершенствование органов чувств, особенно глаз, достигающих в ряде
случаев высокой дифференцировки и даже способности к аккомодации
(например, у Pterotrachea).
Идет все возрастающее освоение этими моллюсками Мирового океа-
на и его разнообразных биотопов, наблюдаются переходы от донной
жизни к пелагической, с чем связаны коренные изменения почти всех
черт организации (например, надсемейства Atlantacea, или Heteropoda).
Наряду с этим среди низших групп Taenioglossa происходит освоение
пресных вод и суши в масштабах, далеко превосходящих то, что наблю-
дается среди Diotocardia. Некоторые семейства — Cyclophoridae из
Cyclophoracea, Pomatiasidae и Acmidae из Littorinacea, частично Hydrobi-
idae из Rissoacea-—перешли от'водного к наземному (сухопутному) об-
разу жизни.
Зоологи еще в конце XIX столетия (Plate, 1895, и др.) видели в се-
мействе Trochidae группу, от которой произошли остальные переднежа-
берные. К этому в основном приходят и палеонтологи. К Trochacea по
уровню строения и ряду общих его признаков ближе всего стоят Archi-
taenioglossa — Cyclophoracea, Viviparacea и Ampullariacea, а также
Rissoacea, Littorinacea и, видимо, Valvatacea, обладающие рядом при-
митивных черт строения, в особенности нервной системы с ее длин-
ными педальными тяжами и следами лестничного строения (А. В. Ива-
нов, 1940а, 19406; Remane, 1959). Остальные надсемейства Taenioglos-
sa, как и Stenoglossa, относящиеся к высшим Monofocardia, в, отно-
шении, строения, поведения и физиологии составляют наиболее много-
численную группу Streptoneura современных морей. В основном им
принадлежит большая часть из 100 современных семейств этого под-
класса.
Некоторые палеонтологи придают важное филогенетическое значе-
ние Trochonematacea как группе, давшей начало ряду семейств Diotocar-
dia и Monotocardia. Так, И. А. Коробков (1960) из Trochonematacea (си-
лур— карбон) выводит Cyclophoracea (известны с раннего карбона) и
Littorinacea (с юры), а не от Trochacea (юра — ныне), которых он сог-
ласно с Венцем производит от Trochonematacea. Однако строение Trocho-
nematacea неизвестно, анатомические же данные связывают Trochacea с
Pleurotomariacea; поскольку и геологически они не разорваны, правиль-
нее Trochacea выводить непосредственно из Pleurotomariacea, а от
Trochacea — Cyclophoracea и Littorinacea. Stenoglossa — группа высших
Streptoneura, представлена преимущественно хищными, наиболее высо-
ко организованными, с сильно концентрированной нервной системой и
наиболее многочисленными по числу видов представителями переднежа-
берных брюхоногих. Относящиеся сюда надсемейства Muricacea, Bucci-
nacea, Volutacea и Conacea связаны близким родством, по признанию
1Q Д. М. Федотов
145
зоологов, друг с другом и с высшими представителями Taenioglossa —
Топпасеа.
В последнее время некоторые палеонтологи, на основании данных
конхиологии (Knight a. all., 1954), в эту цельную, естественную группу
брюхоногих вводят вымершие надсемейства Subulitacea (ордовик — мел)
и Nerineacea (триас — мел) и считают последних родоначальниками под-
отряда Stenoglossa (Neogastropoda), а родословную Taenioglossa ведут
от Loxonematacea (ордовик —-юра), разъединяя, таким образом, Steno-
glossa и высших Taenioglossa, близко родственных между собой. Мне
представляется такой взгляд необоснованным.
По данным сравнительной морфологии, преимущественно анатомии,
конца XIX столетия (Plate, 1895; Pelseneer, 1891), и последнего времени
(Graham, 1949; Boettger, 1952, 1954, и др.) заднежаберные и легочные
моллюски считаются потомками сравнительно примитивных Streptoneura,
претерпевших раскручивание, или деторсион, и в связи с этим утратив-
ших хиастоневрию, т. е. ставших Euthyneura. Известно, что у Actaeonidae
(Opisthobranchia) и среди Ellobiidae и Chilinidae (Pulmonata) имеется
хиастоневрия, у ряда низших представителей обеих групп еще сохраня-
ются сравнительно длинные коннективы и комиссуры центральной нерв-
ной системы. У .большинства же происходит концентрация ганглиев
нервной системы в голове вокруг пищевода, доходящая в отдельных слу-
чаях (Tethys, Nudibranchia) до полного слияния их в одну общую массу
(Беклемишев, 1952, 1964). Процесс раскручивания, затрагивающий в
меньшей мере более консервативных легочных моллюсков, вызывает
у заднежаберных смещение направо и назад мантийной полости и кте-
нидия, лежащего позади сердца, раскручивание петли кишечника и ряд
других изменений организации (см. Morton, 1958)..
В ходе эволюции идет освоение заднежаберными разных глубин моря.
Оно достигалось сначала придонными формами, сохраняющими в
остальном типичное строение раковинных брюхоногих, представленными
по Бёттгеру (Boettger, 1954), 21 семейством, среди которых низшими яв-
ляются Actaeonidae, Pyramidellidae. Преобладающая масса заднежа-
берных (не менее 55 семейств), осваивая разные биотопы моря, эволю-
ирует в сторону облегчения веса раковины, ослабления ее значения как
органа пассивной защиты, обрастания ее мантией, регресса до ее пол-
ного исчезновения; происходит утрата высокого внутренностного мешка.
В связи с этими изменениями происходит регресс: уменьшение мантий-
ной полости, первичного органа дыхания — ктенидия, развитие на зад-
нем конце тела вторичных жабр, а у ряда форм и вторичного кожно-
мускульного мешка, приобретение двустороннесимметричного червеоб-
разного тела и т. д. Такие характерные для высших современных задне-
жаберных изменения исходного типичного строения брюхоногого мол-
люска наблюдаются у Nudibranchia, Aplysiacea, Saccoglossa (Limapori-
tidae и др.), Rhodopidae, Pseudovermidae, Phyllirhoidae и т. д.
Яркими примерами высокой пластичности организации заднежабер-
ных являются такие формы из Saccoglossa, как Berthelinia (рис. 36)
(Тamanovalva, Edentellina} и Midorigai, обладающие подобно Bivalvia
двустворчатой, порой с радиальной ребристостью, раковиной, с муску-
лом-аддуктором, живущие на зеленых водорослях Caulerpa. По виду
они сходны с двустворчатыми, но обладают, однако, организацией, ти-
пичной для заднежаберных (Kawaguti a. Baba, 1959; Kawaguti, 1959;
Сох, 1960; Burn, 1960; Feliksiak, 1961; Boettger, 1962a, 19626).
При всем богатстве и разнообразии строения, способов передвиже-
ния, вплоть до реактивного, биотипов, при большом количестве видов
(10000, Kaestner, 1954/1955) в преобладающем большинстве заднежа-
берные являются типичными морскими обитателями. Все же некоторые-
из них встречаются в пресных водах, а семейства Oncidiidae, Rothouisi-
146
idae и Veronicellidae, которых раньше считали легочными моллюсками —
Stylorrnnatophora, являются амфибиотическими и сухопутными (Boet-
Сравнительно с заднежаберными, легочные моллюски, примитив-
ные формы которых, как и заднежаберных, обладают перекрещенной
нервной системой, характеризуются меньшей степенью деторсиона. При
сближении ганглиев путем укорачивания, а не раскручивания, у них
исчезает перекрест плевровисцеральных коннективов. На очень ранних
этапах своей эволюции, приспособляясь к жизни в береговой полосе
моря, древние формы легочных моллюсков начали использовать для ды-
хания атмосферный воздух. Легочные моллюски, стоящие по своей фи-
зиологии выше заднежаберных, с выходом из морской среды в пресные
воды, а в основном на сушу широко распространились по всей Земле
и в настоящее время представлены более чем 20 000 видов.
Распространившись по всем областям Земли,, включая тропики и
Арктику, приспособившись к перенесению жаркого, засушливого и хо-
лодного сезонов года переходом в спячку, или анабиоз, легочные мол-
люски 'в основном остаются растительноядными, но не микрофагами, а
откусывающими куски растений. Однако среди наземных форм немало
хищников, поедающих дождевых червей и других наземных улиток (Бла-
стов, 1948). ’
Характерно, что и Basommatophora и Stylommatophora, несмотря на
обилие видов, родов и семейств, сохраняют основные черты строения
раковины. Именно на них приходится убеждаться в малом таксономи-
ческом значении раковины сравнительно с анатомией (особенно половой
системы). Освоив условия наземной жизни, стебельчатоглазые не воз-
вращаются для размножения в воду и откладывают яйца, нередко оде-
тые известковой скорлупой, во влажную землю.
Как отмечалось, одни авторы (например, Plate, 1895) выводили зад-
нежаберных и легочных моллюсков от общего предка — из семейства
Actaeonidae, другие (например, Pelseneer, 1898—1899), а позже и Плате
(Plate, 1922) выводили заднежаберных от Actaeonidae, а легочных от
Ellobiidae при общем родоначальнике из семейства Trochidae. Авторы
10s
147
новых сравнительно-анатомических исследований (Fretter a. Graham,
1949а, б, 1962; Boettger, 1955) подчеркивают более близкое родство
заднежаберных и легочных и возвращаются к старой трактовке о
возможности их происхождения от Actaeonidae. По мнению Бёттгера,
от Actaeonidae (учитывая данные палеонтологии, следует сказать
от надсемейства Actaeonacea, в которое входит семейство Actaeonidae,
карбон — ныне), произошли многочисленные (21) семейства надсемей-
ства Cephalaspidea — раковинных заднежаберных. Из разных семейств
Cephalaspidea путем приспособления к жизни в разных биотопах моря
дифференцировались остальные группы заднежаберных — Saccoglossa,
Aplysiacea, Nudibranchia и, судя по сравнительно-анатомическим данным
(Fretter a. Graham, 1954; Boettger, 1955), группа наземных семейств
Oncidiidae, Rathouissidae и Veronicellidae.
По данным Бёттгера, Basommatophora охватывают морские семей-
ства Siphonariidae, Gadiniidae, пресноводные Chilinidae и наземные ЕПо-
biidae, причем с хилинидами связывается происхождение всех осталь-
ных Basommatophora, ' а с элобиидами — всех Stylommatophora. Таким
образом, Плате (Plate, 1895) был ближе к истине, выводя заднежабер-
ных и легочных от общего предка, чем Пельзенер (Pelseneer, 1894), пред-
полагавший происхождение их от разных предков.
Палеонтологическая летопись заднежаберных и легочных брюхоно-
гих бедна, так как несомненные представители этих групп известны
лишь с карбона (Actaeonidae из Opisthobranchia, Siphonariidae и Ellobi-
idae из Pulmonata). Палеонтологи не признают необходимости выделения
надсемейства Cephalaspidea, значительную часть семейств этой группы
они включают в надсемейство Actaeonacea и создают семейство Actaeoni-
dae (карбон — ныне). В. Ф. Пчелинцев и И. А. Коробков (1960) сужи-
вали содержание семейства Actaeonidae и его геологическое распростра-
нение (юра — ныне).
Если в отношении Cephalaspidea палеонтологи вряд ли правы, то в
отношении семейства Actaeonidae, может быть, надо согласиться с такой
перестройкой.
Неправы палеонтологи, создавая по особенностям раковины гетеро-
генное надсемейство Pyramidellacea и выводя Actaeonidae из Руга-
midellidae, а не наоборот. Также мало убедительны положения палеон-
тологов (например, Knight a. all., 1954) о происхождении заднежабер-
ных и легочных моллюсков от ископаемых форм, объединяемых в над-
семейство Subulitacea, на положение которого в системе палеонтологи
смотрят по-разному.
Двустворчатые моллюски
Двустороннесимметричное строение (ротовое отверстие на переднем,
анальное на заднем конце тела, пара ктенидиев в мантийной полости,
целомический перикардий с сердцем, состоящим из желудочка и двух
предсердий, пара выделительных органов, пара гонад, а также раковина
и нога двустворчатых) — основная черта типа моллюсков. Жизнь в тол-
ще ила или песка на дне моря в течение геологической истории с верхов
кембрия, вызвала у двустворчатых появление специфических черт, ха-
рактерных для этого класса. Тело сдавливается с боков, мантия стано-
вится двулопастной, а раковина двустворчатой, нога сдавливается с бо-
ков и теряет ползательную подошву, первичные гребенчатые ктенидии
становятся нитевидными или пластинчатыми. Скрытый, малоподвижный,
нередко прикрепленный образ жизни и пассивный способ питания1 дон-
ным детритом и мелкими организмами ведут к редукции головы и орга-
нов чувств, глотки с радулой и железами. Несмотря на такой образ жиз-
148
ни (с которым связаны стабильность и постоянство главнейших групп
двустворчатых), эти моллюски, благодаря очень высокой плодовитости,
широко распространились (преимущественно с помощью пелагических
личинок, трохофоры и велигера) и заселили Мировой океан от прибреж-
ной, береговой зоны до сверхабиссальных глубин в 10 415—10 687 м (Бе-
ляев и др., 1960). В известной мере они освоили также солоноватые
и пресные воды.
Двустворчатые являются вторым по числу видов (25 000) классом
моллюсков (Kaestner, 1954/1955). Благодаря необычайно тесным связям
их со средой, с толщами дна моря, не только условия жизни оказывают,
правда, скорее сдерживающее, чем стимулирующее, воздействие на дву-
створчатых, но и они нередко становятся мощными биологическими фак-
торами, обусловливающими своей жизнедеятельностью очень сильные
изменения окружающей среды (Воскресенский, 1948).
Жизнь в толще грунта, а также малоподвижный, нередко прикреп-
ленный образ жизни на дне моря очень сильно сказывается на двуствор-
чатых. Приспособление к такому специфическому образу жизни, есте-
ственно, прежде всего отразилось на органе защиты мягкого тела этих
моллюсков — на раковине, а именно на ее форме, строении ее стенок,
замке, лигаменте, а также на строении и функции органов дыхания —
жабрах, которые стали выполнять, помимо функции дыхания, преимуще-
ственно роль вододвигательного и фильтрующего аппарата для ловли
пищи. /'
Это давно было подмечено Неймайром (Neumayr, 1883, 1892, и др.),
построившим классификацию двустворчатых по строению раковины, и
Пельзенером, положившим в основу системы этих моллюсков строение
жабр. Естественной классификации двустворчатых пока нет. Неймайр
'(1883) на основании строения замка делил этих моллюсков на Palaeo-
concha (Ciyptodonta); Desmodonta, Taxodonta, Heterodonta и Dysodonta,
а Пельзеиер (Pelseneer, 1891) по строению жабр — на Protobranchia,
Filibranchia, Pseudolamellibranchia, Eulamellibranchia и Septibran-
chia.
Еще в конце XIX столетия Гертвиг (Hertwig, 1897) осудил односто-
ронний учет лишь некоторых черт для классификации двустворчатых и
подчеркнул необходимость для построения их естественной системы по
возможности одинакового использования различных аппаратов. Он под-
разделил класс на два отряда: Protoconchia и Heteroconchia, но допустил
неправильное объединение в первый отряд Protobranchia и Filibranchia
( = Anisomyaria Тиле и Хааса). Односторонность классификаций дву-
створчатых подчеркивал и А. А. Борисяк (1899).
Последующими работами.палеонтологов в классификацию Неймайра
вносились дополнения, исправления и подкрепления, на основании ряда
других черт строения скелета — лигамента, строения стенок раковины,
отпечатков, оставляемых мускулами-аддукторами, мантийной линии
и т. д. Работы зоологов, среди которых выделяется английская школа,
о строении, развитии, физиологии и экологии современных двустворчатых
дополняли и исправляли классификацию Пельзенера данными, касающи-
мися строения жабр, ноги, сифонов, выделительной, половой, кровенос-
ной и отчасти нервной систем.
В настоящее время распространена компромиссная классификация
Тиле (Thiele, 1935) и Хааса (F. Haas, 1929—1934, 1937—1956), в которой
один отряд — Taxodonta — охарактеризован строением замка, другой —
Anisomyaria — особенностями мускулов-аддукторов, третий, наиболь-
ший — Eulamellibranchia— строением жабр. Последний отряд по осо-
бенностям строения замочного края и зубов подразделен на подотряды:
Schizodonta, Heterodonta, Adapedonta и Anomalodesmata. Эта классифи-
кация, далеко нас не удовлетворяющая, однако широко используется
149
и зоологами и палеонтологами. К сожалению, некоторые палеонтологи
применяют классификацию Неймайра, внося в нее известные исправле-
ния и изменения (Эберзин, 1960), но допускают при этом крупные недо-
четы. Так, прежний отряд Пельзенера Protobranchia, который был цель-
ным и состоял по совокупности признаков из бесспорно наиболее прими-
тивных двустворчатых — Nuculidae, Ledidae и Solenomyidae, так назы-
ваемых Palaeoconcha Неймайра, заменен отрядом Taxodonta. Посколь-
ку в него включены Агсасеа, этот отряд является гетерогенным,
так как современные Arcidae имеют не двоякоперистые гребенчатые кте-
нидии, а нитевидные жабры и ряд черт строения, далеких от организа-
ции Nuculidae, Ledidae и Solenomyidae. Напротив, примитивное и близ-
кое по своей организации к Nuculidae и Ledidae семейство Solenomyidae
выделено из этой группы по отсутствию зубов. Перенесен в конец си-
стемы и отряд Desmodonta вместе с такими высоко специализированны-
ми молодыми семействами как Gastrochaenidae, Clavagellidae, Cuspida-
riidae. Следовательно, отряд Protobranchia в трактовке зоологов обосно-
ван лучше, чем отряд Taxodonta палеонтологов, и его надо сохранить.
Второй отряд современной системы — Anisomyaria, фигурировавший
уже в классификации Гертвига (Hertwig, 1897), включает Filibranchia,
Pseudolamellibranchia и часть Eulamellibranchia Пельзенера и Dysodonta
Неймайра. Позднейшими 'морфологическими, физиологическими и эко-
логическими исследованиями ряда английских зоологов, особенно Йон-
га и его школы, сильно углублены наши знания этой большой группы
двустворчатых и выправлены недочеты в классификации Пельзенера.
После этих исправлений была достигнута значительная цельность и
естественность системы наиболее обширной, разнообразной и содержа-
щей наиболее высоко организованные формы современных двустворча-
тых группы Eulamellibranchia Пельзенера. В основном этот отряд Bival-
via объединяет неймайровские отряды — Schizodonta, Heterodonta и Des-
modonta.
Принимая отряд Eulamellibranchia с включением в его состав этих
подотрядов, надо отказаться от недавно установленных подотрядов
Adapedonta и Anomalodesmata, так как подотряд Desmodonta и без них
объединяет такие семейства, как Solenidae, Myidae, Gastrochaenidae,
Pholadidae, Teredinidae, Pandoridae, Clavagellidae, Cuspidariidae и т. д„
которые размещались между Adapedonta и Anomalodesmata. Нет также
необходимости сохранения отряда Septibranchia Пельзенера, согласно
результатам последних работ Ионга (Yonge, 1954, 1957а, 1959а, 19596).
Нет договоренности о положении в системе рудистов или отряда
Pachyodonta, которых, как и Chamidae, Пельзенер относил к Eulamelli-
branchia.
В самое последнее время Кокс (Сох, 19606) сделал новую попытку
дать классификацию двустворчатых. Учитывая опыт многих предыдущих
авторов, в том числе и Гертвига (Hertwig, 1897), Кокс внес ряд значи-
тельных изменений в существующую и дал основы новой классифика-
ции двустворчатых.
Класс двустворчатых Кокс делит на 3 подкласса: Pro'tobranchia, Ptc-
riomorphia и Heteroconchia, из которых первый делится на 3 отряда, вто-
рой— на 5, третий — на 8. Названия отрядов (прежние и новые) Кокса
отражают преимущественно особенности замка, реже—строение жабр.
В подкласс Protobranchia включены наиболее примитивные двустворча-
тые надсемейства Nuculacea, Solemyacea и (вымершие) Cardiolacea.
В подкласс Pteriomorphia входят надсемейства: Cyrtodontacea, Агсасеа,
Pterinacea, Pinnacea, Pectinacea, Limacea, Anomiacea, Ostreacea, Myti-
lacea и Conocardiacea. Данные анатомии, особенности раковины и эколо-
гии говорят о том, что оба эти подкласса в таком составе являются ра-
циональными объединениями входящих в них надсемейств.
150
Третий, наибольший подкласс — Heteroconchia (название Гертвига,
1897) охватывает главную массу надсемейств двустворчатых: Lyrodes-
macea, Allerismatacea, Laternulacea, Clavagellacea, Pholadacea, Myacea,
Trigoniacea, Megalodontacea, Diceratacea, Hippuritacea, Chamacea, Anthra-
cosiacea, Unionacea, Cyrenacea, Isocardiacea, Arcticacea, Veneracea, Luci-
nacea, Erycinacea, Tellinacea, Solenacea, Mactracea, Cardiacea, Tridac-
nacea, Astartacea, Carditacea, Poromyacea.
Цельность этого подкласса значительно меньшая, чем предыдущих.
За исключением ряда ископаемых надсемейств, подкласс Heteroconchia,
как отмечает Кокс, отвечает отряду Eulamellibranchia Пельзенера. К со-
жалению, это не совсем точно, так как в него включены некоторые се-
мейства, например, Trigoniidae с нитевидными жабрами. Нельзя также
согласиться к Коксом, поставившим высокоспециализированные семей-
ства, вроде Clavagellacea и Pholadacea, в основании системы, ранее мно-
гих других современных семейств Eulamellibranchia.
В целом классификация Кокса производит впечатление более есте-
ственной, чем другие системы.
Значительные затруднения для понимания природы тех или иных
двустворчатых и для построения естественной, филогенетической класси-
фикации доставляют явления конвергентного развития, которое затраги-
вает разные стороны организации. Конвергентное развитие близких по
строению жабр у разных групп (Kaestner, 1954/1955) не позволяет де-
лить большие группы двустворчатых на основании лишь строения жабр
(нитевидные, ложнонитевидные, пластинчатые жабры), хотя в извест-
ной мере эволюция и филогения двустворчатых шла по пути развития
жабр (Protobranchia — Filibranchia — Eulamellibranchia).
Явления конвергенции затрагивают и строение замка. Между тем на
основании наличия таксодонтного замка, как отмечает Нэвил (Newell,
1954), некоторые считают Arcidae близкими к Nuculidae и объединяют
их в отряд Taxodonta. Но еще Дувилл (цит. по Newell, 1954) показал,
что палеозойские и мезозойские предковые Arcidae имели зубы, харак-
терные для многих Pterinacea и Mytilacea из другого отряда — ныне под-
класса Pteriomorphia.
Как известно, для понимания соотношений разных систематических
категорий в системе очень важны численность групп, и их геологическое
распределение.
Из 128 (по не совсем точному подсчету) семейств двустворчатых
8 семейств принадлежит Protobranchia, 30 — Pteriomorphia и 90 — Hetero-
conchia. Из 8 семейств Protobranchia 4 вымерли в палеозое и мезозое,
3 палеозойских существуют ныне и лишь 1 является новым, кайнозой-
ским. Из 30 семейств Pteriomorphia 18 семейств ведет начало с палеозоя,
причем 4 из них вымерло тогда же, 7 — существовали с палеозоя до мезо-
зоя, 7 — с палеозоя до кайнозоя включительно; из 11 семейств, извест-
ных с мезозоя, 9 существует ныне, 2 жило только в мезозое, 1 семейство
возникло в кайнозое. Из 90 семейств Heteroconchia 14 семейств ведет на-
чало -с палеозоя, причем 4 вымерло в палеозое, 5 — в мезозое, 5 семейств
продолжает существовать; 59 семейств известно с мезозоя, из них 21
вымерло в мезозое, 38 существует ныне. Кроме того, 17 семейств извест-
ны лишь в кайнозое. Таким образом, половина семейств Protobranchia
вымерла, половина существует. Среди Pteriomorphia 13 семейств вымер-
ло, 17 — существует. Из Heteroconchia 30 семейств вымерло, 60 — суще-
ствует. Следовательно, наиболее древним, преимущественно палеозой-
ским, и наиболее бедным по числу семейств в течение всей своей исто-
рии вплоть до настоящего времени является подкласс Protobranchia.
Подкласс Pteriomorphia включает значительно большее число семейств,
больше половины которых ведет начало с палеозоя; этот подкласс бога-
то представлен и в современной фауне. В подклассе Heteroconchia число
151
семейств наибольшее, причем только небольшое их число известно с па-
леозоя, преобладающей же массой являются мезозойские и современные
семейства.
Подкласс Protobranchia, как известно,— небольшая группа самых
древних, начиная с позднего, а по данным Мюллера (А. Н. Muller, 1958)
с середины кембрия, и самых примитивных двустворчатых. Состоящая
только из перламутра раковина, нога с ползательной подошвой, неболь-
шие гребенчатые ктенидии, несущие лишь дыхательную функцию
(рис. 37), ток воды для дыхания от переднего к заднему концу, открытая
мантийная полость, улавливание пищи (донного детрита) придатками
околоротовых лопастей, наличие плевральных ганглиев и плевропедаль-
ных комиссур, сердце над кишечником, простые выделительные протоки
для выведения половых продуктов, и образ жизни—все это свидетель-
ствует о примитивности Protobranchia.
Рис. 37. Схема строения жабр у различных групп пластинчатожаберных (из Hennig)
A—Protobranchia с ктенидиями; Б—Filibranchia с нитевидными жабрами; В—Eulamellibranchia с пла-
стинчатыми жабрами; Г—Septibranchia с жаберным аппаратом в виде пористой перегородки, делящей
мантийную полость
Примитивность организации и большая древность Protobranchia сви-
детельствуют об исходном их положении в эволюции двустворчатых как
родоначальной группы. Это давнее предположение Пельзенера (Pelse-
neer, 1891) подтверждено позднейшими .работами зоологов, особенно
Ионга и его школы, и ряда палеонтологов (см. Сох, 1960).
Относящиеся к Protobranchia семейства Nuculidae и Solemyidae свя-
заны многими общими чертами организации (Yonge, 1941). Различаются
главным образом тем, что у Solemyidae выделительные органы располо-
жены позади перикардия, а не впереди его, как у Nuculidae (Odhner,
1912), ктенидии значительно увеличены и участвуют в привлечении
пищи; эти моллюски переходят к роющему образу жизни в мягком
грунте (Yonge, 1941).
Основной группой Protobranchia и по строению, и по образу жизни
являются, без сомнения, Nuculacea (Nuculidae), a Solemyidae — боковая
ветвь; не случайно еще старые авторы связывали филогенетически Nucu-
lidae с Arcidae, хотя резонно систематически их не объединяли.
Подкласс Pteriomorphia (отряд Anisomyaria в классификации многих
авторов) характеризуется формой асимметричных правой и левой ство-
рок, поскольку эти двустворчатые лежат на одной из них. Подкласс
включает формы со всеми переходами от двустороннесимметричных к
асимметричным (по строению тела и раковины), от димиарии через гете-
ро- или анизомиарию к мономиарии аддукторов. Сравнительно-анатомш
чески, онтогенетически и исторически этот путь эволюции Pteriomorphia.
хорошо изучен исследователями (особенно Йонгом и его школой),
вскрывшими перестройку всей организации многих представителей Pte-
riomorphia.
152
Жизнь этих моллюсков на твердом грунте, особенно в литоральной
зоне, лежащих на левой створке (нередко с прикреплением нитями бис-
суса, или с помощью цемента, порой с цементированием раковин особей,
друг с другом), вызывает многие изменения ряда органов этих высоко-
пластичных двустворчатых.
Ряд форм (многие из семейств Pectinidae, Limidae) приобретает спо-
собность прыгать и плавать при помощи быстрого хлопания створками,.,
благодаря развитию поперечнополосатой структуры мускулов-аддукто-
ров, а некоторые, например Lima, плавают реактивным способом (Stud-
nitz, 1931). У таких форм развиваются на краях мантии многочислен-
ные, высокодифференцированные глаза, объединенные общей иннерва-
цией. У Pteriomorphia, особенно у малоподвижных, прикрепленных форм,,
наблюдается эволюция жабр, которые увеличиваются в размерах, диф-
ференцировка их строения от нитевидных к пластинчатым (Ostreacea,.
Pectinacea) и уподобление по строению жабр Heteroconchia, или Eula-
mellibranchia. Возрастает роль жабр, исходных органов дыхания, как
органов захвата пищи путем фильтрации громадных масс воды. Наблю-
дается значительная концентрация ганглиев нервной системы, разви-
ваются различные 'взаимоотношения между перикардием, почками, го-
надами и их протоками. Чрезвычайно возрастает производительность
яичников. Самка устриц, например, за сезон производит от 16 до 60*'
миллионов яиц (Storer a. Usinger, 1957). Благодаря большой жизнен-
ности и громадной плодовитости устрицы, как и ряд других форм Pterio-
morphia (например, Mytilidae), образуют банки из многих миллиардов
особей, расположенные вдоль берегов моря, тянущиеся порой на десят-
ки и более километров. Громадные скопления известны и у ископаемых
форм. Так,.мощные отложения из меловых Exogyra тянутся на протяже-
нии более 2500 миль от Нью-Джерси в США до Мексики. Широко из-
вестны и ныне мощные, преобразующие природу среды . (физико-хими-
ческий состав воды, рельеф дна, изменяющие направление течений и
т. д.) воздействия массовых поселений этих моллюсков, например, мидий.
Признание Pteriomorphia (Anisomyaria) не отрядом, а подклассом-
кажется обоснованным, так как ряд их надсемейств значительно отли-
чается друг от друга. Большинство типичных Pteriomorphia лежат или
прикрепляются на субстрате левой створкой, в отличие от Ostreacea,
прикрепляющихся правой створкой; одни семейства имеют нитевидные-
жабры, другие (Pectinacea, Limacea, Ostreacea, Pinnacea)—пластинча-
тые. У Pterinacea, Mytilacea и Pinnacea раковина преимущественно пер-,
ламутровая, у Pectinacea, Limacea и Ostreacea — кальцитовая (Newell,.
1954; А. Н. Muller, 1958). У большинства выделительные органы лежат
впереди перикардия, а у Arcidae и Pinna,— позади него (Odhner, 1912).
Зоологи, начиная с Пельзенера (1891), и ряд палеонтологов (Newell,
1954, 1960; Сох, 19606) по праву считают Arcacea (Arcidae) родоначаль-
ной группой всех Pteriomorphia. Сравнительно-анатомически, конхиоло-
гически и экологически эволюция Pteriomorphia идет, как было сказано
выше, от симметричных форм (вроде Area) путем ряда изменений к ти-
пичным асимметричным Pteriomorphia (вроде Ostrea и др-). Этому пред-
положению не противоречат и данные хронологии, если действительно
палеонтологическая летопись Arcacea начинается с ордовика (Moore а.
all., 1952), а не с девона, как обычно считают.
Двустворчатые подкласса Heteroconchia (или Eulamellibranchia) яв-
ляются наиболее многочисленными по количеству семейств и видов, наи-
более пластичными и разнообразными по внутренней организации, строе-
нию замочного края раковины, физиологии, образу жизни и экологии.
Они являются преобладающими формами среди современных двуствор-
чатых, заселившими самые разнообразные биотопы Мирового океана
вплоть до сверхабиссальных глубин. Это единственная группа двуствор-
153.
чатых моллюсков, широко проникшая в пресные воды и в целом наибо-
лее процветающая.
Heteroconchia чрезвычайно разнообразны по величине, длина их от
0,5 мм до 120 см, вес до 220 кг (Tridacna). Они чрезвычайно различны
и по своей экологии. Наряду с преобладанием свободноподвижных дом?
вых форм эпи- и инфауны, среди них имеются сверлящие каменистый
грунт, древоточцы, коммспсалисты некоторых иглокожих. Среди осталь-
ных двустворчатых они выделяются также и разнообразием питания.
Преобладающим способом добычи пищи является улавливание нанно-
планктона путем фильтрации масс воды через пластинчатые жабры.
Ресничная мантия, края которой срастаются, образует, помимо отвер-
стия для выхода ноги на заднем конце тела, вводное нижнее и выводное
верхнее отверстия. Жабры и мантия образуют, по выражению В. Н. Бек-
лемишева (1952, 1964), вододвижущий аппарат, 'вызывающий ток воды
V-образной формы и достигающий значительно большего совершенства
сравнительно с таковым у Pteriomorphia. В этой группе, помимо питаю-
щихся наннопланктоном, встречаются ксилофаги (например, корабель-
ный червь) и хищники (Рогошуасеа), а также вторичные детритояды
(Tellinacea). Вместе с тем среди Heteroconchia у мелких форм (Erycina-
сеа, или Leptonacea) (Popham, 1940) наблюдается сходство с Protobran-
chia — Nuculidae и Solemyidae (ползание с помощью ноги с ползательной
подошвой, прямой ток воды в мантийной полости, идущий от переднего
к заднему концу тела, и др.).
У ряда форм из Erycinacea, например Chlamy do concha, Entovalva и
других, раковина обрастается мантией, становясь внутренней. У некото-
рых форм, буравящих и сверлящих твердые породы и дерево (Pholadi-
dae, Teredinidae, а также Clavagellidae), образуется вторичный скелет —
пластинки или трубка, одевающая тело моллюска, и происходят руди-
ментация раковины и в связи с этим изменение формы тела и ряда его
мягких частей.
Большое разнообразие наблюдается и в строении замочного края
раковины: от сложного замка с мощными зубами до замочного края со
слабыми зубами, вплоть до их редукции, что было подмечено и исполь-
зовано для классификации всего класса Неймайром (Neumayr, 1883).
Особенно велики разнообразие и необычайность строения и размеров
раковины, как известно, в группе рудистов, среди вымерших надсе-
мейств— Megalodontacea, Diceratacea и Hippuritacea, которые вместе с
современными Chamacea (Сох, 1960в) объединяются в отряд Pachyodon-
tida подкласса Heteroconchia.
Будучи свободноподвижными формами (кроме Chamacea, прикреп-
ляющихся ко дну обычно левой створкой), Heteroconchia активно рас-
пространяются передвижением взрослых особей и пассивно при помощи
переносимых течениями пелагических личинок.
Как было отмечено выше (стр. 151), Heteroconchia сравнительно с
другими подклассами содержат небольшое число древних семейств, появ-
ляющихся не раньше силура, главным образом это молодые мезозой-
ские и кайнозойские семейства.
Палеонтологии пока неизвестны предковые формы Heteroconchia. Это
могли быть симметричные, димиарные Protobranchia. Сохранение у боль-
шинства современных Protobranchia жабр в виде гребенчатых ктенидиев
заставляет производить Heteroconchia не прямо от Protobranchia, а от
их потомков с нитевидными жабрами. Эти предковые формы были свя-
заны не с Агсасеа, обладающими рядом специфических для них черт
анатомии (семейство Arcidae), а скорее, как думал еще Пельзеиер
(1891), с Mytilacea.
Что касается филогенетических отношений между отрядами или
надсемействами Heterodonta, то пока еще трудно их конкретизировать
.154
и можно лишь отметить довольно обоснованные представления о фило-
генетических связях семейств Heteroconchia (или Eulamellibranchia)
Тиле (Thiele, 1935).
ЛОПАТОНОГИЕ МОЛЛЮСКИ
Лопатоногие (класс Scap.hopoda), обладающие признаками как брю-
хоногих, так и двустворчатых,— своеобразные, исключительно морские
моллюски, с трубчатой, узкой, открытой на обоих концах раковиной; жи-
вут в грунте, выставляя из него задний конец раковины; питаются мел-
кими организмами (корненожками, молодыми моллюсками), захватывая
добычу тонкими щупальцами или усиками; дышат с помощью стенок
мантии; во время развития проходят стадии трохофоры и велигера.
Лопатоногие в основном мелкие моллюски от 3 мм до 13 см, в сред-
нем в 2—4 см, если не считать вымершего Antalis raymondi Young из
верхнего карбона Техаса, которая имела почти 60 см в длину.
Лопатоногие распространены (Беляев и др., 1960) в Мировом океане,
особенно в его субтропической и тропической областях, от литорали до
6930—7000 м глубины; и в настоящем, и прошлом — это малочисленная
группа, более частая, начиная с кайнозоя. Класс состоит из двух се-
мейств: наиболее примитивного, древнего Dentaliidae (ордовик или си-
лур— ныне) и более молодого Siphonodentaliidae (мел — ныне), всего
из 5 современных родов, содержащих примерно 200 современных и более
300 ископаемых видов.
Ведя, подобно многим двустворчатым, скрытый, роющий образ жиз-
ни в грунте моря уже с ордовика или силура, лопатоногйе, однако, су-
щественно отличаются от двустворчатых. Они характеризуются головой
со щупальцами, с помощью которых (а не с помощью ресничных токов)
ловят добычу, ротовой полостью с радулой и челюстью, субрадулярным
органом, положением перикардия с сердцем над кишечником (точнее,
позади него), а также положением печени и непарной гонады, открываю-
щейся в правую почку (как у примитивных Diotocardia) в верхней зад-
ней части вытянутого внутренностного мешка. Центральная нервная си-
стема их сходна с таковой низших двустворчатых — Nuculidae, но обла-
дает буккальными ганглиями. В целом организация лопатоногих ближе
к таковой брюхоногих, чем двустворчатых.
Однако под воздействием условий жизни в грунте моря лопатоногие
в отличие от брюхоногих претерпели регрессивные изменения и приоб-
рели ряд специфических особенностей. К первым относятся некоторый
регресс головного отдела, утрата глаз, отсутствие ктенидиев, лакунар-
ный характер кровеносной системы, отсутствие предсердия. Специфиче-
скими особенностями лопатоногих являются трубчатая узкая форма ра-
ковины и мантии, открытых на противоположных концах, но в онтогенезе
временно покрывающих-тело с боков [как в дефинитивном состоянии у
псаммофильных заднежаберных Berthelinia (Tamanovalva)], и затем
срастающихся своими брюшными краями. Для них характерны специа-
лизированная нога в виде копательного органа с конусом и двумя боко-
выми лопастями на конце, щупальцевый аппарат головного отдела,
состоящий из двух пучков тонких щупалец или усиков, и некоторые дру-
гие особенности. Раковина лопатоногих настолько своеобразна, что по
признанию палеонтологов, не будь современных форм, по ней лопато-
ногих нельзя было бы отнести к моллюскам.
Палеонтологическая летопись лопатоногих, несмотря на наличие у
них прочной раковины, очень бедна — это редкие ископаемые.
Лопатоногие обладают рядом черт организации, в том числе двусто-
ронней симметрией, присущих предкам брюхоногих, двустворчатых и
155
лопатоногих — Praerhipidoglossa Плате и Prorhipidoglossa Пельзенера,
Согласно с последними авторами, а также Гроббеном (Grobben,.
1894) и В. А. Догелем (1940а, б), надо признать, что у лопатоногих
больше родства с брюхоногими, чем с двустворчатыми. Противополож-
ного мнения придерживаются Тиле (Thiele, 1935), Ремане (Remane,.
1954, 1959) и Маркус (Marcus, 1958). Однако для решения вопроса о
родстве лопатоногих с этими классами необходимо в помощь морфоло-
гии привлечь методы химии, физиологии и биохимии, прежде всего се-
рологии.
Головоногие моллюски
Головоногие — двустороннесимметричные, исключительно морские,
придонные, чаще пелагические, очень подвижные, хищные моллюски.
Тело их состоит из головы с парой больших глаз и туловища, одетого
мантией, ограничивающей на брюшной стороне мантийную полость, ко-
торая воронкой сообщается с наружной средой. На голове имеется венец
чаще из десяти или восьми рук, обычно с присосками, который иногда
замещается двумя венцами из многочисленных тонких щупалец. Руки
и воронка в онтогенезе развиваются за счет ноги и иннервируются нож-
ными ганглиями. Известковая раковина у огромного большинства ис-
копаемых форм — наружная, многокамерная, с сифоном, а у небольшой
части ископаемых и у современных форм — внутренняя, на разных ста-
диях регресса вплоть до полного исчезновения. В полости мантии на-
ходится симметричный мантийный комплекс органов: две или одна пара
ктенидиев, медиальное анальное отверстие и парные половые и почеч-
ные отверстия. Высокоразвитая нервная система состоит из сложного
головного мозга, заключенного в хрящевой череп, системы нервов, глаз
сложного строения и других рецепторов. У них имеется хорошо диффе-
ренцированный кишечник с мускулистой глоткой и роговыми челюстями
и радулой, слюнные железы, печень и поджелудочная железа, а также
чернильный мешок. Кровеносная система почти замкнутая и состоит из
сердца с желудочком и четырьмя или чаще с двумя предсердиями, на-
ходящегося в целомическом перикардии, и из системы артерий, вен и
капилляров. Выделительная система представлена реже четырьмя, чаще
двумя почками (целомодуктами). Целомическая полость тела обшир-
ная, реже единая, чаще подразделена на перикардий и полость, содер-
жащую гонады. Головоногие — раздельнополы, нередко с половым ди-
морфизмом. Яйца очень богаты желтком, дробление дискоидальное, раз-
витие прямое, реже со свободной личинкой. Головоногие — очень под-
вижные животные, быстро плавающие реактивным способом при помо-
щи воронки или также благодаря плавникам, могут ползать и с помощью
рук. Современных видов — 400—700, ископаемых (начиная с кембрия)
примерно 13 000. Современные головоногие по строению, физиологии и
поведению относятся к наиболее высокоразвитым беспозвоночным.
Класс состоит из двух подклассов — Ectocochlia (наружнораковин-
ные) и Endocochlia (внутреннераковинные). Сохранение для Ectocochlia
названия Tetrabranchia по наличию двух пар ктенидиев у современного-
Nautilus (реликтовой формы некогда мощного ствола наутилид) неосто-
рожно, так как число ктенидиев у другого мощного ствола аммоноидей
неизвестно. Нет основания сохранять название Dibranchiata для белем-
нитов и современных головоногих (Endocochlia), так как, по-видимому,
ряд палеозойских Ectocochlia, возможных предков Endocochlia, имел
пару жабр. Классификаций наружнораковинных, как известно, много-
(Moore a. all., 1952; Kummel, 1954, 1955; Miller a. Furnish, 1954; Schin-
dewolf, 1954; Руженцев, 1949, 1962; Шиманский, 1957, 1960, 1962; Ши-
манский и Журавлева, 1961). В общем классификация основана на осо-
бенностях строения раковины и сифона. Я придерживаюсь классифика-
156
ции, принятой в работах советских палеонтологов, главным образом
В. Е. Руженцева и В. Н. Шиманского, стоящих на строго материалисти-
ческой концепции развития и наиболее последовательно и полно исполь-
зующих результаты изучения взаимозависимости между онтогенезом
.и филогенезом. Ими разработана следующая система.
1-й надотряд Nautiloidea, кембрий — ныне, включает около 600 родов
и около 2500 видов. Раковина — от прямой до спиральносвернутой и сфе-
рической. Первая камера обычно колпачковидная, сифон у большин-
ства— узкий, реже широкий с внутрисифонными отложениями, началь-
ная часть сифона узкая. Надотряд состоит из 8 отрядов: Volborthellidа,
Ellesmeroceratida, Oncoceratida, Discosorida, Nautilida, Orthoceratida,
Ascoceratida, Tarphyceratida.
2-й надотряд'—Endoceratoidea, ордовик — возможно, силур, около
70 родов. Раковина чаще совершенно прямая, иногда гигантских разме-
щав. Первая камера колпачковидная. Сифон очень широкий, видимо,
является выростом тела, включающим сифон и некоторые органы (в ис-
копаемом состоянии заполнен известковыми эндоконами). Начальная
его часть, как правило, широкая, иногда он занимает всю начальную
.камеру.
3-й надотряд — Actinoceratoidea, ордовик — карбон, около 500 родов.
.Раковина чаще прямая, иногда немного согнутая, может достигать зна-
чительных размеров. Первая камера колпачковидная. Сифон чаще ши-
рокий, с системой из продольных и радиальных «сосудов». Начальная
часть сифона широкая.
4-й надотряд — Bactritoidea, ? силур, девон —пермь, около 20 родов.
Раковина прямая, реже согнутая, небольшая. Первая камера сфериче-
ская, возможно, иногда колпачковидная. Сифон узкий от начала до
конца.
5-й надотряд — Ammonoidea, девон — мел, около 1600 родов (примерно
9000 видов). Раковина чаще спирально свернутая, реже вторично непра-
вильная или прямая. Первая камера сферическая. Сифон узкий от само-
го начала и до конца. В нем различают пять отрядов; Agoniatitida, Goni-
-atitida, Clymeniida, Ceratitida, Ammonitida. Отряд Agoniatitida (поздний
девон — поздний триас) включает 96 родов; раковина плоскоспиральная,
разнообразной формы, сифон вентрально-краевой, лопастная линия аго-
-ниатитового типа. У гониатитов (Goniatitida, средний девон — поздняя
пермь) 183 рода, раковина плоскоспиральная, разнообразной формы,
сифон вентрально-краевой, лопастная линия гониатитового типа. У Cly-
meniida (поздний девон — основание раннего карбона) 33 рода, раковина
плоскоспиральная, преимущественно эволютная, сифон — дорсально-
.краевой. У Ceratitida (ранняя пермь —поздний триас) 395 родов, сифон
на ранних стадиях роста может быть центральным, у взрослых — вент-
рально-краевой. У Ammonitida (юра — мел) 981 род, раковина разнооб-
разной формы, от эволютной до инвалютной, сифон на ранних стадиях
.роста центральный, у взрослых — вентрально-краевой.
Подкласс Endocochlia, к организации которого относится большая
часть данной выше характеристики, делится на надотряд Decabrachia с
•четырьмя отрядами — Belemnoida, Teuthoida, Vampyromorpha и Sepio-
ida — и надотряд Octobrachia с отрядом Octopoda. У белемнитов, ранний
карбон — эоцен?, рук было десять, раковина часто прямая, сильно обыз-
вествленная, с сифоном, состоит из фрагмакона, разделенного септами
на камеры, простракума и рострума.
У Teuthoida, известных с верхней перми, наиболее многочисленной
труппы современных головоногих, раковина, простракум, или гладиус в
виде тонкой роговой пластинки, руки с присосками и крючьями.
У Sepioida, известных с ранней юры, известковая раковина, иногда эндо-
.тастрически завернутая, чаше в виде овальной пластинки, состоящая из
157
Рис. 38. Продольный разрез эмбриональ-
ной камеры и первого оборота раковины
Medlicottia orbignyl (по Шульга — Не-
стеренко).
известкового фрагмакона, простракума и рострума. Руки только с
присосками. У Vampyromorpha, несомненные ископаемые представители
которых пока не установлены, раковина в виде тонкой, почти прозрач-
ной плоской пластинки без извести. Одна пара рук редуцирована до
нитевидных придатков. Отсутствует чернильный мешок. У осьминогов,,
известных с раннего мела, обладающих восемью руками с присосками,
раковина в виде двух тонких конхиолиновых палочек, чаще отсутствует
вовсе, кроме самок Argonauta с вто-
ричной, спирально свернутой рако-
виной.
Наиболее долговечной, древней
(ранний кембрий — ныне), прими-
тивной и разнообразной группой го-
ловоногих являются наутилоидеи, от
которых произошли все остальные
головоногие. Самые древние наути-
лоидеи (ранний кембрий, а по
Л. Ш. Давиташвили, 1958, и сред-
ний) — это мелкие, около 1 см, с
прямой раковиной Volborthella.
Большой перерыв в нахождении
Volborthella и ранних представите-
лей отряда Ellesmeroceratida не по-
зволяет связать филогенетически эти два низших отряда наутилоидей.
Поэтому пока приходится считать отряд Volborthellida боковой вымер-
шей ветвью родоначального ствола головоногих, а отряды слабо согну-
тых и свернутых наутилоидей—потомками примитивного, обширного,,
мощно развитого в ордовике отряда Ellesmeroceratida.
Направление эволюции наутилоидей — от слабо изогнутых, к сверну-
тым формам с несоприкасающимися оборотами эволютной раковины и
к плотно спирально свернутым, инволютным раковинам — протекало не-
просто от слабо изогнутых к спирально свернутым формам. Исторически
процесс увеличения инволютности раковины, по В. Н. Шиманскому
(Руженцев и Шиманский, 1954), сопровождался изменением онтогенеза, .
и в том числе изменением начальной части раковины наутилоидей, на-
зываемой этими авторами у наутилоидей эмбриональной раковиной, и
уменьшением умбонального отверстия. Следует подчеркнуть, что под
термином эмбриональная раковина у моллюсков подразумевается кути-
кулярная или конхиолиновая, в той или иной степени обызвествленная
раковина личинки велигера или соответствующей зародышевой стадии
брюхоногих и двустворчатых моллюсков. Она находится на спинной
стороне личинки и ей гомологичен протоконх ископаемых головоногих..
Начальная же часть раковины с жилой камерой (рис. 38) и несколькими
воздушными камерами молодой особи наутилоидей и аммоноидей гомо-
логична раковине молодого брюхоногого и двустворчатого моллюска, в.
ней помещается их тело. Различие заключается в том, что с ростом мо-
лодых особей брюхоногих и двустворчатых моллюсков, как правило,
раковина растет, не изменяясь в форме. У прямых наутилоидей рост ра-
ковины сопровождается увеличением числа воздушных камер, а у спи-
рально свернутых наутилид и аммоноидей происходят не только рост
раковины и увеличение числа воздушных камер, но и изменение строения •
(формы) раковины. Замечу, что у одних современных форм, например
брюхоногих, продиссоконх сохраняется, у других — отпадает, подобно-
тому, что наблюдается и у ископаемых головоногих.
Прямые и слегка изогнутые формы, известные с кембрия, достигшие
уже в ордовике расцвета, были и абсолютно и относительно господствую-
щими среди моллюсков того времени (ордовик, силур — девон). Это
158
были обитатели прибрежных небольших глубин моря, сравнительно под-
вижные, хотя и не очень поворотливые хищники, нередко достигавшие
громадных размеров, 4—4,5 м в длину. С эволюцией других беспозвоноч-
ных обитателей моря и с изменениями условий-жизни в. палеозое числен-
ность прямых и согнутых наутилоидей стала уменьшаться, и в середине
триаса вымер отряд Orthoceratida (=Michelinoceratida Flower). Начиная
Рис. 39. Схема примерного количест-
венного распределения наутилоидей
(из A. Miiller)
Рис. 40. Схема примерного количе-
ственного распределения аммонои-
дей— 1485 родов (из, A. Miiller)
с ордовика, вместе с прямыми и согнутыми формами продолжается раз-
витие спирально свернутых наутилоидей, которые с правом считают
также происшедшими от Ellesmeroceratida. Нет единого мнения относи-
тельно происхождения отряда Nautilida. По одним авторам (Moore а.
all., 1952, и др.), отряд Nautilida возник в ордовике, т. е. существует-
9 геологических периодов или около 440 миллионов лет, по другим—•
с девона (Шиманский, 1957), т. е. 7 геологических периодов или около
320 миллионов лет. Во всяком случае, это самый долговечный отряд
наутилоидей, проделавший эволюцию от кривых, спирально свернутых
эволютных до инволютных форм. Эта эволюция раковин сопровождалась
превращением простых изогнутых перегородочных линий примитивных
форм '.в извилистые, с лопастями и седлами, что необходимо для боль-
•шей прочности раковины и большей подвижности наутилид, У поздних
(мезозойских) наутилид образуются перегородочные, как отмечает
В. Н Шиманский, почти аммонитовые линии.
Эти важные приспособительные изменения раковин, очевидно, свя-
занные с изменениями строения свернутых наутилоидей отряда Nautili-
da, происходили -в условиях изменения среды во второй половине палео-,
зоя и в мезозое. Как известно, расцвет этого отряда, включающего более-
трех десятков семейств, наблюдался в каменноугольном периоде, значи-
тельного наибольшего разнообразия он достигал в триасе, но из четырех
позднейших семейств — Hercoglossidae, Cymatoceratidae, Aturidae и Nauti-
159-
lidae — лишь последнее выживает и ныне представлено реликтовым ро-
дом Nautilus — единственным современным наружнораковинным голо-
воногим (рис. 39).
Возникновение аммоноидей из бактритоидей сопровождалось свер-
тыванием раковины, вывести же их от наутилоидей требует признания
изменения ряда особенностей строения последних. Эволюция аммоноидей
(Руженцев, 1957а, 19576, 1960, 1962), живших в течение 6 геологических
периодов (рис. 40), начало девона — конец мела, т. е. 240 миллионов лет,
проходила несколькими этапами, включающими подъем, процветание,
связанное с совершенствованием строения, разнообразием экологии и
распространением по всему Мировому океану, кризисы и вымирание
разных отрядов. В общем периоды процветания и кризисов аммоноидей
-отчетливо обусловлены особенностями геологической истории Земли и
связаны с изменениями палеогеографии, температуры, сменами морских
трансгрессий и регрессий, и т. д. Внезапное исчезновение аммонитов в
конце мезозоя связывают с глубокими общими изменениями условий
жизни на Земле. Периоды упадка и вымирания аммоноидей в разное вре-
мя были настолько глубокими, что, например, в конце перми, в связи с
изменением климата, вымирают все, кроме 7 родов аммоноидей, в конце
триаса сохраняются, по одним авторам, всего несколько видов рода
Phylloceras (точнее — Monophyllites'), по другим — 5 родов и 5 видов,
тогда как остальные аммониты вымирают. Эти периоды упадка и выми-
рания, как известно, чередуются с длительными периодами расцвета.
Так, один из 7 переживших пермь родов в благоприятных условиях
триаса дает начало 370 родам, а из единственного рода аммонитов
(Monophyllites), относящегося к семейству Phylloceratidae, развилось
735 юрских и 493 меловых родов аммонитов. Часть этих семейств и родов
аммонитов существовала относительно недолго — лишь в юре, другие
продолжали существовать, эволюировать и дожили до конца мела, до-
стигнув максимального размаха в смысле прогрессивного развития вели-
чины, строения, разнообразия и численности форм и регресса, выражен-
ного в упрощении формы раковины, скульптуры последней, формы лопа-
- стной линии и т. д. (A. Muller, 1961).
Характерными чертами эволюции аммоноидей являются следующие:
постепенное развитие и усложнение лопастных линий от простых у ниж-
недевонских агониатитов до крайне сложных у мезозойских форм, упро-
щение и регресс их в периоды кризисов, сменявших периоды процвета-
ний, а также филетическое увеличение размеров раковин. Последнее,
как указывает Мюллер (A. Muller, 1958), выражается в том, что размеры
большинства девонских и каменноугольных аммоноидей лишь редко
превышают 7,5 см в диаметре, при относительной редкости крупных
форм (35 см в диаметре), тогда как в конце палеозоя увеличиваются
•средние размеры и относительное число крупных форм. Увеличение раз-
меров раковины продолжается, особенно в триасе, например, Pinacoceras
'из верхнего триаса имел 80 см в диаметре, а в юре и мелу достигал еще
-большего размера; в мелу ,в ряде семейств (Desmoceratidae, Diplocerati-
dae) многие виды достигали 120 см в диаметре, a Pachydiscus? seppenra-
densis даже 255—300 см.
Развитие лопастных линий, точнее, перегородок между камерами у
аммоноидей в геологической истории их имеет не отвлеченное, но очень
важное конкретное, жизненное значение, как и сама форма их раковины.
Считают, что форма спирали раковины аммоноидей причинно связана
с характером их передвижения (Чернов, 1922). Плоская форма ракови-
ны лучше всего приспособлена для плавания, раздутая-—для ползания.
Перегородки служили для укрепления раковины и для более прочного
-прикрепления к раковине тела, что особенно было важно для крупных
форм. При быстрых движениях аммоноидей и в известной мере и наути-
160
лоицей — все это вместе с наличием в камерах раковины газа, имело
важное приспособительное значение для этих хищных моллюсков.
По соображениям В. Е. Руженцева (1957а, 19576, 1960, 1962), во всех
отношениях обоснованных гораздо лучше, чем таковые зарубежных па-
леонтологов, эволюция и филогения аммоноидей начинается в нижнем
девоне, когда от наутилоидей или бактритид произошли низшие прими-
тивные агониатитиды (ранний девон — поздний триас). Последние про-
делали довольно сложную эволюцию (в девоне и перми) и, достигнув
триаса в лице немногих высоко организованных форм, вымерли в верх-
нем триасе. Гониатитиды, происшедшие в среднем девоне от агониатитид,
обладавшие простой, но все же более сложной организацией, чем аго-
ниатитиды, претерпели значительную эволюцию и, дав много форм в
карбоне и перми, вымерли в верхней перми. Отряд климений, происшед-
ший в девоне также от агониатитид, достиг значительного развития в
верхнем девоне и в конце девона вымер, кроме двух .семейств, просуще-
ствовавших недолгое время в основании нижнего карбона. Древнейшие
цератитиды с простой организацией произошли в конце перми от аго-
ниатитид.
По сравнению со всеми предыдущими отрядами цератитиды значи-
тельно эволюировали, в преобладающей массе они были представлены
высоко организованными формами с очень сложной лопастной линией и
с плоскоспиральной разнообразной формы раковиной. Цератитиды про-
цветали в триасе и вымерли на границе триаса и юры. Отряд аммонитов,
на раннем этапе своей истории очень близкий к цератитидам, произошел
от примитивных, нижнетриасовых цератитидов. Аммониты, во время
расцвета цератитидов игравшие в триасовых морях незаметную роль,
постепенно эволюировали. Они, как отмечают палеонтологи, проделали
в юре, особенно в мелу, блестящую эволюцию, трагически закончившую-
ся их полным вымиранием в конце мела.
Геологическая история головоногих говорит о наутилоидеях как о
родоначальной группе всех остальных вымерших и современных форм.
Хотя промежуточные формы, связывающие аммоноидей с наутилоидея-
ми, пока не найдены, первые могли произойти только от вторых. В поль-
зу этого говорит то, что древние аммоноидей обладали рядом важных
черт, общих с наутилоидеями: так перегородки у них вогнуты вперед, си-
фонные трубки обращены назад, как у наутилоидей. Характерные черты,
определяющие надотряд аммоноидей и отличающие его от надотряда
наутилоидей, обладающих своими характерными чертами строения и
развития, сформировались в обоих надотрядах в ходе эволюции. Долгое
время самым важным признаком отличия аммоноидей от наутилоидей
считалось сохранение эмбриональной раковины, или протоконха, однако
теперь наличие протоконха установлено и у наутилоидей (Orthoce-
ratida.).
•Одни палеонтологи считают, что аммоноидей, число описанных видов
которых достигает 9000, произошли от свернутых наутилоидей с волни-
стыми перегородками и краевым сифоном, другие производят их от
бактритоидей, полагая, что промежуточным звеном между Bactrites и
аммоноидеями был первый представитель аммоноидей Anetoceras из ниж-
него девона, имевший неплотно свернутую на всем протяжении раковину
(Moore a. all., 1952; Руженцев, 1960). Наличие протоконха у аммоноидей
и большинства бактритоидей и большое сходство перегородочной линии
ряда девонских бактритоидей и наиболее древних аммоноидей, наличие
'вентральной лопасти, строение и положение сифонных дудок и, по-вици-
мому, отсутствие соединительных колец считаются общими чертами бак-
тритоидей и древних аммоноидей. «Узловой группой форм» являются,
по В. Н. Шиманскому, Lobobactrites, Cyrtobactrites, Anetoceras, Teicherti-
ceras, Gyroceratites.
11 Д. M. Федотов
161
Однако и эти палеонтологи (например, В. Н. Шиманский), считают
бактритоидей происшедшими от наутилоидей (Ellesmeroceratida). Пере-
числяя характерные черты строения бактритоидей (прямая или согнутая
раковина, сферический или конический протоконх, почти прямая или
с двумя лопастями перегородочная линия, небольшая лопасть на сифо-
нальной стороне, сифонные дудки, обращенные назад, краевой сифон
только с органической тонкой оболочкой, без спикулярной части), Ши-
манский считает .бактритоидей самостоятельным надотрядом, от кото-
рого он и выводит аммоноидей. Наличие протоконха, краевое положение
сифона и вентральная лопасть являются уертами бактритоидей, общими
с таковыми аммоноидей, раковина же их имеет внешнее сходство с рако-
виной наутилоидей. В. Н. Шиманский подчеркивает экологическую важ-
ность для эволюции аммоноидей образования из прямой раковины бак-
тритоидей спирально свернутой раковины.
По мнению ряда наших и зарубежных палеонтологов, бактритоидеи
дали начало и белемноидеям, имеющим с ними ряд общих черт
(В. Н. Шиманский), по мнению других, белемноидеи произошли от
Orthoceratida.
По этому поводу следует, однако, сказать, что невероятно, чтобы
спирально свернутые, проделавшие блестящую эволюцию и полностью
вымершие в конце мела аммоноидей и прямые внутреннераковинные,
сравнительно малочисленные в прошлой истории Земли белемноидеи и
их потомки — современные головоногие, наиболее высоко организован-
ные из всех групп беспозвоночных животных, были связаны близким
родством друг с другом.
Мне представляется более вероятной мысль о том, что белемноидеи
связаны родством с ортоцератидами, как думали некоторые палеонто-
логи, а может быть и с бактритоидеями. В истории всех этих групп не
было этапа развития спирально свернутой раковины. Наутилоидеи на-
чали свою эволюцию в виде форм с прямой или согнутой раковиной, а
затем в процессе эволюции в разных ветвях возникали формы согнутые,
свернутые в свободную или плотную спираль, иногда вторично развер-
нутые раковины. Аммоноидей начали свою эволюцию значительно позже
наутилоидей и имели исходно спиральную, хотя и неплотно свернутую
раковину, т. е. стояли на более высоком уровне развития, чем наутилои-
деи; в дальнейшем стали преобладать формы с компактно спирально
свернутой раковиной, а с триасй появились формы с вторичнопрямой
раковиной.
Вся геологическая история наружнораковинных головоногих говорит,
следовательно, о том, что спирально свернутая форма наружной рако-
вины и, очевидно, связанная с ней внутренняя организация оказа-
лись нежизненными. Из громадного, относительно разнообразного мира
наружнораковинных головоногих с их блестящей историей в прош-
лом от наутилоидей, как известно, сохранился один современный ре-
ликтовый род Nautilus, а все аммоноидей вымерли. Недавние находки
белемноидей в нижнем карбоне, а если был прав Конинк, то уже и в
девоне (Flower a. Gordon, 1959; Шиманский, 1960), заставляют ду-
мать, что они возникли от неспециализованных форм прямораковин-
ных примитивных наутилоидей и в близком родстве с аммоноидеями
не были.
Сравнительно с длинными прямыми и кривыми раковинами палео-
зойских наутилоидей развитие у наутилид и аммоноидей компактных
спирально свернутых раковин было важным приспособительным дости-
жением. Но эксцессивное развитие наружной раковины у позднейших
наутилид и, особенно, у аммонитов (вызванное появлением более конт-
растных условий жизни в конце мезозоя, в том числе изменением темпе-
ратуры, и обогащением фауны моря тоже хищными, но гораздо более
162
подвижными формами) сыграло известную роль в вымирании этих групп.
Тщательные палеогеографические, син- и аутопалеоэкологические иссле-
дования смогут пролить известный свет на причину вымирания аммони-
тов в верхнем мелу.
Однако надо иметь в виду, что в этот период наблюдалось вымирание
многих других групп, не только морских, но и наземных животных и это
является одной из проблем науки о Вселенной и Земле, а не только био-
логии.
Нижнекаменноугольные белемноидеи,
появившиеся, очевидно, еще ранее, судя
по работе Фл'ауера и Гордона (Flower а.
Gordon, 1959), обладали типичной для
этого отряда раковиной. Перестройка на-
ружнораковинных предков белемноидей,
по-видимому, путем обрастания ракови-
ны мантией, с превращением ее во внут-
реннюю, а в ходе эволюции и облегчение
раковины, не является чем-то необычным
среди моллюсков. У современных хитонов,
двустворчатых и, особенно, у брюхоногих
обрастание раковины мантией, превраще-
ние ее во внутренний скелет и разная
Рис. 41. Схема примерного коли-
чественного распределения иско-
паемых двужаберных моллюсков
(из A. Miiller) ,
степень ее редукции наблюдаются доста-
точно часто. Облегчение раковины, несом-
ненно, происходило и у белемноидей, что
способствовало увеличению их подвиж-
ности, дававшей им большие преимуще-
ства перед их современниками наутилоидеями и аммоноидеями. Мало-
численность палеозойских белемноидей и меньшая их привязанность к
отложениям осадков делают геологическую историю белемноидей очень
неполной. Будучи в палеозое мало распространенными, они стали мас-
совыми формами в мезозое, особенно в юре и мелу, достигая громадных
размеров (2—2,5 м в длину); вымерли они в эоцене. Большинство из
2500 видов белемнитов приходится на долю мезозоя.
Благодаря наличию хорошо развитой раковины геологическая исто-
рия белемнитов выяснена довольно полно (рис. 41). Значительно беднее
фактами геологическая история остальных внутреннераковинных, так
как вследствие облегчения и регресса раковины ископаемые их остатки
неполны и редки. И все же эволюция и филогения современных Endoco-
chlia, особенно благодаря работам Нэфа (Naef, 1931), в основном выяс-
нены главным образом именно по особенностям строения раковины.
У сепий (Sepioida), известных с нижней юры, в редуцированной раковине
сохраняются части известкового фрагмакона с септами, простракума и
рострума, позволяющие сравнивать ее с раковиной белемнитов. Исклю-
чением является Spirilla со спирально свернутой, хорошо развитой рако-
виной. Раковина Teuthoida, наиболее богатой в современной фауне груп-
пы, подвергается еще большему регрессу и представлена тонкой пластин-
кой (гладиусом), соответствующей простракуму белемнитов и, кроме
того, рудиментом фрагмакона и рострума. У некоторых форм, например
Moroteuthis (Кандаков, 1940; Акимушкин, 1963а, 19636; Кабанов, 1964),
рострум достигает значительной величины. Эта группа головоногих изве-
стна с верхней перми. Для Vampyromorpha несомненных представителей
в ископаемом состоянии пока не обнаружено. Рудимент раковины — гла-
диус в этой группе представляет собой тонкую почти совершенно про-
зрачную плоскую без извести пластинку. У осьминогов, наконец, от рако-
вины остаются лишь две тонкие конхиолиновые палочки, чаще же рако-
вина исчезает нацело. Ископаемые осьминоги известны с верхнего мела.
11*
163
По состоянию раковины к белемнитам наиболее близки Sepioida.
У Teuthoida и Vampyromorpha раковины претерпели более значительные
изменения, хотя по общей организации Vampyromorpha боле.е примитив-
ны. Обособление Teuthoida и Vampyromorpha от общего родоначального
ствола Endocochlia, видимо, исторически произошло раньше обособления
Sepioida.
Данные о строении раковины и положении в жилой камере животного
у Nautilus в основном отвечают тому, что наблюдалось у ископаемых
Nautiloidea и Ammonoidea. Все они, как головоногие, очевидно, имели
воронку и венец тонких, сократимых щупалец вокруг рта, сформировав-
шихся за счет ноги, и были свободноподвижными, придонными и пелаги-
ческими животными. С помощью воронки они могли относительно' быст-
ро, насколько позволяли форма и вес раковины, облегчаемой, правда,
газовыми камерами, двигаться реактивным способом в придонных слоях
воды, тогда как для передвижения в вертикальном направлении в толще
воды они пользовались раковиной как поплавком, регулируя количество
газа в камерах. Судя по наличию у современного наутилуса воронки из
двух половинок, складывающихся для движения в коническую трубку,
такая же воронка, вероятно, была у большинства вымерших Ecdocochlia.
Считают (Donovan, 1964), что /древние Ecdocochlia с известковой рако-
виной, имеющей многие газовые камеры, и с сифоном были хорошими
пловцами и, очевидно, самыми древними плавающими из Metazoa.
Так как у некоторых вымерших белемнитов обнаружены остатки
10 рук, то следует предположить, что у них были руки как и у современ-
ных Endocochlia. Вполне вероятно, что белемниты обладали цельной ко-
нусовидной воронкой, более совершенной по строению и функционирова-
нию, чем воронка из двух несросшихся половинок их предковых форм.
Они пользовались воронкой для 'реактивного движения и (по аналогии
с современными головоногими) с помощью плавников двигались голо-
вой вперед. Слабая привязанность ископаемых белемнитов к осадкам
говорит о том, что они были уже хорошими пловцами, более активными
и быстрыми, чем вымершие Ectocochlia, хотя прямая, тяжелая раковина
их, сравнительно с раковиной современных Endocochlia, вероятно, огра-
ничивала подвижность этих моллюсков.
В течение всей своей геологической истории, начиная со времени их
нахождения в раннем кембрии, головоногие были свободноподвижными
придонными, плавающими животными; как крайне редкое исключение
встречалось прикрепление к субстрату, вроде верхнемелового Nipponites.
Они были хищными, активными животными. Просуществовав не менее
десяти геологических периодов, они, вероятно, жили уже в докембрии,
т. е. более 500 миллионов лет, большая часть головоногих сошла со сце-
ны, хотя и дала начало очень высоко организованным современным
формам.
Для наутилоидей и аммоноидей не приходится сомневаться в важ-
ном приспособительном значении их наружной раковины как Органа
пассивной защиты. Большое разнообразие формы и строения раковины
говорит о значительной пластичности и хорошей приспособленности к
условиям жизни этих групп, составлявших важный и многочисленный
элемент морской фауны палеозоя и мезозоя. Сравнительно малоподвиж-
ные прямые и согнутые и даже спирально свернутые толстораковинные
наутилоидеи были в известной степени защищены от врагов-хищников.
Важным шагом в прогрессивном развитии аммоноидей было развитие
инволютной тонкостенной раковины со сложными перегородками, более
легкой и более совершенной, чем раковина наутилоидей, допускавших
значительно большую подвижность. В ходе геологической истории Зем-
ли, с эволюцией и усложнением животного мира в позднем палеозое и
.164
Рис. 42. Схема эволюции раковины у Ecto-
cochlia (из Hennig)
А, Б—Belemniies (ископаемые); В—Bellosepia (ис-
копаемые); Г—Sepia’, Д—Ommastrephes', В—Loligoni-
dae: 1—проостракум; 2—фрагмокон; 5—рострум;
4—сифон; 5—септы
особенно в мезозое, фауна позвоночных моря обогащалась хищниками,
которые, несомненно, уничтожали тогдашних головоногих.
Крупнейшим шагом в эволюции головоногих (легко заметным уже в
конце мезозоя и достигшим апогея в настоящее время) была посте-
пенная редукция раковины, ставшей внутренней уже у белемноидей. Как
показано Г. В. Зуевым, по существу, это были изменения формы строе-
ния и функции раковины (ее эволюция), связанные с образом жизни
придонно-пелагических ка-
ракатиц (Sepioida), нектон-
ных кальмаров (Teuthoida)
и придонных осьминогов
(Optopoda). Округло-оваль-
ная, плоская, значительных
размеров раковина карака-
тиц является щитом, а жид-
кость и газ между известко-
выми пластинками ракови-
ны придают ей значение гид-
ростатического аппарата. У
нектонных, стремительно
плавающих кальмаров рако-
вина редуцируется (рис. 42),
превращаясь в тонкую уз-
кую хитиновую пластинку,
не стесняющую гибкость те-
ла кальмара и его подвиж-
ность. Наконец, у придон-
ных осьминогов от раковины
остаются две тонких палоч-
ки или чаще она исчезает,
что позволяет осьминогам
забираться в узкие щели
между камнями: спасаться
от врагов и подстерегать до-
бычу.
Все это способствовало прогрессивному развитию головного мозга,
органов чувств и рецепторов. Современные головоногие—активные хищ-
ники, они хорошо ориентируются в обстановке, быстро реагируют и на
добычу, и на врага (Акимушкин, 1963а, 19636). Основы всего этого были
подготовлены в течение длительной геологической истории предков сов-
ременных головоногих, эволюционное развитие которых в настоящее
время достигает необычайно высокого уровня. У современных головоно-
гих, очень подвижных, активных животных, разнообразятся способы
движения и изменяются связанные с этим органы. Руки становятся много-
функциональными органами и употребляются для ловли добычи, схва-
тывания врага, передвижения по дну моря и как щупальца, а у осьми-
ногов— и для перетаскивания камней при постройке жилища. У всех,
кроме Vampyromorpha (Pickford, 1939), одна из рук (гектокотиль) слу-
жит для оплодотворения самки. Для плавания используются плавники.
Орган основного, реактивного, способа плавания — воронка использует-
ся и для дыхания при смене воды в мантийной полости. В связи с пре-
обладанием того или иного способа передвижения и в связи с условиями
жизни происходит мощное развитие пары ловчих щупалец, сильная
редукция остальных восьми рук, образование мембраны, объединяющей
руки, и т. д. (Jaeckel, 1955). Прогрессивно развивающиеся подвижность
и активность головоногих, замещение наружной раковины предков внут- '
р'енней раковиной, становящейся своего рода внутренним скелетом, .
165
делают современных головоногих в прямом смысле мягкотелыми/ Их
покровы испытывают важные приспособительные изменения защитного
характера. Головоногие, особенно осьминоги, приобретают свойство
быстро изменять окраску тела путем сильных сокращений и растяже-
ний хроматофоров кожи. Приблизительно у 25% всех видов головоногих
Рис. 43. Схема центрального нервного аппа-
рата различных двужаберных головоногих
(вид сбоку)
A—Omastrephes-, Б—Pepiola; B-—Lollgo\ Г—Sepia,
Octopoda; Д—Octopus', Е—Argonauta (из Беклеми-
шева); 1—церебральный ганглий, 2—педальный
ганглий; 3—висцеральный ганглий; 4—брахиаль-
ный ганглий; 5—лабиальный или верхний буккаль-
ный ганглий; 6—нерв воронки; 7—висцеральный
нерв; 8—зрительный нерв; 9—мантийный нерв; 10—
нервы рук; 11—глотка; 12—пищевод
развиваются светящиеся орга-
ны, расположенные на разных
,частях тела. Они освещают до-
бычу и окружающую среду,
служат светящейся приманкой
для добычи, а слизь, выделяе-
мая этими органами, образует
огневую завесу от врага и т. д.
(Акимушкин, 1963а, 19636).
Судя по низкой организа-
ции центральной нервной си-,
стемы у реликтового предста-
вителя наружнораковинных го-
ловоногих Nautilus, нервные
стволы которого не дифферен-
цировались на ганглии и кон-
нективы, а глаза сохранили
примитивное бокаловидное
строение без хрусталика, эво-
люция нервной системы проис-
ходила лишь у белемнитов и
была, вероятно, незначитель-
ной у наружнораковинных.
У современных головоно-
гих, вследствие укорочения кон-
недтивов и комиссур, происхо-
дит' сближение церебральных,
педальных и висцеральных ган-
глиев и их высокая дифферен-
цировка. В результате у них
образуется громадного объема
головной мозг очень сложного
строения. По совершенству
строения и физиологии голов-
ного мозга головоногие стоят
выше всех беспозвоночных и приближаются к позвоночным (рис. 43).
Двигательные нервы головоногих гигантской длины, огромного диаметра
(до 18 мм), с большой быстротой проведения, почти равной таковой
у позвоночных (Беклемишев, 1964). Мозг заключен в хрящевую кап-
сулу— череп; образуются глазные хрящи для больших глаз, дости-
гающих у крупнейших головоногих до 40 см в диаметре, со своеобраз-
ным приспособлением к аккомодации путем приближения и удаления
хрусталика от ретины. Глаза по своему строению не только не уступают
зрительному аппарату позвоночных, но даже лучше приспособлены к
водному образу жизни (например, напомним наличие отверстия в перед-
ней камере глаза). Зрительные палочки глаз достигают 42 мм в длину
и являются самыми крупными в. животном мире. Высокую гистологиче-
скую дифференцировку оптических центров головоногих и сходство их
с таковыми не только насекомых, но и позвоночных не раз отмечал
А. А. Заварзин (1913, 1950). Характерно, что более новые исследова-
ния показали еще большую сложность и тонкость отправлений оптиче-
ских долей и сетчатки глаз головоногих, наличие у них предметного
166
зрения, близкого к зрению позвоночных (Kalmus, 1959; Young, 1960;
Moody a. Parriss, 1960; .Southerland, 1960), Но глаза головоногих в онто-
генезе возникают не как у позвоночных в виде выпячивания стенки моз-
говых пузырей, а как впячивания эктодермальных -покровов зародыша
(Kaestner, 1954/1955).
В- соответствии с высоким развитием головного мозга и всей нервной
системы, необычайной скорости и точности движений очень высоко раз-
виты рецепторы -—химические, механические, слуховые, равновесия,
обоняния, а также органы движения, особенно плавания. Известно, что
Рис. 44. Схема центральной кровеносной
системы Octopus (из Johansen a. Martin)
1 — аорта; 2 — выносящий жаберный сосуд; 3 —
приносящий жаберный сосуд; 4 — жаберное серд-
це; 5 — желудочек; 6 — жабры; 7—боковая полая
вена; 8 — звездчатый ганглий; 9 — головная
полая вена; 10 — почечная вена
Рис. 45. Схема расположения целома н
мочеполовой системы у головоногих мол-
люсков (из Hennig)
А —четырехжаберные (Ectocochlia); Б — дву-
жаберные (Endocochlia); 1 — гонады; 2 — поч-
ки; 3 — половой проток; 4 — перикардий
ряд форм из отряда Teuthoida по скорости движения не уступают самым
быстрым рыбам, а крупные представители головоногих вступают в борь-
бу даже с морскими млекопитающими.
Редукция раковины у головоногих стоит в связи с развитием внутрен-
него хрящевого скелета, состоящего из головной капсулы, глазных, за-
тылочного и плавниковых хрящей, хрящей основания рук, расположен-
ных позади воронки. Из всех беспозвоночных у головоногих хрящ до-
стигает наивысшего развития и очень сходен с хрящом позвоночных (За-
варзин, 1945, 1947).
Высокого совершенства достигают и другие органы и системы голо-
воногих, в частности кровеносная системы (рис. 44) и кровообращение, с
мускулистым, чисто артериальным сердцем, с артериями и венами, свя-.
занными друг с другом сетью настоящих кровеносных капилляров и об-
ладающими строением, подобно артериям и венам позвоночных. Крове-
носные сосуды настолько прочны, что даже у сравнительно небольшого
осьминога приходится применять напряжение, чтобы перерезать их нож-
ницами. По строению и физиологии кровеносная система достигает очень
высокой степени совершенства (например, у Octopus') и во многом упо-
добляется кровеносной системе позвоночных (Johansen a. Martin, 1962).
Высоко дифференцирован и кишечный тракт, имеются сильные че-
люсти, которые пускаются в ход при защите и для отгрызания куског
от пойманной добычи. Полостное пищеварение (а не внутриклеточное,
обычно распространенное среди многих других моллюсков), протекает
под воздействием секретов и набора ряда ферментов печени и поджелу-
дочной железы. С задней кишкой связан очень своеобразный орган
167
защиты — чернильный мешок, отсутствующий лишь у глубоководных
форм, выпрыскивающий черную жидкость, которая в случае опасности
образует не только дымовую завесу, но и защиту, так как обладает ядо-
витыми и наркотизирующими свойствами. Защитное значение, а не пище-
варительное, имеют и слюнные железы (их задняя пара), секрет которых
ядовит для многих животных (Акимушкин, 1963а, 19636). Они несут
также эндокринную функцию, выделяя тирамин, действующий на тонус
мышц, нервной системы, и на активность хроматофоров (W. Wells а.
J. Wells, 1959 — цит. по Gersch, 1964). Лишь в отношении целомической
полости тела, как было упомянуто раньше, головоногие сохраняют более
примитивные, вернее исходные, отношения сравнительно с большинством
других классов моллюсков, кроме соленогастров. У них сохраняется об-
ширная целомическая полость (рис. 45) в виде сообщающихся друг с
другом перикардиальной и половой полостей или отделов. В первом ле-
жат сердце, жаберные сердца и перикардиальные железы, во втором —
желудок, часть кишечника и гонады.
Наиболее, примитивное состояние целома сохраняется у Vdmpyroteu-
this infernalis, у которого имеется единая обширная полость тела без
перетяжки на половую и перикардиальную, и перикардиальный отдел,
сообщающийся при помощи тонкого канала с апикальным целомом.
У Nautilus половой отдел сохраняется в виде полости тела, в котором
находятся гонады, желудок и петля кишечника и который продолжается
в сифон раковины; перикардиальный отдел, содержащий сердце, обши-
рен. У Sepioida и Teuthoida половой отдел достаточно обширный, в нем
находятся гонада и желудок, в перикардиальном, менее значительном
отделе лежит сердце, жаберные сердца и. перикардиальные железы.
У осьминогов целом сильно редуцирован, сердце лежит в паренхиме, но
полость полового отдела все же довольно обширна, и сообщение между
перикардиальным и половым отделами сохраняется. Я не останавли-
ваюсь на связях между целомом, целомодуктами, почками и половыми
протоками — это хорошо установлено.
Напомню, что крупные, богатые желтком яйца головоногих претер-
певают поверхностное, неполное дробление, без следов спирального де-
терминированного, существование которого у них допускается лишь из
принадлежности их к типу моллюсков и из филетического единства этого
типа. Наряду с преобладающим и характерным прямым развитием, от-
сутствующим у всех других классов моллюсков, у части головоногих,
как было сказано раньше, имеются мелкие яйца и развитие протекает
с помощью личиночных пелагических стадий.
Число современных видов головоногих всего 400—700; это очень мало
против 2500 видов белемнитов, особенно сравнительно с числом видов
современных двустворчатых и брюхоногих, которые исчисляются нес-
колькими десятками тысяч, и лишь немного превосходит численность
бедного видами класса лопатоногих. И вместе с тем во всех отношениях,
кроме строения целомической полости, головоногие, как известно, дости-
гают наивысшего уровня организации и физиологии не только в преде-
лах моллюсков, но и среди всех других типов беспозвоночных.
Современные головоногие по. размерам тела (от 1 см в длину у Idio-
sepius pygmaeus Steenstrup до 20—22 м, считая с руками, у Architeuthis}
стоят на одном из первых мест в животном мире.
Заключительная часть
Основными чертами строения родоначальной формы моллюсков яв-
ляются следующие: двусторонняя симметрия, нежные, мягкие покровы
тела со слизистыми клетками, ортогональный характер нервной систе-
мы, целомического происхождения перикардий с сердцем, кишечник с
168
радулой и парной печенью, пара ктенидиев, отверстия: анальное, пары
гонад и пары выделительных органов, расположенные в заднем конце-
тела и тесно связанные друг сдругом. Эта характеристика близка к той,,
которую дал в свое время Шпенгель (Spengel, 1881). Наличие большин-
ства этих черт, не считая раковины, у всех классов моллюсков говорит
бесспорно о монофилии данного типа..
Предковые формы моллюсков, надо думать, были придонными мор-
скими червеобразно вытянутыми животными с нежной кожей, выделяв-
шей слизь для защиты мягкого тела. Они имели в покровах известковые
спикулы, двигались при помощи сокращения кожно-мускульного мешка
и слизи, жили в спокойных мелких водах и питались мелкими организ-
мами. Спиральное, детерминированное дробление яиц и пелагическая ли-
чинка-трохофора— были основными чертами их индивидуального раз-
вития. Это были вторичнополостные животные, но в отличие от аннелид,
ab origine нечленистые и не обладавшие бесполым размножением и ре-
генерацией. Примерно на таком уровне внешнего строения и внутренней
организации стоят соленогастры, в онтогенезе которых нет признаков
вторичного упрощения, поэтому их и.надо считать наиболее близко стоя-
щими в родословном древе моллюсков к исходной родоначальной форме
этого типа. К сожалению, из-за отсутствия у них раковины, пока ничего
не известно о прошлом вымерших соленогастров. Не исключено, что их
жизнь на колониях полипов является древним признаком.
В ходе эволюции червеобразных придонных предковых форм мол-
люсков, при ползании на брюшной стороне по дну -моря последняя в ре-
зультате формативных раздражений становилась более сократимой, чем
спинная сторона тела. Происходило1 усиление мускулатуры кожно-мус-
кульного мешка брюшной стороны тела: она дифференцировалась и обо-
собилась от мускулатуры спинной стороны тела. Брюшная сторона ста-
новилась толще, усиливалась ее роль >в движении животного. Внутрен-
ние органы, также испытавшие прогрессивное развитие, оттеснялись к
верхней спинной стороне тела, становившейся выше. Жизнь в прибреж-
ной полосе моря вызывала необходимость защиты внутренних органов
спинной стороны, в результате чего развились складки покровов — ман-
тия, далее, вследствие нежности и мягкости покровов, лишенных прочной
кутикулы, стали усиленно размножаться клетки, образующие известко-
вые спикулы, что в дальнейшем привело к развитию пока еще тонкой
раковины. Вероятно, на этом этапе эволюции возникли хитоны. Подобно
соленогастрам, у них имеется одна, низшая для моллюсков личиночная
стадия — трохофора, и развитие тела происходит путем линейного роста
по горизонтальной, передне-задней оси. По многим чертам внутренней
организации хитоны, как известно, еще очень близки к соленогастрам.
Хитоны мало пластичны по строению, развитию и экологии и в настоя-
щее время, и в прошлые геологические эпохи. К сожалению, отсутствие
палеонтологических данных о соленогастрах сильно затрудняет досто-
верное установление путей эволюции хитонов и соленогастров.
После ответвления от общего родословного ствола моллюсков соле-
ногастров и хитонов продолжалось прогрессивное развитие всей органи-
зации примитивных раковинных моллюсков: формирование головного
отдела или головы с ее рецепторами, туловища, одетого на спинной сто-
роне мантией, защищенной раковиной, и сильно мускулистой, сократи-
мой брюшной стороны — будущей ноги. Появляется разнообразие и в
экологии. Надо думать, что на этом раннем этапе исторического разви-
тия примитивных моллюсков, до сформирования за счет части брюшной
стороны тела типичной ноги с ползательной поверхностью, произошла
дивергентная эволюция остальных классов моллюсков. Часть примитив-
ных моллюсков стала приспосабливаться к подвижному образу жизни
и к активной ловле добычи.
169
Как в процессе исторического развития происходило формирование
за счет части мускулистой стенки брюшной стороны тела у исходных
форм головоногих щупалец и воронки,— нам неизвестно; мы знаем лишь,
что щупальца (или руки) и воронка у современных головоногих обра-
зуются из части зародыша, гомологичной зачатку ноги остальных мол-
люсков. Ни в онтогенезе, ни в строении головоногих нет следов того, что
они когда-либо имели ползательную ногу. По-видимому, на очень раннем
этапе эволюции у примитивных головоногих формирование головных
щупалец и воронки было связано с облегчением веса раковины, с обра-
зованием газа, что способствовало подвижному образу жизни, плаванию,
возникновению у них хищничества, активной ловли крупной добычи, а
также развитию, наряду с радулой, сильных челюстей. Известно, что
самое древнее нижнекембрийское головоногое—Volborthella — было
уже типичным головоногим.
Судя по всему, головоногие в самом начале своей эволюции перешли
от обычного для остальных моллюсков (и для большинства групп Spi-
ralia) спирального, детерминированного дробления и образования пела-
гических личинок к неполному, дискоидальному дроблению богатых
желтком крупных яиц и прямому развитию без метаморфоза. Историче-
ски длительный, на протяжении всех геологических периодов подвижный
образ жизни (плавание и активная, охота за добычей) хищных голово-
ногих способствовал их высокому прогрессивному развитию.
Все это говорит об очень раннем обособлении головоногих от родо-
словного древа моллюсков, более раннем, чем ответвления от него брю-
хоногих, двустворчатых и лопатоногих. Общий предок последних, как
предполагали многие зоологи, приспосабливался к жизни в прибрежной
полосе моря с малым количеством осадков, к ползанию по субстрату.
Это способствовало развитию ноги с ползательной подошвой, такие
формы вначале, подобно хитонам, вели малоподвижный образ жизни,
питались соскабливанием преимущественно водорослей и корненожек
с помощью радулы. Развились примитивные брюхоногие.
Брюхоногие, двустворчатые и лопатоногие, обладающие основной
организацией, которая отвечает строению их предполагаемого общего
предка — Prorhipidoglossa Пельзенера или Praerhipidoglossa Плате, а
также индивидуальным развитием со спиральным детерминированным
дроблением, метаморфозом с двумя личинками — трохофорой и велиге-
ром, связаны друг с другом близким родством.
Основным, исходным органом движения этих моллюсков стала нога
с ползательной подошвой: форма, связанная с характером ее использо-
вания, как известно,-характеризует названия классов — брюхоногие, ло-
патоногие, топороногие (Pelecypoda). Брюхоногие, сильно отличающиеся
вследствие скручивания (или торсиона), от двустворчатых и лопатоно-
гих своей асимметрией и оставившие последних далеко позади по свое-
му морфологическому, физиологическому и биологическому прогрессу,
вместе с тем своими низшими, примитивными представителями стоят
ближе к общему родоначальнику, чем двустворчатые и лопатоногие.
Под воздействием условий жизни в иле и песке лопатоногие и еще более
двустворчатые уклонились от исходного плана строения, таким образом,
брюхоногие стоят ближе всего к родоначальной форме, затем следуют
лопатоногие, тогда как двустворчатые — наиболее отклонившаяся и
специализированная группа.
К 'Сожалению, серологических исследований о степенях родства
между классами моллюсков очень мало. Это работы Курамото (Kura-
moto, 1933), Макино (Makino, 1934), Честнут (Chestnut, 1943) и Леоне
г Прайором (Leone а. Ргуог, 1954), причем, судя по работе последних
авторов, полученные результаты противоречивы.
Аннелиды (Annelida)
Центральной группой для всех кольчатых, вернее, вторичнополост-
ных червей, для членистоногих и таких обособленных групп, как онихо-
форы, пентастомиды, и тардиграды, несомненно, являются полихеты.
Сохраняя основной план строения многочленистого метамерного хищ-
ного подвижного животного, полихеты обнаруживают, в связи с различ-
ными условиями жизни, большое разнообразие во многих чертах строе-
ния и отчасти развития.
Внешнее строение тела полихет характеризуют: неметамерный по
природе производный верхнего полушария трохофоры, небольшой про-
стомий, несущий головной мозг и рецепторы осязания, зрения; обоняния
и вкуса, нечленистый задний отдел, или пигидий, с анальным отверстием
и нефиксированное (порой превышающее 800) число сегментов метамер-
ного членистого тела с параподиями.— органами движения, действую-
щими по принципу простого рычага и несущими жабры и цирри. Наруж-
ной членистости тела отвечает метамерное строение и расположение
почти всех частей внутренней организации: вторичной полости тела с ее
диссепиментами и мезентериями, брюшной ганглиозной нервной цепоч-
ки с невросомитами, несущими по 3 пары периферических нервов, замк-
нутой кровеносной системы с ее ангиосомитами, с несколькими дыха-
тельными пигментами в виде гемоглобина, гемэритрина и хлорокруорина,
кожно-мускульного мешка, дифференцированного на миосомиты с пере-
хватами, органов выделения (нефридиев), метамерных зачатков в стен-
ках целома половых клеток, не оформленных в половые железы, и цело-
модуктов, выводящих половые элементы. Половое размножение, спи-
ральное детерминированное гомо- или гетероквадрантное дробление,
телобластическое развитие целома за счет бластомера 4d, свободная тро-
хофорная личинка; бесполое размножение, нередко эпитокия и правиль-
ное чередование полового и бесполого размножения, широкое распро-
странение большой способности к регенерации дополняют основные
черты полихет — типичных морских червей. Полихеты, несомненно, по-
явились в докембрии: уже в среднем кембрии они представлены разно-
образными формами (Walcott, 1911).
Подвижный образ жизни и активное хищничество бродячих полихет
(Errantia), пассивное питание мелким планктоном и детритом сидячих
полихет (Sedentaria), жизнь в песке и толще ила ч разнообразные фор-
мы движения, вплоть до быстрого бегания по дну моря, плавание, движе-
ния. ограниченные внутри временных и постоянных трубок, движения в
толще грунта по гидравлическому типу (по Л. А. Зенкевичу, 1944), ком-
менсализм, отчасти экто- и эндопаразитизм,— в некоторой степени ри-
суют экологию многощетинковых червей.
Будучи наиболее обширным классом аннелид (свыше 5000 видов),
полихеты очень широко распространены в Мировом океане от полосы от-
лива до ультраабиссали— 10 630—10 700 м (см. Беляев и др., 1960),
•171
нередко образуя поселения очень большой плотности, например, до
90 000 экземпляров Maldane sarsi на 1 м2 Баренцева моря (Зенкевич,
1963) и до 40 000 экземпляров Fabricia rivularis на 1. м2 Охотского моря
(Геккер и Ушаков, 1962; Ушаков, 1955). Наряду с чисто глубоководными
имеются эврибатные виды. Широк диапазон у полихет и в отношении
требований вида к температурным условиям и к солености воды. Будучи
в основном аэробами, некоторые полихеты живут в водах с большим де-
фицитом кислорода, в гниющих водах со значительным количеством
сероводорода и даже нефти. Как редкие явления наблюдаются жизнь в
пресной воде, распространение по эстуариям в реки, жизнь в замкнутых
пресных водоемах и даже на суше, среди наземных членистоногих и они-
хофор.
Все это важно для понимания разнообразия строения и отчасти раз-
вития полихет. У хищных, подвижных полихет наблюдается развитие го-
ловного отдела, когда к простомию примыкают 1—4 сегмента тела, на
которых первый, .перистомий, несущий ротовое отверстие, лишен парапо-
дий и Щетинок, но снабжен усиками (cirri), причем у ряда форм не толь-
ко он, но и следующие три передних туловищных сегмента могут сли-
ваться с простомием и друг с другом в головной отдел. Простомий у них
несет упомянутые выше рецепторы, тентакулы (или антенны) и пальпы,
а головной мозг бывает дифференцирован на три хорошо выраженных
отдела: передний (пальпарный), средний (антеннально-глазной), и зад-
ний (нухальный, или затылочный). В мозгу.находится сложный центр
с ассоциативными элементами, ассоциативные клетки собраны в особые
группы (глобулы, гломерулы, стебельчатые тела, особые зрительные
центры) и т. д.
Встречаются высоко дифференцированные глаза с диоптрическим
аппаратом, способные к аккомодации, дающие картинное зрение. Все
эти части простомия, кроме примыкающих к нему цирри метастомия, яв-
ляются неметамерными образованиями, продуктами развития и диффе-
ренцировки простомия, возникающего из верхнего полушария трохофоры
червя. Их развитие связано с очень подвижным образом жизни и хищни-
чеством этих червей и наиболее ярко выражено у семейства Eunicidae.
Перистомий образуется слиянием простомия и ротовой области, воз-
никающих за счет эписферы трохофоры, с первым туловищным сегмен-
том (Anderson, 1959). Данные некоторых авторов о наличии целома в-
головном отделе Protodrilus и Saccocirrus подтвердились и для ряда ти-
пичных полихет. В онтогенезе Scoloplos armiger (Anderson, 1959) просле-
жено наличие целомических полостей или захождение их в головной
отдел, что связано с развитием там мускулатуры и кровеносных сосудов,
и в известной мере с растяжимостью (подвижностью) простомиальных
придатков (пальп и тентакулов). Однако это не дает оснований призна-
вать, как делают Бинар и Жене (Binar et Jeener, 1927), простомий паль-
парным сегментом или насчитывать у полихет несколько головных сег-
ментов.
Сидячий образ жизни в трубках и пассивный способ питания мелкой
пищей, характерные для полихет Sedentaria, ведут к редукции простомия
и головного отдела, к утрате отмеченных выше черт строения головного’
мозга и органов чувств и к'мощному развитию венчика щупалец из пальп
или тентакулов, типичных для подвижных предковых форм. Щупальца'
становятся органами дыхания, захвата добычи, органами чувств, а порой
и выводковыми камерами для развивающейся молоди. Вместе с тем про-
исходит дифференцировка тела на грудной отдел с его кроной щупалец
и брюшной отдел и изменение сегментов. Происходит перестройка пара-
подий и их щетинок, обеспечивающих движение червя внутри трубки.
Дифференцировка тела на грудной и брюшной отделы сопровождается
перестройкой внутренней организации. В грудном отделе полость тела
172
теряет метамерное строение, редуцируются диссепименты, кроме так на-
зываемой диафрагмы, и мезентерии, происходит перестройка и усложне-
ние строения кровеносной системы, ограничение в этом отделе числа
миксонефридиев с выделительной функцией, иногда до одной пары, до-
стигающих большой величины и сложности строения, и т. д. Возникает
гетерономная метамерия, отличная от расчленения тела на ларвальные
и постлар'вальные сегменты, очень сильно выраженная у некоторых форм
(например, Chaetopterus, Sternaspis, и др.).
Не менее ярка зависимость 'между строением и условиями жизни у
полихет, живущих в песке и в толще ила, причем изменения внешнего
строения и внутренней организации порой настолько велики, что в ряде
случаев нелегко решить, что у таких форм, как например Polygordiidae
и Protodrilidae, является результатом вторичного упрощения и что —
примитивными чертами. Головной отдел у них не дифференцирован,
почти нет головных рецепторов, могут редуцироваться параподии, жаб-
ры, щетинки, целом превращается в единую общую полость, диссепимен-
ты и мезентерии выпадают, уменьшается число сегментов тела и т. д.
Такие изменения организации наиболее выражены у полихет, живущих
в толще ила и двигающихся «гидравлическим», по выражению Л. А. Зен-
кевича (1944), способом.
Значительные изменения формы тела, наружного и внутреннего
строения возникают у полихет также под влиянием паразитического об-
раза жизни, что, однако, в основном относится к мизостомидам и в не-
значительной мере к остальным группам полихет. Приближение к ра-
диальной симметрии выражено в развитии округлой формы плоского
тела, в расположении ножек, боковых органов и усиков, в концентрации
брюшной мускулатуры тела и брюшной нервной системы и в отхождении
от нее нервов. К этому добавляется зарастание целома соединительной
тканью, формирование сложного полового аппарата, состоящего из се-
менников, яичников и половых протоков, и ряд других черт взрослых
мизостомид; эти аннелиды, несомненно, близко родственные полихетам,
дают яркий пример мощного формирующего воздействия на организм
паразитического образа жизни, продолжающегося геологически длитель-
ное время — с древнего палеозоя (Федотов, 1937).
Характерные изменения происходят также в строении и экологии
размножения и развития тех немногих форм полихет, которые переходят
от водного (морского) к сухопутному (наземному) образу жизни, о чем
будет сказано ниже.
Такие сильные изменения во внешней и внутренней организации по-
лихет, происходящие под воздействием условий жизни, позволяют счи-
тать эхиурид и сипункулоид потомками древних полихетообразных пред-
ков. Здесь эти классы аннелид я не рассматриваю, поскольку они неве-
лики и являются слепыми ветвями аннелидного типа. Надо, однако, от-
метить, что они служат отличным примером сохранения онтогенезом,
происходящим в условиях открытой жизни, основных типичных черт раз-
вития морских аннелид и преобразующего воздействия условий жизни на
организм червя с переходом к жизни в толще грунта. У эхиурид в более
полном, у сипункулид в менее сохранившемся виде онтогенез характери-
зуется спиральным детерминированным дроблением, трохофорной ли-
чинкой, телобластической закладкой мезодермальных полосок, заклад-
кой сегментов тела, метамерных нефридиев, и т. д.
Родство этих классов с аннелидами выступает отчетливо, хотя в то
же время жизнь этих червей в толще грунта, связанная с передвижением
и рытьем ходов в нем, привела к утрате наружной и внутренней мета-
мерии, приобретению общей полости тела, утрате параподий с их компо-
нентами, смещению порошицы на передний конец тела, удлинению
средней кишки и спиральному ее скручиванию, к утрате кровеносной
173
и дыхательной системами типичного строения и т. д. Такие радикальные
изменения аннелидного типа строения понятны, если вспомнить, что
эхиуриды и сипункулиды найдены в среднем кембрии и что эти черви,
вероятно, жили значительно раньше.
Надо подчеркнуть, что сравнительно с большим разнообразием во
внешней и внутренней организации полихет, связанным с различным об-
разом жизни, их индивидуальное развитие, происходящее в общем в
условиях открытой жизни, протекает более однотипно. Для них характер-
но спиральное детерминированное дробление, гомо- и гетероквадрант-
ность микромеров и макромеров, розетка, крест (рис. 46), клетки прото-
троха, трохофора с теменной пластинкой и венчиками ресниц, телобла-
стическая закладка мезодермальных
полосок, протонефридии, формиро-
вание ларвальных и постларваль-
ных сегментов (рис. 47). Такие осо-
Рис. 46. Крест и розетка трохофоры
полихет
/ — крест; 2 — розетка
Рис. 47. Схема развития лар
вальных и постларвальных сег-
ментов у полихет (из Беклеми-
шева)
1— ларвальные сегменты; 2 — пост-
ларвальные сегменты
бенности типичны для огромного большинства полихет. Ларвальные сег-
менты, возникая в большом числе (от 3 до 13) на оральной стороне
трохофоры, составляя на стадии метатрохофоры и нектохеты тело личин-
ки, позже входят в состав членистого тела червя (Р. Iwanoff, 1928).
После некоторой паузы в зоне роста (около анального отверстия) начи-
нают развиваться постларвальные сегменты, из которых в основном и со-
стоит тело червя.
Такое развитие со свободными пелагическими личинками — атрохной,
монотрохной, дитрохной и политрохной (Свешников, 1961), протекающее
в толще воды, является наиболее распространенным среди полихет
(рис. 48).
В связи с условиями жизни у ряда полихет развитие до стадии некто-
хеты протекает в слизистых кладках или в капсулах. Наконец, у неко-
торых полихет выпадает свободное развитие и возникает живорождение.
Все сказанное подтверждает представление о полихетах как основной
группе кольчатых червей, от которой, очевидно, произошли олигохеты
и пиявки.
174
Рис. 48. Типы пелагических личинок полихет (по Свешникову)
А—протрохофора Chone infundibuliformis (Sabellfdae); Б—метатрохофора Апа-
Hides citrina-, В—нектохета Eteone longa (Phyllodocidae); Г—метатрохофора Наг-
mothoe imbricata; Д—трохофора Castalia punctata (Hesionidae); Е—метатрохофора
Nereis virens (Nereidae); Ж—трохофора Heteromastus filiformis (Capitellidae); 3—
нектохета Heterofnastus filiformis-, И—митрария Myriochele oculata (Oweniidae)
Приходится все-таки отметить, что до сих пор нет общепризнанной
системы полихет, несмотря на большое число работ и сравнительно не-
большое число форм (более 5000 видов). Остановлюсь на двух приме-
рах. По системе Кестнера (Kaestner, 1954/1955), в значительной мере
опирающейся- на работы немецких зоологов, класс полихет делится
на три отряда: Archiannelida, Errantia и Sedentaria. Отряд Errantia со-
стоит из двух подотрядов: Amphinomorpha с семейством Amphinomidae
и Nereimorpha, куда отнесены все другие семейства бродячих полихет..
Отряд Sedentaria составляют пять подотрядов: Spiomorpha, Drilo-
morpha, Terebellomorpha, Hermellimorpha и Serpulimorpha. Sipunculida
считаются вторым классом аннелид.
По Н. А. Ливанову (1940) класс полихет состоит из трех подклассов:
Errantia, Sedentaria и Myzostomida. В подкласс Errantia Н. А. Ливанов
175
включает три отряда: Phyllodocemorpha, Nereimorpha и Eunicemorpha,
а в подкласс Sedentaria 3 отряда — Spiomorpha, Terebellomorpha и Ser-
pulimorpha.
Из сравнения классификаций Н. А. Ливанова и Кестнера нетрудно
видеть, что они в основном сходятся. Непринципиальное различие сво-
дится к тому, что группы бродячих и сидячих полихет Ливанов возводит
в ранг подклассов, а Кестнер считает отрядами. Надо сказать, что в
системе Кестнера выделение семейства Amphinomidae как подотряда
наравне с Nereimorpha и признание его наиболее примитивной группой
бродячих полихет необоснованны. Они имеют плоское, широкое эллип-
соидальное тело, покрытое своеобразными спинными пластинками, ради-
альное расположение и укорочение передних и задних сегментов, т. е. гете-
рономную метамерию, параподии в виде кольчатых ножек с сильно загну-
тыми щетинками, ветвистый кишечник, заросший соединительной тканью
целом и т. д. Графф (Graff, 1888) говорит о том, что Spinther и, следо-
вательно, все семейство Amphinomidae, обладающее также миксонефри-
диями, весьма специализированные, отнюдь не примитивные полихеты;.
но ни Euphrosyne, ни тем более Spinther не могут считаться низшими
полихетами. К сожалению, неизвестно развитие этих форм, а это имело
бы важное значение. Однако и приведенные данные заставляют считать
более естественным положение семейства Amphinomidae, как это делает
Н. А. Ливанов (1940), среди Nereimorpha, а не вместе с Phyllodocidae.
Принципиально, пожалуй, неважно, объединять ли 'всех остальных бро-
дячих полихет, за исключением Amphinomidae, в один подотряд Nerei-
morpha, как делает Кестнер, или делить их на три отряда — Phyllodo-
cemorpha, Nereimorpha и Eunicemorpha, как делает Н. А. Ливанов. Раз-
личия между ними в строении и в известной мере в развитии таковы, что
разделение их не на один, а на три отряда более оправдано. Затруднение
в этом случае заключается, однако, в том, что Eunicemorpha тогда, ока-
зывается, состоят всего из одного типичного семейства Eunicidae, осталь-
ные же семейства представлены или паразитически измененными фор-
мами Ichthyotomidae и Histriobdellidae, или примитивными и в известной
мере вторично упрощенными формами, вроде Protodrilidae, Polygordi-
idae, Saccocirridae, Nerillidae и Dinophilidae. В таком составе отряд
Eunicemorpha недостаточно обоснован, так как близкая родственная
связь Eunicidae с пятью последними семействами довольно проблема-
тична.
Неудовлетворительность существующих классификаций класса поли-
хет и их несогласованность вызваны не только различием точек зрения
авторов, но и самой природой полихет, очевидно, еще недостаточно выяс-
ненной. Судя по последней работе Гудрича (Goodrich, 1945, см. гл. «Мол-
люски»), известного знатока полихет, основа приведенных систем поли-
хет находит и поддержку, и возражения, если исходить из данных о раз-
витии и взаимозависимости между нефридиями и целомодуктами. Прото-
нефридии и протонефромиксии наблюдаются у семейств Phyllodocidae,
Tomopteridae, Nephthydidae и Glyceridae. Метанефридии, становящиеся
при половой зрелости метанефромиксиями и половыми протоками, име-
ются у семейств Sylliidae, Hesionidae и Nereidae. Видимо, эти отношения
наблюдаются у ряда семейств и сидячих полихет — Spionidae, Ariciidae,
Disomidae, Paraonidae, Chaetopteridae и у семейств бродячих полихет —
Chrysopetalidae, Pisionomidae, Thyphloscolecidae, Sphaerodriidae и
Ichtyotomidae. Наконец, миксонефридии наблюдаются у семейств
Eunicidae, Amphinomidae и Aphroditidae из бродячих и у остальных
сидячих полихет — Arenicolidae, Maldanidae, Oweniidae, Cirratulidae,
Opheliidae, Chloremidae, Sternaspidae, Amphictenidae, Ampharetidae, Te-
rebellidae, Sabellariidae, Sabellidae, Serpulidae, и лишь у одного семей-
ства Capitellidae сохраняются самостоятельные и полные как метане-
176
фридии, так и целомодукты. Характерно, что из всей массы полихет
самостоятельностью метанефридиев и целомодуктов выделяется семей-
ство Capitellidae, которое некоторые зоологи рассматривают как родо-
начальное семейство олигохет,' а последние, как и пиявки, также обла-
дают самостоятельными метанефридиями и целомодуктами.
Таким образом, если у примитивных семейств Phyllodocidae и Тогпо-
pteridae имеются примитивные протонефридии и протонефромиксии, то
у семейств Amphinomidae и Aphroditidae, занимающих, например, в си-
стеме Кестнера, место среди примитивных форм, наблюдаются миксо-
нефридии, наиболее распространенные среди высших семейств полихет.
Рассмотренные классификации, основанные на особенностях строения
и экологии .взрослых форм, и подтверждаются, и опровергаются данными
по онтогенезу (Свешников, 1958а, 1960, 1961). Онтогенез отрядов Phyl-
lodocemorpha и Nereimorpha характеризуется монотрохными пелагиче-
скими личинками. Хотя Phyllodocidae и Aphroditidae считаются очень
примитивными полихетами, однако экология взрослых и характер онто-
генеза ряда форм (Nereimorpha и главным образом Eunicemorpha) обна-
руживают наибольшее разнообразие и наибольшую пластичность срав-
нительно со значительной ограниченностью во всем семейств Phyllodoci-
dae и Aphroditidae. В то время как для семейств Phyllodocidae и
Aphroditidae характерны монотрохные личинки и отсутствие эпитокии,
у Eunicemorpha наблюдаются половое и бесполое размножение, часто с
правильным их чередованием, эпитокия, свободное пелагическое, несво-
бодное пелагическое и прямое развитие и живорождение. Кроме того, у
Eunicemorpha имеются атрохная (или прототрохная), монотрохная и по-
литрохная личинки, причем широко распространена личинка наиболее
примитивного типа (атрохная) со сплошным ресничным покровом. В со-
гласии с общими основами классификации данные об индивидуальном
развитии говорят о том, что по сравнению с бродячими, сидячие полихеты
обладают рядом существенных отличий в строении (см. например, Мейер,
1893, 1898), вызванных сидячим образом жизни (головной отдел с коро-
ной щупалец, дифференцировка на грудной и брюшной отделы). Они
далеко не все имеют развитие с пелагическими личинками (у 7 семейств
из 11), причем личинки их гораздо более сложного строения, чем личин-
ки бродячих полихет (Свешников, 1961).
Все это говорит о том, что Eunicemorpha и в меньшей мере Nerei-
morpha являются основной группой полихет.
Надо думать, что примитивные древние представители этой группы,
остатки челюстей и, вероятно, ходов, а также следы ползания которых
известны из докембрийских отложений, дали начало всем остальным
группам бродячих и сидячих полихет и мизостомидам. Несомненно, это
были ползающие донные хищные небольшие полихеты, возможно, как
считает Уолкот (Walcott, 1911), с одноветвистыми небольшими парапо-
диями.
Классификация любой группы животных, не преследуя филогенети-
ческих задач, должна приближаться к естественному порядку. Хотя мно-
гие десятки лет описательным, сравнительным и экспериментальным
методами изучается строение, развитие, в меньшей мере экология и фи-
зиология полихет, тем не менее их систематика остается, как мы видели,
неудовлетворительной.
Это относится, в частности, и к положению в системе полихет так
называемых архианнелид. Они фигурируют то как отряд Archiannelida,
равный отряду Errantia и Sedentaria, несмотря на признание его сбор-
ным, искусственным объединением (Kaestner, 1954/1955), то как отдель-
ные семейства размещаются в отряде Eunicemorpha (Ливанов, .1955).
Несомненно, что семейства Polygordiidae, Protodrilidae, Saccocirridae,
Nerillidae и Dinophilidae имеют общие примитивные черты, причем
12 Д. М. Федотов
177
Polygordiidae, Protodrilidae и Nerillidae, а также Saccocirridae— это од-
новременно и самые примитивные по строению и упростившиеся и отчасти
специализированные бродячие полихеты. Вероятно, это древние потомки
вымерших, еще слабо дифференцированных полихет.
Dinophilidae по своему строению не полихеты, но они могут быть
неотеническими потомками каких-либо примитивных форм Errantia, об-
ладающими одними ларвальными сегментами, как это думал еще
П. П. Иванов. Все это делает более рациональным на данном уровне
знаний рассматривать эти группы червей вслед за Н. А. Ливановым
(1940) как отдельные семейства, не связанные вместе.
Более определенное мнение можно высказать о положении в системе
группы мизостомид, которых одни авторы низводят до положения семей-
ства среди Nereimorpha (Grobben u. Kuhn, 1932), другие считают под-
классом,. как Errantia и Sedentaria (Ливанов, 1940), третьи возводят
мизостомид в ранг класса, приравнивая классу Polychaeta (Kaestner,
1954/1955), наконец, Йегерстен (Jagersten, 1934, 1939а, 19396, 1940а) без
достаточных оснований отрицает за ними природу высших червей. При-
знавая у мизостомид, как и у динофилид, наличие лишь ларвальных сег-
ментов (Федотов, 1937), мы в строении и развитии их видим резкие
отличия от динофилид и сохранение ряда полихетных черт. Почти все
признаки строения мизостомид мы находим у полихет, несмотря на
влияние на организацию этих червей паразитического образа жизни с
древнего палеозоя (Готланда). Характерно, что, несмотря на эндопара-
зитизм и наличие ряда регрессивных изменений, обусловленных подобным
образом жизни, Protomyzostomum сохраняет метамерное строение в рас-
положении параподий, расположении и числе боковых органов, в строе-
нии центральной лестничной нервной системы, отчасти в строении сред-
ней кишки и числе целомодуктов, что еще более связывает мизостомид
с полихетами. Поэтому я считаю более правильным рассматривать мизо-
стомид как группу полйхет, пока на правах третьего подкласса. Мне
представляется, что серологическим методом можно будет уточнить при-
роду и положение мизостомид в системе полихет. Этот метод необходимо
использовать и для исследования всех полихет, поскольку классифика-
ция этого класса нас не удовлетворяет.
Филогенетические отношения групп полихет до сих пор остаются не-
уточненными. Без сомнения, данные о современных полихетах говорят
о более примитивном положении бродячих полихет (Errantia), по срав-
нению с сидячими (Sedentaria), хотя благодаря хищному подвижному
образу жизни некоторые черты их организации (прежде всего развитие
головного отдела с разнообразными рецепторами и головным мозгом)
у них более совершенны, чем очевидно вторично упрощенные, даже реду-
цированные соответственные части сидячих полихет. Наличие уже в сред-
нем кембрии значительного числа родов бродячих полихет (Errantia,
Walcott, 1911), находки в низах кембрия среди остатков медуз, видимо,
пелагической полихеты — Spriggina ploundersi, вероятно, близкой семей-
ству Tomopteridae (Glaesner, 1958), находки ходов, следов и остатков
челюстей и щетинок бродячих полихет, более поздние находки (со сред-
него кембрия) трубок сидячих полихет, несмотря на бедность палеонто-
логических данных (Беккер и Ушаков, 1962), свидетельствуют о более
раннем, по сравнению с Sedentaria, историческом развитии и эволюции
группы Errantia. Бесспорно также, что сидячий образ жизни полихет,
особенно в прикрепленных к субстрату трубках,— явление более позднее,
чем подвижный образ жизни. В связи с этим произошел переход от
активного к пассивному способу питания мелким планктоном или детри-
том при помощи ловчего аппарата в виде венчика щупалец или клейких
длинных нитевидных щупалец. По данным о строении и развитии можно-
со значительной долей правды представить себе пути эволюции от более
178
примитивной к более высокой ступени и пути специализации в обеих
главных группах полихет (бродячих и сидячих). Однако взгляды на при-
роду и состав так называемых архианнелид, которых одни авторы, как
было отмечено выше, рассматривают как отдельные семейства Eunice-
morpha, другие — как отряд (царавне с отрядами Errantia и Sedentaria),
различны. Большинство «архианнелид», если из них выделить семейства
Nerillidae и Dinophilidae (последние живут среди водорослей), являются:
обитателями мезопсаммона и представляют собой гомономно членистых
полихет с примитивными чертами строения (остатки ресничного покрова
в виде поясков и полосок, эпидермальное положение нервной системы,
простота кожно-мускульного мешка, состоящего из продольных мышц и
кровеносной системы, выделительные органы, непосредственно служа-
щие для выведения половых продуктов, и т. д.). Такие черты организа-
ции, как отсутствие или слабое развитие параподий, отсутствие щетинок
или их простота, отсутствие жабр,— имеют характер упрощения. Однако
нахождение в мезопсаммоне примитивных по организации представите-
лей не только полихет, но и других групп, например ракообразных, гово-
рит о мезопсаммоне как о среде, благоприятствующей сохранению при-
митивных организмов, а не их регрессу. Многие черты строения «архи-
аннелид», даже считая ряд черт упрощенными, свидетельствуют о том,
что они являются потомками вымерших полихет, не достигавших высоты
организации, свойственной даже наиболее примитивным представителям
современных Errantia, со слабо дифференцированными параподиями.
Несмотря на простоту, в меньшей мере упрощение в организации Рго-
todrilidae, Polygordiidae и Saccocirridae (последние, конечно, имеют ряд
специализированных черт, например, пенисы), это полихеты гомономно-
и многочленистые, вопрос о ларвальных сегментах которых еще не решен
(строение Chaetogordius мне остается неизвестным). Иное дело семей-
ство Dinophilidae. Хотя некоторые авторы отрицают ларвальную природу
сегментов тела Dinophilidae (Jagersten, 19406, 1947, 1952), тем не менее
трактовка динофилид, данная П. П. Ивановым и В. Н. Беклемишевым,
как неотенических форм полихет, состоящих лишь из ларвальных сегмен-
тов, наиболее обоснована. Помещение динофилид, не находящихся в
родстве с Rotatoria й Gastrotricha, в системе^ рядом с Polygordiidae, Рго-
todrilidae и Saccocirridae, однако, явно искусственно. К сожалению, я не
знаю строения Diurodritus и лучше других динофилид сегментированного
Trilobodrilus (Remane, 1959).
Думаю, что Ремане прав, считая семейство Nerillidae близким к ти-
пичным полихетам, но мне неизвестны работа Делашо о Troglochaetus
из пресных карстовых вод Средней Европы и работа Ремане с описанием
Nerillidium.
Как известно, особых противоречий и трудностей в толковании мор-
фологии олигохет почти нет, по своему характеру они образуют одно-
родную группу гомономно метамерных кольчатых червей, которые срав-
нительно с полихетами несут черты лишь известного упрощения, а не
прогрессивного развития или специализации. Как будет отмечено ниже,
индивидуальное развитие олигохет, во всяком случае на ранних стадиях,
также сохраняет ряд характерных черт, свойственных, развитию полихет.
Как и среди последних, эта сравнительно большая группа кольчатых
червей (2700 видов) включает формы длиной от нескольких миллиметров
с небольшим (5—7) числом сегментов до гигантов в 3 м длиной, обла-
дающих 700—800 сегментами. В целом, сохраняя наружную и внутрен-
нюю метамерию, олигохеты под воздействием условий жизни в почве
утрачивают обособленный головной отдел с придатками (рецепторами
органов чувств) и параподии (кроме щетинок), претерпевают известные
нарушения метамерной организации в виде редукции спинного, нередко
и брюшного мезентериев, частично диссепиментов, испытывают извест-
12*
179
ный регресс частей кровеносной системы, порой некоторое подавление
целома вследствие паренхиматизации и т. д. Наиболее характерной чер-
той олихогет, отличающей их от полихет, является ограничение числа
яичников и семенников в передней трети тела и образование там покро-
вами немногих сегментов тела пояска, спаривание при его помощи и раз-
витие яиц в коконах, которые обычно откладываются в землю. Все это
(обособление половых клеток в семенники и яичники, формирование
компактного полового аппарата, функ-
ционирование которого обеспечивает по-
ловую деятельность олигохет) является
результатом приспособления к жизни в
почве. Это известное прогрессивное раз-
витие олигохет сравнительно с полихета-
ми, Указанными чертами и, пожалуй,
только ими, исключая Acanthobdella с ее
щетинками на первых пяти сегментах
тела (рис. 49), целомической полостью
вокруг кишечника и настоящей крове-
носной системой (Ливанов, 1905) и
ограничивается сходство строения олиго-
хет и пиявок. Порой ограничение разви-
тия половых клеток наблюдается и у по-
лихет, однако открытая жизнь в море
обычно не требует развития у них осо-
бых, специфических приспособлений в их
строении для отправления половой жиз-
ни. Половые клетки развиваются непо-
Рис. 49. Acanthobdella peledina,
передний конец тела со щетин-
ками на первых сегментах
(из Ковалевского)
средственно в полости тела, не оформляются в локализованные половые
железы, обычно не развиваются и сложные выводные половые протоки
и копулятивные органы. Благодаря жизни большими пятнами (массами)
с громадным количеством особей, полихеты, выводя половые продукты
прямо в воду, имеют достаточно шансов для развития потомства и сохра-
нения вида. Реже, подобно знаменитому червю палоло, ряд полихет, как
известно, ко времени размножения переходит от жизни в бентосе к жиз-
ни в поверхностных слоях моря, где и происходит оплодотворение яиц и
начинается развитие. Естественно, что у полихет, особенно у бродячих,
половые клетки развиваются в задней половине тела, что благоприят-
ствует при движении выходу из тела яиц и спермы и их рассеиванию.
Это обратно тому, что имеет место у почвенных обитателей — олигохет,
у которых при затруднении встреч особей и частых повреждениях- более
тонкого заднего отрезка тела половой аппарат располагается в передней
трети тела.
При достаточно согласованном толковании строения олигохет родст-
венные отношения их друг с другом, однако, далеко не выяснены. Одни
считают наиболее примитивным и давшим начало всем группам олигохет
семейство Aeolosomatidae, по мнению других, основной группой этого
класса является семейство Lumbriculidae (Stephenson, 1930) или Phrae-
oryctidae (Ливанов, 1940), семейства же Aeolosomatidae, Naididae и Tu-
bificidae считаются боковой ветвью. Все же не случайно система класса
начинается с семейства Aeolosomatidae, у которого Маркус (Marcus,
1944а, 19446). отмечал ряд примитивных черт. К ним относятся реснич-
ный локомоторный простомий, ресничные головные ямки, положение
головного мозга в простомии, эпидермальное положение нервной системы
лестничного типа с расставленными друг от друга стволами, спинной
кровеносный сосуд, обособленный лишь в передней части тела, а в сред-
ней и задней частях слитый с околокишечным синусом, ресничный кишеч-
180
ник, наличие нефридиев почти во всех сегментах, отсутствие мужских
половых протоков, каковыми неросредственно служат неизмененные не-
фридии, нелокализованное образование половых клеток. Вместе с тем
наличие ряда регрессивных черт не позволяет Маркусу выводить осталь-
ных олигохет от Aeolosomatidae, заставляет считать их примитивной
боковой ветвью (Marcus, 1944а, 19446).
Наряду с некоторыми чертами упрощения в организации других се-
мейств олихогет (Naididae, Tubificidae) они также по сравнению с типич-
ными почвенными олигохетами в известной мере примитивны и просты,
что связано и с более примитивным типом индивидуального развития.
Дробление яиц сохраняет исходный, типичный для полихет спиральный
детерминированный характер. Так, образуется четыре эктодермальных
квартета, пятый участвует в образовании кишечника, выражены крест
и розетка, но гетероквадрантность значительно более резкая, чем у поли-
хет, и начинается уже на двуклеточной стадии дробления. Зачатки экто-
мезенхимы и мезодермы радиально симметричны.
У более высокоорганизованных и специализированных, благодаря
жизни в почве, Lumbricidae спиральное дробление сохраняется лишь на
ранних стадиях, в расположении бластомеров рано проявляется била-
теральная симметрия. Квартеты микромеров, в том числе и их производ-
ные — бластомеры 2d и 4d, играющие большую роль в образовании тела
и органов трохофоры полихет, у олигохет делятся мало. У всех олигохет
в результате их деления зародыш получает билатеральносимметричный
характер делением 2d. У всех олигохет эктодермальные телобласты
являются зачатками наружных покровов, кольцевой мускулатуры, нерв-
ной системы и нефридиев. Делением бластомера 4d, как у полихет, обра-
зуются 2 телобластические полоски целомической мезодермы, за счет
которой образуются мезодермальные сомиты.
У большинства семейств малощетинковых червей — Tubificidae, Lum-
briculidae, Discodrilidae, вероятно также Aeolosomatidae, Phraeoryctidae,
составляющие группу микродрили (Шмидт, 1925; Schmidt, 1924), про-
цессы эмбрионального развития протекают сходно и имеют примитивный
характер. Так, по данным Вейдовского (Vejdovsky, 1888—1892),
П. Г. Светлова (1923а, 19236), Танрейтер (Tannreuther, 1915), Пеннерса
(Penners, 1922, 1926а, 19266), у этих олигохет в результате спирального
дробления образуется пять квартетов микромеров, иногда имеется ро-
зетка и крест, как у полихет; возникает четыре пары эктодермальных
телобластов и одна пара крупных первичных мезобластов, из которых
образуется на каждой стороне зародыша по зародышевой полоске,-позже
срастающихся по длине в плоскости симметрии зародыша. Развитие про-
исходит за счет желтка яйца и является типом неличиночного эмбрио-
генеза (Шмидт, 1925).
Под воздействием особых условий жизни, например вмерзания в лед
(Светлов, 1926; Dawydoff, 1959), происходят вторичные изменения
эмбрионального развития: образуется провизорная защитная эктодерма,
органогенез становится анархическим, изолированные бластомеры обра-
зуют плотную массу клеток, в которых происходит органогенез.
У более высокоорганизованных и специализированных, благодаря
жизни в почве, дождевых червей (мегадрилы, семейство Lumbricidae) тип
онтогенеза сильно изменен. У громадного их большинства спиральное
дробление сохраняется лишь на ранних стадиях, зародыш образуется из
бластомера CD двуклеточной стадии, бластомер АВ дает три бластомера
А, В7, В", имеющих значение осморегуляторов. У Eisenia fetida, обитаю-
щей в навозе, потребности в осморегуляции нет, и бластомер АВ участ-
вует в образовании личиночной эктодермы. Образуется три пары экто-
дермальных телобластов (Светлов, 1928), а не четыре пары, как у низ-
ших пресноводных олигохет. Из одной пары крупных мезобластов
181
возникаем целомическая мезодерма. Очень рано формируется личиночная
глотка, личиночный протонефридий. Зародыш становится инкапсулиро-
ванной личинкой, заглатываемый ею белок обеспечивает дальнейшее раз-
витие червя. У низших олигохет описана редуцированная ларвальная
мезодерма (П. П. Иванов, 1944). По мнению одних эмбриологов
(П. П. Иванов, 1937, 1945), данные онтогенеза не свидетельствуют о про-
исхождении дождевых червей от водных форм; по мнению Г; А. Шмидта
(1951) и отчасти П. Г. Светлова (1928), особенности органогенеза дож-
девых червей истолковываются как изменения хода развития водных
олигохет, почему Шмидт считает возможным выводить первых от вто-
рых. Совокупность данных морфологии и экологии олигохет скорее гово-
рит о том, что древние олигохеты как потомки морских полихет в своей
массе проделали эволюцию путем перехода от жизни в придонном песке
и иле моря к жизни во влажной почве морского прибрежья и распростра-
нения по почве.в глубь материка. Меньшая же часть, исторически быстро
выйдя к более открытой Жизни в пресных водоемах, вторично стала вод-
ными обитателями, в силу чего ни их организация, ни характер онтогене-
за не претерпели тех резких изменений, которые возникли у почвенных
олигохет под воздействием жизни в почве. К сожалению, для подтверж-
дения этого нужно знать индивидуальное развитие Lumbriculidae, Phraeo-
ryctidae и Enchytraeidae, стоящих по уровню организации значительно
ближе к почвенным олигохетам, чем семейства Aeolosomatidae, Naididae
и Tubificidae. Известны случаи вторичного переселения пресноводных
олигохет в почву, например Enchytraeidae, и почвенных олигохет
{Criddrilus lacuum) в пресные воды, что, по мнению Г. А. Шмидта, под-
черкивает контакт сред обитания (пресные воды и почва) и возможности
перехода в них олигохет.
Пиявки морфологически и экологически представляют цельную, мало-
численную (примерно 300 видов), компактную по составу и типу строе-
ния группу хищных и кровососущих червей, ярко демонстрирующую'
глубокую зависимость между строением и отправлениями организма и
условиями его жизни. Для них характерно следующее: удлиненно-яйце-
видная форма обычно'сплюснутого членистого тела (от. 1 до 20 см дли-
ной), постоянное небольшое (33, кроме 30 у Acanthobdella) число сег-
ментов, имеющих вторичную кольчатость, от 2 до 14 колечек в каждом',
передняя и задняя присоски, образованные слиянием 4 и 7-го сегментов,
соответственно прочные упругие, но мягкие покровы тела с кутикулой,
сбрасываемой при линьке целиком, отсутствие придатков тела, кроме
редких исключений в виде наружных жабр, мощно развитая мускулатура
тела, по массе которой пиявки занимают первое место среди всех беспо-
звоночных (Зенкевич, 1944), передняя кишка с хоботом или челюстями
и слюнными железами, секрет которых препятствует свертыванию крови
добычи, парные слепые мешки средней кишки для крови и соков добычи,
переваривание которых длится от 6 до 18 месяцев, отсутствие, кроме
паренхимных глаз, моделированных рецепторов, слияние четырех перед-
них метамер брюшной нервной цепочки в подглоточную массу и семи
последних — в анальную нервную массу, органы выделения — метане-
фридии, половые, протоки — целомодукты, гермафродитизм, анатоми-
чески цельный половой аппарат из половых желез и протоков, с непар-
ными половыми отверстиями в 10—12 сегментах тела, образующих поя-
сок, исключительно половое размножение, развитие в коконах, на ран-
них стадиях — спиральное детерминированное дробление, отсутствие
бесполого размножения и регенерации, в основном пресноводная, реже
морская, как исключение наземная, среда обитания.
Вся организация пиявок очень четко связана с экологическими осо-
бенностями этих червей как кровососов (примерно три четверти видов)
и хищников (остальная четверть видов). Они передвигаются обычно пу-
182
тем попеременного прикрепления к субстрату или телу хозяина задней
и передней присосок,,но могут и плавать.
При отсутствии морфологически дифференцированных органов
чувств, если не считать бокаловидных органов и паренхимных глаз, со-
гласно недавним экспериментальным исследованиям (Kaestner, 1954/
/1955), пиявки тем не менее обладают необычайно тонким, острым вос-
приятием различных внешних раздражений, обеспечивающих их суще-
ствование: колебаний водной среды, вызываемых движением добычи или
врага, незначительных колебаний температуры воды, химических раз-
дражений, исходящих от тела добычи или жертвы, и т. д.
Отсутствие у пиявок способности к регенерации, если не считать зале-
чивания ран, связано со строго фиксированным числом сегментов, с диф-
ференцировкой на обоих концах тела присосок, с компактным типом
внутреннего строения, в том числе сильно сократимой мощной интегри-
рованной мускулатурой тела. При таких особенностях строения сильное
повреждение тела приводит к гибели пиявки. Если у полихет и олигохет
с их бесполым размножением и огромной регенеративной способностью
края раны быстро заживают, легко происходит дедифференцировка всех
тканей и регенерация утраченных сегментов тела, у пиявок даже первый
этап регенерации — заживление незначительного повреждения тела —
оказывается затрудненным.
В индивидуальном развитии пиявок, исходя главным образом из ра-
бот Г. А. Шмидта (ряд работ с 1917 до 1951 г.), можно отметить некото-
рые основные моменты.
В развитии примитивных хоботных пиявок Glossosyphonidae наблю-
дается спиральное детерминированное гетероквадрантное многоклеточ-
ное синхронное дробление, ведущее к образованию пяти квартетов —
трех эктодермальных и двух эктодермальных, образование неполных кре-
ста и розетки, четырехлучевая радиальная закладка эктомезенхимы
и т. д. Цитотипический период, как и общий тип нёлйчиночного развития
этих пиявок, обычно сходен с таковым низших водных олигохет.
У ихтиобделлид, челюстных и глоточных пиявок спиральное дроблен
ние на очень ранних стадиях сменяется билатеральносимметричным и
уклоняется от такового полихет еще в большей степени, чем у дождевых
червей.
После образования пяти квартетов бластомеров у низших хоботных
пиявок, а у остальных без их образования начинается телобластическая
закладка четырех пар эктодермальных полосок, дающих покровы, коль-
цевую мускулатуру, центральную брюшную нервную систему и нефри-
дии, а также закладка одной пары мезодермальных полосок, что опреде-
ляет билатеральносимметричное строение дальнейших стадий развития
пиявок. Инкапсулированное личиночное развитие рыбьих, челюстных и
глоточных пиявок характеризуется образованием в коконе личинки,
заглатывающей белок и снабженной рядом личиночных органов; превра-
щение в червя поэтому естественно связано с образованием, а затем раз-
рушением провизорных органов личинки. Важной чертой онтогенеза всех
пиявок (кроме пока необъясненного исключения — ихтиобделлид), об-
щей с онтогенезом полихет, является развитие надглоточного ганглия,
независимо от зачатка брюшной нервной цепочки, из частей зародыша,
гомологичных верхнему полушарию трохофоры полихет, а также появле-
ние у пиявок головной лопасти.
В онтогенезе всех пиявок происходит развитие за счет мезодермаль-
ных полосок целома и целленхимы, которая у низших в меньшей степе-
ни, а у высших в сильной степени подавляет целом. Происходит морфо-
логическая и физиологическая субституция кровеносной системы лаку-
нарной (Заленский, 1915), состоящей из целомических сосудов и дости-
гающей наивысшей степени развития у челюстных и глоточных пиявок.
183
Простомий, полное расчленение переднего конца тела, слабое обособле-
ние сравнительно небольшой передней присоски, положение головного
мозга над глоткой, хорошо выраженное сегментарное строение кровенос-
ной системы (на примере Proioclepsis, по Ливанову, 1955) и типичное
строение метанефридиев характеризуют хищных и паразитических пло-
ских пиявок Glossosyphonidae как низших, наиболее примитивных пред-
ставителей хоботных пиявок. Это положение подтверждается и данными
онтогенеза (Шмидт, 1941а), в котором у хоботных пиявок сохранилось
больше черт спирального детерминированного гетероквадрантного дроб-
ления, характерного для онтогенеза полихет, чем в развитии какой-либо-
другой группы пиявок.
Ряд специфических черт строения ихтиобделлид, в особенности их он-
тогенеза, сравнительно с таковыми глоссосифонид заставляет считать
ихтиобделлид боковой ветвью, произошедшей от низших представите-
лей— Rhynchobdellea.. Данные строения и онтогенеза челюстных и гло-
точных пиявок позволяют выводить их от низших представителей хобот-
ных пиявок. Акантобделлид, сохранивших среди всех пиявок в наиболее
полном виде некоторые черты строения олигохет, о чем было сказано
выше, обычно не ставят в филогенетический ряд олигохеты — пиявки, а
рассматривают как боковую ветвь.
Нейросекреторный аппарат аннелид, особенно у полихет, развит хо-
рошо, что связано с высокой дифференцировкой нервной системы и об-
щим высоким уровнем жизнедеятельности кольчатых червей. Нейро-’
секреторный аппарат у них намного совершеннее, чем у плоских и пер-
вичнополостных червей.
" В головном мозгу и в ганглиях брюшной нервной цепочки бродячих и
сидячих полихет установлено несколько типов нейросекреторных кле-
ток, секрет которых распространяется по аксонам и выделяется в кровь;
мозг также является источником образования гормонов, действующих
тормозяще на половое созревание и на сопровождающие последнее сома-
тические изменения (эпитокия, столонизация). Другие продукты нейро-
секреторных клеток оказываются стимуляторами регенерационных про-
цессов (Arvy, 1954; Clark, 1959; Clark a. Bonney, 1960; Durchon et Kloot,
1960, 1962; Kloot, 1962; цит. no Gersch, 1964, и сводке, любезно сделанной
А. А. Пановым).
У олигохет нейросекреторные клетки обнаружены в надглоточном
ганглии и ганглиях брюшной нервной цепочки, а у ряда форм установ-
лен вывод их продуктов в кровеносные сосуды. Судя по результатам ис-
следований и опытов, нейросекреторный аппарат олигохет играет зна-
чительную роль в размножении и в процессах регенерации (Harms, 1948;
Hubl, 1953; Michon, 1959; Deuse-Zimmermann, 1960, Juberthie и Mestrov,
1962; Herlant-A'Ieewis, 1956, 1962, цит.. no Gersch, 1964).
У пиявок, так же как у выше рассмотренных групп аннелид, нейро-
секреторные клетки находятся во всех отделах центральной нервной си-
стемы; продукты их секреции, связанные с размножением пиявок, по ак-
сонам попадают в нейропиль (Scharer, 1937; Hagadorn, 1958, 1962; На-
gadorn, Bern and Nishioka, 1962; Legendre, 1959, цит. no Gersch, 1964).
Анализ метамерии аннелид полностью еще не проведен: для очень
многих семейств ни анатомически, ни эмбриологически, ни эксперимен-
тально не установлено число ларвальных сегментов. По имеющимся
данным, число это различно и колеблется между 3 и 13. У семейства
Phyllodocidae ларвальных сегментов 7—8, у Aphroditidae — 4—8 (у под-
семейства Polynoinae — 7—8, Sygalioninae — 4—5?), Glyceridae — 9, Syl-
liidae — 3, Nereidae — 4, Spionidae— 9—13, Capitellidae — 9—13 и у Ser-’
pulidae — 3—7 (7?). По мнению автора теории ларвальных и постлар-
вальных сегментов П. П. Иванова, семейства Polygordiidae, Protodrilidae
и Saccocirridae не имеют ларвальных сегментов, ларвальным отделом у
184
Polygordiidae П. П. Иванов считал лишь головную лопасть. По мнению
В. Н. Беклемишева, трактующего работу Йегерстена (Jagersten, 1952)
по развитию Protodrilus, у этого семейства имеется 6 ларвальных сег-
ментов. Тело Dinophilidae и Myzostomiida состоит из 5 ларвальных сег-
ментов и постларвального нечленистого отдела.
Что касается олигохет, то у семейства. Naididae число1 ларвальных.
сегментов 5—7, у Tubificidae—-5—7, Phraeorycfidae — 5—6?, Lumbriculi-
dae — 7, Enchytraeidae — 3, Lumbricidae — 5. Число ларвальных сегмен-
тов у олигохет, как видно, варьирует значительно меньше, чем у поли-
хет. У пиявок, в основном по анатомическим данным, считается 4—5 лар-
вальных сегментов, что близко к наиболее часто встречающемуся среди
олигохет числу 5 и, видимо, постоянно, поскольку число туловищных сег-
ментов в теле пиявок невелико (33) и вся организация пиявок весьма1
компактна, особенно если сравнить ее с таковой олигохет.
Большое разнообразие в количестве ларвальных сегментов у полихет'
может быть понятно, если вспомнить, что число сегментов у взрослых
червей не фиксировано, оно велико и часто колеблется в очень больших
пределах. Однако у нас до сих пор нет объяснений ряда случаев выпаде-
ния развития ларвальных сегментов. Так, остается непонятным отсут-
ствие ларвальных сегментов, например', у Polygordiidae, скорее более
примитивных, чем вторично упрощенных полихет, имеющих типичное для
полихет развитие с трохофорой. Мало убедительно утверждение
П. П. Иванова, что единственной ларвальной частью у Polygordius яв-
ляется простомий. По данным работы последнего времени (Ander-
son, 1959), у полихеты Scoloplos artniger все сегменты тела разви-
ваются совершенно одинаково (сомиты целома, параподии, щетинки, му-
скулатура). Имеются лишь небольшие особенности в развитии парапо-
дий и щетинок у первого и второго сегментов зародыша. Главное разли-
чие сегментов у Scoloplos заключается в том, что первые 12 сегментов-
образуются у зародыша внутри плотного кокона; это, по выражению
Андерсона, эмбриональные сегменты, а следующие 12 — постэмбрио-
нальные возникают после паузы по вылуплении личинки из кокона.
Лишь по этой.особенности можно различать первые 12 сегментов от сле-
дующих 12 и называть их ларвальными и постларвальными. Однако раз-
личий между ними, по существу, нет. В. А. Свешников (19'59а, 19596)
также не мог ничего сказать о наличии ларвальных и постларвальных
сегментов при развитии этого вида, но он не видел и приостановки в
развитии сегментов, описанной Андерсоном. Характерно, что в западно-
европейских водах развитие трохофоры у Scoloplos armiger протекает'
внутри кокона (Anderson, 1959), а в Белом море (Свешников, 1958а,.
19586) яйцо развивается в поверхностных слоях воды, имеется свободно-
живущая трохофора, далее метатрохофора, которая превращается в.
нектохету с 12 сегментами, становящуюся 24-сегментным червем.
Надо думать, что отмеченные выше (и ряд других) нарушения в раз-
витии сегментов двух категорий у полихет найдут свое объяснение. Боль-
шие различия в числе ларвальных сегментов у полихет, как было сказа-
но выше, могут объясняться большими различиями в числе сегментов
тела взрослых форм и большим разнообразием в их строении и биологии
развития. В силу этого число ларвальных сегментов у современных по-
лихет имеет небольшое филогенетическое значение, но, будучи постоян-
ным в разных семействах полихет, оно становится более важным для
вопроса о происхождении других классов кольчатых червей, потомков
полихет и артропод.
Полихеты и олигохеты, будучи массовыми обитателями моря, почвы
и пресных вод, играют и играли в прежние эпохи важную роль в форми-
ровании и изменении биоценозов и в пищевых связях. Иловые и песчаные
полихеты и дождевые черви, пропуская через кишечник массы грунта,
185,
химически и механически его изменяя, проделывают и проделывали
большую геологическую работу. Так, 10 экземпляров Arenicola (Свеш-
ников, 1963а, 19636) в год пропускают 146 кг грунта; при численности
этих червей 40 экземпляров на 1 м2 (Linke, 1939) за год они практиче-
ски переворачивают весь слой грунта, в котором живут.
Как известно (Гиляров, 1949а, 19496), громадную роль в почвообра-
зовании среди почвенных беспозвоночных играют олигохеты. С их жизне-
деятельностью связаны водопроницаемость, структура почвы, гумифика-
ция, развитие аккумулятивного горизонта, плодородие и т. д. Значитель-
на была роль и вымерших полихет; так, из известковых трубок серпулид
сложены пласты горных пород, например нижнемеловой серпулит мощ-
ностью в 50 м, рифовые известняки среднего миоцена и т. д. (Давиташви-
ли, 1958).
Отмечая особенности биоэкологии аннелид и возможности путей эво-
люции основных их классов, следует иметь в виду, что почва как среда
-обитания во многих отношениях является промежуточной между водной
и наземной средами (Гиляров, 1949а, 19496). При переходе к жизни на
суше организмы должны приспосабливаться к защите организма и орга-
нов дыхания прежде всего от высыхания, хотя процесс газообмена в ор-
ганах дыхания при кожном дыхании возможен и в воздушной среде.
В связи с этим случаи перехода водных (морских) червей непосредствен-
но к наземной жизни очень редки. Известно, что по своим свойствам дно
водоемов является специфической почвой, а почвенная вода — специ-
фическим водоемом (Гиляров, 1949), поэтому в эволюции кольчатых чер-
вей гораздо большее значение имеет переход их от жизни на дне моря
к жизни в почве. Угроза высыхания всего организма, особенно его орга-
нов дыхания, при таком переходе значительно снижается, а следователь-
но, увеличиваются шансы приспособления к жизни в почве — среде, близ-
кой по ряду свойств к водной. Именно такой экологический путь
эволюции кольчатых червей вполне обоснованно рисует М. С. Гиляров
(1949).
Богатство и разнообразие форм в прибрежной зоне как более прими-
-тивных бродячих полихет, так и более подвижных сидячих, живущих во
временных трубках, наличие среди них значительного числа обитателей
грунта являются косвенным обоснованием взгляда на такой экологиче-
'ский путь эволюции групп аннелид. Полихеты — типичные обита-
тели моря в современной фауне — дают отдельные примеры (семейство
Nereidae) перехода через солоноватоводную среду в пресные воды, с
уходом на сотни километров от берега моря, например Lycastis гапаиеп-
sis Feuerborn, Lycastopsis catarractus Feuerborn. Известны и отдельные
примеры перехода полихет к наземному образу жизни в подстилке меж-
ду влажными листьями кокосовых пальм — Nereis vi.tabun.da, Lycastis
vivax, Lycastopsis amboinensis Pflugfelder (последняя живет вместе с
перипатусами, сколопендрами и скорпионами). В связи с таким образом
жизни у них сильно развивается кутикула, образуется своего рода голов-
ная капсула как у сколопендры, некоторые из них могут даже бегать
на параподиях с намечающейся членистостью и поставленных перпенди-
кулярно к почве, как ноги членистоногих. Для них характерно дыхание
всей поверхностью тела, образование углублением покровов зачатков
трахееподобных трубок, развитие, протекающее в яйцах, которые при-
крепляются к спинным циррам параподий, и т. д. Наряду с этим обычная
морская форма Nereis diversicolor, заходящая в солоноватоводную сре-
ду, а также далеко в пресные воды рек, в сухих местах переходит к жиз-
ни в почве, делая ходы, подобно дождевым червям.
Это говорит о том, что полихеты являются наиболее пластичной груп-
пой кольчатых червей не только по многообразию внешнего и внутрен-
него строения, но и по экологическим особенностям, оставаясь в целом
Я 86
морскими обитателями. Это действительно центральная группа аннелид,
указывающая пути, эволюции и происхождения не только олигохет и
пиявок, но и членистоногих, онихофор, тардиград и пентастомид. Можно
пожалеть, что физиологически они остаются слабо изученными.
И морфологическими экологически олигохеты и пиявки значительно
менее пластичны, чем полихеты. Олигохеты, являясь преимущественно
типичными почвенными обитателями (прежнее деление — Terricola, или
Megadrili), представлены меньшей по объему группой пресноводных
форм (Limicola, или Microdrili). Характерно, что среди последних семей-
ство Enchytraeidae, представленное в основном почвенными обитателями,
включает экологически различные формы — амфибиотические, иловые,
а также литоральные морские. Отдельные случаи приспособления к жиз
ни в почве были установлены для водного обитателя Tubifex, а среди
типичных почвенных форм наиболее богатого видами (почти 300 видов)
рода Pheretima Kinberg встречаются виды, ведущие во влажном воздухе
открытый наземный образ жизни и ползающие по деревьям.
Семейства Aeolosomatidae, Naididae. и' Tubificidae (почти исключи-
тельно пресноводные обитатели), Phraeodrilidae и Enchytraeidae обла-
дают рядом примитивных черт в строении и эмбриональном развитии в
большей степени, чем. другие олигохеты, сохранившие спиральное гетеро-
квадрантное дробление с чертами креста и розетки. Эти семейства, оче-
видно, надо считать исходной группой олигохет, которая могла произой-
ти от полихетных морских предков, перешедших от придонной жизни
в береговой полосе моря в-донном иле и песке к жизни в солоноватовод-
ных условиях и позже в пресных водах. В этом отношении не хватает
сведений о строении и развитии тех немногих видов семейств Aeolosoma-
tidae, Naididae и Tubificidae, которые живут в море, тем более, что неко-
торые роды — Heterodrilus, Limnbdrlloides — являются стеногалинными,
чисто морскими обитателями.
Характерно, что и многие пресноводные, и все морские олигохеты
являются обитателями грунта. Эта особенность экологии, дает возмож-
ность предположить, что огромная часть олигохет, так называемых дож-
девых червей (прежние Terricola, или Megadrili), произошла от прими-
тивных олигохет путем их перехода от донной жизни в водной среде к
жизни в почве. Это почти несомненно, тем более, что влажная почва, в
которой проходит активная жизнь, размножение и развитие земляных
червей, по своим основным свойствам очень близка к водной среде и тем
более к условиям дна водоемов (Гиляров, 1949).
Для большинства олигохет характерно и наиболее типично питание
•отмершими частями растений, с чем связана при колоссальных количе-
ствах особей, населяйщих почву, их громадная роль в почвообразовании,
в частности в образовании ее плодородного слоя. Некоторую роль в пита-
нии олигохет играют при этом животные остатки. Лишь немногие олиго-
хеты, например Agriodrilus, Chaetogaster, Eiseniella, являются хищни-
ками. И, наконец, как исключение, среди олигохет наблюдается эктопа-
разитизм (Branchiobdellidae).
Основные данные строения, развития и экологии полихет, олигохет и
пиявок позволяют представить общий ход эволюции аннелид. Очевидно,
эволюция этой группы происходила путем перехода древних (ископае-
мых) примитивных бродячих полихет от придонной жизни, а также жиз-
ни в песке и толще ила литорали и сублиторали моря сначала к жизни
во влажной почве побережья моря, эстуарий рек, солоноватых и опрес-
ненных прибрежных водоемов, а через нее к жизни в почве, что постепен-
но стало характерным для преобладающего большинства (примерно 4/5
видов) современных олигохет. Во многом примитивное строение с рядом,
однако, черт вторичного упрощения, сохранение, особенно в раннем онто-
генезе, некоторых черт, общих с развитием полихет, а также сохранение
187
исходного водного, хотя уж не морского образа жизни, определяют каю
низших олигохет такие семейства, как Aeolosomatidae, Naididae и Tubi-
ficidae. И вместе с тем среди них нет групп, которые по своим особенно-
стям позволяли бы связывать олигохет с какой-либо определенной груп-
пой современных полихет. Единственно, что можно сказать, это что
предков олигохет надо искать среди вымерших полихет отряда Eunice-
morpha или Nereimorpha. Если согласиться с Раймондом, что его силу-
рийская Protoscolex batheri — действительно ископаемый малощетинко-
вый червь, то возникновение класса олигохет относится к раннему па-
леозою.
Вопрос о филогенетических связях олигохет и пиявок в общей'форме
также ясен. Взгляд Вейдовского (Vejdovsky, 1888—1892) о происхожде-
нии пиявок от олигохет был развит в последующих работах многих ав-
торов: Апати, Михаэльсона, Стефенсона, Скрибана, Аутрума и особенно
Ливанова, Шмидта и Изосимова. Наиболее верны взгляды на филогению
олигохет и пиявок Ливанова и Шмидта, основанные на данных анатомии,
эмбриологии и экологии. Многие зарубежные зоологи на основании неко-
торых черт строения и хищного образа жизни ряда форм Lumbriculidae,
и особенно байкальского Agriodrilus, искали родоначальников пиявок в
этом семействе. Тщательное изучение наружного строения и внутренней
организации Agriodrilus и сопоставление их с таковыми всего семейства
Lumbriculidae и пиявок, проделанное В. В. Изосимовым (1940, 1934),
привело последнего к отрицанию значения Agriodrilus как переходной
формы между олигохетами и пиявками, что подкрепило взгляд Н. А. Ли-
ванова, вообще отрицающего значение Lumbriculidae как родоначальной
группы пиявок.
Известное сходство в строении переднего конца тела Agriodrilus и пи-
явок является результатом конвергенции. Целом здесь редуцируется не
путем разрастания соединительной ткани, как у пиявок, а вследствие
мощного развития мускулатуры переднего отдела кишечника, что связа-
но с хищничеством Agriodrilus. Сходство в строении семенных мешков с
таковыми пиявок является чисто внешним, половые отверстия, как и у
остальных олигохет, парные (у пиявок они непарные). Если некоторые.
особенности Agriodrilus имеют конвергентное сходство с таковыми пия-
вок, а другие являются своеобразными, свойственными лишь этой группе,
например, сильное утолщение диагональных- и кольцевых мышц кожно-
мускульного мешка, то в целом основные черты внешнего строения и
внутренней организации Agriodrilus присущи семейству Lumbriculidae.
Кроме других, указанных выше, черт строения Lumbriculidae, говорящих
против возможности выводить пиявок от них, по мнению Н. А. Лива-
нова, уже наличие у них четырех пар нервов в невросомите не позволяет
производить пиявок от Lumbriculidae, так как нет никаких оснований
допускать редукцию четвертой пары нервов.
Более обоснованно считать, согласно Ливанову, что филогенетически
пиявки связаны с олигохетами через семейство Phraeoryctidae (Haplo-
taxidae).
Отсутствие диссепиментов в первых пяти сегментах тела, пять пар
кольцевых кровеносных сосудов, наличие нефридиев, начиная лишь с
6—7-го сегмента (т. е. пяти ларвальных сегментов), метамерные клапаны
в спинном кровеносном сосуде, сравнимые с многоклеточными грозде-
видными метамерными образованиями в спинном кровеносном сосуде
Acanthobdella и низших хоботных пиявок, строение переднего отдела
кишечника с мускулистой глоткой, коротким пищеводом, переходящим
в 6-м сегменте в среднюю кишку, вторичная членистость семенников —
все это является характерными чертами строения Phraeoryctes, близкими
к таковым пиявок. Н. А. Ливанов, кроме этих черт, отмечает также сход-
ство между Phraeoryctes и пиявками в наличии в стенках средней кишки
188
наружных кольцевых и внутренних продольных мышц (у семейства Lum-
briculidae и остальных олигохет расположение этих мышц обратное)
и невросомита с тремя парами нервов (у Lumbriculidae четыре пары нер-
вов). Все это позволяет видеть в Phraeoryctidae группу, которая могла
дать начало пиявкам.
В филогенетическом ряду олигохеты — Phraeoryctidae — пиявки
Н. А. Ливанов правильно помещает паразитических олигохет Branchi-
obdellidae, обладающих типичными чертами олигохет и некоторыми чер-
тами, общими с пиявками.
К олигохетным чертам строения Branchiobdellidae относятся следую-
щие: хорошо развитый целом, отсутствие мезенхимы, строение головного
мозга, брюшной нервной цепочки с тремя парами нервов в невросомите,
строение средней кишки, наличие сердечного тела в спинном кровенос-
ном сосуде, особенности строения полового аппарата. Наряду с этим под
влиянием паразитического образа жизни у Branchiobdellidae возникают
черты, конвергентные с пиявками: так, число сегментов тела постоян-
но (15), передние сегменты сливаются вместе, из трех сегментов обра-
зуется задняя присоска, в передних сегментах частично редуцируется
целом, в стенках средней кишки кольцевые мышцы расположены сна-
ружи, продольные — внутри, мужское половое отверстие непарное,
анальное отверстие лежит на спинной стороне, передвижение при по-
мощи задней присоски, паразитизм. Пример олигохеты Branchiobde.Ha
показывает возможность приобретения олигохетами признаков пиявок
под воздействием условий жизни.
Тардиграды (Tardigrada)
Tardigrada, открытые в 1776 г. Спалланцани, которому они обязаны'
своим названием, широко известны своим анабиозом, асфиксией и инци-
стированием. Это обитатели прибрежной полосы моря, пресных вод и
сырых наземных биотопов; по особенностям строения и развития они.
представляют и большой интерес и вызывают большие трудности при их.
толковании (рис. 50). .
Графф (Graff, 1877) создал для Tardigrada, Myzostomida и Реп-
tastomida отряд кольчатых червей Stelechopoda, объединив их на осно-
вании ряда некоторых общих признаков, прежде всего на основании
наличия нечленистых ножек. Эту точку зрения поддержал в свое время
Геккель (Haeckel, 1896), а позже и Ганстрем (Hanstrom, 1928), который,
анализируя строение нервной системы, признал полихетную природу тар-
диград. Примерно одновременно с Граффом и Геккелем Плате (Plate,
1888) и Эрлангер (Erlanger, 1894, 1895), исходя из результатов собствен-
ных исследований строения и развития тардиград, относили их к типу
членистоногих. В последнее время одни ученые (Беклемишев, 1944, 1952,.
1964; Cuenot, 1949) считают тардиград подтипом Articulata, другие (на-
пример, Kaestner, 1954/1955)—самостоятельным типом, наряду с
Onychophora и Linguatulida, располагающимся между типами Annelida
ri Arthropoda.
Как у пентастомид и онихофор, у тардиград имеются в строении и в
индивидуальном развитии черты аннелидные (полихетные), артроподные
и специфические, своеобразные.
К чертам аннелидным (полихетным) относятся отсутствие обособлен-
ной головы, простомий, представляющий собой передний конец тела, не-
хитиновая кутикула покровов, нечленистые и не сочлененные с телом
конечности, кожные железы ног, секреторной деятельности которых
обязано образование новых коготков взамен изношенных старых, наличие
головного (церебрального) и подглоточного ганглиев, соединенных с
каждой стороны наружной, и внутренней коннективами с первым ганг-
лием брюшной нервной цепочки, иннервация головным ганглием чувстви-
тельных придатков головы, наличие в каждом брюшном ганглии трех
пар периферических нервов, не образующих, однако, кольца, а также-
наличие рта без ротовых органов — измененных конечностей тела —
и гладкой мускулатуры.
Артроподными признаками тардиград являются наличие линек, от-
сутствие ресничек, образование конечностей в виде полых выростов сте-
нок тела, система отдельных специализированных мышечных пучков
вместо кожно-мускульного мешка, слюнные железы и мальпигиевы
сосуды эктодермального происхождения, наличие клеток в полости тела,
соответствующих клеткам жирового тела, редукция вторичной полости
тела, дефинитивная полость тела, образованная остатками целома и схи-
зоцельных полостей, целомическая природа полостей половых желез,.
190
Рис. 50. Тихоходка (Echl--
noscoides sigismundi),
ползающая по водоросли
(из Kaestner по Maijcus)-
образование мужских и женских выводных
протоков целомодуктами, недетерминирован-
ное, неспиральное дробление.
Наряду с признаками аннелидного и артро-
цодного характера, у тардиград имеются свое-
черты. Среди них можно отметить
олигомерную метамерию тела, внешне не сег-
ментированного и состоящего из простомия и
пяти сегментов, наличие четырех пар нечлени-
стых ног, отсутствие органов дыхания и кро-
вообращения, сообщение половых протоков с
клоакой, постоянство клеточного состава ряда
органов, очень сильное укорочение хода ранне-
го эмбриогенеза (деляминационная целоблас-
тула), возникновение ануса на месте замкнув-
шегося бласт,опора, энтероцельное образова-
ние пяти пар целомических мешков.
Простое перечисление черт аннелидного
(полихетного), артроподного и своеобразного
характера в строении и развитии тардиград го-
ворит совершенно ясно об изолированном си-
стематическом и филогенетическом положении
этой группы. Олигомерная метамерия (голов-
ной отдел и пять туловищных сегментов, кото-
рые надо считать ларвальными) и малые раз-
меры тела позволяют признать, аналогично ди-
нофилам и мизостомидам, неотеническую при-
роду тардиград, что уже отмечалось в литера-
туре. При таком толковании отсутствие на-
стоящих ротовых органов (производных конеч-
ностей тела) у тардиград является первичным,
свойственным членистоногим на ранних эта-
пах развития. Это скорее результат остановки
развития, закрепленный исторически, а не рег-
ресса, поскольку тихоходки — свободноживу-
щие формы. Неразрывно с этим и с малыми
размерами связано также отсутствие у них ор-
ганов дыхания и кровообращения. Очевидно, несмотря на сравнительно-
толстую кутикулу покровов, дыхание тардиград, в основном водных ор-
ганизмов, обеспечивается через покровы тела; малая величина тела, очень
сильная его сократимость и отсутствие больших тканевых масс в орга-
нах тихоходок, несомненно, способствуют как легкому газообмену всего
тела, так и циркуляции полостной жидкости, что заменяет функцию кро-
веносной системы. Тот факт,-что мальпигиевы сосуды имеются не у всех
тихоходок, говорит о том, что функция выделения у них в основном осу-
ществляется кишечником. Факт развития органов выделения у трохо-
форных личин,ок полихет, наличие их у взрослых представителей неоте-
нических олигомерных групп кольчатых червей, отсутствие типичных ор-
ганов выделения и наличие мальпигиевых сосудов у тихоходок являются
чертами значительного сходства их с членистоногими. В сущности го-
воря, природа любой из упомянутых выше черт тихоходок, аннелидных
или артроподных, не вызывает сомнения; вопрос заключается в том, ка-
кой категории придавать решающее значение при определении приро-
ды этой группы животных? Однако в организации тардиград признаки
обеих групп находятся в очень сложном противоречивом единстве. Так.
кожно-мускульного мешка, признака кольчатых червей, у тардиград нет,,
мускулатура тела у них представлена отдельными, индивидуализиро-
Рис. 51. Эмбриональное развитие тихоходки Hypsi-
bius convergers (по Mancus из Pflugfelder)
Л — стадия 4 бластомеров; Б — целобластула; В — двуслой-
ный зародыш, образовавшийся деламинацией; Г — образо-
вание ануса на месте замкнувшегося бластопора; Д — энте-
роцельное образование пяти пар целомических мешков:
/ — анус; 2 — целомические мешки
ванными пучками
мышц, что типично
для членистоногих.
Но вместе с тем эти
быстро и энергично
сокращаю щ и е с я
мышцы лишены по-
перечнополоса т о й
структуры, харак-
терной не только
для всей мускулату-
ры членистоногих,
но и для некоторых
мышц других бес-
позвоночных, в том
числе и для анне-
лид. Таким образом,
по одним суще-
ственным ' чертам
тихоходки близки к
аннелидам, по дру-
гим — к членистоно-
гим, но в сочетании,
не свойственном ни
тем, ни другим.
Крайне своеоб-
разными являются
некоторые очень су-
щественные черты
онтогенетическ ого
развития. Образова-
ние рта независимо
от бластопора, су-
ществование которого Маркус (Marcus, 1919, 1958) отрицает, возникнем
венпе ануса на месте бластопора, эктодермы деляминацией, энтероцель-
ное возникновение пяти пар целомов (рис. 51), не образующих вторич-
ной полости тела,— все эти черты являются особенностями онтогенеза
тардиград, выделяющими их из всей группы типов Protostomia, а не
только аннелид и артропод. Нам хорошо известны уклонения от .общего
основного хода индивидуального развития сврбодноживущих форм пара-
зитических форм данного типа. Чаще, как известно, именно в ходе онто-
генеза паразитических форм, утерявших в своем внешнем строении и
внутренней организации признаки класса и даже типа, мы находим со-
хранение черт, общих с таковыми сврбодноживущих родственных форм.
Тихоходки и в этом отношении представляют необычную особенность.
Если в их организации, как отмечалось, наряду со своеобразными, име-
ются аннелидные и артроподные черты, то ряд основных черт их инди-
видуального развития чужд и тем, и другим и вообще всем первично-
ротым.
В целом весь ход индивидуального развития тихоходок носит про-
-стой, примитивный характер сравнительно с развитием артропод и анне-
лид. Давно высказывалось резонное мнение (например, Шимкевич)
о том, что недетерминированное, равномерное дробление и энтероцель-
ное образование целомов является более примитивным, чем детермини-
рованное по спиральному типу развитие и образование целомов из тело-
бластов, дающих начало мезодермальным полоскам, расчленяющихся на
сомиты, в которых образуются целомические полости. Считаясь с этим.
.192
в энтероцельном образовании целомов у тихоходок можно видеть древ-
нюю, примитивную черту онтогенеза.
Таким образом, ряд черт строения тихоходок являются сходными с
таковыми аннелид и членистоногих, и значение их можно понять путем
сравнения с теми и другими. Ряд своеобразных особенностей их орга-
низации также становится понятным, если считать тихоходок неотени-
ческими олигомерно членистыми формами, у которых все же имеется
больше сходства с членистоногими, чем с кольчатыми червями. Однако,
[ , наличие в их онтогенезе черт, чуждых и аннелидам, и членистоногим,
I о которых только что было сказано, подчеркивает своеобразие этой
I группы животных и заставляет считать их самостоятельным типом, по-
добно онихофорам и пентастомидам. Без этого тихоходок можно было
бы признать классом неотенических олигомерных членистоногих, тем
более, что меловые ископаемые тихоходки (Beorn leggi Cooper, 1964)
найдены в янтаре вместе с разными насекомыми, пауками и клещами).
Однако они не связаны ни с одним из подтипов или классов Arthropoda.
I Происхождение тардиград остается пока совершенно загадочным, как
1; и неопределенным положение их в системе.
13 Д' М. Федотов
Пентастомиды (Pentastomida)
Эндопаразитизм и отсутствие среди пентастомид свободноживущих
форм крайне затрудняют выяснение их природы и положения в системе,
между тем известно, насколько в таких случаях важна роль экологиче-
ских особенностей изучаемой группы животных. Обращает на себя вни-
мание очень слабое распространение среди полихет, родство с которыми
находят у пентастомид, эктопаразитических и еще в меньшей степени
эндопаразитических форм. Характерно, что паразиты при этом сохра-
няют и во внешней, и во внутренней организации полихетный тип строе-
ния. Уклонения от типичного строения у паразитических полихет затра-
гивают отдельные черты организации и незначительно превышают осо-
бенности, которые возникают под влиянием условий жизни и у ряда сво-
бодноживущих полихет. Исключением являются мизостомиды с их очень
древним паразитическим образом жизни, который привел их к обособле-
нию в самостоятельную, хотя и близко родственную полихетам группу
(Федотов, 1.915, 1937, см. гл. «Аннелиды»), Напротив, судя по всем при-
знакам и особенностям типичных паразитических полихет, они перешли
к паразитическому образу жизни исторически недавно.
В отличие от полихет у членистоногих при широком распространении
среди них паразитизма наблюдаются, порой даже у эктопаразитических
форм, очень сильные регрессивные изменения, стирающие черты класса
и даже типа.
Для пентастомид — паразитов исключительно позвоночных животных,
рептилии являются основными, вероятно наиболее древними, первичными
их хозяевами, тогда как другие позвоночные — немногие млекопитающие
и совсем единичные виды птиц и амфибий — стали их хозяевами много
позже. Можно думать, что паразитизм пентастомид очень древний
и возник среди водных, вероятно, еще морских представителей рептилий,
которые по условиям жизни могли иметь контакт с полихетными пред-
ками пентастомид. Наиболее примитивные пентастомиды являются пара-
зитами рептилий, а наиболее измененные — паразитируют в теплокров-
ных (рис. 52).
Как известно, Графф (Graff, 1877, см. гл. «Тардиграды») и Геккель
(Haeckel, 1896), признавая аннелидную природу пентастомид, помещали
их вместе с мизостомидами и тардиградами в класс Stelechopoda, как
регрессировавших под влиянием паразитизма аннелид. Более широко
среди зоологов распространен взгляд о принадлежности пентастомид к
артроподам и о близости их к паукообразным (даже клещам). Доводами
в пользу родства пентастомид с членистоногими были следующие дан-
ные, несколько дополненные аргументами и с моей стороны: слоистое
строение хитина, устойчивого к действию едкой щелочи, легко раство-
ряющей кутикулу полихет, линьки, отсутствие ресничек, гистологические
особенности мускулатуры — поперечная полосатость мышц с прикрепле-
нием миофибрилл не прямо к покровам, как у полихет, а с помощью
194
тонофибрилл, проникающих в плазму покровных клеток, гроздевидные
яичники, сложное строение мужских и женских выводных частей половой
системы (рис. 53), коадаптация строения циррусов самцов и вагины са-
мок, внутреннее оплодотворение, характер процессов созревания яиц, яй-
цевые оболочки, дорсальный орган, зародышевые полоски впереди рас-
ширенные в две лопасти и расположенные набрюшной стороне зародыша.
В той или иной мере эти черты пентастомид сближают их с артропо-
дами, тем более, что в различии переднего и заднего отделов тела пента-
стомид также имеется известное сходство с отделами тела членистоногих.
Рис. 52. Внешний вид
пентастомид (из Hennig)
А — Cephalobaena tetrapoda;
Б — Armillifer armillatus
Рис. 53. Схема организации пентастомид (из Hennig)
А—самец Raittietlella mediterranean Б—самка R.. boulengeri', 1—се
менник; 2—семенной пузырек; <3—выносящий, проток;. 4—пенне;.
5—яичник; 6—яйцевод; 7—матка; 8—вагина; 9—семяприемник;
10—половое отверстие
Однако в работах 20-х годов и позже .(Heymons, 1926, 1935; Haffner,
1922, 1924, 1926; Hanstrom, 1928; Ежиков, 1939, 1951) найдены и под-
черкнуты важные черты организации пентастомид аннелидного (полихет-
ного) характера (Jeschikov, 1940). Эти авторы установили ряд фактов,
позволяющих считать пентастомид не мезомерными формами, подобно
паукообразным (в частности, клещам), а измененными полимерными
членистыми животными, подобно кольчатым червям и многоножкам,
число сегментов которых может, превышать сотню. Дольчатость покровов
у взрослых пентастомид, которую необоснованно сравнивали с кольча-
тостью червеобразно удлиненного тела таких клещей, как Eriophyes или.
Demodex, на самом деле является внешним выражением внутренней сег-
ментации, или метамерии. Ей соответствует деление на отрезки продоль-
ных мускулов кожно-мускульного мешка, метамерное расположение ко-
сых (или дорсо-вентральных) мышц, боковых органов и отхождение
по три пары боковых нервов от двух продольных нервных стволов. Это
позволяет считать мнение Геймонса (Heymons, 1926), Гаффнера (Haff-
ner, 1926)'и И. И. Ежикова (1939, 1951) о метамерном строении пента-
стомид обоснованным. Далее, благодаря данным Геймонса, Гаффнера и
Ганстрёма, мы можем считать, что концентрированная короткая брюш-
ная нервная система с пятью парами боковых нервов и с окологлоточным
кольцом, без головного ганглия у более измененных пентастомид, вроде
13:
195
Petitastomum, является результатом вторичного изменения первона-
чально более примитивного состояния лестничной брюшной нервной
системы, связанной окологлоточным кольцом с головными ганглиями.
Согласно данным Гаффнера (Haffner, 1924), мы имеем основания счи-
тать, что два продольных брюшных нервных ствола — это не два задних
нерва, отходящих от последней пары ганглиев, но судя по развитию их из
зачатков, состоящих из ганглиозных клеток, это брюшные стволы, уте-
рявшие ганглионарный характер, но сохранившие метамерно отходящие,
боковые нервы, т. е. элемент невросомитов. Поэтому прав Ганстрём
(Hanstrom, 1928), проводя гомологию между надглоточным ганглием
пентастомид и надглоточным ганглием, или мозгом, полихет.
При таком толковании строения нервной системы более примитивных
пентастомид и полихет очень большое сходство в строении боковых орга-
нов полихет и некоторых пентастомид (Haffner, 1926) позволяет допу-
стить их гомологию. Таким образом, можно считать боковые органы
пентастомид сохранившимся наследием полихетных предков, а не ре-
зультатом конвергенции, так как условия жизни пентастомид слишком
уж отличны от условий жизни типичных полихет или паразитических
мизостомид, также имеющих боковые органы. При этом можно отметить,
что боковые органы пентастомид меньше отличаются от таковых типич-
ных полихет, чем боковые органы мизостомид. Замечу, однако, что такие
зоологи, как В. Н. Беклемишев и Кестнер, видимо, не согласны с по-
добной трактовкой, так как они вообще умалчивают о боковых органах
пентастомид.
Наличие у пентастомид кожно-мускульного мешка, состоящего из
слоя кольцевых мышц, слоя продольных мышц, поделенного на две спин-
ных и' две брюшных ленты, из косых, дорсо-вентральных или диагональ-
ных мышц, также является аннелидной чертой строения. Приходится,
однако, решать, чему придавать большее значение: общему плану строе-
ния мускулатуры тела пентастомид, сходному с таковым полихет, или
тонкому гистологическому строению (поперечной полосатости мышц и
способу прикрепления мышечных волокон тонкими фибриллами, прони-
кающими в плазму эпителиальных клеток), характерному для членисто-
ногих.
Такое смешение в одной системе различных черт, свойственных двум
сильно отличающимся друг от друга группам (аннелидам и артроподам),
заставляет поставить трудный для разрешения вопрос, что же является
в организации пентастомид первичным и что вторичным? Сторонники
взгляда об аннелидной (полихетной) природе пентастомид не считают
наличие поперечнополосатой мускулатуры достаточной аргументацией
противоположной точки зрения, так как поперечнополосатая мускула-
тура встречается у ряда полихет. По этому поводу надо сказать, что по-
перечнополосатые мышцы вообще встречаются не только у полихет, но
и у очень различных групп животных, например у низших червей, мол-
люсков и даже кишечнополостных, причем такое строение свойственно
отдельным группам быстро и сильно сокращающихся мышц. По моим
исследованиям (1937), поперечнополосатая мускулатура наблюдается у
очень подвижных, эктопаразитических мизостомид, но насколько функ-
ционально необходима быстрая сократимость мускулатуры тела у эндо-
паразитов— пентастомид — нам неизвестно. Очень возможно, что она
связана с постоянными и сильными изменениями емкости легких, в ко-
торых живет большинство пентастомид. Однако мышечные волокна пен-
тастомид со слабо дифференцированной поперечной полосатостью мио-
фибрилл, расположенных в один ряд по периферии саркоплазмы, не-
сравнимы со сложными полиэнергидными мышечными волокнами члени-
стоногих, как несравнимы с последними и поперечнополосатые мышцы
других беспозвоночных, в том числе и кольчатых червей, с которыми
196
сходны мышцы пентастомид. Если только Гаффнер прав (Haffner, 1924),
сходство их с мышцами членистоногих заключается лишь в том, что
миофибриллы сначала переходят в сухожильные волокна (тонофиб-
риллы), а затем в клетки гиподермы, а не прямо в последние, как у
червей.
Нарушение вернее выпадение, у пентастомид ряда других сущест-
венных черт строения сегмента, или метамера (конечностей, диссипи-
ментов, выделительных органов, ангиосомитов) вызвано, очевидно, как
и отсутствие ресниц, общим регрессом организации пентастомид, которое
в значительной мере типично и для членистоногих, у которых это в из-
вестной степени обусловлено интеграцией отделов тела, т. е. прогрессив-
ным развитием сравнительно с уровнем организации полихет. Имеется,
несомненно, сходство, хотя скорее внешнее, в строении полости тела пен-
тасдомид и частей целома полихет (Haffner, 1926; Ежиков, 1939). Однако
эпителиальной выстилки стенки полости тела пентастомид не имеют,
кроме того, у нас, к сожалению, нет решающих данных по отногенезу
полости тела у пентастомид и мы не можем установить, является ли по-
лость тела взрослых форм остатками целома или миксоцелем.
К аннелидным особенностям пентастомид относится также отсутствие
и в онтогенезе, и во взрослом состоянии ротовых органов — измененных
конечностей тела, а также нечленистый характер двух или трех пар
личиночных или дефинитивных придатков, хитиновые крючки которых
Гаффнер с некоторым основанием уподобляет щетинкам ножек мизо-
стомид.
Возвращаясь к рассмотрению еще некоторых артроподоподобных черт
пентастомид, можно отметить, что устойчивость покровов тела против воз-
действия едкой щелочи можно объяснить не родством пентастомид с чле-
нистоногими, а результатом воздействия на них организма хозяина. Осо-
бенностями строения пентастомид, сближающими их с членистоногими
вообще (но не с какой-либо определенной группой), являются следую-
щие: сложность строения мужских и женских выводных частей половой
системы (например, наличие длинных тонких, закрученных хитинозных
циррусов), сложность копулятивного аппарата и строения половых же-
лез и цитологические особенности развития половых клеток и особен-
ности раннего онтогенеза.
Следует, однако, отметить, что, подобно многим эндопаразитам, у
пентастомид единственной прогрессивно развитой системой органов яв-
ляется половая система. Если действительно в ряде отношений эта систе-
ма органов пентастомид и сходна с таковой членистоногих, то сущест-
венное отличие между ними заключается в том, что части копулятивного
аппарата у членистоногих, особенно у наземных, развиваются за счет
туловищных конечностей, тогда как у пентастомид они образуются гипо-
дермой тела, как обычно у кольчатых червей.
Мы можем считать, что в основе организации пентастомид лежит
гетерономная и притом не олигомерная, как думают некоторые авторы
(Беклемишев, 1952, 1964), но полимерная метамерия,,как у аннелид и
некоторых классов артропод, выраженная гораздо резче, чем у аннелид.
Исходя из теории ларвальных'и постларвальных сегментов П. П. Ива-
нова, у пентастомид, особенно более примитивных (таких, как Cephalo-
baenidae), можно различать ларвальные сегменты, составляющие тело
личинки или передний отдел тела, и постларвальные — многочисленные
сегменты тела взрослой пентастомиды. Личиночное тело (рис. 54) со-
ставляют две или три пары придатков с крючьями, надглоточный ганглий
и членистая брюшная лестничная нервная система из подглоточного и
четырех брюшных ганглиев с 4 парами боковых нервов, кишечник с ро-
товым отверстием и со слепой средней кишкой. На заднем конце его
после некоторой паузы начинают формироваться постларвальные
197
сегменты, составляющие многочленистое собственно тело половозрелой
пентастомиды.
К сожалению, процессы формирования и роста этих сегментов в онто-
генезе' неизвестны, но природа передних сегментов как ларвальных и ту-
ловищных, так и постларвальных вряд ли вызывает сомнение.
Пробелы в данных о ранних стадиях развития пентастомид не позво-
ляют выяснить ряда важных черт их онтогенеза и сравнить его с тако-
вым полихет и артропод. Отличия от хода развития полихет должны
быть значительными, но они вызваны условиями, в которых проходит
развитие у пентастомид: сначала внутри материнского организма, затем
в теле промежуточного хозяина и, наконец, в дефинитивном хозяине. Все
же наличие дорсального органа (особенно по описанию Оше — Osche,
1959) и стадии зародышевой полоски с двумя передними лопастями у
эмбриона пентастомид напоминают развитие членистоногих.
Рис. 54. Развитие Reighardia sternae' (по Osche из Pfiugfelder)
А — зародыш с брюшной стороны; Б — зародыш сбоку; 1 — антенны; 2 — зачатки
ног; 3 — дорсальный орган
Ввиду их образа жизни физиологических исследований по пентасто-
мидам не имеется, так же как нет по этой группе биохимических, имму-
нологических (в том числе и преципитиновых) и серологических работ.
Морфологические данные (анатомия и эмбриология), как было сказано,
тоже далеко не полны, а сведения по экологии очень ограниченны. Все это
затрудняет достоверное выяснение природы пентастомид и заставляет,
подобно ряду авторов последнего времени (Беклемишев, 1952, 1964;
Haffner, 1922—1926; Nicoli, 1963), рассматривать их как самостоятельный
тип полимерных метамерных беспозвоночных, морфологически, видимо,
и филогенетически близкий не к артроподам, а к аннелидам. Однако не-
смотря на это, вся совокупность данных строения, развития и экологии
пентастомид настолько своеобразна и отлична от таковой другой поли-
мерной метамерной группы — Onychophora, что сравнивать их не прихо-
дится. Нечего говорить о том, что еще меньше общего наблюдается у пен-
тастомид и тихоходок (Tardigrada), хотя ряд авторов все же это находит.
Онихофоры (Onychophora)
Надо сказать, что ни название Onychophora, ни название Protracheata
для этой группы не являются удачными. Первое название кажется не-
сколько лучше второго, которым Геккель (Haeckel, 1896) предложил
заменить первое. Название Onychophora лучше потому, что оно отмечает
характерную черту этих животных, отличающую их от аннелид, тогда как
со вторым — первичнотрахейные — связывается едва ли правильное
представление об этой группе как о членистоногих.
Вопрос о положении онихофор в системе решается пока что методами
анатомии и эмбриологии, причем в трактовке тех или иных черт допу-
скается значительный субъективизм. Онихофоры, на мой взгляд, ближе
к кольчатым червям, чем к членистоногим, о чем свидетельствуют сле-
дующие черты: 1) гомономная метамерия тела, 2) голова без головной
капсулы и внутреннего скелета, не сочлененная с туловищем и слабо от
него обособленная, 3) пара пузыревидных глаз, сходных с глазами поли-
хет по строению и развитию, 4) пара усиков, нечленистых, не сочленен-
ных с головой и возникающих преорально и дорсально на головных ло-
пастях эмбриона, 5) отсутствие наружного скелета тела в виде тергитов
и стернитов как в индивидуальном развитии, так и у взрослых животных;
6) отсутствие сочленений между сегментами тела; 7) наличие слоя плот-
ной фиброзной ткани под покровным эпителием; 8) нечленистые и не
сочлененные с телом конечности; 9) полихетное строение головного моз-
га; 10) наличие в невросомитах кольцевых периферических нервов и сое-
динение друг с другом брюшных нервных стволов над задней кишкой;
11) кожно-мускульный мешок из гладких мышц; 12) метамерно распо-
ложенные выделительные органы типа целомодуктов с мерцательным
эпителием; 13) наличие ресничного эпителия в средней кишке и в поло-
вых протоках — производных выделительных органов, 14) отсутствие
мальпигиевых сосудов.
Некоторые черты онихофор считаются общими с членистоногими: это
хитиновая кутикула, линьки, лапки с коготками на ножках и щетинки на
теле, образование ножек в онтогенезе в виде полых выростов сомитов
тела, трахеи со спиральной нитью, миксоцель как дефинитивная полость
тела, образованная частями целома и первичной полости тела, сердце в
виде спинного кровеносного сосуда, без артериальных и венозных сосу-
дов, с сегментально расположенными остиями с клапанами, лежащее в
перикардии, который отделен диафрагмой от общей полости тела, утрата
полостью тела опорной функции, каковую несут покровы тела, жировая
ткань, нефроциты, артроциты, превращение пары выделительных органов
в слюнные железы, анатомия и развитие половых органов, поверхностное
дробление, наличие бластодермы, бластокинез, форма зародышевой по-
лоски.
Считая, что аннелидные черты онихофор в большей части бесспорны,
а многие черты, по которым онихофор сближают с членистоногими, вызы-
вают сомнение, я разберу главным образом последние.
199
Наличие хитиновой кутикулы, несомненно, признак, сближающий бни-
хофор с членистоногими; но пока нет данных о тонком строении и био-
химии их хитина, нельзя установить, насколько действительно существен-
но сходство хитина, онихофор и членистоногих. Появление хитина у они-
хофор, безусловно, связано с наземным образом жизни, так как хитин
до известной степени предохраняет организм от потери воды. Онихофоры
теряют вдвое меньше воды, чем находящиеся в тех же условиях дожде-
вые черви, однако вдвое больше, чем многоножки, и в 80 раз больше, чем
тараканы, и погибают в сухом воздухе в течение нескольких часов. Живя
в сильно увлажненной среде, онихофоры выходят из-под укрытий в основ-
ном во время дождя, так что и по этому признаку они ближе к кольчатым
червям, чем наземные членистоногие.
Линька онихофор, очевидно, связана с приобретением хитиновой ку-
тикулы и, действительно, характерна для членистоногих. Однако сбра-
сывание кожицы, в сущности та же линька, наблюдается и у аннелид.
Лапки с подошвой и коготками на ножках онихофор имеют значи-
тельное сходство с таковыми ряда трахейных членистоногих, особенно
насекомых, причем ножки онихофор, как и конечности членистоногих,
в отличие от параподий полихет, возникают в онтогенезе как полые вы-
росты тела. Онихофоры используют при движении не червеобразные со-
кращения мускулатуры тела, а, подобно членистоногим, работу мышц
ножек, причем у онихофор ножки одной стороны тела работают асин-
хронно с ножками другой стороны. По новым наблюдениям, онихофоры
двигаются подобно слизнякам, за которых их принял еще Гильдинг, от-
крывший этих животных в 1825 г.
Принципиальным отличием конечностей онихофор от ножек членисто-
ногих является нечленистость ножек онихофор и отсутствие их сочлене-
ний с туловищем. В этом отношении онихофоры ближе к аннелидам, чем
к членистоногим.
Конечно, вряд ли кому-нибудь придет в голову сравнивать ножки они-
хофор с параподиями полихет, но можно вспомнить, как параподии у не-
которых паразитических кольчатых червей, например, у Myzostomida,
преобразовываются в нечленистые ножки с крючками-щетинками, распо-
ложенными на брюшной стороне тела, с помощью которых эти черви не
только ходят, но буквально бегают и по хозяину, и по субстрату. Если
такое превращение параподий в ножки могло иметь место у морских ан-
нелид, то нет ничего удивительного в развитии у онихофор в связи с на-
земным образом жизни ножек и в формировании на концах ножек
подошвы с коготками, похожими на лапки ног наземных членистоногих,
при использовании ножек для передвижения по земле. Поэтому мне не
представляется обоснованным, как это делал Снодграсс (Snodgrass,
1938), по характеру строения ножек создавать для онихофор таксономи-
ческую группу Lobopoda.
Значение кожно-мышечного мешка, составленного гладкой мускула-
турой, как морфологической черты бесспорно аннелидного характера,
усиливается и тем, что мышцы туловища онихофор (Коштоянц, 1957),
подобно мышцам аннелид, очень чувствительны к действию ацетилхолина,
на который мышцы членистоногих не реагируют, и, так же как мышцы
аннелид, не отвечают на действие адреналина. По мнению X. С. Кошто-
янца, различное отношение мышц кольчатых червей и онихофор к атро-
пину, кураре и эзерину требует дополнительной проверки.
Трахеи — особенность, характерная для большой группы членистоно-
гих, но отсутствие у онихофор стигмального аппарата, положение стигм,
находящихся в огромном количестве на голове, около рта, на спине, от-
части боках, не позволяет гомологизировать трахейную систему онихо-
фор и членистоногих, хотя обычно для последних признают независимое
возникновение трахей у разных групп. Несомненно, что трахейная систе-
200
ма у членистоногих развилась, после формирования отделов тела (голо-
‘вы, груди и брюшка) и наружного скелета, после перехода предковых
форм от водного образа жизни к наземному. Метамерия в расположении
стигм трахейной системы связана с наличием у членистоногих наружного
членистого скелета и с возможным возникновением стигм в более тонких
межсегментных перепонках. Факт сохранения онихофорами и до настоя-
щего времени мягкого кожного покрова аннелидных предков объясняет
возникновение у них (независимо от наземных членистоногих) из кож-
ных желез многочисленных, беспорядочно разбросанных трахейных пуч-
ков как приспособление к воздушному дыханию. Иными словами, трахей-
ная система онихофор функционально сходна с таковой членистоногих, но
не гомологична ей.
Характер дефинитивной полости тела и ее физиологическая специфи-
ка как гемоцеля, строение и отправления сердца без сосудов, с остиями,
нефроциты и артроциты у онихофор очень близки к таковым членисто-
ногих, особенно наземных (трахейных и хелицеровых). Однако случаи
сильной редукции вторичной полости тела и распад системы кровеносных
сосудов наблюдаются и у современных групп кольчатых червей, хотя они
и не заходят так далеко, как у членистоногих и онихофор. Следователь-
но, до сих пор неясно, являются ли эти особенности полости тела и кро-
веносной системы онихофор и членистоногих признаками их родства или
результатом приспособления к наземному образу жизни. В пользу пер-
вого предположения говорит то, что и у ракообразных наблюдаются
сходные отношения. В то же время установлено что образование, счи-
тавшееся у онихофор жировым телом, таковым не оказалось, так как в
клетках этой ткани не найдено включений, характеризующих жировое
тело трахейных членистоногих.
Анатомия и развитие половых органов онихофор очень сходны с тако-
выми членистоногих, однако следует напомнить, что у ряда групп коль-
чатых червей также наблюдаются очень сильные отклонения в строении
половой системы. Так, у мизостомид, некоторых паразитических типич-
ных полихет и пиявок происходит обособление половых желез (яичников
и семенников) от вторичной полости тела и соединение половых желез
с половыми протоками в одно целое, а у ряда форм червей половые от-
верстия могут располагаться на заднем конце тела, например, у Myzo-
stomida (женское половое отверстие).
При большом сходстве в анатомии и отногенезе половых органов у
онихофор и членистоногих приходится подчеркнуть, что у онихофор нет
спаривания, которое очень широко распространено у членистоногих.
У онихофор оплодотворение происходит путем проникновения спермато-
зоидов из сперматофор через кожу тела самки (Whitman, 1891; Milne et
Milne, 1954), что напоминает оплодотворение через кожу у ряда пиявок
и, как правило, чуждо членистоногим.
Существенной особенностью онихофор, также не свойственной члени-
стоногим, является то, что они, достигнув половой зрелости к концу пер-
вого— второго года жизни, продолжают расти и живут 6—7 лет, причем
самка дает несколько поколений молоди, которая рождается сформиро-
ванной, а развитие эмбриона в теле матери продолжается от 9 до 13 ме-
сяцев. Лишь немногие виды откладывают яйца в землю.
Как отмечалось выше, в индивидуальном развитии онихофор есть
много общего с членистоногими, причем Коршельт и Гейдер (Korschelt
u. Heider, 1909) находили, что в нем больше артроподных черт, чем
аннелидных. Эти авторы особенно подчеркивали отсутствие в онтогенезе
онихофор детерминированного и спирального дробления, столь харак-
терного для развития аннелид. Однако, если эти черты аннелидных пред-
ков лишь частично сохранились у некоторых водных артропод — низших
ракообразных, то нет ничего удивительного, что они утрачены онихофо-
201
рами в силу их наземного образа жизни и условий развития внутри ма-
теринского организма. П. П. Иванов (1940; см. гл. «Членистоногие»)
почти полностью отождествлял процессы эмбрионального развития они-
хофор и. многоножек (Chilopoda), подчеркивая одинаковое количество у
них (5) передних сомитов. Я не могу согласиться с П. П. Ивановым и
отмечу, что если у трахеат ларвальных сомитов 5, а у хилопод, по Ива-
нову, даже 7, то у онихофор этих сомитов всего лишь 2 — челюстной и
папиллярный.
Я думаю, что многие черты сходства в онтогенезе онихофор и члени-
стоногих вызваны условиями жизни, так типичное для их развития обо-
гащение яиц желтком и поверхностное дробление наблюдаются среди
многих групп наземных животных, а отнюдь не только среди членисто-
ногих. Живорождение и питание эмбрионов за счет организма матери
наблюдается у скорпионов, далеко отстоящих от онихофор, и встречает-
ся„у многоножек, с которыми онихофор обычно сравнивают. Бластоки-
нез, подчеркиваемый в развитии у онихофор Пфлюгфельдером (Pflugfel-
der, 1948) как одно из сходств в развитии онихофор и насекомых, трудно
считать признаком родства между ними.
В способах формирования зародышевой полоски, дифференцировки
зачатков энтодермы и мезодермы, в сегментации мезодермальных полосок
и развитии целомов сходство у онихофор и членистоногих действительно
большое.
Однако в положении широко расставленных половинок зародышевой
полоски у онихофор, как давно отмечалось в литературе, есть сходство
с эмбриональным развитием ряда олигохет и пиявок. В самом существе
процесса сегментации мезодермальных полосок и развития метамерно
расположенных целомических полостей онихофоры сходны с аннелидами.
Временное развитие обширных метамерных целомических полостей до
формирования миксоцеля, особенно у некоторых представителей онихо-
фор, на известной стадии приближается к дефинитивному состоянию вто-
ричной полости тела аннелид в такой мере, в какой оно не наблюдается
в онтогенезе членистоногих.
Если в аннелидной организации взрослых онихофор наземные усло-
вия жизни вызвали значительные изменения, то, естественно, что они еще
сильнее сказались на ходе их онтогенеза и привели к сходству ряда черт
онтогенеза у онихофор и членистоногих, скорее всего конвергентного ха-
рактера.
Многие из указанных выше аннелидных черт онихофор не вызывают
сомнения у большинства зоологов. Часть этих черт свойственна и члени-
стоногим, что понятно, поскольку обе эти.группы обладают рядом приз-
наков, позволяющих объединить их в Articulata. Другая часть исходно,
видимо, аннелидных черт обязана своим своеобразием воздействию ус-
ловий жизни этих животных. Частично это уже было отмечено при рас-
смотрении природы ряда артроподных особенностей.
Гомономность метамерии внешнего строения и внутренней организа-
ции онихофор в своей примитивности имеет аннелидный характер. Она
нарушена преобразованием двух первых пар послеротовых конечностей
в челюсти и ротовые сосочки и сближением их сегментов с головой, т. е.
эволюцией в сторону цефализации, однако голова не обособлена от тела
и не обладает ни головной капсулой, ни эндоскелетом, а усики, на чем
я остановлюсь ниже, являются придатками головных лопастей, а не из-
мененными конечностями туловища. Вряд ли стоит спорить о том, что
мягкость, гибкость покровов и потеря онихофорами внешней сегментации
тела являются результатом приспособления к движению по субстрату;
это могло произойти тем более легко, так как исходно онихофоры не
обладали наружным скелетом, характерным для сегментов тела члени-
стоногих. ’[
202
Нервная система онихофор — лестничного типа, с широко расстав-
ленными брюшными стволами, с многочисленными поперечными, коммис-
сурами в каждом сегменте, без дифференцированных парных ганглиев.
Известно, что находили сходство нервной системы онихофор с таковой
хитонов, соленогастров, архианнелид и до некоторой степени хиллопод.
Если в общем подобное строение нервной системы указывает на ее при-
митивность и на исходно примитивное положение этих групп, то у они-
хофор положение иное. На определенной стадии онтогенеза их брюшная
нервная система представлена зачатками головного мозга, парными
ганглиями («вентральные органы») в каждом сомите и сближенными
друг с другом двумя продольными нервными стволами. В ходе онтогене-
за ганглии дедифференцируются, их ганглиозные клетки распределяются
по продольным стволам, которые расходятся и занимают латеральное
положение по бокам туловища. Таким образом, из ганглионарной брюш-
ной цепочки эмбриона возникает лестничная нервная система взрослого
животного, упрощение которой, несомненно, связано с малоподвижным
образом жизни и очень медлительными движениями онихофор. Чтобы
подчеркнуть аннелидный тип строения нервной системы онихофор, на-
помню еще о принципиальном сходстве их невросомитов, имеющих пе-
риферические кольцевые нервы, с невросомитами аннелид, сохранивши-
мися у онихофор, очевидно, благодаря наличию кожно-мускульного меш-
ка. К этому добавлю и полихетный характер строения их головного
мозга.
Известно, что очень важной особенностью членистоногих, сравнитель-
но с аннелидами, является участие передних сомитов туловища в обра-
зовании головы и головного мозга и преобразование конечностей этих
сомитов в органы чувств и ротовые органы. В связи с этим понятно, что
и сегментацией или составом головы у онихофор также занимались мно-
гие зоологи. И все же единого мнения о природе элементов головы они-
хофор, о числе сомитов, участвующих в ее образовании, у нас нет.
По сравнительным гистологическим исследованиям Гольмгрена
(Holmgren, 1916), строение мозга Peripatus и полихет (Nereis) очень
сходно и вместе с тем существенно отличается, как это подтверждает и
Ганстрём (Hanstrom, 1928, 1935), от мозга членистоногих. Однако в этих
исследованиях оба автора, правильно гомологизируя антенны онихофор
с головными придатками полихет, ошибочно искали гомолога антенн они-
хофор в пальпах, а не в тентакулах полихет, неудачно названных неко-
торыми авторами антеннами и находящихся на дорсальной стороне про-
стомия.
Характерно, что Б. Г. Федоров, также гистологически изучавший нерв-
ную систему онихофор и нашедший принципиальное сходство в строении
невросомита онихофор и полихет (Fedorov, 1926, 1929), о чем было ска-
зано выше, отмечает, напротив, существенное различие в строении го-
ловного мозга онихофор и полихет, причем антенны онихофор он счита-
ет измененными конечностями (1929). Из неметамерной части головного
мозга онихофор (архицеребрума), развивающейся в головной лопасти
и гомологичной мозгу полихет, он исключает зачатки ганглиозных цент-
ров антенны. По его мнению, ганглии антеннального сомита участвуют
в образовании остальной части головного мозга вместе с ганглиями пред-
челюстного, челюстного и папиллярного сомитов. Иными словами, по
Б. Г. Федорову, головной мозг онихофор построен принципиально сходно
с таковым членистоногих и отлично от головного мозга аннелид, с чем
согласен и Н. А. Ливанов (1955).
Б. Г. Федоров (1929) в результатах исследования головного мозга
снихофор во многом расходится с Гольмгреном и считает необходимым
внести серьезные исправления в его схемы строения головного мозга
полихет и онихофор, о которых было упомянуто выше. Так, он отмечает,
203
что антеннальный нерв проходит в мозгу онихофор дорсо-латерально и
дорсально над гломерулосом; он явно обособлен и связан с последним-
лишь некоторым числом волокон, тогда как пальпарный нерв у Nereis
отходит вентрально, пронизывая гломерулус. Иными словами, положение
антеннального нерва у онихофор резко отлично от положения пальпар-
ного нерва у полихет.
Я считаю, что схемы Гольмгрена бесспорно говорят о действительно
общей основе строения головного мозга полихет и онихофор, и ряд воз-
ражений Федорова отпал бы, если бы Гольмгрен уподоблял антенны
онихофор не пальпам аннелид, а тентакулам, которые располагаются у
полихет на спинной стороне простомия и отличаются от пальп по поло-
жению, строению и функции (Haffner, 1959, так называемые антенны).
Известное различие в строении головного мозга полихет и онихофор-
должно быть также и потому, что единственная пара головных органов
чувств — антенны (не считая органов зрения) у онихофор развита гораз-
до сильнее тентакул полихет, так как по сравнению с ними они несут
гораздо большую функциональную нагрузку.
Данные некоторых авторов (Kennel, 1884, 1888; Sedgwick, 1885, 1886,
1888, 1909; Scheldon, 1888а, 18886) говорят о преоральном и дорсальном
положении зачатков антенн у ранних эмбрионов онихофор, о том, что
нервы антенн возникают как прямое продолжение ганглиозной массы
головных лопастей, и о наличии поперечного желобка, отделяющего го-
ловные лопасти с зачатками антенн от зачатков двух продольных стволов
нервной системы, начинающихся зачатками ганглиев челюстей. Все это
указывает на развитие антенн онихофор как придатков акрона (просто-
мия аннелид), а не как конечностей туловища. К сожалению, четких но-
вых исследований по эмбриогенезу нервных центров головных лопастей
и ганглиозных центров антенн онихофор все же нет.
По распространенному взгляду, лишь архицеребрум онихофор при-
надлежит акрону, тогда как остальные части головного мозга образуют-
ся ганглиями сегментов антенн, челюстей и оральных папилл, а по мне-
нию немногих (Fedorov, 1929; Pflugfelder, 1948), признающих существо-
вание и премандибулярного сегмента,— ганглиями четырех посторальных
сегментов. По мнению этих авторов, мозг онихофор, подобно мозгу чле-
нистоногих, состоит из прото-, дейто- и тритоцеребрума и их антенны,
имеющие посторальное происхождение, вторично заняли преоральное и
дорсальное положение, т. е. гомологичны конечностям тела. Однако дан-
ные онтогенеза, отмеченные выше, говорят об антеннах как придатках
головных лопастей эмбриона.
Останавливаясь еще на некоторых данных онтогенеза онихофор, от-
мечу, что некоторые авторы, например Пфлюгфельдер (Pflugfelder,
1948) и Мэнтон (Manton, 1937, 1949, 1958), находили целомические меш-
ки в акроне онихофор и считали их головными целомами, дающими на-
чало целомам антеннального сомита. В этих целомах находили каналы,
якобы зачатки нефридиев, идущие к эктодерме. По классическим, по-
устаревшим представлениям целомы у полихет закладываются лишь В;
сомитах туловища. Однако по отчетливым данным Андерсона (Anderson,
1959; см. гл. «Аннелиды») и Гаффнера (Haffner, 1959), целомические
мешки у исследованных ими полихет развиваются и в простомий. В этом
отношении онихоформы также сближаются с полихетами, и образова-
ние у них целомов антенн из головного целома эмбриона дает лишнее
основание отрицать наличие у онихофор антеннального сомита как та-
кового.
Таким образом, надо считать антенны онихофор гомологами не ко-
нечностей, а простомиальных придатков, а их нервные центры, разви-
вающиеся в акроне, принадлежащими к неметамерной части головного-
мозга. Если у онихофор действительно существует премандибулярный
204
сомит (Fedorov, 1929; Pflugfelder, 1948), лишенный конечностей, то его
надо было бы гомологизировать не с премандибулярным (йнтеркаляр-
ным) сомитом членистоногих, а с буккальным (= перистомиальным) со-
митом аннелид, с которым он сходен и отсутствием конечностей, и отно-
шением его нервных центров к головному мозгу.
В основном я разделяю представление Гольмгрена о том, что голов-
ной мозг онихофор состоит из переднего простомиального мозга, который
включает зрительные и антеннальные нервные центры, и посторального
заднего мозга, содержащего центры задних стомодеальных нервов и
челюстей. Я не вижу оснований для того, чтобы считать головной мозг
онихофор соответствующим прото-, дейто-тритоцеребрумам членис-
тоногих. Передний мозг онихофор гомологичен мозгу аннелид и архи-
церебруму членистоногих, а задний мозг их отвечает метамерной части
мозга членистоногих тем, что его составляют ганглиозные центры челю-
стей и оральных папилл, измененных посторальных конечностей тела
(как у членистоногих ганглии антенн), и других посторальных конеч-
ностей.
Вместе с тем я не согласен с выводами, которые делает Ганстрём
(Hanstrom, 1935), так как принимая положение Гольмгрена о полихетном
характере мозга онихофор, он далеко уклоняется не только от взглядов
этого автора, но, по моему мнению, и от истины. Ганстрём, наряду с поли-
хетными чертами мозга онихофор в архицеребруме, антеннальных гло-
мерулах, служащих еще как стилгломерулы, в стебельчатых телах, гла-
зах и зрительных ганглиях, находит в мозгу онихофор черты сходства
с хелицеровыми в строении центрального тела и симпатической нервной
системы и с ракообразными в дифференцировке центров антеннальных
нервов, строении, форме и положении антеннальных гломерул. Мало
вероятно, чтобы в строении мозга онихофор действительно были особен-
ности, присущие столь отличным от них и по организации, и по образу
жизни группам, как хелицеровые и ракообразные. Мне представляется
такое толкование Ганстрёма неверным и его вывод, будто бы онихофоры
являются промежуточной ступенью между артроподами и аннелидами,
ошибочным.
Превращение двух передних пар посторальных конечностей тела в
ротовые органы (челюсти и оральные папиллы) является очень важной
чертой, сближающей онихофор с членистоногими. Этот шаг в их эволю-
ции, столь характерный для членистоногих, привел к образованию у они-
хофор головы с комплексом органов чувств (антенн и ротовых органов,
челюстей и максилл). Это направление в эволюции онихофор поднимает
их-выше аннелид, но оно возникает скорее всего у онихофор независимо
от членистоногих.
При такой общности направления в эволюции онихофор и членисто-
ногих приходится подчеркнуть отсутствие гомологии между челюстями
онихофор и мандибулами трахейных членистоногих. Челюсти онихофор
возникают из нечленистых конечностей и при развитии лишены члени-
стости, присущей мандибулам. Гомологию этих органов резонно отверга-
ли некоторые как старые (Korschelt u. Heider, 1890—1893), так и новые
авторы (Plate, 1922; Butt, 1957, 1959, 1960). Да и функционируют эти ор-
ганы по-разному (Kaestner, 1954/1955). Челюсти у членистоногих, рас-
положенные поперек к длинной оси тела, работают как два лезвия
кусачек, тогда как у онихофор челюсти, расположенные по продольной
оси тела, двигаются вперед и лежат внутри ротовой полости, а не снару-
жи, на нижней стороне головы, как жвалы членистоногих.
Парные, метамерно расположенные и находящиеся почти во всех со-
митах тела выделительные органы типа целомодуктов относятся к основ-
ным аннелидным чертам организации онихофор. Они сохраняют реснич-
ный эпителей, свойственный нефридиям аннелид. С ними приобрели
205
сходное строение антеннальные, максиллярные и коксальные железы
разных групп членистоногих; целомодукты онихофор в известной степени
участвуют в удалении мочи, но в отличие от энергично функционирующих
антеннальных желез раков и крабов (Picken, 1936) они функционируют
очень замедленно. Перестройка строения и функции исходного типа не-
фридиев водных аннелид в выделительные органы онихофор связаны
с наземными условиями жизни, необходимостью экономии воды организ-
мом, к дефициту которой онихофоры так чувствительны.
Наряду с сохранением почти в каждом сегменте тела пары выдели-
тельных органов, подобно аннелидам онихофоры действительно прояв-
ляют сходство с членистоногими, выражающееся в том, что выделитель-
ные органы второго посторального сегмента становятся слюнными желе-
зами. Однако не следует забывать, что слюнные железы большинства
членистоногих являются эктодермальными производными, а не изменен-
ными выделительными органами.
Эколого-биологическое своеобразие онихофор очень велико. Они
крайне влаголюбивы, живут в среде, насыщенной влагой, выходят, как
известно, из под укрытий наружу после дождя; виды, живущие в более
сухих местах, в сухой период года (Гиляров, 1958) зарываются на это
время в землю, переходя в пассивное состояние. К сожалению, имеется
очень мало эколого-физиологических исследований (они проведены
главным образом Ментон, 1958). Благодаря ей мы знаем, что онихофоры
теряют воду вдвое медленнее, чем дождевой червь, в 40 раз скорее, чем
гладкокожая гусеница бабочки, и в 80 раз быстрее таракана. Это говорит
о малой приспособленности онихофор к жизни в относительно сухой, на-
земной среде. Будем ли мы это приписывать слабой водонепроницаемо-
сти тонких покровов, или особенностям трахейной системы с многочис-
ленными стигмами без замыкательного аппарата, или и тому и другому,
отсутствие контроля над расходом воды у онихофор бросается в глаза,
а это чуждо наземным членистоногим. Казалось бы, будь онихофоры
членистоногими, у них должно быть развитое жировое тело, резервные
вещества которого могли бы давать метаболическую воду для попол-
нения потерянной воды организмом. Однако настоящее жировое тело
у онихофор, как уже отмечено, отсутствует. Известно, что редукция чис-
ла и регрессивный характер органов выделения типа нефридиев отчетли-
во выражены у членистоногих, особенно у наземных форм. Нефридии
остались у них в ничтожном числе, сохранив лишь частично функцию
выделения, перешедшую в основном к мальпигиевым сосудам. Удаление
продуктов распада в виде мочевины непосредственно в заднюю кишку
обеспечивает у наземных членистоногих экономию потребляемой воды.
У онихофор конечные продукты распада отличаются от таковых ан-
нелид, но выведение их с помощью сегментарных выделительных орга-
нов сохраняется: экономия воды достигается крайне замедленным тем-,
пом опорожнения нефридиев, то есть иначе, чем у членистоногих (Man-
ton, 1937). Следует обратить внимание на то, что лишь у одного вида
онихофор яйца, по-видимому, неклейдонические и подобно яйцам боль-
шинства наземных членистоногих откладываются в землю. У большин-
ства же онихофор развитие яиц протекает в теле матери, что, как и на-
ружное оплодотворение импрегнацией через кожу, подчеркивает своеоб-
разие онихофор.
Анализ строения и развития онихофор, на мой взгляд, говорит о том,
что в основном они построены по аннелидному типу, хотя некоторые чер-
ты строения и сближают их с членистоногими. В эмбриональном разви-
тии, хотя оно протекает внутри материнского организма, т. е. в условиях,
совершенно чуждых для онтогенеза аннелид, сохраняются черты онтоге-
неза аннелид и приобретен ряд черт артроподного типа развития. Эколо-
го-биологические особенности онихофор с их крайней зависимостью от
206
влажной среды сближают онихофор с кольчатыми червями и делают их
резко отличными от наземных членистоногих. Все это заставляет видеть
в онихофорах потомков аннелидных выходцев на сушу в среднем кемб-
рии, которые в ходе длительного исторического развития как наземные
животные приобрели ряд черт, сходных с членистоногими, но в целом
получили настолько своеобразное сочетание черт строения, развития и
экологии, что составляют особую самостоятельную группу — тип чле-
нистых беспозвоночных, не входящий ни в тип аннелид, ни в тип артро-
под (Fedotov, 1925; см. гд. «Членистоногие»), Конечно, для подтвержде-
ния такого взгляда на онихофор нужны еще данные физиологии, биохи-
мии и серологии, которых у нас нет.
Мнения о положении онихофор в системе, их значении и эволюции
и филогении различны (Кузнецов, 1951). Вкратце они сводятся к сле-
дующему. Некоторые авторы, например Боас (Boas, 1922), считают они-
хофор аннелидами и как подкласс последних помещают их между Chae-
topoda и Hirudinea, не придавая существенного значения чертам, сход-
ным с артроподными. В настоящее время ясна крайность такого взгляда.
Большинство советских авторов (Догель, 1938; Ливанов, 1955, и др.)
считает онихофор классом примитивных членистоногих — Protracheata,
причем нередко рассматривают их как звено, связывающее аннелид и
артропод, как первый этап в эволюции последних (например, Remane,
1954). Думаю, что и такая точка зрения неверна.
Основываясь на том, что при всем своеобразии строения и развития
онихофор аннелидные черты для них более характерны, чем артропод-
ные, надо считать их хотя и близкой по происхождению к кольчатым чер-
вям (как думали Гольмгрён, Кестнер и другие), но самостоятельной
ветвью, возникшей от аннелидоподобных предков — живыми ископае-.
мыми, а не звеном, связывающим аннелид и членистоногих.
Онихофоры, по-видимому, геологически древняя группа. Хотя несом-
ненных ископаемых онихофор неизвестно, однако среднекембрийская
Aysheala pedunculata (Walcott, 1911) и в значительно меньшей степени
докембрийский Xenusion aucrswaldae Pomp. (Heyman, 1928) имеют не-
которые признаки онихофор.
Членистоногие (Arthropoda)
Хотя значительно (продвинуто решение ряда вопросов филогении чле-
нистоногих, однако все же остается еще много опорного и несогласован-
ного. Так, нет единого мнения о морфологическом значении придатков
и числе .метамеров головы, числе ларвальных сегментов, филогенети-
ческих отношениях четырех ветвей членистоногих, полифилии и моно-
филии членистоногих.
Многие советские (среди них П. П. Иванов и В. Н. Беклемишев) ч
зарубежные зоологи (например, Vandel, 1949; Butt, 1960) гомологизи-
руют антеннулы ракообразных, антенны трахейных и трилобитов с при-
датками простомиума полихет, но не с конечностями их тела.. Большую
роль в этом сыграли данные Гольмгрена (Holmgren, 1916) и Ганстрёма
(Hanstrom, 1928) о нервной системе беспозвоночных. Так, П. П. Ива-
нов не придает значения посторальной закладке антенн; части эмбриона,
на которых закладываются предантенны и антенны, он не признает сег-
ментами и считает, что целомические мешочки не только предантенн, но
и антенн, когда они имеются, возникают как выросты премандибул яр-
кого (интеркалярного) сегмента.
Бат (Butt, 1960), на основании данных эмбриологических работ по-
следних лет и отчасти ряда старых, подчеркивает образование антенн
как выростов головной лопасти (акрона) эмбриона, отрицает существо-
вание предантеннального сегмента и счет сегментов тела, участвующих
в образовании головы членистоногих, начинает с интеркалярного сегмен-
та. В этом Бат подтверждает взгляд П. П. Иванова, который еще в ра-
боте над развитием яванской многоножки Rhysida (1940) считал первы-
ми истинными сегментами, входящими в состав головы трахейных, ин-
теркалярный и мандибулярный.
В. М. Шимкевич, К. Н. Давыдов, Н. А. Ливанов и ряд других наших
и зарубежных зоологов (Plate, 1922; Kaestner, 1956; Slewing, 1956, и др.)
гомологизируют антеннулы ракообразных и антенны трахейных с конеч-
ностями туловищных сегментов аннелид. В. М. Шимкевич (1923) на ос-
новании совокупности данных строения и развития считает усики чле-
нистоногих гомологами конечностей, а не антенн —осязательных при-
датков головной лопасти или верхней части трохофоры аннелид. На
голове кольчатых червей находятся "органы чувств, но нет придатков —
видоизмененных конечностей, или параподий. Две пары антенн у рако-
образных, одна пара антенн у многоножек и насекомых и ротовые орга-
ны всех членистоногих, по его мнению, являются видоизменными конеч-
ностями передних (ларвальных) сегментов тела, слившихся с головной
лопастью при образовании головы. В ходе онтогенеза зачатки антенн
и других придатков головы, закладывающиеся позади рта, перемещают-
ся (вперед, а рот отодвигается назад.
По существу, это же, хотя и без упоминания положений В. М. Шим-
кевича, выражено в трактовке головного мозга полихет и членистоногих
Б. Г. Федорова (1931) и Н. А. Ливанова (1955), которые, однако, вносят
208
существенные изменения в представления Гольмгрена (Holmgern, 1916)
и Ганстрёма (Hanstrom, 1928; см. гл. «Онихофоры») о головном мозге
членистоногих. Головной мозг полихет — неметамерное образование с
палыпарной, антеннально-зрительной и
гичными метамерам брюшной нерв-
ной цепочки. У членистоногих го-
ловной . мозг образован: 1) неме-
тамерным отделом, соответствую-
щим головному мозгу полихет-,'—
это архицеребрум, отвечающий про-
тоцеребруму и гломерулам Гольм-
грена, 2) ганглиями первого сегмен-
та — дейтоцеребрум Гольмгрена, но
без гломерул, 3) ганглиями второго
сегмента — тритоцеребрум, которые
входят в состав головы, причем по-
сторальная комиссура антенн из по-
сторальной становится преоральной.
Поскольку из всей массы совре-
меных членистоногих с антеннами
лишь одни ракообразные сохранили
водный образ жизни, в их органи-
зации и развитии встречаются важ-
ные для решения этого вопроса при-
митивные черты, утраченные боль-
шинством членистоногих в связи с
переходом к наземной жизни. На-
помню, что точные безукоризненные
исследования по развитию паразити-
ческого рачка Lernaea branchialis'L.
Д. Д. Педашенко (1898) показали
гомологию зачатков передних ан-
тенн (рис. 55) и их ганглиев с за-
чатками следующих пар конечно-
стей и их ганглиев науплиуса и не-
зависимое от головного мозга с за-
чатками органов чувств положение
на брюшной стороне зачатков ган-
глиев антеннул.
Примитивные отношения,' под-
тверждающие природу ганглиев ан-
теннул как первой пары ганглиев
брюшной цепочки, а самих антен-
нул — как конечностей, имеются в центральной нервной системе низ-
ших раков, у которых эти ганглии сливаются с протоцеребрумом, возни-
кающим из ганглиозного материала головных лопастей, или акрона
(Plate, 1922; Kaestner, 1956).
Естественно, что от исходных, примитивных отношений, характер-
ных для низших ipaiKOB, у большинства членистоногих, в связи с высоким
уровнем развития головы и интеграции ее составляющих частей — посто-
ральных сегментов тела, сохраняется немного.
Я разделяю мнение о том, что антеннулы ракообразных, как и антен-
ны других членистоногих — гомологи конечностей туловищных сегмен-
тов, а мозг головы членистоногих образуется из неметамерного' от при-
роды зачатка головной лопасти (или акрона) эмбриона и ганглиев пер-
вых двух сегментов тела, участвующих в образовании головы взрослого
членистоногого.
нухальнои ооластями, не гомоло-
Рис. 55. Схема развития составных
частей нервной системы Lernaea
branchialis L. (из Педашенко)
/ — зачаток головного мозга; // — зачаток
брюшной нервной цепочки; 1 — ганглни ан-
тенн I; 2 — ганглни антенн II; 3 — ганглии
мандибул; 4—-ганглии максилл; 5, 6 — ган-
глии максиллипед; 7, 8 — ганглни конеч-
ностей
14 Д- М. Федотов
209
Переходя к вопросу о числе ларвальных сегментов, близко связанно-
му с предыдущим, напомню, что теория метамерии с ларвальными и
постларвальными сегментами П. П. Иванова (1940, 1944, 1945; Iwanoff,
1933) получила широкое, хотя и не общее признание не только у нас, но
и за рубежом и считается классической (Vandel, 1949). Она прямо свя-
зана с вопросом о составе головы и морфологическом значении ее при-
датков. Кроме того, П. П. Иванов и я (Fedotov, 1925) придавали важ-
ное значение числу ларвальных сегментов для установления филогенети-
ских групп членистоногих.
И. И. Иванов (1945), как известно, признавал у ракообразных два,,
у трилобитов, меростомовых и паукообразных —четыре, у многоножек и
насекомых — пять ларвальных сегментов. Однако при их подсчете у него
были известные колебания. Так, отмечая, что три сегмента у науплиуса
закладываются одновременно как типичные ларвальные сегменты, он го-
ворит (И. П. Иванов, 1944) о трех ларвальных сегментах у раков, хотя
в других работах обычно признает у ракообразных два ларвальных сег-
мента, отрицает гомологию антеннул с конечностями и сравнивает их с
пальпами аннелид. У протаопис трилобитов Иванов признавал наличие-
четырех ларвальных сегментов (1933, 1944, 1945), не считая антенны ко-
нечностями и относя затылочное кольцо к головным сегментам. Призна-
вая у мечехвостов и паукообразных наличие четырех ларвальных сег-
ментов, П. П. Иванов отмечал, что данных о развитии ларвальных
сегментов у последних мало. Некоторая непоследовательность наблю-
дается у него и в отношении трахейных. Говоря о наличии у последних,
пяти сегментов головы, Иванов отмечал, что антенны не конечности,
а сегмент ногочелюстей у Chilopoda —• пятый сегмент — является первым.,
грудным сегментом (1944).
iB. Н. Беклемишев (1944, 1952) более строг в счете ларвальных сег-
ментов. Отвергая гомологию антеннул раков и антенн других членисто-
ногих с конечностями, он признает у раков два, у трилобитов, .мечехво-
стов, паукообразных и трахейных и с некоторой оговоркой у пантопод
по четыре ларвальных сегмента. Он находит привлекательной, но необос-
нованной, попытку Солло (Sollaud, 1923) считать, что и ракообразные-
,имеют четыре ларвальных сегмента.
И в отдельных работах и в обзорах зоологов вопрос о числе сегмен-
тов, участвующих вместе с акроном в образовании головы членистоногих,,
решается по-разному, однако нередко говорится о шести сегментах.
В этих случаях счет ведется с предантеннального сегмента, впервые
установленного Геймонсом (Heymons, 1891) у зародыша сколопендры,
работа которого повлияла и на палеонтологов. Иекель (Jaekel, 1909),.
Роу (Raw, 1957), Штёрмер (Stormer, 1942, 1959) насчитывали в голове
трилобитов шесть, а некоторые палеонтологи (Walcott, 1911; см. гл. «Ан-
нелиды»; Нире, 1953 и др.) семь-восемь сегментов. Эти соображения опи-
рались также и на особенности верхней стороны головы наиболее древ-
них нижнекембрийских трилобитов (Нире, 1953), причем палеонтологи
говорят о предантеннальном сегменте, утрате первой пары антенн, мик-
росомитах и ординарных сомитах, видимо, не отдавая себе ясного от-
чета о природе настоящих сегментов. Следует заметить, что Штёрмер
(Stormer, 1942, 1959), говоря о шести сегментах головы трилобитов,,
различает из них четыре посторальных.
Отмечу, что Вандель (Vandel, 1949) и Ват (Butt, 1957, 1960) не счи-
тают сегментами предантеннальный и антеннальный сегменты. По Ван-
делю, это — части акрона; в образовании головы членистоногих участ-
вуют, кроме него, четыре первичных, или ларвальных сегмента, из ко-
торых первый он называет тритоцеребральным, или антеннальным, несу-
щим вторую пару антенн у раков и лишенным придатков вследствие их
редукции у трахейных. По Бату, это — интеркалярный сегмент, парные.
210
придатки его, считающиеся лредантеннами, у трахейных дают верхнюю
губу, а у хелицеровых — рострум, что, мне кажется, требует еще дока-
зательств. Он, как и Вандель, 'признает участие в образовании головы
членистоногих всего четырех сегментов: интеркалярногб, мандибулярно-
го и двух максиллярных.
Таким образом, П. П. Иванов, В. Н. Беклемишев, Вандель и Бат счи-
тают, что в образовании головы трилобитов, меростомовых, паукообраз--
ных и трахейных участвуют, кроме акрона, четыре сегмента.
Несомненно, данные о развитии предантеннального сегмента говорят
против него как сегмента: кроме слабо выраженных придатков и иногда
наличия целомических полостей, других признаков сегмента нет. Данные
о строении нервной системы и развитии антеннулярного сегмента у
низших ракообразных, наличие посторальных придатков, целомических
мешочков и парных зачатков ганглиев у ряда трахейных заставляют
считать сегмент антеннул раков и антенн трахейных первым постораль-
ным сегментом.
Нет также оснований сегмент головы трилобитов, несущий пару ан-
тенн, исключать из серии сегментов, несущих головные придатки, по
одному лишь признаку одноветвистости антенн. Надо думать, что изме-
нение функции, превращение первой пары головных придатков в органы
чувств, а в связи с этим и изменение строения были причинами такого
резкого различия между первой и последующими парами головных ко-
нечностей артропод.
Близость зачатков антенн от головной лопасти у трахейных мне
представляется явлением вторичным, а утверждение Бата и некоторых
других эмбриологов о развитии антенн как непосредственных выростов
акрона есть, видимо, результат недостаточности исследования.
Авторы, отрицающие значение антенн насекомых как конечностей,
ссылаются на работы некоторых экспериментальных эмбриологов (Sei-
del, 1935; Breuning, 1957). Хотя данные последних стоят в противоре-
чии с. работами других, например, Д. М. Штейнберга, получившего путем
регенерации на месте антенны конечность.
В итоге я считаю антеннальный сегмент трилобитов и трахейных на-
стоящим сегментом, а число ларвальных сегментов у них — равным пяти.
На иных позициях стоят Я- А. Бирштейн (1948) и ряд зарубежных
авторов, насчитывающих у всех членистоногих в голове, кроме акрона,
одинаковое число сегментов — .шесть. Как известно, еще в конце прош-
лого столетия Геймонс • (Heymons, 1891) в крупном исследовании по
эмбриологии сколопендры установил наличие у нее шести метамеров го-
ловы; это было принято рядом палеонтологов (Jaekel, 1909, Raw, 1957;
Stormer, 1942, 1959), которые стали признавать и в голове трилобитов
шесть сегментов. В последние годы распространился взгляд, что у всех
членистоногих голова состоит из акрона и шести сегментов (см. Heega-
ard, 1945; Legendre, 1959; Siewing, 1963). Если в будущем эти данные
подтвердятся, они будут свидетельствовать в пользу взгляда о монофи-
лии членистоногих.
Переходя к филогении главнейших групп членистоногих — трилоби-
тов, хелицеровых, ракообразных и трахейных, напомню, что остатки чле-
нистоногих из докембрийских отложений, как и почти все ископаемые
того времени, признаются теперь остатками неорганического происхож-
дения. Из докембрийских отложений известны преимущественно лишь
следы жизни, .следы ползания червей и каких-то членистоногих. Нижне-
кембрийские трилобиты, вероятно, были потомками этих докембрийских
членистоногих.
Характерно, что уже ранние нижнекембрийские трилобиты обладали
всеми типичными признаками этой группы: большим головным щитом
с парой сложных глаз, парой антенн и четырьмя парами двуветвистых
14*
211
ног с жевательными лопастями, сдавленным в дорсо-вентральном на-
правлении телом, со срединной осевой и плевральными частями, состоя-
щими из головы, туловища и пигидия, с большим, непостоянным числом
однородных сегментов тела, обладающими парой однородных конечно-
стей на каждом сегменте, иногда с ясной зоной роста на конце пигидия.
Эволюция этого многочисленного класса (в настоящее время уже изве-
стно 1500 родов и 10 000 видов трилобитов) за период от раннего кембрия
до перми включительйо хорошо известна (см. Stubblefield, 1959; Whit-
tington, 1957, 19(ИЬ исключение представляют мелкие древнепалеозой-
ские малоч^рлен^ые агностиды и эодисциды, в онтогенезе которых
произошла остановка в развитии сегментов туловища. Хотя, особенно в
прежнее время, ряд зоологов и палеонтологов относил трилобитов к ра-
кообразным, на всем громадном протяжении своей истории трилобиты
не проявляют никаких эволюционных направлений, сближающих их с
ракообразными. Важными особенностями экологии, эволюции и филоге-
нии трилобитов является следующее. Для трилобитов характерны жизнь
лишь в море, придонный способ передвижения. Главную массу трилоби-
тов, за исключением лишь немногих форм, составляли животные с тяже-
лым наружным скелетом, передвигавшиеся по дну моря и зарывавшиеся
в ил или песок. Активное и пассивное плавание в толще воды среди три-
лобитов не было распространенным. Приуроченность их к придонным
условиям жизни сказалась на их строении. Во время своей геологической
истории от докембрия до верхней перми трилобиты были членистоноги-
ми, сплющенными в дорсо-вентральном направлении, с большим голов-
ным щитом, туловищем из подвижно сочлененных сегментов и пигидием,
имевшим однородные членистые конечности. Уклонения от этого общего
типа строения сводятся к немногому. Видимо, у некоторых форм к пяти
основным сегментам головы добавлялись один или два грудных, как бы
ассимилируемых головой (более мощно функционируемым отделом
тела). Варьирует число сегментов туловища и пигидия. У группы нижне-
кембрийских— ордовикских мелких Micromera (агностиды и эодисциды)
оно сокращалось до двух, а у большинства оно было непостоянно и до-
стигало 44 (например, у Pedeumias из Olenellidae). Очень разнообразно
было число сегментов и пигидия, от одного (Olenellidae) до 30 (у Aniso-
pyge из Phillipsidae.) При этом наиболее существенным изменением в
пигидии было слияние спинных тергитов его сегментов в неподвижный
щит. С придонным, ползающим образом жизни трилобитов были связаны
способ питания и характер пищи. Они, считается, питались мягкой пи-
щей— разлагавшимися остатками животных и растений. Их ротовые
органы служили для подачи ко рту и размельчения мягкой пищи, а не
для схватывания подвижной добычи.
Кроме наличия антенн, многие из перечисленных черт строения три-
лобитов, к которым надо добавить тип строения конечностей с пателлой
и боковой ветвью — преэииподитом, по толкованию Щтермера (Stormer,
1959, 1963) и Снодграсса (Snodgrass, 1938), сближают трилобитов с хе-
лицеровыми (рис. 56). И эти же черты строения древнейших трилобитов,
не говоря о более поздних, отличают их от самых низко организован-
ных ракообразных.
В богатых трилобитами морских нижнекембрийских отложениях на-
ходятся остатки примитивных меростомовых, вроде недавно описанного
Штёрмером (Stormer, 1956) Paleomerus, а также Aglaspis, Strabops и др.
Как не раз отмечалось в нашей (Fedotov, 1925; Р. Р. Iwanoff, 1933; Бек-
лемишев, 1944, 1952, 1964) и иностранной (особенно работы Stormer,
1942, 1944, 1955а, 19556, 1959, 1963) литературе, общие черты строения
древних меростомовых связаны с таковыми примитивных трилобитов.
Древнейшие хелицеровые, унаследовавшие от трилобитов дорсо-вент-
ральную сплющенность тела и большой головной щит, в начале геоло-
212
гичеюкой истории также были придонными животными береговой полосы
моря. Насколько известна наружная организация древнейших меросто-
мовых, например, Paleomerus (Starmer, 1956), немногим отличалась
от таковой трилобитов. Они уже обладали головогрудным (головным)
Рис. 56. Гомология элементов конечностей трилобитов (А, 5),
скорпионов (В—Д), эуриптерид (£. Ж) и мечехвостов (3)
(по Sterner)
отделом, мало дифференцированным туловищем и мало специализиро-
ванными передними конечностями (рис. 57). Почти несомненно это были
придонные, ползающие формы. Переход меростомовых к хищничеству
был связан с развитием одной из передних пар головных конечностей
Рис. 57. Сравнение метамерии некоторых низших Chelicerata
и Merostomoidea (по Storrner, из Беклемишева)
A—Sidneya ine'xpectans (Merostomoidea, средний кембрий); Б—Strabops
thacheri (верхний кембрий); В—Aglaspides sculptilis (верхний кембрий); Г—
Palaeomerus hamiltonl (нижний кембрий, отряд Agljspida); Д—Hughmitteria
norvegica (верхний силур', Eurypteroidea)
в хватательные хелицеры, с усилением развития и дифференцировкой
на основных члениках ряда головных конечностей жевательных лопас-
тей, в слабой форме имевшихся у трилобитов. В связи с прогрессировав-
шим усилением хищничества шла дифференцировка отделов гела меро-
стомовых и их конечностей. Они становились способными и к плаванию,
к охоте за крупной подвижной добычей. Живя .в береговой зоне моря
при трансгрессиях и регрессиях палеозойских морей, меростомовые по-
213
Рис. 58. Схема примерного количественного распределения 1395 родов
трилобитов (из A. Muller)
падали в особые условия жизни в солоноватоводной среде, эстуариях
рек и во время сильных отливов могли находиться и на обсохших участ-
ках литорали.
В период наибольшего расцвета трилобитов (рис. 58) в позднем
кембрии и раннем силуре (ордовике) и заселения ими всех вод Мирового
океана, из хелицеровых встречаются лишь примитивные Xiphosura и
Eurypterida. Xiphosura в древнем палеозое не ипрали существенной роли
в фауне моря и на протяжении всей своей истории от палеозоя до наше-
го времени продолжали оставаться малочисленными формами. Зато
эуриптериды, уже в силуре и девоне достигшие расцвета, были пред-
ставлены разнообразными, порой гигантскими высоко дифференцирован-
ными многочисленными формами. В блестящих работах Штёрмера
(Stormer, 1942, 1944, 1955а, 19556, 1959, 1963) хорошо изучена природа
эуриптерид, а благодаря Кестнеру (Kaestner, 1931) и главным образом
В. Б. Дубинину (1959, 1962), усовершенствована система, уточнены род-
ственные отношения хелицеровых и определено место эуриптерид в си-
стеме.
Эволюция трилобитов идет от форм многочленистых, с не фиксиро-
ванным числом сегментов тела, примитивных по многим признакам, к
формам с резко дифференцированными отделами тела и, как отмечалось,
214
Рис. 59. Гомология сегментов тела скорпионов и эуриптерид (по Stormer)
Л — Д — скорпионы, Е — 3 — эуриптериды; Г —крышечка; 2 — гребенчатый орган; 3 - легкие;
4 — жабры; 5 — тельсон; римскими цифрами обозначены сегменты тела
на всем протяжении истории от низов кембрия до перми протекала в
условиях чисто морского режима. Ранние этапы эволюции хелицеровых,
именно эуриптерид, протекали в условиях прибрежной жизни, а затем в
солоноватой воде и пресных водах эстуарий и рек. Надо напомнить, что
древнейшие силурийские скорпионы были связаны близким родством
с эуриптеридами (Stormer, 1963) и были первыми среди членистоногих
вышедшими из воды на сушу; поэтому анализ строения скорпионов и
эуриптерид (рис. 59), сделанный в последние годы В. Б. Дубининым
.(выделение им эуриптерид из меростомовых и объединение их вместе
со скорпионами в класс Scorpiomorpha) является обоснованным. Возводя
отряды паукообразных в классы, Дубинин устанавливает классы Scor-
piomorpha, Arachnida, Acaromorpha и Solifugomorpha и объединяет их
с классом Merostomata в надкласс Chelicerata.
С выходом на сушу и освоением условий наземной жизни начинает-
ся новый этап в эволюции и филогении большой ветви хелицвровых, со-
ставляющей прежний класс Arachnoidea. Известно, что уже в карбоне
многие современные и вымершие классы и отряды арахнид были хо-
рошо дифференцированы, следовательно, их эволюция протекала в ран-
нем палеозое задолго до карбона, но пока она нам мало1 известна.
В настоящее же время они представлены четырьмя классами и пример-
но 350 000 видов.
Эта большая ветвь членистоногих включает малочисленных л»еро-
стомовых, морских в течение всей их' истории (хотя некоторые совре-
менные мечехвосты заходят в реки), сохранивших примитивные черты
строения, с отчетливым проявлением в онтогенезе четырех ларвальных
сегментов, и многочисленных хелицеровых, представленных в основном
наземными, далеко отклонившимися от исходного типа по условиям
жизни, по строению и индивидуальному развитию, в том числе и по
(плохо выраженной) закладке ларвальных сегментов. И вместе с тем бес-
спорно, что вся эта ветвь членистоногих через примитивных кембрий-
ских представителей связана родством с примитивными трилобитами.
Признавая большую морфологическую близость позднейших нижне-
силурийских эуриптерид и древнейших скорпионов (Stormer, .1963),
215
надо подчеркнуть экологические факторы, сделавшие возможным пе-
реход их от водного к наземному образу жизни. Во всяком случае у
позднейших эуриптерид жаберный аппарат был хорошо дифференци-
рован, а жаберная полость с ее крышками была настолько изолирована
от внешней среды, что это позволяло им жить не только в морской
среде. При сменах среды у них, очевидно, происходили изменения в фи-
зиологии дыхания. Можно напомнить, что современные скорпионы и
представители многих классов арахнид приспособлены к биотопам
с влажной средой, к дыханию влажным воздухом, они живут нередко в
'непосредственной близости от воды. Экологически наиболее пластичны
клещи, часть которых вполне освоила водную среду (гидракарины).
В разнообразных экологических условиях, включая и паразитизм, среди
арахноморф клещи обнаруживают наибольшее разнообразие как во
внешней морфологии, так и во внутреннем строении и индивидуальном
развитии. И вместе с тем ряд клещей сохраняет древние черты в расчле-
нении тела (Р. Schulze, 1936).
Менее отчетлива родственная связь хелицеровых с панто-подами или
пикногонидами, которых В. Б. Дубинин, а до него ряд зарубежных
авторов, сближают друг с другом. В. Б. Дубинин, правильно, по-нойо-
му истолковав строение палеозойских и отметив ряд общих черт в стро-
ении современных пантапод и хелицеровых (например, хелицеры, боко-
вые ветви кишечника, видимо, четыре ларвальных сегмента) включил
пантопод в эту группу членистоногих, названную им подтипом Chelice-
romorpha.
После работ Д. М. Федотова (Fedotov, 1925), П. П. Иванова (Iwa-
noff, 1933) и особенно Штёрмера (Stormer, 1942, 1944, 1955а, 19556)
трилобитов, за редкими исключениями (Sanders, 1957, 1959; Butt, 1960),
не связывают с ракообразными. Их и ракообразных считают различными
ветвями членистоногих, так как в течение геологической истории они не
сближаются друг с другом. Не случайно древние, наиболее примитивные
раки имеют тело, сдавленное с боков, а не в дорсо-вентральном направ-
лении, что встречается лишь как исключение среди Notostraca и ряда
высших раков. -Головного щита, подобного трилобитному, у ракообраз-
ных нет. Исходный тип состава головы (акрон и три ларвальных сег-
мента) у раков не отвечает тому, что мы знаем о голове трилобитов.
Не случайно также, что при крайне примитивном строении, небольшом
головном отделе, почти гомономией членистости аннелидообразного-
тела, например, у девонской Lepidocaris или современной Hutchinso-
nlella, ряд члеников, обычно брюшка, не имеет конечностей. Различия
в типе строения и онтогенезе, несмотря на сходство экологии (обитание
в водной среде, преимущественно в море), говорят об отсутствии близ-
кого родства между раками и трилобитами.
Если картина исторического развития трилобитов и хелицеровых
достаточно ясна, то этого нельзя сказать о ракообразных, хотя сравни-
тельно прочные покровы тела у большинства их и водный образ жизни
являются важными условиями сохранения последних в ископаемом со-
стоянии. Число видов ракообразных (20 000) почти вдвое меньше, чем
трилобитов и хелицеровых (45 000), но геологическая история и пути
эволюции ракообразных с их необычайным разнообразием строения
очень сложны, причем положение усугубляется еще тем, что для ряда
древнейших кембрийских ракообразных конечности неизвестны, а сле-
довательно, их природа остается спорной. В частности, природа самых
примитивных ископаемых ракообразных Notostraca и Anostraca до сих
пор не выяснена. Некоторые авторы, в том числе и я (Fedotov, 1925),
склонны видеть в среднекембрийской Бургесской фауне представителей
и Anostraca (Opabinia и Yohoia) и Notostraca (Burgessia), относя сюда
и Protocaris из нижнего кембрия. Я допускал также, что Wapiia отно-
216
сится .к Leptostraca или является личиночной стадией Malacostraca.
Однако единого мнения о 'природе этих членистоногих нет (Henriksen,
1928, 1929); Штёрмер, например (Stormer, 1942, 1959), не считает их
раками и в качестве класса Pseudocrustacea относит в свой подтип
Trilobitomorpha вместе с классом трилобитов, находя между ними неко-
торое сходство в строении конечностей.
Но не частности в строении конечностей, а признаки общей органи-
зации говорят о принадлежности этих форм скорее.к Crustacea, чем к
Trilobitomorpha или к труппе членистоногих, с которыми связано про-
исхождение ракообразных, как допускал Ремане (Remane, 1954). Еще
менее оснований у Сандерса (Sanders, 1957, 1959) видеть признаки
трилобитов у Hutchinsoniella в наличии конечностей из семи члеников.
Это несомненно конвергенция, так как слишком уж велико различие в
общей организации этого примитивнейшего ракообразного и трило-
битов.
По-видимому, Сандерс прав (Sanders, 1957), переводя среднедевон-
скую Leptocaris, считавшуюся представителем Notostraca, в отряд (под-
класс) Cephalocarida.
Для экологии ракообразных характерен почти исключительно вод-
ный образ жизни. В отличие от арахнид и трахеат свою эволюцию они
проделали и продолжают ее в море, частично в пресных водах, заселив
все биотопы от прибрежной, литоральной полосы до супраабиссали —
10710 м (Беляев и др., 1960). Они представлены ползающими донными,
порой самыми крупными формами из современных членистоногих, мел-
кими планктонными и нектонными формами, свободноживущими, не-
редко прикрепленными, порой обитателями инфауны, а также много-
численными эктопаразитами и эндопаразитами почти всех групп жи-
вотных. Естественно, что ракообразные при этом претерпевают необы-
чайное разнообразие в строении, физиологии и онтогенезе. Известны
фундаментальные изменения (до полной утраты исходного строения и
исчезновения конечностей, особенно головных), сегментов головы и ту-
ловища, изменения внутренней организации, в том числе фильтрующих
аппаратов. У ряда эндопаразитических форм, как известно, происходит
утрата почти всех признаков не только класса, но и типа членистоно-
гих, и лишь анализ онтогенеза позволяет определить природу таких
ракообразных. Как хищники, питающиеся крупной добычей, планкто-
ноеды, трупоеды и сапрофиты, ракообразные играют громадную роль в
санитарии моря.
Что касается до геологической истории ракообразных, то остатки
Notostraca отмечают в перми, бесспорные же их остатки известны из
триаса; причем некоторые из них (например, Triops cancriformis minor
Irusheim) лишь мелкими признаками отличаются от современных видов.
В нижней юре, по данным Н. И. Новожилова (Novojilov, 1957, 1959),
встречаются весьма своеобразные, напоминающие Notostraca низшие
ракообразные (новый отряд Kazacharthra), нахождение которых еще бо-
лее подчеркивает богатство формообразования среди низших раков.
Наряду с этим, начиная с кембрия, идет развитие (в основном, в мо-
ре) многочисленных и разнообразных двустворчатых раковинных остра-
код, конечности которых неизвестны; с девона развиваются (преимуще-
ственно в пресной воде) Conchostraca, которые, во всяком случае по
внешней форме, мало отличаются от современных представителей и
также не имеют ничего общего с трилобитами.
Некоторые палеонтологи (например, Чернышев, 1934; Roger, 1953)
признают отряд высших раков Archaeosraca Claus, представленный не-
сколькими десятками родов, порой очень крупных (до 16 см длиной),
весьма разнообразных и необычных по форме, существовавших с ран-
него кембрия до триаса. Многие из этих форм (строение конечностей
217
их неизвестно) напоминают представителей отряда Kazacharthra
Н. И. .Новожилова и. вероятно, будут исключены из группы высших
раков. Некоторые формы, вроде нижнедевонокого Nahecaris, по внеш-
нему строению, характеру конечностей близки к современным реликто-
вым Leptostraca. Известный знаток, морфологии высших раков Зивинг
(Slewing, 1956, 1960а, 19606) из всех Archaeostraca лишь эту форму
считает принадлежащей к Malacostraca. Титс и Ментон (Tiegs a. Man-
ton, 1958) также считают Nahecaris самым ранним бесспорным пред-
ставителем Malacostraca. С девона встречаются и редкие остатки неко-
торых других отрядов высших раков, сходные с современными пред-
ставителями.
В основном фауна членистоногих палеозоя была представлена три-
лобитами, развитие же главной массы ракообразных, кроме упомянутых
выше некоторых отрядов (или подклассов), проходило в мезозое, в тре-
тичное время и в кайнозое. Если трилобиты в течение всей своей исто-
рии оставались обитателями морской среды, в основном как придонные,
ползающие, отчасти зарывающиеся формы, то ракообразные после па-
леозоя заселили не только все воды Мирового океана в горизонтальном
и вертикальном направлениях, но и пресные воды (донные, плактонные,
нектонные формы), а недавно, хотя и в незначительной мере, стали и
обитателями суши. В отличие от трилобитов с их ограниченностью в
морфологии и экологии, ракообразные являются ветвью членистоногих
с громадным диапазоном различий в морфологическом и экологическом
отношениях, от крайне примитивных современных форм, вроде Hutchin-
soniella, до высоко организованных высших раков, от активных хищ-
ников и трупоедов до узко специализированных, морфологически край-
не упрощенных эндопаразитов, потерявших .признаки даже типа членис-
тоногих, от типичных водных обитателей до наземных форм.
Последняя, самая большая ветвь членистоногих — многоножки н
насекомые, как известно, фигурирует под разными названиями: Trachea-
ta, Atelocerata, Antennata, Mandibulata. Мне кажется, что трахейная си-
стема, поскольку с ней органически связаны многие основные черты
строения многоножек и насекомых, настолько важна и характерна для
этих групп и для их родства, что следует оставить для них название
Tracheata. Правда, трахеи имеются и у других членистоногих, но счи-
тают, что они возникли в разных группах независимо, конвергентно,
а трахеи, например, паукообразных, кроме того, имеют совсем другое
морфологическое значение, чем у насекомых и многоножек. Основными
органами дыхания паукообразных являются легкие, а трахеи нередко
возникают из последних.
Название Atelocerata неудачно, так как еще надо доказать, что мно-
гоножки и насекомые произошли от предков с двумя парами антенн и
утеряли одну пару. Название Antennata более приемлемо, но при сопо-
ставлении с Diantennata, как некоторые называют ракообразных,, оно
так же неудачно, как и Atelocerata.
В отличие от ветви ракообразных, вся эволюция которой неразрывно
связана с морской и пресноводной средой, эволюция трахейных шла в
основном в условиях наземной жизни. В связи с этим имеется много про-
белов в палеонтологии трахеат: не найдено каких-либо вымерших форм,
которые выявили бы родственные отношения между трахейными и коль-
чатыми червями, нет ископаемых, поясняющих филогенетические отно-
шения между многоножками и насекомыми и разными группами мно-
гоножек. Исключением являются данные палеонтологии о высших кры-
латых насекомых, и то лишь начиная со сравнительно позднего периода
их истории: с карбона и перми. Родственные отношения между много-
ножками и насекомыми в основном приходится выяснять с помощью
данных систематики, анатомии и эмбриологии современных форм. Как
218
известно, начальные данные палеонтологии относятся для многоножек
к силуру, а для насекомых —к девону.
Несмотря на очень древний наземный образ жизни трахейных, бро-
сается, в глаза то, что современные трахейные (многоножки во взрослом
состоянии и в онтогенезе и насекомые в основном в онтогенезе) -обла-
дают многими примитивными аннелидными чертами. По основным чер-
там строения многоножки группы Chilopoda сохраняют большее сход-
ство с аннелидами, чем любой, даже самый примитивный представитель
хелицеровых или ракообразных. Вместе с тем мало обоснованы как
утверждение Бата (Butt, 1957) . о наличии в развитии насекомых три-
лобитной стадии, так и допущения некоторых авторов (Handlirsch, 1926;
Мартынов, 1934, и др.) о происхождении насекомых от трилобитного
предка. Все-таки реальное сходство с трилобитами проявляют лишь
древнейшие морские, хелицеровые, а проявление примитивных черт ме-
тамерии у представителей всех ветвей членистоногих, включая «трило-
битную» стадию развития насекомых, говорит лишь об их отдаленном
аннелидном предке, но никак не о происхождении трахеат от трилоби-
тов.
Я разделяю взгляд Бёттгера (Boettger, 1958), согласно которому в
-основе ствола многоножки — насекомые стоят Chilopoda, наиболее при-
митивные как по строению (однородный характер сегментов тела и ног,
наличие стернитов, артериальных сосудов сердца, положение полового
отверстия на заднем конце тела), так и по типу онтогенеза. Diplopoda
(утерявшие первую пару челюстей, с передним положением половых от-
верстий и имеющие сегменты тела с двумя конечностями каждый, кроме
безногого шейного и трех следующих простых сегментов, несущих по
одной паре ног) и Pauropoda, обладающие рядом регрессивных черт
во внутренней организации, являются вторично (сравнительно с Chilo-
poda) измененными многоножками. По наличию шестиногой стадии,
вылупившейся из яйца, и по некоторым чертам эмбриогенеза (Лигнау,
1911) они отчасти сближаются с насекомыми. Однако другие авторы,
например, П. П. Иванов, в эмбриологии и особенно в развитии лар-
вальных сегментов видят больше сходства с насекомыми у Chilopoda,
чем у Diplopoda. Наконец, в последнее время А. Г. Шаров (1953) под-
черкнул общие черты в положении зародышевой полоски и в ее пере-
мещениях внутри яйца у Pauropoda, Diplopoda и Chilopoda в отличие от
Symphyla, Thysanura и Pterygota, обладающих в этих отношениях своей
спецификой.
Палеонтологи считают, что из трахейных многоножки были первы-
ми выходцами на сушу, подобно скорпионам из хелицеровых. К сожа-
лению, палеонтологические данные о многоножках отрывочны и мало
дают как для выяснения филогенетических отношений в пределах груп-
пы многоножек, так и в отношениях их с насекомыми. Кроме того,
остатки частью недостоверны, частью недостаточно изучены. Последнее
относится к родам Diplopoda Archidesmus Peach из верхнего силура и
девона, Kampecaris Page из древнего красного песчаника и особенно
к родам семейства Euphoberidae Acantherpestes, Euphoberia Meek et Wor-
ten и Palaeosoma Jackson et Brade-Brike из верхнего карбона. У пред-
ставителей этого семейства были сложные и простые глаза, «плава-
тельные» ножки у одних и подобие жабр в виде пластинок и пузырьков
па конечностях — у других (Мартынов, 1934). Думают, что эти древ-
нейшие многоножки вели водный образ жизни.
Данные палеонтологии по наиболее важной в филогенетическом от-
ношении группе многоножек — Symphyla начинаются с олигоцена.
К сожалению, мелкие размеры и нежные покровы этих многоножек не
благоприятствуют их сохранению в ископаемом состоянии. Мало дает па-
леонтология и для выяснения филогенетических отношений между Chi-
219
lopoda и Diplopoda, так как многочисленные и достоверные остатки ти-
пичных Diplopoda известны лишь из карбона — перми. Систематическое
положение Chilopoda (остатков из верхневестфальских отложений) не
достоверно. Несомненные остатки Chilopoda известны лишь из ранне-
третичных отложений (в янтаре).
В организации и отчасти в развитии Symphyla давно отмечалось,
сходство с насекомыми. Анализом строения наружного скелета тела,
а также мускулатуры тела Symphyla, Chilopoda и сопоставлением их с.
таковыми низших насекомых (например, Ephemeroptera) Э. Г. Бек-
кер (1949, 1950) показал, что Symphyla обладают рядом черт, которые
позволяют видеть в них или, вернее, в симфилоподобных многоножках
возможных предков насекомых. В этом отношении А. Г. Шаров, подчер-
кивающий сходство онтогенеза Symphyla с таковым Thysanura и Ptery-
gota, приводит дополнительные доводы в пользу этого взгляда.
В филогении насекомых остро стоит вопрос об отношениях между
первичнобескрылыми и крылатыми насекомыми (Федотов, 1962). Неко-
торые авторы, в том числе и А. Г. Шаров (1959), отказываются считать.
Protura, Collembola и Diplura насекомыми. Они оставляют в классе на-
секомых, кроме крылатых, лишь Thysanura, придавая неоправданно
большое значение эктогнатному типу ротовых органов, в чем я вполне
согласен с Мэнтон (Manton, 1958, 1964). Кроме того, А. Г. Шаров отме-
чает сходство в положении и перемещениях зародышевой полоски у
Thysanura и Pterygota и подчеркивает отличие в этом отношении от
Diplura и Collembola. Сторонники этого взгляда приводят различия этих
трех групп по числу сегментов, в деталях строения трахейной системы,
мальпигиевых сосудов, сердца и подчеркивают почти полное соответствие-
в строении Thysanura и Pterygota. Несомненно, в деталях многих систем,
органов у Protura, Collembola и Diplura -имеются отличия от Thysanura и
Pterygota, однако общими для всех этих пяти групп является следующее:
наличие трех отделов тела —голова, грудь и брюшко, трех грудных сег-
ментов с тремя парами ходильных ножек и связанное с этим значитель-
ное развитие мускулатуры в грудных сегментах, дифференцировка ки-
шечника на тонкий пищевод, проходящий по грудным сегментам, и сред-
нюю кишку, находящуюся в брюшке, где помещаются и половые желе-
зы. Такой тип строения является основным, объединяющим Protura,.
Collembola, Diplura, Thysanura и Pterygota воедино в Hexapoda, или In-
secta, и отделяющим их от других классов трахейных. При наличии этих
основных черт строения особенности ротовых органов (эктогнатный или
энтогнатный тип) не имеют кардинального значения, как не имеют его
и различия в деталях строения тех или иных систем органов. В связи
с мелкими размерами и жизнью в почве или подстилке большинство,
особенностей в строении Protura, Collembola и Diplura носит черты
упрощения и редукции на фоне общего более низкого уровня их орга-
низации сравнительно с Thysanura и Pterygota.
Все это заставляет считать шестиногими, или насекомыми, не только-
Thysanura и Pterygota, но и Protura, Collembola и Diplura; однако для
того, чтобы подчеркнуть большую обособленность последних трех групп
от первых двух, следует класс Hexapoda, или Insecta возвести в ранг
надкласса с двумя классами: класс Apterygota с двумя подклассами
(Ectognatha и Thysanura) и класс Pterygota с двумя подклассами (Ра-
laeoptera и Neoptera). Систему, предложенную А. Г. Шаровым (1959),
делящую Atelocerata, или Tracheata, на восемь классов: Pauropoda, Sym-
phyla, Diplopoda, Chilopoda, Protura, Collembola, Diplura и Insecta c
двумя подклассами: Apterygota, или первичнобескрылые (Monura, Thy-
sanura), и Pterygota, надо отвергнуть как необоснованную. Однако
никто не станет отрицать, что по всем признакам Thysanura ближе •<
Pterygota, чем другие Apterygota и если они и раньше рассматрива-
220
лись многими как группа,' близкая предкам насекомых, то после работ
А. Г. Шарова о развитии лепизмы (1953) и описания им строения Dasy-
leptus (Monura) из нижней перми, мы можем ставить, как это делает
Бёттгер (Boettger, 1957), Thysanura между Symphyla и Pterygota в фи-
логении многоножки — насекомые, а остальные три группы бескрылых
насекомых считать боковыми ветвями, отходящими в сторону.
Что касается филогенетических отношений групп крылатых насеко-
мых, то за последние три десятилетия имеются значительные достиже-
ния в отношении таких групп, как Ephemeroptera, Blattodea, Orthoptera
и др. Основы филогении Pterygota заложены работами А. В. Марты-
нова. Его Palaeoptera и Neoptera, которые надо считать подклассами,
и распределение между ними высших и низших отрядов крылатых на-
секомых, как ископаемых, так и современных, деление Neoptera на Poly-
neoptera, Oligoneoptera и Paraneoptera, как показал сам А. В. Марты-
нов (1938), является, в сущности говоря,'естественной системой Pterygo-
ta и выражением их филогенетических отношений. Естественность систе-
мы Мартынова была подтверждена рядом работ по анатомии высших
‘насекомых. Были подтверждены и его филогенетические соображения,
хотя в них и были внесены известные уточнения и некоторые исправле-
ния, касающиеся, например, мекоптероидного комплекса высших насе-
комых, объединяющего Mecoptera, Lepidoptera, Trichoptera, Diptera и Hy-
menoptera, которые были сделаны одновременно и независимо друг от
друга Хинтоном (Hinton, 1958) и О. М. Мартыновой (1959). В настоя-
щее время с учетом новых данных (см. Родендорф, 1962) Palaeoptera
и Neoptera А. В. Мартынова поделены на Palaeoptera и Archaeoneoptera,
Polyneoptera, Paraneoptera и Oligoneoptera, составляющих .Neoptera, при
этом даны филогенетические отношения между всеми группами насеко-
мых, как современными, так и вымершими.
Пока палеонтология ставит нас перед фактом неожиданного появле-
ния в карбоне — перми отрядов Palaeoptera (вымерших, кроме Odonata
и Ephemeroptera) и некоторых (частью вымерших) отрядов, родо-
начальных для многих современных Neoptera, например, Protoblattodea,
Paraplecoptera, и др. Данные говорят об историческом развитии сначала
преимущественно отрядов с неполным, а затем с полным превращением.
Расцвет последних, как известно, начался в мезозое и происходил в ос-
новном в кайнозое. Это неожиданное появление такого разнообразия и та-
кого числа отрядов насекомых в карбоне-перми, как правильно думает
Б. Б. Родендорф (1947), заставляет считать, что развитие крыла и фор-
мирование отрядов крылатых насекомых происходило задолго до карбо-
на, в девоне или даже до девона. Вследствие трудности нахождения в
континентальных отложениях Ископаемых насекомых, мы пока не знаем
древнепалеозойского этапа эволюции этого класса. Недавнее описание
крылатого насекомого Eopterum devonicum Rohdendorf Б. Б. Родеядор-
форм (1961) оправдало его предположение о существовании в раннем
палеозое крылатых насекомых; но достаточно ли данных, чтобы устано-
вить для этой формы новый отряд Archaeoneoptera, для меня неясно.
Выше было сказано, что сторонники полифилии и монофилии чле-
нистоногих в пользу своих взглядов используют,'помимо других данных,
особенности эмбрионального развития головы и число ее сегментов.
Отмечалось также, что в самое последнее время усилилось течение в
пользу взгляда на монофилию членистоногих, в целом же спор
продолжается. Можно отметить, что сторонниками монофилии членисто-
ногих являются Рай Ланкастер (Ray Lankester, 1904), Геймонс (Неу-
mons, 1901), Карпентер (Carpenter, 1903, 1906), Гандлирш (Handlirsch,
1926), Снодграсс (F. Е. Snodgrass, 1938, I960), Геегард (Heegard, 1945),
Я- А. Бирштейн (1948), В. Н. Беклемишев (1952, 1964), Н. А. Ливанов
(1955), Бат (Butt, 1960), Poo (Raw, 1957), Лежандре (Legendre, 1959)
221
и др. Напротив, мнение о полифилии членистоногих разделяют Кингсли
(Kingsley, 1897), И. Кеннель (1891), Паккард (Packard, 1903), Плате
(Plate, 1922), Франц (Franz, 1924), Д. М. Федотов (Fedotov, 1925),
П. П. Иванов (Iwanoff, 1933), Штёрмер (Stormer, 1942, 1944, 1955а,.
19556, 1956), Шульце (Р. Schulze, 1936, 1939), Вандель (Vandel, 1949),
Тйгс и Ментон (Tiegs a. Manton, 1958), Ментон (Manton, 1964), Вебер
(Weber, 1954) и ряд других авторов. Уже одно .перечисление этих имен
говорит о трудности поднятого вопроса; правильно отмечал В. Н. Бек-
лемишев, что для окончательного решения его недостает фактических
данных.
Однако из всего сказаннного выше видно, что' трилобиты (или три-
лобитоморфные), хелицеровые, ракообразные и трахейные являются
группами, примитивные древние формы которых появлялись в разное
геологическое время; эволюция каждой из этих групп была различна
вследствие особенностей строения, индивидуального развития и эколо-
гии. Родоначальниками трилобитов, хелицеровых, ракообразных и тра-
хейных, очевидно, были морские неспециализированные аннелиды, воз-
никавшие из разных семейств древних полихет и развивавшиеся в эко-
логически разных условиях.
Форониды (Phoronida)
i
Двустороннесимметричные, вторичнополостные, одиночные, червеоб-
разные, живущие в трубках обитатели моря. Верхняя часть цилиндриче-
ского тела несет лофофор со щупальцами, сближенные отверстия петле-
образного кишечника, ротовое — внутри, анальное — вне кроны щупа-
лец. Кожно-мускульный мешок состоит из наружных кольцевых и внут-
ренних продольных мышц. Примитивная нервная система в основном
состоит из диффузного сплетения в эпидермисе, из преорального нерв-
ного поля, кольца и левого нерва или гигантского волокна. Диффе-
ренцированных органов чувств нет. Вторичная полость тела более или
менее подразделена на три отдела. Замкнутая кровеносная система,,
органов дыхания нет. Органы выделения и выведения половых продук-
тов в виде пары целомодуктов. Полы обычно не разделены, яичники
и семенники непарные. Дробление радиальное, иногда спиральное, раз-
витие с хорошо выраженными тремя зародышевыми листками, с пелаги-
ческой личинкой и метаморфозом. Обитатели бентоса мелководья, пи-
тающиеся наннопланктоном, размножающиеся почти исключительно по-
ловым путем.
Форониды — набольшая реликтовая группа, включающая два рода
и несколько видов, судя по косвенным данным существуют с древнего
палеозоя.
Среди исследователей этой группы надо отметить А. О. Ковалевского
(1867), И. И. Мечникова (Metschnikoff, 1871), Колдуэлла (Caldwell,
1883), Икеду (Ikeda, 1901), Кори (Cori, 1891, 1930, 1939а, 19396), Сели
Лоншана (Selys Longchamps, 1907), Е. А. Шульца (1897, Schultz, 1903),
Бэте (Bethe, 1927), Силена (Silen, 1952, 1954а, 19546), Раттенбера
(Rattenbury, 1954) и ряд других (см. Hyman, 1959). Лучшая сводка по-
этой группе сделана Гаймен.
Тело у форонид червеобразное, двусторо'ннеоимметричное, цилиндри-
ческое, нечленистое, со щупальцевым аппаратом на переднем верхнем
конце, колбовидно вздутое на заднем, прозрачное, красное, оранжевое,
или зеленое, реже бесцветное. Сагиттальная плоскость симметрии делит
щупальцевый аппарат и тело червя пополам, по линии, соединяющей
ротовое и анальное отверстия. Эпидермис секретирует цилиндрическую,,
хитиновую, по Гаймен (Hyman, 1958), трубку, которую форониды не по-
кидают в течение всей жизни, а лишь высовывают из нее переднюю часть
тела. При выделении трубки к ней пристают песчинки, кусочки раковин,,
иглы губок и т. д. Форонисы обычно прикрепляются (стелятся) к мяг-
кому, илисто-песчаному или твердому грунту, реже буравят раковины
моллюсков или твердый известковый грунт, делая в последнем вер-
тикальные ходы.
Форониды — небольшие животные. Самым мелким является Phoro-
nis avails — 6 мм в длину, самым крупным —• Phoronopsis californlca —
30—37 см.
223-
Верхняя часть тела — щупальцевый аппарат, или лофофор с щу-
пальцами, скрывающий эпистом, прикрывающий рот; вблизи рта вне
лофофора лежит на спинной стороне анус. Эпистом онтогенетически
принадлежит преоральной лопасти личинки (просоме), лофофор со щу-
пальцами— посторальному ее отделу (мезосоме), а тело форониса —
туловищному отделу (или метасоме). Однако в итоге онтогенеза это
расчленение тела у взрослого червя выражено неясно.
Щупальцевый аппарат состоит из подковообразного лофофора,
у некоторых для экономии места свернутого спиралью и несущего на-
ружный и внутренний ряды многочисленных (порой более тысячи) рес-
ничных полых щупалец. Толстая базальная мембрана под эпидермисом
служит как бы скелетом щупальцевого аппарата. Контакт с внешней
средой обеспечивается этим аппаратом благодаря многочисленным чув-
ствительным клеткам сильно сократимых щупалец и парного лофофор-
ного органа, легко реагирующего на внешние раздражения. Ресничные
клетки эпидермиса щупалец обуславливают вводный ток воды, который
гонит взвесь и мелкие одноклеточные организмы (наннопланктон), об-
волакиваемые слизью, выделяемой железистыми клетками эпидермиса
щупалец и лофофорного органа, к ротовому отверстию. Выводным то-
ком воды, идущим между основаниями щупалец, гонятся из ануса и
нефропоров фекалии и продукты распада. Таким образом, щупальцевый
аппарат выполняет вододвигательную для улавливания пищи и сани-
тарную функции. Наличие в целоме щупалец кровеносных сосудов
с гемоглобином, очевидно, позволяет им выполнять дыхательную
функцию.
Стенка тела туловища состоит из слоя эпидермиса, одетого тонкой
кутикулой, выделяемой эпидермисом базальной мембраны, кожно-мус-
кульного мешка и перитонеума. Среди высоких и узких эпидермальных
клеток разбросаны 'Чувствительные клетки, связанные с базиэпителиаль-
ным нервным сплетением, железистые ацидофильные и базофильные
клетки и 'клетки с гомогенными гранулами. Вещество трубки выделяет-
ся последними (Silen, 1952). Кожно-мускульный мешок состоит из тон-
кого, наружного кольцевого слоя мышц и сильного внутреннего слоя
пучков продольных мышц, образующих синцитий. Мышцы гладкие.
Кожно-мускульный мешок неоднороден. Передняя узкая часть тела наи-
более мускулиста и сократима. У ряда форм кольцевые мышцы, обра-
зующие три сфинктора, отсутствуют в задней, ампуловидно вздутой час-
ти, продольные же мышцы тянутся по всей длине тела.
Представление о нервной системе форонид, приводимое в моногра- -
фиях и сводках (Hyman, 1959), после исследования Силена (Silen, 1954а,
19546) приходится существенно пересмотреть (рис. 60); остаются без
изменений некоторые частные соображения, например, о положении
нервной системы в эпидермисе, наличии подкожного нервного сплетения
п некоторые другие. По-видимому, справедливы также данные Шульца
(Е. A. Schulz, 1903) о развитии при регенерации ганглия путем впячива-
ния эктодермы, протекающем так же, как при развитии его у пресновод-
ных мшанок (Nitsche, 1675; см. гл. «Мшанки»; Safftigen, 1888; Gerwerz-
hagen, 1913).
Главную часть нервной системы, по Силену, составляет диффузное
подкожное сплетение из волокон и уни- и биполярных нервных клеток,
лежащее в эпителии над его базальной мембраной. С ним связаны чув-
ствительные клетки эпидермиса, особенно многочисленные в щупальцах
и лофофорном органе. Волокна же двигательных клеток, иннервирую-
щих. мускулатуру, пронизывают базальную мембрану.
В мезосоме, между эпистомом и анусом, находится нервный центр —
преоральное нервное поле Силена, или церебральный ганглий (по дан-
ным прежних авторов), без ясных границ переходящий в эпидермис
224
Он образует вдоль наружного гребня лофофора нервное кольцо, дающее
нервы щупалец наружного ряда. Нервы щупалец внутреннего ряда от-
ходят непосредственно от нервного центра. Справа от последнего берет
начало боковой нерв, вернее, гигантское волокно, переходящее на левую
сторону метасомы, нс не заходящее в ампулу туловища. Обычно имеется
левое, реже правое гигантское волокно, в наиболее примитивном случае,
сохраняются оба волокна.
Рис. 60. Схема строения нервной системы форониса
> (по Silen)
1 — преоральное нервное поле; 2 — нервное кольцо; 3 — боковой
нерв; 4 — нервное щупальце наружного ряда лофофора; 5 — нервное
щупальце внутреннего ряда лофофора
В мезосоме находится, таким образом, часть нервной системы с бо-
лее многочисленными и сконцентрированными нервными элементами, в
метасоме нервная система вся диффузна. Чувствительная, рецепторная
часть нервной системы, судя по опытам Силена (Silen, 1954; Коштоянц,
1957), находится в мезосоме, в области щупальцевого аппарата, а эффек-
торная— в мускулистой части метасомы. Раздражение щупалец вызы-
вает быстрое синхронное сокращение продольных мышц, приводящее к
сокращению всего тела. Возбуждение проходит по гигантскому нервному
волокну, являющемуся эффекторной частью нервной системы.
15 Д. М. Федотов
225
Диффузный характер раздражения и сокращения, идущих в обоих
направлениях, установлены для нервной сети туловища форонид. Их
нервная система примитивна как по строению и расположению в эпи-
дермисе; так и по физиологии; нервная регуляция как отмечает
X. С. Коштоянц (1957), протекает по типу диффузной нервной сети ки-
шечнополостных.
Кишечник, как выше отмечено, сквозной, петлеобразный со сближен-
ными ротовым и анальным отверстиями. Щелевидный, ресничный рот,
прикрытый эпистомом, ведет в ресничную ротовую полость, .переходя-
щую в пищевод. Последний состоит из ресничных клеток, железистых
клеток, базиэпителиального нервного сплетения, базальной мембраны и
кольцевых мышц. Радиальные мышечные пучки прикрепляются к стен-
кам пищевода и к стенкам тела. Эктодермальный пищевод ведет в экто-
дермальный преджелудок, расширяющийся в желудок, который переги-
баясь переходит в тонкую (эктодермальную) кишку, поднимающуюся
кверху и открывающуюся анальным отверстием на спинной стороне те-
ла. Стенка желудка составлена кубическими ресничными эпителиальны-
ми-.клетками, железистыми клетками, базальной мембраной, кровенос-
ной сетью и мускульными волокнами. Вдоль спинной стенки преджелуд-
ка и желудка проходит полоса ресничных клеток. Пища (наннопланк-
тон), двигающаяся благодаря работе ресничек, в желудке склеивается
секретом желез в шнур, вращающийся под ударами ресничек. Пищева-
рение, как и у личинок, внеклеточное в полости желудка и внутрикле-
точное (Silen, см. Беклемишев, 1952) в синцитиальных выпячиваниях,,
образованных эпителиальными стенками желудка. Тонкая кишка обра-
зована кубическим слабо ресничным эпителием, базальной мембраной
и редкими мышечными волокнами. Кишечник одет перитонеумом, желу-
док подвешен брюшным, а порой и вторичным спинным мезентериями,
а тонкая кишка подвешена правым и левым латеральными мезентерия-
ми. Как ни странно, пищеварение прослежено лучше у личинки акгино-
трохи (Becker, 1937), чем у взрослых форм.
Целом состоит из полости эпистома, полости щупальцевого аппара-
та— подковообразного целомического канала, дающего каналы в щу-
пальца и общей полости тела, отделенной от последнего септой, или диа-
фрагмой. Как и некоторые авторы, я считаю, что эти полости соответ-
ствуют протоцелю, мезоцелю и метацелю ряда других вторичнополост-
ных групп нечленистых. Многие авторы правда признают существование
лишь двух отделов целома. Нет согласованного мнения и о числе мезен-
териев форонид, что отчасти связано с непостоянством их числа у разных
форм. Обычно имеются, как было сказано, брюшной правый и левый ла-
теральные мезентерии, причем брюшной мезентерий форониса отвечает
таковому личинки. У Phoronis avails латеральных мезентериев нет, НО'
зато имеются спинной и брюшной мезентерии (Silen, 1952). Перитоне-
альная выстилка представлена синцитием с ядрами. В полостной жидко-
сти находятся клетки крови с гемоглобином и несколько форм цело-
моцитов.
С брюшным и левым латеральным мезентериями связано непарное
жировое тело, по А. О. Ковалевскому (1867), или вазоперитонеальная
ткань, по Кори (Cori, 1891).
Выделительные органы представлены двумя V-образной формы цело-
модуктами, ресничные воронки которых открываются в метацель по
сторонам пищевода, а выводные отверстия — по бокам порошицы. Цело-
модукт имеет или одну воронку или две воронки — большую и малую.
Онтогенетических доказательств о происхождении таких целомодуктов
путем слияния каждого из двух, как думал Сели Лоншан, однако, нет
(Silen, 1952). В каналах целомодуктов, служащих также и выводными
половыми протоками, находятся экскреторные гранулы. Исследований и
226
экспериментов по экскреторной деятельности целомодуктов очень мало
(Ohuye, 1942, цитировано по Hyman, 1959).
Кровеносная система состоит из продольного спинного сосуда, пере-
ходящего .в целоме лофофора в кольцевой, точнее, подковообразный со-
суд, дающий ветвь в каждое щупальце. Сосуды щупалец переходят в
другой подковообразный сосуд, тесно прилегающий снизу к первому,
двумя ветвями, пронизав септу он переходит в продольный латераль-
ный сосуд туловища. Последний идет вдоль левой стороны нисходя-
щей петли кишечника и образует многочисленные, короткие дивертику-
лы, слепые сократимые капилляры, часть которых открывается в крове-
носный синус в стенке желудка. В этом месте кровеносная система не-
замкнута, и отсюда отходит спинной, или медиальный кровеносный со-
суд. Циркуляция крови достигается ритмическими сокращениями
продольных сосудов, особенно спинного; сокращения латерального сосу-
да и сосудов лофофора слабее и менее правильные; сосуды щупалец и
слепые капилляры сокращаются автономно, независимо от главных
(Bethe, 1927).
Венозная кровь по спинному сосуду движется вперед в верхний этаж
подковообразного сосуда лофофора, артериальная кровь из нижнего
этажа его движется назад по латеральному сосуду (Cori, 1939а, 19396;
Dawydoff, 1960). Однако ток крови может быть и в обратном направле-
нии (Bethe, 1927).
Большинство форонид гермафродиты, раздельнополы лишь некото-
рые представители. Непарные, крупные половые железы расположены в
ампуле туловища, на левой стороне, яичники на спинной, семенники на
брюшной стороне латеральнее кровеносного сосуда. Предполагают, что
жировое тело, или, пожалуй лучше, вазоперитонеальная ткань, служит
для питания гонад. Половые продукты выводятся наружу целомодукта-
ми. Яйца (стр. 230) откладываются в воду или в крону щупалец, реже в
трубку материнского организма. Размножение форонид почти исключи-
тельно половое. Лишь у единичных видов (iBrattstrom, 1943) наблю-
дается поперечное деление; последнее приводит к образованию перепле-
тающихся трубочек, вроде колонии. У Phoronis ovalis, кроме попереч-
ного деления — архитомии (Harmer, 1917), наблюдается и почкование.
(Marcus, 1949); почки образуются как выпячивания стенки тела ампулы,,
реже мускулистого отдела, в которые врастает часть петли кишечника,
кровеносные сосуды и т. д., образуются рот и щупальца, затем почка от-
шнуровывается. Дробление полное, почти равномерное, рано утрачиваю-
щее синхронность, до недавнего времени было плохо изучено. Оно счи-
талось или неправильным, или радиальным, некоторые намеки на спи-
ральное дробление у них отмечали Икеда (Ikeda, 1901) и Коршельт и
Гейдер (Korschelt u. Heider, 1936). У Phoronopsis viridis Раттенбури
(Rattenhury, 1954) описывает спиральное дробление, хотя по ряду осо-
бенностей отличающееся от спирального дробления аннелид, моллюс-
ков и других групп. Из целобластулы инвагинацией образуется гасгрула
с энтодермальной кишкой, или архентероном. Остаток переднего конца
очень большого бластопора, замыкающегося как у полихет с боков и
сзади наперед, становится ртом. Позже, на месте, отвечающем заднему
концу замкнувшегося бластопора (рис. 61), возникает, так же как у по-
лихет, анальное отверстие; задняя кишка образуется за счет конца экто-
дермального кишечника. Эктодермального проктодеума нет. Возникно-
вение мезодермы точно не прослежено. Обычно отмечается, что от
стенок эктодермального архентерона вычленяются клетки мезодермы
(целенхима по Давыдову, 1914; энтомезенхима по Г. А. Шмидту, 1953),
дающие целом. Клетки мезодермы у Phoronopsis viridis лежат у перед-'
ней и боковой губ бластопора (а не позади задней губы, как
у аннелид).
15*
227
Рис. 61. Раннее развитие Phoronopsis viridis (из Беклемишева)
А— стадия 16 бластомер; Б— гаструла; В — стадия роста в длину наружного полуша-
рия зародыша; 1 — анимальный полюс с султаном ресничек (2); 3 — растущая в длину
спинная сторона личинки
У форонид нет образования эктомезодермы из производных микроме-
ра 2d, и энтомезодёрмы — за счет 4d; впячивающаяся при гаструляции
стенка зародыша, образующая архентерон, у губ которого возникают
клетки мезодермы, образована, видимо, более крупными клетками,
производными крупных бластомеров (5а—5d и 5А—5D), и мелкими
клетками, производными четвертого квартета микромеров (Rattenbury,
1954).
Верхнее полушарие зародыша, имеющее теменную пластинку с нерв-
ным утолщением, иногда с 4 глазными пятнами, с султаном длинных
ресниц, покрытое ресничками, становится большой преоральной ло-
пастью в форме капюшона с преоральным венчиком ресниц по краю
(прототрох). Позади рта проходит венчик ресниц (метатрох) и возни-
кает подковообразное утолщение эктодермы, расположенное наискось,
на котором постепенно возникают личиночные щупальца, составляющие
вместе лофофор (рис. 62). Нижнее полушарие (а с ним и кишечник)
растет в длину; на его конце вокруг анального отверстия образуется
анальный венчик ресниц (телотрох). За счет эктодермы возникает пара
личиночных протонефридиев, типичных для трохофорных личинок,
с соленоцитами, парой нефропоров, открывающихся по сторонам по-
рошицы.
Формирование целома, образующегося схизоцельным путем, точно не
прослежено.. Большинство авторов находит два отдела целома—туло-
вищный целом с брюшным мезентерием и отделяющийся от него саптой
(или диафрагмой) пресептальный целом. По данным Икеда (Ikeda,
1901), автора одной из лучших эмбриологических работ по форонисам,
228
5 . J
Рис. 62. Развитие Plwronis (из Беклемишева)
А — замыкание бластопора; Б — трохофора; В — актинотроха; Г — то же, выпячивание зачатка
метасомы; Д — метаморфоз актинотрохи; 1 — преоральная лопасть; 2 — бластопор; 3 — теменная
пластинка; 4— кишечник; 5 — рот; 6 — личиночные щупальцы; 7 — анус; 8—метасома; 9 — зачаток
дефинитивных щупалец
от пресептального целома неполной септой отделяется еще преоральный
целом. Ряд авторов, начиная с Колдуэлла (Caldwell, 1883), принимает
не два, а три отдела целома у актинотрохи и у взрослых форонид. Более
того, Колдуэлл (1883) и К. Н. Давыдов (Dawydoff, 1960) говорят о пар-
ных целомах, которые большинство авторов считает непарными.
К сожалению, даже в работе последних лет (Rattenbury, 1954) этот
вопрос не выяснен. Однако кажется вполне возможным в соответствии с
тремя отделами тела зародыша и личинки (преоральной лопастью, пос-
торальной частью с лофофором и туловищем) принимать наличие и трех
отделов целома.
У актинотрохи развивается кровеносная система и жировое тело, или
вазоперитонеальная ткань. Кишечник образован большой ротовой по-
лостью, эктодермальным пищеводом и эктодермальным желудком с пар-
ными дивертикулами и задней или тонкой кишкой и анальным отвер-
стием; кишечник подвешен брюшным мезентерием. Зачатки половых
желез у актинотрохи неизвестны.
На брюшной стороне, несколько слева, под лофофором, образуется
мешковидное впячивание эктодермы, увеличивающееся по мере развития
личинки — зачаток метасомы.
Сформированная прозрачная личинка — актинотроха, двигательным
аппаратом которой служит телотрох, имеет от 1 до 5 мм в длину, ведет
пелагический образ жизни, питаясь взвесью и одноклеточными расти-
тельными (диатомовыми) и животными организмами, которые улавли-
229
Баются ресничными токами лофофора. Передвижение пищи в кишечнике
обеспечивается главным образом деятельностью ресничного эпителия и
лишь' отчасти сокращением мускулатуры кишечника. В эпителии пище-
вода находятся слизистые клетки, в эпителии желудка всасывающие
клетки и клетки, выделяющие слюну, пищеварение внутриклеточное
(Becker, 1937). Существует зависимость между количеством желтка в
яйцах и характером развития (Silen, 1954). У форонисов (например,
Phoronis miilleri), у которых яйца бедны желтком, все развитие проте-
кает в воде и приводит к образованию пелагических, планктонных личи-
нок — актинотрох, с продолжительной свободной жизнью. У форонисов
с яйцами, относительно богатыми желтком (например, Ph. hippocrepia),
последние сначала развиваются под защитой щупалец лофофора, а за-
тем актинотроха ведет пелагический образ жизни. Наконец, у форм с
крупными, очень богатыми желтком яйцами (Ph. avails') они отклады-
ваются в полость трубки, выходя через отверстие на переднем конце ма-
теринского организма после сбрасывания последним щупалец. Развитие
почти прямое и протекает значительно быстрее, чем у предыдущих видов.
Образуется лецитотрофная (термин Thorson, 1950) личинка, не получаю-
щая строения актинотрохи. Актинотрохи разных видов форонисов осе-
дают при метаморфозе не на любой попавшийся субстрат, но в соответ-
ствии с требованиями вида. Одни прикрепляются к мягкому грунту,
другие — к твердому субстрату — известковый грунт, раковины моллюс-
ков (Silen, 1954а, 19546).
При опускании актинотрохи на дно и прикреплении к субстрату,
а иногда еще до этого, мешковидное впячивание эктодермы на брюш-
ной стороне усиленно растет, выпячивается наружу и свешивается в ви-
де большого цилиндрического придатка почти перпендикулярно к про-
дольной оси актинотрохи. Выпячиваясь он втягивает кишечник и ста-
новится туловищем форониса; спинная сторона актинотрохи, растущая
слабо, превращается в участок переднего конца тела червя, между ро-
товым и анальным отверстиями.
При 'метаморфозе прикрепившейся актинотрохи щупальца сбрасыва-
ются и заменяются двумя рядами дефинитивных щупалец, зачатки ко-
торых появляются еще до метаморфоза у оснований личиночных щупа-
лец. Судьба предротовой лопасти и отчасти щупалец актинотрохи про-
текает или описывается по-разному. Предротовая лопасть и многие
щупальца актинотрохи заглатываются пищеводом (Metschnikoff, 1871;
Caldwell, 1883; Selys Longshamps, 1907) и перевариваются, а потому
эпистом и так называемый церебральный ганглий развиваются заново.
На этом основании Сели Лоншан считает эти образования не гомоло-
гичными таковым других первичноротых. Это утверждение ошибочно
(Cori, 1939а, 19396; Беклемишев, 1952), так как у актинотрохи и у фо-
ронисов при автотомии и регенерации переднего отдела тела восста-
навливаются сброшенные или удаленные части. По данным ряда других
авторов (Caldwell, 1883; Давыдов, 1914; П. П. Иванов, 1937), во время
метаморфоза актинотрохи предротовая лопасть редуцируется, превра-
щаясь в небольшой эпистом, а щупальца дегенерируют. Благодаря из-
менениям .преоральной лопасти у форониса на верхнем конце тела ока-
зывается подковообразный лофофор с двойным рядом щупалец. Клетки
туловища выделяют трубку.
Превращение актинотрохи в червя происходит очень быстро, в тече-
ние четверти часа, как отмечал еще И. И. Мечников (Metschnikoff, 1871).
Изменения внутренней организации ее сравнительно небольшие. В туло- -
вище, образованном выпяченным и выросшим в длину эктодермальным
мешочком, впячивается кишечник, становящийся петлевидным, диффе-
ренцируется кожно-муюкульный мешок из кольцевых и продольных
мышц, происходит образование сфинкторов (по Silen, 1952) и т. д.
230
Ганглий актинотрохи замещается дефинитивным нервным центром,
подробно изученным Силеном (Silen, 1954); кишечник изменяется незна-
чительно, дегенерируют парные дивертикулы желудка, последний диффе-
ренцируется на преджелудок и желудок. Пресептальный целом превра-
щается в подковообразный канал лофофора, дающий ветви в щупальца.
Туловищный целом, отделенный септой, или диафрагмой, от пресепталь-
ного, становится полостью тела форониса; сохраняется брюшной мезен-
терий, на котором подвешен кишечник, но формируются и латеральные
мезентерии. Ив пресептальной кровеносной полости образуется подково-
образный кровеносный сосуд лофофора с сосудами щупалец, располо-
женный в целоме лофофора. Главные продольные кровеносные сосуды
актинотрохи переходят в таковые форониса (Metschnikoff, 1871). Форми-
рование всей кровеносной системы было прослежено Кори (Cori, 1939а,
193916). Протонефридии занимают то же самое положение, что и у акти-
нотрохи, они не обнаруживают признаков дегенерации, но соленоциты
исчезают. Открытые ресничные воронки образуются, очевидно, за счет
целома (Goodrich, 1903).
Процессы регенерации (Шульц, 1897; Е. A. Schulz, 1903) у актино-
трохи (Actinotrocha brachialis) и форониса (Ph. miilleri) поясняют орга-
ногенез, протекающий как обычно. Пищевод у форониса восстанавлива-
ется впячиванием эктодермы, проктодеум не возникает, задняя кишка
восстанавливается из энтодермы, нефридии»— из мезодермы, их ворон-
ки— из целотелия, т. е. это метанефридии или, точнее целомодукты. Пре-
септальный целом образуется из полости тела и дает целом эпистома,
возникает три отдела целома. Головной ганглий возникает впячиванием
эктодермы. Щупальцевый аппарат также восстанавливается иначе, чем
в онтогенезе: на переднем конце образуются две самостоятельные лопас-
ти, позже составляющие единый лофофор, сначала образуются наруж-
ный, а затем внутренний ряды щупалец.
При регенерации актинотрохи, протекающей в пять раз медленнее,
чем у форониса, позади зачатка ганглия личинки впячиванием образует-
ся зачаток будущего ганглия форониса. Полость тела, как при регене-
рации у форониса, образуется в виде трехчленистого целома. Нефридии
восстанавливаются как парные образования из эктодермы, как настоя-
щие протонефридии. Щупальцевый аппарат регенерирует сходно с его
образованием в онтогенезе. Шульц (Е. A. Schulz, 1903), с известным
правом отмечает ряд сходных черт у актинотрохи и трохофоры и фило-
генетический характер способов восстановления у форониса головного
ганглия, щупальцевого аппарата и трех отделов целома у форониса и
актинотрохи, однако его филогенетические соображения о Triarticulata
В. М. Шимкевича (1893) и Trimetamera Мастермана (Mastermann, 1897,
1900) имеют уже исторический интерес.
У форонид широко распространена автотомия переднего конца тела,
при которой отбрасываются лофофор со щупальцами, целомодукты, ро-
товое и анальное отверстия с частями кишечника и септа, затем регене-
рирующие. Регенерация происходит одинаково хорошо в природе и
эксперименте; отрезанная передняя часть тела, как и автотомировавная,
обычно не восстанавливает целое, части же туловища, даже его кусочки
в 1 мм длиной, регенерируют целого червя (Hyman, 1959).
Форонисы — морские, живущие в трубочке, сидячие обитатели мелко-
водья бентоса, литоральной зоны до 50 м глубиной, тропической и уме-
ренной зон всех океанов. Они живут небольшими колониями (сообще-
ствами), порой переплетаясь трубками в виде неправильной сети. Многие
из них, живя .на песчаном или илистом грунте, делают в нем верти-
кальные прямые или извилистые ходы. Некоторые виды буравят извест-
ковый грунт или пустые раковины двустворчатых и брюхоногих мол-
люсков. Как было сказано раньше, форонисы не покидают своих тру-
231
бак, выдаваясь из них над дном моря лишь своим передним концом
с щупальцевым аппаратом.
Будучи небольшой группой по числу видов и особей, форониды игра-
ют незначительную роль в экономике моря. Они входят в состав био-
ценозов со Многими беспозвоночными литоральной .полосы до 50 м глу-
биной. Некоторые, например, Phoronis australis, живут как комменсалис-
ты в трубках Cerianthus. Паразитов у форонид известно мало — это гре-
гарины и метацеркарии трематод, кроме того, мелкие рыбы и десятино-
гие раки повреждают верхние части форонид.
Геологическая история форонид неизвестна. Однако ряд палеонтоло-
гов (Геюкер, 1960) считает возможным приписывать форонидам верти-
кальные ходы в песчаниках древнего палеозоя (кембрий — девон), опи-
санные как Scolithus, которые встречаются, но гораздо реже и в мелу,
палеогене и неогене. Палеонтологи считают, что условия жизни вымер-
ших форонисов были теми же, что и у современных, а самих форо-
нид — древней группой.
Как у представителей любой группы, у форонид в организации
взрослых форм и в ходе онтогенеза имеются черты примитивные и
вторичные, причем первых много. К примитивным особенностям относят-
ся следующие: червеобразная форма тела с тремя отделами — эпистом
(протосома), посторальный щупальцевый аппарат (мезосома) и туло-
вище (метасома), кожно-мускульный мешок из кольцевых и продоль-
ных мышц, эпидермальное, над базальной мембраной, положение нерв-
ной системы, состоящей из сплетения волокон и нервных клеток, с мало
дифференцированным нервным центром, не обособленным от эпидерми-
са, с нервной регуляцией по типу диффузной нервной сети кишечнопо-
лостных," отсутствие органов чувств, наличие пары целомодуктов, внут-
риклеточное пищеварение, размножение одиночных особей почти исклю-
чительно половым путем.
К примитивным чертам относятся также не установившийся тип
дробления (неправильное, радиальное, спиральное), наличие хорошо вы-
раженных трех зародышевых листков, очень длинный бластопор, спо-
соб его смыкания и возникновение рта из остатков бластопора и аналь-
ного отверстия на месте, отвечающем замкнувшемуся заднему его кон-
цу, сходно с аннелидами, наличие у актинотрохи некоторых сходных
черт с трохофорной личинкой — прототрох, метатрох, телотрох, пара
протонефридиев, наличие трех отделов целома (протоцель, мезоцель и
метацель) , метаморфоз, при котором без большой ломки анатомия акти-
нотрохи перестраивается в анатомию взрослого.
Вторичными чертами форонид надо считать нарушение двусторон-
ней.симметрии тела, вызванное наличием левого продольного нервного
тяжа, точнее, гигантского.вол окна и исчезновением правого, положение
на левой стороне гонад и их непарность, асимметричное строение крове-
носной системы, сильное развитие сложного спирально свернутого щу-
пальцевого аппарата, сближенное положение ротового и анального от-
верстий и сведение исходной спинной стороны к небольшому участку
между ними; подразделение кожно-мускульного мешка сфинктерами,
почти полное исчезновение целома в сильно редуцированном эпис-
томе.
Ко вторичным особенностям хода онтогенеза относятся закладка
туловища червя в виде мешочка эктодермы несколько левее от середины
брюшной стороны личинки с превращением его в ножку, направленную
перпендикулярно к длинной оси личинки; и непарный характер цело-
мов.
Прогрессивными чертами являются следующие: мощное развитие и
разнообразие щупальцевого аппарата, замкнутый характер кровенос-
ной системы, кроме небольшого участка в виде синусов в стенке желуд-
232
ка, высокая дифф.еренщиров1ка подковообразного отдела кровеносной
системы на верхнюю (венозную) и нижнюю (артериальную) части, или
этажи, правильная ритмичная пульсация продольных кровеносных со-
судов и некоторые другие. Эти особенности, как и ряд вторичных, выше
перечисленных регрессивных особенностей, связаны с сидячим, доннымг
образом жизни форонисов; они компенсируют затруднения, вызываемые-
таким образом жизни.
Бесспорно, среди группы целомических, нечленистых беспозвоночных,
форониды обладают наибольшим числом черт червеобразных животных,,
очевидно, сохранивших многое от своих предков. По совокупности дан-
ных строения и развития форониды более примитивны, чем мшанки и
брахиоподы. Известные общие черты анатомии и эмбриологии этих трех
групп отмечали многие, поэтому я их не разбираю.
Для мшанок характерно прочное постоянное прирастание к субстра-
ту, размножение, помимо полового, преимущественно почкованием, ве-
дущее к образованию настоящих колоний, связанная с этим утрата ряда
анатомических, функциональных и физиологических черт индивидуума,,
становящегося элементом колониального организма, развитие защитных
приспособлений в виде органического и неорганического скелета, харак-
терны значительные нарушения судьбы зародышевых листков. Эти важ-
ные приспособления к условиям жизни, позволяющие полнее использо-
вать среду обитания, привели вместе с тем к утрате мшанками многих:
исходных черт, сохранившихся у форонид и свойственных отдаленным,
червеобразным предкам. Однако этот путь эволюции биологически ока-
зался очень выгодным. В то время как форониды остались небольшой,,
реликтовой группой, мшанки, став колониальными и утратив ряд черт
червеобразных организмов, широко- приспособились к различным усло-
виям жизни и дали большое разнообразие систематических групп, оби-
лие видов и особей и в прошлой истории -земли и в настоящее время.
Для брахиопод характерно (при прочном постоянном прирастании к
субстрату) развитие наружного скелета в виде двустворчатой раковины,
отсутствие бесполого размножения, почкования и регенерации, размно-
жение исключительно -половым путем и мощное развитие щупальцевого
аппарата, одиночность.
Вся организация брахиопод в целом отлична от таковой форонид и
мшанок. Отмечая известное сходство между актинотрохой и цифонауте-
сом с трохофорой (большее у актинотрохи и меньшее у цифонаутеса),
можно говорить лишь об очень отдаленном сходстве личинок брахиопод.
с трохофорой, сильно отличающихся от личинок форонид и мшанок. Эн-
тероцельное образование целома, отсутствие хитина в скелете, удаленное-
положение рта от ануса, или отсутствие последнего, участие креатино-
фосфата в энергии мышечного сокращения также значительно отличают
брахиопод от мшанок и форонид.
Общими чертами для всех являются крона щупалец, более или ме-
нее выраженные три отдела целома (протоцель, мезоцель, метацель)
и три отдела тела.
Колдуэлл (Caldwell, 1883), давший исключительно точное, кроме не-
которых частностей, описание и истолкование строения и развития форо-
ниса, отмечал сходство и родство между форонидами, мшанками и бра-
хиоподами. Следуя Рай Ленке-стеру (Ray Lankester, 1873), Гатчек (Hats-
chek, 1888), а за ним многие, в том числе Д. М. Федотов (1935, 1936,
1938), объединяли эти группы в тип Tentaculata или, как делает Гаймен
(Hyman, 1959), в Lophophorata. Беклемишев (1952, 1964) сохраняет тип
Podaxonia, куда относит форонид и мшанок, -подчеркивая образование
у них между ртом и анусом личинки ножки и видя в поперечном делении
у некоторых форонид начало возникновения колониальности подакссний.
Наконец, некоторые, в том числе Термье (Termier et Termier, I960),.
23&
придерживаются понятия Trimetamera, предложенного ранее В. М. Шим-
кевичем (1893, 1905) и позже Мастермаиом (Mastermann, 1897) как
Triarticulata. Однако при этом допускается ошибка, когда сюда вклю-
чаются такие отдаленные группы, как иглокожие, гемихордовые, птёро-
бранхии, форониды, мшанки и брахиоподы.
Все эти вопросы вновь рассматривались некоторыми авторами, в том
числе Маркусом (Marcus, 1954, 1958), Сенном (Senn, 1934), К. Н. Да-
выдовым (Dawydoff, 1960) и другими, но в общем не в пользу родства
между всеми этими группами. Нет также оснований говорить о при-
надлежности форонид к аннелидам, как думает К. Н. Давыдов (1960),
пак как никаких намеков на следы метамерии или утраты ее у форо-
нид нет ни в строении, ни в развитии. А. С. Щепотьев (1907), объединяя
в тип Oligomera, предложенный Д. Д. Педашенко (1898), Phoronida,
Ectoprocta и Brachiopoda как модифицированных Trimetamera, Ptero-
branchia, Enteropneusta и Graptolitha как типичных Trimetamera, а также
Endoprocta и Chaetognatha, конечно создал искусственный тип, о чем я
скажу ниже. Здесь лишь отмечу, что А. С. Щепотьев правильно подчер-
кивав общие черты у Phoronida и Ectoprocta и черты Brachiopoda, отли-
чающие их от двух первых.
По морфологическим данным мшанки близки к форонидам, ряд осо-
бенностей в строении у них связан с образованием колоний, что, конечно,
особенно сильно сказалось и на их онтогенезе, хотя спиральное дробле-
ние наблюдается и у тех и у других. Сравнительно с мшанками форо-
ниды сохранили много черт, присущих одиночным, нечленистым вто-
ричнополостным червям. Брахиоподы при наличии некоторых сход-
ных черт в строении и развитии сильно отличаются от форонид и мша-
нок. Поэтому, на мой взгляд, вернее не объединять эти три группы в
Tentaculata, или в Lophophorata, а считать, что они стоят у корня рас-
хождения двух основных стволов — Protostomia и Deuterostomia, причем
'форониды и мшанки ближе к стволу первичноротых, а брахиоподы —
к стволу вторичноротых.
Мшанки (Bryozoa)
Мшанки — двустороннесимметричные, микроскопические (около
1 мм), вторичнополостные червеобразные организмы-зооиды, обра-
зующие почкованием прикрепленные колонии, обитающие преимуще-
ственно в море. Верхняя, выпячивающаяся и втягивающаяся часть зоои-
да— полипид на свободном конце несет венчик щупалец, окружающий
рот петлеобразного сквозного кишечника с анальным отверстием вне
щупалец; нижняя часть зооида — цистид, в котором помещается полипид,
имеет прочные стенки с хитиновой кутикулой, чаще инкрустированной
известью. Мускулатура в виде отдельных пучков мышц. Надглоточный
ганглий лежит между ртом и анусом и дает нервы к щупальцам и внут-
ренним органам. Дифференцированных органов чувств нет. Перито-
неальная (целомическая) полост^ь тела поделена на три, чаще на два От-
дела. Органы кровообращения, дыхания, большей частью и выделения
отсутствуют. Имеются примитивные яичники и семенники без протоков;
особи обычно гермафродитные. Развитие с личинками и некробиотиче-
ским метаморфозом. Закладка зародышевых листков крайне нарушена.
Образование колоний происходит путем почкования зооидов.
Мшанки'морские, реже пресноводные формы бентоса, питаются мик-
роскопически мелкой пищей и детритом; они обладают резко выражен-
ными способностями к регенерации и дегенерации зооидов.
Насчитывается 4000 современных и 15 000 вымерших видов мшанок,
которые известны с верхнего кембрия. Мшанки — тип с двумя подклас-
сами или классами — Phylactolaemata (пресноводные мшанки) и Gymno-
laemata (морские мшанки). Последние обычно делятся на пять отрядов:
Cyclostomata, Trepostomata, Cryptostomata, Ctenostomata и Cheilosto-
mata (Астрова, 1964, 1965). В настоящее время следует различать шесть
отрядов морских мшанок: Cyclostomata, Cystoporata, Trepostomata,
Cryptostomata, Ctenostomata и Cheilostomata (Астрова, 1964, 1965).
По ряду черт строения, в том числе наличию щупалец, начиная с
Гатчека (Hatschek, 1888), некоторые авторы, среди них и Д. М. Федотов
(1933, 1935), объединяли мшанок с форонидами и плеченогими в тип
Tentaculata, однако, в настоящее время такая установка устарела.
Роль зоологии и палеонтологии в изучении мшанок отражена Гай-
мен (Hyman, 1959), Брианом (Brien, 1953, 1960), Астровой (1960, 1964,
1965). Мной использованы, кроме этих источников, также работы
Г. Г. Абрикосова (1936, 1948, 1955), В. Н. Беклемишева (1950, 1952,
1964), В. А. Догеля (1938, 1940), Н. А. Ливанова (1936), Ю. И. Полян-
ского (1946), Мура (Moore, 1952, 1953), Гармера (Harmer, 1930, 1931),
Силена (Silen, 1935, 1942, 1944а—г, 1947, 1950, 1954), Маркуса (Mareus,
1925, 1926, 1934, 1954, 1958), Борга (Borg, 1926, 1930, 1944, 1953, 1956),
Базлера (Basler, 1953), Нитше (Nitsche, 1875), и ряд др.
. Очень характерную черту строения мшанок составляет венчик щупа-
лец на подковообразном (Phylactolaemata) или на круговом (Gymno-
235
laemata) лофофоре на верхней части полипидов. Реснички щупалец вы-
зывают вводной и выводной токи воды, служащие для привлечения пи-
щи и удаления фекалий.
Стенка тела зооида у пресноводных мшанок (Phylactolaemata) со-
стоит из эпидермиса с тонкой хитиновой кутикулой, слоя кольцевых и
продольных мышц и перитонеального эпителия. Часть эпидермальных
клеток высокие, цилиндрические или столбчатые, часть пузырчатые с
вакуолями, содержащими тонко гранулированные включения. Стенка
тела зооидов морских мшанок имеет более упрощенное строение и со-
стоит из хитиновой кутикулы, полигональных эпидермальных клеток и
слоя перитонеума. Такое строение наблюдается у части отряда Ctenosto-
mata; у других его предствителей путем абсорбции воды стенками тела
(их ослизнением) возникает желатинозный, или студенистый, слой не-
известного химического состава, в который погружены зооиды колоний..
У большинства мшанок из современных отрядов Cheilostomata и Cyc-
lostomata, ископаемых Cystoporata, Trepostomata и Cryptostomata имеет-
ся известковый скелет. Между хитиновой кутикулой и эпидермисом вы-
деляется неразделимая смесь арагонита и кальцита с лабильной при-
месью карбоната магния. Сухой остаток состоит из хитина и карбонатов
кальция (Виноградов, 1944; Vinogradov, 1953). Неорганический скелет
мшанок очень лабилен. Его количество изменяется в зависимости от
условий жизни. В солоноватых и пресных водах мшанки теряют СаСОз.
и становятся хитиновыми, как и все Phylactolaemata.
У пресноводных мшанок под эпителием тела, как было отмечено, ле-
жит слой кольцевых и продольных мышц, отсутствующих, вернее утра-
ченных морскими мшанками. Надо считать, что Phylactolaemata обла-
дают более примитивным, чем их древние червеобразные, подвижные
предки, строением стенок тела, сохраняя остаток кожно-мускульного
мешка. У их подвижных колоний (Cristatella) кольцевая и продольная
мускулатура тела зооидов составляет единую мускулатуру всей колонии,
что обеспечивает ее передвижение.
Вряд ли можно сомневаться в том, что морские мшанки, благодаря
развитию неорганического твердого скелета и прикрепленному образу
жизни, утратили мускулатуру стенок тела, то, что составляет. кожно-
мускульный мешок червеобразных организмов.
Остальная мускулатура тела мшанок, не считая продольной.муску-
латуры щупалец, представлена отдельными специализированными мыш-
цами (рис. 63). Дифференцировка и число мышц у пресноводных мша-
нок невелики: это — ретракторы, протракторы, дилататоры полипида,
лофофора и вестибулюма. У морских мшанок эта часть мускулатуры
претерпевает значительные изменения и приобретает гораздо большую
специфичность, характерную для морских мшанок, что в значительной
степени связано с развитием скелета, неподвижностью основной части
зооида — цистида и большой подвижностью нежного полипида. Эта мус-
кулатура обеспечивает высовывание полипидов для ловли добычи, для
дыхания, быстрое втягивание в случае опасности и изоляцию зооида
от среды при наступлении неблагоприятных условий. В этом прини-
мают участие'и гидростатические аппараты — так называемый компен-
сационный мешок некоторых Cheilostomata и перепончатый мешок
Cyclostomata.
Развитие приспособлений для закрывания отверстия у цистида в ви-
де крышечки, воротничка с венчиком щетинок и т. д. привело к образо-
ванию у морских мшанок малочисленных специализированных отдель-
ных мышц, пересекающих полость целома, как и у пресноводных форм.
Это многочисленные париетальные мышцы, мышцы крышечки, ворот-
ника, лофофора и т. д. Специфически развиваются' мышцы у видоизме-
ненных зооидов, например, авикулярий, вибракулярий, необходимые для
236
8
Рис. 63. Схема строения двух зооидов Gymnolaemata
(из Догеля)
/ — рот; 2 — щупальце; 3 — целом; 4 — нервный узел; 5 —анус; 6 — ки-
шечник; 7 — яичник; 8 — семенник; 9— мускул-ретрактор; 10— канатик
их быстрых движений. Как известно, во многих случаях мышцы мшанрк
обладают поперечнополосатой структурой. Естественно, что у хорошо
отграниченных друг от друга зооидов морских мшанок их мускулатура
-строго индивидуализована, в отличие от таковой ряда пресноводных
‘форм.
Нервная система мшанок, как в общем и вся их анатомия, однообраз-
на. Она состоит из надглоточного ганглия, лежащего в целоме лофофора
или в кольцевом целоме между ртом и анусом (на спинной стороне глот-
ки), окологлоточного кольца, чувствительных и двигательных нервов,
идущих от ганглия к лофофору и щупальцам, от нервных тяжей к задне-
му концу полипида, к кишечнику и из субэпителиального сплетения.
"Специальных органов чувств у мшанок нет, имеются лишь чувствитель-
ные нервные клетки, преимущественно на щупальцах (Gerwerzhagen,
1913 а, 1913 6; Saftigen, 1888).
237
Своеобразие Пресноводных мшанок состоит в том, что надглоточный
ганглий и, надо думать, нервные тяжи лофофора, развиваются впячи-
ванием эктодермы с замыканием его краев, благодаря чему обособляет-
ся участок иелома. Вследствие этого надглоточный ганглий и нервные
стволы лофофора, отходящие от него, содержат полость целома. Полу-
чается подобие образования нервной системы вторичноротых, с той суще-
ственной разницей, что у мшанок обособляется участок целомической
полости, прилегающий к надглоточному ганглию, а не эпиневральный
эктодермальный канал, как у иглокожих.
Петлеобразный кишечник (со ртом, окруженным венчиком щупалец
подковообразного или круглого лофофора, с анусом вне венчика на
спинной стороне зооида) несколько проще у пресноводных, чем у мор-
ских мшанок. Кишечник состоит из ротовой полости, глотки, пищевода
(передней кишки) кардиальной части, слепого мешка и пилоруса (же-
лудка, или средней кишки), узкой трубки с анусом (задней, или прямой
кишки). По наблюдениям Силена (Silen, 1944а—г) над морскими мшан-
ками, движением ресничек рта и дистальной части глотки, а затем резки-
ми сокращениями и расширениями кардиальной части желудка пища
гонится в его центральную часть. Здесь она переваривается. Фекалии вы-
брасываются наружу благодаря перистальтике прямой кишки. В стенках
кишечника имеются кольцевые и продольные мышечные волокна. Ре-
зорбирующей частью кишечного тракта является средняя кишка —
центральная часть желудка с базофильными и ацидофильными клетка-
ми эпителия, причем она, как и остальные отделы кишечника, образова-
на эктодермой. Пищеварение для белков и углеводов является по-
лостным, для жиров — внутриклеточным. У пресноводных мшанок (Mar-
cus, 1925, 1926, 1934; Becker, 1937, и др.— см. Hyman, 1959) пища,,
поступающая из рта в глотку, вращается движением ее ресничек и попа-
дает в пищевод, перистальтикой которого гонится в желудок. Здесь она
переваривается под воздействием пищеварительных энзимов, выделяе-
мых эпителием. Пищеварение полностью полостное. Ацидофильные клет-
ки желудка абсорбируют жиры, протеины и экскреторные гранулы. Весь
процесс, включая дефекацию, занимает 1 час по Маркусу, 2—3 часа
по другим авторам. Пищей мшанкам служат мелкие организмы — водо-
росли (диатомовые и др.), простейшие (радиолярии), коловратки, дет-
рит. В жизненном цикле мшанок происходит повторяющийся распад час-
тей кишечника (как и всех других органов зооидов) и их регенерация..
У пресноводных мшанок целомическая полость тела поделена непол-
но на три, а у морских, в связи с отсутствием (утратой) эпистома, на два
отдела. Целом лофофора продолжается в щупальца. У Cyclostomata
полость тела, так называемый перепончатый мешок, участвует во втяги-
вании и выдвижении лофофора. У пресноводных мшанок (Cristatelld)
целомы отдельных зооидов не обособлены и образуют общую полость
тела колонии. Но и у ряда морских мшанок полости тела зооидов коло-
нии 'сообщаются порами. Среди перитонеальных клеток имеются клетки
с пучком ресничек, играющие роль в движении полостной жидкости..
Последняя содержит протеины и свободные клетки — целомоциты. По-
лость целома прорезана группами мышц, более многочисленными у
морских, чем у пресноводных мшанок, и канатиком, с помощью которо-
го слепой конец желудка прикрепляется к стенке тела.
У пресноводных мшанок имеется два ресничных канала (целомодук-
ты), сообщающихся через, ресничные воронки с целомом лофофора и'
общим непарным выводным каналом с наружной средой. Через этот ка-
нал выводятся наполненные экскретом амёбоциты. Предполагают, что
существующий у некоторых из Cheilostomata интертентакулярный орган-
имеет такое же морфологическое значение (Harmer, 1931; см. Hyman,
1959). В процессе выделения участвуют, кроме того, стенки кишечника и
238
бурые тела, последние установлены и у ископаемых форм (Астрова, 1964,
1965).
Несмотря на наличие целома, у мшанок нет кровеносной системы,
что, очевидно, связано с микроскопически малыми размерами зооидов.
У них нет также органов дыхания. Газовый обмен осуществляется через
тонкие стенки щупалец лофофора. Способность тонкостенного полипида
высовываться и втягиваться с лофофором, обширная полость тела, бога-
тая полостной жидкостью, подвижность короткого, петлеобр-азного ки-
шечника являются особенностями мшанок, благоприятствующими газо-
вому обмену, движению полостной жидкости, энергичному пищеварению,
и удалению фекалий. Эти особенности говорят о высоком уровне обмена
веществ и, кроме того, компенсируют биоэкологические трудности, свя-
занные с прикрепленным образом жизни.
Половая система очень проста. Яичники и семенники диффузного,
типа (Догель, 1940); они представлены небольшими скоплениями оого-
ниев и сперматогониев и не имеют выводных половых протоков. Яичники-
возникают у пресноводных форм на брюшной стороне, у морских — на
спинной стороне зооида, на перитонеальной стенке переднего отдела
последнего. Семенники возникают на канатике и на стенке тела в зад-
нем отделе зооида. Мшанки обычна гермафродиты. Оплодотворение у-
них внутреннее.
Лишь у части морских мшанок из Ctenostomata и Cheilostomata яйца
выводятся в воду; у большинства они развиваются до личинки в вывод-
ковых камерах (овицеллах) или в полости тела автозооида. У Cyclosto-
mata и пресноводных мшанок имеет место живорождение. Дробление-
(см. Hyman, 1959) равномерное, нёравномерное, радиальное, бирадиаль-
ное или двулучевое. У Flustrella (Ctenostomata) дробление спиральное с
образованием квартета макромеров и нескольких квартетов микромеров,
хотя автор (Расе, 1906) дробление спиральным почему то не называет.
На стадии 32 бластомеров впячиванием внутрь четырех макромеров
образуется гаструла, макромеры являются мезэнтодермой. После замы-
кания бластопора, очевидно, .деляминацией отделяются клетки мезодер-
мы, окружающие эктодермальный зачаток кишечника (архентерона).
Впячиванием эктодермы возникает глотка. Образуется личинка, которая-
у многих морских мшанок свободноплавающая. Таким образом, у мор-
ских мшанок могут быть выражены все три зародышевых листка — экто-
дерма, энтодерма и мезодерма. У пресноводных форм, развитие которых
идет иначе (стр. 240), наружу выходят молодые личинки.
Свободноплавающая личинка Cheilostomata — цифонаутес имеет те-
менную пластинку, кольцо ресничных клеток вблизи экватора, петлеоб-
разный сквозной кишечник со ртом, глоткой, желудком, задней кишкой-
и анусом, развитую поперечнополосатую мускулатуру и нервную си-
стему. На переднем конце оральной стороны личинки находится груше-
видный орган, на заднем конце — присоска. Двустворчатая кутикуляр-
ная раковинка одевает тело цифонаутеса.
У Ctenostomata и Cheilostomata наблюдаются разные степени упро-
щения внешнего строения и внутренней организации личинок, вызван-
ные условиями жизни.
Для Cyclostomata характерным является полиэмбриония и живорож-
дение. Лишь в некоторых зооидах, так называемых гонозооидах, проис-
ходит дробление яйца в части полости тела, так называемом перепонча-
том мешке (membranacoeus Sack) (Borg, 1926; Silen, 1944 a) и проте-
кает в питающей ткани. Морулоподобный первичный зародыш разра-
стается в колонию, от которой отделяются почки — вторичные двухслой-
ные зародыши. Из каждого такого зародыша образуется свободнопла-
вающая личинка, удлиненной овальной формы, но лишенная многих
органов, имеющихся у личинок Ctenostomata и Cheilostomata.
239
После прикрепления личинки морских мшанок к субстрату происхо-
дит некробиотический метаморфоз, сопровождающийся дегенерацией и
гистолизом органов и частей личинки. Тело ее принимает форму мешоч-
ка — первичного цистида, прикрепленного к субстрату. На его свободной
поверхности впячиванием эктодермы образуется перитонеумом— зача-
ток первичного полипида, одетый клетками мезенхимы. Возникает пер-
вичный зооид, или анцеструла, лишенная половых желез, дающая почко-
ванием начало зооидам колонии мшанки. По мнению Цшише, мезенхи-
ма анцеструлы возникает из эктодермального зачатка первичного
полипида, а не из мезенхимы личинки. Зачаток первичного полипида у
Ctenostomata и Cheilostomata сначала располагается параллельно по-
верхности цистида, затем вращением на 90° получает окончательное
положение, тогда как у Cyclostomata он сразу занимает дефинитивное
положение (Hyman, 1959).
Индивидуальное развитие пресноводных мшанок оказывается гораз-
до сильнее вторично измененным — упрощенным сравнительно с онто-
генезом морских мшанок. В связи с живорождением у пресноводных
•мшанок >все развитие, вплоть до образования зародыша молодой коло-
нии, протекает внутри материнского организма. Это обеспечивает безо-
пасность молоди, но вызывает также подавление личиночной стадии и
нарушение закладки зародышевых листков до полного выпадения энто-
дермы на эмбриональной стадии с превращением зародыша в двуслой-
ный мешок из эктодермы и мезодермы. Этот мешок, минуя стадию сво-
бодноживущей личинки, превращается в молодую дочернюю колонию —
•цистид с двумя полипидами. Такая молодая колония, как ее правильно
называл Harmer (1931), после непродолжительной свободной жизни при-
крепляется к субстрату или оседает на него и дает начало колонии.
При размножении с помощью покоящихся почек — статобластов,
которые способны без вреда переносить длительное время воздействие
крайне неблагоприятных условий — низких температур и высыхания
водоемов, из них возникает один зооид. В образовании зооидов из ста-
тобластов, развивающихся на канатике, участвуют также только эпи-
дермальные и перитонеальные клетки материнского зооида, т. е. те же
элементы, что и при эмбриональном развитии.
В онтогенезе мшанок обращает на себя внимание большая зависи-
мость высоты организации и функциональной активности личинок (и от-
дельных органов) от продолжительности самостоятельной жизни. Ли-
чинки цифонаутес, питающиеся мелкими планктонными организмами и
живущие от двух месяцев до года (Hyman, 1959), являются наиболее
организованными личинками мшанок. Они имеют ряд сходных черт не
только с личинками Kamptozoa, но и с трохофорой полихет (например,
-митрарией). В связи с укорочением продолжительности самостоятельной
жизни у личинок ряда отрядов морских мшанок происходит упрощение
-внешнего строения и в разной степени регресс кишечника, теряющего
пищевод, среднюю и заднюю кишку, так как эти личинки не питаются.
У пресноводных мшанок подавление развития личинки идет еще
дальше. Эмбриональное развитие внутри материнского организма, как
мы видели, приводит к образованию двуслойного зародыша, дающего
начало цистиду с двумя полипидами.
Очень характерной особенностью индивидуального развития морских
мшанок является также сохранение в онтогенезе (и то неполное) лишь
двух зародышевых листков — эктодермы и мезодермы. Энтодерма, за-
кладывающаяся на стадии гаструлы, лишь изредка сохраняется до кон-
ца личиночной жизни, участвуя в образовании сквозного кишечника
личинки цифонаутеса. У пресноводных мшанок, как мы видели, энтодер-
ма как зародышевый листок не закладывается вовсе и на Стадии раннего
онтогенеза существуют лишь эктодерма и мезодерма.
:240
Рис. 64. Monobryozoon limicola (по Тгапёп)
Характерно, что и при метаморфозе, после полного распада орга-
нов личинок морских мшанок при участии (как и у зооидов пресновод-
ных мшанок) лишь эктодермы и мезодермы, у анцеструлы образуется
щупальцевый аппарат, нервная система, весь кишечник, мускулатура,
половые железы, целомодукты и целомическая полость тела. Иными сло-
вами, мшанки обладают всеми органами и тканями (кроме кровеносной
системы), которые у других целомических нечленистых первичноротых
развиваются из эктодермы, энтодермы и мезодермы.
Это говорит или о несостоятельности теории зародышевых листков,
или' о недостаточности обычных морфологических методов при изучении
онтогенеза мшанок и необходимости применения цитохимических иссле-
дований.
Обращает также на себя внимание, что при большем, чем у морских
мшанок, вторичном изменении и упрощении онтогенеза взрослые зоои-
ды пресноводных мшанок в общем близки к Phoronoidea и находятся на
более высоком уровне организации, чем морские формы.
16 Д. М. Федотов
241
Характерной чертой мшанок является их колониальность, причинно
связанная с прикрепленным образом жизни и с сохранением способ-
ности к бесполому размножению путем почкования зооидов. Исключе-
ниями являются недавно открытые Ремане (Remane, 1936, 1938, по, Hy-
man, 1959) и Франценом (Franzen, 1958—1962) псаммофильные мшанки
из Ctenostomata—Monobryozoon ambulans и М. limtcola, вторичный,
характер одиночного образа жизни которых общепризнан (рис. 64).
Как отмечалось (Беклемишев, 1952, 1964), почки у пресноводных мша-
нок возникают на оральной (брюшной) стороне материнского полипида,
а у морских на спинной стороне.
Высшей степени интеграции среди мшанок достигает колония Crista-
tella (Svetlov, 1935; Беклемишев, 1950, 1952, 1964), характеризующаяся
двулучевым строением, ползанием, размножением делением поперек,
способностью в случае повреждения регулировать форму.
Лишенная половых желез анцеструла морских мшанок почкованием
разными путями (Cumings, 1904, 1905, 1912; Borg, 1926, см. Беклемишев,
1951, 1952, 1964; Hyman, 1959; Brien, 1960) дает начало колонии, автозои-
ды которой становятся половыми особями. Путем моно-, би- и полисе-
риального почкования анцеструлы /Hyman, 1959) идет развитие колоний,
имеющих у ряда Ctenostomata й части Cheilostomata форму непра-
вильную (со стелющимися столонами) или правильную, вследствие
почкования зооидов в определенном направлении (как у многих Chei-
lostomata) или благодаря наличию общеколониальной зоны роста, за
счет роста которой идет увеличение размеров колонии, как у Cyclosto-
mata. Здесь нет еще сформированных зооидов и отдельных цистидов, а
имеется лишь общая целомическая полость. С образованием сначала
цистида, затем полипида в этой общеколониальной зоне роста происхо-
дит образование новых зооидов (Borg 1930; Беклемишев, 1952, 1964).
Во время роста колоний, естественно, происходит формирование
автозооидов и гетерозооидов, приводящее к полиморфизму колоний, в
разной степени выраженному у различных отрядов Gymnolaemata. Фор-
ма и характер колоний мшанок чрезвычайно разнообразны: встречаются
стелющиеся, сетчатые, корковые, пластинчатые, веерообразные, листо-
видные, кустистые, древовидные, а также тонкие, стройные, массивные и
тяжелые колонии. Нередко наблюдаются правильно радиально симмет-
ричные колонии, например, из современных у Defrancia (Cyclostomata)
и Kinetoskias (Cheilostomata) и из вымершего отряда Cryptostomata —
у Patellipora. При громадном разнообразии колоний мшанок все же в
каждом отряде, как отмечает Г. Г. Астрова (1960), преобладают опреде-
ленные типы.
Колонии пресноводных мшанок — мономорфны; эмбриональный ме-
шок (термин Маркуса), или оэций, не является измененным зооидом.
Колонии морских мшанок — полиморфны и состоят из автозооидов и
гетерозооидов. Наиболее однородны колонии Ctenostomata, у которых,
кроме автозооидов, гетерозооиды представлены лишь столонами и ризои-
дами. У Cyclostomata полиморфизм выражен значительно сильнее. Кроме
автозооидов — основной массы зооидов, — имеются гетерозооиды в ви-
де крупных гонозооидов, в которых происходит развитие зародышей пу-
тем полиэмбрионии, кенозооиды и наннозооиды. Следует заметить, что-
иглы у Gymnolaemata бывают двух родов, одни являются гетеорозооида-
ми, другие — простыми выростами стенок зооидов (Silen, 1947). Точно
также ризоиды некоторых Cheilostomata (Borg, 1930) являются вы-
ростами стенок зооидов, т. е. они не гомологичны ризоидам Ctenostoma-
ta. Наибольший полиморфизм особей колонии наблюдается у Cheilosto-
mata, молодого, самого обширного отряда из современных морских мша-
нок, У них гетер-озооиды представлены гонозооидами, авикуляриями,
вибракуляриями, иглами, кенозооидами и наннозооидами тогда как ови-
242
целлы, будучи выростами стенки автозооидов, к гетерозооидам не отно-
сятся.
Гетерозооиды обнаруживают разные степени регресса присущей ти-
пичному зооиду организации и обладают специализацией строения, свя-
занной с их функцией в колонии; примером могут служить авикулярии,
функционирующие подобно педицелляриям иглокожих (Hyman, 1959).
Колонии современных морских мшанок сравнительно небольших
размеров, но встречаются колонии Ctenostomata в 45 см (Zoobothrion
pellucidum) и в 60—90 см и более длинные (Alcyonidium) и колонии
Cyclostomata (Flustra.) длиной в 30 см. До 60 см в длину достигали раз-
меры колоний и некоторых ископаемых мшанок. Число зооидов у совре-
менной Membranipora membranacea при длине колонии 152 см и ширине
в 20 см доходит до 2 300 000 (Hyman, 1959). Продолжительность жизни
колоний морских мшанок различна. В холодной зоне гибель мшанок вы-
зывается замерзанием моря, в южных морях мшанки активны круглый
год, там колонии их более долговечны.
Пресноводные мшанки, как правило, недолговечны; холодное время
года в умеренной и холодной зонах и засушливый сезон с высыханием
водоемов, например, в тропической Индии, они переносят в виде стато-
бластов, так как колонии погибают (Harmer, 1931).
Рост колоний, происходящий путем почкования новых зооидов, за-
медляется или прекращается неблагоприятными условиями жизни, свя-
занными с сезонами года (это не относится к районам Мирового океана
в южных широтах).' Однако во всех районах обитания морским мшан-
кам, согласно наблюдениям ряда зоологов (Беклемишев, 1952, 1964;
Hyman, 1959; Brien, 1960, и др.), свойственны необычно сильно выражен-
ные явления дегенерации и регенерации полипидов, поскольку продол-
жительность жизни отдельного полипида невелика., и может сильно
сокращаться под влиянием неблагоприятных условий жизни. Полипиды
'дегенерируют, превращаясь в бурые тела и замещаются новыми полипи-
дами, возникающими путем почкования. У новых полипидов образуются
половые железы и начинается половое размножение. Бурые тела (масса
из дегенерирующих органов и тканей) попадают в кишечник вновь
образующихся полипидов, перевариваются, а остатки выкидываются
через анус. Кроме того, обычно полипиды после размножения деге-
нерируют.
Широко распространены явления дегенерации и регенерации и у
пресноводных мшанок. Их полипиды, как отмечает Гаймен (Hyman,
1959), дегенерируют в результате старения, после полового размножения,
под влиянием неблагоприятных условий жизни, но без образования бу-
рых тел и путем образования нового полипида не в цистиде, а на новом
месте колонии.
Морские мшанки — типичные животные бентоса, прикрепленные к
субстрату; их распространение, естественно, происходит с помощью ли-
чинок и подвижных стадий развития. Требования к условиям жизни со-
временных и ископаемых морских мшанок, по-видимому, сходны (Не-
хорошее, 1934; Duncan, 1957). Как прикрепленные животные, они нуж-
даются в твердом, скалистом грунте или в другом плотном субстрате
вроде вдорослей, раковин моллюсков, брахиопод, трубок кольчатых чер-
вей И т. д. Мшанки требуют чистоты, подвижности и нормальной соле-
ности воды. На их распространении естественно сказывается климат,
температура воды, течения, и т. д.
Среди Gymnolaemata — типичных морских обитателей—существуют
редкие случаи выхода в пресную воду. Так, известны единичные
пресноводные формы Paludicella и Pottsiella (Hyman, 1959) из отряда
Ctenostomata, а также ряд обитателей эстуарий и солоноватоводной
среды.
16;
243
Морские мшанки широко распространены в Мировом океане холод-
ной, умеренной и теплой зон, заселяя главным образом материковые от-
мели (шельф) от полосы прибоя до глубин в 200—300 м. Отдельные ви-
ды спускаются до глубин в 6000 м.
Хотя число видов пресноводных мшанок невелико (50—100), они так-
же распространены по всему земному шару в чистых текучих и спокой-
ных водах; встречаются в больших озерах на глубине до 170 м, напри-
мер, Fredericella sultana (Hyman, 1959), и поднимаются до 3950 —
Plumatelia repens — и даже до 4150 м над уровнем моря — Stolella agi-
lis (Marcus, цит. no Hyman, 1959).
Жизнь в биотопах с чистой, лишенной мути, подвижной водой, может
быть, и мелкие размеры зооидов оберегают мшанок от заражения пара-
зитами. Зараженность мшанок паразитами ничтожна. На Phylactolaemata
паразитируют Nosema и червеобразные Buddenbrockia, неизвестного си-
стематического положения, на Gymnolaemata — грегарины и споры. Сре-
ди мшанок нет паразитических форм. Врагами Phylactolaemata, поедаю-
щими части полипоидов, являются турбеллярий, кольчатые черви, водя-
ные клещи и личинки насекомых.
Следуя тенденции последнего времени, многие считают мшанок са-
мостоятельным типом, состоящим из двух классов — Phylactolaemata и
Gymnolaemata. Теперь, однако, часто вместо типов как крупных объеди-
нений, связанных между собой групп животных, в ранг типов возводят-
ся группы, гораздо меньшие по числу форм, чем мшанки. Я не останав-
ливаюсь на этом вопросе относительно мшанок, поскольку он хорошо
освещен у Гаймен (Hyman, 1959), Г. Г. Астровой (1960, 1964, 1965) и
Бриена (Brien, 1960).
Мшанки, несмотря на значительное число современных (4000) и боль-
шое количество ископаемых видов (15 000), по своему строению состав-
ляют однородную группу животных, с различием главным образом в
деталях внешней морфологии, но объединенных общностью анатомии,
онтогенеза и бесполого размножения. Характерно, что вопросы класси-
фикации или системы мшанок самым интимным образом связаны с во-
просами их эволюции и филогении.
В сущности говоря, принципиально неважно считать ли Phylactolae-
mata и Gymnolaemata подклассами или классами. Отличаясь друг от
друга, они вместе с тем так Сходны, что их близость и родство ни у кого
не вызывают сомнения.
Наличие эпистома, мускулатуры в стенках тела, трех отделов це-
ломической полости, органов выделения, отсутствие известкового ске-
лета, образование почек на брюшной стороне зооидов, жизнь в пресных
водах, реликтовый характер являются достаточными основаниями для
противопоставления Phylactolaemata как класса остальным мшанкам,
если считать мшанок типом.
Отсутствие эпистома, мускулатуры в стенках цистидов, почти исклю-
чительно наличие известкового скелета, двух отделов целомической по-
лости, образование почек на спинной стороне зооидов, жизнь в море
объединяют всех Gymnolaemata как другой класс мшанок. Анатомия
автозооидов, полиморфизм членов колонии, способы развития колоний,
развитие при помощи личинок со сменой поколений — бесполого (анце-
струлы) и полового (автозооидов), жизнь в море достаточно тесно объе-
диняют всех морских мшанок. Соображения Гармера (Harmer, 1930) о
близости Phylactolaemata и Cyclostomata имеют для нас лишь истори-
ческое значение.
Общность анатомического строения современных Ctenostomata, Chei-
lostomata и Cyclostomata, на мой взгляд, делают излишним разбивать,
как делает Борг (Borg, 1926, 1930, 1944), морских мшанок на Gymnolae-
mata (Ctenostomata и Cheilostomata) и Stenolaemata (Cyclostomata и
:244
Trepostomata) или, как делает Маркус (Marcus, 1958),— на Eurystoma-
ta (Ctenostomata и Cheilostomata) и Cyclostamata.
Длинные трубчатые зооиды, терминальные устья, полиэмбриония, поч-
кование от общей зоны роста колонии — признаки, отличающие Cyclos-
tomata от Ctenostomata и Cheilostomata. Первые два являются прими-
тивными чертами, вероятно, утраченными остальными отрядами; полиэм-
бриония, несомненно, вторичная особенность, возникшая из обычного
онтогенеза мшанок; вторичной чертой является и рост колонии из общей
зоны роста, сравнительно с почкованием у остальных отрядов. Однако,
все это не признаки ранга надотряда, и нет оснований противопоставлять
Cyclostomata двум другим современным отрядам Gymnolaemata.
Несмотря на большое разнообразие внешней морфологии, на разную
степень развития полиморфизма зооидов, разнообразия формы колоний,
отчасти химического состава скелета стенок цистидов, анатомическое
строение зооидов, как нередко отмечалось и раньше (например, Silen,
1942), однотипно. Очевидно, у морских мшанок очень давно выработался
тот уровень строения, физиологии и экологии, который обеспечивал в те-
чение нескольких сот миллионов лет существование разных групп Gymno-
laemata. Эволюция этих прикрепленных животных бентоса шла под воз-
действием условий жизни по пути выработки разных форм колоний со
столонами, без столонов, стелящихся, корковых, древовидных, кустистых,
ветвящихся, сетчатых, компактных, обычно сильно обызвествленных, ка-
менистых в ископаемом состоянии, слизистых. Одновременно с этим про-
текала известная эволюция зооидов колоний, шло развитие автозоои'дов
и гетерозооидов — авикулярий, вибракулярий, гонозоойдов, а также эво-
люция строения верхней части зооидов — формы и положения отверстий,
цистидов, скелета, окружающего отверстия, крышечки, механизмов за-
мыкания и втягивания и выдвижения верхней части полипида с его
щупальцами. Происходило также изменение общей формы зооидов, об-
разовывалась удлиненная, трубчатая, укороченная, яйцевидная и другие
формы.
Изучение представителей ископаемых вымерших отрядов Cystopo-
rata, Trepostomata, Chyptostomata и ископаемых форм современных отря-
дов— Ctenostomata, Cyclostomata и Cheilostomata позволяет приписы-
вать всем морским мшанкам сходное анатомическое строение и, вероят-
но, сходный тип онтогенеза с свободно плавающими личинками. Обилие
ископаемых морских мшанок (до 15 000 видов) говорит о приспособлен-
ности их к условиям жизни и широком распространении в морях прошлой
геологической истории Земли и о хорошей приспособленности ныне
существующих морских мшанок к современным, условиям. Совершенно
ясна также роль в распространении и заселении моря у колониальных,
прикрепленных морских мшанок их пелагических личинок.
Как высокую приспособительную особенность мшанок надо считать
огромное распространение у них бесполого размножения путем почкова-
ния, ведущего к росту колоний, оказавшихся в благоприятных условиях
жизни, дегенерации полипидов, превращающихся в бурое тело в течение
естественного хода жизни и под воздействием наступающих неблагопри-
ятных случайных или сезонных факторов и их регенерацию. Сюда же
относится использование бурых тел для регенерации и необычайно силь-
ная регенеративная способность полипидов после дегенерации. Иными
словами, как стадии онтогенеза, так и взрослые особи морских и пресно-
водных мшанок оказываются необычайно морфо-физиологически лабиль-
ными: у них легко выпадает закладка зародышевых листков и утрачи-
вается в онтогенезе стадия личинки; многие части и органы зооидов
легко разрушаются и вновь восстанавливаются. В этом отношении мшан-
ки являются одной из наиболее замечательных и притом далеко не при-
митивных групп животного царства.
245
Немало существует соображений о филогенетических отношениях
между пресноводными и морскими мшанками, между отдельными отря-
дами последних, а также о родстве между Bryozoa Ectoprocta, Kamptozoa
( = Endoprocta) и Phoronida. Здесь я коснусь лишь филогении Phylacto-
laemata и Gymnolaemata, так как на отношениях мшанок к другим груп-
пам я останавливаюсь в другом месте. Из более новых авторов филоге-
нию мшанок рассматривали В. Н. Беклемишев (1952, 1964), Маркус
(Marcus, 1958), Ремане (1954, 1959), Гаймен (Hyman, 1959) и Г. Г. Аст-
рова (1960, 1964, 1965). Я считаю, что соображения Гаймен (1959) наи-
более обоснованы. Не разбирая их, я остановлюсь на своих представлен
ниях о родственных отношениях отрядов и классов.
Строение колоний со столонами, большая обособленность, самостоя-
тельность зооидов, наличие органического скелета обычно лишь в виде
хитиновой кутикулы, слабое развитие полиморфизма (только зооиды^
столоны и ризоиды) надо считать примитивными чертами Ctenostomata
сравнительно с остальными морскими мшанками. У последних колони-
альность достигает высокой степени развития. Основная масса скелета
становится неорганической, индивидуальность даже автозооидов как
самостоятельных организмов значительно подавлена, а гетерозооиды
становятся органами или частями колонии, почти или совсем утратив-
шими анатомические черты, присущие организму. Силен (Silen,
1944а — г), детально исследовавший представителей Ctenostomata и
Cheilostomata и установивший подотряд Cheilostomata — Protocheilosto-
mata с признаками обоих отрядов, правильно подчеркивает близкое род-
ство между этими отрядами. Я не согласен только с проведенной им и
Маркусом (Marcus, 1958) перестройкой классификации мшанок.
Высокая степень развития колоний с известковым скелетом и сильно
выраженный полиморфизм зооидов, являющихся лишь как бы ячейками
колонии, подчеркивают отличие и самостоятельность Cheilostomata как
отряда. Существование в современной фауне таких форм, как Labido-
stomella, говорит лишь о возможности происхождения Cheilostomata от
примитивных форм Ctenostomata. К сожалению, палеонтология не дает
нам документов, подтверждающих этот, взгляд. Палеонтологические
сведения о Ctenostomata, хотя и имеются, начиная с нижнего ордовика и
до нашего времени, но очень бедны; Cheilostomata же известны лишь с
юры, хотя, казалось бы, с их известковым скелетом они должны были
хорошо сохраняться. Остается предположить, что Cheilostomata эволюи-
ровали недавно и их прочный известковый скелет — приобретение исто-
рически позднее. Если так, тогда среди морских мшанок Cheilostomata —
самый новый отряд, но, как думают в настоящее время многие зоологи и
часть палеонтологов, филогенетически он связан с Ctenostomata. Отмечу,
что не случайно такой знаток систематики современных мшанок, каким
был Г. А. Клюге (1896, 1961, 1962), начинает Gymnolaemata с Ctenosto-
mata, а за ними уже ставит Cheilostomata,
Наконец, возможность вторичного перехода Ctenostomata к одиноч-
ному существованию, в известной мере реализованная Monobryozoon
ambulans (Remane, 1936, 1938; см. Hyman, 1959) и М. limicola (Franzen,
1958—1962), бесспорно также надо рассматривать как сохранение пла-
стичности, утерянной всеми другими типично колониальными отрядами
морских мшанок.
Ctenostomata выделялись за всю свою прошлую геологическую исто-
рию медленными, спокойными темпами эволюции; они и сейчас являются
самым малочисленным отрядом морских мшанок. В отличие от них Chei-
lostomata, появившись в юре в виде редких ископаемых, быстро эволюи-
ровали в условиях мягкого режима моря мелового периода настолько,
что были участниками рифообразования в третичном периоде. Они при-
способились к контрастным условиям жизни в Мировом океане умерен-
246
ной, тропической и холодной зон и в настоящее время являются наиболее
обширным, процветающим отрядом мшанок.
В современной фауне не найдены формы, которые по своей организа-
ции или по онтогенезу связывали бы Cyclostomata с Ctenostomata и Chei-
lostomata. Оговорюсь, что несмотря на ряд отличий в частностях анато-
мического строения зооидов, в характере колоний (которые позволили
еще старым зоологам . середины
XIX столетия обосновать деление
современных морских мшанок на
перечисленные отряды и устано-
вить вымершие отряды Cystopo-
rata, Trepostomata и Cryptosto-
mata) принципиальных отличий
(ранга выше отряда)- у Cyclosto-
mata сравнительно с Ctenostoma-
ta и Cheilostomata, как было ска-
зано стр. 244), я не вижу.
Полиморфизм Cyclostomata
гораздо значительнее, чем у'Cte-
nostomata, но слабее выражен,
чем у Cheilostomata. И по поли-
морфизму, и по характеру скеле-
та они эволюировали значительно,
сильнее, чем Ctenostomata. 1
В своем -строении Cyclostoma-
ta не примитивны, из примитив-
ных черт можно отметить только
более вытянутую форму авто-
Рис. 65. Схема примерного количественного
распределения 900 родов ископаемых мша-
нок (по A. Muller)
зооидов и терминальное положе-
ние устья (сравнительно с более короткой, овальной или яйцевидной их
формой у Ctenostomata и Cheilostomata и фронтальным положением
устьев у последних). Дифференцировка вторичной полости тела, вернее,
специфическая функция так называемого перепончатого мешка, отсут-
ствие всяких следов целомодуктов, которые частично все же сохраняют-
ся у некоторых Cheilostomata (интертентакулярный орган), имеют чер-
ты вторичного характера. К вторичным же особенностям относятся у
Cyclostomata полиэмбриония, живорождение и строение их личинок,
измененных сильнее даже наиболее уклоняющихся от типа Cyphonanies,
личинок Ctenostomata и Cheilostomata.
Важным моментом является наибольшая среди мшанок древность
Cyclostomata, представители которых (с оговоркой, однако, что настоя-
щая природа их не установлена) были обнаружены (Buge, I960), в позд-
нем кембрии, когда другие мшанки известны еще не были (М. Fritz,
1947). В течение палеозоя Cyclostomata успели приобрести высокую
дифференцировку колоний, известковый скелет, т. е. типичные черты
Cymnolaemata, и сохранили важный исходный признак — длинную, труб-
чатую форму зооидов. Вероятно, последнее свидетельствует об удлинен-
ной, трубчатой форме червеобразного, одиночного предка мшанок. В те-
чение своей длительной геологической истории Cyclostomata, начиная
с позднего кембрия (рис. 65), развивались постепенно; будучи более
многочисленными в раннем палеозое (ордовик, силур, девон), они все
реже встречаются в верхнем палеозое и становятся редкими в триасе.
С вымиранием отрядов Cystoporata, Trepostomata и Cryptostomata
Cyclostomata начинают быстро эволюировать в юре, достигнув расцвета
в морях мелового периода, мало уступая по численности и многообра-
зию Cheilostomata. Однако далее, в отличие от последних, численность.
Cyclostomata быстро и сильно , стала сокращаться: уже в третичный
247
период и в настоящее время по количеству видов они во много раз
уступают Cheilostomata.
Cystoporata, Trepostomata и Cryptostomata существовали, главным
образом, в палеозое, начиная с ордовика. Cystoporata — небольшая груп-
па примитивных палеозойских мшанок, как самостоятельный отряд была
обоснована Г. Г. Астровой (1964, 1965). Этот отряд вымер в конце перми,
на границе палеозоя и мезозоя, когда вымерли и многие другие группы
беспозвоночных — трилобиты, целые классы и отряды Pelmatozoa и т. д.
Наибольшее распространение Trepostomata наблюдалось несколько
раньше, чем у Cryptostomata (в ордовике), затем их численность стала
постепенно сокращаться в девоне, карбоне и особенно в перми, когда
распространение Cryptostomata, бывших важными рифообразователями,
наоборот, сильно возрастало. Эти мощные в палеозое отряды мшанок
(Шульга-Нестеренко, 1941, 1949) к концу палеозоя вымирают и лишь
в виде отдельных видов доживают до верхнего триаса.
При выяснении филогении отрядов морских мшанок часто приходится
сталкиваться с противоречивостью данных о современных и ископаемых
формах и различно трактующихся палеонтологами. Строение скелета
вымерших отрядов Cystoporata, Trepostomata и Cryptostomata во мно-
гом своеобразно и специализировано. Их филогенетические связи и отно-
шения к другим отрядам морских мшанок удается выяснить комплекс-
ным методом изучения — морфогенетическим, морфо-функциональным,
онтогенетическим и экологическим (Астрова, 1964, 1965).
В настоящее время (см. Астрова, 1964, 1965) можно считать бесспор-
ным, что Cyclostomata являются родоначальным стволом, от которого
отделился отряд Cystoporata, обладающий чертами, сближающими их с
Cyclostomata и Trepostomata. Трубчатой формой ячеек с терминальным
устьем без крышечки и некоторыми другими признаками Cystoporata
выделяются и как древняя ветвь ствола циклостомат, отделившаяся ра-
нее более, высоко организованных Trepostomata, также возникших от
ствола Cyclostomata.
В раннем палеозое Cryptostomata была настолько массовой группой,
что стала важным рифообразователем. В позднем палеозое (карбон и
пермь) они были наиболее распространенными мшанками, а в триасе
совершенно вымерли. Вопрос о филогенетических связях между Cteno-
stomata, Cheilostomata и Cryptostomata довольно сложен. Палеонтологи
находят ряд сходных черт у Cryptostomata и Cheilostomata: грушевид-
ную или коротко-трубчатую форму ячеек, расположение устья на фрон-
тальной стороне, образование начальных стадий колоний анцеструлой
и т. д. Вместе с тем, они отмечают и ряд отличий между этими группами:
своеобразную микроструктуру стенок ячеек (цистид) и наличие системы
капиллярных каналов (Астрова, 1960).
Близкое родство, установленное зрологами между Ctenostomata и
Cheilostomata, и наличие ряда сходных черт между Cheilostomata и
Cryptostomata позволяют считать их родственными группами, составляю-
щими одну филогенетическую ветвь. Вопрос заключается в том, вести ли
происхождение Cheilostomata от Ctenostomata или от Cryptostomata.
По данным зоологии, Ctenostomata и Cheilostomata близко родственны,
причем первые продолжали существовать, когда появились вторые, тогда
как между Cheilostomata и Cryptostomata имеется перерыв во времени
существования: первые встречаются в юре, вторые вымирают в триасе.
Мне представляется более правильным выводить Cheilostomata от Cte-
nostomata, a Cryptostomata считать боковой, слепо закончившейся
ветвью.
Наиболее примитивной группой можно считать Ctenostomata, к ним
близки Cheilostomata, и от обоих удалены Cyclostomata. Вместе с пале-
онтологами можно считать, что Ctenostomata, Cheilostomata и Cryptosto-
248
mata составляют одну филогенетическую ветвь, a Cyclostomata, Cystopo-
rata и Trepostomata—другую.
Судя по внешним признакам и скелету вымерших представителей
Gymnolaemata, их отряды эволюировали уже давно. Отмеченные выше
вторичные особенности Cyclostomata не позволяют выводить из них Cte-
nostomata и Cheilostomata, Очевидно, были две филогенетические ветви,
которые произошли от исходной примитивной группы морских мшанок.
Вероятно, это были мшанки с органическим (хитиновым) скелетом, боль-
шой самостоятельностью зооидов, составлявшие слабо интегрированные
колонии.
В то же время, в то время как Gymnolaemata, продолжая (как и их
предки) жить в море и делая очень слабые попытки выйти в пресную
воду, претерпевают значительные упрощения организации зооидов, у
Phylactolaemata этого не наблюдается. Их организация, как было ска-
зано выше, несмотря на жизнь в пресных водах и на сильные вторичные
изменения индивидуального развития и на реликтовый характер, отме-
ченный Ремане (Remane, 1959), сохраняет почти исходное строение, при-
сущее морским с трехчленистым целомом, червеобразным формам
(вроде Phoronida) и отчасти плеченогим. 'Их правильно считают потом-
ками морских форм, давших начало обеим группам мшанок. Черты
строения одиночных морских форм свидетельствуют об очень раннем
ответвлении от общего ствола предков пресноводных, мшанок, до ответ-
вления отрядов морских мшанок. Однако отсутствие палеонтологических
данных делает соображение о происхождении Phylactolaemata гада-
тельным.
Брахиоподы (Brachiopoda)
Двустороннесимметричное, сдавленное в дорсо-вентральном направ-
лении тело плеченогих, или брахиопод, обычно ориентированное при жиз-
ни передним концом кверху, заключено в двустворчатую раковину, кото-
рая выделяется спинной и брюшной лопастями мантии. У них имеются
эпистом над ротовым отверстием, туловищный отдел со спирально свер-
нутым лофофором, содержащий внутренние органы, и ножка — орган
прикрепления к субстрату. Нервная система состоит из надпищеводного
(церебрального) и подпищеводного ганглиев или только из последнего,
кольца и нервов внутренних органов, лофофора и ножки. Органы чувств
и мускулатура стенок тела развиты слабо; сильно развиты мускулы, от-
крывающие и закрывающие створки раковины и поворачивающие ее.
Внутренние органы представлены неправильно петлеобразным кишечни-
ком с парными пищеварительными железами, реже сквозным, чаще без
ануса, незамкнутой кровеносной системой, целомодуктами (обычно од-
ной парой) как органами экскреции и выведения половых продуктов и
целомической полостью тела, состоящей из трех отделов. Лофофор как
орган улавливания микроскопической пищи, главным образом диатомей,
простейших, мелких личинок беспозвоночных, вместе со стенками мантии
служит и для дыхания. Дыхательным пигментом является гемеритрин.
Полы разделены, половые железы возникают как набухание перитоне-
альных клеток. Размножение исключительно половое. Низшие формы —
яйцекладущие, развиваются без метаморфоза. Высшие — живородящие
со свободноплавающей личинкой и с метаморфозом. Онтогенез характе-
ризуется хорошо выраженной закладкой эктодермы, энтодермы и мезо-
дермы, энтероцельным образованием целома и длительным постэмбрио-
нальным развитием.
Брахиоподы — морские, обычно прикрепленные донные формы пре-
имущественно мелководья или сублиторали (30—200 л*). Размеры совре-
менных брахиопод — от 5 мм до 8 см (Magellania vetwsa достигает
8,4 см) в длину. Размеры ископаемых — в пределах 5—8 см, хотя камен-
ноугольный Gigantoproductus достигал 40 см в длину. Современные бра-
хиоподы являются реликтовой фауной, представленной 260 видами, тогда
как ископаемых около 30 000.
Брахиоподы делятся на беззамковых — Inarticulata, малочисленных
почти на протяжении всей геологической истории (известно — 92 рода), и
на замковых — Articulata, массовых особенно в раннем палеозое, но
сильно обедненных в современных морях (всего известно 1377 родов).
Со времен Гатчека (Hatschek, 1888) многие рассматривали и рас-
сматривают брахиопод как класс типа Tentaculata вместе с классами
Phoronida и Bryozoa. По наличию более или менее выраженного рас-
членения тела и целома на три отдела их относят и к Trimetamera (по
Мастерману). Создавая искусственную группировку, Термье (Termier
et Termier, 1960) объединяют брахиопод вместе с Enteropneusta, Echino-
dermata, Pterobranchia, Graptolitha, Phoronida, Bryozoa. В последнее
250
время брахиоподы считаются самостоятельным типом вторичнополост-
ных нечленистых беспозвоночных, стоящим между первично- и вторично-
ротыми.
За исключением единичных работ нынешнего столетия, исследования
анатомии и общей организации современных брахиопод относятся к кон-
цу прошлого — началу нынешнего столетия. Примерно столетнюю (Laca-
ze-Duthiers, 1859; Morse, 1871, 1873а, 18736;' Ковалевский, 1874) или
полувековую (Yatsu, 1902; Conklin, 1902) давность имеют работы по
эмбриологии этой группы. Современных работ очень немного, причем они
касаются только некоторых вопросов — гистологии (Schaeffer, 1926;
Senn, 1934; Chuang, I960), функции некоторых органов и физиологии
(Richards, 1952; Chuang, 1956, 1959; Atkins, 1958, 1959а, 19596, 1960,
1961; Heller, 1931), постэмбрионального развития лофофора и ручного
\ аппарата (Thomson, 1927; Конжукова, 1948, 1957; Atkins, 1958, 1959,
1961) . То же можно сказать и о работах по эволюции и филогении совре-
менных брахиопод.
К сожалению, гистология, физиология, органогенез и филогения со-
временных брахиопод изучены далеко недостаточно несмотря на сравни-
тельно ничтожное число родов (68) и видов (260).
Палеонтологам наука обязана многими сотнями работ, нередко круп-
ных монографий, по ископаемым брахиоподам, описанию примерно
30 000 видов, установлению многих отрядов, более двух десятков надсе-
мейств, выяснению геологического распространения брахиопод, начиная
•с нижнего кембрия (см., например, Лихарев, 1960; J. Roger, 1960, и т. д.),
выяснению динамики численности, характеристике расцвета, упадка и
вымирания больших групп брахиопод (А. Н. Muller, 1958, 1961).
К этому надо добавить работы о постэмбриональном развитии рако-
вины, ручном аппарате, замке, отпечатках мускулатуры, мантийных си-
нусах и гонадах и т. д. (например, из старых работ Beecher, 1891, 1892;
Muir-Wood, 1955, 1959; из новых Иванова, 1949а, 19496; G. Biernat, 1959,
и некоторые др.). Лишь немногие исследования (например, Иванова,
1962; Ржонсницкая, 1959; Сарычева; 1949, 1960; Сокольская, 1948, 1950,
1954; Muir-Wood, Cooper, 1960; Kozlowski, 1929; Havlicek, 1959; Elliot,
1955, 1957, и др.) охватывают совокупность черт строения раковины,
реже ее онтогенез, а также экологию, эволюцию и филогению брахиопод.
Только такие исследования, когда группа брахиопод изучается с са-
мых ее истоков, начиная с древнейших примитивных представителей и
кончая последними представителями при вымирании группы, позволят
выяснить ход эволюции и филогенетические взаимоотношения надсе-
мейств и семейств. Именно этим путем, как сделано для ряда классов и
типов беспозвоночных и классов позвоночных, палеонтологи смогут вос-
становить филогению вымерших брахиопод, без чего. нельзя выяснить
филогенетические связи современных групп.
До сих пор совершенно неоправданно палеонтологи, да и ряд зооло-
гов, продолжают изучать и рисовать раковины брахиопод, ориентируя
вниз верхнюю (переднюю) часть, а замочную (заднюю) нижнюю часть —
вверх, как делали это старые авторы, считавшие брахиопод моллюсками.
Сплошь и рядом, даже Lingula, изображается стебельком кверху. Мне
представляется необходимым вести описания, исходя из природы брахио-
под, как это делает Бошамп (Beauchamp, 1960).
По основным чертам своей организации и экологии брахиоподы —
однообразная группа морских животных с большим разнообразием лишь
особенностей строения раковины, лофофора и ручного аппарата, что от-
носится почти исключительно к ископаемым формам, о чем будет ска-
зано дальше.
Стенка тела,, где нет раковины, состоит из цилиндрических эпидер-
мальных клеток, среди которых рассеяны железистые клетки, соедини-
251
тельнотканного слоя, слоя мускульных волокон и перитонеального эпите-
лия. Эпидермальные клетки одеты кутикулой и во многих местах тела
имеют реснички. Последние длиннее и многочисленнее на поверхности-
лофофора, его щупалец и на свободной поверхности мантии, что связано
с функцией этих органов.
Наружный эпителий стенки брюшной и спинной лопастей мантии об-
разует двустороннесимметричные, но неодинаковые брюшную и спинную
створки раковины. Предполагают, что в выделении и росте раковины
принимают участие также клетки, находящиеся в папиллах, образован-
ных выростами мантийного эпителия, пронизывающих и стенку ракови-
ны (Borradaile, Potts a. all., 1958). Створки, покрытые снаружи тонким
слоем органического вещества — периостракумом, у большинства без-
замковых брахиопод (например, у Lingula) образованы чередующимися
слоями органического вещества — протеина (Vinogradov, 1953), а не хи- .
тина, как обычно ошибочно считают, и фосфата кальция; у замковых
брахиопод под периостракумом находится тонкий пластинчатый слой из
известковых пластинок, расположенных параллельно поверхности рако-
вины, и толстый внутренний призматический слой с примесью органиче-
ского вещества, причем отдельные призмы направлены под некоторым
углом к поверхности раковины. Это так называемые первичный и вторич-
ный слои раковины брахиопод по Вильямсу (Williams, 1955, 1956).
Эпителий мантии образует и все другие части скелета, элементы зам-
ка, ручной аппарат, отдельные известковые спикулы, а также кутикуляр-
ные щетинки на краях мантии, особенно сильно развитые у беззамковых
брахиопод. Эти щетинки у Lingula служат для защиты мантийной поло-
сти от загрязнения илом и песком (Morse, 1902). Пучки щетинок прини- ,
мают активное участие в движении личинок замковых брахиопод (Кова-
левский, 1874).
Как известно, химический состав раковины беззамковых и замковых
брахиопод различен. У лингулид и дисцинид (Vinogradov, 1953), у кото-
рых неорганическую часть скелета образует фосфат кальция й магния, ’
количество органического, протеинового вещества в раковине велико »
колеблется у Lingula anatina от 25—45 до 52,10% (47,90% составляет
зола). У замковых брахиопод, как и у Craniidae, при различном трое- !
нии в раковине почти нет фосфата кальция (всего 0,5%); он замещается
карбонатом кальция в форме кальцита с небольшой примесью углекис- ,
лого магния. На долю органического вещества приходится всего 2%.
Развитию раковины в онтогенезе предшествует образование органи- i
ческой (из периостракума) эмбриональной двустворчатой раковины —
протегулума (Yatsu, 1902), которая у взрослых обычно не сохраняется. >
Развитие раковины происходит довольно медленно, в связи с ростом жи-
вотного, причем с возрастом.изменяется форма раковины, ее скульптура,. ' ,
развиваются замок, септа и т. д.; постепенное развитие и усложнение в (
ходе онтогенеза проделывает и ручной аппарат, функционально тесно
связанный с лофофором и с ходом его онтогенеза. Раковина редко оста-
ется гладкой (такая раковина наблюдается у примитивных, а также
древних вымерших форм), обычно на ней развивается концентрическая ,
скульптура, в том числе резкие линии нарастания, и радиальная скуль-
птура в виде складок и ребер, а порой и игл. Большое разнообразие в.
этом отношении наблюдалось у ископаемых брахиопод, у которых неред-
ко при массовом, скученном образе жизни возникала также асимметрия,
раковин. Некоторые ископаемые брахиоподы приобрели, в связи с осо- !
бым ростом раковины, с образованием шлейфов, длинную трубчатую '
форму (например, Proboscidella proboscidea, Лихарев, 1960) и т. д. Ряд ;
особенностей формы раковин ископаемых брахиопод, видимо, и трубча- ;
тую форму палеонтологи рассматривают как возрастное, старческое из-
менение раковины.
252
Раковины современных замковых форм, имеющие различную окра-
ску— оранжевую, красновато-коричневую, серую, темно-зеленую и бе-
лую— по форме довольно однообразны, хотя и сейчас встречаются, прав-
да единичные виды, с иглами на раковине (см. Hyman, 1959).
При прикреплении брюшной створкой к субстрату у современных
Crania, Lacazella и у ряда ископаемых брахиопод эта . створка сильно
редуцируется, а порой вовсе атрофируется.
У беззамковых брахиопод, не имеющих замка ручного аппарата,
створки раковины соединены друг с другом мускулами, ножка, или сте-
белек, имеющий собственную мускулатуру, выходит между створками
раковины. Один непарный и пять парных мускулов открывают и закры-
вают створки раковины и двигают спереди назад одну створку относи-
тельно другой и каждую створку в отдельности в бок.
У замковых брахиопод створки, устойчиво соединенные друг с другом
замком, или шарниром, открываются и закрываются действием двух
групп парных мускулов (дедукторов и аддукторов); ножка, выходящая
через отверстие в брюшной створке, лишена собственной мускулатуры.
Движение раковины — повороты вокруг оси, наклон к субстрату и воз-
вращение к вертикальному положению, обеспечивается мышцами, при-
крепляющимися к стенкам раковины и к ножке (так называемые ножные
мускулы).
В физиологическом отношении мускулатура изучена слабо. Наличие
поперечнополосатых мышц среди преобладающего большинства гладких
мускулов створки (задние аддукторы, Blochmann, 1933), по аналогии
с тем, что известно относительно мускулов аддукторов у двустворчатых
моллюсков, позволяет думать, что существуют различия в характере со-
кращения отдельных мышц. Характерно также, что, как и у ряда групп
вторичноротых, источником энергии мышечного сокращения у брахиопод,
как известно (Вержбинская, 1936), служит креатин-фосфат.
В ходе онтогенеза мускулатура раковины брахиопод (Ковалевский,
1874; Yatsu, 1902) возникает рано. По данным А. О. Ковалевского, мно-
гие мышцы личинки переходят в мышцы взрослой брахиоподы.
Орган прикрепления брахиопод — ножка, у беззамковых (лингулид
и дисцинид) является выростом тела, куда продолжается полость тела,
стенки которой состоят из эпидермиса, покрытого толстой кутикулой,
соединительной ткани, слоя мышц и перитонеального эпителия. Ножка,
или стебелек, сильно сократима. У краниид она полностью утеряна. Нож-
ка у замковых брахиопод, которая также редуцируется у некоторых
современных и довольно часто у ископаемых форм, является коротким
плотным выростом тела, без мускулатуры и целомической полости. Она
состоит из эпидермиса, одетого также толстой кутикулой, и соединитель-
ной ткани. У многих современных форм ножка на конце имеет корневид-
ные выросты, или нити, с помощью которых животное прочно прикреп-
ляется к субстрату (Th. Ekman, 1896).
Двустороннесимметричное тело брахиопод, занимающее меньшую,
нижнюю часть мантийной полости, на верхнем, переднем конце несет
небольшой эпистом и сильно вытянутые, часто спирально свернутые руки
лофофора — губные придатки, возникающие вблизи рта. Лофофор зани-
мает большую часть мантийной полости, эпителий его и щупальца, рас-
положенные в чередующемся порядке, несут реснички и являются водо-
двигающим, сортирующим и улавливающим пищу органом, несущим
также дыхательную функцию. Реснички щупалец вызывают два вводных
тока воды к передне-боковым краям раковины, с которыми поступает
пища (рис. 66). Движением ресничек мантии, создающим выводной ток,
вода выносится наружу. Мелкие, легкие частицы — диатомовые, пери-
динеи, взвесь вместе со слизью — гонятся ресничками щупалец по рес-
ничному желобку ко рту, а также падают на дно брюшной лопасти ман-
253
Рис. 66. Lingula '(по Chuang из Grasse). Токи, создаваемые ресничками
в палиальной полости:
А — с вентральной стороны; Б — с дорзальной стороны
тии и выбрасываются наружу выводным током, вызываемым ресничками
мантии и хлопанием створок раковины (Orton, 1914).
В последние годы ряд зоологов и палеонтологов — Ричардс (Richards,
1952), Чуанг (Chuang, 1956), Аткинс (Atkins, 1958, 1959а, 19596, 1960,
1961), Рудуик (Rudwick, 1960), Роуэлл (Rowell, 1960, 1961), Вильямс и
Райт (Williams a. Wright, 1961) —занимается изучением морфологии
частей и функции лофофора брахиопод. Расходясь в некоторых частно-
стях, в целом они подтверждают данные Ортона.
Сравнения фильтрующего аппарата брахиопод и двустворчатых мол-
люсков, частично затрагивающие и роль кишечника, даны Ортоном и Чу-
ангом. Надо сказать, что приспособления к фильтрации воды довольно
примитивны и не затрагивают кишечник. Среди современных биофиль-
траторов брахиоподы играют совершенно незначительную роль в освет-
лении и физико-химическом изменении морской воды, сравнительно, на-
пример, с двустворчатыми моллюсками. Однако роль брахиопод как
биофильтраторов, особенно в периоды их расцвета (когда они жили
громадными сообществами и были широко распространены в морях
палеозоя, достигая порой крупных размеров), несомненно, была гораздо'
значительнее, чем ныне. Лофофор с его функцией фильтрации, ловли
пищи и дыхания является органом, обеспечивающим непосредственную
связь с окружающей средой прикрепленных, донных, заключенных в ра-
ковину животных. Несомненно с помощью лофофора косвенным образом
они реагируют на воздействие среды, на изменение условий жизни, тем
254
более, что других рецепторов, кроме статоцистов (лингулиды и дисцини-
ды), брахиоподы не имеют.
Бичером (Beecher, 1891, 1892) и рядом последующих авторов — Том-
соном (Thomson, 1927), Стели (Stehli, 1956), Е. Д. Конжуковой (1948,
1957) —установлены разные типы лофофора, их развитие и значение для
системы и выяснения родственных связей, о чем говорится ниже.
Лофофор поддерживается известковым ручным аппаратом, развиваю-
щимся за счет эпителиальных стенок спинной лопасти мантии. Лофофор
и ручной аппарат коррелятивно связаны друг с другом, однако считают
(Hyman, 1959), что их отношения изучены недостаточно. У ряда совре-
менных и ископаемых брахиопод прослежены стадии постепенного онто-
генетического развития ручного аппарата, достигавшего у многих иско-
паемых брахиопод, особенно в периоды их прогрессивного развития,
сложного строения и больших размеров (например, у спириферид, назва-
ние которых вызвано спирально закрученной формой ручного аппарата).
Анатомия брахиопод проста и однообразна. В основании лофофора,
между эпистомом и щупальцами лофофора, по середине в плоскости сим-
метрии находится рот, ведущий в глотку, переходящую в пищевод, рас-
ширяющийся в желудок, в который отдельными протоками справа и сле-
ва открывается несколько пищеварительных желез, называвшихся ста-
рыми авторами печенью. Число желез и их протоков различно у разных
групп беззамковых и замковых брахиопод. Желудок переходит в длинную
заднюю кишку, с анальным отверстием, располагающимся в плоскости
симметрии у Crania и справо от нее у остальных беззамковых брахио-
под. У замковых задняя кишка слепая, без ануса. Однако некоторыми
зоологами (Бобрецкий, 1887) плотный тяж, или шнурок, которым кон-
чается кишечник у ряда брахиопод, рассматривается как рудимент
задней, кишки. Наличие же у ряда ископаемых брахиопод отверстия в
замочной 'пластинке, по мнению палеонтологов, например, Бичера
(Beecher, 1892, см. Morse, 1902), позволяет думать, что замковые бра-
хибподы ранее имели анальное отверстие.
Кишечник петлеобразный и подвешен на спинном и брюшном мезен-
териях и на поперечных желудочнопариетальной и кишечнопариетальной
связках.
Гистология кишечника и пищеварение брахиопод изучены недоста-
точно, причем старые авторы (Blochmann, 1933, он же 1892, 1900, по
Hyman, 1959) и новые (Chuang, 1959, 1960) ограничивались изучением
беззамковых брахиопод; анатомия и гистология замковых брахиопод
описаны только А. А. Стрелковым (1946).
Кишечник беззамковых брахиопод (Lingula, Crania) образован столб-
чатым эпителием с ресничками (кроме пищеварительных желез), разбро-
санными железистыми, слизистыми клетками, базальной мембраной,
соединительнотканой стромой, внутренним кольцевым и наружным про-
дольным слоями гладких мышц и перитонеальным эпителием. В цито-
плазме эпителиальных клеток найдены пигментные гранулы, глыбки гли-
когена и липохондрин, в железистых клетках — муко- и гликопротеины.
Пищеварение (Lingula) главным образом, если не исключительно, внут-
риклеточное, но крахмал и гликоген перевариваются внеклеточно.
Колебания ресничек вызывают движение пищи по кишечнику, лишь
несколько напоминающее таковое в кишечнике двустворчатых моллю-
сков. По наблюдениям Морзе (Morse, 1902), происходят перистальтика
пищевода и желудка и постоянные расширения и сокращения пищева-
рительных желез. Это вместе с действием ресничек эпителия кишечника
вызывает передвижение пищевой массы по кишечнику. Выводной ток в
задней кишке гонит фекалии к анусу.
Антиперистальтикой стенок кишечника у замковых брахиопод непе-
реваренные частицы (движением ресничек) выбрасываются изо рта
255
и падают в полость мантии (Richards, 1952), откуда удаляются наружу
выносяийим током.
В то время как замковые брахиоподы питаются лишь диатомовыми
водорослями, перидинеями и взвесью, пищей лингулид служат не толь-
ко микроскопические одноклеточные растительные организмы, но и кор-
неножки, радиолярии, личинки моллюсков и иглокожих, мелкие ракооб-
разные и другие организмы (Hyman, 1959).
Нервная система брахиопод проста, но современными методами
гистологии не изучена. Кольцо вокруг пищевода у беззамковых брахио-
под имеет лишь субэзофагеальный, или подглоточный ганглий, у замко-
вых, кроме того, надпищеводный, или церебральный, ганглий, который
однако значительно меньше первого. Надпищеводный ганглий посылает
пару сильных нервов в руки лофофора, подпищеводный ганглий посы-
лает два спинных и два брюшных мантийных нерва, два нерва ножки и
нервы к мускулам-аддукторам. Нервная система у беззамковых занимает
примитивное расположение в основании эпидермиса, у замковых, по
старым данным Ван Беммелен (см. Hyman, 1959), она лежит в соедини-
тельнотканом слое.
Органов чувств у брахиопод нет, кроме статоцистов (Yatsu, 1902),
которые сохраняются и у взрослых Lingula, со статолитами, находящи-
мися в постоянной вибрации (Morse, 1902). Брахиоподы, хотя у них
не найдены, а вернее не изучены, чувствительные клетки, реагируют на
свет и тактильное раздражение.
Трехчленистый характер целома сохраняется лишь у беззамковых
брахиопод (Hyman, 1959). В их эпистоме находится целом (сравнимый
с протоцелем), заполняющийся соединительной тканью у других брахио-
под. Большой и малый каналы лофофора, и полость тела вокруг пищевода
составляют второй отдел целома — мезоцель, который (среди брахио-
под) вполне отделен септой от метацеля лишь у Crania. Метацель являет-
ся общей полостью тела, в которой находится почти весь кишечник, неф-
ридии и мышцы; лишь у Crania септа полностью разделяет мезоцель и
метацель и сохраняются полные спинной и брюшной мезентерии кишеч-
ника. У остальных брахиопод они неполные или отсутствуют. Поперечные
мезентерии, или связки (желудочно-париетальная и кишечно-париеталь-
ная), пересекают метацель поперек; без особых оснований они рассмат-
риваются некоторыми зоологами, в том числе Бючли (Biitschli, 1924),
как настоящие диссепименты. Лишь у лингулид и дисцинид метацель
продолжается в стебель, или ножку, у остальных брахиопод, имеющих
ножку, последняя плотная без полости целома (стр. 253). У всех брахио-
под -метацель продолжается в лопасти мантии, образуя там мантийные
синусы, с которыми связано развитие и положение половых желез и от-
печатки которых сохраняются у многих ископаемых брахиопод. Целоми-
ческие полости наполнены полостной жидкостью с целомоцитами не-
скольких категорий, среди которых имеются гранулоциты с дыхательным
пигментом — гемеретином, близким по эффекту Бора и другим особенно-
стям не к гемеретрину сипункулид и аннелид, а к гемоглобину позвоноч-
ных (Manwell, 1960). Циркуляция полостной жидкости, вызываемая
ресничками перитонеума, и наличие дыхательного пигмента обеспечи-
вают дыхательную функцию.
Кровеносная система развита слабо, что объясняется наличием об-
ширной целомической полости, с хорошей циркуляцией полостной жидко-
сти и присутствием в ней клеток с дыхательным пигментом, со слабым
развитием соединительной ткани и с хорошей аэрацией всех частей тела
брахиопод. Кровеносная система (Hyman, 1959) изучена недостаточно:
по одним авторам она незамкнутая, по другим — замкнутая.
Находящийся в спинном мезентерии спинной кровеносный сосуд
с сократимым пузырьком, так называемым сердцем, гонит кровь вперед;
256
охватывая пищевод, он входит в лофофор и посылает сосуды в щупальца.
Сзади или внизу спинной, сосуд, разветвляясь, образует сосуды спинной и
брюшной складок мантии: Кровеносная система у Lingula (Schaeffer,
1926) незамкнутая, она образует синусы в гонадах, целомодуктах и
кишечно-париетальной связке; эндотелиальной выстилки в сосудах
нет.
Органы выделения (обычно в числе одной, реже двух пар — у ринхо-
неллид) являются типичными целомодуктами, выводящими половые
клетки и несущими экскреторную функцию, которая экспериментально
доказана советским зоологом М. Геллером (Heller, 1931).
Брахиоподы, как правило, раздельнополы. Известны только единич-
ные гермафродитные виды: три средиземноморских вида Argyrotheca
(Senn, 1934) и австралийский Pumilus antiquatus (Atkins, 1958). Две
(реже одна) пары половых желез находятся в половых синусах мантии.
Последние целомического происхождения и выстланы перитонеальным
эпителием, в связи с которым развиваются половые клетки. Созревшие
половые клетки выпадают в полость синуса и выводятся целомодуктами
(Schaeffer, 1926; Senn, 1934). Исследования последних авторов подтвер-
дили данные А. О. Ковалевского (1874), .нашедшего сходные черты в
этом отношении у брахиопод и аннелид, вернее общие черты со многи-
ми вторичнополостными животными.
Беззамковые брахиоподы — яйцекладущие, замковые — живородя-
щие. Во время размножения самцы выбрасывают сперму в воду. У без-
замковых брахиопод яйца поступают в воду, где происходит их оплодот-
ворение и развитие; у замковых брахиопод яйца поступают в выводко-
вые мешки или в мантийную полость, где происходит дробление и разви-
тие до сформированной личинки.
Дробление недетерминированное, полное, почти равномерное, ради-
ального типа, приводит к образованию целобластулы, которая обычно
путем инвагинации или как исключение (Lacazella.) мультиполярной
иммиграцией превращается в гаструлу. Целом образуется энтероцель-
ным путем от первичного кишечника в виде карманов, превращаясь в два
целомических мешочка; бластопор, щелевидно вытягиваясь, замыкается
от заднего к переднему концу; первичный кишечник теряет сообщение
с наружной средой, превращаясь в замкнутый мешок. Реже {Lingula,
Lacazella) целом (подобно тому, как у некоторых кишечнодышащих)
возникает схизоцельным путем, причем замкнувшийся первичный кишеч-
ник является зачатком мезоэнтодермы, средняя его часть становится
зачатком энтодермальной кишки, а из его боковых частей, отделяющихся
в виде двух плотных клеточных масс, схизоцельным путем возникает
пара целомических мешочков.
Мезенхима не образуется ни у беззамковых, ни. у замковых бра-
хиопод.
Эмбрион растет в длину, становится двучленистым (головной и ту-
ловищный отделы у беззамковых брахиопод — Lingula) и трехчленистым
(головной, туловищный и хвостовой отделы) —у замковых брахиопод.
Внешнее расчленение зародыша не распространяется на его внутреннюю
организацию. Целом расчленен чаще на две пары мешочков, реже на
три пары, например, у Terebratulina (Conklin, 1902) и у Terebratella
4(Percival, 1944, 1960).
На месте бывшего бластопора образуется рот и передняя эктодер-
мальная кишка, возникающие у беззамковых, еще у эмбриона, а у зам-
ковых брахиопод после прикрепления личинки к субстрату.
Круговой складкой эктодермы туловищного отдела у зародыша обра-
зуется мантия, которая у беззамковых брахиопод сразу направлена сво-
им краем вверх, тогда как у замковых брахиопод она направлена вниз
и лишь затем (после прикрепления личинки к субстрату) переворачи-
17 Д. М. Федотов
257
вается вверх. Мантия подразделяется на спинную и брюшную лопасти,
которые образуют соответственно спинную и брюшную створки ракови-
ны. До их образования у беззамковых и некоторых замковых брахиопод
образуется кутикулярная полукруглая раковинка — протегулум.
Вылупившаяся из яйца личинка беззамковых брахиопод (Lingula')
обладает уже строением взрослых форм; после прикрепления, с развити-
ем в перитонеальном эпителии половых клеток, прямо переходит во
взрослый организм. Она имеет раковину, нервную систему, органы чувств
(глазки и статоцисты), сквозной кишечник с ротовым и анальным отвер-
стиями, целомодукты. Развитие заканчивается превращением головного
отдела в эпистом и формированием лофофора, который, будучи лишь
органом движения свободно плавающей личинки, у замковых брахиопод
становится органом улавливания добычи и дыхания. У беззамковых,
кроме того, сохраняется его значение и как локомоторного органа;
непарное щупальце исчезает, статоцисты сохраняются.
Развитие замковых брахиопод сопровождается метаморфозом с ре-
грессом ряда органов личинки, ведущей короткое время свободный при-
донный образ жизни. Утрачиваются органы чувств личинки, происходит
редукция ее головного, или предротового, отдела, становящегося в ре-
зультате упрощения и уменьшения небольшим эпистомом взрослой бра-
хиоподы.
Туловищный отдел личинки и зародыша образует тело брахиоподы.
Четыре пучка временных щетинок утрачиваются, но у беззамковых и
ряда замковых образуются на краях мантии многочисленные щетинки.
Происходит развитие лофофора, который закладывается в виде парных
зачатков щупалец, а затем проходит последовательные стадии усложне-
ния, становясь венчиком, окружающим ротовое отверстие. Происходит
образование спинной и брюшной лопастями мантии — раковины, а на
спинной створке — ручного аппарата. У замковых брахиопод развивают-
ся замок с зубами и зубными (замочными) ямками и идет развитие
мускулов раковины и мышц, обеспечивающих движение раковины.
На месте замкнувшегося бластопора образуется эктодермальная перед-
няя кишка с ротовым отверстием. Задняя кишка и анус не образуются.
Происходит развитие целомодуктов и половых желез.
Хвостовой отдел, хорошо выраженный у личинок замковых брахио-
под,. становится маленькой ножкой, без собственной мускулатуры и цело-
мической полости, тогда как у зародыша беззамковых — у Lingula,
имеющей длинный стебель с собственной мускулатурой и целомической
полостью, внешне он выражен слабо.
Руководствуясь главным образом работой А. О. Ковалевского
(1874), следует отметить ряд особенностей индивидуального развития
Lacazella ( = Thecidium) сравнительно с таковым других замковых бра-
хиопод. Бластула этой брахиоподы сильно удлиненной формы, целом,
как было отмечено, образуется'схизоцельным путем, четырех пучков ще-
тинок не образуется, очень слабо развита брюшная сторона туловищного
отдела, мантия образует только спинную створку раковины, брюшная
образуется частью мантийной спинной стороны туловищного сегмента и,
как пишет А. О. Ковалевский (1874), покровами хвостового сегмента,
стебелек исчезает сразу после превращения.
По толкованию палеонтологов, давно подметивших особенности позд-
него онтогенеза этой брахиоподы по работе А. О. Ковалевского (напри-
мер, Beecher, см. Korschelt u. Heider, 1890—1893; Иванова, 1949а), ман-
тия образует одну спинную створку, на прилежащей к ней части хвосто-
вого отдела личинки образуется пластинка — первичный дельтидиум,
который причленяется к несколько позже возникающей брюшной створ-
ке раковины. Онтогенез Lacazella необходимо дополнительно исследо-
вать.
258
Как известно, по многим чертам строения беззамковые брахиоподы
более примитивны, чем замковые, но вместе с тем некоторые черты их
онтогенеза изменены сравнительно с развитием замковых брахиопод.
Среди современных форм с энтероцельным развитием целома (Entero-
pneusta, Echinodermata) схизоцельное образование целома явно вторич-
ное явление, очевидно, так же, как схизоцельное развитие целома у
Lingula.
Вторичным и пока необъяснимым является расчленение зародыша
беззамковых брахиопод (даже у Lingula с её сильно развитым муску-
листым с целомической полостью стебельком) на два отдела, или сегмен-
та, а не на три, как у замковых брахиопод. Подавление развития треть-
его, хвостового отдела, а по некоторым авторам (Yatsu, 1902) и развитие
стебелька у Lingula не как продолжения конца тела зародыша, а за счет
брюшной лопасти мантии приходится рассматривать как вторичные из-
менения общего хода онтогенеза беззамковых брахиопод. Во всяком слу-
чае стебелек Lingula анатомически является прямым продолжением
тела, а ножка большинства замковых форм — производной части тела,
лишенной почти всех его признаков, кроме эпидермиса с кутикулой, т. е.
данные анатомии стоят в противоречии с данными эмбриологии, но, на
мой взгляд, первые более важны.
Следует еще отметить, что у Crania, не имеющей ножки, последняя
и в онтогенезе полностью отсутствует (Rowell, 1957а). Молодая особь,
прикрепившаяся плоской брюшной'створкой к субстрату и прикрытая
выпуклой спинной створкой, вполне двустороннесимметрична. По длин-
ной оси тела тянется кишечник с ротовым отверстием на переднем и
анальным на заднем конце тела. Это наиболее примитивные, наименее
измененные организация и ориентировка организма во всем типе бра-
хиопод. Личинки, выйдя из материнского организма, по-видимому, пла-
вают поблизости от места отрождения и тут же и оседают, так как на
раковинах крупных брахиопод постоянно находят раковинки различных
размеров, начиная от микроскопически малых.
Наблюдений над жизнью личинок замковых брахиопод очень мало.
Считают, что они быстро прикрепляются к субстрату. По данным Пер-
сиваля (Percival, 1944), личинка Terebratella inconspicua сво'боднопод-
вижной бывает 30 часов, половой зрелости эта брахиопода достигает
через два года, а продолжительность ее жизни — четыре года.
Для замковых брахиопод характерен сложный путь постэмбрио-
нального развития — формирование раковин, лофофора, ручного изве-
сткового аппарата, замка и т. д., что, как известно, позволяет
выяснить природу изучаемых форм и их родственные отношения (Bee-
cher, 1891, 1892, см. Muir-Wood, 1955; Thomson, 1927; Конжукова, 1948,
1957; Stehli, 1956; Atkins, 1958, и Др.). Так, у Laqueus californicus (Кон-
жукова, 1957) в течение постэмбрионального развития раковина прохо-
дит следующие стадии: гвинииформ, цистеллиформ, исмениформ, френу-
лиформ, теребрателлиформ, строение ручного аппарата которых отве-
чает таковому взрослых особей родов Gwynia, Cistella, Ismenia, Frenu-
la, Terebratella.
Ряд последовательных стадий онтогенеза проделывает и лофофор
современных родов замковых брахиопод; у них различают стадии —
таксолофус, трохолофус, схизолофус, спиролофус, птихолофус, циголо-
фус и плектолофус, характеризующиеся определенными чертами строе-
ния (Muir-Wood, 1955; Stehli, 1956).
Многочисленные наблюдения палеонтологов над возрастными изме-
нениями и эволюцией ручного аппарата у ископаемых родов брахиопод
и данные зоологов играют важную роль в выяснении эволюции и фи-
логенетических связей, в построении рациональной классификации
крупных систематических категорий вымерших брахиопод.
17*
259.
Вымершие брахиоподы, очевидно, обладали онтогенезом, сходным с
таковым современных форм, во всяком случае — длинным этапом пост-
эмбрионального развития. Ряд данных о постэмбриональном развитии
вымерших форм был получен палеонтологами до изучения онтогенеза
современных брахиопод. Значение таких исследований для выяснения
родственных отношений вымерших форм подчеркивается рядом недав-
них работ палеонтологов (Е. А. Иванова, 1949а, 19496; Сокольская,
1948, 1950, 1954; Biernat, 1959).
И все же приходится сказать, что органогенез и возрастные пост-
эмбриональные изменения внутренней организации современных бра-
хиопод изучены недостаточно.
Брахиоподы — типичные формы бентоса скалистого грунта и изве-
сткового ила моря, как исключение (лингулиды) ила и песка в солоно-
ватых и эстуарных водах. Населяя преимущественно сублитораль
(на глубинах между 30 и 200 м), они живут небольшими сообществами
или пятнами. Распространенные довольно широко по Мировому океану
брахиоподы наиболее многочисленны около Японии, Южной Австралии
и Новой Зеландии (Hyman, 1959). Являясь реликтовыми, малочислен-
ными формами, брахиоподы играют незначительную роль в жизни сов-
ременного моря, в отличие от ископаемых форм, населявших большие
площади дна палеозойских морей и принимавших участие в рифообра-
зовании. Будучи. доминирующими формами фауны бентоса в палеозое,
брахиоподы в палеозое и мезозое (Лихарев, 1960) населяли мелководье,
в отличие от современных брахиопод, часть которых распространилась
до абиссальных глубин в 5430—5458 м (Беляев и др., 1960), проявляя
лишь в этом прогрессивную особенность сравнительно с вымершими
формами.
Паразитов у брахиопод известно очень мало; врагами их считаются
рыбы; возможно, они уничтожались также головоногими (Сарычева,
1949) и хищными брюхоногими моллюсками (Иванова, 1958).
Исходное состояние организации — тело из трех отделов (эпистома,
туловища со сквозным кишечником и ножки с целомической полостью
и собственной мускулатурой) и трехчленистый целом, как известно, более
или менее полно сохраняются лишь у беззамковых брахиопод, разви-
вающихся без метаморфоза. Сюда еще относится протеиново-фосфат-
ная богатая органическим веществом раковина, без замка.
Примитивное' строение наиболее полно сохраняется у Crania, кото-
рая имеет несколько удлиненное тело, горизонтально лежащее на дне,
с лофофором и ртом на переднем конце и медиальным анусом — на зад-
нем, а также сплошные спинной и брюшной мезентерии. Однако Crania
не имеет статоцистов, утратила ножку и прикрепляется к субстрату
всей брюшной створкой, раковина известковая, бедна органическим ве-
ществом и почти лишена фосфата кальция.
У лингулид с их примитивной по составу раковиной (протеиново-
фосфатной с большим содержанием органического вещества) и сохра-
няющих статоцисты, внутренняя организация несколько менее примитив-
на, чем у краниид. Задняя кишка с анусом повернута вправо, мезентерии
неполные, однако ножка сохраняется вследствие приспособления к жиз-
ни в толще ила или песка, превращаясь в длинный, очень подвижный
стебель, она сильно уклоняется от строения, типичного для ножки других
беззамковых и всех замковых брахиопод. И вместе с тем стебелек лин-
гулид сохраняет исходные черты строения отдела тела — собственную
мускулатуру стенок и целомическую полость.
Прогрессивными особенностями лингулид, сравнительно с другими
беззамковыми и всеми замковыми брахиоподами, питающимися микро-
скопической пищей — водорослями, является питание разнообразными
беспозвоночными и приспособление к жизни в меняющихся усло-
260
виях полосы отлива, в солоноватых и даже в опресненных водах эстуа-
рий.
Представление о беззамковых брахиоподах как о примитивных фор-
мах подтверждается палеонтологическими данными. В нижнем кемб-
рии, вообще бедном морской фауной беспозвоночных, было уже до де-
сятка родов беззамковых брахиопод: Obolus, Obolopsis, Lingulella,
Delgadella, Bicia, Dicellomus, Obolella, Bortsfordia, Mickwitzla, Acro-
treta (Termier et Termier, I960), тогда как из замковых брахиопод были
найдены лишь Nisusia и Billingsella (отряд Orthida), и если не считать
Rustella и Kutorgina, принадлежность которых к Articulata считается
спорной.
Эти мелкие брахиоподы, преимущественно с округлой или несколько
овальной раковинкой, имели короткие ножки, которыми прикреплялись
к субстрату. Очевидно, среди этой фауны беззамковых брахиопод были
формы, позже давшие начало, с одной стороны, формам, приспособив-
шимся к жизни в толще ила или песка, ножка которых развилась в
длинный, гибкий стебелек, и сохранившим такой образ жизни до на-
стоящего времени, подобно современным лингулидам. С другой сторо-
ны, из этой группы древнейших беззамковых брахиопод произошли
формы, лежащие или прикрепленные к субстрату всей брюшной створ-
кой (с чем была связана редукция и даже полная утрата ножки и
брюшной створки), и давшие начало существующим доныне краниидам
и дисцинидам.
Более полутора десятка остальных семейств беззамковых брахиопод,
обладавших короткими ножками и нередко довольно выпуклыми ра-
ковинами, просуществовало в основном только в кембрии, в кембрии и
ордовике или только в ордовике. Среди них, как будет показано ниже,
очевидно, были формы, давшие начало замковым брахиоподам.
Судя по строению раковины ряда современных и ископаемых без-
замковых брахиопод, их скелет и организация были примитивными и
эволюировали за всю свою длинную геологическую историю, начиная ’
с нижнего кембрия до настоящего времени, очень мало. Крайне медлен-
ный темп эволюции и небольшое количество видов и особей характерны
для беззамковых брахиопод, обычно мелких или небольших форм.
Известно всего 92 рода этих брахиопод, из которых до настоящего вре-
мени дошло всего семь родов и три семейства (лингулиды, дисциниды
и кранииды). Беззамковые брахиоподы все же испытали некоторый рас-
цвет в нижнем отделе ордовика, но уже в силуре их численность па-
дает, снижаясь еще сильнее в девоне, позже они становятся малозамет-
ной частью фауны брахиопод (А. Н. Muller, 1958, 1961), типичными
реликтами настоящего времени.
Жизнь лингулид в толще ила и песка и образ жизни дисцинид и
краниид, тесно прижатых к субстрату или прикрепленных к нему це-
ментом всей брюшной створкой, приводили к крайней изоляции организ-
ма от окружающей среды, задержке в появлении эволюционных изме-
нений в строении и в физиологии, но одновременно способствовали и вы-
живанию этих брахиопод.
Палеобиология и палеоэкология большинства других семейств без-
замковых брахиопод известны мало, но, несомненно, в них лежали
причины вымирания большинства брахиопод, прикреплявшихся к суб-
страту ножкой и существовавших, как было сказано раньше, геологиче-
ски недолгое время (как впрочем и большая часть семейств замковых
брахиопод).
Среди всех вымерших надсемейств замковых брахиопод преобла-
дающее большинство семейств просуществовало на протяжении лишь
одного,, двух, реже трех периодов; десятки семейств их известны лишь
из части одного периода, преимущественно ордовика. Только около
261
Рис. 67. Схема примерного количественного распределения (из A. Muller) ископаемых
брахиопод — 956 родов (А), и ископаемых'Inarticulata — 92 рода (Б)
десятка семейств замковых брахиопод (из общего числа 133, см. Лиха-
рев, 1960) существовало на протяжении большей части палеозоя
(рис. 67).
Для преобладающей массы замковых брахиопод характерно при-
крепление ножкой-к субстрату и вертикальное положение раковины.
Фигурально выражаясь, можно сказать, что эволюция замковых
брахиопод шла под знаком эволюции раковины — ее формы, скульпту-
ры, строения и возрастания ее защитной роли. Организм брахиопод
оказался в значительной мере изолированным от непосредственных воз-
действий внешней среды.
Будучи прикрепленными, не передвигающимися обитателями бенто-
са, брахиоподы по необходимости должны были приспособиться легко
реагировать на воздействие условий жизни, прежде всего, изменениями
в строении и форме своих раковин, являющихся буфером между средой
и мягким телом брахиопод. В ходе геологической истории, распространя-
ясь в Мировом океане, правда больше по горизонтали, чем по вертикали,
брахиоподы оказывались под воздействием различных условий среды,
что отражалось на разнообразии формы, скульптуры, строении и' вели-
чине раковины. В периоды расцвета (нарастания численности видов и
количества особей), в ордовике, силуре, особенно в девоне и далее в
карбоне и перми, разнообразие раковин замковых брахиопод было
необычайно велико. Значительные приспособительные изменения в ходе
геологической истории происходили в образе жизни брахиопод. У них
существовали разные этологические типы: якорный, зарывающийся, сво-
боднолежащий и прирастающий (Иванова, 1949а, 19496, 1962),
262
Преобладающим у вымерших и современных замковых брахиопод
(как и для многих кембрийских беззамковых) был якорный тип при-
крепления к субстрату, скалам, твердому грунту моря и к поверхности
различных донных морских животных при помощи короткой ножки,
при этом’ раковина имела вертикальное положение. Ножка, представ-
ляющая собой плотный стебелек с толстой кутикулой без мускулатуры
и полости тела, у замковых брахиопод претерпевала разнообразные
изменения. У ряда форм устойчивость и вертикальное положение ство-
рок выпуклой раковины, опиравшейся на макушечную часть брюшной
створки, достигались при помощи раздваивания ножки и, вероятно
(по аналогии с немногими современными глубоководными формами,
вроде Chlidonophora), при помощи нитей, которые прикреплялись к
субстрату.
Таким образом возникали приспособительные изменения в строении
ножки и приобретение ею добавочной функции. Прирастание к суб-
страту нитями ножки наблюдается у многих современных форм замко-
вых брахиопод.
У ряда палеозойских форм в добавление к прикреплению ножкой
наблюдалось прикрепление к субстрату макушечной частью брюшной
створки — иглами, поверхностью ареи.
Многие палеозойские брахиоподы на раннем этапе жизни прикреп-
лялись ножкой, а затем происходила утрата ее, причем некоторые фор-
мы прирастали к субстрату всей поверхностью брюшной створки.
Наконец, в ходе геологической истории брахиопод наблюдается
также переход от прикрепленного образа жизни к свободному лежа-
нию на дне моря. Это достигалось или развитием длинных игл, или пла-
стинчатых выростов — шлейфов на поверхности раковины, на которые
брахиоподы опирались, или развитием такой формы толстостенной рако-
вины, при которой центр тяжести ее приходился на макушку брюшной
створки. В таких случаях раковина устойчиво лежала на дне моря.
Судя по данным ряда авторов, особенно Е. А. Ивановой, анализ па-
леоэкологии брахиопод позволяет рассматривать разнообразные изме-
нения в строении, форме, скульптуре раковин, в развитии, редукции
ножки, как приспособления, вызванные особенностями условий и образа
жизни брахиопод.
В анатомии современных замковых брахиопод некоторые особен-
ности, сравнительно со строением беззамковых, являются вторичными
регрессивными чертами, это отсутствие органов чувств — статоцистов,
задней кишки и ануса, утрата целомической полости в эпистоме и нож-
ке, утрата последней собственной мускулатуры. В их геологической
истории эволюция мало затрагивала основы анатомического строения.
Прогрессивной чертой замковых брахиопод явилось приспособление их
обмена веществ в сторону все большего использования для построения
раковины солей углекислого кальция моря и все меньшей траты орга-
нического вещества — протеина (Виноградов, 1953). Вместо тонкостен-
ных, богатых органическим веществом раковин беззамковых брахио-
под у них стали формироваться сравнительно толстостенные известковые
раковины с замком, или шарниром, что позволило прочно сочленяться
спинной и брюшной створкам друг с другом. В связи с этим произош-
ло усовершенствование мускулатуры, обеспечивающей четкие движе-
ния створок, раскрывание и плотное их закрывание. У ряда палеозой-
ских замковых брахиопод, помимо этого, развивался сложный внут-
ренний скелет, имевший, по-видимому, различную функцию (Сары-
чева, 1949).
Все это вместе взятое и вертикальное положение раковины, при-
крепленной ножкой, благоприятствовало газовому обмену, улавлива-
нию пищи, повышению жизнедеятельности замковых брахиопод
263
сравнительно с беззамковыми, и способствовало процветанию многих
групп палеозойских замковых брахиопод.
Утрата ножки, ее рудиментация, цементация брюшной створки к
субстрату у палеозойских замковых брахиопод были вторичными явле-
ниями, ограничивавшими жизнеспособность ряда вымерших палеозой-
ских групп этого класса.
Судя по эволюции ручного аппарата, отсутствовавшего среди зам-
ковых лишь у строфоменид и продуктид, у палеозойских брахиопод шло
усложнение и усиление функции лофофора. Значительная эволюция
наблюдалась и в развитии мантийных синусов, т. е. происходило повы-
шение дыхательной функции мантии и нарастание половой продуктив-
ности брахиопод. Со всем этим тесно было связано увеличение размеров
тела брахиопод, которые, как известно, в прошлые геологические перио-
ды достигали размеров, во много раз превосходивших величину совре-
менных, так каменноугольный Gigantoproductus достигал 40 см.
Таким образом, в эволюции брахиопод значительные изменения пре-
терпевал и лофофор, роль которого с увеличением размеров тела и с
массовым плотным заселением небольших пространств дна -моря сильно
возрастала как аппарата дыхания и ловли пищи.
Исходя из однообразия общей организации современных брахиопод
(ринхонеллид и теребратулид), прослеживая их эволюцию и соответ-
ственно филогенетические отношения в этих группах (Ржонсницкая,
1959; Stehli, 1956), начиная с ордовика и силура, можно думать, что
внутреннее, анатомическое строение замковых брахиопод в течение их
геологической истории эволюировало незначительно. Это, видимо, было
также одной из причин вымирания брахиопод в мезозое в борьбе за
существование с другими животными бентоса — подвижными хищными
брюхоногими моллюсками и двустворчатыми, хотя и малоподвижными,
но обладавшими энергично работающим фильтрационным аппаратом.
Совершенствование фильтрующего аппарата брахиопод намного отста-
вало от того, чего достигли двустворчатые моллюски, ставшие мощны-
ми биофильтраторами моря.
Геологическая история брахиопод очень продолжительна и известна
с низов кембрия. Судя по наличию в низах кембрия представителей
беззамковых и замковых брахиопод, ее надо вести, видимо, с докемб-
рия. Однако принадлежность к брахиоподам ископаемых организмов
из протерозоя, описанных как брахиопода, в настоящее время на
основании. всестороннего анализа отрицается (Seilacher, 1956; Wil-
son, 1957). Отложения, в которых найдены Lingulelta и Obolella, отно-
сятся теперь не к протерозою, а к нижнему кембрию (Termier et Тег-
mier, 1960). Ископаемых видов известно около 30 000, современных —
около 260, из более чем 1377 родов в настоящее время сохранилось
68—70, из 35—40 надсемейств — всего 7. Уже эти цифры говорят о том,
что эволюция брахиопод протекала в прошлые геологические эпохи и,
как подчеркивают палеонтологи, в обоих классах она проходила по-раз-
ному. Крайне медленный темп эволюции и бедность количества видов и,
большею частью, особей характерны, как было отмечено раньше
(стр. 250), для беззамковых, обычно небольших или мелких форм.
На долю их приходится ничтожная часть (всего 92 рода) описанных
родов и видов брахиопод. В противоположность этому эволюция пре-
обладающего числа надсемейств замковых брахиопод протекала бы-
стрыми темпами и сопровождалась появлением огромного количества
разнообразных форм. Собственно геологическая история брахиопод за-
полнена летописью замковых брахиопод.
В нижнем кембрии замковые брахиоподы, как говорилось раньше
(стр. 261), были представлены значительно 'безраэд! беззамковых, причем
Среди последних, как предполагают (Termier et Termier, 1960), ,встре-
264
---4 ЪЕ^ЙСС
чались формы вроде Obolus, которые могли быть родоначальниками
замковых брахиопод. . Расцвет большинства надсемейств замковых
брахиопод начинается в ордовике, нарастает в силуре и достигает мак-
симума в девоне, несколько уменьшаясь в карбоне. В это время наблю-
дается громадное разнообразие форм, обилие семейств, родов и видов.
Замковые брахиоподы составляли главную массу прикрепленных форм
бентоса палеозойских морей, подобно ряду классов стебельчатых игло-
кожих. Из более двух десятков надсемейств этого времени большин-
ство вымирает в конце палеозоя, несколько надсемейств переживает
палеозой и заходит в триас и юру, где и вымирает.
Физико-химические условия морей палеозоя с господствовавшим
тогда (ордовик — карбон, Stirton, 1959) мягким, теплым климатом бла-
гоприятствовали массовому развитию брахиопод того времени, остав-
шихся не передвигающимися донными животными даже при переходе
(с утратой ножки) к свободному лежанию на дне моря. Похолодание,
наступившее в конце палеозоя, и тектонические процессы того времени,
очевидно, вызвали вымирание большинства отрядов и надсемейств
замковых брахиопод.
В мезозое произошло, как выражаются палеонтологи (Борисяк,
1905), обновление фауны брахиопод. Слабо представленные в морях
палеозоя надсемейства Rhynchonellacea (с середины ордовика) иТегеЬга-
tulacea (с середины силура) и надсемейства Terebratellacea и Thecidea-
сеа (известные с триаса) эволюировали и составили затем фауну бра-
хиопод мезозоя, третичного и настоящего времени. Для нас, однако,
пока неизвестно, какие особенности, возникшие у них, позволили им
пережить изменения условий среды конца палеозоя — начала мезозоя.
Семейства ринхонеллид, теребрателлид и тецидеид вместе с беззамко-
выми лингулидами, дисцинидами и краниидами составляют современ-
ную реликтовую фауну брахиопод. В бентосе Мирового океана с мезо-
зоя место остальных групп брахиопод, занимают моллюски, процветаю-
щие в настоящее время.
Приходится отметить,, что несмотря на громадное число работ по па-
леонтологии в XX столетии и успехи палеоэкологии, несмотря на массу
описанных хорошей сохранности ископаемых видов, большие успехи
в изучении ручного аппарата, возрастных изменений раковины и этого
аппарата, все же знание о филогенетических связях между большими
группами брахиопод остаются почти на уровне начала нашего столетия.
А. А. Борисяк полвека назад (1905) отмечал, что большие группы бра-
хиопод остаются филогенетически не связанными, друг с другом, что
справедливо и для настоящего времени. Состояние классификации бра-
хиопод подтверждает это положение. Достаточно сравнить классифика-
цию брахиопод, принятую в «Основах палеонтологии» (Орлов, 1960),
Бошампа (Beauchamp, 1960) и Роже (Roger, 1960). В «Основах палеон-
тологии!» б’еззамковые брахиоподы поделены на 6 отрядов с 11 надсе-
мействами, а замковые на 8 отрядов с 29 надсемействами. Роже (I960),'
следуя Вилльямсу (Williams, 1956), делит беззамковых брахиопод на
два отряда (как раньше делало большинство авторов)—Atremata и-
Neotremata с 10 надсемействами, а замковых — на 6 групп: Orthis, Реп-
tamerus, Strophomena, Rhynchonella, Spirtfer, Terebratula, сохраняя
надсемейства и опуская отряды и подотряды.
Таким образом, естественной системы брахиопод мы не имеем и в
настоящее время, как не имеем и упорядоченной классификации бра-
хиопод. Это вызвано широко распространенным среди них явлением
параллельного и конвергентного (или гомеоморфии) развития, что
подчеркивают Б. R. Лихарев (1960) и многие другие авторы, в том чис-
ле и Клоуд (Cloud, 1941, 1948), приведшие поразительные примеры
уподобления родов друг другу, например, Tetractinetta (из среднего
265
триаса), и Cheirothepris (из верхней юры), причем первый род относится
к отряду Spiriferida, а второй — к Terebratulida.
Этим объясняется и отношение к классификации брахиопод зооло-
гов, которые не рискуют строить систему, охватывающую всех брахио-
под современных и ископаемых, тем более, что современные формы
крайне малочисленны .и составляют ничтожную часть (стр. 264) общего
числа описанных видов. Так, Гаймен (Hyman, 1959) для беззамковых
брахиопод сохраняет деление на два отряда (Atremata и Neotremata),
а для замковых приводит лишь надсемейства Thecideacea, Rhynchonel-
lacea, Terebratulacea и Terebratellacea, опуская все ископаемые формы.
Лишь изредка делаются попытки дать картину филогении всех брахио-
под (Kuhn, 1939; Moore, 1952; Beurlen, 1952). Бейрлен, анализируя
отряды замковых брахиопод, перестраивает их систему и высказывает
соображения об их филогении, однако и эта попытка не удовлетворила
палеонтологов. Имеются попытки разобрать филогенетические связи для
некоторых отрядов, например, для строфоменид (Сокольская, 1954);
спириферид (Havlicek, 1959; Иванова, 1959), для рпнхонеллид (Ржонс-
ницкой, 1959), теребратулид (Stehli 1956), продуктйд (Muir-Wood а.
Cooper 1960), однако родоначальные группы этих отрядов указываются
предположительно или не приводятся вовсе.
Таким образом, у нас нет достаточных материалов о филогенетиче-
ских связях обоих отрядов ныне существующих замковых брахиопод,
если не считать их геологической истории и данных об эволюции ряда
черт их организации в ходе исторического развития. Однако не выясне-
ны окончательно положение в системе и родственные связи с отрядом
теребратулид современного семейства тецидеид, отличающегося рядом
признаков строения и некоторыми особенностями онтогенеза (схизолоф-
ный лофофор, прикрепление к субстрату не ножкой, а брюшной створ-
кой, схизоцельное образование целома, развитие раковины и т. д.).
Как известно, одни авторы (Roger, 1960) относят тецидеид к вымерше-
му отряду строфоменид, другие (Лихарев, 1960), как и зоологи,—
к отряду теребратулид. Первое предположение, видимо, вернее, однако
нужны дополнительные исследования.
Хотя современных беззамковых брахиопод вполне можно считать
реликтовыми формами, причем число всех известных ископаемых
форм их вообще очень мало (всего 92 рода против 1377 ископаемых
родов замковых брахиопод), тем не менее родственные отношения их
также недостаточно выяснены. По ряду признаков, хотя бы по наличию
ножки с собственной мускулатурой и целомом и протеино-фосфатного
состава раковины, лингулиды и дисциниды ближе друг к другу, чем к
краниидам, у которых раковина известковая, что является позднейший/
приобретением. Наряду с этим было отмечено, что по многим чертам
организации кранииды, если они имеют и некоторые регрессивные чер
ты (утрата ножки), являются наиболее примитивными формами среди
беззамковых и замковых брахиопод. Среди древнейших нижнекембрий-
ских беззамковых брахиопод (см. стр. 261) имеются формы с разным
составом раковины и разной ее формой. Они, несомненно, дали начало
всем трем существующим семействам беззамковых брахиопод. Среди
них имеются формы, вроде Obolus, которые могли дать начало замко-
вым брахиоподам. Знание впервые появляющихся представителей раз-
личных отрядов ископаемых замковых брахиопод дает возможность
судить об эволюции и филогении этих отрядов, но построение филоге-
нии брахиопод это еще дело будущего.
Кишечнодышащие (Enteropneusta)
Мягкое двустороннесимметричное, длинное червеобразное тело ки-
шечнодышащих покрыто ресничками, лишено наружных придатков,
расчленено на три отдела и состоит из короткого или удлиненного же-
лудеобразного хобота — протосомы, короткого воротника — мезосомы и
длинного туловища — метасомы с целомическими полостями. Вентраль-
ное ротовое отверстие лежит у основания хобота, анальное отверстие —
на конце туловища. Хобот обычно имеет левую, воротник-—правую и
левую целомопоры. В туловище в соответствии с продольными нервны-
ми тяжами и кровеносными сосудами выражены среднее спинное и
брюшное продольные утолщения (ребра); в нем заметны и часто разли-
чимы по окраске жаберный, половой, печеночный и хвостовой отделы.
Окраска тела различна, она может быть серовато-белой, бледно-желтой,
оранжевой, кирпично-красной, темно-коричневой, желтовато-фиолетовой
и черно-фиолетовой.
Размеры тела обычно колеблются от 10 до 50 см, мелкие формы
(например, Sacco gloss us pygmaeus) могут быть 2—3 см длиной, круп-
ные (Balanoglossus gigas} — 1,8—2,5 м длиной.
Кишечнодышащие малоактивные обитатели моря, преимущественно
полосы приливов и отливов, обычно живущие в ходах V-образной
формы в глинистом или песчаном грунте, в которых они двигаются впе-
ред и назад и выкидывают на поверхность грунта кучки фекалий. Ходы
они выстилают слизью желез кожи. Реже они живут под камнями или
среди корневищ водорослей (Kirk, 1938). Кишечнодышащие обычно не
спускаются глубже 350 м, но один вид—Glandiceps abyssicota известен
с глубины в 4500 м. Некоторые виды люминесцируют, некоторые издают
резкий запах йодоформа.
Кишечнодышащие являются небольшой, реликтовой группой очень
примитивных вторичноротых червеобразных животных, состоящей из
четырех семейств — Protoglossidae, Harrimaniidae, Spengelidae и Pty-
choderidae и не более чем из 100 видов. Следы ископаемых пока не обна-
ружены. Анатомия баланоглосса впервые была описана А. О. Ковалев-
ским (Kowalevsky, 1866).
Поверхность всего тела покрыта ресничками, стенки состоят из
ресничного эпидермиса с большим количеством одноклеточных желез
(ретикулярных, бокаловидных, гранулярных) и чувствительных нервных
клеток. Эпидермис туловища выделяет слизь. Слизистых клеток очень
много на хоботе и воротнике. Благодаря работе ресничек слизь движет-
ся по телу и предохраняет его от повреждений при образовании ходов
в грунте. У некоторых форм в эпидермисе имеются железистые клетки;
с деятельностью которых связана люминесценция этих видов (Ptycho-
dera bahamensis, Hyman, 1959).
Между основаниями эпидермальных клеток и базальной мембра-
ной лежит нервное сплетение из волокон и нервных клеток. Под ба-
зальной мембраной находится мускулатура тела, кутиса, или соедини-
тельнотканного слоя, нет. За исключением ProtoglosSus, у многих
(большинства) кишечнодышащих выстилка целома в виде перитонеаль-
267
Рис. 69. Схема строения Balanoglossus (из Hennig)
I— протосома; II— мезосома; /// — метасома; / — стомохорда; 2 — рот;
3— кишечник; 4— жаберные щели; 5 — гонада; 6—печеночные придатки;
7 — анус; 8 и 9 — соответственно брюшной и спинной нервные тяжн; 10 —
протоцель; I/ —мезоцель; 12 — метацель; 13 — гломерулус; 14 — кардно-
пернкарднЙ
кого эпителия, существующая в онтогенезе, не сохраняется, она расхо-
дуется на образование мускулатуры.
У кишечнодышащих как в стенках тела, так и всюду на границах
двух соприкасающихся тканей, в том числе и целомической стенки, каж-
дой тканью выделяется базальная мембрана. За счет базальных мем-
268
бран образуется скелет хобота различной формы в разных семействах,
часто в виде прямоугольной продолговатой пластинки с двумя перед-
ними короткими ушками и двумя длинными задними рожками. Этот
скелет находится -между основанием хобота и выростом ротовой поло-
сти (так называемым стомохордом, или нотохордом). Скелет жаберных
щелей — такого же происхождения (рис. 68).
Нервная система состоит из диффузной интраэпидермальной нерв-
ной сети, одевающей все тело, интраэпителиального нервного сплетения
всего кишечника и брюшного мезентерия, из спинного и более мощного
брюшного нервных тяжей (рис. 69), соединяющихся передней кольцевой
комиссурой на границе хобота и воротника, и пребранхиальной комис-
сурой на границе воротника и туловища. Все эти части нервной сис-
темы лежат поверхностно, интраэпителиально, тонкие концы (основа-
ния) эпителиальных клеток (Hyman, 1959) проникают через нервное
сплетение, прикрепляясь к базальной мембране. Лишь в воротнике
спинной нервный тяж погружен под кожу. Эта часть его закладывается
в онтогенезе в виде желобка, погружающегося под эктодерму и прев-
ращающегося в нервную трубку, передним и задним невропорами
открывающуюся наружу. У большинства кишечнодышащих нервная
трубка становится плотным нервным тяжом, но у многих представите-
лей семейства Ptychoderidae и некоторых из семейства Spengelidae
сохраняется просвет трубки, а у примитивных форм — и оба невропора.
В нервной системе воротника, или нейрохорда по термину Найт-Джон-
са, лежат гигантские нервные клетки, дающие направо и налево гигант-
ские нервные волокна (Bullock, 1944). Нервное сплетение состоит из
нервных волокон, чувствительных и двигательных нейронов и синапсов.
Анатомически и гистологически и спинной, и брюшной нервные тяжи яв-
ляются лишь сгущениями диффузной нервной сети, одевающей все
тело, без дифференцированных органов чувств. Замечу, что основы
строения нервной системы кишечнодышащих были выяснены еще
В. М. Шимкевичем (1889; Schimkewitsch, 1888).
Современными представлениями о строении и физиологии нервной
системы кишечнодышащих мы обязаны новым исследованиям ряда
авторов (Bullock, 1944, 1945; Knight-Jones, 1953; Mookerjee a. all., 1955;
Silen, 1950) и интерпретациям X. С. Коштоянца (1957).
Хобот с его многочисленными чувствительными клетками является
наиболее чувствительным и активным отделом тела кишечнодышащих.
Он играет важную роль в установлении связи между организмом и
окружающей средой. Хобот служит вместе с тем главным органом для
рытья грунта, в котором преимущественно живут кишечнодышащие.
Сокращения при этом начинаются с его кончика. Остановка движений
туловища при удалении хобота указывает на его роль как отдела тела,
интегрирующего движение животного в целом (Коштоянц, 1957). Рез-
кие двигательные акты (Bullock, 1944, 1945) связаны с деятельностью
гигантских нервных клеток и волокон нейрохорда.
В общем, нервная система кишечнодышащих является диффузной,
поверхностной, аганглионарной, не имеющей дифференцированных
центра и нервов. Гистологически и физиологически их нервная система
стоит ниже таковой чуть ли не всех других групп животных, кроме
гребневиков и кишечнополостных, причем нет доказательства, что эта
примитивность вторичная, а не первичная (Bullock, 1945). Все это, оче-
видно, причинно связано с условиями жизни кишечнодышащих, искони
ведущих пассивный образ жизни в толще грунта.
Как отмечалось, оформленных органов чувств нет. Среди эпидер-
мальных разбросаны отдельные, связанные с нервным сплетением пер-
вичные чувствительные клетки, особенно многочисленные в хоботе.
Некоторые из этих клеток являются фоторецепторами с высокой
269
светочувствительностью. Значение недавно описанного преорального
органа чувств (Brambell a. Cole, 1939) новыми данными (Bullock, 1944,
1945; Silen, 1950) отрицается. Образующие его клетки не нервные, а же-
лезистые.
По мнению В. М. Шимкевича (1889, 1892, 1907; Schimkewitsch, 1888,
1892), кишечнодышащие имели раньше щупальца на хоботе, о чем го-
ворит наличие щупалец у некоторых торнарий (Weldon, 1887). На-
сколько это верно — сказать трудно, но своеобразие строения хобота
современных кишечнодышащих, несомненно, результат роющего обра-
за жизни.
Мускулатура тела в общем сохраняет примитивные черты кожно-
мускульного мешка длинных червеобразных животных — это кольцевые
наружные и продольные внутренние гладкие мышечные волокна, возни-
кающие лишь за счет стенок целома. Вместе с тем она обнаруживает
далеко идущие приспособления к роющему образу жизни в грунте.
Как аппарат движения, по В. Н. Беклемишеву (1952, 1964), она нахо-
дится в точном соответствии с образом жизни. Рытье в грунте произво-
дится главным образом хоботом. Вытягиваясь и сохраняя упругость
благодаря заполнению его целома жидкостью, хобот проникает в грунт;
укорачиваясь, он расширяет ход, в который проникает удлинившийся и
сузившийся воротник, куда затем втягивается и туловище. Общая му-
скулатура тела используется при перистальтических плавающих движе-
ниях, когда животные в известные периоды жизни покидают грунт и
выходят на его поверхность (Ikeda, 1908, см. Hyman, 1959).
В связи с таким образом жизни в хоботе кишечнодышащих имеется
тонкий слой кольцевых мышц, масса продольных мышц, почти запол-
няющих целом хобота и дорсо-вентральные мышечные волокна. Мус-
кулатура воротника у разных представителей довольно разнообразна,
в основном она состоит из слоя продольных мышц, располагающихся
между стенкой тела и глоткой, и двух Систем волокон, расходящихся
веерообразно от передней стенки воротника к шейке хобота. Мускула-
тура туловища более однообразна и представлена слабым наружным
слоем кольцевых и сильным внутренним слоем продольных мышц; по-
следние более мощные на брюшной стороне, поделены на два латеро-
вентральных и два латеро-дорсальных тяжа. Кроме того, имеются ра-
диальные мышцы, идущие от стенок кишечника к стенкам тела,
предохраняющие кишечник от сдавливания.
Кишечник в соответствии с длинным червеобразным телом вытянут
в длину, с ротовым отверстием на брюшной стороне, на границе между
хоботом и воротником, и с терминальным анусом. Кишечник обладает
двумя очень характерными особенностями. Он имеет жаберные щели и
соответственно служит для дыхания, в связи с чем вся группа и названа
кишечнодышащими или Enteropneusta. Другой особенностью, менее важ-
ной, хотя сильно переоцененной некоторыми исследователями, но также
очень характерной, является образование слепого преорального выроста,
вдающегося в хобот. Предполагалось, что это нотохорда, которая являет-
ся гомологом хорды хордовых (Bateson, 1884, 1886); отсюда и название
для Enteropneusta и Pterobranchia — Hemichordata. По Уиллею (Willey,
1899), это образование вряд ли гомологично нотохорде позвоночных, и
называется стомохордой. Позже К- Н. Давыдову (Dawydoff, 1948) этот
термин послужил для объединения кишечнодышащих и птеробранхий
в Stomochorda. По мнению Шпенгеля (Spengel, 1893, 1913) и ряда
других зоологов, это не гомолог хорды, а лишь вырост глотки или кишки
хобота.
В последние годы ряд авторов (Silen, 1950, 1954; Newell, 1951), как
отмечает Гаймен, отрицает значение выроста ротовой полости как стомо-
хорды или нотохорды и дает ему другое толкование. Силен, не зная дав-
нишнего высказывания В. М. Шимкевича (Шимкевич, 1889; Schimke-
witsch, 1888, 1892), считает стомохорду лишь преоральным продолжени-
ем кишечника. Шимкевич-,' присоединившийся позже (1907) к взгляду на
гомологию этого органа с хордой, раньше допускал, что предковые фор-
мы кишечнодышащих имели рот на верхнем конце хобота, затем сме-
стившийся к его основанию, й что нотохорда — лишь преоральная часть,
или вырост кишечника. Теперь я также, в отличие от общепринятого и
своего старого взгляда (Федотов, 1923), считаю такую трактовку пра-
вильной, тем более, что у ряда кишечнодышащих, например в семействе
Spengelidae (Догель, 1938), так называемая стомохорда сохраняет строе-
ние отдела кишечника. Гаймен (Hyman, 1959) называет нотохорду бук-
вальным дивертикулом.
За ротовой полостью следует эктодермальная часть пищеварительно-
го тракта, начинающегося глоткой, пронизанной жаберными щелями и
обычно дифференцированной в связи с этим на спинной — дыхательный
отдел и брюшной — пищепроводящий с эпибранхиальным и гипобранхи-
альным желобками, которые Шимкевич описал еще в 1888 г. За глоткой
следует пищевод (дыхательный отдел кишечника), спинная и брюшная
его стенки сплошные, а боковые пронизаны двумя рядами жаберных ще-
лей. В пищеводе семейств Harrimaniidae и Spengelidae имеются кишечно-
кожные каналы, морфологическое значение и функция которых не выяс-
нены; по мнению В. М. Шимкевича (Шимкевич, 1889; Schimkewitsch,
1888) они являются рудиментами жаберных щелей. Пищевод переходит
в печеночную область собственно кишки с многочисленными боковыми
печеночными мешочками с большим количеством железистых клеток.
У рода Glandiceps в этом отделе кишечника на его спинной стороне
находится сифон, сходный с аналогичными образованиями у ряда бес-
позвоночных — капителлид, эхиурид, гефирей, некоторых морских ежей.
За печеночным следует нерезко отграниченный абдоминальный отдел
кишки, на брюшной стороне которого у Glos sob alarms находится задний
сифон, или пигихорд. Пищеварительный тракт кончается прямой кишкой.
Анус снабжен сфинктором. Весь кишечник выстлан ресничками и почти
лишен мускулатуры, если не считать жаберного отдела и пищевода,
имеющих кольцевые мышцы. Пища в виде детрита и мелких организмов
поступает вместе с илом и песком. Для кишечнодышащих характерно
пассивное питание фильтраторов (Knight-Jones, 1953; Ливанов, 1955).
Реснички хобота гонят слизь вместе с частицами пищи ко рту, откуда,
благодаря обилию слизистых клеток и ресничек в передней части пище-
варительного тракта, они направляются дальше и превращаются в пище-
вой тяж во втором отделе пищевода. Стенки последнего очень богаты
железами и, по допущению . Найт-Джонса, выделяют пищеварительные
энзимы. В следующем отделе кишечника происходит всасывание пищи.
К сожалению, процессы пищеварения у кишечнодышащих изучены недо-
статочно;
Жаберный аппарат состоит из метамерно расположенных жаберных
щелей1, имеющих форму подковы или буквы V, обращенной ветвями к
спинной стороне животного. Обычно каждая ветвь подковы подразде
ляется синаптикулами. Жаберные щели на боковых стенках тела наруж
ными жаберными отверстиями сообщаются с наружной средой. У неко-
торых форм, например Stereobalanus canadensis (см. Hyman, 1959), все
жаберные мешки каждой стороны сливаются и открываются в общую
продольную щель. Стенки жаберных щелей поддерживаются скелетом,
образованным утолщениями базальной мембраны. В этом отделе кишеч-
ника, имеющего в поперечном разрезе форму восьмерки, верхний этаж
служит для прохождения воды в жаберные щели, нижний — для прохож-
дения пищи. Число жаберных щелей увеличивается с возрастом. Оно
колеблется от 10 до 250 и более пар. Жаберные щели (Bateson, 1884,
271
1886; Шимкевич, 1889; Schimkewitsch, 1888) в виде отверстий возникают
на месте соприкосновения выпячивания стенки кишечника с эктодермой.
Действием ресничек вызывается ток воды, входящий в рот и выходящий
через жабры, обильно снабженные кровеносными капиллярами, что обес-
печивает газовый обмен в жаберном аппарате.
Сходство в строении и отправлении жаберного аппарата кишечноды-
шащих и низших хордовых (ланцетник) было отмечено уже первыми
исследователями этой группы (Bateson, 1884; Шимкевич, 1889; Schim-
kewitsch, 1888), причем В. М. Шимкевич находил, что строение жаберных
щелей кишечнодышащих напоминает таковое молодого ланцетника.
Целомическая ресничная полость тела энтероцельного происхожде-
ния в соответствии с тремя отделами (хоботом, воротником и туловищем)
состоит из непарного протоцеля и парных мезоцеля и метацеля, дающих
начало мускулатуре и соединительной ткани.
Полость целома хобота занимает центральное положение и сильно
уменьшена развитием его мускулатуры и соединительной ткани. Она
открывается наружу порой на левой стороне. Целом хобота, возникаю-
щий обычно как непарная полость от вершины архентерона, обнаружи-
вает билатеральное строение; у ряда форм он бывает поделен спинным
и брюшным мезентериями. Вентро-латеральные выросты его у Ptycho-
dera и некоторых видов Glossobalanus симметрично вдаются в стебель
хобота. Дорсо-латеральные выросты хоботного целома у ряда форм
(Harrimania, Stereobalanus canadensis, некоторые Ptychoderidae) откры-
ваются порами (Dawydoff, 1948). У некоторых форм, как исключение у
отдельных представителей, встречаются парные поры целома хобота
(Шимкевич, 1907) или они возникают как атавизм при регенерации (Да-
выдов, 1908). Производным целома хобота является перикардий, о кото-
ром будет сказано несколько дальше.
Спинной и брюшной мезентерии парного целома воротника у прими-
тивного рода Protoglossus цельные. У остальных кишечнодышащих они
неполны и полости целома воротника сливаются одна с другой и в зна-
чительной мере заполнены мышцами и соединительной тканью. С внеш-
ней средой сообщаются двумя порами.
Поры целомов хобота и воротника являются наружными отверстиями
каналов, образованных ресничными эпидермальными клетками.
В. М. Шимкевичем (Шимкевич, 1889; Schimkewitsch, 1888) была уста-
новлена их природа как сегментальных органов, которые позже стали
называть целомодуктами, а их наружные отверстия — целомопорами.
Однако В. Н. Беклемишев (1952, 1964) отмечает, что в отличие от типич-
ных целомодуктов у них нет мерцательной воронки и они не выводят
ни половых продуктов, ни экскрета. В. М. Шимкевич (Шимкевич, 1889;
Schimkewitsch, 1888) указал, что поры воротника открываются в первую
жаберную щель. Через пору хобота выходит избыток жидкости, а через
поры воротника происходит наполнение его целомов водой и регулиро-
вание упругости и объема хобота и воротника, необходимое при рытье
грунта.
Спинной и брюшной мезентерии парного целома туловища поддер-
живают кишечник, причем спинной мезентерий часто бывает прерван,
благодаря чему целомические полости туловища сообщаются. Целомы
туловища’ вдаются в воротник в виде парных выростов, найденных и
названных Бэтсоном (Bateson, 1884) перифарингеальными и периге-
мальными целомами; у примитивного Protoglossus они зачаточные (Hy-
man, 1959). Обычно их просвет почти заполнен развивающимися за их
''чет продольными пучками мышц.
Целотелий ресничный и выражен только в туловищном целоме, содер-
жащем амебоидные целомиты, в других отделах он почти нацело расхо-
дуется на развитие мускулатуры и соединительной ткани.
272
Кровеносная система своеобразна (рис. 70).
В связи с червеобразной формой тела животного
она состоит из двух главных продольных сокра-
тимых сосудов — спинного в спинном и брюшного
в брюшном мезентерии. В спинном сосуде кровь
движется сзади наперед, а в брюшном —спереди
назад, как у аннелид. Имеется пропульзаторный
орган — кардио-перикардий, или «головное серд-
це», находящийся в основной (базальной) части
хобота. Движение крови вызывается ритмически-
ми сокращениями мускулатуры перикардия.
Окисление крови происходит благодаря обогаще-
нию кислородом в жаберных щелях — органах
газового обмена. Кровеносные сосуды образова-
ны тканью основного вещества, выделяемого ба-
зальными мембранами мезентериев и других ча-
стей тела. По свбему происхождению онй, подоб-
но кровеносным сосудам беспозвоночных, не дол-
жны иметь эндотелия, хотя некоторые авторы
(Spengel, ом. Hyman, 1959; Давыдов, 1908), ви-
димо, ошибочно описывают эндотелий на мышеч-
ных стенках в спинном и брюшном сосудах.
Кровь, двигающаяся из спинного сосуда впе-
ред, заполняет центральную лакуну, или сердце,
находящееся в хоботе между преоральным вы-
ростом кишечника и перикардием; из хобота в
сердце кровь поступает по двум сосудам.. Пери-
кардий является мешочком, возникающим за
счет целома хобота и состоящим из целотелия и
мышц. На его нижней стенке, как показал
В. М. Шимкевич (Шимкевич, 1889; Schimkewitsch,
1888), имеется желобок, охватывающий цент-
ральную лакуну, или сердце. Ее разветвления за-
ходят в гломерулус, образованный складками
перитонеальной стенки и возникающий у личин-
ки. В нем, по-видимому, происходит очищение
крови от продуктов распада, что предполагали
Бэтсон, Келер и Шимкевич (Hyman, 1959) по на-
личию там желтых и коричневых гранул. Здесь
находятся нефрофагоцитарные клетки. Разруша-
ющиеся амебоциты с заглоченными продуктами
распада выводятся через пору хобота. Следует
отметить, что Бури (Bury, 1895), позже Джемил
(Gemmill, 1914, 1916, 1920; см. гл. «Иглокожие»),
затем Д. М. Федотов (Fedotov, 1924) предполо-
Рис. 70. Схема строения
кровеносной системы Еп-
teropneusta (из Hyman)
1 — хобот; 2 — гломерулус;
3 — кардио-перикардий; 4 —
центральный синус; 5— спин-
ной сосуд; 6 — спинной
нервный тяж; 7 — брюшной
сосуд; 8 — брюшной нервный
тяж; 9 — ла куна рна я сеть
кишечника
жили гомологию перикардия кишечнодышаших и так называемого мад-
репорсвого пузырька личинок иглокожих. Федотов (1924) провел срав-
нение между гломерулусом и осевым органом иглокожих и установил,
что оба образования как производные стенок первого отдела целома име-
ют функцию кровеносной, экскреторной и лимфоидной систем и, может
быть, кроме того, органа внутренней секреции. К сожалению, для окон-
чательного решения этого вопроса
них (см: гл. «Иглокожие»).
Кровь из гломерулуса по двум
брюшной кровеносный сосуд, а из
в жабры; из жабр по выносящим
сосуд. Из брюшного сосуда кровь
не хватает экспериментальных дан-
околоротовым сосудам поступает в
последнего по приносящим сосудам
сосудам кровь поступает в спинной
возвращается в спинной через сеть
18 Д. М. Федотов
273
околокишечных сосудов. В стенках тела и кишечника находятся крове*
носные синусы. Полное описание кровеносной системы дано Горстом
(Horst, 1927—1939).
Кишечнодышащие раздельнополы. Полы могут различаться по ок-
раске. Половые железы представлены небольшими метамерно располо-
женными мешочками, развивающимися за счет целомического эпителия
(Schimkewitsch, 1888; Morgan, 1891, 1894), но лежащими ретроперито-
неально. Короткими каналами гонады открываются наружу. Половые
продукты выводятся в воду, где протекает развитие. Живородящим яв-
ляется лишь Xenopleura vivipara (Gilchrist, 1925); Balanoglossus proli-
fefans (Cap.) размножается делением поперек, причем происходит смена
двух поколений — полового и бесполого (см. Dawydoff, 1948).
Рис. 71. Стадии развития Balanoglossus
(из Heider)
А — гаструла; Б — отшнуровывание протоцеля;
В— образование целомопора протоцеля; Г — тор-
нария; 1 — бластопор; 2 — ротовое впячнванио,
превращающееся в рот; 3 — протоцель; 4 — це-
ломопора протоцеля; 5— мезоцель; 6 — метацель;
7 — ресничный шнур; 8 — теменная пластинка
с султаном ресниц
ч Дробление яиц полное, недетерминированное, слегка неравномерное,
радиального, реже билатерального типа, приводит к образованию жгути-
коносной целобластулы, которая инвагинацией на одиннадцатом или две-
надцатом делении превращается в гаструлу (рис. 71). Широкий бласто-
пор скоро замыкается, но архентерон сохраняет связь с эктодермой.
Энтомезодермы (мезенхимы) не образуется, мезодерма — целобластиче-
ская. Целомы обычно образуются энтероцельным путем, выпячиванием
стенок архентерона в виде непарного переднего (протоцеля) и двух пар-
ных— среднего и заднего целомов (мезо- и метацеля). У некоторых
форм отделившийся от архентерона непарный целом образует по выступу
по сторонам архентерона, которые перетягиваются на средние и задние
целомы. Чаще последние в виде карманов или плотных зачатков возни-
кают из стенок архентерона независимо от переднего целома. Иногда
274
Рис. 72. Торнария (по
Morgan из Heider)
1 — рот; 2 — анус; 3 —про-
тоцель; 4 — целомопора; 5 —
мезоцель; 6 — целомопора;
7 —метацель; « — первая
жаберная щель
Рис. 73. Слегка схематизированное прямое
развитие Saccoglossus horsti (из Kaestner)
А—Е — последовательные стадии развития, закап-
чивающиеся формированием молодого балано-
глосса; 1 — хобот; 2 — воротник; 3 — туловище
с первой жаберной щелью (4); 5 — постанальный
хвост
второй и третий целомы возникают схизоцельным путем из групп мезо-
дермальных клеток.
• Передний целом (протоцель)' мышечным тяжем связан с теменной
пластинкой. На его спинной стороне образуется вырост, который, соеди-
няясь с впячиванием эктодермы, открывается порой наружу. Таким обра-
зом, целомодукт состоит из мезодермального канала и дистальной экто-
дермальной части. За счет спинной стенки переднего целома образуется
перикардий, ритмически сокращающийся у личинки, подобно так назы-
ваемому мадрепоровому пузырьку личинок иглокожих.
Архентерон весь эктодермальный (эктодермальных стомодеума и
проктодеума — нет). Двумя перехватами он разделен на глотку (по-ста-
рому— пищевод), желудок и кишку. Дистальный конец архентерона
направляется к наружной стенке зародыша. Здесь образуются рот и ро-
товая полость эктодермального происхождения. На месте замкнувшегося
18*
275
бластопора, на конце, противоположном теменной пластинке, где архен-
терон сохраняет связь с энтодермой, образуется анальное отверстие. Гра-
ница между эктодермой и энтодермой неясно выражена, но она должна
находиться в самой глубине ротовой полости, где последняя переходит
в эктодермальную глотку.
Как и у ряда других групп, например иглокожих, форонид и т. д.,
у кишечнодышащих существует связь между количеством желтка в яйцах
и типом личиночного развития.
У ряда кишечнодышащих с яйцом; относительно бедным желтком,
образуются цланктотрофные пелагические личинки — типичные торна-
рии (рис. 72), активно питающиеся планктоном, и лецитотрофные' ли-
чинки, развивающиеся за счет желтка яйца. Обычно говорится о прямом
развитии и непрямом с метаморфозом, так как пелагическая торнария по
величине, форме, наличию сложного мерцательного шнура по краю ши-
рокой околоротовой впадины, наличию глазных пятен на теменной пла-
стинке значительно отличается от небольшой в общем овальной формы,
лишь с задним мерцательным венчиком, лецитотрофной личинки. Вскоре
по выходе из яйца такая личинка опускается на дно, плавает у дна с по-
мощью этого венчика, а затем начинает ползать по дну. Метаморфоз
заключается в том, что появляются две поперечные борозды, возникают
хобот, воротник и туловище, молодой червь начинает закапываться в
грунт (рис. 73). Если не считать некоторых частностей, органогенез и ста-
новление взрослой организации при обоих типах развития протекают
сходно, поэтому вместе с В. Н. Беклемишевым (1952, 1964) надо считать,,
что различия между ними второстепенны.
Торнарии, превращение которых в кишечнодышащих впервые было
установлено И. И. Мечниковым (Metschnikoff, 1869, 1881), обычно дли-
ной 2—3 мм, но иногда достигают и 9 мм. Видовая принадлежность боль-
шинства из 62 известных торнарий не установлена.
Торнарии нацело лишены нефридиев. Питаются наннопланктоном,
который ловят с помощью ресничек преоральной лопасти и ротовой по-
лости и находятся, по-видимому, постоянно в движении, вращаясь по
спирали. Главным двигательным аппаратом служит околоротовой венчик
ресничек. Торнарии растут в длину. При опускании на дно они теряют
ресничный шнур, двигаясь с помощью околоанального венчика.
Как и при развитии лецитотрофной личинки, у торнарии перетяжкой
сначала обособляется хобот, причем сначала длина воротника и туло-
вища почти одинаковы. Ползая по дну, молодой баланоглосс концом
хобота ощупывает дно, вытягивает хобот, внедряется им в грунт, под-
тягивая воротник, а затем и туловище.
По отрывочным данным эмбриологии конца XIX века (Balfour, 1880а,
18806), превращение длится немногие часы и состоит в утрате реснич-
ных шнуров и в некотором изменении внешней формы.
Действительно, цланктотрофные и лецитотрофные личинки вытяги-
ваются в длину, главная ось гаструлы становится длинной осью личинки
и взрослого баланоглосса. Внешняя форма и внутренняя организация
личинки двустороннесимметричные, эта симметрия нарушена лишь при-
сутствием у большинства левой поры переднего целома. Сагиттальная
плоскость личинки, делящая ее на правую и левую стороны, становится
таковой баланоглосса. Личинка является типичной диплеврулой. Архен-
терон удлиняется, спинным и брюшным мезентериями он подвешен к
стенкам тела. Сначала закладывается одна пара простых жаберных
щелей (стадия, открытая Bateson, 1884), затем появляется еще несколько
их пар. Вторые целомы образуют два выроста — канала, которые откры-
ваются в первую пару жаберных щелей (Шимкевич, 1889; Schimkewitsch,
1888). Они становятся парой целомодуктов взрослого животного. В них
различаются мезодермальный канал и. эктодермальная наружная конеч-
276
ная часть. За счет стенок целома развиваются мускулатура всех отделов
тела взрослого животного и соединительная ткань.
За счет целомических стенок, как впервые показал В. М. Шимкевич
(Шимкевич, 1889; Schimkewitsch, 1888), а затем и другие авторы, обра-
зуются зачатки гонад, становящиеся полыми мешочками.
У некоторых форм, например, Saccoglossus kowalevskii (Bateson,
1884) и Saccoglossus horsti (Burdon-Jones, 1952), на заднем конце обра-
зуется сильно сократимый, липкий постанальный хвост, которым моло-
дой червь закрепляется, начиная буравить грунт. Позднее хвост отбра-
сывается.
По мере роста тела в длину увеличивается число жаберных щелей
и гонад, продолжается дифференцировка глотки, пищевода, желудка
и кишки. Среди ресничных клеток эпидермиса развиваются несколько
типов железистых клеток, особенно много слизистых. Интраэпителиально
развивается нервное сплетение, одевающее все тело баланоглосса, а так-
же нервная сеть в стенке кишечника и брюшном мезентерии, которые
возникают, вероятно, еще у личинок.
Большая регенеративная способность кишечнодышащих известна по
отдельным наблюдениям различных авторов и по экспериментальным
исследованиям главным образом К. Н. Давыдова (1908), а также Рао
(Rao, 1953). Регенерация из частей, отрезанных на разных уровнях жи-
вотного, приводит в зависимости от уровня разреза или к восстановлению
целого организма или лишь к заживлению раны. Как правило, при реге-
нерации органы и ткани формируются как и при онтогенезе из элемен-
тов зародышевого пласта. Так перикардий возникает отшнурованием
части дорсальной стенки целома хобота, гломерулус — путем образова-
ния складок на стенках целома хобота, прикрывающих перикардий и
сердце, полость последнего является продолжением спинного кровенос-
ного сосуда. Целомодукт (нефридий — по Давыдову, 1908) воротника
возникает из двух зачатков — целомического выроста и впятившегося
участка эктодермы и т. д. Вместе с тем широко распространен и морфо-
лаксис (передифференцировка тканей), который комбинируется с ти-
пичной регенерацией.
Установлены также случаи атавистической регенераций (Давыдов,
1908), когда при регенерации хобота возникает пара целомов и пара
целомодуктов, два перикардия, когда регенерирующий предротовой
вырост кишечника (стомохорда или нотохорда) открывался наружу на
дистальном конце хобота, что указывает на то, что раньше рот нахо-
дился на верхнем конце хобота.
Географическое распространение кишечнодышащих (Hyman, 1959)
слабо изучено. Известно, что они встречаются в тропических, умеренных
и холодных водах. Большинство видов (60 из 100) живет в основном
в теплых водах верхней зоны литорали. В наших водах найдено пока
всего три вида: Saccoglossus (Dolichoglossus) mereschkowskii (N. Wag-
ner) — в Белом и Баренцевом морях, Harrlmania maculosa Rither и Ва-
lanoglossus proterogonius Belichov — в Японском море (Horst, 1934).
Из них S. mereschkowskii, по-видимому, циркумполярная форма.
Данных о паразитах кишечнодышащих мало. В разных органах от-
мечены паразитические простейшие, флагелляты, грегарины, кокцидии,
Haplosporidia, метацеркарии трематод, паразитические веслоногие раки
(Hyman, 1959),
Вся группа кишечнодышащих, если следовать Бурдон-Джонсу (Bur-
don-Jones, 1956), включившему небольшое семейство Protoglossidae
Dawydoff в семейство Harrimaniidae, состоит из трех семейств, а именно
Harrimaniidae Ritter, Spengelidae Willey и Ptychoderidae Spengel. По
простоте строения и дифференцировки отделов кишечника, жаберного
аппарата, скелета хобота, отделов целома, наличию у некоторых пред-
277
ставителей двух пор хобота и т. д. семейство Harrimaniidae наиболее
примитивно. Его организация ближе всего к обычно приводимой схеме
строения кишечнодышащих. Яйца богаты желтком, с чем связано раз-
витие'лецитотрофных личинок с коротким периодом планктонного и при-
донного существования. Это преимущественно мелкие виды, например,
Saccoglossus pygthaeus — 2—3 см. Семейство Phychoderidae является
по организации наиболее высоко дифференцированным и обширным;
яйца бедны желтком, личинки планктотрофные — торнарии. Размеры
очень различны — от 3—5 см (Glossobalanus crozieri) до 2,5 м (Bala-
noglossus gigas).
Примитивными чертами кишечножаберных являются:
1) червеобразная форма тела, главная ось и сагиттальная плоскость
которого совпадает с таковыми гаструлы;
2) превращение личинки во взрослого баланоглосса путем роста
в длину, перехода преоральной лопасти в хобот, а протоцеля — в целом
хобота, рост из посторального отдела в длину воротника и главным
образом туловища и переход мезоцеля в целом воротника, метацеля —
в целом туловища и т. д.;
3) ресничный покров тела и отсутствие под ним кутиса, или дермиса;
4) отсутствие наружного скелета и внутреннего скелета, соединитель-
нотканого происхождения;
5) диффузный, аганглионарный характер нервной системы в виде
интраэпителиальной нервной сети, одевающей все тело, сгущения кото-
рой образуют спинной и брюшной тяжи, и отсутствие дифференциро-
ванных органов чувств;
6) кожно-мускульный мешок из наружных кольцевых и внутренних
продольных гладких волокон;
7) пассивный реснично-мукозный способ принятия пищи;
8) аннелидный тип кровеносной системы, состоящей в основном из
спинного и брюшного сосудов, с движением крови сзади наперед в пер-
вом и спереди назад во втором;
9) отсутствие дифференцированных органов выделения;
10) мешковидные гонады, возникающие за счет целомического эпи-
телия и открывающиеся каждая протоком наружу.
По этим чертам кишечнодышащие — самые примитивные среди не
только вторичноротых, но и многих первичноротых. О кишечнодышащих
нет ни косвенных, ни прямых палеонтологических данных, но несомнен-
но палеоэкология установит следы их ходов в грунте моря, а следова-
тельно, и их возраст. Родство птеробранхий с граптолитами дает осно-
вание предположительно считать, что кишечнодышащие существуют с
древнего палеозоя в толще грунта моря и благодаря такому образу
жизни они сохранили примитивное строение. .
При преобладающем числе примитивных признаков и простоте всей
организации кишечнодышащих они обладают рядом вторичных особен-
ностей, вызванных условиями жизни. К числу таковых относятся: отсут-
ствие наружных придатков тела, положение рта у основания хобота, за-
щищенного воротником, форма хобота, развитие в нем мощной муску-
латуры и опорного образования в виде скелета, производного базальной
мембраны, преорального выроста кишечника и центрального, базального
положения целома — словом, прочность хобота как органа рытья грунта,
положение в хоботе пропульзаторного аппарата кровеносной системы.
Далее к вторичным особенностям относятся превращение воротника,
в связи с участием в рытье ходов в грунте, в крайне мускулистый отдел
тела, в который входят выросты целома туловища (перигемальные и
перифарингеальные), почти нацело заполненные развивающимися за их
счет продольными мышечными пучками, впячивание под кожу спинного
нервного тяжа воротника. К вторичным особенностям относятся также
278
преобладающее развитие брюшных продольных мышц кожно-мускуль-
ного мешка туловища и радиальных мускульных пучков, натянутых
между стенками кишечника и тела. Очевидно, что с жизнью в толще
грунта связаны также наружное оплодотворение и наличие свободнопла-
вающих, преимущественно планктотрофных личинок.
Характерными важными чертами кишечнодышащих, подчеркивающи-
ми родство и принадлежность их к вторичноротым, конечно, являются:
образование рта независимо от бластопора, отсутствие спирального типа
дробления, длительный период клеточных делений, гаструляция, инва-
гинационное образование гаструлы и энтероцельное целома, состоящего
из переднего, среднего и хорошо развитого заднего отделов, не ганглиоз-
ный, а в виде тяжей характер нервной системы, незначительное участие
эктодермы в образовании кишечника, присутствие перикардия целомиче-
ского происхождения, слабое развитие выделительной системы и ряд
других черт (Федотов, 1938).
После работ И. И. Мечникова, первым нашедшего энтероцельное
образование целомов у кишечнодышащих и подчеркнувшего сходство
в развитии и родство иглокожих и кишечнодышащих объединением их
в Ambiilacralia, последующие авторы продолжали находить общие черты
онтогенеза до начала метаморфоза этих групп и резкое различие между
ними на поздних этапах развития. К сходным чертам онтогенеза кишеч-
нодышащих и иглокожих относятся радиальный тип дробления, образо-
вание ресничной целобластулы после десятого деления, инвагинационная
гаструла, монаксонность и гетерополярность зародышей на стадии дроб-
ления с анимальным и вегетативным полюсами, энтероцельное образо-
вание целома, состоящего из протоцеля, мезоцеля и метацеля. К ним
также относятся: образование анального отверстия из бластопора и рта
(независимо от последнего), отсутствие выделительных органов у личи-
нок, двустороннесимметричные форма и внутренняя организация личи-
нок, построенных по типу диплеврулы, в типичном случае с широкой
околоротовой впадиной с волнистым мерцательным шнуром по ее краю
.(торнария, биппинария, аурикулярия) и некоторые другие.
Однако при наличии многих сходных черт в раннем онтогенезе этих
групп в нем имеются и различия. Дробление, например, у морских ежей,
в значительной, степени детерминированное. Различна судьба так назы-
ваемых микромеров, вернее, между последними нет гомологии. У игло-
кожих они, действительно, по размерам микромеры и дают начало ске-
лету личинок. У кишечнодышащих же (Colwin a. Colwin, 1953) эти бла-
стомеры, наоборот, крупные и участвуют в гаструляции. У большинства
личинок иглокожих имеется личиночный известковый скелет.
Несмотря на отсутствие перестройки и на постепенное становление
из организации торнарии взрослого организма и коренной ломки всей
•организации личинок большинства иглокожих, в строении кишечноды-
шащих и иглокожих (несмотря на все своеобразие организации послед-
них) имеются некоторые общие черты. Я имею в виду, что комплекс
органов — перикардий, сердце или кровеносная лакуна, целом хобота
с порой, гломерулус кишечнодышащих (Fedotov, 1924) и осевой комп-
лекс органов (левый и правый осевые синусы, осевой орган и пора каме-
нистого канала) —органы гомологичные, одного морфологического зна-
чения и в известной мере сходные по физиологии, но по-разному сформи-
рованные у взрослых. Вполне сравнимы также ретроперитонеально
расположенные, многочисленные гонады в виде мешочков с эктодермаль-
ными протоками, открывающимися наружу в жаберные карманы у ки-
шечнодышащих, и гонады, сохраняющиеся среди иглокожих в таком
состоянии, например, у офиур. К этому можно добавить еще, что источ-
ником энергии мышечного сокращения у иглокожих и кишечнодышащих
служат аргинин-фосфат и креатин-фосфат.
279
Я считаю, что вряд ли имеются самые отдаленные основания видеть
что-то общее между производным стенок целома хобота, так незадачливо
названным гломерусом, и гломусом предпочки позвоночных.
Родство между кишечнодышащими и иглокожими бесспорно, но не
столь близко, чтобы можно было их объединять в тип Ambulacralia, или
Coelomopora, как по наличию пор целома делал И. И. Мечников.
Значительно популярнее мысли о близком родстве кишечнодышащих
с иглокожими является идея о принадлежности их и всех полухордовых
к типу хордовых на том основании, что у них имеется стомохорда, или
нотохорда, нервный тяж на спинной стороне воротника и жаберного
аппарата. Эти черты безусловно, как отмечает большинство зоологов,
имеют важное филогенетическое значение для многих вторичноротых,
но это значение переоценено. Выше были приведены особенности строе-
ния так называемой стомохорды, или нотохорды, как выроста ротовой
полости, возникающей впячиванием эктодермы. Гаймен (Hyman, 1959),
отрицая гомологию этого образования с хордой, подчеркивает при этом
отсутствие в ротовой полости ясной границы между эктодермой и энто-
дермой, что, может быть, и позволяет считать стомохорду, или нотохорду
энтодермальной. Во всяком случае это — преоральный вырост уже функ-
ционирующего кишечника, без какого-либо отношения к бластопору, в от-
личие от того, что характерно для закладки хорды позвоночных.
Закладка части нервной системы на спинной стороне воротника впя-
чиванием стенки тела — важная черта, сближающая кишечнодышащих'
с хордовыми. Однако сходным образом развивается эктоневральная
нервная система и у ряда классов иглокожих.
Сходство в строении и отправлении жаберного аппарата кишечноды-
шащих и ланцетника (особенно на стадии молодого ланцетника) без-
условно велико. Однако напомню,, что ряд авторов, в том числе и
Д. М. Федотов (1923), признавали существование жаберных щелей и
у некоторых ,вымерших примитивных иглокожих.
Таким образом, нельзя отрицать наличия ряда общих черт в орга-
низации и развитии у кишечнодышащих, низших хордовых и иглокожих.
Вместе с тем уровень организации кишечнодышащих настолько низок,
примитивные черты у них настолько превалируют над прочими, что от-
носить кишечнодышащих к типу хордовых — нет оснований. Из всего
сказанного выше следует, что нет также оснований сохранять тип или
подтип Hemichorda.
Птеробранхии (Pterobranchia)
Тело птеробранхий компактное, овальное, яйцевидно-мешковидное,
мягкое, покрытое ресничками, с наружными придатками в виде рук со
щупальцами. Двусторонняя симметрия может быть нарушена сильным
разрастанием брюшной и правой сторон тела. Тело состоит из протосо-
мы, или головного щита, мезосомы, шейки, или воротника, на спинной
стороне которого имеется от 2 до 18 рук со щупальцами, и из метасо-
мы — короткого мешковидного туловища со стебельком, или ножкой,
отходящей от его брюшной стороны. На границе головного щита и мезо-
сомы, на брюшной стороне, находится рот, на спинной стороне у осно-
вания рук — анальное отверстие. Протосоме, мезосоме и метасоме отве-
чает расчленение полости тела (целома) на непарный протоцель с парой
целомопор, парный мезоцель с парой целомопор и парный метацель без
пор. Расчленение тела на три отдела, ясно заметное при развитии, во
взрослом состоянии у птеробранхий не выражено. Головной щит без
стебелька непосредственно переходит в шейку, или мезосому. Лофо-
фор — руки со щупальцами является полым выростом мезосомы. Раз-
меры тела — мелкие, большинство видов Rhabdopleura в длину менее
0,5 мм\ размеры Cephatodiscus колеблются от 4 до 7 мм с максимумом-
в 14 мм у Cephatodiscus densus.
Птеробранхии — реликтовая группа вторичноротых мшанкоподобных
мелких морских организмов, представленная в настоящее время двумя
семействами (Cephalodiscidae и Rhabdopleuridae), тремя родами (Се-
phalodlscus, Atubaria и Rhabdopleura) и несколькими видами. Ископае-
мые известны с ордовика.
Все птеробранхии, кроме одиночной свободноживущей Atubaria, си-
дячие или полусидячие представители эпифауны, образующие настоящие
колонии, зооиды которых связаны тесно друг с другом (Rhabdopleura'),
или ложные колонии — сообщества самостоятельных зооидов, могущих
выходить наружу из ценоциев и ползать; их стебельки, или ножки, сво-
бодны. Зооиды или автозооиды трех родов (Rhabdopleura) — женские,,
мужские и бесполые; кроме того, часть почек, не развиваясь, становится
зимующими зооидами, которые заполняются запасными веществами.
Колонии могут быть женскими или мужскими. Зооиды Cephatodiscus и
Rhabdopleura заключены в общий домик, или ценоций, неизвестной хи-
мической природы, но состоящий, как подчеркивает Гаймен (Hyman,
1959), не из хитина. Ценоций является продуктом выделения желези-
стых клеток головного щита, который, двигаясь то вправо, то влево,
образует очень характерные веретеновидные отдельности, слагающие
стенки ценоция. Как известно, подобное строение ценоция в животном
царстве наблюдается еще лишь у граптолитов (см. стр. 288). В связи с та-
кой ролью в построении трубок головной щит хорошо развит, его упру-
гость, благодаря жидкому содержимому целома, велика (Ливанов,
1955); несомненно, что в связи с этим головной щит лишен наружных
придатков. Форма ценоция разнообразна: это — вертикально стоящие
281
трубочки, базально связанные друг с другом анастомозами, разветвлен-
ные или неразветвленные, стелющиеся или приподнимающиеся тяжи,
губчатая масса (разные формы Cephalodiscus), или стелющиеся, непра-
вильно дихотомически ветвящиеся столоны с поднимающимися кверху
трубками (Ridewood, 1920). Ценоции и тела животных могут быть окра-
шены в темно-коричневый, красный, оранжевый и черный цвета.
Стенка тела, как руки и щупальца, поскольку они являются полыми
выростами мезосомы, состоит из ресничного эпителия, интраэпителиаль-
ной нервной сети или сплетения, базальной мембраны, продольной мус-
кулатуры и перитонеального эпителия. В центральной части брюшной
стенки головного щита среди эпидермальных клеток находится большое
количество железистых клеток, из секрета которых образуется ценоций.
Нервная система состоит из интраэпидермального нервного сплете-
ния с дифференцированным на спинной стороне мезосомы (или ворот-
ника) спинным ганглием, от которого отходят спинной и брюшной про-
дольные нервы или, точнее, сгущения нервного сплетения. Последнее,
как предполагает Гаймен (Hyman, 1959), развито на поверхности всего
тела. В отличие от нервной системы кишечнодышащих часть нервной
системы на спинной стороне мезосомы (ганглий) не погружена под эпи-
дермис и возникает без впячивания внутрь эктодермальной пластинки.
От ганглия, помимо продольного спинного тяжа, отходят спинной тяж
рук и два более слабых вентральных нервных тяжа рук, медиальный
тяж головного щита и пара глоточных или околокишечных коннективов,
которые, соединяясь на брюшной стороне, переходят в брюшной про-
дольный нервный тяж. Органы чувств отсутствуют. Эксперименталь-
ных исследований по физиологии нервной системы нет.
Мускулатура птеробранхий, как и у кишечнодышащих, целомическо-
го происхождения. Она не имеет или, вернее, утеряла черты кожно-мус-
кульного мешка вследствие сидячего образа жизни животных. Она силь-
нее развита у Cephalodiscus — подвижных организмов, способных по-
кидать ценоций и ползать с помощью головного щита, с использованием
его мускулатуры, и сильно редуцирована у колониальных Rhabdopleura.
Пара продольных мышечных пучков в стебельке продолжается по брюш-
ной стороне туловища, охватывает пищевод и переходит в мезосому; при
их сокращении животное втягивается в трубку. В мезосоме находятся
еще ретракторы рук, при сокращении которых они втягиваются в цено-
ций, глоточные мышцы и два пучка мощных мышц, идущих к головному
щиту и веерообразно прикрепляющихся к его брюшной стенке. Кроме
того, имеются продольные мышцы стенки мезосомы (Щепотьев, 1907,
см. Беклемишев, 1952, 1964). Выпячивание рук достигается повышением
тургора в целомах мезосомы при замкнутых порах и сокращением мышц
его стенок. Мышечные волокна находятся еще в стенке перикардия и
в мезентерии.
Кишечник петлеобразный со сближенными ротовым и анальным от-
верстиями. Рот находится на брюшной стороне, на границе головного
щита и мезосомы, анальное отверстие лежит на спинной стороне позади
оснований рук. Рот ведет в ротовую полость, богатую железистыми клет-
ками. Верхняя стенка ее образует преоральный вырост кишечника,
(Щепотьев, 1907), буккальный дивертикул (Hyman, 1959), нотохорду
или стомохорду прежних авторов, гомологичный таковому кишечноды-
шащих, но не заходящий в протосому. Эпителий преорального выроста
кишечника большею частью вакуолизирован и гистологически отли-
чается от других отделов кишечника. Ротовая полость переходит в глот-
ку, выстланную ресничным эпителием с железистыми клетками. В глотке
у Cephalodiscus и Atubaria имеется пара жаберных щелей, а у Rhabdo-
pleura— пара глубоких дорсо-латеральных желобков (Щепотьев, 1907),
которые, может быть, являются жаберными карманами (Hyman, 1959).
282
Глотка переходит в пищевод, последний — в объемистый, мешковидный
желудок, почти заполняющий туловище, гистологически отличающийся
от других отделов кишечника; его эпителий наполнен зернами секрета.
От желудка отходит узкая кишка, которая загибаясь дорсально, откры-
вается анусом на верхнем конце тела. Этим устраняется загрязнение
фекалиями ценоция, в котором живут птеробранхии.
В отличие от кишечнодышащих у птеробранхии печеночных придат-
ков кишечника нет, жаберные щели не имеют язычков и скелета.
Питание у птеробранхий пассивное. Пища поступает в рот с помощью
движения ресничек рук и состоит из микроорганизмов — диатомей, ра-
диолярий, личинок ракообразных. Процессы пищеварения не изучены.
С пищеварительным трактом функционально связаны руки со щупаль-
цами, одетые ресничным эпителием. На брюшной стороне каждой руки
проходит мерцательная бороздка ко рту, по которой током воды к нему
гонится пища. Сильно сократимые руки и щупальца, способные высовы-
ваться из трубок ценоция и втягиваться внутрь, являются вместе с тем
органами связи или контакта организма с внешней средой. Они являются
также жаберным аппаратом птеробранхий. Считают, однако, что жа-
берные щели Cephalodiscus и Atubaria служат не для дыхания, а для
оттока воды, несущей пищевые частицы из кишечника (Ливанов, 1955).
Надо думать, что такое состояние щелей кишечника и их функции у
птеробранхий являются не примитивными, а результатом деградации
этого аппарата кишечнодышащих и может быть смены их функций.
Полость тела, как было сказано выше, состоит из непарного прото-
целя с парой целомодуктов в головном щите, парного мезоцеля с парой
целомодуктов в мезосоме и парного метацеля в туловище. Протоцель
септой отделен от мезоцеля, последний другой септой отделен от мета-
целя. Мезоцель и метацель продольными спинным и брюшным мезенте-
риями разделены на правую и левую части. У ряда видов Cephalodiscus
в верхней части туловища имеются еще два латеральных мезентерия,
с которыми связаны гонады. У Rhabdopleura нет брюшного мезентерия.
Мезоцель продолжается в руки и щупальца, метацель, почти совсем
заполненный кишечником и гонадами, продолжается в стебель, или нож-
ку. Образования на целомодуктах мезоцеля, считавшиеся А. С. Ще-
потьевым (1907) соленоцитами, по его же признанию (Schepotieff, 1910),
оказались мышечными фибриллами. Перикардий является производным
стенки протоцеля.
Клетки перитонеального эпителия (Щепотьев, 1907) образуют в про-
свет целома протоплазматические выросты; в целоме находятся свобод-
ноплавающие клетки, но и те и другие изучены недостаточно.
Кровеносная система птеробранхий, как и у многих групп животных
малых размеров, в значительной степени упрощена, особенно сильно
у микроскопически мелких, настоящих колониальных Rhabdopleura.
Кроме того, как отмечает Гаймен (Hyman, 1959), вследствие мелких раз-
меров птеробранхий она и недостаточно изучена. Сосуды, еще в боль-
шей степени, чем у кишечнодышащих, носят характер синусов или лакун.
В спинном мезентерии находится спинной синус, переходящий вблизи
спинного ганглия в центральный синус; последний охвачен впяченной
стенкой мускулистого перикардия; вместе они, как у кишечнодышащих,
составляют кардио-перикардий — центральный пропульзаторный орган
кровеносной системы, но он лежит не над выростом ротовой полости,
а спереди, на септе, отделяющей целом головного щита от мезоцеля.
Скопление перитонеальных клеток над центральным синусом с зерни-
стыми включениями, видимо, экскреторного характера, является гломе-
рулусом, однако менее дифференцированным, чем таковой кишечноды-
шащих. -Его строение у Rhabdopleura сильно упрощено сравнительно
с таковым у Cephalodiscus — это отдельные веретеновидные перито-
283
неальные клетки, прилегающие к поверхности центрального кровенос-
ного синуса.
От центрального синуса отходит пара околоротовых синусов, от ко-
торых берет начало продольный брюшной синус, проходящий в брюш-
ном мезентерии и идущий в стебелек. Предполагают, но это эксперимен-
тально не доказано, что, как у кишечнодышащих, в спинном синусе кровь
движется сзади наперед, в брюшном — спереди назад.
Процессы экскреции почти не изучены, к ним имеет, по-видимому,
отношение гломерулус. Функция целомодуктов с их порами связана
с регулированием тургора внутри целомов головного щита и мезоцеля,
в особенности целома рук и щупалец.
Птеробранхии раздельнополы, хотя имеются и гермафродитные виды.
Полы внешне или не различимы (Cephatodiscus) или отличаются друг
от друга (Rhabdopleura). У Cephatodiscus имеется пара мешковидных
гонад, симметрично расположенных в передней части целома туловища,
каждая гонада коротким протоком открывается наружу на спинной
стороне туловища позади границы между ним и мезосомой. У Rhabdo-
pleura имеется лишь одна гонада (семенник или яичник), лежащая в пра-
вом метацеле, открывающаяся половым отверстием справа и позади
ануса.
Размножение происходит половым путем, причем свободноплаваю-
щие личинки служат для распространения вида, а также бесполым пу-
тем— с образованием наружных почек, чем достигается рост колоний
и псевдоколоний.
Эмбриональное развитие изучено во многих отношениях недостаточ-
но, даже в новых работах, например, Джона (John, 1932), и лишь у
Cephatodiscus. Дробление полное, почти равномерное; бластула полая
(целобластула) или плотная (морула); гаструла, видимо, образуется
инвагинацией, деляминацией и полярной иммиграцией. Анальное отвер-
стие возникает на месте бластопора. Покрытый ресничками зародыш
плавает внутри яйцевой оболочки и вылупляется в виде овальной плану-
лоподобной личинки в полость ценоция, становясь свободноплавающей
личинкой с теменной пластинкой на переднем конце. Личинка у Cepha-
lodiscus nlgrescens (John, 1932) червеобразная с прямым кишечником,
со. ртом и терминальным анальным отверстием, похожа на молодого'
баланоглосса, но с парой зачатков рук и без жаберных щелей. Целом
образуется энтероцельно в виде непарного или парного протоцеля и пар-
ных мезоцеля и метацеля. Данные о протоцеле и перикардии противо-
речивы. Превращение личинки во взрослое животное изучено плохо.
Преоральный отдел развивается в головной щит, на спинной стороне
мезосомы закладываются руки, сначала в виде пары зачатков.
У 'Cephatodiscus indicus (Schepotieff, 1910) первая планулообразная
личинка превращается во вторую личинку, настолько похожую на личи-
нок мшанок — Bugula или Alcyonidium, что авторы считали, что
А. С. Щепотьев ошибочно приписал ее Cephatodiscus. Однако, судя по
его описанию и рисункам срезов, органогенез у С. indicus является ти-
пичным для птеробранхий (энтероцельное образование трех отделов
целома, зачаток архентерона вначале лишенный ротового и анального
отверстий и т. д.), т. е. он резко отличен от такового мшанок. Я считаю,
что ошибка Щепотьева заключалась в том, что вторую личинку С. indicus
он реконструировал неверно и отсюда ошибочно придал ей сходство
с личинкой мшанок.
Как было сказано, увеличение числа особей и рост колоний и псевдо-
колоний происходит путем почкования на свободных верхних концах
столона, на концах боковых ветвей колоний (Rhabdopleura) и на конце
стебелька зооидов (Cephatodiscus). Существенной особенностью почко-
вания Rhabdopleura, является то, что зооиды, возникающие на некото-
284
ром расстоянии от активного конца столона, продырявливают стенку
Трубки (перфорирующее почкование, встречающееся еще у граптоли-
тов). Почкование изучено много лучше эмбрионального развития, так
как оно очень распространено. И все же образование колонии первой
особью, возникающей, очевидно, из личинки при половом размножении,
*еще не прослежено. Несмотря на малое количество видов птеробранхий,
Рис. 74. Почка Cephatodiscus,
внешний вид (по Шепотьеву)
•1 — протосома; II — мезосома; III—ме-
тасома; / — лофофор; 2 — ножка
Рис. 75. Почка Cephatodiscus в
разрезе (по Шепотьеву)
I — протосома; II — мезосома; III—
метасома; 1 — кишечник; 2 — прото-
цель; 3— кардио-перикардий; 4 —
мезоцель; 5 — метацель
обилие почек на разных стадиях развития позволило неплохо просле-
дить закладку почек и морфогенез до сформирования взрослых особей,
хотя, в отличие от развития из яйца, для развития при почковании ис-
пользуется главным образом тканевой материал сформированных осо-
бей, однако последние, несомненно, оказывают морфогенетическое воз-
действие на развитие почек. В морфогенезе почек проявляются некоторые
более примитивные черты, не сохранившиеся при эмбриональном разви-
тии. Вместе с тем наблюдаются особенности, связанные с возникновением
нового организма за счет частей и тканей старого.
Почки возникают в виде пузырька; кольцевым перехватом сначала
обособляется большая передняя часть, зачаток головного щита, и мень-
шая задняя, которая затем вторым перехватом делится на меньшую,
шейную часть — зачаток мезосомы — и большую — зачаток тела или
метасомы и стебелька (рис. 74, 75). В полость почки продолжается це-
лом материнского стебелька, который затем соответственно делится
двумя септами на протоцель, мезоцель и метацель. Таким образом, при
почковании на известной стадии развития ясно выражены двусторонняя
симметрия тела, наружное расчленение его на- три .отдела и трехчлени-
стость целома. По некоторым данным, например А. С. Щепотьева (1907),
целом почки поделен на правую и левую половины продольной перего-
родкой, т. е. протоцель является парным; правая часть уменьшается и
становится перикардием; по другим сведениям, — протоцель непарный,
а перикардий возникает за счет его стенки. Разноречивы также данные
о происхождении кишечника; наиболее трудно разрешим вопрос о раз-
витии кишечника у Rhabdopleura-, по А. С. Щепотьеву, он возникает из
мезодермы, и нет никаких признаков участия наружной стенки тела
почки в его образовании. У Cephatodiscus, по ряду авторов, в том числе
по Щепотьеву (1907), средняя кишка почки образуется из эктодермы
впячиванием стенки тела почки. Джон (John, 1932) не согласен с этим
285
и, не приводя доказательств, признает энтодермальное ее образование.
Большинство авторов считает преоральный вырост кишки эктодермаль-
ным, но ротовая полость при почковании, как и при эмбриональном раз-
витий, образуется впячиванием эктодермального стомодеума, выростом
которого, как известно (Fowler, 1905), является этот орган.
Филогенетически важными моментами в развитии птеробранхий яв-
ляются: наличие стадий с двусторонней симметрией, ясное наружное
расчленение тела на три отдела, трехчленистость энтероцельного цело-
ма, наличие червеобразной личинки с длинным прямым кишечником,
с ротовым отверстием на переднем конце брюшной стороны мезосомы
и с терминальным анальным отверстием. Это говорит о происхождении
птеробранхий от двустороннесимметричных червеобразных свободно-
подвижных форм.
Птеробранхий живут на глубинах от 5 до 650 м. (Hyman, 1959), селясь
на различных сидячих животных — гидроидах, кораллах, мшанках, губ-
ках, раковинах и т. д., встречаясь в холодных, теплых и тропических
областях Мирового океана, в том числе и в Антарктике. Данных об их
паразитах мало. Класс состоит из трех родов — Cephalodiscus, Atubaria
и Rhabdopleura, объединяемых одними исследователями (например, Ще-
потьев, 1907) в два семейства, а другими (Dawydoff, 1948; Беклемишев,
1951, 1952, 1964) в два отряда. Действительно, различия между этими
формами таковы, что по аналогии с другими группами, пожалуй, пра-
вильнее считать их отрядами. Между тем некоторые зоологи (Horst,
1927—1939; Hyman, 1959) различают лишь роды.
Данные о строении и отчасти развитии говорят о более примитивном
состоянии Cephalodiscus и Atubaria и о более вторичных изменениях и
деградации Rhabdopleura. Уже одна настоящая колониальность Rhab-
dopleura говорит о них как о наиболее измененных формах, так как сре-
ди всех вторичноротых колониальность исключительное и несомненно-
позднейшее явление, да и среди целомических первичноротых колониаль-
ность также является вторичной.
Птеробранхий реликтовая группа (Родендорф, 1959), она малочислен-
на, ее остатки известны с ордовика. Относясь скептически к данным
Козловского (Kozlowski, 1949) о принадлежности Eocephalodiscus к пте-
робранхиям, я должен отметить, что Rhabdopleuroides, описанный им
(Kozlowski, 1956, 1961) также из ордовика Польши, несомненно, предста-
витель птеробранхий, судя по форме и строению ценоция.
Как известно, несмотря на бросающееся в глаза несходство птеро-
бранхий и кишечнодышащих, такие черты строения, как состав тела из
трех отделов, пять целомических полостей энтероцельного происхожде-
ния с целомодуктами и целомопорами, общий план строения интраэпи-
дермальной нервной системы, преоральный вырост кишечника, пропуль-
заторный в виде кардио-перикардия аппарат кровеносной системы со
спинным и брюшным продольными кровеносными сосудами, с движе-
нием крови сзади наперед в первом и спереди назад во втором, отсут-
ствие типичных органов выделения, являются главнейшими общими чер-
тами птеробранхий и кишечнодышащих.
Хотя онтогенез птеробранхий изучен недостаточно, все же ясны так-
же и существенные различия в строении и онтогенезе птеробранхий и
кишечнодышащих. Разрастанием брюшной стороны главная ось зароды-
ша изогнута так, что кишечник становится петлеобразным, со сближен-
ными ротовым и анальным отверстиями на верхнем конце тела. Тело
становится коротко-овально-мешковидным, лишь временно в ходе онто-
генеза наружно расчлененным. Сильным ростом -брюшной стороны и
правой половины сравнительно с левой и наличием непарной правой го-
нады (Rhabdopleura) двусторонне-симметричное строение нарушается.
Руки и щупальца, производные мезосомы, являются органами дыхания
286
и подгона пищи. Мускулатура тела в основном представлена системой
отдельных мышечных пучков. В мезосоме концентрируется спинной
ганглий, без следов впячивания при развитии. Жаберный аппарат ки-
шечника отсутствует, пара жаберных щелей (Cephalodiscus, Atubaria)
служит лишь для оттока воды. Гонад одна пара или гонада непарная,
половые отверстия — на верхнем конце тела. Птеробранхий почти исклю-
чительно мелкие колониальные и псевдоколониальные, с сильно разви-
тым бесполым размножением путем наружного почкования, придонные,
сидячие или полусидячие организмы эпифауны, живущие в ценоциях.
В отличие от птеробранхий главная ось гаструлы у кишечнодыша-
щих переходит в главную ось длинного, червеобразного, двусторонне-
симметричного тела взрослого животного, с терминальным анальным
отверстием и без наружных придатков. Сохраняется наружное расчле-
нение тела на три отдела и двустороннесимметричная внутренняя орга-
низация. Достаточно полно сохраняется характер мускулатуры тела
в виде кожно-мускульного мешка. Нервная система аганглионарная с
неврохордом в воротнике, возникающем впячиванием эктодермы. Ки-
шечник с метамерными — настоящим жаберным аппаратом и печоноч-
ными придатками. Гонады многочисленные, метамерные, каждая с по-
ловым протоком. Одиночные формы от средней до очень крупной вели-
чины, размножающиеся половым путем червеобразные организмы
инфауны, живущие преимущественно в грунте моря, роющие в нем ходы.
Несомненно, что микроскопические, сидячие или полусидячие, коло-
ниальные, мшанкоподобные с петлеобразным кишечником, со щупаль-
цевым аппаратом, почкующиеся, живущие в ценоциях птеробранхий яв-
ляются вторично измененными сравнительно с кишечнодышащими. Они
занимают в отношении последних такое же положение, какое существует
между мшанками и червеобразными форонидами, утратившими ряд при-
знаков червеобразных предков и претерпевшими ряд регрессивных из-
менений в связи с прикрепленным образом жизни.
Надо напомнить, что и у всех вторичнополостных, двустороннесим-
метричных первичноротых животных, ставших колониальными, сидячими
формами, наблюдается укорочение тела, щупальцевый аппарат, петле-
видный кишечник, дифференцировка кожно-мускуЛьного мешка на от-
дельные специализированные мышечные пучки, утрата или неполнота
мезентериев и т. д,; и все это является вторичными, исторически поздней-
шими изменениями организации и органов червеобразных свободнопод-
вижных предковых форм. При этом, у ряда групп подобные изменения
организации происходили в исторически отдаленное время, в кембрии.
Сидячий образ жизни в твердых трубках вызвал утрату большинства
черт червеобразной организации кишечнодышащих и выработку у пте-
робранхий организации, напоминающей таковую мшанок, однако ни в
каком, даже отдаленном родстве, вопреки мнению Брэма (Вгает, 1911),
птеробранхий и мшанки не состоят.
Специфической чертой, вероятно, не существовавшей и у древнейших,
вымерших представителей кишечнодышащих, является щупальцевый
аппарат, так же как и образованные из спаянных веретенообразных от-
дельностей прочные ценоции. Подобный скелет не свойствен воо'бще
вторичноротым, если не считать трубок погонофор. Позднейшей, также
специфической особенностью птеробранхий является бесполое размно-
жение путем почкования — особенность, широко распространенная сре-
ди групп первичноротых и несвойственная или очень редкая среди вто-
ричноротых (исключение составляют лишь оболочники).
Важным вопросом являются отношения между птеробранхиями и
граптолитами, о родстве которых высказывался, как известно, А. С. Ще-
потьев (1907), вопрос, к кому ближе птеробранхий: к кишечнодышащим
или к граптолитам, будет рассмотрен ниже (см. стр. 289).
Граптолиты (Graptolitha)
Граптолиты — морские, колониальные формы с твердым ценоцием,
широко распространенные в древнем палеозое, известные со среднего
кембрия, многочисленные и разнообразные в ордовике и раннем силуре
и вымершие в конце карбона. Они населяли мелководье палеозойских
морей и сохранились преимущественно в битуминозных, глинистых, от-
части кремнистых сланцах, а также в известняках геосинклинальных
областей. Эта группа чаще представлена лишь отпечатками на породе,
реже остатками тел, в основном скелета, и очень важна для страти-
графии.
Колонии (рабдосомы) с двустороннесимметричными ячейками, или
теками, были разнообразными. Известны колонии древовидноразветвлен-
ные, кустистые, сетчатые со многими ветвями или из одной или двух
ветвей, прямых, изогнутых или спирально свернутых. Ячейки распола-
гались на ветвях правильно, триадами, четырех-, двух- и однорядно или
беспорядочно. Эволюция граптолитов, как и у многих других групп жи-
вотных, согласно идее В. А. Догеля (1954), шла от полимеризации к оли-
гомеризации, от древовидных форм (Dendroidea) класса Stereostolonata
Obut к одноветвистым формам класса Graptoloidea Lapworth. Соответ-
ственно с этим у более древних граптолитов класса Stereostolonata, из-
вестных со среднего кембрия, ячейки зооидов были различными, т. е.
колонии полиморфными. У ряда таких колоний возникали узлы ветвле-
ния. В каждом узле столон давал три ветви: более длинную в столотеку
и две боковых коротких — одну в автотеку, другую — в битеку. Автотеку
строил автозооид, считающийся женской особью, битеку — бизооид,
считающийся мужской особью; концевой зооид (столозооид) строил
столотеку. У геологически более новых граптолитов (класс Graptoloidea),
известных с ордовика, колонии были мономорфными, ячейки, или теки
зооидов были однородными.
Стенки ценоциев и ячеек, или тек состояли из наружного кортикаль-
ного пластинчатого слоя и внутреннего фузеллярного, образованного
спаянными друг с другом веретеновидными с зигзагообразным швом
отдельностями из хитиноподобного вещества.
По аналогии с другими группами в ячейках, несомненно, находились
зооиды, связанные друг с другом полым столоном; строение их неизве-
стно. Рост колоний происходил апертурным почкованием концевых зо-
оидов— столозооидов, т. е. не почкующиеся зооиды, как было сказано
выше (согласно Козловскому), были половыми индивидуумами. Вероят-
но, они же улавливали пищу и ее переваривали.
Размножение было, очевидно, половое и бесполое — почкованием.
По данным А. М. Обута (1953, 1964), существовали мужские и женские
половые клетки и личинки; однако, просмотрев препараты А. М. Обута,
я не смог согласиться с таким их толкованием.
Первый зосид, дававший начало колонии, образовывал конической
формы сикулу с длинной, полой нитью — немой, почкованием он давал
второго зооида, который, продырявив стенку сикулы (перфорирующее
почкование), образовывал первую ячейку первой ветви колонии. Осталь-
288
ные зооиды возникали простым чашечным или апертурным почкованием
столозооида. ‘
Экология граптолитов была довольно разнообразной. Одни колонии
были бентосными формами, прикреплявшимися ко дну моря. Другие,
прикрепившиеся к подвижным плавающим водорослям, были псевдо-
планктонными формами. Третьи, обладавшие пневматофором, или по-
плавком, находились в свободном, взвешенном состоянии и были планк-
тонными организмами. Как в отношении морфологии (см. выше), так
и в экологическом отношении, эволюция граптолитов шла от прикреплен-
ного, бентосного образа жизни (ряд родов класса Stereostolonata)
к псевдопланктонному (ряд родов этого класса и Graptoloidea), а за-
тем к планктонному — ряд представителей последнего класса.
В результате изучения хорошо сохранившихся граптолитов и особен-
но применения метода, разработанного Виманом (Wiman, 1895) —выде-
ление граптолитов путем растворения породы кислотами, заключение
их в парафин и изготовление срезов на микротоме (см. также Обут,
1953),— рядом авторов, особенно польским палеонтологом Козловским
(Kozlowski, 1938, 1947, 1949, 1962) установлены важные черты строения
граптолитов, поясняющие их положение в системе. И все же нет единого
мнения о родстве граптолитов с другими группами.
Согласно взгляду исследователей конца прошлого столетия, к кото-
рому вновь возвращаются некоторые американские зоологи и палеонто-
логи (см. Hyman, 1959; Decker a. Gold, 1957), граптолиты считались ки-
шечнополостными.
Однако в настоящее время мысль А. С. Щепотьева (1907, см. гл.
«Птеробранхии») о родстве граптолитов с птеробранхиями была под-
креплена и развита Козловским (Kozlowski, 1938, 1947, 1949, 1962), уста-
новившим у граптолитов ряд важных черт, общих с птеробранхиями. Он
не только помещает граптолитов в группу полухордовых, но и допускает
возможность, что их предками могли быть птеробранхии, что, однако,
на мой взгляд, малб вероятно. Взгляды Козловского разделяет В. Н. Бе-
клемишев (1951, 1952, 1964); идя еще дальше, последний включает птеро-
бранхий прямо в класс граптолитов как два отряда — Rhabdopleuroi-
dea и Cephalodiscoidea, но, на мой взгляд, без достаточных оснований.
А. М. Обут (1953, 1964), исходя из собственных исследований грапто-
литов, занимает более правильную позицию. .Признавая общие черты
птеробранхий и граптолитов, он выделяет- последних в подтип Grapto-
lithina (1964).
Отсутствие данных о строении зооидов граптолитов крайне затруд-
няет решение вопроса: с какой же из существующих групп животных
они связаны? Почти все черты родства их с птеробранхиями, приводимые
Козловским (Kozlowski, 1938, 1947, 1949, 1962),. Гаймен (Hyman, 1959)
оспаривает и считает включение граптолитов в группу полухордовых не-
достаточно обоснованным. А. М. Обут (1964) в известной мере сходится
с ней в своей критике доводов Козловского и В. Н. Беклемишева и, как
было сказано, в подтип граптолитов птеробранхий не включает.
Виман (Wiman, 1895) считал скелет граптолитов нехитиновым, но
в литературе его называют таковым (см. Hyman, 1958). В последнее
время С. М. Манской и Т. В. Дроздовой (1962) приводятся данные о хи-
тиновой природе скелета этих организмов. У Rhabdopleura скелет нехи-
тиновый. Стенки скелета у граптолитов состоят из наружного кортикаль-
ного и внутреннего слоев из твердых полуколец. У Rhabdopleura он со-
стоит из одного слоя, причем скелет стелящихся трубок состоит из полу-
колец, а у поднимающихся — из полных колец; у Cephalodiscus строение
скелета другое. У граптолитов при развитии имеется1 сикула, а также
нема, или виргула, чего нет у Rhabdopleura. Колонии граптолитов поли-
морфные, а рост их симподиальный, у Rhabdopleura полиморфизма нет,
19 Д- М. Федотов
28!)
а рост колоний моноподиальный. К этому Гаймен (Hyman, 1959} еще
добавляет, что недавно у граптолитов якобы найдены гонотеки и немато-
теки, сходные с таковыми кишечнополостных. Лишь столон связывает
зооидов колоний как у граптолитов, так и у Rhabdopleura.
В. Н. Беклемишев (1951, 1952, 1964), перечисляя довольно много-
численные общие признаки граптолитов и птеробранхий, особо выделяет
фузеллярную структуру скелета, столоны, соединяющие зооиды и пер-
форирующее почкование, и считает эти группы очень близкими. Однако
согласно электронномикроскопическому изучению Ветцеля (Wetzel, 1958;
см. Bulman, 1955, 1963) микроструктура скелета граптолитов и птеро-
бранхий различна.
Рис. 76. Схема примерного количественного распреде-
ления во времени 128 родов граптолитов (из.
A. Muller)
Соглашаясь с В. Н. Беклемишевым о родстве граптолитов и птеро-
бранхий, я не могу, однако, их объединять, как он, в один таксон не толь-
ко ранга класса, но даже и подтипа. Тем более, что нет оснований сохра-
нять название ни Hemichordata, ни Stomochordata, поскольку гомологии
между стомохордой (или нотохордой) и хордой нет.
На данном этапе наших знаний и методики исследования, по моему
мнению, надо считать Enteropneusta, Pterobranchia и Graptozoa (как
называют их Шрок и Твенгофель) (Shrock and Twenhofel, 1953) отдель-
ными типами с общими отдаленными предками в группе вторичноротых,
стоящими друг к другу ближе, чем к остальным типам — Chordata, Ро-
gonophora и Echinodermata.
После работ Козловского и Обута ясно отсутствие сходных черт в
строении граптолитов и колониальных гидрополипов. Резко отличаются
они и по геологической истории. Граптолиты — древнепалеозойские орга-
низмы, процветавшие в ордовике и нижнем силуре (рис. 76). Геологи-
ческая история гидроидных полипов развертывается в настоящую эпоху.
Природа и систематическая принадлежность около десятка форм из:
кембрия, силура и ордовика, с оговоркой отнесенных к граптолитам
(Орлов, 1960—1964), далеко неясны. С ними пока считаться не прихо-
дится.
Иглокожие (Echinodermata)
Иглокожие — морские, прикрепленные и свободноподвижные, донные
исключительно свободноживущие животные, пассивные микрофаги и ак-
тивные хищники-макрофаги, пятилучевой радиальной симметрии с при-
знаками исходной и вторичной двусторонней симметрии,'размерами от
нескольких частей миллиметра до 2—5 м. Известковый скелет, нередко
в виде наружных игл, мезодермального происхождения и коррелятивно-
связанный со многими внутренними органами, из которых очень харак-
терна присущая лишь этому типу амбулакральная (воднососудистая)
система. Кишечник со ртом и анальным отверстием, почти нацело экто-
дермального происхождения, лежит в обширной вторичной полости тела,
энтероцельного происхождения. Нервная система неганглиозного типа,
состоит из трех систем, возникает за счет эктодермы, энтодермы и мезо-
дермы. Кровеносная, дыхательная и выделительная системы —• на низ-
ком уровне организации и физиологии. В плоскости исходной двусторон-
ней симметрии лежат гидропора, или мадрепорит, анальное отверстие,
половое отверстие и части или весь осевой комплекс органов как анато-
мическое целое; элементы последнего отвечают перикардию и сердцу ки-
шечнодышащих.
Общий уровень организации невысок, многие органы многофункцио-
нальны. Обычно полы разделены, размножение половое, реже бесполое,,
делением; регенерация и автотомия очень распространены. Развитие со
сложным метаморфозом, с очень характерными, чаще планктотрофными,
личинками, реже вторично упрощенное.
Современные иглокожие широко распространены в водах нормальной
солености Мирового океана от береговой зоны до сверхабиссальных глу-
бин— 7250—10710 м (Беляев и др., 1960). Ископаемые иглокожие изве-
стны с нижнего кембрия.
Иглокожие — сравнительно малочисленный тип животных. Современ-
ных видов насчитывается 5000—5970 (Kaestner, 1963), ископаемых—•
около 13 000 видов. По разным источникам карпоидей 24 рода, примерно
70—80 видов; цистоидей 65 родов, 360 видов; бластоидей — 350 видов;
эокриноидей —15 родов, 22 вида; паракриноидей 13 родов, вероятно,
16—18 видов; офиоцистий — 5 родов, 7 видов; голотурий—1100 видов;
ископаемых морских лилий 5000 видов, современных— 1000; эдриоасте-
роидей — 22 рода, 101 вид; морских ежей ископаемых — 7200 видов,
современных 800—860 видов, сомастерид—6 родов; морских звезд
ископаемых — 300, современных — 2000 видов; офиур ископаемых—180,.
современных — 2000 видов.
Иглокожие, в общем,— животные средней величины, но диапазон,
размеров значительный не только среди современных, но и среди иско-.
паемых форм. Так, диаметр диска палеозойских морских микролилий;
(Lageniocrinus, Embryocrinus') измерялся десятыми долями миллиметра,.
одним и немногим более одного миллиметра, гигантский мезозойский'
Pentacrinus имел руки длиной до 0,5 м, а стебель — до 21 м. У совреэдрн.-.
19* 2_9Т
кого Antedon диаметр диска всего 6—7 мм,, a Metacrinus superbus имеет
руки до 20 см, стебель до 2 ж длиной, нижнекаменноугольный морской
еж Fournier echinus имел до 35 см в диаметре. Диаметр ряда ископаемых
видов семейства Saleniidae равнялся 3—5 м-, из современных клипеасте-
роид Echinocyamus pusillus имеет всего 6 см в длину, Meoma ventricosa
(из спатангоид) — 18 см, a Sperosoma giganteum (из Echinothuriidae) —
32 см. Из голотурий Leptosynapta minuta измеряется 3—5 мм, a Synapta
maculata более 2 м в длину. Asterina имеет 2,5 см в диаметре, Freyella
remex — 1 м с руками в 45 см. У Nannophiura stimpsoni диаметр диска
0,5 мм, у Gorgonocephalus stimpsoni —14—20 см, а руки — более
0,5 м.
Своеобразие иглокожих, животных, по выражению Грегори
(W. К- Gregory, 1951), «без головы и мозга», выделяющее их среди всех
типов (рис. 77), привлекало внимание многих исследователей. Вместе с
тем их изучение представляло большие трудности, так как для иглокожих
характерны, по выражению В. Н. Беклемишева (1952), «неисчерпаемые
преобразования, непрерывные перестройки, громадное многообразие на-
правлений развития и почти полное отсутствие прогресса». Всем этим
вызвано такое обилие работ по иглокожим, что нет возможности, делать
их обзор, тем более, что литература по иглокожим очень полно исполь-
зована Гаймен (Hyman, 1955). Работ русских и советских зоологов по
иглокожим мало, к ним относятся работы И. И. Мечникова, А. О. Кова-
левского, К. К. Сент-Илера, Д. К. Третьякова, Е. А. Шульц, К- Н. Да-
выдова, И. И. Соколова, Д. А. Ласточкина, П. Г. Светлова, А. М. Дьяко-
нова, А. А. Шорыгина, Г. М. Беляева, Н. А. Вержбинской, Т. С. Савелье-
вой, И. Н. Верховской, М. Н. Зарха, 3. И. Барановой, Д. М. Федотова.
Это исследования по эмбриологии, анатомии, филогении, регенерации,
обмену веществ, систематике и зоогеографии.
Учитывая работы Регнеля (Regnell, 1945 и др.), можно принять сле-
дующую классификацию иглокожих: подтип Pelmatozoa с классами
Carpoidea, Cystoidea, Blastoidea, Eocrinoidea, Paracrinoidea, Crinoidea,
Edrioasteroidea и подтип Eleutherozoa с классами Ophiocistia, Holothu-
rioidea, Somasteroidea, Asteroidea, Echinoidea и Ophiuroidea.
Критика Беклемишевым (1952), Гисленом (Gislen, 1947) и Регнелем
(Regnell, 1948а, 1948в) толкования Уайтхаузом (Whitehouse, 1941) Peri-
dionites и Cymbionites как иглокожих обоснована. Я также не считаю
этих ископаемых иглокожими и думаю, что ни новых классов Cyamoidea
и Cycloidea, ни новых подтипов Haplozoa и Homalozoa, принятых Шро-
ком и Твенгофелем (Shrock a. Twenhofel, 1953), сохранять нельзя. Нет
оснований считать иглокожими и Macheridia Виздерса (Withers, 1926).
Не приходится считаться и с классификацией Спенсера (Spencer, 1938),
предложившего вместо Pelmatozoa и Eleutherozoa подтипы Dactylozoa
(иглокожие без ножек, с пальцами-брахиолами) и Podozoa (иглокожие,
со щупальцами) и включившего в Podozoa морских лилий, поскольку
она построена не на совокупности черт организации, а лишь на некоторых
из них.
Радиальная симметрия внешней формы и внутреннего строения, резко
выделяющая иглокожих среди всех типов двустороннесимметричных жи-
вотных, не только вторичноротых, но и первичноротых, обычно пятилу-
чевая. Однако довольно широко распространены полимеризация амбу-
лакров, или рук.
. Среди Eleutherozoa увеличение числа рук наблюдается у морских
звезд и очень редко у офиур. Полимеризация числа рук довольно часто
встречается у палеозойских, более примитивных морских звезд, в гораз-
до большей степени, чем у позднейших и современных форм. Известны
13-лучевые ископаемые морские звезды, Lepidaster Forbes, Lepidactis
Spencer из семейства Lepidactinidae из Готланда. Многолучевость, от 13
292
до 29 рук, была распространена среди девонских морских звезд (Heptan-
thaster Roenur, Lepidastrella Schuchert; Palaeosolaster Sturtz из семей-
ства Taeniactinidae, Palasterinidae (из вымершего отряда Hemizonida
Spencer). Среди современных морских звезд многолучевость обычно воз-
никает не у примитвных форм, а у более высоко организованных.
Рис. 77. Схема организации иглокожих (из Hennig)
А— морские лилии; Б — морские звезды; В — офиуры; /’ — морские
ежи; Д — голотурий; 1 — анус (целом и нервная система не пока-
заны)
Многие роды семейства Solasteridae — многолучевые, особенно же силь-
ная полимеризация рук наблюдается в семействе Brisingidae, Heliasteri-
dae, Asteriidae отряда Forcipulata, у которых число рук может доходить
до 50, например, Labidiaster из семейства Asteriidae и т. д.
В отличие от морских звезд у офиур увеличение числа рук наблю-
дается очень редко. Известны лишь единичные представители ископае-
мых офиуроподобныхя форм в верхнем девоне — Lepidasterina. Среди
современных типичных офиур имеется всего несколько видов рода
Ophiacantha самого обширного семейства офиур — Ophiacanthidae, вклю-
293
чающего свыше 300 родов. Так, Ophiacantha nutrix Baranova имеет 8 рук,
Ophiacantha cuneactis Clark—9, Oph. aff. cuneactis Clark (из сборов «'Ви-
тязя», по устному сообщению Г. М. Беляева) — 10 рук.
Полимеризация пятилучевой симметрии может идти и путем дихото-
мического разделения или ветвления дистальных частей амбулакров,
остающихся в числе пяти, например, среди ископаемых стебельчатых
морских лилий и современных бесстебельчатых и доходящее, например,
у семейства Comasteridae до 200. Такая же полимеризация наблюдается
у многоветвистых офиур группы Euryalae, у которых, по-видимому, в те-
чение всей жизни особи идет образование громадного числа тончайших
веточек рук.
Полимеризация имеет биологический смысл, так как большое число
амбулакров и ветвистость прежде всего значительно увеличивает поступ-
ление пищи по амбулакральным желобкам (морские лилии и звезды),
усиливает роль многоветвистых рук как ловчего аппарата (Gorgonoce-
phalidae). В ходе эволюции пять простых рук офиур с их мускулатурой
(см. ниже) превратились в наиболее совершенный среди иглокожих ап-
парат передвижения, но дает ли функциональные преимущества увели-
чение числа неразветвленных рук у офиур — неясно. Изредка у современ-
ных офиур и морских звезд наблюдается явление, обратное полимериза-
ции, уменьшение числа рук до четырех.
Известно, что исторически (данные палеонтологии), а также онтоге-
нетически, радиальная симметрия иглокожих возникает из двусторонней
симметрии. Древнепалеозойские Carpoidea, асимметричные вследствие
лежачего образа жизни на дне моря, были иглокожими без следов ра-
диальной симметрии (Bather, 1929). Среди древнепалеозойских прикреп-
ленных или стебельчатых иглокожих, мешкообразных цистоидей, были
формы с переходом или возникновением пятилучевой симметрии из трех-
лучевой. Однако у цистоидей это мог быть и регресс пятилучевой сим-
метрии, так как у гораздо более древних форм, у нижнекембрийских
эдриоастероидей (Stromatocystis), пентамерия была уже вполне выра-
жена. Становление пятилучевой симметрии через трехлучевую наблю-
дается и сейчас в онтогенезе амбулакральной системы у некоторых
современных голотурий.
В общем ходе геологической истории иглокожих идет эволюция ра-
диальной симметрии, однако последняя нарушена или, вернее, не пре-
одолена у большинства Pelmatozoa положением в пятом интеррадиусе
анального отверстия, порового канала или мадрепорита и полового
отверстия. У морских лилий радиальная симметрия нарушается положе-
нием в интеррадиусе лишь анального отверстия, так как поровые отвер-
стия могут'быть в каждом интеррадиусе, а половые железы становятся
многочисленными и перемещаются в руки и пиннулы. У большинства
современных иглокожих (морские лилии, ежи, голотурии), и насколько
известно, у ряда вымерших Pelmatozoa трубчатый кишечник обладает
разной степенью спиральной свернутости, что нарушает радиальную
симметрию общей организации. Лишь у морских звезд и офиур мешко-
видный кишечник приобретает радиальную симметрию. Наибольшей сте-
пени законченности, почти геометрической правильности, достигает
радиальная симметрия у некоторых представителей офиур группы Eurya-
lae, у которых в каждом интеррадиусе образуется по осевому комплексу
органов и притом симметричного строения. Уклонения от радиальносим-
метричного строения, наблюдавшиеся в некоторой степени уже у древ-
них ископаемых форм разных классов, и возникновение наиболее сильно
выраженной двусторонней симметрии у голотурий и неправильных мор-
ских ежей явно обусловлены образом жизни. Однако закономерности,
вызвавшие развитие именно пятилучевой радиальной симметрии,
появление которой из двустороннего строения предковых форм иглоко-
294
жих объясняют переходом к сидячему, прикрепленному образу жизни,
еще требуют глубокого анализа, как, впрочем, и вся проблема формы в
органическом мире.
Следуя Бэзеру (Bather, 1915) и Иекелю (Jaekel, 1918), римскими циф-
рами (I—V) обозначаем амбулакры, арабскими (1—5)—интерамбулак-
ры; плоскость симметрии иглокожих проводим через III амбулакр и 5-й
интерамбулакр, между I и V амбулакрами, где в примитивном случае
лежат гидропора или мадрепорит, половое и анальное отверстия. Более
полно эти исходные отношения сохраняются у Pelmatozoa; у Eleutherozoa
они сохраняются частично или проявляются лишь в онтогенезе (см. Фе-
дотов, .1951) и сильно изменяются с развитием позднейшей вторичной
двусторонней симметрии. '
По морфологии, тесным коррелятивным связям со многими системами
органов и происхождению скелет иглокожих—очень характерная осо-
бенность этого типа. Он внутренний, мезодермальный по происхождению
и возникает интрацеллюлярно в соединительнотканых клетках в виде
зернышек кальцита, развиваясь затем в многоядерном синцитии. Далее
зачаток скелета, проходя стадию или трехногого стульчика или первично-
го креста, превращается в решетчатую пластинку с шестиугольными
ячеями, из которых образуются разнообразные сложные элементы по-
верхностного и внутреннего скелета взрослых особей в виде пластинок,
позвонков, педицеллярий, игл и т. д. грубоячеистого, вплоть до плотного,
строения. У голотурий, кроме того, происходит развитие зачатка скеле-
та в форме пластинки с шестью отверстиями, так называемой пряжки
(Schmidt, 1930). Как известно, личиночный скелет у всех взрослых иг-
локожих, кроме голотурий, исчезает. У последних он сохраняется и обыч-
но, за исключением окологлоточного скелетного кольца, состоит из от-
дельных известковых телец разной формы.
Скелетный элемент иглокожих (Schmidt, 1930; A. Schulz, 1935; Kir-
chner, 1929а, 19296) по фигуре вытравливания, излому, по плоскости спай-
ности и двоякому лучепреломлению подобен одному кристаллу кальцита.
Оптическая ось у элементов типа пряжки расположена тангенциально,
у остальных, развивающихся из решетчатой пластинки,— перпендикуляр-
но к плоскости последней. Будучи производным плазмы соединительно-
тканой клетки, кристалл скелета является биокристаллом (Schmidt,
1930), тесно связанным со строением и функцией частей тела иглокожего
данного вида.
Считают, что на положении оптической оси в биокристалле сказы-
вается его органическая субстанция. При общей филогенетической ста-
•бильности скелета предполагают, что изменение положения оптической
оси скелета может происходить под воздействием чувствительности игло-
кожих к свету или под влиянием кривизны, которую претерпевают ске-
летные части в теле иглокожих (Raup, 1962а, 19626). Тем не менее оп-
тические особенности скелета играют важную роль как в выяснении мор-
фологии скелета, например, скелета рта морских звезд и офиур, так и в
установлении филогенетических отношений в классах иглокожих.
В основном скелет состоит из СаСО3 в форме кальцита; количество
.MgSO4 колеблется между 5 и 15% зольного остатка и в значительной
степени зависит от температуры и ряда свойств морской воды; фосфаты
и сульфаты составляют примерно 1—1,7%; SiO2 и органическая субстан-
ция—сотые части процента (Виноградов, 1937, 1953; Cuenot, 1948). Ске-
лет современных иглокожих, кроме голотурий, составляет большую часть
их веса и по своему химическому составу различен у каждого класса
(Виноградов, 1937, 1953).
У наиболее древних вымерших иглокожих (ряд карпоидей, цистоидей,
некоторые эокриноидеи) скелет состоит из большого числа пластинок не-
правильной многоугольной формы, неправильно расположенных. Упоря-
295
дочение формы и расположения пластинок сначала происходит по ходу
амбулакральных желобков, возрастает по мере их развития и распро-
странения по телу (цистоидеи, эдриоастероидеи) и далее распространя-
ется на скелет всего тела. В ходе эволюции этих классов происходит оли-
гомеризация элементов скелета (отряд Regularia из цистоидеи, бласто-
идеи, часть эокриноидей); ограниченное число венчиков пластинок тела
устанавливается и у морских лилий.
В общем, у Pelmatozoa (Догель, 1954) эволюция скелета идет от
полимеризации к олигомеризации, нарушаясь существованием некото-
рых древних групп с олигомерным скелетом (например, эокриноидеи);
но эволюция этих групп заканчивалась слепо.
То же наблюдается в развитии скелета и у Eleutherozoa. Упорядоче-
ние формы и расположения и уменьшение числа пластинок интерамбу-
лакров у ряда палеозойских морских ежей, установление небольшого чи-
сла рядов пластинок — по два в каждом из амбулакров и интерамбулак-
ров — имеет место у большинства морских ежей. Множественное число
элементов скелета актинальной стороны дискасомастерид и ограничение
числа рядов пластинок скелета амбулакров наблюдается у морских
звезд. Все это и ряд'других примеров говорит о ходе эволюции скелета
амбулакров и интерамбулакров у многих Eleutherozoa от множественного
к ограниченному, олигомерному. Но, как и у Pelmatozoa, здесь олигоме-
ризация встречается в боковых, слепо кончавшихся ветвях, вроде ботрио-
цидарид, ряда офиоцистий, некоторых семейств палеозойских морских
звезд (Hudsonasteridae).
Как известно, современные голотурии имеют лишь микроскопически
мелкие, важные в систематическом отношении, известковые тельца,
очень разнообразной, характерной формы, и крупные части околорото-
вого скелетного кольца. В общем, у голотурий идет процесс редукции
скелета, полностью исчезающего у пелагических Pelagothuriidae. Лишь
как редкое исключение скелет из пластинок встречается у взрослых
Sphaerothuria, Psolus и у молодых особей Cucumaria georgiana и С. stei-
neri (Ekman, 1927; см. Федотов, 1951). Часть известковых телец голоту-
рий является сохранившимся скелетом личинок, остальные образуются
при превращении личинки во взрослую форму и продолжают возникать
и дальше. Вероятно, склериты личинок являются производными истин-
ной пряжки, а склериты взрослых голотурий возникают из решетчатых
пластинок.
Все, что известно относительно современных и ископаемых голоту-
рий, заставляет думать, что у них не было прочного скелета, подобного
панцирю морских ежей. Если скелет у древних ископаемых голотурий
был, то он состоял из тонких пластинок, не мешавших гибкости тела.
При захоронении он, естественно, не мог сохраниться целиком. Очевид-
но, скелет был утрачен голотуриями в отдаленном геологическом прош-
лом, и их эволюция шла и идет по пути прогрессивного развития гибко-
сти, сократимости и подвижности мягкого, нередко червеобразного гела.
Основой первичного абактинального скелета являются центральная
пластинка и правильные венчики, каждый из пяти радиально и интер-
радиально расположенных пластинок. У всех современных иглокожих,
кроме голотурий, они закладываются на абактинальной стороне заро-
дыша и в той или иной мере сохраняются у имаго.
У морских лилий, ископаемых и современных, скелет чашечки обра-
зуется у моноциклических — пятью радиальными и пятью базальными
пластинками (интеррадиально расположенными), у дициклических —
добавляется третий венчик инфрабазальных пластинок, расположенных
радиально. У позднейших форм — псевдомоноциклических (например,
Antedon) инфрабазальные пластинки вовсе выпадают из онтогенеза или
к его концу сливаются с верхним члеником стебля, образуя луковицеоб-
296
разный центральный конус. В ходе онтогенеза скелет чашечки утрачива-
ется, она становится мягкой, например, у Comatulidae, радиальные пла-
стинки становятся прикрытыми центральным конусом, базальные, сли-
ваясь вместе, образуют розетку, уходящую внутрь и становящуюся
крышей камерного органа.
В отличие от современных бесстебельчатых лилий у ряда ископаемых,,
например, нижнекаменноугольных микрокриноидей (Strimple a. Koenig,
1956), базальные пластинки сливаются и образуют центральную пла-
стинку абактинальной части чашечки, подобно центральной пластинке
Eleutherozoa. В ходе геологической истории морских лилий элементы
абактинального скелета претерпевали различные изменения формы, чи-
сла и расположения, наблюдались переходы от радиального к двусто-
роннесимметричному расположению в связи с образом жизни. При ре-
дукции рук или их атрофии рудиментарными становились радиальные-
пластинки, являющиеся опорой рук и т. д. И у современных бесстебель-
чатых лилий в ходе онтогенеза порой наблюдаются уклонения от обычно
пентамерного числа абактинальных пластинок (Mortensen, 1920).
У ряда цистоидей основание чашечки нередко образовано четырьмя
венчиками пластинок (например, отряд Regul'aria), у бластоидей — всего»
двумя венчиками. У цистоидей, как и у морских лилий, также наблю-
даются изменения в числе и положении элементов абактинального ске-
лета. , !
Членистый стебель морских лилий, достигающий у современных форм
свыше 2 ж, а у некоторых мезозойских — до 20 м в длину при кроне в lw
в диаметре, в геологической истории этого класса не раз утрачивался,
происходили переходы к свободному образу жизни, с возвратами вновь
к прикрепленному, (очевидно, вызванные изменениями условий сущест-
вования).
Наличие таких ископаемых форм, как Acanthocystites, Ascocystite^
(Jaekel, 1918), особенности строения стебля современных морских лилий
и характер онтогенеза антедона позволяют думать, что стебель морских
лилий является измененной абактинальной частью тела, прикрепившей-
ся к субстрату.
У морских звезд, ежей и офиур первичный абактинальный скелет со-
стоит из центральной или дорсо-центральной пластинки, венчика из пяти
терминальных, расположенных радиально пластинок, и венчика из пяти
базальных пластинок, расположенных интеррадиально. У морских звезд
терминальные пластинки прикрывают терминальное щупальце и глазок,
у морских ежей они несут отверстие для редуцированного терминального'
щупальца, скопления пигмента и называются глазными пластинками;
на базальных пластинках у морских ежей возникают половые отверстия,
в результате чего они становятся половыми пластинками. Зачаток одной
из базальных пластинок, окружая гидропору, становится у морских звезд
и ежей мадрепоритом, хотя у некоторых морских звезд мадрепорит об-
разуется из особого зачатка еще до появления базальных пластинок.
У офиур с развитием диска базальные пластинки смещаются на акти-
нальную (оральную) сторону диска и становятся оральными пластинка-
ми или ротовыми щитками, один из которых является мадрепоритом.
У многих современных молодых звезд абактинальный скелет состоит из
трех венчиков пластинок, так как развиваются еще зачатки пяти ради-
альных пластинок между базальными и терминальными пластинками.
У некоторых морских звезд, наоборот, происходит выпадение закладок
ряда элементов абактинального скелета и даже отсутствует правильность
в его развитии.
Первичный абактинальный скелет хорошо выражен у ряда палео-
зойских морских звезд, например, Hudsonasteridae и некоторых совре-
менных Pentagonaster, Tosia. У большинства современных форм, у кото-
297-
рых он наблюдается лишь в онтогенезе, их абактинальная сторона по-
крыта вторичным скелетом, который у высоко организованных морских
•звезд, например, Asteriidae, состоит из гибкой скелетной сети. Несомнен-
но, первичный абактинальный скелет будет установлен и в онтогенезе по
мнению Фелла, живой ископаемой сомастериды — Plalasterias latiradiata
(Fell, 1962а — в).
У морских ежей, у которых тело образовано актинальным диском,
•зародыша, а абактинальный диск представлен небольшим апикальным
полем панциря, абактинальный скелет ограничен этим полем. В ходе
эволюции морских ежей, особенно в связи с развитием двусторонней сим-'
метрии, абактинальный скелет претерпевает сильные изменения (см. Фе-
дотов, 1951).
^Общим для большинства классов иглокожих — цистоидей, бластои-
,дей, морских лилий, вероятно, эокриноидей и паракриноидей, офиоци-
стий, поскольку у последних имеется центральная пластинка и венчики
радиально и интеррадиально расположенных пластинок (Volchovia —
Regnell, 1948а; Fedotov, 1926), морских звезд, ежей и офиур—является
наличие радиальносимметричного абактинального скелета. Он также ха-
рактеризует радиальную симметрию иглокожих, как и амбулакральные
желобки на актинальной стороне тела. К сожалению, остается неизвест-
ным абактинальный скелет эдриоастероидей, обладающих амбулакрами,
близкими таковым Eleutherozoa. Можно считать, что в целом абакти-
нальный скелет представителей всех классов иглокожих — гомологичное
образование, но гомологизировать отдельные элементы этого скелета
разных классов все же нельзя. Во всяком случае, по данным онтогенеза
и филогенеза, дорсо-центральная пластинка морских звезд, ежей и офиур
не гомологична ни центральной пластинке ряда палеозойских морских
лилий, ни центральному конусу бесстебельчатых морских лилий. Наличие
абактинального скелета у морских звезд, ежей и офиур, возможно, гово-
рит об их сидячих прикрепленных предках.
Первичный актинальный скелет чашечки морских лилий, цистоидей
и бластоидей состоит из пяти оральных или дельтоидных пластинок и
двойного ряда кроющих и боковых пластинок амбулакральных желоб-
ков; у морских лилий последние продолжаются в руки, абактинальный
скелет которых составляют брахиальные пластинки. На одной или на
всех оральных пластинках находится по одному или по многу поровых
отверстий. Дельтоидные пластинки, расположенные интеррадиально и
прикрывающие рот, хорошо развиты у большинства ископаемых и у не-
многих современных морских лилий, у которых чаще они существуют в
онтогенезе, но исчезают у имаго.
Актинальный скелет рук морских звезд в основном состоит из двойно-
го ряда амбулакральных пластинок, образующих свод амбулакрального
желобка, и двойного ряда адамбулакральных пластинок, расположен-
ных кнаружи и книзу от амбулакральных. Амбулакральные и адамбу-
.лакральные пластинки лежат или друг против друга, или в чередующему-
ся порядке. Неподвижный околоротовой скелет образован первой
парой адамбулакральных и двумя парами амбулакральных пласти-
нок.
У Platasterias latiradiata, несмотря на ряд крупных особенностей ак-
тинального скелета и исходя из описания самого автора (Fell, 1962а — в,
см. его рисунки), у этой звезды, как и у ряда современных форм, надо
признать наличие амбулакральных, адамбулакральных, инфра- и супра-
маргинальных пластинок.
У палеозойских офиур первичный актинальный скелет с открытыми
амбулакральными желобками состоял из двойного ряда амбулакральных
пластинок, расположенных в чередующемся порядке или друг против
друга, и с каждой наружной стороны из ряда адамбулакральных пласти-
.298
вок, часто именуемых латеральными. У мезозойских и современных
офиур, по мере усиления их подвижности, амбулакральные пластинки,
попарно сливаясь, образуют подвижно сочлененные позвонки рук. Адам-
булакральные пластинки превращаются в боковые, прикрывающие руки
с боков; возникающие у них спинные и брюшные пластинки прикрыва-
ют руки сверху и снизу.' Первая пара адамбулакральных и две пары
амбулакральных пластинок образуют подвижный, жующего типа рото-
вой скелет.
Можно согласиться, что у сомастерид амбулакральные пластинки и
виргалии, прилегающие к ним с каждой стороны (Spencer, 1951), гомоло-
гичны основным элементам актинального скелета амбулакров морских
звезд и офиур, но прямо вывести строение их ротового скелета из таково-
го сомастерид нельзя.
У морских ежей весь панцирь, кроме небольшого апикального поля, и
перистомальный скелет образованы актинальным скелетом: амбулак-
ры— амбулакральными, интерамбулакры — интерамбулакральными пла-
стинками. Вопрос о гомологии этих пластинок морских ежей со ске-
летом морских звезд и офиур не решен. Считают (например, Беклеми-
шев, 1952), что амбулакральные пластинки морских ежей не гомологичны
таковым морских звезд и офиур, а гомологичны брюшным пластинкам
последних и даже кроющим пластинкам Pelmatozoa.
Эволюция скелета у морских ежей в палеозое, как отмечалось выше
(стр. 295), шла по пути упорядочения формы, расположения и ограниче-
ния числа интерамбулакральных пластинок, формирования интерамбу-
лакров и амбулакров из нескольких или многих рядов простых пласти-
нок (лепидоцентроиды, мелонехиниды), реже из двух рядов. Лишь у
цидароид уже в палеозое, а у остальных начиная с мезозоя, устанавли-
вается по два ряда пластинок в амбулакрах и интерамбулакрах. Вместе
с тем идет развитие амбулакральных пластинок от простых к сложным,
образующихся из нескольких первичных пластинок. Правильные мор-
ские ежи сохраняют в известной мере однообразие в строении амбулак-
ров и пластинок панциря. С возрастающей дифференцировкой двусто-
роннесимметричного панциря неправильных морских ежей, с разделе-
нием его на верхнюю и нижнюю половины, изменяется функция их ча-
стей, изменяются фррма амбулакров и интерамбулакров, строение их
пластинок, амбулакральных ножек, игл и т. д., достигающих максимума
у ряда современных спатангоид (например, бутылкообразные Pourtalesia,
Echinosigra и др.).
Поверхностный скелет характерен лишь для Eleutherozoa и развит
преимущественно у морских звезд и ежей в виде игл и их производ-
ных— паксилл, педициллярий, сферидий. Иглы (от микроскопических
размеров до 30 см длиной) несут функции зашиты и органов движения —
у морских ежей. Педициллярии, наиболее развитые у морских ежей
(глобиферные, трезубые, офицефальные, трифильные, клювовидные) и
сравнительно однообразные и простые у морских звезд, служат для очи-
стки поверхности тела, для захвата добычи и защиты от врагов. Имея
большое биологическое значение, они важны также и для систематики.
У ряда морских звезд, и особенно ежей, педициллярии (и отчасти иглы)
снабжены сильно действующими ядовитыми железами.
Стенка тела состоит из эпидермиса, соединительнотканого слоя (ку-
тис, кориум, дермис) и перитонеального эпителия. У-голотурий, морских
звезд, у Echinothuriidae из морских ежей и офиур группы Euryalae* между
кутисом и перитонеальным эпителием находится мышечной слой. Эпи-
дермис у голотурий не ресничный, у остальных — с ресничками или жгу-
тиками, которые не разбросаны равномерно, но концентрируются в оп-
ределенных местах тела, преимущественно на абактинальной стороне и в
.амбулакральных желобках. Реснички вызывают на теле иглокожих токи
299
воды, о чем будет сказано ниже. Слабо кутикуляризованный эпидермис
нередко синцитиальный. У офиур, за исключением группы Euryalae, и у
некоторых морских лилий эпидермис может отсутствовать, отчего ске-
лет почти обнажен. Среди эпидермальных клеток находятся чувствитель-
ные, нервные и железистые клетки.
Среди эпидермальных клеток могут быть пигментные. Гранулы пиг-
мента и пигментные клетки встречаются у многих иглокожих. У некото-
рых морских звезд (Brisinga и др.), офиур (Ophiacantha bidentata, Ophio-
scolex glacialis и Др.) имеются крупные фотогенные клетки, глубоко
вдающиеся в кутис, вызывающие свечение. Слизь, выделяемая базофиль-
ными клетками, обильно смачивает тело иглокожих и имеет защитное:
значение.
Жгутики распределены неравномерно, но концентрируясь в опреде-
ленных местах на поверхности тела у представителей всех групп, кроме
голотурий, они создают токи воды, служащие для дыхания, очистки тела1
от грязи и для подачи пищи (Gislen, 1924) (см. ниже).
Очень характерной чертой иглокожих является наличие в стенке тела
соединительнотканого слоя, или кутиса, с коллагеновыми волокнами
того же гистохимического характера, что и коллаген позвоночных (Fer-
guson, 1960). В нем, достигающем наибольшей мощности у голотурий
(чем компенсируется отсутствие у них скелета и обусловливается гиб-
кость их кожистых стенок тела), различают три слоя. В наружном слое
кутиса лежат скелетогенные клетки, образующие скелет, пигментные
клетки и гранулы, обусловливающие цвет тела. Иглокожие бывают крас-
ными, оранжевыми, желтыми, голубыми, пурпуровыми, фиолетовыми, ко-
ричневыми, черноватыми. Наиболее яркими и разнообразно окрашенны-
ми являются морские звезды некоторых отрядов. Окраска тела иглоко-
жих зависит от нескольких пигментов, изученных разными авторами
(Hyman, 1955; Nichols, 1964),— меланина, рибофлавина, других флуорес-
цирующих пигментов, а также — от каротиноидных пигментов: ксанто-
фила, каротина и других от красного до желтого цветов.
Толщина стенок обусловлена мощностью кутиса, который наиболее
сильно развит у голотурий, хотя у некоторых из них, например, синап-
тид, стенки тела и кутис очень тонки. У морских звезд кутис выражен хо-
рошо; у иглокожих с сильно развитым скелетом тела, у морских ежей,
морских лилий и неветвистых офиур главная толща стенок тела состоит
из скелетных элементов.
Мускулатура из гладких мышц в стенке тела развита у голотурий,
морских звезд, морских ежей—эхинотуриид, и у многоветвистых офиур
Euryalae. У голотурий с их мягкими кожистыми стенками тела во внут-
реннем слое кутиса лежат кольцевые мышцы, прерванные пятью орди-
нарными (Apoda, Elasipoda, Dendrochirota) или двойными (Aspidochiro-
ta, Molpadonia) продольными мускульными лентами, которые проходят
по внутренней стороне амбулакров.
Кроме мышц стенок тела, существуют отдельные мышцы, несущие
разнообразные функции. У морских лилий мышцы между брахиальными
пластинками рук и скелетными частями пиннул, сокращаясь, вызывают
их движения, обусловливающие передвижение и плавание морских ли-
лий. У голотурий развиты мускулы-ретракторы, втягивающие переднюю
часть тела со ртом и щупальцами, и протракторы, ее расправляющие,
дилататоры глотки, дилятаторы клоаки.
Системы отдельных мышц между амбулакральными и адамбулакраль-
ными пластинками рук морских звезд, между позвонкам® рук офиур,
мышцы педицеллярий, игл, амбулакральных ножек, целая систем^ из 60
различных мышц в аристотелевом фонаре морских ежей и ряд других
обеспечивают движение животных, сокращение и расправление отдель-
ных их частей и. органов.
300
Для иглокожих, как и для кишечнодышащих, характерным является
участие в энергии мышечного сокращения не только аргинин-фосфагена,
но и креатин-фосфагена. Данные о распределении их в настоящее время
противоречивы. По одним авторам (Judkin, 1954; Prosser a. Brown, 1961),
аргинин-фоофаген имеется у морских звезд, голотурий и морских лилий,
у офиур.— только креатин-фоофаген, у одних морских ежей — только ар-
гинин-фосфаген, у других — оба. По другим авторам (Kerkut, 1960; Roche
a. others, 1960) оба фосфагена найдены у морских ежей, звезд, голотурий
:и офиур. По-видимому, распределение фосфагенов не имеет филогенети-
ческого характера для определенных классов иглокожих.
Нервная система иглокожих чрезвычайно любопытна по строению,
происхождению и физиологии. Она состоит, как известно, из эктоневраль-
:ной чувствительной системы, гипоневральной двигательной, энтонев-
ральной (апикальной, или аборальной) двигательной систем и слабораз-
витых органов чувств. В соответствии с радиальной симметрией всей ор-
ганизации каждая из этих систем представлена кольцом и пятью ра-
диальными нервами, но с известными различиями в разных классах.
Чувствительная эктоневральная нервная система, сплетение волокон
которой сильно развито в покровном эпителии тела и эктодермальном
эпителии кишечника, является производной эктодермы и энтодермы, по-
добно нервной системе кишечнополостных. Невриты чувствительных кле-
ток и ассоциативные клетки расположены более поверхностно — в ба-
зальной части эпителия, двигательные клетки лежат глубже (Smith,
1937, 1947, 1950, см. Беклемишев, 1952, 1964). Гипоневральная и энтонев-
ральная двигательные системы, тесно связанные с эпителиальными
клетками целома, имеют мезодермальное целомическое происхождение.
Однако.это недостаточно точно установлено и требует дополнительных
исследований (Хлопин, 1946).
У морских звезд, ежей, офиур и голотурий эктоневральная чувстви-
тельная нервная система представлена околоротовым кольцом, пятью ра-
диальными нервами и субэпидермальным сплетением; у морских лилий
юколоротового кольца, по-видимому, нет. У морских лилий и звезд эк-
тоневральная нервная система лежит под эпидермисом открытых ам-
булакральных борозд. У морских ежей, офиур и голотурий, в связи с за-
мыканием амбулакральных борозд в эпиневральные каналы, соединен-
ные околоротовым эпиневральным кольцом, эктоневральная нервная си-
стема, образованная дном этих полостей, заключена в них. Однако у го-
лотурий околоротового эпиневрального кольцевого канала нет, а онто-
генез эктоневральной нервной системы, как отмечает В. Н. Беклемишев
(1952), сильно изменен. На границе рта нервное сплетение от околорото-
вого кольца переходит в субэпителиальное сплетение пищеварительного
аппарата. ;Этой части эктоневральной системы приписывают эктодер-
мальное происхождение.
Гипоневральная двигательная нервная система у морских лилий,
звезд и офиур представлена околоротовым кольцом и пятью радиальны-
ми нервами. У голотурий нет околоротового нервного кольца, но имеется
пять радиальных гипоневральных нервов, хорошо развитых в связи с
сильной сократимостью мягких стенок тела. У морских ежей, заключен-
ных в твердый панцирь, двигательные гипоневральные нервы отсут-
ствуют, причем у правильных морских ежей, имеющих аристотелев фо-
нарь, сохраняется пять узелков гипоневрального кольца, у неправиль-
ных ежей гипоневральная нервная система отсутствует.
Энтоневральная двигательная нервная система, как известно, наибо-
лее развита у морских лилий, даже бесстебельчатых; она находится на
дне чашечки, возникая за счет целомического эпителия пятикамерного
органа, и посылает мощные двигательные радиальные нервы в руки, со-
ставляя, по В. Н. Беклемишеву (4952), анатомически и физиологически
301
центральную часть нервной системы морских лилий. Движение рук и
пиннул и координация движений зависят от этой нервной системы; у
морских ежей и офиур она развита слабо; у голотурий отсутствует, что-
связано с иным способом движения и иннервацией органов движения ги-
поневральной нервной системой.
У морских звезд в целомическом эпителии абактинальной (верхней)
стенки диска находится пять радиальных нервных тяжей, сходящихся
друг с другом; у морских ежей и офиур энтоневральная нервная система
представлена нервным кольцом, находящимся в кольцевом аборальном
синусе; она посылает нервы к гонадам.
Эктоневральная нервная система иннервирует амбулакральные нож-
ки щупальца, глазки, сферидии, педицеллярии, иглы и обеспечивает их
целесообразную рефлекторную деятельность. Гипоневральная нервная
система посылает нервы к мускулатуре рук и стенок тела. Поворачива-
ние тела, бегство и другие сложные рефлекторные акты протекают под
координирующим воздействием центрального отдела нервной системы
(Беклемишев, 1952, 1964).
Хотя диффузное нервное сплетение эктоневральной нервной системы
иглокожих напоминает нервную систему кишечнополостных, между ними'
имеется очень большое различие по существу. Так, движения громадного
числа амбулакральных ножек морской звезды, служащих для ловли до-
бычи и для передвижения этих животных, строго координируются дея-
тельностью эктоневральной нервной системы. По образному выражению
Смита (Smith, 1947), деятельность этого аппарата сложнее системы теле-
фонной сети Лондона.
Физиологи (Smith, 1950; Коштоянц-, 1957) допускают у иглокожих на-
личие сложного нейронального комплекса, который обеспечивает возбуж-
дение нерва,.передающееся мышцам (например, у голотурий), и находят
у них в этом сходство с тем, что происходит у позвоночных при перехо-
де возбуждения на мотонейроны в спинном мозгу и в так называемом
мионевральном синапсе.
В последнее время у морских звезд, как и у хордовых, установлено
существование двух главных типов нервных стимуляторов мышечных
сокращений — ацетилхолина и адреналина (точнее норадреналина низ-
ших позвоночных), действующих на нервные окончания взаимопротиво-
положно. В частности, эти стимуляторы вызывают раскрывание и за-
мыкание рта, расслабление и сокращение стенок желудка (Anderson,
1954). Найдены также в моторной нервной системе морских звезд (Unger,
1960 — Marthasterias glacialis) и офиур (Fontaine, 1962 — Ophiopholis-
aculeata) нейросекреторные клетки, давно известные для представителей
многих групп беспозвоночных.
Органы чувств у иглокожих, как животных, медленно двигающихся,.,
живущих нередко в толще грунта моря и в норах, развиты слабо,
в соответствии с низким уровнем развития нервной системы. Слабое раз-
витие органов чувств связано, несомненно, со всей геологической историей
иглокожих, которые изначально были придонными, преимущественно
прикрепленными организмами. Органы чувств наиболее слабо развиты у
морских лилий и офиур, хотя среди них имеются наиболее быстро двига-
ющиеся иглокожие.
Единственными оформленными органами чувств у иглокожих явля-
ются статоцисты синаптид, элазипод и мольпадоний из голотурий и сфе-
ридии морских ежей как орган равновесия, глазки на терминальных щу-
пальцах большинства морских звезд, похожие на глазки, пигментные
пятна глазных пластинок и терминальных щупалец морских ежей, и, на-,
конец, пигментные пятна на основаниях щупалец некоторых синаптид.
Глаза морских звезд как органы зрения трактуются различно (Hyman,
1955). Роль их как органов восприятия при общей значительной свето-
302
чувствительности покровов морских звезд невелика. Однако разная (от
пигментных пятен до сформированных глаз у высших морских звезд—•
семейство Asteriidae) степень развития глазков у морских звезд различ-
ных групп, а также отсутствие их у ряда глубоководных форм говорят о
1 значении их как органов восприятия света. Пигментные пятна глазных
пластинок морских ежей лишены светочувствительных палочек (Догель,.
1940), но положение их, как и глазков морских звезд, у основания терми-
нального щупальца позволяет считать глазные пятна морских ежей по-
! терявшими функцию редуцированными глазками.
С субэпителиальным нервным сплетением связана общая чувстви-
тельность покровов 'иглокожих, в которых находятся как отдельные чув-
ствительные клетки, так и многоклеточные сенсиллы. Последние особен-
но многочисленны на амбулакральных придатках — щупальцах и амбу-
( лакральных ножках морских лилий, звезд, ежей, голотурий, офиур. Ор-
| ганами осязания считаются также терминальные щупальца на конце
I; амбулакров. Как было сказано, покровы иглокожих обладают светочув-
L ствительностью; у одних — отрицательной, у других — положительной,
I что обусловливает перемещения иглокожих из освещенных мест дна
! моря в затемненные и наоборот.
. Оформленных органов обоняния и вкуса у иглокожих также нет, но
многочисленные чувствительные клетки на их амбулакральных ножках
и щупальцах, на ротовых щупальцах, на иглах и в педицелляриях вос-
принимают на значительных расстояниях вкусовые раздражения.
Несмотря на большое число наблюдений за поведением иглокожих и
исследований их органов чувств (Hyman, 1955), точных данных о физио-
логии органов чувств еще мало, что в известной мере вызвано низким
уровнем как их строения, так и отправления.
Как ископаемые иглокожие, не исключая по последним Данным и
Carpoidea, так и современные обладают кишечником с ротовым и аналь-
ным отверстиями, за исключением немногих морских звезд (Porcellana-
steridae, Astropectinidae и Ludiidae) и всех офиур. В связи с планом стро-
ения и в известной мере с различиями в характере пищи наблюдается два
, типа строения кишечника — длийный трубчатый и короткий мешковид- _
ный. Исходным, во всяком случае широко распространенным, является
трубчатый кишечник; им, надо думать, обладали карпоидеи, цистоидеи,
бластоидеи, эдриоастероидеи и вымершие морские лилии; им обладают
современные морские лилии, голотурии и морские ежи. Возможно, что
офиоцистии имели, подобно морским звездам и офиурам, мешковидный
кишечник.
При сидячем образе жизни кишечник становится петлеобразным,
анальное отверстие сближается с ротовым, располагаясь на верхней,,
оральной (актинальной) стороне тела. С этим связана микрофагия, пита-
ние мелкой пищей, поступающей в рот по амбулакральным желобкам
с помощью токов воды, вызываемых ресничками, и обволакиваемой
слизью. Такой способ питания наблюдается у современных морских ли-
лий, у некоторых Eleutherozoa он был свойствен их ископаемым 'пред-
ставителям, а также цистоидеям, бластоидеям и эдриоастероидеям. Боль-
шинством современных свободноподвижных иглокожих добыча ловится
активно.
По аналогии с другими группами говорят о передней, средней и зад-
ней кишке иглокожих, что не обосновано анатомически и эмбриологи-
чески. У личинок всех иглокожих кишечник образован энтодермой, лишь
у личинок некоторых иглокожих (Leptosynapta) имеется эктодермаль-
ный стомодеум. Также и у взрослых иглокожих кишечник почти целиком
' эктодермальный. Эктодерма принимает незначительное участие в обра-
зований переднего отдела кишечника, ближайшего ко рту, и при возник-
новении порошицы.
303
За ротовым отверстием начинается пищевод, переднюю часть которо-
го нередко называют глоткой (например, у голотурий). У форм с труб-
чатым кишечником пищевод развит хорошо, у форм с мешковидным—
почти не выражен. За пищеводом следуют желудок и кишка, которые
у Морских лилий, ежей и голотурий почти не обособлены друг от друга.
Конечную часть кишечника называют прямой кишкой или просто киш-
кой; у многих голотурий она дифференцирована в виде сильно сократи-
мой клоаки. У большинства современных морских лилий рот распола-
гается почти в центре диска, а порошица — эксцентрично в заднем интер-
радиусе, у немногих форм (Actinometridae) рот лежит у периферии, а
анальное отверстие— в центре диска. Трубчатый кишечник, начинаясь
ротовым отверстием, загибается по часовой стрелке и открывается на
конце анального конуса или образует четыре оборота, по длине во много
раз превышая диаметр диска. Характерно, что у морских лилий, как и у
Eleutherozoa, кишечник делает поворот по часовой стрелке, а у многих
ископаемых Pelmatozoa — в обратном направлении (Jaeckel, 1897).
У голотурий рот, активно собирающий пищу с помощью Щупалец,
чаще находится на переднем конце тела, анальное отверстие — на зад-
нем. Их положение может меняться в связи с образом жизни, вплоть
до сближения друг с другом на вытянутом переднем конце тела. Узкий
трубчатый кишечник (в 3—7 раз больше длины тела), прикрепляющийся
к стенкам тела мезентериями, у разных отрядов образует определенное
число петель. У голотурий Apoda, обладающих примитивными и вторич-
ными чертами, кишечник может быть почти прямой трубкой (например,
у Leptosynapta). Как у морских лилий и ежей, кишечник голотурий дела-
ет поворот вокруг продольной оси тела по направлению движения часо-
вой стрелки. У многих голотурий, кроме синаптид и некоторых глубоко-
водных форм, в клоаку открываются водные легкие, а у различных As-
pidochirota органы защиты — кювьеровы органы.
У правильных морских ежей рот находится в центре перистома, на ак-
тинальной стороне, анальное отверстие—в перипрокте, на абактиналь-
ной стороне; у двустороннесимметричных спатангоид (например, семей-
ство Pourtalesiidae) рот и порошица становятся почти терминальными,
смещаясь на передний и задний конец нижней стороны панциря. Рот ак-
тивный и у большинства снабжен аристотелевым фонарем, зубы которо-
го схватывают и пережевывают пищу. Число скелетных элементов ари-
стотелева фонаря доходит до 40, а их мышц — до 60. Некоторые элемен-
ты, например, дужки или компасы, и их мускулатура у большинства пра-
вильных морских ежей участвуют в дыхании, наполняя жидкостью пе-
ристомальные жабры и опорожняя их. У неправильных морских ежей в
связи с характером пищи происходят в разной степени упрощения ари-
стотелева фонаря, например, у клипеастероид. У голостомоид и касси-
дулоид он сохраняется лишь у молодых особей, а у спатангоид наблю-
дается его полная редукция с выпадением закладки в онтогенезе.
На основании ряда данных думают, что части аристотелева фонаря
гомологичны амбулакральным и адамбулакральным элементам, ротово-
го скелета морских звезд.
Узкий трубчатый кишечник правильных морских ежей, выходя из
аристотелева фонаря, поднимается кверху, изгибаясь по направлению
часовой стрелки, и образует круг, потом, поворачиваясь в обратном на-
правлении, делает второй круг. Затем он поднимается кверху и анусом
•открывается в перипрокте на абактинальной стороне панциря. У одних
неправильных морских ежей (спатангоид) кишечник делает два оборо-
та, у других (клипеастероид) —один, но зато широкий.
У большинства правильных морских ежей, кроме цидароид, и части
неправильных ежей кишечник снабжен «сифоном», у некоторых форм
двумя. Кишечник наружным мезентерием прикреплен к стенке панциря.
304
Кишечник морских звезд и офиур, как было сказано выше (стр. 302),
мешковидный, короткий. Рот находится в центре перистома, ведет в ко-
роткий (морские звезды), почти не выраженный (офиуры) пищевод,
переходящий в желудок. Последний у морских звезд (Anderson, 1954)
кольцевой перетяжкой подразделен на нижнюю, кардиальную часть —
объемистый мешок, закрепленный в каждом луче двумя гастральными
лигаментами, возникающими из мезентериев личинки, и на верхнюю,
или пилорическую часть. Последняя образует пять пар длинных желе-
зистых, так называемых печеночных, придатков, прикрепленных парны-
ми мезентериями к верхней стенке рук. Очень короткая кишка, идущая
от пилорического отдела, образует слепые интеррадиальные дивертику-
лы и открывается несколько эксцентрично расположенной порошицей
на абактинальной стороне тела. Неподвижность ротового скелета ком-
пенсируется способностью желудка выворачиваться и охватывать добы-
чу и сильным развитием его пищеварительных желез. Непереваривае-
мые крупные остатки пищи выбрасываются через рот.
У офиур, не имеющих, вернее, утративших заднюю кишку и пороши-
цу, развит мешковидный, сплющенный дорсо-вентрально желудок с
широкими, короткими радиальными и интеррадиальными выростами.
Ротовой скелет, благодаря мускулатуре челюстей и зубов, является под-
вижным жевательным аппаратом. Если ротовой скелет сомастерид обра-
зован тремя парами ближайших ко рту амбулакральных пластинок
(Spencer, 1951), то он не гомологичен ротовому скелету морских звезд
и офиур, образованному двумя парами амбулакральных и одной парой
адамбулакральных. Мало того, на основании данных новых исследова-
ний (например, A. Schulz, 1935; Horstadius, 1939), приходится пересмат-
ривать и гомологию частей ротового аппарата морских звезд и офиур,
установленную когда-то Людвигом (Ludwig u. Hamann, 1889—1907).
Как будет видно ниже, иглокожие преимущественно хищники —
макрофаги. Однако среди всех современных классов распространена и
микрофагия, питание микроорганизмами и детритом реснично-мукозным
способом — путем обволакивания слизью пищевых частиц и их передви-
жения ко рту с помощью ресничек. Последние исследования (Nichols,
1964) говорят, что один и тот же вид,-например, офиур (Ophiolomina
nigra) может быть и микрофагом, и макрофагом.
Пищей морских лилий служат диатомовые, одноклеточные и нитча-
тые зеленые водоросли, различные Protozoa, мелкие ракообразные, ли-
чинки, поступающие с детритом. Микрофагами по Спенсеру (Spencer,
1951) были также и древнепалеозойские сомастериды. Морские звезды
с остроконечными амбулакральными ножками — Porania, Ctenodiscus,
Pontasier, Astropecten и другие из подотрядов Phanerozonia (Gislen,
1924) —также микрофаги, питающиеся мелкими организмами, поступа-
ющими с детритом в пассивный рот с токами <воды, вызываемыми дви-
жениями ресничек на поверхности тела и амбулакральных желобков.
Таким же способом питаются и некоторые офиуры. Микрофагами яв-
ляются и морские ежи из клипеастероид (Hyman, 1955; Goodbody, 1960).
Большинство морских звезд — хищники, питающиеся крупной добы-
чей: двустворчатыми и другими моллюсками, полихетами, морскими
ежами, голотуриями, ракообразными, рыбами. Добычу они ловят педи-
целляриями, амбулакральными ножками и руками, порой выкапывают
(например, Smith, 1961) двустворок из грунта амбулакральными нож-
ками. Мелкая добыча глотается целиком, крупная обволакивается выво-
роченным желудком. Открывание створок моллюсков достигается нарко-
зом (под действием токсинов), силой тяги и просовыванием в щель
между створками стенок желудка (см. Nichols, 1964).
Большинство офиур питается полихетами, ракообразными, особенно
мелкими моллюсками, которых они ловят ножками и руками.
20 Д. М. Федотов
305
Пища морских ежей сравнительно с другими иглокожими гораздо
разнообразнее. Правильные ежи почти всеядны — диатомовые, придон-
ные водоросли, губки, гидроиды, мшанки, сидячие полихеты, ракооб-
разные, иглокожие, падаль идут в пищу плотоядных и растительноядных
морских ежей, которые играют большую роль в очистке дна моря от
органических отбросов. Echinothuriidae могут питаться остатками на-
земной древесины (Mortensen, 1938). Пища неправильных ежей более
однообразна. Многие, роясь в грунте, питаются одноклеточными водо-
рослями, фораминиферами, кусками полихет, моллюсков, иглокожих.
Голотурии ловят планктонные организмы клейкими разветвленными
щупальцами или набивают кишечник илом и песком с водорослями, фо-
раминиферами, радиоляриями, органическими остатками.
В целом, при наличии известных частных особенностей, стенки ки-
шечника иглокожих состоят из жгутиковых, очень узких, высоких клеток
эпителия, субэпителиального нервного слоя, слоя соединительной ткани
с кровеносными лакунами, слоя продольных и кольцевых мышц, слоя
соединительной ткани и ресничного перитонеума. В эпителии разброса-
ны железистые клетки — слизистые, зернистые, секреторные клетки, со-
держащие гликоген, липиды, полисахариды. Гистологическое строение
кишечника исследовалось многими старыми авторами, а в последнее
время у морских звезд — Гайаши (Hayashi, 1935), Андерсоном (Ander-
son, 1953, 1954, 1960), у морских ежей — рядом авторов (см. Nichols,
1964). Новыми исследованиями почти не затронуто изучение пищеварег
ния у голотурий, офиур и морских лилий. В распространении питатель-
ных веществ амбулакральная и кровеносная системы не имеют суще-
ственного значения; оно обеспечивается целомической жидкостью и аме-
боцитами (Nichols, 1964).
Пищеварение лучше известно у голотурий и морских звезд, меньше
у морских ежей и мало у морских лилий и офиур. В кишечном соке и
экстрактах кишечника у морских звезд и голотурий (Федотов, 1951;
Hyman, 1955) найдены липаза, амилаза, инвертаза, триптаза, бутираза,
эриптаза, протеиназа и другие ферменты. Считают, что пищеварение
внеклеточное, кроме внутриклеточного переваривания жиров. В стенках
кишечника и его эпителии находится много амебоцитов, участвующих в
пищеварении.
Общая полость тела, в которой находятся половые железы и кишеч-
ник, является единой полостью, частично поделенной неполными мезен-
териями. Она образуется левым и правым задними целомами личинки;
в ходе метаморфоза левый перемещается на актинальную, правый — на
абактинальную сторону иглокожего и дают гипогастральный и эпигаст-
ральный целомы. У большинства иглокожих (голотурии, морские звезды
и большая часть морских ежей) полость тела велика, но у морских ли-
лий она очень незначительна, так как зарастает соединительнотканными
тяжами, сохраняющими, однако, ресничную перитонеальную выстилку;
у некоторых морских лилий полость тела практически отсутствует.
У ряда морских ежей (отряда Clypeasteroida) мощно развитый внутрен-
ний известковый скелет сильно уменьшает объем полости тела. У типич-
ных офиур полость тела диска занята объемистым желудком, бурсами
и половыми железами. У офиур группы Euryalae целомическая полость
тела морфологически и физиологически замещена эктодермальной, так
называемой третичной полостью тела (Федотов, 1916; Fedotov, 1926),
образованной бурсами. У морских звезд полость тела продолжается в
руки в виде объемистых полостей; у офиур тело дифференцировано на
диск и хорошо обсоблены руки, просвет которых почти заполнен позвон-
ками, полость тела представлена в руках узким спинным каналом. У мор-
ских лилий полость тела чашечки заходит в руки и далее в пиннулы в
виде спинного, или апикального, целомического канала, проходящего в.
306
желобке брахиальных пластинок, и орального, или субтентакулярного,
канала, поделенного вертикальной септой на два.
Часть общей целомической полости тела у иглокожих, за исключе-
нием морских лилий, обособлена около пищевода в виде кольцевого
перифарингеального целома (перифарингеальный синус, оральный целом,
периэзофагеальный синус, или целом, перибуккальный синус). Он обра-
зуется за счет заднего левого целома личинки. У голотурий и морских
звезд он широко сообщается с общей полостью тела, у правильных мор*
ских ежей — с полостью, окружающей аристотелев фонарь, у офиур он
вполне замкнут. Кроме того, от общей полости тела у голотурий отделен
перианальный синус, а у правильных морских ежей на аборальном по-
люсе отделены два синуса — перипроктальный и перианальный.
Общая полость тела выстлана ресничным (жгутиковым) перитоне-
альным эпителием, одевающим также и внутренние органы, лежащие в
ней; как правило, она не сообщается с наружной средой, однако у мор-
ских лилий вторичные поровые каналы открываются непосредственно в.
полость тела, а у некоторых голотурий, например, Paracaudina chilensis
(Yazaki, 1930), полость тела пятью анальными папиллами на заднем
конце сообщается с наружной средой. В спинном целомическом канале
рук у морских лилий находятся группы из 2—6 мерцательных мешоч-
ков или ямок, служащих, видимо, для движения полостной жидкости.
У ряда голотурий Apoda имеются ресничные воронки — урночки, вы-
лавливающие продукты распада из полости тела (Е. Schulz, 1895;
Ласточкин, 1914, и др.); у некоторых голотурий того же отряда
наблюдаются pulsatile rosatte и vibratila clubs французских авторов,
функция которых, однако, не выяснена (Cuenot, 1948).
,В полости тела находится большое количество целомоцитов, по преж-
ним исследованиям, двух типов у морских лилий, звезд и офиур и шести
типов у голотурий и морских ежей (Hyman, 1955). По новым исследо-
ваниям с фазово-контрастным микроскопом (Boolotian, 1962), число
типов клеток достигает 14. Целомоциты, находящиеся и в других полос-
тях целомического происхождения, играют важную роль в обмене ве-
ществ — в пищеварении, дыхании, выделении и в защите организма от
вредного влияния бактерий и паразитов. Известно, что теория фагоцито-
за И. И. Мечникова возникла из его классических опытов с личинками
морских звезд.
Иглокожие не обладают осморегуляцией, их перивисцеральная жид-
кость, наполняющая полость тела и содержащая примесь протеинов и
большое число целомитов, сходна с морской водой, причем в большей
степени, чем полостные жидкости в любом другом типе животных
(Prosser a. Brown, 1961). Жидкость полости тела иглокожих, как и
жидкость амбулакральной системы, обладает определенной забуферен-
ностью (Крепе, см. Федотов, 1951), удерживающей ее реакцию при на-
коплении кислых или щелочных продуктов обмена веществ. Полостная
жидкость по составу мало отличается от жидкостей амбулакральной и
кровеносной систем.
Жгутики перитонеального эпителия, как показано для ряда иглоко-
жих, например, у голотурий Paracaudina chilensis (Yazaki, 1930), Acte-
rias forbesi (Budington, 1942, cm. Hyman, 1955), вызывают движение
полостной жидкости в определенных направлениях, что способствует
обмену веществ в их теле.
Полость тела с ее элементами, входящая в систему тканей внутрен-
ней среды (Заварзин, 1945, 1947), как и многие другие органы иглоко-
жих, несет несколько функций. В известной мере это связано с отсут-
ствием специальных органов выделения, дыхания и слабым развитием
кровеносной системы. Как и у многих других вторичнополостных жи-
вотных, полость тела несет трофическую функцию, которая считается ее
20*
307
первичной функцией (Заварзин, 1945; Ливанов, 1955). Известную роль
в этом играют амебоциты, проникающие в стенки кишечника и выходя-
щие в. полость тела.
Наполненная громадным количеством жидкости (например, у Sphae-
rechinus gratiularis при общем объеме в 225 см3 количество полостной
жидкости достигает 175 см3) полость тела выполняет механическую
роль опоры для мягких стенок тела у голотурий, абактинальной стороны
морских звезд и мягких стенок перистома морских ежей.
Значительна роль полости тела и в дыхании. Газовый обмен полости
тела со средой кожным диффузным дыханием достигается с помощью
щупалец у многих голотурий, папул — у морских звезд и перистомаль-
ных жабер — у большинства правильных ежей. Дыхательная функция
полостной жидкости усиливается наличием дыхательных пигментов —
гемоглобина у разных голотурий, эхинохрома и спинохрома у морских
ежей (Коржуев, 1949, 1964; Prosser, 1952; Hyman, 1955).
Велико значение полости тела и целомоцитов и в процессах выделе-
ния. Защитная роль амебоцитов в борьбе с микробами и продуктами
выделения паразитов была установлена еще И. И. Мечниковым (1892),
участие целомоцитов в удалении продуктов распада — А. О. Ковалев-
ским (1889а, 18896). После их работ выяснением роли целомоцитов за-
нимались многие исследователи, в том числе и Сент-Илер (1897). Фаго-
циты, нагруженные продуктами распада, удаляются наружу, например,
через папулы у морских звезд, собираются у голотурий в урнах
(Е. Schultz, 1895 и последующие авторы); и все же природа включений
в фагоцитах гистохимически еще не установлена (Hyman, 1955).
Кроме общей полости тела, целомическими являются также каналы
амбулакральной системы, перигемальные каналы, осевые синусы и
ампулы осевого комплекса, составляющие своеобразные особенности
организации, свойственные лишь иглокожим.
Амбулакральная, или воднососудистая, система, возникающая из ле-
вого среднего целома (гидроцеля), достигает (сравнительно с другими
системами органов) наибольшего совершенства. В основном она состоит
из околоротового кольца и отходящих от него пяти радиальных каналов
с амбулакральными ножками или щупальцами, каменистого канала,
открывающегося в ампулу; последняя или через гидропору (когда имеет-
ся один поровый канал), или через множество их в мадрепорите сооб-
щается с наружной средой. Сохраняя основной тип строения, амбулак-
ральная система в своих деталях обнаруживает значительные разнооб-
разия, связанные с условиями и образом жизни, с функцией, выполняе-
мой разными ее частями у представителей разных классов. Амбулакраль-
ная система лежит (над эктоневральной и гипоневральной нервными сис-
темами и перигемальной с кровеносной системами) очень поверхно-
стно у морских лилий, гораздо глубже у остальных классов. В основном
она образована хорошо дифференцированными целомическими канала-
ми, стенки которых состоят из жгутикового эпителия,, соединительной
ткани, мускульного слоя и наружного ресничного перитонеального эпи-
телия. Амбулакральная система наполнена жидкостью, едва отличаю-
щейся от морской воды и жидкостей полости тела и кровеносной
системы.
Почти у всех вымерших и современных классов амбулакральная си-
стема сообщается с наружной средой через гидропору или через поры
ситовидно продырявленного мадрепорита, и лишь у современных морских
лилий это сообщение, возникая в онтогенезе, утрачивается у взрослых
животных.
У форм с большим числом рук (например, у ряда морских звезд)
соответственно увеличивается и число радиальных амбулакральных ка-
налов; у многих голотурий (Apoda), живущих в грунте, они, напротив,
308
могут редуцироваться, даже до исчезновения закладки в онтогенезе
(Synapta inhaerens). Радиальные каналы дают альтернирующие (мор-
ские лилии, ежи, голотурии) или расположенные попарно (морские
звезды, офиуры) боковые каналы в амбулакральные ножки, отсутству-
ющие при регрессе радиальных каналов. Амбулакральные ножки в соот-
ветствии с положением амбулакральных каналов находятся на верхней
стороне тела (морские лилии), на нижней (морские звезды и офиуры)
или распространены по всему телу (голотурии, морские ежи). Обычно
расположенные в амбулакрах, они у некоторых современных морских
ежей и голотурий находятся и в интерамбулакрах и даже разбросаны
по всему телу. Амбулакральные ножки, которые по мнению Нихольса
(Nichols, 1962, 1964) являются главными органами дыхания иглокожих,
выполняют разные функции, с чем связано различие в их строении.
Ножки, служащие для передвижения (голотурии, морские звезды и
ежи), кончаются подошвой и снабжены ампулой; ножки или щупальца,
несущие чувствительную и дыхательную функции (офиуры, морские ли-
лии, некоторые морские звезды и голотурии),— остроконечны, без ампул;
правда, офиуры, прилипая щупальцами к отвесным скалам, могут по ним
ползать. У многих лилий, у которых амбулакральные щупальца отходят
от радиального амбулакрального канала по три с каждой стороны, как
и у других прикрепленных вымерших иглокожих, не могли быть орга-
нами движения и служат для привлечения пищи и дыхания. Помимо
отмеченных функций, амбулакральные ножки и щупальца у морских
звезд и офиур и сейчас также служат для удержания и направления ко
рту захваченной добычи.
Две пары щупалец или ножек, ближайших ко рту (морские лилии,
офиуры, многие морские ежи), являются ротовыми щупальцами.
У офиур они, сохраняя значение органов осязания и удержания добы-
чи, приобретают дополнительную функцию — выбрасывают изо рта непе-
реваренные частицы и экскременты. У большинства морских ежей
10 амбулакральных ножек на перистоме утрачивают ампулы и несут,
видимо, функцию органов вкуса. С утратой двигательной функции амбу-
лакральные ножки тривиума у ряда голотурий упрощаются и становят-
ся выростами стенки тела вроде бородавок с чувствительной и дыха-
тельной функциями.
Очень яркие примеры взаимосвязи формы и функции дают у голоту-
рий амбулакральные ножки, ближайшие к ротовому отверстию. Неред-
ко, сохраняя ампулы и отходя от радиальных каналов или непосред-
ственно от амбулакрального кольца, они становятся древовидно раз-
ветвленными, пальчатыми, перистыми ротовыми щупальцами и служат
для дыхания, передвижения, ловли добычи и осязания.
В выполнении основной у современных иглокожих (кроме морских
лилий) двигательной функции амбулакральной системы принимают
участие поровый и каменистый каналы, поскольку через них происходит
как пополнение амбулакральной системы жидкостью за счет морской
воды, так и выведение излишка жидкости наружу. Исходным положе-
нием, учитывая ископаемые формы и состояние в онтогенезе, надо счи-
тать сообщение каменистого канала с амбулакральным кольцом в зад-
нем непарном интеррадиусе (между I и V амбулакрами) и нахождение
в нем порового канала, сообщающегося через участок полости тела с ка-
менистым каналом. У ряда голотурий эти отношения сохраняются,
у других, как и у современных морских лилий, вторично нарушаются.
У всех морских звезд, ежей и офиур происходят прогрессивные измене-
ния, связанные с образованием осевого комплекса органов, что приводит
к увеличению числа функций и возрастанию значения амбулакральной
системы в отправлениях иглокожих.
С двигательной функцией амбулакральной системы связаны полисвы
309
пузыри голотурий, морских звезд и офиур. Это резервуары жидкости —
тонкостенные, сильно растяжимые'пузыри, отходящие в интерамбулакре
от амбулакрального кольца. Функция тидемановых телец — слепых вет-
вящихся выростов амбулакрального кольца, одетых перитонеумом, у
морских звезд (Светлов, 1916), морских ежей (Светлов, 1924) и у офиур
группы Euryalae (Fedotov, 1926) точно не установлена.
Наблюдаемые уменьшение количества гидропор, поровых каналов,'
каменистых каналов и полиевых пузырей до одного и увеличение их до
нескольких десятков и сотен — принципы олигомеризации и полимери-
зации (Догель, 1954).
Основным способом передвижения современных иглокожих, как дон-
ных животных, является ползание, которое обеспечивается амбулак-
ральной системой (голотурии, морские ежи и звезды) или движениями
рук (бесстебельчатые морские лилии и офиуры). Голотурии в этом от-
ношении наименее специализированы. Характерно, что в их передвиже-
нии участвуют щупальца — околоротовые придатки амбулакральной
системы, вероятно, исторически наиболее древние ее дериваты, суще-
ствовавшие у многих наиболее примитивных ископаемых форм. Голоту-
рии ползают при помощи амбулакральных ножек, сокращений мускула-
туры стенок тела и при помощи склеритов, на которые голотурия, напри-
мер, Synaptidae, опирается как змея на ребра и которые принадлежат к
наиболее древнему примитивному скелету Eleutherozoa. Голотурии
являются наиболее медленно передвигающимися свободноподвижными
иглокожими, делающими 6—7 см в минуту. Вместе с тем ряд голотурий
Pelagothuriidae из глубоководного отряда Elasipoda и Galatheathuria из
Aspidochirota приспособились к пелагическому образу жизни.
Более специализированы органы движения у остальных современ-
ных иглокожих. Морские звезды и ежи в известной мере примыкают к
голотуриям, так как у них амбулакральные ножки являются органами
движения, но у морских ежей эту функцию приобретают также иглы
панциря, становящиеся в ходе эволюции даже основными органами
движения: морские ежи опираются на иглы, как на ходули (цидариды и
ряд других правильных морских ежей). Скорость движения морских
ежей зависит от длины игл. Так, Centrostephanus longispinus с очень
длинными иглами в секунду проходит 3,5 см, Arbacia с короткими игла-
ми это расстояние одолевает в течение минуты (см. Kaestner, 1963).
Скорость передвижения морских звезд вполне сравнима с таковой
морских ежей; так, Asterina gibbosa за минуту проходит 2,5 см, Aste-
rias— 5—8 см, Astropecten — 30—60 см. Очень подвижным оказывается
Solaster papposus, который за это же время проходит 200 см.
Более совершенный способ движения выработался у офиур и бессте-
бельчатых морских лилий, которые являются наиболее быстро двигаю-
щимися иглокожими. Движение у этих животных происходит с помощью
рук, благодаря сокращению мышц между их члениками. Сокращением
этой мускулатуры офиуры быстро передвигаются прыжками или толчка-
ми, успевая за минуту пройти 180 см. Кроме того, офиуры передвигаются
змеевидными движениями рук, а по вертикальной поверхности они дви-
жутся, прилипая клейкими амбулакральными ножками.
Бесстебельчатые лилии ползают и плавают также при помощи рук,
при плавании попеременно поднимая и опуская руки. В ходе эволюции,
став из прикрепленных стебельчатых форм свободноподвижными,
морские лилии начали использовать для передвижения членистые
руки с мускулатурой между члениками. Таким образом, у двух резко
различных классов независимо возник сходный способ передвиже-
ния— применение для этого мускулатуры рук. Морские лилии при
плавании в минуту перемещаются на 5 м, при ползании — в час примерно
на 40 м (Hyman, 1955).
310
При поступательном движении иглокожие двигаются как двусто-
роннесимметричные животные, что наиболее четко выражено у голоту-
рий, которые двигаются вперед только по продольной оси тела. При пе-
ремещении других иглокожих также имеется ось движения, связанная
с определенным амбулакром. Однако нередко направление движения
определяется реакцией пятилучевого иглокожего на действие раздражи-
теля.
У морских лилий и голотурий имеются отдельные части, которые
у морских звезд, ежей и офиур анатомически и функционально состав-
ляют единое целое, так называемый осевой комплекс органов. В его
образовании участвуют амбулакральная, кровеносная, половая систе-
мы, передние, средний и задний левый целомы (Fedotov, 1924; Федо-
тов, 1951; Reichensperger, 1934; Smith, 1940).
Этот комплекс состоит из каменистого канала (отходящего от
амбулакрального кольца и открывающегося в ампулу, сообщающую-
ся одним или многочисленными поровыми каналами с наружной сре-
дой), оральной части осевого органа (отходящей от околоротового
кровеносного кольца и заключенной в левый синус, который идет от
перигемального околоротового кольца и открывается в ампулу) и або-
ральной части осевого органа, находящейся в правом осевом синусе
и переходящей в половой тяж, который заключен в половом или або-
ральном синусе. Осевой орган, оральная и аборальная части которого
гистологически несколько отличны, является мощным сплетением кро-
веносных сосудов или лакун в соединительнотканной строме с целомо-
цитами. От аборальной части осевого органа отходят также, двумя
ветвями (например, у морских звезд) кровеносные сосуды кишеч-
ника.
Функция осевого комплекса многообразна. Осевой орган, соединя-
ющий околоротовое и аборальное кровеносные кольца, участвует в
трофической функции; ритмические сокращения правого осевого си-
нуса вызывают движение жидкости в кровеносной системе. Правый
осевой синус и аборальная часть осевого органа (Fedotov, 1924; Федо-
тов, 1951) являются перикардием и сердцем и отвечают кардио-пери-
кардию кишечнодышащих. Осевой орган, видимо, участвует как лим-
фоидная железа в кроветворении и, судя по наличию в нем клеток с
продуктами распада, в процессах выделения. Связь осевого органа
через половой тяж с половыми железами позволяет считать его эндо-
кринной железой. Через гидропору и поровые каналы мадрепорита
происходит выведение излишка жидкости амбулакральной системы и
удаление амебоцитов с продуктами распада. Эти соображения о функ-
циях осевого органа подтверждены экспериментами над морскими
ежами (Boolotian a. Campbell, 1964; Millot, 1964; Burton, 1964).
В осевом комплексе морских звезд,- ежей и офиур существуют
известные различия, например, у морских ежей оральная часть осе-
вого органа обрастает левый осевой синус, у ряда офиур группы Eurya-
lae есть по осевому комплексу в каждом интеррадиусе и т. д. (Федо-
тов, 1951).
У голотурий имеются только каменистый и поровый каналы, из-
редка полость, куда, открываются эти каналы, отвечающая ампуле
осевого синуса; в их онтогенезе нет указаний на закладку и последу-
ющую редукцию частей осевого комплекса. При наличии у голотурий
ряда примитивных черт можно думать,1 что в отличие от морских звезд,
ежей и офиур они вообще не имели осевого комплекса, и положение
у них мадрепорита и полового отверстия вблизи рта, как у древних
вымерших Pelmatozoa, является сохранением примитивного состояния.
У морских лилий имеются вторичные поровые и каменистые кана-
лы и осевая железа, однако в онтогенезе у бесстебельчатых морских
311
лилий в анальном интеррадиусе на прикрепленной стадии развития
временно имеются почти все элементы осевого комплекса органов.
Там находятся пора порового канала, открывающегося в участок
переднего целома личинки — ампулу, каменистый канал, открываю-
щийся в амбулакральное кольцо'и в ампулу, пучок кровеносных сосу-
дов, отходящий от околоротового кровеносного кольца, половой зача-
ток и зачаток осевой железы. Все это позволяет считать пучок
кровеносных сосудов гомологом оральной части осевого органа, а же-
лезистую часть, или осевую, железу,— гомологом аборальной части
осевого органа большинства современных классов (Fedotov, 1930; Фе-
дотов, 1931, 1951; Reichensperger, 1934), с чем, однако, не все согласны
(например, Tuzet et Manier, 1961). При таком толковании у морских
лилий временно имеются в близком соседстве почти все элементы
осевого комплекса, но эти первичные отношения с переходом во взрос-
лое состояние очень сильно изменяются.
Положение с осевым комплексом у иглокожих позволяет считать,
что у голотурий взаимоотношения порового канала, или мадрепорита,
половой железы с ее протоком и кровеносной системы наиболее при-
митивны. У современных морских лилий они вторично нарушены срав-
нительно с голотуриями и большинством вымерших Pelmatozoa.
У морских звезд, ежей и офиур в результате прогрессивного развития
произошла интеграция частей в анатомическое целое — осевой комп-
лекс органов и сильно возросла его функциональная значимость в
жизненных отправлениях высших иглокожих.
Специализированных органов дыхания у иглокожих в соответствии
с общим невысоким уровнем их организации нет. Исключение как
органы газообмена представляют сильно сократимые водные легкие с
сетью кровеносных капилляров голотурий Dendrochirota, Aspidochi-
rota и Molpadonia, которые вентилируются морской водой благодаря
ритмическим сокращениям клоаки. Но как и многие другие органы
иглокожих, водные легкие вместе с другими частями тела играют
важную роль в изменении тургора, в увеличении длины и объема тела
голотурий при поступлении морской воды извне и в укорачивании
тела и уменьшении его объема при выбрасывании воды наружу (Сиё-
not, 1948; Федотов, 1951; Hyman, 1955).
Примитивными органами дыхания, производными стенками тела,
являются папулы большинства морских звезд, отсутствующие у Рог-
cellanasteridae и Brisingidae. Это тонкостенные выпячивания стенки
тела, перистомальные жабры правильных морских ежей, кроме цида-
роид, кустистые выпячивания стенок перистома и некоторые другие.
У голотурий газообмен со средой происходит непосредственно через
тонкие стенки тела. Такими же производными стенки тела, исторически
древними органами дыхания, были у ряда вымерших классов иглоко-
жих ромбовые поры и диплопоры цистоидей, спир акулы и гидроспиры
бластоидей и некоторые другие.
Ряд систем органов (общая полость тела, амбулакральные ножки и
щупальца, околоротовые щупальца голотурий), сохраняя свои основные
функции, участвует в процессах дыхания. У ряда форм в этих полостях
находятся красные кровяные клетки с дыхательным пигментом — гемо-
глобином (Коржуев, 1949, 1964). Участие в дыхании принимают бурсы
офиур, задняя кишка морских лилий, наполняющаяся морской водой и
опорожняющаяся благодаря ритмическим сокращениям анального кону-
са; некоторые приписывают дыхательную функцию также сифонам киш-
ки морских ежей.
Токи морской воды на поверхности соответствующих органов, дви-
жение жидкостей внутри их, вызываемое действием ресничек, ..вме-
сте с сократимостью органов, несомненно, способствует газообмену..
12
В общем, однако, физиология газообмена изучена все же недоста-
точно.
Основу кровеносной системы составляют околоротовое кольцо,
пять радиальных сосудов, отходящих от него в амбулакры, сосуды ки-
шечника и гонад. У морских звезд, ежей и офиур имеется аборальное
кровеносное кольцо, соединенное с оральным осевым органом и даю-
щее ветвь к половым железам. «Осевой» орган у морских лилий со-
ставляет часть кровеносной системы. Оральное и аборальное кольца,
радиальные сосуды и сосуды в стенках кишечника анатомически не
сосуды, а синусы или лакуны. Они представлены тяжами соединитель-
ной ткани с полостями, наполненными полостной жидкостью и цело-
моцитами. Лишь спинной и брюшной кровеносные сосуды кишечника
голотурий и морских ежей, отходящие от околоротового кольца, имеют
характер кровеносных сосудов, при этом у ряда голотурий они соеди-
нены сетью капилляров, а спинной образует чудесную сеть сосудов
водного легкого.
Лишь спинной и брюшной сосуды голотурий и морских ежей нахо-
дятся в общей полости тела. Кольцевые кровеносные сосуды, точнее,
сплетения находятся в оральном и аборальном, радиальные — в ра-
диальных, половые — в половых каналах перигемальной системы цело-
мического происхождения. Перигемальные каналы в виде кольцевого ка-
нала лежат между нервным и амбулакральным кольцами, между
радиальными нервами и амбулакральными каналами. У морских
звезд околоротовое перигемальное кольцо поделено септой' на наруж-
ное и внутреннее. Перигемальное околоротовое кольцо образуется в
четырех интеррадиусах за счет левого переднего целома, в интерра-
диусе с осевым комплексом — за счет левого заднего целома; радиаль-
ные перигемальные каналы являются выростами околоротового кольца.
Аборальное перигемальное кольцо образуется также за счет левого
заднего целома. У морских лилий перигемальная система представле-
на лишь радиальными каналами, идущими в руки и в пиннулы, около-
ротового кольца у них, как и у морских ежей, видимо, нет. У голо-
турий и морских ежей нет аборального перигемального. кольца.
По своему строению кровеносная система иглокожих, как и многие
другие системы, стоит на низком уровне развития, хотя в связи с раз-
витием радиальной симметрии она далеко отошла от исходного состоя-
ния. Можно думать, что продольные кровеносные сосуды кишечника
голотурий и морских ежей являются сохранившимися примитивными
чертами, утерянными морскими лилиями, как и морскими звездами и
офиурами, в связи с вторичным подавлением у них общей полости
тела: кишечник стал мешковидным, коротким, радиально симметрич-
ным. Отсутствие аборального кровеносного кольца у морских лилий
и голотурий надо считать явлением первичным, связанным у первых
с общим строением, а у голотурий — со строением собственно поло-
вой системы, оставшейся в виде непарной половой железы, не подчи-
нившейся радиальной симметрии. Прогрессивному развитию радиаль-
ной симметрии у морских звезд, ежей и офиур подчинился половой
тяж или столон, ставший пентагональным, вследствие чего половой
синус стал также кольцевым на аборальном конце тела, как и заклю-
ченная в него часть кровеносной системы. Иными словами, абораль-
ное кровеносное кольцо этих классов является приобретением, не су-
ществовавшим у голотурий, а не утраченным ими.
Функция и физиология кровеносной системы изучены недостаточно.
Кровеносные сосуды не сократимы, лишь у голотурий и морских ежей
наблюдаются спонтанные сокращения кровеносных сосудов кишеч-
ника. У голотурий кровь движется по спинному сосуду восходящей
петли кишечника, переходит через стенки кишечника в брюшной сосуд
313:
^нисходящей петли кишечника, оттуда в околоротовое кровеносное
кольцо, а из него по радиальным сосудам к стенкам тела (Kawamoto,
1927, см. Hyman, 1955).
Можно думать, что у морских звезд сокращения правого осевого
-синуса — перикардия вызывают слабое движение крови (Федотов,
1951). Хилус из стенок кишечника собирается в кровеносные сосуды
(лакуны) последнего и поступает в аборальную часть осевого органа,
•откуда, благодаря сокращению перикардия, перемещается через
оральную часть осевого органа в оральное кровеносное кольцо и рас-
пространяется по радиальным кровеносным сосудам; часть крови из
аборальной части осевого органа идет в аборальное кровеносное коль-
цо к половым железам.
В основном кровеносная система иглокожих вместе с перигемаль-
ной, служащей для нее опорой, сравнима с лимфатической системой и
несет трофическую функцию; лишь кровеносная система у голотурий
выполняет функцию газообмена.
Кроме урночек некоторых голотурий (Apoda), агглютинирующих
продукты распада и введенные посторонние вещества, органов выде-
ления у иглокожих нет. Главную роль в выделении играют целомо-
циты общей полости тела, амбулакральной системы, осевого органа,
кровеносной системы и ресничные перитонеальные клетки (Cuenot,
1948; Федотов, 1951; Hyman, 1955). Твердые продукты распада с по-
мощью фагоцитов попадают в стенки тела, папул, перистомальных
жабр, в придатки амбулакральной системы (ножки, полиевы пузыри),
в стенки бурс и их производной — третичной полости тела офиур груп-
пы Euryalae, в стенки водных легких голотурий, кишечника, мезенте-
риев и т. д. Часть продуктов распада откладывается в соединительной
ткани стенок тела, обычно же продукты распада выбрасываются раз-
рывом стенок тела наружу. Химическая природа жидких и твердых
продуктов распада изучена недостаточно. Существенным является
установление у иглокожих креатина и его производного — креатинина,
характерных, кроме них, только для позвоночных животных (Baldwin,
1957; Prosser, 1952; Hyman, 1955).
Регенерация широко распространена среди иглокожих, отчасти она
связана с бесполым размножением, а главным образом с частыми
повреждениями и потерями частей тела (особенно рук) при наступле-
нии неблагоприятных условий или при нападении врагов. Степень ре-
генеративной способности различна, она наиболее слабо выражена у
морских ежей, панцирь которых защищает их от повреждений. У них
регенерируют иглы, амбулакральные ножки, педицеллярии и исправ-
ляются части панциря при незначительных повреждениях. Автотомия
и регенерация наиболее развились у голотурий, которые с их мягким,
без скелета телом уязвимы более, чем другие иглокожие, защищенные
скелетом. Однако и у ряда офиур и морских лилий после автотомии
или утраты внутренние органы хорошо регенерируют. Явление регене-
рации достаточно полно изучено и в экспериментальных условиях.
Случаи регенерации очень редко встречаются у ископаемых форм
морских звезд, даже находимых в массовом количестве, но среди со-
временных явление регенерации очень широко распространено у более
высоко организованных морских звезд (отряд Forcipulata), с облегчен-
ным скелетом чаще, чем у более примитивных с более тяжелым скелетом
(отряд Phanerozonia). Есть основания считать, что регенерация разви-
валась как явление приспособительное.
Как и у ряда других типов, например, у кольчатых червей, личинки
иглокожих почти не способны к регенерации.
Основу половой системы составляют половой тяж и развивающиеся
из него гонады, обычно открывающиеся наружу половыми протоками.
314
В онтогенезе непарный половой зачаток вначале лежит в интеррадиусе
каменистого канала, в области левого заднего целома. У морских ли- .
-лий во время метаморфоза половой зачаток перемещается и вокруг
пищевода образует половой тяж, дающий ветви в руки и пиннулы;
в последних за его счет развиваются гонады, продукты которых выво-
дятся наружу разрывом стенок пиннул.
Среди современных иглокожих наименьшие изменения во время
онтогенеза претерпевает половой зачаток у голотурий; он превраща-
ется в гонаду с протоком, открывающимся наружу на переднем конце
тела. У морских звезд, ежей и офиур половой зачаток соединяется с
аборальной частью осевого органа, вырастает в длинный половой тяж,
образующий пятиугольное сильно волнистое кольцо, дающее начало
мешковидным или гроздевидным половым железам. Половые протоки
пяти гонад морских ежей открываются наружу отверстиями половых
пластинок на апикальном поле панциря. У морских звезд парные го-
нады открываются наружу у основания рук, на их абактинальной сто-
роне. У офиур обычно многочисленные мелкие гонады открываются
в полость бурс.
Половой тяж иглокожих заключен в аборальное кровеносное кольцо,
находящееся в половом синусе, производном правого заднего целома.
У большинства ископаемых классов иглокожих, судя по наличию
полового отверстия между ртом и порошицей, вблизи гидропоры, по-
ловая железа, как у голотурий, была непарная.
Полы разделены, гермафродитизм встречается редко. Размножение
происходит половым путем, бесполым — делением тела обычно надвое
(у некоторых офиур, морских лилий, ежей и звезд, особенно в Антарк-
тике), имеет место и живорождение с прямым развитием; наблюдается
также забота о потомстве, вынашивание молоди на теле или в теле
матери.
Размножение происходит в определенные сезоны года; особи дан-
ного вида мигрируют, собираются массами. Яйца выводятся в воду,
где и происходит их осеменение. Как исключение у некоторых морских
звезд и офиур с резко выраженным половым диморфизмом происходит
спаривание, например, у Amphilicus androphorus (Mortensen, 1933).
У ряда иглокожих в яйце до дробления установлены полярность
и симметрия, локализация зачатков будущих частей зародыша, даже
выраженная внешне. Дробление может быть детерминированным, мо-
заичным.
Преобладает радиальное, полное, равномерное или неравномерное
дробление, встречается двусимметричное (некоторые голотурии), по-
верхностное, частичное (например, у живородящих форм). Дробящее-
ся яйцо обладает четырех- или восьмилучевой симметрией. Зародыш —
моноаксонный, гетерополярный, с анимальным и вегетативным полюса-
ми. Дробление после 10-го деления приводит к образованию бластулы,
состоящей из жгутиконосных клеток с кинетидами, подобно киноцитам
бластул губок и кишечнополостных и особей Protomonadina (Захваткин,
1949).
При мозаичном (детерминированном) дроблении (Paracentrotus
lividus) на стадии 16 бластомеров анимальная часть зародыша состоит
из 8 мезомеров, позже дающих эктодерму, вегетативная часть — из
8 макромеров (зачаток энтодермы) и 4 микромеров, дающих начало
первичной мезенхиме.
У большинства иглокожих (без детерминированного дробления)
на месте будущего впячивания в бластоцель уходят отдельные клетки
первичной мезенхимы, из которых образуется известковый скелет ли-
чинок.
315
Рис. 78. Схема развития целомов у личинки иглокожих (из Pflugfelder)'
А, Б — отшнуровывание целома от архентерона; В — деление целома на правый и ле-
вый мешки; Г — выпячивание целомических мешков вдоль кишечника; Д—Ж— даль-
нейшие преобразования; / — аксоцель; 2 — гидроцель; 3 — гидропора; 4 — личиночный
рот; 5 — кишечник
Впячиванием вегетативной, наибольшей части бластулы образу-
ется гаструла с узким эктодермальным архентероном из пищевода,
желудка и задней кишки. На конце, противоположном бластопору,
образуется теменная пластинка с султаном ресниц (морские ежи),
от верхушки архентерона в бластоцель мигрируют отдельные амебо-
идные клетки вторичной мезенхимы, дающей нежную соединительную-
ткань.
Бластопор, или первичный рот, обычно смещается с вегетативного
полюса на брюшную сторону и переходит в анальное отверстие или
замыкается, и тогда на его месте впячиванием эктодермы образуется-
анус.
316
Независимо от бластопора, позже, несколько отступя от переднего
конца личинки, впячиванием эктодермы образуются рот, стомодеум и
часть кишечника. Эта личинка — диплеврула (Semon, 1888), подобно
торнарии кишечнодышащих, овальной формы, с седловидно вогнутой
брюшной стороной, с ресничным шнуром по ее краю и с выпуклой
спинной стороной, с энтероцельными целомами и без органов выделе-
ния. До образование рта у диплеврулы от архентерона энтероцельным
путем возникает целом. В типичном случае верхушка архентерона вы-
пячивается и отшнуровывается в виде непарного целомического ме-
шочка. Он делится на два — левый и правый целомы, которые распо-
лагаются по бокам архентерона. Далее каждый делится на передний
•и задний целомы, затем передние делятся на два. Образуется три пары
целомов — передние, средние и задние (или аксоцель, гидроцель и со-
матоцель немецких авторов). Двусторонняя симметрия личинки нару-
шается неоднородным расчленением правых и левых целомов и мень-
шими размерами редуцированных правых целомов (рис. 78).
Левый передний целом открывается наружу гидропорой на спин
ной стороне личинки. Правый передний целом, как правило, не делит-
ся на передний и средний целомы. Как исключение наблюдается его
сегментация на два целома, а также образование передним целомом
гидропоры. Левый средний целом (гидроцель) соединяется с передним
каналом, будущим каменистым каналом; позже, с ходом развития ли-
чинок, он дугой охватывает пищевод и дает начало амбулакральной
системе. Левый задний целом значительно больше правого заднего;
они дают общую полость тела и, сходясь, в сагиттальной плоскости
образуют первичный мезентерий.
Асимметрия в развитии целомов личинок иглокожих исторически
обусловлена ходом эволюции — развитием из двусторонней симметрии
древних форм радиальной симметрии.
Экспериментальным путем удается получить развитие всех трех
отделов целома, левых и правых, хотя первые больше вторых, что
встречается также и в природных условиях.
Приходится отметить, что наиболее полное расчленение и наимень-
шая асимметрия црломов наблюдается у личинок типа плутеуса мор-
ских ежей и офиур. У морских звезд, у личинок которых разнообра-
зие в ходе развития целомов гораздо больше, чем у офиур, расчлене-
ние целомов на три пары далеко неполное, а у некоторых, например,
Astropecten, нет и этого расчленения: имеются два мешковидных це-
лома, возникающие как два выпячивания верхушки архентерона; от
левого отделяются более крупный левый задний целом, несколько поз-
же— от правого — правый задний, У некоторых морских звезд архен-
терон перетягивается на три части: верхнюю, большую — зачаток
передних и средних целомов и правого заднего, среднюю — зачаток
кишечника — и нижнюю, ближайшую к бластопору,— зачаток левого
заднего целома.
Более чем у морских звезд изменено образование целомов у голо-
турий, хотя личинки их одного типа — аурикулярии и бипиннарии.
В ряде случаев архентерон делится почти на две равные части: верх-
нюю— зачаток всех целомов, нижнюю с бластопором — зачаток ки-
шечника. Непарный целом ложится на левую сторону кишечника и,
делясь, дает верхний мешочек — зачаток левого переднего целома и
гидроцеля — и нижний мешочек, из которого образуются левый и пра-
вый задние целомы, ложащиеся по сторонам кишечника и образующие
спинной и брюшной его мезентерии.
Еще более изменено образование целомов у бесстебельчатых мор-
ских лилий (эмбриология стебельчатых лилий не изучена). После
замыкания бластопора архентерон превращается в слепой мешок,
317
лежащий в заднем конце личинки, который делится на два пузырька:
верхний — зачаток кишечника, переднего и среднего целомов — и
нижний (или задний)—зачаток левого и правого задних целомов.
Однако экспериментально у Antedon (Runnstrom, 1925, 1930) удается
вызвать появление двух передних и двух средних целомов, т. е. обра-
зование всех трех пар целомических мешков.
Общим для всех упомянутых классов иглокожих является развитие
кишечника и целомов за счет архентерона — впячивания вегетативной
части жгутиконосной бластулы. Различия в соотношениях и располо-
жении зачатков кишечника и отделов целома сводится к явлению
меторизиса, смещению границ зачатков. В ходе развития у разных
классов, даже в пределах одного класса, наблюдаются различия в
полноте сегментации целомов и степени подавления развития их на
правой стороне личинки, вплоть до видимого полного исчезновения
правых целомов. Наибольшее отклонение от типичного расчленения
целомов на три пары у морских лилий, очевидно, связано с появлением
у них лецитотрофных личинок — долиолярий, которые сравнительно
с планктотрофными (аурикуляриями, бипиннариями, офиоплутеусами
и эхиноплутеусами), несомненно, являются вторично измененной,
исторически более поздней формой личинок иглокожих. Надо при этом
отметить, что при всем разнообразии хода онтогенеза целомов, в итоге
у всех иглокожих левый второй (средний) целом дает амбулакральную
систему, левый и правый третьи (задние) целомы дают общую по-
лость тела; с передними целомами связано в большей или меньшей
степени развитие осевого комплекса органов как целого или его ком-
понентов.
Вследствие очень кратковременного существования жгутиконосные
бластулы не могут быть расселительной стадией — бродяжками, как
думает А. А. Захваткин (1949); на самом деле расселение вида обеспе-
чивается в основном не ими, а личинками, особенно планктотрофны-
ми — аурикуляриями, бипиннариями, эхиноплутеусами и офиоплу-
теусами, живущими месяцами и питающимися одноклеточными водо-
рослями (особенно диатомовыми).
У иглокожих с яйцами, богатыми желтком (к ним относятся Ante-
don и другие морские лилии), образуются лецитотрофные личинки. При
живорождении (некоторые морские звезды, ежи, лилии, офиуры и голо-
турии) развитие сокращается, типичные личинки обоих типов не обра-
зуются.
Личинки свободноподвижных иглокожих характерны настолько, что
параллельно с классификацией взрослых существует классификация
(Mortensen, 1921) личинок с.подразделением их на отряды, семейства,
роды и виды, совпадающая с ней и в ряде случаев позволяющая разре-
шать сомнения, возникающие в работе систематиков. Недавним иссле-
дованием эта классификация в общем была подтверждена с некоторыми?
поправками и объяснениями причин, по которым иногда близкие семей-
ства все же имеют резко отличные типы личинок.
Развитие иглокожих сопровождается сложным метаморфозом. Дву-
сторонняя симметрия диплеврулы нарушается, как было отмечено выше,
сначала асимметрией или лучше сказать дисимметрией зачатков целома.
Далее в онтогенезе происходит перестройка всей организации личинок:
и развитие радиально симметричного тела имаго с остатками черт пер-
вичной двусторонней симметрии и с развитием у голотурий и морских
ежей вторичной двусторонней симметрии. Такой ход метаморфоза игло-
кожих обусловлен геологической историей этого типа — переходом древ-
нейших вымерших форм от лежания на дне моря, от малоподвижного
образа жизни, к геологически длительному прикрепленному образу жиз-
ни, а затем снова переходом к свободноподвижному существованию.
318
В онтогенезе метаморфоз у морских лилий и некоторых мор-
ских звезд (например, Asterias rubens — Gemmill, 1912, 1914, 1916,.
1920) начинается после прикрепления преоральной частью личинки,,
опустившейся на дно. У остальных иглокожих, в том числе у большин-
ства эмбриологически изученных морских звезд, развитие радиаль-
носимметричного тела происходит у личинок, продолжающих пелагиче-
ский образ жизни.
Характерно, что в ходе развития аурикулярия голотурий (синаптиды)
превращается в бочонкообразную личинку (долиолярию или куколку),
перестающую питаться, похожую на личинку антедона. Более суще-
ственным является то, что при развитии морских лилий и голотурий,
тело их личинок переходит в тело имаго. Но у морских лилий формиро-
вание имаго сопровождается коренной, чрезвычайно сложной, перест-
ройкой всей внутренней организации личинки долиолярии и прохожде-
нием еще двух стебельчатых прикрепленных личиночных стадий —
цистоидеи и пентакринуса. У голотурий не происходит резких перестроек
организации личинки — при формировании из нее имаго атрофируется
лишь небольшая преоральная часть личинки.
У остальных иглокожих радиально симметричный зародыш, будущее-
имаго, развивается лишь из части личинки, другая часть резорбируется
при участии фагоцитоза, что впервые отметил И. И. Мечников (Metsch-
nikoff, 1869), или отбрасывается.
При метаморфозе морских звезд и ежей радиальносимметричный
зародыш возникает из двух пятилучевых зачатков, или дисков ораль-
ного (или актинального), развивающегося на левой стороне личинки, и
аборального (или абактинального), развивающегося на ее правой сто-
роне; будущая вертикальная ось имаго перпендикулярна к длинной оси.
личинки; резорбируется или отбрасывается передняя часть личинки..
Но в то время как у морских звезд актинальный и абактинальный за-
чатки принимают равное участие в образовании тела звезды, у морских
ежей актинальный зачаток сильно растет и образует'тело морского ежа,
тогда как абактинальный диск образует лишь апикальное поле.
При метаморфозе офиур радиальносимметричный зародыш, буду-
щая офиура, образуется из пятилучевых зачатков или дисков (актиналь-
ного и абактинального), возникающих на спинной и брюшной сторонах,
офиоплутеуса. Вертикальная ось имаго с длинной осью личинки обра-
зует гораздо меньший угол, чем у морских звезд и ежей. Однако у офиур,,
как и у морских звезд, в отличие от морских ежей актинальный и абак-
тинальный зачатки принимают равное участие в образовании тела у
имаго.
Общей особенностью метаморфоза морских звезд и офиур является
развитие зачатков тела имаго на поверхности личинки в отличие от мор-
ских ежей, у которых эти зачатки ежа образуются внутри личинки.
Таким образом, при очень большом сходстве личинок и полнот ;
сегментации целомов морских ежей и офиур, развитие их различно по
положению в теле личинки и по форме ее частей, в чем развитие морских,
ежей близко к таковому морских звезд. Это сходство было причиной
того, что прежние авторы (например, MacBride, 1914; см. гл. «Гребневи-
ки») говорили о стадии звезды при развитии морских ежей.
В ходе метаморфоза, вслед за асимметрией, вызванной преоблада-
нием в размерах и более полной сегментацией целомов левой стороны
личинки, а. у ряда форм вызванной редукцией переднего и среднего пра-
вых целомов, наступает развитие радиальной симметрии. Характерно,
что она начинается с образования левым средним целомом (гидроцелем)
амбулакрального кольца, охватывающего пищевод и дающего пять ра-
диальных выростов. В ходе метаморфизма с ними связано образова-
ние радиальных амбулакральных каналов, терминальных щупалец
319
(первичных ножек), амбулакральных ножек, ротовых щупалец голоту-
рий и, наконец, формирование амбулакров имаго. Этот момент онтоге-
неза иглокожих имеет филогенетическое значение, так как и в геологиче-
ской истории иглокожих, в основном Pelmatozoa, формирование радиаль-
ной симметрии начиналось с развития амбулакральных желобков,
идущих от ротового отверстия, служивших, как и сейчас у ряда совре-
менных форм, для приведения пищи ко рту.
Формирование пятилучевой симметрии идет не сразу, что особенно
ясно у голотурий. У некоторых из них сначала появляются лишь три
выроста (зачатки щупалец) на амбулакральном кольце, а затем два
следующих. Возможно, что это также имеет филогенетическое значение,
поскольку у некоторых цистоидей вначале имеется три амбулакральных
желобка, затем один из них разветвляется на два. Формирование пяти-
лучевой симметрии выражается также на абактинальной стороне за-
кладки скелета (абактинального) в виде центральной пластинки и венчи-
ков радиально и интеррадиально расположенных пластинок.
Перестройка двустороннесимметричной организации диплеврулы в
радиальносимметричное имаго сопровождается перемещением ряда за-
чатков и органов, абсорбцией части их, формированием из них имаги-
нальных органов и изменением образа жизни.
Прикрепление к субстрату брюшной стороной первично переднего
конца личинки антедона вызывает переход его в абактинальную сто-
рону будущей лилии, абсорбцию теменной пластинки с султаном ресниц
и превращение первично заднего, или бластопорального, конца личинки
в актинальную сторону имаго. В связи с этим прикрепление вызывает
поворот внутренних органов (зачатка кишечника и целомов личинки)
почти на 90°, их перемещение и перестройку всей организации. Из пе-
реднего конца личинки образуется стебель с известковыми пластинка-
ми, задний конец развивается в чашечку.
Образуется рот и пищевод, открывающийся в преддверие, мешковид-
ный желудок вытягивается в трубчатый кишечник, загибающийся на-
лево в горизонтальной плоскости и образующий спиральный ход. Позже
в интеррадиусе, в котором расположена гидропора, образуется аналь-
ное отверстие. Таким образом, у антедона рот и анальное отверстие
возникают заново.
Наличие в онтогенезе антедона в одном интеррадиусе порового
канала с гидропорой, каменистого канала, ампулы, железистого орга-
на в вертикальном мезентерии с кровеносными сосудами, вблизи
него полового зачатка и анального отверстия подчеркивает общность
черт строения современных морских лилий, цистоидей и ряда других
ископаемых.
У голотурий, морских ежей и офиур метаморфоз происходит у сво-
бодноподвижных личинок, а у ряда морских звезд — после прикрепле-
ния личинки к субстрату с помощью преоральной стебельчатой части
или брахиолами и присоской между ними, расположенными на перед-
нем конце личинки. Итак развитие радиальной симметрии начинается
с образования зачатка амбулакрального кольца. Вслед за тем происхо-
дит вращение по часовой стрелке кишечника личинки, затрагивающее
у голотурий лишь его передний отдел, а у остальных весь кишечник
с вертикальным спинным и брюшным мезентериями, образованными
правым и левым задними целомами. Кишечник, поворачиваясь, занимает
горизонтальное положение. У голотурий, претерпевающих незначитель-
ные изменения организации двустороннесимметричной личинки, при пе-
реходе в имаго кишечник удлиняется и образует нисходящие и восходя-
щую петли. Спинной и брюшной мезентерии левого и правого задних
целомов личинки образуют продольные мезентерии голотурии, но в
брюшном мезентерии возникают разрывы, отчего полость тела у голо-
320
турий становится сплошной. Рот, бывший на брюшной стороне личинки,
смещается на. передний конец, становясь ртом голотурии, бластопор
или переходит в анальное отверстие, или замыкается, и анальное отвер-
стие голотурии образуется заново без проктодеального впячивания.
Личинка превращается в голотурию (Давыдов, 1914; Dawydoff, 1948)
путем эволютивного метаморфоза.
При метаморфозе морских звезд и ежей происходит коренная пере-
стройка личиночного кишечника. Личиночный рот исчезает, пищевод
абсорбируется; у морских ежей дефинитивный рот и пищевод обра-'
зуются впячиванием эктодермы, у морских звезд пищевод возникает
впячиванием стенки эктодермального желудка. Личиночный анус и ко-
нечная часть задней кишки личинки также абсорбируются. Дефинитив-
ное анальное отверстие образуется заново, причем у морских звезд
впячиванием эктодермы возникает проктодеум. Остающаяся часть эн-
тодермальной кишки личинки подвергается гистологической передиф-
ференцировке, сопровождающейся дегенерацией при участии фагоцитов.
У морских ежей желудок личинки удлиняется, образует первый обо-
рот кишечника против часовой стрелки, а личиночная кишка дает второй
поворот по часовой стрелке, в результате чего образуется кишечник
имаго. Прежние вертикальные продольные мезентерии задних целомов
становятся горизонтальными.
У морских звезд выступ на левой стороне личиночного желудка пре-
вращается в желудок имаго, а личиночный желудок — в пилорические
придатки желудка морской звезды; из вертикальных мезентериев, обра-
зованных левым и правым задними целомами личинки, ставшими гори-
зонтальными при метаморфозе, возникают тяжи, прикрепляющие желу-
док морской звезды к стенкам тела. ,
Судьба кишечника у офиур значительно проще, чем у морских звезд
и ежей. Личиночный рот замещается дефинитивным ртом; часть личи-
ночного пищевода абсорбируется, остальная часть дает имагинальный
пищевод, желудок личинки — желудок имаго; задняя личиночная кишка
и анальное отверстие абсорбируются.
В отличие от эволютивного метаморфоза голотурий метаморфоз
морских лилий, звезд, ежей и офиур — некробиотический.
Клетки первичной мезенхимы образуют скелет личинок, а вторичной
мезенхимы — нежную соединительную ткань бластоцеля. За счет клеток
соединительнотканного слоя, или кутиса, возникающего из стенок задних
целомов (соматоплевры), образуется скелет имаго. Эктоневральная
нервная система возникает в виде утолщения эктодермы амбулакраль-
ных желобков; у морских звезд и лилий она остается в их наружном
эпителии, у голотурий, морских ежей и офиур она лежит в эпиневраль-
ных каналах (кольцевом и радиальных), возникающих погружением
стенки амбулакральных желобков внутрь и их смыканием. Развитие
нервной системы в стенке эктодермального кишечника — гипоневральной
и энтоневральной — точно не прослежено, но очевидно, что первая це-
ломического происхождения.
В общем, судьба целомов личинки следующая. Из части переднего
левого целома образуются левый осевой синус с ампулой, поровый ка-
нал и перигемальное кольцо; из части правого переднего целома обра-
зуется правый осевой синус (мадрепоровый пузырек, перикардий), со
стенками левого и правого осевых синусов связано развитие оральной
и аборальной частей осевого органа.
Левый передний целом (гидроцель) дает начало амбулакральной
системе с ее придатками и каменистому каналу. В природе, а также в
эксперименте, например, под воздействием гипотоничной морской воды,
правый йередний целом также образует поровый канал с гидропорой,
а правый средний целом становится правым гидроцелем; они в по-
21 Д. М. Федотов
321
следствии атрофируются. Левый и правый задние целомы дают дефини-
тивную вторичную полость тела.^Из соматической их стенки (сомато-
плевры) развивается слой кутиса или кориума — соединительнотканный
слой стенки тела и ее мускулатура, перитонеальная выстилка полости
тела и мезенхиматозные клетки, дающие скелет имаго. Спланхноплевра
дает начало соединительной ткани, мускулатуре кишечника и перито-
неальному эпителию.
Кроме общей полости тела, из левого заднего целома возникают
перигемальное кольцо с радиальными каналами (у морских звезд и
офиур), перифарингеальное и перианальное кольцо (у голотурий и мор-
ских ежей), перипроктальное кольцо (у морских ежей) и аборальный
кольцевой, или половой, синус (у морских звезд, ежей и офиур).
Из филогенетически важных черт онтогенеза иглокожих можно от-
метить следующие: 1) независимое от бластопора образование рта, 2) пе-
реход бластопора в анальное отверстие, 3) энтероцельное образование
целомов и расчленение их на три пары мешочков, 4) эктодермальное
происхождение мезенхимы, развитие скелета из мезодермы, 5) двусто-
роннесимметричное строение личинок, нарушаемое в ходе онтогенеза
асимметрией в развитии целомов левой и правой стороны и преоблада-
нием в развитии органов левой стороны, 6) поворот рта и пищевода
влево, задней кишки — вправо, 7) развитие радиальной пятилучевой
симметрии, начинающейся с образования гидроцеля, подковообразно
охватывающего пищевод амбулакрального кольца; 8) наличие стадии,
когда закладка у морских звезд, ежей и офиур абактинального скелета
из центральной пластинки и венчиков радиальных и интеррадиально
расположенных пластинок, 9) наличие стадии, когда в интеррадиусе ле-
жат половой зачаток, поровый канал с гидропорой, каменистый канал,
зачаток осевого органа, а в ряде случаев и анальное отверстие.
В развитии представителей разных классов проявляются филогене-
тические черты более частного характера. В онтогенезе бесстебельчатых
морских лилий такое значение имеют стебельчатая стадия, сходная с
цистоидеями и с некоторыми палеозойскими стебельчатыми лилиями.
(Embryocrinus, Hemistreptacrori), и более поздняя пентакринусовая
стадия, сходная с вымершим Pentacrinites. Филогенетическое значение
имеют также стадия дициклического состава скелета чашечки, как и
строение последнего у ряда стебельчатых морских лилий, наличие пяти
поровых и каменистых каналов, так же как у современного Rhizocrinus
во взрослом состоянии и т. д.
Ряд морских звезд, например, Asterias rubens (см. Gemmil, 1914;
Spencer, 1914—1940), в онтогенезе имеет прикрепленную стадию, на ко-
торой положение рта и анального отверстия (до образования дефини-
тивных) сходно, с таковым у сидячих ископаемых иглокожих. Филогене-
тическое значение в онтогенезе типичных офиур имеет стадия с парными
амбулакральными пластинками, наблюдаемая также и у многих палео-
зойских форм. Многоветвистые офиуры Euryalae проходят стадии офиу-
ры и трихастера-— с пятью руками, с системой пластинок на абактиналь-
ной стороне диска, с большой вторичной полостью тела, с мешковидным
кишечником, с асимметричным осевым комплексом, с пятью парами меш-
ковидных бурс и с рядом других черт, характерных для типичных офиур.
Филогенетическое значение имеет закладка аристотелева фонаря у ряда
неправильных морских ежей (спатангоид), лишенных фонаря во взрос-
лом состоянии, и многое другое.
Современные иглокожие, как и ископаемые, исключительно морские
животные бентоса, его эпифауны, реже инфауны и в виде исключения —
пелагические формы (немногие ископаемые'морские лилии, из голоту-
рий— Pelagothuria, Planctothuria, Galatheathuria и некоторые Др.).
В настоящее время иглокожие, особенно офиуры (Hyman, 1955) , освоили
322
все типы дна, моря всех широт и глубин — от литоральной зоны до
сверхабиссальных: морские лилии до 9735 ж, голотурии до 10 710 ж, мор-
ские звезды до ’7614 ж, офиуры до 8006 ж, морские ежи до 7290 ж (Беляев
и др., 1960). Температура, солевой состав воды, особенно ее соленость,
аэрация, глубина, характер грунта, течения и ряд биотических факторов
оказывают сильное влияние на распространение иглокожих. Наряду с
этим на их современном распространении, формировании и характере
фаун разных зоогеографических областей сказались результаты подня-
тий и опусканий земной коры, палеогеографические условия морей преж-
них геологических эпох, и т. д.
Важнейшим абиотическим фактором как в жизни взрослых форм,
так и за время индивидуального развития иглокожих является соле-
ность морской воды. Иглокожие — стеногалинные животные и лишь
немногие формы переносят соленость ниже ЗО%о. Эвригалинных форм
немного, к ним относятся тропические голотурии из Apoda, живущие в
солоноватой воде эстуарий, в мангровых зарослях; Asterias rubens, So-
laster papposus, Henrtcia sanguinolenta живут в Балтийском море при
солености воды 8—15%о (Kaestner, 1963); офиура Ophiophragmus.fi-
lograneus выносит 7,7°/0а солености (Thomas, 1961, цит. по Nichols,
1964).
Подчиняясь, как и все организмы, влиянию окружающей среды, иг-
локожие, образуя нередко громадные скопления, оказывают со
своей стороны сильное преобразующее воздействие на среду. Пропуская
через кишечник массы детрита, ила, песка, иглокожие участвуют в фор-
мировании осадочных пород, играют значительную роль в продуктивно-
сти моря. Они поедают огромное количество разнообразных животных
(Asterias, например, опустошают устричные банки) и вместе с тем мно-
гие иглокожие сами служат пищей для других обитателей моря, особенно
рыб, птиц, некоторых млекопитающих и даже человека.
В прежние геологические эпохи массовыми сообществами, нередко
на больших пространствах, жили некоторые эдриоастероидеи (Cyatho-
cystis), ряд цистоидей в ордовике, особенно многие морские лилии в
палеозое и мезозое, неправильные морские ежи в мезозое, в третичное
время и т. д. Это были породообразующие организмы, результатом жиз-
недеятельности которых и их остатками явились, например, дербошир-
ский мрамор, криноидный известняк и другие строительные материалы.
В настоящее время и, по-видимому, в прежние геологические эпохи
мелководье с его разнообразными условиями заселено фауной иглоко-
жих, количественно и качественно наиболее богатой. Необычайным бо-
гатством иглокожих выделяется Индо-Тихоокеанская провийция со
своим центром — Малайским архипелагом. Фауна иглокожих абиссаль-
ных глубин с их более однообразными и вместе с тем особыми условиями
бедна, но чрезвычайно интересна, так как в ней находятся древние при-
митивные формы, а также крайне измененные под воздействием абис-
сальных глубин. О времени перехода иглокожих к жизни в абиссали
данных пока мало. Есть основания думать, что приспособление к жизни
на больших глубинах среди стебельчатых морских лилий началось в
мезозое.
Если не считать жизни Gorgonocephalus на ранних стадиях на Ger-
semia, а на более поздних — молодых особей внутри взрослых (что при-
носит вред хозяевам), настоящих паразитов среди иглокожих нет.
Сравнительно редки и случаи комменсализма иглокожих с другими жи-
вотными; так, крохотная офиурка Nannophiura lagani (0,5 мм в диамет-
ре диска) йсивет между иглами морского ежа Clypeaster latissimus
(Mortensen, 1933).
Среди паразитов из разных групп беспозвоночных — простейших,
плоских, круглых и кольчатых червей, моллюсков и ракообразных (Ну-
* 323
21
man, 1955) —наиболее характерными являются мизостомиды, некоторые
ракообразные, брюхоногие моллюски и рыбка Fierasfer. Специфические
паразиты иглокожих — мизостомиды, мелкие,, значительно деградиро-
вавшие под влиянием паразитизма аннелид-ы, экто- и эндопаразиты,
главным образом морских лилий, редко морских звезд и офиур (Федо-
тов, 1915; Fedotov, 1931).
. Очень характерными • эндопаразитами морских звезд являются
Ascothoracida, утратившие почти все черты организации ракообразных
(Вагин, см. Федотов, 1960). Своеобразными и сильно измененными ока-
зываются некоторые веслоногие рачки, паразиты офиур и морских ежей,
в том числе и ископаемых (Соловьев, 1961).
Не менее характерными чем мизостомиды паразитами иглокожих
(голотурий, морских лилий, звезд и ежей) являются семейства-брюхо-
ногих моллюсков: Melanellidae, Stiliferidae, Entoconchidae. Среди них
можно проследить цепь переходов от комменсализма к экто- и эндопара-
зитизму, от типичной организации брюхоногого до полной деградации
с утратой черт класса и всего типа моллюсков (А. В. Иванов, 1933—
1953, см. Федотов, 1960).
Судя по сильным регрессивным изменениям организации и развития
этих паразитов и деформациям, вызываемым ими в органах и скелете
иглокожих, например найденных у морских лилий палеозоя (Федотов,
1938), паразитизм ракообразных, брюхоногих моллюсков и особенно ми-
зостомид — геологически древнего происхождения.
Согласно новым исследованиям (Kaestner, 1963), Fierasfer ( = Cara-
pus) не симбионт, а паразит голотурий в личиночном и взрослом со-
стоянии.
Характерно также для эктопаразитических мизостомид (Федотов,
1938) и ряда ракообразных, живущих на морских лилиях (Hyman, 1955),
приобретение покровительственной окраски, сходной с таковой их
хозяев.
Двусторонняя симметрия личинок, вторичное образование рта, закла-
дывающегося независимо от первичного рта, энтероцельное образование
целомов и три их пары — характерные черты иглокожих. К важным чер-
там их онтогенеза относятся также поздняя детерминация, зависимый
характер развития, эпигенетические черты в нем, длительный переход
многочисленных (десять) делений бластомеров, сильная . регулятивная
способность стадий доличиночного развития, инвагинационное обра-
зование гаструлы и эпителиальное построение зародышевых пластов и
органов. Характерными чертами являются также энтодермальное-проис-
хождение мезенхимы, почти нацело энтодермальное происхождение ки-
шечника, образование скелета клетками соединительной ткани, не ган-
глиозный характер нервной системы, закладка ее нередко происходит в
онтогенезе и при регенерации впячиванием эктодермы внутрь, сильное
развитие вторичной полости тела, слабое развитие-кровеносной системы,
отсутствие органов выделения, наличие перикардия, участие в энергии
мышечного сокращения не только .аргинина, но и креатии-фоофата.
Многие из перечисленных черт в той или иной мере присущи кишеч-
нодышащим, птеробранхиям и хордовым. Можно еще добавить гомоло-
гию двух передних пар целомов иглокожих и кишечнодышащих
премандибулярному и мандибулярному целомам хордовых (Федотов,
1923; П. П. Иванов, 1944).
Словом, радиальносимметричные иглокожие имели двусторонне-
симметричных предков, наряду с кишечнодышащими, птеробранхиями,
и хордовыми, и радиальную симметрию приобрели в результате при-
крепленного образа жизни. z • •
В самых общих чертах ход эволюции иглокожих был следующий.
Исходными формами были удлиненные, червеобразные, придонные, ле-
324
жачие, медленно передвигающиеся по дну моря организмы. От них про-
изошли, или были близки к ним, известные пока из среднего кембрия
карпоидеи — животные без радиальной симметрии, ставшие под воздей-
ствием такого образа жизни с течением времени в большей или меньшей
степени асимметричными. Они обладали скелетом из тонких известковых
пластинок, допускавших известную подвижность тела. Очевидно, что на
этом этапе эволюции предковые формы иглокожих были близки к при-
митивным, двустороннесимметричным формам,-давшим начало другим
типам вторичноротых, при этом можно согласиться с тем, что отдален-
ные предковые формы иглокожих были более близкими к предковым
формам хордовых, чем к предкам кишечнодышащих и птеробранхий,
вовсе не имеющих известкового скелета. В пользу такого взгляда свиде-
тельствует: развитие внутреннего скелета у иглокожих и хордовых
(позвоночных) за счет мезодермы (соединительной ткани), нали-
чие настоящих коллагеновых-волокон в кутисе тех и других, образование
у них каротина и каротинина, наличие сложного нейронального ком-
плекса, который обеспечивает возбуждение нерва, передающееся мыш-
цам, сходно с тем, что происходит у позвоночных при переходе,
возбуждения на. мотонейроны в спинном мозгу и в так называемом мио-
невральном синапсе. И что- особенно важно — это обратно противопо-
ложное действие на нервные окончания ацетилхолина и адреналина,
вызывающее расслабление и сокращение мышц, что наблюдается лишь
у позвоночных и иглокожих. Но В ОСНОВНОМ ЭВОЛЮЦИЯ иглокожих и
позвоночных шла различными путями, они разошлись- так давно
и рано, что нет никакой надобности допускать спекуляции с неотенией
и педоморфозом и выведением позвоночных от иглокожих, как думают,
представители английской школы Гарстанга, де Бира и др. .
Двусторонняя симметрия в общем мешковидного, сдавленного тела,
отсутствие радиальной симметрии, .асимметрия в положении отверстий
кишечника сближают типичных карпоидей с кишечнодышащими и Се-
phalochorda (ланцетником). Возможно, что на этом этапе эволюции у
наиболее примитивных иглокожих произошли смена функций вводного
и выводного отверстий кишечника и нарушение двусторонней симметрии
тела, вызванное характером образа жизни, еще до появления радиальной'
симметрии. Однако каких-либо переходных ископаемых форм между
карпоидеями и другими классами иглокожих палеонтология пока не
дала. Хотя в последнее время увеличивается число находок новых форм
карпоидей, класс этот малочислен и, видимо, не является систематиче-
ски единой группой, как отмечал Спенсер (Spencer, 1938). Карпоидей, из-
вестные со среднего кембрия, вымерли в нижнем девоне; их расцвет
наблюдался в кембрии и ордовике (Беккер, 1964).
Эволюция и всех других Pelmatozoa, за исключением Articulata —
подкласса морских лилий, прошла в палеозое. Находки в нижнем кем-
брии представителей эдриоастероидей, эокриноидей, пластинок скелета
цистоидей (Termier et Termier, 1960) и существование в нижнем — сред-
нем ордовике цистоидей, бластоидей, трех надклассов криноидей, а так-
же всех классов Eleutherozoa говорят о том, что иглокожие существовали
еще в докембрии. Климатические и физико-географические условия моря
того времени, очевидно, благоприятствовали эволюции иглокожих, да-
же прикрепленных. В течение палеозоя шло развитие и расцвет всех
классов Pelmatozoa, среди которых доминирующими, наиболее богатыми
численно и по разнообразию форм были стебельчатые морские лилии.
Малочисленные, геологически недолговечные классы — эокриноидей
(нижний кембрий—средний ордовик) и паракриноидей (средний ордо-
вик), обладавшие мало дифференцированным строением и признаками,
общими для цистодей и морских лилий (Regnell, 1960а, 19606),— надо
бесспорно считать близкими к родоначальным формам этих двух классов.
325
По одним авторам (Jaekel, 1918), морские лилии — родоначальная
группа для большинства Pelmatozoa, в том числе и цистоидей, по другим
(Bather, 1915; Яковлев, 1918, и Др.),— более примитивны цистоидеи, и от
них путем утраты поровых ромбов, упорядочением расположения пла-
стинок чашечки, уменьшением их числа могли произойти морские лилии.
Несомненно, что древние Pelmatozoa, прикреплявшиеся к грунту непо-
средственно абактинальным конусом мешковидного тела, одетого боль-
шим числом пластинок неправильной формы и без правильного
расположения, наиболее близки к родоначальным формам иглокожих.
От них недалеки некоторые роды классов эокриноидей и паракриноидей.
Вернее считать, что ни морские лилии не дали начало цистоидеям, ни
цистоидеи — морским лилиям.
Геологически цистоидеи были более древними формами. Они были
и более примитивными. У них, как и у ряда других Pelmatozoa (эдрио-
астероидеи, эокриноидеи, паракриноидеи), между ротовым и анальным
отверстиями лежат гидропора и половое отверстие. У ряда палеозойских
морских лилий гидропора также лежала й анальном интеррадиусе. Как
известно, современные морские лилии проходят в онтогенезе стадию
цистоидеи, не имеющую рук, с венчиками пластинок, отвечающими, та-
ковым цистоидей. Временно на этой стадии между ротовым и анальным
отверстиями лежат гидропора и половой зачаток (Fedotov, 1930). Стадия
цистоидеи бесстебельчатых морских лилий имеет общее с палеозойскими
микрокриноидеями, существовавшими от среднего ордовика до верхней
перми (подкласс Inadunata) и с формами Hyocrinoida того же подклас-
са, имевшими размеры часто меньше 1 мм и нередко лишенными рук
(Weller, 1930; Kallimorphocrinus, Lageniocrinus — Strimple and Koenig,
1956). Вероятно, у цистоидей и близко связанных с ними бластоидей
ромбопоры, диплопоры и гидроспиры возникли для усиления дыхания
в обедненных кислородом придонных слоях воды, где жили эти иглоко-
жие. Развитие рук и длинного гибкого стебля, приподнимавшего чашеч-
ку морских лилий высоко над дном моря, ставило морских лилий
в гораздо более благоприятные условия для улавливания пищи и исполь-
зования кислорода морской воды сравнительно с цистоидеями. Уже эти
обстоятельства могли способствовать появлению новых форм, возникно-
вению разнообразия в строении палеозойских морских лилий, нараста-
нию их численности и вообще высокой морфологической и экологической
пластичности морских лилий Camerata, Inadunata и Flexibilia. В итоге
стебельчатые морские лилии хорошо приспособились к условиям жизни
моря палеозоя. Однако процессы горообразований и отступания бере-
говой линии моря, изменение климата и исчезновение привычной пищи,
как отмечают палеонтологи, были причинами почти полного к концу
перми вымирания стебельчатых морских лилий. Лишь ничтожная часть
подкласса Inadunata (семейство Encrinidae) из трех тогда существовав-
ших подклассов перешла в триас. Несомненно, среди Inadunata были
более стойкие жизненные формы, давшие начало морским лилиям под-
класса Articulata, ныне процветающим бесстебельчатым формам.
Хотя безрукие морские лилии в настоящее время не считаются при-
митивными, так как у ряда этих форм установлена редукция числа рук
и утрата последних, тем не менее в сходстве их с цистоидеями надо ви-
деть черты атавизма, вторичного возврата к более примитивному
состоянию.
К сожалению, пока не найдены низкоорганизованные, примитивные
морские лилии. Самая древняя морская лилия — locrinus ( = Democrinus)
cambriensis (Hicks) оказалась не кембрийской, а нижнеордовикской и
притом уже достаточно типичным представителем Inadunata с хорошо
развитыми руками. Эта форма не проливает света на происхождение
морских лилий. К сожалению, нет ясности и в филогенетических отноше-
326
Рис. 79. Схема примерного количественного распределения во вре-
мени 446 родов ископаемых иглокожих (из A. Muller)
ниях между подклассами морских лилий — Camerata, Inadunata, Flexi-
bilia и Articulata. Первый подкласс, несомненно, более обособлен от
остальных, чем остальные друг от друга; не без основания Иекель
(Jaekel, 1918) выделял его как подкласс Cladocrinoidea и противопостав-
лял подклассу Pentacrinoidea, отвечающему трем подклассам Inadunata,
Flexibilia и Articulata;
Принятая в настоящее время классификация морских лилий рацио-
нальна; она приближается к естественной, хотя и содержит немало сла-
бых мест. Так, подкласс Articulata не обладает ни одним признаком,
который был бы характерен только для него и не встречался бы в других
327
подклассах. И в остальных подклассах имеются формы, у которых приз-
наки больших групп перемешаны (Wanner, 1934). Все это очень затруд-
няет понимание филогенетических отношений между подклассами мор-
ских, лилий.
Как известно, морские лилии среди всех Pelmatozoa были самым
многочисленным, прогрессивным, пластичным и долговечным классом.
Достаточно напомнить, что в настоящее время известно уже 5000 иско-
паемых видов, тогда как следующие, гораздо менее многочисленные
классы, хорошо, однако, сохранившиеся, благодаря прочному скелету
(цистоидеи и бластоидеи), состоят всего примерно из 300—350 видов
каждый. Общеизвестно громадное разнообразие форм и строения палео-
зойских морских лилий и развитие у них вторичной двусторонней сим-
метрии разной степени (Moore, 1952—1964), нарушение пятилучевой
симметрии (Wanner, 1934), переход к свободноподвижному образу жиз-
ни и вторичный переход к прикрепленному образу жизни, но с прикреп-
лением не стебельком, а абактинальной частью чашечки, и т. д.
Как и у других Pelmatozoa, геологическая история морских лилий
развертывалась и протекала в палеозое (рис. 79).;Подкласс Camerata,
состоящий из 32 семейств (около 2500 видов), просуществовал от сред-
него ордовика до верхней перми, где он вымер. Его расцвет наблюдался
в нижнем карбоне.
Следующий по численности подкласс Inadunata (55 семейств, не ме-
нее 1750 видов) существовал с нижнего ордовика до начала триаса.
В перми вымирают все семейства, кроме Encrinidae, которое вымерло в
конце триаса. Данные о сохранении представителей Inadunata в совре-
менной фауне не подтвердились, они оказались принадлежащими к под-
классу Articulata. Расцвет Inadunata происходил в нижнем карбоне.
Цодкласс Flexibilia небольшой (61 семейство, до 300 видов), просуще-
ствовал с ордовика до перми, где он вымер. Представленные довольно
богато уже в ордовике подклассы Camerata, Inadunata и Flexibilia, про-
должая эволюировать в силуре, достигают расцвета в нижнем карбоне,
становясь господствующими иглокожими. Уже в верхнем карбоне чис-
ленность этих морских лилий снижается, а в конце перми, как было, ска-
зано, они вымирают.
Наиболее высоко организованные морские лилии, подкласс Articu-
lata, становятся известными (в виде нескольких семейств) в триасе,
большинство же семейств появляется в юрский и меловой периоды.
Наряду со стебельчатыми, достигавшими порой гигантских размеров
(с кроной до 1 л в диаметре и со стеблем в 21 л длиной — Pentacrinus
sp.), появляются бесстебельчатые формы и даже пелагические. Стебель-
чатые морские лилии представлены примерно всего 80 современными
видами, принадлежащими 6—7 семействам. Это реликты когда-то обиль-
ной фауны стебельчатых морских лилий, ископаемых видов которых
известно не менее 5000, принадлежавших примерно 100 семействам. Гос-
подствующими в настоящее время являются бесстебельчатые ф.ормы
(20 семейств, около 1000 видов), преимущественно мелководные, жи-
вущие громадными сообществами. Это новый значительный расцвет
морских лилий, но уже иного строения и меньшего разнообразия срав-
нительно с палеозойскими и мезозойскими, новый этап их развития, сво-
боднодвигающихся, а не прикрепленных иглокожих. При этом, подоб-
но офиурам и в отличие от остальных Eleutherozoa, для передвижения
они пользуются сокращениями мускулатуры рук, а не амбулакральной
системой.
Несмотря на обилие палеонтологического материала филогенетиче-
ские отношения между подклассами морских лилий не выяснены, так
как не установлена историческая последовательность в появлении раз-
ных таксономических групп, нет определенных направлений в их разви-
328
тии, наоборот, наблюдается большое разнообразие направлений, неред-
ко пестрота в сочетании признаков строения крупных таксонов. По-ви-
димому, большая податливость на воздействия условий жизни вызывала
у морских лилий частые явления конвергенции.
Родственные отношения между Pelmatozoa и Eleutherozoa, как будто
более или менее выясненные по данным палеонтологии, в последнее
время вновь становятся предметом обсуждений.
Со времени работ Бэзера (Bather, 1915) многие, в том числе и я
(Fedotov, 1928; Федотов, 1951), видели в $дриоастероидеях предков мор-
ских звезд, ежей, офиур. Морских звезд они выводили из форм типа
Edrioaster, морских ежей — от Stromatocystis, при' этом эдриоастерои-
деи, как .отмечает Регнель (Regnell, 1960а, 19606), единственные Pelma-
tozoa с амбулакральными порами и, видимо, с ампулами.
Другие авторы, например Убаке (Ubaghs, 1953), сходство в строении
этих классов считали конвергенцией и близкого родства, во всяком
случае между эцриоастероидеями и морскими звездами, не видели. Опи-
раясь на данные Спенсера (Spencer, 1951), предков морских звезд и
офиур Убаке видит в сомастеридах.
. Эокриноидеи, цистоидеи (нижний кембрий), эдриоастероидеи, отно-
сящиеся к немногим наиболее древним из известных иглокожих, являют-
ся среди них тем не менее формами с наиболее высоко развитыми пятью
амбулакрами, например, Stromathocystis (Cabiel, Termier et Termier,
1959);, и без венчиков из пластинок на абактинальной стороне тела.
В этом эдриоастероидеи ближе стоят к Eleutherozoa, чем к Pelmatozoa,
тем более, что у них нет стебелька, а некоторые из них не были при-
крепленными формами. К тому же, по данным Фея (Fay, 1962), стебель-
чатый Steganoblastres (=Astrocystites) оказался не эдриоастероидеей,
как думал Бэзер (Bather, 1915), а представителем, нового класса, как
думает Фей, Edrtoblastoidea с. признаками эдриоастероидей.
Ордовикские сомастериды Спенсера с их легким, подвижным. скеле-
том бесспорно подрывают мнение об эдриоастероидеях (обладающих в
основном прочным, твердым скелетом тела) как родоначальниках мор-
ских звезд и офиур.. Скелет ранних нижнекембрийских эдриоастероидей
(Stromatocystis') был хорошо дифференцирован, основной тип строе-
ния — архитектоника класса была у них так выработана, что свести или
вывести из эдриоастероидей морских звезд, офиур и морских ежей, и
особенно такие формы, как сомастериды, крайне трудно.
Вряд ли надо останавливаться на идее о филогенетических связях
между морскими лилиями и звездами. Кроме наличия рук, в организа-
ции морских лилий ни в прошлом, ни в настоящем нет ничего, что могло
бы поддержать мысль Фелла (Fell, 19626, 1962в, 1963) о близких род-
ственных связях между морскими лилиями, звездами и офиурами; его
толкования морфологии этих групп не обоснованы. Если и наблюдается
некоторое сходство в форме скелета сомастерид и морских лилий, то оно
чисто поверхностное и имеется только у ордовикских сомастерид и сов-
ременных, наиболее эволюировавших бесстебельчатых морских лилий.
Оно полностью отсутствует у сомастерид и древнепалео’зойских морских
лилий. Сомастериды могут быть предковыми формами для морских звезд
и офиур, но и они своей организацией доставляют ряд трудностей для
понимания общего хода эволюции морских звезд, так как древнепалео-
зойские Asteroidea имели тяжелый абактинальный и актинальный ске-
лет. Эволюция морских звезд в целом шла в направлении облегчения
скелета и развития все большей гибкости и подвижности рук.
Иглокожие всех 13 (стр. 292) Классов этого типа на всех уровнях
строения и эволюции имеют солидное мешковидное тело, порой уплощен-
ное, порой, наоборот, удлиненное; лишь у позднейших, начиная с мезо-
зоя, и у современных морских лилий тело приобретает форму цветка.
329
Спенсер (Spencer, 1951), описавший строение Villebrunaster и Chiniana-
ster, говорил о них как о морских звездах, у которых руки находятся в
процессе дифференциации. Это вполне справедливо по отношению
к Archegonaster, общая морфология которой близка к примитивным
эдриоастероидеям, но это неверно для Villebrutiaster и Chinlanas'ter.
По их реконструкции, данной Спенсером (1951), у этих форм имеются
огромной величины пятиугольный рот и преддверие, или vestibulum, и
плоские, широкие руки и нет выраженного диска. Я сомневаюсь в пра-
вильности такой реконструкция и в том, что у них были лишь руки и не
было центрального диска, в отличие от другого семейства сомастерои-
дей — Archegonasteridae, у которого руки, действительно, находились в
процессе дифференциации, а тело было без выраженных рук.
Не обосновано отнесение Спенсером (Spencer, 1951) к сомастеридам
семейства Archophiactidae с родами Archophlactis Spencer, Sturtzura
Gregory и Lepidasterirta Ruedernann, составленного из неродственных
форм, далеко не столь примитивных, как, например, Lepidasterirta, чтобы
их можно ставить у корня, давшего начало морским звездам и офиурам.
Кроме того, сам Спенсер (Spencer, 1914—1940) , устанавливая эти семей-
ства, отмечал бедность материала и ошибки в толковании его строения.
По Спенсеру, Sturt-zura Gregory из силура Австралии по строению
рук, амбулакральных и адамбулакральных пластинок скорее принад-
лежит к вымершему отряду морских звезд Hemizonida (Spencer^ 1951)
и ближе к семейству Tacniactinidae, чем к офиуроподобным формам.
Lepidasterirta Ruedernann из. верхнего девона Северной Америки,
недавно найденная и у нас (Ухткомбинат, ижемская свита, фаменский
ярус), по строению рук, амбулакральным пластинкам в форме сапога,
по чередующемуся расположению этих и адамбулакральных пластинок, '
сходящихся друг с другом на тонких окончаниях рук,— это типичная
палеозойская офиуроподобная форма из Oegophiuroidea.
Природа Archophiactis grayae из верхнего ордовика Шотландии не-
ясна, несмотря на хорошие рисунки автора. Может быть это примитив-
ная форма, но амбулакральные пластинки на рисунках не похожи на
таковые на фото (Spencer, 1951, табл. XXIV, фиг. 4), последние же
имеют форму сапога и расположены в чередующемся порядке.
Судя по палеонтологическим данным последних лет (например,
(Spencer, 1951; Lhemann, 1957), эволюция морских звезд шла более
сложным путем, чем предполагали раньше (Fedotov, 1928; Федотов, 1934, •
1951, и Др.). Прежде всего, исходными, близкими к родоначальным фор-
мам для морских звезд были сомастериды Спенсера, плоские, коротко
пятиугольные астероидного вида иглокожие, с легким актинальным и ;
абактинальным скелетом, почти без диска, со слабо дифференцированны-
ми плоскими руками, медленно двигающиеся микрофаги. Если Спенсер j
прав, эта группа объединяла также формы с исходными чертами строе- j
ния и офиур. Нужно ждать дальнейших находок представителей сома-
стерид из кембрийских отложений, которые позволят пополнить наши
знания о строении этой группы, которую я рассматриваю как класс.
В ордовике протекала значительная эволюция трех ветвей морских >
звезд, сопровождавшаяся развитием рук, их осевого скелета, амбулак-, (
ральных пластинок, дифференцировкой адамбулакральных и маргиналь- . 1
ных пластинок, а также развитием абактинального скелета диска и рук.
Наиболее близкой к сомастеридам была небольшая ветвь морских
звезд, объединенных Спенсером (Spencer, 1951) в отряд Platyasterida,
с плоским телом, широкими руками, сравнительно короткими, с широко
открытыми амбулакральными желобками, с плоско расположенными :J
амбулакральными и адамбулакральными пластинками (Platanaster,
Palasterlscus). На мой взгляд, если принять данные Фелла (Fell, 1962а)
о строении современной Platasterias, она напоминает формы отряда
330
Platyasterida, представители которого найдены в среднем ордовике и
нижнем девоне.
Значительно более мощной и более высоко дифференцированной
ветвью древних морских звезд являются формы отряда Спенсера Hemi-
zonida, с глубокими амбулакральными желобками, свод которых обра-
зован амбулакральными и адамбулакральными пластинками, образую-
щими главную часть актинального скелета рук. Абактинальный скелет
диска и рук состоит из многих пластинок. Морские звезды этого отряда
•более дифференцированы сравнительно с таковыми отряда Platyasterida
и дальше их отошли от сомастерид; они существовали со среднего ордо-
вика до среднего карбона.
Однако филогенетические отношения в этом отряде неясны, уже по-
тому, что подотряды GnathoMerina (семейства Taeniactinidae, Palaste-
rinidae, Schuchertiidae и Lepidactinidae) и Urasterina (семейства Cne-
midactinidae, Urasterellidae и Arthrasteridae) очень различны пр строе-
нию отнесенных сюда Спенсером форм. Природа первых четырех
семейств выяснена еще недостаточно; по форме тела и строению скеле-
та рук они сильно отличаются от семейства подотряда Urasterina.
Последние имеют развитые, очень длинные, узкие руки с хорошо диф-
ференцированным, прочным скелетом, даже если взять наиболее древ-
ние роды, как, например, Cnemidactis, известный из среднего ордовика
нашего Союза.
Следует отметить, что формы отчасти отряда Platyasterida, а главным
образом Hemizonida богато представлены, особенно в нижнем девоне
Зап. Европы. Они достигали очень крупных размеров, более 14 см в диа-
метре, с руками около 10 см длиной, нередко многочисленными, от 13
до 29 (как у Palaeosolaster). Кстати, надо отметить, что многочисленные
жившие с ними офиуроподобные формы (класса офиур) также нередко
отличались гигантской величиной сравнительно с позднейшими предста-
вителями этого класса, например, Encrinaster giganteus, Mastigophiura
grandis (Lehmann, 1957). Это говорит о благоприятных условиях нижне-
девонского моря Зап. Европы и. о достижении известного расцвета этой
группы морских звезд и офиур (точнее, офиуроподобных форм), за ко-
торым последовал упадок в развитии, кончившийся в нижнем девоне их
вымиранием.
В ордовике же началось развитие и третьей ветви морских звезд, та-
ких семейств, как Hudsonasteridae, Xenasteridae, Petrasteridae, Protopa-
laeasteridae и др. Эти морские звезды, объединенные Спенсером в под-
отряд Pustulosa отряда Phanerozonia, обладали тяжелым абактиналь-
ным и актинальным скелетом из небольшого числа пластинок. Очень
рано у них наметилось становление организации настоящих морских
звезд типа Phanerozonia, с хорошо развитыми маргинальными пластин-
ками, образующими вертикальный край диска и рук. Морские звезды
этого подотряда являются очень древней ветвью класса морских звезд.
Они существовали С нижнего ордовика до пермокарбона и, несомненно,
филогенетически связаны с подотрядами Valvata и Paxillosa, которые
известны с юры. Они проделывают известную эволюцию в меловом пе-
риоде, становясь разнообразными, довольно многочисленными и порой
крупными формами. В настоящее время они представлены многочислен-
ными семействами (преимущественно в тропических морях), обладаю-
щими, однако, пятью обычно несгибающимися руками. Более примитив-
ные из них остаются микрофагами с пассивным ртом, питающимися при
помощи ресничных токов; многие сохраняют остроконечные амбулак-
ральные ножки. Некоторые (например, Pentagonaster, Tosia) во взрос-
лом состоянии сохраняют почти исходное строение абактинального ске-
лета в виде строго дифференцированных венчиков радиальных и интер-
радиальных пластинок.
331
В мезозое становятся известными представители важнейших групп
современных морских звезд, которых одни авторы, рассматривают как
два самостоятельных отряда — Spinulosa и Forcipulata, другие—как
подотряды Cryptozonia. Это наиболее высокоорганизованные, почти
исключительно хищные, активно захватывающие нередко очень крупную,
до рыб включительно, добычу, разнообразные, вплоть до шарообразной
формы [современная Podosphaeraster (Clark a. Wright, 1962), ископае-
мые Sphaeraster], достигающие крупных размеров. Более высокооргани-
зованными являются морские звезды Forcipulata, с гибкими длинными
руками, с четырьмя рядами амбулакральных ножек, с облегченным
абактинальным скелетом, нередко в форме ячеистой сети скелетных
перекладин и т. д. Среди морских звезд Spinulosa и особенно Forcipulata
распространена многолучевость, достигающая, как было отмечено рань-
ше (стр. 293), ДО 50 рук у одной особи. Филогенетически эти группы звёзд
близки, причем по ряду признаков Spinulosa сближаются с Pha'nerozohia.
Косвенные данные — распространение Spinulosa по всем морям, a For-
cipulata преимущественно в арктических и антарктических областях —
свидетельствуют о позднейшем происхождении последних. О большой
пластичности обеих групп, достигнутой в ходе эволюции, говорит также
выживаемость ряда их форм (Solaster, Asterias) в опресненных водах,
с соленостью 15—8%о (Дьяконов, 1955).
Эволюция морских звезд, в общем, проходила в направлении диффе-
ренцировки и увеличения рук, удлинения последних, облегчения скелета,
замены тяжелого абактинального скелета диска и рук более легким и
гибким, усиления гибкости рук, перехода от пассивного питания (мйкро-
фагии) к хищничеству и ловле крупной добычи, в увеличении размеров
тела, и т. д. Боковые ветви вымирали в палеозое, основной ствол морских
звезд постепенно эволюировал и разрастался.
Известное совершенствование организации, достижение очень круп-
ных размеров, некоторый расцвет испытали в нижнем девоне примитив-
ные морские звезды отряда Hemizonida и офиуроподобные формы, после
чего наблюдался упадок в их эволюции и вымирание. В мезозое, в мело-
вом периоде, начинается известный расцвет, некоторая эволюция пред-
ставителей уже настоящих морских звезд отряда Phanerozonia,
собственно, его низших подотрядов, преимущественно Valvata, и форми-
рование отрядов Spinulosa и Forcipulata — более высоко организован-
ных морских звезд.
Вследствие особенностей скелета, состоящего из многих отдельных
элементов, морские звезды в ископаемом состоянии сохраняются плохо,
поэтому их геологическая история полна пробелами. Однако из того, что
за громадный промежуток времени (с нижнего ордовика до кайнозоя)
известно немногим более 300 ископаемых видов против примерно
2000 современных, ясно, что эволюция морских звезд шла медленнцми
темпами в палеозое и мезозое, но подъем, начавшийся в конце мезозоя,
быстро нарастал и в настоящее время этот класс находится в расцвете.5
Как известно, морские звезды часто активные хищники, широко распро-
странены в водах Мирового океана, от береговой зоны до ультраабис-
сальных глубин — 7614 м (Беляев и др., 1960).
Из работ Спенсера (Spencer, 1914—1940) было .особенно ясно, что
древние представители класса офиур и морские звезды обладали рядом:
существенных общих черт и имели общих,предков (Федотов, 1951). Од-
нако новую классификацию офиур, которую дал Спенсер- (1951), пол-
ностью принять нельзя. Отмечу, что созданный им отряд Stenurida не-
приемлем , так как туда отнесены такие разнородные формы, как Stena-
ster Billings со звездообразным телом, не обособленным на диск и руки,
и Eophiura Jaekel emend. Schuchert, Palaeura Jaekel emend. Schuchert,.
Pradesura Spencer и другие с хорошо выраженными, дифференцирован-
3.32
ными диском и руками. Возможно -Спенсер прав, находя у них общее в
скелете амбулакров, однако строго противоположное расположение ам-
булакральных пластинок у Stenaster и чередующееся у офиуроподобных
форм других членов этого отряда и.общая морфология тела- последних
в целом настолько различны, что. семейство Stenasteridae нельзя объеди-
нять с другими семействами этого отряда, т. е. весь отряд создан искус-
ственно. ' - .
Мне представляется возможным сохранить отряд Stenurida, но лишь
с одним своеобразным семейством Stenasteridae, обладающим чертами
офиур и морских звезд (Fedotov, 1936), положение в. системе которого
требует еще' тщательного изучения, поэтому его пока приходится игно-
рировать. Тем более, что его значение в эволюции морских звезд и офиур
ничтожно. Остальные семейства этого отряда с диском, руками с чере-
дующимися амбулакральными пластинками явно офиуроподобные фор-
мы и отнести их . надо к Oegophiuroida Матцумото. На этом основании
нужно вернуться к классификации офиур этого автора (Федотов, 1951)
и класс офиур делить на два подкласса — Oegophiuroida и Myophiuroida.
Офиуроподобные формы, геологическая история которых длилась с ниж-
него ордовика до верхнего карбона, обладали рядом общих черт с древ-
ними морскими звездами и не имели многих характерных черт настоя-
щих офиур; они, несомненно, имели общих предков с морскими звездами.
В древнем палеозое они были малочисленными, хотя достаточно широко
распространенными формами. В нижнем девоне наблюдается некоторый
их расцвет, среди них имелись виды (Cheiropteraster giganteus, Masti-
gophiura grandis и.другие), которые были гигантами даже по сравнению
с позднейшими типичными офиурами.
Рано,-уже в нижнем ордовике, началась эволюция класса офиур: на-
метился иной принцип движения, чем у морских звезд — движение с
помощью подвижных рук, снабженных сегментарной мускулатурой,
вместо движения при помощи амбулакральных ножек. С этим было свя-
зано обособление рук от диска, массивный характер амбулакральных
пластинок, между которыми находились мышцы рук. Амбулакральные
желобки были еще открытыми, амбулакральные пластинки — парными,
руки лишь с боков.были защищены адамбулакральными или интерам-
бу'лакральными пластинками, руки были еще слабо дифференцированы.
Такие офиуроподобные формы (Oegophiuroida) вымерли в верхнем
карбоне.
В процессе эволюции офиур происходило замыкание амбулакраль-
ных желобков, превращение парных амбулакральных пластинок в целые
(слитые) позвонки, прогрессивное развитие межпозвоночных мышц, обо-
собление-рук от диска и-превращение их в органы передвижения с по-
мощью мышечных сокращений, более совершенное, чем у морских звезд
и других классов Eleutherozoa, развитие системы наружных пластинок
рук. Становление подкласса Myophiuroida намечалось в нижнем девоне,
откуда известны Aganaster и Ophiurina (Lehmann, 1957), обладавшие
уже позвонками. Число видов типичных офиур несколько возрастает в
карбоне, но заметная эволюция их как самых быстро передвигающихся
иглокожих начинается в мезозое и протекает позже, вплоть до настоя-
щего времени.
В современных морях офиуры наиболее многочисленны по количеству
видов (2000) и по числу особей. Они живут огромными скоплениями,
стадами, являясь руководящими формами ряда обширных биоценозов.
Эти формы, обычно мелкие или небольшие, составляют.отряд Ophiu-
гае, являясь типичными офиурами с пятью простыми руками, хорошо
развитым поверхностным скелетом диска и рук, обладающих ограничен-
ной подвижностью — изгибанием в горизонтальной плоскости, с пятью
парами бурс в форме мешков. Если не считать Astrophiura, все они по-
333
строены однообразно, сравнительно с морскими звездами. У них наблю-
дается бесконечное разнообразие в деталях, связанное с условиями
жизни, но эволюционной направленности в этом разнообразии не видно.
Они широко распространены в Мировом океане от береговой линии до
сверхабиссальных глубин — до 8006 м (Беляев и др., 1960), но по эколо-
гии также довольно однообразны.
В отличие от отряда Ophiurae Euryalae — небольшая группа почти
исключительно многоветвистых, крупных, нередко очень крупных (10—
20 см в диаметре диска) офиур. Они утратили типичный поверхностный
скелет диска и рук, обладающих, благодаря особенностям сочленения
позвонков, способностью искривляться в горизонтальной и свертываться
в вертикальной плоскости. Вторичная полость тела их замещена третич-
ной полостью, возникающей из разросшихся и слившихся вместе бурс.
Euryalae в отногенезе, однако, проходят стадию с организацией и скеле-
том, свойственными представителям типичных офиур.
, Этот отряд — несомненно более поздняя ветвь Myophiuroida,'обла-
дает рядом примитивных черт, отчасти общих с морскими звездами
(Fedotov, 1926), вместе с тем некоторые его представители достигают
максимального среди всех иглокожих совершенства — пятилучевой сим-
метрии. Видимо, филогенетически многоветвистые офиуры Euryalae свя-
заны с нижнекаменноугольными Onychaster и Catyptactis, обладавши-
ми свертывавшимися руками, но с пластинками поверхностного скелета
(Spencer, 1914—1940).
По современным данным, история эволюции морских ежей начинает-
ся, сравнительно с историей морских звезд и офиур, несколько позже
(если не считать Bothriocidaris) — в верхнем ордовике, когда становятся
известными примитивные формы — Aulechinus, Ectinechinus, Myriasti-
ches — с гибким панцирем, с двумя рядами амбулакральных пластинок
и с многочисленными беспорядочно расположенными интерамбулакраль-
ными пластинками. Ими начинается эволюция отряда Lepidocentroida,
продолжавшаяся в силуре, девоне, карбоне и до конца перми. Происхо-
дило увеличение размеров сравнительно гибкого панциря (например,
диаметр нижнекарбонового Fournierechinus достигал 35 см} и числа
амбулакральных пластинок от 2 до 20 в амбулакре, упорядочение распо-
ложения интерамбулакральных пластинок в ряды от 3 до 40 в каждом
интерамбулакре и т. д. Эта группа была наиболее многочисленной среди
палеозойских морских ежей, достигшая расцвета в нижнем карбоне и
вымершая в конце перми. Правда, по ряду существенных признаков к
ним настолько близки так называемые мягкие ежи — Echinothuriidae,
что их относят к отряду Lepidocentroida (Мортенсен, см. Федотов, 1951),
но ископаемые Echinothuriidae пока известны лишь из триаса.
Очень рано, в среднем ордовике, становится известна маленькая
ветвь общего ствола морских ежей — отряд Rothriocidaroida (из од-
ного рода Bothriocidaris с несколькими видами), природа которого как
морского ежа окончательно установлена Р. М. Мяннилем (1962), нашед-
шим у него челюстной аппарат. Этот отряд быстро вымер, не оставив
потомства.
Благодаря примитивности ряда форм, большому разнообразию и не-
устойчивости строения амбулакров и интерамбулакров отряд Lepidocen-
troida является основной группой ствола морских ежей, от которой могли
произойти все остальные. В верхнем силуре появляется ветвь этого
ствола — отряд палеозойских морских ежей Melonechinoida с прочным
панцирем, с двумя и более рядами пластинок в амбулакрах, с одним и
четырьмя в интерамбулакрах, очевидно, взявшая начало от Lepidocent-
roida и просуществовавшая до верхней перми. В силуре же (Regnell,
1959) становятся известными представители новой ветви ствола морских
ежей, отряда Cidaroida, со строго фиксированным числом простых узких
'334
амбулакральных и очень широких интерамбулакральных пластинок
(по два ряда пластинок в очень узких амбулакрах и в очень широких
интерамбулакрах) с крупными бугорками и с сильными иглами, с двумя
типами педицеллярий и без наружных жабр. Отряд Cidaroida филоге-
нетически также выводится из лепидоцентроид, хотя непосредственно
связать цидароид с какой-либо формой последних пока нельзя. Цидарои-
ды,— малочисленные в палеозое, численно возрастают в мезозое и в
настоящее время представлены небольшим числом видов, преимущест-
венно глубоководных, наиболее распространенных в Индо-Тихоокеан-
ской провинции. В течение всей геологической истории морских ежей
цидароиды были незначительной и в общем мало пластичной группой,
у которой эволюировали иглы, как органы движения, в связи с чем раз-
вивались интерамбулакры и мало эволюировали амбулакры.
Все же организация цидароид, сравнительно с таковой лепидоцентро-
ид и мелонехиноид, была более собранной, олигомерной и пока по цеиз-
вестным нам причинам оказалась более жизненной, поскольку все, кро-
ме цидароид, так называемые палеозойские морские ежи, или Perischo-
, echinoidea A'lcGoy, вымерли к концу палеозоя.
По современным данным в мезозое сохраняются лишь цидароиды,
и с того времени начинается новый этап в эволюции морских ежей,
в котором цидароиды играют незаметную роль; в юре начинается эво-
люция остальных правильных, а несколько позже так называемых
неправильных морских ежей.
Вопрос о филогенетических связях цидароид с другими правильными
морскими ежами далеко не решен. Однако трудно представить себе,
.чтобы последние возникли из цидароид с их угнетенными узкими амбу-
лакрами, со слабыми, малочисленными амбулакральными ножками,
играющими у цидароид незначительную роль в передвижении, в отличие
от большинства морских ежей. К реже распространенному взгляду
о происхождении остальных правильных ежей от цидароид (Федотов,
1934) вновь вернулись американские палеонтологи. Одним основанием
для этого является то, что в триасе (откуда пока известны морские ежи
отряда Aulodonta) цидароиды существовали, а лепидоцентроид, если не
считать Echinothuriidae, уже не было. Кроме того, эти авторы думают,
что все последующие группы морских ежей произошли от общего пред-
ка— Mlocidaris нового семейства Miocidaridae. Из морфологически раз-
нообразной, не специализированной группы лепидоцентроид, однако,
гораздо легче вывести формы отряда Aulodonta,’ чем из специализиро-
ванной группы цидароид, тем более, что наиболее древние представители
отряда Aulodonta (Геккер, 1964) имели тонкий, нередко гибкий панцирь
и тонкие иглы, подобно лепидоцентроидам.
В отличие от цидароид эволюция главной ветви родословного ствола
остальных правильных морских ежей шла (что замечалось уже у ряда
палеозойских морских ежей) по пути развития амбулакров, усиления
амбулакральных ножек как органов движения, увеличения их числа и
в связи с этим усложнения амбулакральных пластинок путем образова-
ния из простых сложных пластинок со многими порами. Образуются
перистомальные жабры. Иглы, органы защиты, порой с ядовитыми же-
лезами,. достигают большого разнообразия; иглы используются также
для передвижения, не подменяя в этом амбулакральных ножек; обра-
зуются сферидии, эволюируют педицеллярии (как органы защиты, сани-
тарии, и захвата пищи), которых становится четыре типа и т. д.
Из трех отрядов этой группы морских ежей — Aulodonta, Stirodonta
и Camarodonta [хорошо обоснованной классификации морских ежей
Мортенсена (Mortensen, 1928—1952), лучшей чем новая классификация
Дарема и Мельвиля (Durham a. Melville, 1957)], отряд Aulodonta
становится известен в среднем триасе, отряд Stirodonta — в верх-
335
нем триасе, отряд Camarodonta— в верхней юре. Эти отряды мор-
фологически настолько связаны друг с другом, что их нередко объеди-
няли в один надотряд или как подотряды одного отряда Diadematoida.
Отряды Aulodonta и Stirodonta, испытав значительный расцвет в юре
и особенно в мелу, затем пошли на убыль и являются небольшой частью
современной фауны так называемых правильных морских ежей. Самым
пластичным, достигшим наибольшей высоты организации, распростра-
нившимся по всему Мировому океану, хотя более многочисленным в
тропических водах, является отряд Camarodonta. Надо думать, что
и филогенетически все эти отряды морских ежей связаны друг с другом;
предполагают, что отряд Camarodonta произошел от предков из отряда
Stirodonta, связанного происхождением с отрядом Aulodonta.
Мощной ветвью ствола морских ежей являются четыре отряда — Ho-
lectypoida, Cassiduloida, Glypeasteroida и Spatangoida. Эти отряды, по
классификации Мортенсена (Mortensen, 1928—1952), самостоятельны.
Дарем и .Мельвиль (Durham a. Melville, 1957), вернувшись отчасти к
старой классификации, объединяют Holectypoida и Clypeasteroida в над-
отряд Gnathostomata, a Cassiduloida и Spatangoida в надотряд Atelo-
stomata, куда отнесены два новых отряда Nucleolitoida и Holasteroida,
установленных этими авторами. Классификация Мортенсена хорошо
обоснована и пока нет оснований ее отвергать. Считают, что Holectypoi-
da, Cassiduloida, Clypeasteroida и Spatangoida являются потомками
каких-то примитивных представителей Aulodonta или Stirodonta. В ниж-
ней юре становятся известными представители отрядов Holectypoida,
Cassiduloida и Spatangoida, в верхнем мелу — Clypeasteroida.
В ходе эволюции этих групп морских ежей шло развитие двусторон-
ней симметрии, связанное с переходом от жизни на поверхности грунта
к роющему образу жизни. Панцирь удлиняется, перистом передвигается
вперед, помещаясь иногда на переднем конце панциря; апикальное поле
удлиняется, перипрокт выходит -из него и помещается на заднем конце
тела; соответственно изменяется длина разных амбулакров и интерам-
булакров. В амбулакрах происходит также дифференцировка на верхние
петалоидные части (их амбулакральные ножки становятся жабрами) и
на нижние части, где основными органами движения являются иглы.
Изменяется также и строение интерамбулакров, особенно нижней сто-
роны панциря (например, образуются за счет их пластинок стернум и
нижняя губа).. Иногда панцирь принимает форму бутылки (Pourtalesii-
dae).
Происходит изменение во внутренней организации, а также в фор-
ме и строении аристотелева фонаря — происходит его регресс и исчезно-
вение у взрослых особей (Spatangoida, Cassiduloida). Наиболее совер-
шенной степени двусторонней симметрии достигают морские Spatangoi-
dea, которые в онтогенезе, однако, проходят стадию радиального сим-
метричного морского ежа (Беклемишев, 1952).
Среди этой группы морских ежей Holectypoida, как известно, по сте-
пени сохранения радиальной симметрии'в форме панциря и по строению
амбулакров и интерамбулакров, а также аристотелева фонаря с зубами
и килем на внутренней стороне и по другим чертам близки к формам
отряда Stirodonta или Aulodonta и могли быть родоначальной группой
для Cassiduloida, Spatangoida и Clypeasteroida. По-видимому, по ряду
признаков, -в том числе наличию аристотелева фонаря, Clypeasteroida
сближаются с Holectypoida, по другим чертам и редукции аристотелева
фонаря — с Spatangoida. Возможно, что это сходство обусловлено и
филогенетическими связями. Это основное, что следует взять из цитиро-
ванной выше работы Дарема и Мельвиля (1957), в которой приводятся
детальные филогенетические отношения всех групп класса морских ежей,
в известной степени подтвержденные кристаллографическими особенно-
336
стями скелета — перпендикулярным и тангенциальным положением .оп-
тической оси в скелетных частях (Raup, 1962а, 19626).
Благоприятные условия морей юрского и мелового периодов способ-
ствовали эволюции класса морских ежей (в том числе отрядов Holecty-
poida, Cassiduloida, Clypeasteroida и Spatangoida), расцвету, продолжав-
шемуся после небольшого спада в палеогене и третичном времени
(А. Н. Muller, 1961). Достаточно упомянуть, что именно на эти периоды
расцвета приходится главная масса из 7200 описанных, ископаемых
видов.
В настоящее время фауна морских ежей богата и представлена мно-
гочисленными (860 видов, Storer a. Usinger, 1957) видами и родами,
главным образом из отрядов Camarodonta, Clypeasteroida и Spatangoi-
da. Это преимущественно массовые обитатели полосы от верхней грани-
цы прилива до конца континентального шельфа, особенно в Индо-Тихо-
океанской провинции. Однако среди них имеются и глубоководные
ультраабиссальные формы.
Организация голотурий изучена хорошо, а классификация, построен-
ная на совокупности данных внутренней организации и скелета, рацио-
нальнее классификации любого другого класса иглокожих. Геологиче-
ская история голотурий, вследствие отсутствия у них сплошного скелета,
известна мало. В настоящее время, к сожалению, ни одну из среднекем-
брийских форм Уолкотта (Walcott, 1911; см. гл. «Аннелиды»). нельзя
считать голотурией (Баранова и Геккер, 1964), но Palaeocucumaria из
нижнего девона Зап. Европы (Lehman, 1957; Баранова и Геккер, 1964),
гозможно, голотурия.
Находки отдельных известковых телец ископаемых голотурий отры-
вочны и мало дают для выяснения хода эволюции голотурий. Все же,
даже не помещая верхнеордовикскую Eothuria в класс голотурий, по-
скольку эта форма почти по всем основным признакам, кроме предпола-
гаемого наличия одной пластинки — мадрепоровой с половой порой,
нр укладывается в класс голотурий, это — древняя группа, так как скле-
риты ископаемых голотурий известны из среднего ордовика (Gutchik,
1954).
Родственные отношения голотурий, вследствие отсутствия палеонто-
логических данных, приходится строить лишь на данных сравнительной
анатомии. Можно вместе с Кюено (Cuenot, 1948) считать основной груп-
пой, обладающей наиболее полным комплексом черт организации и ра-
диальной симметрией, отряд Dendrochirota; Aspidochirota, Elasipoda и
Molpadonia можно вывести из отряда Dendrochirota. Затруднение пред-
ставляет отряд Apoda, у представителей которого и в онтогенезе, и у
взрослых имеются примитивные черты, наряду с чертами организации
вторичного характера, связанными или обусловленными жизнью в толще
грунта и являющимися измененными чертами строения, характерными
для класса голотурий, например, редукция амбулакральных радиальных
каналов; амбулакральных ножек (Runnstrom, 1928) и т. д. Все это не
позволяет' вывести отряд Apoda из какого-либо существующего отряда;
скорее всего эти голотурии отошли от общего родоначального ствола
класса до дифференциации его на отряды Dendrochirota, Aspidochirota,
Elasipoda и Molpadonia.
Известно давно, что взгляды зоологов и палеонтологов на родствен-
ные, филогенетические связи классов современных иглокожих противо-
речивы, что в известной мере вызвано особенностями онтогенеза, напри-
мер, офиур, морских звезд й ежей. Напомню, что личинки с их скелетом
и ход сегментации целомов морских ежей и офиур сходны, но у офиур
радиальносимметричные актинальные и абактинальные зачатки тела
• будущего имаго возникают на спинной и брюшной сторонах диплеврулы,
причем отбрасывается задняя часть личинки. При различиях в типе ли-
22 Д- М. Федотов
337
чинок и в ходе сегментации целомов у морских звезд и ежей пятилуче-
вые зачатки тела возникают на левой и правой сторонах личинок, они
сходны друг с другом; отбрасываются передние части личинок; кроме
того, как давно отмечалось, морские ежи в онтогенезе проходят стадию
морской звезды. Между тем личинки морских звезд — бипиннарии — по
своей организации относятся к примитивным личинкам, они близки к
аурикуляриям голотурий и очень далеки от эхиноплутеусов, но у морских
звезд метаморфоз, как и у морских ежей, резко некробиотический, а у
голотурий — эволютивный.
Исторических данных о филогенетической связи морских звезд и ежей
нет, но ее можно допустить на основании характера их морфогенеза.
С другой стороны, при значительном различии хода морфогенеза у мор-
ских звезд и офиур, наоборот, история развития древних палеозойских
морских звезд и офиур говорит об их филогенетической близости, общ-
ности их происхождения. На известном этапе исторического развития,
в древнем палеозое, морские звезды и офиуры легко объединялись ря-
дом крупных общих черт строения в Astefozoa не как классы, а как
более мелкие таксоны. Никаких намеков на такую какую-либо близость
морских звезд и ежей палеонтология не дает. Поэтому ряд общих черт
в морфогенезе морских звезд и ежей надо рассматривать лишь как ука-
зание на развитие от общего отдаленного примитивного пятилучевого
предка до дифференцировки класса морских ежей и Asterozoa, давших
классы Somasteroidea, Asteroidea и Ophiuroidea.
Нет общего взгляда и на филогенетические связи голотурий с други-
ми классами. Если не признавать среднеордовикскую Eothuria голоту-
рией, почти по всем основным чертам отличную от голотурий, то в своей
геологической истории голотурии не имели сплошного наружного скеле-
та, но обладали кожистыми стенками тела с типичными склеритами и
были червеобразными удлиненными формами.
Радиальная симметрия у голотурий не достигает того завершения,
которое характерно для морских ежей, звезд и офиур. Непарная половая
железа, происходящая непосредственно из первичной гонады, половое
отверстие, гидропора, или мадрепорит на переднем конце тела, отсут-
ствие осевого комплекса органов — все это является примитивными
чертами, благодаря которым голотурии резко отличаются от указанных
выше классов. Резко отличается примитивностью и онтогенез голотурий.
Аурикулярии, являющиеся наиболее примитивными личинками иглоко-
жих, сравнимые с торнарией кишечнодышащих, без особой ломки пере-
ходят к имаго, возникающему путем эволютивного метаморфоза, с со-
хранением личиночного скелета, исчезающего у других классов.
К примитивным чертам относится также возникновение пятилучевой
симметрии амбулакральной системы у ряда голотурий через трехлу-
чевую.
Новые черты организации голотурий — кожно-мускульный мешок,
водные легкие с чудесной сетью, кювьеровы органы ряда форм и неко-
торые другие подчеркивают обособленное положение этого класса.
В известной мере сближает голотурий и морских ежей вторичная
двусторонняя симметрия, затрагивающая форму тела и органы амбулак-
ральной системы, но она возникает в обоих классах независимо. Удли-
ненная форма тела и мягкие покровы тела некоторых морских ежей —
также лишь внешнее сходство ежей с голотуриями. Важными общими
чертами в организации этих классов являются положение эктоневраль-
ной нервной системы в эпиневральных каналах и в меньшей степени
удлиненная трубчатая форма кишечника, а также спинной и брюшной
кровеносные сосуды. Однако у голотурий нет полового тяжа, половая
железа непарная. У морских ежей, как и у морских звезд, имеется поло-
вой тяж и пятилучевого строения половые железы.
338
Если считать, что строение эктоневральной нервной системы надо
объяснить происхождением голотурий и морских ежей от общего предка,
тогда приходится допустить, что примитивные древние формы голотурий
утратили периферический скелет, приобрели мягкие гибкие стенки и
червеобразное тело, утратили элементы осевого комплекса органов
и т.д. Тогда надо допустить, что из, онтогенеза голотурий (по неизвест-
ным причинам) выпали основные черты морфогенеза, которые четко
выражены у морских ежей в их некробиотическом метаморфозе, хотя
весь онтогенез голотурий сохраняет примитивный характер.
Вместе с тем приходится подчеркнуть, что у морских звезд и голо-
турий, классов, отстоящих друг от друга дальше, чем морские ежи и
звезды или морские звезды и офиуры, личинки одного типа и притом
более примитивного, чем эхино- и офиоплутеусы. Далее, в биохимии мы-
шечного сокращения у морских звезд и голотурий играет роль аргинин-
фосфаген, тогда как у морских ежей и офиур — креатин-фосфаген; био-
химия первых характеризуется стеллостеролом, а вторых (и морских
лилий) — холестеролом.
Следовательно, общий тип строения, кроме некоторых анатомических
черт, эмбриология, биохимия и пока данные палеонтологии не позво-
ляют сближать голотурий и морских ежей, хотя ранее и нередко сейчас
их объединяют вместе в надкласс Echinozoa.
План строения вновь установленного класса Helicoplacoidea по иско-
паемым веретеновидной формы из нижнего кембрия Калифорнии
(Durham a. Castor, 1963) крайне своеобразен. По-видимому, это иглоко-
жие, но как-либо связать их с каким-нибудь из известных, даже самых
древних классов этого типа нельзя, настолько эти ископаемые своеоб-
разны. Предполагавшегося сходства их с голотуриями и морскими ежа-
ми не оказалось вовсе.. В силу этого приходится по-прежнему допускать
независимое происхождение морских ежей от морских звезд и офиур и
происхождение голотурий, независимое от морских ежей, т. е. полимор-
физм классов Eleutherozoa.
22*
Погонофоры (Pogonophora)
Двустороннесимметричные животные, червеобразные, нитевидные,
морские, непаразитические, преимущественно глубоководные, живущие в
трубках. Тело состоит из трех отделов («сегментов»), передний отдел с
щупальцевым аппаратом для улавливания добычи, переваривания ее и
дыхания, третий отдел сегментированный на конце, со щетинками на каж-
дом сегменте. Эпидермис с хитиновой кутикулой, кожно-мускульный
мешок из кольцевых и продольных волокон, нервная система в виде ин-
траэпителиального сплетения нервных волокон и клеток, образующего
спинной нервный тяж, кишечник с ротовым и анальным отверстиями от-
сутствует. Целомическая полость состоит из непарного протоцеля с парой
экскреторных целомодуктов, парного мезоцеля, парного метацеля с парой
целомодуктов — выводных протоков, пары половых желез, и из метамер-
нс расположенных полостей сегментов членистого конца тела. Кровенос-
ная система замкнутая с главными спинными и брюшными сосудами.
Полы разделены, размножение только половое. Дробление яиц полное,
неравномерное, двустороннесимметричное (или радиальное). Развитие
проходит в материнской трубке без метаморфоза. Погонофоры — недав-
но открытая, пока небольшая группа узко специализированных с чер-
тами регресса животных. Трубки погонофор найдены в среднем кембрии.
Основная масса работ, наиболее полно раскрывших строение, отчасти
развитие и положение погонофор в системе этих поразительно своеоб-
разных животных, принадлежит А. В. Иванову (1949, 1955, 1957, 1959,
1960, 1962, 1964а, 19646, Ivanov, 1963) и лишь отдельные небольшие
работы выполнены зарубежными зоологами (Caullery, 1914, 1948; Jo-
hansson, 1937, 1939; Jagersten, 1956, 1957; Southward a. Southward, 1963;
Webb, 1964a—r).
Иогансон признал описанных Коллери (Caullery, 1914) и П. В. Уша-
ковым (Uschakov, 1933) животных представителями самостоятельного
класса Pogonophora. Сначала Рейзингер (Reisinger, 1938), затем
В. Н. Беклемишев (1944) по очень неполным по тому времени исследо-
ваниям отнесли погонофор к вторичноротым, причем В. Н. Беклемишев
предложил для них новый тип Pogonophora. Близость погонофор к He-
michorda признавали также Коллери (Caullery, 1948) и К. Н. Давыдов
(Dawydoff, 1948). Позже В. Н. Беклемишев (1952) перенес погонофор
как надкласс в тип Hemichorda. А. В. Иванов, подробно изучивший
строение и развитие погонофор и предложивший (1955) для них тип
Brachiata, затем вернулся (1960) к наименованию типа Pogonophora
Beklemischew с одним классом Pogonophora и двумя отрядами — Athe-
canephria Ivanov, 1955 и Thecanephria Ivanov, 1955.
Длина погонофор во много раз превышает ширину, у Siboglinum
caulleryi, например, отношение этих величин равно 600 : 1 (Иванов, 1960).
На переднем конце находится щупальцевый аппарат, на заднем конце
дорсо-вентральное углубление, присоска по Веббу (Webb, 1964а — г).
340
Нет ни рта, ни порошицы, так как нет и кишечника. Спинная сторона
более выпуклая, с ресничной полоской, брюшная более плоская, с
брюшным желобком, находящимся в преаннулярной части метасомы.
Парные выводные отверстия целомодуктов в протосоме и парные поло-
вые женские и мужские отверстия расположены симметрично. Двусто-
ронняя симметрия у Spirobrachiidae нарушена спиральной свернутостью
щупальцев, у Siboglinum— олигомеризацией их до одного.
Размеры тела сравнительно небольшие, мелководные Siboglinum
minutum измеряются 5,5 см в длину, наиболее крупная форма Zenkewit-
chiana longissitna достигает 36 см. В поперечном разрезе тело погонофор
имеет всего 0,13—2,5 мм.
Многие погонофоры полупрозрачны, беловатые, некоторые крупные
виды окрашены, так, Spirobrachia grandis коричнево-красная. Обычно
через стенки тела просвеч'ивает красная кровь сосудов.
Погонофоры всегда живут в трубках из хитина и протеина (Brunet
a-. Carlisle, 1958; Southward a. Southward, 1963), которые образуются их
тубипарными железами; Трубки имеют слоистое строение и бывают
однообразными или состоящими из более плотных темных колец, чере-
дующихся со светлыми эластическими кольцами. У большинства пого-
нофор передний участок трубки более тонкий, нежный, легко спадаю-
щийся, благодаря чему он хорошо защищает животное (Иванов, 1960).
Длина Трубки в несколько раз больше длины погонофоры: у мелких ви-
дов Siboglinum она измеряется 15—20 см, у Zenkewitchiana longissitna.
трубки достигают 1,5 м длины.
До недавнего времени у погонофор различали три отдела тела (или
так называемых сегмента): переднюю, короткую протосому на спинной
стороне с головной лопастью конусовидной или треугольной формы и
на брюшной стороне со щупальцевым аппаратом; более длинную, ци-
линдрическую мезосому и очень длинную метасому, или туловище; по-
следняя двумя-тремя полосками расчленяется на переднюю и. заднюю
области,— преаннулярную и постаннулярную. Протосома обычно отде-
лена от мезосомы кольцевидной бороздкой. Головная лопасть редко
является простым продолжением протосомы, она обычно дифференциро-
вана, у ряда форм она отграничена от протосомы предщупальцевой коль-
цевой бороздкой.
Характерными особенностями мезосомы являются косые и кутику-
лярные кили-уздечки, неизвестного функционального значения. У многих
форм на спинной стороне находятся метамерно расположенные желези-
стые участки эпидермиса, или щитки, а на брюшной — прикрепительные
папиллы с кутикулярной пластинкой каждая, с помощью которых, как
предполагают, погонофоры цепляются за стенку трубки при передви-
жении в ней. У низших форм папиллы в передней части метасомы рас-
положены беспорядочно; у большинства — сначала метамерно, далее
неправильно. В начальной части преаннулярной области находится на
спине дорсальная ресничная полоска, а на брюшной — вентральный
желобок. Две или три (упомянутых выше) полоски образованы хити-
ноподобными, овальными пластиночками, покрытыми зубчиками; каж-
дая пластиночка является наружной головкой маленькой зубчатой ще-
тинки, погруженной в толщу эпидермиса. Зубчатые щетинки, похожие
на uncini сидячих полихет, у погонофор служат- для прикрепления
к стенкам трубки, при передвижении в трубке они открепляются.
Недавние исследования Вебба (Webb, 1964а — г), установившего у
погонофор наличие заднего членистого отдела тела с терминальной при-
соской (рис. 80), соединенного узким стебельком с постаннулярным
отделом метасомы, меняют трактовку сегментации тела, а работа
А. В. Иванова (1964а, 19646) вносит много нового в понимание всей'их
организации и положения в системе.
341
Поэтому к сказанному выше надо добавить следующее: задний, очень
короткий отдел тела погонофор, обычно обрывавшийся при сборах, от-
делен от постаннулярной области коротким плотным цилиндрическим
участком, и состоит из 16—17 сформированных сегментов, каждый из
которых несет метамерно расположенные щетинки, по 4 или по 6 в каж-
дом сегменте. Сегменты расчленены кольцевыми бороздками, имеют
Целомическую полость, поделенную поперечными мускульными септа-
ми— по терминологии А. В. Иванова (1964а, 19646) —диссепиментами.
По данным А. В. Иванова, на заднем конце тела находится зона роста
сегментов, за счет которой образуются новые сегменты. Сегментация в
отличии от таковой у аннелид начинается
с образования мешочков ще-
тинок, затем самих щетинок,
септ (диссепиментов) и кон-
чается появлением наруж-
ных границ сегментов. Как
было отмечено выше, на са-
мом заднем конце находит-
ся терминальное, дорсо-вен-
тральное углубление, при-
соска по Веббу (Webb,
1964).
В соответствии с разде-
лением тела погонофор на
протосому, мезосому и мета-
сому происходит и расчлене-
ние обширной целомической
полости тела, выстланной
перитонеальным эпителием,
на протоцель, мезоцель и
метацель; в ней находятся
кровеносная система, цело-
модукты, половые железы и
многоклеточные тубипарные
железы.
Первый целом (прото-
цель)— непарный, очень не-
большой, в него открывается
первая пара целомодуктов,
он посылает целомические
каналы в щупальца и захо-
Рис. 80. Siboglinum caulleryi (по А. Иванову)
А—внешний вид заднего отдела тела, состоящего из
23 сегментов; Б — продольный разрез заднего конца
тела; / — присоска; // — диссепименты; /// — зона
роста
дит в головную лопасть
(Jagersten, 1956). Возмож-
но, что производным прото-
целя является и перикар-
диальный мешочек у Atheca-
nephria. Сравнительное изу-
чение многих видов погоно-
фор из отрядов Athecanephria (с мешкообразным протоцелем) и Theca-
nephria- (с подковообразным протоцелем), произведенное А. В. Ивано-
вым, позволяет отвергать трактовку Иогансона (Johansson, 1937, 1939)
и Иетерстена (Jagersten, 1956) частей протоцеля.
Мезоцель является парным, разделенным тонким мезентерием, под-
держивающим спинной и брюшной кровеносные сосуды; он не имеет
целомодуктов и от протоцеля отделен массой из соединительной ткани
и мускульных волокон. Мускульная диафрагма, хорошо развитая, отде-
ляет полости тела мезосомы от таковой метасомы.
Метацель — парные обширные целомические мешки с мезентерием,
342
тянутся на всем протяжении метасомы, до сегментированного отдела; в
них находится пара гонад (яичников — у самок, семенников — у самцов)
с целомодуктами, функционирующими в качестве половых протоков.
В заднем сегментированном отделе обширный целом септами (дис-
сепиментами) поделен на сегменты.
Взгляды Вебба и А. В. Иванова на морфологическое значение отде-
лов, или сегментов тела погонофор различны. Вебб (Webb, 1964) считает
мезосомой отдел тела, располагающийся позади протосомы вплоть до
заднего, им открытого, короткого членистого отдела, а последний — ме-
тасомой. А. В. Иванов (1964а, 19646) сохраняет свое понимание отделов
тела и к метасоме относит сегментированный задний отдел. Как будто
прав А. В. Иванов. Все, что известно
относительно так называемых олиго-
мерных и вторичноротых целомических
группах, говорит о наличии половых
желез в туловище или метасоме, а не в
мезосоме. Мы знаем разные пути воз-
никновения метамерии — аннелиды,:
бесчерепные, позвоночные; погонофоры
дают новый пример развития настоя-
щего метамерного строения заднего
отдела тела, при наличии в переднем
участке метасомы лишь метамерного
расположения спинных щитков и при-
крепленных папилл.
Щупальцевый аппарат, детальны-
ми, точными данными о строении, раз-
витии и функции которого мы обяза-
ны А. В. Иванову (Ivanov, 1960), яв-
ляется ведущей, по словам последнего,
системой органов в эволюции погоно-
фор и выполняет жизненно важные
функции, имеет также существенное
значение в систематике этих живот-
ных. Этот аппарат возникает в виде,
непарного правого выроста централы
ного основания головной лопасти, на
переднем крае протосомы и разви-
вается в крону щупалец (обычно от
шести до 100—230). Они, хотя не всег-
да, несут на внутренней стороне ните-
видные пиннулы, или ворсинки. Более
примитивные формы имеют небольшое
число щупалец; в ходе эволюции число
1 — эпителиальная клетка; 2 — пиииула;
3 — выносящий сосуд; 4 — приносящий
сосуд; 5 — выносящий сосуд пиннулы;
6—приносящий сосуд пиннулы; 7*—
ресничная клетка
щупалец возрастает, увеличивается
также на них и число пиннул. У высших форм погонофор щупальца спа-
иваются друг с другом. Идет повышение функционального значения щу-
пальцевого аппарата и его роли в жизни и эволюции погонофор.
В каждое щупальце, образованное цилиндрическим эпителием, под
базальной мембраной которого находится ряд тонких мышц (рис. 81),
от протоцеля заходит целомический канал, в котором расположены при-
носящий и выносящий кровеносные сосуды. Последние дают в каждую
пиннулу, представляющую вытянутую клетку, внутриклеточные прино-
сящие и выносящие .капилляры. На щупальцах, у основания пиннул
расположен ряд крупных мерцательных клеток, вызывающих движение
окружающей воды. Все это говорит о дыхательной функции щупальце-
вого аппарата. Вместе с тем он является и фильтрующим аппаратом:
343
в густой сети пиннул застревают микроскопические организмы и детрит.
Отсутствие рта и кишечника позволяет допускать, что в межщупальце-
вое пространство поступают пищеварительные ферменты, выделяемые,
вероятно, клетками эпителия щупалец, происходит переваривание пищи,
которая всасывается пиннулами (Ivanov, 1960).
Предположения об улавливании пищи, ее характере и переваривании
при участии щупалец высказывались также Иегерстеном (Jagersten,
1956, 1957) и Киркегардом (Kirkegaard, 1961, цит. по Ivanov, 1963).
Критика их предположений и ряд соображений об этом дана так-
же Саусвордами (Southward a. Southward, 1963). Прямые наблюдения
над питанием и пищеварением погонофор подтвердили сообра-
жения А. И. Иванова. Таким образом, как писал и Акс (Ах, 1960), эти
свободноживущие, многоклеточные вторичнопол.остные животные
не имеют кишечника, принимают и переваривают пищу щупальцевым
аппаратом.
Стенка тела состоит из эпидермиса, в толще которого лежит сеть
нервных волокон и клеток, кожно-мускульного мешка и перитонеального
эпителия. Эпидермальные клетки узкие, цилиндрические, на всей поверх-
ности тела одеты кутикулой. Среди клеток эпидеомиса находятся желе-
зистые, нервные и чувствительные клетки. Среди железистых различа-
ются: слизистые, тубипарные и белковые. Зубчатая пластинка образуется
внутри одной эпидермальной клетки. У эмбрионов Oligobrachia dogieli,
молодых особей Siboglinum caulleryi, находящихся еще в материнской
трубке, и в заднем членистом отделе взрослых особей каждая щетинка
развивается в мешочке; на срезах он выглядит впячиванием эпидермиса,
и непонятно, как его возникновение может сначала не зависеть от эпи-
дермиса, каю предполагает А. В. Иванов' (1964).
Тубипарные одноклеточные и многоклеточные железы, находящиеся
в мезосоме, метасоме и членистом заднем отделе, образуют вещество
трубок. Многоклеточные тубипарные железы вдаются в целом метасомы,
они редки у Athecanephria и особенно многочисленны у Thecanbphria.
Пока не ясно, как сказываются различия в отношениях тубипарных же-
лез у обоих отрядов на особенностях их трубок. Не ясно также; какова
функция тубипарных желез в заднем отделе тела.
Эпидермальные клетки спинной ресничной полоски покрыты очень
тонкой кутикулой и снабжены густыми ресничками; функция полоски
не выяснена. В щупальцах, как указывалось выше, у основания пиннул
с каждой стороны один ряд эпидермальных клеток имеет ' длинные
ресницы, очевидно, способствующие циркуляции воды между щу-
палец.
К хорошо развитой базальной мембране, подстилающей эпидермис,
прилегают наружные кольцевые и внутренние продольные мышцы кож-
но-мускульного мешка. Кольцевой слой очень тонок, продольный, осо-
бенно у крупных форм, обладает значительной толщиной и у различных
форм обнаруживает разную степень развития. Пояски зубчатых щети-
нок обладают мускулатурой, видимо, производной кожно-мускульного
мешка, сокращением ее поясок впячивается внутрь и зубчатые щетинки
открепляются от стенок трубки, что способствуют передвижению пого-
нофор в трубке. Прикрепительные паппилы, особенно крупные, обладают
хорошо развитой мускулатурой, формирующейся за счет кольцевых
и продольных мышц кожно-мускульного мешка.
В протосоме развиты дорсо-вентральные пучки мышц, впервые опи-
санные Коллери (Caullery, 1914) и Иегерстеном (Jagersten, 1956),. най-
денные затем у всех погонофор (А. В. Иванов, 1960). Они вызывают
сокращение головной лопасти, протекающее ритмически и, видимо, вы-
зывают проталкивание крови из головных сосудов в спинной. Присоска
на заднем конце тела снабжена ретракторами в виде парных пучков
344
мышц, расходящихся веерообразно
от ее дна (А. В. Иванов, 1964а,
19646).
Кожно-мускульный мешок, мно-
гоклеточные тубипарные железы,
мезентерии, кровеносные сосуды и
пблойые железы одеты 'перито-
неальным, нересничным эпителием.
Предположительно за счет на-
ружного листка целома (сомато-
плевры) развивается кожно-мус-
кульный мешок, а за счет внутрен-
него (спланхноплевры)— мускула-
тура внутренних органов. Клетки
перитонеума нередко бывают круп-
ными, высокими и содержат зерна
экскрета. У ряда форм {Polybrachia
annulata, Lamellisabella zachsi)
спланхнойлевра в средней части
тела разрастается, становясь Губча-
той тканью, заполняющей полость
целома.
Нервная система лежит интра-
эпителиально в базальной части
эпидермиса в виде, очевидно, сплош-
ного ’нервного сплетения из волокон
и клеток (рис. 82), образующего на
сИинйбй1 стороне тела ’ сгущение —
спинной' нервный тяж. Он тянется
до кбнца постаннулярного отдела
метасомы. В протосомё он образует
объемистую нервную массу из кле-
ток и волокон (так называемый
мозг)', частично заходящую в голов-
ную лопасть; на уровне щупальце-
вого’аппарата от нее отходит пара
коннектив, образующих на брюшной
стороне нервное кольцо; нервы, по
одному в каждом щупальце, отходят
от боковых участков кольца. Спин-
ной нервный тяж состоит из про-
дольных нервных волокон; узкий в
Рис. 82. Нервная система Polybrachi.a
annulata (по А. Иванову)
А —вид со спинной стороны; Б —вид сбо-
ку; 7, II н III -прото-, мезо- и метасома:
1 — спинной нервный ствол; 2 — головная
лопасть; 3—нервное кольцо протосомы;
4 — прикрепительные папиллы
протосоме, он расширяется в мета-
соме'на'уровне спинной ресничной полоски, позади которой он вновь
становится узким. Лишь у Lamellisabella, Polybrachia и Spirobrachia
в спинном нервном тяже преаннулярного отдела туловища и части ме-
зосомы установлены неврохорды, гигантские нервные волокна — аксоны
двигательных нервных клеток, характерные для нервной системы мно-
гих многоклеточных.
Если не считать дорсальную ресничную полоску, которая может быть
хеморецептором, дифференцированных рецепторов у погонофор нет, име-
ются лишь одиночные чувствительные клетки в головной лопасти. Фи-
зиология нервной системы погонофор не изучена. Это может быть сде-
лано в будущем на мелководных формах этих животных, доступных для
такого изучения.
Кишечник у погонофор, как известно, отсутствует, но у зародыша он
представлен эпителизованным первичным кишечником с очень узкой
345
полостью (А. В. Иванов, 1960). Пищеварительную функцию несет щу-
пальцевый аппарат.
Кровеносная система замкнутая, хороша развита; она состоит из бо-
лее мощного спинного и более слабого брюшного сосудов, лежащих в
передней половине тела в мезентерии, а далее в стенке тела. В головной
лопасти спинной и брюшной сосуды соединяются сосудами щупальце-
вого аппарата, в задней половине туловища — боковыми комиссураль-
ными сосудами. Спинной и брюшной сосуды находятся и в заднем чле-
нистом отделе тела, но пока неизвестно переходят ли они один в другой
на конце тела.
Спинной сосуд посылает три сообщающиеся друг с другом слепые
ветви в головную лопасть;, в метасоме боковые ветви спинного сосуда
отходят к стенкам яичников у самок и к внутренним поверхностям семе-
проводов у самцов. Брюшной сосуд сильно расширенный лишь в перед-
ней части протосомы, делится на два сосуда щупальцевого аппарата,
которые дают приносящие сосуды щупалец, переходящие в пару выно-
сящих сосудов щупальцевого аппарата, сливающихся в медианный со-
суд, впадающий в спинной кровеносный сосуд. Кровеносные сосуды оче-
видно сильно сократимы, так как снаружи имеют кольцевой слой попе-
речнополосатых мышц; внутренняя стенка их состоит из бесструктурной
мембраны-интимы, без эндотелия.
Участок брюшного сосуда в протосоме, так называемое сердце, обла-
дает толстой мембраной, одет рыхлой соединительной тканью с гладкой
мускулатурой из нескольких кольцевых слоев, чередующихся с диаго-
нальными, и снаружи — продольными мышцами. У форм отряда Atheca-
nephria к спинной стороне сердца прилегает эпителиальный мешочек,
описанный как перикардий (А. В. Иванов, 1955), предположительно
возникающий из протоцеля. Морфологическое значение и функция «пе-
рикардия» не выяснены, непонятно также, если это перикардий, его от-
сутствие у более высокоорганизованных крупных форм Thecanephria с
.сильно развитыми щупальцами. Непонятно также строение сильно со-
кратимого сердца из гладких мышц в отличие от строения спинного
и брюшного сосудов с их поперечнополосатой мускулатурой.
Неясна также природа «сердечного тела» — плотной массы непра-
вильной полигональной формы клеток, описанных Иогансоном (lohans-
son, 1939), а затем А. В. Ивановым (1960). Его видимо надо сравнивать
с сердечным телом полихет.
Красный цвет крови погонофор вызван наличием в их крови гемо-
глобина в растворенном состоянии (Southward a. Southward, 1963).
Прямые наблюдения, сделанные .Карлайлем на живых погонофорах
(см. Ivanov, 1963), подтвердили его предположения о сокращении сердца
и движении крови вперед по брюшному сосуду и назад по спинному.
Кровь проталкивается из брюшного сосуда в щупальцевый аппарат и,
обогащенная в нем кислородом и питательными веществами, по спинно-
му сосуду движется к внутренним органам, яичникам и, очевидно, к се-
менникам.
В’ протосоме находится пара целомодуктов — эпителиальных, реснич-
ных каналов, небольшого размера у. Athecanephria и крупных у Theca-
nephria. Целомодукт состоит из внутреннего канала, без ресничной во-
ронки, сообщающегося с протоцелом, V-образно изогнутого экскретор-
ного отдела и выводного протока, открывающегося наружу у основания
щупальцевого аппарата на брюшной стороне тела. Внутренние каналы
соединяются друг с другом соединительным протоком. Целомодукты
плотно прилегают к головным кровеносным сосудам. Стенка внутрен-
него канала образована кубическими, мерцательными клетками, стенка
экскреторного отдела — крупными переполненными буроватыми зерна-
ми экскрета нефроцитами, с пучками длинных ресниц, выводной проток
346
состоит из невысоких мерцательных клеток. Экскреторную функцию не^
сут также значительные участки внутренних каналов и соединительный
проток. Экскреты, по-видимому, извлекаются целомодуктами из крове-
носных сосудов, допускают также регулирование целомодуктами коли-
чества полостной жидкости. В мезосоме выделительную функцию несут
клетки спланхноплевры. Экспериментальных данных о функционирова-
нии целомодуктов пока нет.
Погонофоры раздельнополы, полового диморфизма у них нет, полы
различаются лишь положением половых отверстий. У самок в передней
, половине метасомы лежит пара длинных яичников, передними концами
прикрепленных к мезентерию позади мускулистой диафрагмы, отделяю-
щей мезоцель от метацеля. Зачатки гонад возникают на боковых по-
верхностях спинного сосуда, под перитонеумом; стенка яичников обра-
зуется перитонеумом, от спинного сосуда в яичник врастают ветви для
питания половых клеток. Фолликулярных клеток нет. Яйцеводами слу-
жит пара эпителиальных V-образно согнутых трубчатых целомодуктов.
В широком колене каждого целомодукта, в воронке, лежит задний'конец
. яичника, узкое колено служит яйцеводом. Яйцеводы открываются на-
ружу на границе между мезосомой и метасомой вентро-латерально.
Эпителий воронок без ресниц. Стенки воронки и яйцеводов снабжены
мускулатурой.
У самцов семенные мешки, являющиеся частью целома, тянутся от
середины до заднего конца метасомы (но не заднего сегментированного
отдела). Спинной кровеносный сосуд, находящийся между семенными
мешками, обеспечивает питание развивающихся половых клеток. Перед-
ние концы семенных мешков сообщаются с семепроводами. Последние—
ресничные эпителиальные каналы, одетые тонким слоем мускульных во-
локон и перитонеумом, являются целомодуктами. Они прикрепляются
к брюшной стенке тела, образуют петлю, направляющуюся назад, а
затем тянутся вперед и открываются наружу на брюшной стороне
двумя половыми папиллами. В семепроводах формируются спермато-
форы.
Погонофоры размножаются половым путем, яйца, очень богатые
желтком, откладываются в полость трубки, где и происходит развитие.
У Siboglinum caulleryi яйцо сильно вытянутое, двустороннесимметричное;
его передний полюс более плотный, гомогенный полупрозрачный отвечает
переднему концу, а задний полюс, более светлый и зернистый,— заднему
концу тела погойофоры; вогнутая поверхность яйца соответствует спин-
ной стороне, выпуклая — брюшной. Анимально-вегетативная ось яйца
отвечает главной, передне-задней оси взрослых погонофор, являющихся
протаксонными животными.
Некоторые данные по онтогенезу погонофор принадлежат Коллери
(Caullery, 1944), Иегерстену (Jagersten, 1957), Саусвордам (Southward
a. Southward, 1963), Веббу (Webb, 1964а — г), капитальными являются
исследования А. В. Иванова (1957, 1963, 1964а, б; Ivanov, 1963).
Дробление яйца у Siboglinum caulleryi полное, неравномерное; без
следов радиального и спирального типов, позже становящееся двусто-
роннесимметричным; целобластулы нет. На стадии 80—85 бластомеров
(рис. 83) на плоской стороне плотного зародыша бластомеры в передней
половине мельче, в задней все более увеличивающиеся, на выпуклой
стороне, на которой бластомеры в среднем крупнее, наиболее крупные
также располагаются в задней части. Кроме того, пара очень крупных
бластомеров лежит у переднего конца зародыша. На стадии приблизи-
тельно 130—140 бластомеров на плоской стороне передние (мелкие)
резко отличаются от задних (крупных) клеток, на выпуклой стороне
разница между клетками менее резка, но пара крупных клеток на пе-
реднем конце на этой стадии всегда имеется. Мелкие клетки наружного
347
стопора, эпителизированного первичного
в в
Рис. 83. Эмбриональное развитие Siboglinum caul-
leryi (по А. 'Иванову)
А — стадия четырех бластомеров; Б, В —ранняя гастру-
ла из 80—85 клеток с плоской (В) и выпуклой (В)
сторонами
слоя являются эктодермой, три большие внутренние клетки представ-
ляют зачаток первичной кишки, две группы внутренних мелких клеток,
располагающиеся между наружными и внутренними клетками, вероятно,
являются мезенхимой. На этой стадии развития нет бластоцеля, бла-
ечника, однако А. В. Ива-
нов считает ее гаструлой.
Далее зародыш не-
сколько увеличивается
(рис. 84), вся внутренняя
часть его заполнена плот-
ной массой клеток -архен-
терона, но без эпители-
ального расположения,
без рта и анального от-
верстия; на его переднем
конце образуется зачаток
переднего непарцого про-
тоцеля и общие зачатки
мезоцеля и метацеля.
После их отделения цент-
ральная клеточная масса
зародыша является, плот-
ным эктодермальным за-
чатком кишечника, точ-
нее, его рудиментом.
После этого- зародыш
Oligobrachia dogieli -силь-
но вытягивается и подраз-
деляется на небольшую
протосому с вытянутой го-
ловной лопастью, на кото-
рой сначала образуется
правое, затем левое щупальце, на сильно удлиненную мезосому, состав-
ляющую большую часть тела зародыша, и очень короткую метасому.
На протосоме и метасоме образуются два провизорных ресничных пояска
и спинная ресничная полоска метасомы. Рот и анальное отверстие отсут-
ствуют. Происходит гистологическая дифференцировка — образуется ку-
тикула, в эпидермисе развиваются одноклеточные железы, под ним —
кольцевые и продольные волокна кожно-мускульного мешка, образуются
спинной и брюшной кровеносные сосуды, развиваются щетинки и т. д.
Эта стадия развития близка к двум последним личиночным стадиям как
О. dogieli, так (по работам Вебба, 1964а—,г) и Siboglinum fiordicum.
У последней формы Веббом прослежены семь стадий. Такая личинка
плавает в материнской трубке с помощью переднего кольца ресниц, вра-
щаясь вокруг продольной оси; заднее ресничное кольцо и спинная рес-
ничная полоска в движении не играют роли. О развитии мезосомы и ме-
тасомы данные Иванова и Вебба расходятся. По Веббу, короткая мета-
сома дает начало членистому, короткому заднему отделу погонофоры,
длинное же тело взрослого животного возникает из длинной мезосомы
личинки (рис. 85). По А. В. Иванову мезосома личинки также длинная,
а метасома очень короткая. Остается неясным, как из короткой мета-
сомы образуется метасомальная, большая часть тела погонофоры и ко-
роткий метамерный задний отдел.
Погонофоры обитатели бентоса, преимущественно абиссали, а неко-
торые, даже сверхабиссали (9000 м), меньшая часть принадлежит к оби-
тателям мелководья и встречается на глубинах от 90 до 200 м: Пока из-
вестно немного видов: 80 описано, 70 — ждет описания, всего же, пр
348
0,1мм
Рис. 84.
I — целом
Продольный разрез зародыша Siboglinum caulleryi (-4) и схема его строения
(5) (по А. Иванову)
протоцеля- // и ///-целом мезо- и метацеля; I-эктодерма; 2- эндодермальный
1 ’ зачаток кишечника
Рис. 85. Три личиночные стадии развития
Siboglinum caulleryi (по Webb)
I, II, ///-прото-, мезо- и метасома; / — перед-
нее кольцо ресничек; 2 — заднее кольцо ресничек
349
мнению А. В. Иванова, может быть до 1000 видов. Несомненно, что даль-
нейшие сборы сильно увеличат число известных видов погонофор и рас-
ширят наши знания об ареале их распространения, так как в настоящее
время большой материал по этой группе собран в морях Дальнего Вос-
тока и прилегающих водах Тихого'океана советскими экспедициями.
И все же скорее всего это небольшая непроцветающая, а узко специа-
лизированная, с упрощенными строением и онтогенезом группа живот-
ных. Погонофоры в ряде случаев являются многочисленными, характер-
ными сочленами биоценозов, организмами, не покидающими своих тру-
бок; они держатся преимущественно вертикально, реже лежат на
субстрате (Webb, 1964а — г). На твердых трубках ряда погонофор по-
селяются фораминиферы, кремнероговые губки, гидроиды, мшанки, аль-
ционарии, серпулиды, морские лилии, асцидии (Иванов, 1960).
Погонофоры, как известно, составляют самостоятельный тип Pogo-
nophora Beklemischew, 1944 с одним классом Pogonophora lohansson,
1937 и с двумя отрядами — Athecanephria Ivanov, 1955 и Theeanephria
Ivanov, 1955.
У отряда Athecanephria (семейства Oligobrachiidae и Siboglinidae)
щупальца всегда разделены, иногда имеется всего одно щупальце. Про-
тосома отделена от мезосомы. Постаннулярная область метасомы с мета-
мерно расположенными на спинной стороне железистыми участками эпи-
дермиса или с обособленными железистыми щитками, против которых
на брюшной стороне может быть по одной большой папилле. Целом про-
тосомы мешкообразный, а целомодукты широко раздвинуты; имеется
перикардий. Сперматофоры веретеновидные. Трубки чаще кольчатые.
У отряда Theeanephria (семейства Polybrachiidae, Lamellisabellidae, Spi-
robrachiidae) щупальца иногда спаяны друг с другом, они могут быть
спирально свернуты; протосома иногда не отделена (слита) от мезосо-
мы. Постаннулярная область метасомы без спинных железистых щит-
ков, с папиллами на брюшной стороне. Целом протосомы подковооб-
разный, ее целомодукты своими экскреторными отделами сближены,
перикардия нет. Сперматофоры более или менее листовидные. Трубки
кольчатые или сегментированные.
Судя по этим данным, различия между отрядами невелики, и фило-
генетические отношения между ними должны быть сравнительно про-
стыми, так как класс погонофор состоит из близкородственных форм.
Можно согласиться с А. В. Ивановым (1960) в том, что строение щу-
пальцевого, ведущего аппарата, и степень внешней метамеризации туло-
вища позволяют считать более мелкие погонофоры Athecanephria при-
митивнее, чем преимущественно более крупные погонофоры Thecaneph-
ria. То, что у многих представителей Athecanephria и низших
Theeanephria (например, Heptabranchia) щупальца свободны, число их
невелико, метамеризация папилл едва выражена или слабая говорит
о вероятном общем предке обоих отрядов. У обширного, более высоко
организованного рода Sibogli.rtu.rn, сравнительно с более примитивными
родами Oligobrachia и Birsteinia отряда Athecanephria, происходит на-
растание метамеризации туловища и вместе с тем упрощение путем
олигомеризации щупальцевого аппарата, несущего одно щупальце и
утратившего у ряда видов пиннулы. Олигомеризация числа щупалец до
двух происходит и у рода Diplobrachia отряда Theeanephria. Наряду
с примитивным родом Heptabrachia основная масса обширного семей-
ства Polybrachiidae (Theeanephria) значительно эволюирует (нарастает
число щупалец до 100, увеличивается число рядов пиннул, у некоторых
родов основание щупалец становится круглым и т. д.). У двух послед-
них, наиболее прогрессивных семейств Theeanephria — Lamellisabellidae
и Spirobrachiidae — протосома слита с мезосомой; соседние щупальца
прочно соединены друг с другом, у Lamellisabellidae образуется полный
350
щупальцевый цилиндр, а у Spirobrachiidae— спирально свернутый щу-
пальцевый аппарат.
Если впоследствии окажутся обоснованным установление Веббом
(Webb, 1964а — г) нового отряда погонофор для рода Sclerolinutn, а
также его примитивность, придется пересмотреть всю родословную по-
гонофор А. В. Иванова (Ivanov, 1960, 1963) и в ее основании поместить
формы не с венчиком из шести щупалец, а всего с двумя щупальцами.
Предположение Иванова о большой древности погонофор оправда-
лось, так как в нижнем и среднем кембрии недавно найдены остатки по-
гонофор (Соколов, 1965).
Из сказанного о строении, развитии и поведении (физиологии) по-
гонофор, основанного почти нацело на данных А. В. Иванова (1949, 1955,
1957, 1959, 1960, 1962, 1963, 1964; Ivanov, 1960), видно необычайное свое-
образие этих животных, не укладывающихся в принятую систему вто-
ричноротых.
Исходя из положения о неразрывности организма.и условий его жиз-
ни, их взаимной обусловленности, сравнительно не трудно для большин-
ства групп (типов, классов) расшифровать планы их строения, а также
найти исходные, примитивные черты и вторичные особенности, прогрес-
сивного и регрессивного характера.
Совершенно необъяснимо, учитывая свободный, непаразитический
образ жизни как мелководных, так и абиссальных погонофор, отсут-
ствие у них кишечника с ротовым и анальным отверстиями, что, очевидно,
вызвало передачу функции пищеварения щупальцевому аппарату. По-
следний поэтому сильно развивается и становится многофункциональным
аппаратом. Хотя данных по онтогенезу пока мало, но и имеющиеся сви-
детельствуют о своеобразии развития погонофор: нет следов бластопора,
полой бластулы, гаструляции, рта, анального отверстия, нет архентерона,
а имеется лишь бесформенная группа клеток, по характеру и положению
в зародыше отвечающая клеткам архентерона. Жизнь погонофор в труб-
ках, прочных, но легко проницаемых для воды (Southward a. Southward,
1963), без оставления их хотя бы на время, очевидно, не может быть при-
чиной редукции кишечника, поскольку у многих других групп, в том
числе и у полихет, этого при сходном образе жизни не наблюдается.
Слабая гистологическая дифференцировка покровов, рецепторов,
низкий уровень дифференцировки нервной системы, кожно-мускульного
мешка, простота кровеносной системы являются как чертами исходного
примитивного строения, так и результатом упрощения, вызванным мало-
подвижной жизнью в прочных трубках.
Раннее эмбриональное развитие погонофор своеобразно. Следует от-
метить двустороннесимметричное дробление яйца с детерминированными
главной осью, передним и задним концами, спинной и брюшной сторо-
нами, без следов спирального дробления. Казалось бы, развитие яйца
в материнской трубке, а не свободно в воде, не может быть причиной ряда
вторичных изменений раннего онтогенеза погонофор, сравнительно с
онтогенезом как представителей вторичноротых животных, так и анне-
лид. Несвободное, инкапсулированное развитие у аннелид, немертин
и других вносит, как мы видели, существенные изменения в онтогенез
этих червей, но тип онтогенеза у них сохраняется. Известное сходство в
онтогенезе погонофор и кишечнодышащих в отношении закладки цело-
мов, как отмечает А. В. Иванов, конечно, имеется, но начальные стадии
их развития кардинально различны, мы до сих пор не знаем происходило
ли у погонофор образование дефинитивного рта, независимого от пер-
вичного рта или бластопора. Кишечнодышащим и иглокожим, особенно
последним, присущ сложный метаморфоз с перестройкой организации
личинки, чего у погонофор нет. Можно также думать, что у них на пе-
реднем конце зародыша, отвечающему переднему полюсу яйца, на кото-
351
ром развивается щупальцевый аппарат, находился рот, а на противо-
положном— анус, т..е. погонофоры не были вторичноротыми животными.
Наличие трех отделов тела с непарным протоцелем и парными, мезо-
целем и метацелем, парных целомодуктов, неганглионарного характера
центральной нервной системы, представленной спинным нервным тяжом,
без затруднения ставило бы погонофор не будь у них сегментированного
заднего отдела, в ряд с кишечнодышащими, как новый тип вторич-
норотых. Но следует отметить, что отношение так называемого сердца-
перикардия к брюшному кровеносному сосуду у Athecanephria говорит
о том, что настоящего кардио-перикардия кишечнодышащих (.стр. 273)
и иглокожих (стр. 310) у погонофор нет. Далее, у низших вторичноро-
тых— кишечнодышащих, птеробранхий и иглокожих — строение половой
системы простое, она состоит у большинства из многочисленных мелких
мешотчатых половых желез с простыми выводными протоками, тогда
как у погонофор имеется пара половых желез, которые являются частью
целомической полости й анатомически не связаны с половыми прото-
ками; половые продукты выводятся парой целомодуктов, ставших по-
ловыми протоками. Такие отношения половой системы близки к тому,
что наблюдается у аннелид. Чертами сходства погонофор с аннелидами
являются также хитиновая природа кутикулы эпидермиса, зубчатые
пластинки с зубчатыми щетинками, сегменты (с 6 щетинками каждый)
.членистого заднего отдела с зоной роста, наличие сердечного тела в
брюшном сосуде, зарастание части целома тканью, наличие цитофоров
у самцов; эти черты погонофор чужды кишечнодышащим, из вторично-
ротых, казалось бы, наиболее близким к погонофорам. Нет сомнения,
что открытие членистого заднего отдела у погонофор еще более укрепит
позиции сторонников взгляда о принадлежности погонофор к полихетам
(Hartmann, 1951, 1954; Ливанов и Порфирьева, 1962, 1965).
Н. А. Ливанов и Н. А. Порфирьева в последней своей статье (1965)
еще категоричнее стремятся найти сходство в строении и развитии у по-
лихет (на примере Owenia) и погонофор, не замечая крайнего своеобра-
зия, присущего всей группе погонофор в целом. Защищая точку зрения
о происхождении погонофор от аннелид (1962, 1965), они сравнивают
планы строения погонофор и Owenia fusiformis (Oweniidae, Polychaeta),
считая брюшной стороной тела ту, вдоль которой проходит нервный
ствол и на которой оказывается тогда и головной мозг. При таком срав-
нении надо было ожидать параллели в онтогенезе погонофор и Oweni-
idae. Однако, как известно (Wilson, 1932; Свешников, 1961), семейству
Oweniidae свойственны специфичные пелагические личинки типа митра-
рия. По исследованиям В. А. Свешникова (1963 и более ранние работы;
см. гл. «Аннелиды»), отдельные семейства полихет имеют строго спе-
цифичные морфологические типы личинок. Сближая погонофор с Owe-
niidae, надо ожидать и некоторого сходства их эмбрионального и личи-
ночного развития. Работ по дроблению Oweniidae нет, но типы личинок
Oweniidae и погонофор совершенно различны и ничего общего друг
с другом не имеют.
Однако полное отсутствие и в заднем членистом отделе следов пара-
подий, ганглионарной нервной цепочки, половых желез в виде набуха-
ний первичных половых клеток, а также образование щетинок внутри
эпителиальных мешочков и порядок формирования новых сегментов го-
ворят скорее о конвергентном развитии членистости погонофор и анне-
лид. Сегменты тела погонофор и аннелид не гомологичные образования
(А. В. Иванов, 1964а, 19646), появление членистости в заднем отделе
погонофор явление вторичное, вероятно, связанное с функцией якоря
при крайне удлиненном теле этих животных.
Мы знаем, что погонофоры, по словам А. В. Иванова (1960), отлича-
ются от других вторичноротых развитием крайне своеобразного щупаль-
352
цевого аппарата на первом сегменте, наличием целомодуктов в первом
и третьем сегментах, примитивной связью гонад с целомом, развитием
своеобразного сердца в брюшном сосуде, полной редукцией кишечника
у взрослых, сильной кутикуляризацией покровов, и резкими вторичными
изменениями эмбрионального развития. На самом деле и строение, и
развитие погонофор значительно сложнее и своеобразнее, чем оно
представлялось сравнительно с таковыми вторичноротых в 1964 г. У по-
гонофор оказалась смесь признаков низших вторичноротых и аннелид.
Данные эмбриологии, которыми мы пока располагаем, говорят о том,
что щупальцевый аппарат образуется на оральном конце эмбриона, где
должен быть блаетопор или первичный, рот, т. е. это животные не вторич-
норотые, а просто утратившие кишечник и рот. Следовательно, погоно-
форы не только самостоятельный тип, как признают В. Н. Беклемишев
(1944, 1946) и А. В. Иванов (1960), но тип, который не укладывается
в группе целомических животных ни в Protostomia, ни в Deuterostomia.
23 Д- Федотов
Филогенетические отношения
типов животных
Как известно, работ по эволюции и филогении животных много, за
последние годы можно отметить следующие: Н. А. Ливанова (1955),
Г. А. Шмидта (1953), В. А. Догеля (1959), В. Н. Беклемишева (1964),
А. П. Маркевича (1964), Гаймен (Hyman, 1940—-1959), Франца (Franz,
1943), Проссера (Prosser, 1952), Ремане (Remane, 1954, 1959), Маркуса
(Marcus, 1958), Дотерти и др. (Dougnerty a. all., 1963), Хаджи (Hadzi,
1963). Мнения об исходной родоначальной форме одноклеточных расхо-
дятся: одни (большинство) видят ее среди биченосцев, другие (меньшин-
ство) родоначальной формой считают амебоидный организм. Теперь я
придерживаюсь последнего взгляда, находя, что идеи А. И. Опарина
(1957, 1964) о гетеротрофности первичных организмов вполне обоснованы.
Отсутствие оболочки, сохраняющейся при фоссилизации, у этих организ-
мов вызвало естественный пробел в их геологической истории. Древ-
нейшие амебоидные организмы были донными формами, и их строение
является исходным типом. В этом я согласен с А. П. Маркевичем (1964)
и Ш. Д. Мошковским (1957). Экологическим моментом, вызвавшим воз-
никновение многоклеточности, вероятно, было объединение нескольких
амебоидных организмов для овладения добычей, что в ходе эволюции из
временного перешло в постоянное. Как видно из сказанного выше
(гл. «Губки»), губки и в настоящее время сохраняют поклеточную диф-
ференцировку, клетки их не образуют тканей и органов; их клетки легко
возвращаются к состоянию амебоцитов и вновь дифференцируются, при-
обретая характер пинакоцитов, хоаноцитов и т. д. На известной стадии
онтогенеза у губок образуются жгутиковые клетки, деятельностью кото-
рых обеспечивается распространение развивающихся губок в море.
В результате дифференцировки, сопровождающейся утратой жгутов, воз-
никают все клетки тела губки. Я отказываюсь от прежнего взгляда на
губок, как на деградированных многоклеточных, родственных кишечно-
полостным, хотя у губок черты упрощения в результате^юдвижного
прикрепленного образа жизни, несомненно, имеются. Надо признать, что
губки — это Parazoa, которые в самом отдаленном геологически прош-
лом (в докембрии) отошли от родоначального ствола, давшего все типы
многоклеточных животных. Вследствие отсутствия у губок общей для
всего типа формы тела и фиксированного определенного числа элемен-
тов радиальной симметрии, их нельзя считать радиальносимметричными
животными.
Родоначальный ствол (родословное древо) многоклеточных начи-
нается кишечнополостными, тело которых состоит из двух клеточных,
слоев •— эктодермы и энтодермы, их' производной — мезоглеи — и при-
митивной, диффузной нервной системы; для кишечнополостных харак-
терна радиальная симметрия, но с элементами первичной двусторонней
и двулучевой симметрии (рис. 86). Из совокупности данных онтогенеза
и строения исходной формой кишечнополостных является полип — при-
крепленный, реже ползающий организм; медузоидная форма является.
354
производной полипа, ее появление связано с формированием гонад и она'
характеризует высших сцифомедуз. Для онтогенеза кишечнополостных
характерно возникновение двух зародышевых листков (эктодермы и
энтодермы), неустановившиеся различные способы их образования
и наличие стадии двуслойной личинки, планулы, видимо, перво-
начально придонной, ползающей, перешедшей затем к плавающему
образу жизни. Наличием стрекательных капсул обусловлено название
кишечнополостных — Cnidaria. Ископаемые кишечнополостные известны
из докембрия, особенно многочисленны, сохранившиеся благодаря из-
вестковому скелету, разные группы коралловых полипов, богато пред-
ставленные и современными шестилучевыми формами.
Объединение с книдариями гребневиков в тип кишечнополостных надо
считать необоснованным; на поверхностном сходстве основано также
сведение гребневика к медузе и предположение о происхождении греб-
невиков от медуз. Некоторое сходство между'ними возникло конвергент-
но, благодаря пелагическому образу жизни. Отсутствие стрекательных
клеток, наличие клейких клеток и общий тип строения резко отличают
гребневиков от кишечнополостных. Не менее характерен» тип онтогенеза
гребневиков, а именно детерминированное дробление, образование мик-
ромеров (эктодермы), макромеров (энтодермы) и мезомеров (среднего
зародышевого пласта) и образование свободноплавающей цидиппидной.
личиночной формы. Широка морфологическая и экологическая пластич-
ность гребневиков: от типичных, грушевидной формы, представителей
отряда Cydippida до плоской планариеобразной, паренхиматозной, пол-
зающей формы — платиктенид, конвергентно уподобляющихся турбел-
ляриям. Однако внутренняя организация ползающих гребневиков на-
столько отлична от организации плоских червей, что их нельзя считать-
родственными формами. Наиболее правильно смотреть на гребневиков
как на слепо кончающуюся ветвь низших многоклеточных животных,,
отошедшую от общего родословного древа.
Двустороннесимметричные многоклеточные (как это принято очень
многими исследователями и как об этом и я писал много раз) образуют
две расходящиеся группы — протостомий и дейтеростомий.
Основными чертами онтогенеза протостомий являются следующие:
образование дефинитивного рта за счет бластопора, а анального'
отверстия — на заднем конце зародыша, спирально детерминированное
дробление, ограниченное число клеточных делений, завершение органо-
генеза шестью синхронными делениями бластомеров, начало образова-
ния зародышевых листков на стадии 128 бластомеров, телобластическая
закладка органов и мезодермальных полосок, образующих парные це-
ломы сегментов тела, участие эктодермы в развитии передней и задней
кишки, неспособность к регенерации ранних стадий развития. Взрослые
протостомий характеризуются наличием наружного скелета (кутикула,,
хитин, раковина), ганглиозным характером центральной нервной систе-
мы, паренхиматизацией, значительным подавлением целома и кровенос-
ной системы. Наряду с морфологическими чертами протостомий .харак-
теризуются аргининфосфорной кислотой, как главным источником мы-
шечного сокращения (за исключением высших классов членистоногих) _
Они обладают большой стойкостью к наркозу, четырьмя дыхательными
пигментами крови (гемоглобин,ом, гемэритрином, гемоцианином и хлор-
окруорином). Широко распространен среди них паразитизм (более двух
третей классов протостомий имеют многочисленных паразитических
представителей, а некоторые классы являются чисто паразитическими).
К первичноротым относятся Plathelminthes, Askelminthes, Nemertini,
Mollusca, Annelida, Tardigrada, Pentastomida, Onychophora, Arthropoda.
Дейтеростомии характеризуются следующими чертами. Дефинитив-
ный рот образуется заново, независимо от бластопора, который часто
35.5
становится анусом, нет спирального дробления, нередка поздняя детер-
минация, период клеточных делений длительный, образование зароды-
шевых листков и начало гаструляции происходят на стадии 1000 и более
бластомеров, гаструла инвагинационная. Кроме того характерно: эпи-
телиальное построение зародышевых листков и зачатков органов, за-
кладка нервной системы впячиванием эктодермальной пластинки, экто-
дермальное происхождение мезенхимы, наличие внутреннего скелета
мезодермального происхождения, энтероцельное образование целомов,
наличие трех пар целомов как постоянное или временное состояние,
неучастие, или слабое участие эктодермы в развитии кишечника, строе-
ние центральной нервной системы в виде тяжа или трубки, слабая
паренхиматизация, сильное развитие целома, обычно наличие перикар-
дия целомического происхождения и сердца. Дейтеростомии очень чув-
ствительны к наркозу, главным источником их мышечного сокращения
является креатинфосфорная кислота, гемоглобин — единственный у них
дыхательный пигмент крови. Среди дейтеростомий отсутствует парази-
тизм, а паразитофауна их специфична (например, у иглокожих — тур-
беллярии и гастроподы). К вторичноротым относятся Enteropneusta,
Pterobranchia и, видимо, Graptozoa, Echinodermata, Chaetognatha,
Chordata.
Можно думать, что филогенетически первичноротые и вторичноротые
взяли свое начало от каких-то примитивных кишечнополостных. Основа-
нием к этому является то, что даже и на данном периоде эволюции у ки-
шечнополостных наблюдаются двулучевая и двусторонняя симметрия
у взрослых форм и в онтогенезе, большое разнообразие типов дробления
яиц и способов развития зародышевых листков. Следовательно, и исто-
рически возникновение первичноротых и вторичноротых очень древнее.
Внизу от ствола первичноротых отходят плоские черви, низшие и,
вероятно, наиболее древние двустороннесимметричные формы, обладаю-
щие, подобно кишечнополостным, целентерическим кишечником и имев-
шими планулообразных предков, ставших придонными животными, дав-
шими начало современным турбелляриям. Возможно, что развитие
сложного гермафродитного аппарата у древних плоских червей было
связано с подвижным образом жизни и сравнительной малочисленностью
популяций, такой половой аппарат обеспечивал необходимое воспроиз-
водство. Наиболее высоко организованными являются обладающие ти-
пичным спиральным дроблением Polyclada, а упростившимися, потеряв-
шими кишечник, с вторично измененным ходом дробления (дуэты вместо
квартетов бластомеров), но с чертами примитивности являются Pseudo-
acoela. От основной группы турбеллярий (рабдоцелид) происходят
остальные турбеллярий: рабдоцелидные предки дали начало ставшим
наружными и внутренними паразитами дигенетическим и Моногенети-
ческим трематодам, удонеллидам и цестодам.
Другую крупную ветвь первичноротых, отошедшую от основного
ствола, составляют первичнополостные черви, имевшие рабдоцелидных
предков (Gastrotricha, Rotatoria, Kinorhyncha, Nematoda, по-видимому,
и Acanthocephala). Возможно, что их рабдоцелидные предки также в от-
даленном прошлом произошли от планулообразных форм. Сравнительно
с плоскими червями эволюция этой ветви червей характеризуется при-
обретением новых черт строения — сквозного кишечника со ртом и ану-
сом, более дифференцированной нервной системы, развитием в парен-
химе щелей, давших начало первичной полости тела, преобладающим
развитием отдельных пучков мышц тела и т. д. Приходится допустить,
что рабдоцелидные предки первичнополостных червей или утратили гер-
мафродитизм, или были раздельнополыми со сравнительно простой по-
ловой системой; от спирального дробления предков сохранились лишь
его следы у ряда классов первичнополостных червей — гастротрих, кино-
:356
апич-
И'ого
. . iiy и,
гется
| них
I 1,ета-
атод
этом
Chor хих
)Д и
: атод
аль-
влей
' нние
£с1м - Пка,
при
1эод-
|езо-
шой
I Со
п )тин
Enieropi |j. и
PieroBr. 5СО.
UrcLploi ’eso-
жно
й —
зую
ом-
А
по-
аб«
по-
ких
ер-
:ых
по
эго
за,
)М-
1ИИ
ге-
ка
-та-
И
я»
ые
яс-
ми
ле
ся
хе-
|1С-
-кЧх—____________________________________________________________, —х-кткэтпхтГ’ЕГ
~ 1 I _______ - --- '
екая история соленогастров неизвестна.
357
Chordata.
Echinoderm at а
Enteropneusta
PieroSranchia
graptozoa
Pogonophora
Brachiopoda
. Bryozoa
Phoronida
Arihropoda
Annelida
Tar dig r a da
Pentaslomida
Ongchophora
Mollusca
Nemertini
AsKelminthes
& £
I
. §
I4
H
Протерозойская эра
700 млн. лет
5л
Cruslaceomorpha Crustacea
Tracheaia \Myriapoda
\jnsecta .
{Polychaeta
\Olcgochaeta
'Hirudinea
q лет
1J
< г
^4 .
|Й>
7-5 I
дк z
sV
Рис. 86. Филогенетическое древо беспозвоночных (по автору)
Для протерозойской, палеозойской и мезозойской эры: сплошными линиями обозначены группы,
имеющие ископаемых предков, пунктиром — не имеющие ископаемых. Для кайнозойской эры: сплош-
ными линиями обозначены ныне существующие группы, пунктиром — вымершие
Ophiurqidea
Asieroidea,
Eleutherozoal Spmqsieroidea
Ecninotdea
Holothurioidea.
\.Ophiocistia
Thecoidea
Crinoidea
„ , Eocrinoidea
'Pelmatozoa {Paracrinoidea
XBlasioidea
J Cystoidea
'Earpoidea
'TriloBitomorpha TriloBHa —
. [Tiphosura.
Eurypterida
Cheliceraia^^0^
SoliFugomorph,
Acaromorpha
Biiralma --------------
Scaphopoda -ту.------y
„ , , \Pulmonaia
gastropoda \OpisthoBranchia
\ProsoBranchia
Cephalopoda iEytgcochlia
, , \Ectocochlia
Loricata -----------------
Solenogastres-------------
Plathyhelminihes
[Kinorhyncha
'Rotatoria
\Nematoda
\Rastrotrioha
[Pseudoacoela
\Monoqenea
Cestoda
Trematoda
\Poluclada
Cienophora
Coelenterata
Spongia
' I
Annetid!h--j!'$ I /
/ i
^'1 ’ 1 1
' /' /'
;
Л'' I I
I /
§
ЬА
•S
§
I
,g
стан . .. . .
мина
шевр
блас*
тели :.
клад '
дерм
мезо. .
Пали
неуч. .
ние
паре
дия '
СТВ1-Г
явля
дыха
тизм
белл.
Ptero
Chor
ВЗЯЛ}
нием
шечн
У взр
яиц 1
риче<
В
веро;
щие,
ШИМ! 1
'ШИМ!
слож
связг
попу,
воде:
пичн
ШИМ!
квар-
acoel
оста.)
пару
ческ!
д
CTBOJ
пред:
и Ас;
дале:
с пл<
обре'
сом, ,
хи-ме
разв!
что р
мафц
ЛОВО]
его следы у ряда классов первичнополостных червей — гастритрик, кино-
.356
ринх, коловраток и нематод, а также, вероятно, акантоцефалов. Типич-
ная организация Askelminthes, надо думать, сформировалась у этого
типа давно, до перехода к паразитическому образу жизни, поэтому и,
вероятно, в силу каких-то особенностей условий жизни она сохраняется
и у паразитических нематод — зоонематод. В сущности говоря, у них
наблюдаются лишь многочисленные и разнообразные различия в дета-
лях, не нарушающих общий тип строения. Как известно, класс нематод
является одним из самых многочисленных классов, уступающий в этом
лишь насекомым. Вместе с тем достаточно ясно происхождение двух, их
подклассов — Adenophorea и Secernenta, в частности, фитонематод и
зоонематод от общих примитивных форм свободноживущих нематод
с чертами организации рабдоцелид.
Появление немертин, паренхиматозных, сдавленных в дорсо-вентраль-
ном направлении червей, обусловлено активным хищничеством (ловлей
крупной добычи с помощью хобота), с.чем было связано формирование
сквозного кишечника и сильное развитие кожно-мускульного мешка,
обеспечившего большую подвижность, необходимую немертинам при
ловле подвижной добычи. Прогрессивное развитие немертин как свобод-
ноживущих червей, привело также к формированию у них за счет мезо-
дермы и возникающей из нее вторичной полости тела — кровеносной
системы, впервые возникшей среди первичноротых в этой группе. Со
свободным существованием в море связано у большинства немертин
сохранение спирального детерминированного типа дробления яиц и
обычно1 развитие пелагических личинок. По совокупности данных, осо-
бенно анатомии, филогенетические связи между Protonemertini, Meso-
nemertini, Metanemertini и Heteronemertini достаточно ясны. Можно
считать, что немертины возникли от примитивных форм, турбеллярий —
-далиеллид, ставших раздельнополыми и утративших сложную половую
гермафродитную систему, и являются слепой ветвью, не давшей потом-
ков. Можно дум-ать, что паразитические плоские черви, первичнопо-
лостные и немертины произошли от примитивных турбеллярий (раб"
доцелид) и взяли свое начало от планулоподобных форм кишечнопо-
лостных.
К сожалению, вследствие отсутствия скелета, нет палеонтологических
(исторических) данных о плоских червях, первичнополостных и немер-
тинах; лишь для двуусток, моногенетических сосальщиков и ленточных
червей имеются косвенные данные об их хронологии.
Следующей ветвью ствола первичноротых являются моллюски, по
своей ^организации, онтогенезу, физиологии и экологии стоящие много
выше предыдущих групп червей и обладающие рядом черт онтогенеза,
общих с онтогенезом аннелид и полихет. Однако в соответствии с ком-
пактным типом организации и -полным отсутствием истинной метамерии
(членистости) онтогенез моллюсков принципиально отличен от онтоге-
неза аннелид (отсутствие закладки метамерии, в том числе и зачатка
постларвальных сегментов). Все, что известно о повторности ряда на-
ружных и некоторых внутренних органов у моллюсков, в том числе и
у Neopilina, относится к так называемой «метамерии упорядочения»
В. Н. Беклемишева.
Предковыми формами моллюсков, надо думать, были придонные
морские червеобразные животные, двустороннесимметричные по наруж-
ной форме и внутреннему строению, без раковины, обладавшие яйцами
со спиральным детерминированным дроблением и исторически вначале
одной пелагической личинкой — трохофорой. На этом уровне находятся
соленогастры, в онтогенезе которых нет признаков вторичного упроще-
ния; в родословном древе моллюсков их располагают ближе всего к ис-
ходной родоначальной их форме. Из-за отсутствия раковины геологиче-
ская история соленогастров неизвестна.
357
В ходе эволюции червеобразных донных ползающих предковых форм
моллюсков усилением развития мускулатуры брюшная сторона тела
становилась толще, а прогрессивно развивавшиеся внутренние органы
на спинной, стороне тела приобрели мантию, образовавшую раковину.
Последняя у хитонов, близких по ряду черт организации к соленога-
страм, состояла из восьми подвижно соединенных пластинок, что было
связано с ползанием по твердому субстрату дна моря (скалам). Хитоны,
подобно соленогастрам, имеют одну личинку (трохофору) и развиваются
путем линейного роста по горизонтальной передне-задней оси. От родо-
словного древа моллюсков хитонов, вероятно, можно отводить вблизи
соленогастров, однако их филогения недостаточно выяснена.
На этом раннем этапе исторического развития примитивных мол-
люсков произошла дивергентная эволюция остальных их классов. Часть
'примитивных моллюсков стала приспосабливаться к подвижному образу
жизни и к активной ловле добычи. С этим было связано развитие щупа-
лец и воронки, сильных челюстей, пары головных глаз. В этом направле-
нии шло становление класса головоногих моллюсков, которые в самом
начале своей эволюции перешли от типичного для моллюсков спираль-
ного, детерминированного дробления яиц и образования пелагических
.личинок к неполному дискоидальному дроблению яиц и кщрямому раз-
витию. Нйжнекембрийский Volborthella был уже типичным головоногим.
Исторически длительный на протяжении всех геологических периодов
подвижный образ жизни хищных головоногих способствовал прогрес-
сивному развитию этого класса. Известно, что многие современные го-
ловоногие по уровню организации и физиологии стоят выше всех осталь-
ных типов беспозвоночных и приближаются к позвоночным, а иные и не
уступают им. История развития головоногих данными палеонтоло-
гии освещена лучше, чем история развития всех остальных беспозво-
ночных. •>
Все говорит об очень раннем обособлении головоногих от родослов-
ного древа моллюсков, более раннем, чем ответвление от него брюхо-
ногих, двустворчатых й лопатоногих. Общий предок последних, так назы-
ваемый Prorhipidoglossum (или Praerhipidoglossum), обладал основной
организацией и цндивидуальным развитием со спиральным, детермини-
рованным дроблением яиц и метаморфозом с двумя личинками (трохо-
форой и велигером). Этот общий предок стал приспосабливаться к жизни
в прибрежной полосе моря с малым количеством осадков, к ползанию
и прикреплению к твердому субстрату. Это способствовало развитию
ноги с подошвой, малоподвижному образу жизни и питанию соскабли-
ванием преимущественно водорослей и корненожек радулой.
Основным органом движения этих моллюсков стала нога с полза-
тельной подошвой; форма ноги, связанная с характером ее использова-
ния, характеризует классы брюхоногих, лопатоногих и топороногих
(Pelecypoda). Брюхоногие, отличающиеся от двустворчатых и лопатоно-
гих асимметрией (вызванной скручиванием или торзионом), благодаря
морфологическому и биологическому прогрессу ставшие процветающим
ныне классом, своими низшими примитивными представителями стоят
ближе к общему родоначальнику, чем двустворчатые и лопатоногие. Под
' воздействием условий жизни в иле и песке лопатоногие и еще более
двустворчатые уклонились от исходного плана строения, стали более
специализированными группами раковинных моллюсков.
История развития брюхоногих и двустворчатых известна достаточно
полно; пока брюхоногие найдены в низах кембрия, а двустворчатые —
начиная с верхов или с середины кембрия; малочисленные современные
лопатоногие и в ископаемом состоянии редко встречаются, начиная с
ордовика или силура, более частыми формами они становятся лишь
в кайнозое.
358
Наличие в нижнем кембрии головоногих и брюхоногих моллюсков
заставляет ожидать их находок и в докембрии, но эти древние предко-
вые формы моллюсков, близко родственные примитивным плоским чер-
вям, плохо сохранились в ископаемом состоянии.
Другой ветвью, произошедшей также от примитивных плоских, па-
ренхиматозных предков, являются кольчатые черви, прежде всего поли-
хеты. Эволюция этих придонных морских червей была связана с хищ-
ничеством и в связи с этим с большой подвижностью. Открытый образ
жизни вызвал развитие многочленистости, охватившей внешнее строение
и внутреннюю организацию, развитие параподий, кутикулярного покрова
и т. д.; все это привело в ходе длительной эволюции полихет (начиная
с докембрия) к становлению у них многих систем органов, типичных для
вторичнополостных членистых первичноротых. Естественно, что шло и
становление типичного для них онтогенеза со спиральным детерминиро-
ванным дроблением, с мезодермальными полосками телобластического
происхождения, с планктонной трохофорой, проходящей несколько ста-
дий, начиная с атрохной, и с закладкой ларвальных и постларвальных
сегментов. Происхождение олигохет от полихет, а от последних пиявок
связано с переходом от жизни в море к жизни в пресной воде и почве,
а затём к хищническому паразитизму. История развития полихет, бла-
годаря хитиновым челюстям,’ трубкам, особенно известковым, 'ходам -и
другим следам жизни, прослежена неплохо, начиная с докембрия; па-
леонтологических данных об олигохетах и пиявках мало.
Говоря о тардиградах, пентастомидах и даже онихофорах, прихо-
дится лишь отметить их своеобразную природу с чертами аннелид и от-
части членистоногих; в системе они все же занимают положение ближе
к аннелидам, чем к членистоногим, но их филогенетические отношения
пока не выяснены и друг с другом они не связаны. Для выяснения при-
роды тардиград, пентастомид и онихофор необходимы новые, более со-
вершенные методики изучения.
Как известно, филогенетические связи между аннелидами, полихе-
тами и членистоногими давно установлены. Разногласие во мнениях
заключается в вопросе — одинаковое или разное число сегментов уча-
ствует в образовании головы членистоногих и монофилетичен или поли-
филетичён этот тип. Согласно многим исследованиям, в головном отделе
разных групп членистоногих участвует разное число сегментов. Трило-
биты’ (или трилобитоморфные), хелицеровые, ракообразные и трахей-
ные, различные по своему строению, индивидуальному развитию й эко-
логии, имевшие родоначальниками морских неспециализированных по-
лихет, развивались в экологически разных условиях и появились в раз-
ное геологическое время. В этом отношении тип членистоногих является
полифил етичным.
При филогенетических построениях не следует переоценивать значе-
ния классификации, не следует забывать ее относительность, недостаточ-
ность знаний и исторической обусловленности многих групп, а также
большую лабильность организмов. Признавая обоснованность и фило-
генетическое значение подразделений двустороннесимметричных живот-
ных на первичноротых и вторичноротых, фактически мы сталкиваемся
с тем, что отдельные признаки встречаются иногда не в той группе, для
которой в норме они характерны. В значительной мере это относится и
к форонидам, мшанкам и брахиоподам, которых раньше хотя и объеди-
няли в тин Tentaculata, но резонно помещали в филогенетическом древе
между ветвями первичноротых и вторичноротых. Если судить по поло-
жению бластопора и рта, — это первичноротые, если же судить по ха-
рактеру расчлененности целома на три отдела, энтероцельному образо-
ванию целома и наличию креатинфосфата у брахиопод, то они имеют
общее с рядом типов вторичноротых. Значительные различия в общей
359
организации и типе онтогенеза говорят об отсутствии близко родствен-
ных связей между форонидами, мшанками и брахиоподами. Геологиче-
ская история форонид, небольшой реликтовой группы современных жи-
вотных (несколько видов), почти неизвестна. Возможно, что они суще-
ствовали уже в кембрии—девоне. Геологическая история обширного
типа — мшанок (4000 современных и 15 000 ископаемых видов) раскрыта
хорошо; филогенетические отношения группы Gymnolaemata выяснены,
но родственные отношения ее и Phylactolaemata еще требуют серьезных
исследований. Геологическая история брахиопод, обширного типа в
прошлом (около 30 000 ископаемых видов) и представленного ныне
всего 260 видами, изучена хорошо, но филогенетические отношения от-
рядов замковых брахиопод еще требуют исследований, и это особенно
относится к выяснению родственных отношений между беззамковыми
и замковыми брахиоподами.
Филогенетические отношения между типами вторичноротых доста-
точно ясны, хотя нотохорду, или стомахорду, кишечнодышащих надо
считать лишь преоральный выростом эктодермальной части кишечника,
а не гомологом хорды. Можно считать, что примитивность червеобразной
формы тела со ртом у основания хобота и анусом на конце туловища
и всей организации двустороннесимм'етричных кишечнодышащих сохра-
нились благодаря свободному существованию в толще грунта, без пере-
хода к прикрепленному образу жизни. Для большинства двусторонне-
симметричных животных, и первичноротых и вторичноротых, исходным
был свободноподвижный образ жизни, и вторичным, подобно птеробран-
хиям, — неподвижное прикрепление и связанное с этим возникновение
петлеобразного кишечника со сближенными ртом и анусом. Надо счи-
тать, что кишечнодышащие также и исторически более древние формы
сравнительно с птеробранхиями, у которых ряд органов, общих с кишеч-
нодышащими, сильно редуцирован или утрачен и развились ценоции
характерного строения, колониальность и, кроме полового, бесполое
размножение путем образования наружных почек. Эта реликтовая груп-
па, представленная почти двадцатью современными видами и немногими
ископаемыми, известна с ордовика. Родственными с птеробранхиями,
согласно исследованиям последних лет, являются граптолиты, обладав-
шие ценоцием, по строению сходным с ценоцием птеробранхий, суще-
ствовавшие со среднего кембрия до конца карбона и во множестве насе-
лявшие палеозойские моря. На данном этапе наших знаний приходится
считать кишечнодышащих, птеробранхий и граптолитов отдельными ти-
пами с отдаленными общими предками в группе вторичноротых, стоящих
друг к другу ближе, чем к иглокожим и хордовым.
Несмотря на бросающиеся в глаза различия между кишечнодыша-
щими, иглокожими и хордовыми, родство между ними доказано рядом
существенных черт онтогенеза, морфологии и физиологии. Не прихо-
дится сомневаться в происхождении иглокожих от двустороннесиммет-
ричных червеобразноудлиненных, придонных, медленно передвигав-
шихся по дну моря, форм. Некоторые из карпоидей, пока известных со
среднего кембрия, в некоторой степени близки к исходным предковым
формам иглокожих. Надо, однако, подчеркнуть, что нет серьезных дан-
ных ни для допущения происхождения хордовых от иглокожих, ни для
объединения последних и кишечнодышащих в тип Ambulacralia. Вернее
всего, что кишечнодышащие, иглокожие и хордовые имели древних, об-
щих предков, эволюция которых, в связи с различиями в образе и усло-
виях жизни, рано разошлась в разных направлениях. Тесная корреляция
между скелетом и многими внутренними органами у иглокожих (доступ-
ная для изучения организации и у большинства ископаемых форм) и
богатство материала по ископаемым иглокожим многих классов позво-
лили установить филогенетические связи между классами этого типа
360
(см. гл. «Иглокожие»), но необоснованным надо признать утверждение
о филогенетических связях морских звезд и офиур с морскими лилиями,
а морских ежей и голотурий — с цистридеями. Тем не менее многое го-
ворит о полифилии классов Eleutherozoa, хотя и происшедших от близко
родственных форм.
Для всех вторичноротых, как известно, характерно наличие в онто-
генезе (хордовые), а у остальных и во взрослом состоянии, трех отделов
(сегментов) целома и обычно целомодуктов в первых двух. Погонофоры
в этом отношении сходны с большинством типов вторичноротых, хотя
они и обладают рядом признаков, общих с аннелидами (полихетами),
однако дажег наличие у них заднего членистого отдела тела не дает
оснований считать погонофор аберрантными полихетами. Даже если
при дальнейшем изучении эмбриогенеза не оправдается природа пого-
нофор как вторичноротых, по большинству основных черт организации
и развития погонофоры, несомненно, наиболее близки к типам вторично-
ротых, и вместе с тем обладают рядом отдельных черт аннелид (поли-
хет). Если у кишечнодышащих и большинства хордовых туловище при-
обретает метамерное строение, каковым оказывается и задний отдел
тела погонофор, то это говорит о независимом развитии членистости
у этих групп животных (как у аннелид). Следовательно, надо считать,
погонофор типом, который не укладывается в группе целомических жи-
вотных ни в Deuterostomia, ни в Protostomia и в известной мере нару-
шает принятую систему, что говорит о том, что ее нельзя абсолютизиро-
вать, поскольку в ней мы имеем дело не с абстрактными величинами, а
с живыми организмами.
Л ИТЕРАТУРА1
ОБЩАЯ
Амалицкий В. П. 1896. О геологическом развитии организмов и земного рельефа.
Варшава, стр. 1—31.
Беклемишев В. Н. 1944. Основы сравнительной анатомии беспозвоночных. Изд. 1.
М., стр. 1—492.
Беклемишев В. Н. 1951. К построению системы животных.— Усп. соврем, биол.,
32, 2: 256—270.
Беклемишев В. Н. 1952. Основы сравнительной анатомии беспозвоночных. Изд. 2.
Изд-во «Сов. Наука», стр. 1—698.
Беклемишев В. Н. 1958. К вопросу о ранней эволюции моллюсков. Сравнительно-
анатомическое значение й функционально-морфологическое толкование организа-
ции Neopilina galathea Lemche.— Зоол. ж., 37, 4: 518—522.
Беклемишев В. Н. 1964. Основы сравнительной анатомии беспозвоночных, т. 1.
Изд. 3. М,, Изд-во «Наука», стр. 1—432; т. 2, стр. 1—446.
Беляев Г. М., Виноградова Н. Г., Филатова 3. А. 1960. Исследование дон-
ной фауны глубоководных впадин южной части Тихого океана.— Тр. Ин-та океанол.
АН СССР, 41: 106—122.
Борисяк А. А. 1905—1906. Курс палеонтологии, т. 1. Беспозвоночные. СПб., стр.
8—368.
Вернадский В. И. 1935, 1939. Проблемы биохимии, т. I (1935) и т. II (1939). М.— Л.,
Изд-во АН СССР.
Виноградов А. П. 1930. Химический элементарный состав организмов моря.
Сообщ. 1.— Тр. Биогеохим. лабор. АН СССР, т. I, стр. 65—278.
В и н о г р а д о в. А. П. 1937. Химический элементарный состав организмов моря.
Сообщ. 2.— Тр. Биогеохим. лабор. АН СССР, т. 4.
Виноградов А. П. 1944. Химический элементарный состав организмов моря.— Тр.
Биогеохим. лабор. АН СССР, т. 6: 5—273.
Воскресенский К. А. 1948. Пояс фильтраторов как биогидрологическая система
моря.— Тр. Гос. океаногр. ин-та, 6(18): 55—120.
Геккер Р. Ф. (ред.). 1947. Развитие жизни на земле. М., Госкультпросветиздат,
стр. 1—46.
Геккер Р. Ф., Осипова А. И., Бельская Т. Н. 1962. Ферганский залив Палео-
генового моря Средней Азии.— Изд. Палеонтол. ин-та АН СССР, т. 1, стр. 1—335,
т. 2, стр. 1—332.
Гиляров М. С. 1944. О функциональном значении симметрии организмов.— Зоол. ж.,
23, 5: 213—215.
Гиляров М. С. 1949а. Роль почвы в филогенезе наземных беспозвоночных.— Усп.
соврем, биол., 27, 3: 325—341.
Гиляров М. С. 19496. Особенности почвы, как среды обитания, и ее значение в эво-
люции насекомых. М.— Л., Изд-во АН СССР, стр. 1—280.
Гиляров М. С. 1958. Эволюция характера оплодотворения наземных членистоно-
гих.— Зоол. ж., 38, 5: 707—735.
Гиляров М. С. 1959. Закономерности изменений морфологических и физиологиче-
ских особенностей членистоногих при переходе к наземному образу жизни.— Тр.
Ин-та морфол. животных АН СССР, 27; 118—133.
Гинецинский А. Г. 1945. Основные направления в физиологии центральной нерв-
ной системы. В сб. «Успехи биол. наук в СССР». М.— Л., Изд-во АН СССР,
стр. 27—34.
Грузов Е. Н. 1963. Строение и систематическое положение эндопаразитического
моллюска Asterophila japonica Randall et Heath.— Зоол. ин-т АН СССР. Л.,
стр. 1—20.
Давиташвили Л. Ш. 1940. Развитие идей и методов в палеонтологии после Дар-
вина. М.— Л., Изд-во АН СССР, стр. 1—263.
1 В общий список литературы включены работы, цитируемые в различных главах
настоящей книги. Специальная литература приводится в списках для каждой группы.
362
.Давиташвили Л. Ш. 1948. История эволюционной палеонтологии от Дарвина до
наших дней. М.— Л., Изд-во АН СССР, стр. 1—575.
Давиташвили Л. Ш. 1958. Краткий курс палеонтологии. М, стр. 1—544.
Давыдов К. Н. 1908. Наблюдения над процессом регенерации у Enteropneusta.—
Зап. Импер. АН, физ.-мат. отд. (СПб.), 20, 10: 1—120.
.Давыдов К. Н. 1914. Курс эмбриологии беспозвоночных. Изд-во «Сотрудник», Пг,—
Киев, стр. XII+502.
Догель В А. 1930. Эволюция беспозвоночных животных.— Человек и природа, 5:
41—75. - '
Догель В. А. 1938, 1940. Сравнительная анатомия беспозвоночных, т. 1 (1938),
стр. 1—600; т. II (1960), стр. 1—495.
Догель В. А. 1951. Общая протистология. М, Изд-во «Сов. Наука», стр. 1—604.
Догель В. А. 1954. Олигомеризация гомологических органов. Изд-во ЛГУ,
стр. 1—368.
Догель В. А. 1959а. Зоология беспозвоночных. Изд. 5. М, Изд-во «Сов. наука»,
стр. ХП + 502.
Догель В. А. 19596. Зоология беспозвоночных. М, Изд-во «Сов. наука», стр. 1—511.
Догель В. А. 1962. Общая паразитология. Изд-во ЛГУ, стр. 1—464.
Ежиков И. И. 1939. Соотношение онтогенеза и филогенеза.— Усп. соврем, биол., П,
2; 217—240.
Ежиков И. И. 1940. Учение о рекапитуляции и его критики. В кн. Мюллер Ф. и
Геккель Э. «Основной биогенетический закон». М.— Л., Изд-во АН СССР,
стр. 7—42.
.Заварзин А., А. 1945. Очерки эволюционной гистологии крови и соединительной
ткани. М, Медгиз, стр. 1—210.
Заварзин А. А. 1947. Очерки эволюционной гистологии крови и соединительной
ткани, т. 2. М.— Л., Медгиз, стр. 1—273.
Заварзин А. А. 1950. Очерки по эволюционной гистологии нервной системы. Избр.
тр., т. III. М.— Л., Изд-во АН СССР, стр. 1—419.
Заленский В. В. 1915. Эмбриология и филогения.— Природа, апрель: 666—691.
Захваткин А. А. 1949. Сравнительная эмбриология низших беспозвоночных. М,
Изд-во «Сов. Наука», стр. 1—395.
Зенкевич Л. А. 1944. Очерки по эволюции двигательного аппарата животных.—
Ж. общ. биол., 5, 3: 129—171.
Зенкевич Л. А. 1951. Моря СССР, их фауна и флора. М, Учпедгиз, стр. 1—368.
Зенкевич Л. А. 1963. Биология морей СССР. М, Изд-во АН СССР, стр. 1—738.
/Иванов А. В. 1937. Морфологические адаптации к паразитическому образу жизни —
Уч. зап. ЛГУ (биол.), 13, 4: 53—94.
Иванов А. В. 1939. Биогенетический закон и исследование филогенеза в палеонто-
логии.— Уч. зап. каф. геол. Моск. гос. пед. ин-та, 1: 175—196.
Иванов А. В. 1949. Строение и развитие эндопаразитического брюхоногого моллюс-
ка Parenteroxenos dogieli A. Ivanov [(семейство Entoconchidae).— Изв. АН СССР,
сер. биол, № 2: 109—134.
Иванов А. В. 1957. Материалы по эмбриональному развитию Pogonophora.— Зоол.
ж, 36, 8: 1127—1144.
Иванов П. П. 1937. Общая и сравнительная эмбриология. М.— Л, Биомедгиз,
стр. 1—809.
Иванов П. П. 1944. Первичная и вторичная метамерия тела.— Ж. общ. биол, 5, 2:
61—95.
Иванов П. П. 1945. Руководство по общей и сравнительной эмбриологии. Л, Учпед-
гиз, стр. 1—351.
Иофф Н. А. 1962. Курс эмбриологии беспозвоночных. М,'Изд-во «Высшая школа»,
стр. 1—266.
К а р п е в и ч А. Ф. 1955. Некоторые данные о формообразовании у двустворчатых
моллюсков.— Зоол. ж, 34, 1: 46—67.
Кнорре А. Г. 1937а. Распространение паразитизма в животном царстве.— Уч. зап.
ЛГУ, сер. биол, 3, 4: 21—88.
Кнорре А. Г. 19376. Распространение паразитов в хозяевах и проблема специфич-
ности.— Уч. зап. ЛГУ, сер. биол, 3, 4: 108—118.
К о р ж у е в П. А. 1949. Эволюция дыхательной функции крови. М.— Л, Изд-во
АН СССР, стр. 1—180.
Коржу ев П. А. 1964. Гемоглобин. Сравнительная физиология и биохимия. М,
Изд-во «Наука», стр. 1—287.
Коштоянц X. С. 1957. Основы сравнительной физиологии, II. Сравнительная физио-
логия нервной системы. М, Изд-во АН СССР, стр. 1—635.
Коштоянц X. С. 1963. Проблемы энзимохимии процессов возбуждения и торможе-
ния и эволюции функций нервной системы.— Баховские чтения, XVII, М, Изд-во
АН СССР: 1—31.
Крепе Е. М. 1933. Сравнительная биохимия мышцы и эволюционное учение.— При-
рода, № 8—9: 65—74.
363
Крепе Е. М. 1963, Исследование фосфолипидов нервной системы в эволюционном
ряду животных.— Биохимия, 28, 6: 978—986.
Ливанов Н. А. 1940. Класс полихет (Polychaeta). В кн. «Руководство по зоологии»,,
т. II. Беспозвоночные. М.— Л., Изд-во АН СССР, стр. ПО—136.
Ливанов Н. А. 1945. Пути эволюции животного мира. Анализ организации типов.—
Уч. зап. Казанок, гос. ун-та, 105, 3: 1—95.
Ливанов Н. А. 1955. Пути эволюции животного мира. М., Изд-во «Сов. Наука»,
стр. 1—400.
Ливанов Н. А. 1960. О закономерностях живой природы.— Уч. зап. Казанск. гос;
ун-та, 120, 6 : 3—32.
Личков Б. Л. 1945. Геологические периоды и эволюция живого вещества.— Ж. общ.
биол., 6, 3: 1—157.
Личков Б. Л. 1954. Об энергетике Земли и причинах тектонических явлений.— Геол,
сб. Львовск. геол, об-ва, т. 1.
Личков Б. Л. 1956. О связи между изменениями структуры Земли и изменениями
климата. В кн. «Доклады на ежегодных чтениях памяти.Л. С. Берга», т. I—III,
М.— Л., Изд-во АН СССР, стр. 192—211.
АГ а к а р о в П. В. 1938. Проблема общего и клеточного наркоза.— Арх. анатомии,
гистол. и эмбриол., 19, № 1—2: 5—104.
А4 а р к е в и ч О. П. 1964. Фелогения тваринного свету. Киев, Изд-во АН УССР,,
200 стр.
Матвеев Б. С. 1937. Задачи проблемы и соотношения онтогенеза и филогенеза,—
Изв. АН СССР, сер. биол., Г: 3—42.
Матвеев Б. С. 1947. Роль эмбриологии в изучении закономерностей эволюции.—
Зоол. ж., 26, 5: 389—402.
Матвеев Б. С. 1957. О преобразовании функции в индивидуальном развитии живот-
ных. (Значение учения А. Н. Северцова о типах филогенетических изменений орга-
нов при изучении индивидуального развития).— Зоол. ж., 36, 1: 4—25.
Мечников И. И. 1892. Лекции о сравнительной патологии воспаления.— Избр. биол..
произведения. А4., Изд-во АН СССР, 1950, стр. 473—651.
Мошковский Ш. Д. 1957. О природе простейших (Protozoa) и границах прото-
зоологии.— Тр. Ленингр. об-ва естествоисп., 73, 4: 129—135.
Мур Р. 1957. Беспозвоночные животные и геохронологическая шкала. В кн. «Земная
кора». ИЛ, стр. 1—787.
Опарин А. И. 1957. Возникновение жизни на Земле. М. Изд-во АН СССР.
О п а р и н А. И. 1965. Происхождение жизни. Изд-во «Медицина».
Орлов Ю. А. 1949. Палеоневрология как один из разделов палеонтологии позвоноч-
ных.— Тр. Палеонтол. ин-та АН СССР, 20: 57—66.
Орлов Ю. А. (ред.) 1960—1964. Основы палеонтологии, 16. т. М„ Изд-во АН СССР.
Павловский Е. И. 1937. Руководство по зоологии, т. I. Беспозвоночные. М.
Парамонов А. А. 1958. Главные направления эволюции фитонематод отрядов раб-
дитид и тиленхид (Rhabditida et Tylenchida) — Зоол. ж., 37, 5: 736—749.
Парамонов А. А. 1962, 1964. Основы фитогельминтологии. М., Изд-во АН СССР,
1962, I: 1—479; 1964, II: 1—443.
Роден дорф Б. Б. 1959. Филогенетические реликты.— Тр. Ин-та морфол. животных
АН СССР, 27: 41—51.
Руженцев В. Е. 1939. Значение онтогенеза'для естественной систематики аммони-
тов.— Изв. АН СССР, сер. биол., № 1: 13—82.
Руженцев В. Е. 1940. Опыт естественной систематики некоторых верхнепалеозой-
ских аммонитов.— Тр. Палеонтол. ин-та АН СССР, 11, 3: 1—134.
Северцов А. Н. 1939. Морфологические закономерности эволюции. М.— Л., Изд-во
АН СССР, стр. 1—610.
Скрябин К. И. и др. 1949—1954. Определитель паразитических нематод. М., Изд-во
АН СССР, т. I (1949), стр. 1—519; т. II (1951), стр. 1—631; т. III (1952),
стр. 1—890; т. IV (1954), стр. 1—987.
Скрябин К. И. (ред.) 1949—1964. Основы нематодологии, 13. т. М., Изд-во
АН СССР.
Соболев Д. Н. 1924. Начала исторической биогенетики. Киев, Гос. изд-во Украины,
стр. 1—205.
Стовас М. В. 1951. Критические параллели земного эллипсоида. Изд. ЛГУ.
С т р а х о в И. М. 1948. Основы исторической геологии. М.— Л., Госгеолиздат.
Страхов Н. М. 1954. Образование осадков в современных водоемах. М.
Талиев Д. Н., Базикалова А. Я. 1934. Предварительные результаты сравнения-
фауны Байкала и Каспия при помощи реакции преципитации.— Докл. АН СССР,
2, 8: 512—515.
Федотов Д. М. 1933. Происхождение беспозвоночных животных.— Вести, знания,
7—8: 239—245.
Федотов Д. М. 1935. Очерк эволюции животного мира.— Тр. Палеозоол. ин-та
АН СССР, 4: 309—324.
Федотов Д. М. 1936. Морфологические закономерности эволюции в применении
к беспозвоночным,— Изв, АН СССР, сер, биол., 5: 1615—1631.
364
Федотов Д. М. 1937. Основные достижения в области морфологии беспозвоночных
в СССР.— Усп. соврем, биол., 6, 3: 471—504.
Федотов Д. М. 1938. Филогения беспозвоночных в СССР за 20 лет.— Зоол. ж., 17,
1: 3—30.
Федотов Д М. 1945. Эволюционное значение регрессивных изменений онтогенеза.—
Бюлл. Моск, об-ва испыт. природы, 50, 3—4: 18—30.
Федотов Д. М. 1946. Эволюционное значение регрессивных изменений онтогенеза.—
Реф. работ учрежд. АН СССР, отд. биол. наук за 1'945 г., стр. 231—232.
Федотов Д. М. .1949. Тип Echinodermata, иглокожие; класс Echinoidea, морские
ежи.— Атлас руководящих форм ископаемой фауны СССР, т. 13. М.
Федотов Д. М. 1954. XIV Международный зоологический конгресс.— Зоол. ж., 33,
2: 468—493.
Федотов Д. М. 1955. Критический разбор основных и некоторых частных обобще-
ний А. Н. Северцова и задачи современной морфологии животных.— Зоол. ж., 34,
1: 216—233.
Федотов Д. М. 1959. Современный подход к разработке проблемы эволюции и фи-
логении животных.— Зоол. ж., 38, 10: 1443—1445.
Федотов Д. М. 1960. Обзор достижений в области морфологии и филогении. бес-
позвоночных в СССР с 1917 по 1947 г.— Тр. Ин-та истории естествозн. и техн.
АН СССР, История биол. наук, 31, 6: 135—246.
Филатов Д. И. 1937. Значение опыта для морфологической характеристики орга-
нов и их гомологизирования.— Изв. АН СССР,; отд матем. и естеств. наук, 6:
955—976.
X л опин И. Г. 1946. Общебиологические и экспериментальные основы гистологии.
Изд-во АН СССР, стр. 1—491.
Циттель К. 1934. Основы палеонтологии, т. I. Беспозвоночные. Горгеонефтеиздат,
стр. 1—1056.
Шванвич Б. Н. 1917. Наблюдения над самкой и рудиментарным самцом Entocolax
ludwigi Vogt.— Зоол. вести., 2: 1—147.
Шимкевич В. М. 1905. Курс сравнительной анатомии позвоночных животных. СПб.,
Изд-во Вольфа.
Шимкевич В. М. 1908. Меторизис как эмбриологический принцип.— Изв. Имп. АН
(СПб.), VI, 2: 997—1008.
Шимкевич В. М. 1918. Биологические основы зоологии, т. I. Изд. 4. Пг., стр VII +
+ 403.
Шимкевич В. М. 1923. Биологические основы зоологии, т. I. Изд. 5. М.— Л., Гос-
издат, стр. 1—384
Шимкевич В. М. 1940. Об основных свойствах живых организмов.— Природа, № 1:
99—105.
Шмальгаузен И. И. 1947. Основы сравнительной анатомии позвоночных живот-
ных. Изд. 4. М., Изд-во «Сов. наука», стр. 1—540.
Шмальгаузен И. И. 1958. Регулирующие механизмы эволюции.— Зоол. ж., 38, 9:
1291—1306.
Шмидт Г. А. 1938. Филогенетическая дегенерация способов развития органов.—
Зоол. ж., 17, 1: 44—74.
Шмидт Г. А. 1951. Эмбриология животных, т. 1. В кн. «Общая эмбриология». М.,
Изд-вб «Сов. наука», стр. 1—355.
Шмидт Г. А. 1953. Эмбриология животных, т. 2. М., Изд-во «Сов. наука», стр.
1—404.
Ще р б ин о в с кий Н. С. 1952. Пустынная саранча шистоцерка. М., Гос. изд-во с.-х.
лит-ры, стр. 1—416.
Эйгенсон М. С. 1954. Твердая Земля и солнечная активность.— Геол. сб. Львовск.
геол, об-ва, 1: 252—270.
Яковлев И. И. 1925. Учебник палеонтологии. Изд. 3. Л.— М., Гос. изд-во, стр. 1—462.
Яковлев Н. И. 1937. Учебник палеонтологии. Изд. 4. Л.— М., ОНТИ НК ТП СССР,
стр. 1—512.
Яковлев Н. Н. 1964. Организм и среда. М.— Л., Изд-во «Наука», стр. 1—148.
А х Р. 1960. Die Entdeckung neuer Organisationstypen im Tierreich.— Die neue Brem-
Bucherei. Wittenberg. Lutherstadt, S. 1—116.
Baldwin E. 1957. Biochemie. Einfuhrung in ihre Dynamik. Verlag Chemie, Weinheim,
S. 1—356.
Baldwin E. and Y u d k i n W. H. 1950. Phosphogens of invertebrates and protochorda-
tes.— Proc. Roy. Soc., B, 136, 885: 614—631.
Balfour F. M. 4880а., A Treatise of Comparative Embryology, v. I, VIE London,
p. 1—1580.
Balfour F. M. 18806 Handbuch der vergleichende Embryologie, Bd. II. Jena,
S. XI+ 980.
«Beer G. R. de. 1954. The evolution of metazoa. In: J. Huxley et all. «Evolution as a pro-
cess». London, p. 24—33.
365
Beklemischew W.-N. 1958. Grundlageii def vergleichende Anatomie der Wirbello-
sen, Bd. I: Berlin, S. VIII + 441; Bd. II. Berlin, S. VII+403.
В о a s J. E. V. 1922. Lehrbuch der Zoologie. Jena, S. XI+ 735.
Boettger C. R. 1952. Die Stamme des Tierreichs in ihrer systematischen Gliederung.______
Abharidl. Braunschweig, wiss. Ges., 4: 238—300.
Boettger C. R. 1954. Die Stamme des Tierreichs in ihrer systematischen Gliederuna —
Kosmos, 50: 68—72.
Borradaile L. A., Potts F. A. and all. 1959. The Invertebrate. Cambridge Univ
Press, p. 1—795.
В u t s c h 1 i 0. 1924. Vorlesungen fiber vergleichende Anatomie. Lief. 4: IV+311.
Dawydoff C. 1928. Traite d’embryologie comparee des invertebres. Paris Masson
p. XIV+930.
Dougherty E. C. (Ed.). 1(963. A review of the lower Metazoa. In «Comparative Biology
and Phylogeny». Univ. California Press, Berkely, p. XI+478.
Easton W. H. 1960. Invertebrate paleontology. N. Y., Harpers Brothers, p. XII+701.
E sco la P. 1956. On the geological eras and the factors controlling organic evolution —
Verhandl. Koninkl. nederl. geol. Injbouwk. Genotsch. Deel. 16.
Fedotov D. M. 1956. Les phenomenes de regression dans le developpement des cer-
tains insectes superieurs.—Proc. XIV Internal. Congr. Zool., Copenhagen, 1953-
493—496.
Fedotov D. M. 1959. Modern methods of development the problems of animal evolu-
tion and phylogeny,— Proc. XV Internat. Congr. Zool., London, 1958, p. 1—3.
F r a n z V. 1924. Geschichte der Organismen. Jena, S. XII + 948.
Franz V. 1943. Die Geschichte der Tiere. In: «Die Evolution der Organismen», G. He-
berer (Hsgb.). Jena, G. Fischer, S. 219—296.
Gersch M. 1964. Vergleichende Endokrinologie der Wirbellosen Tiere. Leipzig, S. 1—535..
Grasse P. P., Poisson R. A. et Tuzet O. 1961. Zoologie, v. I. Invertebres. Paris,’
p. VIII + 919.
Grobben K. 1908. Die systematische Einteilung der Tierreichs.— Verhandl. zool.-bot.
Ges. Wien, 58.
Grobben K. und Kuhn A. 1932. Lehrbuch der Zoologie. Berlin und-Wien, S. XI + 1123.
Grobben K. und Kuhn A. 1963. Lehrbuch der Zoologie, Berlin und Wien, S. XII +
1123.
Hadzi J. 1963. The evolution of the Metazoa. Oxford — London — New York — Paris.
Pergamon Press, p. XII+499.
Haeckel E. 1874. Die Gastrea-Theorie. Jenaische Z. Naturwiss., 8.
Haeckel E. 1896. Systematische Phylogenie der wirbellosen Tiere. Bd. 2, Berlin,
S. XVIII+ 720.
Handlirsch A. 1926. In W. Kiikenthal. «Handbuch der Zoologie». Bd. III,.
S. 1—1158.
Hanson E. D. 1958. On the orgin of the Metazoa.— System. Zool., 7, 1: 16—47.
Hanstrom B. 1928. Vergleichende Anatomie des Nervensystems der wirbellosen Tiere.
Berlin, S. 1—628.
H atschek B. 1888. Lehrbuch der Zoologie. I. Jena, S. 1—432.
HatschekB. 1911. Das neue zoologische System. Leipzig.
Heegaard P. 1945. Remarks on the phylogeny of the Arthropoda.— Arkiv zool., 37 A,
3: 1—15.
H eider K- 1914. Phylogenie der Wirbellosen. Kulture der Gegenwart., 3. Teil., Abt. 4,.
S. 453—529.
Hennig W. 1959. Taschenbuch der Zoologie. H. 3. Wirbellose, II Gliedertiere. Leipzig,.
S. 1—170.
Hennig W. 1963. Taschenbuch der Zoologie. Bd. 2. Wirbellose. I. Ausgenommen • Glie-
dertiere. 2-te Aufl., S. 1—178.
H e r t w i g R. 1897. Lehrbuch der Zoologie, 4-te Aufl., Jena, S. XII+ 612.
HymanL. H. 1940—1959. The invertebrates. N. Y'., 1940, I, p. 1—726; 1951, II, p. 1—550;
1952, III, p. 1—572; 1955, IV, p. 1—763; 1959, V: 1—783.
Jagersten G. 1955. On the early phylogeny of the Metazoa.— Zool. Bidr., Uppsala,.
30: 321—354.
Jagersten G. 1959. Further remarks on the early phylogeny of the /Metazoa.— Zool.
Bidr., Uppsala, 33: 79—108.
Kaestner A. 1954/1|955. Lehrbuch der speziellen Zoologie. Wirbellose, Teil I. Jena,
Fischer, S. XIV+483.
Kaestner A. 1956. Lehrbuch der speziellen Zoologie, I. Wirbellose, Jena.
Kaestner A. 1963. Lehrbuch der speziellen Zoologie, Bd. I. Wirbellose. Jena.
Kerkut G. A. 1960. Implications of evolution. Pergamon Press, p. X+174.
Korsch elt E. 1913. Entwicklungsgeschichte und Morphologie der Wirbellosen. Die-
Kultur der Gegenwart. Ill, 4. Abt., II: 176—332.
Korschelt E. und Heid er K. 1890—1893. Lehrbuch der-vergleichen.d’e. Entwicklungs-
geschichte der wirbellosen Tiere. Spezieller Teil. Jena.
366
Korschelt E. und Heid er K. 1902, 1903, 1909, 1910. Lehrbuch der vergleichende
Entwicklungsgeschichte der wirbellosen Tiere. Allgemeiner Teil. Jena.
Korschelt E. und H eider K. 1936. Vergleichende Entwicklungsgeschichte der Tiere.
I: 1—536; II: 537—1314.
Kreps E. M. 1958. Brain metabolism in the evolution of vertebrates.— Proc. XV Inter-
nal. Congr. Zool., London: 510—512.
Kuhn A. 1955. Vorlesung fiber Entwicklungsphysiologie. Berlin, Springer Verl., S. IX+
506.
Kfimmel B. 1954. Status of invertebrate paleontology, 1953. V. Mollusca: Cephalopo-
da.—-Bull. Mus. compar. zool., Harvard coll., 112, 3: 181—192.
Lemche H. 1957. A new living deep-sea mollusc of the Cambro-Devonian class Mono-
placophora.— Nature, 179, 4556: 413—416.
MacBrideE. W. 1914. Text-book of embryology. London, p. XXXII+ 692.
M a ni M. S. 1953. Organic evolution.— Agra Univ. J. Res. Sci., 2: 83—112.
Mani M. S. 1954. Organic evolution. Some additional.— Agra Univ. J. Res. Sci., 3:
95—108.
Marcus E. 1958. On the evolution of the animal phyla.— Quart. Rev. Biol., 33, 1:
24—58.
Moore R. C. (Ed.). .1952—1964. Treatise on invertebrate paleontology. New York.
Muller A. H. 1958. Lehrbuch der Palaozoologie. II. Invertebrata. Jena, G. Fischer,
S. XV+566.
Mfiller A. H. 1961. Grossablaufe der Stammesgeschichte. Erscheinungen und Proble-
me. Jena, G. Fischer, S. 1—116.
M ii 11 e г F. 1864. Fur Darwin. Leipzig, S. 1—91.
N a ef A. 1931. Phylogenie der Tiere. Berlin, S. 1—200.
О p a r i n A. I. 1964. The chemical origin of life. Illinois, p. IX+ 124.
Pflugfelder O. 1962. Lehrbuch der Entwicklungsgeschichte und Entwicklungsphy-
siologie der Tiere. Jena, G. Fischer, S. XIH + 347.
Picken L. E. R. 1936. The mechanism of urine formation in invertebrates.—'J. Exptl,
Biol., 13.
Plate L. 1922. Allgemeine Zoologie und Abstammungslehre. Jena. G. Fischer, S. VI +
+ 629.
Prosser C. L. (Ed.) 1952. Comparative animal physiology. Philadelphia and London,
p. 1—888.
Prosser C. L. and Brown F. A. 1961. Comparative animal physiology. 2-d ed.
Ray Lankester E. 1873. On the primitive cell layers of the embryo as the basis of
genealogical classification of animals and on the origin vascular and lymph sys-
tems.— Ann. and Mag. Natur. History, 11, 65: 321—338.
R e m a n e A. 1954. Die Geschichte der Tiere. In «Die Evolution der Organismen». 2,
Aufl., G. Heberer (Hrsg.), 2: 340.
Remane A. 1958. Zur Verwandschaft und Ableitung der niederen Metazoen.— Ver-
handl. Dtsch. zool. Ges.; Zool. Anz.: 179—^196.
R e m a n e A. 1959. Die Geschichte der Tiere. In «Die Evolution der Organismen».
G. Heberer (Hrsb.). Bd. 1: 340—422.
R e n s c h B. 1954. Die Entstehung des Lebens auf der Erde.— Kosmos, 50.
Roche J. and all. 1960. Distribution of various phosphagens in invertebrates.— Corn-
par. Bipchem. and Physiol., 1: 44—55.
Runnstrom J. 1925. Zur experimentellen Analyse der Entstehung von Antedon.—
Arch. Entwicklungsmech., 105: 63—113.
Schepotieff A. 1913. Uber die Bedeutung der wassermanischen Reaktion fur die
biologische Forschung.— Zool. Anz., 4, I, 2: 49—54.
Schindewolf О. H. 1950. Grundlagen der Palaontologie. Stuttgart, S. 1—505.
Schindewolf О. H. 1954. Uber die moglichen Ursachen der grossen erdgeschichtli-
chen Faunenschnitte.— Neues Jahrb. Geol. und Palaontol., Monatshefte.
Schindewolf О. H. 1956. Tektonische Triebkrafte der Lebensentwicklung.— Geol.
Rundschau, 45, I: 1—17.
Schindewolf О. H. 1962. Neokatastrophismus? — Z. Dtsch. Geol. Ges., 114: 430—436.
Sedgwick A. 1884. On the origin of metameric segmentation and some other morpho-
logical questions.— Quart. J. Microscop. Sci., N. S., 24: 413—82.
Seidel Fr. 1960. Korpergestalt und Keimstruktur. Eine Erorterung fiber die Grundlagen
der vergleichenden und experimentellen Embrydlogie und deren Giiltigkeit bei pny-
logenetischen Uberlegungen.— Zool. Anz., 164, 7—10: 245—305.
Shrock R. R. and Twenhofel W. H. 1953. Principles of invertebrate paleontology.
2-d ed., N. Y„ X+816.
Snodgrass R. E. 1938. Evolution of the Annelida, Onychophora and Arthropoda.—
Smithsonian Misci Collect., 97, 6: 1—159.
SnodgrassR. E. 1958. Evolution of Arthropod mechanisms.—Smithsonian Misc. Col-
lect., 138, 2: 1—77.
S t i r t о n R. A. 1959. Time, life and man. N. Y., p. VIII+558.
Storer T. I. and U sin ger R. L. 1957. General zoology. N. Y. and London, p. VI+664.
367-
Termier H. et Termier G. 1960. Paleontologie stratigraphique. Paris, Masson,
p. 1—515.
Thiel M. E. 1959. Semaeostomae. Physiologie. In: «Bronns Klassen und Ordnungen
Tierreichs». Bd. II, Abt. 2, Lief. 6, S. 840—1072.
Vinogradov A. P. 1953. The elementary chemical composition of marine ogranisms.—
Mem. Sears Foundat. Marine Res. New Haven Gale Univ.: XVI+647.
WenzW. 1938. Handbuch der Palaozoologie, Bd. 6. Berlin, S. 1—480.
ГУБКИ (PORIFERA)
Митрополитанская P. Л. 1941. О содержании ацетилхолина и холинэстеразы у
простейших, губок и кишечнополостных животных.— Докл. АН СССР, 31, 7: 717—
718.
Резвой П. Д. 1934. Porifera. Губки. В кн. Циттель. «Основы палеонтологии», т. I.
Беспозвоночные, стр. 103—140.
Резвой П. Д. 1937. Тип губок. В кн. «Руководство по зоологии», т. I. Беспозвоноч-
ные. Под ред. Л. А. Зенкевича. М., стр. 228—267.
Р о скин Г. И. 1957. Нервная система губок.— Усп. соврем, биол., 43, 2; 199—207.
Соколов Б. С. (ред.) 1962. Основы палеонтологии. Губки, археоциаты, кишечнопо-
лостные, черви. М., Изд-во АН СССР, стр. 1—485.
A n k е 1 W. Е. und W.i ntermann-Kiiian G. 1952. Eine bei Ephydatia flaviatilis neu
gefundene hochdifferenzierte Zellart und die Struktur der Doppelepithelien.— Z. Na-
turforsch., 7b, 8: 475—481.
Arndt W. 1933. Die biologischen Beziehungen zwischen Schwommen und Krebsen —
Mitt. Zool. Museum. Berlin, 19: 221—305.
Arndt W. 1937. Die Poriferen vom Standpunkt der Strahlungsbiologie.— Sitzungsber.
Ges. naturforsch. Freunde, Berlin, 8—10: 315—355.
Atzeiius B. A. 1961. Flimmer-flagellum of the sponge.— Nature, 191, 4795: 1318—1319.
Burton M. 1949. Observations on littoral sponges including supposed swarming of
larvae, movement and coalescence in mature individuals, longevity and death.— Proc.
Zool. Soc. London, 118, pt. IV: 893—915.
Burton M. 1963. A revision of the classification of the calcareous sponges.— Brit. Mu-
seum Natur. History: 1—693.
В iitschli O. 1884. Bemerkungen zur Gastraeatheorie.— Jahrb. Morphol., 9: 416—427.
Delage Y. 1898. Sur la place des spongiaires dans la classification.— C. r. Acad. sci.
Paris, 126, 7: 545—548.
D uboscq O. et O. Tuzet. 1937. L’ovogenese, la fecondation et les premiers stades du
developpement des eponges calcaires.— Arch. zool. exptl. et gen., 79: 157—316.
Farkas B. 1929. Beitrage zur Kenntnis des feinerem Baues und der Entwicklung der
Spongien. X. Congr. internal, zool. Budapest: 933—941,
Galtsoff P. S. 1923. The ameboid movement of dissociated sponge cells.— Biol. Bull.,
45: 153—161.
Galtsoff P. S. 1929. Heteroagglutination of dissociated sponge cells.— Biol. Bull., 57:
250—260.
Heider K. 1886. Zur Metamorphose der Oscarella lobularis.— Arb. Zool. Inst. Wien, 6.
Jones W. C. 1962. Is there a nervous system in sponges? — Biol. Rev.- Cambridge, 37,
1: 1—50.
Korotkova G. P. 1962. Behaviour of the cellular elements in the calcareous sponge
Leucosolenia. complicata Mont, during regeneration.— Acta biol. Acad, scient. hung.,
XIII, I: 1—30.
Korotkova G. P. 1963. On the types of restoration processes in sponges.— Acta biol.
Acad, scient. hung., XIII, 4: 389—406.
Lackey J. B. 1959. Morphology and biology of a species of Protospongia.— Trans.
Amer. Microscop. Soc., 78, 2: 202—206.
Laubenfels M. W. de. 1955, Are Coelenterates degenerate? — System. Zool., 4, 1:
43—45.
Levi Cl. 19'56. Etude des Halisarca de Roscoff. Embryology et Systematique des De-
mosponges.— Arch. zool. exptl. et gen., 93, L 1—181.
Levi Cl. 1957. Ontogeny and Systematic in Sponges.— System. Zool., 6, 4: 174—183.
Maas O. 1892. Die Metamorphose von Esperia lorenzi O. S.— Mitt. Zool. Stat. Neapel,
10: 408—440.
Maas O. 1893. Embryonal-Entwicklung und Metamorphose der Cornaenspongien.— Zool.
Jahrb., Abt. 2, 7: 331—448.
Maas O. 1898. Die Entwicklung des Spongien.— Zool. СЫ., 5: 581—599.
Maas O. 1900. Die Weiterentwicklung der Syconen nach der Metamorphose.— Z. wiss.
Zool., 67: 2'15—240.
Maas O. 1909. Ueber die Wirkung des Hungers und der Kalkentziehung bei Kalkschwam-
men und anderen kalkausscheidenden Organismen.— Proc. VII Internat. Zool. Congr.
Cambridge: 1—7.
368
Metschnikoff E. 1874. Zur Entwicklungsgeschichte der Kalksschwamme.— Z. wiss.
Zool., 24: 1 —14.
Metschnikoff E. 1876. Beitrage zur Morphologie der Spongien.— Z. wiss. Zool., 27:
274—286.
Metschnikoff E. 1879. Spongiologische Studien.— Z. wiss. Zool., 32: 358—362.
M i n c h i n E. A. 1896. Note on the larva and the post-larval development of Leucosolenia
variabilis n. sp., with remarks on the development of other Asconidae.—-Proc. Roy
Soc., B, 60: 42—57.
Okada Y. 1928. On the development of a Hexactinellide sponge Farrea sollasii.— J. Fac.
Sci. Univ. Tokyo, 2.
Okulitch J. 1960. The Lower Cambrian fauna. In T. W. M. Cameron (Ed.) «Evolu-
tion: Its science and doctrine». Toronto, p. 12—21.
Paschma M. 1958. О systemie nerwowym u gabek.—Przegl. zool., 2, 1: 114—125.
Wrozlaw Polsk. Tow. Zool.
Pavans de Ceccatty M. 1955. Le systeme nerveux des eponges calcaires et siliceu-
ses.— Ann. sci. natur. Zool. et biol. anim. Ser. 11, 17 : 203—288.
Pavans de Ceccatty M. 1957. La nature secretoire des lophocytes de I’eponge siliceu-
se Chondrosia reniformis Nardo.— C. r. Acad. sci. Paris, 244: 2103—2105.
Pavans de Ceccatty M. 1959. Les structures celluliares de type nerveux chez Hip-
pospongia communis Lmk.— Ann. sci. natur. zool. et biol. anim. Ser. 12, 1: 105—112.
Polej aef f N. 1884. Report on the Calcarea dredged by H. M. S. Challenger.— Rept.
Challenger, 8: 1—76.
Rasmont R. 1959. Ultra-structure des choanocytes d’eponges.— Proc. XV Internal.
Congr. Zool. London, 16—23. July 1958: 707—708.
R e i d R. E. H. 1963. Hexactinellida or Hyalospongea.— J. Paleontol., 37, 1: 232—243.
Schulze F. E. 1875—1881. Untersuchungen fiber den Bau und die Entwicklung der
Spongien,— Z. wiss. Zool., 25, 28, 30, 31, 32, 34, 35.
S о 11 as W. J. 1884. On the development of Halisarca lobularis O. S. •—Quart. J. Micros-
cop. Sci., 24: 603—621.
S о 11 a s W. J. 1888. Report on the Tetractinellidae collected bv H. M. S. Challenger, 25,
63, 166: 1—458. . ,
Tuzet O. 1948. Les premiers stades du dfeveloppement de Leucosolenia botryoides Ellis
et Solander et de Claihrina (Lecosolenia) coriacea Mont.— Ann. sci. natur. zool. et
biol. anim. Paris. Ser. 11, 10: 103—114.
Tuzet O. 1963. Presidentiel address.— Bull. Soc. zool. France, 88, 1: 46—53.
Tuzet О., В es si ere Cl. et Paris J. 1961. Recherches serologiques sur les spongiai-
res et les Cnidaires.— Bull. biol. France et Belgique, 45, 1: 150—154.
Tuzet O., Loubatieres E., Pavans de Ceccatty M. 1952. Les cellules nerveu-
ses de I’eponge Sycon raphanus.— C. r. Acad. sci. Paris, 234: 1941—1396.
Tuzet O., Pavans de Ceccatty M. 4953. Les cellules nerveuses de I’eponge calcai-
re homocoele Leucandra johnstoni. C. r. Acad. sci. Paris, 236, 23: 1447—1449.
Tuzet O. et Pavans de Cecc at ty M. 1955. La mobilisation en amoebocytes des cel-
lules des Halisarca (eponges siliceuses). Les polyblastes chez les eponges. Compt.
rend. Soc. biol., 149, 7—8: 799—801.
Tuzet O., Pavans de Ceccatty M. et Paris J. 1963. Les sponges sonttelles des
colonies?—Arch. zool. exptl. et gen., 102, 1: 14—19.
Warburton Fr. E. 1961. Inclusion of parental somatic cells in sponge larve.— Nature,
191, 1794: 1317.
Williams M. Y. 1960. Evolution of Paleozoic life: Ordovician et Permian. In «Evo-
lution: Its science and doctrine». W. M. Cameron (Ed.). Toronto, p. 22—24.
Wilson H. V. 1907. On some phenomens of coalescence and regeneration in sponges.—
J. Exptl Zool., 5: 245—258.
W i 1 s о n FI. V. 1911. Development of sponges from dissociated tissue cells.— U. S. Bur.
Fisheries Bull., 30.
КИШЕЧНОПОЛОСТНЫЕ (COELENTERATA)
Белоусов Л. В. 1960a. Прижизненные наблюдения над клеточными перемещениями
у гидроидного полипа Obelia flexuosa.— Докл. АН СССР, 136, 6: 1490—4494.
Белоусов Л. В. 19606. Клеточные процессы при морфогенезе гидроидных полипов.
Автореф. канд. дисс. М., стр. 1—12.
Белоусов Л. В. 1960в. Клеточные перемещения при морфогенезе гидроидного поли-
па Campanularia Integra.— Ж. общ. биол., 21, 4: 279—288.
Белоусов Л. В. 1960г. Химическая регуляция морфогенеза гидроидных полипов.—
Докл. АН СССР, 130, 5: 1165—1168.
Белоусов Л. В. 19Ь1. Клеточное размножение и рост гидроидных полипов.— Ж. общ.
биол., 22, 4: 281—291.
Белоусов Л. В. 1963. Образование избыточных интерстициальных клеток как пока-
затель неопределенной интенсивности клеточного размножения в морфогенезе Cam-
panularia Integra и Campanularia lacerata.— Докл. АН СССР, 150, 3: 690—694.
24 Д- М. Федотов
369
Бляхер Л. Я, 1963. Константин Николаевич Давыдов. М., Изд-во АН СССР, стр. 1—
243.
Вержбинская Н. А. 1935. К биохимии мышечного сокращения у медуз.— Физиол.
ж. .СССР, 18, 4: 636—677.
Ветохин И. А. 1926а. О работе мерцательного эпителия гастроваскулярной системы
медузы Aurelia aurlta (L.) Lam.— Работы Мурманск, биол. ст. (Мурманск), 11:
107—120.
Ветохин И. А. 19266. О процессах возбуждения в колоколе медузы Aurelea aurlta и
регуляции движений этого животного в морской воде.— Русск. физиол. ж., 9: 517—
536.
Владимирский А. П. 1913. Наблюдения над регенерацией у планул Campanularia
flexuosa Hiks.— Тр. СПб. об-ва естествоисп., 44, 1,' 4—6: 177—191.
Владимирский А. П. 1915. Влияние некоторых факторов на регенеративные про-
цессы у гидроидов.— Тр. Петрогр. об-ва естествоисп., 44, 4: 47—107.
Добролюбова Т. А. 1948. Изменчивость кораллов филогенетического ряда Dibu-
nophullum. bipartltum (McCoy)—Caninia okensis Stuck.— Изв. АН СССР, cep.
биол,, 2: 149—168.
Добролюбова T. A. 1952. Формообразование нижнекаменноугольных кораллов Li-
thostrotion и Lansdaleia в свете мичуринского учения.— Изв АН СССР, сер. биол.,
6:97—110.
Добролюбова Т. А. 1958. Нижнекаменноугольные колониальные четырехлучевые
кораллы Русской платформы.— Тр. Палеонтол. ин-та, 70: 1—224.
Добролюбова Т. А. 1960. Историческое развитие некоторых четыр.ехлучевых ко-
раллов в нижнем карбоне Подмосковного бассейна.— Сб. тр. по геол, и палеонтол.
АН СССР, Коми филиал, Сыктывкар, стр. 330—331.
Заварзин А. А., Стрелин Г. С. 1928. К вопросу о биологическом действии рентге-
новых лучей. Рентгенологические опыты над гидрами.— Вести, рентгенол. и радиол.,
VI, 3: 183—204.
Зенкевич Л. А. 1940. Распространение пресноводных и солоноватоводных кишечно-
полостных.— Зоол. ж., 19, 4: 580—602.
Иванова К. П. 1949. Дробление яйца Aurelia aurlta из Белого моря.— Докл. АН
СССР, 56, 8: 885—887.
Иванова-Казас О. М. 1959. К вопросу о происхождении и эволюции спирально
го дробления.— Вести. ЛГУ, сер. биол., 9, 2: 56—67.
Ильина Т. Г. 1963а. Новые данные о происхождении шестилучевых кораллов.— Докл.
АН СССР, 148, 1: 193—196.
Ильина Т. Г. 19636. Позднепермские и триасовые четырехлучевые кораллы Закав-
казья. Палеонтол. ин-т АН СССР. Автореф. канд. дисс. М., стр. 1—12.
Исаев В. М. 1925. Этюды об органических регуляциях.— Тр. Ленингр. об-ва естество-
исп., отд. зоол. и физиол., 55, 2: 3—183.
Канаев И. И. 1949. Общая схема регуляции нормальной гидры.— Докл. АН СССР,
65, 4: 573—575.
К а н а е в И. И. 1952. Гидра. Очерки по биологии пресноводных полипов. М., Изд-во АН
СССР, стр. 1—370.
Канаев И. И. 1953. К вопросу об «однощупальцевых полипах».— Зоол. ж., 32, 2:
212—214.
Канаев И. И. 1954. К вопросу о «текучести» клеточного состава гидры.— Зоол. ж.,
33, 1: 26—29.
Ковалевский А. О. 1873. Наблюдения над развитием Coelenterata.— Изв. Об-ва
любит, естествозн., антропол. и этногр.: 1—36.
К о р д э К. Б. 1963. Hydroconozoa — новый класс кишечнополостных животных.— Па-
леонтол. ж., 2: 20—25.
Корсакова Г. 1949. Ранние стадии дробления у Tiaropsis.—-Докл. АН СССР, 65, 3:
413—415.
Коштоянц X. С. 1957. Сравнительные данные о строении и функциональных особен-
ностях нервной системы кишечнополостных. В кн. «Основы сравнительной физиоло-
гии», т. II Сравнительная физиология нервной системы. Изд-во АН СССР, стр. 171—
225.
Мечников И. И. 1950 (1886). Эмбриологические исследования над медузами. Избр.
биол. произвед. Изд-во АН СССР, стр. 273—471.
Наумов Д. В. 1953. Общие вопросы метагенеза в связи с установлением первичного
поколения у метагенетических гидроидов.— Тр. Зоол. ин-та АН СССР, 13: 70—90.
Наумов Д. В. 1957. Роль процессов олигомеризации и полимеризации в эволюции
колониальных гидрозоев.— Тр. Ленингр. об-ва естествоисп., 73, 4: 38—42.
Наумов Д. В. 1960. Гидроиды и гидромедузы морских, солоноватоводных и пресно-
водных бассейнов СССР. Определитель по фауне СССР, т. 70. Изд-во АН СССР,
стр. 1-—626.
Наумов Д. В. 1961. Сцифоидные медузы морей СССР. Определитель по фауне СССР,
т. 75. Изд-во АН СССР, стр. 1—97.
Остроумова М. В. 1937. Значение организационного центра гидроида Moerisia in-
kermanica в процессе его регенерации, регуляции и бесполого размножения. Сбор-
370
ник, посвященный акад. Н. В. Насонову к 80-летию со дня рождения и 60-летию
научной деятельности. Изд-во АН СССР, стр. 295—352.
Полянский Ю. И. 1958. Тип Coelenterata. См. А. В. Иванов, Ю. И. Полян-
ский, А. А. Стрелков. «Большой практикум по зоологии беспозвоночных», т. I.
Изд. 2. Изд-во «Сов. Наука».
Соколов Б. С. 1955. Табуляты палеозоя в Европейской части СССР. Введение. Л.—
М., Гостехиздат, стр. 1—525.
Соколов Б. С. (ред.) 1962'. Основы палеонтологии. Губки, археоциаты, кишечнополо-
стные, черви. М., Изд-во АН СССР, стр. 1—464.
Сошкина Е. Д. 1960. Турнейские кораллы Rugosa и их взаимоотношения с девон-
скими.— Со. тр. по геол, и палеонтол. АН СССР, Коми филиал, Сыктывкар, сто.
272—329.
Сошкина Е. Д., Добролюбова Т. А. 1941. Эволюция кораллов Rugosa.— Изв.
АН СССР, отд. биол. наук, 1: 152—162.
Тихомиров А. А. 1887. К истории развития гидроидов.— Изв. Об-ва любит, есте-
ствозн., антропол. и этногр., 50, 2: 1—69 {приложение первое].
Ульянин В. Н. 1876. О происхождении кунин, почкующихся в желудке герионид.—
Изв. Об-ва любит, естествозн., антропол. и этнограф., 24: 1—16.
Хилл Д. 1963. Возможные промежуточные формы между альционариями и табуля-
тами, табулятами и ругозами, а также ругозами и шестилучевыми кораллами.— Тр.
21-го Междун. геол, конгр., т. 1. М., стр. 517—530.
Щелкановцев Я. П. 1905. Наблюдения над строением и развитием Coelenterata.—
Изд. Моск, об-ва любит, естестйозн., VIII; 103.
Яковлев Н. Н. 1928. Наследственность приобретенных свойств и палеозойские ко-
ралллы Rugosa.— Докл. АН СССР, 16—17: 311—312.
А b о п у i А. 1929. Uber Protohydra leuckarti Greet und Hire geographische Verbreitung.—
X Congr. internal, zool. Budapest, 2: 1452—1460.
Aurich H. und Werner B. Uber die Entwicklung der Polypen von Ectopleura dumer-
tieri van Beneden und die Verbreitung der planktischen Stadien in das Siidlichen Nord-
see (Athecatae — Anthomedusae).— Ber. Dtsch. wiss. Kommiss. Meeresforsch., 4:
234—257.
В err ill N. J. 1949. Form and growth in the development of a Scyphomedusa.— Biol.
Bull., 96: 283—292.
Brien P. 1951. Contribution a I’etude des hydres d’eau douce.— Bull. Soc. zool. France,
76: 277—296.
Chapman G. 1953. Studies of the Mesoglea of Coelenterates.— Quart. J. Microscop.
Sci., 94: 155—176.
Dahl E., F a 1 с k В., M e c k 1 e n b u г g Cl. and Myhrberg H. 1963. An adrenergic ner-
vous system in sea anemons.— Quart. J. Microscop. Sci., 104, 4 : 531—534.
Dawydoff C. 1933. Quelques observations sur la morphologie externe et la biologie
des Ctenoplana.— Arch. Zool. exptl et gen., 75, 6: 103—128.
Garstang W. 1946. The morphology and relations of the Siphonophora.— Quart. J.
Microscop. Sci., 87; 103—193..
Glaessner M. F. and Daily B. -1959. The geology and late precambrian fauna of the
Ediacaran fossil reserve.— Rec. South austral, museum, 13, 3: 369—401.
Greenberg M: J. 1959. Ancestors, embryos and symmetry.— System, zool. 8, 4: 212—
221.
G r i m s t о m A. V., Horne R. W., P a n t i n C. F. A. and Robson E. A. 1958. The fi-
ne structure of the Mesenteries of the sea-anemone Mitridium senile.— Quart. J. Mic-
roscop. Sci., 99: 523—540.
Hadzi J. 1918. Shvaeanje Sifonofora.— Rad Jugoslav. Akad. Znan. in Umetn., 219. Razpr.
Mat.—Prirod., 63: 195—277.
Hadzi J. Ш44. Turbeiarijska teorija knidarijev.— Razpr. Slov. Akad. Znan. in Umetn.
Ljubljano Mat.— Prirod. Raz. Dela, 3: 1—238.
Hadzi J. 1951. Die Ableitung der Knidarien von den Turbellarien und einige Folgerun-
gen dieser Ableitung.— Razpr. Slov. Akad. Znan. in Umetn. Cl. IV, Hist, natur. med.,
5: 109—126.
Hadzi J. 1954. Die morphologische Bedeutung der Pneumatophore bei Siphonophoren.—
Razpr. Slov. Akad. Znan in Umetn. Cl. IV, Hist, natur. med., 5: 122—136.
Hadzi J. 1955 Odnos med Scitopolipom in Scifomedusa. —Razpr. (Dissert.) Slov. Akad.
Znan in Umetn. Cl. IV, Hist. Natur. Ljubljana, III: 69—124.
Hadzi J. 1958. Zur Disskussion uber die Abstammung der Eumetazoen.— Zool. Anz,
Suppl., Bd. 21: 169—179.
Hadzi J. 1959. Zum Problem der Knidarien Abstammung.— Bull. Scient. Zagreb, 4, 4:
. 105.
Hand C. 1959. On the origin and phylogeny of the Coelenterates.— System. Zool., 8,
4: 191—202.
Hanitzsch P. 192b. Uber die Eigenart und Entstehung der Vermehrungsweise durch
aborale und orale Proliferation bei Narcopolypen und Scyphopolypen.— Zool. Jahrb.,
Abt. 2, 42, 3: 363—411.
24*
371
Haynes J. and Burnetl A. L. 1963. Dedifferentiation and redifferentiation of cells
in Hydra viridis.— Science, 142, 3598: 1481 —1483.
Hess А., С о h e n A. I. and Robson E. A. 1957. Observations on the structure of Hyd-
ra as seen with the electron and light microscopes.— Quart. J. Microscop. Sci., 98:
315—326.
H о r r i d g e G. A., Chapman D. M. and MacKay B. 1962. Naked axons and sym-
metrical synapses in an elementary nervous system.— Nature, 193, 4818: 899—900.
Horridge G. A. and MacKay B. 1962. Naked axons and symmetrical synapses in
Coelenterates.— Quart. J. Microscop. Sci., 103: 531—541.
Iwantzow N. A. 1928. Beitrage zur Kenntnis der Histologie der Siphonophoren.—
Bull. Soc. Nat. Moscou, 37, 1—2: 1—36.
Jacobs W. 1962. Siphonophore structures help colonies maintain specific depth.— Na-
ture History, 71, 7: 23—27.
К a n a j e w J. 1930. Zur Frage der Bedeutung der interstiziellen Zellen bei Hydra.— Arch.
Entwicklungsmech., 122: 736—759.
Komai T. -1922. Studies on two aberrant Ctenophores, Coeloplana and Gastrodes.. Kyoto,
p. 1—102.
Komai T. 1935. On Stephanoscyphus and НсишИюё.— Mem. Coll. sci. Kyoto univ. Ser-.
В. X, 5: 289—339.
Kramp P. L. i!961. Synopsis of the medusae of the world.—J. Marine Biol. Assoc. U. K.,
40: 1—469.
LaubenfelsM. W. de 1955. Are Coelenterates degenerate? — System. Zool., 4, 1: 43—45.
Lipin A. N. 1926. Geschlechtliche Form. Phylogenie und systematische Stellung von
Polypodium hydrtforme Uss.— Zool. Jahrb., Abt. 2, 47, 4: 541—635.
Mackie G. O. 1959. The evolution of the Chondrophora (Siphonophora — Disconan-
thae). New evidence from behavioural studies.— Trans. Roy. Soc. Canada, Ser. 3, 53:
7—20.
Mackie G. O. 1960. The structure of the nervous system in Velella.— Quart. J. Micro-
scop. Sci., 101: 119—131.
Mergner H. 1957. Die Ei-und Embryonalentwicklung von Eudendrium racemosum Ca-
volini.— Zool. Jahrb., Abt. 2, 76: 63—164.
Mergner H. 1964. Unsere Kenntnisse uber Bau und Funktion der Nesselzellen bei Hyd-
roiden.— Natur und Museum, 94. 1: 15—32.
Metschnikoff E. 1874. Studien fiber die Entwicklung der Medusen und Siphonopho-
ren.— Z. wiss. Zool., 24: 15—83,
Metschnikoff E. 1882. Vergleichende embryologische Studien.— Z. wiss. Zool., 36;
433—444.
Metschnikoff E. 1885. Vergleichende embryologische Studien.— Z. wiss. Zool., 42:
648— 673.
Metschnikoff E. 1886. Embryologische Studien an Medusen. Wien, S. 1—159.
M о о r e R. 19512. Coelenterates. In: R. С. M о о r e, C. G. L a 1 i с к e r and A. G. F i s c h e r.
Invertebrates fossils. N. Y., p. 99—105.
N a g a о Z. 1963. The early development of the Anthomedusa, Polyorchis karajutoensis
Kishinouye.— Annat. zool. japon., 36, 4: 187—193.
Nyholm K. G. 1942—1944. Zur Entwicklung und Entwicklungsbiologie der Ceriantha-
rien und Aktinien.— Zool. Bidrag. Uppsala, 22: 87^-248.
Nyholm К G. 1949. On the development and dispersal of Athenaria Actinia with spe-
cial reference to Halcampa duodecimcirrata M. Sars.— Zool. Bidrag Uppsala, 27:
466—505.
Nyholm K- G. 1958—1960. On the development of the primitive Actinian Protantheu
simplex Calgren.— Zool. Bidrag. Uppsala, 33: 69—77.
Pant in C. F. A. 1960. Diploblastic animals.— Proc. Linnean Soc. London, 171, 1:
1—13.
Pax F. ,1918. Die Antipatharien.— Zool. Jahrb. Syst., 41: 419—478.
Percival E. 1923. On the strobilization of Aurelia.— Quart. J. Microscop. Sci., 67: 85—•
100.
P i c a r d J. 1955. Les hydroides Pteronematidae, origine des «Siphonophores» Chondropho-
ridae.— Bull. Inst, oceanogr. (Fondation Albert I, Prince Monaco), 1059: 1—8.
Rees W. J. 1957. Evolutionary trends in the classification of capitate hydroids and me-
dusae.— Bull. Brit. Mus. (Natur. Hist.) London, 4, 9: 455—534.
Reisinger E. 1957. Zur Entwicklungsgeschichte und Entwicklungsmechanik von Cras-
pedacusta (Hydrozoa, Limnotrachylina).— Z. Morphol. und Okol. Tiere, 45: 656—698.
Robson E. A. 1957. The structure and hydromechanics of the musculo-epithelium in
Metridium.— Quart. J. Microscop. Sci., 98: 265—278.
Robson E. A. 1963. The nerve-net of a swimming anemone, Stomphia cocinea.— Quart.
J. Microscop. Sci., 104, 4: 535—549.
Roch J. and Robin Y. 1954. Sur les phosphagenes des eponges.— Compt. rend. Soc.
biol., 148.
Roch J. and Robin Y. 1957. Sur la presence de creatine chez les invertebrates.— Bio-
chim. et biophys. acta. 24.
372
Russel F. S. and Rees W. J; 1960. The viviparus scyphomedusa Stygiotnedusa fabu-
losa Russel.— J. Marine Biol. Assoc. U. K., 39, 2: 303—317.
Schaeppi Th. 1898. Untersuchungen fiber das Nervensystem der Siphonophoren.— Je-
na. Z. Naturwiss., 32: 483—550.
Schewiakoff W. 1889. Beitrage zur Kenntnis des Acalephenauges.— Morphol. Jahrb.,
'15.
Schindewolf О. H. 1939. Stammesgeschichtliche Ergebnisse an Korallen.— Palaon-
tol. Z„ 21, 4: 321—340.
Schindewolf О. H. 1942. Zur Kenntnis der Polycoelien und Plerophyllen.— AbhandL
Reichsamst. Bodenforsch., Berlin, N. F., 204: 1—324.
Southward A. J. 1955. Observations on the ciliary current of the jelly-fish Aurelia auri-
ta L.—J. Marine Biol. Assoc. U. K., 34: 201—216.
Steinberg S. N. 1963. The regeneration of whole polyps from ectodermal fragments oi
Scyphystoma larvae of Aurelia aurita.— Biol. Bull., 124, 3: 337—343.
Su.giura Y. 1963. On the life-history of Rhizostome Medusae I. Mastigias papua L.
Agassiz.— Annot. zool. japon., 36, 4: 194—202.
S wed mar к В. et Teissier G. 1959. Halammohydra et Otohydra, Hydrozoaires de la
microphaune des sables et d’ordre des Actinulides.— Proc. XVth Internal. Congr.
Zool.: 320—332.
Thiel H. 1962. Clavopsella quadranularia nov. spec. (Clavopsellidae nov. fam.), ein
neuer Hydropolip aus der Ostsee und seine phylogenetische Bedeutung.— Z. Morphol.
und Okol. Tiere, 51: 227—260.
Totton A. K. 1954a. Siphonophora on the Indian Ocean.— Discovery Rept., 27.
To tt о n A. K. 19546. Egg-laying in Ctenophora.— Nature, 174.
Totton A. K. 1960. Studies on Physalia physalis (L.). Part I. Natural history and
morphology.— Discovery Rept., 30.
Uchida T. 1963. The systematic position of the Hydrozoa.— Japan. J. Zool., 14, 1:
1—19.
Uchida T. and N a g a о Z. 1963. The metamorphosis of the Scyphomedusa Aurelia lim-
bata (Brandt).— Annot. zool. japon., 36, 2: 83—91.
Ulrich W. 1949. Uber die systematische Stellung einer neuer Tierklasse (Pogonopho-
ra, Johansson), den Begriff der Archicoelomaten und die Einteilung der Bilate-
rien.— Sitzungsber. Dtsch. Akad. Wiss., 2: 1—25.
Ulrich W. 1951. Vorschlage zu .einer Revision der Grosseinteilung des Tierreiches.—
Verhandl' Dtsch. Zool. Ges., Zool. Anz. Suppl. Bd., 15: 244—271.
Werner B. 1963. Experimented Beobachtungen uber die Wirksamkeit von Aufienfakto-
' ren in der Entwicklung der Hydrozoen und Erorterung ihrer Bedeutung fiir die Evolu-
tion.— Veroffentl. Inst. Meeresforsch. Bremenhaven Sonderband. Bremen: 153—177.
Westblad E. 1935. Neue Beobachtungen fiber Protohydra.— Zool. Anz., Ill: 152—158.
Westblad E. 1947. Notes on hydroids.— Arkiv zool., A, 39, 5: 1—23.
Z w i 11 i n g E. 1963. Formation of endoderm from ectoderm in Cordylophora.— Biol.
Bull., 124, 3: 368—378.
s ГРЕБНЕВИКИ (CTENOPHORA)
Бляхер Л Я. 1963. Константин Николаевич Давыдов. М., Изд-во АН СССР:
1—244.
Камшилов М. М. 1955. Питание гребневика Вегоё cucumis Fab.— Докл. АН СССР,
102, 2 : 399—402.
Камшилов М. М. 1960. Питание гребневика Вегоё cucumis Fab.— Докл. АН СССР,
103,5:1138—1140. .... ,
Ковалевский А. О. 1873. Наблюдения над развитием Coelenterata.—Изв. Об-ва
любит, естествозн., анатропол. и этногр., 10, 2: 1—38. . ..
Мечников И. И. 1955 (1885). Сравнительно-эмбриологические исследования. IV. Об
образовании гаструлы и мезодермы у гребневиков, стр. 152—158. Статьи по срав-
нительной эмбриологии. Академическое издание сочинений, III. М., Гос. изд. медиц.
лит., стр. 42.
Роек ин Г. И. 1917. Строение некоторых сократимых элементов клетки.— Арх. анат.
гистол. и эмбирол., 2, 1: 39—102.
Роек ин Г. И. 4923 Мускульная клетка ктенофор.— Русск. зоол. ж., 3, 3—4: 436—
449.
Третьяков Д. К. 1936. Неотенический характер ктенофор.— Зоол. ж., 15: 252—258.
Chun С. 1880. Die Ctenophoren des Golfes von Neapel und der angrenzenden Meeres ab-
schnitte. In «Fauna und Flora des Golfes von Neapel». I. Monographie. Leipzig.
XVIII + 313.
Dawydoff C. 1933. Quelques observations sur la morphologie externe et la biologie des
Ctenoplana.— Arch. zool. exptl et gen., 75: 103—128.
Dawydoff C. 1936. Les Ctenoplanides des eaux de 1’Indochine frangaise.—Bull. biol.
France et Belgique., 70: 456—486.
Dawydoff C. 1938. Les Coeloplanides indochinoises.— Arch. zool. exptl et gen., 80:
125—162.
373
Dawydoff С. 1941a. Sur 1’embryologie de Ctenoplana.— C. r. Acad. sci. Paris, 212:
653—655.
Dawydoff C. 1.9416. Deux formes larvaires de Ctenoplanidae du plancton des Cotes
d’Annam.— C. r. Acad. sci. Paris, 212, 9: 607—510.
Dawydoff C. 1946. Contribution a la connaissance des Ctenophores pelagiques des
eaux de 1’Indochine.— Bull. biol. France et Belgique, 80: 113—170.
Farfaglio G. 1963. Experiments on the formation of combs in the Ctenophores.— Expe-
rience, 19, 5: 303—304.
Fisch el A. 1887—1889. Experimented Untersuchungen an Ctenophoren.—Arch. Ent-
wicklungsmech., 6—7.
Hadzi J. 1929. Einige allgemein wichtige Resultate meiner Untersuchungen iiber Coe-
lenterata (s. ampl.).— X Congr. internal. Zool. Budapest: 927—931.
Hadzi J. 4957. Die morphologische Bedeutung der Wimperrosetten der Ktenophoren.—
J. Fac. Sci. Hokkaido Univ., Ser. Ill, Zool., 13, 1—4 : 32—36.
Hadzi J. 1958. Zur Diskussion fiber die Abstammung der Eumetazoen.— Zool. Anz.,
Suppl., Bd. 21: 169—179.
Han-strom B. 1928. Vergleichende Anatomie des Nervensystems der wirbellosen Tiere
unter Beriicksichtigung seiner Funktion. Berlin, S. XI+ 628.
Heider K. 1927. Vom Nervensystem der Ctenophoren.— Z. Morphol. und Okol. Tiere, 9:
638—678.
Jagersten G. 1955. On the early phylogeny of the Metazoa. The Bilaterogastraea theo-
ry.— Zool. Bidrag Uppsala, 30: 321—354.
Kerkut G. L. 1960. Implications of evolution.— London. Pergamon Press, p. 1—174.
Komai T. 1922. Studies on two aberrant Ctenophores, Coeloplana and Fastrodes. Kyoto,
p. 1—102.
Korn H. 1959. Zum Nervensystem der Ctenophore Pleurobrachia pileus O. Muller.—
Zool. Anz., 163, 11—12 : 351—359.
Korotneff A. 1886. Ctenoplana kowalevskii.— Z. wiss. Zool., 43: 242—251.
Kowalevsky A. 1880. Uber Coeloplana metschnikowii.— Zool. Anz., 3: 140.
Kowalevsky A. 1886. Entwicklungsgeschichte der Rippenqualen.— Mem. Acad., St.
Petersb., Ser. 7, 10, 4: 1—50.
L a n g A. 1884. Die Polycladen des Golfes von Neapel und der angrenzenden Meeresabsch-
nitte.— Fauna und Flora des Golfes von Neapel. 11. Monogr. Leipzig.
Metschnikoff J. 1885. Vergleichende-embryologische Studien. I. Uber die Gastrulation
und Mesodermbildung der Ctenophoren.— Z. wiss. Zool., 42 : 648—656.
Moore R. C. 1936. Reciprocal inhibition and its reversal strychnine in the modified
Ctenophore, Coeloplana bockii.— Physiol. Zool., 9; 240—245.
Mortensen Th. 1927. Two new Ctenophores.— Vid. medd. Dansk naturhistor. foren.
Kjabenhavn, 83: 277—288.
Pavans de Ceccatty M., H er n and er M. L. et Thiney Y. 1962. Systeme ner-
veux et musculatur chez Beroe ferskali.— C. r. Acad. sci. Paris, 254, 18: 3241—3243.
Prenant M. 1922. Recherches sur le parenchyme des Plathelmintes. Essai d’histologie
comparee.— Arch, morphol. gen. et exptl. Paris, p. 1—1,74.
Remane A. 1956. Zur Biologie des Jugendstadiums der Ctenophore Pleurobrachia pileus
O. Muller.— Kieler Meeresforsch., 12: 72—75.
Spek J. 1926. Uber gesetzmassige Substanzverteilungen bei der Furchung des Ctenopho-
reneies und ihre Beziehungen zu den Determinationsproblemen.— Arch. Entwicklungs-
mech., 107: 54—73.
Tretjakoff O. 1930. Die Gallerte der Rippenquallen.— Anat. Anz., 70: 293—308.
U s P. 1932. Embryona'ni razvoj Ktenofor (Development of Ctenophora).— Razpr. prirod.
muz. Drystva Ljubljana, 1: 155—178.
Yatsu N. 1912a. Observations and experiments on the Ctenophora.— Annot. zool.
japan. Tokyo, 8: 5—13.
Yatsu N. 19126. Experiments on germinal localization of the egg of Bero'e ovata.—J.
Coll. Sci. Japan, Tokyo, 32, 3: 1—21.
Ziegler H. 1898. Experimented Studien fiber Zellteilung. 3. Die Furchungszellen von
Beroe ovata.— Arch. Entwicklungsmech., 7: 34—64.
ПЛОСКИЕ ЧЕРВИ (PLATHELMINTHES)
Беклемишев В. H. 1915. О паразитных турбелляриях Мурманского моря. I. Acoela.—
Тр. Петроград, об-ва естествоисп., 43, 4: 103—172.
Беклемишев В. Н. 1916. О паразитных турбелляриях Мурманского моря. II. Rhab-
docoela.— Тр. Петроград, об-ва естествоисп., 45, 4: 3—78.
Беклемишев В. Н. 1925. Морфологическая проблема животных структур.— Изв.
Биол. н.-и. ин-та Пермск. гос. ун-та, 3, I: 1—74.
Беклемишев В. Н. 1937. Класс ресничных червей (Turbellaria). В кн. «Руководство
по зоологии», т. I. Беспозвоночные. Под ред. Л. А. Зенкевича. М., стр. 386—457.
Богомолов С. И. 1949. К вопросу о типе дробления Rhabdocoela.— Уч. Зап. ЛГУ
сер. биол., 20: 128—142.
374
Богомолов С. И. 1960. История развития Convoluta в связи с морфологией реснич-
ных червей.— Тр. об-ва естествоисп., 63: 155—208. (Уч. зап. Казанск. ун-та, 120,
кн. 6).
Богомолов С. И. 1964. Очерки по сравнительной эмбриологии ресничных червей.
Автореф. канд. дисс. Казань, стр. 1—19.
Быховский Б. Е. 1937. Онтогенез и филогенетические взаимоотношения плоских
паразитических червей.— Изв. АН СССР, Отд. матем. и естеств. наук, сер. биол.,
4: 1353—1383.
Быховский Б. Е. 1957. Моногенетические сосальщики, их система и филогения.
М.— Л., Изд-во АН СССР, стр. 1—509.
Вейдман В. Р. 1939. Развитие и редукция женских половых органов свиного соли-
тера.— Докл. АН СССР, 22, 9: 657—660.
ВейцманВ Р. 1940. Различия в способе полового размножения у ленточных гли-
стов—Докл. АН СССР, 24, 2: 199—201.
Вейцман В. Р. 1953а. Развитие и редукция женской половой системы кошачьего цеп-
ня Taenia crassimllis.— Тр. Ин-та морфол. животных АН СССР, 8: 205—215.
ВейцманВ. Р. 19536. Развитие и редукция женской половой системы Taenia solium.—-
Тр. Ин-та морфол. животных АН СССР, 8: 173—204.
Г и л я р о в М. С. 1964. Определитель обитающих в почве личинок насекомых. М., Изд-
во «Наука», стр. 1—919.
.Иванов А. В. 1952а. Бескишечные турбеллярий (Acoela) южного побережья Сахали-
на.— Тр. Зоол. ин-та АН СССР, 12: 40—132.
И в а н о в А. В. 19526. Строение Udonella caligorum Johnston, 1935 и положение Udone-
idae в системе плоских червей.— Паразитол. сб. Зоол, ин-та АН СССР, 14: ГТ2—
163.
Коштоянц X. С. 1957. Особенности функции нервной системы низших червей. В кн.
«Основы сравнительной физиологии. II. Сравнительная физиология нервной си-
стемы». М., Изд-во АН СССР, стр. 226—230.
Логачев Е. Д. 1957а. Материалы к вопросу о взаимоотношениях тканей внутренней
среды и пограничных тканей у плоских червей в онто- и филогенезе.— Всес. науч,
об-во анатом., гистол. и эмбриол. (Кемерово), стр. 1—22.
Логачев Е. Д. 19576. Микроморфология и эволюция тканей плоских червей (Plathel-
minthes). Автореф. док. дисс. Всес. ин-т гельминтол. им. акад. К. И. Скрябина,
стр. 1—31.
Логачев Е. Д. 1961. Материала к вопросу о способах возникновения клеток у лен-
точных гельминтов. Докл. к годичной научной конференции 1961 г.— Всес. научн.
об-во анатом., гистол. и эмбриол. (Кемерово), стр. 95—109.
Мамкаев Ю. В. 1961. Новый представитель гнатостомулид — Gnathostomula тигта-
nica sp. п.— Докл. АН СССР, 141, 6: 1501—1504.
Павловский Е. Н. 1937. Класс сосальщиков (Trematoda), стр. 461—501; класс лен-
точных червей (Cestoda), стр. 502—555. В кн. «Руководство по зоологии», т. I. Бес-
позвоночные. Под ред. Л. А. Зенкевича. М.— Л.
Переяславцева С. М. 1892. Monographic des Turbellaries de la mer Noire.— Зап.
Новорос. об-ва естествоисп., 17, 3.
Пигулевский С. В. 1958. К вопросу о филогении плоских червей. В кн. «Работы
по гельминтологии. К 80-летию акад. К. И. Скрябина», АН СССР. М., стр. 263—269.
Синицын Д. Ф. 1905. Материалы по естественной истории трематод. Дистомы рыб и
лягушек окрестностей Варшавы. Варшава, стр. 1—210.
Синицын Д. Ф. 1911. Партеногенетическое поколение трематод и его потомство в
черноморских моллюсках.— Зап. СПб. Академии наук, 8, 30, 5: 1—127.
Скрябин К. И. 1947—1962. Трематоды животных и человека. Основы трематодологии,
т. 1—20 [с участием Е. А. Башкировой, Л. X. Гушанской, С. В. Пигулевского,
Ф. Н. Морозова, А. А. Соболева, Е. Г. Евранова, М. М. Белопольской, А. М. Пет-
рова, Т. С. Скрябиловича, Д. Н. Антипина, В. Е. Сударикова, В. П. Коваль].
Федотов Д. М. 1915. Protomyzostomum polynephris и его отношение к Gorgonocep-
halus euchnemis М. et Тг. Дисс. Пг: 1—240.
Фридман Г. М., 1933. Анатомическое строение Baicalarctia gulo Fr. и положение ее в
системе Turbellaria.— Тр. Байкал, лимнол. ст., 5: 179—256.
Шаров И. Н. 1937. К вопросу о гистогенезе при регенерации у планарий., Сб. «Акад.
Н. В. Насонову к 80-летию со дня рождения и 60-летию научной деятельности».
Изд-во АН СССР, стр. 249—293.
Шевченко Н. 1937. Взаимодействие участков различной физиологической активно-
сти у планарий (О роли скачка в градиенте-системе) — Биол. ж., 6, 3: 581—588.
Шмидт Г. А. 1953. Эмбриология животных, т. II. Частная эмбриология. Развитие
плоских червей, стр. 45—48. Развитие низших червей, связанных филогенетическими
отношениями с прямокишечными турбелляриями, стр. 5'9'—72. М., изд-во «Сов.
Наука».
Ш у л ь ц Е. А. 1904. Наблюдения над регенерацией у червей.— Тр. СПб. об-ва естество-
исп., 34, 4: 1—'137.
Шульц Е. А. 1908. Наблюдения над обратимыми процессами развития.— Тр. СПб.
об-ва естествоисп., 38, 4: 1 —134.
375
A х Р. 1961. Verwandschaftsbeziehungen und Phylogenie der Turbellarien.— Ergebn. Biol.,
24: 1—68.
Beklemischew W. 1915. Uber einige acole Turbellarien des Kaspischen Meeres.—
Zooi. Anz., 55: 1—7.
Bennetts H. W. 1928. Life history of Cotylophorum cotylophorum. Illinois Biol. Mo-
nogr., 14, 4: 1—119.
В о a s J. E. V. 1917. Zur Auffassung der Verwaridschaftverhaltnisse der Tiere. I. Kopen-
hagen.
Boettger C. R. 1957. Stammesgeschichte und Verbreitung der Turbellariengruppe Tem-
nocephala.— Abhandl. Braunschweig, wiss. Ges., 9: 26—35.
Bresslau E. 1909. Die Entwicklung der Acolen.— Verhandl. Dtsch. zool. Ges., 19:
314—323.
Bresslau E. 1928. Allgemeine Einleitung zur Naturgeschichte der Vermes Amera.
In: W. К iiken t ha 1. Handbuch der Zool. Bd. 2, 1. Berlin und Leipzig, S. 19—33.
Bresslau E. 1933. Turbellaria. Irt: W. К ii к e n t h a 1. Handbuch der Zoologie, S. 52—320.
Child С. M. 1906. The development of germ cells from differentiated somatic cells in
Moniezia.— Anat. Anz., 29: 592—597.
Clarke A. S. 1954. Studies on the life cycle of the pseudephallidean cestode Schistoce-
phalus solidus.— Proc. Zool. Soc. London, 124: 247—302.
E i s e n b r a n d t L. L. 1936. Precipitin reactions to helminth extracts.— Proc. Soc. Exptl
Biol, and Med., 35: 322—325.
Eisen brand! L. L. 1938. On the serological relationship of some helmints.— Amer. J.
Hyg„ 27: 117.
Enigk K. und Sticinsky E. 1957. Uber die Bohrdriisen der Oncosphare von Davai-
nes.— Z. Parasitenkunde, 18, 1: 48—54.
Euzet L. 1956. Suggestions pour une nouvelle classification des cestodes Tetraphylli-
des.— Proc. 14 Internal. Congr. Zool., 1953: 347—349.
Fuhrmann O. 1928—1933. Trematode. In: W. Kiikenthal. Handbuch der Zoologie,
Bd. 2, I. Berlin — Leipzig, S. 1—140.
Fuhrmann O. 1930—1931. Cestoidea. In: W. Kiikenthal. Handbuch der Zoologie, '
Bd. 2, I. Berlin — Leipzig, S. 141—416.
Hadzi J. 1944. Turbelarijska teorija knidarijev.— Slov. Akad. Zhan in Umetn. i Ljublani
Dela, 3: 238. 1
Hadzi 1958. Zur Diskussion iiber die Abstammung der Eumetazoa.— Verhandl. Dtsch.
Zool. Ges., Zool. Anz., Suppl. Bd., 21: 169—179.
I s h i i Y. 1934. Development of Fasciolopsis buski.— J. Med. Assoc. Formosa, 33: 349—412. ।
Janicki C. 1921. Grundlinien einer «Cercomer» Theorie. Zur Morphologie der Tremato-
den und Cestoden.— Festschrift Zschokke, 30. 1: 22.
Karl in g T. G. 1940. Zur Morphologie und Systematik der Alloecoela Cumulata und ‘
Rhabdocoela Lecithophora (Turbellaria).— Acta zool. fennica, 26: 1—260.
Karling T. G. 1947. Studien fiber Kalyptorhynchien (Turbellaria). I. Die Familien Pla-
corhynchidae und Gnathorhynchidae.— Acta zool. fennica, 50: 1—64.
Karling T. G. 1949. II. Die Familien Karkinorhynchidae und Drasiorhynchidae.— Acta
zool. fennica, 58: 3—42.
К a r 1 i n g T. G. 1950. III. Die Familie Schizorhynchidae.— Acta zool. fennica, 59: 3—33.
К a r 1 i n g T. G. 1952. IV. Einige Eukalyptorhynchia.— Acta zool fennica, 69: 3—49.
К a r 1 i n g T. G. 1955. V. Der Verwandtschaftskreis von Gyratrix Ernenberg.— Acta zool.
fennica, 88: 3—39.
Karling T. G. 1956. Alexlutheria acrosiphoniae n. gen,, n. sp., ein bemerkenswerter
mariner Vertreter der Familie Dalyellidae (Turbellaria).— Arkiv. zool., Ser. 2, 10:
3311—345.
Kiim me 1 G. 1958. Das Terminalorgan der Protonephridien. Feinstruktur und Deutung
der Funktion.— Z. Naturforsch., 13 В, 1: 677—679. j
К ii m m e 1 G. 1959. Feinstrukture der Wimperflamme in den Protonephridien.— Proto- ,
plasma, 1, 3: 371—376.
Lang A. 1884. Die Polycladen des Golfes von Neapol und der angrenzenden Meeresab- I
schnitte. (Fauna und Flora des Golfes von Neapel und der angrenzendem Meers).
11. Leipzig, S. 688. I
La Rue G. 1957. The Classification of digenetic Trematoda: a review and a new sys-
tem.— Exptl Parasitol.,.6 : 306—349.
Luther A. 1955. Die Dalyelliden.— Acta zool. fennica, 87: XI + 337. ’ .
Luther A. 1960. The Turbellarien Ostfennoskandiens I. Acoela. Catenulida Macrosto-
mida, Lecithoepitheliata, Erolicithophora and Proseriata.— Fauna fennica, 7: 1,—155.
Mattes O. 1936, Der Entwicklungsgang des Lanzettegels Dicrocoelium lanceatum.—
Z. Parasitenkunde, 8: 371—430.
Meixner J. 1926. Beitrag zur Morphologie und zum System der Turbellaria-Rhab-
docoela.— Z. Morphol. und Okol. Tiere, 5: 577—624.
Nassonov N. 1932. Zur Morphologie der Turbellaria Rhabdocoelida des Japanischen
Meeres.— Тр. Лабор. экп. зоол. и морф, животных АН СССР, 2: 1—115.
О g г е n R. Е. 1956. Development and morphology of the oncosphere of Mesocestoides
corti, a tapeworm of mammals.— J. Parasitol., 43, 3: 414—426.
376
Ogren R, E. .1957. Morphology and development of oncospheres of the Cestode Oocho-
ristica symmetrica Baylis, 1927.— J. Parasitol., 43, 5: 505—510.
Perejaslavceva С. M. 1885. Sur la developpement des Turbellaries.— Zool. Anz.,
8: 269—271.
Prenant M. 1922. Recherches sur le parenchyme des Plathelmintes. Essai d’histologie
comparee.— Arch, morphol. gen. et exptl. Paris, 5; 1—474.
Rees G. 1940. Studien on the germ cycle of the digenetic trematode Parorchis acanthus
Nicoll.— J. Parasitol., 32: 372—391.
Reisinger E. 1960. Was ist Xenoturbella?— Z. wiss. Zool. 164: 188—198.
Reisinger E. 1961. Allgemeine Morphologie der Metazoen. Morphologie der Coelente-
rata, acoelomaten und pseudocoelomaten Wiirmern.— Fortschr. Zool., 13: 1—82.
Repj a chow W. 1885. Nachtrag zur vorstender Mitteilungen.— Zool. Anz., 8: 272—273.
Riedl A. 1959. Turbellarien aus submarinen Hohlen. 1. Archeophora. 2. Prolecithopho-
ra.— Publ. Staz. Zool. Napoli, 30 Suppl.: 178—208, 209—304.
Salensky W. 1907. Morphogenetische Studien an Wiirmern.— Mem. Acad. Sci. St. Pe-
tersb., Ser. VIII, 19, II: 104—451.
Seiler п-A spang Fr. 1956. Friihentwichlung einer marinen Tricklade (Procerodes lo-
bata O. Schmidt).— Arch, entwicklungsmech., 148: 589—595.
S e i 1 e r n-.A s p a n g Fr. 1957. Die Entwicklung von Macrostomum appendiculatum (Fab-
ricius).— Zool. Jahrb. Anat., 76, 3: 311—330.
Silverman P. H. 1*954. Studies on the biology of some tapeworms of genus Taenia.—
Ann. Trop. Med. and Parasitol., 48, 3: 356—366.
Spatlich W. 1926. Die Furchung und Embryonalhiillenbildung des Eies von Dior-
chis.— Zool. Jahrb. Anat., 47: 101—;112.
Steinbock O. 1958. Zur Phylogenie der Gastrotrichen.— Verhandl. Dtsch. zool. Ges.,
Zool. Anz., Suppl. Bd., 21: 128—169.
Threadgold L. T. 1963. The tegument and associated structures of Fasciola hepati-
ca.— Quart. J. Microscop. Sci., 104, 4: 505—512.
Westblad E. 1950. Xenoturbella bocki n. g., n. sp. a peculiar primitive Turbellarian
type.— Arkiv zool., Ser., 2, 1: 11—29.
W i 1 h e 1 m i R. W. 1940. Serological reactions and species specifity in some helmints.—
Biol. Bull., 79, 1: 64—90.
ПЕРВИЧНОПОЛОСТНЫЕ ЧЕРВИ (ASKELMINTHES)
Богоявленский Ю. К. 1959. Тонкое строение гиподермы некоторых представите-
лей аскаридат.— Acta Parasitol. lithuanica, 2: 43—50.
Богоявленский Ю. К. 1961. Обзор работ по гистологическому строению муску-
латуры некоторых аскаридат.— Тр. Гельминтол. лабор. АН СССР, 11: 38—49.
Богоявленский Ю. К. и Балагана Г. М. 1962. Микроскопическое исследование
кутикулы и гиподермы нематоды Mecristocirrus digitatus (Linstion, 1906) Raillet et
Henry, 1912.— Тр. Гельминтол. лабор. АН СССР, 12: 19—21.
Бластов Б. В. 1953. Европейские и североамериканские коловратки из семейства
нотомматид — сожители Cladocera и их видовая специфичность.— Зоол. ж., 32, 6:
1100—1112.
Бластов Б. В. 1954. Морфология и систематика низших коловраток из отряда
Monogononta. Proales daphnicola — комменсалист дафний и близкие формы.—
Зоол. ж., 33, 1: 50—64.
Бластов Б. В. 1955. К морфологии самцов коловраток отряды Monogononta.— Зоол.
ж., 34, 1: 80—84. ,
Бластов Б. В. 1959. Функциональные модификации формы тела у моногононтных
коловраток (Proalinae) и филогенетические преобразования формы тела в этой
группе.— Тр. Ин-та морфол. животных АН СССР, 27: 101—116.
Ганин М. С. 1877. Наблюдения над эмбриональным развитием Peloderateres.— Тр.
V съезда русск. естествоисп., 11: 35.
Головин Е. П. 1901. Наблюдения над нематодами, т. I. Фагоцитарные органы. Ка-
зань, стр. 1—149.
Головин Е. П. 1902. Наблюдения над нематодами, т. II. Экскреторные органы. Ка-
зань, стр. 1—120.
Губанов Н. М. 1951. Гигантская нематода плаценты китообразных — Placentonema
gigantissima nov. gen., nov. sp.— Докл. АН СССР, 77, 6: 1123—1125.
Дейнека Д. И. 1912. Нервная система аскариды (Ascaris megalocephala Clag.).
Гистологическое исследование.— Тр. СПб. об-ва естествоисп., 12: 101'—359.
Зенкевич Л. А., Филипьев И. Н. 1937. Класс круглых червей. В кн. «Руковод-
ство по зоологии», т. I. Беспозвоночные, стр. 557—627.
Ивашкин В. М. 1962. Проблема парабронематозов животных и перестройка систе-
мы спирурат на основе анализа их онтогенеза. Автореф. док. дисс. М.
377
Мозговой А. А. 1953а. К изучению филогенетических связей и путей эволюции аска-
ридат. Работы по гельминтологии. Сб. «К 75-летию акад. К. И. Скрябина». М.,
Изд-во АН СССР, стр. 422—431.
Мозговой А. А. 19536. Аскаридаты животных и человека и вызываемые ими забо-
левания. В кн. «Основы нематодологии», т. I. Под ред. К. И. Скрябина. М.,
Изд-во АН СССР, стр. 1—351.
Мозговой А. А. 1953в. Аскаридаты животных и человека и вызываемые ими забо-
левания. В кн. «Основы нематодологии», т. II. Под ред. К. И. Скрябина. М.,
Изд-во АН СССР, стр. 1—616.
Муравейский С. Д. 1937. Класс коловраток. В кн. «Руководство по зоологии»,
т. I. Беспозвоночные, стр. 706—749.
Мюге С. Г. 1958а. Сравнительный анализ физиологических адаптаций фитонематод.
В кн. «Сб. работ молодых фитогельминтологов». М., стр. 59—79.
Мюге С. Г. 19586. Эволюция физиологических адаптаций фитонематод к питанию.
Автореф. канд. дисс. М., стр. 13.
Парамонов А. А. 1937. Положение Nematodes в системе.— Зап. Болш. биол. ст.,
10; 97—117.
Парамонов А. А. 1952. Опыт экологической классификации фитонематод.— Тр.
Гельминтол. лабор. АН СССР, 6: 338—368.
Парамонов А. А. 1954а. Специфичность фитогельминтов и ее значение в сельско-
хозяйственной практике.— Зоол. ж., 33, 5: 1002—4024.
Параманов А. А. 19546. Строение и функция фазмид.— Тр. Гельминтол. лабор.
АН СССР, 7: 19—49.
Парамонов А. А. 1957. О принципах таксономической дифференцировки в немато-
дологии.— Зоол. ж., 36, 5: 641—653.
Переяславцева С. 1885. О развитии коловраток.— Зап. Новорос. об-ва естестве-
нен., 9, 4: 1—19.
Попова Т. Н. 1955. Стронгилоидеи животных и человека. Стронгилиды. В кн. «Осно-
вы нематодологии», т. V. Под ред. К. И. Скрябина. М., Изд-во АН СССР, стр.
1—223.
Попова Т. Н. 1958. Стронгилоидеи животных и человека. Трихонематиды. В кн.
«Основы нематодологии», т. VII. Под ред. К. И. Скрябина. М., Изд-во АН СССР,
стр. 1—424.
Попова Т. Н. 1960. Стронгилоидеи животных и человека. Клоациниды, стефануриды,
диафаноцефалидьт В кн. «Основы нематодологии», т. IX. Под ред. К. И. Скряби-
на. М., Изд-во АН СССР, стр. 1—244.
Рыжиков К. М. 1949. Сингамиды домашних и диких животных. В кн. «Основы не-
матодологии», т. I. Под ред. К. И. Скрябина. М.—Л., Изд-во АН СССР, стр.
1—164.
Скрябин К. И. 1946. Строительство советской гельминтологии. М.— Л., Изд-во АН
СССР, стр. 1—211.
Скрябин К. И., Шихобалова Н. П., Соболев А. А. 1949. Спирураты и филя-
риаты. В кн. «Определитель паразитических нематод», т. I. Изд-во АН СССР,
стр. 1—519.
Скрябин К. И., Шихобалова Н. П., Мозговой А. А. 1951. Оксиураты и
аскаридаты. В кн. «Определитель паразитических нематод», т. II. Изд-во АН
СССР, стр. 1—631.
Скрябин К. И., Шихобалова Н. П., Шульц Р. С., Попова Т. И., Боев С.,
Делямуре С. Л. 1952. Стронгиляты. В кн. «Определитель паразитических не-
матод», т. III. Изд-во АН СССР, стр. 1—890.
Скрябин К. И., Шихобалова Н. П., Соболев А. А., Парамонов А. А.,
Судариков В. Е. 1954. Камалланаты, рабдитаты, тиленхаты, трихоцефаляты,
диоктофиматы. В кн. «Определитель паразитических нематод», т. IV. Изд-во АН
СССР, стр. 1—927.
Скрябин К. И., Шихобалова Н. П., Шульц Р. С. 1954а. Трихостронгилиды
животных и человека. В кн. «Основы нематодологии», т. III. Под ред. К. И. Скря-
бина. М., Изд-во АН СССР, стр. 1—683.
Скрябин К. И., Шихобалова Н. П., Шульц Р. С. 19546. Диктиокаулиды,
гелигмозоматиды и оллуланиды животных. В кн. «Основы нематодологии»,
т. IV. Под ред. К. И. Скрябина. М., Изд-во АН СССР, стр. 1'—323.
Скрябин К. И., Шихобалова Н. П., Орлов И. В. 1957. Трихоцефалиды и ка-
пиллярииды животных и человека и вызываемые ими заболевания. В кн. «Осно-
вы нематодологии», т. VI. Под ред. К. И. Скрябина. М., Изд-во АН СССР,
стр. 1—587.
Скрябин К. И., Шихобалова Н. П., Лагодовская Е. А. 1960. Оксиуараты
животных и. человека. В кн. «Основы нематодологии», т. VIII. Под ред. К. И. Скря-
бина. М„ Изд-во АН СССР, стр. 1—557.
Скрябин К. И., Шихобалова Н. П., Лагодовская Е. А. 1961. Оксиураты
животных и человека. В кн. «Основы нематодологий», т. X, 2. Под ред.
К. И. Скрябина. М., Изд-во АН СССР, стр. 1—499.
Скрябин К. И., Шихобалова Н. П., Петров А. М., Левашов М. М. 1962.
378
Строительство гельминтологической науки и практики в СССР, т. 1. Изд-во АН
СССР, стр. 1—296.
Скрябин К. И., Соболев А. А. 1963. Спирураты животных и человека и вызывае-
мые ими заболевания. В кн. «Основы нематодологии», т. XI, 1. Под ред. К. И. Скря-
бина. М., Изд-во АН СССР, стр. 1—541.
Соболев А. А. 1952. Филогенетические отношения и система камалланат,—Тр..
Гельминтол. лабор. АН СССР, 6: 296—301.
Т а у с о н А. О. 1923а. Влияние внешних условий на пол коловратки Asplanchna interme-
dia Huds.— Изв. Биол. н.-и. ин-та Пермск. ун-та, I, 7—8: 129—139.
Таусон А. О. 19236. Влияние ионов Na, К, Са и Mg на пол и жизненные процессы
у коловратки Asplanchna intermedia Hubs.— Изв. Биол. н.-и. ин-та Пермск. ун-та,.
2, 3: 121—133.
Филипьев И. Н. 1912. К вопросу о строении нервной системы у свободноживущих
нематод.— Тр. СПб. об-ва естествоисп., 43, 1; 205—216.
Филипьев И. Н. 1918—1921. Свободноживущие морские нематоды окрестностей Се-
вастополя.— Тр. Особой зоол. лабор. и Севастопол. биол., ст.,.4: 1—614.
Щепотьев А. 1908. О нематодах и близких к ним группах. СПб., стр. 1—90.
Beauchamp Р. 1929. Le developpement des Gastrotriches.—Bull. Soc. zool. Fran-
ce, 35.
Behrenz W. 1956. Vergleichende physiologische Untersuchungen liber die Exkretion pa-
rasitischen Nematoden mit Hilfe der Fluoreszenz mikroskopie.— Z. wiss. Zool., 159:
129—164.
Boveri T. 1892. Uber die Entwicklung der Gegensatz zwischen den Geschlechtszel-
len und den somatischen Zellen bei Ascaris megalocephala nebst Bemerkungen zur
Entwicklungsgeschichte der Nematoden.— Sitzungsber. Ges. Morphol., 8:
lil4—125.
Boveri T. 1899. Die Entwicklung von Ascaris megalocephala mit besonderer Beriick-
sichtigung der Kernverhaltnisse.— Fesrschr. Kupfer. Jena: 383—430.
Brunson R. B. 1949. The life history and ecology of two North American gastrotrichs.—
Trans. Amer. Microscop. Soc., 68: 1—20.
Budde E. 1934. Neue Beobachtungen an parasitischen Rotatorien.— Microkosmos, 27:
65—68.
Chitwood B. G. 1959. The classification on the phylum Kinoryncha.— Proc. XV-th
Internal. Congr. Zool. London, 1958 : 941—943.
Chitwood B. G. and Chitwood M. B. 1950. An introduction to nematology. Sec. I.
Anatomy, p. I—213.
^Chitwood M. B. 1957. Intraspecific variation in parasitic nematodes.— Syst. zool.,
6, 1: 19—23.
Coninck de L. 1962. Problems of systematics and taxonomy in nematology to day.—.
Nematologica, 7, I: 1—7.
Deineka D. M. 1908. Das Nervensystem von Ascaris.— Z. wiss. Zool., 89: 242—307.
G a n i n M. 1877. The development of Pelodera teres Confer.— Zool. Rec., 14.
Gerlach S. A. 1952. Diurodrilus benazzii, ein neue Archiannelidae aus dem Kiisten-
grundwasser des Mittelmeeres.— Zool. Anz., 149: 185—188.
Gerlach S. A., 1963. Die Gattung Haliplectus (Chromadorida, Leptolaimidae), zu-
gleich ein Beitrag zur Morphologie und Phylogenie der Nematoden.— Zool. Anz.,
171, 1—4: 96—113.
Goette A. 1881. Zur Entwicklungsgeschichte der Phabditis nigrovenosa.— Abhandl.
Entwicklungsgesch. Tiere: 59—92.
Goldschmidt R. 1,908. Das Nervensystem von Ascaris lumbrlcoides und Ascaris
megalocephala. I.—Z. wiss. Zool., 90: 73—136.
Goldschmidt R. 1909. Das Nervensystem von Ascaris lumbrlcoides und Ascaris
megalocephala. II.—Z. wiss. Zool., 92: 306—357.
Goldschmidt R. 1910. Das Nervensystem von Ascaris lumbrlcoides und Ascaris
megalocephala. III.— Festschr. zum 60. Geburstag R. Hertwigs, 2: 253—354.
G о о d e у T. 1963. Soil and freshwater nematodes. 2 ed. rev. and newritten by Dr.
J. B. Goodey. London: XVI—544.
Hartwich G. 1952. Vergleichende microskopisch-anatomische Untersuchungen uber;
den Kopfbau einiger Ascarieden.—Wiss. Zool. Univ. Halle, math.—naturwiss. Reihe,
3: 71—83.
Hartwich G. 1957. Zur Systematik der Nematoden — Superfamilie Ascaridoidea.—
Zool. Jahrb., Abt. 3, 85, 3: 211—252.
Higgins R. P. 1962. Morphological, larval and systematic studies in the Kinorhyncha.
Dissert. Abstr. XXII, 10: 3785, Michigan.
Hyman L. G. 1951. The invertebrates: Acanthocephala, Aschelminthes and Endoprocta.
The Pseudocoelomata Bilateria, v. Ill : VI+1572.
Illgen H. 1916. Zur Kenntnis der Biologie und Anatomie der parasitischen Rotato-
rienfamilien —Zool. Anz., 68: 1—9.
Martini E. 1903. Uber Fiirchung und Gastrulation bei Cuculanus elegans.— Z wiss.
Zool., 74: 501—556.
379
Martini E. 1906, 1907. Uber Subcuticula und Seitenfeldern einiger Nematoden. I,
II.—Z. wiss. Zool., 81: 699—766; 86: 1—54.
Mulvey R. H. 1961. The Mononchidae: A family of predaceous nematodes. (Genus
Mylonchulus) Enoplida: Mononchidae).— Canad. J. zool., 39: 665—696.
Murphy D. G. 1963. A note on the structure of Nematode ocelli.— Proc. Helmintol.
soc. Washington, 30: 25—26.
Nachtwey R. 1925. Untersuchungen fiber die Keimbahn, Organogenese und Anato-
mie von Asplanchna priodonta Gosse.— Z. wiss. Zool., 126: 239—492.
Nassonov N. 1897. Sur les organs du systeme excreteur des Ascarides et des Oxyu-
rides.— Zool. Anz., 20: 202—205.
Nassonov N. W. 1900. Zur Kenntnis der phagocytaren Organen bei den parasitischen
Nematoden.— Arch. Mikroscop. Anat., 40: 488—513.
Nyholm K. G. 1947. Studies in the Echinoderida.— Arkiv zool. Uppsala, 39, A, 14:
1—36.
Osche G. 1957. Beitrage zur Morphologie, Okologie und Phylogenie des Ascaroidea
(Nematoda). Parallelen in der Evolution von Parasit und Wirt.— Z. Parasitenkun-
de, 18: 479—572.
P a i S. 1928. Die Phasen des Lebenscyclus der Anguilula aceti Ehrbg. und ihre expe-
rimentel-morphologische Beeinflussung.— Z. wiss. Zool., 131: 299—314. .
P e j 1 e r B. 1S62. Morphological studies on the genera Notholca, K/ellicottia and Kera-
tella (Rotatoria).— Zool. Bidr., Uppsala, 33, 295—309.
Petrov A. M. 1930. Zur Charakteristik des Nematoden aus Kamtschatkaer' Zobeln
Soboliphyme baturini nov. gen., nov. sp.— Zool. Anz., 86: 265—271. .
Raski D. J. 1950. The life history and morphology of the sugar-beet nematode, Hete-
rodera schachti Schmidt.— Phytopatology, 40: 135—152.
Remane A. 1929—1933. Roratoria. In: Bronn’s «Klassen und Ordnungen des Tierreichs»,
Bd. 4, Abt. 2, Buch 1, Teil 1, Lief. 4: 1—576.
Remane A. 1935—1986. Gastrotricha und Kinorhyncha. In Bronn’s «Klassen und
Ordnungen des Tierreichs», Bd. 4, Abt. 3, Buch 1, Teil 2, Lief. 1: 1—385.
Remane A. 1959. Aschelminthes. In: G. Heber er. «Die Evolution der Organismen»,
S. 361—363.
Rothstein M. 1963. Nematode biochemistry. III. Excretion products.— Compar. Bio-
chem. and Physiol., 9, 1: 51—59.
Sacks M. 1955. Observations on the embryology of an aquatic Gastrotrich, Lepido-
dermella squammata.— J. Morphol., 96: 473—495.
Schulte H. 1954. Beitrage zur Okologie und Systematik der Bodenrotatorien.—
Zool. Jahrb., Abt. 3, 82, 6: 551—617.
Schuurmans Stekhoven J. H. 1959. Nematodes . In Bronn’s «Glassen und
Ordnungen des Tierreichs», Bd. 4, Abt. 2, Buch 3, Lief 7: 661—879.
Steinbock О. I. 1957. Zur Phylogenie der Gastrotrichen.— Verhandl. Dtsch. zool.
Ges.: 128—169.
Steiner G. 1917. Uber die Verwandschaftsverhaltnisse und die systematische Stel-
lung der Mermitiden.— Zool. Anz., 48: 363—367.
Steiner G. 1956. The zoological and agricultural status of plant nematodes.— Proc.
XIV Internal. Congr. Zool. Copenhagen: 368—371.
Swedmark B. 1955. Developpement d’un Gastrotriche Macrodasyoide, Macrodu-
sys affinls Remane.— C. r. Acad. sci. Paris, 240, 18: 1812—1814.
Tannreuther G. W. 1920. The development of Asplanchna ebbersbornii (Rotifera).—
J. Morphol., 33, 1: 389—422.
T a u s о n A. 1924. Reifungsprozesse der parthenogenetischen Eier von Asplanchna inter-
media Huds.— Z. Zellforsch. und Gewebelehre, I, I: 56—84.
Tauson A. 1925. Wirkung des Mediums auf das Geschlecht des Rotators Asplanchna
intermedia Huds.— Rev. Hydrobiol. und Hydrogr., 13; 3/4: 130—170; 5/6: 282—325.
Tauson A. 1926. Uber die Wirkung des Mediums auf das Geschlecht der Rotators
Asplanchna intermedia Huds.— Arch. Entwicklungsmech., 107, 2: 355—391.
Tauson A. 1927. Uber die Wirkung des Mediums auf das Geschlecht des Rotators
Asplanchna intermedia Huds.— Arch. Entwicklungsmech., 109, 3: 342—361.
Thorne G. 1961. Principles of nematology. N. Y., McGraw-Hill Book Co., p. XII+553.
Voltzenlogel S. 1902. Untersuchungen fiber den anatomischen und histologischen Bau
des Hinterendes von Ascaris megalocephala und Ascaris lumbricoides.— Zool. Jahrb.
mabt, 2, 16: 487—510.
Wieser W. 1953. Die Beziehungen zwischen Mundholengestalt, Ernarungsweise und
Vorkommen bei freilebenden marinen Nematoden.— Arkiv zool., 4, 26: 439—484.
Wieser W. 1958. Reports of the Lund University Chile Expedition 1948—49. 34. Free-
living marine nematodes. IV General part.— Lunds Univ. Arsskr. N. F. Avd. 2 55,
5: 1—HI.
Wilke U. 1954. Mediterrane Gastrotrichen.— Zool. Jahrb. Abt. 3, 82, 6: 497—550.
Ziegler H. E. 1895. Untersuchungen fiber die ersten Entwicklungsgeschichte der
Nematoden, zugleich ein Beitrag zur Zellenlehre.—Z. wiss. Zool., 60: 351—410.
Zur-Strassen I. 1896. Embryonalentwicklung der Ascaris megalocephala.— Arch.
Entwicklungsmech., 3: 27—105; 133—190.
380
Zur-Strassen О. 1959. Neue Beitrage zur Entwicklungsmechanik der Nematoden.—
Zoologica, 38, 3, 107: 1—142.
НЕМЕРТИНЫ (NEMERTINI)
Беклемишев К. В. 1955. Хищничество у немертин.— Природа, 9: 108—109.
Давыдов К. Н. 1910. Реституция у немертин в связи с вопросом о проспективной
способности зародышевых пластов.— Изв. СПб. АН, 4, 8: 645—'657.
Давыдов К. Н. 1915. Реституция у немертин.— Тр. Особ. зоол. лабор. и Севастоп.
биол. ст. АН СССР, 11, 1: IV+290.
Кайзер Н. А. 1925. Заметка об инцистировании Lineus gesserensis О. F. Muller.—
Б юл л Среднеаз. ун-та, 9: 63—66.
Короткевич В. С. 1955. Пелагические .немертины дальневосточных морей СССР.
«Определитель по фауне СССР», т. 58. Изд-во АН СССР, стр. 1—131.
Ливанов Н. А. 1937.. Тип Немертины. В кн. «Руководство по зоологии», т. 1,
стр. 656—705.
Шмидт Г. А. 1946. Родственные и экологические отношения литоральных немертин
рода Lineus.— Докл. АН СССР, 51, 5: 401—403.
Шмидт Г. А. 1956. Дезоровская фаза в зародышевом развитии немертины Lineus
.ruber (О. Р. Mil Iler, 1774, G. A. Schmidt, 1945).— Докл. АН СССР, 106, 4: 749—
752.
Ш мидт Г. А. 1958. О различиях эмбриональных приспособлений географических рас
немертины Linens ruber (О. F. Muller, 1774, G. A. Schmidt, 1945).— Folia bid.,
6, 4: .265—285.
Шмидт Г. A. 1962a. Изменение экологических отношений у взрослых особей и эво-
люция эмбриогенеза на примере литоральных немертин Lineus desori (mihi sp. n.)
и Lineus ruber (O. F. Muller, 1774, G. A. Schmidt, 1945).— Зоол. ж., 41, 2: 168—
192.
Шмидт Г. А. 19626. Корреляция между закладками хоботка и глотки у немертин
Lineus desori (mihi, sp. п.) и Lineus ruber (О. F. Muller, 1774,
G. A. Schmidt, 1945).—Докл. АН СССР, 147, 6: 1512—1515.
Bohmig L. 1898. Beitrage zur Anatomie und Histologie der Nemertinen.— Z. wiss.
Zool., 64: 478—564.
Bohmig L. 4929. Nemertini. In W. Kiikenthal «Handbuch der Zoologie», Bd. II,
I. H. S. 1—110.
Bresslau E. 1932. Plathelminthes.— Handworterbuch der Naturwissenschaften. Jena.
G. Fischer, Bd. 7, S. 1105—1138.
Brinkmann A. 1917. Die pelagischen Nemertinen.— Bergens Mus. skr., 3, N 1:
1—194.
Burger O. 1897—1907. Nemertini. In Bronn’s «Klassen und Ordnungen des Tierreichs»,
IV Suppl., S. VIII+542.
Coe W.R. 1943. Biology of the Nemerteans of the Atlantic Coast of North America.—
Trans. Cann. Acad. Art and Sci., 35: 129—328.
Dawydoff K. N. 1928. Sur 1’embryologie des Protonemertes.— C. r. Acad. sci. Paris,
186: 173—175.
Gontcharoff M. 1948. Note on 1’alimenfation de quelques nemertes.— Ann. sci.
natur. zool. et biol. anim., Paris, ser., 11, 10: 75—78.
Gontscharoff M. 1960. Le developpement post-embryonnaire et la croissance chez
Lineus ruber et Lineus viridis (Nemertes Lineidae).— Ann. sci. natur. zool. et biol.
anim., Paris, Ser., 12, 2: 225—279.
Hammarsten O. D. 4918. Beitrag zur Embryonalentwicklung der Malacobdella gros-
ser (Muller).— Arbeiten zool. Inst. Stockholm. 1: 1—96.
Horstadius S. 1937. Experiments an determination on the early development of
Cerebratulus lactueus.— Biol. Bull. Woods Hole, 73, 2: 317—342.
H u b r e c h t A. A. W. 1887. The relation of the Nemertea to the vertebrata.— Quart.
J. Microscop. Sci.. 27: 605—644.
Iwata F. 1057. On the early development of the Nemertine, Lineus iorcuatus Cie.— J.
Fac. Sci. Hokkaido Univ., Ser. VI. Zool., 13, 1—4: 54—58.
Iwata F. 1958, On the development of the Nemertean Micrura akkeshiensis.— Embryo-
logia, 4, 2: 103—131.
Iwata F. 1960. Studies on the comparative embryology of nemerteans with spe-
cial reference to their interrelationships.— Pubis. Akkeshi. Marine Biol. Stat, 10:
1—51.
Nussbaum J. und. Oxner M. 1912. Fortgesetzte Studien fiber die Regeneration
der Nemertinen. II. Regeneration des Lineus lacteus Rathke.— Arch. Entwicklungs-
mech., 35, 2: 236—308.
Nussbaum J. und Oxner M. 1913. Die Entwicklung des Lineus ruber Mall. Ein
Beitrag zur Entwicklungsgeschichte Nemertinen.—Z. wiss. Zool., 107: 78—197.
Budemans A. C. 1885. Circulatory and nephridial apparatus of Nemertea.— Quart.
J. Microscop. Sci., 25: 1—80.
381
Salensky W. 1912. Entwicklungsgeschichte der Nemertine im Inneren des Pilidiums.—
Mem. Acad. Sci. St. Petersbourg., Ser. 8, 30, 10: 1—74.
Salensky W. 1914\ Entwicklungsgeschichte der Prosorochmus viviparus.— Mem. Acad.
Sci. St. Petersbourg, ser 8, 33, 2: 1—39.
Schepotieff A. 1913. Uber die Bedeutung der wassermannischen Reaktion fur die-
biologische Forschung.— Zool. Anz., 4, 1, 2: 49—54.
Schmidt G. A. 1937. Vergleichend-embryologische Studien uber die Typen der Embryo-
nalanpassungen bei Hirudineen und Nemertinen.— Z. Morphol. und Okol. Tiere, 32,.
7: 650—671.
Schmidt G. A. 1963. Embryonic development of the Nemerteans Lineus desori sp. n.
and Lineus ruber (O. F. Muller, 1774, G. A. Schmidt, 1945) correlated with the-
ecology of mature individuals and their speciation.— Zool. Polon., Vol. 13.
МОЛЛЮСКИ (MOLLUSCA)
Акимушкин И. И. 1963a. Головоногие моллюски морей СССР. М., стр. 1—234.
Акимушкин И. И. 19636. Приматы моря. М., Изд-во геогр. лит-ры, стр. 1—158.
Беклемишев В. Н. 1959. К вопросу о ранней эволюции моллюсков. Сравнительно-
анатомическое значение и функционально-морфологическое толкование организа-
ции Neopilina.— Зоол. ж., 38, 4: 518—522.
Борисяк А. А. 1899. Введение в изучение ископаемых пелеципод (пластинчато-
жаберных).— Зап. СПб. минерал, об-ва, 2, 37, 1: 1—144.
Властов Б. В. 1940а. Класс лопатоногих моллюсков. В кн. «Руководство по зоо-
логии», т. II. Беспозвоночные, стр. 456—465.
Властов Б. В. 19406. Класс панцирных моллюсков. В кн. «Руководство по зоо-
гии», т. II. Беспозвоночные, стр. 286—322.
Властов Б. В. 1948. Тип моллюски. В кн. «Жизнь животных по А. Брему», т. I. М.,
Госучпедгиз, стр. 1—643.
Воскресенский К. А. 1948. Пояс фильтраторов как биогидрологическая система
моря.— Тр. Океаногр. ин-та, 6(18): 55—120.
Грузов Е. Н. 1957. Новый эндопаразитический моллюск Molpadicola orientalis, gen.
п., sp. п. (семейство Paedophoropodidae).— Зоол. ж., 36, 6: 852—862.
Грузов Е. Н. 1965а. Организация эндопаразитического моллюска Asterophila japoni-
са Randall et Heath. 1. Организация взрослого животного.— Зоол. ж., 44, 8: 1152—
1164.
Грузов Е. Н. 19656 Приспособление брюхоногих моллюсков к паразитизму.— Зоол.
ж., 44, И: 1620—1630.
Ду ев Г. В. 1965. Основные черты и адаптивное отношение эволюции раковины голо-
воногих моллюсков.— Зоол. ж., 44, 2 : 284—286.
Догель В. А. 1940а. Класс двустворчатых моллюсков. В кн. «Руководство по зооло-
гии», т. II Беспозвоночные, стр. 466—547.
Догель В. А. 19406. Филогения типа моллюсков. В кн. «Руководство по зоологии»,
т. II. Беспозвоночные, стр. 610—617.
Журавлева Ф. А. 1955. Прямые и согнутые наутилоидеи ордовика и силура Под-
каменной Тунгуски и Курейки. Автореф. канд. дисс. Палеонтол. ин-т АН СССР,
стр. 1—16.
Заварзин А. А. 1913. Гистологические исследования чувствительной нервной си-
стемы оптических нервных ганглиев насекомых. Дисс. СПб., стр. 1—192.
И в а н о в А. В. 1937. Новый эктопаразитический моллюск из рода Megademus Rosen.—
Докл. АН СССР, 14, 7: 467—470.
Иванов А. В. 1940а. Класс брюхоногих моллюсков. В кн. «Руководство по зооло-
гии», т. II. Беспозвоночные, стр. 323—455.
Иванов А. В. 19406. Половые отношения у паразитических Gastropoda (Явления
раздельнополости, полового диморфизма и гермафродитизма).— Тр. Ленингр. об-ва-
естествоисп., 68, 4: 61—80.
•Иванов А. В. 1945а. Entocolax rimsky-korsakovi nov. sp., новый моллюск, парази-
тирующий в голотурии,—Докл. АН СССР, 49, 7: 553—555.
Иванов А. В. 19456. Морфологические адаптации пищеварительной системы у пара-
зитических Gastropoda.—Уч. зап. ЛГУ (биол.), 75, 15: 112—119.
Иванов А. В., 1945в. Новый эндопаразитический моллюск. Parenteroxenos dogieli nov.
gen., nov. sp.—Докл. АН СССР, 48, 8: 477—480.
Иванов А. В. 1946a. Редукция организации личинок у паразитических брюхоногих.—
Тр. Ленингр. об-ва естествоисп., 69, 4: 93—100.
Иванов А. В. 19466. О путях проникновения эндопаразитических брюхоногих в хо-
зяина.— Тр. Ленингр. об-ва естествоисп., 69, 4: 88—99.
Иванов А. В. 1947.. Строение и развитие эндопаразитического брюхоногого моллюска
Parenteroxenos dogieli A. Ivanov (семейство Entoconchidae). Сообщение II Органи-
зация взрослого животного —Изв. АН СССР, сер. биол., 1: 3—28.
И в а н о в А. В. 1948. О метаморфозе паразитического моллюска Parenteroxenos: dogieli
A. Ivanov.—Докл. АН СССР, 61, 4: 765—768.
382
Иванов А. В. 1949. Строение и развитие эндопаразитического брюхоногого моллюска
Parenteroxenos dogieli A. Ivanov (семейство Entoconchidae). Сообщение II. Органи-
зация личинки и постларвальный метаморфоз.— Изв. АН СССР, сер. биол., 2: 109—
133. Сообщение III. О взаимоотношении между родами Entoconchidae. Там же,
стр. 135—139.
Иванов А. В. 1952. Строение эктопаразитических брюхоногих Stiliferidae, как ре-
зультат их образа жизни.—Тр. Ленингр. об-ва, естествоисп., 71, 4: 86—140.
Иванов А. В. 1953. Строение эндопаразитического брюхоногого моллюска Entoco-
lax rimsky-korsakovi A. Ivanov.— Тр. Зоол. ин-та АН СССР, 13: 250—270.
Кабанов Г. К. 1964. Морфофункциональный и биологический анализ Belemnitida.
Автореф. канд. дисс. Палеонтол. и-f АН СССР, стр. 1—21.
Карпевич А. Ф. 1955. Некоторые данные о формообразовании у двустворчатых
моллюсков.— Зоол. ж., 34, 1: 46—67.
Карпинский А. 1903. О нижнекембрийском роде цефалопод Volborthella
Schmidt.—Изв. АН СССР, 18, 4: 147—153.
Ковалевский А. О. 1879. О строении двух новых форм из рода Neomenia. Речи
и протоколы VI съезда русских естестВоиспыт. и врачей (с 20 по 30 декабря
1879 г.), СПб., стр. 85—86.
Кондаков Н. Н. 1940. Класс головоногих моллюсков. В кн. «Руководство по
зоологии», т. II. Беспозвоночные, стр. 548—609.
Коробков И. А. 1960. Введение в изучение ископаемых моллюсков. Изд. ЛГУ,
стр. 1—279.
Луппов Н. П., Друшиц В. В. |(ред.) 1958. Моллюски головоногие. В кн. «Осно-
вы палеонтологии», т. II. Аммоноидей (цератиды и аммониты). Внутрираковин-
ные. М., АН СССР, стр. 1—359.
Пчелинцев В. Ф., Коробков И. А. (ред.). 1960. Моллюски — брюхоногие.
В кн. «Основы палеонтологии». М., АН СССР, стр. 1—360.
Руженцев В. Е. 1930. Значение онтогенеза для естественной систематики аммо-
нитов.— Изв. АН СССР, сер. биол., 1: 13—32.
Руженцев В. Е. 1940. Опыт естественной систематики некоторых верхнепалеозой-
ских аммонитов.— Тр. Палеонтол. ин-та АН СССР, II, 3: 1—134.
Руженцев В. Е. 1949. Систематика и эволюция семейств Pronoritidae Freeh u.
Medliocottidae Karpinsky.— Тр. Палеонтол. ин-та АН СССР, 19: 1—199.
Руженцев В. Е. 1957а. Филогенетическая система палеозойских аммоноидей.—
Бюлл. Моск, об-ва испыт. природы; отд. геол., 32 (2): 49—64.
Руженцев В. Е. 19576. Процветание и кризисы в истории аммоноидей.— Докл. АН
СССР, 115, 4: 791—794.
Руженцев В. Е. 1960. Принципы систематики, система и филогения палеозойских
аммоноидей.— Тр. Палеонтол. ин-та АН СССР, 53: 1—331.
Руженцев В. Е. (ред.) 1962. Моллюски — Головоногие. I. Наутилоидеи эндоцера-
тоидеи, актиноцератоидеи, бактритоидеи, аммоноидей. В кн. «Основы палеонто-
логии». М., АН СССР, стр. 1—438.
Руженцев В. Е., Шиманский В. Н. 1954. Нижнепермские свернутые и со-
гнутые наутилоидеи Южного Урала.— Тр. Палеонтол. ин-та АН СССР, 50:
1—752.
Федотов Д. М. 1933. Об изменчивости современных пелеципод в связи с изучением
ископаемых форм.— Тр. Палеозоол. ин-та АН СССР, И: 1—16.
Чернов А. 1922. Основные черты в эволюции аммонитов.— Бюлл. Моск, об-ва
испыт. природы, сер. геол., 3 (1): 149—185.
Шиманский В. Н. 1957. Систематика и филогения отряда Nautilida. Бюлл. Моск,
об-ва испыт. природы, сер. геол., 32 (4): 105—120.
Шиманский В. Н. 1960. Новые миссисипские белемниты. Флауер, Гордон. 1959.—
Палеонтол. ж., 2: 158—162.
Шиманский В. Н., Журавлева Ф. А. 1961. Основные вопросы систематики
наутилид и родственных им групп,—Тр. Палеонтол. ин-та, АН СССР, 90: 1—175.
Эберзин А. Г. (ред.) 1960. Моллюски — панцирные, двустворчатые, лопатоногие.
В кн. «Основы палеонтологии». М., АН СССР, стр. 1—300.
Baba К- 1938. The later development of a Solenogastre,, Epimenia verrucosa (Nierst-
rasz).— J. Dept. Agric. Fukuoka, 6.
Baba K. 1940a. Nahrungsaufnahme, Verdauung und Excretion —J. Dept. Agric. Fukuo-
ka, 6.
Baba K. 19406. The early development of a solenogastre Epimenia verrucosa (Nier-
strasz).— Annot. zool. japon., 19: 107—113.
Baba K. 1961a. The shells and radulae in Berthelinia, a bivalved Saccoglossan genus.—
Venus, Japan. J. Malacol., 21, 4: 389—401.
Baba K- 19616. On the identification and affinity of Tamanovalva Umax, a bivalved
Saccoglossan mollusc, in Japan.— Publ. Seto, Marine biol. lab., 9, 1 : 3762, Sirahama.
Boettger C. R. 1955. Die Systematik der euthyneuren Schnecken.— Verhandl. Dtsch.
zool. Ges. Zool. Anz. Suppl., 18: 253—280.
Boettger C. R. 1956. Beitrage zur Systematik der Urmollusken (Amphineura).—
Zool. Anz., Suppl. Bd. 19: 233—256.
383
Boettger C. R. 1962a. Die Verwandschaftsbeziehungen der Kraken — Gattung Argo-
nauts.— Arch. Molluskenkunde, 91, 1/3: 53—55.
Boettger C. R. 19626. Gastropoden mit zwei Schalenklappen.— Verhandl. Dtsch. zooL
Ges. Wien, 2: 403—439.
Brookeb J., Knight J. B. and Yochelson E. L. 1.959. Occurrence and classifica-
tion of fossil Monoplacophora (Mollusca).— Proc'. XV Internet. Congr. Zool. Lon-
don, 16—28 July 1958: 373—376.
Burn R. 1960. Australian bivalve gastropods.— Nature, 187, 4731: 44—46.
Cameron T. W. M. 1960. Evolution: Its science and doctrine. Symposium Roy. Soc.
Canada in 1959: XI+ 242.
Chestnut A. F. 1943. Precipitin studies of certain Mollusca, with special reference to
the cyster. Master’s Thesis, Rutgers Univ. Library. Unpublished.
Clarke A. H., and R. J. Menzies, 1959. Neopilina (Vema) ewingi a second living,
species of the Paleosoic — Devonian class Monoplacophora.— Science, 1'29, 3355:
1026—1027.
Cox L. R. 1960a. A Bivalve Gastropod.— Nature, 185, 4715: 749—751.
С о x L. R. 19606. Thoughts on the classification of the gastropoda.— Proc. Malacol. Soc.
London, 33: 239—261.
Cox L. R. 1960b. Thoughts on the classification of the bivalvia.— Proc. Malacol. Soc.
London, 34: 60—88.
Cox L. R. and Knight J. B. 1960. Suborders of Archeogastropoda.— Proc. Malacol.
Soc. London, 34 : 262—264.
Crofts D. R. 1955. Muscle morphogenesis in primitive gastropods and its relation to-
torsion.— Proc. Zool. Soc. London, 125: 71J—750.
Dahl E. 1954. The question of larval segments and brain structure and their bearing
upon Crustacean phylogeny.— KgL fysiogr. Sallskap. Lund forhandl., 24, 6: 1—9..
Dechaseaux C. 1960. Bivalvea fossiles. In Grasse P. P. «Traite Zool.», V, fasc. 2,
p. 2134—2164.
Donovan D. T. 1964. Cephalopod phylogeny and classification.— Biol. Rev., 39: 295—
287.
Er ben H. K. 1964. Die Evolution der altesten Ammonoidea.— N. Jahrb. Geol. Palaontol.
Abhandl., 120, 2: 107—212
Feliksiak S. 1959. Position of the Monoplacophora in the phylogeny of the mollus-
ca.— Bull. Acad, polon. sci., Cl. II, 7: 427—431.
Feliksiak S. 1961. Slimaki tylosktzelne (Opisthobranchia) о malsowej musli.— Kos-
mos, A, X, 1: 59—64,
Flower R. H. and Cordon AL 1959. More Mississippian belemnites.— J. Paleontol.,
33, 5: 809—842.
Fretter V. and Graham A. 1949a. The structure and mode of life of the Pyramidel-
lidae, parasitic Opisthobranchs,— J. Marine biol. Assoc. Plymouth, 28, 2: 493—532.
Fretter V. and Graham A. 19496. Feeding and reproduction in the Pyramidellids.—
’ Nature, 163, 4140: 361—362.
Fretter V. and Graham A. 1954. Observations on the opistobranch mollusc Actaeon
tornatilis (L.).— J. Marine biol. Assoc. U. K., 33: 565—585.
Garstrang W. 1,896. The morphology of the Mollusca.— Sci. Progr., 5: 38—59.
Garstrang W. 1928. Origin and evolution of larval forms.— Nature, London, 122: 366.
Gegenbauer C. 1898, 1901. Vergleichende Anatomie der Wirbeltiere mit Beriicksichti-
gung der Wirbellosen, Bd. 1, S. 1—978; Bd. 2, S. 1—696. Leipzig.
Goodrich E. C. 1945, 1946. The study of nephridia and general ducts since 1895, I,-
II.— Quart. J. Microscop. Sci., 86: 324—336; 87: 1,13—392.
Graham A. 1949. The molluscan stomach.— Trans. Roy. Soc. Edinburgh, 61, 3: 737—778.
Graham A. 1953. Form and function in the molluscs.— Proc. Linnean Soc. London,.
164, 2: 213—217. .
Graham A. 1057. The molluscan skin with special reference to Prosobranchs (Pru-
dential Adress).— Proc. Malacol. Soc. London, 32: 135—144.
Grasse P. P. (Ed.). 1960. Traite Zoologie, v. 5. Paris.
Grenacher H. 1874. Zur Entwicklungsgeschichte der Cephalopoden.— Z. wiss. Zool.,
24: 419—498.
Grobben K. 1883. Morphologische Studien uber' den Harn- und Geschlechtsapparat,.
sowie die Leibeshohle der Cephalopoden.— Arbeit. Zool. Inst. Wien. 5.
Grobben K. 1894. Zur Kenntnis der Morphologie, die Verwandtschaftsverhaltnisse und'
des Systems der Mollusken.— Sitzungsber. Akad. Wiss. Wien, 103: 61—86.
G г о b b en K. 1900. Einige Betrachtungen Ober die phylogenetische Entstehung der Dre-
hung und die assymetrische Aufrollung bei den Gastropoden.— Arbeit. Zool. Inst.
Wien, 12: 25—44.
Haas F. 1929—1934. Bivalvia. In Bronn’s «Klassen und Ordnungen des Tierreichs».
Bd. III. Abt. 3, 1—5: 1—704.
Haas F. 1937—1956. Bivalvia. In Bronn’s «Klassen und Ordnungen des Tierreichs»..
Bd. 3, Abt. 3, 2, 1—4: 1—923; 5: 1—148.
Hammersten O. D. u. Runnstrom J. 1926. Zur Embryologie von Acanthochiton.
discrepans Brown.— Zool. Jahrb. Abt. 2, 47: 261—318.
384
Harrrmersten О. D. und Run n strom J. 1|926. Ein Beitrag zur Diskussion uber die-
Verwandtschaftsbeziehungen der Mollusken.— Acta zool., Stockholm, 7: 1—67.
Han strom B. 1928. Vergleichende Anatomie des Nervensystems der wirbellosen Tiere
unter Beriicksichtigung seiner Funktion. Berlin, S. XII-1-628.
Hess O. 4957. .Entwicklungsphysiologische Untersuchungen an dem Siisswasser-Proso-.
branchier Bithynia tentaculata.— Zool. Anz., Suppl. Bd., 20: 265—271.
Hoffmann H. 1937. Uber die Stammesgeschichte der Weichtiere.— Zool. Anz., Suppl.
Bd. 10: 33—69. I
Hoffmann S. 1949. Studien uber das Integument der Solenogastren nebst Bemerkun-
gen fiber die Verwandtschaft zwischen den Solenogastren und Placophoren.— Zool.
Bidr., Uppsala, 27: 293—427.
Ihering H. 1877. Vergleichende Anatomie des Nervensystems und Phylogenie der Mol-
iusken.— Z. Naturwiss., 11, N. F. 4. Jena: X+290.
Iwanoff P. P. 1928. Die Entwicklung der Larvalsegmente bei den Anneliden.— Z. Mor-
phol. und Okol. Tiere, 10, 1\: 62—161.
Ivanov A. W. 1933. Ein neues endoparasitisches Mollusk Paedophorus dicoelobius n..
gen., n. sp.— Zool. Anz., 104: 161—165.
Ivanov A. W. 1937. Die Organisation und die Lebensweise der parasitischen Mol-
luske Paedophorus dicoelobius.— Acta zool., 18: 111—208.
Jaeckel S. H. 1955. Bau und Lebenswiese der. Tiefseemollusken.— Neue Brehm-
Bticherei, S. 1—7-0.
Johansson J. 1952. On the phylogeny of the Mollusca.— Zool. Bidr., Uppsala, 29:.
277—292.
Johansen K- and Huston M. J. 1962. Effects of some drugs on the circulatory
system of the intact, nonanesthetized cephalopod Octopus dofleini.— Compar. Bio-
chem. and Physiol., 5: 177—184.
Johansen K- and Martin A. W. 1962. Circulation in the Cephalopod, Octopus dof-
leini.— Compar. Biochem. and Physiol., 5: 161—176.
Kai m us H. 1959. Orientation of animals to polarized light.— Nature, 184, 4682:
228—230.
Kavaguti S. 1959. Formation of the bivalve shell in a gastropod Tamanovalva
Umax.— Proc. Japan Acad., 35, 10: 607—611.
Kawaguti S. and Bab.a K. 1959. A preliminary note on at twovalved sacoglossan
gastropod, Tamanovalva Umax, n. gen., n. sp. from Tammano, Japan.— Biol. J.
Okayama Univ., 5, 3—4: 177—184.
Knight J. B., Batten R. L. and Yochelson E. L. 1953. Status of invertebrate-
paleontology. V. Mollusca: Gastropoda.— Bull. Museum Compar.'Zool., 112, 3: 173—
179.
Knight J. B. and Yochelson E. L. 1958. A reconsideration of the relationships of
the Monoplacophora and the primitive Gastropoda.— Proc. Malacol. Soc. London,.
33, 1: 37—48.
Kiimmel B. 1955. The ancestory of the family Nautilidae.— Breviora, 21c 1—7.
Kuramoto M. 1933. Uber die Antigenitat der Fischweichtier- und Schalfischeiweis-
se.— Arbeit. Med. Fak. Okayama, 4 : 249—261.
L eb our M. 1937. The eggs and larvae of the British Prosobranchs with special refe-
rence to those living in the Plancton.— J. Marine Biol. Assoc. U. K- Cambridge,
22: 105—166.
Lemche H. 1957. A new living deep-sea mollusc of the Cambro-Devonian class Mo-
noplacophora.— Nature, 179, 4556: 413—416.
Lemche H. 1959a. Prostomian interrelations in the light of Neopilina.— Proc. XV-th
. : Internal. Zool. Congr. London: 381—389.
Lemche H. 19596. Neopilina.— Rev. Univ. Madrid, VIII, 29—31i: 411—442.
Lemche H. 1959b. Molluscan phylogeny in the light of Neopilina.— Proc. XV Internat,
Congr. Zool. London, 16—28 July, 1958: 380—381.
Lemche H. and Wingstrand K. G. 1959. The anatomy of Neopilina galatheae
Lemche, 1957 (Mollusca Tryblidiacea).— Galathea Rept. 3: 1—70.
Lemche H. und Wingstrand K. G. 1960. Classe der Monoplacophorus. Monopla-
cophora. In Grasse P. P. «Traite de zoologie», t. V, 2. p. 1787—1821.
Leone Ch. and Pryor C. W. 1954. Serological comparison among four classes of Mol-
lusca,— Biol. Bull, 107: 411—412.
Ma kino K. 1934. Beobachtungen fiber die Immunitatsreaktionen bei. Molluskenarten.—
Z. Immunitatsforsch. und exptl, XX, 81: 316—335.
Menzies R. J., Ewing M., Worzel J. L. and Clarke A. H. 1959. Ecology of
the recent Monoplacophora.— Oikos, 10, 2: 168—182.
Miller A. K. and Furnish W. M. 1954. The classification of the Paleozoic Ammo-
noids.— J. Paleontol., 28, 5: 685—692.
Moody M. F. and Parriss Y. R. 1960. Discrimination of polarized light by Octo-
pus.— Nature, 186, 4728 : 839—840.
Mo’ore R. C. (Ed.), 1957, 1960, 1964. Treatise on invertebrate paleontology, N. Y.
Morton J. E. 1951. The ecology and digestive systems of the Struthiolariidae (Gastro-
poda).— Quart. J. Microscop. Sci., 92: 1—24.
385
Morton J. E. 1953. The function of the gastropod stomach.— Proc. Linnean Soc.
London, 164, 2: 225—235.
Morton J. E. 1955. The evolution of the Ellobiidae with a discussion on the origin
of Pulmonates.— Proc. Zool. Soc. London, 125: Г27—168.
Morton J. E. 1958. Torsion and the adult snail. A re-evaluation.— Proc. Malacol. Soc.
London, 33, L: 2—10.
Neumayr M. 1883. Zur Morphologie des Bivalven-Schlosses.— Sitzungsber. Akad.
Wiss. Wien, 88: 385—419.
Neumayr M. 1892. Beitrage zur einer morphologischen Einteilung der Bivalven.—
Denkschr. Acad. Wien, Math.— Naturwiss. Cl., 58: 701—801.
Newell N. D. 1953. Status of invertebrate paleontology. V. Mollusca: Pelecypoda.—
Bull. Museum Compar. Zool., 112, 3: 161—172.
Newell N. D. 1960. The origin of the Oysters.— Rep. Internat. Geol. Congr. XXL
Nierstrasz H. F. 1909. Die Amphineura I. Solenogastres.— Ergebn. und Fortschr.
Zool., Jena, I.
Nierstrasz H. F. 1910. Die Amphineura 2. Die Chitonea.— Ergbn. und Fortschr. Zool.,
Jena, 2: 367—430.
О d h n e r N. H. 191i2. Morphologische und phylogenetische Untersuchungen fiber die
Nephridien der Lamellibranchien.— Z. wiss. Zool., 100: 287—391.
Odhner N. H. 1921. Norvegian Solenogastres.— Bergens Museum Aarbok 1918—1919,
I, 3: I.—86.
Odhner N. H. 1961. Some notes on the classification of Gastropoda — Proc. Mala-
col. Soc. London, 34, 5: 250—254.
Owen G. 1958. Shell form, pallial attachment and the ligament in the Bivalvia.— Proc.
Zool. Soc. London, 131, 4: 637—648.
Pelseneer P. 1890. Sur le pied de Chitonellus et des Aplacophora.— Bull. sci. Fran-
ce Belg., 22: 489—495.
Pelseneer P. il891. Contribution a 1’etude des Lamellibranches.— Arch, biol., -11:
147—312.
Pelseneer P. 1898—1899. Recherches morphologiques et phylogenetiques sur les mol-
lusques archaiques:—Mem. Cour. Acad. roy. Belgieque, 57. Bruxelles: 1—112.
P e 1 s en e e-' P. 1906. Molluska. In E. Ray Lankester «А Treatise on Zoology», v. 5. Lon-
don, p. 1—355.
Pickford G. E. 1939. The Vampyromorpha. A new order of Dibranchiate Cephalo-
poda.— Vest. Ceskosl. spolec. zool., 6—7: 346—358.
Pickford G. E. 1940. The Vampyromorpha, living fossil Cephalopoda.— Trans. N. Y.
Acad. Sci., ser. 2, 2, 7: 1—13.
Pickford G. E. 1949. Vampyroteuthis infernalis Chun. An archaic dibranchiate Ce-
phalopod. II. External anatomy.— Dana. Rept. Copenhagen, № 82: 1—132.
Pickford G. E. 1950. The Vampyromorpha (Cephalopoda) of the Bermuda Oceano-
graphic Expedition.— Zoologica, 35: 87—95.
Plate L. 1895. Bemerkungen fiber die Phylogenie und Entstehung der Asymmetrie der
Mollusken — Zool. Jahrb. Anat, 9: 162—206.
Plate L. 1897—1901. Die Anatomie und Phylogenie der Chitonen (Fauna Chilensis).—
Zool. Jahrb. Suppl., 4, 5.
Plate L. 1922. Allgemeine Zoologie und Abstammungslehre. I. Das Nervensystem der
Mollusken: G. Fischer, Jena, S. VI+629.
Popham M. L. 1940. The mantle cavity of some of the Erycinidae, Montacutidae and
Galeommatidae with special reference to the ciliary mechanisms.— J. Marine Biol.
Assoc. U. K., 24: 549—587.
Pruvot G. 1890. Sur le developpement d’un Solenogastre.— C. r. Acad sci. Paris, 111:
689—692.
Pruvot G. 1892. Sur les affinities et la classement des Neomeniens.— Arch. Zool. exptl
et gen., 9: 699—805.
Raven Ch. P. 1958. Morphogenesis. The analysis of molluscan development. London,
Pergamon Press, p. XII + 311.
Sachwatkin A. A. 1920. Das Urogenitalsystem von Ampullaria gigas Spix — Acta
zool., 1: 67—130.
Schindewolf О. H. 1954. On development, evolution and terminology of Ammonoid
Suture Lines.— Bull. Museum Compar. Zool., 112, 3: 217—237.
Southerland N. S. 1960. Theories of shape discrimination in Octopus.— Nature,
186, 4728: 820—844.
S pen gel J. W. 1881.. Die Geruchsorgane und das Nervensystem der Mollusken.— Z.
wiss. Zool., 35: 333—383. 7
Stasek C. R. 1961. The ciliation and function of the labial palps of Acila castrensis
(Protobranchia, Nuculidae), with an evaluation of the role of the protobranch organs
of feeding in the evolution of the Bivalvia.—Proc. Zool. Soc. London, 137, 4: 511—
538.
Stenrel H. B. 1957. Nautilus. Treatise on marine ecology and palaeontology. I.— Mem.
Geol. Soc. America, 67: 1135—1141.
386
Storer T. I. and Us in ger R. L. 1957. Phylum Mollusca. General Zoology, N. Y.,
p. 351—367.
Studnitz G. 1931. Die Morphologie und Anatomie von Lima inflata, der Feilenmu-
schel, nebst biologischen Untersuchungen an Lima hians Gmel.— Zool. Jahrb., Abt.
2, 53: 119—316.
SweetW. C. 1959. Muscleattachment impression in some Paleozoic Nautiloid Cephalo-
pods.— J. Paleontol., 33, 2: 293—304.
Termier G. et Termier H. 1952. Classe des Gasteropodes. Dans: «Traite de paleonto-.
logie», Piveteau J. (ed.), v. II, p. 365—460. Paris.
Thiele J. 1891. Die Stammesverwandschaft der Mollusken. Jena.— Z. Naturwiss., 25:
480—543.
Thiele J. 1895. Uber die Verwandtschaftsbeziehungen der Amphineuren.— Biol. СЫ.,
15: 859—889.
Thiele J. 1902.'I. Die systematische Stellung der Solenogastren und die Phylogenie
der Mollusken.— Z. wiss. Zool., 72 : 249—466.
Thiele J. 1929, 1931, 1935. Handbuch der systematischen Weichtierkunde. Jena.
Thompson T. E. 1960. The development of Neomenia carinata Tullborg (Mollusca,
Aplacophora).— Proc. Roy. Soc. London, B, 153, 951: 263—278.
Thorson G. 1946. Reproduction and larval development of Danish marine bottom
invertebrates with special reference to the planktonic larvae in the Sound (0re-
sund).— Medd. Komm. Danmarks Fisk. Havundersogelser, Kjabenhavn, Ser. Plan-
kton, 4, I: 1—523.
Ulrich W. 1959. Neu entdeckte Tiere von allgemeiner zoologischer Bedeutung.— Der
mathematische und naturwissenschaftliche Unterricht, XII, 6: 262—273.
Wenz W. 1938. Gastropoda. In: Schindewolf О. H. «Handbuch der Palaozoologie»,
Bd. IV. Berlin.
Wenz W. 1940. Ursprung und Stammesgeschichte der Gastropoden.— Arch. Mollusken-
kunde, 72, Г: 1—10.
Wilbur К. M- and Yonge С. M. 1964. Physiology of Mollusca, v. I. N. Y. and Lon-
don, Acad. Press, p. ХШ+473.
Wilhelmi R. W. 1944. Serological relationships between the Mollusca and other
invertebrates.— Biol. Bull., 87: 96—105.
Wiren A. 1892. Studien fiber die Solenogastres. I. Monographie der Chaetoderma niti-
dulum Loven.— Kgl. svenska vetenskaps akad. handl., 24, 12: 1—66.
Wiren A. 1893. Studien fiber die Solenogastres. 2. Monographie der' Chaetoderma pro-
duction, Neomenia, Proneomenia acuminata.— Kgl. svenska vetenskaps akad. handl.,
25, 6: 1—100.
Yonge С. M. 1941. The Protobranchiate Mollusca, a functional interpretation of their
structure and evolution.— Philos. Trans. Roy. Soc. London, Ser. B, 230: 79—147.
Yonge С. M. 1947. The pallial organs in the Aspidobranch Gastropoda and their
evolution throughout the Mollusca.— Phil. Trans. Roy. Soc. London, Ser.. B, 232:
443—518.
Yonge С. M. 1949. On the structure and adaptations of the Tellinacea deposit-feeding
Eulamellibranchia.— Philos. Trans. Roy. Soc. London, Ser. B. 234: 29—76.
Yonge С. M. 1953. The Monomyarian in the Lamellibranchia.— Trans. Roy. Soc. Edin-
burgh, 62, 443—478.
Yonge С. M. 1957a. Mantle fusion in the Lamellibranchia.— Pubbl. Staz. Zool. Na-
poli, 29: 151—171.
Yonge С. M. 1'9576. Reflexions on the Monoplacophoran, Neopilina galatheae Lemche.—
Nature, 179, 4561: 672—673.
Yonge С. M. 1959a. The status of the Protobranchia in the bivalve Mollusca.— Proc.
Malacol. Soc. London, 33: 210—214.
Yonge С. M. 19596. Evolution within the bivalve Mollusca.— Proc. XVth 'Internal.
Zool. Congr. London: 367—370. “i
Young J. Z. 1960. Regulation in the-retina and optic lobes of Octopus in relation to
form discrimination.— Nature, 186, 4728: 836—839.
АННЕЛИДЫ (ANNELIDA)
Вагин В. Л. 1954. Asteromyzostomum n. gen.— новый представитель класса мизосто-
мид (Annelides).— Тр. Ленингр. об-ва естествоисп., 72, 4: 16—36.
Геккер Р. Ф., Ушаков П. В. 1962. Vermes. В кн. «Основы палеонтологии». Губ-
ки, археоциаты, кишечнополостные, черви. Под ред. Б. С. Соколова. М., АН
СССР, стр. 435—464.
Заленский В. Д. 1915. Исследования по морфологии и систематике Hirudinea. I
Организация Ichthyobdellidae, т. II, стр. VIII+256.
Иванов П. П. 1944. Первичная и вторичная метамерия тела.— Ж. общ. биол., 5,
2: 61—95.
Изосимов В. В. 1934. Agriodrilus vermivorus Michaelsen и его отношение к фило-
гении пиявок.-— Уч. зап. Казанск. ун-та, Зоология, 2: 5—66.
387
Изосимов В. В. 1940. Класс малощетинковых (Oligochaeta). В кн. «Руководство
по зоологии», т. II. Беспозвоночные. М.— Л., Изд-во АН СССР, стр. 157—204.
Ласточкин Д. А. 1922. Регенеративные явления у Naididae.— Изв. Иваново-Воз-
несенск. политехи, ин-та, 6: 1—74.
Ливанов Н А. 1905. Acanthobdella peledina Grube, 1851.— Уч. зап. Казанск. ун-та,
72: 1—267,
Ливанов Н. А. 1914. Пограничные образования Polychaeta и общее морфологическое
значение таких образований.— Тр. Об-ва естествоисп. Казанск. ун-та, 4.6, 2: 1—286.
Ливанов Н. А. 1940. Класс пиявок (Hirudinea). В кн. «Руководство по зоологии»,
т. II. Беспозвоночные. М.— Л., Изд-во АН СССР, стр. 205—257.
Мейер Е. 1893. Организация трубчатых червей семейства Serpulidae и Hermellidae
как результат их сидячего образа жизни.— Тр. Об-ва естествоисп. Казанск. ун-та,
26, 3: 1—131.
Мейер Е. 1898. Исследования над развитием кольчатых червей.— Тр. об-ва естество-
исп. Казанск. ун-та, 31, 4: 1—356.
Светлов П. Г. 1923а. Ранние стадии Rhynchelmis limosella Hoffmeister.— Изв. Биол.
н.-и. ин-та Пермск. ун-та, 2, 4: 141—152.
Светлов П. Г. 19236. Ранние стадии развития Bimastus constrictus R. (сем. Lumbri-
cidae).— Изв. Биол. н.-и. ин-та Пермск. ун-та, 1, 7—8: 101—НО.
Светлов П. Г. 1926. Эмбриональное развитие в сем. Naididae.— Изв. Биол. н.-и.
ин;та Пермск. ун-та, 4, 8: 359—372.
Светлов П. Г. 1928. Исследования над развитием дождевых червей.— Тр. Особ. зоол.
лабор. и Севастоп. биол. ст. АН СССР, сер. II, 13: 95—329.
Свешников В. А. 1958а. Морфология личинок некоторых Eunicemorpha (Poly-
chaeta).—Докл. АН СССР, 121, 3: 565—568.
Свешников В. А. 19586. Новые для Белого моря виды полихет.— Зоол. ж., 37, 1:
20—26.
Свешников В. А. 1959а. Типы размножения и развития многощетинковых червей
в связи с их географическим распространением.— Зоол. ж., 38, 6: 829—841.
Свешников В. А. 19596. Биология размножения и развития полихет Белого моря.
Автореф. канд. дисс. МГУ, стр. 1—14.
Свешников В. А. 1960. Пелагические личинки некоторых полихет Белого моря.—
Зоол. ж., 38, 6: 829—841.
Свешников В. А. 1961. Пелагические личинки полихет Белого моря — Зоол. ж,
40,2:164—177.
Свешников В. А. 1963а. О типах личинок полихет (Polychaeta).— Докл. АН СССР,
150, 6: 1393—1396.
Свешников В. А. 19636. Биоценотические связи и условия существования некото-
рых кормовых беспозвоночных инфауны литорали Кандалакшского залива Белого
моря.— Тр. Кандалакшей, гос. заповедника, 4. Тр. Беломорск, биол. ст. МГУ 2:
114—134.
Ушаков П. В. 1955. Многощетинковые черви Дальневосточных морей СССР. Опре-
делители по фауне СССР. ЗИН АН СССР, М.— Л., т. 56: 1—455.
Федотов Д. М. 1915. Protomyzostomum polynephris и его отношение к Gorgonocepha-
lus eucnemis. Пг., стр. VIII + 240.
Федотов Д. М. 1937. Специализация и деградация в строении мизостомид под влия-
нием образа жизни. В сб.: «Акад. Н. В. Насонову к 80-летию со дня рождения
и 60-летию научной деятельности». М., Изд-во АН СССР, стр. 52—81.
Чекановская О. В. 1960. Дождевые черви и почвообразование. М.— Л., Изд-во
АН СССР, научно-популяр. серия, стр. 1—206.
Шмидт Г. А. 1917. К развитию энтодермы у Protoclepsis tesselata (Muller).— Дневн
Зоол. отд. об-ва любит, естествозн. антропол. и этногр., новая серия, 4, 1: 1—22.
Шмидт Г. А. 1922. Исследования по эмбриологии кольчатых червей. К вопросу о раз-
витии энтодермы у Rhynchelmis limosella Hoffm.— Русск. зоол. ж., 3, 1—2: 74—93.
Шмидт Г. А. 1923. Исследования по эмбриологии кольчатых червей. К зародышево-
му развитию Archaeobdella esinontii О. Grimm.— Русск. зоол. ж., 3, 3—4: 427—435.
Шмидт Г. А. 1925. Исследования по эмбриологии кольчатых червей. Полярные плаз-
матические массы в яйце Protoclepsis tesselata.— Русск. зоол. ж., 5, 1—2: 138—164.
Шмидт Г. А. 1941а. Дробление яйца Protoclepsis tesselata О. F. Muller.— Тр. Ин-та
цитол., гистол. и эмбриол. АН СССР, 1, 1: 145—182.
Шмидт Г. А. 1:9416. Исследования по сравнительной эмбриологии, пиявок. Сообще-
ние 10. Ранние стадии зародышевого развития рыбьих пиявок (Ichthyobdellidae).—
Сб. «Памяти акад. А. Н. Северцова», 2, 2: 357—489.
Ш умки на О. Б. 1953. Эмбриональное развитие медицинской пиявки.— Тр. Ин-та
морфол. животных АН СССР, 8 : 216—254.
Anderson D. Т. 1959. The embryology of the Polychaete Scoloplos armiger.— Quart.
J. Microscop. Sci., 100, 1: 89.
В i n a r d A. et R. J e e n e r. 1927. Sur 1’existence de cavites coelomiques dans le segment
palpaire des Polychetes.— Rec. Inst. Zool. Torley-Rons' Sean, I, fasc. I: 173—178.
Dales R. Ph. 1950. The reproduction and larval development of the Nereis diversicolor
O. F. Miiller.— J. Marine biol. Assoc. U. K., 29: 321—360.
388
Dales R. Ph. 1952. The development and structure of the anterior region of the body
in the Sabellariidae, with special reference to Phragmatopoma californica.— Quart.
J. 7Aicroscop. Sci., 93 : 435—452.
Dales R. Ph. 1962. The Polychaete stomodeum and the interrelationships of the fami-
lies of Polychaeta.— Proc. Zool. Soc. London. 139, 3: 389—428.
Dawydoff C. 1959. Ontogenese des Annelides. Dans: Gr asse P. P. «Traite de Zoolo-
gie», t. V, I, p. 594—686.
Fedotov D. M. 4929. Beitrage zur Kenntnis der Morphologie der Myzostomiden.— Z.
Morphol. und Okol. Tiere, 15: 156—191.
Ferris G. F. 1953. On the comparative morphology of the Annulata.— Microentomo-
logy, 18, 1: 2—15.
Glaesner M. F. 1958. New fossils from the base of the Cambrian in South Australia.—
Trans. Roy. Soc. South Australia, 81: 185—188.
Graff L. 1888. Die Annelidengattung Spinther.— Z. wiss. Zool., 46: 1—66.
Haffner K. von. 1959. Untersuchungen und Gedanken liber das Gehirn und Kopfende
von Polychaeten and ein Vergleich mit der Arthropoden.— Mitt. Hamburg. Zool
Museum und Inst., 57: 1—29.
Henry L. M. 1947. The nervous system and the segmentation of the head in the An-
nulata.— Microentomology, 12: 65—82.
Henry L. M. 1948. The nervous system and the segmentation of the head in the An-
nulata. Section Onychophore.— Microentomology, 13: 27—64.
Iwanoff P. P. 1928. Die Entwicklung’der Larvalsegmenten bei den Anneliden.— Z.
Morphol. und Okol. Tiere, 10: 62—161.
Jagersten G. 1934. Studien fiber den histologischen Bau der mannlichen Geschlecht-
sorgane und die Ausbildung des Spermium.bei Myzostomum.— Zool. Bidr., Uppsa-
la, 15: 1—22.
Jagersten G. 1939a. Zur Kenntnis der Larvenentwicklung bei Myzostomum.— Arkiv
zool., 31 A, 11: 1—21.
Jagersten G. 19396. Ueber die Morphologie und Physiologic des Geschlechtsapparats
und den Kopulalionsmccanismus der Myzostomum.— Zool. bidrag. Uppsala, 15: 163—
242.
Jagersten G. 1940a. Zur Kenntnis der Morphologie, Entwicklung und Taxonomie der
Myzostomida.— Nova acta Soc. Scient. Uppsala, 11, 8: 1—84.
Jagersten G. 19406. Zur Kenntnis der ausseren Morphologie, Entwicklung und Oko-
logie von Protodrilus rubropharyngeus n. sp.— Arkiv. zool., Stockholm, 32, A, 16:
1—19.
Jagersten G. 1947. On the structure of the pharynx of the Archiannelida with special
reference to there-occuring muscle cells of aberrant type.— Zool. Bidr., Uppsala,
' 25: 551—570.
Jagersten G. .1952. Studies on the morphology, larval development and biology Pro-
todrilus.— Zool. Bidr., Uppsala, 29: 425—512.
Kaiser F. 1954. Beitrage zur Bewegungsphysiologie der Hirudineen.— Zool. Jahrb.
Abt. 1|, 65: 59—90.
L i п к e O. 4939. Die Biota des Jadebusenwates.— Helgol. wiss. Meeresuntersuch., 1:
201—348.
Livanow N. 1931. Die Organisation der Hirudineen und die Beziehungen dieser Grup-
pe zu den Oligochaten.— Ergebn. und Fortschr. Zool., 7: 378—484j.
Marcus E. 1944a. Notes on fresh-water Oligochaeta from Brazil.— Comun. Zool. Mu-
seum Hist. Nat. Montevideo, I, 20: 1—8.
Marcus E. 19446. Sobre oligochaeta limnicos do Brasil.— Bol. Fac. Fil. Cient. Letr.
Univ. S. Paulo, 43, Zool., 8: 5—135.
Penners A. 4922. Die Furchung von Tubifex rivulorum.— Zool. Jahrb. Anat., 43: 323—
368.
Penners A. 1924. Experimentelle Untersuchungen zum Determinationproblem am Keim
von Tubifex.— Arch, microscop, anal., 102: 51—100.
Penners A. 1926a. Die Entwicklung des Keimstreifs und Organbildung bei Tubifex
rivulorum.— Zool. Jahrb. Anat., 45; 251—308.
Penners A. 19266. Experimental^ Untersuchungen zum Determinationsproblem am
Keim von Tubifex. II.— Z. wiss. Zool., 127: 1—140.
Schmidt G. A. 1922. Die Embryonalentwicklung von Piscicola geometra — Zool. Anz.,
53: 123—127.
Schmidt G. A. 1924. Untersuchungen liber Embryologie der Anneliden. 2. Die Beson-
derheiten der Embryonalentwicklung der Ichtyobdellidae und Hire Entstehung.—
Zool. Jahrb. Anat., 46: 199—244.
Schmidt G. A. 1925. Untersuchungen fiber die Embryologie der Annelliden. Die Em-
bryonalentwicklung von Piscicola geometra.— Zool. Jahrb. Anat., 47: 319—428.
Schmidt G. A. 1937. Vergleichende embryologische Studien fiber die Typen der Em-
bryonalanpassungen bei Hirudineen und Nemertinen,— Z. Morphol. Okol. Tiere, 32:
650—671.
Slewing' R. 1953. Morphologische Untersuchungen am «Kopf» der Pisioniden (Pisione
puzae n. sp., Annelidae, Polychaeta).— Zool. Anz., 150, 1—2: 298—313.
25' Д. M. Федотов 389
Stephenson J. 1930. The Oligochaeta. Oxlord, p. XVI+978.
S w e d m a г к В. 1965. Recherches sur la morphologie, le deveioppement et la biofogre
de Psammodrilus balanoglossoides.— Arch. Zool. exptl et gen., 92, 8: 141—219.
Tannreuter G. 1915. The embryology of Bdellodrilus philadelphicus.— J. Morphol., 26..
VejddvskyF. 1888—1892. Entwicklungsgeschichtliche Untersuchungen. Prag.
Walcott C. D. 1911. Cambrian geology and paleontology. Middle Cambrian annelids.—
Smiths. Misc. Collect., 57: 109—144.
ТАРДИГРАДЫ (TARDIGRADA)
Cooper K- W. 1964. The first fossil Tardigrade: Beorn leggi Copper, from Cretaceous
Amber.— Psyche., 71, 2: 41—48.
Cue not L. 1949. Tardigrada. Dans: Grasse P. P. «Traite de zoologie», t. VI, p. 39—59.
Erlanger R. 1894, 1895. Zur Morphologie und Embryologie eines Tardigraden (Mac-
robiotus macronyx).— Biol. Zbl., 14: 582—585; 15: 772—777.
Erlanger R. .1895. Beitrag zur Morphologie der Tardigraden I. Zur Embryologie eines
Tardigraden, Macrobiotus macronyx.— Morphol. Jahrb., 22: 491—513.
Graff L. 1877.. Das Genus Myzostoma (F. S. Leuckart). Leipzig, S. VIII+ 82. .
Hanstrom B. 1928. Tardigrada. In «Vergleichende Anatomie der Nervensystems der
wirbellosen Tiere unter Berficksichtigung seiner Funktion». Berlin, S. 334—338.
Marcus E. 1929. Tardigrada. In Bronn’s «Klassen und Ordnungen des Tierreichs»,.
Bd. 5, Abt. 4, Buch 3, S. VIII+608.
Marcus E. 1958. On the Evolution of the animal Phyla.—Quart. Rev. Biol., 33: 24—58.
Plate L. 1888. Beitrage zur Naturgeschichte der Tardigraden.— Zool. Jahrb., Abt 2,
3: 487—550.
ПЕНТА СТОИМ ДЫ (PENTASTOMIDA)
Ежиков И. И. 1939. К морфологии Pentastomum denticulatum.— Тр. Ин-та морфол.
животных АН СССР, 2, 5: 3—22.
ёжиков И. И. 1951. Класс пятиусток (Pentastomida). В кн. «Руководство по зооло-
гии», III, 2: 9—22.
Haffner К. von. 1922—1925. Beitrage zur Kenntnis der Linguatuliden. I. Ovarium
und Eibildung von Porocephalus armillatus (Wyman).— Zool. Anz., 54: 162—170.
II—1922. Zur Eireifung von Porocephalus armillatus (Wyman).— Zool. Anz., 54:
170—177.
Haffner K. von. 1924. Beitrage zur Kenntnis der Linguatuliden. III. Uber der Hoden
und die Spermatocystenbildung von Porocephalus moniliformis.— Zool. Anz., 58;
'285—290. IV. 1924. Die Korpermuskulatur von Porocephalus armillatus.— Zool. Anz.,
59: 270—276. V. 1924. Drusen von Porocephalus armillatus.— Zool. Anz., 60: 126—136.
VI. 1925. Uber das Nervensystem von Porocephalus armillatus.-.— Zool. Anz., 6h
183—186. VII. 1925. Die systematische Stellung der Linguatuliden.— Zool. Anz., 61:
209—218.
Haffner K. von. 1926. Die Sinnesorgane der Linguatuliden nebst einer Betrachtung
fiber die systematische Stellung dieser Gruppe.— Z. wiss. Zool., 128, 2: 201—252.
Heymons R. 1926. Pentastomida. In W. Kiikenthal. «Handbuch der Zoologie»,
Bd. Ill, 1, S. 69—131.
Heymons R. 1935. Pentastomida. In Bronn’s «Klassen und Ordnungen des Tierreichs»,
Bd. 5, Abt. 4, Buch 1, Teil. 2, S. 1—268.
Jeschikov J. 1940. Zur Morphologie von Pentastomum denticulatum.— Zool. Anz., 132,
11/12: 263—279.
Nicoli R. M. 1963. Phylogenese et systematique de Phylum des Pentastomida.— Ann.
parasitol. humaine et comparee, 38, N 2: 483—516.
Osche G. 1959. «Arthropodencharactere» bei einem Pentastomiden Embryo {Reighardia
sternae).— Zool. Anz., 163, 5/6: 169—178.
Spencer W. B. 1893. The anatomy of Pentastomum teretiusculum (Baird).— Quart. J.
Microscop. Sci., 34; 1—73.
ОНИХОФОРЫ (ON l/CHOPHORA)
Кузнец.ов H. Я. 1951. Подтип трахейнодышащих членистоногих. В кн. «Руководство
по зоологии», III, 2: 107—123.
Butt F. Н. 1957. The role of premandibular or intercalary segment in head segmentation
of the insects and other Arthropods.— Trans. Amer. Entomol. Soc., 83: 1—30.
Butt F. H. 1959. The structure and some aspects of development of the onychophoran
head. Smithsonian Misc. Collect., 137: 46—60.
В u 11 F. H. 1960. Head development in the Arthropo'ds.— Biol. Revs, 35, 1: 43—9'1.
Cuenot L. 1926. L’entonnoir. vibratile de la nephridie des Peripatus. Raports des Pe-
ripates et des Arthropodes.—Ann. Soc. roy. zool. Belgique, 56: 13—17.
E i s i g H. 1887. Monographic der Capitelliden des Golfes von Neapel. Fauna und Flora
des Golfes von Neapel. Berlin, S. XXV+906.
Fahlander K. 1938. Beitrage zur Anatomie und systematischen Einteilung der Chilo-
poden.— Zool. Bidr., Uppsala, 17: 1—148.
390
Fedorov В. 1926. Zur Anatomie des Nefvensvstems von Peripatus.— Zool. Jahrb.,
Abt. 2, 48: 273—332.
Fedorov B. 1929. Zur Anatomie des Nervensystems von Peripatus..— Zool. Jahrb.,
Abt. 2, 50: 279—352.
Haeckel E. 1896. Systematische Phylogenie der wirbellosen Tiere, Bd. 2. Berlin.
S. XVIII+ 7209.
Haffner K. von. 1959. Untersuchungen und Gedanken fiber das Gehirn und Kopfende
von Polychaeten und ein Vergleich mit den Arthropoden.— Mitt. Hamburg. Zool.
Museum und Inst., 57: 1—29.
Hanstrom B. 1928. Vergleichende Anatomie des Nervensystems der wirbellosen Tiere
unter Berficksichtigung seiner Funktion. Berlin.
Hanstrom B. 1935. Bemerkungen fiber das Gehirn und die Sinnesorgane der Onycho-
phoren.— Lunds Univ. Arsskr. Avd. 2, Bd. 31, 5: 3—37.
Heymons R. <1928. Uber Morphologie und verwandtschaftliche Beziehungen des Xe-
nusion auerswaldae Pomp, aus dem Algonkium.— Z. Morphol. OkoL Tiere, 10: 307—
329.
Holmgren N. 1916. Zur vergleichenden Anatomie des Gehirns von Polychaeten, Ony-
chophoren, Xiphosuren, Arachniden, Crustaceen, Myriapoden und Insekten.— Kgl.
Svensk. vetenskapsakad. handl., N. F., 56, I: 1—303.
Kennel J. 1884, 1888. Entwicklungsgeschichte von Peripatus edwardsii- Blanch, und
Peripatus torquatus n. sp.— Arb. Zool. Inst. Wiirzburg, 7: 95—229; 8: 1—93.
Korschelt E. und H eider K. 1909. Lehrbuch der vergleichenden Entwicklungsge-
schichte der Wirbellosen Tiere. Jena. Allg. Teil, IV. S. 167—470.
Manton S. M. 1937. The feeding, digestion, excretion and food storage of Peripa-
topsis.— Philos. Trans.,,B, 227: 411—464,
Manton S. M. 1949. Studies on the Onychophora, VII. The early embryonic stages of
Peripatopsis, and some general considerations concerning the morphology and phy-
logeny of the Arthropoda.— Philos trans. B, 233: 483—580.
Manton S. M. 1958. Habits of life and evolution of body design in Arthropoda.— J.
Linnean Soc. London (Zool.), 44, 295: 58—72.
Milne L. J. and Milne M. J. 1954. The worm that didn’t turn — into anything. (Peri-
patus).— Natur. History, N. Y„ 63: 182—187.
Pflugfelder O. 1948. Entwicklung von Paraperipatus ambionensis.— Zool. Jahrb.,
Abt. 2, 69.
Sedgwick A. 1885. The development of Peripatus capensis. Pt 1.— Quart. J. Micros-
cop. Sci., 25. 449—468.
Sedgwick A. 1886. The development of the Cape species of Peripatus, Pt II.— Quart.
J. Microscop. Sci., 26: 175—212.
Sedgwick A. 1888. A monograph on the species and distribution of the genus Peripatus
(Guibling).— Quart. J. Microscop. Sci., 28; 431—493.
Sedgwick A. 1909. Peripatus. Nat. History, Cambridge, 5: 3—26.
Sheldon L. 1888a. On the development of Peripatus novaezealandiae.— Quart. J. Mic-
roscop. Sci., 28: 205—237.
Sheldon L. 18886. Notes on the anatomy of Peripatus capensis and Peripatus novae-
zealandiae.— Quart. J. Microscop. Sci., 28: 496—499.
Snodgrass R. E. 1938. Evolution of the Annelida, Onychophora, and Arthropoda.—
Smithsonian Misc. Collect., 97, 6: 1—159.
Walcott C. D. 1911. Cambrian geology and paleontology. II, 5. Middle Cambrian an-
nelids. Smithsonian Misc. Collect., 57: 109—144.
Whitman С. O. 1891. Spermatophores as a means of hypodermic impregnation.— J.
Morphol., 4: 361—406.
ЧЛЕНИСТОНОГИЕ (ARTHROPODA)
Беккер Э. Г. 1949. К эволюции наружного скелета и мускулатуры Tracheata (Atelo-
cerata). Сообщение 2. Плейральный и стернальный скелет и мускулатура Chilopoda
. Epimorpha.— Зоол. ж., 28, 1: 39—58.
Беккер Э. Г. 1950. Локомоторные органы и эволюция трахейнодышащих членисто-
ногих (Tracheata).— Вести. Моск, ун-та, 10: 91—106.
Б и р ш т е й н Я. А. 1948. Новые взгляды на филогению членистоногих.— Усп. соврем,
биол., 25, 1: 143—149.
Гиляров М. С. 1956. Роль почвы в происхождении и эволюции насекомых.— Энто-
мол. обозрение, 35, 3: 487—494.
Дубинин В. Б. 1959. О филогенезе хелицеровых животных подтипа Chelicerophora
W. Dub. и о родстве Chelicerata с Pycnogonides.— Тр. Ин-та морфол. животных
АН СССР, 27; 134—150.
Дубинин В. Б. 1962. Надкласс Chelicerata. В кн. «Основы палеонтологии». Члени-
стоногие. Трахейные и хелицеровые. Ред. Б. Б. Родендорф. М., Изд-во АН СССР,
1962, стр. 375—524.
Иванов П. П. 1940. Эмбриональное развитие сколопендры в связи с эмбриологией
и морфологией Tracheata.— Изв. АН СССР, отд. биол. наук, 6: 831—860.
25* 391
Иванов П. П. 1944. Первичная и вторичная метамерия тела.— Ж- общей биол., 5:
61—95.
Кеннель И. 1891. Родственные отношения членистоногих.— Зап. об-ва естествоисп.
(Юрьев) 7.
Л и гн а у Н. Г. 1911. История эмбрионального развития Polydesmus abchasius Attems.
Одесса, стр. 1—249.
М а р т ы н о в А. В. 1934. Myriopoda. В кн. В. К. Ц и т т е л ь. «Основы палеонтологии », т. I.
Беспозвоночные, Изд-во АН СССР, стр. 984—986.
Мартынов А. В. 1938. Очерки геологической истории и филогении отрядов насеко-
мых (Pt'erygota), I. Palaeoptera и Neoptera-Polyneoptera.— Тр. Палеонтол. ин-та
АН СССР, 7, 4: 4—149.
Мартынова О. М. 1959. Филогенетические взаимоотношения насекомых мекопте-
роидного комплекса.— Тр. Ин-та морфол. животных АН СССР, 27: 221—230.
Педашенко Д. Д. 1898. Эмбриональное развитие и метаморфоз Lernaea branchialis
L.— Тр. СПб об-ва естествоисп., 26, 4: 1—307.
Родендорф Б. Б. 1947. Родословное древо беспозвоночных. В кн. «Развитие жизни
на земле», вып. 51. Под ред. Р. Ф. Геккера. М., Госкультпросветиздат, стр. 27—32.
Родендорф Б. Б. 1961. Описание первого крылатого насекомого из девонских от-
ложений Тимана (Insecta, Pterygota).— Энтомол. обозрение, 40, 3: 485—489.
Родендорф Б. Б. (ред.) 1962. Основы палеонтологии. Членистоногие, трахейные и
хелицеровые. М., Изд-во АН СССР, стр. I1—560.
Федоров Б. Г. 1931. Общая морфология головного мозга метамерных беспозвоноч-
ных.— Тр. 4-го всем, съезда зоол., анатом, и гистол. в Киеве, 6—12 мая 1930,
. стр. 100—101.
Федотов Д. М. 1962. Филогенетические отношения членистоногих.— Сб. «Вопр. об-
щей зоол. и мед. паразитол.». Медгиз, стр. 256—277.
Чернышев Б. Н. 1934. Crustacea. Archaeostraca. В кн. В. К. Циттель «Основы па-
леонтологии», т. I. Беспозвоночные. Изд-во АН СССР, стр. 896—899.
Чернышева Н. Е. (ред.) 1960. Класс Trilobita. В кн. «Основы палеонтологии».
Изд-во АН СССР, стр. 1—515.
Ш а р о в А. Г. 1953. Развитие щетинкохвосток (Thysanura Apterygota) в связи с пробле-
мой филогении насекомых.— Тр. Ин-та морфол. животных АН СССР, 8: 63—127.
Шаров А. Г. 1959. О системе первичнобескрылых насекомых,—Тр. Ин-та морфол.
животных АН СССР, 27: 175—186.
Boettger С. Е. 1958. Die systematische Stellung der Apterygota.— Proc. Xth Internal.
Congr. Entomol., Montreal, 1956: 509—516.
Breuning S. 1957. Entwicklungsgeschichtliche Untersuchungen liber die Segmentierung
der Keimanlage am lebendigen Ei von Calliphora erithrocephala Meig.— Zool. Jahrb.,
Abt. 2, 75: 501—580.
Butt F. H. 1957. The role of the premandibular or intercalary segment in head segmen-
tation of the insects and other Arthropods.— Trans. Amer. Entomol. Soc., 73: 1—30.
В u 11 F. H. 1960. Head development in the Arthropods.— Biol. Revs., 35, 1: 43—91.
Carpenter G. H. 1903. On the relationships between the classes of the Arthropoda.—
Proc. Roy. Irish Acad., 24, Sect. B.
Carpenter G. H. 1906. Notes on the segmentation and phylogeny of the Arthropoda,
with an account of Maxillae in Polyxenus lagurus.— Quart. J. Microscop. Sci., 49:
469—491.
Crampton G. C. 1919. The evolution of Arthropoda and their relations with especial
reference to insects.— Amer. Naturalist, 53: 143—179.
E vans H. E. 1959. Some comments on the evolution of the Arthropoda.— Evolution, 13:
147—149.
Fedotov D. M. 1924. On the relation between the Crustacea, Trilobita, Merostomat"
and Arachnida.— Bull. Acad. Sci., Leningrad: 383—408.
Henriksen K. L. 1928. Critical notes upon some Cambrian arthropods described h-
D. Walcott.— Vid. medd. Dansk naturhistor. foren, 86. 1—20.
Henriksen K- L. 1929. Contribution to the interpretation of the cephalic segment of
Arthropoda.— Proc. IVth Internal. Congr. Entomol., Ithaca, 1928, 2: 589—593.
Heymons R. 1891. Die Entwicklungsgeschichte der Scolopender.— Bibl. Zool., 33 Stutt-
gart: 1—244.
Hinton H. E. 1958. The phylogeny of the panorpoid orders.— Annual Rev. Entomol.,
3: 181—200.
Holmgren N. 1916. Zur vergleichenden Anatomie des Gehirns von Polychaeten, Ony-
chophoren, Xiphosuren, Arachniden, Cfustaceen, Myriapoden und Insekten.— Kgl.
svenska vetenskapsakad. handl., 56: 1—303.'
Hupe P. 1953. Classification des Trilobites.— Ann. paleontol. Paris, 39: 61—110.
Iwanov P. 1933. Die embryonale Entwicklung von Limulus moluccanus.— Zool. Jahrb.
Abt. 2, 56: 163—348.
J a e k e 1 O. 1909. Ueber die Agnostiden.— Z. Dtsch. geol. Ges. Abh., 61, 3: 380—400.
Kaestner A. 1931. Die Hiifte und ihre Umformung zu Mundwerkzeugen bei den Arach-
niden.— Z. Morphol. und Okol. Tiere, 22, 4: 721—752.
Kingsley I. S. 1897. The systematic position of the Trilobites.— Amer. Geologist, 20.
392
Legendre R. 1959. Contribution a Tetude du systeme nerveux des Araneides.—Ann.
Sci. nat. (Zool.) (12), Paris, p. 339—473.
Man ton S. M. 1958. Habits of life and evolution of body design in Arthropoda.— J.
Linnean Soc. London (Zool.), 44, 295: 58—72.
Manton S. M. 1964. Mandibular mechanism and the evolution of Arthropods.— Philos.
Trans. Roy. Soc. London, Ser. B, 247: 11—183.
Novojilov N. 1957. Un nouvel ordre d’Arthropodes particuliers: Kazacharthra du Lias
,des Monts Ketmen (Kazakhstan SE).— Bull. Soc. geol. France, 6 ser, 7: 471—-185.
Novojilov N. 1959. Position systematique des Kazacharthra (Arthropodes) d’apres
du nouveaux materiaux des monies Ketmen et Sajkan (Kazakhstan SE et NE).—
Bull. Soc. geol. France, 7 ser., 1: 265—269.
Packard A. S. 1903. Hints of the classification of the Arthropoda; the group of poly-~
phyletic one.— Proc. Amer. Philos. Soc., 42: <142—161.
Plate L. 1922. Allgemeine Zoologie und Abstammungslehre. Jena, I, S. 1;—629.
R a w F. 1957. Origin of Chelicerates.— J. Paleontol., 31, 1: 139—192.
Ray Lankester E. 1904. The structure and classification of the Arthropoda.— Quart.
J. Microscop. Sci., 47: 523—576.
Raymond P. E. 1935. Leanchoilia and other Mid-Cambrian Arthropoda.— Bull. Mu-
seum Compar. Zool. Harvard Coll., 76, 6: 205—230.
R о g e r J. 1953. Sous-classe des Malacostraces. Dans: Traite de Paleontologie., 3, p. 309—
378.
Sanders H. L. 1957. Cephalocarida and Crustacean phylogeny.—System. Zool., 6, 3:
112—128.
Sanders H. L. 1959. The significance of the Cephalocarida in Crustacean phylogeny.—
Proc. XVth Internal. Congr. Zool., London, 16—23 July, 1958: 337—340.
Seidel F. 1935. Der Anlagenplan im Libellenei, zugleich eine Untersuchung fiber die
allgemeinen Bedingungen ffir defekte Entwicklung und Regulation bei dotterreichen
Eiern.— Arch, entwicklungsmech. org., 132: 671—751.
Schulze P. 1936. Trilobita, Xiphosura, Acarina.— Z. Morphol. und Okol. Tiere, 32, 2:
181—226.
Schulze P. 1939. Bemerkenswerte paleozoische Arthropoden.— Z. Morphol. und OkoL,
Tiere, 35: 169-M82.
Siewing R. 1956. Untersuchungen zur Morphologie der Malacostraca (Crustacea).—
Zool. Jahrb. Abt. 2, 75; Jena<
Siewing R. 1960a. Zum Problem der Polyphylie der Arthropoden.— Z. wiss. Zool., 164,
3/4 : 238—270.
Siewing R. 19606. Neue Ergebnisse der Verwandtschaftsforschung bei den Crusta-
ceen.— Wiss. Z. Univ. Rostock, Math.-naturwiss. Reihe, 3: 343—358.
Siewing R. 1963. Zum Problem der Arthropodenkopfsegmentierung.— Zool. Anz., 170,
11/12:429—468.
Snodgrass R. E. 1938. Evolution of the Annelida, Onychophora and Arthropoda.—
Smithsonian Misc. Collect., 96, 6: 1—159.
Snodgrass R. E. 1960. Facts and theories concerning the insect head.— Smithsonian
Misc. Collect., 142, 1: 1—61.
S о 11 a u d E. 1923. Recherches sur Г embryogenie des Crustaces Decapods de la sous-fa-
mille des Palaemoninae.— Bull. biol. France et Belgique. Suppl., 5, 5: 1—234.
Storm er L. 1942. Studies on trilobite morphology. II.— Norsk geol. tidsskr., 21: 49—
164.
St ormer L. 1944. On the relationships and phylogeny of fossil and recent Arachnomor-
pha. Skr. norske Vidensk — Akad. Oslo, 5: 1—158.
St ormer L. 1955a. Chelicerata. In: Treatise on invertebrate paleontology. Moore R. C.
(ed.). Pt. P. Arthropoda, 2.
S former L. 19556. Merostomata. In: Treatise on invertebrate paleontology. Moore
R. C. (Ed.), Pt. P„ Arthropoda, 2.
Stormer L. 1956. A Lower Cambrian Merostome from Sweden.—Arkiv zool., 9, 25:
507—514.
Stormer L. 1959. Arthropoda. General features. In: Treatise on invertebrate paleonto-
logy. Moore R. C. (Ed.). Pt. O, Arthropoda.
Stormer L. 1963. Glgantoscorpio willsi, A new scorpion from the lower Carbonife-
rous of Scotland and its associated preying microorganism. Oslo. Univ, press, p. 1—
170.
Stubblefield C. J. 1959. Evolution in trilobites.— Quart. J. Geol. Soc. London, 115,
.2: 145,—162.
T i e g s O. W. and M a n t о n S. M. 1958. The evolution of the Arthropoda.— Biol. Revs.,
' 33: 255—337.
Van del A. 1949. Generalites de la composition de I’embranchement. Dans: Gras-
se P. P. «Traite de zoologie», 6, p. 79—158.
Weber H. 1954. Grundriss der Insektenkunde. Stuttgart.
Weygoldt P. 1963. Grundorganisation und Primit iv entwicklung bei Articulaten.—
Zool. Anz, 171: 363—376.
393
Whittington H. В. 1957. The ontogeny of trilobites.— Biol. Revs., 32; 421—469.
Whittington H. B. 1961. A natural history of trilobites.— Natur. History, 70, 7: 8—
16.
ФОРОНИДЫ (PHORONIDA)
Геккер P. Ф. 1960. Класс Phoronoidea. В кн. «Основы палеонтологии». Под ред.
Т. Г. Сарычевой. Изд-во АН СССР, стр. 325—326.
Ковалевский А. О. 1867. Анатомия и история развития Phoronis.— Приложение к
XI тому Зап. СПб АН, I: 1—41.
Ливанов Н. А. 1963. Форониды, мшанки и. брахиоподы.'—Тр. об-ва естествоисп. при
Казанск. ун-те, 64: 55—82, или Уч. зап. Казанск. ун-та, 123, И: 55—82.
Педашенко Д. Д. 1898. Эмбриональное развитие и метаморфоз Lernaea branchia-
lis L—-Тр. СПб об-ва естествоисп., 26, 4: 1—246.
Шимкевич В. М. 1893. О первых стадиях развития Notopteropherus.— Вести, естест-
возн., год четвертый: 1—200.
Шульц Е. А. 1897. Образование мезодермы у Phoronis.— Тр. СПб об-ва естествоисп.,
28, I: 31—34.
Щепотьев А. С. 1907. Pterobranchia. СПб, Диссертация: 1.—248.
Becker G. 1937. Untersuchungen uber den Darm und die Verdauung von Kamptozoen,
Bryozoen und Phoroniden.— Z. Morphol. und Okol. Tiere, 33: 72—127.
Bethe A. 1927. Eigentiimliche Formen und Mittel der Blutbewegung.— Z. vergl. Physiol.,
5. ggg_____57g,
Brattstrom H. 1943. Phoronis avails.— Lunds. Univ. Aarskr., Avd., 2, 39, 2: 3—1(7.
Caldwell W. H. 1883. Preliminary note on the structure, development. and affinities
of Phoronis.— Proc. Roy. Soc. London, 34: 371—382.
Cori C. J. '1891. Untersuchungen fiber die Anatomie und Histologie der Gattung P/ioro-
nis.— Biol. Z., 11: 478—481.
С о r i C. J. 1930. Phoronoidea.— Tierwelt Deutschlands, 17: 14—24.
Cori C. J. 1939a. Phoronoidea, Entoprocta und Brachiopoda. In Bronn’s «Klassen und
Ordnungen des Tierreichs», Bd. 4, Abt. 13: 1—183. . .
С о r i C. J. 19396. Tentaculata, Phoronoidea. In: W. Kiikenthal und Th. Krumbach
«Handbuch der Zoologie», III, 2: 67—138.
Dawydoff G. 1960. Classe des Phoronidiens. In: Grasse. P. P. «Traite de Zoologie»,
X, I: 1008—1053.
Gerwerzhagen A. 1913. Das Nervensystem von Cristatella mucedo.— Z. wiss. Zool.,
107: 309—345.
Goodrich E. S. 1903. On the body-cavities and nephridies of the Actinotrocha larva.—
Quart. J. Microscop. Sci., 47: 103—121.
Harmer S. F. 1917. On Phoronis avails Strethill.— Quart. J. Microscop. Sci. 62. London,
Hyman L. H. 1958. The occuremce of chitin in the Lophophorate Phyla.— Biol. Bull.,
114: 106—112.
Hyman L. H. 1959. The Invertebrates, v. V. The Lophophorate Coelomates — Phylum
Phoronida, 228—274.
Ikeda I. 1901. Observations on the development, structure and metamorphosis of Actino-
trocha.— J. Coll. Sci. Tokyo Univ., 13: 507—592.
M a r c u s E. 1949. Phoronis avails from Brazil.— Zoologica, 14.
Mastermann A. T. 1897. On the Diplochorda, 2. The Structure of Cephalodiscus.—
Quart. J. Microscop. Sci., 40: 281—366.
M a s t e r m a n n A. T. 4900. On the Diplochorda. 3. The early development and anato-
my of Phoronis buskii.— Quart. J. Microscop. Sci., 43.
Metschnikoff I. 1871. Uber die Metamorphose einiger Seetiere, 3. Uber. Actinotrocha
Larva.— Z. wiss. Zool., 21: 233—251.
Rattembury J. C. 1954. The embryology of Phoronopsis viridis.— J. Morphol., 95:
289—333.
Safftigen A. 1888. Das Nervensystem der phylactolamen Siisswasserbryozoen.—
Zool. Anz., XI: 96—99..
Schulz E. 1903. Uber die Regenerationserscheinungen bei Phoronis.— Z. wiss Zool.,
75: 391—420, 473—494.
Selys Longschamps M. 1907. Phoronis.— Fauna und Flora .des Golfes von Nea-
pel. Monographic, 30, Berlin, p. 1—280.
Senn E. 1934. Die Geschlechtsverhaltni'sse der Brachiopoden in besonderen die Sperma-
to- und Oogenese der Gattung Lingula.— Acta zool., 15: 1—152.
Silen L. 1952. Researches on Phoronoidea of the Gullmar Fiord area (West Coast of
Sweden).— Arkiv zool., Ser. 2, 4, 4: 95—140. . .
S i 1 c n L. 1954a. Developmental biology of Phoronoidea of the Gullmar Fiord area (West
Coast of Sweden).— Acta zool., 35: 215—257.
Silen L. 19546. On the nervous system of Phoronis.— Arkiv zool., 6, 1: 1—40.
Thorson G. 1950. Reproductive and larval ecology of marine bottom invertebrates.—
Biol. Revs. Cambridge Philos. Soc., 25.
394
МШАНКИ (BRYOZOA)
Абрикосов Г. Г. 1936. О географическом распространении Lophopus cristalliniis
(Pall.) (Bryozoa, Phylactolaemata).— Тр. Зоол. музея Моск, ун-та, III: 245—249.
Абрикосов Г. Г. 1948. Класс Bryozoa. В кн. «Определитель фауны и флоры север-
ных морей СССР», М., стр. 451—461.
Абрикосов Г. Г. 1955. Мшанки. В кн. «Курс зоологии беспозвоночных», т. 1. Изд-во
«Сов. Наука», стр. 262—267.
Астрова Г. Г. 1960. Мшанки. В кн. «Основы палеонтологии». Под ред. Т. Г. Сарыче-
вой. Изд-во АН СССР, стр. 15—1'12.
Астрова Г. Г. 1964. Морфология, история развития и система ордовикских и силурий-
ских мшанок. Автореф. докт. дисс. Палеонтол. ин-т АН СССР, стр. 1—32.
Астрова Г. Т. 1965. Морфология, история развития и система ордовикских и силу-
рийских мшанок.— Тр. Палеонтол. ин-та, 106: 1—428.
Беклемишев В. Н. 1950. К проблеме индивидуальности в биологии. Колонии дву-
сторонне-симметричных животных.— Усп. соврем, биол., 29: 91—120.
Клюге Г. А. 1896. Очерк естественной истории пресноводных мшанок окрестности
г. Казани.— Уч. зап. Казанск. ун-та, 3, 4: 91—145, 146—190.
Клюге Г. А. 1961. Список видов мшанок (Bryozoa) Дальневосточных морей СССР.—
Исследования дальневосточных морей, VII. Изд-во АН СССР, стр. 118—143.
Клюге Г. А. 1962. Мшанки северных морей СССР. «Определитель по фауне СССР»,
т. 76. М,— Л., Изд-во АН СССР, стр. 1—584.
Нехорошее В. П. 1934. Bryozoa. В кн. К. Циттель «Основы палеонтологии»,
т. I. Беспозвоночные. Изд-во АН СССР, стр. 408^458.
Полянский Ю. И. 1946. Класс Bryozoa — мшанки. В кн. «Большой практикум по
зоологии беспозвоночных», т. II. Под ред. Ю. И. Полянского, М., Изд-во «Сов.
Наука», стр. 7—40.
Шульга-Нестеренко М. И. 1941. Нижнепермские мшанки Урала.— Палеонтоло-
гия СССР, V, 5, 1: .1—276.
Шульга-Нестеренко М. И. 1949. Функциональное, филогенетическое и стратигра-
фическое значение микроструктуры скелета тканей мшаиок.— Тр. Палеонтол. ин-та
АН СССР, 23: 1—66.
Basler R. S. 1953. Bryozoa. Part 9. Treatise on invertebrate paleontology.— Geol, Soc.
America and univ. Kansas Press, p. 1—253.
Becker G. 1937. Untersuchungen uber dem Darm und die Verdauung von Kamptozoen,
Bryozoen und Phoroniden.— Z. Morphol. und Okol. Tiere, 33: 72—127.
Borg F. 1926. Studies in recent Cyclostomatous Bryozoa.— Zool. Bidr., Uppsala, 10.
181—507.
Borg F. 1930. Moostierchen oder Bryozoen (Ectoprocta).— Tierwelt Deutschlands, 17:
25—142.
Borg F. 1944. Stenolaematous Bryozoa.— Further Zool. Res. Swedish Antarct. Exped.
1901—1903. Stockholm: 1—276.
В о r g F. 1953/1956. A revision of the recent Heteroporidae (Bryozoa).— Zool. Bidr., Up-
psala, 30: 253—394.
Brien P. 1953. Etude sur les Phylactolemates.— Ann. Soc. roy. zool. Belgique, 84, II:
301—444.
Brien P. 1960. Classe des Bryozoaires. Dans: Grasse P. P. «Traite de zoologie», t. V,
2, p. 1053—1335.
BugeE 1960. Bryozoaires fossiles. Dans: Grasse P. P. «Traite de zoologie», t. V, 2,
p. 1336—1379.
Duncan H. 1957. Bryozoans. In Ladd H. S. (ed.) «Treatise in marine ecology and
paleoecology»,— Mem. geol. Soc. America, 67: 783—800.
Franzen A. 1958—1962. Monobryozoon limicola n. sp., a ctenostomatous Bryozoan
from the detritus layer of soft sediment.— Zool. Bidr., Uppsala, 33: 135—148.
Fritz M. 1947. Cambrian Bryozoa.— J. Paleontol., 21: 434—435.
Gerwerzhagen 1913a. Das Nervensystem von Cristatella.— Z. wiss. Zool., 107: 309—
345.
Gerwerzhagen A. 1913. Beitrage zur Kenntnis der Bryozoen. I. Das Nervensystem
von Cristatella mucedo Cuv.— Z. wiss. Zool., 107 : 309—345.
Harmer S. F. 1930. Polyzoa.— Proc: Linnean Soc. London, 141: 68—118.
Harmer S. F. 1931. Recent work on Polyzoa.— Proc. Linnean Soc. London, 143:
113-168.
Lemche H. 1963. The origin of the Bryozoa Phylactolaemata.— Vid. medd. Dansk. na-
turhistor. foren, 125: 171—193.
Marcus E. 1925. Tentaculata, In «Biology der Tiere Deutschlands», Teil 47: 1—46.
Marcus E. 1926. Beobachtungen und Versuche an lebenden Siisswasserbryozoen.— Zool.
Jahrb., Abt. 3, 52, 4: 279—368.
Marcus E. 1934. Uber Lophopus cristallinus (Pall.).— Zool. Jahrb. Abt. 2, 58: 501—•
606.
Moore R. C. (Ed.). 1952. Invertebrate Fossils. Bryozoans, p. 156—196.
Nitsche.H. 1875.'Beitrage zur Kenntniss der Bryozoen.— Zool. Anz., XI: 96—99.
395
Pace R. M. 1906. On the early stages in the development of Flustrella hlsplda (Fabn-
cius) and on the existence of a «yolk nucleus in the egg of this form».— Quart. J.
Microscop. Sci., 50: 435—478.
Saftigen A. 1888. Das Nervensystem der phylactolamen Stisswasserbryozoen.— Zool.
Anz., 11: 96—99.
S i 1 e n L. 1935. Zur Kenntnis des Polymorphysmus der Bryozoen. Die Avicularien der
Cheilostomata Anasca.— Zool. Bidr., Uppsala, 17: 149—366.
S i 1 ё n L. 1942. Origin and development of the Cheilo-Ctenostomations stem of Bryo-
zoa.— Zool. Bidr., Uppsala, 22: 1—59.
S i 1 e n L. 1944a. The main feature of the development of the ovum, embryo and ooecium
in the ooeciferous Bryozoa Gymnolaemata.— Arkiv zool., Stockholm, 35, A, 17: 1—34.
S i 1 ё n L. 19446. On the division and movements of the alimentary canal of the Bryozoa.—
Arkiv zool., Stockholm, 35 A, 12: 1—40.
S i 1 ё n L. Ц944в. The anatomy of Labidostomella gisleni SiKn (Bryozoa Protocheilosto-
mata).— Kgl. svenska vetenskapsakad. handl., 21, 6: 1—111.
S i 1 ё n L. 1944г. On the formation of the interzoidal communications of the Bryozoa.—
Zool. Bidr., Uppsala, 22: 433—488.
Sildn L. 1947. On the spines of Flustrella (Bryozoa).— Zool. Bidr., Uppsala, 35: 134—
140.
Silen L. 1950. On the inobility of entire zoids in Bryozoa.— Acta zool., 31: 349—386.
Sildn.L. 1954. Bryozoa and Endoprocta.— Acta Univ. Lund. N. F. Avd. 2, 50, 17:
,1—41.
Svetlov P. 1935. Regulationserscheinungen an Cris/ateZ/a-Kolonien. Zum Individuali-
tatsproblem.— Z. wiss. Zool., 147: 263—274.
БРАХИОПОДЫ (BRACHIOPODA)
Бобре цк ий H. 1887. Основание зоологии. Вып. второй. II класс — Руконогие, Вга-
chiopoda, Киев, стр. 701—707.
Вержбинская Н. А. 1936. Природа фосфагена в мускулатуре щетинкочелюстных и
брахиопод и филогенетическое положение этих животных.— Физиол. ж., СССР, 18,
3: 360—365.
Иванова Е. А. 1949а. Онтогенез некоторых каменноугольных брахиопод. Сб. памяти
акад. А. А. Борисяка.— Тр. Палеонтол. ин-та АН СССР, 20: 243—267.
Иванова Е. А. 19496. Условия существования, образ жизни и история развития не-
которых брахиопод среднего и верхнего карбона Подмосковной котловины.— Тр.
Палеонтол. ин-та АН СССР, 21: ХХ+152.
Иванова Е. А. 1958. Развитие фауны в связи с условиями существования.— Тр. Па-,
леонтол. ин-та АН СССР, 59, 3: 1—300.
Иванова Е. А. 1959. К систематике и эволюции спириферид (Brachiopoda).— Па-
леонтол. ж. АН СССР, 4: 47—64.
Иванова Е. А. 1962. Экология и развитие брахиопод силура и девона Кузнецкого,
Минусинского и Тувинского бассейнов.— Тр. Палеонтол. ин-та АН СССР, 88: 1—
152.
Ковалевский А. О. 1874. Наблюдения над развитием Brachiopoda.— Изв. Об-ва
люб. естествозн. антропол. и этногр., 14: 1—40.
Конжукова Е. Д. 1948. О постэмбриональном развитии раковины и брахиального
аппарата некоторых тихоокеанских брахиопод.— Изв. АН СССР, сер. биол., 2:
213—224.
Конжукова Е. Д. 1957. Плеченогие (Brachiopoda) Дальневосточных морей СССР.—
Исследования Дальневосточных морей СССР, 4: 5—84.
Лихарев Б. К. I960. Мшанки, брахиоподы. В кн. «Основы палеонтологии». Под ред.
Т. Г. Сарычева. М., Изд-во АН СССР, стр. 115—324.
Ржонсницкая М. А. 1959. К систематике ринхонеллид.—Палеонтол. ж., 1: 25—35.
Сарычева Т. Г. 1949. Морфология, экология и эволюция подмосковных каменно-
угольных продуктид (роды Dictyoclatus, Pugilis и Antiquatonia).— Тр. Палеонтол.
ин-та АН СССР, 18: 3—304.
Сарычева Т. Г. (ред.). 1960. Мшанки, брахиоподы, форониды. В кн. «Основы палеон-
тологии». М., Изд-во АН СССР.
Сокольская А. Н. 1948. Эволюция рода Productella Hall, и смежных с ним форм
в палеозое Подмосковной котловины.— Тр. Палеонтол. ин-та АН СССР, 14, 3: 1—
168.
Сокольская А. Н. 1950. Chonetidae Русской платформы.— Тр. Палеонтол. ии-та АН
СССР, 27: 5—107.
Сокольская А. Н. 1954. Строфомениды Русской платформы.— Тр. Палеонтол. ин-
та АН СССР, 51: 1—191.
Стрелков А. А. 1946. Класс Brachiopoda — плеченогие. В кн. «Большой практикум
по зоологии беспозвоночных». Под ред. Ю. И. Полянского. М., изд-во «Сов, нау-
ка», стр. 40—56.
Atkins D. 1958. A new species and genus Kranssinidae (Brachiopoda) with a note on
feeding.— Proc. Zool. Soc. London, 131, 3: 559—587.
396
Atkins D. 1959a. The early growth stages and adult structure of the lophophore of.
Macanclrevia cranium (Muller) (Brachiopoda, Dallinida).— J. Marine Biol. Assoc.
U. K., 39: 336—350.
Atkins D. 19596. The growth stages of the lophophore and loop of the Brachiopod Te-
rebratalia transversa (Sowerby).— J. Morphol., 105, 3: 401—426.
Atkins D. 1960. The ciliary feeding mechanism of the Megathyridae (Brachiopoda) and'
the growth stages of the lophophore.— J. Marine Biol, assoc. U. K., 39, 3: 459—479.
Atkins D. 1961. A note on the growth stages and structure of the adult lophopore of
the Brachiopod Terebratella (Waltonia) inconspicua (G. B. Sowerby).— Proc. Zooi.
Soc. London, 136, 2: 255—271.
Beauchamp P. de. 1960. Classe des Brachiopods. Formes actuelles. Dans: Grasse
P. P. «Traitfe de Zoologie», V, 2: 1380—1430.
Beecher С. E. 1891. Development of the Brachiopoda. I. Introduction. Amer. J. Sci.,.
41: 343.
Beecher С. E. 1892. Development of the Brachiopoda. II. Classification of the stages
of growth and decline.— Amer. J. Sci., 44: 133.
Beurlen K. 1952. Phylogenie und System Brachiopoda Articulata.— Neues Jahrb. Geol.
und Palaontol. Monatsh., 3: 111—125.
Biernat G. 1959. Middle Devonian Orthoidea of the Holy Cross Mountains and their
ontogeny.— Paleontol. polon., 10: 1—78.
Blochmann F. 1933. Brachiopoda. In «Handworterbuch der Naturwissenschaften», 2,.
S. 189—201.
Borradaile L. A., Potts T. A., Eastham L. E. E. and Saunders J. T. 1958.
The invertebrata. Cambridge Univ. Press: XVI+796.
Chuang S. H. 1956. The ciliare feeding mechanisms of Lingula unguis (L.) (Brachio-
poda).— Proc. Zool. Soc. London, 127, 1—4: 167—189.
Chuang S. H. 1959. The structure and function of the alimentary canal in Lingula un-
guis (L.) (Brachiopoda).— Proc. Zool. Soc. London, 132, 1/2: 283—311.
Chuang S. H. 1960. An anatomical, histological and histochemical study of the gut of.
the Brachiopod, Crania anomala.— Quart, J. Microscop. Sci., 100, 1:9—18.
Cloud P. J. E. 11941. Homeomorphy and a remarkable illustration.— Amer. J. Sci., 239;
899—964.
Cloud P. E. 1948. Notes on the recent brachiopods.— Amer. J. Sci., 246, 4: 241—250.
Conklin E. G 1902. The embryology of a Brachiopod, Terebratulina sepienirionalis
(Conthony).— Proc. Amer. Philos. Soc., 41: 41—72.
Ekman Th. 1896. Beitrage zur Kenntnis des Stieles der Brachiopoden.— Z. wiss. Zool.,.
62: 169—249.
Elliot G. F. 1955. Shell-structure of Thecidean Brachiopods.— Nature, 175, 4469:
1169.
Elliot G. F. 1957. The origin of the Terebratellacea (Brachiopoda).— Ann. and Mag, Na-
tur. History, Ser. 12, 10 : 251—264.
Havlicek V. 1959. Spiriferidae v ceskem siluri a devonu (Bohemia).— Rozpr. Ust-
redn. ustavu geol., 25: 1—275.
Heller M. 1931. Uber die exkretorische Tatigkeit der Brachiopoden.— Z. Morphol. und
Okol. Tiere, 24: 238—258.
Kozlowski R. 1929. Les Brachiopodes gothlandiens de la Podolie Polonaise.— Paleon-
tol. polon., 1: 1—254.
Kuhn О. 1939. Die Stammesgeschichte der wirbellosen Tiere im Lichte der Palaontolo-
gie. Jena, S. VI+ 150.
Lacaze-Duthiers H. 1859. Memoire sur le systeme nerveux de 1’Haliotide (H. tu-
berculata et H. lamellosa Lam.).— Ann. sci. natur zool. et biol. anim., ser. 4, 12.
Manwell Cl. 1960. Oxygen equilibrium of Brachiopod Lingula hemerythrina.— Science,.
132, 3426 : 550—551.
M о о r e R. C. (Ed.) 1952. Invertebrates fossils. Brachiopoda. P. .197—267. New York.
Morse E. $. 1871. On the early stages of Terebratulina septentrionalis (Conthony).—
Mem. Boston Soc. Natur. History, 2, 29.
Morse E. S. 1873a. Embryology of Terebratulina,— Mem. Boston Soc. Natur. History,
2, 249.
Morse E. S. 18736. On the systematic position of the Brachiopoda.— Proc. Boston. Soc.
Natur. History, 16: 1—60.
Morse E. S. 1902. Observations on living Brachiopoda.— Mem. Boston. Soc. Natur. His-
tory, 5, 8: 316— 386.
Muir-Wood H. M. 1955. A history of the classification of the phylum Brachiopoda.—
Brit. Museum (Natur. History), London, VII+ 124. - .
Muir-Wood H. M. 1959. Report of the Brachiopoda of the John Murray Expedition.—
Brit. Museum (Natur. History), 6: 283—317.
Muir-Wood H. M. and Cooper G. A. I960. Morphology, classification and life ha-
bits of the Productoidea (Brachiopoda).— Mem. Geol. Soc. America, 81: XI+ 447.
Orton J. H. 1914. On ciliary mechanisms in brachiopods.— J. Marine Biol. Assoc. U. K.,
n. ser., X, 2: 283—311.
Percival E. 1944., A contribution to the life-history of the Brachiopod Terebratella in-
conspicua Sowerby.— Trans. Roy. Soc. N. Z. 74, 1: 1—23.
397
Percival E. 1960. A contribution to the life-history of the Brachiopod Tegulorhynchlu
nigricans.— Quart. J. Microscop. Sci., 101, 3—4: 439—457.
Richards J. R. 1952. The ciliary feeding mechanism of Neothyris lenticularis (Deshay-
es).— J. Morphol., 90, 1/3: 65—91.
Roger J. 1960. Brachiopodes fossiles, Dans: Grasse P. P. «Traite de zoologie», t. V,
2: 1431—1499.
Rowell A. J. 1957a. Some early stages in the development of the Brachiopod Crania
anomala (Miiller).— Ann. and Mag. Natur. History, Ser. 13, 9: 35—52.
Rowell A. J. 19576. The genera of the Brachiopod superfamilies Obolellacea and Si-
phonotretacea.— J. Paleontol., 36, 1: 136—152.
.'R о w e 11 A. J. 1960. Some early stages in the development of the Brachiopod Crania ano-
mala (Muller).— Ann. and Mag. Natur History, Ser. 13, 3: 35.
.Rowell A. J. 1961. Inhalant and exhalant feeding current systems in recent Bfachio-
pods.— Geol. Mag., 48, 3: 261—263.
.Rud wick M. J. S. 1960. The feeding mechanisms of spirebearing fossil Brachiopod.—
Geol. Mag., 47, 5: 369—383.
Schaeffer C. 1926. Untersuchungen zur vergleichenden Anatomie und Histologie der
Brachiopodengattung Lingula.— Viertel jahresschr. naturforsch. Ges. Zurich, 72:
186—196.
Sei la ch er A. 1956. Der Beginn des Kambriums als biologische Wende.— Neues Jahrb.
Geol. und Palaontol. Monatsh. Abh. 103, 1/2: 155—180.
Senn E. 1934. Die Geschlechtsverhaltnisse der Brachiopoden im besonderen die Sperma-
to- und Oogenes der Gattung Lingula.— Acta zool., 15: 1—152.
S t e h I i F. G. 1956. Evolution of the loop and lophophore in Terebratuloid Brachiopods.—
Evolution, X, 2: 187—200.
Thomson T. A. 1927. Brachiopod morphology and genera (Recent and Tertiary).— Ma-
nual New Zeal. Board. Sci. Art. Wellington, p. 1—338.
W i 11 i a m s A. 1955. Shell-structure of the Brachiopod Lacazella mediterranea (Risso).—
Nature, 175, 4469: 1123—1124.
Williams A. 1956. The calcareous shell of Brachiopoda and its importance to their
classification.— Biol. Revs., 31: 243—287.
'Williams A.'and Wright A. D. 1961. The origin of the loop in articulate Brachio-
pods.—Paleontol. London, 4, 2: 149—176.
Wilson A. E. 1957. Life in the Proterozoic.— Proterozoic Canada Roy. Soc. Canada
Spec. Pap., 2: 155—180.
Yatsu N. 1902. On the development of Lingula anatina,— J. Coll. Sci. Japan, 17, 4.
КИШЕЧНОДЫШАЩИЕ (ENTEROPNEUSTA)
Давыдов К. H. 1908. Наблюдений над процессом регенерации у Enteropneusta.— Зап.
Имп. акад, наук, 8, 22, 10: 1—120.
Ковалевский А. О. 1867. Анатомия Balanoglossus delle Chiaje.— Зап. Имп. акад,
наук, 10, 3: I—32.
Коштоянц X. С. 1957. Данные о физиологии нервной системы кишечнодышащих.
В кн. «Основы сравнительной физиологии», т. II. Сравнительная физиология нерв-
ной системы. Изд-во АН СССР, стр. 388—397.
Федотов Д. М. 1923. К вопросу о гомологии целомов иглокожих, кишечножаберных
и хордовых.— Изв. Биол. н.-и. ин-та Пермск. ун-та, 2, 1: 1—11.
Шимкевич В. М. 1889. Наблюдения над фауной Белого моря. I. Balanoglossus те-
reschkowskii Wagner.— Тр. СПб об-ва естествоисп., 20, 2: 1—74.
Шимкевич В. М. 1892. О соотношении Enteropneusta и Acrania.— Вести, естествозн.
СПб, 3, 3—4: 92—109.
Шимкевич В. М. 1907. О соотношении между Bilateria и Radiata.— Тр. СПб об-ва
естествоисп., 36, 4: 115—169.
Bateson W. 1884. The early stages in the development of Balanoglossus.— Quart. J.
Microscop. Sci., 26.
Bateson W. 1886. Lower stages in the development of Balanoglossus kowalevskii, and
morphology of the Enteropneusta.— Quart. J. Microscop. Sci., 38.
В rambell F. W. and Cole H. A. 1939. Saccoglossus cambrensis, sp. n., an Enterop-
neust occuring in Walles.— Proc. Zool. Soc. London, 109, pt. II: 211—236.
В u 11 о c k Th. H. 1944. The giant nerve fiber system in Balanoglossus.— J. Compar. Neu-
rol., 80: 355—367.
Bullock Th. H. 1945. The anatomical organisation of the nervous system of Entero-
pneusta.— Quart. J. Microscop. Sci., 86: 55—11.
Burdon-Jones C. 1952. Development and biology of the larva of Saccoglossus hors-
ti (Enteropneusta).— Phil. Trans. Roy. Soc., London, Ser. B, 236, 631: 563—590.
Burdon-Jones C. 1956. Observations on the Enteropneusta, Protoglossus hochleri
(Caullery a. Mesnil).— Proc. Zool. Soc.. London, 127: 35—58.
•Col win A. L. and Col win L. H. 1953. The normal embryology of Saccoglossus kowa-
levskii (Enteropneusta).— J. Adorphol., 92: 401—453.
.398
Fedotov D. M. 1924. Zur Morphologie des axialen Organkomplexes der Echinoder-
men.— Z. wiss. Zool., 123: 209—304.
Horst C. J. van der. 1927—1939. Hemichordata. In Bronn’s «Klassen und Ordnungen
des Tierreichs.», Bd. 4, Abt. 8, Buch 2, Teil 2, Lief. 1—6: 1—737.
Horst G. J. van der. 1934. Die Enteropneusten aus den Meeren der UdSSR.—Zool.
Jahrb. Abt. 2, 58: 61—100.
Kirk H. 1938. Breeding habits and early development of Dolichoglossus otagoensis.—
Trans, and Proc. Roy. Soc. N. Z., 68.
Knight-Jones E. W. 1953. Feeding in Saccoglossus (Enteropneusta).— Proc. Zool.
Soc., London, 123: 637—654.
Kowalevsky A. 1866. Anatomia des Balanoglossus delle Chiaje.— Mem. Acad. Imp.
Sci. St. Petersburg, 7, Ser. X, 3: 18.
Metschnikow I. 1869. Untersuchungen fiber die Metamorphose einiger Seetiere ’
(I. Tornaria).— Z. wiss. Zool., 20: 31—144.
Metschnikow I. 1881. Uber die systematische Stellung von Balanoglossus.— Zool.
Anz., 4, 78:. 139—143: Ibid., 4, 79: 153—157.
Mookerj ее H. K., Ganguly D. N. and G u p t a G. D. 1955. On the study of the cent-
ral nervous system, axial skeleton and anterior metamerism of the Balanoglossid
Ptychodera sp., and their bearing on the phylogeny of chordates.— Anat. Anz., 102,
9/14: 153—164.
Morgan T, H. 1891. The growth and metamorphosis of Tornaria.— J. Morphol., 5: 407—
458.
Morgan Т. Н. 11894. The development of Balanoglossus.— J. Morphol., 9: 1—84.
Newell G. E. 1951. The homology of the Stomochord of the Enteropneusta.— Proc.
Zool. Soc., London, 121: 741—746.
R а о K- R- 1953. The development of Glandiceps (Enteropneusta; Spengelidae).— J. Mor-
phol. 93: 1—13.
Schimkewitsch W. M. '1888. Uber Balanoglossus mereschkowskli Wagner.— Zool.
Anz., 11: 280—283.'
Schimkewitsch W. M. 1892. Sur les relations genetiques de Metazoaires.— Congr.
internal, zool., 2 Sess. Moscou: 215—252.
Silen L. 1950. On the nervous system of Glossobalanus marglnatus Meek (Enteropneus-
ta).—Acta zool., 31, 1: 149—175. ।
Silen L. 1954. Reflections concerning the «Stomochord» of the Enteropneusta.— Proc.
Zool. Soc., London, 124: 63—67.
'S pen gel J. W. 1893. Enteropneusten.— Fauna und Flora des Golfes von Neapel. 18,
Monographie: 1—757.
S pen gel J. W. 1913. Enteropneusta.'—Handworterbuch der Naturwissenschaften, Bd.
3, S. 527—542.
Weldon W. F. R. 1887. Note on Balanoglossus larva from the Batavia.— Proc. Roy.
Soc., London, 42: 146—150.
Willey A. 1899. Remarks on some recent work on the Protochorda with condensed ac-
count of some observations on the Enteropneusta.— Quart. J. Microscop. Sci., 42:
223—244.
ПТЕРОБРАНХИИ (PTEROBRANCHIA)
Щепотьев A. C. 1907. Pterobranchia. СПб. Диссертация, стр. V+248.
Br aem F. 1911. Pterobranchier und Bryozoen.— Zool. Ann., 38: 546—551.
Fowler G. H. 4905. Notes on Rhabdopleura normani Allman.— Quart. J. Microscop.
Sci., 48: 23—31.
G i 1 c h r i s t J. D. F. 1915. Observations on the Cape Cephalodiscus gilchristi (Ridewood)
and some of its early stages.— Ann. and Mag. Natur. History, ser. 8, 16: 233—246.
Horst C. J. van der. 1927—1939. Hemichordata. In Bronn’s «Klassen und Ordnungen
des Tierreichs». Bd. 4, Abt. 4, Buch 2, Teil 2, Lief. 4, 5. Cephalodiscus und Phabdop-
leura: 416—589.
John С. C. 1932. On the development Cephalodiscus. Discovery Repts, Cambridge, 6:
191—204.
Kozlowski R. 1949. Decouverte du Pterobranche Rhabdopleura a 1’etat fossile dans
le Cretace superieur en Pologne.— C. r. Acad. sci. Paris, 228: 1505—1507.
Kozlowski R. 1956. Sur Rhabdopleura du osanich de Pologne (Rhabdopleura z danu
Polski).— Acta paleontol. polon., 1, 1: 3—21.
Kozlowski R. 1961. Decouverte d’un Rhabdopleuride (Pterobranchia) Ordovicien.—
Acta paleontol. polon., 6: 1: 3—16'.
Ride wood W. G. 1920. Cephalodiscus. Brit. Antarct. (Terra Nova). Exped. 1910. Zoo’,.,
IV: 14—81. Brit. Museum (Natur. History). /
S ato F. 1938. Vorlaufige Mitteilungen fiber Atubaria heterolopa gen., nov., sp. nov., einer
irn freiem Zustand aufgefundenen Pterobranchier aus dem Stillen Ozean.— Zool. Anz.,
115: 97—-106.
ocnepotieff A. 1910. Die Pterobranchien des Indischen Ozeans.— Zool. Jahrb. Abt. 3,
28: 429—448.
399
Thomas H. D. and Davis A. G. 1949. The Pterobranch Rhabdopleura in the English
Eocene.— Bull. Brit. Museum (Natur. History), Geology, I, I: I—19.
ГРАПТОЛИТЫ (GRAPTOZOA)
Манская С. M., Дроздова T. B. 1962. Превращение органических соединений в
осадочных породах и органическое вещество граптолитов диктионемовых сланцев.
Геохимия, т. II, стр. 953—962.
Обут А. М. 1953. О морфологии и систематическом положении граптолитов,—Еже-
годн. Всес. палеонтол. об-ва, 14: 91—105.
Обут А. М. 1964. Подтип Graptolithina. В кн. «Основы палеонтологии». Иглокожие,,
гемихордовые, погонофоры и щетиночелюстные. Под. ред. Г. Ф. Геккера. М.,
Изд-во АН СССР, стр. 284—337.
В ulman О. М. В. 11955. Graptolithina with sections on Enteropneusta and Pterobran-
chia. In R. C. Moore «Treatise on invertebrate paleontology». London, p. XVII + 101.
Bulman О. M. B. 1963. The evolution and classification of the Graptoloidea.— Quart.
J. Geol. Soc. London, 119, pt 4, N 476: 401—418.
Decker Ch. E. and J. B. Gold. 1957. Bithecae, gonothecae and nematothecae in Grap-
toloidea.— J. Paleontol., 31: ill54'—1158.
Hyman L. H. 1958. The occurence of chitin in Lophophorata.— Biol. Bull., 114: 106—
112.
Kozlowski R. 1938. Tymczasowe wiadomosci о graptolitach z tremadoku Polski i ich-
teoretycznym znaczeniu.— Ann. Museum Zool. polon., 13, 16: 183—196.
Kozlowski R. 1947. Les affinities des Graptolithes.— Biol. Revs. Cambridge Philos.
Soc., 22: 93—108,
Kozlowski R. 1949. Les Graptolithes et quelques nouveaux groupes d’animaux du
Tremator de la Pologne.— Paleont. Polon., 3: 1—235.
Kozlowski R. 1962. Crustoidea — nouveau groupe de Graptolites.— Acta paleont. po-
lon., 7: 3—52.
Wetzel W. 1958. Graptolithen und ihre fraglichen Verwandten in elektronenmicroskopi-
schen Vergleich.— Neues Jahrb. Geol. und Palaontol. Monatsh., 7: 301—312.
Wiman C. 1895. Uber die Graptoliten.— Bull. Geol. Inst. Univ. Uppsala, 11: 239—316.
ИГЛОКОЖИЕ (ECHINODERMATA)
Баранова 3. И. 1957. Иглокожие Берингова моря.— Исследования дальневосточных
морей СССР, 4: 149—266.
Баранова 3. И., Гек к ер Р. Ф. 1964. Голотурии. В кн. «Основы палеонтологии».
Изд-во АН СССР, стр. 192—199.
Беляев Г. М. 1947. Возрастные изменения силы и скорости движения у некоторых
иглокожих Белого моря.— Зоол. ж., 26, 1: 41—46.
Гек к ер Р. Ф. (ред.) 1964. Иглокожие, гемихордовые, погонофоры, щетинкочелюстные.
В кн. «Основы палеонтологии». М., Изд-во АН СССР, стр. 1—383.
Дьяконов А. М. 1923. Морские ежи. В кн. «Фауна России». Изд. Зоол. музея АН.,
I, 1: 1-261.
Дьяконов А. М. 1933. Иглокожие Северных морей. В кн. «Определитель по фауне
СССР», т. 8. Изд. Зоол. ин-та АН СССР. Л., стр. 1—166.
Дьяконов А. М. 1946. Индивидуальная изменчивость и возрастные изменения у не-
которых групп иглокожих.— Тр. Зоол. ин-та АН СССР, т. 8, 1: 145—193.
Дьяконов А. М. 1949. Определитель иглокожих Дальневосточных морей (Беринго-
ва, Охотского и Японского).— Изв. Тихоокеанск. н.-и. ин-та рыбн. х-ва и океаногр.,
30: 1—130.
Дьяконов А. М. 1950а. Глубоководный элемент в фауне морских звезд Охотского
моря.— Исследования дальневост, морей СССР, 2: 28—57.
Дьяконов А. М. 19506. Морские звезды морей СССР. В кн. «Определитель по
фауне СССР», т. 34. Изд. Зоол. ин-та АН СССР. Л., стр. 1—202.
ДьяконовА. М. 1950в. Фауна иглокожих Малайского архипелага и ее связь с прош-
лым и настоящим Мирового океана.— Тр. Ленингр. об-ва естествоисп., 70, 4: 109—126.
Дьяконов А. М. 1950 г. Монографический очерк морских звезд северо-западной ча-
сти Тихого океана (Echinodermata, Asteroidea).— Исследования дальневосточных
морей СССР, II—IV: 58—139.
Дьяконов А. М. 1952. Иглокожие абиссальных глубин прикамчатских вод.— Иссле-
дования дальневосточных морей СССР, III: 116—130.
Дьяконов А. М. 1954. Офиуры (змеехвостки) морей СССР. В кн. «Определитель
по фауне СССР», т. 55. Изд. Зоол. ин-та АН СССР. Л., стр. 1—135.
Дьяконов А. М. 1955. О способности иглокожих выдерживать понижение нормаль-
ной океанической солености.— Докл. АН СССР, 102, 2: 373—374.
Дьяконов А. М. 1961. Род Henricia Gray из северозападной части Тихого океана.—
Исследования Дальневосточных морей СССР. Обзор видов морских звезд, VII:
Ковалевский А. О. 1889а. О выделительных органах некоторых насекомых, пауков
и многоножек.— Зап. Новоросс. об-ва естествоисп., 14, 2: 201—205.
400
Ковалевский А. О. 18896. О выделительных органах беспозвоночных животных.—
Зап. Новоросс. об-ва естествоисп., 14, 1: 79—84.
Ласточкин Д. А. 1914. К анатомии и физиологии синаптид (Synaptidae).— Тр. СПб
об-ва естествоисп., 45, 5: 1—2, 1—17.
Ласточкин Д. А. 1915. К познанию амбулакральной системы Synaptidae.— Тр. Пет-
рогр. об-ва естественен.,46, 1, 1; 23—39.
.Ливанов Н. А. 1955. Тип иглокожих — Echinodermata. В кн. «Пути эволюции жи-
вотного мира». М., Изд-во «Сов. Наука», стр. 244—288.
Мянниль Р. М. 1962. Таксономия и морфология Bothriocidaris (Echinoidea).— Тр.
Ии-та геол. АН Эстонск. ССР, 9: 143—190.
Светлов П. Г. 1916. К строению Тидемановых телец у Asteroidea.— Тр. Петрогр.
об-ва естествоисп., 45, 4: 83—106.
Светлов П. Г. 1924. О придатках амбулакрального кольца морских ежей (Тидемано-
вы тельца Echinoidea).— Русск. зоол. ж., 4, 1—2: 262—282.
Сент-Илер К. К. 1897. О блуждающих клетках в кишечнике морских ежей.— Тр.
СПб. об-ва естествоисп., 27, 3: 221—248.
Соловьев А. М. 1961. Паразит Canceripustula nocens у позднеюрского морского
ежа.— Палеонтол. ж., 4: 115—119.
Третьяков Д. 1932. Эволюция голкошкурых.— Работы зоол. биол. исслед. ин-та
(Одесса), 1: 7—38.
Федотов Д. М. 1915. Protomyzostomum polynephris Fedotov и его отношение к Gor-
gonocephalus eucnemis М. et. Тг. Пг., стр. V+240.
Федотов Д. М: 1916. Паразитизм Protomyzostomum в Gorgonocephalus.— Зоол.
вести., 1: 162—218.
Федотов Д. М. 1923. К вопросу о гомологии целомов у иглокожих, кишечножабер-
ных и хордовых.— Изв. Пермск. н.-и. биол. ин-та, 2, 1: 1—11.
Федотов Д. М. 1931. Об основном плане строения иглокожих.— Вести. АН СССР,
6: 44—47.
Федотов Д. М. 1934. Иглокожие (кроме морских лилий), В кн. Циттель «Основы
палеонтологии», 1с Беспозвоночные, стр. 216—267, 309—552.
Федотов Д. М. 1939. О субституциях вторичной полости тела у офиур.— Зоол. ж.,
18, 6: 943—959.
Федотов Д. М. 1951. Тип иглокожих. В кн. «Руководство по зоологии», т. III, 2. М.,
Изд-во «Сов. наука», стр. 460—591.
Федотов Д. М. 1960. Обзор достижений в области морфологии и филогении беспоз-
воночных в СССР с 1917 по 1957 г.— Тр Ин-та истории естествозн. и техн. АН
СССР, 31, 6: 135—246.
Яковлев Н. Н. 1918. Новые данные о род$ Cryptocrinus и связь морских лилий с цис-
тоидеями.— Ежегодн. Русск. палеонтол. об-ва. Пг., II: 7—26.
Anderson I. М. 1953. Structure and function in the pyloric caeca of Asterias forbesi.—
Biol. Bull., 105, 1: 47—61.
Anderson I. M. 1954. Studies on the cardiac stomach of the Starfish, Asterias for-
besi.— Biol. Bull., 107, 2: 157—173.
Anderson I. M. 1960. Histological studies on the digestive system of a Starfish, Hen-
ricia, with notes on Tiedemanns pouches in Starfishes.— Biol. Bull., 119, 3: 371—394.
В a t h e r F. A. 1915. Studies in Edriosteroiden. I—IX, p. 1—403.
Boolotian R. A. 1962. The perivesceral elements of echinoderms body fluids.—Amer.
Zool., 2, 2: 275—284.
Boolotian R. A. and Campbell J. L. 1964. A primitive heart in the echinoid Stron-
gylocentrotus purpuratus.— Science, 145, 3623: 173—175.
Burton M. P. 1964. Haemal system of regular Echinoids.— Nature, 204, 4964: 1218.
Cabiel J., Termier H. et Termier G. 1959. Les Echinoderpies Mesocambriens de
la Mongagne Noire (Sud de la France).— Ann. Paleontol., 44: 281—293.
Clark A. M. and Wright C. W. 1962. A new genus and species of recent starfishes
belonging to the aberrant family Sphaerasteridae, with notes on the possible origin
and affinities of the family.— Ann. and Mag. Natur. History, Ser. 13, V: 243—251.
Cuenot L. 1948. Anatomie, ethologie et systematique des Dchinodermes. Dans G r a s-
s ё P. P. «Traite de zoologie», XI, p. 3—275.
Dawydoff C. 1948. Embriologie des Echinodermes. Dans Grasse P. P. «Traite de
- zoologie», XI, p. 277—363.
Durham J. W., Castor К. E. 1963. Helicoplacoidea: A new class of Echinodermes.—
Science, 140, 3668: 820—822.
Durmam J. W. and M e 1 v i 11 e R. V. 1957. A classification of Echinoids.— J. Paleontol.,
31: 242—272.
Ekm an S. 1927. Holothurien der Deutschen Sudpolar-Expedition 1901—11903 aus der
Ostantarktis und von den Kerguelen. Deutsche Siidpolar Exped. 1901—1903. Berlin,
19, Zoologie, 11: 358—419.
Fay R. O. 1962. Edrioblastoidea, a new class of Echinodermata.— J. Paleontol., 36, 2:
201—205.
Fedotov D. M.' 1924. Zur Morphologie des axialen Organkomplexes der Echinoder-
men.— Z. wiss. Zool., 123: 209—304.
401
Fedotov D. M. 1926. Die Morphologie der Euryalae.— Z. wiss. Zool., 127: 404—528.
Fedotov D. M. 1928. Uber die Beziehungen der Echinodermenklassen zueinander (Auf
Grund der Ergebnisse der Zoologie und Palaeontologie).— Тр. Особой зоол. лабор.
СПб. АН, Ser. 2, 12: 31—94.
Fedotov D. M. 1930. Uber die vergleichende Morphologie der Crinoiden.— Zool. Anz.,
89, 11/12: 304—309.
Fedotov D. M. 1931. Uber eigenartigen Parasitismus bei Stachelhautern.— Z. Morphol.
und O,kol. Tiere, 22: 408—415.
Fedotov D. M. 1936. Zur Morphologie und Evolution der Seesterne und Ophiuren des
unteren Silur.— Тр. Палеозоол. ин-та АН СССР, V: 3—32.
Fell Н. В. 1962а. A living somasteroid Platasterias latiradiata Gray.— Univ. Kansas
Paleontol. Contrib. Echinodermata. Art., 6: 1—16.
Fell H. B. 19626. A classification of Echinoderms.— Tuatara, 10, 3: 138—140.
Fell H. B. 1962b. The evolution of the Echinoderms.— Annual Rept. Smithsonian Inst,
4518: 457—490.
Fell H. B. 1963. The phylogeny of Sea-Stars.— Philos. Trans. Roy. Soc., London,
B, 246: 381—435.
Ferguson J. G. 1960. The nature of the connective tissue of the body wall, retractor
hardness and cardiac stomach of the starfish Asterias forbesi.— Anat. Rec., 138, 3:
346.
Fontaine A. R. 1962. Neurosecretion in the Ophiuroid Ophiopholis aculeata.— Science,
138, 3543; 908—909.
Gemmill J. F. 1912. The development of the starfish Solaster endeca Fobes.— Trans.
Zool. Soc. London, 20, 1: 1—71.
Gemmill J. F. 1914. The development and certain points in the adult structure of the
starfish Asterias rubens L.— Philos. Trans. Roy. Soc. London, B, 205: 213—294.
Gem mill J. F. 1916. Double Hydrocoele in the development and metamorphosis in the
larva of Asterias rubens L.— Quart. J. Microscop. Sci., 61: 51—80.
Gem mi 11 J. F. 1920. The development of the starfish Crossaster papposus Muller and
Troschel.— Quart. J. Microscop. Sci., 64, 2: 155—189.
G i s 1 ё n T. 1924. Echinoderm studies.— Zool. Bidr., Uppsala, 9: 1—316.
G i s 1 e n F. 1947. Haplozoa and the interpretation of Perodionites — Zool. Bidr., Uppsala,
25: 402—408.
Goodbody I. 1960. The feeding mechanism in the sand dollar Mellila sexiesperforaia
(Leske).— Biol. Bull., 119, 1: 80—86.
GregoryW. K. 1951. Evolution emerging, v. I, P- XXVI+736. New York.
Gutchik R. C. 1954. Holothurian sclerites from Middle Ordovician of Northern Illi-
nois.— J. Paleontol., 28, 6: 827—829.
Hayashi R. 1935. Studies on the morphology of Japanese sea-stars. I. Anatomy of
Henricia sanguinolenta var. ohshiman, var. n.— J. Fac. Sci., Hokkaido Imp. Univ.,
Ser. VI, Zool, 4, 1: 1—>26.
Horstadius S. 1939. Uber die Entwicklung von Astropecten garanciacus L.— Pubbl.
Staz. zool. Napoli, 17: 221—312.
Hyman L. H. 1955. The invertebrates. Echinodermata, v. IV, p. VII+763.
Jaekel O. 1897. Ueber des Darmsystem der Pelmatozoen.— Sitzungsber. Ges. natur.
Freunde, Berlin, 3: 29—35.
Jaekel O. 1918. Phylogenie und System der Pelmatozoen.—Palaontol. Z, 3: 1—128. .
Jud kin W. H. 1954. Transphosphorylation in echinoderms.— J. Cell, and Compar. Phy-
siol, 44, 3: 507—518.
Kerkut C. A. 1960. Implications of evolution. In «Biochemical studies on Phylogeny».
Pergamon Press: 112—>133.
Kirchner G. 1929a. Uber die Lage der optischen Achse in Skeletteilen der Seeigeln.—
Zool. Jahrb. Abt. 2, 51: 299—312.
Kirchner G. 19296. Die Optik des Crinoidenskelettes.— Zool. Jahrb. Abt. 1, 46:357—510.
Lehmann W. M. 1957. Die Asterozoen in der Dachschiefern des rheinischen Unter-
devons.— Abhandl. Hess. Landesamt. Bodenforsch, 21: 1—160.
Ludwig H. und Hamann O. 1889—1907. Echinodermen. In Bronn’s «Klassen und
Ordnungen des Tierreiches», II, 3. Leipzig.
Metschnikoff I. I. 1869. Studien uber die Entwicklung der Echinodermen und Ne-
mertinen.— Mem. Acad. Sci. St. Petersburg, XIV, 8.
Mil lot N. 1964. Axial organ and fluid circulation in Echinoids.— Nature, 204, 4964:
1216—1217.
Mortensen Th. 1920. Studies in the development of Crinoids.— Papers Dept. Marine
Biol. Carnegie Inst. Washington, 16: 1—94.
Mortensen Th. 1921. Studies of the development and larval forms of Echinoderms.—
Copenhagen, G. E. C. Gad, p. 1—261.
Mortensen Th. 1928, 1935, 1940, 1943, 1948, 1950, 1951. A monograph of the Echi-
noidea. I—V. Copenhagen — London.
Mortensen Th. 1933. Biological observations on Ophiuras, with descriptions of two»
new genera and four new species.— Vid. medd. Dansk. naturhistor-foren. Kabenha-
ven, 93: 171—194.
402
Mortensen Th. 1938. On.the vegetarian diet of some deep-sea Echijioids.—Annot.
Zool. Japon., 17, 3, 4: 225—228.
N i c h о 1 s D. 1962. Echinoderms. London, p. 1—200.
Nichols D. 1964. Echinoderms: experimental and ecological.— Oceanogr. marine biol.
Annual Rev., 2: 293—429.
Raup D. M. 1962a. Crystallographic data in Echinoderm classification.— System, Zool.,
11, 3: 97—108.
Raup D. M. 19626. The phylogeny of calcite crystallographic in Echinoids.—J. Paleon-
tol., 36, 4: 793—810.
Regnell G. 1945. Non-crinoid Pelmatozoa from the Paleozoic of Sweden.— AAedd.
Lunds geol.-mineral Inst., 108: VIII+255.
Regnell G. 1948a. Echinoderms (Hydrophoridea, Ophicistia) from the Orodovician
(Upper Skiddavain, 3 cfi) of the Oslo region.— Norsk geol. tidsskr. Oslo, 27: 14—58.
Regnell G. 19486. An outline of the succession and migration of non-crinoid Pelmato-
zoa faunas in the Lower Paleozoic of Scandinavia.—Arkiv mineral, geol. Kgl. svenska
vetenskaps. akad., 26, A, 13: 1—55.
Regn"ell G. 1959. Donnees concernant de developement ontogenetique des Pelmato-
zoares du Paleozoique (Echinodermes).— Bull. Soc. geol. France, 7 ser., 1: 773—783.
Regnell G. 1960a. Tne Lower Paleozoic Echinoderm faunas of the British Isles and
Balto-Scandia.—Paleontology, 2, 2: 161—179.
Regnell G. 19606. Intermediate forms in early Paleozoic Echinoderms.— Rept. 21st.
Internal. Geol. Congr. Copenhagen, p. 71—80.
Reichensperger. A. 1934. Stachelhauter.— In «Handworterbuch der Naturwissen-
schaften». Jena, Bd. IX: 431—484.
Runnstrom J. 1925. Zur experimentelle Analyse der Entwicklung von Antedon.—
Arch. Entwicklungsmech., 105 : 63—113.
Runnstrom J. 1930. Weitere Beitrage zur Analyse der Metamorphose von Antedon.—
Arch. Entwicklungsmech., 121: 714—725.
Runnstrom S. V. 1928. Entwicklung von Leptosynapta inhaerens.— Bergens Mus..
Aarbok, 1927, 1: 5—80.
Schmidt W. J. 1930. Die Skeletstiicke der Stachelhauter als Biocristalle.— Zool. Jahrb.
Jena, 47, 3:’.357—510.
Schulz A. 1935. Optik der Skeletstiicke der Seesterne.— Zool. Jahrb. Abt. 2, 60:
101—132.
Schulz E. 1895. Uber den Process der Exkretion bei Holothurien.— Biol, Zbl., 15.
Semon R. 1888. Die Entwicklung der Synapta digitata and ihre Bedeutung file die-
Phylogenie der Echinodermen, Jena.— Z. Naturforsch., 22: 175—309.
S m i t h J. E. 1937. On the nervous system of the starfish Marthasterias glacialis (L.).—
Philos. Trans. Roy. Soc. London, B. 22, 3—7: 111—173.
Smith J. E.1940. The reproductive system and associated organs of the Brittle-star;
Ophiothrix. iragilis.— Quart. J. Microscop. Sci., 82: 267—309.
Smith J. E. 1947. The activities of the tube feet of Asterias rubens. L.— Quart. J. Mic-
roscop. Sci., 88: 1—14.'
Smith J. E. 1950. The motor nervous system of Astropecten irregularis.— Philos. Trans..
Roy. Soc. London, 234B: 521—558.
Smith L. S. 1961. Clam-digging behaviour in.the starfish Pisaster breoispinus Stim-
pson 1851.— Behaviour, 18, 1—2: 148—153.
Spencer W. K. 1914—1940. A monograph of the British Paleozoic Asterozoa.— Pale-
ontol. Soc. London, 67, 69, 70, 71, 74, 76, 79, 82, 87, 94.
Spencer W. K. 1938. Some aspects of evolution in Echinodermata. In «Evolution» G. R.
de Beer (Ed.). Oxford, p. 287—303.
Spencer W. K. 1951. Early Palaeozoic starfish.— Philos. Trans. Roy. Soc., London,.
235B, 87—129.
S trimpie H. L. and Koenig J. W. .1956. Mississippian microcrinoids from Okla-
homa and New Mexico.— J. Paleontol., 30, 5: 12S8—1247.
Tuzet O., Manier J. F. 1961. Recherches sur I’organe axial de la- comatule: Antedon.'
mediterranea Link (Echinoiderme-Pelmatozoaire).—Bull. Assoc. Anat., 109: 775—780.
U b a g h s G. 1953. Classe des stelleroides. Dans «Traite de paleontologie» Piveteau J.
(ed.), t. Ill: 774—842.
U-nger H. 1960. Neurohormone bei Seesternen (Marthasterias glacialis).— Symposia.',
Biol. Hungarica, 1: 203—207.
Walcott C. D. 1911. Middle Cambrian holoturians.— Smithsonian Misc. Collect., 57.
Wanner J. 1934. Stachelhauter (Palaontologie). In «Handworterbuch der Naturwissen-
schaften». Jena, Bd. IX: 484—516.
Weller J. M. 1(930. A group of larviform Crinoids from Lower Pennsylvanian Strata of
the Eastern inferior Basin. III.— Stat. Geol. Surv. Rept Invest.,.21.
Whitehouse F. 1941. Early Cambrian echinoderms similar to the larval st-age of re-
cent forms.— Mem. Queensland Museum, 12: 1—28.
Withers Th. 1926. Catalogue of the Machaeridia (Turrilepas and'its. allies) in the De-
partment of Geology. Brit. Museum (Natur. History), XVE+99.
404:
Yazaki M. 1930. On the circulation of the perivisceral fluid in Caudina chilensis
(J. Muller).— Sci. Rept Tohoku Imp. Univ. Sendai, 4 ser., 5: 403—414.
ПОГОНОФОРЫ (POGONOPHORA)
Иванов A. B. 1949. Новый представитель класса Pogonophora.— Зоол. ж., 28, 1.
79—84.
Иванов А. В. 1955. Основные черты организации Pogonophora.— Докл, АН СССР,
100, 1: 175—177.
Иванов А. В. 1957. Материалы по эмбриональному развитию Pogonophora.— Зоол.
ж., 36(8): 1127—1144.
Иванов А. В. 1959. Пути эволюционного развития в классе Pogonophora.— Тр. Ин-та
морфол. животных АН СССР, 27: 274—278.
Иванов А. В. 1960. Погонофоры. Фауна СССР.— Зоол. ин-т АН СССР, новая сер.,
75: 1—271.
Иванов А. В. 1962. Живые ископаемые. В кн. «Наука и человеыество». Изд-во «Зна-
ние», стр. 170—175.
Иванов А. В. 1964а. Тип Pogonophora. В кн. «Основы палеонтологии». Под ред.
Р. Ф. Геккера. Иглокожие, гемихордовые, погонофоры и щетинкочелюстные. Изд-во
АН СССР, стр. 359—367.
Иванов А. В. 19646. О строении заднего отдела тела у Pogonophora.— Зоол. ж., 43,
4: 581—589.
Ливанов Н. А. и Порфирьева Н. А. 1962. Организация погонофор.— Тр. Об-ва
естествоисп. при Казанск. ун-те, 65: 102—119.
Ливанов Н. А., Порфирьева Н. А. 1965. Об «аннелидной» гипотезе происхожде-
ния погонофор.— Зоол. ж., 44, 2: 161 —168.
Свешников В. А. 1961. Пелагические личинки полихет Белого моря.— Зоол. ж., 40,
2: 164—177.
Свешников В. А. 1963. О типах личинок полихет.— Докл. АН СССР, 150, 6:
1393—1396.
‘Соколов Б. С. 1965. Древнейшие отложения раннего кембрия и сабеллитида.— Тез.
докл. Всесоюзный симп. по палеонт. докембрия и раннего кембрия 25—30 октября
1965 г., Новосибирск, стр. 78—91.
Brunet R. С. and Carlisle D. В. 1958. Chitin in Pogonophora.— Natuie. 182, 4650:
1689.
Caullery M. 1914. Sur les Siboglinidae, type nouveau d’invertebres. recueilli par
I’expedition du Siboga.— C. r. Acad. sci. Paris, 158: 2014—2017.
Caullery M. 1944. Siboglinum. Caullery 1914, type nouveau d’lnvertebres, d’affinites
a preciser.— Siboga Exped., Monogr., 25 bis., 26 p.
Caullery M. 1948. Le genre Siboglinum Caullery 1914. Dans Grasse P. P. «Traite
de zoologie», XI, p. 495—499.
Dawydoff C. 1948. Contribution a la connaissence de Siboglinum Caullery.— Bull. biol.
France et Belgique, 82: 141 —163.
Hartmann O. 1951. Fabricinae in the Pacific.— Pacif. Sci., 5 : 379—391.
Hartmann O. 1954. Pogonophora Johansson, 1(937.— System. Zool., 3, 4: 183—185.
Ivanov A. V. 1960. Embranchement des Pogonophores. Dans Grasse P. P. «Traite
de zoologie», \r, I, p. 1521—1622.
Ivanov A. V. 1963. Pogonophora. London, Acad. Press, p. XVI+479.
...Jagersten G. 1956. Investigations on Siboglinum ekmani, n. sp. encountered in the
Skagerrak.— Zool. Bidr., Uppsala, 31: 211—252.
Jagersten G. 1957. On the larva of Siboglinum with some remarks on the nutrition
problem of the Pogonophora.— Zool. Bidr., Uppsala, 32: 67—79.
Johansson К. E. 1937. Uber Lamellisabella zachsi und ihre systematische Stellung.— -
Zool. Anz., 117: 23—26.
Johansson К. E. 1939. Lamellisabella zachsi Uschakow, ein Vertreter einer neuen
Tier-Klasse Pogonophora.— Zool. Bidr., Uppsala, 18: 253—268.
Reisinger E. 1938. Pogonophora.— Fortschr. Zool., 3: 35—54.
Southward A. J. and Southward E. C. 1963. Notes on the biology of some Pogo-
nophora.— J. Marine Biol. Assoc. U. K-, 43: 57—64.
Uschakov P. 1933. Eine neue.F°rm der Familie Sabeliidae.— Zool. Anz., 104.
Webb M. 1964a. The posterior extremity of Siboglinum fiordicum (Pogonophora).— Sar-
'sia, 15: 57—68.
Webb M. 19646. The larvae of Siboglinum fiordicum and a reconsideration of the adult
body regions (Pogonophora).— Sarsia, 15: 57—68.
Webb M. 1964b. Evolutionary paths within the phylum Pogonophora.— Sarsia, 16:
59—64.
Webb H. 1964 r. Additional notes on Sclerolinum bratstromi (Pogonophora) and the
establishment of a new family Sclerolinidae.— Sarsia, 16: 47—58.
Wilson D. P. 1932. On the Mitraria larvae of Owenia fusiformis Delle Chiaje.— Philos.
Trans. Roy. Soc' London, Ser. B, 221: 231—334.
ОПЕЧАТКИ И ИСПРАВЛЕНИЯ
Страница Строка Напечатано Должно быть
С 63 19 СИ. гаструлярных гастральных
67 2 св. многочисленность многочленистость
67 26 сн. более не более
138 13 сн. двубокой двубоковой
212 10 св. малочисленные малочленистые
264 2 сн. богаче беднее
354 16 сн. подвижного неподвижного
Д. М. Федотов