/







































































































































































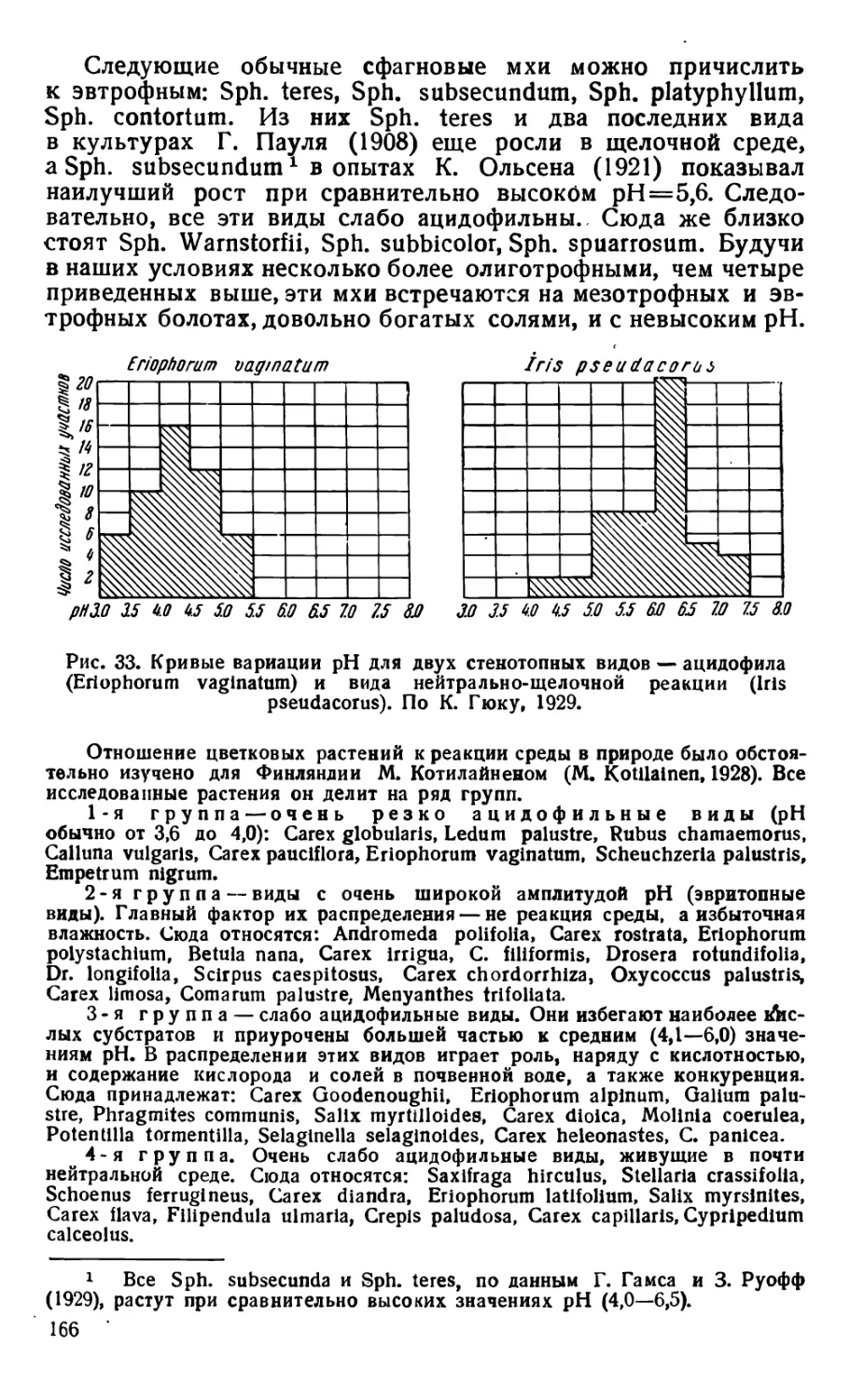



















































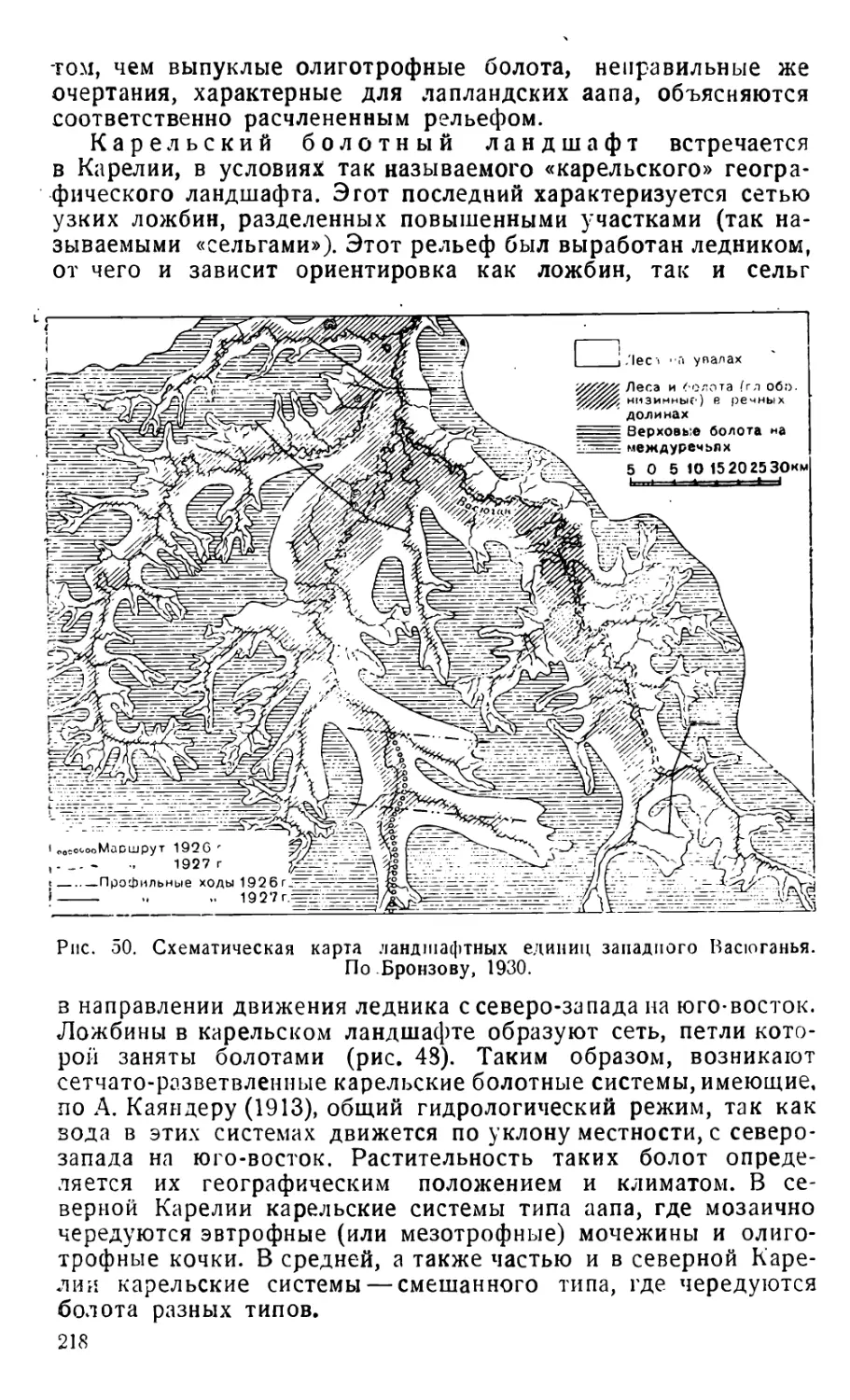



























































































































































































Text
Н.ЯКАЦ
БОЛОТА
И ТОРФЯНИКИ
УЧПЕДГИЗ
4941
Проф. Н. Я. КАЦ
БОЛОТА
И ТОРФЯНИКИ
ПОСОБИЕ ДЛЯ УНИВЕРСИТЕТОВ
*
Утверждено Маркомпросом РСФСР
ГОСУДАРСТВЕННОЕ
> ЧЕБНО-ПЕДАГОГИЧЕСКОЕ ИЗДАТЕЛЬСТВО
НАРКОМПРОСА РСФСР
МОСКВА - 2941
/ ^551
К 30
•доб. 58
ПРЕДИСЛОВИЕ.
В основу книги .Болота и торфяники" положен курс лекций по болото¬
ведению, который автор читал в продолжение 10 лет в Московском, а ча¬
стично и в Горьковском, университетах. Книга может быть использована
в качестве учебного пособия в первую очередь студентами университе¬
тов, а затем студентами сельскохозяйственных вузов и торфяных втузов.
Кроме того, предлагаемое пособие будет полезно географам, почво¬
ведам и, вообще, всем лицам, интересующимся болотоведением, имеющим
достаточную биологическую подготовку.
Болотоведение имеет огромное значение не только при практическом
освоении природных богатств нашей страны, но и в их теоретическом по¬
знании. Поэтому перед автором стояла задача, кроме учебных целей, дать
сводку по теоретическому болотоведению в его современном состоянии.
Этим книга отличается от существующих учебных руководств по болото¬
ведению.
Эти руководства, в том числе и прекрасная книга В. Сукачева, заду¬
маны ^же, чем настоящая книга, соответственно прежнему состоянию болото¬
ведения, и не охватывают предмета в его современном объеме. Сведёние
воедино обширного и весьма разнородного материала, из которого сла¬
гается современное болотоведение, находящееся в периоде бурного роста,
представляло не легкую задачу, и уже поэтому книга не может быть сво¬
бодна от недостатков и притом, вероятно, существенных. Кроме того,
рамки книги заставляли порой суживать объем материала: так, пришлось
опустить весьма важный отдел — описание зарубежных торфяников.
По всем вопросам, возникшим при пользовании этой книгой, автор
просит обращаться по адресу: Москва, Моховая, 11, Государственный, уни¬
верситет, Ботанический институт.
Автор.
Июль 1940 г.
Редактор К. И. Ченалова.
Тираж 3000 экз. Подписано к печати 29/1 1941 г. А 32282.
Печатных листов 25+1 вклейка 0,75. Учетно-издат. листов 30+вклейка 0,19.
Типографских знаков в 1 печ. листе 7.328. Заказ № 722.
Цена без переплета 6 р. Переплет 1 р. 25 к.
Лгниздат. Типография № 3 имени Коминтерна, Ленинград, Красная ул, 1.
ГЛАВА I.
СОДЕРЖАНИЕ НАУКИ О БОЛОТЕ.
В отличие от многих других естественных наук болотове¬
дение не представляет собой единой и совершенно самостоя¬
тельной научной дисциплины. Содержание его складывается
из различных отраслей знания, необходимых для изучения
болот.
Однако научные дисциплины, входящие в состав болотове¬
дения в качестве его разделов, служат только для выявления
разносторонних и сложных особенностей болот как природ¬
ного явления. Объектом болотоведения является, с одной сто¬
роны, взаимодействие почвенных и надпочвенных грунтовых
вод и всех географическо-исторических факторов и, с другой
стороны, растительность, остатки которой (торф и седименты)
имеют самостоятельный научный интерес и хозяйственную
ценность. Отсюда болота как естественно-историческое явле¬
ние и торфяники, прямой их продукт, составляют единство
научно-теоретического болотоведения, исследующего эти слож¬
нейшие закономерности в их многообразных проявлениях.
Соответственно основным объектам исследования болотоведе¬
ние состоит из двух главных разделов: 1) из учения о современ¬
ной болотной растительности и 2) из учения о торфе и водных
отложениях (седиментах) (схема 1). Из этих двух разделов пер¬
вый— учение о растительности—менее разработан, чем второй.
Однако болотоведение и в теперешнем состоянии имеет уси¬
ленно разрабатываемые отделы, характеризующие его много¬
гранные связи с разнообразными областями естественных наук.
Так, один из важных отделов болотоведения, экологиче¬
ское изучение болотной растительности, входит
в область экологии растений и частью соприкасается с физио¬
логией растений. Сюда входят вопросы о ксероморфизме бо¬
лотных растений, о росте болотных растений, в том числе
сфагнов, в зависимости от условий среды, о взаимоотноше¬
ниях между сфагновыми мхами и другими растениями торфя¬
ных болот и т. д. Этот отдел болотоведения довольно широко
разрабатывается в Советском Союзе.
Географическое, или региональное, болото¬
ведение изучает распределение болот и болотных типов как
3
в отдельных странах, так и на земном шаре, картирует бо
лота, учитывает их площадь и районирует их, выделяет
торфяные и болотные районы, области, провинции и зоны. Это
молодой отдел болотоведения. Географическое болотоведение
развивается в Скандинавии, Центральной Европе, а за послед¬
нее время и у нас в Союзе. Оно входит частью в отдел хо¬
рологической географии растений, изучающей распределение
растительных ценозов на земле.
Значительное место в болотоведении занимает отдел, из¬
учающий смены бол отных фитоценозов во вре¬
мени (так называемые сукцессии), их динамику, эволюцию,
превращение. Так как смены болотной растительности, проис¬
ходившие раньше, запечатлеваются в строении торфа, то этот
отдел особенно тесно связан с учением о торфе. Ботаник,
изучающий сукцессии болотной растительности, поставлен
в необычайно выгодные условия. Он судит о сменах непосред¬
ственно на основании строения торфяной залежи, в то время
как при изучении смен в лесах, на лугах и т. д. приходится
прибегать к косвенным методам или к длительным стацио¬
нарным наблюдениям. Этот отдел входит в генетическую
географию растений. Ряд ценных работ в этом направлении
принадлежит шведам.
Более разработано учение о торфе. Здесь выделяются на¬
правления, из которых некоторые представляют самостоятель¬
ные отрасли знания с особым объектом изучения и особой
методикой. В учение о торфе входят:
1. Стратиграфия. Изучает ботанический состав торфа
и водных органических соединений (седименты, сапропели —
см. далее) и устанавливает слои торфа разного ботанического
состава и их смену как в вертикальном, так и в горизонталь¬
ном направлении, т. е., иначе говоря, описывает строение
торфяника.
Стратиграфический метод в основном заключается в микро¬
скопическом анализе остатков растений (частью животных),
живших ранее на самом торфянике (или в водоеме). Страти¬
графия занимается также вопросами возникновения и развития
торфяников, нарастания торфа, скорости этого нарастания
и пр. Изучение стратиграфии, построение стратиграфического
профиля торфяника является сейчас во всех странах необхо¬
димым условием не только научного, но и практического ис¬
следования торфяников.
2. Изучение истории флоры и растительности
местности также базируется на изучении строения торфа.
Однако в отличие от стратиграфии этот отдел изучает исто¬
рию не болотной, а окружающей болото раститель¬
ности и флоры и пользуется для этой цели по преимуществу
остатками растительности, росшей не на самом торфянике,
а вокруг него, а именно: пыльцой древесных, а частью и тра¬
вянистых растений. Сущность этого „метода пыльцевого ана¬
лиза“ и его применение изложены ниже. Этот отдел учения
1
Схема 1
Отделы учения о болоте и связь их с смежными дисциплинами
Экологии растений Хорологическая (пространственная) Генетическая география растений
I 1еография растений |
II 1
Экология болотной География болотной растительности Динамика болотной растительности
растительности | |
г I I
Учение о болотной растительности
Г
Учение о болоте
Учение о торфе
Стратиграфия
торфа
1
I
I
Стратиграфия
водных отложений
I
I
Изучение истории
четвертичной
флоры и раститель¬
ное! н методом
пыльцы
1
I
I
Четвертичная
палеоклиматология
на основе
изучения
торфяников
Четвертичная
палеогеография
на основе
изучения
торфяников
Технология торфа
с рядом отделов
I
Геология
Лимнология
1
Историческая
Палеоклимато¬
Палеогеография
Химия и физика
география
растений
логия
торфа
о торфе в настоящее время наиболее усиленно разрабаты¬
вается. Каждый год в этой области выходят десятки работ.
Хотя по своей цели этот раздел болотоведения и входит це¬
ликом в отдел исторической географии растений, однако
специфичность метода и самого объекта (торф), а также то,
что эти вопросы трактуются почти исключительно в болото¬
ведческих работах и притом почти всегда бок о бок с вопро¬
сами стратиграфии и другими отделами болотоведения,
позволяет отнести этот раздел и к болотоведению, так же как
и два дальнейших раздела. Метод анализа пыльцы в торфе
в настоящее время основной метод изучения послеледниковой
и частью межледниковой истории флоры и раститель¬
ности.
3. Палеоклиматология, учение об изменениях после¬
ледникового климата — это отдел болотоведения и одновре¬
менно общей палеоклиматологии. Он также опирается на ме¬
тод анализа пыльцы в торфе, а равным образом на данные
стратиграфического исследования. Вопрос о послеледниковых
изменениях климата оживленно обсуждается в работах главным
образом скандинавских и среднеевропейских болотоведов.
4. Послеледниковая палеогеография, т. е. изме¬
нение очертаний моря и суши, — особый отдел болотоведения.
Изучение стратиграфии и смены пыльцы в торфяниках по
побережьям Балтийского и Немецкого морей дает ценнейший
материал по истории морских трансгрессий.
5. Технология торфа (и водных отложений — седи-
ментов) может быть разбита на несколько отраслей, рассма¬
тривать которые мы здесь не будем. Этот отдел учения
о торфе имеет многообразные связи с физикой, химией и пр.
Все перечисленные отделы учения о болоте тесно сопри¬
касаются друг с другом и смежными отделами знания. Обычно
научные работы по болотоведению носят типичный ком¬
плексный характер. Например, при описании торфяника,
помимо описания растительности, ее смен во времени, приво¬
дится также и место болота в ряду географических типов
болотных образований (региональный момент), строение тор¬
фяника, данные пыльцевого анализа и т. д. Однако отделы бо¬
лотоведения можно вполне отделить друг от друга не только
мысленно, но и практически. В научных работах очень часто
выдвигается на первый план какая-либо одна сторона — или
стратиграфия, или история послеледниковой флоры и т. д.,
остальные же стороны или затрагиваются слабее, или вовсе не
затрагиваются. Таким образом, можно говорить о работах
преимущественно стратиграфического направления, региональ¬
ного и т. д.
Из предыдущего обзора видны многообразие и многосто¬
ронность учения о болотах и многочисленность корней, связы¬
вающих это учение с другими отделами знания. Это много¬
образие явлений, которые охватывает болотоведение, а также
исключительный общенаучный интерес фактов из области
6
прошлого, с которыми имеет дело учение о торфе, и объяс¬
няют интерес предмета и его большое образовательное зна¬
чение. В частности, болотоведение несравненно шире, глубже,
многограннее и имеет большее образовательное значение, чем
другие специальные предметы по геоботанике, читаемые на
биофаках: степеведение, лесоведение и т. д. Научно-теоре¬
тическое и практическое значение болотоведения заключается
в свойствах самого объекта — болота, представляющего исклю¬
чительные преимущества для комплексного его изучения и
для установления многообразной причинной связи между
самыми разнородными явлениями.
Понятие „болото“ и „торфяник" у разных авторов.
Болотоведение изучает болото. Вследствие многогранности
болотоведения исследователи выдвигали на первый план
разные свойства болота и сообразно с этим по-разному опре¬
деляли самое понятие „болото“. Поэтому естественно, что
единого, общего определения болота не существует, и опре¬
деление ботаника будет отличаться от определения, например,
торфогеолога и т. д. Мы можем различить несколько точек
зрения на сущность болота и соответственно несколько опре¬
делений. Именно, болото можно понимать как бо¬
таническое, геологическое или географическое
явление.
1. Болото—чисто ботаническое понятие. Харак¬
терный признак болота—влаголюбивая растительность, а не
свойства субстрата. Последний может быть или минеральным,
или торфяным. Подобное ботаническое определение болота
дал А. Флеров (1914): „Болотом называется растительное со¬
общество, господствующую роль в котором играют водно¬
болотные, болотные и прибрежноводные растения, требующие
'для своего развития максимальной или избыточной влажности
грунта или даже водной поверхности, независимо от при¬
сутствия или отсутствия слоя из отмерших болотных и водно¬
болотных растений (торфяной слой)“1. В Германии и Сканди¬
навии первые исследователи болот были ботаники. Для них
руководящим признаком болота была именно растительность.
О. Зендтнер (О. Sendtner, 1854), И. Лоренц (J. Lorenz, 1858),
чьи имена одинаково известны и в ботанической географии,
и в болотоведении, считали болотом участки с определенной
растительностью как с торфом, так и без торфа. Лишь для
дальнейших подразделений понятия „болото" ими выдвигался
как решающий признак субстрат. Соответственно этому, оба
названные ученые различали „торфяное болото“ (Torfmoor)
и „минеральное болото“ (Mineralmoor) 1 2.
1 По В. Сукачеву, „Болота, их строение, развитие и свойства", 1926 г.
2 По Г. Шрейберу (Н. Schreiber, 1927).
7
2. Болото — чисто геологическое понятие. Бо¬
лото— это место, где отлагается особого рода масса, обра¬
зованная слабо разложившимися органическими остатками и
называемая торфом. Следовательно, без торфа не может быть
болота. Совершенно очевидно, что растительный покров при
таком понимании болота никакой роли не играет, и понятие
„болото“ более или менее приближается к понятию „торфяное
болото“, или „торфяник“. Еще И. Дау (J. Dau) в 1829 г. опре¬
делил болото как „всякое место, где отлагается торфяная
масса в своем естественном состоянии *.
Е. Вольни (Е. Wollny, 1897), стоя в общем на точке зрения
Дау, уточняет его определение. Он пишет: „Болото — это место,
где, благодаря гниению под водой отмерших частей растений,
возникают более или менее мощные слои торфа“1 2 3.
К. Вебер (С. Weber, 1907) также придерживается геологи¬
ческой точки зрения на болото и, что особенно важно, дает
в своем определении минимальную глубину торфа, которая
позволяет еще считать определенный участок болотом, а также
дает конкретные указания, что нужно считать торфом. Он
пишет: „Болото есть часть земной поверхности, покрытая
слоем торфа толщиной не менее 20 см в осушенном состоянии
и не содержащая в себе заметных на-глаз или наощупь в зна¬
чительном количестве минеральных частиц“8.
Развернутое геологическое определение болот дает также
шведский торфогеолог Л. Пост (L. v. Post, 1916). Болото, по
его формулировке, есть напластование, лежащее над уровнем
моря, где накопляются под водой или на сырой почве био¬
генные (органические) отложения, причем эти отложения дошли
до стадии торфа.
Придерживается геологической точки зрения на болото
и де Лаппаран (De Lapparent, 1900), определение которого
близко к определению Вольни (см. выше), а также И. Фрю
и К. Шретер (1904), примыкающие в своем монументальном
труде к определению К. Вебера (1902), а из более поздних
ученых — издатель „Handbuch der Moorkunde“ и автор I тома
этой серии „Allgemeine Moorgeologie“ (1929) К. Бюлов
(К. V. Bülow).
3. Болото есть понятие географическое, опреде¬
ленный ландшафт. Он складывается из целого ряда фак¬
торов: гидрологического порядка (избыточное увлажнение),
климатических, геологических, топографических, почвенных.
Географическим ландшафтом, „закономерно складывающимся“
и „живущим“, считает болото Р. Аболин. Он проводит этот
взгляд в своей ранней (1914) и более поздней работе (1928).
Ландшафтного понимания болота придерживается и В. Су¬
качев (1926).
1 Цит. по И. Фрю п К. Шретер (J. Früh und С. Schroter, 1904).
2 Цит. по И. Фрю и К. Шретер (J. Früh und С. Schröter, 1904).
3 Цит. по В. Сукачеву (1926).
При определении понятия „болото“ возникает еще вот
какое затруднение. Болота вообще чрезвычайно разнообразны
по своему характеру, особенно, если включать в это понятие,
как это делают некоторые—в том числе и мы, водоемы
с водной растительностью, тундровые болота, засоленные
болота полупустынной полосы и ряд других типов. При таком
расширении понятия формулировка его поневоле делается
весьма общей. Мы определяем болото следующим образом.
Болото — это участок территории, обычно избы¬
точно увлажненный пресной или соленой водой,
стоящей над поверхностью почвы или ниже ее.
застойной или более или менее проточной. Он
может быть с торфом или без торфа. Раститель¬
ность большей частью вод о- или влаголюбивая,
реже мезофильная, а иногда физически или
физиологически ксерофитная. В некоторых типах
болот наряду с болотной большую роль играет и мезофитная
растительность, а иногда и ксерофитная. Примером болот
с мезофитной растительностью могут служить некоторые типы
лесных болот — например, ельники на торфе. Лишайниковые
фитоценозы на высоких, подверженных ветровой эрозии
торфяных кочках-буграх в лесотундре и тундре, служат
примером болот с более или менее ксерофитной раститель¬
ностью.
Болота разделяются на пять основных групп с дальнейшими
подразделениями \
1. Торфяники с мощным слоем торфа, свыше
50 см в неосушенном или 30 см в осушенном состоянии.
2. Минеральные болота пресноводного пи¬
тания с торфом, мощностью в неосушенном со¬
стоянии меньше 50 см или без торфа с субстра¬
том иного рода.
3. Заболоченные земли. Заболоченные земли —
это участки земной поверхности с маломощным
слоем торфа (менее 50 см) или с субстратом
иного рода (гумус, торфянисто-глеевые почвы и пр.).
Растительность носит переходный характер
между болотной (гелофитной) растительностью
и растительностью сухих, незаболоченных мест.
Фито ценозы, покрывающие заболоченные земли,
отличаются обычно малой устойчивостью. Сюда
входят:
а) заболоченные луга: растительный покров образован
болотными и луговыми (мезофитными) видами;
б) заболоченные леса: в растительном покрове сме¬
шаны болотные виды и лесные мезофиты;
в) заболоченные кустарники;
г) заболоченные тундры; наряду с тундровыми мезо- 11 См. Н. Кац (1936).
9
фитами, растительный покров образуют и болотные влаголю¬
бивые виды.
4. Зарастающие водоемы — пространства, по¬
крытые обычно в течение всего вегетационного
периода слоем воды с отложениями минераль¬
ного или органического ила (седименты), а
иногда подводного (лимнического) торфа, и засе¬
ленные погруженной, плавающей или возвышаю¬
щейся над поверхностью воды растительностью.
Сюда относятся, например, озера с зарослями водных расте¬
ний, тростниковые плавни и др.
5. Засоленные болота. Это — пространства, за¬
селенные влаголюбивыми галофитами с посто¬
янным или периодическим избыточным увлаж¬
нением, с субстратом, богатым хлоридами и дру¬
гими соединениями. Сюда относятся, например, со¬
лончаки.
Указанное деление, конечно, довольно грубо и схематично.
Оно не претендует на полную научность и допускает иногда
условные рубрики для разграничения отдельных подразде¬
лений друг от друга (такова, например, условная глубина торфа
в 50 см, отделяющая торфяники от иных типов). Разграни¬
чение некоторых типов болот в природе вызывает в ряде слу¬
чаев затруднения. Тем не менее в общем приведенная класси¬
фикация достаточно проста и удобна, а выделенные типы
достаточно естественны и в общем хорошо один от другого
отграничены. Эти типы естественных угодий имеют еще то
преимущество, что они хорошо «укладываются в географиче¬
скую схему распределения болотных образований. Так, на¬
пример, тундровая зона Европейской части Союза отличается
широким развитием заболоченных земель типа „г“ (заболочен¬
ные тундры) и болот типа 2 (т. е. с маломощным торфом)
и сравнительно слабым развитием торфяников. Полупустынная
зона характеризуется по преимуществу заболоченными водое¬
мами и солончаками. Другие же типы болот представлены
здесь слабо.
Статистика болот.
Учет болот и торфяников. Учесть болота и тор¬
фяники —чрезвычайно важно. Болота разных типов представ¬
ляют в общем неудобные земли, подлежащие мелиорации, и
составляют мелиорационный фонд. Торфяники образуют так
называемый торфяной фонд. Учет инвентаря болот и торфя¬
ников, приведение в наличность имеющихся запасов их —
необходимое условие для планирования мелиораци¬
онных мероприятий и торфодобычи. Только зная
характер болот и торфяников, их распределение по админи¬
стративным областям и районам, можно выработать план
болотно-торфяного хозяйства страны на будущее время,
ю
При том значении, какое имеют для Союза болота и торфяники,
легко себе представить значение учета болот и торфяников,
особенно в условиях планового советского хозяйства. Формы
учета могут быть различные в зависимости от
цели. Важно, прежде всего, учесть площадь земель, подле¬
жащих осушке или требующих других видов мелиорации.
Для торфяного хозяйства важно учесть площадь торфяных
болот. Однако это далеко не достаточно для торфяной про¬
мышленности. Если два района обладают одинаковой пло¬
щадью торфяников, но средняя глубина торфа для одного
района будет, положим, 1,8 м, а для другого — 2,2 м, то второй
район будет богаче торфом, чем первый. Следовательно, для
более правильной оценки нужно иметь сведения не только
о площади торфяников, но и о глубине их. Площадь и глу¬
бина дают уже возможность вычислить запасы торфа в кубо¬
метрах. Сведения о запасах сырого торфа в кубо¬
метрах дают уже лучшее представление о богатстве торфяным
топливом, чем одна только площадь торфяников. Однако и
этих сведений недостаточно для суждения о богатстве торфом.
Сырой торф в естественном состоянии содержит неодина¬
ковое количество воды. В дело идет воздушносухой торф.
Вес воздушносухого торфа, получаемого из определенного
объема торфа-сырца, будет неодинаков. Он зависит от со¬
держания воды в сыром торфе. Следовательно, для прак¬
тики важнее выразить торфяной запас в единицах веса
воздушносухого торфа, а не в единицах объема всей сырой
массы.
Для удобства сравнения с другими видами топлива запасы
торфа и других горючих веществ выражают в тоннах так
называемого условного топлива. Добавим, что важно также
знать, насколько раздроблен торфяной фонд, т. е. сколько
имеется торфяников на той или иной площади. Резюмируем
Сказанное здесь о формах учета болот и торфяников.
Абсолютную площадь болот всех типов в ме¬
рах поверхности важно знать, прежде всего,
мелиоратору.
Для представления о природе края, о ланд¬
шафте, важно знать также, какой процент пло¬
щади занят всеми типами болот вместе, разными
типами болот порознь (торфяники, болота, забо¬
лоченные земли ипр.) и, кроме того, соотношение
площадей разных типов заболоченных про¬
странств.
Для торфяной промышленности важно знать,
прежде всего, абсолютную площадь торфяников.
Более полное представление о запасах торфа
дает объем сырого торфа. Наиболее же важны
сведения о запасах воздушносухого торфа в ве¬
совых единицах, причем эти запасы удобно выра¬
жать в тоннах условного топлива.
11
Состояние учета болот и торфяников. Методы учета.
С учетом площади болот дело обстоит хуже, чем с учетом
торфяников. Можно сказать, что даже для Европейской части
Союза, не говоря уже о менее исследованной Азиатской,
точного учета всех пригодных для мелиорации
заболоченных пространств не имеется. Не уста¬
новлена вполне точно даже площадь, занятая всеми типами
болотных угодий вместе (заболоченные земли, болота без торфа,
торфяники и пр.), не говоря уже о площади, занятой отдель¬
ными типами порознь1.
Лучше обстоит дело с учетом площади торфяников, хотя
и здесь точные цифры как для Советского Союза в целом,
так и для Европейской его части, а равно и для других стран
отсутствуют. Это зависит прежде всего от того, что не все
существующие торфяные болота исследованы в
достаточной мере. Вторая причина неточности сведений
заключается в самой методике учета. Она не одина¬
кова в разных странах, и это приводит к тому, что сведения
по площади торфяников в разных странах часто не вполне
сравнимы между собою. Г. Шрейбер (1927) приводит пример
того, какое значение имеет методика учета торфяников.
В Богемских горах (Вб1ипег\уа1<1) был проделан такой подсчет.
В одном случае минимальная глубина торфа, которая позво¬
ляла считать болото торфяником, была принята за 20 см, и на
определенной территории было установлено всего одно болото,
площадью в 1050 га. При минимальной же глубине торфа в
50 см на той же самой территории было зарегистрировано
36 болот с общей площадью всего в 76,4 га. Добавим, что и
запас сырого торфа во втором случае должен быть значительно
меньше, чем в первом.
Из этого примера видно, какое громадное значение при
учете площади и числа болот имеет минимальная глубина
торфа, т. е. такая, при которой заболоченная площадь еще
считается торфяником. Между тем, минимальная глубина
торфяника, как видно из табл. 1, составленной по данным
того же Г. Шрейбера (1927), различна в разных странах и
у разных исследователей.
Если учесть сказанное выше, то очевидно, что сведения
о площади и числе торфяников, основанные на
разной предельной минимальной мощности
торфа, не могут быть вполне сравнимы. По указа¬
нию Г. Шрейбера сравнение затрудняется тем, что при учете
предельная минимальная глубина, допускаемая для торфяника,
не приводится вовсе.
1 В качестве примера достаточно привести, что в БССР, по Дубаху
(1927), имеется 2318679 га болот, а по Бахтееву (1929) всего 460677
(из С. Тюремнова, 1931). Не лучше обстоит дело и с зарубежными странами.
Для Галиции разные исследователи оценивают площадь болот то в 489100 га,
то в 150000 га, то в 11125 го (!!) (по Н. Schreiber, 1. с.).
12
Таблица 1
Минимальная глубина торфяников в разных странах и у разных
исследователей
(по Г. Шрейберу, 1927)
Г. Шрейбер
Г. Штангеланд (О. 51а1^е1апё), Норвегия
Бременская болотная опытная станция и венские
болотоведы *
Датская комиссия: 1888 г
„ * 1901 г.
, * с 1908 г
Прусский ботанический ферейн
Хаглунд (На£1ипс1), Швеция
Саксонские геологи (сначала)
. „ (позднее)
Советский Союз1
50 см.
40—60 см.
20 см.
40—50 см.
1 фут.
30 см.
70 .
20 „
50 .
Немалое значение при учете болот имеет минималь¬
ная площадь отдельных торфяников, принимаемая при их
картировании. Эта площадь различна в разных странах и у
разных исследователей, что видно из приведенных в книге
Г. Шрейбер (/. с.) данных:
Размер минимальной
площади (в га)
Германско-австрийский болотный
ферейн 0,5
Г. Штангеланд (Норвегия) . . . 0,8—1,0
Геологическое общество Швеции Свыше 5,0
В торфяных фондах, издаваемых Наркомземом на десяти¬
верстке, наносятся лишь торфяники площадью 10 га; торфя¬
ники меньшего размера не входят в подсчет торфяной пло¬
щади и торфяных запасов.
Минимальная площадь торфяников, подлежащих картиро¬
ванию, определяется, прежде всего, масштабом карт, на кото¬
рые наносят торфяники. Кроме того, имеет значение и богат¬
ство страны торфяными фондами, и степень нужды в них.
Недостаточная исследованность торфяных
фондов, с одной стороны, и отсутствие твердо
установленной и единообразной методики учета
их, с другой, служат причиной отсутствия
вполне точных сведений о числе торфяников,
общей площади их, а следовательно, и о запасах
торфа. Эти же причины отчасти затрудняют
сравнение данных по разным странам.
Тем не менее, мы все же приведем сведения о площади
болот и торфяников по разным странам, по отдельным мате¬
рикам и по всему земному шару.
1 По определению Всесоюзной конференции по болотному кадастру
с 1934 г. минимальная глубина болота (торфяника)—30 см в неосушенном
п 20 см в осушенном виде.
13
Таблица 21
Области, республики
Общая площадь в тые.
кв, км
Болот, болотной 1
тундры и плавней а |
Заболоченных лесов г &
§2
» 2
Заболоченных лугов к £
£2
• »
Всего болот и забо« £
лоченных земель
Процент заболоченных
земель к общей площади
Из общей площади заболочен¬
ных земель торфяных болот
в млн. га
Процент торфяных болот
к общей площади
Запасы торфа в млн.
тонн условного топлива2
Северная обл. . .
1119,0
33,5
14,5
0,4
48,4
43,3
15,0(12,35)8
1
13,4
16500»
Ленингр. с Мурм.
районом ....
- (3,6)
Ленинградская с
Карреспубл. . .
484,0
15,0
5,2
0,4
20,6
42,5
6,°
! 12,3
6600
Карельск. АССР .
—
■
—
—
—
—
- (2,5)
1
Западная . . . •
165,1
1,32
0,46
0,52
2,3
13,9
0,8 (0,75)
5,0
880
Московская обл. .
157,9
1,0
0,27
0,45
1,82
11,5
0,60
3,8
664
(Моек, и Калинин¬
ская области) .
—
—
—
—
—
—
(0,65)
Ивановская обл. .
124,4
1.1
0,4
0,4
1,9
15,3
0,6 (0,70)
4,8
660
Кировская и Горь¬
ковская обл.. .
—
—
—
—
—
\ —
(1.2)
(Татарск. АССР) .
—
—
—
—
—
—
(0,042)
Нижегородец обл.
(с Татреспубл.) .
339,0
1,2
0,4
1,2
2,8*
8,3
0,85
| 2,5
935
(Урал с Свердл. и
Челяб. обл.) . .
—
—
—
—
—
—
(2,6)
Уральская (с Баш-
республ.) . . .
984,4
2,5
1,2
0,8
4,5
4,7
1,4
1,5
1540
(Башкир. АССР) .
—
—
—
—
—
—
(0,08)
ЦЧО
192,0
0,55
—
—
0,55
2,8
0,06
0,3
66
Курск, и Воронеж.
обл
—
—
—
—
—
—
(0,09)
Средневолжск.. .
235,3
0,2
—
—
0,2
0,8
0,04
0,2
44
(Саратов, обл.) .
—
—
—
—
—
—
(0,005)
Нижневолжск. . .
458,0
—
—
—
—
—
0,01
0,02
11
(Куйбыш. обл.) .
—
—
—
—
—
—
(0,042)
Северокавказ. . .
305,6
0,6
—
—
0,6
2,0
—
—
— .
Закавказье и
Крым
211,7
0,2
—
—
0,2
0,9
—
—
—
БССР
126,8
2,3
1,0
0,9
4,2
33,1
1.4 (1,5)
11,0
1540
УССР
451,8
1,75
0,2
1,25
3,2
7,1
0,8 (1.5)
1,7
880
Всего . . .
5319,4161,22123,63
6,42191,27
17,1
1 27560
1 5,18
303204
1
1
| (27760)
1
1 Таблица взята из книги „Торф, его происхождение, залегание и распро¬
странение“, 1932 г., с некоторыми изменениями.
2 Средняя глубина залежи принята в 2,13 м. Запас торфа с 1 га 1100
тонн условного топлива. Переводный коэфициент наусловное топливо всюду 0,5.
3 Цифры в скобках — площадь торфяников в млн. га по данным Л. Ле¬
пина (1936).
4 По другим данным свыше 50 млрд. тонн.
14
Площадь болот и торфяников по странам и континентам'.
Заболоченную площадь Европейской части СССР можно оце¬
нить громадной цифрой в 90 млн. га слишним, а площадь
торфяников в круглых цифрах в 27,5 млн. га, запас же торфа —
свыше 50 тыс. млн. тонн условного топлива. Заболоченные
площади и торфяники распределены по Союзу неравномерно.
На нервом месте по общей площади заболоченных земель,,
торфяников и по запасам торфа стоит Северный край. На его
долю приходится больше половины всех болот, тор¬
фяников и запасов торфа Европейской части Союза.
На втором месте стоит Ленинградская область (с Карелией)
(свыше 20% всей площади болот и торфяников). Далее идут
Белорусская ССР и Уральская область (значительная часть ее
находится, впрочем, за Уральским хребтом), а затем следуют
Западная область, Украина, Нижегородская область и централь¬
ные (Московская и Ивановская) области. На долю южных и
юго-восточных областей приходится ничтожная часть общей
площади болот. Однако следует иметь в виду, что наиболь¬
шее значение имеют наиболее доступные и удобно размещен¬
ные запасы торфа в бывшей Ленинградской губернии, в цен¬
тральных областях Союза, БССР и некоторых других районах.
В этих областях и ведется наиболее интенсивная торфодо¬
быча. Громадные же залежи торфа Северного края пока пред¬
ставляют в значительной степени мертвый запас.
В отношении процента заболоченности и заторфованности
(т. е. процента торфяников к общей площади) впереди стоят
также Северный край и Ленинградская область. Заболочен¬
ность здесь в круглых числах 43°/о, заторфованность—12—13%.
Далее следует БССР с 33% заболоченности и 11% заторфо¬
ванности, затем Западная, Московская, Ивановская и Нижего¬
родская области, где заболоченность колеблется от 13,9 до
8,3°/о, а заторфованность от 5,0 до 2,5%. Украинская ССР,
в_ частности ее южная половина, южные и юго-восточные об¬
ласти имеют ничтожный процент заболоченности и заторфо¬
ванности (взято из книги Д. А. Герасимова „Торф, его проис¬
хождение, залегание и распространение“, 1932 г.).
Площадь болот вообще, и в частности торфяников, для Си¬
бири достоверно не известна. В то время как Наркомзем опреде¬
ляет площадь торфяников здесь не менее 100 млн. га и запас,
торфа в 110 млрд, т условного топлива, Госплан опреде¬
ляет общую площадь торфяников Сибири всего в 30 млн. га,
из которых на долю Западной Сибири приходится 7 млн. га,
Восточной Сибири —13 млн. и Дальневосточного края —
10 млн. га1. Это расхождение объясняется тем, что Госплан
имеет в виду лишь более легко доступные в отношении тор¬
фодобычи площади, вне зоны вечной мерзлоты. Общая же пло¬
щадь торфяников Сибири, включая и вечномерзлые районы, 11 Л. Лепин, 1936, а по последним данным Наркомзема РСФСР, на пер¬
вое января 1938 г. площадь торфяников всей Сибири — около 40 млн, га.
15
несомненно, гораздо больше и, возможно, приближается к
указанной выше цифре в 100 млн. га.
Ввиду скудности сведений по площади болот в разных
странах мы приведем лишь площадь торфяников.
Таблица 3
Площадь торфяников и запасы торфа в разных странах
(по данным 1922—29 гг.)
Запасы торфа
в млн. тонн
условного
топлива
Запасы торфа
в процентах
к итогу
Площадь торфяников
в млн. га
Европейская часть СССР . .
30 320
15,7
27,5
Азиатская , „ . .
110000
57,0
100
Весь СССР в целом
140 320
72,7
127,5
Финляндия
13 2001
6,8
12,01 (10,04)
Канада
104501
5,4
9,51 9,5—8,54
США •
88001
. 4,6
8,01 4,0—3,0
Швеция
60501
зд
5,5! 10,0 — 8,04
Польша
30802
1,6
2,82
Германия
2 7501
1,4
2.51 св. 3f04
Англия
2 4201
1,3
2,21 2,54
(без Ир- (вместе с
ландии)Ирландией)
Норвегия
1 3201
0,7
1,21 1,6-1,04
Ирландия
12101
0,6
/’¿1св.0,09*
Голландская Индия
1 1001 2 3 40,6
Эстония
7701
0,4
ОД»
Латвия
6051
0,3
0,551
Япония ....
0,25
Литва
1981
0,1
0,181
Венгрия
165®
0,09
0,15» о,15*
Дания 1
1 143г
0,07
0,13» 0.1*
Италия . . 1
! 1Ю1
0,06
0,101
Голландия
100
0,05
0,031
Новая Зеландия
! ббз
0,03
0,0603
Франция
1 421
0,02
0,0381 0,040*
Австрия
373
0,02
0.0343
Югославия
16,53
0009
0,0153
Чехо-Словакпя
14,51
0,007
0,0131
Швейцария
61
0,003
0,00541 0,0054*
Итого без СССР
52653
27,28
47856
Всего
192973
100
' 175,431
1
1. Hjerstedt, 1928. (Привожу по книге „Торф, его залегание и распро¬
странение*, 1932.)
2. Pfaszicki, М. 1927. (Привожу по тому же источнику.)
3. Reischle und Wächter, 1£22. (По тому же источнику.)
4. Привожу по К. Бюлову (1929). Цифра 1 млн. га для Голландской
Индии более правильная.
Площадь торфяников весьма неравномерно распределена
между отдельными странами. Цифра в 175 с лишним миллио¬
нов гектаров, к которой мы в дальнейшем придем на осно¬
вании других источников, довольно близко характеризует ми¬
ровую площадь, занятую торфяниками. Из этой цифры 72,6%,
т. е. почти %, падает на Советский Союз, причем на его Евро¬
пейскую часть приходится 15,5%, или немного менее % всей
мировой площади торфяников. Соответственно площади на
долю Союза приходится почти % мирового
запаса торфа. Если даже оставить в стороне трудно дося¬
гаемые запасы торфа в Сибири, то все же придется признать,
что Советский Союз по торфяным богатствам далеко опере¬
жает все другие страны как европейские, так и внеевропей¬
ские. Площадь и запасы торфа только Европейской части
Союза почти равны площади и запасам во всех
остальных европейских странах, вместе взятых.
За СССР по торфяной площади и запасам торфа следует Фин¬
ляндия, затем Канада и дальше в меняющемся порядке, в за¬
висимости от того, какие цифры мы примем, США и Швеция.
В общем, СССР и эти четыре страны вместе оказываются без¬
раздельными монополистами мировых запасов торфа, обладая
157—163 млн. га мировой торфяной площади, т. е. в круг¬
лых числах 9/м всей площади торфяников.
Из остальных стран хотя и маленькую долю, но еще за¬
метную в мировом торфяном фонде(от 1,5 до 0,3%) имеют: Гер¬
мания, Англия, Норвегия, Ирландия, Голландская Индия, пло¬
щадь же остальных измеряется большей частью лишь десят¬
ками тысяч гектаров и сотыми и даже тысячными долями
процента мировой площади.
Отметим резкое расхождение в цифрах для некоторых стран,
например для США и Швеции, по различным источникам. Это
свидетельствует, что эти сведения не всегда имеют необходи¬
мую достоверность.
Площадь торфяников по отдельным континентам в точно¬
сти не известна. Приводим имеющиеся сведения (табл. 4).
Таблица 4
Площадь торфяников (в миллионах гектаров)
Азия свыше 100
Европа приблизительно 57
Сев. Америка . . „ 17,5
Южная Америка . не известна
Африка незначительна, точно не известна
Австралия . . . • ' „ . „
Общая площадь торфяников на земном шаре — свыше
175 млн. га. Торфяная площадь Азид складывается целиком
за счет торфяников Сибири; площадь торфяников Японии
увеличивает эту цифру всего на 0,25 млн. га. Если добавить
сюда торфяники островов Голландской Индии, то прибавится
еще всего только 1 млн. га.
2-722
17
Из остальных континентов, кроме Северной Америки (тор¬
фяники США и Канады) и Европы, Африка и Австралия со¬
вершенно не исследованы в отношении своих торфяных пло¬
щадей. Эти площади, однако, несомненно незначительны и не
могут сколько-нибудь заметно изменить мировой итог. Торфя¬
ники Южной Америки не учтены; значительные площади их
имеются здесь в Патагонии и на Огненной Земле. Эти торфя¬
ники увеличили бы весьма немного мировой итог. Таким об¬
разом, мировую площадь торфяников можно оценить прибли¬
зительно цифрой свыше 175 млн. га. Из отдельных материков
впереди стоит Азия, дающая свыше половины миро¬
вого итога.
Европа стоит на втором месте (около трети мировой
площади). Однако по заторфованности (процент торфяников
к общей площади) Европу придется безусловно поставить
на первое место. При этом надо учесть, что тор¬
фяной фонд Европы хорошего качества, находится в
основном в условиях, благоприятных для торфодобывания
(умеренный климат) и удобно размещен в обжитых районах.
Поэтому понятно, что по добыче торфа Европа стоит на
первом месте.
На третьем месте стоит Северная Америка,
имеющая, примерно, 10°/о мировой площади торфа.
Четвертое место следует отвести Южной Америке. Австра¬
лия и Африка имеют ничтожную долю в мировом запасе.
Какова же общая заболоченная площадь земного шара,
считая торфяники и другие типы болот? Сколько-нибудь точно
определить эту площадь невозможно. Если принять, что площадь
болот и заболоченных земель (кроме торфяников) равна пло¬
щади торфяников, то ошибка будет скорее в сторону пре¬
уменьшения, а не преувеличения. Таким образом, если мы
примем площадь болот без торфа и заболочен¬
ных земель в 175 млн. га, то общая заболоченная
площадь земного шара выразится в круглых
числах в 350 млн. га. Отдельные торфяные массивы бы¬
вают весьма обширны. Даже в средней полосе Европейской
части СССР встречаются торфяники до 60 тыс. га. Так
например, если растянуть площадь промышленной залежи1
болота Оршинский Мох вдоль всей длины Октябрьской ж. д.
от Москвы до Ленинграда, то мы будем иметь по обе сто¬
роны железнодорожного полотна непрерывную полосу тор¬
фяника немного уже полукилометра.
Эти размеры—не предельные даже для средних широт
Европейской части Союза. Так, знаменитый Полистовский бо¬
лотный массив на водоразделе рек Ловати, Полисти, Шелони
и Великой имеет около 100 тыс. га („Торфяное дело“ № 1,1929).
1 Промышленная залежь — это части торфяника, имеющие глубину от
0,5 м и более.
18
В межречье Мологи и Шексны есть болотные системы, по-
видимому, еще большего размера. На севере Европейской
части Союза (например Беломорское побережье), где болота
зачастую в результате роста вширь сливаются друг с другом,
площади таких болотных систем бывают еще больше. Наи¬
больших размеров достигают системы торфяников в Западной
Сибири. „Великое Остяцкое болото“ на Обь-Иртышском во¬
доразделе имеет около 1000 км в длину. Площадь его изме¬
ряется многими тысячами квадратных километров (А. Бронзов,
1930).
Важность изучения болот.
Изучение болот не только важно практически, но имеет
и большой научный интерес.
Болото — это прежде всего элемент ландшафта. Всякий,
изучающий ландшафт северной или умеренной зоны — будь
то почвовед, географ, геоботаник, — непременно должен
иметь дело с болотом. Учесть болота, дать их описание, на¬
нести их на карту бывает необходимо при геоботанических
исследованиях. В Европейской части Союза около 17% всей
территории занято болотами различного рода, в том числе и
торфяными. Это очень высокая цифра, особенно если принять
во внимание, что болота распределены далеко не равномерно
и что на юге и юго-востоке их очень мало. Эта цифра пока¬
зывает, что во всей Европейской части Союза в среднем из
каждых 6 км2 поверхности примерно 1 км2 занят болотами.
Иначе говоря, если бы болота были распределены равномерно
и имели одинаковую площадь, то на каждом квадратном
километре поверхности располагалось бы около 17 отдельных
болот, площадью в 1 га каждое. Особенно широко распростра¬
нены болота в северной части Союза, в хвойной зоне и тундре.
Заболоченность целых ботанико-географических зон достигает
50% (Н. Кац, 1936), а в отдельных районах Колы и Беломор¬
ского побережья болота занимают до 90% всей »площади.
Следовательно, на севере болота зачастую определяют ланд¬
шафт. Прав был П. Крылов, когда назвал хвойную зону
Западной Сибири — болотно-хвойной. И в отношении Евро¬
пейской части Союза ботчнико-географы должны после-
довать^его примеру, так как и здесь площадь, занятая
болотами, в хвойной зоне громадна.
Болота имеют большую потенциальную ценность,
реализуемую после осушки и других мероприятий. Всем из¬
вестно, что торфяное топливо имеет большое значение в про¬
мышленности Союза, что торф находит широкое применение
в строительстве, в сельском хозяйстве — в качестве .подстилки
для скота, удобрения и пр. Меньшее значение в настоящее
время имеет использование болот под сельскохозяйственные
культуры. Однако с течением времени по мере использования
свободных земель значение болот и здесь будет возрастать.
Наконец, большое значение имеет превращение болот и
2’ 19
заболоченных пространств в сенокосные, лесные и пастбищ¬
ные угодья после их осушки.
Известную пользу приносят болота и в своем естествен¬
ном состоянии. Некоторые типы болот используются как
сенокосы. Если в более южных районах (например в Полесье)
орошение и осушка изменяют характер и улучшают состояние
этих сенокосных болот, то на севере, в тундре и лесотундре,
некоторые типы болот в их естественном виде сохранят свое
значение как оленьи пастбища еще на значительное время.
Ягодные ресурсы болот как естественных угодий также
имеют значение. Главное место занимает клюква. Сбор ее ягод
на продажу во многих местностях служит важным подспорьем
для населения. Клюква служит также предметом экспорта.
Из других болотных ягод заслуживают упоминания голубика,
или пьяница (Vaccinium uliginosum L.), и морошка. Эти ягоды
служат для потребления на месте, а также и для сбыта на
рынке. Водяника (Empetrura nigrum) на севере также употре¬
бляется в пищу. Если к этому прибавить, что сфагновый мох
идет на утепление строений и пр., то этим почти и ограни¬
чивается польза от болот в их природном состоянии.
Вред, приносимый болотами в их естественном виде, зна¬
чительно превышает пользу от них. Во-первых, болота там,
где они занимают большую площадь, охлаждают местность,
сокращая вегетационный период, что неблагоприятно для
земледелия. Осушка болот отепляет прилегающие к ним
минеральные почвы и делает их более пригодными для зем¬
леделия, что было учтено при проведении больших осушитель¬
ных работ в Швеции К Во-вторых, болота, повышая при своем
росте грунтовые воды, заболачивают окружающие леса и
другие угодья и, разрастаясь вширь, захватывают соседние
участки минеральной почвы. Заболачивание лесов широко
распространено на севере Союза. Оно ухудшает рост деревьев,
сводит на-нет продукцию древесины и превращает обширные
площади, где при иных условиях мог бы расти высокостволь¬
ный строевой лес, в мелколесье, годное в лучшем случае лишь
на дрова. Осушка болот улучшает, следовательно, состояние
наших лесов. Наконец, болота вредны в гигиеническом отно¬
шении. Они способствуют развитию ряда болезней у человека
(ревматические заболевания, малярия, колтун) и у домашних
животных (ящур и др.). Однако главный вред болот тот, что
они без применения специальных мероприятий или вовсе про¬
падают для человека (бросовые земли), или могут быть исполь¬
зованы в крайне ограниченном объеме.
В СССР, как уже говорилось, болота занимают громадную
площадь, а по запасам торфа СССР намного богаче всех
других стран, вместе взятых. Отсюда громадное значение болот
и торфа в экономике Союза. Уже в дореволюционное время
имелись научно-исследовательские и опытные учреждения 11 В. Сукачев, 1926 г.
20
(правда, влачившие жалкое существование) с целью специаль¬
ного изучения торфа, болот и приемов их культуры. Со
времени советской власти интенсивность использования болот
возросла во много раз, а вместе с тем выросли и исследова¬
тельские кадры и опытно-научные учреждения по болотам и
торфу. Важнейшими научно-исследовательскими учреждениями
по изучению болот являются в настоящее время Центральная
торфяная опытная станция Наркомзема, работающая главным
образом по использованию торфа в сельском хозяйстве,
и Институт торфяной промышленности (Инсторф) Наркомтяж-
прома, изучающий применение торфа как топлива. Кроме того,
Белорусская (б. Минская) болотная опытная станция, а также
некоторые областные станции работают по вопросам мелиорации
болот. Особый отдел по изучению торфа имеется в системе
Белорусской академии наук.
Из сказанного видно, что болота заслуживают самого
серьезного отношения к ним и самого глубокого исследования
в целях, прежде всего, их рационального хозяйственного
использования. Одновременно с практическим значением болота
имеют и глубокий теоретический интерес. Болото — совершенно
своеобразный комплекс явлений как неорганического, так и
биологического порядка. Эти явления находятся в своеобраз¬
ной и глубокой причинной связи, для познания которой
болото представляет собой необычайно удобный объект.
ГЛАВА И.
ТОРФ И СЕДИМЕНТЫ.
Органогенные образования, которые мы рассмотрим в этой
главе, делятся на две большие группы — торф и седименты
(гиттии). Эти группы настолько отличны по способу образо¬
вания, ботаническому и химическому составу, физическим
свойствам и пр., что их приходится рассматривать отдельно.
Торф как органогенная порода.
Определение понятия „торф* и место торфа
среди других органогенных образований.
К. Вебер (1908) относит торф к гумусовым породам. Для них
характерно присутствие продуктов неполного разложения
растительных и животных остатков. Торф вместе с перегноем
(Moder)1 относится, по Веберу, к гумусовым породам, наи¬
более бедным золой, — менее 40% золы в воздушносухой
массе (по В. Сукачеву, 1926). Следовательно, в торфе, как
и в перегное, органические вещества преобладают над мине¬
ральными. Однако их происхождение и свойства резко раз¬
личны.
Торф
1. Возникает вообще при избытке
почвенной влаги и при отсут¬
ствии или слабом доступе кисло¬
рода.
2. Растительные остатки в торфе
обычно не разрушаются до конца,
но хорошо сохраняют свою струк¬
туру и различимы простым глазом.
3. В сухом виде имеет связную
структуру, которую сохраняет и
после размачивания в воде.
Перегной
Образуется при умеренной почвен¬
ной влажности и при достаточном
доступе кислорода.
Бесструктурная масса, где раститель¬
ные остатки на-глаз не различимы или
плохо различимы.
Положенный в воду превращается
в землистую массу.
1 Примером перегноя может служить лесная подстилка из перегнив¬
ших листьев и древесины деревьев, напочвенных лесных мхов и т. д.
22
Торф, как было сказано, — продукт разложения органи¬
ческих остатков, главным образом растительных. Сущность
химических изменений последних при пере¬
ходе в торф сводится к обогащению (в процент¬
ном отношении) углеродом и к обеднению
кислородом и водородом.
С оторфовыванием растительных остатков целлюлоза,
клетчатка и другие углеводы, где процент содержания водо¬
рода и кислорода относительно велик, а углерода мал, раз¬
рушаются, лигнин же, более богатый углеродом и бедный
водородом и кислородом, наоборот, накопляется.
Торф обнаруживает несомненное родство
с углем. Торфообразование — процесс современный. Но
процесс этот имел место не только в четвертичный период,
но и в третичный, на протяжении мезозоя и палеозоя и даже
в^ докембрийский период (К. Бюлов, 1929). Но более древние
торфа (до третичного периода включительно) претерпели после
своего образования своеобразные изменения: они прошли
различные стадии обугливания (стадию бурого угля, камен¬
ного угля, антрацита, графита) и называются уже не торфом,
а углями. Сущность процесса обугливания состояла в обога¬
щении углеродом (К. Бюлов). Таким образом, обогащение
углеродом и обеднение водородом и кислородом при отор-
фовании можно проследить и далее в ряде обугливания вплоть
до продуктов, представляющих чистый углерод. Этот гене¬
тический ряд, ведущий от неизменного растительного мате¬
риала до графита, выглядит так (табл. 5):
Таблица 5 1 2В 1С‘ частях вещества
Углерод
Водород
Кислород
Азот
Сфагнум^
50,0
6,5
42,4
Дерево2
50,0
6,0
43,0
1,0
Торф2
58,0
5,5
34,5
2,0
Торф с поверхности1 • .
57,7
5,4
36 0
Торф с глубины 2,5 м1
62,0
5,2
30,6
Торф с глубины 4,6 л«1
64,07
5,0
26,8
Бурый уголь2 •
70,0
5,0
24,0
1,0
Каменный уголь2
82,0
5,0
12,0
1,0
Антрацит2
94,0
3,0
3,0
следы
Г рафит2
100,0
—
Резюмируем сущность процесса оторфования и обугливания.
1. При переходе растительных остатков в
торф увеличивается содержание углеродам
1 По В. Сукачеву (1926). Содержание азота не приведено.
2 По Бюлову (1929). Содержание в обеззоленном органическом веще
стве.
23
уменьшается содержание кислорода и обычно
водорода. Этот процесс продолжается и дальше
по мере дальнейшего изменения торфяной
массы.
2. Образование каменного угля, с точки зре¬
ния химических изменений вещества, есть про¬
должение оторфования. Чем старше уголь, чем
большим изменениям он подвергся, тем богаче
он углеродом и тем сильнее отличается от
торфа.
3. Торф — это наиболее молодой член ряда
обугливания, первая ступень в ряде превра¬
щений растительного вещества в уголь.
До конца XVIII в. представления о торфе носили в общем
фантастический характер и имеют лишь исторический интерес.
Приведем некоторые из этих определений (по И. Макарову и М. Ней-
штадту, 1930).
И. Шейхцер (Scheuchzer, 1706) и некоторые другие считали торф
землей, пропитанной горючими веществами — земляным маслом, горной
смолой или серой. Растительные же остатки считались несущественной
частью торфа.
Патридофилус (Patridophllus) считал, что торфяные залежи образовались
из растворенных и раздробленных водами всемирного потопа органических
веществ, которые затем вновь осели на землю.
И. Фойгт (Voigt) считал торф особой корневой тканью, способной к
росту, но не образующей листьев и стеблей.
Плот (Plot) и Андерссон (Andersson) в 1799 г. высказывали мнение, что
торф сам по себе представляет особое специфическое растение, а не про¬
дукт растительного происхождения.
К концу XVIII в. появляются уже правильные представления о торфе.
Абильгаард С. (Abilgaard S.), Эйзелен (Eiselen, 1795), де Люк (de Luc, 1782)
считали, что торф образовался из растительных остатков и образуется и в
настоящее время.
Одно из первых правильных определений торфа, замеча¬
тельное по своей близости к современному научному опреде¬
лению, принадлежит Риму (Riem, 1794; цит. по Г. Шрейберу,
1927). „Торф — это горючая, растительная, болотная земля
коричневого или черного цвета, которая связана волок¬
нами и корешками разложившихся растений, а частью
также и минеральными частицами, и при горении в боль¬
шей или меньшей степени издает особый запах“. В этом
определении правильно выдвинуты все основные признаки
торфа.
Приведем некоторые из позднейших определений.
Шрейбер (1927): „Торф — это вид почвы, состоящий глав¬
ным образом из связанных друг с другом, более или менее
разложившихся растительных остатков. Эти остатки в свежем
состоянии богаты водой, в сухом — способны к горению. Торф
образуется на протяжении четвертичного периода во влаж¬
ных местах".
Бюлов К. (1929): „Торф — это смесь специфически и не
до конца разложившихся частей растений, окрашенная в более
или менее темный цвет, богатая углеродом и имеющая более
24
или менее кислую реакцию. Торф в истории земли является
наиболее молодым членом генетического ряда углей, образо¬
вание его происходит еще и в настоящее время“.
В обоих определениях не отражена важная особенность
торфа, отличающая его от гумуса, — различаемость раститель¬
ных остатков невооруженным глазом. Кислая реакция,
в определении Бюлова, — не необходимый признак торфа. Зато
здесь особенно резко выдвинут геологический момент — гене¬
тическая связь торфа с углями. Основной морфологический
признак — структурность торфа — ясно подчеркнут в опреде¬
лении Е. Раманна (Е. Кашапп, 1911): »Торф — это образующее
почву скопление более или менее гумифицированных расти¬
тельных остатков, сохранивших растительную структуру".
Раманн добавляет еще: „Для торфа характерна наличность
макроскопически распознаваемых растительных остатков,
часто почти нацело слагающих всю массу".
Более точно определение Г. Стадникова (1930): „Торф есть
сильно обводненный конгломерат битумов г, гуминовых кислот,
их солей, различных других продуктов разложения раститель¬
ного материала без доступа воздуха1 2 и не успевших еще раз¬
ложиться форменных элементов растений (листья, стебельки,,
корни)“. В этом определении очень полно указаны химические
признаки торфа, но не выявлен его генезис и геологическое
прошлое, а также физическая структура — связанность аморф¬
ной гумусовой массы не разложившимися частями растений.
Виды торфа.
Значение „вида“ торфа. Понятие о виде торфа в настоящее-
время не может считаться установленным. Объем и содержа¬
ние этого понятия сильно различаются у разных авторов, как
и самый принцип выделения видов торфа. Однако почти иге
торфоведы признают, что основанием при определе¬
нии торфа или отнесении его к тому или иному
виду служит его ботанический состав. Это зави¬
сит от двух причин. Во-первых, ботанический состав, как и
степень гумификации торфа, служит важнейшим признаком,
который в значительной степени определяет физические,
химические и технические свойства торфа. Во-вторых, оба
эти признака могут быть легко и быстро определены — частью
уже в поле, тогда как другие признаки (зольность, тепло¬
творная способность и пр.) требуют для определения значи¬
тельной затраты труда, времени и средств.
Номенклатура торфов производится обычно от
названий растений, образующих торф. Чаще всего название
1 Битум — смесь восков, жирных кислот, смол и углеводородов. Битумы
получаются из абсолютно сухого торфа действием смеси спирта и бензола
(по Г. Стадникову, 1930).
2 Во 2-м издании 1932 г. слова „без доступа воздуха“ опущены.
25
вида торфа строится из латинских или русских обиходных
названий растений, преобладающих в торфе, например Phrag-
mit es-торф, или тростниковый торф.
Так, финский исследователь Г. Варен (Н. Waren, 1924) в своей класси¬
фикации, которую мы рассмотрим позднее, не только последовательно
придерживается обозначения торфа именем господ¬
ствующего растения, но вдобавок это правило и декла¬
рирует. Тот же автор указывает, как составлять названия торфа, когда
в нем в значительном количестве встречается не один, а два или более видов.
Он рекомендует в этом случае ставить в конце название наи¬
более распространенного компонента. Например, в осоково¬
тростниковом торфе тростник — более важный компонент, осока же образует
лишь примесь.
М. Кириллович и М. Короткина (1933), следуя практике
болотоведов Торфяной опытной станции Наркомзема, дают
правила номенклатуры торфов. Следуя в общем Г. Варену,
они уточняют его правила и составляют названия
торфа на основе глазомерного определения под
микроскопом процента покрытия растительных
остатков в торфе. Правила эти сводятся к следующему:
1. Виды, имеющие менее 15% покрытия \ не входят в на¬
звание торфа.
2. Растения, имеющие покрытие в 15 и более процентов,
но менее 20®/о, входят в название торфа в виде существитель¬
ного.
3. Растения, дающие от 20 и более процентов покрытия,
входят в название в виде прилагательного.
4. Если торфообразователей, дающих не менее 20%,
несколько, то преобладающий ставится на конце.
Пример
Состав торфа
Осока • ... 35%
Гипнум 30о/о
Травянистые остатки .... 20%
Тростник 15%
Хвощ единично
Такой способ обозначения весьма удобен. Лучше, однако,
следуя предложений) 3. Ф. Руофф, вычислять видовой состав
торфа, принимая за сто всю массу, т. е. гумус+растительные
остатки.
Практически мы поступаем при этом так. Возьмем торф, ботанический
состав которого приведен выше. Мы определяем сначала степень его
гумификации (или разложения) по глазомерному микроскопическому методу
П. Варлыгина 1 2.
Предположим, что степень разложения этого торфа равна 50°/0, т. е. что
одна половина (по площади) частиц под микроскопом приходится на гумус,
-а другая половина — на форменные растительные остатки. Удаляем затем
гумус отмучиванием через сито (см. ниже) и в отмученной части определяем
процентный состав торфа. Результаты этого определения приведены выше.
1 Принимаются во внимание лишь форменные остатки. Гумусовая масса
отмучивается и в расчет не принимается.
2 Описание этого метода см. ниже.
Название торфа
Травяно-гипново-осоковый
с тростником
26
Затем делаем пересчет на неотмученный торф и соответственно изменяем
цифры. В название же вводим характеристику степени разложения. Тогда
для приведенного выше торфа мы получаем:
Состав торфа Название торфа
(считая гумифицированную массу)
Г умус 50 %
Осока 17,5 %
•Гипнум 15,0 %
Травянистые остатки 10,0 %
Тростник 7,5 %
Хвощ единично
Травяно-гипново-осоковый с тростни¬
ком со степенью гумификации в 50%
Всего . . 100 %
Это изменение способа М. Кирилловича и М. Короткиной
полезно, так как, вводя процент гумификации, мы даем
более полную характеристику торфа, а количе¬
ственные соотношения компонентов — в их дей¬
ствительном, а не в искаженном виде.
Понятно, что самый способ составления названия торфа
■остается без изменений.
Несколько иной точки зрения держатся торфоведы Инсти¬
тута торфяной промышленности С. Тюремнов и др. В пред¬
лагаемой ими „Классификации видов торфа“ (1935) они зача¬
стую обозначают торф не по господствующим ком¬
понентам, а по тем растениям, которые встре¬
чаются в основном в каждом образчике данного вида торфа,
причем эти растения могут и не составлять главной массы
торфа. Этот принцип нуждается в дальнейшей разработке
и обосновании.
Так, ОЬ 1ц вит-т орф характеризуется постоянным при¬
сутствием ЭрЬ. оМиэиш, хотя процент последнего в массе торфа
иногда не превышает 15.
Эта система обозначения связана с тем, что авторы ее
за основу видов торфа принимают—по крайней
мере, декларативно — не доминирующие виды,
а сочетание остатков характерных видов.
Предложениеназывать торфа на основании
процентного соотношения их главнейших ком¬
понентов следует считать наиболее целесооб¬
разным.
Классификация торфов и седиментов.
Существующие классификации торфов и водных органиче¬
ских отложений (седиментов) можно разделить на две группы:
1) классификации, основанные на принципе
строения торфа, т. е. его ботанического состава;
2) классификации, базирующиеся на генезисе торфа, или
генетические классификации.
27
Эти две точки зрения отмечены отчасти уже Г. Потонье
(Н. Potonié, 1911). Он пишет, что название „беломоховый торф“
(немецкое „Weissmoostorf") употребляется тогда, когда хотят
показать, что это „Sphagnetum-Torf", т. е. торф, продуци¬
рованный сфагновым болотом, сфагнетумом. Когда
же говорят сфагновый торф („Sphagnum-Tori"), то это указы¬
вает, что торф содержит лишь остатки сфагнума.
Подобный торф может образоваться и не на сфагнетуме.
Ниже мы рассмотрим классификации как собственно тор¬
фов, так торфов и седиментов. Значительное большинства
имеющихся классификаций относится к первой группе, т. е.
основывается на составе торфа, причем поэтому
принципу строятся как высшие, так и низшие классификацион¬
ные единицы.
Г. Варен (1924) дал классификацию торфов Финляндии.
Он различал свыше двадцати видов торфа. Эти виды он, сле¬
дуя Г. Андерссону (G. Andersson, 1928), сгруппировал в три
большие группы.
Приведем классификацию Варена.
А. Торфа аз остатков сосудистых растений. В этой группе торфов
преобладают сосудистые растения.
1. Тростниковый торфе преобладанием тростника.
2. Хвощевый торф. Важнейший торфообразователь — хвощ.
(Equisetum limosum, var. fluviatile).
3. Осоковый торф. Здесь преобладают виды Сурегасеае, особенно
осоки. Кроме них, встречаются и широколистные болотные травы, часто
их семена (например, Comarum palustre, Menyanthes tnfoliata). В зависимо¬
сти от второстепенных компонентов различаются такие виды осокового
торфа:
а) тростниково-осоковый торф1,
б) хвощево-осоковый торф,
в) пуши цево-осоков ый торф,
г) шейхцериево-осоковый торф,
Д) буромохово-осоковый торф1 2,
е) сфагново-осоковый торф,
ж) осоковый торф с остатками лиственных пород.
4. Шейхцериевый торф.
5. Пушицевый торф. В чистом виде редок.
6. Кустарничков ы и торф. Низкорослые болотные кустарнички
обычно образуют примесь в осоковых и сфагновых торфах и редко пре¬
обладают в торфе.
7. Древесный торф и лесной торф. Автор называет торфа с
преобладанием древесных остатков „древесными* (немецкое Holztorf). Для
торфов, образовавшихся в лесу, он предлагает название „лесной торф"
(нем. Waldtorf), вводя, таким образом, момент происхождения. Понятия,
„лесной" и „древесный" торф не вполне совпадают. Как указывает Г. Варен,
в древесном торфе древесные остатки не всегда преобладают.
В. Моховые торфа. Главную массу торфа в этой группе образуют мхи.
1. Сфагновый торф. Преобладают торфяные мхи. По преобладанию
того или иного вида сфагнума и по примеси к сфагнуму других видов
различаются:
1 Относительно правила составления названий торфов у Г. Варена
см. выше.
2 Бурые мхи (немецкое название ВгаипшооБе) — это болотные мхи из
порядка Вгуа1ез.
28
а) чистый с ф а г н о в ы й т о р ф (Sph. fuscum-торф, Sph. magellanicum-
торф, Sph. cuspidata-торф идр.),
б) пушицево-сфагновый торф. Sph. fuscum-торф, Sph. magel-
lanicum-торф, Sph. angustifolium-торф),
в) шеихцериево-сфагновый торф (Sph. cuspidata-торф),
г) осоково-сфагновый торф (Sph. cuspidata-торф, Sph. palu-
stria-торф, Sph. papillosum-торф, кустарничково-осоково-сфагновый
торф),
д) кустари ичково-сфагновый торф, сфагновые остатки
более или менее сильно разложились, обильна примесь кустарничков,
е) лесной сфагновый торф: сфагновая масса сильно разложи¬
лась, аморфная, часто мажущаяся, с микроскопическими остатками
древесины; различаются: сосново-сфагновый и березово-сфагновый
торф.
2. Торф из лиственных мхов, не сфагновых.
а) буромо ховый торф; важнейшие компоненты—мхи, виды рода
Drepanocladus, Paludella squarrosa, Meesea,
б) торф из к у к > шкина льна образован видами рода Polytrichum;
важнейший из них Polytrichum commune. Сюда же относится и не
имеющий значения в Финляндии дикрановый торф.
С. Лишайниковые торфа. Они состоят из черных мажущихся остат¬
ков лишайников (Cladonia rangiferina, Cetraría islándica) и образуются на
сухих болотах под лишайниковым покровом. Они имеют лишь теоретиче¬
ский интерес.
Виды торфа, различаемые Г. Вареном, несмотря на тринадцати летнюю
давность его классификации, являются единицами узкого объема и вполне
определенного содержания. Выше отмечалось, что Г. Варен весьма последо¬
вателен в правилах выделения видов торфа и их номенклатуры, базируясь
на преобладающих в торфе растениях. Важно отметить, что в тексте своей
статьи Г. Варен отмечает еще более мелкие единицы торфа, чем в приведен¬
ном списке. Особенно детализированы сфагновые торфа благодаря тому,
что определение сфагнума в торфе часто доводится до вида. Любопытно,
что автор дает результат ботанического анализа торфа в виде формулы.
Цифрами, следуя Е. Мелин (Е. Melin, 1917), он обозначает количественные
соотношения между растениями в торфе:
5—данное растение образует главную массу торфа1
4— !Д —Vo всей массы торфа
з- V* -v; .
2 Vi 6 Ve » » »
1— менее—Vio »
Названия растений в торфе сокращенно обозначаются начальными бук¬
вами латинского родового имени, например, A-Amblystegium, Al-Alnus и т. д.;
части растения обозначаются тоже буквами: R (Radizellen, т. е. тонкие кореш¬
ки), S (Samen, семена) и т. д.
Гумозность (степень разложения) обозначается по десятибалльной шкале
Л. Поста (L. Post). Приведем пример подобной сокращенной формулы торфа:
Нб С5 R В1 Fr S3 (teres) А2. Она показывает, что гумозность (Н) торфа
равна 5, что главную массу“ торфа образуют Сурегасеае (С5), встречающиеся
в виде корешков (R-Radizellen), листьев (Bl-Blatter) и плодов (Fr-Früchte).
Далее, значительна примесь сфагнов (S), именно Sphagnum teres; она равна 3,
т. е. этот вид сфагнума дает от */4 до Vg всей массы торфа. Далее, в незна¬
чительном количестве (цифра 2, т. е. г/1в — У8 всей массы торфа) встречается
мох Amblystegium (А).
Выделение видов торфа у Г. Варена проведено последова¬
тельно и логично. Однако три главные группы торфов вряд
ли удачны. Во-первых, они крайне не равноценны
по своему значению. Так, последняя группа (лишайни-
1 Эта торфяная шкала в точности соответствует известной шкале Гульт-
Сернандера (НиИ-Бегпапёег), выражающей покрытие растений в живом
растительном покрове.
29
новые торфа) имеет ничтожное значение по сравнению с пер¬
выми и дальше не подразделяется. Во-вторых,—и это самое
важное—три группы крайне разнородны. Часто
близкие по ботаническому составу торфа находятся в разных
группах и, наоборот, очень далекие—в одной группе. Так,
буромоховые торфа ближе к буромохово-осоковым, находя¬
щимся в другой группе, чем к сфагновым, с которыми они
соединены. Классификация Г. Варена в общем снизу доверху
проведена по ботанико-систематическому прин¬
ципу, так как в основу не только низших, но и высших
групп торфа положены систематические единицы (группы)
растений.
15Ш1ИИЖ1Р1
■ИиШШШг
1
у//
:■ ' 1Л
их ■<М . .Ж'Я^лъи’. ¿2
Рис. 1. Мало разложившийся тростниковый торф
в естественную величину. По Бюлову, 1929.
Кроме Варена, классификации, базирующиеся в основном
на принципе строения (не генезиса) торфа, дают советские
болотоведы. Таковы вошедшие в учебники классификации
В. Сукачева (1926) и В. Доктуровского (1932). Ниже мы изла¬
гаем в несколько сокращенном виде классификацию В. Сукачева
(1926). Он, следуя К. Веберу, разделяет торфа на три группы
по содержанию в них питательных веществ,
точнее—зольных элементов питания растений: 1 группа—
эвтрофные торфа, богатые питательными веществами;
II группа—мезотрофные торфа, более бедные питатель¬
ными веществами; III группа— торфа олиготрофные,
бедные питательными веществами. В пределах этих групп
В. Сукачев различает девять видов торфа и четыре разновид¬
ности. Эти виды приведены ниже.
I. Торфа эвтрофные. Сюда входят:
1. Тростниковый торф. Состоит из стеблей, корне¬
вищ и листьев тростника. Малоразложившиеся его части в
свежем торфе характерного желтого цвета; они легко отличимы
простым глазом. Торф многозольный, поэтому дает плохое
топливо. Может итти на подстилку скоту, но подстилку дает
тоже плохую (рис. 1).
зо
2. Камышевый торф. Образован стеблями и корневи¬
щами камыша (Scirpus lacustris). По внешнему виду и свой¬
ствам сходен с тростниковым торфом. Строение же стеблей
камыша отличается от тростниковых.
3. Хвощевый торф. Образован стеблями, корневищами
и ветвями хвоща. Все эти части хвоща в торфе весьма сходны
с живыми и хорошо узнаются по черному цвету и зубчатым
листовым влагалищам. Нередко к хвощу примешаны осоки и
другие растения. Для топлива мало пригоден вследствие вы¬
сокой зольности; не годится и для подстилки.
4. Осоковый торф. Состоит из корней и корневищ
осок и представляет в свежем состоянии буроватожелтую
волокнистую массу, темнею¬
щую на воздухе. Этот торф
в зависимости от видов осок,
входящих в него, сильно ва-
риирует в свойствах. Если золы
содержится немного, то идет
на топливо.
5. Г ипновый торф. Об¬
разуется из мхов порядка
Bryales. Цвет темный, часто
почти черный. Консистенция
мажущаяся. Высохший торф
очень легок и дает удовле¬
творительное топливо.
6. Ольшанниковый
торф. В нем обильны остатки рис. 2. Молодой сфагновый торф под
черной Ольхи (Alnus glutinosa) микроскопом (увелич. в 25 р.). По
очень мягкой консистенции. Бюлову, 1929.
Этот торф имеет вид темной,
мажущейся массы. Мало пригоден как топливо. На подстилку
вовсе не годится.
II. Торфа мезотрофные.
7. Лесной торф—сборное понятие. В нем преобладает
то сосна, то береза и встречаются остатки многих травянистых
растений (осоки и др.), а также и мхов. Техническое значе¬
ние торфа невелико.
8. Торф из миа кукушкин лен (Polytrichum) обра¬
зует довольно плотные темные слои небольшой мощности с
хорошо различимыми стебельками мха. Встречается редко.
III. Торфа олиготрофные.
9. Сфагновый торф сложен в основной массе сфагно¬
выми мхами. Имеет две разновидности, образующие мощные,
хорошо видные на разрезах торфяников слои.
Молодой сфагновый торф; образует верхнюю
толщу торфа сфагновых торфяников й состоит из хорошо
сохранившихся, мало разложившихся листочков и стеблей
сфагнума (рис. 2); на топливо не годится, но идет на приго-
31
товление строительного материала (изоплиты) и дает прекрас¬
ную подстилку для скота.
Старый сфагновый торф (рис. 3) слагает более
нижние слои сфагновой толщи; образован также остатками
сфагновых мхов, но .более сильно разложившимися, темнее,
чем предыдущий. В зависимости от примеси к сфагнуму дру¬
гих растений различаются четыре сорта сфагнового торфа:
а) Ш е й х це р и е в о-с ф а г н о в ы й торф; к сфагнуму
»примешиваются стебли, корневища и листья шейхцерии;
б) Пуишцево-сфагновый торф; к сфагнуму примеши¬
ваются остатки пушицы (Eriophorum vaginatum) в виде листо¬
вых влагалищ и корешков; последние имеют вид узких, длин¬
ных, желтоватобурых или более
темных лент.
в) Собственно-сфагно¬
вый торф, часто являющийся
сфагново-сосновым, так как
среди сфагнума в нем часты
остатки древесины и коры бо¬
лотной сосны.
г) Смолистый торф (смо¬
ляк)—по Танфильеву, один из
конечных продуктов гумифика¬
ции болотных растений; имеет
вид черной аморфной массы,
в изломе блестящей.
Классификация В. Сукачева
имеет в низших звеньях виды
торфа, основанные на принципе
его ботанического строения.
Кое-где, однако, выдвигается, хотя и слабо, генетический мо¬
мент, например, для ольшанникового торфа как существенный
признак указано происхождение из черноольхового леса. Выс¬
шие единицы—группы торфов—образованы не попринципу
ботанического состава торфа, а по содержанию
в них элементов питания растений, как и в класси¬
фикации К. Вебера. Таким образом, в отличие от классифи¬
кации Г. Варена принципы образования низших
и высших единиц здесь разные. Надо сказать, что
классификация В. Сукачева еще мало детализирована. Коли¬
чество выделяемых им видов торфа очень ограничено, объем
их велик и нередко недостаточно ясен. В характеристике
видов торфа Сукачева важно описание макроскопических
признаков, физических свойств и применения.
Классификация В. Доктуровского (1932) напоминает клас¬
сификацию В. Сукачева (1926). Здесь различаются почти те
же торфа. Классификация также основана на ботаническом
составе торфа, а не на его генезисе. Однако в отличие от
В. Сукачева разделение у В. Доктуровского одноступенное,
так как приняты единицы лишь одного ранга. Это делает ее
32
Рис. 3. Старый сфагновый торф
лод микроскопом (увелич. в 25 р.).
По Бюлову, 1929.
менее удобной, так как объединение видов торфа в три есте¬
ственные группы—олиготрофную, мезотрофную и эвтрофную
(как у Сукачева) — весьма облегчает ориентировку в разно¬
образии торфов 1.
Замечательную для своего времени классификацию торфов
дал К. Вебер (1908). Он различает три группы торфов по
содержанию в них питательных веществ — олиготрофную, ме¬
зотрофную и эвтрофную. Как отмечалось выше, делением тор¬
фов на эти группы воспользовался и В. Сукачев (1926). Боль¬
шую ценность представляет то, что К. Вебер дает очень
подробную характеристику видов торфа, их ботанического
состава, микроскопических признаков, физических свойств,
консистенции, химизма, применения, а также, что особенно
важно и что обычно отсутствует даже в более поздних клас¬
сификациях,— сведения по географическому распространению
отдельных видов торфа в Германии. Описания видов торфа
у К. Вебера могут и до сих пор служить примером образцо¬
вых описаний.
Схема классификации К. Вебера1 2:
a) Богатые питательными ве’ществами (эвтрофные)
торфа; сюда входят тростниковый, осоковый, камышевын (Cladium maris-
cus), гипновый, трясинный (или ольховый), лесной торфа.
b) Более бедные питательными веществами торфа
(м е з о т р о ф н ы е): торф из мха Polytrichum, торф из соснового леса, вере¬
сковый торф.
c) Бедные питательными веществами (ол и'готрофные)
торфа: пушицевый, шейхцериевый, сфагновый.
Таким образом, классификация К. Вебера так же, как и В. Сукачева,
оказывается двуступенной, причем низшие и высшие единицы осно¬
ваны на разных принципах: первые — на принципе строения (бота¬
нического состава торфа), а вторые — на содержании питательных
веществ. Хотя генетический момент не руководящий для К. Вебера,
однако в некоторых случаях он останавливается и на материнских фитоцено¬
зах, продуцирующих тот или иной вид торфа: так, он указывает, что
ольховый торф образовался из заболоченных лесов с ольхой, лесной же
торф возник из смешанного леса и пр. Надо отметить, что в Центральной
Европе уже сравнительно давнр стали различать довольно много видов торфа
(например, Г. Потонье, 1911). Уже в 1907 г. Цайлер и Вильк (Zailer V. und
Wilk L.) различали тростниковый, осоковый, гипновый, ольховый, шейхце¬
риевый, пушицевый и другие виды торфа, для которых эти авторы приво¬
дят многочисленные, до сих пор фигурирующие в руководствах данные хи¬
мического анализа.
Из более поздних классификаций торфа центрально-европейских торфо-
ведов нужно остановиться еще на классификации Г. Шрейбера (1927). Она
в основном также построена на ботаническом составе торфа 3.
1. Беломоховые торфа:
Сфагновый торф с подразделением на молодой и старый
сфагновый торф.
1 Позднее (1936) В. Доктуровский несколько детализировал свою класси¬
фикацию. Однако разделение на главные группы и здесь четко не проведено.
2 Вебер классифицирует здесь не только торфа, но и водные органоген¬
ные отложения — .мудды" (седименты).. Мы приводим здесь только перечень
видов торфа.
3 Кроме этой, Г. Шрейбер дает тут же и другую систему, по принципу
Л. Поста.
3—722
33
Пушицевый торф.
2. Буромоховые торфа:
Сюда входит гипновый торф из мхов порядка Bryales.
За. Торфа болотистого леса (Bruchwaldtori):
Betula alba-торф (березовый торф).
Quercus-торф (дубовый торф).
Ainus glutinosa-торф (олыпанниковый торф)»
Pinus silvestris-торф (сосновый торф).
Picea excelsa-торф (еловый торф).
ЗЬ. Кустарничковые торфа:
Pinus montana-торф (торф из горной сосны).
Cal luna-торф (вересковый торф).
Oxycoccus-торф (торф из клюквы).
Betula nana-торф (торф из стланика).
Andromeda-торф (торф из подбела).
Salix-торф (ивовый торф).
4. Травянистые торфа:
Осоковый торф.
Cladium-торф (камышевый торф).
Scirpus caespitosus-торф.
Narthecium-торф.
Juncus-торф (ситниковый торф).
Scheuchzeria-торф (шейхецерневый торф)*
Phragmites-торф (тростниковый торф).
Scirpus lacustris -торф (камышевый торф из озерного камыша^
Equise tum-торф (хвощевый торф).
Menyanthes-торф (торф из вахты).
5. Гиттиевые торфа (т. е. торфа, отложившиеся под водой).
Всего К. Шрейбер различает 24 вида торфа. Он дает подробное опи¬
сание структуры, особенно же условий образования торфа в настоящем
и прошлом, местонахождения в торфяной залежи по отношению к другим
видам торфа, географического распространения, применения и пр.
Шрейбер выделяет торфа в основном согласно структурн о-б о тани¬
не с к о м у принципу. Высшие единицы (.роды торфа") построены частью
по генетическому принципу. Например, группа Bruchwaldtorf, как показы¬
вает самое название, имеет материнским фитоценозом заболоченный лес.
Однако генетический принцип проведен недостаточно отчетливо и не везде^
Так, группа кустарничковых торфов основана не на генезисе, а на сложении
торфа из остатков кустарничков. В классификации Г. Шрейбера крайне сум¬
марно разделены моховые и буромоховые торфа, играющие, например, у нас
в Союзе большую роль и хорошо отличающиеся друг от друга по господ¬
ствующему мху. Это особенность многих центральноевропейских торфове-
дов в противоположность скандинавским (сравнить, например, детальное
разделение сфагновых торфов у Г. Варева) и советским. Зато у Шрейбера по¬
дробно разделены древесные торфа, торфа травянистых растений (группа 4)
и, наконец, кустарничковые торфа. Между тем последние весьма незначи¬
тельно распространены, а для некоторых видов этой группы (например,
торф из клюквы) выделение их в самостоятельные единицы возбуждает даже
некоторые сомнения.
В общем, разделение Шрейбера—п римердвуступеннойкласси-
ф и к а ц и и. Нижние ступени выделены по принципу ботанической структуры
торфа так же, как и в классификациях Сукачева, Варена и других, верхние
же частью основаны на том же принципе, частью — на принципе генети¬
ческом.
Чисто структурную классификацию торфов с довольно детальным раз¬
делением на виды дает К. Бюлов (1929) в своем новом Handbuch по боло¬
товедению. Она выглядит в сокращенном виде так:
I. Чист о-м оховые торфа.
1. Сфагновый торф с двумя подразделениями:
а) молодой сфагновый торф,
б) старый сфагновый торф.
2. Буромоховые торфа.
34
II. Смешанные моховые торфа.
3. Пушицево-моховой торф.
4. Шейхцериево-моховой торф.
5. Моховой торф с древесными остатками.
6. Кустарничково-моховойторф, где к сфагнуму примеши¬
ваются остатки кустарничков (Heide-Moostorf).
7. Смешанные торфа из сфагновых мхов и представителей Glumi-
florae (главным образом Сурегасеае).
III. Древесные торфа.
8. Торфа из остатков хвойных пород:
а) торф из соснового леса,
б) торф из горной сосны,
в) торф из елового леса.
9. Торфа из остатков лиственных пррод:
а) березовый торф,
б) ольховый торф.
10. Кустарничков ые торфа.
IV. Торфа из остатков Glumiflorae (злаки и Сурегасеае).
11. Торфа из Сурегасеае с такими подразделениями:
а) осоковые торфа:
1. крупноосоковый торф;
2. мелкоосоковый торф.
б) Торф из камыша (Scirpus lacustris, Scirpus caespitosus'.
в) Ринхоспоровый торф.
г) Торф из Cladium mariscus.
12. Торфа из злаков:
а) тростниковый торф,
б) торф из манника (Glyceria-торф),
в) молиниевый (Molinia-торф).
V. Другие торфа.
13. Шейхцериевый торф.
14. Хвощевый торф.
15. Narthecium-торф.
16. Iris-торф.
17. Menyanthes-торф.
18. Торф из Aspidium Thelypteris.
Классификация К. Бюлова многоступенная. Она основана сверху
донизу иа принципе ботанического состава торфа. Как
указывает сам автор, виды торфа устанавливались им на основании глав¬
ных компонентов, низшие же подразделения — на основании второстепен¬
ных. Отметим крайнюю неравноценность единиц одного м того же ранга.
Иногда единицы низшего порядка оказываются более широкими, чем выс¬
шего. Достаточно указать, что такая важная, большая и разнородная группа,
как осоковый торф, стоит наряду с ринхоспоровым торфом, торфом из ман¬
ника и пр., а пушицево-моховой (сфагновый) торф представляет единицу
более высокого ранга, чем осоковый. Хотя в тексте и выделяются виды
сфагнового торфа по преобладающему виду сфагнума (Fuscuni-торф и пр.),
но в классификации это не отражено. Здесь, как и в других центрально-
европейских классификациях, сфагновые торфа понимаются весьма суммарно,
так же как и буромоховые торфа; торфа же из остатков травянистых ра¬
стений подразделяются в общем весьма дробно, причем здесь выделяются
такие редкие и слабо распространенные торфа, как Iris-торф, ринхоспоровый
торф и др.
Рассмотрим теперь генетические классификации, т. е. основанные на гене¬
зисе, происхождении торфа. Л. Пост, как показывает самое название его статьи
»Das genetische System der organogenen Bildungen Schwedens*, дает генети¬
ческую систему торфов и водных отложений Швеции. Л. Пост считает,
чго при классификации этих образований нужно пользоваться »материн¬
скими формациями, т. е. типами растительности, которые
продуцируют виды торфа. При выделении видов торфа Л. Пост рекомендует
пользоваться не наиболее бросающимися в глаза расти¬
тельными остатками в торфе, а всей совокупностью
3* 35
характерных торфообразующих р а с т е н и й. Именно эта сово¬
купность позволяет определить „материнские растительные формации*. Сле¬
довательно, генетическая система отличается от структурных (см. выше) не
только принципом к л ас с иф и к а ци и, н о и признаками, по кото¬
рым выделяются виды торфа. В сокращенном виде она выглядит
следующим образом:
/. Седименты. Эти органогенные образования отлагаются не на том месте,
где они возникли, но приносятся водой и другими агентами. Это так назы¬
ваемые аллохтонные образования. Сюда относятся:
A. Гитти и (швед. Gyttja); возникают благодаря гниению или брожению;
щелочная вытяжка имеет зеленоватый цвет. Различаются такие виды гиттнй:
Глубоководная гиттия (Profundalgyttja) с вариантами.
Береговая гиттия (Litoralgyttja).
Водорослевая гиттия (Algengyttja).
Известковая гиттия и некоторые другие.
B. Д ю (шведское Dy). Разложение при ослабленном доступе воздуха
дает коллоидные гумусовые вещества. Щелочная вытяжка бурого цвета.
Здесь различаются:
Озерная дю; образуется из продуктов распада планктона в озерах.
Мочежинная дю; возникает в мочежинах сфагновых болот из про¬
дуктов распада сфагновых мхов.
II. Торфа (аутохтонные образования). Они отлагаются на том самом
месте, где образовались. Делятся на два класса, или серии, по потребности
в питательных веществах материнских формаций. Эти серии — ряд эвтроф-
ных торфов (Sumpftorfreihe) и ряд олиготрофных торфов (Moortorfreihe).
Эти два ряда, по мнению Л. Поста, лучше всего отличаются по содержанию
азота. В эвтрофном ряду содержание азота редко меньше 1%, чаще же
2,0—2,5, даже 3,0%. В олиготрофном ряду это содержание большей частью
меньше 1%. Оба ряда делятся на ступени по степени увлажнения материн¬
ских формаций.
A. Ряд эвтрофных торфов.
1. Лимническая (водная) ступень, место отложения торфа по¬
стоянно находится под водой; пример торфов этой ступени — тростнико¬
вый торф.
2. Лимническо-тельматическая ступень (водно-болотная
ступень) носит промежуточный характер между лимнической и болотной
ступенью; сюда относятся хвощевый торф, торф из CladiumMa-
т i s с и s и более редкие виды торфа: Thelypteris-торф, Iris-торф, Glyceria-торф.
3. Тельматическая (болотная) ступень; место образовании
торфа лишь периодически находится под водой; сюда входят: крупноосо¬
ковый торф, буромоховой торф, болотная дю (Sumpfdy).
4. Наземная ступень; место образования торфа находится выше
уровня воды; здесь различаются: мелкоосоковый торф, торф боло¬
тистого леса, лесной перегной.
Аналогичное разделение на ступени дается и для торфов олиготрофного
ряда.
B. Ряд олиготрофных торфов.
1. Лимническая и тельматическая ступени:
Cuspidatum-торф (торф из сфагновых мхов секции Cuspidata), Scheuch-
zeria-торф, осоковый торф, Vaginatum-торф (из пушицы, Sph. magellanicum,
Sph. cuspidatum).
2. Наземная и омброгенная1 ступени с такими видами
торфа: кустарничковыйторф, торф болотистого березо¬
вого лес а, торф болотистого соснового леса, торф боло¬
тистого елового леса, Sph. fuse и m-торф, Sph. mag ell a ni-
cum-торф, вересковый торф (Heidemoder).
Классификация Л. Поста четырехступенная, причем в основу каждой
ступени положен особый принцип:
1 Омброгенная ступень, или ступень атмосферного питания, различается
лишь в серии олиготрофных торфов.
36
1. Отложение органического вещества на месте его
образования или принос со стороны.
2. Содержание питательных веществ определяет разделение
торфов на серии.
3. Степень влажности места, где отлагается торф, по¬
зволяет выделить ряд ступеней.
4. Ботанический состав дает воз но жность выделить
внутриступенивидыторфа.
Нельзя считать эту классификацию вполне генетической, как это делает
Л. Пост. Он не дает описания торфообразующих материнских формаций,
ва которых должна основываться такая классификация, а приводит
данные лишь об условиях существования. Эту классификацию можно назы¬
вать »генетической* скорее с точки зрения генезиса условий, в которых
образовался данный вид торфа.
В классификации, разработанной Институтом торфа, вво¬
дится момент генезиса, устанавливается, из какого типа
растительности возник какой-либо вид торфа,
или, иначе говоря, какова его родоначальная,
„материнская“ растительная труп пировка. При¬
нимается, что все последующие изменения, которым подвер¬
гается торф после своего образования, определяются его ма¬
теринским ценозом* Это и дает возможность установить по¬
следний даже для сильно измененных, сильно гумифицирован¬
ных торфов.
Генетическая классификация различает два типа торфа:
1) низинный тип и 2) верховой тип. Эти типы под¬
разделяются на группы и затем на виды торфа. Приведем ниже
сокращенную характеристику видов торфа. При этом мы сопо¬
ставим виды торфа по генетической классификации с видами
В. Сукачева (1926), где это возможно.
I. Торфа низинного типа.
1. Лесная группа с такими видами торфа:
Торф ольхово-лесной. Образуется в ольховых боло¬
тах и в общем соответствует ольшанниковому торфу В. Сукачева.
Торф березов о-л е с и о й. Образуется в березовых бо¬
лотах с слабо развитой травянистой растительностью.
Елово-согровыйторф. Богат древесными остатками,
образуется в болотистых лесах из ели, березы и кедра на
Урале и в Западной Сибири в так называемых сограх.
О с о к о в о-л есной торф. Преобладают остатки осок;
постоянна кора березы; образуется в осоково-березовых бо¬
лотах.
2. Топяная группа, куда входят:
Тростниково-т опяной торф. Образуется из трост¬
никово-осоковых ценозов типа топи. Название „топяной“
прибавляется по генетической классификации ко всем видам
торфа, образующимся на мокрых безлесных болотах — топях.
Этот вид торфа более или менее соответствует тростниковому
торфу В. Сукачева.
Хвощев о-т опяной торф. Образуется из мокрых хво-
щево-осоковых фитоценозов — топей; он близок к хвощевому
торфу В. Сукачева.
37
О с о к о в о-т опяной торф. Образуется на безлесных
осоковых топях, лишенных мохового покрова. Этот вид торфа
входит в сборный осоковый тип торфа В. Сукачева.
Г и п н о в о-т опяной торф. Образуется на подобных
же топях, но с сплошным ковром из влаголюбивых болотных
мхов (Meesea, Drepanocladus, Calliergon). Отчасти совпадает с
гипновым торфом В. Сукачева.
С ф а г н о в о-т опяной торф. Материнские ценозы',
образующие этот торф, имеют сплошной покров эвтрофных
(требовательных) видов сфагнума (Sph. subsecundum, Sph. ob-
tusum). В. Сукачевым этот вид торфа не различается.
Шейхцериев о-т опяной торф. Этот вид торфа
также относится к эвтрофному (низинному) типу. В нем много
шейхцерии, и он не соответствует шейхцериево-сфагновому
виду торфа В. Сукачева.
3. Переходная группа с такими видами торфа:
Комплексный переходный торф образуется
на болотах, где мозаично чередуются кочки с олиготрофной
растительностью и мокрые межкочечные понижения с эвтроф-
ной растительностью (комплекс).
Осоковый переходный торф образуется на болотах
с осоковым ярусом (Carex lasiocarpa) и с ковром мезотрофных
сфагновых мхов (Sph. Warnstorfii).
Сфагновый переходный торф. Материнский фито¬
ценоз— дальнейшая стадия эволюции переходного болота в
сторону олиготрофного. Здесь уже развивается ковер олиго-
трофного Sph. medium и др. с травостоем из мезотрофной
Carex lasiocarpa и негустым ярусом сосны и березы.
II. Торфа верхового типа.
Сюда входят такие виды торфа: F u s с и m-т о р ф. Обра¬
зуется на верховых болотах с сплошным ковром Sph. fuscum.
Medium-торф, наиболее обычный, в Европейской части
Союза на верховых болотах вид торфа. Материнские ценозы
этого торфа имеют сплошной покров из Sph. medium и Sph.
parvifolium, с пушицей, болотными кустарничками из семейства
вересковых и болотной сосной разной мощности и густоты.
Комплексный верховой торф отлагается в ком¬
плексах верховых болот, где мозаично чередуются торфяные
гряды—бугры из Sph. fuscum (или реже из Sph. medium), с
пушицей и кустарничками и мокрые мочежины с рыхлым по¬
кровом из влаголюбивых сфагновых мхов и шейхцерии.
Parvifolium-торф образуется на сильно обводненных боло¬
тах с рыхлым ковром Sph. parvifolium и обильной шейхцерией.
Шейхцериев ый верховой торф образуется на
подобных -же болотах. Сфагновый ковер образован, однако,
другими видами — Sph. balticum и Sph. Dusenii. Этот вид
торфа в общем близок к шейхцериево-сфагновому торфу
В. Сукачева.
Пушицевый торф. Совершенно гумифицированная
масса, обильно пронизанная волокнами пушицы. Он образо-
38
■зался из зарослей пушицы (Erlophorum vaginatum) с слабо
развитым сфагновым ковром.
С о с н о во-ку стар н и чк о в ы й торф. Сильно гуми¬
фицированный торф с обильными остатками,- кустарничков,
корой сосны и обильными сосновыми пнями. Он образуется
на болотах с хорошо развитой сосной, зарослями кустарнич¬
ков и моховым покровом из Sph. medium й Sph. parvifolium.
Авторы генетической классификации (коллектив сотрудни¬
ков Института торфа) устанавливают виды торфа на основа¬
нии сведений по среднерусским и уральским болотам. Они
считают свою классификацию предварительной. О призна¬
ках, на основании которых образец относится
к тому или иному виду торфа, т. е. о принципах
выделения видов торфа, они не говорят. Мало того,
для большинства видов торфа приводится лишь крат¬
кое описание материнского фитоценоза и не
дается описания сам ого торфа. Поэтомусоставить
ясное представление о том или ином видеторфа
подчас невозможно. Также остается открытым и вопрос,
насколько тот или иной вид торфа действительно связан с
-определенным ценозом. Позднее С. Тюремнов и другие боло¬
товеды Института торфа составили „Классификацию видов
торфа (проект стандарта)*, где уже дана характеристика бо¬
танического состава отдельных видов торфа и устанавливаются
принципы их выделения.
Виды торфа выделены на основании сведений по Москов¬
ской, Ивановской и Горьковской областям. Кроме того,
благодаря большому количеству образцов для каждого вида
торфа, авторы имели возможность обработать некото¬
рые количественные признаки торфов вариа¬
ционно статистически, чего до сих пор, как известно,
не делалось. Большое внимание обращено на технические свой¬
ства отдельных видов торфа.
При наиболее дробной разбивке видов торфа авторы стан¬
дарта различают уже 36 видов против 20 приведенных в более
ранней классификации.
Авторы стандарта классификации объединяют выделенные
ими виды торфа в четырехступенную систему (схема 2). При
этом они вводят в классификацию новый момент: единицы
второго и третьего порядка выделены по при¬
знаку степени разложения и зольности.
Характеризуя современное состояние классификаций торфа,
можно сделать следующие выводы:
1. В большинстве существующих классифика¬
ций торфа низшие единицы (виды торфа) осно¬
ваны на структурном признаке — ботаническом
составе торфа.
2. Одни классификации снизу доверху по¬
строены на структурном признаке. Примером мо¬
гут служить классификации Г. Варена, К. Б ю л о в а.
39
Классификация торфов
(по Инсторфу)
Торфа
Схема 2
1 1
Верховые Переходные
торфа
] Г
<0
* Л
£ с
Л с
<0
со
ас
л х
5 с
2 с
<и и
х
о
из
торфа
СС
5
5 с
2 =
££
о
с:
1
с
& 8
ГГ» л
СО
ос
«о а>
X О)
ос X
сс а»
О 2
ё-
III П1
Низинные торфа
Топяная
группа
более зольная
Топяно¬
лесная
группа
>>
о»
а»
о«
ос
V
ос
3
»
х
X
о
СО
о
е;
СО
с
>>
о.
а:
се
Э
со
X
*
о
<Ы
си
и
с.
3
в
X
*
о
ес
го
се
а,
о
3
о
си
о
X
се се
с с
с с
>» >»
си си
вс ж
Я 5
се д
3 э
3 в
3 *
5 *
3 2
5 ст
22 50
се с-
си &
<и
О)
о.
и
с £
си
и к
X X
ё I
се 3
X ж
х О
2 Л
5 Л
СО ей
се
си а)
2 *=С
2 *>
5 си
2 и
с
с
>*
си
и
X
о
к
X
в
в
X
*
о
>»
о*
X
о
X
<и
X
а>
си
а
с
>»
си
X
се
в
Сй
X
я
о
X
со
X
си
о
в
о
си
о
X
/К/1\/1\ Л Л
Виды торфа Д
с
>»
си
ш
X
я
о
ч
о
се
Л
>»
си
X
Я
о
я
се
си
о»
си
о
се
С
>>
си
В
о
X
£
о
о
2
о
си
о
X
с
с
>»
си.
а
со
X
я
о
вг
го
X
о*
о
а
о
си
о
X
АЛЛ
3. Другие классификации- строят низшие под¬
разделения на структурном признаке, а выс¬
шие — на иных признаках, например на содер¬
жании питательных веществ (Вебер).
4. Л. Пост дал своеобразную классификацию,
ступени которой построены на разных принци¬
пах, как-то: способ отложения органического ве¬
щества, содержание питательных веществ, сте¬
пень увлажнения и, наконец, структура торфа.
Генетической в том смысле, что она основана на
материнских ценозах, продуцирующих торф,
эту классификацию назвать, однако, нельзя.
5. Структурным классификациям можно про¬
тивопоставить генетические классификации,
40
где виды торфа выделены на основании их мате-
ринских фитоценозов. Однако вполне разрабо¬
танных генетических классификаций пока нет.
6. Заслуживают внимания классификации, в
которых основанием для выделения системати¬
ческих единиц служат, кроме ботанического
состава, и другие свойства торфа — как степень
гумификации и зольность (классификации Инс-
торфа).
Химические свойства торфа.
Химизм торфообразователей и торфа. В образовании торфа
участвуют не все составные части растений. По степени уча¬
стия в торфообразовании соединения, входящие в состав расте¬
ний, делятся на три группы:
1. Углеводы, пектиновые вещества и белки при оторфовыва-
нии растений разрушаются, образуют газообразные и раство¬
римые в воде вещества и вымываются из торфа. Только
небольшая часть продуктов распада этих соединений—альдо-
сахара и аминокислоты—превращаются в устойчивые, содержа¬
щие азот гуминовые кислоты.
2. Углеводороды, предельные кислоты жирного ряда и воски,
наоборот, представляют очень устойчивые соединения. Они не
изменяются при образовании как торфа, так и бурых углей.
3. Непредельные кислоты жирного ряда, смолы и лигнин
образуют третью группу. Они не разрушаются, но переходят
в еще более устойчивые соединения. При этом непредельные
жирные кислоты и смолы полимеризуются, причем первые
образуют многоосновные циклические кислоты, а вторые пре¬
вращаются в высокоплавкие и трудно растворимые вещества;
лигнин же под влиянием грибков превращается в гуминовые
кислоты.
Таким образом, торфообразователем оказы¬
вается не все растение в целом, а лишь опреде¬
ленные входящие в его состав химические со¬
единения — углеводороды, воски, смолы, жирные
кислоты и лигнин и, кроме того, иногда альдо-
сахара и аминокислоты.
При переходе растительных остатков в торф изменяется и
состав золы. Хотя сведений подобного рода и немного, однако
сопоставление состава золы торфообразователей и соответствую¬
щих торфов все же позволяет сделать некоторые выводы.
Из таблиц 6 и 7 видно следующее: ’
1) процентное содержание органических веществ в сухой
массе при переходе в торф уменьшается;
2) соответственно этому увеличивается содержание золы;
3) процентное содержание извести, калия, магнезии и в ряде
случаев фосфора при переходе в торф уменьшается;
4) нерастворимые в соляной кислоте зольные элементы, крем¬
незем, алюминий обнаруживают тенденцию к накоплению.
41
Таблица 6
Процентный состав сухого вещества торфообразователей и торфов
(по Е, Гулли, 1913)
Органическое
вещество
Несгораемый С
остаток ?
Растворимых в соляной кислоте
Нераствор.
в соляной
кислоте
Азот
чистом
золы
извести
магнезии
калия
фосфора
Олиготрофные виды сфагно-
СЫгпБеешоог
вых мхов с выпуклого боло-
та (среднее)
97,570
2,430
1,916
0,258
0,128
0,458
0,072
0,514
0,759
в Баварии
Состав торфа в тех же пунктах
95,79
4,21
1,21
0,151
—
0,0363
0,151
3,00
2,080
Мезотрофные виды сфагновых
БбгсЬепэее
мхов с лесного болота (сред¬
и У/еИшооБ
нее)
96,634
3,366
2,460
0,350
0,140
0,846
0,22
0,906
1,524
в Баварии
Состав торфа в том же пункте
89,33
10,67
4,77
0,347
—
0,046
0,260
5,90
2,680
•
Эвтрофные виды сфагновых
мхов с низинного болота
84,495
15,505
5,045
1,336
0,507
0,495
0,166
10,460
1,170
\Vaizernhop-
БрЬ. рЫурЬуПит (среднее)
1егаи
• Состав торфа в том же пункте
71,06
28,94
6,30
0,951
0,343 ;
0,273
0,243
22,64
1,59
в Баварии
Эвтрофный БрИ. соп1огШт,
оттуда же (среднее) ....
88,400
11,630
6,310
2,847 !
0,860 !
0,453
0,145
5,320
1,235
Состав торфа в том же пункте
87,99
12,01
4,20
1,540
0,286
0,111
0,156
7,81
2,050
Надо иметь в виду, что указанные изменения в составе золы
имеют место лишь при нормальном ходе оторфования. В слу¬
чае приноса зольных элементов извне картина резко изменяется,
и состав золы уже зависит от химизма притекающих вод.
Таблица 7
Процентный состав золы растений сфагнового болота и сфагнового торфа
НераствО'
римый
остаток в
НС1
СаО |
Реа08
А1,0,
МяО
к2о
Мл О
рао5
Сфагновые мхи
29
24
7
7,5
11
4,5
Пушица
27
13
5
7
23
10
Шейхцерия
10
22
22
7
19
7
Кустарнички
23
20
3
8,5
15
1,6
6
Верховой сфагновый торф
37
15
17
5
1,5
1,1
2,7
Химические соединения торфа. Если удалить из торфа
воду, не меняя его органической массы, получится абсолютно
сухой торф1. Если подействовать на этот последний смесью
спирта с бензолом, то мы получим так называемый биту м—
смесь восков, жирных кислот, смол и углеводо¬
родов. Г орячая вода извлекает из битума ряд раствори¬
мых в воде веществ. Действуя на нерастворимый вводе
остаток водной щелочью, мы извлекаем растворимые в щелочах
гуминовые кислоты.
Нерастворимый остаток представляет волокнистую массу,
состоящую из клетчатки и неосахаривающихся веществ —
лигнинов.
Таким образом, торф имеет следующий состав:
A. Вода.
B. Сухой остаток.
1. Битумы.
2. Растворимые в воде вещества.
3. Гуминовые кислоты и их соли.
4. Органические кислоты и их соли.
о. Клетчатка.
6. Лигнин.
Минеральная часть торфа, дающая при его сжигании не¬
сгораемый остаток—золу, при указанном способе обработки
входит в состав перечисленных выше компонентов торфа.
Основные органические вещества торфа. Из важнейших
органических соединений, входящих в состав торфа, опустим
битумы, охарактеризованные уже раньше, и остановимся
в дальнейшем на гуминовых веществах.
Гуминовые вещества, по Свену Одену, представляют
аморфные образования с невыясненной структурой от светло-
бурого до темнобурого цвета. Они образуются в природе при
1 См. Г. Стадников, 1932.
43
разложении растительного материала, нерастворимы в бензоле
и обнаруживают ясное сродство к воде, в которой они рас¬
творяются или, по крайней мере, сильно набухают Ч
Гуминовые вещества, способные диссоциировать с отщепле¬
нием водородного иона и давать с сильными основаниями типич¬
ные соли и воду, Свен Оден называет гуминовыми кис¬
лотами.
Гуминовые вещества, нерастворимые в щелочах и лишенные
кислых свойств, он называет гумусовым углем, или
г у м нно м.
Гуминовые вещества различаются в торфе уже простым
глазом в виде бесструктурной массы бурого цвета, вместе с
форменными растительными остатками. Количество этой массы
с возрастом торфа в общем увеличивается. Гумусовая кислота,,
составляющая главную массу гуминовых кислот торфа, имеет
различный элементарный состав. Количество углерода в ней
колеблется от 57,5 до 64,2%, водорода — от 4,3 до 5,4%.
(по Г. Стадникову).
Вода в торфе. Торф в естественном состоянии содержит
очень много воды. В неосушенной. торфяной залежи воды
имеется до 91%. а в осушенной—85% (по Г. Стадникову),
верхний же неразложившийся слой сфагновых торфяников
содержит в естественном состоянии до 94—95% воды Ч Часть-
воды, содержащейся в торфе, можно удалить простым от¬
жиманием без особых потерь органического вещества. Это
так называемая отжимаемая вода. Слабо разложившиеся
торфа содержат м н о г о отжимаемой воды, сильно же
разложившиеся торфа ее не содержат вовсе. При отжи¬
мании таких торфов вместе с водой проходит киселеобразная
масса, содержащая гуминовые кислоты.
Вода, остающаяся в торфе после отжимания, называется
коллоидно-связанной водой. Эта вода прочно удержи¬
вается з торфе коллоидными гуминовыми веществами, и уда¬
лить е<= трудно. При обычной сушке торфа на воздухе эта
вода удаляется не полностью3. Избыток коллоидно-связанной
воды необходимо удалять, чтобы сделать возможным сжига¬
ние торфа в топке. Это происходит обычно на воздухе в
умеренных широтах в течение вегетационного периода. Таким
образом, сушка торфа носит сезонный характер, что
весьма невыгодно для торфяной промышленности. Поэтому
искусственное обезвоживание торфа, т. е. удаление коллоидно¬
связанной воды, представляет очень важную проблему. Ввиду
указанных свойств коллоидов искусственное, механическое
обезвоживание торфа до сих пор считалось невозможным.
В самое последнее время проблема искусственного обезвожива- 11 По Г. Стадникову, 1932.
2 По книге „Торф, его происхождение, залегание и распространение“,
1932.
3 Содержание воды в воздушносухом торфе—25—30%.
44
ния торфа термомеханическим способом в заводском масштабе
разрешена у нас в СССР Главторфом.
Зола в торфе. Путем прокаливания сухого торфа на воз¬
духе получают несгораемый остаток, называемый золой.
Содержание золы в торфе сильно колеблется и зависит от
следующих причин:
1. От содержания золы в самих торфообразо-
вателях, которое у разных торфообразователей не одина¬
ково, например у сфагновых мхов, с одной стороны, и растений
низинных болот—с другой. Верховые торфа, состоящие из
остатков сфагновых мхов, бедных золой, сами малозольны;
низинные же торфа из остатков высокозольных растений
в общем высокозольны.
2.0 т степени гумификации (разложения
торфа). Торфа одинакового ботанического состава в общем
тем богаче золой, чем выше степень разложения торфа.
3. От приноса зольных соединений извне
внешними агентами—водой, ветром. Растения-торфообразова-
тели могут дать предположительно не более 8—10®/« золы
в торфе, минеральное же вещество свыше этого количества—
наносное. Не надо думать, что принос этот просто сво¬
дится к отложению и аккумуляции в торфе нерастворимых
солей. Г. Стадников считает, что „состав золы в торфе
меняется в зависимости от качественного и количественного
состава солей, растворенных в воде, которая приходит в со¬
прикосновение с торфяником".
4. От химических изменений (помимо гумифи¬
кации), происходящих в самом торфянике. Эти изменения
состоят в том, что в верхних горизонтах торфяника образуются
органические кислоты; дающие соли с щелочными металлами.
Эти соли растворимы, и дождевые воды вымывают их из
верхних горизонтов в нижние, где они вступают в обменную
реакцию с солями щелочно-земельных и тяжелых металлов.
Таким образом, происходит обеднение щелочными окислами
золы верхних горизонтов и обогащение ими нижних (Г. Стад¬
ников).
Содержание золы в торфе сильно колеблется в зависи¬
мости от ботанического состава торфа и изменений орга¬
нического вещества растений при переходе в торф, т. е.
в зависимости от степени разложения и других процессов.
Процент содержания золы сильно разнится в низинных, пере¬
ходных и олиготрофных торфах, как это показывают, напри¬
мер, данные Е. Гулли для южнобаварских болот (табл! 10).
Процентное содержание золы для среднерусских болот колеблется в та¬
ких пределах
Низинные болота . . . 6—10%
Переходные болота . . 5—6 .
Верховые болота . . . 3,5—5,0 „ 11 См. „Торф, его происхождение, залегание и распространение“.
45
»Ib
ф
Содержится в 100 частях вовдушносухого торфа
(по Цайлеру и Вильку)
Таблица 8
Органиче¬
ского
вещества
Чистой
золы
Калия
Извести
Окиси
железа и
глинозема
Фосфорной
кислоты
Серной
кислоты
Кремне-
кислоты и
нераствор.
веществ
Азота
Тростниковый торф
Неразложившийся
85,35
14,65
0,244
0,945
4,173
0,167
1,188
7,511
1,86
Мало разложившийся
88,20
11,80
0,853
3,021
1,104
0,169
1,980
5,229
2,29
Сильно разложившийся
89,54
10,46
0,035
4,998
0,496
0,176
1,329
3,162
2,07
Совсем разложившийся ......
87,15
12,85
0,262
0,456
2,573
0,092
0,789
8,497
1,88
Осоковый торф
Неразложившийся
96,16
3,84
0,061
1,774
0,424
0,063
0,761
0,594
2.19
Мало разложившийся
96,03
3,97
0,048
0,507
1,405
0,071
0,284
1,565
1,63
Сильно разложившийся .
96,49
3,51
0,042
1,522
0,999
0,059
0,468
0,296
2,10
Совершенно разложившийся ....
94,32
5,68
0,035
2,538
1,470
0,049
0,287
1,092
1,32
Гипновый торф
Неразложившийся
92,39
7,61
0,128
3,001
0,370 *
0,077
2,254
1,462
2,06
Мало разложившийся
94,27
5,73
0,088
0,432
1,780
0,С89
0,426
2,739
2 25
Сильно разложившийся
96,68
3,22
0,058
1,145
1,292
0,053
0,426
0,268
2,08
Ольховый торф
98,40
1.60
0,055
0,543
0,347
0,046
0,320
0,208
1,37
Торф из шейхцерии
2,62
Мало разложившийся
96,20
3,80
0,048
0,241
0,776
0,319
0,110
2,195
С небольшим количеством волокон
97.76
2,24
0,146
0,330
0,722
0,255
0,148
0,494
2,15
Торф из пушицы
*
0,124
0,85
Мало разложившийся
99,41
0,59
0,057
0,116
0,162
0,027
0,058
С небольшим количеством волокон
99,47
0,53
0,038
0,069
0,134
0,044
0,078
0,141
1,26
Сфагновый торф
0,89
Неразложившийся
98,07
1,93
0,119
0,288
0,275
0,(66
0,150
0,946
Мало разложившийся
99,36
0,64
0,062
0,120
0,070
0,055
0,088
0,186
. 0,79
Сильно разложившийся
96,79
3,21
0.С52
1,789
0,337
0,058
0;305
0,491
1,35
Совершенно разложившийся . . ,
96,08
3,92
0,104
0,089
0,443
0,043
0,111
3,047
0,88
Таблица 9
Химический состав советских торфов (на 100 частей сухого торфа)
(по В. Сукачеву, 1926)
Виды торфа
Гигроско¬
пическая
вода
Потеря
от прокали¬
вания
Зола
Азот
Вещества, перешедшие в раствор 10°!0
НС1
Сфагновый торф близ Ленин¬
града
17,469
96,001
3,998
1,405
0,С64
0,260
0,234
0,161
Травяной торф е древесными
остатками близ Ленинграда . . .
17,660
86,057
13,942
2,325
0,263
—
0,243
0,129
0,154
Сфагновый торф из б. Ленин¬
градской губ
13,82
94,29
5,71
3,11
0,137
0,150
0,669
0,030
0,030
Осоковый торф из б. Мин¬
ской губ
9,094
92,30
7,7С0
3,088
0,195
0,192
2,804
0,052
—
Осоковый торф из б. Мин¬
ской губ
11,290
70,757
29,243
2,831
0,130
0,206
0,612
0,036
—
Ольхово-ивняковый торф
оттуда же
17,500
76,811
23,189
3,888
0,462
0,277
4,500
0,С05
—
Сфагновый торф из б. Мин¬
ской губ., с глубины 0—20 см .
13,72
95,857
4,143
0,480
0,103
0,114
0,364
0,080
—
Сфагновый торф, оттуда же,
с глубины 20—60 см
12,28
93,351
6,649
1,157
0,033
0,108
0,317
0,06
—
В той же книге, по данным западноевропейских исследователей, даются
для болот Западной Европы такие колебания зольности:
Низинные болота . . 8—10%
Переходные болота до 5 ,
Верховые до 3,0»
Олиготрофные болота (верховые) беднее
всего золой, эвтрофные (низинные) наиболее
богаты ею, мезотрофные же занимают среднее
положение. То же надо сказать и относительно отдельных
зольных элементов. Разные виды торфа тоже сильно отличаются
друг от друга по содержанию золы и отдельных ее элементов,
что видно из помещенной выше табл. 8, составленной Цайлером
и Вильком (1907) на основании многочисленных анализов бо¬
лот Западной Европы.
Таблица 10
Содержится на 100 частей воздушносухого торфа
Потеря от
прокалива¬
ния
Торфа сфагнового болота
(олиготрофные) . . . 94,39
Торфа травяного болота
(лугового) эвтрофные . 77,62
Торфа лесного болота (пере¬
ходного) мезотрофные . 86,26
Зольные
вещества
5,61
22,38
13,73
Из приведенных таблиц видно большое разнообразие как
о общем содержании золы, так и отдельных минеральных
соединений в торфах разного ботанического состава. Рассмот¬
рим теперь отдельные минеральные соединения в торфе, их
происхождение и значение.
Кальций содержится в торфе в виде углекислой извести,
а также сернокислых и органических соединений. Происхожде¬
ние его в торфе двоякое: с одной стороны, он попадает туда
из растений, растущих на болоте, с другой стороны — приносится
в растворе почвенных и грунтовых вод. По Б. Оношко (1931),
ключевые во/ о! содержат иногда в растворе громадные коли¬
чества окиси кальция (до половины всех растворенных в воде
минеральных веществ). Эта известь выпадает из раствора в
форме нерастворимых соединений, частью органических, и мо¬
жет накопляться в виде целых прослоек — так называемого
лугового известняка, в виде белесоватой, мажущейся массы.
В 100 частях сухогоЗ торфа содержится ча¬
стей окиси кальция:
В болотах СССР, максимум
* . „ минимум
* , . среднее
„ „ Германии . , 1 2Таблица
Болота
верховые
низинные
1,402
36,902
0,242
3,342
0,652
3,482
0,25^
4,001
11
1 По Флейшеру (1891), для северной Германии.
2 По Б. Оношко (1931).
48
Она бурно вскипает с соляной кислотой и содержит 80—90%
и более золы, в том числе 40—60% окиси кальция.
Содержание кальция в олиготрофных и э в-
трофных торфах резко отлично (табл. 11).
Отсюда следует, что олиготрофные болота беднее известью,
чем эвтрофные; для мезотрофных естественно ожидать про¬
межуточного положения. Приведенные цифры дают основание
думать, что олиготрофные болота Германии беднее известью,
чем болота Союза.
Значение кальция в торфе для растений. Кальций—необхо¬
димый элемент питания растений. При разведении культур¬
ных растений на эвтрофных (низинных) болотах дополнитель¬
ного внесения извести (известкования) не требуется, при
культуре олиготрофных (верховых) торфяников, бедных изве¬
стью, оно необходимо. Значение известкования сводится к
следующему (Г. Пухнер, 1929): известь повышает разложение
торфа, нейтрализует кислоты, усиливает деятельность нитри¬
фицирующих бактерий, способствует поселению клубеньковых
бактерий, усвояющих свободный азот, и тем самым поселению
бобовых растений и, наконец, устраняет вредное влияние на
растения сернистого железа, содержащегося в торфе.
Калий (К) в торфе содержится в очень незначительных
количествах — в десятых и сотых долях процента, считая на
сухое вещество торфа (табл. 12).
Таблица 12
Болота
верховые нвзинные
В 100 частях сухого торфа содержится ча¬
стей КаО:
В болотах СССР, максимум 0,112 0,602
„ , „ минимум 0,012 0,012
, » „ среднее 0.052 0.142
„ » Германии 0,03* 0,10*
Мы видим, что калия в олиготрофных болотах меньше,
чем в эвтрофных, и что данные для северной Германии и для
наших болот весьма близки между собой.
Калий, необходимый элемент питания расте¬
ний, находится в торфе в минимуме. Болота мезо-
трофные и особенно олиготрофные всегда требуют внесения
калийных удобрений. Они нужны и для большинства эвтроф¬
ных торфяников (Б. Оношко, вып. II, 1931). Таблица 13 пока¬
зывает, как реагируют травы на болотах на внесение калий¬
ного удобрения.
Однако в некоторых случаях (например низинное болото
Архангельского опытного поля, по Б. Оношко) эвтрофные
болота сравнительно обеспечены калием, и внесение его дает
лишь очень небольшие прибавки. 1 21 Цифры Флейшера (Fleischer, 1891) для северной Германии.
2 По Б. Оношко (1931).
4-722
49
Таблица 13
Прибавка урожая
от калийного
удобрения
Ч- 219%
1С6%
-г 68%
61%
Фосфорная кислота в торфе имеет различное происхо¬
ждение: в одних случаях она поступает в торф вместе с почвен¬
ными водами, притекающими на болото из окружающих мест
(Б. Оношко, 1931); при этом вымывание фосфорной кислоты
особенно интенсивно, если прилегающие к болоту почвы
распаханы. Если же водосборы покрыты луговой и лесной
растительностью, то вынос фосфора незначителен. В других
случаях фосфорная кислота образуется при выщелачивании
содержащих фосфор горных пород (например апатита), обра¬
зующих дно болота. Отсюда она поступает в торф (Г. Шрей¬
бер, 1927). Наконец, источником фосфора в торфе могут быть
и торфообразующие растения (Г. Пухнер, 1929). Содержание
органически связанного фосфора в торфообразующих расте¬
ниях несравненно больше, чем в торфе, и при оторфовани;'
этот фосфор переходит в растворимые соединения, которые
потом уходят из торфа (Г. Пухнер, 1929).
Эти растворимые соединения, встречаясь в торфе с желе¬
зом, которого здесь иногда бывает очень много, закрепляются
и, соединяясь с железом, образуют трудно растворимую фос¬
форнокислую закись железа, так называемый вивианит, со¬
держащий 28% и более фосфорной кислоты (Б. Оношко, 1931).
Вивианит в залети или бесцветен, или имеет белесоватую
окраску; при окислении на воздухе он становится голубым.
Вивианит располагается в торфянике в виде тонких про¬
слоек, или гнезд, причем больше всего его в периферических
частях болот; к центру же его становится все меньше, что
видно из табл. 14.
Низинное болото
Рудни-Радовельской
Низинное болото
на Москва-реке
При внесении 90 кг К20 ....
То же + фосфорно-кислое удоб¬
рение
При внесении 108 кг К20 ....
То же -f- фосфорно-кислое удоб¬
рение
Таблица 14
Содержание вивианита в торфе в процентах
(по Б. Оношко)
В непосредственной близости коренного берега до-
лины 20,7
В 300 м от коренного берега • 13,0
„ 580 . „ , 11,4
Относительное содержание фосфорной кис¬
лоты в торфе невелико. Таблица 15 показывает процент
фосфорной кислоты в торфе разных типов болот.
50
Таблица 15
(по Б. Оношко)
Болота
верховые низинные
В 100 частях сухого торфа содержится частей фосфор¬
ной кислоты:
В болотах СССР, максимум 0,98 1,42
, „ минимум 0,01 0,11
» „ „ среднее 0,23 0,43
„ , северной Германии (по Флейшеру, 1891) . . . 0,05 0,25
, * Германии 0,04 0,25
Эти сведения, во-первых, показывают, что э в -
трофные болота богаче фосфорной кислотой,
чем олиготрофные, и, во-вторых, что наши бо¬
лота богаче фосфором, чем германские. Вообще,
повидимому, наши олиготрофные болота по содер¬
жанию как золы, так и отдельных зольных эле¬
ментов — кальция, калия и фосфора —б огаче гер¬
манских. Вероятно, объяснить это можно тем, что в Герма¬
нии выпадает больше осадков, чем в СССР, и поэтому
вымывание зольных элементов из торфа идет более энергично.
Фосфорная кислота в торфе—необходимый
элемент питания растений. При этом в вивианите
фосфор доступен для культурных растений. Необходимо,
однако, иметь в виду (Б. Оношко, вып. II, 1931), что в олиго-
трофном, а также часто и в эвтрофном торфе, фосфор на¬
ходится в миним уме для к у л ь т у р н ы х р а с те ний.
Чаще всего он бывает во втором минимуме, после калия, и дает
несколько меньшие прибавки урожая, чем К20. Таким обра¬
зом, при культуре олиготрофных, а также большей частью и
эвтрофных болот требуется внесение фосфорного
удобрения. При этом получаются такие прибавки урожая
сена (по Б. Оношко, вып. II, 1931).
Таблица 16
Прибавка
урожая
в %
Низинные болота 1
Карелии (Лоухи) |
( При внесении 80 кг Р2Ог> (томасшлак)
То же совместно с калийным удобре-
1 нием
-ИЮ
-¡-694
Низинное болото [
Архангельского опыт- <
ного поля 1
При внесении 58 кг Р20* (супер-
фосфат) 2. . . . . .
То же + калийное удобрение ....
+183
-¡-220
Низинные болота Г
б. Владимирской губ. (
При внесении 58 кг Р205 (томасшлак)
То же калийное удобрение ....
+ 26
+ 37
Низинные болота 1
Москва-реки |
[ При внесении 54 кг Р205 (супер¬
фосфат) ’
[ То же “(• калийное удобрение ....
-4- 23
+ 16
Очень резко реагируют на прибавку фосфора эвтрофные
торфяники севера Европейской части Союза. Более слабую
прибавку урожая дали торфяники в б. Московской и Влади-
4* 51
мирской губерниях. Однако, по Б. Оношко (1. с.), существуют
торфяники, настолько обеспеченные фосфо¬
ром, что внесение его не дает прибавки уро¬
жая, а иногда даже снижает последний. Сюда
принадлежат, например, торфяники в долинах Яхромы, Дубны
и других рек Московской области. Эти торфяники весьма
ценны в сельскохозяйственном отношении, так как культура
их не требует искусственных удобрений.
Положительное влияние калийно-фосфорного удобрения
сказывается не только в прибавках урожая сена на низинных
торфяниках, но и в улучшении его качественного состава
(табл. 17).
Таблица 17
(по Б. Оношко, вып. III, 1932)
Группы травостоя
Весовой процент участия
в травостое низинных болот
без удобрений ) удобрено РК
Злаки
58,4
64,6
Бобовые
* 2,6
17,8
Разнотравье
34,4
12,3
Осоки
4,3
5,0
Из таблицы видно, что прибавка калифосфатного
удобрения улучшает качество травостоя, уси¬
ливая весовое участие бобовых за счет разно-
травия.
Азот в торфе. Торф весьма богат азотом. По данным
Флейшера (1891), для северной Германии в 100 частях воздушно¬
сухого торфа содержится (в среднем) азота:
В сфагновых (олиготрофных) торфах 0,8%
В переходных (мезотрофных) торфах 2,0%
В низинных (эвтрофных) торфах 2,5%
Если принять во внимание, что такая богатейшая почва,
как русский чернозем, содержит всего 0,2—0,5°/о азота (дан¬
ные П. Коссовича, 1911), то станет ясно, насколько богат
азотом торф. Однако этот запас азота для культурных
растений лишь потенциальный, так как азот в торфе
в основном находится в форме органоминеральных соединений,
недоступных для культурных растений \ При осушке органиче¬
ский азот минерализуется, т. е. переходит в доступные для
растений нитраты и аммиачные соединения. Таким образом, 11 Есть, однако, указания, что, в то время как эвтрофные торфа содержат
почти весь азот в форме неусвояемых органоминеральных соединений, в
олиготрофных торфах до 20% азота может находиться в аммиачной форме.
Поэтому в первое время после осушки олиготрофные торфа богаче минераль¬
ным усвояемым азотом, чем низинные.
52
осушка и последующее повышение разложения торфа
необходимы для минерализации органического
азота. Поэтому олиготрофные (верховые) торфа, которые
содержат меньше азота и труднее разлагаются, сильно
реагируют на внесение азотистых удобрений
(Б. Оношко, вып. II, 1931). Что касается эвтрофных торфяни¬
ков, то, по исследованию Б. Оношко, они в некоторых слу¬
чаях также обнаруживают определенный азотный минимум.
Таковы болота северных широт (приполярные), где вслед¬
ствие низкой температуры биологические процессы в почве
и нитрификация подавлены. Поэтому для подобных болот
нужно или искусственное внесение азота, или освобождение
его путем обжига. Вообще же эвтрофные торфяники уже с
первого года культуры обеспечены усвояемым азотом и имеют
лишь переходящий азотный минимум, который устраняется
без внесения азотистых удобрений, одними лишь техниче¬
скими приемами.
Сера в торфе, повидимому, входит главным образом в
состав органических соединений (Г. Стадников, 1932). По
Г. Пухнеру (1929), при разложении содержащих серу органи¬
ческих веществ в присутствии богатой железом воды может
образоваться путем обменной реакции кислого углекислого
железа с гипсом двусернистое железо БеБг, которое может за счет
кислорода воздуха окисляться в железный купорос и серную
кислоту; поэтому сернистое железо относится к вредным при¬
месям в торфе. Есть указания, что сернистые соединения
могут приноситься в торфяник ключевыми водами (Г. Шрей¬
бер, 1927). В результате окисления Н2Б в торфе иногда отла¬
гается и чистая сера. Содержание серы в торфах олиготроф-
ных болот не превышает 1,0%> в большинстве же случаев
около 0,5%. В низинных торфах содержание серы выше и
колеблется около 1,5% (Г. Стадников, 1932).
Серная кислота в торфе связана с различными осно¬
ваниями. Г. Шрейбер (1927) указывает, что минеральные источ¬
ники могут приносить в торфяник ряд солей серной кислоты:
гипс, сернокислое железо (Ре$04 + 7ая), сернокислый магний,
глауберову соль, соли калия и алюминия: А12(504)4-К2504+
+24 aq. Эти соли не отлагаются в торфе в виде про¬
слоек, а пропитывают его и при высыхании выкристаллизовы¬
ваются в виде светлого налета, так что торф кажется как
будто посыпанным сахаром. При соприкосновении с сырым
воздухом эти торфа притягивают воду (действие серной
кислоты), причем кристаллический налег расплывается.
Характерен особый запах, который издают эти торфа, и их
горько-соленый вкус. Эти торфа не годятся ни на топливо,
ни под культуру, но служат для устройства лечебных ванн,
применяющихся в Западной Европе уже более 100 лет про¬
тив суставного ревматизма, экссудатов, женских болезней.
На изучение и использование подобных торфов последнее
время обращено внимание и у нас в СССР.
53
Железо встречается в торфе в различных соединениях и
модификациях. Двусернистое и фосфорное железо было уже
рассмотрено выше, когда речь шла о сере и вивианите. Очень
обычное соединение железа в торфе — бурый железняк
(лимонит). По В. Сукачеву (1926), он встречается в двух
модификациях: а) в виде железной охры, иначе желтая
железная руда, болотная железная руда, озерная руда, ключе¬
вая железная руда; б) в виде бурого железняка, иначе
дерновой руды, луговой руды, триплита, бобовой руды.
Железная охра—аморфная, землистая желтая или желто-корич¬
невая масса, очень распространенная на болотах. Она состоит
из гидрата окиси железа Fe2(OH)6 с примесью FeC08, СаС08,
окиси марганца и растительных остатков; образуется или на
поверхности болота, или на небольшой глубине на болотах.
Бурый железняк состоит из гидрата окиси железа FeCOe,
Fe8 (POt)2, крем некисл оты, песка, глины и растительных остат¬
ков. Это аморфная пористая губчатая масса в виде желваков.
Она отлагается в глубине торфяников или в подпочве. Встре¬
чается довольно редко. Лимонит по Г. Шрейберу (1927) обра¬
зуется из закисных соединений железа FeH2(C08)2, которые
возникают при растворении содержащих железо пород водой,
богатой углекислотой. Эти соединения приносятся в торф
железистыми ключами. При соприкосновении с кислородом
воздуха кислое углекислое железо окисляется, выделяется
углекислота и выпадает гидп*т окиси железа вместе с Si02,
Р205 и другими соединениями. Кроме химического
способа образования бурого железняка, окисление желе¬
за происходит и благодаря деятельности бактерий
(Г. Шрейбер).
Железная охра применяется для очистки светильного
газа от серы, а также как желтая и красная краска. Мощные
слои идут для добывания железа (Г. Шрейбер). Лимонит
вредно действует на сельскохозяйственные культуры. Иногда
железная охра выделяется в таком количестве, что заполняет
осушительные канавы и закупоривает дренажные трубы.
Из других соединений железа упомянем о сиде¬
рите, углекислом железе—FeC03. Он встречается в торфе
или в коллоидном виде, или в форме мелких ромбоэдриче¬
ских кристаллов. Аморфная модификация весьма неустойчива,
на воздухе легко окисляется, превращаясь в гидрат окиси
железа; кристаллическая же форма, наоборот, весьма устой¬
чива. Сидерит приносится в торфяник ключами или текучей
водой. Встречается он в виде жил или гнезд, диаметром до
1,5 м и 10—30 см толщины.
Двусернистое железо FeS2 встречается в виде двух
кристаллических модификаций — пирита и марказита
(Шрейбер). Происхождение их, как и многих других минераль¬
ных соединений, в торфе двоякое—или из животных и раститель¬
ных остатков (см. выше), или из ключевой воды, проникающей
в торф. Двусернистое железо может окисляться в железный
54
купорос и серную кислоту. Применения двусернистое железо
не находит, так как встречается обычно в небольших коли¬
чествах, но действует вредно на растения, как и другие
соединения серы.
Содержание железа в торфе вообще не ве¬
лико. По исследованиям Цайлера и Вилька (1907), содержа¬
ние окиси железа и глинозема вместе колеблется от 5,48 до
0,18% на воздушносухое вещество торфа. Однако в случае
приноса железа ключевыми водами количество его настолько
возрастает, что можно говорить о настоящем минеральном
торфе. Пример описания ключевых болот, сложенных целиком
железисто-древесным торфом, можно найти у И. Богданов-
ской-Гиенэф (1926).
Поваренная соль накопляется в торфе в большом количестве
при затоплении торфяников морскими водами. Куски затоп¬
ленного торфа волны могут выбрасывать на берег (так назы¬
ваемый „морской торф“). Он пропитан поваренной солью и
при высыхании покрывается соленой коркой. „Морской торф“
встречается у берегов Голландии и северной Германии, где
торфяники в результате морской трансгрессии были затоплены
морем (Г. Шрейбер, 1927).
Примеси в торфе.
Глина, песок, щебень приносятся в торф извне водой и
ветром и отлагаются в виде примеси к торфу или образуют
целые прослойки. Отложение прослоек морского ила в
толще болот наблюдается в торфяниках, затопляемых мор¬
скими водами, например, в северной Германии. Эти затопле¬
ния— следствие вековых колебаний морского уровня.
Засыпание торфяников делювиальными сно¬
сами обычно в южнорусской лесостепи. Овражные торфяники
часто покрыты здесь минеральными наносами до 70 см мощ¬
ности (В. Матюшенко, 1925). Минеральные прослойки всякого
рода сильно понижают топливные качества торфа и затруд¬
няют торфодобычу.
Вулканический пепел попадает в торф при вулканических
извержениях. Болото, засыпанное пеплом, восстанавливает свой
растительный покров, и торфообразование идет дальше. Слои
пепла оказываются погребенными в торфе. Прослойки вул¬
канического пепла обычны в торфяниках Огненной Земли
(Ауэр, V. Auer, 1933) и восточной Камчатки (Нейштадт М.
и Короткина М., 1936). Из других составных частей торфя¬
ников органического порядка рассмотрим допплерит (иначе
торфяной смолистый уголь, или торфяная печень). Он темно¬
бурого или черного цвета и напоминает то деготь, то каучук;
нерастворим в воде, спирте и эфире, при высыхании тверд,
блестящ, с раковистым изломом, способен гореть и оставляет
богатую кальцием золу (Г. Пухнер, 1929). Допплерит обра¬
зуется всегда в глубоких слоях торфа, в пустотах всякого
55
рода, в трещинах, на месте древесины, корней и стеблей, т. е.
тех растительных остатков, которые с самого начала содержат
полость или делаются полыми при оторфовании. Отложение
допплерита, возможно, происходит при соприкосновении воды,
содержащей гумусовые коллоиды, с грунтовой водой, богатой
солями (Г. Шрейбер, 1827). Химическая природа допплерита еще
недостаточно ясна. По мнению одних — это кальциевая
соль чистой гумусовой кислоты, по мнению других —
гумусовый коллоидальный комплекс, абсорб-
ционно насыщенный известью (Г. Пухнер). Доппле-
рит нередко встречается в торфе.
Физические свойства торфа.
Влагоемкость торфа — его способность поглощать капель¬
ножидкую воду—весьма велика. Она гораздо выше
влагоемкости минеральных веществ. Достаточно
указать, что неразложившийся сфагновый торф может погло¬
тить воды в 15 и более раз по сравнению со своим воздушно¬
сухим весом, минеральная же почва (суглинок) впитывает
воды всего половину своего сухого веса (по Б. Оношко, 1931).
Большая влагоемкость торфа объясняется его капиллярной
структурой — присутствием большого количества пустот
малого диаметра. Эти пустоты зависят от сложения торфа из
растительных тканей. Влагоемкость торфа усиливается еще и
тем, что частицы гумусовых коллоидов обволакиваются тон¬
ким слоем воды (Г. Пухнер, 1929).
Влагоемкость торфа, как и его другие свойства, не одина¬
кова и зависит от его ботанического состава. Особенно
влагоемки сфагновые торфа, что обусловлено строением самого
сфагнума. Прежде всего стебли живого сфагнума с приле¬
гающими к нему (так называемыми поникшими) веточками и
самые веточки с налегающими друг на друга листочками
представляют системы капилляров. Затем водоносные (или
гиалиновые) клетки сфагнума с порами и спиральными утолще¬
ниями также представляют микроскопические капилляры,
наполненные водой. Эти анатомические приспособления еще
более увеличивают влагоемкость сфагновых мхов.
Осоковые, гипновые и другие виды эвтрофных (низинных)
торфов менее влагоемки, чем сфагновые. Приведем ниже
сведения по поглощению воды разными видами сфагнума и
другими растениями (табл. 18).
Сфагновые мхи поглощают значительно больше воды, чем
другие растения, а именно в 15—20 раз и более своего воз¬
душносухого веса. Мхи порядка Вгуакэ поглощают значитель¬
но меньше воды, чем сфагны. Болотные представители этого
порядка мхов (Аи1асопшшт, СНтасшт, ТЬШсИит гесо§пИит,
Нурпит э^аттеит, Н. зсогрю^беэ, Ро1уШсЬит эШ^ит) погло¬
щают воды всего в 17,5—3,7 раза больше своего веса, травя¬
нистые же торфообразователи всего в 3 раза и даже меньше.
56
(из В. Сукачева, 1926)
Таблица 16-
Поглощено воды во столько раз
больше
воздушносухого
абсолютно сухого
веса
веса
БрЬадпцш йдесит1 . . .
23,1
25 Л
гиЬе11ит1 . .
21,0—26,5
24,8—33,7
асиШоИит1 . .
18,5
21,8
п
squaгrosшn1 . .
27,5
32,6
99
ЭизепН1 ....
25,8
30,5
оЫиэшп1 . . .
18,8
22,3
99
атЫурйуИит1
20,5
25,4
гесигуига1 . . .
20,1
25,5
о^ёаШт 1 . .
26,1
31,4
99
тоНивсит1 . .
25,3
29,7
99
БиЬзесипёит1 .
23,5
26,7
9
соп(ог1ит1 . .
16,7
19,8
99
рЫурЬуИит 1 .
19,6
23,5
9
сутЫ^Шит1 .
31,0
38,8
99
рарШоБит1 . .
22,5
27,8
99
виЬЫсЫог1 . .
22,2—26,2
26,4—32,&
99
тесИит1 . . .
26,0
30,7
Нурпит БсЬгеЬей1 2 . . .
9,09
Ну1осопИшп зр1еп(1еп82 .
7,56
ТИШсНит гесодпИит2 . .
5,66
я
аЫеИпит2 . .
•
4,16
СНтасшт (1епс1го1(1е82. .
5,01
Аи1асоттит ра1из1ге 2 . .
10,37
8р11адпит асиМоИит 3 . .
20,10
п
сутЫЬИит3 .
20,16
п
си$р!ёа1ит3 . .
18,77
Нурпит 51гат1пешп3 . .
17,51
„
8согр1о1ёеБ3 . . .
15,21
Ро1уШсЬит эШсШт8 . .
3,71
Сагех 3
3,00
РИ^тЦез соттшИБ3 . .
2,35
Водоцоглотительная способность торфа зависит, кроме
ботанического состава, и от степени его гумификации. Чем
больше гумификация торфа, тем меньше (притом,
же ботаническом составе) его влагоемкость (табл. 19).
Из таблицы 19 хорошо видно, что с увеличением разложения
влагоемкость торфа того же ботанического состава падает.
Таблица 20 показывает то же самое на торфах, слагающих Шува¬
ловский торфяник, сверху вниз. В молодом сфагновом торфе
степень разложения увеличивается сверху вниз с приближе¬
нием к пограничному, сильно разложившемуся горизонту. С
увеличением степени разложения и вместе с тем с глубиной,
1 Данные 3. Руофф (1918).
2 Данные В. Афанасьевой.
3 Данные 2а11ег и. \Vilk (1907).
57
Таблица 19
(по В. Сукачеву, 1926)
Поглощено воды
по отношению
к сухому весу во
столько раз
Сфагновый торф неразложившийся 16,04
„ * мало разложившийся 16,53
„ „ сильно разложившийся . . . 5,23
Осоковый торф мало разложившийся 13,79
„ „ сильно разложившийся 11,22
Гипновый торф мало разложившийся 15,06
„ „ сильно разложившийся 8,87
Тростниковый торф мало разложившийся 8,88
„ „ сильно разложившийся 8,71
Таблица 20
(по В. Сукачеву, 1926)
Поглощено воды
во столько раз
больше абсолютно¬
сухого веса
1. Молодой сфагновый торф:
а) самый верхний слой 14,59
б) самый нижний слой 3,46
2. Пограничный горизонт (сфагновый сильно разложившийся
торф) 1,38
3. Старый сфагновый торф:
а) самый верхний слой 2,82
б) самый нижний слой 7,20
4. Чистый гипновый торф 10,08
5. Тростниковый торф 2,47
6. Осоково-гипновый торф 7,60
уменьшается влагоемкость молодого сфагнового торфа. Старый
сфагновый торф сильнее разложился сверху, ближе к погра¬
ничному горизонту, с глубиной же разложение его падает, а
влагоемкость растет. Наименее влагоемок во всей толще
Шуваловского торфяника сильно разложившийся пограничный
горизонт. Из обеих таблиц видно, что мало разложившиеся
сфагновые торфа имеют наибольшую влагоемкость.
Высокая влагоемкость торфа обуславливает большую
влажность торфяников. Это особенно относится к сфагновым
торфяникам. Большая влагоемкость мало разложившихся
сфагновых торфов используется при употреблении их
на подстилку в скотных дворах. Сфагновая под¬
стилка по своей влагоемкости и другим качествам значительно
превосходит соломенную.
Капиллярность торфа — способность проводить воду по
капиллярам, мельчайшим пустотам — скважинам, пронизываю¬
щим в различных направлениях торфяную массу. В явлении
капиллярности нужно различать скорость и высоту поднятия
(Г. Пухнер, 1929). В сухих крупнозернистых почвах капилляр¬
ный подъем воды происходит быстро (например в гравии
максимум поднятия достигается в несколько секунд), но высота
подъема весьма невелика. В сухих же мелкозернистых почвах,
а также в разложившемся торфе капиллярное поднятие про¬
ба
исходит весьма медленно, но, в конце концов, достигает
большой высоты (Г. Пухнер. 1. с.). Б. Оношко (1931) дает
такие сведения по испытаниям на Новгородской болотной
опытной станции. За 40 суток по этим сведениям вода успела
подняться в болотной минеральной почве на 51 см, в торфе
низинного болота — на 15 см и в торфе верхового болота —
на 5 см.
Э. Гранлунд (Е. Огап1ип<1, 1932) для воздушносухого торфа
нашел иные цифры. Средняя высота капиллярного поднятия
воды в сфагновом слабо разложившемся торфе оказалась
равна 46 см. Она была достигнута через 30—50 дней опыта.
Это капиллярное поднятие соответствует диаметру капилляров
около 0,03 мм. Во всяком случае и мало разложившийся торф
по своим капиллярным свойствам резко отличается от крупно¬
зернистых почв, так как капиллярное поднятие в нем мед¬
леннее, но достигает большой высоты.
Эту особенность капиллярности торфа необ¬
ходимо учитывать при осушке болот, предназна¬
ченных под культуру. Понижение уровня грунтовых вод ниже
70 см для полевых культур и 40—60 см для лугов уже опасно
для растительности, так как поднятие грунтовых вод снизу
идет, как мы видели, весьма медленно (Г. Пухнер).
Кроме того, высота капиллярного поднятия в сфагновом
торфе, как это думает Э. Гранлунд (1932), определяет верхний
предел роста кочек в естественных условиях на неосушен-
ном сфагновом болоте. Это соображение, повидимому, пра¬
вильно и для наших сфагновых болот средних широт. Высота
сфагновых кочек здесь обычно не более 50—60 см, что, при¬
мерно, соответствует высоте капиллярного поднятия в сфагно¬
вых торфах, по данным Э. Гранлунда.
Водопроницаемость торфа зависит от его состояния. Сухой
торф естественной структуры почти водонепроницаем,
потому что воздух между прочно соединенными нераздвигае-
мыми частицами торфа задерживает воду. Наоборот, торфяной
порошок весьма водопроницаем, так как вода, проникая в него
под напором или под влиянием силы тяжести, раздвигает
слабо связанные частицы с заключенным между ними воздухом.
Торф естественной структуры, если он не¬
сколько увлажнен, лишь очень мало проницаем для
воды, так как пузыри воздуха в нем, окруженные тонким
слоем воды, почти неподвижны. Насыщенный же водой
торф пропускает воду весьма быстро (Г. Пухнер).
Водопроницаемость естественного торфа зависит также от его
свойств. Чем больше влагоемкость торфа, тем легче просачи¬
вается через него вода. Увеличение же степени разложения
уменьшает водопроницаемость торфа (В. Сукачев, 1926).
Из сказанного видно, что нельзя считать торф и торфяники
почти водонепроницаемыми. При оценке водопро¬
ницаемости надо принимать во внимание неоднородность
торфяников. Сильно разложившиеся слои торфа, вроде так
59
называемого „пограничного горизонта", почти водонепро¬
ницаемы. Они играют роль водоупорных слоев, поверх кото¬
рых собирается вода. С этими водными горизонтами приходится
считаться при осушке. Наоборот, слои сильно обводненного
торфа, залегающие сверху или в толще торфяника, могут, по-
видимому, в некоторых случаях играть роль водопроводящих
путей, по которым движется вода в толще торфяника.
Водопроводим ость торфа, различаемая некоторыми авто¬
рами (например, В. Сукачев, 1926), тесно связана с его водо¬
проницаемостью. Она определяется быстротой передвижения
воды в торфянике. Эта быстрота по Оппокову (см. по
В. Сукачеву) сравнительно мала—всего 1—2 км в год. Однако
следует принять во внимание неодинаковую водопроводимость
разных слоев торфяника (см. выше). Рассмотренные гидро¬
логические свойства торфа — его большая влагоемкость,
слабая в общем водопроводимость и водопроницаемость —
объясняют важные природные свойства торфяников — в част¬
ности, возможность существования выпуклых сфагновых бо¬
лот. Известно, что олиготрофные сфагновые болота умерен¬
ных широт обычно выпуклы в центре, причем превышение сере¬
дины над краями болот достигает в Европейской части Союза
и в Западной Сибири 5 — 6, а в некоторых случаях даже 8 м.
Так как сфагновые болота в их естественном состоянии напитаны
водой почти до полного насыщения,то понятно, что вся эта масса
воды может быть удержана на выпуклой поверхности торфяника
только благодаря громадной влагоемкости торфа. Сток же и
просачивание воды внутрь толщи. торфяника затруднены
вследствие слабой водопроводимости и водопроницаемости
торфа. Таким образом, самое существование
выпуклых обводненных сфагновых торфяников
объясняется гидрологическими свойствами
торфа. Те же свойства объясняют и ограниченность
сферы действия осушительных канав на болоте.
Особенно ограничено действие канав на сфагновых (верховых)
болотах. По В. Горячкину (1925) (см. по В. Сукачеву), кривая
депрессии грунтовых вод на этих болотах, очень круто под¬
нимаясь от канавы, становится на расстоянии 6 м от нее почти
горизонтальной. Иначе говоря, на этом расстоянии
водоудерживающая способность сфагнового
торфа уже парализует действие осушки. Водо¬
удерживающая способность эвтрофных торфов меньше. По¬
этому на низинных торфяниках канавы дей¬
ствуют сильнее (по Б. Оношко, вып. II, 1931), при глу¬
боких канавах их действие в каждую сторону достигает 100 м
и более.
При закладке канав на болотах для целей культуры сле¬
дует учитывать, помимо характера болота, еще: 1) характер
предполагаемых культур и 2) свойства климата, количество
выпадающих осадков (см. В. Берш, 1909). Полевые культуры
требуют более сухих болот, чем луговые.
60
Несмачиваемость — неспособность сухих торфов впитывать
влагу. Этим свойством обладают отчасти и сухие торфа с
ненарушенной структурой. Сухой торф, хотя вещество его и
тяжелее воды благодаря заключенному в нем воздуху, как
пемза, долго плавает по поверхности воды, не намокая. Осо¬
бенно же несмачиваем порошкообразный торф (Г. Пухиер, 1929).
В случае чрезмерной осушки и распыления верхнего слоя
торф может дойти до состояния несмачиваемости. Такой торф
увлажняется с большим трудом и становится бесплодным.
Пересушка болот поэтому очень опасна (Б. Оношко, 1931).
Торф, раз высохнув, уже не возвращается
к прежнему состоянию набухания, даже при дли¬
тельном смачивании. Это важное свойство делает возможным
сушку торфа на открытом воздухе (см. „Торф, его происхо¬
ждение, залегание и распространение”). Это свойство при¬
суще вообще коллоидам, в том числе и гуминовым кислотам
торфа.
Гигроскопичность торфа — способность его поглощать из
воздуха водяные пары. Гигроскопичность зависит от содержа¬
ния в торфе влаги. Чем меньше в нем влаги, тем быстрее и
больше поглощает торф водяного пара (В. Сукачев). Цайлер
и Вильк изучали увеличение содержания воды в торфе, поме¬
щая его в насыщенную парами атмосферу при 75° (табл. 21).
Таблица 21
Поглощение водяных паров торфом
Содержание воды
Увеличение воды
в воздушносухом
торфе
в °/0 за 3 недели
Сфагновый неразложившийся торф . . .
16,51
-16,00
„ мало разложившийся торф . .
15,10
[-15,60
» сильно разложившийся торф .
13.25
-14,60
Осоковый мало разложившийся торф . . .
13,25
42,40
„ сильно разложившийся торф .
14,07
- 9,20
Гипновый мало разложившийся торф . .
17,77
-12,40
„ сильно разложившийся торф .
15,49
-11,10
Тростниковый мало разложившийся торф
11,87
- 9,90
„ сильно разложившийся торф
14,89
- 5,60
Из таблицы видно, что сфагновые торфа наиболее гигро¬
скопичны, за ними следуют осоковые и гипновые торфа;
тростниковые же стоят на последнем месте. Вышеприведен¬
ные сведения относятся к насыщенной водяными парами
атмосфере; при обыкновенном содержании водяных паров в
воздухе гигроскопичность будет ниже (В. Сукачев).
Абсорбция торфом газов. Кроме водяных паров, торф по¬
глощает разные газы. Большое значение имеет абсорбция
аммиака, выделяющегося при гниении навоза на скотных
дворах. Аммиак особенно хорошо поглощается мало разложив¬
61
шейся сфагновой подстилкой, которая, как мы видели, весьма
влагоемка. Эти два свойства и определяют применение слабо
разложившегося сфагнового торфа как высококачественной
подстилки на скотных дворах.
Испарение воды торфом зависит от ряда факторов. Во-
первых, на испарение воды и торфом, и водной поверхностью,
и растительностью влияют метеорологические факторы —
дефицит влажности воздуха, движение его (ветер) и пр. Во-
вторых, играет роль состояние самого торфа (или вообще
почвы) — именно, степень его влажности, капиллярная водо-
проводимость, обуславливающая подачу воды снизу взамен
испарившейся, и ряд других моментов. Торф в общем испаряет
много больше, чем, например, песок, и почти столько же,
сколько глина (табл. 22).
Таблица 22
(по Д. Оношко)
Количество
испарившейся
воды
(в граммах)
Песок 3328
Глина * 4172
Торф . . . * 4442
При культуре болот, практикующейся в Германии (способ Римпау —
Штраи), осушенное болото покрывают слоем песка в 10—12 см толщиной»
который только и обрабатывается. Слой же торфа под песком служит сре¬
дой для корневых систем культурных растений. При этом покровный песок
служит, с одной стороны, ложем для семян, с другой — улучшает физиче¬
ские свойства ниже лежащей торфяной почвы. Так как песок уменьшает
испарение с поверхности болота, то при способе Римпау необходима более
интенсивная, чем обычно, осушка болот (В. Берш, 1909). Влияние покров¬
ного песка на испарение видно из табл. 23.
5 Т а б л и ц а 23
(по В. Бершу)
Количество
испарившейся
за 2 года воды
(в граммах)
Неизмененный торф 14 526
Торф, смешанный с песком ... 11 609
Торф со слоем песка сверху тол¬
щиной 10 см 8 430
Торф испаряет воды тем больше, чем боль¬
ше он ее содержит. Это видно из исследования Габерланда
(НаЬег1ап<К) (привожу по Г. Пухнеру): болотная земля со
100%-ным содержанием влаги испарила воды больше, чем торф,
где содержалось 50% влаги.
Таким образом, если открытая поверхность торфа и на
осушенном болоте много испаряет, то тем больше испаряет
она на неосушенных.
Болота в естественном состоянии покрыты растительностью.
Как же влияет растительность на испарение с
62
поверхности болота? Уже старые исследования Е. Вольни
(1888) показали, что поверхность торфа, не покрытая расти*
тельностью, испарила за вегетационный период 302,9 мм воды,
а торфа, покрытого травой, — 556,3 мм (по Г. Пухнеру).
Б. Оношко (1931) приводит для новгородских болот такие
цифры:
Таблица 23г
Количество
испарившейся
РОДЫ
Торф без растительности .... 281 мм
Верховые болота с естественной
растительностью . ..... 323 ,
Низинные болота 360 „
Открытая поверхность торфа испаряет мень¬
ше воды, чем болото с естественной расти¬
тельностью. Есть сведения, что культурная растительность
на болоте испаряет особенно сильно.
Для оценки гидрологической роли болот сравним испа¬
рение с поверхности болота и со свободной
поверхности водоема.
С. Тюремнов (1928) установил, что сфагновый покров за
вегетационный период испарил на 11—21% больше, чем сво¬
бодная водная поверхность. Среднее же за 3 года отношение
количества воды, испарившейся с свободной водной поверх¬
ности, к воде, испарившейся с сфагнового покрова, равно 1,16,
Таким образом, сфагновый покров испаряет
больше, чем открытая водная поверхность.
При оценке роли болот как регуляторов питания рек нужно учесть испа¬
рение воды с поверхности болот и условия передвижения воды в торфянике
и отдачи ее торфом. По мнению В. Сукачева (1926). болота не имеют боль¬
шого значения в питании рек. Поэтому осушка болот, расположенных в бас¬
сейне реки, не может вызвать ее обмеления. Сукачев ссылается на мнение
Е. Оипокова, который считает, что болота не только не регулируют питание
рек, но оказывают как раз обратное действие. Дело в том, что болота
обычно насыщены и перенасыщены водой не только летом, но и весной. По¬
этому они не могут задержать и сберечь излишки воды, которые в них по¬
ступают во время весенних разливов и летних дождей. Эти излишки быстро
поступают в реки и расходуются. С другой стороны, болота в обычное время
вследствие известных уже нам гидрологических свойств торфа лишь с тру¬
дом отдают воду рекам, но зато испаряют, как мы видим, много воды. Эта
вода, следовательно, с точки зрения питания рек, пропадает. Таким образом,
болота в их естественном состоянии едва ли служат регуляторами питания
рек. Действительными регуляторами питания рек болота могут стать только
при их канализации, шлюзовании и т. п.
Истинный удельный вес торфа — это вес его абсолютно
сухого вещества при абсолютно плотном заполнении им еди¬
ницы объема (см. „Торф, его происхождение и т. д.а). Следо¬
вательно, для определения истинного удельного веса торф
берется измельченный, не содержащий воздуха. Величина
истинного удельного веса зависит от состава торфа и его
гумификации и колеблется от 1,3 до 1,6. Минеральные почвы
в общем тяжелее торфа: их удельный вес от 2,4 до 2,7 (Пухнер).
63.
Объемный вес, или кажущийся удельный вес торфа —
это вес определенного объема сырого или сухого торфа не¬
нарушенной структуры (т. е. с содержащимися в нем порами
и промежутками, заполненными воздухом или водой) по от¬
ношению к весу того же объема воды. Кажущийся удельный
вес колеблется от 0,2 для сухого мало гумифицирован¬
ного сфагнового торфа до 1,0 или несколько более для насы¬
щенного водой. Дальнейшее увеличение объемного веса торфа
зависит от минеральных примесей.
Выход сухого вещества — это вес воздушносухого торфа,
содержащегося в единице объема сырого торфа (или сырца).
В отличие от каменного и бурого угля, горючих сланцев выход
сухого торфа — величина переменная. Это объясняется прежде
всего тем, что количество воздушносухого вещества в опре¬
деленном объеме торфа зависит от содержания в нем воды.
Обычно выход сухого вощества выражают в килограммах воз¬
душносухого торфа, содержащегося в 1 м3 сырца. По сведе¬
ниям Бременской болотной станции, разные виды торфа дали
из 1 м8 сырца такой выход сухого вещества в килограммах:
Таблица 24
(по В. Доктуровскому, 1932)
Выход сухого
вещества
из 1 м3 сырца
(в килограммах)
Сфагновый торф мало разложив¬
шийся 90
Сфагновый торф хорошо разло¬
жившийся 120
Переходный торф 180
Низинный торф 250
В таблице 25 приведены цифры для Мезиновского болота Московской
области по выходу сухого вещества и по степени гумификации (1932).
Таблица 25
(по В. Доктуровскому, 1932)
Выход воз¬
душносухого
вещества
в кг
Верховой, мало разложившийся
торф 78
Годный на изоплиты 86
Более разложившийся топливный
верховой торф 102
Степень
разложения
(в процентах)
15
15
26
Из таблицы видно, что с увеличением разложения
выход сухого вещества увеличивается.
Усадка торфа — это способность его умень¬
шаться в объеме при потере воды. Для слабо гуми¬
фицированного торфа усадка происходит за счет уплотнения
растительной массы, для сильно гумифицированного большое
значение имеет свойство гумусовых коллоидов сокращаться
в объеме при высыхании. Степень усадки зависит как от
64
способа выработки, так и от качеств торфа. Кирпичный торф
сокращает объем в 2,7 — 7,2, резной — в 1,9 — 6,3 раза. Эвтроф-
ные болота дают усадку залежи в 10—15°/о, олиготрофные в
20—30% первоначальной глубины за 1—3. года осушки
(В. Доктуровский, 1932).
Степень гумификации торфа и методы ее определения.
Торф состоит из растительных остатков, сохранивших клеточ¬
ную структуру, и из аморфной, бесструктурной гумусовой
массы. Отношения между обеими составными частями торфа
колеблются. Чем больше гумусовой массы в торфе,
тем он сильнее разложился или, как теперь говорят,
тем он более гумифицирован. Как будет видно из
дальнейшего, степень гумификации — весьма важное свойство
торфа, от которого зависят многие другие.
Оценка степени гумификации—далеко не простое дело,
чем и объясняется разнообразие методов ее определения. Су¬
ществуют химические и физические методы определения сте¬
пени разложения. Химических методов несколько. Не входя
в подробности, укажем, что они основаны или на учете гуми-
новых веществ путем извлечения их разными растворителями
(например, различными щелочами), или на определении остав¬
шихся в торфе неразложившихся продуктов, например клет¬
чатки (В. Сукачев, 1926). Химические методы громоздки, до¬
роги и к тому же не вполне безупречны. Гораздо проще и де¬
шевле физические методы определения гумификации. Поэтому
именно эти методы весьма широко распространены не только
при технических, но и при научных исследованиях. Различают
три группы физических методов: глазомерные макроскопиче¬
ские, глазомерные микроскопические и, наконец, количествен¬
ные методы. Из глазомерно-макроскопических полевых мето¬
дов заслуживает внимания метод шведского инженера
Вальгрена. Он предложил шкалу определения гумификации,
основанную на макроскопической структуре торфа, его конси¬
стенции при сжатии в руке, цвете воды, выделяемой из
торфа при его выжимании, и пр. (табл. 26).
Л. Пост воспользовался вальгреновскими принципами опре¬
деления гумозности торфа и переработал его шкалу.
Десятибалльная шкала Л. Поста (Л. Пост и Е. Граилунд, 1925) выглядит
следующим образом:
Н 1. Совершенно иегумифицированный и лишенный дю1 торф. При сжа¬
тии в руке между пальцами проходит бесцветная, чистая вода.
Н 2. Почти совершенно кегумпфицированный торф, лишенный дю. При
сжатии его вытекает почти чистая, но окрашенная в желто-коричневый цвет
вода.
Н 3. Очень слабо гумифицированный торф, содержащий очень мало дю.
При сжатии его проходит отчетливо мутная вода. Однако торфяная масса не
просачивается. Остаток (после сжатия) не имеет кашеобразного характера.
Н 4. Слабо гумифицированный торф, иначе говоря, содержащий некото¬
рое количество дю. При сжатии вытекает очень мутная вода. Остаток не¬
сколько кашеобразный.
1 Дю—совершенно разложившаяся бесструктурная масса торфа.
5—722 65
Таблица 26
Шкала разложения Вальгрена
А4- Торф
д сильно
разложив-
А — шийся
Растительные
остатки не раз¬
личимы про¬
стым глазом
Вода при 1
сдавливании
торфа не
выделяется
Масса при сжи¬
мании хороша
проходит через
пальцы, пачкая
руку
АВ-{- Хорошо
дд разложив-
п шийся
АВ-
Заметны лишь
некоторые
растительные
остатки
Вода не
выделяется
или ее выде¬
ляется очень
немного
Выделяю¬
щаяся вода
темно-
коричневая
Масса продав¬
ливается, не¬
много пачкая
руку
В+ Мало
п разложив¬
шийся
В—
Остатки расте¬
ний заметны
Вода выде¬
ляется не¬
много
Вода корич¬
невая
Масса почти не
продавливается,
рук не пачкает.
После сжатия
поверхность
торфа шерохо¬
ватая от расти¬
тельных остат¬
ков
ВС-(- Плохо
пГ разложив-
шийся
вс¬
Так же, как и
в предыдущем
Вода выде¬
ляется
в большом
количестве
Вода желтая
Масса продав¬
ливается очень
мало
ей- Неразло-
£ жившийся
С—
Растительные
остатки ясно
различимы про¬
стым глазом
Вода выде¬
ляется, как
и в преды¬
дущем
Вода совсем
почти
не окраши¬
вается
Масса не про¬
давливается
Н 5. Довольно гумифицированный торф, содержащий довольно много
дю. Растительная структура различима, но не вполне отчетливо. При сжатии
вытекает очень мутная вода и проходит некоторое количество торфяной массы.
Остаток сильно кашеобразный.
Н 6. Довольно гумифицированный торф, содержащий довольно много
дю. Растительная структура неясна. До 1/з всей массы торфа проходит при
сжатии между пальцами. Остаток сильно кашеобразный, но с более ясной
растительной структурой, чем неотжатый торф.
Н 7. Довольно сильно гумифицированный торф, т. е. содержащий зна¬
чительное количество дю. Растительная структура еще довольно хорошо
различима. При сжатии проходит до половины торфяной массы. Эта масса
тонко-кашеобразная и интенсивно темная.
Н 8. Хорошо гумифицированный торф, содержащий много дю, с очень
неясно видимой растительной структурой. При сжатии проходит до 2/3 тор¬
фяной массы. Иногда еще выделяется некоторое количество кашеобразной
воды. Остаток главным образом из более стойких в отношении разложения
корешков и пр.
Н 9. Почти совершенно гумифицированный торф, иначе говоря, почти
нацело состоящий из дю, в котором почти не видна растительная структура.
При сжатии почти вся масса торфа в виде гомогенной кашицы проходит
через пальцы.
66
Н 10. Совершенно гумифицированный торф, нацело состоящий из дю;
при сжатии проходит вся масса торфа, свободной же воды не выделяется1.
Шкала Л. Поста более удобна и представляет несомненные преимуще¬
ства перед шкалой Вальгрена. Соотношение между обеими шкалами таково:
Н 1)
Н 31
Н 5
= с
= ВС;
Н 2)
Н 4)
Н 6
Н 71
= В; =АВ;
Н 8)
9Ц.
Н ку
П. Варлыгин и В. Чеботарев (1929) предлагают придать
шкале Л. Поста количественный характер (табл. 27).
Таблица 27
Ступень шкалы
Приблизительное отжимаемое количество
коллоида
Н 1
От 0 до */к от взятого объема сырца
Н 2
п 5 » Б » » » »
НЗ
» 5 ** 5 ** » » **
Н 4
» ^/5 »> */б » » » •>
Н 5
„ */5 » полного объема
По мнению этих авторов, такое изменение шкалы Л. Поста
уточнит и упорядочит работу. Однако ввиду трудности учета
количества отжимаемого коллоида авторы предлагают свой
особый глазомерный полевой метод определе¬
ния степени гумификации в раскрытом челноке непо¬
средственно после взятия пробы. При этом объем гумусового
коллоида учитывается на-глаз. После тренировки можно на¬
учиться отсчитывать: 7ю> 7#> 7в> 7г. 7в> V* объема, что соответ¬
ствует 10, 20,... 50,65 и 75%. Если отсчет фиксируется между
указанными процентами, то мы получаем, например: между
0 и 7м объема — 5%, для интервала 7н> и 7б — 15°/о, для ин¬
тервала между 7-1 и 7» приблизительно 30% и т. д. Все числа
через 10% также отсчитываются. Таким образом, достигается
желаемая точность отсчета.
Кроме полевых глазомерных макроскопических методов,
существует лабораторный, микроскопический гла¬
зомерный. Этот метод впервые предложен П. Варлыгиным
(1924).. Сущность его состоит в том, что под микроскопом
с небольшим увеличением (не более 140 раз) учитывается на-
глаз в поле зрения процент площади, занятой гумусовыми
частицами, по отношению ко всей площади (гумусовые ча¬
стицы-{-растительные остатки); площадь тех и других вместе
принимается за 100. Для каждого образца торфа, в зависимо¬
сти от требуемой точности, производится от 10 до 30 подсче-
1 Шкала Л. Поста в приведенном здесь виде представляет перевод не¬
мецкого текста О. ЕгсКтап (1921). Немецкий текст этой шкалы имеется еще
у К. Ви1о\у (1929). П. Варлыгин (1929) и В. Доктуровский (1932) дают не¬
сколько сокращенный и измененный текст.
5*
67
тов (или полей зрения), из которых берется среднее. Этот
метод, по мнению П. Варлыгина и В. Чеботарева (1929), дает
близкие к описанному полевому глазомерному методу цифры,
лишь несколько преуменьшенные (в среднем на 5%).
К лабораторным количественным методам от¬
носится метод В. Кудряшева (1920). Это метод весо¬
вой, основанный на отмучивании. Образец торфа, предвари¬
тельно размятый и перемешанный (чтобы достичь большей
однородности), делится на две равные по весу части. Одна вы¬
сушивается до воздушносухого состояния и потом взвешивается;
другая отмучивается через ситечко с отверстиями в */4 мм.
При этом удаляется гумус и остаются форменные раститель¬
ные остатки. Отмученная часть образца высушивается и взве¬
шивается; вес ее делится на вес неотмученной: частное, выра¬
женное в процентах, представляет весовой процент неразло-
жившихся в общей массе растительных остатков торфа. Вы¬
читая это частное из 100, получаем процент гумуса по отно¬
шению к этой массе.
Автор считает, что этот метод дает несколько преувеличенные цифры
разложения, так-как сквозь сито проходят, кроме гумуса, и форменные ча¬
стицы. Как отмечают П. Варлыгин и В. Чеботарев (1. с.), отмучивание дает
в общем параллельные результаты с микроскопическим и полевым процент¬
ным методом П. Варлыгина, но преувеличивает степень гумификации.
Советские торфоведы в последнее время особенно усиленно
разрабатывают физические методы определения гумификации
и стремятся выработать стандартные образцы торфа опреде¬
ленной степени разложения. Получение таких стандартов очень
важно для согласования методов определения гумификации.
За границей вопросы методики определения степени гумифи¬
кации далеко не пользуются таким вниманием, как в СССР.
У нас же из лабораторных методов наиболее широко приме¬
няется микроскопический метод П. Варлыгина.
Тепловые свойства торфа. Теплопроводность торфа. Теп¬
лопроводность вещества характеризуется количеством тепла
(в калориях), проходящим в секунду через пластинку того или
другого вещества площадью в 1 см2 и толщиной в 1 см при
разности температур по обе стороны пластинки в 1°. Так как
торф неоднороден, то его теплопроводность зависит от того,
сколько в нем по объему твердого вещества, воды и воздуха
(Г. Пухнер, 1929). Г. Карстен (Н. Karsten)1 нашел, что тепло¬
проводность сухого торфа равна 0,00027, а насыщенного водой —
0,0011. Следовательно, теплопроводность торфа
значительно повышается с увеличением содер¬
жания в нем воды. Зависит это от того, что воздух
обладает меньшей теплопроводностью, чем вода, а торф,
чем богаче водой, тем беднее воздухом. Торф вообще пло¬
хой проводник тепла. На этом основано его примене¬
ние как изолятора. Если принять изоляционную способность
1 По Г. Пухнеру (1928).
68
воздуха равной 1,00, то для
обыкновенного торфа она бу¬
дет 1,38, для торфяной муки —
3,32, для пробковой муки —
3,90 [данные Бирка из работы
В. Вальяжникова (1924), по
В. Сукачеву (1926)]. Таким
образом, изоляционная спо¬
собность торфа еще выше,
чем такого хорошего изоля¬
тора, как воздух, причем осо¬
бенно хороший изолятор —
измельченный торф.
Торф, даже влажный, плохо
проводит тепло. Иначе говоря,
амплитуда колебаний темпе¬
ратуры торфа быстро зату¬
хает с глубиной. По К. Гюк’у
(К. Ниеск, 1929), в различных
ассоциациях сфагнового бо¬
лота РЮ^хепШеЬе! в восточной
Пруссии суточные колебания
температуры на глубине 25 см
в июле обычно не превыша¬
ли 0,5°. При этом температура
днем здесь была значительно
ниже, чем на воздухе и на по¬
верхности мохового ковра, а
ночью, наоборот, выше. Ам¬
плитуда же суточных колеба¬
ний на воздухе и на поверх¬
ности сфагнового ковра была
13,5 и 22°. Подобный же ре¬
зультат дали измерения и на
сфагновом болоте близ Лахты
Ленинградской области (по В.
Сукач.еву, 1926). Здесь суточ¬
ные колебания температуры
за 15—16 июля на поверхно¬
сти сфагнума достигали 26,4°,
на глубине же 25 см всего 0,3°.
Приводим ниже сведения для
сфагнового болота близ Лахты
по затуханию суточных коле¬
баний с глубиной (по В. Сука¬
чеву, 1926).
Годичные колебания темпе¬
ратуры в торфе, понятно, зату¬
хают на большей глубине, чем
суточные. По В.Сукачеву(1926)
оо
см
се
Я
к
4
ХО
св
Н
>*
н
«е
о.
о
5 ^
« №
н <и
СП
К *0
в *
в >>
се О
хо
о ^
с; Ю
§ О
од
3
х
у
о
Г-
СО О 00 Ю 00
СО СМ тГ ТГ ^ Г"-“ ю
юля
ю
10 ^10 010 «о оо
»-Г см 1-4 |С ио
16 И
СО
ГГ ^0)0^^Ю^оо^
О оо 05 ^
-
ОСО0ОО
СО СО тГ Е* ьо
го
с<
оо 0_ СП О^ со^ ^ 00^
ос" Г'-" *-н тг ^ С-Г 10
^
сч
Т-Н ^^^1000
^ ГГ со ^ ю
о>
^ ю оо ^ оо
СО Ю (О со ю
№
О 10 оо
О со оо ТГ ^ с-Г Ю
2
5
го
00 СМ^ОО^ го^тг оо
о^оГсо
ГО
го
Ю оо^со 05 см^^оо
05 0*0 еогсю
^ со см ^ ^
-
ОС^СО^ГГ 05_ ^ т* 00^
оо О ОС СО
*-н СО 1-1 ^ ^
0>
05 О, эо^
юс0о ~10
»-1 см ^ —• —•
05 о С*. тГ °о
СМ со Е* ^ о с>Г ю
1—« 1—1 1—« 1—<
2
^ та
и
то
У
<и я
н >>
о в
о и
з 5
в у" ^ »»вс
се ^
=£28888
% о
>»* ее
э £я
8§£ ■ ■ ■ ■
со Ей
69
годичная амплитуда температуры в торфяниках на глубине
130—150 см равна всего 4—6° и даже меньше. По Танфильеву
на приленинградских торфяниках на глубине 4,2—5,6 м тем¬
пература постоянна в течение всего лета. Она близка к 6°.
Такая же температура была в торфе на глубине 6,4 м в на¬
чале марта, откуда следует, что на этой глубине тем¬
пература постоянна в течение всего года. Таким
образом, в достаточно глубоких торфяниках средней полосы
у дна можно найти слой торфа с постоянной годичной тем¬
пературой.
Плохая теплопроводность торфа имеет большое значение
для жизни растений. Летом, в солнечные дневные часы, поверх¬
ность болота сильно нагревается, так же как и воздух, окру¬
жающий надземные части растений, в то время как на глубине
10—20 см, в ярусе корней, держится сравнительно низкая
температура. Ф. Фирбас (Б. НгЬав, 1931) дает сводку по макси¬
мальным температурам на поверхности мохового покрова
сфагновых болот. Здесь летние температуры часто бывают
выше 4-30°, в отдельных же случаях выше 409, доходя даже
до 47° тепла. По тем же сведениям, температуры в торфе
на глубине корневых систем за вегетационный период лишь
редко достигают 20°, обычно же не превышают 16—17°. Таким
образом, для местного климата (микроклимата) сфагновых
болот характерен резкий контраст условий на поверхности
мохового ковра (и воздуха над ним) и на глубине корневых
систем. Вместе с тем плохая теплопроводность торфа легко
вызывает ночные заморозки. Это происходит потому, что при
охлаждении поверхности болота ночью выравнивание темпе¬
ратуры между нижними, более теплыми, слоями торфа и верх¬
ними, остывшими, идет очень медленно, так как торф плохо
проводит тепло. Благодаря малой теплопроводности торфа
болота по сравнению с минеральной почвой медленно про¬
мерзают и также медленно оттаивают. Поэтому мерзлота
торфа весной держится долго, особенно в сухих сфагновых
кочках. Указывают, что в подмосковных осушенных торфяни¬
ках мерзлота сохраняется до середины июня (В. Сукачев, 1926).
Таким образом, вегетационный период на болотах
вследствие поздних весенних и ранних осенних
заморозков, а также вследствие долго держа¬
щейся почвенной мерзлоты оказывается более
коротким, чем на минеральной почве.
Слабая теплопроводность торфа объясняет, почему болота
представляют „холодные местообитания“ по сравнению с ми¬
неральными почвами. Эта „холодность“—свойство преиму¬
щественно сфагновых торфяников, почему сфагновые болота
средних широт и служат убежищем для более северных
тундровых растений.
Медленное оттаивание торфяников ведет к тому, что, пере¬
двигаясь на север, мы встречаем вечную мерзлоту сначала
в торфяниках, а потом уже в минеральных почвах. При этом
70
она раньше появляется на повышениях микро- (или мезо*)
рельефа, на торфяных буграх, которые медленнее, но глубже
промерзают и медленнее оттаивают, чем мочежины. Лишь
дальше к северу вечная мерзлота появляется и в мочежинах,
так что торфяники покрываются непрерывным вечномерзлым
слоем неглубоко под поверхностью.
Теплотворная способность торфа измеряется количеством
тепла, выделяющимся при сгорании. Тепло это измеряется
калориями. Различают малые калории и большие калории.
Малая калория — это количество тепла, нагревающее 1 г воды
с 0° до 1° С. Большая калория поднимает температуру 1 кг
воды с 0° до 1° С. Различают калориметрическую и
полезную теплотворную способность. Первая опре¬
деляется сжиганием торфа в герметически закрытой металли¬
ческой бомбе в. кислороде под давлением. В этом случае
определяют теплотворную способность органического вещества
торфа (беззольного и безводного) или абсолютносухого веще¬
ства (безводного, но с золой).
Полезную теплотворную способность определяют, сжигая
воздушносухой торф (т. е. торф с 30% влаги) в топке. Калори¬
метрическая теплотворная способность значительно выше
полезной (табл. 29).
Таблица 29
Полезная теплотворная способность торфа и других
веществ
(по Г. Шрейберу, 1927)
(в калориях)
Воздушносухое дерево 2 400—3 800
Землистый бурый уголь 1 500—3 400
Богемский бурый уголь • 4 300—5500
Каменный уголь 5 500—8 100
Каменноугольный кокс 5 900—7 500
Древесный уголь 6 900—7 500
Торф лучшего качества 3 500—4 200
Торф среднего качества 2800—3500
Торф низкого качества 2 000—2800
Торфяной кокс 7 300—7 600
Торф обладает высокой теплотворной способ¬
ностью и, будучи даже среднего качества, дает в среднем
не меньше тепла, чем дрова. Торф хорошего качества значи¬
тельно превосходит дерево и низшие сорта бурого угля,
уступая лучшим его сортам и каменному углю. Продукт же
переработки торфа — торфяной кокс—не уступает и лучшим
видам топлива.
Полезная теплотворная способность торфов разного ботани¬
ческого состава видна из табл. 30.
Из таблицы 30 видно прежде всего, что теплотворная способ¬
ность торфов одного и того же ботанического состава зависит
от степени гумификации и что чем выше степень разло¬
жения, тем больше теплотворная способность.
71
(по Г. Шрейберу)
Таблица 3©
Теплотворная
способность
Содержание
воздушно¬
сухого торфа
золы (в °/0)
(в калориях)
Тростниковый торф, слабо разложившийся . . .
3 286
14,0
Тростниковый торф, сильнее разложившийся . .
3 368
13,3
Осоковый торф, слабо разложившийся
3 770
1.7
Осоковый торф, сильнее разложившийся ....
3792
4.0
Буромоховый торф, слабо разложившийся . . .
3441
7,1
Буромоховый торф, сильнее разложившийся . .
3 631
' 5,7 .
Олыианниковый торф
3 777
1.7
Березовый торф * , . . .
3 958
2,1
Пушицевый торф
3 741
0,5
Сфагновый торф, слабо разложившийся «...
3 179
0,6
Сфагновый торф, сильно разложившийся . • .
3591
4,1
Гумус верещатников
3 374
10,0
Кроме того, высокозольные торфа имеют малую
теплотворную способность. Можно сказать, что для
олиготрофных (верховых) торфов с малой зольностью, которая
притом колеблется в небольших пределах, содержание золы
практически не влияет на теплотворную способность. Для
верховых торфов эта способность определяется другими
физическими свойствами торфа (см. ниже). Рабочая тепло¬
творная способность эвтрофных торфов с большой зольностью
зависит от содержания золы. Особенно резко падает тепло¬
творная способность высокозольных низинных торфов.
Зависимость теплотворной способности от
степени разложения лучше всего можно проследить
путем калориметрического определения (исключив влияние
золы и воды,) на беззольном и безводном органическом веще¬
стве торфа. Л. Кузнецова (1935) установила, что для олиго¬
трофных торфов теплотворная способность органического
вещества в общем растет с увеличением степени разложения.
При этом для интервала степени разложения от 0 до 40°/»
теплотворная способность непрерывно растет с увеличением
степени разложения и притом настолько правильно, что за¬
висимость между общими свойствами можно выразить форму¬
лой. Таким образом, теплотворную способность можно вычи¬
слить для данного вида торфа теоретически с достаточной
степенью точности, исходя из степени его разложения. В ин*
тервале степени гумификации от 40 до 60% теплотворная
способность торфа почти не меняется. При дальнейшем повы¬
шении гумификации теплотворная способность вновь начинает
расти.
Подобную зависимость теплотворной способности от гумификации
Л. Кузнецова (I. с.) объясняет тем, что на первых стадиях оторфования не¬
прерывно нарастают высококалорийные соединения за счет низкокалорийных
углеводов и сходного растительного материала. В средне разложившихся
72
верховых торфах распад низкокалорийных легко разлагающихся соединений*
в основном, вероятно, уже заканчивается, и начинается разложение таких
компонентов, как лигнин. Эти компоненты по калорийности близки уже
к конечному продукту распада — гуминовым веществам. Поэтому при даль¬
нейшем оторфовании их переход в гуминовые вещества уже не сопрово¬
ждается увеличением теплотворной способности торфа, и она остается более
или менее неизменной.
Эвтрофные торфа, по исследованиям Л. Кузнецовой, связь между тепло¬
творной способностью и степенью гумификации обнаруживают в более слабой
степени, чем верховые торфа, причем теплотворная способность растет лишь
в интервалах степени разложения от 0 до 10 и от 23 до 40.
Теплотворная способность органического вещества сфагно¬
вых торфов в общем увеличивается со степенью разложения.
Она для мало разложившихся сфагновых торфов равна 4700—
5100, а хорошо разложившиеся выделяют до 6100 калорий..
Большая теплотворная способность сильно разложившегося
торфа зависит от того, что по мере разложения в торфе уве¬
личивается содержание высококалорийных соединений—смол,
воскообразных и гуминовых веществ. В мало же разложившемся
торфе, наоборот, большую роль играют низкокалорийные
соединения — клетчатка, пектиновые вещества и пр. В общем,
теплотворная способность органического вещества наших
олиготрофных торфов колеблется между 4700 и 6000, а для
эвтрофных от 5500 до 6200 калорий.
Рабочая (полезная)теплотворная способность
торфа зависит от содержания в нем влаги: чем больше влаги,,
тем ниже теплотворная способность. При 30% влажности
рабочая теплотворная способность верховых торфов колеб¬
лется от 3400 до 3900 калорий и практически зависит от содер¬
жания не золы, а влаги и, кроме того, в большой мере от
степени гумификации. Полезная способность эвтрофных тор¬
фов зависит также от степени гумификации и, кроме того,
от влаги и содержания золы. При малой зольности тепло¬
творная способность близка к верховым торфам, у много¬
зольных же эвтрофных торфов она падает до 2800 калорий
и ниже.
Сухие торфа.
Название „сухой торф“ (Trockentorf) целесообразно удер¬
жать (по Е. Раманну, 1911) для органогенной породы, состоя¬
щей из растительных остатков с более или менее сохранив¬
шейся структурой. Сухой торф имеет вид спутанной, плотной
волокнистой массы. При таком определении сухого торфа
ему противопоставляется как особое понятие „перегной"
(„Moder“). В перегное растительные остатки хуже различимы
и более измельчены, чем в торфе. При таком понимании на¬
звание „грубый гумус“ (Rohhumus) приобретает более широкое
значение и объединяет сухой торф и перегной. Грубый лесной
гумус обычно делится (по Е. Раманну) на два слоя — верхний,
имеющий характер сухого торфа, и нижний — перегноя. В этом
понимании „сухой торф" отличается от обычного торфа и тем,
73
что он образуется в общем без участия почвенной воды, но
в условиях влажного воздуха. Поэтому „сухой торф" в зна¬
чительном количестве образуется лишь в особенно богатых
осадками частях влажных климатических зон — в северных и
горных областях, где содержание зольных почвенных элементов
невелико, а температура воздуха и почвы низка. Все это замед¬
ляет разложение растительных остатков и способствует обра¬
зованию сухого торфа и гумуса (К. Бюлов, 1929).
Можно различить следующие виды „сухих торфов“ (в ос¬
новном по Е. Раманну, частью по Бюлову):
А. Лесные сухие торфа.
1. Буковый торф. Состоит из листьев, частью из других остатков
бука, причем в верхних слоях листья хорошо различимы. Этот торф обра¬
зует в странах с морским климатом (между прочим, в Германии)
мощные слои.
2. Дубовый торф из остатков листьев и плодов дуба. По Раманну
встречается редко.
3. Еловый торф. Еловые леса, даже в молодом возрасте, легко
образуют значительные отложения сухого торфа, состоящего из хвои и
шишечных чешуи.
4. Сосновый торф. В этих торфах, достигающих иногда 30—40 см
мощности, большое участие принимают лесные мхи — спутники сосны,
а также и цветковые растения; в зависимости от сопутствующих видов
сосновые торфа носят различный характер.
5. Пихтовый торф встречается обычно лишь в виде тонких слоев
б. Сухие торфа из невысоких деревянистых растений
(кустарников и кустарничков).
6. Торф из горной сосны (Latschentrockentorf) образует в горах
на болотах и вне их нередко слои значительной мощности.
7. Торф из падуба (Ilex aquifolium).
8. Торф из рододендрона. Rhododendron fernigineum, реже
Я. hirsutum обильно отлагают торф в альпийском горном поясе.
9. Торф из Azalea procumbens встречается там же и достигает
40 см мощности.
10. Торф из Empetrum nigrum встречается в тундровой зоне.
А. Чильман (A. Kihlman, 1890) отмечает чистый торф из водяники мощ¬
ностью 0,1— 0,3 м на восточном побережье Колы (Орлов, Три острова).
Из других кустарничковых торфообразователей в образовании сухого
торфа участвуют: вереск, Erica tetralix, брусника, черника. Первые два вида
образуют сухой торф атлантических вересковых пустошей.
С. Травянистые сухие торфа.
Carex firma и Сагех curvula — важные образователи сухого торфа в вы¬
сокогорной области Альп.
Полуторфа.
Полуторф (На1Мог{) по И. Лоренцу (1858)х— это торф, бо¬
гатый минеральными примесями—глиной, песком и пр.
При дальнейшем обогащении торфа примесями мы имеем уже
минеральные почвы, содержащие гумус. Таким образом, полу-
торфа — промежуточное звено ,между крайними членами 11 По Г. Потонье (1911).
74
ряда — собственно торфом и минеральной почвой. Минеральные
примеси в полуторфах могут быть различны (по К. Бюлову).
1. Кремнеземистые примеси: песок речного, озерного или
эолового происхождения или диатомовые панцыри.
2. Глинистые примеси. Они характерны для слоев торфа на
границе с речными и другими седиментами. Подобные полу-
торфа представляют глины с более или менее обильным
содержанием гумуса и с растительными остатками.
3. Различные примеси, вносимые в торф грунтовыми или
ключевыми водами и отлагающиеся здесь в виде солей, про¬
питывающих торф (например вивианит, сернокислое и угле¬
кислое железо и пр.). При значительном содержании их мы
имеем полуторфа.
Водные отложения (седименты).
Седиментами называют аллохтонные водные отложения,
осаждающиеся не на том месте, где они образовались. Так, на¬
пример, планктон, состоящий из микроскопических организмов,
взвешен в толще воды. Отмирая, он отлагается уже на дне
водоема в виде аллохтонного образования — седимента. Седи-
ментам Л. Пост (1. с.) противопоставляет торфа — автохтонные
«образования, отлагающиеся на месте их материнского ценоза.
Седименты и торф. Свойства седиментов. Седименты,
богатые органическим веществом (так называемые гиттии,
см. ниже), существенно отличаются от торфа по составу и
свойствам. Это видно из табл. 31.
Таблица 31
(в основном по Г. Потонье)
Г И Т т и и
1. Образуются из настоящих вод¬
ных организмов, главным обра¬
зом планктона
2. Исходный материал богаче жиром
и восками, что влияет и на со¬
став продукта (гиттии)
3. Смолы отсутствуют или их не¬
много
4. Беднее углеродом
5. Беднее углеводами
6. Богаче азотом и кислородом
7. При образовании гиттии нака¬
пливается водород
8. Гумусовые кислоты отсутствуют
9. Реакция воды щелочная
10. Образуются в результате восста¬
новительного процесса (гниение,
битуминизация) 1111. Обладают восстановительной спо¬
собностью (очищают железо от
ржавчины)
Торфа
1. Образуются в основном из водно¬
болотных растений
2. Исходный материал беднее жи¬
ром и восками
3. Смолообразные вещества обиль¬
ны
4. Богаче углеродом
5. Богаче углеводами
6. Беднее азотом и кислородом
7. При образовании торфа содержа¬
ние водорода чаще уменьшается
8. Гумусовые кислоты имеются
9. Реакция воды большей частью
кислая.
10. Оторфование носит промежуточ¬
ный характер между образова¬
нием гумуса (окисление) и гит¬
тии (восстановление)
11. Не обладают восстановительной
способностью
75
12. В свежем состоянии предста¬
вляют студенисто-желеобразную
массу
13. В сухом состоянии весьма тверды
14. Окраска от желтой до черной
15. После высушивания уже не раз¬
мокают в воде
12. В свежем состоянии предста¬
вляют связную, волокнистую
массу
13. В сухом состоянии легко разла¬
мываются
14. Окраска более светлая
15. После высыхания обычно размо¬
кают вновь
Образование гиттий сопровождается изменением хими¬
ческого состава исходного материала планктона. При этом,
как и при образовании торфа, растет содержание углерода
и золы, а количество кислорода и азота уменьшается (см. М. Со¬
ловьев, 1932). Сднако водород ведет себя при оторфовывании
и битуминизации не совсем одинаково (табл. 32). При обра¬
зовании гиттии содержание его во всяком случае не умень¬
шается; оно или остается постоянным, или растет вместе
с углеродом за счет других элементов. При образовании же
торфа содержание водорода чаще убывает.
Таблица 32
Состав гиттии и гумуса (в процентах)
(по Г. Потонье, 1908)
Возраст
г
и т т и и
Гумусовые породы
с
Н
Н : С
С
н
Н :С
Четвертичный
50-57
6-7
12
50—СО
5-6
10
Третичный
65
8,5-9
13
60—75
4—6
7,5
Мезозойский
69,5—76
8,5-12
14
75-87
4-5
5,5
Палеозойский
75-83
7,5-10
11
80-95
1,5-6
4
Особенно характерно изменение отношения водорода к уг¬
лероду (последние колонки). В то время как это отношение
в ряду гиттий остается постоянным, в ряду гумусовых пород
оно резко падает. Это показывает, что при битуминизации
водород и углерод ведут себя более или менее одинаково.
Виды седиментов, классификация их. Е. Науман (Е. Nau¬
mann, 1930) делит седименты следующим образом:
a) Седименты, состоящие из органического материала
(илистые отложения). Сюда относятся:
1. Гиттия (частично совпадает с понятием сапропель).
2. Дю.
b) Седименты, состоящие главным образом из минераль¬
ных (неорганических) отложений.
Здесь различаются пять типов седиментов:
1. Глинистые отложения.
2. Кремнеземистые отложения.
3. Известковые отложения.
4. Железистые отложения.
5. Марганцовые отложения.
76
В группе органических водных отложений два основных
типа — гиттия и дю, выделенные впервые Л. Постом (1862),
отличаются следующими признаками (табл. 33).
Таблица 33
(по Е. Науману)
Гиттия
Дю
Материнская формация
Эвтрофные водоемы
Богатые гумусом оли-
Количество планктона
Вода богаче планкгоном
готрофные водоемы
Вода беднее планкто¬
в воде
Окраска воды, завися¬
Менее выражена
ном
Более выражена
щая от присутствия
гумуса
Деятельность фауны
Более интенсивная
Менее интенсивная
Цвет
Серый
Коричневый
Микроскопическая
Характерны плактонные
Характерны хлопья гу¬
структура
организмы
муса
Гиттия образуется из отмирающих организмов, населяющих
водоем, и состоит в основном из их остатков, дю же — в более
бедных жизнью дистрофных озерах с большим содержанием
гумуса. Этот гумус вымывается в озеро из окружающих мест
и отлагается в виде хлопьев на дне, образуя дю. Обычно
к этому гумусу всегда примешиваются и остатки водных
организмов, так что отложения приобретают характер дю —
гиттии. В чистом же виде отложения типа дю не встречаются
(Е. Науман, 1. с.).
Наиболее разработана классификация седиментов Г. Лунд-
квиста (О. ЬипсЦшз!, 1928). Она основана на макроскопических
признаках. Приводим ее с некоторыми сокращениями и до¬
полнениями:
A. Песчанистые гиттии1 с различным содержанием
песка. При содержании его от 50®/о и более он заметен
на-глаз; при большом содержании песка говорят о г и т-
тиевидном песке.
B. Суглинистые гиттии (Ье1ш1(гуи]а) содержат от
20% и более частиц глины и песка; остальное прихо¬
дится на грубый и тонкий детрит, остатки водорослей
и пр.
C. Отложения с заметным содержанием из¬
вести. Детрит под микроскопом стекловидный. В за¬
висимости от содержания извести различаются:
1. Озерный мергель. Отложения с преобладанием
извести или почти нацело из нее состоящие.
1 Эги типы гиттий не входят в классификацию Лундквиста, поскольку
они не всегда различимы макроскопически.
77
2. Известковые гиттиис обильным содержанием
органического вещества.
O. Раковистые гиттии с заметными н а - г л а $
раковинами моллюсков. Основная масса без-
извести.
Е. Отложения с заметной на-глаз глиной.
Детрит серый или почти стекловидный. В зависимости
от содержания глины различаются:
1. Гиттиевидные глины.
2. Глинистые гиттии.
P. Грубодетритовые гиттии1. Основная масса боль¬
шей частью определима под микроскопом, грубая, жел¬
товатая или другого цвета — в зависимости от проис¬
хождения. Характерны ткани высших растений, мхов и
остатки крупных водорослей.
О. Водорослевые гиттии1. Основная масса тоже
большей частью определима под микроскопом, состоит
из хорошо сохранившихся мелких водорослей и пр.
Детрит стекловидный.
Н. Тонкодетритовые гитти и1. Основная масса обычно
неопределима под микроскопом, тонкой структуры или
бесструктурная, стекловидная или желтоватая.
Л. Диатомовая охра. На-глаз заметна примесь окислов
железа. Детрит от этой примеси желто-коричневый или
красный.
К. Озерное дю. Основная масса во влажном и сухом
состоянии темная, часто грубой структуры, перемешана
с хлопьями гумуса.
Ь. Пыльцевая гиттия (ИттепМ, ЬеисЫой) состоит
в своей массе из пыльцы (в виде экзины древесных
пород, принесенной в водоем ветрами и здесь отло¬
жившейся). Особенно много пыльцы ольхи, встречается
также пыльца хвойных. Фимменит светлой окраски
в воздушносухом состоянии легок, весьма горюч. Мощ¬
ность отложений фимменита достигает 0,6—1,8 м, занимая
площадь до 20 га (данные для Германии — Г. Потонье,
т. III, 1912).
М. Намывной торф (8с1шетт1ог1). Этот вид седиментов
состоит из остатков растений (листья, ветви, куски древе¬
сины и т. д.) как водно-болотных, так и наземных, а
также из торфяного детрита. Намывной торф образуется
из приносимого водой органического материала, отлагающе¬
гося на дне водоема или у его берегов. Намывной торф
по Г. Лундквисту (1927)—крайний вариант грубодетрито-
вой гиттии. Он имеет промежуточный характер между
седиментами и болотными торфами.
1Все эти виды гиттий не обнаруживают макроскопической примеси
глины, извести и железа.
78
Еще в большей степени это относится к так называемым
гиттиевым (сапропелевым) торфам. Последние
образованы остатками как крупных водных цветковых расте¬
ний (корневища ЫутрЬаеа и Мирйаг, рдесты), так и планктон¬
ных организмов. Характер разложения органического вещества,
а также его химический состав—промежуточные между гит-
тией и торфом. Материнскими ценозами этих отложений яв¬
ляются заросли рдестов и водяных лилий (В. Сукачев, 1926).
Седименты СССР1 пока еще изучены недостаточно. При¬
ведем некоторые сведения о гиттиях Союза (по В. Алабышеву,
19281 2 * и другим источникам).
Гиттиевидные пески и глины плотностуденистой
консистенции, окраска—от желтой до серой; они оказываются
первыми членами озерных отложений. Из водорослей обычны
остатки диатомовых и зеленых (всепебеэтиэ и Ре&аэ^ит).
Обычны и остатки зоопланктона. Мощность этих седиментов
невелика—до 10—25 см. Они известны для ряда болот
б. Ленинградской губ. (Толполовского и других) и залегают
непосредственно на позднеледниковых и послеледниковых
отложениях, выше переходят в гиттиевые торфа и образова¬
лись в некоторых случаях в субарктический период вслед
за отступанием ледника. На это указывают попадающиеся
здесь иногда остатки арктической флоры (например Огуаэ).
Практического значения они не имеют.
.Озернуга“ (местное название) из торфяников Свердловского района,
близкая к гиттиевидным пескам, имела такой химический состав (В. Толмачев,.
1915; привожу по В. Сукачеву, 1926):
Процент V
Кремнезем (Si02) 38,04
Окись железа (Ре9Оз) . . . .... 4,63
Глинозем (А1203) 5,27
Известь (СаО) 2,72
Магнезия (MgO) 0,92
Окись марганца (МпО) Следы
Серная кислота (Si02) 2,80
Органические вещества и вода . . 45,48
Данные С. Орловой (1932) по химическому составу гиттий из ряда озер
б. Тверской и Костромской губерний и б. области Коми показывают, что
эти гиттии в громадном большинстве случаев относятся к рассматриваемой
группе и часто близки по химическому составу к „озернуге“ из Свердловского
района. Содержание в них золы редко менее 20%, часто близко к 50% и
нередко доходит до 60% и более. Состав золы близок к „озернуге4*. В золе
много кремнезема (обычно более 50% по отношению к весу всей золы,
а нередко и до 75% последней).
Седименты, богатые известью. Сюда относятся
пресноводные мергели (иначе озерные мергели), образованные
в своей основе углекислой известью. Они бывают белыми,
желтыми, кремовыми, розоватыми и более темных оттенков.
1 Литературу по гиттиям СССР см. Известия Сапропелевого комитета,
вып. 1—6, 1928—1932.
2 Терминология употребляется мной применительно к Г. Лундквисту
(1927).
79
При высыхании белеют, становятся хрупкими. Растительных
остатков мало (хары, диатомовые), иногда встречаются массами
части раковин пресноводных моллюсков. Озерные мергели
близки по характеру к известковым туфам в торфе (см. выше)
и связаны переходами с гиттиями, бедными известью. Озерные
мергели — ранние члены свиты озерных отложений. Их нахо¬
дят в виде мощных слоев от 0,5 до 4—5 м толщиной в глу¬
боких частях болот. Они залегают здесь на ледниковых и
послеледниковых аллювиальных наносах и иногда слагают
■целиком всю толщу озерных отложений. Выше их залегает
торф (погребенные озерные мергели), отделенный от них
иногда еще слоем гиттиевого торфа. Озерные мергели найдены
в торфяниках Ленинградской области (Парголовское болото,
•близ Петергофа и др.), в пойме Дубны (Московская область)
и в ряде других районов. Имеют значение как удобрительные
туки при известковании почв. По своему происхождению
озерный мергель—выпавшая из воды углекислая известь. Вы¬
падение извести происходит при потере двууглекислой известью
растворенной в воде углекислоты и воды1. Озерный мергель
часто слоист. В нем, и вообще в сапропелях, различаются тем¬
ные, богатые органическими веществами прослойки, отложив¬
шиеся летом, и более светлые, бедные гумусом зимние. Темная
и соседняя светлая прослойки образуются в течение одного
года. Годичные слои „микрозоны“ обычно не толще листа
писчей бумаги. Сосчитав их, можно определить время отложе¬
ния озерного мергеля. Методика определения возраста слои¬
стых отложений разработана Б. Перфильевым (1927). Химиче¬
ский состав сухого вещества седиментов, богатых известью,
виден из табл. 34.
Таблица 34
(по В. Сукачеву, 1926)
* Состав известковой гиттии
Торфяник Сверд¬
ловского района
Озеро Перегород-
ное б. Тверской губ.
в процентах
Кремнезем (Sl02) . . .
Окись железа (Fe2Os) .
Глинозем (А120,) . . .
Известь (СаО)
Магнезия (MgO). . . .
Окись марганца (МпО) .
Серная кислота (SO3) .
Органические вещества
I
2,56
8,8
0,51
3,4
1,52
40,64
725
1,17
2.4
50,26
3.2
51 81
—
1 Ф. Мюнихсдорфер (Р. MUnlchsdorfer, 1928) различает по способу обра¬
зования такие типы известковых гиттий: 1) альм (Alm)— углекислую известь,
оседающую в результате неорганического процесса под действием гумусовых
коллоидов; 2) озерный мергель (Seekreide), образующийся преимущественно
благодаря деятельности водорослей; 3) известковый туф, образующийся под
водой или вне ее, при участии или без участия растений и без участия
гумусовых веществ.
80
Из седиментов типа гиттии В. Алабышев (1928) приво¬
дит данные по „сапроколлам“, т. е. более старым, уплотнен¬
ным, обычно прикрытым торфом, полуископаемым гиттиям.
Они имеют плотно-желеобразную консистенцию, легко рас¬
слаиваются, имеют в изломе „резиновый" характер и тусклый
маслянистый блеск. При высыхании и промораживании они
расслаиваются на тонкие слои.
Из растительных остатков в сапроколлах обычны: древес¬
ная пыльца, диатомовые и десмидиевые водоросли, реже сине-
зеленые и зеленые (из зеленых иногда Botryococcus Braunii).
Крупные растительные остатки (плоды и ткани рдестов, листья
деревьев, иногда шишки хвойных) в сапроколлах встречаются
лишь рассеянно. Здесь же иногда находят Trapa natans. Из
животного мира попадаются остатки зоопланктонных организ¬
мов (Copepoda, Cladocera), реже иглы губок. Сапроколлы в
болотах (ископаемые) найдены во многих пунктах Ленинград¬
ской области, в б. Тверской губ.
В незаросших озерах гиттиевые отложения, особенно в
верхних слоях, отличаются от болотных гиттий, погребенных
и спресованных налегающим сверху торфом. Эти отложения
очень жидки, типа расплывающегося студня. Лишь с глубины
8—9 м озерные гиттии делаются более густыми и отчасти
приближаются к сапроколлам. По мере отложения торфа по¬
верх гиттии и налегания сплавин они превращаются в иско¬
паемые гиттии (сапроколл). В озерных гиттиях среди раститель¬
ных остатков преобладают сине-зеленые,. диатомовые, реже
зеленые водоросли, а из животных—Cladocera, Copepoda,
Spongiae. Подсплавинные гиттии отмечены на Кудиновских
озерах (Маслово болото, в 30 км от Москвы по Горьков¬
ской ж. д.) и в некоторых других местах. Попадаются также
панцыри ракообразных, коловраток,, иглы губок. Мощность
гиттиевого торфа невелика, обычно всего около 0,5 м. Он
найден в некоторых болотах Ленинградской области.
Торфяные илы близки к намывному торфу (см. выше).
Образуются на дне дистрофных (гумусовых) озер, среди
торфяников вследствие размывания торфяных берегов и ниж¬
них частей сплавин и отложения торфяного ила на дне водоема.
Состав торфяных илов крайне непостоянен. Форменных остат¬
ков прибрежно-водных растений и планктонных организмов
здесь сравнительно немного. Торфяные илы найдены в озерах
б. Ленинградской, Новгородской и других губерний. Несо¬
мненно, что это весьма широко распространенный тип отложе¬
ний в озерах с торфяным дном и берегами, где имеет место
размыв и отложение торфа.
Соленоводные седименты выделяются в особую
группу. В. Алабышев (1932) выделяет в Европейской части
СССР зону солоноватых и соленых озер, где отлагаются эти
седименты. Они характерны для засушливого и пустынного
климата. Этот автор выделяет два типа соленых озер с раз¬
личными отложениями:
6-722
81
1) озера с илами черного или серо-черного цвета, богатыми
сернистым железом; это лечебные грязи,
2) самосадочные озера с отложениями преимущественно
минерального характера.
Минеральные грязи во многих случаях встречаются азо¬
нально и в подзолисто-таежной области—в местах выхода соле¬
носных ключей разного химического состава, вытекающих' из
соленосных пластов коренных пород. Сюда относятся: 1) Старо-
русские соленые источники, вытекающие из известково¬
глинистых девонских слоев, 2) минеральные грязи и источники
Сольвычегодска, вытекающие из пермских горизонтов и др.
Ископаемые гиттии. Отложение гиттий происходило
и в очень древние эпохи.
Горючие сланцы в ряде случаев, несомненно, принадлежат
к гиттиям. Так, прибалтийские горючие силлурийские сланцы,
так называемые кукерситы, возникли, по М. Залесскому, из
планктонной сине-зеленой водоросли Gleocapsomorpha prisca,
близкой к современным Gleocapsa.
Горючие сланцы нижневолжского яруса в Поволжье с остат¬
ками зеленых и сине-зеленых водорослей, червей и корнено¬
жек тоже относятся к гиттиям. По Бертрану и Рено, сапро¬
пелитовые угли — богхеды — образованы студенистыми слое¬
вищами водоросли Pila, а несколько сходная с нею водоросль
Reinschia образует горючие пермокарбоновые сланцы в Южном
Уэльсе. К битуминозным горючим сланцам близка нефть.
Гкттии, образовавшиеся в древние эпохи в водоемах1, претер¬
певали различные изменения, спрессовывались под давлением
отлагавшихся сверху минеральных наносов. Если в водоем
попадали гумусовые вещества, то они под действием этого
давления утрачивали штриховатость и сплавлялись с гиттией
в однородную массу, сначала бурую, потом черную, блестящую,
с раковистым изломом (так называемые витритовые угли).
К таким сапропелитово-гумусовым углям принадлежат, напри¬
мер, черемхиты Кузбасса, причем они относятся как к бурым,
так и к каменным углям. В углях может преобладать то гуму¬
совая, то сапропелитовая часть.
Известны и межледниковые водные отложения, например,
известковые седименты рисс-вюрмской эпохи в южной Баварии
(Ф. Мюнихсдорфер, 1928).
Седименты и геолого-петрографическое строение местности.
География седиментов.
Характер седиментов, как это показали прекрасные
работы шведских лимнологов, тесно связан с химическим
составом воды в водоемах. Характер же водоемов зависит в
свою очередь от геолого-петрографического строения мест-
1 Об условиях образования седиментов в прежние эпохи см. Г. Стадии-
ков (1932), М. Соловьев (1932).
82
Рис. 4. Карта региональной лимнологии южной^ и средней Швеции.
По Е. Науману, 1930.
1 — озерная руда; 2 — болотная руда; 3 — известковые горы;
4 — богатые известью почвы; 5 — граница морских отложений.
ности. Е. Науман (1930) выделяет в Швеции такие геолого-
петрографические районы с определенными типами озер
(рис. 4).
1. Область послеледниковой морской трансгрессии с глини¬
стыми отложениями, богатыми питательными веществами.
Озера в этой области эвтрофного типа.
2. Возвышенности, сложенные известняками (Kalkgebirge).
Почвы здесь богаты известью, однако часто довольно бес¬
плодны. Озера здесь при избытке извести в воде бедны азотом
6* 83
Рис. 5. Карта областей седиментов южной Швеции. Из Е. Наумана, 1930.
1 — области железистых седиментов; 2 — области седиментов
типа дю; 3 — области водорослевой гиттии; 4 и б — области
известковых седиментов; 6 - области глинистых седиментов.
и фосфором. Они принадлежат к известковому типу (der
alkalitrophe Typus).
3. Области морен, богатых известью, имеют, подобно
области морской трансгрессии, плодородные почвы. Водоемы
сходны с водоемами первой области.
4. Область коренных пород, прикрытых мореной (кроме
богатых известью морен),—это область бедных почв. Водоемы
здесь также олиготрофны. Они бедны и известью, и фосфором,
и азотом. Здесь в водоемах по преимуществу содержится
гумус. Обычны также отложения железа (железная руда) в
болотах и озерах. Озера здесь олиготрофного, дистрофного
84
солонцово-солончаковая с самосадочными озерами; ь — тундровая зона пресноводных мало¬
мощных сапропелей; У — горная зона пресноводных сапропелем; 10 — северная граница вы¬
щелоченных черноземов.
п — пресноводные сапропели; и — известковые сапропели; к кремнеземистые сапропели;
м — минеральные грязи; г — глауберова соль; пс — поваренная соль; с — соли магния;
ж — железные руды. (По районам, не заштрихованным на карте, автор книги точных данных
не имеет).
и ацидотрофного (т. е. с резко кислой реакцией воды), а также
сидеротрофного типа (т. е. богаты железом).
Озерные отложения в описанных выше областях различны.
В областях с глинистыми почвами и известковой мореной (области 1 и 3)
дно озер образуют различные типы гиттий.
В области коренных пород (область 4) широко распространены богатые
гумусом отложения типа дю, а также болотные и озерные отложения железа.
В области известняков (область 2) распространены главным образом из¬
вестковые гиттии и озерная известь.
Сравнение карты региональной лимнологии, приведенной выше, с кар¬
той областей сециментов Г. Лундквиста (рис. 5) показывает, что области
геолого-петрографические довольно хорошо совпадают с областями седиментов.
Мощные отложения озерной гиттии нашел М. Нейштадг (1936) в озере
Сомино Переславль-Залесского района Ивановской области. Профиль
озерных отложений Сомина дает такую картину сверху вниз.
О—3,5 м. Озерное дю, в основном из хлопьев гумуса с небольшой при¬
месью диатомовых.
3,5—10,0 м. Тонкодетритовая гиттия. Основная масса неопре¬
делима под микроскопом.
10,0—31,0 м. Диатомовая гиттия. Состоит почти нацело из диато-
мей — Melosira, Navícula, Synedra, Cyclotella, Campilodiscus и других.
Мощность пробуренной не до конца толщи озерных отложений озера
Сомино—31 м. М. Нейштадт (1. с.) считает, что эта мощность наибольшая.
Гиттиевые торфа — конечная стадия озерных отложе¬
ний, знаменующая исчезновение водоема и наступление болот¬
ной стадии. Эти торфа являются верхним членом ряда седи¬
ментов, залегая и непосредственно под сплавинами, и между
озерными и болотными отложениями. Макроскопические ос-
Схе м а 3
Практическое использование гиттий
(по В. Алабышеву, 1928)
Г и тт и я
I
Смола
(деготь)
1
Под¬
смольная
вода
1 1
Газы Кокс
Метило¬
вый
спирт
1
Уксусная
кислота
1
Водный
аммиак
i
Легкие
масла
бензин
1
Средние
масла
керосин
1
Пек
Лак
1
Тяжелые
масла
1
Газо¬
i
Вазе¬
1
Мотор¬
i
Лампо¬
I
Соля¬
1
Пара¬
вое
лино¬
ное
вое
ровое
фино¬
масло
вое
масло
масло
масло
вое
масло
масло
(сапро-
воск)
гзо-
^•верный^гропи^/
.60-
\
^ • Экватор
у
4.. 1
Южный тропик_\ -r- ~- - 7
30-
^Ожны^
ьляриый
1500
Масштаб по экватору
0 1500 3000
45рО*<Н
к востоку от Гриивича
^ /
> в ^
1
Ш
2
■
И
3
Рис. 7. Схематическая карта зон озерных отложений земного шара. По В. Алабышеву. 1932 г. 1—маломощные озерные сапропели тундры; 2 —мощные (до 30 м) пресноводные сапропели таежно¬
подзолистой зоны; 3—пресноводные сапропели субтропической и тропической зон; 4 — зона соленоводных сапропелитов, минеральных грязей, самосадочных озер и солончаков; 5—горная зона
пресноводных сапропелей (до 10 м).
татки растений составляют в гиттиевых торфах половину и
более всей массы. Обычны осоки и тростник, иногда кувшинки
и рдесты. Встречаются и семена этих растений, а также семена
Nuphar, Scirpus lacustris, Najas и изредка плоды Trapa natans.
Присутствуют аморфные торфяные частицы, что сближает
эти отложения с болотным торфом так же, как и остатки
высших растений и волокнистая структура. Из водорослей
встречаются десмидиевые, зеленые, реже диатомеи.
География гиттий в CCfcP. Характер седиментов
тесно связан как с петрографическим составом пород, так
одновременно и с почвами. В стране с зональным расположе¬
нием почвенных типов, как Европейская часть СССР, седименты
располагаются также зонально (рис. 6). Рис. 7 дает схему
зон озерных отложений для земного шара (см. ниже). Исполь¬
зование гиттий дает схема 3.
ГЛАВА III.
ОБРАЗОВАНИЕ БОЛОТ НА МЕСТЕ ВОДОЕМОВ.
Интенсивность заболачивания.
Болота возникают двумя путями: 1) на месте суши в резуль¬
тате ее заболачивания и 2) в результате заселения водоемов
болотной растительностью.
Превращение водоема в болото может итти по-разному,
при этом роль растительности бывает различной. Заселение
водоема растительностью — процесс прежде всего биологиче¬
ский. Отсюда понятно, как важно учесть роль растительности
и внешних факторов в процессе заболачивания.
Быстрота размножения водных растений весьма велика,
что видно из приводимых ниже цифр о длине годичных по¬
бегов у некоторых водно-болотных растений одного из фин¬
ских озер.
Таблица 35
Длина годичных побегов водно¬
болотных растений (в см)
(по Л. Аарио, 1933)
Equlsetum fluvlatile 110—130
Typha latifolla, иногда свыше . 200
Typha angustliolia до 130
Phragmites communis 215—245
Scirpus lacuster 150
Эти цифры характеризуют довольно значительную скорость
поступательного движения зарослей водно-болотных растений.
Л. Аарио (1. с.) нашел, что заросль Scirpus lacuster продвину¬
лась за один год на 1—1,5 м. Заросли Equisetum и Typha,
как видно из рис. 8 и 9, полученных в результате повторных
съемок растительности, продвигаются значительно быстрее
зарослей Scirpus.
Увеличение площади, занятой зарослями водно-болотных
растений, позволяет оценить их роль в заболачивании водое¬
мов. Typha latifolia занял в течение 9 лет площадь свыше 15 га (!),
Sparganium ramosum заполнил за то же время речное русло
на протяжении нескольких сот метров, a Scirpus lacuster за
88
Рис. 8. Разрастание зарослей
Бскриэ ио^ег и ТурЬа на одном
из озер Финляндии. По Л. Аарио,
1933.
1 — АПвта; 2 — ТурЪа в 1931 г.; 3 — при¬
рост ТурИа за 1931/32 г.; 4 — БагриБ в
1931 г.; 5 — прирост Загрив за 1931/32 г.
9 лет образовал круглые заросли радиусом в 12 м (Л. Аарио,.
1. с.). Т. Менкель-Щапова (1930), сравнивая современную
картину зарастания Белого озера (из группы Косинских озер
под Москвой) с прежними кар¬
тами, нашла, что Acorus Calamus,
встречавшийся в 1911 г. на озере
лишь в нескольких пунктах, за
15 лет занял большую часть
береговой линии этого озера,
общим протяжением свыше 2 «ж
С громадной быстротой раз¬
множается, как известно, Elodea
canadensis. По сведениям П.
Гальцова (по Т. Менкель-Щапо¬
вой, 1. с. ), Elodea canadensis,
впервые появившись в Белом
озере в 1905 г., через несколько
лет размножилась так,
что местами сделала не¬
возможным передвижение на лодке, а уже в 1911 г.
ее масса значительно уменьшилась.
Весьма быстрым темпом заполняет водоемы и моховая
растительность. В б. Богородском у. Московскбй губернии я
неоднократно наблюдал торфяные выемки, вырытые 40—50 лет
назад и превратившиеся
в мелкие озерки. Во время
моего посещения эти ка¬
рьеры, зачастую площадью
(ТТЛ/ в тысячи квадратных ме-
г™ трое, представляли часто
■ЗГГ сплошные сфагновые (или
буромоховые) зыбуны, силь¬
но обводненные, но ме¬
стами уже выдерживающие
человека (Н. Кац, 1922).
ф ¡о ¡2 ',о 50м Быстрота размножения
водно-болотных растений
в умеренном климате дает
лишь слабое представле¬
ние об интенсивности этого
процесса в более южных
широтах, где раститель¬
ность достигает роскошного
развития. По Л. Шапошникову и Н. Верещагину (1931) зара¬
стание озер, образовавшихся благодаря разливам Сыр-Дарьи,
идет весьма быстро. Уже на второй год после образования
такого озера оно покрывается по краям густой тростниковой
зарослью. Через несколько лет тростник образует уже мощ¬
ные заросли, до 8 м высотой, причем на 1 мг приходится
150 живых стеблей. С каждым годом тростник продвигается
89
Рис. 9. Разрастание зарослей Scirpus
lacuster и Equisetum limosum. По Л.
Аарио, 1933.
1 — Alisraa; 2 — Equisetum в 1931 г.; 3 — при¬
рост Equisetum за 1931/32 г.; 4 — Scirpus в
1931 г.; 5 — прирост Scirpus за 1931—1932 гг.
в глубь озера, образуя на более глубоких местах зыбуны сво¬
ими сплетающимися корневищами.
Продукция надземной растительной массы—весьма важ¬
ный момент в процессе заболачивания водоемов. Ведь распро¬
странение зарослей растений по зеркалу водоема еще не
означает превращения его в болото. Чтобы судить о быстроте
последнего процесса, важно учесть продукцию растительной
массы и условия ее сохранения. Цифр по величине продукции
водно-болотных фитоценозов очень мало. Приведем некото¬
рые (табл. 36).
Таблица 36
(из Л. Аарио, 1933)
Урожай надземной массы
в тоннах
на 1 га
свежий вес
сухой вес
Ассоциация AUsma plantago ....
17,8
3,0
?»
Sparganium ramosura . *
50,8
10,6
ь
Phragmites communis . .
. 25,8
10,4
■
Scirpus lacuster ....
24,5
8,1
т
Typha latifolia
72,8
17,5
»
Equlsetum fluviatile . . .
35,2
6,1
»
Phragmites-Equisetum. .
31,7
9,5
„
Scirpus-Equisetum . . .
28,4
6,1
»
крупных OCOK
15,4-10,1
о
со
1
ос
со
При оценке этих цифр надо учесть, что они получены
путем пересчета на гектар урожая с маленьких проб¬
ных площадок в 0,25 м2. Этот способ, видимо, несколько
преувеличивает результат. Тем не менее, и с этой поправкой
надо признать, что продукция массы водно-болотной раститель¬
ности весьма велика.
Приводим для сравнения цифры по урожайности сена
заливных лугов низкого уровня приильменского района (из
Г. Ануфриева, 1925) и лугов среднего уровня волжской поймы
(А. Шенников, 1930) (табл. 37).
Таблица 37
Урожай сена на заливных лугах (в тоннах на 1 га)
(по А. Шенникову, 1930, и Г. Ануфриеву, 1925)
Cariceta gracilis
Carlceta vulpinae
Phalaris+Carex gracilis
Glyceriaeta aquaticae
Некоторые типы Plialarideta до
Волжские заливные луга среднего уровня
3.0
4,4
3,15
3,75
6.0
4,0—5,0
Мы видим из сравнения с табл. 36, что ассоциации водных
и водно-болотных растений далеко превосходят по продук¬
90
тивности даже наиболее урожайные заливные луга. Лишь
наиболее производительные канареечниковые луга несколько
приближаются к ним по урожайности. Крупноосоковые же
болотные ассоциации по продуктивности близки к заливным
лугам, причем эти ассоциации в Финляндии и приильменском
районе дают весьма близкие цифры.
Материалы по другим озерам значительно отличаются от
материалов Л. Аарио. Так, Г. Рике (Н. ЮскеЪ 1922) для
озера МесЫе в США нашел для глубинной зоны от 0 до 1 м
вдвое меньшую продукцию свежей массы, чем для той же
зоны озера ЫигтЦат,—16 т против 32 т на 1 га. Для другого
озера США тот же автор нашел еще значительно меньшую
продуктивность. Несомненно, что производительная способ¬
ность растительной массы в более южных широтах, например
в тростниковых плавнях дельт южнорусских рек, гораздо
выше, чем в Финляндии.
Масса подземных частей растений при образо¬
вании донных отложений играет гораздо большую роль, чем
надземная, сильнее подверженная разрушению. Поэтому учесть
подземную массу весьма важно. Однако такие сведения весьма
скудны. Соотношение веса подземных и надземных органов
у водно-болотных растений видно из табл. 38.
Таблица 38
Отношение веса подземных орга¬
нов к весу надземных у водно¬
болотных растений
(по Л. Аарио, 1933)
Бс^рив 1аси5(ег 9:1
РЬодтйез соттигш 5:1
Equisetuш Пиу1аШе 5:2
ТурЬа !аШо11а 1:1
АПэша р1ап(а£о 2:3
Бра^апШт гапкиит 1:2
Учитывая, что подземная масса у этих растений многолет¬
няя, важно знать ее годовой прирост. У ТурЬа около поло¬
вины подземной массы отрастает за один вегетационный
период, у Бйгриз 1аси$1ег—гораздо меньшая часть, точно не
установленная. Для других растений сведений нет (Л. Аарио, 1. с.)
Даже из этих отрывочных сведений видна значительность
продукции подземной массы водных растений, особенно важ¬
ной при образовании донных отложений.
Скорость отложения седиментов дает нам представление
о быстроте заполнения водоема растительными остатками.
Г. Лундквист (1925) считает, что эта скорость отложения зави¬
сит прежде всего от внешних факторов. Один и тот же вид
седимента может в соседних пунктах нарастать с различной
скоростью. В общем известковые и глинистые седименты,
водорослевая, а также грубодетритовая гиттия литоральной
зоны нарастают сравнительно быстро. Значительно медленнее
91
отлагается тонкодетритовая и глубоководная грубодетритовая
гиттия (Г. Гаме — Gams, 1927).
Абсолютные цифры скорости нарастания седиментов можно
вычислить, только когда определен возраст отдельных слоев
при помощи пыльцевых диаграмм или археологических нахо¬
док. Понятно, что и в этом случае скорость нарастания опре¬
деляется лишь приблизительно, поскольку и самая датировка
не может быть точной. Г. Гаме (1. с.) вычисляет скорость
отложения седиментов в различные климатические периоды
послеледниковой эпохи (табл. 39) для озер Лунца (б. Австрия).
Таблица 39
Скорость отложения седиментсв в различные климатические периоды
послеледниковой эпохи
(по Г. Гам су, 1927)
Untersee u. Rehbergmoor
Obersee u. Rotmoos
Хронология по I
Максимальная седиментация
Седиментация
Общая мощ¬
ность отложе¬
ний
За 1000 лет
отложилось
Общая мощ¬
ность отложе¬
нии
За 1000 лет
отложилось
Блитт-Сернандеру
j 1 оды
Около 3 М
Около 2 м
Около 1 Чг М
Около \Чг М
Субатлантиче¬
ский
1900
1000
Около 4 м
Около И м
Суббореальный
— moo
— 2000
— 3000
Около 1 Чг М
Около 1 м
Около 1 М
Около 2 м
Атлантический
— 4000
— 5000
Около 1 м
М
Около 1 м
Чг М
Около 30 см
Около Чг М
Около 25 см
Бореальный
— 00.0
1 м
Около Чг М
30 см
15 см
Инфрабореаль-
ный
Субарктиче¬
ский
— 7000
— 8С00
40 см
Около 35 см
Арктический
— 9000j
—10000
Из таблицы видно, что скорость отложения седиментов неодинакова
в различные климатические периоды1. В добореальный холодный период*
отлагалось всего 0,15—0,5 мм в год. В бореальный период отлагалось
0,5—1,0 мм. В теплый же послеледниковый (атлантическое и суббореальное
время) седиментация достигла максимума —1—2 мм в год. При вычислении
скорости отложения седиментов надо иметь в виду, погребены ли они под
толщей торфа и спрессованы, или же лежат свободно. Приведенные здесь
цифры далеко не максимальны. М. Нейштадт (1936) для озера Сомино Пе-
реславль-Залесского района б. Ивановской области определил возраст толщи
седиментов в 31 м в 10 000 лет, что дает скорость отложения в 3,1 м
в тысячелетие. Автор объясняет это тем, что седименты здесь сносились
водой в сравнительно узкую воронку и быстро заполняли ее.
1 Известную роль играло спрессовывание нижних слоев.
92
Из приведенных вычислений видно, что скорость на¬
копления седиментов во всяком случае не
меньше, а иногда и больше, чем скорость нара¬
стания торфа.
Сведения по быстроте исчезновения озер и сокращения
их площади чрезвычайно скудны. Приведу некоторые из них
(из И. Фрю и К. Шретер, 1904). Ф. Рупрехт путем сравнения
с старыми картами установил, что для превращения озер
окрестностей Tocho под Ленинградом в болота потребовалось
300 лет и, как минимум, —180 лет. А. Корнгубер (А. Когп-
huber, 1886) установил, что озеро Kônigsee в Венгрии, пло¬
щадью в 8 га, совершенно заросло в течение 30 лет. Цеппе¬
лин (Zeppelin) сообщает, что осоковый берег Боденского озера
с 1824/25 года переместился в глубь озера на 120 м. Р. Спарро
и А. Дубах (1918) исследовали зарастание озера Святого
в б. Бежецком у. Тверской губернии. За 400 лет поверхность
озера уменьшилась с 310 до 166 га. При этом озеро потеряло
при заболачивании около 37 млн. кубометров воды (по В. Док-
туровскому, 1932).
Экологические факторы заболачивания.
Влияние глубины водоема на распределение фитоценозов.
Не все водные ценозы одинаково зависят от глубины водоема.
Так, планктон и сплавины, или зыбуны, т. е. растительные
ковры, плавающие на воде и связанные с берегом, в общем
не зависят от глубины и образуются как в мелких, так и глу¬
боких водоемах. То же надо сказать отчасти и относительно
свободно плавающих растений. Зато укореняющаяся водная
растительность зависит самым тесным образом от толщины
водного слоя. Эта толщина прежде всего определяет необ¬
ходимый для растений минимум света. Количество проникаю¬
щего в воду света непосредственно зависит от прозрачности
воды. Чем прозрачнее вода, тем глубже идет водная
растительность. Между обоими факторами существует прямая
взаимозависимость (рис. 10).
В общем, для видов, индиферентных к температуре (напри¬
мер, Elodea), или видов, приспособленных к низкой темпера¬
туре воды (Fontinalis), основной фактор, ограничивающий их
распространение в глубь водоема, — это баланс ассимиляции.
Для видов теплолюбивых (Myriophyllum, Potamogetón) лими¬
тирующий фактор — это понижение температуры с увеличи¬
вающейся глубиной (Ф. Руттнер—F. Ruttner),
В. Сукачев (1926) указывает, что меньше всего света нужно
низшим споровым, поэтому они идут глубже других растений.
На большой глубине дно водоема покрывают именно эти ор¬
ганизмы, главным образом водоросли сине-зеленые, а также
немногие диатомовые и зеленые. Глубоко спускаются также
некоторые мхи, например в Женевском озере Thamnium до
глубины в 660 м. На значительно меньших глубинах посе-
93
ляются цветковые растения. В общем, фитоценозы в зарастаю¬
щих водоемах распределяются соответственно глубинам, об¬
разуя концентрические пояса.
В. Сукачев (1926) дает схему. распределения раститель¬
ности на зарастающих озерах от центра к периферии для
Средней России.
1. Зона микро фи тов — наиболее глубоководная. Она
образована микроскопическими донными водорослями, глав¬
ным образом сине-
JLJL.JL.JL м
1П"А]*
Ё
зелеными, немногими
зелеными и диатомо¬
выми. На несколько
более мелких местах
встречаются более
крупные водоросли —
Vaucheria, Cladophora и
др. Следовательно, эта
зона исключительно
споровых растений.
2. Зона макро¬
фитов состоит ча¬
стью из крупных во¬
дорослей (Chara, Nitel-
1а), главным же обра¬
зом из цветковых;
роголистника (Cerato-
phyllum demersum),
узколистных рдестов
(Potamogetón obtusifo-
lius, P. mucronatus), a
также из некоторых
мхов (Calliergon gigan-
teum). Все они своими
вегетативными органа¬
ми целиком погруже¬
ны в воду.
3. Зона широко¬
лиственных рде¬
стов состоит из ви¬
дов Potamogetón с широкими листьями (Potamogetón perfo-
liatus, Р. lucens, Р. praelongus). Встречаются также Myriophyl-
lum, Sparganium natans.H заходят из предыдущей зоны рого¬
листник и хары. Эта зона также состоит целиком из погружен¬
ных растений.
Толщина слоя воды здесь не превышает 4—5 м, обычно
же меньше. В этой зоне изменяется и характер донных отло¬
жений. Они состоят из смеси остатков планктонных организ¬
мов и преобладающих там остатков высших растений (гиттие-
вый торф). В предыдущих зонах отлагается гиттия, образо¬
ванная планктоном.
94
м
j I i I I || IU-J.
6 ■ Ё I I 1 I “ +?
«f I I -1 1 15
w\ ill ^ *^0
12
n
16
18
го
гг
2ч
20
28
30
Рис. 10. Соотношение между глубиной види¬
мости и границей глубинного распространения
литоральной растительности в озерах Альп и
северной Германии. Из Тинемана.
Заштрихованные столбики — максимальная
глубина видимости; белые столбики — граница
глубинного распространения растительности.
1 — Женевское озеро; 2 — Боденское озеро; 3 — Мориц-
ское озеро; 4 — Галльштеттское озеро; б — Шаальское
озеро; 6 — Большое Пленское озеро; 7 — Цклейское
озеро; 8 — Большое Эйтинское озеро.
4. Зона водяных лилий. Преобладают здесь виды
Nymphaea, Nuphar и плавающий рдест. Все они, наряду с по¬
груженными, имеют и плавающие на воде листья.
5. Зона камышей состоит из растений, погруженных
в воду лишь основаниями стеблей. Это будут тростник (Phrag-
mites communis), камыши (Scirpus lacuster), хвощ (Equisetum
limosum) и др. Сюда заходят также растения из более глубо¬
ководной зоны: очень часто в зарослях тростника плавают на
воде между его стеблями листья водяной лилии. Толщина
водного слоя в этой зоне не более 2—3 м. Отложения имеют
уже характер лимнического торфа.
6. Мелководная зона с непостоянным составом рас¬
тительности: Carex gracilis, Heleocharis palustris, Sagittaria sagit-
tifolia, Alisma Plantago, Polygonum amphybium, Naumburgia
thyrsiflora, Potamogetón gramineus, водные лютики, Hippurls
vulgaris и др. Эта зона располагается между зоной камы¬
шей и урезом воды. Глубина редко больше метра.
Вышеприведенная схема распределения растительности по
глубинным зонам вообще типична для прохладно-умеренного
климата.
Сходные пояса выделяют на заболачивающихся озерах и американские
исследователи (И. Уивер и Ф. Клементс — J. Weaver and Т. Clements, 1929).
Они считают их последовательными стадиями водной серии, т. е. суксцессии,
наблюдаемой при заболачивании водоемов. Различаются такие стадии (stage):
1. Стадия погруженных растений (Submerged Stage). Из от¬
дельных видов здесь Elodea, рдесты, Myriophyllum, Najas, водные лютики,
Utrlcularla, Vallisneria, водоросли микроскопические и крупные (Chara).
2. Стадия растений с плавающими листьями (Floating
Stage). Здесь, наряду с среднеевропейскими родами, представленными частью
викарирующими видами (например Nymphaea polysephala, Potamogetón,
Polygonum), растут также Nelumbo, Castalia. Глубина воды здесь от 1,8 до
2,5 лс.
3. Стадия тростникового болота (Reed-swamp Stage). Предста¬
вители последнего частью те же виды, что и у нас (Typha la tifo lia, Phrag-
mites communis), частью викарирующие американские (Scirpus validus, заме¬
няющий европейский Scirpus lacuster). Глубина воды здесь от 0,3 до 1,2 м.
Лишь Scirpus validus растет на глубине 1,8 м и более.
4. Осоково-луговая стадия (Sedge-meadow Stage).
Здесь растут виды Carex, Juncus и Heleocharis (palustris). Весной
и в начале лета почва здесь весьма влажная и иногда покрыта слоем воды
в несколько дюймов.
5. Лесная заключительная стадия (Climax Forest).
В табл. 40 сопоставлены зоны зарастания озер по иссле¬
дованиям разных авторов для различных областей. Эти зоны
сопоставлены с лимническими, а также и гидрографическими
границами. Легко видеть большое сходство корреспондирую¬
щих растительных зон в разных областях. Сходны не только
жизненные формы растений, характерные для определенной
глубинной зоны, но и порядок смены поясов растительности
в пространстве и во времени, а также видовой состав расти¬
тельных поясов. Последний в основном сходен в Восточной
и Центральной Европе. Даже в Северной Америке мы имеем
обычно или те же самые виды, или викарирующие виды из
Типы и пояса растительности
Тип органогенных
образовании
Г идрографическпе
границы
Лимнологические зоны
по Г. Лундквисту
(1927) и Е. Науману
(1930)
]
Супралитораль
Никогда, длитель¬
но не заливается
Типы и зоны
Г. Лундквист (1927)
Наземный (тер-
рестриче-
ский) торф
Болотный (тель-
матический)
торф
Низкое стояние
воды
Эулитораль
Зона амплитуды
колебаний уровня
Ма§посаг1сеинп
воды
Лимнический
(водный)
тростнико¬
вый, камыше-
вый и прочие
торфа
Высокое стояние
воды
3
•=*
о
09
СО
о
о*
*
о
X
о
а 1 2Рагуосаг1се1ит
А
Ч 1
«в
о!
н
X
.5
Верхняя субли¬
тораль
!
Тростниковые за¬
росли: РБ^тИез,
Б^гриэ, ТурЬа,
иногда Ма£посаг1-
сЫа
о
и
Нижняя субли-
Растения с плаваю¬
тораль
щими листьями
(ЫутрБавасвае)
Верхняя элито-
Погруженные
раль (макроэли-
макрофиты (Шо-
■А
Ч
Ч
тораль)
геНа и др.)
о.
о
н
X
ч
СТ)
Нижняя элито-
Погруженные ми¬
раль(микроэли-
.тораль)
крофиты
Глубоководная
зона
Бактерии
1 Главным образом в горах.
2 Цветение также над водой.
Таблица 40
в заболачивающихся водоемах
.растительное!
ч
i В. Сукачев (1926)
i
Г. Стеффен (1931)
Г- Вальтер (1927)
Ф. Клементс и Ф. Уи
Восточная Пруссия
Германия
вер (1929)—CL11CA
1 Эвтрофное болото
Заключительная
лесная стадия
(Climax Forest)
Мелководная зона: | Зона заливания: ,
Осоково-луговая
Сагех gracilis, Hel- 1 низкорослая расти-,1
стадия (Sedge-mea¬
eocharis, Sagittarla, | тельность из пест-1
dow Stage): Сагех,
Alisma и многие | рой смеси болот-'
Juncus, Heleocha-
другие водно-бо- ! ных и сорно-луго-
ris (palustris)
лотные виды. Глу* i вых видов |
бина но 1 м
Зона камышей:
Phragmites, Scirpus
laoustris, Scolo-
chloa festucacea
(глубина 2—3 м),
Equisetum limosum
И др.)
Комплекс тростни¬
кового болота:
1. Зона Л^поса-
йсеШт
2. Зона 01усепа
aquatica
3. Зона Phгagmites
до 1,5 м глубины
4. Зона БагриБ 1а-
сиз1ег
Формация болот¬
ных растений:
Scirpus lacuster,
Phragmites, Typha,
Glyceria aquatica,
Equisetum limosum,
виды Сагех и Jun-
cus 1
Стадия тростнико¬
вого болота (Red-
swamp StageJrScir-
pus validus, Typha,
Phragmites commu¬
nis. Глубина
0,3—1,2 m
| Зона водяных ли-
] лнй: Nymphaea,
¡ Nuphar, Potamo¬
getón natans
i
i
1
§
о
! м
¡Зона нимфейных:
¡Nymphaea, Nu-
¡phar, Limnanthe-
mum, Potamogetón
natans.. Polygonum
amphibium, иногда
Trapa. Глуб.1 —3jk
Фитоценозы с пла¬
вающими листьями:
Nymphaeaceae, Po¬
tamogetón natans,
виды лютиков,
Limnanthemum,
Trapa natans
Стадия растений с
плавающими ли¬
стьями: Nymphaea
(polisepala), Nelum-
bo, Cas talia ,Po tam o-
geton, Polygonum.
Глубина 1,8—2,4 м
; Зона широколист-
1 венных рдестов:
Potamogetón perfo-
! liatus, P. praelon-
gus, P. lucens, My-
: riophyllum, Sparga-
. nium natans. Глу¬
бина не более“
i 4—5 м \
1 о ¡Зона погужеиных
1 S ¡рдестов: К crispus,
! S"| P. perfoliatus,
g i P. lucens, Myrio-
! j Iphyllum spicatum
gl и др. Глубина
g i 3—7 м
А !
1 =5 .
! <U
Формация погруженных растений |
Фитоценозы це¬
ликом погру¬
женных расте¬
ний с соцветия¬
ми над водой:
виды рдестов,
Myriophyllum
и др.
Стадия погруженных растений
(Submerged Stage): Elodea, Po¬
tamogetón, Myriophyllum, Na¬
jas, Ütricularia, Vallisneria, во¬
доросли макроскопические
(Chara) и микроскопические.
Глубина менее 6 м
1 Зона макрофитов:
узколистные
рдесты: Р. obtusi-
íolius, ,Р. mucrona-
tus, Ceratophyllum,
Calliergon gigante-
i um, Chara, Nitella
н
н
и
ез
Он
Зона Nitella. Глу¬
бина свыше 7 м
Фитоценозы це¬
ликом погру¬
женные2 Najas,
Zannichellia,
Ruppía, Cerato-
phyllum, xapo-
вые
Зона микрофитов:
сине-зеленые, частью
зеленые и диатомовые
водоросли и пр.
7—722
тех же родов. Сходство процессов зарастания водоемов,
порядка смены растительных зон и их видового состава зави¬
сит от нивелирующего влияния водной среды и широкого
распространения водных растений.
Конечная судьба водоема и заключительная стадия водной
растительности различны и сильно зависят от климата,
а также от характера окружающей водоем местности. Так,
например, в южной части лесной зоны США конечной стадией
растительности заболачивающегося водоема будет лес (Climax
Forest).
В хвойной зоне Европейской части Союза такой стадией
будет торфяник и т. д.
Влияние глубины водоема на его заболачивание. В очень
глубоких водоемах возможность развития донной флоры, осо¬
бенно высшей, крайне ограничена. Повышение дна здесь
происходит исключительно за счет отложения остатков планк¬
тона. Образование же сплавин происходит в водоемах лишь
в том случае, если они защищены от ветра и притом неве¬
лики по площади, так как сплавины образуются лишь в спо¬
койной воде. Следовательно, зарастание водоема высшей
укореняющейся растительностью может начаться лишь тогда,
когда водоем значительно обмелел. Обмеление водоема, от
чего бы оно ни зависело, стимулирует развитие водной расти¬
тельности и ускоряет темп зарастания. Понижение уровня
часто вызывается искусственными причинами. В других слу¬
чаях это понижение зависит от естественных причин—напри¬
мер, от изменения рельефа в связи с развитием речной сети.
Результатом этого является самоосушка озер, спуск их и
заболачивание.
Зарастание водоема вследствие понижения его уровня не
всегда сводится лишь к поселению в нем водно-болотной
растительности и к его заполнению растительностью, а про¬
цесс заболачивания приобретает более сложный характер.
Это бывает, например, при одновременном отложении гиттии
и образовании сплавины. Одновременное образование зыбунов
и обильная продукция планктона с интенсивным накоплением
гиттии, по мнению Г. Гамса (1927), обычны на мезо- и эвтроф-
ных озерах. Оба процесса часто протекали особенно интен¬
сивно в сухой (суббореальный) послеледниковый климатиче¬
ский период. Заболачивание водоема при одновременном
надвигании сплавины и отложении гиттии сводится к следую¬
щему (см. Г. Гаме, 1. с.). Рост сплавины ведет к увеличению
ее мощности. При этом она начинает давить на рыхлую, легко
подвижную массу гиттии и выжимает ее внутрь озера в более
глубокие зоны. Продвигание же гиттии внутрь озера, в свою
очередь, благоприятно для одновременного роста сплавин
по направлению к центру озера, так как отложение гиттии
в центральных частях озера создает здесь береговые условия.
При этом комбинированном процессе заболачивание идет
особенно быстро.
9S
Смены растительных зон при зарастании водоемов —
один из лучших примеров эндодинамических и вместе с тем
экогенетических смен (В. Сукачев, 1926 г.). Экогенетическими
мы называем смены, происходящие под влиянием изменения
экологических факторов. Смены эти в той мере эндодинами-
ческие, поскольку они вызываются не одними внешними фак¬
торами, а главным образом самой растительностью, изменяю¬
щей в результате своей жизнедеятельности обстановку суще¬
ствования и условия для следующего звена сукцессии. Веду¬
щий экологический фактор смены фитоценозов на зарастаю¬
щем водоеме — уменьшение глубины вследствие отложения
органического вещества и постепенного повышения дна во¬
доема. Каждый фитоценоз, отлагая на дне водоема орга¬
нические вещества, вызывает обмеление, благоприятное для
нового, более мелководного фитоценоза. Более глубоководные
фитоценозы все время подвигаются в глубь водоема, а их
место занимают более мелководные.
Однако смены при зарастании водоемов объясняются не
только изменением экологических факторов самой раститель¬
ностью. Прежде всего, в смене растительности на зарастаю¬
щем озере существенное значение имеет конкуренция между
растениями. Глубины, занимаемые фитоценозами, вариируют
в довольно широких пределах. Верхняя и нижняя границы
любого пояса зарастания в большой степени зависят от кон¬
куренции. Например, тростник без более мелководного пояса
осок может продвигаться на меньшие глубины, а при отсут¬
ствии более глубоководного конкурента ЭНгриэ ^оЫег может
занимать и большие глубины. Пояс тростника при отсутствии
конкурентов изнутри и извне (по отношению к водоему)
расширяется и растет при большой амплитуде глубины. Кон¬
куренты суживают пояс тростника и амплитуду его глубин.
Хотя значение конкуренции при зарастании неоспоримо, но
мы почти не знаем, в каких формах она происходит. При
смене глубоко погруженных фитоценозов ассоциациями с пла¬
вающими листьями большую роль играет отнятие последними
света.
Смены растительности при зарастании являются эндодина-
мическими только с большими ограничениями. Глубина водо¬
ема далеко не всегда изменяется лишь от деятельности расте¬
ний. Она зависит также и от колебаний уровня водоема. Эти
колебания вызываются внешними факторами — искусственными
(осушка) или естественными (самоосушка озер, климатические
изменения). Внешние факторы, если они понижают уровень
водоема, содействуют процессу зарастания. Они, так сказать,
подгоняют эндодинамический процесс растительности. В слу¬
чае же, если внешние факторы вызывают повышение уровня
водоема, то они могут или вовсе остановить процесс зара¬
стания или даже направить его в обратную сторону. В этом
случае более мелководные пояса растительности могут сме¬
ниться более глубоководными.
7*
99
Озеро Ребегеее в Верхней Швабии представляет исключительно счаст¬
ливый объект для изучения взаимодействия факторов экзодинамических,
т. е. внешних, условий и эндодинамических, внутренних;условий на протяжении
всей его многовековой истории. История Рейегвее может быть хорошо дати¬
рована благодаря остаткам доисторических построек по его берегам. Можно
установить такие этапы этой истории (см. Г. Вальтер, 1927):
1- й период: 15000—2500 лет до нашей эры. Озеро сохраняло в те¬
чение всего этого периода б. или м. неизменную площадь —примерно 27 км2.
2- й период: 2500—1000 лет до нашей эры. Озеро сократило свою
площадь почти вдвое—с 27 до 14 кмР. Это сокращение произошло благодаря
интенсивному зарастанию озера, вероятно, вследствие наступления сухого
(суббореального) периода
3- й период: 1000 лет (до нашей эры) —1787 год (после нашей эры).
Уровень озера более или менее постоянен и заметного зарастания не проис¬
ходит. Временами за этот период уровень озера поднимался даже выше
уровня во второй период. Небольшое уменьшение площади озера с 14
до 11 км2 объясняется, вероятно, зарастанием в результате частичного
спуска озера вследствие самоуглубления стока.
4- й период: 1787—1911 гг. Площадь озера резко сократилась с 11 км2
(1787) до 7 км2 (1808), а в 1911 г. до 1,5 км2. Это сокращение площади
было вызвано зарастанием озера вследствие искусственного углубления
стока на 1 м в 1787/88 г. и настолько же в 1808 г. Зарастание пошло
быстрым темпом и продолжается и теперь.
Анализ приведенных сведений показывает, что во 2-й период благопри¬
ятные климатические условия, повидимому, содействовали эндодинамиче-
скому процессу зарастания и ускоряли его. То же имело место и в 4-й
период, когда процесс зарастания стимулировался понижением уровня озера,
но уже в силу искусственных причин.
В 3-й период процесс зарастания был более или менее приостанов¬
лен действием внешних факторов, поддерживавших высокий уровень озера.
Не исключается, что временами в этот период обычный ход смены фитоце¬
нозов шел в обратном направлении, а именно — менее глубоководные пояса
растительности сменялись более глубоководными.
Соотношение между растительностью и внешними факто¬
рами среды в разных случаях бывает неодинаковым. Зарастание
крупных водоемов с большой площадью водосбора и не слиш¬
ком мелких направляется в основном внешними факторами-
колебаниями уровня водоема. Эндодинамический процесс
смены фитоценозов развертывается лишь тогда, когда этому
содействуют внешние факторы, т. е. когда уровень озера в силу
тех или иных причин понижается. В этих случаях большую
роль играют вековые изменения климата, особенно еще потому,
что зарастание таких водоемов длится тысячелетиями (при¬
мер— Реёегэее). Если водоем невелик и неглубок, а площадь
водосбора незначительна, то перевешивают эндодинамические
процессы. Смена более глубоководных фитоценозов мелковод¬
ными протекает в этих случаях более или менее неуклонно.
Внешние факторы, может быть, замедлят или ускорят процесс,
но едва ли остановят на длительное время, а тем более не
смогут повернуть в противоположную сторону. Можно ду¬
мать, что особенно энергично протекает зарастание неболь¬
ших мелких водоемов, находящихся в стадии угасания. Самая
быстрота зарастания подобных водоемов (иногда немногие
десятилетия) исключает влияние климатических колебаний, по 11 Последний как раз падает на этот промежуток времени.
100
крайней мере — длительных. Кроме того, следует отметить,
что не все водоемы могут заболачиваться, даже если они
находятся в подходящих климатических условиях.
Быстрота течения и зарастание. Быстрое течение препят¬
ствует зарастанию водоемов. Всем известно, что реки зара¬
стают гораздо медленнее, чем пруды и озера. Притом при
зарастании рек дело обычно не доходит до образования бо¬
лота. Случай превращения русла реки в болото, наблюдав¬
шийся мной в б. Богородском у. Московской губернии
(Н. Кац, 1922), несомненно, весьма редкий. Лишь только в том
случае, когда участок живого русла отрезается от реки и пре¬
вращается в стоячий водоем — старицу, он начинает быстро
заболачиваться.
Разные растения неодинаково приспособлены к существо¬
ванию в быстро текущей воде. На деревянных мельничных
водосливах, где вода падает со скоростью водопада, высшая
растительность отсутствует. Здесь селятся лишь более при¬
способленные к быстрому течению водоросли и мхи. То же
наблюдается и в водопадах (Г. Вальтер, 1. с.). Вообще в быстро
текущих ручьях обычны ассоциации нереид — мхов и водорос¬
лей, прикрепляющихся к твердому субстрату ризоидами. Так
как в таких ручьях каменистое дно не дает укрепиться мхам
и корням цветковых растений, требующим рыхлого грунта,
здесь берут перевес плотно прирастающие к твердому суб¬
страту нереиды: водоросли—Cyanophyceae, диатомеи, неко¬
торые зеленые (Cladophora, Vaucheria), отчасти багрянки
и, наконец, некоторые мхи (Fontinalis, Cinclidotus) (Г. Вальтер).
При еще большей силе течения, когда дно ручья образовано
подвижным щебнем, водная растительность вообще развивается
слабо ввиду механического повреждения растений щебнем, пе¬
редвигаемым водой. Лишь когда сила течения допускает
отложение песка, поселяется высшая водная растительность-
виды рдестов, водных лютиков и т. д. Таким образом, сила
течения действует на распределение растительности не только
непосредственно, но и косвенно через характер субстрата. Сила
течения сказывается и на зарастании реки.
П. Никитин (1925) отмечает слабое развитие водной и прибрежно¬
водной растительности реки Дона в средней части ЦЧО и связывает это
слабое развитие с многоводностью реки, большой скоростью течения, мощ¬
ным половодьем и открытостью ее ветрам. На реке же Воронеж, где эти
факторы действуют лишь в слабой степени, водная растительность (по тому
же автору) развита чрезвычайно мощно. В дельте Волги, как мне пришлось
лично убедиться в 1936 г., большие многоводные протоки с сильным тече¬
нием почти лишены водной растительности и окаймлены лишь мощной сте¬
ной тростника, который после спада весенней воды оказывается на сухом бе¬
регу. Берега же менее мощных протоков с слабым течением окаймлены часто
почти непрерывной лентой зарослей плавающих растений, главным образом
Limnanthemum nymphaeoides.
Действие ветра и волн на зарастание. В областях, где не
бывает особенно сильных, продолжительных и постоянных вет¬
ров, ветры влияют на зарастание водоемов главным образом
косвенно, вызывая волнение, препятствующее зарастанию.
101
Если ветры, даже сами по себе не сильные, дуют преимуще-
ственно в одном направлении, то зарастание водоема проис¬
ходит главным образом с подветреной стороны. Противо¬
положный наветреный берег не зарастает или даже эроди¬
руется.
В Прибалтике, где господствуют северо-западные ветры, зарастают обычно
северо-западные берега, юго-восточные же, куда ударяют ветер и волны,
бывают лишены водной растительности. Эта закономерность носит название
„закона Клинге* (1890). Она
наблюдается и дальше на
восток в средней части'
Союза, а также и в Герма¬
нии (Г. Вальтер, 1927).
На рис. 11 изображена
схема зарастания озера по
Клинге. Кроме северо-за¬
падного, заболачивается и
западный берег. Здесь забо¬
лачивание вызывается впа¬
дающей речкой. Понятно,что
волны и ветер развивают
достаточную силу только на
более или менее крупных
И открытых озерах, где и
бывает выражен закон Клин¬
ге. На малых же и мелких
озерках зарастание не обна¬
руживает подобной законо¬
мерности. В частности не¬
большие вторичные озерки,
обычные на выпуклых тор¬
фяниках, зарастают с бере¬
гов, обращенных вверх по
склону болота, более или
менее независимо от их
ориентировки по странам
света. Нижние же берега
этих озерков не зарастают
и даже эродируются (см.
ниже). Все, что уменьшает
силу ветра н волн, благо¬
приятствует зарастанию.
Так, постройка купален на
Белом озере (из группы Косинских озер), создав защиту для водно-болот¬
ной растительности от ветров, способствовала ее развитию. Наоборот, раз¬
рушение купальни оказалось неблагоприятным для зарастания (Т. Мень-
кель-Шапова, 1930).
Рис. 11. Влияние господствующих ветров на
зарастание водоема (закон Клинге). По
К. Гюку, 1929.
1 — лес; 2 — ассоциации пушицы; 3 — сплавина; 4 — ас¬
социации мочежин; 5 — водяные лилии.
Типы заболачивания.
Лимнологические типы озер, их растительность и зара¬
стание. А. Тинеманн (А. ТЫпешапп, 1921, 1923) выделяет следую¬
щие лимнологические типы озер1:
1) эвтрофные озера,
2) олиготрофные озера,
3) дистрофные озера.
1 Помещенная ниже характеристика типа озер приведена по Г. Вальтеру
(1927).
102
Эвтрофные озера, богатые питательными веществами, имеют
богатое водное население и богатый количественно планктон.
Последний делает воду несколько мутной и зеленоватой. Оли-
готрофные озера бедны питанием. Живое население здесь го¬
раздо беднее. Вода прозрачная, голубая. Дистрофные водоемы
бедны солями вообще и всегда бедны известью. Поэтому гумус
не выпадает из воды, а остается в ней в коллоидальном со¬
стоянии, сообщая воде цвет крепкого чая. Вода кислой реакции,
что зависит от растворенных гуминовых кислот. Три указан¬
ных типа основаны на общем характере биологии водоема.
Естественно, что высшая растительность этих трех типов озер
неодинакова. Г. Самюэльссон (G. Samuelsson, 1925) исследовал
высшую растительность 81 озера в Dalarne (Швеция). Он раз¬
личает следующие четыре типа озер: 1) Dyseen, 2) Lobelia-Seen,
3) Lagunen-Seen и 4) Potamogeton-Seen.
Первый тип соответствует эвтрофным озерам. Берега этих
озер поросли Equisetum, Phragmites, Carex lasiocarpa, Menyan-
thes, иногда Calla, Cicuta и др. Большей частью здесь обычны
Potamogetón natans, Nymphaea candida, Nuphar luteum и некото¬
рые другие. Погруженные растения с укороченными стеблями
(типа Isoetes) почти вовсе отсутствуют. Нехарактерны и погру¬
женные растения с длинными стеблями. е
Lobpl ia-See п (озера с лобелией). Для этих озер осо¬
бенно характерны погруженные растения с укороченным сте¬
блем— два вида Isoetes (особенно много I. lacustris), Lobelia
(часто массами) и некоторые другие виды. В этих озерах почти
всегда обильны растения с плавающими листьями — Nymphaea,
Nuphar, Potamogetón natans. Береговые заросли образованы
хвощом, тростником и камышом. Из погруженных растений
всюду Myriophyllum alternifolium, из болотных типичны Alisma,
Scirpus palustris, Carex lasiocarpa, C. rostrata, Menyanthes. Эти
озера относятся к олиготрофному типу Тинеманна.
Lagunen-Seen (лагунные озера). Это мелкие озера с пес¬
чано-иловатым дном, сильно заливаемые полыми водами,
с сильно колеблющимся уровнем, с довольно прозрачной водой.
Обычны погруженные растения с укороченным стеблем (глав¬
ным образом Isoetes echinospora, Scirpus acicularis, Subularia
и др.). Однако Lobelia и Isoetes lacustris, характерные для озер
типа Lobelia, или отсутствуют, или встречаются в небольшом
количестве. Характерны Myriophyllum alternifolium, Potamogetón
perfoliatus, иногда Elodea. Отношение лагунного типа к озерным
типам лимнологов неясно.
Potamogeton-Sееп (рдестовые озера) имеют мощные
•отложения гиттии. Здесь наблюдается цветение воды и наибо¬
лее сильно развита болотно-водная растительность: густые
прибрежные заросли тростника, камыша и хвоща, заросли
Sparganium simplex и Sagittaria. Nuphar luteum образует часто
непрерывный пояс впереди тростников. Здесь массами растут
Potamogetón natans, Nymphaea candida, а местами Polygonum
amphibium. Большей частью обильны и погруженные рдесты,
103
Ceratophyllum, Utricularia vulgaris. Погруженные короткосте¬
бельные растения (кроме Scirpus acicularis) в типичных озерах
отсутствуют вовсе, вероятно, вследствие конкуренции с более
высокорослыми растениями. Эти озера принадлежат к эвтроф-
ному типу лимнологов.
А. Донат (Donat, 1926) устанавливает сходные для Германии типы озер.
Так, тип Lobelia-lsoetes близок к типу Lobelia Самюэльссона. Сюда отно¬
сятся также олиготрофные озера. К ним принадлежат и глубокие альпийские
озера, а в Германии также озера бесплодных песчаных местностей.
Подобные озера атлантических пустошей (Heideseen des atlantischen Gebie-
tes), по Г. Вальтер (1927), характеризуются бедной солями, слабокислой водой.
Растительность бедная: Litorella uniflora, Lobelia Dortmanna, Isoetes lacustrls,
а кроме того, еще Subularia aquatica, Myriophyllum alternifolium и др. Все эти
озера можно назвать физиологически олиготрофными. Береговая раститель¬
ность развита здесь слабо, литоральная водная растительность также весьма
умеренно. Планктон также беден количественно. Зарастание подобных водо¬
емов, повидимому, идет медленно. Эвтрофные озера с плоскими берегами имеют
богатую водную растительность и обильный планктон. То и другое связано
с богатством солей в воде. В этих озерах хорошо выражены глубинные
пояса растительности: пояс тростников и Scirpus, пояс Nymphaea, пояс рде¬
стов и, наконец, пояс хар. Эти озера можно отнести к типу Pbragmites —
Potamogetón. Они близки к Potamogeton-Seen Г. Самюэльссона. Этот тип
характерен для северогерманских озер и, особенно, для балтийской низмен¬
ности. Эвтрофные озера этого типа по всем признакам энергично зарастают.
Другой характер носят эвтрофные водоемы с крутыми берегами. Несмотря
на значительный запас солей, они неблагоприятны для развития высшей ра¬
стительности благодаря крутизне берегов. Видовой состав иногда близок
к эвтрофным озерам с плоскими берегами, и некоторые из этих озер можно
причислить к типу Phragmites-Potamogeton. Однако количественно эта ра¬
стительность развита слабо; растительные зоны вследствие крутизны берегов
узки. Вследствие слабого развития цветковой растительности эти озера бедны
фосфором и калием. Зарастают они слабо. Несмотря на эвтрофную раститель¬
ность, эти озера, видимо, относятся к олиготрофному типу лимнологов. В про¬
тивоположность физиологически олиготрофным озерам это физически олиго¬
трофные водоемы.
Зарастание в разных климатических зонах. Несмотря на недостаточ¬
ную изученность растительности водоемов тундры, можно сказать, что она
скудна, слабо развита. Видовой состав водной флоры тундровой зоны, в общем,
беден и однообразен. Б. Городков (1935) считает причинами слабого развития
высшей растительности в водоемах северной тундры низкую температуру
воды и краткость вегетации. Лишь на юге зоны, в мелких и лучше прогре¬
ваемых водоемах встречаются заросли из немногих видов: Ranunculus hypcr-
boreus, Hippuris vulgaris и Potamogetón alpinus. В лесотундре зарастание на¬
блюдается чаще, и видовой состав водной флоры богаче: кроме названных
видов, еще Ranunculus Gmelini. Nuphar pumilum, Nymphaea tetragona, Pota¬
mogetón natans, Sparganium hyperboreum и немногие другие. Подобно тунд¬
ровым и озера альпийской ступени на севере Скандинавии, как правило,,
вовсе лишены водной растительности (Т. Фриз—Th. Fries, 1913). В субальпий¬
ской области северной Швеции водная растительность уже богаче. Здесь
образуют заросли Alopecurus fulvus f. natans, Sparganum submuticum, Batracnium
peltatum, Ranunculus hyperboreus (изредка), Potamogetón gramineus и P. alpinus.
По берегам субальпийских водоемов нередко кайма Carex aquatilis впере¬
межку с зарослями Equisetum fluviatile, а впереди этого пояса заросли Hip¬
puris vulgaris. Таким образом, многие растительные виды тундр Союза встре¬
чаются и здесь.
Водоемы севера и особенно северо-востока Колы (Ю. Цинзерлинг, 1934)
также бедны водной флорой. Здесь встречаются: Caiex nmpullacea, Carex
aquatilis, Carex gracilis, Hippuris vulgaris, Callitriche, Equisetum limosum, виды
Potamogetón (редко), Arctophila fulva, водная форма Alopecurus fulvus, Spar¬
ganium submuticum; последние три вида лишь на севере. Значительное боль¬
шинство этих видов свойственно и тундрам вообще, а также и горным озерам
104
Северной Швеции. На северном Канине зарастание водоемов описывает
В. Андреев (1931). Большие озера зарастают здесь Сагех aquatilis, а местам»
Equisetum Heleocharis limosum. В зарастании малых озер в крупнобугристой
тундре участвуют, наряду с цветковыми, также и мхи — сфагновые и гип-
новые. Зарастание сфагнумом бывает или глубинное (Spli. riparium, Sph. obtu-
sum), или сфагнум (Sph. Lindbergh) образует ковер на воде. С сфагнами на¬
двигается и Сагех aquatilis. При зарастании водоемов гипновыми мхами уча¬
ствуют и цветковые (главным образом та же Сагех aquatilis). В этом случае
впереди идут обычно подводные заросли Hippuris vulgaris maritima с примесью
гипновых мхов—Drepanocladus exannulatus, Dr. aduncus. По мере повышения
дна надвигается Сагех aquatilis уже с другими мхами. Эти мхи, образуя
сплошной ковер, сменяют друг друга в таком порядке: Calliergon cordi-
folium, С. cordifolium и Drepanocladus uncinnatus, Paludella squarrosa. Стадия за¬
рослей Сагех aquatilis c Paludella характеризует уже болото с мощным торфом-
Дальнейшее накопление торфа на зарастающем водоеме приводит к вытесне¬
нию Сагех aquatilis менее влаголюбивыми осоками—сначала Сагех rariflora,,
а потом С. rotundata и к образованию политрихового ковра. На Колгуеве
в зарастающих озерах сначала появляется Hippuris, затем Сагех aquatilis
с гипновыми мхами и т. д. (И. Перфильев, 1928). Весь процесс зарастания
напоминает только что описанный. В лесной зоне количество видов, уча¬
ствующих в зарастании, сильно увеличивается, так же как количество цено¬
зов и количество глубинных поясов растительности.
Схема зарастания В. Сукачева (1926) более или менее типична для хвой¬
ной лесной полосы Европы. В этой зоне появляется целый ряд новых по
сравнению с тундровой зоной видов, в том числе и среди господствующих
в зарастании. Так, в Фенноскандии, по данным Г. Самюэльссона (1934), на¬
ходят свою северную границу между прочим Sagittaria natans, Scolochloa festu-
cacea, Nymphaea alba, Trapa natans, Limnanthemum nymphaeoides, Potamo¬
getón crispus, Acorus Calamus, Ranunculus circinnatus, Sium latifolium, Nastur¬
tium amphibium, Typha angustifolia, Oenanthe aquatica, Hottonia palustris (три
последних вида не переходят в северную Карелию и онежскую Карелию), Typha
Iatifolia, Elodea canadensis, Hydrocharis morsus ranae, Spirodela polyrrhiza, Spar*
ganium ramosum, S. glomeratum, Potamogetón lucens, Alisma Plantago — aqua¬
tica, Sagittaria sagittifolia, Butomus umbellatus, Stratiotes aloides, Scirpus la-
custer, Calla palustris, Lemna minor, L. trisulca, Iris pseudacorus, Nuphar luteum,
Ceratophyllum demersum, Myriophyllum verticillatum, Cicuta virosa и другие.
Большинство названных видов играет большую роль в зарастании водоемов
лесной зоны Европейской части СССР. Северные границы их здесь неизвест¬
ны, но можно уверенно сказать, что в зарастании водоемов тундры, по
крайней мере типичной, эти виды участия не принимают. Также и микро¬
флора в водоемах лесной полосы, несомненно, более обильна, чем в водоемах
тундры. Более богатая растительная жизнь приводит к более быстрому на¬
коплению в лесной зоне гиттии и торфа и к более интенсивному темпу за¬
растания, по сравнению с тундрой. Гиттии тундровых водоемов, повидимому»
недостаточно мощны (см. В. Алабышев, 1932), так же как и торфа. В лесостеп¬
ной и степной зонах Европейской части Союза в зарастающих водоемах по¬
являются растения, несвойственные лесной зоне, в частности третичные ре¬
ликты— Salvinia natans, CaJdesia parnassifolia, Aldrovanda vesiculosa, Trapa na¬
tans (данные для Украины E. Лавренко, 1927). С другой стороны, некоторые
северные виды здесь выпадают. Олиготрофные виды сфагновых болот севера,
из которых некоторые участвуют и в зарастании водоемов, в общем не до¬
ходят до степной зоны. В зарастании степных водоемов большую роль иг¬
рают Сагех stricta, Phragmites communis, Typha angustifolia и др. Два послед¬
них вида занимают громадные площади в дельтах южнорусских рек. Это
важнейшие компоненты плавней.
В дельте Волги, по моим наблюдениям (1936), громадную роль играют,
кроме тростника, Sparganium ramosum, Limnanthemum nymphaeoides, Trapa,
Najas marina, Potamogetón perfoliatus. Много здесь Butomus umbellatus, Salvi¬
nia natans. Изредка встречается и каспийская роза (Nelumbo caspica).
В лесостепи при зарастании сфагновые и гипновые мхи образуют спла¬
вины и отлагают гиттии как минеральные, так и органические (см. Е. Лав¬
ренко, 1927а и 1927b). Водные отложения здесь часто прикрыты сверху
105
осоковыми, гипновымн, а также и сфагновыми торфами. В степных водоемах
мхи уже не заметны, а седименты сильно минерализованы. Они типа солоно¬
вато-водной гиттии или минеральной грязи (В. Алабышев, 1932).
Для полупустынь характерны засоленные грунты и засушливый климат.
Здесь обычны пересыхающие соленые водоемы, связанные переходами 'с со¬
лончаками. В связи с этим органогенные отложения не имеют никакого значе¬
ния, так как растительные остатки подвергаются здесь более или менее нацело
разложению. Следовательно, роль растительности в зарастании незначительна,
несмотря на весьма мощное в ряде случаев ее развитие. Господствующий
тип седиментов здесь—это различные соли в самосадочных озерах (В. Ала¬
бышев, 1. с.). Видовой состав водной растительности беднее, чем в более се¬
верных широтах. Е. Коровин (1934) для растительности водоемов Средней
Азии отмечает бедность как количественную, так и видовую.
Ничтожное развитие водной растительности в реках он объясняет их
быстрым течением и большим количеством взвешенных частиц. Развитию ра¬
стительности в озерах мешает их быстрое осолонение. Из широко распро¬
страненных в Средней Азии и Казахстане видов Е. Коровин называет Ruppia
marítima, Zannichellia palustris, рдесты, Najas marina, Utricularia vulgaris, Sal-
vi nia natans, Myriophyllum spicatum, Cerataphyllum demersum и др. Водных
.растений, свойственных лишь водоемам равнины Средней Азии, не имеется.
Севернее, в Казахстане, водная флора уже богаче и приближается к более
умеренным широтам. Так, Н. Павлов (1928) приводит для центрального Ка¬
захстана как повсеместные виды, обычные и на севере, Potamogetón natans,
Р. perfoliatus, Р. lucens (из обычных у нас рдестов Р. praelongus и Р. crispus
здесь редки), Butomus umbellatus, Allsma Plantago, Sagittaria sagittlfolia, Najas
marina, Typha angustifolia, Phragmites и др. Из обычных наших видов не
заходят в Центральный Казахстан или имеют здесь южную границу: Equlse-
tum Heleocharis, Hydrochaiis morsus ranae, Stratiotes aloides.
Типы заболачивания водоемов. Заболачивание водоемов
происходит различными путями. Необходимо различать:
1) зарастание водоемов,
2) нарастание,
3) механическое заполнение.
1. Зарастание — процесс, связанный с поселением на дне
водоема укореняющейся растительности, или погруженной,
или имеющей плавающие листья, или, наконец, возвышаю¬
щейся над водой. Зарастание состоит в надвигании укореняю¬
щейся растительности из периферических, более мелководных
участков водоема на его центральную часть, причем, как
говорилось выше, различные растительные пояса сменяют
друг друга в определенном порядке в пространстве и времени.
О зарастании было подробно сказано выше.
2. Нарастание — это процесс надвигания на водоем расти¬
тельного ковра, лежащего на воде и обычно связанного с бе¬
регом. Этот ковер называется „зыбуном“, или сплавиной.
Зыбуны (см. В. Сукачев) часто образуются растениями с длин¬
ными корневищами, горизонтально стелющимися по воде, —
Calla, Menyanthes, Comarum. Эти корневища образуют основу
сплавины. На ней затем поселяются другие растения — цвет¬
ковые и мхи. Их остатки заполняют промежутки между кор¬
невищами и придают сплавине крепость. Одновременно спла¬
вина растет и в толщину и по площади—к центру водоема.
От нижней поверхности сплавины отрываются частицы торфа,
которые оседают на дно и способствуют обмелению водоема.
При понижении уровня озера сплавина, оседая вниз, оказывает
106
давление на лежащую под ней гиттию и выжимает ее в цен¬
тральную часть озера, способствуя, таким образом, его запол¬
нению.
Нередко образование зыбуна начинают не цветковые рас¬
тения, а мхи — Bryales (чаще виды Drepanocladus или Spha-
gnales). На таком еще не прочном зыбуне селятся позднее
цветковые растения, скрепляющие зыбун своими корневищами.
На сфагновых зыбунах чаще поселяется шейхцерия, Сагех
limosa. На гипновых зыбунах видовой состав цветковой флоры
богаче. Сфагновые сплавины образуются обычно в более бед¬
ных солями, олиготрофных водоемах, гипновые же зыбуны —
в более минерализованной воде. В природе весьма часто
наблюдается на одном и том же водоеме комбинированное
заболачивание — путем зарастания и нарастания. Пре¬
обладание того или иного процесса зависит от [физико-геогра¬
фических условий водоема. Процесс нарастания в противопо¬
ложность зарастанию более или менее не зависит от глубины
водоема.
Любопытный пример нарастания сплавины на очень глубокий водоем
описывает К. Скоттсберг (С. Scottsberg). Озеро в кратере вулкана на остро¬
вах Пасхи имело поперечник в 600—700 м при глубине, вероятно, не менее
100 jk (!). В настоящее время оно покрыто толстым слоем торфа, выдержи¬
вающим человека. Лишь окна свободной воды указывают, что здесь раньше
была водная поверхность. Теперь же поверх воды — мощный моховой зыбун,
образованный Campylopus turflcola.
Образование сплавины мало зависит от глубины, зато
в сильной степени — от ветра и волнения. Зыбуны, особенно
моховые, развиваются преимущественно в защищенных от
ветра водоемах. Сильное волнение может разрушить сплавину
и нередко отрывает от нее куски. Эти плавающие острова
держатся на воде вместе с покрывающей их растительностью,
в том числе и древесной. Они могут прибиваться к берегу
и там закрепляться.
Третий способ заболачивания водоемов — механическое
заполнение. В. Ауэр (V. Auer, 1927а, 1933) называет его „me¬
chanische Verwachsung“, или „mechanical Alling“. Этим путем
заболачиваются вторичные озерки с торфяным дном и берегами,
лежащие среди сфагновых болот. Этот тип заболачивания по
В. Ауэру широко распространен в Северной Европе (Лаплан¬
дии) и, вероятно, также и в Канаде. Растительность при ме¬
ханическом заполнении водоема, по крайней мере сначала, не
играет никакой роли. Главная роль в процессе принадлежит
эродирующей деятельности воды. Она размывает торфяные
берега водоема и откладывает на его дне измельченный
(„намывной") торф. При этом площадь озера растет прямо на
глазах местных жителей (лопарей в финской Лапландии).
Это увеличение отмечается на протяжении немногих десяти¬
летий (В. Ауэр, 1927). Одновременно вследствие отложения
торфяного детрита на дне озеро мелеет. Таким образом,
маленький, но сравнительно глубокий водоем превращается
в более обширный, но мелкий (рис. 12).
107
Обмеление водоема подготовляет его заселение осоковой
растительностью или сфагнумом и окончательное превращение
в болото. Процесс, аналогичный описанному, неоднократно
приходилось наблюдать и мне на озерах среди сфагновых
болот в средней части Союза. Эти озера, если они не слиш-
/ Стадия
Рис. 12. Две стадии механического заполнения
озера в финской Лапландии. По В. Ауэру, 1927.
ком малы и дают простор ветру и волнам, как правило,
имеют сильно эродируемые торфяные берега и, возможно,
увеличивают свою площадь. В случаях измерения глубины даже
наиболее значительные из этих озер оказывались весьма мел¬
кими, иногда не более 1—2 м. Дно таких озер зачастую по¬
крыто мощным слоем торфяного ила. Зарастания таких озер
обычно не наблюдается. Это зависит от характера дна, небла-
Рис. 13. Время заболачивания первичных озер и образования вторичных
на торфяниках Лапландии, датированное на пыльце. По В. Ауэру, 1927.
гоприятного для укоренения растений, а также отчасти от
кислой реакции воды и бедности ее солями. Развитию же
сфагновых зыбунов мешает ветер и волнение.
Заболачивание водоемов прежде и теперь.
По затрагиваемому здесь вопросу весьма мало сведений. Тем
не менее, ввиду особого интереса, который он представляет,
целесообразно привести и то немногое, что имеется. В. Ауэр
(1927а) отчетливо устанавливает разницу между характером
зарастания озер в настоящее время и в более раннюю эпоху
для севера Фенноскандии. В настоящее время в Лапландии на
108
/
¡1
Сфтобый торф
р
соснового леса
березового леса
Дт
Торф из тростника
оШйшт
Гиттия
Песок
Сфагновый торф
Внизу с линзами
пушицевого торфа
¡орф болотистого
сосновоголеса
Р
1орф болотистого
березового леса
ОлыианникобыО торф
береговой торф слан
зам и осокоВого торта и
торфа из тростника и
СТаФит
Песок
Ш Субатлантическое зарастание
Р- граница еловой пыльцы
Рис. 14. Типы зарастания водоемов в различные периоды послеледниковой эпохи. По Л. Посту, 1926.
вторичных озерах торфяных болот господствует описанный
процесс механического заполнения, проходящий без участия
растительности. Соответственно этому дно таких озер обра¬
зует не гиттия, а намывной торф. Образование этих озер
и их механическое заполнение могут быть датированы по
пыльце (рис. 13). Оно произошло сравнительно недавно, когда
в Лапландии впервые появилась ель и когда пыльца сосны,
достигнувшая в предшествующий период своего максимума,
начала уже убывать, а пыльца березы увеличиваться. Это
произошло, примерно, 2500 лет назад.
Древние озера в Лапландии, в настоящее время уже исчез¬
нувшие и погребенные торфом, зарастали иначе. В типичных
случаях на дне таких погребенных озер залегает гиттия, над
нейдю, затем хвощевый и, наконец, осоковый торф (рис. 13).
Таким образом, эти озера в прошлом заболачивались благо¬
даря зарастанию их укореняющейся раститель¬
ностью (осока, хвощ). В настоящее время зарастание вто¬
ричных озер укореняющейся растительностью наблюдается
(по Ауэру) чрезвычайно редко.
В. Ауэр (1927) для юго-восточной Канады устанавливает,
что при заболачивании озер в более ранний послеледниковый
период преобладающая роль принадлежала бурым мхам (Нур-
пасеае), а в настоящее время — сфагнумам. В прежнее же
время сфагновое заболачивание почти не имело значения.
Подобное же явление В. Ауэр (1933) отмечает и для Огнен¬
ной Земли. Хотя в общем зарастание озер в прошлом здесь
шло по типу современного, однако есть и различия. С одной
стороны, прежние извержения вулканов способствовали обра¬
зованию вторичных озер, заболачивание которых шло несколько
иначе. С другой стороны, прежде большую роль при зараста¬
нии озер играли, так же как и в Канаде, бурые мхи и, кроме
того, осоки. В области летнезеленых лесов Огненной Земли
особенно резко выступает увеличение роли сфагнума в зара¬
стании озер в позднейшее время, тогда как раньше на мощных
отложениях гиттии поселялась богатая цветковая раститель¬
ность — Hippuris, виды Carex, Marsippospermum и бурые мхи.
Вообще, по мнению Ауэра (1933), флористический состав вод¬
ной растительности прежде был богаче, чем теперь. Так на юго-
востоке Канады в зарастании озер принимали участие разнооб¬
разные виды: Typha, Scirpus, Decodon, Najas, Ceratophyllum, My-
riophvllum и др. Это обстоятельство указывает на более благо¬
приятные климатические условия в прежнее время. Л. Пост (1926)
уже 25 лет назад сделал замечательную попытку установить
типы зарастания озер в южной Швеции в разные климатические
периоды послеледниковой эпохи. Он выделяет три типа смены
седиментов и торфов при зарастании водоемов (рис. 14).
Объясняются эти типы зарастания водным режимом озер,
периодическими колебаниями их уровня. В разные климатиче¬
ские периоды этот режим был неодинаковым.
ГЛАВА IV.
ЗАБОЛАЧИВАНИЕ СУШИ.
Роль физико-географических факторов и растительности.
Заболачивание суши—сложный процесс, возникший в на¬
чале послеледниковой истории и продолжающийся до наших
дней. Несмотря на связанность процессов заболачивания в про¬
шлом и настоящем, характер и темп заболачивания были
неодинаковыми в разные моменты послеледниковой истории.
Не вполне одинаковой была и смена растительности. Поэтому
в интересах изложения полезно в известной мере разделить
вопросы заболачивания в настоящем и прошлом, тем более
что по заболачиванию в прошлом за последние годы нако¬
пился весьма интересный фактический материал.
Заболачивание суши в прошлом, когда образовывались со¬
временные торфяники, происходило в грандиозном масштабе.
Этот процесс, в общем, был гораздо более распространен, чем
заболачивание водоемов. К этому выводу приводит изучение
строения торфяников. Приведем лишь некоторые цифры. По
исследованиям А. Бакмана (A. Backman, 1919), в одной из
финских провинций 95°/« современной площади болот было
занято раньше лесами (по В. Ауэру, 1927). Несомненно,
что и в хвойной зоне Европейской части СССР торфяники
сухопутного происхождения далеко превосходят по площади
торфяники, возникшие на месте водоемов. Дальше к югу
роль водного заболачивания возрастает. Во всяком случае,
если принять во внимание громадную площадь торфяников
в СССР, то грандиозная роль заболачивания суши в прошлом
станет совершенно очевидной.
Заболачивание суши может происходить или в связи
с существующими болотами, озерами и реками, или вне связи
с ними. Рассмотрим сначала первые случаи.
Заболачивание в связи с уже существующими болотами
подробно описано А. Каяндером (1913). Он устанавливает
здесь разные случаи в зависимости от характера почв. Все
же они в целом объясняются одной общей гидрологической
причиной—способностью торфяников при нарастании повышать
ill
уровень своих вод в торфе. Если торфяник окружен легко
водопроницаемыми (например песчаными) почвами, то повы¬
шение уровня воды в торфянике вызывает подъем грунто¬
вых вод в прилегающей к болоту минеральной почве; он
сказывается, прежде всего, в непосредственной близости
к болоту, но захватывает иногда больший радиус. Если окру¬
жающая болото местность ровная и низменная, и сток
затруднен, то заболачивание может итти весьма быстро.
'С подъемом грунтовой воды почва делается постоянно сырой,
появляются сфагновые кочки, смыкающиеся затем в сплошной
ковер, погребающий прежнюю растительность. Так заболачи¬
ваются леса верескового типа на бедных песчаных водопро¬
ницаемых почвах.
Если почвы водонепроницаемы,заболачивание идет по-иному.
В этом случае особое значение приобретает рельеф. При зна¬
чительной расчлененности рельефа в Финляндии болота часто
располагаются выше окружающей местности. Если такое бо¬
лото целиком заполнило котловину до ее краев, то перепол¬
няющая торфяник вода начинает стекать вниз по склону.
Дальнейшее заболачивание склона в большой степени зависит
от его крутизны. По крутому склону вода стекает быстро,
ручейками, и заболачивания не происходит. Пологий и с запа¬
дниками склон при водонепроницаемой почве заболачивается,
так как здесь застаивается вода. Если склон имеет террасовид¬
ные уступы, то особенно легко заболачиваются террасы.
Заболачивание усиливается, если стекающая с болота вода,
уходя вглубь, просачивается вновь на поверхность склона,
постоянно и равномерно его увлажняя. Таким путем забола¬
чиваются даже крутые склоны. В результате этого процесса
длинный склон, разделяющий нередко два расположенных на
разных уровнях торфяника, оказывается сплошь заболочен¬
ным, и соседние болота сливаются.
Наблюдаются случаи и комбинированного заболачивания
под действием воды, стекающей поверхностно и одновременно
просачивающейся снизу грунтовой. В этих случаях сравни¬
тельно водопроницаемая почва допускает все же заболачива¬
ние, если весной и осенью она бывает обильно пропитана
водой, стекающей с выше расположенного болота. Много
интересных сведений по заболачиванию сообщает и Л. Аарио
<1932).
Заболачивание прибрежной полосы озера не следует сме¬
шивать с зарастанием самого водоема. Это заболачивание наблю¬
дается, когда озеро в силу различных причин повышает уро¬
вень на длительный период и затопляет берега, причем
последние заболачиваются. Это повышение уровня может
возникнуть, когда перестает действовать сток из озера
вследствие его закупорки торфяными отложениями (рис. 15).
В других случаях трансгрессия озера вызывается зарастанием
протока водно-болотной растительностью. Аарио (1932) отме¬
чает это явление при зарастании озер в Финляндии. Часто
112
бывает, что одновременно с выступлением озера из берегов
происходит отложение гиттии на дне, и водоем мелеет. Это
обмеление способствует зарастанию озера укореняющейся
растительностью. Таким образом, болото растет сразу с двух
сторон: заболачиваются берега и зарастает водное зеркало.
Надо добавить, что увели¬
чение поверхности озера
усиливает расход воды ис¬
парением. Это также уско¬
ряет зарастание водоема.
Трансгрессия озера не
всегда сопровождается за¬
болачиванием его берегов;
Так болото, прилегающее
к озеру Нурмиярви в Фин¬
ляндии, после трансгрессии
этого озера вовсе прекра¬
тило свой рост там, где
поверх сфагнового торфа
отложился слой гиттии.
Даже в местах болота, не
бывших под водой, рост
его почти прекратился (Л.
Аарио, 1933). Задержка
в развитии сфагновых болот
в результате поднятия уров¬
ня в соседнем озере—до¬
вольно обычное явление.
Особые условия наблюда¬
ются иногда во вторичных
озерах, окруженных сфаг¬
новыми болотами. В этом
случае отложение седимен-
тов на дне часто не влечет
за собой обмеления водо¬
ема и его трансгрессии, так
как уровень воды в озере
непрерывно повышается с
ростом окружающего бо¬
лота вверх (Л. Аарио, 1932).
Заболачивание речных
пойм зависит от двух фак¬
торов — от деятельности
реки и ключей, выходящих по склонам коренных берегов.
Здесь мы рассмотрим только заболачивание, зависящее
от деятельности самой реки. Поймы больших рек
с сильной аллювиальной деятельностью в общем мало под¬
вержены заболачиванию, так как этому препятствует большая
амплитуда колебаний водного уровня и интенсивная седимен¬
тация.
8—722
Рис. 15. Схема заболачивания береговой
полосы озера при выступании последнего
из берегов. По К. Мальмстрем, 1923.
I — профиль и план озера в исходной стадии;
II — то же после закупорки стока торфяными
отложениями.
113
Лишь там, где ослабевают сила течения и аллювиальный
процесс и где полые (или дождевые) воды надолго застаи¬
ваются и медленно спадают, создаются благоприятные усло¬
вия для заболачивания пойм. Эти условия следующие: 1) мед¬
ленность течения, 2) меандрирование русла, 3) низкие берега.
К этим особенностям поймы присоединяются внешние причины,
замедляющие течение реки, поднимающие ее уровень и вы¬
зывающие затопление. К ним принадлежат факторы как свя¬
занные с деятельностью человека (плотины, заколы для рыб¬
ной ловли), так и не связанные с ней (предметы, упавшие
в воду и перегородившие русло, как деревья и пр.). Извест¬
ную роль в заболачивании поймы может играть еще прирус¬
ловый вал: он служит как бы барьером, задерживающим
стенание полых вод в реку.
Некоторые случаи заболачивания заслуживают специального упоминания
(Г. Шлаффнер — Н. Schlaffner, 1921). Особый тип долинных болот возникает
у слияния двух рек благодаря тому, что одна река подпирает воду другой.
Это происходит или потому, что одна река имеет больший уклон, чем дру¬
гая, или вследствие непостоянного уровня воды в одной из рек. Заболачи¬
вание долины может вызывать просачивание в пойму вод, пропитывающих
песчаные прибрежные наносы (Infiltrationsmoore). Это просачивание происхо¬
дит тогда, когда ложе реки благодаря отложению приносимого ею материала
расположено выше прилегающей части поймы. Если просачивающиеся через
,прирусловые пески воды нодпруживает другая река, текущая параллельно пер¬
вой, опять-таки возникает заболачивание. Такого происхождения болота по
Дунаю у Нейбурга. Эти болота образовались просачиванием вод Дуная через
прирусловые аллювиальные наносы и поцпорой со стороны его альпийских
приюков.
Гидрологические условия заболачивания. Обильное увлаж-
некие почвы необходимое условие заболачивания и торфо-
образования. Это увлажнение имеет разные причины. Его
источниками могут быть различные стоячие водоемы, ручьи,
постоянный ток поверхностных вод, действие ключей или,
наконец, высокое стояние грунтовых вод, если оно оказы¬
вается длительным. К. Мальм стрем (1931) для двух пунктов
северной Швеции установил, что болотистые еловые леса
и болотные фитоценозы распространены лишь там, где глу¬
бина грунтовых вод в течение вегетационного
периода в среднем равна 10—30 см. Появление
сфагновых мхов и Polytrichum, предвестников заболачивания,
на границе между минеральной почвой и торфяником, по
этому автору, происходит лишь при немного более низком
уровне грунтовых вод. Обильное увлажнение почвы создает
фон заболачивания. Все остальные факторы заболачивания
(см. ниже) действуют лишь на этом фоне и сами по себе
обычно не имеют решающего значения.
Рельеф и заболачивание. Рельеф определяет совместно
с Почвой высоту стояния грунтовых вод, сток воды и дренаж.
В общем, чем выше расположена местность, тем глубже уро¬
вень грунтовых вод и тем менее благоприятны условия для
заболачивания. Низменный же рельеф с близким уровнем
грунтовых вод благоприятен для заболачивания. Едва ли не
114
большее значение для заболачивания имеет дренирую¬
щая способность рельефа. Легко заболачиваются такие фор¬
мы рельефа, где дренаж незначителен и сток воды затруднен,
особенно если это — низина с высоким стоянием грунто¬
вых вод. Эта комбинация условий — отсутствие стока и бли¬
зость грунтовых вод—осуществляется в бессточных котлови¬
нах рельефа. Эти котловины, особенно в условиях сравни¬
тельно континентального климата большей части Союза (см.
ниже „Климат и заболачивание“), являются преобладающим
местом болотообразования. Г. Шлаффнер (1920) также и для
Германии считает чашевидные формы рельефа наиболее бла¬
гоприятными для поселения здесь болотной растительности
и образования торфа. Эти котловины не всегда имеют посто¬
янное водное зеркало и вмещают озера. Они могут или вре¬
менно покрываться водой, или иметь лишь высокий уровень
грунтовых вод. Особенно большое значение для заболачива¬
ния имели в прошлом отрицательные формы рельефа, выра¬
ботанные ледником. В настоящее время здесь часто распола¬
гаются болота, возникшие после отступания ледника.
Если бессточные замкнутые формы рельефа стимулируют
заболачивание, то расчлененный рельеф с хорошо развитой
речной сетью и хорошим дренажем оказывается его антагони¬
стом. Необходимо иметь в виду, что, при оценке тех или иных
физико-географических факторов в заболачивании, мы встре¬
чаемся с явлением взаимной компенсации факторов.
Например, крутые склоны по условиям рельефа неблагоприятны
для заболачивания. Однако эти неблагоприятные условия
могут быть компенсированы или ключевым увлажнением, или
увлажнением поверхностными водами, или, наконец, климати¬
ческими условиями.
Роль растительности в заболачивании. Сфагнум может
впитывать большие количества капельножидкой воды. Эта
способность зависит от анатомического строения (присутствие
водоносных клеток) и его морфологии. Самая форма листоч¬
ков, их положение на веточках стебля (они черепитчато
покрывают друг друга), а также облегание стебля так называе¬
мыми поникшими веточками—все это создает систему капил¬
лярных ходов, по которым передвигается и удерживается
вода. Благодаря этому капиллярные свойства, зависящие от
особенностей клеточного строения, и влагоемкость сфагнума
возрастают.
В природе растеньица сфагнума почти всегда образуют
скопления, так называемые дернины. Соседние растеньица
в дернине тесно переплетаются друг с другом, и вся дернина
в целом представляет сложную систему капилляров, причем
влагоемкость дернины повышается по сравнению с влагоем-
костью отдельных растений, так как к капиллярности внутри¬
клеточной и капиллярности морфологической прибавляется
еще капиллярность групповая, зависящая от совместного роста
многих особей. При оценке роли сфагнума в заболачивании
8* 115
надо считаться именно с влагоемкостью всей дернины в целом.
Большая влагоемкость сфагновой дернины позволяет ожидать,
что сфагнум, поселяясь, например, в лесу, может вызвать
заболачивание. Этому как будто противоречит то обстоятель¬
ство, что сфагновая дернина не только поглощает много воды,
но и много ее испаряет (см. выше). Однако при этом нужно
еще учесть, что в лесу, особенно еловом, испарение с поверх¬
ности почвы сильно ослаблено (см. В. Сукачев, 1926). Кроме
того, сфагновая дернина высыхает обычно лишь сверху,
нижние слои и слой торфа под ней остаются обычно
влажными. Таким образом, возможно постепенное накопление
влаги в почве и дальнейшее заболачивание. Дренирующее
влияние леса ослабляется тем, что значительная часть атмо¬
сферной влаги поглощается моховой дерниной и не достигает
корней. Влияние других мхов, растущих дернинами, подобно
влиянию сфагновой дернины, но более слабое. Особенное
значение имеют мощные дернины кукушкина льна (Polytrichum
commune). Поселяясь в лесу, он нередко накапливает на
минеральной почве слой политрихового торфа. Аккумуляция
влаги в дернине и торфянистом горизонте происходит так же,
как и в случае поселения сфагнума. Кукушкин лен очень
часто бывает предвестником сфагнума. Подушки сфагнума
среди дернины кукушкина льна обычны в сырых лесах.
В дальнейшем эти подушки могут смыкаться; кукушкин лен
погребается сфагнумом, и заболачивание идет дальше.
Кроме кукушкина льна, накоплению влаги в почве способ*
ствуют и обычные лесные мхи, образующие дернины,—Pleuro-
zium Schreberi и Hylocomium splendens. В этих случаях сфаг¬
нум поселяется прямо среди этих мхов, и стадия кукушкина
льна выпадает. Роль мхов, подготовляющих условия для
сфагнового заболачивания, вероятно, не ограничивается
только накоплением почвенной влаги. Эти мхи образуют слой,
изолирующий сфагнум от минерализующего действия грун¬
товых вод.
Это имеет особое значение для поселения более олиготроф-
ных видов сфагнума (например Sph. recurvum s. 1.), которые
к тому же оказываются энергичными торфообразователями.
Часто заболачивание леса происходит за счет не сфагно¬
вых мхов, а травянистых растений—широколистных (Calla,
Menyanthes, Comarum и др.) и осок (Carex laevirostris, Сагех
vesicaria и др.). Этот тип заболачивания связан с более бога¬
той, чем сфагновый тип, а вместе с тем с более влажной
почвой. При этом типе заболачивания роль гидрологических
факторов больше, а самой растительности, по крайней мере
на первых стадиях, меньше, чем при сфагновом заболачивании.
Кроме того, при травянистом заболачивании роль живого
растительного покрова как собирателя влаги во всяком случае
меньше, чем при моховом заболачивании, а во многих слу¬
чаях и вовсе незначительна. Здесь особое значение в собира¬
нии влаги и в дальнейшем заболачивании приобретает именно
116
накопление мертвых растительных остатков (см. дальше),
которое в свою очередь требует уже с самого начала доста¬
точно влажной почвы. Эта влажность зависит как от близости
грунтовых вод, так и от скопления поверхностных вод при
водонепроницаемой подпочве. Однако иногда цветковая расти¬
тельность и при жизни все же может задерживать влагу и
способствовать заболачиванию (например плотнокустовые
злаки—Deschampsia caespitosa, нередко сильно разрастающиеся
в сырых вторичных березняках, северная Deschampsia flexuosa
и др.).
Мертвый растительный покров и заболачивание. Мертвый
растительный покров обладает большой влагоемкостью.
Накопляя почвенную влагу, вследствие капиллярности, он
способствует заболачиванию. Это действие мертвого покрова
проявляется при заболачивании как лесов (см. выше), так и
открытых пространств—лугов и пр.
Приведу яркий пример влияния мертвого покрова на раститель¬
ность луга из своей практики. Участок сырого луга с господством
Deschampsia caespitosa (щучка) в долине р. Яхромы (близ г. Дмитрова, Москов¬
ской области) в 1922 г. был огорожен, и на нем была прекращена пастьба
скота и покос. Окружающий щучковый луг выпасался и выкашивался,
как обычно. Уже через три года опыта огороженный участок резко изменил
свою физиономию — фон травостоя создавала здесь уже не щучка, а осока
(Carex gracilis) (Н. Я. Кац, 1926). При этом урожай осоки возрос по сравне¬
нию с 1922 г. почти в 17 раз, а количество стеблей почти в 9 раз. Почти
все злаки, наоборот, уменьшили свой урожай, в том числе и господствую¬
щая щучка — последняя более, чем в П/г раза. Эта замена луга болотом
произошла вследствие накопления мертвой подстилки, которая на огорожен¬
ном участке не удалялась ни сенокошением, ни пастьбой.
Теория болотообразовательного процесса В. Р. Виль¬
ямса (1931) в основном построена на представлении о постепенном нако¬
плении мертвой растительной массы. При ее накоплении изменяются условия
существования, и начинается смена одних типов растительности другими,
которые характеризуются определенными биологическими типами растений,
для которых существенным является характер подземных органов. Смена
растительности, ведущая в конце концов к образованию болота, проходит
по В. Вильямсу следующим образом. В дерново-подзолистой зоне на лесных
прогалинах цервая стадия луговой растительности — это стадия корневищ¬
ных злаков (вейники и др.). Более плотная, по сравнению с лесной, мертвая
подстилка из остатков этих злаков препятствует семенному возобновлению
леса. Она труднее разлагается, в ней создаются анаэробные условия, неблаго¬
приятные для корневищных злаков. Они сменяются следующей стадией
рыхлокустовых злаков (Dactylis glomerata, Роа nemoralis), для которой харак¬
терны бобовые. Последние снабжают азотом рыхлокустовые злаки, неспособ¬
ные усвоить органический азот. С дальнейшим накоплением мертвой под¬
стилки, увеличением гуминовых (креновых) кислот и анаэробных условий
рыхлокустовые злаки изживают себя, наступает голод, вызванный недостат¬
ком в почве усвояемых соединений фосфора, азота и серы. Наступает послед¬
няя стадия лугового периода—господство плотнокустовых злаков. Эта стадия
неизбежно сменяется болотом вследствие дальнейшего накопления мертвой
подстилки и сопутствующих изменений почвенных процессов. Стройная
картина естественного развития луговой растительности, нарисованная
В. Р. Вильямсом, неоднократно подвергалась критике. Не оспаривая ее в
целом, отметим только, что представлять этот процесс как чисто эндоди-
намический, зависящий от самой растительности, как это делает В. Р. Вильямс,
нельзя. Внешние факторы играют при этом громадную роль. Выше было
отмечено, что заболачивание идет лишь при определенных гидрологических
условиях, т. е. если почва в силу тех или иных причин уже достаточно
117
увлажнена. Далеко не всякий луг, следовательно, превра¬
щается в болото. В лесной же зоне, которую имеет в виду
В. Р. Вильямс, суходольный луг, предоставленный самому себе, чаще зара¬
стает лесом. Необходимо помнить, что суходольные луга в лесной зоне
сплошь вторичные и обязаны своим возникновением деятельности человека.
Следовательно, они не могут быть звеном в естественном процессе смены
фитоценозов, как это думает В. Р. Вильямс.
Заболачивание лесосек. Заболачиванием лесосек интересо¬
вался еще Г. Танфильев (1889). Он отметил роль кукушкина
льна при заболачивании вырубки из-под соснового леса.
Уже на второй год на такой вырубке сильно разрастается ку¬
кушкин лен; он заглушает лесные мхи и погребает бруснику.
Через три года среди его сплошного ковра появляется
сфагнум, а на лесосеках 18-летнего возраста, где появилась
сосна и ель, имеется уже много сфагнума, клюквы, Сотагит
и появился торфяной слой в несколько дюймов толщиной.
Дальнейшее заболачивание, по мнению Г. Танфильева, может
быть приостановлено лишь осушкой. В заболачивании лист¬
венного и елового леса, по Г. Танфильеву (1. с.), большую
роль играет Са1ап^гозй'з, развивающийся на второй год после
вырубки. Его листья, плотно прикрывая почву, способствуют
в дальнейшем поселению уже настоящих болотных расте¬
ний — Сотагит, Сагех атри11асеа и даже сфагнума—и нако¬
плению торфа. В общем, Г. Танфильев, повидимому, считает,
что главной причиной заболачивания лесосек оказывается
сама растительность.
B. Сукачев (1926), придавая вообще большое значение
гидрологическим факторам, считает, что заболачивание
лесосек начинается лишь в случае, если и первоначально
почва была влажной. Он отмечает и разницу в почвенных
условиях между лесом и лесосекой,—последняя подвергается
действию солнца и ветра и имеет иную растительность, что
сказывается и на ходе заболачивания. Многие авторы считают,
что основная причина заболачивания лесосек—подъем грун¬
товых вод после удаления леса, транспирация которого дей¬
ствует как мощный дренирующий фактор. Так Эбермайер
(ЕЬегтауег, 1920) указал, что лес осушает сырые местности,
которые после вырубки заболачиваются вновь, и связал это
с дренирующей ролью леса. П. Отоцкий (1905) установил,
что грунтовые воды под лесом стоят глубже, чем на безлес¬
ном пространстве при прочих равных условиях, а С. Яков¬
лев (1925) указал, что грунтовые воды под лесом падают
как раз во время вегетационного периода (время транспирации),
а с начала октября начинают подниматься, и что ход годич¬
ной кривой уровня грунтовых вод под лесом иной, чем вне
его. Также и В. Сукачев считает, что уничтожение леса
уменьшает испарение воды из почвы и поэтому вызывает
заболачивание. Растительность лишь содействует этому
процессу.
C. Архипов (1932) также считает, что заболачивание лесо¬
сек зависит от уничтожения естественного водного насоса—
118
леса, причем травянистая растительность содействует этому
процессу. Этот автор, кроме того, подчеркивает и значение
естественного возобновления при заболачивании лесосек, а
также значение величины лесосек и их концентрации. Увели¬
чение размеров лесосек и концентрация рубок, затрудняя
естественное возобновление, способствует тем самым забола¬
чиванию.
Специальные исследования над зависимостью заболачива¬
ния от возобновления леса на вырубках были произведены
в Ленинградской области В. Буренковым, А. Кащеевым и др.
(1934). Исследованы были лесосеки еловых, сосновых и елово¬
сосновых лесов с черникой и брусникой. Установлено, что
при вырубке этих лесов наблюдается заболачивание. Оно
происходит в этом случае за счет увлажнения почвы поверх¬
ностной водой — верховодкой — без участия грунтовых вод,
которые здесь стоят глубоко, ниже 1,5 м. Заболачивание
идет более или менее энергично в первые 15—20 лет, причем
молодое насаждение, вырастающее' на лесосеке, заметно
угнетается. Если возобновление достаточное, молодняк более
или менее сомкнут и равномерно распределен, то к 20—35 го¬
дам заболачивание ослабевает и даже вовсе затухает.
К 40—45 годам заболачивание оставляет следы лишь в лесной
подстилке, а к 50—60 годам такие следы остаются лишь
в почве. Затухание заболачивания зависит от дренирующего
действия леса на поверхностные слои почвы, а также и от
затенения и засыпания продуктами лесного отпада мхов и
травянистой растительности. На основании современных
сведений можно утверждать, что лесосеки могут заболачиваться
при соответствующих физико-географических условиях.
1. Заболачивание происходит: а) при высоком стоянии
грунтовых вод за счет их подъема вследствие удаления
леса или б) вследствие верховодки, которая поддерживает
верхний слой почвы и накопляющиеся растительные остатки
во влажном состоянии.
2. Растительный покров—мхи (особенно сфагновые) и
травянистые растения—могут лишь содействовать заболачи¬
ванию.
3. Возобновление леса на лесосеках задерживает заболачи¬
вание. Если это возобновление идет энергично, а обводнение
почвы невелико (например в случае верховодки), то заболачи¬
вание с течением времени затухает совершенно. Если вырубка
леса вызывает высокое стояние грунтовых вод, то процесс
заболачивания, повидимому, становится необратимым, и лес,
в конце концов, превращается в болото.
Заболачивание лесных пожарищ во многих отношениях
сходно с заболачиванием лесосек. Оно зависит в основном
тоже от повышения уровня грунтовых вод вследствие удале¬
ния леса. Это заболачивание лесных гарей может итти на¬
столько интенсивно, что уже на следующий год после пожара
появляются озера, иногда в несколько гектаров, как это
П9
наблюдал еще Докучаев (по В. Сукачеву). В этих случаях
появление озер есть результат прекращения дренирующего
влияния леса, так как за такой промежуток времени влияние
травянистой растительности на заболачивание не может еще
сказаться. Впоследствии, когда на гарях появляется травяни¬
стая растительность и мхи, они могут играть известную роль.
Г. Танфильев (1889) придает большое значение мхам — мар¬
шанции и Fuñaría hygrometrica. Талломы маршанции иногда
появляются на гарях в большом количестве. Они плотно
прижаты к земле и защищают ее от высыхания. Типичный
мох гарей Fuñaría обычно обильно плодоносит. Ножки густо
стоящих спорогониев этого мха хорошо задерживают дож¬
девые капли и передают их вниз моховым дернинкам.
Последние низки и плотно прилегают к земле в виде корочки,
препятствующей высыханию почвы. Поселяющаяся вслед за
мхами на гарях Deschampsia flexuosa также хорошо задержи¬
вает влагу в плотных дерновинках между щетиновидными
листьями. Г. Танфильев отмечает также роль дернин кукуш¬
кина льна и сфагнума в заболачивании пожарищ. По мнению
некоторых исследователей (А. Флеров, 1899) пожары оказы¬
вают еще специфическое воздействие на лесную почву, так
как при этом происходит сильное ее уплотнение, а образо¬
вавшаяся зола притягивает влагу. Н. Степанов (1925) устано¬
вил, что лесные пожары сильно увеличивают содержание в
почве солей щелочных и щелочноземельных металлов, особенно
сернокислых и углекислых, вследствие чего почва делается
щелочной. Кроме того, разложение алюмосиликатов под
влиянием пожара дает растворимые полуторные окислы же¬
леза и алюминия, которые делают почвы непроницаемыми
для воздуха и воды и к тому же весьма влагоемкими. Влагоем-
кость почвы увеличивается еще за счет изменений ее физиче¬
ских свойств. Щелочная реакция почвы и ее высокая влагоем-
кость неблагоприятны для поселения древесной раститель¬
ности (особенно хвойных, чувствительных к щелочной реакции)
на гарях, что, как мы знаем, может служить причиной забола¬
чивания. Водонепроницаемость почв гарей действует в том
же смысле.
Значение подобных изменений почвенных свойств пожарами оспаривает
В. Сукачев. Многие авторы придают большое значение в образовании болот
лесным пожарам. Так, швед Э. Хаглунд (Е. Haglund) считал, что образование
сфагновых болот в Швеции происходило в результате пожаров („теория
пожаров" — Brandtheorie). На это указывает нахождение в Швеции под
сфагновой толщей угля с мощными стволами деревьев. Эти деревья росли
в эвтрофном торфе, подстилающем сфагновый, и после пожара уменьшили
свой годичный прирост вследствие ухудшения условий существования.
Ухудшение роста было вызвано появлением сфагнума. Эти лесные пожары,
способствовавшие поселению сфагнума, по мнению Хаглуида, объясняются
широким распространением в прошлом огневой культуры (по В. Докту-
ровскому, 1932). Решающее значение придавал лесным палам в образовании
болот в Дальневосточном крае и А. Левицкий (1910). Он указывает, что
уничтожение леса огнем повышает уровень грунтовых вод, а также вызы¬
вает более глубокое промерзание почвы зимой. Оба момента благоприятны
120
для развития болот. Значения лесных палов в болотообразовании нельзя
отрицать, но и нельзя придавать этому фактору такое исключительное зна¬
чение, как это делает Хаглунд.
Смена фитоценозов,
Заболачивание лесов. Заболачивание лесов в настоящее
время происходит в связи с болотами и на их окраинах,,
(рис. 16). Ход заболачивания и смена растительности при этом
протекают различно в зависимости от следующих условий:
1) от состава древесного яруса,
2) от богатства почвы,
3) от степени увлажнения окраин,
4) от крутизны склона минерального берега к болоту.
Рис. 16. Карта растительности на заболачивающейся окраине торфяника,
переходной от леса к болоту. По Л. Аарио, 1932,
Состав древесного яруса заболачивающегося леса влияет
следующим образом. В хвойной полосе Европейской части СССР
при заболачивании сфагнумом сосняков смены пород обычно
не происходит: сосна продолжает оставаться господствующей
породой на всех стадиях заболачивания; она лишь постепенно
мельчает; форма сосны минеральной почвы уступает место
сначала Pinus silvestris uliginosa, затем уже типичной болот¬
ной угнетенной форме Р. silvestris fo Litwinowii. Таким обра¬
зом, сосновый бор переходит постепенно в сосново-сфагновое
болото (схема 4).
Заболачивание ельников происходит иначе. Ель, чувстви¬
тельная к сфагновому заболачиванию, резко ухудшает рост,
постепенно выпадает из состава древостоя и, не давая возоб¬
новления, заменяется березой. Стадия березового болота
121
Схема 4
Смена растительности при переходе от соснового леса с черникой
к сфагновому болоту
(укороченный тип заболачивания при меньшем количестве влаги (по Н. Кацу,
1927; несколько переработано)
Сосновый лес (иногда с елью) с господством черники и лесных НуЬсопиа
I
Заболоченный сосновый лес с господством сфагнума (Sph. recurvum, s. 1.
и др.) и черники
1
Сосново-сфагновое болото с господством черники и болотных вересковых
Сосново-кустарничковое сфагновое болото с господством болотных вереско¬
вых (Ledum, Cassandra)
Схема 5
Смена растительности при переходе от елового леса к сфагновому
болоту в средней части СССР
(удлиненный тип заболачивания при большем количестве почвенной влаги
и в условиях более богатой почвы) (по Н. Кацу, 1926, частью переработано)
Еловый лес на минеральной почве
I i
Заболоченный елово-березовый Заболоченный елово-березовый лес
лес с господством сфагнума и с господством сфагнума и Calla
Calamagrostis lanceolata j
I Березово-сфагновое болото с гос¬
подством вахты
Березово-сфагновое болото ^
с господством Сагех lasiocarpa <
Сосново-сфагновое болото с гос¬
подством Сагех laslocarpa
Сосново-сфагновое болото с бо¬
лотными вересковыми и пушицей
I — более обычный путь заболачивания
II — более редкий путь заболачивания
весьма характерна при заболачивании ельников (схема 5).
В некоторых случаях, когда к ели была примешана сосна,
при заболачивании ель пропадает, сосна же остается и по¬
степенно достигает господства. Березовая стадия при этом
122
Смена растительности при переходе от леса на более бедной почве к сфагновому болоту
(по И. Паасио, 1933; несколько переработано)
Схеме 6
Вересковый сосновый бор
III»
Заболоченный вересковый бор
с Сагех globtilaris
1 I
Заболоченный вересковый бор
с лесными Hylocomia, Polytrl-
chum commune исфагнами
1
U
Сосново-сфагновое болото
с ярусом различных болотных
вересковых
II
„ I 1
Сосново-вересковое сфагновое
болото
III
Брусничный сосновый бор
|
Заболоченный брусничный сосновый бор
с господством Polytrichum commune
i ♦
Заболоченный вересковый сосновый бор
с господством Polytrichum commune
III
Заболоченный лес с господством Polytrlchum
commune, с вереском и осоками между кочками
i
Фитоценозы мокрой окраины сфагнового болота
I—Заболачивание при крутом склоне минерального берега к болоту
II и III — заболачивание в случае пологого склона
II — заболачивание при меньшем количестве влаги
III —заболачивание при большем увлажнении
выпадает. Заболачивание ельников нередко идет при боль¬
шем увлажнении, чем сосняков, может быть потому, что
древесный ярус в этом случае более изрежен, и его роль
как насоса, удаляющего почвенную воду, меньше.
Обводненность окраин весьма сильно влияет на заболачи¬
вание леса. Она зависит от количества воды, притекающей
к окраине со стороны болота и с его берегов, затем от про¬
ницаемости почвы, т. е. насколько задерживается или про¬
сачивается вода (см. Л. Аарио, 1932). При обильном увлаж¬
нении окраин растительность проходит часто стадию безлес¬
ного мокрого осоково-сфагнового болота, как это часто
можно наблюдать на наших болотах. Сходное явление отме¬
чается и для Финляндии (схема 6, путь III и схема 7, путь II).
При меньшем увлажнении стадия безлесного болота выпадает,
и заболачивание с начала до конца проходит по лесному
типу (схема 6, путь II и схема 7, путь I). Крутизна склона
минерального берега имеет существенное значение (Л. Аарио,
1932; И. Паасио, 1933). При крутом склоне полоса заболачи¬
вания узкая, ряд сменяющих друг друга типов короткий
(схема 6, путь 1). При пологом склоне эта полоса широка
и число промежуточных звеньев велико (схема 6, путь Ни III).
При заболачивании богатых почв число промежуточных
звеньев также увеличивается, и наступление стадии сфагнового
олиготрофного болота отодвигается. При заболачивании
бедных почв наблюдается обратное.
В зависимости от комбинации указанных факторов можно
проследить разные пути заболачивания лесов. Они в общем
сходны для Финляндии и хвойной полосы Европейской части
СССР. При заболачивании вересковых боров на бедной почве
в Финляндии, если склон к болоту крутой, вересковый бор
сразу же переходит в сосново-сфагновое болото с тем же
вереском. При более пологом склоне выдвигаются обычно
промежуточные стадии—например сфагновое болото с раз¬
личными вересковыми, которое уже потом сменяется вереско¬
вым сфагновым болотом (схема 6). В средней полосе Европей¬
ской части СССР вересковые, а также черничные боры, быстро
переходят в сосново-сфагновое болото, но уже не с вереском, а
с другими Ericaceae (Ledum, Cassandra—схема 4). При несколько
более влажном типе заболачивания брусничных (а частью и
вересковых) боров часто характерно господство Polylrichum
commune. Конечная стадия может быть та же (схема 6).
На несколько более богатых почвах мы имеем леса с гос¬
подством черники, сосны и ели. Здесь (для Финляндии) ве¬
ресковое сфагновое болото уже не сменяет лес непосред¬
ственно, как это часто бывает при заболачивании бедных
почв (см. выше), а только через стадии лесного болота с
различными Ericaceae (кроме вереска) или с господством
кукушкина льна. Кроме того, здесь появляются новые про¬
межуточные стадии (например с Equisetum silvaticum). Особенно
же характерно участие березы, даже березовой стадии осоково-
124
Смена растительности при переходе от леса на более богатой почве
(по И. Паасио, 1933, несколько переработано)
Черничный сосново-еловый лес
I1
Заболоченный черничный
березово-еловый лес
I
Лесное (обычно с преобладанием сосны)
болото с господством сфагнов, черники
и участием болотных вересковых
I
. I
Сосново-сфагновое болото с ярусом
различных болотных вересковых
1 1
Сосново-вересковое сфагновое болото
1
Заболоченный березово-еловый
лес с господством Ро1у1г1сЬиш
сошлите, черники (иногда брус¬
ники)
Березово-еловое болото с гос¬
подством сфагнов (частью Poly-
trichum commune) и черники или
морошки, а иногда Equiselum SJ1-
vatlcum
г1
Березовое (иногда с участием
ели и сосны) осоково-сфагновое
болото с примесью кустарничков
I11
Фитоненозы мокрой окраины
сфагнового болота
Схема 1
к сфагновому болоту
Чернично*кисличный и кислично*
майниковый еловый лес
I
Крупнопапоротниковый
еловый лес с сфагнами
I I
Еловое болото с господством
широко- и узколистных гелофитов
с участием сфагнов и других мхов
I —заболачивание при меньшем количестве влаги
II — заболачивание при большем увлажнении
сфагнового болота (схема 7). Подобную стадию, как отмеча¬
лось выше, проходят при заболачивании и наши еловые
леса средней полосы (схема 5). Еловые леса с кислицей
и майником на богатой почве заболачиваются иначе (схема 7).
В заболачивании их большую роль играют травянистые ра¬
стения—широколистные (в том числе часто и папоротники)
•и осоки. Роль сфагновых мхов на первых стадиях заболачи¬
вания обычно невелика. Заболачивание проходит чаще по
влажному типу, так как обычная здесь глинистая почва
способствует накоплению воды. Как в Финляндии, так и у
нас (схема 5) характерна стадия мокрого березово-осоково¬
сфагнового болота.
На севере лесной зоны (Печорский край) сосновые боры
заболачиваются или за счет поверхностной влаги (верховодки;
схема 8), или за счет подъема грунтовой воды (схема 9),
что может происходить вследствие роста окружающих болот1.
В первом случае сфагновая стадия наступает особенно быстро.
Сфагнум или сразу внедряется в кладониевый ковер бора,
что бывает при наиболее бедной почве (схема 8, ряд 1), или
же при более богатой почве (черничный бор; схема 8, ряд И);
сфагновой стадии предшествует стадия Polytrichum. Во вто¬
ром случае при медленном подъеме грунтовой воды сфагно¬
вую стадию предваряет гилокомиевая (схема 9). Особенно
широко распространено на севере заболачивание ельников.
По тем же материалам приводим ряд ельников по увеличению
мощности торфяного слоя: 1. Сфагновые ельники с Betula
nana. 2. Сфагновые ельники с багульником. 3. Осоково¬
сфагновые ельники (чаще с Carex globularis). 4. Сфагновые
ельники с Equisetum silvaticum. 5. Сфагновые ельники с мо¬
рошкой.
Заболачивание лесов вне связи с болотами наблюдается
гораздо реже. Здесь в общем наблюдается сходная смена
растительности. В средней полосе на более бедных почвах
заболачивание начинается прямо с поселения сфагнума и
Polytrichum, причем травянистый покров долгое время со¬
храняет обычный для лесов состав. При наличии более бога¬
той почвы заболачивание происходит при участии болотной
травянистой растительности—осоки широкотравных гелофитов
(в том числе и папоротников). Роль мхов, в том числе сфагнов,
в этом случае первоначально незначительна.
Заболачивание лугов. Заболачиваться могут как суходоль¬
ные, так и заливные луга. Рассмотрим сначала заболачивание
суходольных лугов. Я. Гетманов (1925) указывает, что забола¬
чивание суходольных лугов в Заплюсском районе, Ленинград¬
ской обл., происходит только по соседству с торфяниками
вследствие подъема уровня грунтовых вод при росте болот
вверх. В других же условиях суходольные луга, вопреки
взглядам В. Вильямса, могут существовать неопределенно
1 См. Труды Ботанического музея Академии наук СССР, вып. XXIV, 1932.
126
Схема 8
Смена растительности при заболачивании сосновых боров в Печорском
крае под влиянием верховодки 1
(Труды Ботанического музея Академии наук, т. 24, 1932)
Черничный кладо-
ниевый бор
1 и
Заболоченный кла-
дониево-политрихо-
вый бор с преоблада¬
нием лесных вереско¬
вых
I
Брусничный кладониевый Водяничный к ладо-
бор ниевый бор
II I
Заболоченный кладониево-
сфагновый бор с лесными
и болотными вересковыми
Сосново-кустарничковый сфагновый
бор
1
Сфагновое болото
Схема 9
Смена растительности при заболачивании сосновых боров в Печорском,
крае под влиянием подъема грунтовой воды
(Труды Ботанического музея Академии наук, т. 24, 1932)
Кладониевый сосновый бор
I
Кладонпево-гилокомиевый сосновый бор
I
Гилокомиевый сосновый бор
1
Сфагново-гилокомиевыи сосновый бор
I
Сфагновый сосновый бор
долго, не превращаясь в болота. В. Сукачев (1926) считает,
что утаптывание суходольных лугов при пастьбе, а также
их истощение при сенокошении и стравливании скотом может
повести к их заболачиванию. При неумеренной пастьбе, особенно
весной, когда луг еще влажен, на лугу уплотняется почва и
застаивается вода. Вслед за этим селятся мхи и начинается забо¬
лачивание, особенно при кочковатом микрорельефе, который
нередко создается на сырых лугах под влиянием пастьбы.
Между кочками легко застаивается вода, сток же ее бывает
затруднен; создается избыточное увлажнение, и происходит
заболачивание. Неумеренное стравливание и сенокошение могут
повести (по В. Сукачеву) к истощению почвы и вырождению
луга, т. е. изреживакию злакового травостоя и развитию мхов —
1 Изменена номенклатура лесных типов.
127
Hylocomium squarrosum, Thuidium abietinum и даже кукушкина
льна. Моховой покров проявляет при этом свое обычное дей¬
ствие— накапливает влагу, что иногда ведет к заболачиванию.
Однако следует отметить, что в некоторых случаях вытаптыва¬
ние и стравливание лугов оказывают противоположное действие
(см. А. Шенников и Р. Бологовская, 1927). Выпас формирует
менее пестрый, более однородный и ценный травостой с пре¬
обладанием довольно ценных в кормовом отношении злаков,
моховый же покров угнетается и иногда вовсе исчезает, задер-
нованность уменьшается, а микрорельеф выравнивается. Так
бывает с сильно выродившимися замшенными разнотравными
и белоусниковыми лугами на бедной и плотной минеральной
почве. Для подобных лугов, суходольных и заливных (высокого
уровня), выпас полезен и предотвращает их вырождение.
В частности, весенний выпас, которому приписывается особенно
вредное влияние на мокрую почву, на лугах с сухой почвой
не ведет к заболачиванию, а, наоборот, улучшает травостой.
Следовательно, представление о том, что выпас и утаптывание
ведут к заболачиванию лугов, надо сильно ограничить. На
заливных лугах седиментация ила в общем препятствует
заболачиванию, так как она постепенно повышает уровень
поймы и способствует ее обсыханию. Этот процесс иногда,
повидимому, ведет к тому, что на месте пойменных водоемов
при зарастании их возникают мокрые болотистые луга, которые
в дальнейшем переходят в более сухие луга среднего уровня,
а затем даже в луга высокого уровня. Эта сукцессия, т. е.
ряд „разболачивания*, в пойме Волхова проходит так (схема 10):
Схема 10
Смена растительности в приматериковой части
р. Волхова, связанная с уменьшением увлажнения
(по П. Овчинникову, 1926; схематизировано)
Заросли водных растений (камыша, кувшинки и др.)
Caricetum aquaiills
Caricetum gracills
1*
Glycerlaetum aquatlcae, Sie tum
Alopecuretum pratensis
1
Descbampsietum caespitosae _
Nardetum strictae Hleracletum umbellatl.
Однако, несмотря на „разболачивающее“ действие седиментации,
в поймах может итти и обратный процесс — переход менее
влажных ассоциаций в более увлажненные *. При этом луга
превращаются в болота (схема 11). 11 О причинах заболачивания пойм см. на стр. 113.
128
Схема 11
Смена растительности в приматериковой части
поймы Волхова, связанная с заболачиванием
(по В. Овчинникову, 1926; схематизиро¬
вано)
Hieracietum umbellati
1
Phalaridetum arundinaceae
1
Caricetum gracilis
Caricetum filiformis
i
Sphagnettttn
В поймах даже крупных рек на месте лугов могут возни¬
кать болота, причем в некоторых случаях конечным членом
ряда бывает сфагновое болото. Последняя стадия наступает
лишь с затуханием процесса седиментации и с выходом со¬
ответствующей территории из-под влияния речных разливов.
Заболачивание тундр. Заболачивание—широко распростра¬
ненный процесс в тундре. В Малоземельской тундре, по данным
некоторых исследователей1, все типы тундр, без вмешатель¬
ства человека, в конце концов заболачиваются. Торфянистая
стадия в тундре наиболее продолжительна. Мы отметим,
однако, что хотя заболачивание в тундре и протекает в
широких размерах, но превращение этих заболоченных
земель в торфяники с мощным слоем торфа происходит
в современный период лишь медленно вследствие не¬
благоприятного для торфонакопления климата. В север¬
ной же части тундровой зоны заболоченные тундры с мало¬
мощным слоем торфа оказываются весьма длительной,
пожалуй, заключительной стадией (клаймаксом), так как
торфяники здесь в современный период не образуются. Глав¬
ные причины заболоченности тундры — это низкая испаряющая
сила климата и вечная мерзлота. Песчаные лишайниковые
тундры заболачиваются следующим образом. Талая весенняя
вода, просачиваясь вглубь, встречает близкий уровень вечной
мерзлоты и начинает двигаться по уклону вечномерзлого
слоя вниз. Эта движущаяся вода размывает песок под поверх¬
ностью и создает первоначальную диференцировку повышений
и западинок. В западинках застаивается вода, и поселяются
сфагновые мхи, вытесняющие остальную растительность, в том
числе и лишайники. С поселением сфагнума диференцировка
микрорельефа идет усиленным темпом, уровень мерзлоты по¬
вышается, и начинается торфонакопление; возникают торфя¬
ные кочки, и лишайниковая песчаная тундра сменяется торфя¬
нистым кочкарником. Подобная смена идет сначала в пони¬
жениях рельефа, а затем и на склонах. Эти кочкарники
в конце концов затягиваются сфагнумом. В сфагновую тундру 11 См. сборник «Оленьи пастбища Северного края», Л. 1936.
9—722
129
переходят и другие типы незаболоченных тундр, например
мохово-пятнистые и ерниково-моховые тундры. Так, на по¬
бережье Голодной губы (устье Печоры) слой торфа из зеле¬
ных мхов в 20—30 см толщиной прикрыт сверху сфагнумом.
Таким образом, сфагновое заболачивание — это современный
процесс. Оно характерно, однако, лишь для юга тундровой
зоны; к северу роль сфагнов в заболачивании незначительна
(Б. Городков, 1935).
На Новой Земле (А. Зубков, 1932; в результате заболачивания пятнистой
тундры возникает торфяно-суглинистая тундра. Повышения микрорельефа
с суглинистой почвой заняты здесь Nardosmla frígida, Deschampsia árctica,
Camptothecium trichoides. В понижениях — торф мощностью до 30 см, с
ассоциацией Nardosmia — Calllergon sarmentosum. На дальнейшей стадии
заболачивания повышения покрываются торфом, мощностью в 20—25 см к
ковром Sph. fimbriatum, Polytrichum и другими мхами с арктическими
цветковыми. В мочежинах господство сохраняет тот же Calliergon sarmen¬
tosum, к которому присоединяются Cal. Rlchardsonii, Cal. straminem.
Nardosmla по мере накопления торфа уступает здесь свое место Сагех
stans, Dupontia Fischer!, Eriophorum medium.
В связи с заболачиванием суши стоит и тундровое почво¬
образование. В противоположность лесной зоне подзолообра¬
зование здесь слабо выражено. Зато широко развиты процессы
оглеения. В типичной тундре и лесотундре Северного края
почвы главным образом слабо- и скрытоподзолисто-глеевые
(Б. Городков, 1932). Это зона скрытоподзолистая с широким
распространением торфяно-глеевых почв. Севернее располо¬
жена зона арктического почвообразования, также с большим
развитием торфяно-глеевых почв.
Заболачивание суши прежде и теперь.
Вопрос о ходе заболачивания в разные периоды послеледни¬
ковой эпохи имеет не только громадный теоретический
интерес, но и большое практическое значение. Выяснение
интенсивности заболачивания прежде и теперь тесно связано
с вопросом об опасности заболачивания и о мерах борьбы
с ним. Ряд недавних работ по этому вопросу проливает не¬
который свет на заболачивание в прошлом и настоящем.
Современные торфяники часто начинали развиваться на
месте мелких водоемов, оставленных ледником. К. Мальм-
стрем (1923) излагает историю возникновения одного торфя¬
ника в северной Швеции. Мелкие послеледниковые озера на
месте современного торфяника начали отлагать на дне гиттию
и повышать свой уровень. К этому присоединилось зарастание
стоков из озер водно-болотной растительностью. Озера
выступили из берегов, и началось заболачивание последних.
Зарастание речек и ручьев в районе современного болота
вызвало их запруживание, растекание, увлажнение прилегающей
местности и затем заболачивание и торфообразование. Так
возникли первые отложения торфа. Вместе с заболачиванием
суши началось и зарастание ледниковых озер. В. Ауэр (1933)
130
ис. 17. Ход заболачивания, температура, осадки,
ыльцевые зоны, стратиграфия торфяников и хроно-
огия в послеледниковый период в южной Швеции.
По Е. Гранлунду, 1932.
§
§
§
, т* 7 т* ... . 7* , Q .с-2 .1.1
3№£?ив Щ?мное всеш\ Атлантическое время \ Суббореаль- -||f Cuiamammeme
^"чТТ ^ & i ног время §| время
шеро то?0 Акриловоеозеро \ Литориковое море \Mope L/macea
Климатические условия
в Швеции по сернанВеру
Стадии балтийского
моря по мин те
Пыльцевые зоны Готланда
по шкале поста
fts¡ N
^ I ^
Поверхности обратного
развития и пыльцевые зоны
установленные автором
+100 ММ
Современ¬
ные осадки
-ЮОмм
ход изменения
осадков
+Ход изменения средней
температуры
ход заболачивания
для Огненной Земли также указывает, что послеледниковые
озера были обычно первыми очагами торфообразования. Он
также связывает заболачивание суши с изменением уровня
этих озер. Это заболачивание захватило сначала берега водое¬
мов, а затем, с повышением уровня, и соседние участки
суши. В других случаях очагами заболачивания были не
озера, а участки в котловинах, а также и на склонах, ока¬
завшиеся после отступания ледника в условиях избыточ¬
ного увлажнения. Подобный тип послеледникового забо¬
лачивания отмечен Мальмстремом для северной Швеции
(1931).
После отступания ледника заболачивание суши приняло
широкие размеры, и весьма вероятно, что это заболачивание
началось уже вскоре после начала отступания льда (см. В. Ауэр,
1927; Е. Гранлунд, 1932). Последующий ход заболачивания
достоверно неизвестен. Однако некоторые авторы предполагают,
что заболачивание суши шло неравномерно и что периоды
более интенсивного заболачивания чередовались с периодами
покоя (Е. Гранлунд, 1932). Рисунок 17 изображает вероятный
ход заболачивания в южной Швеции после отступания ледника,
причем приведен также вероятный ход изменения температуры
и осадков за тот же период. Намечаются четыре „взрыва"
заболачивания. Эти периоды заболачивания датированы хро¬
нологически. Кривая хода заболачивания хорошо совпадает
с кривой осадков, причем усиление заболачивания совпадает
с резким повышением количества осадков. Два последних
„взрыва" заболачивания сопровождались, кроме того, пони¬
жением температуры. Прерывистость хода послеледникового
заболачивания суши, по мнению Е. Гранлунда, — следствие
изменения климата. Приведенные сведения — лишь первая
попытка дать картину послеледникового заболачивания. По¬
следний период заболачивания в южной Швеции падает по
схеме Е. Гранлунда на первую половину субатлантического
периода. Также и В. Ауэр (1927) считает, что в Лапландии
заболачивание суши в субатлантический период (кроме вто¬
рой его части) протекало весьма интенсивно, а для области
дождливых лесов Огненной Земли тот же автор уста¬
навливает (1933) повышение интенсивности заболачивания
во вторую половину последнего периода послеледниковой
эпохи.
Заболачивание в ближайший к нам период обстоятельно
исследовал К. Мальмстрем (1931). Еще ранее он установил
(1923) для торфяника Ое§его в северной Швеции, что в эпоху
до появления ели это болото за 3—4 тысячи лет увеличило
свою площадь на 51% (по отношению к современной его
площади), тогда как в последующую эпоху ели за 4 тысячи
лег прирост площади равнялся всего 20°/». Эти цифры ука¬
зывают на значительное сокращение интенсивности заболачи¬
вания за последние тысячелетия. Специальное изучение воз¬
раста заболоченных земель в северной Швеции, а также
132
наблюдения над изменением растительности за ряд лет при¬
вели того же автора к таким выводам:
1. Процесс заболачивания в настоящее время
фактически замер.
2. Громадная часть площадей современных
торфяников и заболоченных земель возникла
еще в доеловое время или в начале елового
периода. Позднее заболотились лишь весьма
незначительные площади.
К. Мальмстрем указывает, что заболоченные земли, которым
обычно приписывают недавнее происхождение в результате
вторичного заболачивания, в действительности образовались
зачастую уже в весьма давний период в результате первичного
заболачивания оставленных ледником мокрых площадей. Того
же мнения держится и Гранлунд (1932). К. Мальмстрем считает,
что заболачивание суши вообще идет сначала весьма энергично,
но постепенно, если не изменяется климат, ослабевает и за¬
тухает по мере зарастания озер, установления гидростатиче¬
ского равновесия и окончательного формирования рельефа.
Это не мешает возникновению местных очагов заболачивания
вследствие запруживания озер при их зарастании, лесных
пожаров, вырубок и пр. Последний тип заболачивания носит,
впрочем, временный характер, и заболачивание прекращается,
как только здесь вновь поселяется лес. Автор считает, что
его выводы имеют значение для всей провинции Могг1апс1 и
что предупредительная осушка еще здоровых
лесов в том объеме, в котором она производится
в Швеции, излишняя.
Выводы Мальмстрема о затухании заболачивания в современный период
находят подтверждение. В. Ауэр (1927) отмечает ослабление заболачивания
в Лапландии в современный период, а также остановку и замедление
в расширении площади торфяников в некоторых областях Огненной Земли
(В. Ауэр, 1933). Е. Гранлунд (1932) считает, что в южной Швеции в настоящее
время заболачивание идет очень слабо и носит лишь местный характер.
Г. Бооберг (в. ВооЬе^, 1930) установил, что торфяник в провинции ЛатИаш!
за все еловое время расширил площадь всего на 4% (по отношению к
современной площади болота). К. Пфаффенберг -(К. ИайепЬе^, 1934) для
района Бремена установил, что после интенсивного заболачивания в субатлан¬
тический период оно остановилось и что в настоящее время нет оснований
опасаться его дальнейшего развития. За все буковое время (т. е.за 5000 лет)
здесь накопилось всего 70—60 см торфа, тогда как за предшествующие
4000 дет торфа наросло в 3—4 раза больше. К. Бюлов (1932) установил
интенсивное заболачивание лесов в Гарце в субатлантический период.
В геологически молодых местностях, где не установилось еще гидростатическое
равновесие, заболачивание и торфообразование может итти усиленным
темпом и в современный период (К. Мальмстрем 1931). Так, Л. Аарио (1932)
для морского побережья Финляндии в провинции 8а1акипО указал, что
в районе, освободившемся от моря всего 600—700 лет назад, успели
образоваться болота мощностью свыше 1 м.
ГЛАВА V.
СТРОЕНИЕ ТОРФЯНИКОВ.
Толща торфяника, или торфяная залежь, неоднородна.
Она слагается из слоев торфа разного ботанического состава.
Эти слои залегают не случайно, а в более или менее опреде¬
ленном порядке. Эта смена торфов отражает сь&ну болотных
материнских фитоценозов, происходившую в прошлом, в период
развития торфяника. Эта смена зависела в значительной
степени от изменений условий существования самой раститель¬
ностью, т. е. была эндодинамической. С другой стороны, сюда
врывались и действовали нарушающим образом разные внеш¬
ние факторы.
В качестве конкретного примера смены фитоценозов в про¬
цессе развития болота приведем один из торфяников под
Ленинградом. По Г. Ануфриеву (1925), этот торфяник образо¬
вался на месте озера (рис. 18). Заболачивание последнего
началось с отложений, где преобладали минеральные частицы.
Позднее преобладание перешло к планктонным организмам,
сначала диатомовым, образовавшим нижний слой гиттии (на
рисунке оливково-бурый сапроколл), а затем десмидиевым. По¬
следние господствуют в вышележащих слоях гиттии. Эти
слои характеризуют уже сильно обмелевший водоем. Когда
глубина водоема уменьшилась до 0,5 м, на озеро надвинулись
с берега тростниковые заросли, сначала в виде отдельных
островов, а позднее образовавшие тростниковый зыбун над
слоем гиттии. В этот период отложения приобретают смешан¬
ный характер из остатков планктонных организмов и тростника
(так называемый сапропелевый торф), причем с течением
времени гидробиологические процессы затухают, а роль
тростника возрастает. Сапропель и сапропелевый торф отно¬
сятся к лимнической стадии развития торфяника. Со смыканием
тростниковых зарослей начинает отлагаться чисто тростниковый
торф, и образуется тростниковое болото, относящееся уже к
тельматической (т. е. болотной) стадии развития.
Поселение плавающих дернин сфагнума среди зарослей
тростника ведет к постепенному исчезновению этих зарослей.
На месте тростника поселяются осоки, и образуется мокрое
с редкой древесной растительностью переходное травяно-сфаг-
134
А
в
! . Сфагновый очес
Пушицево-Сфогновыл торф
Ш$Ш Пограничный горизонт
Старый сфагновый торф
11|1!1|Щ;1Н!!1| Кустарниково-древесный торф
Горизонтальный —51—!
саж
1::"В8с1 Древесно-травяной торф
Тростниковый
1-^'аалН Гипновый
ШШШ Сапропелевый
1111111Ш111Д Красновато-бурый сопроколл
Масштаб
1 Вертикальный2 ' 0
- метр
ил и6ко б о -бурый сопроколл
Песок
Зеленовато-серая глина
Желто-бурый суглинок
Нивелир отм
Оь
саж
Рис. 18. Строение выпуклого сфагнового болота Ладожско-Ильменско-Западнодвинской области.
По Г. Ануфриеву, 1929.
новое болото, представляющее мезотрофную стадию развития
торфяника (на рисунке эта стадия не представлена соответ¬
ствующими отложениями). Уплотнение сфагнового торфа при¬
водит к вытеснению осок и других эвтрофных видов спутни¬
ками сфагнума — главным образом пушицей. Возникает сфаг¬
новое болото, отлагающее сфагновый торф (старый сфагновый
торф на рис. 18), и торфяник переходит в олиготрофную
стадию. С этого момента развитие торфяника направляется
уже иными факторами. Это были, с одной стороны, факторы
климатические, вызывавшие смену климатического порядка,
запечатлевшуюся в отложении пограничного горизонта.
С другой стороны, изменения влажности на болоте в про¬
цессе роста самого торфяника вызвали на олиготрофной
стадии смену более влажных редколесных фитоценозов
(сфагново-пушицевое болото) менее влаголюбивыми сосново-
сфагновыми. Большую роль на болоте играли и антропоген¬
ные факторы, в частности пожары, неоднократно уничто¬
жавшие естественную растительность. Более детальное изуче¬
ние верхней пушицево-сфагновой толщи торфяника, несом¬
ненно, установило бы там следы упомянутых смен расти¬
тельности, зависевших от изменения влажности, а также смен
антропогенного характера.
Залежь как отражение эндодинамических смен раститель¬
ности.
Рассмотрим идеальную схему строения торфяника с наиболее
полной серией слоев торфа и с законченным развитием. Такая
схема будет характерна для торфяников в наиболее благо¬
приятном для торфообразования климате и в условиях более
или менее богатой почвы. Пусть это будет притом торфяник,
возникший в результате заболачивания водоема.
Все органогенные образования (рис. 19) можно разделить
на четыре группы по условиям существования их материнских
фитоценозов: 1) седименты, 2) лимнические (водные) торфа,
материнские ценозы которых росли при постоянном зеркале
воды; 3) тельматические (болотные) торфа, материнские
ценозы которых питались грунтовыми водами и или вовсе не
были покрыты водой, или заливались лишь периодически;
4) омброгенные торфа, ценозы которых питались главным
образом за счет атмосферных осадков, причем они водой не
заливались.
Причины смены седиментов в вертикальном направлении
были указаны выше (глава III), так же как и факторы, напра¬
вляющие смену лимнических торфов. Фактор—уменьше¬
ние глубины — с переходом к тельматической группе фито¬
ценозов перестает действовать. .Смены фитоценозов этой
последней группы, сохраняя характер эндодинамических,
направляются уже другим ведущим фактором, постепен¬
ным обеднением минеральными солями. Это
136
Торта лимничесной Торта тельмати-. Гл___ „„и.,и„„^л-л „
Седаменты мч* стадии ияпкпГ, Гтпг)„п ТоРФа минимального питания
чесной стадии
$1
<\> 5^
$
£:
£ с:
«Ч) ^
11
3
3
с:
I
-1
5*1
а 1
к ^
*§>
р
*§■
п>
$
Г
V
О)
I
>
§*
6.
3
*
а
§>
5^
*
§ь
3 <?
**
§>
$>
0>
$
§>
й.
5
I
&
<Ъ
оь
8
3§
§ 7
^ 3
I
4
•§|
С*
I
^ £5
**§
I?
С)
<\>
щ
I'
щ
1Г=
7 >
!! !Ж1 () (/Т';<! (д*
)§/г
, ГП 1(ПЧ I
||
|§>
;§*
'в-з
О
<>»
^-§!| г?
|1| ¡11
£§>
Эвтрофна я
Ч/'
стад ил
.л
/Цезбтроф- Олиготрофная стадия
стадия
Рис. 19. Идеальный профиль торфяника с полной серией слоев торфа и законченным развитием. Орнг.
обеднение происходит вследствие роста по¬
верхности болота и удаления грунтовых вод
от корневых систем растений. Несомненно, что и
другие экологические факторы тоже изменялись с нарастанием
болота вверх, но обеднение солями все же ведущий фактор.
Следовательно, в тельматическом отрезке нашего профиля
(рис. 19) фитоценозы сменяли друг друга снизу вверх, от
более требовательных к менее требовательным.
Специальных сведений по изменению содержания зольных элементов в
залежи снизу вверх почти не имеется. Приведем поэтому лишь средние
данные зольности для некоторых торфов.
Таблица 41
Средняя зольность некоторых видов торфа Ленинградской области
(составлено по Г. Ануфриеву, 1931)
Процент зольности
Олиготрофные Сфагновые торфа 1,48—1,68
торфа Сфагново-пушицевый торф .... 1,54—2,05
Мезотрофные Сфагново-шейхцериевый торф . . . 1,23
торфа Сфагново-осоковый торф . ...... 2,68—3,91
Сфагново-древесный торф 3,75
Эвтрофный Осоково-гипновый торф 6,75—7,68
торф
Содержание золы в этих трех группах торфа в общем со¬
ответствует их положению в залежи (рис. 19).
Убывание минеральных соединений определяет смену мате¬
ринских фитоценозов лишь в пределах эвтрофной тельматиче-
ской и мезотрофной групп торфов (рис. 20). С переходом
в олиготрофную (омброгенную стадию) влияние грунтовых
вод, вследствие удаленности их от корней, ослабевает, и э т и
воды перестают быть ведущим фактором смены.
Сумма зольных элементов на этой стадии остается более или
менее постоянным общим запасом, так как переменное слагае¬
мое—зольные элементы, поступающие снизу, — невелико. Кон¬
центрация же зольных элементов хотя и меняется, но в основ¬
ном только в зависимости от влажности и степени разложе¬
ния торфа, причем эта концентрация колеблется то в ту, то
в другую сторону. Из приведенной выше таблицы и рисунка 20
видно относительное постоянство содержания зольных элемен¬
тов на олиготрофнойстадии. Ведущим фактором смены
в этой стадии торфяника становится, можно ду¬
мать, влажность. На олиготрофной стадии она не оста¬
ется постоянной, но колеблется частью вследствие
деятельности самой растительности, частью под влиянием внеш¬
них факторов. На сфагново-шейхцериевой стадии (рис. 19) мы
имеем избыточно увлажненную поверхность бо¬
лота, причем влага при ровном микрорельефе
распределена более или менее равномерно. На
следующей пушицево-сфагновой стадии, при о б-
438
щем о’б с ы х а н и и поверхности болота вслед¬
ствие его роста вверх, намечается неравно¬
мерное распределение влаги в связи с начав¬
шейся д и ф ере нц иров к ой микрорельефа (кочки
»пушицы).
Рис. 20. Распределение зольных элементов в торфянике в вер¬
тикальном направлении. По Л. Посту, 1926.
С переходом в заключительную сфагновую стадию микро¬
рельеф резко диференцируется на большие
кочки и мочежины. В связи с этим влага распреде¬
ляется неравномерно на площади болота и,
139
кроме того, в каждой точке поверхности тор¬
фяника нередко степень увлажнения изменя¬
ется и во времени. В результате этого отлагается
комплексный, разнородный торф, где в общую
массу сфагнового торфа, отложившегося в более сухих усло¬
виях, включены линзы шейхцериево-сфагнового торфа, обра¬
зовавшиеся в мокрых мочежинах (рис. 19).
Общий ход смены торфов и материнских фитоценозов
в залежи с. изменением экологических условий представлен
такими стадиями торфообразования:
1- я стадия. Материнский фитоценоз-планк¬
тон. Отложения типа седиментов.
2- я ст а д и я—л и м и и ч е с кого торфообразования.
Ведущий фактор смены фитоценозов — умень¬
шение глубины слоя воды (см. выше).
3- я стадия—тел ьматичес кого (болотного) тор¬
фообразования. Ведущий фактор—уменьшение
содержания зольных элементов, в связи с рос¬
том торфяника вверх.
4- я с т а д и я—о мброгенного торфообразования.
Ведущий фактор—колебания влажности. На пер¬
вых ступенях стади и — равномерное распреде¬
ление влажности, позднее оно делается нерав¬
номерным как на площади болота, так часто
и во времени.
Роль физико-географических факторов в строении
торфяников.
Строение торфяников и климат. Строение торфяников
в разных климатических зонах резко различно. Торфяники
хвойной зоны имеют в типичных случаях такие особенности:
1) наиболее полную серию слоев торфа; 2) законченное раз¬
витие. Конечный член серии — более или менее мощный слой
сфагнового торфа.
Надо отметить, что эти свойства выражены не везде, а лишь
в определенных условиях почв и рельефа. Эти торфяники
оптимального развития отличаются еще тем, что эндодинами-
ческий фактор — растительность — играет в их развитии осо¬
бенно большую роль, часто заслоняя влияние внешних факто¬
ров. Это зависит от того, что торфонакопление в этой клима¬
тической зоне особенно интенсивно, а следовательно, ярко вы¬
ражено и нарастание поверхности торфяника и энцодинамиче-
ский характер смен материнских фитоценозов.
Торфяники к северу и к югу от зоны оптимального разви¬
тия, т. е. в зоне тундр и в зоне степей, имеют такие особен¬
ности: 1) серия слоев торфа неполна, число слоев невелико;
2) конечная олиготрофная стадия развития не выражена, что
целиком подходит к степным торфяникам и, повидимому,
и к арктическим торфяникам. Порядок залегания отдельных
140
слоев торфа менее определенен и фиксирован, чем в торфя¬
никах оптимального развития. Зависит это от того, что в раз¬
витии этих торфяников внешние факторы (морозы, почвен¬
ная мерзлота, ветер, водная эрозия и пр.) зачастую преобла¬
дают над эндодинамическим фактором— растительностью.
Последняя лишь в слабой степени определяет развитие тор¬
фяника, так как торфонакопление и в условиях степного
и тундрового климата ослаблено, и во многих случаях берет
верх разрушение торфяника внешними агентами.
Рассмотрим более детально зависимость строения торфяников от климата
по данным Л. Поста (1926) для южной Швеции.
Таблица 42
Распределение разных видов торфа по отдельным торфяным областям
Южной Швеции
(по Л. Посту, 1926).
Торфяные провинции южной Швеции
(№ провинций см. карту)
Общее количество
торфа (в млн. кубо¬
метров)
Сфагновый торф
Торф заболочен¬
ного леса
•о*
о.
о
н
•Я
2
со
о
я
о
о
О
Г иттия
в процентах
I. Область сухопутного заболачивания
Геталанда
7366
63,9
23,8
9,3
3,0
II. Область Венер
3410
69,7
14,2
9,5
6,6
III. Область сухопутного заболачивания
Свеланда
4002
70,8
8,8
12,5
7,9
IV. Повышенная область Геталанда с тор¬
фяниками, образовавшимися на месте
озер
3303
44,7
25,2
17,3
12,8
Va. и Vb. Повышенные области Све¬
ланда с торфяниками на месте быв¬
ших озер
2282
47,7
13,8
23,4
15,1
VI. Пониженная область Геталанда с тор¬
фяниками на месте озер
2641
20,0
15,6
23,8
40,6
VII. Пониженная область Свеланда с тор¬
фяниками на месте бывших озер . .
1479
16,1
17 4
26,8
39,9
VIII, IX и X. Известковые области Шоне-
на, западного и восточного Геталанда
1071
16,6
26,0
32,7
24,7
, Сравнение рис. 21 и рис. 22 показывает прежде всего, что процент
площади сфагновых торфяников хорошо отражает распределение осадков
и что местности, богатые осадками, наиболее богаты сфагновыми торфяниками.
Это будут как раз провинции I—III, где сфагновые торфа наиболее распро¬
странены по сравнению с другими типами торфов (табл. 42). Вместе с тем
эти провинции — по преимуществу районы торфяников сухопутного проис¬
хождения (рис. 23). Провинции IV—VII менее богаты осадками. Процент
площади сфагновых торфяников и объем сфагновых торфов здесь меньше,
чем в провинциях первой группы. В этих провинциях многие торфяники
произошли путем заболачивания озер, соответственно чему седименты (гнттии)
занимают по объему до 40% всех органогенных образований (табл. 42), а
участие сухопутных торфяников в общей площади последних падает (рис. 23).
141
Рис. 21. Распределение годового количества осадков в южной Швеции.
По Л. Посту, 1926.
Наконец, группа известковых провинций (VIH—X) отличается сравнительно
малым количеством осадков, слабым развитием сфагновых торфяников,
малой кубатурой сфагновых торфов н преобладанием эвтрофных. Впрочем,
для этой группы провинций большое значение, кроме климата, имеет и
петрографический фактор — известковая подпочва.
Из сказанного ясно, что для южной Швеции сфагно¬
вый тип строения залежи связан с районами, богатыми
осадками, и что при уменьшении количества послед¬
них усиливается роль эвтрофных торфов в торфяной
залежи.
На связь количества осад¬
ков и мощности сфагнового
торфа указывают для Баварии
также Г. Пауль и 3. Руофф.
Здесь более близкие к краю
Альп торфяники, где выпадает
до 1400 мм осадков в год,
имеют более мощный торф, чем
торфяники, удаленные от Альп,
где количество осадков не
более 900 —1000. И у нас в
Европейской части СССР тор¬
фяники, лежащие ближе к Бал¬
тийскому морю, имеют в об¬
щем более мощный слой сфаг¬
нового торфа, чем расположен¬
ные дальше в глубь конти¬
нента.
Геолого - петрографи¬
ческий и почвенный ха¬
рактер местности и
строение торфяников.
Химический состав ко¬
ренных пород и почв
определяет богатство
грунтовых и поверхност¬
ных вод зольными эле¬
ментами и этим самым
также и строение тор¬
фяников.
1. В богатых зольными элементами районах
в залежи преобладают эвтрофные торфа, и
эвтрофная стадия развития торфяника
длительна. Конечная ж е, о л и г о т р о ф н а я, ста¬
дия задерживается на долгое время, так
как поселению сфагновых мхов мешает
богатый солями субстрат.
2. В бедных минеральными солями районах
эвтрофная стадия торфяника сокращается,
серия эвтрофных торфов немногочленна, олиго-
трофная стадия наступает рано, так как сфагновые
мхи легко поселяются на бедном субстрате.
Пример первого рода—области известняков (см. К. Бюлов,
1929). Кроме богатого солями, в частности известью, субстрата,
для таких областей характерен карстовый ландшафт с глубо¬
143
Рис. 22. Сфагновые торфяники южной'
Швеции в процентах к общей площади.
По Л. Посту, 1926.
ким и непостоянным уровнем грунтовых вод. Торфяников
здесь мало, они часто озерного происхождения. В залежи
преобладают эвтрофные торфа, лежащие часто на мощных,
богатых известью седиментах. Сфагновые торфа встречаются
обычно лишь в центре обширных эвтрофных торфяников,
где минерализующее влияние грунтовых вод проявляется
слабо. К подобным областям принадлежат торфяные провин¬
ции в известняковых районах южной Швеции.
Районы кристалличе¬
ских пород по условиям
торфообразования в общем
противоположны известко¬
вым районам (см. К. Бюлов,
1929). Почвы бедны мине¬
ральными солями; легко
образуется кислый гумус.
На нем часто непосредст¬
венно поселяется сфагнум.
Широко распространены
торфяники, образовавшие¬
ся путем заболачивания су¬
ши. Понятно, что сказанное
относится лишь к тем
олиготрофным областям,
которые лежат в пределах
умеренно прохладного и
влажного климата, допу¬
скающего образование оли-
готрофных сфагновых тор¬
фов. Пример таких обла¬
стей — фенноскандийский
гранитный щит.
Г. Пауль и 3. Руофф (1932)
устанавливают для Баварии связь
между строением торфяников и
формами рельефа, характером
дна и некоторыми другими мо¬
ментами. Они различают ряд се¬
рий (рис. 24).
I. Нормальная полная серия сфагнового болота
характеризуется такой серией слоев (снизу вверх): эвтрофный осоковый
торф — переходный сфагновый торф. При этом переходный торф — сфагново¬
осоковый и в противоположность иным сериям не носит характера лесного
торфа. Этот переходный торф весьма маломощный, в то время как олиго¬
трофный торф имеет большую толщину, чем эвтрофный и переходный вместе.
Подобное строение имеют юрфяники на глинистых почвах Альпенфорланда
в условиях волнистого рельефа, затем в котловинах главного ледника
^аттЬескеп) и вообще в условиях ландшафта основной морены.
II. Неполная серия сфагнового болота. Здесь отсутствует
нижний член серии — эвтрофный торф,—и развитие начинается с переходного
торфа. В зависимости от характера последнего здесь два варианта: 1) сфаг¬
ново-осоковый переходный торф—олиготрофный сфагновый торф и 2) лесной
переходный торф — олиготрофный сфагновый торф. Первый вариант свой¬
ствен центральным частям больших болотных областей. Подобные торфяники
244
Рис. 23. Торфяники сухопутного проис¬
хождения в процентах к общей площади
торфяников в различных торфяных про¬
винциях южной Швеции. По Л. Посту,
1926.
I — X —^торфяные провинции (см. габл. 42).
развивались в котловинах центральной части ледников, особенно там, где
долгое время стояли ледниковые озера. Бедное известью дно этих болот
объясняет укороченный тип их развития: эвтрофная стадия выпадает, и
развитие начинается с переходной стадии. Характер переходного торфа
(сфагново-осоковый) указывает на влажные условия, в которых развивались
эти болота. Во втором варианте развитие начинается с лесного переходного
торфа. Этот тип развития наблюдается на небольших болотах и на окраинах
более крупных. Первый член серии
здесь — лесной торф, что указывает,
повидимому, на развитие в более
сухих условиях.
III. Серия сфагнового
болота со стадией эвтроф-
ноготорфа болотистого
леса (Bruchwaldtorf). Здесь
такой порядок слоев: 1) торф эвтроф-
ноге болотистого леса — олиготроф-
ный сфагновый торф или 2) осоковый
торф—торф эвтрофного болотистого
леса — олиготрофный сфагновый
торф. Более обычен первый ряд,
когда заболачивается лес. Серия
с участием Bruchwaldtorf связана
с текучей водой или с высоким и
колеблющимся уровнем грунтовых
вод. В том и другом случае имел
место постоянный приток питатель¬
ных веществ и, вероятно, частичная
аэрация, что благоприятно для раз¬
вития болотистого леса.
IV.Серия переходного бо¬
лота представлена обычно целиком1
переходными сфагново-осоковыми и
лесными торфами. Предполагают, что
1 Иногда наблюдается еще ниж¬
ний слой осокового и тонкий верх¬
ний слой сфагнового торфа.
L L L
L L
L L I
VTTT>
глина
с (раеновы и тору
осоковый тордз
торф болотистого леса
Осоково - гипновый торф
Рис. 24. Различные серии смены торфов. По Г. Пауль
и 3. Руофф, 1932.
10—722
145
наступление сфагновой стадии в этой серии задерживалось характером
минерального окружения торфяников (влияние береговых вод).
V. Обращенная, или нарушенная серия. В этом случае
естественная смена прерывалась, иногда неоднократно, заносом торфяника
илом. В некоторых случаях смена осокового эвтрофного торфа переходным
повторялась трижды, прежде чем поверхность болота поднялась над уров¬
нем разливов и образовался сфагновый торф.
Строение торфяников и возраст местности. Различные
точки земной поверхности освободились от ледяного покрова
или от моря не одновременно. Торфяники в разных пунктах
тоже начали свое развитие не одновременно и имеют неодина¬
ковый возраст. В некоторых случаях более молодые торфя¬
ники более просто построены и сложены из меньшего числа
слоев.
Рис. 25. Схема возникновения и развития торфяников в полосе отступания
моря в зависимости от возраста местности. По Л. Аарио, 1932.
В приморской полосе Финляндии (провинция Сатакунта —
Satakunta), где суша в настоящее время поднимается, Л. Аарио
установил, что строение торфяной залежи изменяется от
берега моря в глубь континента (рис. 25). Болота, наиболее
молодые, лежащие ближе всего к морю, носят характер
болот без торфа (Sumpf). Эти болота „в состоянии возникно¬
вения" лежат прямо на гиттии и образуются в результате
заболачивания морских заливов. На более высоком уровне,
дальше от берега, на участках, раньше освободившихся от
моря, встречаются уже болота иного типа. Это будут осоко¬
вые болота, или болотистые эвтрофные леса (Bruchmoore)
с двух- или трехчленной серией слоев, где поверх гиттии
лежит травянистый торф, а выше — слой лесного торфа. Еще
выше и дальше в глубь континента идут пушицевые болота.
Они еще более старшего возраста и в типичных случаях
с четырехчленной свитой торфов, так как торф болотистого
леса здесь уже прикрыт тонким слоем пушицевого торфа.
Еще дальше от моря встречаются выпуклые торфяники
с полночленной серией торфов и конечной сфагновой стадией
146
развития. Сверху здесь залегает слой сфагнового торфа, при¬
чем-его мощность и возраст увеличиваются по мере продвиже¬
ния в глубь континента. Так называемая „граница еловой
пыльцы в торфе" (РЮМепроНепдгепге)1 позволяет установить,
что осоковые болота, болотистые эвтрофные леса и пушице-
вые торфяники в приморской полосе образовались уже после
появления ели, тогда как выпуклые сфагновые болота внизу
не содержат еловой пыльцы и, следовательно, обычно доело-
вого, т. е. старшего возраста. Лишь более' близкие к морю
выпуклые торфяники — послеелового происхождения (рис. 25).
Они имеют притом менее мощный слой сфагнового торфа.
Любопытно, что типичная для выпуклых болот Сатакунты
серия: гиттия — травяной торф — осоковый торф (или торф
болотистого леса) — пушицевый торф — сфагновый торф —
повторяет в вертикальном направлении смену типов болот
и типов строения залежи, которая наблюдается при движе¬
нии от моря в глубь континента. Отсюда важный в-ывод,
что развитие торфяников, т. е. смена материн¬
ских фитоценозов и торфов, в различные пе¬
риоды послеледниковой и стории происходи ло
по одному типу. В горах наблюдается упрощение строе¬
ния торфяников снизу вверх.
Г. Шрейбер (1927) указывает, что торфяники больших высот, поздно
освободившиеся от льда, сложены немногими слоями торфа (всего 1—3),
причем число их в общем убывает с высотой, так же как большей частью
и мощность. Болота Зальцбурга, расположенные на высоте 1600—1900 м,
по Шрейберу, имеют всего один слой торфа, мощностью большей частью
всего в 1 м. Этот торф отложился в течение одного климатического периода,
поэтому не отражает послеледниковых изменений климата. Подобные тор¬
фяники сложены травянистыми растениями (Юе<Иог1) и лишь иногда при¬
крыты сфагновым торфом мощностью всего в 10—20 см (рис. 28, IV и V). На
молодой в общем субатлантический возраст альпийских торфяников, хотя
и не во всех случаях, указывает и Г. Гаме (1932/33). Он отмечает также и
малую мощность этих торфяников.
Строение залежи и движения земной поверхности.
Движения земной поверхности отражаются на сменах торфо¬
образующих ( фитоценозов и производят глубокие и много¬
образные нарушения в порядке залегания слоев. Эти движе¬
ния бывают различными, и их влияние на строение торфяников
сказывается неодинаково. Торфяники прибрежной полосы
морей, где в послеледниковую эпоху происходили поднятия
и опускания суши, имеют совершенно необычный порядок
слоев. Эта нарушенность напластований зависела от общего
подъема или опускания уровня грунтовых вод в связи с транс¬
грессией или регрессией моря, причем менялся режим мине¬
рального питания торфяника. В этом случае воздействие моря
носило косвенный характер. В других случаях море непосред¬
ственно покрывало торфяник и отлагало на нем слой ила.
После отступания моря прерванное развитие торфяника могло
1 Ниже этой границы пыльца ели в торфе не встречается, что указывает
на отсутствие ели в прежнее время.
10*
147
возобновляться. Как прямое, так и косвенное влияние моря
вело к инверсии (нарушению) слоев. В случае косвенного
влияния поверх олиготрофного торфа откладывались эвтроф-
ные (например тростниковые) торфа. Подобные |Торфяники
в области морских трансгрессий (рис. 26) дают ценнейший
материал для изучения этих трансгрессий (см. ниже). Вековые
движения суши иногда сказываются на строении торфяников
и внутри континента. В таком случае на это строение влияют
общие изменения уровня грунтовых вод, воздействие же моря,
прямое или косвенное, не имеет места.
П. Томсон (Р. ТЬотэоп, 1933) указывает, что торфяники
северной прибрежной Эстонии имеют слой осокового торфа
лишь редко более метра. Южнее же расположенные торфя¬
ники имеют обычно мощный слой этого торфа. Причина этого,
по мнению Томсона,—неравномерное вековое поднятие Эсто¬
нии. В северной прибрежной полосе Эстонии, где общее на¬
правление стока в послеледниковое время не менялось, подъема
грунтовых вод не было, и торфяники скоро вышли из осоко¬
вой стадии грунтового питания. Дальше на юг вследствие
более быстрого поднятия суши на северо-западе Эстонии водо¬
раздел между Финским заливом и бассейном Эмбаха посте¬
пенно передвигался на юг. Эта область ранее имела сток
к северу, впоследствии же с поднятием суши сформировался
сток к югу, в реки Эмбах и Пернаву. В связи с этим на юге
страны имел место длительный подъем уровня грунтовых вод,
и образовались мощные слои низинного торфа, а наступление
сфагновой стадии задержалось на долгое время. В частности,
уровень озера Чудского (в устье Эмбаха) повысился с подня¬
тием суши против прежнего на 6 м, а сток из него через
Нарву уменьшился.
Нарушение (инверсия) порядка слоев торфа в залежи проис¬
ходит и при резких, подчас катастрофических движениях
земной коры, например, при опускании слоев торфа в кар¬
стовые воронки в районах распространения известняков.
При этом опускании резко изменяются условия увлажнения
и минерального питания, что нарушает обычный ход смены
торфообразующих фитоценозов и вызывает инверсию слоев.
С. Толпа (5. То1ра, 1932) в карстовом торфянике западного Полесья
установил такую смену торфов снизу вверх: лесной торф—гиттия — сфаг¬
новый торф — переходный и эвтрофный торф — сфагновый торф. Мощность
торфяника в пункте бурения была 7,3 м.
Вулканические извержения и строение торфяников. Выше
(глава II) отмечалось нахождение в торфяниках слоев пепла.
Эти слои наблюдаются в торфяниках юго-восточной Камчатки
и Огненной Земли, где имеются вулканы. В последней стране
В. Ауэру (1933) удалось, изучая торфяники, установить пять
вулканических извержений в послеледниковую эпоху, оставив¬
ших след в торфяной залежи в виде слоев пепла \ Засыпание 11 Этими слоями пользуются для датировки торфяников.
148
пеплом, несомненно, влияло на ход развития торфяника,
однако, не столь сильно, как можно было бы ожидать, и
притом, повидимому, не всегда отрицательно (В. Ауэр, 1. с.).
|: :\ Морскойип ЩЩПесон ЩЩ Леской торф
¿glllllllllllSphagnum \ÍK*>\Eriophorum ШШйогел ЩШРШдт/tes
Betula Pinus Ainus Fagus Carpinus Picea
■—
Corylus
йиегш Tilia lilmas
Рис. 26. Профиль торфяника в северо-западной Германии с нарушенной под
влиянием морских трансгрессий серией слоев. Справа — кривые пыльны
Ericaceae и Gramineae -f- Сурегасеае. По Е. Шуберту, 1933.
149
Большей частью отложения пепла даже способствовали росту
сфагнового ковра, что, однако, не совсем понятно. В некото¬
рых случаях пепел защищал поверхность болота от высыха¬
ния или же способствовал отложению специфического, свой¬
ственного этой области, торфа из Машррозрегтит. В торфя¬
никах юго-восточной Камчатки можно установить две основные
прослойки вулканического песка, мощностью от 2 до 25 см
Рис. 27. Строение торфяников на юго-восточной Камчатке с прослойками
вулканического пепла. По М. Нейштадту и М. Короткиной, 1936.
а — торф; Ь — слой вулканического пепла.
(М. Нейштадт и М. Короткина, 1936; см. рис. 27). Слои пепла
сильно увеличивают зольность торфа и понижают его топлив¬
ные качества.
Строение залежи и деятельность рек. В болотах, распо¬
ложенных в речных поймах, обычны слои речного ила в тор¬
фяной толще. Влияние этих слоев на смену торфообразующих
фитоценозов несомненно, хотя сведений об этом мало.
В некоторых случаях, как это постоянно имеет место в дель¬
тах южнорусских рек, воздействие реки носит катастро¬
фический характер, и торфяник погребается под мощным
150
слоем аллювия. Такое чередование торфа (обычно тростнико¬
вого) со слоями речного ила может повторяться в залежи
не один раз (В. Матюшенко, 1925).
Строение залежи и история климатов.
Климат в послеледниковую эпоху (см. об этом гл. Х)пне
оставался неизменным. Колебания климата нашли свое
отражение в строении торфяников. Роль этих колебаний
Атлантический
бореалоный
Ли^иглациольноя
стадия
Готская стадия
Датская стадия
Собрепенный
лесной торф
Молодой
сфагнобый
торф
молодой
лесной
торф
Старый
сфагнобый
торф
Старый
лесной
Старый
осокобый
торф
i
I
В
ш ш
1о§у>?1 Морена
V//////X Осокобый торф
[л :: | Сфагнобый торф
| - -- - - 1 Торф болотистого
леса
Рис. 28. Число слоев торфа, время образования торфяников и смена
климатов. По Г. Шрейбер, 1927.
I — сфагновые болота в пологих впадинах рельефа.
11 — сфагновые болота на месте озер.
Ш — сфагновые болота в депрессиях.
IV — эвтрофные торфяники в Альпах и на севере.
V — осоковые болота на месте озер.
в развитии торфяников недостаточно изучена и оценивается
по-разному. Г. Шрейбер (1927) дает схему строения торфяни¬
ков Средней Европы в зависимости от климатических периодов
Блитт-Сернандера (рис. 28). Каждый слой торфа по этой схеме
(см. профиль 1) более или менее соответствует определенному
периоду. В частности, оба слоя сфагнового'торфа отложились
в прохладном и богатом осадками климате, причем верхний
молодой сфагновый торф образовался в субатлантический
период, а нижний, старый сфагновый торф в атлантический
период Блитт-Сернандера. Оба слоя лесного торфа образова¬
лись в континентальном климате с малым количеством осадков
151
более молодой
сфагновый торф
и жарким летом. При этом верхний (молодой) лесной торф
между обоими слоями сфагнового падает на суббореальный
период, а нижний лесной торф—на конец бореального пе¬
риода, когда наступил температурный оптимум.
Наконец, нижний слой древнего травянистого торфа относится
к более ранней, также сухой и теплой части бореального
периода. Г. Пауль и 3. Руофф (1932) при исследовании бавар¬
ских торфяников не
ОснГ“““““““ ^ установили столь тес¬
ной связи между перио¬
дами Блитт-Сернан-
дера (и, следователь¬
но, климатом) и строе¬
нием залежи, как
Шрейбер, и отводят
в смене торфов важную
роль эндодинамиче-
ским факторам (т. е. ра¬
стительности), а также
условиям, в которых
развивается торфяник
(см. стр. 144). Эту точку
зрения следует считать
правильной, хотя в не¬
которых случаях влия¬
ние изменений климата
на строение торфяни¬
ков проявляется весь¬
ма резко.
К числу подобных
явлений относится и
образование так назы¬
ваемого погранич¬
ного горизонт а—
слоя сильно разложив¬
шегося торфа, образо¬
вавшегося, как пред¬
полагают, в сухой (ксе-
ротермический) период (рис. 29). Помимо этого мощного
горизонта высыхания во многих местах в залежи обнару¬
жены мелкие прослойки сильно разложившегося торфа, также
климатогенного происхождения. Подобные прослойки в СССР
были впервые установлены на болоте Галицкий Мох б. Твер¬
ской губернии1.
Здесь, кроме пограничного горизонта, как ниже, так и выше
его отмечено еще несколько маломощных прослоек сильно
гумифицированного торфа. В средней части Союза эти доба¬
вочные прослойки, вероятно, широко распространены, хотя
^-1-1 Пограничный горизонт
более старый
сфагновый торф
260 см
272 см
282 см
ГипкоВыйторф
ТростникоВыи торф
Осоко-гилнобый торф с
дребе слыл и остаткам
Шок и глина
Рис. 29. Схема строения торфяника под Ленин
градом. Из В. Сукачева, 1926.
См. Труды Опытной торфяной станции, вып. I, 1923.
152
степень выраженно¬
сти и число их варьи¬
рует (см. 3. Руофф,
1934). Далее на се¬
веро-запад число их
сокращается, и в
Шуваловском тор¬
фянике выше погра¬
ничного горизонта
остается лишь одна
разложившаяся про¬
слойка. На Галицком
Мхе, где эти про¬
слойки особенно вы¬
ражены, их морфо-"
логия и генезис были
обстоятельно изуче¬
ны 3. Руофф (1. с.).
Здесь выше погра¬
ничного горизонта
наблюдается иногда
до семи прослоек
(рис. 30).
Торф этих про¬
слоек разложился
сильнее, чем в про¬
межуточных слоях.
В торфяной массе
прослоек часто
встречаются пни
сосны, нередки
остатки пуши¬
цы, несколько
иной в них и со¬
став сфагновых
мхов. Разложение
прослоек колеблет¬
ся от 20—30% (в
верхних) до 30—40%
(в нижних прослой¬
ках), в то время как
степень разложения
остального сфагно¬
вого торфа не выше
15%- Прослойки
отм ечаются так-
шШ
1
Л>/М
^¡¡¡'
же повышенной
з о л ь н о ст ь ю: от
$
§ К»
1
к
с\, С\|
К
*
О
е?
О
X
л
«=:
5
£
о»
ч
X О
«л
о —
8 I
О. с
С
& «=
^ 'Л 4
<ц 05 «п
С/Т О«
22 >> г. V
§Ч |§
£ ® с«
- С Ьл£
О «о X
£ а
О X 1 X
и о с5 5
рг Н
2 к со
? О е
К 3
д 0> -
со Э «2
о* —
>
О «
С о.
о
РЗ
1 т
I :
3 Е
•О*
о
о*
С
О*
со
я
си
I
5
Со
1,88 до 9,26%
шихся слоях.
золы против 0,91—5,17% в менее разложив-
153
Исследования Д. Бегака и Н. Беликовой (1934) показали,
что прослойки богаче микроорганизмами, чем соседние
с ними слои слабо разложившегося торфа. При общей кон¬
центрации микроорганизмов в верхнем слое торфяника (при¬
мерно от 20 до 60 см от поверхности) наибольшее количество
их (до 500 млн. бактерий на 1 г естественного влажного торфа)
приходится на первую прослойку. Ниже расположенные
прослойки гораздо беднее микроорганизмами, чем верхняя,
но все же их здесь обычно больше, чем в соседних слоях
менее разложившегося торфа. Повышенное разложение торфа,
особенности ботанического состава и большее содержание
микроорганизмов, вызывающих распад растительных остат¬
ков, —все это заставляет предполагать, что образование
прослоек происходило в более сухие климатиче¬
ские периоды. В это время поверхность болота несколько
обсыхала, улучшалась аэрация, усиливалась деятельность торфо¬
генных микроорганизмов, и на болоте создавались лучшие усло¬
вия для роста сосны. 3. Руофф(1. с.) определяла возраст про¬
слоек на Галицком Мхе методом пыльцы и пришла к выводу,
что прослойки образовались в сухие и теплые климатические
периоды, типа тридцати пятилетних периодов Брюкнера.
В XIX столетии было три таких периода: 20-е, 60-е и 80-е годы.
Им соответствуют две ясные разложившиеся прослойки.
В прежние столетия прослойки образовывались, видимо, реже,
может быть, потому, что влияние коротких теплых и сухих
периодов за последние столетия сказывалось особенно резко
вследствие уничтожения лесов и прямой осушки болот. Как бы
то ни было, мелкие разложившиеся слои в тор¬
фяниках (микростратиграфия) представляют
след климатогенных см ен растите л ьности, з ав и-
се в ш их, в о т л и ч и е от пограничного горизонта,
от климатических смен мелкого масштаба. Про¬
слойки разложившегося торфа отмечались во многих обла¬
стях различными исследователями (см. литературу в книге
3. Руофф, 1. с.). Их образование связывают обычно с климати¬
ческими изменениями. В последнее время Е. Гранлунд (1932)
в торфяниках южной Швеции установил наличность прослоек
климатогенного происхождения. Образование гумифицирован¬
ных прослоек, их число тесно связаны с климатом. Макси¬
мум развития наблюдается в континентальном климате сред¬
ней части Союза. В более влажном климате Ленинградской
области их не более трех (в верхней субатлантической толще),
в юго-западной Швеции их нет вовсе. Вообще при осадках
свыше 600 мм в год, если климат прохладный, прослоек
обычно не образуется. Однако в некоторых областях (Ирлан¬
дия) даже при осадках свыше 1000 мм вся залежь оказы¬
вается сильно разложившейся, причем в ней уже имеются
слабо разложившиеся прослойки. В этих случаях комбинация
довольно высокой температуры и большого количества осад¬
ков, вероятно, стимулирует разложение; слабо же разло-
154
шившиеся прослойки образовались в более сухие периоды
(3. Руофф, 1. с.).
При увеличении осадков выше известного предела их
влияние на залежь делается прямо противоположным (инверсия
факторов).
Наличием разложившихся прослоек не исчерпывается
микростратиграфия торфяника. В верхней олиготрофной толще
встречаются еще мелкие, а иногда и значительной мощности,
слои, образующиеся в основном благодаря эндодинамическим
сменам растительности.
Основные положения о строении торфяников можно
формулировать следующим образом:
1. Смена слоев торфа в торфянике в течение
его развития складывается из двух моментов,
часто противоположных: первый момент —
эндодинамический — сводится к торфонакопле-
нию, повышению поверхности торфяника, уда¬
лению ее от уровня грунтовых вод и к посте¬
пенному изменению питательного режима в
сторону ол иг от р о ф н о с т и; второй момент — со¬
вокупность различных внешних факторов. По¬
следние часто действуют в противоположном
направлении: они могут или задерживать эндо-
динамическую смену, или вовсе останавливать
ее, причем после остановки, развитие начи¬
нается снова (например, при заносе пойменного
болота аллювием), или направлять ее в обрат¬
ную сторону, или, наконец, ускорять ее, содей¬
ствовать ей.
2. Ведущая роль растительности в смене
слоев торфа, т. е. наибольшая „эндодинамич¬
ность“ смен, наблюдается в общем в зоне,
оптимальной для торф о н а к о п л е ния. С замедле¬
нием торфообразования как к северу, так и
к югу от оптимальной зоны возрастает роль
внешних факторов в смене торфяных слоев,
значение же растительности соответственно
ослаблено.
ГЛАВА VI.
ФЛОРА И РАСТИТЕЛЬНОСТЬ БОЛОТ.
Болотная флора, особенно сфагновых болот, — северного
происхождения. Еще Миддендорф установил, что форпо¬
стами северной флоры на юге являются сфагновые болота.
Гильберт (Hilbert) нашел, что из 125 видов, растущих в
северной Германии на сфагновых болотах, не менее 106, т. е. 85°/о*
распространены в Лапландии, Исландии, Сибири и в
Альпах (по Г. Шрейберу, 1927). Это вполне справедливо
и в отношении флоры болот Союза. Замечательно, что многие
виды, растущие в умеренных широтах исключительно или
преимущественно на торфяниках, на равнинах севера расширяют
свою экологическую амплитуду и встречаются на разнообразных
субстратах. Таковы, например, Betula папа, Empetrum nigrum,
Ledum palustre. Также ведут себя болотные виды и в горах.
Еще Р. Тольф (R. Tolf, 1897) отметил, что в высокогорной
области видовой состав на болотах и на минеральной почве
почти один и тот же; с другой стороны, по Клеве (А. Cleve,
1901), в высокогорной области Скандинавии Azalea procumbens,
Empetrum nigrum, Vaccinium uliginosum, Betula nana и др.
большей частью не встречаются на болотах, но растут на
минеральной почве (по Шрейберу). Соединяя болотные
виды по их ареалам, мы получаем такие группы (Г. Вальтер,
1927):
Настоящие бореальные растения ограничены в
своем распространении в основном хвойной зоной Европы
хотя некоторые из них заходят частью и в тундру. К ним
относятся: Scheuchzeria palustris, Drosera anglica, Cassandra
calyculata, Ledum palustre, многие виды осок и др.
Арктическо-бореальные виды широко распростра¬
нены в хвойной зоне, но заходят далеко и в тундру: Eriophorum
alpinum, Е. Scheuchzeri, Е. polystachium, Scirpus caespitosus, Carex
chordorrhiza, Betula nana, Drosera rotundifolia, Saxifraga hirculus,
Rubus chamaemorus, Empetrum nigrum, Vaccinium oxycoccus,
V. uliginosum, Andromeda polifolla. 11 Все перечисленные здесь настоящие бореальные и арктическо-бореаль-
ные виды встречаются и в Сибири.
156
Европейские виды распространены по всей Европе,
кроме крайнего севера и крайнего юга. Виды этой группы,
встречающиеся только или главным образом в Европе, лишены
индекса; виды, встречающиеся, кроме Европы, также и в Азии,
обозначены euroas; виды, встречающиеся, кроме того, еще и
в Америке, обозначены hol: Butomus umbellatus (euroas), Iris
pseudacorus, Filipéndula ulmaria (euroas), Comarum palustre
(hol), Impatiens noli tangere (euroas), Epilobium palustre (hol),
Calluna vulgaris, Menyanthes trifoliata (hol).
Флора болота по своей экологии отличается от флоры
минеральных почв. Особенно своеобразны и интересны в эколо¬
гическом отношении растения олиготрофных выпуклых болот.
В дальнейшем мы будем иметь в виду главным образом
именно эту группу растений.
Влажность болотного субстрата и распределение видов.
Хотя в общем принято считать, что болотная флора растет
на избыточно увлажненном субстрате, однако, в о-п е р в ы х,
болотные растения неодинаковы в своих требо¬
ваниях к почвенной влажности, и, в о-в т о р ы х, не¬
которые из них постоянно растути на сухом
•субстрате. Можно выделить такие группы
растений по фактору почвенного увлажнения:
1. Настоящие водные растения.
2. Растения очень мокрых местообитаний,
сильно обводненных мочежин, где вода тонким слоем в несколько
сантиметров стоит на поверхности или под поверхностью
болота: Scheuchzeria palustris, Carex limosa, C. rariflora, Rhyn-
chospora alba, Drosera anglica; из мхов: Cephalozia flúitans,
Sphagnum Dusenii, Sphagnum cuspidatum, Sphagnum balti-
cum и др.
3. Растения влажных местообитаний, где почвен¬
ная вода постоянно стоит ниже поверхности: Eriophorum
vaginatum, Andrómeda polifolia, Carex pauciflora (Scirpus caespi-
tosus). Эти растения имеют оптимум или в умеренно влажных
мочежинах, или в переходной полосе от мочежин к кочкам,
или на низких, плоских кочках.
4. Растения сухих местообитаний, более или менее
высоких кочек, где вода стоит на 20 см и более ниже поверх¬
ности: Ledum palustre, Cassandra calyculata, Rubus chamaemorus,
Oxycoccus microcarpa, Sph. medium, Sph. fuscum, Mylia
anómala и другие.
5. Растения очень сухих местообитаний, верху¬
шек высоких кочек, особенно торфяных бугров в тундре, откры¬
тых для иссушающих зимних ветров. К ним относятся главным
образом лишайники—виды кладоний (Cladonia silvática,
Cladonia mitis, Cladonia rangiferina, Cladonia alpestris и др.),
цетрарий (Cetraria nivalis), Ochrolechia tartarea, далее некото¬
рые мхи — тундровые Dicrana, Polytricha,
157
Выше шла речь о видах олиготрофных болот. Виды
эвтрофных болот очень трудно разделить на подобные
группы, потому что на не слишком влажных местообитаниях
на эвтрофных болотах появляются обычные на минеральной
почве луговые и лесные мезофиты. Это зависит от того, что
богатый солями торф и отчасти гидрологические его свойства
(часто сильная степень разложения, малая влагоемкость) до¬
пускают поселение мезофитов на повышениях микрорельефа.
На олиготрофных болотах бедность солями и другие свой¬
ства субстрата более или менее исключают возможность
поселения видов минеральной почвы, и даже сравнительно
сухие местообитания здесь бывают заняты специфическими
видами олиготрофных болот. Наглядное представление об от¬
ношении растений к влажности субстрата дают так называемые
экологические ряды. Обычные цветковые растения олиготроф¬
ных болот дают такой экологический ряд (схема 12):
Схе’ма 12
Экологический ряд важнейших цветковых рас¬
тений олиготрофных болот средней части Союза
по возрастанию увлажнения
Vaccinium uliginosum
Ledum palustre
i
Cassandra calyculata
Andromeda polifolia
Erlophorum vaglnatum
Rhynchospora alba
Scheuchzeria palustris
Сфагновые мхи дают такой ряд (схема 13):
Схема 13
Экологический ряд важнейших олиготрофных сфагновых мхов для
средней части Союза по убыванию увлажнения
Глубина грунтовых вод в см (приведена по А. Гребенщиковой, 1933)
от
до
средняя
0,5
Sph. cuspidatum
5
14
3
12
4
13,0
Sph. Dusenii
Sph. baltlcum
\ Мхи мочежин
—
—
14,0
Sph. paplüosum
J
35
25
30
Sph. angustifolium
> Мхи кочек
15
15
15
Sph. magellanicum
32
30
31
Sph. fusctun
)
158
Графическое вычерчивание амплитуд растений в отношении
факторов среды (в том числе и увлажнения) дает более полное
представление об экологии растения, чем экологические ряды.
Эти амплитуды могут быть вычерчены или в виде прямых
линий (рис. 31), или—что лучше — в виде кривых (рис. 32).
Глубина стояния воды в см. '§
, Empetrum nigrum |
CaUuna vulgaris. j
Sphagnum fuscum. \
Sphagnum magellanlcum. j
Erlophorum vaginatum. \
Sphagnum ru6ellum. j
Andromeda polifolla j
Scheuchzeria palustns j
Sphagnum Baltlcum !
Car ex limosa !
Rhgnchospora alda |
SphagnumDusenji.
Sphagnum cusp/datum. \.
Sphagnum angust/folium..\
Sphagnum papillosum j
Sphagnum apiculatum. j
0 5 10 15 W 25 ЗП 35 40 45 50 55
I I I * I■i 4 |
I I I
T-h !
i
•v
I
,1
J
'T
T
• •• Y««**
0 5 /0 15 20 25 30 35 40 45 50 55'
Олиготрофные сфагны более збтрофные сфагны
Высшие растения
Рис. 31. Амплитуда колебаний грунтовых вод, при которой доминируют
болотные растения. По И. Богдановской-Гиенэф, 1928.
P/nus silvestris- Pinus silvestr/s- Carex limosa- Scheuchzer/a pair
Ledum palustre- Eriophorum oagin-r Sphaanaceae Sphagnaceae
Cassandra calycr Sphagnaceae
Sphagnaceae
Рис. 32. Встречаемость вересковых в четырех ассоциациях сфагновых
болот, расположенных по возрастанию увлажнения. По Н. Кацу, 1926.
Питательный режим и распределение растений.
Издавна вошло во всеобщее употребление разделение болот¬
ных растений по их требовательности к элементам питания на
три группы: 1) олиготрофные, 2) мезотрофные и 3) эвтрофные.
1. Олиготрофные растения довольствуются мини¬
мумом зольных элементов. Раньше предполагалось, что
единственный источник питания этих растений — атмосферная
вода. Это, однако, не совсем правильно, так как влияние
грунтовой воды все же отчасти сказывается даже на раститель¬
ности выпуклых олиготрофных болот. Эти растения можно
назвать олиготрофными. К этой группе относятся
растения олиготрофных болот, корневые си¬
стемы которых полностью залегают вслоеоли-
готрофного торфа. Содержание золы в послед¬
нем невелико, в среднем от 2,5 до 3,4% в воз¬
душно-сухом торфе (данные для центральной части Союза).
2. Мезотрофные растения довольствуются умерен¬
ным содержанием зольных элементов. Их корневые си¬
стемы лежат вмезотрофном торфе с содержа¬
нием золы на воздушно-сухое вещество в пре¬
делах 4—б°/0.
3. Эвтрофные растения требуют высокого содержа¬
ния зольных элементов. Сюда принадлежат растения эвтрофных
болот. Корни их лежат в эвтрофном торфе с
содержанием золы свыше 5°/о-
Надо иметь в виду, что вообще не существует растений, при¬
уроченных лишь к одному типу болот. Особенно это относится к
цветковым растениям мезотрофной и дажеолиготрофной группы.
Почти все цветковые олиготрофные виды встречаются и на
болотах, богатых зольными элементами, а иногда даже доми¬
нируют там вместе с эвтрофными видами. Олиготрофные
цветковые не встречаются обычно на богатых солями субстра¬
тах лишь потому, что не могут здесь конкурировать с эвтроф¬
ными видами. Последние же, встречаясь на мезотрофных
болотах, реже заходят на олиготрофные, чем олиготрофные
виды на эвтрофные болота. Многие эвтрофные виды даже совсем
не встречаются на олиготрофных болотах и свойственны исклю¬
чительно мезотрофным и эвтрофным местообитаниям 1. Ниже
мы приводим списки растений трех указанных выше групп, при¬
чем растения каждой группы, встречаясь и в других группах, наи¬
более распространены в пределах своей группы местообитаний.
Олиготрофные виды: Calluna vulgaris, Empetrum nigrum, Vac-
cinium uliginosum, Ledum palustre, Cassandra calyculata, Andromeda pollfolia,
Oxycoccus microcarpa, Rubus chamaemorus, Erlophorum vaginatum. Drosera
rotundifolia, Dr. obovata, Dr. angllca, Rhynchospora alba, Scheuchzeria palustris, 11 В некоторых случаях появление эвтрофных растений на торфянике
с олиготрофной растительностью объясняется тем, что слой олиготрофного
торфа мелок и, следовательно, корни растений находятся в условиях более
богатого питания.
160
Сагех pauciflora, Sph. cuspidatum, Sph. Dusenii, Sph. balticum, Sph. ru bell am,
Sph. medium, Sph. angustifolium, Sph. fuscum, некоторые виды кладоний и
некоторые другие.
Мезотрофные виды1: Сагех lasiocarpa, Sph. amblyphyllum, Sph.
aplculatum.
Эвтрофные виды. Ainus glutinosa, Betula humllis, Сагех diandra,
C. dioica, C. caespitosa, C. paradoxa, C. gracilis, C. vesicarla и многие дру-
гие виды Сагех, Epipactis palustris, виды Orchis, Ophrys muscifera, Stellaria
crassifolia, Comarum palustre, Equlsetum, Heleocharis, Phragmites communis,
Menyanthes trifoliata, Galium palustre, C. uliginosum, Solanum Dulcamara,
Lycopus europaeus, Lysimachla vulgaris, Drepanocladns vernicosus, D. Sendtneri,
Calllergon giganteum, Camptotheclum nitens, Paludella squarrosa, Meesea
triquetra, Acrocladium cuspidatum и многие другие цветковые растения и
мхи. Сюда относится громадное большинство болотных видов; здесь при¬
ведена лишь незначительная часть их.
Отношение сфагнов к минеральным солям. Действие солей
кальция и некоторых других соединений на сфагновые мхи
было предметом довольно многочисленных специальных ис¬
следований.
Уже О. Зендтнер, И. Мильде (J. Milde, 1861) установили, что сфагны не
выносят оснований и в том числе извести. В. Ельман (V. Oehlmann, 1898)
установил, что чрезвычайно вредное действие на сфагновые мхи оказывают
не все соединения кальция, но главным образом углекислые, а затем серно- и
азотнокислые. М. Дюггели (М. Düggeli, 1903), поливая сфагновые мхи на их
естественных местообитаниях солевыми растворами, установил, что сфагны чув¬
ствительны вообще к минеральным солям, причем наиболее вредна для них
углекислая известь. При этом он нашел, что разные виды неодинаково
чувствительны к поливке солевым раствором. Основной вывод М. Дюггели,
что сфагновые мхи чувствительны к минеральным солям вообще. В этом он
примыкает к точке зрения П. Гребнера (Р. Graebner, 1898, 1901, 1904).
К. Вебер пришел на основании опытов к иному выводу, а именно, что
сфагны не чувствительны к извести. Несмотря на довольно длинную историю
вопроса, решить его в общих чертах удалось лишь Г. Паулю (1908). Он
установил прежде всего, что сфагновые мхи чувствительны к
углекислой извести и притом в неодинаковой степени.
Приведем результаты действия различных солей по данным Г. Пауля
(1. с.) на Sph. medium (табл. 43).
Опыты Г. Пауля показали, что сфагнум хорошо переносит нейтральные
соли самых различных кислот, например, CaSOé, Са(ИОз)2. K2S04, Na2S04,
KCl, NaCl, CaCl2, MgS04, а также слабокислые соли (KHS04,“NaHSO*). Он
может расти при больших концентрациях этих солей. Щелочные соли гораздо
хуже им переносятся и действуют гибельно при гораздо более слабых кон¬
центрациях. Вредно действуют уже такие щелочные соли, как КНСОз,—
МаНСОз, К2НРО4. Еще сильнее действие сильно щелочных К2С03 и Na2C03.
Очень сильно действуют щелочи. Таким образом, rio мнению Пауля,
сфагны чувствительны не к извести или каким-либо
другим соединениям, а к щелочной реакции среды. Он
объясняет это тем, что органические кислоты, вырабатываемые сфагнумом и
служащие для растворения твердых минеральных частиц, которыми он пи¬
тается, связываются щелочными соединениями, и растение таким образом ли¬
шается питания. Сфагнумы выпуклых болот в своем питании целиком зависят
не от растворенных в воде солей, а от твердых мельчайших пылевых частиц,
1 Выделение этой группы весьма затруднительно, так как видов, ясно
приуроченных к мезотрофным болотам, весьма мало. Многие из эвтрофных
видов весьма обычны и на мезотрофных болотах. Отметим, что центрально-
европейские болотоведы, например, Г. Пауль (Paul, 1910), под растениями
переходных болот подразумевают нечто иное по сравнению с мезотрофными
видами в нашем понимании. Равным образом, их понимание переходного
болота расходится с нашим.
11—722
161
которые доставляет им из воздуха атмосферная вода. Эти виды имеют наи¬
большую кислотность (см. ниже) и вследствие особенностей своего питания
очень чувствительны к связыванию своих кислот и вместе с тем к концен¬
трации щелочных соединений (табл. 44). Виды более богатых солями место¬
обитаний — эвтрофных болот и пр.—питаются в значительной мере растворен¬
ными солями, обладают меньшей кислотностью и менее чувствительны к ще¬
лочной реакции (табл. 44а).
Таблица 43
Действие различных солей на Sphagnum medium
Соединения
Содержание
в мг на 1 л
Окислы
Содержание
в мг на 1 л
Реакция мха
СаСО,
134
СаО
1
75
Отмирает
СаБ04
Са (N63)2
Концентр.
—
—
Рост продолжается
966
СаО
330
Отмирает
КНС03
ИаНСОз
240
к2о
225
170
Na20
125
К2СОд
ИазСОз
149
к2о
100
V
107
Na20
62.5
я
кнбо4
720
к26
52,0
я
ИаН504
340
Na20
к26
175
»
к2бо4
6480
3500
я
5725
Na20
2500
»
ИаС1
292
—
—
Сильно поврежден, но
не отмирает
КС1
372
—
—
Угнетается, но не
СаС12
кн2£о4
1100
.
отмирает
Не окончательно погиб
50
—
—
Доза выше смертельной
К2НР04
46
К20
25
То же
М£804
2400
—
—
Еще растет
н3ро4
—
25
Отмирает
Н2804
147
120
Предел жизни
шо3
82
—
Отмирает
КОН
56
—
—
я
ЫаОН
40
—
—
я
Таблица 44
•ЩГ
Виды сфагновых мхов
Sph. rubellum . .
Sph. papillosum .
Sph. molluscum
Sph. medium . . .
Sph. Dusenii. . .
Sph. angustifolium
Sph. platyphyllum
Sph. recurvum . .
Отмирание
произошло при
содержании мг
на 1 л воды
СаСОэ
СаО
77
43
89
50
89
50
134
75
178
100
223
125
223
125
312
175
Местообитания сфагновых мхов
Кочки на выпуклом болоте
Мочежины выпуклого болота
Болотистый лес
Низинное болото
На всех болотах
162
Таблица 44а
Кислотность сфагновых мхов с раз¬
личных местообитаний
(по Гулли)
Sph. rubelJum . . 100 1
Sph. medium . . . 86,7 I Выпуклое
Sph. papillosum . 84,2 | болото
Sph. molluscum . 81,2 I
fSh ¡¡^!!Г • Лесное болото
Sph. recurvum , . 63,3)
Sph. platyphyllum 50,0 Низинное болото
Исследование К. Ольсен (С. Olsen, 1921) и В. Мевиуса
(W. Mevius, 1921) подтвердили большое значение реакции среды
для сфагновых мхов и различие в амплитудах pH у различных
видов. И. Стоклаза (J. Stoklasa, 1922), культивируя сфагны,
нашел, что в сфагновых мхах содержатся свободные водород¬
ные ионы и что ионы натрия, калия, кальция и фосфора вред¬
но действуют на сфагновые мхи. Интересно, что некоторые
соединения алюминия, задерживая поступление в растение пе¬
речисленных ионов, ослабляют их вредное влияние на мхи.
Факт вредного действия щелочных соедине¬
ний на сфагновые мхи не подлежит сомнению
так же, как и необходимость кислой реакции
для роста ацидофильных видов сфагнов. Одна¬
ко вопрос о причинах чувствительности их
к щелочам весьма сложен и в настоящее вре¬
мя еще не разрешен. Объяснение Пауля не может быть
принято. Прежде всего сфагнум долгое время растет и в ди¬
стиллированной воде, вовсе обходясь без минеральных солей.
Кроме toro, наиболее олиготрофные виды, имеющие и наи¬
большую кислотность, должны были бы выносить наиболь¬
шую концентрацию щелочных соединений, а между тем они
оказываются наиболее чувствительными к последним. Кон¬
центрации солей, которые сфагновые мхи выносят в культуре,
не дают никакого представления о концентрациях солей
в естественных местообитаниях (табл. 45).
Предельные концентрации солей для Sphagnum medium
в культуре и содержание их в воде сфагновых
(выпуклых) болот на 1 млн. частей воды
Таблица 45
В культуре 1 В болотной воде
Na20 „
СаО в разных соединениях
к2о „ « п
62,5 —2500 1,8— 4,7
75—330 0,7-11,7
25-3500 2,2— 5,95
»
1 По Г. Паулю (1908).
2 По Рамзнну (1911).
11*
163
Концентрации солей на природных местообитаниях, таким
образом, во много раз меньше, чем в культуре. Выдерживать
столь высокие концентрация сфагнум может лишь при отсут¬
ствии конкуренции со стороны других растений. Несо¬
мненно, что в природе многие виды сфагнума
могут служить надежными индикаторами бед¬
ных солями, и в частности известью, местооби¬
таний.
Отношение цветковых растений 6oaoï к минеральным
солям. Простые наблюдения в нашей полосе показывают, что
цветковые олиготрофных болот чаще встречаются на эвтроф-
ных болотах, чем олиготрофные сфагновые мхи. Из этого
следует, что в природе они легче приспособляются
к более высоким концентрациям солей, чем
сфагновые мхи. Специальному исследованию подверга¬
лись лишь немногие растения олиготрофных болот, в частно¬
сти Calluna vulgaris. Этот типичный олиготрофный вид в при¬
роде встречается почти исключительно на бедных, с низким со¬
держанием извести, почвах. Лишь редко Calluna встречается
на почвах, содержащих до 3,4% карбонатов (по Г. Краусу,
1911), т. е. относящихся по Контежану (С. Conte jean, 1881) уже
к известковым.
Бюсген (М. Büsgen, 1913)1 культивировал вереск на грядках ра¬
ковистого известняка в течение пяти лет. Растение росло в высоту, но
было примерно вдвое ниже и менее обильно цвело, чем на песчаных грядках.
Обстоятельные исследования над влиянием извести при культуре вереска
в лаборатории, в почве и в водных растворах произвела М. Rayner (1913,
1921)1 2 В почве с содержанием СаС03 41,3% Calluna показал понижен¬
ную и замедленную всхожесть семян, остановку в росте и деформацию
главного корня и вообще всей корневой системы, а также отсутствие
роста наземного побега. В водных культурах кальция и магния Calluna рос
лишь при очень слабых концентрациях. Оптимальная концентрация для
него была 0,05%, при более же высоких всходы развивали всего одну-две
пары листьев и через два месяца большая часть их погибла. Любопытно,
что вересковые, как это давно известно, растут на кислых почвах,
как и сфагновые мхи. Между прочим известно, что вереск содержит щаве¬
левокислую известь и может адсорбировать значительные количества каль¬
ция. Возможно, что вредное действие извести зависит от изменения реакции
клеточного сока. Несомненно, что чувствительность вереска к извести не
подлежит сомнению, хотя причины этого до сих пор не выяснены. Воз¬
можно, что вереск и другие болотные Ericaceae близки в этом отношении
к сфагновым мхам.
Отношение болотных растений к реакции среды. Об отно¬
шении сфагновых мхов к реакции среды в условиях культуры
говорилось выше. И в природе разные виды сфагнов встре¬
чаются при различных pH. Р. Нордхаген (R. Nordhagen, 1928)
для норвежской области Sylene (Силене) приводит та¬
кие значения pH и делит сфагновые мхи по их кислотности
так:
1 По реферату 3. Руофф.
2 По реферату 3. Руофф.
164
1. Наиболее ацидофильные виды:
Sph. acutifolium .
Sph. fuscuin . . .
Sph. angustifolium
Sph. Kussowii . .
Sph. medium . .
Sph. Dusenii • . .
Sph. Jensenii . .
Sph. Lindbergii .
Sph. molluscum .
Sph. rubellum . .
Sph. compactum .
Sph. papillosum .
Sph. Girgensohnii
Sph. riparlum . .
pH
3.6
3.6
3.6
3.6
3.6
3.6
3,8; 3,9; 4,0; 4,1; 4,4
3,6; 3,7; 3,8; 3,9; 4,0; 4,1; 5,2; 5,3
3,8; 3,9; 4,0; 4,1
3,8; 3,9; 4,0; 4,1
3,8; 3,9; 4,0; 4,1; 5,2; 5,3
3,8; 3,9; 4,0; 4,1; 5,2; 5,3
3,6; 4,1; 4,2; 4,3; 4,4; 4,6; 4,8; 5,0; 5,2
4,0; 4,1; 4,8
2. Ацидофильность от сильной до умеренной:
Sph. Angstroemil... « 4,1; 4,8; 5,2; 5,3-
Sph. squarrosum 4,1; 4,3; 4,7; 4,8; 5,0; 5,1; 5;2
Sph. platyphyllum 4,7; 4,8; 6,1; 6,3
Sph. plumulosum 5,2; 5,3
3. Ацидофильность от умеренной до слабой:
Sph. contortum 16,0; 6,1; 6,2; 6,3; 6,7
Sph. subsecundum Lr
Sph. teres *
Sph. Warnstorfii 5,2; 6,0; 6,2; 6,3; 6,4: 6,5; 6,7
Sph. centrale 6,0—6,7
В.общем, для сфагновых мхов так же, как и для цветковых
растений, можно принять, что мхи, растущие в сильнокислой
среде, приурочены и к минимальному содержанию солей,
в частности, извести и являются олиготрофными. Виды же,
растущие в среде слабокислой или более или менее нейтраль¬
ной, более требовательны к минеральным солям и являются
более или менее эвтрофными. Выше были перечислены наи¬
более распространенные сфагновые мхи олиготрофных болот
нашей полосы: Sph. cuspidatum, Sph. Dusenii, Sph. balticum, Sph.
rubellum, Sph. medium, Sph. angustifolium, Sph. fuscum. Все эти
виды—резкие ацидофилы (табл. 46).
Таблица 46'
pH торфяной воды на олиготрофных болотах
средней части Союза для разных видов сфаг¬
новых мхов
(по А. Гребенщиковой, 1933) (средние цифры)
Sph. fuscum h 2 3,61—4,03
Sph. medium 2 3,48—4,22
Sph. angustifolium 2 3,60—4,30
Sph. Dusenii i» 2 3,44—3,98
Sph. cuspidatum 3,88—4,06
Sph. balticum 3,41—4,0
Sph. rubellum 1 3,64—3,71 1 21 Stenooxyphile Arten, по Г. Гаме и 3. Руофф (1929), имеющие pH = 3—4.
2 Наиболее ацидофильные виды, также по Р. Нордхагену (1928).
165'
Следующие обычные сфагновые мхи можно причислить
к эвтрофным: Sph. teres, Sph. subsecundum, Sph. platyphyllum,
Sph. contortum. Из них Sph. teres и два последних вида
в культурах Г. Пауля (1908) еще росли в щелочной среде,
a Sph. subsecundum1 в опытах К. Ольсена (1921) показывал
наилучший рост при сравнительно высоком pH=5,6. Следо¬
вательно, все эти виды слабо ацидофильны. Сюда же близко
стоят Sph. Warnstorfii, Sph. subbicolor, Sph. spuarrosum. Будучи
в наших условиях несколько более олиготрофными, чем четыре
приведенных выше, эти мхи встречаются на мезотрофных и эв-
трофных болотах, довольно богатых солями, и с невысоким pH.
Iris pseudacorùô
Рис. 33. Кривые вариации pH для двух стенотопных видов — ацидофила
(Eriophorum vaginatum) и вида нейтрально-щелочной реакции (Iris
pseudacorus). По К. Гюку, 1929.
Отношение цветковых растений к реакции среды в природе было обстоя¬
тельно изучено для Финляндии М. Котилайненом (М. Kotllainen, 1928). Все
исследованные растения он делит на ряд групп.
1- я группа — очень резко ацидофильные виды (pH
обычно от 3,6 до 4,0): Carex globularis, Ledum palustre, Rubus chamaemorus,
Galluna vulgaris, Carex pauciflora, Eriophorum vaginatum, Scheuchzeria palustris,
Empetrum nigrum.
2- я группа — виды с очень широкой амплитудой pH (эвритопные
виды). Главный фактор их распределения — не реакция среды, а избыточная
влажность. Сюда относятся: Andromeda polifolia, Carex rostrata, Eriophorum
polystachlum, Betula nana, Carex irrigua, C. filiformls, Drosera rotundifolla,
Dr. longifolia, Sclrpus caespitosus, Carex chordorrhiza, Oxycoccus palustris,
Carex limosa, Comarum palustre, Menyanthes trifoliata.
3- я группа — слабо ацидофильные виды. Они избегают наиболее khc-
лых субстратов и приурочены большей частью к средним (4,1—6,0) значе¬
ниям pH. В распределении этих видов играет роль, наряду с кислотностью,
и содержание кислорода и солей в почвенной воде, а также конкуренция.
Сюда принадлежат: Carex Goodenoughil, Eriophorum alpinum, Galium palu¬
stre, Phragmites communis, Salix myrtilloides, Carex dioica, Mollnia coerulea,
Poten tilla tormentilla, Selaginella selaglnoides, Carex heleonastes, C. panicea.
4- я группа. Очень слабо ацидофильные виды, живущие в почти
нейтральной среде. Сюда относятся: Saxifraga hirculus, Stellaria crassifoiia,
Schoenus ferrugineus, Carex diandra, Eriophorum latlfolium, Sallx myrslnites,
Carex flava, Filipéndula ulmaria, Crepls paludosa, Carex capillaris, Cypripedium
calceolus.
1 Все Sph. subsecunda и Sph. teres, по данным Г. Гамса и 3. Руофф
(1929), растут при сравнительно высоких значениях pH (4,0—6,5).
166 ’
Важно отметить, что, наряду с видами с очень широкой
амплитудой pH (2-я группа М. Котилайнена), есть растения
с узкой амплитудой, причем они приурочены или к низким
значениям pH (сильно ацидофильные виды 1-й группы), или,
наоборот, к высоким значениям pH (4-я группа) (рис. 33). Для
растений с узким значением pH (стенотоп-
ные виды) реакция среды — важный фактор их
рас[п ределения в природе1. Эти виды могут
служить поэтому индикаторами реакции среды
Carex Goodenougiw
Molima coerulea
Hola palustris.
Peucedanum palustre....
Callana uulgaris
Eriophorum uaginatum..
Car ex pañi cea
Vaccinium oxycoccus
Carex rostrata
Salix re pens
Comarum palustre
Geum riuale
Menganthes trilolíala..
Carex filiform is....
Galium palustre
Filipéndula almario
Phragmites communis...
Carex paradoxa
Cardamine pratensis
Pamassia palustris....
Рн =
¿t.o 5.0 So to
So
Рис. 34. Амплитуды pH различных болотных
видов Дании. По К. Ольсену.
при геоботанических исследованиях. В общем
можно сказать, что многие виды, изученные М. Котилайне-
ном, подобным же образом ведут себя и на наших болотах.
Наглядное представление о pH различных болотных растений
дает графическое изображение их амплитуд (рис. 34).
Распределение растений на болоте иногда хорошо соответ¬
ствует реакции среды. Рис. 35 показывает распределение
кислотности на одном болоте в Пруссии, а рис. 36—распре¬
деление четырех видов в окраинной зоне болота: Hottonia
palustris, Iris pseudacorus, Chrysosplenium alternifolium, Ranuncu¬
lus ficaria—виды нейтральных или слабощелочных местооби¬
1 Это не противоречит тому, что сама реакция среды в свою очередь
зависит от других факторов — содержания С02, карбонатов, гуминовых кис¬
лот и пр. (С Скадовский).
167
таний. Все они характерно приурочены к участкам с высоким
значением pH , встречаясь только на южном и западном бе¬
регу болота.
На выпуклых олиготрофных болотах pH так же, как и
другие сопутствующие факторы, изменяется в общем от пе¬
риферии к центру. Соответственно этому растительность здесь
также располагается концентрическими поясами.
Нарастание мха на болоте как экологический фактор.
Благодаря нарастанию мха и торфа поверхность сфагнового
болота медленно, но неуклонно растет вверх. Болотные растения
выработали различные приспособления против погребения их
нарастающим мхом. Можно различить здесь такие типы при¬
способлений:
168
1. Корневища, растущие вверх наклонно ил»
вертикально. Подобные корневища прирастают ежегодно
на известную величину прямо вверх или вкось, и таким обра¬
зом, следуют за нарастающим мхом. Вертикально восходящее
корневище имеют виды Drosera (росянка, рис. 37), наклонное—
Trichophorum alpinum (рис. 38). Впрочем, как показали обстоя¬
тельные исследования К. Метсевайнио (К. Metsevainio, 1931),
изучившего корневые системы 117 растений, часто один и
тот же вид в разных условиях дает то вертикальное, то более
или менее горизонтальное корневище (например, Scirpus caes-
pitosus, многие осоки, например, Carex rostrata, С. pauciflora).
При этом вертикальные корневища образуются в быстро рас-
Рис. 36. Распределение растений на болоте Плетцендибель в зависимости
от реакции среды. По К. Гюку, 1929.
Четыре вида слева направо: Hottonia palustris, Iris pseudacorus, Ranunculus ficaria, Chryso-
splenium alternifolium связаны с высоким pH.
тущем сфагновом ковре, а горизонтальные — в уплотненном,
сильно разложившемся и медленно нарастающем торфе. Кроме
того, длина междоузлий и подземных побегов больше в быстро
растущих моховых коврах.
К. Бертш (К. Bertsch, 1925) наблюдал вертикальные корне¬
вища на сфагновом болоте Brunnenholzried у многих растений:
Oxalis acetosella, Majanthemum bifolium, Knautia silvatica, Gen¬
tiana asclepiadea, Crepis paludosa, Valeriana dioica, Phyteuma
orbiculare, Polygonum Bistorta, Potentilla tormentilla, Luzula
pilosa, Carex fusca, Molinia coerulea, Listera ovata, Epipactis la-
tifolia, Lysimachia vulgaris, Menyanthes trifoliata, Equisetum
limosum. Многие из этих растений свойственны минеральной
почве, другие, если и встречаются на болотах, то обычно не
на сфагновых быстро растущих торфяниках. Из многочислен¬
ных, тщательных рисунков К. Бертша. мы приводим лишь
два — Oxalis и Majanthemum (рис. 39). Замечательно, что оба
они так же, как и все другие в приведенном списке, вырабо¬
тали на быстро нарастающем с ф а г н о в ом ко вре
типичные восходящие корневища, как у растений
169
быстро растущих сфагновых торфяников1. Этот пример лиш¬
ний раз подчеркивает большую пластичность растений.
2. Растущие вверх боковые по¬
беги корневища выполняют одну
и ту же задачу. Так, Сагех dioica
в быстро растущих болотах имеет
корневище, разветвленное на гори¬
зонтальные побеги, и восходящие
вверх ветви (К. Метсевайнио, 1. с.).
Вероятно, у ряда растений сфагно-
/Л
№г
№2г.
Рис. 37. Вертикально нарастаю¬
щее корневище росянки (Dro¬
sera rotundifolia) в торфе. Ви¬
ден годичный прирост. По В.
Сукачеву, 1926.
8,5 см
1325г,
Рис. 38. Косо нарастающее корневище
Trichophorum alplnum в торфе. По Метсе¬
вайнио, 1931.
вых торфяников с более или ме¬
нее горизонтальной главной осью
корневища (например, шейхцерия,
ср. К. Метсевайнио, 1. с.) подобные
ветви и побеги и являются приспо¬
соблением, выносящим ассимили¬
рующую систему кверху. Lysimachia
vulgaris, например, дает ежегодно
побег из боковых почек корневища,
растущий прямо вверх, причем эти
почки закладываются каждый год
все выше. Lycopodium annotinum,
Rubus saxatilis, Fragaria vesca и от¬
части Ajuga reptans pa сфагновом
ковре обычно избегают обрастания
при помощи своих надземных по¬
бегов. Наконец, некоторые растения
1 На болотах иных типов; имеющих медленный прирост, те же растения
развивают короткие, совершенно иного характера корневища.
170
избегают обрастания благодаря сильному вытягиванию гори¬
зонтального корневища, которое пронизывает сфагновую кочку
и достигает ее края. Так, корневище Paris quadrifolia прохо¬
дит в сфагновой кочке до 40 см, достигая ее поверхности.
Подобным же образом ведет себя и Anemone nemorosa (К.
Бертш, 1. с.).
Рис. 39. Вертикальное корневище ОхаПв и МаЗаШЬешит в сфагновой кочке.
Виден годичный прирост. По К. Бертшу.
3. Образование придаточных корней у дере¬
вянистых растений. Низкорослые деревянистые растения
сфагновых болот способны образовывать придаточные корни
на лежащем во мху и торфе корневище, которое сильно вет¬
вится, достигает значительной длины и образует по мере об¬
растания торфом и мхом многочисленные придаточные корни.
Таким образом, деятельность корневой системы все время со¬
средоточивается в верхних горизонтах. Старые же корневища
постепенно отмирают, так что части одной особи могут
171
обособиться \ На рис. 40 ясно видны более глубокие части кор¬
невища с слабо разветвленными, вероятно, уже недеятельными
корнями и более поверхностные, молодые части с густыми
сильно и тонко разветвленными придаточными корешками.
Широко развитая способность образовывать придаточные корни
свойственна всем обычным кустарничкам сфагновых болот,,
а также и лесным Ericaceae, попадающим на болото (см. К. Мет-
севайнио, 1. с.). Сюда относятся: Empetrum nigrum, Ledum ра-
lustre, Cassandra calyculata, Andromeda polifolia, Vaccinium Mvr-
tillus, V. vitis idaea, V. uliginosum, Oxycoccus palustris, O. mi-
Рис. 40. Корневая система Cassandra calyculata в сфагновом торфе.
По К. Метсевайнио, 1931.
crocarpa, Calluna vulgaris, Betula папа. У всех них главную роль-
играют придаточные корни. Главный корень не играет роли
даже тогда, когда он имеет длительное существование, как
это иногда бывает у Ledum и Calluna. Обычно он бывает не¬
долговечным, а придаточные корни, развивающиеся из корне¬
вищ, возникают уже в первое лето. Образовывать придаточ¬
ные корни в торфе могут и некоторые кустарники (Rhamnus
frángula), а также травянистые растения (Rubus chamaemorus,
Comarum palustre, Cornus suecica, Piróla minor, P. rotundifolia,
P. secunda, Viola palustris, V. epipsila (К. Метсевайнио, 1. с.).
Корневые системы болотных растений. Моховой ковер и
торф на болотах, особенно на сфагновых, представляют со¬
вершенно ' особую среду. Кроме способности торфа и мха
к нарастанию, весьма большое значение для растений имеют
еще три свойства болотной среды:
1) низкая температура уже на небольшой глубине под по¬
верхностью; 11 Для Cassandra calyculata отмечена длина корневища в 2,5 м (Метсе¬
вайнио, 1. с.), для Andromeda polifolia свыше 1,5 м, считая надземный стебель
(ibidem), для Betula папа-2л(В. Сукачев, 1926).
172
2) обильное содержание воды, зависящее от большой вла-
гоемкости торфа;
3) бедность кислородом.
Рост корневища вверх у болотных растений и способность
давать придаточные корни не служат только приспособлением
против затягивания мхом. Болотные растения таким образом
выносят подземные органы в верхний слой субстрата,
где тепловой и кислородный режим более бла¬
гоприятны. От этого зависит поверхностное
залегание подземных органов у болотных расте¬
ний; корни растений минеральной почвы идут
в общем гораздо глубже. У большинства болотных
растений корни проникают на глубину всего 10—15 см, иногда
же всего на глубину максимум 5 см. Лишь у немногих видов
корни идут глубже 50 см (К. Метсевайнио, 1. с., табл. 47).
Таблица 47
Глубина корневых систем у болотных растений
(по К. Метсевайнио, 1. с.)
Drosera longifolia и др
Rhynchospora alba, Eriophorum gracile, Drosera rotundifolia, Em-
petrum nigrum, Oxycoccus microcarpa, Galium palustre и др.
Scheuchzeria palustris, Carex vesicaria, Epilobium palustre, Oxy¬
coccus palustris, Crépis paludosa и др
Eriophorum alpinüm, Carex aquatilis, C. teretiuscula, C. loliacea,
Filipéndula ulmaria, Calluna vulgaris н др
Carex chordorrhiza, C. flava, С. pauciflora, Rubus arcticus, Peuce-
danum palustre, Vaccinium uliginosum и др .
Equisetum palustre, Betula nana, Cassandra, Andromeda и др. . .
Eriophorum latlfolium, E. vaginatum, Scirpus caespitosus, Carex
limosa, C. canescens, C. dioica, Ledum palustre, Carex Goo-
denoughii, C. caespitosa, C. globularis, Rubus chamae-
morus 35—60 см
Наиболее глубокая корневая система у Equisetum limosum fluviatlle (до
80 см), Phragmites communis (100 см), Carex filiformis (75 см), Eriophorum po¬
ly stachium (до 80 см\ обычно 25—30 см), Calla palustris (70 см).
Приведенные цифры не имеют абсолютного значения, так как глубина
корней у одного и того же вида значительно колеблется
в зависимости от условий (табл. 48).
Чем глубже грунтовые воды, тем ниже кор¬
невая система. Можно думать, что корни опускаются в
поисках за водой. Однако, так как глубина стояния грунто¬
вых вод несколько ниже глубины проникновения корней1,
можно предполагать, что растения избегают водоносного слоя
и держат свои корневые системы в общем несколько выше1 2 * *1 Особенно, если принять во внимание, что для корней даны максималь¬
ные глубины, а не средние, которые должны быть еще меньше.
2 С этим хорошо согласуется то обстоятельство, что свыше 9/10 всех
живых корней находится выше уровня грунтовых вод и лишь# около у10
ниже этого уровня (данные К. Метсевайнио).
Глубина
корней
не более:
5 см
Ю .
15 .
20 .
25 .
30 .
173
Таблица 48
Средняя максимальная глубина корней и высота грунтовых вод
(по К. Метсевайнио)
Тип болота
Средняя
высота
стояния
грунтовых
вод в см
Средняя
максимальная
глубина
корней
Кустарниковые болота
57,5
37,6
Лесные болота
28,8
22,0
Сфагновые болота
24.0
19,2
Буромоховые болота
17,1
13,5
в погоне за кислородом, содержание которого возрастает
только выше уровня воды. М. Котилайнен (1927) правильно'
считает недостаток кислорода одним из важнейших факторов
распределения растений на болоте. Влажность, как не раз
указывалось, — другой важнейший фактор, действие кото¬
рого подчиняется своеобразному »закону мак¬
симума". В общем—за сравнительно редкими исклю¬
чениями—почвенной воды растениям на болоте хватает, и при¬
том с избытком. Этот избыток и действует как ограничиваю¬
щий фактор. Багульник и подбел, например, всюду находят
на сфагновом болоте достаточное количество воды. Ограни¬
чивает их распространение не недостаток
воды, а ее избыток: в избыточно увлажненных
мочежинах они, как правило, не встречаются.
Одна из причин »ограничивающего“ влияния избытка влаги,
вероятно, бедность воды кислородом.
Анатомическое строение корней болотных расте¬
ний достаточно своеобразно. Как и у водных растений, их
корни, подобно надземным органам, имеют обычно хорошо
развитую систему воздухоносных полостей.
Значение этих полостей в значительной степени сводится
к снабжению подземных органов кислородом, содержание кото¬
рого в торфе, как отмечалось, весьма не велико. В общем
связь между глубиной проникновения корней и их строением
несомненна. У растений с поверхностной корневой системой
воздухоносные полости и межклетники развиты слабо. У бо¬
лотных же растений, корни которых проникают туда, где
кислорода особенно мало, аэренхима развита особенно сильно
(К. Метсевайнио).
Широко распространена у болотных растений микориз а—
мицелий гриба, симбиотирующего на корневых системах.
Двадцать два вида болотных растений—настоящие мико-
трофы, т. е. зависят в своем питании от грибных гиф, по¬
стоянно арисутствующих на них. Эти растения лишены
корневых волосков, которые заменяются, повидимому,
174
внутриклеточной микоризой. К микотрофам относятся все
обычные кустарнички сфагновых болот: Betula nana, Empetrum
nigrum, Ledum palustre, Cassandra calyculata, Andrómeda polifo-
lia, Vaccinium uliginosum, Oxycoccus palustris, O. microcarpa,.
Calluna vulgaris и некоторые другие растения. Относительно
некоторых из них установлено, что они облигатные (обязатель¬
ные) микотрофы и для развития требуют заражения грибом
(опыты М. Рейнер с Calluna, 1913) 7 Кроме отмеченных настоя¬
щих микотрофов, микориза обнаружена еще у 23 растений —
между прочим, у многих орхидей. Эти растения имеют, кроме
гиф, и корневые волоски.
Особенности древесной растительности на болотах.
Из древесных пород и кустарников на болотах Союза наи¬
более обычны: Pinus silvestris, Р. sibirica, Larix dahurica, береза,
(Betula pubescens), виды ольхи (Alnus glutinosa, A. incana,.
Alnaster fruticosus), Pinus pumita, Rhamnus frángula, виды ив,
ель и некоторые другие виды.
Pinus silvestris, —самое обычное дерево на сфагно¬
вых торфяниках Европейской части Союза и Западной Сибири.
Она растет здесь почти всюду группами на повышениях микро¬
рельефа, избегая мокрых понижений (мочежин), а на более
сухих болотах образует насаждения, иногда довольно густые.
Отличительная особенность сосны—неспособ¬
ность давать придаточные корни. Поэтому она на
торфяниках сильно угнетается сфагновым мхом и образует ряд,
экологических форм разной степени угнетения. Привожу описа¬
ние этих форм, данное Р. Аболиным (1915) по В. Сукачеву (1926).
1. Pinus silvestris L. f. uliginosa Abolin. Дерево
10—12 м высотой. Ствол более или менее прямой, стройный,
с шарообразной кроной, занимающей лишь треть и даже чет¬
верть длины ствола и состоящей из массы толстых корявых
ветвей. Обильно плодоносит. Предельный возраст 120—140
лет. Эта сосна растет на молодых неглубоких или старых су¬
хих торфяниках. Благодаря медленному нарастанию мха и тор¬
фа корневая шейка1 2 к концу жизни дерева погружена в торф
всего на 10—15 см. Это — наименее угнетенная форма сосны,
наиболее близкая к сосне минеральных почв.
2. Pinus silvestris L. f. Litwinowii Sukacz.
(рис. 41). Высота дерева 2 4 м. Ствол более или менее
прямой с овальной, яйцевидной, реже шаровидной кроной,
занимающей */г—7з ствола. Ветви довольно мелкие, отходят
от ствола под прямым углом. Предельный возраст 80—100 лет.
Произрастает на старых умеренно сырых болотах, где мох
растет быстрее, чем на торфяниках, на которых обитает пре¬
дыдущая форма; поэтому к 60—70 годам жизни дерева кор-
1 Последние исследования (М. Молл и ар — М. Molliard, 1937) показали
впрочем, что Calluna не оказывается облигатным микотрофом.
2 Корневая шейка — место отхождения главного корня от стебля.
175
■левая шейка погружается на глубину 30—35 см от поверх¬
ности. Вследствие более быстрого нарастания мха угнетение
сосны выражено гораздо сильнее, чем у Pinus silvestris f. uligi¬
nosa.
.Pinus silvestris L. f.
Wilkomii Sukacz. Де¬
рево 1—3 м высоты, ствол
прямой с тупоконусовидной
или цилиндрической кро¬
ной. Ветвление густое, все¬
стороннее, ветви отходят от
ствола под прямым углом,
начиная почти от корне¬
вой шейки. Плодоношение
обильное. Предельный воз¬
раст 70— 80 лет. Растет на
таких же торфяниках, как
и предыдущая, но более
светолюбива (рис. 42).
Pinus silvestris L. f.
p u m i 1 a A b o 1 i n. Куст
высотой 75—150 см, считая
от корневой шейки. Глав¬
ный ствол почти отсут¬
ствует или имеет всего 10—
30 см, так что густое вет¬
вление начинается почти от
корневой шейки. Шишки
редки. Предел жизни 60—
70 лет. Растет на очень
мокрых быстро растущих
болотах. Глубина погруже¬
ния корневой шейки 40—
60 см. Над поверхностью
мха поднимаются лишь от¬
дельные ветви; вся нижняя
часть погружена в торф.
Это наиболее сильно угне¬
тенная форма, найденная
Р. Аболиным на торфяниках
б. Псковской губернии
(рис. 43). Основной фактор,
вырабатывающий формы
Рис. 41. Pinus silvestris fo Litwinowii болотной СОСНЫ, вероятно,
Suk. Из Сукачева, 1926. быстрота нарастания торфа
и мха.
Сосна весьма нетребовательна в отношении содержания
минеральных солей и потому растет на олиготрофных место¬
обитаниях— как на песках, так и сфагновых торфяниках, куда
она вытесняется более сильными и требовательными конкурен-
176
тамн — елью и другими породами. Неспособность развивать
придаточные корни одна из главных причин ее угнетения на
сфагновых торфяниках. Повидимому, та же причина в значи¬
тельной мере объясняет и чувствительность сосны к
вечномерзлым торфяным грунтам. Можно сказать,
что сосна, за редкими исключениями, на таких торфяниках не
растет. Поднимающаяся с ростом торфа вечная мерзлота по¬
нижает температуру в области корневой шейки и быстро гу¬
бит дерево (Б. Городков)1. В Западной Сибири в области
Пура торфяники на
водоразделах делают¬
ся (по Б. Городкову),
(1. с.) безлесными там,
где глубина вечной
мерзлоты летом менее
1 м. Сосна пропадает
совершенно на торфя¬
никах севернее 64-й
параллели. Восточнее,
в Вилюйском районе
(по Р. Аболину, 1929)
сосна еще растет на
сфагновых торфяниках
с мерзлотой на глу¬
бине 25—45 см. Это,
вероятно, зависит от
того, что благодаря
теплому и сухому лету
в Якутии торф сверху
настолько нагревается,
что корни сосны функ¬
ционируют даже при
близкой вечной мерз¬
лоте. Также ведет се¬
бя сосна и на запад
от Урала. Б. Городков
(1932) считает присут¬
ствие соснового на¬
саждения на сфагновых торфяниках Северного края надеж¬
ным признаком отсутствия вечной мерзлоты. Крупнобугри¬
стые торфяники за редкими исключениями вовсе лишены
сосны. На северных сфагновых торфяниках, где еще нет веч¬
ной мерзлоты, сосна растет сравнительно хорошо и не обра¬
зует форм крайнего угнетения (вроде 1. рит11а), как на бо¬
лее южных и вместе с тем тяготеющих к Балтийскому морю
торфяниках. Причина этого по Б. Городкову — замедление
Рис. 42. Ршиэ БНуевЩв {о \УПокттИ Бик.
Из Сукачева, 1926.
1 Вечная мерзлота и растительность. Материалы КЕПС, № 80, Академии
наук СССР. Здесь же см. обстоятельный разбор распределения древесных
пород на вечномерзлых, в том числе и торфяных, грунтах.
12-722
177
к северу прироста сфагновых мхов, что позволяет сосне успеш¬
нее расти на торфяниках, там где не сказывается еще решаю¬
щим образом влияние низких температур.
Кедр сибирский, Ппиэ эИппса (кедровая сосна), растет
в Западной Сибири и на минеральных почвах, и на торфяни¬
ках. Интересные сведения об ее произрастании здесь приво¬
дит Б. Городков (1916). На южной границе хвойной зоны
Западной Сибири
кедровая сосна со¬
вершенно отсутст¬
вует на торфяниках,
заменяясь здесь сос¬
ной. Севернее кедр
начинает попадаться
на болотах, но в юж¬
ной половине этой
зоны его меньше,
чем сосны. Здесь он
растет в виде фор¬
мы, вполне аналогич¬
ной рямовой сосне
(f. папа РЫ1), лишь
более рослой Веч¬
ная мерзлота задер¬
живает распростра¬
нение сосны как на
минеральном грунте,
так и на торфяниках:
ее северная граница
в Западной Сибири
близка к южной гра¬
нице вечной мерзло¬
ты. Зато кедр растет
здесь сравнительно
хорошо не только на
мерзлых минераль-
Рис. 43. РИшэ вНчевЩв 1о ршп11а АЬоПп. ных почвах, но И на
Из В. Сукачева, 1926. вечномерзлых тор¬
фяниках, так как
образование придаточных корней выше слоя мерзлоты по¬
зволяет ему приспособиться к этому фактору, на что не¬
способна сосна, лишенная этого приспособления. Здесь на
вечномерзлых торфяниках кедр достигает даже 15 м высоты
при почтенном возрасте 250 лет. Хороший рост кедра здесь
объясняется медленным нарастанием сфагнового ковра, что
благоприятно для роста кедра, весьма чувствительного к бы¬
строму нарастанию сфагнума. На более южных болотах, где
нарастание сфагнума идет быстрее, кедр, несмотря на отсут¬
ствие здесь вечной мерзлоты, растет в виде более низкорос¬
лой и угнетенной рямовой формы.
178
Распространение сосны и кедра в бассейне Полуя. притока Оби, изучал
А. Лесков (1935). Он приходит к тем же выводам, как и Б. Городков
Сосна здесь встречается только на талых грунтах — в том числе и на лишен¬
ных мерзлоты торфяниках, как на мелких, так и на глубоких, сильно обвод¬
ненных. Кедр же хорошо растет здесь как на мерзлых торфяниках, где он дости¬
гает 6-8 л высоты и успешно возобновляется, так и на глубоких талых
сфагновых болотах, где он представлен более угнетенной болотной формой.
Подтверждая в общем данные Б. Городкова по экологии кедра и сосны на
торфяниках, А, Лесков (1. с.) указывает, что кедр избегает сухих с близком
вечной мерзлотой торфяников так же, как и вообще сухих субстратов, на
что в свое время указал еще Р. Поле.
Таким образом, основные черты экологии кедра следую¬
щие:
1. Стойкость в отношении вечно мерзлых тор¬
фяных грунтов, зависящая от способности да¬
вать придаточные корни.
2. Чувствительность к нарастанию сфагно¬
вого мха, вследствие чего на быстро растущих
торфяник а,х он вытесняется сосной.
3. Чувствительность к сухости местообита¬
ний, благодаря чему он избегает на севере
сухих торфяников.
Даурская лиственница (Ьапх баЬипеа) также характерна ллч
сфагновых торфяников Восточной Сибири и Дальневосточного
края, как сосна для Европейской части Союза. Она образует
на торфяниках сравнительно редкое насаждение. Даурская
лиственница чрезвычайно холодоустойчивая порода, образую¬
щая северную границу леса в Восточной Сибири в наиболее
суровой в климатическом отношении части лесной зоны всего
земного шара. Своей холодоустойчивостью лиственница обя¬
зана, вместе с другими признаками, крайне поверхностной
и пластичной корневой системе, почему она и растет на веч¬
номерзлых торфяниках. Кроме способности листвен¬
ницы переносить низкие почвенные темпера¬
туры, большую роль играет ее способность, как
и кедра, развивать придаточные корни выше
уровня мерзлоты.
Даурская лиственница свойственна не только вечномерзлым
торфяникам Восточной Сибири и Дальневосточного края.
В Биробиджане она постоянно встречается на торфяниках,
лишенных вечной мерзлоты, на сфагновых кочках, а местами
образует, по наблюдениям автора, на небольших пространствах
негустые насаждения. Негустое стояние, по сравнению с сос¬
ной, вообще свойственно лиственнице на торфяниках.
Болота с ярусом лиственницы наблюдаются и дальше на з.'пад, с районе
Хабаровска (Ц. Минкина и Р. Федорова, 1936).
В общем даурская лиственница, приспособленная к вечномерзлому тор¬
фяному субстрату, достигает на торфяниках значительного долголетия по
сравнению с болотной сосной (на Буреинском хребте, по данным В. Сочавы,
(1934) до 200 лет, большей же частью 150 лет). Тем не менее, даурская
лиственница все же образует на торфяниках угнетенные формы, более или
менее корреспондирующие болотным формам сосны. Эти формы, как и формы
сосны, характерны для определенных типов лиственничных болот. Так, в Vдеком
12 * 179
округе Дальневосточного края заболоченные сфагновые лиственничники
с рослой лиственницей (Laricetum sphagnosum) соответствуют Pinetum sphag-
nosum ленинградских ботаников. На торфяниках здесь наиболее обычен тип
Sphagnetum magno-laricosum, соответствующий Sphagnetum magno-pinosum.
Средняя высота лиственницы в этом типе около 2 м при диаметре у корне¬
вой шейки 4—5 сщ средний возраст 60—80 лет. Эта форма лиственницы
отчасти соответствует Pinus silvestris fo Lltwinowii. Более редок тип Sphag¬
netum nano-laricosum, аналогичный европейскому Sphagnetum nano-pi-
nosum. Лиственница здесь возвышается над мхом всего на 40—100 см, а ниж¬
ние части стволиков погребены во мху и торфе (А. Гожев, 1934). Эта форма
лиственницы более или менее аналогична сильно угнетенной Pinus silvestris ío
Lltwinowii. В Биробиджане, по моим личным наблюдениям, лиственница на сфаг¬
новых болотах обычно соответствует сосне, промежуточной между ío uliginosa
и fo Litwinowil. Встречается и сильно угнетенная форма, вроде fo Lltwinowii.
Е л ь в средней полосе Европейской части Союза довольно
обычна на безмоховых болотах намывного питания, где она
имеет хороший рост и нередко образует насаждения, часто
в смеси с березой и черной ольхой. Она встречается здесь и на
буромохобых болотах грунтового питания, хотя обычно еди¬
ничными деревьями. Будучи требовательной породой, на мезо-
трофных болотах ель обычно не растет и совсем отсутствует
в центральных частях олиготрофных сфагновых болот на мощ¬
ном сфагновом торфе. К северной границе лесной зоны ель
расширяет свою экологическую амплитуду, встречаясь в лишай¬
никовых редколесьях и заходя на сфагновые торфяники.
Эти торфяники относятся или к мезотрофным, или к олиготрофным
однако с примесью в растительном покрове эвтрофных видов. Залежь же
сложена сверху обычно мезотрофными (сфагново-осоковыми и др.) торфами,
так что корни ели все же располагаются в сравнительно богатом солями
торфе. Подобные олиготрофные торфяники с елью описаны для Колы
(К. Маляревский, 1926; Л. Соколова, 19361). Ель здесь образует редкий ярус,
часто с примесью сосны, и сильно угнетена. В Центральной Европе известны
случаи нахождения ели на выпуклых сфагновых торфяниках на мощном
олиготрофном сфагновом торфе. Например, она довольно обычна на болотах
Тюрингенского леса в Германии, на высоте 800—1000 м (К. Гюк, 1929).
Во всяком случае, ель как требовательная порода непри-
способлена к условиям бедных солями олиготрофных сфагно¬
вых торфяников с энергично нарастающим сфагновым ковром
и, за редкими исключениями, не встречается там. Притом на
сфагновых болотах она бывает резко угнетена. К. Бертш
(1925) описывает интересный случай обитания ели на быстро
растущем сфагновом торфянике. Деревцо в 12 лет имело
всего 40 см с небольшим в высоту, причем из мха выста¬
влялись лишь веточки длиной в 15—20 см, стволик же целиком
был в торфе. Ежегодный прирост дерева вверх равнялся всего
3,3 см, т. е. в десять раз меньше прироста на минеральной
почве. Интересно, что на месте отмерших мутовок ветвей
в торфе из ствола возникли придаточные корни. На способ¬
ность ели образовывать на болоте придаточные корни указы¬
вает и В. Гр. Лейнинген (W. Gr. zu Leiningen, 1907)1 2.
1 В Приуральской низменности (Северный Урал) единичная ель обычна
на олиготрофных сфагновых торфяниках.
2 Эта книжка специально посвящена вопросам распространения древес¬
ных пород на болоте и их экологии,
180
Из хвойных пород, свойственных только Западной Европе, заслуживает
внимания горная сосна — Pinus montana, растущая в горах Средней
Европы, на торфяниках и на минеральной почве. Все четыре расы этой гор¬
ной сосны встречаются на болотах. Горная сосна образует две характерные
формы роста, являющиеся, может быть, устойчивыми: одну — низкорослую,
кустарниковидную, типа криволесья (так называемую Latsche, Kuschel)
и другую — высокорослую прямоствольную (Splrke). Обе встречаются
на горных выпуклых болотах. В некоторых случаях распределение этих
двух форм по высотным поясам весьма характерно, и они оказываются
руководящими для определенных высотных типов болот: прямостволь¬
ная—для нижнего, а низкорослая—для верхнего (см. ниже). В общем
горная сосна — это экологический тип, несомненно, более приспособленный,
к существованию на сфагновых торфяниках, чем Pinus slJvestris. Об этом
свидетельствуют ее более высокий рост1 и способность образовывать очень
частое насаждение на 6оло1е, развивать густую крону и угнетать вследствие
эгого растительность, в том числе и сфагновые мхи (см. К. Рудольф, Ф. Фир-
бас и Г. Зигмонд —К. Rudolf, F. Flrbas u. H. Sigmond, 1S28).
Вне Европы из хвойных встречается на болотах кедровник
Pinus pumila Rgl. Он в виде стланца растет на маломощных
сфагновых торфяниках Восточной Сибири и Дальневосточного
края и способен образовывать придаточные корни1 2 3 *. Из лист*
венных пород на болотах обычны береза и ольха. Береза (обычно
Betula pubescens) встречается и часто образует насаждение на
эвтрофных болотах. Береза и черная ольха—наиболее влаговы¬
носливые из наших древесных пород. Береза может расти, хотя и
в сильно угнетенном виде, на очень сильно обводненных болотах
и притом с застойной водой, куда не заходит ни одна другая
древесная порода. Зато она более требовательна, чем сосна, и
не растет на неосушенных олиготрофных сфагновых болотах.
Alnus glutinosa (черная ольха) — типична для эвтроф¬
ных безмоховых болот намывного питания, где она часто
образует насаждения. Черная ольха чувствительна к застой¬
ной бескислородной воде и, вероятно, поэтому плохо мирится с
сплошным покровом болотных мхов. Поэтому она нехарактерна
для моховых эвтрофных болот, несмотря на богатство их со¬
лями, не говоря уже о сфагновых болотах. Интересно присут¬
ствие на корнях черной ольхи, а также у Alnus incana, A. vi¬
ridis и некоторых других видов 8 клубеньков, образование кото¬
рых вызывается микроорганизмами, относимыми в настоящее
время к актиномицетам. Эти организмы симбиотируют с ольхой
и способны усваивать атмосферный азот. Таким образом,
ольха оказывается собирателем азота в почве, что объясняют,
впрочем, не только деятельностью клубеньковых микроорга¬
низмов, но и богатством продуктов ольхового отпада азотом.
Во всяком случае, заслуживает внимания взгляд, что богатство
и нитрофильность травянистой флоры ольшанников зависят
1 Прямоствольная форма горной сосны может достигать на болоте
20 м в высоту (В. Лейнинген, 1907), кустарниковидная же на субальпийских
болотах — всего 20—100 см (К. Рудольф, Ф. Фирбас и Р. Зигмонд, 1928).
2 О нахождении кедрового стланца на болотах см. А. Миддендорф (1867),
В. Сукачев (1910), Б. Городков, Б. Тихомиров (1933).
3 См. Т. Работнов и Я. Меднис (1936); там же и история изучения
симбионтов ольхи.
181
от деятельности самой ольхи. В связи с этим заслуживает
серьезного внимания вопрос об использовании ольхи (главным
образом серой) для улучшения бедных лесных и луговых почв.
Измерение прироста мохового ковра и торфа.
На особенностях приспособления растений к обрастанию мхом
и торфом основан учет прироста торфа и мха на болоте. Этот
учет производится различным о б р а з о м; и результат,
и значение получаемых цифр также различны в зависимости
от метода измерения прироста1. Существует несколько мето¬
дов измерения прироста.
1. Непосредственно по самому сфагновому
мху измеряют прирост в тех случаях, когда граница между
участками мохового стебля прошлого и текущего года отме¬
чается исчезновением хлорофилла в прошлогоднем участке:
зеленая часть соответствует приросту текущего года. Прирост
сфагнума за год особенно легко измерить, когда он плодоно¬
сит год за годом, и на стебле остаются старые спорогонии
(В. Сукачев, 1926). Однако это бывает редко, и практически
этим методом не пользуются. Зато определение прироста по
зеленой части сфагнума, введенное в практику Д. Бегаком
(1928), в настоящее время широко применяется советскими
болотоведами. Этим методом мы определяем линейный
прирост отдельных растеньиц сфагнума за те¬
кущий год. Величина линейного прироста зависит (Д. Бегак,
1928): а) от вида сфагнума, б) для одного и того же вида —
от его местообитания (главным образом от степени увлажне¬
ния и климатических особенностей места и года). Достаточно
указать, что в Архангельском районе Брк ¡иэсиш
за 1934 г. показал прирост от 11 до 16 мм в год, а БрЬ. Ьа1-
Исит в том же районе — от 29 до 54 мм в год (Н. Кац,
М. Кириллович и Н. Лебедева, 1936). Для одного и того же
вида прирост тем больше, чем больше влажность. По тем же
данным, Брй. ЬаШсит в межкочиях дал прирост в 29 мм,
а рядом, в более влажных условиях, в мокрых мочежинах —
45—54 мм. Климат тоже влияет на линейный прирост: чем
дальше на север, тем прирост в общем меньше. Определение
линейного прироста важно еще потому, что дает исходный
материал для определения годичного урожая сфагновой массы
на торфянике. Этот урожай дает понятие об ежегодном накоп¬
лении органического вещества. Д. Бегак (1927) нашел, что на
верховом болоте Калининской области ежегодно прирастает
в среднем 2 тонны органического вещества на 1 га. Впрочем, ме¬
тодика подсчета урожая пока еще окончательно не выработана.
2. Определения прироста по травянистым ра¬
стениям производится по тем видам, которые образуют
1 Обзор исследований по приросту торфа см. Н. Кац, М. Кириллович
и Н. Лебедева (1936).
182
в торфе вертикальное корневище (см. выше). У нас в СССР для
этой цели пользуются исключительно росянкой (Drosera); за гра¬
ницей, кроме последней, также Eriophorum vaginatum и Scirpus
caespitosus. У росянки в торфе сохраняются вертикальные кор¬
невища с розетками листьев. Участок корневища между двумя
соседними розетками отрастает в один год. Благодаря этому
приспособлению, росянка избегает погребения нарастающим
мхом. Можно считать, что длина корневища между соседними
розетками близка к нарастанию сфагновой дернины за год.
Следовательно, по росянке мы измеряем уже не прирост отдель¬
ных растеньиц сфагнума, а годичный прирост всей
сфагновой дернины. При этом получаются величины
прироста, несколько отличные от получаемых первым методом,
большей частью меньшие, иногда гораздо меньшие (Н. Кац,
М. Кириллович, Н. Лебедева, 1936). Величина прироста, опреде¬
ленного по росянке, также зависит от вида мха, условий
местообитания (влажности), от ассоциации, в которой изме¬
ряется прирост, и от климатических условий—прирост тем
больше, чем южнее местность.
Так, для б. Орловской и Курской губерний был найден годичный прирост
в 48 и 45 мм, для Полесья и б. Псковской и Новгородской губерний—30,5 мм,
для района Ленинграда—28,6—30,8 мм, а для Колы—всего 8 мм (Д. Бегак, 1926).
3. Определение прироста по сосне. Прораста¬
ние сосны из семени на сфагновом болоте происходит в общем
на поверхности мохового ковра. С течением времени с нара¬
станием поверхности болота корневая шейка, т. е. место
отхождения от стебля главного корня, постепенно все глубже
погружается в мох и торф. Принимая корневую шейку не¬
подвижной, измеряют расстояние по вертикали от поверхно¬
сти болота до корневой шейки, т. е. глубину ее погружения.
Примем ее равной А см. Спиливая сосну у корневой шейки,
определяем ее возраст по числу годичных колец. Пусть этот
возраст равен а годам. Очевидно, это будет время, за кото¬
рое поверхность болота наросла на А см. Отсюда линейное
нарастание поверхности болота за год равняется í=^-cm.
Эта величина дает в отличие от предыдущих мето¬
дов нарастание сфагновой дернины и торфа вместе.
Так как сфагновая дернина с течением времени спрессовывается и
разлагается, то величина прироста по сосне будет меньше определенной
двумя первыми методами. Кроме того, чем старше сосны, по которым
определяется прирост, тем в общем меньше годичный прирост, так как
чем старше слой торфа над корневой шейкой, тем он сильнее спрессовался
и разложился (В. Сукачев, 1926). Следовательно, чем старше сосны, тем
ближе найденная величина характеризует прирост торфа, а чем сосны
моложе, тем ближе эта величина к приросту сфагновой дернины. Определе¬
ние прироста по сосне широко применяется в СССР и за границей.
Другие методы определения прироста основаны не на ра¬
стениях, живущих на торфянике, и здесь не рассматри¬
ваются1. Упомянем лишь, что прирост собственно
1 Обзор этих методов см. Н. Кац. М. Кириллович и Н Лебедева (1936).
183
торфа лучше определять за большой промежу¬
ток времени. Это возможно, когда удается хронологиче¬
ски датировать возраст определенного слоя торфа. Зная
его мощность, можно вычислить средний годичный прирост
за многовековой период. Однако датировка слоев торфа не
бывает абсолютно точной, и поэтому годичный прирост торфа
определяется лишь приблизительно.
Водный баланс растений выпуклых болот и вопрос об их
ксероморфизме.
Большинство растений на выпуклых олиготрофных торфяни¬
ках обладает рядом анатомических и морфологических призна¬
ков, свойственных растениям сухих местообитаний, т. е. ксеро¬
фитам. Поэтому можно говорить о ксероморфизме этих растений.
Обстоятельные исследования по вопросу о ксероморфизме бо¬
лотных растений произвел Ф. Фирбас (1931). Морфологическими
признаками ксероморфизма болотных растений служат:
1. Присутствие, на нижней стороне листьев волосков
(Calluna vulgaris, Ledum palustre), чешуек (Cassandra caly culata),
воскового налета (клюква, Andromeda polifolia, Vaccinium
uliginosum).
2. Свернутый нижней стороной внутрь (эрикоидный) лист
с устьицами, расположенными лишь на нижней стороне, и
притом в отчасти замкнутом пространстве, что уменьшает
испарение. В сильной степени свернут лист у Calluna и
Empetrum, а также у Ledum palustre, в меньшей — у Andromeda,
Oxycoccus. Ксероморфные черты в анатомическом строении
листа таковы (Ф. Фирбас, 1931):
1. Сильно кутинизированный эпидермис с одной стороны
или с обеих сторон листа, большей частью с сильно разви¬
той кутикулой, свойственен большинству растений выпуклых
болот: всем зимнезеленым и летнезеленым кустарникам, а так¬
же Scirpus caespitosus, Eriophorum vaginatum, Rubus chamae-
morus, Carex pauciflora и другим.
2. Погружение устьица ниже уровня эпидермиса, или вы¬
пячивание над устьицами соседних клеток. У Calluna, например,
устьица погружены в бороздку, и вход в нее закрыт волосками.
3. Уменьшение размеров отдельных листьев (Calluna,
Empetrum).
4. Образование водоносной ткани, например, больших бес¬
цветных клеток в губчатой паренхиме молодых листьев Ledum.
На основании этих морфолого-анатомических особенностей
Ф. Фирбас (I. с.) приходит к выводу, что вечнозеленые
Ericaceae и Empetrum имеют резко выражен¬
ный ксероморфизм и что у большинства летне¬
зеленых растений отчетливо выражены от¬
дельные ксероморфные признаки (особенно у
Eriophorum vaginatum, Trichophorum caespitosum, Carex pauci¬
flora, также у Vaccinium uliginosum, Myrica gale, Betula nana»
184
и в меньшей степени у Rubus chamaemorus). Из обычных у
нас видов выпуклых болот лишь Scheuchzeria и Rhynchos-
рога ксероморфны в слабой степени, а виды Drosera гигро-
мезоморфны, т. е. вовсе лишены ксероморфных признаков. На
этом основании Ф. Фирбас (1. с.) опровергает утверждение
К. Монфора (С. Monfort, 1918), который считал, что флора
выпуклых болот в основном состоит из гигроморфных видов
с примесью лишь немногих ксероморфных. Причина этого*
расхождения—слишком широкое понимание флоры выпуклых
болот К. Монфором, который включил сюда виды, не харак¬
терные для этих местообитаний.
Причиной ксероморфизма растений выпуклых болот давно*
считали затрудненную подачу воды, зависящую от „физио¬
логической“ сухости субстрата. „Физиологически“ сухим
в отличие от физически сухого называют субстрат, где
воды достаточно, но из которого она, в силу некоторых
свойств его, не может в достаточном количестве поступать
в растение.
Разные авторы выдвигали различные причины физиологической**
сухости торфяного субстрата (обзор взглядов по этому вопросу см. Г. Потонье*
г. 3, 1912 и Ф. Фирбас, 1939):
1. Одни видели в ксероморфизме результат задержки всасывания воды
корнями и причину этой задержки искали в различных факторах, как-то:
a) в низкой температуре торфяного субстрата (А, Чильман, 1890j.
К. Гебель —К. Gôbel, 1891 и мн. др.);
b) во временном высыхании почвы (С. Швендер — S. Schwender, 1882
и др., в последнее время также М. Котилайнен, 1928);
c) в высокой водопоглощающей способности торфа (Е. Варминг —
Е. Warming, 1896 и др.; позднее также Г. Потонье, 1912);
d) в слабой водопроводимости торфа (Оден — Oden и др.),
e) в кислотности торфа (гумусовые кислоты) (Шимпер— A. Scliimper,
1898 и др.; последнее время R. Japp, 1912);
i) в незначительном содержании кислорода (Е. Трансо—Е. Transeau,
1906; А. Дахновский — A. Dachnowski, 1908, 1909 и др.); *
g) в токсическом действии некоторых соединений в торфе (продукты
выделения корней, продукты жизнедеятельности бактерий) (Б. Ливингстон —
В. Livingston, 1905; Дахновский, 1908, 1909, 1910; Г. Ригг —G. Rlgg, 1913
и мн. др.);
h) в высоком осмотическом давлении почвенного раствора.
2. Ряд исследователей считали причиной ксероморфизма повышенное
испарение, зависящее от разных факторов, как-то:
a) от внешних — от временно наступающего высокого дефицита влажно¬
сти (К. Монфор, 1917), ветра (R. Japp, 1912) и т. д.;
b) от внутренних причин, например, от неспособности устьиц закрываться;
c) от сравнительно высокой транспирации, зависящей от недостаточного
развития корневой системы (И. Фрю и К. Шретер, 1904).
3. Фосс и Цигеншпек (Voss und Zlegenspeck, 1929) и В. Гаугер*
(Gauger, 1929) видят причину физиологической сухости в микотрофии
болотных растений (т. е. присутствии микоризы).
4. И. Пристли (I. Priestley, 1924) считает, что интенсивная выработка
жиров (при кутинизации и пр.) вызывает ксероморфную структуру.
5. А. Кернер (А. Кегпег, 1887) и некоторые другие до настоящего вре¬
мени считают некоторые анатомические признаки приспособлением против
смачивания водой.
6. Е. Варминг (1888) и др., а в последнее время и К. Монфор (1918)
объясняют особенности строения растении выпуклых болот историко¬
филогенетическими факторами.
185
Ксероморфно-морфолого-анатомические признаки, по данным Ф. Фирбаса
'(1931) такие:
1) сильно развитая сосудистая система (больше, чем у степных ксеро¬
фитов), особенно у растений мокрых сильно растущих сфагновых ков¬
ров;
2) уменьшение отношения поверхности ассимилирующих органов (если
принять во внимание воздухоносные полости) к живому весу (меньше
^ем у мезоморфных солнечных растений);
3) большое число устьиц.
У растений мокрых сфагновых ковров ксероморфные
признаки особенно резки. Вечнозеленые вересковые, растущие
в подобных условиях, отличаются от экземпляров более сухих
'болотных кочек повышенной ксероморфностью, а именно:
•более мощным эпидермисом, ббльшим числом устьиц, большим
развитием проводящей системы. Таким образом, мокрые
растущие сфагновые ковры особенно способ¬
ствуют выработке ксероморфных приспосо¬
блений у растений в той же степени, как и
солнечные минеральные почвы.
Из других ксероморфных признаков, уже физиологиче¬
ских, важно отметить высокое осмотическое дав¬
ление в тех же пределах, как у настоящих
ксерофитов при полном насыщении субстрата
водой.
Транспирация у болотных растений сксеро-
морфными признаками слабая, слабее, чем у мезо¬
морфных солнечных растений. Однако если учесть малое
содержание воды в растении, то обмен ее все же значи¬
телен. Интересно, что болотные растения приспособлены
к воспринятию воды корнями при низких темпе¬
ратурах. Так у Calluna, Andromeda, Oxycoccus перенос из
воды с температурой в 20° в воду с температурой в +1°
лишь незначительно замедляет поглощение воды кор¬
нями.
Между прочим, Ф. Фирбас установил, что испаряющая
сила воздуха на поверхности болота, измеряемая количеством
воды, испаряющейся на единицу свободной поверхности,
велика и лишь немного меньше испаряющей силы ксерофит-
ных местообитаний. Если еще предусмотреть высокую тем¬
пературу у поверхности болота в жаркий летний день и,
наоборот, низкую температуру в то же самое время в обла¬
сти корней, то микроклимат болота, особенно растущих
сфагновых ковров, следует признать неблагоприятным
для растений.
Из исследований Ф. Фирбаса можно сделать такие выводы: 11. Большая часть типичных растений выпук¬
лых болот (кроме Drosera, шейхцерии, очерет-
ника) несомненно, ксероморфны, обладая ря¬
дом м о рф о л о го - а н а т о м и ч е ск и х признаков на¬
стоящих ксерофитов.
186
2. Ксероморфные признаки растений выпук¬
лых болот особенно выражены на растущих, мо¬
крых сфагновых коврах. На таких местообитаниях у ра¬
стений наблюдается стимуляция устьичного испарения и уси¬
ление проводящей системы. Ксероморфные признаки у растений
этих местообитаний не зависят, таким образом, от затруднен¬
ности водного баланса растений Ч
3. На высоких кочках, где растения иногда
испытывают недостаток воды, вследствие сла¬
бой водопроводимости торфа, ксероморфные при¬
знаки (например, высокое осмотическое давление) можно
поставить в связь с затрудненным водным балан¬
сом, так как растения понижают в этих усло¬
виях свою транспирацию.
4. Растения , выпуклых болот представляют
своеобразную экологическую группу, по ряду
признаков близкую к ксерофитам, а по другим
от них отличную.
Надо иметь в виду, что для вечнозеленых вересковых
имеет значение выработка ксероморфных признаков в качестве
приспособлений против испарения в зимних условиях.
Болотные фитоценозы.
Классификация болотных фитоценозов. Необходимо
различать классификацию болот и болотных фитоценозов.
Болото характеризуется рядом признаков — растительности,
субстрата, формы поверхности и др. Описывая типы болот
(и торфяников) и классифицируя их, необходимо принимать
во внимание все эти признаки, выдвигая на первый план те
или другие из них. При классификации фитоценозов примени¬
тельно к современным воззрениям в фитоценологии за основу
берут растительность. Это, конечно, никоим образом не исклю¬
чает желательности при характеристике раститель¬
ных типов указывать и основные черты место¬
обитания. Эта точка зрения проведена и при классификации
болотных фитоценозов, которая приводится ниже. Следует
отметить, что между классификацией болотных типов и
фитоценозов часто даже в учебных руководствах не проводится
четкой границы. Так, В. Доктуровский (1935) под видом
классификации болот приводит в сущности классификацию
болотных фитоценозов (стр. 32). Ю. Цинзерлинг (1932), клас¬
сифицируя болотную растительность, всюду вместо „фитоце-
нозы" применяет слово „болота". Ниже мы приводим класси¬
фикацию и краткое описание болотных фитоценозов на основе
исследований по Европейской части СССР1 2.
1 Эти ксероморфные признаки, по мнению автора, зависят от химической
природы субстрата, что, однако, мало понятно и недостаточно обосновано.
2 Н. Кац, 1936; там же см. и литературу.
187
A. Эвтрофные фито ценозы намывного питания1.
I. Лесные фитоценозы намывного питания:
1. Ельники.
2. Березняки.
3. Черноольшанники.
4. Сероольшанники.
5. Сосняки.
II. Кустарниковые и кустарничковые фитоценозы намыв¬
ного питания:
1. Ивняки кустарниковые (лесной зоны).
2. Ивняки тундровые.
3. Кустарничковые фитоценозы Betula nana.
III. Травянистые фитоценозы намывного питания:
1. Фитоценозы с постоянным слоем воды на поверхно¬
сти (тростниковые, камышевые, рогозовые и др.).
2. Фитоценозы, периодически заливаемые водой (Cariceta
gracilis, С. vesicariae и др.).
B. Эвтрофные моховые фитоценозы грунтового
питания.
I. Лесные моховые фитоценозы грунтового питания:
1. Ельники моховые.
2. Березняки моховые:
a) буромоховые.
b) сфагновые.
3. Сосняки моховые:
a) буромоховые.
b) сфагновые.
II. Кустарничковые моховые фитоценозы грунтового питания:.
1. Буромоховые.
2. Сфагновые.
III. Травянистые фитоценозы грунтового питания:
1. Сфагновые.
2. Буромоховые.
3. Безмоховые.
C. Мезотрофные сфагновые фитоценозы.
I. Лесные мезотрофные сфагновые фитоценозы:
1. Ельники сфагновые.
2. Березняки сфагновые.
3. Сосняки сфагновые.
II. Кустарничковые мезотрофные сфагновые фитоценозы:
1. Фитоценозы Betula nana.
2. Фитоценозы Betula humilis.
1 Эту группу фитоценозов, соответствующею „заливаемым болотам
намывного питания* Р. Аболина (1929), мы, следуя этому автору, отделяем
от фитоценозов грунтового гитання.
188
3. Ивняки.
4. Фитоценозы болотных вересковых.
III. Травянистые мезотрофные фитоценозы:
1. Сфагновые.
2. Безмоховые.
D. Олиготрофные сфагновые фитоценозы.
I. Лесные олиготрофные сфагновые фитоценозы.
II. Кустарничковые олиготрофные сфагновые фитоценозы.
III. Травянистые олиготрофные фитоценозы:
1. Сфагновые.
2. Безмоховые.
E. Олиготрофные лишайниковые фитоценозы.
I. Ягельные фитоценозы с Cladonia (лесной зоны).
II. Ягельные фитоценозы с Cetraria и Cladonia (тундры и
лесотундры).
III. Корково-лишайниковые фитоценозы с Ochrolechia
(тундра).
F. Олиготрофные л и ш а й н и к о в о-д и к р а н о в ы е фи¬
тоценозы.
G. Олиготрофные политриховые фитоценозы.
H. Олиготрофные фитоценозы печеночников.
I. Олиготрофные фитоценозы P.leurozium Schreberi.
J. Фитоценозы водорослей на обнаженном торфе с слабым
развитием высшей растительности.
Описание болотных фитоценозов. А. Эвтрофные фито¬
ценозы намывного питания. Основной экологический признак
этих фитоценозов—принос элементов питания со стороны в рас¬
творенном состоянии и в виде ила речными, озерными, а иногда
и делювиальными водами. Субстрат—торф, перемешанный в раз¬
ной пропорции с илом, иногда ил. Местообитания этих фито¬
ценозов—берега и долины озер, рек, ручьев. Обычно богатое
питание и приток кислорода обуславливают пышное раз¬
витие травянистой растительности. Фитоценозы
этой группы дают в общем наибольшую продукцию
травянистой массы. Мхи, в частности болотные,
развиты слабой обычно не образуют сплошного
ковра. Постоянный глубокий слой воды или, в случае
временного заливания, сильно колеблющийся ее уровень
неблагоприятны для развития мхов, так же как и седиментация
ила. Приводим описание фитоценозов заиливаемых болот.
I. Лесные фитоценозы намывного питан и я.
Общий признак этих фитоценозов — древесный ярус. Травяни¬
стый покров развит хорошо и изреживается лишь там, где
имеется сомкнутый древесный полог. По видовому составу
это одна из наиболее богатых групп болотных фитоценозов.
Характерно постоянное участие мезофитов, и в
частности лесных видов, причем они часто бывают
189
доминантами. Это объясняется тем, что летом, после спада
воды, уровень ее опускается глубоко и местообитания оказы¬
ваются сравнительно сухими. Особенно это относится к кочкам
у стволов деревьев, часто довольно высоким. Эти кочки по
преимуществу и дают приют лесным видам. Эти фитоценозы
частью относятся к заболоченным землям с маломощным
торфом, частью к торфяникам. По преобладанию той или иной
древесной породы в этой группе различаются ельники, берез¬
няки, черноольшанники, сероольшанники и сосняки.
1. Ельники. Эта группа ценозов в общем наименее гигрофильна,
наиболее богата мезофитами и лесными таежными видами. По флористи¬
ческому составу она стоит наиболее близко к ельникам на минеральной
почве, с которыми связана и генетически. Мы различаем такие типы ельни¬
ков, расположенные, примерно, в порядке убывания влажности.
a) Ельники с господством настоящих болотных видов в понижениях
между стволами (Calla, Menyanthes, Сагех laevirostris и др.).
b) Ельник и-к очкарники с Сагех caespitosa.
c) Ельники с примесью березы и значительным развитием Filipén¬
dula ulmaria.
d) Елово-березовые леса с Filipéndula и крупными папоротниками.
e) Ельники с Filipéndula, крупными папоротниками и лесным широко-
травьем (Athyrium fllix íemina, Aconitum, Cirslum oleraceum, Oxalis и др.).
f) Ельники c Filipéndula, Equlsetum pratense и лесным широкотравьем,
близкие по составу к предыдущему типу.
g) Ельники с господством широкотравья и примесью Filipéndula.
Из первых — Aconitum, Geranium silvaticum, Veratrum lobelianum. Северный тип.
h) Ельники с папоротниками и лесным широкотравьем с участием
дубравных видов, а также широколиственных пород в подлеске. Южный тип.
2. Березняки. Многие из типов березняков, описанных ниже, анало¬
гичны типам ельников.
a) Березняки с редким покровом из мезофитов и болотных видов
(Betuletum turfosum 3. Смирновой, 1928).
b) Березняк и-к очкарники с Сагех caespitosa и часто еще с
Сагех paradoxa. Они широко распространены в средней части Союза. Глубина
торфа до 4,5 м.
c) Березняки с Сагех caespitosa, Filipéndula и широкотравьем. Имеют
аналога в ряду ельников.
Во всех трех первых типах доминирует Betula pubescens, в четвертом
же — Betula Kusmischeffli.
d) Березовое криволесье с ярусом кустарниковидной Betula
Kusmischeffii и широкотравьем (Veratrum lobelianum, Aconitum septentrionale
Geranium silvaticum, также Calamagrostis phragrmtoides и др.). Это березняки
Кольской лесотундры.
3. Черноольшанники подразделяются на такие группы (частично
по 3. Смирновой, 1928 г.).
a) Черноольшанники и березово-ольховые леса с
господством в понижениях между стволами деревьев болотных видов (Calla,
Menyanthes, Iris pseudacorus), крупных осок (Сагех rostrata, С. riparia). Виды
ельников на минеральной почве развиты слабо, лишь на кочках. Это наиболее
мокрые черноольшанники, обычно с глубоким торфом.
b) Ч е р н о о л ь ш а н н и к и-к о ч к а р н и к и с Сагех caespitosa. Более
дренированы, чем предыдущий тип. Обычны в средней части Союза.
' с) Черноольшанники с значительным развитием или
даже господством Filipéndula ulmaria. Обычны папоротники (Athyrium
filix femina), типичные для ольшанников виды (Scutellaria galericulata, Viola
epjpsila, Lycopus europaeus, Lysimachia vulgaris и др.), виды же ельников
представлены слабо. Эти ольшанники весной сильно заливаются, летом же
более или менее сухи. Торф —до 1 м и меньше.
d) Черноольшанники и березов о-о льховые лесас Filipén¬
dula и крупными папоротниками. Значение видов ельников здесь возрастает»
190
e) Березов о- и елово-черноольховые леса с сильным раз¬
витием на кочках лесного широкотравья, в том числе и дубравных видов.
В подлеске Tilia, иногда Ulmus, Acer. Олыианниковые мезофиты главным
образом в понижениях между стволами, болотные же виды представлены
слабо. Это наиболее эвтрофные и наиболее дренированные ольшанники
с мелким торфом. По флористическому составу и генетически они ближе к
еловым лесам.
f) Черноольшанники с господством Dryoptens thelypteris и других
болотных растений (Calla) в межкочечных западинах с стоящей на поверх¬
ности водой.
g) Черноольшанники с бедным травянистым покровом. Сильно
затопляются весной и слабо дренированы летом. Бедность травяного покрова —
следствие затенения ольхой, которая достигает здесь максимальной сомкну¬
тости крон и исключительного господства в деревостое.
h) Черноольхов о-б ерезовые леса с сфагновым покровом из
Sph. squarrosum, Sph. teres, Sph. riparium и др. Это наименее эвтрофная,
почти незаливаемая группа ольшанников. Она ближе к мезотрофным, чем к
эвтрофным болотам.
4. Сероольшанники. В некоторых случаях болотистые серооль-
шанники представляют, несомненно, вторичный тип; в других же случаях
вопрос о генезисе их не ясен. Ввиду почти полной неизученности серсоль-
шанников они здесь не рассматриваются.
5. Сосняки. Заиливаемые сосняки в общем редки. Из этой группы
известны сисняки-кочкарники с Сагех caespltosa.
И. Кустарниковые и кустарничковые фитоце^
нозы намывного питания..Сюда относятся фитоценозьГ
без древесного яруса, но с верхним ярусом кустарников или
низкорослых кустарничков (не выше 1 м). Под кустарниковым
бывает иногда еще травяно-осоковый ярус. Моховой покров
слабо развит, как и вообще на заиливаемых болотах. Здесь
различаются:
1. Кустарниковые ивняки из Salix cinerea, S. pentandra с примесью
кустарниковидной березы, ольхи и др. Эти ивняки в лесной зоне возникают
как дериваты лесных заиливаемых болот.
2. Кустарниковые или кустарничков ые тундровые
ивняки на местах, орошаемых проточной водой, часто с травянистым
ярусом (Сагех aquatihs и др.) под ивами. Из ив обычны Salix glauca, S. lap-
pon um, S. phyllcifolia, S. lanata, S. stipullfera, S. myrsmites, S. hastata. Эти
ивняки — излюбленные оленьи пастбища.
3. Кустарничковые фитоценозы с Betula nana. Встречаются
в тундровой зоне в тех же условиях, что ивняки.
III. Травянистые фитоценозы намывного пита¬
ния. В фитоценозах этой группы верхний ярус образуют
растения злакового типа или, реже, широкотравье. Эти ценозы
наиболее просты морфологически, часто одноярусны, с резким
преобладанием одного вида вплоть до чистых зарослей.
1. Фитоценозы с постоянным слоем воды, в которую по¬
гружены нижние части стеблей. В этих фитоценозах нередки настоящие
водные растения, и их можно отнести как к болотным, так и к
водным фитоценозам. Многие близкие ценозы с одним и тем же доминирую¬
щим видом могут быть отнесены как к этой, так и к следующей группе,
гак что резкой границы между обеими группами в сущности нет. В эту
группу входят: Phragmiteta communis, Typhaeta latifoliae, T. angustifoliae,
Scirpeta lacustrls, Scolochloaeta festucaceae, Equiseteta limosi, Saglttariaeta
sagittifoliae, Alismaeta Plantago, Acoreta Calami, Irideta pseudacorus, многие
ассоциации крупных осок (Сагех rostrata, С. gracilis, С. aquatilis) и другие.
Эти фитоценозы развиваются в зарастающих водоемах на месте ценозов
водных растений, с которыми они генетически связаны.
101
2. Фитоценоз ы, периодически заливаемые водой. Сюда
•относятся некоторые magno-Cariceta (С. gracilis, С. rostratae, С. vesicarlae,
С. vulplnae, С. strictae, некоторые С. pseudocyperus) и ряд других ассоциаций.
В. Эвтрофные моховые фитоценозы грунтовою питания
Характерные признаки этого типа:
1. Более или менее сомкнутый ковер эвтрофных болотных
Bryales (бурые мхи, или Braunmoose) или некоторых эвтрофных
сфагнов.
2. Питательные вещества доставляются грунтовой водой,
•а не приносятся в заметном количестве извне.
3. Избыточное или достаточное увлажнение и более или
менее устойчивый уровень грунтовых вод. Отсутствие седимен¬
тации и устойчивый водный режим благоприятны для болот¬
ных мхов. Избыточное и вместе с тем устойчивое увлажнение
сказывается в слабом участии в этих фитоценозах мезофитов,
в частности растений минеральных почв, и в резком преобла¬
дании болотных видов.
4. Продукция древесной и травянистой растительной массы
здесь сильно понижена по сравнению с фитоценозами намыв¬
ного питания вследствие менее благоприятного питательного
и кислородного режима.
Этот тип разделяется на лесную, кустарничковую и травяни¬
стую группы.
1. Лесные моховые фитоценозы грунтового
питания. Основной признак этого типа древесный ярус.
Под ним или ярус кустарников (иногда с нижележащими
травянистыми ярусами), или, же один, два и даже три яруса
травянистых растений. Общее число ярусов может достигать
пяти. Морфологическая структура лесной группы сложнее,
экологическая диференцировка компонентов резче, а видовой
состав богаче, чем в кустарничковой и травянистой группах.
Это следствие лучшего дренажа в лесной группе, допускаю¬
щего участие в составе растительности, кроме гигрофитов,
также и менее влаголюбивых растений.
Лесные фитоценозы делятся на такие группы.
1. Ельники моховые. Здесь пока известны три группы фитоценозов:
a) ценозы с господством болотного и лесного широкотравья и осок,
сюда относится Piceetum fontinale некоторых авторов с преобладанием
в напочвенном покрове Sph. teres и Mnium affine;
b) ценозы с господством на кочках у стволов лесных Hylocomia и лесных
и болотных вересковых; в понижениях доминируют болотные мхи (Sph.
Wamstorfii) и травянистые виды (Equlsetum heleocharis и др.).
c) ценозы с господством Equisetum silvaticum и сфагнов.
2. Березняки моховые. Это самые обычные лесные болота
грунтового питания. По преобладанию сфагновых мхов, или Bryales, мы
различаем здесь:
а) Березняки буромоховые с более или менее сомкнутым ков¬
ром болотных Bryales с двумя группами:
а—1. Березняки буромоховые кустарничковыес господ¬
ством под древесным ярусом кустарничков (например, Betula humills);
а—2. Березняки буромоховые травянистые. Это главная
группа буромоховых березняков; в травянистом ярусе господствуют: Сагех
paradoxa, Dryopteris thelypteris (часто в обоих случаях еще с нижним ярусом
192
Menyanthes и иногда с верхним ярусом из тростника), Сагех lasiocarpa
(иногда в комбинации с теми же ярусами), Menyanthes. Из мхов преобладают
Camptothecium nitens и Aulacomnium palustre.
b. Березняки c Sph. Warnstorfil, также с двумя группами
фитоценозов — кустарничковой и травянистой:
Ь—1. Кустари и чковые березняки с Sph. Warnstorfiiu
обычно с господством Betula humllis.
b—2. Травянистые березняки с Sph. Warnstorfil гораздо
шире распространены, чем кустарничковые. При господстве в моховом
покрове Sph. Warnstorfil здесь доминируют: Сагех paradoxa, Aspidium thelyp-
terls, Menyanthes, иногда Сагех caespitosa и др.
с) Березняки без моховые. Моховой покров разорванный. Эта
группа близка флористически к моховым березнякам и связана с ними
генетически. Деградация мохового ковра, вероятно, вторичное явление и
зависит от конкуренции с травянистым ярусом и некоторых других причин. По¬
этому мы относим эту группу к моховым березнякам. Распространена она слабо
3. Сосняки моховые делятся на группы, аналогичные березнякам.
a) Сосняки буромоховые. Под сосной господствуют или кустар¬
нички, или травянистые растения, а моховой покров образуют бурые мхи.
Сюда входят:
а-Л Сосняки буромоховые кустарничковые. Кустар-
ннчковый ярус образует Betula humllis, моховой покров — Aulacomnium pa¬
lustre и Camptothecium nitens. К северу Betula humills заменяется Betula nana.
а—2. Сосняки буромоховые травянистые. В травянистом
ярусе доминируют: Сагех caespitosa, С. paradoxa, Menyanthes, Сагех lasiocarpa
и другие виды. Моховой покров из тех же Aulacomnium и Camptothecium.
b) Сосняки с Sphagnum Warnstorfil делятся так же, как
—Кустарничковые сосняки с Sphagnum Warnstorfil.
Кустарничковый ярус образует Betula humilis (на севере). На средней
Печоре под Betula папа иногда второй ярус Cassandra и Andromeda, а
иногда еще и третий ярус Menyanthes. Эти фитоценозы в классификации
занимают положение между мезотрофными и эвтрофными;
Ъ—2. Травянистые сосняки с Sphagnum Warnstorfil.
В травянистом покрове господствуют осоки (Сагех caespitosa, С. lasiocarpa
и др.) или широкотравье (Menyanthes trifoliate), в моховом доминирует Sph.
Warnstorfil. II.II. Кустарничковые моховые фитоценозы
грунтового питания. При отсутствии древесного
насаждения верхний ярус образуют кустарнички, т. е. деревяни¬
стые растения не выше 0,8—1,0 м. Количество ярусов здесь
меньше, видовой состав беднее, экологическая диференцировка
видов слабее, чем в лесных фитоценозах грунтового питания,
что объясняется большей в общем влажностью субстрата,
чем в лесной группе. В связи с этим суживается число видов,
растущих на подобных местообитаниях, причем выпадают
менее влаголюбивые, настоящие же болотные виды решительно
преобладают так же, как и болотные мхи. Роль мхов усили¬
вается за счет других ярусов, так же как и влияние их на
остальную растительность. Необходимо различать по моховому
покрову две группы кустарничковых фитоценозов, с дальней¬
шими подразделениями внутри каждой группы.
Ь Кустарничковые буромоховые фитоценозы по
господствующему кустарничку делятся:
а) Ценозы с господством Betula humilis, иногда с ярусом
вахты или сабельника под березкой. В моховом ковре чаще всего господ¬
ствуют Camptothe.eium и Aulacomnium palustre.
и омоховые, на две группы:
13-722
193
b) Ценозы c Betula папа, обычно с господством Camptothecium mtens,
покрывают гряды на эвтрофных болотах на севере лесной зоны. Иногда
Betula папа образует ценозы с другими мхами — Paludella squarrosa и даже
с гигрофильными Drepanocladus (D. vernlcosus), в последнем случае совместно
с осоками.
c) Ценозы с преобладанием низкорослых ив (Salix lanata, S. glauca)
и мхов (Philonotis fontana, Mnium, Aulacomnlum идр.). Встречаются на Коле.
2. Кустарничковые сфагновые фитоценозы. В напочвен¬
ном покрове доминируют эвтрофные мхи (Sph. Warnstorfli, Sph. subbicolor).
Здесь различаются:
a) Ценозы с ярусом Betula humilis ис ковром Sph. W arnstorfii.
b) Ценозы с Betula папа и сфагновыми мхами — тем же Sph»
Warnstorfli, а иногда с Sph. subsecundum и Sph. obtusum.
c) Ценозы с экологически гетерогенными доминантами разных ярусов —
например, в верхнем Betula папа, в следующем—Cassandra или Andromeda,
еще ниже—иногда ярус вахты и моховой ковер из Sph. subbicolor.
d) Подобные же фитоценозы с верхним ярусом Cassandra (или Andromeda)
н нижним из вахты при господстве Sph. Warnstorfli.
III. Травянистые фитоценозы грунтового пи¬
тания. Верхний ярус здесь образуют травянистые растения;
кустарничковый и древесный ярусы отсутствуют. Основные
признаки этого типа следующие:
1. Безусловное господство типичных гигрофитов и почти
полное отсутствие мезофитов.
2. Экологическая недиференцированность отдельных членов
фитоценоза—все они весьма требовательны к влажности
субстрата.
3. Простота сложения ценозов—большей частью из двух
ярусов.
4. Сильное развитие мхов, определяющих условия жизни
верхнего яруса.
5. Бедный видовой состав, особенно сравнительно с лесной
группой.
Все эти признаки зависят от избыточного увлажнения.
Оно в общем выше, чем в древесной и кустарничковой
группах. Здесь следует различать буромоховые, сфагновые
и безмоховые фитоценозы (без сплошного мохового ковра).
1.Травянистые буромоховые фитоценозы с рядом
подразделений.
a) Осоковые фитоценозы с Calliergon giganteum.
Этот мох образует часто полупогруженный, залитый водой, трудно доступ¬
ный ковер. Из цветковых доминируют Carex diandra, С. rostrata, Menyanthes.
На севере (Канин) в подобных условиях встречаются Calliergon cordifohum
с Carex aquatilis.
b) Фитоценозы с господством мхов из рода Drepanocladus
(D. vernicosus, D. lycopodioides, D. Sendtneri, также D. intermedlus, D. exan-
nulatus идр.), Acrocladlum cuspldatum, Scorpidium scorpioides, Meesea triquetra,
Philonotis fontana, Calliergon, tnfarium, иногда Paludella squarrosa, болотные
виды Mnium и др. В травянистом ярусе господствуют Сагех, а также
Menyanthes, Coma rum, Equisetum Heleocharls, Dryopterls thelvpteris и другие
виды. Эта группа развивается при меньшем увлажнении, чем предыдущая,
но все же грунтовые воды лишь на несколько сантиметров ниже поверхности.
c) Ценозы с господством мхов Aulacomnium и Camp¬
tothecium с Сагех rostrata, С. limosa, С. laslocarpa, Menyanthes и другими
в травянистом ярусе. Уровень грунтовых вод здесь нередко на несколько
десятков сантиметров ниже поверхности.
194
2. Травянистые сфагновые фитоценозы имеют сплош¬
ной ковер эвтрофных сфагнов и разделяются так:
a) Осоково-сфагновые ценозы с господством Sph. teres и Sph. subsecun-
dum, реже Sph. platyphyllum, Sph. obtusum и с ярусом из различных Carex.
Это сильно увлажненные фитоценозы.
b) Осоково-сфагновые ценозы с господством Sph. Warnstorfii (уровень
грунтовых вод здесь ниже).
3. Травянистые безмоховые фитоценозы. Сплошной
моховой ковер отсутствует, мхи встречаются отдельными дернинами и обычно
угнетены. Это вызывается длительным застаиванием здесь бедной кислородом
воды и часто еще отравлением растительности газообразными продуктами
разложения торфа. Безмоховые фитоценозы возникают по тем же причинам
и на мезо- и олиготрофных травянистых болотах. Всюду они занимают на
болотах понижения — мочежины — и называются „римпи" \
Подобные „римпи" очень обычны в Карелии и на Коле. В них домини¬
руют из цветковых главным образом осоковые (Carex lasiocarpa, С. limosa,
С. lívida, С. rotundata, С. rariflora и др.) Erlophorum angustifolium, Е. russeolum,
Trichophorum caespitosum, иногда Menyanthes, Equisetum heleocharis и др.
Многие из этих растений образуют ярус не только в эвтрофных, но и в ме-
зотрофных и олиготрофных римпи, причем сопровождающие растения в
этих случаях различны.
С. Мезотрофные сфагновые фитоценозы. Основные при¬
знаки этого типа:
1) обычно сплошной ковер мхов, преимущественно сфаг¬
новых,
2) питание за счет вод, бедных солями и в частности из¬
вестью,
3) избыточное или достаточное и притом обычно устойчи¬
вое увлажнение, благоприятное для развития мхов.
В мезотрофных фитоценозах доминируют или мезотрофные
растения, оптимум которых лежит в мезотрофных условиях,
или же олиготрофные и эвтрофные виды доминируют в одном
и том же фитоценозе, часто в разных ярусах.
Мезотрофные фитоценозы делятся на лесную, кустарничко-
вую и травянистую группы с дальнейшими подразделениями,
аналогичными фитоценозам грунтового питания.
I. Лесные мезотрофные сфагновые фитоце¬
нозы. Основной признак этой группы ■ древесный ярус. Под
ним или кустарничковый ярус, иногда еще с нижним травяни¬
стым, или травянистый. Последний бывает иногда расчленен
на два, редко на три яруса. Лесная группа мезотрофных фи¬
тоценозов сложнее по своей морфологии и богаче по видо¬
вому составу, чем кустарничковая и травянистая мезотрофные
группы, вследствие лучшего дренажа. По сравнению же с лес¬
ными фитоценозами грунтового питания лесные мезотрофные
фитоценозы проще устроены и беднее видами, что зависит
от худшего питания. Лесные мезотрофные фитоценозы де¬
лятся по признаку господства древесной породы.
1. Сосняки сфагновые мезотрофные с рядом групп:
а) Сосняки сфагновые кустарничковые. В этой группе
олиготрофные кустарнички, в том числе вересковые, господствуют или во 11 Финское слово.
13*
195
всяком случае очень обильны. Моховый ковер обычно сплошной. В нем
иногда участвуют олиготрофные сфагновые мхи (например, Sph. médium),
но постоянны и более требовательные—Sph. recurvum s. 1., Sph. Russown,
Sph. Warnstoríü, Sph. subblcolor, иногда Sph. acutifoilum и др. Иногда зна¬
чительна роль лесных мхов. Кроме состава мохового ковра, мезотрофныи
характер этим ценозам придает в одних случаях примесь, иногда значитель¬
ная, мезотрофных видов (например Сагех lasiocarpa), а также участие
эвтрофных цветковых. Эти фитоценозы в значительной части относятся
к торфяникам. В других кустарничковых мезотрофных сосняках постоянна
примесь лесных видов. Эти сосняки с господством то Ledum, то Cassandra, то
Vaccinium ullginosum, то Betula nana со сфагновым покровом из Sph. acuti-
folium, Sph. angustifolium, Sph. Russowii и др. относятся к заболоченным
лесам обычно с мелким торфом.
Ь) Сосняки сфагновые травянистые. Это широко распро¬
страненная группа. Сюда относятся, прежде всего, типичные мезотрофные
фитоценозы с господством в травянистом покрове Сагех lasiocarpa и в мо¬
ховом—Sph. recurvum. Эти фитоценозы играют наибольшую роль в группе.
Иногда фитоценозы с Сагех lasiocarpa имеют до шести ярусов: 1) сосна,
2) тростник, 3) Сагех lasiocarpa, 4) Dryopteris thelypteris, 5) Menyanthes,
6) Sphagnum subblcolor + Sph. recurvum. Эти фитоценозы носят мезоэвтроф-
ный характер. К этой же группе относятся сфагновые фитоценозы с Moli-
nla coerulea (в Карелии), заболоченные сфагновые сосняки с Calamagrostis
lanceolata, с Сагех globulans и некоторые другие.
2. Березняки сфагновые мезотрофны е—широко распростра¬
ненная группа. Здесь можно выделить:
Березняки сфагновые травянистые. Сюда относятся ти¬
пичные, большей частью трехъярусные мезотрофные березняки на более или
менее глубоком торфе с господством Сагех lasiocarpa и сфагнов (Sph. recur¬
vum, иногда Sph. subbicolor и некоторых других видов). Иногда к осоке
присоединяется еще верхний ярус тростника и нижние ярусы из Dryopteris
thelypteris и иногда еще Menyanthes. Эти фитоценозы более эвтрофны и
аналогичны описанным выше фитоценозам сосновых болот. Аналогичные
сосновой группе фитоценозы образует под березой и Calamagrostis lanceo-
lata. Во всех этих фитоценозах доминирует Betula pubescens. Березовое криво-
лесье на Коле с ярусом Betula Kusmischeffii и с обильной морошкой также
относится к мезотрофным березнякам.
3. Ельники сфагновые мезотрофные В эту слабо изученную
группу входят главным образом заболоченные ельники и частью фитоценозы,
переходные от торфяников к заболоченным лесам, реже фитоценозы с мощ¬
ным торфом. Следует различать кустарничковые и травянистые фитоценозы:
a) Ельники сфагновые кустарничковые. Из них известны
ельники с ярусом Ledum, с ярусом Betula папа (в этом случае нередко еще
нижний ярус из вересковых, а иногда еще, кроме того, и яр>с морошки)
и некоторые другие.
b) Ельники сфагновые травянистые, куда относятся, между
прочим, заболоченные леса с Сагех globularls, фитоценозы С ярусом Сагех
lasiocarpa и фитоценозы с ярусом Rubus chamaemorus— обе последние группы
с более глубоким торфом. II.II. Кустарниковые мезотрофные сфагновые
фитоценозы с верхним ярусом из низкорослых деревяни¬
стых растений — кустарничков или невысоких кустарников.
Выше было отмечено, что эти фитоценозы беднее видами и
проще устроены, чем мезотрофные лесные. Для кустарничко-
вых фитоценозов лесной зоны это зависит от большего в
общем увлажнения, для кустарничковых же фитоценозов
тундры, кроме того, и от климатических условий. И по сравне¬
нию с кустарничковой группой грунтового питания мезотроф¬
ные кустарничковые ценозы также беднее видами, что зави¬
сит от худшего питания.
196
По доминирующему к\старничку различаются:
a) Це н о з ы с В е t u fa humilis.
b) Различные сфагновые фитоценоэы с Betula папа
в лесной зоне и за ее пределами.
c) Ценозы кустарничковых ив (Salix repens, S. myrtilloides,
S. lapponum), встречающиеся как в лесной зоне, так и в тундре, например
Кольской.
d) Ценозы с господством олиготрофных вересковых
(например, Andromeda) со Sph. recurvum и с участием эвтрофных видов.
Сюда же относятся и многие типы заболоченной кустарничковой тундры.
JII. Травянистые мезотрофные фитоценозы.
Верхний ярус здесь образуют травянистые растения, чаще
узколистные (преимущественноосоковые), реже широколистные.
Видовой состав и структура фитоценозов этой группы в общем
беднее и проще, чем в кустарничковой мезотрофной группе,
вследствие большего увлажнения. От травянистых моховых
эвтрофных фитоценозов эта группа отличается более бедным
солевым режимом и более бедным видовым составом.
1. Травянистые сфагновые фитоценозы с такими группами:
a) Ценозы с Сагех Jasiocarpa с господством Sph. recurvum, Sph.
obtusum, Sph. papillosum, иногда Sph. riparium, Sph. Dusenii, Sph. subbico¬
lor, Sph. subsecundum, Sph. Wamstorfii и некоторых других. Широко
распространенная группа.
b) Ценозы с Сагех rostrata с господством Sph. recurvum, Sph.
obtusum также широко распространены.
c) Ценозы с Trichophorum caespitosum, чаще с ковром Sph.
papillosum. Обычны в Карелии.
d) Ценозы с преобладанием Сагех chordorrhiza и С. limosa
с Sph. recurvum.
e) UeH03biMenyanthes с Sph. Jensenii, Sph. riparium, Sph. recurvum,
Sph. subbicolor.
f) Ценозы Scheuchzeria с некоторыми видами Drepanocladus (D.
exannulatus и др.), а также c Sph. papillosum и ряд ценозов других цветко¬
вых растений,
2. Травянистые безмоховые фитоценозы. Отсутствие
сплошного мохового покрова в этой группе объясняется так же, как и
в безмоховых эвтрофных фитоценозах, длительным вымоканием, отсутствием
кислорода, иногда отравлением мхов газообразными продуктами разложения
торфа. »Голые“ (безмоховые) мезотрофные фитоценозы распространены там
же, где и эвтрофные без мхов (ем.-выше). Изучена -оте-сяябе.-
Олиготрофные фитоценозы. Общий признак этих фито-
цейозов — питание сильно обедненными водами, в значитель¬
ной части атмосферными. По напочвенному покрову здесь
различается ряд типов, описанных ниже.
D. Олиготрофные сфагновые фитоценозы. Основные
признаки их следующие.
1. Сомкнутый ковер сфагновых мхов. Он резко выделяется
физиономически и в сильной степени определяет существова¬
ние остальной растительности.
2. Бедность видами и простота морфологической структуры
фитоценозов.
3. Достаточное или избыточное и вместе с тем устойчивое
увлажнение бедной солями водой.
Здесь различаются лесная, кустарничковая и травянистая
группы.
197
I. Лесные олиготрофные сфагновые ф и т о ц е :
нозы. Так как здесь доминирует почти всегда одна сосна,
то можно называть этот тип:
1. О л и г отр о фн ы м и с о сн о в о-сф а г н о в ы м и фито¬
ценозами. Это наиболее сложный по структуре тип ол иготроф-
ных сфагновых болот, где число ярусов иногда доходит до
пяти. Также и видовой состав здесь богаче, чем в кустарнич-
ковых и травянистых олиготрофных фитоценозах, вследствие
меньшей влажности субстрата. Однако по сравнению с лес¬
ными эвтрофными и мезотрофными фитоценозами эти фито¬
ценозы проще устроены и беднее видами вследствие более
бедного солевого режима. Под сосновым насаждением в этой
группе располагается или кустарничковый, или травянистый
ярус, в зависимости от чего здесь различаются два подраз¬
деления:
a) Кустарничковые сосново-сфагновые фитоценозы
с ярусом олиготрофных кустарничков под сосной. Здесь такие группы:
я—/. Наиболеесложные (4—5 ярусов) фитоценозы. Верхний ярус
образует Betula nana, под ней более низкие болотные Ericaceae, еще ниже
ярус морошки и, наконец, сфагновый ковер (Sph. angustifolium, Sph. fuscum)
Эти ценозы обычны на севере лесной зоны.
я—2. Ценозы с ярусом болотных Ericaceae (Ledum, Cas¬
sandra и др.) и ярусом морошки под ним. Обычны в средней и
северной полосе тайги.
я—3. Ценозы с ярусом карликовых кустарничков во¬
дяники и вереска, большей частью с господством Sph. fuscum. Они встре¬
чаются в Европейской части Союза в ее западной и северо-западной частях.
а—4. Ценозы сярусом болотных Ericaceae (Ledum, Cassandra
и др.), без яруса морошки, следовательно, трехъярусные. Эти ценозы особенно
распространены в южной части хвойной зоны.
b) Травянистые сосново-сфагновые фитоценозы с тра¬
вянистым ярусом под сосной. Здесь различаются:
Ь—/. С осново-пу шицево-сф агновые фитоценоз ы. Они
трехъярусные (считая древесный ярус). В травянистом ярусе доминирует
пушица—Erlophorum vaginatum, в моховом обычны Sph. medium, Sph. angu¬
stifolium, а иногда Sph. recurvum. Эти фитоценозы широко распространены.
b—2. Ценозы с ярусом низкорослых кустарничков
(Empetrum, Andromeda, Calluna) под пушицевым ярусом. В моховом покрове
доминируют Sph. fuscum, Sph. angustifolium.
2. Безмоховые соснов о-кустарничковые фито-,
ценозы возникают из сосново-кустарничково-сфагновых,
благодаря деградации сфагнового ковра, частью вследствие
конкуренции с верхними ярусами. Большой роли эти фитоце¬
нозы не играют.
II. Кустарничковые олиготрофные сфагновые
.фитоценозы. Древесный ярус здесь отсутствует и верхний
ярус образован кустарничками. Морфологическая структура
этих фитоценозов проще, роль сфагнума выступает резче,
и компоненты растительного покрова более однородны эко¬
логически, чем в лесных сфагновых фитоценозах. Последнее
зависит от того, что менее влаговыносливые лесные виды, еще
играющие некоторую роль в лесных фитоценозах, выпадают
здесь вовсе, и видовой состав почти нацело слагается из уме¬
ренно влаголюбивых растений олиготрофных болот. Выпадение
198
древесного яруса и замена лесных фитоценозов кустарничко-
выми наблюдается или в более влажном климате (например,
в Прибалтике и Прибеломорье), где быстрый рост сфагновых
мхов сильно угнетает древесную растительность, или на севере,
где последняя не растет на торфяниках вследствие низкой
температуры.
Различаются две группы кустарничковых фитоценозов:
1. Кустарничково-сфагновые фитоценозы с мощным
сфагновым ковром. Они удобно делятся дальше по господствующим видам
сфагнов, которые, резко выступая физиономически, к тому же в большой
степени определяют и остальной флористический состав. Здесь можно раз¬
личить:
a) Sphagneta fusel. При господстве Sph. fuscum число ярусов бывает
различно: 1) четыре (это максимум), например:—1-й Betula nana, 2-й—болотные
Ericaceae (Cassandra, Andromeda и др.), 3*й —Rubus chamaemorus, иногда
Empetrum, 4-й — мох; 2) три яруса — те же, что и выше, но без Betula nana;
3) два яруса: верхний образован или более высокорослыми кустарничками
{Ledum, Cassandra), или карликовыми (Calluna, Empetrum), нижний — мхом.
Sphagneta fusel — важнейшая группа кустарничковых фитоценозов.
b) Sphagneta angustlfolli. Доминирует Sph. angustlfolium. Из
цветковых здесь реже доминируют (по сравнению с Sphagneta fusel) Cal¬
luna, Empetrum и Rubus chamaemorus, Ledum же и Cassandra чаще. Структура
фитоценозов, как и в предыдущей группе.
c) Sphagneta magellanicl. Структура фитоценозов и доминирую¬
щие цветковые близки к предыдущей группе. Обе последние группы широко
распространены.
d) Sphagnet^ rubelli с господством Sph. rubellum. Из кустарнич¬
ков чаще доминируют Andromeda и Calluna. Распространена эта группа
слабо. Кустарничковые ценозы с другими сфагнами вовсе редки.
2. Кустарничков ые фитоценозы с разреженным сфаг¬
новым покровом. Сюда относятся ценозы повышенного микрорельефа
торфяников севера Колы. Сфагновый покров развит слабо; из кустарничков
доминируют Betula nana, Empetrum nigrum или Ericaceae (Ю. Цинзерлинг, 1934). III.III. Травянистые олиготрофные сфагновые
фитоценозы. Верхний ярус этих фитоценозов из узко¬
листных (злакового типа) травянистых растений. Морфологи¬
ческая структура весьма проста, — обычно всего два яруса.
Видовой состав беднее всех других болотных фитоценозов —
часто всего 2—3 вида на 1 м2. Причина этого — большая влаж¬
ность и, кроме того, бедность субстрата. Хотя влияние здесь
сфагновых мхов на остальную растительность и не так велико,
как в кустарничковых фитоценозах, но все же оно весьма
значительно. Особенно же резко в типичных случаях высту¬
пает влияние избыточного увлажнения.
По доминирующим сфагнам мы выделяем такие группы фитоценозов:
a) Sphagneta cuspidatl. Из цветковых доминирует чаще всего
шейхцерия. Это наиболее гидрофильная группа, где вода нередко стоит на
поверхности.
b) Sphagneta Dusenli. Редкий верхний ярус из шейхцерии, иногда
Carex limosa, Rhynchospora alba, Eriophorum vaginatum. Эта группа обычна
в глубоких мокрых мочежинах сфагновых болот.
c) Sphagneta Jensenii с шейхцерией и Carex limosa развиваются
в сходных условиях так же, как и группа d.
d) Shagneta Ltndbergii с теми же цветковыми, что и предыду¬
щие группы, а на севере, кроме того, с Scirpus caespitosus, Eriophorum
nisseolum, Carex rotundata и другими.
199
e) Sphagneta baltici c Eriophorum vaginatum, Scheuchzeria palustris
и др„ а на севере c Scirpus caespitosus, Eriophorum russeolum, Carex rotundata,
C. rarlflora. Это менее гидрофильная группа, чем предыдущие. Широко
распространенная на болотах лесной зоны ассоциация Eriophorum vaginatum —
Sph. baltlcum занимает сравнительно сухие мочежины.
f) Sphagneta papillosi с шейхцерией,Eriophorum vaginatum,Scirpus
caespitosus и другими цветковыми часто образуют мокрые ровные ковры.
g) Sphagneta aplculati. Из цветковых доминирует шейхцерия и
другие виды.
h) Sphagneta rubelli, чаще с ярусом Eriophorum vaginatum, встре¬
чаются чаще на западе Европейской части Союза в переходной полосе между
кочками и мочежинами.
i) Sphagneta magellanici. Эти фитоценозы характерны для повы¬
шений микрорельефа — кочек. Особенно часто верхний ярус образует Erio¬
phorum vaginatum.
j) Sphagneta angustifolil. Также по преимуществу фнтоценозы
кочек. И здесь очень часто верхний ярус из той же пушицы.
k) Sphagneta Russowll. Олиготрофные фитоценозы этой группы
очень часто имеют ярус из Eriophorum vaginatum.
l) Sphagneta compacti распространены слабо. Верхний ярус обра¬
зует Carex pauciflora, шейхцерия, Scirpus caespitosus.
m) Sphagneta fuse). Из фитоценозов этой группы особенно широко
распространены Sphagneta с верхним ярусом Eriophorum vaginatum и ниж¬
ним из Andromeda, Calluna и Empetrum.
Довольно обычны также ассоциации с ярусом морошки, В общем Sphag¬
neta fusel широко распространены.
Олиготрофные безмоховые фитоценозы
(„р им пи“) с деградированным, разорванным и угнетенным
моховым ковром возникают часто вследствие отмирания
сфагнового ковра в мочежинах сфагновых болот. Подобные
римпи особенно обычны на болотах Прибалтики, Прибело-
морья, в Карелии и некоторых других районах. Они связаны
переходами с олиготрофными юнгерманниевыми ценозами.
Наиболее обычны римпи с ярусом Rhynchospora alba и
Scirpus caespitosus. Иногда в травянистом ярусе римпи доми¬
нируют также Eriophorum vaginatum, шейхцерия и некоторые
другие виды.
Е. Олиготрофные лишайниковые фитоценозыХарактер¬
ный признак этих фитоценозов более или менее сплош¬
ной покров лишайников. Деревья или вовсе отсутствуют,
или, если встречаются, не образуют яруса. Верхний ярус
состоит обычно из кустарничков (из травянистых растений
часто лишь Rubus chamaemorus), но очень часто они растут
лишь отдельными группами. Причина плохого роста сосны
на лишайниковом покрове торфяников севера объясняется
прежде всего климатическими условиями; южнее же, в лес¬
ной зоне, повидимому, сказывается специфическое влияние
лишайникового покрова на деревья. Оно, вероятно, сво¬
дится к затруднению возобновления. Разреженный рост
кустарничков на лишайниковом ковре также объясняется
угнетающим влиянием лишайников. К нему на севере при¬
соединяются суровые климатические условия, особенно резко
выраженные на кочках, открытых действию морозо^ и ветров.
Лишайниковые фитоценозы — по преимуществу недостаточна
увлажненные фитоценозы.
200
Мы выделяем здесь три группы:
I. Ягельные фитоценозы с Cladonia лесной зоны. В
напочвенном покрове доминируют виды Cladonia — главным образом С. alpe-
stris, С. rangiferina, С. silvática. Кустарнички — Callana (на западе), Ledum,
Cassandra, V. uliginosum—или растут разреженно, или образуют ярус. Эти
фитоценозы широко распространены на кочках и грядах торфяников При-
беломорья и Прибалтики, причем небольшие пятна лишайников часто мозаично
чередуются со сфагновыми пятнами.
II. Ягельные ценозы тундры и лесотундры с видами
Cladonia (С. uncialis, С. gracilis, С. mJtis), Cetraria (С. nivalis) и
арктическими кустарничками. Эти ценозы покрывают кочки с неглубокой
вечной мерзлотой.
III. Корково-лишайниковые фитоценозы с Осhrole-
chia tartarea широко распространены в лесотундре и тундре на высоких
торфяных буграх с близкой вечной мерзлотой, подверженных сильным мороз¬
ным зимним ветрам. Ветровая эрозия нередко разрушает здесь лишайнико¬
вый покров. На лишайниках кустарнички и морошка образуют редкий ярус
или отдельные группы.
F. Олиготрофные лишайниково-дикрановые фитоценозы.
В этой группе напочвенный покров образуют тундровые
мхи — Dicranum (D. elongatum, D. fuscescens и др.). и ли¬
шайники, накипные (Ochrolechia, Pertusaria), а иногда и<
ягели. При этом лишайники, особенно корковые, поселяются
на мхе и сильно угнетают его. Древесная растительность,
так же, как и в предыдущей группе, отсутствует. Кустарнич-
ковая же и травянистая сильно угнетена и часто не образует
яруса. Наиболее обычны Betula nana, Empetrura и другие ку¬
старнички, а также Rubus chamaemorus. Неглубокая вечная
мерзлота, сухость субстрата, крайние климатические условия—
действие морозных ветров, ветровая эрозия—характерны для
этих фитоценозов, обычных на торфяных буграх тундры и
лесотундры.
G. Олиготрофные политриховые фитоценазы.. Сюда от¬
носятся тундровые фитоиенозы с неглубокой вечной мерз¬
лотой.
Здесь пока намечаются две группы
1. Достаточно увлажненные ценозы понижений моче-
жинных торфяников тундры с господством осок (С. rariflora)
и Polytrichum Jensenii.
2. Ценозы кочек-бугров недостаточного увлажнения
с господством Polytnchum strictum и куста рничковым ярусом.
При мелкокочковатом микрорельефе больших бугров здесь чередуются
небольшие пятна политриховых, корковых и кустисто-лишайниковых, дикра-
новых, а иногда и сфагновых ассоциаций (микрокомплекс В. Андреева, 1931V
H. Олиготрофные фитоценозы печеночников. . Эти ценозы
обычны среди сфагновых, в результате деградации которых
они и возникают.
Здесь различаются:
I. Му Пае ta anomalaec господством Mylia anómala. Они характе¬
ризуются умеренным увлажнением, встречаются на кочках и возникают чаще^
на месте дернин Sph. fuscum.
II. Ценозы сильно увлажненных мочежин и межкочий
образуются вследствие дегенерации гидрофильных сфагновых ценозов и свя¬
заны переходами с римпи. Из печеночников здесь доминируют чаще виды
Cephalozia (С. fiuitans), нередко с ярусом Rhynchospora, или Gymnocolea
201
inflata с юй же Rhynchospora и Scirpus caespitosus, или Ptilidium ciliare
с ярусом Scheuchzerla, Erlophorum vaginatum и Andromeda. Иногда верхний
ярус в юнгерманниевых фитоценозах не развивается. Эти ценозы близки
к следующей группе.
I. „Голые" болота. Сюда относятся мочежины на торфя¬
никах, где цветковая растительность и мхи не образуют яруса
и встречаются отдельными экземплярами и дернинками. Поверх¬
ность торфа здесь или более или менее обнажена, или покрыта
слизистым налетом микроскопических водорослей.
К. Олиготрофные фитоценозы Pleurozium Schreberi.
Они распространены слабо, встречаясь небольшими участ¬
ками в хорошо дренированных частях болот, или оде¬
вают иногда гряды мочежинных комплексов. Они могут
быть или с ярусом сосны и господством кустарничков (Ledum,
Cassandra, Betula nana), или без древесного яруса, также
-с кустарничками (Betula nana, Cassandra, Calluna, Empetrum).
ГЛАВА VII.
ФИЗИКО-ГЕОГРАФИЧЕСКИЕ УСЛОВИЯ И РАСПРО¬
СТРАНЕНИЕ БОЛОТ.
Болота и климат.
От климата зависит распространение болотных растений
и, кроме того, самый процесс торфообразования. Размещение
болот по земной поверхности и основные признаки их в об¬
щих чертах, так же как и растительности, определяются
климатом.
При изучении связи болот с климатом влияние его на
заболоченность и на торфообразовательный процесс удобно
рассматривать отдельно.
Климат и заболоченность. Заболоченность измеряется
отношением площади болот ко всей площади административ¬
ной единицы (например области) или естественного природ¬
ного района. Можно сопоставить заболоченность с разными
элементами климата—температурой, количеством осадков
и т. д.; особенно же удобно пользоваться комбинацией разных
климатических элементов, или так называемыми комплексными
факторами, например климатическим коэфициентом. Климати¬
ческий коэфициент — это отношение количества
выпавших осадков к количеству испарив¬
шейся за тот же период влаги. Связь климатиче¬
ского коэфициента и отдельных элементов климата с заболо¬
ченностью особенно ясна в странах со спокойным рельефом,
например, в Европейской части СССР, где климат изменяется
на территории постепенно, без резких скачков. Сравнение
рис. 44 и 45 позволяет сделать такие выводы:
1. Изолинии климатического коэфициента в общем парал¬
лельны друг другу и идут с юго-запада на северо-восток,
так же как и полосы с одинаковым процентом заболоченности.
Это указывает на связь климатического коэфициента с заболо¬
ченностью.
2. Заболоченность в общем возрастает с юго-востока на
северо-запад, как и климатический коэфициент. Значит, между
этим последним и заболоченностью существует
прямая зависимость, т. е. чем больше климати-
203
Рис. 44. Карта заболоченности Европейской части СССР. Цифры указывают
процент заболоченности. По Б. Оношко, 1931.
ческий коэфициент, тем больше заболочен¬
ность.
В Европейской части СССР наиболее заболочены ее се¬
веро-западная (Ленинградская область) и северная части.
Здесь климатический коэфициент достигает своего макси¬
мума 1,50—1,75. На юго-востоке заболоченность падает до
минимума так же, как и климатический коэфициент (0,25—
0,50). Легко понять, что величина климатического коэфициента
количество выпадающих осадков
количество испаряющихся осадков
зависит
от
величины как числи¬
теля, так и знаменателя. На севере, в тундре осадков сравни¬
тельно немного, но из-за низкой температуры мало испарение.
В результате климатический коэфициент высок, и велика
заболоченность. На северо-западе (Ленинградская область}
выпадает много осадков, и поэтому, хотя при сравнительно
высокой температуре испарение довольно велико, климатиче¬
ский коэфициент так же высок, как и заболоченность. А. Костя¬
ков, по Б. Оношко (1931), выделяет в Европейской части Союза
три области с различными климатическими коэфициентами:
1. Область избыточного увлажнения, где клима¬
тический коэфициент постоянно выше единицы. Сюда входят
Ленинградская область, север Союза, Белоруссия. Это область
наибольшего развития болот.
2. Область неустойчивого увлажненияс клима¬
тическим коэфициентом от 0,5 до 1,3. Сюда относятся: Украин¬
ская ССР, центральная часть РСФСР (Московская, Иванов¬
ская области, Татреспублика, Удмуртская и Марийская авто¬
номные республики и некоторые другие). Заболоченность
здесь гораздо меньше, но все же довольно велика.
3. Область недостаточного увлажнения с по¬
стоянным превышением испарения над осадками. Климатиче¬
ский коэфициент здесь постоянно меньше единицы. Сюда
принадлежит юго-восток Европейской части Союза. Заболо¬
ченность здесь очень мала. Сосредоточение болот по морским
побережьям с их влажным климатом хорошо видно на при¬
мере США.
Температурные границы существования болот. Низкая
температура, свойственная тундровой зоне, не препятствует,
а, наоборот, способствует образованию болот — тундра сильно
заболочена. Нижняя температурная граница существова¬
ния болота та же, что и высших' растений: болота отсутствуют
лишь в областях вечных снегов и льдов (нивальная зона)
и на снежных вершинах гор. Верхнего температурного пре¬
дела для болот на земном шаре не существует вовсе. Болота
встречаются—и нередко в значительном количестве — даже
и на равнинах экваториального пояса. Наименее благо¬
приятны условия для болот в областях с высокой температу¬
рой и малым количеством осадков. В таких местностях(полу¬
пустыня) встречаются лишь засоленные болота и водная
растительность.
206
Климат, распространение торфяников и торфообразование.
Распространение торфяников, как и болот, связано с клима¬
том. При этом климатические границы сущест¬
вования торфяников более узки, чем болот.
Нижний температурный предел существования торфяники
находят в климате арктической тундры. В настоящее время
мощные торфяники здесь не образуются из-за низкой темпе¬
ратуры. Встречающиеся в арктической тундре торфяники,
несомненно, реликтового характера. Они образовались в более
теплом периоде, чем современный, и в настоящее время не
только не нарастают, но зачастую разрушаются, деградируют.
Однако торфообразование в арктической тундре происходит
и в современный период. Накопление торфа, хотя и медленно,,
но происходит, главным образом, на низких уровнях. При
этом торф достигает лишь незначительной мощности (А. Зуб¬
ков, 1931). Верхней температурной границы для торфяников
не существует. Мощные торфяники имеются не только на
Батумском побережье с его теплым климатом, приближаю¬
щимся к субтропическому, но и в экваториальной полосе —
например, на Зондских островах, где они занимают весьма
значительные площади. Необходимо, однако, учесть, что выпа¬
дающее здесь большое количество осадков компенсирует не¬
благоприятное влияние высокой температуры на торфообразо¬
вание. Обильные осадки не только способствуют громадной
продукции растительной массы, образующей торф, но и создают
главным образом условия повышенной влажности в почве
и насыщенную водными парами атмосферу, что понижает
испарение и способствует торфообразованию.
Иные условия в засушливых областях с высокой темпе¬
ратурой. В жарких пустынях, например, наземное торфообра¬
зование невозможно, и торфяники отсутствуют.
Таким образом, мм встречаемся с явлением компен¬
сации метеорологических факторов: высокая тем¬
пература сама по себе неблагоприятна для торфообразования,
но это влияние компенсируется обильными осадками. При
малом количестве осадков в засушливом жарком климате влия¬
ние высокой температуры прекращает торфообразование.
Осадки и торфообразование. Осадки благоприятны для
торфообразования, однако до известного предела, выше ко¬
торого они не усиливают, а задерживают этот процесс. Так,
в богатой осадками северо-западной Германии олиготрофные
болота зарастают кустарничками и лишайниками; рост тор¬
фяников замирает, и растительность приобретает характер
пустоши. При дальнейшем увеличении осадков начинает
преобладать разрушение, размыв торфа. Это наблюдается
в тех местностях Европы, где выпадает свыше 1000 мм
годовых осадков (Англия, Богемские горы — Г. Освальд,
1925) при одновременном действии сравнительно низкой тем¬
пературы. В богатых осадками областях иногда проис¬
ходит выливание торфяных масс — „болотные извержения“.
207
Особенно резко эрозия и разрушение торфяников прояв¬
ляются в высокогорных областях (например, в субаль¬
пийском поясе Исполиновых гор; см. К. Рудольф, Ф. Фирбас
и Г. Зигмонд, 1928), где к низкой температуре и обильным
осадкам присоединяется еще сила воды на горных склонах.
Совершенно очевидно, что эти торфяники возникли и разви¬
вались в более теплом и сухом климате, чем современный.
Торфяники могут существовать даже при гораздо большем, чем
1000 мм, количестве осадков, но в более теплом климате. Таким
•образом, не только осадки компенсируют неблагоприятное
влияние температуры на рост и распространение торфяников, но
к обратно: второй фактор может компенсировать первый. Т а-
ким образом, существует взаимная компенса¬
ция факторов (в этом случае температуры и осадков).
Климат и распространение болотных типов. Климати¬
ческие амплитуды распространения болот, как мы видели,
шире, чем торфяников. При этом эвтрофные торфяники
имеют более широкую амплитуду, идут дальше на север
■в области холодного климата в Европейской части Союза
.и дальше спускаются на юг в область континентального
теплого и засушливого климата, чем выпуклые олиготрофные.
Типичные выпуклые болота (Hochmoore) свойственны пре¬
имущественно умеренному морскому климату. В США этот тип
отчетливо приурочен к морским побережьям умеренных широт.
Особенно много занимались исследователи изучением зави¬
симости от климата именно этих болот. Г. Освальд (1923)
считает, что северо-восточная граница выпуклых болот
в Европе хорошо совпадает с январской изотермой —10'
(рис. 46), юго-восточная с июльской изотермой + 20° (рис. 47).
Примерно так же дело обстоит, по Г. Освальду (1. с.), и в Се¬
верной Америке1. К. Бюлов (1929) дает, вероятно, следуя
Г. Освальду, границы для выпуклых болот—январскую изо¬
терму в —10° и июльскую в + 20°, так же как и Г. Гросс
(Н. Gross, 1930) (минимальная температура — январь — 9°,
максимальная — июль от +18 до +20°). Надо сказать, что
значение подобных сопоставлений не особенно велико: во-пер¬
вых, понятие „выпуклое болото* довольно неопределенно и
различно у различных авторов; во-вторых, географические
границы выпуклых болот установлены недостаточно точно;
в-третьих, и это самое главное, как мы видели выше, темпе¬
ратурные границы -чрезвычайно изменяются в зависимости от
других климатических факторов (от количества осадков и пр.).
Достаточно указать, что в Западной Сибири олиготрофные
торфяники с резко выпуклым рельефом поверхности и рядом
черт типичных выпуклых болот приморских областей идут
1 В Европейской части СССР Hochmoore с менее выпуклым рельефом
широко распространены и далеко на северо-восток от январской изотермы
(—10°); кроме того, в громадной части Украины и далеко на восток
в пределах области, очерченной этой изотермой, Hochmoore отсутствуют
совершенно.
203
еще далеко к северу от Нарыма, имеющего среднюю январскую
температуру в —27,5° (!).
Осадки и распространение НосЬтооге. Г. Гросс (1930)
для Восточной Пруссии считает, что верхняя и нижняя гра¬
ницы осадков, допускающие существование типичных выпук¬
лых болот, лежат между 475 и 775 мм (в год). Е. Гран-
лунд (1932) для южной Швеции дает нижнюю границу осадков
для выпуклых болот в 460 мм, а верхнюю—в 1000 мм. И эти
границы также весьма условны. Годовое количество осадков
в Нарымском крае всего 486 мм (Васюганье, по А. Бронзову,
1930), тем не менее, здесь, как говорилось выше, широко
распространены выпуклые болота.
Коэфициенты водного баланса и распространение болот.
Как указывалось выше, связь между распространением болот
и климатом лучше устанавливать с помощью коэфициентов,
образованных двумя или несколькими факторами. Кроме кли¬
матического коэфициента (см. выше), пользуются и другими.
Рассмотрим некоторые из них. Г. Раббов (Н. РаЬЬо\у, 1925)
предложил в качестве коэфициента водного баланса отноше¬
ние количества осадков в год (в сантиметрах)
к дефициту влажности. Он нашел, что при значении
коэфициента ниже 32 образование выпуклых болот невозможно
(по Г. Пауль и 3. Руофф, 1932). Для областей Германии,
богатых выпуклыми болотами, этот коэфициент значительно
выше: для Бремена — 41, для Мемеля — 54, для Кенигсберга —
45. Приведем еще комплексный коэфициент П. Варлыгина (1922),
по его мнению, пропорциональный болотистости:
оса тки X облачность X влажность воздуха
глубина грунтовых вод X температура X скорость ветра
В этот коэфициент, кроме климатических факторов, входит
еще глубина грунтовых вод. Однако против этого коэфициента
можно привести между прочим и то, что сюда не вошли такие
важные топографические факторы, как рельеф местности и др.
Болота и рельеф.
Если климат определяет распространение и характер болот
в общих чертах, то рельеф обуславливает их распределение
в деталях. Поскольку самое возникновение болот зачастую
зависит от рельефа, то и самое их современное распростра¬
нение тесно связано с этим фактором в большей степени,
чем многих других „зональных“ типов растительности, как
лес, степь. Влияние рельефа на болота сказывается различным
образом.
Рельеф и конфигурация болот. Формы рельефа, в частно¬
сти депрессий, в большей мере обуславливают очертания бо¬
лотных массивов. Приведем некоторые примеры. В западно¬
сибирской Барабинской лесостепи олиготрофные сфагновые
болота (по-местному — „рямы“) обычно весьма правильной
210
округлой или овальной формы, что зависит от очертаний за¬
падин, в которых образовались рямы. Эти западины в прош¬
лом были по большей части озерами правильной формы.
Болотные массивы в Балахнинской низменности между Волгой
и Окой к западу от Горького вытянуты в общем параллельно
Волге с запада-юго-запада на восток-хеверо-восток, отражая на¬
правление ложбин флювио-гляциального ландшафта. В Карелии
болота часто имеют форму длинных полос, вытянутых с севе¬
ро-запада на юго-восток и образующих разветвленную сеть
(рис. 48). Форма этих болот соответствует ложбинам рельефа.
Глубина депрессий и характер болот. Глубина котло¬
вины, в которой лежит болото, влияет на его водный ре¬
жим и питание и тем самым на его растительность. Близость
грунтовых вод от дна депрессии и количество верховодки,
притекающей из окружающих мест, зависят от глубины
котловины. В флювио-гляциальной приволжской полосе,
в низовьях реки Керженца, в глубоких междюнных котлови¬
нах, по моим наблюдениям, обычны сильно обводненные,
нередко почти лишенные древесной растительности мезотроф-
ные болота, а в более плоских и сухих — чаще олиготрофные
болота. Т. Работнов (1930) для приархангельского района уста¬
новил разницу в растительности и характере болот в котловинах
разной глубины. В депрессиях, глубиной на 10—12 м, где уже
сказывается влияние грунтовых вод, растительность олиго-
трофных болот приобретает несколько мезотрофный оттенок.
Вместе с тем это торфяники сильно обводненные, тогда как
в менее глубоких котловинах торфяники чисто олиготрофные
и менее обводнены.
Величина депрессий и характер болот. Уже раньше неоднократно
отмечалось (Н. Кац, 1928), что крупные олиготрофные торфяники об¬
ширных депрессий отличаются от торфяников в небольших котловинах,
расположенных по соседству. Первые обычно сильнее обводнены, менее
облесены и отличаются некоторыми другими признаками (см. об этом
дальше). Большая обводненность крупных торфяников—следствие застаи-
нанпя воды в их центральных частях. Это зависит от затрудненности стока
воды из середины болота вследствие удаленности окраин. Малые торфяники
обычно облесены и сравнительно суше. На зависимость типа болота от его
размера указывали и другие авторы (А. Бронзов, 1930; Ю. Ципзерлинг, 1934).
Формы рельефа, размещение болот и заболоченность.
Выше было отмечено влияние рельефа на заболачивание.
Тесная связь существует и между рельефом и современным рас¬
пространением болот. Равнинный рельеф и слабо раз¬
витая речная сеть благоприятны для распро¬
странения болот; возвышенный и сильно расчле¬
ненный рельеф с хорошо развитой речной сетью
действует обратно. В средней полосе (лесная зона)
Европейской части СССР, где климат не оптимальный для суще¬
ствования болот, они сильно зависят от рельефа. Это сказывается
не только в том, что болота развиваются главным образом
в понижениях рельефа, но и в большой заболоченности
равнинных и низменных районов, которые оказыва-
14* 211
Рис. 48. Карельский болотный ландшафт. По К. Маляревскому, 1926.
ются настоящими болотными очагами. Примерами таких сильно
заболоченных районов служат: Полесская котловина (забо¬
лоченность местами достигает до 45%)* Мещерская низмен¬
ность между Окой и Клязьмой, Ильменско-Волховская впадина,
Молого-Судский песчаный район по Мологе, Суде, Чагодоще,
Полистовский район водоразделов Ловати, Полисти, Шелони,
Великой (площадь болот до 90 тыс. га) и многие другие.
Дальше на юг (зона широколиственных лесов, лесостепь и т. д.)
зависимость болот от топографии выражена еще резче. Здесь
болота расположены только во впадинах рельефа, вполне зави¬
сят в своем питании от грунтовых вод, а дальше на юг
постоянно связаны с зеркалом воды. В более богатых осадками
областях болота меньше зависят от рельефа. Уже в приморской
части Ленинградской области сфагновые торфяники имеют
часто слабоволнистое и плоское, нередко лишь незначительно
вогнутое дно. Зависит это от того, что при большом коли¬
честве осадков торфяники могут расти и в таких формах
рельефа, где грунтовые воды более или менее глубоки и где
условия для застаивания воды неблагоприятны. При еще боль¬
шем количестве осадков торфяники располагаются во всех
условиях рельефа, следовательно, зависимость их от рельефа
минимальна. В областях, где осадков свыше 1000 мм (Англия,
окраинные горы Богемии), торфяники покрывают как впадины,
плато, так и склоны и даже гребни холмов непрерывным
торфяным плащом, следующим в общем за рельефом дна
(„Terrainbedeckende Hochmoore“ — Г. Освальд, 1925; рис. 49, см.
описание этого типа — стр. 248). Интересно положение гор¬
ных болот по отношению к рельефу. В Богемском лесу
(К. Рудольф, 1928) выпуклые торфяники холмистых предгорий
и нижней горной ступени, где выпадает 700—800лш осадков,
приурочены к отрицательным формам рельефа. Выше, где
осадков более 1000 мм, торфяники размещаются обычно на
выпуклых формах рельефа — седловинах, склонах, гребнях гор.
Размещение торфяников при обильных осадках на склонах и
на выпуклых формах рельефа объясняется не только прямым,
но и косвенным влиянием осадков. В подобном климате верх¬
ние слои почвы интенсивно промываются и выщелачиваются,
что создает благоприятные условия для поселения олиготроф-
ных болотных растений, особенно сфагновых мхов, и для
роста сфагновых торфяников не только вниз, но и вверх по
склонам.
Почвы и распространение болот.
Связь между распространением типов болот и почвообразо¬
ванием несомненна. Зона подзолистых почв (в Европейской
части СССР) в общем совпадает с зоной максимального рас¬
пространения торфяников вообще и сфагновых в частности.
Сравнительно влажный климат здесь ведет к выщелачиванию,
обеднению солями верхних почвенных горизонтов и образо-
ванию подзолов, что способствует поселению сфагновых мхов
и образованию сфагновых торфяников. В зоне деградированных
и мощных черноземов количество торфяников резко падает.
Почвы здесь, вследствие более сухого и теплого климата,
богаче солями. Поэтому торфяники этой зоны эвтрофные.
Лагг
Логг
Ложбина Пожбинп
с
Эрозионный
ручей
Рис. 49. Схематические профили различных типов выпуклых болот Европы.
Г1о Г. Освальду, 1925.
л—лесное выпуклое болото; ¿г—настоящее выпуклое болого; с—плоско-выпуклое болото;
и—болою, покрывающее рельеф; е—болото аапа (островковый тип); /—болою ааиа
(кольцевой тип).
214
В зонах каштановых, светлобурых почв, сероземов и солонцово-
солончаковых комплексов происходит дальнейшее повышение
концентрации солей в почвенном растворе. Торфяники (над¬
водные) здесь в общем отсутствуют, и их место занимают
солончаки и другие типы засоленных болот. Сказанное можно
иллюстрировать следующей схемой:
Схема 14
Связь почвенных типов с типами болот
Почвенные типы
Подзолистые почвы
Черноземы: деградированные, мощ¬
ные
Каштановые, светлобурые почвы,
сероземы, солонцово-солончаковые
комплексы
Типы болот
Торфяники, особенно сфагновые
(Hochmoore ), широко распространены
Торфяники эвтрофные. Количество их
резко уменьшается
Засоленные болота (солончаки и дру¬
гие типы). Особенно распространены
в области солоицовосолончаковых
комплексов
Геологическое прошлое страны и распространение болот
и торфяников.
В областях, подвергавшихся оледенению, как Европейская
часть Союза, Западная Сибирь, легко установить роль леднико¬
вого периода в распространении болот и торфяников. Значение
периода оледенения заключается, прежде всего, в выработке
определенного рельефа и затем в создании гидрологических
и климатических условий, влияющих на современное распро¬
странение болот. Проследим соотношение между современным
распространением торфяников и границей максимального
оледенения.
В Европейской части СССР к западу от Волги, к югу от
границы максимального (рисского) оледенения торфяники
в общем встречаются лишь спорадически1. Широкое распро¬
странение они получают лишь значительно севернее этой гра¬
ницы. При этом выпуклые сфагновые торфяники почти отсут¬
ствуют южнее границы рисского оледенения. Восточнее Волги
граница рисского оледенения уходит далеко на север, и
южная граница торфяников оказывается значительно южнее
границы оледенения, хотя широкое распространение торфяники
получают и здесь лишь к северу от этой границы. Сфагновые
же торфяники, нередко довольно обширные, в бассейне Камы
встречаются значительно южнее упомянутой границы. Расхо¬
ждение границы максимального оледенения и южных границ
распространения торфяников вообще и сфагновых в частности
1 Лишь к западу от долготы Киева сплошь заболоченный район южного
Полесья выдвигается несколько к югу от южной границы этого оледенения,
так же как и олиготрофные сфагновые торфяники.
особенно резко выражено в Западной Сибири. Здесь область
сплошного распространения громадных и типичных выпуклых
болот лежит, повидимому, в общем к югу от границы оледе¬
нения1. Спорадически же еще вполне типичные резко выпук¬
лые сфагновые болота встречаются километров на 500 к югу
от этой границы. В США соотношения, повидимому, ближе
к европейским. Здесь главная масса торфяников лежит в части
страны, покрытой раньше ледником. Южная граница распро¬
странения торфяников довольно близко совпадает с линией
конечной морены. Вне ее торфяники встречаются лишь рас¬
сеянно (А. Дахновский, 1912). Из сказанного можно сделать
следующий вывод:
1. В Европейской части СССР торфяные бо¬
лота так же, повидимому, как и в Северной
Америке, занимают главным образом террито¬
рию, подвергшуюся наибольшему оледенению.
К югу от границы последнего торфяники в об¬
щем встречаются рассеянно и обычно невелики.
Южная же граница олиготрофных сфагновых
торфяников в общем близка к южной границе
оледенения.
2. В Западной Сибири сфагновые олиготроф-
ные торфяники продвигаются далеко к югу от
южной границы оледенения. Мало того, гро¬
мадные скопления выпуклых торфяников на¬
блюдаются также к югу от этой границы.
Болотные ландшафты.
Как мы видели выше, между характером рельефа и рас¬
пространением, а также характером болот существует опре¬
деленная зависимость. Кроме того, на характер болот влияет
и их гидрология, следовательно, режим водного питания.
В определенной области, более или менее однородной
климатически, при сходных условиях рельефа и водного ре¬
жима повторяются одни и те же типы болот, притом в опре¬
деленном сочетании с участками незаболоченной территории.
Иначе говоря, определенному ландшафту соответствует и тип
болот. Ландшафты с широким развитием болот мы называем
болотными ландшафтами. Они еще слабо изучены,
но некоторые их типы можно выделить уже и теперь.
Нарымский ландшафт со сплошным разви¬
тием водораздельных болотных систем. Подобный
ландшафт имеет громадное распространение на Обь-Иртыш-
ском водоразделе Западно-Сибирской равнины. Описание его
дает А. Бронзов (1930). При крайне равнинном рельефе Запад¬
ной Сибири, в частности бассейна Васюгана (приток Оби),
1 По новейшим сведениям эта граница в Западной Сибири проходит
между 59 и 60° северной широты (Б. Земляков и С. Эпштейн, 1936).
216
болота занимают громадные площади. При этом они в бассейне
Васюгана и соседних к западу притоков Иртыша (Демьянка,
Б. Юган) представляют не отдельные отграниченные друг
от друга площади (или болотные массивы), а большей частью
громадные болотные системы, возникшие из слившихся друг
с другом болот. Подобная система на Обь-Иртышском водо¬
разделе посылает ветви длиной до нескольких сот километров
на водоразделы притоков Оби и Иртыша, а от этих ветвей
отходят в свою очередь ответвления на водоразделы второго
порядка между более мелкими реками. Эти водораздельные'
болота — типичные выпуклые олиготрофные сфагновые тор¬
фяники. Эти торфяники представляют здесь господствующую^
ландшафтную единицу и оттесняют леса к самым речным до¬
линам (рис. 50), где остаются лишь узкие незаболоченные
полосы. Ландшафт дополняется обширными долинными эвтроф-
ными болотами, которые часто отделяются от водораздельных
болот лишь сравнительно узкой полосой прибрежных увалов,
занятых лесами.
Молого-судский ландшафт с обширными, преиму¬
щественно сфагновыми болотами на песчаных межречьях.
Мологи, Суды и Чагодощи. Он сходен во многом с нарымским
ландшафтом благодаря широкому развитию сфагновых болот,
занимающих в некоторых случаях большую часть водораздела
и почти вплотную подходящих к рекам. Однако здесь забо¬
лоченность менее выражена, размеры болот мельче, и характер
их иной.
Болотный ландшафт Прибеломорской низмен¬
ности возник в результате морской трансгрессии. Эта низ¬
менность прерывается многочисленными, но невысокими ска¬
листыми островками гранитов и гнейсов. Она заболочена
местами на 80—90%, причем олиготрофные торфяники имеют
особый приморский характер, отличаясь обилием лишайников
и некоторыми другими признаками от торфяников, располо¬
женных в глубине континента (Ю. Цинзерлинг, 1934). Болота
здесь так же, как и в нарымском ландшафте, имеют характер
систем. Они занимают нередко водоразделы почти сплошь,,
кроме упомянутых выше отростков и узких полос вдоль речек.
Последние во многих случаях служат естественными границами
этих болотных систем.
Болотный ландшафт с лапландскими аапа
свойственен равнинным частям Кольского полуострова. Забо¬
лоченность здесь громадна, иногда до 80—90%. рельеф равнин¬
ный, причем участки незаболоченной суши в виде небольших,
островков, а иногда отдельных, большей частью небольших
и невысоких горных массивов и изолированных возвышен¬
ностей разбросаны среди громадных, неправильных очертаний
болотных систем. Они имеют характер уже не олиготрофных
торфяников, а болот с мозаичным чередованием участков
с олиготрофной и эвтрофной растительностью (так называемые
аапа). Тип «аапа» связан с более холодным и влажным клима¬
217
том, чем выпуклые олиготрофные болота, неправильные же
очертания, характерные для лапландских аапа, объясняются
соответственно расчлененным рельефом.
Карельский болотный ландшафт встречается
в Карелии, в условиях так называемого «карельского» геогра¬
фического ландшафта. Этот последний характеризуется сетью
узких ложбин, разделенных повышенными участками (так на¬
зываемыми «сельгами»). Этот рельеф был выработан ледником,
от чего и зависит ориентировка как ложбин, так и сельг
* оос^соо Маршрут 1920 '
1927 г
I Профильные ходы 192 6 г
Рис. 50. Схематическая карта ландшафтных единиц западного Васюганья.
По.Бронзову, 1930.
в направлении движения ледника с северо-запада на юго-восток.
Ложбины в карельском ландшафте образуют сеть, петли кото¬
рой заняты болотами (рис. 48). Таким образом, возникают
сетчато-разветвленные карельские болотные системы, имеющие,
по А. Каяндеру (1913), общий гидрологический режим, так как
вода в этих системах движется по уклону местности, с северо-
запада на юго-восток. Растительность таких болот опреде¬
ляется их географическим положением и климатом. В се¬
верной Карелии карельские системы типа аапа, где мозаично
чередуются эвтрофные (или мезотрофные) мочежины и олиго¬
трофные кочки. В средней, а также частью и в северной Каре¬
лии карельские системы — смешанного типа, где чередуются
болота разных типов.
218
Припятский болотный ландшафт свойственен бас¬
сейну реки Припяти. Это ландшафт с широким развитием
различного типа эвтрофных болот и в речных долинах и
в межречьях. Сильная заболоченность и преобладание эв¬
трофных болот зависят от гидрологических условий. Медлен¬
ное течение полесских рек и низкие берега способствуют
длительному заливанию речными водами не только речных
долин, но и низких ровных межречий Полесья.
Песчано-дюнный болотный ландшафт обычен
в зандровых областях, по притеррасным песчаным районам,
например, по левобережью Волги в области нижних течений
Ветлуги и Керженца, а также и ниже — в районе Козьмодемь-
янска и Казани. Для этих ландшафтов характерно чередование
эоловых дюн, покрытых сосновыми борами, и понижений
с торфяными болотами. Болота эти обычно олиготрофного
или мезотрофногр характера, что зависит от бедности песчаных
почв минеральными соединениями. Размер торфяников, степень
их облесенности и обводненности зависят от размера меж¬
дюнных котловин, их глубины и пр.
Конечно-моренные ландшафты также имеют характерные
черты в расположении и особенностях болот. Так, Опочецко-
Невельский район (Ю. Цинзерлинг, 1934) местами имеет все
черты конечно-моренного (Вюрмского)'ландшафта с болотами,
большей частью небольшого размера, разбросанными среди
моренных холмов часто вместе с ледниковыми озерами.
ГЛАВА VIII.
ТИПЫ БОЛОТ.
В настоящее время весьма часто болота делят на три
группы, которые характеризуются растительностью, условиями
питания и другими признаками.
1. Низинные болота, питающиеся богатыми солями,
грунтовыми или речными водами, располагаются в понижениях
рельефа, имеют плоскую или вогнутую поверхность. Залежь этих
болот образована эвтрофными (богатыми питательными веще¬
ствами) торфами, или же преобладает минеральная почва.
Растительный покров слагается главным образом изэвтрофных,
требовательных видов. Сюда относятся, например, тростни¬
ковые, осоково-гипновые болота, некоторые типы лесных болот
(ольшанники и др.).
2. Выпуклые болота, питающиеся водами, бедными
минеральными солями, в значительной части атмосферного
происхождения \ располагаются обычно на водоразделах и
имеют выпуклую в центре поверхность.
Верхняя часть торфяной толщи этих болот (а иногда и вся
залежь) сложена олиготрофным торфом, а растительность
центральной части болота слагается из олиготрофных растений,
мало требовательных к содержанию питательных веществ.
Сюда относятся болота, где преобладающая роль принад¬
лежит сфагновым (торфяным) мхам.
3. Переходные болота с мезотрофцой раститель¬
ностью имеют по своим признакам промежуточный характер
между выпуклыми и низинными болотами. В некоторых случаях
эти болота и генетически представляют стадию эволюции
от низинного болота к выпуклому.
Указанное разделение достаточно обосновано, так как ба¬
зируется на существенных признаках, а именно:
1) на характере растительности,
2) на характере субстрата (торфа),
3) на форме поверхности болота, 11 Считать, как это обычно делается еще и теперь, что выпуклые болота
питаются исключительно атмосферными осадками, неправильно. Поэтому и
называть их болотами атмосферного питания тоже нельзя.
220
4) на характере питания,
5) на положении по отношению к рельефу.
Из сказанного видно, что для того чтобы установить при¬
надлежность болота к одному из этих трех типов, нужно,
в сущности, иметь по крайней мере данные нивелировки,
чтобы судить о форме поверхности, изучить состав торфа —
его химизм, не говоря уже о растительности. Однако на
практике обычно достаточно лишь изучения
одной растительности для отнесения болота
к тому или иному типу, так как растительность
хорошо характеризует другие свойства болота.
Главные подразделения болот и соответствующие названия
имеют значительную давность (см. Г. Шрейбер, 1927).
Термин „гвдтзсЬептоог“ (иеЬе^апдзтоог) более позднего
происхождения. Русские названия: выпуклое, переходное и
низинное болото взяты с немецкого. Приведенное выше раз¬
деление не охватывает всего многообразия известных в на¬
стоящее время болотных типов. Поэтому мы приведем ниже
другое разделение. Теперь же рассмотрим сначала то, что
называют выпуклым болотом, а затем типы болот, замещающие
к северу выпуклые болота1.
Выпуклые болота.
Выпуклые болота давно привлекали внимание как уче¬
ных исследователей, так и практиков, первых—исключительным
своеобразием своей растительности и строения, вторых — топ¬
ливными запасами. Это одна из причин большой изученности
выпуклых болот. Другая причина — это сравнительная простота
их строения, в частности их растительности, что облегчало
и разделение выпуклых болот на более мелкие единицы —
географические типы. Поэтому классификация выпуклых болот
разработана лучше, чем классификация других* типов. Тем
не менее до сих пор определение понятия „выпуклое болото",
его объем и дальнейшее подразделение неодинаковы у раз¬
личных авторов.
Определение понятия выпуклое болото. И. Фрю и К. Шретер (1904)
считают существенным для выпуклых болот:
1) бедность воды минеральными соединениями, особенно известью;
2) питание за счет атмосферных осадков;
3) распространение лишь в богатых осадками умеренных и холодных
областях;
4) выпуклую центоальную часть, способность расти от центра к пери¬
ферии, благодаря чему наиболее старые (по возрасту) части лежат в центре
болота;
5) своеобразную растительность с рядом экологических особенностей.
Среди этих признаков, между прочим, отсутствует указание на особен¬
ности субстрата (олиготрофный торф). 11 Типы мезо- и эвтрофных болот умеренной климатической зоны здесь
не рассматриваются.
221
Примерно также характеризует выпуклое болото и Г. Потонье (190S г.,
т. I, стр. 38—41). И он также ничего не говорит о строении торфа.
В. Вангерин (W. Wangerin, 1926) считает, что выпуклые болота имеют
такие признаки: атмосферное питание, выпуклая поверхность, господство
в напочвенном покрове сфагновых мхов, центральное возвышенное безлесное
„плато*, характерный »регенеративный* комплекс ассоциаций, характеризую¬
щийся образованием мочежин и их зарастанием (цит. по И. Богдановской—
Гиенэф, 1928). Определение выпуклого болота у этого автора более узкое,
чем у предыдущих.
Ф. Könne (F. Koppe, 1926) относит выпуклые болота к олиготрофньш,
считая, что последнее понятие шире первого. Г. Освальд (1923), не давая
определения понятия „выпуклое болото", указывает, что он понимает по¬
следнее как географическо-топографическую единицу и притом широкого
объема, что видно и из его подразделения выпуклых болот на географиче¬
ские типы.
А. Каяндер (1913) понимает выпуклое болото (sensu stricto) достаточно
узко. Здесь имеется более или менее горизонтальное, округленных очертаний
центральное возвышенное плато („Hochfläche*) с бесчисленными мочежинами
или озерками. Это плато через более или менее отчетливый краевой склон
переходит в окраинный пояс низинного болота. В Северной России, даже
в ее западной, смежной с Финляндией части, типичные выпуклые болота,
по мнению А. Каяндера, отсутствуют (1913 г.).
Е. Гранлунд (1932) определяет выпуклое болото как геолого-топографи¬
ческое понятие: выпуклое болото—это омброгенный (т. е. атмосферного
питания) торфяник, возвышающийся над минеральным берегом, сверху со
слоем сфагнового торфа, мощностью не менее 40 см. В этом определении
важно указание на строение, однако минимальная глубина сфагнового торфа,
может быть, слишком низка. Кроме того, отсутствует указание на раститель¬
ность, и, как и в большинстве определений, имеется признак атмосферного
питания, который нельзя принять. К вышеприведенному определению при¬
мыкает и Г. Гросс (1933).
Целесообразно характеризовать выпуклое болото такими
чертами морфологии, строения и растительности:
1. Выпуклая в центре поверхность, причем эта выпуклость
не зависит от рельефа дна.
2. Более или менее мощный верхний слой олиготрофного
торфа, в котором располагаются корни современной раститель¬
ности.
3. Питание водами, бедными минеральными соединениями,
кислой реакции.
4. Олиготрофная (нетребовательная к содержанию мине¬
ральных соединений) растительность.
Все эти признаки не обуславливают непременно друг друга,
и поэтому их следует оговорить отдельно. Рассмотрим теперь
свойства выпуклых болот более подробно.
Форма поверхности выпуклых болот. Различаются три
типа равнинных выпуклых болот (рис. 51—53) по форме их по¬
верхности:
a) болота с центральной возвышенной, более или менее
горизонтальной частью (центральное плато) и краевыми скло¬
нами значительного падения (1);
b) болота с поверхностью, более или менее равномерно
повышающейся от периферии к центру (2);
c) болота с краевыми склонами и несколько выпуклым
центральным плато (3).
°22
Форма поверхности болота в одних случаях более или ме¬
нее симметрична, причем наивысшая точка располагается при¬
мерно в центре болота, а краевые склоны развиты более или
менее равномерно. В других случаях выпуклость эксцентрична*
краевые склоны, если они есть, развиты неравномерно, и
профиль поверхности асимметричен. Первый случай часто
наблюдается там, где дно котловины, занятой болотом, более
или менее равномерно понижается от краев к середине и имеет
в общем симметричную форму. Второй случай наблюдается
тогда, когда котловина, занятая болотом, асимметрична и
имеет неровное дно. Особенно заметна асимметрия профиля
у болот горных склонов (рис, 54). В областях, богатых
осадками, выпуклые болота не¬
редко развиваются почти на ров¬
ном месте, в очень плоских депрес¬
сиях. Такие торфяники получают
возможность равномерно расти
во все стороны и часто имеют
весьма симметричный профиль.
Подобные выпуклые болота с бо¬
лее или менее ровным дном и
симметричным профилем попада¬
ются и в Советской Прибалтике.
Сложный рельеф поверхности вы¬
рабатывается иногда на болотах,
образовавшихся в результате слия¬
ния прежде самостоятельных бо¬
лот. В этом случае обособляются
отдельные выпуклые участки („шап¬
ки“) на поверхности болота, раз¬
деленные сильно обводненными
Рис. 51—53. Формы поверхности
выпуклых болот. По И. Паа-
сио, 1933.
1—болото с горизонтальным цешраль-
ным плато; 2 — равномерно-выпуклое
болото; 3—промежуточным тип меж¬
ду первым и вторым.
понижениями, представляющими более молодые участки на
месте слияния соседних болот (рис. 55).
Рельеф выпуклого болота зависит также от его вели¬
чины и возраста. Крупные торфяники Нарымского края (пс
А. Бронзову, 1930) имеют хорошо развитое центральное
плато, занимающее до 50% всего поперечника (рис. 56).
У болот меньшего размера краевые склоны развиты сильнее
центра.
Наконец, небольшие болота вовсе лишены центрального
плато и имеют равномерно повышающиеся от периферии к
центру края. Нарымские болота в громадном большинстве
случаев уже прошли эту стадию, зато выпуклые болота Ба-
рабы сплошь этого типа (рис. 57). На форму поверхности
болота сильно влияют и окружающие условия, которые встре¬
чает торфяник при разрастании по периферии. Торфяники
Барабы, окруженные засоленными почвами, неблагоприятными
для сфагнума, показывают лишь слабый периферический рост;
рост же в высоту у них идет беспрепятственно, благодаря
чему вырабатывается резко выпуклая поверхность.
Морфологическое расчленение поверхности выпуклого
болота. На болотах с равномерно выпуклой поверхностью
часто вовсе нельзя выделить различные части морфологиче¬
ской структуры. Болото же с центральным плоским или
слабо выпуклым плато в типичных случаях имеет такие части
(рис. 51—53):
1) центральное, более или менее ровное, возвышенное
„плато* (Hochfläche);
.X 2) краевой склон (Randgehänge), т. е. более или менее кру¬
той уступ, ограничивающий центральное плато;
224
профилем; С — высокогорное болото на гребне.
Рис. 55. Профиль болота, образовавшегося путем слияния двух соседних болот. По А. Бронзову, 1920.
I — торф из Sph. fuscura; II — торф из Sph. medium; III — торф из Sph. Dusenil; IV — торф из Sph. parvifolium; V — пушицевый торф; VI — шейх-
цериевый торф; VII — древесный торф; VIII — торф из Sph. papillosum; IX — торф из Sph. balticum; X — осоковый торф.
п — пушица; ш — шейхцерия; б — береза; с — сосна; к — кустарнички; 2 — сосново-кустарничково-пушицевый комплекс; [3, б, 7, 10 — сосново-куетар-
ничковый сфагновый комплекс; 4, 6, 8, 9 — мочежинный комплекс; II — сосново«пушицевый сфагновый комплекс.
’Глубина торфа
гПикеты
*?!«•» ШШ *1ЛШ ШШ т4)б шг№ М'¡02 500Ш ^ 750.6 м-750.7М*Мб Ы*5и ш 575 1,751/\Ж*Ш ш ^м*т^4}.7 **г и ?
0.7 3.0 3.9 ¿0 6.5 3.7 5.9 ¿3 8.5 7.7 8.3 8.7 ЯЛ Яч Я* 77 Я.9 67 72 7.4 73 86 7.9 7.4 О г, Яп Оп 6п\ ¿с. Л £ ч 4а ~Го 7л \ ±.. -¿г-
А ЧГ _ чиА° - ■ ™-г ■ ~ зиг ” ~ 4*ь ииь — ои- — щ •" ' 5П | 50.8 ^' Ш ^ ШI ^ ® 45 7 и ? **/ /л
0,7 3.0 3.9 60 6.5 5.7 5.9 6.3 8.5 7.7 5.3 87 8.5 8.5 82 77 8$ 67 7.2 7.4 75 8.5 7.3 7.3 84 до Зо 8.0^ 85 77 6.5 ¿6 "Г$ ¿8 \ 43 2.7 2; ?7 ру
П8 /20 122 ' № \ 186 ¡28 ' 130 132 № 136 ¿8 \ Щ М /44 ‘ '"/42 "" 150 ‘ */52 ' к? 75б~
Ш^сфагм.
3§ пушиц
ш
-С(рагн: е
\ пушиц осон\
Ш пушиц, -сфогн срагн -пушиц.- осок У//Л осоковый
дно торфяника- синяя глина 0
МАСШТАбЫ горизонтальный
0г 0.4 0.6 08 юно
-I -г-1 I I I > НЧ
древесно - осоковый пушиц
’ Ю0.5 О Ю 2.0 3.0 4.0М
вертикальный
Рис. 56. Профиль и растительность болота с хорошо развитым центральным плато в Западной Сибири.
По А. Бронзову, 1930.
Буквы обозначают небольшую примесь: п — пушицы, о —: осоки, и/ — шейхцерии, г — гнпнема, •* — хвоща, ку — кустарничков, древесины
кедра, е — ели, л* —пихты, и — ивы, во - вода. 1, 19, ?5 — гадья (мокрое безлесное сфагновое болото); 2, 3, 4, 6. 8, 10, 12, 14, 17, 21. 23 —
сосново-кустарничковый комплекс; 16, 18, 20, 22 — озерный комплекс; 5, 7, 9, 11, 13, 15 _ мочежннный комплекс; 24 — че!
ного п еосново«-кустарничкового комплексов; 27 — заболоченный сосновый комплекс.
чередование мочежин-
Сое ново -кустарничтыи комплекс
Условные отметки
лоееръноат
11^
ч
ч
ч
8?
%
51
70
80
Глубина торта
в м
%
ч
$
Пикеты
14
¡3
12
11
10
0
8
7
6
5
4
3 2 1
Расстолние в м
86 14
¡00100 ¡00 /00 100 100
' ЮО
' 100
ЮО 4060 100 ЮО ЮО
Горизонтальный
вертикальный
О О/ О г 03 04 -05 0.6 0.7 0.0 09 10 КМ
Масштабы
О 05 /О /5 го 25 во 3 4.0 45 5.0 КМ
11орф
|~~~| •
из ЗрОадпит ГизсипА
рог ш /ас
Т
примесь
тростника
1егеь.
0
*
осока
осиШо1%
г
11
гипнов
Х7777Л -
осоковый
д
N
древесина березы
СИ-£3 ■
древесный
с
и
- • ^ сосны
шшпи •
тростниковый
уголь
"
Рис» 57. Растительность и строение болота с равномерно выпуклым
профилем в Барабе. По А. Бронзову, 1930.
3) мокрая окраина — понижение между краевым склоном и
склоном минерального берега к болоту. Эта окраина называется
Ьадд (шведское слово, вошедшее в обиход и других языков).
Некоторые авторы (Л. Аарио, 1932) на надвигающихся на
сушу болотах (трансгредирующих) выделяют еще четвертую
часть — „заболачи¬
вающуюся окраину*
(Тгаг^геввШпзгапё).
Это зона без торфа,
где происходит вне¬
дрение болотной ра¬
стительности на ме¬
сто растительности
минеральной почвы.
— это запади¬
на по краю боло¬
та, обычно сильно
увлажненная благо¬
даря воде, стекаю¬
щей сюда по крае¬
вому склону из цен¬
тра болота, а от¬
части и с его бере¬
га. Таким образом,
Lagg важен гидро¬
логически, удаляя
излишнюю воду из
центра болота. В не¬
которых случаях пе¬
реполняющая Lagg
вода образует здесь
маленькую речку
(рис. 58) и вызы¬
вает эрозию краево¬
го склона (рис. 59).
Lagg в общем обра¬
зуется лишь там,
где минеральный бе¬
рег наклонен к боло¬
ту и где поверхность
болота выпукла. Он
образуется различ¬
ными путями.
1л^’и в типичных случаях безлесны и заняты, как указы¬
валось, гидрофильными осоковыми фитоценозами, нередко
с сплошным сфагновым ковром. Краевые склоны обычно срав¬
нительно сухи, благодаря стоку воды, и нередко покрыты
довольно рослым сосновым насаждением (рис. 59).
Центральное плато, краевой склон и Lagg бывают отчет¬
ливо выражены в областях с значительным количеством осад-
228
Рис. 58. План части лагга на болоте Комоссе
(Швеция). По Г. Освальду, 1923.
1 — окраинный комплекс; 2 — осоково-злаковое болото; 3 —
ассоциация Eriophorum polystachium — Sph. cuspldatum;
4—ассоциация Carex limosa без мохового покрова; б—вода;
6 — сосны; 7—ели; S—березы.
ков. В Европейской части Союза выпуклые болота с упомя¬
нутыми морфологическими частями встречаются в Прибалтике
и примыкающей части Ленинградской области. Хорошо раз¬
виты эти части на торфяниках приленинградской Прибалтики,
в юго-западной Финляндии, в южной Швеции и на побережье
Рижского залива. В средней части Союза болота слабо вы¬
пуклы, и их части здесь обычно ясно не обособлены.
Гидрологическая система выпуклого болота. Как уже
говорилось в главе о торфе, сфагновые болота поглощают
громадные количества воды. Однако там, где выпадает много
Рис. 59. Профиль лагга и краевого склона на болоте Комоссе (Швеция).
По Г. Освальду, 1923.
Вверху — неэродированный, внизу — эродированный краевой склон.
1 — эвтрофный торф, 2 — сфагновый торф, 3 — вода.
атмосферных осадков, болото не может удержать их целиком.
Избыток воды уходит или поверхностным стоком, или под
поверхностью торфа. В первом случае мы говорим о поверх¬
ностной системе дренажа болота, во втором — о подземной.
Формы поверхностного стока лучше изучены. Сюда относятся
Ь১, ложбины стока и эрозионные ручьи.
1. Lagg, как уже указывалось, отводит излишнюю воду,
притекающую по склону из центра болота.
2. Ложбины стока (нем. ИйНеп, швед. отводят воду
из центра болота к периферии. Они имеют в одних случаях
отчетливое русло, ограниченное довольно отчетливыми скло¬
нами, нередко облесенными, причем здесь иногда растут не¬
обычные для олиготрофного болота древесные породы—ель,
береза. В других случаях ложбины стока вовсе плоские,
229
и склоны незаметны на-глаз (рис. 60). Растительность ложбин
разнообразна и резко выделяется на фоне окружающего вы¬
пуклого болота: в одних случаях это пушицево-сфагновое
болото, в других осоково-сфагновое; иногда здесь поселяется
тростник, иногда же образуется сильно обводненный зыбун
с шейхцерией или вахтой. Часто здесь наблюдаются скопле¬
ния воды, причем вода то течет сверху постоянно или перио¬
дически, то скрывается под торфом в виде подземного ручья.
К. Вебер (1902) подробно описывает ложбины стока на
Аугустумальском болоте и их происхождениеоно бывает
различным. Ложбины стока представляют или заросшие ручьи,
протекавшие раньше по минеральному дну болота, или пони¬
жение между двумя росшими навстречу и не вполне сливши¬
мися болотами, или образуются, когда трансгредирующее бо¬
лото обтекает минеральный остров и образует по другую его
сторону два языка, которые постепенно сближаются и обра¬
зуют борозду (ложбину стока), упирающуюся в минеральный
остров на его противоположной (от болота) стороне. Г. Ос¬
вальд (1923) описывает случай образования ложбин стока,
близкий ко второму способу Вебера; в этом случае ИйНе пред¬
ставляет остаток ровного (осоково-сфагнового) болота, на
котором начали расти навстречу друг другу две выпуклые
шапки выпуклого болота, причем осталась лишь узкая, непо¬
крытая ими полоса прежнего болота—теперешняя ложбина
стока. Явления влажного климата—ложбины стока—образуются
там, где выпуклые болота не могут удержать всех выпадаю¬
щих на их поверхность осадков и где избыток их стекает.
В СССР плоские ложбины стока с недиференцированным рус¬
лом я наблюдал в Ленинградской области.
Третий тип поверхностного стока—эрозионные ручьи. Это
ложбины в торфе, с хорошо оформленным руслом, с ясной
эрозией и с более или менее быстрым течением (Г. Освальд,
1923) (рис. 61). Эрозионные ручьи и вообще эрозия—явления
еще более резко выраженного морского климата, где выпадает
очень большое количество осадков (южная Швеция, Шотлан¬
дия). В СССР до последнего времени эти образования не были
известны. Крайне любопытные эрозионные торфяные овраги
были обнаружены в 1933 г. Л. Кудряшевым и В. Эпштейном
на островных торфяниках дельты Северной Двины под Архан¬
гельском (Н. Кац, Л. Кудряшев и В. Эпштейн, 1935). Размыва¬
ние краевых частей этих торфяников, подходящих часто к
самой воде, ведет к дренажу и оседанию таких участков. Обра¬
зуется зачаточная ложбина, в дальнейшем углубляемая сте¬
кающими по ней дождевыми и снеговыми водами. В дальней¬
шем овраг начинает расти своей вершиной, как растут овраги
в черноземной полосе, и постепенно подвигается к центру
болота. В некоторых случаях такие овраги вдаются довольно
далеко в глубь последнего и спускают даже находящиеся там 11 Подобное описание ложбин стока см. также Reimers и К. Hueck (1929).
230
Рис. 60 Ложбина стока с плоским руслом (ИйИе) на болоте Комоссе
(Швеция). По Г. Освальду, 1925.
Рис. 61. Эрозионный ручей на болоте Комоссе (Швеция).
По Г. Освальду, 1925.
вторичные озерки, большей же частью оканчиваются в пери¬
ферическом поясе болота. Периферическая старая и глубокая
часть торфяного оврага зарастает на склонах совершенно не¬
обычной для сфагновых болот растительностью; здесь встре¬
чаются: береза, черемуха, рябина, шиповник, крушина. К центру
болота овраг становится все мельче и уже.
Воронки. К формам, вырабатываемым эрозией, относятся
воронки. Воронки, повидимому, двоякого происхождения
(Г. Освальд, 1925). В одних случаях они образуются вследствие
оседания и обвалов торфа под действием ручьев, текущих под
поверхностью болота. В других случаях они возникают в ре¬
зультате зарастания текущего в торфе ручья и представляют
его остатки. Форма этих воронок различна: то более или ме¬
нее округлая, то вытянутая. Размеры нередко весьма значи¬
тельны— одна воронка на болоте Комоссе в Швеции имела до
1,5 м в ширину (Г. Освальд, 1923). Обычны меньшие размеры..
Глубина может достигать нескольких метров (Г. Освальд,
1925). На дне, как правило, имеется вода. Нередко несколько
воронок располагается в ряд, обозначающий течение подзем¬
ного ручья. Свежие воронки, недавно образовавшиеся вслед¬
ствие обвала, имеют более или менее вертикальные стенки
с голой поверхностью торфа (рис. 62). Старые, давно образо¬
вавшиеся воронки имеют более пологие склоны, заросшие
кустарниковидной березой Betula odorata (в океанической Нор¬
вегии). Их растительность в верхней части воронки постепен¬
но переходит в растительный покров окружающего болота
(рис. 63).
Воронки обычны там, где болота эродируются — в Европе в приморских
областях с большим количеством осадков: в юго-западной Швеции (Г. Дю
Риэ, G. Du Rietz, 1925), в океанической Норвегии (Г. Освальд, 1925),—а также
в богатых осадками горных областях [в Гарце они часты на высоте 700 ж
и более (К. Гюк — Hueck, 1928)]. В Европейской части Союза воронки
отмечены лишь в приморской Прибалтике, да и то чрезвычайно редко
(И. Богдаиовская-Гиенэф, 1928). Дальше на восток они, повидимому, от¬
сутствуют совершенно. Есть указание, что образования, похожие на воронки,
имеются на торфяниках западного побережья Камчатки (В. Тюшев, 1906).
Повидимому, они более или менее соответствуют воронкам европейских
болот (устное сообщение Е. Любимовой).
Озерки. Стоячие водоемы довольно обычны на выпуклых
болотах. По своему происхождению они бывают первичными
и вторичными.
Первичные озера представляют остатки от прежних более
обширных озер и или имеют еще зеркало свободной воды
(Restseen К. Вебера, 1902, стр. 78), или они нацело заросли
и погребены под поверхностью болота (так называемые Vor¬
seen). Иногда первичные озера—это остатки заросших болот¬
ных речек или прежних Lagg (см. выше) (Г. Гаме и 3. Руофф,
1929).
Вторичные озера образуются на поверхности болота. Они
особенно характерны для определенных типов выпуклых бо¬
лот. Величина их сильно колеблется, начиная от нескольких
232
Рис* 62. Недавно образовавшаяся провальная воронка на океаническом
болоте Норвегии. По Г. Освальду, 1925.
Рис. 63. Старая заросшая воронка на океаническом болоте Норвегии.
По Г. Освальду, 1925.
квадратных метров. В общем величина вторичных озерков
увеличивается с размерами самого болота. На громадных тор¬
фяниках Васюганья (Обь-Иртышский водораздел) площадь
озер достигает несколь¬
ких гектаров (А. Брон¬
зов, 1930). Форма озерков
различна: то более или
менее округлая, как
обычно на болотах Во¬
сточной Пруссии (К. Ве¬
бер, 1902; Г. Гросс, 1912),
то вытянутая. Округлые
озерки образуются на го¬
ризонтальной поверхно¬
сти болота, вытянутые —
на его склонах. На при¬
морских болотах Прибал¬
тики (И. Богдановская-
Гиенэф, 1928)озерки обычно вытянуты перпендикулярно нукло¬
ну болота. Длина озерков до 200 м, ширина обычно 10—20 м,
редко больше. Любопытно, что берега этих озерков неоди¬
наковы. Обращенный вниз по склону болота берег, на который
Рис. 64. Грядово-озерньш комплекс на
болотах Советской Прибалтики. По И.Бог-
дановском-Гненэф. Стрелка показывает
уклон.
Рис. 65. Вторичные озерки с островами на выпуклом болоте в Восточной
Пруссии. По Г. Гроссу, 1922.
давит вода и особенно лед (весной), более извилист, отвесен
и эродирован (рис. 64). От него иногда отходят мочежины, кото¬
рые под действием эрозии могут расширяться вниз по склону,
благодаря чему соседние параллельные друг другу озера соеди¬
няются поперечной перемычкой. Обращенный вверх по склону
234
берег озера—прямой, спускается к воде наклонно, не эроди¬
руется и нередко зарастает с краю сфагнумом. Глубина озер¬
ков от 1,5 до 4,5 ж (И. Богдановская-Гиенэф, 1928; Г. Гросс,
1912; К. Вебер, 1902; Н. Кац, Л. Кудряшев, В. Эпштейн, 1935).
На озерках встречаются острова, в некоторых случаях с дере¬
вьями (береза, сосна) до 10 м высоты (Г. Гросс, 1912) (рис. 65).
Растительность озерков обычно развита слабо и бедна видами:
преимущественно сфагновые мхи и их обычные спутники
(шейхцерия), реже другие цветковые (например, Nymphaea
candida Presl на болоте Оршинский Мох близ Калинина).
Озерки то встречаются поодиночке, то сгруппированы помногу
вместе и образуют „озерные комплексы“. Последние характерны
для выпуклых болот более или менее влажного климата. Они
обычны в северо-западной Европе: в юго-западной Финлян¬
дии, южной Швеции и Советской Прибалтике. Кроме того,
эти комплексы обычны и на севере Европы, в том числе и
в Европейской части СССР. Здесь они встречаются на боло-
Рис. 66. Комплекс с большими кочками и мочежинами на выпуклом
болоте. По Г. Потонье, 1911.
тах типа аапа, а также на бугристых болотах тундры и лесо¬
тундры.
Микрорельеф сфагновых болот. Можно различить такие
типы микрорельефа выпуклых болот:
I. Ровный микрорельеф. Повышения если и есть, то
235
невелики и едва приподнимаются над окружающим ровным
болотом. Растительность сравнительно однородная.
II. Мелкокочковатый микрорельеф. Повышения
в виде сфагновых кочек, высотой до 30 см и более, однако
Рис. 67. Карта комплекса с параллельными грядами и мочежинами. По Г. Гаме
и 3. Руофф, 1929.
площадь их невелика, так же как и лежащих между ними
западинок. Последние не являются настоящими мочежинами.
Растительность кочек и западинок может довольно резко раз¬
личаться.
III. Мозаичный микрорельеф с повышениями разной
формы и мочежинами. Повышения более или менее сухие, раз¬
личной формы, высотой до 50,
реже до 70 см и более, чере¬
дуются с мокрыми, иногда
с водой на поверхности, по¬
нижениями— мочежинами. Те
или другие, или же обе формы
этого микрорельефа занимают
значительные площади. Расти¬
тельность повышений и моче-
жин резко отлична и при пе¬
стром чередовании тех и дру¬
гих дает впечатление мозаич¬
ного комплекса.
Можно различить:
1. Микрорельеф с больши¬
ми кочками и мочежинами
(рис. 66). Кочки более или
менее округлые или лопаст¬
ные и разбросаны более или
менее изолированно среди
мочежин.
2. Микрорельеф с парал¬
лельными грядами и мочежинами. Те и другие в этом случае
вытянуты и ориентированы более или менее параллельно друг
другу (рис. 67). При этом мочежины или изолированы друг
от друга и более или менее замкнуты грядами, или же
обособленные гряды разбросаны на фоне мочежин.
236
Рис. 68. Сетчатый грядово-мочежин-
ный микрорельеф на болоте в при-
ленинградской Прибалтике. По И.Бог-
дановской.
1 — гряды; 2 — ассоциации мочежин с Ш1уп-
сЬоврога и юнгерманниями; 3 — мочежины
со сфагнумом.
3. Сетчатый грядово-мочежинный микрорельеф. Узкие
гряды сливаются друг с другом, образуя как бы неправильную
сеть с мочежинами в ее петлях (рис. 68). В других случаях
сеть образуют мочежиьы, а в петлях располагаются повыше¬
ния микрорельефа. Параллельности элементов в этом типе
микрорельефа не наблюдается.
4. Микрорельеф с врезанными кочками и мочежинами. На
более или менее ровном, часто мокром болоте располагаются
отдельные кочки и мочежины; уровень тех и других незначи¬
тельно отличается от уровня окружающего болота.
IV. Мозаичный микрорельеф с повышениями и озерками
(а часто вместе с тем и с мочежинами). Этот микрорельеф
напоминает микрорельеф с мочежинами, но место последних
занимают озерки с водой.
Пока здесь можно различить:
1. Микрорельеф с параллельными вытянутыми озерками и
грядами (рис. 64).
2. Микрорельеф с более или менее округлыми озерками.
3. Микрорельеф с ровными „площадями", мочежинами и
озерками. Здесь чередуются более или менее ровные про¬
странства с лишайниково-сфагновым покровом („площади“), мо¬
чежины и озерки неправильной формы (Н. Кац, Л. Кудряшев,
В. Эпштейн, 1935).
V. Эрозионный микрорельеф возникает в результате вод¬
ной эрозии на болоте. Эрозионный микрорельеф в Европей¬
ской части СССР не получает заметного развития. Эрозионные
явления свойственны преимущественно западным зарубежным
болотам.
Формирование микрорельефа выпуклых болот. Централь¬
ные части крупных выпуклых болот очень часто имеют совер¬
шенно своеобразный ландшафт благодаря развитию мозаичного
микрорельефа, с мозаичным распределением растительности.
Происхождение этого микрорельефа уже давно привлекало
внимание исследователей, и было высказано много взглядов
для объяснения этого явленияг. В формировании микро¬
рельефа надо различать две группы факторов — биогенные и
физиогенные.
Биогенные причины образования кочек. Сюда относится
кочкообразующая деятельность растений. На кочкообразую¬
щую роль вересковых кустарников указывали еще А. Гризе-
бах (А. Grisebach, 1845) и Лекере (Lesquereux, 1847), а затем
и многие другие: Филарский (Filarszky, 1893), Ситенский (Sy-
tenski, 1891), Зальфельд (Salfeld, 1883), К. Вебер (С. Weber,
1902), М. Дюггели (М. Düggeli, 1903). К. Вебер всюду на¬
ходил в сфагновых кочках остатки древесной растительно¬
сти и считает, что все кочки образуются или вокруг кустар¬
ничков или около стволов сосны. В. Сукачев (1906) также 11 Обзор исследований по формированию микрорельефа см. Н. Кац,
М. Кириллович и Н. Лебедева (1936).
237
отмечает роль деревьев — сосны и березы: их корни удер¬
живают моховую дернину и не дают ей спадаться, что спо¬
собствует диференцировке микрорельефа.
Из травянистых растений кочки образуют обычные виды
сфагновых болот—ЕйорЬогит уа£та1ит и Багрив саезриозиэ
(О. Зендтнер, 1854; Г. Фрю и К. Шретер, 1904).
Важную роль в образовании кочек играют сфагновые мхи.
В общем можно сделать такие заключения:
1. Сфагновые мхи, несомненно, способны уси¬
ленно нарастать около поселяющихся на болоте
деревьев и кустарничков, а отчасти и около коч¬
кообразующих травянистых растений (пушица
и др.).
2. Э та с п ос о б н о с ть создает начальную дифе-
ренцировку микрорельефа и обособление кочек.
3. Роль цветковых растений при образовании
сфагновых кочек сводится в основном к сти¬
муляции роста сфагновых мхов и к созданию
опоры для растущих кочек.
Биогенные причины образования мочежин. Сфагновые
мхи, как мы видели, усиленным ростом способствуют образо¬
ванию кочек. Другие же растения нарастают, наоборот, мед¬
ленно, и участки, ими занятые, обгоняются соседними участ¬
ками, покрытыми сфагнумом, и с течением времени оказываются
ниже их. Таким образом, возникают депрессии, мочежины. За¬
медление или остановка роста болота чаще вызывается л и-
шайниками, печеночниками и водорослями.
Роль лишайников в образовании мочежин отмечена рядом
исследователей (Ш. Ganong, 1897; К. Вебер, 1902; Купфер,
1913; Р. Аболин, 1914; В. Вильямс, 1922; А. Бронзов, 1930).
Особенно подробно описали образование мочежин на месте
лишайниковых кочек скандинавские авторы — А. Каяндер (1913),
Л. Пост и Р. Сернандер (1910). А. Каяндер указывает, что кочки
БрЬ. ¡иэсит по мере роста вверх делаются все суше, что дает
возможность поселиться здесь лишайникам. Последние посте¬
пенно вытесняют сфагнум, и на вершине кочки образуется
лишайниковый покров. С его образованием рост кочки в вы¬
соту более или менее прекращается, соседние же сфагновые
мочежины продолжают расти вверх. При этом уровень грун¬
товых вод поднимается, и края кочки, где сохранился сфаг¬
новый покров, продолжают расти вверх, обгоняют централь¬
ную часть кочки и обрастают ее в виде валика. Вершина же
кочки делается кратерообразным углублением, где скопляется
вода, стекающая с краев. Таким образом возникает маленькая
мочежина, или углубление с водой на вершине прежней кочки.
Этот способ образования мочежин отмечен также для Гарца
(К. Гюк, 1928) и приленинградской Прибалтики (Богдановская,.
1928). Старые лишайниковые кочки в результате этого процесса
могут оказаться на уровне дна мочежин. Сфагновая стадия
жизни кочки, таким образом, сменяется лишайниковой, оста-
238
вляющей след в кочке в виде узкой прослойки сильно раз¬
ложившегося торфа (густая штриховка на рис. 69). В даль¬
нейшем лишайниковая стадия иногда опять сменяется сфагновой*
и поверх сильно разложившегося торфа вновь отлагается сла-
Рис. 09. Профиль кочки на сфагновом болоте с чередованием более и
менее разложившихся слоев. По А. Каяндеру, 1913.
бее разложившийся сфагновыйГторф (более редкая штриховка).
Такая смена иногда повторяется несколько раз, в результате
чего торф кочки приобретает неоднородную структуру (рис. 69).
Описанное Каяндером превращение кочки в понижение часто.
Асе-Под о то гопд1/еппо
‘ А с с —С о Пип а—Sph. tmbri cotum
I гт=
i
I
1
1
Асе.—Са Ни no—C lodo то rangifenna-silvoti со
Acc-Collona—Sph. mflgellomcum
t'—
л
Acc—Crimmio hypnotdes
Асе— Colluno—tiglocomíum
_L t
Acc—Colluno—Sph tenellum
Acc—triophorum wginotum—Jph. magellomcum Acc-Scirpus oustrtacus—Sph
i i
ПочежОны, частью с водной поверхностью
торе И от сит
Рис. 70. Схема смены ассоциаций в регенеративном комплексе на
болоте Комоссе (Швеция). По Г. Освальду, 1923.
на этом не останавливается. Часто в мокрых понижениях noce-
ляется сфагнум, и здесь восстанавливается (регенерирует) сфаг¬
новый покров. Возникают гидрофильные сфагновые ассоциации,
из которых по мере дальнейшего нарастания формируется
239
сфагновая кочка, покрывающаяся затем снова лишайниками.
Эта смена мочежины лишайниковой кочкой и обратно иногда
повторяется, т. е. приобретает циклический характер и носит
название регенерации *. Регенеративный процесс протекает
указанным образом на богатых лишайниками болотах. Он
описан для болот Швеции Г. Освальдом (1923) и для приленин-
градской Прибалтики И. Богдановской-Гиенэф (1928) (рис. 70).
Кроме лишайников, задерживают рост болота и мелкие эпифитные пече¬
ночные мхи, поселяющиеся на сфагнуме (Л. Пост и Р. Сернандер, 1910;
Р. Аболин, 1914; А. Какс, 1914; Г. Жозефи, 1920). М. Юрьев (1935) выделяет
даже особый тип мочежин, обязанных своим происхождением печеночникам,
и называет их гепатогенными. Мочежины с печеночниками возникают и вслед¬
ствие вымокания лишайниковых кочек. В дальнейшем они или зарастают
сфагнумом, или превращаются в озерки (Г. Освальд, 1923; И. Богдановская-
Гиенэф, 1928). Водоросли играют большую роль главным образом в Западной
Европе. Их роль в образовании мочежин отмечает К. Вебер (1902), Г. Жо¬
зефи (1920), П. Томсон (Р. ТЬотэоп, 1924), а в СССР, между прочим,
Р. Аболин (1914). Мочежины с водорослью 2у£0£оп1иш широко распростра¬
нены на болоте Комоссе в Швеции (Г. Освальд, 1923). На западе эта водо¬
росль развивается иногда в таком количестве, что болото приобретает
красноватый оттенок—Красное болото („па сгегтчопет*) в 6. Польше. Это
болото посетила пятая международная ботанико-географическая экскурсия
(В. Шафер — 82а(ег, 1928).
Мочежины возникают вследствие затенения сфагнума более
рослыми растениями (яBeschattungsschlenken“ Р. Сернандера),
кустистыми лишайниками, некоторыми лиственными мхами,
кустарничками или сосной. Сосна, поселяясь на сфагновых
кочках, задерживает рост сфагнума и вызывает образование
кратерообразных углублений на верхушке кочек. Появление
этих воронок объясняют или затенением сфагнума сосной
(Лекере, 1847), или боковым обрастанием сфагнумом (К. Ве¬
бер, 1902), или образованием льда (А. Не1аакозк1, 1912)х, или
стенанием дождевой воды по стволу вниз, что неблагоприятно
для таких мало влаголюбивых мхов кочек, как БрЬ. {ивсит
и Ро1у1псЬит эШсШт (А. Каяндер, 1913).
Во всех рассмотренных случаях причина образования моче¬
жин—сама растительность (так называемые фитогенные моче¬
жины). В других случаях мочежины возникают благодаря
деятельности животных (зоогенные мочежины).
Физиогенные способы образования микрорельефа. Сюда
относится образование микрорельефа под влиянием физических
факторов. Хотя эти факторы очень часто действуют совместно,
но для удобства изложения их лучше разделить на группы.
К механическим факторам относится прежде всего действие
силы тяжести, вызывающее смещение торфяных масс. Послед¬
нее проявляется в разном масштабе и в разных формах.
К. Вебер (1910) предполагает, что торфяник под давлением
собственной массы обводненного торфа изменяет степень вы¬
пуклости и, растекаясь, увеличивает свою поверхность, причем
образуются разрывы, в общем параллельные краям торфяника. 11 По А. Каяидеру (1913).
240
По линиям разрыва обособляются мочежины и озерки, разде-
ленные повышениями (грядами). Болота горных склонов часто
расчленены на террасы. Эти террасы состоят из более или
менее обводненной горизонтальной части (плато), где лежат
обширные мочежины, иногда разделенные грядами, и уступы
с менее влаголюбивой раститель¬
ностью типа гряд (Г. Андерссон
и Г. Гессельман — й. Апбегевоп
и Н. Невве^ап, 1908; К. Рудольф,
1928). Причины этой диференци-
ровки — оползание торфяных
масс вниз по склону. По сход¬
ным причинам происходят и бо¬
лее мелкие расчленения микро¬
рельефа на вытянутые и парал¬
лельные друг другу гряды и
мочежины. Это расчленение (гря-
дово-мочежинный микрорельеф)
широко распространено на север¬
ных болотах. При рассматрива¬
нии подобных болот с высоты или при картировании их (рис. 71)
видно расчленение на вытянутые мочежины (или озерки) и гря¬
ды, расположенные концентрически и параллельно друг другу и
берегу болота. Ближайшее рассмотрение показывает, что
мочежины и гряды располагаются на разных уровнях, в общем
параллельно высотным уровням и перпендикулярно к склону
Рис. 71. Концентрическое распо¬
ложение озерков (черное) на вы¬
пуклом болоте (Швеция). По
Е. Гранлунду, 1932.
Рис. 72. Расположение кочек и мочежин на склоне болота.
По К. Гюку, 1928.
болота (рис. 72). Такое расположение встречается всюду, где
при достаточной влажности болота имеется наклон его по¬
верхности. Сморщиванию торфа в складки и его скольжению
вниз по склону сопутствует давление снега, снежной каши
весной и расширение воды при замерзании (Тантту — ТапИи,
1915).Снег,поТантту, особенно действует тогда,когда его много
и когда он лег на незамерзшее болото, замерзание воды — в хо¬
лодные осени и бесснежные зимы, снежная каша — при быстром
таянии после снежной зимы. По В. Ауэру (1920) давление
талых вод весной сдвигает по склону болота кочки в ряды,
потом обрастающие сфагнумом и превращающиеся в гряды.
16-722 241
Оба последних автора особенно выдвигают роль термических
факторов вместе с механическими в выработке упомянутого
микрорельефа. Большое значение имеет неравномерное замер¬
зание и оттаивание. На склонах болот торф оттаивает полосами.
Полосы, дольше остающиеся мерзлыми, оказываются припод¬
нятыми, заселяются сфагнумом и превращаются в гряды;
промежутки между ними, оттаивающие раньше, оседают, ока¬
зываются пониженными и превращаются в мочежины. Сильно
замерзающие участки, кроме того, выпирают соседние, слабо
замерзающие. Таким образом, грядово-мочежинный
микрорельеф формируется при сложном взаимодей¬
ствии механических и термических факторов.
Гидрологические причины. В этой группе факторов
большую роль играют изменения влажности. Эти изменения
захватывают большие участки болота, когда оно обводняется
со стороны другого, выше лежащего и с ним соединенного,
или когда болото самоосушается при образовании естествен¬
ного стока (А. Каяндер, 1913). Уже при слабой первоначальной
диференцировке микрорельефа влага распределяется неравно¬
мерно и стимулирует дальнейшую диференцировку, причем
повышения растут дальше, а понижения, наполняясь водой,
делаются мочежинами. При местном повышении влажности
происходит так называемое „регрессивное“ образование моче-
жин (А. Нильссон — A. Nilsson, 1890; А. Каяндер, 1902—1903,
1913; Е. Мелин — Е. Melin, 1917; К. Рудольф, 1928). При высы¬
хании мокрого болота на нем обособляются гряды, а увели¬
чение влажности на более или менее сухом болоте вызывает
образование мочежин и гряд, представляющих остатки преж¬
ней поверхности болота (В. Ауэр, 1920). При чередовании
обводнения и высыхания мочежин сфагновый ковер в них не
развивается, растительность отмирает, разложение торфа уси¬
ливается, образовавшийся торфяной ил уплотняется и оседает,
а мочежины углубляются (К. Рудольф, 1928). С появлением
воды начинается эрозия мочежин. Они при этом углубляются,
расширяются и превращаются в озерки (Р. Аболин, 1914;
В. Вильямс, 1922; Раманн, 1911; М. Юрьев и Б. Полынов, 1925).
Изменение увлажнения еще специфически воздействует
на микрорельеф. При усилении притока воды с выше распо¬
ложенного участка болота или в силу ухудшения стока зна¬
чительные участки болота иногда обводняются, причем гряды
Sph. fuscum затопляются и гибнут (А. Бронзов, 1930). Ново-
образовавшиеся гряды могут запруживать расположенную
выше по склону мочежину, которая начинает расти в сторону,
или способствовать образованию новой мочежины выше
по склону (К. Мальмстрем, 1922; Л. Аарио, 1932). Мочежины,
как отмечалось, могут возникать в связи со вторичными
озерами, вследствие затопления берега озера, направленного
вниз по склону болота.
Влияние вертикального давления на микро¬
рельеф. Разные причины, как местные дожди, замерзание
242
болота, могут усилить давление на верхние слои торфа. При
этом вода с растворенными газами поднимается вверх и вы¬
ходит на поверхность там, где растительная дернина слабо
развита (Л. Аарио, 1932; П. Томсон, 1924; Я. Гетманов, 1925).
Нередко места выхода газов слегка приподняты над мочежиной
в виде очень плоских грязевых холмиков. Растительность
здесь часто отмирает в результате отравления газами, и по¬
верхность торфа обнажается.
Уплотнение и оседание торфа постоянно сопут¬
ствуют другим факторам при формировании рельефа. Они
вызывают образование или содействуют развитию мочежин
(Раманн, 1911; В. Сукачев, 1906; М. Юрьев, 1925, 1925—1927;
Д. Бегак, 1928).
Неодинаковое разложение торфа. Этот фактор
действует вместе с уплотнением и оседанием. Его значение
отмечают Б. Полынов и М. Юрьев (1925), О. Кузенева (1911)
и другие.
Мочежины могут появляться на мокрой поверхности болота,
вследствие возникновения на ней гряд, разделяющих эту по¬
верхность на отдельные участки — мочежины. Эти мочежины
называются первичными („primäre Flarken“ у Г. Ранкен —
Н. Rancken, 1912 и Е. Мелин, 1917) в отличие от вторично
образующихся на более сухом болоте.
Типы выпуклых болот Западной Европы Ч
Г. Освальд (1925), развивая разделение, предложенное Г. По-
тонье (1910), дает классификацию типов выпуклых болот и
близких к ним типов для всей Европы. Он выделяет четыре
типа Hochmoore и устанавливает их связь с климатом. Эти
типы в порядке возрастания влажности климата следующие
(рис. 49):
1. Лесное выпуклое болото (»Waldhochmoor").
2. Настоящее выпуклое болото („eigentliches Hochmoor“).
3. Плоско-выпуклое болото („Flach-Hochmoor*).
4. Болото, покрывающее рельеф („Terrainbedeckendes Hoch¬
moor“).
Первый тип Г. Освальда (1925) в общем соответствует
континентальным выпуклым болотам Г. Потонье. Эти болота
выпуклы, имеют Lagg, покрыты сравнительно редким сосновым
насаждением и зарослями кустарничков, скрывающих сфаг¬
новый ковер (рис. 73). Этот континентальный тип распростра¬
нен на территории б. Польши, Финляндии, Восточной Швеции
и в Советском Союзе.
Настоящее выпуклое болото также имеет хорошо выражен¬
ную выпуклость в центре и Lagg. На больших болотах раз¬
виваются ложбины стока (Rfillen — см. выше). Краевой склон
болота нередко довольно хорошо облесен, в центре же в типич- 11 По Г. Освальду (1925).
16*
243
ных случаях — угнетенные деревья лишь кое-где по кочкам.
Растительность центральной части болот не всюду одинакова.
В центральных частях области распространения этих болот
в Европе большую роль играет „регенерационный“ комплекс
ассоциаций, где постоянно происходит зарастание мочежин
сфагнумом и развитие на них кочек. Следовательно, здесь
происходит энергичное нарастание торфа и болота.
Рис. 73. Общий вид лесного сфагнового болота.]
По Г. Освальду, 1925.
Смена ассоциаций при зарастании мочежин идет все время от гидро¬
фильных ассоциаций к менее увлажненным фис. 70). Мочежины, частью
с водой, с гидрофильными сфагновыми мхами, юнгерманниями и водорослями
(Яубо^опШт на Комоссе), зарастают менее влаголюбивыми сфагнами и пу¬
шицей. В дальнейшем пушицу заменяет вереск и возникают вересково¬
сфагновые ассоциации. С дальнейшим ростом кочек и их обсыханием
сфагновые мхи заменяются лишайниками (вересково-лишайниковые ассо¬
циации). Образование лишайникового ковра ведет к остановке роста кочки.
Это поворотный пункт сюкцессии. В дальнейшем кочка делается мочежиной,
последняя вновь зарастает и т. Д-’
244
Таким образом, вся смена принадлежит к круговым или циклическим
как бы повторяющимся сменам. В регенерационном комплексе преобладают
интенсивно растущие вверх сфагновые ассоциации. Процент площади, занятой
важнейшими ассоциациями, в этом комплексе следующий (табл. 49).
Таблица 49
Процентный состав регенерационного комплекса на болоте Комоссе
в Швеции
(по Г. Освальду, 1923, схематизировано)
Вересково-сфагновые ассоциации • • 63,6 48,6
Лишайниковые ассоциации главным образом с верескбм . . 5,6 10,6
Мочежины главным образом смешаннсго типа с сфагнумом,
печеночниками и водорослью 2уяо§оп1иш 17,2 24,2
Сфагновые ассоциации с Сурегасеае 13,5 15,7
Озерки, эродированные площади и др 0,1 0,9
Сумма .... 100,0 100,0
Рис. 74. Спокойный комплекс с преобладанием лишайников, где не
происходит нарастания торфа, на болотах Гарца. По К. Гюку, 1928.
В областях, более богатых осадками, чем те, где распро¬
странены болота с регенерационными комплексами, например
в юго-западной Швеции, на более значительных высотах в го¬
рах Средней Европы, например в Гарце, настоящие выпуклые
болота носят несколько иной характер (Г. Освальд, 1923).
Сфагнум здесь во многих случаях отступает на задний план,
и господство принадлежит так называемым «спокойным» ком¬
плексам ассоциаций (ЗШ^апбэкотр^хе) (рис. 74), т. е. таким,
245
где нарастание торфа если и происходит, то незначительно,
и где главную роль играют ассоциации без сфагнового покрова.
Мочежины здесь главным образом с водорослями Zygogonium,
а на кочках много лишайников (табл. 50).
Рис. 75. Общий вид солигенного болота в Шотландии.
По Г. Самюэльссону, 1910.
Таблица 50
Процентный состав »спокойного* комплекса (БШЫапсккотрих)
на Комоссе
(по Г. Освальду, 1923, схематизировано) I II
Лишайниковые ассоциации главным образом с вереском . . 23,2 34,8
Сфагновые ассоциации с вереском 53,3 18.4
Сфагновые ассоциации с Сурегасеае 4,3 1,0
Чисто водорослевые мочежины 11,1 44,3
Смешанные мочежины с сфагнумом, водброслями и юнгер-
манниями 7,9 0,6
Чисто сфагновые мочежины 0,2 0,9
Сумма • . . . 100,0 100,0
Подобные комплексы на более восточных выпуклых болотах, как Комоссе,
имеют лишь подчиненное значение. Крол/е «спокойного» комплекса на запад¬
ношведских болотах, начинает играть роль эрозия, — возникает эрозионный
комплекс, где до 40% площади в той или иной степени эродировано
Вследствие весьма влажного климата болота здесь образуются даже на слабо
выпуклых поверхностях, от чего зависит отчасти и слабое развитие Lagg.
246
Рис. 76. Строение торфяника в Англии на высоте 235 м над уровнем моря.
По Г. Эрдтману, 1928.
Черное — лесной торф; штриховка — торф из ЕпорЬогшп уа^гЫит; белое — глинистые
прослойки; А, В, С, И — пункты бурения.
10
53
52
Рис. 77. Распределение типов болот и пунктов болотных
извержений в Ирландии. По Е. Гранлунду, 1932.
Сплошные линии — контуры выпуклых болот; пунктирные —
* -содигенных болот; черные точки — пункты болотных извержений.
Климат этих мест лежит уже за пределами оптимума, для развития выпуклых
болот—он здесь слишком мягок и богат осадками. Чем дальше на восток
(в Швеции), к Балтийскому морю, тем энергичнее нарастание поверхности
болота, тем больше роль регенерационных комплексов и сфагнума. В при¬
балтийской восточной Швеции на выпуклых болотах начинает играть неко¬
торую роль древесная растительность.
Область распространения настоящих выпуклых болот,
по Освальду, — южная и средняя Швеция, юго-западная Фин¬
ляндия, южное побережье Финского и Рижского заливов,
северная и северо-западная Германия, горы Средней Европы
(например Альпы). Этот тип Освальда в общем соответствует
Рис. 78. Ландшафт субальпийского болота в Исполиновых горах (сред¬
няя Европа) с криволесьем Plnus montana. По К. Рудольфу, 1928.
«Seeklima - Hochmoore* Г. Потонье. Он свойственен областям
с приморским климатом, более влажным, чем климат лесных
выпуклых болот.
В областях с наиболее резко выраженным морским клима¬
том 1 (Англия, Шотландия), отчасти в горных местностях
с обильными осадками (окраинные горы Богемии), болота
резко выраженного атлантического характера. Они грязносеро¬
зеленой окраски, почти лишены сфагнума и покрыты главным
образом Eriophorum vaginatum и Scirpus caespitosus. Их по¬
верхность следует в общем за конфигурацией рельефа, покры¬
вая его неровности как бы плащом торфа. Эта [особенность
1 От 1000 мм и более осадков в год.
248
подобных болот оправдывает их на¬
звание: Terrainbedeckende Hochmoore1
(рис. 75, 76, 77, 78, 79). Это тре¬
тий тип Г. Освальда. Водная
эрозия здесь достигает максимальной
силы. Многометровая толща торфа
иногда до дна прорезается мощными
эрозионными оврагами естественного
происхождения. Эрозия приводит
иногда к полному сносу торфа на
больших площадях, причем в резуль¬
тате деградации торфяника возникают
белоусовые луга.
Некоторые особенности этих болот наме¬
чены уже у настоящих выпуклых болот край¬
него приморского варианта, а именно: отступа¬
ние сфагнума на задний план, остановка торфо-
образования, расположение (иногда) на выпук¬
лых формах рельефа, намечающаяся эрозия.
При дальнейшем увеличении осадков эрозия
уже определяет существование болота. Несо¬
мненно, что Terrainbedeckende Hochmoore об¬
разовались при ином климате, чем современ¬
ный. В настоящее время они разрушаются.
Между этим типом и настоящими выпуклыми
болотами стоит четвертый тип Г. Освальда —
плоско-выпуклые болота (Flachhochmoore).
Этот тип очерчен Г. Освальдом (1925) весьма
схематично. По рельефу поверхности и расти¬
тельности (три комплекса: регенерационный,
эрозионный и спокойный) он напоминает на¬
стоящее выпуклое болото. Однако здесь не
выражен краевой склон (отсюда и самое на¬
звание) и обычно не развивается и Lagg. К
этому типу принадлежат частью болота юго-
запада Швеции, далее приморские болота запад¬
ней Норвегии, болота североскандинавских
гор и некоторые другие. Позднее Г. Освальд
(1925) подробно описал приморские плоско-
выпуклые болота западной Норвегии. Некото¬
рые из них весьма своеобразны—роль лишай¬
ников в «спокойных» комплексах замещает
здесь мох Grjmmia hypnoides1 2 (рис. 80). В дру-
1 Болота, покрывающие'местность (как бы
торфяным плащом).
2 Дернины этого мха, серопушистые от бе¬
лых верхушек листочков, покрывают в некото¬
рых областях целые квадратные километры. На
торфяниках Сг1шт1а 11урпо1без распространена
главным образом на островах Англии, Фа¬
рерских, в Исландии, Норвегии, на Земле
Франца-Иосифа, а также в горах. Торф, обра¬
зованный этим мхом, большей частью не пре¬
вышает 0,5 м (см. Г. Гаме, 1930/31; там же
интересные сведения об экологии, географии
этого мха и ближайших к нему видов).
гих районах этой области эти болота сильным развитием сфа! нового покрова
напоминают выпуклые сфагновые болота. Мочежины в приморской Норвегии
развиты слабо вследствие мягкого климата. Несомненно, что Р1асЬЬосЬшооге
Освальда — весьма сборное понятие, которое в дальнейшем будет разбито
на отдельные типы.
Рис 80. Комплекс с СгИпппа 11урпо1беэ (белые кочки) и озерками
на океаническом болоте Норвегии. По Г. Освальду, 1925.
Типы выпуклых болот СССР.
Из четырех типов Г. Освальда плоско-выпуклые болота и
болота, покрывающие рельеф, неизвестны в СССР и вряд ли
здесь существуют вообще. У нас встречаются лишь болота
лесного выпуклого типа Г. Освальда и болота с грядово-мо-
чежинным микрорельефом, во многом сходные с его настоя¬
щими выпуклыми болотами. Однако на наших болотах обычно
отсутствует циклический регенеративный процесс, почти не
выражены явления эрозии и т. д. В общем выпуклые болота
СССР не умещаются в рамки типов Освальда и в дальней¬
шем классифицируются по схеме, предложенной автором этой
книги в его прежних работах. Мы выделяем три большие
группы выпуклых болот:
О лесные олиготрофные сфагновые болота;
2) олиготрофные болота с комплексами гряд и мочежин,
в иногда и с озерками;
3) безлесные олиготрофные болота без мочежин.
Последняя группа слабо представлена в СССР и почти не
изучена. Сюда относятся болота севера лесной зоны с слабо
выраженной выпуклостью и едва намеченными другими при¬
знаками Hochmoor. Этих дегенерирующих форм выпуклых бо¬
лот мы касаться не будем и остановимся на первых двух
группах.
1250
Лесные олиготрофные сфагновые болота. Основные при¬
знаки этих болот такие:
1. Диффузное, т. е. более или менее равномерное, как в
лесу (а не групповое), расположение деревьев, образующих
ярус по всему болоту. Древесный ярус, хотя сравнительно
редкий и низкорослый, придает, особенно издали, болоту сход¬
ство с лесом.
2. Сплошной сфагновый ковер при различном составе тра-
вяно-кустарничковоге яруса.
3. Отсутствие мочежин и озерков, мелкокочковатый ми¬
крорельеф.
В зависимости от древесного яруса и других признаков
в этой группе различается несколько типов болот, являющихся
вместе с тем и региональными типами.
1. Сосново-кустарничковые сфагновые бо¬
лота— восточноевропейский тип.
Древесный ярус из сосны, под ней губтые заросли болот¬
ных вересковых кустарничков, багульника, Кассандры, голубики
и других, высотой до метра. Подними скрыт сплошной сфаг¬
новый покров (Sph. medium и Sph. angustifolium). Поверх¬
ность болота лишь слабо выпуклая (обычно от 1 м, и меньше),
сравнительно сухая, Lagg обычно не выражен, озерки и мо-
чежины отсутствуют. Верхний сфагновый слой торфяной
залежи в южной части области распространения этого типа
более или менее сильно гумифицирован, часто с многочислен¬
ными погребенными пнями. В северной части лесной зоны
к вересковым часто присоединяется Betula папа, а под кустар¬
ничками образует ярус морошка, а иногда водяника (Empe¬
lrum) (северный вариант сосново-кустарничковых болот). На
юге хвойной зоны эти северные виды отсутствуют (южный
вариант). Этот тип широко распространен в хвойной полосе
Европейской части СССР. Он в общем близок к лесным вы¬
пуклым болотам Освальда. По преобладанию того или иного
кустарничка здесь можно выделить багульниковые, кассан-
дровые и другие болота.
2. Сое но в о-пу ш и ц ев о-ку ста рн и ч ко в ы е сфагно¬
вые болота. Восточноевропейский тип. Это тоже лесные
болота, но сосновый ярус в общем реже и ниже, чем на сосново-
кустарничковых болотах, вследствие большей их влажности.
Однако мочежин и озерков нет и здесь. Микрорельеф, морфо¬
логия поверхности и строение в общем, как у предыдущего
типа. Под древесным ярусом здесь чередуются участки ассо¬
циаций пушицы (Eriophorum vaginatum), приуроченные к не¬
большим межкочечным понижениям, и участки ассоциаций
кустарничков на кочках у стволов сосны. По господствующему
кустарничку можно различать багульниково-пушицевые, кас-
сандрово-пушицевые и другие болота. Этот тип обычен в
Европейской части СССР, в центральной и западной частях
хвойной зоны.
251
3. Сосново-пушицевые сфагновые болота
(рис. 81). Восточноевропейский тип. В отличие от предыдущего
большая часть поверхности болота в центре занята сфагно¬
выми ассоциациями пушицы. Среди них кочки с зарослями
кустарничков встречаются лишь редкими и небольшими пят¬
нами. Распространены эти болота там же, где и сосново-пу-
шицево-кустарничковые.
В рассмотренных трех типах черты типичных Hochmoor
зыражены слабо: выпуклость незначительна, морфологическая
диференцировка болота неотчетлива, настоящих Lagg обычно*
нет, естественная дренажная система не развита, водных ре-
Рис. 81. Сосново-пушицевое сфагновое болото. По Г. Потонье, 1912.
зервуаров нет, нет часто и характерного мощного верхнего
слоя неразложившегося сфагнового торфа. Сфагнум физионо-
мически подчинен цветковой растительности, роль его в со¬
здании экологических условий болота не так исключительна.
Добавим, что растительность этих болот, если не считать крае¬
вой эвтрофный пояс, более или менее однородна, и резко
обособленные концентрические растительные пояса здесь
обычно не выражены. Эти неразвитые лесные выпуклые бо¬
лота на юге хвойной зоны—конечный, более или менее устой¬
чивый климатический тип, а в средней ее полосе обычно яв¬
ляются лишь ранней стадией развития.
4. Сосново-ку старничковые болота сЗрЬЛиэсиш.
Западносибирский тип. Эти болота свойственны преимуще¬
ственно болотно-березовой зоне и лесостепи Западной Сибири.
По растительности они близки к восточноевропейскому типу,.
252
отличаясь главным образом сплошным ковром Sph. fuse um.
Однако они настолько отличны по своей морфологии, строе¬
нию и окружающей растительности, что выделяются в особый
региональный тип.
Особенности их следующие:
1. Сильно выпуклая поверхность Ч
2. Мощный слой слабо разложившегося торфа из Sph. fuscum.
3. Наличность в зоне лесостепи по окраине торфяников
засоленных почв. Смена центрального олиготрофного болота
растительностью засоленных почв проходит через ряд весьма
характерных растительных поясов.
Сильная выпуклость этих торфяников (по-местному «ря-
мов») объясняется тем, что они при беспрепятственном росте
в высоту лишены возможности расти вширь вследствие харак¬
тера окружающих почв. Мощный же слой слабо разложивше¬
гося сфагнового торфа указывает, что развитие их в прошлом
шло в условиях иного климата, чем современный засушливый
климат западносибирской лесостепи. Во всяком случае, на¬
личность подобных выпуклых болот совершенно необычна
для европейской лесостепи, где выпуклые торфяники вообще
отсутствуют. Столь же необычно и проникновение далеко на
юг в область лесостепи на рямы таких северных болотных
растений, как Rubus chamaemorus, Betula папа1 2.
Описание растительности этих болот см. у А. Генкеля(1933),
а залежи — у М. Нейштадта (1936).
5. Лиственничные кустарничковые сфагновые
болота (приморско-нижнеамурский тип).
В Восточной Сибири и на Дальнем Востоке весьма обычны
плоские с неглубокой залежью и близкой вечной мерзлотой
сфагновые болота с ярусом даурской лиственницы. С при¬
ближением к Хабаровску, где уже действует смягчающее
влияние моря, вечная мерзлота в торфе исчезает (N. Katz, 1932).
Болота, примерно с долготы Хабаровска, во многих случаях
приобретают слабо выпуклую поверхность, торфяная залежь
делается более глубокой, при средней глубине в 1,0—1.5 м,
причем сверху залегает слой сфагнового торфа, хотя и неболь¬
шой мощности; остальная же залежь сложена мезотрофным и
эвтрофиым торфом3. Растительность этих болот в общем того
же типа, что и мерзлых лиственничных болот; здесь тоже
довольно разреженный ярус даурской лиственницы, в среднем
5—6 м высоты, при сомкнутости крон 0,3—0,5. Эти болота
1 Выпуклость достигает иногда 2 м на протяжении 25 ж (А. Бронзов,
1930), а по профилю превышение середины над окраиной может доходить
до 3,5 ж.
2 Betula nana и Vaccinium uliglnosum встречаются еще в Курганском
округе Западной Сибири, хотя лишь редко и только в северной части
(А. Генкель, 1935).
* Описание этих болот см. Н. Кац, Ц. Минкина, Р. Федорова, Г. Эйдель¬
ман (рукопись).
253
можно назвать лиственничными кустарничково-сфагновыми со
слабо развитыми чертами выпуклых болот вследствие суро¬
вого климата с холодной зимой. Признаки Hochmoor чем бли¬
же к морю, тем лучше выражены. Сплошной сфагновый ковер
этих болот часто образован Sph. medium, нередко вместе с Sph.
fuscum. Под лиственницей ярус кустарничков: чаще домини¬
рует Кассандра, часто много также Betula ovalifolia. Наряду с
нашими видами встречаются и дальневосточные Scolochloa
spiculosa, Smilacina triflora, Triadenum asiaticum (во встречаю¬
щихся иногда мочежинах) и др. (см. Н. Кац).
Олиготрофные болота с комплексами гряд и мочежин.
Микрорельеф и растительность в центральной части этих болот
весьма своеобразны и носят характер мозаичного комплекса.
Повышения микрорельефа—то длинные, во много десятков
метров, параллельные или анастомозирующие гряды, то боль¬
шие лопастные или более или менее округлые кочки, то пло¬
щади, часто мелкокочковатые — возвышаются над соседними
мочежинами обычно не больше, чем на 70 см. Разделяющие
их мокрые понижения—мочежины—часто с водой на поверхно¬
сти, тогда как на грядах вода стоит на глубине 30—40 см.
Соотношение площади гряд и мочежин в комплексе различно:
преобладают то те, то другие. Растительность повышений
резко отлична от мочежин. Она менее гидрофильна. Здесь часто
встречается сосна то группами, то отдельными деревьями, а
в травяно-кустарничковом ярусе доминируют то Ericaceae
(чаще Cassandra, Andromeda, на западе Calluna), то Eriophorum
vaginatum.
В напочвенном покрове господствуют сфагны (главным
образом Sph. medium или Sph. fuscum), а иногда и лишайники.
В мочежинах доминируют влаголюбивые виды из секции Cuspi¬
date— Sph. balticum, Sph. Dusenii, Sph. cuspidatum, а из цвет¬
ковых редкий ярус образуют Eriophorum vaginatum (иногда
Scirpus caespitosus), а в сильнее обводненных местах Rhyncho-
spora и Scheuchzeria. Избыточное увлажнение мочежин — при¬
чина крайне бедного видового состава и простоты структуры
фитоценозов — обычно всего два яруса — травянистый и мохо¬
вой, а в наиболее мокрых местах нередко один сфагновый
ковер с редкими стеблями шейхцерии. Нередко сфагнум в мо¬
чежинах деградирует и заменяется желтоватобурой коркой
печеночников (юнгерманниевые мочежины), или напочвенный
покров вообще слабо развит и выступает обнаженный торф.
Среди этих мозаичных комплексов нередко разбросаны озерки,
а в некоторых случаях целые скопления их—озерные ком¬
плексы. Центральная часть болота с грядами и мочежинами
окаймлена олиготрофными комплексами с слабо диференци-
рованным микрорельефом (без мочежин). Эти комплексы бы¬
вают облесенные или безлесные, кустарничковые или пуши-
цевые, и в виде концентрических поясов в определенном по¬
рядке располагаются вокруг центральной части, часто в свою
очередь замыкаясь окраинным кольцом мезотрофных или
254
эвтрофных болот. Концентрическое расположение поясов
зависит от концентрического распределения увлажнения, мощ¬
ности торфа и пр., что в свою очередь зависит от выпук¬
лого рельефа болота. Характер поясов, их ширина неодинаковы
в разных районах. Диференцировка растительности на концен¬
трические пояса отличает эти болота от слабо диференциро-
ванных лесных сфагновых болот (см. выше). В районах, где
грядово-мочежинные и лесные сфагновые болота встречаются
вместе, первые—обычно крупные по площади болотные мас¬
сивы конечной стадии развития, а последние—небольшие
болота начальной стадии. Важно отметить, что перифериче¬
ские комплексы грядово-мочежинных болот в общем повто¬
ряют растительность малых сосново-сфагновых болот. Выпук¬
лость грядово-мочежинных болот обычно больше, чем сосново-
Рис. 82. Профиль торфяника советско-прибалтийского типа с распределе¬
нием растительных комплексов. По И. Богдановской-Гиенэф, 1928.
сфагновых, и они более обводнены. Верхняя толща залежи
состоит из более или менее мощного слоя сфагнового торфа,,
часто мало разложившегося (подстилка) и почти без пней.
1. Болота с комплексами вересково-лишайни¬
ково-сфагновых гряд, мочежин и озерков (совет¬
ско-прибалтийский тип). Болота этого типа встре¬
чаются в приморской полосе Прибалтики (рис. 82). Они
подробно описаны И. Богдановской-Гиенэф (1928). Кроме
обычных для более восточных болот комплексов с сфагно¬
выми грядами и мочежинами, здесь широко распространены
комплексы с сетчатым грядово-мочежинным микрорельефом
(см. выше описание и рис. 68), где на грядах, кроме сфагно¬
вых мхов, весьма обильны лишайники, занимающие иногда
50—80% всей площади гряд. Мочежины между грядами в зна¬
чительной части тоже лишены сфагнума и покрыты коркой
печеночников. Таким образом, нарастание болота в этих ком¬
плексах незначительно. И. Богдановская называет эти ком¬
плексы регрессивными. На болотах прибалтийского типа часты
озерки, нередко сгруппированные в обширные грядово-озер¬
ные комплексы.
255
Из флористических особенностей болот отметим господство вереска
(СаПипа) на грядах в центре болот, а также почти полное отсутствие здесь
древесной растительности. Болота эти принадлежат к типично выпуклым
(выпуклость иногда до 7 м) и обычно сверху имеют мощный слой мало
разложившегося сфагнового торфа из Брй. {иэсит (Г. Чернова-Лепилова,
1928). Торфяники советско-прибалтийского типа довольно близки к настоя¬
щим выпуклым болотам Г. Освальда (1925). Их сближает со шведскими
болотами этого типа обилие лишайников (а с западношведскими, кроме
того, и обилие озерков). Однако И. Богдановская (1928) все же выделяет их
в особый географический вариант.
2. Болота с комплексами вересково-лишайни-
к о в о-с фагновых гряд, мочежин и с озерками.
Южно-прибеломорский тип. Эти болота известны
из прибеломорской Карелии (Кемь) («поморский тип» Ю. Цин-
зерлинга, 1934) и из приархангельского района Северной Двины.
Описаны они в первом районе К. Маляревским (1926) и Ю. Цин-
зерлингом (1934), во втором — А. Шенниковым и М. Голу¬
бевой (1930), Д. Бегаком (1932) и в последнее время Н. Ка¬
цем, Л. Кудряшевым и В. Эпштейном (1935). В последней
работе дана характеристика этого типа. Он во многом сходен
с советско-прибалтийскими торфяниками, а именно:
1. Сильно развиты лишайники на повышениях в центре
болот 1.
2. Обилен вереск, он является главным доминирующим ра¬
стением гряд. ,
3. Имеются озерки, собранные в комплексы, например, на
болотах Двинской дельты.
4. Сфагнум в мочежинах деградирует, и образуются моче-
жины без сфагнума, с печеночниками и пр. („римпи“), осо¬
бенно на Карельском побережье.
5. Центральная часть болота безлесна. Верхний слой за¬
лежи, как и на прибалтийских болотах, сложен мало разло¬
жившимся сфагновым торфом.
Оба рассмотренных типа отличаются от других восточно¬
европейских НосЬшоог’ов рядом особенностей, зависящих от
приморского положения в сравнительно влажном климате. Эти
особенности следующие:
1. Широкое развитие лишайников, частично заменяющих
сфагнум на повышениях микрорельефа. 2. Деградация сфаг¬
нума в мочежинах и образование „римпи“. 3. Обилие вереска
в глубь континента, быстро .исчезающего с болот.\4. Безлесье
в центре торфяников. 5. Присутствие, часто обилие озерков.
Можно думать, что в эпоху, предшествовавшую современной,
климат был для развития сфагнума оптимальным. В это время от¬
ложились мощные слои мало разложившегося сфагнового торфа.
Позднее климат (вероятно, вследствие понижения температуры)
стал менее благоприятен для сфагнума. Сфагновый ковер дегра¬
дировал и был частично заменен лишайниками, а в мочежинах
1 На ровных повышенных участках в центре болота, имеющих характер
«площадей», на двинских дельтовых болотах лишайники занимают 50 и
более процентов площади повышений.
*256
печеночниками. При значительном сходстве оба типа имеют и
отличия.
Более северный прибеломорский тип отличается массо¬
вым развитием Scirpus caespitosus, который частично заменяет
здесь в мочежинах обычную у нас шейхцерию. Местами (напри¬
мер в дельте Северной Двины) весьма обилен арктический Sph.
Lindbergii. Оба эти растения — редкость на прибалтийских бо¬
лотах. Зато широко распространены здесь, как и в средней
части СССР, Eriophorum vaglnatura и Sph. medium. Эти ра¬
стения на прибеломорских болотах играют значительно мень¬
шую, а в приархангельском районе и вовсе небольшую роль.
Необходимо также иметь в виду, что на обширных прибело¬
морских низменностях, частью образовавшихся после отступ¬
ления моря, торфяники в равнинных условиях занимают гро¬
мадные площади и сливаются друг с другом, образуя болот¬
ные системы (прибеломорский болотный ландшафт), прибал¬
тийские же торфяники встречаются в виде обособленных
пятен, т. е. отдельных болотных массивов.
3. Болота с комплексами пушицево-сфагновых
гряд с Sphagnum fuscum и мочежин — ильменско-
западнодвинский тип. В отличие от предыдущих эти
болота—чисто сфагновые; сфагнум почти безраздельно
господствует здесь как на повышениях, так и в мочежинах.
В центре болота большую площадь (иногда, по моим данным,
свыше 50% всей площади) занимают грядово - мочежинные
комплексы, часто с параллельной ориентировкой элементов
микрорельефа, направленных обычно перпендикулярно к склону
болота.
Гряды или лишены вовсе деревьев, или лишь с еди¬
ничными сильно угнетенными соснами, относящимися к fo
Litwinowii, часто очень угнетенной и близкой к fo pumila, или
же с этой последней. Верхний ярус гряд образует чаще пу¬
шица, реже—Ericaceae. Под пушицей обычен ярус кустарнич¬
ков—Andromeda, иногда Empetrum, а ближе к морю и Са1-
luna.
В грядово-мочежинном комплексе гряды с пушицей из¬
дали заслоняют мочежины и дают впечатление однотонного
седоватого пушицевого болота. Этот оттенок весьма харак¬
терен для ильменско-западнодвинских болот. В мочежинах —
редкий ярус шейхцерии, пушицы, а часто и очеретника (Rhyn-
chospora alba) и рыхлый мокрый ковер из Sph. cuspidatum,
Sph. Dusenii и Sph. balticum. Грядово-мочежинцые комплексы
центра окаймлены олиготрофным поясом без мочежин. На бо¬
лее крутых склонах болота встречаются кустариичковые или
сосново-кустарничковые комплексы (нередко с ассоциациями
вереска и водяники), а также пушицево-кустарничковые ком¬
плексы, а на более пологих склонах, особенно в их нижних
частях, куда стекает вода, располагается безлесное пушицево-
сфагновое болото. Эти болота типичные выпуклые (до 8,5 м,
считая максимальное превышение середины над краями), сверху
17—722 257
с мощным слоем сфагнового торфа, часто слабо разложивше¬
гося. Эти торфяники описаны мной для бассейна озера Иль¬
мень и верховьев Западной Двины, где климат менее влажен,
чем в Прибеломорье и в Прибалтике. Климат этот в настоя¬
щее время (и был, повидимому, в прошлом) наиболее благо¬
приятен для роста сфагновых мхов и образования сфагнового
торфа. В приморских районах в современном климате, как мы
видели, сфагновый ковер деградирует, а к юго-востоку и во¬
стоку от области распространения описываемого типа, пови¬
димому, уменьшается выпуклость и мощность сфагнового
верхнего слоя торфа и увеличивается его разложение.
4. Болота с комплексами кустарничково-сфаг-
новыхгряд и мочежин. Среднепечорский тип.
Эти болота в центре также с сплошным сфагновым покровом
и комплексами с грядами и мочежинами. На сфагновых гря¬
дах, сложенных чаще из Sph. fuscum, а иногда из Sph. medium
и Sph. angustifolium, бросается в глаза кустарничковый ярус
из Cassandra и Andromeda, а под ним обычно ярус морошки.
Пушица же здесь чаще не образует яруса, встречаясь обычно
лишь единично. Сосны на грядах то совсем отсутствуют, то
растут единично, иногда же группами. Pinus silvestris fo pu-
mila здесь, повидимому, не встречается. В мочежинах доми¬
нируют: шейхцерия, пушица, Sph. balticum, Sph. Dusenii; Sph.
cuspidatum и Rhynchospora alba, обычные на ильменско-западно-
двинских болотах, вовсе не характерны для среднепечорских.
Грядово-мочежинные комплексы центра опоясаны, как обычно,
кольцом олиготрофных комплексов без мочежин. Среди
них преобладают сосново-кустарничково-сфагновые, а часто и
сосново-кустарничково-пушицевые сфагновые комплексы. Без¬
лесные же с ровным микрорельефом пушицевые болота не
получают здесь такого развития, как на ильменско-западно¬
двинских болотах. Выпуклость печорских болот, судя по немно¬
гим сведениям, невелика, а разложение верхнего слоя торфа
значительно1. Эти болота отличаются от ильменско-западно¬
двинских меньшей выпуклостью, вероятно, более разложив¬
шимся верхним слоем торфа, большой облесенностью, более
бедным видовым составом (редкость Rhynchospora и Sph. cuspi¬
datum), обилием морошки и важным физиономическим при¬
знаком: кустарнички на грядах замещают здесь пушицу. Этот
тип установлен мной для средней Печоры и, вероятно, свой¬
ственен всему бассейну верхней и средней Печоры и Онеги.
5. Болота васюганского (нарымского) типа
с комплексами гряд, мочежин и озерков. Болота
этого типа хорошо изучены А. Бронзовым (1930) на Обь-
Иртышском водоразделе в бассейне Васюгана. Выше был
описан нарымский болотный ландшафт. Крупные болота с
1 По профилю одного болота на водоразделе Вычегды и Печоры
превышение середины над краями всего около 2 м (Н. Кац и Ц. Минкина,
1936).
258
хорошо выраженной плоской центральной частью („плато“),
занимающей до 50% длины профиля, с пологими длинными
склонами и мокрой ровной окраиной—„гальей", соответствую¬
щей Lagg, из которой может вытекать речка (рис. 56). Выпук¬
лость в некоторых случаях может достигать 10 м. Эта цифра,
вероятно, наибольшая из всех известных в настоящее время.
В центре наиболее крупных болотных систем громадные пло¬
щади занимают озерные комплексы (они часто господствуют
по площади) и грядово-мочежинные. На меньших болотах
склоны занимают относительно большую площадь, центральная
же часть менее развита. Она здесь не так обводнена и занята
грядово-мочежинными, а также сосново-кустарничковыми ком¬
плексами, без озерков. Гряды обычно с сосной, образующей
нередко ярус. Сосново-сфагновые ассоциации занимают также
значительные участки на более крутых склонах болот. Из
мхов на грядах доминирует Sph. fuscum, часто с большой
примесью лишайников. Они обильны также и в сосново-сфаг¬
новых ассоциациях склонов болот. Из цветковых на грядах доми¬
нируют Andromeda, Cassandra, а также Eriophorum vaginatum.
Rubus chamaemorus заметной роли не играет. В мочежинах
хорошо развит сфагновый покров, чаще из Sph. Dusenii и Sph.
baiticum с редким ярусом из шейхцерии или пушицы. Длин¬
ные склоны крупных болот тоже с грядово-мочежинными
комплексами, примерно того же типа, что и в центре, причем
эти комплексы вытянуты не перпендикулярно к склону, а
чаще идут параллельно ему. Таким образом мочежины здесь
могут служить водостоками, по которым стекает из центра
лишняя вода. На склонах меньших болот располагаются гря¬
дово-мочежинные комплексы вперемежку с сосново-кустарнич¬
ковыми, т. е. периферический пояс этих болот по раститель¬
ности мало отличается от центральной части. Залежь этих
болот сложена в большей своей части мощным слабо разло¬
жившимся, а вверху почти совсем не разложившимся сфагно¬
вым торфом. У дна почти всюду древесный торф, указываю¬
щий, что торфяники эти возникли вследствие лесного забола¬
чивания. Болота нарымского типа достаточно резко отличны
от соседних с ними поту сторону Урала—печорских и, как ни
странно, по своей морфологии и гидрологии ближе к более запад¬
ным болотам, тяготеющим к Прибалтике. Они в общем близки
к советско-прибалтийскому и ильменско-западнодвинскому
типу и представляют, как и оба последних типа, сильно выпук¬
лые Hochmoore с поверхностью, часто расчлененной на части,
свойственные типичным выпуклым болотам, и с мощным слоем
слабо разложившегося сфагнового торфа. По обилию озерков
они сходны с советско-прибалтийскими, но регрессивные явле¬
ния здесь слабее и лишайников меньше, хотя последние здесь
все же обильнее, чем на ильменско-западнодвинских болотах.
Зато более резко выраженное, чем на балтийских болотах,
господство сфагнума сближает их с ильменским типом. Резко
отличаясь по морфологии и строению от печорских, нарымские
17* 259
болота флористически все же ближе к ним, чем к более
западным. Кроме отсутствия вереска, малая роль Rhynchospora
alba и Sph. cuspidatum сближает эти болота с печорскими
и отличает их от более западных. Присутствие типичных
выпуклых болот в лесной зоне Западной Сибири, вопреки
прежнему мнению Миддендорфа и А. Каяндера (1913), которое
повторяет и Г. Освальд (1923), как будто не вяжется с конти¬
нентальным климатом этой области. Однако надо учесть прежде
всего, что количество летних осадков, особенно важных для
существования выпуклых болот, в Нарымском крае довольно
велико и близко к тому, что выпадает в Ленинграде. С дру¬
гой стороны, физико-географическая обстановка после отступа¬
ния ледника, именно наличие громадных пресноводных водое¬
мов, подпертых ледником и создавших особый местный кли¬
мат, была весьма благоприятна для грандиозных процессов
заболачивания. Наконец, сами болота, вследствие крайней
равнинности рельефа, беспрепятственно разраставшиеся вширь,
заняли громадные площади и сами изменили климат в благо¬
приятную сторону — именно в сторону большей влажности.
Наконец, надо иметь в виду, что при громадном размере
болот в центре их всегда создаются отличные от окружаю¬
щих условия, в том числе и климатические, причем централь¬
ная часть их приобретает признаки болот более северных
и влажных областей. В свете сказанного здесь типично выпук¬
лый характер васюганских болот не должен нас особенно
удивлять.
6. Болота западнокамчатского типа с ком¬
плексами кочек и мочежин. Некоторые интересные
особенности западнокамчатских Hochmoor были отмечены еще
В. Тюшевым (1908). Однако представление об их раститель¬
ности дают лишь работы последних лет (М. Нейштадт, 1936;
Е. Любимова, 1937), на основании которых эти болота и опи¬
саны. Основные черты западнокамчатских выпуклых болот
следующие:
1) выпуклость поверхности, причем эта выпуклость иногда
заметна на-глаз, так как она не скрадывается древесной расти¬
тельностью;
2) безлесье и даже полное отсутствие древесной расти¬
тельности, кроме Alnus hirsuta, встречающейся кое-где в неболь¬
шом количестве по долинам рек и ручьев, прорезающих
болота;
3) массрвое развитие осок не только в мочежинах, но
и на повышениях микрорельефа. Эти осоки определяют уны¬
лый буроватозеленый фон болота. Особенно много Сагех
Middendorfii Fr. Schmid и С. Lyngbyei Hornem. Эти виды
не только образуют фон в окраинных комплексах олиготроф*
ных болот с мелкокочковагым лишайниковым и сфагновым
покровом, но частью образуют редкий ярус (например Сагех
Lyngbyei Hornera) и на кочках мочежинных комплексов
в центре болота;
2С0
4) плохое развитие и низкий рост болотных кустарничков.
Так, Ledum, Cassandra, Vaccinium uliginosum, Andromeda poli-
folia редко превышают 25—30 см, обычно бывают значи¬
тельно ниже. Как правило, они обычно ниже осокового яруса;
5) присутствие Myrica tomentosa;
6) мочежинные комплексы в центре этих болот носят такой
характер. В мочежинах, частью сливающихся друг с другом,
растут Сагех rotundata, С. rariflora, Sph. Lindbergii. Однако
сфагновый покров часто вовсе отсутствует, тогда мы имеем
мочежины типа „римпи“. Кочки среди мочежин — большей
частью высокие, сложенные Sph. fuscum с редким ярусом
Сагех Lyngbyei и кустарничков. При дальнейшем развитии
мочежин за счет кочек мы получаем комплекс с резким пре¬
обладанием „римпи“, с голой или покрытой печеночниками
поверхностью торфа с редким ярусом Сагех rariflora. Среди
этих римпи встречаются лишь маленькие кочки с Trichopho-
rum caespitosum, Сагех pauciflora, иногда с дернинками сфаг¬
нума, реже лишайников. Иногда мочежины наполнены водой
и имеют характер озер (озерно-мочежинный комплекс);
7) на болотах встречаются долинообразные понижения
с сильным уклоном, с направлением с востока на запад (как
и реки, текущие среди болот), с небольшими овальными или
круглыми озерами, диаметром в 5—10—20 м, значительной
глубины (свыше Н/г—2 м), с обрывистыми берегами. Эти озерки
напоминают карстовые воронки \ Можно считать эти воронки
следствием оседания и провалов толщи торфа под влиянием
подмывания их текущими под поверхностью торфа ручьями.
Подобные воронки, как известно (см. выше), встречаются на
болотах морского климата ( в Скандинавии);
8) болота занимают местами громадные площади, покры¬
вая водоразделы, спускаясь по пологим склонам и занимая
невыработанные долины маленьких речек и ручейков, проре¬
зающих болота;
9) торфяная залежь имеет большую мощность—до 7, а
в приморской полосе даже до 8 м (М. Нейштадт, 1936). В ее
сложении принимают значительное участие сфагновые, частью
мало разложившиеся торфа. Замечательная особенность строе¬
ния торфяной залежи этих болот—„инверсия“, нарушение обыч¬
ного порядка смены слоев торфа (рис. 83 и 84). Очень часто
на поверхности лежит осоково-сфагновый, довольно сильно
гумифицированный торф, мощностью в 0,5—0,75 м. Очевидно,
что материнский фитоценоз этого торфа—современная расти¬
тельность болота, где сильно представлены осоки. Под этим
слоем залегает слой неразложившегося сфагнового торфа,
чаще мощностью около 1 м. Ниже обычно залегает слой эвтроф-
ного торфа, из осок, сфагновых и гипновых мхов. Таким обра¬
зом, на торфяниках Камчатки поверх олиготрофного торфа
залегает торф более мезотрофный и вместе с тем происходит 11 Подобные образования описывает и В. Тюшев (1908).
2G1
изменение растительного покрова в мезотрофном направлении.
Возможная причина этого — приближение уровня грунтовых
вод к поверхности болота. Это приближение может вызы¬
ваться причинами общего характера, например, общим подъ¬
емом грунтовых вод благодаря общему опусканию местности.
Рис. 83. Строение торфяника на побережье Камчатки с нарушен¬
ным порядком залегания слоев торфа. По М. Нейштадту, 1936.
1 — чисто сфагновый торф; 2 — гипново-сфагновый торф;
3 — гипново-осоковый торф; 4 — хорошо разложившийся торф*
Рис. 84. Строение торфяника Камчатки с инверсией слоев
торфа. По М. Нейштадту, 1936.
1 — морская галька; 2 — глина, 3 *- ил; 4 — гипново-осоковый
торф; 5 — сфагновый торф; 6 — Sph. fuscum; 7 — Sph. medium;
8 - Sph. Lindbergii; b — осоковый торф; 10 — гипновый торф.
Вышеприведенное описание относится к болотам полого¬
волнистой равнины, тянущейся вдоль морского побережья Кам¬
чатки (данные Е. Любимовой). Болота приморской равнины,
тянущейся полосой у самого берега моря, отличаются, судя по
описаниям М. Нейштадта (1. с.), от болот, расположенных в глубь
полуострова, более развитым сфагновым покровом, большей
ролью лишайников. Обилие озерков, однообразие растительного
покрова и олиготрофная растительность (много Empetrum
nigrum) дополняют отличия приморских болот от континен¬
тальных.
В общем болота западного побережья Камчатки отличаются
своеобразной смесью признаков, с одной стороны, приморских
262
Hochmoor, а с другой — арктических болот, имея, кроме того,
в своем составе дальневосточные виды.
Приморские черты западнокамчатских болот следующие:
1) мощная торфяная залежь, где значительную роль играют
мало разложившиеся сфагновые торфа;
2) деградация сфагнового покрова (образование римпи),
размывание торфа с образованием воронок, обилие озерков
и ручьев на болоте;
3) распространение, подчас сплошное, болот в разных
условиях рельефа — на водоразделах, склонах, в долинах;
4) безлесье болот;
5) некоторые черты флористического состава—например,
* присутствие Myrica tomentosa, близкой к Myrica gale, харак¬
терной для расположенных далеко на запад болот примор¬
ской Западной Европы. Арктические черты проявляются осо¬
бенно резко в растительности — присутствие и часто большое
обилие арктических видов: Carex rariflora, С. rotundata, Sph.
Lindbergii, Erapetrum nigrum.
Наиболее распространены и обильны на болотах дальне¬
восточные виды осок — Carex Middendorffii и С. Lyngbyei.
7. Карельские болотные системы смешан¬
ного типа. Этот тип болот связан с карельским ландшафтом
(см. выше). Эти сетчатые системы болот, описанные выше,—
„смешанные" (карельский тип — Ю. Цинзерлинга), так как
здесь чередуются разные типы болот. Среди них преобладают
олиготрофные сосново-сфагновые с пушицей (Sph. médium
и Sph. angustifolium), частью грядово-мочежинные болота, но
встречаются также вперемежку с ними мезотрофные ценозы
с сосной и без нее, эвтрофные ценозы, а также и заболочен¬
ные леса. Все эти типы сменяют друг друга на первый взгляд
беспорядочно, без концентрической поясности растительности,
характерной вообще для выпуклых олиготрофных болот. По
А. Каяндеру, причина такого чередования — разные условия
питания, что в свою очередь зависит от степени наклона дна
ложбин. Там, где наклон сильнее, количество протекающих
солей бывает больше и развивается более требовательная
растительность.
8. Болота среднерусского типа с сосново-
кустарничково-пушицевыми и мочежинными
комплексами. Эти болота носят переходный характер
между облесенными кустарничково-пушицевыми сфагновыми
болотами и болотами с грядово-мочежинными комплексами.
Последние не занимают сплошь центральной части массива,
как, например, на ильменско-западнодвинских Hochmoor, а пере¬
межаются с сосново-сфагновыми комплексами и часто вкрап¬
лены в них в виде островков. Эти же сосново-сфагновые ком¬
плексы образуют также широкое кольцо и вокруг комплексов
с мочежинами. Таким образом, на долю последних комплексов
приходится значительно меньше половины всей поверхности
болота. В комплексах с участием мочежин последние обычно
263
мельче и занимают меньшую часть комплекса, чем на болотах
более северных районов, повышения же микрорельефа — чаще
не гряды, а большие кочки, то более или менее изодиаметри-
ческие, то лопастные. В общем эти болота сравнительно мало
обводнены, с чем связана и их облесенность и, вероятно, ука¬
занная выше форма кочек, так как сравнительно сухой и
плотный торф, особенно при малом угле падения склонов
болота, не образует параллельных гряд-складок. Раститель¬
ность комплексов кочек и мочежин имеет такие особенности:
1) на болотных кочках с ненарушенной растительностью
особенно характерны Sph. medium, Eriophorum vaginatum,
редкий ярус сосны (ассоциация Pinus-Eriophomm-Sphagnum
medium); большую роль играют также ассоциации Кассандры
с тем же мхом и сосновым ярусом;
2) в мочежинах доминируют шейхцерия, пущица, очеретник,
Sph. balticum, Sph. Dusenii и Sph. cuspidatum. Обилие очерет-
ника и Sph. cuspidatum — признаки, общие с ильменско-западно¬
двинскими Hochmoor.
Сосново-сфагновые комплексы, покрывающие ] большую
часть болота, главным образом сосново-кустарничково-пуши-
цевого типа. Из кустарников доминируют Cassandra, Andromeda,
ближе к периферии Ledum. Кроме того, большие площади
занимают и сосново-кустарничково-сфагновые комплексы. Хоро¬
ший пример болот среднерусского типа — Писцовское и
Сахтыш-Рубское болота Ивановской области (Н. Кац, 1923).
Северные болота с мозаичным растительным покровом.
Болота, расположенные севернее области выпуклых болот, резко
отличаются от них по своему характеру. Часто эти северные
болота нельзя отнести ни к одному из трех основных типов
болот более южных районов. Северные болота характеризуются
пестрым, мозаичным и обычно беспорядочным чередованием
участков с различной растительностью, то олиготрофной, то
эвтрофной. Это чередование происходит как в мелкомозаичных
комплексах, где олиготрофные ассоциации занимают повы¬
шения микрорельефа, а более эвтрофные — понижения (моче-
жины), так и на значительных площадях. При этом участки
торфяника, занятые олиготрофной, мезотрофной и эвтрофной
растительностью, чередуются друг с другом, а также с заболо¬
ченными участками без торфа более или менее беспорядочно,
часто без центро-периферической поясности, характерной для
выпуклых болот. Торфяная залежь этих болот в основном
сложена эвтрофными торфами, причем олиготрофные торфа
залегают обычно более или менее тонким слоем — отдельными
островками — кочками или грядами, а также более значитель¬
ными участками. Эти особенности мозаичных болот зависят
от климатических условий, в которых они развиваются. Су¬
ровый климат задерживает нарастание торфа и выход болота
из сферы влияния грунтовых вод. Таким образом, формиро¬
вание выпуклой поверхности с концентрическим распреде¬
лением растительных комплексов оказывается в этом климате
264
невозможным. Очень часто при сравнительно небольшой мощ¬
ности залежи форма поверхности болота определяется рельефом.
Она более или менее следует за ним. Северные болота неспо¬
собны создавать свой собственный рельеф, свой гидрологи¬
ческий режим и олиготрофный режим питания и сильно
зависят от окружающих физико-географических условий, как
рельеф дна, режим грунтового питания, минеральное окру¬
жение и пр. Растительность этих северных болот не так резко
отличается от растительности минеральных почв, как расти¬
тельность выпуклых олиготрофных болот. Флористически
состав северных болот и даже зачастую доминирующие виды
в общем те же, что и на заболоченных минеральных почвах.
Северные мозаичные болота по характеру растительности
распадаются на ряд типов, являющихся вместе с тем регио¬
нальными. Они описаны ниже.
ЁЗ Аморфный, торф дна мочетины '&PZÜ Олиготрофный ptoptp
Звтрофный торф
Рис. 85. Схема строения болота типа аапа (а) и выпуклого болота (Ъ).
Болота типа аапа (Аарашооге). Слово «аапа» — финский
народный термин, обозначающий определенный тип болот и
вошедший в другие языки. В научной литературе его впервые
употребил А. Каяндер (1913). Болота «аапа» представляют
мозаику элементов олиготрофных (выпуклых) и эвтрофных
болот. Последние имеют характер травянистых (чаще осоковых)
болот с бурыми или сфагновыми мхами, а иногда и типа
римпи, и образуют фон, по которому разбросаны кочки из оли-
готрофного торфа, покрытые олигогрофной растительностью.
Если кочки эти сближены, то создается впечатление олиготроф-
ного болота с повышениями и мочежинами. Однако это сход¬
ство— лишь кажущееся (рис. 85), так как в основе болот аапа
лежит эвтрофный торф, на котором располагаются отдельные
олиготрофные кочки.. Мочежины здесь, таким образом, пер¬
вичные. Выпуклые же болота имеют сверху сплошной слой
сфагнового торфа, и мочежины здесь вторичные. Аарашооге
нельзя считать лишь стадией развития выпуклых болот. Регу¬
лярное затопление мочежин весной, ведущее, между прочим,
к образованию римпи, препятствует смыканию сфагновых кочек
и превращению аапа в Hochmoor. Так характеризует аапа
Г. Дю Риэ, следуя Г. Ранкену (1912).
265
Г. Освальд (192о) в общем дает то же определение аапа. Он подчерки¬
вает момент устойчивости сфагновых кочек и эвтрофных мочежин и отсут¬
ствие между ними скжцессионной1 связи. Расширению сфагновых кочек
мешает достаточно богатая солями вода, затопляющая болото весной. Он
считает аапа климатическим типом, замещающим в северной Европе выпук¬
лые болота, а в Альпах — сменяющим их при поднятии вверх. А. Каяндер
(1913) характеризует Ааратооге несколько иначе. Они свойственны северной
Финляндии и Лапландии и представляют сильно разветвленные болотные
системы нередко громадных размеров. Эти болота в основном безлесны.
А. Каяндер подчеркивает, что эти болота изобилуют римпи с слабо разви¬
тым, вследствие длительного застаивания воды, и угнетенным покровом мхов.
Эги римпи чередуются с повышениями характера гряд, покрытыми то сфаг¬
новыми, то бурыми мхами. Подобно предыдущим авторам, А. Каяндер
•считает аапа устойчивыми во времени. В противоположность выпуклым
болотам, имеющим центрифугальный сток, в аапа вода течет в одном на¬
правлении. Аапа в понимании А. Каяндера значительно отличаются от аапа
в понимании предыдущих авторов и представляют более гетерогенные
образования. В тех случаях, когда гряды с бурыми мхами чередуются
о эвтрофными мочежинами типа римпи, это будут чисто эвтрофные ком¬
плексы, а не мозаика олиготрофных и эвтрофных участков, характерная для
аапа в понимании Г. Дю Риэ и Г. Освальда. Болота типа аапа для смежных
с СССР частей Финляндии были описаны А. Каяндером (I. с.). Для Карелии
конкретное описание ассоциаций карельских аапа мы находим у К. Маля-
ревского (1926). Типы карельских аапа описаны Ю. Цинзерлингом (1934).
В последнее время довольно подробное описание болот этого типа в се¬
верной Карелии дал Г. Благовещенский (1936)1 2. Для средней Печоры
болота аапа описаны Н. Кац (1928) и Ю. Цинзерлингом (1929).
Следующие черты характерны для советских аапа:
1) чередование эвтрофных мочежин и повышений с олиго-
трофными, а иногда и мезотрофными ассоциациями;
2) обилие „римпи", т. е. мочежин с угнетенным и разре¬
женным моховым покровом и обнаженным торфом;
3) вогнутая, горизонтальная или наклонная, но не выпуклая
поверхность;
4) преобладание эвтрофных торфов в залежи, причем оли-
готрофные встречаются тонким слоем и отдельными участками,
слагая кочки;
5) отсутствие (так же как у Hochmoore) вечной мерзлоты
в торфе.
Аапа в Европейской части СССР — климатический тип бо¬
лот. Они располагаются в основном к северу от выпуклых бо¬
лот. Несмотря на Недостаточную изученность их, можно выде¬
лить несколько типов аапа, имеющих региональное значение.
1. Тип припечорских аапа. Эти аапа исследованы
в придолинной полосе средней Печоры (N. Katz, 1930). Они
встречаются в виде обособленных площадей (массивов). В цен¬
тре этих болот широко распространены комплексы аапа, часто
с обильными вторичными озерками. На грядах этих комплексов
обычно образуют ярус Ericaceae (Cassandra, Andromeda), а
часто и Betula nana, моховой же ковер — Sph. medium и Sph.
angustifolium. Здесь обычна примесь некоторых эвтрофных
видов — иногда даже ярус из вахты (Menyanthes). Мокрые, часто
1 Сюкцессия—смена фитоценозов во времени.
2 См. также другие статьи в сборнике «Геоботаника», вып. 3, Л. 1936.
26G
колышущиеся при ходьбе и с трудом проходимые мочежины
с разорванным моховым ковром из угнетенных сфагнов (Sph.
Jensenii) и Drepanocladus exannulatus, местами с обнаженным
торфом и коркой печеночников. Редкий ярус образуют здесь
шейхцерия, а также Carex limosa и Menyanthes. Эти аапа-ком-
плексы бывают окаймлены или прямо узким поясом олиготроф-
ного кустарничкового болота, или широким кольцом олиго-
трофных грядово-мочежинных комплексов. В последнем случае
болота носят смешанный характер аапа-олиготрофных болот.
Подобные смешанные болота широко распространены в При-
печорье.
2. Тип карельских аапа (см. Ю. Цинзерлинг, 1934). Эти
аапа обычны в северной Карелии в условиях карельского ланд¬
шафта (см. выше). Они представлены уже не отдельными бо¬
лотами, а большей частью сетчато разветвленными болотными
системами с ориентировкой гряд и мочежин перпендикулярно
к длинной оси болота и общему наклону местности с северо-
запада на юго-восток. Гряды образованы часто Sph. fuscum,
а мочежины эвтрофные, с преобладанием Equisetum limosum,
Carex lívida, C. lasiocarpa, C. limosa, Menyanthes и с слабо раз¬
витым моховым покровом. Нередко на таких болотах — скоп¬
ления (комплексы) озерков. Аапа-комплексы нередко окаймлены
олиготрофными ценозами. Карельские аапа богаче флористи¬
чески, чем припечорские, и ассоциации мочежин здесь более
разнообразны. По строению эти болота эвтрофны.
3. Тип лапландских аапа. Эти болота встречаются
на Кольском полуострове в условиях равнинного „лапланд¬
ского“ ландшафта. Они занимают сплошь большие простран¬
ства в виде болотных систем неправильных очертаний. Повы¬
шения микрорельефа в аапа-комплексах—в виде островков или
гряд.
Кайма из олиготрофных фитоценозов может присутство¬
вать или же не развивается. К западу и к востоку от Имандры
лапландские аапа не одинаковы. К западу они, вследствие
сильно расчлененного рельефа, имеют меньшие размеры.
В мочежинах здесь господствуют Carex ampullacea, Scirpus
caespitosus, Eriophorum angustifolium. Арктические Cyperaceae
роли здесь не играют, и по растительности эти аапа напоми¬
нают более южные карельские.
К востоку от Имандры в условиях равнинного рельефа аапа
занимают громадные площади, а растительность их имеет от¬
части арктический характер: в гмочежинах, наряду с Erio¬
phorum angustifolium и другими, распространены арктические
Carex rotundata и С.-rariflora.
Крупнобугристые болота. Для этих болот характерны
крупные возвышения бугрообразной или вытянутой (грядооб¬
разной) формы с залегающей неглубоко в торфе вечной
мерзлотой (рис. 86). Подобные бугры чередуются с понижениями
обычно в виде мочежин. Торф этих мочежин, в противопо¬
ложность буграм, оттаивает к концу лета обычно до дна.
267
Это мозаичный комплекс крупных бугров и мочежин. Высота
бугров различна. Т. Фриз (1913) в северной Швеции находил
бу'-ры до 7 мвысоты. На р. Полуе (приток Нижней Оби)
высота бугров 4—6 м (Н. Властова, 1936), в Енисей¬
ской лесотундре бугры до 8—10 м при средней высоте
3—4 м (Г. Эйдельман, 1936). Для Колы обычна высота бугров
3—3,5, иногда до 4 м(А. Чильман). В. Андреев (1931) дает
для северного Канина наибольшую высоту 3—4 м. Форма бугров
более или менее округлая, лопастная или вытянутая. Еще
А. Чильман отметил (1. с.), что наиболее высокие бугры
Рис. 86. Крупиобугристое болото. По Р. Поле.
связаны всеми переходами с расположенными рядом с ними
маленькими кочками обычного для кочек сфагновых болот
типа. Еще Р. Поле (R. Pohle, 1908) отметил, что в крупнобуг¬
ристых комплексах сочетаются в пространстве резко ксеро-
фитные ценозы крупных бугров и гидрофильные ассоциации
понижений. И действительно, растительность бугров — яркий
пример ксерофитных болотных фитоценозов. Приведем опи¬
сание растительности бугров по Г. Ануфриеву (1922). Вершины
высоких бугров зимой не покрываются снегом. Здесь обычны
пятна обнаженного, подвергающегося морозному выветриванию,
торфа. Эти пятна часто разбиты неглубокими трещинами на
маленькие ячейки. Кроме участков голого торфа, вершины
бугров покрыты лишайниковым ковром (главным образом
корковые лишайники Icmadophila и Ochrolechia tartarea), при¬
дающим буграм издали серую окраску. Цветковая раститель¬
ность развита на верхушках бугров слабо. Она ютится по глу¬
боким трещинам, часто пронизывающим поверхность бугров,
268
'-HtjcmapHi
cmpetrum-Dimm elongatum-Ass
Empetram-Sphagmim fasern-Ass
или образует маленькие стелющиеся куртинки высотой 2—3,
максимум 10 см по небольшим западинкам. Среди цветковых
главную роль играют крайне угнетенные арктические кустар¬
нички— Empetrum nigrum, Betula nana, а также Ericaceae
(Ledum palustre, Vaccinium ullginosum, V. vitis idaea и др.)
и. кроме того, Rubus chamaemorus. Условия существования
для растительности крайне неблагоприятны: иссушающее дей¬
ствие морозных зимних ветров при отсутствии снегового по¬
крова и, как и всюду на буграх, близкая вечная мерзлота.
На склонах бугров, где действие морозных ветров не так
сильно, рост кустарничков лучше, а в нижней части склонов
они образуют уже со¬
мкнутые заросли, высотой
иногда до 1 м, причем
здесь часто преобладает
полярная березка. Пони¬
жения между буграми
заняты мокрыми осоко¬
выми фитоценозами сков-
ром то сфагновых, то бу¬
рых мхов, а иногда и
с слабо развитым мохо- ^Exrostrata-uErlophaiwivbgiiiatuiii ассоциации ит.д.
вым покровом и участ-
ками голого торфа. В по¬
нижениях играют боль¬
шую роль, наряду с арк¬
тическими, Carex rotun-
data, С. rariflora, Eriopho-
rum russeolum, иногда
также Eriophorum angustifolium и другие виды осок — на ниж¬
ней Печоре Carex chordorrhiza, С. limosa и некоторые другие.
Среди понижений между буграми постоянно встречаются за¬
чатки будущих бугров в виде сфагновых кочек, часто из Sph.
fuscum, а иногда в виде участков дернины мочежин, припод¬
нятых действием мерзлоты. Развитие бугров и их постепенный
рост сопровождаются сменой фитоценозов от влаголюбивых
к все более ксерофитным (рис. 87). По мере роста бугров
начинает преобладать морозная ветровая эрозия, уничтожающая
растительный покров. 1Рис. 87. Сюкцессия ассоциаций на крупно-
° бугристом болоте в Лапландии.
По Г. Дю Риэ, 1925.
Для объяснения происхождения бугров высказывались различные
взгляды !. Они сводятся к следующему:
). Бугры — это останцы прежнего торфяника и возникают вследствие
размыва сплошного ранее торфяника водой (А. Чильман, 1890). Это объяс¬
нение, несомненно, приложимо лишь к отдельным случаям.
2. Бугры образуются вследствие биогенного процесса — нарастания
сфагнума вверх — и представляют в сущности маленькие выпуклые торфя-
нички, возникшие на минералыой почве. Промежужи между буграми—это
естественные мочежииы (Г. Танфильев, 1911). Однако рост сфагнума может
вызвать лишь первоначальную диференцировку микрорельефа, но не объ¬
ясняет дальнейшего роста бугров в высоту. Кроме того, нередко зарождаю-
1 Обзор их дает Г. Ануфриев (1922).
2С9
щиеся бугры образованы осоковым торфом—отложением мочежин, приподня¬
тым вверх механической силой (см. ниже).
3. Бугры образуются благодаря накоплению торфа кругом стволов и
корней деревьев (Р. Поле, 1903). Этот способ происхождения бугров вряд
ли, однако, играет заметную роль.
4. Большинство исследователей выдвигает как главный фактор бугро-
образования влияние вечной мерзлоты и периодическое замерзание и оттаива¬
ние избыточно увлажненного грунта. Этот фактор проявляет себя различно.
При замерзании верхнего слоя болот (на Алтае), лежащих на насыщенных
водой щебнистых моренах, притекающая вода, не находя выхода, разрывает
замерзший слой, и в образовавшуюся трещину внедряется снизу жидкий
торф. Последний, замерзая, выпирает поверхность болота около трещины.
Образуются бугры до 1,5 М высоты (С. Яковлев, 1911). Подземная вода,
замерзая осенью, расширяется, образуя подземною наледь. Последняя при¬
поднимает дернину, и возникает бугор (Р. Аболин, 1913). Подобное выпу¬
чивание вызывается иногда почвенным горизонтом в болоте, если он ока¬
зывается сжатым между двумя замерзающими осенью почвенными слоями
(В. Сукачев, 1911).
Несомненно, что в разных случаях действуют различные из указанных
причин. Взгляды С. Яковлева (1911), Р. Аболина (1913) и В. Сукачева (1911)
относятся к северным и восточным частям Сибири, где типичные крупно¬
бугристые торфяники отсутствуют.
Происхождение крупных бугров выясняется при изучении
их строения на разрезах. Это строение и, следовательно,
происхождение не везде одни и те же. Весьма часто в осно¬
вании торфяных бугров находится минеральное ядро, т. е.
выпуклина минеральной почвы. Иначе говоря, минеральная
почва под буграми выше, чем в мокрых понижениях между
буграми. Это установлено для Колы (А. Чильман), для нижней
Печоры (наши наблюдения 1932 г.). Минеральное ядро занимает
нередко меньшую часть бугра, как это часто имеет место
в европейской бугристой тундре. Такие бугры сложены пре¬
имущественно или же даже нацело из торфа \ В западносибир¬
ской лесотундре высокие до 6 м бугры почти нацело мине¬
ральные— слой торфа не более 0,6 м (Н. Властова, 1932),
Это зависит от сурового и континентального сибирского кли¬
мата, неблагоприятного для торфообразования. За Енисеем
климат еще суровее, крупные торфяные бугры сходят на-нет,
и в северо-восточной Сибири слой торфа на болотах вообще
лишь незначительной толщины1 2. Минеральное ядро внутри
бугров указывает, что они образуются путем выпирания грунта.
Это действие производит образующаяся летом талая вода,
когда она замерзает и расширяется. Значение этого фактора
при образовании бугров отмечалось, как мы видели, разными
авторами. В частности, для крупных бугров этот способ воз¬
никновения отмечает для северной Швеции Т. Фриз. Несо¬
мненно, что это весьма обычный тип бугрообразования и у нас.
Давление замерзающей воды вызывает особенность строе¬
ния торфа бугров. Слои осокового и других видов торфа,
1 Чисто торфяные бугры без минерального ядра наблюдались в север¬
ной Швеции (Т. Фриз).
2 О распространении крупнобугристых торфяников см. Б. Городков
(1935) и другие работы того же автора.
270
образующиеся в мокрых ровных понижениях болот, в буграк
образуют складки, приподнятые над соседними ровными мок¬
рыми западинами (рис. 88). Эти слои могли приподняться лишь
после своего образования под действием замерзающей воды.
Этот способ возникновения бугров наблюдается и в настоящее
время. На нижней Печоре мы наблюдали эмбриональные бугры
в виде невысоких возвышений беловатого цвета, издали напо¬
минающих камни. Эти бугорки были нацело сложены тем же
видом сфагнума, что и окружающая мокрая сфагновая дернина.
Приподнятый мерзлотой сфагнум высох и побелел. Он смерзся
в плотную массу, но вместо льда, как внутри сформировав¬
шихся торфяных бугров, между стеблями сфагнума образо¬
валась мерзлота в виде плотного кристаллического снега..
Не всегда, однако, торфяные
бугры образуются воздей¬
ствием мерзлоты. Эмбриональ¬
ные торфяные бугры, вполне
сходные по внешнему виду
с обычными большими сфагно¬
выми кочками мочежинных
болот более южных широт, Рис. 88. Строение торфяного бугра
НО С вечномерзлым ядром в северной Швеции. По Т.Фризу, 1913
внутри, обычны на нижней Пе- Штриховк%Тло! -ПкГы"РтГрТГ' Т°РФ
чоре. Вероятно, эти бугры воз¬
никают так же, как и кочки сфагновых торфяников, вне
зоны вечной мерзлоты. В такой сфагновой кочке, вследствие
ее плохой теплопроводности, быстро появляется вечная мерз¬
лота, способствующая затем дальнейшему росту кочек и пре¬
вращению их в настоящие бугры. А. Чильман выдвинул теорию
образования бугров путем эрозии поверхности болота текучей
водой. Бугры в этом случае являются останцами сплошной
поверхности торфяника. Значение этого способа образования
бугров преувеличено, но в некоторых случаях он, несомненно,
имеет место. В Малоземельской тундре описан «эрозионный
комплекс» бугристых болот, возникший указанным способом
(сборник «Оленьи пастбища Северного края», 1933). Эти бо¬
лота легко отличаются по ряду признаков от крупнобугристых
болот неэрозионного происхождения. Мочежины здесь совсем
особого характера: в них массами развивается Сагех aquatШs.
Из сказанного следует, что крупнобугристые болота обра¬
зуются различным путем.
Крупные торфяные бугры имеют различный ботанический
состав. В верхней части бугров обычны торфа, образовавшиеся
в сравнительно сухих условиях. Здесь большую роль играет
сфагнум; на Коле (по А. Чильману, Г. Ануфриеву, 1922) бугры
одеты мощным слоем чисто сфагнового торфа (55рЬ. ^эсит),
то же на нижней Печоре, а в северной Швеции — сфагново-
поЛитриховым торфом (Т. Фриз). Сверху эти торфа выветри¬
ваются, трескаются, покрываются коркой лишайников, зеленых
мхов (Шсгапит). Ниже сфагнового торфа залегают торфа более
271
влажных и эвтрофных местообитаний. Здесь обильны остатки
осок — чисто осоковые торфа (в северной Швеции Сагех гоШп-
<Ша, С. ациаиНэ — Т. Фриз), осоково-сфагновые, иногда осо-
ково-гипновые. В основании бугра, как указывалось, залегает
обычно минеральное ядро. В западносибирской и енисейской
лесотундре ботанический состав торфа иной. Олиготрофные
виды сфагнума, роль которых здесь в современной раститель¬
ности невелика, и в торфе играют небольшую роль. Иногда
Рис. 89. Строение торфяников Енисейской лесотундры.
По Г. Эйдельману, 1936.
1 — сфагновый торф; 2 — осоковый; 3 — гипновый; 4 — древес¬
ный; б — хвощевый; 6 — из остатков вересковых.
они образуют лишь тонкий поверхностный слой сфагнового
торфа, в других случаях весь торфяной бугор целиком сло¬
жен из эвтрофных торфов (рис. 89).
!".■ Крупные бугры — важнейший член комплексов крупнобуг¬
ристых болот. Видимые издалека в виде возвышений уже
не микро-, а мезорельефа, эти бугры создают совершенно
своеобразный ландшафт. Надо иметь в виду, что остальные
члены растительного покрова этих болот весьма разнообразны.
Не говоря о том, что крупные бугры сочетаются то с олиго-
трофными, то с мезотрофными, то с эвтрофными мочежинами
272
с сфагновым или с гипновым ковром, или же с слабо развитым
моховым покровом, в сложении крупнобугристых комплексов
участвуют ассоциации олиготрофных болот, болот аапа и других
болотных типов. Так, в Кольской лесотундре крупные бугры
часто разбросаны среди элементов аапа. К последним относятся
мезотрофные и эвтрофные мочежины, частью без сплошного
мохового покрова (римпи), и кустарничковые фитоценозы кочек
с слабо развитым сфагновым ковром. Смешанные аапа-буг-
ристые болота широко распространены в Кольской лесотундре.
В Кольской тундре структура этих комплексов еще сложнее —
сюда присоединяются еще плоские дикрановые бугры — элемент
более северных тундровых болот. На нижней Печоре бугры
разбросаны обычно среди невысоких олиготрофных сфагновых
и лишайниковых кочек или гряд, т. е. элементов более южных
олиготрофных болот. Эти болота промежуточного характера
между крупнобугристыми и олиготрофными. Сложность расти¬
тельного покрова крупнобугристых болот увеличивается еще
тем, что комплексы с крупными буграми не покрывают обычно
всего болота, а чередуются с различными комплексами, ли¬
шенными этих бугров, например, олиготрофными грядово-моче-
жинными, комплексами типа аапа и другими.
Болота с комплексами мерзлых дикраново-лишайниковых
бугров и мочежин. На поверхности этих болот мозаично
чередуются мочежины и бугры. Первые в общем весьма похожи
на мочежины крупнобугристых болот, вторые же значительно
отличаются от крупных бугров,—прежде всего меньшей высо¬
той. Наиболее типичны описания этих болот в Большеземель-
ской тундре (В. Андреев, 1933, 1935)L Высота бугров здесь
всего до 0,6 м и ниже, т. е. бугры гораздо ниже, чем
на крупнобугристых болотах. Эти бугры имеют пло¬
скую верхушку и занимают значительную площадь (от 5ХЮл*2
до 15X25 м2). Они различной формы: то овальные, то в виде вы¬
тянутых узких гряд. На западе Большой Земли они оттаивают
всего на 15—20 см. Напочвенный покров бугров из мхов (глав¬
ным образом тундровых видов Dieran um — D. angustum, D. fus-
cescens, D. elongatum, часто также Polytrichum strictum) и
лишайников (на западе Большой Земли — Cetraria nivalis, С. cu-
cullata, Sphaerophorus fragilis, Cladonia mitis, Ochrolechia frigida).
При этом лишайники, особенно корковые, образуют верхний
ярус, поселяясь на моховой дернине. Она образует нижний
ярус напочвенного покрова и частично отмирает. Верхний
ярус бугров в общем слабо развит и сильно угнетен. Здесь
встречаются Betula nana, Ledum palustre, Rubus chamaemorus,
Empetrum nigrum, Vaccinium uliginosum, V. vitis idaea, Andro¬
meda polifolia и некоторые другие (в Малоземельской тундре—
Н. Кац и Ц. Минкина, 1936). Растительность мочежин очень
бедна видами и однообразна в разных районах. Здесь редкий 11 В. Андреев (1935) называет их плоскобугристыми (термин Б. Город
нова).
18—722
273
ярус арктических осок; особенно широко распространены
Carex rotundata и С. rariflora. Моховой покров обычно сом¬
кнут— преобладают то сфагновые мхи (Sph. Lindbergii, Sph.
balticum, иногда Sph. riparium), то Bryales — главным образом
виды Drepanocladus (D. revolvens и др.). В мочежинах мерз¬
лота оттаивает глубже, чем на буграх, — в Малоземельной
тундре в сентябре она была на 0,45 — 0,70 см ниже поверх¬
ности и, как исключение, на 1,0 м. Иногда мочежины оттаивают
до дна (Н. Кац и Ц. Минкина). Эти болота в общем соответ¬
ствуют „плоскобугристым болотам“ Б. Городкова 1. Под этим
названием они часто и описываются в литературе. В Евро¬
пейской части СССР они распространены в области тундры.
Это более северный тин по сравнению с крупнобугристыми
болотами. Строение этих болот изучено весьма слабо. Причины
образования бугров в некоторых случаях, повидимому, те же,
что и крупных бугров, — действие мерзлоты. В западной части
Большеземельской тундры эти бугры располагаются на выпу¬
клинах минеральной почвы (В. Андреев, 1932). Другие типы
болот описаны при описании отдельных торфяноболотных
областей. 11 Конкретное описание этих болот см. Б. Городков (1932), а общее опи¬
сание их и данные о географическом распространении — он же (1935).
ГЛАВА IX.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ БОЛОТ
И ТОРФЯНИКОВ.
Болота Европейской части СССР.
Распространение болот Европейской части Союза зависит
от климатических и других физико-географических факторов.
На территории Восточно-Европейской равнины они постепенно
изменяются с севера на юг и с запада на восток. Соответ¬
ственно этому изменяется количество болот и их характер.
Растительность Европейской части Союза имеет достаточно
определенно выраженную зональность; существуют подобные
же зоны и болот. Описание болот удобно вести по выделен¬
ным болотным зонам (Н. Кац, 1936). В пределах зон выделены
уже более дробные болотные области \ Следует различать
такие болотные зоны в Европейской части СССР (с севера
на юг, рис. 90).
I зона — минеральные болота и заболоченные земли арктиче¬
ской Большеземельской тундры.
II зона — торфяники типичной тундры с мерзлыми ди-
краново-лишайниковыми буграми и мочежинами.
III зона — торфяники с крупными мерзлыми буграми и
мочежинами.
IV зона — аапа-торфяники северной тайги.
V зона — олиготрофные торфяники таежной полосы с ком¬
плексами гряд и мочежинами.
VI зона — эвтрофные и олиготрофные сосново-сфагновые
торфяники юга хвойной зоны.
VII зона — эвтрофные осоковые торфяники и тростниковые
болота полосы широколиственных лесов и лесостепи.
VIII зона — прибрежно-водные тростниковые болота и за¬
соленные болота степной полосы.
I зона. Минеральные болота (без торфа) и заболоченные
земли арктической Большеземельской тундры. Эта зона
занимает так называемую арктическую тундру — крайний
северо-восток Европы — Югорское и Карское побережья 11 Описание областей приведено лишь для зоны V.
18* 275
Рис. 90. Карта торфяно-болотных зон и областей Европейской части СССР-
По Н. Капу, 1936.
Объяснение см. в тексте.
Северного полярного моря и хребет Пай-Хой, а, кроме того,
остров Вайгач и южный остров Новой Земли. Слабая испа¬
ряемость и повсеместная мерзлота вызывают здесь громадную
поверхностную заболоченность. На болота здесь приходится
от 51,5 до 66,3%'» если же прибавить заболоченные моховые
тундры, то 70°/о и более всей площади (В. Андреев, 1935).
Суровый арктический климат препятствует отложению торфа.
Болотные пространства арктической тундры — не торфяники,
а минеральные болота и заболоченные земли. Эти болота имеют
вид ровных пространств с торфом всего в несколько санти¬
метров, лежащим на вечномерзлом оглеенном субстрате. Они
поросли главным образом северной Сагех адиабПз и близким
арктическим видом С. в1апз. Торфяники с мощным слоем
торфа здесь редки, невелики, и торф в них в настоящее
время не нарастает. Это реликтовые образования ксеро-
термического периода, когда климат в тундре был теплее
современного и накопление торфа шло более быстрым темпом.
Для этой зоны, следовательно, характерны: 1) громадная за¬
болоченность, 2) преобладание осоковых болот на неглубоком
торфе, 3) слабое распространение настоящих торфяников и
реликтовый их характер.
И зона. Торфяники типичной тундры с мерзлыми дикра-
ново-лишайниковыми буграми и мочежинами. Эта зона за¬
нимает в Большой Земле моховую и ерниковую подзоны
тундры и северную лесотундру, а в Малой Земле — мохово-
ерниковую и моховую тундровую подзоны В. Андреева (1932).
Кроме того, сюда относится Северный Канин и остров Колгуев \
Следовательно, сюда входит типичная тундра Л. Берга
(1931) и его южная тундра. На водоразделах этой болотной
зоны широко распространены торфяники с дикраново-лишай-
никовыми грядами и мочежинами. При ровном низменном
рельефе эти торфяники делаются господствующим элементом
ландшафта. Например, в Вангурейской лапте1 2 (Большеземель-
ская тундра) они занимают более 80% всей территории
(В. Андреев). Заболоченность в этой зоне, так же как и в
предыдущей, очень велика, что объясняется теми же причинами.
Болота в Большеземельской тундре занимают на западе ее
от 8,9 до 52,6%, а в восточной части — от 19,3 до 45,6%
(В. Андреев, 1936). Если учесть еще заболоченные тундры,
то цифры будут выше. Общий процент заболоченности
в Большеземельской тундре, таким образом, вероятно, близок
к 50. Эту цифру мы пока примем и для всей зоны в целом.
В отличие от более северной зоны мы имеем здесь значитель¬
ную заторфованность, т. е. большой процент площади при¬
ходится на долю торфяников. Так, для некоторых районов
севера Малоземельской тундры при общей заболоченности
1 Положение Колгуева с точки зрения зональности неясно; может быть,
его следует отнести к I зоне.
2 Низменность по побережью Северного Полярного моря.
277
в 60—80% на торфяники приходится от 10 до 40% всей
исследованной площади (Н. Кац и Ц. Минкина). Широкое разви¬
тие торфяников показывает, что климат здесь благоприятнее
для торфообразования, чем в более северной зоне. В связи
с этим торфяники сравнительно глубокие — в Малой Земле
у озер, где залежь наибольшая, — до 2,5—3,0 м; при удалении
от озер залежь быстро выклинивается (Н. Кац и Ц. Минкина).
Кроме характерного для
зоны типа—торфяников с ди-
краново-лишайниковыми буг¬
рами,— здесь встречаются и
другие типы болот. На границе
с предыдущей зоной арктиче¬
ские осочники с Carex aqua-
tilis, о которых говорилось
выше, становятся преобладаю¬
щими; к югу они постепенно
редеют. Встречаются и крупно-
бугристые болота.
В Малой Земле они пред¬
ставлены: 1) комплексами
крупных бугров и озерков,
2) комплексами эродируемых
бугров и мочежин cCarex aqua-
tilis (эрозионный комплекс см.
„Оленьи пастбища Северного
края“, 1933). Из заболоченных
земель отметим: 1) кустарни¬
ковые ивняки с ярусом Salix
lapponum, S. phylicifolia, S.
lanata и др.; это ценные оленьи
пастбища; 2) сфагновые тунд¬
ры, особенно распространен¬
ные на юге зоны; в них господ¬
ствуют то кустарнички — Be-
tula nana, Ledum, то травяни¬
стая растительность (Luzula
Wahlenbergii). На болотах зоны
всецело преобладают арктиче¬
ские виды: Empetrum nigrum,
Betula папа, Rubus chamaemorus, Carex rotundata, C. rariflora,
Eriophorum russeolum и др. Обычные же для болот нашей лес¬
ной зоны растения—Cassandra calyculata, Scheuchzeria и другие
вовсе не характерны для здешних болот. Болота зоны
в естественном виде — оленьи пастбища средних бонитетов.
Торфа северной части Малой Земли обнаружили удовлетвори¬
тельные топливные показатели. Однако торфодобыча в тундре
наталкивается на неблагоприятные для сушки торфа климатиче¬
ские условия и вечную мерзлоту. Подстилочные торфа тундры
найдут применение в сельском хозяйстве. Ботаническое строе-
278
Рис. 91. Строение торфяников Мало-
земельской тундры. По Н. Кацу н
Ц. Минкиной. 1936.
1 - сфагновый торф; 2—гнпново-сфагновый;
3 - осоково-сфагновый; 4—гипново-осоковый;
5 — осоково-древесный; 6 —гипновый; 7—ди-
крановый; 8—кустарничковый (вересковые
и Betula nana); 9 — осоково-кустарничково-
дрсвеоным; 10 — хвощ; 11—травяно- (вахта)
осоковый, 12 — осоковый.
ние торфяников в Малоземельской тундре (более сухие участки)
таково (см. Н. Кац и Ц. Минкина, 1936, рис. 91): сверху торф
сильно выветрившийся, часто минерализованный дикрановый
или из кустарников; ниже идет нередко мало разложившийся
сфагновый торф; еще ниже — эвтрофные или смешанные торфа,
нередко с прослойкой древесного торфа. Иногда книзу опять
встречаются сфагновые торфа. В общем для тундровых
торфяников характерен верхний слой дикранового (тундрового)
торфа и слабое участие олигогрофных сфагновых торфов.
Однако все же роль сфагнов сильно возрастает здесь и
в растительности, и в торфе по сравнению с более северной
зоной.
Ш зона. Торфяники с крупными мерзлыми буграми и
мочежинами. Эта зона в Европейской части СССР тянется
с перерывом у Белого моря от Урала до Финляндии. На вос¬
ток она продолжается в Западную Сибирь, а на запад —
в Финляндию и далее в северную Скандинавию. Она зани¬
мает южную лесотундру и север лесной зоны.
Северная граница зоны образована или морем, или северной
границей лесов. Южная граница к востоку1 и западу от Печоры
идет в общем по 66-й параллели, а ближе к Мезени и дальше
на запад о? нее идет в общем по южной границе лесотундры,
как и на Кольском полуострове. Характерный болотный
тип на водоразделах зоны — крупнобугристые болота. Ком¬
плексы с крупными буграми, весьма пестро сложенные,
чередуются с комплексами без бугров, повидимому, часто
занимая лишь подчиненное положение. Заболоченность зоны
велика. По исследованиям почвоведов (Л. Прасолов, 1927;
А. Красюк, 1933), при приближении с юга к южной границе
болотной зоны заболоченность резко возрастает, а на карте
Л. Прасолова соответствующая полоса показана сплошь
заболоченной.
Определяя заболоченность по растительности, а не по почве,
нельзя принять болотистость нашей зоны близкой к 100°/о,
как это показано на карте Л. Прасолова. Все же она очень
зысока, вероятно, не ниже 50%- Для отдельных районов
ваболоченность, несомненно, гораздо выше. Значительная
часть болот зоны приходится на заболоченные земли — в этом
сходство с поверхностно заболоченными северными зонами.
Однако здесь, благодаря более благоприятному для торфо-
образования климату, участие торфяников в общей
заболоченной площади больше, чем на севере.
Глубина торфяников здесь также более значительна. Средние
глубины на нижней Печоре по нашим данным были от 1,13 до
1,5 м (Н. Кац и Ц. Минкина, 1936). Крупные бугры предста¬
вляют характернейший ландшафтный признак зоны. Высота
торфяных бугров здесь максимальная — к северу они заме¬
няются сравнительно низкими дикраново-лишайниковыми
1 Ближе к Уралу она, видимо, спускается несколько южнее.
279
буграми, к югу низкими же, уже лишенными вечной мерзлоты
грядами и кочками сфагновых болот. Благодаря сравнительно
теплому лету здесь образуется значительное количество талой
воды, которая, замерзая, производит сильное действие, проя-
вляющееся в образовании громадных бугров.К северу, благодаря
короткому лету, талой воды бывает меньше,' эффект от ее
замерзания небольшой, и высота бугров невелика. Накопление
торфа или торфогенный процесс в нашей зоне идет не только
в мочежинах, как в двух северных зонах, но и на повышениях
микрорельефа на первых стадиях их формирования. Невысокие
кочки одеты здесь часто торфогенным сфагновым покровом.
Дальнейший рост их идет уже главным образом под влиянием
мерзлоты, причем торфогенный покров по мере роста кочки
вверх и ее обсыхания заменяется лишайниковым, почти не
образующим торфа. С дальнейшим ростом и выходом из-под
защитного снегового покрова начинается разрушение торфа
на вершинах высоких бугров. Роль сфагновых мхов в этой
зоне больше, чем на севере. Они играют не только большую
роль в мочежинах, но часто образуют и кочки. Олиготроф-
ные сфагновые торфа здесь сильно распространены, слагая
часто невысокие кочки и образуя иногда довольно мощный
слой в мочежинах и в некоторых случаях главную массу
крупных бугров. Возрастание роли сфагновых мхов и сфагно¬
вых торфов — следствие более мягкого климата этой зоны.
Кроме бугристых болот, в этой зоне встречаются и другие
болотные типы. Из более южной зоны заходят сюда олиготроф-
ные сфагновые болота разных типов. Они иногда встречаются
в виде обособленных массивов, часто же образуют смешанные
массивы с крупнобугристыми участками. Встречаются в этой
зоне также различные эвтрофные и мезотрофные болота, а
в западной ее части и комплексы типа аапа. Из типов
заболоченных земель здесь обычны заболоченные леса со
сфагнумом и ярусом Betula nana, осок и других растений.
IV зона. Торфяники аапа северной тайги. Эта зона, ве¬
роятно, имеет прерывистое распространение и представлена,
повидимому, лишь в северном Приуралье, с одной стороны,
и на Коле и в северной Карелии — с другой, откуда она про¬
должается в Финляндию. Мало вероятно, как это предполагают
некоторые, что болота типа аапа и переходные между ними
и крупнобугристыми обычны на севере всей лесной зоны
Северного края. Кроме болот аапа, описанных выше и пере¬
ходных между ними и другими типами, здесь распространены
различные олиготрофные, а также мезотрофные и эвтрофные
болота. Растительность болот этой зоны уже близка к болотам
более южной зоны: здесь всюду широко распространены
болотные Ericaceae (в числе них на востоке и Кассандра),
Scheuchzeria, Scirpus caespitosus (в Карелии). Из арктических
видов морошка всюду встречается массами, водяника же,
Betula nana, Carex rotundata, C. rariflora имеют массовое распро¬
странение главным образом на Кольских аапа.
280
V зона. Олиготрофные торфяники тайги с комплексами
гряд и мочежин. Эта зона тянется непрерывно, переходя на
запад за пределы СССР и через Урал далее на восток в Запад¬
ную Сибирь. Ширина зоны вдвое с лишком больше ширины
всех расположенных к северу болотных зон, вместе взятых.
В Карелии и в северном Приуралье эта зона граничит с зоной
аапа, а на остальном протяжении ее северная
граница там, где она не образована морем, идет
по южной границе лесотундры и лишь к востоку
от Печоры, подходя к 66-й параллели, глубоко
вдается в таежную зону.
Интересно проследить смещение болотных типов и зон относительно
ботанико-географических зон с запада на восток. Крупнобугристые болота
на Коле, свойственные лесотундре, заходят в тундру и почти не заходят
в лесную зону. К востоку от Печоры эти болота тоже занимают лесотундру,
но южная их граница уже глубоко смещается в лесную зону. В Западной
Сибири этот тип болот встречается главным образом уже в лесной зоне,
в лесотундре же на водоразделах здесь господствуют плоскобугристые бо¬
лота (Б. Городков, 1928). Смещение крупнобугристых болот относительно
лесной зоны к югу объясняется тем, что сухой континентальный климат
Западной Сибири, несмотря на низкую температуру, способствует продви¬
жению леса на север и, наоборот, неблагоприятен для торфонакопления и
для образования крупных бугров. Понятно, что, вместе с зоной крупнобуг¬
ристых болот, смещается к югу и зона выпуклых олиготрофных мочежинных
болот.
В целом широкое распространение этого типа болот опре¬
деляется климатом. К северу от этой зоны с ухудшением
климата появление местной вечной мерзлоты вызывает обра¬
зование крупнобугристых болот; к югу с уменьшением клима¬
тического коэфициента начинают преобладать эвтрофные
болота, олиготрофные же приобретают характер лесных болот,
уже без мочежин. Однако распространение грядово-мочежин-
ных торфяников обуславливается и рельефом. Как отмечалось,
этот тип представлен обычно сравнительно обширными боло¬
тами. Условие образования таких болот — равнинный рельеф
с большими депрессиями. Даже в зоне наиболее благоприят¬
ного для мочежинных болот климата эти торфяники развиваются
на водоразделах в условиях равнинного рельефа. Там же, где
рельеф расчленен, в олиготрофных условиях встречаются пре¬
имущественно сосново-сфагновые болота. К югу же от зоны гря-
дово-мочежинных болот сосново-сфагновые болота
уже повсюду начинают преобладать в олиго¬
трофных условиях. Рельеф имеет значение не только
для распределения олиготрофных типов внутри рассматри¬
ваемой зоны, но и в большой степени определяет южную
границу зоны. Так, от Камы до Вятки эта граница идет по
южной окраине низменных болотистых районов (А. Фокин, 1930),.
Дальше на запад от р. Вятки до р. Унжи эта граница идет
по северной окраине Волжско-Двинского водораздела (А. Шен-
ников, 1938), также по границе обширных болотистых низмен¬
ностей, к югу от которых лежат районы с расчлененным
рельефом, относящиеся уже к следующей болотной зоне.
281
Наконец, от Волги (у Калягина) до Смоленска эта граница
идет по северной окраине возвышенностей, причем зона
захватывает лежащие к северу низменности, может быть,
предледниковые вюрмские, с крупными олиготрофными
болотами. По Мещерской и Волжско-Окской предледниковым
низменностям мочежинные комплексы, хотя и несовершенно
выраженные, идут далеко на юг, разрезая следующую болот¬
ную зону и вызывая смещение болотных зон. На остальном
протяжении южная граница зоны установлена весьма прибли¬
зительно и во многих пунктах вообще проведена лишь условно.
Грядово-мочежинные торфяники господствуют на низменных
ровных водоразделах этой зоны. Районы с расчлененным
рельефом дают приют сосново-сфагновым торфяникам. Наконец,
в отдельных районах с особыми почвенно-геологическими
условиями преобладают и эвтрофные болота. Все же во всей
зоне в целом безусловно преобладают олиготрофные
болота над эвтрофными в среднем примерно
в отношении 3,8:1,0. Это зона климатического
оптимума для выпуклых сфагновых торфяников.
Вместе с тем сфагновые мхи и сфагновые торфа здесь макси¬
мально распространены.
Сфагновые мхи на олиготрофных болотах большей частью
господствуют безраздельно1, одевая сплошь повышения микро¬
рельефа и выстилая сомкнутым ковром мочежины. На крупных
болотах на протяжении целых километров сфагнум опреде¬
ляет условия существования и физиономию растительного
покрова, причем остальная растительность отступает на задний
план. Помимо большой роли сфагновых торфов, важно отметить,
что олиготрофные виды сфагнов часто почти
нацело слагают торф, образуя чисто сфагновые
торфа лишь с незначительной примесью цветковых растений.
Эта болотная зона вместе с тем — зона оптимального торфо-
образования как в настоящее время, так, вероятно, и в течение
всей послеледниковой истории болот. Это видно из того,
что торфяники достигают здесь наибольшей мощности1 2,
а процент площади торфяников по отношению к общей
заболоченной площади достигает максимума3. Зато процент
заболоченности значительно падает по сравнению с северными
болотными зонами, и заболоченные земли с неглубоким торфом
абсолютно и по отношению ко всей площади болот играют
меньшую роль, чем на севере. Меньшее, чем на севере, развитие
процессов заболачивания зависит здесь от климатических
условий, большей испаряемости и отсутствия вечной мерзлоты.
Цифры заболоченности относятся к прежним административным
районам, не совпадающим с природными. Заболоченность
1 Исключение составляют лишь приморские районы, где на повышениях
микрорельефа и в мочежинах сфагны частью заменяются другими растениями.
2 Средняя глубина торфа в этой зоне принята в 2,62 м.
3 Это отношение приблизительно равно 50%.
в общем повышается к северу, доходя в б. Ленинградской
области и Карелии до 42,5%. На юге зоны она в общем
падает: в центральной части Московская, Ивановская области
заболочены на 16 —11,5%, на юго-западе зоны (БССР) за¬
болоченность выше 20 — 30%.
Как уже отмечалось выше, характерный тип болот на
равнинных водоразделах зоны — выпуклые грядово-мочежин-
Рис. 92. Карта растительности мезотрофного болота Камское, Горьковского
края. По Н. Властовой, 1933.
1 — осока нитевидная, Sph. subbicolor и береза; 2 — осока нитевидная, Sph. recurvum^ бе¬
реза; 3 — осока нитевидная, Sph. recurvum и сосна; 4 — Sph. recurvum, сосна и кустарнички;
5 — Soh. recurvum, шейхцерпя и сосна; 6—Sph. recurvum, пушица, сосна; 7—грядово-мочежин-
ный комплекс с пушицей и сосной; 8 — грядово-мочежинный комплекс с осокой нитевидной;
9 — березовый лес с осоками и Sph. subbicolor; 10 —березовый лес с папоротниками и зеле¬
ными мхами; 11 — береза и ольха с папоротниками и Sph. subbicolor; 12 — Sph. papillosum.
ные торфяники. В разных частях зоны эти торфяники не¬
одинаковы и образуют особые региональные типы. Отметим,
что в грядово-мочежинных болотах мозаично чередуются
в горизонтальном направлении, а иногда и в вертикальном,
т. е. сверху вниз (в глубь залежи), торфа, сложенные моче-
жинной растительностью, и торфа, сложенные растительностью
кочек. Первые торфа образуют дно мочежин, вторые —
повышения микрорельефа. Кроме грядово-мочежинных олиго-
283
трофных болот, всюду в соответствующих усло¬
виях рельефа встречаются сосново-сфагновые
болота. Они принадлежат к трем основным типам: сосново-
кустарничковому сфагновому, сосново-пушицевому сфагновому
и сосново-кустарничково-пушицевому сфагновому. Эти типы
сосново-сфагновых болот распределены неодинаково на про¬
тяжении болотной зоны. Кроме олиготрофных, встречаются
здесь также мезотрофные и эвтрофные болота. Последние
в отдельных районах с особыми почвенно-геологическими
условиями могут даже преобладать. Мезотрофные болота
главным образом двух типов:
1) сфагновые, обычно облесенные сосной или
березой с преобладанием осок, чаще Сагех
1аз1осагра, образующей верхний ярус травяного покрова;
Рис. 93. Профиль строения Шатурского торфяника. Работы Торфяной
академии „Шатурская болотная система*, I, 1921.
1 — сфагново-пушицевый торф; 2 — пушицевый; 3 — осоковый;
4 — включения пушицы; 5 — пни; 6 — гипново-осоковын торф;
7 — сфагново-осоковый; 8 — включения шейхцерии.
2) лесные болота с сосной и участием ели,
с ярусом кустарничков, преимущественно
болотных вересковых, и моховым покровом
из сфагновых мхов и лесных Ну1осош1а.
Болота первого типа чаще невелики по площади. Однако
встречаются и крупные болота, где олиготрофная раститель¬
ность встречается лишь в виде островов на переходном болоте.
Таково, например: переходное Камское болото Горьковской
области, где широко распространены БрЬ. гесигуига, БрИ. виЬЫ-
со!ог, Сагех 1азюсагра (рис. 92). Прекрасный пример мезо-
трофного болота с небольшими линзами сфагново-пушицевого
торфа на мощной основе осокового торфа представляет
известная Шатурская система (рис. 93.).
Болота второго типа описаны под Архангельском. Они
имеют часто глубокую (около 4 м) залежь и развиваются
путем заболачивания елового леса (А. Шенников и М. Голу¬
бова, 1930). Вероятно подобные болота обычны на севере
зоны. Типы эвтрофных болот весьма разнообразны. Рассмотрим
лишь некоторые из них. Особенно интересны гипновые болота.
Своеобразные по видовому составу, они дают приют некоторым
интересным флористическим элементам. К ним близки по видо-
284
вому составу эвтрофные сфагновые болота с массо¬
вым развитием эвтрофных сфагновых мхов, особенно БрЬ.
\№агп81огШ. Очень часты смешанные эвтрофные болота с гипно-
выми и сфагновыми мхами. Эти гипновые, сфагновые, а также
смешанные болота1 бывают двух типов:
1. С комплексами гряд и мочежин в центре.
Микрорельеф напоминает олиготрофные грядово-мочежинные
комплексы. Мочежины выстилают влаголюбивые виды Бгера-
пос1абиз, некоторые СаШе^а (С. ЦИанит), Меезеа Шцие1га и др.
Здесь редкий ярус осок, древесная и кустарничковая расти¬
тельность отсутствует. Повышения в виде кочек или гряд
Рис. 94. Строение эвтрофного болота. По Н. Кацу и^Ц. Минкиной, 1936.
1 — гипново-осоковый торф; 2 — осоково-шейхцериевый с гип-
нумом; 3 — древесный; 4 — осоково-сфагновый; б — хвощ.
образованы менее влаголюбивыми мхами, главным образом
Sph. Warnstorfii, Camptothecium nitens. Верхний ярус гряд
образуют низкорослые кустарнички — Betula папа (на севере),
и В. humilis (на юге зоны). Встречаются и деревья (сосна
и береза), иногда образующие насаждение. К периферии эти
комплексы обычно переходят в более сухое облесенное
гипновое болото.
Хороший пример представляют болота из более северной части зоны
(район южнее Архангельска) (Т. Работнов, 1930). Южный тип подобных
болот (с Betula humilis) встречается в Московской области. Залежь этих болот
сложена нацело эвтрофными торфами, иногда сверху донизу осоковыми
и осоково-гипновыми. На средней и нижней Печоре верхняя осоковая
и осоково-гипновая толща обычно подстилается торфами, богатыми древес¬
ными остатками — осоково-древесными, а иногда и чисто древесными. Эти
болота В центре с грядово-мочежинными эвтрофными комплексами (иногда
здесь частью смешанные комплексы с мезотрофными кочками и эвтроф¬
ными мочежинами). По периферии они облесены сосной, с болотными Erica¬
ceae и сплошным сфагновым покровом (главным образом Sph. Warnstorfii)
г1Лх можно объединить под названием эвтрофные моховые болота.
285
и носят мезотрофный или даже иногда олиготрофный характер (Н. Кац и
Ц. Минкина) (рис. 94).
2. Второй тип моховых эвтрофы ых болот
с ровным или мелкокочковатым микрорельефом,
без мочежин. По своему флористическому составу и фито¬
ценозам болота этого типа близки к предыдущим. Здесь
следует различать:
a) Безлесные мокрые болота. Моховой ковер обычно гипно-
вый—главным образом виды Drepanocladus. Травянистый
ярус часто редкий и низкий, из осок.
В районе станции Плесецкой, к югу от Архангельска, не¬
глубокая залежь этих болот сложена обычно гипново-осоко-
вым торфом. Болота эти, видимо, с самого начала были
безлесны (Т. Работнов, 1930).
b) Облесенные то сосной, то березой, более сухие болота.
Эти болота имеют чаще смешанный моховой ковер из гипно-
вых и сфагновых мхов —часто последние преобладают. Иэ
Bryales на облесенных болотах доминируют менее влаго¬
любивые мхи — Camptothecium nitens, Aulacomnium palustre.
Из сфагнов преимущественно Sph. Warnstorfii. Подобные
болота встречаются, например, в Калининской области (Торо-
пецкий район).
Эвтрофные моховые болота большей частью жестководные.
Они питаются водами, богатыми солями (в том числе и кальцием)
и встречаются обычно в местах выхода ключей на надпойменных
речных террасах, куда не достигают речные разливы (напри¬
мер, на нижней Печоре, по течению ее притока Ижмы и на
впадающей в последнюю р. Ухте). В районах с близкими
известняками моховые эвтрофные болота заходят и на водо¬
разделы, как к югу от станции Плесецкой (южнее Архангельска)
(Т. Работнов, 1930). Можно думать, что в западной части зоны
(Ленинградская область) эти болота менее распространены,
чем дальше на восток. Может быть, это зависит от большей
выщелоченности грунтов в более богатой осадками западной
части Союза.
Из других эвтрофных типов широко распространены
лесные заиливаемые болота, питающиеся речными
разливами. Эти болота обычно без сплошного мохового
покрова. Зато травянистый покров богат видами и обычно
хорошо развит, причем заметна роль широкотравья. Последнее
иногда доминирует, и тогда осоки отходят на задний план.
Широкотравье в значительной части слагается из лесных видов.
Лесные заиливаемые болота различны по характеру раститель¬
ности. Опишем вкратце главнейшие типы их.
1. Лесные кочкарники с Сагех caespitosa,
с которой иногда согосподствует С. paradoxa. Древесный ярус
образован березой, черной ольхой, редко сосной. При вырубке
деревьев возникают травянистые кочкарники. Эти кочкарники
как безлесные, так и лесные, часто относятся к заболоченным
землям, так как растут на мелком торфе или на торфянисто-
286
глеевой почве, например в пойме р. Волхова \ Лесные коч¬
карники широко распространены в Московской области1 2, где
они хорошо изучены (Н. Кац, 1929). В юго-восточной Мещере
(Рязанский район) это торфяники с мощностью торфа
в IV2 — 2 м (Д. Мещеряков, 1930).
Здесь остатки кочкообразующих осок встречаются лишь в самом верхнем
слое торфа; ниже идут березово-осоковые торфа с менее требовательными
осоками (Carex rostrata, С. lasiocarpa). Следовательно, генетически березовые
кочкарники—тип более молодой, развившийся из менее богатых солями типов,
эвтроф но го болота. Эту смену Д. Мещеряков ставит в связь с улучшением
питательного режима болот вследствие распашки окружающих пространств
и вырубки лесов.
2. Черноол ьшанни ки. Сюда относятся лесные болота
с ярусом черной ольхи и разнообразным и богатым видами
травянистым покровом. По растительности черноольшанники
можно разбить на несколько типов; они различны и по.
характеру субстрата (3. Смирнова, 1928).
1. Ольшанники на заболоченной минеральной‘
почве, иногда с небольшим слоем торфа. Эти ольшанники
во многих случаях развились на месте еловых лесов в результате
их заболачивания. '
2. Ольшанники с мощным (до 4 ж) торфяным
слоем, причем под древесным торфом залегает слой осоко¬
вого или осоково-тростникового. Эти ольшанники развились
из осоковых болот.
В юго-восточной Мещере черноольшанники также растут и на совсем
мелком торфе (заболоченные леса), и на торфе свыше 3 м (торфяники^. Те
и другие отличаются по растительности. Кроме того, последняя зависит
и от питательного режима черноольшанника. Здесь, как и в Приозерной
Мещере, черноольшанники—образование молодое. Тонкий слой чернооль¬
хового торфа подстилается здесь торфами иного состава — осоковыми, бе¬
резовыми, березово-осоковыми. Следовательно, улучшение питательного
режима привело в этом случае к смене менее требовательных типов расти¬
тельности более требовательным (ольшанники). Причина этого, по Д. Меще¬
рякову (1930), та же, что и при развитии осоковых кочкарников (см. выше).
Широко распространены в рассматриваемой зоне смешан¬
ные эвтрофные болота, представляющие комбинацию моховых
эвтрофных и лесных заливаемых болот. При расположении
этих болот в долинах, параллельных реке, растительность
распределяется на них полосами, в общем, параллельными
реке. Болото со стороны реки затопляется полыми водами
и носит характер лесного заиливаемого болота—чаще кочкар¬
ника с березой или ольхой. Более удаленная часть питается
грунтовыми водами и занята моховым эвтрофным болотом.
Если прилегающее к коренному берегу болото находится под
минерализующим влиянием делювиальных вод, здесь также
развивается осоковый кочкарник. В этом случае центральное
моховое болото, следовательно, окаймлено с двух сторон
1 Материалы по исследованию реки Волхова и его бассейна, вып. IX,
Л. 1926.
2 Эти кочкарники, а также кочкарники Рязанского района относятся,
собственно, уже к следующей зоне.
287
осоковыми кочкарниками — и со стороны реки, и со стороны
коренного берега. Смешанные эвтрофные болота описаны
в Московской области (Н. Кац, 1929). Заслуживают упоминания
еще крупноосоковые болота, также заиливаемые. Эти
болота с ярусом крупных осок (Carex gracilis, С. vesicaria,
С. vulpina) образовались из лесных заиливаемых болот. Они
образовались после вырубки леса и поддерживаются сено¬
кошением, при котором уничтожается поросль. Торф сильно
минерализованный. Эти болота используются как сенокосы.
Болота этой зоны резко отличны от болот трех северных
зон. К отличиям, отмеченным выше, надо еще добавить сле¬
дующие: болота всех типов, особенно олиготрофные, резко
отличны по своему видовому составу, растительности, эко¬
логическим условиям и субстрату от окружающих минераль¬
ных почв. Северные болота, как указывалось, обособлены от
окружения не так резко. Болота рассматриваемой зоны — и
олиготрофные, и эвтрофные — имеют обычно однородную
растительность на значительных площадях в том смысле, что
она принадлежит к одному какому-либо типу питания. Расти¬
тельность северных болот, как отмечалось выше, мозаична,
т. е. слагается из пестрого чередования больших или меньших
участков с растительностью различного типа (например,
олиготрофных и эвтрофных). По флористическому составу
и доминирующим в растительном покрове видам болота зоны
резко отличны от северных болот. Из арктических цветковых
на болотах доминируют лишь Empetrum nigrum и Rubus
chamaemorus и притом лишь в северной части рассматриваемой
зоны. В общем же господствующее значение на олиготрофных
болотах приобретают Scheuchzeria palustris, Eriophorum vagi-
natum, Cassandra calyculata, Andromeda polifolia. Все эти виды
повсюду — главные образователи ассоциаций. В северных
болотных зонах (кроме аапа) значение их невелико, и некоторые
.из них находят там северную флористическую границу. Также
Rhynchospora alba, широко распространенная в центре и на
западе зоны, почти не перешагивает ее северной границы.
Резко изменяется при движении на север видовой состав
и ассоциации также и эвтрофных болот. Например, многие
широколиственные лесные мезофиты (особенно дубравные
виды) совсем не встречаются в северных зонах.
Несмотря на скудность наших знаний о географическом
распространении типов болот, все же можно сказать, что
ряд болотных типов, свойственных северным
зонам, не встречается в зоне грядово-мочежин-
ных торфяников, и обратно. Все типы болот с вечной
мерзлотой отсутствуют в нашей зоне. Наоборот, болота
с Betula humilis и черноольшанники, хотя и распространены
здесь не везде (см. ниже), вовсе отсутствуют в северных
болотных зонах. Болота зоны имеют исключительное эконо¬
мическое значение. Торфяные запасы зоны — главный
торфяной топливный фонд СССР, как в количе-
288
ственном, так и в качественном отношении.
Кроме того, надо отметить значительную обжитость (кроме
северных частей), доступность торфяных запасов, а также
благоприятные климатические условия для добычи и сушки
торфа. Зона разделяется на шесть болотных областей, описы¬
ваемых ниже.
V—1. П е ч о р с к о-О н е ж с к а я область олиготрофных гря-
дово-мочежинных торфяников. От Верхней Уижи до Урала эта
область граничит со следующей зоной, причем граница в значительной сте¬
пени топографическая. От Унжи до Вологды граница с Среднерусской обла¬
стью условно проходит по северной окраине Волжско-Двинского водораздела.
От Вологды на запад граница идет условно по Шексне, Белоозеру и Ковже.
От Онежского озера до низовья Северной Двины область V—1 граничит
с областью смешанных болот карельского типа по проведенной условно гра¬
нице карельского ландшафта.
На низких ровных водоразделах этой области широко распространены
грядово-мочежинные торфяники среднепечорского типа (см. выше).
Кроме этого руководящего типа олиготрофных болот, в области обычны
и сосново-сфагновые болота. Это небольшие, но нередко и крупные по раз¬
мерам болота. Сосново-сфагновые болота главным образом сосново-кустар-
яичково-сфагиового типа, с ярусом морошки под кустарничками. Сосново-
пушицево-кустарничковые болота в общем более редки. Более распространены
они на юго-западе области, а к северо-востоку редеют. Из мезотрофных
болот обычны лесные сфагновые болота с Сагех ^Шогпш. Из эвтрофных
интересны большие моховые торфяники с комплексами гряд и мочежин,
обычные на древних террасах нижней Печоры, Ижмы и Ухты. Всюду встре¬
чаются лесные эвтрофные болота с березой и елью. Широко распространены,
особенно на севере, заболоченные сфагнумом сосновые и еловые леса. Забо¬
лоченность области сильно возрастает к северу, где, кроме климата, этому
способствует и рельеф. Обширные низменности этой части области — Моше-
Онежская, Лаче-Кубинская, Средне- и отчасти Верхне-Сухонская (А. Шен-
ников, 1930), далее Кулойский долинный район, водораздел Пезы и Мезени
заболочены чрезвычайно сильно.
V — 2. Прибеломорская область олиготрофных торфя¬
ников с л и ш а й н и к о в о-с ф а гн о в ы м и г ря д а м и, м о ч е ж и на м и
и озерками окаймляет юго-западный, южный и юго-восточный берега
Белого моря.
Характерный тип олиготрофных болот здесь прибеломорский с лишай¬
никовым покровом на грядах, с деградацией сфагнового покрова, с безлес¬
ной центральной частью и другими признаками болот приморского климата.
Возможно, что островное нахождение черной ольхи на южном побережье
Белого моря1, присутствие тростниковых торфов в донном слое приархан-
гельских болот и тростниково-осоковых болот на побережье Онежской и
Северо-Двинской губы также отражают влияние морского климата. Забо¬
лоченность области весьма велика, особенно в области беломорской транс¬
грессии со сглаженным рельефом, где болота, почти целиком олиготрофные,
иногда занимают до 70% и более всей площади (Шурецко-Сорокинский район).
Сплошное распространение обширных болотных систем создает подчас
особый болотный ландшафт Прибеломорской низменности (см. выше опи*
сание и рис. 51).
V — 3. Об.ласть карельских болотных массивов сме¬
шанного типа. Границы этой области определяются распространением
карельского ландшафта (см. выше). Степень заболоченности в общем зна¬
чительна. Болотные массивы носят смешанный характер с преобладанием
олиготрофных типов болот (см. выше описание этого типа). Конкретных
1 Далее на восток северная граница черноольшанников этого южного
типа спускается далеко на юг. Между прочим черноольшанники отсутствуют
в большей северо-восточной части Печорско-Онежской области.
19—722
289
описаний этих болот весьма мало. Имеются сведения по району средней
Карелии, прилегающему к Заонежскому полуострову (Солоневич, 1936).
Впрочем, здесь болота невелики и обычно представляют изолированные
болотные массивы. Торфяники этого района по строению в основном эв-
трофные. Сильно распространены осоковые торфа (иногда осоково-гипновые,
осоково-сфагновые); встречаются (обычно у дна) и лесные торфа, слагающие
на краях болот иногда залежь сверху донизу. Слой олиготрофного сфагно-
во-пушицевого (а иногда и пушицевого) торфа встречается лишь на участ¬
ках болот с олиготрофной растительностью.
Незначительное участие сфагновых торфов, вероятно, характерно для
всей области.
V — 4. Прибалтийская область олиготрофных торфяни¬
ков с комплексами лишайниково-сфагновых гряд, моче*
жин и озерков занимает узкую полосу на советском побережье Фин¬
ского залива (район VI Ю. Цинзерлинга, 1934). Олиготрофные болота
советско-прибалтийского типа. Эти болота, как отмечалось, носят ряд черт
приморского климата и близки к прибеломорским, от которых они отличаются
некоторыми признаками вследствие своего более южного положения.
V — 5. Ладожско-Ильменско-Западнодвинская область
олиготрофных грядово-мочежинных торфяников занимает
бассейн озер: Ильмень, Ладожского и Чудского, бассейн верхней и частью
средней Западной Двины и частью верхнее правобережье Днепра с притоками.
Олиготрофные болота преобладают над мезотрофными и эвтрофными. Круп¬
ные и среднего размера болотные массивы относятся к типу, хорошо выра¬
женному в бассейне оз. Ильмень и Западной Двины и свойственному, ве¬
роятно, и остальной части области. Этот тип мы назвали ильменско-западно¬
двинским. Физиономическая особенность грядово-мочежинного комплекса
в центре этих болот—господство в верхнем ярусе пушицы.
Она маскирует расположенные под ней кустарнички1
и создает впечатление однотонного седоватого пушицевого болота. Господ¬
ство пушицы на грядах, слабое развитие морошки, обилие в мочежинах,
наряду с обычными шейхцерией, пушицей, Sph. balticum, Sph. Dusenii,
тгкже очеретника и Sph. cuspidatum отличает эти Hochmoor от средне¬
печорских болот, так же как и более значительная выпуклость и мощная толща
сфагнового торфа, имеющая часто характер подстилки. Эти болота находятся
в климатических условиях, наиболее благоприятных для торфообразования
вообще и, вероятно, для сфагнового в частности. Сфагновый ковер здесь
столь же мощный, как и на среднепечорских болотах, так как современные
условия для произрастания сфагнума весьма благоприятны как там, так и
здесь. Однако большая выпуклость и большая мощность субатлантической
сфагновой толщи торфа указывает на лучшие условия торфообразования
на западе. Еще западнее—на Финском побережье — климатический оптимум
сфагновых мхов уже превышен, и начинается деградация сфагнового по¬
крова. Вместе с тем в современный период ослабевает и торфообразование.
Выпуклость Hochmoor, повидимому, опять понижается в приморской полосе,
что, может быть, указывает на менее благоприятные за время развития болот
условия торфообразования. Болота западной части нашей области, где часто
обилен вереск и усиливается роль лишайников, по растительности напоми¬
нают советско-прибалтийский тип.
Небольшие олиготрофные торфяники области сосново-сфагнового типа,
чаще сосново-кустарничково-пушицево-сфагновые.
В общем, для болотной области (по крайней мере, для части, входящей
в состав Ленинградской административной области) характерны обширные
олиготрофные торфяники с мощным верхним слоем мало разложившегося
(степень разложения 5—15%) сфагнового торфа, с почти беспнистой централь¬
ной частью и слабопнистым торфом на окраинах. Нижняя половина залежи
хорошо разложившаяся (40—50%). Пограничный горизонт—мощный на окраи¬
нах и почти не выражен в центре. В нем обычны мощные пни сосны, часто
не уступающие по своему развитию соснам на минеральной почве.
1 Здесь образуют ярус под пушицей Andromeda, а иногда Empetrum и
Calluna.
290
Другие типы болот изучены недостаточно. Из обычных лесных мезотроф-
ных болот здесь встречаются болота с Сагех ЙШогпИз. Обширные мезотрофные
болота с большим участием этой осоки и сфагновых мхов описаны из района
оз. Ильмень (Г. Ануфриев). Эти болота иногда сплошь мезотрофные или же
содержат в центре участки олиготрофной растительности.
Из эвтрофных типов в области встречаются гипновые и лесные заили¬
ваемые болота. Особенно надо подчеркнуть значительное распространение
черноолынанников во всей области.
Заболоченность сильно варьирует в зависимости от рельефа. Выделяются
громадной заболоченностью следующие низменные районы (Ю. Цинзерлинг,
1934): 1 2 3Вверху одно из болот б. Иваново-Вознесенской губ., внизу Мезинское болото б. Влади¬
мирской губ.
1 — осоково-гипновый торф; 2 — осоковый; 3 — древесный; 4 — гипновый; б — сфагново-осоко¬
вый; 6 — сфагновый; 7 — сфагновый (БрИ. йксит); 8 — пни сосны; у — березы; 10— суглинок
11 — песок.
1. Ильменско-волховская впадина с громадными олиготрофными и долин¬
ными эвтрофными болотами.
2. Молого-судский равнинный песчаный район по Мологе, Суде, Чаго-
доще. Олиготрофные болота здесь часто почти сплошь занимают водоразделы.
3. Полистовский район равнинных водоразделов Ловати, Полисти, Ше-
лонн, Великой, где расположен громадный Полистовский болотный массив,
частью олиготрофного типа. Другие районы, наоборот, бедны болотами или
вследствие особенностей геологии (Силурийское известковое плато), или
вследствие расчлененного рельефа (Валдайская возвышенность, Опочко-Не-
вельский район конечных морен)»
V — 6. Среднерусская область олиготрофных сфагно¬
вых болот. Крупные олиготрофные болота относятся к среднерусскому
типу. Комплексы кочек и мочежин на этих болотах обычно вкраплены в виде
островов в основной фон сосново-сфагнового болота. Олиготрофные болота
меньшего размера, а иногда и крупные, относятся к сосново-кустарничково-
пушицевому сфагновому типу. Небольшие болота обычно сосново-кустарнич-
19* 291
ковые сфагновые. Крупные
олиготрофные болота по дан¬
ным для восточной части б
Центрально-промышленной об¬
ласти (С. Тюремнов,1928) имеют
в центральной части такое
строение снизу вверх (рис. 95).
1. Осоково-гипновый торф,
мощностью 0,5—1,0 м.
2. Древесно-осоковый, хо¬
рошо разложившийся.
3. Сфагновый торф, средне
разложившийся из Sph. medium
и Sph. parvifolium.
4. Мало разложившийся
торф из Sph. fuscum (мощность
до 2,5 м).
5. Средне разложившийся
с участием Sph. medium, Sph.
parvifolium.
6. Пограничный j горизонт,
иногда до 40 — 50 см мощ¬
ностью.
7. Слабо или средне разло¬
жившийся торф из Sph. medium
и Sph. parvifolium с прослойка¬
ми хорошо разложившегося
торфа. Эти болота в отличие от
северо-западных имеют более
гумифицированный и пнистый
торф, подстилка же выражена
слабо.
На менее крупных болотах
и окраинах больших, по сведе¬
ниям для того же района, сфаг¬
новый торф менее диференци-
рован на слои различного со¬
става, сильнее гумифицирован
и богат пнями.
Болота с преобладанием
мезотрофной растительности
довольно часты, но обычно не¬
велики. В естественном виде
они облесены березой или сос¬
ной, с сплошным сфагновым
покровом и с ярусом Сагех
lasiocarpa. Встречаются и круп¬
ные болота с преобладанием
мезотрофной растительности и
островами олиготрофной. При¬
меры — Шатурская система и
Камское болото Горьковского
края (Н. Властова, 1936)х, о
которых говорилось выше.
Из эвтрофных болот здесь
встречаются все типы, перечи¬
сленные при описании болот- 11 И то и другое болото
располагаются на границе с сле¬
дующей болотной зоной.
292
вой зоны* Лишь моховые эвтрофные торфяники с грядами и мочежинами
в общем нехарактерны для области. Если они и встречаются, то в виде
смешанного типа: грядово-мочежинные комплексы вкраплены в лесные ком¬
плексы без грядово-мочежинного микрорельефа. Обычно же моховые болота
более или менее ровны иди мелкокочковаты. Строение эвтрофных болот
крайне разнообразно. Общее описание растительности и строения эвтрофных
болот юга области (Мещера) можно найти у Д. Мещерякова (1930).
С. Тюремнов (1928) дает краткое описание строения эвтрофных залежей
восточной части б. ЦПО. Как здесь, так и там часто наблюдается, что на без¬
лесных болотах нижняя часть залежи богата древесными остатками. Здесь
залегают осоково-древесные, а иногда чисто древесные торфа. Верхняя
часть — осоковая, иногда осоково-гипновая, без древесных остатков. При
значительной мощности верхних бездревесных торфов можно думать, что
переход облесенного болота в открытое произошел давно в силу изменения
естественных условий — увеличения увлажнения. Во многих случаях, как
это обычно имеет место в Мещере, верхний осоковый или гипново-осоковый
торф имеет всего 20—50 см и подстилается снизу осоково-древесным. В этих
случаях, вероятно, влияли антропогенные факторы — уничтожение древесного
яруса, которое сопровождалось обводнением болота.
Болота, теперь облесенные, были часто такими с момента их возникно¬
вения. Об этом свидетельствует залегающий сверху донизу осоково-древесный
торф (Мещера). Но бывают и обратные случаи: открытые болота переходят
в лесные после частичной их осушки. Особый тип строения имеют торфяники
в районах, богатых известью и питаемых известковыми ключами (рис. 96).
Так, на Уткинском болоте (Середской район, Ивановской области) в торфе
много извести то в виде отдельных прослоек, то в виде мощных пластов.
Извести много и в озерных отложениях под торфяником. Залежь Уткин-
ского болота во многих местах развивалась по безлесному типу под влия¬
нием избыточного ключевого увлажнения, и лишь в самое последнее время
наступила стадия лесного эвтрофного болота. В других пунктах болото
было облесено в течение всего своего развития.
Из флористических особенностей болот области отметим, что Empetrunr
nigrum вообще редок и лишь в редких случаях выступает как доминант.
Морошка встречается чаще, и, мало того, местами определяет своим массо¬
вым развитием северный „морошковый" вариант олиготрофных болот
(N. Katz, 1927). В общем же роль этих двух арктических видов сильно умень¬
шается по сравнению с печорско-онежской и ильменско-западнодвинской
областями.
Заболоченность среднерусской области значительно меньше, чем двух
последних* областей. Заболоченность областей Московской и Ивановской,
входящих большей частью в рассматриваемую болотную область, равна 11,5
и 15,3%.
Эти цифры дают представление и о заболоченности всей среднерус¬
ской области. Наиболее заболоченные районы (б. Орехово-Зуевский и
Егорьевский уезды) дают 22 и 17% заболоченности (Л. Хаустов, 1931). Рас¬
пределение болот, как и всюду, зависит от рельефа.
VI зона. Эвтрофные и олиготрофные сосново-сфагновые
торфяники юга хвойной зоны. Эта зона расположена южнее
предыдущей, главным образом в пределах полосы хвойных
лесов с широколиственными породами. Северная граница ее
с предыдущей зоной в значительной части обусловлена релье¬
фом.
Южная граница почти совпадает с северной границей
широколиственных лесов. Для зоны в целом харак¬
терно преобладание эвтрофных торфяников
над олиготрофными, в среднем (по всей зоне)
в отношении 1:0,5.
В предыдущей зоне отношение было обрат¬
ное. Следовательно, климатический оптимум
293
развития олиго трофных сфагновых болот
в этой зоне уже превышен. Вместе с тем здесь
уменьшается средняя мощность торфа (2,02 м
п ротив 2,62 м в предыдущей зоне), интенсив¬
ность торфообразования (вследствие повышения
темпа разложения растительных остатков), роль сфагно¬
вых торфов и роль сфагновых мхов в совре¬
менном покрове болот. Все эти черты зависят
от увеличения испаряемости, от уменьшения
отношения количества выпадающих осадков
к испаряющимся. Уменьшение роли сфагнума в современ¬
ном покрове сказывается не только в уменьшении площади
олиготрофных болот, но и в том, что физиономическая
роль сфагнума на болотах выступает не столь
резко, как в предыдущей зоне, а кроме того,
он не в такой степени, как там, определяет
условия существования остальной раститель¬
ности. Вспомним, что на лесных сфагновых болотах, характер¬
ных для этой зоны, сравнительно мощно развитой ярус сосны
маскирует моховой ковер, и при рассматривании издалека подоб¬
ное болото напоминает лес, и что, кроме того, сфагновый ковер
иногда бывает почти совсем закрыт густыми кустарничками.
Заболоченность зоны резко падает по ср а вне¬
нию с предыдущей. Максимум заболоченности (40°/о)
дают наиболее заболоченные равнинные районы Белорусско-
Украинского Полесья (Е. Лавренко, 1927; С. Тюремнов, 1931).
В общем заболоченность больше всего на западе зоны,
в Полесье, к востоку же падает. Отношение площади торфя¬
ников ко всей заболоченной площади высоко и, вероятно,
близко к предыдущей зоне. Олиготрофные болота зоны отно¬
сятся к сосново-сфагновому типу, что отражено и в названии
зоны. Сосново-сфагновые болота были описаны выше. Необхо¬
димо иметь в виду, что распространение этих болот, как мы
видели выше, не ограничено одной рассматриваемой зоной.
В частности, они встречаются и в зоне грядово-мочежинных
торфяников. Однако в той и в этой болотной зоне они пред¬
ставляют различные стадии развития. Там они представляют
начальную стадию развития и свойственны в основном неболь¬
шим, „эмбриональным" болотам. Здесь, в более южной зоне,
этот тип представляет устойчивую, климатически обусловлен¬
ную стадию развития: он свойственен здесь как небольшим, так и
более значительным по площади болотным массивам. При этом
сосново-сфагновые болота грядово-мочежин-
ной зоны, с одной стороны, и рассматриваемой
зоны—с другой, неодинаковы по раститель¬
ности и строению. На юге эти болота бедны видами. Здесь
выпадают как образователи ассоциаций Empetrum nigrum и
Betula nana, a Rubus charaaemorus и Sph. fuscum лишь спора¬
дически определяют физиономию болота. Что касается торфя¬
ной залежи, то она на болотах этой зоны обычно значительно
294
гумифицирована и богата пнями. В более северной зоне, кроме
этого типа залежи, встречается и другой—с мало разложив¬
шимся и мало пнистым торфом. Примером строения сосново¬
сфагновых болот в рассматриваемой зоне могут служить
торфяники в б. Медынском уезде, Калужской губернии \ Здесь
наблюдаются два слоя олиготрофного торфа: верхний—хо¬
рошо или средне разложившегося сфагнового торфа и ниж¬
ний— хорошо разложившегося пушицево-сфагнового торфа.
Залежь богата сосновыми пнями. Выпуклость невелика. По¬
добные болота с указанным строением встречаются на южной
окраине распространенных сфагновых болот, по линии от
Казани на юг б. Минской губернии2, т. е. в пределах рас¬
сматриваемой зоны. По характеру растительности, как указы¬
валось, сосново-сфагновые болота можно подразделить на
сосново-кустарничково-сфагновые, сосново-кустарничково-пу-
Рнс. 97. Профиль болота по р. Припяти. По С. Тюремнову, 1931.
Л — сфагново-осоковый торф; 2 — лесной (ольховый); 3 — гипново-топяной; 4 — осоково-лесной;
5 — осоково-топяной; 6 — хвощево-топяной; 7 — тростниково-топяной.
шицевые сфагновые и сосново-пушицевые сфагновые. Все эти
три типа встречаются в нашей зоне, но распространены не
везде одинаково. Мезотрофные болота рассматриваемой зоны
в общем напоминают соответствующий тип северной зоны.
И здесь весьма характерны сфагновые болота с Carex lasio-
сагра, в ненарушенном виде с сосновым или березовым
ярусом. Эвтрофные болота в общем тех же типов, что и в
соседней к северу зоне. Черноольшанники весьма обычны
уже во всей зоне. Здесь и в следующей зоне они достигают
максимума своего распространения. Г ипновые торфяники
здесь несколько иного характера, чем в зоне грядово-моче-
жинных торфяников. Мочежинный микрорельеф
угасает на них так же, какинаолиготрофных
болотах, встречаясь, впрочем, еще кое-где на эвтроф-
ных болотах. (Строение см. рис. 97.)
Изменяется несколько и ф л ори стичес к ий
состав гипновых торфяников. Например, Betula
nana, широко распространенная на севере предыдущей зоны,
заменяется здесь Betula humilis. 11 •,Торфяное дело*, 1924.
s Там же.
295
VII зона. Эвтрофные осоковые торфяники и тростнико¬
вые болота полосы широколиственных лесов и лесостепи.
Эта болотная зона занимает в общем зону широколиственных
лесов и лесостепи (северная степь В. Алехина). Северная
граница в общем совпадает с северной границей широколи¬
ственных лесов1, местами отступая то к северу, то к югу от
нее. Южная граница в общем совпадает с южной границей
лесостепи. Основные черты этой зоны: 1) господство эвтроф-
ных болот и отсутствие выпуклых олиготрофных1 2; 2) большое
участие заболоченных водоемов в общей площади болот.
Отношение площади надводных торфяников к общей площади
болот для зоны в целом неизвестно. Для отдельных областей
это отношение, вычисленное на основании данных книги „Торф,
его происхождение и т. д.“ (1932), таково: УССР — 0,25%
Средневолжская область — 0,20; ЦЧО—0,11. Эти цифры,
несмотря на их неточность, показывают, что указанное отно¬
шение невелико. Относительно большое распространение
зарастающих водоемов—следствие неблагоприятного для
торфообразования климата с высокой испаряемостью. В север¬
ных подзонах тундры заболоченные пространства также сильно
преобладают над торфяниками. Однако это будут уже не водо¬
емы, а суша (заболоченная тундра и болота без торфа), и
причина преобладания будет другая: низкая температура и
мерзлота, препятствующие торфообразованию и способствую¬
щие заболачиванию. Есть и еще разница между тундрой и
лесостепью: заболоченность тундры очень велика, а лесостепи
незначительна. Это потому, что климат тундры способствует
заболачиванию, а климат лесостепи нет. Заболачивание суши
в лесостепи не получает развития. Оно здесь идет лишь при
наличии постоянных свободных запасов воды (т. е. в водоемах)
или при заливании полыми аллювиальными водами. Заболочен¬
ность зоны невелика и абсолютно, и по сравнению с соседней
к северу зоной. Наиболее заболочена северо-западная часть,
прилегающая к Полесью.
Замедленность торфообразования сказывается не только
в малой площади торфяников, но и в небольшой их средней
глубине 1,76 м против 2,02 м в предыдущей зоне. Эта малая
глубина может быть скорее следствие замедленности торфо-
накопления, чем молодости этих болот. Резюмируем вкратце
характеристику зоны: 1) заболоченность зоны невелика абсо¬
лютно, невысок и процент заболоченности;
2) заторфованность зоны весьма невелика абсолютно и по
сравнению с общей заболоченной площадью, а равным образом
и по сравнению с заторфованностью двух следующих к северу
болотных зон;
1 Согласно Е. Лавренко, это отношение можно принять и для восточной,
почти неисследованной части зоны, как это сделано и здесь.
2 Островное нахождение их в пределах зоны не исключено.
8 Если исключить Украинское Полесье, относящееся к предыдущей
зоне, то это отношение будет меньше.
296
3) средняя глубина торфяников также невелика;
4) широко распространенный тип болот—зарастающие
водоемы;
5) все указанные особенности торфо* и болотообразования
зависят от климата—в первую очередь, от невысокого клима¬
тического коэфициента, т. е. от большой испаряемости.
Характеристика растительности зоны в дальнейшем приве¬
дена для ее западной части (Украина); восточнее Украины
данных почти нет. Можно различить здесь (по Е. Лавренко)
два основных типа эвтрофных болот: 1) прибрежно-водные,,
приречные и приозерные болота с господством тростника
(Phragmites), отчасти камыша (Scirpus lacustris) и рогозов
(Typha latifolia, часто Т. angustifolia); эти болота частью без
торфа, частью с лимническими (водными) отложениями разной
мощности; 2) осоковые болота, которые бывают двух типов:
первый тип—аллювиальные, крупно-осоковые болота с пре¬
обладанием Carex gracilis, иногда С. stricta, С. riparia, С. acuti-
formis, а иногда и Glyceria aquatica; второй тип—осоково-
гипновые болота, более или менее свободные от влияния
аллювиальных вод, преимущественно на надлуговых террасах.
Здесь доминируют: Carex lasiocarpa, С. teretiuscula, С. limosa,.
С. stricta, С. paradoxa, часто с хорошо развитым моховым
ковром из Drepanocladus aduncus, D. vernicosus, Acrocladium
cuspidatura, Calliergon giganteum, C. cordifolium (E. Лавренко,
1927; В. Матюшенко, 1928). Иногда осокоьо-гипновые болота
бывают облесены зарослями ив (Salix rosmarinifolia, S. aurita,
S. cinerea), Betula pubescens, B. humilis (лесные болота). Черно-
ольшанники были широко распространены в лесостепи и
севернее. В настоящее время здесь их площадь сильно-
сократилась вследствие вырубок. Осоково-гипновые торфяники
зоны с гладкой поверхностью—грядово-мочежинный микро¬
рельеф здесь совсем отсутствует. ¿
Другие отличия этих и осоковых торфяников от более северных—
флористические. Они сводятся к следующему1: Обеднение видового со¬
става. Многие массовые виды северных болот сходят здесь на-нет. Роль их,
несомненно, меньше, чем на севере (например Carex rostrata). То же отно¬
сится и к мхам. Например, широко распространенные на северных болотах
Camptbotheclum nitens, Meesea triquetra не отмечены в районах северо-
западного правобережного Полесья (Д. Зеров, 1934), первый же мох редок
в Черниговском Полесье (Ф. Левина, 1937), т. е. даже к северу от зоны 1 2„
Ряд видов (например С. riparia, С. acutiformis, С. stricta), играющих на
здешних болотах большую роль, встречаются значительно реже в хвойной
зоне, а в северной ее части, повидимому, и вовсе сходят на-нет. Многие
на севере типично мезотрофные и олиготрофные виды, как, например,
Rhynchospora alba, Carex lasiocarpa, переходят здесь на эвтрофные болота.
Строение эвтрофных болот Украины изучено В. Матюшенко
(1925, 1928) и Е. Лавренко (1927) (рис. 98). Примером строения
1 На севере правобережного Полесья.
2 Правда, в верховьях Трубежа, т. е. уже в пределах рассматриваемой'
зоны, этот мох отмечен еще на осоково-гипновом болоте (В. Матюшенко,
1928).
297
осоково-гипновых болот может служить торфяник по Трубежу
(рис. 99).
Мезотрофные болота распространены слабо. Они
встречаются по пескам надлуговых террас, иногда и вне их
на тех же почвах, и невелики по площади. Это или: 1) сухие,
облесенные березой и сосной сфагновые торфяники суходоль¬
ного происхождения, часто особенно богатые мезотрофными
и олиготрофными болотными видами, в частности Ericaceae,
и иногда даже напоминающими северные болота и своими ассо-
Рис. ?8. Строение торфяников восточной Украины.
По Е. Лавренко.
I — сфагново-осоковый мезотрофныи торфяник; II — осоковый
торфяник межречья; III — пойменный гипново-осоковый торфя¬
ник; IV — притеррасный гипново-осоковый торфяник; V — сфаг¬
новый сплавинный торфяник.
1 — сфагновый торф; 2 — осоковый; 3 ~ осоковый со сфагну¬
мом; 4 — гипновый, 5 — гипновый со сфагнумом; Ь—гипново-
осоковый; 7 — тростниковый; 8 — тростниково - осоковый;
9 — минерализованная прослойка; 10 — гиттия; 11—гиттие-
вый торф; 12 — черный ил; 13 — ил с раковинами; 14 — песок;
15 — озерный мергель; 16 — вода.
циациями (В. Доктуровский, 1925; Н. Савич, 1928) \ но обычно
лишь с тонким слоем сфагнового торфа поверх эвтрофных
торфов1 2 или 2) сфагновые сплавины водного происхождения
1 Одно из болот, описанное Н. Савич в б. Раненбургском уезде, по
своей растительности может быть причислено к олиготрофным.
2 В некоторых случаях, как на Леонидовском сфагновом болоте под
Пензой, мало разложившийся сфагновый торф достигает 2,5 м толщины,
298
в западинах надлуговых террас (например, в б. Харьковской
губернии; см. Е. Лавренко, 1927). Все они водного происхожде¬
ния и представляли раньше мелкие водоемы, обычно типа
кочкарников с Сагех stricta и водной растительностью (Pota¬
mogetón natans и др.).
Заболачивание этих водоемов шло при помощи сфагново¬
осоковой сплавины.
В строении торфа можно наблюдать обычно такую после¬
довательность: над минеральным дном залегают водные отло¬
жения (гиттия), выше идет осоковый торф, над ним сфагно¬
вый торф. Иногда встречаются еще слои осоково-гипнового
торфа, а в некоторых случаях еще и слой воды или очень
Рис. 99. Строение торфяников Украины. По В. Матюшенко, 1925.
I. II — пойменный торфяник; III — овражный торфяник; IV — плавневый торфяник.
1 — сфагново-осоковый торф; 2—гипново-осоковый; 3 — осоковый; 4— тростниково-осоковый;
5 — тростниковый; 6 — ольшанниковый; 7 — ил с раковинами; 8 — овражный делювий; У—су¬
глинок; 10 — песок; 11 — черный ил; 12 — вода; 13 —прослойка.
жидкого торфа, на котором покоится верхний слой сфагно¬
вого торфа.
Современный растительный покров этих торфяников до¬
вольно разнообразен. В центре обыкновенно осоково-сфаг¬
новые ассоциации, часто зыбунного характера со сплошным
сфагновым покровом и осоками (преобладают Сагех lasiocarpa,
С. teretiuscula и др., часто также много Calamagrostis lanceolata).
Нередко выделяются концентрические пояса растительности.
Иногда по периферии сохранилась полоса осокового кочкар¬
ника из Сагех stricta, на которую надвигается осоково-сфагновая
сплавина (рис. 100).
В описываемой зоне встречаются еще особые осоковые
болота, обычно с Сагех stricta и иногда еще с тростником
в западинах древних лёссовых террас. Слой торфа здесь
невелик, водный режим непостоянен—к концу лета эти болота
299
!
часто пересыхают. Эти болота развиваются на месте дегради¬
рованных солонцов и часто связаны пространственно и гене¬
тически с осиновыми кустами.
Из более южной зоны заходят сюда засоленные болота.
Местами, например в Киевской лесостепи, в блюдцах древних
террас они даже весьма часты. Процессы торфообразования
в описываемой зоне ослаблены. В современный период, не
говоря уже об осушке и эксплоатации торфяников, преобла¬
дают даже естественные процессы их разрушения.
Уничтожение лесов вызы¬
вает быстрое таяние снегов,
сильные паводки рек и заносы
пойменных торфяников илом.
Сильное развитие оврагов и
их быстрое углубление вызы¬
вает опять-таки погребение
долинных торфяников илом,
их осушку, отмирание болот¬
ной растительности и прекра¬
щение торфообразования (ри¬
сунок 99).
Отметим общие закономер¬
ности в распространении бо¬
лотных типов. Выше указы¬
валось, что на эвтрофных
торфяниках зоны многие се¬
верные виды сходят на-нет, и,
наоборот, большую роль игра¬
ют виды, слабее распростра¬
ненные на севере. Одни и те
же типы болот севернее рас¬
сматриваемой зоны изменяют
свой характер. Например, юж¬
ные черноольшанники отлича¬
ются от более северных при¬
террасных, во-первых, своей
длительной поемностью, что зависит от их низкого положения
над уровнем весенних разливов (указание Р. Еленовского),
во-вторых, отсутствием ели и ее спутников, которыми насыщены
северные черноольшанники. Не только ассоциации, но и неко¬
торые типы северных болот отсутствуют в этой зоне, например,
заиливаемые болота с ярусом ели, болота с массовым разви¬
тием Sphagnum Warnstorfli.
Среди эвтрофных болот, как уже указывалось, большую
роль играют осоково-гипновые торфяники. Роль их не везде
одинакова. Осоково-гипновые болота широко распространены
в Полесье, т. е. уже за пределами рассматриваемой зоны, и
обильны еще на севере последней. Южнее они широко распро¬
странены еще в низменном районе слабо расчлененных древних
террас Днепра (Полтавский район), а в других районах в общем,
зоо
Sphognetum
^can cosum
о о Corex stncta
°о о Good
••яУ Сыроватый луе по дерееу
Рис.
100. Растительность сфагнового
болота в восточной Украине
(б. Харьковская губ.). По Е. Давренко.
слабо. Роль их, следовательно, уменьшается в пре¬
делах зоны с севера на юг, и в степной полосе,
уже за ее пределами, они сходят на-нет. По всей
вероятности, также распространены и осоково-гипновые торфа.
Для всей зоны характерно расположение болот
и особенно торфяников в придолинных частях
рек, в поймах, надпойменных террасах, на дне
и склонах балок. Водораздельные болота в степных блюд¬
цах редки, невелики и со слабым торфообразованием.
VIII зона. Прибрежно-водные тростниковые болота и
засоленные болота степной полосы. Эта зона занимает
в Европейской части СССР полосу ковыльных и полупустынных
степей. Характерный признак — отсутствие здесь
за редкими исключениями настоящих надвод¬
ных торфяников. Климатический коэфициент (отношение
выпадающих осадков к испаряющейся влаге) здесь невелик,
и условия торфообразования еще более неблагоприятны, чем
в предыдущей зоне. Торфообразование, если и происходит, то
в очень ограниченном размере и в связи с речками и озерами.
Торфа обычно лимнические (водные) или лимническо-тельма-
тические (водно-болотные). Надводные торфа не образуются.
Вторая особенность зоны—широкое развитие
засоленных болот, куда относятся настоящие солончаки,
солоноватые болота и прибрежно-водные ценозы соленых
водоемов. Солончаки (и солонцы) попадаются уже в зоне
мощных черноземов (лесостепь). Здесь и южнее, в зоне обык¬
новенных черноземов, их немного. В каштановой зоне их
больше. Местами в солонцовых комплексах оба типа засолен¬
ных грунтов преобладают. Здесь, а также севернее солончаки
занимают долины, заливаемые и древние поймы, верховья и
склоны балок. Южнее каштановой зоны, в зоне солонцовых
комплексов на соленосных глинах Каспия уже сплошное
распространение этих комплексов—они занимают до 75% всей
поверхности. Преобладают солонцы, но много и солончаков.
Последние занимают здесь уже водораздельные впадины,
берега и днища соленых озер. Еще южнее, в зоне сероземов
и светлобурых почв, в Прикаспийской низменности, в низовьях
Урала, Волги, Кумы, Терека, площадь солончаков возрастает
за счет солонцов, и они занимают все понижения рельефа.
Это—область сплошного распространения засоленных болот
(солончаков). Можно различить такие типы засоленных болот
(по Е. Лавренко):
1. Мокрые солончаки переменного увлажнения с Salicornia,
Halocnemum, Obione, Salsola, Suaeda.
2. Солоноватые болота с более постоянным увлажнением
с Scirpus Tabernaemontani, S. maritimus, Juncus maritimus.
3. Тростниковые (иногда Scirpus lacustris или Typha, при засо¬
лении—T. Laxmanii), заросли в озерах разной степени засоления.
Для солончаков характерно, кроме избытка солей, непо¬
стоянство водного режима. Весной они нередко представляют
301
соленые озера, позднее—соленые грязи и, наконец, пухлые
солончаки со сплошной коркой солей. Засоление и увлажнение
меняются и по годам. С этим связан и неустойчивый характер
растительности. В области подвижных песков засоленные
болота могут подвергаться засыпанию песком.
Кроме засоленных болот, в рассматриваемой
зоне широко распространены плавни. Это при¬
брежно-водные заросли, где господствует обычно тростник;
нередко громадные площади занимает Турйа angustifoИa.
Кроме зарослей надводных растений, в плавнях обычны ценозы
водных прикрепленных и плавающих растений. А. Флеров
(1930) описывает плавни р. Кубани и выделяет здесь ряд
типов. Органические отложения плавней в виде то ила, то
тростникового торфа, или вернее — мощной, частью живой
тростниковой дернины. Настоящие тростниковые торфа обычно
погребены под мощными речными наносами (рис. 99).
Своеобразны встречающиеся в западинах степных водоразде¬
лов и древних террас, в так называемых подах, подовые
болота. Растительность здесь лугово-болотная, прибрежно¬
водная или солончаковая.
К северу, с выщелачиванием дна подов и оподзоливанием
подовые болота заменяются осоковыми кочкарниками с Сагех
вШ^а и осиновыми кустами. Водный режим подов не¬
постоянен, как и растительность. Это эфемерные болота-
При распашке степей они заносятся илом и постепенно
исчезают.
Из северных типов болот черноольшанники распространены
здесь лишь спорадически.
Осоково-гипновые болота в степной зоне исчезают. Виды
северных олиготрофных болот, кроме немногих, не доходят
до степей так же, как большинство эвтрофных болотных
северных видов.
Болота Западной Сибири.
Западно-Сибирская низменность — равнина с еще более
сглаженным рельефом, чем Восточно-Европейская. Климат
и здесь — основной фактор, определяющий в общих чертах
распространение болотных типов. Изменяясь зонально, климат
обуславливает, как и в Европейской части СССР, зональное
размещение болот. Поэтому при дальнейшем описании удобно'
держаться болотных зон, как это было сделано при первых
попытках районирования болот Западной Сибири (Н. Кац,
1929; А. Бронзов, 1930). Накопившийся с тех пор новый
материал позволяет углубить и расширить прежнее райониро¬
вание (Н. Кац) и описание болотных типов Западной Сибири.
В настоящее время можно различить такие болотные зоны.
I зона. Заболоченные земли и минеральные болота
арктической западносибирской тундры. В Западной Сибири
эта болотная зона занимает арктическую и мохово-лишайнико-
302
вую подзоны тундры и отчасти заходит в подзону кустарнико¬
вых тундр1. Эта зона — продолжение той же болотной зоны
Европейской части СССР и сходна с нею по основным при¬
знакам. И здесь, так же как и там, основной тип болот—
осоковые болота с ровным или мелкокочко¬
ватым микрорельефом, без бугров и мочежин
и с маломощным'слоем торфа. Торфяники здесь
редки и носят характер реликтов ксеротерми-
ческого периода. Растительность болот весьма одно¬
образна в арктической зоне и Европы, и Сибири. Господ¬
ствуют Сурегасеае, главным образом Сагех chordorrhiza,
С. rariflora, С. stans. Eriophorum russeolum, а в моховом покрове
Calliergon Richardsonii, С. sarmentosum, С. stramineum, Drepa-
nocladus exannulatus, D. vernicosus, иногда эвтрофные сфагны
(Sph. obtusum, Sph. subsecundum). Однако последних гораздо
меньше, чем гипновых1 2.
Как известно, роль сфагнов в северной тундре вообще сильно падает
за счет гипновых мхов при заболачивании как суши, так и водоемов.
Иногда среди осоковых пространств — невысокие (0,5—0,75 м) бугры, зани¬
мающие в комплексе с мочежинами не более 20% площади, причем моче-
жины сходны по растительности с арктическими осоковыми болотами. Эти
комплексы Б. Городков называет мелкобугристыми. Они представляют
как бы слабый арктический отголосок плоскобугристых комплексов. Впро¬
чем, по сравнению с осоковыми болотами площадь мелкобугристых ком¬
плексов в северных подзонах тундры невелика. В. Андреев (1934) в Запад¬
ной Сибири выделяет два типа тундровых осоковых болот:
1. Осоково-приозерный тип. Образует пояс заболачивания вокруг тунд¬
ровых озер. Господствует Сагех stans.
2. Осоково-водораздельный тип. Занимает громадные площади на водо¬
разделах. Господствуют из осок Сагех stans, С. rotundata, С. chordorrhiza,
из мхов Drepanociadus revolvens, Sph. squarrosum и др. Торф осоково-гип-
новый, слой его всего в 10—20 см. Мерзлота оттаивает на 50—60 см.
Совершенно своеобразный тип болот, самой северной подзоны аркти¬
ческих тундр — это „полигональные болота“, аналог сухих полигональных
тундр (Б. Городков, 1935). Дернина этих болот растрескивается на много¬
угольники, диаметром 5—20 м. Трещины окаймлены грядками субстрата
до 0,5 м высотой. Растительность полигонов обычного типа арктических
осоково-гипновых болот. На грядках мощность торфа всего 0,2—0,3 м; под
ним выпяченный минеральный субстрат. Основу грядок образуют сфагны
(Sph. lenense, Sph. Warnstorfii, Sph. fimbriatum), на вершинах их тундровые
виды Dicranum и Polytrichum, Сагех stans и другие арктические цветковые.
Для восточной части Обско-Тазовского полуострова В. Говорухин (1933)
описывает такие типы болот низинной тундры: 1) Magnocaricetum с двумя
ассоциациями: a) Magnocaricetum drepanocladosum и b) Magnocaricetum
sphagnosum; этот тип близок к осоково-приозерному типу В. Андреева;
2) Nanocaricetum с господством осок (Сагех chordorrhiza, С. rotundata,
С. rariflora) и гипновых мхов. Этот тип близок к осоково-водораздельному
типу В. Андреева. Однако у В. Говорухина эти болота — мозаичный ком¬
плекс, по мокрому осоковому фону проходят мерзлые гряды и встречаются
плоские возвышения.
Эти полигональные арктические болота образуются в крайне суровом
арктическом климате. Полигональные трещины — результат морозобоя.
1 Терминология подзон тундры по Б. Городкову (1935).
2 Описание Б. Городкова (1935), касающееся арктических осоковых
болот вообще.
303
Этим далеко не исчерпываются типы болот арктической тундры, где
широко развиты заболоченные осоково-кочковатые тундры разных типов
(8. Андреев, 1934).
Заболоченность зоны так же, как и в Европейской части
•СССР, весьма велика. При этом на севере зоны (подзона
арктических тундр) заболоченность больше; на юге она падает.
Б. Городков (1935) оценивает (для всей Евразии) процент
площади болот (и озер) для первой подзоны в 60, а для
•подзоны лишайниково-моховых тундр в 25. Однако сюда,
вероятно, не входят заболоченные тундры. Таким образом,
общая болотистость еще выше. Для значительной части
интересующей нас зоны площадь болот составляет 25—35°;0
(В. Андреев). Если прибавить сюда заболоченные тундры, то
общую заболоченность западносибирской
зоны арктических болот придется оценить
цифрой, близкой к 50°/0, при этом скорей в сто¬
рону преуменьшения.
Болота рассматриваемой зоны имеют громад¬
ное значение как кормовая база оленеводства;
с точки же зрения торфодобычи эти болота не играют ни¬
какой роли.
II зона. Торфяники южной тундры и северной лесотундры
с мерзлыми буграми и мочежинами. Характерный и
распространенный тип болот этой зоны —
торфяники с хорошо выраженным мозаичным
комплексом, где чередуются мочежины и по¬
вышения в виде гряд или невысоких бугров.
Эти торфяники, несомненно, весьма близки к торфяникам
с мерзлыми дикраново-лишайниковыми грядами и мочежинами
европейской тундры, несколько отличаясь от них по своей
растительности. Высота бугров сравнительно невелика — чаще
0,5—0,7 м. Б. Городков называет эти торфяники плоскобугри¬
стыми. Этот тип в Западной Сибири широко распространен
в кустарниковой подзоне тундры и северной лесотундре.
Обе эти подзоны и входят в состав болотной зоны. Б. Городков
(1928) считает, что плоскобугристые торфяники замещают
крупнобугристые к северу от лесной зоны. При этом с повыше¬
нием уровня вечной мерзлоты и уменьшением мощности
талого слоя высота бугров на водоразделах уменьшается,
и крупнобугристые торфяники делаются плоскобугристыми.
Вечная мерзлота залегает здесь уже сплошным панцырем и
в буграх, и под мочежинами, тогда как в крупнобугристых
торфяниках мочежины оттаивают обычно до дна. Уступая
водоразделы плоскобугристым торфяникам, крупнобугристые
торфяники на север дальше всего продвигаются по краям
водоразделов вдоль лесных островов, где мерзлота, промы¬
ваемая талыми водами с лесных островов, частично исчезает.
В Западной Сибири сплошь мерзлые торфяники идут с севера
примерно до 64-й параллели. Южнее, между 64 и. 62° север¬
ной широты мерзлота уже распадается на отдельные участки.
.304
Это главным образом область крупнобугристых торфяников
с местной (спорадической) мерзлотой.
Б. Городков (1935), описывая плоскобугристые болота вообще, отме¬
чает, что по сравнению с крупнобугристыми болотами на плоских буграх
цветковая растительность более угнетена, разрежена и низкоросла, зеле¬
ные же мхи и лишайники лучше развиты; голых пятен, денудированного
торфа меньше, так как зимой невысокие бугры лучше защищены снегом,
чем крупные бугры более южных широт, и разрушение торфа здесь слабее.
Мокрые мочежины между буграми имеют весьма сходную с мочежинами
крупнобугристых болот растительность. Однако здесь чаще преобладают
зеленые мхи, а не сфагны, и эвтрофные ассоциации более распространены,
чем олиготрофные; флористический же состав несколько беднее. Господ¬
ствующие виды мочежин по Б. Городкову: Sph. balticum, Sph. fimbriatum,
Sph. Lindbergli, Calliergon cordifolium, C. sarmentosum, Drepanocladus exan-
nulatus, D. vernicosus, Polytrichum Jensenii и другие мхи, а из цветковых
обычные арктические — Сагех rariflora, С. rotundata, С stans. Отличия от
плоскобугристых торфяников на запад от Урала установить трудно. Веро¬
ятно, бугры богаче мхами (в частности Dlcranum) и беднее лишайниками,
чем на западе, по крайней мере на зональных и плакорных водораздельных
торфяниках К Собственно для Западной Сибири сведения по интересую¬
щему нас типу болот скудны. В. Андреев (1934) выделяет четыре типа
плоскобугристых болот, различающихся по условиям рельефа, раститель¬
ности и глубине мерзлого слоя, и дает краткое описание их. Он различает:
1) торфяно-плоскобугристую водораздельную раз¬
ность; на буграх мхи (Aulacomnium turgidum, Dicranum fuscescens), лишай¬
ников мало; вечная мерзлота в мочежинах на глубине 0,5—0,7 м\
2) т о р ф я н о-п л о с к о б у г р и с ту ю котловинную разность,
в котловинах с топкими мочежинами с Сагех aquatllls, где мерзлота лежит
глубоко, и с сфагновыми буграми;
3) т о р ф я н о-п л о с к о б у г р и с ту ю долинную разность, тоже
с сфагновыми буграми; встречается в притеррасных частях долин;
4) торфяно-плоскобугристую деградирующую раз¬
ность в котловинах и депрессиях на толщах погребенных торфяников.
Торфяники прорезаны ложбинками, спинки бугров обрушиваются. На буг¬
рах обильны лишайники. Зонально и широко распространена, повидимому,
первая разность.
Для лесотундры левобережья Оби описаны1 2 плоскобугристые комплексы
с торфом, мощностью обычно всего 1—1,5 м с трещиноватыми разрушаю¬
щимися буграми, высотой около 1 м.
На буграх деградирующий сухой ковер Polytrichum strlctum и Dicra¬
num и пятна Ochrolechia. Иногда на буграх обильны лишайники. Мочежины
в этих комплексах с редким ярусом обычных арктических Сурегасеае и
черным налетом юнгерманний.
Из других типов болот в этой зоне надо выделить арктиче¬
ские осоковые болота с мелким торфом. Эти болота весьма
близки по растительности к мочежинам плоскобугристых
комплексов. Площадь их в описываемой зоне постепенно
возрастает к северу за счет плоскобугристых, и в зоне аркти¬
ческой тундры, как мы видели, они господствуют. Заболочен¬
ные тундры и здесь, как и всюду, широко распространены.
Они встречаются среди всех типов тундры, но роль их среди
последних не везде одинакова.
1 В. Андреев (1935) отмечает, что на буграх плоскобугристых болот на
востоке Большеземельской тундры роль лишайников меньше, чем на западе
той же области.
2 В. Андреев, Формации равнин левобережья, „Советское оленевод¬
ство", вып. 5, 1935.
20—722
305
В целом эта болотная зона с ее неглубокими1 торфяниками,
где широко развиты комплексы мочежин и невысоких бугров,
и с большими площадями заболоченных тундр, несомненно,
продолжается и на запад от Урала, составляя одно неразрыв¬
ное целое с зоной болот с дикраново-лишайниковыми буграми
и мочежинами в Европейской части СССР.
III зона. Крупнобугристые торфяники южной лесотундры
и севера лесной зоны Западной Сибири. Эта зона занимает
юг западносибирской лесотундры и север лесной зоны.
Характерный „зональный" тип на водоразделах — это торфя¬
ники с комплексами крупных вечномерзлых торфяных бугров
и мочежин. Эти торфяники в общем близки по характеру
растительности к европейским крупнобугристым торфяникам.
Впрочем, сведения Об этих торфяниках крайне скудны.
В. Андреев (1934) различает два типа водораздельных торфяников на
северной окраине лесной зоны Западной Сибири, давая, впрочем, лишь
самую общую характеристику.
1. Водораздельная торфяно-сфагновая разность. Здесь
крупные бугры влажные, сфагновые; мокрые осоковые мочежины часто от¬
таивают до дна. Часты небольшие озерки.
2. Водораздельная торфяно-лишайниковая разность.
Бугры сплошь покрыты лишайниками, часто сливаются. Площадь мочежин
меньше площади бугров.
Обе разности широко распространены на водоразделах севера лесной
зоны. На буграх встречаются нередко угнетенные сосны и кедры. Мощ¬
ность торфа'3—4 м.
На незаливаемой террасе Полуя крупнобугристые болота
весьма бедны торфом (Н. Властова, 1936)1 2. Бугры имеют высоту
4—6 м, однако слой торфа большей частью всего в 5—10 см,
а остальной бугор состоит из минерального грунта. Максималь¬
ная мощность торфа здесь всего 0,6 м. Торф на буграх сильно
выветривается, сверху сфагновый, под ним слой осоково¬
древесного или осоково-гипнового торфа. В мокрых пониже¬
ниях между буграми торф залегает лишь пятнами. Он сфагно¬
вый или осоково-сфагновый, тоже неглубокий (0,6 м, редко
1,0 м). Заболоченность зоны весьма велика: для всей зоны
(в Европе и в Азии вместе) некоторые исследователи оценивают
ее в 60% (Б. Городков, 1935). Почти та же цифра (59%) при-
водится и для севера таежной зоны Западной Сибири (для
еловолиственничной подзоны). В южной лесотундре болот
меньше — около 40%, не считая заболоченных тундр (В. Ан¬
дреев, 1934). Таким образом, общая болотистость
этой зоны по скромному расчету 50%, а ве¬
роятно, и выше. Из этой площади значительная часть
приходится на заболоченные пространства. Они различны
в разных частях зоны. На севере ее, в лесотундре, эти про¬
странства представляют заболоченные тундры и частью
1 Мощность торфа 0,5—2,0 м (В. Андреев, 1934).
2 Эти болота представляют не типичные крупнобугристые торфяники,
а лишь комплекс заболоченных земель.
306
лиственничные редколесья, на юге (лесная зона) это заболо¬
ченные леса.
IV зона. Олиготрофные грядово-мочежинные торфяники
лесной зоны Западной Сибири. Эта зона имеет громадную
протяженность с севера на юг. Ее ширина с севера на юг
не меньше ширины трех первых зон, вместе взятых. Север¬
ную границу зоны за недостатком сведений провести трудно.
Южную границу А. Бронзов (1930) проводит несколько север-
1 — болотно-хвойная зона; 2 — болотно-березовая зона; 3 — лесостепная зона;
4 — южная граница зоны верховых болот.
нее южной границы хвойной зоны (рис. 101). Таким образом,
эта болотная зона лишь немного уже зоны хвойных лесов.
Она служит продолжением европейской зоны грядово-моче-
жинных торфяников. Для всей зоны характерно следующее:
1. Господство (по площади) принадлежит
олиготрофным выпуклым торфяникам, эвтроф-
ные же отступают на второй план.
2. Торфообразование здесь наиболее ин¬
тенсивно, площадь торфяников по отноше¬
нию к общей площади болот (т. е. заторфован-
20* 307
ность) наибольшая, так же как и глубина
торфа.
¿.Сфагновые торфа имеют здесь макси¬
мальное распространение, так же как и
сф агновые мхи в современном растительном
покрове болот.
Характерный тип торфяников в центральной части этой
зоны — это торфяники с комплексами гряд, мочежин и озерков
(васюганско-нарымский тип). Мы охарактеризовали его как
тип оптимального развития, который можно поставить наряду
с выпуклыми болотами северо-запада Европейской части
СССР, находящимися в приморском климате. Именно торфя¬
ники этого типа чрезвычайно распространены на равнинных
пространствах в центре зоны, покрывая почти сплошь водо¬
разделы; они создают характерный для этих районов нарымский
болотный ландшафт. Растительность торфяника зависит от
его размеров, формы поверхности и, ближайшим образом, от
его водного режима.
К северу с ухудшением климата выпуклые болота изменяют
свой характер. О болотах, расположенных между Самаровым
и Березовым вдоль по Оби, имеются данные Н. Властовой
(1936). По характеру растительности здесь выделяются три
типа олиготрофных болот, представляющие различные стадии
развития.
1. Сосново-сфагновые болота с довольно рослой сос¬
ной (Pinus silvestris fo Litwinowii), Кассандрой, пушицей, Sph.
medium, Sph. recurvum. Комплекс с мочежинами имеется
лишь в центре болота и на незначительной площади. Однако,
несмотря на олиготрофную растительность, торфа, сла¬
гающие эти болота, не чисто олиготрофные. Они сфагно¬
вые, но с примесью хвоща, осок, древесных остатков
(сосна, береза) и имеют, кроме того, повышенную золь¬
ность сравнительно с нормальной для олиготрофных тор¬
фов (в среднем 6ф/0)- Глубина торфа незначительна — от 1,5
до 1,75 м.
2. Второй тип болот характеризуется комплексами с пре-,
обладанием шейхцериевых мочежин и с угнетенной сосной.
Они более глубокие—залежь от 2 до 4 м, однако она сложена,
несмотря на олиготрофную современную растительность, не
сфагновыми, а шейхцериевыми и осоковыми или вахтовыми
торфами. Это—болота склонов вторых террас.
3. Третий тип—это болота с обширными комплексами
гряд и мочежин. Последние сильно преобладают по площади
над грядами с Sph. fuscum, где часто сфагнум отмирает и
поселяются лишайники. Деградация сфагнового ковра в моче-
жинах приводит к возникновению пятен голого торфа и
образованию скоплений воды. Глубина залежи тоже сравни¬
тельно невелика—2 м, лишь местами до 2,5—3 м. Торфяная
залежь целиком сфагновая. Сверху — до глубины 1,5—2 м
торф подстилочный со степенью разложения ниже 25°/о. Ниже—
308
торф тоже сфагновый, но более разложившийся. Зольность
очень низкая—в среднем всего 2,09%>
Болота второго и третьего типа напоми¬
нают по растительности болота нарымско-васюганского
типа:
1) широко распространен грядово-мочежинный комплекс;
2) явления деграда¬
ции сфагновой дернины
и регрессивного вымо¬
кания (лужицы свобод¬
ной воды); эти явления,
может быть, слабее вы¬
ражены, чем в Васю-
ганье. Отличия этих
болот от нарымско-
васюганских таковы:
1) меньшая глубина за¬
лежи, 2) преобладание
часто мезотрофныхтор-
фов, а в случае олиго-
трофного типа строе¬
ния залежи сравнитель¬
но малая мощность сфа¬
гнового торфа. Эти
признаки, а также
слабо выпуклая
поверхность
этих торфяников,
вероятно, указы¬
вают на неблаго¬
приятные для раз¬
вития выпуклых
болот климатиче¬
ские условия с при¬
ближением к се¬
верной границе
распространения
этого типа.
Олиготрофные сфаг¬
новые болота в Запад¬
ной Сибири встреча¬
ются еще далеко к северу от Березова, заходя в нижнюю часть
бассейна реки Полуя (А. Лесков, 1935). В верхней и средней ча¬
стях бассейна Полуя олиготрофные сфагновые торфяники с сос¬
ной и кедром. Севернее, в нижней части Полуя эти торфяники
уже только с одной редкой сосной, без кедра. Несмотря на
то, что их растительный покров здесь напоминает более юж¬
ные олиготрофные болота, залежь имеет незначительную
мощность (0,5—0,8 м). Это крайние аванпосты выпуклых
болот к северу, дегенерирующие представители этого типа.
209
Хотя для описываемой зоны весьма характерно резкое
преобладание олиготрофных торфяников над эвтрофными, но
последние также заслуживают описания. Они не одинаковы
в разных районах. Для южных частей зоны (западное Васю*
ганье) весьма характерны обширные, простирающиеся на
тысячи гектар долинные гипновые торфяники, главным образом
в притеррасных частях речных долин (М. Барышников, 1929).
Эти торфяники жестководного питания—болотные воды
богаты окисью железа и солями кальция. На них наблюдаются
такие зоны растительности (от центра к периферии) (рис. 102).
1. Центр: безлесное мокрое осоково-гипновое болото
с периферическим узким поясом комплексного осоково-гип-
нового болота с мочежинами и невысокими и узкими грядами
(так называемые веретья)1.
2. Лесное моховое эвтрофное болото, комплекс кочек из
Sph. Warnstorfii с сосной, иногда кедром и другими породами,
и мокрых гипновых мочежин.
3. Третий пояс (притеррасный) образован осоковыми
кочкарниками (Carex caespitosa, С. paradoxa). Мочежины здесь
пропадают.
В настоящее время намечаются лишь слабые признаки
перехода этих болот в более олиготрофную стадию на грядах
и иногда ближе к окраинам. Глубокая залежь этих болот
почти нацело сложена гипновыми и осоковыми торфами
(рис. 103). У дна обильны древесные остатки, указывающие,
что болота образовались путем заболачивания лесов. Любо¬
пытно, что иногда эти болота имеют выпуклую поверхность
и на них — правда, в виде редкого исключения — попадаются
вторичные озерки. Это придает им некоторое сходство
с выпуклыми болотами.
В Васюганье, как и везде в хвойной зоне Западной
Сибири, кроме ее южной окраины, эвтрофные и в частности
гипновые болота встречаются на речных террасах среди без¬
раздельно господствующих на водоразделах олиготрофных
болот. На восточном склоне северного Урала эвтрофные
болота развиты весьма широко, выходя из долин на водо¬
разделы. Здесь выделяется целый район холмистых восточных
склонов Урала, где обширные эвтрофные моховые торфяники—
преобладающий тип болот.
Эти торфяники иного характера, чем описанные выше1 2.
Громадные болотные массивы площадью (включая и окраины) до
10 000 га на склонах водоразделов рек Турьи, Каква, Лобва (притоки Сось-
вы) в центральных частях безлесны, со сфагновым покровом (главным
образом эвтрофный Sph. Warnstorfii) с преобладанием Betula папа и осок
(главным образом Carex laslocarpa, меньше С. dioica и С. teretiuscula).
Отдельными пятнами, примыкая к суходольным островам, среди втих
болот встречаются выпуклые подушки олиготрофных торфяников, сложен¬
ные торфом из Sph. fuscum. Ближе к краям втих болот — значительные
1 На схеме не изображен.
2 Торфяное дело, № 3, 1926.
310
У///Л Остбьш торф
УШШ ГипноВый торф
Рис. 103. Строение долинного гипнов.ого болота в Васюганье (Западная Сибирь). По М. Барышникову, 1929.
б — примесь вахты; п — пушицы; х — хвоща; г — гипновых мхов; о — осок; с — древесины сосны; к — древесины кедра; б — древесины березы;
пх — древесины пихты.
площади редколесных (сосна и береза) болот с ковром олиготрофных
сфагнов. Болота окаймлены широкой полоской „согр*— болотистых лесов
с сосной, кедром и елью. Залежь сложена осоковыми, сфагново- и гип-
ново-осоковыми торфами средней разложенности. Обычная глубина залежи
3—3,5 м»
Подробное описание целой системы обширных торфяников на восточ¬
ном склоне северного Урала дает Н. Властова (1937). Эти обширные тор¬
фяники (Нижне-Салдинская группа) располагаются на плоском водоразделе
рек Тагила и Салды (притоки Туры). Подробно изученное болото этой
группы, как и вышеописанные болота, имеет центральную глубокую гип-
новую часть и окраинные мелкие участки, сложенные нацело олиготроф-
ным торфом. Сочетание олиготрофных и эвтрофных участков, видимо*
характерно для восточного склона Урала вообще. Мощная шестиметровая
толща в центре болота сложена главным образом осоково-гипновыми
и осоковыми торфами. В центральной части болота громадные площади
занимает эвтрофный грядово-мочежинныи комплекс. В мочежинах в мохо¬
вом покрове доминируют Drepanocladus vernicosus и осоки (Carex diandra,
С. limosa, С. chordorrhiza, С. stricta). Здесь много Triglochin maritima.
На грядах, перпендикулярных к уклону болота, редкий угнетенный cocho»
вый ярус, заросли Betula nana. Моховой ковер гряд образует Camptothe-
cium nitens, а иногда Sphagnum Warnstorfli. Происхождение этих болот
можно связать с близким залеганием соленосных третичных пород.
Заболоченность зоны в целом неизвестна. Если судить по
Васюганью, где водоразделы почти сплошь заболочены и где
полоски суши встречаются лишь вдоль рек, то придется
считать болотистость зоны значительно выше 50%« Торфя¬
ники зоны — колоссальный, в настоящее время
совершенно почти неиспользованный топлив¬
ный фонд, а мощная неразложившаяся верхняя
толща сфагнового торфа болот средней части
зоны является тоже громадным резервом под¬
стилочного и строительного материала.
V зона. Гипновые торфяники юга хвойной и болотно-бере¬
зовой зоны. Эта зона занимает юг хвойной зоны и значи¬
тельную часть болотно-березовой. Эта зона примерно соот¬
ветствует „зоне южных болотных комплексов“ А. Бронзова
(1936). Описание ее приводится по этому автору. Эта зона
неширока — с севера на юг менее 100 км, но протянулась
с запада на восток от Урала до Енисея. Несмотря на
сравнительно южное положение, заболочен¬
ность зоны громадна — свыше 50%. Чрезвычайно
характерными моментами являются: 1) положение болот
как на равнинных водоразделах, так и в доли¬
нах, 2) господство гипновых болот при остров¬
ном распространении олиготрофных. Гипновые
торфяники достигают здесь громадных размеров — например,
южная часть Большого Васюганского болота на Обь-Иртыш-
ском водоразделе, далее болота к востоку от Оби (водораз¬
дел Кети и Чулымы). В южной гипновой части Большого
Васюганского болота1 среди огромных безлесных осоково-
1 Этот колоссальный массив, площадью в 1,5 млн. га, к северу изме¬
няет свой характер. Здесь чередуются слегка выпуклые, широкие (0,5 км)
полосы олиготрофных болот с безлесными мокрыми площадями главным
образом мезотрофного характера.
312
гипновых болот местами встречаются вторичные озерки и
всюду проходят узкие гряды „веретья“ с березой (Betula
pubescens), реже — с сосной. Грядово-мочежинный микрорельеф
характерен (по Бронзову) для осоково-гипновых болот всей
Западной Сибири вообще. Кроме мочежин и веретьев, на
Большом Васюганском болоте имеются еще несколько вытя¬
нутые островки олиготрофного болота в несколько десятков,
метров в длину и ширину (по-местному „шоломочки“).
Для осоково-гипновых мочежин характерен хорошо развитой моховой
покров главным образом из Drepanocladus vernicosus и D. Sendtneri и до¬
вольно редкий ярус осок (господствуют Сагех diandra, С. chordorrhiza,.
С. limosa, С. Hudsonii, местами С. rostrata, С. lasiocarpa). Всего А. Бронзов
выделяет здесь 15 ассоциаций.
Гряды (веретья) обычно с ярусом кустарничков. Чаще всего господ¬
ствует Betula nana, реже Cassandra и Andromeda. Сплошной моховой по¬
кров чаще из Sph. Warnstorfii, реже (на более низких грядах) из Campto-
thecium nitens. Иногда наверетьях ярус Betula pubescens. Около нее олиго-
трофные виды сфагаоз (Sph. parvifolium, Sph. medium, Sph. fuscum) обра¬
зуют кочки.
На олйготрофных островках („шоломочках") хорошо развит ярус
из болотных Ericaceae (главным образом Ledum palustre) и моховой ковер
из Sph. fuscum. Залежь Большого Васюганского болота как в южной, так
и в северной (сфагновой) его части образована в нижней части осоковыми
и осоково-хвощевыми торфами — в северной части с меньшей, в южной
с большей примесью тростника: В южной гипновой части болота донный осо¬
ковый торф, мощностью от 0,5 до 2 м, прикрыт осоково-гипновым торфом.
Сфагновые торфа здесь встречаются лишь островками, слагая упомя¬
нутые выше „шоломочки*. Они, следовательно, недавнего происхождения.
В северной сфагновой части болота верхняя толща из осоково-сфагновых
н сфагновых торфов. Южная часть болота, принадлежащая к рассматри¬
ваемой зоне, возникла, видимо, на месте прежних галоидно-сульфатных
солончаков и солонцов, может быть, с осоково-тростниковыми болотами
(займищами) в понижениях рельефа. Эти водоразделы были, вероятно, до
заболачивания совершенно безлесны. Севернее, где сейчас раскинулась сфаг¬
новая часть Большого Васюганского болота, засоленность была меньше,
грунтовые воды мягче. Поэтому осоковая стадия болота была здесь короче,
и скоро наступила сфагновая стадия. Весьма вероятно, что примерно там,
где теперь идет граница между осоково-гипновой и сфагновой частью
болота, находилась раньше граница степи и лесостепи. Эта граница, может
быть, соответствует и прежней климатической границе, проходившей здесь
прежде, когда климат был суше современного. Вряд ли эта граница обусло¬
влена современным климатом этой полосы, допускающим сфагновое болото-
образование.
Из других районов той же зоны имеются некоторые сведения о торфя¬
никах болотно-березовой подзоны на водоразделах Уй-Тара и Ишим-Иртыш,
а также долины Иртыша (Г. Бронзова, 1932). Торфяники располагаются здесь
и на водоразделах, и в долинах. На водоразделах преобладают осоково-гип-
новые болота, но обычны также осоковые, березово-осоковые и переходные
с участками олйготрофных. Залежь водораздельных болот обычно мелкая,
торфа довольно малозольные. Эти признаки и состав торфов свойственны
также и вышеописанным болотам. Здесь распространены осоковые, тростни¬
ково-осоковые и гипново-осоковые торфа. В долине Иртыша болота смешан¬
ного типа с олиготрофными и низинными участками. Эвтрофные торфа
долины Иртыша того же состава, что и водораздельные. Некоторые долин¬
ные гипновые торфяники Иртыша образовались путем заболачивания стариц
(М. Спиридонов, 1928). Здесь также наблюдается микрорельеф с грядами.
На заливаемых рекой осоковых болотах господствуют Сагех gracilis,
С. intermedia и др. В восточной части зоны в Ачинском районе, где рельеф
расчлененный, почти все болота долинные. Они гипново-осоковые, осоково¬
березовые и елово-согровые (Г. Бронзова, 1932).
313
Как уже указывалось, среди обширных эвтрофных торфя¬
ников нередки островки олиготрофных торфяников. Эти
последние сосново-кустарничково-сфагновые (обычно с гос¬
подством Sph. fuscum), западносибирского типа, по-местному
„рямы“ (см. выше). Олиготрофные торфяники в рассматривае¬
мой зоне изучены весьма слабо по сравнению с теми же
болотами в лесостепи. На рямах б. Ишимского уезда
на западе зоны сильно развиты кустарнички из вере¬
сковых (А. Гордягин, Б. Городков, 1915). В долине Иртыша
(б. Тарский уезд, в лесной зоне) на рямах также обиль¬
ны кустарнички, здесь много Ledum, а в сфагновом
ковре, кроме господствующего Sph. fuscum и Sph. recur-
vum, много Sph. medium. Залежь до глубины 0,7 м сло¬
жена мало разложившимся сфагновый торфом, ниже до
глубины 1,3 м — хорошо разложившийся сфагновый торф,
внизу—с большой примесью осоки. Рям опоясан крупно¬
осоковым болотом. Водораздельные рямы хвойной зоны
окружены лесами, то сосновыми, то смешанными из сосны,
пихты, кедра, ели и березы—„рямовая corpa“ (Б. Городков,
1916).
К северу от рассматриваемой зоны сфагновые болота
совершенно своеобразны (А. Бронзов, 1930). Здесь, как отме¬
чалось выше, слегка выпуклые олиготрофные болота чере¬
дуются в виде полос с широкими, плоскими, весьма мокрыми
безлесными болотами типа аапа, приуроченными к небольшим
понижениям. Эти болота сфагновые. Наряду с олиготроф-
ными видами (Sph. Dusenii, Sph. balticum), здесь большую
роль играют эвтрофные сфагны (Sph. obtusum, Sph. subsecun¬
dum), к которым местами примешан Drepanocladus vernicosus.
Местами сфагновая дернина деградирует и заменяется
печеночниками. Среди таких болот разбросаны вытянутые
кочки Sph. fuscum. Эти своеобразные болота« напоминаю¬
щие в некоторых отношениях болота аапа, сплошной неши¬
рокой полосой тянутся к северу от этой зоны, соединяя
южную, относящуюся к рассматриваемой зоне, часть Боль¬
шого Васюганского болота с северной его частью, которая
уже относится к зоне грядово-мочежинных олиготрофных
болот.
VI зона. Осоково-тростниковые и засоленные болота
лесостепи и степи Западной Сибири. Эта зона занимает
западно-сибирскую лесостепь и степь. Характерные признаки
этой болотной зоны следующие (см. А. Бронзов, 1930):
1. Преобладающий тип болот — это солончаковатые
травяные, т. е. тростниковые, тростниково-осоковые и осоковые
болота и солончаки. 2. Торфа этой зоны (на юге и в центре ее)
засолены галоидно-сульфатными солями; к северу это засоле¬
ние сменяется карбонатным. 3. Болотистость весьма высока,
особенно на севере лесостепи; в некоторых районах болот —
до 50°Л> всей площади. 4. Болота, как и в болотно-березовой
зоне, встречаются как в долинах, так и на водоразделах.
■314
При слабо расчлененном рельефе, особенно на севере лесо¬
степи, болота занимают обширные площади в центре межре-
чий, при более же расчлененном рельефе они встречаются
в межгривных понижениях и в депрессиях на широких гривах
водоразделов. В лучше исследованной Барабинской лесостепи,
где заболочено до 50% всей площади, главный тип водораз¬
дельных болот — травяные болота, так называемые займища
(М. Нейштадт, 1935). Займища — это тростниковые и осоково¬
тростниковые болота. Главную роль играет тростник, некото¬
рые осоки (Сагех riparia, С. caespitosa), много также Scolochloa
festucacea. Глубина торфа часто не более 15 см. Когда есть
торф, мощность его обычно невелика (0,5—1,5 м, редко
больше). Торфа главным образом тростниково-осоковые,
с значительной зольностью1, но вполне годны на топливо.
Для западной части зоны растительность займищ описана для б. Курган¬
ского округа (лесостепь).
А. Генкель (1935) выделяет здесь такие типы заболоченных водных про¬
странств и болот:
1. Плоские мелкие, пересыхающие в сухие годы пресноводные озера-
блюдца с обильными Butomus umbellatus, Typha angustifolia, Сагех intermedia.
Флористический состав этих озер богатый.
2. В разной степени соленые озера с более бедной
флорой. Целый ряд видов пресных озер здесь выпадает. Постоянны
здесь Phragmites, Potamogeton pectinatus, Lemna trisulca 1 2. В наиболее засо¬
ленных озерах отсутствует высшая береговая растительность, а береговая
полоса заселена галофитами.
3. Проточные водоемы. Сюда относятся русла рек с водной
растительностью (Potamogeton perfoliatus, Р. lucens, Lemna trisulca, также
Nymphaea, Nuphar, Stratiotes aloides), старицы и озера поймы Тобола и его
притоков с водной растительностью, тростником, Glyceria aquatica, Сагех
aristata v. ortostachys 3.
4. Тростниковые займищ a—мощные заросли тростника в обшир¬
ных плоских водораздельных впадинах и по краям крупных озер.
5. Осоковые болота. Они неодинаковы на севере и на юге лесо¬
степи. На севере эти болота сильнее и более устойчиво обводнены, чем
на юге, и торф здесь более мощный. В центральных частях северных
осоковых болот мощные кочкарники с Сагех stricta, часто с примесью трост¬
ника. На периферии этих болот в условиях периодического обводнения
доминируют Сагех riparia и Alopecurus ventricosus, а при большем обводнении
Typha latlfolia и Сагех intermedia. Из других особенностей северных коч¬
карников укажем ассоциации с Сагех rostrata, С. lasiocarpa, С. limosa с сфа¬
гнумом и некоторыми северными видами. Эти ассоциации свойственны
участкам, наиболее удаленным от минерализующего влияния берегов.
В южной лесостепи осоковые болота — это мощные кочкарники с Сагех
stricta и Drepanocladus aduncus в межкочечных понижениях. Торфообразование
здесь ослаблено, и в иные годы кочкарники вовсе пересыхают и исполь¬
зуются как пастбище. Северные виды сюда уже не заходят, зато постоянны
степные виды, заходящие из окружающих мест, и. пастбищные формы.
6. Осоковые болота боровых районов. Характерно пре¬
обладание Сагех rostrata, С. limosa, С. lasiocarpa по моховому ковру из
1 10—25% на абсолютно сухое вещество.
2 Оба эти типа озер свойственны лесостепи. Озера боровых районов
более глубокие, отличаются от озер лесостепи осоковыми сплавинами и
видовым составом — в частности, массовым развитием таких редких в
Курганском округе видов, как Chara, Potamogeton flliformis, Najas marina.
3 Оба последних вида свойственны исключительно бассейну Тобола в
пределах Курганского округа.
315
Drepanocladus vernicosus и Camptothecium nitens. Это настоящие осоково-
гипновые болота. Кроме них, в боровых районах встречаются и осоковые
болота такие же, как в лесостепи.
7. Сфагновые торфяники. В более восточном районе описаны
болота и пресноводные озера б. Петропавловского уезда (южндя лесо¬
степь) (М. Спиридонов, 1928). Озера здесь иля в глубоких котловинах,
или в плоских блюдцах; и те, и другие могут полностью пересыхать.
Из господствующих цветковых растений в этих озерах можно отме¬
тить: Carex orthostachys, Phragmites, Scolochloa festucacea, Scirpus lacu-
stris, часто Carex nutans и некоторые, другие. Иногда в озерах — плавающие
ковры Drepanocladus Sendtneri. В высохших озерах и в мелкой воде у бере¬
гов — ковер мха Drepanocladus aduncus. Торфянистые отложения там,
где они есть, лишь ничтожной мощности. М. Спиридонов подтвер¬
ждает обмеление озер в районе
к югу от Сибирской ж. д.,
установленное многими иссле¬
дователями, и приписывает
главную роль в этом явлении
сносу почв с прилегающих
склонов. Этот снос усиливается
с усилением выпаса и особенно
с распашкой склонов озерных
котловин. При частых и силь¬
ных обмелениях на дне озера
поселяется осоковый кочкар¬
ник. За этой фазой умирания
озера следует стадия ивняка,
а затем стадия березового кол¬
ка. Последняя представляет этап
внедрения леса в степь*
Очень интересны оли-
готрофные торфяники.
В описываемой зоне они
представлены сосново¬
сфагновыми болотами, по¬
местному „рямами“. Под
сосновым насаждением густой ярус кустарничков, — следова¬
тельно, эти болота сосново-кустарничково-сфагновые. Рямы
эти в лесостепи в общем хорошо изучены. Они близки к бо¬
лее северным рямам, отличаясь от них в некоторых призна¬
ках. Растительность рямов расчленена на концентрические
пояса. Для Барабы типичны такие пояса от центра к перифе¬
рии ряма (П. Красовский, 1925):
1. Сосново-кустарничково-сфагновое болото.
2. Березово-сфагновое болото. 1Рис. 104. Расположение рямов, займищ и
солончаков (Бараба, Западная Сибирь).
По М. Нейштадту, 1936.
1 — рямы (выпуклые сфагновые болота); 2—займища
(тростниковые болота); 3 — солонцы и солончаки.
3. СаПсеЦип ЬеМоэит.
4. Саг^сеЬ—Р1и^тИ:е1:ит.
Рям окружен солонцами и солончаками, и весь переход
от центрального олиготрофного болота к засоленному мине¬
ральному берегу происходит всего на протяжении 0,5 км. Этот
контакт сфагнового ряма с засоленными почвами — явление
обычное вообще для рямов западно-сибирской лесостепи'.
Рямы таежной полосы имеют иные окраины—они окружены
1 Оно отмечалось еще А. Гордягиныы и другими (например Е. Ники¬
тиной, 1928).
316
лесом. Подобные пояса отмечают и
другие исследователи. Л. Трефилова
(1925) дает для той же Барабы более
расчлененные пояса:
1. Сосново-сфагновое болото.
2. Березово-сфагновое болото.
3. Болото с Сагех caespitosa и бе¬
резовым ярусом.
4. Безлесное болото с Сагех caes¬
pitosa.
5. Phragmitetum (мокрый пояс трост¬
ника).
6. Мокрый осоковый пояс (Sco-
lochloa festucacea, Сагех riparia, Сагех
ortostachys).
7. Alopecuretum на минеральной
почве.
8. Солонец или солончак.
В общем рямы обычно разбросаны
островками среди займищ, или по¬
следние кольцеобразно окружают
рямы (рис. 104)). Рямы обычно округ¬
лые или овальные. Площадь их срав¬
нительно невелика—в Барабе в сред¬
нем от 100 до 300 га, редко — до
1000 га (М. Нейштадт, 1936). В Барабе
в каждом ряме имеется озеро. Рельеф
ряма резко выпуклый (рис. 105). Про¬
исхождение рямов связано с озерами.
В настоящее время они довольно
интенсивно наступают на озера. По
строению они довольно однообразны.
Глубина торфа всюду значительная—
до 6,5 м одного сфагнового торфа.
Этот мало разложившийся сфагновый
торф из Sph. fuscum и Sph. medium об¬
разует верхнюю толщу залежи. Этот
торф в случае наиболее полной серии
слоев подстилается тростниковым или
осоково-тростниковым торфом, под
которым лежит сапропель (первый
тип строения; рис. 106). В других слу¬
чаях сфагновый торф лежит прямо
на сапропеле (второй тип). Наконец,
иногда сапропель отсутствует, и за¬
лежь сложена лишь сфагновым и
осоково-тростниковым торфом. Раз¬
ные типы строения встречаются на
одном и том же ряме.
Рямы с резко выпуклой поверхностью и мощ¬
ной толщей мало разложившегося сфагнового
торфа вовсе не соответствуют континенталь¬
ному климату западно-сибирской лесостепи
и окружающим засоленным почвам. Не менее
резок контраст между растительностью, окру¬
жающей рямы, и северными элементами их фло¬
ры. Захождение последних в глубь западно¬
сибирской лесостепи совершенно необычно
для лесостепи Европейской
части СССР.
К числу таких растений относятся
Sph. fuscum (массовый вид рямов), Rubus
chamaeraorus, Betula nana, Oxycoccus
microcarpa. Из них Rubus chamaemorus
всюду встречается в Барабе и нередко
очень обилен (П. Красовский, Е. Ники¬
тина, М. Нейштадт, 1936), то же и
в б. Ишимском у. (А. Гордягин, Б. Го¬
родков, 1915), а также в б. Курганском
округе (А. Генкель, 1935).
Довольно обычна, хотя и реже,
Betula nana. Она отмечена в Курган¬
ском округе (лесостепь) — правда, на се¬
вере; нередко встречается и в б. Ишим¬
ском у.1 на самой северной границе
лесостепи (севернее Тюкалинска, А.
Гордягин). Встречается она и в Барабе. Интересная особен¬
ность рямов лесостепи — массовое развитие брусники. На юге
б. Тобольского уезда брусника уже не встречается на тор¬
фяниках, и в урманно-болотистой подзоне тайги она встре¬
чается на рямах только на окраинах (Б. Городков, 1915).
Болота Таймыра.
Для полуострова Таймыра так же, как и для более запад¬
ных частей Сибири, хорошо выделяются ботанико-географиче¬
ские подзоны, аналогичные подзонам северной части Западной
Сибири. Эти подзоны- в общем совпадают и с болотными
зонами, по которым мы в дальнейшем будем производить
описание. На Таймыре выделяются такие болотные зоны:
I зона. Арктические осоково-пушицевые бо¬
лота. Эта зона соответствует арктической тундровой подзоне.
Краткое описание Ф. Кьельмана (F. Kjellmann, 1883) отмечает
преобладание осок и разных пушиц (Eriophorum angustifolium,
Е. russeolum, Е. Scheuchzerii). Эти болота широко распростра¬
нены всюду в арктической тундре и почти лишены торфа.
II зона. Плоскобугристые болота. Этот тип широко
распространен не только в южных подзонах тундр, мохово-
1 Б. Городков (1915) отмечает здесь также и Oxycoccus microcarpa.
А 6 8
Ш/ <ШЗ П
Рис. 106. Типы строения
сфагновых болот (рямов)
в Барабе. По М. Ней-
штадту, 1936.
1—сфагновый торф; 2—трост¬
никовый торф; 4 — сапропель;
3 и 5 — глина.
318
пушицевой и мохово-кустарничковой, но и в лесотундре
и даже в лесной зоне (Хетская равнина) вплоть до северного
склона центрально-сибирского плато. Пока мы относим все
эти ботанико-географические подзоны к одной болотной зоне.
В тундре средняя высота бугров 0,7 м. На буграх мо¬
ховой ковер из Aulacomnium turgidum, Camptothecium nitens,
Drepanocladus uncinatus, Ptilidium ciliare; лишайники зани¬
мают всего до 10% площади бугров. Сфагновые мхи встре¬
чаются лишь изредка у основания бугров. Из кустарнич¬
ков на буграх на севере преобладают ивы (Salix reptans,
S. pulchra), на юге Betula exilis, а кроме того, много багуль¬
ника и голубики. В мочежинах доминирует Сагех aquatilis,
С. rotundata, из мхов Drepanocladus revolvens, D. exannulatus.
В лесной зоне бугры лишайниковые, площадь их больше, чем
мочежин. В мочежинах сплошной ковер Сагех rotundata, С.
saxatilis, С. chordorrhiza. Залежь глубже, чем можно ожи¬
дать. 1 Из других типов болот отметим полигональные
болота с повышенными гривками вокруг „полигонов“.
Последние представляют пониженные участки с раститель¬
ностью обычных мочежин арктических болот, распространен¬
ных как в тундре, так и в лесной части зоны. Низинные
пушице-осоковые болота — обычный тип в тундро¬
вой части зоны. Они близки по растительности к осоково-
пушицевым болотам севера Таймыра, которые замещают их
к северу. На юг (Хетская равнина лесной части Таймыра)
они замещаются близким типом — осоковыми эвтрофными
болотами. Глубина торфа в них различна — от 0,1 м и выше.
Под торфом часто каменный лед.
В лесной подзоне встречаются еще ерниковые болота.
Из заболоченных тундр отметим некоторые типы ивняков и
ерников (последние с торфянистым горизонтом и сфагновым
покровом), пушицево-тальниковую моховую тундру и неко¬
торые другие типы. В лесной части Таймыра имеются забо¬
лоченные сфагнумом лиственничные леса. В целом для Тай¬
мыра, как и для севера Сибири вообще, характерна большая
заболоченность. Наличие полигональных болот указывает на
континентальность климата.
Особенность Таймыра — значительное одно¬
образие болотных типов, идущих через тундру
(кроме ее северной, арктической подзоны) в лес¬
ную зону. Внедрение тундровых типов болот
(плоскобугристых) в лесную зону Таймыра
зависит от суровых климатических и почвен¬
ных (вечная мерзлота) условий последней.
Болота тундровой зоны Якутии1.
Болота якутской тундровой зоны весьма близки к болстам
тундр Западной Сибири. Большая поверхностная заболочен-
1 См. В. Сочава, 1933, 1935.
319
'яость, связанная с мерзлотой, замедленное торфообразование
создают и здесь, как и на Таймыре, благоприятные условия
для развития болот без торфа и ограничивают распростране¬
ние торфяников. Характеристика болотных типов может быть
лишь самой общей из-за скудости материала. Можно разли¬
чить такие типы болот:
¡.Арктические болота с осоковыми моче-
жинами и разбросанными среди них торфя¬
ными грядами и буграми. Площадь гряд — около 20%
всей площади. В подзоне арктических тундр в мочежинах
доминирует хорошо поедаемая оленями Carex stans, которая
южнее замещается в этом типе болот непоедаемыми осоками—
Carex rotundata, С. rariflora, С. chordorrhiza. На грядах обычно
преобладает Betula exilis, из мхов — Dicranum и Polytrichum;
лишайников же мало. В бассейне Анабары бугры среди аркти¬
ческих осоковых болот до 1 м высоты, с ледяным ядром,
покрытым осоковым торфом. Чаще здесь не бугры, а узкие
гряды, образующие сетку, разделяющую поверхность осокового
болота на клетки или ячейки.
Эти болота широко распространены в арктической подзоне,
а также и южнее. Они близки к плоскобугристым. В тундрах
Якутии они занимают от 14,5 до 18,3% всей территории.
2. Болота с Arctophila fulva и Carex stans по
зарастающим тундровым озерам.
Остальные типы относятся к заболоченным тундрам и редко¬
лесьям.
В арктической подзоне Якутии сфагны — флористическая
редкость. Южнее, в подзоне лишайниковых тундр, они чаще,
но в общем все же распространены незначительно и на не¬
больших площадях. Значительно больше их в кустарниковой
подзоне. В лесотундре сфагны уже широко распространены в ли¬
ственничных марях, и широко развито сфагновое заболачивание.
О заболоченности тундр Якутии можно получить предста¬
вление, суммируя площади различных заболоченных угодий.
Процент заболоченности, определенный таким путем, по разным
районам от 26 до 40.
Эта заболоченность хотя и велика, но все же несколько
ниже, чем в тундре Западной Сибири, что можно приписать
не только более расчлененному рельефу, но, вероятно, и
более континентальному климату.
Болота тундровой зоны Дальневосточного края1.
Типы болот и заболоченных пространств тундр Дальне¬
восточного края описаны весьма схематично. Ниже приводится
краткая характеристика болот главным образом на основании
более подробно рассмотренного Пенжинского района.
1 В. Сочава (1932), Б. Городков (1933). Здесь описываются главный
образом равнинные болота.
. 320
1. Осоковые эвтрофные болота. Это болота плос¬
ких понижений. Особенно распространены они по берегу
Полярного моря. Здесь господствуют обычно Carex stans,
С. rotundata и С. rariflora; первая — при более проточном, по¬
следние— при застойном увлажнении. Кроме того, здесь много
Carex chordorrhiza, виды Eriophorum и др. Из мхов — Calliergon
stramineum, Drepanocladus vernicosus, иногда сфагновые мхи.
Покрытие осок в одном случае было 60%, а сфагнов — 30%.
Торф иногда до 0,5 м. В лесотундре Carex stans заменяют
плохо поедаемые оленями С. rariflora и С. rotundata, и сфагно¬
вые мхи преобладают над гипновыми. При понижении уровня
мерзлоты преобладание получает Eriophorum vaginatum.
2. Осоковые эвтрофные мочежины в ком¬
плексе с невысокими буграми (мелкобугристый ком¬
плекс Б. Городкова). Мочежины с Carex stans, иногда с другими
осоками и сфагнами представляют описанные выше осоковые
болота. Плоские бугры, высотой в 0,2 — 0,3 м1, иногда сфаг¬
новые. Иногда они вытянуты до 10 м в длину, при 2 — 3 м
ширины. Эти бугры, сверху одетые торфом, довольно обычны
в Пенжинском районе. Крупнобугристые торфяники на водо¬
разделах здесь отсутствуют, а минеральные бугры, свойственные
Забайкалью и смежным частям Якутии и представляющие
более южный тип, встречаются лишь в речных долинах.
3. Прибрежная растительность зарастающих
озер. Озера зарастают сфагновыми мхами и различными
цветковыми — Carex utriculata, Comarum palustre, Equisetum
limosum, Rubus chainaemorus, Menyanthes, Arctophila fulva и др.
Два последних вида составляют хороший корм для оле¬
ней. Этот тип свойственен лесотундре и лесной зоне, в тундре
он отсутствует. Основные особенности болотообразования
в тундре Дальневосточного края следующие:
1. Громадное развитие на равнине, а также
в широких долинах предгорий заболоченных
кочкарников, в частности с Eriophorum vagi¬
natum. Пушицевые кочкарники — наиболее характерный тип
растительности для северо-востока Сибири. Уже в Якутии
роль их уменьшается.
2. Замедленный темп торфообразования,
отсутствие в общем торфяников и сравни¬
тельно слабое распространение болот, в том
числе осоковых. Они уступают свое место
кочкарникам. Причину этого надо искать не
только в рельефе, но, вероятно, и в климате.
Болота Среднесибирского плоскогорья.
Среднесибирское плоскогорье — территория к востоку от
Енисея (вплоть до Алдана и его притока Маи), орошаемая
Леной и ее притоками (Л. Берг, 1931). К северу это плоскогорье
1 По В. Сочава — до 0,5 м.
21—722
321
переходит в более пониженную область, тяготеющую
к Таймыру. Равнинные части этой территории относятся
к лесной зоне. Для южной части Якутии можно выделить
такие типы болот1:
1. Заросли прибрежно-водных растений (Sco-
lochloa festucacea, Phragmites communis, Scirpus Tabernaemontani)
no берегам водораздельных озер (аласов), в долинах рек и пр.
(в долине Лены — Scirpus lacustris, Acorus calamus, Glyceria
aquatica и др.).
2. Осоковые луга часто на торфянисто-глеевых почвах
с признаками засоления, кругом аласов в менее топких местах,
чем фитоценозы первой группы, а также всюду в долинах
рек, логов и всякого рода понижениях. Здесь доминируют
Carex aristata, С. intermedia, С. aquatilis, С. wiluica, С. caespi-
tosa. Эти луга используются как сенокосы. Р. Аболин относит
их к болотистым кочкарникам.
3. Вейниковые луга (Calamagrostis Langsdorfii). Они
встречаются в приаласных котловинах, но там, где поверх
соленосных суглинков залегают органические озерные илы,
а также в разнообразных понижениях рельефа — в логах,
котловинах, долинах рек и пр. Р. Аболин1 2 относит вейниковые
луга к болотистым кочкарникам и считает их самыми распро¬
страненными в этой группе на Лено-Вилюйском плато.
4. Сфагновые торфяники, Обычно небольшие (иногда
с Sph. fuscum, Sph. lenense), на склонах, питающихся почвен-
но-груивовыми водами, а иногда кругом озер и в долинах
речек (в этом случае с Cassandra, Calamagrostis, Sph. obtusum).
5. О со к о в о - г и пно в ые болота, частью ключевые,
встречаются у выходов грунтовых вод, у подножия склонов
в заболоченных понижениях и по берегам озер. Болота эти с
Carex rostrata, С. limosa, Drepanocladus aduncus, D. lycopodioides.
В песчаных районах низовьев Вилюя такие типы эвтрофных
болот в безозерных заболоченных понижениях (Т. Работнов):
a) болотца с кочками, покрытыми Кассандрой и ковром
Sph. subsecundum и Carex chordorrhiza в межкочиях;
b) где менее влажно, к господствующей Кассандре при¬
соединяются Calamagrostis Langsdorfii, Carex wiluica, ковер мха
Drepanocladus uncinatus;
c) болота более богатого грунтового питания с преоблада¬
нием осок (С. Meyeriana; как примесь — С. wiluica) с несплош¬
ным ковром мхов (Drepanocladus intermedius и D. vernicosus)
с примесью к ним Meesea triquetra и Sph. subsecundum,
при большем увлажнении преобладает Carex chordorrhiza, С.
limosa, С. rostrata с теми же мхами. Иногда здесь среди мс-
чежин—повышения с зарослями Кассандры;
d) сфагновые болота грунтового питания с грядово-моче-
жинным микрорельефом; на повышениях преобладают лишай¬
1 А. Каяндер (1902), Р. Аболин (1929), Т. Работнов (1935).
2 Р. Аболин отмечает в некоторых случаях под вейниковыми кочкар¬
никами травяной торф грубой структуры, мощностью до 0,5 м.
322
ники и багульник; в мочежинах — ассоциации с Carex lasiocarpa
или с Eriophorum Chamissonis и с Sph. platyphynura, или
с Carex limosa и С. rostrata и с другим сфагном (Sph. Dusenii).
В центре этих болот интересная ассоциация Carex limosa —
Cratoneuron glaucum. Торфяной слой этих болот весьма мел¬
кий— не более 0,7 м.
Болота долины р. Алдана по среднему течению описал В. Кор-
жевин (1934). Типы болот напоминают только что описанные:
a) моховые болота с Carex wiluica и Camptothecium nitens;
b) осоковые кочкарники чаще тоже с Carex wiluica и осо-
ко-вейниковые болота; заливаются полыми водами;
c) болотистые ивняки;
d) облесенные кочкарники (Carex Shmidtii) на нeзaливáe•
мых террасах; мощность торфа—0,15—0,25 м\
e) сфагновые торфяники с Sph. fuscum, Sph. acutifolium
с пятнами лишайников и с кустарничками. Торф обычно
всего 0,4 м (иногда до 0,85 м).
6. Заболоченные леса. Сюда относятся некоторые
типы сырой тайги, в частности, леса из даурской лиственницы
(Laricetum muscosum) с ковром разнообразных мхов и Laricetum
sphagnosum. Последний тип по растительности напоминает
олиготрофное сфагновое болото. Лиственницы здесь высотой
5—7 м и старше 300 лет.
7. Солончаки встречаются и в долинах, и на водо¬
разделах. Наряду с железисто-карбонатными, имеются и на¬
стоящие галоидно-сульфатные (или черные) солончаки с солян¬
ками (Salicornia, Suaeda и др.).
Характерные особенности болотообразования в Якутии
следующие:
1) довольно значительная заболоченность
при почти полном отсутствии торфяников;
2) преобладание среди болот осоковых и,
особенно, вейниковых кочкарников; эти кочкар¬
ники отличаются по своей растительности от
кочкарников тундр Якутии;
3) наличность засоленных болот.
Болота лесной области Дальневосточного края. Болота
Амурской области.
Амурская область — в основном холмисто-горная страна и
лежит большей частью в области вечной мерзлоты. Оба фак¬
тора накладывают печать на болота этой области. Весьма
характерны олиготрофные сфагновые более или менее плоские
болота с редким ярусом даурской лиственницы и обычно с
кустарничками. Залежь неглубокая с близкой вечной мерзлотой
(этот приморско-нижнеамурский тип болот описан выше).
Кроме того, можно выделить такие типы болот:
1. Вейниковые заливаемые болота речных долин
с Calamagrostis Langsdorfii, иногда с маломощным злаковым
21* 323
торфом. В южном Биробиджане они используются под рисо¬
сеяние.
2. Осоковые эвтрофные болота (с Carex caespitosa, С.
Meyeriana, иногда с С. filiformis).
Болота с С. Meyeriana иногда с хорошо развитым ковром
(Sphagnum subsecundum) или с пятнами мхов. Эти болота
довольно обычные в прилегающей к Амуру южной части
Биробиджана (Н. Кац), и встречаются также и в других рай¬
онах. Мощность торфа здесь наибольшая по сравнению
с другими типами осоковых болот. Если мощность торфа
этих болот приемлема для эксплоатации, то высокая зольность
(19—52°/о на абсолютно сухое вещество) часто препятствует
использованию торфа на топливо. Это возможно лишь для
менее зольных торфяников. Осоковые болота других типов
имеют совсем мелкий и сильно заиленный торф и на топливо
непригодны.
3. О соково-вейниковые и осоково-разно¬
травные болота.
В первом типе доминируют те же осоки, что и на осо¬
ковых болотах, и Calamagrostis Langsdorfii. Залежь мелкая
(0,3—0,4 м); сильно минерализованная, обводненная; золь¬
ность от 40% и выше. Болота эти обычно заливаются по¬
лыми водами. Болота второго типа с теми же осоками и
разнотравьем. Они располагаются в понижениях, между релок
(холмы), в центре обводнены и имеют характер зыбунов.
4. Пушицевые болота (с Eriophorum vaginatum). Они
отмечены О. Кузеневой (1911) и Доктуровским (1911) на севере
области и встречаются также и в Хабаровском районе. Глубина
торфа, повидимому, всюду незначительна (О. Кузенева дает
глубину иловато-болотной или торфяно-болотной почвы всего
в 0,25—0,65 м). Часто к пушице примешаны Cassandra, Ledum
и другие цветковые олиготрофных болот вместе с сфагновыми
мхами, иногда образующими сплошной покров. Эти пуши¬
цевые болота не имеют ничего общего с пушицево-сфагно-
выми торфяниками Европейской части СССР. Эти заболочен¬
ные земли напоминают скорее пушицевые кочкарные тундры.
Заслуживают упоминания еще различные типы заболоченных
земель (мари) то с лиственницей, то с кустарничками. Кроме
этих типов, для Амурской области описаны еще и другие,
приуроченные к наиболее суровому климату — к горным райо¬
нам. Сюда относятся:
5. Наледные болота горных долин с мощным слоем не¬
глубоко лежащего льда1.
6. Сфагновые горные болота с россыпями горных пород1 2.
7. Бугристые мари с мелким торфом и буграми минераль¬
ной почвы, возникшими поД действием мерзлоты. Этот тип
широко распространен и на севере Амурской области.
1 Растительность наледных болот Якутии описал Т. Работной (1937).
2 Горные болота Буреинского хребта подробно описал В. Сочава (1934).
324
В общем Амурская область с широким раз¬
витием заболоченных пространств и слабым
развитием торфяников напоминает соседнее
Среднесибирское плато и тундровую зону
Дальневосточного края. Главное отличие
Амурской области — широкое развитие на водо¬
разделах и пологих склонах заболоченных
сфагнумом лиственничных лесов, переходящих
в плоские, неглубокие, мерзлые торфяники.
Приморская область Дальневосточного края.
При переходе от Амурской области к Приморью климат
делается более морским и мягким. Изменяются и типы болот.
Условия для торфообразования и для образования сфагнового
торфа делаются при приближении к морю более благоприят¬
ными, увеличивается мощность торфа, торфяники делаются
ЕЭ ирагков. (ZZZ3 осоковый 1чо>\ пуш.-трав-осомов.
EZ3 сщ-осот. ЗШ трое- осоте. Ww/л i вой./ ■ трав. ■ осоков.
масштаб * горизонтальный —< ЮОм.
nULUimUUt ВРПтииП Лкикп) г , ]
Рис. 107. Строение одного болота в Биробиджане. По Ц. Минкиной.
выпуклыми. Можно наметить ряд переходных типов от плоских
мерзлых торфяников восточной части Амурской области
к типичным выпуклым болотам Приморья.
На границе Амурской области и Приморья (юг Биробид¬
жана, Хабаровский район) на плоских водоразделах, иногда
на высоких террасах обычны торфяники, нередко весьма об¬
ширные, со смешанной растительностью и довольно мощной
залежью. Последний признак и отсутствие вечной мерзлоты
отличают эти торфяники от мерзлых плоских и мелких лист¬
венничных сфагновых торфяников центра Амурской области.
Растительность этих торфяников — комплекс олиготрофных
кочек или гряд и эвтрофных мочежин.
На повышениях микрорельефа сфагновый покров из Sph.
medium, иногда с примесью Sph. fuscum, ярус вересковых
кустарничков (доминирует обычно Кассандра) и даурская
лиственница, единично или редкими группами. Иногда олиго-
трофные кочки смыкаются в сплошные, значительные по
площади участки. Мочежины, довольно обводненные, эвтроф-
ного или несколько мезотрофного характера. Здесь доминируют
Сагех Meyeriana, Rhynchospora alba. Сплошной ковер образуют
эвтрофные сфагновые мхи — Sph. subsecundum, Sph. obtusum,
иногда с примесью гипновых (Drepanocladus vernicosus). Залежь
этих болот из эвтрофных торфов: осоковых, также травяно-
325
осоковых, иногда с значительным
а участием хвоща (рис. 107). Сфаг-
| новые и осоково-сфагновые торфа
I встречаются островками, слагая
с олиготрофные кочки или покрывая
“ участки с олиготрофной расти-
« тельностью. Мощность их весьма
§ невелика, редко более 0,5 м. Не-
| смотря на довольно высокую золь-
| ность (13,0—19,5% на абсолютно
_ ° сухое вещество), торф этих болот
| имеет приемлемую теплотворную
s | способность и представляет удо-
| * влетворительное топливо. Топлив-
= ные качества повышаются большой
^ | плотностью и сухостью торфа. Пол-
0 §«• ное отсутствие пней облегчает до-
с || бычу.
* 21 Наряду с подобными торфяника-
§ 31 ми начинают попадаться торфяники
о «о преимущественно с олиготрофной
® *| растительностью, с господством
§ Ц сфагновых мхов и кустарничков
о.«У (главным образом Sph. medium
s I® и Cassandra calyculata) и с ред-
Jj* |f ким ярусом лиственницы. Иногда
о« в центре встречаются мочежины, а
“ ¿о местами растительность делается
s эвтрофной.
| s Залежь этих болот в осно-
5. | s ве эвтрофная, по степени раз-
ложения, зольности и ботаниче-
= «- скому составу близка к вышеопи-
5 §= санному типу. Однако более мощ-
g* ный сфагновый торф (до 1—0,75 м),
и i покрывает в виде шапки непре-
§ f рывным слоем большую часть бо-
" | лота, которое принимает слабо
я | выпуклую форму (рис. 108). Эти
1 торфяники уже ближе к типичным
® болотам с выпуклой поверхностью,
J, чем мерзлые лиственнично-сфагно-
ё вые или болота со смешанной Ра-
O' стительностью.
■= Дальше на восток, в Примор-
§ ской полосе, между хребтом Си-
J_ хотэ-Алинь и берегом Японского
у моря, торфяники приобретают ха¬
рактер уже настоящих выпуклых
болот. Так, небольшой торфяник
у реки Лянчихе сверху с довольно мощным слоем сфагнового
торфа и выпуклой поверхностью К Существование подобных
торфяников возможно лишь в сравнительно мягком и бога¬
том осадками климате Приморья. Вероятно, в дальнейшем
можно будет выделить особую область выпуклых болот по
берегу Японского моря.
Ряд типов болот Приморского края и Амурской области
общие, например, вейниковые, осоковые болота, заболочен¬
ные сфагнумом лиственничные леса и др. Если в Южном
Приморье (Ханкайская низменность) болота лишены вечной
мерзлоты с крайне мелкой залежью, вейниковые или осоко¬
вые 1 2, то на севере (Удский край) сфагновые болота с близ¬
кой мерзлотой. Болота здесь, по А. Гожеву (1934), чаще всего
расположены на 2-й и 3-й террасах. Есть они и на пологих
склонах, реже в поймах. Наиболее частый тип — сфагновое
болото с рослой лиственницей (5р1^пе1ит п^по-1аг1созит).
В мочежинах этих болот растительность эвтрофная (Не1оШит
1апа1иш, Огерапос1абиз ехаппиЫиз). Повидимому, они напоми¬
нают вышеописанные болота смешанного типа. Дальнейшая
стадия развития этих болот — торфяники с карликовой лист¬
венницей, возвышающейся над мхом всего на 0,4—1 м. Кроме
того, в Удском крае встречаются еще лесные болота с оль¬
хой и березой и небольшие площади кочкарников (с Сагех
саеврйоза).
В Приморье после громадного перерыва вновь появляются
олиготрофные выпуклые болота, которые в типичном виде не
встречаются на всем пространстве к востоку от Енисея до
западной части Приморья.
Болота Камчатки3.
Камчатка, кроме крайнего севера и горных цепей, отно¬
сится к лесной зоне. Торфяники известны лишь в южной
части полуострова. Болота эти резко отличны от
болот тундровой материковой части Дальне¬
восточного края и болот центральной Сибири.
Южная Камчатка—это область выпуклых оли-
готрофных болот. В этом отношении она сходна
с восточным Приморьем, где, как мы видели, также
встречаются выпуклые болота. Однако на Камчатке они резко
отличны от приморских как по растительности, так и по
строению и представляют совершенно своеобраз-
1 В. Савич говорит об ясно выпуклых сфагновых торфяниках, поросших
лишь редкой лиственницей в бухте Гроссевича.
2 М. Болотова (1930).
3 В. Тюшев (1906), В. Комаров (1919), М. Нейштадт (1936), М. Нейштадт
я М. Короткина (1936), Любимова (1937).
327
ный региональный тип торфяников — описанный
выше под названием западнокамчатского. Бо¬
лота Камчатки вообще, а выпуклые в частности,
настолько своеобразны, что Камчатку следует
выделить в особую болотную провинцию. Нали¬
чие здесь, как и в Приморье, выпуклых болот объясняется
морским климатом Камчатки с более мягкими и снежными
зимами по сравнению с сибирским континентом. Не надо
забывать, что вечная мерзлота, повидимому, отсутствует здесь,
как и в Приморье. Однако более суровый летний климат
Камчатки накладывает отпечаток на торфяники, придавая им
северный характер и вызывая резко выраженную деградацию
сфагнового покрова. Далее, своеобразие камчатских торфяни¬
ков объясняется особенностями их флоры (например, присут¬
ствием Myrica tomentosa). Наконец, ключи к разгадке особен¬
ностей строения торфяной толщи кроются в истории развития
полуострова, а для западной Камчатки, кроме того, в влиянии
вулканических извержений. Особенности распределения забо¬
лоченности и торфяников .Камчатки следующие:
1. Заболоченность Камчатки весьма велика, хотя и неравно¬
мерна. Особенно богато болотами западное побережье Кам¬
чатки. Менее заболочены районы с возвышенным рельефом
(район Петропавловска). Однако и здесь болот достаточно.
2. Заторфованность Камчатки также весьма велика. Торфя¬
ники занимают, вероятно, большую часть заболоченных про¬
странств и весьма глубоки. Широкое распространение и
мощность торфяников резко отличают Камчатку от смежных
частей Азиатского континента, где площадь торфяников и
глубина их незначительны.
Ввиду того что торфяники западного и восточного побе¬
режья Камчатки сильно отличны друг от друга, рассмотрим
их отдельно. Растительность олиготрофных торфяников за¬
паднокамчатского типа была описана выше, так же как и
некоторые черты их строения. К сказанному надо добавить
постоянное присутствие в более старых торфяниках мощного
слоя глины. Этот слой, может быть, отложился в ре¬
зультате местного оледенения Камчатки (М. Нейштадт).
Торфа западной Камчатки имеют несколько повышенную по
сравнению с Европейской частью СССР зольность. Это объяс¬
няется, вероятно, некоторой заиленностью торфов, благодаря
работе рек и моря (особенно на торфяниках морского побе¬
режья) и, кроме того, заносом вулканического песка и пепла
из восточной и центральной Камчатки. Тем не менее, несмотря
на повышенную зольность, теплотворная способность торфа
весьма хороша. Таким образом, западное побережье Камчатки
обладает большими запасами топливного торфа. Мало разло¬
жившиеся сфагновые торфа, довольно обычные здесь, можно
использовать и на подстилку.
Возникновение сплошных торфяников на западном побе¬
режье рисуется в следующем виде (М. Нейштадт): приморская
328
низменность в начале постплиоцена была выше и затем раз¬
мывалась осадками и блуждающими реками, пока не превра¬
тилась в современную низменность с останцами в виде при¬
морских увалов и гряд. Очагами болотообразования послужили
старые русла рек. Блуждая по равнине, эти реки иногда за¬
носили торфяники илом и прерывали их развитие, пока не
отложились значительные толщи торфа. Разрастаясь, торфяники
захватили водоразделы, окружили увалы и заболачивали их.
Другие авторы, впрочем, считают, что западнокамчатская низ¬
менность была раньше дном моря, доходившего до середин¬
ного камчатского хребта и потом отступившего. Интересно
размывание торфа водами Охотского моря и речками в области
их устьев (М. Нейштадт). Причина этого, повидимому,—насту-
пание моря на сушу в современный период. Море, наступая,
заставляет реки менять русла и размывать прибрежные тор¬
фяники. Размытый торф в дальнейшем засыпается морской
галькой.
Торфяники восточного побережья Камчатки исследованы
в районе Петропавловска (М. Нейштадт и М. Короткина, 1936).
По отношению к рельефу различаются:
1) торфяники морских прибрежий, наиболее крупные по
площади;
2) торфяники пологих склонов, занимающие меньшие пло¬
щади;
3) горные торфяники, сравнительно небольшие (100—500 га).
Торфяники морских прибрежий имеют в общем ровное,
плоское дно и слегка выпуклы. Они часто весьма мокрые,
с открытой водой на поверхности. Здесь часты иногда встре¬
чающиеся группами озера. Лишь редко олиготрофная расти¬
тельность преобладает. Эти чисто олиготрофные болота
с сплошным сфагновым ковром (господствует Sph. fuscum) и
по видовому составу напоминают европейские. Новыми видами
будут Myrica tomentosa, Betula exilis, Carex Middendorfii и дру¬
гие. Чаще приморские болота мезотрофные, иногда с остров¬
ками олиготрофной растительности. На мезотрофных болотах
громадным распространением пользуется Myrica tomentosa
(восковник), характернейшее растение торфяников Камчатки,
особенно восточной.
Myrica образует довольно много ассоциаций с разными ви¬
дами осок (Carex Middendorfii, С. filiformis и др.) то с сфаг¬
новым покровом (Sph. médium, Sph. fimbriatum, Sph. Warnstor-
fii), то без него.
Часто на мезотрофных болотах большие пространства за¬
няты эвтрофными ассоциациями. Это смешанные эвтрофно-
мезотрофные болота.
Из эвтрофных видов доминирует чаще Equisetum limosum,
образующий иногда обширные заросли в воде или делящий
господство с осоками (Carex Lyngbiei, С. filiformis, С. vesicaria).
Приморские торфяники восточной Камчатки сложены
в основном эвтрофными и мезотрофными торфами, что соот¬
329
ветствует их современной растительности (рис. 109). Наиболее
распространены осоковые торфа, за ними хвощево-осоковые.
Обычны также осоково-сфагновые. Чисто сфагновые торфа
редки. В общем, по строению залежи торфяники восточного
побережья эвтрофные, реже мезотрофные. Залежь олиготроф-
ного типа встречается небольшими участками. Отличительная
•особенность восточнокамчатских торфяников — прослойка вул¬
канического пепла и высокая зольность, зависящая от при¬
меси вулканического пепла. Эта зольность часто делает торфа
непригодными для топлива. Можно предполагать, что торфя¬
ники прибрежной полосы образовались на местах, где раньше
было море. Следовательно, в противоположность западному
побережью здесь скорее можно предполагать поднятие суши.
Второй тип—торфяники пологих склонов с наклонным
дном и поверхностью.
Третий тип—горные торфяники с резко выраженным па¬
дением дна. За его рельефом следует и рельеф поверхности.
Болота Кавказа.
Торфяники Кавказа1 бывают равнинные и горные.
Равнинные расположены на приморской низменности и тя¬
нутся с перерывами почти от Сухуми до Батуми. Лучше
исследованы Потийские болота в Рионской низменности.
Около рек, прорезающих низменность,—болотистые ольшан-
ники, а между ними обширные безлесные кочковатые осо¬
ково-тростниковые болота. Среди последних разбросаны
•сфагновые торфяники, площадью иногда до 12 км1 2. Сфагно¬
вый торф, всего 0,5 м мощностью, подстилается осоковым.
Такие торфяники встречаются и севернее. Иного типа при¬
морские болота Аджаристана 2 (рис. ПО). Это—настоящие
сфагновые выпуклые торфяники. Сплошной сфагновый покров
образован главным образом Sph. cymbifolium, Sph. acutifolium®.
В начале лета на сфагновом ковре доминируют осоки
(главным образом Carex lasiocarpa), а во вторую половину
фон образует Molinia litoralis Host, и появляются заросли
Rhynchospora alba, Rh. caucásica, Bulbostylis tenerrima Palla.
Интересно присутствие на болотах, наряду с Osmunda
regalis и третичными кустарниками Кавказа,— Rhododendron
ponticura, Vaccinium Arctostaphylos, также и северных ви¬
дов—угнетенной Rhamnus frángula, Drosera rotundifolia, Me-
nyanthes trifollata. Один из сфагновых торфяников (к се¬
веру от Кобулет) имел ясно выпуклую поверхность (мак¬
симальное превышение середины над краем 3,0 л«) и сверху
1 См. О. Зедельмейер (1927).
2 Растительность этих торфяников описана О. Зедельмейер (1927),
а строение В. Доктуровским (1931).
* Из остальных интересны западные атлантические виды Sph. imbricatum
и Sph. crassicladum.
330
р.Тихая
Масштаб
горизонт, омгтоот*
ч'о^гЪ*.
Ё2/ ^Шз и» 15 \ппп\? ЕЕЗ$
ШЁ&2 1°*°\Н I*18 ф Ю
Рис. 109. Строение торфяников восточного побережья Камчатки. По М. Нейштадту и М. Короткипой, 1936.
I — осоковый торф; 2 — гипново-осоковый торф; 3 — сфагново-осоковый торф; 4 — хвощево-осоновый торф; б — травяные остатки; 6 — вахта;
7 — пушица; 8 — древесные остатки; 9 — прослойки вулканического песка; 10 — ольха.
Масштаб
213 026 638м горизонт.
О 21 02 60 л вертин.
Уровень__
р. Черная
Уровень
моря
ЕЭ/ ЕВ? ШЖ'З \ЕШ'4 Г~*~15 Щр Р^\7
уровень
р_Ачиуа_
Уровень
моря
Рис. 110. Профиль строения одного торфяника в Закавказье. По В. Доктуровскому, 1935.
1 — торф сфагновый; 2 — с травяными остатками; 3 — осоковый; 4 — с тростником; б — древесина ольхи; 6 — песок с морской галькой;
7 — глина.
мощный слой мало разложившегося сфагнового торфа.
Под ним осоковые торфа с примесью тростника, травяных и
древесных остатков, иногда внизу осоково-травяно-древесные,,
а еще ниже тростниковые торфа. Эти сфагновые торфяники
близки к настоящим северным Hochmoor. Их на первый взгляд
парадоксальное нахождение в условиях жаркого климата
Черноморского побережья объясняется лишь большим коли¬
чеством осадков, компенсирующих неблагоприятное влияние
высокой температуры.
Отличительные особенности болот Кавказа следующие:
1) южный (по флоре) характер равнинных эвтрофных бо¬
лот с их богатой флорой, с реликтовыми видами и эндемами;
2) наличие на Черноморском побережье выпуклых сфагно¬
вых болот, по строению и рельефу напоминающих северные,
но со своеобразной растительностью;
3) бедность горных болот бореальными и аркто-альпийскими
видами;
4) слабое распространение горных болот на. Кавказе и
отсутствие здесь (в отличие от Альп) выпуклых болот.
ГЛАВА X.
ЧЕТВЕРТИЧНАЯ ИСТОРИЯ РАСТИТЕЛЬНОСТИ ПО ДАННЫМ
ИЗУЧЕНИЯ ТОРФЯНИКОВ.
Пыльцевой метод исследования торфяников.
Метод пыльцевого анализа в настоящее время основной
метод при изучении истории послеледниковой растительности,
почему мы и рассмотрим вначале его сущность, сферу при¬
менения, методологические обоснования и необходимые кор¬
рективы, которые надо иметь в виду при его применении.
Основателем пыльцевого метода следует считать шведа Н. Ла-
герхейма (Nils Lagerheim) (см. Г. Гаме, 1926—1927), который
первый систематически стал его применять, хотя первый
подсчет ископаемой пыльцы в торфе в 90-х годах произвел
К. Вебер. От Лагерхейма воспринял этот метод Л. Пост, после
чего он получил широкое применение сначала в Швеции, а
затем и в других странах.
Техника пыльцевого анализа заключается в следующем *. Пробу торфа,
если он сырой, тщательно перемешивают, берут 1—2 смь торфа и кипятят в
10—15% растворе едкого кали или натра 1 2 1—2 минуты. Если торф сухой
и не раздавливается пальцами, то его наскребают ножом с разных сторон
куска для большей однородности пробы. После кипячения рекомендуют до¬
бавлять воду, чтобы кашица приняла более однородную консистенцию и при
покачивании не оседала на стенках сосуда. Перед взятием пробы для ана¬
лиза жидкость тщательно взбалтывают. Затем капля жидкости наносится
пипеткой (при густой консистенции — стеклянной палочкой) на предметное
стекло, причем полезно удалить оттуда крупные частицы торфа. Подсчет
пыльцы производится под микроскопом с увеличением около 200 раз. При
большом обилии пыльцы употребляют более сильное увеличение (около 250
раз). В этом случае поле зрения меньше, что облегчает подсчет. Лучше вы¬
бирать такую комбинацию объектива и окуляра, чтобы большое увеличе¬
ние сочеталось с большим полем зрения (у Лейтца и Рейхерта это будет
объектив 7 и окуляр 1 или объектив 7 и окуляр 0).
Кипячение в едкой щелочи удаляет из пыльцы гумус и делает отчетли¬
вым строение экзины3. Пыльцу удобнее считать при подвижном столике.
1 Описание этой техники можно найти у русских авторов в статьях
В. Доктуровского (1923), М. Нейштадта (1933) и др.
2 Можно употреблять также щавелевую и азотную кислоту.
3 К. Фегри (К. Faegгi, 1936) применял окраску пыльцы в торфе при по¬
мощи смеси фуксина и метиленовой синьки. Пыльца и основная масса торфа
окрашиваются при этом различно. Этот способ применяется для бедных
пыльцой препаратов и при плохой сохранности пыльцы в торфе.
333
По Л. Посту, достаточно подсчитать для получения надежного среднего IOC
зерен. Г. Эрдтман и К. Рудольф считают, что для получения среднего не¬
обходимо подсчитать не менее 150 пылинок. У нас чаше подсчитывают 100.
В бедных пыльцой торфах для сбережения времени обычно ограничиваются
этой цифрой.
При анализе подсчитывается пыльца древесных пород и
вычисляется процент каждой породы к общему количеству
пылинок. Пыльца орешника не входит в эту сумму. Она
подсчитывается отдельно, а процент ее вычисляется от суммы
пылинок всех древесных пород. Дуб, липу, вяз, а также ясень
и клен, если они встречаются, объединяют обычно вместе под
названием «смешанного дубового леса», а часто, кроме того,
подсчитывают и порознь.
При обычном анализе подсчитывают не виды, а древесные
породы, или, лучше сказать, роды: ольха, береза и т. д. Если
в районе распространены не один, а два или более видов
какого-либо рода, то суммарный подсчет дает схематизиро¬
ванную картину. Подсчитывая, например, суммарно пыльцу
Betula папа и резко отличающихся от нее хотя бы по сво¬
ему распространению В. pubescens и В. verrucosa, мы сильно
уменьшаем ценность выводов. Поэтому теперь все чаще пы-
таются. определить пыльцу до вида (главным образом для
Betula и Pinus). Делается это преимущественно путем изме¬
рения пыльцевых зерен с последующей вариационно-статисти¬
ческой обработкой. Обнаружилось, что разные виды одного
и того же рода дают различные вариационные кривые, что
делает до известной степени возможным их различение. Так,
по П. Штарку (Р. Stark, 1927), средний размер пыльцы Pinus
montana больше, чем Р. silvestres, вариационные кривые обоих
видов различны, что позволило ему различить пыльцу обоих
видов в торфе. Е. Шуберт (1933) и другие исследователи раз¬
личали в торфе Betula nana, В. verrucosa, В. pubescens. Впро¬
чем, Ф. Фирбас (1935) установил уменьшение размеров пыльцы
Betula и Pinus от нижних слоев торфяников к верхним и при¬
шел к выводу, что на размеры ископаемой пыльцы влияет
среда, их заключающая, и что поэтому определение видов
сосны и березы вариационно-статистическим методом невоз¬
можно. Однако для других родов, в частности Picea, для ви¬
дов Picea Omorica, P. omoricoides и P. excelsa, оказалось воз¬
можным применить этот метод. Определение до вида облег¬
чается при наличии морфологических различий Ч В настоящее
время все больше входит в употребление добавочный подсчет
недревесной пыльцы. Эта пыльца подсчитывается по отдель¬
ным систематическим группам: Gramineae, Cyperaceae, Ericales,
Chenopodiaceae, Caryophyllaceae; иногда еще подсчитывают
споры папоротников, например, Ф. Овербек и Г. Шмиц (1931).
Пыльца неизвестных растений, не вошедших ни в одну из
этих групп, относится к группе Varia (разные). В случае оби- 11 См. также П. Чернявский (P. Ceinjavski, 1935).
334
—А —д-
Aù/es (пихта)
—
—X— —О— В et и La (береза!
—V-
—▼— —д-
Carpinus (граб)
—•Я—* XX XX —
-*-х-
—▲—
со-со-со
лия подсчитывается также пыльца СотроэНае и водных расте¬
ний *; иногда, кроме того, споры сфагнума и т. д.2.
Подсчет пыльцы недревесных пород производится разными
способами.
1. Берется про- —□— (ольха)
центное отношение
суммы пыльцы не¬
древесных пород к
сумме пыльцы дре¬
весных пород (т. е.,
как для орешника)8.
2. Берется про¬
цент недревесной
пыльцы по система¬
тическим группам
к сумме пыльцы
древесных пород
(обычный способ).
3. Берется отно¬
шение суммы недре¬
весной пыльцы по
группам к общей
сумме этой недре¬
весной пыльцы (К.
Фегри, 1935).
Каждый из спо¬
собов имеет свои
преимущества. Во¬
обще подсчет не¬
древесной пыльцы
важен для установ¬
ления степени об-
лесенности страны.
Для этой цели луч¬
ше употреблять пер¬
вый способ, дающий
суждение об отно¬
шении количества
древесной пыльцы
к количеству всей
недревесной. Вто¬
рой способ дает
менее наглядные результаты,
третьему способу' позволяет
-ф— —П— i- i-и •- •- CoryLus (орешник)
—— Lonx (лиственница>
— •— Pinas (соснау
—д— Рюеа_ (ель)
—»—•— Quercus (дуб)
В— —□— —Т— —В— Смешанный
т “ дубовый лес
—ф— Sa ¿ix (она)
— Tilia (липа)
Ulmus ¡ваз)
—о— Г NBP( сумма не древесной пыльцы - Nichtbaum-
pollen'-
—— Ericaceae (вересковые)
—О— Сурегасеае (осоковые)
—(8)— Gramineae (злаки)
—о— Водян-растения (WPP- У as serßfLa теп pa lien)
Рис. 111. Таблица сигнатур, употребляемых при
вычерчивании пыльцевых диаграмм. По Г. Гамсу,
1929.
Подсчет пыльцы в торфе по
иногда восстановить историю
растительности для безлесных местностей (К. Фегри, 1935).
1 См., например, К. Фегри (1935).
2 Для внеевропейских стран подсчитываются иногда многие отсутствую¬
щие у нас древесные породы и недревесные виды (см., например, В. Ауэр,
1933).
3 Подсчеты у Овербек и Шмиц (1. с;), диаграммы у К. Фегри (1. с.).
335
Составление пыльцевых диаграмм. Результаты подсчета пыльцы наносят
на диаграммы. Каждая древесная порода имеет свой условный знак. В общее
употребление вошли обозначения Л. Поста (1916). Однако в обозначении
пыльцы смешанного дубового леса, орешника и граба, а также пород, не
встречающихся в Швеции, например пихты, нет единых общепризнанных
знаков (Г. Гаме, 1929). Приводим таблицу сигнатур, употребляемых при вы¬
черчивании пыльцевых диаграмм (рис. 111).
При построении диаграммы на горизонтальной оси откладывается процент
пыльцы (обычно с интервалами через 10%), а на вертикальной оси на каждом
горизонте в соответствующем месте ставится сигнатура данной породы, и со¬
седние по вертикали горизонты соединяются при помощи линий *.
Обоснование пыльцевого метода. Основная цель пыльце¬
вого анализа—установить «пыльцевой спектр», т. е. процент
соотношений пыльцы древесных пород в прошлом, и рекон¬
струировать картину лесов соответствующего периода. Послед¬
нее возможно лишь при допущении, что количественные соот¬
ношения пыльцы в горизонте торфа отражают количественные
отношения древесных пород в период отложения этого гори¬
зонта. Следующие соображения можно привести против воз¬
можности переносить соотношения данных пыльцы и древесных
пород:
1. «Местная» пыльца с деревьев, растущих на самом болоте
и в ближайших окрестностях, затушовывает влияние пыльцы,
приносимой издалека. Если это так, то пыльцевой анализ не
дает общей картины состава лесов, и значение его не велико.
2. Различные породы производят разное количество пыльцы,
например, хвойные больше лиственных. Поэтому породы с вы¬
сокой пыльцевой продукцией преувеличенно представлены
в пыльцевом спектре * 2. Например, сосна, ель, бук, дуб пе¬
речислены в порядке убывания преувеличенности (Е. Васмунд—
Е. Wasmund, 1931).
3. Пыльца разных пород переносится на неодинаковое
расстояние и поэтому не в одинаковой мере попадает в торф.
4. Пыльца разных видов неодинаково сохраняется в торфе.
Второе соображение — неодинаковая продукция пыльцы —
очевидно само собой. О дальности переноса древесной пыльцы
свидетельствует ее нахождение, например, на севере Гусиной
Земли и на Маточкином Шаре (Новая Земля). На основании
этого дальность переноса пыльцы можно оценить в 700—900 км
(Г. Гессельман—Н. Hesselman, 1919). На такое расстояние
заносится наиболее легкая пыльца сосны.
Для нас в этом случае интереснее вопрос о неодинаковой транспорта¬
бельности пыльцы. И. Дьяковская (I. Dyakowska, 1936) экспериментально
определила скорость падения пыльцы разных древесных пород и вычислила
затем среднее расстояние, на которое распространяется пыльца. Приведем
эго расстояние:
Abies pectinata • 0,7 км
Picea excelsa 21,6 „
Plnus silvestris 74,7 „
Ainus glutinosa . . • 132,3 n
Salix caprea . . • 213,2 *
xO различных способах составления диаграммы см. Г. Гаме (1929).
2 См. Ф. Фиргаппер (F. Vierhapper, 1926).
-336
Из приведенных цифр видно, насколько различил транспортабельность
пыльцы у разных пород.
На неодинаковую стойкость пыльцы в торфе указывают
Г. Пауль и 3. Руофф (1927). В эвтрофных и мезотрофных
торфах иногда пыльца очень плохо сохраняется, особенно
пыльца лиственных пород. Последняя может разрушаться до
неузнаваемости (особенно в гипновом торфе), так что подсчеты
дают совершенно неверный результат.
С точки зрения оценки высказанных соображений гораздо большее зна-
чение, чем теоретические рассуждения, имеет практическая проверка пыльце¬
вого метода, предпринятая некоторыми исследователями (Г. Эрдтман, 1921;
К. Рудольф, 1932).
Она производилась путем сравнения современного пыльцевого спектра
(в поверхностном слое торфа) с составом окружающих лесов. К особенно
определенным выводам пришел К. Рудольф (табл. 51).
Таблица 51
Состав лесов Германии по отдельным породам и пыльцевой спектр
(по К. Рудольфу, 1932)
Смешан¬
Сосна
Ель
Пихта
Бук
Граб
ный дубо¬
Береза
Ольха
вый лес
1.
Буковое время.
Средний процент
пыльцы
19
9
8
26
4
8
10
15
2.
Современный пе¬
риод. Средний
процент пыльцы .
Участие отдель¬
46
17
3
9
1
6
9
9
3.
ных пород в со¬
ставе лесов по
данным 1900 г. .
45
20
3
14
—
7
11
11
Таблица показывает хорошее совпадение данных анализа современного
пыльцевого спектра и современной картины лесов, полученной лесоводствен-
ными методами, и дает, таким образом, положительный результат проверки
пыльцевого метода1.
При анализе пыльцы в водных отложениях (гиттиях) присоединяются
еще новые факторы по сравнению с торфами, а именно: дрифт (действие
течения); различная способность пыльцы разных пород держаться на воде
(разная скорость погружения) и пр.1 2. Тем не менее и анализ пыльцы в седи-
ментах передает картину окружающих лесов, т. е. и здесь пыльцевой метод
оправдывает себя. Это видно из следующего:
1. Поверхностный слой гиттии дает такой же пыльцевой спектр, как
и поверхностный слой торфа с соседних болот (Г. Эрдтман, 1921).
1 На гребне Рудных гор в пыльцевом спектре с поверхности болота
преобладали ель и сосна (рис. 112). Первая господствует в настоящее
время в лесах горного гребня, вторая — на поверхности болот, которые были
исследованы. Затем следует пыльца бука и пихты, образующих леса ниже
по склону. Еще меньше в торфе пыльцы дуба, липы и других пород, расту¬
щих еще ниже [К. Рудольф и Ф. Фирбас, 1924; см. еще те же авторы, 1927,
а также П. Мюллер (Р. Müller, 1937)].
2 См. Е. Ьасмунд (1930).
22 — 722
337
Современный
еловый период
$
Ч)
11
£ «* *5) <5-
с§»с:Л)
1и1
^ ^ Г*
Г*
<5>
«ъ
'ё
1
?■§
1|
Рис. 112. Средняя диаграмма пыльцы вершины Рудных гор
как пример диаграммы средней горной ступени Средней
Европы. По К. Рудольфу и Ф. Фирбасу, 1927.
2. Изучение послеледниковой истории лесов по данным пыльцы в гит-
тиях дает ту же картину, что и анализ торфа, а изменение пыльцевой диа¬
граммы при переходе из гиттии в вышележащий торф происходит плавно,
постепенно (по исследованиям П. Штарка, 1927; по Г. Шмицу, 1930).
Пыльцевой анализ (в торфе и седиментах) дает в общем
хорошее представление о составе лесов в отдельные этапы
их послеледниковой истории и оказывается почти един¬
ственным методом их изучения.
Однако часто от результатов пыльцевого анализа прямо
переходят к составу лесов соответствующего периода, игно¬
рируя различную продукцию пыльцы у разных пород и дру¬
гие факторы, рассмотренные выше. Поэтому заслуживает вни¬
мания попытка вывести для отдельных пород поправочные
или переводные коэфициенты, позволяющие судить по коли*
338
о
JO 20 JO 40 50 60 70 80 SO 100%
л
* ь
□
М
>
<d
»—-—*
~\
Г
“1
2
V
L
г
\
у
!
\
\
~у_
ц
Г
\
П А
гу
J
^1
Ï
Q
г
fw
”|
гг
[1
■/À т
X
/
У
7^
~*v.
г
\
\/
АУХУАххУххАхУХЯЯЯЯ**
iïlîiîSïîïîîîîoîïïïï
ïîïîïï5îïiîïiïïîS«î
Iiiïiîiîtïi2i$ïtïî»
í&í¿íííí£?tííÍííííí
íñííáííistíííiiííí
ДХАОДХАААААААХАДАА*Д
y Д ХА XA A XA ХДД ХДД A Д XA X
ДДХДХДХАДДХ0ДДХДХДХ4
д хд уд хд XA X А X Д X Д ХА ХД X
«iíiííSító'ílMxió«
íiííMíííií
Д ХД ХД ХДД ХД х_
А ™Д >&А хд ХДДХДДХ XÄX
ХД Д хд'хд°д *Д ХА ХХА А ХДД
ХХА хдхдхдх
;S£ôo^ïôï|^ît|xJÏÎ
Д хд хд А хд ХДД ХД хДААДХ
..д^ххдххддхдххдхддххд
ХХАДХХДХ1ХХДХДХХДХXX
Л ХХД X Д*Х Д ХДДХД УДДХДХД
XAXXADA/AXAAOXAXAXAX
дхдх хахахдОхадхахахд
а X хд хд ХДХ хд хд ХХД хдх
ХХХХДХХХХ|ХХХХХХХ/,
_i¿ х Д X XQ X X А ХД хд ХД ХД X
хххх/дхдх хахдО ххд хх,
X д хдххдхдххдххдххдхх
Д ХДХ ХДХ ХД ХА ХА ХД ХД ХДД
ХД ХДХ ХДДХХХДХАОДХХХХ
хдх ХДД ХД ХДДОДХД ХДДХД
ДХДДХДОХДДХД ХДХДХДХХ
д д хд хО Д хд ХДХОА ХД А'ХД д
ХХДХХХДХХДХХОДХХХДХД
Д xxOxaxxxxDxxxxxx XXXI
х хд х X хд хдх хдх хд X Д х :д
□ххххдхОхвххххххххдх
хд
(ДХХДХДХХД ХХХдОДЛХ
• - ГДХХД0Д*ДХ4
к ХДД хдд; -=*
Д ДХ хд Х.Д X хд ХД X ХД|
ххддхд х ДОДД ХДД х_
ДА ХДДХД ХД XIAX/ADAXAK
ХДД XAD ХД ХА ХД ХД ХА ХД ХД
Д ХАДДАД хдадОдхд ХД ХДХ
ХДХД ДАДД ХДД ХА ХХА ХА ХД
ДХДОХХДЧХДХД XAAXAXÜX
ХД ХДДДХД ХДДХД ХДХДОДА
да ДА ХДД ДД XАД УД ХД ХА'ХД
ХДДХД ХДДДОДОАХДДДХДХ
ДА’А Д X Д Д Д ХДД Д • д ДА ХД ДД
ХД хд ХД Д Д Д Д ДД хд Д ОД ADA х
AXAAX46AXA¿U4AXAAAXA|
А ХХА XI
□ х ХД ХД ХДХДХОДХД ХДХСМ
Д X Д ХД ХД X ХД ХД ХД XAX ААХ
ХД ХД ХА ХА А ХАОСАХ ХА ХХД
Д XDA X Д хд ХД ХД ХД XAUM«
ХХДХХДХХХДХОХХАХДДХД
ХОХ XX ХХА XX* ХХХХ XX XXX
ХХЛАХОХХОХ/АХОХХАХХО
ХД ХКХХВ ХА ХХвХХХ_ХД ХХА
XXQXXX ХХ*ХХХОХв;ХХХ
■ XX ХХОХДХХХХХХХХХД*'
:ххД ХХХХ ХХВХАXXXAXXJ
AXXXXBXAXXXXXXBXXXXI
ХВХХВХДХВХХВХХХВХХД
ХХХДХВХОХХВХВХХХХВХл
«ïüi?i5.x5it||Wîi?î
ХВХХХХХХХХХХХО
хххоххвхххахехх, _
ХВХХВХХХВХХХХХВХХХХВ
ХХХХХХХХХХХХХХУХХХХХ
XX ххххвххххвхххххвх
■XXXXххххххХХХХХХХХ
ххвххаххвхххв XXХХХХ
ХХХХХХХХХХХХХХ ха XXX
вххххвхххххвхххххох»
х Abi es • Pi nus д Picea и Ainus о Betula Afagus
■я Смешанный дубовый пес * орешник
Рис. 113. Пыльцевая диаграмма болота Etzelwllermoor (слева) и пред-
полагаемый состав лесов (столбик справа). По П. Мюллеру, 1937.
чественным соотношениям пыльцы об участии отдельных дре¬
весных пород в составе лесов.
Попытку вычислить эти коэфициенты сделал П. Мюллер. Понятно, что
породы с большой продукцией пыльцы (например, сосна), преувеличенно
представленные в пыльцевом спектре, имеют переводный коэфициент меньше
единицы, породы же е малой продукцией, наоборот, больше единицы. Рис. 113
дает представление о составе лесов во вторую половину послеледникового
22* 339
периода, который был установлен путем перечисления из пыльцевой диа¬
граммы с помощью поправочных коэфициентов. Рассмотрим влияние на пыль¬
цевой спектр местных условий, на которые приходится вносить поправку.
Это, между прочим, расширяет сферу применения метода. Нередко породы,
растущие на самом болоте, преувеличенно представлены на диаграммах,
особенно ольха (Г. Пауль и 3. Руофф, 1927; О. Эрнст — О. Ernst).
Не всегда преобладание пыльцы в торфе отражает преобладание этой
породы в лесах. Хороший пример в этом отношении — сосна. Благодаря
легкости ее пыльцы для сосны большое значение имеет занос пыльцы изда¬
лека (так называемый Ferntransport). Поэтому * преобладание пыльцы сосны
может указывать на обезлесение местности, когда решающую роль играет
в спектре легкая пыльца сосны, принесенная издалека. Так, по Ф. Овербеку
(1928) возрастание сосновой пыльцы в верхней »культурной“ части пыльцевой
диаграммы объясняется уничтожением лесов человеком. Влияние господству¬
ющих ветров такжё сказывается на пыльцевом спектре. В Эстонии, на запад¬
ной стороне одного болота в торфе содержание пыльцы широколиственных
пород было выше, чем на восточной. П. Томсон (1933) объясняет это влиянием
господствующих западных ветров. Интересный пример влияния преобладающих
ветров и вместе с тем влияния расположенных у самого болота лесов на
пыльцевой спектр дает Ф. Овербек (1928).
Интересны „локальные“ (местные) пыльцевые спектры. Они иногда
хорошо отражают состав и историю лесов однородных в физико-географи¬
ческом (почвенном, геологическом и пр.) отношении районов. Так, Г. Гесмер
(н. Hesmer, 1933) для Бранденбурга установил, что уже со времени теплого
послеледникового периода дубовые и сосновые леса распределялись соот¬
ветственно почвенным условиям примерно так . же, как и теперь, а именно:
дубовые леса (позднее частично вытесненные буковыми) заняли плодородные
почвы морен, а песчаные районы были заселены сосной. Спектры песча¬
ных районов повсюду богаты сосновой пыльцой, причем господство сосны
идет далеко вглубь послеледниковой эпохи. В Эстонии, в области морской
трансгрессии, где почвы выщелочены, сосна была обильна со времени бореаль-
ного периода, когда ее пыльца в спектре доходила до 90%.
Внутри страны на моренных отложениях ее участие всегда было меньше.
Пыльца же широколиственных пород давала обратное соотношение (П. Том¬
сон, 1926). Таким образом, для Эстонии можно установить два »локальных*
спектра: один, богатый сосной, — для береговых, более бедных в почвенном
отношении, районов и другой, более бедный сосновой пыльцой, — для вну¬
тренней части страны. То же имеет место и в Литве (П. Томсон, 1931).
Значение пыльцевого анализа и сфера его применения.
Значение пыльцевого анализа все больше возрастает по мере
расширения его применения и углубления его методики.
Успехи изучения послеледниковой истерии лесов, а также
распространения отдельных древесных пород в прошлом и
путей их расселения стали возможными только с помощью
пыльцевого анализа \
Изучение смены древесных пород по пыльце—до настоя¬
щего времени один из наиболее надежных способов для сужде¬
ния об изменениях климата после отступания ледника: так,
появление пыльцы теплолюбивых древесных пород указывает
на потепление климата и т. д. Пыльцевые диаграммы позво¬
ляют увязывать хронологически более или менее отдаленные
пункты. Если, например, для какого-либо пункта определенный
горизонт пыльцевой диаграммы точно датирован, то горизонт
со сходным пыльцевым спектром в другом пункте можно
отнести примерно к тому же времени. Пыльцевой метод ока- 11 См. Г. Гесмер (Н. Hesmer, 1927).
340
зывает ценные услуги археологии и антропологии, облегчая
датировку находок в торфе *.
Пыльцевой анализ дает возможность установить положение
границы леса в прошлом, т. е. соотношение между лесом и
безлесными формациями, тундрой или альпийской раститель¬
ностью 1 2 или между лесом и атлантическими пустошами типа
Heide 3>4, наконец, между лесом и степью 5. Пыльцевой ме¬
тод дал возможность подойти вплотную к генезису и после¬
ледниковой истории безлесных растительных формаций. Из¬
учение истории безлесных северных и атлантических типов
растительности при помощи пыльцевого метода возможно
главным образом благодаря подсчету недревесной пыльцы.
Ф. Фирбас (1934) считает количественное отношение недревесной и дре¬
весной пыльцы хорошим мерилом степени облесенности. Так, в лесных мест¬
ностях процент недревесной пыльцы по отношению к древесной колеблется
от 16 до 49, в бедных лесом районах — от 77 до 819, а в безлесных—от
438 до 1093. Этот автор различает в торфе пыльцу некоторых кустарничков
(Calluna, Erica tetralix, Ericales, Empetrum), что позволяет различать „спектр*
пыльцы, например, болотами соснового леса с ярусом кустарничков, а также
различные типы безлесных формаций, например, тундру и атлантические
пустоши. В разных случаях преобладает пыльца различных видов кустар¬
ников. Этим методом удалось подойти к разрешению вопроса о генезисе
дустошей6 побережья Немецкого моря, и притом иначе, чем это делали
раньше на основании изучения лишь современной растительности и лесных
архивов. В монографии по истории пустошей Е. Колумбе (Е. Kolumbe, 1934)
пришел к выводу о бифилитичности Heide. В песчаных районах—это древ¬
ний, естественный тип растительности, сменивший в литориновую эпоху
вследствие изменения климата прежние сосновые леса. Пустоши в районе
древних морен антропогенного происхождения: они сменили здесь в течение
немногих последних столетий уничтоженные человеком леса.
Наиболее широко пыльцевой анализ применяется при исследовании
торфов и сециментов. Однако его применяют при анализе минеральных почв —
лёссов, летучих песков 7, а также трепела, лесного гумуса и т. д. Помимо
послеледниковых органогеннных отложений, пыльцевой метод используется
для исследования межледниковых отложений. Попытки анализа третичных
бурых углей предприняты Г. Геком (Н. Heck, 1927)8 и К. Рудольф (1935).
Весьма интересна попытка пыльцевого анализа глетчерного льда В. Ва
реши (V. Vareschi, 1935). Она, несомненно, имеет значение для гляциологи¬
ческих проблем.
Анализ современного „пыльцевого дождя* применяется еще, чтобы
установить растения-возбудители так называемой „сенной лихорадки*
(„Heufieber“)—воспаления слизистых оболочек вследствие раздражения их
вдыхаемой пыльцой (см. В. Люди и В. Вареши — W. Lüdi, V. Vareschi, 1936).
Послеледниковая история растительности тесно связана
с историей климата и суши. Поэтому прежде чем перейти
1 См. Е. Васмунд (1935), В. Риц (W. Rytz, 1930).
2 В. Ауэр (1927).
8 К. Фегри (1935).
* Также и Ф. Ионас (F. Jonas, 1935) считает, что пустоши северо-западной
Германии — доатлантического возраста,
5 В. Ауэр (1933).
6 Пустоши — безлесные пространства с вереском и др. растительностью.
7 Анализ пыльцы в гумусированных летучих песках (braune Flugsande)
дал сходные результаты с анализом торфа (Ф. Ионас, 1934).
8 По Ф. Фирбасу (1929); там же см. о возможности пыльцевого
анализа в отложениях этого возраста.
341
к обзору истории лесов по данным пыльцевого метода,
необходимо остановиться на изменениях климата и суши
в псслеледниковую эпоху.
История Балтики и климатов в послеледниковое время.
Изменения климата и пограничный горизонт. Пользо¬
вавшаяся до самого последнего времени почти всеобщим
признанием теория послеледниковых изменений климата, так
называемая теория Блитт-Сернандера, была впервые ясно сфор¬
мулирована в 1876 г. норвежцем А. Блиттом (A. Blytt). Окон¬
чательный вид эта теория получила в связи с работами Р. Сер-
нандера. Последний удержал большинство названий климати¬
ческих периодов Блитта. Блитт и Сернандер различают такие
климатические периоды (с хронологической датировкой по
де Гееру (de Geer) и другим) *:
1. Новое время (современный период). Последние 1000 лет
до настоящего времени. «
2. Субатлантическое время (500 лет до н. э., 1000 лет после
н. э.). Климат океанический, но в меньшей мере, чем атланти¬
ческий, на что указывает приставка „суб“.
3. Суббореальное время (3000—500 лет до н.э.) Климат конти¬
нентальный, но в меньшей степени, чем в бореальный период.
4. Атлантическое время (50СО—3000 лет до н. э.). Климат
резко океанический.
5. Бореальное время (7000—5000 лет до и. э.). Климат
резко континентальный.
6. Переходное время (арктическое и субарктическое время
Сернандера).
Периоды Блитт-Сернандера до настоящего времени фигу¬
рируют всюду в литературе. Однако в оценке климатических
особенностей этих периодов нет единого мнения, притом эта
оценка далека от первоначальной. Особенно приходится при¬
знать это относительно суббореального периода 2. Р. Сернан¬
дер и его ученики и последователи считали этот период
ксерстермическим, против чего еще в конце XIX века высту¬
пил швед-географ Г. Андерссон (G. Andersson). Он считал, что
послеледниковый максимум тепла наступил гораздо раньше,
в атлантический период. К этому мнению примкнул Л. Пост,
Г. Гаме и ряд других исследователей. Далее весьма вероятно,
что максимум континентальности не совпадал с температур¬
ным максимумом, а имел место гораздо раньше. В пользу
того взгляда, что послеледниковый максимум тепла был до¬
стигнут еще в атлантический период, приводится распростра¬
нение теплолюбивых широколиственных пород по данным
пыльцевых диаграмм 34. Кривая солнечных сияний, вычислен- 1 2 * 41 Привожу по Г. Гамсу и Р. Нордхагену (1923) и Г. Гроссу (Н. Gross, 1931).
2 См. Г. Гаме (1930).
ЗГ. Гаме (1925).
4 К. Б юл08 (1930).
342
ная астрономами (рис. 114), показывает всего один после-
ледниковый максимум тепла в пятом тысячелетии до н. э.,
т. е. в атлантическое время. Во всяком случае теперь все
более склоняются к тому, что суббореальный период, во-пер¬
вых, не был ни самым теплым, ни самым сухим периодом
послеледниковой эпохи и что, во-вторых, в этот период на¬
ступило заметное ухудшение климата по сравнению с атлан¬
тическим. Это ухудшение по Г. Гроссу (1930) сказалось в по¬
нижении температуры и уменьшении количества осадков.
Последних, впрочем, было еще достаточно, и климат оста¬
вался влажным (по Г. Гроссу). Во всяком случае, считать
этот период ксеротермическим, как это делают, вряд ли воз¬
можно.
Рис. 114. Кривая солнечной радиации в четвертичный период (по Милан-
ковичу) и оледенения (по Кёппену) (Кбрреп). Из Г. Гамса, 1930.
Ученик Л. Поста Е. Гранлупд (1932) попытался вычислить количество осад¬
ков с суббореалыюго времени до настоящего момента. Ход кривой подтвер¬
ждает современное представление о значительном увеличении количества осад¬
ков от суббореального периода к субатлантическому. Однако, по исследова¬
ниям Гранлунда, количество осадков в суббореальный период не отличалось
заметно от такового в атлантический (рис. 17). Таким образом, и попытка
эмпирически вычислить ход осадков не дает представления о суббореальном
периоде как особенно сухом. Ход температуры, вычисленный Гранлундом,
показывает в согласии с данными астрономов (см. выше) один температурный
максимум, впрочем, имевший место несколько позднее—от 4009 до
2500 лет до н. э. Резкое падение температуры от атлантического периода
к суббореальному прекрасно заметно у Гранлунда и хорошо согласуется со
сказанным выше об ухудшении климата в суббореальный период. В настоя¬
щее время целесообразно говорить о длительном теплом послеледниковом
периоде, который включает бореальное и атлантическое время (die postglaziale
Warmezelt Л. Поста) и который сменился в суббореальное время более холод¬
ным периодом. Вопрос о послеледниковых климатах, впрочем, далеко еще не
разрешен1. Одна из предполагаемых причин—это, повидимому, изменение
интенсивности солнечного сияния. Солнечная радиация, по вычислениям
астрономов, непрерывно поднималась со времени окончания ледниковой эпохи
и затем достигла максимума, за которым последовал за 5000 лет до н. э.
п максимум температуры. После максимума началось непрерывное падение.
Максимум приходится, вероятно, на атлантический период.
Факторами второго порядка, значение которых оценивается величиной
в 5%, будут: холод непосредственно на самом леднике и влияние ледниковых
антициклонов. Первый фактор имел значение лишь там, где в этот
момент находился ледник, а второй действосал в Средней Европе лишь до
тех нор, пока ледник занимал значительную площадь. Эти сухие ледяные
1 Г. Гаме (1925, 1926), К. Бюлов (1930), Е. Васмунд (1929); там же и
о методах установления климатических изменений.
343
ветры влияли не только на температуру, но и понижали влажность воздуха,
а вместе с тем оказывали вредное влияние на растительность. Влияние лед¬
никовых ветров постепенно падало, по мере того как ледник уходил к се¬
веру. На климат областей, прилегающих к Немецкому и Балтийскому морям,
значительное влияние оказала история этих морей, изменение их уровня
и солености. Можно сказать, что история климата южной Прибалтики скла¬
дывается главным образом из двух факторов: из общего хода температуры,
зависящего от космических причин, и геологических изменений Балтий¬
ского моря — в первую очередь его площади. Внутрь материка значе¬
ние второго фактора падает, и решающее значение приобретает общий ход
температуры. Колебания температуры более мелкого порядка, несомненно,
имевшие место, мы оставляем сейчас в стороне.
С вопросом о послеледниковых климатах самым тесным
образом связан вопрос о так называемом „пограничном гори¬
зонте“ (ОгепгЬойгоп!). Веберовский пограничный горизонт—
это слой сильно разложившегося торфа из остатков пушицы
и вересковых, лежащий между верхним мало разложившимся
„молодым“ сфагновым торфом и нижним сильнее разложив¬
шимся „старым“ сфагновым торфом. К. Вебер, основатель
учения о пограничном горизонте, неизменно в течение многих
лет держался взгляда, что этот слой образовался в течение
тысячелетнего относительно сухого, может быть, и относи¬
тельно теплого климатического периода. Этот период отно¬
сится к суббореальному времени. Пограничный горизонт пред¬
ставляет, таким образом, важнейший руководящий страти¬
графический горизонт и служит существенным доводом
в пользу предполагаемого ксеротермического послеледнико¬
вого периода. Однако в последние годы в связи с критикой
теории Блитт-Сернандера подвергся пересмотру и вопрос
о пограничном горизонте и вызвавших его причинах. Прежде
всего отметим,, что вопрос о пограничном горизонте весьма
сложен и еще очень далек от разрешения. Синхронность
(т. е. единовременность) пограничного горизонта в разных
местностях еще не доказана. Следовательно, не доказан во
всех случаях и его суббореальный возраст. Далее, и страти¬
графически, т. е. по положению в торфяной залежи, погра¬
ничный горизонт не везде равноценен. Так, Г. Гросс (1930)
считает, что „истинный“ пограничный горизонт К. Вебера
всегда разделяет две толщи омброгенного торфа определен¬
ного состава. Нижняя, более гумифицированная, толща—про¬
дукт выпуклого болота теплого океанического климата
с „спокойными комплексами“, эрозией и замедленным торфо-
образованием, верхняя же—продукт болота сравнительно
холодного климата с энергично нарастающими регенерацион¬
ными комплексами. В средней части СССР сфагновый торф
под „пограничным горизонтом“ в понимании советских уче¬
ных носит иной характер — здесь это уже не омброгенный
торф атлантического климата. В связи с этим по Г. Гроссу
пограничный горизонт среднерусских болот не есть погра¬
ничный горизонт в смысле Вебера.
В СССР пограничный горизонт, аналогичный веберовскому,
был впервые установлен В. Сукачевым на Шуваловском тор-
344
фянике под Ленинградом. Однако состав его иной, чем?
в Германии. Здесь это слой в 20 см почти черного, сильно
разложившегося торфа с обильной пушицей и мощными пнями
и стволами сосны. Пни стоят вертикально, по размерам близки
к пням сосен на минеральной почве и далеко превосходят
мелкие болотные сосенки. Несомненно, что во время образо¬
вания пограничного горизонта торфяник был покрыт мощным
сосновым лесом. Пограничный горизонт резко отграничен
от верхнего, слабо разложившегося молодого сфагнового
торфа и, наоборот, постепенно переходит в нижний, сильно
разложившийся „старый" сфагновый торф. Подобный погра¬
ничный горизонт мог отложиться лишь в условиях сильного
высыхания болота, замедления торфообразования и развития
на болоте мощного соснового леса. „Пограничный горизонт"
в виде слоя сильно разложившегося торфа в Европейской
части Союза был обнаружен во многих торфяниках под Ленин¬
градом и в других частях Ленинградской • области (в бывших
Псковской и Новгородской губерниях), а также в средней
части Союза (в б. Тверской, Московской, Владимирской,
Рязанской, Иваново-Вознесенской, Калужской и других губер¬
ниях) (В. Сукачев, 1926). Таким образом, замедление торфо¬
образования не было явлением местным, а более или менее
всеобщим.
Если „пограничный горизонт“ типа Шуваловского торфяника трудно
толковать иначе как следствие более сухого периода, то в других местах
этот горизонт как климатический индикатор не однозначен.
В горах Средней Европы, как мы видели выше, пограничный горизонт
образовался в наиболее благоприятный для торфообразования период.
В бедных осадками областях, как средняя часть СССР, уменьшение их
количества в суббореальный период могло вызвать замедление нарастания
торфа. Вопрос о пограничном горизонте разрабатывался главным образом
в Прибалтийских странах (Швеция, Германия). Как раз здесь за последние
годы ряд исследователей поставил под сомнение значение пограничного гори¬
зонта как индикатора сухого периода, а вместе с тем и сухости суббореаль¬
ного времени. Решительный противник теплого и континентального, суббо-
реального периода в духе теории ¿Злитт-Сернандера— Г. Гросс (1930, 1933).
Суббореальный период, по его мнению, — время постепенного ухудшения кли¬
мата при переходе от влажного и теплого атлантического времени к менее
океаническому, более холодному и более сухому субатлантическому, а суббо-
реально-субатлантический контакт, представленный пограничным горизонтом,—
лишь переход от атлантического типа торфообразования к торфообразованию
субатлантическому (см. выше). Этот переход от атлантического Hochmoor
с ослабленным торфообразованием к субатлантическому выпуклому болоту
с интенсивным торфонакоплением произошел благодаря нескольким факто¬
рам, а именно: общему понижению температуры при сокращении вегетацион¬
ного и удлинении морозного периода и уменьшению количества осадков до
современного. Против существования ксеротермического суббореального
периода Г. Гросс приводит доказательства из разных областей науки. Инте¬
ресно, что микроскопическое исследование пограничного горизонта в северо-
западной Германии (Д. Шредер, 1930) обнаружило там присутствие гидрофиль¬
ных сфагновых мхов (Sph. cuspidatum), а также убывание по сравнению
с ниже лежащим старым сфагновым торфом пыльцы вересковых. Последняя
же в больших количествах встречается как раз в сильно гумифицированном
и выветрившемся торфе. Эти обстоятельства говорят также против образо¬
вания этого горизонта в условиях сухого климата. Также и Ф. Ионас (1933)
считает невозможным отделение суббореального периода от атлантического,
345-
История Балтики» климата, лесов и торфяников в Фенноскан
ч _
о „ —
=■ olS
Sj¡3
?u р-
о» Ó «9
=;=íu
Важнейшие геологические событии
Стадии
Балтики
Периоды
по де-Гееру
Зарубежная
Прибалтика
Район Ленинграда
1
Швеция,
Финляндия
(по Мунте
и Саурамо)
i
Район Ленинграда
(по Маркову,
1934)
Море Му а
1000
Начало
эры
Поднятие. Возникновение
островов Невской дельты.
Облесение дюн
1000
2000
*
Сужение
Датского
2400
! н
и
зунда
3000
к
4000
es
20
i °
4500
! *
Открытие
5000
¡ ®
английского
i 1
канала
! е*
! «
¡ ч
1 О
! ч
1 u
,5600
в
Связь между
ЪЮО
! с
Балтийским и
Немецким мо-
|
рем через
ütOO
Датский зунд
7010
Разделение
7210
i <3
Скандинавско-
i i
го ледника на
-
ч
двое (начало
ев
S СS
Шведской
35
геохронологии)
7800
4 2
Í— н
5 г->
Отделение
8000
X
Балтийского
.8300
5
моря от океана
Образование дюн у Сестро¬
рецка. Образование лагун
на месте морских заливов
(у Сестрорецка). Образова¬
ние здесь торфяников
Море на месте Сестрорецка,
Лахты, Ленинграда, Петер¬
гофа, от устья Невы вверх
морской залив
Максимум поднятия вплоть
до Кронштадта, где впадает
р. Tocho, текущая на месте
Невы
Море Limnaea
Начало солонова¬
той фазы (умень¬
шение солености)
2-я регрессия
И литориновая
трансгрессия
1-я литориновая
регрессия
Максимум лито-
риновой транс¬
грессии
i
«U
о.
о
S
о
о
со
о
S
Я
о.
о
I
Начало литори-
новой трансгрессии
Максимум Анцн-
ловой транс¬
грессии
о
о.
0>
о
о
я
<
Иольдиево море
2-е Иольдиево
море
(Море Eclieneis)
Таблица 52
дии и в районе Ленинграда со времени последнего оледенения
Уровень по
i!
Периоды и климаты
Развитие лесов п
Периоды культуры
отношению
о
к совремсн-
а> сэ
3 2
по Блитт-Сернандеру,
торфяников в районе
в Фенноскандин
ному
s a
Ленинграда
(по Монтелиусу)
в Ленинграде
=* 42
Гроссу и другим
и в СССР
(Марков, 1934)
ч ^
3 о
С S
(по Ануфриеву, 1131)
4-0 Ж
Новое время
Новое время
1 И II
Субатлантическое
В лесах преобладание ели.
Средние века
время» Климат про-
Максимум березы и сосны.
(Время викингов)
хладный и влажный
Сильное убывание ольхи и
Древняя история
4. О
смешанного, дубового леса.
Железный век
О
В торфяниках образуются
мощные толщи сфагнового
х + 1.0 м
о.
торфа
3 лито
моря
III
Суббореальное время.
Климат более холод-
Убывание смешанных дубо¬
вых лесов, особенно ольхи.
Рост торфяников замедля-
Бронзовый век
S
ный и сухой, чем в
о
атлантическое время
ется. Отлагается^ погранич-
О»
и
а»
Ос
ный горизонт
$1е1п1и51епгеЦ
1
■+■ 3.5 м
IV II V
Атлантическое время.
В лесах господство березы
Оаг^гаЬегге'й (время
Климат теплый и
влажный
и сосны. Внедрение ели. К
концу периода она становит¬
ся лесообразующей породой.
Окончательно поселяются
вырытых в земле мо¬
гильников).
Время дольменов,
н В СССР.: неолит.
— 58
- « ¿ s
®*ев х и
сначала ильм и липа, позд¬
нее дуб и лещина. К концу
периода их максимум. Обра¬
* стоянки у Сестро-
3 рецка, на Клязьме,
£ Окская дюнная сто¬
В = н £“ +8 м
зование сфагнового торфа
янка и пр.
Н
в водоемах Trapa u Najas
5
marina
о Кампинийская
2 культура в Европе
£
— 2.4 м
VI
Бореальное время.
В лесах господствуют бере¬
Время Маглемозе.
Климат теплый и
за и сосна. Ель почти исче¬
я Азильская — Тар-
_ о
сухой
зает. Широколиственные по¬
§ деноизская куль-
5 и
г о
VII и
VIII
роды (гл. о. ильм) редки,
« тура
у * я
gog-
~ О
ольхи также мало. Отлага¬
ются преимущественно раз¬
та
с
к
ь- S О
ложившиеся торфа с древес¬
с
СГ>
IIa* *+-4.5 м
ными остатками
*• та
а
2 — 2 jií
IX
Время улучшения
климата
Растительность сначала арк¬
Магдалениевая
ч
тическая, потом распро¬
£ культура
о
X о*
страняются березы, сосна
ч
о
(нижняя), ель, серая ольха.
Начало торфообразования
а*
в О
= S
ч
та
о
(гипновые торфа) и отложе¬
С
<ь>
Q,
ние гиттии
^ 1»
* С.
4
Важнейшие геологические события
Стадии
Балтики
<и £
5 я
2
Швеция,
Район Ленинграда
5 ¿>о
_ ф
3 ф
Зарубежная
Финляндия
(по К. Маркову,
1934)
5 g-s
к g «
°u Р*
и .
S ф
Ф п
Прибалтика
Район Ленинграда
(по Мунте
и Саурамо)
*=:t=tu
Сс
9.00
Край ледника
Субарктическая
у фенноскан-
дийских конеч¬
ных морен
1-й прорыв лед¬
трансгрессия Бал¬
тийского ледни <.
озера (В 11I-G1 IV)
1-е Иольдиево
ников Балтий¬
море
ского озера у
Конец Балтийско¬
10000
Биллинген (Шве¬
го ледникового
№
Оставление Ленинграда лед-
ция)
озера
S
ником
Уровень озера
G 111
ГО
«а
н
11000
а
Начало обра-
Ленинград под льдом, юж-
Плотинные лед¬
зования лен¬
нее плотинные ледниковые
никовые озера
се
точных глин
озера
(GI, ОН, GIII).
го
в Фенноскан-
SÉ
дии
О
н
о
12000
и
Край ледника
в Шонен
(Экапе)
130С0
Изостатич.
поднятие ска-
нодатского
14000
моста суши
15000
16000
Осцилляция
17000
се
К
«3
А11егб<1
180С0
го
н
о
19000
се
го
Край ледника
X
у балтийских
о
конечных мо¬
20000
н
го
рен
(датировка
CÍ
ненадежна)
а вместе с тем и выделение особого пограничного горизонта. Последний,
по его мнению, лишь верхняя часть старого сфагнового торфа. Начало торфо-
образования по субатлантическому типу и переход от атлантического ста¬
рого торфа к молодому сфагновому, образовавшемуся в более прохладном
климате, осуществляется, по Ф. Ионас, через особый слой торфа, так назы¬
ваемый „торфопредшественник* — УогШ^ог^ Этот Уог1аи!з1ог1 (заменяющий
сухой пограничный горизонт), залегающий поверх старого сфагнового торфа,
носит резко гидрофильный характер и образовался, благодаря подъему
грунтовой воды, имевшему место одновременно с субатлантическим пони¬
жением температуры. В выводах Ф. Ионаса, впрочем, не бесспорных, для
Продолжение таблицы 52
Уровень по
отношению
к современ¬
ному
в Ленинграде
(Марков, 1934)
Пыльцевые зоны
(по Маркову, 1934)
Периоды и климаты
по Блитт-Сернандеру,
Гроссу и другим
Развитие лесов и
торфяников в районе
Ленинграда
(по Ануфриеву, 1931)
Периоды культуры
в Фенноскандии
(по Монтелиусу)
и в СССР
+ 31
X
Субарктическое время
+ 23 .и
i
4- 38 м
1
XI
Арктическое время
I
нас важно отрицание особого суббореального климатического периода
и непосредственный контакт атлантического и субатлантического слоев торфа.
Любопытные данные по интересующему нас вопросу получил Е. Гранлунд
(1932). Он установил на шведских болотах ряд горизонтов обратного разви¬
тия (рис. 17) (Rekurrenzflachen). Эти горизонты представляют отчетливый
контакт между сильно разложившимся сфагновым торфом и залегающим
выше, слабо гумифицированным торфом. Гранлунд считает, что выпуклое
болото при постоянном количестве осадков дорастает до определенного
уровня (Grenzfläche), после чего рост прекращается, и на поверхности отла¬
гается сильно гумифицированный торф. Дальнейший рост болота начинается,
когда увеличивается количество осадков, после чего вновь начинает отлагаться
слабо гумифицированный торф. Граница, отделяющая слабо и сильно разло¬
жившийся торф и указывающая на перемену характера торфообразования
(Umschlag der Torfbildung), остается в залежи в виде Rekurrenzflache. Увеличе¬
ние количества осадков, вызывавшее образование этих горизонтов» достигало»
по мнению Гранлунда, 50—100 мм. Пограничный горизонт — одна из таких
Rekurrenzflächen на границе суббореального и субатлантического времени.
Всего на суббореальный период падает по Гранлунду три Rekurrenzflächen.
Для нас важно, что Е. Гранлунд считает пограничный горизонт не слоем,
а лишь контактом между слабо и сильно гумифицированным торфом и по¬
лагает, что этот контакт — следствие увеличения количества осадков. В. Вольф
(W. Wolf, 1930) также отрицает климатическое происхождение старого
сфагнового торфа, а вместе с тем и пограничного горизонта, и приписывает
их образование почвенным процессам *.
Таким образом, для Прибалтики вряд ли есть основания принимать су¬
ществование длительного второго сухого климатического периода и погра¬
ничного горизонта в виде обособленного слоя, носящего ясные следы обра¬
зования в сухом климате. Период, отделяющий атлантическое и субатланти¬
ческое время, характеризовался, видимо, постепенным понижением темпера¬
туры и уменьшением количества осадков и вряд ли был суше, чем субатлан¬
тический.
На болотах Европейской части СССР „пограничный гори¬
зонт“ носит ясные признаки сухости климата. Кроме указан¬
ных особенностей, для него характерно обилие грибных гиф
и иногда увеличение количества пыльцы вересковых. Все это
говорит в пользу сухости климата. Другой вопрос, насколько
велико было уменьшение количества осадков по сравнению
с атлантическим и субатлантическим периодом. Весьма воз¬
можно, что в условиях континентального климата средней
Европейской части СССР, где количество осадков близко
к минимуму, необходимому для существования выпуклых
болот, даже небольшое уменьшение их могло иметь резуль¬
татом замедление торфонакопления и образование погранич¬
ного горизонта 1 2 * * * *.
История Балтийского моря. (Табл. 52.) Послеледниковая
история Балтийского моря тесно связана с историей флоры
и лесов и историей послеледникового климата. Мы рассмот¬
рим здесь эту историю в самых общих чертах 8. В настоящее
время, благодаря работам финских, датских, шведских, частью
немецких геологов, выделяют такие стадии Балтийского моря:
1. Балтийское ледниковое озеро (рис. 115). Это
был замкнутый пресный водоем, северный берег которого был
образован краем отступающего ледника. Датско-шведский
мост суши отделял этот бассейн от Немецкого моря. Берего¬
вая линия (так называемая „балтийская граница“) этого озера
в южной Швеции лежала на 155—161 м, а под Ленинградом
на 100—106 м выше уровня современного Балтийского моря.
В эго время в Прибалтике господствовал суровый субаркти-
1 Критику положений В. Вольфа см. у К. Бюлова (1930).
2 О послеледниковых изменениях климата в северном полушарии (в том
числе и Америки) см. Е. Васмунд (1929); о климатических колебаниях в США
см. А. Дахновский (1922), а в Южной Америке см. В. Ауэр (1933).
п См. сводные обзоры Г. Гамса (1929, 1930), также В. Дееке Оееке»
1930).
350
ческий климат. Прорыв этого озера в области нынешнего«
озера Веттер (южная Швеция) имел место в первой половине
восьмого тысячелетия до н. э. и привел к соединению Бал¬
тийского озера с Немецким морем, его осолонению и освобо¬
ждению больших пространств Финляндии и Швеции от моря.
Образовалось Иольдиево море (2-я стадия).
Рис. 115. Карта Балтийского ледникового озера (Baltische Eissee) во время
I Сальпаусселька, около 8800 л. до н. э. По Рамзаю; из Г. Гаме, 1931.
1 — край ленника; 2 — изобазы Балтийского ледникового озера; 3 — береговая линия Балтий¬
ского озера; 4 — современная береговая линия.
2. Иольдиево море (рис. 116) \ Название его происхо¬
дит от руководящего моллюска — Joldia árctica (= Portlandia).
Его берега образуют так называемую „морскую границу“
(„Marine Grenze“) и лежат ниже берегов Балтийского озера.
Флора и фауна в это время были еще в основном арктические,
но к концу Иольдиевого периода в южной Швеции и под 11 Обзор истории Иольдиевого моря и пыльцевые датировки см. К. Мар¬
ков (1933).
Ленинградом появились уже не только ель и облепиха
(Hippophae), но также ольха, орешник и ряд сравнительно
теплолюбивых водных растений. Иольдиево время было непро¬
должительным (по разным данным 700 лет и меньше) и
окончилось в восьмом тысячелетии до н. э., с началом боре-
Рис. 116. Карта Иольдиевого моря незадолго до его конца, около 7900 л.
до н. э. По М. Саурамо; из Г. Гамса, 1931.
1 — край ледника; 2 — изобазы Иольдиевого моря; 3 — береговая линия_ Иольдиевого моря;
4 — современная береговая линия.
ального периода. Возможно, что к концу этого времени мы
имеем уже послеледниковый температурный максимум.
3. Анциловое озеро (рис. 117). Дальнейшее поднятие
Фенноскандии вновь привело к разъединению Немецкого и
Балтийского морей и образованию замкнутого бассейна —
Анцилового озера. Оно начало постепенно опресняться. Это
352
озеро далеко заливало нынешнюю сушу по обоим берегам
Ботнического залива, осевшую под давлением ледяных масс.
На юге же (например, на дне теперешнего датского Зунда и
к югу от о. Борнгольма) росли леса и были болота там, где
теперь находится море глубиной 35—70 м. По мере поднятия
Фенноскандии море постепенно заливало противоположный
Рис. 117. Анциловое озеро в период своего максимума, около 7400 л. до и. э.
По М. Саурамо; из Г. Гамса, 1931.
Л — край ледника; 2— изобазы Анцилового озера; 3 — береговая линия Анцилового озера;
4 — современная береговая линия.
южный берег (так называемая анциловая трансгрессия):
Время существования Анцилового озера (7800—5600 лет до
н. э.) соответствует теплому бо реальному периоду. В эту
эпоху лед таял, и быстро расселялись теплолюбивые растения.
Название водоема — по имени руководящего моллюска Апсу1из
Аиу1аШ|$.
4. Литориновое море (рис. 118). С повторным возник¬
новением сообщения между Балтийским и Немецким морями
пресное Анциловое озеро сменилось соленым Литориновым
23-722
морем. Для этой стадии характерно наступание моря на
южный берег (так называемая литориновая трансгрессия),
в то время как Фенноскандийский щит, освобождаясь от
давления льда, продолжает непрерывно подниматься. В ли-
ториновую трансгрессию Балтийское море впервые достигло
на севере Германии своих теперешних берегов. Литориновая
Рис. 118. Литорииовое море в период своего максимума, около 4000 л. до
н. э. По М. Саурамо; из Г. Гамса, 1931.
1 — изобазы Литоринового моря; 2 — береговая линия Литоринового моря; 3 — современная
береговая линия.
трансгрессия не была непрерывной, но чередовалась с отсту¬
паниями моря или остановками уровня. Различают три лито-
риновые трансгрессии 1, из которых самую древнюю называют
01уреи5-Тгап5£ге881оп, по названию руководящей диатомеи.
Наиболее высокого уровня и наибольшей солености Лнтори-
новое море достигает около 4600 лет до н. э. Первая половина
литориновой трансгрессии и ее апогей относятся к атланти¬
1 См. Е. Нуурра (1932). Этим трансгрессиям предшествовала еще чет¬
вертая трансгрессия Ма$1о£1о1а (Л. Аарио, 1935).
354
ческому периоду с его теплым и влажным климатом, макси¬
мальным распространением смешанного дубового леса и
интенсивным ростом болот. Конец Литоринового моря отно¬
сится уже к суббореальному периоду.
Море Limnaea и море Му а. Уменьшение солености
Балтийского моря, вероятно, вследствие сужения датского
Зунда и общего понижения уровня, наступило в середине
суббореального периода, около 2000 лет до н. э. В связи
с этим постепенно начала исчезать фауна, в том числе и
моллюски Литоринового моря, среди них и руководящий
вид этого моря Litorina. Их заменили пресноводные мол¬
люски, в числе их Limnaea ovata f. báltica—руководящий
вид новой стадии моря Limnaea. Вторая половина моря
Limnaea1 соответствует началу субатлантического периода,
а первая половина — послеледниковому ухудшению климата
(суббореально-субатлантический контакт). Последняя стадия —
море Муа (моллюск Муа arenaria) — относится уже к истори¬
ческому времени. Она нерезко отграничена от предыдущей и
характеризуется увеличением сосны и ели в пыльцевом
спектре, что зависит от уничтожения лесов человеком. Как
отмечалось, Фенноскандийский щит по мере освобождения от
ледяного покрова со времени анциловой стадии в общем
почти непрерывно поднимался (изостатические движения).
Это видно хотя бы из следующих данных для Финляндии 1 2
(Средняя Усмия), показывающих превышение уровня моря
над современным в метрах:
Трансгрессия Регрессия
1. Фаза Rhabdonema (преданциловая) . . . .
2. Анциловая фаза
3. Фаза Mastogloia (предлиториновая) . . . .
4. Литориновая фаза 1-я стадия
я „ и п П
5. Фаза Ытпаеа
0. Трансгрессия, вероятно, относящаяся к же¬
лезному веку
-60,5
49-50
63
—
42
—
35,5
30
30,5
27
24,2
23,5
17,9
13,5
7,0
—
На южном побережье Балтийского моря большую роль
играли эустатические движения, т. е. колебания уровня моря.
В этой области отмечено наступание моря в анциловую и
литориновую эпохи (анциловая и литориновая трансгрессии).
Соотношение между морем и сушей здесь осложнялось еще
тектоническими явлениями в четвертичную эпоху. Здесь пре¬
обладало во вторую половину послеледниковой эпохи в общем
опускание суши так же, как и на южном берегу Немецкого
моря, где опускание продолжается частью и теперь. Однако
во вторую половину послеледниковой эпохи на побережье
Немецкого моря, повидимому, не было непрерывного опуска-
1 От 500 года до н. э., до времени викингов.
2 Л. Аарио (1935); там же и связь с археологией.
23»
355
ни я, как это думали раньше. Так, между устьями Везера и
Эльбы Г. Шютте установил четыре послеледниковых опуска¬
ния и три поднятия.
Ф. Овербек (1934) подтверждает его выводы но основании исследования
торфяников и датирует отдельные стадии Немецкого моря.
1- я трансгрессия — между 8000 и 7000 лет до н. э. (бореальный период).
2- е поднятие — недостаточно доказано.
2-я трансгрессия—между 6000 и 50и0 лет до н. э. (па границе бореаль-
лого и атлантического периодов).
2- е поднятие — недостаточно доказано.
3- я трансгрессия — начало — 3000 лет и конец между 1000 и 2С00 лет
до н. э.
3- е поднятие — хорошо выражено: его начало 1500 лет до н. э.
4- я трансгрессия — 500-900 лет после н. э.
Послеледниковая история флоры 'и лесов.
Древнейший послеледниковый (добореальный) период.
Первыми поселенцами на освобожденной ледником территории
были арктические, тундровые растения. Это видно из того,
что тундровые растения в ископаемом состоянии были найдены
во многих пунктах далеко к югу от границ современного рас¬
пространения (рис. 119), а именно в южной Швеции, Норве¬
гии, Дании, Англии, Шотландии, в Прибалтике, а также под
Ленинградом, Вологдой и в других местах. Характерные пред¬
ставители этой флоры — арктические виды — низкорослые ивы
Salix reticulata, S. polaris, Dryas octopetala, также Betula nana,
Arctostaphylos alpina и арктические мхи, например, Scorpidium
turgescens (Jensen) (рис. 120). Особенно характерна Dryas.
По ее названию этот период называется временем Dryas, а
флора — флорой Dryas. Остатки этой флоры обычно находятся
не в торфе, а ниже его, в подстилающих торф отложениях.
Это показывает, что торфообразование в значительных раз¬
мерах началось лишь позднее.
Типичное местонахождение флоры Dryas—это глинистые отложения
(Dryas-Tone) мелких водоемов. Они содержат, кроме остатков водных
растений, и обильные остатки сухопутных тундровых растений, в том числе
п листья Dryas. Остатки сухопутных растений, ловидимому, приносились
в эти водоемы водой (см. П. Томсон, 1933).
Иногда различают древний и более молодой периоды Dryas (К. Иессеи —
К. Jessen, 1920) для Дании К Древний характеризуется присутствием Betula
nana, Dryas octopetala, Salix polaris, S. reticulata; новый, кроме тундровых
растений, — Betula nana, Dryas, Salix, Arctostaphylos alpina, еще растениями
более умеренного климата—Caltha palustris, Trlglochin maritima. Между двумя
периодами Dryas вклинивается период потепления и появляется древесная
растительность (см. ниже).
Продолжительность периода Dryas пока не выяснена. Е. Гииппе (1933)
считает этот период кратковременной, обусловленной местными влияниями,
стадией растительности на самой окраине ледника — стадией, которая быстро
сменилась лесом.
К. Марков для Ленинграда установил, чт« среди тундровой раститель¬
ности, появлявшейся всего в 60—80 км от края ледника, через 150—200 лет 11 По Г. Шрейберу (1927).
356
после его отступания уже встречалась ель. Косвенное подтверждение факта
быстрого поселения деревьев после отступания ледника дают наблюдения
над растительностью морен горных ледников в Западной Норвегии (К. Фегри.
1933). После поселения здесь сначала пионерной растительности, а затем
криптогамных ценозов уже через 40—55 лет (иногда же свыше 180 лет)
последние сменяются зарослями кустарничков (Empetrum, Calluna). Низкорос¬
лая же древесная растительность (Alnus incana, Betula tortuosa) поселяется
иногда уже через 20 лет, а через 70 лет (а иногда и более, чем через 180 лет)
образует уже заросли. Однако весьма вероятно, что в других случаях деревья
Рис. 119. Известные до 1895 г. местонахождения ископаемой ледниковой
арктической флоры (черные точки). По А. Натгорсту, 1892;
из Е. Вульфа, 1932.
Двойная линия — граница оледенс'ния.
появлялись после отступания ледника лишь через значительный промежуток
времени. Что касается окончательного образования сомкнутых лесов на
освобожденной от ледника территории, то для этого потребовался весьма
длительный период (см. ниже).
Климат времени БгуаБ, по мнению Г. Андерссона (1910), не был аркти¬
ческим, а глациальным, сухим, холодным и континентальным. Продолжитель¬
ность вегетационного периода была 4—5 месяцев, июльская температура
-I- 9°. Это время соответствует Даниглациалу (Оап^1а21а1\ Готиглациалу
(йо^к^а!)1 и Балтийскому ледниковому озеру. Его продолжительность 11 Разные стадии отст)пания ледника см. рис. 121.
357
{по Г. Гроссу, 1930) 19000—8300 лет до н. э. Громадная длительность периода
Ргуав по этим данным на первый взгляд резко противоречит представлению
о кратковременности этого периода (см. выше). Вряд ли есть основание,
однако, считать этот период совершенно лишенным древесной растительности.
Достаточно указать, что остатки древесной растительности находят даже под
отложениями Балтийского ледникового озера. Приведенные выше сведения
о климате времени Бгуаэ не противоречат возможности произрастания дре¬
весной растительности. Если температура самого теплого месяца здесь
несколько и ниже, чем та,
при которой в настоящее
время произрастают деревья,
то зато сравнительно про¬
должительный вегетацион¬
ный период и сухость воз¬
духа компенсируют небла¬
гоприятную для деревьев
температуру лета. Возможно,
однако, что данные Г. Ан-
дерссона, в случае, если они
правильны, характеризуют
лишь самую раннюю часть
позднеледникового периода.
Е. Гииппе (1933) считает,
что на Карельском пере¬
шейке и в окрестностях Ле¬
нинграда уже в начале позд¬
неледникового времени (т. е.
за 12000—13 000 лет до н. э.)
температура перешагнула за
предел произрастания дре¬
весной растительности (т. е.
+10° в самый теплый месяц),
так как нельзя иначе объяс¬
нить появление здесь в это
время березы и сосны. По
мнению этого автора, уже
в первую половину поздне¬
ледникового времени темпе¬
ратура лета была близка к
современной. Он считает,
что глациальный климат Ан-
дерссона существовал лишь
на самой окраине отступаю¬
щего ледника и что темпе¬
ратура быстро повышалась
по мере его отступания.
Рис. 120. Распространение северных мхов
в настоящее время Зи в дилювии. По Г. Гамсу.
Scorpidium turgescens (Jensen): 1 — ю.-з. граница ареала;
2—эксклавы; 3—отдельные местонахождения; 4—иско¬
паемые местонахождения Thuidium Blandowii (Web et
Mohr); 5 — ю.-з. граница ареала; 6 — современные от¬
дельные местонахождения; 7 — ископаемые местонахож¬
дения.
Время добореаль-
ных лесов. В Сред¬
ней Европе первой
стадией послеледниковых лесов были березово-сосновые леса.
Характер этих лесов выяснен пока недостаточно. Ф. Фирбас
(1930, 1934) считает, что первые леса в Средней Европе
носили субарктический характер. Для них были характерны
такие моменты: 1) бедность древесными породами — только
береза, сосна и ивы являются лесообразующими, широко¬
лиственные же встречаются очень редко; 2) первые бере¬
зовые леса были еще редки, о чем свидетельствует про¬
цент иедревесной пыльцы; 3) остальная растительность
358
сохраняла еще в значительной части характер более ранней
тундровой фазы. Здесь присутствовали еще северные виды
Empetrum, Betula nana, Selaginella selaginoides. Для Средней
Европы первым лесным периодом было время березово¬
сосновых лесов, причем сначала обильнее была береза, а затем
усилилась сосна. Для некоторых областей Средней Европы
удается разделить более раннюю фазу субарктических бере¬
зовых и более позднюю фазу сосновых лесов. Последние были
уже более густыми и сомкнутыми, чем березовые. Selaginella
в этих лесах уже вовсе не встречалась, a Betula nana и Ешре-
ttrum присутствуют еще в более древних отложениях соснового
времени, к концу же его, повидимому, пропадают1.
4 Какой характер носили еловые леса в период „нижнего максимума ели*
в северной половине Европейской части СССР — сказать трудно за недостат¬
ком сведений. Судя по совместному нахождению ели и флоры Dryas1 2, можно
думать, что и эти леса были редколесьями с тундровой растительностью.
Интересно раннее появление теплолюбивых широколиственных пород.
В Средней Европе уже к концу березово-соснового времени орешник
и породы смешанного дубового леса были налицо, хотя и в небольшом
количестве. Мало того, они встречались уже во всей области в границах их
современного ареала (Ф. Фирбас). Также и в Прибалтике эти виды уже
встречались, хотя и в ничтожном количестве, в иольдиево время, когда
ледник еще был в области морен средней Швеции (данные Е. Гииппе, 1933,
Л. Аарио, 1933 и др.) Следовательно, первое появление этих пород
относится — самое позднее — к пребореальному времени. Изучение пребо-
реальных диаграмм, таким образом, дает возможность установить лишь
историю распространения древесных пород, а не историю их первого по¬
явления (Ф. Фирбас). Последнее же отодвигается на еще более древний период.
Какой же промежуток времени отделяет максимум последнего оледенения от
первого лесного периода? К. Рудольф (1930) принимает большую
продолжительность этого переходного времени, даже
большую, чем вся послеледниковая история лесов. Таким
образом, Рудольф предполагает, что исходное положение для этой истории
надо искать не в ледниковых убежищах на юге Европы, а в распределении
древесных пород в указанное переходное время с его ледниковыми стадиями и
интерстадиальными промежутками. Поэтому вряд ли правильно, что это время
было совершенно лишено древесной растительности 3. В связи с осцилляциями
края ледника, повидимому, в Европе происходили временные вспышки раз¬
вития древесной растительности, затем угасавшей и сменявшейся тундровой.
Нахождение под Ленинградом макроскопических древесных остатков
{ель, ольха) под отложениями Балтийского ледяного озера и пыльцевой
анализ (С. Яковлев, 1926) отчасти это подтверждают. П. Томсон (1935) считает,
что лес с сосной, елью, березой, ольхой и даже с ничтожной примесью вяза,
липы и орешника (!) достигал Финского залива еще в конце готиглациала
(Gotlglazial) (рис. 121) в период потепления климата (климатический оптимум
готиглациала). Весьма вероятно, что это продвижение леса соответствует
периоду Alleröd Иессена (Jessen) в Дании. Последующее ухудшение климата
привело к деградации этого леса 4. Другие авторы (К. Фегри, 1935) для запад¬
ной Норвегии устанавливают даже двукратное продвижение и отступание
лесов в позднеледниковый период, причем последнее продвижение, возможно,
соответствует стадии Alleröd.
1 Данные для Рейнского Пфальца (Ф. Фирбас, 1934),
2 Например, под Ленинградом (Г. Ануфриев, 1925).
3 Е. Гииппе (1933) еще более решителен: он отодвигает время первых
лесов еще дальше, к готиглациалу.
4 Другие относят это продвижение лесов в Прибалтике к еще более
(Раннему времени—самое позднее к началу готиглациала (Ф. Фирбас, 1934).
359
В свете современных данных пыльцевого анализа можно
сделать такие выводы о позднеледниковом периоде истории
лесов:
1. Первый лесной период в Средней Европе — березово¬
сосновый, причем сначала максимум дает береза, потом сосна. 2Рис. 121. Стадии отступания ледника в период последнего оледенения. По
М. Саурамо; из Г. Гамса, 1929.
1 — морены датском стадии; 2 — морены готской стадии; 3 — морены финиглациальной стадии;
4 — конечные послеледниковые морены; 5 — годы по шведской геохронологии; 6 — годы но
финской геохронологии.
2. Первые (березовые) леса носят субарктический характер.
Они редки (редколесья), бедны древесными породами; тепло¬
любивые широколиственные породы большей частью совсем
отсутствуют; обильны представители тундровой флоры.
360
3. От максимума последнего оледенения до первого лес¬
ного периода прошло, вероятно, весьма длительное время.
Можно предполагать, что в этот переходный период в Средней
Европе и Прибалтике уже имело место, по крайней мере,
однократное развитие лесов, сменившееся затем периодом
его деградации и наступлением тундровой стадии.
4. Смена леса и тундры в этот переходный период вызыва¬
лась осцилляциями (колебаниями) границы ледника. Эти
колебания сопровождались изменениями климата. Первое
появление лесов, вероятно, совпадает с позднеледниковым
температурным оптимумом, сменившимся затем субарктиче¬
ским похолоданием.
Интересен вопрос о миграциях древесных пород перед облесением
Средней Европы. Весьма вероятно, как это думает К. Рудольф (1930), что
уже задолго до березового времени в Средней Европе шло расселение
древесных пород. Они могли продвигаться во время интерстадиальных про¬
межутков (т. е. между временными продвижками вперед ледника), когда климат
делался более мягким, и пережидать „стадии" (т. е. лродвижки льда) с их
суровым климатом в соответствующих „убежищах1*.
Послеледниковая история лесов и флоры прибалтийских
ч стран.
История растительности Прибалтики особенно хорошо
изучена. Кроме того, здесь особенно хорошо установлена
связь лесных периодов с стадиями Балтийского моря,
а вместе с тем и хронология этих периодов. Поэтому удобно
начать обзор послеледниковой истории1 Европы именно
с Прибалтики. Здесь можно различить такие периоды
1. Время сосны и орешника. Соответствует
бореальному периоду и анциловой стадии Балтики. Хроно¬
логия 7800—5600 лет до н. э. Это время редких парковых
сосновых и березовых лесов с прогалинами—травяными сте¬
пями (лесостепь) — и часто с орешником. Климат был срав¬
нительно континентальным, довольно сухим, с суровыми
зимами, но с летом, сначала умеренно теплым (температура
июля около 12°), к концу же периода на 2,4° теплее нынеш¬
него. Вегетационный период был на 15 дней длиннее, чем
теперь. Бореальное время обозначает наступление теплого
послеледникового периода. Аркто-альпийская и субарктиче¬
ская флора отступает в горы и на север и остается в виде
реликтовых очагов на болотах. Быстро начинают расселяться
теплолюбивые деревья: черная ольха, вяз, липа, дуб, позднее
клен, ясень. Однако роль их в лесах пока еще незначительна.
Равным образом расселяются теплолюбивые травянистые
растения — Cladium Mariscus, виды Najas, Trapa natans. К концу
периода ряд растений расселяется к северу, дальше их тепе¬
решних границ: Quercus, Tilia, Ulmus, Ainus glutinosa, Corylus,
1 Позднеледниковая история здесь не рассматривается.
2 См. сводку Г. Гросса (1930).
30Í
^11
£6
-Рис. 122. Границы распространения теплолюбивых растений в Финляндии теперь
и в теплый послеледниковый период. По М. Саурамо; из Г. Гамса, 1929.
1 — Trapa natans; 2 — Corylus avellana; 3 — Ceratophyllum demersum — прежняя северная гра¬
ница; 4 — Lycopus europaeus — прежняя северная граница; 6 — Сагех pseudocyperus—прежняя
.северная граница; 6 — Ceratophyllum demersum — современная северная граница; 7 — Lycopus
europaeus — современная северная граница; 8 — орешник — современная граница; 9 — Сагех
pseudocyperus — современная граница.
Cladium mariscus, Najas, Carex pseudocyperus, Lycopus, Cerato-
phyllum demersum, Trapa natans (рис. 122)Примером служит
орешник, который с этого времени потерял в Швеции 84000 км2
пространства2. Границы растений в горах южной Норвегии,
по вычислению Рекштадта3, поднимались на 350 — 400 м выше,
чем теперь. Тогдашнее распространение некоторых растений
и животных, а также положение верхней границы деревьев
Tilia Queráis —д—Carpms —д—fagus
i —ь—Picea —•—Corylus —a—AUws
—■ — Смешан. дубоСыйлес (имт+тпю +аиегж) -■
Рис. 123. Основная диаграмма пыльцы для южной
Швеции. По Л. Посту; из Г. Гросса, 1930.
в горах позволили Г. Андерссону* * вычислить, что в теплый
послеледниковый период температура была на 2,7— 2,3° выше,
чем теперь. Другие авторы считают, что превышение летней
температуры против современной равнялось даже 4°3.
1 Водяной орех, в настоящее время совершенно вымерший в Финляндии,
известен здесь в ископаемом состоянии (в болотах) из 35 пунктов (Г. Линд¬
берг — Н. Ыпс1Ьег§).
2 По Г. Андерссону; привожу по К. Бертшу (К. ВеПзс!), 1928).
3 Рекштадт (Рек$1а(И, 1903), привожу по Г. Шрейберу (1927).
* Г. Андерссон (1905), цит. по К. Бертшу (1928).
5 Г. Линдберг (1911); привожу по К. Шрейберу (1927).
303
2. Время смешанного дубового леса (рис. 123).
Соответствует литориновой стадии Балтики и первой половине
моря Limnaea и датируется 5600—500 годами до н. э. Это»
время делится на три периода:
1- й п ер и од. Температурные условия лета близки к преды¬
дущему периоду, но зима делается мягче. Количество же,
осадков значительно возрастает, особенно в горах и на морском
побережье. Этот период, как и следующий, относится к после¬
ледниковому теплому времени. На более богатых почвах
господствуют смешанные дубовые леса с подлеском из ореш¬
ника. Эти леса сменили здесь прежние сосновые леса. На плохих
почвах продолжают существовать березовые и сосновые леса,
а в сухих районах—безлесные участки (степи). Продолжается
расселение Тгара и понтийских видов Adonis vernalis, Oxytropis
pilosa, Stipa pennata *. На востоке (побережье Финского залива)
начинают распространяться ельники. Этот период соответствует
первой части атлантического периода и литориновому времени
до первого максимума.
2- й период. Климат приобретает все черты, присущие
атлантическому времени: теплое лето, обилие осадков, особенно
в приморской полосе, где климат типичный океанический,
теплый. После литоринового максимума температура лета
понижается. Кульминация смешанного дубового леса продол¬
жается. Ель продолжает распространяться в северо-восточной
Прибалтике и появляется под Берлином, в Курляндии и в Вос¬
точной Пруссии, а несколько позднее в северной Швеции.
Теплый климат способствует дальнейшему распространению’
Тгара natans, орешника, а также продвижению на север
и северо-восток Hedera, Taxus, Acer campestre. Зато сухо¬
любивые понтические виды отступают, атлантические же
занимают свой теперешний ареал. Некоторые из них проникают
далеко на север и восток, как Erica, Myrica, Drosera intermedia.
Благодаря влажному климату сосновые леса на песках вытес¬
няются пустошами, образуется ортштейн (также и в первый
период). Под конец литоринового времени бук доходит на
север до южной Швеции, на восток—до восточной Пруссии.
Происходит закрепление и частью облесение дюн. В литори-
новую эпоху образуются омброгенные болота, и отлагается
на западе старый сфагновый торф. Второй период совпадает
с второй половиной атлантикума и временем от литоринового
максимума до конца Литоринового моря. Его конец датируется
2030 лет до н. э.
3- й период. Соответствует первой половине стадии
Limnaea и суббореальному времени. Датируется 2000 — 500
годами до н. э. По Г. Гроссу, климат ухудшается, температура
лета понижается на 2,4° и сокращается вегетационный период.
Осадки также убывают; на востоке значительно, на западе
и юге меньше. В связи с ухудшением климата смешанный 11 Доходит в Швеции до Vastergotland.
364
дубовый лес резко идет на убыль; северные границы тепло¬
любивых растений отодвигаются к югу. Граница леса в горах
понижается. На севере вымирает Trapa natans, сильно отсту¬
пают к югу Najas и Cladium, начавшие отступать еще в преды¬
дущий период. Частично отступают на запад и атлантические
виды. Деградируют также атлантические пустоши. Зато сильно
распространяется ель. В Швеции же начинается ее массовое
развитие. С елью движутся на запад и другие восточные виды,
ее спутники. Одновременно с елью в северной Германии
Рис. 124. Бореадьная фаза орешника в Средней Европе. По К. Рудольфу, 1930.
А орешник; />' Pinus; В - Picea; Г Abies; Д - Fagus; Е— Betula; Ж — Ainus; 3 — сме¬
шанным дубокыи лес; S — Salix; // — остальное. Черная линия — граница распространения
Pinus silvestris.
и южной Скандинавии распространяется и бук, частью дальше
его нынешней северной границы. В западной Прибалтике на
болотах получают преобладание регенера1ивные, растущие
комплексы, и образуется пограничный горизонт. В континен¬
тальных областях последний выражен как горизонт высыхания
с мощными пнями.
•3. Время бука и ели. Соответствует субатлантическому
периоду, второй половине времени Limnaea и времени моря
Муа. Продолжительность от 500 л. до н. э., до наших дней.
Климат прохладный, близкий к климату конца предыдущего
периода. Количество осадков примерно то же. Смешанный
305
дубовый лес деградирует за счет дальнейшего распространения
буковых и еловых лесов, а позднее вследствие вмешательства
человека. Распространение бука и ели идет примерно до
половины периода, после чего под влиянием человека начи¬
нается деградация еловых, особенно же буковых лесов. Бук
отступает в восточной Прибалтике, в южной Швеции и север¬
ной Германии. Ель же продвигается на запад и юго-запад
в южной Швеции и восточной Пруссии. Продолжается начав¬
шееся ранее движение на запад восточных видов области еловых
лесов (в восточную Пруссию и восточную Прибалтику). Аркти¬
ческие и субарктические виды распространяются немного
дальше на юг за пределы своих ареалов и на некоторых
реликтовых местообитаниях, атлантические же виды сокращают
свои реликтовые ареалы. Сильно распространяются сорняки,
рудеральные и адвентивные растения, среди них Elodea cana¬
densis, быстрое распространение которой привело к исчезнове¬
нию некоторых водных растений. В очень влажных областях
идет развитие солигенных, питающихся притекающими со сто¬
роны водами, болот и заболачивание суши; в области омбро-
генных (выпуклых) болот идет сначала рост болот и образование
молодого сфагнового торфа, а позднее торфообразование
местами замедляется, и на поверхности болот получают пре¬
обладание спокойные комплексы и эрозия.
История лесов Средней Европы 1.
1. Время орешника (бореальное время). Максимум
пыльцы орешник дает на западе и в горных районах востока
(наиболее высокие цифры — до 450%) (рис. 124), в то время
как в спектрах Европейской части СССР пыльцы орешника
содержится всего только несколько десятков процентов.
Обилие пыльцы орешника, повидимому, указывает на существо¬
вание густых его зарослей в еще редких сосновых лесах. Это
время называют также сосново-орешниковым2. В этот период
шло массовое распространение смешанного дубового леса, ель
же была лишь в горах (Альпы, Карпаты) и только позднее
появилась в северной предальпийской области. Пихта лишь
начала распространяться в горах на западе. Береза и сосна
идут на убыль. Время орешника—это послеледниковый мак¬
симум тепла. На это указывает и более высокое положение
границ древесной растительности в горах Средней Европы.
Так, в Рудных горах граница массового распространения орешника была
тогда на 400 м выше, чем теперь (К. Рудольф и Ф. Фирбас, 1927), так же как
и в Vorarlberg’e (Ф. Фирбас, 1925/26). Эти цифры весьма близки к цифрам,
полученным в Скандинавии. В отличие от добореального лесного периода
болотообразовапие идет в широком масштабе. Болота, главным образом
травяные (Богемия)®, частью переходят в омброгенную стадию (Бавария) * *.•
1 Периоды лесов горных местностей, несколько отличные, см. дальше.
- К. Рудольф и Ф. Фирбас (1927), Г. Пауль и 3. Руофф (1932).
* К. Рудольф (1928).
* Г. Пауль и 3. Руофф (1932).
366
Хронология этого периода 7800—5600 лет до и. эЛ Время
мезолита.
2. Время смешанного дубового леса (атлантиче¬
ское время). Смешанный дубовый лес господствует в нынешней
области лиственных лесов Средней Европы. Сначала в нем
много липы и вяза (иногда доминируют), позднее господствует
дуб. В горах уже рано начинается вытеснение дуба хвойными,
приводящее, в конце концов, к господству пихты на западе,
в среднем поясе гор, и ели на востоке, а на западе в более
высоком поясе. Орешник не образует самостоятельных зарослей
и встречается лишь в подлеске. К концу атлантикума—массо¬
вое распространение бука в сопровождении граба. На равнинах
в торфяниках отлагаются древесные торфа (Богемия), выше
в горах образуется старый сфагновый торф (Богемия, Бавария).
Хронология—5600—2500 лет до н. э., начало расцвета неоли¬
тической культуры.
3. Буковое время (суббореально-субатлантическое).
Бук достигает максимума в области лиственных лесов, где он
часто доминирует, и в области хвойных, где он делит господ¬
ство с пихтой. В первую половину периода в Богемии на
болотах образуется пограничный горизонт с мощными пнями,
прекращается торфообразование на малых болотах и на окраи¬
нах больших. В высокогорных же областях (Исполиновые горы)
рост болот резко усиливается. Во вторую половину периода
усиливается рост болот на равнинах, и отлагается молодой
сфагновый торф, в высокогорных же болотах торфообразование
прекращается. Хронология 2500 до н. э. — 15G0 н. э. Первая поло¬
вина букового времени соответствует позднему неолиту и брон¬
зовому времени, вторая—железному и историческому времени.
4. Культурное время. Уничтожение лесов и создание
искусственных насаждений. В области лиственных лесов при
содействии человека хвойные породы вытесняют широколист¬
венные; то же идет и в горных местностях. Это время назы¬
вают также елово-сосновым, так как в спектре господствует
пыльца хвойных. В Богемии на высокогорных болотах торфяники
начинают разрушаться, болота горной ступени зарастаю.т лесом
или криволесьем. В эпоху бука климат ухудшается, граница
леса в горах понижается и начинается вымирание растений,
появившихся в Средней Европе в теплое послеледниковое
время. Так обстоит дело в Швабии по К. Бертшу (1928) с
Trapa natans и Najas flexilis, N. marina. Исчезновение их, впро¬
чем, можно объяснить уничтожением подходящих местообита¬
ний (заболачивание водоемов).
Ленинградская Прибалтика.
История лесов Ленинградского района в общем была изучена
хорошо, а за самое последнее время особенно детально благо¬
даря работам К. Маркова (1934, 1935). Кроме того, для этой 11 Здесь и всюду дальше хронология по Г. Пауль и 3. Руофф.
ЗЬ
области хронологические датировки и увязка со стадиями Бал¬
тийского моря наиболее надежны, а за последнее время пыль¬
цевые диаграммы Ленинграда удалось хорошо увязать с де¬
тально разработанной пыльцевой шкалой Л. Поста. В общем,
з настоящее время история лесов и датировка пыльцевых
диаграмм Ленинграда являются исходными для остальных об¬
ластей Европейской части СССР. Поэтому рассмотрение истории
лесов Европейской части СССР мы начнем с ленинградской При¬
балтики. Ниже сопоставим лесные периоды для ленинградского
района с периодами Блитт-Сернандера и стадиями Балтики 1.
XI зона — арктическое время1 2. Господствует пыльца
сосны и березы (последняя дает здесь нижний максимум); кроме
того, немного пыльцы ели, ивы и ольхи. Встречаются остатки
флоры Dryas. Соответствует Балтийскому ледниковому озеру.
Хзона — субарктическое время. При господстве
березы и сосны последняя дает здесь свой нижний максимум.
Кроме того, здесь нижний максимум ели. Соответствует транс¬
грессии и последней стадии ледникового Балтийского озера
(В III по Рамзаю).
Эти две зоны соответствуют добореальному периоду с гос¬
подством пыльцы сосны и березы, с максимумом ели и ивы.
В это время лесов не было, а имелись лишь отдельные деревья,
з начале периода росшие среди тундровой растительности.
Торфообразование (гипновый торф) и отложение гиттии в водое¬
мах лишь начинались.
IX зона — время улучшения климата. Продолжаю¬
щееся господство пыльцы сосны и березы, причем последняя
дает второй максимум. Ель сходит на-нет. Зона соответствует
Иольдиевому морю.
VIII и VII зоны — бореальный период. Господство
пыльцы березы и сосны и второй максимум последней. В верх¬
ней части периода начинается непрерывная кривая смешанного
дубового леса и орешника, которая поднимается так же, как
и кривая ольхи. Кривая ивы сходит на-нет; ель местами почти
пропадает. Этот период соответствует максимуму анциловой
трансгрессии. Благодаря сухому и теплому климату отлагаются
разложившиеся торфа с древесными остатками.
VI зона—переходный период. Береза дает третий
максимум (более низкий, чем второй), количество пыльцы
сосны резко падает; увеличение широколиственных пород, ореш¬
ника и ольхи продолжается.
V и IV зоны — атлантический период. При пре¬
обладании березы над сосной смешанный дубовый лес, ольха
и орешник дают максимум. Ель образует непрерывную кривую
и идет на подъем, в конце периода делаясь лесообразующей
породой. Литориновое море достигает максимума стояния и
1 Г. Ануфриев (1931), К. Марков (1935).
2 Пыльцевые зоны от более древних вверх, в порядке убывания, приве¬
дены по К. Маркову (1935). Границы их, впрочем, весьма субъективны.
•солености. Происходит интенсивное отложение сфагнового
торфа и зарастание водоемов тростником. В гиттии встречаются
остатки теплолюбивых растений—Trapa natans и Najas marina.
III зона — суббореальный период. Количество
пыльцы ольхи, орешника и смешанного дубового леса падает,
ель дает максимум (нижний максимум верхней ели). Березы
меньше, чем в предыдущий период, сосна на прежнем уровне.
К концу периода на¬
растание торфа заме- о ю го зо w so ео% ю го%
дляется, и образуется g|re 1 1 ? If» 1 1 1 |,, | |g
пограничный горизонт.
Литориновое море от¬
ступает и опресняется.
II и I зоны — суб¬
атлантический
период. Количество
пыльцы ели достигает
наивысшего за всю по¬
слеледниковую исто¬
рию максимума (верх-
fir
Е
Г
—
;
г
—
Л
~2
х
Г
ЙП
л
]г
в
тт.
то
У_
W
а
Л
X
1
/
Г
X
X
Г
JT
Т
г—^
Ж
р
h
1
щ
\
1
—о—
- titula
Salix
—О—
Pí ñus
ели) и затем начинает
падать. Пыльца оль¬
хи после небольшого
подъема также резко
убывает. Сплошная
кривая смешанного ду¬
бового леса уже в на¬
чале периода преры¬
вается, и широколист¬
венные породы встре¬
чаются лишь отдель¬
ными зернами. Балтий¬
ское море к началу пе¬
риода почти достига¬
ет современного уров¬
ня. Это время наибо¬
лее благоприятно для сфагнового торфообразования и сфа¬
гновых болот с господством Sphagnum fusctim. Максимум ели
совпадает, примерно, с прослойкой разложившегося торфа,
образовавшейся около середины субатлантического времени,
повидимому, вследствие непродолжительного сухого периода.
—д—
Picea
—я—
Corylus
виегсешп mixtura Que rcus tíl mus
Pinus
Tilia
Рис. 125. Средняя диаграмма пыльцы Пере-
славского района как пример пыльцевой диа¬
граммы центральной части СССР. По М.
штадту, 1928.
Ней-
Центральная часть СССР.
В средней части СССР отсутствовали стадии Балтийского
моря. Следовательно, здесь, как и дальше на восток, хроно¬
логическое соответствие данных пыльцевых диаграмм и отне¬
сение их к периодам Блитт-Сернандера возможны лишь при
сравнении с пыльцевыми диаграммами Прибалтики. Средняя
диаграмма пыльцы Переславского района Ярославской области
24—722 369
Таблица 53
Лесные периоды в разных частях Европейской СССР в послеледниковое время
Типы
Периоды '
по Блитт-Сернандеру
Северный тип
(Шенкурск.)
(по М. Ней-
штадту, 1930)
Северо-запад¬
ный (Ленинград-
ский) (по
Г. Ануфриеву
и К. Маркову,
1935)
Западный
(Западная
область)
Юго-западный
(С.-З. Украина)
(по Д. Зерову,
1934)
Среднерусский
Восточный
(Казанский)
Уральский
(по Д. Гераси¬
мову),
6.
Субатлантический период
Время сосны,
ели и березы
Время сосны,
ели и березы
Время сосны,
березы и ели
Время сосны
и максимума
граба
Время сосны,
березы и ели
Время сосны
Время сосны,
березы и ели
5.
Суббореальный — Атлантический
Время сосны,
березы и ели,
с максимумом
смешан. дубо>
вого леса
(и ольхи)
Время сосны,
березы и ма¬
ксимума сме¬
шанного дубо¬
вого леса и
ольхи
Время
смешанного
дубового леса
и ольхи
Время сосны
с участием
смешанного
дубового леса
Время сосны,
березы и ма¬
ксимума ольхи
и смешанного
дубового леса
Время сосны
и максимума
смешанного
дубового леса
и ольхи
Время березы,
сосны, макси¬
мума смешан¬
ного дубового
леса и появле¬
ния пихты
4.
Атлантический
Время березы
(преобладает)
и сосны
Время березы
(преобладает)
и сосны
Время березы
(преобладает)
и сосны
То же
Время березы
(преобладает)
и сосны
Время сосны
и березы
Время сосны
и березы
3.
Бореальный
Время сосны
(преобладает)
и березы
Время сосны
и березы
Время сосны
и березы
Время сосны
Время сосны
(преобладает)
и березы
Время сосны
(преобладает)
и березы
То же
«
2
1
«в
2. Субарктический
Время
нижней ели
Время
сосны, березы
и максимума
нижней ели
Время
нижней ели
(максимум)
сосны и ивы
Время
сосны, березы
и ивы
Время
нижней ели
(преобладает)
сосны, березы
и ивы
(максимум)
Время
нижней ели,
березы и ивы
Время
лиственницы,
ивы и нижнего
максимума ели
в>
а
о
ю
о
е*
1. (Арктический)
Время
сосново-бере¬
зовых редко¬
лесий с флорой
Огуаз
Время
сосны и березы
(редколесья?)
По 3» Руофф (1936) с дополнениями и некоторыми изменениями.
характерна для центра Европейской части СССР. Намечаются
такие периоды послеледниковой истории лесов1:
1. Максимум ивы и нижний максимум ели
(зоны 1 и 2; рис. 125). Соответствует добореальному (суб¬
арктическому) более или менее холодному периоду. Возможно,
что »нижняя“ ель—это северная Picea obóvata.
2. Максимум сосны и березы (зоны 3,4,5).Соответ¬
ствует бореальному периоду.
3. Максимум смешанного дубового леса и
ольхи (зоны 6, 7, 8). Соответствует атлантическому периоду1 2.
4. Верхний максимум ели и абсолютный мини¬
мум березы (зоны 9, 10, 11). Субатлантический период.
5. Современный период (зона 12).
" Эти периоды выделяются на диаграмме по стратиграфическим уровням,
представляющим максимум кривых отдельных пород или точки пересечения
этих кривых. Эти уровни следующие (рис. 125):
a) пересечение кривых березы и ели,
b) максимум березы,
c) максимум ольхи,
d) верхний максимум ели.
Кроме уровней, выделяются еще 12 зон.
Запад Европейской части СССР.
Для западной области 3. Руофф (1936), по данным, главным
образом С. Тюремнова, устанавливает такие периоды:
1. Березово-сосновый период. Этот древнейший
период, повидимому, не установленный пока для центра Евро¬
пейской части СССР, относится или к арктическому или к суб¬
арктическому времени.
2. Период нижней ели, сосны и ивы (субаркти¬
ческий). Соответствует периоду с максимумом ели и ивы
в центре СССР.
3. Березово-сосновый период.
4. Березовый период.
5. Период ольхи и смешанного дубового леса.
Максимум последнего выражен здесь весьма резко (до 35°/о).
6. Сосновый период с верхним максимумом ели.
Восток Европейской части СССР (Казань).
Здесь выделяются такие периоды (3. Руофф 1. с.)3.
1. Период нижней ели и ивы (добореальный).
2. Сосновый период.
3. Березовый период.
4. Сосновый период с максимумом смешанного дубового
леса и ольхи. Максимум смешанного дубового леса здесь неве¬
лик и не превышает 20°/«.
5. Сосновый период.
1 М. Нейштадт (1929, 1933), М. Neustadt (1928).
2 Суббореальный период не имеет определенного пыльцевого спектра.
Контакт же между суббореальный и субатлантическнм периодом обозначается
на диаграмме близким схождением кривых всех пород.
3 По Д. Герасимову.
24*
371
Урал Ч
Для диаграмм Урала западного и восточного характерно
присутствие в нижних слоях, кроме ели и ивы, значитель¬
ных количеств пыльцы лиственницы. Таким образом, самый
древний период уральских болот — это время нижней ели,
лиственницы и ивы (3. Руофф, 1. с.). Этот период отмечен на
болотах как среднего, так и южного Урала. Другая особен¬
ность болот Урала — это то, что ель здесь не исчезает в атланти¬
ческий период, а дает непрерывную кривую, лишь несколько
снижаясь перед суббореальным подъемом. Третья особен¬
ность-присутствие пыльцы пихты, она появляется на Урале
поздно и в древних горизонтах отсутствует.
Западная Сибирь.
Немногочисленные пыльцевые диаграммы с юга хвой¬
ной зоны Западной Сибири (Васюганье1 2) во многом сходны
с уральскими. И здесь в нижних горизонтах торфяников
есть лиственница, что, может быть, дает возможность син¬
хронизировать их с уральскими. Однако пихта и кедр по¬
явились здесь рано — их пыльца встречается уже в ниж¬
них слоях. Притом процент кедра довольно высок (до 40°/©).
Раннее по сравнению с Уралом появление пихты и кедра в За¬
падной Сибири указывает на их миграцию с востока. Пыль¬
цевые диаграммы из Барабинской лесостепи3, как васюганские
и уральские, имеют пыльцу пихты и кедра. Первая отмечена
лишь у самой поверхности, вторая небольшим процентом
идет до дна. Интересно, что ель встречается лишь внизу за¬
лежи, в середине же исчезает совершенно, не встречаясь уже
до самой поверхности, где ее заменяет пихта.
Украина.
Для северо-западной Украины установлены такие лесные
периоды4:
1. Время ивы, березы, сосны. Из остальных древес¬
ных пород встречается лишь ольха и очень немного дуба (1,5е/©).
Вероятно, это время первого поселения древесной раститель¬
ности, когда настоящих сомкнутых лесов еще не было. Болота
этой стадии неизвестны. Отложения с пыльцой относятся
кседиментам. Предположительно этот период относится к суб¬
арктическому времени.
2. Время сосны. Резко преобладает сосна (80—93°/©)-
Кроме нее, встречаются те же породы, что и раньше. Вероятно,
в это время уже образовались настоящие леса, но они еще
не покрывали всей страны. В это время началось торфообра-
зование и отложился гипновый и гипново-осоковый торф.
Этот период, вероятно, относится к бореальному времени.
1 См. Известия Главного ботанического сада, 1930.
2 А. Бронзов (1930).
8 М. Нейштадт (1936).
4 Р. 2егош (Д. Зеров, 1934).
372
3. Время дуба и сосны. В этот период появляются и
начинают распространяться широколиственные породы—дуб,
вяз, липа, а также орешник. Все они в этом периоде образуют
уже непрерывную кривую. Появляется граб. В это время пре¬
обладали тростниковые и тростниково-осоковые болота. Атлан¬
тический период.
4. Время граба, бука и сосны. Характерен максимум
граба (иногда свыше 10%). Впервые начинается отложение сфаг¬
нового олиготрофного торфа на ранее образовавшихся эвтроф-
ных болотах. Олиготрофные сфагновые болота возникают
также и вновь, благодаря заболачиванию минеральной почвы.
Характерные отличия пыльцевых диаграмм Украины от
западноевропейских—это, во-первых, отсутствие времени
орешника и, во-вторых, резкое преобладание пыльцы сосны во
всех горизонтах. Первое представляет географическое явление,
повидимому, общего порядка, так как время орешника вообще
отсутствует на диаграммах Европейской части СССР, по край¬
ней мере хвойной зоны. Объясняется это климатическими усло¬
виями. Преобладание или большое содержание сосновой пыльцы
является, повидимому, особенностью зандровых областей, к ко¬
торым относится и северо-запад Украинского Полесья. В подоб¬
ных областях „сосновый” тип диаграммы, как мы это видели
выше, хорошо выражен и весьма устойчив на протяжении длин¬
ного послеледникового промежутка времени. Значительное уча¬
стие граба в последнем периоде отличает украинские диаграммы
от более восточных и сближает с польскими и восточнопрусски¬
ми. Обилие граба в пыльцевых диаграммах понятно, так как
изученный район входит в область его современного распростра¬
нения *. Ель в этих диаграммах отсутствует. Это тоже понятно,
так как район лежит в области островного распространения ели,
к югу от границы ее сплошного распространения. Следова¬
тельно, ель и раньше там не встречалась. Крайне слабо уча¬
стие ели и на других, пока немногочисленных, диаграммах
для южной окраины лесной зоны, например, района Пензы 2.
Северный край (западная часть).
Для Шенкурского района б. Архангельской губ. установлены
такие этапы истории лесов* * * * 8 (начиная от более древних).
1. Время сосны. Сосна преобладает над другими по¬
родами. На втором месте стоит береза, на третьем—ель.
Породы смешанного дубового леса вовсе отсутствуют.
2. Время березы. Береза абсолютно преобладает над
другими породами, за ней идет сосна, затем ель. К концу
периода — отдельные зерна пыльцы смешанного дубового леса,
главным образом, вяза.
1 О распространении граба теперь и раньше (по пыльце в торфе)
см. Г. Эрдтман (1934); карту о пыльце граба в торфе советских болот — см.
Известия Главного ботанического сада за 1930 г.
2 См. В. Доктуровский (1925).
8 М. Нейштадт (1930).
373
3. Время сосны, березы и ели. Все три породы
примерно в одинаковом количестве. На это время прихо¬
дится максимум дубового леса, правда, незначительный, не
свыше 3%- Много также пыльцы ольхи, иногда дающей
здесь максимум.
4. Время сосны и ели. Ель дает максимум, после чего
идет на убыль. Следующая по преобладанию порода — сосна.
5. Время сосны. Породы смешанного дубового леса
почти исчезают.
Пыльцевые диаграммы из района Онежского озера1 в не¬
скольких сотнях километров к западу от Шенкурска сходны
с только что описанными. За исключением наиболее древнего
соснового периода, не выраженного на онежских диаграммах,
вероятно, вследствие молодости болот, там можно выделить
все периоды, отмеченные для Шенкурского района. Максимум
ольхи и небольшой максимум смешанного дубового леса
и здесь приходится на период сосны, березы и ели. Последний
период выделяется и на диаграммах Карельского перешейка 1 2,
где на его начало также падает максимум широколиственных
пород. Этот период Е. Гииппе датирует литориновым временем.
Общий обзор послеледниковой истории лесов и расселение
древесных пород в Европейской части СССР.
Из предыдущего обзора мы могли убедиться, что отдельным
областям Европейской части СССР свойственны свои особые
„региональные“ пыльцевые спектры. Прежде всего, следует
отметить, что современный состав лесов опреде¬
ленной области в значительной степени имеет
след и в послеледниковой ее истории. Так,
в Украинском Полесье, где ель в настоящее время встречается
спорадически, эта порода и в прошлом принимала лишь
ничтожное участие в составе лесов.1 Сибирские породы — кедр,
лиственница и пихта, характерные только для диаграмм
Урала,—в настоящее время встречаются лишь на востоке
Европейской части СССР.
Из таблицы 53 видно, что послеледниковая история
лесов Восточноевропейской равнины от Урала до западной
границы Союза и от Украины до Ленинграда имеет много
общего. Первый добореальный сосново-березовый лесной
период (или арктическо-субарктический) изучен еще очень
слабо. Вторая же половина добореального периода на
востоке и в средней части СССР характери¬
зуется временем нижней ели. Так называемая „нижняя
ель“ (возможно, северная Picea obovata) дает здесь нижний
высокий максимум—обычно 30 — 40°/о> на Урале совместно
с лиственницей, и затем количество ее пыльцы резко падает.
1 К. Марков, В. Порецкий и Е. Шляпина (1934).
2 Е. Гииппе (1933).
374
Этот нижний выступ ели заметен и на диаграммах западной
части СССР (Ленинградская область, БССР), но здесь он ниже,
и ель подчинена сосне и березе, тогда как восточнее ель
часто абсолютно преобладает. Еще дальше на запад, вне
СССР, нижний максимум ели или едва намечен1, или вовсе
отсутствует*. Таким образом, „нижняя“ ель была широко
распространена в добореальный период в средней и северной
части Европейской СССР. Интересно отметить, что в этих
областях ель встречалась в лесах в течение всей после¬
ледниковой истории, и между нижней и верхней елью вовсе
или почти не было перерыва1 2 3. На западе (Ленинградская и б.
Западная области) верхняя и нижняя ель отделены друг от
друга промежутком, когда ель почти совсем сходила со сцены.
Следующий сосновый период всюду харак¬
теризуется господством сосны, часто дающей абсо¬
лютный максимум. Иногда этот период, повидимому, не от¬
граничен от следующего периода березы. Так, на Урале
различают период сосны и березы. В этот период появляется
смешанный дубовый лес и орешник, и кривая их пыльцы
делается непрерывной. На севере (Шенкурский район
б. Архангельской губ.) пыльца широколиственных пород в это
время еще совершенно отсутствует.
Следующий березовый период датируют
обычно атлантическим временем. Он характеризуется максиму*
мом березы, обычно абсолютным. Содержание березовой пыльцы
в период максимума — 50 — 65%. в районе Ленинграда—72е/#4-
Березовый максимум особенно отчетлив на северо-западе Евро¬
пейской части СССР (Ленинград)5. В березовое время продол¬
жается подъем кривой пыльцы смешанного дубового леса и ольхи.
На севере (Шенкурский район) широколиственные породы, глав¬
ным образом вяз, появляются лишь к концу березового периода.
Период максимума смешанного дубового
леса и ольхи относится к суббореальному, а по некоторым
авторам — к атлантическому времени. В это время широко¬
лиственные породы, ольха и орешник достигают своего
максимума за всю послеледниковую историю лесов. Максимум
смешанного дубового леса не везде одинаково выражен.
Абсолютного преобладания этот лес достигает в западном
типе пыльцевой диаграммы, где содержание пыльцы равно 35°/«-
Если учесть меньшую продукцию пыльцы дубового леса
1 В торфяниках Готланда нижняя ель дает всего до 5% [Г. Мунте,
И. Геде и Л. Пост (Н. Munthe, J. Hede och L. Post), 1927]; привожу по К. Мар¬
кову (1934).
2 В Средней Европе (К. Рудольф, 1930) нижний максимум ели отсут¬
ствует и начинает обозначаться лишь в восточных Альпах и Карпатах.
8 М. Нейштадт (1930).
8 Среднее, выведенное мной из очень большого количества диаграмм
Г. Ануфриева и В. Доктуровского.
8 Еще западнее (южная Швеция, Англия) мы имеем пыльцевые спектры
с еще более резким преобладанием березы. Однако этот березовый макси¬
мум вряд ли синхроничен с нашим.
375
по сравнению с хвойными породами, то это соотношение,,
несомненно, указывает на существование здесь широколиствен¬
ных лесов с примесью хвойных, и этот период можно назвать
временем господства широколиственного леса. В диаграммах
в средней части Европейской СССР, где средний максимум
широколиственного леса не превышает 23в/о» обычно же бывает
20% и ниже, где абсолютно преобладают береза и сосна,
лучше говорить о максимуме, а не о господстве смешанного
дубового леса. Слабо выражен этот максимум на Урале, где
он к тому же сильно запаздывает и примерно совпадает с по¬
явлением пихты, как и на севере (Шенкурский район), где
пыльца дуба и его спутников достигает самое большее 5%.
Период хвойных (сосново-еловый) соответствует суб¬
атлантическому времени. Этот период выражен на всех типах
диаграмм, кроме Украины, где он заменяется грабово-дубово¬
сосновым. Преобладание сосны или ели объясняется частью
географическим положением, частично характером почв района,
к которому относится диаграмма. Так, диаграммы юго-запад¬
ного типа (Украина) показывают почти полное отсутствие
еловой пыльцы; они относятся к району вне сплошного рас¬
пространения ели. Вообще бедные елью и очень богатые
сосной диаграммы Минского Полесья — пример диаграмм зан-
дровых песчаных областей, где сосна господствовала в течение
чуть ли не всей второй половины послеледникового периода.
Смешанный дубовый лес, ольха и орешник в этот период
идут на убыль, причем на севере (Шенкурский район) пыльца
смешанного дубового леса почти исчезает.
Послеледниковая история растительности СССР.
Освобожденная от льда территория Восточной Европы за¬
селилась сначала тундровой флорой примерно того же состава,
что и в Западной Европе. Под Ленинградом в ископаемом виде
были найдены относящиеся к этому периоду тундровые цвет¬
ковые растения и мхи1; карликовые ивы: Salix polaris, S. reti¬
culata, S. arbuscula, S. myrsinites, S. herbacea, Dryas octopetala,
Betula nana, Polytrichum Jensenii, Aulacomnium turgidum, Drepa-
nocladus latifolius, Calliergon turgescens и др. Betula папа и Dryas
были найдены также под Вологдой1 2 3. Недалеко от Вологды,
под Тотьмой, наряду с тундровыми видами, были найдены два
новых — Thalictrum alpinum и Armeria sibirica, последняя —
на расстоянии 650—350 км от ближайших современных место¬
нахождений 8. Почти все перечисленные растения встречаются
в настоящее время намного севернее их ископаемых место¬
нахождений, что, несомненно, указывает на холодный, аркти¬
ческий климат, существовавший на широте Ленинград—Вологда
1 Г. Ануфриев (1925), С. Яковлев (1926) и Ю. Цинзерлинг (1934), где
дан перечень находок тундровой флоры в окрестностях Ленинграда.
2 И. Перфильев и Г. Ширяев (1915).
3 К. Солоневнч и А. Корчагин (1934).
376
вскоре после отступания ледника. То же было и в Сибири.
Так, на Иртыше — в лесной зоне были найдены в торфе остатки
цветковой тундровой и альпийской флоры: Dryas, Salix polaris,
S. herbácea, Betula nana, Pachypleurum alpinum. Это указы¬
вает на то, что граница тундры в Сибири спускалась значи¬
тельно южнее, чем теперь. Очень рано среди тундры появились
первые представители .древесной растительности, главным
образом береза, сосна, ива, несколько позднее — ель. Эти
деревья росли разреженно, лесов еще не было. На месте
современного Балтийского моря в это время существовало
так называемое Балтийское пресное ледниковое озеро, питав¬
шееся талыми водами ледника и отделенное от океана.
С отступанием ледника температура начала повышаться,
и наступил длительный теплый период, захватывающий боре-
альное и атлантическое время. Произошли важные изменения
в растительности, указывающие на потепление климата. Так,
теплолюбивые растения, например Trapa natans, продвигаются
далеко на север по сравнению с нынешним их распространением
(рис. 122); в Ленинградской области массами произрастает
теплолюбивая Najas marina. Najas minor продвигается до-
Вологды километров на 500 севернее современной границы
(В. Кудряшев, 1923). Несомненно, что и широколиственные
древесные породы шли дальше на север, чем теперь. На это
указывают хотя бы находки дуба в аллювиальных отложениях
района Вологды1 и на южном берегу Ладожского озера1 2 3, где
дуб сейчас не растет.
Для Шенкурского района (б. Архангельской губ.) пыльце¬
вой анализ тоже как будто показывает, что широколиственные
породы — дуб, клен, вяз, липа, а также орешник—в более
раннее время шли дальше на север, чем теперь (М. Нейштадт,
1930).
В пользу существования теплого послеледникового периода
как в Европейской части СССР, так и в Сибири, можно
привести ряд фактов из различных областей. Сюда относятся:
1. Находки остатков древесной раститель¬
ности в тундре иногда далеко за пределами
современных лесов. Так, Сукачев (1922)* нашел в Кар¬
ской тундре в торфе остатки лиственницы, ели, березы (Betula
pubescens), ольхи (Ainus fruticosa), а также остатки сравнительно
теплолюбивых растений — малины и роголистника, растущих
теперь значительно южнее.
А. Зубков (1931) нашел в арктической тундре Западной Сибири, наряду
с видами Equisetura (Е. silvatlcum и Е. limosum), и Lycopodium (L. annotinum),.
которые теперь там не растут, также остатки березы (Betula alba).
2. Существование торфяников, иногда доста¬
точно глубоких, в высоких широтах — там,
1 М. Нейштадт (1930).
2 А. Иностранцев (1892).
3 Там же смотри и соответствующую литературу.
377
где торфообразования в настоящее время
почти не происходит. Образование мощных слоев торфа
могло происходить лишь в условиях более теплого климата,
чем современный. Подобные реликтовые торфяники в тундре
описывались неоднократно: торфяники на Новой Земле \ в За¬
падносибирской тундре1 2 3 4, в Карской тундре8 и других пунктах,
-а вне СССР А. Натгорстом в Северной Гренландии, где торфя¬
ники сейчас покрыты слоем льда, и Г. Андерссоном (1910) на
Шпицбергене, где они достигают 1,8 — 2,4 м мощности и где
в настоящее время торф не образуется. Эти торфяники часто
содержат остатки древесной растительности, и ботанический со¬
став торфа говорит за их образование в более теплом климате.
В современном климате обычно идет их эрозия и разрушение.
Немногочисленные пыльцевые анализы тундровых торфяников
подтверждают факт значительного продвижения в минувший
теплый период древесной растительности к северу. А. Егорова
(1930) по пыльце установила, что древесная растительность
в Карской тундре была распространена дальше на север, чем
теперь, в частности, сосна примерно на 200 км.
3. Реликтовые северные нахождения тепло¬
любивой растительности. Изолированные место¬
нахождения ильма (Ulmus montana) на полуострове Заонежье,
орешника в южной части Лодейнопольского округа, черной
ольхи на Беломорском побережье у Сороки — во всех случаях
много севернее их современного сплошного распространения—
указывают на их более широкое прежнее распространение,
как и островные северные местонахождения в Ленинградской
области Asperula odorata, Asarum europaeum и некоторых
.других (Ю. Цинзерлинг, 1934).
Несмотря на слабую изученность вопроса, в настоящее
время все же можно собрать довольно много фактов из раз¬
ных областей в пользу сухого и теплого послеледникового
времени. Эти факты следующие:
1. Строение торфяников. Существование погранич¬
ного горизонта в торфяниках СССР трудно объяснить иначе,
как наличием сравнительно сухого климата в послеледниковое
время, хотя возможно, что в это время (суббореальный период)
температура по сравнению с атлантическим периодом уже пони¬
зилась и что теплый послеледниковый период уже закончился.
2. Современное распространение степных
растений. Некоторые степные виды, например, Carex supina,
Sisimbrium junceum, Agroporum cristatum, Linum perenne входят
изолированными вкраплениями в Якутскую тайгу и тундру
северо-востока Сибири*. Проникли они на север, вероятно,
в теплое послеледниковое время. Большое значение тундрово-
степному элементу во флоре Арктики придает А. Толмачев
1 В. Кудряшев (1925).
г А. Зубков (1931).
3 В. Сукачев (1922).
4 Е. Лавренко (1933); привожу по Е. Вульфу (1936).
.378
(1927)'. Он считает, что в теплый и сухой период степные
»иды далеко распространились на север и проникли в тундру.
Признаки остепнения есть и в лесной зоне. Так, на верхней
Волге (б. Тверская губ.) А. П. Ильинский (1922) отмечает
■степные виды: Festuca sulcata, Clematis recta, Anthyllus poly-
phylla, Phleum Boehtneri, Verónica spicata, может быть, относя¬
щиеся к последней теплой и сухой эпохе, так же как степняки
в таежной части Горьковской области (б. Нижегородской губ.).
В Ленинградской области (в б. Псковской и б. Новгород¬
ской губерниях) реликтами ксеротермического периода явля¬
ются Anemone patens, А. pratensis, Astragalus arenarius, Koeleria
glauca, К. granáis, Filipéndula hexapetala и некоторые другие
(Ю. Цинзерлинг, 1934). Труднее связать с климатом нахожде¬
ние в Ленинградской области некоторых восточных (сибирских)
видов, а также находки в торфе ископаемой лиственницы
приблизительно в 600 км к западу от ее современной границы,
в б. Псковской губ.1 2.
3. Факты из области почвоведения и физи¬
ческой географии также говорят в пользу сухого периода.
Приведем некоторые3.
Ископаемые черноземовидные почвы, принадлежащие
прежним степям и покрытые сверху подзолистой почвой
и тайгой, найдены в Нарыме Д. Драницыным (1914). Это
указывает как на сухой климат в прошлом, так и на последо¬
вавшую позднее смену его более влажным, благодаря чему на
степь надвинулся лес.
Согласно Д. Виленскому (1924) и К. Гедройцу (1926) осоло¬
девшие почвы на юге Европейской части СССР, в чернозем¬
ной полосе Западной Сибири и в Якутии произошли из
солонцов, а эти в свою очередь — из солончаков в результате
выщелачивания, связанного с увеличением влажности климата.
Таким образом, сухая послеледниковая эпоха была солонча¬
ковой, а современная, более влажная, характеризуется дегра¬
дацией (осолодением) солонцов.
Нахождение в южнорусских степях на лёссе чернозема
указывает также, что климат раньше был суше, а теперь сде-
. л алея более влажным. Деградация черноземов указывает на
надвигание леса на степь и, следовательно, на смену климата.
О том же говорит и увеличение площади осиновых кустов
в Хреновской степи (б. Воронежской губ.) за последнее время.
Возникновение дюнных ландшафтов (например, около Се-
•строрецка) также связывают с сухим климатом (С. Яковлев, 1926).
В отношении истории лесов теплая эпоха характеризовалась
-сначала господством сосны (бореальный период), а затем
березы (атлантический период). В это время в заметном коли¬
честве появились широколиственные породы и орешник, и
1 По Е. Вульфу (1936).
2 Б. Арапов (1914), Н. Кузнецова (1927).
» См. Л. Берг (1931).
379
количество их, постепенно возрастая, достигло вместе с ольхой
максимума распространения.
Климат с наступлением суббореального периода начал
ухудшаться. Наступил прохладный и сравнительно влажный
субатлантический период, повлекший за собой изменения
растительности в сторону современных условий. С ухудшением
климата теплолюбивые травянистые растения, например, Trapa
natans, и широколиственные породы начали отступать к югу
от современной границы их распространения. Острова степной
растительности на севере сократились, лес начал наступать
на степь, и произошла деградация чернозема. Некоторые ис¬
следователи (Л. Берг, 1931) считают, что в настоящее время
с ухудшением климата происходит общее смещение ботанико¬
географических зон к югу, т. е. тундра надвигается на лес,
лес — на степь и т. д. В умеренной климатической зоне про¬
исходит интенсивное накопление сфагнового (молодого) торфа,
а на севере торфообразование прекращается, и во многих
случаях начинается разрушение образовавшихся ранее торфя¬
ников. В лесах увеличивается роль хвойных — сосны и ели,
широколиственные же породы идут на убыль.
Послеледниковая история лесов Северной Америки.
Лучше изучена история лесов юго-восточной Канады и сосед¬
ней части США в районе Великих Озер. Картина смены дре¬
весных пород всюду весьма сходная. Намечаются три периода
развития лесов, начиная с более древнего:
1. Время хвойных лесов. Господствуют и особенно
характерны ель и пихта, а также сосна.
2; Время максимума широколиственного
леса. В районе озера Мичигана этот период характеризуется
господством (относительным) леса этого типа. Главный ком¬
понент здесь дуб; кроме того, встречаются клен, вяз. Коли¬
чество пихты и ели резко падает, местами они почти исчезают.
К северу в общем их количество возрастает и, например, уже
для Северного Мичигана можно говорить о развитии в этот
период хвойно-широколиственного леса.
3. Время возрастания количества хвойных
пород и убывания широколиственных.
Порядок появления древесных пород в Канаде такой:
раньше всего появились ель, сосна, пихта й береза. Позднее
пришли вяз, орешник (или сходная с ним по пыльце Ostrya),
ольха, дуб и Сагуа; еще позднее Tsuga, Tilia, Juglans. Наконец,
самые поздние пришельцы — Fagus и Пех — появились в конце
периода, соответствующего, может быть, нашему бореальному,
или в начале атлантического. На юге (район Мичигана) Tsuga
также появилась сравнительно поздно (в середине второго
лесного периода), одновременно с грабом. Порядок расселения
пород во многом напоминает Европу. И здесь первыми
поселились сосна и береза, а на востоке рано появилась и ель-
380
Широколиственные породы и в Европе пришли позднее, а бук
и граб — последними. Однако пихта в Европе и на Урале
в отличие от Северной Америки появилась поздно.
Американские исследователи, например Р. Sears (1935), признают суще¬
ствование для Северной Америки, подобно Европе, следующих периодов:
периода возрастания температуры, теплого послеледникового периода
и периода ухудшения климата. Здесь главное значение придается темпе¬
ратуре. Г. Фуллер (G. Fuller, 1935), выделяя те же климатические периоды,
соответствующие лесным, особенно подчеркивает, что в области Мичигана
не было смены влажных и сухих периодов, а было изменение температуры.
Вопрос о послеледниковых климатах, впрочем, далек от разрешения, как
мы видим, даже в Европе, а тем более в Америке.
История растительности четвертичного периода.
Граница между четвертичным периодом и плиоценом до сих
пор геологами еще не установлена (А. Жирмунский, 1936).
Вместе с тем до сих пор не только не решен вопрос о числе
оледенений, новее еще имеются „моноглациолисты", т.е. сто¬
ронники одного оледенения. Громадное большинство гео¬
логов признает теперь множественность оледенений (от двух
до шести, даже до семи). Разногласия зависят во многом от не¬
определенности самого понятия „оледенение". А. Жирмунский
различает шесть оледенений и четыре межледниковые эпохи.
Приводим их ниже с датировкой Миланковича. Эти оледенения и меж¬
ледниковые эпохи вместе составляют четвертичный период К
1. Предгюнцкая ледниковая эпоха .... 682—832 тысяч лет 2 * * * * *.
2. Гюнцкая ледниковая эпоха 546—682 * *
3. Межледниковая эпоха 477—546 * ,
4. Миндельская ледниковая эпоха .... 430—477 * ,
5. 2-я межледниковая эпоха 234—430 * ,
6. Рисская ледниковая эпоха 185—234 ^ „
7. 3-я межледниковая эпоха 118—185 „ „
8. Вюрмская ледниковая эпоха 113—118 » .
9. 4-я межледниковая эпоха 74—113 „ „
10. Неовюрмская ледниковая эпоха .... 74— 9 „ „
11. Послеледниковая (холоден) 9— 0 , ,
Чередование ледниковых и межледниковых эпох довольно хорошо от¬
ражает минимум и максимум на вычисленной Милакковичем для четвертич¬
ного периода кривой солнечной радиации.
При чередовании оледенений и межледниковых эпох арктическая расти¬
тельность сменялась лесной с рядом теплолюбивых видов. При этом опре¬
деленные стадии развития растительности в общем повторялись в после¬
дующие оледенения и межледниковые эпохи, как это можно видеть из при¬
водимой ниже таблички 8.
1 Некоторые авторы, как П. Бек (Р. Beck, 1933) и Г. Гаме (1935), от¬
носят древнейшие оледенения к плиоцену.
2 Другие авторы (Г. Гаме, 1933) сокращают продолжительность четвер¬
тичного времени до 300 тысяч лет и, вдвигая в этот промежуток времени
все оледенения, начиная с Миндельского, сокращают продолжительность
ледниковых эпох.
8 Не надо думать, что это сходство было полным. Отдельные эпохи
четвертичного периода сильно отличались по продолжительности, темпера¬
туре и, несомненно, по растительности (Г. Гаме, 1935).
381
Стратиграфия польского дилювия на флористической основе*
(по В. Шаферу).
s
<и
а
&\
и
>
И«
с g
■§8
> с
S »
2
ш
JQ О
as я
с я
U
*
О)
I. Наступание ледников (трансгрессия С).
Стадия кустарниковой тундры (флора пока неизвестна).
II. Оледенение С: флора Dryas.
III. Отступание ледников (регрессия С): стадия кустар¬
никовой тундры. I. II. III. IV. V. VI. VII.I. Стадия лесотундры.
II. Стадия сосново-дубовых лесов.
III. Стадия смешанных лесов с сосной, пихтой, грабом и
другими широколиственными породами.
IV. Стадия [широколиственных лесов — оптимум темпе¬
ратуры.
V. Стадия смешанных лесов, в общем напоминающая III.
VI. Стадия бореальных хвойных лесов с участием еще
широколиственных пород.
VII. Стадия субарктических хвойных лесов уже без
широколиственных пород.
Е.
V (9
Ж О.
S <и
з с
о S
>
I. Трансгрессия Vi: стадия кустарниковой тундры.
II. Оледенение Vx: флора Dryas в средней и северной
Польше, кустарниковая тундра и лесотундра в южной.
III. Регрессия V* (флора пока неизвестна).
Межледниковая ступень VjV2 (Masovien II): леса с уча¬
стием граба, бука, тисса (изучены недостаточно).
с
0>
I
со
**
(С
>
I. Трангсрессия V2 (растительность неизвестна).
И. Оледенение V^ флора Dryas.
III. Регрессия V2: кустарниковая тундра.
Послеледниковая ступень.
и
Э
3
п
О
С
[туры | температуры
Мы видим, таким образом, что на территории б. Польского государства
кульминационные точки трех оледенений — С, Vlf V2 — имели сходную расти¬
тельность (флора Dryas). Температура в это время упала до минимума,
и климат был арктический. С приближением межледниковой эпохи темпе¬
ратура начинала повышаться, и флора Dryas уступала место кустарниковой',
тундре (с Betula nana и другими кустарничками) с первыми деревьями—
Betula tortuosa, Ainus viridis и др. С дальнейшим повышением температуры
среди тундр формируются первые леса (стадия лесотундры).
С наступлением межледниковой эпохи лесотундра сме¬
нилась лесом. Примерно те же стадии, но в обратном порядке
проходила растительность с приближением нового оледенения
и понижением температуры. В течение межледниковой эпохи
растительность тоже изменялась. Лесные стадии сменялись
по мере приближения температуры к максимуму в середине
межледниковой эпохи. Это хорошо видно на интерглациале
c/v. Температурный оптимум этой эпохи характеризовался
широколиственными лесами богатого видового состава из липы,,
кленов (Acer tataricum, A. campestre), ясеня, орешника, граба
и других пород. Хвойные в этих лесах заметной роли не
играли. Богата была и водная флора этого периода. На тер¬
ритории б. Польши, кроме обычных водных растений,
в это время встречались третичные виды, в настоящее
время вообще в Европе1 частью вымершие: Nvmphaea
lotos, Tsuga (из древесных пород), Brasenia, Dulichium
spathaceum, частью вымирающие в Европе и на терри¬
тории б. Польши, — Aldrovanda vesiculosa, Najas flexilis, виды
Trapa и др. С приближением оледенения v2 климат ухудшается,,
увеличивается роль хвойных, сосны, пихты, Tsuga—стадия
смешанных лесов. Широколиственные породы — Acer tataricum
и Brasenia начинают вымирать. С дальнейшим ухудшением-
климата доминируют хвойные леса с елью, сосной, листвен¬
ницей. Теплолюбивые водные растения вымирают. Развиваются
выпуклые торфяники. Следующая стадия — субарктические
чисто хвойные леса из сосны, лиственницы, без широколиствен¬
ных пород. Начинается расселение тундровых видов (Betula,
nana) из реликтовых местонахождений. С началом оледенения
наступает тундровая стадия. Сходные лесные периоды наблю¬
даются и в первой половине интерглациала c/v при повы¬
шении температуры от оледенения С к максимуму интергла¬
циала c/v.
На пыльцевых диаграммах из межледниковых эпох удается
выделить лесные периоды. Предпоследний интерглациал в
б. Польше дает такую картину (К. Piech, 1932) (рис. 126).
1. Фаза сосново-березовых лесов
2. „
сосново-ду б овых
3. „
дубовых
4. „
дубово-липовых
5. „
липово-грабовых
6. „
грабово-еловых
3 См. В. Шафер (1930).
383
7. Фаза пихтовых
лесов.
8. Фаза пихтово¬
еловых лесов.
9. Фаза сосновых
лесов с тремя под¬
фазами.
Мы видим, что
смена лесов напо¬
минает в общих чер¬
тах послеледнико¬
вую. И там первая
после отступания
ледника стадия бе¬
резово-сосновых ле¬
сов сменяется широ¬
колиственными ле¬
сами, за ними сле¬
дуют хвойные—пих¬
товые или еловые
леса.
Хотя флористиче¬
ский состав четвер¬
тичного периода и
был близок к совре¬
менному, однако,как
уже говорилось, в
это время существо¬
вали также и те ви¬
ды, которые в совре¬
менную эпоху в Ев¬
ропе вымирают или
вовсе вымерли. Глав¬
ным образом, это
были третичные ви¬
ды. Проследим
вкратце историю не¬
которых из них в
Европе (см. Г. Гамс,
1935).
1. Виды, вымер¬
шие раньше всего,
которых в послед¬
нюю межледниковую
эпоху уже не было
в Европе: Tsuga (два вида—Т. diversifolia и Т. canadensis), Euryale
ferox, Dulichium vespiforme. Из них Tsuga живет в современную
эпоху в Америке и Японии, Euryale ferox—в Маньчжурии и южнее.
2. Виды, вымершие в Европе уже в последнюю межлед¬
никовую эпоху: Picea omorikoides, Brasenia purpurea, Dulichium
384
spathaceum. Picea omorikoides имеет близкий вид Picea omorlka,
живущий еще и теперь на Балканах, Dulichium spathaceum
живет еще в Северной Америке, a Brasenia, кроме того,
в Африке и Восточной Азии.
Кроме названных видов, до последней межледниковой
эпохи в Тюрингии удержались Fraxinus americana, Thuja ос-
cidentalis, Pavia и Juglans.
Из слонов в миндель-рисскую межледниковую и рисскую
ледниковую эпоху жил Elephas meridionalis, от него произошли
позднейшие виды: Е. antiquus, Е. trogontherii и Е. primigenius.
Первый из них вымер в Средней Европе в начале последнего
оледенения, Е. primigenius — еще раньше, а от Е. trogontherii
произошел мамонт, просуществовавший до конца плейстоцена.
Из носорогов Rhinoceros etruscus, современник Elephas meridio¬
nalis, вымер немного позднее его — в рисс-вюрмскую меж¬
ледниковую эпоху. Из позднейших видов Rhinoceros, сменив¬
ший R. etruscus, R. Merckii вымер в начале последнего оледе¬
нения, a Rhynoceros trichorhinus, современник мамонта, вымер
вместе с последним.
Из предков человека — питекантроп жил в середине
миндель-рисской эпохи, гейдельбергский же человек жил
в конце ее, а также в рисскую эпоху. Неандертальская
раса существовала в рисскую (позднее гейдельбергского чело¬
века), рисс-вюрмскую эпохи и захватила также последний
интерглациал. Ориньякская и кроманьонская расы жили уже
в последнем оледенении.
В СССР первые сведения о межледниковой флоре отно¬
сятся еще к 1890 г. (Н. Криштофович). С 1904 г., со времени
описания Н. Боголюбовым Лихвинского отложения в 1904 г.,
интерес к этой области значительно возрос. Наиболее же
интенсивно эта область развилась лишь за последние 12 лет.
Сводка современных знаний по межледниковой растительности
СССР составлена В. Сукачевым (1936). В середине плиоцена,
сведения о котором вообще весьма скудны, флора Европейской
части СССР, судя по находкам П. Никитина (1928)1 в Воронеж¬
ской области, имела еще много азиатских и североамери¬
канских видов (Azolla, Brasenia, Decodon и др.). К концу плио¬
цена, по данным П. Никитина (1933) для Сызрани и И. Пали-
бина (1936) для Азербайджана, флора была в общем близка
современной, хотя и встречались вовсе вымершие или нерасту¬
щие здесь в настоящее время виды. Древнейшая межледниковая
эпоха, изученная в СССР, это миндель-рисская, куда относится
знаменитое Лихвинское отложение, давшее главный материал
по этой эпохе. Флора нижних озерных отложений Лихвинского
местонахождения носила северный характер. Пыльца принад¬
лежала сосне, ели, пихте и березе. Пыльцы широколиственных
пород и теплолюбивых водных растений здесь не найдено.
Это указывает на то, что климат был во всяком случае не ii Привожу по В. Сукачеву (1936).
25-722 v385
теплее современного. Выше начинает встречаться пыльца
широколиственных пород, что указывает на потепление. На¬
хождение здесь пыльцы падуба (Ilex), семян Najas, Euryale,
спор Salvinia указывает на климат более теплый, чем совре¬
менный. В суглинках и супесях, поверх озерных отложений,
встречена лишь пыльца сосны, ели, ивы и березы. Это может
указывать на похолодание, зависящее от приближения оледе¬
нения. Таким образом, и здесь намечается закономерность
в смене растительности, отмеченная выше для польского ди¬
лювия,— переход от более северной растительности к более
теплолюбивой и затем опять к более северной. Впрочем, для
достоверных заключений пока еще мало данных. Несомненно
одно, что в эту межледниковую эпоху одно время климат был
теплее и влажнее, чем теперь, и имелись виды, к настоящему
времени уже вымершие. Несравненно лучше исследованы рисс-
вюрмские межледниковые отложения, известные из 46 пунктов.
Список флоры этой эпохи содержит свыше 125 видов цветковых
растений и более 25 видов мхов. Эта флора близка к совре¬
менной, вымерших видов вовсе нет, но есть виды, которых
нет в современной флоре. Особенно интересен американский
вид Brasenia purpurea, указывающий на климат, более теплый,
чем современный. О том же говорит нахождение в средних
широтах Европейской части СССР Salvinia natans, Aldrovanda
vesiculosa, Trapa natans, Najas marina, Najas flexilis. Все они
были шире распространены, чем теперь. В середине этой эпохи
южные древесные породы продвигались на север и частью
на восток дальше, чем теперь. Так, Acer tataricum и Tilia pla-
typhyllos доходили до Москвы, а граб—до Галича или, может
быть, еще севернее. Ilex же заходил на запад Евро¬
пейской части СССР. Интересно отметить, что северная Sela-
ginella selaginoides, а также Picea obovata и Abies доходили
в эту эпоху до Москвы, т. е. южнее, чем теперь. Подобные
находки говорят за то, что временами климат рисс-вюрмского
интерглациала был холоднее современного. Пыльцевой анализ
рисс-вюрмских отложений (В. Доктуровский, 1931; И. Покров¬
ская, 1936) показал, что здесь имеются три стадии лесной
растительности:
1) Н ижняя (ближайшая к рисскому оледенению) с преоб¬
ладанием или ели, или березы, сосны и ивы, или сосны;
2) Средняя, где, кроме пыльцы этих пород, встречается,
а иногда преобладает пыльца смешанного дубового леса и
орешника;
3) верхняя (ближайшая к вюрмскому оледенению) —
с преобладанием сосны и ели.
Нижняя стадия, вероятно, соответствует холодному климату
после рисского оледенения, верхняя —также холодному кли¬
мату перед вюрмским оледенением, а средняя—климатическому
оптимуму в середине интерглациала.
ГЛАВА XI.
ИСПОЛЬЗОВАНИЕ БОЛОТ.
Болота в своем естественном состоянии, как это указы¬
валось в начале этой книги, могут быть использованы чело¬
веком лишь с весьма малой эффективностью. Польза, которую
при этом извлекают из болот, обычно далеко не покрывает
вреда, приносимого ими. Использование болот в их естест¬
венном виде без вложения в них труда и капитала называется
экстенсивным. Наиболее экстенсивные виды использо¬
вания— сбор ягод, грибов, ловля рыбы в болотных озерах
и т. д. Большой эффект дает рубка леса на лесных болотах,
далее сенокошение и пастьба скота. Сенокошение широко
практикуется на неосушенных болотах, в том числе и на тор¬
фяных, и на заболоченных землях. Наибольшее значение
в этом отношении имеют открытые (безлесные) травяные
болота, где главную массу травостоя образуют осоки, реже
злаки. Еще до самого последнего времени в северной части
Украины, отчасти и севернее широко практиковалось сено¬
кошение даже на болотах, где травостой образуют жесткие,
узколистные осоки, имеющие весьма ограниченную кормовую
ценность. Гораздо большую ценность имеют так называемые
крупно-осоковые заливаемые болота в речных поймах. Эти
болота, из которых наибольшее хозяйственное значение, по¬
жалуй, принадлежит осочникам из хорошо поедаемой скотом
в молодом состоянии Сагех gracilis, луговеды относят отчасти
к заливным лугам низкого уровня. Это едва ли не единствен¬
ный тип болот, использование которых и без особых мелио¬
ративных предприятий дает недурной хозяйственный эффект.
Интересно отметить, что крупно-осоковые болота средней
полосы Европейской части СССР в большей своей части воз¬
никли из лесных, эвтрофных, менее ценных в хозяйственном
отношении болот в результате вырубания леса и ежегодного
сенокошения.
Однако все же в целом болота в их естественном виде
являются балластом для народного хозяйства и имеют лишь
потенциальную ценность, которая реализуется лишь после
вложения в них значительного труда и капитала. Вопрос
об интенсивном использовании болот не ставится в настоящий
387
момент, по крайней мере в широких размерах, лишь для слабо
обжитых районов на северной границе земледелия — в лесо¬
тундре и тундре. Здесь, на ближайший отрезок времени,
использование болот следует вести по линии организации на
них упорядоченной пастьбы скота, в частности оленей. Не
исключается впрочем и использование торфа в ограниченном
размере как топлива и как удобрения.
В более же обжитых южных районах вопрос об интенсив¬
ном использовании болот уже давно поставлен на очередь
самой жизнью и в особенно широких масштабах осущест¬
вляется Советской властью. Интенсивное использование болот
в СССР и за границей ведется в основном по двум линиям:
по линии с.-х. мелиорации и по линии торфодобычи.
Хотя из двух упомянутых направлений эксплоатации у нас
в СССР гораздо большее значение имеет торфодобыча, однако,
необходимо упомянуть и о грандиозных мероприятиях Совет¬
ской власти по осушке с мелиоративной целью огромных
заболоченных пространств (в Колхиде, Белоруссии, Западных
областях, в Московской области (по реке Дубне) и в других
районах).
В СССР вопросы культуры болот разрабатывались и раз¬
рабатываются в основном опытными учреждениями (Минской
болотной опытной станцией, Новгородской болотной стан¬
цией и др.). В Карелии, Полесье и некоторых других районах
культура болот внедряется уже в практику в довольно широ¬
ких размерах. При культуре болот последние превращаются
в полевые или в луговые угодья. Основные мероприятия по
культуре болот заключаются прежде всего в осушке, затем
в раскорчевке леса, кустарника и пней, удалении кочек с по¬
следующей вспашкой и боронованием. Интенсивность осушки
(частота канав) варьирует в зависимости от предполагаемой
культуры и влажности климата. Для определенных геогра¬
фических районов установлено опытным путем расстояние
между осушительными канавами. У нас культивируются
эвтрофные (низинные) болота, за границей также и олиго-
трофные (верховые). Не только олиготрофные, но в боль¬
шинстве случаев и эвтрофные нуждаются в искусственном
удобрении, последние, главным образом, во внесении фосфора
и калия.
Следует особо упомянуть об орошении болот. Оно обога¬
щает торфяник минеральными солями, улучшает физические
свойства почвы и приносит вместе с водой семена ценных
кормовых трав. В результате урожай трав сильно повышается
и улучшается качество травостоя.
В последние годы орошение болот изучалось на Яхром¬
ской болотной опытной станции под Москвой.
Гораздо большее значение в СССР имеет эксплоатация
торфа. При наличии громадной площади болот Советский
Союз обладает колоссальными запасами торфа. Эти за¬
пасы составляют около 3,5% суммы всех топливных ре-
388
сурсов СССР, причем торф стоит на втором месте после
каменного угля. Эти громадные запасы составляют по неко¬
торым расчетам свыше 62°/« мировых запасов торфа. Соответ¬
ственно той роли, которую играет торф в Советском Союзе,
его добыча приобрела при Советской власти гигантский раз¬
мах. К началу третьей пятилетки (1938 г.) торфодобыча у нас
увеличилась против довоенного времени более, чем в 16 раз.
Торфяная промышленность СССР занимает теперь первое место.
Громадный рост торфодобычи тесно связан с переходом
к механизированным методам добычи. Вместо полуручного
элеваторного способа в дореволюционной России, теперь торф
добывается механизированными способами — гидроторф, бо-
иерный, фрезерный. При этом первые два способа дают куско¬
вой торф, последний — фрезерную крошку. В настоящее
время можно наметить такие основные линии использования
торфа:
1. На топливо, частью после соответствующей переработки
(коксования).
2. Для получения технических продуктов, в результате
переработки на химических заводах.
3. Для целей строительства.
4. Для нужд сельского хозяйства.
5. Для нужд медицины и санитарии.
Подавляющее количество добываемого у нас торфа идет
на топливо. Основные потребители торфа — электростанции.
В дореволюционное время на торфе работала всего одна не¬
большая электростанция—электропередача под Москвой, мощ¬
ностью в 15 тыс. кет. В настоящее время на торфе рабо¬
тают девять мощных районных электростанций общесоюзного
значения (НИГРЭС, Шатурская и другие), с общей мощ¬
ностью в сотни тыс. кет. Эти электростанции дают энергию
крупным промышленным центрам — Москве, Ленинграду,
Горькому, Иваново, Ярославлю и др. Кроме того, на торфе
работает ряд электростанций местного значения. Торф упо¬
требляется как бытовое топливо. Из торфа (фрезерная крошка)
на специальных заводах (в Орехове Московской обл.) изго¬
товляются плотные, не крошащиеся кирпичи-брикеты, которые
хорошо горят в печах. Из кускового торфа после обжига
получается высококачественное топливо — торфяной кокс,
который по своей теплотворной способности лишь немного
уступает лучшим сортам каменного угля.
Кроме использования на топливо непосредственно или
после соответствующей обработки, торф дает ряд других
ценных технических продуктов. Так, из него добывается газ,
а получающаяся при газофикации и коксовании торфяная
смола при дальнейшей переработке на специальных торфо¬
химических заводах дает ряд высокоценных продуктов —
воск, пек, креолин, фенолы и пр.
Применяется торф и для нужд строительства. Из мало
разложившегося сфагнового торфа приготовляются так назы¬
389
ваемые изоплиты, употребляющиеся в строительстве и пред¬
ставляющие прекрасный утепляющий материал. В СССР
имеется четыре специальных торфоизоляционных фабрики.
В сельском хозяйстве торф находит многообразное приме¬
нение. Мало разложившиеся сфагновые торфа дают высоко¬
качественную подстилку для скотных дворов. По энергии
поглощения жидких и газообразных выделений эта подстилка
значительно превосходит соломенную. Кроме подстилки, торф
представляет хорошее удобрение. Торф можно использовать
на удобрение как в виде торфяного навоза, так и непосред¬
ственно. В первом случае торфяная подстилка пропускается
через скотные дворы и затем в виде торфяного навоза выво¬
зится в поле. Торфяной навоз дает большие прибавки уро¬
жая, чем соломенный; Непосредственно на удобрение с боль¬
шим успехом, особенно на песчаных почвах применяются
низинные торфа. При этом хорошо разложившиеся торфа
следует предварительно проветрить и несколько подсушить.
Слабо разложившиеся торфа складываются сначала в кучи
с известью или золой и навозом — так называемый торфяной
компост. Верховые торфа во всяком случае следует про¬
пустить через компостные кучи, где торф переслаивается из¬
вестью. Торфофекалин представляет ценное удобрение для
огородов. Кроме удобрения сфагновый торф (в виде порошка)
в смеси с патокой служит для приготовления торфомеляссо-
вого корма для скота. Западноевропейская практика давно
уже дала положительную оценку этого корма. К тому же
выводу привели и опыты, производившиеся у нас. В торфя¬
ном порошке хорошо сохраняются овощи и фрукты. Следует
упомянуть еще об изготовлении из торфа цветочных горшков.
Торф применяется для санитарных и медицинских целей.
Сфагновой подстилкой засыпают выгребные ямы, чем создается
среда малоблагоприятная для развития бактерий. Торф при¬
меняется и для бальнеологических целей (лечебные грязи).
ЛИТЕРАТУРА.
Аарио Л. (Aario L.), Pflanzentopographische und paläogeographische Moor¬
untersuchungen in N — Satakunta, Fennia, 55, Nr. I, 1932.
— Vegetation und postglaziale Geschichte des Nurmijärvi-Sees, Ann. Bot. Soc.
Zool. — Bot. Fenn. Vanamo, T. 3, Nr. 2, 1933.
Аболин P., Постоянная мерзлота грунтов и ископаемый каменный лед,
Записки Читинского отделения Приамур. отдела Русск. геогр. об-ва,
вып. IX, 1913.
— Опыт эпигенологической классификации болот, Болотоведение, 1914.
— Болотные формы Pinus silvestrb, Тр. Бот. музея Росс, академии наук,
XIV, 1915.
— К вопросу о классификации болот Северо-западной области, Изд. Гос.
ин-та с.-х. мелиораций, М. 1928.
Алабышев В., Краткая программа по исследованию болот и торфяников
как месторождений сапропелитов и родственных им отложений, Изв.
Сапропел. комитета, вып. 4, Л. 1928.
— Зональность озерных отложений, там же, вып. 6, 1932.
Андерссон Г. и Гессельман Г. (Andersson G. u. Hessel-
man Н.), Vegetation und Flora im Staatsforst „Harania-Kronopark*. Mitt.
der staatl. forstlich. Versuchsanst. Schwedens, H. 4, 1908.
Андреев В.. Растительность тундры Северного Канина, сборн. „Оленьи
пастбища Северного края*, Архангельск 193L
— Кормовые ресурсы оленеводства в западной части Большеземельской
тундры, сборн. „Оленьи пастбища Северного края", Л. 1933.
— Растительность и природные районы восточной части Большеземельской
тундры, Тр. полярной комиссии Акад. наук, вып. 22, Л. 1935.
Ануфриев Г., О болотах Кольского полуострова, Работы организован¬
ного в 1920 г. Геогр. ин-том Кольского почвенно-ботанического отряда,
вып. III, Петроград 1922.
Архипов С., Заболачивание и типы леса в Котласском леспромхозе, 1932.
Ауэр В. (Auer V.), Über die Entstehung der Stränge auf den Torfmooren,
Acta Forest. Fennica, 12, 1920.
БакманА. (ßackman A.), Torvmarksundersökningar i mellersta Osterbotten,
Acta forestalia Fennica, 12, 1919.
Б e гак Д., К вопросу о приросте верховых торфяников, Торф, дело, № 9/
1926.
— О приросте торфяников, Торф, дело, № 11 —12, 1927.
— Прирост торфяника Галицкий Мох.,Тр. Научно-исслед. торф, ин-та, вып. I,
1928.
— Перспективы торфодобычи на Севере, Гос. геол. изд., 1932.
БегакД. и Беликова Н., Количество и распределение микроорганизмов
в верховых торфяниках, Тр. Научно-исслед. торф, ин-та, вып. 14, 1934.
Берш В. (Bersch W.), Handbuch der Moorkultur, Wien und Leipzig 1909.
Бертш К. (Bertsch К.), Das Brunnenholzried, Veröffentl. der Staatl. Stelle
für Naturschutz bei Württ., Landesamt für Denkmalpflege, H. 2, 1925.
Благовещенский Г., Эволюция растительного покрова болотного
массива 10Э7 км у ст. Лоухи (Карелия). Тр. Бот. ин-та Акад. наук СССР,
сер. III, сборн. „Геоботаника", вып. 3, 1936.
391
Богдановская-Г иенэф И., Ключевые болота Кингисеппского у. Ле¬
нинградской губ., Журнал Русск. бот, об-ва, т. И, 1926.
— Растительный покров верховых болот русской Прибалтики, Труды Петер-
гофского ест.-науч. ин-та, М® 5, 1928.
— Образование и развитие гряд и мочежин на болотах, Сов. ботаника»
№ б, 1936.
Бооберг Г. (Booberg G.), Gisselasmyren, Norrländskt Handbibliotek. Bd.
XII, Uppsala 1930.
Бронзов А., Верховые болота Нарымского края, Тр. Научно-исслед.
торф, ин-та, вып. 3, М. 1930.
Буренков В., Кащеев А., Мальчевская Н.9 Материалы по изуче¬
нию процессов заболачивания сплошных лесосек в Лисинском леспром¬
хозе, Тр. Лесотехнической академии, Nk 4 (42), 1934.
Бюлов К. (Вü 1 о w К.), Allgemeine Moorgeologie,Handbuch d. Moorkunde,I»
Berlin 1929.
— Die Oberharzer Vermoorung als geologisches Phänomen, Cbl. f. Miner, u.
Geol., Abt. B, Nr. 2, 1932.
Бюсген M. (Büsgen M.), Kieselpflanzen auf Kalkboden, Englers»Fes»band
der Bot Jahrb. f. System etc., Bd. 50, Suppl., 1914.
Вальяжников В., Торф как изоляционный и консервирующий материал,
изд. Инсторфа, 1924.
Вангерин В. (Wangerln W.), Über die Anwendung der Bezeichnung
Hochmoor in der Pflanzengeographie, Bot. Arch., Bd. XV, 1926.
Варен Г. (Waren H.), Untersuchungen über die botanische Entwicklung der
Moore, Wissensch. Veröffentl. d. Finnischen Moorkulturvereins, Nr. 5,
1924.
Варлыгин П., К вопросу о схеме отношений между образованием болот
в Европейской России и физико-географическими условиями среды,
Вести, торф, дела, Nfc 1 — 2, М. 1932.
— Определение степени разложения торфа, Торф, дело, № 9, 1925. -
Варлыгин П. и Чеботарев В., Методика практического полевого
определения степени разложения торфа, Вести, торф, дела, № 3, 1929.
Варминг Е. (Warming ЕА Om Grönlands Vegetation, Meddel. от Grön¬
land, 12, 1888.
— Lehrbuch der ökologischen Pflanzengeographie, Berlin 1896.
Вебер К. (Weber C.), Über die Moore, mit besonderer Berücksichtigung
der zwischen Unterweser und Unterelbe liegenden, Jahresbericht der Män¬
ner vom Morgenstern. Heimatband an Elb-und Wesermündung, H. 3, 1900.
— Über die Vegetation und Entstehung des Hochmoores von Augstumal im
Memeldelta, Berlin 1902.
— Главнейшие виды гумуса и торфа и их участие в строении северогер¬
манских болот. Почвоведение, № 4, 1908.
— Was lehrt der Aufbau der Moore Norddeutschlands über den Wechsel des
Klimas in postglazialer Zeit, Zschr. d. Dtsch. Geol. Ges., 62, 1910.
Вильямс В., Общее земледелие, ч. II, М. 1922.
— Земледелие с основами почвоведения, 2-е изд., М. 1931.
Властова Н., Торфяные болота нижнего течения реки Оби, Тр. Центр.
торф, опытной станции НКЗ, т. I, М. 1937.
Вольни Е. (Wollny Е.), Untersuchung über den Einfluss der Pflanzendecke
und der Beschattung auf die physikalischen Eigenschaften des Bodens,
Forschungen aus dem Gebiete der Agrikulturphysik, 10, 1888.
— Die Zersetzung der organischen Stoffe und Humusbildungen mit Rücksicht
auf die Bodenkultur, Heidelberg 1897.
Гаме Г. (Gams H.), Das Graumoos (Racomitrium lanuginosum) und seine
Begleiter als Torfbildner, Mikrokosmos, 24 Jahrg., H. 10, 1930/31.
— Die höhere Wasservegetation. Handbuch der biologischen Arbeitsmethoden
(методика и общие вопросы изучения водной растительности).
— Die Geschichte der Lunzer Seen, Moore und Wälder, Intern. Revue der
gesamten Hydrobiologie und Hydrographie, Bd. XVIII, H. 5/6, 1927.
— Von den Follateres zur Dent de Mordes, Pflanzeng. Kom. d. Schweiz.
Naturf.-Ges., Beitr. z. geobot. Landesaufn., 15, 1927.
Гаме Г. и Нордхаген (Gams H. und Nordhagen R.), Postglaziale
392
Klimaänderungen und Erdkrustenbewegungen in Mitteleuropa, Landeskund¬
liche Forschungen, H. 25, 1923.
Ганс Г. и Руофф 3. (Gams Н. und Ruoff S.), Geschichte, Aufbau und
Pflanzendecke des Zehlaubruches, Schrift, der Phys.-Ökon. Ges. zu Königs¬
berg i. Pr., Bd. LXV1, H. 1, 1929.
ГанонгВ. (Ganong W.), Upon raised peat Bogs in the province of New
Brunswick, Transact. roy. Soc. of Canade, II ser., v. III, Sect. IV, 1897.
Гау rep B. (Gauger W.), Untersuchungen über die Biozönose und die Phy¬
siognomie eines ostpreussischen Hochmoores (Zehlau) im Jahresprofil, Diss.,
Königsberg 1929.
Гебель К. (Goebel KO, Pflanzenbiologische Schilderungen, Marburg, 1889u.
1891.
Генкель А., Водная и болотная флора б. Курганского округа западно¬
сибирской лесостепи, Уч. зап. Перм. гос. ун-та, т. I, вып. 4, 1935.
Г еоботаника (сборник), Тр. Бот. ин*та Акад. наук СССР, серия III, вып. 3,
Л. 1936.
Геринг П. (Ноеring Р.), Moornutzung und Torfverwertung, Berlin 1915.
Гетманов Я-, К вопросу об эволюции лугов и болот, Л. 1925.
Городков Б., Наблюдения над жизнью кедра (Pinus sibirica Mayr.) в За¬
падной Сибири, Тр. Бот. музея Акад. наук, вып. XVJ.
— Вечная мерзлота и растительность. Материалы Кепс. Акад. наук СССР,
№ 80 (отдельный оттиск).
— Крупнобугристые торфяники и их географическое распространение,.
Природа, № 6, 1928.
— Вечная мерзлота в Северном крае, Тр. Совета по изучению производи¬
тельных сил, серия северная, вып. I, изд. Акад. наук, Л. 1932.
Растительность тундровой зоны СССР, изд. Акад. наук СССР, 1935.
Гранлунд Э. (Granlund Е.), De svenska Högmossarnas Geologi. Sveri-
ges Geologiska Undersöknings Arsbok, 26, 1932.
Гребне p П. (Graebner P.), Bildung natürlicher Vegetationsformatione«
im norddeutschen Flachlande, Arch. der Brandenburgia, IV, 1898.
— Die Heide Norddeutschlands, Leipzig 1901.
— Handbuch der Heidekultur, Leipzig 1904.
Г ризебах A. (Griesebach A.), Ober die Bildung des Torfes in de«
Emsmooren, 1845.
Гросс Г. (Gross H.), Ostpreussens Moore mit besonderer Berücksichtigung
ihrer Vegetation, Schriften der Physik.-ökon. Ges., Jg. LII1, 1912.
Гулли E. (Gully E.), Ober die Beziehungen zwischen vegetationchemischer
Zusammensetzung und Düngebedürfnis der Moore, Mitteil, der K. Bayer.
Moorkulturanstalt, H. 3, 1909.
— Untersuchungen über die Humussäuren, Mitteilungen der K. Bayer. Moor¬
kulturanstalt, H. 5, 1913.
Гюк К. (Hueck KO. Die Vegetation und Oberflächengestaltung der Ober¬
harzer Hochmoore, Beitr. z. Naturdenkmalpflege, Bd. XII, H. 2, Berlin 1928.
— Die Vegetation und die Entwicklungsgeschichte des Plötzendiebels, Beitr.
z. Naturdenkmalpflege, Bd. XIII, Berlin 1929.
Дау И. (D a u J.), Über die Torfmoore vSeelands, Kopenhagen u. Leipzig 1829.
Дахновский A. (Dachnovski A.), The toxic propertyofbog-water and
bog-soil, Bot. Gaz., 46, 1908.
— Bog toxins and their effect upon solls, Bot. Gaz., 47, 1909.
— Physiologically arid habitats and drought resistance in plants, Bot. Gaz.,
— Peat deposits of Ohio, their origin, formation and uses, Geol. Survey of
Ohio, 4 th ser., bull. 16, 1912.
Де Лаппаран (De Lapparent A.), Traité de Géologie, 4-me ed., 1900.
Д e Л io к И. (De Luc J.), Physicalische und moralische Briefe über die Ge¬
schichte der Erde und des Menschen, Leipzig 1782.
Доктуровский В., Торфяные болота, 1936.
Донат A. (Donat А.), Die Vegetation unserer Seen und die biologischen
Seetypen, Ber. d. Dtsch. Bot. Ges., 44, 1926.
Драни цын Д., Вторичные подзолы и перемещение подзолистой зоны на
севере Обь-Иртышского водораздела. Изв. Док. почв, комитета, II, №2,1914.
393
Дюгелли М. (M. D ü g g е I i). Pflanzengeographische und wirtschaftliche Mo¬
nographie des Sihltales bei Einsiedeln. Vierteljahrsschr. der Naturf.-Ges. in
Zürich, Jg. 48, H. 1 u. 2, 1933.
Д ю P и э Г. (du Rietz G.), Die regionale Gliederung der skandinavischen
Vegetation. Svenska Växtsoc. Sällskapets Handl., VIII, Uppsala 1925.
Ельман В. (Öhlmann V.). Vegetative Fortpflanzung der Sphagnaceen
nebst ihrem Verhalten gegen Kalk, Diss., Freiburg 1898.
Жозефи Г. (Josephy G.), Pflanzengeographische Beobachtungen auf eini¬
gen schweizerischen Hochmooren, Diss., Milt. Bot. Mus. Univ. Zürich, Wien
1920.
Зальфельд (S а 1 f e 1 d), Geographische Beschreibung der Moore des nord¬
westlichen Deutschlands, Landw. Jahrb., XII, 1883.
3 e и д t he p O. (Sen.dtner O.), Die Vegetationsverhältnisse Südbayerns,
München, 1854.
3 у б ко в А., Тундры гусиной земли, Tp. Бот. музея Акад. наук, вып. XXV,
1932.
Известия сапропелевого комитета, вып. 1 —6, 1923 —1932.
Какс А., Болота окрестностей озера Дулова, Мат. по изуч. восточного
болотного района Псковской губ, III, 1914.
Кац Н., Отчет об исследовании болот Богородского у. Московской губ.,
Материалы по опытному делу Моек, губ., вып. 16, М. 1922.
Katz N., Sphagnum Bogs oi Central Russia, Phytosociology, Ecology and
Succession, J. Ecol., v. XIV, Nr. 2, 1926.
— Sphagnummoore im nördlichen Teile des Moskauer Gouvernements, Бюлл.
Моек, об-ва испыт. .природы, т. XXXVI, вып, 3 —4, 1927.
— Покос и пастьба как факторы, регулирующие жизнь сырых лугов,
Научно-агрон. журн., № 1, 1926.
— О типах олиготрофных болот Европейской части СССР и их широтной
и меридиональной зональности, То. Научно-исслед. ин-та при физ.-мат.
фак. МГУ, М. 1928.
— Zur Kenntnis der Moore Nordosteuropas, Beili. z. Bot. Cbl., Bd. XLVI,
Abt. II, 1930.
— Zur Kenntnis der Moore des Fernen Ostens, Ber. Dtsch. Bot. Ges., Bd. L,
H. 6, 1932*
— К а ц С., Атлас растительных остатков в торфе, Сельхозгиз, 1933.
— Кудряшев Л., Эпштейн В., О торфяниках дельты Северной
Двины, Землеведение, т. XXXVII, вып. 4, 1935.
— О торфяно-болотных образованиях Европейской части Союза и их гео¬
графическом распространении, Бот. журн. СССР, 1936.
— Минкина Ц., Торфяные болота северной части бассейна реки Пе¬
чоры, Сборн. „Торфяные болота Коайнего Севера и Азиатской части
СССР“, ОНТИ, 1936.
— Кириллович М., Лебедева Н., Движение поверхности сфагновых
болот и формирование их микрорельефа, Землеведение, т. XXXVIII,
вып. 1, 1936.
К а ян дер A. (Cajander A.), Die Alluvionen des unteren Lena-Tales, Acta
Soc. Sc. Fenn. XXXII, Nr. 1, 19027
— Ein Beitrag zur Entwicklungsgeschichte der Nordfinnischen Moore, Fennia,
20, Nr. 6, 1902— 1S03.
Кернер A. (Kerner A.), Pflanzenleben, I Aufl., 1887.
Кириллович M. и Короткина M., Инструкция по определению
ботанического состава торфа. Сборник инструкций по исследованию
торфа и торфяных залежей, изд. Всес. ин-та торфа, М. 1933.
Клеве А. (Cleve A.), Zum Pflanzenleben in nordschwedischen Hochgebir
gen, 1901.
Контежан K. (Contejean C), Géographie botanique. Influence du ter¬
rain sur la végétation, Paris 1881.
Könne Ф. (Koppe F.), Die biologischen Moortypen Norddeutschlands, Ber.
Dtsch. Bot. Ges., Bd. 44, 1926.
Корнгубер A. (Kornhuber A.), Botanische Ausflüge in die Sumpfniede-
rung des „Waser“, Verh. der k.-k. zool.-bot. Ges., Bd. XXV, Wien 1886.
Костяков А., Перспективы мелиорации в СССР, Госплан СССР, М. 1925.
394
Коссович П., Основы учения о почве, ч. II, вып. 1, СПБ 1911.
Котилайнен М. (Kotilainen М.), Untersuchungen über die Beziehungen
zwischen der Pflanzendecke der Moore und der Beschaffenheit, besonders
der Reaktion des Torfbodens, Finska mosskulturforeningen, Nr. 7, 1928.
Краус Г. (Kraus G.), Boden und Klima auf kleinstem Raum, Jena 1911.
Кудряшев В., К вопросу о пограничном горизонте среднерусских торфя¬
ников, Вести, торф, дела, № 4, 1918, М. 1920.
Купфер, Лекции о растительности болот для слушателей курсов по куль¬
туре болот и луговодству при Рижском политех, ин-те, 1913.
Левицкий К., К вопросу об эволюции болот Амурской области, Почво¬
ведение, № 2, 1910.
Лекере (Lesquereux), Untersuchungen über die Torfmoore im Allgemei¬
nen, Berlin 1847.
Лепин Л., Торфяной фонд на начало 1936 г.
Лесков А., Северный предел распространения кедра (Pinus slbirica Mayr.)
и сосны (Pinus silvestrls L.) в бассейне реки Полуя, Бот. журн. СССР,
№ 1. 1935.
Лейнинген В. Гр. (W. Gr. zu L е i n i n g е n), Die Waldvegetation praealpiner
bayerischer Moore, Naturwlss. Zschr. f. Land- und Forstwirtschaft, Mün¬
chen 1907.
Ливингстон Б. (Livingston B.), Physiologlcal propertfes of bog water
Bot. Gaz., 39, 1905.
Лоренц И. (Lorenz J.), Allgemeine Resultate aus der pflanzengeographischen
Untersuchung der Moore des praealpinen Hügellandes Salzburg, Regens¬
burg 1858.
Луидквист Г. (Lundquist G.), Bodenablagerungen und Entwickelungs¬
typen der Seen, Die Binnengewässer, Bd. II, Stuttgart 1927.
Макаров И. и Нейштадт М., К истории литературы по торфу, Торф,
№ 3—4, 1930.
Маляревский К., Колонизационное обследование территории, отведен¬
ной Мурманской ж. д., Сборн. »Второй год колонизационной работы
Мурманской ж. д.", Л. 1926.
Мальмстрем К. (Malm ström С.), Degerö Storniyr, Meddel. fran Statens
Skogsförsöksanstalt, H. 20, Nr. 1, 1923.
— Über die Gefahr der Versumpfung des Waldbodens in Norrland (Nord¬
schweden), Meddel. fran. Statens Skogsförsöksanstalt, H. 26, Nr. 1, 1931.
МевиусВ. (Mewius W.\ Beiträge zur Physiologie „kalkfeindlicher* Ge¬
wächse, Jahrb. f. wissensch. Bot., Bd. 60, 1921.
Мелин E. (Melin E,), Studier över de Norrländska myrmarkernas Vegetation,
Norrlänkst Handbibliotek, VII, Uppsala, 1917.
Метсевайнио К. (Metsävainio К.), Untersuchungen über das Wurzelsy¬
stem der Moorpflanzen, Ann. Bot. Soc. Zoologicae-Botanicae Fennicae Va-
namo, T. I, Nr. 1# 1931.
Миддендорф А., Путешествие на крайний север и восток Сибири, т. I,
ч. IV, Растительность Сибири, СПБ 1867.
Мильде И. (Milde I.) Übersicht über die Schlesische Laubmoosflora, Bot.
Zeit., 1861.
Монфор К. (Mon fort C.), Die Xeromorphie der Hochmoorpflanzen als Vor¬
aussetzung der physiologischen Trockenheit der Moore, Zschr. f. Bot., 10,
Мюнихсдорфер Ф. (Münichsdorfer F.), Über Almbildung und einen
interglazialen Alm in Südbayern, Geognost. Jahresheft, 1927, XL. Jg., Mün¬
chen 1928.
Науман E. (Nauman E.), Einführung ln die Bodenkunde der Seen. Die
Binnengewässer, Bd. IX, Stuttgart 1930.
Нейштадт M. и Эйдельман Г., Геобэтанический анализ торфа, Минск
1935.
К истории развития озер в послеледниковое время, Почвоведение, № 2,1936.
— Торфяные болота Барабинской лесостепи, Тр. Центр, торф, станции, т. I,
1936.
— Короткина М., Торфяные болота юго-восточной Камчатки, Тр.
Центр, торф, ст., т. I, 1936.
393
Нильсон A. (Nllsson A.), Nagra drog ur de svenska Växtsarnhällenos
utvecklingshistoria, Botanlska Notiser, 1899.
Нордхаген P. (Nordhagen R.). Kalktuffstudien aus dem zentralen Norwe¬
gen, Naturwiss. Wschr., Nr. 10, 1922.
— Die Vegetation und Flora des Sylenegebietes. I. Die Vegetation, Norske Vi-
denskaps-Academi in Oslo, IMatem.-Naturvidensk. Klasse, 1927, Nr. I, 1928..
Овчинников П., Очерк растительности поймы реки Волхова от дер.
Завижье до с. Пчевы, Мат. по исследованию реки Волхова и его бас¬
сейна, вып. IX, Л. 1926.
Ольсен К. (Olsen С.), Studier over Jordbundens Brintlonkoncentration ög.
dens Betydnlng for Vegetationen saerlig for Plantefordelnlngen 1 Nafuren,
Meddeleiser ira Carlsberg Laboratoriet, 16 Bd., Nr.l, Kobenhavn, 1921.
— Studles on the hydragenion concentration of the soil and its signiflcance
to the végétation, especially to the natural distribution of plants, С.—г.,
trav. Labor. Carlsberg, 15, Nr. 1, Copenhague, 1923.
О н о ш к о Б., Болотоведение, Сельхозгиз, 1931.
— Культура болот с основами болотоведения, вып. II и III, М., Сельколхоз-
гиз, 1931 и 1932.
Освальд Г. (Oswald Н.), Die Vegetation des Hochmoores Komosse, Sven-
ska Växtsoclologiska Sällskapets Handlingar, I, Uppsala 1923.
— Die Hochmoortypen Europas, Festschrift, C. Schröter, Veröfientl. des Geog..
Institutes Rübel in Zürich, H. 3, 1925.
— Zur Vegetation der ozeanischen Hochmoore in Norwegen, Svenska Växtso-
ciol. Sällskapets Handlingar, VII, Uppsala 1925.
Отоцкий П,, Грунтовые воды, Труды опыт, леснич., 1905.
Паасио И. (P a a s 1 о I.), Über die Vegetation der Hochmoore Finnland«,.
Acta Forestalia Fennica, 39, 1933.
Павлов H., Флора Центрального Казахстана, ч. 1, 1928.
Патридофилус (Patridophilus), Danske Ekonom. Mag., Nr. XIV.
Пауль Г. (Paul H.), Die Kalkfeindlichkeit der Sphagna und ihre Ursache,.
Mitteil. Bayer. Moorkulturanstalt, H. 2, 1908.
— Руофф 3. (PaulH. u. Ru off S.), Pollenstatlsli sehe und stratigraphische
Mooruntersuchungen im südlichen Bayern, I u. II Ber. Bayer. Bot. Ges.,.
Bd. XIX u. XX, 1927-1930.
Перфильев И., К методике изучения иловых отложений. Тр. Бородинской
преснов. биол. станции в Карелии, т. V, Л. 1927.
Пирсэлл В. (Pearsall W.), The aquatic végétation of the englisch lakes,.
Journ. of Ecology, v. VIII, Nr. 3, 1920.
Поле P. (Pohle R.), Pflanzengeographische Studien über die Halbinsel Kanis
und das angrenzende Waldgebiet, T. I, Acta Horti Petro poli ta ni, T. XXI,
Fase. 1, Petersb. 1903.
— К биологии сибирского кедра, Йзв. Бог. сада, т. XIII.
Пол ынов Б. и Юрьев М., Лахтинская впадина, Изв. Научно-мелиор.
ин-та НКЗ, вып. 8—9, 1925.
Пост Л. (Post L.), Das genetische System der organogenen Bildungen Schwe¬
dens, Comité internat, de Pédologie, IV Comiss., Nr. 22.
Пост Л. и СернандерР. (Post L. u. Sernander R.), Pflanzenphy-
siognomische Studien auf den Torfmooren im Närke, Stockholm 1910.
— Die stratigraphische Zweigliederung schwedischer Hochmoore, Sverig. Geo-
logiska Undersökning, Arsb. 6, Nr. 2, 1912.
— Einige südschwedische Quellmoore, Uppsala 1916.
— Гранлунд E. (Post L. och Granlund E.), Södra Sveriges Torvtillgon-
gar I., Sver. Geol. Unders., Arsb. 19, 1925.
— Einige Aufgaben der regionalen Moorforschung, Sveriges Geologiska Under¬
sökning. Ser. C., Nr. 337, Stockholm 1926.
Потонье Г. (H. Potonié), Die Entstehung der Steinkohle und der Kaust o-
biolithe überhaupt, Ber.in 1910.
— Die rezenten Kaustoblolithe und ihre Lagerstätten, Bd. II, Die Humus-Bil¬
dungen, Abh. Preuss. Geol. Landesanst., N. F., H. 55, II, Berlin 1911.
Прасолов Л. (Prasolow L.), On the distribution of bogs and bogs soils
over the northwestein lake regfon of the USSR, Sec. internat. Soil Science
Congress, 1930.
396
Пристли И. (Priestley L), Ecology ofMoorland Plants, Nature, 114,1924.
Пухнер Г.. Торф, перев. с нем. под ред. И. Вихляева и П. Танеева, Но¬
вая деревня, М. 1929.
ПфаффенбергК. и ГассенкампВ. (Pfaffenberg К. u. Has¬
se n к а m р W.), Über die Versumpfungsgefahr des Waldbodens im Syker
Flottsandgebiet, Abh. Naturwiss. Ver., Bremen, Bd. XXIX, H. 1/2, 1934.
Раббов Г. (Rabbow G.), Beitrag z. Kenntnis der Vegetationsverhältnisse des
Kieshofer Moores. Mltt. Natur. Ver. f. Neuvorpommern und Rügen, 50/51,
Greifswald, 1925.
Работнов T., О болотах Архангельского округа, Торф, дело, JSfe 5, 1930.
Работнов Т. и Меднис Я., Ольха как азотособиратель, Природа, № 6,
1936.
Раманн Е. (Ramann Е.), Bodenkunde, 3, Auf)., Berlin 1911.
Р а н к е н Г. (Rancken Н.), Torfmarkernas utweckling i Lappland, Finska Moss-
kulturfören. Arsbok., 1911, Heisingf. 1912.
P e й н e p M. (Rayner M.), The ecology of Calluna vulgaris, p. I а. II, The
New Phytologist, v. XII а. IX, 1913 a. 1921.
Риг г Г. (Rigg G.), The effect of some puget sound bog waters on the root
hatrs of Tradescantia, Bot. Gaz., 55, 1913.
Рикет Г. (Ricket H.), A quantitative Study of the larger aquatic Plants of
Lake Medote, Transact. Wisc. Acad. of Sei., Arts et Leiters, 20, 1922.
Рудольф К., Bisherige Ergebnisse der botanischen Mooruntersuchungen in
Böhmen, Belh. z. Bot. Cbl., Bd. XLV, Abt. II, H. 1, 1928.
— Фирбас Ф. иЗигмондГ. (Rudolf К., F i r b a s F. und Sigmond H.).
Das Koppenplanmoor im Riesengebirge, Naturwiss. Zschr. «Lotos», Bd. 76,1928.
Рупрехт Ф. (Ruprecht F.), Ein Beitrag zur Frage über die Zeitdauer, welche für
Sumpf- und Torfbildung notwendig ist, Btill. Acad. Sc., St. Petersbourg, t. VII.
Руттнер Ф. (Ruttner F.)t Über die Kohlensäureassimilation einiger Wasser¬
pflanzen in verschiedenen Tiefen des Lunzer Untersees, Intern. Revue der
ges. Hydrobiol. u. Hydrographie, Bd. XV, H. 1/2.
P у о ф ф 3., Die Aufsaugungsvermögen des Torfmooses und seine Verwend¬
barkeit als Verbandstoff, Mitt. d. Vereins zur Förder. der Moorkultur im
Deutschen Reiche, H. 14—15, 1918.
— Морфология и возраст прослоек в верхней толще сфагнового торфа
среднерусских болот, Труды Научно-исслед. торф, ин-та, вып. 14, 1934.
Обзор русской ботанической литературы по болотам и торфу за 1925—
1934 гг., Сов. бот., № 4. 1936.
С*амюэльссон Г. (Samuelsson G.), Scottisch peat mosses, Bull. Geol. Inst.,
Uppsala, X, 1910.
— Untersuchungen über die höhere Wasserflora von Dalarne, Svenska Växtso-
ciol. Sällskapets Handlingar, IX, Uppsala 1925.
— Die Verbreitung der höheren Wasserpflanzen in Nordeuropa (Fennoskandien
und Dänemark), Acta Phytogeographica Suecica, VI, Uppsala 1934.
Ситенский (Sitensky), Ober die Torfmoore Böhmens, 1891.
Скин M. (Skene M.), The Acidlty of Sphagnum and its Relation to Chalk
and mineral Salts, Ann. Bot., v. XXIX, London 1915.
СкоттсбергК. (Skottsberg C.), The Vegetation of Easter Island, The
natural Hlstory of Juan Fernandez and Eastern Island, v. II.
Соловьев M., Проблема сапропеля в СССР, изд. Акад. наук, 1932.
Стадии ков Г., Химия торфа, Госиздат., М.—Л. 1930 (1-е изд.), 1932
(2-е изд.).
Степанов С., Физико-химические особенности почв лесных гарей, Тр. по
лесному опытному делу, вып. II (LXV1), 1925.
Стоклаза И. (Stoklasa I.), Über die Verbreitung des Aluminiums in der
Natur und seine Bedeutung beim Bau und Betriebsstoffwechsel der Pflan¬
zen, Jena 1922.
Сукачев В., Материалы к изучению болот и торфяников озерной области,
Тр. преснов, био л. стан. СПБ о-ва естествоисп., т. II, 1906.
— К вопросу о влиянии вечной мерзлоты на почву. Изв. Росс. Акад. наук, 1911.
— Болота, их образование, развитие и свойства, 3-е изд., Л. 1926.
Тантту А. (Tanttu А.), Über die Entstehung der Bülten und Stränge der
Moore, Acta Foiestalia Fennica, 4, 1915.
397
Танфильев Г., О болотах Петербургской губ., Труды Вольного экономи¬
ческого об-ва, № 5, 1889.
ТинеманА. (Thinemann А.), Seetypen, Die Naturwissenschaften, 192L
— Die Binnengewässer Mitteleuropas. Eine limnologische Einführung, Die Bin¬
nengewässer, 1, Stuttgart 1926.
Тихомиров Б., Пожары зарослей кедрового стланика в Пенжинском
крае, Бот. журн. СССР, т. 18, № 6, 1933.
Тольф Р. (То 1 f R.), Svenska Mosskulturförenlngens Tidskriit, 1897.
Томсон П. (Thomson Р.), Der Einfluss der üaseruption auf die Oberilä-
chenformen der Hochmoore, Bot. Arcb., 8, 1924.
Транзо E. (Transeau E.), The bogs and bog flora of the Huron River Val¬
ley, IV P., Bot. Gaz., 41, 1906.
T у н м а p к C. (Thunmark Sven), Bidrag tili kännedomen om rechta kalk-
tuffer, Geoi. Förening. i Stockh. Förhandlingar, Now.-Dec., 1926.
Тюремнов С., Микроклиматические исследования на верховом болоте.
Тр. Научно-исслед. торф, ин-та, вып. 1,М. 1928.
— Истомина Е., Овсянникова М. и др., Классификация видов
торфа (проект стандарта), М. 1935.
Т ю шов В., Изв. Русск. географ, об-ва, т. XXXVII, № 2, 1906.
Т о л п а С. (Т о 1 р а S.), Krasowe torfoviska kolo Rözanyn a Polesu (Карстовые
болота у Рожана в Полесье), Acta Soc. Bot. Polon., v. IX, Suppl., War¬
szawa 1932.
Филарский Ф. (Filarsky F.), Von den Torflagern am Fusse der Hohen
Tatra, Jahrb. d. Karpat. Vereins, XX, 1893.
Фирбас Ф. (Firbas F.), Untersuchungen über den Wasserhaushalt der
Hochmoorpflanzen, Jahrb. f. wissensch. Bot., Bd. 74, H. 4/5, 1931.
Флеров А., Изучение и исследование болот, Вести, торф, дела, № 1, 1914.
Флейшер (Fleischer), Die Eigenschaften des Hochmoorbodens als land¬
wirtschaftliches Kulturmedium, Landw. Jahrb., 1891.
Фосс и Цигеншпек (Foss und Z i e g e n s p e c k), Zur Biozönose des
Moorwaldes, Bot. Arch., 25,1929.
Фойгт И. (Voigt I.), Versuch einer Geschichte der Steinkohlen, der Braun¬
kohlen und des Torfes, Weimar 1802 (1782?).
Фриз T. (Fries Th.), Botanische Untersuchungen im nördlichsten Schweden,,
Akad. Abh., Uppsala u. Stockholm 1913.
Фрю И. иШретер К. (Früh J. und Schröter C.), Die Moore der
Schweiz, Bern 1904.
Хаглунд (Haglund), Brandtheorie, реф. в журн. Болотоведение, 1912.
Цайлер В. и Вильк Л. (Z aller V. und W i 1 k L.), Über den Einfluss der
Pflanzenkonstituenten auf die physikalischen und chemischen Eigenschaften
des Torfes, Zs ehr. f. Moorkultur u. Torfverwendung, 1907.
Цинзерлинг Ю., Очерк растительности болот по среднему течению
р. Печоры, Изв. Главн. бот. сада СССР, т. XXVIII, вып. 1—2, 1929.
Чернова -Лепилова Г., Верховые торфяники Куровицкого плато, Тр.
Петерб. естеств. научн. ин-та, JN® 5, 1928.
Чильман А. (Kihlmann А.), Pflanzenbiologische Studien aus Russisch-
Lappland, Acta Soc. Fauna et Flora Fennica, 6, 1890.
Чильман Ф. (Kjelmann F.), Über den Pflanzenwuchs an der Nordküste
Sibiriens, Die wissenschaftlichen Ergebnisse der Vega-Expedition, 1883.
Швенденер C. (Sch wendener S.), Die Schutzscheiden und ihre Ver¬
stärkungen, Abh. Akad. Wiss., Physik. Kl., Abh. 3, Berlin 1882.
Шеин и ков А. и Болотовская Р., Введение в геоботаническое обос¬
нование организации пастбищ на севере, Труды Вологодской обл. с.-х.
опытной станции, вып. 1, Вологда 1927.
— Волжские луга Средне волжской области, Л. 1930.
— Голубева М., Растительность болотного участка Архангельского
болотного опытного поля, Тр. Арханг. болотн. опытн. поля, вып. 1, Ар¬
хангельск 1930.
Шейхцер И. (Scheuchzer J.), Von dem Züricher Torf, Beschreibung der
Naturgeschichte des Schweizerlandes, T. I, Zürich 1706.
Шимпер A. (Schlmper A), Pflanzengeographie auf physiologischer Grundr
läge, Jena 1898.
398
Шлаффнер Г. (Schlaffner Н.), Die geographischen Bedingungen der
Moorbildung in Deutschland, Diss., 1921.
Шрейбер Г. (Schreiber H.), Moorkunde, Berlin 1927.
Шуберт E. (Schubert E.), Zur Geschichte der Moore, Marschen und Wäl¬
der Nord Westdeutschlands, Mitt. der Provinzialstelle für Naturdenkmal¬
pflege, H. 4, 1933.
Эбермайер (Ebermayer), Einfluss der Wälder auf die Bodenfeuchtigkeit, 1920.
Эйдельман Г., Торфяные болота Дальне-Восточного края, Торфяные
болота Крайнего Севера и Азиатской части СССР, 1р. Центр, торф,
опытной станции, т. I, 1936.
— Торфяные болота Енисейского севера, Тр. Центр, торф, опытн. станции
НКЗ, т. I, М. 1936.
Эрдтман Г., Literature on pollen-statistics published before 1927. Geol.
Förening. i Stockh. Förh. Mars—Apr., 1927.—Literature on poll, statistics
publish, during thé Jears 1927—1929. Ibidem, Mars—April, 1931. —Liter,
on poll, statist, and related topics publish. 1930 and 1931. Ibidem,
Nov.— Dec., 1932. — Liter, on poll, statist, and related topics, publish,
1932 and 1933. Ibidem, Mai — Okt. 1934 (Библиография литературы по
пыльцевому анализу по отдельным странам).
Эрдтман Г. (ErdtmanG. a. Erdtman Н.), The improvement of Pollen¬
analysis Technique, Sv. Botanisk Tidskrift, Bd. 27, H. 3, 1933.
Эй зелен И. (Eiselen J.), Handbuch zur höheren Kenntnis des Torfwe.
sens 1795.
Юрьев M., Шуваловский торфяник .Черная гора", Изв. Научн. мелиор. ин-та,
вып. 11—12, 1925.
Яковлев С., Наблюдение над влиянием леса на грунтовые воды в тече¬
ние 1904 г., Изв. Гос. ин-та опыт, агрон., т. III, № 2—4, 1925.
Япп Р. (Japp R.), On the stratification of a marsh, and its relation to evapo¬
ration and temperature, Ann. Bot., 23, 1909.
— Spiraea Ulmaria L. and its bearing in the problem of xeromorphy in marsh
plants, Ann. Bot., 26, 1912.
Каяндер A. (Cajander A), Studien über die Moore Finnlands, Helsing¬
fors 1913.
Никитин П., О речной растительности рр. Воронежа, Дона и Усмани,
Народное хозяйство, 1925.
Ши м пер A. (Schimper A.), Pflanzengeographie auf physiologischer
Grundlage, 3 Aufl., herausgegeben von D-r F. Faber, Bd. I и il, 1935.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
А. Бронзов, .Почвоведение" N® 2, 1936.
Е. Вульф, Введение в историческую географию растений, Сельколхозгиз,
1932.
Б. Городков, Труды Дальневосточного филиала Академии наук.
A. Жирмунский, Материалы по четвертичному периоду СССР, 1936.
Е. Лавренко, В1стник природознавства, Ns 3—4, 1927.
К. Марков, Труды комиссии по изучению четвертичного периода, т. IV,
вып. 1, 1934.
B. Матюшенко, Труды Научно-исследовательского Торфяного Инсти¬
тута, в. 1, 1928.
В. Матюшенко, Торфяные болота Украины, Торфяное дело, 1925.
Д. Мещеряков, Сб. Мещерский край, Изд. о-ва исследов. Рязанского края.
М. Нейштадт, Труды Центр. Торф. Станции, вып. 3, 1930.
М. Нейштадт, Сборник инструкций по исследованию торфа и торфяных
залежей, ч. 1, Москва 1933.
В. С о ч а в а, Известия Государ. Географического Общества, т. LXV, вып. 4,
1933.
В. С о ч а в а, Амгунь-Селемджинская Академия наук СССР, ч. I, Л. 1934.
В. Сукачев, Метеорологический вестник, т. XXXII, Nfe 1—4, 1922.
B. Сукачев, Основные черты развития растительности СССР во время»
плейстоцена, Материалы по четвертичному периоду СССР, 1936.
C. Тюремнов, Торфяное дело, № 7, 1928.
С. Тюремнов, Торфяное дело, № 1, 1931.
ОГЛАВЛЕНИЕ
Стр.
предисловие 2
Глава I. Содержание науки о бо¬
лоте з
Понятия „болото* и „торфяник“ у разных
авторов 7
Статистика болот 10
Площадь болот и торфяников по странам
и континентам 15
Важность изучения болот ... * 10
Глава II. Торф и седименты. ... 22
Торф как органогенная порода —
Виды торфа 25
Классификация торфов и седиментов ... 27
Химические свойства торфа 41
Примеси в торфе 55
Физические свойства торфа 56
Сухие торфа 73
Полуторфа 74
Водные отложения (седименты) 75
Седименты и геолого-петрографическое
строение местности. География седи¬
ментов 82
Глава III. Образование болот на
месте водоемов 88
Интенсивность заболачивания . —
Экологические факторы заболачивания . • 93
Типы заболачивания 102
Заболачивание водоемов прежде и теперь . 108
Глава IV. Заболачивание суши . , ш
Роль физико-географических факторов и
растительности ..•• —
Смена фитоценозов * 121
Заболачивание суши прежде и теперь . . . 130
Глава V. Строение торфяников . . 134
Залежь как отражение эндодинамических
смен растительности 136
.Роль физико-географических факторов в
строении торфяников 140
Строение залежи и история климатов ... 161
Глава VI. Флора и растительность
болот • 156
Влажность болотного субстрата и распре¬
деление видов 157
Питагельный режим и распределение ра¬
стений 160
Нарастание мха на болоте как экологиче¬
ский фактор 168
Особенности древесной растительности на
болотах 175
Измерение прироста мохового ковра и
торфа Г 182
Водный баланс растений выпуклых болот
и вопрос об их ксероморфизме .... 184
Болотные фитоценозы 187
бтр.
Глава VII. Физико-географические
условия и распространение бо¬
лот 208
Болота и климат —
Болота и рельеф 210
Почвы и распространение болот 213
Геологическое прошлое страны и распро¬
странение болот и торфяников .... 215
Болотные ландшафты 216
Глава VIII. Типы болот. ...... 220
Выпуклые болота 221
Типы выпуклых болот Западной Европы . 243
Типы выпуклых болот СССР 250
Глава IX. Географ и ческоё распро¬
странение бОЛОТ И ТорфЯНИКОВ . 275
Болота Европейской части СССР 275
Болота Западной Сибири 302
Болота Таймыра 318
Болота тундровой зоны Якутии 319
Болота тундровой зоны Дальне-восточного
края 320
Болота Среднесибирского плоскогорья . . 221
Болота лесной области Дальневосточного
края. Болота Амурской области . . . 323
Приморская область Дальневосточного
края 325
Болота Камчатки 327
Болота Кавказа ♦ . . . . 330
Глава X. Четвертичная история ра¬
стительности по данным из¬
учения торфяников ззз
Пыльцевой метод исследования торфяни¬
ков —
История Балтики и климатов в послеледни¬
ковое время 342
Послеледниковая история флоры и лесов. 356
Послеледниковая история лесов и флоры
прибалтийских стран ......... 361
История лесов Средней Европы 366
Ленинградская Прибалтика 367
Центральная часть СССР 369
Запад Европейской части СССР 371
Восток Европейской части СССР —
Урал ... 372
Западная Сибирь ... * —
Украина -
Северный край (западная часть) 373
Общий обзор послеледниковой истории ле¬
сов и расселение древесных пород
в Европейской части СССР 374
Послеледниковая история растительности
СССР 376
Послеледниковая история лесов Северной
Америки . • 380
История растительности четвертичного пе¬
риода 381
Глава XI. Использование болот. . 387
Указатель литературы 39.1
ИСПРАВЛЕНИЕ
Напечатано на стр. 2 (выходные данные/ Цена без переплета 6 р.
Переплет 1 р. 25 к.
Должно быть: Цена без переплета 6 р., в переплете 6 р. 60 к.
Н. Я« К а ц. Болота и торфяники. Зак. М 722.