/



























































































































































































































































































































































































































































































































































































































































































































Author: Верещагин Н.К.
Tags: биология млекопитающие




Text
Главный редактор
Директор Зоологического института АН СССР акад. Я. Я. ПАВЛОВСК ИЙ
Редактор издания проф. А. А, СТРЕЛКОВ
-^***«*WR
Л’ 'ТЕК
ПУШЮ...
ПРЕДИСЛОВИЕ
Книга Н. К. Верещагина — труд не совсем обычный. Хотя он и не претендует на полноту ни в одном из составляющих его разделов, все же подчинение исследований одной главной цели — прослеживанию прошлых и современных путей формирования фауны своеобразной и сложной страны — позволило автору достигнуть выдающихся результатов. В сущности, это первая в отечественной зоологии попытка разработки обобщенной и цельной картины развития наземной фауны Кавказа с неогена до наших дней на фоне геологической истории и смен ландшафтов.
Для нее был привлечен весь арсенал современной биологической науки— от палеонтологических документов, с анализом отдельных филогенетических этапов четвертичных млекопитающих, до массовых фактов современной зоогеографии и экологии ныне живущих форм.
По мнению автора, восточное Средиземноморье (включая Кавказ) в неогене было захвачено единым процессом последовательных смен териокомплексов, общим для всей территории современной Голарктики.
Корни четвертичной териофауны Кавказского перешейка уходят глубоко в плиоцен. Собственно кавказский — горно-лесной — териокомплекс развивался преимущественно на месте по крайней мере с верхнего миоцена. Наряду с этим утверждается, что формирование териофауны в четвертичном периоде (антропогене) всего Кавказского перешейка происходило главным образом за счет вселений и вымираний видов.
Местный морфогенез, т. е. видообразование в пределах перешейка, имел по сравнению с другими типами формирования фауны подчиненное значение.
Высказывания автора об отсутствии видимых качественных скачков в морфологической эволюции четвертичных млекопитающих еще нуждаются в дополнительных исследованиях и подтверждениях.
Особо подчеркнутая разрушительная (деструктивная) деятельность человека, лавинообразно нараставшая на Кавказе с верхнего палеолита вплоть до современности, обрывала историю целого ряда охотничье-про-мысловых зверей. Приведено много и современных примеров коренных перестроек экологических группировок и изменения образа жизни (экогенеза) уцелевших млекопитающих в различных зонах.
Оригинальная постановка и методика разработки большого ряда во-прдЬов фаунологии, затронутых в этой книге, несомненно, будет способствовать организации дальнейших углубленных палеонтологических и зоологических исследований не только на Кавказе, но и на других территориях нашей Родины. Эту методическую значимость книги следует отметить особо.
1*
4 Предисловие
Имеются в книге и важные для текущей практики сведения о промысловом значении местных и акклиматизированных на Кавказе видов зверей, а также материалы для дела охраны природы и заповедников.
Нет сомнения в том, что история формирования фауны млекопитающих Кавказа будет полезна и интересна не только для зоологов, но и для широкого круга людей других специальностей — ботаников, геологов, археологов и географов.
Академик, генерал-лейтенант медицинской службы
Е. Н. Павловский
ВВЕДЕНИЕ
Выяснение исторических закономерностей в геохронологическом и пространственном распределении организмов в связи с их эволюцией — интересная и трудная проблема, полная диалектических взаимосвязей и противоречий.
Ее разработка необходима как для научного обоснования практических работ в области реконструкции фауны, так и для дальнейшего целеустремленного развития фау но логических исследований.
Проблема эта особенно сложна для Кавказского перешейка, развитие ландшафтов которого на протяжении всего кайнозоя было обусловлено появлением и исчезновением прилежащих морей, горообразованиями и оледенениями. Следствием упомянутых процессов является и быстрая смена современных климатов и ландшафтов на небольших пространствах.
Кавказ — большая и сложная в широком физико-географическом смысле страна. Исследованиям ее геологического строения, флоры, фауны и истории населявших ее народов посвящены многочисленные труды выдающихся ученых почти за 250-летний период.
Исследования по фауне млекопитающих Кавказа касались главным образом выявления видового состава, географического распределения и систематических особенностей видов, в меньшей степени — вопросов экологии и еще меньше — истории фауны. Главнейшие исследования принадлежат К. А. Сатунину, Н. Я. Диннику, Н. А. Смирнову, М. В. Шидлов-скому, 3. С. Родионову, П. А. Свириденко, С. И. Огневу, Л. Б. Бёме, С. С. Турову, В. Г. Гептнеру, А. И. Аргиропуло, А. А. Насимовичу, С. К. Далю, И. В. Жаркову, П. П. Гамбаряну и многим другим.
Труд, проделанный отечественными зоологами и коллекторами по сбору и накоплению документальных материалов о кавказских зверях, неоценим. Мы можем лишь частично отразить его в библиграфическом списке.
В результате всех этих исследований выявлен, по-видимому почти полностью, современный видовой состав млекопитающих страны и создана возможность установления основных закономерностей в распределении видов.
Большая палеонтологическая работа по сбору и описанию остатков ископаемых млекопитающих Кавказа проделана В. В. Богачевым, А. А. Борисяком, В. И. Громовым, Е. И. Беляевой, В. И. Громовой, Р. Д. Джафаровым, Л. К. Габунией, Н. О. Бурчак-Абрамовичем, мною и другими. Неоценимы и остеологические сборы археологов, особенно С. Н. Замятнина, Г. К. Ниорадзе, А. П. Круглова, Е. И. Крупнова
'6
Введение
Все упомянутые работы и накопленные материалы, в сущности, и позволили мне сделать попытку разработки самостоятельной проблемы происхождения и истории формирования фауны млекопитающих Кавказа.
История формирования фаун млекопитающих на Кавказе неразрывно связана с историей развития Кавказской суши, ее флоры, ландшафтов, с развитием жизненных форм и ареалов отдельных видов зверей.
Появление и исчезновение морей третичного периода в восточной части Средиземноморской геосинклинали, т. е. на территории современного Черного моря, Кавказа и Каспия, их очертания, характеристика и смены фаун моллюсков изучены к настоящему времени с большими подробностями геологами — академиками Андрусовым (1888) и Архангельским (1934), Архангельским и Страховым (1938), а также Жижченко, Колесниковым, Эберзиным (1940) и другими, главным образом в связи с запросами нефтяной промышленности. Особенно детальны эти работы для неогена. Подробная зоогеографическая сводка, посвященная фаунам Каспия и Черного моря, выполнена еще Совинским (1904).
Геоботаниками и палеоботаниками также опубликован ряд очерков истории наземной флоры прикаспийских стран и особенно Кавказа, основанных на флористическом анализе и на палеонтологических документах (Н. И. Кузнецов, 1909; Палибин, 1936; Гроссгейм, 1936, 1948; Малеев, 1941, и др.).
История наземной фауны Кавказа до сих пор была освещена лишь в самых общих чертах, преимущественно на материале по ископаемым млекопитающим: в очерке истории фауны Европейской части СССР академика Мензбира (1934), в очерке истории фауны СССР Серебровского (1935), в обзоре Богачева (1938) для третичного периода Кавказа, в обзоре местонахождений ископаемых остатков третичных млекопитающих Борисяка и Беляевой (1948), по истории четвертичной фауны — в работах Громова (1939, 1948), Громовой (1948), Бурчак-Абрамовича (1951в), Пидопличко (1951, 1954).
История происхождения фауны наземных зверей Кавказа разбиралась и на современном зоогеографическом материале Сатуниным (1896, 1901а, 19016, 1904, 1909, 1913), Динником (1911), Шидловским (1940а, 19416, 1945, 1947), а в последнее время Б. А. Кузнецовым (1949, 1950).
Слабее освещено на основе данных зоогеографии происхождение фаун других классов наземных позвоночных: птиц — Пузановым (19386), рептилий, и амфибий — Никольским (1913), Соболевским (1929), Ляйстером (1931), Черновым (1939).
История фауны наземных беспозвоночных Кавказа изложена в зоогеографических очерках по скорпионам Бялыницким-Бирулей (1917), по стрекозам, а также млекопитающим, рептилиям и амфибиям — Бартеневым (1933, 1934), по жесткокрылым — Семеновым-Тян-Шанским (1936), А. В. Богачевым (1947) и по прямокрылым — Уваровым (Uvarov, 1921).
Наиболее характерная черта большинства упомянутых работ — одностороннее освещение вопросов истории формирования фаун. Эти вопросы обсуждались либо с палеонтологических, либо с чисто зоогеографических позиций, без попыток синтеза, а также без учета экологических и морфофизиологических материалов и фактов.
Несомненно, что подобный подход к делу не может дать истинного представления о таком сложном историческом процессе, каким является
Введение
7
формирование фаунистического комплекса в тот или иной геологический отрезок времени. Правильнее в этом едином процессе различать три главнейших и взаимосвязанных направления:
1) эволюцию среды и условий существования отдельных видов живот-' ных под влиянием вековых колебаний климата, поднятий и опусканий земной коры и жизнедеятельности самих биоценозов; на позднейших стадиях приобретает важнейшее значение еще антропогенное влияние;
2) морфологическую эволюцию отдельных видов под влиянием внешней среды и внутренних закономерностей развития организмов;
3) экологическую эволюцию отдельных видов и биоценозов в целом, в связи с непрерывными изменениями условий существования, изменениями видового состава биоценозов под влиянием вымирания одних, трансформации других, вселения или выселения третьих видов.
Не все перечисленные направления процесса формирования фауны одинаково доступны для изучения палеонтологам, зоогеографам и экологам, но синтез материалов и выводов этих исследователей необходим.
Кавказ имел сложную геологическую историю, не менее сложную историю развития почвенного и растительного покрова, длинный ряд смен человеческих культур. По мере приближения к современности нарастали, взаимно комбинируясь, различные воздействия человека на ландшафт и фауну. Поэтому понять и установить главнейшие черты формирования современной фауны Кавказа можно лишь на основе комплексного освоения того научного наследства, которое было накоплено для этой территории в области геологии, геоморфологии, геоботаники, археологии, палеонтологии, систематики, фаунистики, экологии и зоогеографии.
Обширность подобной задачи вполне естественно заставила ограничить пределы наших личных исследований и подразделить их на три взаимосвязанных цикла.
1. Палеонтологическое и археологическое обоснование истории формирования фауны.
2. Анализ ареалов и происхождения четвертичных млекопитающих в связи с их экологией и морфогенезом.
3. Териогеографическое районирование Кавказского перешейка и пути новейшей эволюции фауны.
По каждому из этих разделов исследования развивались в зависимости от наличия материалов и технических возможностей.
Материалы, маршруты, методологические и методические замечания
Основными палеонтологическими материалами по третичным и плейстоценовым млекопитающим послужили коллекции, собранные палеонтологами, геологами, археологами и хранящиеся в: Естественпоистори-ческом музее Академии наук Азербайджанской ССР, Зоологическом (ЗИН) и Палеонтологическом (ПИН) институтах Академии наук СССР, Всесоюзном геологическом институте, Институте геологии Академии наук Грузинской ССР, Институте геологии Академии наук Армянской ССР и в городских музеях Пятигорска, Ставрополя, Краснодара, Темрюка, Новочеркасска, Ростова, Астрахани. Наши личные сборы и тафономические наблюдения при изучении третичных млекопитающих производились на миоценовом захоронении у Беломечетской в центральном Предкавказье, на плиоценовом захоронении Косякин карьер близ Ставрополя
Введение
и на верхнеплиоценовых — нижнечетвертичных захоронениях по Псе-купсу и Таманскому полуострову (рис. 1).
Такие же наблюдения и сборы остатков плейстоценовых млекопитающих и растений производились в плейстоценовых речных песках, в долинах Кубани и Кумы, близ Крапоткина и Георгиевска в Предкавказье, а в Закавказье в битуминозных слоях Ашперонского полуострова (рис. 2).
Небольшие сборы костей, кремневых орудий и дополнительные наблюдения были сделаны нами и на старых раскопах палеолитических пещер
ад
ад
ад
Верхний I плиоцен ' 1
& БАТУМИ
I Продуктивная толща “моцен Понтический 50 0 SO ТОО 150 200км
КРАС
ОРДЖОНИКИДЗЕ
ТБ
ад
ЕРЕВАН
&
3£
\1евенцовка£——' 4 1 нрвоче^асск ксай
РОСТОВ
ф Верхний Гх Меотический плиоцен Сарматский
® миоцен Чокраксний
о Олигоцен
Петровское Благодарное z
"осякинКа Беломечётска
ад 44 .46 48________________________________
иняя Балка
Рис. 1.
Каоьео Нимб,
МАХАЧКАЛА
МЕСТА НАХОДОК ОСТАТКОВ ЗВЕРЕЙ
В ОТЛОЖЕНИЯХ ТРЕТИЧНОГО ПЕРИОДА Датировка по фауне позвоночных и морских моллюсков
Ярусы - куяльникский I Апшеронский +Акчагыльский
Средний киммерийский омоцен ]1 Продуктивная
СУХУМИ
Кушчи
АКУ к- Батан
" е
Черноморского побережья и бассейна Риона: Ахштырской, Сакажиа, Гварджилас-Клде, а также в недавно раскопанной пещере Кударо.
В итоге было исследовано более 1500 фрагментов костей третичного возраста и более 55 000 плейстоценовых.
Для палеонтологического обоснования изменений ареалов промысловых млекопитающих и мелких грызунов в голоцене было предпринято изучение костных остатков из различных типов захоронений. Исследовались следующие категории остатков.
1. Пищевые и промышленные отбросы человека в слоях стоянок, поселений, городищ от мезолита до последних веков нашей эры (Саркел, Семибратнее городище, Елизаветинская, Кепы, Фанагория, Тамань, Геленджик, Анаклия, Калакент, Мингечаур, Баку и др.).
2. Костные поделки, украшения, черепа и скелеты диких и домашних животных (уложенные согласно погребальному ритуалу) из могильников (Нальчик, Самтавро, Триалети, Севан и др.).
Введение
&
3. Коллекции черепов парнокопытных в горных святилищах — пещерах, навесах, где они накопились на протяжении столетий в связи с обрядовыми празднествами и поклонением богу охоты (ряд ущелий в Осетии).
4. Костные остатки грызунов и насекомоядных в пещерах, навесах и мелких пустотах скал, натасканные в течение последних столетий филинами и сычами (предгорные и горные районы Предкавказья и Закавказья).
-----МЕСТА находок остатков
йб
ю
38
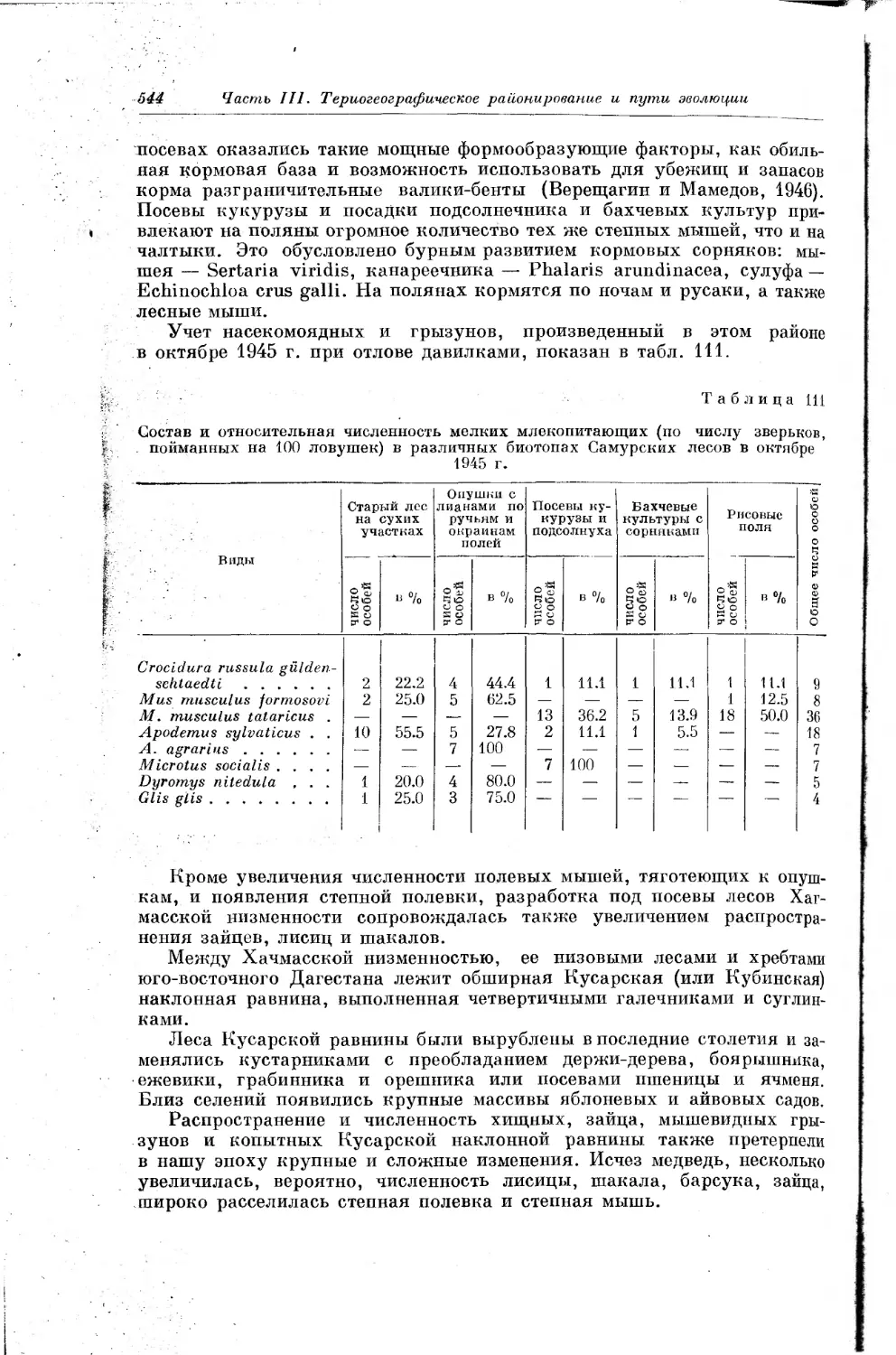
РОСТОВ
ИННуЛЬ
44
Датировка ло кремневым изделиям и фауне А Верхний палеолит А Средний палеолит нижний палеолит о Верхний плейстоцен(хвапынский ярус) • Средний плейстоцен (хазарский ярус) & Нижний плейстоцен (бакинский ярус узунларские слои)
S3 О so 100 150 200 км
ЕРЕВАН
Эилас
&
&
. рV?
Карьер' Цимбал
Воронцовым
ыДаховская
Ахштырская
МАХАЧКАЛА
ЗВЕРЕЙ В ПЛЕЙСТОЦЕНОВЫЕ ОТЛОЖЕНИЯХ
Типы захоронений
д Пещеры
* Открытые палеолитические стоянки в делювии Озерные и озерно -речные
& отложения
Предгорный и овражный делювий
± Асфальтированные делюви-альные и озерно речные пески
X Травертины
|-е- Прибрежноморские отложения
Кнр-Ману
БАТУМИ
ЛШакет
БАКУ Вннагады
Тамтама А
Рис. 2.
5. Кости и рота утонувших животных и кухонные отбросы племенв живших на берегах и в свайных постройках быстро мелеющего ныне озера Севан.
6. Единичные находки костей в голоценовых суглинках.
Всего, таким образом, было исследовано в различных ландшафтах более 70 голоценовых захоронений (рис. 3), содержащих около 52 000 определимых костей и их фрагментов.
Выбор микрорайонов для исследований голоценовых захоронений, особенно при поисках скоплений остатков пищи филинов и сычей, производился с расчетом проверки теоретических соображений о путях проникновения видов и истории послеледниковш! эволюци ареалов фаунистических комплексов. В качестве примеров укажем на исследования по плейстоценовым реликтам Апшеронского полуострова, Ставропольской возвышенности и Пятигорья (Верещагин, 1949в, 1953а).
Изучение и картирование местонахождений ископаемых остатков, а также просмотр обширной литературы позволили составить общую схему
' IQ
Введение
распространения на Кавказском перешейке костеносных отложений третичного и четвертичного периодов.
Подъемы кавказских хребтов в кайнозое обусловливали вынос на предгорные равнины огромных масс галечников, песков и ила, образовывавших слоистые толщи. Этот терригенный материал перерабатывался волнами в прибойных участках или вступал в спокойный контакт с морскими отложениями в тихих бухтах окружающих морских бассейнов.
46
АСТРАХАНЬ
РОСТОВ
СТАВРОПОЛЬ
42
Севан
Мерагз
50
46
Лй1
- Датировка по керамике и кремневым изделиям
• XV-XIX ев. н.з
О ГХ1УВВ.Н.З
о Скифская эпоха ([тысячелетие до н.з.)
о Поздняя бронза
© Ранняя бронза
с Энеолит
е Неолит
<5 Голоцен, точнее не датированный и мезолит
50 О 50 100 1S0 200км
льчик
& А л хаете ОВДЖО'НИКИДЗЕ
(КРАСНОДАРУ \ЖИК
МАХАЧКАЛА
К алане нт
рн
СУХУМИ
Самтавр
Атбилиси
& БАТУМИ
МЕСТА НАХОДОК ОСТАТКОВ ЗВЕРЕЙ В ГОЛОЦЕНОВЫХ
ОТЛОЖЕНИЯХ
Типы остатков и ЗАХоронений
। Кухонные отбросы в слоях открытых поселений
+„ Священные” коллекции
в капищах и в могильниках х Остатки пищи сов в скалах - Остатки смешанного пропет хождения в озерно-речных и пещерных слоях
АКУ
Рис. 3.
Накопление и захоронение скелетов шло, как правило, в дельтовых участках рек и в морских заливах в результате выноса трупов погибших зверей постоянными и временными потоками. Песчано-гравийные костеносные линзы различных эпох кайнозоя заполняют здесь обычно ложбины стока или эрозионные русла, врезанные в более древние слои. Таковы местонахождения третичных и раннечетвертичных захоронений остатков млекопитающих на наклонных предгорных равнинах Предкавказья, Ставрополья и в широких межгорных долинах Закавказья. Вскрытие этих слоев и находки костей происходят обычно в результате новейшей эрозии и при искусственных выемках грунта.
До сих пор на Кавказе неизвестны пещерные местонахождения костных остатков третичного возраста. Объясняется это, по-видимому, крупными перестройками рельефа и гидрографической сети, которые уничтожили древний карст.
Районы развития карста четвертичного возраста были использованы археологами для поисков палеолитических стоянок. Находки палеолита
Введение
11
сопровождались обычно находками и сбором «фауны», т. е. более или менее -обильных костных остатков животных, оставленных в пещерах первобытными охотниками или четвероногими и пернатыми хищниками. Позднейшие, послепалеолитические (постплейстоценовые) захоронения кост-* пых остатков залегают преимущественно в покровных суглинках, делювиальных овражных смывах, аллювии первой надпойменной террасы рек, также в пещерах и под навесами скал.
Большинство захоронений остатков голоценового возраста, доступных для раскопок и изучения, было обусловлено охотничьим промыслом человеческих племен и деятельностью хищных птиц. Однако стихийная, массовая гибель диких животных и захоронения их остатков наблюдаются та Кавказе и в настоящее время (Верещагин, 19516, см. также рис. 4).
Уже в результате рассмотрения изложенной схемы становится ясно, то палеонтологическое обоснование истории происхождения фауны кавказских высокогорий не может быть полным.
Причины этому тафономические — отсутствие открытого осадконакопления в высокогорье и отсутствие находок пещер с остатками животных плиоценовой эпохи.
Критическое освоение накопленного палеонтологического материала могло быть произведено лишь при всестороннем учете особенностей захоронений, типов сохранности костных остатков и костного вещества. В связи с этим каждому региональному обзору захоронений остатков по геологическим (стратиграфическим) слоям в работе предпосланы разделы, поясняющие основные пути и характер накопления и захоронения остатков на том или ином этапе (см. «Тафономию» Ефремова, 1950).
Правильное освещение характера захоронений — их природы — позволяет сделать ряд важных заключений о древних ландшафтах, условиях жизни и гибели организмов. Между тем природа захоронений далеко не всегда учитывалась палеонтологами, что приводило нередко к ложным выводам, особенно в тех случаях, когда захоронения имели разновозрастные костные остатки.
В большинстве случаев палеонтолог, исследуя захоронения, имеет дело со случайным набором остатков животных, отражающим лишь особенности образования данного захоронения, а отнюдь не состав экологических группировок древнего района.
Чем больше видовой состав животных (и растений), остатки которых образуют захоронения, и чем больше состав остатков отражает природные соотношения численности видов и особей в популяциях, тем больше «универсальность» захоронения.
В конечном счете степень универсальности захоронений зависит от трех факторов: 1) условий первоначальной гибели животных; 2) условий накопления остатков во вмещающей породе; 3) условий консервации и «поведения» остатков в образующемся слое.
Понятно, что характер и особенности каждой группы факторов определяют в совокупности и возможность реконструкции древних биоценозов и ландшафтов на основе изучения обнаруженных костных остатков.
В качестве высокоуниверсального захоронения упомянем Бипагадин-ское (глава П). Толстые пластины асфальта улавливали и консервировали на плейстоценовом Апшероне почти одинаково успешно как растения, так и насекомых, птиц, тушканчиков и носорогов. В результате представилась возможность создания весьма достоверных ландшафтных картин прошлого. - ;
12 Введение
Иное положение создается при обсуждении результатов сбора и исследовании костных остатков в осадках делювиального, речного, озерного и морского происхождения, а также на палеолитических стоянках: и особенно в пещерах.
Накопление костных материалов в речных отложениях происходит, как это было показано нами (Верещагин, 1953в), в результате размыва рекой костеносных слоев, а также при гибели животных в русле потока,, в пойменных озерах и на прирусловых отложениях. Попадая в действующее русло, костные остатки, подобно валунам и галечникам, подвергаются жесткой обработке, сортировке и распределению по фациям в; соответствии со своим удельным весом и «парусностью».
Таким образом, более правильное представление о комплексе погибавших в долинах рек животных может быть получено лишь, при сборе костных остатков по фациям, т. е. в галечниках, песках и илах.
Такие сборы, как правило, прежними исследователями не производились, почему имеющиеся коллекции отражают нередко лишь последний этап выборочности — методику раскопок.
Фациальное распределение костных остатков имеет место и при выносе^ трупов животных речкой в озеро и в морские бухты. Чаще захоронение скелетов идет в толще дельтовых отложений, но плавающие трупы могут переноситься далеко течениями и, опускаясь на дно, захороняться в толщах ила и ракушечника спокойных бухт.
Накопление костей в палеолитических и послепалеолитических слоях стоянок, поселений, могильников зависело прежде всего от охотничьих навыков и обычаев разных племен. В пещерных же стоянках остатки пищи человека перемешивались нередко с остатками пищи хищных зверей (медведей, волков, гиен, леопардов) и хищных птиц (филинов и сычей), реже имело место и замывание костей в пещерные слои водой потоков. Ясно, что без учета навыков охотников и образа жизни самих животных возможны грубые ошибки в выводах, которые и делались исследователями, механически подсчитывавшими число остатков домашних и диких видов в слоях поселений различного типа и возраста. Известно, например, что современные кавказские охотники нередко переносят по горным тронам на расстояние 20—25 км по 30—40 кг мяса диких коз, косуль, свиней. Кавказские козлы же совершают суточные переходы на 1500—2000 м по вертикали. Филин, по нашим наблюдениям в пещерном районе Имеретии, нередко летает кормиться из пещер предгорий на субальпийские луга, поднимаясь на 1000 м и более по вертикали.
Поэтому строить предположения о палеоландшафтах и понижении или повышении снеговой линии на основе находок в пещерах остатков козлов и прометеевых полевок, живущих в нашу эпоху в высокогорье, можно лишь после обсуждения особенностей всех возможных факторов накопления костеносных слоев.
Точно так же нельзя сравнивать и состав костных остатков из помоек греческих городов на Таманском полуострове (Фанагория, Кепы) с составом кухонных остатков из мелкого рыбачье-охотничьего поселка римского времени в устье Кубани (Семибратнее городище) для выводов о деградации или развитии охотничье-промысловой фауны района. В данном случае будет иметь значение прежде всегоимевно величина и характер населенного пункта, его микротопографическое положение, а отнюдь не появление и вымирание охотничье-промысловых животных на окружающих пространствах.
Рис. 4. Преобладающие типы гибели зверей и захоронений их остатков в кайнозое Кавказского перешейка.
14 Введение
Везде, где позволял материал, в книге приведены численные соотношения костей и особей в захоронениях, а также видовой состав домашних животных. Такие списки даются для Кавказа впервые, помогая судить по составу домашних животных о развитии культуры, направлении хозяйства племен и о характере ландшафта. Так, отсутствие остатков лошади и обилие остатков свиньи в поселениях равнинной Колхиды указывают, что ландшафтные черты этого района (повсеместная заболоченность) в эпоху энеолита и бронзы были очень сходны с существовавшими здесь еще лет 50 назад. В связи с этим и соображения некоторых геологов о наличии в Колхиде в I тысячелетии до н. э. обширных сухих равнин, основанные на непроверенных сообщениях древних авторов, не подтверждаются.
Для заключений о видовом составе и распространении охотничье-про-мысловых животных в век бронзы и железа были привлечены также рисунок и скульптура древних мастеров. Особого внимания заслуживали бронзовые, серебряные и золотые вещи с изображениями животных из курганов Армянского нагорья, долины Куры и с Закубанской наклонной равнины — Цалка, Самтавро, Майкоп, Келлермес и др. При комплексном учете данных о вероятном месте производства вещей, художественных вкусов заказчиков — урартов, кобанцев, скифов — и экологии изображенных животных реалистическое искусство древних мастеров служит отличным материалом к истории былого распространения и постепенного вымирания некоторых крупных животных.
Дело в том, что на протяжении нашей эры численность многих крупных зверей настолько понизилась, что вероятность попадания их остатков в какое-нибудь захоронение была совершенно ничтожна. Тем менее вероятна находка этих остатков. Поэтому так называемый «зверовый стиль» искусства в I тысячелетии до н. э. может дать ряд ценных указаний зоологу. Зоолог должен, конечно, и помочь археологам правильно определить видовую принадлежность изображений животных на основе морфологических и зоогеографических данных.
Большинство драгоценных вещей зверового стиля из кавказских могильников кобанской, ходжалинской, скифосарматской и других культур было просмотрено и обсуждено заново не только по печатным изданиям (Толстой и Кондаков, 1889; Уварова, 1900; Куфтин, 1940, и др.), но ив натуре в особых кладовых Государственного Эрмитажа (Ленинград), Музея Грузии (Тбилиси) и Исторического музея (Москва). Результаты использованы при обзоре видов и анализе их ареалов (глава III). Зверовый стиль в кавказских древностях заслуживает, в сущности, и специального подробного зоолого-этнографического исследования.
При изучении кайнозойских фаун Кавказа мы придерживались геохронологических схем подразделения неогена, предложенных для восточного Средиземноморья (Черного моря, Кавказа и Каспия) академиками Андрусовым (1918), Павловым (1925), с теми уточнениями, которые были внесены в последние десятилетия Ковалевским (1933, 1936), Жижченко, Колесниковым и Эберзиным (1940).
Переход от плиоцена к плейстоцену в области Кавказского перешейка признается нами в согласии с Павловым (1936) и Варданянцем (1948) в конце апшеронского века. Этой переходной эпохе, видимо, соответствует последний этап существования в Предкавказье таманско-псекупского фаунистического комплекса, установленного впервые Громовым (1948, 1950) и охарактеризованного нами во II главе.
Введение
Подразделение четвертичного периода Кавказа принято двучленное — на плейстоцен и голоцен. Под плейстоценом здесь всюду подразумевается эпоха довольно длительного развития фаунистических комплексов — от верхнего апшерона до окончания последнего крупного оледенения Кав-* казн и наступления ксеротермической эпохи, знаменующей начало голоцена. Плейстоцен подразделен на нижний, средний и верхний, чему по каспийской стратиграфии условно соответствуют бакинский, хазарский и хвалынский ярусы с общеизвестными для Русской равнины фаунистическими комплексами млекопитающих. Расчленение четвертичной толщи предгорных равнин Кавказского перешейка и сопоставление речных террас с пещерными слоями и морскими террасами, в связи с находками остатков позвоночных, приняты с учетом работ Мирчинка (19376), Громова (1948), Варданянца (1948) и Николаева (1953).
Археологическая хронология Кавказа при сопоставлении с указанными подразделениями четвертичного периода не несет принципиальных отличий от европейских (западносредиземноморских) схем, разработанных Обермайером (1913), Осборном (1924) и Пенком (Penk, 1939). Особенности датировок кавказского палеолита в связи с обнаруженными следами экологических группировок млекопитающих учтены по работам Замятина (19506, 1957) и других кавказских археологов.
Весьма важной задачей было установление критериев относительного геологического возраста изучаемых костных остатков.
Проблема определения относительного и абсолютного геологического возраста ископаемых костей приобретает все большее значение в геологии, палеонтологии и археологии. Установление древности ископаемой кости, как известно, производится по: 1) физико-химическому составу (метод прокаливания и радиологический); 2) систематическим признакам (родовой, видовой и подвидовой принадлежности); 3) принадлежности к тому или иному фаунистическому комплексу (по сопровождающей «фауне»); 4) сопровождающим остаткам человеческой культуры (каменным орудиям, керамике и т. п.); 5) вмещавшему кости геологическому слою после установления возраста этого слоя по фауне беспозвоночных и по косвенным геоморфологическим данным (например, по возрасту морских или речных террас); 6) типу сохранности костного вещества.
Каждый из названных методов и критериев обладает своими недостатками. Так, методы физико-химические, в частности метод прокаливания и радиоактивных изотопов, дают сравнимые результаты лишь при изучении костей, захоронявшихся в совершенно одинаковых условиях и не долее полумиллиона лет. Решающее значение здесь имеет состав вмещающей природы, ее радиоактивность, водный и солевой режим грунтов в период захоронения и т. п.
Впрочем, метод прокаливания костных образцов и сравнения коэффициентов остаточного органического вещества, применяемый Пидопличко (1952), безусловно может давать пригодные показатели абсолютного и относительного возраста при условии разработки собственных шкал для различных физико-географических областей и типов захоронений.
Более сложен и слабо разработан метод определения абсолютного* возраста по изотопам радия, урана, тория (Чердынцев, 1955), дающий сбивчивые показатели.
Определение геологического возраста кости по видовым и фаунистическим признакам имеет смысл при наличии детально разработанных схем эволюции морфологических признаков для данного рода или вида)
Л6
Введение
и смен фаунистических комплексов для данной территории. Необходима, кроме того, хорошая сохранность остатков (наличие черепов).
Каменные орудия, а также керамика и другие «следы» человека дают ценные показатели относительного возраста костеносных горизонтов лишь при наличии разработанных хронологических шкал смены человеческих культур для различных географических областей и районов. Между тем теперь становится все очевиднее, что становление палеолитических культур в Западной Европе протекало в несколько иные геологические отрезки времени, нежели в Восточной. Кроме того, необходимо, чтобы изделия и кости залегали «in situ».
Определение возраста кости по вмещавшему ее геологическому слою мало пригодно для континентальных четвертичных слоев в связи с нередкой переотложенностью остатков. Кроме того, эти горизонты больше всего и нуждаются в расшифровке возраста на основе находок остатков позвоночных. Последний способ, учитывающий органолептически тип сохранности костного вещества, требует значительного опыта и простейших приемов для объективной оценки. Он дает также показатели лишь относительного возраста.
Весь кавказский палеонтологический материал по четвертичным видам потребовал сплошного нового просмотра, так как одностороннее применение морфологического — морфометрического критерия и обращение с костью как со своего рода зоологическим фетишем приводило прежних исследователей к курьезам и засорению списков видов.
Однако при внимательном разборе и учете признаков сохранности костного вещества, а также при учете условий захоронения вполне возможно различать древность кости в пределах основных подразделений четвертичного периода. Для этого следует учитывать степень обеднения органическим веществом (фоссилизацию) и вторичную минерализацию, т. е. новое насыщение костного вещества солями, по цвету в свежем разломе, степени и глубине прокраски от поверхности вглубь, наличию запаха сырой кости при поскабливании ножом и т. п. Таблица для определения относительного возраста ископаемых костей четвертичного периода дана нами (Верещагин и Громов, 1953а).
Для обзора и анализа ареалов ныне живущих видов были использованы сборы и материалы наших полевых исследований, проводившиеся с перерывами с 1935 по 1941 г. в Азербайджане, отчасти в Армении, а с 1945 по 1952 г. — в Азербайджане, Грузии, Северной Осетии, Кабарде, Сванетии, Дагестане. Наши маршруты показаны на карте (рис. 5).
Были использованы также превосходные коллекции шкур, спиртовых и сухих тушек, черепов млекопитающих, собранных многочисленными зоологами и натуралистами, хранящиеся в ЗИНе, Зоологическом музее Московского университета, Музее Грузии и Институте зоологии Академии наук Грузинской ССР (сборы до 1944 г.), Институте зоологии Академии наук Армянской ССР (сборы до 1943 г.), Институте зоологии Академии наук Азербайджанской ССР (сборы до 1948 г.), на Ставропольской и Бакинской противочумной станциях (сборы 1945 г.). Общее число пунктов сбора различных видов зверей превышает 3300, а количество коллекционных экземпляров достигает 13 500.
Выводы о новейших изменениях относительной численности и ареалов 23 видов пушных зверей сделаны по материалам государственных заготовительных пунктов пушнины в Предкавказье и Закавказье за 25-летний период — с 1925 по 1950 г. Ареалы этих же видов были уточнены
Введение 7 7
по данным 257 кавказских пунктов, принимавших пушнину в 1945— 1948 гг. (рис. 5). Динамика заготовок шкур пушных видов отражена на графиках, отдельно для Предкавказья и Закавказья, а также по республикам и краям. По Астраханской области таких данных у нас не было.
Пункты находок ископаемых остатков, поимки мелких зверьков и добычи промысловых видов наносились различными знаками на специально изготовленную и изданную для этой работы бланковую карту масштаба 1:5 000 000. Для пушных видов показана малыми точками и относительная плотность популяций на отдельных участках ареала по данным заготовок. Для этого среднее ежегодное число шкур каждого вида, принимавшееся по каждому пункту, разносилось в пределах ареала вида в своем условном точечном масштабе по сантиметровым клеткам стандартной сетки нашей карты. Учитывался также район (радиус) действия заготовительных пунктов. Общее число шкур пушных видов, послужившее для отметок на картах, превышает 3 665 000 штук. Контуры ареалов вычерчивались преимущественно для стенотопных видов. Для обычных л широко распространенных видов копытных и хищных, например для лбана, косули, оленя, сайги, козлов, выдры показано распространение, известное нам по личным наблюдениям в природе.
? Непроверенные мною литературные сведения, но заслуживающие внимания, поставлены всюду со знаком вопроса.
— Обзор истории развития и современного состояния ареалов выполнен J* не для всех видов. Для этого были выбраны преимущественно виды узко распространенные (стенотопные), а также те, для которых имелось больше палеонтологических фактов.
Расшифровка природы и истории ареалов кавказских зверей оказалась наиболее легкой при учете «борьбы» ксерофитного, пустынного и степного ландшафтов с лесным и вообще с мезофитным, а также при учете климатических смен в четвертичном периоде. Эта история ареалов в связи с историей ландшафтов более или менее ясна теперь лишь для четвертичного периода. О более глубоких, древних связях можно высказывать лишь вероятные предположения. Понятно также, что чем древнее и прочнее связи какого-либо зверя с тем или иным типом ландшафта, чем меньше число занимаемых им биотопов, тем более легки и тем более достоверны выводы.
Разрывы ареалов и «экологические барьеры» обсуждаются здесь только по линии связи вида с тем или иным типом ландшафта, особенно фитоландшафта, а также с точки зрения наличия физических и климатических границ. Несомненно, что экологические барьеры нуждаются в более обстоятельной расшифровке, поскольку они определяются нормами физиологических реакций, морфологическими возможностями и жизненными привычками («производственными показателями») видов. В правильности же наших теоретических предпосылок убеждало уже то, что специальные поиски индикаторных видов в соответствующих ландшафтах, организованные при работах по уточнению границ их ареалов, неизменно увенчивались успехом.
Специальные морфологические исследования применялись преимущественно к ископаемому материалу по четвертичным видам для выяснения происхождения последних, их стратиграфической изменяемости п быстроты эволюционного процесса. Они проведены преимущественно на хищных (Верещагин, 19516), отчасти на копытных и грызунах (глава IV). Географическая изменяемость на современном геологическом этапе была также прослежена на некоторых видах хищных, грызунов и копыт-
2 Н. К. Верещагин
18
Введение
ных й использована для выяснения происхождения и геологического возраста видов и подвидов. Учитывалось, что морфо-функциональный анализ организмов должен быть увязан с историей ландшафтов.
Б При описаниях и перечислениях по фрагментарному материалу иско-паемых четвертичных млекопитающих возникали значительные затруднения с наименованием видов и подвидов. Дело в том, что одни млекопитающие образовывали с среднего плейстоцена до наших дней последовательные ряды форм, слабо отличающихся друг от друга по остеологическим признакам, другие же давали на различных этапах ряды потомков более или менее отличных, которых можно считать либо подвидами живу-Я щих, либо подвидами ранее описанных ископаемых видов, либо особыми видами.
Л Как правило, палеонтологи считают возможным называть ископаемых | верхнечетвертичных млекопитающих именами современных видов, либо f) описывать их в качестве подвидов от современных на основе правила приоритета, рассчитанного на географическую изменяемость. Между тем
J обозначение предковой формы в качестве подвида современной противо-f речит здравому смыслу, тем более, что критерий вида и подвида часто ;
г условен и субъективен. !
г Считая эту практику порочной, мы все же оставили в сводных табли-I цах №№ 62 и 103 наименования описанных подвидов, прибегая для обо-k значения плохо изученных в смысле стратиграфической изменяемости j . форм к наименованиям conformis — сходный и affinis — родственный.
В существующих Правилах палеозоологической номенклатуры (1932) ; эти вопросы, к сожалению, не разобраны.
Возможность экстраполяции данных экологии современных млекопитающих на ископаемые формы была для нас достаточно бесспорна и подтверждена изучением «универсального» захоронения в Бинагадах.
Важность этих выводов для палеоэкологов и палеогеографов, занимающихся реконструкцией ландшафтов, очевидна.
Тем не менее, признавая известное постоянство ведущих экологических черт организмов в четвертичном периоде, мы отнюдь не отрицаем возможности и вероятности довольно быстрой и глубокой эволюции их физиологии и вообще изменения образа жизни при сохранении прежних морфологических черт.
Зоогеографический анализ всей голоценовой териофауны и районирование Кавказского перешейка являются завершением исследований по истории происхождения современных фаунистических комплексов. В основу анализа положен не только генетический, но и возрастной (стратиграфический) принцип. В отличие от формальных схем, основанных, как правило, на геоморфологическом или фитоландшафтном подразделении и приспосабливании к нему зоогеографических категорий, здесь выявляется историческая сущность ареалов видов и ареалов фаунисти- > ческих комплексов. *
Вопрос о родине — происхождении того или иного вида решается нами, помимо соображений, высказанных в III главе, еще на основе критериев, предложенных Арльдтом (Arldt, 1919).
Остается далее уяснение перспектив ближайшей эволюции ареалов видов и фаунистических комплексов. Решение этих вопросов лежит, несомненно, в исследованиях, раскрывающих сущность процесса формирования новейших экологических черт отдельных видов и их группировок. Ведущим фактором в этом процессе на данном геологическим этапе является человек. В связи с этим экологические исследования и состояли |
Введение
19
в наблюдениях над численностью и поведением зверей в первобытных и в созданных человеком ландшафтах и биотопах.
Эти исследования помогают уяснить новейший ход развития или деградации численности и ареалов, а также направления и пути формирования фауны в целом. Они производились по преимуществу в восточном Закавказье в различных ландшафтных зонах.
Количественные учеты зверей в различного типа угодьях производились по общепринятой, реже по особо разработанной методике, во время возглавлявшихся мною экспедиций Азербайджанской противочумной станции и Института зоологии Академии наук Азербайджанской ССР с 1938 по 1948 г. при изучении опасных в эпидемиологическом отношении грызунов и запасов ценных промысловых животных.
Новейший этап целенаправленных воздействий человека на природу степей изучался нами преимущественно в пределах Азербайджана. Подобное исследование должно быть в далънейягем поставлено в Предкавказье, где развито уже весьма широко устройство лесополос.
Преднамеренное введение в фауну новых видов зверей, которое столь широко вошло в практику в советское время, в этой книге подробно не освещено.
Акклиматизация новых видов и вообще биотехнические мероприятия в охотничьем хозяйстве — это целая самостоятельная проблема, над которой работают крупные научные силы. Тем не менее некоторые примеры темпов и характера формирования ареалов чужеземных видов хищных и грызунов в пределах Кавказа использованы для характеристики процесса развития ареала вида.1
При изучении новейших преобразований экологических группировок млекопитающих учитывалось, что ландшафтные изменения, вносимые человеком, накладываются на естественный процесс эволюции ландшафта и сложно взаимодействуют с ним, нередко целиком подавляя его. Так, например, развитие формации угнетенных колючих кустарников в предгорьях восточного Предкавказья на месте бывшего леса происходит не только под влиянием рубок и выпаса скота, но и под влиянием современной климатической тенденции. Наоборот, осушение центральных частей Колхидской низменности и насаждение там дренирующих пород деревьев совершенно подавляет естественные процессы заболачивания почвы и развития гидрофильной растительности. В обоих случаях первобытные исходные группировки крупных и мелких млекопитающих разрушаются нацело.
В этой книге приняты следующие экологические и зоогеографические понятия и термины.
Экологическая группировка млекопитающих — понятие, под которым разумеется сочетание видов, связанных в пределах биотопа общностью экологических условий, отвечающих на данном историческом этапе выработавшимся морфо-физиологическим особенностям видов.
Под фаунистическим комплексом млекопитающих разумеется сложившееся на данном историческом этапе сочетание ряда экологических группировок млекопитающих, связанных в пределах ландшафтной зоны общностью территории и общностью физико-географических факторов, воздействовавших на тот или иной географический район.
1 Сам я принимал непосредственное участие в работах по введению в гфауну Кавказа болотного бобра, енота-полоскуна, американской норки и консультировал ряд других опытов.
2*
20
Введение
Например, фаунистический комплекс сухих низменностей восточного Закавказья включал в плейстоцене и ныне включает экологические группировки открытой полупустыни, тугайных зарослей, тростниковых озер и болот и т. д. Этот комплекс развивался и исторически складывался с плиоцена до наших дней в условиях сухого и умеренно теплого климата.
Термин фауна употребляется нами для обозначения серии фаунистических комплексов, свойственных многозональным территориям.
Термин «фауна», применяемый большинством современных геологов, палеонтологов и археологов для любого набора встреченных в слоях ископаемых остатков животных, нами не применяется или берется в кавычки, так как отождествлять выборочный, а нередко и разновозрастный набор остатков животных в захоронениях с фауной или фаунистическим комплексом данного района или целой страны нет никаких оснований.
Наши понятия и термины вполне сохраняют свое значение и в палеогеографическом смысле, т. е. для различных стратиграфических этапов прошлого. Фаунистический комплекс может существовать на протяжении целой геологической эпохи, постепенно трансформируясь в отношении состава и этапов морфогенеза составляющих его элехментов. На практике восстановление полного состава экологических группировок и фаунистических комплексов прошлого по ископаемым остаткам достижимо с раз-. личной степенью приближения, зависящей, как уже было показано, от «неполноты геологической летописи». Именно поэтому для палеореконструкций приходится нередко пользоваться лишь индикаторными, массовыми видами, остатки которых встречаются в определенных стратиграфических горизонтах наиболее постоянно и в большом числе.
Находки остатков таких индикаторных видов плейстоценовых степей Евразии, как мамонт, лошадь, бизон, сайгак и другие, позволяют предполагать одновременное существование на подходящих участках и сопутствующих им сусликов, сурков, корсаков, степных хорьков и т. п. Сходные основания для экстраполяций могут быть и при находках остатков индикаторных горных видов.
Степень освоения литературных данных по различным разделам неодинакова. Для выполнения задуманного пришлось оперировать с обширной палеонтологической, зоологической, ботанической, геологической и археологической литературой — более чем 3200 названий. В библиографическом списке показаны лишь важнейшие использованные и цитированные работы.
Печатные свидетельства древнегреческих, римских, арабских и древнеармянских натуралистов, историков и географов о животных Кавказа использованы как по полным переводам, так и по обзорам Гана (1884— 1890), Латышева (1893—1904, 1947, 1948) и Караулова (1901).
Огромная литература по синонимике и вообще по формальной систематике видов приведена только для кавказских форм, и лишь наиболее существенная.
Кавказ — Кавказский перешеек — рассматривается в этой книге в следующих пределах: с севера — Кумо-Манычский водный рубеж, с юга — государственная граница закавказских республик.
Совершенно естественно, что в процессе полевых исследований и работы над книгой возникло и возникает много дополнительных проблем и вопросов. Наиболее важные из них — это вопросы стратиграфической параллелизации костеносных местонахождений и палеолитических па
Введение
21
мятников в условиях горной страны, лежащей на рубеже нескольких климатических, флористических и фаунистических провинций.
Осуществление этой работы могло быть сделано лишь при всемерном содействии дирекции Зоологического института Академии наук СССР и Президиума Академии наук Азербайджанской ССР. Б. С. Виноградову и А. А. Стрелкову я обязан весьма ценными редакционными замечаниями и советами.
Таблицы штриховых рисунков сделаны художником В. Н. Ляховым, а рисунки зверей—Е. Я. Захаровым и ироф. К. К. Флеровым. Все ландшафтные снимки сделаны мною.
На местах — в Азербайджане, Армении, Грузии, Дагестане, Северной Осетии, в Ставрополье и Краснодарском крае — я встречал всегда живейший интерес и содействие в работе со стороны всех советских учреждений и отдельных лиц, как специалистов, так и рядовых рабочих на новостройках и колхозников в глухих горных и степных селениях.
Пользуюсь случаем выразить благодарность всем лицам, так или иначе содействовавшим успеху дела.
Первый, более подробный вариант нашей работы был выполнен еще в 1954 г. При подготовке к печати были произведены большие сокращения текста, а также внесены новые материалы.
ЧАСТЬ II ЕР В А Я
ПАЛЕОНТОЛОГИЧЕСКОЕ
И АРХЕОЛОГИЧЕСКОЕ ОБОСНОВАНИЕ ИСТОРИИ ФОРМИРОВАНИЯ ФАУНЫ
Глава 1
РАЗВИТИЕ ЛАНДШАФТОВ И ФАУН МЛЕКОПИТАЮЩИХ КАВКАЗА В ТРЕТИЧНОМ ПЕРИОДЕ
ОБЗОР ЗАХОРОНЕНИЙ КОСТНЫХ ОСТАТКОВ В ТРЕТИЧНЫХ СЛОЯХ
На месте современного Кавказа суша была уже в палеозое, о чем свидетельствуют обильные остатки растений каменноугольной флоры, обнаруженные в сланцах и песчаниках северного склона Главного хребта.
После мощных трансгрессий моря отдельные острова возникают и
Рис. 6. Морской бассейн и суша (заштрихована) в Восточной Европе и Передней Азии в верхнем эопене — начале олигоцена. (По Архангельскому и Страхову, 1938). Точкой отмечено верхнеолигоценовое захоронение у Бенара.
существуют здесь вновь в мезозое.
Уже в юре кавказские острова были покрыты папоротниками, гинкго, саговниками, но палеоботаники начинают историю флоры прикаспийских стран с мелового периода, когда на новых островах появляется древесная растительность, содержавшая лиственные виды — платан, тополь, дриандру, мирику — наряду с гигантскими хвойными — араукариями, секвойями и др.
О наземных позвоночных этих островов, сливавшихся постепенно в общую сушу, можно судить лишь по следам игуанодонов, открытым на натеках травертинов у пещеры
Сатаплия близ Кутаиси (Канделаки и Дзоценидзе, 1937; Габуния, 19566).
По представлениям Павлова (1936), обширная суша в начале палеоцена тянулась от Передней Азии до Балтики. В районе Кавказа имелись невысокие складки юрских и меловых слоев.
Затем воды европейского Средиземного моря (Тетиса) залили южную Европу, Украину, Крым и частично Кавказ, сливаясь с Азиатским морем,, уходившим далее на юго-восток.
Таким образом, оформление суши и ее рельефа в области Кавказа, предопределивших развитие и формирование здесь фаун четвертичного периода, происходило целиком в кайнозое.
г.
Часть I. Обоснование истории формирования фауны
В эоцене в области северного побережья Малой Азии, а также современного Сурама, Казбека и Эльбруса начались мощные вулканические проявления, в результате которых Кавказский остров (рис. 6), поднимавшийся среди Тетиса, приобрел, по-видимому, горный рельеф. Фактических данных об эоценовых флорах нет.
В море, омывавшем Кавказский остров, обитали небольшие киты рода Zeuglodon из подотряда древних китов Archeoceti. Их остатки найдены в отложениях коунской свиты на Апшеронском полуострове и в районе речки Сумгаит.
Остатки наземных позвоночных обнаруживаются впервые на Кавказе лишь в олигоценовых толщах,
ОЛИГОЦЕН
В олигоцене тропический характер природы Кавказского острова сохраняется. Из туфо-песчаников нижнеолигоценового возраста горы Дарры-Даг в долине Аракса описаны остатки папоротников — Blechnum Brauni, Pteris oeningensis, пальм — Sabal haeringiana, хвойных — Podocarpus, лавровых — Cinnamomum, мирики, зелква, мелких злаков — Panicum miocenica и др. Палибип (1936) сопоставлял дарры-дагскую олиго-ценовую флору с нижнео лигоценовыми флорами Балкан и средней Европы, считая, что она свидетельствует о наличии влажных тропических лесов, покрывавших вулканические острова.
Костных остатков из нижнеолигоценовых слоев пока неизвестно, но весьма вероятно, что Кавказский остров и прилежащие к нему с юга участки суши были заселены в это время представителями той разнообразной фауны из примитивных копытных и хищных, которая хорошо известна из эоценовых и олигоценовых отложений Западной Европы.
Богачев (1938г) все же упоминал о находке отпечатка лапы пятипалого хищника из подотряда креодонтов, достигавшего размера крупной собаки, на слое окремнелого вулканического пепла Дарры-Дага близ города Джульфы.
В морских — среднеолигоценовых — майкопских отложениях Предкавказья и Закавказья богато представлены остатки тресковых и сельдевых рыб. Известны также находки остатков морских коров — II ahi he-rium cf. schinzi Каир, и других, ближе не определенных видов, но возможных предков современных дюгоней, по южному берегу Черного моря — близ Платана, в Чиатурском марганцевом бассейне Грузии и в майкопской свите Ашперонского полуострова в урочище Ахзы-Хазры (Богачев, 1938в). По-видимому, из таких же слоев восточного Закавказья Лидеккером (Lydekker, 1892) описаны остатки мелких китов Zeuglodon (Microzeuglodon) caucasicum Lyd. и Iniops caucasicum Lyd.
Остатки Microzeuglodon aff. caucasicum Lyd. описаны также из отложений, вскрытых речкой Сумгаит в 20 км к северу от Баку (Рябинин, 1938).
Ряд находок остатков китообразных родов Microzeuglodon, Iniops, Delphinus (sensu lato), Zeuglodon перечисляется и Богачевым (1938в, 1939а) для майкопской свиты Ашперонского полуострова.
В майкопских же слоях на речке Сумгаит найдены остатки листьев вечнозеленых видов растений из семейств Combretaceae, Sapotaceae, Ternstomiaceae, с примесью тропических хвойных пород.
Все это обилие наземной растительности и морских млекопитающих рисует девственную тропическую природу Кавказского острова и саргассовый тип лагун его южного побережья. На Малоазиатской суше, судя
Г I
г
Гл. I. Развитие ландшафтов и фаун млекопитающих в третичном периоде 27
по находкам с южного склона Тавра, в олпгоцепе также существовала растительность, содержавшая подокарпус, мирику, дуб, циннамон, андро-меду, эвкалипт и другие виды.
Захоронение остатков наземных олигоценовых зверей было открыто геологом М. Ф. Хучуа в 1948 г., в местности Бенара Ахалцихского района южной Грузии. Немногочисленные и сильно разрушенные фрагменты трубчатых костей и зубов обнаружены в так называемой верхней
Рис. 7. Остатки олигоценовых копытных из Бенара.
j 4 — и os lunatum. BenaraHierium kaUistrati Gab.; 2 — M3 Acertitherium ct‘. filholi Osb.; 3 — метатарзальная кость Schis3th.erium chufiu-ъе; 5 — фрагмент лопатки мелкого парнокопытного.
пестроцветной песчаниковой толще, согласно залегающей над лигнито-носноп свитой. Условия захоронения изучены слабо. Мелкие обломки костей приурочены главным образом к конкрециям сильно уплотненного песчаника. Отсюда определены и описаны Габунией (1951а, 19516, 1953. 1955а, 19556) остатки грызунов и копытных:
Rodcntia Fam. gen.
Perissodactyla
Schizotherium chucuae Gab.
Benaralheriuni callistrati Gab.
Artiodactyla
Anlhracotherium sp.
Lophiomeryx benarensis Gab.
28
Часть I. Обоснование истории формирования фауны
Габуния проводит стратиграфические параллели бенарского местонахождения с богатой «фауной» олигоценовых фосфоритов Кверси во-Франции и со среднеолигоценовыми местонахождениями остатков зверей в Монголии и в Казахстане, где кости схизотериев и лофиомериксов известны пз верхнеолпгоценовых слоев (рис. 7).
Фаунистические находки близ Ахалциха подтверждают соображения палеоботаников об олигоценовых ландшафтах в области Кавказа и свидетельствуют о существовании прерывающихся временами континентальных связей Малокавказской суши в олигоцене со Средней и Центральной Азией, а также с Западной Европой.
МИОЦЕН
В нижнем миоцене Кавказский остров тянулся примерно от Анапы на западе до верховьев Самура на востоке. В районе современного Дзи-рульского кристаллического массива существовал другой остров мень-
Рис. 8. Морские бассейны и суша в среднем миоцене. (По Жижченко, — см. Жижченко и др., 1940). Точкой отмечено местонахождение остатков наземных позвоночных у Бсломечетской.
шего размера. В районе Трпалети—Ахалциха и Боржоми в нижнем миоцене было море и отлагались морские пески, по в конце нижнего миоцена суша появилась и здесь (Жижченко, Колесников, Эберзип, 1940). В слоях этой эпохи известны лишь остатки морских млекопитающих.
Из миоценовых отложений Локбатапа, южнее Баку, описаны остатки тюленя — Phoca sp., близкого к Р. vindobonensis Toula (Богачев, 19276). Остатки тюленя Phoca sp. обнаружены также в известняках Килязин-ской косы к северу от Баку, а остатки китов цетотериев — Cetotherium meyeri. Brandt в Локбатапе и у сел. Джорат на Апшеронском полуострове (Богачев, 1938в).
Гл. 1. Развитие ландшафтов и фаун млекопитающих в третичном периоде 29
В начале среднего миоцена в так называемом гельветском ярусе Кавказский остров сохранял прежние, нижнемиоценовые очертания. Море, отложившее осадки тархапского и чокракского ярусов, расстилалось от северного побережья Кавказского острова до Ростова на северо-за-’ [аде и Степного (бывш. Элиста) на северо-востоке. Южнее Сурамского острова к этому времени появляется, по-видимому, крупный массив Малокавказской суши, а сами кавказские острова сильно увеличиваются.
Растительность островов в среднем миоцене, судя по находкам в Предкавказье и в Закавказье, еще полностью сохраняла субтропический характер (Палибпн, 1936), но уже появляется небольшая примесь видов с опадающей листвой — Castanea sativa и хвойных северного корня — Pinas neptuni. В среднем же миоцене, в чокракский век, Кавказская •сvina постепенно поднимается и, разрастаясь, становится полуостровом, црпчлененным к Переднеазиатскому массиву (рис. 8).
Из чокракского яруса в Предкавказье известны и обильные остатки наземных млекопитающих.
Центральное Предкавказье
Бел оме четска я. Местонахождение это, обнаруженное в 1926 г. А. В. Дановым, находится у станицы Беломечетской, на правом берегу Кубани, севернее Черкесска (рис. 1).
Чокракский горизонт достигает на этом участке плоскогорья 60 м мощности и выражен преимущественно крупнозернистыми песчаниками я песчано-известняково-глинистымн отложениями. Современная Кубань, подмывая эродированную толщу миоценовых всхолмлений, вскрыла в ряде мест костепосные песчаные линзы, которые находятся на высоте иколо 50 м над меженным уровнем реки. Скорее всего здесь протекала река, прорезавшая чокракские слои и размывшая какое-то первоначальное озерное пли болотное захоронение трупов. Костеносная линза у южной окраины Беломечетской состоит из хрящеватого кварцевого песка с зернами 0.5—1.0 мм, слабо цементированными иловатыми частицами. .Местами на разрезе песчаная толща прокрашена окисью железа в виде вертикальных или косых полос. Резко выраженной речной слоистости пег, что возможно объясняется метаморфизацией отложений. В толще хрящеватого песка заметны участки иловатого, более плотно сцементированного песка сероватого цвета, свидетельствующие об изменении режима течения. Видимая мощность уцелевшего костеносного слоя на этом участке достигает 3.5—4.0 м. Сверху он перекрыт довольно плотной, почти горизонтальной известняковой плитой 30—40 см толщины. Эта прослойка, вероятно, свидетельствует о быстром наступлении моря и перекрывании речных отложений морскими. Выше ее пески вновь чередуются с известняками и прикрываются местами четвертичными суглинками (рис. 9).
Остатки наземных и морских животных — целые трубчатые кости, их обломки, фрагменты черепов и изолированные зубы — залегают разбросанно: или непосредственно в хрящеватом песке, пли чаще в скоплениях сизоватого, плотно цементированного иловатого песка, прокрашенного местами охрой. Кости с поверхности палевого, светло-коричневого или темно-бурого цвета. В свежем разломе вещество диафизов светло-коричневое, трещины и поры эпифизов прокрашены окисью, реже — закисью железа. Так же сильно прокрашен дентин зубов. Эмаль их блестящая, темно-сизая, иногда черная у основания коронок. Древние раз
30
Часть I. Оиоснование истории формирования фауны
ломы трубчатых костей обычно заметно затерты, «закатаны» водой, а полости костей забиты песком.
Погрызы костей хищниками не подмечены. Восстановить сезон и причины первоначальной гибели животных трудно. Вряд ли эта гибель была единовременной и катастрофической. Разрозненность и фрагментарность
Рис. 9. Костеносная толща у Беломечетской.
Фот. автора, 1950.
остатков, обилие изолированных зубов мелких копытных, их положение в толще песка в горизонтальном, а иногда и в вертикальном положениях свидетельствуют о неоднократном переотложении. Гибли как растениеядные, так и хищные формы во взрослом и в молодом возрасте. Остатки болотного мастодонта — платибелодона состоят из фрагмента лицевой части черепа и нижних челюстей, принадлежавших одному взрослому и одному молодому индивиду.
В захоронении присутствуют остатки наземных п водных — морских млекопитающих, а также водных черепах — триониксов. Характер сохранности и степень минерализации костей у тех и других оказались
Гл. 1. Развитие ландшафтов и фаун млекопитающих в третичном периоде 3J
почти одинаковыми. Это подтверждает вторичную аккумуляцию остатков животных в толще аллювия. Кости, очевидно, вымывались из первичных скелетоносиых линз олигоцепо-миоцеиового возраста речным потоком или, что менее вероятно, морскими волнами. При этом кости, подходящие * ио удельному весу и «парусности» для захоронения, оказались перераспределенными в толще отложений реки. Первоначальные захоронения трупов наземных животных, судя по прокраске костей солями железа, происходили в застойном пресноводном водоеме, вероятно пойменного типа. Таким образом, следует признать, что эпоха существования большей части животных беломечетской «фауны» более древняя, нежели эпоха образования дошедшего до нас костеносного слоя. Другая, меньшая часть животных гибла, несомненно, и во время образования беломечетской аллювиальной толщи. Здесь могли мацерироваться трупы наземных зверей, приносившиеся речкой, и трупы заплывавших в русло морских млекопитающих.
В коллекциях, хранящихся ныне в ПИНе, ЗИНе и Институте палеонтологии АН Грузинской ССР, имеются остатки видов и групп млекопитающих, включенпых в табл. 1.
Таблица 1
Видовой состав и численность остатков млекопитающих в сборах из среднемиоцеповых слоев Беломечетской 1
Отряды и виды Число костей Отряды и виды Число костей
Carniv ora Anchitherium aurelianense Cuv. 4
\niphicyon sp Hyaena sp Ursavus sp Dicerorhinus caucasicus Boris. 172
9 6 4 Chalicolheriidae gen Fam. gen 3 1
Tubulidentata Artiodactyla
Orycteropus sp A Kubanochoerus robustus Gab. . 1
1 Hippopolamidae (?) gen. . . M icromeryx jlourensianus Lar- 1
Rodentia
tet 7
Palaeocricelus caucasicus Arg. 6 Dicrocerus elegans Lartet . . 3
Proboscidea Paratragocerus caucasicus Soc. Eotragus cf. martinianus Lar- 8
Platybelodon danovi Boris. . . 4 tet 12
Mastodon sp 131 Hypsodontus miocenicus Soc. . 1
Perissodactyla Fam. gen 52
Paranchiiherium karpinskii Bo- Cetacea
ris 20 Fam. gen 34
Имеются также в коллекциях остатки рыб Pisces и водных черепах — Trionyx. Общее количество известных теперь форм миоценовых млекопитающих Беломечетской достигает 20, ио систематическая принадлежность некоторых из них пока еще загадочна (рис. 10—15).
Стратиграфическими, руководящими формами Борисяк (1937, 19386) считал паранхитерия, анхптерия, носорога и платибелодона. Беломечет-ский парапхитерий характеризуется той же степенью специализации,
1 Определения даны по печатным работам Борисяка (1928а, 1943), Аргиропуло (1938, 1940в), Соколова (1949), Габунии (1955г, 1956а), а также по неопубликованным данным К. К. Флерова и моим.
'32
Часть I. Обоснование истории формирования фауны
что и Parahippus из миоцена Северной Америки. Анхитерий — вероятно, более ранняя филогенетически форма, характерная для миоцена Евразии.
Беломечетского носорога Борисяк считал одним из наиболее примитивных представителей Dicerorhinae, с зубами, имеющими ряд примитивных и в то же время специализированных черт. Возможно, что его следует рассматривать как древнейшего представителя ветви Rhinoceros schleier macheri.
Своеобразный мастодонт — платибелодон наиболее близок к представителям рода Phiomia и Palaeomastodon из олигоцена Африки, объединяю-
Рис. 10. Остатки хищных из Беломечетской.
2, 2 — Hyaena sp.; 3 — Amphicyon sp.
щих возможных предков длиниорылых мастодонтов. Нижние, уплощенные дорзо-вентралыю и плотно соединенные вместе бивни платибелодона образуют вытянутый ковш, как и у Ambeloclon jricJti Barb, из плиоцена Небраски. По предположениям Борисяка (19286) и Осборна (Osborn, 1936), эти мастодонты не имели длинного хобота. Они жили и питались по берегам в заводях рек, озер, доставая сочные водные растения и их корневища нижними уплощенными бивнями.
-% Мелкие дикроцерусы, микромериксы и эотрагусы — руководящие формы средне- и верхнемиоценовых слоев Евразии. Вообще ряд признаков говорит о смешении в костепосном слое Беломечетской стратиграфически различных форм. В связи с этим и выделить группу индикаторных видов фаунистического комплекса среднемиоценового возраста трудно.
Гл. I. Развитие ландшафтов и фаун млекопитающих в третичном периоде 33
Специфичность беломечетской «фауны» не позволяет пока провести ближайшие параллели с другими миоценовыми фаунами Евразии, и ее зоогеографические связи остаются недостаточно ясными, но при ма-
Рнс. 11. Черен п зуб Platybelodon danovi. (По Борвсяку, 19286).
лой вероятности ее самобытного развития в пределах небольшой Кавказской суши широкие связи последней с южным континентальным массивом несомненны.
При попытке воссоздания вероятных ландшафтов эпохи обитания и накопления трупов беломечетских животных возникает мысль, что наряду с горными цепями па Кавказской суше должны были существовать
3 H. К. Верещагин
34
Часть 1. Обоснование истории формирования фауны
и широкие участки спокойных равнин с хорошо развитыми лесами, кустарниками, с озерами и тихими заводями рек, заросшими обильной болотной растительностью. В этих заводях и озерах, очевидно, и кормились платибелодоны.
Охарактеризованные литологически и морской фауной моллюсков последующие горизонты среднего миоцена, известные под именем кара-ганского и конкского ярусов, сохранили и ряд местонахождений остатков насекомых и растений.
В карагаиских слоях Дагестана обнаружены остатки тополя — Populus mutabilis, саппндуса — Sapindus fulcifolius и вечнозеленых цин-
Pirc. 12. Череп п зубы Paranchitherium karpinskii. (По Борпсяку, 1937).
намонов — Cinnamomum sclieuclizeri. Из песчано-глинистых отложении прибрежной зоны спаниодоптеллового моря к юго-западу от Дербента определены листья вечнозеленых пород, характерных для верхнеолиго-цеповых и иижнемпоценовых отложений Европы: мирики — Myrica lia-keaefolia, лавра — Laurus primigenia, ардпзип — Ardisia cf. oceanica,. хурмы— Diospyros paradisiaca и др. Судя по фауне морских моллюсков, в это время возможно наступление значительного опреснения миоценового бассейна как следствие его изоляции от Тортоиского бассейна Европы в районе Балкан и вероятное изменение климата в сторону большей влажности и большего приноса воды реками (Жижченко, Колесников,. Эберзин, 1940).
Богатейшая «фауна» насекомых была обнаружена в слоистых мергелях карагапского яруса к западу от Ставрополя. Насекомые сносились,, вероятно, небольшими ручьями в мелководные опресненные лиманы се1-
Гл. I. Развитие ландшафтов и фаун млекопитающих в третичном периоде 33
верного побережья и захоронялись в тонких илах. Определено до 90 видов насекомых. Из водных форм здесь обычны поденки — Ephemeroptera, стрекозы — Odonata, клопы-водомерки — Gerridae, водяные жуки — , Dytiscidae, Hydrophilidae, ручейники — Trichoptera, комары — Dip-tera. Из сухопутных растениеядных видов, сапрофагов, хищников и паразитов жили прямокрылые — Ortoptera, клопы — Hemiptera, тли, тараканы, термиты, цикады, бабочки — Lepidoptera, двукрылые, перепончатокрылые — Himenoptera и жуки (Родендорф, 1939).
Рпс. 13. Верхние резцы гш антского непарпопалого из Беломечетской. (0pm .).
Рис. 14. к. 1ык неизвестного парнокопытного (Hip-popotatnidae?) из Беломечетской. (Ориг.).
Этот набор жизненных форм свидетельствует о субтропическом климате и об обильной травянистой растительности на Кавказской суше в конце среднего миоцена.
Северное побережье Конского бассейна, судя по находкам многочисленных остатков растений в долине р. Крынки в Донбассе (Кришто-фовпч, 1930), обладало, по-видимому, растительностью умеренного типа, резко отличной от субтропической растительности кавказского неогена.
В верхнем миоцене море по-прежпему широко заливает юг Украины, равнинный Крым, Предкавказье, значительную часть Закаспия, а возможно, и Прана.
Фауна Сарматского моря отражает дальнейшее опреснение и значительно меняется по фацпям и возрасту.
В нижнем сармате Кавказская суша увеличивалась за счет прироста с юга. С^дя по мощностям отложений Сарматского моря, рельеф «Яфе-тиды» п прибрежных участков сохранял в основном тот же характер, что и в среднем миоцене. Наибольшие горные поднятия намечались в нижнем сармате в области Гурийского хребта и Хевсуретин, а в среднем сармате еще и во внутреннем Дагестане. Северо-западное побережье продол
36
Часть I. Обоснование истории формирования фауны
жало оставаться равнинным. Море здесь мелело в связи с поднятием Ставропольского массива.
В верхнем сармате море сильно сокращалось в размерах, оставляя северную часть современного Каспия и большую часть Закаспия. Ставро
Рис. 15.
1 — челюсть Kubanochoerus robustus (по Габуния, 1955а); роговые стержни: 2 — 1>ага-tragocerus caucasicus; 3 — Eotragus cf. martinianzis', 4 — зубы Hypsodontus miocenicus (по Соколову, 1949).
1 1
1 ] j
1
польская возвышенность полностью осушалась и выдвигалась па север в Предкавказскип пролив. На юге от Кутаиси и до Телава разрастался широкий перешеек, соединявший «Яфстиду» с малокавказской сушей. В восточном Закавказье образовался узкий Куринский залив (Жижченко, Колесников, Эберзпп, 1940).
Ландшафты Кавказа сарматского времени могут быть восстановлены со значительной степенью правдоподобия по находкам остатков растений и животных.
гл ч< с сс
C1 П( ВС иг лг В£
зи от ци Са sts Ph
Гл. I. Развитие ландшафтов и фаун млекопитающих в третичном периоде 37
Прибрежноморская растительность мелководий состояла из фукусов п цистозейр, остатки которых обнаружены в долинах Сунжи и Сулака и на Малокабардинском хребте.
Сарматское море было обильно заселено тюленями — Phocidae и мелкими беззубыми китами рода Cetotherium. Остатки цетотериев найдены в прибрежных сарматских ракушечниках и илах близ Дербента, Махачкалы, в синих сарматских глинах близ Майкопа (Спасский, 1939). Остатки тюленей обнаружены в белых морских песках близ Ставрополя, в синих
Рис. 1G. Морские бассейны и суша в верхнем миоцене. Верхний сармат. (По Колесникову,—см. Жижченкои др., 1940). Точками отмечены главнейшие местонахождения остатков наземных позвоночных.
глинах близ Майкопа и Горячего Ключа и в ряде других мест. Встречаются они в смешанных захоронениях в пределах Ставрополья вместе с остатками сухопутных третичных млекопитающих. Все это были, не-? сомиепно, теплолюбивые формы, вряд ли знакомые со льдами.
Наземная растительность Кавказского полуострова в сармате состояла из хвойных, вечнозеленых и листопадных видов. Из обнажений по р. Супсе известны остатки ели — Abies sp. сосны — Piiius sp., секвойи — Sequoia sp. С Кахетинского хребта и с Закубанской равнины пз низов среднего сармата определены остатки магнолий — Magnolia, лавров — Laurus primigenius, цинпамопов — Cinnamon шп polimorphum, вязов, ив и груш.
Захоронения в слоях среднего сармата ряда пунктов Восточной Гру-; зин, содержат около 30 определенных видов деревьев, до 70°о которых относится к породам с опадающей листвой. Кроме вечнозеленых лавров, цинпамопов, мирик и магнолий, здесь уже росли третичные виды граба — Carpinus, дуба — Quercus, ивы — Salix, клена — Acer, каштана — Castanea, дзельквы — Zelkova Ungeri и кустарники: кутра — Аросупо-phyllum ibericum, пузырник — Colutea Salieri и др. По мнению Палибина
38
Часть I. Обоснование истории формирования фауны
(1936), вечнозеленые представители этой растительности, «одевавшей всю поверхность Яфетиды», были в сармате последним остатком тропической и субтропической флоры, свойственной Кавказу с начала третичного периода. Этот факт начавшегося процесса «бореализации» флоры важен и для понимания дальнейшей эволюции фауны. Сопоставляя с этой флорой верхнемиоценовые флоры Малой Азии, Гроссгенм (1936) приходит к выводу, что они обнаруживают большую близость и характеризуются той же смесью бореальных и субтропических элементов.
С точки зрения эколога и фауниста было бы важно знать тафономию этих захоронеппй, так как несомненно, что при наличии горного и даже высокогорного рельефа, в среднем миоцене могла уже выработаться высотная зональность растительности. Именно этим-то, быть может, и объясняется в ряде случаев «смешанный» характер миоценовой флоры, отмечаемый ботаниками.
В сармате на Кавказском полуострове появляются представители гиппарионовой фауны. Это проникновение шло с юга по широкому закавказскому перешейку (рис. 16).
Первое открытие захоронений остатков животных гиппарионовой фауны в Закавказье принадлежит, по-видимому, Рябинину (1913).
В урочище Кясаман, на левом берегу р. Поры, в красных глинах с'прослойками песчаников, содержащих остатки водорослей — Chara cf. Escheri и моллюсков — Planorbis sp., Рябинин обнаружил кости и зубы Hipparion gracile Каир и обломки зубов представителей Rhino-cerotidae. Остатки тех же зверей Рябипип отмечает в глипах с гипсом, залегающих под акчагыльскими слоями в обнажениях хребта Кацаху-р ис-Кеды.
Южнее, на правом берегу Иоры, в обрывах хребтов Эйляр-Оуги и Палан-Тикян, геолог Домбровский (1914) обнаружил в 1913 г. большое захоронение остатков третичных млекопитающих. Оно известно под именем эльдарского.
Карталинские плоскогорья
Эльдар. По описаниям Домбровского (1914), Андрианова и Ларина (1935), костеиосный горизонт прослеживается на протяжении 6 км от ущелья, соединяющего степь Эльдар с поймой Иоры, превращаясь по мере следования с запада на восток из конгломерата ракушечника сначала в песчаник, затем в известняк, снова в песчаник, переходящий в глины, и, наконец, теряется в мощной толще глин верхнего сармата. Скопления костей — части скелетов — встречаются как бы гнездами, с промежутками через 10, 20 и несколько сот метров. На поверхности слоя костеносного песчаника видны местами знаки водной ряби.
Характер захоронения свидетельствует о наличии в этом районе низкого побережья и илистого залива. В последний и сносились трупы животных, перемещавшиеся незначительными течениями и слабым прибоем в пункты затпшья. Здесь они погребались в песчано-глиппстых осадках. Причину массовой гибели сухопутных млекопитающих авторы видят в степных пожарах, землетрясениях или паводках, при которых погибавшие животные выносились в залив рекой или селевыми потоками.
По нашим наблюдениям, все уцелевшие полые кости, даже нижние челюсти п ребра, оказались сплющенными последующим давлением отложившихся толщ. Некоторые кости ломались в процессе захоронения, благодаря чему их полости, забитые песком и илом, сохранили первоначальную форму.
Гл. I. Развитие ландшафтов и фаун млекопитающих в третичном периоде 39
С поверхности кости ровного каштанового или шоколадного цвета, .а в свежем разломе более светлых, серых тонов. Вторичная минерализация и вообще метаморфизация костного вещества невелика. Сохранились и добыты копролиты.
Кости затертые прибоем, единичны. Имеется много сочлененных костей конечностей. Вообще ряд фактов позволяет согласиться с заключениями прежних исследователей о природе этого захоронения.
В коллекциях, хранящихся в Москве и Баку, по определениям Богачева (1927а), Алексеева (1930), Борисяка и Беляевой (1948), имеются виды, указанные в табл. 2 (рис. 17 и 18).
Таблица 2
Видовой состав и численность остатков млекопитающих в сборах из верхнемиоценовых слоев Эльдара
Отряды п виды
Число костей
Отряды и виды
Число костей
Primates
Tam gen......................
Carnivora
Crocuta eldarica Bog.........
Hyaena cf. eximia Gaudry . .
Proboscidea
Mastodon longirostris Каир . M. pentelici Gaudry..........
Mastodon sp..................
Dinotherium giganteum Каир.
Perissodactyla
Hipparion gracile Каир . . .
Chalicotherium sp...........
Aceratherium transcaucasi-cum Bog......................
Picerorhinus aff. orienialis Schloss......................
1
4
6
24
3
164
2
6
65
Artiodactyla
Sus erymanthius Roth, et
Wagn.....................
Achtiaria borissiakii Alex. . .
Camelopardalis (Helladotheri-um) sp.....................
Tragocerus aff. leskevitschi
Boris....................
Tragocerus sp. Nr. 1.......
Tragocerus sp. Nr. 2.......
Gasella sp..........
Pinnipedia Phoca sp............. . . .
Cetacea Delphinus sp.................
Cetotherium sp..............
5
69
159
6
Отсутствие переотложения остатков и безусловное единовременное существование большинства видов перечисленного списка позволяют рассматривать набор эльдарских наземных животных как часть единого фаунистического комплекса верхнемиоценового Закавказья. Наиболее массовыми ландшафтными видами в этом комплексе, судя по сборам, были гпппарпон, козлорогп, носороги и жирафы.
У д а б н о. В 1931 г. на водоразделе рек Куры и Иоры, в Гареджий-ской степи, близ сел. Удабно, Н. А. Гедройцем было обнаружено другое крупное захоронение остатков млекопитающих фауны гиппариона. Позднее Музеи Грузии организовал здесь раскопки.
Это захоронение находится в сарматских глинах, переслаивающихся песчаниками ширакской свиты. Его тип и характер, по-видимому, близки к эльдарскому. Так же близок и видовой состав остатков. По исследованиям Бурчак-Абрамовича и Габашвили (1945, 1950), Церетели (1942),
Рис. 17. Остатки хищных и копытных из Эльдара.
7, 3— череп Hyaena sp.; 2— копролнт гиены; 4, 5 — скакательный сустав и роговой стержень Т гадости* sp.; 6’, 7 — роговой стержень и челюсть Gazella sp.
Гл. I. Развитие ландшафтов и фаун млекопитающих в третичном периоде 41
Борисяка.и Беляевой (1948), предварительный список форм в материалах, хранящихся в ПИНе и Музее Грузии, таков:
Primates Perissodactyla
’ Udabnopithecus garedziensis et Gab. Carnivora Hyaena sp. Rodentia Hystricidae gen. Proboscidea Mastodon sp. Dinolherium sp. Burtsch. Hipparion gracile Каир Aceratherium sp. Artiodactyla Sus sp. Cervidae gen. Achtiaria sp. Tragocerus sp. Gazella sp.
Особый- интерес для разработки вопроса происхождения человека представляет здесь находка зубов крупной человекообразной обезьяны — гареджийского удобнопитека (рис. 19).
Рис. 18. Остатки хищных и копытных из Эльдара.
1 — челюсть Crocuta eldarica; 2 — резец Acerat helium, transcaucasicum’, 3—5 — зубы Tragocerus leshevitschi (по Богачеву, 19276).
A p к ii e т п. На самом западном участке Закавказского залива Сарматского моря костоносный участок обнаружен в 1.5 км к востоку от сел. Аркпети Юго-Осетпи. Здесь обнажаются серовато-желтые суглинки мощностью в 1.5—2.0 м с раздробленными костями млекопитающих. Костеносная линза длиной 3 м и мощностью 50—70 см содержит пеокатанпые-п нередко сочлененные трубчатые кости и черепа млекопитающих. В собранных коллекциях Габуния (19526, 1955в) определил остатки непарно-и парнокопытных:
Perissodactyla
Hipparion sp. (cf. garedzicum n. sp.)
Artiodactyla
Sus sp.
Eostylocerus sp.
Dicrocerus salomeae Gab.
Tragocerus ex gr. leskevitschi Boris.
Tragocerus sp.
Gazella cf. gaudryi Sclilos.
Gazella sp.
Giratfidae gen.
Phronetragus arknelhensis Gab.
42
Часть I. Обоснование истории формирования фауны
Захоронение это, по мнению Габунии, весьма близко по возрасту к Удабно и может быть датировано верхним сарматом — меотисом.
Состав наземных млекопитающих сармата Закавказья свидетельст вует о наличии значительных предгорных равнин, покрытых растительностью типа саванны и гилей, в которых паслись миоценовые жирафы и козлороги. Речные потоки, стекавшие с оформившегося Кавказского хребта, были, очевидно, достаточно
мощны для того, чтобы выносить в бухты Сарматского моря трупы мастодонтов и носорогов.
В целом фаунистический комплекс млекопитающих кавказского сармата подтверждает характер ландшафта, известный по палеоботаническим данным, особенно если учесть, что остатки растений в захоронениях часто свидетельствуют не только о «смешанности» сарматской флоры, но и о высотной дифференцировке растительности.
garedziensis.
, наружной,
Udabnopitecus { .») с передней, н жевательной (x2l/J поверхно-
Рис. 19. Зубы 1—4 — Рт2 (х 1 внутренней стей; 5—7 — М1 (X I1/») с передней, наружной и жевательной поверхностей. (По Бурчан-Абра-мовичу и Габашвили, 1950).
СМ
Ставрополье
Гиппарионовый комплекс появился в сармате и в Предкавказье, проникнув частью в обход поднявшихся горных складок — по побережьям, частью непосредственно через плоскогорья и хребты центрального участка полуострова. Известны находки остатков гиппариона — Hipparion sp. и динотерия — Dinotherium gigant-heum Каир в среднесарматских известняках близ села Бурлацкое (Хоменко, 1913а) и мелкой жирафы — Camelopardalis parva Weith. в верхнесарматских отложениях близ села Благодарное Ставропольского края (Павлова, 1933а).
Гора Куцай. Северо-восточнее Ставрополя, близ села Петровское, на горе Куцай А. А. Иванчин-Писарев обнаружил в 1915 г. более крупное захоронение остатков позвоночных. Подобно закавказским сарматским захоронениям оно содержит как морские, так и наземные формы.
По описаниям Иванова (1916), костеносная толща горы Куцай сложена песками и глинами, залегает между подстилающими ее песчанистыми и вязкими глинами среднего сармата и прикрывающими известковыми рыхлыми песчаниками. Остатки млекопитающих концентрируются в среднезернистых морских кварцевых песках с прослойками крупнозернистых. Характерна плохая сохранность остатков: найдены по преимуществу изолированные зубы, обломки позвонков, метаподий, фаланг. Кости тюленей и китов находятся в нижнем 2—-3-метровом слое глин и песков верхнего сармата, остатки же наземных зверей лежат несколько выше, по все же в нижних 3—5-метровых слоях песчано-глинистой толщи.
Набор сухопутных видов в определениях Иванова (1916) говорит ю том, что здесь захоронены остатки представителей фауны гиппариона:
Гл; I. Развитие ландшафтов и фаун млекопитающих в третичном периоде 43
Perissodactyla
Hipparion gracile Каир
Hipparion sp.
Aceratherium sp.
»Rhinoceros sp.
Artiodactyla
Listriodon sp.
Sus sp.
Gazella sp.
Fam. gen.
Pinnipedia
Phoca cf. pontica Eichw.
Cetacea
Cetolherium priscum Brandt
Cetotherium (?) sp.
Найдены также остатки птиц, черепах, рыб.
Единичные находки остатков верхнемиоценовых носорогов родов Dicerorhinus и Aceratherium известны близ Майкопа в слоях зеленоватого ила и песка, а также в морских песках близ Бешпагира и Ставрополя. В целом состав видов зверей из предкавказского сармата весьма близок к составу закавказского.
В поисках истоков гиппарионового комплекса, появившегося на Кавказе, уместно сравнить состав зверей из сарматских местонахождений Кавказа с другими миоценовыми «фаунами» Евразии. Для оценки фаунистических влияний в миоцене представляют интерес южноазиатские миоценовые «фауны» в Пенджабе, у южного подножья Гималаев, в области Салт — в бассейне Инда, Сиваликских холмов в бассейне Инда и Ганга, среднесарматская «фауна» Крыма, верхнемиоценовая «фауна» Ирана с восточного побережья оз. Урмия и ряд местонахождений в Малой Азии и на Балканах.
«Фауны» Сиваликских холмов и Салта, по недавней сводке Колберта (Colbert, 1935), захоронены в колоссальной толще континентальных отложений, достигающей мощности 7000 м. Эта толща содержит слои от верхнего миоцена до нижнего плейстоцена и образовалась в результате сноса обломочного материала с непрерывно поднимающихся хребтов в предгорную область опускания, на которой временами существовали пресноводные стоячие бассейны. В этих слоях захоронялись и трупы погибших зверей. Отсюда известно теперь до 288 видов млекопитающих.
В нашей палеонтологической литературе и у геологов укоренилось мнение, ведущее начало, вероятно, еще от Осборна (Osborn, 1921), что «гиппарионовая фауна» Южной Азии — Сиваликских холмов — «проникла» в Восточную Европу через Кавказ.
По мнению Андрусова (1918), фауна пикермийского типа распространялась в Европе с востока, следуя двумя путями: по северному побережью постепенно сокращавшегося Сарматского моря и через Малую Азию.
Пикермийский тип гиппарионовой фауны в среднесарматских пластах русской территории, как думал Андрусов, выражен еще неполностью, но усиливается в верхнем сармате и достигает полного развития в мэо-тических пластах, оканчивая, по-видимому, существование в юго-восточной Европе, в самых низах понта.
Борисяк (1928а, стр. 376) полагал, что гиппарионовая фауна проникла в Восточную Европу из Передней Азии через Черноморскую сушу и Кавказ.
По мнению Богачева (1938 г, стр. 36—37), «Вторжение африканоси-валикской фауны через Иран и Закавказье на еще неразвившуюся окончательно зону Главного Закавказского (sic!) хребта, а оттуда через Ставропольское плато дальше — в южнорусские и украинские степи, отмечается поэтапно находками остатков млекопитающих».
Часть I. Обоснование истории формирования фауны
Между тем фаунистические списки Колберта из верхнемиоценовых слоев Сиваликов (Lower Siwaliks, Kamlial) показывают, что сходство их со списками фауны кавказского сармата практически отсутствует.
В нижних слоях Сивалика нет ряда групп зверей, характерных для сармата прикаспийских стран, и прежде всего самих гиппарионов.
Более поздние слои нижнего Сивалика в области Салт (Chinji) содержат до 85 описанных к 1935 г. видов из отрядов приматов, грызунов,, хищных, хоботных, непарнокопытных (гиппарионов, носорогов), парнокопытных (свиней, трагулид, оленей, жираф). Важной особенностью этих горизонтов является отсутствие представителей семейства полорогих. Общими с фаунистическим комплексом кавказского сармата являются здесь лошадиные, а также безрогие носороги, жирафиды и приматы.
Севастопольская «фауна» в Крыму, которой придают среднемиоценовый возраст, содержала следующие формы (Борисяк и Беляева, 1948)1
Carnivora
Ictitherium tauricum Boris.
Perissodactyla
Hipparion gracile var. sebastopolianum
Boris.
Aceratherium zernovi Boris. A. zernovi var. asiaticum Boris.
Het сомнения, что здесь присутствовал фаунистический комплекс, весьма близкий к эльдарскому, но существование прямых зоогеографических связей недостаточно ясно.
Судя по современным геологическим данным, появление гиппарионо-вого комплекса в Крыму могло произойти лишь с юго-запада в конце среднего миоцена и начале сармата, так как позднее, вплоть до плиоцена, Кавказская суша была, по-видимому, отрезана с севера от Русской равнины и от Крыма широкими морскими разъединениями (Андрусов, 1918, 1926; Муратов, 1951).
Особый интерес для воссоздания миоценовых ландшафтов и фаунистических связей Передней Азии с Кавказом представляет «фауна» намытых глин и песков южного склона горы Сехенд у Мерагэ.
По определениям Родлера и Вейтхофера (Rodler und Weithofer, 1890), Меквенема (Mecquenem, 1924), Богачева (1928) и Бурчак-Абрамовича (19526), марагинская «фауна» состояла из следующих форм:
Artiodactyla
Achtiaria expectans Boris.
Tragocerus leckevitschi Boris.
Tragocerus sp.
Gazella sp.
Reptilia
Testudo sp.
Trionyx sp.
Primates
Mesopithecus orientalis Kittl M. pentelici Gaudry
Carnivora
Hyaenarctos maraghanus Mecq. Ictitherium, hipparionum Gaudry I. robustum Gaudry Hyaena eximia Wagner Meles polaki Kittl M. maraghanus Kittl
Felis brevirostris Groiz. et Job.
F. attica Wagner Muchairodus aphanistus Каир M. orientalis Kittl
Tubulidentata
Orycteropus gaudryi Major
Proboscidea
Mastodon pentelici Gaudry
Perissodactyla
Aceratherium persia Pohlig
Rhinoceros morgani Mecq.
Chalicotherium pentelici Gaudry
Hipparion mediterraneum Hensel
Artiodactyla
Sus (Microstonyx) erymanthius Rotlu et Wagner
Тл.-I- Развитие ландшафтов и фаун млекопитающих в третичном периоде 45
Helladotherium gaudry Mecq.
Alcicephalus neumayri Rodl. et Wei th. Camelopardalis attica Gaudry Urmiabos aserbaidzanicus Burtsch.
Gazella gaudry Schlosser
*d. brevicornis Gaudry
G. capricornis RodL et Weith.
Palaeoryx pallasi Gaudry Prot oryx caroli пае Major
Antilopinae gen.?, мелкая форма Tragocerus rugosifron Schlos. Protragelaphus scozesi Dames. Tragelaphus hontom schindleri Rodl. et Weith.
Helicophora rotundocornis Weith. Oiceros rothi Wagner
O. atropatanes Rodl. et Weith.
O. boulei Mecq.
Известны также остатки птиц: страуса — Struthio sp. и урмиорниса — Urmiornis maraghanus Mecq.
Богатство видов в марагинской «фауне» свидетельствует о высокой степени универсальности этого захоронения. Хотя тафономические черты его в деталях и неизвестны, можно думать, что на протяжении столетий в верхнем миоцене существовала однообразная причина гибели животных и последующий снос их остатков к участку захоронения. Такой причиной могли быть ядовитые газы (фумарол) и селевые потоки из пеплов и глин. Сносившийся материал отлагался в прибрежной зоне предгорного озерного бассейна. По мнению Полига (Pohlig, 1886), захоронение это озерно-речного происхождения, сходное с Пикермийским близ Афин ] Вальд-Арно близ Флоренции, и связано с береговой полосой озера кра-Урмия, уровень которого был тогда значительно выше, нежели теперь. Высокая комплексность видна и по наличию в захоронении остатков животных различных эколого-морфологических типов, например приспособленных к древесной среде мезопитеков, к обитанию на открытых пространствах саванны или даже степей — жираф, газелей, страусов к различному питанию — хищников, всеядных, травоядных форм Явное преобладание обитателей открытых пространств при наличии форм обитающих и в субтропических лесах, позволяет говорить о смешанных саванново-гилейных ландшафтах, свойственных в миоцене северным частям современного Иранского плоскогорья. F
Марагинская «фауна» носит «африканский» облик в нашем современном его понимании, она была довольно близка, хотя и не идентична более древней фауне закавказского сармата. ’
пт ВнУт₽®н™е современного Ирана и Анатолии, по взглядам Шталя (Stahl, 1907) и Освальда (1915—-1916), оформились как суша в верхнем миоцене, причем в замкнутых бассейнах — остатках миоценовых морей отлагались мощные толщи соленосных и гипсоносных глин. Богачев и Шишкина (1915) дали картинные представления этих процессов, важных и для развития фауны млекопитающих и для возникновения костеносных захоронений. По их мнению, после поднятия страны в ерещем миоцене и образования складок, разделивших морской бассейн на ряд бессточных засоленных впадин, здесь имел место сухой пустын-ный режим внутри страны.
Эти представления о сухом и пустынном режиме страны не вяжутся с экологическим анализом верхнемиоценовых млекопитающих Мерагэ. Вернее, на фоне сухой и жаркой обстановки здесь существовали и участки тропических лесов, приуроченные к береговым линиям остаточных бассейнов.
Фаунистический комплекс марагинского типа был, вероятно, характерен и для всей Передней Азии. Подтверждением этому служит генезис ландшафтов Малой Азии, известный по многочисленным работам геоло-
га (см. Фюрон, 1955).
' 46 Часть I. Обоснование истории формирования фауны
В Малой Азии остатки верхнемиоценовых млекопитающих обнаружены в ряде местонахождений под Стамбулом, Верхним Гедизом, Муглы, в Галатии и Каппадокии. В сборах преобладают остатки иктитериев, куниц и махайродов из хищных, мастодонтов — из хоботных, гиппариона и носорогов — из непарнокопытных, жираф, газелей и антилоп — из парнокопытных. Такого же типа «фауна» прослежена и на Самосе, и на Балканах у Афин (Пикерми).
Мощные вулканические проявления, известные по разрезам в Каппадокии, сопровождавшиеся выпадением пепла и образованием грязевых потоков, обусловливали гибель животных и накопление их остатков в формировавшихся туфах.
В целом верхнемиоценовая фауна зверей Передней Азии, Кавказа и Крыма, а также Балкан обнаруживает большие черты сходства, но установить очередность появления и развития гиппарионового комплекса на этих территориях до просмотра и изучения всех материалов одним исследователем невозможно.
В конце миоцена — в мэотический и понтический века — море, омывавшее Кавказ, стало мелеть и еще более опресняться. Ставропольское поднятие вновь стало расширяться и выдвигаться на север. Сильно выдвигалась также на северо-восток суша современного горного Дагестана и почти исчезал Восточнозакавказский залив. В области Маныча, вероятно, существовал временами неширокий пролив.
Крупные поднятия кавказских хребтов третичного периода к этому времени были, по-видимому, завершены, и формирование биокомплексов высокогорного кавказского типа шло уже полным ходом в условиях высотнозональных климатов и вероятных местных оледенений.
Нет, однако, никаких оснований относить к эпохе верхнего миоцена начало гипотетического ряда грандиозных покровных оледенений Кавказа, о которых писал Ковалевский (1936), изучивший континентальные толщи Аджиноура.
Предположения Ковалевского и Гроссгейма (1936) об истреблении верхнемиоценовым оледенением всей тропической флоры и фауны Кавказской суши не подкреплены палеонтологическими документами. Совершенно не доказано также и существование в гюнце и минделе столь мощных ледников, которые могли бы выполнять и «выпахивать» Алазано-Агричайскую долину в восточном Закавказье, тем более, что ее прогиб, несомненно, более позднего возраста.
Гроссгейм (1936, 1948) считал мэотис временем широкого проникновения на Кавказ южной ксерофильной флоры, но в ископаемом состоянии известна лишь мезофильная лесная флора из Гурии, состоящая из смеси вечнозеленых пород субтропического типа Rhododendron ponti-cum с явным преобладанием листопадных — Fagus orientals, Acer tri-lobatum и с некоторой примесью хвойных — Taxus grandis. Остатки столь же мезофильных растений: ольхи, черешни, хмелеграба, ивы, осок и тростника — обнаружены в Нахичеванском ниже епл и оцепов ом месторождении каменной соли и листоватых глин (Палибин, 1936; Богачев и Шишкина, 1915).
Ископаемые остатки высокогорных растений и животных этой эпохи нам неизвестны, но можно предполагать, что, помимо их автохтонного развития, на Кавказе уже имели место и широкие миграции относительно холодолюбивых элементов флоры и фауны по цепям горных хребтов альпийской системы. i
Гл. I; Развитие ландшафтов и фаун млекопитающих в третичном периоде 41
В качестве современных «живых ископаемых», возникших в эту эпоху, можно считать малоазийских хомяков рода Mesocricetus, прометееву полевку — Prometheomys и кавказских козлов.
Наземная фауна предгорных равнин самого верхнего миоцена в области Кавказского перешейка до сих пор также плохо известна и плохо-датирована стратиграфически. Гиппарионовый комплекс, вряд ли обедненный, продолжал существовать по равнинным окраинам кавказской, горной системы, особенно в ее юго-восточной части. Небольшие захоронения остатков зверей известны в восточном Закавказье и западном Предкавказье. Одно из них было обнаружено в 1935 г. Н. А. Кудрявцевым: в овраге на северном отроге Кахетинского хребта в Закавказье.
Стратиграфическое положение этого захоронения недостаточно выяснено, но условно относится к верхнему миоцену или нижнему плиоцену. По описанию Орлова (19386), костеносный слой просматривается на левом склоне глубокого оврага, проходящего через сел. Джапаридзе (близ Цители-Цхаро) в сторону р. Алазани. Разрозненные и разломанные трубчатые кости, челюсти и зубы млекопитающих залегают здесь в слоях морских (?) глин, перекрытых континентальными глинистыми слоями, чередующимися со слоями погребенной почвы. Здесь обнаружены, остатки видов:
Proboscidea Fam. gen.
Perissodactyla Hipparion gracile Каир Rhinocerotidae gen.
Artiodactyla
Sus sp.
Cervidae gen.
Giraffidae gen.
Gazella sp.
В Предкавказье сборы остатков млекопитающих верхнемиоценового > возраста были сделаны еще в 1940 г. Я. М. Эглоном в глинах мэотического (?) яруса на правом берегу Кубани, близ Армавира. Единичные фрагменты сильно минерализованных костей принадлежат гиппариону — Hippa-rion sp., антилопе величиной с сайгу — Antilopinae gen., мелкому бобру — Castoridae gen. и мелкому тюленю — Phoca sp., саблезубому тигру — Machairodus sp.
Обломки коренных мышевидного грызуна величиной с домовую мышь (Rodentia, Muridae) были обнаружены А. В. Сизовым в миоценовых отложениях ущелья р. Аксая в Грозненской области, на границе отложений верхнего сармата и мэотиса в желтовато-зеленых глинах. Там же найдены остатки травянистых растений, пресноводные остракоды и отолиты рыб — бычков — Gobius.
На юге Русской равнины к мэотическому ярусу условно относятся остатки позвоночных в континентальных отложениях под Новочеркасском.
Трупы, по-видимому, сносились в Танаисский залив Мэотического моря пра-Доном и захоронялись в косо слоистых белых песках и песчаниках, залегающих теперь «между понтом и размытым сарматом» (Жиж-ченко, Колесников и Эберзин, 1940). Эти пески достигают мощности 12 м и содержат очень тонкие прослои голубовато-серой глины с диатомовыми водорослями, куски окремневших стволов деревьев и зубы хоботных.
У хутора Янова в таких песках, вместе с моллюсками родов Congeria, Neritina найдены кости окуня — Perea sp. Остатки млекопитающих, добытые К. Лисицыным и Быкодоровым у хутора Янова, хутора Поповка ив карьерах по р. Тузловке, по определениям Хоменко, Богачева, а также* Соколова (1954), принадлежат следующим четырем видам:
48
Часть I. Обоснование истории формирования фауны
Proboscidea Dinolherium. gigantheuni Каир
Mastodon borsoni Hays. Artiodactyla
M. cf. tapiroides Cuv. ' Palaeoryx longicephalus Soc.
Итак, конец миоцена на Кавказе может быть охарактеризован как эпоха дальнейшего расширения площади суши, возникновения и развития процесса «бореализации» флоры, становления горной зональности растительности под воздействием тектоники и изменения климата в сторону похолодания. Те же процессы вместе взятые явились толчком к прогрессивному развитию группы лошадей, полорогих жвачных и оленей при одновременном затухании жирафид.
ПЛИОЦЕН
Фаунистические «следы» плиоценовых бассейнов в Каспийской области дали основание геологам выделить понтический ярус, балаханскую продуктивную толщу, акчагыльский и апшеронский ярусы. Для плиоцена Черноморской области, кроме понта, выделены киммерийский и куяльницкий ярусы, гурийские слои и слои Чауды, соответствующие трем последним ярусам Каспийской.
В нижнем плиоцене—начале понта Кавказский полуостров имел очертания, близкие к верхнемиоценовым и омывался слабосоленым озером-морем, не соединявшимся с океаном (Андрусов, 1918). По северному побережью имелись большие открытые заливы: Кубанский — на северо-западе и Терский — на северо-востоке. С запада в современную Колхиду вдавался до долготы Кутаиси Рионский залив, а на востоке — Самур-ский и Куринский заливы. Судя по мощности понтических отложений и по прибрежной зоне конгломератов (Колесников, Жижченко и Эберзин, 1940, стр. 402, карта), последние четыре залива отличались большой глубиной, что свидетельствует о значительных горных поднятиях побережья в области современной Абхазии и северного Азербайджана.
Широкие континентальные связи с Передней Азией допускали проникновение на Кавказ южных растений и животных.
Ряд геологов отмечал похолодание в начале понтической эпохи с вероятным появлением ледоставов на реках Русской равнины (например, Соколов, 1904).
Климат Кавказской суши между тем оставался довольно теплым в начале плиоцена. Многочисленные остатки растений, происходящие из песков и песчаников ергенинской свиты в 50 км к северу от Сталинграда, в бассейне р. Иловли, дают, по определениям Баранова (1952), представление хотя и о листопадной, но все же относительно теплолюбивой флоре верхнего миоцена—нижнего плиоцена. Там росли Corilus fossilis, Alnus incana, Quercus sp., Castanea sp., Parrotia persica, Araliacea.
Показателем относительно теплого климата в нижнеплиоценовую эпоху на Кавказе служит произрастание в Ергенях железняка — Parrotia persica, пережившего плейстоцен лишь на юге Каспия, т. е. на 10 географических градусов южнее.
Начиная с среднего понта, на Кавказе наступает новое потепление климата, что доказано послойными сборами остатков растений в понтических отложениях на мысе Пицунда (Мчедлишвили, 19546).
В конце понтического века море вовсе отступает из Предкавказья. В области Каспия водоем сохраняется лишь на месте его южной котловины (рис. 20). В восточном Предкавказье и в восточном Закавказье, вероятно, оформились обширные пространства полупустынь.
Гл. J. Развитие ландшафтов и фаун млекопитающих в третичном периоде 49
Позднее — в киммерийском веке — климат стал близким к тропическому, на что указывает обилие железнорудных прослоек на Таманском и Керченском полуостровах и в предгорьях Северного Кавказа. В западном Закавказье такие условия подтверждены и остатками тропических растений в прибрежно-морских слоях (Мчедлишвили, 1954а)
Рис. 20. Киммерийский и Балаханский бассейны. (По Колесникову, — см. Жижченко и др., 1940). Точками отмечены главнейшие местонахождения остатков наземных позвоночных.
и в современных им вулканических туфах Годердзи на Аджаро-Имеретинском хребте, где обнаружены фикусы, араукарии, пальмы (Палибин, 1936).
Широкое материковое соединение, впервые появляющееся в эту эпоху между Кавказским полуостровом и Русской равниной, создавало предпосылки к обмену флористическими и фаунистическими элементами этих территорий.
Именно в эту эпоху могло уже идти проникновение с северо-востока некоторых древних элементов фауны среднеазиатских пустынь далеко на запад — в равнины восточной части Кавказского перешейка.
На месте Черного и Азовского морей в киммерийском веке остается водоем, границы которого близко совпадают с современными береговыми линиями этих бассейнов.
Захоронения остатков наземных позвоночных эпохи нижнего и среднего плиоцена известны на Кавказском перешейке преимущественно из континентальных захоронений.
Ставропольская возвышенность
Наиболее крупное и комплексное из плиоценовых захоронений находится в 10 км к западу от Ставрополя, в толще песчаных отложений, вскрытых Косякинским карьером. Плиоценовая река и ее про-
4 Н. К. Верещагин
№
Часть I. Обоснование истории формирования фауны
ток прорезали сарматские известняки на западной окраине Ставрополь
ского плато, оставив после своего постепенного угасания костеносную линзу мощностью до 7—9 м и шириной около 90 м (Каспиев, 1939; Гни-ловской и Егоров, 1955).
В древней аллювиальной толще наблюдается до семи комплексов отложений с различным петрографическим составом — от мелкозернистого серовато-белого песка до гравия с глинистыми прослоями и глинистыми гальками до 1.5 см диаметром (рис. 21 и 22).
Рис. 21. Разрез Косякинского карьера.
а, б — гумусированные суглинки; в—ж — костеносные речные пески косой и иной слоистости; справа - - мощность в метрах.
По нашим исследованиям, прослои гальки в нижних горизонтах толщи1
состоят из местных сарматских пород Ставрополья и из истертых обломков костей морских сарматских и чаще плиоценовых наземных зверей. Кости животных, попавшие в русло, расчленялись и, неоднократно переотлагаясь, захоронялись преимущественно в однообразных песчанохрящеватых фациях, отложенных при скоростях от 1.3 (нижний горизонт) до 0.1 м/сек. (верхний горизонт).
В разрезах можно наблюдать все степени истирания кости от совершенно целой до кусочков, затертых в виде костяной гальки.
Большинство (до 92%) собранных в карьере костей при полной потере органического вещества вторично почти не минерализовано. Кости напоминают рыхлый мергель или мел. С поверхности они кремового или сероватого цвета, довольно легкие и слегка «звенят», как фаянс. Все трещины костей этого типа сохранности прокрашены охрой. Марганцевожелезистые дендриты на этих костях редки, но часто заметны извилистые черточки — следы корней растений новейшего происхождения. Встречаются иногда и лунки неправильной формы — следы древнего воздействия воды и водных беспозвоночных.
Кости этой группы большей частью целы и разломы их — результат небрежных сборов при добыче песка. Меньшая часть костей (8%) сероватой, пятнистой окраски, созданной обилием дендритов. Эти кости, как
к
CI п.
Л1
6i ВС
щ в тр та
эт; кл
ЧТ<
Гл. 1. Развитие ландшафтов и фаун млекопитающих в третичном периоде 51
правило, минерализованы сильнее и значительно тяжелее костей мелового типа сохранности. В свежем разломе их вещество напоминает сливной песчаник, и лишь принадлежность их к тем же видам да «меловое» ^стояние поверхностного слоя в некоторых местах заставляют призна-। ть их геологическую одновозрастность с костями первой группы. Эмаль joob во всех случаях отличной сохранности и либо натурального, либо
iветло-горохового и розоватого цветов.
Следы погрызов хищниками редки, но все же имеется 9 костей с явно с рызеипыми эпифизами. Изредка попадающиеся позвонки сарматских
Рис. 22. Косякпнскии карьер.
Фот. автора, 1952.
китов и тюленей, вымытые из сарматских известняков, легко отличаются сильной вторичной минерализацией и буроватой окраской от остатков плиоценовых наземных позвоночных.
Разнотипность в сохранности плиоценовых костей объясняется различными особенностями условий первичного захоронения, сроками пребывания па отмелях и косах реки при вторичном их отложении и разпо-возрастностью в пределах эпохи существования долины и русла реки.
Весьма вероятно, что в долине и русле ставропольской палеореки были примерно те же условия для накопления костей, что существовали и в четвертичном периоде в среднем течении рек Допа, Волги, Урала. Часть трупов накоплялась в старицах, а часть непосредственно в русле и омутах.
Размывая собственные и более древние морские отложения различных этапов накоплении, река переносила на небольшое расстояние и переоткладывала вымываемые кости.
Направление течения реки пока не установлено, по пе исключено, что опа текла и не с Кавказа, а с Русской равнины, например с Ергеней,
Z *
52
Часть I. Обоснование истории формирования фауны
в эпоху низкого стояния моря и слабых поднятий в области Ставрополья.
Несмотря на известную разновозрастность остатков, подавляющее большинство плиоценовых видов зверей этого захоронения, несомненно, можно рассматривать в качестве фаунистического комплекса, свойственного той эпохе плиоцена, на протяжении которой существовала река.
Фрагментарность остатков и специфичность ставропольского комплекса затрудняет определения, и большинство форм подробно еще не описано.
Предварительный список видов из сборов, хранящихся в ПИНе и ЗИНе, по определениям Беляевой (19406, 1944), Борисяка и Беляевой (1948), Аргиропуло (19396, 1940в) и моим (Верещагин, 1954), и численность остатков приведены в табл. 3.
Таблица 3
Видовой состав и численность костей млекопитающих из плиоценовой толщи Косякинского карьера
I Отряды и виды ! Число костей Отряды и виды Число костей
Insectivora Proboscidea
* Crocidura sp J Anancus arvernensis Cruiz.
* Talpa sp 5 et Job 21
Desman a sp Carnivora 1 Dinotherium sp Perissodactyia Hipparion sp 12 3
Ursus cf. arvernensis Cruiz. 12 Hipparion gracile Каир . . 18
Dinocyon cf. thenardi Jo- Tapirus cf. arvernensis Dev. 3
urdan 4 et ВоиШ
* Canidae gen Fells cf. issiodorensls Croiz. 3 Dicerorhinus oriental is
Schlos 72
et Job 1 Aceratherium cf. incisivum
Каир 8
Lagomorpha Rhinocerotidae gen. (<f. Chilo-therium) 1
* I epus sp 25 Rhinocerotidae gen 33
* Ochotona cf. antique. .... •Э Artiodactyla
Rodentia Propotaniochoerus provinci-
* * Amblicastor caucasicus Arg. Steneo fiber sp C rice Lus sp Mus sp 1 •> 37 1 alls Gerv Procapreolus sp Pliocervus sp Pseudalces sp Giraffidae gen. (cf. Swathe- 10 7 9 2
rium) 2
Gaze ll a sp 5
При м с а а н я е. Формы, отмеченные звездочкой, обнаружены и собраны мною в 1 952 г.
Кроме остатков перечисленных сухопутных зверей, в этом захоронении найдены также остатки птиц — Charadrius cf. morinellus L., водных черепах — Тrionychiidae (14 обломков костяных щитков), нижняя челюсть мелкой ящерицы — Lacerlilia, кости мелких амфибий — Anura — лягушек или жаб (рис. 23—25).
При признании стратиграфической одновозрастности большинства остатков фаунистический комплекс древнего Ставрополья следует считать весьма богат.ым. Ставропольские мастодонты, тапиры, носороги, бородавочники обитали, несомненно, в субтропическом ландшафте с ме-
Гл. I. Развитие ландшафтов и фаун млекопитающих в третичном периоде 53
зофптной растительностью. Здесь имелись, очевидно, приречные заросли тугайного типа, чередующиеся с открытыми луговыми полянами, населенными кротами, хомяками, пищухами.
Преобладание остатков носорогов, свиней (рис. 26, 27) — животных, тяготеющих к зарослям речных долин, наличие остатков бобров, тапиров, водных черепах как обптателей болот, стариц и протоков, прокраска остатков окисью железа являются известным подтверждением данных выше ландшафтных картин и тафономических заключений. Обитание
Рис. 23. Остатки мелких позвоночных из Косякинского карьера.
1 — челюсть ящерицы сем. Lacerlidae; 2 — плечевая Talpa sp.; 3 — челюсть Desmana sp.; 4, 5 — плечевая и пяточная Lepus sp.; 6, 7 — верхние коренные и бедренная Steneofiber sp.; 8 — челюсть Cricetus sp.
мелкой выхухоли в плиоценовой речке указывает также на наличие в плиоценовом Ставрополье весьма древних эрозионных долин и рек с устойчивым гидрологическим режимом, близким к режиму Дона или Волги.
В стратиграфическом смысле ставропольский комплекс не верхнеплиоценового возраста. Борисяк (1943) датировал его понтом. На нижнеплиоценовый возраст указывает отсутствие здесь настоящих слонов и лошадей. Аргиропуло (1932, 1940в) сближал кавказского амбликастора с верхнемиоцеповыми представителями того же рода из Монголии и Северной Америки. Олени ставропольского комплекса нижпеплиоценовых и средпеплиоценовых типов.
Сопоставляя ряд известных геологических фактов и состав ставропольского фаунистического комплекса, можно пока условно датировать
Рис. 24. Остатки хищных и хоботных из Косякииского карьера.
J,3 — челюсти Ursus cf. arvernensis; 2 — челюсть Dinocyon cf. thenar di (X1/*); 4 — преднореином Canidae gen. (x2); 5,6 — челюсть и плечевая Felis cf. issiodoiensis; 7 — Anancus arvernensis.
1, 2
4 — 41
IHOK lets.
Рис. 25. Остатки копытных из Косякинского карьера.
1, 2 — метаподии Hipparion gracile и Hipparion sp.; 3— челюсть Tapirus cf. arvernensis; /—челюсть Propotamochoerus provincialis; 5, 6 — рог и челюсть Procapreolu^ sp.; 7 — челюсть Cervidae gen.
об
Часть I. Обоснование истории формирования фауны
последний первой половиной плиоцена, считая, что он стратиграфически моложе тараклийского комплекса Молдавии, описанного Хоменко (19146), павлодарского — на Иртыше и довольно близок к комплексу Малуштени в Румынии (Simionescu, 1930).
В предгорном Предкавказье аналогами морских киммерийских отложений некоторые геологи склонны считать обломочную континентальную
Рис. 26. Челюсть Dicerorhinus orientalis из Косякинского карьера.
толщу мощностью до 700—800 м, описанную Швецовым (1928), в Кабарде и Северной Осетии. Эта толща, состоящая из суглинков, песков, туфогенных брекчий, содержит местами крупные валуны из изверженных
Рис. 27. Клык (?) Rhinocerotidae gen. из Косякииского карьера.
пород и напоминает до некоторой степени морену. Она образует местами хорошо выраженный предгорный барьер на северном склоне Черных гор и слагает Змейский хребет, залегая непосредственно па сарматских глинах. Эти отложения были образованы, вероятно, мощными грязевыми потоками, сносившими массу плохо сортированного материала с незакрепленных горных склонов во время катастрофических Ливией. В толще обнаружены наземные и пресноводные моллюски — Helix sp., Paludina sp.,
Гл. /. Развитие ландгиафтов и фаун млекопитающих в третичном периоде 57
.Xeritina sp., травянистые растения и листья деревьев, а также кости наземных позвоночных. М. В. Павлова определила в сборах Швецова рог оленя — Cervus (?) и «челюсть крысы».
В восточном Закавказье в киммерийском веке накапливались колоссальные толщи песчано-глинистых отложений мощностью до 1000—1400 м, которые известны в области Апшеронского полуострова под названием балаханской, или продуктивной, свиты. Эти толщи послужили затем крупнейшим резервуаром для нефти.
Генезис продуктивной толщи, как известно, и до сих пор является предметом горячей дискуссии геологов. Колесников (Жижченко, Колесников и Эберзии, 1940), излагая разнообразные воззрения по этому поводу, приходит к выводу, что эти слои отлагались в области «развивающейся тектонической ванны». Другими словами, толща образовалась в мелководных бассейнах с прогибающимся дном за счет сноса терригенного материала с молодых хребтов, поднимавшихся по восточной оконечности Большого Кавказа.
Остатки наземных позвоночных в этой толще встречаются чрезвычайно редко.
В 1932 г. В. Подгорнова обнаружила остатки млекопитающих в светлых мелкозернистых песчаниках продуктивной толщи с восточной стороны вулкана Лок-Батан, расположенного к югу от Баку. По описаниям Богачева (19386, 1941) и нашим данным, здесь имеются следующие виды:
Carnivora
Vulpes khomenkoi Bog.
Artiodactyla
Cervus (cf. Eucladocerus) sp. Gazella sp.
Найдены также остатки сома — S iluris giants L.
Остатки лисицы представлены двумя полными рядами верхних коренных и фрагментами челюстей, запрессованными в светло-серый и мелкозернистый песчаник. Вторичная минерализация остатков значительна. Эмаль зубов блестящая и совершенно черная, а вещество, заместившее кость, в свежем разломе палевого или розоватого цвета. Остатки газели представлены фрагментом осевого черепа с полными рядами верхних коренных. Остатки оленей состоят из двух полных метакарпальных костей. Отождествлять локбатанские среднеплиоценовые формы с четвертичными — благородным оленем и джейраном, — как это делал Богачев, нет никаких оснований. Захоронение это было связано, конечно, с пресноводным потоком, скорее всего с дельтовым участком одной из рек, стекавших с восточного Кавказа, похожей на современный Сумгаит или Пирсагат.
Смежные горизонты продуктивной толщи на Апшеропе характеризуются, по Голубятникову (1925), пресноводной фауной моллюсков, например Planorbis cornu var. manteli Dunk., P. costatus Klein, рядом лимпеид-—Limnaea armanensis Noul. и др., Unto jasamalicus D. Golub., Melania glacilicosta Sandb. Известны также харовые водоросли —Chara sp. С пашей точки зрения, такой набор видов вряд ли мог бы существовать в условиях пустынного ландшафта и в пересыхающих засоленных речках позерах. Западнее, из балаханскойтолщи Ширакской степи, известны остатки древесины бобового растения — Albizzioxylon hyrcanum, характеризующего субтропические условия.
58
Часть I. Обоснование истории формирования фауны
Немногочисленные остатки зверей, найденные в отложениях века балаханской толщи, свидетельствуют лишь о вероятной дифференцировке фаунистических комплексов по географическим участкам Кавказского перешейка в связи с дальнейшим формированием различного типа климатов и ландшафтов — от субтропического лесного до жаркого и сухого ландшафта полупустыни.
На юго-западе Русской равнины в эпоху среднего плиоцена обитали млекопитающие, эволюция которых была связана еще с миоцена с различными участками суши восточного Средиземноморья. Особый интерес для понимания истории фауны Предкавказья представляют поэтому средиеплиоценовые высокоунпверсальные захоронения остатков зверей, известные в Молдавии из галечно-песчаных вееров древних потоков в бассейнах рек Сальчи, Кагула, Прута, а на Украине столь же универсальные захоронения в промоинах (катакомбах) одесских известняков, заполненных красной глиной.
Средпеплиоцеповая «фауна» Молдавии содержала обезьян, грызунов, хищных, носорогов, гиппарионов, гиппопотамов, верблюдов, оленей и антилоп. Зайцеобразные и грызуны здесь были представлены уже современными родами зайцев, сеноставок, белок, бобров, дикобразов, слепышей (Хоменко, 1914а, 1915; Борисяк, 1943).
В «фауне катакомб» Одессы определено до 30 видов млекопитающих из отрядов насекомоядных, грызунов, хищных, хоботных и парнокопытных, а также остатки страусов (Грицай, 1938, 1939; Борисяк, 1943; Пидопличко, 1954).
Фаунистические комплексы северного Причерноморья в среднем плиоцене отражают мозаичность ландшафтов: наличие рек и ручьев с развитыми поймами и древесными зарослями, а также и большую остепнеппость водоразделов. Возможные пути развития ареалов этих комплексов или их отдельных представителей лежали и в юго-восточпом направлении — в Предкавказье.
Как молдавская (руссильонская), так и одесская средпеплиоцеповая «фауна» имеют еще ряд теплолюбивых, по крайней мере субтропических форм.
Наличие в руссильонском ярусе восточного Средиземноморья обезьян и гиппопотамов ярко свидетельствует об этапе послепонтпческого потепления. Можно думать, что климатические условия северного побережья Киммерийского бассейна в конце среднего плиоцена были сходны с существующими ныне в южной Италии, Испании, Франции. В Предкавказье климат был, вероятно, несколько более суровым и континентальным.
На протяжении верхнего плиоцена Кавказская суша существовала в виде обширного перешейка в очертаниях, близких к современным. Временами она отрезалась от севера в области Мапыча неширокими проливами. Области развития морей — Каспийской и Черноморской котловин — оказывались в такие эпохи более или менее разъединенными.
В Черноморской области большинство местонахождений остатков верхнеплиоценовых млекопитающих приурочено к окраинным участкам древних морей, омывавших северо-западное побережье Кавказа и южные границы Русской равнины. Эти захоронения расположены, очевидно, по зоне контакта дельтовых участков и галечных вееров палео-Дона. палео-Донца, палео-Днепра, а также древних речек западной оконечности Кавказа с Куяльницкпм и послекуяльницкими бассейнами (рис. 28).
Га. Развитие ландшафтов и фаун млекопитающих в третичном периоде 59
Ряд авторов прошлого и начала нашего столетия — Фишер фон Вальд-гейм (1809), Эйхвальд (1850), Нордман (1858—1860), Павлова (1895), Богачев (1923—1924, 1938 г), Шерстюков (1926, 1927), Григор (1929) — упоминали, реже описывали обычно не датированные стратиграфически
Рис. 28. Куяльницкий и Акчагыльский бассейны. (По Колесникову, — см. Жиж-ченко и др., 1940). Точками показаны пункты главнейших захоронений.
единичные находки следующих форм млекопитающих верхнеплиоценового Приазовья и Прикубанья:
Carnivora
Hyaena sp.
Meles sp.
Rodentia
Trogontherium cuvieri Fisch.
Proboscidea
Mastodon arvernensis Croiz. et Job.
Elephas lyrodon Weithofer
E. meridionalis Nesti
E. antiquus Falc.
Perissodactyla
Equus stenonis Coccbi
Artiodactyla
Camelus sp.
Cervus sp.
Bos sp.
Специальные поиски и сборы остатков верхнеплиоценовых зверей в Приазовье были организованы Громовым (1933, 1936) в 1933—1936 гг. в хрящевато-гравийных песках у станиц Морской, Мержаповки, Хапры н в Воловой балке под Таганрогом. Сходные остатки собирались мною в карьерах Левенцовки под Ростовом в 1954 г. Фрагментарные, нередко сильно окатанные кости, прокрашенные солями железа из хапровских песков, говорят о различных условиях и этапах их захоронений в аллювиальных выносах и в зоне морской абразии и аккумуляции осадков. О пребывании костей в морской воде свидетельствуют, в частности, следы сверлящих и минирующих морских беспозвоночных на их поверхностях (рис. 29).
60
Часть I. Обоснование истории формирования фауны
По определениям Громова (1948), Борисяка и Беляевой (1948) и нашим, из Приазовья имеются остатки следующих видов:
Carnivora
Canis cf. lupus L.
Cants sp.
Ursus sp.
Hyaena sp.
Ma chair odus sp.
Rodentia
Trogontherium cuvieri Fisch.
Proboscidea
Mastodon sp.
Elephas planifrons (?) Falc.
E. nieridionalis Nesti
Perissodactyla
Ilipparion sp.
Equus stenonis Nesti (cf. major Boule)
Rhinoceros sp.
Elasmotherium sp.
Artiodactyla
Sus sp.
Catnelus (P aracamelus) cf. kujalni-kensis Chom.
Eucladocerus pliotarandoides Alles.
Cervus cf. elaphus L.
Cervus sp.
Известны также кости страусов — Slrutio sp.
При известной стратиграфической разновозрастное™ остатков, отмечавшейся еще Богачевым (1923—1924), этот набор видов, прослеженный
Рис. 29. Челюсть Elephas nieridionalis из хапровских песков правобережья Дона, источенная беспозвоночными. (Ориг.).
и западнее в ряде мест, может рассматриваться все же в качестве руководящих форм фаунистического комплекса, свойственного в верхнем плиоцене широким пространствам юга Русской равнины.
3 а к у б а п с к а я равнина
В пределах Кавказского перешейка североазовским местонахождениям больше всего отвечают захоронения в галечниковой толще Закубаиской наклонной равнины по р. Псе к у псу. Основные сборы были сделаны еще в 1930—1932 гг. Громовым (19356, 1937, 1948) в районе станиц Бакинской и Саратовской, па левом берегу Псекупса, где различались две надпойменных террасы: нижняя — вюрмского и верхняя — рисского возраста.
Геологические разрезы, приведенные Громовым (1948), свидетельствуют о существовании в западнокавказских предгорьях пресных водо
1 j <
I
а
Д
Г L { п к о У Р1
1« ш 6t М( ТВ НС ле
ра не
Гл. I.- Развитие ландшафтов и фаун млекопитающих в третичном периоде 61
емов, менявших свой режим от текущего со средней скоростью потока до озерно-болотного, застойного водоема.
По Громову (1948, стр. 56), в нижнем и среднем комплексе отложений были собраны остатки:
Proboscidea
Elephas meridionalis Nesti — зубы и кости конечностей.
Perissodactyla
Rhinoceros etruscus Falc. — почти полный череп и части скелета.
Equus stenonis Cocchi — метаподии, фаланги.
Eguus sp. (aff. stenonis) — кости конечностей.
Artiodactyla
Bison sp. _ — разбитый череп.
Bos sp. (Leptobos) — разбитый череп.
Eucladocerus pliotarandoides Aless. — череп с рогом и обломки рогов.
Cervus sp. (Rusa) — обломки рогов.
Cervus sp. (ex. gr. polycladus?) — обломки костей конечностей очень
Cervus sp. крупного вида (крупнее лося!) оленя.
Известны также остатки страуса — Strutio sp., нижняя часть голени.
Упомянутые находки с Псекупса остались неописанными и были утрачены во время войпы.
В сохранившихся в ПИНе сборах имеется сложно разветвленный рог оленя — Eucladocerus (рис. 30) и череп этрусского носорога. Костное вещество этих остатков сильно ожелезиепо и палевой окраски.
Судя по наличию целых черепов и рогов в этом захоронении, следует предполагать наличие в верхиеплиоценовом Закубанье равнинных пространств, пересеченных речками с относительно спокойным течением, впадающими в обширные стоячие водоемы. Такие условия могли существовать лишь при значительно Л1епьших поднятиях или при большой пенепленизации западного Кавказа в верхнем плиоцепе.
Восточнее Псекупса единичные остатки крупных верхнеплиоценовых млекопитающих прослеживаются в ряде мест галечниковой толщи, входившей в контакт с предгорными водоемами. Таковы местонахождения разрозненных костей и зубов слонов — Elephas cf. planifrons Falc., £. meridionalis Nesti по рекам Лабе, Кубани, в районе Тщикских плавней (Навозова, 1951) и у станиц Ново-Лабинской, Некрасовской, Григоро-полисской (Громов, 1937) и южнее Армавира. Более часты находки остатков южных слонов, носорогов — Rhinoceros cf. etruscus Jaeg., а также оленей — Eucladocerus pliotarandoides Aless. в ожелезненной толще песка у станицы Воскресенской, западнее Ставрополя, и в нижних горизонтах речных песков карьера Гирей у станции Крапоткин (рис. 31).
Экологическая обстановка в эпоху существования этого комплекса, судя по обилию остатков оленей, характеризовалась относительно мягким, умеренным климатом и наличием в западном Предкавказье развитых широколиственных лесов типа колхидских или гиркаиских. Появление быков, лошадей и страусов свидетельствует о продолжающемся процессе местного остепнения и развития на водоразделах открытых пространств типа саванн. Животные, по-видимому, гибли довольно часто па предгорной равнине и в ущельях от селевых потоков, погребавших трупы и скелеты в выносившейся массе галечников.
В других случаях накопление костей в галечниках было следствием размыва озерно-болотных костеноспых линз позднейшими потоками при перестройке речной сети. В связи с этим эпоху существования животных
62
Часть I. Обоснование истории формирования фауны
Рис. 30. Рог Eucladocerus pliotarandoides пз галечников Псекупса.
псеку некого и описанного ниже таманского комплексов следует, вероятно, считать более древней, нежели эпоху образования костеносных галечников предкавказских равнин.
Стратиграфическое сопоставление большинства псекупских, Лабинских и кубанских костеносных горизонтов проводится обычно с акчагыль-ским, реже — с апшеронским ярусами морских отложений каспийского типа.
Таманский полуостров
Несколько более поздние, чем нижние горизонты Псекупса, костеносные местонахождения прослеживаются в ряде мест па Таманском полуострове (рис. 32): у Синей Балки, Сенной, Фонталовской, Кучугур, Капустиной Балки. Костеносная брекчия у Сине й Балки была открыта еще в 1912 г. Губкиным (1914), который отнес пресноводные песчаные толщи северного берега полуострова к самым низам плейстоцена. Как правило, в этих толщах обильны пресноводные моллюски родов Vivipara, Bythinia, Dreis-sensia, Uni о и др. (рис. 33). Разрозненные, иногда окатанные обломки костей сильно минерализованы, особенно в местонахождениях у Сенной и Ф о н т а л о в с к о й. В песчано-гравий -ных слоях у Сенной весьма характерно также наличие сильно минерализованных экскрементов гиен, свиней и жвачных. По коллекциям, хранящимся в ЗИНе и ПИНе, из упомянутых местонахождений описаны и определены (Борисяк, 1914; Беляева, 1925, 1933а, 19336; Верещагин, 1951а,
1957а; Бурчак-Абрамович, 1952а) виды зверей, указанные в табл. 4 п па рис. 34.
Близ Сенной в о желез пенно м песке найден и позвонок крупной рыбы. Фаланга тюленя, которого Кирпичников (1953) сближает с каспийским Phoca caspica Ginel., вряд ли происходит из тех же слоев, что и остатки наземных видов в Синей Балке. В результате тафономического обзора и экологического анализа перечисленных форм следует признать большинство видов этого списка частью единого фаунистического комплекса. Этот комплекс существовал в какую-то фазу регрессии, вероятно, послекуяльпицкого (Апшерон-ского) бассейна па обширной суше, тянувшейся от Предкавказья до Дуная. Наличие весьма разнообразных животных, приспособленных к жизни в степи (лошади, зубры), в древесно-кустарппковых зарослях (носороги, лесные антилопы, олени, слоны) и в озерах и протоках (бобр), указывает
Гл. J, Развитие ландгиафтов и (фаун млекопитающих в третичном периоде 63
па вероятное существование в западном Предкавказье умеренно теплого климата и открытых равнин с протоками, окаймленными густыми зарослями лиственных пород деревьев п кустарников. По составу и этапу морфогенеза ряда форм этот комплекс стоит ближе всего к виллафранкскому
Рве. 31. Чаудинский и Апшеронский бассейны. (По Колесникову, — см. Жижченко н др., 1940). Точками отмечены главнейшие местонахождения остатков наземных позвоночных.
нЗападной Европе и к ниховапскому в Юго-Восточной Азии. В то же время ряд видов из таманских местонахождений практически неотличим от форм хапровского комплекса (бобр трогонтерий, южный слои, верблюд;
Таблица 4
Видовой состав и численность костных остатков верхнеплиоценовых млекопитающих из пяти местонахождений на Таманском полуострове
Отряды и виды Число иостей Отряды и виды Число костей
Carnivora
Cards tamanensis N. Лег. . . 2
Panthera sp 1
Kodentia
Castor tamanensis N. Ver. . . 2
Trogontherium cuiieri Fisch. . Proboscidea 5
Elephas mer idio nails ft esti1 . .
E. trogontherli Pohl f antiquus Falc. Perissodactyla Elasmolherium caucasicum По- | 803
ris 66
Rhinoceros cf. etruscus Falc. 5
Equus aff. sussenbornensis
Wiist 54
Artiodactyla
Sus tamanensis N. Ver. . . . 2
Camelus cf. kujalnikensis Chom. 1
Eucladocerus sp 22
Megaceros cf. euryceros Al dr. . 1
Tamanalces caucasicus N. Ver. 3
Cervidae gen 41
Gazella sp 2
Tragelaphus sp 2
Strepsicerotini gen. (cf. Tau-
rotragus) 18
Bison cf. schoetensacki Freud. 12
Bison sp 2
1 По мнению Гарутта (1958), это особый слон — Phanagoroloxodon mammontoides.
64
Часть 1. Обоснование истории формирования фауны
Верещагин, 1957а). Весьма сходные виды прослеживаются в отложениях апшеронского яруса в восточном Предкавказье и в Закавказье.
В Каспийской области развитие фаунистических комплексов в верхнем плиоцене и физико-географические условия прослеживаются по захоронениям остатков млекопитающих и растений по окраинам Акчагыль-ского и Апшеронского бассейнов, связанных с котловиной современного Каспия.
В акчагыле депрессия земной коры приобрела в области северного Каспия меридианное направление в противоположность прежним, ши-
Рис. 32. Обрывы и оползни северного берега Таманского п-ва у мыса Литвинова.
Фот. автора, 1954.
ротным. Осадки Акчагыльского моря прослежены от восточного Закавказья и предгорий Копет-Дага на юго-востоке до Казани и Уфы на севере [Архангельский, 1934; Ковалевский, 1933; Колесников (Жижченко, Колесников и Эберзин, 1940)].
В акчагыле существовал узкий Куринский залив, доходивший почти до Тбилиси, и широкий Терский залив, доходивший до Моздока. В них стекали реки Кавказского хребта, выносившие в морские бухты и на предгорную равнину трупы млекопитающих.
В период максимума трансгрессии Акчагыльскпй бассейн переливал часть своих вод в Куяльницкий по узкому проливу, который пролегал будто бы пе по Мапычу, а южнее Ставропольской возвышенности (Гату ев, 1932).
Эго новое развитие моря из остаточного бассейна века продуктивной толщи Ковалевский (1933) объясняет вторжением в область Каспия океанических вод, но гипотеза того же автора о бореальной трансгрессии,
Гл. I. Развитие ландгиафтов и фаун млекопитающих в третичном периоде 65
т. е. о сообщении с Северным океаном, отвергается современными данными изучения фауны моллюсков в северных частях бассейна.
Климатические смены, не носящие катастрофического характера, столь же вероятны на протяжении акчагыла, как и для других этапов плиоцена. Однако возникновение покровных оледенений Большого Кавказа, как это предполагал Ковалевский (1936), вряд ли могло иметь мести. Для акчагыла характерно оживление вулканической деятель-
5 Н. К. Верещагин
Часть I. Обоснование истории формирования фауны
пости, о чем свидетельствуют прослойки пеплов и пемзы в толщах суглинков и галечников предгорий. Перегораживая на огромном пространстве Русскую равнину, Акчагыльский бассейн ограничивал возможности миграций фаунистических элементов из Средней Азии в Предкавказье и обратно (рис. 28).
Наземная флора эпохи Акчагыльского моря известна по находкам с окраин Курипского залива, близ Нафталана, в Кахетии и в степи Шираки. Отсюда определены остатки кавказского бука — Fagus orientalis, дуба — Quercus sp., ивы — Salix alba, гранатника — Punica granalum, дзельквы — Zelcova crenata, липы — Tilia platyphyllus, ольхи — Alnus glutinosa, шелковицы — Morns Andrussovi, пицундской сосны — Pinus pithyusa, секвойи — Sequoia Langsdorlii и многих других (Палнбнн, 1936).
Судя по этому набору лиственных и хвойных видов, условия существования и характер растительности окраин Курипского залива были крайне близки к существующим и в наши дни в предгорьях восточного Закавказья.
Палибин (1936) и Гроссгейм (1936, 1948) считали, что тропический характер растительности Кавказа в акчагыле уже не возвратился. Последние вечнозеленые секвойи и лавровишни в Шираках существовали, вероятно, во влажном климате с умеренно холодными зимами.
Остатки наземных позвоночных в акчагыльских слоях Кавказа редки. Известны преимущественно находки зубов мастодонтов, плосколобых слонов и стеноновых лошадей. Последние два вида появляются на Кавказе впервые именно в акчагыльском веке.
В Предкавказье зубы Mastodon arvernensis Croiz. eL Job. и Elephas planijrons Falc. найдены на хребте Малгобек, в 40 км южнее Моздока. Зубы и кости плосколобого (?) слона найдены также в овраге Соленом, близ Грозного (Павлова, 1931).
В восточном Закавказье кости крупных позвоночных залегают вместе с остатками наземных п пресноводных моллюсков — Helix s#., Planorbis, Ciclostoma и растений современных видов в конгломератах акчагыльского яруса в междуречье Алазапи и Куры (Кудрявцев, 1932). Из акча-гыла западного Азербайджана известен также рог оленя — Cervus sp.
Фрагментарные остатки млекопитающих, среди которых определены зубы Elephas all. planifrons Falc. и Equus stenonis Cocchi, обнаружены в грубообломочной брекчии хребта Бедепи в Цалкинском районе Грузии. Эта брекчия подстилается слоем долеритовой лавы и перекрыта песчаниками и глинами озерного происхождения. Озерные осадки в свою очередь перекрыты пластом долерита (Заридзе и Татришвплн, 1948). Таким образом, звери существовали и погибли на этом участке в эпоху перерыва деятельности вулканов Малого Кавказа.
Новая, развивающаяся трансгрессия в области Каспийской котловины, но не достигающая уже прежних размеров, известна под именем Апшершь ского моря.
Куринский залив Апшеронского моря доходил лишь до долготы Киров- | абада, а вероятное временное сообщение Терского залива с Эвксипскпм бассейном происходило в области Маныча. Па север это морс доходило лишь до широты Сарепты и Ипдера. 2 _ ч
Климат и ландшафты Кавказа в апшеропский век вряд ли заметно I няя че отличались от акчагыльских. Вулканические явления в эту эпоху про- | должались с той же силой, что и в акчагыле. Катастрофические ливни создавали временами сели, выносившие с гор массы галечников и руупные
н-ве )а-но
ам lulls, ш, 1US [1US 6). де-лли ого
кий По-1ЛИ,
цш. бых казе
phas ока. ном,
юсте irbis, ыль-акча-s sp. лены жены узпи. чани-вредь обра-а дея-
»вины, перон-
^пров-гнским ходило
аметно у проливни
Рис. 34.
I — челюсть Cants tamanensis;
няя челюсть Trogonlherium cuvieri; 5 и пясть Equus aff. sussenbornensis
10 — пенек и рог Т amanalces sp.; ц
Остатки зверей таманского комплекса.
2 локтевая Panthera sp.; 3 — череп С astor tamanensis* 4 — вепх-- верхняя челюсть и зубы Elephas meridionalis; О, 7_________ Pin
челюсть Sus tamanensis; 9 — рог Eucladocerus sp •* . , f — стержень рога bison sp.
уупные
5*
68
Часть 1. Обоснование истории формирования фауны
валуны, которые встречаются иногда на равнинах Кахетии (Кудрявцев,
1933).
Наземная растительность, отложении Ширакской степи,
Рис. 35. Разрез костсносиых песков у Георгиевска.
а—в — галечники выгоном террасы с обломками обсидиана; г—и — светлые и о?келезненные песни с окатышами пепла, костями слонов и копытных; справа — мощность в метрах.
известная по находкам из апшеронскпх содержала ель — Picea orientalis и ряд лиственных форм современного типа: бук, дуб, осину, яблоню, иву, лещину, медвежий орех, грецкий орех — Juglans regia, дзелькву, жимолость — и «гиркан-ские» формы: дуб — Quercus castaneifolia, ольху — Alnus subcordata, клен — Acer ibericum (Палибин, 1936). Окончательное исчезновение в Закавказье вечнозеленых видов субтропических растений происходило в апшеропском веке в условиях дальнейшего охлаждения климата.
Лесные ландшафты побережий Ку-рипского залива Апшеронского моря, по-видимому, были сходны с современными ландшафтами Колхиды и Гиркаяии, ие измененными человеком, но при отступлениях вод Акчагыл некого и Апшеронского морей па восточных участках Кавказской суши, несомненно, возникали аридные условия с образованием прибрежных степей или даже полупустынь. Эти условия определили и возможность развитя ареалов степных и пустынных форм млекопитающих.
Остатки млекопитающих в отложениях апшеронского яруса встречаются чаще, чем в акчагыльском, по представлены также единичными трубчатыми костями и зубами хоботных и непарнокопытных. Эти захоронения приурочены большей частью к морским слоям, главным образом к прибрежным фациям спокойных бухт, и соответствуют различным уровням бассейна. Захоронения происходили и в толщах континентальных отложений — в зоне накопления предгорного «барьера», а также в травертинах Пятигорья.
В восточном Предкавказье «следы» фаунистического комплекса, близкого по составу к псекунекому и таманскому, прослеживаются в своеобразных натеках из
вестняка — травертинах и суглинках гори Машук, а восточнее — в суглинках побережий Апшеронского бассейна. Кости слонов и копытных, встречающиеся в травертинах, хрупки,
легко крошатся. Наблюдаются случаи полного растворения кости и заполнения образовавшейся полости кристаллами кальцита. Из наиболее древнего, верхпеплноцепового «комплекса травертинов» горы Машук описаны и упоминались остатки: Elephas meridionalis Nesti, Е. cf. anti-
Гл* I. Развитие ландшафтов и фаун млекопитающих в третичном периоде 69
ши/s Falc.,-Е. aif. trogontherii Wiist, Bovinae, Cervinae (Егоров, 1932; Иванова, 1948; Громов, 1948).
Близ станицы Виноградной и у Георгиевска в аллювиальной толще , серых песков верхней террасы Кумы нами собраны ниже слоя перемытых обломков базальта и вулканических пеплов (рис. 35) зубы овернского мастодонта — Mastodon arvernensis Croiz. et Job., южного слона — Elephas nieridionalis Nesti, лошади — Equus cl. stenonis и трубчатые кости оленей — Eucladocerus sp. (рис. 36).
На склоне горы Таш-Кала, близ Грозного, в апшеронских суглинках найдены остатки южного слона — Elephas nieridionalis Nesti и лошади — Equus sp. (Павлова, 1931). По-видимому, из апшеронских слоев происходят и два зуба лошади Стенопа — Equus stenonis Cocchi, найденные близ верхних Ачалук на Сунженском хребте, а также стержень рога крупной антилопы — Bubalis sp., описанный Бурчак-Абрамовичем (1952а).
Из захоронения у села Винодельного, к северо-востоку от Ставрополя, Ворисяк и Беляева (1948) определили следующие формы по материалу, хранящемуся в ПИНе:
Proboscidea
Elephas nieridionalis Nesti
Perissodactyla
Equus (stenonis?) sp.
Artiodactyla
Paracamelus gigas Schloss.
Нижняя челюсть верблюда из этого захоронения была описана особо Хавесоном [(19546) (рис. 37)].
Единичные находки зубов южных слонов отмечались и в галечниках предгорном равнины у Махачкалы.
Апшеронский полуостров
Особенно многочисленны находки морских и наземных млекопитающих в известняках апшеронского яруса на Ашперонском полуострове близ Бинагады, Шихово, Баку.
Здесь, по определениям Богачева (1923, 1938а, 1940 г), были найдены остатки дельфина — Delphinus delphis L., своеобразного морского хищника—иекромита из ластоногих, близкого к выдро-тюлеиям — Necromites nestoris Bog., и непарнокопытных — Hipparion crassum Gerv., Hipparion^. Под 22-метровым слоем апшеронского известняка близ Баладжар найдена и плечевая кость тюленя, неотличимая от таковой Phoca caspica GmeL Расшифровывая палеозоогеографическую загадку находки черноморского Delphinus delphis L., Богачев (1939а) приходит к выводу, что дельфин проник в Каспий из Средиземного моря в акчагыле, одновременно с иммиграцией моллюсков — Mactra, Cardluni, Potamides. Вымирание же дельфина в Каспии совпадает, по мнению Богачева, с концом апшеронского пека в связи с наступлением ледниковой эпохи и появлением льдов.
То же самое следует сказать п по поводу недостаточно изученных остатков иекромита, которого Богачев (1940 г) сближал с семантором — Scmantor macrurus Orl., описанным из плиоценовых отложений Западной Сибири.
Западнее находки приурочены к глинистым хребтам Кабристаиа и к так называемым «боздагам», образующим два-три ряда параллельных
Рис. 36. Остатки зверей из речных песков Георгиевского карьера.
П1
J, 2 — остаток М5 Elephas meiidionahs и его стиравшаяся поверхность; 3, 4 — стиравшаяся
и язычная поверхность 1*П13 Equus stenonis-, 5 — пясть Е иг la docent s sp. ПЛ
п<
do
Гл, I. Развитие ландшафтов и фаун млекопитающих в третичном периоде 71
всхолмлений, протянувшихся от Иорского и Карталпиского плоскогорий на восток до ущелья Гардыман-Чая. Эти всхолмления сложены толщей глин, суглинков с прослоями галечников мощностью до 400 м и прослеживаются также под Малым Кавказом от Кировабада до Нафталана.
Находки, сделанные геологами преимущественно в толщах хребтов Копджашен, Палан-Тпкян, Караджа, Боз-Даг, Чузгуп-Тапа и других, определявшиеся Богачевым (1938в), Бурчак-Абрамовичем (1951а, 1952в)
Рис. 37. Челюсть Paracamelus gigas из Винодельного.
к мною, позволяют дать следующий список видов наземных позвоночных из толщ апшеронского яруса западнее Апшеронского полуострова:
Rodentia
Trogontheriuni cuvieri Fisch.
Muridae gen.
Proboscidea
Aiiancus arvernensis Croiz. et Job.
Elephas nieridionalis Nesti
Perissodactyla Equus sp.
Rhinoceros cf. etruscus Falc.
Rhinoceros sp.
Artiodactyla
Cervus sp.
Eucladocerus sp.
Bison sp.
He исключена возможность, что и верхняя челюсть стегодона—Stego-don sp., хранящаяся в Естественноисторическом музее в Баку, происходит из каких-то верхнеплпоценовых отложений восточного Закавказья (Богачев, 1935). В целом видовой состав руководящих форм из апшеронских, верхнеплпоценовых слоев Закавказья весьма напоминает таковой пз посткуяльницкпх горизонтов западного Предкавказья. Мастодонты апшеронского яруса были либо последними экземплярами, дожившими в восточном Средиземноморье до эпохи, переходной к плейстоцену, либо их остатки попали в эти горизонты из более древних слоев.
В южном Закавказье, на Армяио-Аиатолийском нагорье, находки остатков индикаторных для верхнего плиоцена форм зверей известны из-под Ленинакана и Эрзурума и связаны, вероятно, с отложениями галечников и гравийных песков, сносимых потоками во внутренние небольшие водоемы с отложениями озерных пресноводных бассейнов.
Еще Абих (1899) писал о скоплении окаменелых костей в верхнетре-тпчных известняковых мергелях и конгломератах, залегающих под пластом туфовой лавы на склонах строящейся крепости у Ленинакана. По определениям академика И. Ф. Брандта, здесь были остатки «Maslo-imtidae, Cervidae и Bovidae».
' 72
Часть I. Обоснование истории формирования фауны
Схема стратиграфического распределения главнейших местонахождений (захо
pout
Эпохи Отделы Ярусы Местонахождения: Черноморская область и Предкавказье
- Верхний Чаудинские слои Гурийские слои 7 Таманский полуостров: Синяя; Балка, Сенная, Кучугуры Ставрополье: Воскресенская Закубанская равнина: Псекупс,: Бакинская ; 1 i Am
Плиоцен Куяльницкий Е Таманский полуостров: Капустина Балка 1 Акч
Средний Киммерийский > Змейский хребет: Дарг-Кох Пре
- Нижний Понтический । Ставропольская возвышенность; i Косякинский карьер Пон
Миоцен Верхний г ч Мэотический Сарматский Север Азовского побережья: Новочеркасск, Туз ловка ' -- 4 Ставрополье: Бурлацкое, Благодарное, Петровское | Мао* Caps
* .1 Средний Конкский Караганский Чокракский Гельветский Ставрополье: Беломечетская К он] Kapi Чок] Гель
* ’ • Нижний
Олиго -цен 'if*-*'- ’ - -ч \ -
Кг
I
i Гл. /. Развитие ландшафтов и фаун млекопитающих в третичном периоде 73
: . Таблица 5
i- ронсний) остатков третичных млекопитающих в пределах Кавказского перешейка
1 Ярусы Местонахождении: Каспийская область и Закавказье Преобладающие группы видов
Апшеронский Терский хребет: Малгобек, Грозный, Ачалуки Апшеронский полуостров: Бинагады, Хурдалан, Ши-хово Армянское нагорье: Ленинакан, Нурнус Таманские волки Южные слоны Бобры трогонтерии Стен о новы лошади Сложнорогие олени Таманские зубры
i ккчагыльский Иорское плоскогорье: Эльдар, Джейран-Чели Кировобадское плоскогорье: Карасахкал, Нафталан Иорское плоскогорье: Палап-Тикян Овернские мастодонты Ацератерии Гиппарионы Сложнорогие олени
г Продуктивная толща Апшеронский полуостров*. Локбатан Лисицы Олени Газели
j j Понтический < * . s < Диноционы Овернские мастодонты Хомяки Двурогие носороги Тапиры Гиппарионы Косули
к ; Мэотический . Сарматский i Карталинское плоскогорье: Джапаридзе Горийская котловина: Арк-нети Иорское плоскогорье: Эльдар, Удабно Карталинское плоскогорье: Кясаман, Коцахурис-Кеды Гиены Длинно рылые мастодонты Ацератерии Гиппарионы Жирафы, кизлороги Газели
1 1 Конкский Караганский Чокракский Гельветский Амфиционы Болотные мастодонты Паранхитерии Анхитории Г ига нтс кие с в инь и Антилопы
Джавахетия: Ахалцых Бенаратерии Халикотерим Ан трак отер ии Трагулиды
Часть I. Обоснование истории формирования фауны
Богачев (1923—1924) упоминал о находке в серых вулканических песках близ Ленинакана остатков армянского слона — Elephas armeniacus Falc., по-видимому идентичного с Е. meridionalis Nesti.
В позднейших сборах из того же района Бурчак-Абрамовичем были определены остатки Elephas sp., Rhinoceros mercki Jaeger, Equus stenonis Cocchi (Авакян, 1948).
Для верхнеплиоценовых отложений диатомита близ села Нурнус на р. Занге Богачев (1938в) приводил следующий список остатков млекопитающих. несомненно нуждавшийся в уточнении:
Carnivora
Muslela filholi Gaudry Putorius sp.
- >> Lagomorpha Lepus sp.
Rodent ia Gerbillus sp.
Perissodactyhi
Hippar ion sp.
Rhinoceros etruscus Falc.
Artiodactyla
Tragocerus sp.
Гамбарян (1934), описавший этот пласт диатомита, отметил, что он достигает мощности 7 м и залегает на андезитовых лавах, а сверху перекрыт базальтом. Другими словами, какой-то обширный пресноводный бассейн существовал в районе средней Занги в эпоху между двумя крупными извержениями лав. Прослои пемзы и пеплов в глинах этого пласта свидетельствуют о том, что вулканическая деятельность не прерывалась и в период накопления осадков.
Находка костей песчанки — «Gerbillus» должна указывать на известную ксерофитность ландшафтов южных окраин Армянского нагорья в верхнем плиоцене.
Особенности захоронений и видовой состав верхнеплиоценовых зверей Закавказья подтверждают наличие там в апшеронском веке умеренного, но не холодного климата, бурной вулканической деятельности и отражают до известной степени аридные или семиаридные условия в восточной и южной частях страны.
В составе верхнеплиоцёнового комплекса Кавказа, особенно по захоронениям на Тамани, уже прослеживается переход к следующей послетретич-ной эпохе — плейстоцену.
В зоогеографическом j смысле фаунистические комплексы наземных млекопитающих неогена Кавказской суши не являются самобытными. Олигоценовая «фауна» Малого Кавказа обнаруживает родственные связи с центральноазиатскими и европейскими «фаунами» этой эпохи.
Среднемиоценовая «фауна» Предкавказья, известная из-под Беломечетской, — это кавказский вариант миоценовых «фаун», общих для Северной Америки и Евразии. Эта «фауна» появляется здесь в результате оформления временных континентальных связей с северными и южными массивами суши, но южное влияние было более вероятно.
В верхнем миоцене — сармате — южное влияние сказывается в проникновении на Кавказскую сушу гиппариопового фаунистического комплекса в результате широкого слияния Кавказского острова с Малокавказской сушей.
С миоцена же, очевидно, развивалась и местная горная фауна. Нет сомнения, что некоторые из современных горных эндемиков Кавказа являются потомками представителей местной верхнемиоценовой горной фауны. j
Гл. / Развитие ландшафтов и фаун млекопитающих в третичном периоде 75
Плиоценовые фаунистические комплексы предгорных равнин и плоскогорий развивались на Кавказе главным образом в результате миграций с юга и с севера. При этом южное фаунистическое влияние преобладало, являясь следствием развития жарких и сухих эпох плиоцена.
Северное фаунистическое влияние начало сказываться главным образом лишь в самом конце плиоцена при дальнейшем похолодании климата и появлении так называемых плейстоценовых родов и видов млекопитающих.
В плиоцене же продолжало идти и самобытное видообразование, особенно в сохранившихся на фоне общей пенепленизации высокогорных ландшафтах.
Палеонтологические факты, касающиеся наземных млекопитающих Кавказского перешейка, не дают оснований поддерживать гипотезу о континентальных связях западного Кавказа, Крыма и Малой Азии через посредство «Понтической суши», занимавшей будто бы котловину Черного моря (Андрусов, 1918; Вульф, 1944; Пузанов, 1949). Эта гипотеза отвергнута теперь и геологами (Муратов, 1951).
Однако тесные связи западного Предкавказья с Крымом вследствие отсутствия в верхнем плиоцене Азовского моря более чем вероятны (Верещагин, 1957а).
Современные взгляды на стратиграфическое положение захоронений остатков третичных наземных млекопитающих Кавказского перешейка даны в виде обзорной схемы (табл. 5).
Нет сомнения, что представители многих родов и виды верхнеплиоценового —• «таманского комплекса» продолжали существовать на Кавказском перешейке и трансформироваться позднее в нижнем и среднем плейстоцене. - • .
РАЗВИТИЕ ЛАНДШАФТОВ И ФАУНИСТИЧЕСКИХ КОМПЛЕКСОВ КАВКАЗА В ЧЕТВЕРТИЧНОМ
ПЕРИОДЕ
О ГРАНИЦЕ ПЛИОЦЕНА И ПЛЕЙСТОЦЕНА В ПРЕДЕЛАХ J
КАВКАЗСКОГО ПЕРЕШЕЙКА
В применении к Кавказу решение вопроса о нижней границе четвертичного периода крайне осложнено специфичностью генезиса всех ландшафтных элементов этой страны, лежащей между двумя древними материками. Несомненно, что разнообразие ландшафтных зон фаунистических комплексов и причудливость биогеографических границ в верхнем плиоцене были здесь уже не меньшими, чем в наши дни.
Поэтому геоморфологический анализ, так же как и единичные находки ископаемых растений и животных в континентальных отложениях, может отражать здесь нередко лишь узкоместные явления.
Геологами и биологами этот вопрос чрезвычайно запутан даже для Европы, что видно хотя бы по обзорам Жирмунского (1936) и Николаева (1947, 1950). В согласии с последним мы считаем, что определение границы третичного и четвертичного периодов должно решаться не изолированно, на основе какого-то одного критерия, а в комплексе, с учетом всех данных , взаимосвязанных дисциплин.
В качестве критериев для установления границы третичного и четвертичного периодов предлагались, как известно, самые разнообразные смены ландшафтной обстановки, идущие под воздействием двух основных факторов — тектоники и климата. Таковы колебания уровня и смены морей, орогенические и эрозионные циклы, оледенения, смены растительного и животного мира, а также этапы морфогенеза ископаемых организмов.
Соответственно субъективным вкусам и нижняя граница четвертичного (ледникового) периода в пределах Кавказа проводилась то с верхнего сармата (геолог Ковалевский, 1936; ботаник Гроссгейм, 1936), то с конца акчагыла (геологи Рейнгард, 1931, 1936а; Герасимов и Марков, 1939; палеонтолог и геолог Громов, 1948), то с верхнего апшерона (геологи Пав-: лов, 1936; Мирчинк, 19366; Варданяпц, 1948).
Отечественные зоогеографы — Мензбир (1934), Серебровский (1935) — j нового решения по сравнению с западноевропейскими схемами в приложе-1 нии к Кавказу не дали. =
В настоящее время взгляды Ковалевского, Гроссгейма, а также Шве- , цова (1928) и других, развивавших гипотезу о плиоценовых покровных f
. Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
77
оледенениях Кавказа, приравнивая последние к гюнцу и минделю европейской гляциологической схемы, могут быть, вероятно, оставлены как не выдерживающие критики.
Основываясь на геоморфологических и палеонтологических фактах, Громов (1948) признавал, что где-то на грани третичного периода и плейстоцена происходило прогрессивное «ухудшение» климатических условий, отнюдь не означающее еще обширного оледенения. Он считал «хапров-ский и псеку некий комплексы» (см. главу I) характерными для переходной эпохи — от плиоцена к плейстоцену, по крайней мере в Предкавказье и на юге Русской равнины.
Начало четвертичного периода велось Громовым с «таманского комплекса», тогда еще мало изученного и мало известного.1
Для низов доледниковья в пределах Кавказского перешейка индикаторными формами признавались: «Elephas trogontherii (=Е. wusti М. Pawl.), Bison schoetensacki Freud., Elasmotherium caucasicum Boris., Cricetus cri-cetus L., Spalax microphtalmus Giild.». Перелом фауны здесь усматривался в исчезновении мастодонта, плосколобого и южного слонов, гиппариона и этрусского носорога, а также в появлении зубра. Таким образом, био-стратиграфический критерий к верхней границе третичного периода у нашего палеонтолога вполне сходен с критериями западноевропейских авторов. Хапровский и псекупский комплексы сопоставляются далее с пойдем и гюнц-минделем, а таманский — с миндельским оледенением западноевропейской гляциологической схемы.
К гюнцу относился и первый эрозионный цикл четвертичного периода, соответствующий отложению галечников 175-метровой террасы р. Кубани.
Сходные палеонтологические материалы и суждения, разработанные Ппдопличко (1940в, 1952, 1954), существуют и для севера Черноморского побережья в пределах Украины.
Новые палеонтологические материалы действительно подтверждают, что качественные изменения в составе растительных и животных группировок на равнинах и в предгорьях Кавказского перешейка произошли лишь в конце апшеронского века. Эти изменения свидетельствуют об установившемся умеренно теплом или даже умеренном климате.
В пресноводных верхнеплиоценовых толщах Таманского полуострова близ Кучугур и в нижних горизонтах аллювиальных песков карьера Гирей на Кубани нами обнаружены слегка окремнелые остатки древесины широколиственных пород: дуба — Quercus sp., бука — Fagus sp. и вяза Ulmus sp.2 Весьма вероятно, что субтропических видов во флоре этой эпохи в Предкавказье уже не было вовсе. Обильные остатки наземных моллюсков — Chondrula, Helix, пресноводных — Corbicula, Anodonta, Bithinia, Micromelania и других из континентальных толщ посткуяль-яицких (?) горизонтов на Тамани и в верхнеапшеронских слоях восточного Закавказья свидетельствуют скорее всего также об умеренном климате.
Именно в верхнеапшеронских горизонтах предгорий Северного Кавказа вовсе исчезают остатки мастодонтов, гиппарионов и появляются остатки лошадей — Equus cl. stenonis, Е. all. sUssenbornensis, эласмотериев — Elasmotherium caucasicum, верблюдов рода Paracamelus и бизонов — Bison sp.
i На самом деле таманские захоронения весьма близки к псекунеким и хапроиским (Верещагин, 1957 а).
- Определения А. И. Зубкова.
78 Часть I. Обоснование истории формирования фауны
По этапу морфогенеза кавказские слоны верхнего апшерона соответствуют переходной стадии — Elephas meridionalis — Е. trogontherii (Верещагин, 1957а).
Аналогичная картина прослеживается и в континентальных отложениях Закавказья, в частности на Армянском нагорье, но абсолютная хронологическая близость этих захоронений, сохранивших следы сходных фаунистических комплексов и этапов морфогенеза, к предкавказским уже вызывает сомнения из-за их значительной географической разобщенности.
Итак, кроме приобретения растительностью умеренного, а местами и ксерофитного облика, появления на обширных, освобожденных морем пространствах ряда «степных» и даже «пустынных» форм копытных, Кавказский перешеек в конце ашперонского века был подготовлен к новым эрозионным циклам и к новому этапу горообразования.
В связи со всем сказанным мы считаем возможным приурочивать начало антропогена в области Кавказа, вслед за Павловым (1936), к концу регрессии Ашперонского моря и началу развития Бакинского моря, учитывая, что эта континентальная фаза была, несомненно, весьма длительна.
ПРОБЛЕМА ГЕНЕЗИСА КЛИМАТОВ И ЛАНДШАФТОВ КАВКАЗА В ПЛЕЙСТОЦЕНЕ В СВЯЗИ С ИЗУЧЕНИЕМ ФОРМИРОВАНИЯ ФАУНИСТИЧЕСКИХ КОМПЛЕКСОВ
Основные проблемы генезиса климата и ландшафтов Кавказа в после-апшеронское время, важность которых для разработки истории фауны неоспорима, следующие.
1. Тектонические циклы и динамика рельефа суши в связи с речной эрозией.
2. Колебания уровня морей и континентальные связи Предкавказья с Русской равниной.
3. Масштабы и число оледенений в связи с тектоникой и колебаниями климата.
4. Смены растительности и формирование растительных зон.
Исследования в области этих проблем на Кавказе велись практически без привлечения палеонтологических материалов по наземным видам животных и растений. Поэтому имеет смысл подвести их итоги с точки зрения значимости для проблемы истории фауны.
Динамика рельефа суши. Картина динамики рельефа Кавказа в плейстоцене весьма сложна. Геоморфологами признается, что к концу плиоцена — ашперонского века — произошло значительное выравнивание древних горных массивов Большого и Малого Кавказа вследствие замирания орогенеза.
Следы древнего выравненного рельефа действительно сохранились по Главному хребту, например в Абхазии, Сванетии, Кабарде, а особенно в Дагестане, где широкие, в настоящее время высокогорные, плато, заболоченные и с дряхлой речной системой, достигали, по-видимому, в конце плиоцена всего 500—800 м абсолютной высоты. Возраст этого выравненного рельефа определяют то ашперонским, то бакинским веком. Мир чинк (1936а) склонен приравнивать его к концу апшерона—началу баку. Отдельные горные гряды в области Водораздельного и Боковых хребтов поднимались будто бы не выше 1000—1500 м над уровнем Ап-шеронского моря. Для воссоздания нижнеплейстоцеповых фаунистических и флористических комплексов находки ископаемых остатков
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
70
зверей и растений на этих высокогорных плато представляли бы чрезвычайный интерес, но возможности образования и сохранения захоронений в условиях энергичного сноса, очевидно, отсутствовали.
По соображениям Николаева (1941, 1949), рельеф внутренних плато Большого Кавказа значительно древнее апшерона. Рельеф Армянского нагорья, по утверждению Паффенгольца {1948, 1950), настолько древен, что обусловил возникновение оз. Севан уже в начале четвертичного периода при запруде глубокой эрозионной долины пра-Занги базальтовыми лавами вулкана Богу-Дага.
На основании наблюдений в Дагестане и Осетии развивалась гипотеза о горстовом поднятии в плейстоцене центральных участков Главного хребта на высоту 1000—1500 м (Рейнгард, 1939; Варданянц, 19336, 1948). Николаев (1941, 1949) признавал сводовое поднятие Большого Кавказа при размахе колебаний тектонических движений в «послеверхнеплио-ценовое время» в 3—4 км. По-видимому, речь шла здесь об амплитуде между поднятиями центральных частей хребта и опусканиями в области котловин Черного и Каспийского морей.
Указывалось и на наличие дифференцированных новейших тектонических движений в области центрального Кавказа, Ставрополья и Таманского полуострова. Лакколиты Пятигорья, резко выделяющиеся на рельефе, «испытывали на общем фоне относительно большие поднятия, чем соседние участки». Подъем Большого Кавказа в плейстоцене до 2 тыс. м признавал и Бонов (1949), работавши!! «объемным методом».
Таким образом, по общепринятым взглядам, в основе развития рельефа горного Кавказа в четвертичном периоде лежит сводовое, а местами и горстовое поднятие с одновременным прогибанием в области предгорных долин и равнин, а также в области котловин Черного и Каспийского морей.
Важным следствием этих колебательных движений было сближение древних денудационных уровней с более молодыми в районе предгорий и последующий уход этих древних уровней (террас) под молодые отложения в районе равнин. Этот процесс перекрывания осадков террас в предгорьях и на равнинах крайне важен для правильного сопоставления возрастов горных террас с террасами равнин и для заключений о возрасте ископаемых фаунистических находок.
Подъем кавказских хребтов неизбежно приводил к оледенениям (об этом см. дальше) и к образованию скального рельефа с обилием россыпей в области сбросов, куэст, и содействовал, следовательно, развитию скальных группировок растений и животных в новых типах местообитаний. Неизбежным следствием было также усиление динамичности вертикальных ландшафтных зон, содействовавшее экологической и морфологической дивергенции, развитию в одних случаях и расчленению в других ареалов видов и их группировок.
При подъеме хребтов на протяжении плейстоцена происходило также энергичное расчленение склонов речной эрозией. Горные речки углубляли свои долины поэтапно, образуя террасы в соответствии с горообразовательными циклами. В то же время они пропиливали лежащие на их пути молодые поднимающиеся хребты, в результате чего образовывались узкие каньоны — щели.
Такие каньоны-щели постплиоценового возраста имеются па северном склоне Большого хребта — па Баксане, Чегеме, Череке, Урухе, в боздагах восточного Закавказья — на Геок-Чае, Турьян-Чае, Алазани, Kvpe, в районе Мцхети, на участке юго-восточных отрогов Карабахских гор на Араксе и на Вазару-Чае в Талыше.
8$ Часть I. Обоснование истории формирования фауны
Это дифференцированное воздымание хребтов в комбинации с речной эрозией приводило к образованию как бы отгороженных экраном участков долин. В результате перехвата осадков экранирующими хребтами высоко в горах возникали своеобразные участки с сухим умеренным климатом, которые послужили своего рода убежищем для сухолюбивых видов растений и животных. Такими участками на протяжении плейстоцена стали внутренний Дагестан, продольные долины горных районов Осетии и Кабарды, Горийская котловина на Куре, верхняя часть долины р. Бамбак на Армянском нагорье и Зувандская котловина в Талыше. Значение своеобразных убежищ для немногих видов мелких сухолюбивых млекопитающих: сусликов, хомяков, степных полевок — эти участки сохранили и до наших дней.
В области известняковых массивов речная эрозия содействовала развитию карста с образованием ниш и пещер, заселявшихся некоторыми хищниками, а позднее и первобытным человеком.
Снос речками обильного обломочного материала с хребтов на предгорные равнины и в долины приводил на протяжении всего плейстоцена к образованию мощной мантии предгорных галечников и суглинков. Такими отложениями выполнены Закубанская и Терско-Сунженская долины, Кусарская и Кировабадская наклонные равнины в Закавказье и террасы Черноморского и Каспийского побережий.
На этих предгорных равнинах формировались ареалы млекопитающих, связанных с предгорным лесостепным ландшафтом и, как правило, с умеренной влажностью. Кроме того, предгорные равнины служили в плейстоцене основными путями расселения для видов (южных — к северу и северных — к югу), тяготеющих по своим адаптациям к среде с небольшими высотами над уровнем моря и к равнинному рельефу.
В высокогорьях, а отчасти и на предгорных равнинах (на Апшерон-ском и Таманском полуостровах) большое значение для развития рельефа и формирования ареалов зверей имели вулканические явления. Особенно крупные масштабы имели эффузии лав на Армянском нагорье, где лавовые потоки перекрывали огромные пространства плоскогорий в различных направлениях, занимая иногда до 50% общей площади (Маруашвили, 1946). Влияние этих процессов на формирование ареалов зверей сказывалось, помимо прямого единовременного воздействия, еще и через длительное косвенное изменение ландшафта. Так, например, современное безлесие западной Армении некоторые геоботаники склонны объяснять именно эффузиями лав, происходившими в плейстоценовой эпохе (П. Ярошенко, 1941).
Горообразовательные циклы и их скорость были, по-видимому, тесно связаны с колебаниями уровня морских бассейнов четвертичного периода.
Колебания уровня морей. Угасшее Апшеронское море в области Каспия сменилось Бакинским, давшим новую обширную трансгрессию с обилием дидакн. На смену Бакинскому пришло Хазарское море с архаичными дидакнами — Didacna surachanica Andr., которому в области Черного моря соответствовало Новоевксинское. После перерыва Хазарское море сменилось сильно опресненным Хвалынским, протянувшимся далеко на север и оставившим осадки с полупресно водной фауной моллюсков даже под Уральском и Казанью. Его осадкам в области Черного моря соответствуют древнечерноморские слои. С последней крупной трансгрессией в области Каспийской котловины — Хвалынским морем — связывается обычно и представление о последней стадии крупного оледенения Кавказского хребта. Отступая, Хвалынское море превращалось
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
81
в послеледниковый Каспий, который давал лишь небольшие трансгрессии, ие выходившие далеко за пределы его современных очертаний (Ковалевский, 1933; рис. 38).
В эпохи крупных плейстоценовых трансгрессий морские береговые » террасы перешейка как на восточном, так и на западном побережьях сильно сокращались.
Моря начинали размывать либо предгорный барьер, например, в восточном Закавказье, либо наклонные равнины, затрудняя тем самым воз-
Рис, 38. Кавказ и Каспий в верхнем плейстоцене.
1 — область древнепресноводных отложений в бассейне Волги;
2 — область арало-каспийсной мореной трансгрессии; 3 — область распространения четвертичного оледенения. (По Ковалевскому, 1933).
мощности поселений и миграций равнинных зверей. Сокращались также площади аллювиальных равнин: Ейско-Кубанской, Кумо-Терской, Рион-ской, Кура-Араксинской, которые также служили участками, прово-t дящими или сохраняющими расселяющиеся фаунистические элементы В различного происхождения и различного возраста.
Образовавшийся при трансгрессии Манычский пролив должен был Г- ограничить проникновение зверей с Русской равнины, по крайней мере зимоспящих и не способных к кочевкам через голые пространства льдов. S Моря в эпохи регрессий оставляли в низменностях обширные засоло-| пенные пространства и болотные низины с остаточными лиманами и t озерами. Эти пространства рассолонялись наиболее быстро в западном £ Предкавказье; где они занимались степями. В восточном Предкавказье • и восточном Закавказье процесс рассолонения шел медленнее, здесь г С Н. К. Верещагин
, 82 Часть Z. Обоснование истории формирования фауны ?
|
возникали полупустыни, заселявшиеся тушканчиками, песчанками, сайгаками и джейранами. Сохранившиеся местами на тех же низменностях | лиманы и озера существовали в виде обширных заболоченных массивов, | II. тростниково-нимфейных и камышовых плавней, а в Колхиде — в виде
В ольхово-нимфейных и осоково-сфагновых топей. Длительность существо-
11: вания таких массивов зависела от гидрологического режима подпиты-
Г. вавших их речек и быстроты заполнения остаточных котловин аллювием.
Они являлись пристанищем таких водолюбов, как выдра, водяная полевка, речной бобр, кабан. ;
; . Масштабы и кратность оледенений. Колебания климата ' г и оледенения горных массивов, происходившие на Кавказе, могли быть J наиболее действенными фаунообразующими факторами в четвертичном i л .. периоде. |
| - Все учение об оледенениях Кавказа и их кратности основано, в сущ- !
I ности, на геоморфологических материалах и фактах, а также на сопостав- |
I ленйи оледенений с орогеническими циклами и колебаниями уровней Г
| морей. Это сопоставление встречало всегда большие затруднения и по- I | Стоянно осложнялось стремлением отечественных геологов подогнать ! I схему оледенений и межледниковых эпох Кавказа к западноевропейским £ | стратиграфическим схемам и терминам, разработанным для Альп. Само- I I; стоятельных стратиграфических исследований и стратиграфических г | схем, свободных от влияния «западноевропейского гляциологического । гипноза», для Кавказа пока почти неизвестно. I
| Между тем по мере углубления геоморфологических исследований I к выяснялось своеобразие развития Кавказа в плейстоцене в зависимости от качественно и количественно различных с Альпами факторов.
Академик Павлов (1925) считал, что конец апшеронского века в Кавказской геоморфологической области совпал с развитием плиоценового = оледенения (гюнцского) и ознаменовался поднятием хребтов, вызвавшим : образование ледников в горах., и усилением отложений галечных выно- г сов, представляющих обычное явление в верхней свите апшеронского I яруса. Признавая многократность оледенений, Павлов приравнивал } миндельское оледенение к верхам бакинского яруса, рисское — к верхам ‘ хазарского, а хвалынский ярус сопоставлял с эпохой таяния вюрмского; ледника. Эта схема удержалась, в сущности, в общих чертах и до настоя-1 щего времени. |
Рейнгард (1937, 19476) признавал для Кавказа четыре или пять круп-| ных оледенений. Из них от первого — гюнцского, — приходящегося^ на перерыв акчагыл—апшерон и захватывавшего предгорья, известны только остатки покровных галечников. При втором же — миндельском, — относимом на перерыв между апшероном и баку, ледники выходили будто бы на 80 км к северу, на предгорную равнину центрального участка Кавказа. Это оледенение сменилось второй, особо продолжительной межледниковой эпохой. В течение последней в предкавказских равнинах шло накопление мощной толщи делювиальных суглинков в резуль-г тате сноса материала с гор. Третье оледенение — рисское — в этой схеме1 считается сильнейшим и оказавшим наибольшее влияние на выработку современного рельефа долин. Последовавшая вслед за тем короткая межледниковая эпоха, при которой ледники сохранялись в вершинах ледниковых долин, вновь сменилась четвертым и последним оледенением, в течение которого ледники нигде не выходили за пределы горных ущелий Позднее были уже только четыре стадии отступания ледников с одной фазой наступления, приходящейся на средние века нашей эры.
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 83
По представлениям Рейнгарда, подъем Каспия начинался в связи с уменьшением испарения вместе с оледенением и вначале проходил медленно, затем, при усилении таяния ледника, шел быстро, а наибольшей высоты достигал к концу второй половины оледенения, когда трансгрессия Черного моря еще далеко не достигала своего наибольшего развития (рис. 39).
По Герасимову и Маркову (1939), вся ледниковая история Кавказа делилась на два периода. В первом из них, падающем на акчагыл—апше-
КАВКАЗ ЛЕДНИКОВЫЕ И МЕЖЛЕДНИКОВЫЕ ЭПОКИ КА С П И Й TPAHCFPECKU И PEEPECUU
Современная эпоха Cardium eduLe Diducna trigonoides Л. surachanica =^Калинская ороеени-^рхческая фаза Л. carditoides '=у\Предрисская орогена ^Рческая фаза В. г и dis, Л. catiltus Лослвгкнщская ^яорогеническая фаза Верх ни и} Бакинские $ [Средне- апшеран\ слои \ ' Каспийские \ С / слои \слои Sea
V-S Стадии отступания
d-fi Стадии отступания<=^Ё.
W R-W Вюрм-оледенение Рисе -вюрмская межледниковая V эпоха
R-II Рисс-оледенение
BIRR Межледниковая фаза
RI Рисе-оледенение
MR Большая миндель-ДллЁ= рисская межлед-р_ :..... никовая эпоха V- -—
М Миндель-оледенение
G-M Гюну-миндельская pdd межледниковая зпохаШ^
G ГюнЦ'-аледенекие
Доледниковое время- нижний апшерон
Рис. 39. Соотношение оледенений и орогенических фаз Кавказа с колебаниями Каспия. (По Рейнгардту из Богачева, 1940).
рои, центральный и восточный Кавказ испытывали будто бы одно- или двухкратное покровное оледенение вследствие поднятия горных хребтов. Во втором же периоде, приходящемся на вторую половину четвертичного периода, весь Большой Кавказ испытывал двухкратное оледенение, развивающееся на «горном остове, подготовленном пижнечет-вертичными тектоническими движениями». При этом авторы сомневались в синхронности этих оледенений с альпийскими, но считали несомненным «коренное неоднократное изменение климатического режима п множественность (3—4) ледниковых эпох Кавказа».
Рекордное число — десять—покровных оледенений Кавказа насчитывалось с верхнего миоцена Ковалевским (1936).
6*
Часть /. Обоснование истории формирования фауны
*
Трех-, четырехкратное оледенение Кавказа признавал и Варданянц (1948), но он же склонен принять и схему Громова (19356) о наличии на Кавказе всего двух оледенений, сопоставляемых с рисским и вюрм-ским оледенениями Европы, и соответствующих: первое — началу хазарского, а второе — началу хвалынского века с последующими стадиями отступания (рис. 40).
Для Черноморской котловины этот согласованный процесс оледене-и трансгрессий мог осложняться периодическим возобновлением
НИИ
контур моря; 2 — контуры площадей оледенения. (По Варданянцу, 1948).
зависящей от опусканий и поднятий в районе Сводная стратиграфическая схема Варданянца
Рис. 40. Оледенение Кавказа в эпоху предхазарскои орогеническои фазы. / — предполагаемый
связи моря с океаном, Босфора и Дарданелл, дана в табл. 6.1
Учитывая появление периодических связей морских бассейнов, Вар* ; данянц дает картину «каскада» от Арала—Каспия—Черного моря в Среди- । земное море с периодическим «промыванием» пресными водами Каспия | и стока этих вод через Манычский пролив после орогенической фазы, | Каскады в восточном направлении с осолонением морей происходят в j послетретичный период в карангатскую и древнечерноморскую эпохи, [ когда воды Средиземного моря будто бы прорывались до Каспия. j
Странно, что гипотеза геологов о покровных оледенениях Кавказа j не нашла возражений у биологов, а даже встретила поддержку. f
Формирование растительных зон. В представле- ? ,ниях кавказоведов: ботаников Кузнецова (1909), Медведева (1915), зоологов: Сатунина (1910), Динника (1911) — существовала одна эпоха ; _____ _______ £
1 Другие геологи, например Понов (1955) сопоставляют, впрочем, хвалынский L ярус с новоэвксинскими слоями, хазарский же — с карангатскими и т. д. J
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 85
Таблица 6
Сводная стратиграфическая таблица послетретичной истории Черного моря, Кавказа и Каспия (по Варданянцу, 1948, стр. 23) с добавлением связей в области
Маныча
Босфор Черное море Кавказ Маныч Каспийское море
Современные отложения (Стадии бюля) Открыт (?) Послехва лынский ярус
Закрыт Опускание, регрессия, опреснение Поднятие, оледенение (бюль или неовюрм) Закрыт Опускание, регрессия
Древнечерномор-ские слои (Стадии вюрма). ! Открыт Хвалынский ярус
Открыт. * Опускание, вторжение соленой воды Поднятие и оледенение (вюрм) Закрыт Опускание и регрессия
% 1 1 Н овоэвксинские слои. (Рисс-вюрм) (?) Хазарский ярус
Закрыт Опускание, регрессия, опреснение- Поднятие и оледенение (рисе) Закрыт Опускание и регрессия .
Карангатские слои Мин де ль-рисс Открыт. Верхний баку (переходные слои)
13 Открыт Опускание, вторжение соленой воды Поднятие Закрыт Регрессия
Узунларские и Древнеэвксин-ские слои Закрыт Средний баку Нижний баку
Чаудинские слои Поднятие и оледенение (мин-дель) Закрыт Опускание, регрессия
Краснодарский горизонт? (Г юнц-минде ль) Закрыт Верхний апшерон
1 Закрыт Регрессия и опреснение (?). Поднятие и оледенение (гюнц) Закрыт Опускание, регрессия
Гурийские слои Открыт Нижний и средний апшерон.
86 Часть I. Обоснование истории формирования фауны
горного покровного оледенения, когда происходило насыщение кавказской флоры и фауны бореальными элементами и полное вытеснение горных животных и растений на предгорные равнины.
На основе анализа современной флоры Кавказа Гроссгейм (1948) признает необходимым допустить ряд жарких межледниковых эпох для объяснения наличия и концентрации в отдельных участках перешейка ксерофильного флористического ядра южного корня. Множественность оледенений и теплых межледниковых эпох на Кавказе признают также геоботаники Малеев (1946) и Колаковский (19476), разбирая вопросы происхождения флоры восточного Средиземноморья, в частности Колхиды. Они прямо связывают с ледниковыми плювиальными периодами значительное распространение мезофильных широколиственных лесов, с интерплювиалами, аридными эпохами — редукцию мезофильных лесов и широкое развитие гемиксерофильной и термофильной растительности. Нетрудно заметить, однако, что эти соображения ботаников основаны либо на анализе дизъюнкций ареалов и флористического состава отдельных районов, либо просто повторяют высказывания геологов. Такие высказывания были и у меня (Верещагин, 1949в). Между тем происхождение разрывов ареалов зависит от большого числа взаимосвязанных явлений, а не только от прямого воздействия климатических смен. Даже факты далекого проникновения на юг настоящих северных растений, например сфагновых мхов, росянки и других на Большой Кавказ, в болотистую Колхиду и в высокогорье Малого Кавказа, можно правильно оценить лишь при точном знании условий и способностей распространения этих видов в прошлом и настоящем. Что же касается использования фитоценотических преобразований для оценки климатических смен, то и они вызывают весьма спорные и противоположные высказывания даже для такого небольшого геологического отрезка, как историческая эпоха (например, Синская, 1933; Федоров, 1952).
Малое число находок макроскопических ископаемых остатков плейстоценовых растений не дает оснований для разработки схемы климатических смен. По имеющимся же палеоботаническим и геоботаническим материалам, все же становится очевидным, что уже с середины плейстоценовой эпохи на Кавказе существовала флора, крайне близкая к современной, а фито ландшафтная зональность приобрела в основном современные черты. В конце ледникового периода древесная растительность была та же, что и сейчас, и существовала почти на тех же местах, что и в наши дни (Гроссгейм, 1936, 1948).
Никто не будет отрицать, конечно, наличия в прошлом грандиозных по сравнению с современными долинных ледников, например на северном склоне западного Кавказа. Троговые участки долин Лабы, Зеленчуков, Кубани, Баксана и других рек, а также гряды конечных морен, бараньи лбы отчетливо видны здесь и для каждого непосвященного (рис. 41). Тем не менее троги занимают все же лишь верхние участки ущелий, далеко не достигая даже Скалистого хребта. При любой попытке воссоздания условий существования растительных и животных группировок в эпоху крупнейших долинных ледников Кавказа становится все же ясно, что никаких принципиальных различий по сравнению с современностью здесь не было. Высокогорная растительность в ледниковую эпоху была сдвинута книзу, но отнюдь не вытеснена целиком на предгорные равнины.1 Для животных горного и скального рельефа в эпоху
1 Это вынужден был признать и Гроссгейм (1936, стр. 203), несмотря на солидарность со взглядами Ковалевского.
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
87
наи( элыпего оледенения было, несомненно, достаточно мест обитания на )бтах меридианного направления и на боковых цепях гор в непосредственной близости к ледникам. Горные — хвойные, лиственные и смешанные — леса, насыщенные горными группировками животных, благополучно существуют и в данный момент в непосредственной близости с ледниками, например на Череке, Урухе, Рионе (рис. 42).
Что касается «явных следов» гипотетических покровных оледенении в впдр галечниковых толщ и даже крупных валунов на предгорных рав-
я
>-;е
>-х
<е й, о-то ie-ая
>н, ия ая
ей-ти— :им то-ре-:ен-сть о и
аых верней-реи, аого зтки
по-?ных ?ано-знию щни-м на поху [ицар*
Рис. 41. Трог и концевая морена, зарастающая сосной, в Цейском ущелье Северной Осетии.
Фот. автора, 1947.
винах, то в большинстве случаев они могут быть объяснены разновременный спонтанными выносами в грязеводяных валах во время паводков и извержений вулканов.
С понятием о воздействии ледниковой эпохи на фаунистические ком-чсксы млекопитающих Кавказа должно быть связано представление снежном факторе, а также о сдвигах широтных ландшафтных зон на русской равнине под воздействием оледенения севера Европы. Между тем глубина и продолжительность снегового покрова в плейстоцене почти не изучены, или о них судят лишь по косвенным примерам распространения п миграций некоторых современных млекопитающих.
По мнению геоботаников (Медведев, 1915; Гроссгейм, 1936, 1948), обилие так называемых бореальных видов растений па Кавказе может соъясняться лишь тем, что «языки рисского ледника подступали достаточно близко к Кавказскому хребту». Другими словами, предполагалось, что в районе Маныча и Сальских степей была и танга, и тундра. Резуль-а ы новейших палинологических исследований так же как и ископа
88
Часть I. Обоснование истории формирования фауны
емых остатков позвоночных, показывают, что между низовьями Дона и Волги в плейстоцене всегда простирались степи.
Оценка зоопалеонтологических и зоогеографических фактов для разработки стратиграфической схемы Кавказа дается после обзора плейстоценовых захоронений. Здесь же следует попутно отметить, что необходимым следствием признания грандиозных покровных оледенений Кавказа было бы сведение всей эволюции высокоспециализированных горных
Рис. 42. Расположение ледника и леса в верховьях Рпоиа.
Фот. автора, 1957.
форм (козлов, снеговых полевок и других) к ничтожно малому отрезку времени — к послеледниковой эпохе.
В итоге вся гляциологическая схема Кавказа и особенно гипотеза покровных оледенений нуждаются в коренном пересмотре с широких биологических позиций.
Абсолютная геохронология четвертичного периода. Попытки определения абсолютной длительности четвертичного периода в применении к Кавказу не имели оригинальных решений. Большинство отечественных авторов следовало в этом отношении Зергелю (Soergel, 1925), Мпланковичу (1939), определяя весь ледниковый период в 600—650 тыс. лет. Ковалевский (1933) относит Арало-Каспийское море за 230 столетий, Хазарское — за 135 столетий и Хвалыпское — за 34 столетия до наших дней, необычайно «омолаживая» таким образом всю группу явлений, связанных с формированием фауны и становлением человечества. Позднейшие исследователи считали, впрочем вполне про-визорно, что важнейшие события четвертичного периода: горообразовательные циклы и оледенения — происходили на Кавказе и на смежных участках за значительно более длинный отрезок времени — на протяже-
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 89
нпи 500—600 тыс. (Варданянц, 1948) и даже 1 млн лет (Николаев, 1949; Ролов, 1949). Дальнейшие исследования физико-химического состава горных пород и костных остатков, а также осадконакоплений еще внесут поправки в эти цифры, крайне важные для учета быстроты ’шолюционного процесса, видообразования и формирования фауны.
По условиям образования захоронений удовлетворительно докумен-фованным палеонтологически оказался на Кавказе лишь верхний 1 [ейстоцеп, т. е. эпоха предположительного наибольшего оледенения и последующие стадии его сокращения.
ОБЩАЯ ХАРАКТЕРИСТИКА УСЛОВИЙ ОБРАЗОВАНИЯ ЗАХОРОНЕНИЙ МЛЕКОПИТАЮЩИХ В ПЛЕЙСТОЦЕНЕ
Природа, происхождение плейстоценовых захоронений, т. е. условия пбелп зверей и погребения их остатков, выясняются, как правило, легче п достовернее, нежели третичных.
Остатки послетретичных млекопитающих встречаются на Кавказе в морских береговых террасах Черноморского, Азовского и Каспий-ского побережий, в речных и озерпых террасах, в покровных суглинках я пещерах.
Захоронений морского и лагунного типа в послеапшеропских слоях ь Кавказе обнаружено мало. Причина этого лежит в особенностях новей-i, ii тектоники, так как по численности особей и по богатству форм эко-т гпческие группировки плейстоценовых зверей вряд ли сильно уступали • рхпетретпчпым. Массовые захоронения остатков плейстоценовых я ркопптающпх, погибших от разных причин, образовывались, несом-в нно, в устьевых пространствах впадавших в морские бухты рек см. «Введение»). Меньшие скопления трупов могли образовываться в облети среднего п нижнего течения крупных рек, где также накапливались Ц 1щп аллювия. Между тем все эти участки, расположенные в низовьях уоапи, Терека, Риона, Куры и перегруженные наносами, находц-шсь в плейстоцене и находятся теперь в зоне тектонических прогибов, следовательно, не имеют сейчас предпосылок к вскрытию естественным порядком. Они могут быть обнаружены лишь при бурении скважин или при крупных выемках грунта.
Исключением из указанной закономерности могут быть захоронения в самых вершинах бухт плейстоценовых морей, если эти участки позднее оказались в зоне предгорных поднятий. Такие участки могут находиться в среднем течении Кубани — Крапоткин—Невинномысская; на Тереке — в районе Моздока и Котляревской; на Рионе — по долготе Кутаиси и Зестафонп; наконец, па Куре — в районе Мцхети—Тбилиси.
Образование захоронений в «зоне качания» долинных прогибов и предгорных поднятий, по-видимому, обусловливало и мпогослойность захоронений, если гидрологические условия сохранялись на протяжении нескольких эпох.
Большинство обнаруженных па перешейке плейстоценовых захоронений остатков зверей представлено единичными, целыми н разрозненными скелетами в континентальных отложениях различного происхождения: озерпых, аллювиальных, делювиальных, эоловых, пещерных, а также в натеках известняковых туфов и в асфальтовых лужах на равнинных и предгорных участках перешейка.
В горах условий для природных захоронений костей было мало, несмотря па постоянную гибель горных зверей от случайных падений'
90
Часть I. Обоснование истории формирования фауны
лавин, камнепадов, гололедиц, голодовок. Мелкозернистый аллювий горных речек в пределах самой горной системы, как правило, маломощен ниу и недолговечен (Шанцер, 1951), а возможности захоронения костей в толще лоз крупного галечника очень невелики. Конечные и боковые морены плейстоценовой эпохи в ущельях, как правило, не содержат остатков позво- теч<
ночных, что также объясняется истиранием костных остатков в толще рей
морены при ее образовании. сте!
Захоронения костных остатков в делювиальных шлейфах склонов, <
расположенных вне влияния крупных рек, не были и не могли быть мае- обл<
совыми, так как единичные (не накопленные по какой-то причине) скелеты тип.
павших зверей обычно разрушались бесследно при медленном накопле- । ]
нии делювия. тич]
Чаще и лучше сохранялись скелеты от трупов зверей, вынесенных на чаю
равнину селевыми потоками—в толще грязи и конгломератов конуса вы- люс
носа. Образование грязевых потоков, наносимые ими разрушения и гибель 1932
в них животных наблюдались на протяжении всего четвертичного пе- щих
риода. О нп весьма картинно описаны Ивановским (1911) для долины и ос Аракса, Муратовым и Гвоздецким (1948) — для долины Хасаута на Се- Bull
верном Кавказе. Известны они и для речек восточного Закавказья, па- I
пример в ущелье Демир-Апаран-Чая. ш стиг
Возникновение этих потоков вызывалось и вызывается извержением слав
вулканов, влекущим растапливание фирна и льда, а также быстротеч- F
ными ливнями. деле
Эти селевые потоки, временами более частые и мощные в плейсто- ( него цене, чем в наши дни, нередко являлись причиной массовой гибели жи- мер,
вотных. prises
Постоянная и катастрофическая гибель зверей, обитавших на равни- Челб
нах Кавказского перешейка, вызывалась, вероятно, в плейстоцене, как водое
и в наши дни, многоснежьем и гололедицей. Захоронение костей на рав- лой
нинах, вне речных долин, происходило редко, лишь в оврагах и покров- йены
ных суглинках делювиального происхождения. . Koctj
Новый тип массового накопления костных остатков возник в плей- Они стоцене в результате деятельности первобытного человека, бросавшего беспо остатки пищи на открытых стоянках и особенно в пещерах. На ли
Универсальность таких захоронений, т. е. накопление остатков многих зарск видов зверей в одном местонахождении, была обусловлена в плейсто- У цене, кроме деятельности первобытного человека, еще и охотой филина, черра а также четвероногих хищников, затаскивавших кости своих жертв в пе- пеще{ щеры. под и
Этот тип захоронений особенно важен, так как он встречается наиболее хорон часто именно в горах, создавая предпосылки для обоснования истории К формирования горной фауны. Главнейшие местонахождения рассматри- аллюг ваются далее по районам. остатв
изучет
ОБЗОР ЗАХОРОНЕНИЙ КОСТНЫХ желез
ОСТАТКОВ В ПЛЕЙСТОЦЕНОВЫХ ОТЛОЖЕНИЯХ Ка
вскры
Западное Предкавказье зелено
залега
После отложения костей верхнеплиоцеповых млекопитающих в га- галечн
лечниках Псекупса и конгломератах Таманского полуострова в западном Ниже] Предкавказье существовал какой-то значительный перерыв в образова- и внов нии массовых и универсальных захоронений. с оСта,
Гл. II, Развитие ландшафтов и фаун в четвертичном периоде
91
Единичные фаунистические находки, которые могут быть датированы нижним и средним плейстоценом, представлены большей частью зубами поздних южных и трогонтериевых слонов.
| В пределах Прикубанской равнины, т. е. по правобережью широтного •пения Кубани, известные захоронения остатков плейстоценовых зве-находятся на значительной глубине, в толщах аллювия древних рек, стекавших с Кавказа.
Эти захоронения образовались, очевидно, за пределами выноса грубо-сломочного материала, на границах водоемов плавневого или лиман-ипа, принимавших мелкозернистые наносы горных потоков.
В глинах п песчанистой толще Прикубанской равнины среднечетвер-т.чного, «хазарского» возраста остатки крупных млекопитающих встре-<ыотся вместе’с остатками древесины и раковинами пресноводных мол-прсков — Unto, Paludina, например в районе Краснодара (Православлев, 1 32). В вышележащей толще песчанистых лёссовидных суглинков, имею-irIX иногда прослои вулканического пепла, кроме костей млекопитающих j остатков растений, встречаются еще и раковины наземных моллюсков — Ihminus tridens Mull., свидетельствующие о степных условиях.
В районе станицы Темижбекской, где мощность этих суглинков до-с нгает 50 м, были обнаружены кости «Bos, Equus, Cervus, Elephas» (Право-4 iaB.ieB и Апшелес, 1930).
Ряд сохранившихся находок из других местонахождений был опре-д»лен памп в Краснодарском музее в 1952 г. Зубы и обломки черепа раннего мамонта у- Elephas trogonlherii — Е, primigenius происходят, например, из станицы Дпнской. Остатки скелета длиннорогого зубра — Bison prisons longicornis обнаружены под станицей Крыловской, на среднем '(елбасе. Скелет был захоронен в тонком иловатом песке какого-то стоячего водоема. По типу сохранности эти кости напоминают алебастр при малой вторичной минерализации. В аналогичных условиях были захороним остатки длиннорогого зубра и мамонта у стапицы Скобелевской. Костя залегали на глубине 16.5 м под лёссом, в илистом болотном слое. Они имеют на поверхности характерные следы каких-то минирующих беспозвоночных (личинки Trichoptera?!) в виде продолговатых насечек. Наличие длиннорогого зубра подтверждает среднеплейстоценовый, «хазарский» возраст этих находок.
У Краснодара же, на глубине 14 м в супесях второй аллювиальной террасы Кубани, было извлечено пять почти целых скелетов малых пещерных медведей (Григор, 1926), описанных Борисяком (1930, 1931) под именем «степной» формы — Spelaearctos rossicus, Гибель зверей и захоронение их трупов Борисяк объяснял действием мощного селевого потока.
Карьер Гирей. Попытка стратиграфического расчленения аллювиальной толщи Прикубанской равнины и захороненных в этой толще остатков млекопитающих была произведена Громовым (1948) па основе изучения «опорного разреза» по песчаным карьерам у поселка Гирей и железнодорожной станции Кавказской (г. Крапоткин).
Карьеры, заложенные в 10—15-метровой вюрмской террасе Кубани, вскрывали здесь слой покровных супесей и песков в 2 м и затем толщу зеленоватого суглинка с вертикальными корнеточинами в 1.5 м. Далее залегала толща косо слоистых галечников с линзами песка, прослоями галечников мощностью до 3. 5 м и с остатками костей мамонта в основании. Ниже шла прослойка глины мощностью 0.8 м с остатками растений (рис. 43) и вновь горизонт песков мощностью до 3 м, подстилаемый галечниками с остатками бизонов, оленей, слонов.
92
Часть J. Обоснование истории формирования фау,ньъ
Из галечниковой толщи, по Громову (1948), извлекались остатки следующих форм: «Elephas wiisti, Е. aff. trogontherii, Е. primigenius, Rhinoceros mercki, R. antiquitatis, Bison priscus cf. longicornis, B. priscus de-minutus».1
Остатки слона Бюста и носорога Мерка, по наблюдениям геолога А. Д. Колбутова, происходят из нижнего горизонта галечников, залегающего под глинами с лигнитизированной древесиной. Громов сближал
Рис. 43. Обнажение галечников и песков в карьере Гирей. Слой покровных суглинков снят.
Фот. автора, 1952.
эти остатки по возрасту с остатками зверей из тираспольского гравия, а остатки мамонтов из более высокого горизонта песков и галечников относил к рисскому времени, синхронизируя их с «фауной» из Ильской палеолитической стоянки (см. дальше). В итоге в сборах выделялись три стратиграфических комплекса фаун: 1) постхазарский с остатками Elephas primigenius, 2) хазарский с остатками Е. trogontherii, 3) дохазар-ский с остатками Е. wiisti.
Возраст большей части костей из карьера Гирей, по мнению Громова, хазарский, т. е. раннерисский или даже миндель-рисский.
Наше изучение особенностей захоронения костных остатков на новых карьерах у поселка Гирей в 1952 г. показало, что толща древнего аллювия в 12—13 м залегает здесь под покровными суглинкамми па сарматских сизых глинах, прорезанных современной Кубанью на глубину 5—6 м (рис. 44). В нижних горизонтах песков и глии, на глубине 10 м, обычны остатки крупных древесных стволов, нередко одетых чехлом из ожелезпенпого ила, напитанных водой и как бы обугленных. По древесине
1 Эти материалы были утрачены во время войны.
• Гл. II,. Развитие ландшафтов и фаун в четвертичном периоде 93
' здесь были определены: дуб — Quercus sp., бук — Fagus sp. и тополь — Populus sp., т. е. породы, характерные для зоны современных предгорий. Костные остатки прослеживаются в галечниках, непосредственно налегающих на пески с древесными стволами. Они большей частью сильно --------------------------------------------------------------^3
окатаны, истерты, вторично минерализованы и прокрашены охрой.
Исследованные нами в Краснодарском музее новые сборы зубов слонов из карье-ра Гирей принадлежат: Elephas meridionals— два коренных с эмалью 3.5—4 мм; £. Irogontherii — три коренных с эмалью 2,4—2.8 мм; Е. trogonlherii primigenius — один зуб с эмалью 1.8—2.2 мм. По типу сохранности костного вещества зубы южного и трогонтериевого слонов из Гирея весьма близки. Цемент их «мергелевого» типа сохранности, розовато-кофейного цвета. Зуб раннего мамонта (8.5 петель эмали на 10 см у М®) имеет более свежий вид. с цементом белым в разломе, с тер-нымн прожилками и со слабой прокраской местами охрой. В сборах имеются также фрагменты костей оленей из группы Cervus elapnus и обломок рогового стержня бизона —Bison cf. schoetensacki.
Сохранность этого обломка «мергелевого» типа. Таким образом, в песчаногравийной толще у Кавказской смешаны к захоронены остатки животных, вымытые и переотложенные из озерных слоев различного возраста. В целом комплекс с южным и трогонтериевым слонами, возможно, несколько более поздний, нежели таманский (см. главу I).
Зубр уже крупнее таманского, меньше разнообразие оленей, по-видимому отсутствуют остатки антилоп.
Кости млекопитающих, изредка находимые в верхних горизонтах лёсса я в подстилающем его горизонте ископаемой почвы водоразделов северо-западного Предкавказья, датируются более поздней эпохой. Исключением могут быть упомянутые захоронения в озерных отложениях под Крыловской и Скобелевской, которые следует датировать средним плейстоценом. Мирчинк (19366) сопоставляет верхние горизонты лёсса Предкавказья с вюрмским оледенением, а горизонты ископаемой почвы — с писс-вюрмским межледниковьем.
На Закубанской наклонной равнине к самым низам четвертичного периода относились средние горизонты галечников Псекупса с остатками южного слона, стеноновой лошади и какого-то быка (Громов 1948). v ’
а <
Ь ййЁЙЙ
8.0
15
ОА ОБ
ОБ
ОБ
Рис. 44. Разрез костеносных галечников и песков карьера Гирей. а — покровные суглинки; б — супесь с прослоями гравия; в — речные песни с прослоями галечников и глин; г — галечники и пески с костями млекопитающих и древесными стволами; 0 — сарматские голубые глины; справа — мощность в метрах.
2.0
1.5
94. Часть I. Обоснование истории формирования фауны
Находки, которые могут быть датированы нижним и средним плейстоценом, представлены обычно остатками южных и трогонтериевых слонов. Они обнаруживаются, как правило, в наиболее высоких террасах левых притоков Кубани.
В окрестностях станицы Преградной на Большом Зеленчуке найден зуб нижней челюсти Elephas trogontherii (коллекция ЗИНа).
В фондах Краснодарского музея с Закубанской равнины хранятся г коренные зубы слонов промежуточного типа между Elephas trogontherii f и Е> primigenius с толщиной эмали 1.8—2.4 мм и имеющие от 7 до 8 эмалевых петель на 10 см стирающейся поверхности. Захоронение остатков слонов происходило, как правило, в мелкозернистом речном или озерном песке в условиях последующего заболачивания, о чем свидетельствует обычная прокраска охрой. По типу сохранности эти зубы выглядят всегда несравненно моложе зубов южных слонов с Псекупса и Тамани. Дентин и цемент плотные, светло-кремового или желтоватого цвета. В щебневом карьере под станицей Некрасовской на Лабе был найден зуб слона промежуточного типа между Elephas meridionalis и Е. trogontherii, нижний, коренной зуб лошади — Equus aff. sussenbornensis, обломок пенька рога оленя — Eucladocerus sp. и обломок рога гигантского оленя — Mega-ceros euryceros. Это местонахождение по видовому составу и по типу сохранности при значительной прокраске охрой костной ткани еще сильно напоминает некоторые захоронения на Таманском полуострове.
В новых сборах костных плейстоценовых остатков, сделанных Краснодарским музеем в гравийном карьере у поселка Ивановского, были обнаружены и коренные зубы настоящих мамонтов с толщиной пластин эмалн 1.4—1.8 мм и с 12 петлями на 10 см. Коренной зуб мамонта найден и на р. Пшиш. В Майкопском музее хранятся зубы южного и трогонтериевого слонов из-под станицы Ново-Лабинской с берега Лабы.
Стратиграфическое расчленение этого музейного материала затруднено не только из-за неудовлетворительной этикетировки, но и из-за особенностей образования захоронений в условиях предгорной равнины (см. выше). Конусы выноса селевых потоков, захоронявших трупы зверей, на равнине могли перекрывать друг друга в разные геологические эпохи, в зависимости от мощности образовавшегося селя. Реки же, врезавшие вслед за тем свои долины в этих галечниках и конгломератах, а также в озерных осадках, заново перераспределяли весь костный материал в аккумулятивных террасах. В связи с этим толщи речных террас содержат весьма разновозрастные фаунистические остатки. Поэтому попытка синтеза геоморфологических и палеофаунистических данных в работах Мирчинка (19366) по сопоставлению речных террас на юге Русской равнины и в Предкавказье может быть принята лишь с большими оговорками. Мирчинк отнес 175-метровую террасу под Черкесском к плиоцену на основе находок зубов Elephas planifrons\ 100—125-метровую террасу, вслед за А. П. Рейнгардом, — к миндельскому оледенению. Последняя в пределах Кубанской степи прикрыта двумя горизонтами лёссовых пород и характеризуется двумя комплексами ископаемых почв, соответ-1 ствуя «палюдиновым» пескам под Таганрогом, Бессергеновкой и Ейском. | 1
Необоснованная палеонтологически 50-метровая терраса под Черкесском 1 связывается Мирчинком с моренными образованиями селения Хумарин- ] ского и вторым сверху горизонтом лёссовых пород водоразделов Запад- s ного Предкавказья, которым он придает рисский возраст. Весьма вероятно, что эти последние отложения Предкавказья синхронны горизонтам галеч- ] ников, залегающим в основании второй надпойменной террасы низового i
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
95
Поволжья и содержащим остатки зверей «хазарского фаунистического комплекса»;
Значительные уточнения в видовой состав фаунистического комплекса равнин западного Предкавказья в верхнем плейстоцене внесло изучение костных остатков, собранных на Ильской палеолитической стоянке.
Илье кая палеолитическая стоянка. «Следы» обитания палеолитического человека в западном Предкавказье прослежены по кремневым орудиям, находимым по отмелям Курджипса и Белой у Майкопа, а также у Фортепьяновки. Здесь было, видимо, много летних открытых стоянок. Пещеры в предгорьях, о которых имеются сообщения в краевой хронике, не исследованы еще палеонтологами. Так называемая Фанагорийская пещера — узкая и холодная щель, промытая в известняковых конгломератах в верховьях речки Аюк, к западу от поселка Горячий Ключ, не была пригодна для обитания крупных зверей.
Ильская палеолитическая стоянка была обнаружена еще в 1898 г. Де Байем (см. Волков, 1899). Она принадлежит к стоянкам открытого типа и находится на правом берегу Иля, левого притока Кубани в 40 км к юго-западу от Краснодара, в зоне современных обезлесенных предгорий. Вторично эту стоянку разыскал в 1925 г. Замятнин (1934). Раскопки стоянки производились в 1927—1928 гг. Замятиным, а в 1930 и 1936— 1937 гг. — Городцовым (1940).
Стоянка располагалась под доломитовыми обрывами, на второй, 10— 14-метровой террасе Иля, сложенной делювиальными суглинками с двумя горизонтами погребенных почв. В нижнем из них и залегают палеолитические остатки.
Многочисленные орудия, описанные упомянутыми археологами, сделаны из доломита, реже из кремня. Замятнин, учитывая небрежную обработку доломитовых орудий, датировал стоянку мустьерской стадией, Городцов же, ссылаясь на более тщательную обработку орудий из кремневой гальки, относил эту стоянку к эпохе солютре. Геологическая датировка по террасам — рисс-вюрм, начало рисса (Громов, 1933, 1948).
Поверхность рисской террасы, как думал Городцов, заболотилась и покрылась ржавчинными лужами с началом делювиальных процессов. К этому времени орда забросила стоянку. Одновременно вследствие подвижек слоев из доломитового массива происходило истечение нефти, вытеснявшей воду и консервировавшей слои стоянки — ее палеолитические орудия и костные остатки.
Костные остатки с Ильской стоянки неоднородны по типу сохранности. Так, одни фрагменты «звенят» и минерализованы настолько сильно, что лишь слабо липнут к мокрым пальцам, а по удельному весу похожи на плиоценовые. Такие кости обычно сильно прокрашены окисью железа, заполняющей поры, или облеплены частицами иловатого мелкозернистого песка, характерного для болотистых источников поймы рек. Эмаль зубов в этом случае окрашена в совершенно черный цвет. Другие же кости, консервированные нефтью, производят впечатление менее древних. Все извлеченные кости представлены «кухонными остатками». Эпифизы трубчатых костей обломаны под самую шейку, а диафизы расколоты на мелкие куски для добычи мозга. Особенно много обломков челюстей и изолированных зубов зубров. Имеются кости полу взрослых мамонтов, обгрызенные крупными хищниками.
В растворенных бензином кусках асфальта, облеплявших кости зубров, нами найдены еще остатки кавказской мышовки, мелкой змеи, жуков и травянистых растений.
.96
Часть /. Обоснование истории формирования фауны
Состав и численность остатков млекопитающих в сборах Замятнина и Городцова, по определениям Громовой (1932а, 1937) и нашим исследованиям, даны в табл. 7 и на рис. 45. Нами учтены и трудно определимые остатки различных видов.
Таблица 7
Видовой состав и численность костных остатков млекопитающих из Ильской стоянки
Виды Число костей Число особей
Ursus spelaeus 2 2
Cants aff. lupus (мелкая форма) 19 4
C. lupus ..... 1 1
Crocuta spelaea . . 24 7
Elephas primigenius 210 5
Lepus aff. europaeus 15 1
Sicista cf. caucasica 2 2
Muridae^ ближе не определен. . 9 2
Equus caballus 13 5
E. cf. hidrunlinus 23 3
Sus scrofa 1 1
Cervus elaphus . 33 4
Megaceros eury ceros 37 4
Bison priscus 2401 43
Saiga tatarica ......... 38 2
Artiodactyla, ближе не определен. 549 —
Всего 3377 86
Отсутствие остатков волосатых носорогов, лосей и первобытных туров, вероятно, не случайно. Эти животные были редки и трудно добываемы. Еще более редки или даже вовсе отсутствовали в ту эпоху на Закубанской равнине эласмотерии и верблюды, характерные виды среднего плейстоцена Поволжья.
Зубр, кости которого составляют основную массу (87%) костных остатков на Ильской стоянке, был представлен, по мнению Громовой, уже измельчавшей формой — Bison prisons deminutus.
На самом деле наравне с относительно мелкими экземплярами коренных зубов и пястных костей (самок?) имеются и неотличимые остатки от крупных костей зубров из хазарского яруса средней и нижней Волги. Процент их, впрочем, невелик, но для заключений об этапе морфогенеза зубров необходимо учитывать и охотничью избирательность древних закубанцев, которые могли убивать преимущественно самок.
В окрестностях стоянки были, вероятно, лугостепи и лесостепи, в которых обитали крупные стада бизонов, лошадей и оленей.
Ландшафтные особенности верхнеплейстоценового Закубанья выявляются лучше по наличию остатков сайги и кабана. Сайга не водилась на левобережье Кубани в историческую эпоху: здесь летом было слишком сыро, а зимой скапливалось много снега. Кабан же был еще недавно весьма многочислен и массами спускался на эту лесистую равнину при многоснежье в горах. Таким образом, судя по наличию сайги, остеп-ненность Закубанья в эпоху обитания человека на стоянке была большей,
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
97
нежели в наши дни. Климат был суше, и катастрофические снегопады отсутствовали. Стада сайгаков могли, впрочем, прикочевывать в предгорья с севера лишь при пурге и гололедице, как это наблюдается и теперь в восточном Предкавказье.
Массивы горного леса были, очевидно, значительно выше, нежели ныне, чем и объясняется ничтожное число остатков кабанов.
Весьма характерно также отсутствие остатков животных скального и горного рельефа, козлов и баранов, а также «северных» млекопитающих.
Отсутствие остатков козлов и серн нельзя использовать, однако, для заключений о датировке стоянки и размерах оледенений Кавказа. Эти высокогорные ныне копытные даже в эпоху наибольшего оледенения могли держаться вдоль нижнего края ледников — в горах, близ вершин Семашхо,~Оплепен, на 60—70 км от стоянки, т. е. все же слишком далеко, чтобы попадать регулярно в сферу действия ильских первобытных охотников. Ближайшие же горы были пологи и низки. Отсутствие остатков песца, северного оленя, овцебыка скорее всего доказывает, что эти виды не проникали на Северный Кавказ вообще.
Остатки слонов на Ильской принадлежат мамонту позднего типа (с эмалью зубов толщиной 1.3—1.4 мм). Выявлено по крайней мере две взрослых особи и три подростка на стадии смены М3 на М4.
Несомненно, что новые поиски и сборы костей мелких видов, захороненных в асфальтовых натеках этой стоянки, могли бы дать много нового для восстановления деталей истории четвертичной фауны Кавказа.
Обломки хитиновых покровов насекомых, пойманных на стоянке нефтяными лужами, были исследованы по нашей просьбе А. В. Богачевым. Определено частью до вида, частью до рода 18 жуков, муравей и оса (?). Среди жуков наиболее обычны водолюбы и плавунцы: Dytiscus sp, n., Agabus sp., Illibius sp., Hydroporus sp., Colymbetes fuscus L., жужелицы: Carabas sp. n., Platysma sp., Amara sp., Bembidium sp.; листоеды: Gastroidea sp., Donacia sp., навозники: Aphodius subterraneus L. Степные формы представлены Tentyria nomas Pall. Много видов, значительно отличающихся от современных, что говорит о древнем возрасте этой фауны.
Остатки растений из кусков битума, взятых С. И. Замятниным, по исследованиям М. Г. Кипиани, содержали до 20представителей травянистой растительности, среди которых преобладали остатки: Cruciferae gen. et sp., Gompositae gen. et sp., Lepidium perfoliatum, Polygonum avi-culare, Euphorbia cf. palustris, cf. Statice, Chenopodium sp. et cf. Atrip-lex и единичные остатки Alisma sp., cf. Carex, Scirpus sp., Plantago sp. и др. Макроскопических остатков древесных пород не обнаружено. Приведенный список видов характеризует группировку выгрева, ее древнюю «засоренность» и присутствие растений небольших луж.
Абсолютная датировка Ильской по радиоактивности костного вещества составляет в определениях В. В. Чердынцева 39 тыс. лет.
По нашему мнению, фаунистические остатки с Ильской стоянки свидетельствуют о начале верхнего плейстоцена и о значительном остепнении предгорий.
В 1957 г. А. А. Формозов организовал разведочные раскопки входа в пещеру в ущелье реки Белой у станицы Даховской. В раскопе на глубине трех метров было собрано до 80 обломков костей плохой сохранности вместе с единичными орудиями мустьерского типа. По нашим определениям, кости принадлежали следующим видам: Canis sp. (10/2), Vulpessp. (17/4),
7 H. К. Верещагин
Рис. 45. Остатки млекопитающих с Ильской стоянки.
1 1 — челюсть Cams lupus; 2 — челюсть Crocuta spelaea; 3,4— М4 Elephas primigenius и его
£ стиравшаяся поверхность; 5 — верхняя челюсть Sicista cf. caucasica (х8); 6 — фаланга
J Equus hidruntinus; 7 — пясть Е. caballus; S — челюсть Megaceros euryeeros; 9 — пясть
Cervus elaphus; 10 — стержень рога Saiga talarica; 11, 12 — M3 и эпифиз голени Bison priscus.
' Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 99
Ursus spelaeus (3/2), Crocuta spelaea (1/1), Meles meles (7/2), Panthera spelaea (4/1), Felis cf. silvestris (2/1), Lepus europaeus (3/1), Cricetus crice-tus (3/2), Elephas cf. primigenius (14/1), Cervus elaphus (5/2), Megaceros ^yceros (5/2), Capra cf. caucasica (2/1), Bison priscus (19/3).1 Этот состав рактеризует экологическую группировку эпохи переходной от среднего
Рис. 45 (продолжение).
к верхнему плейстоцену, в общем довольно близкую к той, что была отмечена в Ильской. Особый интерес представляют какие-то мелкие формы волка и лисицы.
Центральное Предкавказье
В пределах Ставропольской возвышенности и Пятигорья остатки плейстоценовых млекопитающих обнаруживаются чаще всего в толщах делювиальных суглинков, перекрывающих водоразделы и выполняющих
1 В числителе показано число фрагментов, а в знаменателе число возможных осоки -
7*
. 100
Часть I. Обоснование истории формирования фауны
древние овраги. Более древние, нижнеплейстоценовые, захоронения происходили и в отложениях каких-то древних потоков, а также в отложениях морских или озерных заливов на окружающих равнинах. Старые опубликованные находки представлены остатками эласмотерия — Elasmotherium sibiricum из-под Дивного, в области Манычской депрессии (Павлова, 1916). К нижнел1у плейстоцену следует отнести и описанные Хоменко (1913а) sy&biEquuscL stenonis из лёссовидных суглинков близ села Мирного. Средним плейстоценом, хазарским веком могут быть датированы: рог Mega-ceros hibernicus, найденный Хоменко в делювиальных глинах береговых обрывов Калауса; остатки Saiga prisca (=*У. tataricd) — в делювиальных глинах в балке Коронкояр, в 35 км к югу-юго-востоку от Ставрополя; остатки Equus caballus fossilis — в делювиальных глинах Шатыр-Кургана и остатки Bos sp. —• в песках на дне колодца в русле Мутнянки.
Среди зубов слонов, хранящихся в Ставропольском музее, нижнеплейстоценовым, трогонтериевым слонам принадлежат три нижнекоренных зуба с 7 петлями на 10 см и толщиной эмали в 2.5—2.8 мм. В том же музее еще в 1948 г. хранилось 6 фрагментов черепов длиннорогого зубра | и один фрагмент черепа первобытного тура — Bos primigenius, собранных ' на территории Ставрополья. Большинство их происходило из подстилающей лёссовидные суглинки толщи и может быть датировано средним плейстоценом.
Ряд новых находок был сделан В. Г. Гниловским в 1941 г. на трассе Кубано-Егорлыкского канала по западной окраине Ставропольской возвышенности. Два обломка нижних челюстей верблюда — Paracamelus < cf. gigas, нижний эпифиз пясти, пятка, прогрызенная каким-то хищником, были извлечены из котлована консольного сброса на первой террасе Егорлыка с глубины 6 м. Оттуда же происходят весьма фрагментарные остатки костей короткорогого зубра — Bison cf. schoetensakii, состоящие из обломков шейных позвонков, обломка стержня рога и предплечья, обломка правого плеча и астрагала.
Остатки эти «мелового» типа сохранности, легки и слабо прокрашены по трещинам окисью железа.
С глубины 9 м, из слоя аллювия, налегающего на майкопские глины в первой террасе Егорлыка, происходят и обломки массивных верхнекоренных зубов быков — Bos sp. или Bison sp., а также верхний коренной и первая фаланга мелкой лошади — Equus sp. Все эти остатки, фоссили-зованные, прокрашенные охрой и неокатанные, залегающие непосредственно на древнем базисе эрозии, также отмечают, по-видимому, нижний, реже средний этап плейстоцена.
К верхнему плейстоцену, а частью и к послеледниковые относятся находки Рябинина (1918) в Удельной степи в верховьях Калауса. Рябинин обнаружил обильные костные остатки млекопитающих в верхних горизонтах толщи лёссовидных суглинков, залегающих «in situ» в срезах левого берега балки Малый Киянкуль, а также в вершине балки 1-й Терновой и в русле Калауса, близ устья балки Малый Янкуль. В балке Малый Янкуль многочисленные обломки костей зверей были захоронены на глубине 2.5 м в бурой бесструктурной песчанистой глине, с гипсом, и с журавчиками из белой углекислой извести. По определениям Рябинина, здесь оказались остатки: «Equus caballus fossilis; Sus scrofa fossilis; Bos taurus fossilis; Saiga tatarica fossilis и стержень рога, похожий на джей- ( раний — Gazella subgutturosa». Найденные там же мезолитические орудия} («культуры солютре, или мадлена», по Рябинину) подтверждают самый! поздний этап плейстоцена или даже переход к голоцену. i
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 101
Таким образом, на окраинах Ставропольской возвышенности прослеживаются местами «фаунистические следы» основных этапов плейстоцена, которые совершенно не подтверждают вытеснения на равнину горных кавказских млекопитающих в эпоху великого оледенения, как это пред-; ваялось Рябинину и многим фаунистам и зоогеографам. Не обнаружены щсь и северные виды. Значительные уточнения в состав степного и лесостепного комплекса верхнеплейстоценового Ставрополья могли бы внести исследования существующих и погребенных пещер на южных и западных склонах горы Стрижамент (Верещагин и Громов, 19536).
R области лакколитов Пятигорья и на Кабардинской наклонной рав-! отметим старую находку остатков мамонта в гравийных карьерах илпз Ессентуков (Карпинский и Дрейер, 1904). Еще Эйхвальд (1850, нр. 190) писал, что близ Кисловодска им были найдены целые челюсти ,b)i адеи — «Equus caballus fossilis», покрытые известковым туфом.
Травертины Пятигорья. Теперь известно, что накопление травертинов в Пятигорье тянулось с перерывами с плиоцена, причем известковые источники меняли свое расположение на склонах после каждого горообразовательного цикла. Кости зверей, погибгпих от разные причин, скатывались или растаскивались хищниками по склонам и перекрывались делювием, а попадая в струйки известковой воды, консервировались вследствие осаждения на них слоя туфа.
Травертины отлагались почти на всех лакколитах—Лысой, Железной, Змепке, Бештау, но изучены в палеонтологическом отношении лишь травертины на горе Машу к (Иванова, 1948; Мирчинк, 19366; Громов, UK.;),
Иванова насчитывала четыре возрастных комплекса травертинов.
флме 1-го плиоценового комплекса, упомянутого в I главе, остальные относятся к плейстоцену и голоцену.
Травертины 2-го комплекса миндельского и миндельрисского возрастов располагаются небольшими пятнами на северном и восточном скло-вх. В них были найдены зубы и части челюсти Elephas aff. antiquus. Еще обширнее пятно травертинов 3-го комплекса, залегающее северо-восточнее. Его толща сопоставлялась с 55-метровой террасой Подкумка, в которой были обнаружены многочисленные остатки оленей — Cervidae. Упомянутой террасе придают рисский и рисс-вюрмский возраст. По-видимому, из этих же травертинов в карьере Бойченко Кобутовым добыты обломки зубов Lepus sp., Capreolus sp. и мелких полорогих. Сохранность их верхнеплейстоценового облика.
Толщи травертинов 4-го верхнеплейстоценового и современного комплекса залегают на уровне первой и второй террас на южном склоне горы у Пятигорска, вплоть до русла Подкумка. По Громову (1948), из этой толщи происходят зубы Equus sp. позднего типа, а зубы Elephas primi-genius обнаружены в галечниках и суглинках второй — вюрмской террасы.
В Пятигорском музее из находок плейстоценового возраста хранится кусок доломитизированного травертина с кальцитовым «слепком» стержня рога некрупного бизона — Bison sp. Обломок верхней челюсти мамонта (3-м молочным зубом, хранящийся в этом музее, происходит с северного склона Машука, из каменоломни № 1. Свежесть кости этого образца, совершенно необычная для южных находок, создает впечатление, что мамонт в районе Пятигорья вымер лишь в голоцене. Более древнего типа сохранности оказался обломок верхнекоренного зуба мамонта, найденный в травертинах у Медового водопада близ Кисловодска, а также
102 Часть I. Обоснование истории формирования фауны
зубы, мамонтов, извлеченные из толщ песков и суглинков близ станицы Каменской и из обрыва берега р. Юцы на глубине 12 м.
В том же музее хранятся 2 челюсти волосатых носорогов — Rhinoceros tichorhinus и 6 фрагментов черепов первобытных зубров верхнеплейстоценового типа сохранности.
Все эти фаунистические остатки отмечают различные стадии накопления толщ аллювио-делювиальных песков и суглинков на наклонной равнине центрального Предкавказья и дают лишь самое схематичное представление об экологических группировках млекопитающих мамонтовой фауны в природе предгорных ландшафтов. Последние в первой половине плейстоцена были, несомненно, сильно остепнены. Для конца же плейстоцена был, напротив, характерен лесной облик района, во всяком случае в пределах самого Пятигорья. Подтверждением этому служат остатки оленей и косуль в травертинах 3-го комплекса горыМашук и «постплиоценовых флор». В травертинах близ Пятигорска Красновым определены остатки 14 видов травянистых и древесных растений, характерных и для современного широколиственного леса предгорий: бук — Fagus sylvatica, орех — Corylius avellana, граб — Garpinus betulus, вяз — Ulmus campestris, дуб — Quercus robur, платанолистный клен — Acer platanoides и др. Близкая по составу, но несколько более поздняя «флора» с горы Железной в обработке Палибина (1913) содержала: ивы — Salix alba и др., липу — Tilia cordata, ясень — Fraxinus excelsior, мушмулу — Mespilus germanica, «Бореальных» элементов в этих позднеплейстоценовых или раннеголоценовых захоронениях нет. Наоборот, прослеживается скорее «южное» влияние.
ВосточноеПредкавказье
Большая часть равнин восточного Предкавказья лежит в пределах каспийских трансгрессий, и плейстоценовая фауна его остатками млекопитающих представлена очень слабо.
Полупустыни и степи в низовьях Кумы, Терека и Сулака — это область обширного опускания. Они перекрыты толщей послеледниковых морских и речных наносов, лежащих ниже современного базиса эрозии рек, в связи с чем и находки костных остатков плейстоценовых зверей делаются здесь западнее — в высоких террасах, по окраинам заливов плейстоценовых морей, на предгорных равнинах и склонах предгорных всхолмлений.
Наклонные равнины восточного Предкавказья перекрыты делювиальными шлейфами и аллювием Терека, Сунжи, Сулака и других рек. Однако рельеф предгорий по сравнению с западным Предкавказьем осложнен поднятиями третичного возраста — вытянутыми по широте Терским и Сунженским хребтами. Эти пустынные ныне всхолмления, испытывав-, шие при энергично идущей эрозии и пенепленизации поднятия на про-| тяжении плейстоцена, резко отгораживают Владикавказскую и Сунженскую наклонные равнины от аллювиальных равнин западного При-ка спия.
Еще Полигу (Pohlig, 1886) были известны остатки мамонта из долины Сунжи, хранившиеся в Кавказском музее в Тбилиси. Муллаевым (1929) был описан зуб Elephas trogontherii, найденный в послетретичных отложениях Урсдона близ села Карагач, в юго-западном углу наклонной Владикавказской равнины. Обломки лопатки трогонтериевого слона, извлеченные с 5-метровой глубины Моздокского песчаного карьера, хранятся в Пятигорском музее. Зубы трогонтериевых слонов раннего типа,
Гл. JI. Развитие ландшафтов и фаун в четвертичном периоде 103
найденные в пределах Грозненской области, хранятся в краеведческом музее Грозного. Все эти захоронения происходили преимущественно в аллювии нижнечетвертичного возраста.
Громов (1948) упоминал два черепа Bison prisons deminutus и фрагмент черепа Megaceros sp., извлеченные из первой надпойменной террасы Терека под Моздоком. Полости черепов зубров оказались забитыми вулканическим пеплом. Этот факт позволяет синхронизировать находки с отложением вулканических пеплов, обнаруженных под галечниками первой надпойменной террасы, в уступе второй — рисской террасы восточного склона горы Машук.
Возможно, что именно последняя вспышка вулканической деятельности Казбека и Эльбруса в верхнем плейстоцене и содействовала в ряде случаев как гибели зверей, так и захоронению их остатков в толще пеплов, смываемых селевыми потоками. Из находок в долине Кумы ниже Буденновска упомянем о нижнекоренном зубе мамонта (коллекция Пятигорского музея). К находкам верхнеплейстоценового возраста относятся фрагменты нижних челюстей кабанов — Sus scrofa ferns, обнаруженные геологом Л. М. Пламеневским в аллювии первой — вюрмской — террасы правого берега Терека, выше Орджоникидзе.
Вообще следует отметить, что принципиальной разницы в характере формирования ландшафтов и фаунистических комплексов в восточном Предкавказье по сравнению с западным не было. Здесь прослежены остатки тех же крупных зверей нижнего и среднего плейстоцена: трогон-териевых слонов, гигантских оленей, первобытных зубров, а в верхнем плейстоцене — и кабанов.
Лесная зона в верхнем плейстоцене была, по-видимому, отчетливо выражена по нижнему поясу северного склона Кавказского хребта даже в Дагестане. Из туфов предгорий близ Махачкалы Палибин (1913) определил в качестве «посттретичных» видов остатки рябины — Sorbus aria и граба — Carpinus betulus, растущих здесь местами и в наши дни.
Обширная область предгорного и внутреннего Дагестана, а также морские террасы западного побережья Каспия, от Сулака до Апшеронского полуострова, охарактеризованы остатками плейстоценовых млекопитающих чрезвычайно бедно. В музее Махачкалы хранятся обломки зубов Elephas trogontherii ранней формы, с толщиной эмали 2.5—3.0 мм, происходящие из ожелезненных галечников р. Манас. Там же имеется и обломок зуба более позднего слона с признаками Elephas primigenius, и атлант слона, найденный близ сел. Унты в горах.
Поиски палеолита и раскопки пещер и навесов в апшеронском известняке над Махачкалой, а местами и во внутреннем Дагестане могли бы значительно улучшить наши представления об особенностях формирования фауны в среднем и позднем плейстоцене этой части страны.
Западное Закавказье
Остатки плейстоценовых млекопитающих обнаружены в западном Закавказье по преимуществу в пещерных захоронениях в виде «кухонных отбросов» палеолитического человека и остатков пищи хищных животных.
Другие типы захоронений большей частью выпали здесь совершенно из-за отсутствия осадконакопления и быстроты потоков, перетиравших в валунах и галечнике даже крупные кости, из-за большой влажности климата, обусловливавшей быстрое разрушение костного вещества на
104
Часть I. Обоснование истории формирования фауны
поверхности почвы, и быстрого опускания морского дна у побережья, не допускавшего образование дельт.
Небольшим исключением были захоронения в делювии террас и в мелких тростниково-нимфейных и сфагновых болотах и озерцах, сохранившихся и ныне в районе Сочи, Пицунды, Очамчири. Некоторые из них. оказавшиеся в зоне предгорного поднятия, позднее были вскрыты речной эрозией. Крупные по объему и площади ольховые и сфагновые болота » и трясины в низовьях Риона и Супсы (Поти, Кобулети) лежат в области f колхидской депрессии и интенсивно погружаются при загрузке колхидского долинного прогиба наносами рек. Поэтому даже весьма обильные захоронения костей плейстоценовых млекопитающих, которые, вероятно, образовывались здесь, не будут доступны непосредственному наблюдению в ближайшие столетия без искусственных грандиозных выемок грунта. I
В литературе имеется упоминание о зубе Elephas primigenius Blum., | I хранящемся в Сочинском музее (Громов, 1948), но он мог быть и завезенным. | F Рябинин (1937) описал обломок правой ветви нижней челюсти крупной f кошки, близкой к тигрольву — Felis cf. spelaea, найденный в 1935 г. в 6 км. • - к северу от Сочи, в толще галечников третьей морской, карангатской ;
i террасы, содержащей створки Cardium. Tapes, Mactra и датируемой
? в то время рисс-вюрмом, а ныне, по Варданянцу (1948), — началом I рисса. I
х Черноморское побережье Кавказа в плейстоцене было, по-видимому, I весьма удобно для поселений палеолитических охотников. Обилие узких
L долин и ущелий, береговая полоска приморских террас, пригодная для кочевок тяжеловесных копытных как с севера, так и с юга, наконец, обилие пещер, заселенных пещерными медведями, содействовали успеху загон ных охот на крупного зверя.
Подъем отрогов Большого хребта, происходивший в плейстоцене, обеспечивал и сохранение от размыва и вымывания речками отложений на днищах пещер. Следы палеолита окаймляют предгорья от Сочи и Туапсе на севере до Кутаиси и Батуми на востоке и юге. Они продолжаются, вероятно, и на северное побережье Малой Азии.
Еще Замятнин (1937а) описал 43 точки палеолитических местонахождений в Абхазии и дал карту этих находок. Он подразделял их на три хронологические группы: 1) домустьерские, 2) мустьерские, 3) верхнепалеолитические. Большая часть этих местонахождений представлена, впрочем, единичными кремневыми орудиями.
Ефименко и Береговая (1941) перечисляли для западной Грузии 15 стоянок открытого типа и 14 пещерных.
Остатки млекопитающих известны лишь из пещерных стоянок, расположенных в ущельях рек Хосты, Кудепсты, Мзымты, Цебельды, Риона, Квирилы и других, более мелких.
Пещеры Черноморского побережья ।
В 1936—1938 гг. Замятнин (1940) производил раскопки Навалишинской и Ахштырской пещер в Сочинском и Адлерском районах Краснодарского ' края. |
Н а в а лишин с к а я пещера находится близ сел. Навалишиио, | в 12 км от моря на правом берегу Кудепсты. При раскопках были выделены [ два культурных слоя — верхний (верхний палеолит), содержащий кости । хомяка и во вторичном залегании — пещерного медведя, и нижний (мустьег [ ориньяк) — с остатками лося, козла, хомяка, пещерного медведя, волка, t
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 105
Тут же встречались кости птиц, раковины Anodonta и Helix. По мнению Замятина, эта пещера была лишь сезонным пристанищем древнего человека,
Громов (1948) дал список видов зверей по слоям этой пещеры, включенный в табл. 8.
Таблица 8
Видовой состав и численность костных остатков млекопитающих в Навалишинской пещере
Виды Средний палеолит Верхний палеолит
Canis lupus 1 —
Ursus spelaeus ...... ... 165 229
Cricetus cricetus — 1
Alces alces — 1
Capra sp 1 1
Всего 167 232
Громов (1948) упоминал еще о раскопках в Широкопокосской пещере (именуемой им Навалишинской II), расположенной поблизости от Навалишинской I и содержащей орудия верхнего палеолита. Среди костей, извлеченных из этой пещеры, были определены по преимуществу остатки пещерного медведя и единичные кости благородного оленя.
Из Воронцовской пещеры в ущелье Хосты, раскапывавшейся в 1946 г., добыты остатки пещерного и бурого медведей (коллекция Краснодарского музея). Сохранность костей пещерного медведя неоднородна. В свежем разломе костное вещество нижней челюсти буроватое и фоссилизованное. Лучевая и бедренная кости темно-бурой окраски, слабо фоссилизованы, при поскабливании дают запах свежей кости. Создается впечатление, что пещерный медведь существовал здесь и в послеледниковье.
Ахштырская пещера. Пещера находится на правом берегу р. Мзымты, у выхода ее из скалистого каньона, против сел. Ахпттырь, в 15 км от Адлера (рис. 46). Она заселена теперь лишь летучими мышами, преимущественно подковоносами. Устье пещеры обращено к югу и открывается в обрыв высотой около 100 м над руслом речки. В эпоху жизни палеолитических охотников обрыв был, вероятно, значительно ниже. Высота жерла пещеры достигает 4.35 м, ширина ~ 2 м, а длина — более 100 м. Перед жерлом располагается свободный навес с двумя входами и площадками, из которых восточная и была раскопана Замятниным (рис. 47). К пещере подходит с запада только узкая тропка над обрывом, которую было удобно защищать.
Такое положение жилой площадки приводило к тому, что большая часть костных остатков и орудий была выброшена древними обитателями с обрыва и унесена речкой. Замятнин полагал, что пещера заселялась человеком периодически, много тысячелетий подряд. При отсутствии людей в ней жили медведи и летучие мыши. Западнее и ниже в обрыве имеются еще небольшие щели-пещеры, также заселявшиеся медведями.
По мнению Громова (1948), эпоха образования большой пещеры совпадала с возникновением 60—80-метровой террасы Черноморского побережья. испытавшей подъем до 112 м у крыла Ахштырской антиклинали.
106
Часть /. Обоснование истории формирования фауны
Прослеживаются две фазы в геологической истории пещеры. Первая из них заключалась в появлении двух перпендикулярных трещин: одной сбросовой и другой — в плоскости наслоения известняка, в результате чего и при процессе выветривания стала образовываться сталактитовая пещера, на дне которой накапливались щебень и суглинок. Вторая фаза состояла в дальнейшем росте пещеры за счет разрушения потолка и дальнейшего накопления слоев на ее дне в виде щебня и обломков сталактитов.
Рис. 46. Ущелье Мзымты ниже Ахштырской пещеры.
Фот. автора, 1954.
Громов высказал предположение, что накопление щебнистых слоев происходило в условиях общего «ухудшения», т. е. похолодания климата, а разрушение щебня и образование из него суглинков происходило при потеплении и увлажнении. Привлечение для этих же выводов фаунистических данных оказывается мало состоятельным (об этом см. дальше).
В опубликованном разрезе Замятниным (1940) выделено пять культурных слоев — от среднего палеолита (нижнее и верхнее мустье) до наших дней. Слои нижнего мустье Ахштырской Замятнин синхронизировал со слоями Ильской стоянки в Предкавказье и со слоями Киик-Коба в Крыму.
При раскопках было собрано более 3000 определимых обломков костей преимущественно в слоях среднего палеолита. Они принадлежали летучим мышам, хищным, грызунам и копытным, а также птицам.
Остатки крупных зверей — обломки эпифизов и диафизов трубчатых костей — являются типичными кухонными отбросами. Кости летучих мышей и экскременты накапливаются в пещере и в паши дни. Кости хомяка, вероятно, были занесены филином.
Большинство остатков пещерного медведя представлено метаподиями фалангами, изолированными зубами, обломками крупных трубчатых
Гл. 11. Развитие ландгиафпгов и фаун в четвертичном периоде 107
костей. При пробных раскопах на закрытом участке пещеры Замятнину попадались и целые трубчатые кости — бедренные, берцовые, плечевые и кости предплечья, а также обломки черепов медведей, принадлежавшие, вероятно, особям, погибшим естественной смертью.
По типу сохранности костного вещества обломки из слоев мустье и верхнего палеолита на глаз почти неотличимы. Наоборот, в одном и том же слое попадаются остатки различного типа сохранности. Преимуществен-
Рис. 47. План устьевой части Ахштырской пещеры.
пая окраска костей розовато-желтоватая, с разводами светло-коричневого оттенка. Эмаль зубов либо натурального цвета, либо потемневшая, с резко заметными черными трещинками. Корни зубов окрашены в лимонный или в светло-кирпичный и коричневый цвета. Для слоев мустье преобладает более темная окраска костей. Местами по поверхности разбросаны черные пятнышки дендритов, иногда сливающихся и придающих темно-бурую окраску всей кости. В разломе кости преимущественно светло-кремовой окраски. Они относительно легки и слабо «звенят». Вторичная минерализация большей частью отсутствует.
Видовой состав и численность остатков млекопитающих (по нашим исследованиям) показаны в табл. 9 и на рис. 48.1
1 Подсчет числа остатков и особей произведен по коллекциям, хранящимся в ЗИНе. Результаты несколько отличаются от цифр Громовой. Остатки их слоев нижнего и верхнего мустье объединены, так как разницы в составе, численности и типе сохранности костей не намечается.
108
Часть I. Обоснование истории формирования фауны
В мустьерских слоях найдены еще шейный позвонок какого-то орла, а в верхпепалеолитических — кости ястреба — A slur gentilis L., сороки — Pica pica L. и мелких воробьиных — Passeriformes.
Численность костей в верхнепалеолитических слоях резко падает по сравнению с мустьерскими. Наименьшее возможное число особей забитых пещерных медведей и выявленное по первым фалангам — 39.
Рис. 48. Остатки млекопитающих из мустьерских слоев Ахштьгрской пещеры.
1 — пястная Canis lupus; 2 — голень Vulpes vulpes; 3, 4 — клык я астрагал Ursus spelaeus; 5 — бедренная Mai les sp.; 6 — голень Criceius cricetus; 1 — пястная Sus scrofa; 8 — 2-я фаланга Cervus elaphus; 9 — 1-я фаланга Capreolus capreolus 10 — запястная A Ices a Ices.
Это был, очевидно, излюбленный и легко добываемый вид, селившийся по ущельям. От бурого медведя удалось обнаружить лишь коренной зуб в слоях верхнего палеолита.
Следующие места по возможному числу особей занимают: козел, олень и кабан.
Остатки горных и скальных копытных все же сравнительно редки: имеется лишь козел, а серпы, по-видимому, нет вовсе. Некоторое увели-
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
109
Таблица 9
Впювой состав и численность костных остатков млекопитающих в Ахштырской пещере
1 Виды Средний палеолит Верхний палеолит Всего особей
число костей число особей число костей число особей
rmolophus ferrum-equinum .... — — 1 1 I
Jfipes vulpes ........... 9 1 3 1. 2
Cenh' lupus 9 1 — — 1
arctos ............ — — 1 1 1
fwlaeus ............ 2946 32 543 7 39
1 tries cf. foina 1 1 1 1 2
Ms cf. silvesiris I 1 2 1 2
f fains cricetus .......... 1 1 — — 1
V 1 7 3 4
•1. де alces — — 3 1 1
i'caus elaphus 18 3 4 1 4
tftpiceros euryceros 2 1 — — 1
fapreolus capreolus — — 2 I 1
В <on priscus ........... 9 2 — — 2
fiprn caucus tea 6 2 9 4 6
Всего 3004 576 68
viine числа остатков кабана в слоях верхнего палеолита может являться in* только показателем роста численности этого вида в окрестностях пещеры, но и постепенного вымирания пещерных медведей, в связи с чем потникам пришлось переключиться на другие объекты охоты.1
Отсутствие баранов в верхнем плейстоцене Черноморского побережья вполне увязывается с представлениями о древних лесных, мезофитных шдшафтах этого района, полученными на основе общего облика состава фуны» Ахштырской.
Вызывает удивление, что кости зубра и гигантского оленя обнаружены только в мустьерских слоях, хотя первый просуществовал здесь до наших дней. Остатки лося и косули появляются лишь в верхнепалеолитических слоях, но эти виды существовали па Кавказе по крайней мере с нижнего плейстоцена до современности. Таким образом, соображения Громова (1948, стр. 262) о климатических изменениях по наличию или отсутствию на Кавказе этих видов не могут быть приняты. Вообще численность остатков копытных и хищных, кроме пещерного медведя, слишком мала для обоснования заключений о фаунистических и климатических сдвигах.
Для правильной палеогеографической оценки состава видов зверей из раскопа Ахштырской пещеры необходимо представлять действительные условия обитания зверя и условия охоты первобытных племен.
Черноморское побережье с его сильно пересеченным рельефом и большой облесенностыо было, несомненно, мало пригодно для обитания мамонтов, лошадей, гигантских оленей, сайгаков, первобытных туров,
1 Остатки баранов — нижний эпифиз плюсны и вторая фаланга, — особо отмеченные Громовой (1948) как принадлежащие мелкому дикому виду «Ovis cf. ophion), u Hi ближайшем рассмотрении оказались современного, голоценового типа сохранности, и пх принадлежность к слоям мустье ошибочна. Очевидно, это остатки домашней овны, свалившейся в раскоп из послепалеолитических горизонтов.
110
Часть I. Обоснование истории формирования фауны
степных зубров. Охота производилась, конечно, поблизости от жилья па те виды, которые были под рукой.
Между тем топографические условия Черноморского побережья обусловливали па протяжении плейстоцена значительную стабильность вертикальных ландшафтных зон, несмотря на вероятные изменения климата. Этим и объясняется однообразие состава зверей в добыче древних ахштырцев на протяжении среднего и верхнего палеолита.
Несмотря на слабую универсальность захоронений остатков зверей в Навалишинской и Ахштырской пещерах, выявленный частично фаунистический комплекс района отчетливо показывает, что в верхнем плейстоцене на западе Кавказского хребта уже существовал, как и в наши дни, резкий зоогеографический рубеж для степных и лесостепных видов Русской равнины. Этот барьер преодолевался лишь эвритопным обыкновенным хомяком. Настоящие северные виды пе прослеживаются здесь, как и в предгорьях северо-западного Кавказа. Отсутствовало и южное — переднеазиатское — влияние: нет фаунистических элементов сухих нагорных степей Малой Азии.
Южнее по Черноморскому побережью остатки плейстоценовых млекопитающих обнаруживались будто бы в пещерах под Сухуми (Крон-гельм, 1873), а недавно стали известны из пещеры Абласкира, и из гротов по ущелью Цебельды.
Пещера Абласкира находится на западном склоне одного из южных отрогов Панавского (Кодорского) хребта, в 2 км к северо-востоку от сел. Отак и в 35 км от Очамчири. Из устья пещеры па высоте 265 м над ур. м. вытекает речка Ачхитизго. В выемках стен пещеры, недалеко от входа, зарегистрировано наличие палеолита. Описание Аб-ласкиры было дано Сатуниным (1910) и Маруашвпли (19386).
В 1946 г. А. А. Садовский собрал в пещере 75 обломков костей пещерных медведей, принадлежавших двум взрослым и одному молодому экземпляру. Кости были обнаружены в 50 м от входа, под водой, в толще тонкой глины и гравия, и потом собирались на протяжении 350 м по дну ручья. Были найдены целые берцовые и плечевые кости, метаподии, фаланги, а также почти целый осевой череп очень короткомордого взрослого экземпляра. Звери гибли здесь от старости и от других причин, без участия человека, очевидно так же, как бывало временами и на глубинных участках Ахштырской пещеры. В разломе кости беловаты, напоминают мел и легко крошатся при нажиме. Вторичной минерализации нет, дендриты па поверхности отсутствуют. О геологических условиях захоронения, времени образования глинистых отложений дна пещеры и обитания пещеры человеком данных пе имеется.
В ущелье Цебельды шурфовка гротов производилась археологом Л. Н. Соловьевым близ сел. Цебельдипского в 1941 г. Палеолитические орудия и костные остатки были собраны в Холодном Гроте и Кей-Богазе.
В переданной ЗИНу остеологической коллекции в 234 номера оказались фрагменты зубов, трубчатых и плоских костей животных, добытых палеолитическими охотниками и частично филинами. Состав видов показан в табл. 10 и на рис. 49.
Судя по типу сохранности костного вещества, большинство остатков . может быть отнесено к самому верхнему плейстоцену, а небольшая часть, особенно кости грызунов, — даже к голоцену. По составу видов с этих, наиболее южных пунктов вновь прослеживается лишь местный горпо- ] лесной облик фаунистического комплекса. Отмечен и южный предел | проникновения обыкновенного хомяка.
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
111
Таблица 10
Видовой состав и численность остатков млекопитающих на палеолитических стоянках Холодный Грот и Кей-Богаз
Виды Число костей Виды Число костей
чип spelaeus 3 Sus scrofa .......... 75
, (irctos 1 Cervus el a ph us 60
1 dries sp 6 Capra caucasica 58
'dis silvestris 1 Bison pr i sc us 8
iri'icola Urn sir is 2 —
hcelus cricetus ....... 20 Всего 234
Пещеры Имеретии
Северо-восточная окраина Колхидской низменности образована невы-пким плоскогорьем, сложенным толщей меловых известняков, па кото-налегают пески и илы олигоцена.
Рис. 49. Остатки млекопитающих из грота Кей-Богаз.
1 — плечевая Martes sp.; 2 — плечевая Felis silvestris; 3 — плечевая Cricetus cricetus; 4— бедренная Aimcola ten eshis; 5 — астрагал Sus scrofa; 6 — 2-я фаланга Cetvus elaphus; 7 — 1-я фаланга Rupicapra rupicapi a; 8 — эпифиз пясти Capra caucasica.
Плоскогорье расчленено притоками Риона, прорезавшими каньоны I толщах известняков па глубину до 50—60 м. В результате карстовых явлений в стенках известняков па протяжении тысячелетий образовалось я открылось много пещер, заселявшихся первобытными охотниками, пещерными медведями и филинами (датируются верхним палеолитом).
Часть I. Обоснование истории формирования фауны
Открытие палеолита в западном Закавказье относится еще к первой
половине прошлого столетия, причем первые находки каменных орудий были сделаны, по-видимому, в «гроте Язона» (пещера Сакажиа), около Кутаиси, швейцарцем Э. Фавром в 1863 г. (Замятнин, 1957).
В одной из пещер близ сел. Ргани в Чпатурском районе инженер Бернацкий (1884) собрал обожженные и расколотые кости зверей, которые
и направил в Берлин Рудольфу Вирхову, а
Рис. 50. Вход в пещеру Сакажиа.
Фот. автора, 1954.
частично оставил в Тбилиси, где их изучал Анучин (1887). В сборах Бернацкого преобладали будто бы кости пещерного медведя.
Все изученные до сих пор пещерные стоянки Имеретии относятся Замятнипымк верхнему палеолиту. Наиболее । ранние из них открыты в Чи-атурском районе С. А. Кру-ковскпм: пещера Хергулис-Клде у сел. Вачеви и навес Таро-Клде у сел. Шукрутп. В «3-м слое» Хергулис-Клде среди кремневых орудий му-стьерского и верхнепалеолп-тнческого облика оказались единичные расколотые кости I и зубы пещерного медведя — Ursus spelaeus, лошади — Equus caballus, козла — Cap-, га cf. caucasica и зубра—I Bison cf. priscus.
Из пробной траншеи раскопок Круковского в Таро-Клде вместе с верхнепалеоли-тнческими резцами и единичными мустьерскими остроконечниками были извлечены зубы пещерного медведя, I кости малоазийского хомяка — Mesocricetus aff. awra/wd зубы и расколотые трубчатые кости кавказского козла * и зубра.1
Средняя группа палеолитических стоянок включает, по Замятнину. пещеры: Сакажиа, Уварова, Бнеле-Клде, Девнс-Хврели. Сюда же относится и многослойная пещера Сагварджиле на речке Дзеврула, о которой появились лишь предварительные публикации (Киладзе, 1953; Гзели-
швили, 1953).
Пещера Сакажиа (Вирхова), пожалуй, наиболее примечательная находится на левом склоне ущелья речки Цхал-Цитела, правого приток Квприлы, близ монастыря Моцаметн и в 3 км от Кутаиси. Высота ее
1 Определения костей из этих местонахождений сделаны мною по зарисовкам С. Н. Замятнина.
над ;
50 м.
разл< Вход баюп
д
щеры
ника п вать 31 горах.
Фил то лит
Сак; раскош павшим шое чис пещеры
Огр< померо! датпров 1953; 3. печнике
. Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 113
над уровнем моря около 300 м и над современным уровнем ручья около 50 м. Образовалась пещера почти у поверхности древнего расчлененного разломами мелового плато, в выступающей теперь глыбе известняка. Вход высотой около 5 м в виде расширенной вверху арки ведет в заги-"пщйся влево рукав (рис. 50 и 51).
Днище речки с ее каскадами, размытыми скалами и окрестности пе-iPpv даже при теперешних стравленных зарослях самшита и грабин-
ас-ро-ли-ди-
Рис. 51. План пещеры Сакажпа.
щя, )МЯ-itus, бча-
ину.
>тно-орой шли-
зная, ггока га ее
*овкам
;шка поразительно живописны и уютны. Обитатели пещеры могли добыть зверя как на заболоченной равнине Риона, так и в прилежащих прах
(рилины в пещерах, по-видимому, не жили, а медведи если и жили, - шиь короткие сроки.
Сакажиа была заполнена 2-метровой толщей отложений и целиком ^скопана в два приема: в 1914 г. Р. Шмидтом и А. Козловским, проковавшими прямой коридор, а в 1936 г. — Г. К. Ниорадзе. В 1954 г. небольшое число костей и кремневых орудий было собрано мною в дальнем конце пещеры, где осталось еще немного отложений.
Огромная коллекция кремневых орудий и сколов в несколько тысяч номеров с обилием резцов и ножевидных пластинок (рис. 52) позволяет тировать эпоху обитания пещеры солютрейской стадией (Ниорадзе, [953; Замятнин, 1957). Собрано также много поделок — шильев и наконечников дротпков пз расщепленных диафизов метаподий лошадей и зуб-
И. К. Верещагин
114
Часть I. Обоснование истории формирования фауны
ров. При значительной мощности культурного слоя такая однообразная датировка его мало понятна, так как, несомненно, что пещера была чрезвычайно привлекательна и в послесолютрейскую эпоху. В костных сбо
j 1
1 i с р м Л Р
ш
Де
ных не ме круш
Рпс. 52. Сакажиа. Кремневые орудия (по Замятпипу, и костяные изделия (по Ниорадзс, 1953).
1 К очевидно 2 н
казанк предстал коллек
диафиз казан ;
ным ун
рах встречаются остатки домашних животных, происходящие, из верхнего слоя.
Кухонные костные отбросы верхнепалеолптпческой эпохи лены зубами, фалангами, обломками челюстей, эпифизов и трубчатых костей. В коллекции Шмидта их насчитывается 472 номера,
Гл. 11. Развитие ландшафтов и фаун в четвертичном периоде
116
в моих сборах — 130, а у Ниорадзе — свыше 16 000.1 Большинство их, впрочем, неопределимо, так как кости дробились на мелкие осколки. Единичные кости — пятка и локтевая тигрольва оказались обгрызены крупными хищниками. Тип сохранности костного вещества свидетель-’ сгвует о позднем плейстоцене. Обломки большей частью серой окраски, реже палевые, почти без дендритов, фоссилизованпые, но без вторичной минерализации. Эмаль зубов прижизненной окраски, реже потемневшая. Явно более молодыми — голоценового возраста — выглядят клыки бурого медведя.
Беляева (1940в) и Громов (1948), изучившие в сборах Ниорадзе небольшое число зубов и фаланг, опубликовали список в 18 видов зверей. Результаты наших исследований по всем сборам обоих коллекций приведены в табл. 11.
Таблица 11
Видовой состав и численность костных остатков млекопитающих в пещере Сакажиа
Виды Число костей Число особей
Vulpes vulpes .......... 5 9
Canis lupus 13 3
Ursus spelaeus ......... 35 5
U. arctos и 3
Panther a spelaea 12 3
Felis cf. lynx 1 1
Martes cf. foina 3 1
Meles meles .......... 2 1
Chionomys roberli 1 1
Castor fiber2 1 1
Hystrix cf. hirsutiroslris .... 1 1
Equus caballus 102 8
E. hid runt inus2 1 1
Sus scrofa 75 7
Cervus elaphus 140 10
Capreolus capreolus 6 2
Alces alces 5 1
Bison priscus 1488 32
Capra caucasica 3 166 25
Rupicapra rupicapra 8 2
Всего 2076 НО
Кроме фрагментов, отмеченных в таблице, к остаткам копытных и хищны : среднего размера — козлов, кабанов, серп, волков — относится пе менее 8800 осколков диафизов трубчатых костей, а к остаткам зверей к упного размера — зубров, оленей, лошадей, пещерных медведей и тп-
видно,
едстав-афизов юмера,
i Коллекция Шмидта хранится в Одессе, а коллекция Ниорадзе — в Тбилиси.
2 Наличие и численность остатков речного бобра и плейстоценового осла показаны по данным В. II. Громова и Е. И. Беляевой, так как в сохранившихся коллекциях их не обнаружено.
з В этой и следующих таблицах имеретинских местонахождений вид козла показан западпокавказской, а пе восточной формой, в противоположность ошибочном указаниям Громовой (1948) и Громова (1948).
Рис. 53. Остатки млекопитающих из пещеры Сакажиа.
1, 2 — клыки Vulpes vulpes; 3, I — клыки Meles meles; 5, 6 — челюсть Maries sp.; 7 — Pm-j l-'ells lynx; 8 — ь'лык Pells silreshis; 9 — l’ni4 и JO — клык Panthera spelaea; Il — Mt Cants lupus; 12. 13— пястная и плюсневая Ursus spelaeus; 11 — челюсть (X 2) и /5 — 1-й коренной (ХЮ) Microtus cf. roberti; 16, 17 — Pin3 и его стиравшаяся поверхность Equus caballus; 18 — М3 Cervas elaphus; 19 — М3 Rupicapia rupicapra; 20 — M3 Capra сайга-sua: 21 — M3 Rism priscus;
29
30
Рис.
scro/a; 23
Sus
•• - 1-п раланга
2Ь — зи — стер-га и астрагал
53 ( п родолжепие )
ж _______ , 24 — 1-я фаланга и астрагал Cervus ektphus;
рога'"астрагал, эпифизы пясти и плюсны Bison priscus; 26, 27 — 1-л Ф Capra caucasica.
118
Часть /. Обоснование истории формирования фауны
грольвов — не менее 6000. В этой пещере обнаружено и несколько обломков костей человека.
Итак, на первом месте в добыче сакажийцев был зубр, затем шли козел, олень, лошадь кабан, пещерный медведь. Столь значительное разнообразие видового состава объясняется расположением пещеры на рубеже горного и низменного ландшафтов. Сакажийцы охотились в болотистой Рионской долине на кабанов, лосей и бобров, на Имеретинском плато добывали зубров, оленей и лошадей, а козлов и серн — в ближайших скалах по ущелью Риона.
В гро тах Уварова и Бараташвили в том же ущелье Шмидт собрал при закладке шурфов до 30 зубов кавказского козла, 12 зубов _ зубра, 12 обломков костей пещерного медведя, 2 зуба оленя и 1 зуб лошади, 1 пяточную кость волка, 1 обломок таза бобра вместе с серией верхнепалеолитических орудий. Сохранность этих костных материалов вполне сходна с сакажийскощ
Из пещеры Бнеле-Клде имеется небольшое количество обломков костей, которые определялись Смирновым (1923—1924) вместе с костями из поздней стоянки Гварджилас-Клде.
Пещера Девис-Хврели находится в 4 км от станции Харагоули, на правом склоне ущелья речки Чхеримелы, в кристаллических известняках сеномана. Вход в пещеру расположен на высоте 340 м над ур. м. и на высоте 80 м над руслом Чхеримелы. Он обращен на юго-запад. Раскопки велись Г. К. Ниорадзе в 1926 г. Длина исследо-- ванной части пещеры достигает 25 м при ширине 4.5 м у входа и высоте 6.5 м. Ниорадзе (1933) раскопал небольшую часть пещеры, вскрыв 4 слоя: 1) верхний, с современными остатками животных; 2) бурый, содержащий также остатки животных; 3) культурный слой и 4) глинистый слой. По
Таблица 12
Видовой состав и численность костных остатков млекопитающих в пещере Девис-Хврели
ЧИСЛО : костей Р
Виды
Число костей
Виды
Canis lupus . ? Capreolus capreolus 2
Ursus spelaeus ? Alces alces 1
U. aff. arctos ........ 20 Bos или Bison ....... ?
Mesocricetus cf. auratus . . . ? Capra cf. caucasica . ... . 22
Sus scrofa .......... 30 Rupicapra rupicapra 3
Cervus elaphus 1
кремневому инвентарю Ниорадзе относит эту стоянку к верхнему ориньяку, а Замятнин — к позднему отделу мадленской культуры. В остеологических сборах из пещеры преобладают сильно раздробленные кухонные остатки с небольшой примесью костей из погадок филина. Список видов, по определениям Громовой (1929) и М. В. Павловой, дан в таблице 12, Число костей в сборах, оказавшихся у Павловой, осталось неизвестные
В целом состав костных остатков из этой пещеры чрезвычайно напоминает таковой, выявленный для пещеры Сакажиа. Меньшее видовог разнообразие объясняется лишь меньшими месштабами раскопок.
Некоторое недоумение вызывает, впрочем, отсутствие остатков лошади, так как стоянка эта находится как раз в центре сухого плоскогорья. Сл£
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 119
дующие местонахождения средней и поздней групп располагаются уже ближе к отрогам Рачинского хребта.
В 1934 г. Замятнин (19376) обнаружил верхнепалеолитические орудия под известняковыми навесами близ сел. Мгвимеви Чиатурского района, на правом берегу р. Квирилы. В 1940 г. Киладзе (1944) продолжила раскопки под одним из навесов.
Кости из-под навесов Мгвимеви весьма фрагментарны. Они фосси ли-зованы, но вторичной минерализации нет, кроме поверхностных натеков кальцита.
Костные остатки из сборов Замятнина и Киладзе определялись Громовой (1948), а позднее мною. Был выявлен список видов, показанный в табл. 13.
Таблица 13
Видовой состав и численность костных остатков млекопитающих под навесами Мгвимеви1
Виды
i
।
; Число костей
I
।
Cants lupus .........
Ursus spelaeus ........
Panthera spelaea.........
Mesocricetus cf. auratus ....
Equus caballus...........
Bison priscus .........
Capra cf. caucasica......
Ovis cf. ammon ..........
Caprovinae ближе не определен.
Число особей
Всего..............
На первом месте по численности в добыче здесь был также, очевидно, первобытный зубр, а затем козел и баран. Последний представлен крупным видом аргали, что особенно знаменательно. Обильны в Мгвимеви остатки лошади. Наличие остатков пещерного медведя и тигрольва указывает все же на плейстоцен, хотя бы и самый поздний. Отсутствие остатков кабана, оленя, косули объясняется скорее всего малым объемом раскопок и материалов.
К наиболее поздним палеонтологическим памятникам относится пещера Гварджилас-Клде, раскопанная частично в 1915— 1916 гг. инженером Круковским (1916), а в 1953 г. — Каландадзе и Тушаб-рампшвили (1955).
Пещера эта находится в ущелье ручья Чирулы, правого притока Квприлы, близ сел. Ргани. Вход в нее -- горизонтально вытянутый овал — обращен на юг и расположен на выступающем мыске правой стенки каньона, чуть ниже половины ее высоты. Выше и на противоположном берегу имеется еще несколько гротов. Длина пещеры достигает 30м при ширине у порога 19.5 м и высоте около 4 м (рис. 54 и 55).
По описанию Круковского, в истории пещеры было три или четыре, фазы обвалов кальцитового потолка, под обломками которого оказались погребенными два древних культурных слоя.
1 цифры даны с учетом материалов, поступивших в остеологическую лабораторию ЗИНа дополнительно после публикации 1948 г.
120
Часть 1. Обоснование истории формирования фауны
Орудия в этой пещере представлены большим количеством резцов скребков и но/кевпдных пластинок из розового кремня. Имеется также значительное количество поделок из кости — наконечпиков, дротиков и шильев (рис, 56). По каменному инвентарю стоянку относят к самов верхней стадии палеолита — мадлену-азплю.
Рис. 54. Пещера Гварджилас-Кпдс.
Фот, С. II. Замятины;!, 1026.
С. А. Круковский собрал до 150 обломков костей и зубов хищных, копытных и грызунов, а Смирнов (1923—4924) определил среди них 14 видов.
В сборах Каландадзе и Тушабрамшвили насчитывается свыше 4b костных фрагментов, и мною было собрано там же еще 78.
Все кости крупных зверей — это типичные кухонные остатки палеолитических охотников: разбитые эпифизы, диафизы, черепа, изолировав ные зубы. Имеется и небольшое число костей насекомоядных, грызунов! и птиц, происходящих явно из погадок филинов. Сохранность костиог вещества весьма свежая. Кости фоссилизованы слабо, желтовато-кремовые с поверхности. Эмаль зубов нередко прижизненной окраски, реже!
| ' Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
121
е в й
с рыжеватом налетом и мелкими трещинками с синевой. Поскабливание дает ясный запах сырой кости, и по типу сохранности костное вещество не может быть древнее самого конца плейстоцена, а вернее, относится к началу голоцена. Многие кости погрызены лесными мышами.
Результаты определения костных остатков из всех сборов и слоев даны в табл. 14. Объединение сделано с учетом отсутствия разницы в видовом составе зверей в обоих слоях.
К трудно определимым осколкам диафизов костей крупных копытных —
зубров и оленей — может быть отнесено еще до 900 фрагментов из сборов
Тушабрамшвили, а к остаткам козлов, серн, косуль — не менее 550. Кроче того, неопределимых мелких фрагментов диафизов копытных и хищных (рис. 57) насчитывается около 2970 шт.
Большое количество костей из Гварджилас-Клде позволяет довольно уверенно утверждать о значительных преобразованиях в составе экологических группировок млекопитающих района, которые произошли здесь на рубеже плейстоцена и голоцена. На плоскогорье исчезли лошадь, баран, возможно также и дикобраз. Вообще из южных нагорно-степных форм уцелел только малоазийский
Рис. 55. План пещеры Гварджилас-Клде.
хомяк. Так как нет осно-
ваний думать, что резко изменились вкусы и охотничьи навыки первобыт-
ных охотников, то следует предположить, что исчезновение упомянутых видов было обусловлено увеличением облесенности плоскогорья.
Н. А. Смирнов, Громов (1948), Пидопличко (1951) рассматривали список зверей из Гварджилас-Клде как единую в стратиграфическом
и экологическом смысле группировку.
Названные авторы повторяли одни и те же соображения о смещении вертикальных ландшафтных зон книзу под воздействием оледенения Большого Кавказа, что было основано на том, что древние охотники вряд ли добывали козлов и серн далее, чем за суточный переход с тяжелой ношей, т. е. за 15—20 км, а теперешний обитатель альпийских лугов — прометеева полевка — будто бы не могла существовать в окрестностях пещеры в условиях того относительно ксерофитиого ландшафта, который имеется здесь в настоящее время.
Наши наблюдения на месте показали, что рассуждения упомянутых исследователей беспредметны. Распространение промыслового зверя, в частности кавказских козлов, в те времена было более обширным, нежели теперь. Уже ближайшие скалы в верховьях речки Чирулы, в 6 км от пещеры, даже сейчас вполне пригодны для обитания козлов. Что ка-
122
Часть I. Обоснование истории формирования фауны
£•
£-
Таблица 14
Видовой состав и численность костных остатков млекопитающих в пещере Гварджилас-Клде
Виды Число костей Число особей
Talpa aff. caucasica 1
Erinaceus aff. europaeus 2 2
Vulpes vulpes 1 1
Cants lupus 6 2
Ursus arctos nieridionalis .... 6 2
U. arctos caucasicus 5 1
U. spelaeus 6 2
Gulo gulo 1 1
Mustela nivalis caucasica .... J 1
Mesocricetus aff. auratus 17 8
Prometheomys aff. schaposchnicovi . 5 3
Sus scrofa 3 2
Capreolus capreolus 2 1
Cervus elaphus 34 5
Capra cf. caucasica ....... 254 13
Rupicapra rupicapra ...... 1 1
Bison bonasus (caucasicus?) . . . 235 8
Всего 580 54
2л?'
м
& « f
сается филина, то я наблюдал в апреле 1954 г. один экземпляр, регулярно I летавший по вечерам из дневочной каверны, расположенной ниже Гвард- Ц жилас-Клде, на ближайшие субальпийские поляны и в леса Рачинского В хребта. Из сухих окрестностей пещеры филин, очевидно, таскал хомяков и в плейстоцене, а из горно-лесной и субальпийской зон — прометеевых В полевок. Таким образом, для обсуждения состава фаунистических на- i ходок в Гварджилас-Клде не требуется признания ни смещения зон, ни мозаичности ландшафта, ни существования в одной ландшафтной || зоне двух различных экологических типов зверей: высокогорных и мезо- F фильных ныне кавказских козлов и прометеевых полевок и ксерофиль-ных малоазийских хомяков.
Находки остатков прометеевой полевки и малоазийского хомяка в среднем течении Квирилы могут свидетельствовать и о существовании в верхнем плейстоцене фаунистических связей западной части Большого Кавказа с Малым Кавказом, проходящих по Имеретинскому плато и западным склонам Сурамского хребта.
Итак, верхнеплейстоценовый фаунистический комплекс северо-восточной окраины Колхиды, известный по остаткам промысловых видов и жертв филинов из пяти пещерных стоянок, имел ряд общих черт и существенных отличий с комплексом Черноморского побережья. Сходство сказывалось в наличии широко распространенных горно-лесных видов хищных и копытных: волка, лисицы, медведя, лесного кота, куницы, кабана, косули, оленя, лося, зубра при полном отсутствии индикаторных видов евразийских равнин: мамонта и первобытного тура.
В противоположность Причерноморью здесь обитал ряд выходцев из , нагорных степей Передней Азии. Особенно показательны в этом отношении малоазийский хомяк, дикобраз, аргалиобразный баран, лошадь и плейстоценовый осел.
Ь
С
1
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
123
Вселение этих видов в западное Закавказье происходило с юга и было обусловлено преимущественно наличием карста, т. е. природной сухостью грунта и почв, и в меньшей степени влиянием сухой климатической фазы, имевшей место где-то в среднем плейстоцене.
Лесные звери — кабан, лось, косуля — в этом комплексе были довольно многочисленны и связаны главным образом с заболоченной Колхидской низменностью. Жил в ней п такой древний реликт, как речной бобр.
Рис. 56. Кремневые и костяные орудия из пещеры Гварджилас-Клде. (По Замятнину, 1957).
Высокогорные «альпийские» виды были распространены несравненно шире, чем в наши дни.
Район стыка, или «экологический барьер», для некоторых северных (европейских) видов зверей и выходцев из нагорных степей Передней Азии оформился в верхнем плейстоцене по ущельям Кодора и Цебельды.
Общий список видов зверей, известный из верхнеплейстоценового комплекса западного Закавказья по палеолитическим стоянкам и другим типам захоронений, насчитывает до 30 видов, распределяющихся по отрядам таким образом:
Insect ivora.................2
Chiroptera...................1
Carnivora...................13
Lagomorpha...................1
Rodentia ....................7
Perissodactyla...............3
Artiodactyla ...............13
В этом комплексе следует вновь подчеркнуть, кроме отсутствия мамонта и первобытного тура, еще и отсутствие северных форм: овцебыка, северного оленя, песца, зайца-беляка. В то же время здесь совершенно
Рис. 57. Остатки млекопитающих из пещеры Гварджилас-Клде.
1 — челюсть (Х2) Talpa caucasica; 2 — челюсть Guio all. yulo; 3 — бедренная (X 2) Mesoci icetus auratus; 4 — челюсть Prometheomys schaposchnikovi; 5 — челюсть Cervus elaphus; 6—8 — M3, астрагал и эпифиз пясти Capra caucasica; 9—10 — M2 и 1-я фал а ига Bi so a b onasus.
Гл. II. Развитие ландшафтов и фаун е четвертичном периоде
125
нет и теплолюбивых видов более южных широт. От западнопредкавказ-ского плейстоценового комплекса (предгорной равнины) этот комплекс отличается преимущественно присутствием лесных и вообще мезофильных, а также горных форм зверей. Это позволяет утверждать, что основные черты современной фауны и ландшафтов сложились в западном Закавказье уже к среднему плейстоцену.
Переход к голоценовой (послеледниковой) фауне в западном Закавказье совершается, по-видимому, более стремительно, нежели это имело место на протяжении среднего и верхнего плейстоцена. Фаунистические изменения в конце верхнего плейстоцена заключаются в вымирании пещерных медведей, тигрольвов, пещерных гиен; в отступлении к югу ксе-рофильных видов переднеазиатских плоскогорий: малоазийского хомяка, дикобраза,-аргали; в измельчании местных лесных и горно-альпийских видов: зубров, козлов, серн, косуль, при сохранении общего лесного облика фауны млекопитающих района вплоть до наших дней.
По-видимому, и климатические изменения с конца среднего плейстоцена проходили в западном Закавказье несколько иначе, чем в других районах, и были обусловлены узко местными орографическими и климатическими особенностями этой территории.
Дальнейшие спелеологические исследования в районе карста Имеретии, Абхазии и Краснодарского края, несомненно, уточнят существующие на данном этапе палеографические представления.
Должны быть сделаны здесь и специальные поиски пещер нижнего плейстоцена и верхнего плиоцена.
Центральное Закавказье
Район Сурамского водораздела, верховьев Риона, Большой Лиахвы п Горийской котловины представляет для палеонтолога и зоогеографа особый интерес как путь вероятной миграции на Кавказ неогеновых и четвертичных млекопитающих Передней Азии. Остатки плейстоценовых зверей известны из пещерных палеолитических стоянок и из делювия.
В1952 г. Любин (1953) обнаружил впервые ряд древнепалеолитических орудий в долине Большой Лиахвы и Метехской Проне на высотах около [500 м, близ Джиджойты, Морго и Лаше-Балта. Изделия, найденные V Лаше-Балта: ручные рубила, отщепы, нуклеусы, —изготовлены из андезита и окремнелых пород. Все они окатаны в потоках и переотложены после вымывания из какой-то высокой, по-видимому, 60-метровой террасы. Как и в Абхазии, костные остатки животных, добывавшихся первобытным человеком эпохи изготовления этих орудий и залегавшие в террасах, очевидно, не сохранились. Большую ценность приобретают поэтому палеонтологические находки в нижних слоях карстовых пещер.
Пещеры горы Часовали-Хох. Палеолитические и более поздние слои в пещерах урочища Кударо в Южной Осетии были открыты В. П. Любиным в 1955 г. Четыре наиболее примечательные пещеры расположены здесь на правом склоне ущелья речки Джоджоры, пад селениями Кваиси и Часовали, в выдающемся в рельефе склона доломитовом массиве на высоте около 300 м над уровнем речки и на 1700 м над ур. м. (рис. 58). Обильные археологические материалы доставила верхняя пещера Кударо I. Она имеет два входа, обращенные на юг и восток, наиболее сухая, с полого поднимающимся к северу дном (рис. 59 и 60). Раскопками 1955, 1956 и 1957 гг. были вскрыты глинистые слои мощностью до 2.5 м с глыбами известняка, содержащие вверху керамику
126
Часть I. Обоснование истории формирования фауны
и орудия эпохи железа, бронзы, меди, неолита, а глубже — орудия среднего и нижнего палеолита. Отщепы, ручные рубила, остроконечники могут быть отнесены частью к эпохе мустье, частью — к ашелю. Они изготовлены обычно из окремцелого песчаника. Все слои насыщены костными остатками, занесенными в пещеру преимущественно людьми и филинами. Костные остатки из нижнепал еолитических слоев сильно фоссилизованы, светло-каштановой окраски с обильными дендритами. Нередко они окатаны, как и галька, действием небольшого потока, про-
Рнс. 58. Гора
Часова Л! l-Х ох. Стрелками показано
расположение
пещер.
Фот. автора, 1957.
( (.
I С I А Л л 6 р р с ь л/ Р1 л. л/ м л/ Рг и Jih Sl Сс. Га Са Ov От Bit Ale.
текавшего через пещеру во влажные эпохи и сезоны. Края фрагментов заметно «звенят» при их ощупывании. Преобладают обломки разбитых трубчатых костей, зубы, фаланги и мета подии хищных и копытных, трудно определимые до вида.
Глинисто-известняковые слои в шурфе перед входом пещеры Ку-даро III на глубине трех метров содержали орудия верхнепалеолитп-ческого типа и обилие мало поврежденных костей хищных и копытных свежей сохранности и светлой окраски. Видовой состав костных остатков из палеолитических слоев трех пещер весьма разнообразен (табл. 15 и рис. 61). К 1958 г. выявлено до 40 видов зверей плейстоцена.
В палеолитических слоях Кударо 1 обнаружено также 42 кости птиц — Aves\ 3 кости ящериц — Lacerta sp.: 43 кости жаб — Bufo sp.; 22 кости лягушек — Rana sp.; и свыше 30 000 костей рыб — Pisces. преимущественно лососевых — SalmonidaeP Обилие костей рыб в пижпе-
1 В сообщении о сборах 1955 г. (Верещагин, 19576) единственная найденная тогд;, межчелюстная кость лосося была ошибочно указана как кость крупной ящерицу
па/ что Дж
хар вал бар CKIH гор] здес 0ТП( захс нахе
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
127
Таолйца 15
Видовой состав и численность костных остатков млекопитающих из палеолитических слоев пещер Кударо
Виды
| Кударо 1 нижний и средний палеолит
Кударо II
Кударо III верхний палеолит
falpa Sp..............................
Neomys sp.............................
Chiroptera, ближе не определен........
Масаса р..............................
Canis cf. lupus.......................
Сноп sp. -............................
Vulpes cf. vulpes .................. Grocuta speluca.......................
Lrsus spelaeus .................... Martes cf. foina......................
lluslela ci. nivalis..................
Meles cf. meles .................. Gulo cf. gulo.........................
Pant her a spelaea ................... p, pardus ...................... Carnivora, ближе не определен.........
Г pus cf. europaeus ................. Marmot a sp...........................
Hyslrix cf. leuciira ................. Allaclaga cf. williamsi..............
Mesocricetiis aff. auratus...........
Microtus aff. majori.................
Я. aff. gud..........................
Prometheamys aff. tschaposchnicovt...
\luridac, ближе не определен.........
Rhinoceros sp.........................
Sus scrofa...........................
Cervus cf. elap/1us...................
Capreolus aff. capreolus.............
Capra cf. caucasica..................
Ovis cf. ammo и ................... Ovis или Capra, ближе не определен...
Bison prisons .................... Мелкие фрагменты костей копытных и хищных—
Artiodactyla — Carnivora.............
1
1
27 4
27 3
33
1 2979
9
1 И 11
3
9 18 12
10
18
2 56 16
5 13 52
6
9
213 36 55
26 189
5
13013
20
11
11
135
палеолитических слоях обоих входов пещеры Кударо 1 наводит на мысль, что отложения образованы волнами какого-то водоема, возможно пра-Джоджоры, приносившими трупы и истиравшими кости в гальке и песке.
В целом состав животных из палеолитических слоев Кударо I характеризует ландшафт относительно сухих плоскогорий, где существовали широко распространенные виды: волк, лисица, пещерный медведь, зарсук и др.; виды горные, кавказские — барс и сурок; нереднеазиат-ские—средний хомяк, баран и дикобраз. Широкое проникновение на горный Кавказ переднеазиатских иагорио-степпых видов и переживание здесь конца плиоцена обезьянами представляет выдающийся интерес. Об относительно слабо расчлененном рельефе в теплом сухом климате в эпоху захоронения основной массы палеолитических остатков свидетельствуют находки костей дикобраза, барана и малое число костей козлов и серп.
128
Часть I. Обоснование истории формирования фауны
Сравнение составов животных этой стоянки и верхнепалеолитических стоя-
нок Имеретии наводит даже на мысль о
Рис. 59. План пещеры Кударо I. 1, 2— раскопы 1955—1958 гг.
существовании изолированных очагов обитания кавказских козлов и других зверей горного и скального рельефа на дифференцированно поднимавшихся в плейстоцене массивах кавказских хребтов. Эти поднятия в районе верховий Риона с эпохи нижнего палеолита достигали действительно крупных масштабов — порядка 500 и более метров.
Поздние этапы существования плейстоценовой фауны в центральном Закавказье прослежены пока лишь по единичным находкам зубов мамонтов. Один из таких зубов Elephas prirnigenius, происходящий из Душетско-го района, описан Бурчак-Абрамовичем (1946). Зуб мелкого мамонта был найден также в г. Гори, в отложениях первой надпойменной террасы Куры. Габуния (1952а) относит зуб к поздним мамонтам плейстоцена или даже к низам голоцена.
По времени эта находка, вероятно, совпадает с некоторыми остатками поздних мамонтов в Пятигорье.
Обитание мамонтов в верхнем плейстоцене Закавказья вряд ли следует расценивать как продвижение «северных» видов на юг. Скорее всего, что здесь происходила самобытная трансформация хоботных, как и в западном Средиземноморье.
В о с т о ч и о е Закавказье
В послетретпчный период восточное Закавказье было низменной территорией, зажатой с севера, запада и юга тремя обособленными горными системами, а с востока — морем.
Море по крайней мере трижды наступало на сушу в плейстоцене, захватывая восточную половину низменности, сильно сужая тем самым участки обитания равнинных видов зверей.
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
/29
Открытые к настоящему времени захоронения остатков четвертичных млекопитающих расположены здесь па морских террасах Апшеронского полуострова и местами на речных террасах предгории в западных частях пишешюсти.
• Кухонные и ппые костные остатки из плейстоценовых слоев пещер восточном Закавказье пока почти неизвестны.
Остатки плжнеплепсто-1 еловых зверей обнаружены в прибрежных от-. нкениях Бакинского мо-ри па Апшеронском полуострове. Близ станции Кишлы, к северу от Баку, г слое грубого раковиино-известняка со створками / idacna rudis Nal. и D. su~ r chanica Au dr. на глубине ; м были найдены фрагмен-пчерепа и зубов молодого г сорога — Rhinoceros cf. wcki, крупной лошади — / quus sp. и гиены— Сго-i sp. Череп гиены был •.шее отнесен к виду Ну па striata (Богачев, 1925а, ’ »о8б), что совершенно не-кроятио для нижнего ппстоцена. В ПИНе хранится обломок нижней че-юстй и зуб эласмотерия, яапдепные на глубине 12 м в нефтяном колодце № 19, лпз сел. Бинагады. Он происходит из древнекас-шшской террасы с Didacna mchanica Andr., который подстилает на ее южной
Рис. 60. Раскопки южного входа Кударо I. Фот. автора, 1956.
окраине кпровые пласты с остатками зверей бинага-дпиского комплекса (об лои см. дальше).
В пропитанных нефтью галечниках древнекаспийской террасы, на-егающих на нефтеносные пласты продуктивной толщи у Хурдалаиа иаАпшероне, была найдена нижняя челюсть жеребенка (Богачев, 1938в), которую Громова (1949) отнесла к Equus aff. sussenbornensis.
Находки остатков зверей в средне- и верхпепленстоцеповых слоях более часты, нежели в нижнеплейстоцеповых. К сожалению, точная
стратиграфическая датировка их встречает затруднения в связи с малой изученностью и с нередкой дислоцироваиностью каспийских террас.
Большинство находок сделано также па Апшероне.
Образованию захоронений способствовали здесь сильные изменения рельефа под воздействием тектонических, водно-эрозионных и эоловых факторов. Наступавшие и отступавшие воды древнекаспийских бассей-
9 Н К Верещагин
. . см
iiiii___J
/4
Pi c. 61. Остатки позвоночных из палеолитических слоев пещеры Кударо I.
1 — межчелюстная кость (Х2) лосося Salmo sp.; 2 -М3 (X 2) Масаса sp., 3—5 — молочный клык (X 2), М2 и плюсневая Ursus spelaeus; б — верхняя челюсть Panlheia paidus; 7 — челюсть Maimota sp.; 8 — челюсть (X 2)М esocricetus all. awatus; 9 — боновая и стиравшаяся поверхность зуба (X 2) Ilyshix sp.; 10 — копытная фаланга Rhinoceros sp.; 11— рог Capreolus cf. capreolus; 12 — 1-я фаланга Cervus cf. elaphus; 13 —15 — пясть, 1-я фаланга, астрагал Oris cf. ammon.
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
131
нов в одних случаях уничтожали, в других лишь сглаживали или, наоборот, подчеркивали следы воздействия наземных агентов.
После крупных трансгрессий Каспия рельеф Апшерона был осложнен еще местами извержениями грязевых вулканов, образовавших молодые, ’ иные на основной рельеф шапки, или конусы. Как правило, все ввестт.ыс захоронения послебакинского возраста приурочены к выхо-кш окисленной нефти, т. е. битума или асфальта.
Н и фти и жидком асфальте гибли многочисленные животные, а их
и в
статки
консервировались
совершенстве
сохранялись на
долгие ты-
етпя.
Кроме того, пласты битуминозных песков, обладая большой
власти1 костью, предохраняли от размыва и развевания образовавшиеся Блимп захоронения остатков зверей.
В б льшинстве .случаев образование захоронений остатков плейстоце явых животных было связано с пресной водой и одновременными исте-.•илям’ нефти или асфальта, т. е. носило комбинированный характер.
Наиболее известны Бинагадинское и Кир-Макинское захоронения, к лде.|ьные находки костей делались и в других местах.
Так, например, при рытье канавы в Сураханах была найдена челюсть Ярблю, а — Camelus sp., а из битумов на острове Артема добыты черепа Грога благородного оленя — Cervus elaphus (Богачев, 1924, 1932). Р рхпе елюстпые косги лошади «азиатского типа» — Equus cf. caballus .ща )ужепы в глинистой брекчии горы Бог-Бога в Балаханах, 15 км б ро-БОСточпее Баку. Остатки лошади известны также из древнекас-шшскои террасы на третичных всхолмлениях близ сел. Али-Байрамлы, .страгичного всхолмления Баба-Занан близ Сальяп — остатки быка — Б^р. (Богачев, 1938в).
Бинагады
Б разработке истории развития четвертичной фауны Кавказа Бинага-iniicKoe захоронение занимает особое место. Наличие в этом захоронении остатков растений, моллюсков, насекомых, рептилий, птиц и зверей всех «олого-морфологических типов позволяет воссоздать в деталях ландшафт зревпрго Апшерона и изучить важный этап в формировании фауны.
Захоронение это было открыто в 1938 г. студентом А. Мастан-Заде, унимавшимся генезисом апшеронских битумов. Первоначальные рас-гапкп были организованы В. В. Богачевым в 1938 г. (Азербайджанский риал АН СССР) и продолжались при участии С. Касабовой и Р. Сул-тенова вплоть до 1941 г., а затем были возобновлены в 1946 г. под ру-оводством Н. И. Бурчак-Абрамовича.
Планомерности раскопок сильно помешала кустарная добыча кира,1 1роцзводпвшаяся па протяжении многих столетий, в результате которой костсносиая площадь оказалась покрытой ямами и шурфами различии глубины и величины (рис. 62).
Для воссоздания цельной стратиграфической картины при надлежаще давлешшх раскопках необходимо будет прорезать всю площадь за-
Рронеяпя взаимно-перпендикулярными канавами.
Бппагадииское захоронение остатков животных и растений находится йгребне отрога холма в 0.5 км к юго-востоку от сел. Бинагады и в 7 км । северу от Баку. Ближайшие расстояния от морского побережья — tO км па юг и 25 км па север.
1 Кпр или битум, употреблялся и употребляется для заливки плоских крыш-
132
Часть 1. Обоснование истории формирования фауны
Костеносная площадь занимает около Р/2 га поверхности гребня отрога холма, близ бугра Кыррар. Современная высота участка 54—57 м над уровнем океана и 48 м над уровнем озера Беюк-Шор. К северу возвышается остаток конуса древнего грязевого вулкана Кичик-Даг, за которым лежит вытянутая меридианно котловина соленого озера Маса-зырского (Мирдаляби), а северо-восточнее — озера Бинагадинского. На восток, после сильного падения к солончаковой депрессии Кариа-тах-Шор, начинается подъем на Балаханское плато. С юго-востока
из и пл на* В раг В 01 вае
Рис. 62. Бпнагады. Яма от добычи кира на участке находок костей носорогов. ’ Фот. автора, 1941.
к подножью Бинагадинского холма прилегают занефтеванные ныне солончаки и впадина соленого озера Беюк-Шор, уходящая далеко на юго-восток (рис. 63).
Судя по карте Воларовича и Леднева (1913—1929) и по описание Богачева (19406), в основе Бинагадинского холма лежит антиклинальная складка шпротного простирания, образованная слоями майкопскш (олигоценовых) и диатомовых (миоценовых) глин, а также песчаника^ продуктивной толщи среднеплиоценового возраста, окаймленных с юга отложениями древнекаспийской, срсднечетвертичпой террасы. Сглажен’ иый ныне конус грязевого вулкана Беюк-Даг, достигающий 105 м ао солютной высоты, был «насажен» в плейстоцене на северо-восточны’ участок миоценовой складки. Этот конус образовался, по-видимому в два возрастных этапа. Его основание, сложенное лёссовидными пес ками, датируется Q21S, т. е. рисс-вюрмом, а более поздняя увенчивании шапка Q2b относится к началу вюрма.
Свод складки в районе захоронения оказался срезан волнами нг ступавшего моря, в результате чего создались условия для выхода ней
Ю; каспи' раковз An dr., возмог хазаре Сама о зарски очевид] гичной образох как этс В ЭПОХ1 островк Совр ствия т чинк, 1 геологи Ясамаль мому, «I смотря j пяков. £ рола всл в верхне
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 133
нз песков продуктивной толщи в ракушник древнекаспийской террасы и в более поздние четвертичные слои. В соответствии с фазами сжатия пластов продуктивной толщи на поверхности складки образовывались натеки нефти, асфальтирующей покровные наносы различного генезиса.
результате на выравненной поверхности складки появилась серия нзновозрастных асфальтированных слоев, содержащих остатки жи-|мтяых. и растений. Новейший покровный натек асфальта просматри-нется на северо-восточном склоне отрога холма; он образовался, по-ви-
•.юму, частично в результате добычи нефти в последние два столетия.
Рис. 63. Современный рельеф на участке костеносных местонахождений Бпнагады и Кир-Маку.
1
л
Я X
И а ь ь :й
с-
а-ти
£ Южная окраина плейстоценового битумного пятна граничит с древнекаспийской террасой «хазарского» возраста, содержащей индикаторные раковины моллюсков: Dreissensia polymorpha Pall., Didacna surachanica Aiidr., P. kovalevskii V, Bog., Neritina pallasii Lindh. и др. He исключена возможность, что и нижние горизонты костеносных слоев синхронны ! хазарской террасе, т. е. эпохе отложения прибрежного ракушника.
Сама терраса слегка наклонена к югу, что свидетельствует о послеха-зарских сдвигах. На северном склоне холма терраса не прослеживается, очевидно, в хазарский век там была суша, позднее подвергшаяся энергичной эрозии. Вообще, если предположить, что рельеф района в эпоху образования захоронения был в основных чертах сходен с современным, как это признают многие геологи, то тогда нужно признать также, что в эпохи трансгрессий эта местность превращалась в архипелаг мелких островков и полуостровков.
f Современный рельеф Апшерона был создан в результате взаимодействия тектонических, эоловых и водно-эрозионных факторов (М. Мир-ийнк, 1934). Наличие здесь плоских, вытянутых котловин — шоров — геологи склонны объяснять действием ветра. Даже глубокая щель — Ясамальская долина, тянущаяся на юг от Бинагадов до моря, по-види-ному, «выдута» ветрами северных румбов в третичных суглинках, не-; смотря на прикрывающую их местами пластину апшеронских извест-I пяков. В целом следует признать, что важные изменения рельефа Апше-I рона вследствие дислокаций и эпейрогенических колебаний происходили
8верхнем плейстоцене и в голоцене.
134
Часть I. Обоснование истории формирования фауны
По исследованиям Шлепнева (1947), взаимные перекосы отдельных точек Апшерона как следствие эпейрогенических колебаний достигали только за 16 лет — с 1912 по 1928 гг. — 114 см! Таким образом, в связи
с дислокацией четвертичных террас современный рельеф Апшерона в районе Бинагадов не может уже дать солидных отправных точек для воссоздания деталей рельефа и природы за-
Рис. 64. Разрез асфальтированной толщи в Бинагадах.
а — покровные суглинки; б — битуминозные пески с битой раку-шей, костями и древесными остатками; e — слой «жирного» асфальта; г — занефтеванные пески с остатками животных и растений; б — пески продуктивной толщи; справа — мощность в метрах.
хоронения.
Типичный геологический разрез четвертичных слоев в центре костеносной площади показан на рис. 64.
Верхний покровный слой а представлен делювием — суглинками мощностью от 0.2 до 1.0 м. Ниже идет слой б мелкозернистого песка, с довольно крупнобитой раку шей морских моллюсков, пропитанный нефтью. Он достигает мощности от 0.75 до 1.4 м и содержит нередко у южной границы крупные стволы и ветви можжевельника. Изредка здесь встречаются также обломки костей копытных — лошади, оленя и крупных птиц. Далее идет прослойка в из темно-коричневого битума, вязкого и плотного, мощностью от 0.1 до 0.5 м. Она постепенно выклинивается к краям костеносной площади, что создает впечатление линзообразного залегания. В кровле битума бывает местами много мелких птичьих костей, особенно уток, остатков жуков и травянистых растений. Кости крупных животных на границе слоев бив редки и бывают погружены в самый кировый слой.
Битум подстилается вновь мелкозернистым песком — слоем г, содержащим мелкобитую ракушу, а изредка и мелкий галечник. Этот слой мощностью от 0.3 до 0.6 л имеет местами слабые признаки горизонтальной, как бы речной слоистости, а нередко и отпечатки узких листьев болотных растений. Это основной костеносный слой, из которого и была извлечена главная масса костей млекопитающих, птиц, рептилий, насекомых, моллюсков и остатков растений.
Анализ механического состава грунта иа слоев биг показал преобладание мелкозе-
ма — 62.5 и 75.4%, что, по-видимому, свидетельствует о прибрежно-эоловом происхождении песчаных бинагадинских слоев (Верещагин, 19516), Скопления остатков животных и растений залегают в подстилающем кировый слой занефтеванном песке как бы гнездами, или «карманами», насыщенными битумом сильнее, чем окружающая толща.
Костеносный слой на глубине 3 м от поверхности грунта незаметно
переходит в мелкозернистые влажные и местами чуть пропитанные нефтью пески продуктивной толщи. На сохранившихся разрезах некоторых ям, в самой высокой точке бугра, на глубине 0.8—0.1 м, в толще
н л г п 6 л< Тс и
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
135
голоценовых суглинков, прослеживается еще тонкий кировый слой мощностью 10—15 см, который бывает переполнен костями мелких птиц, образующих плотные сплетения. Этот слой свидетельствует о какой-то новейшей фазе выжимания нефти.
Происхождение различных слоев описанного разреза не совсем ясно. Слой б образовался, очевидно, в волноприбойной зоне какого-то стоячего водоема, чем и объясняется наличие грубобитой морской ракуши с диаметром частиц от 2 до 5 мм. Происхождение слоя битума в рассматривается обычно как результат выжимания нефти по линии разлома слоев продуктивной толщи несколько севернее плейстоценовой битумной линзы.
Мастан-Заде (1939) высказал соображение о возможном излиянии нефти при извержении вулкана Кичик-Даг.
Несомненно одно, что процесс битуминизации, т. е. окисления и загустевания нефти, шел в надгрунтовых условиях. Во всяком случае на дне водоема — под водой — битуминизации не происходит, так как нефть поднимается на поверхность воды и сгоняется ветром. Учитывая это, необходимо признать, что слой б образовался в результате трансгрессии какого-то водоема, перекрывшего битумную линзу.
Происхождение костеносного слоя г, вероятно, связано с каким-то временным водоемом, но вряд ли с текучим. Границу костеиосного слоя и песков продуктивной толщи практически трудно и подметить. Граница размыва — базис эрозии — не выражен. Наличие чешуек мелкобитой ракуши диаметром от 2.0 до 3.5 мм с ничтожным процентом встреч (0,2—1%) свидетельствует и здесь об участии, кроме наносного мелкозернистого песка, еще каких-то береговых отложений морского или лиманного типа.
Залегание побегов сухолюбивых травянистых растений, мелких древесных веток, детрита вперемежку с костями животных и остатками жуков свидетельствует о каких-то очень небольших грязе-нефтяных потоках, оставлявших наносы в углублениях рельефа. Разрозненные кости птиц, остатки насекомых и растений, запрессованные в мозговые коробки крупных зверей с битумной массой, также говорят об отсутствии сортировки материала, как это бывает в речном потоке. Скорее подтверждается захоронение на месте в застойном водоеме при небольших подвижках костей в субстрате.
Скелеты животных, за исключением единичных скелетов птиц размером до голубя, в этом захоронении полностью расчленены. Остатки связок п перьев наблюдаются лишь на костях птиц из более молодых слоев, например из битумной прослойки в горизонте б. Целых скелетов крупных зверей при первых раскопках обнаружено не было, но это следует отнести скорее за счет несовершенства раскопок. Кости скелетов крупных и средних зверей как бы разбросаны в асфальтовой толще на некотором удалении друг от друга и залегают большей частью горизонтально. В некоторых раскопках Н. И. Бурчак-Абрамовичу удалось найти и разрозненные на пространстве нескольких квадратных метров кости одного скелета ........... лошади, оленя. Прочные черепа зверей средней величины — волка, гиены, барсука — залегают в битуминозном песке отдельно от шейных позвонков. Даже атлант оказывается в стороне. Известна находка 6 соединенных шейных позвонков осла и нижней челюсти лошади, обломанный передний кусок которой залегал в грунте рядом с горизонтально стоящими «салазками». Обломанные кости крупных видов зверей и птиц встречаются, как правило, в нижнем горизонте слоя б. В слое г
136 Часть I. Обоснование истории формирования фауны
кости обычно целы и нередко сохраняют полуотвалившиеся эпифизы. Целы даже хрупкие трубчатые кости крупных птиц, содержащие в пустой полости блестящие натеки чистого асфальта, образовавшиеся от нефти, просочившейся по тканевым капиллярам.
Кости, погрызенные волками, гиенами, медведями, встречаются сравнительно редко. Описаны обгрызенные плечевые кости медведя, таз плейстоценового осла, бедренная кость носорога (Верещагин, 19516; Бурчак-Абрамович, 1953а). Черепа грызунов не имеют следов ударов клюва или когтей хищных птиц и вряд ли являются погадковыми или попавшими в асфальт из желудка погибших птиц. В носовых и мозговых полостях черепов встречаются, но не часто, скопления ложнококонов мясных мух. Такие скопления найдены в углублениях рельефа плечево!! кости носорога, причем открытые капсулы показывают, что мухи благополучно вывелись. Неизвестно ни одного случая насечек, зарубок и переломов костей, сделанных рукой первобытного человека, но обращает внимание повышенный процент костей с различного рода заболеваниями — остеомиэ литами, рахитами, гельмиптозами (Верещагин, 19466, 19516; Г. В. Гаджиев и Д. В. Гаджиев, 1952). Обтертость, окатан-ность костей в потоке, а также трухлявость, трещиноватость и обтертость песком, появляющиеся при пребывании на поверхности или в хорошо аэрируемом грунте, у бинагадинских костей наблюдается очень редко. Известны всего две кости волка и оленя с заполированными нижними эпифизами.
Минерализация костного вещества сравнительно слабая. При распиле диафизов чувствуется легкий запах сырой кости, чего обычно не бывает у костей среднеплейстоценового возраста. По исследованиям Мамеда-лиева и Каплан (1948), в кости плеча волка были обнаружены белковые вещества, а в древесине можжевельника из слоя б 30.52% целлюлозы.
Окраска просушенных костей темно-бурая, и лишь на гладких поверхностях диафизов просматривается местами темно-коричневый и бронзовый цвета. В свежем разломе кости буровато-коричневого или каштанового цвета. Хрупкость и светло-каштановый цвет костей появлялись лишь после выварки их в щелоке. Эмаль и дентин зубов окрашены обычно в буровато-каштановый цвет, но для крупных коренных зубов гиен, сайгаков, быков крайне характерна голубоватая окраска эмали ; язычной поверхности зуба, особенно яркая при свежей расчистке. j
Крупные раскопки в Бинагадах, проводившиеся с 1938 по 1942 г., j были недостаточно научно документированы, поэтому природа захоро- J нения осталась слабо исследованной. |
Тем не менее вопросу происхождения Бинагадинского захоронения, t т. е. причинам гибели животных и условиям захоронения их остат-1 ков, а также древним ландшафтам Апшерона, уделял внимание почти | каждый исследователь, работавший с тем или иным материалом из г-Бинагадов. f
Богачев (1939в, 19406, 1944) признавал древность современного! рельефа и рисовал картину обширного пресного озера, остатком коего! теперь является Беюк-Шор. В этом озере с плавающей нефтью и завя4 зали старавшиеся добраться до чистой воды носороги, лошади, быки,| олени. Собиравшиеся на их трупы четвероногие и пернатые хищники,| вступая в драку, гибли в свою очередь. Многочисленные ручьи, будто? бы впадавшие в озеро, приносили растительный детрит и стволы можжевельников, плоды груш. Климат района в эпоху захоронения был| более холодным и несколько более влажным, нежели в наши дни. Былв|
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 137
хорошо выражены перелеты птиц. Учитывая все это, Богачев проводил аналогии в типе захоронения с асфальтированными «кладбищами» вСтаруни (Галиция) и в Ранчо ля Бреа (Калифорния). Правильно было рпчеркнуто длительное и постепенное накопление органических остат-(ов без какой-либо единовременной катастрофы, погубившей многие а’ячи разнообразных по морфологии и экологии существ.
Эта стройная картина существенно нарушается рядом фактов. Если ьшзнавать древность современного рельефа, то Бинагадинский холм при высоком стоянии вод озера надо представлять в виде небольшого островка. Тогда не останется места ни для животных, ни для концентрации нефти.
Отсутствие в костеносном слое остатков пресноводных моллюсков ыб, ксерофильность известных из него растений не вяжется с картиной пирного пресного озера. К тому же при обилии ручьев и озерных ыдиемов животным нет необходимости лезть в нефтяную грязь. Гибель носорогов и кабанов от увязания в иле почти исключена. Кроме того, иловатых отложений в бинагадинских слоях нет, а ничтожная мощность к^стеносного мелкозернистого песка и битой ракуши исключает воз-\ кность увязания.
Ботаник Петров (1939) на основе списка определенных из асфальта пиков растений утверждал о существовании на Апшероне ландшафта аяны или аридного редколесья, а на месте захоронения — дельтового астка речки.
Аргпропуло и А. Богачев (1939), основываясь на наличии остатков iccHoii сони, краснохвостой песчанки, серого хомячка, тушканчиков в ряда видов относительно мезофильных жуков, считали, что в районе ахоронения при более влажном, нежели ныне, климате были леса тугайного типа.
Орнитолог Серебровский (1948), анализируя видовой состав птиц, признавал существование перелетов, а местность района захоронения изображал в виде мелководных топей с камышами, располагавшихся ,1 окраине морского залива. Гибель птиц Серебровский объяснял дей-гвием нефтяной пленки.
Геолог Султанов (1947) предполагал, что участок захоронения был берегом морского мелководного залива, до которого слабо докатывалась волна. Что касается гибели носорогов, слепушонок, тушканчиков в волнах наступавшего моря и гибели пролетавших птиц в зоне, отравленной газами фумарол, то подобные курьезные представления этого автора о геологических и биологических событиях не нуждаются в коммента-ПИЯХ.
Ошибочными оказались и утверждения Ефремова (1950) в его превосходной «Тафономии» о речном наносном происхождении костеносного поя <?, который будто бы сохранился от позднейшего размыва лишь под защитой асфальтовых масс.
II. Громов (1952), изучавший грызунов, также объяснял накопление органических остатков в Бииагадах с тафономических позиций И. А. Ефремова — приносом речкой — и признавал существование на верхнеплейстоценовом Апшероне степи при отсутствии настоящей лесной растительности.
В высказываниях всех упомянутых авторов остаются биологически 8 тафономически неувязанными факты совместного захоронения в одном слое и в одной точке остатков растений, моллюсков, насекомых, птиц, мелких грызунов, средней величины хищников и крупных копытных.
138
Часть I. Обоснование истории формирования фауны
Если принос и накопление остатков речкой в ее дельте находят известное подтверждение, то причины гибели, места гибели, характер и дальность транспортировки животных и растений остаются в этих высказываниях без ответа. Между тем анализ всей совокупности геологических и биологических фактов вполне может дать истинное представление о природе Бинагадинского захоронения и ландшафтах плейстоценового Апшерона.
Кроме Бинагадов, нами изучались и примеры современной массовой гибели насекомых, птиц и зверей на Апшероне и в Кабристане, а также условия образования длительных захоронений остатков этих животных.
Уже изучение геологических разрезов в ряде старых и новых раскопок на костеносной площади показало, что озерное, а тем более речное происхождение костеносного слоя г сомнительно. Между тем костеносный слой всегда связан с асфальтовой прослойкой <?, и при ее выклинивании в слое г исчезают и органические остатки. Другими словами, присутствие нефти — асфальта было непременным условием накопления остатков растений и животных.
Наблюдения в Кабристане у Хурдалана, Кир- j кишлага и Бешбармака, показали, что натеки и; толстые скопления асфальта образуются на склонах и в углублениях рельефа близ своеобразных трещин и кратеров, из которых выжимается нефть. Грязе-нефтяные кратеры, или колодцы, подают, кроме асфальта, и газ и солоноватую воду. По краям они нередко зарастают захудалым тростником (найденным и в Б и наг ад ах, рис. 65), зеленые кусты которого составляют весной поразительный контраст с желтым фоном горно- i солянковой полупустыни. Колодцы эти часто | привлекают скот, причем в загустевшей грязи | у Бешбармака вязнут и тонут коровы и ослы.
На поверхности пластин асфальта также скапливается часто дождевая вода, которая при нагреве и разжижении асфальта образует в его толще крупные линзы пресной воды, очень привлекательные для животных в период засухи. Такие асфальтовые лужи в течение года улав-1. ливают огромное количество остатков растений, насекомых, птиц и звс-рей — от мыши до джейрана и ишака. Разлагающиеся остатки тонут в асфальтовой массе, и кости животных проходят в подстилающие иловатые слои вместе с минеральными частицами, приносимыми ветром
СМ
Рис. 65. Бинагады. Верхушка стебля из сближенных междоузлий Phragmites communis из костеносного слоя.
и временными потоками.
Для накопления остатков растений и животных вовсе не обязательно,: чтобы асфальтовые скопления были в крупной водомоине или долине,; овраге. Наоборот, их расположение на пологом склоне или даже гребнеf холма гарантирует более длительное и эффективное действие, так как здесь они дольше не перекрываются наносами (Верещагин, 19516).
Итак, современные примеры говорят за то, что первоначальная гибель к животных могла происходить и в плейстоцене на месте или близ места р захоронения, без дальнего переноса. |
Мелкие и крупные млекопитающие, птицы и насекомые могли гибнуть f и накапливаться на Бипагадинском холме в течение веков вследствие^
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 139
отравляющего и улавливающего действия тяжелой бинагадинской нефти. Выжимание нефти сопровождалось образованием липких пластин и нефтеводяных ванн с пресной водой, располагавшихся, вероятно, близ края морского залива. Они улавливали зайцев, тушканчиков, песчанок, полевок, хомячков и бесчисленное множество птиц, насекомых. Более толстые слои загустевшего асфальта могли удержать и более
крупную добычу — носорога, лошад привлекали хищных млекопитающих лисиц, волков, гиен по сравнению с ничтожным числом их на отмелях рек только и можно объяснить привлечением хищников обильной пищей в какую-то ловушку. Охота лисиц на погибающих в асфальте птиц была шиеана мною (Верещагин, 1946а).
Принос трупов водой и их распад на месте захоронения мог иметь место лишь в небольших масштабах.
Потоки, приносившие трупы, были небольшими и эфемерными, так как в-захоронении нет остатков мезофильных растений и животных лесной и луговой зон, а также пресноводных моллюсков и рыб. Это были, вероятно, кратковременные селевые потоки, зарождавшиеся не далее нескольких километров, а вернее,
►, быка. Трупы этих травоядных и хищных птиц. Обилие остатков
Таблица 16
Возрастной состав волков в Бинага-динском захоронении
Число особей Возраст
абсолютное в процентах
0.8 Около месяца.
20' 16.6 3.5—4 месяца.
15 12.5 7—8 месяцев.
61 50.8 От 1 года до 3 лет.
20 16.6 От 3 до 5 лет, со слабо стертыми зубами.
3 2.5 Более 5 лет, с сильно стертыми зубами.
дан метров от точки захоронения.
Слой б с битым ракушечником и со стволами можжевельника образо-
вался, видимо, в результате наступления новой трансгрессии моря на мелководье лимана. Он перекрыл битум и спрессовал костеносный слой. Обломки костей могли попасть в пего в связи с размыванием южного края основной костеносной линзы и, несомненно, переотложены. Древесные стволы также заносились и выбрасывались волнами из залива В слой б (рис. 64).
Важные указания на природу Бинагадинского захоронения может дать обсуждение и таких биологических факторов, как сезон гибели животных и состав их популяций. Ясно, что насекомые и плоды груш попадали в асфальт лишь в теплый период года, но наличие в составе птиц, прилетавших на гнездовье с юга: колпицы, рыжей цапли, черного аиста и на зимовку с севера — белой совы говорит о гибели их и летом и зимой.
В составе погибавших зверей были все возрастные стадии — от сосунков до глубоких стариков с совершенно стертыми под корень зубами. Особенно эффектно это видно на примере с волками, установление истинной численности остатков которых в связи с их величиной и прочностью меньше зависит от способов извлечения материала, чем у других видов. Так, при подсчете нижних челюстей волков после определения их индивидуального возраста были получены данные, указанные в табл. 16.
Уже на основе этих цифр видно, что в бинагадинскую ловушку
попадали животные всех возрастов, а отнюдь не очень старые и не совсем молодые. Из табл. 16 следует, что основная масса (67.4%) захороненных волков приходится на животных взрослых, достигших полной силы (рис. 66). Другими словами, в захоронении оказалась не какая-то выбо
140
Часть /. Обоснование истории формирования фауны
рочная возрастная группа зверей, а порции популяций, существовавших на протяжении большей части годового цикла.
Гибель животных в течение всего теплого времени года усматривается
и при учете костных материалов по
Рис. 66. Образцы нижних челюстей вол ков различных возрастных групп из Бинагадинского захоронения. (Иллюстрация к табл. 16).
всем другим видам хищных (Верещагин, 19516).
Все эти факты подтверждают еще раз, что для всех — и мелких и крупных — животных существовала единая причина гибели и именно на участке захоронения, а не где-то в другой местности.
Эти предварительные предпосылки позволяют подойти с большей уверенностью п к оценке полноты фаунистического комплекса, отмеченного в Бинагадах, а также к обсуждению экологической обстановки в восточном Закавказье в эпоху образования захоронения.
По подсчетам Бурчак-Абрамовича и Джафарова (1953), количество костей копытных, извлеченных при раскопках 1938—1942 гг., достигает 8213 при наименьшем возможном количестве особей в 412. Было определено также 2043 кости хищных, принадлежащих 11 видам и по крайней мере 440 особям. Зайцеобразным и грызунам принадлежит около 2300 костей, относящихся к 15 видам и минимум 180 особям, и насекомоядным — 3 вида до 50 костей.
Общее число извлеченных птичьих костей не меньше 30 000. Найдено около сотни остатков черепах, ящу-
рок и змей, много тысяч остатков насекомых, преимущественно жуков и сотни раковин наземных моллюсков. Определение и обработка остат-1 ков млекопитающих производились Богачевым (19386, 19406), Аргиропу-j ло (19416), мною (Верещагин, 1948,1 1949в, 19516), Бурчак-Абрамовичем (1951 в, 1952г), Джафаровым (1950, 1955), Алекперовой (1952, 1955), i И. Г ромовым (1952) и др. Было выявлено 39 видов.
Подсчет числа извлеченных осо бей млекопитающих был произведен
нами по нижним челюстям для массовых форм и iro всем костям для единичных. Для копытных приведены цифры но Бу рчак-Абрамовичу и Джафарову (1953). Были учтены все коллекции препарированного материала, хранившегося в 1945 г. в музеях Баку, Ленинграда, Москвы. Результаты
Гл. Л. Развитие ландшафтов и фаун в четвертичном периоде
141
Таблица 17
видовой состав и численность особей млекопитающих в Бинагадинском захоронении
Виды Число особей Виды Число особен
Insectivoi а Mesocricetus auratus planicola 3
fwidura aff. russula . . . . Rtmiechinus aff. aunlus . . . 3 3 Merlon es erythrourus interme-dius Allactaga elater dzhajarovi . . 12 4
^r./niceus aff. euro pe us .... J A. williamsi 10
Итого 7 A. jaculus bogatchevi .... 26
Dyromys nitedula ...... 1
Carnivora Microtus arvalis ....... M. socialis ......... | 70
• «.ms lupus apscheronicus . . Canis sp........... nilpes corsac V. vulpes aff. alpherakyi . . . pvuta spelaea 120 J 85 125 20 M. apscheronicus ....... Ellobius aff. lute seen s .... Arvicola cf. terrestris .... Hystrix vinogradovi ...... 2 10 1 25
arclos binagadensis . . . 4 Итого 177
Wrnclu peregusna 13 Perissodactyla
tides meles aff. minor . . . . 55
Р.1И/Л ra spelaea ....... 11 Equus aff. hidruntinus .... 73
M.< tf. lybica Jtatfiup jubalus z 4 E. cab alius subsp Rhinoceros binagadensis . . . 154 31
Итого ' 440 Итого 258
Lagoinorpha Arliodactyla
l/fiis europaeus gureevi . . . 4 Sus apscheronicus Megaceros cf.euryceros .... 11
2
Итого 4 Cervus elaphus binagadensis 52
Rodentia Saiga tatarica binagadensis . . Ovis cf. ammon 82 1
Hus musculus ........ 2 Bos mastan-zadei 6
Iwdemus sylvaticus 3 Итого
i ricelulus migratorius argiro- s 154
puloi Всего
1040
Таблица 18
Состав птиц по отрядам в
Бинагадинском захоронении
Отряды Число видов Отряды Число ВИДОВ
Limicolae 22 Coraciiformes (совы) . . . G
Anseres 21 Steganopodes 9
Accipitres 17 Lariformcs 2
Gressores 9 Pterocletes 1
Passcres 8 Columbus 1
Palhidicola 7 Podicipites 1
14 2
Часть I. Обоснование истории формирования (фауны
сведены в табл. 17. Цифры числа костей и особей отражают не только действительную численность захороненных животных, но и методику извлечения костей из породы. Численность мелких видов преуменьшена. Характерные остатки показаны на рис. 67, 68, 70, 71.
Общий список птиц, по исследованиям Серебровского (1948) и Бурчак-Абрамовича (1951в), достигает 97 видов (табл. 18).
Из пресмыкающихся известны остатки греческой черепахи — Testudo graeca L. и мелких ящериц, ближе не определенных.
Остатки насекомых очень многочисленны, но хорошо сохранились только жесткокрылые. А. Богачевым (1947) описано 107 видов жуков семейств: 4
Cicindelidae Carabidae Dytiscidae Cyprinidae Staphylinidae Silphidae Histeridae Buprestidae
Dermestidae Hydro ph ilidae Coccinellidae Tenebrionidae Cerambycidae Chrysomelidae Curculionidae Scarabaeidae
A. H. Кириченко определил еще остатки шести видов клопов — мейств Thyreocoridae, Pentatomidae, Gerridae.
I 1 i
Рис. 67. Би па гады.
Челюсти: 1 — Hemiechinus anialvs; 2 — Eiinaceus europaeus.
Остатки мух, их ложнококонов, бабочек, стрекоз, саранчовых, плохо сохранившиеся и трудно определимые, не описаны. Раковины моллюсков принадлежат мелким сухопутным видам Helix, llelicella, Pupilla.
Остатки растений, определенные Петровым (1939) и мною (Верещагин, 1949в), представлены видами:
Pirus salicifolia Pistacia cf. mutic.a Vitis cf. silvestris Zozimia sp. Tragacanthum sp.
J unipcrus polycarpos Punica granatum Allium sp.
Phragmites communis Colchicus sp.
Scirpus sp.
Богачевым (19406) упоминались еще из костеиосного слоя остатки Alhagi, Isatis, Artemisia, Salsola, Tamarix, но подтверждений этих определений не последовало.
Некоторые древесные остатки этого списка происходят и из ракушечника слоя б, будучи, следовательно, моложе основного костеиосного
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 143
захоронения. Но это не исключает факта длительного существования устойчивых ксероморфиых ландшафтов в восточном Закавказье.
Палеонтологические материалы из Бинагадов оказались настолько полными, что позволяют говорить о целых фаунистических комплексах различных систематических групп и о биоценозах, существовавших в восточном Закавказье в плейстоцене.
Ландшафтные, климатические и зоогеографические особенности древнего Апшеропа выявляются с большой полнотой при рассмотрении эволюции экологических группировок млекопитающих. Изменения в видовом составе зверей с плейстоцена до наших дней следует проследить далее по отрядам (табл. 19, 20, 22, 24) с привлечением современной зоогеографии для северо-восточных и юго-восточных окраин Большого хребта.
11 а с е к о м о я д н ы с
Отсутствие на плейстоценовом Апшероне бурозубок и крота свиде-»льствует о ксерофнтном ландшафте. Длиннохвостая белозубка обитает чипе в лесу п в полупустыне, ушастый еж — в степи и в полупустыне, не избегая и аридного редколесья. Обыкновенный еж живет в сырых лесах и в сухих кустарниках полупустыни.
Таблица 19
(равнение битового состава насекомоядных четвертичного периода в предгорьях восточного Кавказа
Плейстоцен Историческая эпоха
Виды Аншерон Апшерон и Кабристан предгорный Дагестан
Crocidura russula С. leucodon ............ Suneus etruscus .......... Ileiniechinus auritus Erinaceus europacus ........ ? । -4-
Всего 3 + 2 ? 4+ ? 4
Примем а и не. Знаком ? в этой и следующих аналогичных таблицах отмечены виды, наличие которых в ту или иную эпоху весьма вероятно.
X п щ н ы е
Отсутствие лесных видов хищных: лесной куницы, лесного кота, рыси говорит о том, что значительных лесов па древнем Апшероне не было; отсутствие выдры и порки — о том, что не было рыбной речки и рыбного камышового озера. Редкие и в наши дни па Апшероне ласка, куница белодушка могли и не попасть в захоронение. Наличие бурого медведя, часто обитающего и ныне в аридном редколесье Закавказья, создает представление о древиелг ландшафте Апшеропа как и сухом плоскогорье с островками арчево-фисташковых лесов в распадках оврагов. О сходстве
144
Часть I. Обоснование истории формирования фауны
того древнего ландшафта с современным свидетельствует и наличие мелкой лисички и корсака, почти неотличимых от современных. Важно отметить отсутствие пещерных медведей, что, вероятно, объясняется чрезмерной сухостью.
Широко распространенные и подвижные хищники: волк, лисица, корсак, пещерная гиена, барсук, тпгролев, гепард, пятнистый кот — могли существовать в таком составе в степи или полупустыне с небольшими участками кустарников п деревьев. Климат района мог быть и более холодным, чем ныне, но прп невысоком снеговом покрове, дававшем возможность охоты пятнистому коту, корсаку, лисице.
Для выяснения природы захоронения п эволюции группировок видов с плейстоцена до наших дней было интересно сравнить относительную численность разных видов хищных, погибавших в Бинагадах, с современной, выявленной при охотничьем промысле в соседнем с Баку Шема-хпнском районе. В результате были получены списки и цифры, подчеркивающие сходства и различия в экологической сущности обеих группировок и перекомбинации численности пяти массовых форм — лисицы, волка, корсака, барсука и шакала (табл. 21).
Сравнение списков видов хищных вновь подтверждает отсутствие связей бинагадпнской ловушки с горным ландшафтом и с настоящими мезофитиыми лесами, из сохранившихся массивов которых в Шемахииском районе поступают в настоящее время шкуры куниц и кошек.
3 а и ц е о б р а з и ы е и i р ы з у н ы
Состав грызунов в плейстоцене (табл. 22) был почти вдвое богаче, нежели па современном Апшеропе, и близок к группировке, существующей ныне в предгорном Дагестане. Бпнагадпиские грызуны в целом характеризуют ландшафт плейстоценового Апшерона как предгорную степь с небольшими участками сухих лесов, а местами и с озерцами, поросшими тростником и рогозом.
Современные черноватые хомяки живут в более мезофптной обстановке, чем существующая на Апшероне в наши дни (см. главу III).
Наличие обыкновенной и апшеронской полевок также говорит о существовании более мезофитных, чем современные, биотопов. Нижняя граница современного распространения обыкновенной полевки проходит теперь в зоне предгорных кустарников близ Шемахи, на высотах около 700 м, а ныне живущей кустарниковой полевки — на высотах около 1000 м.
Остальные виды — заяц, серый хомячок, степная мышь, красно-хвостая песчанка, тушканчики, степная полевка, слепушонка и дикобраз — в таком сочетании характерны теперь для жарких сухих долин северного Ирана и Карабаха, имеющих поросли колючих астрагалов, иволистпых груш, можжевельников и боярышников.
Сравнение относительной численности грызунов, уловленных бина-гадинской ловушкой в плейстоцене и отлавливаемых теперь на Апшероне сычами, ловцами Азербайджанской противочумной станции и нефтяными лужами, показывает, что в голоцене произошло увеличение численности ксеротермофильпых форм за счет сокращения и даже исчезновения мезофильных (табл. 23).
Сильпое обеднение состава грызунов с плейстоцена до наших дней отнюдь нельзя объяснять влиянием человека (об этом см. главу VII),
2 -epscft< spelae. 19, 2t
f" >-
м
в ю Э“ 1-а-п-а»
не ли эм
Рис. 68. Бинагады.
ie, 10-ка-•пь ос-
ке,
су-1ЯЯ Дит оло оло
1К0-лин IOB,
1на-юне &ТЯ-пенсия
Хней •II).
j, ? — череп Vulpes corsac; 3, 4 — череп V. vulpes aff. alpherakyi; 5, 6 — черепа ( anis lupus apscheronicus; 7,8 — клыки Ursus arctos binagadensis; 9—12 — черен, атлант и плечевая Crocuta spelaea; 13—17 — черепа и плечевая Meles meles aff. minor.; 18— череп Vormela peregusna; j(2fl — челюсти молодых Acinonyx aff. jubatus; 21 — плечевая Felis aff. lybica; 22, 23 — челюсти молодых и 24 — взрослого Panthera spelaea.
]Q H. К. Верещагин
Рис. 69. Бицагады.
16, 17 реннап и берпог
1,2 — челюсти Lepus europaeus дитеегг; 3—6 — челюсть, бедренная, плюсневая и берцовая Allactaga elater; 7—10 — челюсть, бедренная, плюсневая, берцовая A. viiliamsi dzjafaroti 11—15 — череп, бедренная, плюсневая и берцовая A. jaculus bogatschevi;
> 15
32
3!
см
рновая zjafarovi',
Рис. 69 (продолжение)
|(ц /; — челюстная и бедренная Cricetulus rnigraloriun ai giro) mini; 18—21— челюстные, бед-реннап и берцовая Mesocricetus а ига/us planicola; 22—26 — челюстные, плечевая, бедренная • и берцовая Mervmes erythrourus intei medins; 27—30— череп, бедренная п берцовая ЬИоЬгик aff. lutescens; 31 — 32— челюстная и лопатка Hyshix innogradovi.
148
Часть I. Обоснование истории формирования фауны
Таблица 20
Изменение видового состава хищных четвертичного периода в предгорьях восточного Кавказа
Виды Плейстоцен Историческая эпоха
Апшерон Апшероп и Кабристан предгорный Дагестан
Canis aureus 1— —
Canis sp C. lupus 1'—~
Vulpes corsac Tz. vulpes —|— —
Hyaena striata Crocuta spelaea .......... —
Ursus arctos 4- —|—
Vormela peregusna ......... Muslela nivalis .......... —
M. lutreola ............ — — —|—
M. eversmanni — —
Lutra lutra ............ — —
Martes foina ............ Meles meles „1 —1—
Panthera spelaea .......... — —.
P. leo ............... —
P. pardus ............. — +
Felis lynx ............. —
F. silvestris — ?
F. lybica ............. + ?
F. chaus .............. Acinonyx jubatus ? "t"
Всего 11 12 + 4? 17-4-3?
Таблица 21
Относительная численность хищных на Апшероне по отлову асфальтовой ловушкой и по данным охотничьего промысла в Шемахинском районе
Апшерон в плейстоцене Шемахинский район, 1930—1940 гг.
виды процент особей виды процент особей
Лисица 28.1 Лисица 75.5
Волк 27.5 Барсук 16.4
Корсак 19.2 Шакал 3.5
Барсук 13.0 Волк 2.3
Пещерная гиена 4.5 Кошка лесная и камышовая 0.8
Перевязка 2.9 Куница лесная и белодушка 0.7
Тигролев 2.5 Медведь 0.2
Медведь 0.9 Рысь 0.2
Гепард 0.6 Ласка 0.1
Пятнистый кот 0.4 Выдра 0.04
Мелкий волк 0.2 Перевязка 0.04
— — Полосатая гиена 0.001
— — Леопард 0.0001
Число видов .... 11 Число видов .... 15
Число особей . . . 440 Число особей . . . 12890
Таблица 22
Изменение видового состава зайцеобразных и грызунов четвертичного периода в предгорьях восточного Кавказа
Виды
Lepus europaeus..........................................
Citellus pygmaeus........................................
Microtnys mi nut us.....................................
][us musculus...........................................
Apodemus sylvaticus.....................................
Л. jlavicollis...........................................
.4. agrarius............................................
(ricelulus migratorius..................................
ytesocricetus auratus planicola 1.......................
\{, auratus nigriculus..................................
Meriones erythrourus....................................
Alactagulus acontion....................................
Mlaclaga elater.........................................
J. williamsi............................................
1, jaculus ..................
Dyromys nitedula........................................
Microlus arvalis........................................
V. social is............................................
y. apscheronicus........................................
Ellobius aff. lulescens.................................
£. talpinus.............................................
Arvicola terrestris.....................................
Ityslrix Vinogradovi....................................
Всего ......................
Плейстоцен
Историческая эпоха
Апшерон
16
Апгперон и предгорный
Кабристан Дагестан
Таблица 23
Изменение видового состава и относительной численности (в о/ ) грызунов и на Аншсроне с плейстоцена до наших дпей Р У
Виды Плейстоцен, отлов асфаль- XVII-XX вв. 1939— 1940 гг. 1947 г., отлов
товыми лужами н. э., отлов сычами отлов капнанамп нефтяными лужами
Степная полевка 23.8 18.1 4.6 Отсутству Отсутству Вымер 85 4 25.0 ет ет | 29.2 1 20.8 ет 12.5-0.3 4.1 ет
Обыкновенная полевка . . Большой тушканчик . . . Бннагадипский дикобраз 2 Краснохвостая песчанка . . . . } 38.6 14.4 13.8 7.1
Сорный тушканчик .... 5.5 9.5 2 0
Горная слепушонка .... >рый хомячок Палый тушканчик .... 5.5 4.4 2.2 42.4 0 2 Этсутству 2.9 0.3 0 6
Заяц-русак * • 4 • * * 2.2
равнинный хомяк Тесная мышь Стенная мышь 1.6 1.6 1.1 } 5‘7 Отсутству 0.2 I Ч 9
Апгаеронская полевка . . . Лесная соня 1.1 0.5 O.Zj | Вымерла 0 1
Серая крыса — — 0.1 —
Число впдов 15 7 9
Число особей 181 347 2473 24
i Возможно, что современный черноватый ? Возможно, что современный индийский
хомяк — потомок бинагадинского. дикобраз — потомок бинагадинского.
150
Часть I. Обоснование истории формирования фауны
Непарнокопытные
Наличие плейстоценового осла, лошади, носорога (табл. 24) характеризует плейстоценовый Апшерон как степную равнину с кустарниками по оврагам и травянистой растительностью, вероятно, выгоравшей к середине лета.
Таблипа 24
Изменение видового состава непарнокопытных четвертичного периода в предгорьях восточного Кавказа
Виды Плейстоцен Историческая эпоха
Апшерон Апшерон и Кабристан предгорный Дагестан
Equus hidrunlinus — —
Е. hernianus — “Г • 1
Е. caballus gtnelini —
E. caballus subsp + — —
Rhinoceros binagadensis — —
13 сего 3 1 4-1 ? 2
П а р н о к о п ы т н ы е
Плейстоценовый кабан древнего Апшерона мог обитать по тростникам I лиманов, а также в фисташково-можжевеловых лесах. Кабаны держатся и сейчас круглый год в аридном редколесье Закавказья с зарослями можжевельника и фисташки. Наличие сайгаков дает представление об обширных открытых пространствах со степной растительностью.
Таблица 25
Изменение видового состава парнокопытных четвертичного периода в предгорьях восточного Кавказа
Плейстоцен
Историческая эпоха
Виды
Апшерон
Апшерон п Кабристан
предгорный Дагестан
Sus apscheronicus ........
S. scrofa 1 ...................
Cervus elaphus .........
M egaceros euryceros...........
Saiga tatarica.................
Gazella subgutlurosa...........
Ovis cf. amnion................
Bos maslan-zadei ........ B. primigenius^................
Bison bonasus...............•
Всего..................
Бинагадинские бык и олень могли заходить па Апшерон во время него весеннего развития трав. Обеднение Апшерона копытными к рической эпохе — результат влияния человека.
пыш исто-1
1 Возможно, что современный кабан — потомок апшеронского.
2 Европейский тур жил на Апшероне позднее существования бинагадинской ло-1 2 вушки.
1— 4 —
Рш3— Д'
£
>в
ам гея [МИ об
пыш-исто-
ло
Рис. 70. Бинагады.
}-4— IP, М1, пястная и копытная фаланга Equus caballus subsp.; 5—9— М3, М3, pm—Mj, пястная и тазовая Equus hidruntinus; 10, 11 — череп Rhinoceros binagadensis (сверху и снизу).
152'
Часть I. Обоснование истории формирования фауны
Плейстоценовые ландшафты Апшерона могут быть характеризованы и по другим группам. Так как птицы в связи с их сезонными пролетами мало показательны, остановимся на жуках. Среди жуков, определенных А. В. Богачевым, оказалось, по нашим подсчетам, такое распределение видов и числа особей по отношению к воде и биотопам суши, как указано в табл. 26.
।
Рис. 71. Бинагады.
1, 2 — эпистрофей и плюсневая Sus apscheronicus; 3,4 — плюсневая и плечевая Ovis aff. аттоп; 5—8 — роговой стержень, атлант, лопатна, пястная Saiga tatarica binagadensis; 9, 10 — череп Bos mastan-zadei сверху и сбоку. (нат. вел. j (по Богачеву, 1940).
Водные жуки сами по себе не являются показателями ландшафта. Известно, что водолюбы и плавунцы, обладающие сильным полетом, часто летят к водоемам издалека даже в больших пустынях. Однако сухопутные формы жуков из бипагадинского костеиосного слоя, по подсчетам А. В. Богачева, содержат 8% (из 75.7) относительно мезофильных форм, свойственных ныне зоне предгорных кустарников у Шемахи и отсутствующих теперь па Апшероне. Другими словами, и на сдвигах ареалов жуков подтверждается общая закономерность — ксерофити-зация ландшафтов в голоцене.
Гл, II. Развитие ландшафтов и фаун в четвертичном периоде
163
Таблица 26
Распределение числа видов и особей жуков, собранных в бииагадин-ском асфальте, по экологическим группам
Экологические группы жуков Число видов Число особей
абсолютное в процентах абсолютное в процентах
Водные 15 14.1 760 21.1
Земноводные 11 19.2 74 2.0
Сухопутные 81 75.7 2758 76.9
Всего 107 100.0 3592 100.0
Итак, весь комплекс фактов, связанных с видовым и численным составом млекопитающих, жуков и растений плейстоценового Апшеропа, говорит о наличии в эпоху накопления остатков менее сухой и теплой, чем ныне, обстановки. Экологические условия здесь были близки к существующим ныне на южных склонах Армянского нагорья, на высотах [500—1600 м, в зоне так называемого аридного редколесья, или в предгорном Дагестане, на высотах от 0 до 500 м.
Универсальность Бинагадинского захоронения позволяет сделать п других выводов, касающихся стратиграфии, палеоэкологии и истории формирования фауны.
В бинагадинском фаунистическом комплексе хорошо выражена группа широко распространенных видов плейстоценовых степей и лесостепи, известных по палеолитическим стоянкам и другим типам захоронений в Европе. К ним относятся:
Erinaceus europaeus Canis lupus Vulpes corsac V. vulpes Crocuta spelaea Ursus arctos Meles meles Panthera spelaea Allactaga jaculus
Apodemus sylvaticus Mus musculus Microtus arvalis Arvicola terrestris Equus caballus Megaceros euryceros Cervus elaphus Saiga talarica
Происхождение большинства этих видов все же недостаточно ясно, и о проникновении в восточное Закавказье с севера можно говорить только в отношении корсака, большого тушканчика, сайги.
В экологическом смысле эта группа неоднородна и включает виды, адаптированные ныне к открытым ландшафтам и биотопам, и лишь отчасти виды, связанные с лесными угодьями и увлажненными участками речных долин в тех же степных равнинах. Настоящих ксерофилов в этой группе нет.
Ряд видов этой группы — лисица, волк, медведь, барсук, большой тушканчик — представлен в Бинагадах южными, местными формами, что доказывает консервативность и древность условий, создавших предпосылки для формирования бинагадинского фаунистического комплекса.
Вымерли из этих видов совершенно пещерная гиена, тигролев, лошадь, гигантский олень, и значительно сдвинулись ареалы лишь у корсака, большого тушканчика и сайги. Сдвиг ареалов обыкновенной полевки и водяной крысы имеет лишь местный характер.
ш
Часть I. Обоснование истории формирования фауны
Весьма значительна группа древних средиземноморцев, адаптированных к ксероморфным ландшафтам — к нагорным степям Передней Азии. Сюда относятся:
Crocidura russula Hemiechinus auritus
V or me la feregusna Felis lybica
Ac inonyx iubatus Lepus europaeus Allactaga williamsi Cricetulus migratorius Mesocricetus auratus
Microtus socialis Ellobius lutescens Meriones erythrourus Hystrix vinogradovi Rhinoceros binagadensis Equus hidruntinus Sus apscheronicus Ovis amnion Bos mastan-zadei
В экологическом смысле эти виды либо широко приспособлены к сме- j шаниым биотопам: длиннохвостая белозубка, ушастый еж, перевязка, ! степной кот, гепард, горный тушканчик, степная полевка, кабан, либо £ специализированы ныне к степным нагорным формациям: дагестанский j хомяк, серый хомячок, горная слепушонка, и к биотопам жарких долин | и низменностей: краснохвостая песчанка и бинагадинский дико- | браз. |
Большинство этих средиземноморцев также представлены плейсто- ! ценовыми формами подвидового значения. Основные массивы их ареалов в плейстоцене явно располагались к югу от Кавказского перешейка. , Такого же южного и юго-восточного происхождения и вымершие: плей- f стоценовый осел, бинагадинский носорог и бинагадинский тур. Из эндемичных кавказских видов можно отметить лишь вымершую апшерон-скую полевку, остатки которой на основе строения нижних коренных | были отнесены к роду Pity ту s европейского типа. Для дагестанского J хомяка характерен сдвиг ареала во внутренний Дагестан, а для горной j слепушонки — в нагорную степь Талыша. I
Итак, самое общее направление эволюции фаунистического комплекса | равнин восточного Закавказья с эпохи бинагадинской ловушки заклю- ;
чалось в исчезновении (вымирании и сдвигах ареалов) относительно ]
мезофильных форм вследствие увеличения сухости климата и ксерофи-тизации ландшафта. j
Общая палеозоогеографическая характеристика бинагадинского ком- i
плекса выявляется при сопоставлении с плейстоценовыми фаунами юга < Русской равнины и Передней Азии. с
В Бинагадах отсутствовали широко распространенные виды среднего и верхнего плейстоцена Восточной Европы: мамонт, волосатый носорог, j первобытный зубр, пещерный и степной медведи; отсутствовали и север- с ные виды: песец, заяц-беляк, лемминг, северный олень, овцебык. с
Этот ряд особенностей может быть объяснен частью ксероморфностью ] ландшафта на востоке Кавказского перешейка, частью геологическим возрастом эпохи захоронения. 0
В эпоху существования бинагадинской ловушки ареалы тундровых в видов, очевидно, далеко не доходили сюда. Не доходили с севера и аре- т
алы мыши-малютки, полевой мыши и степного хоря. Таким образом, г<
Апшерон и Кабристан в плейстоцене являлись районом стыка лишь некоторых фаунистических элементов юга Русской равнины и ирано-мал оазийских плоскогорий. С другой стороны, здесь не было и передне- Т1
Гл, II. Развитие ландшафтов и фаун в четвертичном периоде
155
азиатских видов, обнаруженных в Палестины, Сирии, Ливана:
Herpestes ichneumon L.
Felis chaus Giilci.
Panthera pardus Schreb.
Battus rattus L.
Spalax sp.
Phacochoerus garroda Bate
Hippopotamus amphibius L.
средне- и верхнеплейстоценовых слоях
Duma mesopotamica L.
Gazella arabica L.
Gazella cf. subgutturosa Giild.
Capra primigenia Frass
C, beden Schreb.
Bubalus sp.
Procavia cf. syriaca Schreb.
Учитывая значительную удаленность Сирии и Палестины от Апшерона, можно думать, что эта разница в видовом составе имеет зоогеографическое значение, удержавшееся на разных стратиграфических этапах.
Наконец, по сравнению с современным составом фаунистического комплекса равнины восточного Закавказья бинагадинский характеризовался отсутствием ряда эвритопных видов, что можно объяснить только более поздним проникновением их на Апшерон. К таким видам относятся ныне живущие: S uncus etruscus, С unis aureus, Hyaena hyaena, tfustela nivalis, Felis chaus, F. lynx, Panthera pardus, Rattus norvegicus, Qtrzella subgutturosa, Martes foina, недавно вымершие: Pantheraleo, Equus heraionus.
Таким образом, выясняется интересная деталь, что ряд южных теплолюбивых видов продвинулся в восточное Закавказье, а ряд европейских непных видов отступил в восточное Предкавказье уже после существования бинагадинской ловушки.
Все это, подчеркивая специфичность бинагадинского комплекса, позволяет также утверждать, что асфальтовая ловушка действовала относительно небольшой отрезок плейстоцена.
Стратиграфические сопоставления и возраст бинагадинского комплекса
Установление возраста Бинагадов представляет огромный интерес для разработки вопросов фауногенеза и морфогенеза млекопитающих в четвертичном периоде.
Попытки определения возраста бинагадинского костеносного горизонта й бинагадинского фаунистического комплекса делались на основе учета возраста древнекаспийской террасы, учета вероятных эпох сжатия пластов — орогенических фаз, сопровождавшихся истечением нефти и на основе палеозоогеографических сопоставлений.
Богачев (1939 г, стр. 47—51 и др.) датировал Бинагады рисс-вюрмом, учитывая хазарский возраст террасы, подстилающей закированные пласты. Сжатие пластов продуктивной толщи и истечение нефти он связывал с так называемой «калинской» фазой складчатости, приходящейся, по Рейнгарду (1937), на начало второго, рисского оледенения. По Варданянцу (1948, табл. 3, стр. 104, 125), значительные сжатия, сопровождавшиеся образованием складок на Апшероне, а также пробуждением грязевых вулканов, происходили в предхазарскую и в предхвалыпскую орогени-тяческие фазы, а затем в неоплейстоцене и голоцене. По этой схеме бинагадинский слой мог образоваться и в предхазарскую, и в предхвалыпскую эпоху.
Однако датируя время срезания бинагадинской складки хазарской трансгрессией, Богачев учитывал, что нефть калинской орогенической
156 Часть I. Обоснование истории формирования фауны
фазы не могла бы иметь значения для улавливания и консервации растений и животных, так как ее выжимание предшествовало хазарской трансгрессии. В связи с этим Богачев (19406, стр. 14) прибегал к признанию дислокации слоев с D idacna surachanica и существования двух Явлинских фаз сжатия.
Иная датировка Бинагадов была бы при объяснении срезания бина-гадинской складки водами Бакинского моря. Тогда образование асфальтовой ловушки можно было бы связывать и с предрисской, и с калинской орогенической фазами.
Своеобразие бинагадинского комплекса по сравнению с четвертичными комплексами Русской равнины затрудняет биостратиграфические сопоставления, тем более, что в нем не обнаружено остатков таких руководящих форм,.как слон и зубр.
Громов (1948, стр. 424 и др.) датировал бинагадинский комплекс поздним доледниковьем, проводя хронологические параллели со среднечетвертичной «хазарской фауной Поволжья», отнесенной Николаевым (1937) к миндель-риссу.
По обзору Бурчак-Абрамовича и Джафарова (1955) видно, что большинство палеонтологов, не вдаваясь в самостоятельные исследования, следовало датировке, предложенной Богачевым.
В работе о хищных бинагадинского асфальта (Верещагин, 19516) такой датировке следовал и я с той оговоркой, что эпоха захоронения может быть уточнена при производстве более совершенных раскопок и при установлениии истинного соотношения на Бинагадинском холме морских отложений с континентальными.
Сопоставление состава видов зверей из Бинагадов со списками видов из палеолитических пещер Палестины, Сирии и Ливана (Bate, 1937; Picard, 1937) показывает, что наибольшее число сходных форм из этих двух районов приходится на мустье и ориньяк, т. е. на верхний плейстоцен.
Наши определения остаточного органического вещества в костях бинагадинских лошади и сайги методом прокаливания (применяемом Пидопличко, 1952) показали, что после извлечения нефти горячим щелоком и бензином показатель прокаливания колеблется в пределах от 480 до 520, т. е. оказывается близким к показателям для костей хазарского комплекса. ?
Попытка определения возраста бинагадинских костей радиоактивными методами, произведенная В. В. Чердынцевым, успеха не имела.
На основе обзора всех палеобиологических и геологических фактов эпоху существования бинагадинского фаунистического комплекса и образования захоронения следует относить к началу хазарской трансгрессии, т. е. датировать концом среднего плейстоцена. В схеме Рейнгарда это будет соответствовать низам среднекаспийских слоев или второму рис-скому оледенению — «Вы», а в схеме В. И. Громова рисской стадии -«Q и».
Продолжительность эпохи существования бинагадинского комплекса в описанном составе точно неизвестна. Однако его отдельные элементы существовали здесь и в Хвалынском веке. Другие же заменялись викарными формами. Пример такого более позднего захоронения, в котором азиатский тип быка (быкМастана) заменяется европейским туром, но сайга продолжает существовать, имеется для другого асфальтированного кладбища — Кир-Маку.
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
157
Формирование некоторых морфологических черт у зверей восточного Закавказья на протяжении верхней половины четвертичного периода рассматривается в главе IV.
Подводя итоги тафономическому и зоогеографическому анализу Вина-гадов, приходим к следующим выводам.
1. Бинагадинские млекопитающие существовали в ландшафте сухой степи и аридного редколесья, развитого местами по оврагам и россыпям скал. Климат района был несколько более холодным и влажным, нежели в наши дни.
2. Животные различных систематических и экологических типов гибли в жидкой нефти и в толще вязкого асфальта, частью же тонули в селевых потоках.
3. Захоронение остатков животных и растений происходило на месте или близ места гибели в асфальтированных песках и в мелкобитом ракушечнике. Асфальтированная толща была позднее перекрыта эоловыми песчано-ракушечниковыми и озерно-лиманными отложениями.
4. Залегание костеносного слоя на окраине древнекаспийской террасы с индикаторными моллюсками хазарского яруса позволяет датировать бинагадинский фаунистический комплекс послебакинской эпохой.
Связывая обильное истечение нефти и процесс захоронения с ороге-пическими фазами, можно приурочить бинагадинский фаунистический комплекс либо к началу хазарского, либо к началу хвалынского века.
5. Сопоставление бинагадинского фаунистического комплекса с комплексами, известными из четвертичных отложений ксерофитной зоны Передней Азии (палеолит Сирии, Ливана, Палестины) позволяет датировать эпоху его существования культурой мустье. По показателям остаточного органического вещества кости бинагадинских зверей близки к костям хазарского яруса Поволжья.
6. При 39 обнаруженных видах млекопитающих бинагадинский комплекс содержал до 19 (48.7%) плейстоценовых видов, широко распространенных на равнинах Западной и Восточной Европы. Остальные 19 видов (48.7%)“плейстоценовые переднеазиатского, иранского, происхождения, а лишь один вид (2.6%) может считаться эндемичным кавказским.
В этом комплексе, по-видимому, совершенно отсутствовали арктические виды, горные звери, а также пещерные медведи, слоны и зубры. В эпоху существования бинагадинского комплекса на Апшероне смыкались ареалы некоторых зверей степей Русской равнины и нагорных степей Передней Азии. Более поздние захоронения на Апшеронском полуострове уже ее отличаются такой универсальностью, как Бинагадинское.
Кир-Маку
Захоронение находится в полутора километрах к югу-юго-востоку от сел. Дыгя, в центре Апшеронского полуострова, на склоне бугра Кир-Маку (т. е. «асфальтового»).
Кир-Макинское «кладбище» было открыто еще в 1870 г. А. А. Штукен-сергом, который собрал здесь и передал в Казанский университет заки-рованный череп самки сайги и нижнюю челюсть волка.
По карте Воларовича и Леднева (1913—1929) видно, что холм Кир-Маку достигает 100 м абсолютной высоты при широтном диаметре осно-вашш около 1.5 км и возвышается над окружающей местностью на 60— (О м. Шапка холма сложена древнекаспийскими отложениями, которые подстилаются песками средней свиты продуктивной толщи (средний плиоцен), поставленными почти вертикально. Холм имеет два широких, по-
Ш
Часть I. Обоснование истории формирования фауны
гребенных или по лупогребенных асфальтовых потока на северном склоне и один — на юго-восточном. Костеносный закированный слой расположен здесь несколько ниже вершины холма и залегает на глубине от 2 до 5 м в виде пятна площадью в 1.5—2 га. Обобщенный разрез четвертичных отложений Кир-Маку на участке за-кированных пятен приведен по Султанову (1947) на рис. 72.
По имеющимся наблюдениям, битум встречается здесь часто в виде вертикальных жил диаметром у основания до 20 см при В длине до 1.5—2 м, но основная его масса К представлена все же сплошным слоем скорее В всего натечного происхождения. Кости на- ходятся как в натечном битуме, так и в би- К тумных жилах. По-видимому, нефть, выходя К на поверхность при сжатии пластов, обра- зовывала на склонах липкие натеки, а ме- К стами и лужи, которые затем перекрывались ж песчано-пылеватыми наносами. Ряд живот- В ных, несомненно, погибал, завязая в нате- К ках асфальта. Как правило, здесь обнаружи- В вались лишь разрозненные части скелета ж и черепа. ж
По-видимому, Кир-Макинское захороне- " ние имеет ряд общих черт с травертинами горы Машук на Северном Кавказе. Однако асфальтовые натеки обладали, очевидно, самостоятельной высокой уловистостью.
Уже давно здесь был найден и описан (Богачев, 1924, 19256) череп первобытного тура, не того же равнине
Богачев датировал эпоху образования битуминозного солончака у сел. Дыгя после-бакинским временем, а гибель и захоронение быка связывал с эпохой увлажнения и распространения степной флоры на Annie-ронском полуострове в хвалынский век.
В 1940—1941 гг. в Кир-Маку было извлечено значительное количество костей перво-
бытного тура, сайгака, волка, лисицы, пещерной гиены, грифов, добытых землекопами при разработке битума и перезахороненных в особой яме (Богачев, 1944; Султанов, 1947).
В стратиграфическом смысле кир-макинские слои моложе бинага-динских. Здесь не вызывает сомнений возраст рельефа, так как битуминозный слой залегает соответственно рельефу склона. |
Первобытный тур из Кир-Маку — более поздний тип быка, сменивший бинагадинского тура в верхнем плейстоцене восточного Закавказья, В кир-макинской плейстоценовой группировке, однако, еще существовала сайга, а возможно и гиена бинагадинского типа.
Верхним плейстоценом следует датировать и находки в битумах о. Артема. Месторождение битумов находится в северо-восточной части острова.
Рис. 72. Разрез асфальтированных слоев в Кир-Маку.
а — покровные суглинки; б — песчано-глинистая толща; в—слой «жирного» асфальта; г — пески продуктивной толщи; справа — мощность в метрах.
отличавшийся по типу от особей вида, существовавших на Русской в верхнем плейстоцене и голоцене.
г
л
Д
я
Т(
32
Ж
&
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
159
Отсюда были извлечены два черепа благородного оленя с обломанными кронами рогов, хранящиеся в Естественноисторическом музее в Баку (Богачев, 1938а). Здесь же в старых отвалах в 1944 г. были собраны и другие кости благородного оленя, несколько костей лошади, волка и крупных птиц (Бурчак-Абрамович, 1951в).
Судя по типу сохранности костного вещества, возраст этих островных захоронений близок к кир-макинскому.
Обитание оленей и лошадей на о. Артема имело место, вероятно, в относительно влажную и холодную эпоху самого верхнего плейстоцена,
ИСТОРИЧЕСКАЯ ЭПОХА
Большой
Кавказ ।
Широколиственные
Альпийская зона----j
леса
Нигер но ‘•солянковая полупустыня
Эфемеровая полупустыня1
Винагад
полустепъ
Шемаха
гБзба-Даг
3000 \~2000
-1000
,--т--т-т--т--т--т-, >riW
О 10 20 30 W 50 60 70 80 90 100 НО 120 130 1Ы1150 160 км.
СРЕДНИЙ ПЛЕЙСТОЦЕН
Большой Кавказ
Рис. 73. Изменения и сдвиги ландшафтных зон на Апшеронском полуострове с среднего плейстоцена по данным изучения Бинагадинского захоронения.
когда копытные широко кочевали по Апшерону, перебираясь и на прилежащие острова, особенно в весенние сезоны пышного развития эфе-меретума. Связывать заселение острова с регрессией Каспия нет необходимости, так как узкие проливы звери могли преодолевать вплавь.
История формирования ландшафтов и фаунистических комплексов равнины восточного Закавказья в плейстоцене по имеющимся находкам представляется в следующем виде.
В нижнем плейстоцене здесь преобладали ксероморфные ландшафты, унаследованные, вероятно, еще с среднего плиоцена (века продуктивной толщи). В эпоху бакинскотт трансгрессии по побережьям Куринского залива паслись эласмотерии, носороги, лошади, верблюды. Из хищных жила гиена типа пятнистой или бурой гиены Африки.
HIW
160 Часть /. Обоснование истории формирования фауны
К среднему плейстоцену на равнинах восточного Закавказья оформи- г ровалея комплекс млекопитающих, состоящий из видов, широко распро- [ страненных в плейстоценовых степях умеренного пояса Евразии, и из нагорно-степных видов Передней Азии.
* По аналогии с западным Закавказьем в горах Большого хребта в это же время, вероятно, обитали бурые и пещерные медведи, куницы, рыси, стада козлов и серн.
Оледенение восточных отрогов Большого хребта вряд ли было велико. Тем не менее в верхнем плейстоцене на общем фоне ландшафта аридного редколесья и степи, на равнинах произошел ряд фаунистических преобразований. |
Бык азиатско-индийского типа был замещен европейским туром, ис-чезли, по-видимому, аргали и гигантский олень. В какую-то фазу верхнего j плейстоцена, возможно связанную с наибольшим похолоданием и увлаж- I нением климата, в ^Закавказье, а возможно, и на Иранское плоскогорье { проник ряд видов плейстоценовых степей и лесостепи Восточной Европы. : В эпоху похолодания, вероятно, имело место некоторое смещение высот- । ных ландшафтных зон вниз (рис. 73), не нарушившее тем не менее непре- I рывного существования здесь ксерофильной группировки зверей передне- I азиатского происхождения. г
Отступание ареалов восточноевропейских степных видов: корсака, I большого тушканчика, сайгака —к северу произошло в самом конце верх-1 него плейстоцена при новом этапе потепления и иссушения ландшафтов. :
Наступление сухой и теплой голоценовой эпохи вызвало здесь новую миграционную волну (южных теплолюбивых видов, заселивших Закавказье, а частью проникших и далеко к северу в предкавказские равнины.
Нагорье Малого Кавказа (южное Закавказье)
Высокие плоскогорья южной Грузии, Армении, Карабаха в тафономическом смысле занимали в плейстоцене особое положение. Горный рельеф обусловливал частое возникновение селевых потоков, лавин, камнепадов, вулканических извержений, а следовательно, и гибель животных, но условия для образования захоронений были редки. Известные плейстоцене- j вые захоронения находятся здесь либо на окраинных участках широких речных долин, либо в озерных западинах высокогорных плато. Они образовывались в конусах выносов речек, грязевых потоков, в делювии склонов и в озерных отложениях.
Роль древнего человека в образовании захоронений фаунистических остатков на Армянском плоскогорье оказалась ничтожной, несмотря на обилие палеолитических находок (Замятнин, 1947; Паничкина, 1948, 1950а, 19506; Сардарян, 1954). Причина этому заключается, по-видимому, в разрушении большинства среднепалеолитических пещер и в размыве слоев открытых стоянок.
Исключительный интерес представляет недавно найденное геологом А. К. Векуа захоронение костных остатков нижнеплейстоценовых млекопитающих в древних озерных отложениях близ Ахалкалаки. В находках 1958 г. Векуа определил остатки следующих видов: Canis sp., Crocus spelaea, Meles cf. meles, Marmota sp. (мелкаяформа), Elephas cf. trogontherii} Equus sp., Rhinoceros etruscus, Hippopotamus sp. (крупная форма), Mega-ceros sp., Bos cf. primigenius. Наличие здесь бегемота и какого-то своеоб-г разного сурка придает южный облик этой группировке, свидетельствуя;
CJ б.
TJ 01
Д} K1 то ле
ко ло к а ку
яв в
с I гад Ко OTJ Ав фо< сох
Гл. II, Развитие ландшафтов и фаун в четвертичном периоде
161
о ландшафте низких плоскогорий с развитой гидрографической сетью и теплом климате района на заре антропогена.1
В 1928 г. в песчано-галечниковых карьерах близ Казачьего поста, ца южной окраине Ленинакана, было обнаружено большое скопление костей, до «300 кг» которых было собрано и перевезено в музей Еревана (Богачев, 1938 в, 1938г; Авакян, 1948). Богачевым были определены остатки: Bison prisons, Megaceros sp. (hibernicus?), Corvus elaphus moral, Equus caballus. Остатки слона — Elephas (cf. armeniacus) были найдены 8 более глубоком горизонте, отнесенном к верхнему плиоцену (апшерон-
Рис» 74. Череп Bos primigenius из Баяндура на Армянском нагорье.
сапГярус и его эквиваленты). Остальные находки датировались плейстоценом. Авакян и Бурчак-Абрамович (1945) описали в сохранившихся из пого пункта сборах остатки верблюда типа Camelus knoblochi, а в качестве сопутствующих видов назвали: Equus stenoms, Rhinoceros ci. mercki, Elephas mendionalis, E. trogontherii (2), Cervus sp., Bos sp.
Зубы слонов из-под Ленинакана, изображенные Сардаряном (1954), свидетельствуют о преобладании в сборах остатков Elephas trogontherii, дпзкого по тину к тираспольскому Е. wusti и отчасти к слонам из конгломератов Таманского полуострова. Отсюда же Бурчак-Абрамович (1951 г) описал фрагмент черепа Bos primigenius, хранящийся в Музее Грузии.
фрагмент черепа Bos primigenius (рис. 74), найденный близ сел. Баян-дУр Лепинаканского района, был описан Громовой (1931). Уже этот краткий обзор показывает, что под Ленинаканом имеется аллювиальная пища с весьма разновозрастными остатками, очень напоминающая галечниковые отложения наклонных равнин Предкавказья.
В основной массе сборов все же прослеживается пижиечетвертичный комплекс с такими индикаторными видами, как трогонтериевый слон, лошадь Стенопа и носорог Мерка. Все это позволяет сопоставлять ленина-канскую пресноводную толщу с верхними горизонтами галечников Псекупса и нижними горизонтами карьера Гирей на средней Кубани.
Близкими — нижне- и частью средиечетвертичными — по возрасту являются остатки зверей из песчапо-гравийной толщи близ сел. Эйлас,
12 км к юго-западу от Еревана. Серые мелкозернистые речные пески костями млекопитающих залегают в урочище Аджи-Эйлас под слоями галечников и покровных лёссовидных суглинков па глубине 3—3.5 м. Костеноспые линзы достигают здесь мощности 1 — 1.5 м. Кости частью переотложены, очевидно, из каких-то более древпих захоронений озерного типа. Авакян (1946) датирует эти слои рисской эпохой. Фрагменты сильно фоссплпзованы. Обломки рогов «мелового» пли «мергелевого» типа сохранности. Эмаль зубов оленей буроватая от прокраски дентина. Про
1 Возможно, что возраст этой группировки более древний — верхнецлиоценовый. IJ Н. К. Верещагин
.Часть I. Обоснование истории формирования фауны
смотренные нами костные остатки из этого пункта содержат зубы Ek-phas trogontherii, фрагмент черепа быка типа «дилювиальных туров» — Bos trochoceros, обломки рогов и зубов оленей — Cervus ex. gr. elaphus и лани — Dama cf. mesopotamica (рис. 75).
Более поздние, верхнеплёйстоценовые захоронения, отчасти обуслов- L ленные деятельностью первобытных охотников, обнаружены пока лишь I на северных участках нагорья Малого Кавказа. В литературе имеется f сообщение о находке обломков бивней и костей скелета Elephas primi- | genius, а также зуба Equus caballus на глубине 10 м у Налбанда, в 30 км I
Рис. 75. Аджи-Эйлас.
J — череп Bos trochoceros; 2, 3 — обломок рога и челюсть Dama cf. mesopotamica, к северу от Ленинакана, на левом берегу речки Чичхана, а также костей грызунов и обломка оленьего рога (Каракаш, 1898).
Зуртакетская стоянка. Особое стратиграфическое значение придавалось вначале палеолитической стоянке, открытой Маруапь вили (1946) в южной Грузии на Цалкинском плоскогорье. Она была обнаружена в 1945 г. на правом берегу долины р. Карабулах, правого притока Храма, в 15 км к северу от Башкичети, на высоте около 1320 м. Речка прорезает здесь лавовый покров толщиной 15 м. Следы человеческой деятельности представлены в виде раздробленных костей, орудий из кремния, обсидиана и кости. Преобладают обсидиановые изделия, которых в 10 раз больше, нежели кремневых. Орудия из кости и камня уже прикреплялись к древку, так как напоминают наконечники дротиков и стрел. По каменному инвентарю стоянка датируется эпохой солютре.
Полагая, что лавовый покров перекрыл и обжег культурный слой, вытеснив древнее племя, Маруащвили сильно «омолаживал» возраст
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 163
рельефа нагорья и время интенсивной вулканической деятельности. Эти представления были опровергнуты Паффенгольцем (1951), установившим, что стоянка располагалась в обрушившейся теперь нише стенки дол ери-говоп лавы.
Собранные Маруашвили в 1945 и 1948 гг. остатки млекопитающих состояли из коренных зубов, резцов и эпифиза пясти лошади, из коренных осла, позвонков празубра и фрагмента лучевой барана.
Костное вещество остатков большей частью выветрелое, с небольшим числом дендритов на поверхности. Тип его сохранности говорит о весьма поздней эпохе плейстоцена.
Бурлак-Абрамович (1951а) дал для Зуртакети список материала (табл. 27).
Таблица 27
Видовой состав и численность костных остатков млекопитающих Зуртакетской стоянки
Виды
Число костей Число особей
Equus caballus subsp.............
Е. (Asinus) sp...................
Bison priscus....................
Ovis cf. gmelini.................
Всего..................
Известны также остатки куропатки, близкой к кеклику.
Судя по видовому составу животных в добыче зуртакетцев, верхне-плейстоценовый ландшафт бассейна Храма рисуется в виде плоскогорий увалов с лугово-степной и со слабо развитой лесной растительностью.
Присутствие ослов-куланов, мелких баранов было недавно весьма характерным и для современных ландшафтов более южных безлесных территорий.
Деятельность древних охотников прослеживается и в верховьях Храма, где близ сел. Бармаксыз Куфтин (1941) обнаружил пещерную эшшалеолитическую стоянку на высоте около 8 м над уровнем реки.
Культурный слой с обсидиановыми орудиями достигал не более 40 см, а почвенный покровный слой — до 80 см мощности. Было собрано j)900обламков обсидиановых орудий и 600 кремневых на площади 70 м2 размытого склона.
Костный материал состоял из кусочков разбитых диафизов и из прокрашенных гумусом зубов копытных
По определениям Бурчак-Абрамовича (19516), в сборах оказалось j коренных зубов мелкой лошади и задний нижнекоренной какого-то дикого быка.
Столь же поздние геологические находки были сделаны Далем (19506) и побережье оз. Севан при искусственном понижении уровня вод.
В собранных обширных материалах преобладают фрагменты домашних даотных послепалеолитической эпохи (об этом см. дальше).
Напболее древние по типу сохранности остатки принадлежат карликовому туру— Bos cf. minutus и одногорбому верблюду — Camelus
11*
164 Часть I. Обоснование истории формирования фауны
dromedarius, предположительно отнесенного Хавесоном (19546) к дикой форме.
Несмотря на относительную бедность видового состава зверей в плей-стоценовых слоях южного Закавказья, здесь все же усматривается закономерность в смене фаунистических комплексов, сходная с Предкавказьем.
Интересно отметить, что в плейстоценовых захоронениях, обнаруженных на плоскогорьях Малого Кавказа, прослеживается ряд индикаторных видов зверей ланшафта «холодных степей» Европы: трогонтериевый слон, носорог Мерка, лошадь, верблюд Кноблоха, гигантский олень, благородный олень, первобытный тур. Из специфических видов Передней Азии известны лишь лань и муфлонообразный баран. Южный, восточносредиземноморский характер здешнего фаунистического комплекса был подчеркнут, очевидно, в большей степени среди мелких плейстоценовых форм.
Детального стратиграфического расчленения известного набора видов еще не произведено. Не выявлены и морфологические отличия руководящих форм с учетом стратиграфической и географической изменяемости.
«Следы» высокогорного верхнеплейстоценового комплекса, в противоположность Большому Кавказу, здесь пока не прослежены. Частично это объясняется слабым развитием и исследованностью карста. Бедность палеолитических находок не позволяет судить и о влиянии плейстоценовых похолоданий и оледенений хребтов плоскогорья на формирование фауны. Несомненно, однако, что большая, чем ныне, обводненность и мезофит-ность плоскогорья в нижнем и среднем плейстоцене не препятствовала существованию верблюдов в жарких межгорных долинах. Вертикальные сдвиги ландшафтных зон и фаунистических комплексов при плейстоценовых похолоданиях были здесь, вероятно, еще меньшими, нежели на Большом Кавказе.
Переход к голоцену на Армянском плоскогорье плохо документирован, но совершался, по-видимому, довольно сходно с восточным Закавказьем.
Стратиграфическое и географическое распределение местонахождений остатков плейстоценовых млекопитающих в пределах Кавказа показано на рис. 2 (см. «Введение»).
ФАУНИСТИЧЕСКИЕ СВЯЗИ И ЛАНДШАФТЫ КАВКАЗСКОГО ПЕРЕШЕЙКА В ПЛЕЙСТОЦЕНЕ ПО ПАЛЕОНТОЛОГИЧЕСКИМ
ДАННЫМ
В результате регионального обзора плейстоценовых захоронений и на основе геологических фактов особенности формирования кавказской фауны представляются теперь в следующем виде.
Фаунистический комплекс нижнеплейстоценовых горных поднятий Кавказа, известный пока лишь по раскопкам пещеры Кударо I, был уже в достаточной мере специфичен и представлен прямыми предками современных кавказских видов насекомоядных, грызунов и копытных, характерных для средиземноморской системы альпийской складчатости.; Сильная пенепленизация Кавказа, происшедшая будто бы к началу плейстоцена, которая предполагается некоторыми геологами (см., например, Варданянц, 1948), с точки зрения зоолога, совершенно невероятна. Формирование горной кавказской фауны с такими специали-:
Гл, II, Развитие ландшафтов и фаун в четвертичном периоде
16д
зированными видами, как прометеева и снеговая полевки, кавказские козлы, серна, шло параллельно аналогичным процессам в Альпах, Карпатах, Малой Азии по крайней мере с миоцена из сходного, родственного материала. Развитие и возникновение этих горных видов < лишь на протяжении плейстоцена трудно представить себе уже потому, что в конце среднего плейстоцена это были уже вполне современные виды.
Основные миоценовые поднятия центральных участков Большого ; Кавказа, очевидно, все же сохранялись до плейстоцена, хотя бы и сглаженные, срезанные в той или иной степени эрозией.
! Лучше известные захоронения остатков животных средне- и верхнеплейстоценового возраста отражают в пределах Кавказа дальнейший этап ! общего похолодания климата и модернизации фауны.
К среднему плейстоцену в Предкавказье и в Закавказье исчезли эла-j смотерии и сложнорогие олени* На предгорных равнинах шло развитие I степного и лесостепного ландшафтов, сменявших ландшафты полупустынь | и саванны. Следствием этого было дальнейшее обособление зональных | фаунистических комплексов. В лесостепи предгорий Предкавказья в эту эпоху обитали трогонтериевые слоны, носороги Мерка, благородные олени, длиннорогие зубры. В восточном и южном Закавказье зубры были замещены первобытными турами.
Широкое внедрение некоторых сухолюбивых видов арало-каспийской фауны в восточное Предкавказье, например предков малого суслика, среднеазиатских песчанок и тушканчиков, могло происходить весьма рано, даже в конце апшеронского века. Предки этих видов при трансгрессиях Каспия могли сохраняться по окраинам Восточнопредкавказского залива, либо исчезать здесь нацело и заселять территорию при новых регрессиях моря совершенно заново.
В западном Закавказье по круто обрывающемуся к морю южному склону Большого хребта в среднем плейстоцене существовал горно-лесной фаунистический комплекс, содержавший медведей, кабана, оленя, кавказского козла, зубра.
Фаунистическое влияние степей Русской равнины уже ощущалось здесь по наличию обыкновенного хомяка, проникавшего на юг по крайней мере до Цебельды.
Низкие плоскогорья Имеретии являлись в верхнем плейстоцене пре-I делом распространения на северо-запад сухолюбивых форм нагорий Передней Азии: среднего хомяка, дикобраза, осла, аргалиобразного барана. Наличие лошади и осла свидетельствует о значительном остей-нении в бассейне среднего течения Риона. Уцелели здесь с плиоцена даже обезьяны.
j Аналогичные остепненные ландшафты, усложняемые местами грандиозными поверхностными излияниями лав, существовали и на нагорьях Малого Кавказа, а также в центральном Закавказье.
С концом нижнего плейстоцена связывается на Кавказе и представление о начале заметного непосредственного воздействия палеолити-1 ческого человека па фауну крупных млекопитающих. Это воздействие [ । было более заметно в западной половине перешейка, нежели в восточной.
1 В восточном Закавказье на протяжении среднего плейстоцена устой-
’ чпво держался еще с верхнего плиоцена ландшафт сухих предгорий
г можжевелово-фисташковыми лесами и высыхающим к лету травянистым / покровом степного типа. Фаунистический комплекс района в среднем
~ плейстоцене содержал своеобразную смесь зверей плейстоценовых сте-
пей Русской равнины: корсака, сайгу и др.; переднеазиатских нагорий:
[-
, 166
Часть I. Обоснование истории формирования фауны
»
пятнистого кота, гепарда, слепушонки и др.; пустынь туранского типа: малого тушканчика; наконец, местных кавказцев: апшеронскую полевку.
Характерно было отсутствие здесь пещерных медведей, слонов, зубров. Северное фаунистическое влияние в среднем плейстоцене восточного Закавказья ощущалось лишь в смысле проникновения некоторых видов плейстоценовых степей Русской равнины: корсака, большого тушканчика, сайги (рис. 76).
ВЕРОЯТНЫЕ АРЕАЛЫ ТЕРИОКОМПЛЕКСОВ В КОНЦЕ
Г
<
Рис. 76.
Ареалы степных «северных» и «южных» видов развивались по окраинам перешейка навстречу друг другу, вероятно пульсациями, неравномерно и разновременно. Можно думать, что и «перекрывание» Кавказского перешейка ареалами некоторых других северных плейстоценовых видов, например обыкновенной полевки, первобытного тура, происходило также неодновременно уже в среднем плейстоцене, в соответствии со сменой сухих и влажных климатических фаз.
Фаунистическое влияние Русской равнины прогрессивно нарастало на протяжении плейстоцена и сказывалось сильнее в равнинном Предкавказье, нежели в Закавказье.
В начале верхнего плейстоцена, позднее существования бинагадинского комплекса, в предгорьях западного Предкавказья обитали уже настоящий мамонт и несколько измельчавший зубр.
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 167
Значительная остепненность предгорной Закубанской равнины в начале верхнего плейстоцена выясняется по наличию в Ильской стоянке остатков ряда степных травянистых растений и жуков, лошади, осла, сайги. Остатки зверей из местонахождений верхнеплейстоценового возраста па равнинах Предкавказья показывают лишь все ту же остепнен-рость ландшафтов. Здесь, по-прежнему встречаются лошадь, кабан, бла-родный олень, сайга, зубр, первобытный тур. Такая группировка чкет свидетельствовать о степи, лесостепи, но уж никак не о сплошных « ах, не о мезофитных лугах и тем более не о моховых болотах и тундре. Судя по современным палеофаунистическим данным, влияние оледе-лш Восточной Европы, так же как и горнокавказских, не отражалось четно на составе фаунистических комплексов предгорных равнин, оборот, по массовым сборам костей на отмелях низового Дона легче следить этап или этапы развития степного и даже полупустынного цшафтов, нежели лашдшафтов мезофитных. Наличие верхнеплейсто-, ювых и голоценовых остатков зайца-беляка и северного оленя на ничем Доне следует рассматривать лишь как результат весьма поздних нов развития их ареалов к югу по поймам крупных рек Русской рав-ИЫ, по не как прямого следствия сдвигов их ареалов под влиянием поеданий и оледенений севера Европы. Из предгорных и равнинных плейстоценовых захоронений с самого Кавказского перешейка совершенно неизвестны остатки горных форм, например снеговых полевок, твказских козлов, серн. Это может указывать до известной степени на сутствие покровных оледенений и тех грандиозных сдвигов фитоланд-чЬтных зон (с выходом альпийских группировок на равнину), которые повались некоторым прежним зоологам и современным геоморфологам.
При признании покровных оледенений кавказских хребтов с выходом дников на предгорные равнины необходимо было бы считать, что высоко-.рные ныне козлы, серны, прометеевы полевки либо нацело вытеснялись , предгорные равнины, либо откочевывали на какие-то соседние хребты-убежища, не подвергавшиеся оледенению, либо нацело вымирали.
Похолодание, а следовательно, и общее понижение вертикальных .шдшафтных зон прослеживаются по современным палеонтологическим материалам в западном и восточном Закавказье со среднего плейстоцена. Оно сказывалось в более широком распространении относительно мезофильных зверей и зверей горно-лесной зоны.
Следует отметить также, что Кавказский перешеек в связи с его более южным положением и разнообразным рельефом обладал в плейстоцене, вероятно, более богатым составом зверей, нежели юг Русской равнины. Очевидно также, что в Закавказье существовали убежища — рефугии — ця ряда субтропических и гидрофильных видов зверей на протяжении всего плейстоцена. В Колхиде и Гирканике благодаря близости Черноморского и Каспийского бассейнов могли пережить плейстоцен, например, акне виды, как макак, черная крыса и дикобраз.
Все это довольно хорошо согласуется с результатами изучения обильях палеонтологических материалов в Палестине, Сирии, Ливане (об пом см. далее). Рассмотрение верхнеплейстоценовых фаунистических комплексов Кавказа и сопоставление их состава с сирийско-ливанским убеждают в том, что пути далекого проникновения на Кавказ и вообще к югу лесных европейских форм, как например лесной куницы и других, могли идти в большей степени через Эгейскую сушу, т. е. через Балканы и краевые хребты Малой Азии, чем через юг Русской равнины и равнины Предкавказья.
168 Часть I. Обоснование истории формирования фауны
Вместе с этим Кавказский перешеек на протяжении плейстоцена был, । по-видимому, непреодолимым экологическим барьером для проникнове- { ния ряда степных зверей Русской равнины, например степного хоря, | малого суслика, на юг, в Закавказье, и зверей нагорных степей Передней | Азии — песчанок южного типа, горной слепушонки и других видов — ( в Предкавказье. |
Существование ряда ксеротермических эпох на протяжении плейсто- | цена не улавливается на современном зоопалеонтологическом материале. |
Можно говорить пока только об одной эпохе остепнения в среднем f плейстоцене и о послеледниковой сухой и теплой климатической фазе. J
Потепления климата на протяжении плейстоцена отнюдь не приводили к возврату фауны субтропического типа даже в Закавказье. Фауна пред- f гории и низких плоскогорий оставалась сухолюбивой и лишь относительно t термофильной. В Закавказье происходило продвижение на север по пере- i шейку отдельных видов малоазийско-иранских нагорий, а в Предкав- I казье —• собственно в районе Приманычья — продвижение к западу и юту зверьков пустынь Турана. Никаких новых вселений настоящих суб-1 тропических млекопитающих из южной и Передней Адзии, а тем более! из Африки в теплые (межледниковые?) фазы плейстоцена не было даже -в Закавказье. Аналогичная картина наблюдалась, в сущности, и среди растительных форм.
Пути и этапы формирования плейстоценовой фауны Кавказа выявляются также при сопоставлении ее с плейстоценовыми фаунами смежных территорий: Русской равнины, Крыма, Передней Азии. Возможность сравнения имеется преимущественно для среднего и верхнего отделов плейстоцена, так как данные для нижнего плейстоцена и эпохи, переходной к плиоцену, пока недостаточны.
Общий список млекопитающих среднего и верхнего плейстоцена Русской равнины включает ныне, по Громову (1948) и Пидопличко (1954), с учетом наших неопубликованных данных по Волге и Дону, до 74 видов, Из них общих с Кавказом можно насчитывать до 36, т. е. около 50%. Это сходство было бы, вероятно, еще большим при успешных поисках костных остатков в прирусловом аллювии рек равнинного Предкав-; казья.
Основное отличие кавказской плейстоценовой фауны от фауны Русской, равнины — это уже отмечавшееся отсутствие бореальных видов: песца, зайца-беляка, леммингов, северного оленя, овцебыка — и наличие горных видов: козлов, серн, баранов, а также ряда грызунов передне-азиатских нагорий.
При сопоставлении плейстоценовых фаун Кавказа с фауной Крыма, известной ио исследованиям Бирули (1930а, 19306), Виноградова (19376), Громовой (1935а), Громовой и Громова (1937), Громова (1948), выясняется, что из 58 названных для Крыма видов 35, т. е. до 50%, оказываются общими с Кавказом. Среди общих видов отметим представителей равнинной, степной группировки: ушастого ежа, волка, лисицу, корсака, пещерную гиену, барсука, тигрольва, большого и малого тушканчиков, мамонта.
Основное же отличие сказывается в отсутствии на Кавказе широко распространенных степных форм: малой пищухи, рыжеватого суслику хомячка Эверсманна, желтой пеструшки — и северных видов: песца? зайца-беляка, северного оленя, — столь характерных для слоев верхнего палеолита во многих крымских палеолитических стоянках. Большое сходство состава крупных зверей этой маленькой горной страны с Кавказом позволяет думать, что эти территории унаследовали и получили
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
169
I
й а, те е-
:а
мн
Hi, ую та. жо ка, Да, иго шое давили
плиоценовых и плейстоценовых зверей из одного источника и даже соединялись друг с другом в . нижнем плейстоцене через какую-то сушу на месте Азовского моря. Фаунистическое влияние юга Русской равнины было главнейшим для обеих территорий.
При рассмотрении южных и юго-западных фаунистических связей Кавказа в плейстоцене следовало бы прежде всего охарактеризовать палеонтологически южное побережье Черного моря и внутренней Анатолии. Однако данные по плейстоценовой фауне из этих районов отсутствуют, хотя следы палеолитического человека ашельской стадии и прослежены внутри страны Питтаром (Pittard, 1929), Пфанненштилем (Piannenstiel, 1941), Сотэ (Sauter, 1948).
Судя по распределению находок палеолита, прослеженным Сотэ, человек освоил внутренние части Малой Азии (район оз. Туз и другие) уже на первых стадиях своего развития — шель—клектон—ашель — и продержался здесь до мезолита. Это свидетельствует о благоприятном характере природы и, в частности, об обилии крупного четвертичного зверя на нередко опустыненных и засоленных ныне территориях смежной с Кавказом страны.
Фюрон (1955) также отмечает, что ландшафты внутренней Турции в плейстоцене были менее опустынены, нежели в наши дни. Глубина озер Туз и Будур была на 100 м больше современной. Из плейстоценовых похолоданий здесь оставило следы вюрмское.
Плейстоценовая фауна еще более южных районов Передней Азии и северо-восточной Африки известна главным образом по исследованиям Бланкевхорна (Blanckenhorn, 1901, 1910, 1921 —1922), Вофрея (Vaufrey, 1931), Бейт (Bate, 1937), Пикара (Picard, 1937). Из названных этими авторами 69 достоверных видов 22, т. е. до 31%, являются общими с Кавказом. Если учесть еще сомнительных по определению тигрольва, тура, волосатого носорога, то сходство будет еще большим. Нет, однако, оснований считать, как справедливо и отметил Пикар, что в Сирии и Ливане в плейстоценовую эпоху жили лось и северный олень. То же самое следует сказать и относительно Египта, для которого Бергом (1947, стр. 82), некритически цитировавшим Бланкенхорна, были названы эти северные формы. Для разработки истории развития фауны Кавказа важно учесть наличие в плейстоценовых отложениях Палестины остатков лесной куницы, перевязки, лесного кота, малоазийского хомяка, водяной крысы, косули и пр. Наконец, наличие здесь в верхнем плейстоцене видов южноазиатского происхождения: шакала, полосатой гиены, камышового кота, кулана, джейрана — позволяет проследить и понять поздний, послеледниковый этап вселения на Кавказ этих видов. Основу плейстоценовой палестино-сирийской фауны составляют все же передне-азиатские, североафриканские — средиземноморские элементы, включая вивер, льва, настоящих ослов, бородавочников, бегемотов, синайских козлов, ареалы которых, вероятно, в плейстоцене не подходили близко к Кавказу. Достоверное отсутствие в добытых материалах ! остатков мамонта при наличии трогонтериевого слона в ашеле может I служить хорошим руководящим указанием для выводов о вымирании и характере расширения ареалов отдельных «северных» плейстоценовых видов к югу через Эгейскую сушу и Кавказский перешеек.
Климатические и ландшафтные смены в Палестине выяснены, по данным изучения остатков зверей, несколько полнее, чем на Кавказе. '• Бейт (Bate, 1937) сделала попытку изобразить эти смены графически, г попользовав численность извлеченных при раскопках остатков костей
170 Часть /. Обоснование истории формирования фауны
газелей (ряд видов), животных ксероморфного ландшафта, и лани — Duma mesopothamica, животного, связанного с широколиственными лесами и с более мезофитным ландшафтом. Газели и лани были основным источником питания древних кармелитов, живших в пещерах Вади эль Мугара (Wadi el Mugara) и других.
В результате наибольшая численность остатков лани по отношению к числу остатков газелей оказалась в нижнем ангеле, затем в верхнем
ЭПОХИ И ГОРИЗОНТЫ ПРОЦЕНТЫ И ОБЩЕЕ ЧИСЛО ФРАГМЕНТОВ КОСТЕЙ КЛИМАТ И ФАУНА
100 90 80 70 60' SO 40 30 20 10 0 0 10 20 30 40 50 60 70 80 90 100
Неолит о Мезолит Очень сухой и теплый '
4 ^3102 Ъ>78 CazeLla sp.p31™ ^87 Наступлю ьше арюсти итепли Слабое увлажнение Увеличение сухости Теплый
Б- Мустье Лата s' mesopotamica ^8998 * Влажный, резкая смена фауны.
Мустье С ^23? Сухой и теплый
Мустье Возвращение сухости
< АшельЕ. 1 | ;<а» «VI sw юН" Теплый и сырой,иногда с появлением сухости Смешанная фауна, бегемоты и носороги
Ашель F ** *о Сырой, тра пическии
Рис. 77. Колебания численности костей газели и лани в кухонных отбросах пещер горы Кармел в Палестине на протяжении антропогена. [По раскопкам Гаррод и Бейт (Garrod and Bate, 1937)].
мустье и в верхнем ориньяке. Нижний ашель, по представлениям Бейт, характеризовался теплым и сухим климатом, когда Палестину населяли бегемоты и носороги. В среднем мустье был сухой и теплый климат, а в верхнем — влажный плювиальный, с резкой сменой фауны. Последнее слабое увлажнение усматривается также в верхнем ориньяке (рис. 77).
Эти плювиальные эпохи в Палестине соответствовали, вероятно, каким-то горным оледенениям на Малом и Большом Кавказе. Пикар (Picard, 1937) приходит к выводу, что прежние представления Блапкен-хорна (Blanckenhorn, 1910, 1921) и других о том, что климат Палестины в плейстоцене был сходен с современным климатом Германии, ошибочны.
Гл. II Развитие ландшафтов и фаун в четвертичном периоде 171
Наоборот, зоопалеонто логический материал свидетельствует о существовании здесь еще с плиоцена довольно устойчивого сухого средиземно-морского климата.
О похолоданиях и плювиальных периодах, по его мнению, говорят лишь отложения галечников, возможно, синхронные большим оледенениям севера. При этом главная плювиальная фаза Палестины соответствует наибольшему, рисскому оледенению Европы. Кроме того, с ашеля до мезолита заметны еще также два других плохо выраженных плювиала. В целом здесь, как и ныне, был ландшафт сухой пустыни с островками широколиственных лесов в более влажных ущельях.
На юге и юго-востоке от Кавказа, в пределах северо-западного Ирана, в настоящее время имеется три опорных точки, откуда известны остатки плейстоценовых млекопитающих. Это делювий мергелевых холмов близ г.Мераге, пещеры Тамтама и Бизитуп. В сборах А. Гебеля из-под Мараги (см. Брандт, 1870) в ЗИНс хранятся фрагментарные плейстоценового типа сохранности остатки: Canis lupus, Crocuta spelaea, Rhinoceros cf.
Equus caballus, E. cf. hemionus, Bos. sp., Ovis cf. ammon.
Из палеолитических слоев пещеры Тамтама близ Резайе, западнее оз. Урмия, по раскопкам Куна (Coon, 1951) определены остатки 15 видов зверей, из которых отметим песчанок — Meriones, дикобраза — Hystrix, оленя — Cervus elaphus, джейрана — Gazella subgutturosa.1 Остатки оленей составляют до 66%, а лошадиные — до 12% всех сборов.
В пещере Бизитун у известной скалы Бехистун, на дороге Багдад— Камадан, из верхнепалеолитических и позднейших слоев извлечены остатки 16 видов зверей, среди которых представляют значительный интерес находки шакала — Canis aureus, барса — Panthera pardus, пищухи— (k'hotona sp., джейрана и др. По численности и здесь на первом месте стоят олени — до 50% всех сборов, затем лошадь — 23% и джейран — 12%.
Ошибочно также указываются вновь остатки бобра. Изображен зуб тюленя, занесенный, вероятно, с Каспийского побережья. Эти исключительно интересные материалы свидетельствуют о верхнеплейстоценовых северных границах ареалов некоторых южных видов: шакала, барса, джейрана, — не обнаруженных в Бипагадах. Судя по фаунистическому составу, ландшафт и климат севера Иранского плоскогорья в верхнем плейстоцене были довольно близки к современным. Обилие оленей указываеат, впрочем, на существование хорошо развитых тугайных десов, позднее уничтоженных человеком.
Стратиграфическое и географическое распределение плейстоценовых видов на Кавказе и смежных территориях дано в табл. 62.
При современной изученности плейстоценовых «фаун» Кавказа и рассмотренных выше смежных территорий степень сходства и древность фаунистических связей может быть охарактеризована цифрами, указании в табл. 28.
Приведенные цифры отражают, конечно, и степень изученности плейстоценовых слоев, и их тафономические черты. Тем не мепее подтверждаются и уже высказанные выше соображения о том, что в среднем и верхнем плейстоцене на равнинах и в предгорьях перешейка преобладало фаунистическое влияние с севера и северо-запада (юго-запада) уже в то
1 По определениям Фразера (Fraser), Кун называет остатки речного бобра — ;<tor fiber, за которого и были приняты, очевидно, кости дикобраза.
172
Часть 7. Обоснование истории формирования фауны
1
Яр
Л|<.
время, когда ареалы южных — переднеазиатских и местных — кав-казских — форм нижнего плейстоцена либо отодвинулись на юг, либо приобрели реликтовый характер.
В результате обзора этапов и путей формирования фауны зоогеогра- i фическое районирование Кавказского перешейка может быть дано пока > лишь для верхнего плейстоцена. В эту эпоху здесь выделялись следую- \ шие округа. L
1. Предкавказский степной и лесостепной, охватывающий равнины! Предкавказья с Ставропольской возвышенностью и Таманским полу-Г
? -
Таблица 28
Степень сходства средне- и верхнеплейстоценовых «фаун» млекопитающих Кавказа и смежных территорий1
Кавказский перешеек Русская равнина Крым Передняя Азия
Число видов, известных из средне- и верхнеплейстоценовых слоев 83 74 58 69
Процент видов, общих с средне- и верхнеплейстоценовой «фауной» Кавказского перешейка 100 54 53 33
и й
s'
островом, от Маныча на севере до подножья Апшеронского полуострова на юго-востоке, мамонтов, лошадей, пещерных медведей сайгаков.
2. Кавказский горный, охватывающий горную систему Большого Кавказа. Преобладание мезофильных горно-лесных форм млекопитающих: бурых и пещерных медведей, горных полевок, козлов, серн — при наличии немногих сухолюбивых видов в восточной половине системы, уцелевших и ныне в Дагестане и Кабарде, по происхождению нижнеплейстоценового возраста: средний хомяк, безоаровый козел, малый суслик.
3. Закавказский предгорный, охватывающий предгорья и равнины западного и восточного Закавказья. Преобладание грызунов передне* азиатского происхождения, носорогов южного типа, лошадей и туров -на востоке и зубров — на западе.
4. Малокавказский плоскогорный, охватывающий плоскогорья Малого Кавказа. Обилие лошадей, муфлонов, первобытных туров. В жарких долинах южной окраины нагорья вероятно обитание верблюдов, а по северо-западным склонам — горио-лесных видов: кавказских полевок, козлов, серн.
Большого
Характерно преобладание й, гигантских оленей, зубров,
хребта на юге г
Шс
С01
W
KOI
к
1 Точность этих подсчетов быстро стареет в связи с открытием новых место-.нахождений и поступлением новых сборов. ,j=
Kai лит (Пи раз
г
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
173
ОБЗОР ЗАХОРОНЕНИЙ КОСТНЫХ ОСТАТКОВ В ГОЛОЦЕНОВЫХ ОТЛОЖЕНИЯХ
Условия гибели животных и пути образования захоронений в голоцене
В послеледниковье продолжались те же самые процессы гибели животных и захоронения их остатков, которые протекали и в плейстоцене. Ца новом этапе они имели все же важные отличия. Наступление сухой й теплой эпохи, очевидно, уменьшило масштабы водной эрозии склонов н отложений делювия и аллювия, а также вынос трупов и костей животных в область речных дельт, озерных и морских заливов.
U2
46
44
ОБЛАСТИ |
РАСПРОСТРАНЕНИЯ I КУЛЬТУР ЭПОХИ БРОНЗЫ
/г i
Я
Рис. 78.
ОТ ХУДО * X6S AOh Э доше.
Стало сильнее проявляться прямое воздействие человека на ландшафт: выжигание и вырубка лесов, изменение режима потоков.
Под прямым и косвенным воздействием человека в голоцене быстро сокращалась численность крупного зверя на равнинах, и природные захоронения их остатков становились весьма редкими.
Некоторые случаи природной массовой гибели и захоронения остатков зверей в наши дни были описаны для равнин Кавказского перешейка (Колесников, 1950; Верещагин, 1951г).
Следы человеческих культур послеледниковой эпохи прослежены на Кавказе уже с большой полнотой, причем открыты памятники эпохи мезо-дцта, неолита, меди, бронзы и железа, содержащие костеносные слои (Пиотровский, 1949). Е. И. Крупнов любезно предоставил нам схему размещения посленеолитических культур — рис. 78.
1Т4 Часть I. Обоснование истории формирования фауны
С неолита в корне меняется характер расположения и масштаба человеческих поселений. Они приурочиваются к побережьям рек и озер, а пещерные поселения остаются лишь как пережиток палеолита, развиваясь затем вновь в средневековье вследствие грабительских войн феодалов.
Малый размах тектонических процессов в голоценовую эпоху не создает уже условий для вскрытия морских, озерных и дельтовых захоронений так, как для третичных и плейстоценовых. Значительно возрастают зато возможности находок костных остатков из относительно недолговременных захоронений в делювии и в мелких нишах, пещерках, где эти остатки накапливались в связи с временными поселениями человека и хищников, особенно сыча и филина.
Кухонные костные остатки в слоях голоценовых поселений представлены теми же типами фрагментов, что и в палеолите, но число и разнообразие поделок из кости и рога сильно увеличивается.
Весьма характерно появление погрызов костей, сделанных домашними собаками, но фаланги и метаподии уже не дробятся так тщательно для добычи мозга, как раньше, в палеолите.
С энеолита на Кавказе появляются также погребения, содержащие целые черепа и скелеты диких и домашних млекопитающих, уложенные вместе с покойником в соответствии с погребальным ритуалом.
Наконец, за последние столетия нашей эры на Кавказе накопились и сохранились своеобразные «священные» коллекции черепов домашних и диких животных в результате религиозно-культовых обрядов у ряда горных племен.
Все эти типы захоронений, исследованные в той или иной степени, рассматриваются далее по геоморфологическим районам.
Западное Предкавказье и юг Русской равнины
Окончательное оформление в голоцене широких сухопутных связей предкавказских степей с южнорусскими обеспечивало возможность широкого встречного проникновения млекопитающих этих территорий.
Исследования по плейстоценовой фауне низового Дона показали обилие остатков млекопитающих голоценового типа сохранности, вымываемых из пойменных отложений на песчано-щебнистые косы. Кроме того, обильные сборы костей были сделаны при археологических раскопках поселений и курганов, проводившихся на низовом Дону Институтом истории материальной культуры АН СССР.
Особо ценные костные материалы были добыты В. Гольмстен при раскопках землянковых поселений XV—X вв. до н. э. на р. Царице и при раскопках хазарской крепости Белая Вежа близ станицы Цимлянской (Артамонов, 1952).
В коллекциях из раскопок на низовом Дону и особенно из Белой Вежи (Саркела) среди более чем 30 000 костей нами определено 11 видов домашних и 23 вида диких млекопитающих.1 В составе последних отражен еще богатый комплекс зверей степной зоны и долинных лесов. Группа охот-ничье-промысловых зверей включает 16 видов — от корсака, степного хоря и бобра до медведя, лося и зубра. Эти материалы показывают, что еще в средневековье (VIII—XIII вв. н. э.) ареалы ряда лесных зверей
1 Эти материалы публикуются особо.
Гл, II. Развитие ландшафтов и фаун в четвертичном периоде
175
нежной, и лесостепной зон тянулись по долинам рек к предкавказским равнинам, смыкаясь в пойменных дебрях Ей, Кубани с кавказскими ареалами этих же животных.
Главнейшие археологические памятники в пределах самого Предкавказья представлены слоями неолитических поселений, скифскими курганами, многометровыми культурными толщами древнегреческих колоний и так называемого Боспорского царства (Ростовцев, 1918; Каллистов, 1949) на Таманском полуострове, Прикубанской и Закубанской равнинах.
Из наиболее древних поселений района отметим Геленджикское.’
JL Аханов добыл в 1952 г. значительное число костей из ям землян-кового поселения эпохи позднего неолита — ранней бронзы на берегу Геленджикской бухты. Это обломки диафизов и позвонков, пестрого или желтовато го цвета. Они слегка звенят, как это бывает у обожженных костей. Многие были погрызены после человека какими-то мелкими хищниками. Основная масса костей принадлежала крупному рогатому скоту и дельфинам.
Из диких видов обнаружены еще олень, кабан, рысь, заяц (табл. 29).1 * Найдены также единичные косточки рыб, раковины морских моллюсков 8 виноградной улитки. 5
Таблица 29
Ввдовой состав и численность костных остатков млекопитающих из землянок века ранней бронзы близ Геленджика
Виды
Число костей
Виды
Число костей
Домашние
Лошадь ..................
Свинья........о..........
Крупный рогатый скот . .
Мелкий рогатый скот, преимущественно овцы .
Дикие
Итого .............
2
55
396
106
559
Felis lynx ....... Lepus europaeus . . . . Delphinus delphis . . . Sus scrofa ....... Cervus elaphus..........
Итого...........
2
1
282
2
6
Всего................ 852
м
и и й
и
I-щ т-го го зй
Состав диких видов из геленджикского поселения указывает на важную роль морского зверобойного промысла у древних таманцев или по крайней мере на использование ими выброшенных морем погибших дельфинов. Имелось также поблизости обилие лесных видов: рысей, кабанов, оленей.
Мощные костеносные слои, достигающие 2—3, а местами и 10 м, пробиваются ныне по берегам Таманской бухты на месте древнегреческих городов: Кепы, Фонагории, Тамани. Они энергично подминаются морем вследствие значительного опускания берега за исторический период. Обильные кухонные остатки этих поселений представлены
1 Эта и ряд следующих аналогичных таблиц составлены по нашим неопубли-
кгванным материалам.
176
Часть I. Обоснование истории формирования фауны
костями домашних животных, осетровых рыб и характерными прослоями фиолетовых раковин мидии. Остатков диких зверей в этих слоях практически нет, что вполне соответствует и представлению о большой окульту-ренпости района в далеком прошлом (рис. 79).
Рис. 79. Обрывы культурных слоев Фанагории на южном берегу Таманской бухты.
Фот. автора, 1952.
Сбор и исследование обломков костей на протяжении нескольких сот метров в обрывах культурных слоев этих трех городов дали результат, отраженный в табл. 30.
Таблица 30
Видовой состав и численность костных остатков домашних млекопитающих в слоях Таманских городов
Виды Кепы Фанагория Тамань
Собака 4 10 5
Кошка 2 — 3
Лошадь 11 18 13
Свинья 5 6 2
Крупный рогатый скот 9 68 27
Овца 15 34 32
Коза 3 6 —
Мелкий рогатый скот, ближе не определенный — 4 10
Всего 49 146 92
Гл. II. Развитие ландшафтов и фаун в четвертичном пери д 177
Судя по такому составу домашних животных, ландшафты Таманского полуострова во второй половине I тысячелетия до н. э. были в общем Ьпзкп к современным. Кочевые стада, пригонявшиеся скифами, и мест-< >«р животноводство греков должны были уже в те времена сильно опу-книть полуостров. Обедненная крупными видами группировка млеко-гающих этого района содержала тогда, очевидно, как и теперь, обык-Бшого ежа, лисицу, зайца, степных и лесных мышей, обыкновенную [ 1мевкуг слепца.
Более подробны и интересны данные по С е м и б р а т н е м у г о-тодищу близ станицы Варенпковской, восточнее устья Кубани. Раскопки Анфимова в 1938—1939 и 1951—1952 гг. показали, что здесь с\ в. до и. э. существовал вначале небольшой поселок, затем в III—II вв. w it э. значительный город и вновь, после разрушения и заброса, неболь-д охотничье-рыболовное поселение в I в. н. э. Костный материал, (Ф..«ранный по слоям, вполне подтверждает выводы, сделанные по данным \а<рпальной культуры. Весьма характерно, в частности, большое число с лков диких видов в V в. до н. э. и затем лишь в I в. н. э. (табл. 31).
:их лъ-
Таблица 31
Видовой состав и численность костных остатков млекопитающих в слоях Семибратнего городища
Века паша эра До нашей эры Всего
особей
I I IT III IV 1 V
Домашние
ака ..... 2/2 1 46/32 26/17 25/16 48 31 99
I Лошадь 7/5 10/6 102/52 33/18 74/32 142/75 188
1 яшья 15/8 5/4 64/44 36/16 101/42 103 70 184
ауппый рогами скот . . . 39/14 13/5 153/67 87/37 215/53 303 183 309
•ада 9/6 13/12 72/43 27/11 98/37 152 72 181
Коза Чезкпй рогатый — — 17/8 4/3 32/17 63 28 56
скот, ближе не определенный . — 3/2 — 3 2 — — 4
I "
Итого . . . 1021
Дикие («й lupus . . . — — — 4 4 4
Ш* meles . . . — — 1 — — 2/2 3
lutra ... — — — 1 — — 1
/pus europaeus . — — 1 — 2/2 4/4 7
te scrofa .... 25/7 2/1 4/2 3/3 2/2 16/3 28
Vrws elaphus . . 'tpr olus capreo- 24/7 12/8 6 6 4/3 6/6 6/5 35
lus — 1 • 1 **—- 2
Итого . . . 80
Всего . . . 1101
Примечание. Косая черта разделяет число ноитей и число особей. |2 Н. К. Верещагин
178 Часть 1. Обоснование истории формирования фауны
В слоях этого городища собраны также немногочисленные остатки | птиц: домашней курицы, диких уток, журавля — и многочисленные ? кости осетра, севрюги и карповых, преимущественно сазана.
Уровень древнего боспорского животноводства был явно невысок. | Собаки были весьма однотипны и мелки, размеров шпица. Служили они, | очевидно, лишь для сторожевых целей. Козы, овцы и свиньи поражают I легким костяком и, следовательно, заморенностыо. Коровы были не- I сколько крупнее, нежели северный, «торфяной» скот, и принадлежали I к европейскому короткорогому типу. Наличие остатков выдры, кабана • и оленя позволяет представить окружающую картину зарослей трост- v ников кубанских плавней. Крупных массивов леса поблизости, по-види- | мому, не было. г
Костные материалы из раскопок скифских курганов середины I тысячелетия до н. э. в районе Майкопа известны лишь по записям и зарисов- ? кам Веселовского (1901), который отмечал многие десятки и сотни скеле» | тов лошадей, убитых при похоронах вождей. В
Небольшие раскопки городищ и могильников Прикубанья восточнее I Краснодара, произведенные Покровским и Анфимовым (1937), выявили | в слоях IV в. до н. э.—III в. н. э. кости лошадей, крупного и мелкого Е рогатого скота. Обрезки и амулеты из мощных рогов оленей и из клыков К кабанов, найденные в тех же могильниках, хранятся в Краснодарском музее. U
Исследованные нами 337 костей из могильников, раскопанных Н. В. Ан-ш . фимовым у станицы Уст ь-Л а б и п с к о й, дали только 8 видов • домашних животных, приносившихся в жертву при погребениях и в качестве пищевых запасов покойнику (табл. 32).
Таблица 32
Видовой состав и численность костных остатков млекопитающих в курганах близ Усть-Лабинской .
Вена Виды Наша эра До нашей эры Всего особей
I—II I II Ш IV V VI
Собака Лошадь Свинья Крупный рогатый скот . . . Овца Коза Мелкий рогатый скот, ближе не определенный . 1/1 1/1 32/5 3/1 19/3 18/4 6/2 2/1 17/1 5/5 49/5 1/1 31/2 11/3 90/3 20/6 55/6 6/1 33/3 13/6 1/1 10/1 3/1 1/1 8/1 1/1 3 14 13 18 5 3 1° J
Всего . . . 66 |
Примечание. Косая черта разделяет число костей и число особей.
Собаки этого района были размером с мелкую промысловую лайку. Мелки были также свиньи, коровы, овцы и козы.
Городцов (1935, 1936), производивший в 1934—1935 гг. раскопки хЕлизаветинского городища и курганов левого берега Кубани, в 13 км
нош сып белт
I
пале
Гл. IL Развитие ландшафтов и фаун в четвертичном периоде
119
западнее Краснодара, упоминает, что в слоях первых веков нашей эры он находил «много костей коров двух пород, мелкой свиньи, овцы, козы (?) й костей собак трех пород. Кости диких животных очень редки; оточены лишь рога благородного оленя, которые укладывались в могилу ччте с покойником».
Из степей Приманычья остатки животных века бронзы добыты раскопками Артамонова (1937) в районе Манычстроя. В толще курганов века поздней бронзы с северного берега Маныча, у хутора Спорного, обнаружены кости животных:
Домашние
Лошадь
Свинья
Крупный рогатый скот
Овца
Дикие
Lepus europaeus Marmota bobac
Наибольший интерес для палеогеографа представляла бы находка копей сурка на южном берегу Маныча, так как зоологам XX столетия сурки с предкавказских равнин документально неизвестны. Однако с юж-нет берега Маныча, у поселка Веселого, из курганов середины I тысячелетия до н. э. известны остатки иного состава (табл. 33).
Таблица 33
Видовой состав и численность костных остатков млекопитающих в курганах южного берега Маныча
Виды Число костей Число особей
Домашние
Собака 16 1
Лошадь 12 2
Крупный рогатый скот 4 2
Овца 71 5
Итого 103 10
Дикие
Vulpes vulpes . 38 3
Canis lupus .......... 1 1
Meles meles ....... .... 1 1
Lepus europaeus ......... 2 1
Итого . . . 42 6
Всего 145 16
Если остатки домашних животных здесь — результат жертвенных приношений, то кости лисиц, волка, барсука и даже зайца извлечены из на-сыви кургана; они оказались захороненными в ней после природной ги-чеш зверей в норах.
Полоса предгорий и ущелья речек северо-западного Предкавказья палеонтологически не освещены, хотя интересные результаты, вероятно,
12*
180
Часть I. Обоснование истории формирования фауны
могли быть получены при исследованиях костей из слоев древнеаланских поселений, развалины которых имеются в ущельях по Лабе, Белой, Зеленчуку, а также в сохранившихся от разработок натеках асфальта у Хадыженска (Винда, 1910).
Из случайных находок отсюда известны только скелет длиннохвостой снеговой полевки — Mircrotus (Chionomys) roberti, обнаруженный в могильной камере XIV столетия в ущелье р. Малой Лабы, близ станицы Андрюковской, и рог кавказского лося — Alces alces caucasicus, найденный в 1949 г. близ станицы Отрадной в галечнике р. Уруп.
ЦентральноеПредкавказье
Будучи ныне своеобразными лугово-лесными «мезофитными островами» t среди степных равнин, Ставропольская возвышенность и Пятигорье № могли сохранить до наших дней и ряд мезофильных видов зверей, про- ж двинутых сюда в холодные эпохи плейстоцена частью с севера, частью W с юга — со стороны Большого Кавказа. Ж
По коллекциям Ставропольского музея и по литературным данным * (Динник, 1914а), было известно о недавнем существовании в островных лесах Ставрополя оленей, рысей, позднее исчезнувших совершенно.
Такие недавно исчезнувшие или исчезающие виды могли быть и среди мелких видов зверей. Особый интерес представляло также палеонтологическое решение вопроса о древности изоляции малого суслика во внут-
*•' ренних долинах северного склона Большого Кавказа.
J В связи с этим, кроме исследования костных остатков из слоев древ-
них городищ, здесь были предприняты специальные разведочные раскопки в пещерах и навесах по окраинам Ставропольского плато и по лакколитам Пятигорья, а также по обрывам каньонов речек наклонной Кабардинской равнины.
Район наклонной Кабардинской равнины был, несомненно, широко заселен человеком в раннем голоцене, о чем свидетельствуют находки неолитических орудий, многочисленные курганы и остатки древних поселений. В энеолитических погребениях, раскопанных А. А. Миллером в Нальчике в 1929 г., были обнаружены украшения, состоявшие из зубов домашних и диких животных: кошки, лисицы, кабана, благородного оленя, быка очень крупной формы, домашней или дикой, козы или овцы.
Такие же украшения известны по раскопкам Круглова, Пиотровского Подгаецкого (1941) из неолитических погребений Нальчикского кургана близ Агубековского городища. Они состояли из зубов кабана, оленяД дикого быка, лисицы, медведя.
Из кухонных остатков аланского поселения VI—VIII вв. н. э. близ сел. 3 е ю к о в о, в 53 км от Нальчика, по раскопкам Деген-Ковалевского (1935) известны преимущественно кости домашних видов (табл, 34).?
Остатки быка здесь мелки, их принадлежность к домашней породеJ сомнений уже не вызывает. Характерно появление домашней свиньи! и весьма примитивной породы козы с изгибом рогов типа Capra prison
Еще Кистяковский (1935), исследовавший многочисленные погадки хищных птиц из степей, примыкающих к северной и восточной окраин^ Ставропольского плато, зарегистрировал остатки 15 видов ныне живу! щих грызунов, не обнаружив существенных отклонений от очертании их современных ареалов. f
Небольшое количество костей грызунов и насекомоядных из погй док филина было собрано и Лютым (1940) на западных склонах возвышен
Гл, II. Развитие ландшафтов и фаун в четвертичном периоде
181
Таблица 34
Видовой состав и численность костных остатков млекопитающих из слоев аланского поселения у Зеюково
Виды
Число костей Число особей
Домашние
Собака .....................
Лошадь......................
Свинья........м.............
Крупный рогатый скот........
Мелкий рогатый скот.........
2
8
15
29
10
Итого........................ 64
»
Дикие
Cervus elaphus....................... 1
__________________________. f______
Всего........................ 65
i и в районе Ставрополя, причехМ зоогеографических новостей также чено не было.
Осенью 1950 г. нами была произведена палеонтологическая разведка западной окраине известнякового обрыва горы Стрижа мент южной окраине Ставропольской возвышенности. Наличие здесь лесного месива, ряда источников и «моря скал» позволяло надеяться на находки углей лесных и степных зверей, оставленных древними охотниками и пещерными хищниками.
В четырех небольших пещерках были обнаружены лишь единичные кисти овед и коров, затащенные пастухами и волками. Неустойчивый, обшивающийся потолок пещер и массивные глыбы известняка на их дне затруднили более основательную шурфовку.
В одной из каверн обрыва было собрано значительное количество костей мелких млекопитающих из остатков, принесенных филином на протяжении последнего столетия. Результат их определения дан в табл. 35.
Наиболее интересная находка — остатки кустарниковой полевки, неизвестной ранее в литературе для этого района, но, как оказалось нередкой и ныне на лугах плато. Значительный отрыв ареала кустарниковой полевки от основного его кавказского массива является известным биогеографическим свидетельством наличия в прошлом более влажной, ш ныне, эпохи.
Исследования в Пятигорье по аналогии со Ставропольским плато велись на древних изолированных возвышенностях, вздымающихся среди степи. Особое внимание было обращено на участок вековой мерзлоты горы Развалки, в 5 км к северу от Железноводска, в на каньон речки Березовки, к югу от Кисловодска.
Разведочные раскопки были предприняты на восточном склоне горы Развалки, у пещеры Селитряной. Здесь среди сплошного широколиственного леса подымаются два живописных утёса трахитовых скал, изъеденных ветровой эрозией. Под южным утесом есть небольшая сухая
Часть I. Обоснование истории формирования фауны
Таблица 35
Видовой состав и численность костных остатков млекопитающих в погадках филина у горы Стрижамент
Виды Число костей Число особей
Hemiechinus auritus 13 2
Erinaceus europaeus • . 25 4
Talpa caucasica 7 1
Mustela nivalis caucasica .... 8 2
M. nivalis dinniki 5 1
Lepus europaeus . 3 2
Cricetus cricetus 19 3
Mesocricetus auratus 290 34
Cricetulus migratorius 211 29
Microlus arvalis Л/. majori . } 215 } 32
Spalax microphtalmus 17 3
Apodemus sylvaticus 30 7
Micromys minutus 5 3
Mus musculus 1 1
Rattus norvegicus 5 1
Всего 852 125
пещера> образовавшаяся на месте стыка трех взаимно перпендикулярных вертикальных трещин. Толща конуса отложений под входом в пещеру достигает мощности 8—9 м, они накапливались на протяжении
последних тясячелетии за счет смыва почвы с верхних площадок утеса, непрерывного разрушения его скальной стенки и выброса обломков скал
древними поселенцами из самой пещеры. Развалка использовалась, очевидно, древними поселенцами для стойбищ, так как подножие утесе и конус пещерных выбросов переслоены костями мелких и крупных жи-
вотных и керамикой.
Наша разведочная траншея шириной в 1 м и длиной в 3 м прорезала слой конуса перед входом в пещеру на глубину в 1.50—1.90 м. В сборах оказалась по преимуществу грубая, черная с битой ракушей керамика древнеаланского типа, а керамика из более глубоких слоев имеет ямочный и штриховой орнамент, характерный для глиняных изделий века бронзы. В верхнем 60-сантиметровом слое оказался, кроме того, обломок обсидиана, обломок каменного молотка и ручного точила из черного
сланца.
Обломки костей крупных животных — типичные кухонные отбросы, реже — обломки изделий из рога оленя. Остатки мелких животных — это кости из погадок хищных птиц, преимущественно филинов, садившихся на скалы над пещерой. Здесь имелось очень ценное и редкое сочетание трех типов накопления костных остатков: кости приносили птицы; охотники, использовавшие пещеру в качестве убежища, приносили диких зверей; накоплялись кости при обрядовых пиршествах племен или просто пастухов древних стойбищ.
Собранные послойно костные остатки — всего до 1500 фрагментов -оказались приблизительно одного типа сохранности. Кости домашних и охотничье-промысловых животных мелко раздроблены. Все кости почти без признаков фоссилизации, желтого или белесого цвета.
Гл. II. Развитие ландшафтов и фаун е четвертичном периоде 183
Таблица 36
Видовой состав и численность костных остатков млекопитающих у пещеры Селитряной в Пятигорье
ip-пе-
гии
>са, кал
1СЬ, *еса жи-
;ала .рах [ика иоч-века бло-ного
осы, [X — 1ДИВ-е со-сили )ИНО-
пле-
?ов — шних почти
Глубина (в см) Всего ностей
Виды 0-60 60-120 120-200
г Домашние
Собака 1 1 — 2
Лошадь ' 9 8 2 19
Свинья 34 17 7 58
Крупный рогатый скот Овца 103 103 74 72 39 216 175
I Коза • • ’ Мелкий рогатый скот, ближе не 29 32 гг 77 — 34 176
| определенный 67
J Итого 311 254 115 680
Дикие
} faniiechinus а иг i t u s ........ 2 — — 2
| Erinaceus europaeus ........ — 2 — 9
1 foipfl caucusica .......... 1 1 1 3
Vulpes vulpes ........... * — 1 1
(Jrsus arclos........... 1 — — 1
Mustela nivalis .......... 2 1 — 3
Meles meles ............ 4 — 4
Putorius eversmanni ........ 2 — — 2
Vormela sarmatica ......... 1 — — 1
r ffiis Silvestris ........... 1 — — 1
| Lepus europaeus .......... 17 1 1 19
j Elisglis 10 4 1 15
Criceius cricetus .......... 173 23 1 197
Mesocricetus auratus ........ 159 22 2 183
Cricetulus migratorius ....... 13 4 — 17
Arvicola terrestris 31 10 — 41
)licrotus arvalis 18 — — 18
}1. majori 1 1 — 2
: Rattus norvegicus ......... 2 — — 2
ptpodenuis sylvaiicus ........ 2 2 — 4
\ius musculus ........... 1 —- — 1
Spalax microphthalrnus 1 — —— 1
Sus scro/a 4 8 — 12
Cerras elaphus ........... 1 5 — 5
Cupreolus capreolus 3 1 1 6
Итого 450 85 8 543
Всего 1 761 339 123 1223
184
Часть I. Обоснование истории формирования фауны
Возраст всего исследованного слоя отложений можно определить в общем в пределах последних 2000—2500 лет.
В результате определения костей зарегистрировано 6 видов домашних и 26 видов диких животных (табл. 36).
Отсюда же собраны остатки птиц, жаб, лягушек и раковины клаузи-лий и геликсов.
При обсуждении результатов становится ясно, что наиболее интенсивное накопление остатков диких животных, особенно принесенных фили-
Рис. 80. Пещерка, обитаемая филином на горе Лысой. Сверху лежат трупы 7 ежей и 1 коростеля.
Фот. автора, 1952.
ном, происходило относительно недавно. Быстрое уменьшение числа остатков грызунов с глубиной показы вает, что этот участок был освоен человеком раньше в большей степени, нежели теперь. В эпоху древнеаланских поселений склон Развалки был, вероятно, сильно обезлесен, в связи с его об житостью филины и другие хищники были оттеснены. В последней четверти нашей эпохи Пятигорье и Развалка были вновь облесены и населены крупными лесными охотничье-промысловымп видами: медведем, кабаном, оленем, косулей. Из записей старожилов теперь известно, что олени исчезли здесь в начале нашего столетия, медведь же, косуля и кабан изредка заходят в леса Пяти-горья и в наши дни, но не живут уже здесь постоянно. Охота филина за тот же поздний период — 4—5 последних столетий — дала набор мелких видов, характерных для районов лаколлитов и в наши дни. Основу питания филина составляли три вида хомяков, водяная, кустарниковая и обыкновенная по-
левки, полчок. Остатки снеговой полевки — Chionomys gud были собраны наравне с остатками хомяков и водяных крыс в расщелинах скал над каменистой россыпью участка вековой мерзлоты.
В скалах с «вековым холодильником» горы Развалки удалось добыть и несколько экземпляров кавказской снеговой полевки па высотах около 400—500 м над ур. м. Ближайшее местонахождение зверька было обнаружено лишь в 3 км к югу от Кисловодска, в каньоне речки Березовки, т. е. после перерыва в 50—55 км. Таким образом, наличие кавказской снеговой полевкп на горе Развалке является свидетельством наличия
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
18»
в прошлом холодной эпохи, вероятно максимального оледенения, продвинувшего горных зверьков на эти небольшие возвышенности Пятигорья.
Обппыше остатки насекомоядных, мелких хищников из древних погадок филина были собраны также под навесами скал горы Лысой
Fiic. 81. Расщелина в известняках западного борта каньона речки Березовки у Кисловодска — место дневок филина.
Фот. автора, 1954.
цик. 80), крайнего северо-восточного лакколита, и в каньоне ущелья речки Березовки, южнее Кисловодска (рис. 81). В последнем пункте
Таблица 37
Видпвой состав и численность костных остатков млекопитающих в погадках филина у Кисловодска
Виды Число особей 1 Виды Число особей 1
чпг ага neus I Arvicola lerrestris 51
Talpa соеса . 3 Microtus arvalis . ... . 2
tfustcla nivalis 3 M. gud 4
pulorius eversmauni .... о Apodemus sylvaticus . . . 1
frpus europaeus 1 Mus musculus 2
Dyron js nitedula 1 Rattus norvegicus 3
Cricetus cricetus 45 Spalax microphtalmus . . . 2
Jksocricetus auratus . . . 23 .—
Crictiidus migrate rius . . . 3 Всего 148
i Число особей показано по
подсчету нижних челюстей.
186
Часть J. Обоснование истории формирования фауны
материал более обилен, древен и добыт в полуметровом слое голоценовых пылеватых отложений под вертикальной трещиной стенки известняков нижнего мола. Убежище это было заброшено филином не менее сотни лет тому назад, так как находится всего в километре от южной окраины Кисловодска. Накопление костей происходило на протяжении ряда столетий второй половины нашей эпохи.
Рис. 82. Остатки млекопитающих из погадок филина в каньоне Березовки.
1 — плечевая Talpa caucasica; 2— череп Mustela nivalis; 3 — тазовая Lepus europaeus; 4 — челюсть R a this norvegicus; 3, 6 — череп Cricetus cricetus; 7, 8 — череп Mesocricetus auratus nigiiculus; 9, 10 — череп Arvicola terrestris.
Всего было собрано 1582 косточки. Сохранность их неодинакова. В нижнем слое они приобрели кофейный цвет и имеют даже депдриты в верхнем же — просто желтые.
Видовой состав остатков зверьков свидетельствует о том, что в эпоху их накопления окружающий ландшафт был весьма близок к современному (табл. 37 и рис. 82).
Здесь же собрано 305 костей птиц и 19 экземпляров амфибий. По обилию остатков выделяется, как и па Развалке, группировка относительных сухолюбов — степняков: хомяки, хорек, слепец, — в сущности нигде пе заходящая в таком полном составе так далеко в горы. Обилие
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
187
остатков водяных крыс и появление снеговой полевки объясняются наличием участков влажных лугов и россыпей скал по каньону Березовки.
Хомяков и слепца в этом районе следует рассматривать как авангард-относительно сухолюбивую группировку, продвинувшуюся в послед-овую эпоху с севера, из сухих степей Предкавказья. Это «наступление» мекрыло здесь на десятки километров северные «форпосты» лесных и Ьпшских видов: кустарниковой и снеговой полевок, лесной сони, волчка, — задержавшихся на мезофитных участках лакколитов и ущельях рек с более влажной и холодной эпохи. Пределами для распро-.т нрнпя этой группировки вверх служат влажные луга Пастбищного та, а также устьевые участки узких каньонов, прорезанных речками
м хребте (Верещагин, 1953а).
Подобная картина наблюдается и на западе района, по правобережью I идианного отрезка течения Кубани, но на меньших высотах.
Исследования голоценовых остатков зверей в центральном предгор-iow Предкавказье позволяют сделать следующие выводы.
I . Район интенсивно осваивался человеком с неолита до наших дней, чт вело к быстрому уменьшению площади лесов и исчезновению крупного р ря: зубра, оленя, а позднее косули и кабана.
2 Установлено наличие мезофильных реликтов холодной (ледниковой) г Xi уцелевших среди степи на лакколитах Пятигорья и на Ставро-л ьскоп возвышенности. К ним относятся снеговая и кустарниковая ь ;рВКИ.
3 Преобладающая в районе группировка относительно сухолюби-mi видов грызунов: хомяков, слепушонки и слепца, — характерная пя открытого ландшафта, существовала в предгорьях в районе Кисло-г:ска и несколько южнее, в течение последнего тысячелетия, а быть । кет, и на протяжении всей нашей эры. В течение этого же времени районе Пятигорья отсутствовал малый суслик. Признание проникнове- я этого суслика в продольные долины хребта в голоцене может быть лано лишь после находок его ископаемых остатков в более глубоких „юях, чем раскопанные нами. Такая находка могла бы доказать суще-Ьвание в послеледниковую эпоху более сухой и жаркой климатической А|зы, нежели та, что характерна для наших дней.
Восточное Предкавказье
В области степей восточного Примапычья для палеозоолога представлю г интерес следы микролитической культуры, обычно приравниваемой L к позднему неолиту. Стоянки микролитических племен известны с южной Украины и степного Крыма, а затем прослеживаются по всей северной окраине Каспия и на чинках Усть-Урта, уходя далее в глубь Средней Кип. Особенно постоянно заселялись этими племенами Бэровские бугры, окруженные ильменями, богатыми рыбой и кабанами.
Остатки предметов материальной культуры и фрагменты костей животных этой эпохи находят здесь на выдувах — на участках развеянных курганов и бугристых песков. Из таких стоянок в районе Прикумских теней известны фрагменты костей слепышей — Spalax gigantheus, сайга-^вповец, а на песках черных земель и больших песчанок — Rhombomys рШ. Между низовьями Кумы и Волги археологом И. В. Синицыным Шружепо обилие костей разных животных вокруг остатков древних острпгц и много поселений и могильников эпохи бронзы и средневе-.ОВьЯ.
188
Часть I. Обоснование истории формирования фауны
Для предскифской эпохи из группы курганов Три брата близ Степного известны, по фотографии Синицына (1948, стр. 151), остатки крупного быка со стержнями рогов, близкими по типу к первобытному туру.
Датируя погребения концом II—началом I тысячелетия до и. э, Синицын сближает эту культуру с закавказской культурой Триалети (Куфтин, 1941).
Аналогичные и более поздние памятники предскифской и скифской эпохи были прослежены Крупновым (1947) в степях между Тереком п Кумой. Остеологические материалы отсюда, вероятно, могли бы уточнить наши представления об истории ареалов среднеазиатских пустынных зверей и соображения Крупнова о значительных ландшафтных сдвигах в последнее тысячелетие до нашей эры.
Южнее — с северо-восточных предгорий Дагестана — костные остатки известны из поселений и курганов века бронзы. В районе Дербента, Каякента и Дешлагара примитивные археологические раскопки производились Русовым (1879) и Цилоссапи (1879). В протоколах раскопок у них есть указания на находки в пещерах, склепах, курганных могилах «костей грызунов», «костей животных», «костей крупных и мелких домашних животных» и т. п. В ЗИНе хранятся костные материалы, добытые раскопками Круглова (1946а) в 1941 г. в слоях поселения второй половины II тысячелетия до н. э. у селения К а я к е и т и Д ж е м и к е н т. Фрагменты костей из этих слоев светло-коричневой окраски и слегка звенят. Кости кабана гигроскопичны подобно верхнеплейстоценовым. Определения и подсчет фрагментов даны в табл. 38.
Таблица 38
Видовой состав и численность костных остатков млекопитающих в слоях поселения века бронзы у сел. Джемикент
Виды Число костей Число особей
Домашние
Собака 82 1
Крупный рогатый скот 129 6
Коза 11 4
Овца Мелкий рогатый скот, ближе пе 46 5
определенный 401 15
Итого Дикие 669 31
Phoca caspica 1 1
Ursus arctos 1 1
Vulpes vulpes 1 1
Equus hemionus ......... 6 1
Sus scrofa 3 1
Gazella subgutturosa 6 1
Итого 18 6
Всего 687 37
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
189
Заслуживают внимания факты добычи тюленей и обитания в районе джейранов и куланов (рис. 83).
В разведочных раскопках археологов Дагестанского филиала АН СССР, произведенных в 1954 г. под живописными утесами Капчугая, апа нее Махачкалы, оказались слои конца II—начала I тысячелетия дон. э. В сборах из этих слоев нами определены фрагменты костей: лошади — 3, крупного рогатого скота — 12, овец — 14 и оленей — Cervus elaphus —12. Такое соотношение указывает и на обилие оленей, и наваж-
Рис. 83. Остатки млекопитающих из слоев поселения эпохи бронзы близ Джемикента.
i — челюсть Vulpes vulpes; 2 — заплюсневая Ursus arctos; 3—5 — 1-я "фаланга, копытная фаланга и астрагал Equus hemionus; 6—9 — роговой стержень, головка бедра, 1-я фаланга, берцовая Gazella subgutturosa.
ную роль охотничьего промысла у древнего пастушеского племени. Утесы чокракских песчаников испещрены здесь контурными рисунками животных, сделанных всего лишь несколько столетий тому назад. В некоторых из них можно узнать барса, лошадь, оленя и сайгу.
Остатки зверей более поздних эпох — века железа — известны из раскопок древних поселений в долине Сунжи и ее притоков.
Крупнов (1948) опубликовал результаты раскопок слоев поселения скифского времени — второй половины I тысячелетия до н. э. близ сел. А л х а с т е, на левом берегу р. Ассы (табл. 39).
Сходные отношения чисел остатков домашних животных Крупнов (1949) приводил по определениям В. И. Цалкина для слоев Нестеровского и Лугового поселений VI—V вв. до н. э., находящихся в той же долине Ассы. В последней среди массы костей домашних животных был обнаружен роговой стержень сайги и единичные кости косули.
В восточной части долины Сунжи раскопки поселений и могильников П-Ш вв. н. э. велись в 1938 г. А. Б. Кругловым близ селений И с т и-Су, Ал х а н-К ала, Хор о-Ч а й. Костный материал отсюда, определенный нами (табл. 40), тяжел, прокрашен местами гумусом. Остатков 1иких зверей здесь также мало, но значительный интерес представляет
190
Часть I, Обоснование истории формирования фауны
Таблица 39
Видовой состав и .численность костных остатков млекопитающих в слоях поселения скифской эпохи у сел.
Алхасте
Виды 1 Число костей Число особей
Домашние
Собака 6 3
Лошадь 21 6
Свинья 175 15
Крупный рогатый скот 101 10
Мелкий рогатый скот 54 8
Итого 337 42
Дикие
Vulpes vulpes 1 1
Cricetus cricetus ........ 5 1
Cervus elaphus ......... 2 1
Capreolus capreolus 6 3
Итого 14 6
Всего 371 48
т а б л и ц а 40
Видовой состав и численность костных остатков млекопитающих в слоях поселений и в могильниках долины Сунжи
Виды Число костей Число особей
Домашние
Собака 75 4
Кошка о 1
Лошадь 55 3
Свинья 32 10
Крупный рогатый скот 64 7
Овца 145 12
Коза 6 3
Итого 3X2 40
Дикие
Equus hemionus I 1
Cervus elaphus 1 1
Итого 2 2
Всего 384 42
1 Определения произведены Н. А. Сугробовым. В остатки домашней свиньи вошли, вероятно, и остатки дикого кабана.
Гл. 7/. Развитие ландшафтов и фаун в четвертичном периоде
191
новое подтверждение былого обитания степных видов крупных зверей р предгорьях .восточного Кавказа.
Наши разведочные поиски костей мелких видов зверей района были I ^приняты в 1952 г. в нишах и россыпях сарматских известняков над 'йлачкалой и в кавернах песчаников под Капчугаем. Было собрано зна-штелыюе количество костей: ушастых ежей — Hemiechinus auritus, Еа — Lepus europaeus, малого суслика — Citellus pygmaeus, чернового хомяка — Mesocricetus auratus nigriculus и степной полевки — 1 croZws socialis. Все они происходят из погадок филинов и пролетных Сипиков, присаживавшихся на утесах.
Дальнейшие палеонтологические исследования голоценовых захорони с учетом прежних сведений археологов (Комаров, 1879; Штейн, (7У; Иессеп, 1935) могут, несомненно, значительно уточнить намеченную иую картину появления в районе послеледниковых степных видов нгытвых, деградацию их численности и состава по мере приближения ; ^временности.
Продольные долины северного склона
Большого Кавказа и внутренний Дагестан
Продольные тектонические долины, отгороженные с севера Пастбищ-цм и Скалистым хребтами, хорошо выражены к востоку от Эльбруса. ; iK и внутренний безлесный Дагестан, они были весьма привлекательны ря послепалеолитпческпх племен, селившихся здесь с учетом хорошей Ьпродной защищенности, сухого и относительно теплого климата. Неявно в этих долинах и во внутреннем Дагестане обнаружены мезолитиче-Ьш? стоянки. В век бронзы какие-то племена оставили здесь замечательные погребальные памятники в виде курганов и подземных могильников — ватакомб для коллективных захоронений, а на месте своих поселений — •евольпо мощные слои «кобанской культуры» (Уварова, 1900, 1902).
От аланов осталось здесь за эпоху средневековья множество развалин крепостей п боевых башен, забаррикадированных пещер и индивидуальных боевых ячеек в отвесных стенах скал, а их потомки — осетины накопили уже упомянутые, своеобразные коллекции черепов копытных । святилищах, или так называемых «дзуарах».
В природных нишах и в развалинах крепостей накопилось в ряде мест I и значительное количество костей грызунов пз погадок сычей.
Все эти типы захоронений костей, за исключением кобанских могиль-аиков и поселений, были исследованы нами.
Мезолитические стоянки ущелий Баксана и Аварского Кой-Су
Многометровые накопления делювия, содержащие кремневые орудия мезолитической стадии, были обнаружены и исследованы С. Н. Замятиным в 1955 г. па правом склоне ущелья Баксана (рис. 84), в начале первой теснины, ниже сел. Быллым. Раскопом навеса Сосруко были последовательно вскрыты слои средневековья, раннего железа и пять политических, содержащих характерные кремневые орудия переходно!! йохп. Общая глубина раскопа достигала 6.85 м.
Большинство костей пз всех слоев мелко раздроблено, фоссилизацпя ^агмептов на самой начальной стадии. Определения извлеченных облом-да костей дали результаты, показанные в табл. 41 и на рис. 85.
Рпс. 84. Грот Соеруко в ущелье Баксана.
Фот. С. II. Замятнина, 1956.
Гл. /Г Развитие ландшафтов и фаун в четвертичном периоде
193
Таблица 41
Ыной состав и численность костных остатков млекопитающих в мезолитических слоях навеса Сосруко в ущелье Баксана
Виды Средневековые слои Раннее железо Мезолитические слои
1 2 3 4 5
ипья . . . . • репный рогатый /ют
MKU11 рогатый
Итого • *
Дикие rulpes . .
Irw SP.........
[L n:d I'S . • • iii'ra pardus . gp turt paeus . Бм pygmaeas П/mux sylvati-
iulpirius . i&rtus yud . . tatt/ux capreo-
..............
>v daphits . . rupi-
fyii Л. cattcasi-
^dactyla, бли-’ fc пе опреде-
ляйте . . . ^
Biero . . •
7 4 — — — — —
20 15 7 — — — —
23 28 44 — — — —
50 1 47 51 — — 1 —
1 । 1 о 1) 1 1
1 ' и Z_ — —
— — 1 — — 1 1
— 1 — — —
— — 1 1 1 — —
— — — — 1 — —
— — 6 7 3 1 —
— 4 — — 1
1 1 1 2 12 — —
— — — — — 7 1
— — 4 1 11 — —
7 3 । 1 '3 125 526 198 45
10 9 87 140 556 208 50
60 56 138 140 556 208 50
Кроме того, в мезолитических слоях оказалось до десятка костей куртки, болотной совы, белоголового сипа и мелких воробьиных.
Приведенный состав видов зверей этой стоянки при всем его разио-не дает зоогеографических новостей и был характерен для Слка еще в прошлом столетии, до уничтожения па Скалистом хребте имей, серп, козлов. Особый интерес для решения вопроса о древности игляцяп сусликов в горах Кавказа представляют остатки этого вида «имолптпческпх слоев. Однако эти остатки могли попадать в слои из угадок, сброшенных с утеса не только филином, но и пролетавшими па иг^ицпикамп, п могли быть таким образом занесенными из степных равна Прима ничья.
Мезолитическая стоянка Чох во внутреннем Даге-•пнебыла открыта в 1955 г. В. Г. Котовпчем в 2 км к северу от сел. Чох |3 Н. К. Верещагин
194
Часть I. Обоснование истории формирования фауны
Гунибского района. Стоянка расположена под крутон известняковой скалой (рис. 86) на правом берегу небольшого потока, глубоко разрезавшего горную ложбину, покрытую альпийским лугом. Мощность мезолитического слоя в раскопанном участке превышает 0.5 м. Культурный слой богат кремнем на всех стадиях его обработки и мелкобитыми костями. Среди кремневых орудий особенно характерны орудия с затупленной спинкой, острия, ножевидные пластинки и разнообразные микролиты.
По типу сохранности костного вещества фрагменты близки к обломкам, происходящим из слоев III и II тысячелетий до н. э. в предгорьях Даге-
Рис. 85. Остатки млекопитающих из мезолитических слоев грота Сосруко.
1 — челюсть Panthera pardus; 2 — челюсть (Х2) Citellus pygmaeus; 3 — челюсть (X2j Ellobius lalpinus; 4 — челюсть Cervus elaphus; 5 — астрагал Capra caucasica.
стана. Большинство фрагментов неопределимо. Результаты псследовави костных сборов 1955—1957 гг. даны в табл. 42 и на рис. 87. ОстатЗ грызунов принадлежат видам, характерным для района и в напшдД На стоянку они попали, конечно, из погадок хищных птиц. Среди копы] пых наиболее интересно наличие остатков зубра и мелкого .барана вероятного потомка баранов пижнепалеолитической стоянки .Кудаи
Из кобанских могильников века бронзы в Осетии археологи уже давно извлекались скульптурные изображения местных диких жив^ пых, изготовленных из бронзы, реже— золота и серебра.
Подвески, фибулы, бляхи, пряжки, павершия жезлов часто долам кобапскими мастерами в виде голов кавказских козлов, оленей, лосЛ диких и домашних баранов, быков, коз, лошадей, медведей (Уварова, 1М
Былое наличие на Кавказе некоторых из названных видов было J тверждеио нашими исследованиями в осетинских капищах, где отдм поздний этап существования богатой группировки млекопитающих J северном склоне Большого Кавказа.
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
195
Таблица 42
Видовой состав и численность костных остатков млекопитающих в мезолитических слоях стоянки Чох в Дагестане
Виды
Число ностей
Виды
Число костей
Домашние (?)
Лошадь........
-Дикие
Fells lynx . . . . Lepus europaeus . Mesocricetus aura-tus................
Arvicola terrestris Sus scrofa .... Cervus elaphus . . Ovis sp. (cf.
gmelini) . . . .
1
1
5
5
1
48
302
Capra sp. (cf. aegagrus или C. cylindricor-nis) 8
Ovis или Capra 94
дикие .... Caprovinae, ближе не опреде-
ленные .... Bison bonasus 7202
(caucasicus?) . Мелкие обломки трубчатых костей, преимущественно Аг- 20
tiodaclyla . . 712
Всего . . . 8400
Дзуары Северной Осетии
О священных коллекциях черепов домашних и диких копытных в Кавказе было известно уже давно, еще по заметкам западноевропей-их щчешествепников по Черкессии в XVII столетии, например Жана • Люка (1879), по записям Гюльденштедта конца XVIII столетия (Пал-1831), статьям Диннпка (1890а) и Сатунина (1914). Эпоха накопле-ля черепов в дзуарах приходится на пять последних веков — начиная XIV—XV столетия.
В 1947 и 1948 гг. мы исследовали 18 дзуаров в ущельях Уруха, Ардона, Фиагдона п Гпзельдона (рис. 88).
Коллекции черепов в дзуарах, расположенных па предгорных равни-мх. например в священной роще Хетаг, были вывезены на костеобжига-Ььпые заводы в 30-х годах. Лишилось остеологических коллекций и Ю 30°о дзуаров в горной зоне (Верещагин и Семенов-Тян-Шапский, $8; Верещагин и Наниев, 1949).
Для устройства капищ у адыгейцев, дигорцев, осетин использовались шоппсные рощи на выдающихся отрогах хребтов, навесы скал, пещеры. Ьружалпсь и специальные массивные постройки: церкви, часовни, скрытые массивные сараи — из камня или из толстых бревен.
(зуары посвящались различным божествам и принадлежат отдельным шенш1м, жители которых собираются здесь для пирушек в определенный «пь весны, лета или осени.
Обычаи прирезки животных в дзуарах и пиршества как пережиток даства существовал и в Абхазии. По преданиям абхазов (Вейдепбаум, 59), из пещеры Оггип осенью появлялся белый бык, который и при-। яся в жертву.
13*
196
Часть I. Обоснование истории формирования фауны
Наряду с прирезкой для пиршества домашних копытных у дигорцев п осетин был развит еще обычай приносить к капищам черепа убитых диких копытных. Этот обычай приноса черепов в дзуары связан с верой
Рис. 86. Чохская стоянка на Гуппбс. Фот. В. Г. Котовпч.
в покровителя охотников и охотничьих животных — бога Авсати,которому посвящались черепа. Весьма вероятно, что в прошлые столетия, когда зверя было еще много, артель охотников отправлялась перед праздником
Рис. 87 Остатки млекопитающих из мезолитических слоев Чохской стоянки.
I—з — 1-я фаланга, эпифиз пясти и астра! i i (пн ( Г. gmelini.
слопенпых к степам стволов. Однако
за добычей и притаскивала ту-1 шу и голову убитого оленя, лося или зубра к дзуару. Об этом I свидетельствует и наличие в ; глубинных слоях черепов пеще-1 ры Дигоризед костей ног зубров' с копытами. Черепа диких копытных, вывареппые охотника-} ми, приносились в дзуар и вне! зависимости от празднеств. | Огги бережно укладывались в об-щую кучу или па специальные!
завалинки, полати и полки I или вешались па полуобрублеп-1 ные сучья принесенных и при рассчитать древность накопле
ния коллекций по числу черепов диких животных затруднительно. I Сохранность черепов диких копытных, обусловленная манипуляциями с ними и препаровкой, оказалась неодинаковой в различных святилищах, Ни один ттз черепов не ггмел нижней челюсти. Подавляющее большинство!
Гл. II- Развитие ландшафтов и фаун в четвертичном периоде
197
черепов зубров сохранило лишь наиболее прочную часть лобных и затылочных костей со стержнями рогов, совершенно так же, как это бывает v черепов первобытных зубров с береговых отмелей Дона, Волги и Урала. пишь немногие черепа сохранились целиком с базальной частью, меж-
Рис. 88. Расположение некоторых дзуаров в Северной Осетии.
/ — сохранившиеся леса; 2 — обследованные святилища; з — граница распространения оленя в конце XIX столетня; 4 — участок обит шип последних: зубров и лосей в конце. XVIII сто штня; •> — современные ледники.
челюстными костями и роговыми чехлами. Роговые чехлы удерживались на стержнях железными наконечниками стрел, заткнутыми в прорубленные отверстия, или же намеренно снимались для использования в качестве кубков для вина. Вываренные черепа туров, серн, косуль приносились обычно уже с огрубленной наискось базальной частью. У сырых «реиов самцов оленей и лосей, убитых осенью, рога вырубались вместе с полоской кожи п лобных костей. Мозг доставался через образовавшееся
198
Часть I. Обоснование истории формирования фауны
отверстие. Затем черепа варились, очищались от мяса и костей и водворялись в святилище. Черепа оленух и лосих оказались все без затылочной кости, которая срубалась до половины высоты затылка. Большинство черепов оленей не имело ростральных частей, отвалившихся преимущественно при варке.
Наиболее примечательны во всех отношениях дзуары: Дигоризед, или Олисай-Дон, Лесгор I и Лесгор II, Реком.
Рис. 89. Пещера Дигоризед в стене юрских сланцев ущелья Уруха. Фот. автора, 1947.
Пещера Дигоризед расположена на правой стороне ущелья Уруха, в отвесной южной стене Скалистого хребта. Она образовалась в результате выветривания юрских известняков. Этот гигантский обрыв имеет местами уступы, живописно задрапированные порослью сосны, можжевельника, шиповника.
Высота пещеры над уровнем моря около 1200 м. Принадлежит она сел. Заделеск, расположенному в 1.5 км южнее.
Устье пещеры треугольной формы, высотой около 12 м и шириной у основания 5.5 м. Снизу оно забрано прочной стенкой высотой 2.5 м, устроенной из обломков известняка па гаже (рис. 89 и 90).
Пиршественный зал шириной около 7 м и длиной до 12 имеет грубо сложенный очаг, узкие скамьи и столы из расколотых сосновых бревен, железную кружку для денежных приношений, два прислоненных к скале сучковатых ствола с подвешенными резными из дерева фетишами и обеденный инвентарь, состоящий из двух огромных деревянных мисок для мяса, деревянной ложки — уполовни — п полутора десятков рогов зубров служащих кубками. Куча черепов и рогов в левом дальнем углу зажата
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
199
Рис. 90. План пещеры Дигорнзед.
1 — обвалившиеся глыбы; 2 — очаг; з — священная посуда; 4 — снамьи; б — полати с рогами оленей; б — каменная стенка; 7 — черепа домашних полорогих: оленей, косуль, серн; л — черепа лосей; з — черепа зубров.
палочки — шомпуры для шашлыка, (рис. 92) и мелкие серебряные монеты.
неЖ Двумя обрушившимися известняковыми глыбами. В передней исти зала на высоте 2.7 м устроены полати из нескольких бревен и жер-
на которых навалена огромная куча оленьих рогов. Число их достигало i —150 пар (рпс. 134). Потолок и стены зала прокопчены смоляной ко-Lbio костров. Отходящий вправо узкий отрог перегорожен при входе иикальным бревном с жестяной иконой св. Николая. Будучи у входа в рост человека, отрог далее понижается и заканчивается слепо I, восьмом метре. Он во всю jpihv заполнен метровым р черепов и рогов парно-|.'1ЫТПЫХ (рис. 91).
Ежегодное пиршество в киризеде происходит в ре, во время сенокоса.
При нашем первом посе-дап пещеры в августе Ц] г. в первых рядах пережатого отрога находились Ьрепа лося, а также до 15 ! .дои зубров. Очевидно, ргойвпдели Гюльденштедт ВЬшнмк. Упомянутых Дии-рш 3 черепов верблюдов Ьружепо не было, что и Ьавляет предполагать, что v нпх былп приняты чере-। лосей. Толща черепов, Едоцих в левом углу пир-Ктвениого зала, была Сильно пересыпана вековы-Lg слоями пыли, а основная реачерепов в правом отроге уя, вероятно, недавно пе-юрошена археологами или Ерпстами. При разборке доя черепов постоянно попадись изогнутые дугой Ю в виде петли прутья — ямполическое ярмо, выданные полу обожженные j вакопечники железных стрел
Основную массу остеологической коллекции Дигоризеда составляли иерепа овец, затем в убывающем порядке шли черепа оленей, крупного жатого скота, домашних коз, зубров, косуль, сери, лосей, дагестанских дав, днких кабанов и, наконец, буйвола. Такой порядок был обусловлен ^только ценностью охотничьих приношений для божества, по и чпслен-кктыо отдельных видов промысловых животных в районе экскурсий охотников сел. Заделеск. Наилучшая сохранность костного вещества наблюдалась у черепов зубров и лосей, лежавших в поверхностном слое iwtoee сухих мест пещеры, например у входа в боковой отрог. Многие взчерепов зубров еще имели на лбу и затылке обрывки связок, сохранившееся из-за отсутствия кожеедов. Один из черепов зубров имел вырезанную на лоу ножом надпись «1833». Того же типа сохранности были и че-
200
Часть 1. Обоснование истории формирования фауны
репа лосей. В лунках их зубов хорошо сохранилась труха от веток деревьев и кустарников. Рогов лосей в пещере не оказалось. У одного пз черепов лося, лежавшего в переднем ряду, рога были срублены вместе со всей лобной п затылочной частью, у другого же, найденного в дальнем углу, они отвалились или были сбиты по венчику. Возможно, что рога лосей были похищены из пещеры или оставлены охотником дома как особо ценная добыча.
Рога оленей, лежащие вперемежку с черепами на участках, не доступных дождю и снегопадам, сохранили натуральный коричневый цвет
Рис. 91. Внутренний вид пещеры Дигори зед.
Фот. автора, 1947.
Лишь единичные экземпляры рогов оленей, убитых в августе, были с* следами линьки кожи. Основная же масса рогов, находившаяся на полатях, под воздействием дождя, снега и солнца равномерно побелелаI с верхней стороны и почернела от копоти с нижней. Черепа серн и косуть встречалась преимущественно в среднем и дальнем участке костиож слоя. По характеру сохранности они не отличались существенно от черепов зубров и лосей.
Дагестанскому козлу принадлежало всего 4 черепа с уцелевшими роговыми чехлами, по типу сохранности они были среднего возраста. 4J репа диких свиней — всего 3 штуки — были обнаружены лишь в поверх-1 постных слоях боковой кучи. Их наличие объясняется, вероятно, утратой религиозных предрассудков дигорцами и резким уменьшением болм ценного зверя в окрестностях. Свежие вываренные черепа и разбитые! топором кости конечностей, ребра принадлежали только крупному рога! тому скоту, козам и овцам. Было ясно заметно, как па смену мощщЛ черепам взрослых быков древности в новейшее время приходят черепа? полу взрослых и мелких особен, которые стали разрубаться вдоль.
Гл. 'П. Развитие ландшафтов и фаун в четвертичном периоде
201
Судя по черепам, породный состав крупного рогатого скота района был • овершепно однотипен в течение всего периода накопления костного слоя. Все мо были крупные животные с вогнутым междурожьем и рогами,
лепными несколько вперед, в стороны и вверх, относившиеся । у ’ ично укрупненному европейскому типу Bos brachyceros. Черепа овец tag принадлежали сравнительно мелким породам со слабо развитыми Бм я\ш рогов и роговыми чехлами. Если считать, что с момента осно-£( святилища ежегодно закалывалось по бычку и 2—3 барашка, то
taw длительность накопления этой коллек-и составит не мнеее пятисот лет.
Ка оказалось, черепа в правом отроге жэл 1 на небольшом слое известняковой Ьуи и вековой пыли, подстилаемой сплош-ЮН плитой. Пробный шурф, забитый в ле-tai ч лу пиршественного зала под грудой лреы на глубину7 60 см до известняко-fro дна, показал отсутствие костей в слое вековоJ пыли.
Ц>..-па зубров и лосей рассматрива-ILft дигорцами как особо ценные припоше-0/; Австатн, п можно думать, что боль-gaHCT O черепов этих зверей, убитых охот-Кш Заделеска, попадало в святилище.
Становится ясно, что численность зубров |i .|. ?1но лосей была уже очень невелика Ktci- XV—XVI вв. Начало накопления черной в пещере вряд ли может быть древнее £|\ XV вв., а прекращение накопления t'4'В зубров и лосей падает на конец \ ЦП начало XIX в. Прекращение массо-
юп) поступления оленьих черепов и рогов Рис. 92. Железные наконечни-Ейцеру, судя но характеру^ их сохранно- кп стрел из пещеры Дигори-
Сгп. ышь незначительно запаздывает по зед'
кшаешпо с черепами п рогами зубров
। досей. Олени все же дожили до появления в руках осетин трехли-luiiKU. Охотники Заделеска убивали в год в среднем до десятка оленей. Особый интерес остеологическая коллекция Дпгоризеда представляет
, отношении учета возрастного п полового состава добывавшихся охотно-промысловых животных, а также сроков добычи но сезонам года по отражено в таол. 43.1 юда,
Главный промысловый зверь — о тень —
* досыпался преимущественно
воссшшо месяцы, т. е. в период гона.
Дзу а р Лесгор! (нижний) устроен на небольшом выступе той же
Скалистого хребта, ио с левого борта ущелья Уруха и почти на Коковой высоте с пещерой Дигоризед. Оп принадлежал заброшенному Ер в пяча ле нашего столетия сел. Лесгор. Коллекция черепов раскол о-в два этажа за небольшой каменной оградой па открытой стенке. Кнпй ряд положен прямо па землю, верхний пристроен па толстом «новом бревне-колоде, укрепленном в расселине скалы и па «вешалке» рцде сучковатого ствола сосны. Вся верхняя куча черепов сильно про-Kbna дымом священного костра, систематически разводившегося под
i Учтены тр черепа, возраст и пол которых удалось установить достоверно.
202
Часть /. Обоснование истории формирования фауны
Таблица 43
Половой п возрастной состав диких копытных в коллекции пещеры Дигоризед
Виды Самки Самцы
полувзрослые взрослые итого молодые полу-взрослые взрослые итого
Sus scroja .... — — — 3 3
Capra cylindri-cornis — 1 1 3 3
Rupicapra rupicapra caucasica . 15 15 -— 10 10
Bison bonasus Caucasians .... 9 26 35 1 8 21 30
Capreolus capreolus 10 10 20 — 2 32 34
C ervus elaphus mar al 10 265 275 35 384 439
Alces alces cauca-sicus — 4 4 — — 2 2
скалой. Большинство черепов и рогов оленей нижнего ряда истлело от сырости. По численности черепа располагались в следующем убывающем | порядке: овца, корова, коза, олень, восточнокавказский козел, косуля, зубр, лось. Совсем свежими казались череп молодого зубренка и совершенно прокопченный череп зубрихи с сохранившимся правым роговым чехлом.
Лесгор II (верхний) расположен метров на 250 выше по обрыву, сбоку от очень крутой тропинки, идущей по живописной щели. Коллекция черепов находится под небольшим навесом скалы и плохо защищена от осадков. Рога оленей лежат здесь на горизонтальных вешалах пз сосновых стволов. Часть черепов, лежащих на земле, «вросла» в почвенный слой и была уже перекрыта дерном. Черепов диких копытных здесь больше, чем в Лесгоре I, так как дзуар расположен ближе к охотничьим участкам и дальше от селения. Порядок преобладания видов такой: овца, восточнокавказский козел, олень, корова, коза, зубр, серна, лось, косуля.
Черепа восточнокавказского козла, как и в Лесгоре I, большей частью без роговых футляров, сбитых и, вероятно, унесенных в поселки (рис. 93).
Для выяснения прежних мест обитания и истории деградации ареалов зубра, лося и оленя эта группа урухскпх дзуаров имеет важное значение. Принос черепов и рогов в дзуары по горным тропам был, конечно, нелегок. Охотники притаскивали черепа и рога преимущественно в наиболее почитаемый дзуар, а также в тот, который был ближе к месту добычи зверя. Судя по расположению древних вьючных трон, связывающих селения I i Заделеск и Лесгор с предгорной равниной через перевалы Скалистого хребта, и по расспросам старожилов сел. Заделеск, удалось установить, что главная масса черепов оленей, лосей п зубров поступала в урухские дзуары снизу, из лесов Черных гор, а также с Терской наклонной рав-* |
нины, особенно из района сел. Ахсарисар. Олени держались еще в 20-х I годах в период гона па верхних широких участках ущелья Уруха, в районе I i селений Стыр-Дигора, Погкау, Камунта, Дупта. Однако последние лоси с и зубры бродили в конце XV11I столетия преимущественно в лесах пред- д горни Осетии. Подтверждением этому служит отсутствие их черепов и в упомянутых верхних святилищах. В самой же продольной долине между! в
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
203
истым и Боковым хребтами олени, лоси и зубры были истреблены и вытеснены, вероятно, еще до дпгорцев, густой сетью поселений и вышкой природных лесных рощ древним населением. Есть основание ду-что ro время путешествия Гюльденштедта единичные зубры и лоси
Сроди.н.г в лесах Осетии.
ГД з у а р Реком наиболее известен в литературе и особенно по-гзется осетинами (Вязпгпн, 1929). Он расположен в Цейском ущелье, [течет приток Ардона, в зоне сосновых лесов Бокового хребта, и прн-Ьежат сея. Цен. Но сюда приходят жители и других селений.
Рис. 93. Черепа восточнокавказских козлов в дзуаре Лссгор II.
Фот. автора, 1947.
ho массивная, из толстых тиссовых и сосновых бревен постройка, хной дверью, без окоп. Живописная поляна, па которой стоит Реком, данная вековым сосновым лесом, скалами, с видом на снежные вер-iu, была издревле культовым местом, использовавшимся еще в век юы. Постройку сарая датируют обычно концом XVII в. (рис. 94). Уцелевшая от расхищения туристами коллекция черепов и рогов ко-цх расположена здесь на завалинках и на специальных полках и ах стен. Она представлена черепами и рогами кавказских козлов, пнпх овец, коров, коз, оленей. Сатушги (19146) упоминал, что в Рс-пмелпсь черепа зубров. Большинство остатков кавказских козлов •тавлено полувыветрелыми и расслоившимися от влаги и солнца пни чехлами и трухлявыми костными стержнями, но было и 6 свеже-донных черепов, принесенных в последние два-три десятка лет раин ближайших ледников. Здесь же над святилищем, в скалах, и и пор живут серны. Олепи и косули, вероятно, отсутствовали в округе яыпо пз-за глубоких зимних снегов. Принос черепов и рогов оленей (омпроизводился главным образом с пизких участков бассейна Ардона.
204
Часть 1. Обоснование истории формирования фауны
Роговые чехлы н осевые черепа кавказских козлов с их костным стержнями принадлежат взрослым и даже старым особям — возраст
от 6 до 17 лет. Наибольшее число случаев приходится на козлов возраст 10—11 лет (рис. 95 п 96).
Всего по 18 дзуарам нами было разобрано до 10 тыс. черепов, рого. п их фрагментов от 8815 особен, принадлежащих И видам копытных Домашние животные оказались представленными быком, буйволом овцой, козой; дикие -
Рве. 94. Дзуар Реком. Фот. автора, 1947.
4. Численность козлов, серп, косуль и
кабаном, зубром, восточ нокавказским козлом, сер ной, лосем, оленем, косу лей (табл. 44).
11 аши исс ледованш коллекций черепов в осе тинских дзуарах при со поставлении литературных сведений и современные фактических данных по зволяют сделать следующие выводы.
1. Полтора столетия тому назад на Тереке-Сунженской наклонно! равнине и в ущельях се верного склона Большого Кавказа обитало 8 видов парнокопытных: кабан, зубр, восточнокавказский и западпокавказский ко,-лы, серна, благородный олень, лось, косуля.
2. Зубр и лось былина грани вымирания уже в конце XVIII в. , когда чис-лепность оленя была еще велика.
3. Окончательное ист-, ребленпе оленя па исследованном участке Большог. Кавказа приходится не второе десятилетие XX в кабанов резко сокращалась
па протяжении прошлого и начала нынешнего столетня.
5. Основной причиной полного вымирания одних и сокращен^ распространения и численности других видов копытных являлось здесц прямое и косвенное воздействие человека. Это воздействие состояли в пеограничейном промысле при помощи совершенствующегося огнестрельного оружия и в обезлесении предгорной наклонной равшим!
Святилища, аналогичные описанным, существуют также в Хевсуре, тин, Тушетии и Дагестане. Их остеологические коллекции состоя преимущественно из черепов и рогов восточнокавказских и безоа ' ровых козлов, оленей (Дппиик, 1914а: Макалатия, 1935; МаруатпвилЛ 1955).
5
и к
я
_ я g ф д ф д
я
KUlLHilkU X
а s
О g ? »
’1«л pvtl< »u К' шытн их и
1 Бассейн «Риардона Е ас •С(‘йн / крдона Кассой и Vpyxa
Виды • о л tef 0J >—< -—к. Итого
Дзнвгпс Табаху Нар Зарамаг нижний Еыз Мизур । верхний Геком Садов in । пnil Эгид Ход Дунта Ногнау с. р t5 !>• Ханаз Сч о Лесгор ] (нижний Лесгор 1 (верхний
Домашние
Буйвол К р у п пы и рога гг ый — — — — — 1 — — — — — — — — — — 1 — — 1
скот 4 118 1 55 166 42 68 39 53 81 73 79 39 3 6 500 492 118 50 2086
Овца 3 30.5 223 134 123 108 72 39 76 8) 62 156 10 11 1000 1396 138 125 4061
Коза 2 60 73 27 41. 17 6 27 56 53 1<8 53 5 6 200 244 73 30 991
Итого 7139
Дикие
Sus scroja — — — — — — — — — — — — — — — 3 — — 3
Capreolus capreolus . Cervus elaphus ma- 9 — — — 2 — — — — — — — — — — 51 3 1 66
ral Alces alces caucasi- 12 з 1 — 1 — 17 — — 76 1 — О 67 661 54 49 945 9
cus — — — — — — — ( — — — — — — 6 1 1
Capra cylin dricor nix C. caucasica .... 1 10 — .) — 5 292 11 5 10 11 21 — 8 — 4 40 86 509
— — — — — — 6 — — — — — — — — — — — 6
Rupicapra rupicapra 58
caucasica Bison bonasus cauca- — — — 7 3 — — 1 10 — — 27) о 3
sicus —— —- — — — — — — — — 1 — — — — 73 2 •) 79
Итого . . 1675
Всего I 1 8814
206
Часть /. Обоснование истории формирования фауны
Остатки мелких видов млекопитающих в погадках филинов и сычей были собраны нами в двух пунктах внутренних долин. Близ сел. Дзивгис
Рис. 95. Соотношение возрастных групп и размеров рогов восточнокавказских козлов в дзуаре Реком. Длина роговых чехлов измерялась по большой кривизне.
в ущелье Фиагдона эти остатки залегали па дне небольших каверн в изве| стняковой скале. Они были совершенно отбелены и хорошей сохранности.
Таблица 45
Видовой состав и численность костных остатков млекопитающих в погадках филина и сыча у сел. Дзивгпс
Виды
Число к остей
Число особей
Vulpes vulpes....................
Mustela nivalis..................
Lepus europaeus ......... Cricetulus migralorius ...... Microtus arvalis.................
Mus musculus.....................
Apodemus sylvaticus..............
jRattus norvegicus...............
139
22
8
15
93
I
1 22 ~4
* 5 л
н
Всего..................
45
Гл. II- Развитие ландшафтов и фаун в четвертичном периоде
207
Древность их накопления не более чем 150—200 лет. Современный ландшафт иа этом участке представлен крутыми задернованными сухими склонами с группировкой трагакантовых астрагалов, россыпями скал. Близ русла речки располагаются огороды, покосы и выгоны. Абсолютная 1 ысота около 1400 м. Состав остатков показан в табл. 45.
Здесь же обнаружено до 40 фрагментов ящериц и жаб.
Все названные виды зверей живут в сухой части долины Фпагдона и настоящее время. Следует только отметить, что существование здесь истока, домовой мыши, а
з вести.
цожет быть, и серого хомячка в значительной степени обусловлено деятельностью «довека, уничтожившего лес о дну ущелья.
Иной состав костей в по-цках, принадлежавших сы-L был установлен в вер-ивьях Донпсер-Дона, пра-и>го притока Уруха, в разницах средневековой крепостцы.
Верхние части продоль-jpil ДОЛИНЫ с пологими склонами покрыты здесь м-зофитпыми лугами и по-’ослью березняка. Абсолютная высота около 1600 м. Видовой состав остатков ж-рьков соответствует современному ландшафту (таблица 46).
Сопоставление этих двух писков при учете развития ландшафтов внутренних дйлпи показывает, что состав к распределение мышевидных во внутренних сухих долинах изменялись па протяжении последних сто-aefiiii в широких преде-.л.
Иссушение долин вследствие вырубки рощ и лесов склонов, а также । |цпаса скота приводило к замене мезофильной экологической группи-меки грызунов, состоящей из Microtus arralis, М. majori, Chionomys rd. группировкой ксерофилыюй: Cricetulus migralorius, Mus tniisculus planus, Lepus europaeus — и внедрением синантропных форм: llallus и Mus musculus musculus.
Фактов недавнего вымирания каких-либо видов мелких зверь-№ па обследованных участках не подмечено. По выявлено дсь и остатков древвих видов: горного суслика — Citellus jnjg-musicus и горпого хомяка — Mesocricetus auratus raddei, — харак-прпых для участков внутренних долин Кабарды на западе и Дагестана на востоке.
< Л;
-
Рис. 96. Роговые чехлы козлов и рога оленей на завалинке Рекома.
Фот. автора, 1947.
208
Часть I. Обоснование истории формирования фауны
Таблица 46
Видовой состав и численность костных остатков млекопитающих в погадках сыча в верховьях Донисер-Дона
Виды
Sorex ar а пей s ..........
Mustela nivalis.......
Microlus gud ..........
Microtinae............
В том числе достоверно определены
Al. arvalis .........
М. majori .........
Ahis musculus ..........
Apodemus sylvalicus...
Число костей Число особей
.) 1
16 1
37 6
/27 80
— (77)
— (3)
43 14
14 8
844 110
Всего .................
Западное Закавказье
Поселения человека в голоцене были приурочены в районе к террасам Черноморского побережья, ущельям горных речек и к берегам водоемов болотистой Колхиды.
Особо интересны фаунистические находки в неолитических и позднейших слоях Ахштырской пещеры близ Адлера, которые налегают непосредственно на палеолитические. Остатки из верхних слоев пещеры в сборах С. Н. Замятнииа трудно, однако, разграничить точнее по послепалеолптпческпм культурам. Кости эти легкие, большей
Таблица 47
Видовой состав и численность костных остатков млекопитающих в послепалеоли-тических слоях пещеры Ахштырской
Виды Число костей Число особей Виды Число i гостей Число особей
Домашние Собака Лошадь ..... Свинья Крупный рогатый скот Коза Мелкий рогатый скот 3 1 8 7 17 25 1 1 4 2 6 4 Ди кие Ursus arctos Canis lupus ...... Meles nicies Lepus europaeus .... Halt us rat I us Sus scrofa Corvus ela pints Capreolus capreolus . . . 4 4 2 J 5 25 о 1 1 1 4 1 л
Итого 6J Итого 71 I(i
Been» 132 34
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
209
частно светло-желтые, но имеют и темную пятнистость. Некоторые обломки слабо фоссилизованы и напоминают кости из верхнепалеолптиче-ских слоев. В определимых остатках преобладают обломки эпифизов. Список видов и численность остатков даны в табл. 471 (рис. 97).
По сравнению с составом кухонных остатков из других послепалео-зитпческих поселений Кавказа здесь выявлен весьма разнообразный
Рис. 97. Остатки млекопитающих из послепалеолитических слоев пещеры Ахштырской.
1 — плитная Ursus arctos; 2 — лучевая Meles meles; 3 — бедренная Lepus europa-eUS; 4 — бедренная Rattus rattus; 5 — плюсневая Capreolus capreolus-, 6 — пястная Capra caucasica.
$став диких форм, так как пещера долго служила убежищем охотникам и пастухам.
Очень показательно исчезновение остатков горных копытных, отмененных, очевидно, в горы, пещерного медведя, лося, гигантского оленя, зубра. Показательно появление черной крысы в слоях с культурой железа.
В оолотпетой Колхиде наиболее древние культурные слои, содержаще остатки животных, известны по раскопкам энеолитичсского селища. Оно помещалось на полузатопленном ныне в болоте искусственном холме, Ьнзсел. Очамчирн, па левом берегу ручья Джикумура (Соловьев, 1939). В костных сборах В. И. Громов определил остатки мелкой коровы, «уашней свиньи, коз и овец. Из диких животных обнаружены остатки меня, косули, дельфина, а также рыб, в частности камбалы. Характерно jjfcyrcTBHP остатков . лошади.
1 Материалы из слоев неолита и железа объединены, так как состояние костей щ^ает уверенность в смешивании их в слоях и при раскопках.
14 Н. К Верещагин
210
Часть I. Обоснование истории формирования фауны
Одно из поселений эпохи ранней бронзы было раскопано археологами Зугдидского музея у Анаклии, близ устья Ипгура. Культурный слой перекрыт здесь новейшим иловатым аллювием. Весьма первобытная черпая керамика, по мнению Б. А. Куфтина, позволяет датировать этот памятник II тысячелетием до п. э. Кости из этого захоронения блестящие, каштанового цвета, слабо дробленные, что указывает на обилие животной пищи у древних обитателей побережья. Видовой состав остатков свидетельствует о примитивном «болотном» животноводстве и развитом охотничьем промысле (табл. 48).
Таблица 48
Видовой состав и численность костных остатков млекопитающих в слоях поселения эпохи бронзы близ Анаклии
Виды Число ностей Число особей
Домашние
Свинья 103 17
Крупный рогатый скот 258 17
Коза 47 5
Итого 408 39
Дикие
С an is aureus 1 1
Delphinus sp 4 3
Sus scrofa 15 3
Cervus elaphus 4 1
Capreolus capreolus 36 4
Итого 60 12
Всего 468 51
Отсюда же известны единичные кости птиц и рыб.
В настоящее время оленя здесь поблизости нет, но живет масса шака-лов, и изредка встречаются кабаны и косули.
Характерно отсутствие остатков лошади и малое число остатков дельфинов по сравнению с геленджикской стояпкой.
Раскопки многочисленных курганов и остатков поселений древнегреческих и более поздних времен в Колхидской низменности, вдоль среднего и нижнего течения Риона, дали обильный костный материал, позднее, видимо, утраченный. По фотографиям, относящимся к раскопкам кургана Наохваму, предоставленным мне археологом Б. А. Куфтиным, определены остатки домашней козы, дикого кабана, оленя и косули.
Ряд оболомков рогов благородного оленя, найденных в торфяниках Колхиды при рытье дренажных каналов, хранится в музее г. Поти.
На восточной окраине Колхидской низменности весьма важное захоронение кухонных костных остатков вскрыто раскопками голоценовыхI (неолитических и позднейших) слоев стоянки Сагварджплс (Киладзе! 1953).
Гл, II, Развитие ландшафтов и фаун в четвертичном периоде
211
И
е-я
(К
о-
*И т-м
В просмотренных мною костных сборах в этих слоях присутствуют в порядке преобладания остатки следующих диких животных:1
Sus scrofa
Census elaphus Capreolus capreolus Bison bonasus
Capra caucasica Rupicapra rupicapra
Ursus arctos Equus caballus Fells silvestris Panthera pardus Castor fiber
Находка кости бобра хорошо подтверждает переживание в Колхиде ним видом плейстоцена Кавказа.
Среди многочисленных памятников древнего зодчества и скульптуры с анималистическим орнаментом, сохранившихся в западном Закавказье,
i ока-
дел ь-
зегре-щиего зднее, ргапа елены
пиках
ое за-•новых шадзе,
Рис. 98. Горельеф на капители колонны храма Баграта. Барсы, терзающие козла.
Фот. автора, 1956.
услуживает упоминания храм Баграта (постройка 1003 г.) в Кутаиси. На япптелях колонн там изображены гепарды или барсы, терзающие лошадь ййкулана и козла. Учитывая широкие культурные связи средневековой Грузии, трудно, конечно, утверждать на основе этих изображений, что тланп пли дикие лошади обитали в Имеретии в X в. (рис. 98).
Самые поздние «следы» формирования экологической группировки иедкпх млекопитающих Имеретии прослеживаются по костным остаткам, натасканным филинами в небольшие каверны в стейках каньонов, рассекающих Имеретинское плоскогорье. Наши сборы были сделаны в 1952 г. только аз п е щ е р ы С а к а ж и а в ущелье Цхал-Цителы. Результаты опре-«енпй показаны в табл. 49.
Наиболее интересный факт — это появление сразу четырех видов: зайца, полчка и серой крысы, не обнаруженных в палеолити-taux слоях имеретинских пещер.
В горной части Западного Закавказья, в ущельях Иигура, Риопа и jjjeinic-Цхалн, населенной сванами, до наших дней не сохранилось та-х коллекции черепов охотничьих животных, как на восточном Кав-
i Судя по типу сохранности, в этих материалах, изучаемых II. И. Бурчак-Абра-г чем, присутствуют, несомненно, и кости палеолитической эпохи.
14*
• 212
Часть I. Обоснование истории формирования фауны
Видовой состав и численность в голоценовых погадках филина Таблица 49 остатков млекопитающих в ущелье Цхал-Цителы
Виды Число костей Число особей
Erinaceus europaeus ....... 37 4
Cams aureus 1 1
Lepus europaeus 2 1
Glis glis ............ 12 3
Rattus norvegicus 74 12
Всего 126 21
казе. Радде (1866), правда, писал, что он видел в 1864 г. груду рогов кавказских козлов, сложенных будто бы охотниками близ часовни сел. Чу-биани (Ушкуль) в истоках Ингура. Их было там «несколько тысяч». Однако в 1948 г. мне удалось разыскать только 7 пар рогов и роговых стержней козлов: 6 — Capra caucasica и 1 — С. cylindricornis, — находившихся в алтаре часовни сел. Жабеши и в алтаре монастыря Ламар-дия, выше Ушкуля. Подтверждения сведений о существовании в прошлом «тысяч турьих рогов» не удалось получить пи от стариков, ни от местных органов власти. Как оказалось, охотники этого селения храпят рога и лопатки кавказских козлов и серн на чердаках своих боевых башен. К 1948 г. эти коллекции были уже заброшены и раздарены, а новых пополнений не поступало из-за истребления зверя.
Сваны, в противоположность осетинам, отрубают рога козлов лишь с небольшой перемычкой лобной кости. Рога козлов и лапы медведей прибиваются здесь и теперь к верандам домов или вмазываются в стены каменных жилищ, сделанных из шиферного сланца.
Итак, исследования голоценовых костных остатков в западном Закавказье показали обширное распространение в неолите и позднее крупных промысловых животных: кабана, оленя, косули, зубра, медведя,—а также наличие речного бобра в болотистой Колхиде. Крайне характерно также появление остатков новых видов, неизвестных из плейстоценовых | (палеолитических) слоев: шакала, пасюка и черной крысы. Судя по составу домашних животных и характеру захоронений из остатков, голоценовые ландшафты Колхиды и вообще Черноморского побережья были крайне близки к существовавшим здесь в начале нашего столетия. I
Центральное Закавказье
По разведочным раскопкам и сборам В. П. Любина в 1952 г. над навесами урочищ Сароты-Ор и Шаршиат-Кадзах, в окрестностях Стали-нири, из неолитических и позднейших слоев мною определены обломки I костей малорослых домашних свиной, бычков, коз и овец, а также зубы [ бурых медведей и лисиц. Обильные материалы доставили также в 1955— I 1957 гг. голоценовые слои пещер Кударо в верховьях Риона.
В долине Куры и Арагвы обильные костные материалы были извле-1 чены археологами Кавказского музея на многослойных могильных по-
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
213
лях и городищах в Самтавро и Армази, близ Мцхети.1 По архивным фотоснимкам и выставочному материалу этой эпохи поздней бронзы- — IX— VIII вп. до н. э. — можно судить, что здесь были обильны остатки лота-«4. г ров, овец, коз, кабанов, амулеты из клыков бурого медведя и рез-« . 5ров. Обнаружены недавно и скелеты погибших позднее куниц —
foina, а также степных полевок — Microtus socialis. Наиболее Kiuu пзделпя из рогов благородного оленя, представленные так назы-Кнлш «выпрямителями копий». Изящные бронзовые бляхи и безде-1\нпс этой эпохи из Самтавро и Армази нередко изображали собак, лсиц, львов, пантер, зайцев, оленей, джейранов, баранов, кавказских Квлов и первобытных туров.
При сопоставлении палеонтологических фактов, известных для Закавказья в целом, эти изображения подтверждают существование па Кавказе в I тысячелетии до н. э. ряда крупных промысловых животных, вымерших лишь в средних веках нашей эры.
Восточное Закавказье
В пределах Куринской низменности и в предгорьях Большого Кавказа, Карабаха и Талыша голоценовые захоронения костных остатков пбиаружепы в делювии, аллювии, пещерах, асфальтовых натеках и в культурных слоях древних поселений.
I Б литературе упомянута находка рога благородного олепя в делю-вш склона близ сел. Алты-Агач, у восточной оконечности Большого хребта (Деминский, 1901). Значительное число находок сосредоточено 13 Ашиеропском полуострове.
С. А. Ковалевский обнаружил в 1941 г. скелет полосатой гиены — Hyaena hyaena па глубине 4 м в толще голоценовых лёссовидных суглинков в низовьях речкп Сумгаит, севернее Баку. Скелет залегал под культурным слоем со средневековой керамикой и под костным слоем другого, ►1олее древнего поселения, перекрытого отложениями новейшего Каспия. Учитывая горизонт захоронения и сохранность костного вещества, Ьзраст этой находки можно датировать ранним послеледниковьем.
Осевой череп очень старой полосатой гиены, кости лошади и каспийского тюленя — Phoca caspica были найдены С. А. Ковалевским в Баилов-скоя пещере, на южной окраине Баку (Богачев, 19386).
В Бинагадах из отвалов асфальтированных покровных суглинков вад средпечетвертичными слоями нами извлечены две кости ног джейрана — Gazella siibgiitturosa, явно более свежие и слабее прокрашенные нефтью, чем кости плейстоценового комплекса. В асфальтовых натеках горы Киркпшлаг нами обнаружены еще кости зайца русака и волка. Г Скелетики мелких грызунов: тушканчиков — Allaclaga elater, хомячив __ Cricetulus migratorius, полевок — Microtus socialis, —погибавших в асфальте, обнаруживались, кроме Киркишлага, и на горе Зигиль-Ппря (Верещагин, 19516).
Обилие в Азербайджане поселений и могильников голоценовой эпохи (Пассок и Латынин, 1926) обусловило и получение значительных кост-k сборов из культурных слоев.
Гуммель (1948), копавший курганы века бронзы на наклонной равнине западнее Хаплара (бывш. Еленендорф), упоминает о многочислен-
1 Этп материалы остались не определенными и были утрачены.
214
Часть I. Обоснование истории формирования фауны
пых находках быков, овец, коз и домашней свиньи. Ничтожны находки остатков лошади.
Раскопки поселений и погребений древнего Мингечаура на правом берегу Куры, близ современной ГЭС, вскрыли культурные слои, датируемые в довольно широких пределах — от II тысячелетия до н. э. до раннего средневековья. Костные материалы отсюда весьма велики, но еще не все определены. При предварительном просмотре нами этих материалов здесь отмечены многочисленные остатки лошадей, ослов, коров, овец, коз, реже домашних свиней. Остатки диких животных представлены здесь костями благородного оленя — Cervus elaphus, населявшего еще сравнительно недавно куринские галерейные леса этого района, джейрана — Gazella subgutturosa и кабанов — Sus scrofa. Последние два вида были обычны в районе и до недавнего времени.
Особый интерес представила наша находка в Мингечауре среди кувшинных погребений костей черной крысы — Rattus rattus вместе с остатками лошади, осла, крупного рогатого скота и кабана. Погребения в кувшинах, сопровождавшиеся жертвоприношениями домашних козлят и запасов зерна, датируются серединой I тысячелетия до п. э. В связи с тем, что могильное поле было занято уже много столетий солянковой группировкой полупустыни, а крысы могли обитать па нем либо в эпоху существования поселения с оросительной системой, либо в эпоху культа массовых могильников, эта находка отлично документирует наши соображения о древности существования черных крыс в Закавказье (Верещагин, 1949в).
Весьма древнее поселение было обнаружено в голоценовых суглинках уустья речки Сумгаит в 1941 г. С. Л. Ковалевским. Кухонные костные остатки залегали на глубине 3 м под культурным слоем с средневековой керамикой и хорошо сохранились в условиях большой сухости. Состав обнаруженных видов дан в табл. 50.
Таблица -50
Видовой состав и численность костных остатков млекопитающих в слоях древнего поселения у Сумгаита
Виды Число ностей Число особей
Домашние
Лошадь 6 1
Крупный рогатый скот И 3
Овца 12 3
Итого 29 7
Дикие
Lepus europaeus 2 1
Gazella subgutturosa 3 1
Итого 5 2
Всего 34 9
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
215
Здесь же найдено 10 фрагментов костей лебедя-шипуна — Cygnus и гуся — A user anser L., всего от 3 особей.
По заключению С. А. Ковалевского, костеносный слой перекрыт 1гдолениями современного Каспия и поэтому может быть датирован лег ‘ предшествовавшей последней в голоцене трансгресии Каспия — Н ле I тысячелетия до н. э.
Обильные сборы костей сделаны также в 1953, 1954 гг. археологами лбайджапской Академии наук при раскопках крепости Урень-Кала ча ан), в центре Мильской степи. По нашим наблюдениям, в средне-девых слоях здесь преобладали остатки овец, крупного рогатого скота, пощади, с небольшой примесью костей джейранов — Gazella sub-
УГОИ.
Из (лоев средневековья костные материалы были добыты также рас-р taJ- I В. Н. Левиатова во дворе дворца Ширван-Шахов в Баку. Рас-рмки скрыли остатки небольших поселении, последовательно располагав п я друг над другом па холме современной Бакинской крепости.
ученные слои датируются по керамике IX—XIII вв. н. э. Это эпоха
он1,а арабского халифата, век Низами и начала монгольских прорывов воск иное Закавказье. Часть костных материалов захоронена здесь,
го в результате падежа скота, так как весьма обычны целые лопатки
пдалечья овец и коз, но большинство фрагментов не что иное, как
с сохранившиеся в сухом грунте кухонные отбросы.
Таблица 51
Видовой состав и численность костных остатков млекопитающих в слоях средневекового поселения на холме Бакинской крепости
Виды Число костей Число особей
Домашние
Собака 3 3
Кошка 1 1
Лошадь 39 10
Осел 15 5
Верблюд 3 2
Крупный рогатый скот 111 14
Овца 202 29
Коза 3 2
Мелкий рогатый скот, ближе
не определенный 223 15
Итого 600 81
Дикие
Phoca caspica 1 1
Vulpes vulpes 1 1
Equus hemionus 1 1
Gazella subgulturusa 48 18
Итого 51 21
Bcei о * . . 651 102
216
Часть I. Обоснование истории формирования фауны
Характерно преобладание костей домашних животных: овец, коров, лошадей, ослов. Остатки диких видов: джейрана, кулана, лисицы и тюленя — немногочисленны (табл. 51).
Отсюда же известны остатки птиц: Cygnus sp., Anser sp., Otis tarda L., O. tetrax L., курицы,—всего 21 фрагмент от 10 особ<&, Кроме того, остатки рыб: Acipenseridae—25 фрагментов от 4 особен к Cyprinidae — 58 фрагментов от 8 особей.
Рис. 99. Развалины крепости Чухур-Кабала.
Фот. автора, 1951.
Наибольшее число остатков джейранов приходится на слои X—XI вв. Плюсна кулана обнаружена в слое XIII в.
Находки в древнем Баку хорошо подтверждают указания об обилии джейранов и куланов в восточном Закавказье. Установлено также существование промысла на тюленя, водоплавающую птицу и рыбу в Бакинской бухте.
В более высоких слоях, датируемых XV—XVI вв. п. э. по материалам тех же раскопок, обычны кости и целые черепа серых крыс—Rattus norvegicus. Однако одновременность этих остатков данным слоям не доказана, так как крысы нередко самостоятельно проникают в глубокие слои по расщелинам кладки и норам.
На северной окраипе Курипскоп низменности и в Алазапо-Агричап-ской долине нами исследовались костные материалы в слоях древнем крепости Чухур-Кабала (рис. 99), разрушенной монголами в XIV в. (Крымский, 1934). Остатки крепости и города расположены между двумя левыми притоками Турьяи-Чая, среди сплошного широколиственного леса. В эпоху расцвета города эти леса были, очевидно, сильно вырублены, так как обломки квадратных кирпичей и керамики
Гл. II» Развитие ландшафтов и фаун в четвертичном периоде
217
встречаются в окрестностях и особенно по дну ключей Кара-Су, протекающих через ольхово-лапиновые джунгли.
Кости диких и домашних животных вперемежку с керамикой, кпр-жми, золой п землей заполняют в крепости глубокие колодцы, энер-» Шо подмываемые теперь с востока речкой Кара-Чай. Состав костных ков домашних и диких видов, собранных нами в отвалах обрыва j I русле речки, указан в табл. 52.
Таблица 52
Видовой состав и численность костных остатков млекопитающих в средневековых слоях крепости Чухур-Кабала
Виды
Число костей
Число особей
Домашние
Осел........................
Лошадь......................
Крупный рогатый скот........
Буйвол .....................
Мелкий рогатый скот.........
1
3
13
1
38
1
2
2
1
5
Итого...................... 56 11
Дикие
Cervus elaphus...................... 1
Capreolus capreolus................. 1
Итого........................ 2
Всего
При специальных раскопках видовой состав диких видов был бы, ко-( по, сильно расширен.
Сбор остатков мелких зверьков из погадок сычей был предпринят нами в 1946—1947 гг. на северной окраине Куринской низменности и Апшероне. Такие сборы были сделаны между Алд жиган- Чаем иГеок-Чаем в недолговечных пещерках по обрывам третичных всхолмлений, „целяющих Алазано-Агричайскую долину от Куринской низменности. Следование остатков из этого района подтвердило видовой состав совре-I ой группировки насекомоядопых и грызунов, известной по матс-рлам отловов капканчиками (табл. 53).
Более подробные материалы были получены при изучении прежнего и современного распространения мелких млекопитающих па Апшерон-дом полуострове, особенно в связи с возможностью сравнения современно состава видов с плейстоценовым.
В известняковой скале восточнее мыса Б и л ь г я, на северном |>егу Апшероиа, в 1944 г. Н. И. Бурча к- Абрамович обнаружил узкую ведь (рис. ЮО), занимавшуюся па протяжении нескольких столетий подвид домовыми сычами. Косточки насекомоядных и грызунов, птиц, реп-Ьи, амфибий, а также хитин жуков из распавшихся погадок образовали |гд шве каверны слой толщиной в 10—12 см. В верхних слоях косточки
218
Часть I, Обоснование истории формирования фауны
Таблица 53
Видовой состав и численность костных остатков мелких млекопитающих в погадках сычей из Геокчайского боздага
Виды Число костей Число особей
Hemiechinus auritus 2 1
Crocidura russula 33 7
Dyromys nitedula 4 ' 1
Cricetulus migratorius 5 2
Microtus socialis ........ 106 19
Mus musculus 16 6
Apodemus sylvaticus 36 9
Meriones erythrourus 15 1
Всего 217 46
были коричневые, в нижних — уже совершенно побуревшие. Отсюда было извлечено И. М. Громовым в 1944 г. и мною в 1945 г. до 5500 костей,
Рис* 100. Убежище домового сыча в скалах мыса Бильгя.
принадлежащих млекопитающим. Кости птиц, ящериц, жаб составляли не меньшую цифру.
Состав видов зверьков, поедавшихся сычом, оказался очень близким к составу зверьков, обитающих и ныне на северном Апшероне и известных нам по массовым отловам капканчиками (табл. 54).
Несмотря на обилие и относительную древность накопления погадок сычей, в их «сборах» оказался, в сущности, вполне современный состав видов.
Отсутствие сходства в отношении численности различных грызунов при охоте сычей и при отлове ловушками отражают лишь избирательную способность птиц и требования, предъявляемые к ловцам противочумной станции, — ловить преимущественно тот или иной вид грызуна.
Относительная стабильность экологической группировки мелких млекопитающих Апшерон
ского полуострова на протяжении последнего отрезка голоцена несомненна. Вместе с тем изучение распространения современных млекопитающих показывает, что обыкновенный еж, лесная соня, лесная
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
219
Таблица 54
Видовой состав и численность мелких млекопитающих на Апшеронском полуострове
Виды Пойманные сычом за последние 3—4 столетия Добытые капканами в 1939—1940 г.
Erinaceus europaeus - Редок.
Hemiechinus auritus — Обычен.
Crocidura russula . . 8 Обычна.
Pachyura etrusca 2 Не добыта.
Dyroniys nitedula ........ — Редка.
Cricelulus migrator ins 147 73
Microtus socialis ........ 83 115
Meriones erythrourus 63 2123
Allactaga elater 1 8
A. williamsi 33 50
Afus musculus 1 90 81
Apodenius sylvaticus J ZU 6
Rattus norvegicus — 2
Всего 3571 2458
рпь и степная полевка являются реликтами, уцелевшими с какой-p. более влажной эпохи на немногих участках полуострова. Отметим, L релпктовость в распространении была подмечена и для ряда других Ьоночиых, в частности для озерной лягушки — Rana ridibunda L. । болотной черепахи — Emys orbicularis L. (Верещагин, 1949в) (рис. 101).
Косвенные указания на обитание некоторых крупных зверей в историчную эпоху в восточном Закавказье даются и в изобразительном ис-цсстве кочевых племен.
В Кабристапе, к юго-западу от Баку, под скалами апшеронских из-истняков расположен ряд древних стоянок кочевников. Живописные егалы, навесы, использовавшиеся, вероятно, и для религиозных обря-wBf имеются у гор Кызыл-Кум, Беюк-Даш, Джангир-Даг. Древние кдожнпки изобразили на скалах контурные рисунки львов, благородных оленей, безоаровых и домашних козлов, домашних быков и лошадей {Верещагин п Бурчак-Абрамович, 1948). Некоторые из этих рисунков уносятся, вероятно, к неолиту, другие — к веку бронзы и нашей эпохе (pc. W2).
Подводя итоги исследованиям голоцеповых захоронений в восточном Закавказье, приходим к выводу, что в послеледниковую эпоху в восточном Ькавказье появляются: из хищных — полосатая гиена, а возможно и кг из грызунов — черпая и серая крысы; из копытных — кулан и джей-Несколько ранее здесь, очевидно, исчезает ряд крупных видов зве-мц, характерных для среднего и верхнего плейстоцена.
Следует отметить также, что распространение ряда копытных, например yvM и джейрана, было здесь еще недавно несравненно большим, причем уеии жили в тугайных лесах среднего течения Куры.
1 Число особей показано по подсчету нижних челюстей.
Рис. 101.
Рис. 102. Рисунки козлов и других животных на скалах Джингир-Дага.
.Гл. 11. Развитие ландшафтов и фаун в четвертичном периоде 221
Распространение обыкновенного ежа, лесной сони, леспой мыши, верной лягушки и болотной черепахи на Апшеронском полуострове приобрело в нашу эпоху реликтовый характер, свидетельствуя об отно-KiibLio устойчивом климатическом режиме на протяжении исторического вЧода и о новейших изменениях ландшафта под влиянием человека.
Нагорье Малого Кавказа
статкп млекопитающих послеледниковой эпохи на Армянском плоско-цры обнаружены в делювиальных отложениях, туфоконгломератах, [верных отложениях, пещерах, культурных слоях поселений п могиль-
Находки в делювии и на берегах оз. Севан.
Ли» н 1926 г. А. Б. Шелковников доставил в ЗИН окатанные фрагменты а зубра — Bison bonasus subsp., извлеченные из послеледниковых I конгломератов у истока Занги из оз. Севан (Громова, 1935в).
По типу сохранности костного вещества эти фрагменты могут быть юваны эпохой бронзы.
Крупный черен оленя с отлично сохранившимися рогами был найден • я₽ юв1ш склона близ сел. Кабахлу Азизбековского района Армении |>. 1947а).
I аллювиальных суглинках правобережья Занги, в окрестностях Ряи на глубине 6.5—7.0 м от поверхности земли был обнаружен фраг-м черепа суслика — Citellus cf. xanthoprymnus. Суглинки залегают авовом покрове, излившемся, как предполагают, в III—II тысяче-L| до н. э., и эпоха былого обитания сусликов в этом районе оценивается полько более поздним периодом (Даль, 1949а). В настоящее время . »чпая граница распространения суслика проходит па 40 км западнее.
•начптелыюе количество костей зверей захоропялось в голоцене в при-допой полосе оз. Севан. Эти кости принадлежали частью провалив-1пся под лед озера и утонувшим животным, частью животным, погибам в горных потоках. Значительное число костей — кухонные остатки донпх охотников и скотоводов, селившихся на побережьях в свайных Ьройках. Близ остатков таких построек у истока Занги были извлечены ре пары мощных рогов благородных оленей, хранящиеся в Кавказском да (Баперн, 1871). Свайные постройки существовали также близ Но-Ыаязета и у оз. Гилли (Лалаян, 1929).
g 1947—1948 гг. при начавшемся спуске вод озера Даль (19506) г^рал па обнажившейся полосе северного берега много обломков костей, «еди которых определил остатки оленя, лесного кота, кабана и медведя. J3 четководной бухты Артанишского залива с глубины 1.5 м была изучена значительная коллекция обломков рогов оленя, обломков че-кабанов и быков.
В серых речных песках побережья Севана: в низовьях речки Гава-гет в окрестностях речки Агбулах и у мыса Сары-Кая — были пай-деы более древние по типу сохранности обломки рогов оленей, обломки tpfnoP П позвонки крупных быков, близких по размерам к первобытному пли зубру. Под мысом Сары-Кая в этих же отложениях найден об-Ьк черепа карликового тура — Bos ci. minutus (Даль, 1950а).
Некоторое число костей собрал также на берегах Севапа по нашему -учению Я. И. Хавесон. Этп сборы были сделаны преимущественно в северо-западном берегу у Шорджи, а на южном — близ мыса Нора-ф и Сары-Кая.
222
Часть I. Обоснование истории формирования фауны
Обе эти коллекции были исследованы нами.
Большинство собранных С. К. Далем обломков коричневого и светло-коричневого цвета, а в сборах Я. И. Хавесона — преобладают светлые. Такие кости лишь кажутся тяжелыми и минерализованными из-за отложившегося известнякового осадка. Кости, собранные в волноприбои-нон зоне, сильно обтерты, кремового цвета, а затонувшие в местах затишья и захороненные в иле целы и темно окрашены. От оленей уцелело больше всего обломков рогов, часто полуразрушенных от пребывания в воде и последующего выветривания на берегу. Имеются целые метаподии и челюсти домашних быков и буйволов. По степени прокраски костного вещества и фоссилизации материал разбит на две возрастные группы. Видовой состав и численные соотношения даны в табл. 55.
Таблица 55 Видовой состав и численность костных остатков млекопитающих на побережьях оз. Севан
Виды Полуископаемые ч Современные Итого
Домашние
Собака 12 8 20
Кошка 1 ’ 1
Лошадь 61 23 84
Свинья 6 2 8
Верблюд 3 3 6
Крупный рогатый скот 70 67 137
Буйвол 5 2 7
Мелкий рогатый скот, ближе не определенный 13 14 27
Итого 171 119 290
Дикие
Vulpes vulpes 1 1 2
Ursus arctos 1 — 1
Meles meles — 1 1
Sus scrofa 5 — 5
Cervus elaphus 19 33 52
Итого 26 35 61
Всего 197 154 351
Особый интерес в этих сборах представляют остатки одногорбого верблюда. Характер сохранности костей предплечья, лопатки и нижней] челюсти (рис. 103), вещество которых прокрашено в светло-кофсшшйj цвет на глубину 4—5 мм, говорит о значительной древности этих остатков-3.5—4 тысячи лет.
Хавесон (19546), изучив индексы зубного ряда, относил челюсть верблюда к дикому дромадеру — Camelus dromedarius dahli. Подтвержде-пне этого диагноза на более полноценных находках свидетельствовало! бы о весьма большой ксероморфизации ландшафтов Армянского нагорья! в какую-то раннюю фазу голоцена.
]
1 с г б с
с 6l
Э1 Э1 к г К( НС Да по Ис д* 30< и пр ДИ]
Тр дат рог
сел рен
нар бли: оста кра] По • роли Зате хоро га на львы
Т ров coxpf пше стади пзобр TCKI1N
Тл. II» Развитие ландшафтов и фаун в четвертичном периоде
223
Самостоятельный заход диких верблюдов в мезофитный ландшафт современного типа присеванского высокогорья весьма сомнителен. Другие дикие виды: медведь, кабан и олень — характерны скорее для мезона — луговой и лесной — обстановки, чем для нагорной степи и
Находки в могильниках и слоях поселений. , ? точно датированы костные остатки, добытые из могильников и до древних городищ с побережий оз. Севан, с Карабахского плоско-ю 1, с берегов Запги, из ущелья Дебеда-Чая и из верховьев Храма, Б Цалки на Трпалет-
хребте. Ж
] о мнению Пиотров- ДЖ
(1949, стр. 92), наи-древней стоянкой
U1 голоцена является ДЯ
।птпческая стоян-h Iа берегу Зан-
• I . сел. Шенгавит, рас-
ЬвДОЯ Е. А. Байбуртя-
Гв 1936—1938 гг. Ее Jgg»
Jjn’уют второй ПОЛОВИ-
j 11 тысячелетия до н э. ч.
I леев стоянки С. К. ' ’
см
%- I определил кости ое- t < i.»а111 i
,9'1)вых козлов, лошадей
’ ' *-* В—4 ЕВ ЦТ Л ТТ TZ^Z'^i'T. Е"*^ л 71 / л Л* Г nZ w л 1'11 л гя м» 1 я гя ГТ Г'\ £
Рис. 103. Челюсть Camelus cf. dromedarius с побережья Севана.
। шб. Остатки лошадей ыадлежалй, вероятно,
м особям.
амечательные находки были сделаны Куфтиным (1939) в 1930 г. на □стеком хребте в верховьях Храма. В курганных погребениях, руемых энеолитом — III тысячелетием до н. э., — здесь обнаружены ... благородных оленей.
Пз одного индивидуального позднеэнеолитического погребения близ до, Озни Цалкинского района в коллекции Куфтина имеется нижнеко-инои зуб медведя — Ursus arctos meridionalis.
В более поздних курганах — середины II тысячелетия до н. э. — об-рружсны обломки черепов, метаподии и фаланги огромных быков, Низких по величине к первобытному туру — Bos primigeniuSj а также доки собаки, лошади, кабана, овцы, козы. Все эти остатки про-гдаы гумусом, темно-бурого цвета, частью сильно истлевшие, [io представлениям Куфтина, почетные покойники подвозились к ог-Ьрным погребальным ямам на колесницах, запряженных быками, м быкл убивались, съедались, а шкуры с черепами и костями ног Емпплись с умершим. На серебряном ведерке, найденном в этих курсах, изображены благородные олени и серны, а па золотом кубке — щи.
Такой же тип могилы с остатками быка был обнаружен в 1948 г. в К и-«пзкане (Пиотровский, 1950). Более полная, чем в Триалети, ' ирапность черепа позволяет утверждать, что древние племена, осгавпв-Еатоттпп погребений, имели дело с первобытным туром па первых дцпях одомашнивания. На золотой чаше, найденной в этой могиле, ’ । ржепа пара рычащих львов, весьма близких по стилизации к хет-। мш рисункам этого вида.
224
Часть I. Обоснование истории формирования фауны
Еще по описаниям Реслера (1896) было известно, что курганы и могильники в Арчадзоре и у сел. Ходжалы на Карабахском плоскогорье прикрывают «кострища с массой золы и огромными залежами пережженных костей животных: лошадей, баранов, собак, кабанов, буйволов и быков*, Эти памятники датируют теперь началом I тысячелетия до н. э. и связывают с ассирийской культурой (Пиотровский, 1949). В каменных ящи-
Рис. 104.
Черепа: 1 — Vulpes vulpes; 2 — Maries foina; 3 — Lepus europaeus из урартских могильников побережий Севана.
ках — дольменах, разрытых Бельком и Ивановским в Карабахе н под Кадабеком, также находили черепа лошадей, кости барана, верблюдов и остатки диких животных (Сысоев, 1925).
Из раскопок урартской крепости Цовипар с южного берега оз. Севан, по Пиотровскому (1944), известны остатки домашней свиньи, домашнего быка, овцы. Обширные могильные поля, оставленные ура рта ми на во-сточном берегу оз. Севан у сел. Загалу, раскапывались с 1905 по 1908г. Лалаяном (1929). Здесь и около селений Нор-Баязет, Адиаман былв большие земляные курганы трех типов, перекрывавшие каменные) склепы с коллективными захоронениями. По инвентарю переходно! эпохи от бронзы к железу эти погребения датируют началом I тысячи летия до н. э. (Пиотровский, 1949, стр. 86). Из этих могильников были
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
225
описаны (Сатунин, 19076) черепа куниц — Martes latijrons (=Martes /ш) п барсуков — Meles urartuorum (=Meles meles minor), а в 1930г. \ Б. Шелковппков доставил в ЗИН нижнюю челюсть лошади, череп
щсепка, два черепа лисиц — Vulpes vulpes, четыре черепа куниц — irtes foina и череп зайца — Lepus europaeus.
Укладка в могилу трупов диких и домашних животных имела у урар-в важное ритуальное значение. Сам Лалаян указывает, что он постоянно нодпл в могилах скелеты лошадей, крупного рогатого скота, овец и pa, которые и выбрасывал на месте. Закладывались ли трупы диких I орков целиком или только их черепа (рпс. 104), осталось неизвестным.
Украшения, добытые в этих могилах, изображают обычно благородна оленей. На сердоликовой печатке, найденной в одной из могил, пзо-| клен бегущий медведь.
Раскопки урартской крепости К а р м и р - Б л у р (Тейше-| |!ш) на р. Запге, близ Еревана (Пиотровский, 1944, 1949), доставили гнып материал, освещающий ряд черт животноводства и фаунистиче-v \ особенностей Армянского нагорья в середине I тысячелетия до н. э.
Тепшебаини — крупный административный центр государства ) фту — был взят приступом в VII в. до н. э. прорвавшимися в Закав-1же скифами. При штурме в горящих зданиях крепости были погре-ыпод обвалившимися потолками и стенами домашние и дикие живот-। латакже разнообразная утварь. Даль (1952), исследовавший 4667 кост-I осгатков, извлеченных при раскопках Тейшебаиии, привел список р.шв (таол. 56).
Т а б л п ц а 56
Видовой состав и численность костных остатков млекопитающих урартской крепости Тейшебаишг
Виды
Число особей
Виды
Число особей
Собака...................
Лошадь...................
Осел.....................
Свинья...................
Крупный рогатый скот Буйвол ..................
Овца.....................
К-'за....................
3
8
6
6
22
2
25
7
Итого.............
Martes foina............
Felis lybica............
Cricetulus migratorius . Mus musculus............
Apodemus sylvaticus . . M er tones sp...........
Capra aegagrus..........
Ovis gmelini............
Gazella subgutturosa . . Cervus elaphus..........
Итого..............
1
1 2
2 4
1 t
1 2
4
Дикие
Сг ddura russula . . . ii miechinus auritus . .
Всего ............
Знание своеобразных условий единовременного захоронения и личные рвдеиия па раскопках позволили С. К. Далю считать, что песчанка [рдестыи еж были позднейшими пришельцами на развалины крепости, впита белодуптка и степная кошка были, возможно, прирученными >1ькамп, служившими для ловли грызунов. Длиннохвостая белозубка,
15 Н. К. Верещагин
226
Часть I, Обоснование истории формирования фауны
хомячок, лесная и домовая мыши относятся к местным синантропам, заселявшим в те времена жилые постройки. Кости и поделки из рогов безо-арового козла, барана, оленя, джейрана дают некоторое представление об охотничьем промысле на северных окраинах Урарту. Находки трубчатых костей джейрана — первое документальное свидетельство обитания этой газели в средней части долины Аракса.
Находки в убежищах хищников. Новые палеонтологические материалы по последниковой эпохе нагорья Малого Кавказа были собраны зоологами при изучении остатков пищи хищников, накоплявшихся в пещерах и мелких кавернах.
Еще Поляков (1882), обследуя пещеры по берегам Севана в 1879 г., обнаружил в пещере Песчаной горы череп ласки, челюсти полевок, мышей и уток.
Обследуя современную фауну позвоночных Сарайбулахского (Урц-ского) хребта, Даль (19406, 1954а) собрали определил остатки животных, скопившиеся в четырех карстовых пещерах — промоинах — в результате хищничества филина, волка, а также самостоятельного захода некоторых зверьков. Пещера, давшая наибольший материал, находится в третьем с запада ущелье, на западной стороне хребта, над сел. П1а-гаплу. Входное отверстие ее расположено на правом склоне каньона, на высоте 11 м от дна ущелья (абсолютная высота 1750 м). Общая длина исследованных ходов пещеры составляла 41.5 м. Кости животных были разбросаны преимущественно в понижениях и камерах и в большом количестве содержались в глинистых наносах. В этой пещере собрано 1850 костей, принадлежащих по меньшей мерс 660 особям земноводных, птпц и зверей.
Сохранность и возраст извлеченных костей различны. Кости некоторых домашних животных, лежавшие на поверхности, сохранились лучше, | чем замытые в грунт. Они серо-желтоватой окраски. На некоторых кос- I тях успела образоваться известковая корка толщиной до 1.3 мм или выро-ели сталагмиты до 5 мм высоты. Наиболее старые кости растворяются в соляной кислоте полностью. Их возраст оценивался С. К. Далем от 2 до I 3—4 тыс. лет. К таким древним остаткам относились кости шакала, домашней собаки, лисицы, каменной куницы, перевязки, барсука, зайца, I осла, лошади, оленя, быка, безоарового козла, арменийского муфлона. I
Судя по тому, что у изученных мною (Верещагин, 19516) черепов I барсуков из этой пещеры признаки барсука урартской формы выражены I более резко, чем у черепов из могильников Севана, можно думать, I возраст некоторых костных остатков более древний, нежели его определил Даль. К наиболее древним остаткам относятся и челюсти крупной сеноставки, обнаруженные в дальнем конце пещеры. Современного типа I оказались остатки белозубой, ночниц, лисиц, ласок, куниц и грызунов. I Они большей частью отпрепарированы водой из погадок филина.
В результате определения всех остатков из четырех пещер был полу-1 чен большой список видов, приведенный в табл. 57.
Извлечены также остатки жаворонков, клушиц, голубей, серой ц| каменной куропаток, желтопузика, озерной лягушки.
Учитывая расположение пещер в переходной ландшафтной зопе ц| особенности поступления остатков зверей в пещеры, здесь пет основании I делать далеко идущие выводы о ландшафтно-климатических сменах в го|
л оцепе.
В настоящее время из названных в списке диких видов па Сарайбу-
лахском хребте совершенно
отсутствуют шакал,
медведь, пищуха, кулан
• Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
227
Таблица 57
Видовой состав и численность костных остатков млекопитающих в пещерах Урцского хребта
Виды Число костей Виды Число костей
Домашние Meles meles ...... 14
Сооака 11 Felis lybica 1
Лошадь 8 Lepus europaeus .... 98
Осел 11 Ochotona sp 3
Крупный рогатый скот 15 Allactaga williamsi . . 9
Овца 28 Meriones persicus . . . 135
Коза 26 Cricetulus migratorius . Mesocricetus auratus . . Microtus socialis .... 135 794 140
Итого 99
M. nivalis ....... 6
Дикие Arvicola terrestris . . . 2
Ellobius lutescens . . . 323
Erinaceus europaeus . . 1 Equus hemionus .... 4
Crocidura russula . . . 1 Cervus elaphus . 24
Myotis oxygnathus , . . 3 Capra aegagrus 85
Canis aureus 3 Ovis gmelini 76
C lupus ........ 4
Vulpes vulpes I'rsus arctos 35 16 Итого 1931
Xlustela nivalis .... 3
lories foina 15 Всего
Vormela peregusna . . . 1 2030
1епь, причем исчезновение крупных видов — несомненно, результат , ;еисгвпя человека.
Цццпй облик фаунистического комплекса района, даже в эпоху нале! ня наиболее древнпх остатков был все же весьма близок к совре-рьому, характеризуя ландшафт сухих плоскогорий Передней Азии.
В юго-восточной и центральной Армении исследования костей из по-филина производились весьма тщательно в 1946—1949 гг. Сосни-v 11 (1947, 1948) в различных вертикальных зонах и ландшафтах: 1|зопе колючих трагакантнпков (1500—1600 м над ур. м.); 2) в зоне намой типчаковой степи и аридного редколесья (2100—2200 мнад ур. м.) । Дзпзбековском районе и у оз. Севан; 3) в зоне горных лугов (2600 м цур. м.) в Нахичеванской АССР. Эти данные позволяют хорошо уяснить । большом материале (в несколько тысяч костей) характер распреде-г! мелких млекопитающих в условиях высоких плоскогорий и воз-Ьпую роль климатических и ландшафтных сдвигов для формирования Езкологическпх группировок. В табл. 58 эти данные обобщены.
Большинство исследованных костей было накоплено, очевидно, в саде* последние столетия и даже в последние десятки лет.
Основные группировки мелких зверьков — ксерофильпая и мезофиль-де — либо замещают, либо дополняют здесь друг друга, характеризуя ^•i ioландшафт, а частью ареал и особенности охоты популяций филинов.
данные показывают также биологические черты филипа: его зпа-дедьную оседлость, оставление погадок преимущественно близ места детЫг использование большого набора видов и способность охоты в разио-Апннх биотопах. Наряду с этим в зимнее время часть филинов, оче-откочевывает из высокогорий в долины, а при отсутствии убежищ г 15*
228
Часть Z. Обоснование истории формирования фауны
Таблица 58
Видовой состав и относительная численность (в °/0) остатков ^млекопитающих в погадках филина на Армянском плоскогорье
Виды Ущелье Чн-рахана (1500—1600 м над ур. м.) Южный склон Севанского хребта (2000—2100 м над ур. м.) Ущелье Джа г рп (2200 м над УР- м.) Склоны it оз. Анна-лич (2600 и над ур. м.)
Erinaceus europaeus 0.7 2.4
Crocidura russula — — 1.5 —
C. leucodon — — 1.1 —
Sorex minutus — —— 0.3 —
Neotnys fodiens — — 0.3 —
Myotis oxygnathus — " 0.3 —
Mustela nivalis caucasica .... — — 0.3 1.2
Lepus europaeus 4.5 2.5 0.7 —
Ochotona sp 4 5 — — 4.7
Allactaga williamsi — — 1.2 —
Dyromys nitedula • — 0.3 —
Apodernus sylvaticus — 2.5 5.0 —
Mer tones sp 31.9 •— — —
Cricetulus migratorius 9.1 2.5 9.6 —
Mesocricetus auratus 18.2 82.5 22.5 17.3
Microtus arvalis ........ M. socialis ........... ! 9.1 10.0 . 44.2 62.5
M. majori — — 4.9 2.4
M. nivalis 4.5 — 4.9 2.4
Arvicola lerrestris ........ — — 0.7 4.7
Ellobius lute see ns 18.2 — 2.0 2.4
в охотничьей зоне может улетать на дневку в другую, богатую такими убежищами.
Интересен факт находки остатков пищухи, по-видимому, вымершей в нашу эпоху на Армянском плоскогорье.
В зоне нагорной полупустыни долины Аракса сборы костей пз погадок сычен п филинов производились Соснихиной (19506) в пещерке лавовой горы Кархапа (900 м над ур. м.) близ сел. Новый Шахварут, к западу от Еревана. Восточнее аналогичные сборы производились памп в 1947 г. в пещерах под навесами скал у подножий своеобразных скал Алинджа-Даг, Иляплу-Даг и Дарры-Даг и у пещер близ сел. Клпт, восточнее Ор-дубада, в начале теснины Аракса.
Современный ландшафт в районе Абракуппса представлен платооб-разными увалами, покрытыми кустиками колючих астрагалов, горных I солянок, со слабо развитым злаковым эфемеретумом. Остатки кустарнике-1 вой растительности, сохранявшиеся ио расщелинам, состоят из миндаля, I крушины, эфедры и свидетельствуют о большой ксерофптпости района I даже до развития скотоводства. Увалы прорезаны сетью сухих каньонов с россыпями скал и пещерами, в которых отдыхают филины и сычи. Одна I из таких пещер была обследована в «ущелье Гиен» — Кяфтар-Дара| (рис. 105).
На левом склоне теснины Аракса мы обследовали 3 карстовых пещеры. расположенные между станциями Ордубад и Мигры, у сел. Клпт.
Эти пещеры были посещены еще в 1879 г. Поляковым (1882), разыски-! вавпшм палеолитические стоянки. В раскопе, заложенном в первом зале
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
229
ц а 58
Щ1Х
ДОНЫ . Аь-на-(2600 м ур. м.)
средней пещеры па глубине 1.5 м, И. С. Поляковым были найдены две тазовых кости безоарового козла — Capra aegagrus; кости ног бурого медведя — Ursus arctos; несколько костей зайца — Lepus europaeus; бедренная небольшого хищника вроде собаки» и кости голубя. Раскопом оылп вскрыты сверху: слон пушистой глины и толща помета летучих мышей в 35 см, а далее глинистые слои с пометом и костями летучих мышей.
2.4
1.2
4.7
17.3
62.5
2.4
2.4
4.7
2.4
о такими
зымершой
t из noi'a-*рке лаво-, к западу t в 1947 г. Алвнджа-сочнее Ор-
I платооб->в, горных устариико-з миндаля, стп района х каньонов сычи. Одна яфтар-Дара
Рис. 105. Устье пещеры, заселенной филином, в ущелье Кяфтар-Дара.
Фот. автора, 1947.
ых пещеры, шт.
>), разыски-первом зале
Все 3 пещеры открываются в вертикальной стенке доломитизирован-його известняка па склон восточной экспозиции одного из распадков ущелья Аракса. В этом же массиве известняка имеются и другие пещеры, >1 их отверстия недоступны для посещения без специальных прпспособ-ленпи. Наиболее крупная — средняя сталактитовая пещера «Длинно-Крылов», о которой писал еще Сатупии (1915а). Ход в ее первый зал идет наклонно вниз, затем с середины зала начинает повышаться п после не-оильшого перевала переходит в длинный и узкий отрог с натеками кальцита. Влево от задней части зала уходит вниз другой отрог в 20 м длиной, идущий в нижележащую камеру шириной в 15 м с 50-метровым отрогом. Общая длина зала и длинного отрога до 250 м.
230
Часть I. Обоснование истории формирования фауны
Высота первого зала от 12 до 18 м, но есть и более высокие купола. В обвалах раскопа И. С. Полякова виднелись кости длиннокрылов — Miniopterus schreibersit. В левом углу зала, близ лежки какого-то хищника (барса?), нами было найдено 14 обглоданных костей современного типа сохранности; лошади, осла, овцы, быка, несколько покрытых кальцитовой корочкой костей длиннокрылов. В средней части пещеры на россыпи камней найдены кости молодого барса — Panthera pardus и куницы белодушки — Martes foina, также современного типа сохранности.
Из живых обитателей пещеры был замечен и добыт один большой подковонос — Rhinolophus ferrum equinum и насчитано с десяток летавших длиннокрылов. Между тем в 1893 г. К. А. Сатунин насчитывал их тысячами. На вековой пыли были замечены следы барса, куниц и белозубой.
На устьевых участках двух крайних пещер были собраны и немногочисленные остатки грызунов из погадок филина.
Сопоставление материалов Т. М. Соснихиной по мелким видам с нашими показывает заметное увеличение ксерофильности и видовое обеднение группировки мелких млекопитающих с запада на восток (табл. 59),
Таблица 59
Видовой состав и относительная численность (в %) остатков млекопитающих в погадках сычей и филинов из пещер долины среднего Аракса
Виды Новый Шах-варут, гора Нарахана Абрдкунис, пещера Кпфтар-Дара Орлубад, пещера «Длиннокрылое»
Erinaceus europaeus — 3.1 —
Hemiechinus auritus 25.2 1
Crocidura russula Г.2 3.1 —
Miniopterus schreiber sit 0.6 —
Mustela nivalis 0.6 — —
Lepus europaeus 3.7 6.4 6.7
Allactaga williamsi . . 4.9 6.4 —.
Allactaga elater 14.2 — —
Mus musculus 4.3 3.1 6.7
Apodemus sylvaticus 0.6 6.4 —
Meriones blackleri M. persicus 14.2 | 40.0 | 80.0
Mesocricelus auratus ...... 0.6 — 6.7
Cricetulus migrator ius ...... 4.3 6.4 —
Calomyscus bailwardi ...... * 6.4 —
Arvicola terrestris 1.2 3.1 —
Microtus arvalis ........ 17.8 —
M. socialis — 6.4 —
Ellobius lutescens ........ 0.6 9.2 —
Весь этот материал — до 1500 костей из погадок сов в долине Аракса, в общем современного типа сохранности, — собран с поверхности и не дает указаний на появление или исчезновение каких-либо зверьков
в нашу эпоху в зоне полупустыни. Аналогичный состав был установлен
и при отлове капканчиками. Отметим уточнение распространения мышевидного хомячка у горы Дарры-Даг, открытого близ Илянлу-Дага лишь в 1938 г., и случаи находок костей малоазийского хомяка, не живущего ныне в зоне нагорной полупустыни.
Гл. II • Развитие ландшафтов и фаун в четвертичном периоде 231
Нагорье Талыша и Эльбурса
еверных предгорий Талыша и из Ленкоранской низменности кост-латков послеледниковой эпохи пока неизвестно, хотя о древности ля человеком южной Мугани и свидетельствуют древние курганы, е затронутые археологами. Неизвестны кухонные остатки костей d l аагорной степи Талыша.
Сборы костей грызунов были сделаны мною в 1945 г. по мелким пещеркам скал, освоенным сычами на трех участках злаковой нагорной степи .. е.^ючих астрагалов в верховьях Виляж-Чая, у поста Дыман, и в вер-. < Вашару-Чая, у Келаханы и Кельвяза.
явность этих костей не более 2—3 столетий. В большинстве случаев защищены грунтом и постепенно выветриваются. В табл. 60 дан-
!Я всех трех участков объединены.
Таблица 60
Видовой состав и численность костных остатков млекопитающих в погадках сычей из нагорной степи Талыша
Виды Число костей Число особей
Hemiechinus auritus 2 1
Lepus europaeus 1 1
Allactaga williamsi ....... 53 9
M er tones persicus ........ 2 1
Cricetulus migratorius 35 14
Mesocricetus auratus 111 17
Microtus socialis ........ 76 15
M. arvalis ........... 1 1
Ellobius lutescens ........ 13 7
Итого 294 66
Все перечисленные мелкие зверьки существуют в этом районе и в на-поящее время, составляя современную группировку, характерную для игорной степи северо-запада плоскогорий Ирана и Армянского нагорья. 3 этом списке нет лишь обыкновеного ежа, длиннохвостой белозубки, цчшой мыши и снеговой полевки. Эти виды здесь крайне редки, а рас-хранение снеговой полевки реликтовое и приурочено лишь к более лш вершинам. Сходство видового состава нашего списка со списками (|шхиной (1947, 1948), данными для степных — трагакантовых — участ-
Дрмянского нагорья, доказывает если не тождество, то большую Диетическую близость фауны и ландшафтов обеих территорий.
С смежных участков Иранского плоскогорья остатки голоценовых иекопитаюпщх добыты Куном (Сооп, 1951) на восточной оконечности щбгрсского хребта, в 50 км от Астрабада, в мезолитических и неоли-веских слоях пещеры Белт. Среди 1170 фрагментов определены остатки к видов. Преобладают кости джейрана — Gazella subgutturosa (67%), следует первобытный тур — Bos primigenius (18.8%), козел — Capra (5.7%), баран — Ovis sp. (4.4%), каспийский тюлень — Phoca caspica ,= 1?ф и собака — Canis sp. (1.7%). Упомянуты также единичные остатки dpessp., Felis sp., Viverridae, Citellus sp., Castor fiber, Hystrix sp., Mus .’„Equus asinus, Sus sp., Cervus elaphus, Cervidae. Определения остатков
232 Часть I. Обоснование истории формирования фауны
вивер, речного бобра и первобытного тура нуждаются в проверке. Остальные виды до известной степени характеризуют близкие к современным группировки млекопитающих жарких плоскогорий северо-западного Ирана и влажной лесной Гирканики.
В целом исследования различных типов захоронений костных остатков в голоценовых слоях Армянского нагорья, долины Аракса, нагорного Талыша и Эльбурса показали существование в недавнем прошлом в этих районах ряда видов крупных зверей: зубра, первобытного тура, кулана, оленя, джейрана — и недавние сдвиги ареалов некоторых мелких видов: малоазийского суслика и малоазийского хомяка.
Обзор исследований голоценовых захоронений остатков млекопитающих на Кавказском перешейке позволяет суммировать достигнутые результаты в сводных таблицах 62—63.
Наши исследования сборов костей из послепалеолитических культурных слоев и из природного делювия дали возможность проследить появление на Кавказском перешейке в послеледниковую эпоху ряда видов крупных зверей, проникших сюда со смежных территорий, а также вымирание многих плейстоценовых видов. Прослежены быстрое развитие животноводства и его характерные черты на различных участках, а также прогрессивное увеличение относительной численности остатков домашних животных по сравнению с дикими в кухонных отбросах с неолита до современности. Исследования костных материалов из мест дневок филина и сыча позволили значительно уточнить былое и современное распространение многих насекомоядных и грызунов. Редкость находок многослойных голоценовых захоронений мелких костей при массовых встречах поздних костных материалов в небольших пещерках и навесах создает впечатление недавнего существования какого-то перерыва в образовании убежищ и в накоплении остатков пищи сов. Однако это явление правильнее объяснить относительно быстрым механическим выветриванием открыто лежащих костей грызунов и частым разрушением мелких каверн,
В изученном материале голоценовой эпохи представлено до 77 видов млекопитающих, обитавших на Кавказе в конце эпохи верхнего палеолита и частично вымерших к XX столетию.
Сравнение списков зверей из голоценовых и плейстоценовых захоронений показывает крупные преобразования в составе и распространении млекопитающих на Кавказском перешейке, происшедшие преимущественно на рубеже плейстоцена и голоцена (табл. 61).
Итак, в голоценовую эпоху на Кавказе совершенно исчезают хоботные1 и обезьяны и катастрофически сокращается число видов непарно-и парнокопытных. Появление в голоценовых слоях большого числа остатков видов грызунов объясняется преимущественно разницей в природе вскрытых и обнаруженных захоронений.
Категорические выводы о появлении или исчезновении тех или шшх видов в голоцене могут быть сделаны, таким образом, лишь в отношении массовых видов крупных животных. Состав видов зверей из захоронений голоценовой эпохи и их местонахождения дают представление о почта современной фауне и о почти современном типе распространения видов и фаунистических комплексов.
1 Возможное переживание плейстоцена на Кавказе мелкими мамонтами требует проверки и новых материалов в связи с находками геологически молодых остатков мамонтов в районе Пятигорья и в Горийской котловине.
Гл, II. Развитие ландшафтов и фаун в четвертичном периоде 233
Таблица 61
Численность видов млекопитающих, остатки которых известны из плейстоценовых и голоценовых отложений Кавказа
Отряды Плейстоцен Голоцен
Insectivora 6 9
Chi гор ter а 2 3
Primates 1 —
Carnivora 21 14
Lagomorpha 1 2
Rodentia 26 33
Proboscidea 2 —
Perissodactyla < 6 2
Artiodactyla . 18 14
Всего 83 77
Рассмотрение постплейстоценовых фаунистических преобразований , отрядам на основе палеонтологических материалов дает следующую фтину.
Насекомоядные. Из плейстоцена в послеледпиковье переходит по •нишей мере пять видов: крот, длиннохвостая белозубка, водяная ку-фа, ушастый и обыкновенный ежи. Кроме них, в голоценовых захо-юнениях обнаружены остатки белозубок: малютки, белобрюхой и ма-юй — вероятно обитавших на Кавказском перешейке и в плейстоцене.
Рукокрылые. Из плейстоценовых слоев известны остатки большого* подковоноса. В голоценовых слоях пещер нагорья Малого Кавказа шычны остатки больших ночниц и ДЛИННОКРЫЛОЕ.
Обезьяны. Данных об обитании макаков в голоцене пока нет.
Хищные. При 21 виде, известных из плейстоценовых слоев, в голосовых по костным остаткам обнаружено только 15. Выясняется отсутствие в голоценовых захоронениях Кавказа остатков пещерного и степного медведей, пещерной гиены, гепарда, тигрольва и, возможно, росо-нахи- НаРяДУ с этим, в голоценовых отложениях впервые появляются остатки шакала, полосатой гиены. Вероятно также проникновение й Кавказ в голоцене ряда других видов хищных южного происхождения, остатков которых в голоценовых захоронениях пока не обнаружено, например камышового кота, льва и тигра. Отсутствие находок впослеледниковых захоронениях остатков льва, тигра и гепарда может ейть объяснено и малой численностью этих зверей в историческую эпоху.
Зайцеобразные. Из плейстоценовых отложений известен один вид — прусак. В голоценовых отложениях обнаружены остатки зайца-русака и крупной пищухи, очевидно существовавшей па Малом Кавказе 8 в плейстоцене.
Грызуны. Выясняются крупные изменения состава и ареалов ряда происходившие в разные этапы голоцена. Характерно выми-дикобраза и апшеропской полевки, отступление малоазийского голяка с Апшерона и из Имеретии, уменьшение ареалов малоазий-суслика на Армянском нагорье, кавказских полевок в Пред-а3ье и т. п. Наличие остатков черных крыс в неолитических слоях ш Черноморском побережье и в слоях середины I тысячелетия до н. э.
ш
Часть /. Обоснование истории формирования фауны
Таблица 62
Видовой состав и стратиграфическое положение находок костных остатков млекопитающих. обнаруженных в четвертичных слоях Кавказского перешейка
Отряды и виды
Insectivora
Erinaceus europaeus L. (s. lato) . . Hemiechinus auritus Gmel. (s. lato) Sorex minutus L......................
S. araneus L.........................
Neomys fodiens Schr. (s. lato) . . . Crocidura leucodon Herm..............
C, russula Pall. (s. lato)...........
S uncus etruscus Savi ...............
Talpa caucasica Sat..................
T sp...............................
Chiroptera
Rhinolophus jerrum equinum Schr. . . . Myotis oxygnatus Mont....................
Miniopterus schreibersii Kuhl............
Chiroptera gen..........................
Primates Macaca sp. ..............................
Carnivora Canis aureus L...........................
C. lupus L. (s. lato)....................
C. lupus apscheronicus N. Ver............
* C. tamanensis N. Ver.1...................
Canis sp.................................
Cuon sp..................................
Vulpes corsac Pall.......................
V. vulpes L. (s. lato)...................
V. vulpes aff. alpherakyi Sat............
Ursus arctos L...........................
Z7. arctos binagadensis N. Ver...........
U. spelaeus Rosm (s. lato)..............
/7. rossicus Boris.......................
Hyaena hyaena L. . ......................
•Crocuta spelaea Goldf...................
Martes foina Erxl........................
M. foina latifrons Sat...................
M. cf. foina Erxl........................
M. cf. martes L..........................
Vormela peregusna Guld...................
1 Звездочкой отмечены формы переходной верхнеплиоценовой—нижнеплейсто. ценовой эпохи, обнаруженные на Таманском полуострове.
Гл, II- Развитие ландшафтов и фаун в четвертичном периоде
235
Таблица 62 (лродолжение)
Отряды п виды
Плейстоцен
Голоцен
ность
lilra lutra L,....................................
MusUle nivalis L. (s. la to)......................
у tits meles L. (s. la to)........................
V. meles urartu orum Sat..........................
mdes aff. minor Sat............................
ihks sp. * * * *..................................
Cdo aff. gulo L...................................
I s Iveslris Schr. (s. la to).....................
f tyhica Forst....................................
f aff lybica Forst................................
f, lynx L. (s. lato)..............................
falhtra pardus L............................ . .
p. spelaea Goldf.................................
•Paithera sp.......................................
*jtinonyx aff. jubatus Schr........................
-Lagomorpha
Otkotuna sp............•........................
kpus europaeus Pall............................
L. europaeus gureevi I. Grom...................
L tf. europaeus................................
Rodeiitia
Jlcwiotfl sp..................................
pUfKih? pijgmaeus Pall........................
C aff. citellus xanthoprimnus Benn. . № fiber L.....................................
•CJgnianensis N. Ver. ........................
Wiyontherium cuvier I Fisch.................
jtylrix cf. leucura Sykes.....................
Я, vinogradovi Arg............................
hrwys nitedula Pall..........................
(llii glis L.................................
iidsla subtilis Pall.........................
5 cf, caucasica Vin...........................
illuctaga jaculus Pall.......................
X jaculus bogatschevi Arg....................
L|, wdliamsi Thos............................
williamsi dzhafarovi 1. Grom. . . . J. cf. uilliainsi.........................
J. Hater Licht...............................
^ы1ах microphtalmus G uJ d...................
p [ioanlheiis Nehr...........................
j|;^uniys min ulus Pall......................
musculus L. (s. lato)....................
edemas agrarius Pall.........................
д. syl^ticus L. (s. lato)....................
flatus rattus L..............................
I’tori-tgicus Berk..........................
236
Часть 7. Обоснование истории формирования фауны
Таблица 62 {продолжен^}
Отряды и виды Плейстоцен Голоцен
нижний средний верхний мезолит неолит бронза железо _1 средневековье совремеи-1ТОС*Г1.
Cricetus cricetus L.......................
Mesocricetus auratus W ater...............
Л/. aff. auratus Water....................
M. auratus raddei Nehr....................
Л7. auratus planicola Arg.................
Cricetulus migratorius Pall...............
C. migratorius argiropuloi I. Grom. . . . Calomyscus bailwardi Thos.................
Meriones erythrourus Gray.................
M. erythrourus intermedins I. Grom. . . M. blackleri Thos.........................
M, persicus Blanf.........................
Lagurus lagurus Pall......................
Arvicola terrestris L. (s. lato)..........
Microtus arvalis Pall.....................
71/. socialis Pall........................
71/. cf. roberti Thos.....................
M. nivalis Mart. . .......................
71/. aff. gud Sat.........................
Л/. aff. majori Thos......................
Л/. apscheronicus Arg.....................
Prometheomys aff. schaposchnikovi Sat. Ellobius talpinus Pall....................
E. lutescens Thos.........................
E. aff. lutescens Thos....................
Muridae gon...............................
Proboscidea
* Elephas meridionalis Aesti..............
*Z7. antiquus Falc........................
E. trogontherii Pohl......................
E. primigenius Blum.......................
Perissodactyla
Equus hemionus Pall.......................
E. aff. hidruntinus Reg...................
E. caballus L. (s. lato) .................
E. caballus gmelini Ant...................
*£'. aff. sussenbornensis Wiist...........
* E. stenonis Cocchi......................
Rhinoceros t ichor hi nus Fisch...........
Rh. cf. mercki Jaeger.....................
Rh. binagadensis Dzhaf....................
Rh. sp....................................
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
237
Отряды п ВИЦЫ
Таблица 62 (продолжение)
до, cf. etruscus Falc.
fIjsmolherium sibiricum Pcuicasicum Boris. .
Fisch..........
Artiodactyia
?l,. sc r of a L. (s. lato).................
I, dpscheronicus Bur. e! Dzhaf..............
5 hmanensis N. Ver..........................
ujppopotamus sp. ...........................
Cmdus dromedarius dahli Chav, (ferus?) J knoblochi Nehr.............................
piracanelus gigas Schloss....................
P. cf. kujalnikensis Chom....................
Cavus elapkus L. (s. lato)...................
licladocerus sp..............................
! /fri’idflc gen.............................
httr.d cf. niesopotamica Brooke..............
’/rgdceros euryceros Al dr...................
^[qaeeros sp.................................
jllcej alces caucasicus N. Ver...............
д aff, alces L...............................
ij (mania Ices sp • • •••••••••*•• »apreo/«s capreolus L. (s. lato).............
,kuigd lalarica L............................
fatlla subgutturosa Giild....................
sp....................................
• T^laphus sp...............................
• ^rtpsicerolini gen. et sp.................
Rupicapra rupicapra caucasica Lyd. • . . Capra cylindricornis Blyth...................
f. caucasica Giild..........................
aff. caucasica Giild......................
L^gdgrus Gin el............................
vitielini Blyth..........................
0. cf. gnielini Blyth......................
p. cf. ammon L.............................
ininutus Malsh...........................
if. primigenius Boj........................
p irochoceros Leitn........................
Ц. maslan-zadei В arts ch..................
Jis,n bonasus caucasicus Sat...............
B. pr scus Boj. (s. lato)..................
•B rf. schoetensackl Freud...................
В sp.......................................
Ляп или Pos................................
238
Часть I. Обоснование истории формирования фауны
Сводная стратигра главнейших местонахождений остатков индикаторных млекопитающих чет
фич вер’
Черноморская область
западное Предкавказье и Русская равнина
западное Закавказье
Климат и лавд-шафты по состав? териофауны '
CJ м< CI от же
Слон морских отложений
Подразделения четвертичного периода
Крепость С арке л, IX—XIII вв.
бурый медведь речной бобр кулан лось
благородный олень сайга зубр
ПК — 2841
Семибратнее городище у устья Кубани волк выдра кабан олень
Шалаши колхидян у Анаклин н Очамчири шакал кабан косуля благородный олень
Современный климат и рез-! кое усиление | антропогенных воздействий иа ландшафты.
Развитие широко-лиственных лесов в горьях.
пред.
Землянковое поселение у Геленджика рысь заяц-русак благородный олень дельфин
ПК — 417
Пещера Ахштырская, верхние слои бурый медведь барсук черная крыса КОСУЛЯ
ПК —460
Сильное л онио и остеп-пение предгорий. Сокращение долинных ледников.
потеп-
1 Показатели прокаливания костей (НК) даны по коллагеновому методу (Индо- клип к
. Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
23<J
Табл и ц а 63
кая таблица
периода па Кавказском перешейке и смежных территориях
Каспийская область
восточное II ред кав к а а ье п Русская равнина
восточное Закавказье
Нагорье Малого Кавказа н север Иранского плоскогорья
Дзуары Северной Осетии ‘ XV—XIX вв. благородный олень косуля кавказский лось восточнокавказскпй козел серна кавказский зубр
Поселение IX—XIII вв. у Баку лисица кулан джейран тюлень
Поселения долины Сунжи, Пстп-су, Алхан-Кала кулан благородный олень
Поселения у Каякента и Джемикента лпспца медведь кулан джейран
ПК — 275
।
| Навес Сосруко на Баксане заяц-русак малый суслик слепушонка кабан
| благородный олень кавказский козел
ПК — 241.
I
I
Кувшинные погребения у Мипгечаура и поселение у Сумгаита полосатая гиена черная крыса кабан благородный олень джейран
Крепость Кармир-Блур куница бел о душка благородный олень арменийский муфлон бсзоаровый козел джейран
Свайные поселения и могильники побережий Севан а
куница белодушка барсук
кабан благородный олень карликовый тур
ПК—207
Пещера Белт у Астрабада дикобраз благородный олень джейран первобытный тур тюлень
! Стоянка Чох в Дагестане
• дагестанский хомяк муфлон
ПК-317
«52). Анализы любезно выполнены в лаборатории И. Г. Пидоплпчко в Клеве.
- 240 Часть /. Обоснование истории формирования фауны
Подразделения четвертичного периода Слои морских отложений Черноморская область Климат и ланд-шафты по составу териофауны “ же
западное Предкавказье и Русская равнина западное Закавказье
X
ф я
Костеносныи аллювии поздних террас долины Дона лисица малая сеноставка малый суслик большой тушканчик желтая пеструшка водяная крыса лошадь лось благородный олень северный олень сайга
В
И
о ь СЗ
к
Св
св
я
(D
Д’
О
К
о Я Я о
’Я я №
св И
Стоянка Пльская у Краснодара пещерная гиена мамонт кабан благородный олень сайга первобытный зубр
ПК—518
Костепоспый аллювий ранних террас долины Дона степной хорек малый суслик тушканчик-прыгун слепыш желтая пеструшка благородный олень
Пещера Гварджилас-Клде крот бурый медведь росомаха малоазийский хомяк прометеева полевка кавказский козел зубр ПК —512 Общее сильное похолодание в вероятное смещение книзу ландшафтных зон при оледенении высоких хребтов.
Пещеры Сакажиа, Дэвис-Хвроли, Мгви-меви пещерный медведь лошадь речной бобр лось серна кавказский козел аргали ПК —580 Ьолыиие подъемы хребтов. •
Пещера А хштырска я. нижние слои волк пещерный медведь обыкновенный хомяк б л ci город! i ый о л е и ь гига втеки ы о л ен ь первобытный зубр ПК — 70G Вертикальная зональность современного i типа. • •
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
241
Таблица 63 {продолжение)
Каспийская область
Сл п морских отложении
восточное Предкавказье и Русская равнина
восточное Закавказье
Нагорье Малого Кавказа и север Иранского плоскогорья
Делювий балки Янку ль в верховьях К а лауса лошадь сайга
Верхний комплекс травертинов горы Машук в Пятигорье мамонт лошадь косуля
Битуминозные суглинки Кир-Маку и о. Артема волк лошадь благородный олень сайга европейский тур
ПК—158—208
Делювиальные суглинки у Мерагэ волк пещерная гиена лошадь носорог аргали
Пещеры Тамтама и Бизи-тун в северном Иране шакал пещерная гиена барс песчанки благородный олень джейран
Зуртакетская стоянка в верховьях Храма лошадь осел муфлон первобытный зубр
Костеносные пески в террасах Терека у Моздока первобытный зубр
Костепосный аллювий древней, j клубленной долины Волги пещерная гиена тигролев мамонт волосатый носорог шпроконалая лошадь кноблохов верблюд гигантский олень длиннорогий зубр ПК —484
Битуминозные пески у Бпнагадов на Апше-ронском полуострове ушастый еж корсак бинагадинский медведь перевязка гепард краснохвостая песчанка дикобраз осел бинагадинский носорог благородный олень сайга бинагадинский тур ПК 450—520
16 Н. К. Верещагин
242
Часть I. Обоснование истории формирования фауны
Слои Черноморская область
Подразделения мор- Климат и ланд.
четвертичного CKI1X in афты по составу
периода отло- западное Предкавказье западное Закавказье териофауны
жений и Русская равнина
Костеносные нески у В Закавказье
Гирея и Краснодара, Пещера Кударо в вер- сильное остеп-
средние слои ховьях Риона, некие плоско-
трогонтериевый слон нижние слои горий, повлек-
длиннорогий зубр макак шее проникно-
степной медведь красный волк вение на Кав-
пещерный медведь каз зверей Не-
барс малоазийский хомяк редней Азии.
Е-ч И а сурок.
Я о о »—< прометеева полевка
ф ЕГ ф 1=5 о кустарниковая по-
о S Сб ф левка
Е-| О а к с а дикобраз
ф к а »я а о носорог
л а а t-f кабан
й а косуля
W аргали
ПК - 914
1 Костеносные пески у Б равнинном
Гирея и Ивановского Предкавказье
на Кубани, нижние климат yjjg.
слои ренный и су-
южный слон хой. в пред.
древний слон горьях лайд-
олень шафт широко-,
короткорогий зубр лиственных ле. сов.
Конгломераты и пески Кучугур, Синей Балки и Цимбала на Таман-
ском полуострове бобр трогонтерий таманский бобр
южный слон
г4 древний слон
а о кавказский эласмо-
го терий Фауна неизвестна.
Рх лошадь
о 1-1 сложнорогие олени
ЕЕ Ф ЕГ О а к х ь а а >а 2 верблюд лесные антилопы
9 ф о О о короткорогий зубр
К и со ПК 913—1203
О а Пески и галечники
-_> а древних террас Ку-
бани у Воскресенской и Псекуиса у Бакин-
ской и Саратовской
южный слои В равнинном
1 степонова лошадь Предкавказье
1 этрусский носорог климат теплый.
сложнорогие олени Ландшафт са-
ПК —1033 ванны и галерейных лесов.
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде
243
Таблица 63 (продолжение)
Каспийская область Нагорье Малого Кавказа
восточное Предкавказье и Русская равнина восточное Закавказье и север Иранского плоскогорья
Костеносные пески верхних террас Кумы и Под-кумка у 1 еоргиевска южный слон стенонова лошадь олени быки ПК — 1073 Морской ракушечник у Хурдалапа на Апше-ропском полуострове лошадь носорог Костеносные пески Аджи-Эйласа у Еревана трогонтериевый слон лань первобытный тур Палеолитические стоянки средней Занги и подножия Алагеза Фауна неизвестна. Костеносные пески Ленинакана, верхние слои трогонтериевый слон лошадь мерков носорог гигантский олень кноблохов верблюд первобытный тур
Подхазарские слои у Бинагадов кавказский эласмо-
Костеносные пески у Подлесного и Дивного близ Маныча южный слон сибирский эласмотерий Таш-Кала у Грозного плосколооый слон стенонова лошадь терии Морской ракушечник у Кишлы на Апше-ронском полуострове гиена стенонова лошадь Делювий у Цинандали этрусский носорог Еникент на Алазанн бобр трогонтерий Костеносные пески Ленинакана, нижние слои овернский мастодонт Нурнус на Занге мелкие куницевые заяц этрусский носорог антилопа орикс
16*
244
Часпгъ I. Обоснование истории формирования фауны
Таблица 63 [окончание]
Подразделения четвертичного периода
Слои морских отложений
Черноморская область
западное Предкавказье и Русская равнина
западное Закавказье
Климат и ланд. шафты по соста >у териофауны
Костеносные пески у Хапров и Ростова на Азовском побережье плосколобый слон овернский мастодонт махайрод гиппарион степонова лошадь верблюды
ПК — 1056—1142
Фауна неизвестна.
Климат теплый. На юге Русеков равнины ландшафт саванны в галерейных лесов.
в Ку римской низменности позволяет говорить о вероятной автохтошюсти вида п переживании им плейстоцена на теплых и влажных низменностях у Черного моря и южпого Каспия. Такой же вывод о пасюке сомнителен. Подтверждено также обитание речных бобров в неолитическую эпоху в Колхиде.
Большинство других находок уточняет данные о распространении ныне живущих видов.
Непарнокопытные. Проработанные материалы не дают оснований говорить о переживании плейстоцена па Кавказе волосатым носорогом и плейстоценовым ослом. Документально подтверждено появление в голоцене в восточном Закавказье и в восточном Предкавказье современного кулана. Несомненно, обитание здесь в голоцене и диких лошадей, но пока в костных материалах не выявлены их отличия от остатков домашних лошадей различного происхождения. Дикие и домашние лошади служат одним из главных объектов жертвоприношений и питания у кавказских племен на протянпжеи большей части исторической эпохи.
Парнокопытные. Прослежено широкое распространение по Кавказскому перешейку дикой свиньи, становящейся в голоцене одним из главных объектов охотничьего промысла, а также первобытного тура на Малом Кавказе. Существование диких верблюдов в голоценовую эпоху на Кавказе пока не доказано.
Прослежено широкое распространение благородного оленя почти по всему Кавказскому перешейку с плейстоцена вплоть до XIX столетия; косуль и лосей, обитавших по долине низового Допа, приречным лесам и камышам Кубани и в лесистых низменностях Предкавказья и в Колхиде, — до конца XVIII столетня. Голоценовых остатков большерогого оленя и лани не обнаружено.
оказано обитание зубров на Малом Кавказе в эпоху бронзы, а на среднем и низовом Дону — в средине века пашей эры. Сайга исчезает из восточного Закавказья на раннем этапе голоценовой эпохи. Остатки джейрана начинают попадаться в слоях, датируемых II тысячелетием до н. э., 43 восточном Предкавказье и в Закавказье. Особенно многочисленны они в средневековых слоях пашей эры на Апшеропском полуострове.
Гл. II. Развитие ландшафтов и фаун в четвертичном периоде 245
Палеонтологические материалы по голоценовым млекопитающим позволяют сделать и ряд общих выводов о фаунистических и ландшафтных изменениях в пределах Кавказского перешейка, происходивших с конца хрхнего плейстоцена. Отсутствие в голоценовых слоях остатков млеко-автающих, существовавших еще в эпоху наибольшего похолодания, г: пдетельствует о каком-то переломе в экологической обстановке, содействовавшем их вымиранию.
Вообще палеонтологические материалы показывают, что паиболь-Ье потрясения испытали в послеледниковую эпоху экологические группировки млекопитающих предгорных равнин. Горная фауна была, веро-«ито, затронута в значительно меньшей степени.
Судя по тому, что такие виды, как пещерный медведь, пещерная гиена, ппролев, плейстоценовый осел, гигантский олень и другие плейстоцено-I. f виды, благополучно существовали в холодном климате верхнего пдйстоцена, можно предполагать, что основной причиной пх вымирания были резкое потепление п иссушение ландшафтов. Это иссушение ландшафтов должно было оказывать на фауну значительно более сильное влияние иа низменностях и в предгорьях восточных районов страны, не-1к^[Н на западе Кавказского перешейка. В этом отношении ряд поучи-ьных примеров отступания ареалов относительно мезофильных видов Си т дан при характеристике Бинагадинского захоронения.
Как и плейстоценовым слоям, голоценовым отложениям Кавказа принце полное отсутствие остатков северных холодолюбивых видов: север-пло оленя, зайца-беляка, песца. Не найдено голоценовых остатков этих з<реп даже в пределах низового Дона. Становится ясным, что пх ареалы в это время отодвинулись еще дальше па север, а пе приблизились к Кавказу. Образование изолированных участков обитания горных мезофильных видов грызунов: кустарниковой полевки — па лугах Ставрополь-4К01 возвышенности, кавказской снеговой полевки — иа горе Развалка । Пятпгорье, малоазийской снеговой полевки — на Келахапской сопке л сухой Зувапдской котловине горного Талыша и степной полевки — на \пп еропском полуострове — также свидетельствует о наступлении эпохи сильного потепления, повлекшего сдвиги ландшафтных зон. Этп и другие примеры разрывов распространения рассматриваются далее особо, в обзоре истории развития и современного состояния ареалов отдельных видов.
Следующим фактом, знаменующим новый этап в формировании верхне-четвертичных фаунистических комплексов, служит появление на Кавказе новых видов зверей, не зарегистрированных «универсальной» палеонтологической летописью в битуминозных плейстоценовых слоях Ап-, шеропского полуострова. Уже упомянутые находки в голоценовых отданиях остатков шакала, полосатой гиены, кулана, джейрана п их современное обитание здесь1 указывают па сравнительную молодость ареалов этих видов на Кавказском перешейке. Расположение основной части ареалов упомянутых видов южнее и восточнее Кавказа, а также то обстоятельство, что этп животные не переносят глубокого и продолжительного снегового покрова, служат доказательствами пх южного п при этом недавнего проникновения па Кавказ. Список этих видов должен быть, вероятно, расширен за счет камышового кота, льва и тигра, ископаемые остатки которых с Кавказа пока неизвестны (подробнее об этом см. в главе III).
I За исключением вымершего кулана.
246 Часть /. Обоснование истории формирования фауны
Наши палеонтологические материалы свидетельствуют лишь об одном цикле потепления и ксерофитизации ландшафтов, наибольшее развитие которого приходится на довольно раннюю фазу послеледниковой эпохи.
Весьма вероятно, что на материале по млекопитающим в условиях такой сложной в геоморфологическом смысле страны, как Кавказ, и вообще трудно подметить более мелкие климатические колебания в пределах голоцена, которые тем не менее могли иметь место.
Ксеротермическую эпоху признавали и геоботаники Малеев (19396, 1946), П. Ярошенко (1956), Гроссгейм (1948), Тахтаджян (1941), объясняя ею узко местное распространение растений-сухолюбов в западном Закавказье и в ряде мест нагорья Малого Кавказа среди более мезофит-ных формаций.
Отмечали ее также почвоведы Яковлев (1914) и Захаров (1935), геоморфолог Щукин (1925), зоолог Свириденко (1927, 1937), геологи Герасимов и Марков (1939).
Прослеживаются на остеологическом материале и ландшафтные смены, протекавшие на последнем этапе исторической эпохи под воздействием человека.
Обильные в кухонных остатках кости кабанов, оленей, лосей, косуль в средневековых слоях Саркела свидетельствуют о мощных пойменных лесах, покрывавших в то время долину Дона. Уничтожение этих лесов к XVIII столетию и привело к отрыву кавказских ареалов ряда видов европейских копытных.
Находки полуископаемых остатков оленей в Ставрополье, находки остатков кабанов и оленей в ряде сухих ныне нагорно-степных участках Армянского нагорья могут подтвердить до известной степени значительно большую облесенность этих стран в позднем голоцене.
Существование значительных массивов леса на безлесных ныне нагорьях западной Грузии, у селений Даби и Гоми, в южной Грузии, близ Цалки, в Азербайджане, на пустынных хребтах близ Кировабада, было доказано недавно Яценко-Хмелевским и Канделаки (1940, 1941а, 19416), исследовавшими угли из культурных слоев поселений II тысячелетия до н. э. (Цалка) и XII в. н. э. (Ганджа).
Наши исследования священных коллекций черепов в горных долинах Северной Осетии указали также (в связи с исчезновением лосей и зубров) па истребление лесов ’ к XVIII в. по предгорным наклонным равнинам восточного Предкавказья.
Изученные палеонтологические материалы дают много и других фактов изменений ареалов, расселения синантропных видов и вымирания промысловых зверей под влиянием непосредственного истребления их человеком и косвенного воздействия через коренное изменение ландшафтов.
^^фд^^И****
Ч АС Т Ь ВТ О Р А я
АНАЛИЗ ПРОИСХОЖДЕНИЯ ЧЕТВЕРТИЧНЫХ МЛЕКОПИТАЮЩИХ КАВКАЗА В СВЯЗИ
С ИХ РАСПРОСТРАНЕНИЕМ, ЭКОЛОГИЕЙ И МОРФОГЕНЕЗОМ
Глава III
ОБЗОР ПРЕЖНЕГО И СОВРЕМЕННОГО РАСПРОСТРАНЕНИЯ
И ЧИСЛЕННОСТИ НЕКОТОРЫХ МЛЕКОПИТАЮЩИХ
Детали и закономерности формирования фаунистических комплексов любой страны прослеживаются и понимаются лучше при изучении истории развития ареалов, их современного состояния и экологии отдельных видов. Хорошо очерченные и сгруппированные ареалы являются и основой зоогеографического районирования.
Следует, однако, учитывать, что современный ареал палеарктического вида может быть лишь остатком древнего очага или его более или менее древним приростком, развившимся в четвертичном периоде. Поэтому история происхождения видов получает наибольшую убедительность лишь при сопоставлении палеогеографических, палеонтологических и современных зоогеографических фактов с данными современной экологии видов и при учете влияния человека на протяжении исторической эпохи.
Факторы, влияющие на распространение расселяющегося или исчезающего вида, многообразны, но среди них имеются главнейшие и подчиненные.
Уже исследования Бинагадинского захоронения (глава II) показали, что особенности прежних и современных ареалов млекопитающих восточного Закавказья зависели преимущественно от эволюции четвертичных ландшафтов этого района. В меньшей степени эти особенности зависели п зависят от эволюции морфологических адаптаций видов и их биоцено-твческпх связей друг с другом. Создается даже впечатление, что при развитии и угасании ареалов растениеядные млекопитающие ведут себя в значительной мере автономно-независимо друг от друга. Подчиненное значение имеет, очевидно, и влияние на распространение травоядных зверей преследующих их хищников.
Исходя из теоретических предпосылок, выяснение происхождения и истории развития ареала наиболее доступно у крупных и массовых видов: хшцных, хоботных, грызунов, копытных, — остатки которых чаще попадают и сохраняются в различного рода отложениях. В то же время важно, чтобы это были относительно степотоппые виды, т. е. такие, которые связаны все же с определенным типом ландшафта и с относительно небольшим набором биотопов.
Такие виды оставляют больше ископаемых и современных «следов» прежних ареалов. Напротив, узко распространенные эндемики с их реликтовым ареалом, как правило, позволяют сделать только тот вывод, что они, вероятно, развивались в данном районе. Эвритопные и массовые 'яквисты дают обычно лишь хорошие палеонтологические факты, особенно
250 Часть II. Анализ происхождения четвертичных млекопитающих
если у исследователя имеется последовательный ряд разновозрастных захоронений.
Для других же видов, в частности большинства насекомоядных и рукокрылых, ископаемые остатки которых редки, при выяснении происхождения приходится ограничиваться лишь зоогеографическим картированием и анализом современных ареалов, проводя экологические аналогии и экстраполяции на основе более изученных примеров по хищным, грызунам и копытным.
Далее сделана попытка проследить прежнее и современное распространение на Кавказе ряда живущих и угасших млекопитающих, факторы, влиявшие и влияющие на их ареал и численность. Сделана также попытка выяснить время появления этих видов на Кавказе и пути вероятного проникновения для несомненных вселенцев. Выбранные примеры охватывают различные отряды, экологические и палеозоогеографические группы видов.
Отряд Н А С F К О М О Я Д Н Ы — IIV S Е С ТI V О В А
Палеонтологическая древность ряда родов этого отряда весьма велика. Ископаемые остатки насекомоядных известны с эоцена. Радиальная адаптация и разнообразие связей с ландшафтами у насекомоядных огромны. Известны подземные, водные и широко специализированные формы. В пределах Кавказского перешейка наиболее древние остатки принадлежат 3 родам: выхухоли — Desmana, кроту — Talpa и землеройке — Crocidura. Обнаружены они в нижнеплиоценовом комплексе Ставрополья (см. главу II). Из плейстоценовых слоев здесь известны остатки 5 видов, относящихся к 5 родам: Erinaceus, Hemiechinus, Crocidura, Neomys, Talpa.
В современной фауне Кавказского перешейка насчитывается 15 видов насекомоядных, относящихся к 8 родам. В экологическом смысле они принадлежат только к 3 жизненным формам: роющей, подземной — 1 род и 3 вида (кроты); земноводной — 2 рода и 2 вида (выхухоль и водяная кутора); остальные — более или менее широкой специализации, связанные с довольно разнообразными типами ландшафтов и биотопов (ежи, землеройки).
Сем. ЕЖОВЫЕ — ЕЕINAШ1) А Е
Обыкновенный еж — Erinaceus europaeus L. (s. slato). Ископаемые остатки видов рода Erinaceus известны в Европе с миоцена. На Кавказе остатки обыкновенных ежей найдены в среднеплейстоценовых слоях Апшеронского полуострова и в верхнеплейстоценовых слоях западного Закавказья. Современное распространение этого ежа на Кавказе весьма обширно (карта 1). Он встречается от уровня Каспия до 2500 м абсолютной высоты по всему перешейку. Отсутствует па альпийских лугах, в ровной открытой пустыне, полупустыне и по л у степи, в постоянных болотах и крайне редок в горных лесах. Наибольшая численность наблюдается в низовых лесах Закавказья и в разреженных лесах и кустарниках наклонных равнин Предкавказья.
В полупустынях восточного Предкавказья и Закавказья этот еж живет в тугайных зарослях, садах и оазисах.
Сокращение ареала, происшедшее с плейстоцена на больших пространствах восточного Закавказья, и реликтовость распространения на Аише-
Ill, Прежнее и современное распространение и численность млекопитающих 251
энском полуострове объясняются послеледниковым иссушением района (Верещагин, 1949в).
Происхождение обыкновенного ежа на Кавказском перешейке недо-, аточно ясно. Ареал современного вида уходит далеко на север и юг от ^вказа. Отсутствие ежа в открытой пустыне, полупустыне и в равнинной степи свидетельствует тем не менее о мезофильности зверька, о его •февних связях с лесными ландшафтами и о вероятном происхождении плиоценовых предков, обитавших в мезофитных ландшафтах восточ-1Г0 Средиземноморья. Находки остатков ежей в среднеплейстоценовых дожениях Закавказья также подтверждают нижнечетвертичную, вер-зе дочетвертичную, древность этого вида в пределах Кавказа.
В настоящее время обыкновенный еж быстро распространяется по рав-чнам в связи с развитием орошения степей и пустынь.
Ушастый еж — Hemiechinus auritus Gmel. Ископаемые остатки этого да найдены в среднеплейстоценовых слоях на Ашперонском полуострове в верхнеплейстоценовых отложениях севера Прикаспийской низмен-остп (Верещагин и Громов, 1952). На юго-западе остатки ежей звестны из палеолитических слоев пещер Палестины. В голоценовых тложениях кости ушастого ежа, натасканные филинами в пещерки, най-ены нами во многих местах Предкавказья и Закавказья.
Современное распространение этого ежа на Кавказе приурочено к сули низменностям Предкавказья, восточного Закавказья и высоким пло-когорьям южных районов (карта 1).
В предгорьях Большого Кавказа ушастый еж распространен от уровня цорядо 300—350 м абсолютной высоты, но в южном Закавказье и Талыше живет в нагорной степи и в полупустыне до высоты 1600—1800 м. В зоне горных и низовых лесов зверька нет. Наибольшая численность вида наблюдается в зоне полупустыни, например в Кура-Араксинской низменности.
Адаптации к ландшафту полупустыни и степи у ушастого ежа ярко выражены в пищевой специализации — питании моллюсками, саранчовыми и жуками, реже мелкими рептилиями и грызунами. Малая потребность в воде, способность переносить высокие температуры и использовать для убежищ норы черепах, песчанок, сусликов, лисиц, барсуков также характеризуют этого ежа как зверька южного ксерофильного типа.
Большое вертикальное распространение в южном Закавказье и уход ареала ушастого ежа на широких пространствах к югу свидетельствуют о древнем, вероятно верхнеплиоценовом, проникновении вида из Передней Азии.
Современный ареал ушастого ежа несколько увеличивается в связи с антропогенным иссушением предгорий.
Сем. КРОТОВЫЕ—ТАЕР1ПАЕ
Кавказские кроты — Talpa caucasica Sat., Т. orieiltalis Ogn. Остатки кротов рода Talpa известны в Европе и Северной Америке с миоцена (Trouessart, 1898—1899а; Simpson, 1945). На Кавказе наиболее древние остатки мелких кротов Тalpa sp. обнаружены в плиоценовых слоях у Ставрополя.
Остатки современного вида 7\ aff. caucasica найдены в плейстоценовых слоях западного Закавказья.
В голоценовых местонахождениях они весьма обычны, особенно в рав-нинном Предкавказье.
252 Часть II. Анализ происхождения четвертичных млекопитающих
Учитывая древние изоляции кавказских кротов от европейского — Т. europaea L. и средиземноморского — Т. соеса Savi (см. карту у Бобринского, Кузнецова, Кузякина, 1944), мы рассматриваем их далее в согласии с Огневым (1926а, 1928) в качестве особых видов. По морфо-физиологическим адаптациям оба кавказских вида настолько близки, что их распространение описано ниже совместно и показано на карте 2 в объединенном ареале. Современная изученность ареалов видов позволяет отметить лишь, что Предкавказье, Большой Кавказ и Малый Кавказ населяет кавказский крот с подвидом Т. caucasica ognevi Str. на Рачинском и Триалетском хребтах, а Черноморское побережье, Колхидскую низменность и Талыш — восточный крот.
Современный ареал кавказских кротов чрезвычайно показателен для понимания истории развития ландшафтов страны.
В западном Предкавказье крот обитает в долинах Дона, Ей, Челбаса, Егорлыка и Кубани.
На Прикубанской равнине крот вымирает в засушливые годы на больших пространствах, сохраняясь лишь в более сырых понижениях.
На лугах и в лесах Ставропольской возвышенности многочисленная популяция крота имеет как бы островное распространение.
В северокавказских предгорьях крот распространен широко, но отсутствует на остепненпом Таманском полуострове, а на остепненной Кабардинской равнине живет по долинам рек. В Пятигорье он обычен в дубовых лесах, а по долине Подкумка — вплоть до Георгиевска. Севернее встречается еще по долине Кумы до Буденновска, а восточнее Орджоникидзе становится уже редким.
В горных лесах северного склона Большого Кавказа крот весьма обычен — от верховьев бассейна Афипса до Аргуни, особенно по влажному хребту Черных гор.
Логинов (1949) получил такие данные относительной плотности населения крота, отлавливая зверьков в июле 1937 г. в различных вертикальных зонах (табл. 64).
Таблица 64
Относительная численность крота в ландшафтах северо-западного Кавказа
Зоны Количество кротов, пойманных за сутки 100 ловушками
Лесостепь предгорий 40
Широколиственные леса 30
Темнохвойные леса 5
Альпийские луга 10
В сухих продольных долинах от Эльбруса до Дагестана крота нет, как и во внутреннем Дагестане.
В западном Закавказье крот обычен по террасам Черноморского побережья и по береговым грядам рек болотистой Колхиды, а на запад распространен по северным склонам восточного Тавра.
По южному склону Большого Кавказа крот прослежен от Геленджика до Закаталы, причем в бассейне Цебельды, Ингура, Цхенис-Цхали он обитает по пологим участкам ущелий, поросших пихтово-кленовым лесом. На Сурамском хребте и в Южной Осетии крот обычен на средних высотах
111. Прежнее и современное распространение и численность млекопитающих 253
Ьоодт, но отсутствует в сухой Горийской котловине. Отсутствие крота в восточной части Большого хребта обусловлено преимущественно большой крутизной склонов, тонкостью слоя почвы, налегающей на коренные ш-р дыв высокогорье, и сильными засухами в нижней части зоны буково-Ьвых лесов, вызывающими неустойчивость кормовой базы. Крота • я в сырой в зимнее время, по сильно пересыхающей летом Алазано-|гатайской низменности.
Неожиданно крот уцелел с какой-то влажной эпохи в зоне полупу-И1Н , в Караязском галерейном лесу, восточнее Тбилиси (Верещагин,
Рис. 106. Местообитание крота (в дубовых рощах) и краев ох вое той песчанки (на полянах с кермеком и солеросом) в Караязском лесу долины Куры.
Фот. автора, 1958.
1940а). В этом реликтовом местообитании крот держится по дубовым жам и на разнотравных лугах вдоль берегов ключевых речек Кара-Су. Рядом же, на солончаковых полянах с кермеком и древовидной солянкой, (рис. 106), обитают степные полевки и краснохвостые песчанки. Таким лбразом, здесь имеется замечательный пример мозаичности биотопов п сосуществования бок о бок влаголюбивых и сухолюбивых зверьков.
На Малом Кавказе крот распространен в зоне широколиственных лесов н субальпийских лугов северных склонов Аджаро-Имеретинского и Трпалетского хребтов в Джавахетии, на Бамбакском и Безобдал неком хр бта\. В верховьях Акстафа-Чай и Дебеда-Чай Даль (1944а) наблюдал наибольшую плотность — 42 крота на гектар в смешанном — буковограбовом — лесу на высотах 1770—1800 м. В окрестностях Севана и на Даралагезском хребте крота нет. Местами он уцелел на северном склоне Севанского хребта в верховьях Дзегама и Шамхора. Па южном склоне Армянского нагорья крот образует ряд изолированных реликтовых популяций по ущелью Занги (Даль, 19506). В Нагорном Карабахе он прослежен мною по лесистому ущелью Тертер-Чая в пределах от 700
254
Часть II. Анализ происхождения четвертичных млекопитающих
до 1600 м, но и здесь весьма редок. В лесном Талыше крот существует на высотах от 400 до 1700 м, но редок из-за чрезвычайной летпей сухости склонов. В ущельях Виляж-Чая, Вазару-Чая, Вашару-Чая в июле 1945 г. па 100 ловушко-суток приходилось всего 3—4 крота.
Далее на юго-восток крот, вероятно, живет п по северному склону Эльбурсского хребта.
Документальных данных об обитании крота на Иранском плоскогорье и в восточной Анатолии нет. Бленфорд (Blanlord, 1876) о кроте ничего не говорил. Радугин (19176) включил крота в список зверей оазиса Хой, лежащего па высоте около 2000 м, возможно приняв копки слепушонок за кротовые.
В результате обзора объединенного ареала кавказских кротов видно,, что распространение их весьма тесно связано с ландшафтом широколиственных лесов, мезофитпых лугов и слабее — с ландшафтом разнотравных степей. Существование крота обусловлено узко определенным!! физическими свойствами почвы, особенно ее влажностью, и наличием запасов пищи — значительной биомассы беспозвоночных. Миграции крота через сухую степь и полупустыню исключены, н, несомненно, что разъединенные ныне участки ареала на Большом Кавказе, Малом Кавказе и в Талыше имели древние ландшафтные связи. Объединенный ареал кротов па Кавказе показывает, таким образом, ту территорию, которая имела в третичном периоде единый тип ландшафта. Тип ареала кавказских кротов — с главным массивом в западных районах страны — сложился, вероятно, уже к верхнему плиоцену в связи с рапним развитием аридных участков на востоке Предкавказья и Закавказья. Современные разрывы ареала кротов в пределах Кавказского перешейка обусловлены всей историей «борьбы» мезофптного п ксерофитного ландшафтов при длительных изменениях климата п особенностей орографии на протяжении плиоцена п плейстоцена.
Миграции кротов с Русской равнины иа Кавказ и обратно, видимо,могли иметь место в неогене и в верхнем плейстоцене через Прикубанскую равнину и Ставрополье. Кавказ был, однако, п самостоятельным очагом видообразования для кротовых.
Разрывы ареала крота в Предкавказье, на Армянском нагорье и в долине Куры у Караяз, вероятно, послеледникового возраста. Разъединение Татыш—Карабах может быть и более древнее, дочетвертичное.
Современный ареал крота на Кавказе сокращается в связи с вырубкой лесов и ксерофптизацией предгорных и горных районов при выпасе скота и распашках. Непосредственное воздействие человека при добывании кротов па шкурку сказывается заметно па численности вида лишь в предгорьях западного Кавказа (график I). Истинного колебания численности промысел не отражает.
Выхухоль — Desmana moschata L. Кости выхухолей описывались из плейстоценовых слоев южной и средней Европы (Trouessart, 1898— 1899 a, Wolf; 1938, 1939) в качестве подвидов к современным D. moschata hungarica Kormos, D. moschata fossilis Larlet, D. moschata magna Owen.
В пределах Кавказа остатки мелкой выхухоли — Desmana sp. (фрагмент ипжней челюсти) обнаружены в аллювии плиоценовой реки близ Ставрополя. Остатки плейстоценового возраста известны из палеолитических стоянок Украины и из четвертичного аллювия Днепра, Дона, Волги.
Современный ареал выхухоли захватывает бассейны Дона, Волги, Урала. К Кавказскому перешейку теперь прилегает лишь небольшой
207-
Гл, III: Прежнее и современное распространение и численность млекопитающих 255
участок ареала, расположенный по долине Допа, от Маныча до устья Дона, где зверек уже давно был редок (карта 2).
Глубокие морфо-фцзпологическпе адаптации выхухоли к земноводному ечцествовапию в пойменных озерах с обильной кормовой базой из бес-/ .воночиых отражают длительную эволюцию ее предков в бассейнах пук с весьма устойчивым гидрологическим режимом, характерным для обширных равнин.
В молодых горных реках плиоценового и плейстоценового Кавказа I и непрерывно менявшимся базисом эрозии и катастрофическими паводками выхухоли вряд ли могли существовать. Поэтому есть основание считать восточноевропейский вид довольно древним, верхнеплиоценовым эснеицем лишь в западном Предкавказье — для рек Ей, Челбаса, вероятных прежних притоков пра-Дона.
В паши дни для выхухоли пригодны некоторые пойменные озера в ни-апйях Кубани, например у станицы Гривенской, и некоторые речки Ко. хиды: Ция, Цива, Пичера.
I Отряд ХИЩНЫЕ—СА RN I Г ORA
Происхождение и возраст ареалов большинства хищных Кавказа кпнавливается более или менее четко, частью по палеонтологическим документам, частью по особенностям ареала. Низкая, как правило, численность особей в популяциях п редкость массовой гибели не создавали условий для образования природных захоронений остатков хищных. Тем не менее остатки хищных встречаются в кухонных остатках палеолитического человека и в слоях поселений позднейших эпох в связи с раз-цитпем добычи этих животных для получения меха. Громов (1948) Линял даже высокую стратиграфическую оценку остаткам четвертичных иицпых. Из кайнозойских отложений Кавказского перешейка известны остатки представителей семейств Canidae, Ursidae, Hyaenidae, Muste-lidtie, Felidae. Виды этих же семейств составляют фауну хищных Кавказа ь наши дни.
Сем. ПСОВЫЕ — CAXIDAE
Из плейстоценовой фауны Кавказа известно 5 видов псовых родов Ca/tisn Vulpes, среди них 2 вымерших. В голоценовой фауне здесь существует 5 видов.
Шакал — Canis aureus L. Ископаемые остатки шакала часто указываются для палеолитических слоев пещер южной Европы (Франции, Италии, Румынии), Севершш Африки и Передней Азии.
Однако трудности диагностики мелких собачьих по скелетам заставляют сомневаться в идентичности мелких плейстоценовых собак Европы современному шакалу Африки и Азии, тем более, что в Восточной Европе иСредней Азии остатков плейстоценовых шакалов не обнаружено. Остатки в слоях ашеля и мустье пещер Палестины, Ливана, Сирии можно считать близкими современному виду (Bate, 1937). В плейстоценовых захоронениях Кавказа остатков шакала не обнаружено. Это и позволило нам указать на недавнее появление шакала па перешейке (Верещагин, 1949в, 19516). Поздние остатки шакала найдены в слоях поселения ранней бронзы близ Анаклии, в пещерах близ Кутаиси и в пещерах Сарайбулахского хребта в Армении (Даль, 19406).
Современное распространение шакала приурочено преимущественно к низменностям (карта 18). В западном Предкавказье шакал случаев
256
Часть II. Анализ происхождения четвертичных млекопитающих
и редок, а в восточном — обитает по долинам Терека и Сунжи от Моздока и Грозного. По Черноморскому побережью Кавказа шакал многочислен от Геленджика до Батуми, но в горы здесь пе поднимается выше 400— 500 м. Особенно много его в ольховых дебрях низовьев рек Колхиды. На восток ареал проходит через Сурам, который изредка переходят одиночные звери. Наибольшая численность в восточном Закавказье наблюдается па низменностях: Закатал о-Исмаиллинской, Хачмасской Ленкоранской. Зимой шакал многочислен по тугаям и окраинам тростниковых болот средней Куры и низового Аракса. Наиболее высокие заходы шакалов в горы Большого Кавказа наблюдаются осенью, например у Закаталы, когда звери, питаясь преимущественно фруктами, особенно плодами мушмулы, встречаются на высотах до 1000 м. Лесную зону Талыша шакал населяет до ее верхней границы — 1700—1800 м. В открытой полупустыне шакал бывает обычен летом на кормежке саранчовыми, а зимой лишь при обилии зимующих стрепетов.
На высоких сухих плоскогорьях севера Ирана и Армении шакал не живет. Редок он здесь и по глубоким ущельям.
Несмотря на кажущуюся приспособленность к умеренному климату, шакал — относительно теплолюбивый хищник, но выносящпй глубоких снегов. Его южное происхождение выявляется при сильных снегопадах в Куринской низменности, когда звери становятся довольно беспомощными при передвижении и добыче пищи и часто гибнут от истощения и холода (Верещагин и Дюнин, 1949). Присутствие шакала в голоценовой фауне южной Европы и отсутствие его па Русской равнине, вероятно, объясняется разницей в тепловом и снеговом режимах.
В новейшую эпоху шакал подвергается всюду энергичному преследованию, но тем не менее численность его еще велика. В 30-х годах на каждые 1000 км2 добывалось в Азербайджане 113 шкур шакалов, в Грузии — 32.4 и в Дагестане — 10 (Верещагин, 1947 г). За последнюю четверть века наблюдался значительный подъем добычи в 1934 г. (график 2). Назначение повышенной премии за шакала, как вредителя животноводства и охотничьего хозяйства, привело к увеличению добычи шкур в 1949 г. в 2х/2 раза, что показало полное благополучие популяции шакала в современном культурном ландшафте Закавказья.
Волк — Canis lupus L. (s. lato). В верхнем плиоцене и нижнем плейстоцене Европы уже существовал ряд мелких волков из полиморфной группы С. lupus. Остатки крупной формы волка С. lupus появляются в слоях эпохи наибольшего оледенения и особенно характерны в Германии и Австрии для вюрма (Freudenberg, 1914).
На Кавказе наиболее древняя находка нижней челюсти мелкого волка — С. tamanensis описана из нижпеплейстоценовых конгломератов Таманского полуострова. Родственные отношения этого вида к современному С. lupus неясны.
Остатки крупного волка начинают встречаться здесь с нижнего плейстоцена и далее уже обычны вплоть до современности (карта 19).
На юг от Кавказа остатки С. cl. lupus обнаружены в мустьерских слоях пещер Палестины (Bate, 1937) и в верхпеплейстоцеповом делювии северо-западного Ирана.
К северу остатки средне- и верхненлейстоценовых крупных волков обычны на отмелях Днепра, Допа, Волги, Урала и в слоях палеолитических стоянок Русской равнины и Крыма.
Современное распространение волка на перешейке повсеместное, но неравномерное.
UHL Прежнее и современное распространение и численность млекопитающих 257
По данным государственных заготовок шкур, наибольшее число волков добывается на единицу площади в западном Предкавказье и в восточном Закавказье (карта 19). На 1000 км2 добыча шкур волка в 30-х гцах составляла: Азербайджан — 11.6, Армения — 9.9, Северный Кй :аз — 6.7, Дагестан — 6.7, Грузия — 0.8.
Большинство шкур, поступающих на заготовительные пункты, проис-пз равнинных районов и принадлежит животным, битым в холодные сезоны года.
Наиболее плотно населены волком низовья Терека, Сулака, Кумы, । в Куринской низменности побережья Кызыл-Агачского залива, где, кроме домашнего скота, этот зверь питается кабанами и рыбой.
В ольховых болотах Колхиды волк обычен в низовьях речек Гагида, о л I, Пичера.
В горных районах Кавказа большая численность волка наблюдается в Ii нзказском, Боржомском и Закатало-Лагодехском заповедниках, лк , как и в камышах Каспийского побережья, держатся оседлые вшуляции волка, которые совершают вертикальные и горизонтальные дерекочевки вслед за дикими копытными. В Кавказском заповеднике воль держится довольно равномерно во всех зонах, причем лишь зимой |ывают заметна несколько большая концентрация этого зверя в поясе Ьнр.жолиственных лесов. В этот период здесь держится большая часть вию.ювья кабана, косули, оленя. Регистрация встреч волка по заповедник) (в процентах от общего количества) дала цифры: альпийская зона — 30% темпохвойиые леса — 32 %; широколиственные леса — 38 % (Теплев. 19386). По' данным Теплова, отход молодняка кабапов, оленей, косуль, сори и кавказских козлов в заповеднике от волка составляет и течение 5 месяцев от 34 до 61%. Кроме диких копытных, волк уничто-Ьетп сотни голов домашнего скота в прилежащих к заповеднику угодьях.
об обилии волка в районе Боржоми писал еще Диппик (1914а). В За-щшьском заповеднике волк живет так же оседло, как и в Кавказском (Ларков и Млокосевпч, 1935; Верещагин, 1938а).
На остальной территории перешейка, особенно в восточной его части, волк совершает теперь правильные перекочевки вслед за стадами домашнего скота, которые выпасаются летом в горах, а зимой — в низменностях. В весеннее время наибольшая концентрация волка наблюдается в сухих предгорьях с оврагами п навесами скал, где звери выводят молодых.
История колебания численности волка па перешейке в голоцене может быть представлена лишь в общих чертах.
По мере истребления человеком диких копытных па равнинах численность волка должна была сокращаться, но с развитием скотоводства постепенно восстанавливалась. Усовершенствование огнестрельного оружия п вообще способов добывания, а также разведение крупных собак для охраны стад повели, вероятно, к новому сокращению численности зверя.
Резкое увеличение добычи волков после второй мировой войны в связи с назначением высоких премий показало полное благополучие кавказских популяций этого зверя (график 3).
Голарктическое распространение волка в четвертичном периоде, Iciaoo выраженная (и изученная) географическая изменяемость и нсдо-।. даочиость палеонтологических материалов крайне затрудняют установление области или района происхождения этого зверя. На Кавказе [ оН рассматривается памп в качестве плиоценового обитателя.
Н. К. Верещагин
258
Часть II. Анализ происхождения четвертичных млекопитающих
Лисица — Vulpes vulpes L. (s. lato). Голарктический род Viilpes верхнемиоценовой древности. Из Восточной Европы известны остатки плиоценовых лисиц V. mericlionalis Nordm., V. moravicus Wold., достигавших величины современной среднерусской.
Не описаны еще остатки примитивных плиоценовых лисиц из одесских катакомб и из верхнеплиоценовых слоев Азовского побережья. В пределах Кавказа остатки V. kchomenkoi с признаками зубов рыжей лисицы и песца обнаружены в продуктивной толще (средний плиоцен) Апшеронского полуострова (Богачев, 1938в, 1944; Верещагин, 19516). Найден изолированный ложнокоренной зуб лисицы и в плиоценовых слоях у Ставрополя.
Остатки лисиц группы V. vulpes в плейстоценовых и голоценовых отложениях Западной и Восточной Европы, Крыма и Кавказа весьма обычны как в палеолитических стоянках, так и в других типах захоронений.
К югу от Кавказа остатки лисиц найдены в палеолитических слоях северного Ирана (Coon, 1951), Сирии, Ливана, Палестины, причем Пикар (Picard, 1937) перечисляет там V. alopex для мустье и орипьяка, V. cf. niloticus Rupp. — для ашеля и мустье и Canis vulpes — для оринь-яка и мезолита.
Современное распространение лисицы на Кавказе повсеместное (карта 20). Наибольшая плотность населения лисиц наблюдается в зоне полупустыни, степи, предгорной лесостепи, нагорной степи и наименьшая — на альпийских лугах. В горных буковых лесах южного склона Большого Кавказа лисицы почти не живут. Указание Дппника (1914а) на обилие лисицы в лесах Кавказа относится к разреженным лесам, например в предгорьях и на плоскогорьях Северного Кавказа. В Кавказском заповеднике наибольшее число лисиц держится в течение всего года в зоне широколиственных лесов — до 43.1% встреч; меньше — в зоне еловопихтовых лесов — до 31.8%; наименьшее — в зоне альпийской и субальпийской— до 25.1% встреч (Хонякина, 1938). Такое распределение соответствует в общем и особенностям размещения здесь кормовой базы: мышевидных грызунов, птиц, насекомых, ягод. Иная картина наблюдается в Закавказье.
По данным маршрутных учетов, проведенных нами в 1935—1936 гг. на южном склоне Большого Кавказа, в Азербайджане, были получены показатели распределения встреч свежих следов, экскрементов и самих л псиц по вертикальным ландшафтным зонам (табл. 65).
В зоне полупустыни плотно заселены весной склоны оврагов, холмов предгорий, а зимой — морское побережье п предгорья.
Из кормов летом здесь имеют основное значение саранчовые, а зимой — личинки жуков, гусеницы, степная полевка и песчанки. Морское побережье привлекает лнспц обилием пищевых выбросов: погибших птиц, рыб, раков. Огромное значение для современного распределения лисиц по биотопам имеет сельскохозяйственная деятельность человека (см. главу VI).
Добыча лисиц па шкурку в течение 1930—1940 гг. показала, что наибольшая плотность лисьего населения наблюдается в равнинном Предкавказье и в восточном Закавказье. Были получены следующие средние цифры выхода шкур на 1000 км2 территории: Северный Кавказ 276, Азербайджан — 268, Армения — 158, Дагестан — 138, Грузия —74.
Обилие лисиц находится в обратной зависимости от степени облесен-ности и гористости страны (Верещагин, 1947 г.). Особенно редка лисица в западном лесистом и гористом Закавказье.
i.i. HL Прежнее и современное распространение и численность млекопитающих 259
Таблица 65
Частота встреч лисиц и их следов в различных зонах восточного Закавказья
Число лисиц и следов, встреченных на маршруте в 20 км
Чипы угодий fосточного Закавказья 1935 г. 1936 г.
июнь январь ИЮНЬ январь
Целинная полупустыня 2 4 Я 12
Низовые леса долины Агри-Чая . . | 4 6
Горные буково-грабовые леса . . . — — 1 2
Альпийские луга — 1 1 2
Скалы и осыпи перевалов .... । — — — —
По учетам II. Н. Бакеева, плотность населения лисицы в 1938—1941 гг. |Спицевском районе Ставропольского края достигала летом 12.5, а в осен-Ь-знмине сезоны 10.3—16.5 особей на 1000 га (Чиркова, 1952).
В течение исторической эпохи ареал и численность лисицы па Кавказе, Кншдпмому, развивались под благоприятным влиянием сельскохозяйственной деятельности человека. Даже весьма интенсивный промысел ю шкурку в пашем столетии не может подавить численности вида.
Материалы заготовок за последнюю четверть века показывают резкие яолебапия числа добываемых шкур в Предкавказье и менее резкие — I Закавказье. Последнее обусловлено большим разнообразием закавказ-квх условий и большей устойчивостью популяций. Судя по заготовкам шкур, с 1925 по 1952 г. в Предкавказье имело место три пика нарастания численности: 1927, 1936 и 1946 гг. — с соответствующими падениями 11931, 1941 и 1951 гг. (график 4 и 5).
Учитывая наличие близкой формы уже в среднем плиоцене Кавказа н поразительную полиморфность современного вида в пределах перешейка [глава IV), лисицу следует относить в группу местных — восточносредп-земноморских — видов верхнетретичного возраста.
Корсак — Vulpes corsac L. Область распространения корсака в плейстоцене тянулась широкой полосой степей от Западной Европы до северного Китая. Остатки этой лисицы найдены в пещерах Швейцарии, Чехословакии. Достоверно известны они из ряда палеолитических стоять Русской равнины, в частности пз Кодака, и из пещер Крыма.
В пределах Кавказского перешейка многочисленные остатки корсака найдены только в среднеплейстоценовых песках па Апшероне. К югу, в частности в Сирии, Палестине, Иране, корсаков, очевидно, пе было. Их замещалп там другие нагорпо-пустыипые формы мелких лисиц.
В XX столетии корсак был уже распространен па Кавказе, преимущественно в северо-восточной половине перешейка (карта 21).
Сокращение западного участка ареала вида началось, очевидно, ше в верхнем плейстоцене и стало резко заметным в последние полтора столетия, когда сильно увеличилось влияние промысла. По западному побережью Каспия ареал корсака отступил к северу со среднего плейстоцена па 700 км.
Причины сокращения ареала в доисторическое время в Европе и н-а Кавказе недостаточно ясны. На примере доступных нам материалов
17*
260
Часть II. Анализ происхождения четвертичных млекопитающих
становится известно, что изменения в структуре группировки мелких стенных хищников происходили уже с среднего плейстоцена. В Западной Европе, например, вымер бесследно ряд мелких волкообразных и шакалообразных собак.
Численность корсаков в биоценозе полустепи восточного Закавказья времен бинагадинской асфальтовой ловушки была еще довольно велика, но затем катастрофически сокращалась. Вычисленная нами по костным остаткам и по добыче шкур относительная численность корсаков по отношению к лисице и волку представлена для четвертичного периода цифрами табл. 66.
Таблица 66
Относительная численность (в °/о) остатков псовых в четвертичном периоде
Датировка и местонахождение Корсак Лисица Волк
Голоцен, XX в. Средняя годовая добыча шкур по Северному Кавказу за 1935— 1940 гг.— 63600 (1.06 97.5 2/i
Голоцен, Саркел—VIII—Х1Г1 вв. л. а. Общее число особей, добытых хазарами и славянами — 44 6.8 77.3 15.9
Средний плейстоцен, Бин ага ды. Общее число особей, пойманных асфальтом—327 . . . 25.6 37.7 36.7
Корсак был, таким образом, уже издавна менее жизненной формой, нежели лисица и волк. Тем не менее в оптимальных для корсака районах численность зверька иногда п теперь еще превышает численность лисиц и волков. Так, например, к югу от Акмолинска в злаково-полынной степи в 1947 г. было найдено 37 нор корсака, 3 норы лисицы и 1 нора волка, а в 1948 г. там же было отмечено 42 поры, пз них 34 корсачьих и 8 лисьих (Чиркова, 1952, стр. 402).
Родина корсака, по-видимому, и лежит где-то в области степей Казахстана. По экологической сущности это животное пустынных степей, питающееся мелкими грызунами, птицами, рептилиями и насекомыми.
Корсак был обычен в степях западного Приманычья и встречался под Сальском еще в первом десятилетии нашего века (Диннпк, 1914а; Богачев, 1918). В настоящее время отсюда поступают лишь единичные шкурки (график 6). Распашка степей и вообще сельскохозяйственная деятельность человека оказывают отрицательное влияние па корсака. Теперь это зверек обычен лишь в районе Терско-Кумскпх песков.
По типу ареала совремепной экологии и палеонтологическим данным корсака с юдуст считать в кавказской фауне типичным вселенцем эпохи среднего плейстоцена со стороны степей Русской равнины. Вселение это произошло, очевидно, при наибольшем развитии к югу ландшафта холодных степей.
Сем. ГЛ 1111ОВЫЕ — 11 1 A ENID А Е
Остатки гиен и иктитериев известны в Евразии с верхнего миоцена (Simpson, 1945). Особенно они характерны для плиоценовых отложений Средиземноморья.
Гл. IIL Прежнее и современное распространение и численность млекопитающих 261
Для Кавказа известно 3—4 вида ископаемых и один живущий вид гиены. Наиболее древние остатки обнаружены в среднемиоценовом захоронении у Беломечетской (карта 21).
Уже с верхнего миоцена наметились две линии специализации: тип /’госн/п, с хищническими зубами, близкими по строению к зубам крупных кошек, и тип Hyaena, более примитивный, с менее мощными зу-'амп.
В четвертичном периоде здесь существовало два вида. Их прямые родственные связи с третичными формами недостаточно ясны.
Пещерная гиена — Crocuta spelaea Gold!. В плейстоцене на Кавказом перешейке обитал, по-видимому, только этот вид, известный по \м местонахождениям: в средпеплейстоцеповых слоях Апшеронского олуострова (Бинагады) и в верхнеплейстоцеповых отложениях Закубаи-ой равнины (Ильская).
К северу от перешейка ареал этой гиены занимал в плейстоцене ог-юмные пространства Евразии (Пидоплпчко, 1951).
К югу остатки этого вида прослежены в верхнем плейстоцене северо-ыпадного Прана (Брандт, 1870; Coon, 1951).
Средпепленстоцоновая пещерная гиена Русской равнины была зпа-Ительно крупнее закавказской (Верещагин, 19516), что соответствует более крупным размерам травоядных тех районов, например зубрам и •шадям.
По особенностям черепа и размерам бинагадипская гиена из Бина-г дон занимала промежуточное место между пещерной гиепой Русской равнины и ныне живущей пятнистой гиеной — С. crocuta L. Африки. Вообще прямые родственные связи этих форм вполне вероятны.
Распространение пещерной гиены быстро сокращалось в верхнем плейстоцене, отодвигаясь на юго-запад, в Африку, по не исключена возможность, что и в голоцене трансформированный вид еще держался южнее перешейка. Так, например, пятнистая гиена жила в Палестине в эпоху мн’тье, в ориньяке и в голоцене «natulian», при новом наступлении сухого и’жаркого климата (Bate, 1937).
Особый интерес пррдставляет вопрос о причинах вымирания этого зверя на огромной территории Евразии. Такой причиной явилось, несомненно, постепенное вымирание хоботных и крупных копытных в верхнем плейстоцене. Палеолитический человек вряд ли мог иметь непосредственное значение для столь раннего исчезновения этой гиены, так как п современному охотнику, при его совершенном оружии, гиена практически не нужна. Мнение Пидоплпчко (1951, стр. 160) о роли «антроппче-I и» причины в исчезновении гиен в Европе и Азии мало обосновано. 1 допой пещерной гиены следует считать Средиземноморье, а огромное разрастание ареала в плейстоцене необходимо рассматривать в совокуи-вости с распространением всей мамонтовой фауны. Наступление ксеротермической послеледниковой эпохи ознаменовалось на Кавказе вселением с юга меиее специализированного вида — полосатой гиены.
Полосатая гиена — Hyaena hyaena L. Остатки полосатой гиены обнаружены в палеолитических пещерах Палестины, Марокко, Алжира н Португалии. Находки во Франции мало достоверны (Wolf, 1938, стр. 59). Пидоплпчко (1951) вслед за Шарффом (1918) считает, что поедая гпепа попала в Европу еще в плиоцене с юго-запада, при наличии плиоценовой суши, простиравшейся от Африки до Британских островов. Однако еще не доказано, что Н. prisca Mars, de Series, H. mons-^ulana Croiz. et Job. из плейстоцена южной! Франции и, наконец,
262
Часть II. Анализ происхождения четвертичных млекопитающих
Н. antiqua Lankest. из плиоцена Англии были предками современного вида.
Важно отметить, что нигде на Русской равнине и в Сибири остатков полосатой гиены не обнаружено. Только в Средней Азии, южнее Самарканда, в среднепалеолитических слоях пещеры Аман-Ку тан найдены обломки костей и зубы этого вида (коллекция ЗИНа).
Отсутствие остатков полосатой гиены в плейстоценовых захоронениях Русской равнины, Крыма, Кавказа, особенно в бинагадинском комплексе,
Рис. 107. Места обитания полосатых гиен в долине среднего Аракса (Абракунпс).
Фот. автора, 1947.
доказывает, что она проникла на перешеек относительно поздно. Остатки полосатой гиены обнаружены на Кавказе только в голоценовых отложениях Апшеронского полуострова. Современный ареал ряда подвидов этой гиены охватывает Среднюю и Переднюю Азию, Африку. Все районы, заселенные ею, характеризуются сухим и жарким климатом. Слабая развитость подшерстка гиены также указывает па ее южное происхождение. На Кавказе она держалась и держится преимущественно в жаркой полупустыне низменностей и межгорных долин (рис. 107) в восточном и южном Закавказье. В конце прошлого столетия гиена встречалась еще и в восточном Предкавказье, между Дербентом и Махачкалой (Дмя-ник, 1914а; Сатупип, 1915а). Отмечены случайные заходы и под Владикавказ (ныне Орджоникидзе) (Богданов, 1873). В 60-х годах прошлого века гиен довольно часто убивали под Тбилиси, Ереваном, Нахичеванью (Радде, 1899). В 30-х годах нашего столетия она обитала в небольшом числе только в малонаселенных уголках западного Азербайджана и в восточных районах Грузии (Верещагин, 19426). За десятилетие — с 1930 по 1940 г. — здесь было добыто всего 26 гиен, а с 1940 по 1950 г. — не более 5—6. Небольшое число гиен еще существует в распадках пус
[л. III.- Прежнее и современное распространение и численность млекопитающих 263
тынных хребтов Карталинского плоскогорья, близ Удабно и Гереджи. Типичные места обитания гиены в восточном Закавказье приурочены к размытым глинистым хребтам (боздагам) с зарослями можжевельни-в и фисташки.
* Распространение и численность гиены сокращались на Кавказе стиль быстро в течение последнего полустолетия главным образом под Ьянпем сознательного истребления из-за случаев нападения на
Д’Н’П.
Сем. ЛВДВЕЖЬИ — UJRSIDAB
Пз кайнозойских слоев Кавказа теперь известны представители 2 poll медведей: Ursavus, Ursus.
Ursavus — среднемиоценовый род, обнаруженный в Беломечетской, Ur is — нижпеплиоценовый становится известен па Кавказе с эпохи плиоценового захоронения у Ставрополя (карты 22 и 23).
Рис. 108. Череп малого пещерного медведя из Краснодара.
В четвертичной фауне Кавказского перешейка существовало три вда медведей рода Ursus, из которых до наших дней дожил лишь один.
Малый пещерный медведь — Ursus (Spelaearctos) rossicus Boris. Описан в 1931 г. по 5 скелетам, извлеченным из песчаных отложений каких-то древних потоков среднеплейстоценового возраста у Красно-дара (рис. 108).
Остатки малого пещерного медведя указаны также из-под Херсона Ьорнсяк, 1931), из бассейна Большого Иргиза (Беляева, 1935, 1939а), с реки Урала (Верещагин и Громов, 1952), а теперь обнаружены и в пе-щерал Северного Урала. В связи с отсутствием остатков этого вида в палеолитических слоях пещер западного и центрального Закавказья, а также в битуминозных слоях Апшеронского полуострова можно думать, что южная граница его распространения проходила где-то в предгорьях Предкавказья. Малый пещерный медведь обитал, по-видимому, по долинам рек, степным балкам, склонам холлов и был, вероятно, менее связан с карстовыми районами, нежели следующая форма. Большая тупобугорчатость коренных указывает па пре-ямпцественную растениеядность зверя.
264
Часть II. Анализ происхождения четвертичных млекопитающих
В плейстоценовой фауне перешейка этого медведя следует считать нпжпеплейстоценовым вселепцем с Русской равнины, вымершим, вероятно, к верхнему плейстоцену.
Большой пещерный медведь — Ursus (Spelaearclos) spelaeus Ro-senin. В фауне Западной Европы и Северной Африки пещерный медведь был одним из ландшафтных видов па протяжении второй половины плейстоцена.
На Кавказе он известен по остаткам из Ильской стоянки у Краснодара и из пещер западного Закавказья (рис. 109). Указания Богачева (1939, 19406) п Громовой (1948) на присутствие этого медведя в бпнага-дипском комплексе ошибочны (карта 22).
Рис. 109. Череп большого пещерного медведя с укороченной лицевой частью из пещеры Абласкпра.
Остатки пещерного медведя из Воронцовской пещеры под Хостоц привлекают внимание относительно слабой фоссилизацией. Возможно, что небольшая популяция пещерных медведей несколько пережила плейстоцен в северо-западном Закавказье.
С Русской равнины, исключая север Черноморского побережья, остатки этого вида неизвестны. Однако в районе Самарской Луки существовал в плейстоцене некрупный пещерный медведь, занимавший по строению черепа и зубов среднее положение между малым и большим пещерными медведями.
На Малом Кавказе, в Талыше и к югу от перешейка, в частности в Курдистане, и в Сирии остатки пещерных медведей не найдены. Поэтому западное Закавказье следует считать пока южной границей вида в восточном Средиземноморье.
Родственные взаимоотношения этого вида с малым пещерным медведем подлежат дальнейшему изучению, по считать этот вид верхпеплел-стоцеповым потомком малого нет оснований, так как имеются близкие по геологическому возрасту находки обеих форм. По происхождению большого пещерного медведя следует считать местным — восточписре-диземно морским — видом.
Бурый медведь — Ursus arctos L. (s. lato). По сравнению с пещерными бурые медведи — более прогрессивная ветвь, сохранившая в то же время ряд примитивных признаков. Из плейстоценовых слоев пещер Западной Европы, Северной Африки и Передней Азии (Сирии) описано не менее десятка форм.
В верхнем плейстоцене бурый медведь был широко распространен на Кавказском перешейке, о чем свидетельствуют единичные находки
Гj. III. Прежнее и сое ременное распространение и численность млекопитающих 265
его остатков в пещерах Имеретии и в делювии Ашперонского полуострова.
На север от перешейка остатки бурых медведей найдены в плепстопе-а жьтх песках и галечниках долин Дона, Волги и Камы, а также в делю-•i ряда палеолитических стоянок Русской равнины. Две формы — U. cf. les п U. syriacus — указывались для палеолита Ливана, Сирип, Од естпны (Picard, 1937).
В галеценовых отложениях Кавказа остатки бурого медведя немпого-щкаепны, но тем не менее закономерно сменяют остатки большого пе-Ijppaoro в многослойных захоронениях, свидетельствуя о широком рас-
Рис. 110.
1 — 2 — бронзовые ггодвестш — фигурки медведя (1:1) из кобаястсих могильников Осетин (по Уваровой, 1900); <7 — изображение бегущего медведя (Х2) на сердоликовой печатне из урартских могильников (ориг.).
птхитрапеппп зверя в послеледниковую жну (карта 23).
В древнем искусстве у народов Кав-изображения медведей сравнительно «m u. В кобанских погребениях в Север-14 Осетии добыты подвески и брелоки г, бронзы, изображающие медведя. Бе-пщ'Ш медведь изображен па сердоликовой печатке (рпс. 110) из урартского могил ника па побережье оз. Севан. Мед-поднимающийся на дерево, изображен ва .-еребряном сосудике из Майкопского кургана.
’ Современное распространение медведя л Кавказе еще довольно обширно. По Ьерпому склону Большого хребта оп оГптает от Новороссийска па западе до «ровьев Дженгн-Чая па востоке, всюду,
XXесть участки леса, горные кустарники и луга. Наибольшая численность ]рря наблюдается теперь в Кавказском заповеднике. На Ставропольской возвышенности медведь был уничтожен уже в прошлом столетии, а в леса Пятигорья заходит теперь лишь изредка. В Кабарде и Северной Осетии ч.цкедя мало. Нет его па большей части центрального Дагестана, но он ^встречается в лесах Дешлагара. В низовых лесах Самура и на Кусар-скоп наклонной равнине медведя истребили около 30 лет тому
назад.
На южном склоне Большого хребта медведя теперь больше, чем на северном. Он обычен в лесистой Абхазии, Имеретии. В болотистой Колхиде пе водится. В Сванетии и Южной Осетии медведь обычен в верховьях Кодора, Пнгура и Риона, несмотря на энергичное истребление местным населением. В Азербайджане он многочислен в бассейнах речек Бслокап-Чай, Катехп-Чай, Кахи-Чай и многих других, до верховьев Гсрдымап-Чая на востоке.
При созревании каштанов и грецких орехов в сентябре—октябре медведи концентрируются в лесах и садах Алазано-Агричайской долины. При обилии буковых орешков и желудей много медведей держится осенью п у верхней опушки леса. На низких хребтах Карталинского, Порского и Аджипоурского плоскогорий оп живет по можжевелово-фисташковому редколесью, где кормится ягодами фисташки, можжевельников п травой, особенно астрагалами. По днищам распадков п вдоль обрывов медведи устраивают здесь торные тропы. Недавно медведи держались и по тугайным лесам Куры, особенно па участке между Упнгечауром и устьем Алазани.
266 Часть II. Анализ происхождения четвертичных млекопитающих
На Малом Кавказе медведь теперь сильно истреблен и редок повсеместно. В безлесной Джавахетии его нет вовсе. На Армянском нагорье он обитает местами по можжевеловому редколесью и в уцелевших дубовых лесах. Весьма редок медведь и в горных лесах Талыша.
Больше всего добывается медведей в восточном Закавказье. На 1000 км2 в среднем за десятилетие — с 1930 по 1940 г. — заготавливалось шкур: Азербайджан — 1.5, Армения — 1.5, Грузия—0.9, Северный Кавказ — 0.9, Дагестан — 0.2, по большая часть добытого «оседала» у местного населения.
Бурый медведь на Кавказе, очевидно, местного — плиоценового — происхождения. Подтверждением этому служат находка остатков в плиоцене под Ставрополем, высокая численность его в стране и значительное разнообразие форм. На различных участках страны могли быть, впрочем, новейшие генетические влияния с севера и юга смежных популяций. Так, например, недавние — голоценовые — связи ареала северных и кавказских рас медведей существовали по долине Дона, степным балкам и камышам западного Предкавказья, что и обусловливало сходство медведей северо-западного Кавказа с европейскими (Смирнов, 1916а).
Влияние человека на численность и ареал медведя на Кавказе сказывалось непосредственно и через изменение ландшафта. Добыча шкур в течение четверти века — с 1925 до 1950 г. — несколько повысилась к 1930 г., затем упала и вновь поднялась к 1941 г. (график 8).
Перспективы дальпейшего существования медведя па Кавказе невелики, особенно в связи с продолжающимся лесоистреблен нем и преследованием его как промыслового зверя и вредителя животноводства.
Сем. КУНЬИ — MUSTJELIUA E
Остатки куньих появляются в Евразии с нижнего олигоцена. Из плейстоценовой фауны Кавказа известно 6 видов куньих, из которых один, по-видимому, вымер к голоцену.
Росомаха — Gulo all. gulo L. Остатки росомахи известны из многих пещер юга и средней полосы Западной Европы, начиная с нижнего плейстоцена. В пределах СССР остатки росомахи найдены в палеолите Крыма, Украины, на отмелях Камы, в пещерах Алтая и Урала.
На Кавказе обломок нижней челюсти росомахи обнаружен в верхнепалеолитическом слое пещеры Гварджилас-Клде и в нижне- и в среднепалеолитических слоях пещеры Кударо I. Это самое юго-восточное местонахождение плейстоценового вида.
Смирнов (1923—1924) считал росомаху горным зверем и объяснял ее вымирание на равнинах Евразии исчезновением в послеледниковье плотных осевших снегов, па которых этот зверь мог догонять копытных средних размеров.
Пидопличко (1951) объяснял быстрое вымирание росомахи и отступление ее ареала к северу в Восточной Европе «антропическими факторами». Интересно, что очень сходное сокращение ареала отмечается для песца, северного оленя и других арктических видов. Однако в отличие от них у росомахи образовалось первоначально значительное разъединение древнего средиземноморского очага и его северного, плейстоценового придатка. Кавказский очаг вида затухал, вероятно, медленно, чему способствовало обилие пищи и разнообразие рельефа горной страны, обеспечивающее всевозможные типы состояния снегов. Существовал, видимо, целый комплекс причин, приведших к вымиранию зверя на Кавказе.
Прежнее и современное распространение и численность млекопитающих 267
Наюольшее значение могло иметь пониженное размножение на современной стадии эволюции вида. По происхождению росомаху следует считать охтоном горных окраин Средиземноморья, откуда опа и распространись на север, сохранившись до наших дней на новом участке ареала.
Время окончательного вымирания росомахи на Кавказе пока не уточнено. Нет, однако, ничего невероятного в том, что она вымерла здесь виздмее, чем на юге Западной Европы, может быть даже в век бронзы.
Куница белодушка — Martes foina Erxl. Род Martes становится тестей в Евразии с нижнего плиоцена. При более широком понимании )да, т. е. включая настоящих куниц в род Mustela, можно думать, что а группа развивалась в Голарктпке уже с верхнего миоцена. На Кав-нзе в плиоценовых отложениях Армянского нагорья были указаны тки Mustela filholi Goudry и Putorius sp. (Богачев, 1938в).
фрагментарные остатки куниц — Martes sp. собраны археологами средне- и верхнеплепстоценовых слоях пещер западного Закавказья: днптырская, Сакажиа. На Русской равнине челюсть куницы Martes sp. Зваружена нами в плейстоценовых песках низовой Камы. Очевидно, Л следует сближать с лесной куницей. По нижнему течению рек Русской 1ВШ1НЫ остатков настоящих куниц не найдено, ио в Крыму они обычны Ь палеолитических Слоев. Из палеолита Палестины оппсапа М. palaeo-гш'а Nehr., близкая к М. foina. Куница урартских времен с побережья »<«нана была описана под именем М. latifrons Sat. без достаточных обо-вапий.
Современный ареал белодушкп огромен: он тянется от берегов Испари полосой средних широт до северного Китая и Маньчжурии. Кавказ-.да подвид — М. foina nehringi Sat. обитает в лесистых предгорьях, горных лесах и на безлесных плоскогорьях (карта 24). На Ставропольской возвышенности и на волнистых равнинах к западу от Ставрополя зна держится по кустарникам балок и по линиям обрывов сарматских звестияков. Шкурки ее ежегодно поступают также из среднего течения Ныв районе Буденновска. Предгорья Большого Кавказа, внутрен-долины и центральный Дагестан прочно заселены белодушкой. Осо-бепно много белодушкп в бассейнах левых притоков Терека, Сунжи, Манаса и в районе Гунпба. Порядочно шкур белодушек поступает из Самарских низовых лесов. Мы добыли эту куницу у Баку, близ основания Апшеронского полуострова.
В галерейных лесах Кура-Араксинской низменности белодушка не водится.
В западном и восточном Закавказье белодушка занимает преимущественно предгорья и нижнюю часть горных лесов. В зоне низовых лесов, например по Алазано-Агрича гской долине, весьма многочисленна, отца в основном и поступают ее шкурки. Па Армянском нагорье и в Та-лыше белодушка встречается всюду, где есть хотя бы небольшие кустар-fliKOBbie заросли и уцелевшие леса. В верхней зоне гор белодушка образует прочные поселения. Однако в пустынных межгорных долинах, ^пример по среднему Араксу, звери держатся лишь но крупным россыпям скал п ущелий обособленных вершин, например Иляплу-Даг, Дарры-Даг и др.
Судя по ареалу и экологии, белодушку па Кавказе следует рассмат-ивать в качестве древнего, плиоценового обитателя, появившегося с юга.
Воздействия человека, в том числе и промысел, сказываются на бело-шке слабее, чем на следующем виде.
268
Часть II, Анализ происхождения четвертичных млекопитающих
Лесная куница — Martes martes L. Остатки лесной куницы обычны в плейстоценовых слоях пещер Западной Европы — от Англии, Франции, Италии до Венгрии. Отмечены они и для мустьерских слоев Палестины (Bate, 1937). Кости этой куницы, трудно отличимые от костей бело-душки, вероятно, имеются в некоторых палеолитических стоянках западного Закавказья.
Современное распространение кавказского подвида — М. martes Lorenzi Ogn. весьма своеобразно. Опа селится преимущественно в высокоствольных буковых лесах северного и южного склонов Большого Кавказа, постепенно исчезая к востоку. На Малом Кавказе она обитает по северному склону Аджаро-Имеретинского хребта, а на Триалетском хребте — от Боржоми и Бакуриани до Белого Ключа. На запад ареал уходит по северному склону Тавра в Малую Азию (карта 25). Восточнее Дебеда-Чая и Акстафа-Чая этой куницы либо пет, либо она уже крайне редка. В лесах Карабаха за последние 25 лет она ни разу не добывалась
Следует отметить, что Сатунин (19076) описал череп молодой куницы из могилышка восточного берега Севана под именем М. martes (?). Диагноз этот ничем, кроме «тонкого сложения» и «малой величины», не подтвержден, и череп этот, хранящийся в Музее Грузии, принадлежит на самом деле белодушке.
Таким образом, ареал лесной куницы на Малом Кавказе не достигал района оз. Севан и в середине I тысячелетия до н. э. Во всяком случае на 7 известных нам черепов куниц из урартских могильников желто-душковых не встретилось ни одного.
Нет лесной куницы и в Талыше, несмотря па обилие леса и полчков— весьма важной пищи. Указание Эллермапа и Моррисон-Скотта (Ellermann and Morrison-Scott, 1951, стр. 245) па наличие желтодушки под Астра-бадом основано, вероятно, иа недоразумении.
Отсутствие лесной куницы в восточной части Малого Кавказа и в Талыше может быть объяснено только относительно поздним появлением вида на Кавказе и наличием древнего безлесного барьера в бассейне Храма. Следует учитывать также и увеличивающуюся конкуренцию со стороны белодушки в юго-восточиых районах страны.
Наибольшая численность лесной куницы наблюдается на западном Кавказе. По данным заготовительных пунктов, в 30-х годах на 1000 км2 добывалось шкурок обеих куниц 1: Грузия — 37.8, Армения — 30.3, Азербайджан — 19.4, Дагестан — 10.3, Северный Кавказ — 7.8.
Высокая численность куниц в Грузии объясняется сравнительно большой еще облесенностью страны. В Армении вся добыча шкур приходится иа куницу белодушку.
По наблюдениям Донаурова (1949), над лесной куницей в Кавказском заповеднике в 1935—1936 гг., процент встреч ее экскрементов летом и следов па снегу зимой распределялся так, как указано в табл, 67.
Такое распределение обусловлено обилием важнейших растительных и животных кормов: летом — мышевидных грызунов и насекомых; осенью и зимой — рябины, черники, тисса, грызунов.
Малая численность лесной куницы в нижней зоне леса обусловлена, вероятно, вытеснением ее более сильной белодушкой. Па южном склепе Большого хребта распределение зверька несколько иное.
В табл. 68, составленной по нашим записям 1935—1936 гг., показана частота встреч следов жизнедеятельности куниц в Заката л иском заповед-
1 При государственных заготовках шкур оба вида куниц записываются совместно.
/!. III. Прежнее и современное распространение и численность млекопитающих 269
Таблица 67
Частота встреч (в °/0) следов лесной куницы на западном Кавказе
Место обитания Лето Зиму
Широколиственные леса 0.7 6.9
Пихтово-буковые леса без ягодников . Пихтово-буковые леса с подлеском и 16.9 9.4
ягодниками 37.8 51.8
Верхняя опушка леса 40.9 30.9
Субальпийские луга 3.7 1.0
Вке и прилежащих угодьях.
Несомненно, что в нижней и средней зонах гор эти данные у нас и «наурова относятся и к кунице белодушке.
По многолетним наблюдениям Млокосевича (Марков и Млокосевич, 1135), в районе Лагодехи и Закаталы лесная куница держится преимуще-с -веяно в верхней зоне горных лесов и не способна конкурировать с более о ьной белодушкой в предгорьях. На безлесных равнинах ее нигде нет.
По морфо-экологической сущности лесная куница — более спецпали-грованпый и молодой филогенетически вид по сравнению с белодушкой.
Гм собленпе лесной куницы, очевидно, произошло в Европе еще в нижнем 1епстоцене, так как к среднему плейстоцену это был уже сформировавшая вид. Особенности кавказского ареала и современной экологии лесной куницы говорят скорее всего о довольно поздней — верхпсплей-пщеповои — миграции из Европы через Балканы и Малую Азию.
Непосредственное и косвенное влияние человека на численность и ,pea.i кавказских куниц стало заметно сказываться в последние столетия.
Промысел лесных кунпц на Кавказе был сильно развит в древней Церкессип. О добыче шкур куниц в XVIII в. в горах западного Кавказа номинал еще Дюбуа де Монпере (изд. 1937). Радде (1866) отмечал, что из верхней Свапетип в середине прошлого века ежегодно вывозилось до 500 шкур кунпц. Хищнический промысел куниц развился на Северном Кавказе с 60-х годов прошлого столетия при появлении новых пересе-енцев (Дипник, 1914а; Туров, 1937). Истребление крупных массивов леса за последние столетня в восточной части Большого хребта, например
Таблица 68
Частота встреч (в %) следов лесной куницы па южном склоне Большого Кавказа
Место обитания Лето Зима
Буково-грабовые леса нижней зоны . Буковые леса с черешней и лещиной 12.5 13.2
средней зоны Дубово-буковые осветленные леса 30.6 47.8
верхней зоны 50.0 39.0
Субальпийские луга 7.9 —
270
Часть II. Анализ происхождения четвертичных млекопитающих
в Дагестане п Азербайджане, также могло сказываться на сокращении ареала (Туров, 1937). Добывание шкур обоих видов куниц в течение последней четверти века уменьшалось в 1930—1931 гг. и увеличивалось в 1937 и 1947 гг. Некоторое падение промысла наблюдалось, в Абхазии в 30-х годах и было будто бы следствием развития промыслу на полчков (Марков, 1938) (график 9).
Сем. КОША ЧЬИ — IELIDA E
Из неогеновых отложений Кавказа и смегкных с ним территорий известно не менее 10 форм кошачьих. Из сарматских отложений близ; Армавира известны остатки Machairodus sp., а из Марагинского верхнеплейстоценового захоронения в северо-западном Иране указывались, остатки Felts sp. средних размеров. Более поздние фрагментарные находки костей кошек Felts, Panthera датируются нижним и верхним плиоценом (Ставрополь, Таманский полуостров).
В плейстоценовой фауне Кавказа открыто 6 видов: тигролев, барс,, рысь, лесной кот, пятнистый кот, гепард. Наконец, в голоцене здесь существовало по крайней мере 9 видов, из которых 3 вымерло в историческую эпоху.
Тигролев — Panthera spelaea Goldf. Остатки крупной плейстоценовой кошки, известной чаще всего под именем пещерного льва, обнаруживались, в средне- и верхнеплейстоценовых слоях Евразии — от Англии до Новосибирских островов (Trouessart, 1904—1905). Тигролев — характерный индикаторный вид в плейстоценовой мамонтовой фауне Русской равнины и Крыма (Громова, 1932а; Громов, 1948; Пидопличко, 1951). Южная граница распространения этой кошки проходила, по-видимому, где-то. в области Средиземноморья. Остатки кошек из палеолита Ливана, Сирии и Палестины, вероятно, принадлежат уже настоящему льву (Picard, 1937). Не найдены остатки тигрольва и в пещерах Ирана. На Кавказе остатки крупных плейстоценовых кошек обнаружены в пещере Сакажиа,. в делювиальных суглинках близ Сочи и в асфальтированных песках Апшеронского полуострова (рпс. 111 и карта 33).
Предположение Громова (1948) о том, что тпгролев вымер па Кавказе позднее, чем в других районах, чуть ли не в век бронзы, вряд ли имеет серьезные основания.
Вероятнее, что он вымер одновременно с пещерным медведем, полосатым носорогом, мамонтом. Родственные ему виды — лев и тигр, появляющиеся иа Кавказе, — это послеледниковые иммигранты с юга.
Лев — Panthera leo L. Вопрос о существовании льва в голоцеповой фауне Кавказа и о родственных взаимоотношениях его с плейстоценовым тигро львом Евразии весьма запутан. Большинство палеозоологов считало, что тигролев — Р. spelaea был непосредственным предком современного льва. Пидопличко (1951), не вдаваясь в детали, называл львом и крупную плейстоценовую котику. Однако, как показали наши исследования (Верещагин, 19516), тигролев обладал более специализированными кошачьими признаками черепа, нежели современный лев Африки.
Некоторые западноевропейские палеонтологи отмечали остатки львов, в ряде пещер Англии, Франции, Испании, Германии, Италии, Польши, Швеции, Чехословакии, Венгрии, Марокко и Родезии, преимущественно, дтя голоценовых слоев (Woli, 1938, 1939).
Га,Ш> Прежнее и современное распространение и численность млекопитающих 271
Остатки льва указывались из слоев ашеля и мустье Палестины (Picard,, 1937).
Громова (1932а) упоминала о зубах и метаподиях льва из древнегреческого склепа в Ольвии, но эти остатки могли происходить от ковра-нурн, привезенной с юга, а не ог зверя, добытого на севере Черноморского побережья.
Великолепные барельефные и горельефные изображения львов, наивные при раскопках древнего Шумера, Вавилона и Ниневии, свидетель-/ч лот о том, что львы были самыми обычными животными в Передней Ач и в Ш—I тысячелетиях до н. э. Ряд новых изображений львов на
Рис. 111. Череп тигрольва (самки) из бшпи адипского асфальта.
цуис, выполненных хеттами, в центральной части Малой Азии был роликовая Остином (Osten, 1929—1930).
На Кавказе лев известен пока лишь по изображениям и древним ли-Киитурньш сведениям. Имеются следующие заслуживающие внимания находки: рельефные изображения двух львов на золотой обкладке ста-к*на из курганов в Триалетп — II тысячелетие до н. э. (Куфтин, 1941): контурные изображения двух львов на золотой чаше из погребения в Ки-ровакане — II тысячелетие до н. э. (Пиотровский, 1949); контурные изображения львов на скалах в Кабристане — I (?) тысячелетие до и. э. (Верещагин п Бурчак-Абрамович, 1948); контурные изображения львов па серебряных сосудах (рис. 112) и скульптурные изображения в виде золотых бляшек (рис. ИЗ) из кургана близ Майкопа — II тысячелетие до н э. (Фармаковскпй, 1914); рельефное изображение льва па золотой Лэдке колчана из курганов Таманского полуострова.
Художественные изображения львов и львиц, терзающих ланей и куланов, имеются на золотой фиале из кургана Солох на южной Украине — VI в. дон. э. Манцевич (1949) пришла к выводу, что эта фиала сделана ионийскими мастерами, но совершенно напрасно называет куланов (вопреки нашему совету) лошадьми, а львиц — леопардами.
* Возможность того, что лев проник в ксеротермическую эпоху в Предкавказье п па Русскую равнину, не исключена. Погребова (1950) с полным основанием указала, что большинство художественных вещей и изображений зверей, найденных в скифских курганах, выполнено на месте — в Скифии — скифскими мастерами, а следовательно, и по местному ани-малпстнческому материалу. В подтверждение этому укажем, что в слоях
272
Часть II. Анализ происхождения четвертичных млекопитающих
древнего Танаиса была обнаружена плита местного грубого известняка (43x56 см) с рисунком льва. Камень этот хранится в Эрмитаже (Книпович, 1949).
Более поздние изображения льва имеются на многих памятниках армянских, арабских и иранских времен в Армении, Азербайджане и Грузии. Отметим изображение головы льва на стенах крепости старого Дербента, сделанное еще при арабском халифате, и львиных голов на воротах крепости в Баку, воздвигнутых в честь шаха Аббаса в XVI] столетии.
В средневековом искусстве горного Дагестана изображение льва встречается весьма часто (Башкиров, 1931). В Иране в эпоху сефевидов охотничьи ковры изготовлялись с рисунками львов (Кверфельд, 1940).
L____I___I____1___L_—
Рис. 1 12. Золотые бляхи, изображающие львов из Майкопского кургана.
Все эти ранние и более поздние изображения львов на Кавказе проще всего объяснить влиянием хеттской, ассирийской, вавилонской, древне-иранской, арабской культур и даже завозом готовых вещей с юга — из Передней Азии. Тем не менее сопоставление рисунков на скалах Кабристана и скифских вещах Предкавказья с историческими справками заставляет смотреть па дело иначе. Следует иметь в виду, что ареал европейско-азиатского льва неудержимо сокращался в историческую эпоху даже в Передней Азии.
Существуют указания Геродота и Аристотеля об обитании львов на Балканах во второй половине I тысячелетия до и. э. Лев фигурирует в поэзии Гомера (Илиада). Отряд греков в V в. до н. э. встретил в верховьях Евфрата фаунистический комплекс саванны Северной Африки со страусами, онаграми, от которых к XIX в. по осталось и следа (Ксенофонт, изд. 1896).
Обзоры прежнего распространения льва на Ближнем Востоке давались уже неоднократно на основе таких литературных справок (Sundevall, 1863; Keller, 1909; Громова, 1928; Korner, 1930).
О какой-то крупной кошке (льве?!), прыгнувшей па лошадь охотника-скифа, имеется сообщение у Лукиана Самосатского (125 г. и. э.) (Латышев, 1947—1948). Вероятность обитания какой-то крупной кошки в южных степях в древиеславяиские времена признавал, например, Житков (1936, стр. 229), разбирая зверовую терминологию — «лютый зверь» — в «Слове о полку Игореве» и в «Поучении» Владимира Мономаха. Следует указать, впрочем, что у Сементовского (1857) под именем «лютого зверя» подразумевается всего навсего волк. Признавая обычную неполноценность переводов, упомяпем о Моисее Каганкатвацп (X в. и. э.; изд. 1861 г., стр. 6), который писал: «Благословенна страна Агвап (т. с. древняя Албания, — Азербайджан Н. В.}. . . есть (в пей) и хищные звери: львы, тигры, барсы, дикие ослы, и множество птиц: орлы, соколы и подобные I
lit. Прежнее и современное распространение и численность млекопитающих 273
цу». Интересно, что упоминание о диких ослах получило подтверждение в виде костей кулана, найденных в 1946 г. при раскопках слоев поселения \]Ц в. в Баку. Отметим, и то восточное Закавказье в эпоху арабского «фата и позднее было вполне пригодно для обитания львов. Тугап \ Куре 11 Араксу, необозримые пространства тростников по разливам вМхтанп были обильно населены кабанами, оленями, а в открытой полу-пстынс паслись стада куланов и джейранов. Всхолмления в предгорьях Ьицамн можжевелово-фисташковых лесов и пышным злаковым покрс-
Рпс. 113. Золотая обкладка колчана из Семибратних курганов. Лев, терзающий оленя.
L также привлекали весной, летом и осенью стада тех же копытных, ц охотничьей оде «Шикария», посвященной сыпу Ширван-хапа Мену-fcnv. поэт Хагапп (Авсараддип), живший с 1120 по 1194 г. в Шемахе, Свевает охоту Ширван-хапа на льва, указывая па местность между Ьуегш Каспия и Шемахой (История Азербайджанской литературы, » 1. 1943. Текст Араслы, па азербайджанском языке). На фарепдеком -до слово «шпр» означает льва, и можно думать, что вся область Шир-Ы получила свое имя не только от правивших ханов, ио и от обитав-«в ней львов. Пизами Гапджеви (XI—XII вв. и. э.) в поэмах «Искан-«Семь красавиц» и других постоянно давал художественные ШИЧ1ПЯ со львом. Усама иби Мункыз (изд. 1922—1923) описы-«охоты на львов в Сирии и Палестине в XI [ в.; причем львы были там обычны, как у пас зайцы.
8конце XIX столетня Блэпфорд (Blandford, 187G) указывал, что львы «вымерли па севере Ирана, по еще живут в Месопотамии, на западных
и Н. К. Верещагин
274
Часть II. Анализ происхождения четвертичных млекопитающих
отрогах Загроса п юго-восточнее Шираза. Отметим, что персидский лев — Р. leo persica Meyer, 1826, был описан по экземпляру из Тегерана, т. е. из северо-западного Ирана.
По сообщению Шардена (Chardin, 1735, стр. 51), львы п тигры еще водились в лесах Мингрелии и Имеретии в начале Х\ II в. Живший в начале прошлого столетия в Армении Шопен (1852) указывал, что в его время львов уже не было, но из-за Аракса изредка забредали тигры. Денфорд и Эльстоп (Daniord a. Alston, 1880) упоминали об истреблении львом лошадей в 1873 или 1874 г. близ Биледжика, на верхнем Евфрате. По сообщению Киннера (Kinnear, 1920), львов уже не было в конце прошлого столетия в Малой Азии, но они еще жили в Месопотамии и Аравин. Однако в результате массового завоза нарезного оружия на Ближний Восток во время первой мировой войны количество азиатских львов быстро сократилось. По мнению Пококка (Рососк, 1930, 1939) и Гарпера (Harper, 1945), быстрое вымирание азиатских львов было всецело обязано европейской колонизации и азартной охоте английских офицеров, убивавших со скуки ио нескольку десятков этих кошек за время пребывания в Азии.
Учитывая все сказанное, можно думать, что лев действительно обитал и в Закавказье, проникнув сюда в голоцене с юга, а к концу средневековья уже исчез, постепенно отступив на юго-восток и юго-запад (карта 33). В этом отношении имелась почти полная аналогия с сокращением численности и ареалов тигра и полосатой гиены, которые продержались здесь до первой половины XX столетия.
Тигр — Panthera tigns L. Ископаемые остатки настоящих тигров известны, по-видимому, лишь из пещер Китая и Явы (Wolt, 1938). Указания на находки во Франции и в Сибири относятся, вероятно, к остаткам тигролова. Не найдены остатки тигров и в среднеазиатских палеолитических пещерах: Тешик-Таш, Аман-Кутап.
Для понимания истории тигра па Кавказе наиболее знаменательно отсутствие его остатков в Бинагадинском захоронении.
Основная часть ареала тигра лежит ныне в пределах юго-восточной Азин. Сопоставляя все это, следует считать современного тигра на Кавказском перешейке таким же послеледниковым вселепцем, как шакала, гиену п льва. Тигр распространялся из юго-восточной Азии в северо-западном направлении, очевидно, лишь в позднем голоцене. Характерно, что в искусстве древних пародов Междуречья он не отражен, но был уже знаком средневековым иранцам, хотя и не пользовался столь большой популярностью, как лев.
Изображения тигра попадаются изредка в древиеиранской художественной миниатюре. В 20-х годах XVII столетия тигра видел в шаховом зверинце в Казвине и красочно описал русский купец Федот Котов (1852).
Обзор и критика сведений о распространении тигра на Кавказе в конце XIX и начале XX столетия были даны Динпиком (1914а), Сатуни-пым (1915а), Огневым (1935).
Мною (Верещагин, 1947) была дана карта ареала тигра в Закавказье и пунктов его добычи в последние десятилетия. Восточное Закавказье было до недавнего времени крайним западным участком ареала тигра.
В 60-х годах прошлого столетия тигр был столь обычен под Ленкоранью, что за год здесь убивали несколько штук (Радде, 1899). Тигр попадался довольно часто и в конце 90-х годов па Ленкоранской низмен-пости и в горных лесах Талыша. Тогда ежегодно убивали здесь по крайней мере одного тигра (Сатунип, 1915а). Но в начале нашего столетиями-
Гл. IIP Прежнее и современное распространение и численность млекопитающих 276
елейность тигра под влиянием постоянного преследования стала быстро падать. Уже в 20-х годах тигры (рис. 114) только заходили в пределы военного Закавказья из лесов южного побережья Каспия, но пе жили здесь
(О'д-Ю.
’ С1920 по 1950 г. отмечено только два случая добычи тигров: в 1922 г. —
, i Тбилиси и в 1932 г. — в Талыше (карта 33).
Советские офицеры, жившие в 1942—1943 гг. в Гнляне и Мазандеране, |йбщалн мне, что им приходилось неоднократно слышать о тиграх и
Рис 114- Тигр в ленкоранских джунглях. Первая половина XX в.
нно наблюдать их следы, растерзанных кабанов и домашних животных • досах северного склона Эльбурсского хребта.
Энергичная вырубка горных и низовых лесов, истребление кабанов — LioBHoii пищи тигра и прямое преследование пе создают условий для су-Л^ств вапия этого зверя на Кавказе и в будущем.
рарС __ Panthera pardus L. (s. lato). Остатки леопарда отмечены для «доо.титическпх слоев многих пещер Африки в Индии.
В пещерах Сирии, Палестины, Ливана остатки этого вида прослежены ।слоях с ашеля до неолита (Picard, 1937; Bale, 1937); в центральном Праве пандепы в палеолитических слоях пещеры Бнзитун (Coon, 1952); юиицер Западной Европы описаны остатки близких форм (Wolf, 1938). В теплые фазы плейстоцена в Западной Европе мог обитать и леопард .^•мешюго африканско-азиатского типа. В плейстоценовых слоях Ьпочной Европы остатков леопарда пе найдено. Это свидетельствует Ьш взгляд о том, что в плейстоцене северная граница распространен « гря проходила южнее, па Кавказе и в Конет-Даге.
Liiniciвеппая находка костей плейстоценового барса па Кавказе Кт в палеолитических слоях пещер Кударо I и III в верховьях репа. „
Из голоценовых захоронении остатки барсов, быть может и завезеп-,л, достоверно известны теперь для греческих и римских слоев Ольвии
'276 Часть II. Анализ происхождения четвертичных млекопитающих
Рис. 115. Бронзовые головки леопардов из курганов Украины.
п древней Феодосии (коллекции Института зоологии АН УССР и ЗИНа), а в пределах Кавказа из мезолита грота Сосруко па Баксане и из пещеры «Длиннокрылой» в теснине Аракса.
Отсутствие барса в среднеплейстоценовой фауне Апшерона позволяет думать, что он появился вначале на центральном Кавказе, проникнув сюда вместе с баранами и дикобразом с юга.
В ранненсторпческую эпоху барс был распространен широко. В равнинном Предкавказье он был известен скифам. Художественные изображения голов барса из бронзы найдены в скифских курганах юга Русской равнины и в Предкавказье (рис. 115). Из Келлермесского кургана извлечена золотая фигурка лежащего барса. Великолепная бронзовая статуэтка барса найдена также недавно в раскопках в Самтавро, западнее Тбилиси (см. репродукцию в Вестнике древней истории, 1948, № 3).
В начале нашего столетия распространение барса было ограничено предгорными и горными районами Большого и Малого Кавказа, а также низменностью восточного Закавказья (Динник, 1914а; Сатунип, 1915в). В последнее полустолетие шло быстрое сокращение числа барсов.
За четыре года—с 1894 по 1898г,— в районе Кубанской охоты егерями было добыто 11 барсов. В 1904г. там же было добыто 2 барса, а Вообще на Главном хребте в нашем
столетии барс был уже очень редок. В 1912 г. 2 барса были убиты в районе Закаталы, а после этого в ущельях левых притоков Алазани наблюдались уже только следы проходных одиночных особей хищника, несмотря па обилие на этом участке козлов, сери, оленей. В Северной Осетии последние 2 барса были убиты в начале 20-х годов, по позднее появились вновь, и в 1949 г. 1 барс был убит в ущелье правого притока Сунжи. В Южной Осетии 1 барс был убит в 1952 г. близ Сталипирц,
В Талыше и Карабахе барсы более часты и живут оседло. В 50-х годах прошлого столетия Радде получил по заказу за 7 педель 12 свежих барсовых шкур в Ленкоранском уезде. С 1930 по 1940 г., т. е. примерно через 70 лет, во всем Азербайджане было принято на пушные базы всего 2 шкуры барса, да небольшое число гпкур было скуплено частными лп-
в 1905 и 1906 гг. — уже ни одного.
цами.
Па всем Кавказе за упомянутое десятилетие было добыто не более 10 барсов, преимущественно па Запгезурском хребте и в Талыше. С 1929 по 1932 г. были известны регулярные заходы барсов в камыши центральной Муганп, например на оз. Махмуд-Чала. Близ сел. Хармандали в 1929 г. жителями была найдена мертвая и будто бы закопанная самцом самка барса. Барс самец, свалившийся в колодец во время снежной бури, был убит близ сел. Маштаги на Апшеропском полуострове 15 февраля 1946 г. (Бурчак-Абрамовпч и Джафаров, 1949). Большие снегопады и метели согнали тогда с гор этого зверя.
рд.'Ш. Прежнее и современное распространение и численность млекопитающих 277
Встречи и добыча барсов неожиданно участились после второй мировой войны. Свежие шкуры барсов начали поступать из Талыша, пз Нахичеванского края л из Карабаха. Так, в 1947 г. в Азербайджане было
принято 5 шкур, в 1948 г. — 36 шкур.
Такое увеличение численности объясняется, очевидно, переходом зверей пз северо-западного Ирана. Современное постоянное распространение барсов па Кавказе ограничено горными лесами Талыша, хребтами
I арабаха, Запгезура и Нахичеванского края (карта 34).
Барсы, обитавшие на Большом Кавказе, относились к бледно окрашенному подвиду Panthera pardus cis-Viucasiciis. Теперь это прак-гически вымершая форма. Закавказские Р. pardus tul-ims Vai. (рис. 116) более яркие, с большей примесью ржавых тонов, имеют низкий мех. К ним принадлежал и барс, убитый на Апшероне. Быстрое исчезновение барсов на Большом Кавказе в пашем столетни является, несомненно, результатом настойчивого преследования с ирезным оружием. При отсутствии запретов па добычу барсов этот зверь в ближайшие десятилетия совершенно кчезнст па Кавказском пе-
Рис. 116. Леопард трехмесячного возраста из Талыша. Типичная пятнистость молодого зверя.
Фот. автора, 1947.
решейке.
Рысь — Felix lynx L. (s. lato). Близкие виды подрода Цпх, Pardina по крайней мере среднеплпоценовой
древности и известны из верхнетретичных отложений южной Европы: Молдавии, одесских катакомб, Франции, Италии. Из нижпеплиоцепо-вых отложений близ Ставрополя извлечена нижняя челюсть кошки,
близкая по структуре зубов к мелкой рыси.
Остатки рыси современного типа весьма характерны для раине- и нозднепалео литических стоянок Западной и Восточной Европы, рас подавшихся в пределах лесной и лесостепной зоны (Wolt, 1938, 1939; Громов, 1948; Ппдопличко, 1951). Па Кавказе они обнаружены в пещере Сакажиа (западное Закавказье).
В палеолите восточного Средиземноморья — в Ливане, Сирии -катков рыси не найдено (Picard, 1937; Bate, 1937), так же как п в Иране iQhmi. 1952). Это дает основание утверждать, что и в плейстоцене распространение рыси в Передней Азии далеко па юг не шло. Современный ареал рыси кругополярпый и связан с равнинными и горными лесами.
В голоцене он был сильно сужен при преследовании зверя человеком, а главное — обезлесением. Интересно, что в приречных лесах степной юны рыси не держатся и к югу далеко не идут и ле шли, несмотря на обн-
278
Часть II. Анализ происхождения четвертичных млекопитающих
лие нищи— зайцев, бобров, косуль. Возможно, что дело здесь в весенних разливах, и именно поэтому остатки рысей крайне редки в отложениях рек низового Урала, Волги, Дона.
Современный ареал рыси — F. lynx orientalis Sat. на Кавказе охватывает всю лесную зону Большого и Малого Кавказа и Талыша (Динник, 1914а; Сатушш, 1915а; Верещагин, 1947г) (карта 35). В смежных районах Турции и Ирана рысь встречается в островках горных лесов. Вообще в горных районах эта кошка держится даже при наличии лишь небольших кустарников. Так, например, на восточной оконечности Большого хребта рысей порядочно в кустарниковых зарослях и мелколесье близ селений Кызыл-Бурун и Хизы, где многочисленны зайцы. Встречается изредка рысь и в безлесном Дагестане, а также в небольших островках дубовых лесов Армянского плоскогорья, например у Шалбуза, в верховьях Нахичевап-Чая. Существует рысь и в почти безлесном ландшафте современной Палестины (Bodenlieimer, 1935). Вообще в южных горных
странах рысь не может служить показателем исключительно лесных ландшафтов, как думает Пидопличко (1951). По происхождению рысь следует относить к древним местным средиземноморцам. В последние десятилетия добыча шкур рыси па Кавказе колебалась в довольно широких пределах как в Предкавказье, так и в Закавказье. Достоверные подъемы численности имели место в 1928—1929, 1932—1933, 1938 и 1950 гг. (график 15). Трудности промысла в горах ограничивают возможности истреб
ления зверя.
Лесной кот — Fells silvestris L. Остатки мелких кошек/1, pygmaea Lartet, F. media Lartet, F. attica Wagner и других близких к лесному коту видов известны из миоценовых и плиоценовых отложений Италии, Австрии, Греции, в частности пз пикермийской фауны. Остатки современного вида обычны в плейстоценовых и голоценовых слоях огромного числа пещер всей Западной Европы, за исключением Скандинавского полуострова (Trouessart, 1898—1899а; Wolf, 1938). Обычны они в слоях пещер Крыма, датируемых средним и верхним палеолитом (Пузанов, 1929; Громов, 1948). На Русской равнине нам известна находка нижней челюсти мелкой кошки плейстоценового типа сохранности в русловых отложениях низового Дона (Цимлянская). На Кавказе найдено несколько костей этого кота в палеолитических слоях Ахштырской и Цебельдинских пещер. На юго-востоке Средиземноморья остатки лесного кота отмечены в слоях пещер Палестины и Сирии — от ашеля до неолита (Picard, 1937* Bate, 1937).
Голоцеповые остатки лесного кота найдены нами только в Пятигорье. Интересно, что в средневековых слоях крепости Саркел на Допу обнаружены остатки только домашних кошек. В связи с этим создается впечатление, что ареал кавказского подвида — F. silvestris caucasicus Sat. не соединялся в голоцене с европейским через Русскую равнину.
Современный ареал лесного кота охватывает Западную Европу, Малую Азию, Кавказ. В Крыму оп вымер, очевидно, в голоцене. В пределах Кавказа распространение лесного кота постепенно уменьшается в восточном направлении (карта 3G). Из Талыша и Эльбруса документальных находок пока нет, а неясные литературные данные о какой-то кошке в лесах Ленкорани, Гиляна и Мазандарапа (Бламберг, 1853; Смирнов, 1922; Бобринский, Кузнецов, Кузякин, 1944) не подтверждены. Связи с лесными биотопами у этого вида прочные, но в Предкавказье дикий кот довольствуется и небольшими уцелевшими участками кустарников, леса и тростников. Такие примеры имеются в равнинной Кабарде, по долине
Ги Ш Прежнее и современное распространение и численность млекопитающих 279
? Кумы, на Ставропольской возвышенности и в дельте Кубани. ! а юге страны он связан с лесными массивами значительно теснее, а па Ьпесепном нагорье Малого Кавказа практически отсутствует. В западни Закавказье кот многочислен в буковых лесах Аджаро-Имеретинского ’ та и в ольховых топях Колхиды. При разнообразном рельефе нашей luuoii страны распространение кота не показывает зависимости от высоты усового покрова, которая была помечена Формозовым (1946) на равни-Ьг 1? подчеркнута Пидопличко (1951).
Наибольшая численность лесного кота наблюдается в нижней трети горного леса. Предпочитаемым типом угодий являются буково-грабовые леса. По учетам в Кавказском заповеднике, из 43 встреч котов приходилось на широколиственные леса, 18.6% — на темнохвойпые "‘о — субальпийскую зону (Теплов, 1938а). Основная масса шкур и вверпом Кавказе добывается в предгорных лесах.
Итак, но палеонтологическим данным лесной кот* автохтон Среди-г । морья, а по особенностям современного ареала и экологии на Кав-и. его следует считать ранним плейстоценовым вселенцем из Европы. Ьдакновепие его на Кавказ происходило скорее всего через Балканы । Чалую Азию.
Промысел лесного кота за последние четверть века при мелких коле-а иях в числе добываемых шкур не оказал пока заметного влияния на Кипе численности вида (график 16).
Камышовый кот — Fells chaus Giild. Ископаемые остатки этого кота пз-। тш из плейстоценовых слоев Индии (Wolt, 1938). В Палестине и Си-и: 1 они обнаружены во всех слоях — с среднего мустье до неолита (Pi-ид!, 1937), предположительно показаны В. И. и В. И. Громовыми (1937, - 86) для азильских и тарденуазских слоев пещер горного Крыма.
азаипяна находку остатков в Западной Европе, например в травер-1ак близ Штуттгарта, вряд ли заслуживают внимания. На Кавказе 1 лютоцеповые и голоценовые остатки этого вида неизвестны. Находки Коцеповых остатков на Русской равнине также недостоверны. В слоях ш-хш столетий крепости Саркел у Цимлянской обнаружены 2 Liinx эпифиза плечевых костей кошки, близкой по размерам к камышовому коту.
U слоях славянского поселения XIV столетия в Полтаве был обнаружен пижнпй эпифиз плеча кошки, близкой по размерам к этому виду 11, Громов, 1948), но достоверность и этого определения невелика. Наши ждайпше исследования ископаемых остатков кошек показали, что етвертнчном периоде в Крыму и на юге Русской равнины жил пятнистый кот — Fells lybica, а не камышовый. В восточное Предкавказье и па юг Русской равнины камышовый кот проник, очевидно, лишь в голоцене.
В настоящее время основной ареал камышового кота находится в южной и юго-восточной Азии, а на Кавказском перешейке оказывается «пе-пзвитым». Камышовый кот обитает в восточном Закавказье от уровня Ьслпя до 900—1000 м абсолютной высоты. Па запад он распространен Jjfcinne Куры до Гори, а за Сурамом уже неизвестен. По долине Аракса тцышовый кот встречается до долготы Алагёза. По побережью Каспия и)ычен, особенно близ Самура и по низовьям Сулака и Терека, проникая далее по камышам вдоль Бэровскпх бугров и в дельту Волги (Доброхотов, 1939).
В восточном Предкавказье этот кот встречается до Грозного — но долине Сунжи, и до Моздока — по долине Терека (карта 37).
2S0
Часть II. Анализ происхождения четвертичных млекопитающих
В западном Предкавказье камышовый кот теперь не водится. Поступление шкур из Краснодарского края объясняется завозом их с юга-востока. В научных коллекциях из этого района камышовый кот неизвестен, и литературные указания, начиная с Богданова (1873), па обитание этого кота в дельте Кубани никем пока пе подтверждены.
Сведения Формозова (1946) об обитании этого кота в камышах низовой Кубани ошибочны, так как шкура и череп из-под станицы Грпвенской. на которые ссылается этот автор, принадлежали на самом деле лесному коту.
Наибольшая численность вида наблюдается в низовых лесах восточного Закавказья. Излюбленные биотопы — это тростниково-рогозовые крепи болот п озер, тугайные заросли вдоль рек и речек, где кот охотится
Рис. 117. Челюсть гепарда из бинагадинского асфальта.
на водяных крыс и болотных птиц. В горных лесах отсутствует, ио предпринимает переходы и по открытым местам закавказской полупустыни.
Проникновению в западное Закавказье мешают какие-то факторы, точнее пока неизвестные. На первый взгляд кажется, что в ландшафтах Колхиды, весьма сходных с гирканскими, при обилии тепла и болот камышовый кот мог бы процветать наравне с шакалом. Одной «молодостью» ареала кота на Кавказе объяснить его отсутствие в Колхиде трудно, так как Сурамский перевал невысок, а расселение хищников может идти весьма быстро. Отсутствие остатков камышового кота в Бинагадинской захоронении, «неразвитость» ареала в пределах перешейка, наконец, несомненная стенотопыость животного и относительиая теплолюбивость позволяют утверждать, что это — последпиковый вселепец южного происхождения. Площадь угодий, где обитает камышовый кот, увеличивалась на низменностях Азербайджана при развитии туземных оросительных систем и сокращалась при их забросе. В последнее время численность и ареал его уменьшаются в связи с уничтоженном тугайных лесов, тростников и интенсивным промыслом. Проследить динамику численности впда по добыче шкур трудно, гак как в сводках объединяются шкуры этого вида и леспого кота.
Назначение в 1948 г. высокой премии за добытого кота — вредителя путриеводства — резко повысило выход шкур, показав тем самым благополучие численности впда.
Гепард — Acinonyx jubalus Schr. Ископаемые остатки гепарда известны из верхнего плиоцена южной Европы и Африки (Simpson, 1945).
Прежнее и современное распространение и численность млекопитающих 281
В четвертичных отложениях остатки гепарда крайне редки и до недавнего времени были известны лишь из верхнего плейстоцена Китая (Pei, 1939). Па Кавказе они (рис. 117) были впервые обнаружены в средиеплей-стоценовых слоях Апшеронского полуострова (карта 38). В ту эпоху ге-[ард (рис. 118) мог охотиться здесь па сайгаков, ослов, лошадей и зайцев. Возможно, что его распространение в плейстоцене простиралось i в восточное Предкавказье. Современный ареал гепарда занимает Африку и южную часть Азии вплоть до Китая. Предпочитаемые биотопы, судя по сведениям Рузвельта, а также Поккока (Рососк, 1939, 1941)
Рис. 118. Гепард в плейстоценовой полупустыне восточного Закавказья.
и других охотников п натуралистов, — равнинные и холмистые саванны, полупустыни п пустыни. Находит приют в скалах, кустарниках. В Туркот и Иране держится в открытой песчаной, глинистой и каменистой пустыне, но все же предпочитает долины рек с тугайными зарослями, пз которых выходит па охоту за газелями и зайцами.
Документальных данных об обитании гепарда на Кавказе в историческое время пет. Изображение этой кошки (рис. 119) имеется на орнаменте серебряного сосудика из Майкопского кургана (Фармаковский, 1914). Наличие ошейника указывает па вероятность приручения гепардов уже в скифскую эпоху. На керамике XII в. и. э. из раскопок Старой Гянджи имеются изображения гепарда, несколько напоминающие древнеегипетские.
Сведения об обитании гепарда в восточной Грузии — Картли — в средине века имеются в грузинских летописях «Картлпс Цховреба». Отсюда эти данные, очевидно, попали в художественную литературу, причем некоторые местные писатели (К. Гамсахурдиа. Десница великого мастера. 1945) обнаруживали весьма курьезные познания об этом звере, приписывая ему обитание в высокогорье. Об охотах на малоазийских га-«мем с гепардами в XII столетии в Сирии и Палестине постоянно упоминал Усама ибн Мункыз (изд. 1922—1923 гг). В русских летописях, а также в «Словно полку Игореве» часто упоминаются «пардусы», т. е.
282
Часть II, Анализ происхождения четвертичных млекопитающих
ловчие «леопарды» — гепарды, которые служили русским князьям в XIXII столетиях для охоты (Житков, 1936). Способ этот был заимствован, по мнению Житкова, у персов или половецких ханов. Все это очень правдоподобно, тем более, что военные, культурные и торговые связи кочевников Русской равнины с Предкавказьем и Закавказьем имели место еще с века бронзы.
Пидоплпчко (1951, стр. 178) вслед за Аристовым (1866) ошибочно приписал все эти древпеславянскпе сообщения о «пардусах» леопарду, который непригоден для охоты. У азербайджанских ханов, армянских и кар
ем
Рис. 119. Изображения гепардов на серебряном сосуде из Майкопского кургана (По Фармаковскому, 1914).
тлийских князей охота с ручными гепардами просуществовала, по-видимому, до XIV столетия. Иосаф Барбаро видел в 1474 г. у армянского князя 100 охотничьих гепардов.
Адам Олеарий (изд. 1870 г.) сообщал, что в его присутствии персидский шах травил гепардами онагров в исфаганском зверинце. В XVII столетии в Гиляие и Мазандарапе гепардов было еще много. По этому поводу Олеарий писал (стр. 935): «Там (в области Гплян, — II. В.] водится много и хищных зверей, как например тигров, леопардов, волков и медведей. Тигров (гепардов, — Н, В.) здесь такое множество, что их привозят стадами на продажу по 10—20 голов. . . Обыкновенно тигров приучают к употреблению на охоте потому, что они чрезвычайно быстро бегают и делаются очень ручными, так что охотник свободно сажает тигра позади себя на лошадь». Имеются позднейшие сообщения о гепарде в Мазандеране и вообще в прикаспийских лесах (Filippi, 1865; Blanford 1876).
В Малой Азии и Аравии гепард был уже редок в конце прошлого и начале текущего столетия (Daniord a. Alston, 1880; Aharoni, 1930; Bodenhcimer, 1935).
Гепард, вероятно, продержался в Кура-Ара ксииской низменности и в долине среднего Аракса до XVIII столетия в диком состоянии. В этом убеждают быстрое сокращение ареала и численности гепарда в новейшее время в Туркмении и его недавнее обитание в северном Иране (южном Азербайджане).
Исчезновение гепарда в Закавказье и в Передней Азии объясняется уменьшением численности степных копытных, а также интенсивным отловом молодых гепардов для дрессировки.
fj, [II: Прежнее и современное распространение и численность млекопитающих 283
Гарпер (Harper, 1945), прослеживая современный ареал гепарда | в Азии и Африке, констатирует повсеместное сокращение ареала и численности этой своеобразной кошки.
I Отряд ЗАЙЦЕОБРАЗНЫЕ — LAGO М ОRP Н А
Удвоешгорезцовые довольно древние. Некоторые роды отряда, ^пример Mytonolagus, Shamolagus, известны уже с эоцена.
В пределах Средиземноморья, Средней Азин и Монголии третичные Ецацеобразные представлены пищухами и зайцами (Trouessart, 1904— 1905: Simpson, 1945; Борпсяк и Беляева, 1948).
На Кавказском перешейке известны остатки представителей только дб-.х родов — Ochotona и Lepus, Они появляются с нижнего плиоцена.
Сем. ЗАЯЧЬИ — ILEPO1HDAE
Заяц-русак — Lepus europaeus (s. lato). Наиболее древние фрагментарные остатки мелких зайцев обнаружены в плиоценовых речных от-лонениях близ Ставрополя, а более поздние — в плиоценовых озерпых Сложениях средней Занги, севернее Еревана.
Многочисленные остатки зайца-русака из среднего плейстоцена ниточного Закавказья (Бинагады) дают уже представление о виде, ) Ссьма близком к современному.
Верхнеплейстоценовые остатки русака обнаружены и в прирусловых сложениях низового Допа. В голоценовых захоронениях Кавказа остатки русака не представляют редкости как в слоях человеческих I мелений, так и в пещерках, обитаемых филинами.
Современный р>сак населяет практически весь Кавказский перешеек, ।ареал его уходит иа север и на юг. Южная граница неясна, но, вероятно, шходит до Междуречья.
Многолетние данные государственных заготовок показывают такое мсп ре деление выхода шкур русака по Кавказу на 1000 км2 (Верещагин, Ц47 г): Северный Кавказ — 3300; Азербайджан — 770; Дагестан — 594; Грузия — 219; Армения — 127.
Действительный убой русаков в Закавказье, особенно в Азербайджане, значительно выше, так как большинство шкурок «оседает» па местах. Добыча иа Северном Кавказе сконцентрирована преимущественно в Краснодарском крае и в равнинном Дагестане. В Грузии почти все зайцы добываются в ее восточной, сухой части (карта 39).
На альпийских и субальпийских лугах Большого Кавказа численность зайцев ничтожна. Более обычны они в сухих внутренних долинах северного склона хребта, а также на остепненпых высокогорных лугах внутреннего Дагестана. Па Гунибском плато русаки залегают на день в уцелев-дих сосновых и березовых рощах.
В сплошных массивах горных буковых, грабовых и пихтовых лесов траков нет совершенно. Правда, они проникают в лесную зону с субаль-шшскпх лугов вниз по обезлесенным склонам и гребням хребтов, а по ^сушенным вырубкой ущельям — снизу вверх — в лесную зону, по >го*происходит большей частью при изменениях ландшафта, вносимых человеком.
Вообще при рассматривании современного распространения русака на широких пространствах Кавказского перешейка становится ясно, что
284 Часть II. Анализ происхождения четвертичных млекопитающих
это — животное сухих нагорий и умеренно сухих равнин с кустарниками или редколесьем.
Что касается распределения русака по биотопам, то здесь следует различать места дневок — лежек — и места жировок. Они совпадают далеко не всегда. Кроме того, природное распределение русака на лежке и жировке повсеместно резко изменено на протяжении тысячелетий деятельностью человека и домашних животных.
В равнинном Предкавказье русаки кормятся на открытой целинной степи лишь весной и в первой половине лета, где и залегают. При высыхании, выгорании степей зверьки концентрируются по долинам речек. В осенне-зимнее время они кормятся на выдувах, по буграм и южным склонам и залегают в балках и кустарниках.
В окультуренных районах и в районах интенсивного выпаса скота размещение русака сильно усложнено. Почти все типы окультуренных угодий в Ставрополье отвлекают зайцев с целины, особенно в тех случаях, когда последняя используется для выпаса скота. Наибольшее значение в качестве кормовых угодий для русаков имеют посевы люцерны и других |трав. Привлекают их также созревшие дыни на бахчах и зерно пшеницы и ячменя на жнивье (Колосов и Бакеев, 1947, стр. 12). Кормежка зерном на жнивье развилась, видимо, недавно в результате крайне небрежной уборки хлебов комбайнами.
Зимой русак кормится на озимях, где и залегает. Часто ложится он также на зяби, обязательно пересекая при преследовании ближайшую лесополосу. Постоянные жировки и залегание русака в самих лесополосах наблюдаются лишь при большой ширине насаждений и при значительном возрасте их. В снежные зимы русаки оказываются серьезными вредителями молодых посадок.
Современное широкое распространение русака в предгорьях, особенно на Закубанской наклонной равнине, обусловлено деятельностью человека, истребившего леса и осушившего болота.
В западном Закавказье русак редок, держится по наиболее сухим участкам черноморских террас, а в сырой Колхиде — лишь по расчищенным полянам предгорий.
В восточном Закавказье наибольшая численность русака наблюдается в сухих предгорьях и по склонам холмов Карталинского, Иор-ского и Аджииоурского плоскогорий в формации де ржи-де рева и бородача. Только появление зимой больших стад овец с овчарками временно вытесняет зайцев из этого типа угодий (Верещагин, 19426). Много русака и по изреженным тугаям, особенно в зоне тамарикса. В Алазано-Агричайской и Хачмасской низменностях русак держится там, где первобытные леса уничтожены вырубкой, залегая на лежку по окраинам лесных полян. В ровной полупустыне Кура-Араксинской низменности русак исключительно редок даже в солянковых и каперцовых группировках. Распашка полупустыни и устройство полей, садов, оазисов сразу же обусловливают появление зайца. Особенно постоянно держится закавказский русак в виноградниках и на старых залежах. Посевы люцерны и хлопка также привлекают русака, исключая время поливов и прополки хлопковых рядов.
На Ашперонском полуострове заяц держится, кроме виноградников, и по дюнным всхолмлениям, залегая среди кустов эфедры, песчаного овса. Зимой при преследовании охотниками и при выпасе овец в виноградниках русак залегает на поднятом паре и на озимых посевах ячменя. Здесь, очевидно, имеет значение микрорельеф, так как спрятаться в бо-
1
I в б
о в м
м
6i of
Л(
щ 19 ка
ту ло и
СО1
НО1
в д
par на к (
Ми
вит
1 Из
< впе] стог пле! У к че в уг кавв стол нахо близ
IV. прос
i. Прежнее и современное распространение и численность млекопитающих 285
I роз чу или среди крупных комков земли легче, чем на озими. В виноград-
никах зайцы охотно поедают созревший виноград и грызут арбузы на г бахчах, утоляя жажду.
> 1ид воздействием охотников за последние десятилетия на Апшероне
' овалась популяция русака, особи которой залегают на день на их каменных заборах или на развалинах домов и уже издали за-|.чечают опасность.
По предгорным равнинам зайцы залегают на сухих руслах рек, маскируясь среди валунов и легко уходя здесь от шакалов и лисиц.
F нагорье Талыша и в долине среднего Аракса основной природный 1 и русака — это днища и склоны ущелий с зарослями тамарикса,
облепихи и крушины. Вообще этот южный мелкий русак весьма на-i поминает повадками среднеазиатского толая.
- j Круглогодичное размножение и наибольшая численность на юге, сокра-, • щенке числа пометов по мере продвижения на север (Верещагин, 19386,
4 ; 19426; Колосов, 1941; Колосов и Бакеев, 1947) характеризуют русака
как весьма пластичный вид южного, средиземноморского происхождения.
1 ’ Колебания численности русака происходят на Кавказе под влиянием
i туляремийных эпизоотий, кокцидиоза и реже ранних заморозков (Ко-
t и Бакеев, 1947). До известной степени эти колебания сказываются
промысле (график 17—18).
1 । к последние годы наибольшее значение для резкого повсеместного
> сокращения численности зайца имеет круглогодичное браконьерство — ночнне охоты с автомашин.
г В культурных ландшафтах Кавказского перешейка русак будет i в дальнейшем одним из наиболее ценных охотничье-промысловых зверьков.
Отряд ГР ЫЗ У НЫ— К О I) JEW TIA
11 Из кайнозойских отложений Кавказского перешейка и с юга Русской равнины теперь известно свыше 40 видов грызунов. В голоценовой фауне ; На этих территориях существовало не менее 50 видов, относящихся j к семействам Scinridae^ Castoridae, Myoxidae, Dipodidae, Spalacidae, ’ tfuridae, Cricetidae, Hystricidae.
L Ниже рассматриваются особенности ареалов их некоторых представителей.
Сем. БЕЛИЧЬИ — SCIUHIDAE
- В четвертичной фауне Кавказа известно всего 4 вида беличьих. ’ f Из них вымер сурок, а живут 2 суслика и кавказская белка.
Сурок— Marmota sp. В 1957 г. по находкам в пещере Кударо I стало впервые известно, что какой-то сурок жил на Большом Кавказе в плей-1 стоцене и голоцене. Затем кости сурков были обнаружены и в верхне-плейстоценовых отложениях в бассейне р. Храми на Малом Кавказе v сел. Орозмап. Несколько довольно свежих черепов сурков, близких | к черепам байбаков, было извлечено геологами из карстовой воронки в ущелье р. Белой (карта 40). Не исключена и возможность того, что кавказский сурок вымер на северо-западном Кавказе в начале нашего столетня (Верещагин, Гептнер, Строганова, 1959). Особняком стоит находка мелкого сурка — Marmota(l) sp. в нижнечетвертичных слоях близ Ахалкалаки.
Малый суслик — Citellus pygmaeus Pall. (s. lato). Изучением pac-L простра нения и происхождения малого суслика на Кавказе занимались j
*
1
286
Часть II, Анализ происхождения четвертичных млекопитающих
фаунисты, зоогеографы, паразитологи, но и до сих пор эти вопросы остаются недостаточно выясненными.
Реальные факты палеонтологии, морфологии, зоогеографии и экологии тем не менее могут продвинуть вперед поставленную задачу.
* Прежде всего род Citellus миоцено-плиоценовой древности для Север-
ной Америки и по крайней мере плиоценовый для Евразии (Simpson, 1945),
Остатки сусликов типа С, pigmaeus были определены Бюхнером, а также Пидоплпчко (1951) из нижнечетвертичных отложений у Но-гайска Запорожской области.
Верхнеплейстоценовые остатки малого суслика достоверно известны теперь из пределов современного ареала на равнине Восточной Европы. Виноградов и Громов (1952) различали более древнюю форму — С. pyg-maeus muslcoides, и более молодую — С. pygmaeus caspicus, подчеркивая, очевидно, сохранение каких-то архаичных черт у горного подвида.
Из раниеголоцеповых, мезолитических слоев остатки суслика из-Г вестны теперь из первой теснины Баксана. Поздние костные остатки | малых сусликов из погадок филина собраны мной на окраинах Кавказ-| ского участка ареала: в верховьях Калауса, в Ставрополье и под Г Махачкалой.
? Современный ареал малого суслика расположен на равнинах Восточ-1 ной Европы, северного Крыма, Предкавказья и северных частей Средней Азии, в степной, полупустынной и пустынной золах.
В связи с неогеновой древностью этой территории есть основание j считать ее родиной вида, не прибегая к гипотезе перекочевки его предков откуда-то с юга через Кавказ, как это сделал Пидопличко (1951).
На равнинах Предкавказья юго-западная граница малого суслика пересекала в начале 50-х годов нашего столетия Манычскую впадину в низовьях Егорлыка и, огибая Ставропольскую возвышенность с севера, вдавалась далеко к юго-востоку, в верховьях Калауса, к сел. Султанскому, не доходя на 15—18 км до станции Курсавка. Далее на восток граница южных колоний выходила на Куму у Воронцово-Александров-ского и шла параллельно широтному течению Терека. От устья Терека на север тянется сплошная область распространения грызуна. Обособленная колония имеется между низовьями Терека и Сулака.
На правобережье Сулака суслик распространен по предгорьям от I Чир-Юрта вплоть до Буйнакска, а по приморским террасам Каспия проникает на юг почти до Каякента.
Более изолированные от основного массива ареала популяции суслика имеются в широтной долине между Сунженским и Терским хребтами, к западу от Грозного, а затем в ущельях верхнего течения рек Кубани, Малки, Баксана, Чегема и Безингийского Черека (карта 41), Территория, заселенная сусликом, между Сунжей и Тереком в конце 30-х годов состояла из четырех участков общей площадью около 1000 га (Огнев, 1947).
Что касается горных колоний малого суслика, то со времени Менетрие (1832), описавшего зверька с северного склона Эльбруса, распространение его прослеживалось и описывалось по крайней мере 14 зоологами в конце прошлого и в первой половине нашего столетия.
По Свириденко (1927, 1937), самые западные колонии горного eye- j лика в 30-х годах нашего столетия были расположены по ущелью Учку- | лапа и его притоков. Самая верхняя колония на Учкулапе пом по го не ! достигала устья Махара и Джалпа-Кола. Книзу суслик не распростра- i нялся далее устья Худеса.
|Ч. HI. Прежнее и современное распространение и численность млекопитающих 287
Наиболее крупный массив поселений находился и находится в верховьях Малки и ее притоков: Хасаут, Гарбази, Большого Талуко. %о — обширное плоскогорье, слабо наклоненное к северу и востоку, •гжит в пределах 1500—2500 м абсолютной высоты. Свиридепко (1937) проследил соединение этого участка ареала с правомалкписким па высоте 3072 м через перевал Бурупташ, где зверьки держатся уже неда-ико от ледников Эльбруса.
Отсюда суслики, вероятно, попали и в бассейн Уч кулана.
Рис. 120. Местообитание малого суслика в ущелье Баксана.
Фот. автора, 1952.
ПоХасауту суслик распространялся в 30-х годах до урочища Нарзан, а также заселял бассейн Талуко и юго-западный склон горы Кинжал, шее книзу по долине Малки не шел.
На восток колонии суслика тяпутся и теперь по ущельям Куртыка в Джоургена, левых притоков Баксана. По ущелью Баксапа суслик встречается в 1.5—2 км выше теснины, прорезанной рекой в Скалистом фбте. Отсюда суслик распространялся в 1952 г. непрерывно по обоим склонам ущелья вплоть до сел. Нижний Баксан (рис. 120). Особенно иного его было в районе сел. Быллым.
Выше Нижнего Баксана идет снова теснина, и колонии суслика после ^^километрового разрыва появляются вновь в изобилии ио левому (Клону ущелья. Проникновение зверька в ущелье Верхнего Баксана произошло, очевидно, по ущелью речки Кыртык, левому притоку Баксана, с запада. В верхнем троге Баксапа суслик в 1952 г. встречался дав рашшо Тегсиекли и даже несколько выше, а во время обследования йиридеико (1937) он не доходил и до устья Адылы-Су.
Пи долине речки Джигиом, правого притока Баксапа, суслик про-инкал и в долину Чегема, в которой живет весьма высоко, особенно по
288 Часть II. Анализ происхождения четвертичных млекопитающих
притоку Баши-Бузу-Су. По северным склонам горы Чегем-Баши его распространение проходит и в ущелье Безингийского Черека.
В Балкарском Череке в 1952 г. суслика еще не было, хотя и здесь есть для него вполне подходящие места, например у селений Купим и Зильги.
Нет его также, да вероятно и никогда не было, в ущелье Уруха. В 1947—1948 гг., пройдя по всему Уруху и его притокам, а также по Балкарскому Череку и через Черекско-Рионский перевал, я нигде не нашел сусликов, а от местных жителей слышал, что на их памяти и раньше сусликов не было. Не нашел их в 1883 г. и Динник (1884а, 1884в). Таким образом, сообщение Россикова (1887, стр. 44) о массе сусликов встреченных им в горной Дигории, — очевидно, чистейшая фантазия. Огнев (1947, стр. 139), цитируя Россикова, внес новую путаницу, вообразив, что гора Кунь-Итыга и хребет Желаноко, т. е. Джинал находятся в Северной Осетии, в то время как они расположены в бассейне Малки иначе в предгорной Кабарде.
Современная географическая изменяемость малого суслика в пределах Кавказского перешейка (глава IV) не дает четкого ответа на древность изоляции отдельных участков кавказского ареала зверька, так как мы попросту не знаем быстроты формирования отдельных признаков вида в различных условиях среды.
При изучении распределения суслика в пределах всего ареала выявляются древние приспособительные черты зверька к биотопам полынной полупустыни. Именно в зоне полынной полупустыни на глинистых почвах при известном разнообразии растительных группировок наблюдается наибольшая плотность населения суслика.
Учитывая неогеновую древность ландшафта полынной полупустыни на северных окраинах Средней Азии (Лавренко, 1938) и нагорных ксерофитов Восточного Средиземноморья (Буш, 1935; Гроссгейм, 1948), можно думать, что возраст указанных адаптаций по крайней мере верхнеплиоценовый.
В настоящую ковыльную степь суслик проникает лишь местами. Факторами, ограничивающими распространение малого суслика, Бируля (1941) считает высокий и сомкнутый травостой, его однообразие, обилие талых вод весной и застойность перегретого воздуха в ковыльниках.
Тот же автор высказал справедливое мнение о том, что в процессе эволюции зверек адаптировался к открытым биотопам. На ровной низкотравной степи были обеспечены быстрое взаимное оповещение об опасности, сложное норное хозяйство и нормальный теплообмен при наличии движения припочвенного слоя воздуха. Важно отметить, что и у горнокавказских сусликов эти древние адаптации не сгладились, а лишь несколько преобразовались.
Суслики, селящиеся в горах на морене или на аллювиальных галечниках, приспособились рыть норы под крупные валуны, предупреждая тем самым обваливание нор в песчаном грунте и используя рыхлые промежутки между булыжниками.
На горных остепнениых лугах суслик устраивает колонии обычно на обнажениях и близ выходов скал и роет норы в вертикальных стенках троп, пробитых скотом на склонах. Эта привычка рыть норы в различного рода вертикальных стенках характерна и для суслика Волго-Донских степей.
В ущельях Баксана, Чегема наиболее постоянно и плотно заселены сусликом сухие и жаркие днища долин, где условия жизни близки
fi, HI.-Прежнее и современное распространение и численность млекопитающих 289
к пустынно-равнинным. По мере поднятия по склонам плотность поседении быстро падает. Так, на Баксане, близ Быллыма, в августе 1952 г. нами при линейном учете были получены данные о числе нор сусликов, нпшые в табл. 69.
Т а б л и ц а 69
Плотность населения сусликов в ущелье Баксана
Число нор на п стар .... Полынно-тимьннная группировка на нижней, 1-й террасе, местами с кучами камней, каменными заборами и сорняками. Высота 1100 м Типчаково-полынная группировка на верхней, 3-й террасе по каменистым россыпям. Высота 1200 м Субальпийский луг с разнотравьем, местами с выходами скал. Экспозиция склона около 30° на юг. Высота 1850 м
76 25 2
казания Свириденко (1937) о том, что суслики хорошо мирятся со (-I ними, покрытыми древесной растительностью, и охотно поселяются \ давая большую плотность нор на полянах среди большого леса, аименыпую — на полынно-злаковых выгревах, можно объяснить । ь стремлением этого автора всемерно подчеркнуть морфо-экологическую обособленность горного суслика от равнинного.
Древность и происхождение изоляции отдельных колоний суслика редкавказье. неодинаковы. Обособление крайних юго-восточных ко-jjnun в предгорном Дагестане друг от друга обусловлено лишь большим ни меньшим постоянством течения нижних участков русел Терека и Гулака. Вследствие частой бифуркации и меандрирования рек появится И исчезают более или менее широкие участки тугайных лесов и камышовых озер, являющиеся экологическими преградами для сусликов. При этом целые популяции зверьков механически переносятся с берега на берег. Было также доказано опытами, что суслики могут переплывать неширокие реки (до 150 м) и переходить их по мостам ^приденко, 1927, 1937). Известны также случаи дальнего переноса единичных недобитых беременных сусликов хищными птицами, например чайками-хохотуньями.
Тем не менее изоляция колоний сусликов, живущих между Сунженским и Терским хребтами, представляется нам весьма древней по ряду соображений. Во-первых, суслики из Алхан-Чуртской долины морфологически уже отличаются от прикумских и дагестанских и описаны в качестве подвида С. pygmaeus boehmii Krass. Во-вторых, прорыв Терека через Терские ворота между Змийским и Терским хребтами по крайней мере среднеплейстоценового возраста. От устья Малки до устья Сунжи Терек является мощной рекой с быстрым и выпрямленным течением, почти не образуюхцим стариц. Вдоль Терека здесь хорошо развиты полосы старых галерейных лесов, непригодных для обитания суслика. Северный склон Терского хребта также покрыт лесами, непроходимыми для сусликов.
Проникновению сусликов в Алхан-Чуртскую долину с востока препятствовали обширная полоса широколиственных и тугайных лесов пи Гудермесской наклонной равнине и реки Аргунь, Гудермес и Аксай.
Поэтому образование терско-сунженских колоний суслика следует, вероятно, сопоставлять со временем образования ландшафта типчаково-19 Н. К. Верещагин
290 Часть II. Анализ происхождения четвертичных млекопитающих
ковыльной и бородачево-ковыльной степи Алхан-Чуртской долины, т. е, по крайней мере с средним плейстоценом.
Наиболее загадочно происхождение горных, эльбрусских колоний сусликов.
По этому поводу до сих пор существовала одна, в той или иной степени видоизмененная гипотеза Щукина (1925) о том, что суслик проник в горы вслед за отступавшим ледником по приледниковой степи, а потом был отрезан с севера образовавшейся полосой леса. Эта гипотеза, возникшая в связи с критикой представлений Кузнецова (1890) и Краснова (1893—1894) о проникновении сухой нагорной растительности по продольным внутренним долинам Северного Кавказа из Дагестана, подразумевала проникновение суслика с равнины по поперечным ущельям. Однако вопрос о происхождении горного суслика не может решаться столь упрощенно, так как зверек заселяет под Эльбрусом не только днища ущелий, но и перевалы меридианных хребтов. Кроме того, колонии, расположенные на увалах в бассейне Малки, вовсе не отрезаны с севера полосой леса, так как предгорья этого района были издревле безлесны.
Еще Свириденко (1927) предполагал, что малый суслик как «выходец из Средней Азии» мог появиться в Волго-Донских степях и в Предкавказье лишь в послехвалынское время. Молодость ареала суслика в предгорном Дагестане и с нашей точки зрения находит подтверждение в отсутствии зверька в центральном Дагестане, где условия для него весьма благоприятны. Позднее, изучив особенности распространения горного суслика, Свириденко (1937) пришел к выводу, что суслики появились в горах центрального Кавказа еще в эпоху максимального оледенения — рисского (т. е. задолго до хвалыиской трансгрессии, -Н. В.), в результате которого «расширились открытые степные про-странства». Это соображение подкреплялось опровергнуто!! уже гипо-тезой Тутковского (1909) об иссушении климата в приледниковой зоне. | Между тем нигде на Кавказе ни ледники, ни фены, дующие с них, не I образуют приледниковых степей. Наоборот, современные ледники не-1 редко врезаются непосредственно в сосновые и смешанные леса, как | например Караугомский, Цейский и ряд других. Формирование сухой | степной зоны внутри горной системы связано с образованием отгора-| живающих краевых хребтов, играющих роль дождевых экранов, а влия-I ние ледников отнюдь не сказывалось. Так, например, в центральном | Дагестане ледников практически нет, а сухость там велика. Важным | соображением П. А. Свириденко является, однако, признание рсликЛ товости горных сусликов со времени большого оледенения Кавказа, | чем он, в сущности, отрицает прежние данные геологов и геоморфологов I о наличии громадных ледников на местах, ныне заселенных сусликами, | Кроме того, зоогеографические построения Свириденко отлично совпа-1 дают и с представлениями Буша (1935) о том, что горно-степная расти-1 тельность продольных долин Северного Кавказа — не что иное, как । дериват древней горноксерофильной растительности Средиземноморской | области в целом. F
Принимая гипотезу Щукина о проникновении суслика в горы вслед за отступавшим ледником, Иофф (1936) был склонен объяснять всю исто- j рию образования разрывов ареала суслика в Предкавказье бурными I колебаниями численности, расселением и вымиранием зверька на боль- [ ших площадях. [
Действительно, в начале 50-х годов нашего столетия разрыв ареала между наиболее южными колониями суслика в верховьях Калауса и на 5
fi. 1П. Прежнее и современное распространение и численность млекопитающих 291
. склонах Бермамыта достигал по прямой всего 92—95 км, а между колониями в Прикумской степи и в ущелье Баксана — 120 км. Естественно возникает мысль, что в близком будущем этот разрыв может быть заполни, тем более, что расселение суслика к югу ускорялось в последние делшлетия влиянием деятельности человека и, быть может, влиянием природного иссушения ландшафтов при вековых колебаниях климата, г. считалось, что массовое вселение суслика в пределы предкавказских
. началось будто бы из бывш. Астраханской губернии около 1900 — Л г. (Пирковский, 1913; Свириденко, 1927). На самом деле суслик, I вероятно, существовал здесь и раньше, но не обращал на себя внимания 1 в связи со слабо развитым земледелием, а главное — в связи с отсутствием организаций, его изучающих. Могли быть и местные колебания числен-костп суслика, как это отметил еще на основе отчетов Лебедева (1912) Сатувпн (1920, стр. 81). Все это не исключает и не опровергает, однако, : доказанного быстрого продвижения суслика к югу в наши дни (Свири-: денко. 1927; Романова, 1936; Бабенышев, Бируля, Беседин, Голосовская, ' Егоров, Корф, Янушко, 1937).
Г В 1952 г. я обнаружил сусликов уже около Султанского, а Миронов, Павлов, Пушница и Ширанович (1952) опубликовали сводную карту, на которой отмечен сдвиг ареала сусликов к югу почти по всей границе на несколько километров.
Тейп расселения к югу в последние десятилетия достигал здесь 2.5— ; год. При таких скоростях равнинные суслики могут достигнуть t юрш> х колоний в верховьях Баксана и Малки в течение каких-нибудь г 30-40 лет!
; Судя по геологическим фактам (Варданянц, 1939, 1948; Николаев, 1941). можно думать, что суслик появился на Большом Кавказе уже вначале четвертичного периода, когда спад вод Ашперонского моря и значительная пенепленизация хребтов создавали предпосылки для ши-j рокого расселения по Кавказскому перешейку степных и пустынных видов
. и фауны. Концентрируясь на участках нагорноксерофильной рас-, пВОСТИ продольных долин и южных склонов квестовых уступов, ; суслик мог пережить в горах и наибольшее оледенение. Однако и более поздние — плейстоценовые и голоценовые — миграции суслика в горы ссевера не менее вероятны.
J При позднем расселении суслика на Кавказ все дело заключалось в том, i насколько был преодолим для него экологический барьер в виде лесных и зтговых формаций Черных гор, Пастбищного и Скалистого хребтов. Про-- викновение сусликов в горы по поперечным ущельям речек центрального Кавказа, как это рисовалось Щукину (1925), с нашей точки зрения, \ было возможно не повсеместно. Ущелья в Скалистом хребте восточнее 1 Чегема имели и имеют узкие сырые теснины, или каньоны, непроходимые на протяжении 8—10 км для суслика. Эти теснины в плейстоцене все более углублялись в соответствии с тектоническими циклами и в послеледни-ковье достигли наибольшего развития.
Вероятно, из-за упомянутого барьера суслика и нет в ксерофитной зоне верховий Балкарского Черека, Уруха, Ардона, Фиагдона, Гизель-допа. Сунжи, Аргуни, а также во всем центральном Дагестане, хотя условия обитания там безусловно не хуже, нежели под Эльбрусом.
•? Особенно показательно сопоставление наличия суслика в Алхан-Чурт-1 да долине и отсутствие его в долинах правых притоков Сунжи.
На относительно недавнее появление суслика в горах косвенно ука-j дает слабое развитие ареала на восток по продольным долинам. Так,
19*
292 4 Часть II. Анализ происхождения четвертичных млекопитающих
например, суслик еще не успел заселить ущелье Балкарского Черека.
В свое время при продвижении суслика с равнины к Эльбрусу путь расселения лежал, очевидно, западнее и восточнее лесистого Пятигорья, которое было своего рода преградой для проникновения зверьков на юг и в пещерах которого могли откладываться кости сусликов, съеденных филинами.
Палеонтологическая разведка, предпринятая нами в 1952 г. в районе Пятигорья, показала, что на протяжении последних полутора-двух тысячелетий кости сусликов в районе Железноводска, Пятигорска и Кисловодска не накапливались.
Однако эта разведка не доказала, что суслика не было в окрестностях Пятигорья в ксеротермическую эпоху послеледниковья или в плейстоцене, особенно в связи с находкой его остатков в раннеголоценовых слоях в теснине Баксана.
Вопрос о происхождении суслика в горах Кавказа может быть разработан далее при организации более глубокой шурфовки навесов и пещер гор Развалки, Верблюда и по ущельям Кич-Малки, Кумы и Подкумка, а также при изучении акклиматизации горных сусликов на равнине, а равнинных — в горах. При этом эксперименте можно было бы проследить и скорость эволюции экологических и морфологических адаптаций у обеих форм. Пока же, оценивая все изложенные факты, следует признать относительно позднее — голоценовое — расселение суслика в горы с равнин Предкавказья.
В новейшую эпоху человек своей деятельностью бессознательно содействовал быстрому расселению суслика как в горах, так и на равнинах (см. главу VI). Истребление и промысел на шкурку (график 20) на первых этапах не могли приостановить этого процесса.
Современные «сплошные очистки» от сусликов и дальнейшая распашка степей, а также развитие лесных полезащитных полос и искусственного орошения вскоре приведут к практическому уничтожению сусликов на предкавказских равнинах.
Малоазийский суслик — Citellus citellus L. Остатки сусликов С. citel-loides Kormos обнаруживались в плейстоценовых слоях пещер Европы (Wolf, 1939), a Citellus sp. — в верхнем палеолите Сирии (Wolf, 1939), На Кавказе известны только поздние — голоценовые — остатки мало-азийского суслика из аллювиальных суглинков в ущелье Занги, к северо-западу от Еревана (Даль, 1949а).
Современное распространение юго-восточного подвида — С. citellus xanthoprymnus Benn, охватывает Армянское нагорье и Малую Азию.
Суслик занимает в Армении довольно большой район долины Аракса, от района станции Кара-Бурун и Алагез до села Илли, к северо-западу от Ленинакана, верхнюю часть долины и бассейна р. Бамбак до станции Амамлы, западные и северные склоны Алагеза (карта 41).
Распространение зверька по вертикали (Даль, 19486; Аветисян, 1950) занимает зону почти в 1.5 км — от 1100 до 2700 м. В долине Аракса он существует в условиях горно-полынной полупустыни, а в горах — в условиях лугостепи и даже субальпийского луга. На настоящих мезофитных лугах этот суслик, однако, не живет, так же как не распространяется и вниз по пустынной долине Аракса.
При значительной экологической изменчивости (Аветисян, 1950) малоазийского суслика следует рассматривать как вид, формировавшийся на фоне предгорного и горного ландшафта аридной и относительно жар
Гл. Ш- Прежнее и современное распространение и численность млекопитающих 293
кой области. Областью развития вида следует считать Малоазийско-Эгей-ско-Балканскую сушу.
Прежнее обитание этого суслика в ущелье Занги свидетельствует о несколько более широком распространении вида в отдельные эпохи четвертичного периода. Современное расселение зверька на подходящие для него участки, например на аридные третичные всхолмления по северной окраине Куринской низменности, возможно лишь в обход с востока Карабахских гор. Главной причиной слабого развития кавказского участка мреала этого вида по сравнению с ареалом степной полевки и малоазий-Ького хомяка следует считать большую консервативность малоазиатского вдслика к месту обитания и меньшие его миграционные способности.
Местные разрывы и сокращения ареала этого суслика на Армянском нагорье можно рассматривать как следствие не только прогенических Iпроцессов, но и обширных излияний лав, перекрывавших большие площади/В эпохи похолоданий и оледенений понижение верхней границы его распространения на Армянском нагорье достигало, вероятно, не менее 400—500 м. Возможно, что суслик заселял в плейстоцене и южную часть Джавахетии.
В новейшую эпоху ареал малоазийского суслика расширяется в верхней зоне и сужается в нижней в результате иссушения склонов под влиянием выпаса скота. Происходит и исчезновение отдельных пупуляций под влиянием промысла на шкурку и истребления химическими средет-лами (Аветисян, 1949).
Сем БОБРОВЫЕ — (ASTOPJOAE
Остатки бобровых известны с нижнего олигоцена Евразии и Северной Америки. По приспособлениям бобровые близки частью к наземным дикобразам, частью к земноводным полевкам и восьмизубым. Остатки бобров миоцено-плиоценовых родов Steneofiber, Amblicastor, Trogontherium и плиоценово-плейстоценового рода Castor многочисленны в пределах широкой полосы северного Средиземноморья и Средней Азии — от Британских островов до Монголии.
На Кавказском перешейке обнаружены остатки бобров 4 родов: Steneo-fiber, Amblicastor, Trogontherium, Castor (карта 42).
Речной бобр — Castor fiber L. Остатки речных бобров на Кавказе появляются впервые в верхнеплиоценовых конгломератах Таманского полуострова. Таманский бобр — С. tamanensis N. Ver. из урочища Синяя Балка являлся, по-видимому, непосредственным предком современного речного бобра (Верещагин, 1957а). Другие плиоценовые представители семейства Castoridae — Steneofiber, Amblicastor и Trogonthe-пит, — известные с Кавказского перешейка, не имеют прямых родственных связей с речными бобрами.
В верхнем плейстоцене и в голоцене на Кавказе существовал речной бобр вполне современного типа. Его остатки к настоящему времени обнаружены в пещерах Сакажиа (Громов, 1948) и Уварова, близ Кутаиси, в неолитических слоях пещеры Сагварджиле и в слоях века поздней бронзы на могильных полях Самтавро, близ Мцхети.1
Таким образом, обитание бобра в голоцене на притоках Риона и Кви-рилы, а также на притоках Куры и Арагвы доказано теперь документально
1 Сведения о находках костей бобра в Сагварджиле и в Самтавро любезно предоставлены доктором биологических наук Н. О. Бурчак-Абрамовичем.
294 Часть II. Анализ происхождения четвертичных млекопитающих
(рис. 121). В четвертичных отложениях на Русской равнине остатки речного бобра весьма обычны, но в Крыму обнаружены только в слоях поздней скифской эпохи. К югу от Кавказа достоверных находок остатков бобра пока нет. В пещерах Палестины, Сирии и Ливана они не найдены (Bate, 1937).
Указания на находки остатков бобра в пещерах Ирана (Coon, 1951) сомнительны, так как, очевидно, их спутали с остатками дикобраза.
Обилие поздних остатков бобра в пойме низового Допа (Саркел у Цимлянской, Новочеркасск) говорит о том, что обитание этих зверей в нашу
Рис. 121. Остатки речного бобра.
1 — бедренная из слоев I тысячелетия до н. э. в Самтавро; 2 — обломок тазовой из неолитических слоев пещеры Сагварджиле; з — обломок тазовой из палеолитических слоев пещеры Уварова*
эпоху на реках Прикубанской и Закубанской равнины было вполне вероятным. До XVIII в. вполне подходящие места обитания бобров имелись по Псекупсу и ряду мелких левых притоков Кубани и даже Терека.
Литературные сведения об обитании бобра на Кавказе в нашу эпоху весьма многочисленны и довольно общеизвестны.
Упоминания о бобрах в Скифии и Колхиде обычны у античных авторов (Геродот^ изд. 1888 г.; Страбон, изд. 1879 г.). Поздние упоминания о бобрах в Мингрелии и вообще в Грузии появляются у Ламберти (1654, изд. 1913 г.) и Шардена (Chardin, 1686), в грузинских летописях «Картлис Цховреба» и в «Географии Грузии» царевича Вахушти (1904), написанной в начале XVIII в. Еще в конце XVIII и начале XIX в. шкуры бобров и «бобровая струя» имели в Грузии, по-видимому, известное экономическое значение как при вывозе, так и при ввозе. Об этом отчетливо говорится в положении о тифлисской таможне 1803 г.
Гюльденштедт (1879) и Паллас (1831) отметили вероятность обитания бобра на Кубани, Сунже, а Менетрие передал в коллекцию акад. Брандта череп бобра, найденный будто бы на берегу Сунжи. Ровинский (1809) указывал, что бобры изредка попадаются на Кубани. Нордманн (1840) сообщил, что несколько штук бобров было убито в 30-х годах XVIII столетия в бассейне речки Нотанеби, в Колхиде, и указал на обитание бобра на Тереке. Гогенакер (Hohenacker, 1837) упоминал об обитании бобра на Араксе.
[{'Прежнее и современное распространение и численность млекопитающих 295
Весьма сомнительные сведения о бобрах и «бобровой струе», добываемой в Армении, Турции и Иране, имеются у Шопена (1852, стр. 807).
Все эти данные повторены и в монографии Брандта (1855).
, Череп бобра, хранящийся в ЗИНе за № 6330 («Menetries, 1831») и изоб-:;еннып у Брандта (1855), не носит следов водной окатки, речного песка I ила, но тщательно препарирован и скоблен ножом. Другими словами,
, хна принадлежать животному, убитому незадолго до приезда Менет-Oii. на Сунжу, если только он не был подобран упомянутым исследова-плен где-либо севернее, по дороге на Кавказ или обратно. Сам Менетрие ' iietries, 1832) не называл бобра среди кавказских зверей (!).
Радде (1866) указал на достоверное обитание бобра в верхней Сванетии V Оссейне Цхенис-Цхали, у Лентехи, в источниках речки Хеледулы. 1 жрадов (1870) писал, что бобры жили в его время в уреме Малого юнчука на Закубанской равнине.
Богданов (1873) утверждал, что бобры водились недавно по речкам За-ь Лиской степи и что в 1864 г. был убит, вероятно, последний бобр ш: Лабе, недалеко от ее устья. Динник (18846) высказал предположение tv обитании бобра в верховьях Лабы. Кеппен (1902) опубликовал сведения ( 2 бобрах, будто бы убитых в 1947 г. на Араксе, и высказал несколько i пересных соображений о возможности обитания бобров в верховьях оакса. Обильные, но непроверенные сведения о бобрах были напечатаны itpm Лшгстовым (Linstow, 1908).
Описывая историю бобра на Кавказе, Сатунин (1920) повторил обзоры йшпена и Линстова, но добавил, что бобр будто бы еще жил в 1909 г. бассейне Су псы, в Колхиде, и, возможно, сохранился где-нибудь, в неисследованных уголках западного Закавказья и северо-западного Кавказа.
Недавний обзор Огнева (1947) о бобре на Кавказе с приложением карты прежних мест его распространения (1?) добавил ряд недоумений для будущих исследователей.
При изучении болот и речек Колхиды в 1931 и 1944 гг. в связи с опытами разведения нутрий я не нашел здесь нигде погрызенных пней и остатков бобровых плотин, но убедился, что бобры могли жить недавно в Рион-скоп долине — от устья Квирилы до берега Черного моря.
Вполне подходящие для бобра речки, но теперь уже с голыми берегами, имеются, например, в районе курорта Цхалтубо и во многих других местах Колхиды.
Точную дату полного исчезновения бобра на Кавказе установить трудно, но, вероятно, оно произошло в середине XIX в.
К югу от Кавказа бобр был известен будто бы в прошлом столетии ({Месопотамии и широко распространен в Иране. В Турции он обитал будто бы в речках системы Кизыл-Ирмака, где его добывалось в год до 2000 штук (!). Во второй половине прошлого столетия бобр будто бы существовал в болотистой местности между Кайсарие и Ипьесу, к югу от среднего течения Кизыл-Ирмака (Danlord a. Alston, 1877, 1880).
Все эти литературные сведения, подробно изложенные Кеппеном (1902), а затем Линстовым (Linstow, 1908), выглядят очень солидно, но ^ело в том, что никто никогда не видел научных документов: рисунков, фотографий, коллекций, подтверждающих эти сообщения. Наряду с этим в качестве примера беззаботной научной фантазии можно указать .на сообщение Радде (1866), который «поселил бобра» на правых притоках Риона в высокогорной Сванетии в узких горных щелях, с бешено мча-щпмися потоками, которые не посещаются даже выдрами.
296 Часть Л. Анализ происхождения четвертичных млекопитающих
Что касается прежнего обитания бобра в центральных частях Малой Азии и Ирана, то трудно представить себе, чтобы при тамошнем безводье и безлесье бобр мог существовать где-нибудь в историческое время. Известно, что бобр питается древесиной тополя, осины, ивы, березы. Крайне сомнительно, чтобы ежевика, облепиха, лох и карагач — основные современные кустарники и деревья, растущие вдоль русел рек в Иране и Турции, были бы для него достаточным кормом. Однако следует, конечно, учитывать и крайне быстрое изменение ландшафтов в последние столетия. Недавнее открытие остатков бобра в слоях Неаполя Скифского в Крыму показало, что бобры жили по речке Салгиру на сухой и совершенно безлесной ныне части полуострова еще в середине I тысячелетия до н. э. (Цалкин, 1947). В связи со старыми сообщениями об обитании бобров на степном юге Украины Пидопличко (1951) справедливо разъяснил, что бобры могли и могут существовать в зоне степей и даже полупустынь по речкам, пересекающим эти зоны, при условии наличия прирусловых лесов. Эти соображения Пидопличко пригодны для подтверждения прежнего распространения бобров в Предкавказье; что же касается нагорий Передней Азии, то там ограничивающими факторами являются крайне неустойчивый режим горных потоков и резко континентальный, засушливый климат.
Бобр не может жить на быстрых горных речках с галечниковым руслом, так как здесь нет ни убежищ, ни пищи. Даже при наличии леса по берегам жизнь бобров на горных речках с большим уклоном невозможна, потому что каждый паводок неминуемо сносит любую построенную ими плотину, а устраивать норы негде.
Тем не менее бобры уцелевали при интенсивных горообразовательных процессах в таких широких межгорных долинах, как Куринская и Рион-ская, и на предгорных равнинах. Чаще же они вторично заселяли уже пенепленизированную страну.
В связи с историей бобра на Кавказе особый интерес представляет вопрос о его родине и о развитии морфо-физиологических приспособлений к подледному образу жизни.
Первоначальной родиной речных бобров были, несомненно, области с теплым климатом, в частности Средиземноморье. Адаптации к жизни в замерзающих водоемах, неоднократно подчеркнутые исследователями (Linstow, 1908; Федюшин, 1935; Верещагин, 1939в; и др.), стали развиваться, очевидно, уже с нижнего плиоцена при начавшихся похолоданиях климата. Процесс приспособления к плаванию подо льдом и к работе резцами под водой был, видимо, значительно ускорен в первой половине плейстоцена.
Быстрота расселения бобра в послеледниковое время по территории, освободившейся от материковых льдов, оказалась поразительной. Учитывая трудности, связанные для бобра с пересечением водоразделов, следует признать, что обилие этого зверя в Скандинавии, на Кольском полуострове и на Карельском перешейке в историческое время могло бы служить отличным подтверждением отсутствия великого оледенения равнинной Европы для антигляциа листов.
Причинами исчезновения бобра на Кавказе были: неумеренный промысел — для первой стадии уменьшения численности и антропогенное изменение ландшафта — для заключительной стадии. Изменение условий обитания состояло в вырубке приречных лесов, в увеличении быстроты течения, эрозии и паводков рек при уничтожении лесов и выпасе скота в их бассейнах. Этот процесс протекал быстрее на речках Предкавказья,
//L Прежнее и современное распространение и численность млекопитающих 297
чем в Закавказье. Дольше всего бобр продержался в глухих болотах и ; протоках Колхидской низменности.
. В 40-х годах нашего столетия несколько пар бобров, завезенных из ^Воронежа в Караязский нутриевый совхоз, размножалось здесь при кле-Еочнсм содержании. При подсадке тополей и ив на берегах речек бобры Помогли бы жить здесь и на воле.
J В итоге нашего обзора приходим к следующим выводам.
I 1. Речные бобры жили на Кавказском перешейке в плиоцене и четвер-
|личном периоде.
I 2. К XVIII в. распространение речного бобра стало здесь реликтовым. L Он уцелел в болотах и на речках западного Закавказья, в Колхиде, а воз-| можно, и на предгорных равнинах Предкавказья по притокам Кубани, | Терека, Сунжи. В горных районах Кавказа бобров в историческое время не было.
.j = кончательное вымирание бобров на Кавказе произошло в середине .YJX а.ш в начале XX в. в Колхиде.
ж 4. При упорядочении водного, лесного и охотничьего хозяйства страны точного бобра можно будет развести в небольшом числе на некоторых Ьевых тритонах Кубани, речках Хачмасской низменности и в Колхиде. Юсобенно пригодными для этих целей могут оказаться речки типа ключей ^кара-су» с постоянным дебитом при посадках на их берегах тополей, ив к карагача. При быстром развитии сельского хозяйства это мероприятие ^ряд ли дает заметный экономический эффект.
Сем МЫШИНЫЕ — М VJRIDAE
Из четвертичных отложений на Кавказском перешейке известны ос-• шки 15 видов этого семейства. В современной фауне Кавказа зарегист-^фовано 29 видов. Особый интерес представляет здесь история появле-Жя рода Rattus.
^Черная крыса — Rattus rattus L. Остатки крыс R. cf. rattus были из-Вкгны уже давно из четвертичных слоев многих пещер Западной Европы — К островов Великобритании до Карпат (Woldrich, 1882; Брандт и Вольд-Жу 1887). Находили их обычно с костями животных верхнего плейсто-Кна: пещерного медведя, волосатого носорога и других — в слоях, датируемых верхним палеолитом, например в ориньякских слоях пещеры Эвелины в Сомерсайтшире (Англия). Большинство находок костей черных крыс в пещерах приходится все же на побережье Италии и вообще Средиземного моря (Wolf, 1939).
В пещерах Сирии, Ливана, Палестины кости черной крысы обнаружены в слоях мустье при наличии в то время носорога Мерка и гиппопотама (!) (Bate, 1937). Как правило, остатки черных крыс одновременны включающему их слою, т. е. залегают «in situ», но, очевидно, имеется много когда косточки современных крыс сваливались на дно раскопов крысы сами залезали по щелям в глубокие слои.
ВТ Тем пе менее нет оснований говорить о проникновении черной крысы Европу в послеледниковое время, так же как и о «завозе ее еще в доисто-ИКеские времена», что нередко переписывается из одних популярных руко-вКдств и научных статей в другие. Видимо, черная крыса в Средиземноморье является плиоценовым реликтом. Плейстоцен она пережила па теплых побережьях юга Европы и севера Африки, а быть может, и Кавказа -Верещагин, 1947г, 1949в).
298 Часть II, Анализ происхождения четвертичных млекопитающих
I
В пределах СССР кости черной крысы найдены в древнегреческих и римских слоях Ольвии, на юге Украины (Пидопличко, 1938в).
На Кавказе находки костей этого вида единичны в неолитических слоях Ахштырской пещеры, на могильном поле с кувшинными погребениями I тысячелетия до н. э. на Куре у Мингечаура (рис. 122).
В настоящее время черная крыса обитает на Кавказе весьма широко как в «дикой» природе, так и в постройках человека (карта 51). Она живет в портовых сооружениях всех более или менее крупных населенных пунктов на побережьях Азовского, Черного и Каспийского морей. В равнинном и горном Предкавказье черной крысы в настоящее время нет и, по-видимому, не было и раньше.
Среди многих тысяч костей из погадок филина в Ставрополье и Пятигорье, собранных Кистяковским (1935) и нами, костей черной крысы также найдено не было.
Широко распространена теперь черпая крыса по Черноморскому побережью — от Батуми до Новороссийска. Здесь она живет в садах, лесу где обычно замещается пасюком. Фле-
Рис. 122. Тазовая кость черной крысы из слоев I тысячелетия до н. э. в Мингечауре.
и реже по берегам водоемов,
ров (1927) наблюдал ее в лесу севернее Батуми, а Шидловский (1947, 1948, 1950) отметил для многих пунктов Абхазии, Мингрелии, Аджарии, ука-
зав, что в первой грызун распространен от уровня моря до 600 м, а в последней — до 1500 м абсолютной высоты. Мною черйая крыса добыта на оз. Бебесыри вместе с пасюками осенью 1946 г., но в болотах, окружающих оз. Палеостом, ни разу не наблюдалась и не вылавливалась (Верещагин, 19416). По Степанову (1931), черная крыса в Батуми уступает в численности пасюку. В восточном Закавказье черная крыса живет «дико» в лесах северной Армении, где встречается и в населенных пунктах. Особенно часта она была в 1937 г. в шахтах Шамлуга и на мельницах селений Ку-лали, Ачаджур и Марц (Шидловский, 1942). В Южной Осетии редка и найдена лишь в населенных пунктах в глубине ущелий. В Кахетии и вообще в Алазано-Агричайской долине живет и в садах, и в постройках, причем особенно часто на чердаках (Аргиропуло, 1940а). Обычна летом в зарослях ежевики по окраинам полей у Куткашена и Варташена. В Куринской низменности держится по оазисам, особенно вдоль оросительных канав, в зарослях ежевики.
В Баку черная крыса обитает в домах прибрежной полосы и, как правило, не встречается далее 100 м от берега бухты. По материалам массовых отловов, на судах бакинского порта она составляет 60% улова, а в портовых постройках — только 9%. Остальное приходится на пасюка (Верещагин, 1942а).
Особенно часта эта крыса в Ленкоранской низменности — от сел. Масаллы до Астары и далее по южному побережью Каспия — в Гиляне и Мазандеране. Здесь она живет в фруктовых садах и держится во всех портах, особенно на рыбных промыслах. Грызун был известен из Гир-кании зоологам еще в начале прошлого столетия (Менетрие, 1832; Радде 1886).
По материалам массовых отловов, в Ленкорани и прилежащих селениях в конце 30-х и начале 40-х годов нашего столетия черная крыса составляла только 0.51% от всех вылавливаемых синантропных грызунов
I Гл. 111- Прежнее и современное распространение и численность млекопитающих 299
^Верещагин, 1949г). Высоко в горы грызун не идет и здесь, а держится Инйптб в низменной части района. Способность черной крысы существо-рать круглогодично в Колхиде и Гирканике в «дикой» природе и устраи-- вать доры под пнями, заборами и убежища в дуплах подтверждает ее < алюценовую автохтонность как для Средиземноморья вообще, так и для
Закавказья в частности.
А. С развитием человеческой культуры, устройством землянок, а позднее ж I надземных строений из сырцового кирпича, камня и дерева, постройкой Т мореходных лодок и кораблей черная крыса быстро приспособилась к по-типу убежищ, используя наличие тепла и пищи. Приспособление Вобитанию в портовых сооружениях и на кораблях было, по нашему мне-лг чпю, связано именно с первоначальным обитанием крысы на теплых по-
s бережьях Средиземноморья, где имелась возможность кормежки на лито-I ради. Позднее эта крыса начала проникать в глубь страны, вдоль рек, JL придерживаясь населенных пунктов, и постепенно заселила зону низовых ж лесов Закавказья.
К. Серая крыса — Rattus norvegtens Berck. Достоверных остатков этого пз плейстоценовых слоев Передней Азии, Кавказа и Западной Ев-Яропы неизвестно. Остатки пасюков, обнаруженные в пещерах Чехосло-Жвакии, Мальты, Гибралтара, Ирландии, залегали вместе с остатками по-Яслеледниковых животных и по типу сохранности не были древними (Wolf, 01939). Кости пасюка на Кавказе известны из слоев позднего средневе-Жковья (XV—XVI вв. и. э.) по раскопкам во дворе дворца Ширван-Шахов В в Баку (Верещагин, 1949в) и из пещерок с погадками филина древностью Игйе более нескольких столетий в Ставрополье, Пятигорье, Северной Осетии I н в Имеретии. Наиболее древние, дошедшие до нас литературные указа-Ания об опасных крупных мышах, вредящих посевам и обгрызающих ветви Икеревьев в области каспиев, принадлежат Клавдию Элиану в III в. н. э. «(Латышев, 1904).
г Арабы, по-видимому, знали серую крысу и имели с ней дело уже Т в раннем средневековье (Житков, 1944).
I В настоящее время пасюк зарегистрирован на Кавказе во всех круп-| № населенных пунктах страны, связанных железными и автомобильными дорогами, а также во всех приморских селениях, на рыбных промыслах п в портах Каспийского и Черноморского побережий, а также на реках: Кубани, Сулака, Терека, Куры, Риона, особенно в их судоходной части (Верещагин, 1947г) (карта 52).
| Пасюк обитает на Кавказе местами и в «дикой» природе. Такие дико-живущие пасюки особенно распространены по влажным предгорьям Кабарды, Северной Осетии, Грозненской области. Здесь пасюк нередко является настоящим вредителем огородов и полевых складов — «сапеток» (Росснков, 1887; Бугданов, 1936). В Кабарде и Северной Осетии пасюк живет в аулах, особенно близ мельниц, почти до верхних участков ущелий Большого хребта (Туров, 1926в).
Дикие популяции пасюков существуют также в болотах Колхиды, (рис. 123) Гирканики и на древних озерах левобережья Куры в Азербайджане (Верещагин, 19416, 19426, 1947г, 1949г; Львов, 1949).
1 Живут пасюки и в россыпях скал пустынного Каспийского побережья, I например на Апшеронском полуострове, подкармливаясь выбросами моря. I Их жилища в болотах Рионской и Куринской низменностей состоят из открытых гнезд — площадок в камышах, а закрытые убежища устраиваются под карчами ольхи. В «дикой» природе пасюк питается в значительном степени животной пищей, но растительная все же преобладает.
300
Часть II. Анализ происхождения четвертичных млекопитающих
При длительных ледоставах и снегопадах пасюки, обитающие в камышовых и ольховых болотах Закавказья, частью вымерзают, частью переживают тяжелый период в прибрежных населенных пунктах (Верещагин и Дюнин, 1949).
Будучи тесно связан с водными биотопами, пасюк, вероятно, не смог бы перенести ледниковую эпоху ни в Предкавказье, ни в Закавказье. Более широкое, нежели у черной крысы, современное распространение пасюка на земном шаре не противоречит соображениям о плиоценовой древности первого и голоценовом возрасте второго вида в Средиземноморье. По срав-
Рис. 123. Места обитания серой крысы в Колхиде.
Фот. автора, 1939.
нению с черной крысой пасюк обладает большей пластичностью физиологических функций (Руттенбург, 1950), которая и позволила ему расселиться вместе с человеком от тропиков до Арктики. Но в «дикой» природе Закавказья пасюк оказывается менее устойчивым в зимнее время из-за своей гидрофильности.
Итак, в развитие взглядов Житкова (1944) и наших первоначальных соображений о плиоценовой релпктовости крыс рода Rattus в Средиземноморье (Верещагин, 1949в) пасюка следует рассматривать здесь в качестве послеледникового иммигранта, прочно вошедшего в фауну болот Закавказья и лесов нижней горной зоны Предкавказья. В связи с этим возникают вопросы о путях и более точных сроках проникновения грызуна на Кавказ.
Пасюк пе мог, конечно, перебраться в Европу из пустынь Средней Азии по той простой причине, что его там пе было. Еще Кащенко (1912) справедливо считал, что пасюк пе мог проникнуть на Кавказ из Индии через сухой и пустынный Иран по караванным путям, как это иногда рисовалось позднейшим популяризаторам. Путь проникновения его был,
>П. Прежнее и современное распространение и численность млекопитающих 301
конечно, прибрежно-морской. Если учесть, что египтяне и финикияне совершали плавания по Красному и Средиземному морям уже во II тысячелетии до н. э., а позднее проникали и на юг Азии (Авдиев, 1953), то ста-т ясным, что пасюк мог быть завезен судами в Средиземноморье еще до ' лпец эры. Первоначально пасюк попал, очевидно, на Черноморское юбережье Кавказа.
Проникновение пасюка на Каспийское побережье произошло позднее, >.)сле заселения Черноморского. Оно могло идти двумя путями: водным — о Манычу и по суше, особенно по влажным наклонным равнинам Пред-авказья.
До бурного развития городов с их многоэтажными домами, бойнями, аналпзацией и развития железнодорожного и автомобильного транс-ина пасюк, видимо, существовал преимущественно в «дикой» природе — о болотам и дельтам рек с тростниковыми зарослями, — как это наблю-ается в настоящее время в устьях и плавнях Дуная, Днестра (Айзен-падт, 19506), в болотах Риона и в дельте Волги.
Во времена Палласа, видевшего каких-то крыс в дельте Волги, пасюк, ероятпо, уже жил там наравне с водяной полевкой.
Новейший этап развития ареала пасюка на Кавказе был связан со гроптельством в XIX и XX вв. шоссейных и железных дорог, крупных пунктов сельского и городского типа. Населенные пункты Куринской ценности осваивались пасюком вначале медленно по водным магистра-т. е. по Куре, Араксу и их притокам. В Алазано-Автаранскую долину ;рысы также проникали вначале по левым притокам Куры: Турьян-Чаю, >ок-Чаю, Алджиган-Чаю. Позднее же, с появлением автомобильного чюбщепия, заселение было неизмеримо ускорено. На сухих плоскогорьях Армении расселение пасюка шло медленнее. В Ереване и Джульфе, например, пасюки появились лишь в 30-х годах нашего столетия, после постройки железной дороги, связавшей названные города с Тбилиси и баку.
В равнинном Предкавказье расселение пасюка пошло интенсивно в 20-х годах нашего столетия с развитием автомобильного транспорта. Появление пасюков в новых населенных пунктах Сальских степей зарегистрировал Губарев (1941).
В Ставрополе пасюк был известен еще Сатунину (1901г), а судя по нашим находкам костей пасюка в погадках филина на горе Стрижамент, ок жил в лесистых ущельях Ставрополья уже в прошлом столетии.
Критический обзор взглядов на происхождение пасюка в северной половине Евразии и на его взаимоотношения с человеком, данный Кузякиным (1951), не прибавил ничего нового для понимания истории появления зверька в СССР.
Полевая мышь — Apodemus agrarius Pall. Ископаемые остатки житника известен из плейстоценовых слоев пещер Чехословакии (Wolf, 1939). На Кавказе они пока неизвестны, но обнаружены в верхнечетвертичных отложениях низового Дона.
Современный ареал житника занимает зону широколиственных лесов и южную часть тайги Евразии с перерывом в области Забайкалья и верхней части бассейна Амура (Бобринский, Кузнецов, Кузякин, 1944). В степ-шшзоне Русской равнины житник обитает лишь по долинам рек. В Крыму отсутствует, но обнаружен в плейстоценовых слоях. Изолированный теперь с севера участок обитания расположен в Предкавказье. Распространение грызуна приурочено здесь преимущественно к влажным предгорьям, а в степи и полупустыне он встречается лишь вдоль русел рек (Свириденко,
302 Часть II. Анализ происхождения четвертичных млекопитающих
1944). По Черноморскому побережью прослежен к югу до Сухуми (Шид-ловский, 1947, 1950).
Живет житник также на Ставропольской возвышенности и по тугайным лесам Кумы, Терека и Сулака. Изолированный участок обитания расположен на юго-востоке в лесах Хачмасской низменности — от низовьев Самура до Хачмаса (Беме, 1928; Верещагин, 1944).
Распространение по вертикали незначительно; наивысшие участки обитаемых мезофитиых лугов и полей на северо-западном Кавказе лежат на отметках 950—1000 м. В районе Нальчика грызун прослежен до высоты 700 м, а под Орджоникидзе и Алагиром — до высоты 800—850 м.
По наблюдениям Свириденко (1944), особенности распространения житника в Предкавказье зависят от количества годовых осадков. Эврп-топность зверька характерна при осадках не менее 500 мм в год. В более засушливых областях зверек обитает лишь во влажных биотопах, как например в низовых лесах Хачмасской низменности у побережья Каспия. Здесь житник держится по сырым днищам лесистых оврагов и берегам ключевых ручьев под защитой зарослей ольхи, ежевики, перевитых лианами, и не вредит посевам зерновых, как это бывает в других местах. На более сухой Кусарской наклонной равнине, как и на всем остальном побережье Каспия, зверек отсутствует.
В целом распространение житника в Предкавказье приурочено преимущественно к районам с густой сетью мелких потоков и ключей, в зоне развития предгорных шлейфов. Характерными биотопами здесь служат кустарники вдоль небольших речек и луговины вдоль заболоченных ручьев.
В результате в предгорьях Кавказа имеется ряд более или менее обособленных друг от друга участков обитания: Кубанский (с Черноморским побережьем), Терско-Сунженский, Самурский (карта 54). Разрывы между участками обитания в Предкавказье обусловлены наличием сухих остеп-ненных водораздельных пространств.
Ранее — в плейстоцене — житник вряд ли проникал на юг далее хачмасских низовых лесов, так как в плейстоценовых битумах Апшеропа его остатки не найдены. Подтверждением тому же служит и отсутствие зверька в Алазано-Агричайской долине, где существуют подходящие для
него условия.
Во влажном западном Закавказье полевая мышь распространена по террасам Черноморского побережья на юг до Сухуми. Она встречается здесь чаще по перелескам, вырубкам, кустарникам и садам.
Распределение житника по биотопам на Черноморском побережье характеризуется отловами и учетами Шидловского (1947) (табл. 70).
Таблица 70
Распределение полевой мыши по биотопам в Абхазии
«Живые изгороди» Огороды Сады Чайные плантации Кукурузные поля Берега водоемов Кустарники Сплошные леса с с о Р5
Число пойман- ( абс. . 1 27 36 12 10 31 135 2 254
пых зверьков ( в % 0.4 10.6 14.1 4.7 3.9 12.2 53.2 0.8 100.0
fo.111. Прежнее и современное распространение и численность млекопитающих 303
В. Эти данные показывают, что антропогенные изменения ландшафта Ярове Черноморского побережья, особенно в смысле обезлесения, являются Янгоприятным фактором для развития здесь ареала и увеличения числен-ТГностп грызуна. Возможно, что значительное распространение житника яа , ио Черноморскому побережью в историческую эпоху обусловлено ландшафтными изменениями, произведенными человеком. Важно отметить, что далее на юг, в Колхиде, имеются биотопы, очень сходные с абхазскими. Отсутствие- в них житника можно объяснить, вероятно, фактором избы-
Д точной влажности.
д Своеобразный неразвитый ареал житника на Кавказе позволяет пред-Яклагать довольно позднее верхнеплейстоценовое проникновение зверька севера. При наступлении послеледниковой эпохи кавказские популяции Яйтника были, видимо, отрезаны развитием широкой полосы осушенных Чптепев
(Ь- утствие зверька в плейстоценовом и современном Крыму также Жлодтверждает позднее развитие ареала в южном направлении.
Б новейшую эпоху распространение полевой мыши на Кавказе уве-f лпчивается при вырубках лесов на участках избыточного увлажнения — > в юго-западных районах — и при организации орошения — в сухих < районах на севере и востоке страны.
JL Маюазийский хомяк — Mesocricetus auratus Water, (s. lato). Bo-kp°c ° мропсхождешш хомяков рода Mesocricetus на Кавказе довольно Тсложен. Впервые остатки хомяков — род Р aleocricetus— появляются на Кавказе уже с среднего миоцена. В нижнем плиоцене Предкавказья жил хомяк, близкий по размерам к средним хомякам восточного Среди-i земноморья. Таким образом, сам Кавказ мог быть территорией развития 1 средних хомяков в такой же степени, как и Передняя Азия.
Остатки ископаемых хомяков рода Mesocricetus известны из среднего пверхнего палеолита Палестины — пещера Этлит (Picard, 1937), а в пре- делах Кавказа из пещер бассейна Риона и Квирилы: Гварджилас-Клде, j Мгвимеви, Кударо — и из плейстоценовых слоев Апшеронского полу-Г острова.
j Важно отметить, что в четвертичных отложениях Русской равнины Г до сих пор обнаруживались остатки только хомяков рода Cricetus.
В послеледниковых пещерных отложениях остатки малоазийского 1 хомяка весьма обычны на Армянском нагорье, в центральном Закавказье д й в центральном Предкавказье.
! В связи с значительной географической изменяемостью средних хо-1 мяков кавказские представители рода рассматриваются обычно система-Г дамп в качестве 2 или 3 самостоятельных видов. Признавая южное — ' переднеазиатское — происхождение ныне живущих на Кавказе средних хомяков, мы рассматриваем далее малоазийского, дагестанского и черноватого хомяков в качестве последовательного генетического ряда под-? видов убывающей древности.
’ Современное распространение этого хомяка охватывает Сирию, Малую j Азию, Армянское нагорье, Талыш, северо-запад Иранского плоскогорья, в восточное Закавказье, центральный Дагестан и равнинное Предкавказье. 1 Обзоры ареала давались Аргиропуло (1935, 1939а), Нейгаузер (Neuhauser, Г 1936), Шидловским (1940в), Эллермаиом (Ellermann, 1948). Современное I вертикальное распространение среднего хомяка па Кавказе лежит в пределах от 0 до 2400 м. Высотный оптимум закавказского подвида — М. aura г № koenigi Sat., по Шидловскому (1940в), находится в пределах 1200— ! 2000 м (карта 58) <
I ’
. *
304 Часть II. Анализ происхождения четвертичных млекопитающих
На Армянском нагорье, в пределах СССР, этот хомяк наиболее распространен на относительно сухих и теплых верхних участках речных долин. В долине речки Бамбак, по окраинам оз. Севан, на Сарайбулахском и Зангезурском хребтах он живет на участках типчаковой нагорной степи. На холодном плоскогорье Джавахетии держится и на остепненных лугах близ оз. Табисцхури.
В Нагорном Карабахе хомяк известен из-под Геворкевана.
В Талыше прослежен от верховьев Виляж-Чая на юг до Кельвяза, в зоне нагорной злаково-полынной степи на высотах от 1600 до 2000 м. В группировке колючих астрагалов и акантолимонов, как и на Армянском нагорье, не живет, но концентрируется на более влажных площадках по днищам ущелий, особенно по окраинам посевов.
Распространение зверька на хребтах севера Иранского плоскогорья, вероятно, разорвано на отдельные участки. *
В восточном Закавказье хомяк обитает в Горийской котловине, занимая участки древнего остепнения, либо площади, где вырублен лес. В плейстоцене и в голоцене он был распространен в Имеретии и в верховьях Риона. Восточнее, на Иорском и Карталинском плоскогорьях, он живет в формации плотнокустового злака бородача на высотах 550— 600 м. Восточнее Алазани — до Шемахи — его нет, ио возможно, что реликтовые колонии будут обнаружены со временем к северо-востоку от Шемахи, например в ущельях Килязи-Чая, Гильгин-Чая и западнее Дербента.
Современное распространение дагестанского подвида — М. auratus raddei Nehr. ограничено внутренним Дагестаном в долинах Аргуни, верхнего Сулака и Самура. В ущелье Аргуни зверек прослежен от аула Ша-тили до Евдокимовского. По Россикову (1887), известен из Андийского Койсу. По данным экспедиции Огнева, по сведениям Беме (1925), Дюкова (1927), хомяк водится по всему Хунзахскому плоскогорью на полях аулов Баитль, Чондотль и др.
Хомяк живет здесь на участках нагорной ксерофитной степи, тяготея к возделанным участкам — зерновым полям. Вертикальное распространение — от 1600 до 2300 м (Гептнер и Формозов, 1941).
В среднем плейстоцене эти участки обитания, очевидно, имели связи с предгорными, апгперонскими.
Эти связи, вероятно, прервались в верхнем плейстоцене при расчленении Дагестана водной эрозией и при образовании лесной зоны на юго-восточной его окраине.
Современный ареал дагестанского хомяка несколько увеличивается под влиянием сельскохозяйственной деятельности человека.
Распространение предкавказской формы — черноватого хомяка (М. auratus nigriculus Nehr.) весьма широко, но все же преимущественно «прижато» к горам (см. карту 58).
Места обитания на севере Предкавказья лежат в зоне ковыльно-типчаковых степей, а на Ставропольском плато и в предгорной Кабарде -на остепненных лугах и культивируемых участках с посевами кукурузы. Держится зверек также в бурьянистом разнотравье долины Терека (Огнев, 1924).
В ряде мест этот хомяк активно проникает в мезофитные предгорья Предкавказья, где вырублены леса, как например на Закубанской, Кабардинской и Терско-Сунженской наклонных равнинах. Вообще в Предкавказье этот хомяк более эвритопен, нежели его закавказский родич, но все же занимает преимущественно сухие выгревы.
[л. /П. Прежнее и современное распространение и численность млекопитающих 305
| По вертикали в Предкавказье хомяк распространен от уровня моря I до 1000 м. Наиболее далеко заходит он в горы в центральном Предкавказье I у Кисловодска и Нальчика, но в сухие, продольные долины северного I шона он не проник. По долинам левых притоков Терека черноватый I хомяк распространен только до каньонов в хребте Черных гор.
1 В последние десятилетия замечено расселение хомяков в пределы )вской области, в 40-х годах они были обнаружены уже северо-за-
1 Маныча.
свете данных о характере современного ареала малоазийского хо-мяк«>. и его трофических связей представляется вероятным, что проник-I мовеяие предка современного вида на Кавказскую сушу произошло с юга с j запада в третичном периоде, скорее всего в какую-то сухую эпоху чего плиоцена.
уть развития ареала хомяка с нагорий Малого Кавказа в восточное dpt ^кавказье, очевидно, проходил по остепненной зоне предгорий вдоль I подножия юго-восточного склона Большого хребта и далее через низкие то время плоскогорья Дагестана. Позднее этот путь был нарушен развитием сноса суглинков и галечников на предгорные заболоченные рав-1 нины, особенно в районе современных боздагов Аджиноура и Кусарской 1 наклонной равнины. В среднем плейстоцене при обширном остепнении I горных склонов ареал хомяка был, очевидно, наибольшим. Последующая 1 мезофитизация ландшафтов и сдвиги высотных зон в ледниковую эпоху приводили к разрывам ареалов и к понижению границ обитания хомяка. I Hoi ое [нарушение ареала произошло при наступлении послеледниковой i ксеротермической эпохи в условиях расчлененного горного рельефа ’ плоскогорий Передней Азии.
j Во влажных горных районах Имеретии и южной Осетии раннего-I доценовое иссушение хребтов и склонов, наоборот, несколько содействовало расширению ареала хомяка, который был вытеснен там лишь позднее, при новом увлажнении климата.
I Иное толкование истории развития ареала хомяка на Большом Кавказе 1 к в Предкавказье может быть дано при признании видовой самостоятельности дагестанской и предкавказской форм с учетом возможности проис-хождения их от местного третичного предка.
В новейшую эпоху сохранившиеся участки ареала в южном Закавказье относительно устойчивы и сокращаются лишь при перевыпасе скота и распашках больших площадей с применением орошения и частичного уничтожения зверя ядами, как вредителя.
1 Малоазийская песчанка — Meriones blackleri Thos. Остатки песчанок рода Meriones указывались из ашельских и мустьерских слоев пещер Палестины (Picard, 1937; Wolf, 1939). Для Кавказа, кроме упоминаний о находке остатков Gerbillus sp. в плиоценовых слоях Армянского нагорья (Богачев, 1938в), известны лишь остатки современного вида в поверхностных слоях пещер и навесов долины Аракса.
Современный ареал малоазийской песчанки расположен в Малой Азии, Сирии, западном Иране и в части Закавказья (Ellermann, 1948; кюринский, Кузнецов, Кузякин, 1944; Погосян, 1949).
В Закавказье эта песчанка обитает в долине среднего Аракса, местами ? б Зангезурской теснине Аракса, по восточным предгорьям Карабаха, вМильской степи, в южной Мугани и на Кировабадской наклонной равнине. По широкой долине Акера-Чая она распространена почти до Ла-до, т. е. до высоты около 1500 м. Численность малоазийской песчанки в долине среднего Аракса на равнинных участках больше, чем других
20 Н. К. Верещагин
306 Часть II. Анализ происхождения четвертичных млекопитающих
видов, но на каменистых склонах гор она быстро уступает персидской. На левобережье Куры песчанки нет, хотя там имеются условия, но отличимые от правобережных. По Кировабадской наклонной равнине ее распространение прослежено до низовьев Храма. По материалам массовых отловов, в предгорьях Малого Кавказа она преобладает численна над краснохвостой песчанкой.
В Мильской степи малоазийская песчанка обитает наряду с краснохвостой почти повсеместно, но в предгорьях Карабаха держится преимущественно по выгревам с группировками полыней до 350—400 м абсолютной высоты. В северной части Муганской степи эта песчанка сильно* вытеснена распашкой и орошением, распространена главным образом в полынной полупустыне вдоль границы — от Карадонлы до Белясувара. На северо-восточных склонах Талыша прослежена до заставы Эшакчи на высотах около 600—700 м (карта 63).
Зверек селится чаще всего на плотных аллювиальных и делювиальных лёссовидных суглинках широких речных долин с формацией полыни и злакового эфемеретума.
Учитывая особенности распространения малоазийской песчанки в восточном Закавказье и, в частности, отсутствие ее на Куринско-Иорском междуречье и в Ширванской степи, можно думать, что она расселилась на север относительно недавно — в плейстоцене, а быть может даже и в голоцене. В южном Закавказье, т. е. в долине среднего Аракса, малоазийская песчанка может рассматриваться как местный вид верхнетретичного возраста.
Персидская песчанка — Meriones persicus Blanf. Ископаемые остатки этого вида из плейстоценовых слоев достоверно неизвестны.
В голоценовых отложениях остатки известны из пещер южных склонов Армянского нагорья и из долины среднего Аракса.
Основная часть ареала персидской песчанки (карта 62) находится в Турции, Иране, Афганистане и на юге Туркмении (Neuhauser, 1936; Гептнер, 1940; Ellermann a. Morrison-Scott, 1951).
На Кавказе она многочисленна по южным склонам Армянского нагорья, а также в нагорном Талыше. В бассейне Алинджа-Чая ее численность, судя по массовым отловам капканами, составляет в среднем 62.5% от численности других видов песчанок. По левобережью теснины Аракса эта песчанка не идет на север далее Минджевани. По ущельям южных склонов Зангезурского хребта распространена до 1800 м в формации трагакантовых астрагалов и известна даже из-под Зангелана, Шах-буза.
На Сарайбулахском хребте, по Далю (19406), песчанка составляет до 17% от численности других видов грызунов в рационе филина. По ущелью Занги прослежена до высоты 1230 м (Даль, 19486) и весьма обычна по склонам северной окраины долины среднего Аракса в формациях горпо- ! солянковой полупустыни (Флеров и Гуреев, 1934; Аргиропуло, 19396;. ; наши данные 1947 г.).
В нагорной степи Талыша эта песчанка обычна в верховьях Виляж- j Чая и в Зуванде на высотах 1500—1800 м. Так, близ поста Дыман в 1945 г, ее остатки в погадках сыча, по нашим сборам, составляли 26% от остат- I ков других грызунов.
Вообще в сухих горных ландшафтах северного Ирана и Малой Азии это — по-видимому, самый многочисленный и широко распространенный вид песчанки. Обычный биотоп персидской песчанки — каменистые склоны ущелий, покрытые местами подушками колючих астрагалов.
Гл> Ш' Прежнее и современное распространение и численность млекопитающих 307
акантолимонов, полынью, а весной еще и эфемерными злачками и маками. По ущельям зверек выходит местами и на водоразделы под границу злаковой нагорной степи.
Привычка рыть норы под камни, под корни кустарников, искусное лазание по камням, голые ступни, длинный хвост с метелкой и длинные вибриссы персидской песчанки — все это, по мнению Аргиропуло (1939а), ie приспособительные признаки, формировавшиеся в условиях гор-аменистой пустыни.
Зверек этот по преимуществу зернояден: обычной его пищей в первую половину лета служат семена мелких злачков, а во вторую — горных солянок.
В целом это — древний индикаторный вид переднеазиатского нагорно-пустынного териокомплекса.
Сельскохозяйственная деятельность человека местами содействует процветанию персидской песчанки, так как близ зерновых посевов и вдоль заборов, устроенных из голышей, численность зверька бывает обычно несколько повышенной. Перегонное скотоводство и неумеренный выпас скота ведущий к расширению формации колючих астрагалов, также вызывает некоторое расширение ареала грызуна.
Краснохвостая песчанка — Meriones erythrourus Gray (s. lato). Ископаемые остатки этой песчанки известны из -среднеплейстоценовых слоев Апшеронского полуострова.
В настоящее время этот полиморфный вид (описано до 23 подвидов), отождествляемый с североафриканской М. libycus Licht., имеет огромный ареал: Северная Африка, Передняя Азия, восточное Закавказье, Средняя Азия до Семиречья. Северная граница на Кавказе проходит на широте Шемахи (Ellermann a. Morrison-Scott, 1951; Бобринский, Кузнецов, Кузякин, 1944).
В Закавказье она распространена по всей Кура-Араксинской низменности и в предгорьях — от Тбилиси на западе до берега Каспия на востоке (карта 61). Особенно многочисленна краснохвостая песчанка на Апшеронском полуострове. На побережье Каспия не наблюдалась севернее Килязи и южнее Кызыл-Агачского залива.
Центральные участки низменности заселены этой песчанкой повсеместно.
В ровной полупустыне Мильской, Ширванской и Муганской равнин большие плотности населения наблюдаются в полынно-солянковой и каперцовой группировках. Зверек селится чаще всего по аллювиальным грядам и по окраинам блюдцеобразных понижений — солончаков, заросших кустами древовидной солянки, австрийской полыни и полыни Совпча. Наибольшие плотности отмечены все же по песчаным береговым грядам Куры, Алазани, Иары, Аракса, особенно в зоне тамариксовых зарослей. Такие же высокие плотности населения бывают и по дюнным всхолмлениям побережья Каспия, там, где песчаные бугры закреплены тамариксом и эфедрой. В таких местах насчитывается до 40 особей на гектар. Огромное значение для поселений этой песчанки имеют биотопы, созданные человеком, особенно виноградники, заброшенная оросительная сеть, стоянки овцеводов в степях и многолетние залежи.
По типу активности это сумеречное и ночное животное летом, сумеречное и дневное — весной и осенью, дневное — зимой. По типу питания краснохвостая песчанка преимущественно зерноядна: весной питается побегами эфемерных однолетников; летом — побегами и семенами эги-лопса, рейграса, костра и насекомыми, в частности нестадными саранчо-20*
308 Часть II. Анализ происхождения четвертичных млекопитающих
выми; осенью — новыми побегами эфемерных злаков; зимой — стеблями полыни, семенами древовидной солянки, шведки и др.
Нередко в поисках семян влезает довольно высоко на кусты солянок. При снегопадах эти песчанки обычно отсиживаются в норах до момента его стаивания, реже пасутся на близлежащих к норе кустах.
Обилие зверьков на песчаных повышенных грядах в ровных степях объясняется легкостью устройства вентилируемых нор и зимней жировки в группировке полыни и солянок. Здесь же устраиваются и стоянки для скота — «ятаги». Сочетание обилия грызунов и скота создает иногда у неопытных наблюдателей впечатление благоприятного воздействия скотоводства на состояние зимующей популяции песчанок (Гладкина, 1952).
По современному ареалу и ландшафтной приуроченности краснохвостая песчанка сформировалась как вид в пределах южного и юго-восточного Средиземноморья. Кавказский перешеек она заселила с юга и юго-востока, вероятно, еще в плиоцене или но крайней мере в нижнем плейстоцене, если учесть, что в среднем плейстоцене она уже жила на Аи-шеронском полуострове.
Эпохи трансгрессий четвертичного Каспия песчанка переживала на третичных всхолмлениях по северной и западной окраинам Куринского залива. В восточной части Кура-Араксинской низменности этот вид является одним из немногих «молодых» колонизаторов территории, освободившейся в голоцене от морских вод.
Местная земледельческая культура сказывалась до сих пор благоприятно на процветании вида. Однако современная инженерная техника орошения и дальнейшее развитие хлопководства с применением обильных поливов будут быстро вытеснять эту песчанку на обширных площадях.
Степная полевка — Mlcrotas socialis Pall. (s. lato). Остатки этого вида найдены в срэднеплейстоценовых слоях Апшеронского полуострова, в голоценовых слоях пещеры Сарайбулахского хребта в Армении и во многих других пунктах, лежащих в пределах современного ареала.
Современный ареал зверька захватывает Киренаику, Балканы, южную Украину, степной Крым, Переднюю Азию, Среднюю Азию, восточное Предкавказье и восточное Закавказье (Минин, 1938; Виноградов и Аргирэпуло, 1941; Ellermann, 1948).
В Предкавказье эта полевка распространена в Сальских степях по долине Маныча, в районе низовий Малки, Баксана, на склонах Терского и Сунженского хребтов и в остепненных предгорьях северного Дагестана. В Терско-Кумский массив песчаной полупустыни зверек проникает по прирусловым полосам протоков и каналов Кумы, Куры и Терека.
На Кусарской наклонной равнине полевка распространена по выгрев-ным участкам до высоты 600 м, где ее ареал соприкасается уже с ареалом обыкновенной полевки. В низовых лесах Хачмасской низменности зве- ; рек обитает лишь на широких распаханных полях.
От западного побережья Каспия степная полевка распространена s ; восточном Закавказье до высоты 700—800 м на предгорных увалах (карта : 72).
В низовые леса Алазано-Агричайской долины полевка проникает I по обезлесенным участкам. На Ашперонском полуострове и в Кабристане ; имеются ныне небольшие реликтовые колонии на участках, имеющих ? всего до 280 мм осадков в год, но при местном дополнительном увлажне- ; нии почвы, подстилаемой водонепроницаемыми плитами апшеронского известняка. Сплошное распространение зверька в Кура-Араксинской низменности начинается в районе, где выпадает не менее 350 мм осадков :
Гл. И/. Прежнее и современное распространение и численность млекопитающих 309
I
a
и я
У )-). j-i-э-
>М гт-
ia го
Ul о-
II-
•о-дх
IX.
Да ja, во
ж-эч-
I и
по эго на.
по
юв-ТОМ nie-
а в рта
сает гапе 1Ц11Х к некого жой JKOB
I р. год. В юго-восточной Мугани предел распространения полевки в Ленко-I раненую низменность ставится большой зимней влажностью почвы при | осадках около 800 мм в год (Верещагин, 1942в, 19466).1
По долине Куры полевка встречается до Мцхети, а теперь и в Мух-’ райской долине, куда проникла лишь несколько десятков лет тому назад 5 в связи с обезлесением этой долины. После перерыва, образованного ле-J спетым узким ущельем Куры у Мцхети, зверек обитает вновь в сухой Го-J дошской котловине на высотах 600—800 м. Здесь обособился слабо отличающийся от типичного подвид М. socialis goriensis Arg. НаКировабад-ской наклонной равнине полевка обитает до современной нижней границы леса, т. е. до высоты 600—700 м, а по днищам некоторых ущелий поднимается и выше. На обезлесенных северо-восточных склонах Карабаха степная полевка распространена до высоты 800 м близ Мадагиса, Марда-! керта, Мартуни и других пунктов, где ее ареал соприкасается с нижней ; границей ареала кустарниковой полевки. По южным склонам Карабаха полевка проникает высоко в горы по долинам Акера и Охчи-Чай. Такая же картина наблюдается по ущельям южного склона Армянского нагорья.
В полупустыне Кура-Араксинской низменности полевка образует неустойчивые поселения в группировке каперцов на каштановых поч-! вахп слабее в полынно-злаковой. Летние условия для зверька взоне полу-{ пустыни песснмальны вследствие «физиологической сухости» корма и высокой температуры почвы. Полевка выдерживает эти условия, переходя отчасти на питание саранчовыми и моллюсками и устраивая глубокие норы (Верещагин, 19466).
Численность полевки в полупустыне ежегодно катастрофически па-Г дает к нюню вследствие прекращения размножения и быстрого вымирания взрослых особей из-за высыхания корма и высоких температур. Отдельные жизнеспособные колонии зверьков уцелевают при этом по не-Г многим ложбинкам, оврагам с зарослями расторопши, в увлажненных поливом садах, под кустами каперцов и т. п.
В Муганской степи, в районе Белясувара и Астрахан-Базара, в годы массового размножения наблюдаются бессистемные массовые передвижения стенных полевок по ночам в первой половине июня вследствие «физиологического беспокойства», вызванного высыханием кормовой растительности и перегревом почвы (Мамедов, I960).2
В связи с своеобразными изменениями суточной и сезонной активности, зависящей от температурных условий и инсоляции, степные полевки в Азербайджане истребляются хищными птицами и млекопитающими преимущественно в зимнее время.
Очень характерные «пульсации» плсшадей распространения зверька прослеживаются по годам близ основания Апшеронского полуострова. В случае ряда последовательных теплых и влажных зим и прохладных летних периодов, как правило, происходят нарастание численности полевок и вспышка массового размножения. При влажной осени в полу-’ мстыне образуются большие запасы корма в виде побегов растений, раз-потравно-злакового эфемеретума. Это позволяет полевкам продвигаться с Кабристанских увалов по долинам пересыхающих речек почти до мор-
------------
1 Наряду с этим рассказ К. Н. Россикова о массовой перекочевке полевок днем в августе в Мильской степи, переписанный Огневым (1950) как достоверное событие, является только вымыслом (Верещагин, 19466).
I 2 Сообщение Огнева (1950) об сбптанвп этой полевки в Ленкоранской низменности |\ основано на путанике в местоположении географических пунктов: пост Кельвяз на-> ходится в высокогорном Талыше, а не в «Талышской низменности».
310 Часть II. Анализ происхождения четвертичных млекопитающих
ских береговых гряд, на 30—40 км от границы постоянного обитания! Между тем первый же жаркий и сухой сезон вновь ведет к вымиранию зверьков и к новому отступлению к северо-западу границы их распространения. Это явление крайне показательно для объяснения характера и быстроты миграций, а также истории развития ареала зверька в результате вековых изменений климата и ландшафта.
В Талыше степная полевка живет на высотах 1500—1800 м в нагорной злаковой степи, но отсутствует в малокормной группировке колючих астрагалов. Большая плотность населения полевки развивается на восточных и северных склонах Талыша, близ верхней опушки леса, которая сильно «спущена» вниз рубкой и выпасом скота. Верхние части ущелий Виляж-Чая в районе Ярдымлы и в верховьях Вазару-Чая близ Лерика, используемые под посевы зерновых, являются ныне настоящими очагами массовых размножений полевки. Возникновение этих очагов обусловлено целиком деятельностью человека. Отсюда полевка проникает вниз до днищ ущелий восточных склонов Талыша в том случае, если лес разрежен или целиком уничтожен человеком.
В долине среднего Аракса полевка встречается лишь в Аралыхе и на прирусловых участках некоторых притоков Аракса, например по Алинджа-Чаю, Нахичеван-Чаю, Арпа-Чаю, и совершенно отсутствует на выжженных солнцем склонах и увалах Зангезурского хребта в зоне каментстой горно-солянковой полупустыни и трагакантовых астрагалов. Эго связано с малэкормяэстью упомянутых группировок. В центральной j части Армянского нагорья полевка проникает по долинахМ рек левобережья f Аракса. В верхней части долины р. Бамбак, в зоне нагорной злаковой степи, на высотах 120Э — 1800 м расположен участок обитания горно-степной полевки, описанной под именем М. socialis schidlovskil. Этот участок отгорожен с запада холодным Ахалкалакским плоскогорьем, а с севера — лесистыми отрогами Бэзобдальского хребта и тесниной ущелья р. Бамбак. Он не имеет никакого отношения к ареалу горийского подвида, в противоположность мнению Огнева (1950). Наоборот, связи этого участка ареала надо искать на юге в районе Ленинакана и Карского нагорья, где в сухих горных долинах, вероятно, имеются популяции, морфологически близкие к полевке Шидловского. Даль и Захарян (1951) показывают распростра- f некие полевки Шлдловского от верховьев Бамбака по долине Аракса до широты Еревана.
Наибольшие плотности населения степной полевки в периоды вспышек массового размножения, при которых количество отверстий нор на гектар доходит до 9Э ЭЭЭ, отмечается на пахотных и природных угодьях в зоне предгорной борэдачевой степи на Аджиноурском и Карталинском плоскогорьях, в восточных предгорьях Карабаха и на северо-западных склонах Талыша. В этом степном ландшафте и находится современный экологиче- L ский оптимум зверька (Сатунин, 1912а; Верещагин, 1942в, 19466). Е
Ж 1знеспособлые даже в периоды сильнейшего угнетения численно- Е сти колонии грызуна прочно связаны здесь с кустами держи-дерева, I дающими надежное убежище от пернатых хищников и перегрева. Грани- F чащие с борэдачевой степью можжевелово-фисташковые леса — так I называемое «аридное редколесье» — заселены полевкой слабо. Она дер- Е жится здесь по осветленным полянам с разнотравьем, устраивая норы г под кустами эфедры. ;
Характер и сроки размножения степной полевки зависят от высоты | и обитаемой зоны. В то время как в зоне полупустыни на низменности | восточного Закавказья ее размножение приурочено преимущественно I
Гл. 1 IL Прежнее и современное распространение и численность млекопитающих 311
к октябрю—ноябрю и к марту—апрелю, а в случае теплой зимы и ко всему периоду вегетации эфемеретума — с октября по май, в зоне нагорной степи Талыша и на Армянском нагорье интенсивное размножение цблюдается в июне, соответственно более позднему выгоранию раститель-* ти.
На богарных посевах зерновых в зоне предгорной степи и на полив-до посевах зерновых (рис. 124) и люцерны в зоне полупустыни восточ-
Рис. 124. Степная (общественная) полевка среди посева ячменя.
Фот. автора, 1939.
кого Закавказья полевка сохраняет дольше, нежели на целине, половую активность и вообще жизнедеятельность.
Основные природные факторы колебания численности и размеров ареала степной полевки — климатические, особенно в восточном Закавказье. Тем не менее эпизоотии, хищные млекопитающие и птицы имеют также огромное значение для снижения численности этого грызуна в годы его массового размножения. Наибольшее влияние пернатых хищников на численность полевки сказывается в полупустыне и в предгорной степи ^ноября по апрель, т. е. во время пролетов и зимовки птиц (Верещагин, 19466).
Сельскохозяйственная деятельность человека содействовала расширению пределов существования этого вида, предоставляя ему новые территории при вырубке низовых лесов и обеспечивая интенсивное «змножение сочным и калорийным кормом иа озимых посевах зерновых.
Основные морфологические черты степной полевки с ее слабо дифференцированным, легко намокающим шерстным покровом, короткими конечностями, мало приспособленными к быстрым и дальним передвижениям уплощенным черепом и удлиненной диастемой, показывающими огромную способность рытья нор в почвах тяжелого и среднетяжелого механического состава, свидетельствуют о формировании вида в условиях относительно ксерофитных ландшафтов восточного Средиземноморья.
Часть ZZ. Анализ происхождения четвертичных млекопитающих
Своеобразное распространение степной полевки на Кавказе и в Средней Азии, нигде не выходящее в настоящую пустыню, но вкрапленное в нее интразонально на размытых останцах, говорит о широких древних связях разобщенных ныне участков степей и полупустынь, которые, очевидно, имели место в отдельные отрезки плейстоцена. Соображения по этому поводу уже были высказаны (Верещагин, 1942в, 19466; Гептнер,1945).
Мнение Пидопличко (1951) о недавнем развитии ареала этой полевки в восточном Средиземноморье не обосновано в связи с фактическими ошибками в описании ареала.
В восточном Средиземноморье, на относительно сухих плиоценовых плоскогорьях. Малой Азии, протекала, по-видимому, первоначальная эволюция целой группы форм южных серых полевок. Одна из них — наша степная полевка — и заселила с нижнего плейстоцена сухие участки побережий восточносредиземноморских бассейнов.
История развития ареала степной полевки на Кавказе в связи с лабильностью ее экологических черт особенно сильно зависела от горообразовательных процессов, колебаний уровня Каспия и смещений ландшафтных зон как по горизонтали, так и по вертикали. Так* например, изоляция горийского подвида была обусловлена подъемом северных отрогов Триа-летского и южных отрогов Кахетинского хребтов, перегородивших долину Куры у Мцхети. Более древнее обособление имелось у полевки Шидловского, которую, вероятно, следует считать уже особым видом. Это обособление было связано с подъемом Армянского нагорья в четвертичном периоде при сохранении ксерофитных группировок на отдельных его участках. Изоляции способствовали, возможно, также обширные излияния лав, перекрывавших в разных направлениях участки древнего плато (Верещагин, 1942в; Шидловский, 1945).
Ареал степной полевки на перешейке, очевидно, неоднократно «пульсировал» на протяжении плейстоцена соответственно развитию и сокращению пригодных для обитания ландшафтов в различные климатические фазы. В нашу эпоху идет энергичное расширение ареала грызуна за счет проникновения его в предгорья и в горы при вырубке лесов и осушении склонов. Особенно хорошо оно заметно на горных склонах в Карабахе и Талыше. Проникновение полевки в полупустыню ив пустыню происходит при проводке каналов и освоении для зерновых посевов новых территорий в восточном Предкавказье, причем современные истребительные работы не в состоянии сократить сколько-нибудь заметно этот процесс.
Обыкновенная полевка — Microtus arvalis Pall. Ископаемые остатка обыкновенной полевки известны из многих плейстоценовых захоронений в пещерах и в аллювиальных отложениях Западной Европы — от Ирландии до Карпат и от Италии до Швеции. В пределах СССР остатки этих полевок обычны в верхнепалеолитических слоях Крыма (Виноградов, 19376) и в средне- и верхнеплейстоценовых отложениях нижнего Дона и Урала (Верещагин и Громов, 1952).
На Кавказе кости и черепа этой полевки обнаружены в среднеплей* стоце новых слоях Апшеронского полуострова вместе с костями степной полевки (Аргиропуло, 19416; Громов, 1952).1
В нашу эпоху эта полевка распространена в таежной, лесостепной и даже пустынной зонах — от Атлантического океана до Хингана и верховьев Амура. Распространение ее на Кавказе своеобразно (карта 71).
1 В 1958 г. нижние челюсти полевок этого вида найдены в нижнеплейстоцено-вых слоях пещеры Кударо I.
Гл. П!..Прежнее и современное распространение и численность млекопитающих 313
В Предкавказье почти сплошной ареал полевки идет из Донских степей через Маныч по побережью Азова, охватывая низовья Кубани и Таманский полуостров.
Широко распространена затем эта полевка на Ставропольской воз-'ценности, где заселяет мезофитные луга и вообще площади, на которых блей лес.
Северо-восточная граница сплошного распространения обыкновенной п:(‘вки в предкавказских степях проходит примерно на долготе Буденновска, совпадая с границей полупустынь в области низовой Кумы, но вдается далеко на восток по долине реки в массив Терско-Кумских песков. По грядами понижениям с луговыми, тугайными и остепененными шировками вдоль русел Терека и Сулака эта полевка обитает вплоть-и побережья Каспия. В сухой степи Предкавказья большое значение ,! обитания полевок имеют увлажненные, но незасоленные понижения ефа (Наумов, 1948). На Большом Кавказе эта полевка живет только , ^кубанской наклонной равнине и в предгорьях до 900—950 м, но выше . сах не водится.
1а горных лугах западной части Большого Кавказа она всюду заме* а кустарниковой полевкой.
h наклонной равнине центрального Предкавказья полевка всюду обыкновенна, но в горах или редка, или отсутствует (Огнев, 1950). На Плесенной Терско-Сунженской наклонной равнине многочисленна деду и является вредителем. Туров (1926в) считал ее довольно обыкно-ч ной только в лесной зоне Северной Осетии. По нашим наблюдениям цельях Фиат-Дона и Гизель-Дона, эта полевка в массе обитает на ; песенных полянах и субальпийских лугах Лесистого и Скалистого
лов. Обычна эта полевка и в ущелье Уруха на высотах 1500—1800 м. це на восток прослежена в бассейнах Аргуни и Сулака на высотах ЯОдо 2700 м (Беме, 1933; Гентнер и Формозов, 1941).
В предгорьях Дагестана от Хасав-Юрта до Дербента, а также южнее,.
। низкой приморской террасе вплоть до Баку, этой полевки нет.
Отсутствует она и в сырых местах Хачмасской низменности, населен-степными полевками, лесными и домовыми мышами. На Кусар-(()й наклонной равнине нижняя граница ареала проходит на высота п.)0—650 м, в зоне стравленного леса — «шибляка», и лишь по луго-s5$ участкам долины Самура спускается до 250—300 м.
Указание Беме (1928, стр. 143) на поимку зверька близ устья Самура сдаивает внимания,' если только не основано на ошибке.
На восточной оконечности Большого Кавказа этот грызун встречается на полях зерновых и на субальпийских лугах в районе Конах-и Хизы. В районе Шемахи нижняя граница ее распространения доходит на высотах около 800 м, но далее к западу на субальпийских лу--пверховъях Гердыман-Чая этот вид замещен кустарниковой полевкой.
В районе обезлесенных человеком участков южного склона Боль-шо хребта, по ущельям Демир-Апа раи-Чая, Бум-Чая, Мазым-Чая шз Куткашена, Варташена, Лагодехи, полевка проникает с горных 5гов по полянам, заросшим папоротником орляком, до самых поднови крутых склонов, в зону низовых лесов Ал аз а но-Авта райской додал На высокогорных лугах восточной половины южного склона Большого Кавказа полевка эта концентрируется обычно по окраинам обильно даоженпых стоянок скота, там, где пышно развивается конский щавель 1 сборная ежа. Плотно заселяет она и субальпийские луговины по обез-десенным гребням боковых хребтов меридианного направления.
314 Часть II. Анализ происхождения четвертичных млекопитающих
Далее полевка обычна на луговых участках Кахетинского хребта и в сильно обезлесенной Южной Осетии, вплоть до верховьев Риона и его притоков, где, как и на северном склоне, целиком замещается кустарниковой полевкой.
Этой полевки нет в сухой Горийской котловине и в сырой Колхиде, по Черноморскому побережью, на южном склоне западного Кавказа и на северных склонах Аджаро-Имеретинского хребта (Сатунин, 1913* Шидловский, 1947, 1948, 1950).
На Триалетском хребте и его отрогах обыкновенная полевка обычна всюду, как и на Мокрых горах и по луговым угодьям Джавахетии.
На Армянском нагорье, широко заселенном обыкновенной полевкой, нижняя граница ее распространения сильно поднимается на востоке и юге. На хребтах, окружающих оз. Севан, эта полевка повсеместна (Туров и Турова-Морозова, 1928), но на северном сильно обезлесенном и относительно сухом склоне она не была обнаружена нами ниже Кода-бека (1400 м).
На горных лугах Карабаха, например в районе Дели-Дага, Сары-Ери, Басаргечара, она очень обычна, а на обезлесенных восточных склонах •спускается до высоты 1200—1300 м.
По южному склону Зангезурского хребта в бассейне Акулис-Чая прослежена нами на мочажинах среди остепненных и сильно засоренных выпасом скота отрогов и ущелий лишь с 1800 м и выше. Вовсе нет этой полевки в зоне трагакантников, т. е. в формации «фриганы» и «гариги», а тем более в полынной и солянковой полупустыне, долине среднего Аракса и Кура-Араксинской низменности. Нет этой полевки и в Ленкоранской низменности, а тем более на Мугани, вопреки утверждениям Огнева (1950, стр. 209) и картам Кузнецова (Бобринский, Кузнецов, Кузякин, 1944). В высокогорном Талыше полевка начинает встречаться лишь с 2000—2100 м по остепненным луговинам, например у постов Кельвяз и Космальян. В лесной зоне Талыша она не живет даже в местах, заселенных кротом. Вообще современное распространение обыкновенной полевки в восточной половине Кавказского перешейка до известной степени противоположно распространению степной полевки. В северо-западном Иране и в северо-восточной Турции обыкновенная полевка распространена на высотах 2000—2500 м, а далее к югу зона обитания, очевидно,, еще более повышается. Бленфорд (Blanford, 1876) отметил эту полевку под именем Arvicola myslacinus De Fil. между Ширазом и Исфаганом, на высоте около 2800 м.1 На восточном Эльбурсе она отмечена Гудвином (Goodwin, 1939). Водится, вероятно, наСавеланеи добыта в районе оз. Урмии экседицией Кавказского музея. В коллекциях Музея Грузии имеются экземпляры из Карса, Ольты, с Большого Арарата и из других мест Турции. Многочисленность этой полевки в высокой Алапь кертской долине — в верховьях Евфрата — объясняется обилием заболоченных мест и ручьев. В центральной Турции немногие уцелевшие места обитания считаются реликтовыми (Neuhauser, 1936; Ellermann, 1948).
В связи с повышением суточных и годовых температур фактор влажности почвы и мезофитности растительности приобретает для грызуна на юге все большее значение. Поэтому зона распространения обыкновенной полевки по вертикали закономерно повышается в южном направлении, а ее ареал рассекается на изолированные реликтовые островки, приуроченные к высоким плоскогорьям и хребтам.
1 В нагорном Талыше и далее в Иране обичает по-видимому, полевка, близ» кая к М. guentheri Danf. et Alst.
1
I к'Ш' Прежнее и современное распространение и численность млекопитающих 315
бта она ку-
да, :аза ИЗ;
шна
кой, се и (Ту-м и 'еда-
Ери, о нах
-Чая эрея-в нет : «га-сред-Лен-
ниям ецов, ггъся остов стах, овен-стной iBepo-левка ания, метил юм и [ечена в рай-t Гру-
И из \лаш->м заевшие 1948). влаж-ызуна новен-равле-ровки,
I, близ»
J Ареал зверька, очевидно, полностью выклинивается на высоких гор-I вых хребтах в среднем течении Тигра и Евфрата.
f Отсутствие зверька в западной части Кавказа и Закавказья уместно Уяснять избыточным увлажнением, а в лесном и низменном Талыше^— *1 летней сухостью почвы и растительности.
По типу питания обыкновенная полевка — типичное травоядное жи-! вогное. Концентрированные корма в ее современном рационе появились ' как следствие влияния человека в историческом периоде.
Основные природные факторы, ограничивающие численность этой по-левкп в горах и на равнинах, — это неблагоприятные климатические g погодные условия. Воздействие четвероногих и пернатых хищников взетшш сезон на эту полевку в условиях Кавказа, значительно больше, : ш на степную.
Судя по современному ареалу и экологии, обыкновенную полевку принято считать европейским мезофильным видом, эволюционировавшим g условиях умеренного климата и влажности на растительных группировках лугового типа.
Согласно этому представлению, появление и полное обособление круп-• йх популяций обыкновенных полевок на хребтах Передней Азии и Кавказа может быть объяснено древним расселением вида в какую-то влаж-! ную эпоху при широком развитии луговых или по крайней мере степных формация в опустыненных теперь жарких долинах и склонах.
При таком толковании особенностей современного ареала и экологи-| веских черт этой полевки появление ее на Кавказе вряд ли могло произойти j ранее середины плейстоцена, в эпоху значительного похолодания и надви-] гааяя мезофитных луговых группировок на зону полупустыни и пустыни. | Местное — высокогорное — происхождение обыкновенной полевки 1 з горах восточного Средиземноморья и последующее расселение ее к се-* веру могли бы получить признание при получении доказательств среди-; коморского происхождения формаций северных мезофитных лугов.
Высокогорное происхождение лугов и возможность спуска их в плей-, стоцене с гор к северу признается теперь некоторыми ботаниками (см. Федоров, 1952).
Кустарниковая полевка — Microtus (Pitymus) та fori Thos (s. lato). ’ Ископаемые остатки полевок, относимых к подроду Pitymys, известны j аз плейстоценовых отложений Евразии и Северной Америки (Wolt, 1939; , Simpson, 1945). На Кавказе остатки особого вида полевки М. apschero-1 nicos Arg. — вероятного предка современного кавказского вида обнару-s жены в среднеплейстоценовых слоях Апшеронского полуострова.
i распространение кавказской кустарниковой полевки захватывает
Вольшой Кавказ, Малый Кавказ, Малую Азию, Талыш и Эльбурс (карта
: Я
В Предкавказье кустарниковая полевка обитает от северной границы предгорных лесов до осыпей перевалов Главного хребта.
На Закубанской равнине зверек распространен местами до высоты ЙО-300 м и обычен от Крымской до Майкопа — на первых увалах • уцелевшими участками дубовых лесов и мезофитных лугов. До етного русла Кубани эта полевка не доходит даже по долинам ле-1 sax притоков и всюду заменена здесь обыкновенной полевкой. На восток граница повышается и проходит южнее Черкесска на высоте около {00 м. В лесной зоне и на альпийских лугах северо-западного Кавказа зверек всюду очень обыкновенен, и обильные коллекционные сборы мются из Кавказского и Тебердинского заповедников.
316 Часть II, Анализ происхождения четвертичных млекопитающих
В центральном Предкавказье зверек обитает по наиболее мезофит-ным участкам ущелий, и на большей части сухой Кабардинской наклонной равнины отсутствует.
Изолированный участок обитания находится в массиве дубовых лесов Пятигорья (Бештау, Железная, Развалка). Еще далее к северу выдвинут обособленный участок обитания на Ставропольской возвышенности. Здесь полевка живет на засоренных выпасом луговых полянах среди остатков широколиственных лесов, например в окрестностях горы Стрижамент.
Разрыв этого участка ареала с основным горнокавказским достигает 50—60 км и занят ныне остепненными предгорьями. Имеются реликтовые пятна обитания и в верховьях Калауса по байрачным лесам балок. На Терской и Сунженской наклонных равнинах она обитает теперь на мезофитных лугах, недавно возникших после вырубки леса.
Вопреки мнению Огнева (1950), этой полевки нет ни под Моздоком, ни под Грозным: здесь для нее слишком сухо. В центральном Дагестан она обитает на луговых участках на высотах 1600—2500 м.
В западном Закавказье кустарниковая полевка обычна всюду от уровня берега Черного моря до осыпей перевалов хребтов. В сырой Колхиде ее нет лишь на участках периодически затапливаемых ольховых дебрей, моховых и тростниковых болот. На альпийских лугах Сванетии полевка весьма многочисленна от опушек березового леса с черничником до самых верхних участков злаково-лютиковых ковриков, вдающихся в осыпи шиферного сланца. Здесь она полностью замещает обыкновенную полевку. На восток зверек прослежен по всему южному склону Большого Кавказа вплоть до верховьев речки Пирсагат в Азербайджане.
На альпийских лугах в районе Нухи, Куткашена зверек бывает замещен в отдельные годы обыкновенной полевкой. Утверждения Арги-ропуло (1939а) о том, что кустарниковая полевка не живет в зоне леса Большого Кавказа, ошибочны, наоборот, на. опушках леса и в разреженных насаждениях дубово-грабовых лесов наблюдается нередко высокая численность зверька.
В Алазано-Агричайской долине, в ущельях Аджиноурского плоскогорья и на Куринской низменности эта полевка не живет нигде. Упоминания о находке зверька в Геок-Тепе, на пустынных всхолмлениях к северу от Евлаха (Шидловский, 1938; Огнев, 1950), ошибочны (Верещагин, 1949в).
В восточном Тавре и по системе хребтов Малого Кавказа зверек распространен преимущественно на северных склонах, а также на высоких влажных плоскогорьях в Джавахетии, Цалке.
Весьма обычен он как на обезлесенных рубкой склонах ущелий Шах-дагского хребта, так и в сохранившихся буковых лесах ущелий Тер-тер-Чая в Карабахе, где спускается до 1000—1200 м абсолютной высоты. В Талыше кустарниковая полевка живет лишь по опушкам у верхней границы леса и чрезвычайно редка вследствие большой сухости в летнее время (Верещагин, 1949д). Самый юго-восточный участок ареала зверька расположен по северному склону Эльбурсского хребта (Ellermann, 1948).
Особенно постоянно заселены этой полевкой разнотравные луга близ верхней опушки леса. На этих участках с наибольшей численностью населения и лежит экологический оптимум зверька.
По типу питания кустарниковая полевка близка к обыкновенной, на большие роющие способности свидетельствуют о бблыпей роли корневищных частей травянистых растений в ее рационе.
Гл. III. Прежнее и современное распространение и численность млекопитающих 317
Динамика численности этой полевки значительно устойчивее, нежели v степной и обыкновенной. Значение пернатых хищников для снижения численности кустарниковой полевки значительно меньше в связи с более юдземным образом ее жизни и обитанием в высокотравье и лесах.
По степени дифференцировки мехового покрова на подпушь и ость она занимает промежуточное место между степной и обыкновенной полевками.
В целом кустарниковая полевка — типичный мезофильный вид восточного Средиземноморья, сформировавшийся на горных лугах и в широколиственных лесах. Это — хороший индикаторный вид, свидетельствующий о длительном существовании мезофитных условий на отдель-г IX участках Малоазийско-Кавказской суши. Наибольшее распростра-t нпе эта полевка, очевидно, приобрела в эпоху самого сильного похо-дания в плейстоцене.
Ставропольско-кавказский разрыв ареала, вероятно, образовался в начале голоцена, малокавказско-гирканский — несомненно значительно з :-евнее.
Кавказская снеговая полевка — Microtus (Chionomys) gud Sat. i lato) (рис. 125). Ископаемые остатки этой полевки обнаружены при j иконках пещер Кударо в верховьях Риона. В настоящее г.-земя зверек распространен в горных районах Большого и Ма-•?}Го Кавказа и восточного Тавра (карта 68). На северо-западном Кавказе эта полевка чаще всего встречается в пределах от 1600 до 2000 м а Перевальном и Скалистом хребтах и отсутствует на передовой цепи [еряых гор (Насимович, 1935).
В ущелье Кубани и Теберды, по нашим наблюдениям, полевка появится на высотах 1500—1700 м по склонам Скалистого хребта и особенно адогочисленяа выше в субальпийской зоне по россыпям крупных скал, поросших сосной, березой, ивой, далее на альпийских лугах, вплоть до □сыпей и ледников.
На Ставропольской возвышенности этой полевки теперь нет (Верещагин и Громов, 19535), но она живет в россыпях трахитовых скал на участке вековой мерзлоты горы Развалка в Пятигорье, на высоте всего около 600м. Эта популяция в настоящее время изолирована от основного ареала ва50 — 60 км к северу, так как ближайшее местонахождение полевок находится в каньоне ручья Березовка, к югу от Кисловодска (Верещагин, 1953а). Выше кавказская снеговая полевка живет по ущельям Малки, Баксана, Чегема, Черека, особенно в россыпях скал в каньонах, проре-I заиных этими речками в Скалистом хребте. Встречается она местами и ! во внутрэнних продольных долинах на высотах 1200—1500 м, там, где ее-тьрэссыпи камней. В Черных горах под Нальчиком эта полевка редка.
В Ардонском ущелье наибольшая численность полевок наблюдалась вамп на боковой морене в сосново-березовом лесу близ конца Цейского ледника, среди гранитных скал, заросших папоротником, черникой, j голубой жимолостью и рододендроном.
I Для ущелья Терека описано обилие этой полевки на склоне горы I Фетхус близ Орджоникидзе (Формозов, 1926).
1 В центральном Дагестане эта полевка весьма многочисленна на Гунибе, | наБогосском хребте, в ущельях притоков Самура на высотах 1300, 1600, | 2100, 2500 м (Гентнер и Формозов, 1941).
1 В августе 1952 г. я наблюдал множество этих полевок в сухой и жар-
•п1 котловине в верховьях ручья Манас среди россыпей скал мергеля
318
Часть II. Анализ происхождения четвертичных млекопитающих
у сел. Лаваши, на высоте около 1000 м. Летние условия для зверька здесь уже близки, по-видимому, к пессимуму.
По южному крутому склону Большого Кавказа в восточном Закавказье эта полевка встречается по ущельям на высотах 1300—1400 м, например в ущелье Гердыман-Чая и другим, ио обычна лишь в амфитеатрах истоков речек в березово-грабовых опушках на высотах 2000—2600 м. Встречается также на гребнях поперечных хребтов у верхней опушкн леса, образованной горным дубом па высотах 2200—2700 м. Далее на
Рис. 125. Кавказская снеговая полевка.
£По акварели К. К. Флерова.
запад полевка прослежена на Кахетинском хребте выше Телави вплоть до снеговой линии (Шидловский, 1951). В относительно сухой и обезлесенной Южной Осетии эта полевка начинает встречаться в скалах с 2000 м (Огнев, 1950).
В верхней Сванетии кавказская снеговая полевка очень многочисленна среди пихтово-клеповых лесов. В сухой долине Ингура она живет в прогреваемых солнцем гнейсовых скалах, заросших желтой азалией, например у сел. Местия. В Абхазпи по крутым юго-западным склонам опа обычна с высоты 500 м до ледников, хотя Шидловский (1950) и указывает ее только для высокогорий в истоках р. Бзыби. На Сурамском хребте эта полевка сейчас, по-видимому, не встречается. В Малой Азии она вероятно, обитает на большей части восточного Тавра, так как найдена на горе Варзамбёг, южнее Ризе, на высоте 3000 м (Neuhauser, 1936). Недавно она была обнаружена Шидловским и в Джавахетии.
Южная часть ареала этой полевки не столь обширна, как на Большом Кавказе.
Наиболее плотно заселяются кавказской снеговой полевкой устоявшиеся осыпи крупных скал, закрепленные березовым и сосновым лесом с черничником, жимолостью, малиной и понтийским рододендроном в подлеске.
Га. Ш* Прежнее и современное распространение и численность млекопитающих 319
Кроме наличия корма, для жизни зверька основное значение имеют, невидимому, структура и расположение камней, скал, обеспечивающие cyMie укрытия, устройство и сохранение запасов сушеного корма. Состав твердого субстрата имеет меньшее значение, так как заселяются извест-•; новые, гнейсовые, гранитные и сланцевые россыпи.
Состав летних и зимних кормов у этой полевки сильно варьирует по участкам обитания. По Насимовичу (1935), основным кормом в Кав-зеком заповеднике являются анемоны, папоротники, малина и кав-к якни рододендрон. В верховьях Теберды, по наблюдениям П. Н. Сте-
Рис. 126. Запасы сена, устроенные кавказской снеговой полевкой в Баксанском ущелье.
Фот. автора, 1952.
Панова, 32 разобранных стожка содержали 47 видов растений, причем наиболее часто встречались папоротники, малина, мхи, рябина, ива, береза,, элакп, земляника, ель, сосна, брусника.
По моим и О. И. Семенова-Тян-Шанского наблюдениям, в Цейском ущелье зверьки запасают преимущественно веточки голубой жимолости, чернику» иван-чай, ветки рябины, кавказский рододендрон, костянику. Они скусывают также веточки березы и кустарников, забираясь для этого* высоко на гладкоствольные деревья. На относительно сухих лугах до лин Донисер-Дона и Баксана запасы сена, устроенные в развалинах крепостей и расщелинах (рис. 126), состоят из вероники, шалфея, крапивы, клевера и т. п.
В целом кавказская снеговая полевка приспособлена к скальному рельефу и избегает рытья. Среди других видов полевок она выделяется поразительными лазательными способностями, близкими к таковым У сонь И белок. u
Крупных колебаний численности у этой полевки нет. Тем не менее в отдельные годы некоторые ущелья бывают почти лишены зверьков,.
. 320
Часть II, Анализ происхождения четвертичных млекопитающих
что может свидетельствовать о наличии местных эпизоотий. Климатические и погодные воздействия не сказываются на этом виде столь неблагоприятно, как на других полевках.
Из хищников заметное значение имеют лишь куницы и ласки, из пернатых опасны в основном крупные совы. Вообще эта полевка может дать довольно устойчивую кормовую базу для нового вселяемого небольшого пушного хищника, например для соболя.
Судя по наибольшей численности и размерам особей, современный экологический оптимум кавказской снеговой полевки расположен в субальпийской зоне западной половины Большого Кавказа.
Для развития ареала этой полевки часто имело большее значение образование скального рельефа при горообразованиях и оледенениях, чем наличие мезофитной растительности. В этом убеждают большие плотности населения зверька в сухих внутренних долинах и в центральном Дагестане, где проявляются черты сходства в экологии этого вида с малоазийской снеговой полевкой. Анализ ареала и распределения по местообитаниям позволяет считать кавказскую снеговую полевку древним местным видом, связанным в своей эволюции с развитием высокогорных растительных формаций миоценового возраста. Оледенения Большого Кавказа в плейстоцене не могли вытеснять популяции этих зверьков на равнину, как это представлялось некоторым фаунистам и палеонтологам, а лишь частично понижали границы их распространения. Для полевок оставалось много мест обитания на хребтах меридианного направления и на передовых продольных цепях даже при депрессии снеговой линии в 1000—1100 м. Тем не менее холодный и влажный климат ледниковой эпохи, приводивший к развитию луговых формаций в предгорных равнинах, создавал условия для проникновения .зверьков в нижележащие области по береговым обрывам некоторых речек. В одну из холодных эпох полевка появилась, очевидно, и в Пя-тигорье, но вряд ли достигала когда-либо Ставропольской возвышенности. Находки ее ископаемых остатков средпечетвертичного возраста в предгорных районах могут уточнить в дальнейшем эти соображения.
В настоящее время выпас скота и вырубка горных лесов до некоторой степени сужают распространение кавказской снеговой полевки в восточной части Кавказа, но расширяют — в западной.
Малоазийская снеговая полевка — Microtus (Chionomys) nivalis Mart. (s. lato). Остатки этого вида указывались для плейстоценовых и голоценовых слоев многих пещер Англии, Франции, Германии, Италии, Австрии, Швеции, Чехословакии, Венгрии (Wolf, 1939).
Из пещер горы Кармел в Палестине описаны остатки особого вида -М, {Chionomys) machintoni Bate, встречавшиеся с ашеля до мезолита (Bate, 1937). У нас известны лишь остатки голоценового возраста на Армянском нагорье. Современный вид широко распространен в горах Средиземноморья — от Пиренеев до Балкан, в Сирии, Палестине, Малой Азии, на Кавказе и в западном Копет-Даге. В верхнем плейстоцене эта полевка, возможно, жила и на Больших Балханах.
Па Большом Кавказе зверек известен лишь с северного склона хребта: с вершин Фишт, Пшекиш, Большой Тхач, Ачешбок, Бамбак, из ущелий Теберды и верховий Черека. В Балкарском Череке и далее к востоку никем не добывалась. Известные точки находок па Большом Кавказе расположены на высотах 1500—2000 м, в субальпийской зоне. Места обитания здесь те же, что и у кавказской снеговой полевки (карта 67).
[Л'1И. Прежнее и современное распространение и численность млекопитающих 321
На Малом Кавказе малоазийская снеговая полевка обитает в лесной п льшшской зонах Триалетского хребта, в Джавахетии, на всех хреб-m Армянского нагорья, Карабаха и в Талыше.
Вертикальное распространение ее в общем повышается в юго-восточном управлении соответственно расположению фито ландшафтных зон, побывают и исключения. На Триалетском хребте она найдена на высотах-ООО м у Бакурьян; на Бамбакском хребте обычна на высотах 1800— jO и. На хребте Шах-Даг прослежена нами по ущелью Шамхор-Чая г 1600 до 2300 м абсолютной высоты. Южнее встречается в зоне нагор-иксерофлтнои степи, иапрпмер у Севана, на Сарайбулахском, Дарала-шском хребтах (Туров и Турова-Морозова, 1928; Даль, 19446, 19496). Лц Ереваном обнаружена на высоте 1200 м в россыпях скал с кустарни-г,\1И (Флеров и Гуреев, 1934).
На южном склоне Зангезурского хребта по ущелью Акулис-Чая уоазпйская снеговая полевка прослежена нами в 1947 г. на высотах 1600 до 2500 м. Зверек держится здесь в россыпях камней, проросших up ей, шиповником, миндалем, боярышником и крушиной, в зоне Еагакаитовых астрагалов.
В буковом лесу по ущелью Тертер-Чая в Карабахе нами пе встречена, выше известна из альпийской зоны у Кельбаджар.
Б Талыше распространение зверька реликтовое, так же как и в Ко-IH-Даге. Небольшая популяция полевок была обнаружена мною в июне «<45 г. на Келахапской сопке в сухой Диабарской котловине на высоте *410 м. Зверьки держались здесь в россыпях тешенитовых скал с бедным редким ксерофитным покровом пз камнеломок, пивяныиков, побегов ра и ксерофильных папоротников среди горио-соляпковой полупу-:Ш1Ш (Верещагин, 19456). Вероятно, эта полевка живет и на Савелане, hi Эльбурсском хребте.
Наибольшая численность зверька наблюдается в зоне остепненного кга иапрпмер у побережья Севана. Скальный субстрат — основа пропитания п этого вида. Привычка к запасанию кормов у малоазийской световой полевки развита не столь сильно, как у кавказской, а способ-кть к обитанию в разнообразных условиях неизмеримо больше. Ее пищевые адаптации п отношение к влажности варьируют в широких уделах. Если в субальпийской зоне западного Кавказа эта полевка Ст бок о бок с кротом и дождевым червем, то в Талыше и на южных клонах Армянского нагорья она обитает в зоне нагорных ксерофитов KfljQ7 с фалангой и скорпионом. Вообще она адаптирована в большей иенени к пониженному давлению и к скальному субстрату, чем к сочной ♦те и низкой температуре.
Измельчание малоазийской снеговой полевки к востоку, а также ис-«зновенпе ее поселений на средних высотах южных склонов Армян-Кр нагорья указывают, что юго-восточные участки ареала находятся 11 области современного пессимума.
Прерывистость в распространении зверька на юго-востоке и горные • дяции в Талыше и Копет-Даге можно объяснить прогрессивным иссу-гнием восточного Средиземноморья после наивысших возможностей ря расселения вида, которые проявились в какую-то плювиальную I холодную эпоху.
Однако далекие и различные по типу обособления участки ареал а сиего-«оиполевки в Средиземноморье в целом могут свидетельствовать и о том, что п-еювые полевки Пиренеев, Альп, Карпат и Кавказа являются особыми ви-1яп. возникшими параллельно из генетически родственного материала.
? Н. К. Верещагин
322
Часть II. Анализ происхождения четвертичных млекопитающих
Послеледниковое потепление и подъем фитоландшафтных зон по вертикали приводили к сужению участков ареала снеговой полевки, к углублению разрывов и к изоляции различных географических рас снеговой полевки.
Ограниченное распространение малоазийской снеговой полевки на Большом Кавказе может объясняться недостаточной приспособляемостью этого южного вида к перенесению большой влажности на юго-западе Большого Кавказа. Труднее объяснить отсутствие этого вида в восточной части Большого хребта, в сухих продольных долинах северного склона и в Дагестане.
Длиннохвостая снеговая полевка — Microtus (Chionomys) roberti Thos. Единственная находка четвертичных остатков известна из верхнего палеолита западного Закавказья — пещеры Сакажиа. Современное распространение этой полевки охватывает северные склоны восточного Тавра в Малой Азии, Аджаро-Имеретинский хребет, Триалетский хребет п Большой Кавказ (карта 69).
На северо-западном Кавказе эта полевка встречается от хребта Черных гор на высотах 800—1000 м и до альпийской зоны. В ущельях центрального Предкавказья редка и отсутствует на Ставропольской возвышенности. Обычна в Северной Осетии па высотах около 800—1000 м.
В Дагестане водится в буковых лесах в верховьях Андийского Койсу у аулов Тахота, Порода, но на остальной территории этой страны, по-видимому, отсутствует (Гептпер и Формозов, 1941). Нет этой полевки и во внутренних продольных долинах Северного Кавказа.
В Азербайджане добыта мною (Верещагин, 1940а) в 1935 г. в ущелье Катехи-Чая, к северо-западу от Закатал, на высоте 1450 м, а восточнее пока неизвестна.
Вертикальное распространение длиннохвостой снеговой полевки на черноморском склоне Большого Кавказа — от уровня моря до верхней границы зоны леса (Шидловский, 1947, 1950). В Аджарии зверек населяет лесную зону от 800 до 1800 м абсолютной высоты (Шидловский, 1948). Распространена и во всей Южной Осетии, кроме сухой Горийской котловины.
Типичные места обитания длиннохвостой снеговой полевки с высокой численностью — это затененные глухне и сырые участки ущелий. В западной части Большого Кавказа и на Малом Кавказе такие биотопы бывают в пихтовых и пихтово-буковых лесах с широколиственным высокотравном из лопухов, борщевника, крапивы, сборной ежи, заполняющим дно распадков. В Предкавказье зверька больше в низкой влажной части Черных гор, чем в верхних частях крупных ущелий. В восточном Закавказье наблюдается обратная картина.
По морфологическому типу, в частности по строению мехового покрова, полевка эта до известной степени близка к водяной крысе. Зверек нередко плавает в горных ручьях и вообще существует в условиях крайней влаж
ности.
Формирование этой полевки как вида было связано с древними мезо-фитиыми и даже гнгрофитными фитолаидшафтамп и биотопами, существовавшими с миоцена в горах, окаймляющих с юго-востока и востока Черноморскую впадину. Сокращение распространения длиннохвост» снеговом полевки в восточном направлении вполне соответствует из. менению типов ландшафта и биотопов горного леса.
Существование изолированной, реликтовой ныне популяции в верховьях Антщйского Койсу в Дагестане можно объяснять расселение!
[л. IH Прежнее и современное распространение и численность млекопитающих 323
зверька с южного склона в какую-то влажную эпоху. Такая же миграция бука в Дагестан в голоцене была описана Тумаджаиовым (1940).
В целом длиннохвостая снеговая полевка — индикаторный и эндемичный вид древнего мезофильного фаунистического комплекса в Кавказском округе.
Рыжая полевка — Cletrionomys glar coins Schreb. Ископаемые ос-гатьп представителей рода известны в Европе с плейстоцена. Основная часть ареала этого вида расположена в таежных и широколиственных аесах Евразии. В зону степи зверек проникает лишь по лесным участкам речных долин. В экологическом смысле рыжая полевка мезофильна и холодолюбива. Па юге восточного Средиземноморья существует только Ьнтппский подвид — С. glareolus ponticus Thos, распространенный по се-кртым склонам Тавра — па южном побережье Черного моря (Thomas, <«<*6: Nculiauser, 1936) и иа Аджаро-Имеретинском хребте (карта 72}.
В Аджарии зверек живет у истоков р. Супсы в еловом лесу (Шпдлов,-Kiiii, 1940а). Эют изолированный ныне Босфором южный участок обширного ареала зверька служит замечательным примером северного фау-иетпческого влияния на Кавказ через Балканы и Малую Азию.
Судя ио отсутствию рыжей полевки в западной части Большого Кав-ц.-а, проникновение ее на южный берег Черного моря произошло, вероятно, весьма поздно, в эпоху наибольшего похолодания в верхнем плейстоцене.
Степная пеструшка — Lagurus lagurus Pall. Ископаемые остатки япго вида верхпсплейстоценовой древности известны из ряда пунктов риской равнины по долинам Дона, Волгп, Урала. Современный ареал шеположеп в степной п полупустынной зонах от низовьев Днепра до вер-зовьев Енисея и границ северо-западного Китая (Виноградов и Аргиро-кцо. 1941). Широко распространена степная пеструшка и па участках., гвежных с Кавказским перешейком по северной окраине Черного моря, \-,иьа и Каспия. В пределах перешейка она живет лишь па северных метках предкавказскпх степей по междуречьям Кумы, Калауса, Ма-бпа, т. е. к северо-востоку от Ставропольского плато (Свириденко, |{28). Кистяковский (1935) обнаружил остатки этого зверька в погад-хищных птиц в окрестностях Ачикулака, Благодатного, Сухой Паяны, Петровского, Камбулата (карта 64).
Степная пеструшка проявляет себя в Предкавказье лишь в периоды I весовых размножений и возникающих при неблагоприятных условиях I верекочевках. Неразвитость ареала пеструшки в предкавказскпх ст£-цх, прочные связи ее с ландшафтом и группировками злаково-разно-твпых, ковыльно-типчаковых и полынных степей умеренней полосы [вразип позволяют думать, что этот вид проник па перешеек недавно., 1верхном плейстоцене пли голоцене со смежных участков степей Рус-I voii равнины.
Такое вселсппе могло происходить в эпохи исчезновения Мапычского I пролива.
Прометеева полевка — PromeLheoniys schaposchnicovi Sal. (рис. 127). Цопаемые остатки этой полевки, занесенные филином, известны « иерхпепалсолитических слоев пещеры Гварджилас-Клдс в ущелье речки Квпрплы в западном Закавказье, па высоте около 450—500 м и и шпкпсналеолптических слоев пещеры Кударо I.
Пещера Гварджплас-Клде лежит вне пределов современного ареала церька, по для того чтобы судить о сдвигах ареала с верхнего плейсто-№ia. следует еще проследить детали современного распространения этой
324
Часть II. Анализ происхождения четвертичных млекопитающих
Рис. 127. Прометеева полевка. По акварели Е. Я. Захарова.
полевки на Рачинском хребте и изучить радиус охотничьих полетов филина в этом районе.
Современное распространение прометеевой полевки приурочено к западной половине Большого Кавказа и Аджаро-Имеретинскому хребту на Малом Кавказе (карта 65). На Большом Кавказе опа встречается от вершин Фишт и Оштен до Казбека — Крестового перевала, Гудаура.
В Кавказском заповеднике прослежена в долине Мзымты, ниже оз. Кар-дывач, па лугах в истоках Курджипса, Пшеки-ша, в бассейне Киши и Уруштеие, в истоках Малой Лабы и блпз Красной Поляны (Туров, 1926а, 19266). В Абхазии зверька ловили в урочище Ават-хара (на высоте около 1500 м). В Сваиетии опа прослежена мною в 1948 г на высокогорных лугах в верховьях Ипгура и Рпо-па, где особенно обильна на перевале Загар Леч-хумского хребта, на высотах 1800—2800 м. В продольной сухой долине Ин-гура этого грызуна нет. В Южной Осетии прометеева полевка добывалась в верховьях Большой Лиах-вы (Шидловский, 1951. На центральном участке Большого хребта она прослежена на горных лугах от Мампссонского перевала до станции Казбек и Гудаур (Туров, 1926а, 19266). В верховьях Балкарского Черека, Донп-сер-Допа и Ардона мне
не удалось разыскать ни зверька, ни следов его жизнедеятельности. Нет данных об обитании этой полевки в верховьях Кубани, Баксана, I Малки. Таким образом, ареал па северном склоне Перевального хребта разорван по крайней мере от бассейна Лабы и Зеленчука до Ардона, Это можно объяснить скорее всего истребительным действием горных лед- I ников.
На лугах Скалистого хребта в Осетии, Кабарде и в Грозненской об- I ласти, местами вполне пригодных ныне для обитания этого грызуна, I он не найден. По-видимому, севернее сухих продольных долин прометеева полевка по распространялась и в плейстоцене.
Восточнее Казбека прометеевой полевки нет и па Перевальном хребте. В тщательно обследованпом мною Закатальском заповеднике ее нет на-1
Гл. III. Прежнее и созрел енное распространение и численность млекопитающих 325
верное, так же как п в центральном Дагестане. Вообще быстрое выклинивание распространения грызуна на восток от Казбека может быть объяснено значительным сужением зоны альпийских лугов и ее ксерофи-пзацней, сказывающейся в развитии типчаковых и белоусовых г рупии-Лвок вместо разнотравной.
На Малом Кавказе прометеева полевка обнаружена пока лишь в вер-к»вьях речки Баквис-Цкали, левого притока Супсы, близ курорта Бах-^ро, на высоте 1600 м (Шидловский, 1940а, 1947). Этот участок субаль-Ескпх лугов расположен западнее вершины Готнмария (Метис-Цкаро,
-3 м). Ареал грызуна здесь, несомненно, значительно обширнее. Судя приведенным данным, вертикальное распространение прометеевой по-звки в настоящее время лежит в пределах 1500—2800 м. Возможны щонахожденпя и на более низком уровне. Экологический оптимум ^ерька в нашу эпоху находится в пределах зон субальпийского и аль-Йского луга с преобладанием манжетки и рыхлокустовых злаков: gfcpiioii ежи, мятлика и др. Плотно заселены также луговины между за-taiiHMH кавказского рододендрона и березы. В рододендрон грызун, пнако, не заходит, так как там нет пищи, но в березовых колках и вообще । верхней опушке леса с высокотравьем столь же обычен, как и па откры-лугах. Обитание этой полевки на луговых полянах среди букового №, описанное Туровым (19266) для Кавказского заповедника, можно сматривать в ряде случаев как следствие наступления лесных формант на зону альпийских лугов в современную эпоху. В сильно затенен-ух буковых и грабовых лесах полевка эта при отсутствии там травяного ярма не живет. В ряде мест зверок держится па полях овса, пшеницы, Кртофеля пли заходит на их окраины, как например в верховьях Иигура.
Строго подземный образ жизни и питание преимущественно прикор-уиыми частями травянистых растений отражены в морфологии зверька: толощонном черепе, большом радиусе кривизны резцов, удлиненной •шастеме, рудиментарных глазах, укороченных рычагах конечностей. Череп с его гребнем, идущим почти до затылка, очень напоминает он-шровый. По слабо дифференцированному и поставленному вертикально росяному покрову, окраске, нередко меланистической, зверек напо-иппает слепушонку.
Вообще в морфо-функциональном смысле прометеева полевка совмещает до известной степени черты ондатры, водяной полевки и слепушонки, ю живет, в противоположность последней, в мезофитных условиях (см. дат Виноградов, 1926; Огнев, 19266).
По наблюдениям Гамбаряна (1952), большие рогсшие способности врометсевоп полевки совмещаются, в противоположность слепушонке, (питанием надземными, а не корневыми частями растений.
Такие адаптации указывают па весьма большую древность вида и его । прение связи с ландшафтом высокогорных мезофитных лугов.
Колебания численности прометеевой полевки связаны с массовой ги-(елью при весенних ливпях и последующих холодах, когда вода замер-I лгт в норах (Туров, 1926а).
Строгая эпдемичность вида указывает на болтшую консервативность «•логических условий на западном Кавказе, на древность и большое постоянство существования ландшафта и зовы мезофитных субальпийски альпийских лугов, по крайней мере с нижнего плиоцена.
Разрыв ареала зверька на два участка — кавказский и малокавказ-сй - мог произойти уже лишь в послеледниковье в связи с иссушением
Сурамского кристаллического массива.
326 Часть II, Анализ происхождения четвертичных млекопитающих
Прометееву полевку следует рассматривать в качестве древнего плио Ценового, эндемичного для Малого и Большого Кавказа вида с ареалсш реликтового типа, несколько увеличивающимся в нашу эпоху вследствие расширения зоны высокогорных лугов под влиянием рубок леса и выпаса скота.
Водяная полевка — ArvLeola terrestris L. (s. lato). Ископаемые остатки этой полевки широко известны из плейстоценовых отложений Ев ропы.
На Русской равнине остатки водяных крыс обычны для верхнеплейсто ценовых и голоценовых отлогкений. К югу от Кавказа остатки этой по левкп найдены в ашельских слоях пещер Сирии. На Кавказском пере Шейке известны единичные находки костей в среднечетвертичпых слоях восточного Закавказья, в верхнем палеолите западного Закавказья и многочисленные находки в голоценовых слоях пещер на местах дневок филинов.
Современное распространение вида охватывает лесную, степную и полупустынную зоны Евразии. На юг от Кавказа обитание прослежено в северном Иране, Малой Азии, вплоть до Палестины.
На Кавказе водяная полевка живет от уровня Каспия до 2800—2800 и абсолютной высоты во всех зонах, исключая горные широколист венные и хвойные леса, а также зоны скал и осыпей перевалов (карта 68). Наибольшая численность наблюдается на пойменных лугах низо вого Дэна, несколько меньшая — в плавнях Кубани.
В Колхидской и Ленкоранской низменностях опа крайне редка, но все жэ живет в непересыхающчх водоемах. В полупустыне восточного Предкавказья и Закавказья она обычна по обширным озерам и прото кам, заросшим тростником и рогозом, где держится летом, не приближаясь к берегам, и устраивает шарообразные гнезда в гуще зарослей.
В предгорьях наибольшая численность водяной полевки паблюдается по ключевым речкам, например на Терско-Сунженскои равнине.
Во внутренних степных долинах северного склона Большого Кавказа и в центральном Дагестане образовалась особая экологическая форма полевки, отличающаяся подземным образом жизни. Зверьки селятся там по днищам ущелий, особенно на орошаемых участках полей. В альпийской зоне Большого Кавказа эта полевка редка и известна лишь из верховий Терека и Арагвы.
На Армянском нагорье, в Малой Азии и в Иране водяная полевка обычна на субальпийских лугах, по самым верховьям речек, например в Карабахе, в верховьях Акстафы и в озерном районе Джавахетии. В долине Аракса много этих полевок на ключевых речках Аралыхской котловины.
Итак, распространение этой полевки всюду узко местное и тяготеет преимущественно к водным биотопам.
Экологический оптимум подземных популяций этой полевки находится на средних высотах в сухой зоне плоскогорий Кавказа, а гидрофильных популяций — в зо la полупустыни при отсутствии избыточной атмосферной влакнэсти. Низкая численность водяной полевки па Черноморском поберэкьэ и в болотах Колхиды и Гпркании свидетельствует до известной степени о сходстве и особенностях распространения с обыкновенной полевкой. Эту низкую численность вида в болотах Колхиды можно объяснить огромной конкуренцией и преследованием со стороны пасюка, а в Гиркании — сильным пересыханием водоемов летом и крайней уплотненностью ленкоранских желтоземов (Верещагин, 19416, 1949г).
1’л. НЕ Прежнее и современное распространение и численность млекопитающих 327
Судя по удлинению диастемы, увеличению радиуса кривизны верхних резцов, укорочению хвоста и т. и., которые выявляются у горных подземных популяций по сравнению с гидрофильными, дивергенция в пределах современного вида намечается по линии вторичной изоляции от водной среды и приобретения черт слепушонки.
В Передней Азии и на Кавказе такие тенденции, вероятно, были обусловлены поднятиями горных систем в четвертичном периоде.
Судя по палеонтологическим фактам, ареалу и экологии, древность водяной полевки па Кавказе по меньшей мере верхнеплиоценовая. В Закавказье численно преобладают ныне южные, относительно ксерофиль-ше популяции.
Устойчивость ареала вида в восточном Средиземноморье обеспечена как природным и искусственным развитием пресноводных биотопов, так я экологической пластичностью зверька.
Отряд НЕ ПАРНОКОПЫТНЫЕ — PERISSOPA CTYLA
В четвертичной фауне .Кавказа существовали представители семейств Equidae и Rhinocerotidae, К голоцену остаются только лошадиные, история появления и исчезновения которых рассматривается ниже.
Сем. Л ОШАДИНЫЕ — EQ XJIJDAE
Наиболее, древние остатки крупных кабаллоидпых лошадей Equus sltfionis Cocchi известны с Кавказа и прилежащих территорий из отло-/Ki'iiiiu верхнеплиоценового возраста. Появление лошадей происходит, ^-видимому, одновременно с южным слоном, знаменуя особый этап мсмене фауны. Позднее, с среднего плейстоцена, на перешейке появляется полуослы.
История настоящих лошадей прослеживается на Кавказе пока еще весьма схематично по ряду разновозрастных захоронений.
Лошадь четвертичного периода — Equus caballus (s. lato). После крупных стеноновых лошадей, остатки которых найдены в акчагыльских в апшеронских отложениях перешейка, наиболее близка к лошадям четвертичного периода таманская верхнеплиоценовая—пижнеплейсто-цеяовая лошадь — Е, aff. sussenbornensis Wust. из конгломератов Таман-да’о полуострова и галечников древнекаспийской террасы у Баку (глава П п карта 79).
Остатки среднеилейстоценовых лошадей, подробно еще не описанные, Сильны в асфальтированных песках у Бинагадов и встречаются единично 5 суглинках хазарского возраста в ряде мест Ставрополья и Краснодарского края. Эти лошади были близки к Е. caballus chosaricus Grom, из ,'редпепленстоцеповых отложений Русской равнины.
Остатки лошадей из Ильской палеолитической стоянки в Предкав-газье п из палеолитических стоянок Имеретии и пагорья Малого Кав-аза, по-видимому, близки к шпрокопалой лошади — Е, caballus latipes вгош. Русской равнины (см. Громова, 1949).
На юг от Кавказа остатки верхнеплейстоценовых лошадей известны аз палеолитических слоев пещер Тамтама п Бизитуп и из делювиальных суглинков близ Мараги в Иране, а па юго-западе — из пещер Палестины
328 Часть II. Анализ происхождения четвертичных млекопитающих
и Сирии. Южные лошади были легче сложены, нежели северные.
Число особей плейстоценовых лошадей, учтенное по костным остаткам в различных типах захоронений, по отношению к другим видам составляет: Ильская стоянка — 6.6%, Бинагады— 5.2%, пещера Сака-жпа — 2.5 °о , навесы Мгвпмеви — 16.5%, Зуртакетская стоянка — 50.0°/0.
Если принимать перечисленные выше формы плейстоценовых лошадей, обитавших на перешейке, за единый генетический ряд, то происхождение лошадей, живших здесь в голоцене, нет надобности связывать с прико-чевкой исходных форм извне.
Что касается диких лошадей, распространенных на Кавказе в после-ледниковье, то выделение их остатков среди остатков одомашненной лошади представляет большие трудности.
Между тем кости лошадей весьма обычны в слоях послепалеолпти-ческпх поселений, где представлены кухонными остатками, а в погребениях — черепами и целыми скелетами. Весьма вероятно, что эти остатки принадлежали в значительной степени тарпанам — Е. caballus gmelini Ant. пли их одомашненным потомкам.
Учет численности остатков лошадей в кухонных отбросах поселений и могилышках Кавказа исторической эпохи показывает, что это животное имело огромное значение в хозяйстве, пищевом рационе и в обрядах различных племен и пародов (табл. 71).
Большое число остатков лошадей известно также из погребений урар-тов X—XII вв. до н. э. в бассейне Севана (Лалаян, 1929) и в культурных слоях поздней бронзы и средневековья близ Самтавро и у Мингечаура.1 Лошадь, в том числе и дикая, служила* одним из излюбленных объектов рисунка и скульптуры эпохи бронзы в Передней Азии и • на Кавказе.
Еще в середине I тысячелетия до н. э. дикие лошади водились в Передней Азии в изобилии. Это были тонконогие животные с небольшими ушами на легкой голове восточного, арабского типа. По сухости конституции и изяществу форм они не имели ничего общего с тарпаном. На них охотились с дротиками и травили крупными догами, как о том свидетельствуют великолепные рельефные изображения на степах ассирийского дворца в Ниневии (Атлас по древней истории Египта, Передней Азия, Индии и Китая, 1937, табл. 156, рис. 2). Все это еще раз подтверждает, что тарпан был аборигенной лошадью Европы и северной половины Азин, а отнюдь не прикочевал туда с юга в конце плейстоцена и тем более голоцена, как это иногда представляется палеонтологам.
В памятниках кобанской культуры в Осетин и Абхазии на бронзовых поясных пряжках и на топорах лошадь изображалась в виде довольно стройного и длиннохвостого существа. В материалах, добытых в Осетии Уваровой (1900), попадаются бронзовые фигурки лошадей с мощной гривой и длинным хвостом. Если не принимать во внимание стилизацию, то по конституционному типу, в частности по легкой голове, эти лошади мало подходят к тарпану, а скорее близки к лошади южного типа.
В крупных скифских курганах Предкавказья и юга Русской равнины при раскопках археологам неоднократно встречались массовые ритуальные захоронения лошадей в несколько десятков и даже сотен (до 300) экземпляров.
В скифских памятниках обычны п реалистические изображения лошадей. Упомянем о контурном рисунке жеребца с короткой стоячей^гри-
1 Фактические материалы из этих мест ие сохранились.
[л. III» Прежнее и современное распространение и численность млекопитающих 329
,т-ю-:а-/о-•и, не ю-
ie-ю-
ги-бе-ки ni
1ИЙ □т-щх
Таблица 7 %
Численность кухонных и ритуальных остатков лошадей по сравнению с другими видами животных в слоях поселений и могильниках Кавказа1
* ——---
Районы и пункты
Датировка
Отношение числа особей лошадей к числу особей других животных (в %)
Предкавказье и Русская равнина Крепость Саркел (славянские и хазарские слои)........................
П-селенде у Цимлянской (хазарские .да)..............................
Поселение Зеюково близ Нальчика. [, родища Исти-Су, Алхан-Кала в долине Сунжи.............................
ьгпльники близ Усть-Лабинской. . .
кредите Алхасте на Ассе...........
Сошбратнее городище на Кубани. . .
Кепы, . . .
Фанагория.
Тамань. . .
VIII—XIII вв. н. э.
VIII—X вв. н. э.
VI—VIII вв. н. э II—III вв. н. э.
VI в. до II. э.—II в. н. э.
VI—V вв. до н. э.
V—I вв. до н. э.
ар-;ых а. 1 тов 1зе> >ед-ши цпц •хо->ст-:ого нт» аету аны тем
U30-[ЬНО тш гри-1ИЮ, 1ади
Древнегреческие города Таманского полуострова
Курганы на Маныче. . .
Цимлянская на Дону
землянка I землянка II
Восточное Закавказье
Крепость Чухур-Кабала...............
Поселение в район крепости Баку. . .
Поселение у Сумгаита................
Западное Закавказье
Пешера Ахштырская близ Адлера. . .
V—I вв. до и. э.
I тысячелетие до н. э.
Половина II тысячелетия до н. э.
XII—XIV вв. н. э. IX—XII вв. н. э.
I тысячелетие до н. э.
Неолитические и более поздние слои.
Ины аль-300)
ло-гри-
10.3
3.7 16.7
‘ 7.1
21.2
13.0
17.0
22.5
12.3
14.1
12.5
13.2
12.0-
16.6
8.1
8.3
2.9
вой, с толстыми ногами, тяжелой головой, с маленькими ушами и тупой уордоп, напоминающего тарпана на серебряном сосуде из Майкопского кургана (Фармаковский, 1914; рис. 128). Общеизвестные изображения доадей на серебряной вазе из кургана Куль-Оба представляют, вероятно, уже облагороженных домашних лошадей. Контурные рисунки лошадей имеются и на скалах в Кабристане, южнее Баку. Они выполнены, дамо, в древнеримские и доримские времена, когда дикие лошади, возможно, водились в прикуринских степях (Верещагин, и Бурчак-Абрамович, 1948).
Со времен Палласа, Гмелина и Гюльденштедта, наблюдавших тарпанов в конце XVIII в. в южнорусских и предкавказских степях, истории окончательного исчезновения этого животного во второй половине XIX в. было посвящено много статей и заметок (Левченко, 1882; Кеппеи, 1896; Анучин, 1896; Гептнер, 19346; Antonins, 1938; и др.). Особенно полный и беспристрастный обзор состояния вопроса о тарпане и задачах его изучения был дан Кеппеном, настаивавшим ла необходимости изу-
1 Неопубликованные материалы авторе.
330 Часть II. Анализ происхождения четвертичных млекопитающих
чения скелетов лошадей из курганов южной России. Но такого изучения еще не производилось.1
В противоположность большинству исследователей, Анучин отрицал существование дикой лошади в степях и пустынях Евразии как вида, считая ее за вторично одичавшую лошадь кочевников. Несомненно, однако, что если бы даже это дичание и имело место в больших масштабах, то эта одичавшая лошадь была бы чрезвычайно близка к исходному материалу — дикой лошади позднего палеолита и раннего неолита. Спе-
Рис. 128. Изображение лошади, первобытных туров, львов и других животных на серебряном сосуде из Майкопского кургана. (По Фармаковскому, 1914).
циально для Кавказа сообщений о тарпанах мало. Ровинский (1809, стр. 131) писал: «Дикие кони, или тарпаны, встречаются косяками своими в Кубанской степи, около устьев Урала, и за оным».
Мне не удалось найти исторических сведений об обитании диких лошадей в Закавказье. Вообще плоскогорья северных частей Передней Азии, издавна густо населенные, сохранили к средневековью, по-видимому, уже только куланов. Лошади же были, вероятно, целиком переловлены и одомашнены хеттами, ассирийцами, урартами еще до нашей эры.
Быструю эволюцию лошадей в четвертичном периоде принято связывать с дальнейшим развитием остепненных ландшафтов и жесткотравных, злаковых группировок, появившихся еще в плиоцене. Адаптивные признаки и современная экология живущих диких и дичающих домашних лошадей подтверждают это. Тем не менее удовлетворительная способ-
1 Черепов и скелетов лошадей из прежних раскопок крупных курганов Украины и Предкавказья нс сохранилось.
Tj. Ш. Прежнее и современное распространение и численность млекопитающих 331
иисть крупных косяков к активной защите от стайпых хищников — волков и способность доставать корм из-под снега позволяли лошадям обитать в лесостепи и даже в лесной зоне Европы, где они кормились на полянах п болотах совершенно так же, как это делают ныне домашние юшади Якутии.
Вообще верхнечетвертичная лошадь всегда напрасно считалась стено-гопным, стопным животным, так как она воплощала в своем конституциям типе некую универсальную форму среди копытных. Доказательством тужит успешное применение домашних лошадей в самых различных ындшафтах: от ледников Гренландии, Антарктики и Альп до тропических аесов, болот и пустынь всех материков.
Неравно Ветуляпп (1952) развил замечание Кеппена о том, что в средой Европе дикие лошади были лесными животными, утверждая далее, •по тарпан приспособился к обитанию в лесах средней полосы Европы I стал лесным животным лишь на последней стадии существования, т. е. в историческую эпоху.
В свете современных наблюдений над парнокопытными причина такой Ьелы биотопов заключается в развитии лесных формаций на месте прежней лесостепи и в преследовании лошади человеком.
Па Кавказе, с его пересеченным рельефом и с относительно густым населением, подобных приспособлений произойти, очевидно, не могло, L(* как крутые лесные и безлесные склоны были все же непригодны для ршадей, а пологие участки не давали убежища.
Окончательное вымирание тарпанов на Русской равнине, как известно, ^тировалось семидесятыми годами прошлого столетия. Недавнее сообщение Гептпера (19346, стр. 482, 1955) о последнем тарпане, будто бы мигаем в Полтавской области в 1918—1920 гг., значительно приближает у полного исчезновения этого животного.
Причины исчезновения диких лошадей па Кавказе состояли в пресле-«вании пх конницей кочевников, в распашках степей, в заселении чело-•екоммс! т водопоев и, наконец, в охоте за ними с огнестрельным оружием (сч. также Кеппен, 1896).
Кулап — Equus hemionus Pall. (s. lato). Ископаемые остатки куланов несения к настоящему времени из плейстоценовых и голоценовых слоев пропчущ^твенно степной и полупустынной зон Евразии. Остатки кулаков из" Западной Европы, описанные как Е. hemionus fossilis Nehr., по мнение Громовой (1949), могут быть отнесены к мелким лошадям, но воз-додакть далекого проникновения куланов па запад пз Азин при после-1ДНИКОВОМ остенненпп равнин не исключена. Фаланги н метанодпи полуслов из верхних стоев стоянки Афонтова гора на Енисее, па песчаных выдувах у Тронцкосавска на Селенге, на Иртыше и в пещерах Алтая (Громов, 1948; Верещагин, 1956) большей частью верхпеплейсто-Ьпового п голоценового тина сохранности. Описаны и остатки из верх-плейстоцена Кптая.
В Средней Азии достоверные остатки кулана известны пз палеолити-шик слоев Ьухтармипской пещеры (Верещагин, 1956) и из слоев века Едая пещеры Каупчп-Тапа. Из древнейших слоев Анау под Ашхаба-относимых к 30Э0—1500 гг. до и. э., Дюрст (Ducrst, 1908) описал ататки тонконогой лошади — Е. caballus pumpellii Duerst, которые, пи соображениям Громовой (1949), могут быть также отнесены к кулану.
На юг от Кавказа кулан жил с нижнего палеолита—ашеля до совре-«остн (Bate, 1937). Определены остатки кулана и в палеолитических Кк пещеры Барда-Балка в южном Курдистане.
332 Часть II. Анализ происхождения четвертичных млекопитающих
Таким образом, по современным данным, кулан (рпс. 129) несомненно южный вид, сформировавшийся на фоне полупустыни и нагорной степи Евразии.
Основным препятствием для правильного решения вопроса о путях и времени появления кулана на Кавказском перешейке является недостаточная ясность родственных взаимоотношений плейстоценового осла — Е.diidruntinus Reg.—с куланом. До сих пор прямая филогенетическая связь этих видов обычно отрицалась на том основании, что кулап имеет более примитивные черты строения зубов (Stelilin et Graziosi, 1935; Громова, 1949). Судя по аналогии с филогенетическими рядами четвертичных слонов, носорогов, лошадей и зубров, возможность того, что кулан — непосредственный потомок плейстоценового осла, не исключена. Отсюда может исходить и утверждение об автохтонном происхождении кулана на Кавказе. Остатки голоценового полуосла — кулана известны из ряда мест Кавказского перешейка и смежных мест Русской равнины (карта 78). Наиболее древние остатки относятся ко второй половине 11 тысячелетия до н. э. пз слоев поселения у Джемикента, южнее Махачкалы, и позднейшие — к слоям XIII в. поселения у Бакинской бухты.
Исторические сведения об обитании кулана на Кавказе и смежных территориях довольно многочисленны. Еще Ксенофонтом (изд. 1896 г.) картинно описаны охоты древних греков на куланов, обитавших вместе со страусами в полынных степях верховьев Евфрата. По свидетельству Моисея Корейского (изд. 1893 г.), армянские цари династии Аршакидов (I—II вв. н. э.) охотились на онагров (кулапов) и кабанов в долине среднего Аракса. Эти сообщения были недавно разобраны Саркисовым (1946а), который ссылался еще па упоминания об онаграх в Армении в «Своде законов армяно-григорианской церкви», в «Географии Армении» Аиапиа Шпракцп (VII в.) и у Григора Нарэкаци (XI в. н. э.).
Под влиянием идей Сатунина Саркисов высказывал и необоснованное предположение о проникновении онагров в Армению из «Арало-Каспийской низменности», ссылаясь на наличие горных барьеров с юга и востока, хотя ясно, что эти животные водились в историческую эпоху в каждой мало-мальски подходящей межгорной долине и на плоскогорьях Передней Азии.
Еще более убедительны примеры, доказывающие существование кулана в Армении, из задачника Анании Шпракцп, разобранные Тер-Погосяном (1947).
Об обитании диких ослов в Бардинском царстве (в современной Миль-ско-Карабахской степи) упоминает армянский историк X в. Моисей Ка-ганкатваци (изд. 1861 г.). Низами Ганджеви (XII в.) воспевает охоты на диких ослов в Кура-Араксипской низменности. Рашид-Ад-Дии (XIV в., изд. 1946 г.) указывал, что в числе жертв грандиозной загонной охоты, устроенной Газан-ханом в Талыше, были и дикие ослы. В начале XVII в. Адам Олеарий (изд. 1870 г.) описал охоту иранского шаха на онагров — «гуров» в одном из отделений огромного зверинца близ столицы Ирана Испагаии.
Позднейшие заметки о кулане принадлежат русским натуралистам и свидетельствуют о том, что это животное уже во времена Палласа было распространено лишь за Волгой.
Хорошее описание повадок кулана для района киргизских степей Эмбенского уезда было дано Леваневским (1894). Из него явствует, что кулан был обычен еще во второй половине прошлого столетия в полупустыне между Эмбой и Уралом. В то время, очевидно, уже шло быстрое сокращение поголовья кулана и смещение ареала вида на восток и юго-
Гл\ IIL Прежнее и современное распространение и численность млекопитающих 333
восток. Трудно предположить, чтобы это смещение и сокращение численности было обусловлено только истреблением при помощи усовершенствованного нарезного оружия или вследствие увеличения численности населения. Основным оружием у ногаев и казахов по-прежнему оставались пистонные или кремневые винтовки, а увеличение числа кочевий ограничивалось кормовой емкостью пастбищ пустыни и полупустыни. Вероятно, были и еще какие-то причины, воздействовавшие на размножение и динамику численности животных.
Уже в первые десятилетия нашего века кулан сохранялся в пределах СССР лишь в полупустынях северо-восточного Казахстана — Бедпак-
Рис. 129. Куланы в плейстоценовой полупустыне восточного Закавказья.
$ала. Семиречье, — где эти звери исчезли в 30-х годах, а также на юге Туркмении — в Бадхизе. Особенности распространения кулана на Иранском плоскогорье, а также в Аравии пока неизвестны.
Учитывая значительную стенотопность и адаптации кулана к пустынно-степному ландшафту, появление его на Кавказском перешейке можно связывать с наступлением ксеротермической эпохи. В Закавказье он мог проникнуть с юга через долины и низкие хребты северо-западного Ирана, а в Предкавказье — с Русской равнины и в обход северного Каспия из Средней Азии.
Наибольшее расширение ареала кулана имело место в век бронзы. Исчезновение куланов в Закавказье возможно совпало с эпохой вторжения монголов в XIII в. и с позднейшим ростом Иранского государства, когда звери полупустыни в массе уничтожались па охотах многотысячной конницей. В северо-восточном Предкавказье куланы продержались, очевидно, до XIX в.
Отряд ПАРНО НО ПЫТПЫР-ARTIODACTYLA
Б четвертичной фауне Кавказского перешейка из этого отряда существовали представители семейств Suidae, Camelidae, Cervidae, Bovi-dae.
334 Часть II. Анализ происхождения четвертичных млекопитающих
До голоцена здесь дожили свиньи, олени и полорогие. Палеонтологические и зоогеографические материалы, позволяющие судить об истории развития ареалов и эволюции численности голоценовых видов парнокопытных, рассматриваются ниже.
Сем. СВШ1ЫЕ — SUIJJAE
Ископаемые остатки бунодоитных нежвачных семейства Suidae известны в Евразии с нижнего олпгоцепа. Род Sus, включая вымерший подрод Microstonyx, нижнеплиоценовой древности. Наибольшее число остатков ископаемых свиней обнаружено в верхнемпоценовых и плиоценовых отложениях Средиземноморья и Юго-Восточной Азии.
Кабан — Sus scrofa L. (s. lato). Остатки свиней полиморфного вида — S. scrofa появляются на Кавказском перешейке с верхнего плиоцена. Предком современного кабана можно, по-видимому, считать крупного таманского кабана — S. tamanensis (Верещагин, 1951г).
Следующим эволюционным этапом является апшеронский кабан — S. apscheronicus Burtsch. et Dzhai. из среднеплейстоценовых суглинков восточного Закавказья, описанный Бурчак-Абрамовичем и Джафаровым (1948).
В Ахштырской пещере на Черноморском побережье остатки кабанов прослеживаются во всех слоях — от мустье до поздиего железа причем намечается возрастание их числа по мере приближения к современности. Так, в мустье они составляли 0.06%, в верхнем палеолите — 1.2% и в послепалеолптических слоях — 20.0% от общего числа костных остатков каждого слоя.
В Западной Европе кабаны существовали как в доледниковую, так и в послеледниковую эпохи (Amon, 1938). К северу от Кавказа — на Русской равнине — имеются единичные находки кабанов в верхнеплпоценовых и среднеплейстоцеповых отложениях среднего Поволжья. Вообще в плейстоцене кабан был, очевидно, исключительно редок даже по долинам рек Русской равнины, проникая сюда спорадически с юга в малоснежные зимы. Из обильных костных сборов с отмелей Дона и Волги нам известен только один фрагмент челюсти кабана плейстоценовой древности (полуостров Тунгус), а из долины Урала — лишь остатки голоценовых кабанов (Верещагин и Громов, 1952). Вообще ряд фактов говорит за то, что широкое расселение кабанов с Кавказа на Русскую равнину произошло лишь в голоцене.
К югу от Кавказа — в северном и центральном Иране, а также в Палестине, Сирпи — остатки кабанов обнаружены в палеолитических и в послепалеолптических слоях поселений, хотя далеко по часто, что соответствует и преобладающим сухим ландшафтам этих стран.
Учитывая современные экологические черты среднеазиатских и кавказских кабанов: питание по преимуществу сочной и концентрированной растительной пищей, добываемой либо па поверхности, либо в почве, корот-коногость, относительно бедный шерстный покров, а в связи с этим п слабую приспособленность к низким температурам и к глубокому снеговому покрову, — следует признать, что именно последние факторы п определяли распространение этого животного в четвертичном периоде. В этом отношении соображения Формозова (1946) о «хиопофобпости» ка-бапов вполне разумны. В условиях такой горной страны, как Кавказ, с его разнообразными климатами и ландшафтами, глубина снегового покрова не имела и не имеет такого решающего значения, как па равна-
Га. III. Прежнее и современное распространение и численность млекопитающих 335>
пах. Именно этим и объясняется благополучное существование кабанов на Кавказе в четвертичном периоде.
В слоях поселений и в погребениях эпохи бронзы и более поздних культур Кавказа остатки кабана присутствуют почти постоянно. Особенно обычны они для древних поселений болотистой Колхиды, как например у Анаклин, и в слоях древних городищ Кубани. Обилие кабанов было характерно в средние века нашей эры, в частности, для долины Дона, так как в славянских и хазарских слоях Саркела у Цимлянской остатки этих зверей составляют до 6% от всех собранных костей. О былом распространении и численности кабанов на Кавказе в историческую эпоху ионию судить до известной степени также по предметам искусства и литературным источникам. Изображения диких свиней в орнаменте и среди погребального инвентаря века бронзы на Кавказе неизвестны и наблюдаются лишь в более позднюю,скифскую эпоху. Эффектные изображения кабанов (рис. 130) имеются в погребальном инвентаре из курганов Прикубанья и Таманского полуострова.
К древнеримским временам относится замечание Моисея Хоренского (ивд. 1893 г.) об охоте царей династии Аршакидов-в I—11 вв. н. э. на кабанов
в долине Аракса, в Армении. В описании царствования Артавадза и его войны с римлянами (кн. 2, гл. 22, стр. 73) говорится: «Он весь был предан яствам и питию; бродил, блуждал по болотам, по чаще тростников, по крутизнам, охотясь иа онагров и кабанов»; и далее (кн. 3, гл. 55): (Другой раз пришлось в камышах зажженных охотиться на кабанов».
Быстрее всего уменьшение численности кабанов и сокращение его ареала происходило на Армянском нагорье при отсутствии религиозного запрета па мясо кабанов и интенсивном процессе обезлесения горных склонов. Доказательством былого обилия кабанов на этом нагорье служат постатки, находимые ныне по обмелевшим берегам оз. Севан. Современная многочисленность кабанов в значительно более сухих районах Ирана н Турции подтверждает основную роль охотничьего промысла в христианской Армении для исчезновения кабанов. Вместе с этим в Грузии обилие горных лесов и труднодоступных болот долго поддерживало численность зверя на высоком уровне, и только в XIX и XX столетиях кабаны стали быстро исчезать.
Наоборот, в мусульманских районах развитие земледелия, в частности культуры кукурузы, риса, бахчевых, в ряде случаев содействовало развитию кормовой базы и процветанию кабанов. Такие примеры наблюдались в Ленкоранской низменности и в Самурских лесах. Вообще распространение ислама в средние века на Кавказе явилось, пожалуй, одной из даных причин, содействовавших расширению ареала и росту численности кабанов в историческое время. Даже в настоящее время в Азербайджане и Дагестане кабанов больше, чем в других местах. Об обилии кабанов па Кавказе в XVIII—XIX столетиях можно судить по многочисленным заметкам кавказских натуралистов, краеведов, путешественников.
Быстрое сокращение численности кабанов началось, очевидно, со второй половины прошлого столетия, после заселения Кавказа русскими..
336
Часть II. Анализ происхождения четвертичных млекопитающих
Первоначально оно проявилось на нижнем Дону, затем на Кубани, позднее — на Тереке и Сулаке.
Кабаны были редки в гирлах Дона и на Азовском побережье уже в начале XIX в. и совершенно исчезли около 1812 г.
В 70-х годах прошлого столетия кабаны еще попадались, по свидетельству Динника (1914а), в лесах под Ставрополем. В XX столетии их здесь уже не было.
На северо-западном Кавказе число кабанов сильно уменьшилось к началу нашего века в связи с развитием там поселков охотников армян, занимавшихся круглый год охотой.
В плавнях Кубани немного кабанов обитало еще в 1932 г. близ станиц Гривенской и Петровской, причем за сезон здесь убивалось местными охотниками не более десятка этих животных.
Небольшое количество кабанов жило в 30-х годах нашего столетия в ольховых болотах низового Риона, между Поти и Самтреди, которые на протяжении многих тысячелетий кишели этими животными. Вообще в западной Грузии, Имеретии, Абхазии, число кабанов к 30-м годам было уже повсеместно невелико.
В Караязском лесу, восточнее Тбилиси, еще в 1909 г. брали за один день охоты до 50 кабанов. В 1911 г. за два дня здесь было взято 120 штук (Калиновский, 1900; Марков, 1931а, 1935). В настоящее время в Караязах вряд ли можно взять за день и 5 кабанов.
В Кура-Араксинской низменности численность кабанов сильно колебалась па протяжении всего четвертичного периода в связи с опусканиями и подъемами уровня Каспия, а также прорывами береговых гряд, при которых на месте полынных степей образовывались обширные тростниково-рогозовые болота и озера.
Случай такого прорыва Аракса был в 1896 г., когда в Мугапской степи образовались обширные разливы Ax-Чала (Шелковнпков, 1907). От размеров паводков на Куре, степени наполнения степных озер и выгорания камышей ;прп засухах зависит благополучие и численность обитающих кабанов и в наше время.
В брошюре «Сельское и лесное хозяйство Ленкоранского уезда» (1914, стр. 21—22) говорилось: «Лет десять тому назад диких свиней было так много, что они являлись настоящим бичом для крестьян, но в последнее время охота на них приняла промышленный характер и количество свиней значительно уменьшилось. Ежегодно зимой наезжают сюда армяие из Елизаветпольской губернии для охоты на диких свиней ради шкуры, которая идет на лапти; местные русские охотятся на свинью ради мяса. За последние два года через Ленкорань было отправлено в Елизавет-польскуто губернию около 5000 шкур».
В 30-х годах нашего столетия кабанов уже пе было в лесах северо-восточного ^Карабаха и по северным склонам Малого Кавказа вплоть до Грузии. Они встречались еще, однако, в долине Акера-Чая и по лесистым северным склонам Зангезурского хребта, где их популяции пополняются особями, перекочевывающими через Араке из можжевеловых лесов северного Ирапа. j
В конце 40-х годов нашего столетия огромный массив тугайного леса, протянувшийся на 70 км по р. Куре — от устьев Иоры п Алазани до Мипгечаура, — был вырублен для подготовки ложа Мппгечаурского водохранилища. При этом были частью истреблены, частью вытеснены и кабаны, населявшие эти тугайные леса.
[л. Ill- Прежнее и современное распространение и численность млекопитающих 337
В 50-х годах, после закрытия Мингечау рекой плотины, стали быстро щсыкать и уничтожаться тугаи ниже Евлаха, а за этим следовало исчезновение и кабанов.
Уменьшение кабанов в последние десятилетия хорошо заметно даже »1лыше, горные леса которого справедливо считались еще недавно од-пз самых богатых кабанами участков.
h целом сокращение участков распространения кабанов на Кавказе иторпческом периоде произошло в результате прямого истребления . человеком как ценного промыслового зверя и вредителя. Антропо-(<й} ю изменения ландшафтов и занос эпизоотий имели все же второстепенное значение.
Современный ареал кабанов на Кавказе еще довольно значителен. Ив встречаются по всему северному склону Большого хребта и пред-ml iM, даже под Кисловодском и Нальчиком. В горном Дагестане каба-К еало. Наибольшая плотность их популяций существует ныне в Кав-им заповеднике, в низовьях Терека п Сулака, на Кусарской наклоп-S равнине, в Алазано-Агрпчайской низменности, местами по ту! аям |м|и Аракса и в Талыше (карта 80). В смежных районах Ирана кабана ftub.не всего по северным склонам Эльбурсского хребта (Саркисов, 1944 0.
По вертикали кабан распространен на Кавказе от уровня моря до аннинской зоны. Наиболее постоянно заселены, одииако, предгорпые тппости, в то время как в горах распределение кабанов сильно за-Ейг от сезонности кормов и снегопадов. Постоянство обитания по-гтно зависит прежде всего от кормовых условии. По учетам Допау-мВ. п Тенлова (1938), процент встреч кабанов в среднем за год в Кавказ-£jv заповеднике распределялся следующим образом: зона широьолп-Бцннх лесов — 48%, темнохвойных, лесов — 39% субальпийская — 1, альпийская — 2%.
1роцепт встреч свежих следов жизнедеятельности кабанов — пороев, twyvi; экскрементов, — по нашим наблюдениям в 1935 и 1936 гг., Екатальском заповеднике и в долине Алазани указан в табл. 72.
Таблица 72
Встречаемость следов кабанов на южном склоне Большого Кавказа в Азербайджане
Зоны и биотопы Процент встреч от общего числа на типовых 20-километровых маршрутах
август 1935 г. октябрь 1935 г. январь 1936 г.
Визовые леса А газани 45 15 52
Предгорья с фрукта ринками 32 35 33
Буковые леса средней зоны 10 32 15
Дубовые опушки субальпийской зоны 10 28 —
Альпийские луга 3 — —
В Талыше и в Ленкоранской низменности при отсутствии альпийской «ни. ио при обилии плодовых у верхней опушки, а также посевов зерно-вуа в зоне нагорной полустепи распределение кабанов несколько иное. К<баны концентрируются у верхней опушки леса в шопе—июле, но осенью Срои живут преимущественно на низменности и в средней зоне леса.
оо Н. К. Верещагин
338
Часть II, Анализ происхождения четвертичных млекопитающих
На суточный цикл активности кабапов и их распределение оказывают влияние не только корма, но и степень и характер преследования человеком. В восточном Предкавказье и Закавказье кабаны залегают па лежку нередко близ полотна железной дороги (Динник, 1914а), а также в садах и по бурьянам окраин полей хлопка (в Ширванской и Муганской степях).
Там, где кабапов не стреляют и вообще не тревожат, они выходят на жировку еще засветло, а в глухих камышах Кызыл-Агачского заповедника бродят и днем. В местах постоянного преследования кабаны выходят на жировку лишь поздней ночью и уходят затемно на лежку в глухие места, иногда за 10—15 км от места жировки.
Излюбленные биотопы кабана в восточной половине Кавказского перешейка — это непролазные чащи тугайных и низовых лесов, состоящие из лоха, карагача, пвы, кустов ежевики и зарослей тростника. Эти дебри и есть первобытный, исходный биотоп, соответствующий морфологическим адаптациям зверя.
Нарушения в закономерностях горизонтальных и вертикальных кочевок в связи с поиском кормных мест и уходом от глубоких снегов приводят теперь к гибели кабанов. Описанная Диипиком (1914а, стр. 47—48) массовая гибель кабанов от многоснежья, холода и голода в горах северо-западного Кавказа, наблюдавшаяся в 80-х годах прошлого столетия и в 1902—1908 гг., объясняется, с нашей точки зрения, исчезновением возможности широких откочевок в теплую низменность Прикубанья.
При наличии массивов леса, или тугаев, по речкам Закубапской равнины кабаны благополучно откочевывали бы в долину Кубани из области многоснежья в горных лесах. В XX столетии инстинкт откочевки уже заглушался воздействием прямого преследования, обилием поселений и наличием открытых, обезлесенных пространств на предгорной равнине.
Общее число добываемых па Кавказе кабанов оценивалось в 40-х годах в 8Э00 голов, пз которых приходилось на Азербайджан до 4000, на Северный Кавказ — до 2000, на Дагестан — до 1500, па Грузию—до 300 и Армению — до 200 (Верещагин, 1947 г.).
Будущее кабанов па Кавказе зависит преимущественно от дальнейших форм развития сельского и лесного хозяйства.
Кабан не терпим как вредитель полей кукурузы, ячменя, пшеницы, риса, бахчевых и в местах развития этих культур будет так или иначе со временем уничтожен или сведен до минимума.
Дальнейшее упорядочение оросительной сети в восточном Закавказье I и регулирование паводков рек с использованием земель под хлопок будет I неизбежно обусловливать быстрое уменьшение численности кабапов I вследствие сокращения защищенных участков.
В горных лесах Большого Кавказа и Талыша при нормированной I добыче кабаны смогут существовать более или менее благополучно не- I ограниченно долгое время.
Сем. ОЛЕНЬИ — CERVIDA Е I
Из плиоценовых слоев Кавказского перешейка известны остатки пред-1 ставнтелей родов Eucladocerus. Procapreolus, Р Uocervus. В послетретичных ! отложениях достоверно известны представители родов Cervus. Damn, I Megaceros. A Ices. Capreolus. Из них до современности дожили лишь! благо родный олень, лось и косуля.
Благородный олень — Cervus elaphus L. (s. ]ato). Остатки оленей I группы C. elaphus известны в Европе с плиоцена. Только пз плейстоцене-1
HI. Прежнее и современное распространение и численность млекопитающих 339
ают эве-кку щах их), г на вед-одят ухие
-кого зтоя-
Эти ЛО-
IL ко-при-—48) щеро-гия п й воз-
й рав->ласти и уже пшй и в ни не.
I 40-х Ю0, па
Э — до
итьпей-
еницы, иначе
авказье к будет <абанов
Эванной
чио пе-
ки и редети ч пых , Da та.
II' лишь
оленей
1стоцепо-
|вых слоев Средиземноморья описано до десятка различных форм. В пределах Кавказа остатки этих оленей прослеживаются с нижнего плейстоцена до современности. Как правило, они относились к ныне живущему С. elaphus maral Ogilby. Местонахождения ископаемых остатков, пока-1 занные на карте 83, свидетельствуют о весьма широком распространении !вмда.
Севернее перешейка благородный олень был таким же обычным обитателем пойм рек и припойменных равнин па протяжении всего плейстоцена, как зубр и лошадь. В Новочеркасском музее его остатки стоят на четвертом месте по численности после остатков слонов, зубра, тура. В прирусловых отложениях низового Дона кости благородного оленя составляли 3.3°о от всех собранных на отмелях костей, а на отмелях низовой Камы и средней Волги — от 0.5 до 5.0%.
К югу от Кавказа — на Иранском нагорье — остатки плейстоценовых благородных оленей обнаружены в палеолитических и послепалео-даческих слоях ряда пещер и в делювии северо-восточнее Мараги. Важно отметить, что в пещерах близ Урмии и Бехистуиа кости оленей составляют до 57 % от общего числа извлеченных костей. В мезолитических и неолитических слоях пещеры Белт у восточной оконечности Эльбурс-ского хреота остатков олепеп оказалось уже немного — 0.8% (Coon, 1951). Еще реже остатки оленей попадались в неолитических слоях городища Апау в западной Туркмении (Duerst, 1908). Благородные олени были cjyoii обычной пищей у палеолитических охотников в Ливане, Сирии, Палестине. Несмотря на столь широкое распространение, не может быть сомнений в том, что в передней Азии оно было узко местным, т. е. приурочено к долинам рек, к лесистым склонам и т. п., по крайней мере с плиоцена.
В послеледниковых отложениях остатки благородного оленя столь $е многочисленны, как и в плейстоценовых. Особенно обычны черепа, зубы, рога олепей в голоценовых суглинках Ставрополья и Пя-дарья, по долинам рек Егорлыка, Кумы, Подкумка, а также в делювии склонов и по берегам Севана на Армянском нагорье.
Обычны они в могильниках, в слоях поселений и городищ Кавказа — I с энеолита до позднего средневековья.
Все эти находки (см. главу II) позволяют думать, что олепи держались ва остспненных равнинах Предкавказья и на плоскогорьях Закавказья повсеместно по крайней мере весной, осенью и в начале зимы.
С этими фактами широкого прежнего распространения олепей в степ-' вых ныне местностях вовсе не следует связывать большую облесенность ^ррпторнп, как это делали Шелковнпков (1930), Даль (1947а) и др. 11Я оленей в весенне-летнее время вовсе пе обязательно наличие крупно леса. Наоборот, они откочевывали из лесов па степные и луговые v-rtCTKii, как это делают местами и сейчас, спасаясь от кровососущих ИСКОМЫХ.
| На протяжении нашей эры олепи обитали по долинам Дона, Волги, । Урала ппо прилежащим участкам степи и лесов; несомненно, что кавказ-дая популяция благородных оленей имела постоянные связи с восточноевропейской. В средневековых слоях крепости Саркел близ Цимлянской j ’(<тК благородных оленей составляли до 4.3 % всех отобранных для I |хлсдовапия костей.
I В течение всей послепалеолитпческой истории благородный олень I ввлялся излюбленным сюжетом для художников и скульпторов Кавказа I щи украшении глиняной посуды, оружия, одежды. Начиная с наскаль-I 22*
340
Часть II. Анализ происхождения четвертичных млекопитающих
ных контурных рисунков (рис. 131) в Кабристане, «олений сюжет» прослеживается в бронзовых подвесках кобапской культуры, в орнаменте ход-жалинских и севанских бронзовых поясных пряжек и особенно в золотых вещах скифо-сарматской культуры Закубанской равнины и южной Украины (рис. 132—133). Видное место благородный олень занимал и в религиозных верованиях, ритуалах. Так, по преданиям абхазцев и имеретин, в день святого Георгия к священным пещерам или часовням приходили олени, которых парод резал для пиршеств (Джапашия, 1915). В ряде дигорских дзуаров жертвенные черепа и рога оленей преобладают
Рис. 131. Скала Беюк-Даш южнее Баку. Изображение охоты на оленя.
Фот. автора, 1946.
над остатками других диких копытных. Все это говорит о том, что числен- I ность оленей в послеледниковую эпоху не снизилась столь быстро и ката- I строфически, как у тура и зубра.
Находки костных остатков оленей в четвертичных отложениях степных J районов нередко используются натуралистами либо в качестве доказатель- к ства о степном образе жизни оленей в прошлом, либо в качестве доказательств прежнего наличия леса в степных районах. Па основе современных наблюдений за поведением и питанием оленей, а также по строению их низких коренных зубов ясно, что благородные олени плейстоцена и исторического времени держались в степях преимущественно весной и осенью. Они паслись на молодой траве степей, а остальное время жили ио пойменным и байрачным лесам. В степи и лесостепи олеин были широко распространены по крайней мере до позднего средневековья. Так, Воплай (изд. 1832 г., стр. 92) в начале XVII столетия застал стада оленей в украинских ' степях: «По степям украинским разгуливают целыми стадами олени, лани, сайги >. Правдивость этого сообщения подтверждается благополучным существованием оленей на подножном корме в степном заповедник Аскания-Пова в наши дни.
fx Ш. Прежнее и современное распространение и численность млекопитающих 341
Впрочем, Скальковский (1885, стр. 189) говорил в XVIII в. по поводу ей й в Запорожье несколько иначе: «Они (т. е. охотники, — Н, В.) иске ловили зайцев, волков и даже оленей, которые до 1760 годов водп-, в прекрасных лесах Заднепровского Запорожья».
I протяжении исторической эпохи происходило интенсивное со-чше численности оленей в связи с развитием кочевого скотоводства, м п земледельческой культуры. Наиболее сильное сокращение по-
Рвс. 132. Северная Осетия. Кобанская культура. (По Уваровой, 1900).
1 — бронзовый ясезл с изображением охоты на оленя; 2 — напершие.
-чляций оленя в степной и лесостепной зонах Восточной Европы и Кав-цза произошло, очевидно, при одомашнив алии лошадей и широком приме-енпп их для устройства загонных охот. Быстрое истребление степных Ьсуляции оленей происходило при военных походах скифов, гуннов, •.таков, а позднее и монголов. Возможно, что именно с этими передви-Ьпямп и коллективными загопными охотами степняков (Малеин, 1911) иасдует связывать первоначальное оттеснение оленей в леса п лесистые гдгорья, где их пе могла преследовать конница. Барбаро (изд. 1836 г.) .дел в начале XV столетия конную охоту — «пагапею» татар па дроф вереи, когда загон охватил 120 миль (!) и «от татар бежали олени и -у и* звери».
Истребление степных популяций благородного оленя явилось, таким |шом, первым этапом в общем сокращении ci о численности в псторпче-, й эпохе.
Второй этап сокращения численности о теней начался с позднего сред-щековья в связи с широким применением подсечной системы земледелие быстрым истреблением лесов. В предгорьях и па высоких плоско-рях Кавказа это уничтожение леса сопровождалось иссушением тер-I рчорпп п, следовательно, сокращением пастбищ.
342 Часть II» Анализ происхождения четвертичных млекопитающих
Литературные сведения о распространении и количестве добывавшихся оленей известны по крайней мере с XVI столетия. Грузинские и иранские летописи, древняя художественная литература Грузии, Азербайджана, Ирана полны сообщениями об охотах на оленей. На загонные охоты в леса Гиляна, Карабаха в позднем средневековье персидские шахи сгоняли тысячи загонщиков — крестьян и воинов. Крупные звери, преимущественно олени, убивались при этом многими сотнями.
О множестве оленей в XVI в. в соседних с Талышом угодьях Мазенда-рана и Гиляиа можно судить по сообщению Адама Олеария (изд. 1870 г., стр. 730) о том, что шах Тахмасп со своей свитой в конце сто-
Рис. 133. Закубанская равнина. Скифская культура. Золотая бляха — лежащий олень из кургана близ станицы Костромской.
летия убил во время одной из охот 2000 оленей и джейранов — «агу» и «повелел выстроить башню из их рогов». Царевич Вахушти сообщал в «Географии Грузии» (1904), что 94-й царь Вахтанг в начале XVIII столетия со своей свитой убил в один день 180 оленей на Триалетском хребте юго-западнее Тбилиси.
Историк Сехнпа Чхеидзе писал: «Царь Вахтанг Леванович (1703—1721) пригласил высокопоставленных отправиться на охоту в Триалетию и на Шанбианп. В первый день убили до 60 оленей и несметное число других зверей» (История Грузии, 1945, стр. 408).
Еще в XVIII в. олени населяли камыши и пойменные леса Дона, Кубани, Маныча, Кумы, Куры, Терека и Сулака.
Исчезновение оленей в Предкавказье в XVII—XVIII вв. происходило вследствие заселения края казаками и массового завоза огнестрельного оружия. Уже Диннпк (1914а, стр. 97) отмечал, что в конце прошлого столетия в лесах под Ставрополем он находил лишь обломки оленьих рогов как доказательство того, что олени водились здесь «лет 60—70 тому назад», т. е. в 50—60-х годах XIX в.
В 10-х годах нашего столетия олени обитали еще близ Краснодара, на правом берегу Кубани. Здесь они уже тогда находились под охраной и были окончательно выбиты во время гражданской войны.
Северную границу ареала оленей иа северо-западном Кавказе Дин-ник (1914а) проводил по станицам Передовой, Каладжинской, Махашев-ской и севернее Майкопа. Наибольшее количество оленей водилось тогда в верховьях Белой, Большой и Малой Лабы, причем за день можно было
fx. ///• Прежнее и современное распространение и численность млекопитающих 343
встретить 2—3 десятка оленей. В зимнее время олепи заходили даже под Новороссийск и Кабардинку. В дальнейшем происходило постепенное смещение к югу границы распространения. В начале 30-х годов северная граница обитания оленя проходила уже в предгорьях по линии селений Соленой, Бугуж, Даховской, Самурской, Нефтяной, Навагпнской. Западными пределами были верховья р. Пшиш (Наспмович, 1936а). Восточнее Кавказского заповедника олени еще встречались в 40-х годах нашего столетня под станицей Преградиой и Микоян-Шахаром.
На предгорной равнине, севернее Нальчика, на оленей охотился еще в70-х годах прошлого столетия Россиков (1887).
Рис. 134. Апофеоз оленьих охот в Дигории. Полати в пещере Дигоризед. Фот. автора, 1947,
В предгорьях Северной Осетии последние олени были выбиты (рис. 134) jконце 20-х годов (Верещагин и Наниев, 1949). Дольше они сохранились ао притокам Сунжи у Гудермеса п уцелели до сих пор в тугайных лесах Стлана и Терека в равнинном Дагестане.
‘ В горном Дагестане олени были многочисленны по ущельям Аварского Койсу. В лиственных и хвойных лесах близ Анцуга и Тлядаля, йтакже в Хевсуретии и Тушетии в капищах находится много оленьих рогов (Динник, 1914а; Маруашвили, 1955). Олени бывают здесь и теперь, но лишь в летнее время, когда откочевывают с южного склона Главного хребта, спасаясь от кровососов. Поздней осенью, в октябре—ноябре, олени возвращаются из Дагестана на зимовку па южный склон. На пере-валах-то их стреляли и стреляют дагестанцы.
В Самурско-Хачмасских низовых лесах олени уже не водились в начале нашего столетия. Выше, в лесах Кубинского района, по свидетельству старожилов сел. Кусары, олепи перевелись в 80-х годах XIX в.
В Куринской низменности олени водились еще в конце прошлого столетия в тугайных лесах и камышах среднего и нижнего течения Куры. Старожилы древнего селения Шильян и Ширвапской степи помнят оле-яец обитавших по окраинам камышей Шильяпского озера, и тропа через камыши и до сих пор носит имя оленя — «марал гёль», «марал ёлу». В Талыше олени были малочисленны уже во время Радде, Сатупипа (19056) |Дпнипка (1914а). По свидетельству охотииспектора Азербайджана В,В. Витовича, последний олень в Талыше был убит в 1912 г.
• 344 астъ II. Анализ происхождения четвертичных млекопитающих
В Карабахе, в ущельях Дзегама и Шамхора, олени исчезли в 20-х годах нашего столетия. По Мелик-Шахпазарову (1898), в 90-х годах в Зап-гезуре было такое обилие оленей и косуль, что некоторые охотники убивали по 150—200 голов за год. Осипов (1898) указывает, что олени были обычны в его время в окрестностях Чаикенда, т. е. южнее современного Кировабада.
В 30—40-х годах несколько оленей еще держалось в Караязском лесу на Куре, где в 90-х годах прошлого столетия пх убивалось по 15— 20 экз. за однодневную охоту.
В западной Грузии олени были истреблены или оттеснены в горы из низовьев Риопа, Кодора и Ингура в начале первой мировой войны при начавшейся рубке леса.
В настоящее время ареал благородного оленя состоит из 4—5 изолированных участков (карта 83). Наибольший из них — северо-западный, охватывающий территорию Кавказского и Тебердипского заповедников, в бассейнах рек Пшиша, Курджипса, Белой, Лабы, Малого и Большого Зеленчуков и Кубани. Наибольшая численность оленей летом — в глубинных участках Кавказского заповедника — в пихтовых лесах с полянами на высотах 1700—2200 м. Зимовки оленей расположены чаще всего на выгревах в нижней части пояса пихты и зоне буковых лесов с глубиной снега от 0.3 до 0.5 м, причем звери держатся как на склонах, так и по долинам. Общее поголовье оленей в заповеднике в 1934 г. определялось примерно в 900 штук (Насимович, 1936а). Вообще после резкого сокращения числа оленей во время гражданской войны поголовье их стало возрастать п к 40-м годам в Кавказском заповеднике составляло не менее 3000 голов.
На северо-востоке Предкавказья олени сохранились в лесах и камышах низового Терека, примерно от станицы Червлегшой до Кизляра, и на Сулаке — от Хасав-Юрта до низовьев реки.
Олени этой популяции держатся круглый год на низменности, в тугаях. Им некуда теперь откочевывать: северные предгорья Дагестана ныне обезлесены и заселены.
В западном Закавказье небольшое число оленей держится еще в лесах по среднему течению Ингура. В буковых лесах Боржомского заповедника, т. е. по восточным отрогам Аджаро-Имеретинского хребта, Эль-Мар [(Е. »Г. Марков), 1939] насчитывал в 1934 г. до 900 голов оленей. Летовкп боржомских оленей были расположены раньше на высоких, безлесных и холодных плато Джавахетии и Ахалкалакского нагорья. В восточном Закавказье олени обитают в Караязском лесу и на южном склоне Большого хребта — от верховьев Алазапи до речки Гердымап-Чай. Наибольшая численность их наблюдается в Закатальском заповеднике, т. е. в бассейнах речек Белапух-Чай и Катехи-Чай. Число оленей в Закатальском заповеднике ориентировочно оценивалось в 40-х годах в 1000 штук. Порядочно оленей и восточнее, в Куткашенском и Исмаиллинском районах, как в горах, так и на низменности. Здесь они держатся близ селений Чухур-Кабала, Исмаиллы, Каланджак, Рушан-Кейд и др.
По нашим наблюдениям в 30-х годах, олени в летнее время держатся на южном склоне хребта, в верхней трети буковых лесов и в субальпийской золе. При отсутствии скота и собак они выходят и на яйлагп. Зимой они живут па средних высотах в буковых лесах по хребтам меридианного направления.
В многоснежные зимы они изредка выходят па открытую холмистую степь южнее Шемахи. Эти факты, а также круглогодичное обитание оле-
Гл. !!!• Прежнее и современное распространение и численность млекопитающих 345
зависеть здесь главным
Рис. 135. Семибратние скифские курганы. Бронзовая головка лани Эрмитаж.
noii в тугаях и камышах Сулака делает понятным и захоронение остатков [•леней вместе с остатками ослов и сайгаков на Аишеропском полуострове ч плейстоцене. Тем не менее современный благородный олень — в массе настоящий горно-лесной вид, привыкший обходиться без дальних кочевок ш равнину. Разница в повадках и в направлении миграций у различных юпуляций этого оленя свидетельствует только о большой пластичности шеря и большой роли человека в воздействии на высшую нервную деятельность в формировании современных привычек.
Ежегодная добыча оленей браконьерами на Кавказе достигала в 40-х годах нашего столетия 200—250 голов (Верещагин, 1947г).
В наши дни Кавказ является основным резерватом благородного меня в СССР. Дальнейшая судьба оленя будет «разом от постановки дела охраны. После организации борьбы с браконьерством оленя можно будет вновь развести в лесах Карбаха, Талыша, Кусарскоп наклонной равнины. Кроме того, кавказские популяции оденя могут быть использованы в будущем 1я заселения пойменных и водораздельных ьтов Русской равнины.
Дань — Dama cf. mesopotamica Brooke. Ископаемые остатки ланей, близких к сре-iL3iiMHOMopcKon D. dama L., описывались из плиоценовых и плейстоценовых захоронении Италии, Англин, Германии.
Обнаруженные на Кавказе в плейстоценовом аллювии Аджи-Эйласа у Еревана фрагмент нижней челюсти и обломок рога иляются пока единственными. Отнесение их к месопотамскому виду сделано здесь условно.
На юге Малой Азии и в Палестине лани были наиболее частой добычей палеолитических летников (Bate, 1937).
Художественные изображения головок средиземноморских ланей, сделанные из бронзы (рис. 135), обнаружены в Семибратппх скифских грганах Таманского полуострова, а также в курганах южной Украины. Все это дает основание думать, что в голоцене ареал обоих видов ланей, г е, средиземноморской и месопотамской, охватывал юг Балканского полуострова, Причерноморье, Малую Азию, Ирак и Иран (карта 82).
Лань — обитатель широколиственных лесов и перелесков, а также прирусловых зарослей речных долин Средиземноморья и более теплолюбива, нежели благородный олень.
К XX столетию лань была почти истреблена в при родном ареале, и позднее искусственно вторично разведена по всей Западной Европе IB Северной Африке (Ellermann a. Morrison-Scott, 1951).
Удачный опыт разведения ланей был произведен перед революцией а на Кавказе близ Боржоми (см. главу VI).
В будущем, при упорядочении охотничьего хозяйства, лапь, вероятно, 1ВПТСЯ наиболее благодарным материалом для заселения зоны предгор-
кустарников и лесов Кавказа. Ею могут быть заселены леса Ставрополья, предгорного Дагестана, восточной Армении, Нахичеванской \ССР и Нагорного Карабаха.
346
Часть II. Анализ происхождения четвертичных млекопитающих
Гигантский олень — Megaceros euryceros Aldr. Остатки этого оленя становятся известными в Евразии с верхнего плиоцена.
Область распространения гигантского оленя в плейстоцене захватывала лесостепную и степную зоны Европы и Азии. Остатки его особенно часты в хазарской толще долины Волги. Там на отмелях они составляют от 2 до 8% всего костного материала. Обычны его остатки и в плейстоценовых слоях Западной Европы. На Британских островах они обнаруживались в пещерах, в делювии и в торфяниках. Судя по рисунку, сде-
Рйс. 136. Золотые пластинки из скифских курганов Закубанья с изображениями, похожими на гигантских оленей. Берлинский музей. (По Bachofen-Echt, 1937).j
данному/в XII столетии, и довольно свежим остаткам в торфяниках, находимым в Ирландии, некоторые палеонтологи полагают, что гигантский олень дожил там и до нашей эры (Mitchell a. Parkes, 1949). Таких свежих остатков в Восточной Европе неизвестно, и, по-видимому, этот олень вымер здесь в верхнем плейстоцене. Тем не менее сведения Пи-допличко (1951) о находках геологически молодых остатков гигантского оленя в СССР, основанные на результатах прокаливания костей, заслуживают внимания и дальнейшей проверки.
В то время как кухонные остатки этого оленя обильны в палеолитических пещерах Крыма, они совсем неизвестны пз пещер Палестины, Сирии, Ливана. На Кавказском перешейке (карта 82) его остатки встречаются с верхнего плиоцена (Таманский полуостров) до мустьерских слоев включительно (пещер Абхазии и стоянок Закубанской равнины).
В Абхазии гигантский олень жил, очевидно, в горной мезофитной обстановке.
В плейстоценовой степи восточного Закавказья этот олень был чрезвычайно редок, так как в битумах Бипагадов обнаружена всего одна кость этого вида на несколько десятков тысяч костей других видов. Ни в одном из голоценовых захоронений, даже в таких массовых, как побережья оз. Севан и средневековые слои Саркела, остатков гигантского оленя пет. Неизвестны и его изображения в рисунке или скульптуре у палеолитических и послепалеолитических племен Кавказа. Стилизованные изображения благородного оленя на золотых пластинках из скифских курганов Закубанья, несколько напоминающие изображения гигантского (рис. 136, 137), дали основание Бахофен-Эхту (Bachofen-
Га. ПГ. Прежнее и современное распространение и численность млекопитающих 347
Edit, 1937) ошибочно утверждать, что гигантский олень якобы существовал на Кавказе в I тысячелетии до н. э.
Обычные захоронения скелетов гигантского оленя, находимые ] поименных отложениях и в торфяниках, свидетельствуют о том, что w экологии он несколько напоминал лося. По конструкции метаподий и особенно копытных фаланг гигантский олень занимал промежуточное .оложенне между лосями и быками. Он, вероятно, выносил довольно ыубокпй снеговой покров, но не обладал столь большой проходимостью w болотам и снегу, как лось. Скорее всего это был обитатель холодной
Рис. 137. Череп гит антского оленя из окрестностей Ростова-на-Дону.
достепн и долин равнинных рек с пойменными лесами и с зарослями кустарников, в которых и обитал вместе с эласмотерием и трогонтериевым лоном.
Кавказский лось — Alces alces caucasicus N. Ver. Остатки лосей группы Л. alces L. известны из плейстоценовых отложений северной юловпны Голарктики. Как правило, точки находок не выходят южнее .J-н параллели. В западном Средиземноморье плейстоценовые остатки зоеей либо редки, либо принадлежат другим видам рода Alces (Wolf, 138; Пидопличко, 1951, на карте — стр. 79 у этого автора все находки внесены к A. alces). Старые сведения о находках остатков лосей в палеолите Палестины, Сирии, Ливана (Blankenhorn, 1910) основаны на ошибках в определении (Picard, 1937). Тем более это относится к Египту. К северу от Кавказа остатки плейстоценовых лосей обнаружены в стоянке Боршево II под Воронежем и в обрывах берега Волги близ сел. Осыпной Бугор, выше Астрахани. В палеолите Крыма лось неизвестен. На отмелях средней Волги и особенно низовой Камы остатки лосей составляют обычно до 5% всех костей крупных видов.
На Кавказском перешейке остатки лосей определены из верхпеплей-Явценовых (верхнепалеолитических) слоев пещер западного Закавказья л Хосты до Кутаиси.
Наряду с этпм остатки лосей не найдены в мустьерских слоях Ахпггыр-пещеры, Ильской стоянки и в асфальтах Апшеронского полуострова. Создается впечатление, что лось как бы заново проник на Кавказ с се-Ка в верхнем плейстоцене, хотя первоначальное развитие предковых км могло происходить и на Кавказе, и вообще в области восточного Средиземноморья. Теперь выяснено, что лось пережил па Кавказе верх-дй плейстоцен и вымер лишь в начале XIX столетия.
Присутствие лося па Кавказе в век бронзы можно было предполагать пи бронзовым пряжкам и подвескам эпохи кобанской культуры из ар
348
Часть II. Анализ происхождения четвертичных млекопитающих
хеологических раскопок в Северной Осетии (Уварова, 1900, стр. 80, рис. 75; Верещагин 1949а). В коллекциях Государственного исторического музея в Москве имеется бронзовая пряжка, изображающая две головы лосих, из раскопок у аула Атаге в долине Сунжи, происходящая из слоев кобанской культуры II периода (рис. 138). Интересное рельефное изображение, напоминающее лося, высечено на камне из Эйюка,
города хеттов, существовавшего
в половине 11 тысячелетия до н. э. в северо-восточной части Малой Азии.
Таким образом, возможно, что лоси жили в историческую эпоху и южнее Кавказа, на лесистых участках Малой Азии.
Изображение лосей па золотых и бронзовых пластинках (рис. 138
Рис. 138. Бронзовая пряжка из аула Атаге на Супже с головами лосих. Исторический музей.
Рис. 139. Бронзовая бляха в виде головы лося (1 : 2). Южная Украина.
и 139) и накладках колчанов известны также пз курганов па Закубанской предгорной равнине и с юга Украины.
Наконец, Келлер (Keller, 1909) упоминал о находках золотых подвесок, изображающих лосей, в древнегреческих слоях поселений северного побережья Черного моря.
Все эти фигуры лосей скифской эпохи имеют большое значение не только для восстановления прежнего ареала лося, но и для правильного толкования происхождения золотых и бронзовых вещей скифских курганов с орнаментом и рельефом анималистических сюжетов. Учитывая, что на Кавказе и в северном Причерноморье проходила в голоцене южная граница распространения лося, становится ясно, что большинство скифских художественных вещей зверового стиля изготовлялось не па юге Балкан, не в Малой Азии и не на островах Эгейского моря, как думают нередко историки и археологи, а па месте — в Скифии.
О кавказских и южноевропейских лосях есть упоминания и в литературных произведениях.
Жан де Люк (изд. 1879 г.) при путешествии в земли «ногайских татар» в начале XVI ] в. отметил наличие у них лосей наравне с дикими лошадьми, оленями п другими животными.
Замечание Кларка (Clarke, 1810, стр. 386) о двух молодых и очень ручных «elks», виденных им под Екатерине даром, относится, конечно,
[л, III. Прежнее и современное распространение и численность млекопитающих 349
h леням, а ие к лосям, поэтому Кприков (1952) напрасно использовал т\ цитату для доказательств былого наличия лосей на Кавказе.
Палаас (1831) в своей «Зоогеографии» указывал, что лось водится It Кавказе. Наконец, в описании верований и обычаев черкесов Люлье |§70) писал, что богом лесов у черкесов являлся Мезитха, разъезжавший на кабане с золотой щетиной. По мановению этого бога «олени и л.»сп собирались в лесах, и тогда девушки доили их самок». Тем не менее вйличпе лося в современной фауне Кавказа отрицалось позднее луч-1и tn зоологами (Дппник, 1896, 1914а; Сатунин, 19036). Сатуннн пи-л: «Таким же манером он (Паллас, — Н. В.) поселил на Кавказе
Рис. 140. Полуископаемый череп лося в музее Ставрополя.
Фот. автора, 1945.
в лося (4. alces L.), относительно которого теперь уже никто более не сомневается, что его не только нет теперь на Кавказе, но и раньше никогда не было». В наше время Кулагин (1932) и Бутурлин (1934) вновь подвергли сомнению наличие лося на Кавказе. Оговорка Кулагина 0 возможности обитания лосей в Закавказье в связи с появлением рассказа Полферова отношения к делу не имеет, так как последний в рассказе «Эзпос», упоминая лосей и глухарей, описал свои впечатления, полученные где-то в зоне тайги, а не на границе Турции, как предста-иилось Кулагину.
К 30-м годам нашего столетия кости лося стали известны из слоев гпродшц VIII—-ХИ вв. и. э., раскопанных археологами у станицы [[шпянской на нижнем Допу. В 1944 г. в коллекциях Ставропольского уузея мною был обнаружен полуископаемый череп старого лося (рис. 140). В 1947 г. при обследовании осетинских святилищ — дзуа-!ш —в бассейне Уруха мы обнаружили там 7 черепов лосей (рис. 141), указав тем самым, что лось жил в предгорьях Северного Кавказа па-иавне с зубром и вымер или был истреблен здесь одновременно с последним в начале XVIII столетия (Верещагин и Семенов-Тян-Шапский, ОЙ* Верещагин, 1949а, 19496, 1955; Верещагин и Паниев, 1949).
Священные коллекции, содержащие остатки лосей, подобно дпгор-скпм, существуют, вероятно, и в других ущельях северного склона Большого Кавказа, в частности па Балкарском Череке.
350
Часть II. Анализ происхождения четвертичных млекопитающих
Во время процветания осетинских дзуаров в Дпгорпи и накопления в них черепов и рогов лось был уже очень редок по сравнению с оленем и даже зубром. 7 обнаруженных черепов лосей пз 16 изученных святилищ составляют всего 10% от численности черепов зубров и 0.8% от
черепов оленей. Вымирание
зверя на центральном участке Кавказа было обусловлено, вероятно, помимо непосредственного истребления, еще и повсеместным уничтожением лесов на Терско-Су ижевской равнине. Возможно, что последние лоси были убиты здесь в самом начале XIX столетия.
При раскопках крепости Саркел на нижнем Дону в 1950—1951 гг., проведенных Государственным Эрмитажем (Артамонов, 1952), было обнаружено много кухонных костных остатков лося вместе с остатками оленей, кабанов, сайгаков, медведей, бобров. Остатки лосей составили там от 0.3 до 0.8% всех костей домашних и диких животных, взятых в коллекцию при раскопках.1
Наконец, остатки лося и северного оленя стали недавно известны также из римских слоев Ольвии (Пидопличко и Топачевский, 1953).
А. А. Садовский (устное сообщение) видел в 1937 г. нижнюю челюсть лося среди большого количества костей диких копытных, в том числе оленя и косули, добытых при раскопках древнего кургана Наохваму в Мингрелии. Уцелевшие ольховые топи в низовьях Риона с их зарослями ивняка и вахты трилистной и сейчас представляют идеальные места для обитания лосей.
Недавнее существование лося в За-кубанье было подтверждено находкой рога (рис. 142) близ станицы Отрадной, в галечнике речки Уруп. Рог хранится в Краснодарском музее. Он выветрел
Рис. 141. Череп лося из пещеры с поверхности, но внутри еще совсем Дигоризед. свежий.
Фот. автора, 1948. Итак, если считать лося па Кавказе
верхнеплейстоценовым вселеицем с севера, то можно думать, что основной путь проникновения этого вида на юг проходил по долинам Дона и Волги. Не исключен путь вселения и с юго-запада через Балканы и Малую Азию. Распространение лося на Кавказе, вероятно, ограничивалось предгорьями (карта 84). Лоси (рис. 143) держались на участках с ключевыми и болотистыми речками Закубанской и Терско-Сунжепской равнин, по приморским
1 Исследование 200.000 фрагментов костей проводилось в Саркеле в 1952— 1953 гг. под моим руководством К. Б. Юрьевым, Г. В. Храбровым и др.
Гл. III. Прежнее и современное распространение и численность млекопитающих 351
11 я ем 1Я-от
<3 а
IMO
и на но, {ОСЬ
на $ен-рта-юго есте ков, >ста-iCTeii итых
того <е из ко и
террасам Абхазии и в Колхидской низменности. В противоположность благородному оленю, лось, по-видимому, не приспособился на Кавказе к горной лесной обстановке.
При упорядочении лесного и охотничьего хозяйства страны лося можно будет поселить снова на Закубанской наклонной равнине по долинам Афипса, Псекупса, Лабы и Зеленчука. Возможно, он прижи
вется местами и в горных лесах с молодыми насаждениями сосны, осины и ивы, например в Тебердинском и Кавказском заповедниках.
ЛОСЯ диких кули, куренные с их астной места
в За-ходкой |адпой, >анится дветрел совсем
Кавказе с севе-го вида ръ все-гранение рта 84). угистыми (морским
j в 1952—
Рис. 142.
Рога лосей: 1 — из слоев крепости Саркел на Дону; 2 — из галечников речки Уруп.
Косуля — Capreolus capreolus L. и С. capreolus pygargus Pall. Род Capreolus среднеплиоценовой древности (Simpson, 1945). Предком современного рода, вероятно, является Procapreolus, известный из нижнего плиоцена Европы и Азии. Флеров (1952, стр. 98—99) считает современную косулю потомком миоплиоценовых Cervulinae.
Немногочисленные остатки мелких европейских косуль известны из различных типов четвертичных захоронений Европы и Азии и почти все в пределах современного ареала (Woli, 1938; Пидоплпчко, 1951). На Кавказе остатки косуль известны из верхпечетвертичпых травер-тяпов горы Машук и из пещерных стоянок Закавказья — с апгеля до современности: Кударо I, Ахштырская, Гварджилас-Клде и др. Крупная форма ail. pygargus отмечена и южнее, в пещерах Палестины, Сирин, Ливана, с ашеля до неолита (Picard, 1937; Bale, 1937).
На Русской равнине остатки крупной косули обнаружены в верхне-палеолптических слоях Костепок иа Дону, в позднем всрхиечетвсртич-
352 Часть II. Анализ происхождения четвертичных млекопитающих
ном аллювии Среднего Поволжья (Хрящевка). Западнее, на Украине и в Крыму, в палеолитических слоях встречаются уже только остатки мелкой европейской косули. Остатки косуль многочисленны и в голоценовых отложениях Кавказа, например в слоях стоянки века меди— бронзы близ Анаклии, в послепалеолитических слоях Ахштырской пещеры и т. д. (карта 85).
В слоях VIII—XIII вв. н. э. остатки крупных, сибирских косуль встречены при раскопках крепости Саркел на Дону. В капищах Северной Осетии, например, в пещере Дигоризед хранятся черепа и рога мелких косуль типа С. capreolus, добывавшихся по Уруху с XV по XX вв.
Рис. 143. Кавказский лось.
Изображения косуль, оставленные древними племенами, на Кавказе редки. Упомянем о контурных рисунках косуль со стрелами у груди на серебряном ведерке из курганов века бронзы на Триалетском хребте (Куфтин, 1941).
Современный ареал косули на Кавказе еще довольно велик, и вообще косуля сохранилась лучше других копытных. Северный Кавказ населяет несколько более крупная форма, близкая к сибирской (Динник, 1914а), но наряду с ней там же встречается и мелкая, распространенная и в Закавказье. В камышах низового Дона, Кубани косуль в настоящее время нет, но несомненно, что кавказский ареал крупной косули связывался по долине Дона с ареалом сибирского подвида еще в позднем средневековье. Наличие крупной косули в Предкавказье, вероятно, объясняется сравнительно поздним, послеледниковым вселением ее с севера.
В островных лесах под Ставрополем косуля окончательно исчезла в 20-х годах нашего столетия. Мало ее и в уцелевших широколиственных лесах Закубанской наклонной равнины — между Хадыженском и Майкопом. В буково-грабовых лесах северо-западных отрогов Боль-
1IL Прежнее и современное распространение и численность млекопитающих 353
Шого Кавказа, например у Горячего Ключа, Крымской, косуль поря-дочно и в настоящее время.
В Кабардино-Балкарии и в Северной Осетии косуля держится по ;<-сам Черных гор, в ущельях Скалистого хребта и по мелколесью северного склона Терского хребта. По долине Сунжи косули теперь нет, о она обитает в тугаях Терека и Сулака между станицей Червленой : Кшяром. Живет косуля и по уцелевшим лесам предгорных увалов эозненекой области, но под Махачкалой бывает как большая редкость, (камышах низовий Сулака и Терека косуля не держится из-за обилия да. В центральном сухом Дагестане ее нет вовсе. В низовых лесах амура косуля обитала еще в начале 20-х годов, но в 40-х годах уже яершэнно исчезла. В лесах Кусарской наклонной равнины ее мало. ; целом современное распространение косули на северном склоне Кав-аза ограничено высотами 1500—2000 м для летнего времени. Зимой Рй катастрофических снегопадах косули спускаются на предгорные (внины, забираясь даже в селения Майкопского района (Оленич-Гне-;да, 1955).
В западном Закавказье косули в нижнем поясе леса мало вслед-да густой сети поселков; больше ее в средней трети зоны леса, jущельях верхней Сванетии, например по Ингуру, ее теперь нет, да < оаиыпе она постоянно вытеснялась отсюда глубокими снегами.
Небольшое количество косуль еще обитало в 40—50-х годах : i колхидских ольховых топях близ станции Чаладиди и в среднем те-1ении речки Пичеры. Потийские охотники убивали их ежегодно до й’штук. Обычна косуля по северным склонам Аджаро-Гурийских хреб-• r0S( а также на Триалетском хребте. На юго-запад ареал ее уходит j ।Малую Азию. В Палестине косуля была еще обычна в прошлом веке ка горе Кармел, но последний экземпляр был убит в 1912 г. (Bodenhei-аег, 1935).
В восточном Закавказье, по южному склону Большого хребта, кода весьма обычна и заселяет преимущественно нижнюю треть зоны торных лесов с зарослями кизила, боярки, грабинника. В буково-грабо-! вых лесах она держится близ полян, заросших гигантским папоротни-j ком орляком и ежевикой.
I В Закатальском заповеднике в 1935 г. мы наблюдали такое распре-косуль по высоте в бассейне Тала-Чая и Катехи-Чая, учтенное vсвежим следам на отмелях ручьев и на снегу (табл. 73).
Таблица 73
Распределение косуль по числу встреченных за день следов летом и зимой в Закатальском заповеднике
Дата учета Южные отроги хребтов. Буково-грабовый лес с полипами и осинником, 700—1800 м Глубинные участки ущелий с буковым лесом, 1200—1600 м Верхняя опушка леса и субальпийские луга, 2000 -2400 м
22—26 VIII 1935 8 2 1
21—26X1 1935 (снег 15— 16 3 —
—30 см) 1 I 1936 (снег 30—50 см 7 1 —
и более)
•п Н. К. Верещагин
354 Часть II. Анализ происхождения четвертичных млекопитающих
Косуля держится постоянно и в дубово-лапинных лесах Алазано-Автаранской низменности, причем зимой ее бывает здесь больше, чем. летом. В тугаях, тростниках и оазисах Кура-Араксинской низменности косуля не живет.
На Малом Кавказе косуля держится всюду, где есть значительные лиственные леса. По ущельям верховий речек Дебеда и Акстафа она живет летом в субальпийском высокотравье, близ уцелевших островков леса, на высотах 1800—2000 м.
В Карабахе косуль много в сохранившихся буковых лесах Тертер-Чая. На южных склонах Армянского нагорья ее нет.
В Талыше и далее в лесах Эльбурсского хребта косули мало. Она держится здесь как бы «пятнами» по некоторым осветленным ущельям. В 1945 г. за июнь—июль нам удалось встретить в ущельях Вазару-Чая и Виляж-Чая лишь единичные следы этого зверя.
В целом на Кавказе косуля нередка, однако ныне она нигде не достигает здесь такой численности, как в предгорьях северо-восточного Тянь-Шаня или в Приморье.
В нагорьях Ирана косуля если и есть, то в ничтожном числе по уцелевшим островкам леса. Бленфорд (Blanford, 1876) указывал косулю лишь для лесов Гирканики. В северных районах Междуречья проходит, видимо, и южная граница распространения этого зверя.
Хотя основной ареал косули и расположен ныне на севере Евразии, тем не менее на Кавказе — это, несомненно, древний, верхнеплиоценовый вид, ближайшие предки которого существовали и обнаружены в нижнем плиоцене Предкавказья и вообще в Средиземноморье.
По адаптациям косуля — лесной, преимущественно листоядный зверь, способный быстро передвигаться высокими скачками в кустарниках и высокотравье, но глубоких снегов косуля не выдерживает.
Быстрое сокращение ареала и численности косули на Кавказе произошло в последнее столетие в результате неограниченного промысла зверя и уничтожения лесов человеком (см. главу VI).
Общая добыча косуль в настоящее время исчисляется приблизительно: на Северном Кавказе — 4600, в Азербайджане — 1500, в Грузии — 700, в Армении — 300, в Дагестане 200 голов (Верещагин 1947г).
Создание полезащитных лесных полос в равнинном Предкавказье, несомненно, откроет широкие возможности для разведения косуль в культурном ландшафте.
Сем. ПОЛОРОГИЕ — BOVIDAE
В четвертичной фауне Кавказского перешейка известны представители подсемейств Bovinae, Antilopinae, Caprinae. В голоценовой фауне здесь существовали представители семи родов: Gazella, Saiga, Rupicapra, Capra, Ovis, Bos и Bison.
Джейран — Gazella subgutturosa Giild. Ископаемые остатки многочисленных видов рода Gazella известны в Евразии с верхнего миоцена — нижнего плиоцена. Вообще это род миоценовой древности. Фрагменты роговых стержней и зубов газелей, частично сходных с средиземноморской плиоценовой G. deperdita Gerv., найдены на Кавказе в сарматских отложениях близ Георгиевска, в среднеплиоценовых песчаниках южнее Баку и в нижнечетвертичной толще Таманского полуострова.
Гл, III, Прежнее и современное распространение и численность млекопитающих 355
Кроме того, в послеледниковых суглинках верховьев Калауса, юго-западнее Ставрополья, был найден обломок стержня рога, похожего на рог джейрана (Рябинин, 1918).
Остатки настоящих джейранов известны по раскопкам Куна (Coon, 1951) из плейстоценовых слоев пещер Тамтама и Визитун в центральном Иране, из мезолитических слоев пещерных навесов близ Джебела и Красноводска (раскопки А. П. Окладникова, 1949 г.), из слоев Анау у Ашхабада (Duerst, 1908).
В Ливане, Палестине, Сирии остатки газелей, близких к С. gazella Pall., G. dorcas L. и G. subgutturosa, зарегистрированы из палеолитических слоев пещер от ашеля до позднего неолита (Picard, 1937; Bate, 1937). На Русской равнине ископаемых остатков джейранов из плейстоценовых отложений неизвестно. Не было джейрана в восточном Закавказье ни в среднем, ни в верхнем плейстоцене. Достоверные остатки джейранов на Кавказском перешейке известны только из голоценовых слоев исторической эпохи как в Предкавказье, так и в Закавказье (карта 87).
Особо многочисленны остатки джейранов в слоях древнего Баку, где они составляют 14.5% от общего числа костей и 39% — от костей баранов и коз.
Главный массив современного ареала джейрана приурочен к северо-западному Ирану и Афганистану, а также к пустыням Средней Азии.
Отсутствие джейрана в бинагадинской среднеплейстоценовой фауне н обилие его остатков в голоценовых слоях восточной части перешейка позволяют утверждать, что джейран появился на Кавказе относительно недавно — в послеледниковое время — и «заместил» в Азербайджане сайгу. Вероятнее всего, что он проник непосредственно с юга через хребты северо-западного Ирана, окаймляющие с юга Муганскую степь. Такое расширение ареала стало возможным при наступлении сухой и теплой фазы голоцена. Не исключена, впрочем, и возможность завоза джейранов в Куринскую низменность древними кочевыми племенами, проводившими лето на нагорьях северного Ирана, а зиму — в Муган-ской степи подобно тому, как это еще недавно проделывали шахсе-ваны.
Скотоводы в Мильскои и Муганской степях и в настоящее время постоянно ловят и держат около кибиток молодых джейранов, легко приручающихся. В Сирии, Палестине, Ливане и вообще в странах Ближнего и Среднего Востока новорожденные газели вылавливались тысячами ежегодно для приручения и просто па мясо. У Усамы ибн Мун-кыза (изд. 1922—1923 г., стр. 204) имеется такое описание этой охоты: «Когда газели приносят детенышей, охотники выходят пешком и за-опрают тех детенышей, которые родились в ночь, в предыдущую ночь уИ за 2—3 ночи раньше. У крепости Джабар собрали 3 тысячи в день». Подросшие газели пересылались вождям и царям в качестве подарков 1 нередко на весьма большие расстояния.
(Преследование джейранов конницей мидян, албанцев, персов, арабов. монголов, вероятно, также было одной из причин расселения этих газелей на плоскогорьях и проникновения их в изолированные долины. П в наши дни можно наблюдать, как под влиянием длительного пресле-1ованпя на лошадях и автомашинах газели забираются в солончаки, I тугая, тростниковые заросли или убегают в распадки и на склоны гли-ввстых увалов. Тем не менее через день-два они выходят на открытую
23*
356 Часть II. Анализ происхождения четвертичных млекопитающих
равнину с твердой почвой, так как среди зарослей и в горах они часто оказываются беспомощными при нападении волков.
Старые литературные сведения свидетельствуют о широком распро- I
странении и большой численности джейрана в восточном и южном За- I
кавказье в средние века нашей эры. В поэзии ганджийского Низами (XIII в. н. э.) образ джейрана очень часто фигурирует в качестве художественного сравнения.
Олеарий (изд. 1870 г.) упоминал о стадах джейранов, замеченных им в 1633 г. у Джевата в Муганской степи, и об обилии этого зверя на смежном Иранском плоскогорье. Соймонов (1763) наблюдал джейранов в 1720—1724 гг. на о. Святом (ныне о. Артема), на который они были либо завезены еще арабами для развода, либо перешли с Апшеронского полуострова во время какого-то послеледникового падения уровня Каспия. Соймонов пишет (1763, стр. 110): «Однако живет на острову много диких коз, которые там плодятся, и неизвестно чем они кормятся, разве растущим на камнях мохом». «Дек. 3 (1724 г.). . . между тем на Святом острову казаки Донские с генералом бывшие стреляли несколько | коз диких». I
Отметим, что у арабов, по свидетельству академика Дорна (1875), | о. Святой был известен под названием острова «степных овец». О наличии I джейранов еще в середине прошлого столетия в долине среднего Аракса I имеются указания у Нордмана (1840) и Шопена (1852), Очевидно, в этой I долине джейраны были уничтожены на загонных охотах лишь во второй I половине прошлого столетия. Джейран приспособлен к обитанию на I ровных площадях полупустыни с твердой почвой (подробнее см. Be- I рещагин, 1937). Большая скорость (до 50—55 км в час), развиваемая I джейраном накоротке, позволяет ему легко спасаться от волков и даже f от гепарда. Он не выдерживает преследования волков и собак на вязкой | почве и поэтому не живет на свежевспаханных и особенно на политых | участках. На Аджиноурском плоскогорье джейраны живут и среди по- I севов ячменя и пшеницы, но только на богаре, где почва всегда более I или менее уплотнена. |
Будучи наиболее многочисленным в ровных степях, джейран держится все же на участках нагорных ксерофитов на пересеченном рельефе по третичным всхолмлениям Джейран-Чель, Боздаг, Турут-Саруджа и Кабристана. Массовая гибель джейранов в Куринской низменности наблюдается при сильных заморозках и обильных снегопадах (Верещагин и Дюнин,4949). Вообще джейран относится к «хионофобам», так как не может доставать корм из-под снега и быстро передвигаться при глубоком снеговом покрове. Эта особенность также подтверждает его недавнее появление в Закавказье с юга.
Площадь распространения джейрана и численность его в степях > восточного Закавказья менялись в историческую эпоху в широких пределах в связи с колебаниями уровня Каспия, прорывами береговых гряд и разливами Куры и Аракса, а также в связи с колебаниями чис- I ленности кочевых и оседлых племен и их земледельческой техники. I
В 20-х годах нашего столетия распространение джейрана в Закав- I ] казье совпадало с площадями полынно-солянковой полупустыни и I бородачевой степи и на западе доходило почти до Тбилиси. I €
Резкое сокращение численности и ареала джейрана в восточном I г
Закавказье началось с 20-х годов нашего столетия и особенно усилилось I р ' в 30-х годах, когда развитие автомобильного транспорта и применение I для охоты автомашин и самолетов сделало доступным для браконьеров! и
| [л.Ш. Прежнее и современное распространение и численность млекопитающих 357
I *1—----------------------------------------------------------------------------
массовое избиение этого зверя. В начале нашего столетия джейраны были вытеснены с Апшеронского полуострова, а к 20-м годам — из степи Шираки.
В 30-х годах охота вдогон на легковых и грузовых машинах при-’ яла такие размеры, что джейраны были почти уничтожены в северной \,сти Ширванской степи. Только у станции Карасу браконьеры добывали на автомашинах более 500 джейранов в год. Браконьеры на автомашинах Мильских хлопковых совхозов добывали в 30-х годах до 500— НО газелей ежегодно. Столько же добывали и пастухи, зимующие в Миль-[\:ой степи. К 1937 г. распространение джейрана в Азербайджане ока-• дось приурочено к 8 изолированным участкам полупустынных и слабо ^военных участков прикуринских степей и увалов Аджиноура и Кабестана общей площадью около 8219 тыс. км2, а численность их дости-, ла в это время 5—6 тыс. голов (Верещагин, 1939а) (карта 87). В по-i едуютцие годы ареал и поголовье продолжали быстро сокращаться 1 связи с бурным развитием автомобильного транспорта, браконьер-f вом и освоением площадей полупустыни под хлопок.
К 40-м годам джейраны были вытеснены с территории севернее речки (Дмгаит. В Кабристане они оттеснялись в глубь полупустынных увалов вникающими нефтяными промыслами. В 1935—1937 гг. стайки джей-рднов еще встречались на Гездекском плато и у горы Кергез, в 10 км к юго-западу от Баку. В 40-х годах отдельные особи попадались не ближе ;5-ЗО км от Баку, в районе Карадага и Шахи-Кая.
Орошение и распашка Кура-Араксинской низменности приведут к ближайшие десятилетия к полному вытеснению джейранов из восточ-: юто Закавказья; если не будут организованы степной заповедник и действенная охрана этого зверя.
; Сайга — Saiga tatarica L. Остатки сайги известны в плейстоценовых ; и голоценовых слоях от Британских островов на западе до Новосибир-j ских островов на севере и до Аляски на востоке (Nehring, 1890; Черский, j 1891; В. И. Громов, 1948; Пидопличко, 1951). Тем не менее устойчивость пластических признаков сайгаков с среднего плейстоцена до наших дней 1 ^ет основание считать, что древность рода по меньшей мере плиоценовая । Ш даже миоценовая. Отсутствие находок близко родственных форм 1 в плиоценовых слоях Палеарктики довольно загадочно, но, учитывая 1 адаптации современного вида, можно думать, что он формировался в ус-1 данях саванн и степей.
j На северном побережье Средиземного моря, т. е. в южной Франции, | Лдаи п Греции, остатки сайги, как правило, отсутствуют (Wolf, 1938, I 1939), несмотря на то, что в остальном плейстоценовая фауна этих рай-I оков была весьма сходна с фауной средней полосы Европы. Наиболее 1 вдае находки плейстоценовых остатков сайгаков имеются в степном I ЦрЫМу и на Кавказском перешейке. Кости сайгаков являются массо-| вкм и надежным индикатором средне- и верхнеплейстоценовых отложе-1 нвп Русской равнины.
На Кавказском перешейке ископаемые остатки сайги известны из Предкавказья и из восточного Закавказья (карта 86).
| К югу от Закавказья сайга, по-видимому, не проникала. Не было 1 ее и в западном Закавказье, что в общем соответствует установив-1дася здесь древней облесенности района и древнему пересеченному рифу*
Появление сайги на Кавказе можно относить по крайней мере к сред-$цу плейстоцену.
368 Часть II. Анализ происхождения четвертичных млекопитающих
Остатки сайгаков, принадлежащие минимум 70 особям, в бинагадин-ских слоях битума на АпшСронском полуострове очень хорошо документируют путь проникновения в восточное Закавказье европейско-азиатских степных форм с севера по западному побережью Каспия. По-видимому, в преДхазарской фазе регрессии древнего Каспия отсутствовал экологический барьер для степных видов, существующий ныне в виде лесов Хачмасской низменности и Кусарской наклонной равнины. В связи с зимними миграциями сайгаков, описанными еще Г личем (1865), возникает вопрос: насколько постоянно было обитание сайгаков в восточном Закавказье. Не были ли это лишь зимние забеги во время суровых зим? Ответ на это дает изучение возрастного состава погибших в асфальте зверей. 60 нижних челюстей сайгаков, сохранивших коренные, дали следующие возрастные группы.
1. Молодые июньские-июльские; стираются молочные коренные, прорезается из челюсти 1-й истинно коренной — 33.3%.
2. Молодые августовские-сентябрьские; 1-й истинно коренной прорезался и стирается—21.7%.
t. 3. Полувзрослые — ноябрьские; 2-й истинно коренной прорезался, молочные сменяются на постоянные — 5.0%.
4. Взрослые, с стирающимися истинно коренными, и старые особи с сильно стертыми зубами — 40.0%.
Таким образом, не подлежит сомнению, что сайгаки размножались на Апшероне и обитали в восточном Закавказье по крайней мере летом, Г осенью и в начале зимы. Сайга продержалась в восточном Закавказье позднее и была, вероятно, вытеснена лишь хвалыпской трансгрессией, сильно сузившей предгорную остепненную полосу суши.
В Предкавказье в верхнем плейстоцене сайга обитала вместе с зубром и гигантским оленем на Закубанской предгорной равнине и, видимо, на Таманском полуострове. Были ли это лишь зимние забеги из Прикубанской равнины и через Керченский пролив из Крыма, по имеющимся материалам судить трудно. Что такие перекочевки по льду были возможны, говорят, например, случаи захода сайгаков на острова Аральского моря (Берг, 1905).
В послеледниковую эпоху ареал сайги был еще огромным и простирался от Карпат до Монголии. В равнинном Предкавказье сайгаки обитали в голоцене от Азовского до Каспийского побережий. Отсюда и особенно с смежных участков на Русской равнине известны многочисленные находки голоценовых остатков сайги. Относительная численность остатков этого вида по отношению ко всем костям животных, извлеченным из культурных слоев, представляется в таком виде: 1) станица Цимлянская, эпоха бронзы, XV—X вв. до и. э. — 5% по одному и 12% по другому участку раскопок; 2) хутор Красный Яр близ Цимлянской, — эпоха поздней бронзы — 4.2%; 3) сел. Луговое в долине Ассы, слои скифской эпохи — 1 стержень рога среди большого числа остатков домашних животных; 4) станица Цимлянская, слои с хазарской культурой VIII—X вв. н. э. — 14.8%; 5) крепость Саркел близ Цимлянской, IX—XI вв. н. э., по раскопкам 1934 г. —6.5% и по раскопкам 1950 г.— 3.3%.
В рисунке, скульптуре и фольклоре кавказских народов сайга неизвестна, если не считать изображения какой-то антилопы на сосуде из Май- j конского кургана (рис. 128) и маловыразительных недавних, почти Современных рисунков на чокракских песчаниках у Капчугая в Дагестане, описанных Марковиным (1953).
Гл, IIL Прежнее и современное распространение и численность млекопитающих 369
На Украине сайга была обычна в позднем средневековье (Боплан, йзд. 1832 г.); Литвин (изд. 1890 г., стр. 48) примерно в 1550 г. писал о сайгаках и косулях Киевской области Великой Литвы: «Дикие козы в таком огромном количестве перебегают зимой из степей в леса и летом обратно в степи, что каждый крестьянин убивает их до тысячи в год».
В прошлом столетии южная граница распространения сайги в Закас-пни проходила по Усть-Урту, Аралу и северным Кызыл-Кумам.
В XIX и начале XX столетия сокращение численности сайги под влиянием прямого истребления шло очень быстро на всем уцелевшем ареале (Глия, 1865; Динник, 1914а; и др.). Имела значение также и распашка степей. Во времена Россикова (1887) сайгаки водились еще в низовьях Малки. В 90-х годах прошлого столетия сайгаки встречались под Ставрополем: их убивали близ Сенгилеевского озера и сел. Старо-Марьев-ского (Динник, 1914а). При Диннике сайгаки были еще обычны около Арзгира, Рагулей, Туркменской Ставки, но уже исчезли на его памяти из окрестностей Большой Кугульты, Джалги, Бурукшуна, Тахты. В западном Приманычье их уже не было в начале нашего столетия (Богачев, 1918). В начале 20-х годов многочисленные вооруженные отряды в степях способствовали катастрофическому сокращению поголовья сайгаков. Сайга уцелела лишь местами на Черных землях между низовьями Кумы, Волги, Урала, Эмбы, в северном Казахстане и в Монголии. В Терско-Кумской полупустыне она совершенно исчезла, и в 30-х годах ее кочующие стайки встречались лишь как большая редкость у Арзгира и Терекли-Мектеба. Тем не менее последующий запрет добычи, изъятие у населения па резного оружия и постепенное обезлюдение полупустыни обусловили восстановление поголовья сайгаков.
По подсчетам Адольф (1950, 1952), в 1939 г. между низовьями Кумы л Волги обитало до 3500 сайгаков, а в 1950 г. встречались стада более 1500 особей; общее количество зверя на правобережье Волги составляло несколько десятков тысяч голов.
После второй мировой войны сайга стала обычна и в Терско-Кумских степях (Бабенышев, 1948).
В суровую зиму 1948/49 гг. тысячи сайгаков погибли в астраханских й предкавказскпх степях от холода и голода. На восточных склонах Ергеней весной 1950 г. трупы сайгаков валялись по степи группами по 5-8 штук (Колесников, 1950). Множество сайгаков устремилось при снегопадах на юг, к Тереку и Сулаку, причем отдельные группы животных достигали предгорий Дагестана. Лавровский (1950) описал также «копления десятков тясяч сайгаков севернее Черных земель на участках развитых бурьянов.
При пурге и холодных бурях в суровые зимы сайгаки, как и джейраны, ищут спасения в тугаях и камышах, поэтому преодоление этими животными тугаев по Тереку и Сулаку не должно удивлять. Наряду с этим бывала гибель сайгаков и при дальних перекочевках в бесснежные холодные зимы, когда они в поисках снега для утоления жажды проходили сотни километров к северу, а затем оказывались застигнуты внезапным бураном. В результате упомянутой откочевки летом 1951 г. значительное число сайгаков паслось на полях кукурузы и пшеницы в Грозненской области.
Судя по современному ареалу и привычкам, сайгу следует считать животным степного ландшафта, но в пустыню она была, конечно, «загнана» человеком. Весьма вероятно, что в плейстоцене и голоцене она жила в лесостепи и во всяком случае держалась в пойменных зарослях и камы-
360
Часть II. Анализ происхождения четвертичных млекопитающих
шах, подобно некоторым африканским антилопам. Существование дальних кочевок из леса в степь и обратно, хотя бы и в необычные бесснежные зимы, подтверждается и упомянутым сообщением Литвина. Вообще современная стенотопность сайги в значительной мере кажущаяся и » вынужденная. Дальнейшие перспективы существования сайги в Предкавказье невелики. Распашки и орошение больших массивов степей и пустынь будут сужать с каждым годом площади, пригодные для ее оби-
тания.
В 1950 г. 15 сайгаков было выпущено на о. Булла, южнее Баку. Вероятно, сайгаки могли бы существовать ныне вновь и во всем полупустынном восточном Закавказье.
Рис. 144. Изображение серны на серебряном ведерке из курганов на Триалетском хребте (По Куфтипу, 1941).
Серна —Rupicapra rupicapra caucasica Lyd. Ископаемые остатки серн обнаруживались неоднократно в плейстоценовых и голоценовых слоях пещер Бельгии, Франции, Германии, Италии, например в гроте Коломби с его чрезвычайно богатым составом остатков животных. Места находок были всегда приурочены к горным или к предгорным районам.
На Кавказе единичные остатки верхнеплейстоценовых серн найдены лишь в пещерных палеолитических стоянках западного Закавказья, а голоценовые средневековые — в святилищах Северной Осетии. Остатки серн в плейсто-
ценовых и голоценовых слоях обширных предгорных равнин Кавказа нигде не обнаружены. В
Иране, Сирии, Палестине, Ливане остатки серн в плейстоценовых отложениях неизвестны. Современное распространение серны приурочено к Пиренеям и Кантабрийским горам, Альпам Франции, Швейцарии, Италии, Апеннинам, Карпатам, Балканам, Малой Азии и к Кавказу (Couturier, 1938).
Таким образом, палеонтологические данные и современная зоогео-
графия дают представление о серне как о горном виде, возникшем на альпийских складчатых горах, окаймляющих с севера западное Средизем-
номорье и с юга — восточное.
Контурные изображения серн с остриями стрел у груди имеются на серебряном ведерке из кургана века ранней бронзы на Триалетском хребте (рис. 144). Если учесть, что серны плохо переносят условия неволи и вряд ли завозились когда-либо древними племенами далеко от мест природного обитания, то эта находка указывает археологам, что ведерко было изготовлено в пределах Армянского нагорья и Малой Азии. Современный ареал серны на Кавказе еще довольно велик (карта 88). На Большом хребте ее распространение начинается на западе от перевала Гойтх и горы Семешхо (1032 м) — в верховьях Пшиша (Дииник, 1896,. 1914а; Насимович, 19496). В 1955 г. несколько серн еще жило даже северо-западнее Туапсе, в скалах района Джубги. Отсюда серна распространена на восток по обеим сторонам Кавказа вплоть до горы Баба-
]
] ]
I
1 г с I
л
В
н в]
[а. Ш. Прежнее и современное распространение и численность млекопитающих 361'
Дат в Азербайджане. На западном Кавказе она живет в пределах от 250— 300 до 3000/м над ур. м. В конце прошлого и начале нашего века серна была наиболее многочисленна в верховьях Белой, Уруштена и Малой Лабы. Здесь за день встречали до 400 серн и попадались стада до 90 голов Дипник, 1914а). В 1926—1937 гг., по мнению Насимовича (19496), в Кавказском заповеднике держалось минимум 4000 серн, а Жарков (1940а) шзывал даже цифру в 12 000. Места зимней концентрации серп приурочены здесь к склонам хребтов преимущественно западной и восточной «позиции.
Наибольшее число серн сохранилось теперь на средних высотах в суб-льпийской зоне, на высоте от 1500 до 1700 м.
В центральной части Северного Кавказа серна обитает на склонах льбруса и в сосновых лесах хребтов, окаймляющих верховья Баксана, •егема, Черека. В передовой цепи Скалистого хребта на этом участке рна не сохранилась и начинает попадаться лишь в Дигории и Осетии. Дагестане она распространена по лесистому хребту Черных гор до вредно течения Аргуни. Здесь серна живет всюду на скалах в весьма влажен обстановке субальпийского высокотравья и буково-грабовых лесов. •> верховьях Уруха и Ардона серны обитают на скалистых участках, оросших сосновым лесом, например около Ногкау и Цея. В централь-ом безлесном Дагестане серны теперь нет, и только на восточном склоне оры Шах-Даг живет небольшое число этого зверя.
По южному склону Большого хребта серны весьма обычны в Абхазии Сванетии. В верхних частях ущелий Кодора, Ингура. Риона они живут о очень крутым участкам склонов, поросших пихтой, елью и кленом. Наиболее богатые сернами места находятся здесь в верховьях Цхенис-[хали. В Южной Осетии серн мало, так же как и в Кахетии. Далее к воску они многочисленны в буковых и березовых лесах ущелий Катехи-[ая, Шин-Чая и бассейна верховий Турьян-Чая в Азербайджане.
В Закатальском заповеднике в августе наблюдаются стада по 50—7О5 серн. Общая численность серн в этом заповеднике определялась в 30-х годах примерно в 1000 голов (Марков и Млокосевич, 1935). Обычные места обитания здесь серны летом — это опушки березовых лесов и очень крутые задернованные склоны субальпийской лугостепи с белоусом, овсяницей, местами с выходами скал черного сланца. На скалистых участках склонов близ Курбан-Эфенд в Исмаиллинском районе серна держится и летом начиная с высот 800—900 м. В зимнее время большинство серн обитает здесь в буковых лесах в пределах 800—1200 м.
На Малом Кавказе ареал и численность серны значительно меньше. Серна обитает еще в бассейне среднего течения Чороха в районе Артвина п далее на запад, по Восточному Тавру, например в окрестностях Тра-пезунда. Новых сведений об обитании серн по северному склону Аджаро-Имеретпнского хребта нет. В буковых лесах и скалах левобережья Куры в районе Боржоми серна была обычна в конце прошлого столетня. На территории Боржомского звериного отвода в 130 га было загорожено в 1890 г. 5пиук (Шильдер, 1892). На Триалетском хребте серна водится в районе Белого Ключа, но, по-видимому, истреблена в других местах. На без-десном плоскогорье Джавахетии серн нет и, вероятно, не было.
Об обитании серны на Алагезе сообщал еще Богданов (1873) со слов Калишевского, который будто бы наблюдал ее на этой горе в 1904 г. Ни Бурчак-Абрамович в 1944 г., ни Даль в 1950 г. серны на Алагезе не нашли. По северо-восточной окраине Армянского нагорья серна жила еще в прошлом столетии. В 60-х годах XIX в. серна водилась в верховьях
362
Часть II. Анализ происхождения четвертичных млекопитающих
речек Дзегам-Чай и Шамхор-Чай до сел. Барсуна (Радде, 1899, стр. 74). Краевед Осипов (1898) называл серну для горных окрестностей сел. Чайкснд. В 30-х годах нашего столетия серн уже не было ни в Карабахе, ни на Шах-Дагском хребте, хотя Саркисов (1944ж) и утверждал, что они перевелись на Мров-Датском хребте только к 1930 г. Вообще сведения о распространении серн на северных отрогах Шах-Дагского хребта, па Дорийском нагорье и в северо-восточном Карабахе, приведенные Саркисовым (1944ж), требуют проверки.
Рис. 145. Лежка серны над обрывом в ущелье Катехи-Чая.
Фот. автора, 1935.
Исчезновение серн в Карабахе и вообще на Малом Кавказе нельзя объяснить только воздействием человека, так как по Мров-Дагскому и другим хребтам еще порядочно глухих лесов, в которых уцелели, например, безоаровые козлы.
В Талыше и на Эльбурсском хребте серны нет и, вероятно, не было, но она, несомненно, могла бы там существовать. К югу от Армянского нагорья серна, по-видимому, еще встречается на участках с сохранившимися лесами. В коллекции ЗИНа имеется рог серны из гор южного побережья оз. Ван.
Приспособленность серны к сильно пересеченному скальному рельефу гор меньше, нежели у козлов. Костяк ее более легкий, копыта менее массивны. Серпы избегают ходить по таким тяжелым скальным местам, как кавказские козлы, хотя некоторые натуралисты придерживаются другого мнения. Главные враги серны — крупные кошки (барс п рысь). От волка и медведя она спасается на «отстойных» скалах и, обладая большой скоростью передвижения в условиях скального рельефа, серны часто пасутся рядом с медведями. Сезонные и суточные кочевки не выражены у серны так отчетливо, как у козлов. Серны не откочевывают •зимой в малоснежный Дагестан и не выходят летом на ледники из леса.
[..HL Прежнее и современное распространение и численность млекопитающих 363
Нападения кровососущих двукрылых они переносят, очевидно, легче, чем козлы. Связи с лесом (рис. 145) и с мезофитной луговой растительностью у серны прочнее, чем у козлов. В целом серна — это вид, приспособленный к мезофитным участкам горных поднятий Средиземноморья. Ъ четвертичном периоде серна, как специализированный горный вид, на равнины, видимо, уже не спускалась. Расселяться с хребта па хребет леко по равнине в это время она не могла, так как, не обладая в этих j данях такими скоростями, как косуля и джейран, она была бы истреб-л па хищниками, прежде всего волками.
Отсутствие остатков серн в равнинных захоронениях и данные по ее с временному распространению и адаптациям свидетельствуют о само-с ятельном происхождении кавказской и альпийской серн от общих п едков существовавших в третичном периоде на складчатых возвышенностях побережий Сарматского моря.
Говорить о расселении серны в четвертичном периоде из малоазийско-ывказских гор в европейские Альпы или, наоборот, из Европы на восток через Балканы и Малую Азию нет оснований. Кавказскую серпу следует, в сущности, считать не подвидом швейцарской, а самостоятельным видом. По сравнению с альпийской у кавказской серпы ются не только известные морфологические отличия, но и иной гс ос — длинный шипящий звук, отмеченный еще Динником (1896).
Отсутствие серн в Талыше и Эльбурсе, быстрое сокращение числен-яо:тп и ареала серны в XIX и XX столетиях на Малом Кавказе — все говорит о том, что современный экологический оптимум зверя и, вероятно, центр возникновения лежат в районе хребтов, окружающих • остока п юга Черное море.
При организации рационального лесного и охотничьего хозяйства гграпьг серну следует развестп вновь в Карабахе, в верховьях Дзегама, Памхора и в Талыше.
Западнокавказский и восточнокавказский козлы — Capra саи-tuica Giild. и С. cylindricornis Blyth. Ископаемые остатки представителей подсемейства Caprini начинают встречаться в Евразии с нижнего ппоцена.
На Кавказе обнаружены пока лишь остатки козлов плейстоценового п голоценового возрастов. Все пункты находок плейстоценовых остатков расположены в западном и центральном Закавказье по окраине границ современного ареала и нередко удалены от мест современного обитания на 50—60 км по прямой (карта 89).
Эти факты можно объяснить не только влиянием оледепеппй, «отжимавших» козлов вниз, а преимущественно большей численностью зверя п более обширным распространением его в плейстоцене (см. главу II).
На юге — в палеолите Ливана, Сирии, Палестины — известны остатки местных, южных форм типа С. sinaitica ЕгепЬ., описанные под пмепем: С. beden Wagn., С. ibex L., С. cf. пиЫапа Cuv. (Picard, 1937). Остатки плейстоценовых козлов известны также из многочисленных горных п предгорных стоянок Западной Европы — от Ирландии и Бель-ш до Болгарии. Эти остатки, иногда даже целые черепа, относимые обычно к нескольким видам современных европейских козлов — С. ibex L., С. aegagrus L., С. pyrenaica Schinz и ископаемым — Ibex priscus Wold., /, cebennarum Gerv. и др., — встречаются там как в плейстоценовых, так и в голоценовых слоях (Woldrich, 1893; Wolf, 1938). Какой-то крупный козел Capra sp. обитал в палеолите и в горах Крыма (Громова и Гронов. 1937).
364
Часть II. Анализ происхождения четвертичных млекопитающих
Рис. 146. Бронзовое на-всршие, изображающее восточнокавказского козла. Северная Осетия. Кобанская культура. (По Уваровой, 1900).
Скульптурные изображения кавказских козлов, преимущественно восточнокавказского тина, изготовлялись нередко в век средней бронзы. Эти небольшие бронзовые фигурки козлов употреблялись древним населением горного Предкавказья в качестве наверший жезлов, подвесок, брелоков (рис. 146).
Несколько напоминают западнокавказских козлов и контурные изображения на серебряных сосудиках из Майкопского кургана (рис. 128).
Палеонтологические и археологические документы не дают пока оснований говорить о прежнем обитании кавказских козлов на Малом Кавказе и в Тавре.
Тем ие менее такое обитание их на Малом Кавказе кажется весьма вероятным, особенно при проведении параллелей с другими эндемиками: кавказской мышовкой, прометеевой, кустарниковой и снеговыми полевками, кавказским тетеревом. Возможность благополучного существования кавказских козлов на Малом Кавказе в наши дни доказана и опытом Боржомского звериного отвода (Шильдер, 1892).
Современное распространение кавказских козлов охватывает почти весь Большой Кавказ на протяжении 9 географических градусов (между 39°45' и 48°30' восточной долготы от Гринича). Козлы обитают преимущественно на склонах Перевального хребта, но держатся и на далеко выступающих к северу и югу параллельных хребтах и отрогах меридианного направления.
На этом огромном прострапстве к нашим дням обособилось два вида: западный — С. caucasica (С. caucasica sevprtzowi) и восточный — С. су~ lindricornis, различающиеся преимущественна структурой рогов самцов (см. главу IV).
Распространение западпокавказского козла (рис. 147) начинается ныне на западе с горы
Чугуш (Насимович, 1949а), но в 80-х годах прошлого столетия, по сведениям Динника (1914а), козлы водились и на 30 км западнее, на вершинах Фишт и Оштен. В Кавказском заповеднике козлы держатся по Водораздельному хребту и по его северным отрогам, причем на горах Большой Бамбак, Джуга, Ахцархва и Ятыргварты участки местообитаний летом до известной степени изолированы друг от друга (Насимович, 1949а). В начале нашего столетия козлы были нередки и на Скалистом хребте — горе Ачешбок. Недолговременные заходы единичных козлов за последнее полустолетие отмечены и на вершины Дудугуш, Чура, Хацавита.
От верховий Белой и Мзымты западный козел распространенно Водораздельному хребту и его северными южным отрогам до верховьев Балкарского Черека. В верховьях Теберды и Кодора козлов теперь крайне мало. В Тебердипском заповеднике летом близ ледников встречаются их группы в 10—15 голов.
В верховьях Ингура их теперь почти пет. Восточнее, в верховьях Мульхры, они еще живут по южным склонам Тетпульды, а единичные особи встречаются и над сел. Ушкуль. Па Шоде и Лайле, вершинах, изолированных от Водораздельного хребта, козлы редки. На северном склоне Эльбруса их еще порядочно по меридианным хребтам. В начале
Гл. I IL Прежнее и современное распространение и численность млекопитающих 365
нашего столетия они жили на горе Кинжал, а в начале прошлого столетия водились даже в Пятигорье на горе Бештау (Vietingholt, 1812). Над селениями Тегенекли и Верхний Баксан в 50-х годах встречались группы । 20—25 козлов. Довольно обычны они и по ущелью Безингийского Че-
Рис. 147. Западнокавказские козлы зимой. По акварели Е. Я. Захарова.
река. Здесь в 40-х годах некоторые пастухи убивали до 15 козлов за осень. Более обычны козлы в верховьях Балкарского Черека па склонах Дых-Тау. В августе 1948 г. мы встретили за день в ущелье Дыхсу 3 самок (Молодыми и 2 молодых самцов. Близ ледников здесь всюду было порядочно следов и объеденная козлами трава.
В верховьях Уруха западнокавказский козел встречается уже очень редко.
366 Часть II, Анализ происхождения четвертичных млекопитающих
Распространение восточнокавказского козла ограничено теперь на западе истоками Ингура и Цхенис-Цхалн на южном склоне, истоками Баксана и Малки — на северном. В ЗИНе имеется череп самки этого вида даже пз Теберды; в Музее Грузии оттуда же хранится череп самца с рогами переходного типа от С. cylindricornis к С. caucasica, а с Баксана — череп 11-летнего самца переходного типа от С. caucasica к С, cylindricornis. 13 черепов с северных отрогов Эльбруса и с горы Кинжал в коллекции ЗИНа принадлежат также восточнокавказской форме. В Ушкуле у истока Ипгура сван В. Г. Хачвани имеет чучело типичного восточного самца 5.5 лет, убитого у вершины Шхара в декабре 1947 г. Из этого же селения Радде вывез в прошлом столетии рога старого козла переходного (гибридного?) типа между западно- и восточнокавказским.
В ущелье Псыгансу Наспмович (1950) видел в 1949 г. 5 брошенных охотниками черепов, принадлежавших восточному козлу, и 1 череп — западному. В верховьях Безингииского Черека 4 черепа имели рога западного или центральнокавказского типа, al — восточного. В ущелье Балкарского Черека 2 найденных черепа принадлежали восточным козлам. В 1887 г. Диннпк (18906) видел здесь же в ауле Кундюм 2 черепа западнокавказского типа и 8 черепов — восточного. А 4G просмотренных нами черепов (ущелья Уруха и Урсдона в селениях Стыр-Дигор и Ногкау) принадлежали восточному козлу.
В Цейском ущелье бассейна Ардона в святилище Реком из 298 пар рогов 292 пары было восточного типа и 6 — западнокавказского.
Отсюда на восток по всему Водораздельному хребту идет сплошная область распространения восточного козла (карта 89).
По сведениям, собранным мною и В. И. Наниевым в 1948 г. в Северной Осетии, восточный козел встречается по всему северному склону Бокового и Перевального хребтов в верховьях и бассейнах речек Урух, Ардои, Фиагдон, Гизельдон. В верховьях речки Архон встречаются стада до 60 голов. Стада до 100 голов встречались изредка и в районе селений Дунта, Архон, Аксау.
В Грозненской области и в центральном Дагестане козлы теперь редки. На Гунибском и Хунзахском плато их, вероятно, не было уже во время кавказской войны в середине XIX в. В среднем течении Кара-Койсу их теперь нет, но несомненно, что в прошлом столетии они водились тут повсеместно. В Самурском и Лакском районах бывают лишь случайно зашедшие сюда звери. В бассейне Аварского Нойсу козлы весьма обыкновенны по Богосскому хребту. В 20-х годах здесь встречались их табунки штук по 20 (Гептнер и Формозов, 1941). Многочисленны козлы и в верховьях Кара-Чая и Кусар-Чая в скалах Шалбуз-Дага и Шах-Дага.
На южном склоне в верховьях Арагвы п Алазани козлов теперь мало. Нет их ныне вовсе на Телавском хребте, хотя в недавнем прошлом они, вероятно, здесь жили. В Лагодехском заповеднике в 30-х годах насчитывалось примерно 200—300 голов (Марков, 1940). После какого-то мора их стало мало в Белапухском и Мазымчайском ущельях (Верещагин 1938а).
Особенно обильны козлы летом и зимой в Катехском ущелье, где встречаются и пыпе стада по 200 и более особей. Общая численность козлов в Закатальском заповеднике определялась в 30-х годах в 3500—4000 голов (Марков и Млокосевич, 1935; Верещагин, 1938а). Многочисленны козлы и в амфитеатрах ущелий Шин-Чая, Демир-Апарап-Чая. Особенно много их в настоящее время на южной окраине Дагестана — в верховьях Самура (рис. 148) и па участке трех высоких вершин Базар-Дюзи, Баба-
1.1. Ill, Прежнее и современное распространение и численность млекопитающих 367
Даг, Шах-Даг, что было отмечено и в 1896 г. Сатуниным (Радде, 1899). В 30-х годах нашего столетия мы наблюдали здесь много стад по 30—40 голов, а иногда и скопления по 250 и более голов. Крайняя восточная гоапица ареала восточнокавказского козла находится у горы Айвазель, Iллометров на 25 восточнее горы Баба-Даг. Группы в 15—20 козлов тречаются и ныне под Конахкендом в верховьях Гильгип-Чая.
Пределы вертикального распространения восточного козла (рис. 1 9) значительны. В труднодоступных с юга ущельях Закатальского за-11 веднпка козлы держатся летом даже на высотах 1200—1300 м. Они
Рис 148. Оз. Халахи в верховьях Самура. Окрестные скалы — любимое местообитание восточнокавказских козлов.
Фот. автора, 1937.
приходят к минеральным источникам и переходят днища ущелий на высотах всего около 800—1000 м. Восточнее, над Варташеиом и Исмаиллы, они живут летом на лесистых склонах меридианных хребтов на высотах I4QQ-15OO м. Зимой нижняя граница опускается па 200—250 м лишь ка заснеженных склонах, но по днищам ущелий остается той же, что и дам Напившись утром у серных источников в глубине ущелий, старые козлы лпбо остаются у русла речки на высотах 1400—1500 м, либо поднимаются до высот 2800—3000 м, где и отдыхают близ завалов снега. Таким образом, размах суточных кочевок достигает 1500—2000 м. В августе они нередко переходят и через перевалы на высотах 3400—3500 м.
На южном склоне от верховьев Алазани до верховьев Пирсагата популяции козлов бывают разделены летом на две группы — перевальную в лесную, так как вся полоса альпийских лугов занимается стоянками скотоводов со стадами овец и коров, охраняемых собаками. Первая группа козлов держится все лето в лесу, отдыхая па осыпях и карнизах скал, другая же — в скалах и на осыпях перевалов, над зоной лугов.
368
Часть II. Анализ происхождения четвертичных млекопитающих
Западнокавказские козлы в нашу эпоху стали весьма мезофильными животными, но вероятнее всего, что их приспособление к влажному типу леса и к альпийским лугам — явление позднее, вторичное.
И сейчас кавказские козлы достигают все же наибольшей численности в восточной, более сухой части Кавказа, а пасутся чаще всего на участках лугостепи, а не па пышных высокогорных лугах. Крайне характерная привычка козлов лесной популяции выходить и отдыхать длительное время на скальных выступах, осыпях и солнцепеках среди леса, возможно, также является отзвуком древних привычек их предков, обитавших в более сухих и безлесных горах.
Степень связи с фирновыми снегами и с ледниками у кавказских козлов наибольшая в летнее время. В августе козлы обычно выходят к фирновым снегам и отдыхают на солнцепеке, пользуясь прохладой и отсутствием мух (Верещагин, 1949а). Заходить на оледенелые склоны и ледники козлы всегда остерегаются. При каждом новом выпадении снега козлы предпочитают ступать на голые камни. Большие площади ледников козлы преодолевают очень редко. При переходе обширных ледников у горы Лаборда и восточнее горы Шоды в августе 1948 г. я нигде не видел козлиных следов на самих ледниках. По наблюдениям егерей (М. Г. Ивкин и др.), в районе Лагодехи и Белоканы большие стада козлов ежегодно откочевывали в ноябре—декабре в Дагестан, пользуясь всегда бесснежными гребнями хребтов.
Общие границы распространения кавказских козлов за последние 150 лет изменились мало. Можно думать, что в период кавказских воин в середине XIX столетия козлов убивалось больше, чем сейчас. Новое падение численности козлов началось при появлении русских поселений в предгорьях Предкавказья и с завозом в горы четырехлинейных винтовок Бердана, а затем с начала нашего столетия при завозе трехлинеек.
В первой половине XX в. поголовье кавказских козлов на большинстве участков обитания постепенно сокращалось. Исключение составляла территория кавказской великокняжеской охоты в бассейне Кубани, охрана зверей на которой была хорошо налажена. С начала 30-х годов при организации действенной охраны происходил рост поголовья козлов на территории Кавказского и Закатальского заповедников.
В 80-х годах козлы были вытеснены с Фишта и Оштена. В 1908 г. они были выбиты на горе Ачишко у Красной Поляны (Касимович, 1949а). В истоках Ингура (сел. Ушкуль) охотники прошлого столетия убивали за 35—40 лет охоты до 200—250 козлов. В первой половине XX столетия сыновья тех же охотников за тот же срок, пользуясь более совершенным нарезным оружием, убивали не более 15—20 козлов.
На центральном участке Северного Кавказа козлы были выбиты на передовой цепи Скалистого хребта, в частности па Кинжале и Бармамыте, в начале нашего столетия (Динник, 1914а). Еще в 80-х годах прошлого века Дропов добыл серию в 13 черепов козлов в верховьях Малки.
В Пятигорье — на Бештау — козлы были выбиты, вероятно, еще в начале прошлого столетия.
В Дигории, судя по черепам в святилищах Лесгор и Дигоризед, восточнокавказские козлы водились еще в начале нашего столетия па Скалистом хребте, отгораживающем с севера продольную долину Донисер-Доиа. В 30—40-х годах козлы здесь уже исчезли, о чем можно было судить и по отсутствию свежих черепов в святилищах.
Общее число добываемых ежегодно в 30-х годах XX в. кавказских козлов было, вероятно, не менее 4000 голов (рис. 150).
III. Прежнее и соеременное распространение и численность млекопитающих 369
Вообще кавказских козлов отнюдь нельзя еще назвать вымирающими, зающими животными, несмотря на то, что охотничий надзор в горах существует. Наличие трех горных заповедников — Кавказского. Те-
рне. 149. Восточиокавказские козлы летом.
По акварели Е. Я. Захарова.
L^некого и Закатало-Лагодехского — служит значительной гаран-1,.я длительного существования многочисленных популяций.
Развитие скотоводства, сопровождавшееся истреблением волков и ... ш барсов, имело для существования кавказских козлов как по-Сельпое, так и отрицательное значение. На первой стадии развития скотоводства влияние волков на кавказских козлов было не-
94 Н. К. Верещагин
370
Часть II. Анализ происхождения четвертичных млекопитающих
сколько ослаблено, так как большая часть этих хищников стала подкармливаться летом в горах домашним скотом, а на зиму оттягивалась вслед за стадами па низменность. Однако по мере развития скотоводства и увеличения нагрузки на пастбища началось заражение козлов паршой, ящуром н другими болезнями (Динник, 1914а; Верещагин, 1938а; Насимо-вич, 1941).
Легкость приручения кавказских козлов, скрещивания с домашними и безоаровыми козами, акклиматизации на равнинах позволяет надеяться, что в дальнейшем распространение обоих видов будет сильно расширено
Рис. 150. Рога восточнокавказских козлов на веранде осетинского дома. Ущелье Уруха.
Фот. автора, 1947.
искусственно. Эти козлы будут, вероятно, разведены в горах Карабаха, на Шах-Дагском, Триалетском и Аджаро-Имеретинском хребтах, а также в Талыше и Эльбурсе.
Значительные хозяйственные результаты могут быть получены и при гибридизации кавказских козлов с домашними (Верещагин, 1938а).
Учитывая эндемизм и большую морфологическую обособленность кавказских козлов от других представителей рода (см. главу IV), их следует считать местными видами, сформировавшимися от кавказских -третичных предков.
Безоаровый козел — Capra aegagrus Erxl. Ископаемые остатки этого козла известны из плейстоценовых и голоценовых слоев пещер Италии. Ливана, Сирии, Палестины (Picard, 1937; Wolf, 1938). Па Кавказе они обнаружены лпшь в послеледниковых слоях пещер теснины Аракса (Поляков, 1882), пещер Сарайбулахского хребта, в слоях века бронзы развалин крепости Тейшсбаини у Еревана (Даль, 19406, 1952) и в капищах Тугаетии п Хевсуретпи.
л. III. Прежнее и соеременное распространение и численность млекопитающих 371
Изображения, напоминающие безоарового козла, сделанные в век бронзы, имеются на скалах известняка в юго-восточном Кабристане, • западу от Баку (рис. 151). Эти рисунки, быть может, указывают на не-двнее обитание козлов в низких хребтах близ Каспия — в Кабристане. чень изящные бронзовые фигурки (рис. 152) безоаровых козлов, отно-(чщиеся к кобаискои и каякентско-хорочаевской культурам (первая । .ювпиа I тысячелетия до н. э.), известны из Дагестана и хранятся I исторических музеях Москвы, Махачкалы и Грозного.
Рш. 151. Изображения козлов на скалах Беюк Даш южпее Баку.
Фот. автора, 1945.
Современное распространение безоарового козла, охватывает, кроме Кавказа, горы Передней и Средней Азии — от Крита до западной Индии
Турцию, Иран, Афганистан. На Кавказском перешейке козлы I >атся пынс па 3 изолированных участках: в восточной части Большого Кагказа, в центральной и восточной частях Малого Кавказа. На Большом Кавказе ареал ограничен верховьями Алазани, Аргуни и верховьями Гердыман-Чая (карта 90), т. е. сухой и безлесной частью горной страны. В 20-х годах нашего столетия безоаровые козлы жили в Лакском округе Ьгестапа. По словам Динника (1914а), этот козел встречался в начале Сщего столетия километрах в 30 от Гуниба.
На южный лесистый и влажный склоп Большого хребта от Лагодех о Шемахи и иа перевалы водораздела безоаровые козлы заходят пз а стана очень редко: раз в два-три десятилетия.
Безоаровый козел обычен в верховьях Аварского Койсу, в районе •еленпн Тлядаль, Кидеро, Чорода, Тахота. Здесь он держится в уцелевши сосновых и буковых лесах. В верховьях Терека и Арагвы безоаровый 24*
372
Часть II. Анализ происхождения четвертичных млекопитающих
I-----1----->-----1—----1----1 см
Рис. 152. Фигурка безоарового козла. Бронза. Музей в Махачкале.
козел не водится п, судя по отсутствию его черепов в осетинских капищах, не водился и раньше.
На Малом Кавказе распространение его более обширно. На юго-западе козел обитает в пихтовых лесах верховьев Чороха, местами на Аджаро-Имеретипском хребте и на западных отрогах Триалетского хребта в районе Боржоми, а затем в юго-восточной части Армянского нагорья. На Шах-Дагском хребте козлы обычны в районе Кедабека и Калакента, например в верховьях Шамхора близ вершины Кашкар (3379 м), на горе Кяпяз — над оз. Гек-Гель и под вершиной Мров-Даг (3740 м).
Небольшое число коз обитало в 1940 г. и в осыпях среди буковых лесов на юго-восточном склоне горы Кечал Даг восточной оконечности Шах-Датского хребта. Сторож лесопильного завода ежегодно убивал здесь 7—8 коз.
Самой восточной вершиной, заселенной в 30-х годах нашего столетия безоаровыми козами, была гора Кире (2743 м) над Степанакертом.
В центральных и южных частях Армянского нагорья безоаровые козы обитают в скалистых горах от долины речки Азат до склонов Занге-зурского, Мегринского и Баргушет-ского хребтов. На севере нагорья пределом их распространения являются теперь южные склоны Гегам-ского и Варденисского хребтов (Даль, 19516). Наиболее часты они на Урцском и Айоцдзорском хребтах, а также на ближайших к ним ска
листых массивах. В 1950 г. Даль (19516) насчитывал на Урцском хребте пять «очагов» площадью в 33.5 км2, на которых держалось 124 козы.
В 70-х годах прошлого столетия козы встречались еще близ Еленовки па Севане, откуда И. С. Поляков вывез один череп.
В Нахичеванской АССР козы держались в 1947 г. на 11золированиых нагорной полупустыней вершинах Алинджа-Даг и Илянлу-Даг близ селений Абракупис, Боянур и Ханагя. На протяжении нашей эры козы, конечно, неоднократно вытеснялись с этих вершин при использования последних древнеармянскими племенами в качестве естественных крепостей. По сравнению с кавказскими безоаровые козы более пластичны, так как нередко легко пересекают значительные пространства пустынных увалов, обладая, следовательно, огромными возможностями расселения.
По Зангезурскому хребту п его южным отрогам козы обитают к востоку вплоть до теснины Аракса. Особенно многочисленны они в Неграм-ских горах между Джульфой и Ордубадом, на высотах 1000—1200 м. На участке теснины Аракса козы распространены по юго-восточным склонам Мегринского хребта вплоть до Охчи-Чая. В 1947 г. небольшое количество зверя держалось еще в пустынных скалах близ Акулиса и Орду-ба да.
Об обитании безоаровых коз в Талыше есть литературные указания XIV столетия, например Рашид-ад-дина. Козы водились в высокогорье Талыша еще в середине прошлого столетия, например па вершине Кыз-Юрды (Радде, 1899, стр. 75), по позднее их там уже никто не находил.
Гл. III. Прежнее и современное распространение и численность млекопитающих 375
На юг ареал этого козла уходит в сухие нагорья Малой Азии и Ирана. Коллекционные экземпляры, добытые в конце прошлого и начале нашего столетия, известны из-под Эрзурума, Вана, Карса, Арарата, Тебриза, Гиелана, Демавенда. В пределах СССР эти козы обитают еще на Копет-|>\ и в Больших Бал ханах. По сообщениям Саркисова (1944д), козы prrpi чаются в Иране во всех горах провинций Азербайджана, Курди-г:ип, Керманшаха и «Нуристана. В южном Иране козы распространены ровпнциях Иезд и Шираз, затем в Мекране, горах Белуджистана и до Сулеймановых гор.
Наиболее обычные местообитания — это сухие скалистые горы с крупной, бояркой, можжевельником и другими ксерофитами. Современнее вертикальное распространение безоаровых коз па Кавказе — от 900 до3500 м, но они отлично уживаются и ниже — на уровне моря.
С< временная приуроченность ареала к восточному Средиземноморью и весь образ жизни безоарового козла свидетельствует о длительной эво-..юцгч вида в сухих, жарких и относительно низких горах со скальным рельефом. На Большой Кавказ безоаровый козел проник, по-видимому, с юга — из Передней Азии — в одну из ксеротермических эпох плиоцена вяп плейстоцена аналогично хомякам рода Mesocricetus. Проникновение ли 1 гло идти как по Сураму, так и по остепнепным предгорьям восточного Закавказья. Последующая изоляция в сухой восточной половине Большого Кавказа может объясняться оледенениями хребтов и развитием широкой мезофитной зоны леса и лугов па западе. Более позднее пичв енпс этого козла в Дагестане, например в раннем голоцене, менее веро тно.
Влияние человека па этот вид стало сильно сказываться лишь в последние десятилетия при развитии перегонного скотоводства и завозе и ы массы дальнобойного огнестрельного оружия. Сокращение ареала и численности коз в результате промысла особенно заметно и в наши
I па юго-востоке Армянского нагорья.
При усовершенствовании охотничьего хозяйства Кавказа безоаровые козы смогут существовать и далее неограниченное время. Ареал этих коз дано расширить, расселив их искусственно по продольным долинам центрального Кавказа, например в ущелье Малки, Баксана, Черека, Уруха, Фпаг-Дона, в которых имеются благоприятные для обитания участки по квестовым уступам Скалистого хребта.
Бараны, аргали — Ovis cf. аттоп L. и арменийский муфлон — 0. pnelini Blyth. В Западной Европе остатки аргалпобразных баранов известны с плиоцена до верхнего плейстоцена, а му флопообразных — I плейстоцена до современности.
На Кавказе остатки крупных баранов типа архара (рис. 153) — очень I редки и встречены лишь в средне- и верхнеплейстоценовых слоях Закавказья как в горах, так и па равнинах. Еще более редки находки остатков иуфлопообразпых баранов, обнаруженных лишь в палеолите па Малом I Кавка, о и в мезолите в Дагестане.
Ближайшие к Кавказу местонахождения остатков плейстоценовых аргалпобразных баранов отмечены в ангельских п орипьякских слоях пещер Крыма. Из ашельско-мустьерских слоев пещеры Киик-Коба и из орипьякских слоев пещеры Аджи-Коба добыты кости муфлопообразпого барана 0. cf. argaloides Nelir. (Громова, 1935а; Громова и Громов, 1937). В Палестине, Ливане и Сирии остатки баранов из палеолитических и пео-лптпческпх слоев пещер неизвестны (Picard, 1937), несмотря па подходящие ландшафты.
314
Часть II. Анализ происхождения четвертичных млекопитающих
В северо-западном Иране в четвертичных суглинках близ Мераге найден зуб iipynnoro архара, а остатки Oris sp. — в пещерах близ Урмии и Астрабадского залива (Соои, 1951).
Остатков архара пз голоценовых слоев на Кавказе нет.
Кости постплейстоценового мелкого барана — О. ci. gmelini, кроме мезолита Дагестана, обнаружены лишь в пределах границ современного обитания этого вида: в развалинах крепости Кармир-Блур близ Еревана (VII—V вв. до н. э.) и в слоях пещер Сарай-Булахского хребта (Даль, 1941, 1952) (карта 91).
Древние литературные данные, а также изображения барана в рисунке и скульптуре у кавказских народов бедны. Наиболее древнп скульп-
Рис. 153. Остатки архаров.
1,3 — пястная гг плечевая, средний плейстоцен, асфальты близ Баку; 2 — М3,верхний плейстоцен, делювий близ Мераге.
турные изображения. Головка муфлонообразного барана на поясной пряжке известна в южной Грузии из Триалетских курганов, относимых к середине II тысячелетия до п. э. (Куфтпн, 1941). Головки баранов с рогами аргалиобразного типа в виде бронзовых подвесок довольно обычны в погребениях и поселениях кобанской культуры. Эти изображения дают, однако, представление скорее о домашнем, нежели о диком животном, так как в них не чувствуется ни мощи, пн экспрессии дикого зверя (рис. 154). По-видимому, аргалиобразные бараны исчезли иа Кавказе к голоцену. Не уцелели архары даже в центральном Дагестане, где обстановка для них была наиболее благоприятна.
Рисунки п рельефные изображения мелкорогих баранов типа муфлона известны по золотым вещам из скифских Курганов западного Предкавказья — Келлермес и других, а из Закавказья — па так называемом «сасапидском металле» (220—657 гг. и. э.). Па серебряных блюдах, хранящихся в Эрмитаже, изображена конная охота Хосрова 1 и Шапура II за мчащимися баранами (Орбели и Тревер, 1935). Район производства
n III. Прежнее и современное распространение и численность млекопитающих 375
серебряных блюд — Ирано-Анатолийские плоскогорья — соответствует современному ареалу переднеазиатского вида барана.
Современный ареал муфлопообразиого барана Гмелпна ограничен S пашей стране южным Закавказьем (карта 91).
I В Музее Грузии хранятся рога и шкуры этого вида из Еревана, Нахи-вани, Пеграма, Джульфы, с горы Дарры-Даг и сел. Пузиан в Занге-Бре. Из ближайших окраин Ирана там же имеются экземпляры с Котур-'.ага, пз окрестностей Хоя и с о. Коюн-Даг на оз. Урмия.
В ЗПНе имеются рога, доставленные акад. Броссе в 1849 г. с Алагеза, Мастарт в 1911 г. Пфицепмайером.
Рис. 154. Бронзовые головки баранов типа архара из кобанских могильников Северной Осетии. (По Уваровой, 1900).
। череп, добытый с той же горы у сел. Имеются также черепа и шкуры из Армении вообще; из Нахичеванской ГТ: окрестностей сел. Шахбуза, Ешалдина, Аразпп; из Турции:
t горы Арарат, окрестностей Бая-а, сел. Гурджи-Булах, горы Пир-Бшида в 60—80 км от оз. Ван. В «веро-западном Иране были добыты jora, черепа и шкуры из Кара-Дага, । югу от Ордубада, Зорских гор, . к Урмии, горы Савелан и пз окрест-тей Таврпза (сборы Мнльковпча, ЕО-г., Брандта, 1879 г., фон Вика, |Ч14 г., Хаиыкова 1851 г., Введен-кого, 1908 г.).
Распространение баранов в южном Закавказье быстро сокращалось < прошлого столетия. Шопен (1852) также указывает на обитание баранов Алагезе, но современные авторы—Саркисов (1941, 1944а, 19446)
п Даль (1945, 19496) — описывали распространение баранов лишь к вос-иК) от р. Занги па Сарай-Булахском и Даралагезском хребтах.
В окрестностях Абракуниса, Джульфы, Неграма, Ордубада и на горах Дарры-Даг и Иляллу-Даг в 1947 г. бараны были уже большой редкостью, но небольшое число их еще держалось па Зангезурском хребте, обешю в окрестностях селений Биченах н Шахбуз.
Распространение муфлона связано ныне в Закавказье с нагорными степями где преобладают группировки ксерофитных плотпокустовых маков и зиббальдии с формацией колючих астрагалов на склонах средней полосы гор и с солянково-полынной группировкой в долине Аракса. В жаркие долины бараны спускаются преимущественно в зимнее время, ; летом держатся в высокогорье.
Судя по общему распространению баранов в Евразии (Насонов. 1923) ь жпо думать, что на Кавказском перешейке и архар и муфлон былп [пришлыми видами, ареал которых развился па протяжении плейстоцена с юга вслед за развитием ксерофитных ландшафтов.
Учитывая древность дивергенции аргалиобразных и муфлолообразных баранов, прослеженной с среднего плейстоцена (Верещагин, 1953в), нет осповапий рассматривать архаров и муфлонов в качестве единого вида, подвергшегося лишь географической трансформации.
Ближайшее будущее диких баранов па Кавказе, как и других кош»[т-1ых, будет зависеть главным образом от охотничьего законодательства.
376
Часть II. Анализ происхождения четвертичных млекопитающих
Аргалиобразными баранами следует заселить центральный Дагестан в районе Гуниба, Ботлиха, Хунзаха.
Первобытные и кавказский зубры — Bison sp., В. cf. schoetensacki Freud., В. priscus Вор, В. bonasus caucasicus Sat. Остатки зубров появляются в Евразии с верхнего плиоцена. На Кавказе наиболее древними находками считаются роговые стержни мелких бизонов — Bison sp., В. cf. schoetensacki в верхнеплиоценовых апшеронских глинах Кабристана (в восточном Закавказье) и в конгломератах Таманского полуострова (Бурчак-Абрамович, 1949; Верещагин, 1957а).
Рис. 155. Кавказский зубр.
Остатки среднеплейстоценовых, наиболее крупных зубров — В. priscus longicornis и измельчавших верхнеплейстоценовых — В. priscus demi-nutus обнаружены во многих местах Предкавказья и Западного Закавказья. Наибольшее число находок сделано в озерно-речных и пещерных слоях (карта 92).
В восточном Закавказье, па Армянском и Иранском нагорьях зубры в плейстоцене были редки пли даже полностью замещались первобытным туром. Особенно загадочно отсутствие зубров в бинагадипском комплексе. Однако к юго-западу — в пещерах Ливана, Сирии, Палестины — остатки зубров становятся вновь обычными II известны с ашеля до неолита (Picard, 1937).
Наметившаяся в конце плейстоцена тенденция к измельчанию зубров Евразии была усилена и закреплена преследованием человека и иссушением ландшафтов. Кавказ по представил в этом исключения. Нет оснований приписывать это измельчание влиянию «ухудшения» климата в верхнем плейстоцене, как это нередко указывается зоологами и палеонтологами, так как именно похолодание и развитие луговых формаций и обусловило с начала плейстоцена поразительное увеличение размеров зубров к среднему плейстоцену. Современный кавказский зубр (рис. 155),
Гд. III. Прежнее и современное распространение и численность млекопитающих 377
несомненно, являлся прямым потомком местных плиоцено-плейстоценовых зубров.
Измельчание зубров в верхнем плейстоцене и в послеледниковье сопровождалось и обособлением их популяций по долинам рек, на высоких ’лскогорьях п в широколиственных лесах горных склонов. Кавказские юбры не сохранили к нашей эпохе степных привычек своих плейстоценовых предков и американских собратий. Они сделались обитателями мезофмтпой лесостепи п пояса горных лесов. Их остатки прослежены остами в западном Закавказье и на Малом Кавказе.
В средине века кавказская популяция зубров еще имела, по-видимому, связи с популяцией зубров Восточной Европы по лесистой долине Дона.
Разница в судьбе евразийских и американских зубров в голоцене объясняется преимущественно различным влиянием человека (Верещагин, 1956). Было бы даже странно, если бы тяжеловесные зубры и туры уце-дйди в степях и лесостепи Восточной Европы и Азии после кочевок по всспстребляющей конницы хазар, половцев, кипчаков, монголов. Одсако еще в XVI в. в Великой Литве и Мазовии на зубров охотились плесах с копьями—«метабуло» (Виженер, изд. 1890 г.), а в Западной Сп-бнрп зубры и туры жили еще в XVIII в. (Белл, 1776). Соображение Пи-допличко (1951) о том, что европейский плсйстоцеповый зубр был постоянным обитателем степи, слабо обосновано.
Странно, что скульптурные и плоские изображения зубров у древних ш иен Кавказа почти неизвестны. Только среди кобанских бронзовых бе. дедушек можно иногда усмотреть маловыразительные фигурки каких-то быков, отдаленно напоминающих зубров. Литературные сведения о порах Кавказа известны нам с средневековья.
Еще Рашид-Ад-Дин (изд. 1946 г.) сообщал,что Абага-хап, зимуя в 1275— 1276 гг. в Аррапе и в 5 фарсангах от Шахруда, стал охотиться в лесу на Ффиых буйволов». При описании охоты Газан-хана в Талыше в 1301/02 г. дот летописец (стр. 188—189) отметил: «После этого воины устроили облаву п гпалп дичь, как-то: горных буйволов, джуров, диких «аз п ослов, шакалов, лисиц, волков, медведей и других всевозможных диких и хищных зверей вовнутрь изгороди, до тех пор, пока они все не Игрались в том загоне». Под джурамп разумеются, вероятно, косули — цжуюр», а под дикими козами — безоаровые козлы, либо джейраны.
Отметим, что слухи о каких-то диких быках в горах Талыша и Эльбурса сохранились и до XIX столетия (Менетрие, 1832). Критический обзор Брандта (1867) по этому поводу новых материалов не дал. Возможно, что вымирание зубров в лесах Эльбурса и Талыша произошло примерно в то же время, что и в лесах центрального Кавказа, т^в^ в XV11I—XIX вв.
Первые литературные указания о зубрах в Черкессии принадлежат, возможно, Жану де Люку (1625, изд. 1879 г.), а в Абхазии— Арканджело Ламберти (1654, изд. 1913 г.).
О зубрах, водящихся в Кабарде, Осетии, Ингушетии, было известно в Москве задолго до материалов академиков Ловича, Гюльдепштедта и других, которых принято считать (Башкиров, 1940) первооткрывателями зубра на Кавказе.
На стр. 7994 X тома «Полного собрания законов Российской империи с 1649 года» говорится: «Именной приказ, данный из кабинета Ея Величества императрицы Анны Иоанновны Астраханскому оберкомепданту — О ловле и присылке ко Двору и в Измайловский зверинец ежегодно рзпых живых зверей. . . Еще известно нам, что в Кабарде есть дикие
375
Часть II. Анализ происхождения четвертичных млекопитающих
быки и кдосы, которые по тамошнему называются домбап, того ради имеете вы всячески стараться, не жалея на то употребить несколько пз казны пашей денег, чтобы тамошние князья оного рода бычков и телок молодых по 5 или по 10 велели ловить и присылали в Кизлярскую крепость. а там оных несколько времени прикармливать к хлебу, а когда привыкнут, то присылать в Астрахань водою, а пз Астрахани отправлять их с прочими зверьми в Москву, и о том же имеете вы от себя писать п к
Рис. 156. Черепа зубров в пещере Дигоризед.
Фот. автора, 1947.
Шамхалу Горскому, также в Кизлярскую крепость к коменданту и к Элмурзе Черкасскому, чтоб в ловле и присылке оных бычков и телок старание приложили».
Возможно, что в Чечне и Северной Осетин зубры водились еще во время путешествия Гюльденштедта, т. е. в J 770—1774 гг.
Динник (1890а), как известно, еще застал стариков сел. Заделеск, рассказывавших об отцах и дедах, стрелявших зубров-до мбаев железными пулями. На одном из черепов пещеры Дигоризед (№ 10, Североосетинский педагогический институт) видна вырезанная надпись «1833», возможно, свидетельствующая о дате отстрела одного из последних в Осетии домбаев (рис. 156).
В середине XIX столетия зубры уцелели лишь в горах и лесах северо-западного Кавказа. Постепенное исчезновение зубров в этом районе распадалось на несколько этапов. Северная граница распространения зубра проходила на шпроте станиц Дагестанской и Абхазской. Восточной границей было, вероятно, долготное течение Кубани, вплоть до устья Малого Зеленчука. На западе зубры доходили до речки Пшиш. Башкиров (1940), сопоставляя краеведческие заметки, считал возможным обитание
[д,///. Прежнее и современное распространение и численность млекопитающих 379
зубра в то время и в верховьях речек Шахэ, Сочи, Мзымты, Псоу, Бзыби, Кодора и Ингура.
Общая численность зубров в 70-х годах прошлого столетия определясь более, чем в 2000 голов.
•Кавказские войны и заселение казаками предгорий постепенно оттес-• г зубров на участок между верховьями Белой и Лабы. Развитие ското-
Рис. 157. Зубр (самец) в пихтовом лесу.
Фот. Д. П. Филатова, 1909.
водпва, рубка и сплав леса, охота непрерывно сокращали численность и ареал зверя.
। К 90-м годам прошлого столетия численность кавказских зубров, яо Сатунпну (1898), определялась в 500—700 голов, а по Кулагину (1918) — в 442 зубра.
Организация Кубанской великокняжеской охоты в 90-х годах на тер-риторпп в 522 тыс десятин в районе обитания зубров и предпринятая борьба с браконьерством до некоторой степени прекратили падение поголовья.
Специальные обследования Филатова (1910, 1912) показали все же альниппее сокращение занятых зубрами участков. На карте, составленной Филатовым, распространение зубров было показано лишь между верховьями Белой и Большой Лабы.
Поголовье зубров Филатов определял в 1910 г. в несколько сот голов. Держались они преимущественно в пихтарниках (рис. 157).
В 1917 г. число зубров, по-видимому, не превышало 500. Появление на территории Кубанской охоты после революции 1917 г. пастухов со стадами скота, лесорубов, дезертиров и охотников, вооруженных трехлинейными винтовками, ознаменовало последний этап существования кавказского зубра. Большая часть зверей была быстро перебита па мясо и шкуры,
380
Часть II. Анализ происхождения четвертичных млекопитающих
а в 1919 г. среди оставшихся вспыхнула будто бы какая-то эпизоотия, занесенная, возможно, в горы домашним скотом.
К 1920 г. все поголовье исчислялось в 50 особей, в том числе в районе Киши-Хамышки не менее 20 зубров. Преследование зубров продолжа-» лось браконьерами и в дальнейшем.
Организация Кавказского заповедника в 1924 г. не спасла зубра в связи с трудностями охраны зверя в горах и обилия у местного населения пастильного оружия.
С 1921 по 1926 г. было отмечено несколько случаев убоя зубров: близ станицы Линейной, в 50 км западнее Майкопа (1921 г.), близ горы Гефо (1923 г.), в верховьях Кодора (1925—1926 гг.), на Алоусе и Мастакане. Здесь на горе Алоусе пастухи имеретины убили в 1926 г. трех зубров, вероятно последних. После этого сведений о зубрах не поступало (Башкиров, 1940).
С 1940 г. на Кавказе начинается уже история гибридного, кавказско-американского зубра (см. главу VI).
Зубробизонами, обладающими повышенным жизненным тонусом, при желании и средствах на охрану можно заселить горные леса всего Северного Кавказа и Закавказья.
Первобытные туры — Bos trochoceros Meyer, В. mastan-zadei Burtsch., В. primigenius Boj., В. minutus Malsb. Историю быков рода Bos принято вести из северной Индии, плиоценовые и плейстоценовые отложения которой сохрапили остатки большого числа угасших видов бычьих.
Как и зубры, туры также дали на Кавказе последовательный генетический ряд родственных форм, по менее четко выраженный. Кроме того, здесь имелись и представители боковых филогенетических ветвей.
История первобытного тура европейского типа — ветви В. primigenius — начинается на Кавказском перешейке с среднего плейстоцена.
Наиболее древней находкой следует считать череп гигантского В. trochoceros в среднеплейстоценовых песках Аджи-Эйласа, южнее Еревана (Авакян, 1946). Более поздние, верхнеплейстоценовые находки известны из битумов Апшеронского полуострова (Богачев, 19256; Бурчак-Абрамо-вич, 1951 г, 1952 г), с Армянского нагорья и из Предкавказья (карта 93). На этом история тура на Кавказе как бы обрывается, но в погребениях II тысячелетия до н. э. [хеттская (?) культура] на Триалетском хребте у Цалкп и Кировакана прослежены черепа, метаподии и фаланги быков, близких по размерам к голоценовым диким турам.
Упомянутые смежные формы туров представлены на Кавказе апшерон-ским средпеплейстоцеиовым быком — В, mastan-zadei из битумов Апшеронского полуострова (Бурчак-Абрамович, 1952 г) и послеледниковым карликовым туром — В, ci. minutus пз голоценовых песков побережья Севана (Даль, 1950а). Местонахождения эти единичны.
Интересно, что, в противоположность зубрам, первобытный тур на Кавказе не сделался обитателем леса и не ушел в горы. Разница в экологии этих четвертичных видов может быть уяснена по распространению их остатков в Восточной Европе и па Кавказе.
Так, на север от Кавказского перешейка остатки первобытных туров широко известны из аллювиальных и делювиальных отложений долин рек Русской равнины.
Численность туров почти повсеместно была ниже численности зубров. При этом относительное падение численности остатков туров особенно хорошо заметно в юго-восточном направлении. Имеются такие показатели
ft. 1П. Прежнее и современное распространение и численность млекопитающих 381
зо достоверно определенным музейным коллекциям, собранным по ире-Ьуществу на отмелях рек (табл. 74).
Таблица 74
Распределение находок черепов первобытных туров и зубров в музеях на Русской равнине
Музеи Зубры Туры
абс. % абс. %
Казанский • .... 23 85.1 4 14.9
Уральский 13 76.5 4 23.5
Куйбышевский 11 84.5 2 15.4
Саратовский 36 81.8 8 18.2
Астраханский 67 95.7 3 4.3
Новочеркасский 38 74.5 13 25.5
Ставропольский 6 8э.7 1 14.3
Такое распределение подтверждают до известной степени соображения идоплячко (1951), что туры были обитателями более влажных ландшафтов и 6 ютопов, нежели зубры.
В Европе тур был преимущественно лесным мезофитным животным, особенно в конце своего существования. На Кавказе и в Передней Азии лого не произошло: туры держались здесь по увлажненным долинам и яа лугах открытых плоскогорий и, конечно, были легко уничтожены. Подтверждением этих соображений может служить современная практика пазведенпя крупного рогатого скота па равнинах Средней Азии. Там животине выпасаются в группировках пустыни или полупустыни лишь в короткий срок развития весенних эфемеров, а позднее, с июня, концентрируются по лугово-болотным формациям окраин озер атмосферного питания, например на озерах Чушка-Куль у города Туркестана и по долинам рек. Такое же положение наблюдается и в жесткотравной степи. По мере огрубения плотнокустовых злаков крупный рогатый скот переходит изстеппв лога, в овраги, в сырые поймы.
Судя по тяготению к луговым формациям, тур получал, очевидно, наибольшие возможности для расселения в южных районах — в Передней Дзпи и на Кавказе — в эпохи похолоданий и увлажнения климата.
В плейстоценовой полупустыне — степи восточного Закавказья — апшеронские туры обитали, видимо, сезонно, во время развития зимне-весеппнх эфемеров. Летом их, наверно, не было здесь вовсе. В лесистом и болотистом западном Закавказье туры не жили, но далее па юг были роду обычны по остеппенным и лесистым плоскогорьям Передней Азии.
Остатки В. primigenius, найденные в мезолитических слоях пещеры йлт близ Астрабадского залива, вместе с остатками джейранов (Сооп, 1951), вероятно, следует относить к потомкам В. mastan-zadei — форме, |цее ксерофильиой, нежели европейский тур. Потомком среднеплейстоценового апшеронского быка является, очевидно, и В. namadicus Falc., остатки которого были описаны для нижних, неолитических слоев городища Анау у Ашхабада (Duerst, 1908).
В конце плейстоцена и в голоцене иа Армянском нагорье существовали, очевидно, две формы туров: мелкая и крупная. Судя по находкам костей
382
Часть II. Анализ происхождения четвертичных млекопитающих
Рис. 158. Золотая бляха, изображающая первобытного тура. Майкопский курган (пат. вел.).
быков в неолитических слоях и в слоях городищ Урарту, можно думать, что была одомашнена и та, и другая форма.
О существовании туров на Кавказе в нашу эпоху документальных данных нет.
Факт поразительной редкости находок кухонных остатков туров в слоях древнпх городищ отмечен и для Русской равнины, но толкование этого в том смысле, что дикий бык был слишком опасным противником для доисторического человека (Громова, 1931, стр. 362) и что при появлении домашних животных человек стал «избегать охотиться за этой опасной добычей», конечно, весьма наивно. Этому явно противоречат сцены охот па барельефах Шумера и Ассирии.
Объяснение заключается в резком сокращении численности зверя к неолиту, во-первых, и, во-вторых, в привычках европейского тура, затруднявших его добывание, так как на зубра, обитавшего в лесу и ничуть не менее опасного, человек продолжал упорно охотиться на протяжении всего голоцена. В средневековье туры встречались уже только на отдельных участках и нередко специально охранялись (Инженер, изд. 1890 г.).
Изображения первобытных быков па Кавказе известны с верхней бронзы и особенно часты для скифской эпохи. Контурные рисунки туров имеются на серебряном сосуде из Майкопского кургана (11 тысячелетие до н. э.), а в скульптуре туры представлены в виде бронзовых объемных фигурок и золотых плоских бляхах, изображающих тура в профиль (рис. 158).
Контурные рисунки быков и коров, имеющиеся на скалах (рис. 159) в Кабристане, южнее Баку, и относящиеся, вероятно, к римским временам, изображают, видимо, уже домашних животных (Верещагин и Бурчак-Абрамович, 1948). Бронзовая фигура мощного быка, близкого по конституции к туру, известна из кобанского могильника в Осетии. Рельефные изображения голов быков имеются на печатках, перстнях пз могильников I тысячелетия до н. э. близ Мпнгечаура (коллекция Музея истории АН Азербайджанской ССР). На золотой печатке из Чертомлык-ского кургана па южной Украпне также имеется великолепное изображение фигуры ревущего тура.
К югу п юго-западу от Кавказа, т. е. в Месопотамии, Сирии, Египте, широко известны великолепные барельефы древнеегипетских, шумерпп-ских, вавилонских, ассирийских, хеттских художников, изображавших тура и охоту на него. Изготовлялись они во II и начале I тысячелетия до н. э., когда дикие быки были еще весьма обычны в Передней Азии п в северо-восточной Африке. Усама ибн Мупкыз (изд. 1922—1923 гг.) охотился па каких-то быков в Сирии еще в ХП в.
Среди исторических записей по Кавказу нам не удалось найти указаний па наличие в этол стране первобытного тура. Таким образом, о времени окончательного исчезновения тура в степях Предкавказья п па плоскогорьях Малого Кавказа можно судить лишь косвенно, проводя параллели с хорошо известными художественными сравнениями и историческими справками в древнерусских летописях, «Слове о полку Игоревен былинами и в «Поучении Владимира Мономаха» (см. Долгих, 1905; Громова, 1931; Колесник, 1936).
Весьма вероятно, что на лесостепных участках ПредкавказьяинаМалом Кавказе полное вымиранпетурапроизошлонесколько раньше,чем в Европе.
[л. III. Прежнее и современное распространение и численность млекопитающих 383
Обзор историй появления, развития и современного состояния рассмотренных четвертичных млекопитающих показал преимущественно трудности, связанные с оценкой фактов палеонтологии, зоогеографии и экология, для выяснения происхождения на Кавказе тех или иных видов и путей формирования фауны.
Эти трудности оказались особенно велики для уяснения происхождения родины эврптопных убиквистов, например обыкновенного ежа,
Рис. 159. Изображения быков на скалах Б шок-Даш южнее Баку.
Фот. автора, 1945.
дисицы, волка, а также для ряда крупных массовых видов четвертичного периода, слабо изменявшихся на свопх обширных ареалах, например оленей, зубров. Тем но менее становится очевидным, что корпи местного ядра четвертичной териофауны Кавказа уходят в плиоцен и даже в верхний миоцен, хотя ископаемые остатки большинства предковых форм четвертичных видов в названных слоях пока и неизвестны.
История развития ареалов многих млекопитающих на Кавказе оказалась большей частью целиком зависящей от формирования четвертичных ландшафтов.
В меньшей степени особенности ареалов зависели в зависят от эволюции морфо-фттзпологическпх адаптаций видов и их биоцепоти-ческих связей друг с другом. Создается даже впечатление, что шрп развитии и угасапип ареалов растениеядные млекопитающие ведут себя в значительной мерс автономно, независимо друг от друга. Подчиненное эначыше имеет, очевидно, и влияние на распространение травоядных зверей преследующих их хищников.
Рассмотренные примеры распространения видов п экологических зависимостей позволяют выделить три основных генетических типа ареалов млекопитающих Кавказского перешейка.
384 Часть II. Анализ происхождения четвертичных млекопитающих
1. Кавказский лесной и высокогорный — связанный с третичными мезофитными лесными и горно-луговыми ландшафтами. Примеры: ареалы кротов, прометеевой и кустарниковой полевок, серны.
2. Переднеазиатский нагорно-пустынный и нагорно-степной — связанный с третичными ксерофитными ландшафтами, развивавшимися и на Кавказской суше в сухие и жаркие эпохи кайнозоя. Примеры: ареалы персидской песчанки, краснохвостой песчанки, малоазийского хомяка, степной полевки, безоарового козла.
3. Европейско-азиатский степной — связанный с развивавшимися на предкавказскпх равнинах в плейстоцене степными ландшафтами. Примеры: ареалы корсака, малого суслика, сайги.
Второстепенные типы ареалов четвертичных млекопитающих развивались на основе трех перечисленных и являются преимущественно следствием расселения по Кавказскому перешейку европейских, среднеазиатских и южноазиатских видов в плейстоцене и голоцене. Наиболее обособлен и, в сущности, чужд Кавказу тип ареала пустынных видов Средней Азии (см. главу V).
По палеонтологическим документам современным ареалам и современной экологии рассмотренных видов зверей можно дать и самую общую хронологическую схему происхождения четвертичной фауны млекопитающих Кавказа. В этой фауне могут быть выделены три возрастных (стратиграфических) группы видов: третичная (плиоценовая), плейстоценовая и голоценовая.
Группа местных третичных форм, существование которых (или их предков) на Кавказе уже в плиоцене не вызывает сомнений, состоит из насекомоядных, рукокрылых, хищных, грызунов, хоботных, непарнопалых и парнокопытных, сформировавших частью и основу голоценовой фауны. По экологии и происхождению эта группа неоднородна и подразделяется на мезофильную и ксерофильную подгруппы.
Виды первой подгруппы — местные, мезофильные, возникшие в обстановке мезофитных восточносредиземноморских ландшафтов, т. е. на фоне широколиственных, горных и низовых лесов, высокогорных лугов. К таким видам относятся, например, кавказские кроты, трогон-терий и речной бобр, черная крыса, прометеева, кавказская снеговая, длиннохвостая снеговая полевки, косуля, отчасти кавказские козлы.
В эту же подгруппу можно отнести, очевидно, и эвритопных убиквистов на том основании, что в плиоценовых отложениях встречены остатки родственных им форм, например: у белозубок, лисиц, волков, медведей, оленей туров и зубров. До наших дней из этой подгруппы дожили по преимуществу широко распространенные виды, игорно-лесные виды и виды, уцелевшие в горно-лесном ландшафте. _ z
Вторая подгруппа — ксерофильная — состоит из видов, свойственных сухим и жарким ландшафтам восточного Средиземноморья, теплолюбивых, известных в зоогеографической литературе под именем ирано-мало-азийских и средиземноморских. На Кавказском перешейке они появились по преимуществу с юга, так как основные массивы их ареалов уходят на юг, а на Кавказе захватывают главным образом южные и юго-восточные районы. Другие уцелели с плиоцена на месте — грузинский макак.
Среди типичных представителей второй подгруппы отметим куницу белодушку, малоазийского суслика, среднего хомяка, иранских песчанок, степную полевку, безоарового козла. Немногие из перечисленных форм распространены ныне на ксероморфных участках Большого Кавказа и его
Гл. Ш* Прежнее и современное распространение и численность млекопитающих 385
предгорий. Другие же заселяют только южные окраины Закавказья. Весьма вероятно, что в эту подгруппу следует отнести и таких относительно эвритопных, но все же сухолюбивых и южных по адаптациям зверьков, как ушастый еж, перевязка, серый хомячок, ареалы которых троходят насквозь по восточной половине Кавказского перешейка — * л нагорных степей и полупустынь севера Ирана до степей Русской рав-цшы(см. карты 1, 26, 60). Ареалы некоторых видов этой подгруппы ши развиты в плейстоцене на Большом Кавказе несколько шире, {ежели в голоцене, как например у среднего хомяка и других передне-азиатских форм. У других же они развивались в северном направлении, е. на равнины Закавказья и даже в Предкавказье, относительно не-(авн0—.в верхнем плейстоцене или даже в голоцене, как например малоазийской песчанки, кулана и джейрана.
Фаунообразующее значение имели на перешейке и европейско-азиат-кие гидрофильные и лесные виды. Появление лесных видов зверей европейского типа из Западной Европы на Кавказе могло происходить и в тре-пчный период, и в плейстоцене, особенно в те эпохи, когда массивы широколиственных лесов Европы и Кавказа получали взаимную связь через 4алую Азию и Балканы или через Русскую равнину. Но нельзя отрицать I параллельного видообразования, особенно у горных форм, например рыси, кавказской снеговой полевки, косули, серны;
К более молодой — плейстоценовой — по возрасту генетической руппе следует отнести виды, проникшие на перешеек с севера, с юго-дада и юго-востока. Древние предпосылки для такого вселения заключались в оформлении северных массивов суши Кавказского перешейка, развитии в эпохи похолоданий лесных и лугово-степных ландшафтов j а степных и полупустынных ландшафтов в предкавказских равнинах I в эпохи потеплений. Весьма вероятно, что для миграций лесных видов на Кавказ и с Кавказа в плейстоцене было два пути: северный — j по пойме Дона и юго-западный — по северному побережью Малой I Азми.
I Такими путями могли попадать на Кавказ лось и бурый медведь но-I дальней расы. Северный путь был возможен также для житника, j крупной азиатской косули и многих других видов. Юго-западный путь • более вероятен для лесной куницы, лесного кота и несомненен для ры-Ш полевки. Время проникновения на Кавказ названных видов было, конечно, различным. Наиболее раннее характерно для обыкновенного j Х0да и наиболее позднее — для европейского медведя и азиатской I косули. Имелись в плейтоценовой группе и репные элементы.
I Раннеплейстоценовыми вселеицами в предкавказские равнины из 1 непной подгруппы можно считать корсака, малого суслика, сайгу и других, более поздними степную пеструшку. Северное степное влияние достигало в плейстоцене даже до восточного Закавказья.
(Развитие на перешейке ареалов южных — переднеазиатских — видов в плейстоцене было, затруднено в связи с прогрессирующим похолоданием.
Так называемые межледниковые — теплые и сухие — эпохи по ареа-«дам млекопитающих прослеживаются все же на перешейке по распространению туранских и отчасти переднеазиатских видов.
Самая поздняя генетическая группа зверей, появившаяся па пере-шейке в голоцене, состояла из южных, теплолюбивых видов, получивших ; возможность продвинуться па Кавказ при послеледниковом потеплении < а иссушении ландшафтов. В нее вошли шакал, полосатая гиена, лев, 25 к- Верещагин
" Часть II. Анализ происхождения четвертичных млекопитающих
тигр, камышовый кот, кулан, джейран и, вероятно, некоторые насекомоядные и рукокрылые. Часть названных видов вымерла на перешейке при содействии человека в последние столетия.
В историческую эпоху началось и проникновение на Кавказ случайных видов: серой крысы и европейских рас домовой мыши, — завозившихся вначале на судах при морских и речных путешествиях древних греков, славян, хазар и варягов, а позднее — по железнодорожным и шоссейным путям.
Изучение прежних и современных особенностей распространения кавказских млекопитающих показало также большую природную динамичность численности и границ ареалов отдельных видов на протяжении четвертичного периода. Выявилась и огромная зависимость численности и ареалов многих видов от влияния человека в исторической эпохе.
Разрывы ареалов относительно стенотопных и стенотопных млекопитающих Кавказского перешейка в большинстве случаев удовлетворительно объясняются чередованием влажных и сухих эпох на протяжении кайнозоя, а также горообразованием и оледенениями хребтов.
Наиболее характерны высокогорные разрывы ареалов кавказских эндемиков: кавказской снеговой и прометеевой полевок, серны в районе Сурамского хребта. Возраст этих разрывов, вероятно, раннеголоценовый. Разъединения ареалов мезофильных и лесных видов, например у крота, кустарниковой полевки, косули, характерны и для центрального Предкавказья, и для восточного Закавказья. Время образования таких разрывов приходится частью на плейстоцен, частью — на после-ледниковье. Известны и разрывы ареалов ксерофильной^руппы видов: у среднего хомяка, степной полевки, безоарового козла\
Анализ таких разрывов, особенно у ареалов грызунов, позволяет проследить наличие на Кавказе по крайней мере двух эпох с ксерофитизи-рующим воздействием: одной весьма древней и другой недавней, молодой. Кроме того, прослеживается воздействие холодной и влажной эпохи, располагающейся между названными сухими.
Попытки уточнения степени и характера фаунистических влияний смежных территорий на основе изучения стратиграфической и географической изменяемости, а также общий анализ ареалов всех голоценовых зверей на основе зоогеографических карт даны в главах IV и V.
Е Глава IV
МАТЕРИАЛЫ ПО СТРАТИГРАФИЧЕСКОЙ И
I ГЕОГРАФИЧЕСКОЙ ИЗМЕНЯЕМОСТИ1 КАВКАЗСКИХ
К МЛЕКОПИТАЮЩИХ ЧЕТВЕРТИЧНОГО ПЕРИОДА
К Разработка вопросов филогенеза и формообразования для различимых филогенетических ветвей четвертичных млекопитающих Кавказа Кв настоящее время осложнена многими трудностями,
И Ископаемые костные материалы из большинства местонахождений '• обычно настолько фрагментарны, что возможности морфометрический •1 сравнений сильно ограничены. Кроме того, ископаемые остатки бывают яередко в различной степени деформированы в процессе фоссилизации, | Коллекции черепов и шкур современных видов мелких зверьков собира-I лись многими коллекторами в разные годы и разные сезоны, применяв-| щими разные способы консервировки. В результате выводы по изменяв-А мости млекопитающих во времени и пространстве бывают недостаточно «убедительны. Так, например, изменчивость роста и веса у популяций гры-Ж-аунов с коротким жизненным циклом (полевки) по отдельным годам и Ж сезонам может значительно перекрывать или затушевывать изменяемость К пространственную — географическую — в связи с колебаниями местных ж факторов среды.
Я Малая пригодность сборов насекомоядных и мышевидных, фиксиро-Ш ванных в жидкостях и сухим способом для тонких систематических ис-:« сзедований, отмечалась, например, Шидловским (1953а, 19536, 1954а, •в 19546). Получение доказательных результатов по географической изменяв-! мости пушных зверей при исследовании и обработке массовых сборов ! шкур пушносырьевых баз также возможно лишь с большими оговорками ! g при достаточной изученности индивидуальной, половой и возрастной изменчивости популяций видов.
j Кроме того, отсутствие четких критериев степени родства морфологи-1 дай близких видов и пределов вида иногда попросту затрудняет состав-j ленпе филогенетических и географических рядов форм.
Считая тем не менее, что сравнительные исследования морфосистема-1 тическпх особенностей млекопитающих Кавказа и Русской равнины могут Я содействовать познанию истории формирования фауны, мы даем здесь
I 1 Считая этимологически более правильным называть эволюционные и географи-I ческие изменения живых организмов изменяемостью, мы оставляем установившийся | термин «изменчивость» для обозначения индивидуальных и возрастных изменений.
I 25*
I
388 Часть II, Анализ происхождения четвертичных млекопитающих
краткий обзор имеющихся материалов по некоторым отрядам. Для прослеживания морфологических изменений во времени в ряде случаев были использованы лишь наиболее массовые фрагменты, пригодные для морфометрического исследования.
Отряд НАСЕКОМОЯДНЫЕ- IТМ S IE С ТI V О Н А
Кавказские материалы по ископаемым представителям отряда ничтожны. Среднеплейстоценовые ежи — Hemiechinus atf. auritus и Erinaceus aff. europaeus из бинагадинского битума Апшеронского полуострова по размерам и конструкции нижних челюстей и зубов практически не отличались от современных.
Географическая изменяемость обычного ежа на Кавказе прослеживается по потемнению окраски игл и меха брюшка у популяций населяющих серые и лесистые низменности. С Черноморского побережья Кавказа и Малой Азии описаны, например, темные формы: Erinaceus europaeus concolor, Е, europaeus ponticus, В Предкавказье обитают восточноевропейские Е, europaeus rumanicus, а в полупустынях Закавказья — светлые особи типа Е. europaeus transcaucasicus.
Современные ушастые ежи изменяются на перешейке слабо. Наиболее малорослые популяции, описанные как Hemiechinus auritus cal!igoni и Н. auritus brachyotis, обитают в полупустынях восточного Предкавказья и долины среднего Аракса.
Стратиграфическая изменяемость кавказских кротов (род Talpa) нам неизвестна. Современные популяции кротов дают здесь отчетливо выраженные географические формы, особенно на давно изолированных друг от друга участках ареала.
Наиболее крупные кавказские кроты — Talpa caucasica ognevi обитают на южных склонах западной половины Большого Кавказа и на Триалет-ском хребте (Огнев, 1926а, 1928; Строганов, 1948).1 В южном и юго-восточном направлениях кроты мельчают. Популяции наиболее мелких кротов — Т. orientalis transcaucasica и Т. orientalis talyschensis обитают на далеко изолированных от основного массива ареала участках — хребтах Армянского нагорья и в лесном Талыше. Впрочем, в последние годы Н. К. Депарма обнаружила на северо-восточном Кавказе популяцию кротов еще более мелких, нежели талышский.
По размерам черепа и плюсны кавказские кроты дают ряд, указанный в табл. 75.
Такое распределение географических форм с измельчанием к юго-востоку объясняется, по-видимому, длительной изоляцией относительно небольших популяций зверьков на горных хребтах в условиях усиливающейся сухости, увеличения годовых температур и обеднения кормовой базы — почвенных беспозвоночных.
Для понимания характера видообразования кавказских бурозубок следует учитывать, что они, как мезофильные жизненные формы, испытывали на Кавказе, подобно кротам, влияние длительной изоляции в горнолесных массивах.
1 В 1944 г. С. У. Строганов провизорно отнес 8 экз. кротов из района Кутаиси и Боржоми к крупному итальянскому кроту — Т, romaпа ognevi. До окончательного выяснения родственных взаимоотношений кавказских кротов с западиосредиземно-морскими мы рассматриваем эту форму в качестве подвида Т. caucasica. Точно также ' форма Т, europaea transcaucasica, описанная Далем (1944а), с Бамбакского хребта условно отнесена к Т, orientalis.
fa IV, Стратиграфическая и географическая изменяемость млекопитающих 388
Таблица 75
Географические изменения размеров (в мм) кавказских кротов1
» Промеры Запад Большого и Малого Кавказа Talpa caucasica ognevi 8 экз. Большой Малый Кавказ Кавказ Talpa саиса- Talpa orien-sica caucasica tails orientalis 240 ЭКЗ. 170 ЭКЗ. Малый Кавказ Talpa orientalis trans-caucasica 62 экз. Талыш Talpa orientalis taly-schensis 3 экз.
Кондилобазальная длина черепа . . . Длина верхнего ряда зубов Длина ступни . . . 35.9 33.9 30.1 32.4 29.6
35.1—37.2 14.5 33.1—35.7 29.8—32.6 13.4 12.2 31.3—33.0 11.2 29.5—29.8 11.1
14.0—15.0 19.6 12.5—13.8 11.9—12.7 18.3 16.9 10.5—11.7 16.5 11.0—11.2 15.5
19.0—20.0 17.5—19.2 16.0—17.9 14.2—17.5 15.0—16.0
Примечание. В числителе указана средняя величина, а в знаменателе — пределы.
Современные бурозубки (род Sorex) также мельчают в пределах Кавказского перешейка с запада на юго-восток, как например Sorex minutus. Кавказская популяция обыкновенной бурозубки рассматривается теперц как подвид от европейской — iV. araneus satunini (Огнев, 1928). Эта бурозубка отличается широкой и короткой носовой частью черепа, будто бы более короткой нижней челюстью и т. п.
В иных случаях на Кавказе формировались более крупные самостоя-|тельные виды бурозубок, например темноокрашенная и длиннохвостая raddei, близко родственная европейской — S. araneus.
I Водяные землеройки Кавказа (род Neomys) — формы, морфологиче* |ски и экологически более специализированные, нежели сухопутные землеройки. Генетические взаимоотношения кавказских представителей рода разработаны слабо, и описанные формы: Neomys leptodactylus, ft, schelkovikovL TV. balcaricus, N. dagestanicus—рассматриваются нами как подвиды от европейской — N. fodiens. Для первых двух из названных форм отмечена еще Сатуниным (1915а) узколапость, а для дагестанского подвида — недоразвитость киля хвоста, бледность окраски и т. п. — признаки, свойственные зверькам, обитающим в странах, испытывающих прогрессивное усыхание гидрографической сети (Испания, Крым),
Среднеплейстоценовые белозубки из Бинагадинского захоронения — Crocidura russula, С. leucodon не отличались заметно по строению черепа и зубов от современных особей, обитающих в восточном Закавказье. Южные по происхождению белозубки распространены на перешейке нередко значительно шире бурозубок. Особенно это относится к С. russula foldenstaedti, обильной как в засушливой, так и во влажной зонах.
Современный вид дает хорошо заметную географическую изменяемость окраски, связанную с обитанием в гидрогенных биотопах жаркой полупустыни и в сырых тенистых лесах Колхиды и Гирканики. По Шидловскому (19536), длиннохвостые белозубки Закавказья мельче и темнее в сырых лесах прибрежных низменностей Черного моря и Каспия, нежели в центре страны и на засушливых участках восточного Закавказья (табл. 76).
измерениям Строганова (1948), Даля (1944 а), Верещагина (1945 6) (Талыш).
390 Часть II. Анализ происхождения четвертичных млекопитающих
Таблица 76
Географические изменения размеров (в мм) и окраски длиннохвостой белозубки в Закавказье1
Промеры и типы окраски Западное Закавказье, Колхида Crocidura russula monacha 150 экз. Центральное Закавказье, долина Куры Crocidura russula guldenstaedti ; 170 кз. Восточное Закавказье, Талыш Crocidura russula caspica 40 экз.
Длина тела .... Длина ступни . . . Кондилобазальная длина черепа . . . Длина верхнего ряда зубов Окраска 70.6 74.1 70.6
60—89 12.7 60-91 13.0 63—80 13.1
11.0—15.0 18.1 11.0—15.0 18.7 12.0—13.6 19.3
16.7—19.4 8.1 17.5—19.8 8.2 18.6—20.2 8.9
7.6—8.8 Все тело темно-каштаново-бурое, хвост очень темный со всех сторон. 7.3—8.9 Верхняя сторона тела мыши но-серо-бурая, нижняя серо-белесая, хвост темный, одноцветный. 8.3—9.4 Все тело темно-каштаново-бурое, хвост темный, одноцветный.
Примечание. В часлдгелз указана средин я величина, в знаменателе — пределы.
Понятно, что если будет доказана видовая обособленность каспийской белозубки, то сходство в окраске следует отнести за счет конвергентного развития признаков на столь сходных экологически участках, как Колхида и Гирканика#
1
I
< I
г
Д о
У в в р к т: н и; п<
С1
Pf П] сс сс С(
Отряд РУКОКРЫЛЫЕ — С ПТКОР Т Е К А
Ископаемые остатки рукокрылых известны на Кавказе, за ничтожными исключениями, лишь из голоценовых слоев, поэтому говорить о филогенетических изменениях представителей этого отряда у нас нет оснований. По литературным данным (Бобринский, Кузнецов, Кузякин, 1944; Кузякин, 1950), у некоторых кавказских представителей Rhino lophidae по мере продвижения с северо-запада на юго-восток современная географическая изменяемость выражается в посветлении окраски и в увеличении размеров по мере подъема над уровнем моря (Rh. hipposideros, Rh, ferrum-equinum).
Широко распространенные виды рода Myotis и Vespertilio иногда занимают по величине и окраске промежуточное положение между европейско-сибирской популяцией и центральноазиатской, например Myotis-mystacinus. В других случаях, например у Vespertilio serotinus — более близки к европейским.
1 Шидловский (19536), давший эти измерения, считал, что по сумме признаков С. russula caspica должна быть выделена в особый вид — С. casplca. Это заслуживает специального исследования, так как длиннохвостая белозубка — вид огромной экологической пластичности и морфологически изменчив.
сл
,l IV. Стратиграфическая и географическая изменяемость млекопитающих 391
Для получения достоверных выводов по географической изменяемости летучих мышей Кавказа необходимо иметь солидные сравнительные материалы из Передней Азии, так как ареалы большинства видов уходят отсюда на юг и юго-запад.
Отряд ХИЩНЫЕ— С Л HXI V О ЕЛ
Обычная для четвертичных захоронении редкость остатков хищных ине затрудняет исследование морфологической эволюции в этом от-ie. Единственное исключение представляют Бинагады, в которых
ЯЕсобо многочисленны остатки псовых.
г У кавказских волков со среднего вгейстоцена несколько сократилась Кина мозговой капсулы, сузились Реальные концы носовых костей, к умении 1лись широкоскулость, дли-fca верхнего ряда коренных и длина Ксочного сужения (рис. 160). Инте-КсСно, что в очертаниях и вообще в Конструкции зубов разницы подменить не удалось. Небольшое сокращение длины верхнего ряда зубов произошло не только за счет более тесной Ксадки зубов, но и в результате по-Еепенного исчезновения М3. У 33 че-К?ов волков бинагадинской серии М3 Е^исутствовал в двух случаях, что вставляет 6 %, а среди 222 черепов ^временных волков из пределов КССРМ3 был встречен лишь в 0.9% ву-лев. В скелете конечностей уда-рзось подметить также сужение лопаток к современности (Верещагин,
Ы9516).
В Современная географическая из-Г пеняемость волков на перешейке j изучена слабо, но тем не менее из-1 вестно, что в равнинном Закавказье
Рис. 160. Изменение височного индекса [длины височного сужения (в %) к длине мозговой части черепа] у волков Кавказа с среднего плейстоцена.
1 — современные Canis lupus cubanensis; 2 — ископаемые С. lupus binagadensis, восточное Закавказье, Бинагады. Средние величины показаны черной точной.
1 возки несколько мельче, чем в Предкавказье. На Армянском нагорье ! животные вновь крупнеют, и, по описанию Даля (1951а), обитающий
i miCanis lupus hafastanicus, не уступая в росте кубанским, отличается 1 мягким и длинным мехом, окрашенным на спине в ярко-белесо-сероватые Bi палевые тона с сильным развитием черноты. Пушники различают на Р^ерешепке также «степных», относительно светлоокрашенных волков | открытых равнин, и «лесных», более темных. В Кабарде встречаются волки j ярко-рыжей окраски, являя довольно редкий пример аберративной из-4 I менчпвости. Три таких шкуры хранятся в ЗИН АН СССР.
I Вероятный среднеплиоценовый предок современной лисицы
I Fulpes khomenkoi из продуктивной толщи восточного Закавказья обладал
относительно мелкими коренными и короткими клыками, напоминая 3 ЭТОМ отношении песца (Богачев, 1938в; Верещагин, 19516). К концу среднего плейстоцена (хазарский ярус) лисица оказалась сформированной
392
Часть II. Анализ происхождения четвертичных млекопитающих
как вполне современный вид, и последующие изменения в строении черепа оказались ничтожными. Среднеплейстоценовые лисицы из восточного Закавказья (Бинагады) имели еще довольно широкие пределы вариаций черепных признаков, свойственных ныне как лисице Куринской низменности — Vulpes vulpes alpherankyi, так и высокогорным — V. vulpes kurdistanica, И. vulpes alticola. Однако в массе по строению и размерам зубов нижней челюсти они почти не отличались от -первой, что можно проследить и на наших графиках (рис. 161). /
Географическая изменяемость черепа и волосяного покрова современных лисиц Кавказа весьма велика. Она была в общих чертах прослежена
Рис. 161. Изменение размеров зубов закавказских лисиц с среднего плейстоцена.
I — длина нижнего ряда коренных; II — длина М, по коронке; III — длина
по коронке. 1 — современные Vulpes vulpes alpherahyi, 2 —• ископаемые V. vulpes aff. alpherakyi; восточное Закавказье, Бинагады. Средние величины показаны черной точкой.
еще Огневым (1931) по единичным черепам и шкурам. Огневым была дана и вполне формальная карта распространения описанных кавказских подвидов лисиц: Vulpes vulpes stepensis, V. vulpes karagan, V. vulpes caucasica, V. vulpes alticola, V. vulpes kurdistanica, V. vulpes alpherakyi, Кузнецов (Бобринский, Кузнецов и Кузякин, 1944) описал весьма близкое к истине распределение географических форм кавказских лисиц по меховым признакам на основе массовых заготовок шкур.
Впервые карта распределения кряжей, т. е. цветовых и иных меховых особенностей, а также размеров и вариаций их у лисиц на Кавказском перешейке, была опубликована нами (Верещагин, 1947 г).
Пушники различают здесь на материале в сотни тысяч ежегодно добываемых шкур шесть кряжей: украинский, донской, кубанский, северо-кавказский, ереванский и закавказский.
Характеристика кавказских лисиц по меховому покрову на основе просмотренных нами более 13 тыс. шкур на Ростовской и Тбилисской пушных базах может быть дана кратко в следующем виде.
Гл. IV. Стратиграфическая и географическая изменяемость млекопитающих 39&
Украинский кряж. Подразделяется еще обычно на североукраинский и центральноукраинский. Это — крупные лисицы с длинной, относительно жесткой остью, создающей на спине рыжевато-белесый фон, н на боках — фон соломенного цвета. Брюхо чаще всего рыжеватое, фронтальная поверхность лап черная. Распространены в предкавказских степях, но здесь не характерны и редки. Популяции этих лисиц как бы зкраплены местами среди типичных северокавказских — от северо-за-ладного Приманычья до равнинной Кабарды. Присутствие лисиц этого ^ряжа в 40-х годах в Предкавказье стало заметным, по мнению пушников,, ишь после второй мировой войны.
Донской кряж. Довольно крупные светлые лисицы, рыжевато-бе-сые, с алым цветом плеч и брюха. Ость по сравнению с украинским ряжем низкая. Наиболее часто встречающиеся типы окраски, связан-ые, вероятно, с полом и возрастом: рыжая, светло-рыжая, рыже-серая; еже встречаются темно-рыжие и серо-рыжие особи.
Эти лисицы обитают на севере Азовского побережья, в области ни-ового Дона, в Сальских степях. На восток распространены до Егор-ыка, а на юг — до Ей.
Кубанский кряж. Лисицы несколько крупнее, чем донские. Характеризуются ярко-рыжей и даже красноватой окраской морды, спины и особенно шеи и плеч, в связи с чем образуется заметный густоокрашенный крест». Ость на спине ниже, чем у украинских лисиц. Наиболее часты )со’би насыщенной рыжей окраски, несколько меньше рыже-серых и еще меньше светло-рыжих.
Распространены в Прикубанских степях, на Закубанской наклонной равнине от Керченского пролива на западе до водораздела Кубани и Кумы на востоке.
Северокавказский кряж. Лисицы несколько меньших размеров, чем: предыдущие. Мех грубый, ость нередко образует на спине «завихрения» — пятна неровно укладывающихся грубых волос. Брюхо черное, темно-серое. Встречаются в двух вариантах — красной и красно-серой, отличающейся более бледной окраской. Красно-серая вариация увеличивается в числе к юго-востоку. К этому кряжу относятся лисицы, описанные под именем Б vulpes caucasica.
Распространение лисиц этого кряжа охватывает центральное и восточное Предкавказье: от Закубанской равнины до побережья Каспия и от низовьев Кумы и Терека до Самура. Шкурки таких лисиц поступают и пз восточного Закавказья, что свидетельствует о большой близости окраски и иных особенностей шкурок лисиц восточного Предкавказья 2 восточного Закавказья.
Закавказский кряж. К этому кряжу принадлежат мелкие лисицы равнин восточного Закавказья, дающие три основных вариации: закавказскую красную, закавказскую красно-серую и закавказскую серую. Они отличаются сравнительно грубым низким мехом, нередко с большой примесью черных остевых волос на крупе и плечах, наличием более или менее широкого желтоватого или рыжеватого «ремня» вдоль спины, особенно хорошо заметного у бледно и грязноокрашенных полувзрослых особей. Серая окраска, как правило, свойственна молодым особям — сеголеткам. К этому кряжу относится мелкая лисица восточного Закавказья, описанная под именем V. vulpes alpherakyi.
Лисицы закавказского кряжа обитают в низовьях Терека и Сулака, вдоль западного побережья Каспия, во всем равнинном восточном Закавказье и в долине среднего Аракса. К этому кряжу в 40-х годах отно
-394
Часть II. Анализ происхождения четвертичных млекопитающих
сили и закавказскую караганку, вкрапленную в популяцию лисиц восточного Закавказья.
Закавказская караганка по государственному стандарту 40-х годов — это мелкая лисица с ровным темно-серым или даже слегка темно-бурым мягким мехом. Ноги до локтевого и коленного сочленений окрашены в черный или черно-бурый цвет. С караганкой Средней Азии эти лисицы имеют мало общего и представляют цветовую аберрацию — темноокра-
46'
54'
56
Темрюк
Махачкала
42‘
АСТРАХАНЬЮ
Элиста
РАСПРЕДЕЛЕНИЕ ТИПОВ ОКРАСКИ ЛИСИЦ ПО ЗАГОТОВКАМ 1945г (в процентах}
1 Темно-рыжая 2 рыжая 3 Светло -рыжа?
4 Рыже-серая
5 Серо-рыжая
6 Темно-серая срыхе^шяреш 7Свепто-сераясрыжеватым^мнеь 8 Гоязно'серая
з Светло-серо-рыжая ж Ь Заготовительные пункты
3 4
икл
44
4Z
40'
ахичЕван
Тавриз
38'
Решт0
оз. Урмия
36'
46°
48'
52’
54'
Казеин
50’
Рис. 162.
КРАСНО Новороссийск Туапсе
Батум ___ Трапезунд
РАСПРЕДЕЛЕНИЕ КРЯЖЕЙ ЛИСИЦ ПО МАТЕРИАЛАМ ЗАГОТОВОК EZZJ Украинский
БАКУ
^А71еморань
ГТПТП1 Северокавказский Кубанский Ереванский
2323 Закавказский с Караганка
385
шейный вариант лисицы Алфераки, аналогично чернобурым особям лисиц севера.
Ереванский кряж. Крупная лисица, но мельче предкавказских, со слабым черепом, резко отличающаяся от других кавказских исключительно густым и пышным мехом. Ость очень длинная, на ощупь шелковистая. Окраска варьирует от светло-лимонной до грязно-буровато-рыжей. К этому кряжу относятся лисицы, описанные под именем V. vulpes allicola и V. vulpes kurdistanica.
Эти лисицы обитают на Армянском нагорье, а далее к югу, вероятно, в хребтах северо-западного Ирана и восточной Турции. Из высокогорий Большого Кавказа, в частности из К)жной Осетии и центрального Дагестана, поступают сходные, но не идентичные шкурки.
Следует попутно отметить, чтопри составлении карты ареалов лисиц различных кряжей (рис. 162) и диаграмм соотношения типов окраски в ряде глубинных пунктов (рис. 163) не могла быть учтена возрастная изменчивость животных. Цифры и проценты выхода кряжей даны в табл. 77.
О шнур (в °/о) лИсин 1 > 1 зличных кряжей но район;; Кавкааского перешейка1
Пункты сбора шнур Унраии-^ ский Донской Кубанский Северо-кавказский. красный Северо-кавказский красносерый Закавказский красный Закавказский красносерый Закавказский серый В ара-ганка Ереванский Сиводушка и крестовка Лисица без кряжа Число исследованных шкур
Сальские степи, хутор Веселый 20 75 5 - 301
Прикубанские степи, Славянская 6.2 47.4 46.0 -— —— — 11" —— 352
Закубанская равнина, Но-волабинская 23 72 5 W— WWW*. — 310
Приманычские степи, Арз-гир 2.3 3.5 0.3 88.5 — 5.2 м. 1* — 1 "1 1250
Терско-Кумские степи, Моздок 0.2 —— 2.5 97.0 0.5 1 1 — —- III" 1 .1» — 1199
Предгорный Дагестан,Буйнакск — — 57.5 3.5 9.0 3.5 32.5 1 IIIMI — 872
Каспийское побережье, Дербент 10 53 "1 '11 17 20 11 1 1 566
Колхидская низменность, Ланчхуты _р —и — 21 — 79 19
Центральное Закавказье, Сталинир — —- —— — —— — 33 1 1 57 76
Центральное Закавказье, Тбилиси 1 —— 1 I— 2 1.5 15.3 24 18 2.5 34.2 2 — 3704
Муганская степь, Сабира-бад — 24 8.5 65.5 2 685
Армянское нагорье, Ереван — — — 0.2 11.6 5.6 — 10.0 72.5 —— 3738
Долина Аракса, Нахичевань — — — 26 10 13 — 23 ' г л 1 i ' 28 — — 127
1 Материалы этой и следующей таблиц собраны автором на Ростовской пушной базе в 1945 г.
396 Часть II. Анализ происхождения четвертичных млекопитающих
Таблица 78
Географическая изменяемость размеров (в см) сырых шкур лисиц на Кавказском перешейке и Украине
Кряжи и пункты сбора шкур Длина от носа до основания хвоста Ширина посередине Площадь шкурки (в см2) Длина хвоста без волос Число промеренных шкур
Центральноукраинский, Сталине 85 79—90 24 23—25 4106 44 42—47 31
Донской, Новочеркасск 76 73—86 22 21—26 3384 42 39—45 34
Кубанский, Славянская ...... 79 71—94 22 19—25 3424 44 37—45 30
Северокавказский, Благодатное . . 76 70—86 21 19—24 3380 41 35—43 28
Без кряжа, Батуми 77 70—88 14 14—17 217^ ) 39 37—43 28
Без кряжа, Богдановка 76 70—84 15 15—17 2312 40 40—44 34
Закавказский, Евлах 68 66—80 16 16—22 2208 38 38-44 38
Закавказский, Баку 66 54—76 15 13—22 2020 34 24—45 65
Ереванский, Цалка 75 70—84 15 13—18 2302 42 35—45 23
Ереванский, Ереван 77 70—90 20 17—21 3048 43 40—48 29
Примечание. В дробях в числителе указана средняя величина, в знаменателе~~пределы.
Таблица 79
Географические изменения размеров черепов (в мм) лисиц Кавказского перешейка1
Районы и пункты сбора черепов Основная длина Ширина между концами слуховых трубок Число исследованных черепов
Западное Предкавказье и Придонье Тарасовский район Азовский район 131.5 47.0 5
125—138 128.0 46—49 46.8 5
125—137 46—48
1 Для таблицы использованы коллекции ЗИНа и Московского университета. Взяты черепа взрослых особей — самцов и самок — со сменившимися и начавшими стираться коренными.
[a. IV. Стратиграфическая и географическая изменяемость млекопитающих 397
0
з
I
Таблица 79 (продолжение)
Районы и пункты сбора черепов Основная длина Ширина между концами слуховых трубок Число исследованных черепов
Сальский район 129.0 47.0 5
123—138 45—49
Закубанская равнина, Майкоп . * 133.5 47.5 10
128—142 46—51
Центральное Предкавказье
Курсавский район ........ 133.0 47.5 37
124—141 46—50
Восточное Предкавказье
Uпатово, Бешпагир 134.5 47.0 7 (
123—150 46—49
Орджоникидзе 132.0 48.5 48
123—141 46—51
Кизляр, Махачкала 129.0 46. И
120—143 46—49
Большой Кавказ
Гвилеты 133.0 47.7 4
122—138 46—50
Черноморское побережье
Туапсе—Гагры 128.0 45.8 6
126—135 43—48
Восточное Закавказье
Кировабад 121.5 44,4 14
115—129 43—46
Малый Кавказ
Боржоми 125.0 45.0 3
118—133 44—46
Елоновка 128.0 47.2 7
119—138 44—50
средняя величина, в зна-
Примечание. В дробях в числителе указана менателе — пределы.
Аберративпая изменивость окраски кавказских лисиц редка. В Предкавказье и Закавказье встречаются изредка сиводушки и крестовки. В Дагестане попадаются и своеобразные чернобурые лисицы с мехом шоко-шно-фполетового оттенка. В Закавказье чернобурые лисицы встречаются в Закатальском и Нухинском районах: одна-две — на 20 000.
Пушники не разделяют на кряжи шкурки лисиц западного низменного Закавказья. Эти лисицы отличаются мелким ростом и рыжеватой окраской «обобщенного» лисьего типа. Судя по заготовкам шкур в Ба-
. ^93 Часть II. Анализ происхождения четвертичных млекопитающих
туми, в Аджарии преобладают рыже-серые особи, составляющие до 71 % всех добываемых здесь лисиц. До 16% приходится на лисиц, близких по окраске и росту к закавказскому кряжу. Наконец, на окраинных участках нагорья Джавахетии (Богдановка, Цалка) обитают лисы переходного типа — от ереванского к закавказскому кряжу.
Размеры шкур и черепов лисиц некоторых из перечисленных кряжей приведены в табл. 78 и 79.
При анализе всех приведенных данных по распределению типов окраски, размеров шкур и черепов выясняется весьма сложная картина
Рис. 164.
географической изменяемости лисиц на перешейке. Несомненно, что лисицы закономерно уменьшаются в размерах в равнинном Предкавказье с запада на восток, а их окраска тускнеет и бледнеет. В Закавказье эта закономерность нарушается фактором высотности, но наименьшие размеры и тусклость окраски характерны все же для самых восточных популяций, населяющих жаркую Кура-Араксинскую низменность. В качестве примера зависимости окраски лисиц от ландшафта приведем еще распределение кряжей лисиц в восточном Закавказье, составленное по данным 41 заготовительного пункта за сезон 1935/36 г. Общее число добытых шкур, использованных для составления картограммы (рис. 164) достигало 28 250 штук.1
1 Цифровая таблица опущена в целях облегчения текста.
Гл. /V. Стратиграфическая и географическая изменяемость млекопитающих 399'
Рассмотрение картограммы приводит к выводу, что наиболее тускло-окрашенные лисицы обитают в зоне полупустыни, а по мере продвижения в предгорья и горы, особенно к западу, увеличивается численность ярко-окрашенных лисиц.
В горнолесном Закатальском районе добывается, до 4.2% чернобурок, 4.2% — сиводушек, 2.1% — крестовок.
Отметим, что на юг и юго-восток от Закавказья расположены ареалы мелких рас той же полиморфной группы черноухих лисиц Палеарктики: F. vulpes flavescens Gray, V. vulpes persicus Blanf., V. vulpes leucopus Blyth.
Учитывая столь пышное формообразование у лисиц Кавказа и следуя общепринятому критерию области происхождения вида, можно при*
Рис. 165. Изменения размеров зубов корсаков со среднего плейстоцена,
I —длина нижнего ряда коренных; II — длина М, по коронке; III — длина М14-М3. 1 — современные Vu/pes corsac, Средняя Азия; 2 — ископаемые V. aff. corsac, восточное Закавказье, Бинагады. Черные точки — средние величины.
знать подобласть восточного Средиземноморья и Передней Азии наиболее вероятной первоначальной родиной голарктической лисицы.
Значительно большие черепные изменения, нежели у лисицы, происходили во времени у корсака. Судя по остаткам из бинагадинского асфальта, I восточнозакавказские среднеплейстоценовые корсаки были несколько* мельче современных предкавказских и среднеазиатских (Верещагин, 19516). Кроме того, со среднего плейстоцена шло заметное сокращение размеров М3, М3 и Даже полное исчезновение М3 (рис. 165,166). Бинагадинский кор-’ сак обладал еще стопроцентным наличием М3 (при 43 случаях), в то время ! как у современных этот зуб присутствует всего у 66% особей (8 из 12 слу* чаев). Современные корсаки, как известно, крупнеют в восточном на-J правлении и на своей вероятной родине — в степях Прибайкалья и За-I баикалья — достигают наибольшей величины.
! Бурые медведи Кавказа с среднего плейстоцена заметно мельчали. 1 [дагадинский медведь из среднеплейстоценовых слоев Ашперонского полуострова был близок к современному большому кавказскому мед-Ursus arctos caucasicus и, очевидно, являлся его предком, но имел' одее крупные коренные зубы хищнического, плотоядного типа. Осо
400
Часть II. Анализ происхождения четвертичных млекопитающих
1
бенно это заметно по остробугорчатости пятки М^. Измельчание зверя и его зубов к голоцену сопровождалось развитием плоскобугорчатости зубов, а следовательно, и развитием растениеядности. Бинагадинский плейстоценовый медведь мог бы быть выделен, в сущности, в самостоятельный вид (Верещагин, 1951 в).
Наибольшей величины достигал среднеплейстоценовый медведь Русской равнины, известный по находкам с низовой Камы (Мысы). Этот медведь, современный бинагадинскому, названный нами камским — U. kamiensis N. Ver. sp. nova, обладал длинным и стройным черепом с низким лбом (рис. 167). По величине — основная длина черепов: 381; 386 мм— он не уступал пещерным медведям плейстоцена. В верхнем плейстоцене медведи Прикамья (Татарии) стали мельчать и приобретать значительную выпуклость лба. Последняя отчетливо видна у черепа медведя — U. karmalkiensis N. Ver. sp. nova (рис. 167,2) из асфальта близ сел. Нижние Кармалки на речке Шешме. В послеледниковую эпоху медведи Русской равнины и Кавказа сильно измельчали, что хорошо прослеживается и по размерам последних коренных. Наш график (рис. 168), совмещающий элементы стратиграфической и географической изменяемости и индивидуальной изменчивости размеров М2 медведей, отлично иллюстрирует отмеченные выше факты.
Современная популяция медведей Кавказа в генетическом смысле оказывается весьма пестрой. По-вп-димому, в этом случае нельзя гово
рить о закономерностях в географической изменяемости единого консолидированного вида, так как, судя по исследованиям Смирнова (1916а), кроме крупного кавказского подвида, здесь обитает некрупная номинальная — северная раса и южная (пли средиземноморская) — мелкая. Несомненная взаимная метизация всех этих форм, северная из которых вселилась, по-видпмому, относительно недавно, крайне затрудняет познание сути современной изменяемости зверя в пространстве.
Черепа бинагадинских барсуков среднего плейстоцена имели широкий диапазон индивидуальной изменчивости. Здесь еще представлены в широких пределах признаки как песчаного — Meles meles leptorhinus} так и обыкновенного барсуков. В массе же эти барсуки стояли блпже всего к современным барсукам восточного Закавказья и северного Ирана — М. meles minor,, М. meles canescens Blanl. Интересно, что объем мозговой части черепа и мозговой полости у современных барсуков биометрически достоверно уменьшился, что указывает и иа уменьшение интенсивности всех физиологических процессов! (рис. 169). Первый верхний моляр барсуков восточного Закавказья заметно сузился (по отношению к своей длине) с среднего плейстоцена почти иа 8?о, ио па Армянском нагорье еще в век бронзы жили барсуки и с более широким М1, нежели древнеапшероиские (Верещагин, 19516).
2
Рис. 166. Нижние челюсти корсаков.
1—современного,Средняя Азия,№ 9470,ЗИН;
2 — ископаемого, восточное Закавказье, Бина гады, № 23674, ЗИН.
].♦ /V. Стратиграфическая и географическая изменяемость млекопитающих 401
Современные барсуки более крупны в Предкавказье и наиболее мелки в неточном Закавказье.
Почти не испытали морфологических изменений с среднего плейсто-I на черепа перевязок восточного Закавказья. Современные перевязки
•о-
1Ы
s, не 1ГО
Рис. 167. Осевые черепа медведей.
j _ современный Ursus u?cfos cancasicus, Большой Кавказ, № 6169, ЗИН; 2—U. harmaikiensis N. Ver. sp. nova, Татарин, Няжние Кармалки, верхний плейстоцен, № 3, Казанский университет; 3 — U. hamiensis N. Ver. sp. nova, Татария, Мысы, средний плейстоцен, № 1, Казанский университет.
.ем но-ше ыи по-яи-
Ьолько мельчают по мере продвижения с запада па восток, в преде-х степной и пустынной зон СССР.
Росомаха эпохи верхнего палеолита западного Закавказья (рис. 57, 2) пилась от современных тундрово-таежных большой высотой нижней * .дтп и крупными размерами коренных. В этом отношении она пре-тпла даже более древних, средпеплепстоцеттовых росомах Русской ванны п Урала (табл. 80). Такое противоречие с правилом Бергмана Кг, впрочем, удовлетворительно объясняться крупной величиной
26 Н. К. Верещагин
'402 Часть 1/, Анализ происхождения четвертичных млекопитающих
местной добычи — кавказских копытных. Самой мелкой, как ока залось, была росомаха в среднем плейстоцене среднего Урала.
Рис. 168. Стратиграфические и географические изменения длины М2 плейстоценовых и современных медведей.
1 — Ursus arctos arctos, Карелия; 2 — U. arctos caucasicus, Большой Кавказ; <3 — U. arctos meridionalis, Малый Кавказ, Талыш; 4 — U. a. arctos (subfoss.), Воронеж, голоцен; 5 — U. karmaikiensis N. Ver. sp. nova, Татарин, Нижние Кармалки, верхний плейстоцен; 6 — U. karniensis N. Ver. sp. nova, Татария, Мысы, средний плейстоцен; 7 — U. a. binagadensis, восточное Закавказье, Бинагады. Черные точки — варианты, за-; .
штрихованные кружки — средние величины.
О морфологических изменениях кавказских куниц рода Martes в чет-
вертичном периоде документальных
Рис. 169. Изменения объема мозга барсуков восточного Закавказья с среднего плейстоцена.
данных не имеется, но современные куницы образовали здесь заметно обособленные формы. Череп лесной куницы Кавказа — М. martes lorenzi в среднем на 2.0—2.5 мм длиннее, чем у среднерусской — М. martes ruthena Ogn., и с более широкой расстановкой скуловых дуг. От шведских — М. martes martes L. кавказские отличаются более развитой лицевой частью черепа при сравнительно небольшой черепной коробке и более узкой за глазничной перетяжке (Кузнецов, 1941). Более резкие отличия наблюдаются в окраске меха. ( )
1 — современные Meles meles minor; 2 — ископаемые М. meles aff, minor, восточное Закавказье, Бинагады. Черные точки — средние величины.
Эту разницу не следует считать значительной даже если учесть, что зверек проник на Кавказ относительно недавно, так как она может быть обусловлена и большой спецификой кавказских
местообитаний. Современная лесная куница мало варьирует по величине и окраске в пределах кавказского ареала. Значительно заметнее индиви-
Гл ‘IV. Стратиграфическая и географическая изменяемость млекопитающих 403
Таблица 80
Стратиграфические и географические изменения размеров (в мм) зубов и нижней челюсти росомах Кавказа и Восточной Европы1
Промеры Верхний плейстоцен Средний плейстоцен Современность
Кавказ, пещера Гварджилас -Клде Музей Грузии Украина, Гонцы Институт зоологии АН УССР Татария, Мысы на Каме Казанский университет Север среднего Урала, пещера Низе л ЗИН 3 экз. Норвегия-Анадырь ЗИН 5 экз.
Высота челюсти позади Mi 30.3 22.0 28.0 20.5 24.0
19.5—21.0 21.0—26.0
Высота челюсти в 23.3 21.5 17.8 19.3
области Рт3 . . . — 17.5—18.0 18.0—21.0
Длина Рю2 7.1 9.0 9.0 7.5 7.0
7.3—7.6 6.0—8.0
nlapniia Рт2 • • - • & 5.0 4.8 5.3
4.5—5.1 4.5—6.0
Длина Ртз 13.2 12.7 13.5 10.5 .11.0
10.0-11.0 11.0—11.1
Ширина Ртз 9.0 6.5 7.4
6.0—7.0 7.0—8.0
Длина Mj 24.0 22.0 24.0 19.5 22.1
19.0—20.0 21.0—23.2
Ширина Mi .... 10.8 9.0 9.8
8.0—9.5 9.0—10.2
Примечание. В дробях в числителе указана средняя величина, в знаменателе—пре-j дам.
§
сальная и возрастная изменчивости, сказывающиеся, например, в форме 1 । даже окраске горлового пятна.
| Эта изменчивость, по данным заготовок Ростовской пушной базы, представлена на диаграмме (рис. 170).
• * Отличия у кавказской популяции, более древней куницы бело душки — V, foina nehringi, от западноевропейских — М. foina foina Erxl. оказа-* гь. каки следовало ожидать, биометрически еще менее существенными.
м не менее черепа этих куниц с Кавказа крупнее (длиннее и шире) ч 2-2.5 мм, чем у швейцарских, и на 5—6 мм, чем у крымских — М. таг-«nori Martino.
Череп кавказской норки— Lutreola lutreola turovi также значительно пшее, нежели среднерусской (Бобринский, Кузнецов, Кузякин, 1944). явление увеличения роста, как известно, бывает вообще довольно v'актерпо для популяций, обитающих на новейших окраинах ареала.
Детальные морфологические и экспериментально-экологические ис-.'цования необходимы для выяснения родственных взаимоотношений i v.joii и большой кавказских ласок — Mustela nivalis caucasica и М. ni~
i Неопубликованные материалы автора.
26*
404 Часть II. Анализ происхождения четвертичных млекопитающих
valis dinniki, которые живут нередко бок о бок, и, следовательно, могут считаться самостоятельными видами.
Среди кошек особый интерес, в смысле выяснения их происхождения на Кавказе и темпа видообразования, представляют барс, гепард, лесной кот, рысь.
Размеры и пропорции зубов (и трубчатых костей) леопардов практически не претерпели изменений с среднего плейстоцена (табл. 81;
Рис. 170. Вариации размеров и формы горлового пятна кавказских лесных куниц (I) и куниц белодушек (2). Показана частота встречаемости.
И"1
рис. 61,6, 85,1) Оказались консервативны и зубы гепардов (Верещагин. 19516).
Современные барсы Кавказа мельче, с менее пушистым мехом и ярче окрашены в Закавказье, чем в Предкавказье (Динник, 1914а; Сатунин, 1915а). По размерам они превосходят южноазиатских и африканских.
Современный лесной кот Кавказа — Felis silvestris caucasicus отличается от европейского — F. s. silvestris несколько более крупным черепом и ростом. Кондилобазалъная длина черепа самцов первого 99.1 (89.8—102) мм, второго — 86—96 мм. Верхний зубной ряд соответственно 33.1(30—34.7) мм и 28—32 мм.
Как правило, коты, обитающие в камышах низовой Кубани, более однотонной окраски, нежели живущие в буковых лесах. В крупных массивах последних у Ну хи, Закатал и Мадагйса ДМалый Кавказ) нередки меланистические особи. Для основательных выводов о масштабах обособления кавказской популяции нужны серии шкур и черепов с севера Малой Азии, с Балкан и Карпат.
Современная кавказская рысь — Felis lynx orientalis обычно рассматривается в качестве подвида от европейской. По величине тела и черепа
Гл. IV. Стратиграфическая и географическая изменяемость млекопитающих 405
опа нс уступает, а иногда и превосходит последнюю. По окраске же кавказские рыси более ярки, нежели северные. Морфологические изменения черепов рысей в пределах кавказского ареала состоят в небольшом меныпении их размеров и в упрощении вырезки хоан у малокавказ-кой популяции.
Таблица 81
Размеры зубов (в мм) современных и ископаемых леопардов Кавказа
Местонахождение и датировка Виды Длина Pm4 по коронке Длина Mi по коронке
аиадный Кавказ, современность. Panthera pardus . . 25.0 18.1
24.5—26.2 17.8—19.3
;;с.1ье Баксана, грот Соеру ко, ранний голоцен (мезолит). Р. aff. pardus . . . 18.8
ерховья'Риона, пещера Кударо, средний плейстоцен (нижний палеолит). Р. cf. pardus . . . 26.0 ——
Примечание. В дробях в числ ителе указана средняя величина, в знаменателе —
ределы.
Отряд ЗА Й ЦЕОБР А ЗНЫЕ — ЕА СО МО1УРНА
Судя по измерениям И. Громова (1952), у закавказских русаков с эпохи отложения бинагадинских битумов произошло сужение лицевой части черепа, появилась угловатость очертаний орбиты, уменьшились в объеме сдаовые пузыри, сузились крылья подвздошной кости. Современные русаки Кавказа дают хорошую иллюстрацию к правилу Бергмана: они крупнеют по мере распространения с юга на север. Закавказские русаки — Lepus europaeus cyrensis имеют общую длину черепа 93—97 мм, J дай вес до 3.5 кг и тусклую палево-буроватую окраску. У предкавказ-скихГ. europaeus caucasicus общая длина черепа колеблется от 95—105 мм, j дан вес достигает 4.5 кг, окраска преимущественно буровато-желто-( вато-серая (Бобринский, Кузнецов, Кузякин, 1944). По нашим измере-I йИЯм, кондплобазальная длина черепа взрослых зайцев (самцов и самок) j из Куринской низменности восточного Закавказья равна 85.3(79—94) мм i (32 экз.); а в западном Предкавказье — 88.4 (83—96) мм (25 экз.). Живой : вес взрослых восточиозакавказских зайцев (самцов и самок) составляет 1 соответственно 3.780 г (14 экз), а предкавказских — 4.760 г. (9 экз). Резкое укрупнение зайцев намечается в районе Хачмасской низменности, j ка наклонной Кусарской равнине и в низовых лесах Самура. Этот дол-1 лнып тип изменяемости подтверждает южное происхождение вида.
Отряд Г Р Ь1 3 УНЫ — Е() J) ENTIA
Только немногие виды кавказских грызунов известны нам с плиоцена,
>1 филогенетические ряды большинства форм оказываются слишком ко-j роткпмп для разработки стратиграфической изменяемости. Однако ши-I рокое распространение и массовость многих видов позволяют легко про-1 даивать изменяемость географическую.
I Географическая (широтная) изменяемость морфо-физиологических осо-| (еииостеп малого суслика сказывается в увеличении зверька и в потем
406 Часть II. Анализ происхождения четвертичных млекопитающих
нении окраски, а также в укорочении сроков летней спячки по мере продвижения из полупустыни в степь.
По измерениям Огнева (1947), черепа подвида Cittelus pygmaeus ka-labuchovi из Сальских степей в среднем почти на 2 мм больше, нежели у подвида С. pygmaeus pallidus из Прикумской полупустыни. Еще заметнее изменения по высотным зонам. Суслики изолированных популяций Предкавказья показывают закономерное увеличение размеров и потемнение окраски по мере продвижения в горы (та£л^ 82).
- Таблица 82
Географические изменения размеров (в мм) малого суслика на Кавказском перешейке
Промеры Низовья Терека CiteUus pygmaeus planicola 25 экз. Предгорья Дагестана CiteUus pygmaeus satunini 40 экз. Терско-Сунткен-ское плоскогорье CiteUus pygmaeus boemii 7 экз. Склоны Эльбруса CiteUus pygmaeus musicus 48 экз.
Длина тела .... Длина хвоста . . . Длина ступни . . . Кондилобазальная длина черепа . . . 206.7 208.5 207.5 220.0
182—230 35.6 182—230 36.3 193—215 37.8 205-240 47.4
30—40 32.1 32—43 32.1 28—40 33.2 45—50 37.4
27—35 40.7 30—36 40.0 28—34 41.0 36-38 43.5
39.5—42.0 37.9—42.6 37.8—41.9 42.5-45.2
Примечание. В числителе указана средняя величина, в знаменателе — пределы.
При распространении от полупустыни в низовьях Терека до верховий Баксана длина тела сусликов увеличивается в среднем на 14 мм, длина хвоста — на 12 мм, ступни — на 5 мм и кондилобазальной длины черепа — на 3 мм (рис. 171).
Однако горные популяции сусликов обнаруживают не меньшую изменяемость в зависимости от обитания либо на сухих и жарких днищах троговых долин (на высотах 900—1200 м), либо на склонах гор — субальпийских лугах (на высотах 2500—2800 м). Как правило, популяции, обитающие по днищам ущелий, наиболее сходны с равнинными, особенно по окраске.
Европейский суслик — Citellus citellus, по утверждению Виноградова и Громова (1952), увеличивается в размерах и по относительной длине хвоста при распространении с востока на запад — с Украины на Балканы. Малоазийский подвид этого суслика проявляет прежде всего хорошо заметную морфо-физиологическую изменяемость в зависимости от обитания в той или иной вертикальной зоне. По Аветисяну (1950), суслики, обитающие в районе Алагеза в зоне полупустыни, пробуждаются в половине марта и залегают в летнюю спячку во второй половине июля, а в нагорной степи пробуждение происходит лишь в начале апреля, С высотой у суслика увеличиваются размеры тела и вес, укорачивается хвост (табл. 83), темнеет окраска.
На примере тушканчиков (род Allactaga) удается проследить некоторые части генерального направления эволюции и быстроту специализации в этой группе.
Гл, IV. Стратиграфическая и географическая изменяемость млекопитающих 407
Таблица 83
Изменения размеров (в мм) и веса (в г) малоазийского суслика (самцов и самок) при обитании в различных высотных зонах
Промеры Зона 1255—1550 м над ур. м. Зона 1550—2190 м вад ур. м.
Длина тела Длина хвоста Вес 203 215
175—230 45 180^90 (43 J
33—37 239 21—"59 278
188-372 184—430
Примечание. В числителе указана средняя величина, в знаменателе — пределы.
Современные тушканчики Предкавказья и Апшеронского полуострова приобрели с среднего плейстоцена (Бинагады) более совершенные прыга-
Рис. 171. Изменения размеров малого суслика в Предкавказье. Прямоугольники показывают возведенную в квадрат длину ступни. Популяция из низовий Терека принята за единицу. Черные кружки — места отлова.
тельные способности. Их голень по отношению к бедру заметно удлинилась, повысив тем самым динамичность зверьков. Наиболее отчетливо это видно на примере малого и малоазийского тушканчиков, у которых
1
408
Часть II, Анализ происхождения четвертичных млекопитающих
z длина голени чу ЛААЧ -
индекс прыжка (~бедра Х *uU) становится особенно высоким (табл. 84).
И. Громов (1952) отметил еще, кроме того, более примитивный но сравнению с современными тип строения метаподип у большого тушканчика из бинагадинского асфальта — Allactaga jaculus bogatschevi, выражающийся в более широкой расстановке дистальных концов плюсны, а у малого тушканчика — в более длинном свободном конце второй метатарзальной кости (mtt2).
Хомяки (подсемейство Cricetinae) обнаружены на Кавказе уже с среднего миоцена. Однако установить прямое родство миоценовых (Palaeo-cricetus, Беломечетка) и плиоценовых (Ставрополь) хомяков с четвертичными родами — Cricetus и Mesocricetus представляется затруднительным. Хомяки из нижнеплиоценового аллювия Ставрополья (Косякин
Таблица 84 Изменения индекса прыжка у тушканчиков с среднего плейстоцена1 карьер) отличались очень массивными коренными. Средний хомяк из бинагадинского асф^бьта — Mesocricetus аига-tus pbanicola занимал по величине среднее место между
Виды Индекс прыжка
бинагадин-ские современные М. auratus raddei и М. аи-ratus nigriculus, отличаясь
Allactaga jaculus . . . A. willtamsi A, elater 125.0 126.1 129.5 127.0 129.5 136.8 от известных в СССР форм Mesocricetus коротким твердым нёбом и короткими резцовыми отверстиями. Популяция верхнеплейстоцено-
вых хомяков Чиатурского марганцевого района, судя по строению деталей посткраниального скелета, была впрочем не менее специфична, нежели бинагадинская.
Географические изменения современных популяций малоазийского хомяка сводятся преимущественно к небольшим различиям в окраске: степени яркости рыжеватого цвета спины, боков и развития черноты брюшка (Аргиропуло, 1937). Кроме того, горные популяции (например, дагестанская), как правило, крупнее предгорных, предкавказских. Экологическая пластичность вида велика. На высоких плоскогорьях Армении хомяк залегает в спячку в конце октября—начале ноября, а на низком плоскогорье в Кахетии — лишь в начале декабря.
Современные серые хомячки Апшеронского полуострова с среднего плейстоцена приобрели более узкое в переднем отделе нёбо, более крупные коренные, круче поставленный венечный отросток, слабо изогнутый резец нижней челюсти и М3 с укороченным талонидом. Вообще функциональное значение этих мелких отличий и направление эволюции большей частью трудно объяснить. Их легче просто зарегистрировать, как это и сделано И. Громовым (1952).
Картина морфо-экологической изменяемости на Кавказе у такого полиморфного вида, как домовая мышь — Mus musculus, весьма сложна.
1 Индексы вычислены нами по средним цифрам абсолютных промеров, приведенных И. Громовым (1952). Попытка Громова использовать морфологические отличия бинагадинских тушканчиков для реконструкции древних ландшафтов Апшеропа, прибегая к допущению то более мягких грунтов, то к мозаичности типов угодий, то к наличию в Бинагадах разнообразного рельефа, мало состоятельна.
Г.1-. 1V. Стратиграфическая и географическая изменяемость млекопитающих 409
В восточном Закавказье этот вид процветал в среднем плейстоцене (Бинагады), как и в настоящее время. Степень современной морфо-экологической дифференцировки у подвидов: белобрюхих и короткохвостых (степных и полупустынных) — М. musculus hortulanus, Al musculus lataricus; серобрюхих и длиннохвостых (горно-лесных) — М. musculus abbotti) М. musculus formosovi и, наконец, у завозных домовых — М, musculus musculus, в сущности, вероятно, не меньше, нежели у лесных мышей Кавказа (см. дальше), но еще менее поддается выражению современными средствами морфологического исследования (см. Гентнер, 1930; Гулий, 1930; Свириденко, 1935а).
Систематика лесных мышей Кавказа (род Apodemus) до сих пор слабо разработана. Учитывая исследования Аргиропуло (1940а, 1946), Свириденко (1936), Кузнецова (Бобринский, Кузнецов, Кузякин, 1944), Шид-довского (1953а), а также собственные наблюдения, мы принимаем здесь (впредь до специального экспериментально-экологического исследования) четыре вида: A. mystacinus, A. flavicollis, A, fulvipectus, A, syl-Mticus (см. также главу V).
Малоазийская мышь — A. mystacinus, распространенная на Балканах, в Малой Азии и в западйдм Закавказье, оказывается заметно мельче на Кавказе, чем на Балканах. По измерениям Шидловского (1953а), кондялобазальная длина черепа балканских особей составляет 28.4 мм (34 экз.), у закавказских 26.5 мм (200 экз.), а длина верхних коренных соответственно 4.9 и 4.5 мм. Зверьки балканских популяций окрашены бледнее малоазийских и закавказских. По мнению Шидловского (1953а), проникновение этого зверька на южный склон Большого Кавказа (см. карту 55) — явление совсем недавнее, последних десятилетий. Таким дазохг в данном случае зверек явно мельчает, распространяясь в новую для него мезофитную страну. Палеонтологические исследования в пещерах Имеретии помогут выяснить в дальнейшем первичность или вторич-востьтакого энергичного вселения, а следовательно, и характер дивергенции этого восточносредиземноморского вида.
Общая картина географических изменений остальных трех видов на Кавказском перешейке значительно сложнее. Желтогорлая мышь европейского типа с ступней в 24—25 мм (близкая к A. flavicollis samarien-ns) встречается только в приречных лесах низового Сулака в вос -i точном Предкавказье.1 На крайнем северо-западе южная граница рас- пространения европейского вида проходит по низовому Дону (карта 55).
Относительно мелкая кавказская желтопятная мышь — A. fulvi-pctus2 с ступней в 22—23 мм длины живет в широколиственных лесах нижнего горного пояса в Краснодарском крае, Абхазии, Аджарии, северо-западном Азербайджане и местами в северной Армении (карта 56). Она довольно легко отличима даже в молодом и юношеском возрасте 1 от лесной мыши крупными размерами, большой длиной (относительно | проченной) ступни и наличием округленного или удлиненного желто-I ватого пятна на груди. В открытые и осветленные места этот зверек за-? бегает нередко на кормежку даже днем. Легко, и охотно лазает на деревья i «часто живет в дуплах. В актографе отличается от лесной мыши слабой реакцией на силу освещения. Весьма вероятно, что относительно корот-; 1 Если только этикетки хранящихся в Московском зоологическом музее экземпля-
] пов не перепутаны. Гептнер и Формозов (1941) называли отсюда 5 экземпляров этой 1 Lmh. Мною изучен один. Происхождение этого вида здесь для нас еще неясно.
-2 К этому виду мы относим кавказские подвиды: A. flavicollis ponticus, A. flavi-parvus, A. sylvaticus fulvipectus прежних авторов.
410 Часть II. Анализ происхождения четвертичных млекопитающих
кая ступня и голень, удлиненный хвост желтопятной мыши и ее относительно узкие хоаны (Аргиропуло, 1946) по сравнению с таковыми лесной мыши — все это приспособления к передвижению менее быстрыми темпами и к более совершенному лазанью.
Популяций этой мыши из различных участков ареала неравноценны. Как правило, они несколько мельчают в южном, но вновь крупнеют в восточном направлении. Морги-левская (1954), не различавшая лесных и желтопятных мышей приводит такие результаты массовых измерений для пяти смешанных популяций. Кондилоба-зальная длина черепа: Сухуми-23.8, Гагры — 22.3, Батуми -23.1, Ахалкалаки — 22.7, Лагоде-хи — 23.7 мм. Длина задней ступни: Сухуми — 22.6, Гагры —21.3, Батуми—21.4, Лагодехи— 22.7 мм.
Популяции крупных желтопятных мышей обычны в ряде ущелий центрального Предкавказья (Уру-ха, Ардона, Терека) и на предгорных равнинах восточного Предкавказья. Здесь они значительно варьируют по величине в разных ущельях, что, несомненно, объясняется как генотипической изолированностью ..популяций, так и отличиями р местообитаниях. Делались дажеАпопытки объяснить более крупный рост мышей в ущелье Армхи (притоке Терека) повышенной ионизацией воздуха в области тектонического разлома (У шатии ская- Дека ленке, 1933). Если это так, то величина мышей в горных популяциях могла бы служить своеобразным индикатором для поисков радиоактивных изотопов.
В восточном Закавказье популяции этой мыши обитают преимущественно в предгорных низменностях. Тщательно промеренные
единым приемом серии сухих тушек одновозрастных кавказских желтопятных и лесных мышей дали вариационные ряды географических изменений длины ступни, показанные на рис. 172.
Единичные остатки лесной мыши — A. sylvaticus subsp. обнаружены в среднеплейстоценовых асфальтах Апшеронского полуострова. Морфологические изменения к современности по этим остаткам не установлены. В Предкавказье эта мышь широко распространена на равнинах, в предгорьях и горах, особенно по участкам с разреженными насаждениям (карта 57). Горные популяции лесной мыши отличаются повышенным
18 19 20 21 22 23 24 25 26 мм
Рис. 172. Изменения длины ступни лесных мышей на Кавказском перешейке.
Apodemus flavicollis: 1 — Ростовская область. A, fulvipecius: 2 — Кавказский заповедник; 3 — Алазано-Агричайская долина (Закавказье); 4 — Малый Кавказ (Делижан); 5 — Талыш. A. syivaticus: 6 — Ростовская область; 7 — Кавказский заповедник; 8 — Северная Осетия (Ларс, Зарамаг); 9 — Алазано-Агричайская долина (Заката лы); J0 — долина среднего Аракса. Слева подошва лесной, справа — желтопятой мыши.
По оси ординат — число случаев.
f Гл. /У. Стратиграфическая и географическая изменяемость млекопитающих 411
I
I
IT
я
Й
в-
1-
,e-y-.3, lM. it-ий зу-ед-ед-эНО [ЫХ »яс-ЛИ-: и Де-[ИТЬ I в ека) ^уха раз-нко, шна огла нди-1ТИВ-
ЕОПу-;ИМУ~ шеи-нные эпят-изме-
жены орфо-лены, пред-1ИЯМИ щным
содержанием эритроцитов крови (Ка лабухов, 1940). В Закавказье лесная мышь несколько крупнее и нередко труднее отличима от желтопят-яой. На груди взрослых экземпляров почти всегда имеется желтый мазок.
1 В целом Кавказ представляет древний, вполне самостоятельный и активный и в наши дни очаг формообразования домовых и лесных мышей, tTO подтверждает и их большую — плиоценовую — древность в этой тпане.
Краснохвостая песчанка из бинагадинского битума — Meriones rythrourus intermedins обладала промежуточными признаками между современными песчанками, обитающими к западу и к востоку от Каспия.
Примитивные черты среднеплейстоценовой песчанки сказываются ; укорененности берцовой кости, составляющей 113.8% от длины бедрен-юй против 125.9% у современных песчанок Апшеропа.1
Учитывая, что удлинение берцовой кости по сравнению с бедренной сарактеризует прыгательный тип передвижения (тушканчики), можно тверждать, что современные песчанки значительно «отушканились» термин Н. А. Смирнова) с среднего плейстоцена, т. е. приобрели большие арыгательные способности.2
Кавказские полевки (роды Prometheomys, Arvicola, Microtus) представляют не менее благодарный материал для изучения изменений во времени и пространстве, нежели мыши.
Зубы и нижние челюсти прометеевой полевки из ашельских слоев Кударо I и верхнепалеолитических слоев пещеры Гварджилас-Клде не несли заметных отличий от современных. Это и понятно, так как консервативность морфологических черт древнего подземного грызуна весьма велика. Изменения в типах окраски заметны у современных популяций на различных участках (табл. 85).
Таблица 85
Изменения численности меланиот^в в пределах ареала прометеевой пдлевКи3
Окраска Верховья Лабы и Белой Верховья Б зыби Верховья Терека
абс. % абс. % абс. %
Серовато-бурая . . 13 52 16 100 15 100
Черная 12 48 — “— — —
? По-видимому, повышенный процент меланистов и на самом деле при-1 прочен лишь к западной части ареала.
Особенно сильно изменяются в пределах Кавказа и прилежащих территорий современные популяции водяной полевки — Arricola terre-jlris. Только с перешейка описано по крайней мере 9 форм, а со смежных I участков Ирана, Турции и Русской равнины — не менее 6. Географиче-1 ский характер морфологических изменений зверька уловить трудно.
j 1 Индексы средних вычислены нами по абсолютным цифрам измерений в ра-j боте И. Громова (1952).
- И. Громов (1952) также указывал, что бинагадинская песчанка была «более бегающей» (т. е. менее прыгающей,—Н. В.}, нежели современная.
| з По коллекции ЗИН АН СССР-
4
Часть ZZ. Анализ происхождения четвертичных млекопитающих
Как правило, северные болотные формы из низовьев Дона, Кубани и Волги немного крупнее горных и южных кавказских (табл. 86).
Популяции этого вида из отдельных участков обладают мелкими морфологическими особенностями, которые свидетельствуют лишь о специфике местообитаний. Горные «подземные» популяции во внутренней горной зоне Балкарии, Осетии и в Дагестане в связи с огромной практикой рытья имеют несколько более «слепушонковый» тип положения верхних резцов, т. е. с большим радиусом кривизны, нежели болотные, и менее дифференцированный мех.
В пределах Передней Азии, Кавказа и Средней Азии известно до 8 подвидов снеговой полевки, распространенных от Палестины до Копет-Дага. Географическая изменяемость малоазийской снеговой полевки от Малого Кавказа до Копет-Дага выражается в посветлении окраски по мере увеличения сухости и инсоляции, в укорочении М3 при распространении с запада на восток и в уменьшении роста (табл. 87).
Таблица 86
Изменения величины черепа (в мм) водяной полевки на Кавказском перешейке 1
Промеры Низовья Дона Arvicola ter-restris tanai-ticus 10 экз. Низовья Кубани Arvicola ter-restris cuba-nensix 11 ЭКЗ. Внутренний Дагестан Arvicola ter-restris kuruschi 32 экз. Закавказье Arvicola ter-restris persi-cus 32 экз.
Кондилобазальная длина черепа . . . Длина верхнего ряда коренных .... 41.2 41.0 38.5 39.6
41.0—41.5 10.2 < 39.5—42.6 9.9 38.2—38.8 9.4 37.3—41.9 10.1
10—lOA- 9.2—10.5 9.6—10.8
Примечание, менателе — пределы.
В дробях в числителе указана средняя величина, в зна-
По промерам черепа наиболее мелка снеговая полевка Большого Кавказа, что как бы указывает на малую пригодность влажных, мезофитных районов для этого зверька. Измельчание этой полевки в юго-восточном направлении, а также исчезновение ее поселений на средних высотах южных склонов Армянского нагорья свидетельствует о том, что юго-восточные участки современного ареала находятся теперь в области большого угнетения (пессимума) и что когда-то была иная, более влажная эпоха, пригодная для расширения ареала на восток.
Географическая изменяемость кавказской снеговой полевки на Большом Кавказе выражается в заметном уменьшении размеров тела и в посветлении окраски при продвижении с запада на восток. Последнее связано, вероятно, с выцветанием меха в безлесном и солнечном Дагестане. Эту картину посветления в широтном направлении затемняет несколько инверсия зон на центральном участке Большого Кавказа.
Серии зверьков из различных участков ареала дают убывающий к востоку ряд цифр (табл. 88).
1 Измерения Огнева (1933).
IV. Стратиграфическая и географическая изменяемость млекопитающих 413
Таблица 87
Географические изменения размеров (в мм) малоазийской снеговой полевки1
Промеры Северо-западный Кавказ, гора Бамбак М icrotus nivalis logi-novi 7 экз. Малый Кавказ, берега оз. Севан Microtus nivalis satunini 8 экз. Талыш, гора Келахан М icrotus nivalis subsp. 1 экз. Колет-Д аг, гора Шах-Шах Microtus nivalis demen-tievi 3 экз.
109.4 121.9 115.0 103.2
Длина тела 99—121 109—142 97—120
Длина хвоста . . . 58.1 54.0 52.0 55.3
52—68 32—90 51—60
Длина ступни . . . 18.8 17.9 18.5 18.5
12.2—20.0 13—20 17—20
Общая длина черепа 26.4 27.8 27.3 27.2
25.0—27.8 26.3—29.6 25.8—28.5
Длина верхнего ряда коренных .... 5.9 6.3 6.3 6.1
5.5—6.2 6.1—6.7 5.5—6.5
яйтрл? ” ппепелы " В ДрОбяХ в числителе указана средняя величина, в знаме-
IjalCJXC 11 1,041X51 •
Таблица 88
Географические изменения размеров (в мм) кавказской снеговой полевки2 !, /'
Промеры Западный Кавказ Microtus gud nenjukovi 128 экз. Центральный Кавказ Microtus gud gud 116 экз. Восточный Кавказ, Дагестан Microtus gud Ighesicus 8 экз.
Длина тела 120—152 102—149 100—121
Длина хвоста 68—106 61.0—85.0 58—78
Длина ступни . . . . 19.0—26.9 18.0—23.0 18.5—21.2
Кондилобазальная дли-
на черепа 26.4—28.6 25.0—29.2 26.4—27.2
Длина верхнего ряда
коренных 6.7—7.8 6.0— 7.0 6.0—6.6
Следует, впрочем, отметить, что изменяемость размеров этой полевки й0 вертикальным и ландшафтным зонам еще не прослежена.
Среднеплейстоценовая апшеронская полевка — Microtus (Pity ту s) timcheronicus отличалась от современной кустарниковой полевки Кавказа крупным ростом, большой длиной зубного ряда (6.3 мм против 5.7 мм для ныне живущего вида) и деталями очертаний нижней челюсти. Возможно также, что она была менее мезофильна, нежели современный вид.
Географическая изменяемость у современной кустарниковой полевки выражена слабо, и описанные в пределах Кавказа подвиды отражают преимущественно субъективизм исследователей или экологические (и
1 Промеры автора по материалам ЗИНа.
2 По измерениям Огнева (1950) и автора (для Дагестана).
414 Часть /Л. Анализ происхождения четвертичных млекопитающих
возрастно-стадиальные) формы зверьков, связанные с условиями обитания и развития в том или ином году. Таковы: ЛГ. majori majori из северо-восточной части Малой Азии, Карского нагорья, Джавахетии, Мингре-лии, центральной и западной части Большого Кавказа; М, majori ciscau-casicus — с северного склона Большого хребта в его западной и центральной части; М. majori suramensis — с центральной части обоих склонов Большого хребта, Су рама и Малого Кавказа; М. majori daghestanicus— из внутреннего Дагестана, Тушетии, северо-западного Азербайджана, северо-восточных склонов Малого Кавказа и Джавахетии; М. majori schelkoimikovi — из лесного Талыша.
Талышские экземпляры отличаются интесивным ржавым оттенком на боках и небольшими размерами. К последнему подвиду должна быть отнесена, вероятно, и М. majori dorothea Ellerm. — с Эльбурсского хребта. Крупные отличия полевки Шелковникова от кустарниковых полевок Малого и Большого Кавказа подтверждают значительную древность изоляции зверька на этом юго-восточном участке ареала. Перекрывающие и заходящие друг за друга ареалы для первых четырех подвидовых категорий говорят сами за себя. О необходимости специальных эколого-морфологических исследований для обоснования географической изменяемости говорят и расплывчатые диагнозы описанных форм.
Обыкновенная полевка — М icrotus arvalis на Русской равнине, как известно, заметно мельчает в направлении с севера на юг. На Кавказском перешейке, с его разнообразным рельефом, при эффекте инверсии зон, Бергмановский тип изменяемости затушевывается. Географические изменения для этого вида должны быть исследованы специально в разрезе высотных и ландшафтных зон, с учетом изменчивости популяций зверьков по сезонам и годам.
Кавказские подвиды дают ряд цифр, указанный в табл. 89.
Таблица 89
Географические изменения размеров в (мм) обыкновенной полевки на Кавказе 4
I )
Промеры Предгорья Северного Кавказа М icrotus arvalis macr ocraninus 162 экз. .. / В нутренние долины центрального Кавказа М icrotus arvalis gudauricus 52 экз. Армянское нагорье М icrotus arval is tran ясаиса-sicus 133 экз. Нагорный Талыш М icrotus arvaiis mystacinus . 4 экз.
Кондилобазальная длина черепа . . . Длина верхнего ряда коренных .... Длина ступни . . . 26.3 23.8—27.3 5.7 25.3 25.2
25.8—27.3 6.0 24.0—26.9 5.9 24.0—26.0 5.5
5.8—6.5 17—18.2 5.3—6.2 15.5—19.6 5.2—6.0 14.1—18.0 5.2—5.7 16.2 15.0—17.0
П р и м е я а н и е. нателе — пределы.
В дробях в числителе указана средняя величина, в знаме-
1 По измерениям Красовского (1930), Огнева (1950) и автора (для Талыша).
/V. Стратиграфическая и географическая изменяемость млекопитающих 415
Юкраска зверьков, обитающих в сухих продольных долинах к востоку от Эльбруса и во внутреннем Дагестане, более светлая, нежели у обитаю-
щих во влажных предгорьях.
Описанный тип географических изменений с учетом характеристики ареала свидетельствует о древнем происхождении зверька на мезофитных Ьднятиях Средиземноморья.
F Средпсплейстоценовая степная полевка Microtus socialis из бинагадин-кого асфальта по строению черепа почти не отличалась от современной Громов, 1952).
По исследованиям Огнева (1950, стр. 400), подвиды этой полевки крупнеют при распространении с севера на юг, увеличиваются также их духовые пузыри и усложняется структура М1 * и М3, но в этом направле-щш необходимы дальнейшие исследования с учетом особенностей географической среды, прежде всего высоты над уровнем моря. При учете огромности ареала эта изменяемость не так уж велика.
Н’? по позволяет считать степную полевку и ее связи с относительно Героф ггными ландшафтами Средиземноморья весьма консервативными древними, по крайней мере нижнеплейстоценовой древности.
тряд ХОБОТНЫЕ — JP R O B €) S С I I) Е А
На юге Русской равнины, на Кавказе и в Передней Азии (как и вообще з ^Средиземноморье) настоящие слоны появляются сразу в качестве высоко-Ойециалпзировапных форм — Elephas (Archidiskodon) planifrons, Е. (Hespe-^nloxodon) antiquus, E. (Archidiskodon) meridionalisс среднего плиоцена (Тго-j uessart, 1898— 1899в; Павлова, 1910a; Богачев, 1923—1924; Osborn, 1942;
Simpson, 1945). Столь же древним следует, по-видимому, считать недавно j описанного Гаруттом (1957а) мелкого слона — Phanagoroloxodon mam-1 обнаруженного в каких-то верхнетретичных слоях западного I Предкавказья.1 Родственные связи этого слона с названными выше недостаточно выяснены, а сам автор описания склбнен сближать его то cE.(J/esperoteodon) antiquus, то с Е. (Parelephas) trogontherii. Несомненно, ошибочно приурочивание этой формы Гаруттом к нижнему плейстоцену.
I Отсутствие остатков прямых предков названных форм в олигоценовых, j Моденовых и нижнеплиоценовых слоях, видимо, не случайно. Напомним I о достоверном отсутствии остатков слонов в низах послепонтических отло-I женин в Предкавказье и в северном Причерноморье, например в аллю-I юнКосякинского карьера у Ставрополя и в глинах промоин понтического | ювестпяка, вскрытых одесскими катакомбами. В обоих захоронениях при 1 jcei их «тафономической универсальности» из хоботных обнаружены I даь остатки мастодонтов (в Косякине и динотериев).
I Эволюция слонов прослеживается у нас преимущественно по находкам I зубов. Именно на последних, в сущности, и основана слабо разработан-* \ гипотеза о существовании прямого генетического ряда: плосколо-,iii <лоп — южный слон — трогоптериевый слон — мамонт. Недоста-i точно ясным бывает обычно систематическое положение остатков, относи-
I 1 Фанагорпйскнй слон описан по сильно минерализованному осевому черепу > «д.гбптыми зубами) неизвестного происхождения, хранящемуся в Краснодарском Фотографии Гарутта (1957а) изображают зубы слонов, найденные в конгломе-... V Сенной и Ахтанизовской на Тамани, и отнесение их к фанагорийскому . слабо обосновано. Нами эти зубы отнесены к южному и древнему слонам (Всре-
; 1957а). Мы не имеем также возможности останавливаться здесь на довольно
| Данных взглядах и рассуждениях Шерстюкова (1954).
416 Часть II. Анализ происхождения четвертичных млекопитающих
.'’'С,*?
мых к древнему слону, которого наши палеонтологи помещали то в группу Loxodonta, то сближали с плоеколобым и южным слонами. Дуброво (1957) отнесла его к роду Palaeoloxodon. Редкость находок остатков древ-* него слона позволяет ограничиться лишь картой их распределения (карта 76). В нашем распоряжении было более 400 эстампов и описаний различных зубов верхнеплиоценовых и четвертичных слонов, собранных на Русской равнине и Кавказе и исследованных нами лично в отечественных музеях и собраниях. Наряду с картами находок слонов на Кавказе (карты 74—76) мы имеем возможность дать здесь лишь самую общую описательную картину различных зубов на отдельных филогенетических этапах и сводную таблицу главнейших морфологических показателей. Зубы и вообще остатки плиоценовых и четвертичных слонов обнаруживаются на Кавказском перешейке, как правило, в переотложенном состоянии. Все они вымыты из первоначальных скелетоносных линз какими-то потоками и перераспределены в толщах конгломератов, галечников и песков. Местонахождения остатков плосколобого и южного слонов приурочены в Предкавказье к зоне дельтовых выносов древних потоков и селей, стекавших с Большого Кавказа к побережьям Акчагыльского и Ашперонского морей. Нередко они залегают и в толщах ожелезненных галечников и песков, высоких (третьих—четвертых) террас рек на предгорных наклонных равнинах. По мнению Богачева (1923—1924), большинство зубов южных слонов на севере Азовского побережья (низовья Дона) также вымыто из среднеплиоценовых слоев и переотложено в гравийных песках верхнеплиоценового.возраста.
В восточном Закавказье эти местонахождения приурочены преимущественно к толщам лёссовых суглинков и галечников, оконтуривающих древние побережья Куринского залива Акчагыльского и Ашперонского морей. На нагорьях Малого Кавказа кости верхнеплиоценовых и нижнечетвертичных слонов обнаруживаются в речных и озерно-речных наносах, например под Ленинаканом и Эрзурумом, а в Цалке даже в конгломератах между слоями долеритовых лав, датируемых гюнцем и гюнц-мин-делем (см. также Бурчак-Абрамович, 1951а).
Зубы плосколобого 1 и южного слонов были относительно небольшой величины, низкие, с короткими и слабыми корнями. Их эмалевые карманы закладывались в виде грубо рифленых, гребневидных пластинок из сросшихся продольно эмалевых цилиндриков. Обрастая дентином и цементом, эти редко посаженные карманы образовывали как бы эмалевую гармонь, а при стирании — сплющенные поперечно зубу и выступающие на 5—6 мм (у взрослых экземпляров) петли толстостенной эмали (рис. 173). Рабочая, стиравшаяся поверхность зуба представляла собой крайне грубую терку — жернов для раздавливания и растирания объемистой растительной пищи. Донные участки карманов эмалевой гармонии, обращенные к пульпарной полости, образовывали ровный закругленный валик с слабо выраженными бугорками — сосочками. Цемент зуба этих ранних слонов обычно метаморфизирован и ослаблен настолько, что сами корни, как правило, бывают разрушены, а остатки пульпарной полости всегда забиты грунтом.2 На Русской равнине остатки верхнеплиоцеповых слонов встречаются преимущественно в южных районах и к северу от широты Москвы и Казани, как правило, не обнаружены.
1 Недавно Гарутт (19576) высказал еще малообоснованное предположение, что плоско л обого слона в пределах СССР не было вовсе, а остатки, отнесенные к этому виду, принадлежат ранней форме южного слона.
2 Цемент и дентин этих зубов напоминают обычно алебастр или мергель.
Гл /V- Стратиграфическая и географическая изменяемость млекопитающих 417
По типу питания плосколобые и южные слоны были, по-видимому, еще довольно близки к мастодонтам. Штрихи и царапины па эмалевых грезах зубов этих слонов весьма напоминают царапины на эмали зубов шастсдоитов. У последних они имеют, однако, преимущественно поперечно ’ правление к челюсти, а у слонов — продольное, что обусловлено, Квдпо, лишь различным типом жевания. Эти царапины были, несомненно, рдпы песчинками и камешками, попадавшими на зубы прп пожирании Ьре:! п корневищ древесных и травянистых растении.
Сезонной жировкой слонов в зарослях болотных растений по окраи-Врм водоемов и в болотах речных долин, несомненно, и объясняются I относительно частые захоронения костей этих толстокожих в гидро-Кр|ых фациях.
[ft 173. Распил AI5 южного слона с севера Азовского побережья по вертикали и горизонтали.
Кры (тонки эмалевых карманов п особенности образовании петли (а — петля, б — карман).
Характер смены и число сменявшихся зубов у верхиеплпоценовых cicioi еще недостаточно изучены на сериях черепов и челюстей. Обычно Ьедполагают, что в каждой челюсти стиралось и сменялось по 5 корен-I <ы\. поочередно выдвигавшихся сзади, и сверху из каждой челюсти, Ьдобио тому, как это происходит у современных индийского и африканского слонов. Последний—6-й — зуб, наиболее крупный и толстоэмальный, отличался отсутствием задней отжимной площадки и функционировал на последнем жизненном этапе зверя. Этот тип смены, сохранившийся, оче-|мдио, и у мамонта, учитывается и нами при дальнейших обозначениях и ^осуждениях.1
К нижнему антропогену уже произошло значительное увеличение ь уела эмалевых карманов, их глубины, а следовательно, и высоты зуба, тюнчепие эмалевой стенки и уменьшение ее извитости — плиссирован-।jocTii. Зубы, особенно нижние, становились относительно уже, длиннее, [побьем и вес увеличивались и относительно, и абсолютно. Этот этап морфогенеза слопов отмечен серией захоронений па более низких (вторых-третьпх) террасах речных долин и известен под именем слона Бюста и тро-гонтерпевого слона — Elephas (Parelephas) wiisti, Е. (Parelephas) trogontherii.2
1 На фрагменте осевого черепа южного слона из Хапров, хранящемся в Институте [ 1ивгическпх наук ЛИ СССР, видно, что замена второго верхнего зуба третьим про-Е жюдила на месте — сверху, как у мастодонтов, а не сверху и сзади, как у мамонтов и I (овремепннх слонов.
2 Последнее наименование мало соответствует действительности, так как । расцвет численности и ареала бобра трогоптерпя был, по-видимому, раньше — в верхнем плиоцене.
27 Н. К. Верещагин
418
Часть Л. Анализ происхождения четвертичных млекопитающих
Местонахождения костей трогонтерневых слонов
наиболее
известны
в галечниках третьей террасы левобережья Днестра у Тирасполя в долине Аракса, у Еревана и Ленинакана (карта 75).
В среднем плейстоцене начался последний,
з ак л ючите л ьпый
существования филогенетического ствола слопов.
Это
этап
совпало с бурным
расцветом численности человеческих орд быстро совершенствовали способы добычи
среднего палеолита,
которые
и слонов
Трогонтериевый слои к этому
крупного зверя,
том
числе
времени
уже испытал
дальнейшее
измельчание и превратился в мамонта — Е. (Mammonteus) primigenius. Облик зверя в целом стал уже совершенно иным: появился обплышн
шерстный покров,1 удлинились бивни и увеличилась их кривизна, про» пзошло упрочнение запястных и заплюсневых суставов. В зубах продол-
жалось сближение эмалевых петель, увеличение их числа на единицу длины зуба, утончалась эмаль, уменьшалась морщинистость ее поверхностей (рис. 174). Днища эмалевых карманов приобрели прпострешшй
край с сидящими по нему остроконечными сосочками, выдающимися в нудь-парную полость. Корни зубов развивались в длинные, изогнутые и прямые полые трубкп. На срезах довольно тонкой эмали трогонтерневых слонов
в
и мамонтов уже редко удается подметить следы раздавливания дресвы, Часто насаженные эмалевые пластины делали жернов менее грубым п, быть может, увеличивали его прочность. Раздавливание и перетирание веточного корма, в достаточной степени грубого, несомненно, продолжало иметь место, по, очевидно, большую роль начало иметь и питание травой, что достоверно известно по вскрытым желудкам замерзших трупов мамоптов Сибири.
Остатки настоящих мамонтов на перешейке более обычны в Предкавказье, чем в Закавказье. Опп залегают как правило, в молодых (первых-вторых) террасах рек. Тип сохранности эмали, дептипа и цемента зубов мамонтов всегда резко отличается от таковых у зубов более ранних слонов. Корневые раструбы с пульпарной нблостыо у зубов мамонтов не
редко сохраняются целиком.
Наиболее ранние остатки слонов мамоптового типа обнаружены на Кавказе, в карьере Гирей — вторая терраса Кубани. Хорошо датированы лишь находки на Ильской стоянке в Закубапье. Отсюда имеются 4 обломка коренных, мпого обломков бивней и до 40 обломков трубчатых костей, костей заплюсны, запястья.
Стратиграфически наиболее поздняя находка — это фрагмент верхней левой челюсти с сильно изношенным 6-м зубом из травертинов северного склона Машука. Беловатая эмаль, светло-коричневый цемент, светло-коричневая в свежем разрезе челюстная кость, дающая при доскабливании характерный запах сырой кости, совершенно необычны для столь южных захоронений. Малые размеры зуба, тонкость эмали (1.0-1.2 мм) при малом диаметре футляра бивня позволяют отнести этот фрагмент к какому-то карликовому и очень позднему мамонту эпохи позднего палеолита или даже неолита.
В Закавказье зубы мамонтов обнаружены пока лишь в Горпиской котловине (Бурчак-Абрамович, 1946; Габуния, 1952а). Зуб, описанный Габунией (1952а), найден в первой надпойменной террасе р. Куры в г. Гори. Это — пестертый М4 нашего счета. Основываясь па мелких размерах зуба п 12 пластинах на 10 см, толщине эмали в 1.5 мм, Габуния сближал свою находку с «карликовыми» мамонтами западного Средиземноморья,
^Сведений о шерстном покрове трогонтерневого слона не имеется.
IV. Стратиграфическая
и
географическая изменяемость млекопитающих
419
ПО
условиям
залегания
датировал началом послевюрмского
времени — от
шпала
мезолита
до неолита.
гавшей
оснований
В отличие от Громовой (1948), предпола-
существование
каких-то «мамонтов в Закавказье
дорнсскую эпоху», и Бурчак-Абрамовича (1946), считавшего мамонта Закавказье северным тундровым представителем, Габуния правильно
1
Рис. 174. Продольные распилы:
1 — Мб; 2 — М6 мамонтов с Русской равнины.
а
подчеркивал вероятность формирования на юге Восточной Европы и в Закавказье мелких местных форм мамонта.
Если северная форма мамонта и была «продвинута» в эпоху наибольшего похолодания на юг до Кавказа, то вряд ли опа распространялась далее Предкавказья. Путь вселения для северных мамонтов в верхнем плейстоцене па западе через лес п кручи скал был неудобен, а па востоке ря mix было слишком сухо и жарко.
27*
420
Часть II. Анализ происхождения четвертичных млекопитающих
Закавказье, по-видимоМу, являлось южной границей распространения мамонтов. Подтверждением этому служит отсутствие достоверных находок остатков мамонтов на Армянском нагорье, в Иране и вообще в Передней Азии. Ксероморфная обстановка при слабо развитой древесной растительности, которая сложилась (по крайней мере в низких долинах этих районов) к среднему плейстоцену, была мало пригодна для слонов позднего и северного типов.
На север от предкавказских равнин остатки мамонтов — наиболее частые палеонтологические находки в долинах Дона, Волги, Урала, и по мере движения на север численность их по сравнению с остатками более древних слонов непрерывно увеличивается. По учету, проведенному нами в 1950—4955 гг., эта численность представлена цифрами, указанными в табл. 90.
Таблица .90
Численность остатков зубов различных ископаемых слонов в краеведческих музеях
М узеи Мамонты Трогонтериевые слоны Южные слоны
абс. % абс. % абс. %
Архангельский 12* 100.0 — - — —
Великого Устюга 33* 100.0 — —— —
Вологодский 23* 100.0 — — -— —
Казанский университет 59 1 1.6 -— —
Уральский 10 'ЗЗ.З7 2 16.7 -— -—
Куйбышевский 10 77.0 3 23.0 — —
Саратовский 21 81.0 5 19.0 — —
Астраханский 51* 43.5 71* 56.5 — —
Новочеркасский 3 18.7 4 25.0 9 56.3
Темрюкский и сборы с Тамани в Пине 3 7.3 8 19.5 30 73.2
Краснодарский 7 33.3 И 52.4 3 14.3
Ставропольский 3 25.0 2 16.7 7 58.3
Пятигорский 4 44.5 2 22.2 3 33.3
Махачкалинский 1 33.3 — -— 2 66.7
Примечание. Звездочкой (*) отмечены случаи учета не только зубов, но и других костей.
Такое распределение указывает не только на уменьшение условий захоронения для мамонтов на юге и не только на увеличение эрозии и вскрытие здесь более древних слоев с остатками южных слонов. Главная причина этого распространения состоит, конечно, в том, что высокая численность мамонтов и основной ареал зверя был на севере, а у трогоитерие-вых и южных слонов — на юге.
Описанные выше морфологические преобразования зубов в предполагаемом генетическом ряду могут быть прослежены на фотографиях их продольных распилов (рис. 173, 174), а также на эстампах и контурах (рис. 175). Практическое значение для стратиграфических сопоставлений может иметь также таблица (91) морфометрических показателей зубов по некоторым «ключевым» захоронениям Кавказа и Русской равнины.1 *
1 Эта таблица дается с учетом того, что прямое родство верхнеилиоценовщ
И четвертичных форм слонов не всегда доказано.
За
Рис. 175. Эстампы стиравшейся поверхности (петель эмали) и боковые контуры зубов
Mg ископаемых слонов Русской равнины и Кавказа (М6— а и Мв— б).
tlepkas (Mammontens) primigenius: la, 16— долина Сухоны, № 10401, № 10870, Музей Тотьмы;
а -торфяник долины Дона, без номера, Музей Воронежа; За — Кирилловская верхнепалео’ литическая стоянка под Киевом, № 302, Инет. зоол. АН УССР;
Рис. 175 (продолжение).
46 — среднепалеолитическая стоянка Чокурча в Крыму, без номера, музей Симферополя; ба — вторая терраса Кубани, карьер Гирей у Кавказской, без номера, Музей Армавира; 6а — долина Терека близ Наурской, без номера, Музей Грузии. 7а, 7б — хазарский ярус долины Волги, Черный Яр, без номера, ПИН. («Поздний» трогонтериевый слон или «ранний» мамонт).
Юа
Рис. 175 (продолжение).
Ftnhas'fParelephas) trogontherii: 8а, 86 — третья терраса Днестра у Тирасполя, без номера, Москов-«йй геолого-разведочный институт. Elephas (Archidiskodon) meiidionalis: 9a—долина Аранса, JL».» Казачий пост, без номера, Геологический институт АН АрмССР; 10at Юб — конгло-JW“wnaB» 1249
мепаты Таманского полуострова, №, ПИН (Фанагорийский (!?) слон).
Рис. 175 (продолжение).
116 — конгломераты Таманского полуострова, № ’232'» ПИН (Фанагорийский (!?) слон);
126 — Обиточное, Украина, № 24239, ЗИН; 13а — Ставрополь, № 25284, ЗИН.
1 а блиц a 91
Изменения основных морфометрических показателей зубов (М| — Mg) слонов из антропогеновых и плиоценовых слоев Кавказа и Русской равнины 1
Эпохи и ярусы Местонахождения (захоронения) Показатели м1 2 м2 Мз М4 Мь ЛТ6 м6
Мамонты
Число исследованных зубов 2 1 2 18
Низкие террасы 1 3 8 22
Северной Двины и Сухоны, Н аиболынее число эмалевых карманов . , 12 — 17 25
музеи: Волог- 12 18 20 29
ды, Тотьмы, . Великого Ус- Число эмалевых карманов на 10 см . . 12 9 9.3 (8.5—10) 8.8(7—12)
га >s тюга, Соль-Вычегодска, 12 8.7 (8—10) 8.9(7.5-10) 8.7(6.2-12)
„ к Я о Ф № га Й о из Г_| М Архангельска. Толщина эмалевых стенок (в мм) 1.0 1.0 1.4 (1.1—1.6) 1.4 (1.0—1.8)
1.0 1.0—1.2 1.6 (1.0-2.0) 1.5 (1.0—2.5)
О Сб га § * Число исследованных зубов . . . Ч /Ч . —— 2 3 4 5
й 1 — 1 3 1 2 -—
я Наибольшее число эма-
Й и ф Верховья Десны, стоянка Ели- левых карманов . . — W 12 14 15 тг "17 23
га сеевичи (верхний палеолит). Число эмалевых карманов на 10 см. . , —— 11.5(11-12) 9.3 (8—11) 8.1 (7—10) 8.2 (7-9)
14 11.3(10—12) 9.5 8.8 (8-9.5) —
Толщина эмалевых стенок (в мм) — 1—1.1 1.5 (1.0—1.9) 1.9 (1.6-2.2) 1.7 (1.2—2.0)
- 0.9 1.3 (“1.1—1.5) 1.8 2.1 (1.9—2.2) —-
1 Неопубликованные материалы автора. Промеры и эстампирование зубов производились на местах раскопов (Мезип) и в музеях РСФСР, Белоруссии, Украины, Крыма и Кавказа. Половая изменчивость не изучена.
Эпохи и ярусы М естонахож дения (захоронения) Показатели м! к,2 м2
хвалынский ярус Средняя Десна,1 стоянка Мезин (верхний палеолит). < Число исследованных зубов Наибольшее число эмалевых карманов . . Число эмалевых карманов на 10 см . . s| । о] । Ч । Т "15 12
Толщина эмалевых стенок (в мм) 0.8
Плейстоцен верхний \ Средний Днепр, Кирилловская стоянка под Киевом (верхний палеолит). Число исследованных зубов Наибольшее число эмалевых карманов . . Число эмалевых карманов на 10 см . . . — -
ярус Толщина эмалевых стенок (в мм) —— —
хазарский Северные предгорья Крыма, пещера Чокур-ча (средний палео лит—мустье) . Число исследованных зубов Наибольшее число эмалевых карманов . . — —
1 Промеры толщины эмали сделаны по эстампам. Подсчет ч как зубы сидели в челюстях.
Таблица 91 (продолжениеj
Мз 4 М4 м5 „6 м6
3 2 4 77 1
1 о 6 - 3
"ПТ — — —
И (10—12) 9(8-10) 9.5(9-10) 8.5
10 10.5 (9—1.2) 9.3(8.5—10) 8.3(8—8.5)
1.2 (1.0—1.4) 1.5—1.7 • 1.6 (1.5—1.7) 1.6
1.1 1,4—1 Л 1.6(1.4—1.8) 1.9(1.8—2.0)
— 3
Т ТЕ 4
— 0 22
"13 s тг .... 22
— — — 8.9 (8—10)
1 11 110-12) 8.9 (7—1.2) 8.5 (8-9) 8.5 (8-9)
— — — 1.9 (1.8—1.9)
1.2 (1.0—1.4) 1.7 (1.2—2.0) 1.9-2.0 2.0 (1.8-2.3)
1 1 4 1 —
т 1 2 Т
18 20 " 1
1 "14 18 1 20 "25
пела эмалевых карманов («петель»)
Мезине был
затруднен, так
ЮПП пип
Таблица 91 (продолжение
1 Промеры тилщнпи* ----
как зубы сидели в челюстях-
Плейстоцен |5 : | нижний —бакинский ярус средний — хазарский ярус il; | . । .;v-1 • и ” ( , , . . : гния) Северные пред- ( горья Крыма, 1 пещера Чокур- < ча (средний палеолит—мустье). Галечники и суглинки низких террас рек Предкавказья и Закавказья: < Ильская, Гирей, Лабинская, Душети и др. Галечники в цоколе высоких террас Дне-стра, Дона, Тирасполь, Ростов (ископаемые долины прарек). it,. । . 11TCJ1 И L Число эмалевых карманов на 10 см . . . Толщина эмалевых стенок (в мм) Число исследованных зубов Наибольшее число эмалевых карманов . . Число эмалевых карманов на 10 см . . Толщина эмалевых стенок (в мм) Число исследованных зубов Наибольшее число эмалевых карманов . . Число ^ма^евых карманов на 10 см . . . Толщина эмалевых стенок (в мм) Тр< ма эгонте^ 10-11 9.8(8.5—11) 10 1.5 (1.3—1.9) 1.4 1 2 9 Я О 11.5 7-8 1.6
1.2 (0.8—1.5) 2 13 10—10 2.1 (1.9—2.2) 5 3 “2Г 8.4(7-10) 7.2 (6.5-8) 1.6(1.5—1.71 1.6 (1.4—1.9) 3 4 тг 6.5 (6-7)
1.3(1.!—1.5) ?иевые слоны 8—10 1.4 2.0-2.1
6.7 (6-8) 2.4 (1.8—3.3) 1.9 (1.8—2.3)
м о
8.0 (7-9.5)
2.0(1.8-2.1;
7
5
24
7.7 (6-9.5)
7.5(7.0—8.б[
1.9 (1.2—2.2)
1.8 (1.5—2.2)
2
•8
18
23
5-6.5
6.8 (5.5-8)
2.9 (2.3—3.4)
2.4 (1.9—3.3)
Та б лица 91 f продолжение)
Эпохи и ярусы Местонахождения (захоронения) Показатели Mi Mg * Мз М4 м| м|
й ярус Галечники в цо- коле высоких Число исследованных зубов ....... — — — Т 4 2 4 5
К к к ф о tr к С S ЕН И £ Я ’S ю ф террас рек Предкавказья: Гирей, Лабинская, Преград- ’ ная, Пяти- Наибольшее число эмалевых карманов . . Число эмалевых карманов на 10 см . . Толщина эмалевых стенок (в мм) — — — — 6.1 (5—8) 26 7.0 (6-8)
Пл нижний — горек, Ставрополь (ископаемые долины прарек). — — — 5.5 T9 6.4 (6.2—6.5) 2.3 (1.7—3.2) 2.4 (2.2—2.5) 7.0 (5.5—8) 2.8 (2.5—3.0) 2.2 (1.4—2.8)
Южные слоны
Q Число исследованных зубов — — . — 1 1 J. 7~
г*^ Он и 5S S о Пески и галечники долины < среднего Аракса. Ленинакан Наибольшее число эмалевых карманов . . Число эмалевых карманов на 10 см . . -— — — — 6.5 25 6.2
ф 2 « S4 (Казачий пост). т 6.4 (5.3—8)
о g к а а « 1 Толщина эмалевых стенок (в мм) 2.8 2.3
к ® 1 1 X 2.1 3.0 (2.7—3.6)
верхний Конгломераты Таманского полуострова: Сенная, Цимбал. 1 Число исследованных зубов Наибольшее число эмалевых карманов . . j j — । “9“ ®| I - 4 5 — 2 3 12 11
Эпохи и ярусы
Местонахождения (захоронения)
Показатели
Конгломераты Таманского полуострова: Сенная, Цимбал.
Плиоцен верхний — акчагыльский ярус
Пески и галечники прарек Предкавказья: Псекупс, Гирей, Воскре- < сенская, Георгиевск, Малго-бек, Грозный и др.
Пески и галечники прарек севера Азовского побережья: Ле-венцовка, Ак- < сай, Хапры, Морская, Таганрог, Мариуполь и др.
Число эмалевых карманов на 10 см . .
Толщина эмалевых стенок (в мм).........
Число исследованных зубов .............
Наибольшее число эмалевых карманов . .
Число эмалевых карманов на 10 см . ,
Толщина эмалевых сте нок (в мм).........
Число исследованных зубов .............
Наибольшее число эмалевых карманов . . Число эмалевых карманов на 10 см . .
Толщина эмалевых стенок (в мм).........
«
1 1
j м3 M4 Ms
- .. _ ’ 7 5.4 (5—5.5) 5.7(5—6.3) 2.8 (2.1—3.8) 5.7(4.8-6.5) . ; < ) 4.7 (4.5—5.0) ; / 2.7—2.9 '•.
8-9 8-8 1.8
gl I ч । ч । н । । । । । s GO '-— 1.6 (1.1—2.0) ~ T 2.9 (2.8—3.5) 2 8 IT 5—6 3.5 (2.9—4.0) 3 ’ 4 12 13 4.8 (4.5-5) 5.0 (4.5—5.5) 3.1 (2.3—3.5) 3.7 (3.0—4.5) 2 “4 16 15 4.8 (4.5-5) 4.6 (4.0—5.0) 3.5(3—3.9) 3.8 (3.5—4.1) J
- 'f-cc cd । 1 । il il 1 1 1 lL° e 1 1H 1 p° 1И 1 ко rn £S 1 ' 1 1 lQ OJ cd 5.3 (5-6) 3.5 (2.8—4.1) 3.2 (2.7—3.9) ТГ 4.4 (4.2—4.5) 3.0 (2.8—3.3)
430
Часть IJ, Анализ происхождения четвертичных млекопитающих
Если учитывать только изменения зубов, то с эпохи появления слонов в Черноморско-Каспийской области до момента их вымирания в раннем
голоцене наибольшее число эмалевых петель у М| увеличилось с до — .
1 о 19
а число петель, приходящихся на 10 см стертой или боковой поверхности
4 1 2 05
увеличилось на Толщина стенок эмали уменьшилась на мм.
4.2 2.50
Описанные столь схематично якобы скачкообразные изменения в строении зубов четвертичных слонов могут объясняться скорее с геологических (тафономических) позиций, нежели с биологических. Известные нам как бы обособленные этапы морфогенеза слонов могут соответствовать всего лишь этапам перестроек гидрографической сети и циклам осадконакоплений, обусловивших возникновение костных захоронений. Это касается, впрочем^ и других зверей, а не только слонов. Инертность эволюции форм, а она почти всегда имела место, приводила к запаздыванию адаптивных изменений при быстро менявшихся факторах среды, от которых зависело, конечно, и осадконакопление. Наоборот, мы склонны рассматривать описанные изменения как непрерывное (но не обязательно скачкообразное) накопление мелких признаков, оборванное эволюцион
ным тупиком и вымиранием.
К сожалению, у нас нет пока никаких основательных данных для утверждений о том, что развитие мамонтовых признаков происходило у слонов вполне одновременно как на Кавказе, так и на Русской равнине, но дальнейшие сопоставления исследований палеонтологов и геологов, несомненно, раскроют истину.
Отряд НЕПАРНОКОПЫТНЫЕ — Р EEI S 8 ODACTY Е А
Филогенетические ряды в этом отряде разработаны слабо. До настоящего времени среди палеонтологов нет установившегося мнения о том, что носороги верхнего плиоцена и антропогена — Rhinoceros etruscus, Rh, mercki, Rh. tichorhinus — представляют единый филогенетический ряд форм. Крупные морфологические отличия этих стратиграфически обособленных видов все же, по-видимому, сглаживались на гранях эпох. Черепа еще плохо изученного носорога Rh. binagadensis из среднего плейстоцена Апшеронского полуострова (Бинагады) отличались от черепов верхнеплейстоценового волосатого носорога сильным прогибом крыши в области височного сужения, слабым развитием второго рога и большой вздутостью мозоли для первого рога, укороченной заглазнич-ной областью черепа, тонкими сосцевидными отростками и небольшими (сферическими) затылочными мыщелками. Зубы этого носорога были значительно крупнее (шире и длиннее), чем у волосатого, и аборальная грань последнего моляра выдвигалась далеко за передний край орбиты (рис. 176, 2). Предглазничные отверстия были весьма малы. Судя по положению сравнительно узкого затылка, голова этого зверя держалась горизонтальнее, чем у волосатого, и, по-видимому, бинагадинский носорог был более веткойдным и менее травоядным, чем волосатый. Большая стройность метаподий, предплечья и голени бинагадинского носорога свидетельствует также, что он был более легкого и подвижного сложения, чем волосатый. Обилие этого зверя в явно ксерофитном ландшафте древнего Апшерона, весьма отличном от ландшафтов, населенных волосатым носорогом, подтверждает его видовую или даже родовую обособлен-ндсть (см. Джафаров, 1955). Все сказанное отнюдь не исключает возмож-
м в
Ч(
К( ш бо (Б
но rei ЛО1 ИЛ] зат
Гл. /V. Стратиграфическая и географическая изменяемость млекопитающих 431
ностп проникновения волосатого носорога с севера в восточное Закавказье п далее в Переднюю Азию позднее, в верхнем плейстоцене. Это тем более вероятно, что смена форм, а именно: проникновение крупных зверей с севера в позднем плейстоцене — прослежена на Апшеронском полу-। траве, например, у первобытных туров.
Фрагментарность находок остатков волосатых и более древних носо-югов на Кавказском перешейке позволяет нам ограничиться лишь де-
Рис. 176.
Черепа носорогов: 1 — Rhinoceros tichorhinus, верхний плейстоцен, Русская равнина, Владимир, № 10699, ЗИН; 2 — Rh. binagadensis, средний плейстоцен, восточное Закавказье, Бинагады, № 24402, ЗИН.
г працпеи карты находок их остатков (см. карту 77), не вдаваясь пока , дталн генезиса форм и стратиграфической изменяемости морфологи-Lkux особенностей в возможных филогенетических рядах.
У эласмотериев {Elasmotherium caucasicum — Е. jiScTieri), эволюция Ьгорых известна нам лишь па их последнем жизненном этапе, с верхнего Ьзцсна до среднего плейстоцена, произошло заметное измельчание зу-Ев прп развитии большей складчатости (извитости) эмалевой пластинки i:> шсяк, 1914).
Столь же сложен, как и для носорогов, вопрос о прямой или косвен-Lj преемственности морфологических признаков у лошадей аитропо-L Кавказа и Русской равнины. Огромные миграционные способности единых прп пх почтп универсальной приспособленности к временному Eji постоянному обитанию во всевозможных типах ландшафтов крайне Ьгдияют удовлетворительное решение происхождения восточноевро
• 432
Часть II. Анализ происхождения четвертичных млекопитающих
пейских и кавказских лошадей четвертичного периода. Так, например, до сих пор неясно, является ли русский тарпан трансформированным потомком местной верхнеплейстоценовой лошади (широкопалой лошади верхнего палеолита Придонья) или прикочевал относительно недавно — в голоцене — с юга. Несомненно все же, что на Кавказском перешейке и на Русской равнине образовывались местные ландшафтные формы лошадей, сменявшие друг друга па различных геологических этапах. Генеральное направление эволюции лошадей проявлялось в развитии крайней гппсодоптнп и однополости, по в различных районах по-своему.
Учитывая все это, пока можно говорить лишь о стратиграфических и географических изменениях послетретичных лошадей пашей страны вообще, не связывая их с формированием какого-то определенного вида. Судя по кавказским и восточноевропейским материалам, эволюция лошадей представляется здесь в следующем виде. Верхнеплиоценовые лошади типа Equus stenonis из Приазовья были крупны, относительно тяжеловесны, с крупными зубами, отличающимися сравнительно тонкой и сильно пзвптой эмалевой пластинкой. Метаподпи их пе несли еще пи черт крайней специализации к быстрому бегу в условиях полупустынного и степного ландшафтов, пи черт шпрокопалости, необходимой для передвижения по луговым и торфяным болотам тайги и тундры.
Лошади самого верхнего плиоцена — нижнего плейстоцена типа Е. sussenbornensis (из конгломератов Таманского полуострова) имели зубы уже вполне «кабаллоидпые»; метаподпи их были более стройны характеризуя эту форму как явно скоростное животное, обитавшее в ландшафте саванны или выгоравшей летом степи. По размерам опп превосходили лошадей четвертичного периода (Верещагин, 1957а).
В среднем плейстоцене в восточном Закавказье обитала менее крупная лошадь, с более уплощенными в передне-заднем направлении диафизами метаподий. Индексы выступания валика нижнего эпифиза плюсны п пясти как показатели костно-хрящевого типа крепления путовых суставов у бинагадинской лошади были уже несколько меньше, нежели у лошадей верхнего плиоцена. Зато, очевидно, усилилось крепление боковых связок. Копытные фаланги этой лошади были относительно невелики, с значительным углом подъема передней стенки, что и обусловливалось ксерофитпым ландшафтом и плотными грунтами того района. Верхпе-плейстоценовые и голоценовые лошади Кавказа по ископаемым материалам известны нам плохо, но несомненно, что в Закавказье обитали лошади южного типа — сухой конституции (см. главу III). Лошадь верхнего плейстоцена Русской равнины характеризовалась необычайной массивностью ног, шпрокопалостыо. Особенно хорошо широкопалость прослеживается на костяках пз верхнепалеолитических слоев Костенок па Дону.
Отличия в величине опорной площади от таковой у лошади ксерофит-ного восточного Закавказья здесь настолько велик и, что сомнений в приспособлениях к иному типу грунта и ландшафта пе вызывают (рис. 177). Лошадям верхнего палеолита Придонья приходилось, очевидно, пастись и вообще передвигаться в условиях зыбких лугов, тундровых и таежпо-моховых болот в поймах рек и на водоразделах. Сходный тип строения копытной фаланги, но все же пе столь широкой, имелся и у лошадей крайнего северо-востока Сибири, обитавших, очевидно, па несколько более плотных грунтах и при менее глубоких снегах.1
1 А. А. Слудский (устное сообщение) связывает, впрочем, птирококопытность передних ног северных и верхнеплейстоцеиовых копытных с потребностями в тебеневке т. е. в разгребании глубокого снега для добычи корма.
Рис. 177. Стратиграфические и географические изменения формы и опорной площади копытных фаланг лошадей Евразии. Контуры фаланг пе-1
родней (а) и задней (б) пог. -ту нат. вел.
1(1 16— Equus caballus gmelini, современный «тарпан», Русская равнина, № 521, ЗИН; 2а, 26 — Е. przewalskii, современная лошадь Пржевальского, Джунгария, № 17591, ЗИН; За, 36 — Е. caballus subsp. (foss.), ранний голоцен, Новосибирские острова, № 4419, ЗИН; 4а, 46 — Е. caballus talipes, верхний плейстоцен (верхний палеолит), Воронеж, Костенки XV, № 25183, ЗИН; 5а, 56 — Е. caballus subsp., средний плейстоцен, восточное Закавказье, Бинагады.
И. К. Верещагин
434
Часть II. Анализ происхождения четвертичных млекопитающих
Размеры площади передних и задних копытных фаланг, различных лошадей (и отношение их друг к другу) даны в табл. 92. Оказывается, что развитие относительно большой площади передней фаланги имело место у степных и южных лошадей.
Таблица 92
Стратиграфические и географические изменения величины опорной площади (в мм2) копытных фаланг лошадей в аптропогене Азии
Промеры Средний плейстоцен Восточное Закавказье, Бинагады Equus caballus subsp. 4 экз. Верхний плейстоцен Верховья Дона, Ко-стенки Equus caballus latipes 4 экз. Верхний плейстоцен Новосибирские острова Equus caballus subsp. 8 энз. Украина Equus caballus gmelini Ant. № 521, ЗИН Джунгария Equus przewalskii Pol. № 17591. ЗИН
1. Передняя нога . . 28.8 ! 43 40 34 37.5
27—32 39—50 27—48
2. Задняя нога . . . 25.5 40.9 37 31.5 33.5
37—46 33.5—40.5
3. Индекс разницы (2 : 1 X 100) . . 88.5 95.0 92.5 92.6 89.3
Примечание. В дробях в числителе указана средняя величина, в знаменателе — пределы.
*
В эпоху наибольшего похолодания и расширения мезофитных ландшафтов широкопалая лошадь распространялась далеко к югу, по-видимому до предгорий Кавказа. Есть основания думать, что ксеротермическая эпоха и широкое остепнение Русской равнины обусловили дальнейшую дивергенцию местной верхнепленстоценовой лошади в направлении развития признаков лесного и степного тарпанов.
Подтверждение такой морфологической эволюции затруднено отсутствием достоверных скелетных материалов от этих столь недавно вымерших лошадей (см. Громова, 1949; Гептнер, 1955).
Отряд ПАРНОИОПЫ Т II ЬГ Е — Л К Т I О II А С Т Г i А
Значительная разработанность филогенетических рядов в этом отряде позволяет довольно хорошо проследить главнейшие направления морфологической эволюции некоторых кавказских представителей.
Свиньи группы Sus scro/a—S. vittatus появляются в пределах Восточной Европы и Кавказа впервые в верхпеплиоценовых слоях как вполне сформировавшиеся в основных морфологических чертах животные.
Еще Амоном (Amon, 1938) была отмечена чрезвычайная изменяемость черепных признаков плиоцепо-плейстоценовых свиней, обитавших на огромных пространствах Евразии. Ареал этой группы видов был настолько обширен уже в нижнем плейстоцене, что имеются основания считать позднейшие стратиграфические находки в наших пределах за реально существовавший преемственный генетический ряд форм.
Таманский кабан — 5. tamanensis из верхнеплпоценовых слоев Таманского полуострова отличался от более поздних кавказских крупной
Тл. /Г. Стратиграфическая и географическая изменяемость млекопитающих 435
елпчиной п необычайной шириной нижней челюсти в области диастемы \ взрослых самцов (Верещагин, 1957а). Кабан из среднеплейстоцеиовых слоев Апшеронского полуострова — S. apscheronicus, не отличаясь от со-виеменпых по величине, обладал сильно оттянутыми вниз слуховыми кап-• амп и слезными костями промежуточного типа между характерными 1й5. scrofa и 5. vittatus (Бурчак-Абрамович и Джафаров, 1948). Промеры Ьрины нижней челюсти сзади клыков у четвертичных кабанов дали я цифр, который мы приводим в табл. 93.
Таблица 93
Изменения величины кабанов по промеру нижней челюсти (в мм)
тонахождения и геологический возраст Виды Ширина шикнем челюсти сзади клыков Число исследованных экземпляров
Кв*каз, современность. Sus scrofa attila 46.0 5
45—55
•г. ни I Дон, X—XIII вв. п. э. S. scrofa attila 59.0 з
55—64
Ц»..1жье, полуостров Тунгус, Кципй плейстоцен. S. cf. scrofa 65.0 1
Звдвказье, Апшеропский нолу-тров, средний плейстоцен. S. apscheronicus 52.0 2
50—54
Ц^дкавьазье, Таманский полу-тров, верхний плиоцен. S. tamanensis 73.0 1
Примечание. В дробях в числителе указана средняя величина, в знаменателе — пре-дач.
Рядншкппх челюстей и черепов кабанов аптропогена показан на рис. I 17S. Наряду со значительными изменениями пропорций отдельных костей левого и висцерального черепа зубы кабанов среднего плейстоцена по выпчине изменились мало. При одинаковой величине у бинагадин-I скпх п современных кавказских кабанов заметно все же, что пяточная кть этого зуба усложнилась к современности за счет развития одного-двух столбиков бугорков пятки (см. рис. 178, I, 4).
Резкое, скачкообразное сокращение размеров зубов, особенно замет-Ijtt на М|, произошло только в неолите у одомашненных популяций.
Достоверно определенные остатки оленей группы благородного — Сег-I-iiselaphus s. lato появляются впервые на территории СССР в слоях амого нижнего аптропогена.1 Олени эти, очевидно, трансформировались из каких-то плиоценовых средиземноморских форм. Павлова описала череп и крайне простые, не ветвящиеся рога оленя — (m'ds tschelekensis пз акчагыльских — верхиеилиоцеиовых — слоев За-йасппя. В иижнеплейстоценовых конгломератах Таманского нолуост-рова встречаются одиночные обломки рогов плиоценовых оленей с пе-I «только отставленным от венчика первым отростком. Их принадлеж-
11 Указания на более ранние находки, например в среднем плиоцене Закавказья | Вогачск, 1938в), недостоверны и относятся, вероятно, к оленям рода Eucladocerus.
436
Часть II. Анализ происхождения четвертичных млекопитающих
ность к группе благородного оленя не выяснена (Верещагин, 1957а), но весьма возможно, что здесь были и предковые формы. По размерам эти олени были еще относительно невелики. Нижнеантропогеновые олени, известные по находкам в Тираспольском гравии (на Днестре), имели уже рога с двумя сближенными и низко посаженными надглазничными отростками и безкронный, примитивный тип концевого ветвления.
Рис. 178. Осевые черепа и нижние челюсти кабанов.
1а, 16 — Sus scrofa attila, современный, Большой Кавказ, № 10485, ЗИН; 2 — S. scrofa attila, X—XIII вв. н. э., долина Дона, Саркел; За, 36 — S. apscheronicus, средний плейстоцен, восточное Закавказье, № 26031, ЗИН; 4 — S. tamanensis, верхний плиоцен, Кавказ, Таманский полуостров
№ 26000, ЗИН.
Тип «тесного» ветвления рога над венчиком оказался далее поразительно устойчивым, сохраняясь у оленей Восточной Европы и Кавказа на протяжении всего антропогена. Но в долине Волги и далее к востоку у средне- и верхпеплейстоценовых оленей имелось более «свободное» размещение первых отростков на стволе, наблюдающееся п у современных сибирских изюбрей, и у североамериканских вапити. Образование более или менее мощной кроны рогов у благородных олепей Европы началось, по-видимому, с среднего плейстоцена. Жидкая крона, вернее ее зачаток, наблюдалась уже у некоторых рогов, вымытых пз хазарского аллювия долины Волги (рис. 179) и Дона, а у некоторых рогов из битумов Ашперонского полуострова опа была резко выражена.
Из весьма обширного описания бинагадииских материалов по оленю (Алекперова, 1952) можно усмотреть все же, что среднеплейстоценовыц олень восточного Закавказья, пе отличаясь заметно по величине, имел
Г /V. Стратиграфическая и географшческая изменяемость млекопитающих 437
более легкие рога, чем современный олень Большого Кавказа. Судя по приведенным Алекперовой рисункам, в битуме встречались рога и со с ибо развитыми и редко посаженными надглазничными отростками. Лее эти материалы не позволяют установить роль миграций и местного формообразования у благородных оленей Восточной Европы и Кавказа в антропогене под влиянием смен и сдвигов ландшафта и зон. Тем не менее ясно, что на протяжении четвертичного периода в Восточной Ев-ьше и на Кавказе шла своего рода «борьба» эляфоидного (Cervus elaphus —
Рис. 179- Рога благородных оленей — Cervus elaphus subsp. из среднеплейстоценового аллювия долины пра-Волги. Куйбышевский музей.
с развитой кроной) и вапитоидного (С. canadensis — без кроны и с раздвинутыми первыми отростками) типа ветвления. Для кавказской популяции это ложно усмотреть из описаний Дипника (1914а) и проследить па таких массовых коллекциях, как пещера Дигоризед в Северной Осетии или мхзей Боржомского заповедника. Эляфоидный тип рогов у современных кавказских оленей, как правило, подавлен. В коллекции Дигоризеда, содержавшей в 19ч8 г. 62 пары и 121 разъединенный рог, оказалось 27% рогов эляфоидного типа (рис. 134) и 73% — вапитоидного.1 Впрочем, дотоверность этого подсчета осложнена тем, что многие рога в этой ^лекции имеют промежуточный тип концевого ветвления, т. е. с зачатком редкой кроны.
Еще Смирнов (1923—1924) указал на измельчание кавказских оленей с верхнего палеолита, основываясь па находках зубов в пещере Гвард-•дас-Клде. Вновь обнаруженные в этой пещере нами и Д. М. Тушаб-ршшвпли коренные зубы действительно поражают своей величиной, вапомипая в этом отношении лосиные.
1 Упоминавпе Динника о 500 парах рогов в Дигоризеде — простое преувеличение.
438
Часть II Анализ происхождения четвертичных млекопитающих
сз
При наличии в коллекциях лишь фрагментарных остатков четвертичных оленей более подробно можно говорить пока только о стратиграфической изменяемости величины рогов. Рога и, по-видимому, сами звери постепенно крупнели с нижнего плейстоцена до верхнего. Наиболее крупные рога встречаются в верхнем палеолите севера Русской равнины (Костен-ки под Воропежом). Позднее — в голоцене — начинается некоторое измельчание, по далеко не столь сильное, как иапрпмер у зубров. Олепи долины среднего Дона в средневековье (крепость Саркел под Цимлянской) были все же еще весьма крупны (рис.180, 181). Интересно, что по сравнению с зубрами (см. дальше) эволюция роста оленей как бы «запаздывает». Кавказская популяция олепей, пережившая четвертичные похолодания в наиболее благоприятных условиях, возможно, и дала особо крупный генетический материал для колонизации Русской равнины в конце верхнего плейстоцена. Расселение кавказских оленей на север шло, очевидно, по долинам Дона и Волги. Во всяком случае утверждение Флерова (1952) о том, что олени достигали наиболее крупного роста в среднем плейстоцене (миндель-рисс) и затем постепенно мельчали, несомненно, ошибочно.
Для современных кавказских оленей характерны промежуточные размеры тела и величины рогов по сравнению с западноевропейскими и азиатско-амерп канскими, правильно отмеченные, и в первоописанип Оджильби.
Географическая изменяемость современного оленя — С. elaphu maral в пределах Кавказа пе про слежена в достаточной мере и, в сущности, «затушевана» индивидуальной и . возрастной изменчивостью. По наблюдениям Дин-
Рис. 181. Изменение мощности рогов благородных оленей группы Cervus elaphus. Полусумма диаметров основания рога под коронкой. Черные кружки—средние величины.
По вертикали — число случаев.
1 — XV—XIX вв. н. э., Северная Осетия, пещера Дигоризед; 2 — X—XIII вв. н. э., долина Дона, Саркел; 3 — 1 тысячелетие до н. э., Кавказ, могильники: Махачкала, Самтавро и побережья озера Севан; 4 — верхний плейстоцен (верхний палеолит), долина Дона, Хостенни XIV; 5 — средний плейстоцен, битумы Апшеронского полуострова, Бинагады; 6 — средний плейстоцен, аллювий пра-Волги; 7 — нижний плейстоцен, аллювий пра-Днестра, Тирасполь; 8 — верхний плиоцен, Кавказ, Таманский полуостров.
440
Часть II. Анализ происхождения четвертичных млекопитающих
ника (1914а), наиболее крупны олени северо-западного Кавказа (Кубанской области). Здесь они приближаются по росту к изюбрю и вапити. В восточном и южном направлениях олени Кавказа мельчают. Особенно мелки олени, живущие в предгорьях Дагестана, по тугаям низовий Терека и Сулака. И в ископаемых материалах, и у современной популяции здесь отмечен легкий тип рогов с небольшим числом (5—6) отростков и почти всегда лишенный кроны. Относительно мелки были и олени, обитавшие на Армянском нагорье и в Талыше. Все это находит объяснение в оптимальных условиях северо-запада: обилии сочных кормов, обширности диких горно-лесных пространств и т. п. — по сравнению с издревле окультуренным жарким и сухим востоком и югом.
Все сказанное объясняет лишь сложность картины формирования морфологического типа и ареала благородных оленей в пределах Восточной Европы и Кавказа в кайнозое, наглядно показывая, что превосходные временами палеозоогеографические экскурсы Гентнера и Цалкина (1947) могут быть подтверждены или отвергнуты именно путем дальнейшего сбора и детальной проработки палеонтологических материалов.
Филогенетическая ветвь современных лосей рода Alces известна пока лишь с среднего плейстоцена. Тем не менее нет сомнения, что Alces alces сформировался по крайней мере в верхнем плиоцене, так как в среднем плейстоцене, это животное уже не отличалось от современного. Лосеподобные плотнорогие из отмерших в нижнем плейстоцене боковых ветвей — Tamanalces и Alces lalifrons уже в верхнем плиоцене обладали некоторыми лосиными признаками черепа и рогов, сформировавшимися, очевидно, задолго до антропогена.
Приспособления современного северного лося к жизни в таежных болотах и прп высоких снегах: крайняя высоконогость, большая опорная площадь копыт, брахиодонтность и веткоядпость — могли возникнуть, конечно, задолго до проникновения лося в тайгу и моховые болота.1 * В этом нас убедили наблюдения над лосями, обитавшими еще недавно (в 1951 г.) в пойменных болотах и уреме Среднего Поволжья. Поэтому утверждения Флерова (1950, 1952) о недавнем появлении этих адаптаций п о питании лосей преимущественно листьями и корой не соответствуют действительности. Как известно, лоси в течение 7—8 месяцев в году питаются преимущественно твердым (замерзшим) веточным кормом.
Черепа и рога среднеплейстоценовых лосей из хазарских слоев Среднего Поволжья были уже практически неотличимы от современных. Кавказский лось — A. alces caucasicus, известный теперь на Кавказе с верхнего плейстоцена, в XVIII столетии был относительно мелким зверем с ярко выраженными признаками европейской формы. Его череп был сравнительно широк, особенно в предглазничной и лобной частях, по сильнее сужен в носовой части, чем у северных лосей. Коренные отличались особой брахиодонтностью с усложненной клиновидными складками лабиальной поверхностью Pm3 и Pm4. Вообще морфологические признаки кавказского лося достаточно велики для того, чтобы датировать его появление и начало обособления на Кавказе по крайней мере средним плейстоценом.
Сайга — Saiga tatarica дает пример значительной морфологической устойчивости со среднего плейстоцена. Правда, в коллекциях, как у нас, так и в Западной Европе, имеются преимущественно зубы и лобно-затьь
1 Следует напомнить, что брахиодонтность (и веткоядпость), крайняя высоконо-
гость и даже развитие большой опорной площади копыта возникали и без снега в тре-
тичном периоде у жираф и некоторых антилоп Средиземноморья и Африки.
Рис. 182. Стратиграфические и географические изменения в строении стержней рогов и лобно-затылочных фрагментов черепов сайгаков.
1
-g- нат. вел.
Вид сбоку и спереди; 1а, 16 — Saiga borealis Tscher., ранний голоцен, Якутия, долина Вилюя, № 13636, ЗИН; 2а, 26— S. tatarica, верхний плейстоцен (хвалынский ярус), аллювий пра-Урала, № 24201 (10), ЗИН; За, Зб — S. talarica binagadensis, средний плейстоцен, битумы Апшеропского полуострова, Бинагады, № 22385, ЗИН; 4а, 46 — Saiga sp. средний плейстоцен (хаварский ярус), аллювий пра-Волги, № 1084, ЗИН.
' -442
Часть II. Анализ происхождения четвертичных млекопитающих
Рис.. 183. Контуры стержней рогов и лобно-затылочного отдела черепа современных сайгаков.
1 — Saiga tatarica, Волго-Уральские степи, № 7064, ЗИН; 2 — S. tatarica mongoiica, северо-западная Монголия, № 8465, ЗИН.
лочные фрагменты со стержнями рогов. Наиболее древний и, по-видимому, примитивный тип лобно-рогового фрагмента обнаружен в хазарских отложениях Среднего Поволжья. Череп средпеплейстоценовой восточноевропейской сайги был сравнительно небольшим, с совершенно прямыми относительно короткими и слабо разваленными стержнями рогов (рис. 182, 4а. 46).
Весьма близок к нему череп сайги из бинагадинского асфальта восточного Закавказья. Не отличаясь от хазарского типом развала рогов, бинагадинские черепа имеют более высокие (длинные) стержни и с заметным изгибом назад в сагиттальной плоскости (рис. 182, За, 36).1
Черепа более позднего времени из хвалынских, т. е. верхнеплейстоценовых, отложений долины Урала (наши сборы 1950 г.) приобрели уже больший развал рогов, нередкую изогнутость стержней в фронтальной плоскости, массивность в одних случаях или еще большую удлиненность в других (рис 182, 2а. 26).
Этот тип строения лобно-затылочных частей черепа сохранялся у сайгаков на огромных пространствах Восточной Европы, Средней Азии и Западной Сибири с верхнего плейстоцена до современности.
Однако в Якутии — на Вилюе и Лене — в позднем плейстоцене и раннем голоцене существовали популяции северной сайги — 5. borealis Tscher., морфологически, несомненно, обособленные. Черепа этой сайги отличались некоторой удлиненностью зароговой части, слабым развалом рогов, прямизной п длиной стержней, сильно сплющенных с боков (рис. 182, 1а, 16).
Современные сайгаки дают весьма широкий диапазон индивидуальной изменчивости лобно-затылочного участка черепа, но практически их черепа уже неотличимы от упомянутых выше фрагментов черепов с среднего Урала (рис. 183, 1).
Географические изменения у пыпе живущих популяций связаны, по-видимому, с обитанием па различных типах пастбищ. В нагорной полупустыне внутренней Монголии обособилась, например, и захудалая по-пулляция — подвид 5. tatarica mongolica Ванн. (рис. 183, 2).
Цифровые показатели размеров и индексы лобно-затылочных фрагментов сайгаков даны в табл. 94.
1 К сожалению, объемистое описание бинагадинской сайги, сделанное Алекперовой (1955), чрезвычайно трудно использовать для каких-либо выводов.
1д. IV. Стратиграфическая и географическая изменяемость млекопитающих 443
Г а б л и ц а 94
Стратиграфические и 1еографические изменения размеров (в uu) и пропорции (в %) лобно-затылочных фрагментов сайгаков1
- « Ь ы их местонахождение и геологический возраст Длина стержня рога по хорде Ин 1,екс массивности (обхват основания стержня в % от его длины) Наибольшая ширина черепа в орбитах Индекс длины зарожья (длина зароговой части в % от ширины в орбитах)
Северо-западная Монголия, современность, Saiga tatarica motigolica, 1 экз. джптарпя, современность, Saiga 1а-2 экз Qi и и Дон, X—XIII вв. н. э., Saiga lata tea, 100 экз Якутия, Оленек и дельта Лены, поздний плейстоцен, Saiga borealis, 2 экз. Долина Урала, поздний плейстоцен, 5 be tatarica, 10 экз Ышеюнскип полуостров, средний плейстоцен, Saiga tatarica blnagaden sis, 2 экз Jo.wia Волги, средний плейстоцен, У i т sp.. 1 экз 99 127 62.7 76.7 106 125 62.0 47.5
124—130 139 76.7—76.9 78.5 125—125 62 2 46.5—49.5 45
110—165 146 71—91 70 130—144 122.0 44.2—47.0 56.1
139—153 156.1 66.7—73.4 77 117—127 134.5 55.0—57.2 51.8
141—179 135.5 65.9—87.1 75 129—142 130 127 49.2—55.6 52.2 57.5
135—136 1 15 74.3—76.0 82.6
Примечание. В дрооих в числителе указана средняя величина, в знаменателе—предо.
В целом видно, что с среднего плейстоцена до голоцена роговые стержни сапгаков постепенно удлинялись, череп делался более широким в орбитах, а зароговая область черепа относительно укорачивалась. Позднее произошло новое уменьшение длины стержней.
В величине и пропорциях трубчатых костей конечностей, в длине зубного ряда и конструкции самих зубов плейстоценовых сайгаков трудно подметить существенные отличия от современных. Так, например, индекс бокового сжатия диафизов метаподпи, т. е. отношение переднезаднего диаметра диафиза пясти и плюсны к поперечному диаметру (посередине кости), у бинагадинской сайги колеблется в пределах: пясть —90—105% [5 экз.), плюсна — 128—140% (3 экз.), а у современной волго-уральской соответственно 90—102 и 114—146% (5 экз.). Индекс прыжка, т. е. отношение длины берцовой кости к длине бедра, было равно у бинагадинской сайги 114.9% (3 экз.), а у современной — 114.2% (5 экз.).
Иными словами, бшгагадинские сайгаки уже обладали пе меньшими, чем в наши дин, скоростями передвижения нижних рычагов конечностей.
j также подтверждает в известной степени уже отмеченную большую древнюю устойчивость морфологической структуры зверя.
Стратиграфическая изменяемость кавказских козлов — Capra caucasica, С. cylindrical*nis представляет особый интерес для выяснения происхождения н быстроты эволюции горных млекопитающих, но, к сожале
1 Неопубликованные материалы автора но коллекции ЗННа. Выбраны промеры, дающие существенные различия. 1 2 р
2 Здесь использовано 4 экз.
444 Часть If. Анализ происхождения четвертичных млекопитающих
нию, до сих пор обнаружены костные остатки преимущественно поздних — верхнеплейстоценовых и голоценовых — особей.
Верхнеплейстоценовые (верхнепалеолитические) козлы западного Кавказа отличались от современных более крупными размерами, о чем свидетельствуют промеры зубов и метаподий (табл. 95). Метаподии козлов из пещеры Гварджилас-Клде (рис. 184) поражают своей величиной.
Рис. 184. Нижние эпифизы пясти (1—3) и плюсны (б—7) кавказских козлов — Capra caucasica (foss.) из пещеры Гварджилас-Клде, верхний палеолит; пястная кость (4) современного 5-летнего С. caucasica.
Что касается конструкции зубов и метаподий, то достоверных отличий, кроме несколько большей эйрисомпости (шпрококостности), верхне-плейстоценовых и раппеголоценовых козлов (рис. 97) подметить не удалось. Вообще создается впечатление, что основные морфо-физиологические черты кавказских козлов, видов, адаптированных к высокогорному ландшафту, сформировались уже к началу аптропогена.
Географическая изменяемость современных кавказских козлов наиболее отчетливо прослеживается по степени специализации рогов самцов в зависимости от долготы. При распространении с запада па восток наблюдается последовательное усложнение строения рогов.
Именно по строению рогов, пли, вернее, роговых футляров, на Кавказе (с запада на восток) различали четыре вида козлов: С. dinniki, С. vertzovi, С. caucasica, С. cylindricornis (см. Динник, 1914а). На самом деле
Гл.- /V. Стратиграфическая и географическая изменяемость млекопитающих 44.5
Таблица 95
Стратиграфические изменения размеров зубов и метаподий (в мм) западнокавказских козлов 1
Промеры зубов и метаподий Верхнеплейстоценовые, пещеры Сакажиа п Гварджилас-Клде Современные, вападный Кавказ
среднее пределы число экз. среднее пределы число экз.
Передне-задний диаметр М3 24.8 24—25 3 23.8 22—26.5 10
Передне-задний диаметр М3 Поперечный диаметр нижнего эпифиза 31.6 25—36 16 26.8 26—29 6
пясти Поперечный диаметр нижнего эпифиза 45.6 42—48 5 38 35—42 3
плюсны 41.7 36—45 3 32 30—36 3
и восточнокавказская.
всего две основные формы: западнокавказская
Западпокавказскнй козел морфологически менее постоянен, чем восточнокавказский, и дает ряд переходов от центральнокавказского — С, caucasica caucasica к западнокавказским — С, caucasica severtzovi и С. caucasica dinniki.
Впрочем, в Сванетии, в верховьях Ингура, встречаются и переходные, очевидно гибридные, особи между восточнокавказскими и кавказскими козлами (Радде, 1899; Верещагин, 1938а).
Наиболее западным козлам свойственны массивные и короткие рога
отли-рхне-
УДа-
>гпче-рному
наи-амцов ок иа-
а Кав-I С. sexi. деле
саолевидпого профиля, со слабым развалом и отклонением концов вперед и наружу, а иногда и со слабым проявлением положительного, т. е. одноименного, перекручивания оси роговых футляров — «гомонимпости» Насонова (1923). Западные козлы (туры Северцова) имеют рога преимущественно нейтрального «ибексовидного» типа с большими буграми на наружной, большой кривизне, с значительным расхождением концов роговых футляров при легком проявлении гомонимности, т. е. со слабым перекручиванием правого футляра вправо и левого — влево. Козлы центрального Кавказа (туры Гюльденштедта) характеризуются рогами с большим развалом п с концами, слабо загнутыми внутрь при легкой «гетеронимности» футляров, т. е. перекручивании правого рога влево и левого — вправо. Наконец, у восточнокавказских козлов (палласовых, или дагестанских туров) рога сильно расходятся в стороны и футляры загибаются у старых особен назад и внутрь, образуя пологую спираль. Наблюдается при этом и небольшая гетеронимность. Эта географическая изменяемость иллюстрируется рис. 185. Кроме того, шов теменной и лобных костей образует узападиокавказских козлов прямую линию, а по мере движения на восток v восточнокавказских популяций этот шов приобретает форму тупого угла, характерную для овечьего черепа.
Если исходной примитивной формой козлиных рогов считать рога саблерогого профиля — ибексовые, безоаровые, — то можно думать, что рога восточиокавказского козла наиболее «молоды» в филогенетическом смысле*
1 Дли промеров были взяты зубы наиболее крупных 8—12-летних и метаподий \-*7-аетних козлов, имевшиеся в коллекции 311 На.
446
Часть II. Анализ происхождения четвертичных млекопитающих
Происхождение и возникновение такой «широтной» изменяемости и вообще морфологическое обоснование обоих форм козлов на Кавказе при наблюдающихся ныне горизонтальных миграциях и скрещиваниях могут быть объяснены весьма быстрым темпом эволюции некоторых признаков у этих козлов. Учитывая современную экологию обоих видов (глава III). нет оснований приписывать пх обособление влиянию ледниковой эпохи. Интересно, что подобный же пример усложнения рогов при распростране-
Рис. 185. Черепа и рога козлов Кавказа.
1 — Qapia caucasica severtzovi, западный Кавказ, № 12325, ЗИН; 2 — С. caucasica caucasica центральный Кавказ, № 697, ЗИН; <? — С. cylindricornis, восточный Кавказ, № 6964, ЗИН.
шш с запада на восток имеется и в роде Ovis, ио уже на обширнейших пространствах Евразии.
Вопрос о происхождении кавказских козлов и их ближайших родственных связях недостаточно ясен. В наши дни они обнаруживают известное сходство в морфологии черепа и рогов, да и в экологии с другими козлами типа козерога, обитающими в Евразии. По строению черепа и рогов они ближе всего к альпийскому ибексу — Capra ibex L., по сильно
Рве. 18G. Стратиграфические и географические изменения в черепах и роговых стержнях зубров Русской равнины и Кавказа. Контуры лобной поверхности „ 1 черепов и стержней, -tf пат. вел.
1 — Bison bonasus caucasicus, XV—XVIII вв. н. э., Северная Осетия, пещера Дигоризед; 2 — В. bonasus bonasus L., X—XIII вв. н. э., долина Дона, Сарксл; 3 — В. bonasus bonasus L., XII в. н. э..Белоруссия, Гродно; 4 — В. priscus (aff. deminutus), верхний плейстоцен (верхний па* леолмт), западное Закавказье, пещера Сакажиа, без номера, Одесский университет; 5 — В. piiscus deminutus, верхний плейстоцен, Татария, Нижние Кармалки, № 1, Казанский университет; 6 — В. longicornis. средний плейстоцен (хазарский ярус), аллювий ира-Волги, Сталинград ГЭС, № 26134 (1), ЗИН, 7 — В. schoetensacki, нижний плейстоцен, аллювий ира-Дпестра, Тирасполь, Московский геолого-разведочный институт; 8—В. cf. schoetensacki нижний плейстоцен, Кавказ, Таманский полуостров, № 26009, 31111; 9 — В. sp. (tamanen-six N. Ver.), верхний плиоцен, Кавказ, Таманский полуостров, № 26010, ЗИН.
А48 Часть II. Анализ происхождения четвертичных млекопитающих
отличаются от наиболее близкого географически синайского — С. si-naitica Ehrenb. и сибирского тека — С. sibirica Meyer. Еще больше морфологические различия в черепе и рогах у кавказских козлов по сравнению о пиренейским — С. pyrenaica Schinz.
Обе кавказские формы — западная и восточная — имеют, конечно, значительно больше общих морфо-экологических черт между собой, нежели с любым другим названным видом козлов. Так, например, строение слезной кости, а частью и зароговой области у восточнокавказского и за-паднокавказского козлов идентично. Поэтому объединение «С. ibex severtzovi Menzb., С. ibex nubiana F. Cuvier, C. ibex sibirica Ра11.»в один вид — C. ibex L., сделанное недавно Эллерманном и Моррисон-Скоттом (Е1-lermann a. Morrison-Scott, 1951), должно рассматривать как пример поспешных и неудачных морфо-систематических экстраполяций.1 Правда, синайский козел и сибирский тек по форме рогов, строению слезной кости и зароговой области напоминают друг друга, однако их объединение -с альпийским и запа дно кавказским козлами не выдерживает критики, тем более, что С. caucasica выделен почему-то в качестве отдельного вида, а С. cylindricornis — в качестве подвида к нему.
Морфологическая обособленность кавказских козлов от других евразийских свидетельствует о древней, среднечетвертичной изоляции их предков на Кавказе.
Среди других полорогих наиболее отчетливый, последовательный ряд морфологических изменений прослеживается с верхнего плиоцена у зубров (род Bison).
Эволюция зубров (как и слонов мамонтового ряда) проходила на широких пространствах Голарктики одинаково как в Азии, так и в Северной Америке. Лишь судьба и история угасания голоценовых популяций бизонов на этих материках оказалась совершенно различной (Верещагин 1956).
Некоторые американские исследователи, например Скиннер и Кай-зен (Skinner a. Kaisen, 1947), рассматривают геологически обособленные этапы морфогенеза зубров в ранге рода, что вряд ли правильно.
На Кавказе и Русской равнине филогенез зубров известен из нескользких геологических этапов пресноводных осадконакоплений на протяжении всей четвертичной системы вплоть до современности: 1 — плоскорогий и небольшой Bison sp. (tamanensis N. Ver.) в апшеронском ярусе западного Предкавказья (Таманский полуостров) и восточного Закавказья (Кабристан); 2 — значительно увеличившийся В. schoetensacki из третьей террасы Днестра (у Тирасполя); 3 — огромный длиннорогий В. longb cornis 2 из аллювия высоких террас Предкавказья и хазарского аллювия долин Волги и Урала; 4 — вновь несколько измельчавший В. priscus с подвидами из покровных делювиальных суглинков и галечников первых-вторых террас Предкавказья (позднемустьерская стоянка Ильская) и пещер западного Закавказья (ориньякская стоянка Сакажиа и др.), а также в Хвалынском ярусе и его аналогах Русской равнины (асфальтированные суглинки Нижних Кармалок в Татарии и др.); 5 — резко измельчавший в послеледниковье В. bonasus исторической эпохи с двумя дожившими до наших дней подвидами: литовским — В. bonasus bonasus и кавказским — В. bonasus caucasicus (см. рис. 186, 187).
1 К аналогичному выводу пришел недавно иЦалкин (1955).
2 Соблюдая последовательность, необходимо рассматривать длиннорогого бизона из хазарского аллювия — более раннего, чем номинальная форма — Bison priscus JBoj., — не в качестве подвида от последнего, а в ранге вида.
Гл. IV.- Стратиграфическая и географическая изменяемость млекопитающих 449
Уже верхнеплиоценовые фрагменты черепов и трубчатых костей зубров дают представление о вполне консолидированных морфологических чертах расцветшего в плейстоцене рода. Во всем последующем ряду,
Рис. 187. Стратиграфические и географические изменения размеров зубов зубров Русской равнины и Кавказа. Передне-задний диаметр на уровне альвеолы (М3 ~а\ М3 — б).
j.j „ Bison bonasus bonasus L., Белоруссия, Белове?к, современность; 2а, 26 — В. bonasus саи-«Gsicus,Кавказ, современность; За, 36 — В. priscus, верхний плейстоцен, западное Закавказье, пещеры Гварджилас-Клде и Сакажиа; 4а, 46 — В. priscus, верхний плейстоцен, За куб анская равнина, стоянка Ильская; За, 56 — В. longicornis, средний плейстоцен (хазарский ярус), аллювий пра-Волгп, Сталинград ГЭС; ба, 66 — В. cf. schoetensacki, нижний плейстоцен, аллювий пра-Днестра, Тирасполь; 7 — В. sp. (tamanensis N. Ver.), верхний плиоцен, Кавказ, Таманский полуостров
по имеющемуся материалу, подмечается скорее эволюция количественных признаков (величина, длина, объем), а не качественных. Изолированные зубы и трубчатые кости длиннорогого зубра хазарского аллювия Волги — в сущности, не более, чем пропорционально увеличенные копии нпжнеплейстоценовых зубров тираспольского гравия и верхпеплиоце-
29 Н. К. Верещагин
450 Часть II. Анализ происхождения четвертичных млекопитающих
Таблица 96
Изменения размеров (в мм) черепных фрагментов зубров в плейстоцене 1
Местонахождения и геологические этапы Длина собственно стержня рога по хорде Передне-задний диаметр основания собственно стержня Ширина лба за глазницами
Кавказ, пещера Дигоризед, XVI—XVIII вв. н. э 128 (18 экз.) 60.1 (63 экз.) 223 (27 экз.)
Кавказ, пещера Сакажиа, верх- 95—185 44—90 186—263
ний палеолит Татария, Нижние Кармалки, 210 85 —
верхний плейстоцен 270 87 291
Поволжье, Сталинградская ГЭС и др., средний плейстоцен . . Молдавия, Тирасполь, нижний плейстоцен Кавказ, Таманский полуостров, 350 (7 экз.) 120 (8 экз.) 328 (7 экз.)
371—560 308 (8 экз.) 290—340 105—140 106 (8 экз.) 85—120 301—352
верхний плиоцен 210—260 (5 экз.) 80—90 (2 экз.) —440
Примечание. В дробях в числителе указана средняя величина, в знаменателе — пределы.
новых зубров таманских конгломератов. Тем не менее даже малые изменения в скелете нередко свидетельствуют о глубоких внешних и физиологических преобразованиях.
Для стратиграфических целей, кроме рис. 186 и 187, имеют значение и размеры черепных фрагментов зубров (табл. 96), особенно при учете половой, возрастной и индивидуальной изменчивости зверя.
Изучение морфогенеза зубров Кавказа и смежных территорий приводит к выводу, что наша небольшая горная страна была в сфере эволюции этих полорогих на протяжении всего антропогена и сохранила в горных лесах до современности замечательны!! измельчавший и угасший реликт плейстоценовой эпохи.
Иной была история первобытного тура (род Bos). так как Кавказ в плейстоцене колонизировали формы различного происхождения (Бурчак-Абрамович, 1957).
Обсуждая результаты краткого обзора примеров морфологической эволюции кавказских и восточноевропейских млекопитающих во времени и пространстве, мы видим, что затронут широкий круг биологических явлений и закономерностей.
Прежде всего становится ясным, что филогенетические ветви индикаторных форм четвертичного периода (хищных, слонов и копытных) развивались на Кавказском перешейке довольно сходно с тем, что имело место на остальных широких пространствах северной части Евразии.
1 В основу этой таблицы положены неопубликованные материалы автора по-промерам коллекций черепов самцов и самок в пещере Дигоризед и взрослых плейстоценовых самцов в коллекциях Казанского геологического музея, Московского геолого-разведочного института, ряда волжских музеев, Одесского палеонтологического музея и ЗИН а.
Гл. IV- Стратиграфическая и географическая изменяемость млекопитающих 45'1
Иными словами, в антропогене Кавказский перешеек был захвачен единым процессом морфологической эволюции млекопитающих, общим для всей Палеарктики или даже Голарктики. Более того, в этой эволюции можно усмотреть те же черты, что были описаны Хойером (Hooijer. 1949) Для млекопитающих юго-восточной Азии. Как правило, большинство видов зверей увеличивалось в размерах на протяжении плейстоцена и резко мельчало в конце его, как бы подтверждая правило Бергмана во времени.
Рассмотрение морфологических изменений в филогенетических ветвях млекопитающих на протяжении антропогена приводит к выводу, что основная эволюция (и трансформация) четвертичных форм шла где-то (и когда-то) гораздо раньше— в дочетвертичные эпохи. Уже в фрагментах костей скелета и черепа нижнеплейстоценовых и даже верхнеплиоценовых млекопитающих (хищных, грызунов, хоботных, копытных) мы узнаем характерные черты строения не только родового ранга верхнеплейстоценовых и голоценовых форм, но и подрода и даже вида, т. е. узнаем лисицу группы красных лисиц, хомяков подродов Cricetus и Mesocricetus, оленя г: пшы Cervus elaphus и т. д.
На отдельных известных нам стратиграфических этапах антропогена ,h следовательные этапы родственных форм млекопитающих (в большинстве генетических рядов) разнятся друг от друга большей частью количественными, нежели качественными признаками. Благородный олень верхнего палеолита и исторической эпохи Придонья — это как бы увеличенная кспия нижнеплейстоценового оленя тираспольского гравия и т. д. Во всем этом можно усматривать и отсутствие видимых (качественных) скачков в течение четвертичной истории млекопитающих. Отсюда, вероятно, вытекает и необходимость пересмотра критерия видов. Не вызывает сомнения., о в антропогене морфологическая эволюция скелета млекопитающих шла преимущественно по линии сравнительно мелких приспособлений [в духе частных идиоадаптаций Северцова (1939)]. Именно поэтому соображения некоторых отечественных палеонтологов, например, Флерова [1952), Громова (1948) о геологической молодости некоторых родов млекопитающих— северного оленя, лося и их морфо-физиологических адап-[щий к ландшафту тайги и тундры — неверны и свидетельствуют лишь о слабости недавних представлений о генезисе и древности ландшафтов северной части Голарктики.1 Для нас же несомненно, что антропоген был преимущественно периодом формирования ареалов и экологических группировок уже вполне сложившихся видов.
При ландшафтных изменениях, за которыми часто не могла поспевать морфологическая эволюция, млекопитающие антропогена использовали преимущественно ту экологическую (физиологическую) пластичность, которая была заложена в них еще в плиоцене. В других случаях они просто вымирали или выселялись — меняли ареал.
Эволюция многих четвертичных видов нередко была оборвана человеком. Воздействием последнего может быть объяснена частично и поразительная незаконченность эволюции многих четвертичных видов и их, казалось бы, беспричинное вымирание.
1 Новейшие исследования геологов и палеонтологов на севере Сибири, особенно := Якутии, показывают, что эта страна испытала в кайнозое преобразования климатов и ландшафтов (и фауны), весьма сходные с теми, которые имели место в Средиземноморье. Следовательно, там можно ожидать находок остатков нижнечетвертичных северных оленей, лосей, сайгаков.
29*
452 Часть II. Анализ происхождения четвертичных млекопитающих
Среди общих выявленных закономерностей следует отметить также неравномерность темпов формообразования и неравноценную пластичность признаков в различных филогенетических рядах, иногда даже систематически близких форм. Укажем на консервативность черепных признаков лисиц по сравнению с признаками корсака, отставание укрупнения и измельчания оленей и лосей по сравнению с зубрами и т. п.
В ряде случаев остается неясным, имеем ли мы дело с мелкими адаптациями к среде обитания и начальными этапами дивергенции, в том смысле, как это было показано Виноградовым (1946) на грызунах, или с последовательным развитием признаков генерального направления эволюции. Трудно, например, решить окончательно, без дополнительных йсследований, является ли удлинение голени и усиление динамичности у современных тушканчиков и песчанок Апшеронского полуострова, по сравнению с плейстоценовыми, следствием ксерофитизации (опусты-нения) ландшафтов или это закономерный результат эволюции их организма в определенном направлении. В пользу последнего говорят как будто описанные случаи специализации зубной системы у волков, корсаков, слонов, идущей до тех пор, пока естественный отбор не приводит к угасанию ту или иную филетическую ветвь.
Основной движущей силой морфологических трансформаций антро-погеновых млекопитающих было, конечно, воздействие изменений Внешней среды. Именно в антропогене эти изменения климатов и ландшафтов были особенно велики.
При всем этом эволюция млекопитающих Кавказского перешейка в антропогене имела, конечно, и свои специфические черты, обусловленные своеобразием экологической обстановки.
Мы видим здесь и самостоятельное древнее развитие многих реликтовых, горно-лесных форм, приводившее к далекой морфологической дивергенции и консерватизму (кавказские полевки, козлы), и бурные начальные этапы видообразования с распадением на локальные экологические формы (кроты, лисицы, лесные мыши, водяные полевки и др.).
Кавказские популяции европейско-азиатских видов зверей, как правило, образовали здесь местные формы подвидового значения, морфологическая обособленность которых позволяет иногда судить не только о происхождении и древности ареалов видов, но и о темпах морфологической дивергенции.
Географические изменения современных млекопитающих в пределах Кавказского перешейка заметны преимущественно по измельчанию и по-светлению окрасок при распространении в юго-восточном направлении (лисицы, снеговые полевки), увеличению размеров тела с высотой обитания (суслики), усложнению строения рогов полорогих в восточной части ареала (кавказские козлы). Эти примеры изменений нередко лишь повторяют в микромасштабах сходные закономерности, наблюдающиеся на обширных пространствах Евразии в долготном и широтном направлениях. Они также способствуют познанию происхождения и истории ареалов видов в этой стране. Становится все более ясным, что в процессе формирования антропогеновой фауны млекопитающих всего Кавказского перешейка основная роль принадлежала вселениям, колебаниям численности и вымираниям организмов, а дивергенция и вообще морфогенез имели подчиненное значение.
ЧАСТЬ ТРЕТЬЯ
ТЕРИОГЕОГРАФИЧЕСКОЕ
РАЙОНИРОВАНИЕ КАВКАЗСКОГО ПЕРЕШЕЙКА И ПУТИ НОВЕЙШЕЙ ЭВОЛЮЦИИ ТЕРИОФАУНЫ
I
I
1
i
$
h
АНАЛИЗ ГОЛОЦЕНОВОЙ ТЕРИОФАУНЫ И СХЕМА РАЙОНИРОВАНИЯ
Исследования истории происхождения и экологии некоторых кавказских млекопитающих (глава III) дали возможность выделить руководящие экологические и географические группы видов, ареалы которых обрисовывают более или менее четко определенные естественные районы различного генезиса и возраста. Несомненно, что рассмотрение распространения всех остальных голоценовых видов зверей с оценкой их происхождения и древности на Кавказе по типу ареала может значительно уточнить полученную териографическую основу упомянутых районов. Смысл такого териогеографического районирования и териогеографиче-ских характеристик заключается в познании закономерностей эволюции фауны в связи с изменениями лика страны и в выявлении перспектив целенаправленной реконструкции фауны.
Следует, однако, учитывать, что ареалы отдельных видов, состав экологических группировок и фаунистических комплексов, а следовательно, и зоогеографические границы не остаются постоянными. Они эволюционируют с тем большей быстротой, чем больше меняется ландшафт страны, границы ландшафтных зон и чем меньше сами живые организмы оказываются способными приспосабливаться к этим изменениям. Поэтому районирование, особенно в пределах подобласти, провинции, можно установить лишь для относительно небольших геологических отрезков времени, что далеко не всегда подчеркивается зоогеографами. Более того, географическая граница ареала «типа фауны», «фаунистического комплекса» и т. п. — понятие чисто условное. Такая граница в виде линии может существовать лишь для каждого вида в отдельности на данное календарное число. В какой-то геологический момент она может лишь приблизительно совпадать для ряда видов, обладающих близкими экологическими показателями.
Современные схемы зоогеографического районирования и характеристики комплексов видов различных зоогеографических категорий даются биогеографами обычно по составу остаточного набора видов, уцелев-ш и х в условиях самых разнообразных антропогенных воздействий на протяжении исторической эпохи. Попытки привлечь некоторые исторические сведения о наличии в недавнем прошлом угасших форм и исторически проанализировать динамику ареалов отдельных видов бывают обычно робки и недостаточны.
Такой подход между тем неправилен уже потому, что при сознательном исключении недавно вымерших видов, тем более вымерших под влиянием
456
Часть III. Т ериогеографическое районирование и пути эволюции
человека, теряется практическая направленность зоогеографии по выяснению перспектив реконструкции фауны.
Начиная со второй половины XIX столетия происхождению фауны и зоогеографическому районированию Кавказского перешейка уделял * внимание почти каждый исследователь, так или иначе занимавшийся зоогеографическим подразделением Евразии. Еще Уоллес (Wallace, 1876) разделил Кавказский перешеек вдоль по Большому хребту, отнеся Предкавказье к Европейской подобласти, а Закавказье — к Средиземноморской.
Гениально просто и фактически почти правильно подразделил Кавказский край Северцов (1877), отнеся Северный Кавказ, западное Закавказье и Кавказский хребет к Средиземно морскому округу, остальное Закавказье — к Западноазиатскому округу, а северо-восточное Предкавказье — к Среднеазиатскому округу. В этом подразделении было уже признание разнородности фауны перешейка.
У. и Ф. Склэтеры (W. a. Ph. Sclater, 1899) объединяли Кавказский перешеек с Малой Азией, относя его к Европейско-Азиатской подобласти.
Наиболее полное признание разнородность происхождения современной фауны Кавказского перешейка получила в работах Сатунина (1901 г, 19036, 1904—1906, 1909—1910, 19126, 1913).
Основными теоретическими предпосылками у Сатунина были преимущественно монофилетическое происхождение видов и «миграционизм» без начала и конца.1 Он, по-видимому, предполагал, что где-то на юге (за Закавказьем) лежал центр видообразования, откуда, как из рога изобилия, расселялись на север через Балканы и Кавказ готовые высоко-и малоподвижные виды. В то же время Сатунин был первым русским зоологом, который сделал попытку комплексного освещения истории развития фауны Кавказской суши на основе зарождавшихся тогда геологических сведений о третичных и послетретичных морях, проливах и гляциологических представлений. Основанием для миграционистской гипотезы Сатунина являлось, видимо, убеждение, что эволюция млекопитающих идет весьма медленно и что Кавказский перешеек значительно 1 моложе в геологическом смысле, нежели прилежащие территории Пе- I редней Азии и южной России, вследствие чего он, т. е. Кавказ, и не может ? быть территорией самостоятельного видообразования. Препятствия для миграций млекопитающих на Кавказ с севера Сатунин видел в Маныч- | ском проливе, а с северо-востока — в Арало-Каспийском бассейне. {
Крайне интересна была его первая умозрительная попытка установления геологического возраста на Кавказе вселявшихся видов млекопитающих. Так, например, Сатунин (1901) считал, что Microtus arvalis. Arvicola amphibius существовали на Кавказе уже и во время ледникового периода. После исчезновения Манычского рукава «пришли на Кавказ с севера только Putorius foetidus, Vulpes corsac, Cricetus vulgaris, Sminthus subtilis, Castor fiber, Alactaga saliens, Lepus europaeus, Saiga tatarica*.
«К позднейшим переселенцам с севера, кроме упомянутого уже Cricetus vulgaris, следует причислить также Мus agrarius и М. minutus»r
Происхождение фауны всего Кавказского края было освещено позднее i (Сатунин, 1909—1910), когда перешеек был разделен на Северный Кавказ,
1 Признавая, подобно Пидопличко (1953), значительную роль местного развития ряда видов, группировок и комплексов млекопитающих в формировании фауны Па- | леарктики и восточного Средиземноморья в частности, я, отнюдь, не отрицаю обоснован' ! ного палеонтологическими фактами расселения видов и фаун на протяжении кайно зоя, особенно в четвертичном периоде. t
Гл. V. Анализ голоценовой териофауны и схема районирования 457
Хребет и Закавказье, а в них намечено 9 округов. В этой работе особо подчеркивалась значительная древность и самобытность альпийской фауны, наличие пришельцев в лесной области по склонам гор и было высказано соображение о выходе предков животных горной фауны Кавказа из гор Передней и Центральной Азии. Указывалось, что арктическая фауна никогда не доходила до Кавказа, так как в высокогорье здесь нет ни бетой куропатки, ни зайца-беляка. Далее утверждалось, что фауна горной страны Передней Азии автохтонная, что африканского влияния на фауну Закавказья нет и что она остается чисто азиатской.
Происхождение на Кавказе «арало-каспийской фауны» объяснялось наступлениями ледников и затоплениями прикаспийских степей, а обособление Аралыхского участка в долине Аракса — вулканическими явлениями в районе юго-восточных отрогов Зангезурского хребта. Фауна лесного округа Талыша рассматривалась как северо-западный аванпост индийской фауны.
Общая характеристика фауны перешейка в этой работе была кратко вьражена в следующих словах: «Мы видим здесь и древнюю автохтонную фауну Большого Кавказа, и остаток средиземноморской фауны в запад-псм Закавказье, и оригинальную, самобытную фауну нагорья восточной Милой Азии, крайний северо-западный аванпост индийской фауны, и, на-д нец, фауны, пришлые из степей южной России и пустынь Средней Азии».
Зоогеографические исследования Сатунина получили завершение в работе «О зоогеографических округах Кавказского края» (19126),. когда было выделено 5 подобластей и 11 округов. Его зоогеографическое Районирование Кавказа пользуется наибольшей популярностью.
Сам Сатунин видел свое основное достижение в том, что его округа и ; (астки совпадали почти полностью с фитогеографическими подразделе-! киями Кавказа. Если учесть, что Сатунин объездил почти весь край, то он придавал большое значение непосредственному впечатлению от естности и что фауна не только зависит от типа ландшафта, но и оставляет его неотъемлемую часть, то в упомянутом совпадении не будет шчего удивительного.
Списки млекопитающих,которыми Сатунин характеризовал выделенные кавказские округа, предельно точны, и только тогдашняя малая изученность особенностей ареалов и фаун смежных территорий, а также предвзятость прежних идей мешали правильному толкованию фактов.
Некоторые ошибки Сатунина были повторены и в этой последней работе. Схема деления Кавказского перешейка на зоогеографические округа осталась по-прежнему «геоморфологична», «фитоландшафтна» и не вытекала из истинных очертаний ареалов видов и их комплексов. Границы его округов шли либо по водоразделам хребтов, либо по подножиям последних, а на юге страны были нанесены чисто провизорно. Аралокаспийская фауна по-прежнему проникала через восточное Закавказье по ущелью и долине Аракса до подножий Арарата и Алагеза, а Лесной округ восточного Закавказья чудесным образом вклинивался на запад до Артвина, захватывая на северо-востоке половину внутреннего безлесного Дагестана.
Из других кавказских фаунистов упомянем Динника (1911), который указал на преимущественно южный облик фауны зверей Кавказа, отметив, что в степях Северного Кавказа имеется 3 вида общеевропейских, 5 видов восточноевропейских, 13 видов азиатских млекопитающих. Дин-ник также считал, что горные животные попали на Кавказ с юга, особенно, с Малой Азии.
458
Часть III. Териогеографическое районирование и пути эволюции
Критикуя через 27 лет Сатунина, Ляйстер (1931) показал, что Аралых-ский участок долины Аракса, отнесенный Сатуниным к округу степей восточного Закавказья, имеет 85% видов млекопитающих общих со смежным Ираном и только 41 % — общих с Закаспием.
» Действительно, из арало-каспийской фауны в долине среднего Аракса обитает всего 2 вида: малый тушканчик и манул, да и те имеют здесь древние связи с Закаспием через северный Иран, а не через восточное Закавказье и Предкавказье.
Однако утверждение Ляйстера о том, что подавляющее большинство животных, характерных для Армянского нагорья, пришло из Ирана, являлось, как и у Сатунина, лишь данью беспочвенному миграционизму, так как наличие здесь суши с олигоцена позволяет считать современное нагорье Малого Кавказа попросту краевым участком древнего видообразовательного очага Передней Азии.
Мензбир (1934), владевший и палеонтологическим материалом, от-I носил все западное равнинное Предкавказье к Понто-Барабинской под-| провинции Европейско-Сибирской подобласти, а восточное Предкав-5 казье — к Киргизской подпровинции Средне-Азиатской подобласти. Кав-< каз и Закавказье были отнесены им к Кавказской подпровинции Восточносредиземноморской провинции Средиземноморской подобласти Палеарктики.
Северная граница Кавказской подпровинции Мензбира проходит в области предгорий Северного Кавказа по линии от Новороссийска на i Черноморском побережье до Хачмаса на Каспийском. Южная же граница проходит от Батуми на Ереван и огибает с юга Талыш. Фауна Кавказской подпровинции, к которой относился и южный Крым, смешанного характера — европейско-азиатская. По мнению Мензбира, Кавказ находился под влиянием двух фауногенных центров — средиземноморского и центральноазиатского.
Границы выделенных Мензбиром подпровинций почти полностью совпадают с современной фитоландшафтной и климатической зональностью, данной в самых общих чертах, и становится ясно, что в цитируемой книге попросту охарактеризованы фаунистически ландшафтные зоны перешейка.
Наиболее четкие воззрения на фауну и природу Кавказского пере^ шейка были кратко, но с широких географических позиций изложены Пузановым (1938а).
Пузанов относит к Средиземноморской подобласти все Закавказье, которое вместе с Греческим архипелагом, Малой Азией, горным Крымом, входит в Восточносредиземноморскую провинцию. Северные склоны Эльбурса и Талыш выделены в Гирканскую провинцию, лежащую на границе с Средне-Азиатской подобластью.
Большой Кавказ Пузанов охарактеризовал как «сложный фаунистический узел, на юге которого преобладают виды средиземноморские, на севере — европейские, на востоке — среднеазиатские, но ядро которого составляет эндемичная альпийская фауна большой древности».
Специальное териогеографическое подразделение Кавказского перешейка было выполнено Кузнецовым (1949, 1950), использовавшим новые обширные коллекции млекопитающих, собранные в советское время. По схеме деления Палеарктики, предложенной Кузнецовым, равнинное Предкавказье и равнинное восточное Закавказье оказались в под-«области степей и пустынь, предгорное Предкавказье — в переходной
Гл. V. Анализ голоценовой териофауны и схема районирования 459
зоне лесостепи, Большой Кавказ и большая часть Закавказья — в Средиземноморской провинции подобласти северных лесов.
Обоснование и характеристика выделенных 6 кавказских районов ! 9 подрайонов были даны на основе списка зверей в 113 видов, с при-' течением фитол андшафтных описаний Буша (1935). Обе статьи написаны - напечатаны Кузнецовым наспех, изобилуют парадоксальными умозаключениями и просто фактическими ошибками. Несмотря на правильное понимание и толкование Средиземноморской подобласти, к ко-орой отнесен почти весь Кавказский перешеек, представления Кузнецова о путях формирования фауны недостаточны, что видно из такой фразы: «Третью группу видов кавказской маммалиофауны составляют ормы явно средиземноморского (!?—Н. В.) происхождения». Выходит, то кавказские, т. е. средиземноморские, виды происходят от самих себя, мутны также представления об Армянском нагорье, так как одна и та не территория называется то Армянским горно-лесным районом, то Чагорно-Армянским районом. Талыш путанно охарактеризован как лрно-лесной район, в то время как еще Сатунин четко подразделял го фауну на горно-лесную индийского типа и на нагорно-степную — райскую.
В списке видов не хватает: кожанка Огнева, кожанка Бобринского, кладчатогуба, малоазийской лесной мыши, кавказской снеговой по-евкп, сайги, джейрана, зубра, не говоря уже о недавно вымерших лосе, улане и тарпане. Имеется также много фактических ошибок в заключе-шях о происхождении фауны, в фаунистических характеристиках районов, особенно в части подразделения Кавказского округа и в описаниях границ.
В качестве примера ограничимся Азербайджанским районом, граница которого (Кузнецов, 1950) охарактеризована следующим образом: «На-пшаясь у г. Астара на Ирано-Советской границе у восточной оконечности (?—Н. В.) Талышского хребта, эта линия идет по предгорьям гряды (!?—Я. В.), по южному пределу Муганской, Мильской и Карабахской степей к озеру Севану (!—Н. В.) и далее через г. Караклис, южнее г, Тбилиси к Сурамскому перевалу. Здесь она резко поворачивает на восток через гг. Гори, Мцхет, Лагодехи, Закаталы, Нуху, к восточной оконечности Главного Кавказского хребта и по Кубинской низменности к побережью Каспийского моря» (стр. 157). В результате такого описания получается совершенно чудовищная путаница, так как в подобласть степей и пустынь попали Ленкоранская низменность, Алазано-Агричай-ская и Хачмасская низменности с их почти субтропической лесной растительностью и буково-грабовые леса северных склонов Шах-Дагского я Триалетского хребтов, ограничивающие оз. Севан.
Окраинные части Кура-Араксинской низменности не только «неоднократно», но и никогда не заливались в четвертичном периоде водами Арало-Каспийского бассейна, поэтому фауна этой территории не только не молода, как представляется Кузнецову, а сохранила даже такие древние элементы, как малоазийский хомяк и крот (!).
В списке млекопитающих этого района, который дальше описан в виде равнины с полынной полупустыней и пойменными лесами, показаны такие виды, как обыкновенная и малая бурозубки, мышь-малютка, лесная полевка (!?—Н. В.), кустарниковая полевка, обыкновенная полевка, безусловно отсутствующие в Кура-Араксинской, Ленкоранской и даже на большей части Алазано-Агричайской низменностей, и не показаны такие характерные виды, как поздний кожан, кожан Огнева, куница белодушка, перевязка, барс и благородный олень.
.460
Часть III. Териогеографическое районирование и пути эволюции
Заслугой Кузнецова является, однако, более правильное, чему прежних авторов, толкование Средиземноморской подобласти.
В последние десятилетия уточнением ареалов грызунов в западном Закавказье занимался Шидловский (1940в, 19416, 1945, 1947, 1948, 1950, 1951). Он признавал большую разнородность фаунистических влияний в фауне грызунов Грузии и занимался преимущественно уточнением границ округов Сатунина (Шидловский, 19416). Судя по тому, что одни виды грызунов, по Шидловскому, проникли в Грузию с запада, с востока, с юга, а другие являются древними «реликтовыми», этот исследователь стоит в общем на правильных зоогеографических позициях, пытаясь иногда выяснить даже геологический возраст и историю ареалов видов, слагающих «родентофауну» Грузии. Так, например, время проникновения малоазийского хомяка на Кавказский перешеек Шидловский относил на конец миоцена — начало плиоцена, а разрыв ареала прометеевой полевки на Большом Кавказе и в Аджарии связывал с иссушением Дзирульского массива в пос ле ледниковье. В противовес Сату-нину, Шидловский предлагал выделить альпийскую зону Большого £ Кавказа в качестве самостоятельной зоогеографической единицы, мотивируя это наличием в ней прометеевой полевки и кавказской мы-шовки.
Бобринский (1951) относил фауну основной части Предкавказья к фауне Европейско-Казахстанских степей, а его северо-восточный учас-b ток — к фауне Арало-Каспийских пустынь. К фаунам Арало-Каспийских <- пустынь и Европейско-Казахстанских степей отнесена и фауна восточ-. ного Закавказья. Вся горно-лесная фауна Большого Кавказа, северных склонов Малого Кавказа и восточных склонов Талыша отнесена к фауне Европейского широколиственного леса. Фауна альпийской зоны Большого хребта и Малого Кавказа отнесена к фауне Гор подобно фаунам Копет-Дага, Тянь-Шаня, Алтая и восточносибирских хребтов. Цитируя Сатунина и приводя его карту, Бобринский повторял и его ошибки, признавая преобладание арало-каспийской фауны в восточном Закавказье. Пестрит ошибками также характеристика фауны нагорий Малого Кавказа (стр. 314), для которых отрицается наличие оленя, лесной куницы, крота, безоарового козла.
Происхождение фауны Кавказа обсуждалось Бобринским на основе современных геологических данных. Отмечено преобладание юго-западного пути заселения Кавказа европейскими животными, через Балканы и Малую Азию. Развитие ареалов европейских обыкновенной и снеговой полевок на Кавказ допусклось на основе признания возможности широчайших миграций.
Кроме этих спорных утверждений, вызывает недоумение предположение о том, что прометеева полевка, кавказский тетерев, козлы и кавказский улар начали обособляться еще в то время, когда Главный Кавказский хребет представлял собой остров, так как островом Кавказ переставал быть и в олигоцене и в нижнем и в верхнем миоцене.
Весьма подробное эколого-ландшафтное районирование территории Армении было выполнено недавно Далем (1954а), который охарактеризовал списками наземных позвоночных естественные (преимущественно фитоландшафтные) участки Армянского нагорья и долины среднего Аракса. Использование для зоогеографической характеристики сразу четырех классов позвоночных и учения о «типах фаун» Евразии крайне затруднило понимание происхождения и состава териофауны Закавказья даже
Гл. V. Анализ голоценовой териофауны и схема районирования 461
для самого автора. При зоогеографическом районировании Закавказья Даль придерживался преимущественно схемы Кузнецова (1949).
В результате этого краткого обзора зоогеографических представлении о современной фауне млекопитающих Кавказского перешейка выясняется, как и на нашем палеонтологическом материале (глава III), что прп наличии древнего местного видообразовательного и фаунообразовательного центра в горных районах эта территория находилась под значительным влиянием териокомплексов смежных территорий.
Для того чтобы правильно оценить эти фаунистические влияния и их распределение во времени и пространстве, необходимо остановиться па древности и происхождении кавказского местного и смежных териокомплексов, а в особенности на происхождении и природе Средиземноморья, в понимании которого нет единого мнения. Палеогеографы подразумевают под Средиземноморьем побережья остаточных бассейнов моря Тетис или по крайней мере побережий Сарматского моря, тянувшегося от Атлантики до Закаспия. В климатологическом смысле к Средиземноморью относят южную Европу, Северную Африку, часть Передней Азии и даже часть Средней Азии (см. Берг, 1938).
Геоботаники Алехин (1938), Вульф (1944) понимали под Средиземномор-ск)й подобластью, или провинцией Голарктики, побережья Средиземного и Черного морей, включая Северную Африку, Испанию, Италию, Грецию, Малую Азию, Крым. Между тем Малеев (1946) и Гроссгейм (1936, 1948) неоднократно указывали на генетическое родство, если не на тождество кавказских формаций маквиса, гариги, фриганы, широколиственных лесов с такими же формациями западного Средиземноморья, явно стремясь к их объединению. В настоящее время Лавренко (1958) считает, что в фитогеографическом смысле лесная область Кавказа по происхождению и экологии (климату) родственна широколиственным лесам 1 Европы, а не Средиземноморью.
В понятиях прежних зоогеографов Средиземноморская подобласть J чаще всего охватывала побережья Средиземного моря, Малую Азию, | северную часть Аравии и Закавказье (Уоллес, Склэтеры, Северцов, \ Кобельт).
- Современные орнитологи, например Серебровский (1928), рассмат-: рпвали Переднюю Азию (Сумерию) как территорию встреч и обмена фау-| мистических элементов Африки, Центральной Азии, Индии, Европы. 1 Штегман (1938) видел средиземноморский тип фауны, кроме побережий, собственно Средиземного моря как раз в Сумерии Серебровского, а в Закавказье и в южном Крыме — лишь оттенок этого типа.
Кашкаров и Коровин (1931) подмечали черты средиземноморского влияния даже на флору и фауну средних зон жизни Тян-Шаня, а ныне проявляются тенденции рассматривать фауну Кавказа и Гирканики как часть провинции широколиственных лесов Европы (Рустамов, 1945; Кузнецов, 1950; Бобринский, 1951).
Анализируя очаги развития пустынно-степной фауны млекопитающих Палеарктики, Гентнер (1939, 1940, 1945) развивает мысль о том, что на юге этой области существовали лишь древние центры развития пустынной фауны, которых он насчитывает четыре. Гептнер (1945) отрицает самобытность средиземноморской фауны и целесообразность выделения соответствующей подобласти на том основании, что в Средиземноморье нет будто бы древних и специфических форм млекопитающих. Побережья севера Африки и юга Европы, по Гептнеру, просто лишь пронизаны выходцами с севера и юга.
462 Часть III. Териогеографическое районирование и пути эволюции
Такие рассуждения неправильны уже потому, что жизненные арены (ландшафты) и разнообразие биотопов пустынь и Средиземноморья несравнимы между собой.
Средиземноморье предоставляло всегда больше возможностей для иммиграции животных и растений, нежели Сахара и Кара-Кумы, но значение его как самостоятельного древнего формообразовательного центра от этого не уменьшалось.
Правильное толкование понятия «Средиземноморья», а следовательно,, и средиземноморского типа фауны может быть достигнуто лишь при комплексном освещении природных преобразований, происходивших в кайнозое в пределах Средиземноморской геосинклинали.
Как известно, Сарматское море, тянувшееся от Гибралтара до Закас-пия, позднее — в плиоцене и плейстоцене — неоднократно распадалось на цепь изолированных бассейнов, имевших временами взаимную связь в эпохи трансгресий. Отдельные участки их побережий развивались, в неодинаковой обстановке. Побережья Арала и северной части Каспия по условиям рельефа и большей континентальности были рано опустынены и выведены из единой системы зональности средиземноморского типа.
Тем не менее остальные участки побережий этой цепи кайнозойских бассейнов, в том числе и Кавказ, унаследовали ряд единых первобытных черт ландшафтов, флоры и фауны, корпи которых уходят в миоцен. Именно поэтому под Средиземноморьем, учитывая его палеогеографическую сущность, следует понимать южные окраины Европы (включая юг Украины и Крым), Северную Африку, Малую Азию, Кавказский перешеек (за исключением Приманычья), Талыш и Эльбурсский хребет.
Различия в физико-географической обстановке обусловили также неоднородность истории формирования ландшафтов и кайнозойской фауны; Средиземноморья в западной — Пиренейско-Балканской и восточной — Балкано-Эльбурсской частях. Еще в олигоцене на южной окраине восточного Средиземноморья сказывалось фаунистическое влияние центрально-и среднеазиатской континентальной индрикотериевой фауны. Позднее западная часть Средиземноморья сохраняла теплолюбивый облик фаунистических комплексов северного и южного побережий вплоть до начала плейстоцена. Эти комплексы содержали приматов, виверр и бегемотов. Только недавно стало известно, что бегемоты и приматы в верхнем плиоцене—нижнем плейстоцене жили и в Закавказье. В это же время и в районе Азовского моря присутствовало еще некоторое количество относительно теплолюбивых форм — слонов и антилоп, но териокомплекс в целом характеризовал здесь лишь ландшафт саванны или лесостепи на грани умеренного и субтропического' поясов.
Западная часть Средиземноморья в плейстоцене была защищена с севера высокими хребтами Альп и Карпат, в то время как восточная была открыта действию масс воздуха, охлаждавшихся в зимние сезоны над Русской равниной. В результате оледенение севера Европы не оказывало такого воздействия на ландшафт и фауну Испании, Италии, Греции,, как на Крым и Кавказ.
Наибольшее, Днепровское оледенение продвинуло на Русской равнине ряд видов таежного или даже тундрового териокомплекса вплоть до южного побережья Крыма. Юго-восточнее, т. е. на Кавказский перешеек,, такого продвижения северных видов не произошло, так как зона илейсто-
Гл. V. Анализ голоценовой териофауны и схема районирования 463’
К ценовых степей на Русской равнине выгибалась в районе северо-восточ-Р ного Азова далеко к северу. Кроме того, имело значение и наличие в эпохи трансгрессий Каспия Манычского пролива (см. главу II).
Под ландшафтом средиземноморского типа биогеографы нередко > понимают современный ландшафт Испании, Сицилии, Палестины, т. е. К выжженные солнцем и объеденные козами каменистые и сухие склоны И гор с плантациями маслины, фиг и винограда на подходящих участках, Я с колониями кроликов, куницами белодушками и уцелевшими кое-где И муфлонами и безоаровыми козлами. На самом же деле как в западной, Ж так и в восточной части Средиземноморья наряду с ксерофитными ланд-т шафтами гариги, фриганы и нагорных степей имеются реликтовые ландшафты третичных хвойных и широколиственных лесов, мезофитных лугов альпийского и субальпийского типов (см. Ильинский, 1937). Именно на этих мезофитных горных участках формировались и сохранялись такие специфические виды млекопитающих, как малоазийская мышь, снеговые полевки, прометеева полевка, серна, пиренейские, альпийские и кавказские козлы, макаки — формы, вряд ли менее древние, чем путорак, тушканчики, песчанки, селевиния и другие зверьки смежных пустынных очагов видообразования.
Высокогорные ландшафты Кавказа, как и других горных систем альпийской складчатости, формировались уже в миоцене. Если в плиоцене Кавказ и был пенепленизирован в какой-то мере, то главные горные | цепи все же оставались.
J Поэтому-то голоценовый горно-лесной териокомплекс Кавказа и имеет i так много сходных и близкородственных форм с горно-лесным териокомплексом альпийских поднятий западного Средиземноморья. На восток й этот тип териофауны прослеживается в лесном Талыше и по южным 1 склонам Эльбурса. Кавказский горно-лесной териокомплекс развивался j преимущественно на месте. Целиком он во всяком случае ниоткуда не | приходил. Этим и объясняется его высокий видовой и подвидовой энде-! мшм. Фаунистические влияния на него шли в жаркие и сухие эпохи | плиоцена с юга, а в плейстоцене — главным образом с северо-востока, I северо-запада и юго-запада.
| Южные фаунистические связи были наиболее широки, древни и по-I СТОЯННЫ.
| На передне-азиатских нагорьях расположен самостоятельный очаг I формообразования двух подтипов териокомлексов плиоценового возраста: нагорно-степной и нагорно-пустынный.
Дифференциация переднеазиатского комплекса на упомянутые подтипы прослеживается почти во всех отрядах млекопитающих. Она свя-I зана с сильной расчлененностью рельефа страны и разнообразием кли-f матов. В то время как в широких межгорных долинах и узких ущельях j вырабатывался биоценоз нагорно-пустынного типа, на смежных высо- них плоскогорьях и хребтах формировались экологические группировки ландшафта нагорных степей и даже лугостепи.
Воздействие этого южного комплекса на Кавказский перешеек рас-J падалось, несомненно, на несколько этапов. Наиболее древним следует 1 считать еще миоцено-плиоценовый, т. е. эпоху расселения по перешейку ; гиппарионовой фауны.
Весьма вероятны значительные фаунистические влияния с юга и в верхнем плиоцене.
В плейстоцене, особенно верхнем, продвижение на север южных форм: было, очевидно, затруднено.
4<64 Часть III. Териогеографическое районирование и пути эволюции
На севере Кавказский перешеек примыкает к Русской равнине, южные части которой в районе водоразделов Днепра, Дона и Волги существуют как суша по крайней мере с нижнего миоцена. Степное флористическое и фаунистическое ядро юга Русской равнины, по данным пыльцевого анализа и палеофаунистики, сформировалось уже в плиоцене.
Заселение предкавказскпх равнин выходцами из восточноевропейских и североказахстанских степей могло начаться уже с среднего плиоцена, но массовое проникновение зверей этого степного териокомплекса на Кавказский перешеек происходило, несомненно, в плейстоцене под влиянием сдвига к югу ландшафтных зон. Во влажные эпохи плейстоцена на Кавказ мог оказывать влияние и териокомплекс широколиственных и таежных лесов Европы и Азии. Лесные мезофильные виды могли расселяться по долинам Дона и Волги, а также через Балканы и по северному берегу Малой Азии.
На востоке, за Каспием, лежат пустыни Средней Азии, териокомплекс которых оформился, по-видимому, в главнейших чертах уже в нижнем плиоцене. Проникновение представителей среднеазиатского пустынного териокомплекса на Кавказский перешеек могло идти с северо-востока и юго-востока, в обход Каспия, а частью и непосредственно с востока.
Плейстоценовые и голоценовые вселенцы (несомненные выходцы из полупустынь и пустынь Турана) — тушканчики и песчанки — появлялись в восточном Предкавказье, вероятно, в эпохи регрессий древнего Каспия. Поздний — голоценовый — этап проникновения на Кавказский перешеек ряда теплолюбивых южных млекопитающих хорошо заметен по ареалам ряда видов.
Переходя к систематическому анализу голоценовой териофауны Кавказского перешейка в целом, отметим, что привлечение для него не только остаточных, ныне живущих видов, но и вымерших в историческую эпоху под прямым и косвенным воздействием человека сделало наши списки зверей Кавказа значительно обширнее, нежели применявшиеся до сих пор другими исследователями.
Учитывалось, как уже было отмечено, что анализ современной фауны должен вскрывать историко-генетическую и экологическую сущность видов, т. е. характеризовать не только происхождение ареала, но и его возраст. В связи с этим соображения о древности того или иного вида на Кавказе, при отсутствии палеонтологических доказательств, строились на основе сопоставлений материалов по древности и эволюции предпочитаемых животными ландшафтов с данными об экологии видов и особенно о их миграционных способностях.
Рассматривая современные и восстановленные ареалы обитавших и обитающих на Кавказском перешейке четвертичных млекопитающих (карты 1—93) и сопоставляя их с данными палеогеографии, оказалось возможным значительно уточнить нашу схему генетического состава голоценовой фауны перешейка, данную в III главе. В полном виде эта схема выглядит следующим образом.
ПАЛЕОГЕОГРАФИЧЕСКАЯ И ЭКОЛОГИЧЕСКАЯ СУЩНОСТЬ КАВКАЗСКИХ МЛЕКОПИТАЮЩИХ
I. Третичные (плиоценовые):
1) кавказские, мезофильные и широко распространенные;
2) переднеазиатские, сухо- и теплолюбивые;
v 3) восточноевропейские, гидрофильные и степные;
Гл. F. Анализ голоценовой териофауны и схема районирования 465
4) туранские, пустынные.
II. Плейстоценовые: , . '
5) европейские, лесные, мезофильные;
6) европейско-азиатские, степные;
* 7) туранские, полупустынные и пустынные.
III. Голоценовые:
8) южноазиатские, теплолюбивые;
9) туранские, пустынные;
10) завозные, случайные.
Основные типы ареалов некоторых индикаторных зверей выделенных оксов показаны на рисунках 188—194.
^^географический и возрастной анализы голоценовой фауны зверей Кавказа ведутся далее по отрядам.
Н А С В К О М О Я Д Н Ы JE — 1 N 8 С ТI V О ВА
Кавказский перешеек — один из самых богатых видами насекомоядных участков СССР. При исследовании 436 местонахождений и просмотре нами 1224 экземпляров здесь насчитано 15 живущих видов (карты 1—6k Современные сводки по Кавказу и смежным территориям (Огнев, 1928: Бобринский, Кузнецов, Кузякин, 1944; Кузнецов, 1950; ЕПег-
I шапп a. Morrison-Scott, 1951, и др.) позволяют считать, что на перешейке I недостает длинноиглых ежей Средней Азии и Ирана, путорака — Dip-j lomesodon и некоторых белозубок западного Средиземноморья.
! Основное ядро насекомоядных на Кавказе составлено местными мезофильными видами. К ним относятся кроты южного, средиземно морского j типа, кавказская и малая бурозубки, серая белозубка. В эту же группу | мы относим такие широко распространенные виды, но связанные все же I с мезофитными ландшафтами, как обыкновенный еж и водяная кутора, ( происхождение которых на перешейке неясно. Второе место по численности принадлежит видам, связанным с третичными ландшафтами степей и полупустынь восточного Средиземноморья, например ушастому ежу, белобрюхой белозубке, этрусской белозубке.
Из третичных реликтовых видов Русской равнины отметим выхухоль, встречающуюся лишь на самом северо-западном участке перешейка. Плейстоценовыми вселенцами на Кавказе, появившимися с севера, можно считать обыкновенную бурозубку, ареал которой недоразвит в того-{ восточном направлении, и малую белозубку, которая в восточном Закав-| казье и Передней Азии крайне редка.1
I Генетический состав насекомоядных Кавказского перешейка пред-I ставлен по нашей схеме в табл. 97.2
i ——----------
I * К этому виду в Закавказье отнесена Crocidura dinniki Ogn., систематическое по-'t ложение которой не совсем ясно.
f 2 Автор не имеет возможности останавливаться здесь на критике взглядов Бобрин-1 ского, Кузнецова, Кузякина (1944) по систематике землероек, столь полно вое-(принятых Эллерманном и Моррисон-Скоттом (Е Hermann a. Morrison-Scott, 1951).
Наиболее курьезные превращения были произведены с серой белозубкой. В свое время I Огнев (1928) ошибочно отнес к этому виду спиртовой экземпляр длиннохвостой бе-! лозубки из северного Ирана (Мала-Кала), хранящийся в коллекции ЗИНа. Бобрин-• да и Кузякин соединили (по созвучию?!), дальневосточную — Crocidura lasiura Dobs. J t понтийской — C. lasia Thos. Получилась неимоверная путаница с наименованием I «распространением обеих видов. 4
30 Н. К. Верещагин
466 Часть III. Териогеографическое районирование и пути эволюции
Таблица 97
Генетический состав насекомоядных Кавказа
Виды Erinaceus europaeus L Hemiechinus auritus Gmel. . . Talpa orientalis Ogn T, caucasica Sat Desmana moschata L Sorex minutus L araneus L S. raddei Sat Neomys jodiens Schr Crocidura svaveolens Pall. . . C. leucodon Herm C\ russula Herm C. lasia Thos C. zarudnyi Ogn Suncus etruscus Savi Третичные (плиоценовые) Плейстоценовые Голоценовые
кавказские, мезо- i 1 ! 1 ।—। 1 т ! "—г 1 "1” фильные и широко р аспр остра ценные , , । . । । I t । , переднеазиатские, + "Г 1 1 т 1 1 1 1 1 1 1 1 + 1 сухо-и теплолюби- вые восточно-европей-1 1 1 1 1 1 1 1 1 1 + 1 1 1 1 ские, гидрофиль- ные и стенные 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 туранские, пустын- I I 1 1 I 1 I 1 ц 1 1 1 I I I европейские, лес- 1 1 1 1 1 1 1 1 г 1 1 1 1 1 1 ные, мезофильные 1 ( । I I । I I I । । I [ 1 I европейско-азиат- 1 1 1 1 1 т 1 1 1 1 1 1 1 1 1 ские, степные туранские, полупу-I 1 I I I | 1 I I I 1 1 | I I стынные и пустын- ные I । । । । I I ] [ I I । 1 । I южно-азиатские, И 1 1 1 1 1 1 1 1 1 | 1 1 1 теплолюбивые . . , । , । ( . I.,. туранские, полупу- 1 1 i 1 1 1 1 1 1 1 1 1 1 1 1 стынные и пустын- ные . | | } | | | | j | j | | J | | завозные, случайные
Р Г КОКРЫЛ ЫН — С Н IBOPTER А
Голоценовая фауна рукокрылых Кавказского перешейка, судя по 419 местонахождениям и 785 просмотренным нами экземплярам, насчитывает 27 видов (карты 7—17). Есть основание думать, что в верхнем плейстоцене она была несколько беднее, так как в связи с похолоданием ареалы многих теплолюбивых видов отступали на юг, а в голоцене восстановились. Известные трудности, связанные с коллекционированием летучих мышей, позволяют считать возможным в будущем новые находки неизвестных ранее для Кавказа форм.
Судя по сводкам Аллена (Allen, 1940), Тета (Tate, 1942), Кузякина (1950), Эллерманна и Моррисон-Скотта (Ellermann a. Morrison-Scott, 1951), на Кавказе нет представителей субтропических семейств Ptero-pidae^ Rhinopomatidae, Embalonuridae, Nycteridae, Hipposiderinae, характерных для южных частей Передней Азии. В остальном по сравнению с Европой и Азией Кавказский перешеек весьма богат видами летучих мышей. Основная масса летучих мышей, обнаруженных на Кавказе, имеет ареалы, приуроченные к Средиземноморью в его широком понимании. В экологическом смысле эти древние, плиоценовые средиземноморцы могут быть подразделены на относительно мезофильных, тяготеющих к массивам широколиственных лесов, нередких дуплогнездников, места летовок и зимовок которых приурочены преимущественно к краевым участкам крупных горных сооружений перешейка, и на более сухолюбивых и теплолюбивых средиземноморцев, предпочитающих ландшафт безлесных низких плоскогорий, предгорий и жарких низменностей.
Гл. V. Анализ голоценовой териофауны и схема районирования
467
В первую группу отнесено большинство ночниц и кожанков. В нее же попал и поздний кожан, живущий на Кавказе чаще в буковых лесах, например, в Талыше, нежели на открытых равнинах.
Во вторую группу входят преимущественно подковоносы, широко-средиземноморский нетопырь. Влияние европейской лесной фауны рукокрылых для нас не совсем ясно, но вряд ли оно было велико, так же как и пустынной среднеазиатской. Среди таких пустынных видов следует упомянуть кожанка Огнева, распространение которого крайне характерно и приурочено к третичным всхолмлениям в Куринской низмен пости восточного Закавказья.
i поздним плейстоценовым вселенцам из Европы могут быть условно отнесены всего два вида: гигантская вечерница и северный кожанок. Возможно, что их появление на Кавказе было вторичным в результате окончания ледникового периода.
Общий список с разноской на генетические группы дается в табл. 98 •I 1 (именованиях русских авторов — Огнева (1928) и Кузякина (1950).
Таблица 98
Генетический состав рукокрылых Кавказа
Третичные (плиоценовые) Плейстоценовые Голоценовые
, ft ф 1 л? Я 3 ° Я Q 1 1 ф £ J к га н • 2 gg > ь со 1 1 н И ф &g ф S >s
Виды Ф ft Й S S Ф -3 И о _ rt a s ft, к й о R ь о СО R S к £0 ф К Ч О И <D ftO га дои ft ft с о >> я аГ 1=3 2 ft’s 2 ft СО Нч 1 га о и X к й ь к & И сГ р-» Д ф 33 и га 2 Я §& ф р4 я о
К a? b S S о CO X О cd Н ф и s ЕЧ £ t- ft К X о о ° .2 со ф ° ? О ф « Ен ф О 3 ф 5 з я ю ? != х ft §й ф S
aS 3 ft и: 1 о й д я ° Q X X га
PQ R О сб К d ft й 2 О ft S ft .Xi &2 п2 & S тур г СТЫ1 ные г-0 Кг £ е ft S 2 >1^3 Ь ft X О га
Квй йод СЙ О X Ь X со. £ ф М £0 X. О о йь СО
nolophus ferrum equinum
ГЛ ^hr — 1 — — — — — — —-г*-»
mehelyi Matsch — + — — — — — — — —
1 atryale Blas -— —р — — —, — — — — —
kipposideros Bechst. . . . — 1 — — —— — — • —
blasii Peters —. — — — — — — —
bechsteinii Kiihl . . . — — — —— — —
if, cmarginatus Geoffroy. . . . — —i—* — — — 1 и™ — — —
V, wygnathus Mont 1 — — — —'—* -— — — —
У, wfitacinus Kiihl + — —— — —. —- — 1
If nattareri Kiihl . . . . * * — — — — — —
у:>>iopterus schreibersii Kiihl -t-1 — — — — — — —
auritus L —р — —- — — —
^rbaslella darjelingensis Dobs. — — —t— — — — •—’ —
1 barbastella Schr * + — — — — — — — —
leisleri Kiihl .... —— ~р — — — — — —
V, siculus Pall — — — -— f — — — — ——-
,V. noctula Schr — ™ 1 — — — —— - —
Vespertilio pipistrellus Schr. 1 — — — ——- — — — — ——
f, natkusii Keys. et. Blas. . . — — ~р — — —
Г. kuhlii Kiihl — “Г — — — — — — —
y, Bonap -р — — — — — — — — —
Г. murinus L —р — — — — — — "— — —
bobr inski i Kuz *" — — _р —" — -— — — —
1, nilssonii Keys. et. Blas. , . — — — " i — — — —
Г. serotinus Schr —1— — — — — — — — —
V. ugnevi Bobr — — — + — — — — —
padarida taeniotis Raf. . . . 1 1 — — — — — . —
30*
4Q8' Часть III. Териогеографическое районирование и. пути эволюции
Для некоторых видов — Vespertilio bobrinskii Kuzyakin и V. ognevi Bobr.— необходимо, очевидно, дальнейшее сравнение: для первого — с Г. (Epte-sicus) nasutus Dobson, 1877, и для второго — с У. (Eptesicus) sodalis Barn Hamilton, 1910.
*
В голоценовой фауне Кавказа насчитывалось 25 видов хищных. Верхнеплейстоценовая фауна вряд ли была значительно богаче. В наши дни уцелело до 22 видов. Обильные коллекции ископаемых остатков (92 местонахождения), черепов и шкур живущих видов (376 местонахождений), просмотренные нами экземпляры (417), визуальные наблюдения и многолетние государственные заготовки шкур (257 пунктов) позволяют подойти |к зоогеографическому анализу этой группы на основе огромного факти-1ческого материала (карты 18—-38). По сравнению с окружающими территориями (Огнев, 1931, 1935; Бобринский, Кузнецов, Кузякин, 1944; ? Ellermann a. Morrison-Scott, 1951) Кавказский перешеек весьма богат хищ-? ными. В кавказской фауне можно отметить отсутствие представителей семейства вивер — Viverridae, северная граница ареала которых проходит несколько южнее, в Ираке, и представителей подсемейства медоедов — Mellivorinae, обитающих также значительно южнее и восточнее. Осталь-। ные палеарктические семейства хищных представлены на Кавказе довольно полно. Из собачьих сюда не заходят ареалы некоторых лисиц Передней Азии: Vulpes ruppeli Schinz., У. сапа Blanf., Fennecus zerda Zimm., а из кошачьих отсутствуют настоящие пустынные виды: Felis margarita Loche, Fells caracal Schr. Большинство кавказских хищников принадлежит к широко распространенным формам с невыясненными путями происхождения — убиквистам, которые тем не менее отнесены нами к группу кавказских средиземноморцев в связи с находками на перешейке ископаемых остатков близких форм в среднем плейстоцене и даже в верхнем плиоцене, таковы: волк, лисица, бурый медведь, барсук, рысь.
Южные — переднеазиатские средиземноморцы — виды степных, и пустынных ландшафтов представлены куницей бе л о душкой, перевязкой, ливийской (пятнистой) кошкой и гепардом.
Плиоценовые восточные — турано-афганские — виды представлены манулом.
Плейстоценовые европейские вселенцы, проникшие с севера и юго-запада, легко узнаются по неразвитости ареалов, которые выклиниваются в юго-восточном направлении. К ним относятся: лесная куница, норка, лесной кот и, может быть, медведь европейского типа на Большом Кавказе.
Хищные голоценовой группы вселенцев все относительно теплолюбивые и южного происхождения. Их позднее появление на перешейке выясняется по сопоставлению фактов современной экологии, состоянию ареалов и отсутствию остатков в битуминозных плейстоценовых захоронениях на Апшероне и в пещерах верховьев Риона. К ним отнесены: шакал, полосатая гиена, тигр, лев и камышовый кот. Происхождение этих видов на Кавказе разъяснено в главе III.
Наименования хищных в табл. 99 даны преимущественно по номенклатуре, принятой у отечественных авторов.
Гл. V. Анализ голоценовой териофауны и схема районирования 469
Таблица 99
Генетический состав хищных Кавказа
Третичные (плиоценовые) Плейстоценовые Голоценовые
Виды кавказские, мезофильные и широко распространенные переднеазиатские, сухо- и теплолюби- ! вые । восточно-европейские, гидрофильные и степные туранские, пустынные европейские, лесные, мезофильные европейско-азиатские, степные туранские, полупу-। с тынные и пустын- ные южно-азиатские, теплолюбивые туранские, полупустынные и пустын- ные завозные, случайные
. Cgpis aureus L (Е lupus L. Vulpes vulpes L V. corsac L Hyaena hyaena L Ursus arctos L Martes martes L M. foina Erxl Vormela peregusna Guld. . . . ? Putorius putorius L . P. eversmanni Less Lutreola lutreola L ; Mustela nivalis L \ IM. erminea L Meles meles L i Lutra lutra L J * Panthera tigris L . *P. Leo L P. pardus L ' Felis lybica Forst | F\ silvestris Schr I F. chaus Giild I F. lynx L 1 Otocolobus manul Pall. . . . | * Acinonyx jubatus Schr. . . . 11+111+11++1+111111 + 11++1 1 1111111++1111111111+1 u 1+ 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 I 1 1 1 1 1 1 1 1 1 +1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 IIII+IIIIII+I+I+II+IIIIII 1 1 1 +1 1 1 1 1 1 +1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 М 1 i 1 М I 1 1 1 +1 1 1++1 11 11 1 1 1 1 1 1 +1 1 1 + 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 + 1 1 1 1 1 1 1 1 1 1 1 1 1
Примечание. ? — отмечены виды, обитание которых в пределах перешейка окончательно не доказано; ♦ виды, вымершие в нашу эпоху.
ЗАЙЦГО БГАЗП ЬГЕ^ LAG О М ОВРНА
В голоцене на Кавказе существовало два вида из этого отряда: заяц-ру-сак и арменийская пищуха. Материалы по ископаемым кайнозойским зайцам и пищухам юга Русской равнины и Предкавказья, данные современной экологии и особенности ареалов обоих видов позволяют рассматривать их в качестве древних плиоценовых (миоценовых) вселенцев перешейка. Эволюция их проходила в третичных ксероморфных ландшафтах восточного Средиземноморья. Из более южных форм на Кавказе отсутствует арабский заяц — Lepus arabicus Ehr., а из западных средиземноморцев— кролик — Oryctolagus cuniculus L.
Проникновение с севера на Кавказ в плейстоцене малой пищухи и зайца-беляка не доказано.
Оба голоценовых кавказских вида относятся к третичным по возрасту, восточносредиземноморским по происхождению и к относительно сухолюбивым по современной экологии зверьков (табл. 100).
• ' 410
Часть III. Териогеографическое районирование и пути эволюции
ГРЫЗЛ НЫ — ROD ENTIА
В голоценовой фауне Кавказа насчитывается не менее 50 видов грызунов. Видовой состав кавказских грызунов можно считать теперь практически исчерпывающим (при 1770 точках сбора ныне живущих форм и при 10 623 просмотренных нами экземплярах). Средне- и верхнеплейстоценовая фауна грызунов вряд ли была значительно богаче послеледниковой.
Пестрота зоогеографического и экологического составов грызунов, их значительная геологическая древность позволяют сделать значительное число достоверных выводов об особенностях и путях формирования этой группы фауны. Учитывая возможность влияний фаунистических комплексов смежных территорий, следует признать, что на перешейке недостает ряда видов грызунов. Для такого обзора и сравнения использованы работы Огнева (1940, 1947, 1948, 1950), Бобринского, Кузнецова, Кузякина (1944), Эллерманна и Моррисон-Скотта (Ellermann a. Morrison-Scott, 1951).
Из восточноевропейского степного комплекса здесь нет: сурка,1 серого, рыжеватого и большого сусликов, эверсманнова хомячка. Из европейского лесного и горно-лесного комплексов не хватает: обыкновенной белки, сонь родов Eliomys, Muscardinus, балканского Dolomys, пашенной полевки, экономки и ряда видов подземных полевок рода Pitymys. Из среднеазиатского пустынного и переднеазиатского нагорно-пустынного комплексов на Кавказе недостает желтого и тонкопалого сусликов, тушканчиков родов Pygeretmus, Jaculus^ мышевидной сони, боялычной сони, многих видов песчанок родов Gerbillus, Psammo-mys, Tatera, земляных крыс Nesokia.
Нет здесь и ряда видов западного Средиземноморья: африканского дикобраза — Hystrix cristata L., берберийской мыши — Lemniscomys Barbaras L., Arvicanthis niloticus Desm, Acomys cahirinus Desm. и др.
Наиболее многочисленную группу здесь составляют плиоценовые виды влажных и лесных ландшафтов восточного Средиземноморья.
Мезофильные грызуны этой группы, связанные по происхождению с верхнетретичными широколиственными лесами и горными лугами, представлены видами:
кавказская белка полчок лесная соня черная крыса домовая мышь кавказская желтопятная мышь лесная мышь
малиазийская мышь прометеева полевка водяная полевка кавказская снеговая полевка длиннохвостая снеговая полевка кустарниковая полевка
Под домовой мышью здесь подразумевается ряд дикоживущих форм мыши, связанных с биотопами зоны горного, предгорного и низменного леса: Mas musculus formosovi Heptn., M. musculus abotti Wat. — и с биотопами степи и полупустыни: М. musculus hortulanus Nordm., M. musculus tataricus Sat. Аналогично с этим крупные кавказские лесные мыши
1 Возможно, что сурок байбак все же жил в историческую эпоху в степях Предкавказья.
Гл. V. Анализ голоценовой териофауны и схема районирования
471
типа Apodemus fulvipectus Ogn.: A. flavicollis ponticus Svirid., A. flavi-collus saturatus Neuh., A, flavicollus argiropuli Vin. et Arg. — лесов Кавказа, образующие на юго-востоке переход к менее крупным, светлым, с слабым горловым мазком лесным мышам зоны полупустыни восточного южного Закавказья типа A. arianus Blanf., рассматриваются в качестве полиморфного вида восточносредиземноморской (кавказской) желто-пятной мыши.
К древним сухолюбивым видам восточносредиземноморского происхождения отнесены:
малоазийский суслик
малоазийский тушканчик
малоазийский хомяк серый хомячок
мышевидный хомячок краснохвостая песчанка персидская песчанка
малоазийская песчанка песчанка Виноградова горная слепушонка малоазийская снеговая полевка горный слепыш дикобраз
Древность появления этих видов на разных участках перешейка и мрактер их расселения неодинаковы, и ряд примеров этого уже был дан I главе III.
Из третичных видов восточноевропейского происхождения, гидрофильных и степных по экологии, можно назвать речного бобра и предкав-казских слепышей. При этом, не предрешая вопроса о первоначальной истинной родине бобра, здесь подразумевается, что он существовал на Кавказском перешейке и Русской равнине еще в плиоцене.
Более сложен вопрос о происхождении и появлении слепышей в Пред-авказье. При отсутствии палеонтологических находок остатков слепышей на Кавказском перешейке и до получения доказательств расселения их из Малой Азии следует считать оба предкавказских вида потомками миоцено-плиоценовых слепышей, обитавших на юге Русской равнины. Вселение обыкновенного и гигантского слепышей на перешеек могло произойти уже в эпоху образования балаханской толщи, т. е. в среднем плиоцене, причем последующие трансгрессии морей они могли переживать и в кавказских предгорьях.
Единственным представителем древних выходцев из пустынь Турана и Афганистана является малый тушканчик. Его ареал, захватывающий восточное Предкавказье, восточное и южное Закавказье, может дать интересный пример влияния «арало-каспийской фауны».
Сопоставление особенностей ареалов и палеогеографических данных позволяет выделить довольно мощную группу плейстоценовых вселен-цев различного возраста и происхождения.
((Европейских» лесных и вообще мезофильных видов насчитывается по крайней мере семь:
лесная мышовка мышь-малютка полевая мышь желтогорлая мышь
обыкновенный хомяк обыкновенная полевка 1 рыжая полевка
Ареалы этих видов па перешейке как бы недоразвиты. В случае далекого проникновения этих видов в Закавказье экологические русла
1 Недавняя находка костей этой полевки в ангельских слоях пещеры Кударо I указывает, что этот вид на Кавказе более древен.
472
Часть III. Териогеографическое районирование и пути эволюции
расселения связаны либо с пойменно-болотными формациями на предгорных равнинах, либо с нагорными мезофитными лугами и горными хвой-. ными лесами. Географически русла проникновения тяготеют к.побережьям Черного моря и Каспия, обтекая Кавказский хребет с запада и востока, как например, у мышки-малютки, полевой мыши, обыкновенного хомяка. Реже они идут по высокогорным мезофитным формациям, как у лесной мышовки и обыкновенной полевки. Ареал европейской желтогорлой мыши типа Apodemus /lavicollis samariensis ограничен тугайными лесами низовьев Терека и Сулака в восточном Предкавказье и тем самым довольно загадочен.
Учитывая, что эта форма вряд ли могла возникнуть здесь самостоятельно, нужно признать, что зверек мог вселиться сюда лишь со стороны низовой Волги скорее всего в верхнем плейстоцене, так как позднее — в послеледниковье — ландшафты северного Прикаспия были опустынены и уже не создавали возможностей для такого проникновения.
К плейстоценовым вселенцам мы относим и восточных сухолюбивых зверьков из южноказахстанского полупустынного и отчасти среднеазиатского пустынного комплекса. Их всего три вида: емуранчик, мохноногий тушканчик, тарбаганчик.
Плейстоценовыми их следует считать по той причине, что на нижнем Дону встречены их остатки верхпеплейстоценового возраста. Другими словами, эти виды проникали из пустынь Закаспия далеко на запад по крайней мере уже в эпоху послехазарской регрессии моря. Хвалын-скую трансгрессию они могли переживать частично и на южном берегу Терского залива этого моря. Процесс нового вселения этих видов, а также пустынных туранских в Терско-Кумскую полупустыню и в Приманычъе мог произойти снова в голоцене. К таким голоценовым вселенцам из пустынь Средней Азии следует отнести песчанок: большую, полуденную и тамариксовую. Основанием этому служат отсутствие их остатков в четвертичном аллювии низового Дона и ограниченность современного ареала. Большая песчанка вымерла в Предкавказье в историческое время.
Кроме такого вторичного проникновения в голоцене пустынных видов на северо-восток, естественно было бы ожидать появления на юго-востоке перешейка и новых южных грызунов в ксеротермическую эпоху послеледниковья. Между тем таких примеров нам неизвестно, за исключением, может быть, расселений малоазийской песчанки в пределах Кура-Араксинской низменности и местных расселений степной полевки, персидской песчанки и некоторых других под влиянием антропогенного изменения ландшафтов.
Из случайных завозных видов, заселявших перешеек уже в историческую эпоху, имеется лишь один — серая крыса. Кроме того, продолжается завоз на Кавказ домовых мышей северного типа, а серые и черные крысы широко расселяются по железным дорогам, водным и воздушным путям.
Общий генетический состав грызунов дан в табл. 100. Наименования грызунов приведены в написаниях, принятых у русских авторов, т. е. большой тушканчик называется Allactaga jaculus Pallas, а не A. major Kerr.; емуранчик — Alactagulus acontion Pallas, а не A. pumilio Kerr.; краснохвостая песчанка по-прежнему названа Мег tones erythrourus Gray, а не M. lybicus Licht.; дикобраз — Hystrix leucura Sykes, а не Hystrix iridica Kerr, и т. д.
।
Ф
।
С7 ।
г< । Г
tn
I '•И
Примечание. Звездочкой (*) отмечены виды, вымершие в историческую эпоху.
I I I I I I I I I I 4—F [ + I Г + I + I I I I I I I
I 4- I I 4-4-4- I 4-4-1 I 4-4- I 4- I I I I I I I I I 4- I I I I I 4-
I I I I I 1 I I I I I I I I I I I I I I I I I I I I++I I I I
I I I I I I I I I I I I I I I I I I i I I I I I I I I I I I I I
I N I I I I I I I I II I 4-1 I I + I ++ I I I I I I I I I |
I I +4- I I I I I I I I I I I I I I I I I | | | | II I I I I I
I I I I I I I I I I I I I I I I I I I I I I I I I I I I 4-4-4- |
I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I |
I I I I I I 4-1 I 4-4- I I I I I I I I I I I I I I I I I I I I
I I I I I I I I I I I I I I .1 I I I I I I | I 4-1 'I I I Г I I I
• 1 Виды i Генетический сос
кавказские, мезофильные и широко распространенные л ч АЗ W 07 АЗ
переднеазиатские, сухо-и теплолюбивые К Л ГЙ Я О Й w Е Дс Я -ф О о\
в осточно-евр опсй-ские, гидрофильные и степные (плиоце. te) я W д Е и
туранские, пустынные и гр
европейские, лесные, мезофильные Я Й сь W Я
европейско-азиатские, степные ДЛСТОЦ' вые о W я
туранские, полупустынные и пустынные CD а о । ДЭ td Л АЗ н
южно-азиатские, теплолюбивые Ч о Й о я cd и о W S3 АЗ О я
туранские, полупустынные и пустынные й д АЗ
завозные, случайные Е CD О р
Анализ голоценовой териофауны и схема районирования 47 &
4Я4 Часть III. Териогеографическое районирование и пути эволюции
Т а б лица 100 (продолжение)
Виды Третичные (плиоценовые) Плейстоценовые Гол о ценовые
кавказские, мезофильные и широко распространенные переднеазиатские, сухо- и теплолюби- ; вые восточно-европейские, гидрофильные и степные туранские, пустынные европейские, лесные, мезофильные 1 европеиско-азиат- 1 ские, степные туранские, полупустынные и пустын- ные южно-азиатские, теплолюбивые туранские, полупустынные и пустын- ные завозные, случайные
Arvicola terrestris L
Мicrotus nivalis Mart *— I- _ — " " — — — —
M. gud Sat JL i — —— — —- — — — -—.
M. roberti Thos — ———• — — — — — * " -
M. majori Thos — — —-— — ——~ — — — ы
M. arvalis Pall — — — Ч- —1 — — —
M. socialis Pall — —|— — — - --- — * ~ —_
Clethrionomys glareolus Schr. — — — -— ——. — —
Hystrix leucura Sykes — — — — — —— — — — —
ЕНАРНОКОПЫТНЫЕ
PEBI88OPACTYLA
Судя по коллекционным материалам и древним историческим справкам, в послеледниковье на Кавказе существовало два или три вида лошадей. В связи с запутанностью вопросов морфологии и генезиса лошадей Старого Света происхождение тарпана не совсем ясно, поэтому он -отнесен в группу степных плейстоценовых вселенцев с севера, согласно лишь общепринятым взглядам. Кулан же показан в качестве голоценового вселенца с юга. Вероятнее же, что послеледниковая история лошадей на севере и юге страны была неодинаковой: в Предкавказье имело место заселение лошадьми восточноевропейской равнины, а в Закавказье — переднеазиатскими.
Как и в случае с некоторыми грызунами и хищными, также имеет место неравнозначность происхождения отдельных видов для различных участков территории Кавказа. Так, кулана и лошадь переднеазиатского типа в южном Закавказье можно рассматривать в качестве местных верхнетретичных видов, а в восточном Закавказье и в Предкавказье они будут уже голоценовыми вселенцами.
В нашей генетической схеме голоценовые непарнопалые Кавказа занимают, следовательно, различные места (табл. 101).
ПАРНОКОПЫТНЫЕ-ARTIODA CTYLA
По числу видов и численности особей парнокопытных на единицу площади Кавказский перешеек занимает первое место в СССР. Это — крупный формообразовательный очаг для парнокопытных.
Из арктических форм сюда, очевидно, не попали северный олень и Тем более овцебык. Из южных теплолюбивых видов к Кавказу далеко
Гл. V. Анализ голоценовой териофауны и схема районирования 47б
Т аб л и ц а 101
Генетический состав непарнокопытных и парнокопытных Кавказа
Виды Третичные (плиоценовые) Плейстоценовые Голоценовые
кавказские, мезофильные и широко распространенные переднеазиатские, сухо- и теплолюби- вые восточно-европейские, гидрофильные и степные туранские, пустынные европейские, лесные, мезофильные европейско-азиатские, степные туранские, полупустынные и пустын- ные южно-азиатские, теплолюбивые туранские, полупустынные и пустын- ные завозные, случайные
Perissodactyl а
* hemionus Pall . .
:: caballus gmelini Ant.
* . caballus L. (subsp.) . Artiodactyla
< ' scroja L. ............
’ -pus elaphus L.........
’ Hees alces L............
’ preolus capreolus L. . . у i'gd tatarica L.........
C>;.Ma subgutturosa Guld. В iptcapra rupicapra L. . C-pra caucasica Ciild. . . C cylindricornis Blyth. . t aegargus Erxl...........
с ‘is gmelini Blyth . . . .
Bison bonasus L.........
Bps primigenius Boj. . .
Примечание. Звездочкой (•) отмечены виды, вымершие в историческую эпоху.
не доходят теперь ареалы малоазийской и месопотамской ланей Dama и ряда видов переднеазиатских газелей: Gazella dorcas L., G. gazella Pall., Cj. leptoceros Cuv.
Из горных форм Средней Азии на Кавказе нет винторогого козла, а из за-падносредиземноморских и западноевропейских — гривистого барана — :\nmotragus lervia Pall, и козлов: Capra sinaitica Ehr., C. ibex L., C. pyre-naica Schinz.
В четвертичном периоде на перешейке происходило постепенное обеднение парнокопытными, несмотря на вселения новых видов со стороны. В среднем плейстоцене на перешейке существовало по крайней мере 15— 16 видов, а в голоцене — 13. В наши дни уцелело 11 видов. Основная группа кавказских парнокопытных представлена местными горно-лесными и нередко эндемичными формами. К эндемикам относятся кавказская серна, западнокавказский и восточнокавказский козлы. Широко распространенные лесные и вообще более или менее мезофильные виды, входящие в эту же группу, представлены кабаном, благородным оленем, косулей, зубром и первобытным туром. Возможно отнесение в эту же группу и лося, когда будет доказано его южное происхождение.
Переднеазиатские сухолюбивые виды, связанные преимущественно с нагорно-степными ландшафтами, представлены безоаровым козлом и малоазийским муфлоном.
476
Часть III. Териогеографическое районирование и пути эволюции
В группу плейстоценовых вселенцев отнесены лось северного типа, появившийся на Кавказе, быть может, уже вторично в качестве лесного зверя, и сайга, наибольшее развитие ареала которой на перешейке приходится на средний плейстоцен. Вероятно, в эту группу должна быть отнесена и крупная форма косули, живущей в Предкавказье.
Из поздних — послеледниковых — вселенцев можно назвать только джейрана, проникшего в восточное Закавказье с юга, а в восточное Предкавказье — возможно, еще и с востока. В целом генетический состав голоценовых парнокопытных Кавказа показан в табл. 101.
В результате палеогеографического и зоогеографического анализов голоценовой териофауны Кавказа можно сделать следующие заключения.
1. Голоценовая фауна Кавказского перешейка содержала до 136 видов млекопитающих из отрядов насекомоядных, рукокрылых, хищных, зайцеобразных, грызунов, непарнокопытных и парнокопытных. В наши дни сохранилось до 124 видов.
2. Основу этой фауны — до 36% видов — составляют местные (преимущественно мезофильные) и широко распространенные формы плиоценовой древности, эволюция которых протекала на Кавказском перешейке и окружающих участках восточного Средиземноморья, сохранявших благодаря особенностям рельефа, близости моря и теплового режима свои первобытные (мезофитные) черты.
3. Следующей по значимости является группа переднеазиатских сухолюбивых вселенцев третичного возраста — до 27% видов.
4. Восточноевропейские степные и гидрофильные звериг проникшие частью в плиоцене и частью в плейстоцене, составляют в сумме до 12.5% видов.
5. Европейские лесные и мезофильные виды, появившиеся на Кавказе главным образом в плейстоцене, составляют до 11% видов.
ft. Та б л и ц а 102
Генетический состав (число видов по типам происхождения) современной (голоценовой) фауны млекопитающих Кавказа по отрядам
Отряды Третичные (плиоценовые) Плейстоценовые Голоценовые Я о м fe. я о о st £Г © Я В о О
кавказские, мезофильные и широко распространенные переднеазиатские, сухо- и теплолюби- вые в осто чн о-ев р о пей-ские, гидрофильные и стенные туранские, пустынные европейские, лесные, мезофильные европейско-азиатские, степные туранские, полупустынные и пустын- ные южно-азиатские, теплолюбивые туранские, полупустынные и пустын- ные । завозные, случайные
Insectivora Chiroptera Carnivora Lagomorpha .... Rodentia Perissodactyla . . . Artiodactyla .... 8 10 8 14 8 4 9 4 2 15 1 2 I I «I I 3 1 1 1 2 4 7 1 1 2 5 1 1 1 1wl 1 1 1 5 1 1 1 11|« 1 1 1 15 27 24 2 52 3. 13.
Всего | • • • 48 35.7 37 27.2 7 5.1 5 3.6 15 11.0 10 7.3 3 2.2 7 5.0 3 2.2 1 0.7 136
Гл. V. Анализ голоценовой териофауны и схема районирования 477
6. Послеледниковые вселенцы — виды, теплолюбивые и не переносящие глубоких снегов, появляющиеся с востока и с юга (из Средней и Передней Азии), составляют до 7% всего состава.
Наконец, завозные, случайные виды, появляющиеся в историческое ' мя, составляют менее 1%.
Все эти данные, существенно отличающиеся от представлений других авторов, объединены в сводной таблице 102.
А Р ЕАЛЫ кавказских лесных и высокогорных видов
Малоазийскаялес-
' нал мышь
\ Кустарниковая по-левка
> Кавказская снеговая полевка
' Длиннохвостая снеговая полевка
Полевка Прометея
& Р
МАХАЧКАЛА
200 км
БАКУ
ЕРЕВАН
Рис. 188.
КРАСНОДАРА-
СУХУМИ
& БАТУМИ
Предлагаемая схема териогеографического районирования Кавказского перешейка основана на результатах зоогеографического картирования и анализа териофауны, изложенных в главе III и в настоящей.
Для выделения округов, районов и участков использованы обобщенные контуры ареалов индикаторных, относительно стенотопных видов млекопитающих кавказского комплекса и комплексов пришлых (рис. 188—194). При районировании учитывалось прежде всего не формальное большинство видов зверей той или иной зоогеографической группы, а палеогеографическая сущность территорий и видов. Так, например, к Кавказскому горно-лесному округу, кроме Большого и Малого Кавказа, отнесены восточные склоны Талыша и северные склоны Эльбурса, хотя от древнего кавказского териокомплекса па последних осталось всего 4 явно автохтонных вида, остальное же принадлежит плейстоценовым и голоценовым наслоениям. Наряду с этим все степное, а частью и лесостепное Предкавказье отнесено к Предкавказскому району степных восточноевропейских плейстоценовых вселенцев, так как
Рис. 190.
АРЕАЛЫ европейско-азиатских мезофильных и лесных видов
СТАВРОПОЛ
ТБИЛИСИ
ЕРЕВАН
Рис. 192.
СУХУМИ
БАТУМИ
Рис. 191.
МАХАЧКАЛА
Полебая мышь
/' Обыкновенный хомяк
Рыжая полевка < Лось (Восстановленный ареал)
200км
БАКУ
Рис. 193.
АРЕАЛЫ
Шакал
видов.
Джейран (босста-нобленный ареал)
Рис. 194.
Гл. V. Анализ голоценовой териофауны и схема районирования 481
голоценовая фауна этой территории уже не содержит заведомо автохтонных кавказских элементов плиоценовой древности, В нашем подразделении нет той дробности, к которой стремились иные зоогеографы, пытаясь w что бы то ни стало разрезать перешеек на возможно большее число *;усков, привлекая для этого, в сущности, больше геоморфологию и геоботанику, нежели конкретный зоологический материал. Тем не менее фито-ландшафтные границы имеют и в нашей схеме ведущую роль, так как связи индикаторных видов млекопитающих с типом фитоландшафта остаточно велики.
Признавая, что большая часть Кавказского перешейка находится у пределах восточной половины Средиземноморской подобласти Палеарктики, или, что тоже, в Восточносредиземноморской провинции, мы относим северную часть равнинного Предкавказья к Предкавказскому району подобласти Восточноевропейских и Североказахстанских степей, а северо-западный угол восточного Предкавказья — к Терек о-Кумск ому гчастку подобласти Среднеазиатских пустынь. Весь Большой Кавказ, северные склоны Малого Кавказа и восточные склоны Талыша объединены в Кавказский округ, в котором выделены Гирканский, или Галышско-Эльбурсский, и Дагестанский районы. Нагорье Малого Кавказа ^Армянское нагорье) отнесено к Севанскому району, а полупустыни и степи восточного Закавказья — к Куринскому району Переднеазиатского округа той же Восточносредиземноморской подобласти. Нагорностепной Талыш и долина среднего Аракса рассматриваются как коренные участки Переднеазиатского округа. Общая схема, историко-геологическая характеристика намеченных округов, районов, участков и их териокомплексов даны ниже и показаны в табл. 103 и на рис. 195.
ВОСТОЧНОСРЕДИЗЕМНОМОРСКАЯ ПОДОБЛАСТЬ
КАВКАЗСКИЙ ОКРУГ
(округ кротов и кустарниковой полевки)
Ареалы некоторых индикаторных видов кавказского териокомплекса занимают почти целиком площади, покрытые лесами предгорных наклон-дых равнин в Предкавказье, низовыми лесами — в Закавказье, горными лесами, высокогорными лугами и скалами перевалов. Они и послужили для выделения Кавказского округа, северная граница которого проходит в области предгорных наклонных равнин.
В степном Предкавказье к описываемому округу относятся два изолированных участка — Ставропольский и Пятигорский (рис. 195). В восточном Закавказье южная граница округа проходит на широте Шемахи, Ивановки, по долине Агри-Чая и Алазани, выходя к Кахетинскому хребту у Телави. В К а рта линии и Южной Осетии граница окаймляет сухую Горийскую котловину с севера и выходит к подножью Су-рэмского хребта. Отсюда она резко заворачивает к востоку по подножьям Триалетского хребта, проходит над Тбилиси, затем в нижней части долины Храма и у подножья Шах-Дагского хребта, южнее Шамхора и Кировабада. На юго-востоке граница окаймляет Карабах на высотах 600— 100 м у Мартуни, Карягино и, захватывая частично бассейн Охчи-Чая, поворачивает на север и северо-запад, огибая с востока и северо-востока высокие степи и лугостепи Армянского (Севанского) нагорья. Южная граница малокавказской части округа идет на запад по водоразделам Шах-Дагского и Бамбакского хребтов, проходит несколько южнее Киров-
31 Н. К. Верещагин , $
482
Часть III. Териогеографическое районирование и пути эволюции
акана и, огибая с севера Джавахетию, уходит в восточную Турцию по водоразделу Аджаро-Имеретинского хребта. Западная граница проходит от Трапезунда по Черноморскому побережью до Сухуми и, отклоняясь несколько к востоку, совпадает затем с границей Предкавказского района степных вселенцев.
На крайнем юго-востоке Закавказья к этому округу относится изолированный Гирканский, илиТалышско-Эльбурсский, район. Его границы почти совпадают с очертаниями лесной зоны восточных склонов Талыша и северных склонов Эльбурса. В пределах перешейка к этому району относится Талышский участок.
В основном массиве Кавказского округа выделен Дагестанский район, зажатый между мезофитными ландшафтами краевых хребтов восточного Кавказа (рис. 195).
Следует отметить, что граница Кавказского округа, проходящая по подножьям хребтов и предгорным низменностям, представляет местами широкую переходную полосу, на которой обитают нередко бок о бок степные звери равнин и звери горно-лесного ландшафта. Тем не менее с лесостепью Русской равнины эта полоса, вопреки мнению Кузнецова (1950), имеет мало общего даже в Предкавказье.
Главный массив округа существовал как суша по крайней мере с олигоцена, постепенно наращивая площадь по периферии. Возникновение крупных горных цепей произошло в миоцене. Центральные части горной системы Большого Кавказа были образованы юрскими сланцами и изверженными породами. Делювиальный чехол и почвенный покров оказались хорошо развитыми лишь на высоких плоскогорьях, в ущельях же коренные породы выходят на поверхность в виде скал и осыпей. Краевые хребты состоят преимущественно из песчаников и известняков, на которые и налегают в предгорьях отложения кайнозойских морей и аллювио-делю-виальные шлейфы.
Сходная картина наблюдается на Малом Кавказе и в Талыше, но меловые отложения встречаются здесь реже. В плиоцене и постплиоцене при замирании орогенеза происходила пенепленизация хребтов, чередующаяся с циклами горообразований и оледенений.
Древность формирования современных типов растительности Кавказского округа — по крайней мере нижнеплиоценовая. Современный тип зональности климата и вертикальных фитоландшафтных зон в пределах этого округа выработались уже к первой половине четвертичного периода.
Первобытные ранне голоценовые ландшафты Кавказского округа неоднородны на его огромной площади. В западной части предгорная Закубанская равнина местами слабо всхолмлена и пересечена многочисленными почти не меандрирующими речками.
В верхней половине эта равнина покрыта дубово-карагачевыми лесами, чередующимися с обширными полянами остепненных лугов. После перерыва в области центрального Предкавказья этот тип ландшафта появляется вновь на Терско-Сунженской наклонной равнине и окончательно исчезает к востоку перед Сулаком, восточнее Гудермеса.
В Закавказье — в Колхиде, в Алазано-Агричайской, Хачмасской и Ленкоранской предгорных низменностях — развиты особого типа низовые леса с преобладанием длиннолистного дуба, кавказской лапины и карагача, увитых колючими лианами. Для заболоченных участков характерна поросль черной ольхи. При обилии орехоплодных кустарников и деревьев на этих низменностях существовали благоприятные условия для обитания лесных грызунов, копытных и хищных.
Гл. V. Анализ голоценовой териофауны и схема районирования 483
Склоны хребта Черных гор, самой северной горной гряды Большого Кавказа, покрыты влажными буково-грабовыми лесами, причем местами на выходах известняковых пород существуют степные реликты травянистой растительности.
"Следующая гряда — Скалистый хребет — имеет на перевалах субальпийские луга, а на южных склонах — горные сосняки, замещающие буковники. По днищам торговых долин Кубани, Теберды и других рек, особенно в продольных широтных долинах от Эльбруса до центрального Дагестана, вклиниваются участки ксерофитной растительности горных склонов.
Большая часть горных склонов нижнего и среднего пояса южного склона Большого Кавказа, северных склонов Малого Кавказа и восточных Талыша покрыты буковыми и грабовыми лесами, создающими сезонное обилие корма для лесных грызунов и копытных.
В верхнем поясе горного леса западного Кавказа развиты пихтовые р и зновые леса, а на южном склоне — пихтово-кленовые. При развитом скальном рельефе они обильно населены снеговыми полевками и сернами. Верхняя опушка леса, как правило, представлена высокогорным дубом и березой. Характерны также заросли кавказского рододендрона, вдающиеся в зону высокогорных лугов.
Сама высокогорная зона альпийского типа, с мезофитными лугами, осыпями и скалами перевалов, с ледниками, расположенными на высотах 2800—3500 м, полностью выражена в западной и центральной частях Большого Кавказа, а также на наиболее высоких хребтах Малого Кавказа. В высокогорном Талыше верхняя опушка леса граничит с нагорной степью и формацией нагорных ксерофитов. В восточной части Большого Кавказа субальпийские луга, как правило, переходят в формацию плотнокустовых злаков, особенно типчака и белоуса, которая является как бы восточным вариантом альпийского луга. На средневысоких плоскогорьях Дагестана этот процесс остепнения при малых осадках и физиологической сухости тонкого слоя почвы пошел еще дальше. Нагорные остепненяые луга и лугостепи с низкими злаками оказались преобладающей формацией. Задернованные участки плато и склонов при обилии травяного корма и быстром скате дождевой и снеговой воды особо благоприятны здесь для подземных грызунов: полевок, хомяков. В этой же зоне находятся основные кормовые ресурсы высокогорных копытных.
Современный комплекс зверей Кавказского округа состоит, при самой общей характеристике, преимущественно из местных третичных видов мезофильного ряда, связанных с высокогорными лугами, горными и низовыми лесами. Формирование этого комплекса и развитие ареалов зверей, входящих ныне в его состав, происходили главным образом за счет местного видообразования и биоценотических процессов, протекавших здесь в неогене и четвертичном периоде.
Вселение в этот округ зверей со смежных территорий происходило на протяжении четвертичного периода, преимущественно в предгорную, переходную полосу.
В результате в териокомплексе Кавказского округа насчитывалось в голоцене до 88 видов, основная масса которых принадлежала I генетической группе (см. стр. 464). Слабо оказалась представленной II и несколько больше III группа; вовсе нет туранских форм, т. е. IV группы; довольно сильно выражено ядро V группы; отсутствуют вовсе представители VII, но имеется несколько видов VIII и X групп (табл. 103).
Индикаторными видами являются местные представители I группы.
484
Часть III. Териогеографическое районирование и пути эволюции
К этим мезофильным плиоценовым видам кавказского фаунистического ядра и широко распространенным принадлежат по крайней мере 48 видов, т. е. более половины общего числа голоценовых зверей округа (табл. 103).
В генетическом и экологическом смыслах эта группа неоднородна. В ней имеются звери, связанные с мезофитными лесами и луговыми формациями, довольно широко распространенные и в широколиственных лесах Европы, как например малая бурозубка, полчок, косуля, благородный олень. Более специфичен набор эндемичных кавказских высокогорных видов, связанных с мезофитными луговыми формациями альпийского типа. Характерные горные ареалы имеют здесь кавказская мышовка, прометеева полевка, длиннохвостая снеговая полевка и кавказская снеговая полевка. Более подвижные и крупные, но тем не менее стенотоп-ные виды, связанные с горным рельефом, осыпями, скалами, альпийскими лугами и горными лесами: кавказские козлы и серна, — как и названные грызуны, являются индикаторами кавказского териокомплекса.
Для Ставропольского, Пятигорского участков и для Дагестанского и Гирканского районов характерна ненасыщенность и значительная обед-ненность видами I группы, вызванная чаще всего антропогенными причинами, комбинировавшимися с послеледниковыми изменениями климата и ландшафта. Так, например, в Ставрополье, Пятигорье и Талыше отсутствуют теперь олень, козлы, серна, прометеева и длиннохвостая снеговая полевка; в центральном Дагестане — прометеева полевка и т. д.
Из недавно вымерших форм этой группы отметим речного бобра и кавказского горного сурка.
Иная судьба была у другого третичного реликта — черной крысы.
Виды плиоценовой древности — сухолюбивые и теплолюбивые (II группа) — представлены здесь рукокрылыми, хищниками, грызунами и копытными, вселившимися на соответствующие ландшафтные участки.
При преодолении экологических барьеров по периферии горных систем ряд видов этой группы, вероятно, мог бы населить и некоторые участки, лежащие внутри горной системы, например продольные долины в Сванетии, Кабарде, Дигории.
Живущих ныне видов не менее 16. Наиболее распространены из них подковоносы, куницы белодушка, заяц-русак, дагестанский хомяк, дикобраз, безоаровый козел.
Плейстоценовые вселенцы (V группа) — лесные и мезофильные по их экологической сущности — северного (европейского) происхождения. Они обитают большей частью только на Большом Кавказе в зонах горного леса и высокогорных лугов. Таковы, например, места обитания обыкновенной землеройки. Другие распространены шире, как например кавказский лесной кот и кавказская лесная куница, но их все же нет в центральном Дагестане и Талыше. Лесной куницы нет и в восточной части Малого Кавказа. Норка и полевая мышь, представленные предкав-казскими формами подвидового значения, заходят в пределы округа лишь в зоне предгорий северо-западного Кавказа. Своеобразны ареалы обыкновенной полевки, связанной преимущественно с луговыми формациями высокогорий, но отсутствующей в западном Закавказье, и рыжей понтийской полевки, часть ареала которой заходит из Малой Азии на северный склон Аджаро-Имеретинского хребта. Распространение кавказского лося, вымершего в прошлом столетии, было ограничено, видимо, и в плейстоцене Предкавказьем и западным Закавказьем.
Гл. V. Анализ голоценовой териофауны и схема районирования 485
Исключительным фактом является присутствие в округе представителя VI генетической группы — малого суслика, обитающего в сухих продольных долинах северного склона Большого хребта рядом с кавказской снеговой полевкой и кавказскими козлами.
*Из VIII группы — южных теплолюбивых форм, вселившихся недавно, отметим шакала, тигра и гиену, обитающих по окраинам округа.
Единственный вид X группы — пасюк — заселяет нижний горнолесной пояс Предкавказья и низовые леса западного и восточного Закавказья. Он особо многочислен в болотах Колхиды и Гирканики. В эту же группу следует отнести, вероятно, и несколько рас домовой мыши — $us musculus subsp., завезенных на Кавказ из более северных районов.
Кавказский округ следует рассматривать не только как климатический и ландшафтный рефугиум для всех мезофильных видов животных вообще, но главным образом как убежище для крупных промысловых млекопитающих, вытесненных человеком из предгорных обезлесенных равнин, низменностей и с высоких безлесных плоскогорий. Это — об-обшпрный природный заповедник, сохраняющий уцелевший запас крупного зверя. Такой фонд еще может послужить в будущем для самых разнообразных научных и практических целей.
Именно защитными особенностями горного рельефа и лесной растительности объясняется то поразительное обилие числа видов и особей крупных хищных и копытных, которое уцелело здесь с плейстоцена по сравнению со смежными равнинными и безлесными территориями.
Несомненно, что современные горно-лесные коты, барсы, горные кабаны, олени, косули, зубры при прекращении преследования человеком и развитии на смежных пространствах подходящих ландшафтов выселились бы на окружающие равнины и Кавказ явился бы, таким образом, своебразным новейшим центром расселения.
Выделение высокогорного, или альпийского, комплекса и района на основе изучения млекопитающих обосновать трудно. Дело в том, что в высокогорной зоне Кавказа нет ни одного млекопитающего, которое бы так или иначе не было связано с субальпийской и с горно-лесной зоной. Так называемые альпийские виды — прометеева полевка, кавказская мБЛповка — обитают, кроме альпийских лугов, еще и па субальпийских п луговых полянах в середине зоны горного леса. Кавказские козлы, серна, кавказская снеговая полевка достигают наибольшей численности именно в лесной зоне. Таким образом, здесь можно говорить лишь об альпийской экологической группировке. Правда, настоящие альпийские виды не распространяются до нижнего горно-лесного пояса и отсутствуют в предгорных низменностях, но этим и обусловлен переходный характер названных фитоландшафтных и геоморфологических категорий.
Экологическая группировка зверей высокогорной зоны Большого Кавказа относительно бедна видами. В ней насчитывается 3 вида насекомоядных, 8 видов хищных, из которых 5 бывают только летом; 1 вид зайцеобразных, 8 видов грызунов и 5 видов копытных, из которых 2 прикочевывают только на летний сезон.
Учитывая миоценовую древность горного рельефа Большого Кавказа п верхнемиоценовую древность кавказских горных растительных формаций, начало обособления этих форм следует относить, вероятно, к нижнему плиоцену. Очевидно, с нижнего плиоцена ведут начало и известные ныне морфо-физиологические адаптации высокогорных млекопитающих, отражающие до известной степени процесс горообразования и сопровождающие его физические явления.
486 Яастъ III, Т ериогеографическое районирование и пути эволюции
У кавказских высокогорных зверей имеются общеизвестные примеры адаптаций к пониженному атмосферному давлению и температуре в виде: повышенного обмена веществ, повышенного количества эритроцитов и гемоглобина крови, увеличенного сердца и т. и. Развитие привычек хождения и лазания по карнизам и наклонным стенкам скал у козлов, серн, снеговых полевок и ослабление привычек к рытью нор у последних явились следствием формирования рваного рельефа и россыпей скал. Суточные и сезонные миграции кабанов, косуль, оленей, козлов, серн по вертикали и горизонтали развивались под воздействием кровососущих двукрылых, распределения минеральных источников, сезонного недостатка кормов при выпадении снега. Сенозаготовительная деятельность у снеговых полевок возникла, несомненно, в связи с обилием укрытий в скалах, обеспечивающих сохранность подсохшего корма, несмотря на обилие осадков.
Возникновение ряда эндемичных высокогорных роющих грызунов — кустарниковой и прометеевой полевок, как видов, питающихся надземными частями травянистых растений, — было обусловлено здесь исчезновением древесно-кустарниковых формаций и развитием пространств увлажненных лугов, лугостепей и оголенного грунта при подъеме хребтов.
Упомянутая относительная бедность видового состава зверей высокогорной зоны Большого Кавказа, хорошо заметная даже при сравнении с. остепненным плоскогорьем смежного Малого Кавказа, была обусловлена, на наш взгляд, четырьмя главными причинами:
1) значительной быстротой горообразования, вследствие чего много специализированных видов миоценового фаунистического комплекса горного леса, очевидно, попросту вымерло;
2) отсутствием других близких центров горного видообразования, т. е. относительно древней изолированностью страны и затрудненностью новых вселений;
3) территориальной ограниченностью жизненной арены высокогорья, обусловленной рваным рельефом: крутизной склонов и отсутствием пригодных для миграций зверей высокогорных плато; именно этим можно объяснить, например, современное отсутствие здесь горных баранов и сурков;
4) слабым влиянием оледенения Европы на продвижение таежноарктических видов, которые не смогли колонизировать Кавказское высокогорье.
В четвертичном периоде происходили относительно небольшие сдвиги вертикальных границ распространения кавказских млекопитающих, обусловленные наступлениями и отступлениями ледников и соответственными сдвигами фито ландшафтных зон. Разрывы ареалов названных выше эндемичных видов на два участка — кавказский и малокавказский — произошли преимущественно в послеледниковье, с развитием ксерофит-ных формаций на Сурамском хребте.
Наивысшее развитие кавказский тип горно-лесного териокомплекса имеет на хребтах и склонах, окаймляющих Колхидскую (Рионскую) низменность. Эту насыщенность кавказскими эндемиками западного Кавказа вряд ли следует объяснять существованием плиоценовой и четвертичной Понтиды на месте современной Черноморской котловины, как это иногда пытаются делать. На восток по Большому и Малому Кавказу эти древние автохтонные элементы и более поздние плейстоценовые фаунистические наслоения, связанные с высокогорными лугами и горными широколист
Гл. У. Анализ голоценовой териофауны и схема районирования 481
венными лесами, постепенно убывают, их распространение становится прерывистым или даже переходит в иные зоны и зоогеографические районы.
Сама Колхидская низменность отличается теперь значительной бедностью видового состава зверей, особенно грызунов и копытных. Для роющих грызунов здесь очень тяжел режим периодически затапливаемых пространств, а многие копытные, судя по палентологическим и историческим фактам, были уничтожены человекам в нашу эпоху.
”аким образом, учитывая насыщенность кавказского териокомплекса, ;азский округ можно было бы разделить на Западный и Восточный районы Большого Кавказа — примерно на долготе Тбилиси и Казбека — н на район Малого Кавказа. Последний несколько обеднен видами по сравнению с Большим Кавказом. Здесь нет теперь кавказских козлов, < ц распространение лесной куницы и т. д.
апболыпим фаунистическим своеобразием отличаются внутренний Дагестан и горно-лесной Талыш. По происхождению они принадлежат, несомненно, к Кавказскому округу. Однако в связи с глубоким преобразованием исходного фаунистического комплекса в четвертичном периоде они заслуживают выделения в особые категории.
Дагестанский район
(район дагестанского хомяка и безоарового козла)
Территория этого района занимает бассейны Аргуни, Андийского Копсу, Аварского Койсу и Самура. Северная граница проходит по хребту Черных гор, по линии Шатой—Буйнакск и выходит на западе к верховьям Поры и Крестовому перевалу. Южная граница идет от верховьев Иоры по водораздельному хребту до горы Баба-Даг. Восточная граница тянется по линии от Буйнакска на гору Баба-Даг. В четвертичном периоде эта страна испытывала поднятие и энергичное расчленение речной эрозией. Судя по наличию фаланг и скорпионов, страна имела сухие ландшафты на протяжении всего кайнозоя и древние фаунистические связи с южными массивами суши. В фитоландшафтном смысле район характеризуется формацией горного остепненного луга и лугостепи, а на высоких плоскогорьях имеются березовые рощи и субальпийские остепненные луга. По днищам и южным склонам ущелий характерны полынные ассоциации с примесью сухолюбивых кустарников: держи-дерева, барбариса. В верховьях Аргуни и Андийского Койсу развиты горные сосняки и буковые леса.
При таком безлесье, сухости и высокогорности Дагестанского района его кавказский териокомплекс сильно обеднен древними лесными и высокогорными видами. Здесь нет кротов; из 22 видов рукокрылых, известных для Кавказского округа, во внутреннем Дагестане документально отмечено только 6, в том числе обе широкоушки, нетопырь-карлик, остроухая и яаттерерова ночницы, поздний кожан. Нет здесь прометеевой полевки, а длиннохвостая снеговая полевка держится только в верховьях Аварского Койсу, нет косули, а олень бывает только на летовках. Мало серны, которая держиться лишь у краевых хребтов, там, где имеются леса. Обилен восточнокавказский козел, и, видимо, недавно вымер муфлон.
В то же время присутствие переднеазиатских горных и сухолюбивых форм —безоарового козла, дагестанского хомяка, образовавшего в изоли-
488 Часть 111. Териогеографическое районирование и пути эволюции
рованных ущельях локальные крупные расы, свидетельствует о древних — верхцёплиоценовых, среднеплейстоценовых — связях с южным Закавказьем. Характерны мелкие расы полевок: кавказской снеговой и водяной, живущей здесь на сухих и поливных участках подобно горной сле-। цушонке.
- Гирканский район
" * -
(район талышского крота и полчка)
Район охватывает низменное побережье юго-западного и южного Каспия, восточные склоны Талыша и северные склоны Эльбурса. В пределах перешейка расположены лишь Ленкоранская низменность и горнолесной Талыш, которые могут быть выделены в Талышский участок этого района.
Северная граница Талышского участка начинается с южной Мугани (от сел. Масаллы) и отрогов Талыша у Астрахан-Базара. Восточная граница идет по берегу моря от устья Виляж-Чая, а западная проходит на высоте 1700—1800 м, совпадая с нижней границей нагорной степи. В геологическом смысле наиболее молода заболоченная зимой Ленкоранская низменность, сложенная выносами речек Талыша и морскими отложениями. Фитоландшафты низменности и горных склонов представлены сплошным массивом широколиственных низовых и горных лесов с небольшим числом теплолюбивых эндемиков третичного возраста, как например шелковой акации, железняка и колючих лиан.
Геоботаники и зоогеографы обычно выделяют южное побережье Каспия в особую — Гирканскую — провинцию в связи с ее современной изолированностью и значительным своеобразием. С точки зрения палео- и териогеографии Талышско-Эльбурсский горно-лесной массив должен быть все же объединен с Кавказским округом по следующим соображениям.
Еще в нижнем миоцене северные отроги Эльбурса, Талыша, Малого Кавказа и Восточного Тавра представляли собой единое южное побережье миоценового бассейна и, следовательно, несмотря на воздействие с востока полтавской флоры, имели много общего в типе ландшафта. Эти общие ландшафтные черты сохранились и до наших дней, хотя известная изоляция упомянутых участков побережий наступила уже в верхнем миоцене—сармате. Так, например, морфологически и по видовому составу болотные ольшаники и горные буков ники Гиркании весьма сходны, а часто и неотличимы от соответствующих формаций Колхиды.
Талышский участок отличается своеобразным климатическим и почвенным режимом. Горные склоны и приморская низменность крайне пересыхают летом, хотя здесь зимой и выпадают обильные осадки. Субальпийской зоны здесь практически нет, а верхняя опушка широколиственного леса, образованная талышским дубом и фруктовыми розоцветными, переходит в ксерофитное высокотравье, а затем в нагорную степь или в формацию колючих астрагалов. В связи с этой летней сухостью, крайней уплотненностью желтоземов и отсутствием луговых формаций на низменности нет ни крота, ни лесной мыши, ни кустарниковой полевки, а водяная крыса водится лишь в немногих непересыхающих болотах — бывших морских заливах (морцах). Обыкновенная и снеговая полевки также не живут здесь в лесной зоне, а встречаются лишь па наиболее высоких участках нагорной степи.
Вообще древних миоцено-плиоценовых млекопитающих Средиземноморья в районе теперь мало, но все же они есть, и здесь лежит крайний юго-восточный предел их распространения.
Гл. V. Анализ голоценовой териофауны и схема районирования 489
Талышский участок характеризуется крайней обедненностью кавказскими млекопитающими. Здесь нет кавказской белки, кавказской мышовки, прометеевой и кавказской снеговой полевок, малоазийской мыши, кавказских козлов, серны. Возможно, впрочем, что некоторые из названных мелких зверьков еще найдутся на Эльбурсе.
Кроме остатков древнего третичного ядра и широко распространенных видов, относимых к I группе нашей схемы, голоценовый териокомплекс района сложен представителями II, VIII и X групп.
Из Г группы широкое распространение в районе имееет темная длиннохвостая белозубка. Крот и бурозубка-малютка обитают в верхней, более влажной части зоны горного леса.
Рукокрылые Талыша весьма обильны. Они преобладают на низмен-I юти, тяготея к постройкам и дуплам. Видовой состав летучих мышей южного побережья Каспия известен, впрочем, недостаточно.
Распространение хищных этой группы не несет в Талыше оригиналь-। ,ix черт. Лиса в горных лесах крайне редка, как и всюду на Кавказе. Ь елкий южный медведь здесь тоже редок. Выдра особенно многочисленна по глухим рыбным речкам.
Грызуны представлены по преимуществу древесными формами. Осо-( чгно многочислен полчок, распространенный от верхней опушки леса ; ) берега моря. Черная крыса живет только на низменности, а лесная ? ышь — только в горных лесах. Кустарниковая полевка живет лишь - верхней трети горного леса и крайне редка. Бобра ныне нет нигде, но воз-ожно, что он существовал на низменности еще в голоцене. Из копытных наиболее многочислен кабан. Олень в Талыше уничтожен в начале ашего столетия, но есть еще в лесах Гиляна и Мазендерана. О возмож-юсти существования зубров и первобытных быков уже говорилось в ; III главе.
Южные— переднеазиатские — виды, связанные с сухими ландшафтами (II группа) в районе малочисленны. К ним отнесены: куница бело-душка,заяц и степная полевка, проникающие из Муганской степи и степного высокогорья, а также дикобраз, распространенный от уровня моря до перевалов Талыша.
Безоарового козла на Талыптском участке теперь нет, но, вероятно, он водился здесь раньше. На Эльбурсе он обитает и сейчас.
В Гирканском районе в силу исторических причин и особенностей экологии нельзя и ожидать присутствия европейских и восточноевропейских гидрофильных, лесных и степных видов зверей плиоценового и плейстоценового возраста, так же как и ту рано-афганских, пустынных.
Второе место по значимости занимают виды, проникшие на перешеек в голоцене (VIII группа).
Среди насекомоядных и рукокрылых таких видов достоверно неизвестно, но, очевидно, они есть, так как Талыш и Эльбурс также испытывали влияние плейстоценового похолодания. Более достоверны представления о недавнем появлении здесь шакала, полосатой гиены, тигра и, возможно, льва.
Из случайных, завозных видов (X группа) укажем на пасюка, живущего теперь в болотах низменности и горных лесах.
Териологическая характеристика Гирканского района ясно показывает, что это — типичный уголок восточного мезофит ног о Средиземноморья, а все «индийское» в нем — это лишь позднее наслоение.
490 Часть III. Териогеографическое районирование и пути эволюции
В связи с установлением древней самобытности развития кавказского горно-лесного териокомплекса возникает вопрос о степени влияния его на териокомплексы смежных территорий, т. е. юга Русской равнины, пустынь Закаспия и нагорий Передней Азии. Характер и степень этого \ влияния определялись большой специализацией представителей кавказского комплекса к горному рельефу и к условиям большой влажности лесных и луговых ландшафтов.
Эти приспособления исключали миграции большинства кавказских зверей на остепненный север и опустыненный северо-восток. Такая закономерность начала проявляться уже с среднего плиоцена, когда широкие пространства суши, обнажившиеся вслед за отступившим Понтическим бассейном, были, вероятно, опустынены при жарком и сухом климате века продуктивной толщи. Даже в ледниковом плейстоцене горнолесные виды выселялись лишь на наклонные предгорные равнины Предкавказья, достигая в лучшем случае Ставропольской возвышен-( нести.
Южный, юго-западный и юго-восточный пути расселения были более *•’ доступны для кавказских млекопитающих. Влияние высокогорной груп-* пировки кавказских млекопитающих сказывалось в альпийской зоне Армянского нагорья и в восточной Турции.
В этих направлениях могли расселяться: обыкновенная полевка, кавказская снеговая полевка, кавказская мышовка, кавказский олень, ; косуля, возможно даже кавказские козлы. Такие расселения на юг могли j: быть значительны в эпохи крупных похолоданий. Позднее — в голоцене — J все эти влияния были сглажены, а местами и уничтожены наступившим потеплением и иссушением ландшафтов.
Более существенна была роль горно-лесной территории Кавказского округа в качестве проводника на восток европейских лесных видов. Расселение из Европы через Балканы и по северным лесистым склонам Тавра ясно намечается для лесной куницы, понтийской рыжей полевки, орешниковой сони и, возможно, для лося.
ПЕРЕДНЕАЗИАТСКИЙ ОКРУГ
(окрух' малоазийского хомяка и песчанок)
Под Переднеазиатским округом здесь подразумевается Восточносредиземноморская провинция других авторов без Большого Кавказа и Крыма: территория Малой Азии, северо-западного Ирана, Армянского нагорья и низменности восточного Закавказья.
Индикаторные для этого округа звери принадлежат к двум подкомплексам — нагорно-пустынному и нагорно-степному, — образуя соответственно два варианта ареалов. Эти ареалы местами мозаично, а местами высотно-зонально сменяют друг друга. Нагорно-степной вариант не дает на перешейке резко выраженных границ, так как его представители: средние хомяки, горный тушканчик, безоаровый козел и другие — более широко специализированные виды, трансформируясь проникали по подходящим ландшафтам далеко к северу (рис. 189).
Нагорно-пустынный вариант включает узко специализированные, пустынные виды и уже поэтому более консервативен, образуя резко , выраженную северную границу ареала.
Наибольшее выполнение северной границы ареала нагорно-пустынного подкомплекса дают песчанки: персидская, малоазийская, Виногра
Гл. V. Анализ голоценовой териофауны и схема районирования
491
дова. Эта граница очерчивается более или менее резко, хотя и она причудливо врезается местами по ущельям и долинам рек в Севанский район в соответствии с экспозицией склонов и развитием ксерофитных формации (рис. 190).
Теологическая древность большей части территории округа по крайней мере олигоценовая. В долине среднего Аракса известно местонахождение остатков олигоценовых растений, близ Ахалциха — захоронение остатков ; олигоценовых млекопитающих, у оз. Урмии — верхнемиоценовых, а по ущелью Занги — плиоценовых млекопитающих. Окраины округа, = например Куринская низменность, значительно моложе — верхнеплио-I ценового и четвертичного возраста.
; Преобладающий современный ландшафт нагорий северо-западного t Ирана, восточной Турции и долины среднего Аракса — это выжженные солнцем хребты из вулканогенных и осадочных толщ, чередующиеся ! с долинами и узкими ущельями. Склоны гор покрыты группировкой [ трагакантовых астрагалов с мелкими сухолюбивыми кустарниками, > крушиной, миндалем, эфедрой. Местами сохранилось можжевеловое редколесье с группами восточного боярышника и иволистной груши. Для жарких широких долин характерна формация горной древовидной солянки и полыни, а для берегов речек — тугайные заросли из разно-листного тополя, лоха, облепихи и тамарикса.
Участки плоскогорий на высотах 1600—1900 м покрыты нагорно-стегиыми группировками с преобладанием овечьей овсяницы, а местами и рыхлокустовых злаков. Луговые формации альпийского типа встре-Е чаются лишь на высотах не ниже 2000 м, например в районе Хоя, Баязета.
В восточной Турции, в районе Вана, Карса, Кагызмана, уцелели еще , и небольшие массивы горных сосновых, реже дубовых лесов. Дубовые ! леса сохранились местами и на Армянском нагорье.
Териокомплекс округа сформирован представителями I, II, IV, V ; и VIII генетических групп.
Следует отметить, что большинство представителей VII группы, являю-J щихся для более северных частей перешейка послеледниковыми вселен-I цами, на территории этого округа — либо аборигены, либо пришельцы ; с юга, значительно более ранние. Другими словами, VIII группа для этого округа может быть частично упразднена и переведена во II.
В пределах нашей страны типичный териокомплекс округа представлен лишь на двух небольших участках. Кроме того, к этому же округу относятся два крупных района с сильно преобразованными териокомплексами.
Общая и териогеографическая характеристика участков и районов этого округа, лежащих в советских пределах, даны ниже и в табл. 103.
Западноиранский район
Араксинекий участок
(участок персидской песчанки и мышевидного хомячка)
Северная граница этого участка начинается на западе в районе Кагызмана и идет по южному склону Армянского нагорья и Карабаха, уходя на восток за Араке, в иранскую Мугань в районе устья Акеры. Южная граница условно проводится по Араксу. Таким образом, в этот участок входит вся левая половина долины среднего Аракса с ее свое
492 Часть III. Териогеографическое районирование и пути эволюции
образными скальными возвышенностями: Алинджа-Даг, Илянлу-Даг, Дарры-Даг, Неграм-Даг. Основной тип растительности — это полынносолянковая группировка и злаковый эфемеретум. По берегам речек в голоцене были развиты тугайные заросли, позднее почти уничтоженные, а местами замененные искусственными рощами из платанов и грецких орехов.
Голоценовый териокомплекс Араксинского участка содержит почти полный набор видов переднеазиатского типа в его нагорно-пустынном варианте.
Представители I группы — все довольно широко распространенные виды, и настоящих мезофильных кавказцев среди них, в сущности, нет. Преобладают рукокрылые: ночницы, кожаны; хищники и мышевидные грызуны.
Большинство видов этой группы тяготеет здесь либо к уцелевшим лесистым участкам, либо к биотопам культурного ландшафта — садам, поселкам.
Представители II и VIII групп (идентичной здесь со II) дают основной фаунистический фон: ушастый еж и мелкие белозубки — из насекомоядных; подковоносы — из рукокрылых; гиена, куница белодушка, перевязка и другие — из хищных; хомяки, песчанки — из грызунов; кулан, безоаровый козел, муфлон, джейран — из копытных. Всего до 34 видов. Индикаторными видами следует считать персидскую песчанку, горную слепушонку, мышевидного хомячка.
Значительно число недавно вымерших видов: кулан, лошадь, олень, джейран, дромадер (?). Их исчезновение объясняется ранним освоением участка скотоводческими племенами и легкостью истребления крупных зверей в условиях межгорной долины. Среднеазиатские, или ту рано-афганские, элементы (IV группа) представлены здесь слабо. К ним принадлежат манул, аралыхский малый тушканчик, и, возможно, араздаян-ская песчанка. В области среднего Аракса лежит их современный северо-западный предел.
В составе послеледниковой (VIII) группы отметим здесь тигра, камышовую кошку и шакала. Тигры изредка забредали сюда еще в прошлом столетии. Камышовый кот живет и сейчас по тугаям и камышам долины Аракса. Шакал в долине среднего Аракса исключительно редок. Весьма возможно, что послеледниковое потепление продвинуло с юга в пределы участка ряд видов рукокрылых, которые признаны нами в качестве древних средиземноморцев. В качестве новейшего завозного вида (X группа) отметим пасюка.
Нагорно —-Талышский степной участок (участок персидской песчанки и серого хомячка)
Участок занимает плоскогорье в верховьях Виляж-Чая и Зувандскую котловину в верховьях Вашару-Чая и Вазару-Чая, близ Кельвяза и Келаханы. Нижние пределы этого участка лежат на высотах 1700—1800 м по верхней границе леса и суховыносливого высокотравья на западных, северных и восточных склонах Талышского хребта. Ландшафт плоскогорья представлен пологими склонами и выравненными площадками с каштановой почвой, чуть прикрытой низкими рыхлокустовыми злаками: костром, заячьим ячменем и другими, — чередующимися с формацией колючих астрагалов и подушек акантолимонов. Местами возвышаются
. Гл. V. Анализ голоценовой териофауны и схема районирования 493
; обособленные вершины Кыз-Юрды, Келаханской сопки и других с россы-; пями скал изверженных пород.
Голоценовый териокомплекс участка сформирован преимущественно видами II и V генетических групп. Из насекомоядных здесь обычны 1 длиннохвостая белозубка, ушастый еж. Из хищных — лиса, волк, Ж барсук. Характерно обилие русака. Из грызунов обитают: малоазийский тушканчик, степная полевка, горная слепушонка, персидская песчанка, 1 серый хомячок и малоазийский хомяк. На Келаханской сопке живет * малоазийская снеговая полевка, распространение которой здесь явно S реликтовое. На возвышенных участках плоскогорья, там, где культи-ж вируется рожь, обитает обыкновенная полевка.
I Копытных теперь нет, но несомненно, что еще недавно здесь жили ® кулан, баран и безоаровый козел.
V Севанский район
ж (район малоазийской снеговой полевки и малоазийского хомяка)
V Севанский район занимает остепненные нагорья Малого Кавказа, у Северная граница этого района проходит по перевалам Аджаро-Имеретинского хребта, огибает с севера нагорье Джавахетии, включая Мокрые горы, Сомхетский и Безобдальский хребты. Далее на восток она идет по водоразделу Бамбакского хребта и окаймляет с севера оз. Севан и часть остепненных нагорий Карабаха. Южная граница проходит на высоте 1500—1700 м на южном склоне Карабахского, Зангезурского # и Даралагезского хребтов; западнее проходит над Ереваном и по южному ® склону Алагеза, а затем уходит в пределы Турции, в районе Кагызмана. Ж По типу ландшафта Севанский район — это высокое плоскогорье т с мягкими очертаниями хребтов и обособленных вершин. На более низких участках расположены степные формации с преобладанием типчака и рыхлокустовых злаков. По ущельям и выгревам, а также по мелкокаменистым россыпям на плоскогорье выходят формация трагакантовых астрагалов и большей частью уничтоженное ныне аридное редколесье с можжевельником, иволистной грушей, восточным боярышником. Изредка по северным склонам ущелий сохранились участки дубовых лесов. Для возвышенных участков, начиная с 2000 м, характерны остепненные, а выше и настоящие мезофитные луга.
«Вообще Севанский район следует рассматривать как район борьбы ксерофитных горно-пустынных ландшафтов центральных частей Передней Азии и мезофитных лесных ландшафтов кавказского типа. Эти черты борьбы ландшафтов отражены и в голоценовом териокомплексе района. Этот комплекс совершенно не несет оригинальных черт, а состоит из смеси кавказских и переднеазиатских видов.
Наибольшее выполнение границ района дают относительно мезофильные или, вернее, относительно суховыносливые виды грызунов, обитающие в ландшафте нагорной степи и остепненного луга. Но и их распространение уходит далее, правда лишь на больших высотах, в Иран и восточную Турцию.
Териокомплекс района сформирован из представителей I, II, V, VIII и X групп, всего до 43 ныне живущих форм.
Наиболее полно представлены I и II группы; отсутствуют представители европейской древней лесной и восточно-европейской степной фауны (III и VI группы); нет здесь и туранцев (IV и IX группы).
494
Часть III. Териогеографическое районирование а пути эволюции
Состав древних кавказских и широко распространенных видов (I группа) по сравнению с таковым Кавказского округа сильно обеднен. Тем не менее здесь обитают: закавказский еж, кавказский крот, кавказская бурозубка, складчатогуб, ночницы, южный медведь, кавказская мышовка, кустарниковая и кавказская снеговая полевки, косуля.
В голоцене, вплоть до наших дней, здесь вымерло 5 видов копытных, частью живших оседло, частью прикочевавших из лесной зоны северных склонов Малого Кавказа: кабан, олень, серна, зубр, первобытный тур.
Перечисленные виды, за исключением рукокрылых и хищников, занимают и занимали наиболее мезофитные, либо даже гигрофитные биотопы на повышенных участках района; на берегах ручьев, родников, заболоченностей и в уцелевших рощах широколиственного леса.
Рысь и медведь живут в остатках дубовых лесов.
Из древних плиоценовых видов ирано-малоазийского или восточносредиземноморского корня (II группа) в Севанском районе живут звери, связанные с биотопами нагорной степи. Нагорно-пустынные виды представлены слабо: ушастый еж, белозубка Зарудного), подковоносы, куница белодуптка, русак и вымершая теперь пищуха, горный тушканчик, малоазийский суслик и горный слепыш, малоазийский хомяк, снеговая полевка Сатунина и горно-степная полевка Шидловского, безоаровый козел и муфлон. В голоцене вымерла лошадь и, возможно, дромадер.
Малоазийский суслик и горный слепыш заселяют теперь лишь западную часть района. Однако распространение на плоскогорье этих видов было подвержено в голоцене значительным колебаниям.
Местами представители I и II групп образуют в районе мозаичную смесь видов, обусловленную горным рельефом.
При полном отсутствии древних европейских лесных, степных и гидрофильных видов, а также среднеазиатских полупустынных и пустынных здесь обитают некоторые виды северного (европейского?) происхождения, распространявшиеся, очевидно, позднее — в плейстоцене (V группа). Таковыми следует считать обыкновенную полевку (?), живущую на субальпийских лугах, и кавказскую лесную кошку, живущую в уцелевших лесах. В качестве послеледниковых вселенцев (VIII группа) трудно теперь назвать для района достоверно какие-либо виды, но, очевидно, что они имеются в различных отрядах. В частности, весьма вероятно, что на Армянском нагорье после верхнеплейстоценового похолодания появились уже вторично некоторые рукокрылые и начали заходить сезонно южные хищники: полосатая гиена, лев, тигр.
Пустынные виды копытных — кулан и джейран, — вероятно, также заходили временами на более или менее пологие, остепненные участки плоскогорья из долины среднего Аракса.
Из новейших синантропных вселенцев укажем на пасюка и домовых мышей.
Формирование экологических группировок и териокомплекса Севанского района в четвертичном периоде шло в условиях энергичного горообразования, сопровождавшегося разломами и обширными излияниями лав, перекрывавших в различных направлениях западную часть страны.
Оледенение хребтов нагорья в плейстоцене не было велико и не играло существенной роли в формировании здешнего териокомплекса. Освоение человеком этой территории, наметившееся уже в ашеле и развившееся в век бронзы и железа, было основной причиной исчезновения здесь крупных млекопитающих.
Гл. V. Анализ голоценовой териофауны и схема районирования 495
В пределах очерченной территории Севанского района можно выделить в качестве обособленного участка холодное Ахалкалахское нагорье— *Округ истоков реки Куры» Сатунина. Отличия этого участка от остальной территории района состоят в присутствии здесь гельского слепыша, а на ограничивающем с северо-востока хребте Мокрых гор — ь кавказской снеговой полевки и кавказской мышовки. Холодный климат f плоскогорья Джавахетии и обилие родниковых вод создали здесь усло-f вия для процветания таких видов, как водяная кутора, обыкновенная . полевка, и обусловили отсутствие степной полевки. Природное безлесье f плоскогорья и его равнинный рельеф привели также к раннему вытес-f нению человеком крупных хищных и копытных.
Куринский район
{(район степной полевки и краснохвостой песчанки)
gg Куринский район зажат между восточными частями Кавказского Ж: округа и довольно полно совпадает с площадями полынно-солянковой ® полупустыни и бородачевой степи в низменностях восточного Зйкав-Я казья С востока район ограничен Каспием и вытянут клином к подножью ЯСурамского хребта.
Между Карабахом и Талышом район переходит в основной массив Я@ападноирайского района Переднеазиатского округа.
ЯЬ Геологическая древность территории района неоднородна. Восточная ^его часть в пределах горизонтали — 20 м заливалась, видимо, морем даже в голоцене, а в пределах горизонтали +30, +40 м заливалась кодами Хвалынского, Хазарского, Бакинского морей. Предгорья и предгорные низменности Малого и Большого Кавказа, и вся долина сред-3№ей Куры от Мингечаура до Гори имеют верхнетретичный возраст.
Hr В ландшафтном и геоботаническом смысле район включает ровные ^пространства солянковой и полынной полупустыни, эфемеретумной и бородачевой степи, тугайные леса по Куре, Араксу и их притокам, камышовые заросли озер и разливов, размытые глинистые хребты — боздаги с можжевелово-фисташковыми лесами по северным склонам. Древность этого ландшафта по крайней мере нижнеплейстоценового возраста, но корни его уходят, вероятно, и в верхний плиоцен.
Куринский район отнюдь нельзя назвать бедным или ненасыщенным видами, как это нередко представляется зоогеографам, ссылающимся па то, что степи Кура-Араксинской низменности молоды и недавно вышли из под вод моря.
Бедной видами можно считать лишь группировку млекопитающих переднеазиатских нагорий, которая составляла древнюю основу формирования здешнего териокомплекса. Кроме того, ненасыщен биотоп открытой ровной полупустыни, особенно в Ширванской степи. По генетическому признаку териокомплекс района формировался из элементов 1. II, IV, VIII и X групп.
Кавказских мезофильных и широко распространенных видов (1 группа) здесь не менее 25. Наиболее характерны: кавказский крот, черная крыса, кабан и олень, тяготеющие к реликтовым и мезофитным местообитаниям (тугаям, оазисам, тростниковым болотам и т. п.). Хищные представлены широко распространенными видами; волком, мелкой лисицей, лаской, барсуком, выдрой и медведем.
. 496 Часть III. Териогеографическое районирование и пути эволюции
До 22 видов плиоценовой древности — II группы — являются основой териокомплекса района.
Большинство видов этой группы имеют здесь ареалы, уходящие основной площадью на юг, юго-восток и юго-запад, подтверждая правильность выделения этого района. Индикаторные насекомоядные — ушастый еж, белобрюхая, длиннохвостая белозубки, белозубка-малютка — характерны для сухих биотопов открытой полупустыни.
До 9 видов индикаторных рукокрылых тяготеют к пещерам, постройкам, а кормятся вдоль тугаев и опушек оазисов. Наиболее часты подковоносы, средиземноморский нетопырь Куля, перевязка и пятнистый кот, ареалы которых заходят на Кавказский перешеек с юга именно в этой части восточного Закавказья. Недавно вымер гепард.
До 6 видов переднеазиатских грызунов имеют в районе либо реликтовые ареалы, как например у малоазийского тушканчика и закавказского хомяка, либо неразвитые ареалы, как у малоазийской песчанки. Наибольшую вагильность и численность имеют здесь серый хомячок, краснохвостая песчанка и степная полевка.
Из непарнокопытных в голоцене здесь, возможно, жила еще лошадь южного типа, а из парнокопытных — аргали.
Представителей III группы — европейской —[ здесь нет.
Виды плиоценовой древности — восточные, Туранские и турано-аф-ганские (IV и IX группы) — немногочисленны. В этом отношении комплекс млекопитающих района, очевидно, существенно разнится от комплекса насекомых, в частности от набора жуков (см. Семенов-Тян-Шанский, 1936). Среди насекомоядных, зайцеобразных, непарнопалых и парнокопытных ни ранних, ни поздних туранцев здесь нет.
Только малый тушканчик, обитающий по преимуществу в эфемере-тумной полупустыне на Апшероне и в Кабристане, может быть отнесен к древним иммигрантам туранского происхождения.
Напомним,что в среднем плейстоцене в этот районраспространялись некоторые степные виды с Русской равииныжорсак,большой тушканчик,сайга.
Южные, теплолюбивые виды, проникшие на перешеек в послеледниковое время (VIII группа), представлены всего 6—7 видами. Возможно, что в эту группу следует отнести еще некоторых мелких насекомоядных и рукокрылых, которых мы считали плиоценовыми вселенцами.
Не возбуждает сомнений позднее появление в районе шакала, полосатой гиены, тигра, льва, кулана, джейрана, из которых тигр, лев и кулан совершенно вымерли. К случайным видам, вселившимся в район в историческую эпоху, относится пасюк, обитающий ныне по озерам, болотам и во всех крупных населенных пунктах.
В настоящее время в результате сельскохозяйственной деятельности человека происходит дальнейшее насыщение района мелкими кавказскими видами.
ПОДОБЛАСТЬ ВОСТОЧНОЕВРОПЕЙСКИХ И СЕВЕРОКАЗАХСТАНСКИХ СТЕПЕЙ
ВОСТОЧНОЕВРОПЕЙСКИЙ ОКРУГ
Предкавказский район
(район обыкновенного хомяка и полевой мыши)
Ареалы индикаторных видов зверей этого района развивались с севера. Поэтому его северная граница проводится чисто условно по Ма-
Гл, V, Анализ голоценовой териофауны и схема районирования
497
нычу. Западная граница проходит по морским побережьям от низовьев Дона до устья Кодора. В западном Закавказье к этому району относится лишь узкая полоса береговых террас, по которым проникает к югу несколько восточноевропейских видов. Южная граница совпадает всюду северной границей Кавказского округа. Восточная граница начинается ! на севере-с низовий Калауса, проходит через среднее течение Кумы, выходит на Терек в районе Моздока и огибает его низовья с севера, дости-? гая Каспийского побережья в районе Махачкалы (рис. 195).
| Геологическая древность суши очерченной территории неодинакова. (Наибольшей молодостью отличаются Прикубанская и Приманычская равнины, низовья Терека и Сулака. Наибольшая древность характерна 1 для Ставропольской возвышенности и наклонных равнин предгорий, 1 сложенных майкопскими (олигоценовыми), верхнесарматскими (миоце-! новыми), апшеронскими (плиоценовыми) и четвертичными отложениями..
Первобытные, послеледниковые ландшафты разнообразны и неод-| дородны по происхождению. На севере — в Приманычье — преобладают 5 типчаковые и полынные степи, в предгорных равнинах западной^ части j округа остепненные луга, мезофитные луга и лесостепь с островками j широколиственных лесов и лесов тугайного типа по поймает рек. | В ксеротермическую фазу голоцена здесь имело место значительное 1 природное остепнение Закубанской и Терско-Сунженской наклонных j равнин, которые начали зарастать лесом в начале нашей эпохи (Захаров, t 1935). На юго-востоке района характерны сухие полынные степи, пере-| ходящие в солянковые полупустыни. По лиманам Азовского и Каспий-| £Кого побережий и разливам в низовьях Кубани, Челбаса, Ей, Терека 1 и Сулака имеются обширные заросли тростника, узколистного рогоза, ! морского камыша. Эти заболоченности разграничены местами косами I песчаных наносов с формациями сухой степи, населенной тушканчиками, . хомяками и зайцами. Тугайные леса, чередующиеся с зарослями эриан-| туса и солодки, сохранились лишь по низовьям Терека и Сулака.
I Терпокомплекс района также неоднороден на этой обширной терри-I тории и сформирован представителями I, II, III, V, VI, VIII генетических групп. Основу его составляют виды V и VI групп.
Кавказская плиоценовая группа обеднена и состоит из обыкновенного ежа, кавказского крота, малой землеройки, вечерниц и кожанов, 3 широко распространенных хищных, нескольких мелких грызунов и ко-пытных. Из последних уцелел, впрочем, лишь кабан. В историческую J эпоху вымерли кавказский медведь, речной бобр, кавказский олень, | кавказский зубр.
I Видов южного — переднеазиатского — происхождения (II группа) | насчитывается до 15: ушастый еж, белобрюхая белозубка, подковоносы; 1 куница белодушка и перевязка — из хищных и ряд суховыносливых 1 грызунов — средний хомяк, степная полевка и др. Большинство видов I этой группы приурочено к ксерофитным биотопам предгорной зоны.
Древние восточноевропейские виды (III группа) редки, это — выху-!холь и слепыши.
Есе остальные живущие виды — более поздние вселенцы, плейсто-| ценового и голоценового возраста, проникшие в Предкавказье преиму-I щественно с севера, из лесов Западной Европы и из степей Восточной I Европы.
В V группе, включающей зверей плейстоценовой древности северного 3 происхождения — лесных и мезофильных по их экологической сущ-| яостп __ можно насчитать 9—11 видов. К ним относятся: кавказская J 32 И. К. Верещагин
' 498 Часть III, Териогеографическое районирование и пути эволюции
Видовой состав и географическое распределение
Восточносредиземномор
Виды Кавказский округ
район Большого Кавказа Дагестанский район Малого Гирканский район
район Кавказа Талышский лесной участок
Обыкновенный еж — Erinaceus europaeus L. Ушастый еж — Hemiechinus auritus Gmel. Восточный крот — Talpa orientalis Ogn. Кавказский крот — Talpa caucasica Sat. Выхухоль — Desmana moschata L. Малая бурозубка — So-rex minutus L. Обыкновенная бурозубка— Sorex araneus L. Кавказская бурозубка — Sorex raddei Sat. Водяная кутора — Ncorny s fodiens Schr. Малая белозубка — Crocidura suaveolens Pall. Белобрюхая белозубка — Crocidura leucodon Herm. Длиннохвостая белозубка — Crocidura russula Herm. Серая белозубка — Crocidura lasia Thos Персидская белозубка — Crocidura zarudnyi Ogn. Белозубка-малютка — S uncus etruscus Savi Большой подковонос — Rhinolophus /er rum-equinum Schr. Подковонос Мегели — Rhinolophus mehelyi Matschi Южный подковонос — Rhinolophus euryale Blas. Малый подковонос — Rhinolophus hipposide-ros Bechst. concolor Martin orientalis Ogn. caucasica Sat., ognevi Strog. balcaricus Ogn., schelkovniko-vi Sat. monacha Thos lasia Thos nordmanni Sat. rumanicus Barr, et Ham. volnuchx caucasicus Sat. raddei Sat. dageslanicus Hept. et Form. dinnicki Ogn. gUldenscht transcauca transcaucasica Dahl ognevi Strog. ni Ogn leptodactylus Sat. aedti Pall. lasia Thos sicus Sat. talyschensis N. Ver. caspica Thos + +
Примечание. Звездочками (•) обозначены виды, вымершие на протяжении последних точнее не названных.
Гл. V. Анализ голоценовой териофауны и схема районирования 49 §
Таблица 103 голоценовых млекопитающих Кавказского перешейка
подобласть Подобласть Восточноевропейских степей Подобласть Среднеазиатских пустынь
Переднеазиатский округ Восточноевропейский округ Западный округ
i Западноиранский район Севанский Куринский Предкавказский район северного Прикаспия
Ьяссинский Г участок Нагорно-Талышский степной участок район район район Терско-Кумский участок
transcaucasicus Sat. rumanicus Barr, et Ham.
calligoni Sat. brachyotis Sat. auritus Gmel.
— " transcaucasica Dahl caucasi cus Sat.
— — caucasicus Sat.
j — — raddei Sat. — —
F — — leptodactylus Sat. balcaricus Ogn.
1 — — * —. dinnicki Ogn. suaveolens Pali
persit ca Thos “F ~F
> guldenschtaedti Pall. i — —
| zarudn li Ogn. — — —
! + — —j— + ~F
j + + £ — — —i— ~F ~—-
[етнй и десятилетий нашей эры. Знаком + отмечено наличие-;номинальных форм и форм
. 500
Часть III. Териогеографическое районирование и пути эволюции
Виды В осточносредиземномор
Кавказский округ
район Большого Кавказа Дагестанский район район Малого Кавказа Гирканский район Талышский лесной участок
Подковонос Блазиуса — Rhinolophus blasii Peters. Ночница Бехштейна — Myotis bechstelnii Kiihl Трехцветная ночница — Myotis emarginatus Geoffr. Остроухая ночница — — — —
Myotis oxygnathus Mont.
Усатая ночница — Myotis mystacinus Kiihl —1— —j— —1~
Ночница Наттерера — Myotis nattereri Kiihl —J— — —
Длиннокрыл — Minwp-terus schreibersii Kiihl —j— —
Ушан — Plecotus auri-tus L. wardi Thos
Азиатская широкоуш-ка — Barbastella darje-.lingensis Dobs. — —
Европейская широкоуш-ка — Barbastella barba-stellus Schr. 1 1 — —j— —
Вечерница Лейслера — Nyctalus leisleri Kiihl —1~ —. — —
Гигантская вечерница — Nyctalus siculus Palumbo ““I"11 — —1“^ —
Рыжая вечерница — Nyctalus noctula Schr. — —[— —
Нетопырь-карлик — V espertilio pipistrellus Schr. — + if
Нетопырь Натузиуса — V espertilio nathusii Keys, et Blas. —|— +
Нетопырь Куля — Vespertilio kiihl ii Kiihl — — — ’
Нетопырь Сави— Vespertilio savii Bonap. caucasicus Sat. — — —
Двуцветный кожан — V espertilio murinus L. — *— —i— +
Кожанок Бобринского — Vespertilio bobrinskii Kuz. —— — — —
Северный кожанок — V espertilio nilssonii Keys, et Blas.
Гл* V. Анализ голоценовой териофауны и схема районирования 501
Таблица 103 (продолжение)
спая подобласть Подобласть Восточноевропейских степей Подобласть Среднеазиатских пустынь
Переднеазиатский округ Восточноевропейский округ Западный округ
. западноиранский район Севанский район Куринский район Предкавказский район район северного Прикаспия
Араксинский i участок Нагорно-Талышский степной участок Терско-К умений участок
i 4- ; 1 О * > i "Т j crfilf nils :| Dahl . i я ,'yt] ж :Я • дД — —1— 1 caucasicus Sat. 1 + + 1 I 1 + + 1 z/“h 1 1 1 1 1 + + II 1 1 Illi 1 II II 1 1 1 1 1 1 1 II 1 ’ / -
$02 Часть III. Териогеографическое районирование и пути эволюции
Восточносредиземномор
ж* ’ 'Ягаь Виды Кавказский округ
Гирканский район
район Большого Дагестанский район Малого
ж-и . # ' 1 £ Кавказа район Кавказа Талышский лесной участок
Поздний кожан— Vespertilio serotinus Schr. Кожан Огнева— Vespertilio ognevi Bobr. Широкоухий складчатогуб — Tadarida tae-niotis Raf. Шакал — Canis aureus L. Волк — Canis lupus L. cubanensis Ogn. | — + + +
• Лисица — V ul pes vul pes L. caucasica Dinn. alticola Ogn. +
К V Г’ Корсак —Vul pes corsac L. Полосатая гиена — Hyaena hyaena L. Бурый медведь — Ursus arctos L. Лесная куница — Martes martes L. Куница белодушка — Martes foina Erxl. Перевязка —• V ormela peregusna Giild. Черный хорек—Putorius putorius L. Степной хорек — Putorius eversmanni Less. Норка — Lutreola lutreola L. Ласка — Mustela nivalis L. Горностай — Mustela ermine a L. Барсук — Meles meles L. Выдра — Lutra lutra L. * Тигр — Panthera tigris L. * Лев—Panthera leo L. Барс — Panthera pardus L. Пятнистый кот — Felis lybica Forst. Лесной кот — Felis silve-stris Schr. Манул — Otocolobus ma-nul Pall. Камышовый кот — Felis chaus Giild. Рысь — Felis lynx L. caucasicu. arctos J lorenzi Ogn. turovi Kuzn. et Nov. caucasic* ciscaucasi s Smirn., nehrir caucasica В is Ogn. cus Sat. caucasicus Sat. orienta meridiona lorenzi Ogn. igi Sat. arr. et Ham. canescen tullianus dis Sat. ills Midd. s Blanf. + septentrionalis Sat. Valenc. +
Гл> V. Анализ голоценовой териофауны и схема районирования 503
Таблица 103 (продолжение)
.сная подобласть Подобласть Восточноевропейских степей Подобласть Среднеазиатских пустынь
ж Переднеазиатский округ Восточноевропейский округ Западный округ
Западной ранений район Севанский район Куринский район Предкавказский район район северного Прикаспия
Ара ксинский ‘ участок Нагорно-Талышский степной участок Терско-Кумский участок
1 т l^dpherakyi 1* Sat. ♦ + 1 ' .1 . + * + + nehrin^ caucasica 1 e min tullianus Ve haj astanicus Dahl kurdistanica Sat. meridiona 'i Sat. Barr, et Ham, 1 ” or Sat. dene. orientalis Sat. alpherakyi Sat. satunini Ma* tschi lis Midd. * । stepensis Braun, caucasica Dinn. kalmykor * * arenas L. nehringi Sat. +? turovi Kuzn. et Nov. dinnil +? caucasicus Ogn * caucasicus Sat. ★orientalis (?) Sat. । “Г karagan Erxl. ит Ogn. ci Sat. +? heptneri Ogn.
onalis Ь
504 Часть III. Териогеографическое районирование и пути эволюции
\ 5 - - -
Восточносредиземномор
Виды г 1' ’ Кавказский округ
район Большого Дагестанский район Малого Гирканский район
Кавказа район Кавказа Талышский лесной участок
* Гепард — Acinonyx fu-bat us Schr. f Заяц-русак — Lepus euro- L paeus L. В *Арменийская пищуха — t Ochotona sp. В; Кавказская белка — f'. Sciurus anomalus Giild. Малый суслик— Citellus 1 ' pygmaeus Pall. Г L Малоазийский суслик — f • Citellus citellus L. f * Речной бобр — Castor fiber L. 1 Полчок — Glis glis L. f Лесная соня — Dyromys nitedula Pall. Южная мышовка — Sici-sta subtilis Pall. Лесная мышовка — Sici-sta betulina Pall. Кавказская мышовка — Sicista caucasica Vi-nogr. Малый тушканчик — Allactaga elater Licht. Большой тушканчик — Allactaga jaculus Pall. Малоазийский тушканчик — Allactaga wil-liamsi Thos Земляной зайчик — Ala-ctagulus acontion Pall. Емуранчик—Scirtopoda telum Licht. Мохноногий тушканчик — Di pus sagitta Pall. Обыкновенный слепыш — Spalax micro-phtalmus Giild. Гигантский слепыш — Spalax giganteus Nehr. Горный слепыш - 6*palax leucodon Nordm. Черная крыса — Rattus rattus Ij. \ caucasici musicus Men. as Ogn. су r ens is Sat.
* tschetschei caucasicus Ogn. et Turov strandi Form. A)- * « licus Sat. ognevi Hept, et Form. pictus 1 _ persius Erxl. Blanf. + i r
Гл. V. Анализ голоценовой териофауны и схема районирования
505
Таблица 103 (продолжение)
Г^‘ екая подобласть Подобласть Восточноевропейских степей Подобласть Среднеази атских пустынь
Переднеазиатский округ Восточноевропейский округ Западный округ
i ападноиранский район Севанский Куринский Предка вказский район северного Прикаспия
А 'ОК А н<сиясний , ;часток । Нагорно-Ta лишений степной участок район район район Терско-Кумский участок
1 1 1 * ' " — * — —
1 !> cyrensis Sat. * — caucasic '.us Ogn.
1 i 1 1 < 1 wnihoprym-nus Benn. — xanthoprymnus Benn. * kalabuchovi Ogn., satunini Svirid., boeh-mi Krass. * planicola Sat. , т - . i
rxl. f pic tus Bh inf. tichomirowi Sat. daghestanicus Ogn. et Turov nordmanni Keys, et Blas. 1 1 1 1
j 'л !} walychen-i sis Sat. williai nsi Thos caucasicus Nehr. schmidti Sat. jaculus Pall., fuscus Ogn> kizljartcus Sat. fuscus О^п.
— —' — -— dinniki Sat. —•
**• 1 — — — turovi Hept.
— 1 — — *— — nogai Sat.
1 — — — —
— |r~ armeniaca Me-hely* nehr i ng i Sat. —- — —,
"4" i f — 4-
«<ИШМШН1
506
Часть Iff. Териогеографическое районирование и пути эволюции
В осточносредиземномор
Кавказский округ
Виды район Большого Дагестанский район Малого Гирканский район
Кавказа район Кавказа Талышский лесной участок
Серая крыса — Rattus norvegicus Berck. 'Степная мышь — М us musculus L. colchicus Lvov formosovi Hf Wat iptn., abbotti erh. abbotti Waterh. tataricus Sat.
Мышь-малютка — Mic-romys minutus Pall. Полевая мышь — Apo-demus agrarius Pall. .Кавказская желтопят-ная мышь — Apodemus fulvipectus Ogn. Желтогорлая мышь — Apodemus jlavicollis Melch. Лесная мышь — Apodemus sylvaticus L. Малоазийская мышь — Apodemus mystacinus Danf. et Alst. Малоазийский хомяк — Mesocricetus auratus Waterh. Обыкновенный хомяк — Cricetus cricetus L. Серый хомячок — Crice-tulus migratorius Pall. Мышевидный хомячок — Calomyscus bailwardi Thos Большая песчанка — Rhombomys opimus Licht. Полуденная песчанка — Meriones meridianus Pail. Краснохвостая песчанка — Meriones ery thro tints Gray .Персидская песчанка — Meriones persicus Blanf. Тамарисковая песчанка — Meriones tamaris-cinus Pall. .Малоазийская песчанка — Meriones blackleri Thos Песчанка Виноградова — Meriones vino-gradovi Heptn. caucasicus Duk. ponticus Svirid. ciscaucasi euxinus G. Allen koenigi Sat. pulchei saxatilis Krass. ,cus Ogn. raddei Nehr., avaricus Ogn. et Heptn. * Ogn. argiropuli Vin. et Arg. arianv euxinus G. Allen brandti Nehr. । s Blanf. r
Гл, V, Анализ голоценовой териофауны и схема районирования 607
Таблица 103 (продолжение)
4 окая подобласть Подобласть Восточноевропейских степей Подобласть Среднеазиатских пустынь
i 1 t' Переднеазиатский округ Восточноевропейский округ Западный округ
4 Западноиранский район Севанский Куринский Предкавказский район северного Прикаспия
ipaKi л некий участок Нагорно-Талышский степной участок район район район Терско-Кумский участок
+
atarici is Sat. аЬбоШ Waterh. tataricus Sat. hortulanus wagneri
Nordm. Eversm.., по-
— — — -f- gaiorum Heptn.
- — — caucasicus Duk. •
- — — — planicola Svi-rid. —
- — — — samar ien sis Ogn.
arianus Blanf. ciscaucasicus - —
* — — Ogn. •—
— brandti Nehr. nigriculus Nehr. —
— — — stavropolicus Sat. —
4“ —|— —j— pulcher Ogn. phaeus Pall.
т —— — — —
— — — —
—- — — — nogaiorum Heptn.
+ — — caucasicus Brandt ~- —
rossicus Heptn. — — —- —
1 — 1 — — — ciscaucasicus
Sat.
bogdanwi Heptn. — bogdanovi Heptn. — 4“
1 +- — — — ——
508
Часть III, Териогеографическое районирование и пути эволюции
i Виды В осточносредиземномор
Кавказский округ
район Большого Кавказа Дагестанский район район Малого Кавказа Гирканский район Талышский лесной участок
Араздаянская песчанка — Meriones sp. — 1 — —
Степная пеструшка — Lagurus lagurus Pall. — — — —
Обыкновенная слепушонка — Ellobius tal-pinus Pall. '“I— L- -IM —
Горная слепушонка — Ellobius lutescens Thos — — — ——
Прометеева полевка — Prometheomys scha- poschnikowi Sat. — —
Водяная полевка — Arvi-cola terrestris L. ognevi Tur., rufescens Sat., turovi Ogn. djukovi Ogn. et Form., kuruschi Heptn. et Form. persicus de Fil.
Малоазийская снеговая полевка — Microtus nivalis Mart. loginovi Ogn. —— trialetcus SchidL ‘——
Кавказская снеговая полевка — Microtus nenjukovi Form., gud Ighesicus SchidL, ose-ticus SchidL -J- —
gud Sat. Sat.
Длиннохвостая снеговая полевка — Microtus ro-berti Thos personatus Ogn. — 1 —
Кустарниковая полев- colchicus daghestanicus intermedius schelkovnikovi
ка — Microtus majori Thos SchidL, cis-caucasicus Ogn. SchidL SchidL Sat.
Обыкновенная полев- macrocranius Ogn., gudauricus tra nscaucasicus —
ка — Microtus arvalis Pall. Ol rn. Ogn.
Степная полевка — Microtus socialis Pall. — +
Рыжая полевка — Cle-thrionomys glareolus Schr. 'ponticus Thos 1 — —
Дикобраз — Hystrix leu-сига Sykes — ; — hirsutirostris Brandt
• Кулан — Equus hemio-nus Pall. — — — —
* Тарпан — Equus caballus gmelini Ant. — — — —
* Переднеазиатская лошадь — Equus caballus L. (subsp.) Кабан — Sus scrofa L. Благородный олень — Cervus elaphus L. attila maral Thos. Ogilby *
* Лось — Alces alces L. * caucasicus N. Ver. — — ——
Гл. V. Анализ голоценовой териофауны и схема районирования 509
Таблица 103 {продолжение)
>р сная подобласть Подобласть Восточноевропейских степей По область Среднеаз иатских пустынь
Переднеазиатский округ Восточноевропейский округ Западный округ
иноиранский район Севанский Куринский Предкавказский район северного При каспия
)К АГ а> синений У 1СТОК Нагорно-Талышскнй стенной участок район район район Терс ко-Ку мский участок
Т —* — — — —
—. —— — — -j-
1г + —!— — — — —
persi cut f de Fil. caucasicus Ogn., cubanensis Ogn.
— satunini Schidl. — — —
kovi 1 — — — —— — —
| mystacinus de Fil. transcaucasicus Ogn. — macro Ogn cranius
1 schidlovskii Arg. binominatus Ellerm. parvu s SaU
t i l I * ♦ * * * «
— — — * *
* ♦ * ? — —
— — attila Thus * maral attila Ogilby Thos
- 1 — — * — * caucasicus N. Ver. —
4 3
Часть III. Териогеографическое районирование и пути эволюции
Виды В осточносредиземномор
Кавказский округ
район Большого Кавказа Дагестанский район район Малого Кавказа Гир канский район Талышский лесной участок
Косуля — Capreolus сар-reolusj,. capreolus L., Pygargus Lyd. capreolus L.
Сайга — Saiga tatarica L. — —
Джейран — Gazella sub-gutturosa Giild. Серна—Rupicapra rupicapra L. caucasica Lyd. —
Западнокавказский козел — Capra caucasica Giild. caucasica Giild., severtzovi Menzb., din-niki Sat.
Восточнокавказский козел — C a pra cylin dri-cornis Blyth —1— — '—
Безоаровый козел — Capra aegagrus Erxl. — 1 —
Арменийский муфлон — Ovis gmelini Blyth — * — —
* Кавказский зубр — Bison bonasus L. caucasicus Sat. * — —
* Первобытный тур — Bos primigenius Boj. —_ “— *—
бурозубка, норка, кавказский лесной кот, мышь-малютка, полевая мышь самарский подвид желтогорлой мыши, обыкновенный хомяк. Из вымер-тих в историческую эпоху форм отметим европейского бурого медведя кавказского лося, кавказскую крупную косулю и, возможно, первобытного тура.
Ареалы большинства этих видов прижаты к кавказским предгорьям. Основными доказательствами того, что эти виды имеют на перешейке плейстоценовый возраст, являются неразвитость их ареалов в юго-восточном направлении, данные геологической истории страны и истории фитоландшафтов. Все они относительно холодолюбивы и вряд ли могли продвигаться на жаркий и сухой юг в добакинское время.
Довольно загадочно здесь распространение восточноевропейского подвида желтогорлой мыши, обнаруженной пока только в тугаях по Сулаку.
Степные виды Восточной Европы (VI группа) заселяли район преимущественно в плейстоцене. Сюда относятся: малая белозубка, корсак и степной хорь, малый суслик, степная мышовка, большой и мохноногий тушканчики, емуранчик, степная пеструшка и обыкновенная слепушонка. Из недавно вымерших форм можно указать на сурка и тарпана.
I
Гл. V. Анализ голоценовой териофауны и схема районирования
511
Р
1
:i-
1
1
I
*
Таблица 103 (окончание)*
скан подобласть i * Подобласть Восточноевропейских степей Подобласть Среднеазиатских пустынь
Переднеазиатский округ Восточноевропейский округ Западный округ
Заладноиранский район Севанский Куринский район Предка вказский район северного’ Прикаспия
Араксинский участок Нагорно-Талыгаский степной участок район район Терско-Кумскнй^ участок
capreolus L. — *pygargus Lyd. —
I - at • — 1 1+1
Г * 1 ч 1 — —
Armenian a N; is. — —
- — * ——
"4F* Некоторые из этих видов (малый суслик) внедрились в нагорные* ! степи северного склона Кавказа, другие (большой тушканчик и сайга) — проникали по побережью Каспия в среднем плейстоцене даже в восточное Закавказье.
Иммигранты послеледникового возраста (VIII группа) имеются в от-рядах хищных, непарно- и парнокопытных. К ним принадлежат шакал, камышовый кот и вымершие недавно полосатая гиена, кулан и джейран. Возможно, здесь обитали в историческую эпоху также лев, леопард и гепард.
Ареалы шакала и камышовой рыси, как известно, вдаются в пределы > района лишь узкой полоской по долинам рек и вдоль предгорий в восточ-
> ное Предкавказье.
К случайным, завозным видам исторической эпохи (X группа) отнесен пасюк, живущий в плавнях Дона, Кубани, Сулака, Терека и быстро-; проникающий теперь во все населенные пункты.
i На востоке и северо-востоке район граничит с участком развития
более суховыносливого териокомплекса — полупустынь и пустынь Средней Азии.
612 Часть III. Териогеографическое районирование и пути эволюции
ПОДОБЛАСТЬ СРЕДНЕАЗИАТСКИХ ПУСТЫНЬ
ЗАПАДНЫЙ ОКРУГ
Район северного Прикаспия
Терско-Кумский участок
(участок тамариксовой и полуденной песчанок)
Этот участок занимает территорию полупустынь и пустынь между низовьями Кумы, Терека и Сулака. Западная граница участка идет с севера от водораздела Маныча и Сарыкамыша к юго-востоку, на устье Сунжи, а южная пересекает низовья Терека и Сулака, выходя к Каспию у Махачкалы. Территория этою участка совпадает на перешейке с пределами суши, заливавшимися водами последней крупной — Хвалыиской трансгрессии. Северная его половина образована морскими отложениями ледниковой эпохи и представляет собой равнинную песчано-глинистую полупустыню. Юго-западная треть покрыта бугристыми песками, а юго-восточная — аллювиальными суглинками и глинами послеледниковой эпохи. Наиболее молоды северная половина и юго-восточная треть участка. Часть их заливалась даже при небольших трансгрессиях Каспия исторической эпохи. Большая часть Терско-Кумской полупустыни и пустыни покрыта полынно-солянковой формацией. Бугристые пески закреплены песчаным овсом, полынью, донником. По лиманному побережью Каспия развиты обширные тростниковые заросли, перемежающиеся с бурьянистым разнотравьем на грядовых повышениях.
Голоценовый териокомплекс участка сформирован представителями I, II, IV, VI, VII, VIII и IX групп. Основу его теперь составляют виды VI и VII группы.
Из I группы здесь имеются лишь палеарктические убиквисты, занимающие интразоналыше биотопы, но отнюдь не мезофильные кавказцы. Обыкновенный еж, вол1<. ласка, барсук, степная мышь, кабан заселяют камыши, межгрядовые понгцкения в бугристых песках, но сами существенной роли в биоценозах участка не играют.
Восточносредиземно морские сухолюбивые млекопитающие (II группа) представлены лишь широко адаптированными видами: ушастым ежом, белобрюхой белозубкой, перевязкой, русаком, серым хомячком, степной полевкой. Обыкновенный слепыш замещен здесь гигантским, образующим реликтовые поселения в понижениях между песками. Из древних среднеазиатских видов (IV группа) живет малый тушканчик.
Восточноевропейские степные (VI группа) и туранские (VII и IX группы) виды, заселявшие участок в эпохи плейстоценовых регрессий древнего Каспия и в голоцене, представлены корсаком, малым сусликом, тушканчиками, полуденной и тамарисковой песчанками. В историческую эпоху здесь вымерли большая песчанка, манул, тарпан, но уцелела сайга.
Среднеазиатский оттенок здешнему териокомплексу придают, в сущности, лишь песчанки. Тарпан продержался здесь дольше, чем в Пред-кавказском районе.
Южные — послеледниковые — виды (VIII группа), по-видимому, проникали на участок как с юга, так и с северо-востока, из Средней Азии. Можно полагать, что здесь в голоцене жили почти все виды VIII группы, названные для Предкавказского района. Из них уцелели лишь шакал и камышовый кот.
Гл. V. Анализ голоценовой териофауны и схема районирования
513
Териологическая характеристика Терско-Кумского участка показывает, что это — в сущности, уже не Кавказ, а уголок арало-каспийских (Среднеазиатских) пустынь. Особенно похожи на среднеазиатские группировки животных на песчаных буграх, например на феноменальном * лрхане Кумторкала в предгорьях Дагестана.
Формирование териокомплекса этого участка происходило под влиянием колебаний уровня четвертичного Каспия в большей степени, нежели ta любой другой прикаспийской территории.
Для расселения на участок восточных зимоспящих видов в четвертом периоде большое значение имели не только трансгрессии и ре-рессии.Каспия, но и бифуркации нижнего отрезка Волги.
Обзор смежных фаунистических влияний и характеристика округов, айонов и участков Кавказского перешейка, сделанные на основе истории аспространения видов и палеогеографических материалов, вскрывают олько небольшую часть биологических процессов, происходивших под влиянием сложных преобразований природной обстановки в кайнозое а участках суши, окружающей Черное и Каспийское моря.
Геохронологическая последовательность фаунистических влияний, амеченных нами, будет, несомненно, уточняться по мере накопления алеоитологических фактов и анализа процесса видообразования. Тем е менее наша схема оправдывает себя на данном этапе зоогеографи-еской науки.
Териологическая характеристика, данная выделенным округам, >айонам и участкам, показывает и степень насыщения их видами, открывая дуги для выводов по реконструкции и обогащению фауны. Полный список ждов (и подвидов) голоценовой териофауны Кавказского перешейка :щн в табл. 103.
Новейший ход эволюции состава экологических группировок млекопитающих перешейка и изменений границ фаунистических комплексов под влиянием неорганизованного и целенаправленного воздействия человека показан в следующей главе.
33 Н. К. Верещагин
ИЗМЕНЕНИЯ ЭКОЛОГИЧЕСКИХ ГРУППИРОВОК И АРЕАЛОВ МЛЕКОПИТАЮЩИХ ПОД ВЛИЯНИЕМ ДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА В РАЗЛИЧНЫХ ЗОНАХ
Воздействие человека иа фауну млекопитающих Кавказа можно проследить по кухонным остаткам первобытных племен уже с ашельской стадии. Нет сомнения, что на Кавказе, как и в других районах, палеолитические племена содействовали вымиранию ряда млекопитающих Или сокращению их ареалов. Степень этого воздействия на различных зверей установить пока трудно. Первобытными охотниками добывались, конечно, преимущественно массовые виды и наиболее доступные. Так, на Закубаиской равнине в охотничьей добыче палеолитических орд преобладал зубр, затем шли лошадь, мамонт, благородный и гигантскшт олени, пещерная гиена. В добыче древних охотников Черноморского побережья и центрального Закавказья преобладал пещерный медведь, а следующие места занимали кабан и благородный олень. На нагорьях Малого Кавказа было вновь велико значение зубра и лошади. Состав и соотношение численности различных зверей в добыче палеолитических охотников, вероятно, сильно менялись по сезонам и на протяжении тысячелетий. Много видов мелких зверей, птиц, рептилий, рыб, употреблявшихся в пищу, видимо, вообще ускользнуло от палеонтологов, так как эти животные могли поедаться на месте поимки и на временных охотничьих стоянках.
Некоторые виды зверей, употреблявшихся в пищу первобытными племенами верхнего плейстоцена на различных участках Кавказа показаны в табл. 104. Правда, в приведенных цифрах легче усмотреть зависимость преобладания того или иного вида от типа ландшафта, чем от возраста стоянки.
Таблица 104
Численность остатков (в %) преобладающих видов зверей в различных палеолитических стоянках Кавказа
Виды Северо-западный Кавказ (Илье к ап) Черноморское побережье (Ахштырская) Колхида (Сакажиа) Нагорье Малого Кавказа (Зурта кетен ан)
Пещерный медведь . . 2.6 57.3 5.0 —
Лошадь . 6.5 * 2.5 50.0
Благородный олень . . 5.2 5.9 7.6 —
Первобытный зубр . . 56.0 2.9 26.5 16.0
1л. VI. Изменения группировок и ареалов под влиянием человека
5 1о
Влияние человека на
послепалеолитпческую
млекопитающих Кавказа и их группировки эпоху было многообразным. Оно складыва-
лось
из:
I) непосредственного
цнической охоты;
истребления промысловых видов вследствие
) косвенного воздействия па отдельные виды, их группировки и налы через изменение ландшафта, макро- и микроклимата, почвы
Рис. 196. Бронзовые наконечники стрел и копни боевого и охотничье] о назначения эпохи поздней бронзы (копья) и скифо-сарматской культуры. Коллекция Кавказского музея.
1 вообще экологических особенностей территории в результате продленной н сельскохозяйственной деятельности;
3) случайного заноса и завоза синантропных видов;
4) сознательного изменения группировок зверей ирн помощи целе-пправленного завоза и разведения в естественных условиях новых пдов, частичной охраны и разведения избранных видов и уничтожения
вредных.
Прямое
влияние
человека на крупных
диких животных
Кавказа
быстро росло в послепалеолитпческую эпоху по мере развития культуры, «елейности п расселения древпих племен.
Появление домашних животных не могло уменьшить этого влияния. Сокращение числа остатков диких животных, которое наблюдается ;культурных слоях послепалеолитнческой эпохи по мере приближения Iсовременности (табл. 105), объясняется, в сущности, развитием ското-
щства и отнюдь не означает уменьшения преследования диких жи-
Ютпых.
516
Часть III. Териогеографическое районирование и пути эволюции
Таблица 105
Численность костей диких п домашних животных в послепалеолптичсских стоянках, поселениях и святилищах Кавказа и Русской равнины1
Памятник и его датировка Процент костных остатков животных Характеристика
диких домашних
Дзуары Северной Осетии. Пещера Дпгоризед, Х1\ — XIX вв и. э Хазарская крепость Белая Вежа (Саркел) на Дону, VIII—ХШ вв. н. э Поселение у Бакинской бухты на Апшероне, IX—ХШ вв. и. э Семпбратяее городище в устье Кубани, IV в. до я. э. — I в. в. э Землянковые поселения у Цимлянской на Дону, I тысячелетие до п. э Поселение у Каякента в Дагестане, II тысячелетие до и. э Землянковэе поселение у 1 е-ленджпкской бухты, III тысячелетие до н. э Пещера Ахштырская, стоянка охотников, неолитические и более поздние слои. . . 18.95 26.0 2 20.6 7.4 ’ 20.0, . 28.9 16.2 34.2 46.3 81.05 74.0 79.4 92.6 71.1, . 80.0 <83.8 65.8 53.7 Развитое скотоводство. Обилие горно-лесных зверей, добывавшихся копьями, стрельбой из лука и огнестрельного оружия. Обилие домашнего скота и диких степных и лесных животных, добывавшихся на коллективных загонных охотах и при индивидуальном промысле самоловами. Обилие домашнего скота, джейранов, тюленей, перелетной птицы. Развитая загонная охота на джейранов, куланов и тюленебойный промысел. Развитое низкопородное скотоводство. Роль охотничьего промысла увеличивалась ври угасании городища в начале пашей эры. Животноводство па низком уровне. Обилие степных копытных. Развитое скотоводство. Роль охотничьего промысла, в том числе на тюленей, значительна. Развитый дельфпнобойный промысел. Наличие скотоводства и охоты на лесных зверей. Обилие крупных лесных зверей в окрестностях пещеры.
Прямое потребление и сокращение численности ряда охотничье-про-мысловых зверей Кавказа было тесно связано в последние тысячелетия с историей кавказских племен, народов и пх переселений, с воинами и развитием материальной культуры, а особенно с совершенствованием оружия.
Несмотря на обилие археологических и летописных материалов по Кавказу, детали эволюции охотничьего промысла с палеолита до наших дней прослеживаются с трудом. Древние рисунки на скалах в Кабристапе, рисунки на сосудах из курганов Триалетп, Майкопа свидетельствуют о широком применении луков во IJ и I тысячелетиях до и. э.
1 Неопубликованные материалы автора.
2 Процент костей диких животных в Саркеле несколько искусственно увеличен при отборе их для окончательного определения.
Гл> 17. Изменения группировок и ареалов под влиянием человека
617'
Копья и луки были основными орудиями добывания крупного зверя как у кобанцев и колхидяп, так и у более поздних племен (рис.196). Поразительное разнообразие типов железных наконечников стрел и копии йоевого и охотничьего назначения характерно, в частности, для средие-яковья Северного Кавказа (рис. 197).
Основным охотничьим приемом добывания крупных зверей кочевыми оеменамп на равнинах перешейка в течение последних трех тысячелетий
Рис. 197. Железные наконечники стрел и копий эпохи средневековья — VII—IX вв. н. э. Коллекция Кавказского музея.
были, несомненно, загонные охоты с помощью конницы. В ряде случаев они дополнялись установкой постоянных пли съемных изгородей и тенет, которыми перегораживались овраги, балки, долины рек.
В горных п лесных условиях, кроме коллективных охот, широко практиковался и индивидуальный промысел.
Переломным этапом для сокращения численности ареалов крупных животных Кавказа была, несомненно, эпоха средневековья. Техника изготовления сложных луков и приемы добычи зверя в лесных и горпо-деспых условиях были в это время весьма высоки.
Наличие большого количества свободных вооруженных людей и феодальные общественные отношения обеспечивали широкие возможности использования огромных людских масс для грандиозных облав. Такие облавы практиковались еще и в позднем средневековье.
Рапшд-Ад-Дин, иранский летописец XIV столетия (изд. 1946 г.), приводил рассказ о походе «государя ислама» Газап-хапа (одного из монгольских правителей Ирана) в Аладаг. Зимой 1301/02 г. Газан-хап остановился вставках в Белясуваре (на юге Кура-Араксинской низменности) и Хама-шахрэ п пошел на охоту в горный район по дороге на владения Талыш л Нспахбад.
518
Часть Iff. Териогеографическое районирование и пути эволюции
«Газан-хан приказал среди тех гор построить из жердей и хвороста две сходящиеся клином изгороди длиною в один день пути, так чтобы между концами изгородей в широкой части клина расстояние было около одного дня пути, а в узкой пятьдесят гязов, и в тупике их сделать из дерева наподобие загона. После этого воины устроили облаву и гнали дичь как-то: горных буйволов, джуров, диких коз и ослов, шакалов, лисиц, волков, медведей и других всевозможных диких и хищных зверей— вовнутрь изгороди до тех пор, пока они все пе собрались в том загоне. Государь ислама с Булуган-хатун восседал на помосте, который построили посередине и любовался на тех животных. Часть их перебили, а часть отпустили на волю» (стр. 188—189).
По позднейшим иранским летописям, исследованным Петрушевским (1949, стр. 292—293), известно, что в XVI — XVII столетиях в Иране существовала охотничья повинность — «шикар». Для нее широко привлекались крестьяне.
«Охотники-эмиры с их нукерами и мулязимами собирали на охоту по приказу шаха иногда до десяти и более тысяч человек, окружали большой район, намеченный для охоты, особой цепью заграждения, дабы не дать зверям возможности вырваться из кольца окружения. Эта линия окружения, которая постепенно суживалась по мере продвижения охотников, называлась „джеркэ", „нергэ“ или „комарга" (стр. 292).
«Во время шахской охоты в Мианкале,в лесах Мазандарана в 1028 г. (1618 г. н. э., — Я. Я) кольцо окружения зверей равнялось 3 фарсангам (18—20 км, — Я. В.) в длину и 2/3 фарсапга (4—4.5 км, — И. В.) в ширину. Перед началом большой охоты, во время которой истреблялись сотни и тысячи крупных зверей, в течение нескольких дней происходила загонка зверя в кольцо окружения. В качестве загонщиков зверя привлекались нередко, помимо конных воинов, также пехотинцы— „пийядэганЧ из рапйятов. Во время большой шахской охоты в лесах и чащах Гилям (южное побережье Каспия, — Я. В.) весной 1002 г. (1564 г. и. э. — Я. В.и для загонки зверя в джеркэ было согнано около 10 тысяч крестьян 3 людей Гиляна» (стр. 293). 1
Аналогичные грандиозные облавы на куланов и джейранов существовали в долине среднего Аракса, а на лесного зверя — в северной Армении! и Грузни на протяжении всего средневековья. Об этом ясно свидетель^ ствуют примеры в «Задачнике» Алании Ширакца (см. Тер-Погосян, 1947)J поэмы Руставели (1937) и Низами (1940), «География» Вахушти (1904)?
Ко времени изобретения, внедрения и быстрого совершенствования огнестрельного оружия на Кавказе уже произошло значительное сокращение численности крупных копытных в результате охот с первобытной техникой.
Наличие мелких, постоянно враждующих друг с другом племен н феодалов, занимавших отдельные ущелья, обычай кровной мести обусловливали обилие оружия в крае и широкую возможность применения его для охотничьих целей в течение последних столетий.
По мере сокращения численности зверя к XVIII—XIX вв. грандиозные загоны с устройством заборов стали нерентабельными, но появление мелкокалиберных кремневых и пистонных гладкостволок и винтовок создало новые широкие возможности индивидуального промысла.
Кавказские войны середины XIX столетия также способствовали резкому сокращению численности крупного зверя.
Заключительным этапом в этом направлении было изобретение четырехлинейной винтовки Бердана, а затем и винтовки Мосина — русской
1л. VI. Изменения группировок и ареалов под влиянием человека 619
(рехлинейки. Именно последняя открыла практически неограниченные возможности отстрела горных копытных на дальних дистанциях. Развитие перегонного скотоводства и вооружение пастухов трехлинейками, гражданская война (1917—1922 гг.), сопровождавшаяся массовым завозом в горы оружия, только довершили разгром кавказских популяций оленей и зубров.
Уже в результате обзора эволюции ареалов отдельных видов кавказских млекопитающих, данного в IV главе, выясняется, что за последнюю тысячу лет на Кавказе вымерло при содействии человека не менее 9 видов крупных млекопитающих (табл. 106).
Т а б л и ц а 106
Последовательность исчезновения (вымирания) на Кавказе некоторых млекопитающих
Виды Вероятная дата г гсчезновег г ня Последний участок обитания
Лев. Первобытный тур. Кулан. Гепард. Бобр. Лось. Тарпан.-Зубр. Тигр. X в. XII в. XIII в. XIII в. Конец XIX в. Начало XIX в. 80-е годы XIX в. 20-е годы XX в. 30-е годы XX в. Восточное Закавказье. Западное Предкавказье. Восточное Закавказье, в восточном Предкавказье до XVIII в. Восточное Закавказье. Колхида и Закубанская равнина. Предкавказье. Восточное Предкавказье. Западный Кавказ. Т а лыш.
Кроме того, были сильно сокращены численность и ареалы оленя, косули, джейрана, сайги и других видов (Верещагин, 1947).
Наиболее сложны и разнообразны косвенные воздействия человека на фауну Кавказского перешейка:
I) уничтожение тугайных, предгорных и горных лесов путем вырубки, выжигания, выпаса скота и разделки их под сады и поля;
2) Распашка CTeneii и полупустынь с последующим культивированием различных растений в комбинации с орошением и устройством искусственных древесных насаждений (рощ и садов, полезащитных лесных полос);
3) развитие перегонного скотоводства, сопровождавшееся широким обезлесением и эрозией горных склонов, а также сменами растительных группировок горных лугов, степи, полупустыни, пустыни;
4) развитие строительно-архитектурной деятельности человека (устройство землянок, поселков сельского типа и обширных городов с постройками различных конструкций);
5) развитие строительства дорог, гужевого, автомобильного и железнодорожного транспорта.
Формирование новых повадок животных в связи с созданием человеком различных типов вторичного, или культурного, ландшафта и формирование новых группировок зверей в этих новых условиях следовали за развитием хозяйственного уклада племен, скотоводства, агротехники и индустриализации страны. Особенно сложными оказывались соотношения численности видов в группировках мелких млекопитающих — грызунов и насекомоядных, — ее динамики в сезонном и многолетнем разрезе в связи со сменой биотопов под влиянием
&20 Часть III. ТериогеографичесЪое районирование и пути эволюции
различных агротехнических приемов, принятого севооборота и т. п. (см. Оболенский, 1935; Поляков, 1950, и др.).
Обзор прямых и косвенных воздействий человека на отдельные виды зверей, их экологические группировки и фаунистические комплексы дается далее на ряде конкретных примеров по геоморфологическим и фитоландшафтным районам.
СТЕПИ И ПОЛУПУСТЫНИ ПРЕДКАВКАЗЬЯ
Первобытный послеледниковый ландшафт Прикубанской равнины, низового Дона, Приманычья и степных окраин Ставрополья был представлен разнотравно-типчаково-ковыльными и типчаково-ковыльными степями северного и южного типа (Шиффере, 1953). По долинам рек, степным балкам были развиты кустарники и лиственные леса.
Надпойменные террасы рек интенсивно заселялись скотоводческими племенами с неолита. В век бронзы и железа землянковые поселения и городища вдоль рек встречались уже довольно часто (см.: Городцов, 1935; Покровский и Анфимов, 1937; Артамонов, 1937; Горецкий, 1948).
Увалы Таманского полуострова широко колонизировались греками уже с VI в. до н. э. (Каллистов, 1949). Состав послеледниковых группировок млекопитающих степей Предкавказья был богат. В лесах пойм и в камышах плавень Дона и Кубани держались волки, медведи, выдры, лесные коты, барсы, бобры, водяные крысы, кабаны, косули, олени, лоси, туры и зубры. Собственно степь была населена тарпанами, куланами, сайгаками, джейранами. В группировку мелких зверьков целинной степи Приманычья входили обыкновенный и ушастый ежи, лисица, корсак, степной хорек, сурок, малый суслик, хомяк, тушканчики и слепыш.
По аналогии с современностью и, судя по историческим сведениям для смежных районов (Барбаро, изд. 1836 г.; Михаил Литвин, 1890; Боплан, 1832), исчезновение и оттеснение диких лошадей, куланов, сайгаков, туров и зубров происходили интенсивно в предкавказских степях уже в средневековье под влиянием выпаса стад домашнего скота и загонных охот многотысячной конницы хазар, половцев, монголов.
Сильное обезлесение нижнего Дона, Кубани, Терека, Сулака, произведенное человеком в последние столетия, привело к вытеснению с этих участков медведей, бобров, кабанов, оленей, лосей, косуль (Кондратьев, 1885—1886; Богачев, 1918; Флеров и Баландин, 1931).
Известно, что значительные леса по правому берегу Кубани существовали еще в XVIII в. (Мищенко, 1928; Роговский, 1928; Захаров, 1935).
На карте, составленной Роговским, по правобережью Кубани отмечено 9 реликтовых островков леса, а на карте Кубанской области 1775—1776 гг. показана почти непрерывная полоса леса и кустарников, тянувшаяся от истока Протоки к устью Лабы.
Заметное влияние на ландшафт оказывали с середины прошлого столетия попытки предотвратить разливы Кубани, Терека, Сулака в районах их дельт путем обвалований берегов и забивки прорывов береговых гряд. Тем самым удавалось лишь временно осушить заболоченности, так как разница уровней реки и окружающих пространств все увеличивалась.
Планомерное осушение кубанских плавней и освоение новых участков под пшеницу и посевы риса началось лишь в 30-х годах нашего столетия. Наиболее крупные изменения при осушке плавней претерпевала группировка грызунов и хищников.
Гл. VI. Изменения группировок и ареалов под влиянием человека
521
На огромных пространствах осушавшихся болот и озер вытеснялись норки, выдры, лесные коты, водяные крысы, а вместо них вселялись хорьки, зайцы, обыкновенные полевки, степные мыши, обыкновенные » хомяки, большие тушканчики.
к Намеренные выжигание и вырубка тугайных лесов по Куме, Тереку, г Сулаку производились первоначально из военных соображений при * устройстве кавказской военной линии уже со второй половины XVIII столетия (Попко, 1880; Потто, 1912). Устройство казачьих поселений и постов вело к быстрому истреблению крупного зверя, так как казаки преимущественно занимались охотой и рыбной ловлей. В царствование Анны Иоановны на гребенских казаков была возложена обязанность [оставлять для зверинцев столицы живых кабанов, оленей, коз, камен-•1ых козлов — «штейнбоков», зубров.
Истребление байрачных лесов, разбросанных по северным склонам олмов, балкам предкавказских равнин шло аналогично тому, как это ыло описано для степей Русской равнины (Кузнецов, 1896; Танфильев, 896; Лавренко, 1939). Завершение процесса вытеснения корсаков, куланов, арпанов, сайгаков из предкавказских степей произошло, однако, лишь ри массовой распашке последних, начавшейся в первой половине прошлого столетия при устройстве казачьих поселений. Скотоводческое и зем-едельческое освоение равнинного Предкавказья имело не менее важное качение и для формирования ареалов, численности и группировок мелких млекопитающих. С уничтожением байрачных лесов сокращались площади своеобразных резерватов, сохранявших мезофильных насекомо-j ярых — землероек, крота; мелких хищных — куницу белодушку, | барсука; лесных грызунов — лесную соню, лесную и полевую мышей, j Распространение перечисленных зверьков приобретало на предкавказ-I ских равнинах все более реликтовый, раздробленный на мелкие очажки вид. I Примеры остаточного, разорванного распространения названных зверьков можно наблюдать и в настоящее время в Петровском и Александровском районах Ставрополья, в среднем и верхнем течении Калауса, а также на самой Ставропольской возвышенности.
С начала XX столетия в степном Предкавказье было предпринято истребление волков, сусликов и мышевидных грызунов, получившее плановое развитие лишь при Советской власти. С XX столетия здесь началась и новейшая фаза развития культурного ландшафта, отраженная е обширных статистических данных сборника «Северный Кавказ после районирования» (1925). Интенсификация земледелия и дальнейшее расширение посевных площадей происходили в последние десятилетия при внедрении новых сельскохозяйственных культур, орошении и мелиорации (при устройстве левобережных каналов Малки, Куры, пропуска кубанской воды в Егорлык и осушении кубанских плавней). В 20-х годах нашего столетия началось устройство лесных полос, первоначально вдоль линий железных дорог, а в 30-х годах — на обширных массивах полей. Наиболее крупные полезащитные полосы имеются ныне в районах Краснодара, Тихорецкой, Сальска, где общая протяженность их достигает уже многих сотен километров.
Характер связей, складывавшихся у млекопитающих с биотопами культурного ландшафта равнинного Предкавказья, отражает большую гамму экологических особенностей и потенциальных возможностей приспособления различных видов к новым условиям.
Влияние скотоводства на животный мир было здесь более длительным, нежели влияние земледелия. Выбивание степного травостоя скотом при-
522 Часть 111. Териогеографическое районирование и пути эволюции
иодило на протяжении столетий к замене разнотравно-типчаково-ковыльных степей типчаково-ковыльными, позднее — типчаково-полынными, а также к развитию сорных группировок на стоянках, у водопоев, вдоль скотопрогонных дорог и вокруг селений. Еще в 80-х годах прошлого столетия развившееся в Предкавказье тонкорунное овцеводство стало все более вытесняться экстенсивным хлебопашеством с целинных разнотравных степей Кубани, Ставрополья и Терека в зону типчаково-ковыльных степей бассейна Кумы (Рождаев, 1925). Стравливание целинного покрова скотом и устройство многочисленных тблоков — выбитых стоянок — содействовало быстрому расширению ареала суслика и тушканчиков. Расселению сусликов к югу способствовала также вытеснявшая их из Приманычья распашка степи, устройство мостов через каналы и речки, скотопрогонных дорог (Свириденко, 1927; Формозов, 1929). Суслики и тушканчики получали на тблоках возможность широкого осмотра, быстрого передвижения, свежий зеленый корм из сорного разнотравья и лучшую защиту от хищников. Заброшенные и действующие стоянки скотоводов служили местами концентрации белобрюхих и малых белозубой, степных полевок.
Современные комплексы местообитаний в культурном ландшафте предкавказских степей — «хозяйственные стации» Оболенского (1935), постоянно или временно заселяемые мелкими млекопитающими, — представлены здесь выгонами, дорогами различных типов, культивируемыми площадями и населенными пунктами.
Каждая из этих категорий содержит набор биотопов, определяемый агрокультурными и строительными приемами человека, а также экологической приспособляемостью видов млекопитающих.
Выгоны. При слабой нагрузке скотом выгоны имеют почти полный набор природных биотопов, сохраняя первобытную экологическую группировку, содержащую малого суслика, малого и большого тушканчиков, земляного зайчика, обыкновенного слепыша, степную полевку, степную пеструшку, слепушонку. Выгоны, расположенные близ селений, особенно при наличии несколько увлажненных почв, зарастают несъедобной для скота растительностью. На таких выгопах селятся суслик, большой тушканчик, степная мышь, обыкновенная полевка и слепыш.
Дороги. Полосы отчуждения по краям железнодорожного полотна и крупных автомобильных дорог имеют группировку млекопитающих, близкую к выгонной.
На этих полосках целины сосредоточиваются малая белозубка, черноватый хомяк, обыкновенный хомяк, большой тушканчик, степная мышовка, слепушонка, слепыш, малый суслик. Бурьяны, канавы и кюветы по сторонам железнодорожного полотна привлекательны для ушастых ежей, перевязок, степных хорьков, ласок, степных мышей, житников, обыкновенных полевок, серых хомячков.
Культивируемые площади имеют группировки млекопитающих, зависящие от принятой агротехники, севооборота, размеров пахотных участков. Обычно крупные массивы зерновых заселены здесь лишь обыкновенными полевками и степными мышами. На окраинах посевов держатся малые суслики, хомячки, серые хомячки, слепушонки, слепыши (Оболенский, 1935). В зависимости от характера обработки и состояния посевов на полях держатся также зайцы, а при наличии и доступности грызунов и насекомых — лисицы. Так, например, в зимнее время при малоснежье русаки кормятся на озимях, но залегают на день на зяблевой пахоте. Лисицы в такие же зимы в массе концентрируются
Гл. VI. Изменения группировок и ареалов под влиянием человека
523
< на заселенных грызунами озимях и стерне. Наиболее пестрый состав | экологических группировок зверьков существует при наличии мозаич-j пости участков посевов различных культур, быстро сменяющихся на небольших пространствах участками целины. Резерватами хомячков, ’ полевок, степных мышей, слепышей являются многолетние посевы люцерны и кормовых смесей. Обширные пахотные площади, используемые юд зерновые и регулярно обрабатываемые, имеют сильно обедненную руппировку, состоящую из лисицы, русака, обыкновенной полевки * степной мыши.
, Видовой состав грызунов закономерно сменяется на залежах разного
i озраста. Стерня первого года бывает заселена степными мышами, обыкно-
, енными полевками, изредка лесными и полевыми мышами, еще реже
I щшовками и серыми хомячками. При «старении» залежи и зарастании
, е сорняками — мышеем, лебедой, полынями — устойчивость поселений
1 рызунов быстро возрастает. Появляется группировка, состоящая из
л шастых и обыкновенных ежей, лисиц, зайцев, обыкновенных хомяков,
тепушонок, слепышей. Многолетние залежи заселены обычно сусликами, •ушканчиками, степными пеструшками, и их группировка уже близка 1е ; первобытной.
), Полевые станы, ссыпные пункты, скирды среди полей заселены бело-
брюхой и малой белозубками, степными мышами и обыкновенными по-> явками.
(Населенные пункты степного Предкавказья содержат довольно большой набор биотопов. Каменные ограды приусадебных участков, устроенные из обломков известняка, канавы, заросшие бурьяном, сады, огороды, хозяйственные постройки и склады топлива из стеблей iii подсолнечника, кукурузы, могара заселены постоянно белозубками,
п- ласками, степными хорьками, степными мышами, серыми крысами,
в, обыкновенными хомяками и полевками. В нежилых постройках посе-
гю ляются серые хомячки, а в разросшихся садиках, особенно примыкающих
но к выходам скал или байрачным и долинным лесочкам, — лесные мыши
ля п лесные сони. Берега подпруженных речек (водохранилищ) заселяются
ой водяной полевкой. Наконец, в жилых постройках и церквах селений
обосновываются колонии рыжих вечерниц, поздних кожанов, реже на двуцветных кожанов и остроухих ночниц. В жилых постройках селений
гх. Ж и в полевых станах обычны всюду домовые мыши аборигенного и завоз-Я| кого происхождения. Все крупные селения, связанные с Доном железной то- ’Ж и автомобильной дорогами, были уже в 30-х годах заселены пасюка, Ж ком.
то- Вообще синантропную группировку мелких млекопитающих в селе-
ей, пнях степей Предкавказья в настоящее время можно считать вполне
но- сформированной, так же как и группировку полевых и домовых вреди-
телей (см. также Стальмакова, 1935).
к*)- Стихийное развитие охотничьего промысла в первой половине XX
ров столетия продолжало подавлять численность сайги, корсака, а позднее
;есъ белого хоря. С 1925—1927 гг. начало быстро развиваться добывание
нах на шкурку так называемых «летних» пушных видов: малого суслика,
:кн, обыкновенного и черноватого хомяков, большого тушканчика (Б. Кузне-
тки 1 доп, 1932).
чии Крупные мероприятия по истреблению вредных грызунов были раз-
шее верну ты в Предкавказье с начала XX в. Истребление малого суслика
^ень : в предкавказских степях проводилось вначале для защиты посевов на этся крестьянских наделах. Позднее была выяснена большая эпидемиологи
524 Часть III. Териогеографическое районирование и пути эволюции
ческая опасность этого вида и очерчены границы эндемического по чуме очага.
Плановые истребительные работы по сусликам и мышевидным грызунам были осуществлены в Предкавказье лишь в советское время. Ежегодно обрабатываемые ядовитыми приманками и хлорпикрином площади возросли с сотни тысяч гектаров в 1923 г. до трех миллионов гектаров к 1935 г. Окопанные скирды, газированные зернохранилища насчитывались, к 1940 г. десятками и сотнями тысяч (Свириденко, 1925; Виноградов и Оболенский, 1926; Фалькешптейн, 1933; Б. Кузнецов, 1932).
Колебания в масштабах истребительных мероприятий в 20—30-х годах зависели здесь лишь от вспышек и падений численности мышевидных грызунов и развития эпизоотий на сусликах. Современные истребительные работы по сусликам строятся в степном Предкавказье с расчетом полной очистки территорий от вида. Несмотря на повторные отработки и затрату громадных средств и труда, уничтожить суслика целиком при современных технических возможностях не удалось даже в пределах эндемического предкавказского очага. Налицо лишь резкое ежегодное снижение численности и сильное разрежение популяций в результате истребительных работ. Не наблюдалось здесь и примеров полного подавления массовых видов мышевидных грызунов. Вспышки численности мышевидных удавалось, однако, предотвращать на небольших опытных участках в соответствии с утверждениями Сатунина (1912а) о важности борьбы с грызунами в годы низкой численности и об истреблении грызунов в стациях переживания, особенно в ранневесеиний период (Наумов, 1946).
Значительно более эффективно и грандиозно по масштабам воздействия на ареалы сусликов и мышевидных грызунов было введение севооборотов и устройство лесных полос, а также вылов сусликов на шкурку с промышленными целями.
Резкое снижение численности мышевидных грызунов последовало на больших площадях в середине 30-х годов при введении зяблевой вспашки. Распашка Сальских степей и устройство лесных полос создали также экологический рубеж для проникновения сусликов в Прикубанскую равнину. Дальнейшее развитие лесных посадок, устройство крупных водохранилищ близ Ставрополя, Ново-Троицкого и других центров будет содействовать новому этапу увлажнения климата. По развившимся лесным посадкам, связывающим байрачные и долинные кустарники и леса, уже идет и будет идти расселение лесных и полевых мышей, лесных сонь. Одновременно, очевидно, будет происходить развитие ареалов степной и лесной мышей, обыкновенной полевки и хомяков в восточном направлении — в зону полупустыни — и сокращение ареалов малого тушканчика, емуранчика, мохноногого тушканчика, земляного зайчика, полуденной песчанки. При развитии культурного охотничьего хозяйства произойдет и увеличение численности русака, лисицы.
Мероприятия по заселению предкавказских равнин новыми видами пушных зверей были связаны с ландшафтами низовий Кубани, Терека и Сулака.
В 1934 г. в тугайные леса по Тереку у Кизляра было выпущено 45 енотовидных собак—Nyctereutes procyonoides Gray (рис. 198). В последующие годы енотки широко расселились по тугаям и камышам Терека, Сулака, а по тростниково-камышовым зарослям лиманного побережья Каспия они проникли далеко к северу. Ареал терской популяции енотов довольно быстро соединился здесь с ареалом волжской, размножившейся
Гл. VI. Изменения группировок и ареалов под влиянием человека
526
от зверьков., выпущенных в низовьях Волги в 1936 и 1939 гг. (карта 94). Промысел еноток в Грозненской области и в северном Дагестане начался с 1943—1944 гг. Уже за 1944—1945 гг. по низовому Тереку было добыто /Ю 1000 еноток (график 7; см. также: Лавров, 1946).
Довольно эффективными оказались и опыты интродукции экзотических рнзунов.
В 1932 г. в плавни низовьев Кубани, близ станицы Гривенской, было напущено 43 южноамериканских болотных бобра — Myocastor coy pus
Рис. 198. Енотовидная собака.
Mol., а на старице Сулака — озеро Шайтан-Казак — в том же году было поселено 22 экземпляра. Успеху интродукции этого вида препятствовало замерзание водоемов в суровые зимы. Погибавшие на льду от холода и голода нутрии становились добычей лисиц, собак, хищных птиц (Верещагин, 1936, 1947г, 1950а). В 50-х годах значительный хозяйственный 1 эффект был получен в этих же районах при развитии полувольного раз-i ведения болотных бобров, т. е. при передержке в клетках части племен-j кого стада в морозный период.
J Более успешным оказалось заселение тростниковых водоемов Кубани, 2 Ей, Терека и Сулака ондатрой — Ondatra zibethica L. (рис. 199).
По данным Всесоюзного научно-исследовательского института охотничьего промысла, первые выпуски ондатр начались в плавнях Кубани 4 в 1944 г., а затем с 1947 г. и в тростниках низового Терека. В последую-: щие годы естественное расселение зверьков было неизмеримо ускорено искусственным расселением при начавшейся плановой эксплуатации (карта 95).
Препятствием к процветанию ондатры в Предкавказье будут лишь ; дальнейшие работы по осушке придаточных водоемов рек и вообще регу-1 дарование стока Кубани, Терека, Сулака при строительстве плотин и водохранилищ. ...........
526 Часть III. Териогеографическое районирование и пути эволюции
t
L
Относительно слабый темп роста численности ондатр в Предкавказье объяснялся до сих пор лишь своеобразным гидрологическим режимом рек, приводившим к гибели молодняк при сильных паводках в июне, а также туляремийными эпизоотиями, возникающими на водяных крысах.
В 1950 г. в тугаи низового Сулака выпустили 23 енотов-полоскунов — Procyon lotor L., отловленных в Азербайджане. Обилие перестойных дуплистых деревьев и кормов в виде лягушек и плодов диких розоцветных создают благоприятные условия для быстрого развития численности
Рис. 199. Ондатра на кормовой площадке.
и ареала этого многоядного хищника по уцелевшим лесам низовий Сулака и Терека. Продвинется он также и по долинам Сунжи, примерно до широты Ачалуки.
Енотом можно быстро заселить и тугайные леса по Куме, в районе Буденновска—Георгиевска, а также массивы леса в Пятигорье и по левым притокам Кубани. Целесообразность такого расширения ареала этого хищника находится под вопросом в связи с вероятным вредом, который он может принести птицеводству и дичному хозяйству осенью и весной (Верещагин, 19536).
Дальнейшие перспективы обогащения экологических группировок млекопитающих предкавказскпх равнин промысловыми видами будут зависеть целиком от характера развития охотничьего хозяйства. При истреблении волка и улучшении дела борьбы с браконьерством районы лесных полос можно будет заселить косулями, оленями и резко увеличить там численность русака.
ПОЛУПУСТЫНЯ ВОСТОЧНОГО ЗАКАВКАЗЬЯ
Центральные части Кура-Араксинской низменности заняты преимущественно своеобразными полынными и полынно-солянковыми, реже каперцевыми и злаковыми группировками. На северных, южных и за-
, Гл. VI. Изменения группировок и ареалов под влиянием человека 527
вад пых окраинах низменности полупустыня замещается бородачевой степью (Гроссгейм, 1932). Полупустынные и степные группировки развиты местами и в Горийской котловине, от Мцхети до Сурама.
Внутри этой зоны уцелели еще местами галерейные леса вдоль Куры, ’икса, Поры, Алазани, тростниково-камышовые и рогозовые водоемы i чожжеве лово-фисташковое редколесье по третичным всхолмлениям.
Первобытные голоценовые группировки зверей описываемых про-страпств были довольно богаты. В полупустыне и в степи паслись стада планов, джейранов. Состав насекомоядных и грызунов открытых пространств был, вероятно, в общих чертах сходен с современным. Здесь обитали: ушастые ежи, длиннохвостые белозубки, краснохвостые пес-чайки, степные полевки, малоазийские и малые тушканчики. В тугайных дебрях и тростниково-рогозовых крепях лиманов и остаточных озер в lihi-рванской, Муганской и Мильской степях держались волки, шакалы, из осатые гиены, львы, тигры, леопарды, гепарды, кабаны, олени, джей-'U hl.
Прирусловые озера Аракса и Куры, заливы Каспия кишели во доп л а-!;i щей птицей на пролетах и зимовках. Здесь же обитали водяные крысы, },,1 :.ры, камышовые кошки.
Влияние человека на группировки зверей этой зоны становится до-кучентально известным с века бронзы, когда запасы крупного зверя степей ст; л и энергично использоваться оседлыми и кочевыми охотниками и скотоводами (Верещагин и Бурчак-Абрамович, 1948; Верещагин, 1949в). Следующий этап воздействия на фаунистический комплекс закавказских степей был связан с быстрым развитием здесь скотоводческих племен и ростом крупных поселений, а также с развитием земледелия и ирри-> ;> ЩИ.
Сельскохозяйственное освоение Ширванской, Мильско-Карабахской 1 Муганской степей имеет многовековую историю, которая еще слабо изу-чиа археологами и историками.
Судя по остаткам древних каналов, крепостей и гигантских курганов г Мильской и Муганской степях, обширные пространства были освоены древней земледельческой культуротт при наличии уже высокоразвитой н’хппки ирригации (Маевский, 1902а, 19026). Изучаемые теперь могильные поля и культурные слои Мингечаура и Урень-Калы свидетельствуют о наличии в центре Кура-Араксинской низменности мощных поселений уже в начале I тысячелетия до н. э. (Сысоев, 1925; Пахомов, 1923; Джафар-Заде, 1946, и др).
Еще более древни, очевидно, небольшие оросительные системы, создававшиеся на притоках Куры и Аракса. Воду этих притоков легче было освоить небольшим общинам, в то время как проводка крупных кана-,юв была под силу местному населению лишь на более высокой ступени развития хозяйства. В этом отношении соображения Латынина (1935) но истории ирригации в Средней Азии целиком применимы и к Азербайджану. Наиболее древними оазисами в Кура-Араксинской низменности являются города и поселки: Барда, Агдаш, Геок-Чай, Шильям и др., расположенные на Тертере, Алджиган-Чае, Геок-Чае, Гердыман-Чае п Ах-Су.
Кроме использования под посевы зерновых, а позднее и хлопка, полупустыня давала еще и запасы зимних кормов для кочевого скотоводства.
Тем не менее основное воздействие человека иа ландшафт и зверей Кура-Араксинской низменности состояло в распашке и орошении откры-
-628
Часть III. Териогеографическое районирование и пути эволюции
тых пространств полупустыни. Истребление тугайных лесов, стравливание растительного покрова скотом в зимний период и осушение речных озер и разливов имели для диких млекопитающих второстепенное значение.
Уже первоначальные наблюдения на территории вновь созданных хлопководческих совхозов в Мильской степи над пресмыкающимися и земноводными показали, что орошение приводит к дифференцированному уничтожению или снижению численности одних и развитию других ви-щов (А. Богачев, 1938). Весьма резкие сдвиги в составе и численности группировок гнездящихся и зимующих птиц происходят при орошении полупустыни (Сатунин, 1912а; Верещагин, 19506). Что касается млекопитающих, то эволюция состава их видов на орошаемых и неорошаемых участках зависит от сезона года и агрономических приемов, т. е. принятой ведущей культуры, очередности обработки посевных клиньев, поливов и развития древесно-кустарниковых зарослей и посадок на межах.
Хороший пример качественных и количественных изменений группировок млекопитающих в результате развития в полупустыне поселений сельскохозяйственного и промышленного типа дает современный Ашперон-ский полуостров с созданными виноградниками, ргородами, промыслами и поселками. На Апшероне насчитывается ныне 18 видов зверей, в то время как на смежных участках полупустыни Кабристана — 15. На окультуренном Апшероне обитает 5 видов зверьков, отсутствующих в смежной целинной полупустыне: пасюк, черная крыса, домовая мышь, средиземноморский нетопырь, обыкновенный еж. Здесь уничтожены джейран и волк, но резко возросла численность зайца, серого хомячка, степной мыши в связи с созданием новой кормовой и защитной растительности (Верещагин, 19386, 1949в).
Весьма показательны примеры влияния деятельности человека на привлечение и распределение рукокрылых, которые появляются и оседают в открытой полупустыне даже при устройстве одиночной водонапорной башни, гробницы, овечьей кошары, а тем более в оазисах со старыми дуплистыми деревьями платана, грецкого ореха. В сельских населенных пунктах зоны полупустыни с 1935 по 1945 г. нами наблюдались и добыты следую-щие виды рукокрылых:
Rhinolophus ferrum-equinum Rh. hipposideros
Myotis mystacinus
Vespertilio kuhlii
V. nathusii
V. pipistrellus
V. serotinus
Nyctalus noctula
Все они используют жилые и служебные постройки в качестве дневных убежищ, а кормятся на окраинах селений, по опушкам садов и парков (Верещагин, 19426; Кузякин, 1950). Наиболее часто используются летучими мышами щели под цоколем крыш, щели по углам оконных косяков станционных построек, школ и других зданий, устроенных из тесаного известняка или кирпича, а также каменные купола мавзолеев и мечетей. Нетопыри, особенно средиземноморский (Куля), приспособились к ловле насекомых близ электрических фонарей. В таком крупном центре, как Баку, обитает преимущественно Vespertilio kuhlii, который был исключительно редок в городе до 30-х годов нашего столетия. Его появление и увеличение численности в последнее десятилетия связано, по нашим наблюдениям, с развитием обширных зеленых насаждений, с увеличением численности летающих насекомых и с устройством пресных бассейнов— водопоев. Так, в весенне-летний период 1935 г. мною насчитывалось
Гл. VI. Изменения группировок и ареалов под влиянием человека 629
па определенном маршруте в центральных кварталах города от 1 до 3 нетопырей, в 1940 г. — от 3 до 5 и летом 1949 г. — от 3 до 7 штук.
Примеры с зверями других отрядов взяты далее для участков, орошаемых туземной сетью каналов, при наличии небольших массивов полей, Чередующихся с целиной.
Изменения состава и относительной численности уцелевших ныне •,декопитающих, происходящие при орошении и посевах, по наблюдении в Ширванской и Муганской степях, показаны в табл. 107.
Здесь отчетливо видно, что наибольшие потрясения исходная груп-[ гровка млекопитающих открытой полупустыни испытывает при создали! обширных посевов хлопка. Но при низкой агротехнике и туземной t ютеме орошения, при нехватке воды для орошения и при наличии пе-; смежающихся пространств целины, залежей, заброшенных канав и разини древесно-кустарниковых зарослей происходит явное увеличение •-тленности охотничье-промысловых зверей.
Наибольшее обогащение видового состава имеет место в древних 11зисах с запущенными садами, особенно в тех случаях, когда они примыкают к тугаям и низовым лесам. Все новые вселенцы проникают на освоенные человеком площади из галерейных и низовых лесов, камышовых болот: обыкновенный еж, шакал, камышовый кот, черная крыса, лесная сыть, лесная соня и кабан.
Иные условия создаются для млекопитающих на обширных массивах полупустыни, орошаемых системой каналов. На территории Мильских ювхозов уже в 1936—1937 гг. можно было проследить полное вытеснение степной полевки и песчанок с обильно и регулярно орошаемых участков 1-го, 2-го и 3-го хуторов. На валах оросителей уцелсвали лишь белозубки, ежи и степные мыши, т. е. зверьки относительно слабо связанные с норами.
В последующие же годы в связи с развитием на валах канав бурьяни-стой растительности и древесных посадок поселения названных зверьков становились более устойчивыми, появлялись также степные полевки, зайцы и лисицы.
Реакции отдельных видов зверей на разнообразные изменения обстановки целинной полупустыни важно дополнить несколькими конкретными примерами.
Насекомоядные и грызуны. Обильные и многократные поливы посевов хлопка полностью вытесняют длиннохвостых белозубок, степных мышей, песчанок и степных полевок. Условия для обитания грызунов в посевах хлопка при прополке рядов отсутствуют по крайней мере до зимы. Двукратный полив, применяемый для посевов зерновых и люцерны, действует слабее. Зверьки спасаются при поливах «на сухих огрехах», межах, валах оросителей и вновь быстро размножаются, заселяя посевы после их обсыхания. Особенно это относится к степной полевке (Родионов, 1924), причем многолетние посевы люцерны являются настоящими накопителями популяций этого грызуна. Развитию численности и вредоносности грызунов в полупустыне содействует также мозаичность распределения посевных участков, т. е. беспорядочность чередования посевов с залежами различного возраста и целиной при низком уровне агротехники (Верещагин, 1942в, 19466).
Большое значение для формирования жизнеспособных группировок мелких млекопитающих в полупустыне имеет сама оросительная сеть. Постоянно действующие оросительные каналы иногда заселяются водяной крысой и служат путями для ее расселения.
34 H. К. Верещагин
. ' дЗО Часть ///. Териогеографическое районирование и пути эволюции । <— — —-——— —_ । Т а б л и ц а 107 | Изменения видового состава и относительной численности млекопитающих при орошении полупустыни в восточном Закавказье
' В иды Целинная полупустыня Посевы Залежь двухлетняя 1 Обочины дорог, канавы с бурьяном Запущенные сады поселков
хлопок ячмень люцерна
? . Insectivora Hemiechinus аи-ritus .... Crocidura rus-g sula giilden- g< • schtaedti . . . > C. leucodon . . W; * Erinaceus euro- | paeus .... r Carnivora F Vulpes vulpes . T.. Canis lupus . . F Meles meles . . |? Vormela peregu- sna 1 Mustela nivalis f. • Canis aureus * Felis chaus . . Lagomorpha Lepus europaeus Rodentia Mus musculus tataricus . ‘ . Cricetulus migratorius . . Microtus socialis M er tones eryth- rourus . . . Allactaga willi-amsi .... A. elater . . . * Mus musculus musculus . . * Apodemus sylva-ticus .... * Rattus rattus . * R. norvegicus . 1 * Dyromys nltedula Artiodactyla Gazella subgutturosa .... * Sus scrofa . . . i X IX 1111 1 XX X ^Х X X 1 i XX XXX 1 XX X — . z xz - XXXXXX — XXXXXXX XX XXX XX XX XX XXX ххххх 1 : XX х ь X j
XX fl XX Н ххх И хх S хххх fl XX fl XX 1 ХХХХХ 1 Из
Hr- 4 1 х 1 ' ХХХХ № X ; к * Х г
.. Всего .... 16 2 i 5 « 1 16 16 1 23
Примечания. 1. Знаками X показано наличие и относительная численность видов, 2. Данные летних и зимних количественных учетов обобщены иля целины, залежей, обочин дорог садов. Наличие и численность зверьков на посевах показаны для озимей и развитых посевов’. Во время полива посевов вытесняются нацело почти все виды. 3. Вселяющиеся из других зон* и биотопов виды отмечены звездочной 4. Не включена в таблицу водяная крыса обнаруженная лишь в оазисе Агдама, но отсутствующая в других оазисах низменно-' сти. b’ Для широко распространенных видов—шакала, лисицы — отмечены те угодья, на которых они встречаются на жировке и лежке.
Гл. VI. Изменения группировок и ареалов под влиянием человека
23
, видов, очин до-ых иосе-
5 Других волянан изменно-ноторых
На нутриевой ферме в Караязах множество водяных полевок и серых крыс продырявило своими ходами стенки водосбросных канав и распределителей, подающих воду в бетонированные клетки болотных бобров. Водяные крысы стали здесь синантропами, поедающими остатки зерна ’’ корнеплодов, вымываемые из клеточных бассейнов.
По наружным откосам и валам оросительных каналов обычно селятся лепные полевки, краснохвостые песчанки и степные мыши. Их привлекают сюда хорошая аэрация грунта и легкость устройства нор, быстрый t :ат дождевой воды и кормовые заросли сорняков.
Остатки древних оросительных канав в степях, заметные часто лишь ! виде небольших ложбинок и низких валиков, бывают неизменно за-( лены степной полевкой и краснохвостой песчанкой. Поросшие зелеными номерами, кустами солянок и каперцев, они служат и важными путями : вселения зверьков при осеннем увеличении их численности и активности.
Аналогичное значение для жизни и расселения грызунов имеет в полупустыне железнодорожное полотно. Его приподнятые участки регулярно выселяются степной полевкой, краснохвостой песчанкой, степной мышью в длиннохвостой белозубкой. Рыхлый, песчаный грунт, пригодный для геройства нор, обильные семенами заросли сорняков — Xanthium stru-marium, Cirsium, Artemisia, Salsola, Sueda — и выбрасываемые из ва-dhob семена арбузов, дынь, хлебные объедки весьма привлекательны для гесчанок.
Песчанки изредка устраивают норы и под шпалами, начиная рытье, t ак под камень, с торца и ведут их в глубину полотна. В таком виде норы одновляются и остаются обитаемыми, несмотря на постоянное беспокойно и засорение выходного отверстия при проходе поездов и подбивании ^брошенного зверьками песка путевыми обходчиками.
Степные полевки селятся в летнее время у основания полотна на северных склонах в связи с некоторым затенением, меньшей прогреваемостью почвы, при наличии увлажнения и более сочной растительности. В зимнее же время полевки заселяют южный склон насыпи, так как здесь пазвитпе эфемерных злаков идет интенсивнее и начинается на 2—3 не-[ели раньше, чем на северном склоне.
Важное значение для расселения и сохранения микропопуляций 1 степных полевок имеют также обочины и канавы грунтовых и шоссейных j дорог. На этих участках сохраняются даже в годы депрессии численности I и размножения жизнеспособные колонии зверьков.
I Весьма замечательные приспособительные особенности оказываются я у малоазийского тушканчика, нередко устраивающего норы в самой тяее грунтовых дорог, несмотря на шум, сотрясение и засорение норы при проезде телег и автомашин.
1 Куринские русаки образуют две различные популяции, одна из ко-|торых может быть названа «домашней», а другая — «дикой». Первая обитает на окраинах селений. Обычно зайцы залегают па день в бурьянах изарослях жесткого злака Imperata cylindrica, нередко всего в 30—50 м перед двориками домов. Около небольших селений, состоящих из* 10—15 глинобитных домов, при тщательных поисках можно обнаружить от 3 до я 5 русаков. Зайцы выносят целыми днями обычный шум селения: передви-| жение людей, домашнего скота и лай собак. На расстоянии 80—100 м 1 отселения и далее на 2—3 км расположена зона, совершенно лишенная 1 зайцев, после чего начинают встречаться уже русаки «дикой» популяции. 1 Как показывают наблюдения, это обитание вблизи человека вырабаты-1 вается у зайцев в условиях слабого преследования их местным населением 1 34*
332 Часть III. Териогеографическое районирование и пути эволюции
и в условиях «охраны» зверьков на дневной лежке от лисиц и шакалов домашними собаками.
Заметное влияние, на распределение грызунов в полупустыне и жизнеспособность их популяций оказывало издревле перегонное скотоводство с зимним выпасом больших стад овец, коров, верблюдов, лошадей, а также стоянки скотоводов. В последние столетия они состояли из нескольких войлочных кибиток для жилья и ряда ям, отгороженных Камышевыми щитами для укрытия молодняка скота. Они располагались обычно по древним аллювиальным грядам, весьма привлекательным для краснохвостых песчанок. Следы заброшенных стоянок остаются на десятки лет в виде углублений и унавоженных тблоков, заросших сорняками: расторопшей, белой полынью, лебедой, — и являются характерным элементом современного ландшафта Кура-Араксинских степей. Именно на этих «поранениях целины» и сохраняются обычно в годы депрессии размножения жизнеспособные популяции степных полевок, краснохвостых песчанок. Высокие сорняки дают затенение летом, спасая полевок от опасного перегрева. Зимой же на таких местах быстро развиваются кормовые эфемеры. Эти же заброшенные ямы используются ушастыми ежами, белозубками, лисицами, барсуками, перевязками, а также красными утками для устройства нор, гнезд и убежищ.
Выедание скотом полынно-злакового покрова полупустыни имело на протяжении тысячелетий огромное значение для местного ограничения численности полевок, песчанок и тушканчиков и перераспределения их популяций.
Синантропная группировка зверей во вновь создаваемых временных и постоянных поселках формируется здесь вначале за счет местных форм: длиннохвостой белозубки, степной полевки, серого хомячка, степной мыши и черной крысы.
Особенно обычны и надоедливы степные мыши и длиннохвостые белозубки в полуземлянках рыбаков и охотников, устроенных по окраинам тростниковых озер низменности. На рыбных промыслах озер низменности всегда обильны пасюки. Степные полевки очень быстро заселяют заново отрытые полуземлянки и установленные кибитки кочевых скотоводов, в Ширванской и Мильской степях. Они таскают там хлеб и другие пищевые продукты, отнюдь не стесняясь присутствия людей. Интересно, что серый хомячок является постоянным обитателем жилых построек преимущественно в южной части низменности, от сел. Пушкино и далее к югу, а также в предгорьях Малого Кавказа и в нагорном Талыше.
По мере освоения населенного пункта, иногда через несколько десятков лет, в него проникают космополитические синантропы: домовая мышь, черная крыса и пасюк. Пасюк, впрочем, переходит в жилище человека и из «диких», природных условий, т. е. из популяций, акклиматизировавшихся в тростниковых болотах Закавказья. То же относится и к черной крысе местного реликтового происхождения.
По материалам нашей экспедиции, в марте 1940 г. состав выловленных давилками 1536 зверьков в домах крупных селений в предгорьях Карабаха оказался в среднем следующим:
Mus musculus musculus — 78.2%
М. musculus abbotti — 12.1%
Л/. musculus tataricus — 2.7%
Microtus socialis — 1-4%
Cricetulus migratorius " * — 1.2%
Crocidura russula giildenschtaedti — 0.3%
Гл. VI. Изменения группировок и ареалов под влиянием человека
533
В качестве примера последовательности формирования синантропной (домовой) и оазисной группировок приводим результаты отлова капканчиками мелких и учета крупных млекопитающих в различных типах поселений человека, расположенных в зоне полупустыни (табл. 108-110). t
Таблица 108
Состав мелких млекопитающих иа зимней стоянке овцеводов среди каперцевой группировки в центре Ширванской степи, (5 апреля 1939 г.)
Виды Число зверьков, добытых на ЮО ловушек по всем местообитаниям Место добычи
Insectivora
Hemiechinus auritus i Яма, заросшая бурьяном. Яма для содержания ягнят, прикрытая тростниковой крышей.
Crocidura leucodon 2
Rodentia
Meriones erythrourus 2 На валиках у заброшенных ям.
Mus musculus tataricus 6 2 экз. — в кибитке и 4 экз. — в кошаре для овец.
Microtus socialis 16 6 экз. —на площадке между кибитками и 10 экз. — в кибитках.
В то же время в окружающей полупустыне отлов давилками дал на 100 ловушко-ночей 15 степных полевок и 2 краснохвостых песчанки. Кроме того, вблизи стоянки были отмечены 2 лисицы, 1 заяц, 5 джейранов.
Таблица 109
Состав мелких млекопитающих в станционном поселке К ер ар Ширванской степи. (10—15 апреля 1939 г.)
Виды Число зверьков, добытых на 100 ловушек по всем местообитаниям Место добычи
Insectivora
Crocidura russula giildenschtaedti 3 В сараях для топлива.
Chiroptera
Vespertilio kiihlii p. serotinus .......... 3 1 Пойманы на чердаке здания станции.
Rodentia
Mus musculus musculus 8 В жилых бараках.
M, musculus tataricus 7 В сараях для топлива и птичниках.
Rattus norvegicus . . 5 В жилых бараках.
Примечание. Поселок состоит из десятка деревянных жилых баранов и каменного здания станции. Несколько молодых деревьев акации.
534
Часть III. Териогеографическое районирование и пути эволюции
Окружающая поселок открытая полынно-солянковая и злаково-эфе-меретовая полупустыня, а также опытные богарные посевы ячменя были населены в те же сроки степной полевкой и краснохвостой песчанкой. Кроме того, в окрестностях поселка добыты и зарегистрированы визуально ушастые ежи, лисицы, ласки, зайцы, джейраны.
Таблица 110
Состав мелких млекопитающих в г. Агдаме (24—30 апреля 1940 г.)
Виды Число зверьков, добытых на 1000 ловушек по всем местообитаниям Место добычи
Insectivora Crocidura russula guldenschtaedti 5 В садах и на люцерне.
C. leucodon 3 На люцерне.
Chiroptera
Vespertilio pipistrellus 1 Убит в саду.
Myotis my st acinus 1 Поймана на чердаке жилого дома.
Carnivora
Mustela nivalis 1 Убита в саду.
Lagomorpha
Lepiis europaeus cyrensis .... 2 Убиты в садах.
Rodentia Mus musculus musculus 29 В складах и в жилых домах.
M. musculus tataricus 36 В садах и скирдах.
Cricetulus migratorius 2 В садах и на межах полей.
Meriones blackleri 1 На выгревном пустыре среди по-
Apodemus sylvaticus aria n us . . 5 селка. В садах по канавам.
Microtus socialis 186 На люцерне и на озими ячменя.
Arvicola terrestris 3 В стенках оросительных канав сре-
ди садов.
Примечание. Городок в Карабахской степи с садами яблонь, винограда, окруженный посевами ячменя, люцерны и участками старых залежей. На облов полей (люцерны и ячменя) и построен приходится по 400 ловушко-ночей. Остальные 200 затрачены на сады, канавы, пустыри
Прогон, устроенный через запущенный сад площадью в 15 га на ок-раине поселка, позволил зарегистрировать еще 3 зайцев, 2 лисиц, 5 шакалов и 1 камышевого кота.
В крупных городах низменности синантропная группировка оказалась уже обедненной за счет исчезновения местных видов грызунов и насекомоядных.
В качестве примера может быть упомянут Баку, в пределах которого ныне живет всего 4 вида зверьков: средиземноморский нетопырь, домовая мышь, пасюк и черная крыса.
Хищные. Появление и увеличение численности лисиц, шакалов, ласок, барсуков, перевязок, камышевых котов в оазисах и среди поселков сельского типа в закавказско!! полупустыне связано с развитием более
Гл. VI. Изменения группировок и ареалов под влиянием человека
535
1
j устойчивой кормовой базы из насекомых, грызунов и домашней птицы, I благоприятного рельефа и защитных зарослей, особенно важных для I вывода молодых. Эти условия сохраняют свое значение даже при разви-| тип охотничьего промысла.
JL ’Копытные. Интересные примеры существования и поведения ж в кульурном ландшафте восточного Закавказья имеются для джейрана S я кабана. В течение последних трех десятилетий джейран находился
Рис. 200.
* ОД формальной охраной, но постепенно истребляется браконьерами с автомашин и пастухами. Он вытеснялся, кроме того, вследствие уничто-{ жения кормовых участков и образования вязкого грунта при распашках ; степей и их орошении. Однако джейраны благополучно существуют среди богарных посевов зерновых на Аджиноурском плоскогорье. В мае—
нюне они держатся там даже в высоком ячмене и пшенице, так как грунт богары остается все же достаточно плотным для того, чтобы газели могли спастись от волков. В Мильской и Ширванской степях джейраны держатся очень упорно среди зарослей солянок и шведки, образовавшихся да залежах двух-трехлетнего возраста. Особенно эффектно это прослеживалось в 1936 г. па землях Мильских совхозов (Верещагин, 19386) (рис. 200).
Заполнение степей отарами овец, стадами коров и верблюдов начинается постепенно с октября и ведет сперва к перераспределению стад джейранов. Газели занимают вначале пустующие пространства худших паст
бищ и солончаков, но при увеличении численности пастушеских стоянок
536 Часть III. Териогеографическое районирование и пути эволюции
и стад, а также при затоплении солончаков водой в середине зимы бывают принуждены пастись вблизи стад баранов и даже вперемежку с последними. Эта близость бывает, впрочем, всегда временной и неустойчивой.
Резкая смена мест обитания у джейранов бывает при длительном пре-следовании их конными охотниками. Джейраны забиваются после этого на два-три дня в тугайные кустарники, камыши. От преследования на автомобилях джейраны спасаются на солончаках и на участках с овражным и кочковатым рельефом.
Развитие оазисов обусловило также выработку у кабанов способности мириться и благополучно сосуществовать в непосредственной близости от населенных пунктов. Кабаны регулярно залегают теперь на дневную лежку в кустах ежевики в садах Агдаша, Карабуджаха и других городков и селений Ширвани и Мугани. Нередко эти лежки устраиваются также на валах водохранилищ и на заросших тростником и эриантусом канавах в непосредственной близости от жилых домов и проезжих дорог.
Второстепенные влияния человека на млекопитающих зоны полупустыни сказывались еще через истребление тугайных лесов по Куре, Араксу | и их притокам.
В восточной Грузии тугайные леса стали быстро исчезать лишь в XIX в., а в середине века они будто бы заботливо охранялись и даже подсаживались для хозяйственных нужд (Кецховели, 1942). В Азербайджане истребление тугайных лесов шло интенсивно еще и в первой половине XX в.
Исчезновению тугаев сопутствовали вытеснение из зоны полупустыни v медведей, барсов, тигров, львов, оленей и значительное сокращение площадей обитания кабанов.
В результате этого краткого обзора прямых и косвенных влияний человека видно, что голоценовый фаунистический комплекс Кура-Араксинской низменности сильно беднел крупными млекопитающими. Некоторое обогащение видового состава происходило за счет появления рукокрылых и расширения площадей обитания мелких зверьков защищенных биотопов: белозубок, обыкновенных ежей, шакалов, зайцев, лесных мышей, черных крыс и т. и.
В связи с уничтожением тугайных предгорных лесов, с развитием посевных площадей и орошения здесь происходило постепенное расширение ареалов мелких зверьков — обитателей полупустыни — на ее окраинах и проникновение в ее центральные участки лесных зверьков.
Новейшие преобразования экологических группировок и фаунистического комплекса млекопитающих полупустыни восточного Закавказья идут под влиянием целенаправленных воздействий человека. Ведется истребление степных полевок, краснохвостых и малоазийских песчанок,, шакалов, волков, камышовых кошек, энергично используются запасы зайца, лисицы, кабана. Под формальной охраной продолжает находиться джейран, но отсутствие степного заповедника и надлежащего охотничьего надзора ведет к его постепенному вымиранию.
Массовая борьба с главным вредителем зерновых культур — стенной полевкой — стала производиться с начала XX в. путем обильного полива полей и применения отравленных приманок (Сатунин, 1912а; Родионов, 1924; Верещагин, 1942в, 19466).
Истребление краснохвостых песчанок затравкой нор хлорпикриноА! и выловом капканами стало проводиться только с 40-х годов нашего столетия на Апшеронском полуострове, отчасти на Мугани и в предгорьях Карабаха. Ни в одном случае сужения общих площадей обитания упомянутых видов истребительными работами не было достигнуто, несмотря
Гл. VI. Изменения группировок и ареалов под влиянием человека
537
на то, что эти работы в годы массовых размножений проводились на площадях многих сотен тысяч гектаров. При современном уровне техники истребительных работ осуществление полного истребления степных полевок
л песчанок в условиях Кура-Араксинской низменности вряд ли эконо- чески целесообразно. Исходя из этого, истребительные мероприятия предлагается строить из расчета ограничения численности зверьков при помощи запахивания стерни и применения отравленных приманок с учетом жизнеспособности (жизненности) популяций и предвидения погодных ч ювий (Поляков, 1950).
Основные вредители животноводства и дичного хозяйства — волк, и нал и камышовый кот, — находящиеся под воздействием обычного
Рис. 201. Одичавшие кролики на о. Булла.
охотничьего промысла, весьма далеки от полного исчезновения. Изучение динамики заготовок шкур названных видов за 1925—1955 гг. показывает лишь колебания в числе добытых шкур, обусловленные активностью заготовительных организаций и величиной премий, назначенных за добытого вредителя. Так, например, добыча шкурок шакала при увеличении заготовительной цены в 5 раз в 1949 г. быстро увеличилась на 500%.
Намеренное введение в экологические группировки зоны полупустыни чужеземных зверей проводилось для увеличения добычи ценной пушнины.
Еще во второй половине прошлого столетия на острова Каспия — (Килой, Нарген, Булла, Обливной, Свиной — капитанами парусников п сторожами маяков были завезены беспородные кролики — Lepus си~ nkulus L., которые здесь одичали и прижились. В 1931—1952 гг. на острова Урунос, Жилой, Булла, Лось и Обливной выпускались небольшие партии венских голубых кроликов для организации островных кроличьих хозяйств.
Не имея практически наземных врагов, эти разномастные одичавшие зверьки (с средним весом около 1.5 кг) обитают и размножаются и ныне на перечисленных островах, благополучно обходясь без пресной воды (рис. 201). На о. Булла в 1940 г. обитало до 500 кроликов (Верещагин, 19426). Росту поголовья кроликов на островах препятствуют неограниченный отстрел, ведущийся сторожами маяков и заезжими охотниками,„
538
Часть 111. Териогеографическое районирование и пути эволюции
а также недостаток убежищ и корма. Легко размываемая зимними дождями затвердевшая вулканическая грязь островов мало пригодна для устройства нор, а редкие кусты древовидной солянки, франкении, эфедры, солероса и выгорающие к июню побеги заячьего ячменя и мятлика не дают запасов корма. Устройство искусственных убежищ и подкормки были бы здесь важной предпосылкой для организации рентабельных кролиководческих хозяйств.
Начиная с 1931 г. в ряде тростниково-рогозовых водоемов низменности был поселен болотный бобр (рис. 202, 203) (карта 95).
Рис. 202. Болотные бобры кормятся на плаву.
Фот. автора, 1948.
Размножение и расселение болотных бобров ограничивалось заморозками, ледоставами и пересыханием водоемов летом, в результате чего множество зверьков гибло от шакалов и собак (Верещагин, 1936, 1939в, 19426, 1950а). Естественное эффективное расселение болотных бобров и формирование ареала шло по системам сообщающихся озер Шильян и Кара-Су со скоростью 15—20 км в год, по популяции и промысел оказались неустойчивыми (график 23).
Болотный бобр образует теперь прочные естественные поселения лишь на участках незамерзающих родниковых речек, например па оз. Хулу-флу в Шамхорском районе и на речках Кара-Су в Караязах. Тем пе менее вольное нутриеводство может иметь в Азербайджане большие перспективы при условии регулярного обводнения водоемов, устройства зимовочных загонов и «катастрофического» запаса зверьков в этих загонах на случай сильного заморозка или засухи.
Значительно более крупные изменения в экологические группировки зверей зоны полупустыни будут внесены в ближайшее время новыми гран-
'Гл. VI, Изменения группировок и ареалов под влиянием человека.
539
дпозными оросительными сооружениями и лесными посадками. Устройство Миигечаурского водохранилища создало уже возможность орошения нового миллиона гектаров степей.
Инженерная оросительная сеть обеспечит мощный многократный поря, дальнейшее внедрение хлопка п травопольных севооборотов. Применение усовершенствовапых истребительных средств на сухих огрехах, мах оросителей и других местах обитания подавит поголовье тушканчиков, песчанок и степных полевок на огромных массивах низменности.
Рис. 203. Заросли рогоза на оз. Ши льни, разреженные болотными бобрами.
Фот. автора, 1948.
Создание лесных полос и развитие кустарниковых зарослей будет способствовать в дальнейшем увеличению площадей обитания белозубок, обыкновенных ежей, шакалов, ласок, камышовых котов, зайцев, степных мышей, отчасти черных крыс.
Соответственно должны быть предусмотрены мероприятия по охотничьему промыслу п предотвращению образования опасной эпидемиологической обстановки.
Дальнейшее обогащение териокомплексов степей и полупустынь Закавказья новыми промысловыми видами в условиях развития культурного ландшафта имеет относительно узкие перспективы.
Промысловые грызуны, как и мелкие вредители, практически нетерпимы иа полях зерновых и технических культур. Мелкие пушные хищники без кормовой базы — грызунов — вымирают или становятся вредителями днчп п домашней птицы. Поэтому, например, проект заселения степным хорем восточного Закавказья прп неустойчивой численности там грызунов мало обоснован и биологически и экономически. Дикие степные копытные смогут существовать преимущественно па участках,
640
Часть III. Териогеографическое районирование и пути эволюции
недоступных орошению и непригодных для посевов, например в Кабристане, степи Джейран-Чели. Кроме джейрана, на этих участках, вероятно, смогли бы существовать еще сайгаки и некоторые африканские антилопы: гну, бубалы, прыгун и др.
Существование сайги в плейстоцене Закавказья и опыт завоза 20 сайгаков на о. Булла в 1950 г. свидетельствует о том, что такая интродукция вполне возможна. Понятно, однако, что при отсутствии действенной охраны в степях новые вселенцы не будут иметь по сравнению с джейраном никаких преимуществ.
ЛЕСОСТЕПНАЯ И ЛЕСНАЯ ЗОНЫ
Воздействие человека на формирование новейших экологических группировок и комплексов млекопитающих в лесостепных и лесных районах Кавказа весьма сложно комбинируется с влиянием многих природных факторов.
Послеледниковые ландшафты горного Кавказа складывались под влиянием общего потепления и усыхания. Освобождавшиеся от льдов огромные площади троговых долин и склонов Перевального хребта на западном Кавказе непосредственно занимались луговой и горно-лесной растительностью, надвигавшейся снизу и по отрогам меридианных хребтов.
В зоне горных лесов формировались биоценотические отношения современного типа, причем развитие лесных формаций приводило к зарастанию субальпийских полян (Тумаджанов, 1947). Дальнейшее потепление и усыхание вело к наступлению степных формаций на лесные.
Зона предгорий лесостепи в ксеротермическую фазу голоцена была поднята местами довольно значительно, в связи с чем на высотах до 1000 и по западным отрогам Большого Кавказа и иод Эльбрусом развивались черноземы (Яковлев, 1914; Захаров, 1935; Берг, 1947; Клопотовский, 1948).
В последующий отрезок голоцена — в начале нашей эпохи, — невидимому, происходило обратное: лесные формации стали наступать на степные. Особенно сильно это было выражено на предгорных равнинах. Ценотическая эволюция лесной растительности Кавказа в четвертичном периоде рассмотрена в работах Синской (1933) и Колаковского (1947а) на основе геоботанического анализа. Однако в нашу эпоху естественные сукцессии древесной растительности сами по себе уже не имели решающего значения для формирования ландшафтов и экологических группировок млекопитающих в связи с мощным влиянием бессистемных рубок, пожаров и выпаса скота, далеко перекрывавших природные процессы. Дубовые, фисташковые и грабинниковые леса предгорий, деградируя под воздействием человека, превращаются в шибляк, а затем на их месте возникает бородачевая степь и дажа фригана. Горные буковые леса при вырубках сменяются грабовыми, при дальнейшем стравливании — березняками и зарослями рододендрона и, наконец, переходят в субальпийские луга (Буш, 1935; Г. Ярошенко, 1940, 1945).
Ботаники и почвоведы признают более широкое, чем ныне, распространение лесов предгорий и плоскогорий Кавказа в историческую эпоху, основываясь главным образом на существовании фрагментов прежних лесов в остепненных местностях, особенно в Предкавказье и Армении (Буш, 1900, 1935; Кузнецов, 1900; Троицкий, 1927, 1939; Тахтаджян, 1941; Гроссгейм, 1948).
Гл. V1. Изменения группировок и ареалов под влиянием человека 541
Несомненно, что развитие иных растительных группировок на месте уничтоженного леса и восстановление последнего происходили в различных районах Кавказа на основе местных закономерностей, обусловленных экологической обстановкой и природными сменами растительности.
* Впрочем, новейшие природные тенденции, идущие под влиянием климата, к замене степных формаций лесными (Захаров, 1935) или наоборот (Ярошенко, 1940, 1956) перекрываются влиянием человека, повсеместно ксерофитизировавшнм растительность (карта 97).
Первоначальные воздействия человека на горные леса выражались главным образом в повышении их нижней и в понижении верхней границы. В последние столетия в связи с промышленными рубками начались широкие обнажения склонов с образованием полян и свободных от леса полос по всей высоте (см. Заклинский, 1931; Солнцев, 1940).
Общие последствия обезлесения на Кавказе состояли в иссушении 1'очвенного слоя и значительной ксерофитизации замещающей растительности, в уменьшении влажности климата и количества осадков при отсутствии конденсации туманов в древесной листве и ветвях, в резком усилении эрозии, т. е. в смыве почвенного слоя, разработке распадков н образовании селевых потоков (Кириллов, 1915; Рощин, 1928, 1930, 1931).
Почти повсеместно обезлесение приводило к вытеснению типично псных копытных — оленя, лося, косули, зубра; лесных хищных — мед-дя, лесной куницы; древесных грызунов —полчка, кавказской белки, оз части желтопятной мыши. На менее зависящих от леса грызунах, например на полевках и лесных мышах, обезлесение сказывалось более сложно. Кроме того, на оголенные вырубкой и иссушенные склоны проникали из смежных зон относительно сухолюбивые грызуны и хищники — обитатели открытого ландшафта. Однако в каждом географическом районе имели место и свои специфические изменения, обусловленные различиями в видовом составе млекопитающих и особенностями среды. Они и рассматриваются ниже по природным зонам.
Лесостепь Предкавказья
Обширная полоса наклонных равнин Предкавказья протянулась от Таманского полуострова на западе до побережья Каспия и горы Бешбармак на востоке. Южной ее границей служат крутые подъемы склонов Большого Кавказа, а на севере — широтные отрезки течения Кубани, Терека и северные склоны Ставропольского поднятия.
В ксеротермическую эпоху послеледниковья эта зона была, вероятно, сильно остепнена, а в субатлантическую эпоху заросла дубовыми лесами, чередующимися с участками лугов и лугостепей на водоразделах. Современный лесостепной ландшафт этой полосы имеет явно вторичное, антропогенное происхождение (Буш, 1900; Тумаджанов, 1947, и др.).
Древние, нижнечетвертичные по происхождению участки сухой степи сохранились лишь между третичными всхолмлениями Терского п Сунженского хребтов, а также на южных обрывах известняковых хребтов предгорий и по северным предгорьям Дагестана.
Как показывают археологические исследования (Герц, 1870; Фар-маковский, 1914; Уварова, 1900; Городцов, 1935, 1936; Анфимов, 1935; Иессен, 1940; Крупнов, 1946, 1947, 1948, 1949; Круглов, 1946а), по этим предгорным равнинам располагались поселения эпохи ранней и поздней бронзы, относящиеся к майкопской, северокавказской, кобанской,
,542 Часть HI. Tериогеографическое районирование и пути эволюции
скифо-сарматской и эллинистической культурам. Позднее — в нашу эпоху — здесь развивались аланские поселения. Аланы постепенно вытеснялись пришлыми мусульманскими племенами. Мищенко (1923, 1928) и Захаров (1,935) считали, что уцелевшие дубово-грабовые леса Заку-банской наклонной равнины обрастали в нашу эпоху скифские курганы, располагавшиеся первоначально в степной местности.
Еще в середине прошлого столетия на равнине во многих местах оставались островки леса и священные рощи адыгейцев, так называемые «кодоши», или «кудоши», как свидетели наличия в прошлом обширных лесных массивов (Вейденбаум, 1878).
Обезлесение этой наклонной равнины было уже почти закончено местным населением к началу крупных переселений казаков в середине XIX столетия (Фелицин, 1884; Щербина, 1913).
Крупные станицы с военным и охотничье-промысловым населением устраивались с середины прошлого столетия по долинам левых притоков Кубани и правых притоков Терека, Сунжи. Между тем эти долины с их тугайными зарослями — уремой — были основными руслами сезонных кочевок крупного зверя (кабанов, косуль, оленей, зубров) из горна равнину, особенно важными во время катастрофических снегопадов в горах. Таким образом, кроме непосредственного преследования и истребления копытных, здесь имело еще место лишение зверей возможности сезонных миграций. Оголенные от леса пространства водоразделов и поселки («заставы») в долинах нарушали установившийся инстинкт кочевок, что вело к гибели копытных от голода, особенно кабанов, принужденных оставаться в горных лесах в снежные зимы. Истребление урем на нижних участках пойм левых притоков Кубани, сопровождавшееся изменением гидрологического режима потоков, привело и к исчезновению речных бобров.
Дальнейшая замена ландшафта лесостепи Закубанской равнины современным культурным ландшафтом с большими площадями пахотных земель, выгонов и поселков сопровождалась оттеснением к предгорьям кустарниковой полевки, сокращением численности и площадей обитания лесной мыши, а также развитием численности обыкновенной полевки, малоазийского и обыкновенного хомяков, серого хомячка, степной мыши, слепыша, зайца и лисицы.
Выпас скота содействовал здесь образованию сухих выгревов по днищам долин и устьевым участкам ущелий, благодаря чему названные зверьки энергично внедрялись и в зону горного леса, например в окрестностях Крымской, Ильской и Горячего Ключа.
На наклонной Кабардинской равнине островки лесов остались лишь кое-где по балкам и долинам рек, а также в районе Пятигорья.
Ширина обезлесенной полосы между Пятигорском и Георгиевском, а также между Нальчиком и Прохладной достигает теперь 30— 40 км.
Эта равнина, еще лет 200 назад населенная косулями, оленями, лосями, зубрами, имеет теперь относительно бедную группировку синантропных видов н мелких сельскохозяйственных вредителей. Продолжающееся уничтожение лесов предгорных увалов в районе Нальчика приводит к пересыханию речек. На иссушенных вырубкой и выпасом скота склонах некоторое время уцелевают заросли орешника, грабинника, постепенно отступающие вверх. Вслед за ними продвигаются заяц, обыкновенный и черноватый хомяки, степная мышь, слепыш, обыкновенная слепушонка.
.Гл. VI. Изменения г руппировок и ареалов под влиянием, человека
543*
В новейшее время водораздел Кубани и Кумы засаживается лесными полосами для уменьшения овражной эрозии и предотвращения летних засух.
Аналогичные преобразования прежних ландшафтов прослеживаются п ha более влажной Терско-Сунженской равнине. Близ Алагира здесь уцелела священная осетинская роща Хетаг, посещенная еще Балл асом и состоящая из трехсотлетних лип, буков й лещин. Она свидетельствует о былом обилии предгорных лесов, заселенных зубрами, лосями, оленями, косулями.
В наши дни последствия обезлесения хорошо заметны по изменению видового состава и ареалов мышевидных грызунов.
Кустарниковая полевка и лесная мышь были оттеснены с равнины к лесистым предгорьям. Их место заняли обыкновенная полевка, хомяк и серый хомячок, ставшие вредителями посевов пшеницы. Обилие огородных культур и посевов кукурузы, а также незамерзающих зимой речек позволило серой крысе стать на этом участке настоящим вредителем, полей и огородов (Бугданов, 1929). Вплоть до лесистых склонов Черных гор со стороны сухого Сунженского хребта сюда продвинулась даже* сухолюбивая перевязка (Туров, 19266).
Дубово-буковые леса предгорной Чечни и северо-западного Дагестана между низовьями Сунжи и ущельем Сулака были заменены в результате рубок и выпаса скота своеобразными кустарниковыми зарослями из орешника — Cory 11 us a veil а па, груши — Pyrus caucasica, алычи — Prunus divaricata, крушины — Rhamnus cathartica, кизила — Coruus mas и др. (Гроссгейм, 1948).
От прежней лесной группировки зверей этих районов сохранились, местами кроты, лесные кошки, рыси, лесные сони, кабаны, косули, но, исчезли медведи, полчки и олени.
Предгорные увалы между Махачкалой и Дербентом на протяжении столетий были разработаны местами под посевы кукурузы и виноградники. От прежних лесов в этих предгорьях осталась ксерофитная формация с преобладанием держи-дерева — Paliurus spina christi и грабинника. Результатом всего этого было широкое внедрение малого суслика, степной полевки и серого хомячка в предгорья. Гептнер и Формозов (1941) отмечают, что проникновение в Дагестан наиболее велика у полевки, однако по аналогии с примерами по восточному Закавказью, можно предположить, что менее многочисленный, но более подвижный и эвритопный хомячок расселяется по формации шибляка в ущельях энергичнее полевки.
Южнее — в низовьях Самура и других речек — на Хачмасской низменности частично сохранились низовые леса. От сплошного ранее массива ныне осталась как бы лесная «сетка», узкие полосы, закрытые лианами и окаймляющие большие поляны, разделанные под посевы кукурузы и овощей. Однако и в таком виде этот участок сохранил небольшое количество кабанов, косуль и обилие полчков, лесных сонь, полевых мышей.
На обезлесенные участки и лесные поляны продвинулась и внедрилась с сухих всхолмлений Кусарской наклонной равнины степная полевка, а на рисовых полях сформировались особые гидрофильные популяции степной мыши — Mus musculus tataricus, сходной с М. mus-cuius hortulanus. Гидрофильные привычки у этих мышей — явление,, конечно, древнее, так как зверьки обитают и по окраинам тростниковорогозовых болот, т. е. вне зависимости от посевов риса. Но на рисовых
544 Часть III. Териогеографическое районирование и пути эволюции
I
посевах оказались такие мощные формообразующие факторы, как обильная кормовая база и возможность использовать для убежищ и запасов корма разграничительные валики-бенты (Верещагин и Мамедов, 1946). Посевы кукурузы и посадки подсолнечника и бахчевых культур привлекают на поляны огромное количество тех же степных мышей, что и на чалтыки. Это обусловлено бурным развитием кормовых сорняков: мышея — Sertaria viridis, канареечника — Phalaris arundinacea, сулуфа — Echinochloa crus galli. На полянах кормятся по ночам и русаки, а также лесные мыши.
Учет насекомоядных и грызунов, произведенный в этом районе в октябре 1945 г. при отлове давилками, показан в табл. 111.
•г. Таблица 111
Состав и относительная численность мелких млекопитающих (по числу зверьков, . пойманных на 100 ловушек) в различных биотопах Самурских лесов в октябре
V 1945 г.
Виды Старый лес на сухих участках Опушки с лианами по ручьям и окраинам полей Посевы кукурузы и подсолнуха Бахчевые культуры с сорняками Рисовые поля 1 1 Общее число особей 1
число | особей 1 В % число I особей В % число особей В % число особей В % число особей В %
Crocidura russula gulden-schtaedti . 2 22.2 4 44.4 1 11.1 1 11.1 1 11.1 9
Mus musculus jormosovi 2 25.0 5 62.5 — — — — 1 12.5 8
M. musculus tataricus . — — — — 13 36.2 5 13.9 18 50.0 36
Apodemus sylvaticus . . 10 55.5 5 27.8 2 11.1 1 5.5 — — 18
A. agrarius ...... " 7 100 — — — —— — — 7
Microtus socialis .... — — ’ * — 7 100 — — — —™ 7
Dyromys nitedula , . . 1 20.0 4 80.0 —.— — — —.— — — 5
Glis glis ........ 1 25.0 3 75.0 — — — — — 4
Кроме увеличения численности полевых мышей, тяготеющих к опушкам, и появления степной полевки, разработка под посевы лесов Хаг-масской низменности сопровождалась также увеличением распространения зайцев, лисиц и шакалов.
Между Хачмасской низменностью, ее низовыми лесами и хребтами юго-восточного Дагестана лежит обширная Кусарская (или Кубинская) наклонная равнина, выполненная четвертичными галечниками и суглинками.
Леса Кусарской равнины были вырублены в последние столетия и заменялись кустарниками с преобладанием держи-дерева, боярышника, ежевики, грабинника и орешника или посевами пшеницы и ячменя. Близ селений появились крупные массивы яблоневых и айвовых садов.
Распространение и численность хищных, зайца, мышевидных грызунов и копытных Кусарской наклонной равнины также претерпели в нашу эпоху крупные и сложные изменения. Исчез медведь, несколько увеличилась, вероятно, численность лисицы, шакала, барсука, зайца, широко расселилась степная полевка и степная мышь.
- Гл. VI. Изменения группировок и ареалов под влиянием человека
545
[ Распространение лесной мыши стало мозаичным в соответствии с уце-| девшими древесно-кустарниковыми зарослями. По заброшенным пашням j и выгонам широко расселились степные полевки, степные мыши, ото-* двинулась несколько вверх нижняя граница ареала обыкновенной полевки.
’Недавняя проводка Самур-Дивичинского канала и устройство рисо-вых полей содействовали и здесь образованию водолюбивых популяций степной мыши.
Полчок сохранился на немногих участках уцелевшего перестойного леса, но лесная сопя распространена еще широко.
Перспективы обогащения териофауны предгорных лесостепных рав-) яин Предкавказья новыми видами невелики.
В ряде пунктов на озерах и водохранилищах выпущена ондатра. 1 Местами на Закубанской и Терско-Сунженской равнинах живут енотовидная собака и енот-полоскун (карта 94).
Ближайшее будущее этой зоны — дальнейшее всемерное развитие ( садоводства и интенсивного земледелия при быстром увеличении поселков и населения. На наиболее сухих участках следует создать лесные полосы. Охотничье хозяйство должно предусматривать здесь умеренное развитие численности русака, увеличение численности лисицы, а мес-тами — норки и степного хоря; сельское хозяйство — борьбу с хомя-; ками и полевками в западной половине и со степной полевкой и мышами — в восточной.
I Заповедных участков в этой зоне не выделялось, а теперь такие мероприятия сильно запоздали.
Весьма сходные с описанными преобразования ландшафтов, а затем I и группировок млекопитающих происходили также и на предгорных I наклонных равнинах в восточном Закавказье.
I
I
Лесные низменности Закавказья
1 Теплые и влажные низменности западного и восточного Закавказья — I Колхидская, Алазано-Автаранская и Ленкоранская — в голоцене были 1 покрыты болотистыми дебрями с преобладанием широколиственных по-I род и с обилием колючих лиан.
| Первобытный — не тронутый человеком тип этих низовых лесов недо-1 статочно известен (Альбов, 1892; Синская, 1933; Малеев, 1938; Гроссгейм, | 1948). Характерные породы уцелевших низовых лесов: длиннолистный i дуб (Quercus longipes), граб, ильм, лапина (Pterocaria caucasica), черная ольха, а в Ленкоранской низменности — железняк (Parrotia persica).
Выборочное истребление низовых лесов человеком повсеместно приводило к появлению в них множества опушек, завешенных лианами: Smilax excelsa, Hedera helix, Clematis vitalba, Periploca graeca. Образующиеся поляны зарастали папоротником-орляком и ежевикой или I держи-дерев ом и колючим ситником.
| Несмотря на однотипность ландшафтов, последствия воздействий человека на формирование группировок млекопитающих названных участков оказались различными из-за топографических условий.
Следы человеческой культуры полнее изучены в западном Закавказье. Цо окраинам Колхиды (Рионской низменности) известны стоянки эпохи среднего и верхнего палеолита, всюду обильны памятники эпохи бронзы (колхидская культура), эллинистических и более поздних времен.
В нашу эпоху в Колхиде были созданы обширные поселения с плантациями кукурузы и плодовых садов. Численность и видовой состав
35 Н. К. Верещагин
646
Часть III. Териогеографическое районирование и пути эволюции
зверей сильно пострадали здесь как от непосредственного преследования, так и от сокращения площадей лесов. В конце XIX в. здесь вымер речной бобр, а позднее исчез и благородный олень. Только в глухих болотах еще сохранились кабан и косуля.
Начиная с 1930 г. болота (рис. 204) по Риону подверглись осушению путем кольматации и прокопки каналов (Зунтурпди, 1931; Шапошников и Верещагин, 1932), а ольховые дебри — вырубке и замене плантациями чая, кукурузы, мандариновых садов, посадками эвкалиптов.
Рис. 204. Ольхово-кувшинковые болота близ Поти — характерное местообитание болотных бобров и пасюков.
Фот. автора, 1931.
Все это приводило к резким сменам в составе и численности млекопитающих.
Развитие птицеводства и скотоводства содействовало поддержанию численности шакала и волка. Развитие садоводства при обилии закрытых лианами опушек по оставленным грядам леса обусловило увеличение численности лесной сони, полчка. На вырубках, сухих олуговевших участках расселялась кустарниковая полевка.
Эволюция группировок млекопитающих в связи с осушением ольховых топей и другими типами хозяйственной деятельности человека показана в табл. 112. Наши наблюдения, учеты и отлов зверьков ловушками и капканами производились осенью и зимой 1938—1939 и 1940 гг. у восточного побережья оз. Палеостом и у оз. Бебесыри.
Исчезновение большинства видов млекопитающих па плантациях чая, мандаринов, лимона и тунга объясняется отсутствием па них убежищ и корма.
Синантропная группировка в поселках сельского типа сформирована теперь в Колхиде из завозных видов — пасюка, домовой мыши; частью из туземных — черной крысы, лесной мыши. Для жилых строений Черноморского побережья Шидловский (1947) приводит такие цифры отлова грызунов на 1000 ловушек: домовая мышь — 158, серая крыса —
Гл. VI. Изменения группировок и ареалов под влиянием человека 547
Таблица 112
Изменение видового состава и относительной численности млекопитающих при осушении и вырубке низовых лесов в западном Закавказье
Виды Лес колхидского типа и ольховые топи Поляны с орляком и болотистые луга — пастбища Посевы кукурузы Посадки чая и цитрусов
Insectivora
Sorex araneus Crocidura russula monacha . Talpa coeca Erinaceus europaeus .... хххх хххх XX 1 X хххх
Carnivora
Canis aureus C. lupus Vulpes vulpes . Mustela nivalis Meles me les Lutra lutra ........ Felis silvestris XXX XX XX XI 1X1 1 1
Lagomorpha
Lepus europaeus — X X
Rodentia
Mus musculus Apodentus sylvaticus .... Rattus rattus * R. norvegicus Microtus majori Arvicola terrestris Dyromys nitedula Glis glis .......... * Myocastor coy pus XX хххххх XX XX XX XX XX XX XXX XXI1X1111
Artiodactyla
Sus scrofa ......... Capreolus caprcoins .... X — — —
Всего 21 15 1 4 10
Примечание. Знаком х показано наличие и относительная численность каждого вида, звездочкой (*) отмечены виды, завезенные случайно и намеренно.
125, черпая крыса — 17, лесная мышь — 13. В крупных населенных пунктах низменности последний вид отсутствует. По 18 летним учетам Степанова (1946), в Батуми было отловлено и исследовано противочумной лабораторией 44 771 мышь, 40 000 пасюков, 3505 черных крыс, 1588 александрийских крыс. По мнению Степанова, количество мышей пкрыс в Батуми примерно одинаково.1 Но мышь живет главным образом в жилых помещениях — квартирах, а основная масса крыс живет и питается впе квартир.
1 Это мнение весьма спорно, мышей, как правило, бывает больше.
35*
. 646
Часть III. Териогеографическое районирование и пути эволюции
Итак, общее направление в смене группировок млекопитающих прп развитии современного типа культурного ландшафта в Колхидской низменности состоит в крайнем их обеднении. Исчезают не только крупные копытные и средние хищники, но сокращаются количество видов и численность большинства местных грызунов.
В Алазано-Автаранской низменности северо-западного Азербайджана, при ее ином топографическом положении, последствия освоения территории человеком для млекопитающих оказались весьма отличными от
Рис. 205. Залитые рисовые поля в долине Агри-Чая — пример полного сезонного вытеснения млекопитающих.
Фот. автора» 1950.
колхидских. Эта долина, достигающая в ширину 20—25 км, вытянута на протяжении почти 300 км, от сел. Ахмети в Грузии до сел. Исмаиллы в Азербайджане. Остатки первобытных лесов из длиннолистного дуба и граба сохранились до наших дней преимущественно на восточном участке. Освоение низменности человеком здесь не менее древнее, нежели в Колхиде. Близ Белокан, Куткашена, Ниджа зарегистрированы остатки селений I тысячелетия до н. э. В настоящее время низменность довольно густо заселена, причем широкие пятна поселений разбросаны либо в центре низменности или сосредоточены в устьях горных ущелий. Характерно сильное увеличение речных протоков и канав при разборе горных речек для полива риса (рис. 205), пшеницы, табака и ореховоплодовых садов. Основное воздействие человека па ландшафт и зверей сказывалось здесь также в расчистках леса, но с последующим устройством обширных плантаций грецкого ореха, лесного ореха, каштана, яблонь, груш, алычи, мушмулы, айвы, винограда и распашкой образующихся полян под ячмень, пшеницу, кукурузу, рис, табак и кунжут.
Гл. VI. Изменения группировок и ареалов под влиянием человека
549
Окраины полей, края оросителей и участки выгонов обильно зарастали бояркой, ежевикой, терном, держи-деревом, эриантусом и тростником. Для окраин селений и опушек крайне характерно мощное развитие сорных однолетников, кустарников и деревьев, особенно черной бузины 'Iайланта.
Процесс восстановления первобытных дебрей на местах древних ^брошенных поселений нередко повторялся, причем на месте произ-застания исконных древесных пород появлялись трансформированные насаждения с примесью завозных видов. Так, на месте города Чухур-Кабала образовались дикие заросли ольхи, кавказской лапины, граба, эполя, клена и вторично одичавших — лесного ореха, грецкого ореха, платана.
Вырубка и выжигание джунглей вытесняли таких крупных живот-пых, как кабан, олень, косуля, медведь, рысь, лесной кот, леопард, ;> также лесную куницу, лесную соню, полчка, белку, но вместе с этим здесь увеличивалась численность лисицы, камышового кота, барсука, зайца, степной, водяной и обыкновенной полевок, черной крысы.
По мере старения плодовых и ореховых садов лесная группировка зверей восстанавливалась почти полностью. Отдельные виды — лесная соня, полчок, кавказская белка, отчасти лесная мышь, закавказский подвид лолтопятной мыши, горная домовая мышь и длиннохвостая белозубка — появлением обильных кормов и убежищ в булыжниковых заборах •ажё увеличивали свою численность. Лесная мышь становилась здесь инантропным домовым грызуном. Смена состава и изменение численности зверей на различных типах угодий этой низменности показаны в табл. ИЗ.
Таким образом, видно, что при обезлесении и разработке земель низменности под зерновые (и технические) культуры в новейшее время исчезает 6 видов промысловых животных: олень, косуля, медведь, рысь, десной кот и белка, — но увеличиваются в числе 2 вида: лисица и заяц. В то же время со степного юга вселяются малоазийский тушканчик, степная мышь и степная полевка, а из зоны горного леса — обыкновенная полевка.
В крупных орехово-плодовых насаждениях почти полностью восста- навливается и группировка промысловых зверей, а численность древесных грызунов даже нарастает. Это происходит, впрочем, главным образом сезонно вследствие осенних миграций этих зверей, обитающих ‘ в горных лесах и в заросших лесом оврагах, на кормные садовые j участки.
Синантропная группировка в обширных садовых поселках этой низменности формируется почти целиком из местных видов: лесной мыши, темнобрюхой домовой мыши, черной крысы, отчасти лесной сони и длин-, нохвостой белозубки.
На чердаках и в щелях домов ютятся остроухие ночницы и малые • нетопыри. Завозные синантропы — домовая мышь и пасюк — обитают J теперь во всех крупных поселках, связанных шоссейной дорогой.
i Значительно более резкие ландшафтные и фаунистические изменения произошли в зоне низовых лесов по северо-восточной окраине Кара-
i баха, в низовьях Тертера, и на юго-востоке Мугани, по северной окраине ; Талыша, где культурный ландшафт начисто сгладил первобытные черты j местности. Эти черты могут быть теперь лишь угаданы. Малое развитие j орехово-плодовых посадок не дало возможности восстановиться хотя бы ; части прежнего комплекса лесных зверей.
550 Часть III, Териогеографическое районирование и пути эволюции
; Т а б л и ц а 113
Изменение видового состава и относительной численности млекопитающих при устройстве посевов и садов в Алазано-Автаранской низменности
Нетрону- Поляны с посевами зерно- Старые наса-
Виды тый низ о- вых и нустар- ждения ореха
' * вый лес никами и плодовых
Insectivora .
Crocidura russula guldenschtaedti . XXX X X хххх
C. leucodon — X —
Sorex araneus . Erinaceus europaeus X XX
Carnivora
Canis aureus X XX
C. lupus Vulpes vulpes у^
Ursus arctos X — X
Mustela nivalis X XX х
/. Martes foina у^ ”— х
Meles meles . . Lutra lutra у^ X (по речкам) XX
Felis chaus F. silvestris ух
F. lynx У( — х
* Procyon lot or — XX
Lagomorpha -
Lepus europaeus , ,- < ..... . — XX х
Rodentia
Mus musculus abbotti XX XXX
* M. musculus tataricus XX —
Apodemus sylvaticus ....... XXX X хххх
A. fulvipectus XX X XXX
Rattus rattus — у^ X (близ селе-
НИЙ)
* Microtus arvalis ......... — XX (близ скло- —
нов гор)
* M. socialis — хххх
Arvicola t err e str is ........ — X (по речкам) —
* Allactaga williamsi ....... — —-—
Dyromys nitedula ........ X х XXX
Glis glis ............ XX хххх
Sciurus anomalus у<\ — XX
Artiodactyla
Sus scrofa ............ Cervus elaphus XX х 1
Capreolus capreolus ....... X —
Всего 24 23 24
Примечание. Знаком X показаны наличие и относительная численность каждого вида, звездочкой (*) отмечены виды, аклиматизированные и вселившиеся из других зон.
Гл. VI. Изменения группировок и ареалов под влиянием человека
551
! ло-
Только недавнее существование в этих районах дикобразов, наличие
местами черных крыс и полчков в изолированных садах свидетельствуют о былых лесных группировках зверей.
На большей части Ленкоранской низменности, представлявшей в пер-' вобытпом виде сплошные дебри, — гирканские леса из дуба, ольхи, железного дерева, перевитых колючими лианами, — произошли пока изменения, сходные с описанными для Алазано-Автаранской долины. Разработка больших полян под культуру риса, а позднее широкие расчистки
для культуры чая и мандаринов в комбинации с ростом поселений и развитием охотничьего промысла сократили численность кабана, дикобраза, нацело вытеснили медведя, тигра, оленя, косулю. По вырубкам и плантациям чая из Мугани сюда стал проникать заяц, но степная полевка не смогла проникнуть ни со стороны нагорной степи, ни из Мугани из-за большой влажности почвы в зимнее время. Лесная мышь здесь совершенно отсутствует, а степная нигде не образует тех гидрофильных популяций на рисовых полях, которые столь характерны для Хачмасской низменности и юго-восточного Карабаха.
Создание обширных поселков, виноградных и плодовых садов,рисовых полей и обилие густых опушек, вырубок, колючих зарослей и изгородей, окаймляющих поля, атакже заросших камышами водо-
!..«.• y-...ц,-т-.«. J.——; X------т-дя»-;—L ... .J
I II III IV V VI VII VIII IX X XI XII
Рис. 206. Динамика численности и отлова
домовых грызунов — домовом мыши, пасюка и черной крысы — в Ленкорани и прилежащих селениях в 1941—1942 гг.
хранилищ содействовали увеличению численности шакала, камышового кота, полчка, черной и серой крыс, домовой мыши. Сохраняется здесь и кабан, кормящийся на посевах зерновых. Шакал и камышовый кот при
ви да.
отсутствии интенсивного промысла увеличиваются в числе, подкармливаясь домашней птицей.
В связи с уменьшением числа перестойных деревьев полчки зале-{ тают теперь нередко в дневную и зимнюю спячку на чердаках : жилых и хозяйственных построек. Чердаки и щели деревянных и каменных построек заселяются также усатыми ночницами. Синантропная, \ домовая группировка вредителей состоит всего из 3 видов грызунов: ? домовой мыши, пасюка и черной крысы. Их относительная численность, 1 по данным массового отлова ловушками, представлена такими цифрами: мышь — 90.0%, пасюк — 9.49%, черная крыса — 0.5% (Верещагин, ; 1949) (рис. 206).
Дальнейшее развитие чайных, тунговых и цитрусовых плантаций неизбежно повлечет вырубку уцелевшего первобытного леса и сокра-щепие посевов риса, что в свою очередь обусловит сильнейшее обеднение всех экологических группировок млекопитающих, очень сходное с колхидским.
652 Часть III. Териогеографическое районирование и пути эволюции
Арчево-фисташковое редколесье восточного Закавказья
По палеонтологическим и археологическим исследованиям (Петров, 1939; Верещагин, 1949в; Гуммель и Яценко-Хмелевскищ 1941) известно, что сухие предгорья на юго-восточной оконечности Большого Кавказа, в Кабристане и бассейпе Гильгин-Чая, на Шекин-ском, Карталинском и Иорском плоскогорьях, по наклонной Киров абадской равнине, юго-восточным склонам Карабаха и в Горийской кот-
Рис. 207. Остатки приречных и можжевеловых лесов в ущелье Турьян-Чая.
Фот. автора, 1949.
ловине были покрыты в голоцене так называемыми «светлыми лесами», состоявшими из рощиц кэвовых деревьев — Phistacea mutica и можже вельников — Juniperus polycarpos, чередовавшихся с участками боро-дачевой степи. От этих лесов остались теперь лишь небольшие участки, значительно поврежденные рубкой, и то только на труднодоступных южных обрывах и северных склонах третичных всхолмлений, в Циви-Цкаройском, Самухском, Аг датском, Геок-Чайском районах и отдельные заповедные и «священные» рощи и деревья, заботливо охраняемые мусульманами, например в Кабристане (Верещагин, 1949в). Как показывают наблюдения в уцелевших лесах этого типа, редколесье было населено медведями, полосатыми гиенами, волками, лисицами, барсуками, куницами белодушками, благородными оленями.
В результате хищнических рубок па топливо и постройки и все увеличивающегося выпаса скота арчево-фисташковые леса заменялись вначале зарослями держи-дерева, а затем пагорио-ксерофитной формацией из колючих астрагалов. С истреблением больших массивов редколесья к XVIII столетию распространение медведей, кабанов, оленей сильно
Гл. VI. Изменения группировок и ареалов под влиянием человека
553 ‘
сократилось. Неизмеримо увеличивалась эрозия склонов и их иссушение, приводившее к оттеснению на северные склоны холмов и плоские участки популяции степных полевок и малоазийских хомяков (рис. R-
' Горные леса Большого Кавказа
Новейшая эволюция ареалов видов и формирования экологических группировок млекопитающих в горно-лесной полосе Большого Кавказа оаввсит главным образом от охотничьего промысла, от промышленных рубок и развития скотоводства.
На протяжении последнего полустолетия в этой зоне был уничтожен зубр, катастрофически сократилось распространение оленя и барса, значительно уменьшилась во многих местах численность медведя, кабана, косули, серны, кавказских козлов.
Истребление лесов нижнего пояса, а затем и пихтовых лесов среднего п верхнего поясов описывалось для северо-западного Кавказа еще Дин-ником (1911). Промышленные рубки пихтовых и буковых лесов в долинах Зеленчуков, Теберды и других речек со сплавом леса начались с 90-х годов прошлого столетия.
Наибольшее обезлесение под влиянием рубок и выпаса скота наблюдается по гребням хребтов, веерообразно спускающихся к Таманскому полуострову. Образование полян с мезофитными лугами содействовало-здесь развитию численности кустарниковых полевок и улучшению кормов л условий для кабанов, косуль и оленей.
В глубине ущелий процесс обезлесения комбинируется с развитием искусственных насаждений, хорошо заметных по притокам Афипса, Псекупса и других речек, где черкесами были издавна насажены сады груш, айвы и мумшулы. После выселения черкесов во второй половине прошлого столетия грушевые насаждения одичали, заполнили днища ущелий и слились с массивами широколиственных лесов склонов, а местами даже внедрились в них. Эти своеобразные грушевые леса, описанные Малеевым (1939а), оказались необычайно привлекательными и кормными, особенно в осенний период, для куниц, барсуков, медведей, полчков, лесных сонь, кабанов, оленей, косуль.
На распределение мышевидных грызунов влияние человека сказывается наиболее сильно по днищам ущелий. Кроме лесостепных грызунов, по днищам ущелий наиболее высоко проникают вслед за человеком полевая мышь, мышь-малютка и степная мышь.
Домовая, синантропная группировка в горных поселениях первоначально формируется из туземных видов, особенно из лесной мыши и длиннохвостой снеговой полевки. Последняя часто встречается в погребах, подвалах и селится в парниках и близ огородов (Жарков, 1949).
В восточном направлении антропогенное обезлесение склонов и его последствия для биоценотических смен быстро нарастали в связи с увеличением природной сухости климата. Широкие расчистки горных склонов от буковых и сосновых лесов в ущельях Черека, Уруха, Ардопа, Фиагдона и Терека (см. Буш, 1931, 1935) приводили на склонах южного освещения к образованию сухой лесостепи и формации трагакантовых астрагаловг населенных зайцем и серым хомячком, а на северных склонах к развитию мезофитных лугов, заселенных кротом, обыкновенной и кустарниковой полевками. i
В Закавказье особенно сильному истреблению подверглись буковограбовые и каштановые леса нижнего пояса по террасам Черноморского*
554
Часть III. Териогеографическое районирование и пути эволюции
побережья, от Батуми до Новороссийска. На больших пространствах от прежних дебрей здесь сохранились лишь небольшие кустарниковые заросли, чередующиеся с папоротником-орляком. Основными причинами обезлесения склонов в Абхазии являются сплошные рубки, выпас коз и пожары, препятствующие лесовозобновлению (Заклинскпй, 1931).
Наряду с истреблением лесов горных склонов и развитием многочисленных поселков создавались плодовые сады, преимущественно из яблонь и груш, а также декоративные посадки и парки с платанами, эвкалиптами, пихтами, бамбуками и другими экзотическими растениями.
В этих условиях происходила выработка бедной видами группировки зверей с заметным синантропным оттенком. Этот оттенок придают упомянутой группировке не только живущие в «дикой» природе пасюки и черные крысы, но и полчки, тяготеющие к плодовым культурам, лесные кошки и шакалы, тяготеющие к курятникам поселков, к стоянкам рыболовецких и дельфинобойных бригад.
Приспособления лесных и желтопятных мышей к кормежке на посадках пробкового дуба (Свириденко, 1940, 1944) и домовых мышей к обитанию и кормежке в парниках (Шидловский, 1947) дополняют разнообразие биологических реакций грызунов на новейшие земледельческие приемы человека.
Численность мышевидных грызунов в горах близ Сочи при оставлении на месте неубранного лесного хлама (па лесосеках военного времени) сильно увеличивается вследствие создания большого количества дополнительных убежищ для лесной мыши, мыши-малютки, кустарниковой полевки. Кроме того, по лесосекам проникает в горы заяц и полевая мышь. В то же время сокращается численность желтопятной мыши. Наоборот, при культурных рубках, с полной уборкой сучьев, численность мышевидных резко падает в первый год существования вырубки, но уже на второй, при зарастании ее луговыми травами и кустарниками, столь же быстро восстанавливается, причем в упомянутую группировку добавляется еще и черная крыса (Свириденко, 1945). При хищнических рубках леса древесные грызуны — лесные сони и полчки — снижают численность, а при культурных рубках вовсе исчезают. В связи с этими наблюдениями приобретает известную достоверность соображение Шидловского (1948), что распространение здесь к югу полевой мыши и мыши-малютки — явление последних столетий, связанное с воздействием человека на ландшафты побережья.
Расчистки от леса горных склонов в Абхазии и Аджарии содействовали местами большому распространению популяций прометеевой полевки, сохранившихся на реликтовых полянах в лесной зоне.
На отрогах Рачинского хребта, на Сурамском, Картлийском, Кахетинском хребтах, особенно в окрестностях Сталипири, Тбилиси и Мцхети, интенсивная вырубка буковых лесов приводила к новейшему расширению ареала малоазийского хомяка и смещению ареалов многих лесных видов. Всего за 50—60 последних лет была обезлесена Мухранская долина, еще в конце прошлого столетия населенная медведями, рысями, лесными кошками, лесными мышами, полчками, кабанами, косулями, оленями. От прежних буково-грабовых лесов здесь остались ныне лишь деревья-«свидетели» (рис. 208). Долина в настоящее время распахана под посевы ячменя и кукурузы, а заболоченные части ее используются в качестве покосов и выгонов. На распаханные участки вселилась с запада горийская степная полевка, появились также заяц-русак, степная
Гл. VI. Изменения группировок и ареалов под влиянием человека
555
мышь и лисица. От прежней лесной группировки млекопитающих осталась лишь эврптопная длиннохвостая белозубка.
Сухость склонов вследствие вырубки леса и последующей пастьбы скота на склонах гор в окрестностях Тбилиси и Мцхети необычайно увеличивается и приводит в конце концов к замене луговых формаций степными.
Лесовозобновление здесь отсутствует. Даже вторичные кустарниковые группировки, образующиеся на северных склонах после вырубки буковых лесов и состоящие из представителей родов: Crataegus, Mespi-
Рпс. 208. Деревья-«свидетели» в обезлесенной и распаханной Мухранской долине, заселенной степной полевкой.
Фот. автора, 1945.
lus, Rosa, Prunus, Jasminum, Cerasus, Cotoneaster, Spiraea, CoryIlus,— истребляются при интенсивном выпасе скота. На южных склонах исчезают даже такие кустарники, как Juniperns и Paliurus. Держи-дерево держится особенно упорно, но в конце концов также постепенно отступает вверх по склонам не только из-за механического уничтожения, но и из-за изменения физико-химических свойств почвы при ее уплотнении, иссушении и прогревании солнцем (Сосповский, 1915). Этот антропогенный процесс иссушения территории содействовал широкому распространению малоазийского хомяка, серого хомячка, степной полевки, степной мыши, зайца и лисицы, сокращая в то же время площади обитания лесных хищных, грызунов и копытных. В более мезофитных условиях — на плоскогорьях Южной Осетии — под влиянием человека происходила замена лесных ландшафтов лугово-лесными и луговыми (Н. А. и Е. А. Буш, 1936): в этом районе резко обеднялась лесная труп-
ооб
Часть III. Териогеографическое районирование и пути эволюции
пировка зверей крупных видов — медведь, олень, косуля, кабан — и увеличивалось распространение обыкновенной и кустарниковой полевок.
Восточнее, по южному склону, процесс обезлесения в результате рубок хорошо заметен в районах Кварели, Закатал, Нухи, Куткашена и Шемахи. Выпас скота в верхней опушке леса мешает возобновлению горного дуба вследствие уплотнения и увеличения физиологической сухости почвы (Ярошенко, 1956). В ряде мест в результате постепенной расчистки от леса были созданы скотопрогонные дороги шириной в пес-
Рис. 209. Иссушенные вырубками склоны в среднем течении Гердыман-Чая, населенные куницей белодушкой и лесными мышами.
Фот. автора, 1949.
колько десятков метров, связывающие низменности с высокогорными пастбищами. Эти оголенные полосы, проходящие, как правило, по греб-ням меридианных хребтов, служат настоящими миграционными руслами для лисиц и зайцев, обитающих на низменности и в высокогорье. Миграции этих зверей легко наблюдать осенью при снегопадах. В районе Кварели, Лагодехи п у Варташена широкое и давнее обезлесение склонов, по-видимому, и обусловило расселение обыкновенной полевки из высокогорий на Алазапо-Агричайскую долину.
Последствия новейшего истребления лесов сказались особенно сильно на составе млекопитающих близ восточной оконечности Большого Кавказа, между Шемахой, Лагичем и Конахкептом. Буково-грабовые и дубовые леса верхнего пояса заменены здесь большей частью мезофит-ными лугами на северных склонах ущелий или же низкорослыми зарослями кустарников.
В средней зоне — на высотах от 700 до 1300 м — наблюдается иссушение склонов и образование остепненных группировок вследствие
Гл. Г/. Изменения группировок и ареалов под влиянием человека
557
вырубки леса и выпаса скота. Карст способствовал усилению этих воздействий (рис. 209).
Из крупных животных в верховьях глухих ущелий уцелели восточно-К1вказские козлы, а в островках дубовых лесов — кабаны и немногочисленные медведи. Вытеснены полностью лесные куницы, олени, ко-<ули.
Вселение сухолюбивых млекопитающих в этот район идет со стороны предгорной степи, от Шемахи.
Б районе сел. Демерчи, на высоте около 1200 м, обитают серые хомячки, степные мыши и зайцы. Граница распространения степной полевки проходит пока все же ниже, на высоте 800—900 м.
В целом здесь наблюдается своеобразное обеднение состава млекопитающих при наличии незаполненных, ненасыщенных видами плоскогорных остепненных ландшафтов.
Подводя итоги, следует отметить, что в результате прямого воздей-ст ия человека и образования в лесной зоне Большого Кавказа антро-покультурных ландшафтов происходит общее для всех районов обеднение угодий крупными промысловыми животными (медведем, кабаном, оленем, косулей, козлом, серной), исчезновение специфически лесных видов (лесной куницы, полчка, белки) и весьма дифференцированное перераспределение мышевидных грызунов.
ПРОДОЛЬНЫЕ ДОЛИНЫ БОЛЬШОГО КАВКАЗА И ВНУТРЕННИЙ ДАГЕСТАН
К востоку от Эльбруса до внутреннего Дагестана между Передовыми хребтами расположены так называемые продольные долины в виде цепи гигантских воронок, разделенных небольшими перевалами. Защищенная с севера как бы экраном хребтов, эта своеобразная ксерофитная и теплая 1 на явилась убежищем ряда сухолюбивых и теплолюбивых растений и животных. Ботаники Краснов (1893—1894) и Кузнецов (1909) уже давно рассматривали эту зону наравне с внутренним Дагестаном как территорию выработки особой ксерофильной флоры. Гроссгейм (1948) относил нагорно-ксерофитную формацию этой зоны к типу трагакантников, характерных для сухих нагорий Передней Азии. Ксерофитные нагорностепные формации с ковылем, полынью и барбарисом прослеживаются, впрочем, местами и западнее Эльбруса — в троговых долинах меридианного направления — по Кубани, Теберде, Малому и Большому Зеленчукам, а также в долине верхнего Ингура в западном Закавказье. Продольные долины были освоены человеком еще с века бронзы и явились колыбелью так называемой кобанской культуры (Уварова, 1900), датируемой первой половиной I тысячелетия до н. э. Население этих долин занималось скотоводством и охотой, а позднее и земледелием. К концу средневековья популяции крупных копытных: кабапов, лосей, оленей, косуль, кавказских козлов, серн, зубров — были сильно разрежены на Передовых хребтах и совершенно вытеснены из описываемых долин.
Во внутреннем Дагестане по ущельям Андийского и Аварского Койсу, а также по Кара-Койсу энергично истреблялись олени, прикочевывающие на лето с южного склона хребта. С большей части хребтов, особенно с плоских возвышенностей, были вытеснены восточнокавказские и безоаровые козлы и истреблены бараны. Несомненно, что последние два вида
558
Часть III. Териогеографическое районирование и пути эволюции
уже отсутствовали на Гупибе в период кавказских войн середины прошлого столетия.
Природная ксероморфность продольных долин и Дагестана была усилена человеком. Вообще современное широкое распространение на-горно-ксерофитной растительности на северном склоне хребта было в значительной мере обязано человеку (Тумаджанов, 1947). Во внутреннем Дагестане, где леса никогда не были сильно развиты, многовековое истребление сосняков в верховьях притоков Сулака приводило к замене их
Рис. 210. Сел. Ушкуль с полями зерновых и картофеля в верховьях Ингура. По окраинам местообитания кустарниковой и прометеевой полевок.
Фот. автора, 1948.
березняками — Betula pendula, В. pubescens и до известной степени улучшало условия обитания и кормежки оленей, безоаровых и восточнокавказских козлов. Древнее население ущелий Кара-Чая, Кабарды, Северной Осетии, внутреннего Дагестана, Свапетпи расчистило от камней под сады и поля значительные площади днищ ущелий, сложив булыжники в круглые кучи и в продолговатые гряды. Были воздвигнуты также нередко циклопические каменные заборы и миниатюрные крепостцы с боевыми башнями из гранитных обломков или шиферных сланцев (рис. 210).
Упомянутые гряды, обрастая кустарниками и травой, напоминают разорванные остатки конечных и боковых морен. Как правило, эти гряды, а также каменные заборы аулов и жилые строения, служат местами концентрации степных мышей, лесных предкавказских мышей и кавказских снеговых полевок. Снеговые полевки пользуются кучами камней, щелями оград и степ для просушивания и устройства запасов сена на зиму, а также для устройства гнезд.
• Гл. VI» Изменения группировок и ареалов под влиянием человека 559*
В Ардонском ущелье близ Зарамага Туров (1926в) наблюдал прочное заселение каменных куч серыми хомячками. По Баксану, Чегему и Череку каменные заборы и заброшенные поля являются излюбленными участка ли поселении горных сусликов — вредителей зерновых и картофеля. В Кабарде, Осетии, Дагестане на поливных посевах зерновых и в садах концентрируются горные водяные полевки, которые приобрели огромную способность к рытью, напоминая в этом отношении земляных крыс Средней Азии. Они оказались здесь вредителями, подгрызающими корни молодых яблонь, груш, урюка. Кроме того, поливная вода, быстро проникая в норы, размывает большие участки склонов.
Во внутреннем Дагестане вредителями зерновых посевов сделались малоазипские хомяки и кустарниковые полевки. Некоторые популяции снеговых полевок па плато Гуниба потеряли свои привычки запасания сена на зиму. Они просто пользуются там запасами сена, которое забивается на зиму местными жителями в ниши и пещеры известняковых стен.
Спнатропная группировка грызунов в глухих горных аулах Дагестана формируется из местных видов — лесных мышей и снеговых полевок.
В ряде ущелий Грозненской области возделывание картофеля, ячменя и пшеницы содействовало привлечению в безлесные горы кабанов и некоторому увеличению численности зайца-русака.
В западном Закавказье, по среднему и верхнему течению Ингура, расчистка склонов от пихтово-кленовых лесов сопровождалась вначале образованием луговых формаций с желтой азалией—Rhododendron 11а-vuin. При выпасе скота и истреблении кустарников на топливо появлялось низкое засоренное разнотравье. На склонах южного освещения росли ксерофилтные виды полыни. Номере обезлесения исчезали на большой территории медведи, рыси, лесные кошки, лесные куницы, серны, козлы.
Прп дальнейшем иссушении склонов вследствие распашек под зерновые вытеснялись снеговые, кустарниковые и прометеевы полевки, кавказские мышовки, лесные мыши. Образовавшиеся открытые пространства выгонов и полей оказывались нередко практически свободными от грызунов, так как типичных полевых вредителей — обыкновенных хомяков, серых полевок — поблизости не было.
Такне изолированные горные долины дают теперь отличные примеры свободных экологических ниш, образовавшихся в результате создания культурного ландшафта.
ВЫСОКОГОРНЫЕ ЛУГА И СКАЛЫ БОЛЬШОГО КАВКАЗА
До возникновения и развития перегонного (кочевого) скотоводства,, воздействие человека на ландшафты и групировки млекопитающих этой зоны практически отсутствовало. В историческую эпоху выпас скота содействовал расширению зоны лугов снизу вследствие понижения верхней опушки леса, но и сокращал ее верхний предел разбиванием дерновых луговин, вклинивающихся в каменистые осыпи перевалов. Выпас приводил также к расширению пепоедаемых скотом зарослей рододендрона — Rhododendron caucasicum, в свою очередь препятствующего возобновлению березы. Понижение верхней границы леса за последнее столетие в результате выпаса скота достигло во многих местах Кавказа 150—200 м по вертикали. Оно усилено также вырубкой леса пастухами на топливо. Расширение зоны альпийских и субальпийских лугов привело к силь
660 Часть III. Териогеографическое районирование и пути эволюции
ному увеличению ареала обыкновенной и кустарниковой полевок, горной популяции зайца и лисицы. Стоянки скотоводов, огороженные каменными завалами, утоптанные и обильно унавоженные, быстро зарастают конским щавелем — Rumex, будяками — Cirsium и др. Они служат своеобразными резерватами обыкновенных полевок, концентрирующихся по окраинам утоптанных площадок под защитой многоярусной растительности. В начале лета — до прихода пастухов — на этик же участках кормятся зеленой травой медведи и копаются, разыскивая личинок жуков и червей, кабаны.
Перевыпас — перегрузка пастбищ — приводит к угнетению растительности лугов. Скот прокладывает по склонам множество троп, образующих как бы ступенчатую сетку. В вертикальные стенки ступенек открываются многочисленные норы обыкновенных, кустарниковых, прометеевых полевок. По этим норам легко проникает и скатывается дождевая и снеговая вода, размывая дерн. Самые верхние языки кавказских лугов, образованные, как правило, дерновинками зиббальдии — Sib-baldia procumbens и манжетки — Potentilla, часто заселенные кустарниковыми полевками, быстро разбиваются и уничтожаются при этом на больших пространствах. В результате выпаса скота и роющей деятельности грызунов сползают целые участки травяных склонов. Образующиеся характерные осыпи скал используются кавказскими козлами главным образом для отдыха и убежищ. Усиление эрозии и смыв склонов во время ливней ведут к развитию селей, т. е. потоков жидкой грязи и камней. В большинстве районов Кавказа сели исторической эпохи — явление недавнее, обязанное быстрому развитию скотоводства в последние два столетия. Так, в Нухе с селями познакомились впервые в 1889 г., в Варташепе — в 1895 г. (Пыльцов, 1904). Для бассейна Демир-Апаран-Чая в Куткашенском районе Азербайджана, где сели в настоящее время особенно страшны и мощны, Пыльцов (1904, стр. 268) приводит такие данные: «. . .лет 25 тому назад (т. е. в 80-х годах XIX столетия, — Н. В.) здесь паслось не более 5000 голов баранов, а в настоящее время более 10 000 голов». Перегрузка горных лугов скотом привела теперь в амфитеатре этого ущелья к обнажению огромных площадей склонов, занятых ранее альпийскими лугами. В результате из амфитеатра были вытеснены почти нацело восточнокавказские козлы, серны, обыкновенные полевки.
Особое значение перегонное скотоводство сыграло для диких копытных: оленей, козлов, серн. Домашний скот заносит в горы различные болезни: сап, ящур, сибирскую язву, бруцеллез, чесотку, железницу, паршу и гельминтозы. Гибель диких копытных, в частности козлов, серн от заболеваний, занесенных скотом, была отмечена в районе Кавказского (Динник, 1914а; Насимович, 1949а, 19496) и Закатальского заповедника (Марков и Млокосевич, 1935; Верещагин, 1938а).
Пребывание пастухов с овчарками, стадами овец, коз на высокогорных лугах в течение трех летних месяцев и прямое преследование оленей, козлов, серн привело к большим изменениям в природных повадках и суточном цикле этих зверей. Олени и серны стали держаться летом только в лесной зоне и в верхней опушке леса, выходя на пастбища лишь ночью. Такая же история произошла на западе с кавказскими козлами (Пасимо-вич, 1949а). В восточной части, па южном склоне восточнокавказскне козлы образуют летом две резко выраженных популяции. Одна из них держится на перевалах и осыпях хребта, не спускаясь в лес; другая — живет в лесу, выходя к верхней опушке леса и на яйлаги лишь по ночам ( Верещагин, 1938а).
Гл. VI. Изменения группировок и ареалов под влиянием человека
561
ЛЕСА СЕВЕРНЫХ СКЛОНОВ МАЛОГО КАВКАЗА
Ландшафтные изменения в историческую эпоху были, по-видимому, » шеныпимп на северных склонах Аджаро-Гурийских хребтов, сохра-ивтпих большие массивы широколиственных и пихтовых лесов, из ко-крых и до сих пор поступают крупные партии шкур медведя и лесной < вицы. Тем не менее и здесь всюду прослеживается энергичное повей-ш е понижение верхней опушки леса, а в районе Ахалциха — надвига-п на зону леса фаунистического комплекса сухих нагорий Передней \ пи. Восточнее новейшие изменения группировок млекопитающих 31 чительно заметнее.
Быстрое вытеснение медведя, оленя, косули, куниц из Атенского а ечья Триалетского хребта по мере беспощадного уничтожения сосно-fl х п буковых лесов, начавшегося с первой мировой войны, было опи-
Беловым (1924).
В Северной Армении, на Сомхетском, Безобдалъском и Бамбакском хребтах, жалкие островки леса уцелели местами в отдельных распадках (Троицкий, 1934). О большой древности освоения человеком этих районе свидетельствуют археологические и палеонтологические находки. Антропогенное иссушение склонов в верхних частях лесной зоны особенно заметно при южной экспозиции в верховьях речек, берущих начало н; самом плоскогорье, например по Дебеда-Чаю. На участке ущелья между станциями Амамлы—Кировакан можно наблюдать характерное взаимное захождение — мозаичность ареалов сухолюбивых видов млекопитающих нагорных степей и мезофильных видов лесной зоны. Деятель-пчль человека содействует здесь всюду наступлению группировки сухолюбивых зверьков на группировку мезофильных, вслед за расширением площадей ксерофитпых формаций. Ареалы малоазпйского хомяка, полевки Шидловского, горной слепушонки имеют явную тенденцию к рас-нрению на север, вниз по долине реки.
В нижнем лесном поясе и отчасти в среднем поясе лесоистреблепие приводило к образованию своеобразных формаций засухоустойчивых кустарников и лугостепи, содействуя продвижению вверх по ущельям сухолюбивых зверьков из полупустыни восточного Закавказья: мало-азпйской песчанки, степной полевки, степной мыши.
Антропокультурпые биотопы днищ ущелий — поселки, сады, виноградники, посевы зерновых — явились здесь основными участками стыка мелких видов зверей — выходцев из степей восточного Закавказья и из агорных степей Армянского нагорья.
Среди довольно большого набора видов, увеличивающих численность в культурном ландшафте ущелий, можно выделить ряд групп. К первой, связанной с постройками и садами, относятся: длиннохвостая белозубка, домовая мышь, отчасти лесная мышь и серый хомячок. Во второй, связанной с пашнями и посевами зерновых, выделяются: лисица, заяц, степная мышь, кустарниковая и обыкновенная полевки, а на границе среднего п нижнего пояса —* степная полевка. На луговых участках вторичного происхождения, используемых как пастбища и сенокосы, в среднем поясе развивается группировка грызунов-вредителей, состоящая из кустарников ой и обыкновенной полевок. В верхнем поясе, среди кустарниковых зарослей дуба и высокотравья, обитают крот, обыкновенная и кустарниковая полевки, обычна косуля. Местами при заболачивании истоков ручьев и развитии гидрофильной растительности среди осветленных
3g Н. К. Верещагин
362
Часть TII. Териогеографическое районирование и пути эволюции
вырубкой леса нагорных долинок увеличивается численность и водяной полевки.
Несмотря на значительную древность населенных пунктов, например г.г Кнровакапа, синантропная группировка грызунов еще представлена по преимуществу туземными видами, которые в 1943 г. составляли: домовая мышь — 86%, лесная мышь— 10.3%, серый хомячок — 3.7% общего улова грызунов (Соснихина, 1946). Интересно, что лесные мыши встречаются только в зпмне-весенпий период на участках, недавно освоенных под строительство. Хомячки держатся в городе зимой и летом, а весной отсутствуют.
Восточнее, в широких амфитеатрах ущелий рек Дзегама и Шамхора, остались ныне лишь небольшие массивы буково-грабовых лесов. Здесь совершенно оголены от леса гребпи хребтов меридианного направления. Замена лесных группировок зверей луговыми особенно велика в зоне от 1000 до 2500 м.
Здесь исчезли серны, кабаны, сократилось распространение безо-аровых козлов, косуль, медведей.
Сильно расширилось распространение и увеличилась численность обыкновенной и кустарниковой полевок. Кустарниковая полевка, обитающая на лугах, вышедших из-под леса, образовала здесь слабо трансформированный подвид — Pitymys daghestanicus nasarovi (Шидловский, 1938; Огнев, 1950).
На восточных склонах Карабаха в связи с ранним освоением гор первичные буковые леса уцелели лишь по ущелью Тертера и по северным и восточным склонам горы Кире. Истребление лесов сопровождалось здесь сужением ареала крота, медведя, кавказской белки, полчка, лесной мыши и кустарниковой полевки, косули.
По склонам, подвергшимся вырубке, распашке и иссушению, распространились из предгорий степная полевка, серый хомячок и степная мышь. В ущелье Тертера по пахотным участкам па высотах 650—700 м уже обитает степная полевка, а кустарниковая полевка еще «цепляется» на этой же высоте за тенистые северные участки россыпей скал и рощицы дубово-грабового мелколесья.
Еще более заметно процесс дальнейшего развития безлесных и иссушенных горных ландшафтов прослеживается на юге Карабаха и на самом Армянском плоскогорье.
АРМЯНСКОЕ НАГОРЬЕ И ДОЛИНА СРЕДНЕГО
ТЕЧЕНИЯ АРАКСА.
На высоких плоскогорьях Малого Кавказа расположены участки буково-грабовых лесов, субальпийского высокотравья и нагорных степей. На южных склонах плоскогорий, к долине Аракса, мезофитные ландшафты граничат с аридным редколесьем и сухой типчаковой степью, переходящей в зону колючих астрагалов и акаптолимонов. По долине среднего Аракса развита нагорная полыпио-солянковая полупустыня.
Человек начал осваивать плоскогорье уже с нижнего палеолита (За-мятпип, 1947; Паничкина, 1948), но иа протяжении второй половины четвертичного периода местные ландшафты сильно менялись преимущественно вследствие тектонических и климатических причин.
Даже в послеледниковую и в собственно историческую эпохи растительные ландшафты плоскогорья претерпевали ряд крупных изменений. Участки нагорных степей, развившихся в послеледниковую ксеротермн-
Гл, VI. Изменения группировок и ареалов под влиянием человека 563
ческую эпоху, сократились позднее под влиянием некоторого увлажнения и наступления лесной фазы. Но с эпохи бронзы здесь началось и заметное воздействие человека на ландшафты, сказавшееся прежде всего на распространении лесов и численности крупных зверей. Значительно 'юлыпее, чем ныне, распространение сосновых и дубово-буковых лесов в Джавахетии в историческую эпоху отметили на основе почвенных исследований Клопотовский (1948), на основе геоботанических данных — Кузнецов (1900) и Троицкий (1927, 1939), данных археологии — Гроссгейм (1948).
Существовало, впрочем, и древнее плейстоценовое безлесие огромных площадей Джавахетии и Армении как следствие образования лавовых покровов (Ярошенко, 1941).
В раннем голоцене плоскогорье было еще богато населено лошадьми, кабанами, оленями, косулями, зубрами, первобытными турами и баранин. Высокие безлесные участки служили тогда летним выгулом для тдагчно лесных копытных, находивших здесь спасение от кровососущих двукрылых лесной зопы.
В самом конце II тысячелетия до н. э. человеческая культура Закавказья перешла в стадию бронзового века с широким развитием земледелия, скотоводства, а позднее — и садоводства (см. Пиотровский, 1947, 1949).
Следы древних оросительных систем обнаружены теперь на склонах Алагеза и в Гехамскпх горах. Из клинописных надписей урартов извергло, что у племен Армянского плоскогорья в первой половине I тысячелетия до н. э/ было много скота. Сар дур, сын Аргишти, угнал из Эрпарха (страны, лежащей к северу от Алагеза) 23 194 головы крупного и 63 420 голов мелкого рогатого скота за два похода (Пиотровский, 1949). Именно -iCBoenne для земледелия и скотоводства окрестностей оз. Севан и положило уже три тысячи лет назад начало истреблению лесов в этом районе.
Тем не менее лиственные леса на Армянском нагорье были уничтожены главным образом в XIX столетии (Н. Б., 1861; Даль, 1947а).
На южных склонах Армянского нагорья, в Армении и Нахичеванской АССР, были уничтожены в историческую эпоху большие площади суховыносливого редколесья, состоящего из древовидного боярышника — Crataegus orientalis, иволистной груши — Pyrus salicilolia, можжевельника — Juniperus polycarpos, миндаля — Amigdalus Fenzliana. В нашу эпоху местами уцелели лишь священные деревья — «свидетели», описанные нами (Бурчак-Абрамович и Верещагин, 1950).
Ксерофитпзацпя южных склонов Зангезурского, Даралагезского хребта и Алагеза была необычайно усилена в последние столетия быстрым развитием перегонного скотоводства с неумеренным выпасом скота, приводившим к выбиванию дерна и засорению природных травянистых группировок.
В связи с иссушением склонов нижняя граница распространения некоторых грызунов — обыкновенной и снеговой полевок — оказалась поднята теперь в Зангезуре на 350—400 м. О недавнем распространении этих зверьков на меньших высотах свидетельствуют их далеко нзоли-I ровапиые ныне колонии в наиболее мезофитных уголках ущелья Акулпс-| Чая, в россыпях скал, поросших крушиной — Rhamnus Pallasii, R. spa-thulaefolia п шиповником, па высотах всего около 1200—1300 м. Здесь эти колонии существуют уже бок о бок с поселениями персидской песчанки. Песчанка же постепенно расселяется вверх, проникая по иссушенным и стравленным овцами каменистым склонам по мере расширения фрига-
ЗК*
564
Часть III. Териогеографическое районирование и пути эволюции
ноидной растительности. Современное распределение группировок мышевидных по вертикали показано на рис. 211.
Такие же примеры взаимного захождения ареалов высокогорных (нагорно-степных и луговых) и полупустынных млекопитающих наблюдаются в верховьях Арпа-Чая на Айоцдзорском хребте. Три вида песчанок — Meriones persicus, М. blackeri, М. vinogradovi — активно проникают там в зону фриганы, а малоазийские хомяки концентрируются по межам и окраинам посевов ячменя, а также на залежах, в пределах горно-степпой зоны. Даже нагорно-степной, относительно ксерофильный
Лесная мышь-8
Степная мышь-9 Персидская песчанка-3 Мал о аз и некая песчанка-1 Песчанка Виноградова-! Степная полевка-12 Серый хомячок- 6 Мышевидный хомячок-2
г. Капу дА их (39! 7 м)
г. Илянлу (2000лП
Лесная мышь-3 Серая полевка - 6 Снеговал полевка-19
Лесная мышь -21 • Степная мышь-24 Персидская песчанка-12 Мышевидный хомячок-3
Лесная мышь-15 Степная мышь-3 Персидская песчанка-8 Серый хомячок-3
р Арале (900mJ
г Дарры -Дог (1911 л)
Рис. 211. Зональное распределение грызунов по данным отлова на 100 ловушко-ночей в октябре 1947 г. на объединенном долготном профиле в районе среднего Аракса.
I — россыпи скал и луговины с сорняками (2200—2300 м); II — колючие астрагалы и россыпи скал с шиповником и бояркой (1600—1700 м); III — россыпи скал с кустами крушины и миндаля (1200—1300 м); IV — виноградники в долинах речек и склоны холмов (1и00—1100 м).
комплекс под влиянием деятельности человека имеет здесь тенденцию к расширению (Даль, 19496).
Одновременно с ксерофитизацией долины среднего Аракса и южных склонов Армянского нагорья здесь происходил и противоположный процесс — создание массивов древесных насаждений из грецкого ореха, платана, миндаля, урюка, яблонь. В качестве примера укажем на ореховые рощи-сады в окрестностях Еревана и на обширные рощи вековых платанов по долинам Нахичеван-Чая, Базар-Чая, вызывающие даже споры ботаников по поводу их антропогенного или природного (автохтонного) происхождения. Обширны и плодовые оазисы долины Алинджа-Чая и Акулис-Чая. Созданный еще в VIII в. до н. э. в рабовладельческом государстве Урарту правобережный канал от р. Занги, проходящий туннелем в несколько сот метров через гранитную скалу, и поныне орошает большие площади нагорной полупустыни. Еще тогда же, согласно клинописным надписям урартов, был устроен и ряд рощ, садов и виноградников (Пиотровский, 1949).
Вообще развитие древесных насаждений и опусты пение долшшых территорий в странах Ближнего Востока и Средней Азии па протяжении исторической эпохи зависело большей частью уже не от колебаний климата, а от характера развития человеческой культуры, прежде всего от состояния ирригации (см. также Берг, 1938). Быстрое исчезновение одного из крупных предгорных оазисов произошло, например, в наши дни в ущелье Акулис-Чая, у города Ордубада, где несколько десятков гектаров мощных плодово-древесных насаждений были истреблены засухой в те-
Гл. VI. Изменения группировок и ареалов под влиянием человека
565
ченпе 10—15 лет после разрушения турками армянского селения Аку-лпс в 1917 г.
В историческую эпоху в долине среднего течения Аракса были истреб-»пы лошади, куланы, кабаны, олени и джейраны. Сильно уменьшилась численность баранов. На орошенных землях в долине Аракса и его притоков увеличились численность и распространение степной полевки, серого хомячка и слепушонки. Поселения песчанок и тушканчиков оттеснялись па верхние террасы. В садах-оазисах долин (рис. 212) и виноградинках с их каменными заборами концентрировались длиннохвостые
Рис. 212. Урюковые садики в долине Алинджа-Чая, заселенные горной слепушонкой и лесной мышью.
белозубки, зайцы, лесные сони, лесные и степные мыши, серые хомячки, горные слепушонки. Увеличивалась и активность горной слепушонки, селившейся по искусственно орошаемым участкам долин Веди-Чая, Арпа-Чая (Даль, 1948а, 19496). Аналогичные факты отмечены и нами в долине Алинджа-Чая.
На рисовых посевах долины Аракса сформировалась гидрофильная популяция степных мышей (Погосян, 1945). Вслед за увеличением численности мышевидных возрастала и численность хищников — лисицы, перевязки, ласки, — несмотря на развитие охотничьего промысла.
В целом воздействие человека иа ландшафт этой сухой долины формировало численно обогащенные группировки мелких зверьков, приспособлявшихся к своеобразному режиму оазисов.
Постройки из сырцового кирпича заселялись серым хомячком, превращавшимся в настоящего домового вредителя, а обилие церквей и монастырей содействовало привлечению летучих мышей, особенно подковоносов, нетопырей, ночниц.
366 Часть III. Териогеографическое районирование и пути эволюции
Первоначальная синантропная группировка грызунов в мелких населённых пунктах формировалась здесь из местных грызунов: степной мыши и серого хомячка. Результаты ночного лова грызунов в октябре 1947 г. в жилых постройках некоторых селений долины Алинджа-Чая показаны в табл. Г14.
Таблица 114
Видовой состав и численность грызунов (по числу зверьков, пойманных на 100 ловушек) в мелких населенных пунктах долины среднего Аракса
Виды Ханагя Абранунис
число особей в % число особей В %
Mus musculus subsp — — 4 11.7
М. musculus talaricus 24 66.6 22 64.7
Cricetulus migratorius 10 27.7 8 23.5
Apodemus sylvaticus 2 5.5 — —
Всего 36 34
В Ленинакане и Ереване, расположенных на плоскогорье и связанных железной дорогой с Баку и Тбилиси, произошло вытеснение аборигенных грызунов завезенными недавно космополитическими видами. Данные вылова грызунов, проведенного в этих центрах в начале 40-х годов, показаны в табл. 115 по публикациям Соснихпной (1944), Шид-ловского и Соснихиной (1944).
Таблица 115
Видовой состав и численность (в °/0) грызунов в городах Армянского нагорья
Виды Ленинакан Ереван
Mus musculus subsp <80 50.5
Cricetulus migratorius 15 32.0
Rattus norvegicus .) 17.5
Такое преобразование первоначальной синантропной группировки связано теперь па плоскогорье и в мелких селениях с прокладкой шоссейных дорог и развитием автотранспорта.
Из мероприятий по обогащению териофауны плоскогорья упомянем весьма успешное разведение болотных бобров на речке Кара-Су (Сев-Джур) близ Эчмиадзина, начатое с 1940 г. (Саркисов, 19466), и мало удачные опыты выпуска 100 енотовидных собак в 1934 г. близ Воскресеповки и 15 самок и 4 самцов пятнистых оленей в лесу Хосрова близ Гарни, проведенные в 1954 г. Здесь проектировалось и разведение ондатры на озерах Ахалкалакского нагорья.
Гji. VI. Изменения группировок и ареалов под влиянием человека
567
ЛЕСНАЯ И НАГОРНО-СТЕПНАЯ ЗОНЫ ТАЛЫША
Весьма сходные с описанными для Малого Кавказа ландшафтные и Фиоцепотпческие изменения происходили в историческую эпоху и в Та-1ЫП1Р ’
Под влиянием прямого преследования с огнестрельным оружием в 20-х годах нашего столетия здесь исчезли тигр и олень, произошло резкое сокращение численности барса и кабана. Значительно раньше, возможно вначале прошлого столетия, исчезли зубр и безоаровый козел, вероятно обитавший в высокогорье. Сельскохозяйственная деятельность человека произвела крупные преобразования в группировках и распространении многих видов зверей.
Вырубка здешних широколиственных лесов и выпас скота шли особенно энергично на северных отрогах этого хребта, где образовались широкие сухие поляны, заросшие по опушкам папоротником, орляком и ежевикой. В верховьях Виляж-Чая южные склоны ущелий ныне распаханы под посевы ячменя и пшеницы. По восточным склонам верхняя опушка леса также искусственно понижена па 250—300 м, а местами и значительно больше. В результате обезлесения и иссушения склонов здесь произошло вытеснение лесных зверей и грандиозное расширение ареалов зайца-русака, степной полевкп и серого хомячка, расселяющихся со стороны нагорной, ксерофитной степи. Проникновение полевки по обезлесенным гребням хребтов и склонам происходит книзу до дна ущелий па 1200— 1500 м, тогда как прежняя — природная граница распространения проходила на высоте 1700—1800 м. Быстрое просыхание склонов позволяет полевке переносить it глубоких ущельях огромную влажность (до 1500 мм осадков в год). Между тем в зоне экологического оптимума этого зверька количество осадков пе превышает 500—550 мм, а высшим пределом на равнинах является 800 мм. Этот пример изменчивости экологической устойчивости мелкого грызуна, зависящей от особенностей ландшафта, пе имеет аналогов ни в одном из районов Кавказского перешейка. Именно здесь проникновение сухолюбивых млекопитающих во влажную горно-лесную зону, прослеженное нами на огромном пространстве предгорного и горного Кавказа, получило свое высшее и рельефное выражение. Только ло вселение идет здесь не снизу, как в большинстве других районов, а сверху. В Лерикском п Ярдымлинском районах Талыша степная полевка почти ежегодно угрожает зерновым посевам вспышками массового размножения. Посевы зерновых привлекают в июне к верхней опушке леса и большое число кабанов.
Одновременно с расширением ареалов сухолюбивых видов здесь происходило сокращение ареалов крота, дикобраза, лесной мыши, кабана, отчасти косули.
Группировка домовых млекопитающих в мелких населенных пунктах, расположенных в горных лесах, до крайности бедна и состоит из темной длиннохвостой белозубки, домовой мыши и пасюка.
Развитие скотоводства и земледелия в нагорной степи Талыша обусловило, как и в Нахичеванском крае, полное уничтожение аридного редколесья. Степное нагорье Талыша на высотах 2000—2500 м в настоящее время — район ненадежного богарного земледелия с посевами ячменя, пшеницы, а на более высоких участках — и ржи. Дикие копытные — кулан, баран, безоаровый козел — вероятно, обитали в этой зоне еще полтора-два века тому назад. Земледельческая культура, развившаяся на плоскогорье в последние столетия, способствовала развитию
568
Часть III. Териогеографическое районирование и пути эволюции
численности мышевидных — степной и обыкновенной полевок, горной слепушонки, черноватого хомяка, — а также зайца-русака. Особенно заметна бурная роющая деятельность горной слепушонки на посевах люцерны в долине Зуванда, на высотах 1800—2000 м. В садах и на посевах люцерны слепушонка выбрасывает кучи земли до 1 м диаметром и высотой до 30 см, в то время как на целине эти кучки не превышают в диаметре 25—30 см.
Группировка домовых грызунов в населенных пунктах этого нагорья очень близка к описанной для долины среднего Аракса. В горных селениях Кельвяз, Дыман при отловах давилками попадаются преимущественно местные виды грызунов. В июне 1945 г. они составляли: серый хомячок — до80°о и домовая мышь — до 20%. Серый хомячок живет даже во вторых этажах жилых построек, а летом и на чердаках, приспосабливаясь к весьма высоким температурам (Верещагин, 1949г). Завозные домовые мыши и серые крысы обычны пока лишь в районных центрах Ярдымлы, Лерик, связанных с Ленкоранью автомобильными дорогами.
МЕРОПРИЯТИЯ ПО ОХРАНЕ И ОБОГАЩЕНИЮ ЛЕСНОЙ II ВЫСОКОГОРНОЙ ТЕРИОФАУНЫ
Известия о существовании охранных мероприятий для охотничье-промысловых зверей Кавказа дошли до нас с раннего средневековья, но можно не сомневаться в том, что эти мероприятия были и в рабовладельческих государствах.
Первоначально охрана сводилась к запретам охоты широким слоям населения иа участках, представлявших особую ценность, например для феодалов в Иране, Армении, Грузии. Такие местные запреты охоты в бекских, ханских, шахских, княжеских и царских владениях подразумевали одновременно и охрану природного ландшафта — рощ, камышовых болот, источников и т. п.
Уже в древней Армении в царствование Хосрова (IV—V вв. н. э.) производилось насаждение и ограждение дубовых лесов и последующее заселение их охотничьими животными в районе Гарни, Двина, по долине Мецамора. «Потом царь приказал набрать всяких диких животных и зверей и пускать их внутрь ограды, чтобы превратить эти места в места царской охоты, гуляний и увеселений» (Бузанд, 1953, стр. 17). В XIX столетии крупные массивы горных лесов отводились или арендовались для охот великих князей. Такова была организация Кубанской охоты на площади около 500 тыс. га в ущельях левых притоков Кубани, Кавказской охоты в Караязском тугайном лесу (рис. 213) на Куре, восточнее Тбилиси, Боржомского звериного отвода близ Боржоми (Шильдер, 1892; Башкиров, 1940).
Наиболее показателен опыт Боржомского звериного отвода, где 500 га горных склонов было огорожено высоким забором, препятствующим выходу и имеющим впускные ловушки для зверей. Мазымчанское и Белокапское ущелья южного склона Большого Кавказа близ Лагодехи арендовались с 1903 по 1917 г. для охоты па горных копытных Демидовым Сан-Донато (Марков и Млокосевич, 1935; Марков, 1935, 1938). Все эти участки при хорошей организации охраны и редкости производившихся охот служили, в сущности, заказниками или даже своего рода заповедниками, сохранявшими большие запасы крупного зверя.
В начале XX столетия идея организации таких охотничьих отводов переросла в идею организации государственных заповедников.
Гл. 1/. Из мнения группировок и ареалов под влиянием человека 5(№
После Октябрьской революции (20—30-е годы) были организованы горные и горно-лесные заповедники: Кавказский, Боржомскпй, Тебер-дпнский, Закатальский, Лагодехский, Гумистииский, Рицский, Телаво-Кварельский, Сарай-Булахский — общей площадью до 550 000 га (Марков, 1935; Макаров, 1940; Гамбарашвили, 1937; Князев, 1946; Вере-шагпн, 1947г).
Кроме того, в 30-х годах был создан ряд временных заказников в пред-о >ьях и горах для охотничье-промысловых животных. Общее число
Рис. 213. Результаты однодневной охоты на кабанов в Караязах в 1909 г. Убито 43 кабана.
Фот. Е. Л. Маркова.
заказников в 30—40-х годах достигало 17 площадью в 200 000 га. Значение этих резерватов оказалось в общем невелико, так как запрет на отстрел и отлов животных не предусматривал охраны лесов от рубок и выпаса скота (карта 97).
Организация кавказских заповедников приостановила падение численности крупного зверя, в частности оленей, козлов, серн, а затем способствовала и постепенному росту численности этих видов.
Учитывая, что в ближайшие десятилетня па Кавказе произойдут дальнейшие грандиозные перестройки ландшафтов, обусловленные промышленной разработкой недр и бурным развитием сельского хозяйства, необходимо всячески развивать здесь и дальше дело охраны природы. Сокращение кавказских заповедников и промышленные рубки леса на их территориях следует рассматривать как проявление величайшей близорукости и крохоборства.
о 7 О
Чаешь III. Териогеографическое районирование и пути эволюции
Некоторые иопытки обогатить горно-лесную териофауну чужеземными видами зверей были сделаны на Кавказе в конце XIX столетия.
По рекомендации Шильдера в 1888 г. в Боржомскнп звериный отвод выпустили 54 европейских ланей — Dama dama L. Животные хорошо прижились, и к 1918 г. их насчитывалось несколько сот голов, причем встречались стада по 50—80 штук. В 1919—1920 гг. лани были перебиты местными жителями. В том же 1888 году в отвод было выпущено 7 европейских оленей — Cervus elaphus L. из Вильно и 58 — из Австрии для «улучшения рогов^> местных оленей. По свидетельству охотоведа Е. Л. Маркова, число оленей к 1918 г. достигло в отводе 1000 голов, но позднее из-за развития браконьерства сократилось до нескольких десятков.
В отвод было подсажено также 10 восточпокавказских и западно-кавказских козлов, которые дали ряд интересных ублюдков между собой и местными безоаровыми козлами. Общая численность козлов будто бы достигала к 1918 г. 500 голов, а в период гражданской войны также сошла на нет. Названные огромные цифры плотности зверя в отводе, даже при учете наличия горного рельефа, вызывали необходимость постоянной подкормки животных. Как бы то пи было опыт Боржомского отвода является блестящим примером огромных возможностей в деле разведения на Кавказе местных и чужеземных копытных.
Более или менее планомерные попытки обогащения горно-лесной фауны Кавказа были предприняты только в советское время. К сожалению, они касались преимущественно пушного зверя.
. Следует отметить, что предпринятые введения новых видов в горнолесную зону Кавказа отличались сугубым эмпиризмом. Теория акклиматизации не разрабатывалась сколько-нибудь серьезно. Осуществлялись лишь отдельные, случайные п более или менее рациональные предложения.
Еще Формозов (1930) рекомендовал заселить пихтовые леса Кавказа обыкновенной белкой; Марков (19316, 1935) предложил заселить леса Талыша пятнистым оленем, а горы и леса Кавказа — кроликом, скунсом, енотовидной собакой, шиншиллой. Мантейфель (1943) писал о желательности поселения на Кавказе горала, винторогого козла, памирского архара, шиншиллы, гималайской панды, австралийской кузулисы. Иные предложения предусматривали поселение в Закавказье американских норок, настоящих енотов, завоз в Закавказье степного хоря п расширение ареалов кавказских козлов и серн на Малый Кавказ (Верещагин, 19396).
Опыты интродукции пушных зверьков в леса Кавказа были освещены в работах зоологов (Шапошников, 1938; П. Степанов, 1939; Куранова и Степанов, 1941; Даль,1941; Верещагин, 19416, 1947в; Буковский, 1950).
Ниже упомянуты лишь некоторые наиболее удавшиеся опыты акклиматизации пушных видов и копытных в лесной и высокогорной зонах Кавказа.
На западном Кавказе проводились опыты акклиматизации белки, енотовидной собаки, пятнистого оленя и зубробизона.
Алтайская белка — Sciurus vulgaris altaicus Serebr. (рис. 214) была выпущена в 1937 г. в числе 134 штук в ущелье Теберды в Тебердипском заповеднике, в 1940 г. их было уже 2400, а в 1941 г. — более 3500 (Кура-пова и Степанов, 1941). В 1944 г. белки заселяли леса в бассейне Аксаута, Кубани, Теберды, обоих Зеленчуков, питаясь семенами пихты, ели, сосны, буковыми орешками. К 1948 г. они уже появились в Кавказском заповеднике и проникли на южный склон хребта к тогу до Сухуми, обогнув, вероятно, ледниковые поля в районе верховий Лабы. Единичные шкурки
• Гл. VI. Изменения группировок и ареалов под влиянием человека
571
начали поступать с осени 1948 г. из лесов бассейна Бзыби и Кодора. Дальнейшее продвижение белок па восток по зоне пихтово-кленовых лесов Сванетии шло поразительно быстро. В 1951 г. стали поступать сведения, чтэ белки появились в бассейне Риона. В 1954 г. белка стала уже приносить вред садовым культурам в районе Гагр (Руковскин, 1956).1 В дальнейшем белка, несомненно, заселит всю лесную зону Закавказья и, вероятно, проникнет по лесам Сурамского хребта в широколиственные и хвойные леса Триалетского и Аджаро-Имеретинского хребтов.
Рис. 214. Алтайская белка в сосновых лесах верхней Кубани.
Целесообразность введения белки в леса Кавказа в настоящее время стала сомнительной, так как зверек вредит культурному садоводству, особенно ореховым плантациям. В настоящее время необходимо развитие промышленной добычи грызуна.
Современный (1956 года) и будущий ареалы обыкновенной белки показаны на карте 95.
Закатало-Исмаиллпнская низменность в восточном Закавказье была использована для опытов акклиматизации енотовидной собаки, енота-полоскуна, американской норки и североамериканского скунса. Успешно прижились здесь только еноты.
Выпущенные в 1941 г. западнее сел. Исмаилльт 22 енота быстро увеличивали численность и ареал, несмотря иа случаи браконьерства и истребления пх в садах селений. Вся долина Агри-Чая оказалась пригодной для этого многоядного хищника (рис, 215, 216). Уже в 1945 г. ареал енота занимал около 250 км2 прп численности в 180—200 штук (Верещагин, 1947в, 19536). К 1949 г. площадь обитания составила при-
1 В этой неточной в географическом смысле статье Руковского белка указана для б ллесных окрестностей Новороссийска, Геленджика и других городов, что не соответствует действительности.
о72 Часть III. Териогеографическое районирование и пути эволюции
близительно 850—900 км2 при численности в 800—850 штук. Скорость эффективного расселения и формирования ареала енота в пределах сырой и лесистой низменности была поразительной и достигала в благоприятных условиях 15—20 км в год. При этом еноты нередко тяготели к окраинам селений, близ которых всегда находили пищу: фрукты, лягушек, домашнюю птицу.
Для ускорения естественного расселения в августе—сентябре 1949 г» было отловлено в Исмаиллинском районе 23 енота, из которых 18 штук
Рис. 215. Енот-полоскун из лесов Агри-Чайской долины.
Фот. автора, 1949.
было выпущено в низовых лесах Самура и Хачмасской низменности (Руковский, 1947, 1950). Последующие отловы зверьков были произведены в Куткашенском и Исмаиллинском районах в 1950 и 1951 гг.,а добытые партии енотов выпущены в галерейные леса Терека и левых притоков Кубани, несмотря на паши предупреждения о возможном вреде для птицеводства и дичпого хозяйства от такого предприятия (Верещагин, 19536).
Интродукция других хищников не увенчалась успехом. Скунсы и американские норки погибли уже через год после выпуска от невыясненных причин, что же касается енотовидных собак, то небольшое число этих животных еще существует в низовых лесах Автаранской долины.
Столь же малоуспешными оказались и опыты акклиматизации енотовидной собаки в горных лесах Тебердинского заповедника в Закавказье, в Гумистинском заповеднике в Абхазии, в Южной Осетии и Карталинии (карта 94). Зверьки, как правило, гибли от невыясненных причин в пер-
Гл. VI. Изменения группировок и ареалов под влиянием человека
573
вне годы выпуска, откочевывая нередко на огромные расстояния (П. Степанов, 1939; Даль, 1941; Верещагин, 1947г).
Олъшатнпковые болота и дебри Колхидской низменности были использованы в последние десятилетия для интродукции болотного бобра. Первый выпуск 48 болотных бобров был произведен в 1932 г. восточнее юз. Палеостом. В 1936 г. нутрии наблюдались в районе Кобулет, Супсы, Ланчхуты, у устья Хопы, в 20—25 км от пункта выпуска. На пятый год размножения общая площадь расселения достигала уже не менее 500 км2,
Рис. 216. Следы енота, черного аиста и турача на отмели речки в районе сел. Нидж.
Фот. автора, 1949.
но фактически заселенные и освоенные зверьками участки вряд ли достигали более 3—4 тыс. га (Верещагин, 19416).
Предпринятое в дальнейшем искусственное расселение акклиматизировавшихся зверьков содействовало быстрому формированию ареала по подходящим участкам болот Черноморского побережья, в частности в Абхазии.
Несовершенство регулирования промысла и отсутствие охраны от врагов при заморозках и обсыхании дебрей приводило местами к истреблению нутрий.
Возможности свободного обитания в водоемах Колхиды болотных юобров будут неуклонно сужаться в связи с осушением и вырубкой ольховых топей, но полуволыюе разведение может успешно развиваться и далее на водоемах озер типа Бебесыри и Никит (Верещагин, 1950а).
Субальпийские луга центрального Дагестана были использованы для опыта разведения алтайского сурка — Marmota baibacina Brandt. В 1934 г. 113 сурков, вывезенных из Кош-Агачского аймака Ойротской автономной области, было выпущено на высокогорном плато к западу от сел.
Ь7 /
Часть III. Териогеографическое районирование и пути эволюции
Гуниб. Зверьки обосновались в россыпях скал известнякового песчаника, частью же самостоятельно отрыли поры под плиты известняка на травяных склонах (рис. 217). В августе 1935 г. было учтено 47 сурков, но выводков замечено не было. Общая площадь расселения достигла уже в это время будто бы 25 км2. В 1936 г. было учтено всего 15 взрослых и 2 молодых сурка (Шапошников и Крушинская, 1939; Лавров, 1946). В 1937 г. было насчитано 18 обитаемых нор с 27 зверьками. Наше обследование в августе 1952 г. показало, что сурки распространились по всему
Рис. 217. Местообитание алтайского сурка на северном обрыве плато Гуниба.
Фот. автора, 1952.
Гунибскому плато па площади около 45 км2. Наибольшая плотность их колоний наблюдалась ио северному обрыву плоскогорья иа высоте 1800 м по линиям разломов пластов (рис. 218). Отдельные выводки встречались даже близ сел. Гуниб на высотах 1300—1400 м. Дальнейшее самостоятельное расселение зверьков намечалось, несмотря на значительное браконьерство, в западном направлении. Судя по распределению колоний на Гунпбе, большая часть низких склонов и днищ ущелий Дагестана с быстро выгорающей пагорпо-степной растительностью непригодна для сурков. Однако высокие плато, подобные Гуппбу и Хуизаху, с обильными россыпями скал по их окраинам весьма перспективны для дальнейшего искусственного расселения зверьков.
Сурок может быть, очевидно, успению расселен и по всей пагорпо-степной зоне внутренних долин северного склона Большого Кавказа вплоть до Эльбруса.
При признании целесообразности дальнейшего разведения па Кавказе алтайских сурков наиболее пригодными участками для новых выпусков можно считать россыпи известняковых скал по основанию квестовых уступов южных обрывов Скалистого хребта в ущельях Аргуни,
Гл. VI. Изменения группировок и ареалов под влиянием человека
575-
Ардона, Черека и Баксана, а также в долине Ингура и Риона. Можно расселить этих сурков и на хребтах Армянского нагорья, например в районе оз. Севан. Современный и возможный будущий ареалы сурка показаны па карте 95.
В 1939 г. в ущелье Теберды было выпущено 59 экз. пятнистого оленя — Cervus nippon Тетт. для развития пантового хозяйства. К 1942 г. численность оленей достигла примерно 150 голов, но позднее, при немецкой оккупации района, упала вдвое. В 1954 г. небольшое стадо пят-шстых оленей было завезено и в Кусарский район Азербайджана.
Рис. 218. Алтайские сурки в Дагестане.
Для восстановления зубра на Кавказе в Кавказский заповедник в 1940 г. было завезено 5 условно чистых (1/16 крови по зубру) зубробизонов — Bison bison L. хВ. bonasus L. из Аскании-Нова. В 1944 г. в заповеднике имелось стадо в 11 голов условно чистых зубробизонов, в 1951 г.— 21 (6 самцов и 15 самок), а в 1956 г.—106 голов. Звери содержатся здесь частью в больших вольерах, частью на свободе под охраной конных пастухов и регулярно откочевывают в летнее время на кормежку в субальпийскую и альпийскую зоны. В настоящее время есть полное основание думать, что зубробизоны окажутся достаточно жизнеустойчивыми для того, чтобы ими можно было заселить всю лесную зону северо-западного Кавказа.
Рассмотренные примеры опытов введения в фауну Кавказа новых видов зверей показывают, в сущности, молниеносное (в геологическом смысле) формирование ареалов при наличии экологических пищ. Формирование новых биоценотических отношений при насыщении вселенными видами различных биотопов должно быть предметом специальных исследований прикладной зоологии.
Дальнейшие работы в области перестройки и обогащения лесного и высокогорного териокомплексов Кавказа должны быть организованы с.учетом природных условий и основных направлений развития деятельности человека в различных участках.
И 6 Часть III. Териогеографическое районирование и пути эволюции
Самые общие теоретические предпосылки для постановки работ по дальнейшему обогащению териокомплекса горно-лесных районов Кавказа (Кавказского округа) заключаются в следующем.
1. Комплекс зверей лесной зоны Кавказа недавно обеднел под влиянием человека крупными видами: зубром, лосем, местами оленем и серной.
2. Высокогорная группировка копытных и грызунов Большого Кавказа бедна видами частью в силу древней изолированности страны, частью из-за резкого рельефа альпийской зоны и тяжелого снегового режима, особенно в западной части хребта.
3. Кормовой режим для грызунов и хищных в лесной зоне западной части Большого Кавказа более устойчив, чем в восточной.
4. Малокавказский район Кавказского округа не имеет ряда ценных видов хищных и копытных, свойственных Большому Кавказу.
Учитывая эти особенности, следует усилить внимание к возможности восстановления зубра (или зубробизона), лося, благородного оленя на Кавказе и поселения в предгорьях лани, пятнистого оленя. Кормовые запасы лесной зоны могут, по-видимому, выдержать еще одного пушного хищника типа соболя. На Малый Кавказ необходимо завести кавказских козлов и серну. Лесную куницу и серну следует развести в Карабахе и Талыше и т. п.
Вообще искусственное преодоление экологических барьеров таит огромные возможности для расширения ареалов местных видов и поселения чужеземных.
Исключительный интерес для проблемы видообразования и изучения типов направленной изменчивости будет представлять опыт разведения восточнокавказского козла на западе, т. е. в системе Аджаро-Имеретинских хребтов, а западнокавказского козла — в Карабахе или в Талыше и Эльбурсе.
Совершенно особое место в деле разведения и сохранения высокогорных копытных должен занять внутренний Дагестан. Горный, скалистый рельеф, бедность растительного покрова не позволяют развиться здесь интенсивному земледелию, садоводству, скотоводству. Именно поэтому, а также по территориальной отграниченности Дагестан может быть использован как огромный садок для разведения высокогорных копытных, не нуждающихся в лесной растительности.
Сюда могут быть подсажены и разведены архары — Ovis аттоп L., куку-яманы, или синие овцы, — О. nahura Hodgson, тары — Hemitragus jemlaicus Smith, винторогие козлы — Capra falconeri Wagner, гривистые бараны — Ammo tragus lervia Pallas.
Должна быть восстановлена и численность безоаровых козлов — Capra aegagrus, заселявших ранее все плоскогорье. Несомненно, что наряду с этими мероприятиями должно быть предусмотрено и всемерное развитие истребления волка.
Сухие нагорья Малого Кавказа и Талыша не таят таких широких перспектив, как Дагестан, для разведения горных копытных, так как зону аридного редколесья и трагакантовых астрагалов целесообразно использовать не только в качестве пастбищ, но и для создания лесосадов миндаля, фисташки и маслины. Тем не менее в широкой долине Аракса должен быть восстановлен джейран, который может заселить неудобные для земледелия участки между Вагаршапатом и Ордубадом.
В зоне долинных лесов Колхиды, Закатало-Исмаиллинской и Ленкоранской низменностей, а также на речках Кара-Су в Армении должно йолучить дальнейшее развитие полувольное разведение болотных бобров.
Гл. VI. Изменения группировок и ареалов под влиянием человека 577
ОСНОВНЫЕ ВЫВОДЫ
Рассмотрение прямых и косвенных влияний человека на фауну млекопитающих Кавказского перешейка в голоценовую эпоху показывает, ([то эта эпоха составляет особый качественный — антропогенный — этап 1 эволюции фауны.
На этом этапе эволюция ареалов видов, экологических группировок и фаунистических комплексов шла преимущественно под прямым и косвенным влиянием хаотичной деятельности человека, нередко далеко перекрывавшей природные процессы эволюции ландшафтов и биоценозов.
Некоторые общие и частные закономерности формирования фауны Ь’ юкопитающих Кавказа на антропогенном этапе могут быть сформулированы следующим образом.
1. Прямое воздействие человека на фауну млекопитающих Кавказа в историческую эпоху состояло преимущественно в бесплановом и ограниченном лишь техническими возможностями охотничьем промысле. Грандиозные загонные охоты феодалов в горах и предгорьях и такие же о оты кочевых племен на равнинах были главной причиной быстрого обеднения Кавказа крупными млекопитающими.
Человек значительно ускорил в одних случаях и целиком обусловил в других вымирание обитавших на перешейке льва, тигра, речного бобра, лошади, кулана, лося, первобытного тура, зубра и содействовал сокращению численности и ареалов медведя, полосатой гиены, барса, кабана, косули, благородного оленя, сайгака, джейрана.
Состав и численность пушных промысловых зверей пострадала за исторический период значительно меньше. Катастрофически сокращались в последние столетия и десятилетия лишь численность и ареалы корсака и степного хоря. По остальным же пушным видам, судя по материалам государственных заготовительных пунктов за последние четверть века, наблюдаются преимущественно многолетние колебания численности, обусловленные различного рода природными факторами.
Предпринятые в советское время плановые мероприятия по истреблению волка, шакала, малого суслика, обыкновенной и степной полевок, зтепной мыши не дали еще ни одного примера уменьшения общих или частных границ ареалов перечисленных видов.
2. Процесс вымирания (истребления) и сокращения ареалов промысловых млекопитающих был повсеместно ускорен косвенной деятельностью человека, преобразующей различные типы ландшафтов. Особенно пагубно сказывалось повсеместное хищническое уничтожение лесов.
3. Новейшая эволюция ареалов и привычек в выборе мест обитания у ряда охотничье-промысловых зверей Кавказа была обусловлена в значительной степени влиянием человека на их высшую нервную деятельность. Это влияние нередко совершенно подавляло древние инстинкты горизонтальных и вертикальных миграций, суточной деятельности и т. и.
Основной реакцией копытных на близость человека и его домашних животных была реакция «убегания» в защищенные биотопы: леса, горы, пустыни. При слабом преследовании и создании новых типов кормовых , угодий ряд видов хищных и копытных — медведь, шакал и кабан — I в Азербайджане стал обитать вблизи человека. Человек способствовал также образованию местных экологических популяций млекопитающих, например «лесных» и «скальных» восточнокавказских козлов па южном склоне восточного Кавказа, «домашних», или «садовых», кабанов и зайцев-русаков в оазисах Азербайджана.
37 H. К. Верещагин
578 Часть III, Териогеографическое районирование и пути эволюции-
Создание новых типов биотопов содействовало также развитию изменений экологических черт и даже формированию и появлению новых черт экологии у мелких зверьков, особенно у грызунов. Об этом см. также пункт 7.
4. Преобразование ландшафтов человеком расшатывало природные^ более или менее устоявшиеся ландшафтно-зоогеографические границы^ содействуя повсеместно в связи с обезлесением наступлению сухолюбивых экологических группировок и фаунистических комплексов и отступлению мезофильных.
Тем самым смещались на юг, в предгорья, границы Предкавказского района степных вселенцев, а в Закавказье, на север, — границы Переднеазиатского нагорно-пустынного округа. Этот процесс наступления группировок и комплексов сухолюбивых млекопитающих был выражен наиболее отчетливо на востоке, юго-востоке и юге Кавказа в соответствии с природной зональностью и большей континентальностью и сухостью-климата. Кавказский горно-лесной и высокогорный округ превращался все более отчетливо в мезофитный рефугиум — убежище мезофильных млекопитающих. Северная граница Арменийского переходного района спускалась по иссушенным человеком амфитеатрам ущелий северного склона Малого Кавказа в связи с продвижением в бывшую лесную зону зверьков нагорно-степного фаунистического комплекса. Такая же картина была выражена еще более резко в Талыше.
5. Формирование новых экологических группировок и фаунистических комплексов млекопитающих на распахиваемых и орошаемых пространствах степей и полупустынь Кавказского перешейка происходило за счет исчезновения (вымирания) крупных промысловых видов копытных (тарпана, кулана, сайги, джейрана) и грызунов (суслика, слепыша,, тушканчиков, песчанок), не выносящих распашки и орошения. Одновременно при орошении и создании культурного ландшафта шло и качественное обогащение комплексов за счет вселения мезофильных насекомоядных и грызунов из интразональных типов угодий: галерейных лесов,, тростниковых зарослей, а также из смежных участков лесов низменностей и предгорий. Параллельно происходило и увеличение численности широко распространенных хищников: лисицы, такалаг ласки, перевязки.
6. Частичное восстановление, а иногда даже и обогащение первобытных группировок лесных млекопитающих происходило при замене предгорных и низовых лесов Предкавказья и Закавказья обширными насаждениями плодовых и ореховых культур при условии смыкания этих насаждений с лесами нижнего горного пояса. Это восстановление и перестройка лесных группировок зверей шли за счет сезонных прикочевок крупных охотничьв'Промысловых зверей и увеличения численности поселявшихся.1 оседло древесно-лесных грызунов и хищников.
7. Формирование группировок вредителей сельскохозяйственных культур и синантропных (домовых) происходило в каждой экологической зоне первоначально на основе туземного фаунистического материала.
В культурном ландшафте создавались широкие предпосылки для развития у зверей новых экологических черт и для формообразования. К наиболее эффектным примерам относится образование гидрофильных популяций степной мыши на рисовых полях и посевных популяций степной полевки на богарных посевах пшеницы и ячменя в восточном Закавказье.
Роль случайных, завозных зверьков в формировании группировок сельскохозяйственных вредителей невелика. В качестве примера имеется!
Гл. VI. Изменения группировок и ареалов под влиянием человека 579
лишь пасюк, обосновавшийся на огородах предгорного Предкавказья и Черноморского побережья.
8. Процесс формирования синантропных группировок зверьков возникал многократно и независимо в различных зоогеографических районах и участках Кавказского перешейка на основе приспособления туземных видов к местным образцам строительной техники, а также в соответствии с развитием морфологических черт, экологических возможностей и потребностей отдельных видов. Синантропные группировки формировались и продолжают формироваться и далее из представителей трех отрядов: грызунов, рукокрылых и насекомоядных. Наиболее полное соприкосновение с человеком происходит у грызунов.
Приспособление кавказских грызунов: степной и горно-лесной форм домовой мыши, лесной мыши, серого хомячка, снеговых полевок — к постройкам человека — явление, несомненно, такое же древнее, как и приспособления космополитических форм домовой мыши и крыс.
При совершенствовании жилищ и увеличении поселений человека в условиях культурного ландшафта всюду происходило вытеснение туземных синантропных группировок космополитической.
Вселение завозных видов — серой крысы и синантропных рас домовой мыши — в культурный ландшафт и поселения человека происходило раньше всего по побережьям Черного и Каспийского морей при посредстве судов. Формирование группировок домовых грызунов в населенных пунктах побережий, в сущности, закончено. В новейшее время происходит быстрое заселение глубинных районов Кавказа завозными синантропными грызунами, обусловленное развитием железнодорожного и автомобильного транспорта. Вытеснение туземных грызунов из домовой группировки наиболее замедлено на высоких плоскогорьях, например в Армении.
9. Планомерная реконструкция социалистического сельского хозяйства, предусматривающая развитие лесных посадок и инженерного орошения, упорядочение лесного и охотничьего хозяйств, а также дальнейшее развитие населенных пунктов будут создавать новые предпосылки для быстрой эволюции экологических группировок и фаунистических комплексов млекопитающих Кавказа.
10. Кроме планомерной переделки ландшафтов, для дальнейшего формирования фауны млекопитающих Кавказа будут иметь также наибольшее значение целенаправленные мероприятия по обогащению рассмотренных выше ландшафтных зон и участков кавказскими и чужеземными промысловыми видами.
Особо интересные и важные практически результаты могут быть получены от заселения внутреннего Дагестана центральноазиатскими высокогорными копытными, а малокавказской части Кавказского округа — козлами Большого Кавказа.
37*
ЗАКЛЮЧЕНИЕ
Рассмотрение основных этапов истории фауны млекопитающих Кавказа по накопленным к данному моменту фауно логическим материалам закончено. Эти этапы лишь намечены в третичном периоде, но вырисовываются все рельефнее на протяжении антропогена по мере приближения к современности. Подводим самые общие итоги. История териофауны Кавказа начинается с олигоцена. Кавказский горно-лесной териокомплекс начал формироваться, по-видимому, с среднего миоцена. Вселение гиппарионового комплекса на равнины и в предгорья Кавказской суши произошло с юга в верхнем миоцене, когда фауна побережий Черного моря, Кавказа и Каспия не отличалась принципиально от фауны западного Средиземноморья. Сходство формирования териофауны Кавказского перешейка проявлялось и далее в среднем плиоцене в условиях усиления климатической и ландшафтной дифференцировки. Исчезали мастодонты, гиппарионы, появлялись лошади, быки и слоны.
Переход к плейстоцену в пределах Кавказа был также сходен с западноевропейским. Ареалы крупных индикаторных форм нижнего плейстоцена — трогонтериевых слонов, лошадей, мерковых носорогов, гигантских оленей и зубров — перекрывали Кавказский перешеек, т. е. уходили с Русской равнины в Переднюю Азию. В дальнейшем в соответствии с новыми этапами расчленения ландшафтов и увеличением роли Большого Кавказа как горного экологического барьера специфичность формирования фауны несколько увеличивалась. В Предкавказье вселялись степные виды Русской равнины и полупустынные виды севера Средней Азии, а в Закавказье — нагорно-степные виды Передней Азии.
Развитие териофауны перешейка в четвертичном периоде происходило значительно энергичнее за счет вселения и вымирания видов, нежели за счет местного видообразования.
Известные факты трансформации видов в течение четвертичного периода — превращения трогонтериевых слонов в мамонтов, первобытных зубров в кавказских зубров и т. п. — отнюдь не вели к качественному обогащению фауны.
Скорость эволюции и становления видов, а также направленность адаптаций кавказских млекопитающих оказывались глубоко различными у представителей одного и того же отряда, что подтверждает необоснованность розыска единых закономерностей на этом пути.
Ледниковая эпоха не оказала заметного северного влияния па состав териокомплексов Кавказа, но, несомненно, способствовала смещению ареалов и ускорению трансформации многих видов.
В раннем голоцене в связи с наступлением ксеротермической эпохи на Кавказский перешеек проник с юга ряд сухо- и теплолюбивых млеко-
Заключение 581
питающих. В позднем голоцене формирование экологических группировок и тёриокомплексов равнин и предгорий стало проходить все более под прямым и косвенным влиянием человека. Наступало сильное обедне-нбние фауны крупными хищными и копытными. Сопоставление всех фактов по угасанию отдельных видов, изменению их ареалов и сменам их комплексов в антропогене Кавказа (главы II, III, VI) не позволяют однако преуменьшать и роли природных факторов в этих изменениях. Особенно это относится к зверям происходящим из степей Центральной Азии.
На новейшем этапе развития планового социалистического хозяйства открылись широкие перспективы целенаправленной реконструкции и обогащения териофауны Кавказа при помощи истребления вредных видов и искусственного введения полезных. При дальнейшем развитии народного хозяйства и эксплуатации природных ресурсов Кавказа должны быть предусмотрены крупные государственные мероприятия по охране образцов природных фаунистических комплексов.
Очередные задачи исследований
ближайшие задачи в области организации и теории исследования териофауны Кавказа и ее истории весьма обширны и разнообразны.
Необходимо, конечно, продолжать всемерное накопление документальных костных материалов из известных и вновь открываемых мест захоронений остатков третичных и четвертичных млекопитающих.
Весьма важные и перспективные сборы костных остатков могут, вероятно, дать в ближайшие годы систематические обследования размываемых третичных и четвертичных слоев береговых уступов Цимлянского, Новотроицкого, Тщикского и Мингечаурского водохранилищ.
Дальнейшему углубленному исследованию должны быть подвергнуты также «ключевые» захоронения в третичных отложениях близ Беломечетской, в Эльдаре и Удабно, на Косякинском карьере близ Ставрополя, на песчаных карьерах севера Азовского побережья, нижнечетвертичные захоронения на Таманском полуострове, средне- и верхнечетвертичные асфальтированные захоронения на Апшероне. Дальнейшие поиски палеолита, предпринятые в районе Пятигорья, в Карабахе и в центральном Закавказье, могут дать ценные палеонтологические находки и /уточнить наши представления о генезисе фауны этих районов в антропогене.
Накопление и хранение палеонтологических коллекций в республиканских, областных и районных музеях должно быть всемерно упорядочено. Систематическая разработка поступающих коллекционных материалов из названных пунктов должна расширить и уточнить наши представления о составе древних териокомплексов и облегчить параллели-зацию их с известными кайнозойскими фаунами западного Средиземноморья, Русской равнины и Средней Азии. Именно расширение видового состава ископаемых фаунистических комплексов при одновременном проведении палеоботанических исследований и может помочь воссозданию целостных палеогеографических картин и разработке стратиграфической схемы континентальных кайнозойских толщ.
Должен получить дальнейшее развитие экологический анализ фактов эндемизма, реликтовости и разрывов ареалов кавказских млекопитающих для правильного развития теорий колебаний климата, уровня морей, орогенеза и оледенений.
682
Заключение
Этот анализ необходимо обставлять и сопровождать палеонтологической документацией. Так, например, уточнение наших представлений о древности полупустынных и пустынных видов Средней Азии в восточном Предкавказье будет возможно лишь при глубоких бурениях в районе Терско-Кумских полупустынь, а также при палеонтологических разведках в северном предгорном Дагестане.
При установлении общих закономерностей формирования фауны Кавказа в кайнозое, особенно в антропогене, при изучении видового состава осталось до сих пор много нерешенных вопросов, связанных, в частности, с географической и внутривидовой изменяемостью.
Таковы родственные и экологические взаимоотношения ряда описанных систематиками форм: из насекомоядных — кротов, водяных кутор и белозубой, из хищных — ласок и медведей, из грызунов — лесных и домовых мышей, из копытных — косуль и т. д.
Думается, что окончательное решение подобных вопросов одним морфологическим (морфометрическим) методом мало перспективно.
При всей тщательности подобных работ, даже при дополнении их обычными полевыми наблюдениями, остается множество неясных вопросов: степень взаимной гибридизации, характер наследования признаков и т. п.
Сложность рельефа, многообразие климатов и быстрые смены ландшафтов на небольших пространствах создают здесь крайне сложную картину географической изменяемости. У мелких млекопитающих, например у грызунов, эта географическая изменяемость связана еще также с изменяемостью, зависящей от погодных условий разных лет и сезонов.
Оперирование с коллекционным материалом, собранным в разные годы, в разные сезоны, без учета фактора высотности, может только затемнить суть географической изменяемости.
В результате исследователь морфолог, фаунист, проделав огромную измерительную работу, оказывается способным высказать лишь более или менее достоверные догадки о природе и темпах формообразования. Нам думается, что на современном этапе науки фаунолог должен уметь и иметь возможность совмещать морфологический анализ не только с полевыми экологическими наблюдениями обычного типа, но и с экспериментом в природных условиях или близких к ним.
Постановка таких исследований трудоемка, но необходима, так как именно она будет содействовать разрешению насущных вопросов эволюции организмов.
Дальнейшее изучение и сопоставление фактов новейшей эволюции экологических и морфо-физиологических особенностей видов с палеонтологическими данными позволит правильно решать вопросы темпов эволюции млекопитающих на отдельных этапах четвертичного периода. В результате будет получена еще более реальная возможность использовать остатки ископаемых млекопитающих для стратиграфического расчленения четвертичной толщи и предвидеть фаунистические изменения в будущем.
ЛИТЕРАТУРА
ЗООЛОГИЧЕСКАЯ ЛИТЕРАТУРА
Авакян Л. А. 1946. Первобытный бык (Bos primigenius Boj.) в Армении. Изв. АН АрмССР, естеств. науки, 9 : 59—61.
А г а к я н Л. А. 1948. История находок ископаемых четвертичных млекопитающих Армении. Изв. АН АрмССР, физ.-мат. и техн, науки, 1, 1 : 71—80.
А акян Л. А. 1950. Остатки Elephas trogontherii Pohl, из четвертичных отложений Армении. Научн. тр. Ереванск. гос. унив., сер. геолог., XXX : 149—160. Авакян Л. А. 1952. Остатки Equus stenonis из четвертичных отложений Армении. Научн. тр. Ереванск. гос. унив., сер. геол., XXXVII : 109—116.
\ акян Л. А. 1955. Rhinoceros mercki Jaeg. из четвертичных отложений Армении. Научн. тр. Ереванск. гос. унив., сер. геол., LII, 2 : 35—41.
Авакян Л. А, и Н. О. Бурчак-Абрамович. 1945. Ископаемый верблюд из четвертичных отложений Армении. Докл. АН АрмССР, III, 4 : 99—102.
Аветисян О. Р. 1949. Малоазийский суслик Citellus xanthoprymnus Bennet как вредитель сельского хозяйства в Армянской ССР и возможности его полного уничтожения. Докл. АН АрмССР, 3 : 163—196.
Аветисян О. Р. 1950. Биологические особенности малоазиатского суслика в Армянской ССР. Изв. АН АрмССР, биол. и сельхоз. науки, III, 1 : 173—184. ;,леро'ерг Г. П. 1930. Предварительный обзор русских и монгольских кабанов. Докл. АН СССР, IV : 91—96.
Адлерберг Г. II. 1935. Медведи Кавказа. Изв. АН СССР, отд. матем. и естеств. наук, 1 : 73 -100.
(ольф Т. А. 1950. Сайга в Астраханских степях правобережья Волги. Охрана природы, 10 : 73—82.
дольф Т. А. 1952. Количественный учет сайгаков в Астраханских степях. Сб. «Методы учета числ. и геогр. распр. наземн. позв.», Изд. АН СССР, М.—Л.: 245—250.
ди з е н ш т а д т Д. С. 1950а. Некоторые причины колебания численности и вопросы расселения серой крысы. 2-я Экол. конф, по проблеме «Массовое размножение животных и их прогнозы» (тезисы докладов), Киев : 3—4.
Айзен шт а дт Д. С. 19506. Гнезда серых крыс, устраиваемые во время половодья. Природа, 6 : 81—82.
А й з е н ш т а д т Д. С. 1950в. Расселение серых крыс вдоль железнодорожного полотна. Природа, 4 : 62—63.
A i зеншт а дт Д. С. 1955. К вопросу об истории и путях расселения пасюков (Ratus norvegicus norvegicus Berken.) в пределах СССР. Зоол. журн., XXXIV, 5 : 1045—1152.
Алекперов А. М. 1958. Герпетогеографическое районирование Азербайджана. Уч. зап. Азерб. гос. унив. им. С. М. Кирова, 1 : 65—82.
Алекперова Н. А. 1950. Описание зубов арверненского мастодонта (Mastodon arvernensis Croiz. et Job.) с Кавказа. Изв. АН АзербССР, 12 : 25—32.
Алекперова Н. А. 1952. Марал бинагадинских Кировых отложений. Бинага-динское местонахождение четвертичной фауны и флоры, II. Тр. Естеств.-ист муз. АН АзербССР, V : 5—181.
Алекперова II. А. 1955. Ископаемая бинагадипская сайга. Бинагадинское местонахождение четвертичной фауны и флоры, IV. Тр. Естеств.-ист. муз. АН АзербССР, 10—64.
Александров В. 1956. Зубры в Кавказском заповеднике. Охота и охотничье хозяйство, 8 : 27.
Алексанян А. Б. 1910. Материалы по изучению грызунов в Армянской ССР. Сб. научн. тр. Ереванск. мед. инет., I, Ереван : 91—98.
684
Л итература
Алексеев А. К. 1915а. Об остатках оленя с о. Челекена. Геол, вести., I : 346— 350.
Алексеев А. К. 19156. Фауна позвоночных деревни Ново-Елизаветовки. Одесса : I—XIV+1—453.
Алексеев А. К. 1930. Верхнесарматская фауна млекопитающих Эльдара, I. Achtiaria borissiaki n. sp. Тр. Геол. муз. АН СССР, VII : 167—204.
Алексеев А. К. 1933. Trogontherium cuvieri Fischer из четвертичных отложений Украины. Тр. Комисе, по изуч. четвертичн. периода, III : 5—22.
Алиев Ф. Ф. 1955. Акклиматизация и хозяйственное освоение енота (Ргосуоп lotor L.) в Азербайджане. Докл. АН АзербССР, XI, 8 : 571—578.
Алиев Ф. Ф. 1956. Некоторые данные по размножению и росту енотов (Ргосуоп lotor L.). Докл. АН АзербССР, XII, 8 : 583—592.
Андрианов К. С. и Н. И. Ларин. 1935. Условия залегания верхнесарматских позвоночных степи Эльдара. Бюлл. Моск. общ. испыт. природы, сер. геол., ХШ, 4 : 559—569.
Андрусов Н. 1888. Очерк истории развития Каспийского моря и его обитателей. Изв. Русск. геогр. общ., 24 : 17—36.
Андрусов Н. И. 1918. О возрасте фауны млекопитающих Пикерми. Изв. Росс. АН, XII : 151—155.
Анисимов А. Ф. 1938. Биология гигантского слепца. Уч. зап. Сев.-Осет. гос. пед. инет., I, Орджоникидзе : 165—176.
(Анучин Д. Н.) Anutschin D. 1887. Uber die Reste des Hohlen-Baren aus Transcaucasien. Bull. Soc. Imp. Nat. Moscou, I : 216—221.
Анучин Д. H. 1896. К вопросу о диких лошадях и об их приручении в России. СПб. : 1—75.
Аргиропуло А. И. 1932. Роды и виды хомяков (Cricetinae) Палеарктики. Тр. Зоол. инет. АН СССР, I : 239—248.
Аргиропуло А. И. 1934. Заметки по систематике и распространению серых полевок группы «so ci al is» Pallas. Тр. Зоол. сектора Грузинок, отд. Закавк. фил. АН СССР, I : 165—174.
Аргиропуло А. И. 1935. Заметки по летней биологии общественной полевки в Закавказье. Тр. Азерб. микробиол. инет., V, 1 : 217—227.
Аргиропуло А. И. 1936. К вопросу о расщюстранении водяной крысы (Аг-vicola terrestris Linn.) на Кавказе. Тр. Азерб. тропич. инет., II, 1 : 287—289.
Аргиропуло А. И. 1937. Каталог грызунов Кавказа. Тр. Азерб. фил. АН СССР, XX : 47—69.
Аргиропуло А. И. 1938. К фауне третичных Cricetidae. Докл. АН СССР, XX, 2—3 : 223—226.
Аргиропуло А. И. 1939а. К распространению и экологии некоторых млекопитающих Армении. Зоол. сб., I. Тр. Биол. инет. Арм. фил. АН СССР, 3 : 27—66.
Аргиропуло А. И. 19396. Остатки бобра (Amblycastor caucasicus sp. п.) из плиоцена Предкавказья. Докл. АН СССР, XXV, 7 : 638—640.
Аргиропуло А. И. 1940а. Сем. Muridae мыши. Фауна СССР. Млекопитающие. III, 5, нов. сер., 21 : 1—169.
Аргиропуло А. И. 19406. О нахождении мышевидного хомячка. Природа, 1 :91—93.
Аргиропуло А. И. 1940в. Обзор находок третичных грызунов на территории СССР и смежных областей Азии. Природа, 12 : 74—82.
Аргиропуло А. И. 1941а. О происхождении мышей. Природа, 2 : 83—87.
Аргиропуло А. И. 19416. Четвертичная фауна грызунов и насекомоядных Бинагадов (Апшеронский полуостров). Природа, 3:88—91.
Аргиропуло А. И. 1946. К вопросу об индивидуальной и географической изменчивости у некоторых видов рода Apodemus Каир. (Mammalia). Тр. Зоол. инет. АН СССР, VIII : 195—220.
Аргиропуло А. И. и А. В. Богачев. 1939. Четвертичная фауна апшерон-ских отложений битума (кира). Природа, 6 : 76—78.
Бабенышев В. П. 1948. Сайга в степях восточного Предкавказья. Природа. 6 : 63—64.
Бабенышев В. П., Н. Б. Б и р у л я, Б. Д. Б е с е д и н, Ф. 3. Г о л о с о в-с к а я, А. Н. Е г о р о в, И. Г. К о р ф, П. А. Я н у ш к о. 1937. Распространение сусликов вида Citellus pygmaeus Pall, в пределах Op джени кидзев-ского края. Зоол. журн., XVI, 4 : 736—739.
Бажанов В. С. 1948. Современное распространение и история большого суслика. Изв. АН КазССР, сер. зоол., 8 : 27—50.
Бартенев А. 1933. О некоторых положениях истории и сложения палеарктической фауны. Зоол. журн., XII, 1 : 101—129.
Зоологическая литература 685
Бартенев А. 1934. К характеристике сложения фауны западного Предкав-L казья. Зоол. журн., XIII, 1 : 139—147.
^Бартенев А. 1935. О понтийской фауне. Зоол. журн., XIV, 4 : 762—784.
Башкиров И. Г. 1940. Кавказский зубр. Изд. Комит. по заповедникам, М. : 3—72.
Б < л о в Н. К. 1924. Кое-что о причинах оскудения дичи в Закавказье. Спорт и охота 1 в Закавказье, 5 : 34—35.
Беляева Е. И. 1925. Elephas trogontherii Pohl. Таманского полуострова. Тр. Геол, и Минерал, муз. АН СССР, V : 1—15.
| Бел яев а Е. И. 1933а. Некоторые данные об ископаемых слонах Таманского по-I луострова. Изв. АН СССР, сер. VII, отд. матем. и естеств. наук, 8 : 1209—1211.
[ Беляева Е. И. 19336. О фауне четвертичных млекопитающих с Таманского полу-L острова. Природа, 10 :63.
(Беляева Е. И. 1935. Некоторые данные о четвертичных млекопитающих из Ниж-f неволжского края по материалам г. Пугачева. Тр. Комисе, по изуч. четвер-I тичн. периода, IV : 303—309.
|Беляева Е. И. 1939а. Заметка об остатках четвертичных млекопитающих полуострова Тунгуза. Бюлл. Моск. общ. испыт, природы, отд. геол., XVII (6) : 85—89.
[Беляева Е. И. 19396. Новая находка остатков эласмотерия. Природа, 10 : 75.
[Беляева Е. И. 1940а. О ворошиловской фауне млекопитающих. Палеонтол. обозрение, 2 : 77.
Беляева Е. И. 19406. О новой верхнетретичной фауне млекопитающих на Северном Кавказе. Природа, 2 :82.
Беляева Е. И. 1940в. О фауне млекопитающих из палеолитической пещеры Вир-[ хова (Сакажиа). Бюлл. Комисе, по изуч. четвертичн. периода, 6—7 : 105—107. Беляева Е. И. 1944. Новые материалы по верхнетретичным млекопитающим Северного Кавказа. Природа, 3 : 76—77.
Беме Л. Б. 1925. К биологии и к распространению некоторых грызунов Северного Кавказа. Владикавказ : 29—41.
Беме Л. Б. 1926. Некоторые наблюдения над распространением и образом жизни Chilotus socialis parvus Sat. и Microtus arvalis Pall, на северо-восточном Пред-. кавказье. Уч. зап. Сев.-Кавк. инет, краевед., 1 : 57—62.
Беме Л. Б. 1928. Результаты обследования охотничьего хозяйства Парабочевского и Самурского заказников НКЗ Дагестанской С. С. Республики и краткий обзор фауны наземных позвоночных их населяющей. Изв. Горского пед. инет., Владикавказ : 115—156.
Беме Л. Б. 1929. Охотничье хозяйство Ингушии и его перспективы. Ингушек, научи о-и сел. инет, краевед., Владикавказ : 131—142.
Беме Л. Б. 1933. По Аварскому и Андийскому округам Дагестанской Республики
! в целях зоологических наблюдений. 2-й Сев.-Кавк. пед. инет., Орджоникидзе i 127—146.
Беме Л. Б. иД. Б. Красовский. 1930. Материалы к познанию экологии ногайского тушканчика (Dipodipus sagitta nogai Sat.), Ежегодн. Зоол. муз.
J; АН СССР, XXXI : 419—430.
Берг Л. 1905. Сайгаки на острове Николая I. Природа и охота, 5 : 28—32.
Бернацкий А. 1884. Ископаемые остатки пещерного человека и пещерных животных на Кавказе. Тифлис : 1—6.
Бибикова В. И. 1950. О некоторых биологических особенностях первобытного J зубра. Бюлл. Моск. общ. испыт. природы, отд. биол., IV (5) : 35—43.
Бируля А. А. 1912. Материалы по систематике и географическому распространен : нию млекопитающих, III. Carnivora, собранные Н. А. Зарудным в Персии
в 1896, 1898, 1900—1901 и 1903—1904 гг. Ежегодн. Зоол. муз. Акад, наук, XVII : 219— 280.
Бируля А. А. 1928. К вопросу о нижнем течении р. Волги как зоогеографической границе. Докл. АН СССР .* 338—340.
Бируля А. А. 1930а. Предварительное сообщение о хищных Carnivora из чет-г ' вертичных отложений Крыма. Докл. АН СССР, 6 : 133—144.
Б и р у л я А. А. 19306. Предварительное сообщение о грызунах из четвертичных отложений Крыма. Докл. АН СССР, 23 : 617—622.
Бируля Н. А. 1941. О природе факторов, ограничивающих численность малого-суслика в ковыльных степях. Зоол. журп., XX, 1 : 135—154.
Бобринский Н, А. 1951. География животных. Учпедгиз, М.—Л. :1—384. Бобринский Н. А., Б. А. К у з н е ц о в, А. П. К у з я к и н. 1944. Определи-[ тель млекопитающих СССР. Изд. «Советская наука», М. : 1—440.
Богачев А. В. 1938. Зоологические наблюдения над пресмыкающимися и земноводными в Мильской степи. Изв. Азерб. фил. АН СССР, 4-—5 : 89—100.
586
Литература
Богачев А. В. 1947. Фауна бинагадинских Кировых пластов. Жуки Coleoptera. Тр. Естеств.-ист. муз. АН АзербССР, I—II : 137—160.
Богачев В.В.1912. Каталог Донского краевого музея в Ростове. Ростов н/Дону : 1—738.
Богачев В. В. 1913а. О фауне соленосных отложений русской Армении. Ежегодн. по геол, и минерал. России, XV, 8-—9 : 213—224.
* Богачев В. В. 19136. Донской музей в Новочеркасске. Ежегодн. по геол, и минерал. России, XV, 8—9 : 233—238.
Богачев В. В. 1918. Отчет об осмотре местонахождения костей на Эльдаре. Изв, Кавк, муз., XII : 16—19.
Богачев В. В. 1923. Кость гиппариона, или первобытной лошади, в Апшеронском известняке. Азерб. нефт. хоз., II : 91—92.
Богачев В. В. 1923—1924. Новые материалы к истории третичных слонов в юго-восточной России. Изв. Азерб. унив., 3:99—121.
Богачев В. В. 1924. Послетретичные млекопитающие на Апшеронском полуострове. Азерб. нефт. хоз., 7—8 : 82—83.
Богачев В. В. 1925а. Остатки носорога в слоях бакинского яруса. Азерб. нефт. хоз., 12:88—89.
Богачев В. В. 19256. Первобытный бык-тур (Bos primigenius Boj.) на Апшероне. Изв. Азерб. политехи, инет., I, Баку : 119—135.
Богачев В. В. 1927а. Палеонтологические заметки о фауне Эльдара. Изв. Общ. обслед. и изуч. Азербайджана, 5 : 21
Богачев В. В. 19276. Палеонтологические заметки о фауне Эльдара. Изв. Общ. : обслед. и изуч. Азербайджана, 5 : 194—214.
Богачев В. В. 1927в. Тюлень в миоцене Каспийского бассейна. Изв. Азерб. гос. унив., отд. естеств. и мед., VI : 135—147.
Богачев В. В. 1935. Достигал ли Stegodon в своем распространении Кавказа. Изв. Азерб. индустр. инет., 8(15) :1—6.
Богачев В. В. 1938а. Дельфин в отложениях Апшеронского яруса. Природа, 3 : 78—85.
* Богачев В. В. 19386. Находка послетретичной фауны млекопитающих в окрест-; ностях Баку. Изв. Азерб. АН СССР, 3 : 107—108.
Богачев В. В. 1938в. Палеонтологические заметки. Тр. Азерб. АН СССР, геол, сер., IX, 39 : 1—96.
Богачев В. В. 1938г. Вопросы зоогеографии Кавказа и его геологическая история. Изв. Азерб. фил. АН СССР, 4—5 : 29—47.
Богачев В. В. 1939а. Об ископаемых китообразных Кавказа. Изв. Азерб. фил. АН СССР, 1—2 : 153—154.
Богачев В. В. 19396. Новое о находке костеносных слоев в окрестностях Баку. Природа, 1 :83.
Богачев В. В. 1939в. Четвертичная (межледниковая) фауна на Апшеронском полуострове. Докл. АН СССР, ХХШ, 3 : 315—318.
Богачев В. В. 1939г. Бинагады — кладбище четвертичной фауны на Апшеронском полуострове. Изд. Азерб. фил. АН СССР, Баку : 3—84.
Богачев В. В. 1939д. Новые данные по флоре и фауне Бинагадинского кирового местонахождения. Изв. Азерб. фил. АН СССР, 6 : 91—99.
Богачев Б. В. 1940а. Апшеронская фауна в бассейне Азовского моря. Природа, 12 : 73—74.
Богачев В. В. 19406. Картины первобытной природы Апшерона (Бинагады). Изд. Азерб. фил. АН СССР, Баку : 1—114.
Богачев В. В. 1940в. Находка четвертичной фауны под Новочеркасском. Природа, 5 : 88.
Богачев В.В. 1940г. Открытие новой формы морского млекопитающего в апшеронском ярусе. Изв. Азерб. фил. АН СССР, 1 : 88—89.
Богачев В. В. 1941. Новые находки динотерия (на Дону). Природа, 2: 95—97.
Богачев В. В. 1944. Новые находки четвертичной фауны па Апшероне. Природа, 2 :68—70.
Богачев В. В. и А. И. Шишкина. 1915. Фауна и флора соленосных отложений русской Армении. I, Кульп. II, Бассейн Олты~Чая. Зап. Кавк, муз., сер. А, 2, Тифлис : 1—76.
Богданов М. Н. 1871. Птицы и звери черноземной полосы Поволжья и долины Средней и Нижней Волги (биогеографические материалы). Тр. Общ. естсство-испыт. Казанск. унив., I, 1 : 3—226.
Богданов М. Н. 1873. Этюды русской охоты. Журн. охоты и коннозаводства, 1:94—95.
Зоологическая литература
587
Борйсяк А. А. 1914. О зубном аппарате Elasmotherium caucasicum n. sp. Изв. Акад, наук, 9 :555—584.
Борис як А. А. 1928а. Очередная задача русской палеонтологии. Природа, 4 : 369—384.
Борисяк А. А. 19286. О Platybelodon Danovi n. g., n. sp. — новом мастодонте из чокракских отложений Кубанской обл. Ежегодн. Русск. палеонтол. общ., ' VII: 105—120.
Борисяк А. А. 1930. Ursus spelaeus rossicus nov. и. Докл. АН СССР, V : 102—105.
Борисяк А. А. 1931. Новая раса пещерного медведя из четвертичных отложений Северного Кавказа. Тр. Палеозоол. инет. АН СССР, I : 137—202.
Борисяк А.А. 1935. Новые материалы к филогении Dicerorhinae. Докл. АН СССР, Ш, 8 : 381—384.
Борисяк А. А. 1937. О Paranchitherium karpinskii. Изв. АН СССР, сер. геол., 4 : 770—789.
Борисяк А. А. 1938а. Новый Dicerorhinus из среднего миоцена на Северном Кавказе. Тр. Палеонтол. инет. АН СССР, VIII, 2 : 1—68.
БорисякА. А. 19386. Об остатках анхитерия из среднемиоценовых отложений Северного Кавказа. Изв. АН СССР, сер. биол., 1 : 207—218.
Борисяк А. А. 1943. Обзор местонахождений третичных наземных млекопитающих Союза ССР. Фрунзе : 1—42.
Борисяк А. А. 1945. Об остатках представителей сем. Equidae из среднего миоцена Северного Кавказа. Тр. Палеонтол. инет. АН СССР, XVIII, 1 : 1—52. ' Борисяк А. А. и Е. И. Б е л я е в а. 1928. Новые материалы по русским мастодонтам. Тр. III Всесоюзн. съезда зоологов, анатомов, гистологов, 14—20 декабря 1927 г., Л. : 1000.
Борисяк А. А. иЕ. И. Беляева. 1948. Местонахождения третичных наземных млекопитающих на территории СССР. Тр. Палеонтол. инет. АН СССР, XV, 3 :1—116.
(Брандт И.) Brandt I. 1834. Mammalium Rodentium exoticorum novorum velminus rite cognitorum. Mus. Acad. Zool. Description et leones : 21—106.
(Брандт И.Ф.) Brandt I. F. 1855. Beitrage zur nahern Kenntniss der Sagethiere Russlands. St. Petersb. : 43—128.
(Брандт И. Ф.) Brandt I. F. 1867. Zoogeographische und paleontologische Beitrage. Verhandl. Mineral. Gesellsch-, 8, St. Petersb. : 1—258.
(Брандт И. Ф.) В r a n d t I. F. 1870. Ueber die von Herrn Magister Adolph Goebel auf seiner persischen reise bei der Stadt Maragha und der provinz Aderbeidjan gefundenen Saugethier-reste. Riga : 2—8.
(Брандт И. Ф. и Ж. Н. Вольдрих) Brandt I. F. u. Woldrich J. N. 1887. Diluviale europaisch-Nord-asiatische Saugethierfauna und ihre dezighmun-gen zum Menschen. Mem. Acad. Imp. Sci, St. Petersb., VII, ser. 6, XXV, 10.
(Б p а н д т И. Ф. и Ф. Ф. P а т ц e б у p г) Brandt I. F. u. R a t z e b u r g F. F. 1829. Monographie des Bibers. Mediz. Zool., Berlin, 1 : 12—30.
Бу г д а н о в Г. В. 1929. Фауна вредителей сельского хозяйства района восточных предгорий Северного Кавказа. Ингушек, научно-иссл. инет, краевед., Владикавказ :112—140.
Бугданов Г. Б. 1936. Вредители сельскохозяйственных культур и продуктов в предгорной полосе Северного Кавказа. Горская с.-х. опытн. ст., сер. 1, 11 : 1—32.
Бурчак-Абрамович Н. И. 1946. Elephas primigenius Blum, из Душетского района. Вести. Гос. муз. Грузии, ХИГА : 303—309.
Бурчак-Абрамович Н. И. 1949. Находка древнего бизона Bison sp. в апше-ронских отложениях (верхний плиоцен) Азербайджана. Докл. АН АзербССР, V, И : 474—479.
Бурчак-Абрамович Н. И. 1950а. Ископаемый трубкозуб Oricteropus gaud-ryi F. M. в пределах СССР и смежных стран. Изв. АН АрмССР, биол. и с.-х. науки, III, 10 : 949—954.
Бурчак-Абрамович Н. И. 19506. Новый представитель ископаемых быков Taurina в гиппарионовой фауне Мара гм — Urmia bos azerbaidzanicus Bur. gen. et sp. nov. Докл. АН СССР, LXX, 5 : 875—878.
Бурчак-Абрамович И. И. 1951a. Древний плосколобый слон Elephas planiIrons Falc. и близкие к ному формы в Закавказье. Изв. АН АзербССР, 2 : 75—81.
Бурчак-Абрамович Н. И. 19516. Фауна эпипалеолитической стоянки в Триалети. Сообщ. АН ГрузССР, XII, 2 : 93—97.
Бурчак-Абрамович Н. И. 1951 в. К истории фауны наземных позвоночных Азербайджана. Животный мир Азербайджана, Баку : 7—33.
688
Литература
Бурч ак-Абрамович Н. И. 1951г. Черен первобытного быка-тура Bos pri-migenius Boj. (из Армении). Изв. АН АрмССР, физ.-мат. и естеств. науки, IV, 2 :135—145.
Бурчак-Абрамович Н. И. 1951д. Материалы к изучению фауны палеолита Закавказья (Зуртакетская стоянка). Изв. АН АзербССР, 9 : 19—31.
Бурчак-Абрамович Н. И 1952а. Находка ископаемого буйвола (Bubalus sp.) на Кавказе. Изв. АН АзербССР. 2 : 63—75.
Бурчак-Абрамович Н. И. 19526. Нижнеплиоценовый бык Urmiabos azer-baidzanicus Bur. из южного Азербайджана. Изв. АН АзербССР, 6 : 15—29.
Бурчак-Абрамович Н. И. 1952в. Находки овернского мастодонта Mastodon arvernensis Grois. et Job. в Азербайджане и восточной Грузии. Изв. АН АзербССР, 9 : 23—30.
Бурчак-Абрамович Н. И. 1952г. Первобытный бык (Bos Mastan—zadei mihi species nova) в четвертичной фауне сел. Бинагады на Апшеронском полуострове. Бинагадинское местонахождение четвертичной фауны и флоры, II. Тр. Естеств.-ист. муз. АН АзербССР, V : 181—203.
Бурчак-Абрамович Н, И. 1953а. Материалы к изучению длинных трубча-- / тых костей (диафизов) молодых особей бинагадинских крупных млекопитающих.
£ Бинагадинское местонахождение четвертичной фауны и флоры, III. Тр. Естеств.-
' ист. муз. АН АзербССР, VI : 210—277.
Бурчак-Абрамович Н. И. 19536. Следы повреждений хищными зверями на костях млекопитающих бинагадинского ископаемого кладбища. Бинагадин-В ское местонахождение четвертичной фауны и флоры, III. Тр. Естеств.-ист. муз.
I АН АзербССР, VI : 278—301.
Бур ч ак-Абрамович Н. И. 1953в. Материалы к изучению болезненных в ? травматических образований на костях бинагадинских ископаемых птиц.
С Бинагадинское местонахождение четвертичной фауны и флоры, III. Тр. Естеств.-
f ист. муз. АН АзербССР, VI : 310—323.
t Бурчак-Абрамович Н. И. 1953 г. Находки эласмотерия на Апшеронском I полуострове и Южном Урале. Изв. АН АзербССР, 6 : 75—90.
,Г. Бур ча к - Абрамович Н. И. 1954. Ископаемый носорог (Rhinoceros cf. et-ruscus Falc.) в долине p. Алазани. Изв. АН АзербССР, 4 : 33—48.
? Бурч ак-Абрамович Н. И. 1957. Ископаемые быки Старого Света, 1. Тр. Естеств.-ист. музея АН АзербССР, XI : 1—263-pXI табл.
Бурчак-Абрамович Н. О., Е. Г. Габашвили. 1945. Высшая челове-! кообразная обезьяна из верхнетретичных отложений восточной Грузии. Сообщ.
АН ГрузССР, VI, 6 : 451—457.
Бурчак-Абрамович Н. И., Е. Г. Габашвили. 1950. Находка ископаемой высшей человекообразной обезьяны в пределах Грузии. Природа, 9 : 70—72.
Бурчак-Абрамович Н. О., Р. Д. Джафаров. 1945. Находка гигантского оленя в Кировых отложениях Апшеронского полуострова. Изв. АН АзербССР, 10 : 71—78.
Бурчак-Абрамович Н. И., Р. Д. Джафаров. 1946. Sus apscheronicus sp. n. в составе бинагадинской фауны. Изв. АН АзербССР, II, 6 : 26—57.
Бурчак-Абрамович Н. И., Р. Д. Джафаров. 1948. Остатки дикого кабана из бинагадинских Кировых отложений (Апшерон). Тр. Естеств.-ист. муз. АН АзербССР, I—II : 26—37.
Бурч ак-Абрамович Н. О., Р. Д. Джафаров. 1949. Леопард (Pardus pardus tullianus Valenc.) на Апшеронском полуострове. Тр. Естеств.-ист. муз. АН АзербССР, III : 86—104.
Бурчак-Абрамович Н. И., Р. Д. Джафаров. 1950. Материалы к изучению верхнетретичных Proboscidea (хоботных) Закавказья. Род Mastodon et Dinotherium. Изв. АН АзербССР, 2 : 38—54.
Бур чак-Абрамович Н. И. иР. Д. Джафаров. 1953. Материалы к количественному учету копытных Бинагадинского местонахождения четвертич-ной фауны и флоры, III. Тр. !Естеств.-ист. муз. АН АзербССР, VI : 157— 209.
Бурчак-Абрамович II. И. и Р. Д. Джафаров. 1955. Бинагадинское местонахождение верхнечетвертичной фауны и флоры на Апшеронском полуострове. Бинагадинское местонахождение четвертичной фауны и флоры, IV. Тр. Естеств.-ист. муз. АН АзербССР, X : 89—145.
Бурчак-Абрамович Н. О., 3. С. Эквтимишви ли. 1953. Находка ископаемого носорога (Rhinoceros cf. etruscus Falc.) из Кахетии (с. Цинандали) Тр. Инет. зоол. АН ГрузССР, XI : 229—236.
Бутурлин С. А. 1934. Лоси. Промысловые животные СССР, КОИЗ, М.—Л.: 2—67.
Зоологическая литература
589
(Бюхнер Е.) Buchner Е. 1887. Zur Geschichte der Kaukasischenture (Capra caucasica Giild. und Capra cylindricornis Blyth). Mem. Acad. Imp. Sci., St. Pe-tersb., VIIE XXXV, 8 :1—27.
Б я л ы н и ц к и й - Б и р у л я А. А. 1917. Скорпионы, 1. Фауна России и сопредельных стран. Паукообразные (Arachnoidea), 1 : 1—224.
В*а р ш а в с к и й С. Н. и К. Т. К рыл о в а. 1939. Экологические особенности популяции малого суслика (Citellus pygmaeus Pall.) в разные периоды жизни. Зоол. журн. XVIII, 6 : 1026—1048.
Васильев Я. 1896. Предельные линии распространения в Кубанско-Черноморском районе красного зверя и птицы. Природа и охота, 2 : 1—35.
Верещагин Н. К. 1936. Опыты акклиматизации нутрии (Myocastor coy pus bo-nariensis Rengger) на юге СССР. Тр. Азерб. фил. АН СССР, XXIX : 1—67.
Верещагин Н. К. 1937. Современное состояние джейрана (Gazella subgutturosa Giild.) в Мильско-Карабахской степи. Изв. Азерб. фил. АН СССР, 2 : 155—174.
Верещагин Н. К. 1938а. Дагестанский тур (Capra cylindricornis Blyth) в Азербайджане. Тр. Зоол. инет. АзербССР, IX/45 : 1—70.
Верещагин Н. К. 19386. Млекопитающие Апшеронского полуострова. Баку : 1—34
Верещагин Н. К. 1939а. Джейран в Азербайджане. Тр. Зоол. инет. Азерб. фил. АН СССР, X : 109—122.
Верещагин Н. К. 19396. К вопросу о реконструкции фауны Азербайджана. Изв. Азерб. фил. АН СССР, 1—2 :161—164.
Верещагин Н. К. 1939в. К вопросу об экологических нишах и морфологических адаптациях. Бюлл. Моск. общ. испыт. природы, XLVIII, 1 : 43—52.
Верещагин Н. К. 1940а. Новые находки ископаемых и современных млекопитающих в Закавказье за период 1935—1940 гг. Изв. Азерб. фил. АН СССР, 6 : 108—115.
Верещагин Н. К. 19406. Перспективы разведения нутрии в Абхазии. . Монадире, 1.
Верещагин Н. К. 1941а. Степной кот (Felis ornata Gray) в Восточном Закавказье. Сб. тр. Зоол. муз. Моск, унив., VI : 305.
Верещагин Н. К. 19416. Акклиматизация нутрии (Myocastor coypus Mol.) в западной Грузии. Тр. Зоол. инет. Груз. фил. АН СССР, IV : 4—42.
Верещагин Н. К. 1942а. Домовые грызуны в Азербайджане и методы борьбы с ними. Изд. АН АзербССР, Баку : 1—37.
Верещагин Н. К. 19426. Каталог зверей Азербайджана. Изд. АН АзербССР, Баку : 1—95.
Верещагин Н. К. Г942в. Условия жизни вредителя полей — общественной полевки в Азербайджане. Изв. Азерб. фил. АН СССР, 2 : 85—89.
Верещагин Н. К. 1944. Находка полевой мыши в Азербайджане. Природа, 3 : 74—75.
Верещагин Н. К. 1945а. Животный мир Азербайджана. В кн.: Физическая география АзербССР, Баку : 233—269.
Верещагин Н. К. 19456. Новые фаунистические находки в Талыше. Природа, 6 : 67—68.
Верещагин Н. К. 1946а. Гибель птиц от нефти в Азербайджане. Зоол. журн., XXV, 1 : 69— 80.
Верещагин Н. К. 19466. Основные экологические черты общественной полевки в полупустынной зоне Азербайджанской ССР. Тр. Зоол. инет. АН АзербССР, XI : 144—182.
Верещагин Н. К. 1947а. Новые находки хищных и копытных в бинагадинском асфальте. Докл. АН СССР, LV, 3 : 247—249.
Верещагин Н. К. 19476. Новая раса бурого медведя из бинагадинского асфальта (Ursus arctos binagadensis subsp. nova). Докл. АП СССР, LV, 4 : 351—353.
Верещагин Н. К. 1947в. Американский енот в лесах Исмаиллинского района Азербайджанской ССР. Изв. АН АзербССР, 5 : 68—73.
Верещагин Н. К. 1947г. Охотничьи и промысловые животные Кавказа. Изд. АН АзербССР, Баку : 3—144.
Верещагин И. К. 1948. Лось Alces sp. как недавно вымершее на Кавказе животное. Докл. АН АзербССР, IV, 3 :124—126.
Верещагин Н. К. 1949а. К истории и систематическому положению лося на Кавказе. Докл. АН СССР, LXVI, 3 : 491—493.
Верещагин Н. К. 19496. Зоогеографические карты. В кн.: Атлас Азербайджан* ской ССР. Изд АН АзербССР, Баку : 44.
Верещагин Н. К. 1949в. Плейстоценовые реликты Кабристана и Апшеронского полуострова. Бюлл. Моск. общ. испыт. природы, отд. биол., LIV (4) : 3—14.
590
Литература
Верещагин Н. К. 1949г. О происхождении крыс рода Rattus в Закавказье. Природа, 11 : 62—63.
Верещагин Н. К. 1949д. К экологии и эпидемиологическому значению грызунов Ленкоранской низменности и горного Талыша. Тр. Зоол. инет. АН АзербССР, XIII : 115—129.
Верещагин Н. К. 1950а. Болотный бобр (нутрия) его разведение и промысел в водоемах Закавказья. Изд. АН АзербССР. Баку : 1—146.
Верещагин Н. К. 19506. Зимовки и промысел водоплавающей птицы в Азербайджане. Тр. Инет, зоол АН АзербССР, XIV : 133—212.
Верещагин Н. К. 1951а. Остатки собаки и бобров (Mammalia, Canis, Castor, Trogontherium) из нижнего плейстоцена западного Кавказа. Докл. АН СССР, LXXX, 5 : 821—824.
Верещагин Н. К. 19516. Хищные (Carnivora) из бинагадинского асфальта. Бинагадинское местонахождение четвертичной фауны и флоры, I. Тр. Естеств. ист. муз. АН АзербССР, IV : 28—126.
Верещагин Н. К. 1951в. Млекопитающие. Животный мир Азербайджана. Изд. АН АзербССР. Баку: 84—251.
Верещагин Н. К. 1951г. Условия массовой гибели наземных позвоночных и захоронения их остатков в Закавказье. Зоол. журн., XXX, 6 : 616—619.
Верещагин Н. К. 1952. Остатки животных и растений в битуминозных отложениях. Природа, 3 : 122—123.
j Вер е щ а г и я Н. К. 1953а. К истории ландшафтов Предкавказья в четвертичном периоде. Изв. Всесоюзн. геогр. общ., 85, 2 : 200—201.
Верещагин Н. К. 19536. Опыты разведения новых видов пушных хищников в Азербайджанской ССР. Тр. Инет. зоол. АН АзербССР, XVI : 150—168.
В ерещагин Н. К. 1953в. Остатки аргалиобразного барана Ovis cf. ammon Pall, из битуминозных слоев Апшеронского полуострова. Бинагадинское место-I- нахождение четвертичной фауны и флоры, III. Тр. Естеств.-ист. муз. АН
f АзербССР, VI : 302—309.
1 ’ Верещагин Н. К. 1953г. Великие «кладбища» животных в долинах рек Русской равнины. Природа, 12 : 60—65.
Верещагин Н. К. 1954. К истории фауны позвоночных и развития ландшафтов Ставрополья в неогене. Матер, по изуч. Ставропольск. края, 6 : 169—175.
Верещагин Н. К. 1955. Кавказский лось (Alces alces caucasicus N. Ver. subsp. nova) и материалы к истории лосей на Кавказе. Зоол. журн., XXXIV, 2 : 460—463.
Верещагин Н. К. 1956. О прежнем распространении некоторых копытных в районе смыкания европейско-казахстанских и центральноазиатских степей. Зоол. журнал, XXXV, 10 : 1541—1553.
Верещагин Н. К. 1957а. Остатки млекопитающих из нижнечетвертичных отложений Таманского полуострова. Тр. Зоол. инет. АН СССР, XXII : 19—74, Верещагин Н. К. 19576. Плейстоценовые позвоночные из пещеры Кударо I в Юго-Осетии и их значение для разработки истории фауны и ландшафтов Кавказа. Докл. АН СССР, 112, 6 : 1347—1349.
Верещагин Н. К. и И. М. Громов. 1952. К истории фауны позвоночных района нижнего течения реки Урал. Тр. Зоол. инет. АН СССР, IX : 1226—1269.
Верещагин Н. К. и И. М. Громов. 1953а. Сбор остатков высших позвоночных четвертичного периода. Изд. АН СССР, М.—Л. : 1—37.
Верещагин Н. К. и И. М. Г р о м о в. 19536. О прошлом фауны и флоры Ставрополья. Природа, 7 : 103—104.
Верещагин Н. К. и С. К. Даль. 1948. О распространении дикобраза в Закавказье. Докл. АН АрмССР, IX, 2 : 85—86.
Верещагин Н. К. и А. Г. Д ю н и н. 1949. Поведение птиц и млекопитающих при сильном заморозке в восточном Закавказье. Изв. АН АзербССР, 7 : 34—38, Верещагин Н. К. и А. Д. К о л б у т о в. 1957. Остатки животных на мустьер-ской стоянке под Сталинградом и стратиграфическое положение палеолитического слоя. Тр. Зоол. инет. All СССР, XXII : 75—89.
Верещагин Н. К. и С. И. Мамедов. 1946. Степная мышь (Mus musculus tataricus Sat.) как вредитель риса в Азербайджане. Докл. АН АзербССР, II, 3 : 113—115.
Верещагин Н. К. и В. И. Н а н и е в. 1949. Прежнее и современное распространение копытных в Северной Осетии. Зоол. журн., XXVIII, 3 : 277—280.
Верещагин Н. К., В. Г. Гептнер, А. С. Строганова. 1959. О времени и причинах вымирания кавказского сурка. Научные докл. высш, школы. Биол. науки, 2 : 36—38.
Верещагин Н. К. и О. И. Семенов-Тян-Шанский. 1948. Остатки копытных в святилищах Северной Осетии. Природа, 9 : 70—71.
f
I '
’ Зоологическая литература
59T
: Ветуляни T. 1952. Проблема тарпана на фоне новейших работ Академии наук СССР по истории лошадей в Старом Свете. Зоол. журн., XXXI, 5 : 727—735.
В иноградов А. Ф. 1870. Охота в уреме Малого Зеленчука. Журн. охоты и коннозаводства, 13 : 101—102.
Виноградов Б. С. 1916. К познанию грызунов Закавказья. Зап. Кавк, муз., 1 * А, 3, Тифлис : 1—27.
(Виноградов Б. С.) V i п о g г a d о v В. S. 1926. Some external and osteolo-Ж gical characters of Prometheomys schaposchnikovi Satunin. Proc. Zool. Soc., g* London, 2 : 401—411.
• Виноградов Б. С. 1937a. Тушканчики. Фауна СССР, III, 4, M.—Л. : 1—196.
Виноградов Б. С. 19376. Материалы по изучению четвертичной фауны Крыма. Тр. Сов. секции Междун. ассоц. по изуч. четвертичн. периода, I, Изд. Геол. К уир., Л.—М. : 115—119.
Виноградов Б. С. 1946. К вопросу о морфологической дивергенции близких Ф°РМ млекопитающих. Тр. Зоол. инет. АН СССР, VIII : 89—104.
Виноградов Б. С. 1948. Влияние человека на животный мир пустыни. Живот- ныи мир СССР, II. Зона пустынь. Изд. АН СССР, М.—Л. : 349—358.
Виноградов Б. С. 1950. Млекопитающие — Mammalia. Животный мир СССР, III. Зона степей. Изд. АН СССР, М.—Л. : 26—77.
Виноградов Б. С. и А. И. Аргиропуло. 1941. Определитель грызунов.
у М.—Л. : 1—241.
Виноградов Б. А. и И. М. Г р о м о в. 1952. Грызуны фауны СССР. Изд. АН СССР, М.—Л. : 1—296.
Виноградов Б. С. и С. И. Оболенский. 1926. Грызуны. Тр. по прикладн. энтомол., ХШ, 3 : 177—221.
Витович В. 1924. Охота в Азербайджане. Эконом, вести., Баку, 1 :4—12; 2 : 3—6; 3 : 4—6; 4 : 6—10; 5 : 5—9; 6 : 5—9; 7 : 3—6; 8 : 3—10.
: Воронов А. Г. 1935. Некоторые наблюдения над деятельностью общественной-полевки (Microtus socialis Pall.) на пастбищах предгорного Дагестана. Бюлл. Моск. общ. испыт. природы, отд. биол., XLIX, 6 : 7—8.
Воронов А. Г. 1940. К экологии и географическому распространению Mesocri-cetus Brandti Nehr. Уч. зап. Молотовского гос. пед. инет., VII, 130—152.
Габуния.Л. К. 1951а. Об остатках млекопитающих из третичных отложений Ахалцихского лйгнитоносного района. Сообщ. АН ГрузССР, XII, 3 : 139—141.
Габуния Л. К. 19516. Об остатках халикотерия из третичных отложений Грузии. Сообщ. АН ГрузССР, XII, 5 : 279—284.
Габуния Л. К. 1952а. О зубе «карликового» мамонта из Гори. Сообщ. АН ГрузССР, XIII, 3 : 153—154.
Габуния Л- К. 19526. О новом местонахождении гиппарионовой фауны в Грузии.. Сообщ. АН ГрузССР, ХШ, 5 : 281—283.
Габуния Л. К. 1953. Олигоценовая фауна млекопитающих Грузии. Природа, 4 : 109—110.
Габуния Л. К. 1955а. Об остатках нового вида Dicrocerus из меотических отложений Восточной Грузии. Докл. АН СССР, нов. сер., 100, 2 : 359—360.
Габуния Л. К. 19556. О своеобразном представителе Indricotheriidae из олигоцена Грузии. Докл. АН СССР, XXI, 4 : 177—188.
Габуния Л. К. 1955в. Об остатках нового представителя Bovinae из верхнемиоценовых отложений Восточной Грузии. Сообщ. АН ГрузССР, XVI, 6 : 459—461.
Габуния Л. К. 1955г. Новый представитель Suidae из среднего миоцена Бело-мечетской (Северный Кавказ). Докл. АН СССР, 102, 6 : 1203—1206.
Габуния Л. К. 1956а. О находке остатков ископаемого трубкозуба (Orycteropus sp.) в среднемиоценовых отложениях Беломечетской (Северный Кавказ). Докл. АН АзербССР, XII, 3 : 203—206.
Габуния Л. К. 19566. Следы динозавров в нижнемеловых отложениях Сатаплиа-(Западной Грузии). Тр. Сект, палеобиол. АН ГрузССР, III : 39—87-
Гаджи е в В. Г. 1953. Бинагадинский ископаемый осел Equus cf. hidruntinus Reg. Бинагадинское местонахождение четвертичной фауны и флоры, III. Тр. Естеств.-ист. муз. АН АзербССР, VI : 5—156.
Гаджиев Г. В. и Д. В. Гаджиев. 1952. Признаки рахитического поражения на костях ископаемых бинагадинских волков. Изв. АП АзербССР, 7 : И—20.
Г амбара шви ли Н. Г. 1937. Лагодехский заповедник (фауна). Заповедники и. памятники природы ГрузССР, I, Тифлис : 3—17.
Гамбарян П. П. 1951. К вопросу о систематическом положении горной слепушонки (Ellobius lutescens Thos). Докл. АН АрмССР, XIV, 2 : 45—48.
Гамбарян П. П. 1952. Особенности строения нор прометеевой полевки. Докл.. АН АрмССР, XV, 3 : 85—88.
i 592
Литература
Гамбарян П. П., В. С. Карапетян, К. А. Айрумяи, К. Г. Казарян, С. К. Межлумян. 1957. К экологии прометеевой полевки Prometheomys schaposchnikovi Sat. Мат. по изуч. фауны АрмССР, III, (зоол. сб. X) : 5—16, Гарутт В. Е. 1951. Изменения строения кисти хоботных (Proboscidea) в связи с условиями их обитания. Докл. АН СССР, LXXVII, 3 : 513—515.
Гарутт В. Е. 1957а. О новом ископаемом слоне — Phanagoroloxodon mammontob des gen. et sp. nov. с Кавказа. Докл. АН СССР, 112, 2 : 333—335.
Гарутт В. Е. 19576. Новые данные о древнейших слонах. Род Protelephas gen. nov. Докл. АН СССР, 114, 1 : 189—191.
Гарутт В. Е. 1958. Фанагорийский слон (Phanagoroloxodon mammontoides, gen. n. et sp. n.) и пути филогении в семействе слоновых. Зоол. журн., XXXVII 10 : 1541—1553.
Гентнер В. Г. 1926. Краткий определитель млекопитающих Дагестана. Махачкала : 1—58. ч
Г е пт не р В. Г. 1927. Заметки о песчанках (Gerbillinae), I. О песчанках, обитающих между реками Урал и Терек. Матер, к позн. фауны Нижневолжск. края. Саратов, 1 : 29—37.
(Гентнер В. Г.) Heptner W. G. 1930. Uber die Rassen von Mus musculus im ostlichen Kaukasus (Zis und Transkaukasien). Zool. Anz., 89, 1/2 : 5—92.
(Гептнер В. Г.) Heptner W. G. 1931. Notizen uber die Gerbillinae (Mammalia, Muriuae), III. Zool. Anz., 94, 3/4.
(Гептнер В. Г.) Heptner W. G. 1934a. Notizen uber die Gerbillidae (Mammalia, Glires), VIII. Gerbillidae der Kaukasuslander und der Kalmukensteppe. Folia Zool. et Hydrobiol., VI, 2 : 179—268.
(Гептнер В. Г.) Heptner W. G. 19346. Notizen fiber den sudrussischen Tarpan. Zeitschr. Saugetierk., 9 : 431—433.
Г e n т н e p В. Г. 1935. К познанию географической изменчивости Scirtopoda telum Licht. (Mammalia, Dipodidae). Сб. тр. Гос. зоол. муз. (при МГУ), II : 17—22.
Гептнер В. Г. 1936. Общая зоогеография. Биомедгиз, М. : 1—548.
Гептнер В. Г. 1939. Зоогеографические особенности фауны песчаных пустынь Туркестана и ее происхождение. Бюлл. Моск. общ. испыт. природы, отд. биол., XLVII, 5—6 : 323—342.
Гептнер В. Г. 1940. Фауна песчанок (Mammalia, Glires) Ирана и зоогеографические особенности малоазиатско-ирано-афганских стран. Новые мем. Моск. общ. испыт. природы, XX : 1—54.
Гептнер В. Г. 1945. Пустынно-степная фауна Палеарктики и очаги ее развития. Бюлл, Моск. общ. испыт. природы, отд. биол., 4 (1—2) : 17—38.
Гептнер В. Г. 1948. О систематических взаимоотношениях некоторых африканских форм песчанок (Gerbillidae, Mammalia, Glires) с азиатскими. Докл. АН СССР, LXIII, 3 : 329—332.
Гептнер В. Г. 1955. Заметки о тарпанах. Зоол. журн., XXXIV, 6 : 1404—1423.
Гептнер В. Г. и А. Н. Формозов. 1941. Млекопитающие Дагестана. Сб. тр. Гос. зоол. муз. Моск. гос. унив., VI : 3—74.
Гептнер В. Г. и В. И. Цалкин. 1947. Олени СССР. Матер, к позн. фауны и флоры СССР, нов. сер., Изд. Моск. общ. испыт. природы, 10 (XXV)j 1—176.
Гладкина Т. С. 1952. Биологические основы борьбы с краснохвостой песчанкой (Meriones erythrourus Gray.) в Азербайджане. Тр. Всесоюзн. инет. защ. раст.. 4 : 122—131.
(Г л и ч Ц.) Glitsch С. 1865. Beitrage zur Naturgeschichte der Antilope Saiga Pall. Bull. Soc. Imp. Natur. Moscou, 1 : 207—245.
Греве К. A. 1895. Географическое распространение ныне живущих и ископаемых Hippopotamidae, Suidae. Дневн. Зоол. отд. Общ. любит, естеств. в Москве, II, 3 : 20.
Григор Г. Г. 1926. Находка ископаемых медведей вблизи Краснодара. Тр. Геол, муз., VIII, М. : 31.
Григор Г. Г. 1929. Находки ископаемых форм Hyaena sp., Meles sp., Elephas meridionalis в р-не ст. Кисляковской. Тр. Кубанск. иод. инет., 2иЗ : 193—202.
Грицай Т. Г. 1938. Палеонтолопчш розкопки в карстових вечерах одеських катакомб. BicTi АН УРСР, 4 : 48—52.
Грицай Т. Г. 1939. Работы Одесской палеонтологической экспедиции Академии наук УССР. Природа, 3 : 90—11.
Громов В. И. 1933. Изучение четвертичной фауны Северного Кавказа. Вестн, АН СССР, 4 : 46.
Громов В. И. 1935. Стратиграфическое значение четвертичных млекопитающих Поволжья. Тр. Комисе, по изуч. четвертичн. периода, IV : 309—324.
Громов В. И. 1936. Некоторые итоги полевых исследований в 1935 г. в районах Сухуми, Пятигорска и Ростова. Тр. Сов. секц. Междун. ассоц. по изуч. четвертичн. периода Европы, II : 15—19.
Зоологическая литература
593
s
а
L.
п
н-
Н
3. б.
и
эй •>
11.
dX
X
»л.
.as
12.
ta-
ии
ГН.
их
tax эр-
Гр о м о в В. И. 1937. Новые данные о четвертичных и верхнечетвертичных млекопитающих Северного Кавказа. Тр. Сов. секц. Междун. ассоц. по изуч. четвертины. периода Европы, I : 32—47.
Громов В. И. 1939. Краткий систематический и стратиграфический обзор четвертичных млекопитающих. Сб. «Академику В. А. Обручеву», М., II : 163— ‘ 223.
Громов В. И. 1948. Палеонтологическое и археологическое обоснование стратиграфии континентальных отложений четвертичного периода на территории СССР. (Млекопитающие, палеолит). Тр. Инет. геол, наук, 64, геол, сер., 17 : 1—520.
I э о м о в В. И. 1949. Основные этапы развития четвертичной фауны СССР. Тр. Инет, геогр., 37 : 359—361.
Громов В. И. 1950. Краткий очерк истории четвертичной фауны СССР. Матер, по четвертичн. периоду СССР, 2 : 37—49.
Громов В. И. и Г. Ф. М и р ч и н к. 1936. Четвертичный период и его фауна. Животный мир СССР, т. I. Изд. АН СССР, М.—Л. : 79—123.
Г э о м о в И. М. 1948 Новые данные о прежнем распространении камышового кота на юге Европейской части СССР. Докл. АН СССР, LXII, 6 : 829—832.
Г ) ом о в И. М. 1952. Фауна грызунов (Rodentia) бинагадинского плейстоцена и его природа. Бинагадинское местонахождение четвертичной фауны и флоры, II. Тр. Естеств.-ист. муз. АН АзербССР, V : 203—349.
Громов И. М. 1957. Ископаемые четвертичные грызуны северной части нижнего Дона и прилежащего участка волго-донского водораздела. Тр. Зоол. инет. АН СССР, XXII : 246—318.
Громова В. И. 1927. Материалы к познанию фауны Трипольской культуры. Ежегодн. Зоол. муз. АН СССР : 83—121.
Громова В. И. 1928. Лев в Европе в историческое время. Природа, 10 : 929—930. Громова В. И. 1929. Об остатках млекопитающих каменного века из Закавказья.
Ежегодн. Зоол. муз. АН СССР, XXX, 3 : 427—446.
Г .1 о м о в а В. И. 1931. Первобытный бык или тур (Bos primigenius Boj.) в СССР. Ежегодн. Зоол. муз. АН СССР, XXXII : 293—364.
Сомова В. И. 1932а. Новые материалы по четвертичной фауне Поволжья и по истории млекопитающих восточной Европы и Северной Азии вообще. Тр. Ко-мисс. по изуч. четвертичн. периода, II : 69—184. —-
Громова В.) Gromova V. 19326. Die Fauna eHier mittelpalaeolithischen Station Ilskaja im nordlichen Kaukasus. Тр. Зоол. инет. АН СССР, I : 305—345.
1 р о м о в а В. И. 1935а. Об остатках диких баранов и козлов в четвертичных отложе-‘ ниях Крыма. Докл. АН СССР, IV (XI), 1—2 (70—71) : 97—100.
'ромов а В. И. 19356. Об остатках носорога Мерка (Rhinoceros Mercki Jaeg ) с нижней Волги. Тр. Палезоол. инет., IV : 91—135.
, Громова В. И. 1935в. Первобытный зубр (Bison priscus Bojanus) в СССР. Тр.
| Зоол. инет. АН СССР : 2—204.
£Гром о в а В. И. 1937. Новые находки позднечетвертичной фауны млекопитающих д' на Северном Кавказе. Докл. АН СССР, XVI, 6 : 351—352.
вромова В. И. 1939. Остатки млекопитающих из Ергеней. Уч. зап. Ленингр. гос. унив., 21, 5 : 200—202.
Ж Громова В. 1940. Скульптурные изображения гигантского оленя в доисториче-Ж ское и историческое время. Палеонтол. обозр., 2 : 59—60.
жГро м о в а В. И. 1948. К истории фауны млекопитающих Кавказа. Изв. АН СССР, Ж сер. биол., 5 : 517—538.
«Гром о в а В. И. 1949. История лошадей (рода Equus) в Старом Свете, ч. I. Обзор и описание форм. ч. II. Эволюция и классификация рода. К; II :5—161.
Ж Тр. Палеонтол инет., XVII, I : 1—373.
Громова Во И. 1950а. К истории рода Equus в Восточной Европе и общий очерк истории рода в Старом Свете. Матер, по четвертичн. периоду СССР, 2 : 70—80.
• Громова В. И. 19506. О нижней границе четвертичного периода в свете палеон-В тологии млекопитающих. Матер, по четвертичн. периоду СССР, 2 : 21—29. Жтр° м о в а В. И. и В. И. Громов. 1937. Материалы к изучению палеолитической В фауны Крыма в связи с некоторыми вопросами четвертичной стратиграфии.
Тр. Сов. секции Междун. ассоц. по изуч четвертичн. периода Европы, 1 : 52—94. «Губарев Л. Д. 1941. Распространение серой крысы (Rattus norvegicus Bore.) S в восточных районах Ростовской области. Тр. Росте вс к. н/Дону гос. научно-иссл. п роти во чу мн. ипст. Паркомз драва СССР, II : 133—144.
ТГГули й Г. И. 1930. Систематические признаки курганчиковой мыши (Mus musculus hortulanus Nordm.). Изв. Сев.-Кавк, краев, ст. защ. раст., 5 : 105—128.
(Г ю л ь д е н ш т е д т А. Я.) Guldenstaedt A. J. 1778. Antilope subgutturosa. Acta Acad. Imp. Sci. Petropoli, II, 1 : 251—274.
38 H, К. Верещагин
594
JI wne рату pa
(Г юльденштедт А. Я.) Guldenstaedt A. J. 1783. Beschreibung des-kaukasische Steinbecks aus dessen hinterlassenen Papieren. Neue Nord Beitrag ' IV : 386—392.
Даль G. К. 1940a. Насекомоядные и рукокрылые Армянской ССР и Нахичеванской АССР. Зоол. со. Арм. фил. АН СССР, II : 5—26.
Даль С. К. 19406. К исследованию вымерших и современных животных Сарай-Булагского хребта. Зоол. сб. Арм. фил. АН СССР, II : 27—35.
Даль С. К. 1941. Данные о результатах акклиматизации енотовидных собак в Армянской ССР. Изв. Арм. фил. АН СССР, I (6) : 121—127.
Даль С. К. 1944а. Мелкие пушные звери Памбакского хребта. Зоол. сб. АН АрмССР, III : 47—69.
Даль С. К. 19446. Позвоночные животные Сарай-Булагского хребта. Зоол. сб. АН АрмССР, III : 5—46.
Даль С. К. 1945. О нахождении подковоноса Мех ели (Rhinolophus Mehelyi Matschie) • в Армянской ССР. Докл. АН АрмССР, III, 4 : 97—98.
Д а лъ С. К. 1946. Новые данные о землеройках-белозубках Армянской ССР. Докл. АрмССР, IV, 5 : 143—147.
Д а л ь С. К. 1947а. Новые биогеографические данные об исторических границах лесов в Армянской ССР. Докл. АН АрмССР, VI, 3 : 83—87.
Даль С. К. 19476. Новый подвид ночницы Наттерера из Вайка (Даралагез). Докл. АН АрмССР, VII, 4 : 173^178.
Даль С. К. 1948а. Позвоночные животные Памбакского хребта. Зоол. сб. АН Арм ССР, V : 5—68.
Даль С. К. 19486. Материалы по вертикальному распространению пресмыкающихся, птиц и млекопитающих в долинах рек Занги и Мисханы. Зоол. сб. АН АрмССР, V : 69—86.
Даль С. К. 1949а. Суслик верхнетретичных отложений юга Закавказья. Докл. АН АрмССР, XI, 2 : 67—71.
Даль С. К. 19496. Очерк позвоночных животных Айоцдзорского хребта. Зоол. сб. АН АрмССР, VI : 5—97.
Даль- С. К 1950а. Карликовый тур донных отложений озера Севан. Докл. АН АрмССР, XI, 4 : 133—138.
Даль С. К. 19506. Позвоночные животные прибрежной полосы озера Севан и изменение их группировок в связи со спуском водоема. Зоол. сб. АН АрмССР, VII : 5—53.
Даль С. К. 1951а. К систематике волков Закавказья. Докл. АН АрмССР, XIV, 3 : 87—92.
Даль С. К. 19516. Данные по биологии, распространению и количественному соотношению в стадах безоаровых коз на Урцском хребте. Изв. АН АрмССР, биол. и с.-х. науки, IV, 1 : 33—40.
Даль С. К. 1952. Результаты изучения млекопитающих из раскопок урартского города Тайшебаини. Изв. АН АрмССР, обществ, науки, 1 : 75—86.
Даль С. К. 1954а. Животный мир Армянской ССР, АН АрмССР : 1—415.
Даль С. К. 19546. Динамика видового состава группировок млекопитающих Терско-Кумских песков. Матер, по изуч. Ставропольск. края, 6 *. 193—208.
Даль С. К. 1954в. Палеофауна наземных позвоночных из пещер Урцского-хребта. Изв. АН Арм. СССР, биОл. и. с.-х науки, VII, 2 : 61—71.
Даль G. К. 1957. Закавказская пищуха. Зоол. сб. АрмССР, X : 17—26.
Даль С. К. иХ. А. Захарян. 1951. Обзор численности грызунов, вредителей сельскохозяйственных культур. Изв. АН АрмССР, биол. и с.-х. науки, IV, 8 : 757—765.
Дементьев Г. И. и А. К. Рустамов. 1946. К вопросу о границах распространения пустынной фауны южного типа в Средней Азии. Изв. Турки, фил, АН СССР, 3—4:147—151.
Джафаров Р. Д. 1949. Ископаемые носороги Rhinocerotidae Кавказа. Изп. АН АзербССР, VIII : 51—59.
Джафаров Р. Д. 1950. Материалы к изучению черепа бинагадинского носорога. Изв. АН АзербССР, 8 : 45—52.
Джафаров Р. Д. 1955. Новый представитель четвертичных носорогов (Rhino-ceros binagadensis species nova) из бинагадинской фауны. Б и нага ди некое местонахождение четвертичной фауны и флоры, IV. Тр. Естеств.-ист. муз. АП АзербССР, X : 65—88.
Динник Н. Я. 1882. Кавказский горный козел. Тр. СПб. общ. естествоиспыт., ХШ: 73—91.
(Динник N.) D i n n i k N. 1887. On the Caucasian Mountain Goat (Capra Cauctv ' sica Giild.) Ann. Mag. Nat. Hist., June : 450—461.
Зоологическая литература
595
Динник Н. Я. 1896. Кавказская серна и ее образ жизни. Природа и охота, 2 : 36—59.
Динник II. Я. 1897. Медведь и его образ жизни на Кавказе. Матер, к позн. фауны и флоры Росс, имп., отд. зоол., III : 125—169.
Динник Н. Я. 1901. Млекопитающие горной полосы Кубанской области. Матер. ' к позн. фауны и флоры Росс, имп., отд. зоол., V : 1—30.
Динник Н. Я. 1902. Кавказский олень (Cervus elaphus maral Ogilby). Матер, к позн. фауны и флоры Росс, имп., отд. зоол., VI, М. : 1—42.
Динник Н. Я. 1909. Кавказские каменные козлы или туры. Матер, к позн. фауны и флоры Росс, имп., отд. зоол., IX : 1—47.
Динник Н. Я. 1911. Общие замечания о фауне Кавказа. Тр. Ставропольск. общ. для изуч. Сев.-Кавк, края в естеств.-ист., географ, и антроп. отнош., 1 : 1 —15..
Динник Н. Я. 1914а. Звери Кавказа. Зап. Кавк. отд. Русск. геогр. общ., XXVII, 1 : 247—536.
Динник Н. Я. 19146. Список трудов. Изв. Кавк. отд. Русск. геогр. общ., XXII, 3 : 230—298.
Динник Н. Я. 1916. Бывшее богатство Кавказа дичью и истребление ее в последнее время. Наша охота, XIX : 13—24.
Доброхотов В. И. 1939. К находке камышового кота [Felis (Chaus) chaus chaus Schr. J в дельте Волги. Научно-метод. зап. Комит. по запов., 3 : 192—193.
Долгих И. 1905. Мнимый единорог риму или реэм Востока, ур и тур Европы, Bos primigenius палеонтологии. Рига : I + 85.
Домбровский Б. С. 191X О находке эльдарской фауны млекопитающих. Ежегодн. по геол, и минералХРоссии, XV, 8—9 : 256.
Донауров С. С. 1949. Некоторые данные по биологии лесной куницы в Кавказском заповеднике. Тр. Кавк. гос. запов., III : 5—38.
Донауров С. С., В. К. Попов иЗ. П. Хонякина. 1938. Соня-полчок в районе Кавказского государственного заповедника. Тр. Кавк. гос. запов., 1 : 227—279.
Донауров С. С. и В. II. Т е п л о в. 1938. Кабан в Кавказском заповеднике. Тр. Кавк. гос. запов., 1 : 191—225.
Донауров С. С., В. П. Т е п л о в и П. А. Ш и ш к и н а. 1938. Питание лесной куницы в условиях Кавказского заповедника. Тр. Кавк. гос. запов., 1 : 281—315.
1 у б р о в о И. А. 1957. К родовой систематике подсем. Elephantinae Gray (Слоны).. Vertebrata Palasiatica, I, 3:223—231.
ю к о в Н. Н. 1927. Дагестанский хомяк. Изд. Даг. научно-иссл. инет, и Дагнар-комзема. Махачкала : 1—8.
Егоров Н. М. 1932. Находка бивня южного слона (Elephas meridional is) на горе Машу к. Сов. краевед, на Сев. Кавк. : 57—59.
/. н у к и д з е Г. П. 1951. Материалы к изучению биоэкологии краснохвостой песчанки (Pallasiomys erythrourus Gray) в условиях Самгорской равнины (на груз., яз. с русск. резюме). Тр. Зоол. инет. АН ГрузССР, X : 75—92.
Ефремов И. А. 1950. Тафономия и геологическая летопись, 1. Тр. Палеонтол. инет. АН СССР, XXIV : 1—176.
Жарков И. В. 1938. Экология и значение лесных мышей в лесах Кавказского заповедника. Тр. Кавк. гос. запов., 1 : 153—488.
Жарков И. В. 1940а. Результаты учета животных в Кавказском заповеднике в 1939 году. Научно-метод. зап. Комит. по запов., VII : 138—150.
Жарков И. В. 19406. Зимний учет копытных прогоном в Кавказском заповеднике. Научно-метод. зап. Комит. по запов., VI : 80—87.
Жарков И. В. 1949. Колебания численности мышевидных грызунов в Кавказском заповеднике. Тр. Кавк. гос. запов., III : 65—80.
Лит к о в Б. М. 1930. О разведении нутрии на юге СССР. Пушное дело, 4—5 : 22—24. ЮК и т к о в Б. М. 1936. К терминологии рода Felis. Бюлл. Моск. общ. испыт. природы, * отд. биол., XLV (3) : 226—232.
(Житков Б. М. 1944. Замечания о крысах и некоторых условиях их исслсдова-f ния. Зоол. журн., XXIII, 2—3 : 91—100.
[Заленский Б. 1903. Остеологические и одонтографические исследования над ! мамонтом (Elephas primigenius Blum.) и слонами (Elephas indicus L. и El. i africanus Blum). Научн. результ. эксп., снаряж. Акад, наук, СПб. : 1 —123. |3аленский Б. 1909. Видовые зоологические признаки мамонта. Научи, результ. [с эксп., снаряж. Акад, наук, СПб. : 36—42.
।Зубарева В. И. 1940. Фауна Киева 1000 лет назад. Природа, 8 : 82—86.
(Иванов А. 11. 1916. Фауна позвоночных в верхнесарматских отложениях Став-Ik ропольской губ. Изв. Акад, наук, физ.-матем. отд. : 195—198.
В Иванов В. В. 1952. Сайгаки в Прикаспийских степях. Природа, 8 : 120—121.
| 38*
596
Литература
Иванова И. К. 1948. Остатки Elephas cf. antiquus Falc. из травертинов горы Машук в окрестностях Пятигорска. Тр. Моск, геол.-развед. инет., XXIII : 126—138.
Илларионов В. Т. 1940.. Мамонт. К истории его изучения в СССР. Горький : 1—96.
И о ф ф И. 1936. О географическом распространении сусликовых блох в связи с историей расселения сусликов. Паразит, сб. Зоол. инет. АН СССР, VI : 313—361.
Казнаков А. Н. 1908. Несколько наблюдений над образом жизни Spalax mic-ropthalmus Giild. и Prometheomys schaposchnikowi Sat. Изв. Кавк, муз., IV, 1—2 : 142—149.
Калабухов Н. И. 1939. О нахождении мышевидного хомячка в Закавказье. Природа, 12 : 85.
Калабухов Н. И. 1940. Некоторые особенности проявления адаптивных признаков у близких видов грызунов. Уч. зап. Ленингр. гос. унив., 59 : 80—101.
Калабухов Н. И. и В. В. Раевский. 1935. Материалы по динамике фауны грызунов в Предкавказских степях. Сб. «Борьба с грызунами в степях Предкавказья», Азово-Черном, краев, книгоизд., Ростов н/Дону : 202—230.
Калан тарян М. А. 1952. Новые данные о дикобразе в Армении (вредитель с.-х. культур). Изв. АН АрмССР, биол. и с.-х. науки, 4 : 103—106.
Калиновский. 1900. Очерк кавказской фауны и кавказских охот. Тифлис : 1—106.
Кар а к а ш Н. И. 1898. О новых находках остатков мамонта па Малом Кавказе. Дневн. X съезда русск. естествоиспыт. и врачей в Киеве : 241.
Карпинский А. П., Дрейер. 1904. Об открытии и раскопках остатков мамонта в Ессентуках. Изв. Акад, наук, XXI : 7—9.
Кашкаров Д. Н. м. Е. Н. Коровин. 1931. Опыт анализа экологических путей расселения фАоры и фауны Средней Азии. Экология и биоценология, 1 : 28—97.
Кащенко Н. Ф. 1912. Крысы и заместители их в Западной Сибири и Туркестане. Ежегодн. Зоол. муз. Акад, наук, XVII, 3—4 : 370—389.
Келлер К. 1913—1914. К зоогеографии Кавказа и истории его домашних животных. Изв. Кавк. отд. Русск. геогр. общ., XXII, 2 : 366—384.
(К е п п е н Ф. П.) К б р р е n Fr. Th. 1883. Die Verbreitung des Elenthere 1ш europaischen Russland, mit besonderer Berucksichtigung einer in den funfziger Jaliren begonnenen Messenwanderung desselbe. Beitr. z. Kenntniss d. Russ. Reich., 6 : 141—243.
Кеппен Ф. П. 1896. К истории тарпана в России. Журн. Министерства нар. проев., январь : 96—171.
Кеппен Ф. П. 1902. О прежнем и нынешнем распространении бобра в пределах
России. СПб. : 81 — 92.^
Кири ков С. В. 1949. Образ жизни и начальные этапы дивергенции высших позвоночных в связи с эколого-географическими факторами. Проб л. физ. геогр., XIV : 115—125.
Кириков С. В. 1952. Исторические изменения животного мира нашей страны в ХШ—XIX вв. Изв. АН СССР, сер. геогр., 6 : 31—48.
Кириченко А. Н. 1910. Отчет по командировке на Талыш в 1909 году для собирания коллекций для Зоологического музея Императорской Академии наук. Ежегодн. Зоол. муз. Акад, наук, XV, 3 : 139—152.
Кирпичников А. А. 1953. Каспийский тюлень из нижнечетвертичных отложений побережья Азовского моря. Докл. АН СССР, ХС, 5 : 883—886.
Кистяков с к и й А. В. 1935. Материалы по распространению грызунов в степях Предкавказья. Сб. «Борьба с грызунами в степях Предкавказья». Азовочерном. краев, книгоизд., Ростов н/Дону : 188—196.
Ковдышев В. В. 1935. К вопросу о массовых размножениях полевок (М. arva-lis Pall, и М. socialis Pall.) в Азербайджане. Тр. Азерб. микробиол. инет,, V, 1.
Кожевников Г. А. и В. II. У в а р о в. 1916. К. А. Сатунин. Некролог п перечень печатных работ и новых форм, описанных покойным Сатуниным (с портретом). Тифлис : 1—31.
Колесник Н. Н. 1936. Происхождение и географическое распространение крупного рогатого скота. Изв. АН СССР, сер. биол., 2—3 : 375—417.
Колесников И. М. 1950. Случаи гибели сайгаков в прикаспийской низменности зимою 1949/50 гг. Природа, 10 : 64—65.
Колесников И. М. 1959. Проникновение сайгаков в Сухапско-Терскую низменность. Зоол. журн., XXXVIII, 1 : 137—140.
Колосов А. М. 1941. Биология размножения зайца-русака (Lepus europaeus ' Pall.). Зоол. журн., XX, 1 : 154—172.
Зоологическая литература
№7
Колосов А. М. и Н. Н. Бакеев. 1947. Биология зайца-русака. Изд. Моск, общ. испыт. природы, М. :1—103.
. [Кондратьев В.] 1885, 1886. Сведения о зверях, птицах, земноводных, рыбах, насекомых и червях, водящихся в области Войска Донского, составленное учителем естественной истории и физики Гимназии сего Войска Василием Конд-
1 ратьевым и расположение по отделениям, к коим систематически каждое из описываемых здесь животных принадлежит. Газ. «Казачий вестник», Новочер-* касск : 44—95; 37—47.
Кондрашкин Г. А. 1949. О серых крысах (Rattus norvegicus Berk.) дельты Волги. Бюлл. Моск. общ. испыт. природы, отд. биол., IV, 1 : 29—33.
Корнеев А. П. и И. В. Марисов а. 1950. Новая находка в СССР широкоухого складчатогуба (Tadarida teniotis Rafin.). Тр. Зоол. муз. Кшвськ. держ. утв., II : 159—160.
[ К о х и я С. С. 1952. Материалы к изучению размножения картлийской полевки | (Microtus socialis goriensis Arg.). Сообщ. АН ГрузССР, XIII, 5 : 307—312.
I Красовский Д. Б. 1930. Материалы к познанию фауны млекопитающих г Горной Ингушии. Изв. Ингушек, научно-иссл. инет, краевед., II—III, : 66— f 92.
I- К р а с о в с к и й Д. В. 1932. Материалы к познанию фауны наземных позвоночных - Ру тульского кантона Дагестанской АССР. Изв. Сев.-Кавк. пед. инет., IX, I 2 : 185—216.
j Кузнецов Б. А. 1932. Роль пушного промысла в деле борьбы с грызунами-I? вредителями. Бюлл. VII Всесоюзн. съезда по защит, раст., 2 : 12—14.
Кузнецов Б. А. 1936. О некоторых закономерностях распространения млеко-О питающих по Европейской части СССР. Зоол. журн., XV, 1 : 165—183.
[Кузнецов Б. А. 1941. Географическая изменчивость соболей и куниц фауны Б СССР. Тр. Моск, зоотехн. ипст., 1 : 113 133.
Кузнецов Б. А. 1949. Опыт зоогеографического районирования Кавказа и
Закавказья. Тр. Моск, пушно-мехювого инет., II : 109—143.
Кузнецов Б. А. 1950. Очерк зоогеографического районирования СССР. М. :
Г . 1—175. V
[Кузнецов И. Я. 1935. Борьба с мышами в Азово-Черноморском и Северокав- казском краях в 1933—1934 гг. Сб. «Борьба с грызунами в степях Предкав-
казья», Азово-Черноморск. краевое книгоизд., Ростов н/Дону : 7—33.
Кузякин А. П. 1950. Летучие мыши. Изд. «Советская наука», М. : 1—443.
Кузякин А. II. 1951. История расселения, современное распространение и Й места обитания пасюка в СССР. Матер, к позн. фауны и флоры СССР, Изд.
Моск. общ. испыт. природы, нов. сер., отд. зоол., 22 : 22—81.
[Кулагин И. М. 1918. К истории распространения зубров в России. Изв. АН I СССР, сер. VI—XII, 15 : 1649—1658.
Кулагин Н. М. 1921. К истории фауны Европейской России. Природа, 7—9 : |. 39—58.
[Кулагин II. М. 1932. Лоси СССР. Тр. Лабор. прикл. зоол., 2 : 1—120.
Куликов М. 1939. Уссурийские еноты в Краснодарском крае. Сов. охотник, 10 : 47.
Куражковский Ю. Н. 1949. Об адаптации диких копытных к изменениям ландшафта, созданным деятельностью человека. Зоол. журн., XXVIII, 2 : 188—190.
Курапова М. В. и П. Н. Степанов. 1940. Акклиматизация алтайских : белок (Sciurus vulgaris altaicus Ser.) в Тебердинском заповеднике. Научно-
[: метод, зап. Главк, упр. по заповедникам, зоопаркам и зоосадам: 104—114.
Курапова М. В. и П. Н. Степанов. 1941. Акклиматизация белок на Кав- казе. Советск, охотник, 6 : 33-
»К у т е п о в Н. 1896. Великокняжеская и царская охота на Руси с X—XVI вв., В 1, СПб. : 1—211.
Лавров Н. П. 1946. Акклиматизация и реакклиматизация пушных зверей ж в СССР. Заготиздат, М : 1—219.
Лавровский А. А. 1950. О влиянии летней засухи и суровой зимы 1949—
Е' 1950 годов на численность и размещение сайгаков. Охрана природы, 10 : 83—
87‘
В Ласка рев В. Л. 1911. Заметка о новых местонахождениях ископаемых млеко* Е питающих в третичных отложениях южной России. Зап. Новоросс. общ. есте-I ствоиспыт., XXXVIII : 393—405.
Лебедев Ф. Н. 1912. О борьбе с сусликами при помощи сернистого углерода в Самарской, Ставропольской, Астраханской и Саратовской губерниях в 1909—1911 гг. Деп. земл., СПб. : 3—56.
.$98 Литература
Лебедев Ф. Н. 1925. Мугоджарский суслик. Биологические наблюдения. Вести, микробиол. и эпидемиол., TV, 3 : 25—32.
Лебедева Л. G. 1956. О составе летних кормов кабана в высокогорной зоне Кавказского заповедника. .Уч. зап. Моск. гор. пед. инет., 61 : 273—278.
Леваневский М. 1894. Очерки Киргизских степей Эмбенского уезда. Землеве-» дение, кн. IV: 121—123.
Левченко М. 1882. Исчезнувшие и исчезающие в южной России животные. Киевская старина, III : 373—379.
Логинов В. В. 1936. Новые данные к познанию биологии кавказского кабана (Sus scrofa attila Thomas). Бюлл. Моск. общ. испыт. природы, XLV (1) : 10—21.
Логинов В. В. 1949. Очерк экологии кавказского крота на западном Кавказе. Тр. Кавк. гос. запов., III : 81—130.
Львов Г. Н. 1949. Колхидский пасюк. Уч. зап. Горьковск. гос. унив., 14 : 39— 51.
Лютый А. М. 1940. К вопросу о сельскохозяйственном значении филина. Тр. Ворошиловск. гос. пед. инет., II : 223—224.
Л я й стер А. Ф. 1916. К. А. Сатунин (некролог). Список трудов К. А. Сатунина. Изв; Кавк. отд. Русск. геогр. общ., XXIII, 3 : 328—343.
Ляйстер А. Ф. 1931. К вопросу об иранском влиянии на фауну южного Закавказья. Закавк. краеведческ. сб., 1 : 76—92.
Максимов А. А. 1948. Об исторических изменениях в распространении серой полевки (Microtus arvalis Pall.). Докл. АН СССР, LX, 4 : 677—680.
f Максимов А. А. 1950. Об исторических факторах в развитии туляремии и ту-? . ляремийных очагов. Журн. микробиол. эпидемиол. и иммунол., 6 : 44—46. : Мамедов С. И. 1950. Некоторые наблюдения над миграцией и гибелью полевок >> (Microtus arvalis Pall.) в юго-восточной Мугани. Изв. АН АзербССР, 1 : 49—
4 55 (на азерб. яз. с русск. резюме).
[ Мантейфель И. А. 1934. Реконструкция фауны СССР и одомашнивание новых L . видов животных. Фронт науки и техники, 9, : 80—90.
f Марков Е. Л. 1931а. Дикиехсвиньи Закавказья. Тифлис.
. Марков Е. Л. 19316. Об акклиматизации в Закавказье уссурийского пятнистого оленя. Природа и социалистич. хоз., 6—8 : 121—123.
Марков Е. Л. 1935. Охотничье хозяйство Закавказья. Тифлис : 1—221.
Марков Е. Л. 1937. Боржомский охотничий заповедник. Запов. и памятники природы ГрузССР, 1, Тифлис : 23—39.
Марков Е. Л. 1938. Охотничье-промысловые животные Лагодехского заповедника. Тбилиси : 1—67.
Марков Е. Л. 1940. Акклиматизация пушных зверей в Грузии. Советск, охотник, 7 : 33—34.
Марков Е. Л. 1951. По вопросу разведения северной белки в лесах Армянской ССР. Изв. АН АрмССР, IV, 4 : 387—388.
Марков Е. Л. и Л. Л. М л о к о с е в и ч. 1935. Закатальский заповедник. Изд. Закавк. фил. АН СССР, Баку : 1—65.
Мартынов А. В. 1934. Роль различных экологических категорий для оценки фактов зоогеографии и для истории фауны. Тр. Первого Бсесоюзн. геогр. съезда, 11—18 апреля 1933 г., 3 : 309.
Маруашвили Л. И. 1955. Оленьи рога хевсурских и тушинских святилищ. Природа, 1 : 117—118.
Маруашвили Л. И. 1956. Целесообразность пересмотра существующих представлений о палеогеографических условиях ледникового времени на Кавказе. Изд. АН ГрузССР : 3—129.
(Мензбир М. А.) М е n z b i е г М. А. 1887. On a new Kaukasian Goat (Capra Severtzowi sp. nova). Proc. Zool. Soc. London, dec. : 618—620.
Мензбир M. A. 1934. Очерк истории фауны Европейской части СССР. М.—Л. : 1—223.
(Менетрие Е.) Menetries Е. 1832. Catalogue raisonne des objets de zool ogie recuellis dans un voyage au Caucase et jusqu’aux frontieres actuelles de la Perse, entrepris par ordre de sa majeste Pempereur. СПб. : I—XXXIX-j-1—271.
Минин H. B. 1938. Эколого-географический очерк грызунов Средней Азии. Изд. Ленингр. гос. унив. : 1—184.
Миронов Н. II. и А. А. Лисицын. 1953. К вопросу экологии и происхождения песчанок северо-западного Прикаспия. Сб. научных работ Приволжской противоэнид. станции, 1 : 110—130.
Миронов ы. П., А. Н. Павлов, Ф. А. П у ш п и ц а и П. И. III и рано в и ч. 1952. Изменение границы ареала малого суслика в Донских и Ставропольских степях. Зоол. журн., XXXI, 5 : 752—760.
Зоологическая литература
599
Моргилевская И. Е. 1949. Морфологическая характеристика лагодехской популяции лесной мыши. Тр. Зоол. инет. АН ГрузССР, VIII : 225—237.
.М о. р г и л е в с к а я И. Е. 1954. Морфологическое описание четырех популяций лесной мыши в Грузии. Тр. Инет. зоол. АН ГрузССР, XIII : 71—81.
Моргилевская И. Е., 3. К. Цителова и А. А. Минасян. 1947. । Грызуны города Тбилиси (фаунистический очерк). Тр. Зоол. инет. АН ГрузССР, VII : 205—220.
Морозов а-Т у р о в а Л. Г. 1939. К систематике млекопитающих Кавказа. Бюлл. Моск. общ. испыт. природы, XLVIII (2—3) : 98—106.
Му л л а е в И. 1929. Elephas trogontheri Pohlig из постплиоцена Терской области. Азерб. нефт. хоз., 4 (88) : 100.
Навозова Ф. В. 1951. Палеонтологические находки в Краснодарском крае. Изв. АН АзербССР, 2 : 67—73.
Насим.ович А. А. 1935. К биологии снежной полевки (Chionomys nivalis ne-njukovi Formos.) на западном Кавказе. Бюлл. Моск. общ. испыт. природы, отд. биол., нов. сер., XLIV, VII—VIII : 386—390.
Басимович А. А. 1936а. Динамика запасов благородного оленя в Кавказском заповеднике. Тр. показат. Кавк. гос. запов., 1 : 3—33.
,Н а с и м о в и ч А. А. 19366. Учет туров в Кавказском заповеднике в июне 1935 года. Бюлл. Кавк. гос. запов., 2 : 18—21.
Басимович А. А. 1936в. О некоторых закономерностях зимнего распространения копытных в горах Западного Кавказа. Бюлл. Моск. общ. испыт. природы, XLV (Г) : 3—9.
Басимович А. А. 1938а. Влияние лавин на растительный и животный мир Кавказского заповедника. Природа, 7—8 : 191.
Касимович А. А. 19386. К познанию минерального питания диких животных Кавказского заповедника. Тр. Кавк. гос. запов., 1 : 103—150.
Касимович А. А. 1939. Зима в жизни копытных Западного Кавказа. Вопросы экол. и биоценол., 7 : 3—91.
Касимович А. А. 1940а. Летние корма благородного оленя на Западном Кавказе. Научно-метод. зап. Комит. по запов. \ VI : 149—150.
Басимович А. А. 19406. К методике количественного учета поголовья туров. Научно-метод. зап. Комит. по запов., VII : 23—28.
Касимович А. А. 1941. К методике количественного учета благородного оленя в Кавказском заповеднике. Научно-метод. зап. Комит. по запов., VIII : 173— 177.
Касимович А. А. 1949а. Очерк экологии западнокавказского тура. Тр. Кавк, гос. запов., III : 5—38.
Касимович А. А. 19496. Новые данные по биологии серны на Западном Кавказе. Тр. Кавк. гос. запов., III : 51—64.
Басимович А. А. 1950. Об организации заповедника высокогорной природы Центрального Кавказа. Охрана природы, 11 : 72—79.
Насонов Н. В. 1923. Географическое распространение диких баранов Старого Света. Игр. : 1—254.
Наумов Н.П. 1937. К вопросу о стационарном распределении мышевидных грызунов (обыкновенной полевки, общественной полевки, степной пеструшки, курганчиковой мыши). Уч. зап. Моск. гос. унив., 13 : 3—38.
Наумов Н.П. 1945. Географическая изменчивость динамики численности животных и эволюция. Журн. общей биол., VI, 1 : 37—52.
Наумов Н.П. 1946. О биологических основах планирования борьбы с мышевидными грызунами. Зоол. журн. XXV, 1 : 81—86.
Наумов Н II. 1948. Очерки сравнительной экологии мышевидных грызунов. Изд. АН СССР, М.—Л. : 1—190.
Наумов Н. П., М. М. ДукельскаяиВ. В. Домбровский. 1951. Новые методы борьбы с обыкновенными полевками. Зоол. журн., XXX, 5 : 466—476.
Никольский А. М. 1891. Позвоночные животные Крыма. Прилож. к LXVIII тому Зап. Акад, наук, СПб. : V—XIX, 1—484,
Никольский А. М. 1913. Пресмыкающиеся и земноводные Кавказа. Изд. Кавк, музея, Тифлис : 1—272.
Никольский А. М. 1947. Роль ледникового периода в истории фауны палеарктической области. Бюлл. Моск. общ. испыт. природы, отд. биол., LII (5) : 3—14.
Никольский Г. В. 1947. О биологической специфике фаунистических комплексов и значении ее анализа для зоогеографии. Зоол. журн., XXVI, 3.
Новиков К.Л. 1935. Систематические особенности хомяка (Cricetus cricetus L.). Бюлл. Моск. общ. испыт. природы, отд. би о л., XLIV : 302—313.
600
Л и те ратура
(Нор д ма н A.) N о г d m а п А. 1840. Voyage dans la Russie Meridionale et la Crimee, par la Hongrie, la Valachie et la Moldavie, execute en 1837, sous la direction de M. Anatole de Demidoff, III. Paris : 1—756.
(H о p д м а н A.) N ord m an A. 1858a. Palaeontologie Suedrusslands, I. Ursus spelaeus (odessanus). Helsingfors : 1—110.
(H ордман A.) N ordman A. 18586. Palaeontologie Suedrusslands, II. Felis, Hyeana, Canis, Thalassictis, Mustela, Lutra, Spermophilus, Arvicola, Spalax, Castor, Lepus und Equus. Helsingfors : 15—190.
(H op дма н A.) N о r d m a n A. 1859. Palaeontologie Suedrusslands, III. Bos, Antilope, Alces, Cervus, Capreolus, Rangifer, Palaecmeryx, Camelus, Sus, Elasmotherium und Rhinoceros. Helsingfors : 25—270.
(H ордман A.) N о rd m a n A. 1860. Palaeontologie Suedrusslands, IV. Elephas, Mastodon, Dinotherium, Phoca, Manatus, Cetotherium, Balaena, Balaenop-tera und Delphinus. Helsingfors : 35—360.
О б ер май ер Г. 1913. Доисторический человек. Перевод П. Ю. Шмидта. Изд. Брокгауз—Ефрон, СПб : 1—686.
Оболенский С. И. 1927. Грызуны правого берега Нижней Волги. Матер» к позн. фауны Нижн. Поволжья, 1, Саратов : 1—25.
Оболенский С. И. 1935. Фауна грызунов степей Предкавказья. Сб. «Борьба с грызунами в степях Предкавказья». Азово-Черном, краев, книгоизд. Ростов н/ Дону : 172—188.
Оболенский С. И. 1950. Влияние травопольной системы земледелия на фауну.. Животный мир СССР, 3. Изд. АН СССР, М.—Л. : 567—585.
Огнев С. И. 1924. Грызуны Северного Кавказа. Ростов н/Дону : 1—64.
Огнев С. И. 1926а. Материалы к познанию млекопитающих Кавказа. Уч. зап. Сев.-Кавк. инет, краев., 1, Владикавказ : 31—56.
(Огнев С. И. ) О g n е v S. I. 19266. Prometheomys, a remarkable rodent from the Caucasus. Journ. Mammol., 7, 3 : 215—220.
Огнев С. И. 1928. Звери Восточной Европы и Северной Азии. Насекомоядные и летучие мыши, I. Гос. изд., М. — Л. : 1—631.
Огнев С. И. 1931. Звери Восточной Европы и Северной Азии. Хищные млекопитающие, II, Гос. изд. М.—Л. : 1—776.
(Огнев С. И.) О g n е v S. I. 1933. Materialien zur Systematik und Geoographie der russischen Wasserratten (Arvicola). Zeitschr. fur Saugetierkunde, B. 8, H. 3 : 156—179.
Огнев С. И. 1935. Звери СССР и прилежащих стран (звери Восточной Европы и Северной Азии). Хищные и ластоногие, III. Гос. изд. биол. и мед. лит., М.— Л. : 1—752.
Огнев С. И. 1940. Звери СССР и прилежащих стран (звери Восточной Европы и Северной Азии). Грызуны, IV. Изд. АН СССР, М.—Л. : 1—615.
Огнев С. И. 1947. Звери СССР и прилежащих стран (звери Восточной Европы и Северной Азии). Грызуны (продолжение), V. Изд. АН СССР, М.—Л. : 1—809.
Огнев С. И. 1948. Звери СССР и прилежащих стран (звери Восточной Европы и Северной Азии). Грызуны (продолжение), VI. Изд. АН СССР, М.—Л. : 1—559.
Огнев С. И. 1950. Звери СССР и прилежащих стран (звери Восточной Европы и Северной Азии). Грызуны (продолжение), VII. Изд. АН СССР, М.—Л. : 1— 706.
(Огнев С. И. и В. Г. Гептнер) OgnevS. I. and W. G. Heptner. 1927. A new subspecies of Hamster from Daghestan (Mesocricetus raddei Nehr.), and some Remarks on the Russian species of Mesocricetus (with a Key to them). Ann. Mag. Mat. Hist., ser. 9, 19 : 141—150-
(Огнев G. И. и A. H. Формозов) Ognev S. I. and A. N. F or m osov. 1927. A new form of Watervole from Daghestan (East Caucasus). Ann. Mag. Nat. Hist., London, ser. 9, 19 : 138—141.
Орлов Е.И. и Б. С. Феню к. 1927. Материалы к познанию фауны позвоночных приморской части Калмыцкой области. Матер, к позн. фауны Нижн. Поволжья, 1, Саратов : 41—87.
Орлов Ю. А. 1933. Ископаемые верблюды Старого Света. Пробл. происхожд. домапш. животн., 1, Изд. АН СССР, М.—Л. : 211—217.
Орлов Ю. А. 1938а. Новые местонахождения третичных млекопитающих на Кавказе. Природа, 1 : 125.
Орлов IO. А. 19386. Местонахождение фауны гиппариона в Джапаридзе (Кахетия). Тр. Палеонтол. инет. АН СССР, VIII, 3 : 99—101.
Осборн Г. Ф. 1924. Человек древнего каменного века. Л. : 1—527.
Павлов А. П. 1922. Ледниковые и межледниковые эпохи Европы в связи с исто-х рией ископаемого человека. Отч. Акад, наук за 1921 г., Игр.
Зоологическая литература
ш
Павлова М. 1895. О русских мастодонтах и их отношении к мастодонтам других стран. Bull. Soc. Imp. Nat. Moscou, VIII : 146—154.
(Павлова M.) Pavlova M. 1908. Les elephants fossiles de la Russie. Nouv, mem. Soc. Natur. Moscou, XVII, .1—2 : 57.
Павлова M. 1910a. Послетретичные слоны из разных местностей России. Ежегодн. » по геол, и минерал. России, XI, 6—7 : 167—174.
Павлова М. В. 19106. Каталог коллекций геологического кабинета император* ского Московского университета. Изд. Моск, унив., 1 : 1—175.
Павлова М. В. 1916. Находки Elasmotherium sibiricum в Ставропольской губ. и в других местах России. Зап. Геол. отд. Общ. любит, естеств., ан троп о л*, и этногр., V : 67—80.
Павлова М. В. 1922. Очерк исторического развития изучения третичных и послетретичных ископаемых млекопитающих, найденных в России. Бюлл. Моск. общ. испыт. природы, сер. геол., XXXI : 117—148.
Павлова М. В. 1925. Ископаемые млекопитающие из Тираспольского гравия Херсонской губ. Мемуары Геол. отд. Общ. любит, естеств., антропол. и этногр., 3 : 1—74.
Павлова М. В. 1926. Cervus tchelekensis n. sp. et Alces maeoticus n. sp. Ежегодн. Русск. палеонтол. общ., IV : 45—71.
(Павлова M.) Pavlova М. 1931. Les elephants fossiles du sud de TUSSR. 36. намят! акад. П. А. Тутковського, 2, Кшв, АН УРСР : 67—85.
Павлова М. 1933а. Camelopardalis parva Weith. из сарматских отложений Кавказа. Тр. Палеонтол. инет., II : 269—273.
Павлова М. 19336. Фауна ископаемых млекопитающих, собранных в б. Саратовской губ. близ г. Хвалынска на берегу р. Волги. Тр. Комисе, по изуч. четвертичн. периода, III, 1 : 167—172.
(П а л л а с И. С.) Pallas Р. S. 1779. Capra caucasica es schedisCcl. A. I. Giil-denstaedt redigente. Acta Acad. Imp. Sci. Petropolis, III, 2 : 273—281.
(П а л л а с П. C.) Pallas P. S. 1831. Zoographia rosso-asiatica, sustems omnium animalium in extenso. Imperio Rossico, Petropoli, I : 1—568.
Пи до и л и ч к о И. Г. 1936. К вопросу о палеонтологических находках в Приазовье. Природа, 12 : 117—120.
Пидопличко И. Г. 1937. Шдсумки дослщження погадок за 1924—1935 рр. Тр. 1нст. зоол., б!ол. АН УРСР, XV-Зб. праць Зоол. муз., 19, Ки1в : 101—170.
Пидопличко И. Г. 1938а. Ельдарская палеонтологическая экспедиция Музея Грузии и АН УССР. Вестн. АН УССР, 8—9 : 44—48.
Пидопличко И. Г. 19386. Матер1али до вивчення минулих фаун УРСР, 1 : Вид. АН УРСР, Ки1в : 3—176.
Пидопличко И. Г. 1938в. Фауна Ольвии. Природа, И—12 : 113—116.
Пидопличко И. Г. 1940а. Краткий обзор фауны палеолита У ССР.Сов. археол.,. 5 : 151—158.
Пидопличко И. Г. 19406. Основные черты фауны и геологии палеолита УССР. Бюлл. Комисе, по изуч. четвертичн. периода, 6—7 : 61—62.
Пидопличко И. Г. 1940в. Граница четвертичного оледенения Европейской части СССР в палеонтологическом освещении. Докл. АН СССР, XVI, 3 : 262— 265.
Пидопличко И. Г. 1946. Очерк четвертичной палеогеографии Украины. Тр. Инет, геогр. АН СССР, 37 : 187—194.
Пидопличко И. Г. 1950. История фауны степей. Животный мир СССР, III. Зона степей. М.—Л. *. 492—526.
Пидопличко И. Г. 1951. О ледниковом периоде, 2. Изд. АН УкрССР. Киев : 1—262.
Пидопличко И. Г. 1953. Амвросиевская палеолитическая стоянка и ее особенности. Кратк. сообщ. Инет, археол., 2 : 65—68.
Пидопличко И. Г. 1954. О ледниковом периоде, 3. Изд. АН УССР, Киев : 1—217.
Пидопличко И. Г., В. А. Т о п а ч е в с к и й. 1953. Лось и северный олень-в доисторическом прошлом. Природа, 7 : 101—103.
Пирковский Г. Я. 1913. Результаты обследования губернии в отношении зараженности ее сусликами. Отчет о деятельн. Ставропольск. энтомол. бюро за 1913 г. : 61—73.
Погосян А. Р. 1945. К экологии степной мыши Mus musculus tataricus Sat. в Архмении. Докл. АН АрмССР, II, 4 : 123—127.
Погосян А. Р. 1948. Об экологии обыкновенной полевки Microtus arvalis Pall, в условиях посевов зерновых культур Армянской ССР. Зоол. сб. АН АрмССР, V: 87—115.
4S02
Литеparnypa
Погосян А. Р. 1949. Экология и биология песчанок в Армянской ССР. Зоол. сб. АН АрмССР, VI : 99—125.
Л одъяпольский Г. Н 1949. Животный мир Кабарды. Нальчик : 1—72.
Лодъяпольский Г. Н. 1950. Звери, птицы Кабардинской АССР и проблема их охраны. Сб. «Охрана природа», II : 67—72.
Поляков И. Я. 1950. Травопольная система земледелия и вредные мышевидные грызуны. Журн. Общ. биол. XI, 1 : 40—52.
Правила палеозоологической номенклатуры. 1932. М.—Л. : 1—33.
Прохоров М. Г. 1932. Работы палеонтологической экспедиции на Северном Кавказе. Природа, 8 : 756—757-
Пузанов И. И. 1929. Дикая кошка в дилювиальной фауне Крыма. Тр. Крымск, научно-испыт. инет., 4 : 107—110.
Пузанов И. И. 1938а. Зоогеография. Учпедгиз. М.—Л. : 1—360.
Пузанов И. И. 19386. Орнитофауна северо-западной Черкессии и некоторые соображения о ее происхождении и связях. Тр. Зоол. сект. Груз. фил. АН СССР, II : 125—180.
Пузанов И. И. 1949. Своеобразие фауны Крыма и ее происхождение. Уч. зап. Горьконек, гос. унив., 14 : 5—32.
(П фи цен ма и р Е. В.) Pfizenmayr Е. W. 1920. В astardi erungen von Cavicorniem in Transkaukasien. Sitz. Ber. Ges. natf. Freunde : 154—167. 3 pls.
Пхакадзе В. A. 1940a. Некоторые данные по систематике и биологии слепца в условиях Грузии (предв. сообщ.). Тр. Груз. с.-х. инет., XII : 113—118.
(Радде Г. И.) Radde G. I. 1866. Berichte fiber die biologischgeographischen Untersuchugen in den Kaukasuslander. Tiflis : 1—295.
(Радде Г. И.) Radde G. I. 1886. Die Fauna und Flora des Slid westlichen Caspi-Gebietes. Leipzig : 1—425.
(Радде Г. И.) Radde G. I. 1893. On the present Range of the European Bison in the Caucasus. Proc. Zool. Soc. London, 1 .* 175—177.
(Радде Г. И.) Radde G. I. 1895. Zum andenken an Maximilian Noska. Zwei monographische studien. I, Capra (Aeguceros) caucasica Giild. II, Capella rupi-capra Keys et Blas. Dresden : 9—42.
Радде Г. И. 1899. Коллекции Кавказского музея, 1. Зоология:!—520.
Радищев А. М. 1926. Материалы к познанию фауны грызунов Кабардино-Балкарской автономной области. Изв. Сев.-Кавк, краев, ст. защ. раст., Ростов н/Дону 2 : 83—89.
Радугин В. И. 1917а. Определитель млекопитающих Кавказского края. Зап. Кавк, муз., сер. А, 4, Тифлис : 1—56.
Радугин В. И. 19176. Из наблюдений над природою Котурско-Урмийского района в Северо-западной Персии. Изв. Кавк. отд. Русск. геогр. общ., XXV, 2—3: 284—300.
Резник П. А. 1951. Кавказская мышовка в горных районах Ставропольского края. Природа, 10 : 68.
Реутт Н. 1841. Джикви, или закавказский тур. Коннозаводство и охота, II: 724—797.
Родендорф Б. Б. 1939. О миоценовой фауне насекомых окрестностей г. Воро-шиловска. Природа, 12 : 85—88.
Родионов 3. С. 1924. Биология общественной полевки и опыты борьбы с нею в Закавказье. Отд. защ. раст, от вредит. НКЗ АзербССР, Л. : 1—191.
Романова В. К. 1936. Распространение сусликов в степях Предкавказья.
В кн.: Итоги научно-иссл. работ. Всесоюзн. инет. защ. раст. за 1935 г. Л.: 43—45.
Россиков К. Н. 1887. Обзор млекопитающих животных долины р. Малки. Зап. Акад, наук, физ.-матем. отд., LIV, СПб. : 1—97.
Рубцов И. А. и Б. С. Виноградов. 1950. Влияние человека на животный мир степной зоны. Животный мир СССР, III. Изд. АН СССР, М.—Л. : 539—566.
Руновский Н. Н. 1947. Американский енот в Закавказье. Природа, 2 : 59— 60.
Руновский II. Н. 1950. Акклиматизация енота (Ргосуоп lotor L.) в Азербайджанской ССР. Тр. Всесоюзн. научно-иссл. инет, охоты, промысла, IX : 171— 186.
Ру конский Н. Н. 1956. Современное распространение алтайской белки на Кавказе. Тр. Всесоюзн. научно-иссл. инет, охоты, промысла, XVI : 157—180.
(Рул ье К Ф.) Rouillier. 1841. Aegoceros Pallasii. Ein neuer russischer Zweihufer. Bull. Soc. Moscou, XIV, XI : 908—923.
Рустамов A. К. 1945. Европейская лесная «Гирканская» фауна в Копет-Даге. , Изв. Тур км. фил. АН СССР, 3—4 : 132—-135.
J
Зоологическая литература
603
1945. Felis ornata Gray из Армении. Докл. АН АрмССР, III, 1946а. О распространении онагра (Equus hemionus) в приарак-Армении. Изв. АН АрмССР, естеств. науки, 3 : 75—81.
19466. Южноамериканский болотный бобр в Армянской ССР.
руттенбург С. О. 1950. Физиологические особенности некоторых видов крыс в связи с их географическим распространением. Афтореф. дисс., Л. :1—20.
Рябинин А. И. 1918. Послетретичные млекопитающие и кремневые орудия ив Удельной степи Ставропольской губ. Ежегодн. русск. палеонтол. общ., II : 33—46.
Рябинин А. Н. 1937. Заметка об остатках пещерного льва из окрестностей Сочи на Черноморском побережье Кавказа. Тр. Сов. секции Междун. ассоц. по изуч. четвертичн. периода Европы, III : 60—62.
Рябинин А. Н. 1938. Microzeuglodon aff. caucasicum Lydek. из верхнего Майкопа Кабристана. Пробл. палеонтол., 4 : 137—186.
ЖТ я б о в Н. И. 1940. Материалы по экологии лесной мыши и кустарниковой полевки ВТ Кавказа. Сб. научн. раб. комсомольцев-биологов АН СССР, М.—Л. : 129—138. ЯЕСаркисов А. А. 1941. О географическом распространении Ovis ophion arme-I niana Was. Сообщ. АН ГрузССР, II, 3 : 277—283.
|Саркисов А. А. 1944а. О миграциях Ovis ophion armeniana Nas. Тр. Ереванск. зоол. парка, 1, 2 : 89—91.
^Саркисов А. А. 19446. Арменийский муфлон Насонова Ovis ophion armeniana Н Nas. Тр. Ереванск. зоол. парка, III, 5—52.
Ц -С а р к и с о в А. А. 1944в. К вопросу географического распространения гельского И слепца (Spalax monticola armeniana Mehely). Тр. Ереванск. зоол. парка, 1, 2 : < 129—130.
МП а р к и с о в А. А. 1944г. Дикий кабан Закавказья и Северного Ирана. Тр. Ере-ванск. зоол. парка, 1, 2 : 61—73.
^Саркисов А. А. 1944д. О копытных (Ungulata) Ирана. Тр. Ереванск. зоол. С парка, 1, 2 : 39—59.
«Саркисов А. А. 1944е. Кавказская серна Rupicapra rupicapra caucasica Ly- dek. в Армении. Тр. Ереванск. зоол. парка, 1, 2 : 139—142.
Вбаркисов А. А. ~
Я 3:85—88.
Вк] а р к и с о в А. А. в сцнскпх степях
;ScаркисовА. А.
Изв. АН АрмССР, естеств. науки, 5 : 87—94.
«(Сатунин К. A.) Satunin К. А. 1895. Sorex Raddei Sat. n. sp. und Meles taxus arenarius Sat. n. suhsp. Arch, fur Naturgesch., Jahrb. 1, 1 : 109—113.
•(Сатуяин К. A.) Satunin K. A. z ...._
Saugethierfauna der Kaukasuslander. Zool. Jahrb., Syst
Сатунин К. A. 1898. Кавказский зубр. Естествозн. и геогр., 2:1—21.
(С а т у ни н К. A.) Satunin К. А. 1901а. On a new Hedgehog from Transcaucasia. Proc. Zool. Soc. London, 11, 19 : 284—291.
(Сатунин К. A. ) Satunin K. A. 19016. Uber ein neues Nager-genus (Pro-metheomys) aus dem Kaukasus. Zool. Anz., XXIV, 653 : 572—575.
iC а т v н и н K. A.) Satunin K. A. 1901b. Zwei neue Saugethicre aus Transcau-casien. Zool. Anz., XXIV, 649:461—469.
«Сатунин К. А. 1901г. О млекопитающих степей северо-восточного Кавказа. ® Изв. Кавк, музея, 1, IV : I—П+1—100.
«Сатунин К. А. 1903а. Фауна северо-восточного Кавказа. Природа и охота, V II : 9—13.
^Сатунин К. А. 19036. Обзор исследования млекопитающих Кавказского края. Зап. Кавк. отд. Русск. геогр. общ., XXIV, 2 : 1—63.
Сатунин К. А. 1903в. Дельфиновый промысел на Черноморском побережье Кавказа. Необходимость упорядочения его. Куний промысел в Абхазии. Истребление в связи с ним горной дичи. Газета «Кавказ», 79.
(Сатунин К. A.) Satunin К. А. 1904. The black wild cat of Transcaucasia. Proc. Zool. Soc. London, II : 162—163.
(Сатунин К. A.) Satunin K. A. 1905a. Uber Kaukasischc Steinbocko. Zool. Anz., XXIX 11 : 336—344.
Сатунин К. A. 19056. Кавказский дикий кст (Felis catus caucasicus Satunin subsp. nova). Природа и охота, VI : 1—2.
Сатунин К. А. 1904—1906. Млекопитающие Талыша и Мугани. Изв. Кавк, муз., II, 2—4 : 87—262.
Сатунин К. А. 1907а. Млекопитающие северо-восточного Предкавказья по сбору экспедиций Кавказского музея летом 1906 г. Изв. Кавк, муз., III, 2—3 : 94— 142.
Сатунин К. А. 19076. Барсук и куница конца бронзового века на Кавказе. Jt Изв. Кавк, муз., Ill, 1 : 1—36.
1896. Vorlaufiger Mittheilungen uber die ' " IX : 277—314.
1901а. On a new Hedgehog from Transcau-
19016. Uber ein neues Nager-genus (Pro-
604
Литература
Сатунин К. А. 1907в. Материалы к познанию млекопитающих Кавказского края и Закаспийской области. I—VII. Млекопитающие, собранные в Ленкоранском уезде А. Б. Шелковниковым в 1907 г. Изв. Кавк, муз., III, 2—3 : 239— 260.
Сатунин К. А. 1909. Материалы к познанию млекопитающих Кавказского края и Закаспийской области. XII. Млекопитающие, собранные А. Н. Казнаковым у Гудаура. XIII. Крот Ленкоранского уезда. XIV. Млекопитающие, собранные К. А. Сатуниным и А. А. Ростовцевым в Бакурианах в 1908 г. XVI. Млекопитающие Кавказа и прилежащих стран, хранящиеся в Зоологическом музее Академии наук. Изв. Кавк, муз., IV, 3—4 : 269—301.
Сатунин К. А. 1909—1910. Некоторые соображения о происхождении фауны Кавказского края. Изв. Кавк. отд. Русск. геогр. общ., XX, 2 : 1—12.
Сатунин К. А. 1912а. Животный мир Мугани. Тифлис : 1—88.
Сатунин К. А. 19126. О зоогеографических округах Кавказского края. Изв. Кавк, муз., VII : 7—55.
Сатунин К. А. 1913. Фауна Черноморского побережья Кавказа. Mammalia, Aves. Reptilia, Amphibia, Mollusca, Vermes. Тр. общ. изуч. Черном, побережья, II, СПб. : 1—248.
Сатунин К. А. 1914. Кавказский зубр. Очерки родиноведения Кавказа, 4, Тифлис : 1—14.
Сатунин К. А. 1915а. Млекопитающие Кавказского края. Зап. Кавк, муз., сер., А, № 1, 1 : 1—410.
Сатунин К. А. 19156. Мертвый сезон в жизни степей Восточного Закавказья. Изв. Кавк. отд. Русск. геогр. общ., XXIII, 2 : 161 —172.
Сатунин К. А. 1915в. Новые млекопитающие из Закавказья. Изв. Кавк, муз., VIII : 89—93.
Сатунин К. А. 1916. Обзор фаунистических исследований Кавказского края за пятилетие 1910—1914 гг. Зап. Кавк. отд. Русск. геогр. общ., XXIX, 3: 1—81.
Сатунин К. А. 1920. Млекопитающие Кавказского края. Тр. музея Грузии, 2, Ы : 1—223.
Свириденко П. А. 1922. К систематическому положению дагестанского суслика (Citellus Satunini sp. п.). Бюлл. муз. Грузии, 1 : 69—77.
Свириденко П. А. 1926. К систематике и биологии суслика нагорной Армении. Уч. зап. Сев.-Кавк. инет, краевед., 1 : 147—175.
Свириденко П. А. 1927. Распространение сусликов в Северо-Кавказском крае и некоторые соображения о происхождении фауны предкавказских и калмыцких степей. Изв. Сев.-Кавк, краев, ст. защ. раст., 3, Ростов-Дон : 123— 171.
Свириденко П. А. 1928. Степная пеструшка (Lagurus lagurus Pall.) на Северном Кавказе и ее систематическое положение. Изв. Сев.-Кавк. ст. защ. раст., 4: 47—61.
Свириденко П. А. 1935а. Северокавказские Mus musculus и их распределение по климатическим зонам. Уч. зап. Моск. гос. унив., IV : 70—87.
Свириденко П. А. 19356. Факторы, ограничивающие численность мышевидных грызунов. Уч. зап. Моск. гос. унив., IV : 44—87.
Свириденко П. А. 1936. Грызуны Северного Кавказа и Предкавказских степей. Сб. Научно-иссл. инет. зоол. Моск. гос. унив., 3 : 86—89.
Свириденко П. А. 1937. Суслик Большого Кавказа Citellus musicus и происхождение горной степи. Зоол. журн., XVI, 3 : 448—482.
Свириденко П. А. 1940. Грызуны-вредители пробкового дуба и роль лесокультурных мероприятий в ограничении их численности. Лесное хоз., 6 : 57— 60.
Свириденко II. А. 1943. Экологические факторы, определяющие географическое распространение и эйритопность полевой мыши. Зоол. журн., 5 : 28— 299.
Свириденко П. А. 1944. Распространение, питание и эпидемиологическое значение полевой мыши Apodem us agrarius Pall. Докл. АН СССР, XLI1, 2 : 96—100.
Свириденко П. А. 1945. Роль деятельности человека в накоплении и ограничении численности лесных мышевидных грызунов. Зоол. журн., XXIV, 6 : 347— 361.
Север нов Н. А. 1877. О зоологических (преимущественно орнитологических), областях внетропических частей нашего материка. Изв. Русск. геогр. общ., XIII, 3 : 1—125.
Сев ер до в А. Н. 1939. Морфологические закономерности эволюции. Изд. АН СССР, 4 М.—Л. : 1—610.
Зоологическая литература
605
Сем ев о в - Т я н - Ш а вс кий А. П. 1936. Пределы и зоогеографические подразделения Палеарктической области для наземных сухопутных животных на основании географического распределения жесткокрылых насекомых. Изд. АН СССР, М.—Л. : 1—15.
Сем v нов-Тян - Шанский А. П. 1937. Основные черты истории развития Альпийских фаун. Изв. АН СССР, сер. биол., 4 *. 1211—1227.
Семе итонский Н. 1857. Сказание о ловах великих князей киевских. СПб. : 1—18.
Серебровский П. В. 1928. К орнитогеографии Передней Азии. Ежегодн. Зоол. муз. АН СССР, 29 : 289—392.
Серебровский П. В. 1935. История животного мира СССР (краткий очерк). Л.: 3—129.
Серебровский II. В. 1936а. Очерк третичной истории наземной фауны СССР. Животный мир СССР, I. Изд. АН СССР, М.—Л. : 11—78.
Серебровский П. В. 19366. Послеледниковое формирование современной фауны и воздействие человека на природу. Животный мир СССР, I. Изд. АН СССР, М.—Л. : 124—157.
Серебровский П. В. 1948. Птицы бинагадинских Кировых отложений. Тр. Естеств.-ист. муз. АН АзербССР, I—II : 21—75.
Симонович и Сорокин. 1874. Заметка об остатках мамонта на Кавказе. Изв. Кавк. отд. Русск. геогр. общ., Ш : 28—29.
Симпсон Д. Г. 1948. Темпы и формы эволюции. Изд. иностр, лит., М. : 1—358. Слодкевич В. С. 1930. К остеологии передней конечности El asm о thorium cauca-sicum Boriss. Тр. Геол. муз. АН СССР, VI : 9—29.
Смирнов Н. А. 1916а. Бурый медведь в коллекциях Кавказского музея. Зап. Кавк, муз., А, 4 : 1—25.
Смирнов Н. А. 19166. Заметка об урмийском диком баране (Ovis orientalis ur-miana Guenther). Изв. Кавк, муз., X : 1 — 16.
Смирн о в Н. 1917а. О находке челюсти росомахи при пещерных раскопках в Кутаисской губернии. Изв. Кавк, муз., XI : 1—2.
Смирнов Н. 19176. О видовой самостоятельности закавказского черного кота. Изв. Кавк, муз., XI : 1—3.
Смирнов Н- А. 1919. Краткий определитель грызунов Кавказского края. Работы Земск. опытн. ст. по борьбе с грызунами в Закавк., 3, Тифлис : 1—21.
Смирнов Н. А. 1922. Felidae Кавказа и сопредельных стран по коллекциям Кавказского музея. Изв. Азерб. гос. унив., 3 : 41—42.
Смирнов Н. А. 1923—1924. О некоторых млекопитающих Западного Закавказья в каменном веке. Изв. Азерб. гос. унив., 3, 141—149,
Соболевский Н. И. 1929. Герпетофауна Талыша и Ленкоранской низменности. Опыт зоогеографической монографии. Мем. Зоол. отд. Общ. любит, естеств., антроп. и этногр., 5 : 1—141.
Сов и некий В. К. 1904. Введение в изучение фауны понто-каспийско-аральского морского бассейна, рассматриваемой с точки зрения самостоятельной зоогеографической провинции. Зап. Киевск. общ. естествоиспыт., XVIII : 1—216.
Сокол о в И. И. 1949. Об остатках полорогих (Bovidae Mammalia) из среднего миоцена Северного Кавказа. Докл. АН СССР, 67, 6 : 1101—1104.
Соколов И. И. 1953. Опыт естественной классификации полорогих (Bovidae). Тр. Зоол. инет. АН СССР, XIV, М,—Л. : 5—295.
Соколов И. И. 1954. Новые виды антилоп третичной фауны юга СССР. Тр. Зоол. инет. АН СССР, XVII : 217—223.
Соку р И. Т. 1950. Влияние молодых полезащитных лесополос на количество и поведение полевки степной (Microtus socialis Pall.) в прилежащих полях. Тр. 1нст. Зоол. АН УССР, 3 : 35—60.
С о с н и х и н а Т. М. 1944. Грызуны города Ленинакана. Изв. АН АрмССР, естеств. науки, III : 53—60.
Сое н и х и н а Т. М. 1946. Грызуны города Кировакана. Изв. АН АрмССР, естеств. науки, 3 : 67—73.
Соснихина Т. М. 1947. О нахождении костей пищухи (Ochotona sp.) в Армянской ССР. Докл. АН АрмССР, VII, 2 : 77—78.
Со с н и х и н а Т. М. 1948. Польза и вред филина в сельском хозяйстве Армянской ССР. Изв. АН АрмССР, биол. и с.-х. науки, 1,3: 271—281.
Соснихина Т, М. 1950а. Серый хомячок (Cricetulus migratorius Pall.) в условиях Армянской ССР. Зоол. сб. Инет, фитопатол. и зоол. АН АрмССР, VII, Ереван : 55—82.
Соснихина Т. М. 19506. Хозяйственное значение домового сыча в условиях полупустыни юга Армянской ССР. Изв. АН АрмССР, III, 1 : 95—100.
60G
Литература
Соснихина Т. М. 1952. Результаты опытов по выяснению возможности аккли-. матизации белки-телеутки в лесах Армянской ССР. Изв. АН АрмССР, биол. и с.-х. науки, 9 : 15—32.
Спангенберг Е. П. 1935. Соня-полчок. В сб.: Н а у м о в С. П. и др. Тонкопалый суслик. Соня-полчок. Слепыш. Бурундук. М. : 36—70.
Спасский П. И. 1939. Находки костей верхнетретичных млекопитающих в окрестностях Дербента. Изв. Азерб. фил. АН СССР, 1—2 : 147—152.
Спасский П. И. 1951. Остатки цетотериев из Северного Кавказа (окр. гор. Майкопа). Изв. АН АзербССР, 2 : 57—65.
С п у л ь с к и й Б. А. 1915. Ископаемые слоны ледниковой эпохи Юго-Западной России. Зап. Киевск. общ. естествоиспыт., XXV, 1—2:153—172.
Стальмакова В. А. 1935. Места обитания грызунов в ставропольских степях. Сб. «Борьба с грызунами в степях Предкавказья» : 196—222.
Станчинский В. В. 1927. Экологическая эволюция и формирование фаун. К вопросу формирования орнитоценозов. Тр. Смоленск, общ. естествоиспыт. и врачей, II : 189—204.
Станчинский В. В. 1939. Экологическое направление в изучении природных комплексов-ландшафтов. Научно-метод. зап. Комит. по запов. 3 : 8—23.
Степанов И. В. 1931. Крысы и мыши Батуми. Вести, микробиол., эпидемиол. и паразитол., X, 1 : 83—92.
Степанов М. В. 1946. Материалы по биологии размножения крыс и мышей в Батуми. Бюлл. Моск. общ. испыт. природы, отд. биол., LI (6) : 29—35.
Степанов П. Н. 1939. Опыт акклиматизации енотовидной собаки (Nyctereutes procyonoides Gray) в Тебердинском заповеднике. Научно-метод. зап. Комит. по запов., III : 164—170.
Строганов С. У. 1948. Систематика кротовых. Тр. Зоол. инет. АН СССР, VIII 2 : 327—330.
Сушкин П. П. 1922. Эволюция наземных позвоночных и роль геологических изменений климата. Природа, 3—5 : 3—31.
Сушкин II. П. 1925. Зоологические области Средней Сибири и близлежащих частей нагорной Азии и опыт истории современной фауны палеарктической Азии. Бюлл. Моск. общ. испыт. природы, нов. сер., 34 : 7—86.
Теп л о в В. П. 1936. Положение с волками в заповеднике и борьба с ними. Бюлл. Кавк. гос. запов., 2 : 14—18.
Теп лов В. П. 1938а. Материалы по биологии дикого кота (Felis silvestris caucasicus Sat.) в условиях Кавказского заповедника. Тр. Кавк. гос. запов., 1 : 331-340.
Т е п л о в В. П. 19386. Волк в Кавказском заповеднике. Тр. Кавк. гос. запов., 1 : 366—443.
Тер-Погосян А. Г. 1947. К вопросу о распространении онагра (Equus hemio-nus) в древней Армении. Изв. АН АрмССР, естеств. науки, 2 : 91—93.
Теряев В. А. 1936. Проблема определения возраста позвоночных в палеонтология. Пробл. палеонтол., 1 : 135—169.
Теряев В. А. 1939. Фетишизм в палеонтологии. Бюлл. Моск. общ. испыт. природы отд. геол., XVII (4—5) : 144—151.
Тимофеев С. 1939. Полевки в восточном Закавказье, их образ жизни, вредная деятельность и условия борьбы с ними. Тифлис : 1—24.
Третьяков Д. К. 1941. Третичная фауна одесских катакомб. Сов. наука, 1 : 104.
т р О ф имев В. А. 1951. Об ископаемых свиньях рода Microstonyx. Докл. АН СССР, LXXVI, 6:881—884.
Туга ринов А. Я. 1936. К вопросу о формировании островных фаун. Изв. АН СССР, сер. биол., 2—3 : 501—523.
Туров С. С. 1926а. Новые данные о распространении прометеевой мыши (Prome-theomys schaposchnikovi Sat.). Изв. Горского пед. инет., III, Владикавказ : 4, Туров С. С. 19266. К биологии и распространению Promcthoomys schaposchnikovi Sat. Уч. зап. Сев.-Кавк. инет, краевед., 1 : 23—30.
Туров С. С. 1926в. Опыт систематического обзора млекопитающих Осетии. Уч, зап. Сев.-Кавк. инет, краевед., 1 : 311—337.
Туров С. С. 1927. Определитель позвоночных (кроме птиц), преимущественно Северного Кавказа. Владикавказ : 1—55.
Туров С. С. 1934. Темная вариация Prometheomys schaposchnikovi Mammalia (Muridae). Сб. тр. Гос. зоол. муз. при Моск. гос. унив., I : 43—44.
Туров С. С. 1937. Заметки о куницах Северного Кавказа. Сб. тр. Гос. зоол. муз. при Моск. гос. унив., IV : 27—34.
Туров С. С. и Д. Б. К р а с о в с к и й. 1937. Очерк фауны Присулакского оленьего ' заповедника. Зоол. журн., XII. 4 : 35—56.
Зоологическая литература 60Т
Иг^
i Т у р о в С. С. и Л. Г. Турова-Морозова. 1928. Материалы по изучению ! млекопитающих Северного Кавказа и Закавказья. Изв. Горского пед. инет., 5 V : 157—186.
^Уваров Б. П. 1917. К фауне позвоночных низовьев р. Кумы. Изв. Кавк, муз., f X : 102.
Уйатинская-Декаленко Р. С. 1933а. К экологии прометеевой мыши. Природа, 11 : 71—72.
Ушатинская-Декаленко Р. С. 19336. Материалы к вопросу о влиянии ионизации воздуха на организм животных. Изв. 2-го Сев.-Кавк. пед. инет., IX : 83—88.
[ Фал ь к е н шт е й н Б. Ю. 1933. Мышевидные грызуны в СССР в 1932—1933 гг. \ Сб. Всесоюзн. инет. защ. раст., 7 : 77—83.
I Федоров С. М. 1954. Млекопитающие (Mammalia) Ставропольского края. Матер. < по изуч. Ставропольем края, 6 : 177—192.
Федосов А. В. 1941. Особенности биологии и экологии кавказского волка и борьба с ним в Кавказском заповеднике. Научно-метод. зап. Комит. по запов., 1 : [ 85—89.
Федюшин А. В. 1935. Речной бобр, его история, жизнь и опыты по размножению, М. : 1—356.
^Филатов Д. П. 1910. Летняя и зимняя поездки в северо-западный Кавказ в 1909 г. f для ознакомления с кавказским зубром. Ежегодн. Зоол. муз. Акад, наук, ; XV : 171—217-
фил ат о в Д. П. 1912. О кавказском зубре. Зап. Акад, наук, сер. VIII, физ.-матем. отд., XXX, 8, СПб. : 1—40.
(фишер-В а л ь дгейм Г. И.) Fischer von Waldgeim. 1809. Sur 1’Elas-K. motherium et le Trogontherium deux animaux fossiles et inconnus de la Russie. r Mem. Soc. Imp. Nat. Moscou, II : 250—268.
Клеров К. К. 1927. Очерк фауны млекопитающих долины реки Пороха. Ежегодн.
1 Зоол. муз. АН СССР : 63—73.
Клеров К. К. 1932. Обзор диагностических признаков беловежского и кавказского & зубров. Изв. АН СССР, сер. VII, отд. матем. и естеств. наук, 10 : 1579— | 1590.
К1 леров К. К. 1934. О географическом распространении и систематике лосей. Докл. £ АН СССР, II, 2 : 1—7.
Кф леров К. К. 1950. Морфология и экология оленеобразных в процессе их эволю-К ции. Матер, по четвертичн. периоду СССР, 2 : 50—69.
Клеров К. К. 1952. Кабарги и олени. Фауна СССР. Млекопитающие. 1, 2. нов. & сер. 55 : 1—255.
К леров К. К. и А. А. Гуреев. 1934. О млекопитающих, собранных армянской Ж паразитологической экспедицией 1931 г. АН СССР. Тр. Совета по изуч. произ-R водит, сил, сер. Закавказская, II : 93—123.
№ ормозов А. Н. 1926. Заметки о млекопитающих Северного Кавказа. Уч. зап.
№ Сев.-Кавк. инет, краевед., 1 : 73—90.
К; ормозов А. Н. 1929. Скотобой, его значение для степной фауны и борьбы с вре-К дителями. Природа, 11 : 389—392.
Кф ормозов А. Н. 1930. О возможности акклиматизации белки в Кубанских лесах К Кавказа. Пушное дело, 6 : 17—19.
шф ормозов А. Н.) Formozo v A. N. 1931. Notizen iiber einige Nager des west-к. lichen Kaucasus mit Beschreibung zweier neuer Formen. Folia Zool. et Hygrobiol., К III, 13, VI, Riga : 79—84.
t ормозов A. H. 1937. Об освоении фауны наземных позвоночных и вопросах ее К реконструкции. Зоол. журн., XVI, 3 : 407—442.
Ьормозов А. Н. 1938. К вопросу о вымирании некоторых степных грызунов в позд-кГ нечетвертичное и историческое время. Зоол. журн., XVII, 2 : 260—272.
ормозов А. Н. 1945. Несколько дополнений к статье В. М. Житкова «Замеча-№ ния о крысах и некоторых условиях их исследования». Зоол. журн., XXIV, К 2:126.
КС’о р мозов А. Н. 1946. Снежный покров как фактор среды, его значение в жизни Ц; млекопитающих и птиц СССР. Материалы к позн. фауны и флоры СССР, Изд. № Моск. общ. испыт. природы, нов. сер., отд. зоол., 5 (XX) : 3—142.
KG ормозов А. Н., И. Б. К и р и с. 1937. Деятельность грызунов па пастбищах К' и сенокосах. III. Влияние общественной полевки (Microtus socialis Pall.) и не-к которых других грызунов на растительность Кизлярского района Дагестан-В ской ССР. Уч. зап. Моск. гос. унив., XIII : 59—70.
Иавесон Я. И. 1954а. Третичные верблюды восточного полушария (род Рагаса-К melus). Тр. Палеонтол. инет. АН СССР, XVII : 100—162.
608
Лите ра ту pa
Хавесон Я. И. 19546. Дикий одногорбый верблюд из отложений озера Севан (Армения). Докл. АН СССР, XCVII1, 3 : 475—478.
Хоменко И. П. 1913а. Материалы по палеонтологии третичных и послетретичных млекопитающих Северного Кавказа. Тр. Ставропольск. общ. изуч. врир. Сев. Кавк., Ill, 1 : 1—39.
Хоменко И. П. 19136. Новейшие успехи изучения гиппарионовой фауны Южной России. Дневн. XIП съезда русск. естествомспыт. и врачей в Тифлисе, 2 : 443— 444.
Хоменко И. П. 1913в. Меотическая фауна с. Тараклии Бендерского уезда. L Предки современных и ископаемых оленей. II. Giraffinae et Cavicornia. Ежегодн. геол, и минерал. России, XV, 4—5 : 107—143.
Хоменко И. II. 1914а. Открытие Руссильонской фауны и другие результаты геологических наблюдений в Южной Бессарабии. Тр. Бессарабск. общ. есте-ствоиспыт. и любит, естеств., VI : 3—11.
Хоменко И. П. 19146. Меотическая фауна с. Тараклии Бендерского уезда. I. Грызуны, носороги, лошади, свиньи, хоботные. Тр. Бессарабск. общ. естество-испыт. и любит, естеств. : V : 1—56.
Хоменко И. 1915. Руссильонский ярус в среднем плиоцене Бессарабии и его значение для познания возраста Балтских песков и Кульяницких отложений. Тр. Бессарабск. общ. естествоиспыт. и любит, естеств., VI : 3—28.
X о н я к и н а 3. П. 1938. Материалы по питанию лисицы в Кавказском заповеднике. Тр. Кавк. гос. запов., 1 : 317—330.
Ц а л к и н В. И. 1947. Новые данные к истории фауны Крыма. Докл. АН СССР, LIX, 3 : 605—607.
Ц а л к и н В. И. 1951а. Новые данные о распространении кулана в историческое время. Докл. АН СССР, LXXXI, 2 : 289—291.
Ц а л к и н В. И. 19516. Горные бараны Европы и Азии. М. : 1—343.
Цадкин В. И. 1955. Изменчивость и систематика туров западного Кавказа (Сар-rinae, Artiodactyla). Бюлл. Моск. общ. испыт. природы, отд. биол., LX(4):17—33.
Церетели Д. В. 1942. Раскопки местонахождения ископаемых неогеновых млекопитающих в гареджийском Удабно. Вести. Гос. муз. Грузии, XI-А : 161— 168 (на груз. яз.).
Чернов С. А. 1939. Герпетологическая фауна Армянской ССР и Нахичеванской АССР. Тр. Биол. инет. Арм. фил. АН СССР, III, зоол. сб., 1 : 77—194.
Чернявская С. И. 1956. Сезонное размещение и кочевки диких копытных и медведя в районе Кавказского заповедника в связи с распределением урожая плодов фруктарников и орехоносов. Бюлл. Моск. общ. испыт. природы, отд. биол., 61, 4 : 7—21.
Черский Н. 1891. Описание коллекций послетретичных млекопитающих животных, собранных новосибирскою экспедициею 1885—1886 гг. Прилож. к LXV тому ((Записок Акад, наук», 1, СПб. : 1—706.
Чиркова А. Ф. 1952. Методика и некоторые результаты учетов численности лисицы и корсака. Методы учета численности и географического распределения наземных позвоночных. Изд. АН СССР, М. : 179—203.
Чхиквишвили И. Д. и В. А. Пхакадзе. 1938. Материалы к познанию вредных для сельского хозяйства грызунов, распространенных в районе Каспи. Вестн. Груз. гос. с.-х. инет., 1 (5) : 72—93 (на груз. яз.).
Шапошников Л. В. 1938. Акклиматизация пушных зверей в СССР. Зоол. журн., XVII, 5 : 939—960.
Ш апо ш ников Л. В. 1940. Интродукция пушных зверей в СССР за 1938 г. Зоол. журн., XIX, 5 : 766—775.
Шапошников Л. В. и Н. К. Верещагин. 1932. К вопросу об акклиматизации нутрии в Закавказье и Предкавказье. Землеведение, XXXIV, 1—2 : 40—78. "
Шап ошников Л. В., Е. С. К р у ши н с к а я. 1939. Алтайский сурок (Маг-mota baibacina baibacina Kastsch.) в Дагестанской АССР. Зоол. журн., XVIII, 6 : 1048—1054.
Ш а р ф ф Р. Ф. 1918. Европейские животные, их геологическая история и географическое распространение. М. : I—VII-}-1—233.
Ш е л к о в п и к о в А. В. 1930. Заметки о новейших флористических и фаунистических находках в Армении. Закавк. краоведческ. сб., сер. А, 1,Тифлис: 142—144.
Ш е р с т ю к о в Н. М. 1926. О новых находках ископаемых слопов в Приазовье. Бюлл. Сев.-Кавк, бюро краевед., 5—7 : 27—28.
Ш е р с т ю к о в Н. М. 1927. Палеонтологические находки остатков млекопитающих третичного и послетретичного периода на Северном Кавказе в 1924—1927 гг. , Бюлл. Сев.-Кавк. бюро краевед., 1—6, Ростов н/Дону : 42—46.
Зоологическая литература
609
Шерстюков Н. М. 1954. Описание остатков ископаемых слонов, найденных в Грозненской области. Тр. Естеств.-ист. музея АН АзербССР, VIII : 227—261.
Шидловский М. В. 1919. Материалы к фауне грызунов Закавказья, ч. 1. Полевки.
1. Подрод Arbusticola. Работы Земск. опытн. ст. по борьбе с грызунами в Закавказье, 2 : 1—>46.
Шидловский М. В. 1919. Материалы к фауне грызунов Закавказья, ч. 1. Полевки. 2. Подрод Chionomys. Работы Земск. опытн. ст. по борьбе с грызунами в Закавказье, 5 : 1—38.
Ш и д л о в с кий М. В. 1938. О полевках группы Arbusticola. Бюлл. Гос. муз. Грузии, IX-А : 80—100.
Шидловский М. В. 1940а. Новые данные по фауне грызунов Грузии. Сообщ. Груз. фил. АН СССР, 1, 2 : 143—148.
Шидловский М. В. 19406. Изучение грызунов в Грузии. Природа, 7 : 80.
Шидловский М. В. 1940в. Об особенностях распространения закавказского хомяка по кавказскому перешейку. Тр. Зоосект. Арм. фил. АН СССР, Зоол. сб., II : 37—48.
Шидловский М. В. 1941а. Определитель грызунов Грузии и сопредельных территорий. Тбилиси : 1—55.
Ш и д л овский М. В. 19416. Характерные черты родентофауны Грузии. Сообщ. АН ГрузССР, II, 1—2 : 137—144.
Шидловский М. В. 1942. Родентофауна лесной зоны северной Армении. Тр. Кироваканской лесн. опытн. ст., Ш : 53—80.
Шидловский М. В. 1945. Реликтовые и эндемичные виды в родентофауне Грузии. Сообщ. АН ГрузССР, VI, 6 : 431—439 (на груз. яз. с русск. резюме).
Шидловский М. В. 1947. Родентофауна Черноморского побережья Грузии. Тр. Зоол. инет. АН ГрузССР, VII : 97—146.
Шидловский М. В. 1948. Грызуны Аджарии. Тр. Зоол. инет. АН ГрузССР, VIII : 187—223.
Шидловский М. Ф. 1950. Грызуны Абхазии. Тр. Зоол. инет. АН ГрузССР, ГХ : 135—161.
Ши д л о в с к и й М. В. 1951. Грызуны Юго-Осетии. Тр. Зоол. инет. АН ГрузССР, X : 187—222.
Шидловский М. В. 1953а. Малоазийская горная мышь (Silvimus mystacinus Danf. et A 1st.) в родентофауне Грузии (Rodentia, Muridae). Тр. Инет. зоол. АН ГрузССР, XI : 135—168.
Шидловский М. В. 19536. Белозубая кутора Crocidura russula Herm. в фауне землероек Грузии. Тр. Инет. зоол. АН ГрузССР, XII : 215—228.
Ши дловский М. В. 1954а. Семейство песчанок в родентофауне Грузии (Roden-tia, Gerbillidae). Тр. Ивет. зоол. АН ГрузССР, XIII : 63—69.
Шидловский М. В. 19546. К систематике и распространению полчка (Glis glis L.) в Грузии. Тр. Инет. зоол. АН ГрузССР, XII : 47—61.
Шидловский М. В. иТ. М. Соснихина. 1944. Грызуны города Еревана. Зоол. сб. АН АрмССР, III : 71—108.
Штегман Б. К. 1938. Основы орнитогеографического деления Палеарктики. Фауна СССР, Птицы 1, 2. Изд. АН СССР, М.~Л. : 1—76.
Э й х в а л ь д Э. 1840. Первобытный мир России. СПб. : 1—73.
Эйхвальд Э. 1850. Палеонтология России. Новый период, СПб. : 1—284.
.9 к в т и ми ш в и л и 3. С. 1947. Материалы к изучению биоэкологии водяной крысы (Arvicola amphihius L.) в условиях Бакуриани. Тр. Зоол. инет. АН ГрузССР, VII : 299—324.
Эль-Мар 1939. Рев оленей в настоящем году. Журн. Монадирс, И.
Януш ко П. А. 1938. Смертность полевок Microtus arvalis в природе в условиях степных районов Предкавказья и влияние на нее хищников. Зоол. журн., XVII, 1 : 102—112.
Я цк о И. 1940. Раскопки верхнеплиоценовой фауны в окрестностях Одессы. Палеонтол. обозр., 2 : 76—77.
Aharoni J. 1930. Die saugetiere Palastinas. Zeitschr. Saugetierkunde, V : 327—343.
Aharoni R. 1932. Die Muriden von Palastina und Syrien. Zeitschr. Saugetierkunde, VII, 1—6 : 166—240.
Allen G. M. 1940. Bats. Cambridge : 1—368.
Amon R. 1938. Abstammung Arten und Rassen der Wildschweine Eurasicus. Zeitschr. fur Zucht, R. B. XL, 1 : 49—88.
Antonins O. 1938. On the geographical distribution in former times and today of the recent Equidae. Proc. Zool. Soc., 107, В : 557—564.
Arldt T. 1919. Handbuch der Palacogt ographie, Leipzig, I : 1—680.
Backer J. 1934. Bemerkungen uber den persischen Lowen. Zeitschr. Saugetierkunde, IX : 439—440.
39 H. К. Верещагин
610
Л ите рату ра
Barclay Е. N. 1934. Notes on the Fallow Deer of Asia Minor. Ann. Mag. Nat.-Hist., Incl. Zoology, Botany and Geology. 10 ser., XIV, London : 157—159.
Bate D. M. 1937. The fossil fauna of the Wady-el-Mughara Caves. In: G а г г о d D. M. and D. M. Bat e. The Stone Age of the Mount Carmel, I, Oxford : 139—235.
Bell W. D. 1923. The Wanderings of an Elephant hunter. London : 1—187.
Bennet E. T. 1895. Observations on several Mammalia from Trebizond and Erzeroum. Proc. Zool. Soc. : 89—90.
Blanford W. T. 1876. Eastern Persia. Zoology, II : 1—516.
Bodenheimer F. S. 1935. Animal life in Palestine. Jerusalem : 1—132.
Bbdenheimer F. S. 1937. Prodromus faunae Palestinae. Mem. Inst. Egypte, 33 : 1—284.
Colbert E. H. 1935. Siwalik Mammals in the American Museum of Natural History. Trans. Amer. Philos. Soc., 26 : X-f-402.
Colbert E. H. 1955. Evolution of the Vertebrates. New York : 1—479.
Couturier M. 1938. Le Chamois. Grenoble, XIV+1—455.
Danford C. and E. Alston. 1877. On the mammals of Asia Minor. Proc. Zool. Soc. : 270—281.
Danford C. and E. Alston. 1880. On the mammals of Asia Minor. Proc. Zool. Soc. : 50—64.
Duerst J. U. 1908. Animal Remains from the Excavations at Anau. In : P u mp el-I у R. Explorations in Turkestan, IL Carnegie Inst. Washington Publ. : 341—442.
E Her ma nn J. R. 1948. Key to the Rodents of South-West Asia in the British Museum Collection. Proc. Zool. Soc., 118, III .* 765—816.
E Hermann J. R. and T. C. Morrison-Scott. 1951. Checklist of Palaearctic and Indian Mammals 1758 to 1946. London : 1—810.
Filippi deF. 1865. Note di un viaggio in Persia, nel 1862. Milano : 1—396.
Freudenberg W. 1914. Die Saugetiere des altcsten Quarters von Mitteleuropa. Geol. und Palaeontol. AbhandL, NF, XII, 4/5 : 1—219.
Goodwin G. G. 1939. Five new rodents from the eastern Elburz Mountains and a new race of har from Teheran. Amer. Mus. Novit., 1050 : 1—5.
Grote H. 1925. Ueber das letzte siidrussische Wildpferd (Equus gmelini). Pallasia, Dresden, 2 : 181—184.
Harper F. 1945. Extinct and vanishing mammals of the Old World. New York: 1—850.
Hohenacker R. 1837. Enumeration animalium quae in provinciis transcaucasicus Karabach, Schirwan et Talysch nec non in territorio Elisabethpolensi observavit. Bull. Soc. Nat. Moscou, X : 136—147.
Hooijer D. A. 1949. Mammalian evolution in the Quaternary of southern and eastern Asia. C. R. XVIHe Congr. Intern. Zool., Paris (1948) : 507—511.
Hooijer D. 1950. The study of subspecific advance in the quaternary. Evolution. IV, 4 : 360—361.
Hopwood A. T. 1935. Fossil Elephants and Man. Proc. Geol. Assoc., 46 : 46—60.
Kinnear N. B. 1920. The past and present distribution of the Lion in South-Easter Asia. Journ. Bombay Nat. Hist. Soc., 27 : 33—39.
Lindner K. 1937. Geschichte des deutschen Weidwerks. I. Die Jagd der Vorzeit. Berlin und Leipzig : 1—435.
Lindner K. 1940. Geschichte des deutschen Weidwerks. IL Die .Jagd im friihen Mit-telalter. Berlin : 1—477.
Linstow O. 1908. Die Verbreitung des Bibers im Quart ar. AbhandL und Ber. Mus. fur Natur- und Heimatk. zu Magdeburg, I, IV : 213—387.
Lydekker R. 1885. Catalogue of the fossil Mammalis in the British Museum, L London : I—XXX4-1—268.
Lydekker R. 1886. On the Fossil Mammalia of Maragha in North-Western Persia. Quart. Journ. Geolog. Soc., XLII : 166.
Lydekker R. 1892. On Zeuglodont and other Cetacean Remains from the Tertiary of the Caucasus. Proc. Zool. Soc. : 558—564.
L у d e к к e r R. 1898. Wild Oxen, Sheep and Goast of all Lands. London : 1—318. Lydekker R. 1913—1916. Catalogue of the ungulate mammals in the British Museum, I—V. London. 1913, I : 1—249; 1914, II : 1—295; III : 1—283; 1915, IV: 1—438; 1916, V : 1—207.
Mecquenem R. 1911. Contribution a I’etude du Gisemcnt des Vertebres de Maragha et de ses Environs. Ann. d’Hist. Nat., Paris, 1 : 81—98.
Mecquenem R. 1924. Contribution a Petude des fossiles de Maragha. Ann. Paleon-tol., XIII : 135—160.
Miller G. S. 1912. Catalogue of the mammals of Western Europe. London : XV-p у.-. , ... . ...
Зоологическая литература $13
Mitchell G. F. and H. M. Parkes. 1949. The Giant Deer in Ireland. Proc. Royal Irish Acad., 52, sect. B, 7 : 291—314.
Mivart G. S. 1890. Dogs, Jackals, Wolves and Foxes, a Monograph of the Canidae, London : I—XXV^-l—216.
M u r i e I. 1870. On the Antelope saiga tatarica (Pall.). Proc. Zool. Soc., London:451— . 503.
N e h r i n g A. 1890. Ueber Tundren und Steppen der Jetzt- und Vorzeit, mit besonderer Beriicksichtigung ihrer Fauna. Berlin, F, DiimmlVIII : 1—258.
Nehring A. 1891. Diluviale Reste von Cuon, Ovis, Saiga, Ibex und Rupicapra aus Mahren. Neuen Jahrbuch Mineral., Geol. und Paleontol : 107—155.
Neuhauser G. 1936. Die Murieden von Kleinasien. Zeitschr. Saugetierkundq, II : 163—236.
Noska M. 1895. Zwei monographische Studien I. Capra (Aegoceros) caucasica Giild. II. Capella rupicapra Keys, et Blas. Dresden : 1—42. . .
Osborn H. F. 1915. Review of the Pleistocene of Europe, Asia and northern Africa. Ann. New York Acad. Sci. 26 : 215—315.
Osborn H. F. 1921. The age of mammals in Europe, Asia and North America. New York: 1—636.
Osborn H. F. 1922. Pliocene (Tertiary) and Early Pleistocene (Quarternary) Mammalia of East Anglia, Great Britain, in Relation to the Appearance of Man. Geol. Mag., LIX, X: 433—441.
Isborn H. F. 1936, 1942. Proboscidea. A monograph of the Discovery, Evolution, Migration and Extinction of the Mastodonts and Elephants of the World. 1936, I : 1—802; 1942, II : 805—1676.
’ci W. C. 1939. An attempted correlation of quarternary geology, palaeontology and prehistory on Europe and China. Geochronological table, I. Univ, of London, lust, of Archeol.
5ic ar d L. 1937. Inferences on the problem of the Pleistocene Climate of Palestina and Syria drawn from Flora, Faune and Stratigraphy. Proc. Prehist. Soc., N. S., Ill, 1:58—70.
Pilgrim G. 1913. Correlation of the Siwaliks with Mammal Horizons of Europe. Rec. Geol. Survey India, XLIII, 1 : 264.
Pilgrim G. E. 1939. The fossil Bovidae in India. Mem. Geol. Survey India, N. S.4 XXVI, 1 : 1—356.
Pilgrim G. E. 1944. The Lower Limit of the Pleistocene in Europe and Asia. Geol. Mag., LXXXI, 1 : 28—38.
Pocock R. I. 1929. Tigers. Journ. Bombay Nat. Hist. Soc., XXXIII, 3 : 505—541.
Pocock R. I. 1930. The Lions of Asia. Journ. Bombay Nat. Hist. Soc., XXXIV :
638—665.
Pocock R. L 1935. The Races of Canis lupus. Proc. Zool. Soc., 4 : 647—686.
Pocock R. I. 1938. The Jackals of S. W. Asia and S. E. Europe. Proc. Zool. Soc.,,
108B : 37—39.
Pocock R. I. 1939, 1941. The fauna of British India, including Ceylon and Burma. Mammalia. London, 1939,1 : 1—503; 1941, II : 1—463.
P о h 1 i g Ht 1886. On the Pliocene of Maragha, Persia Resemblance to that of Pikermi in Greece; on Fossil Elephant Remains of Caucasia and Persia and on the Results of a Monograph of the Fossil Elephants of Germany and Italy (com. Dr. J. Hinde). Quart. Journ. Geolog. Soc., XLII, 166 : 177—182.
P о h 1 i g H. 1889. Monographic des Elephas antiquus Falc. mit Beitragen uber Elephas primigenius Blum, und Elephas meridionalis Nesti. Nova Acta. Acad. Coes. Leop. Corel. Germ. Naturae Curiosorum, Dresden : 1—259.
Rod 1 er A. und K. Weithofer. 1890. Die Widerkauer von Maragha. Denkshr. Akad. Wissensch., Wien, 57 : 752—772—[-6 taf.
Schreuder A.1929. Conodontes (Trogontherium) and Castor from the Tegliap Clay compared with the Castoridae from other localities. Arch. Mus. Teyler, ser. Ill, VI : 99—321.
Sela ter W. L. and L. Ph. 1899. The Geography of Mammals. London : 1—335. \ Scia ter P. L. and O. Thomas. 1894—1900. The book of antelopes. London, 1894, I : I—XXXV+1—220; 1896—1897, II : 1 — 194; 1897—1898, III : 1—245; 1899—1900, IV : 1—242.
Simionescu I. 1930. Vertebratele pliocene de la Malusteni (Covurlui). Aoad\ Romana Publ. Fond. Vas. Adam., IX, XLIX : 83—148.
Simpson G. G. 1941. Range as a zoological character. Amer. Jour. Sci., 239 : 785— 804. \
Simpson G. G. 1945. The principles of classification and a classification of mammals.
Bull. Amer. Mus. Nat. Hist., 85 : I—XVLf-l —350.
39*
612
Литература
Skinner М. F. and О. C. Kais е n. 1947. The fossil Bison of Alaska and preliminary revision of the genus. Bull. Amer. Mus. Nat. Hist., 89, 3 : 127—256.
Soergel W. 1922. Die Jagd der Vorzeit. Jena : 1—149.
Stehlin H. G. et P. Graziosi. 1935. Ricerche Sugli Asinidi fossili d’Europa. Ahh. Schweiz, palaont. Ges., 56, Mem., 3 : 1—73.
Stirton R. A. 1935. A review of the Tertiary beavers. Un. Gal. Publ. Bull. Dep. Geol. Sciences, 23, 13 : 391—458.
Sunde vail C. J. 1863. Die Tierarten des Aristoteles. Stockholm : 1—242.
Tate G. H. 1942. Review of the Vespertilionine bats with special attention to genera and species of the Archbold collections. Bull. Amer. Mus. Nat. Hist., 80 : 221—297.
Thenius E. 1949. Ueber die Saugetierfauna aus dem Unterpliozan von Ilhan bei Ankara (Tiirkei). Sitzungsb. Oester. Akad. Wissensch. Abt. I. Biologie, Bd. 158, Hf. 9/10:656—661.
Thomas O. 1906. On a collection of Mammals from Persia and Armenia preesented to the Brit. Mus. by Col. A. G. Bailward. Proc. Zool. Soc., II, XXXV :509—527.
Trouessart E. L. 1898—1899 a. Catalogus mammalium tarn viventium quam fossilium. I, Primates, Prosimiae, Chiroptera, Insectivora, Carnivora, Rodentia, Pinninedia, Berolini: 1—664.
Trouessart E. L. 1898—1899 6. Catalogus mammalium tarn viventium quam fossilium. II. Tillodontia, Ungulata, Sirenia, Cetacea, Edentata, Marsupialia, Allo-theria, Monotrcmata. Appendix (Addenta et Corrigenda). Index Alphabeticus, Berolini : 665—1469.
Trouessart E. L. 1904—1905. Catalogus mammalium tarn viventium quam fossilium. Quinquennale suppiementum, III, Berolini: 1—929.
Uvarov В. P. 1921. The geographical distribution of Orthopterous insects in the Caucasus and in Western Asia. Proc. Zool. Soc., London, 31 : 447—472.
Vaufrey R. 1931. Paleontologie de 1’Acheleen superienz de la grotte d’Oumm-Quatafa (Palestine). L’Anthropologic, XLI, 3—4:253—262.
(.Viet inghof f]. 1812. Discours sur quelques objcts d’histoire naturelle recueillis au Caucase prononces par le Baron de Vietinghoff. Mem. Soc. Imp. Natur. Moscou, III : 83—96.
Wallace A. R. 1876. The Geographical Distribution of Animals. London, 1:1— - 503; II : 1—607.
Woldrich I. N. 1882. Die diluvialen Faunen Mittel-Europas und eine heutige Sareptaner Steppenfauna in Niederosterreich. Mitth. Wiener Anthrop. Geselsch., XI : 1—26.
Woldrich J. H. 1893. Reste Diluvialer Faunen und des Menschen aus dem Wald-viertel Niederosterreichs. Denkschr. der kais. Acad, der Wissensch., 60, Wien:l—632.
Wolf B. 1938. Fauna fossils cavernorum. I. Fossilium Catalogus. I. Animalia, Pars 82 : 1—192; II. Fossilium Catalogus. I. Animalia, Pars 89 : 1 —196.
Wolf B. 1939. Fauna fossilis cavernorum. I. Fossilium Catalogus. I. Animalia, Pars 82 : 193—240. II. Fossilium Catalogus. I. Animalia, Pars 89 : 97—208.
ПРОЧАЯ ЛИТЕРАТУРА
(ботаника, геология, археология и др.)
(А б и х) A b i с h Н. 1857. Ueber das Steinsalz und seine geologische stellung im Russischen Armenien. Palaeontologischer Theil. Mem. de 1’Academy Imp. des ' Sciences de St. Petersb., VI ser., VII : 1—92.
А б и x Г. В. 1899. Геология Армянского нагорья. Орографическое и географическое описание. Перевод Б. 3. Коленко. Зап. Кавк. отд. Русск. геогр. общ., XXI : 1—202.
Авдиев В. И. 1953. История Древнего Востока. Госполитиздат, 2-е изд., Л.: 1—758.
Алехин В. В. 1938. География растений. Учпедгиз, М. : 1—327.
Ализаде К. А. 1955. Геологическое строение бинагадинского местонахождения ископаемых костей. Бинагадинское местонахождение четвертичной фауны и флоры, IV. Тр. Естеств. ист. муз. АН АзербССР, X : 5—9.
Альбов Н. 1892. Леса Абхазии. Общ. сельск. хоз. Южн. России, 4 : 37—53.
Ай а ной а Е. Н. 1954. Палинологические данные. Бот. Журн. 39:343.
Андрусов Н. И. 1918. Взаимоотношения Эвксинского и Каспийского бассейнов в неогеновую эпоху. Изв. Росс. AH, XII : 749—760.
А н д р у с о в Н. И. 1926. Геологическое строение и история Керченского пролива. Бюлл. Моск. общ. испыт. природы, отд. геол., IV (3—4), нов. сер., XXXIV* 294—330.
Прочая литература
613
Андрусов Н. И. 1928. Очерк истории развития Каспийского моря и его обитателей. Матер, по геол, истории Каспийского моря, I, Баку : 17—36.
Аноним. 1853. Географические известия о древних российских и иностранных писателях. Отеч. зап., LXXVI, 2, И : 123.
А н < н и м. 1855. Ведомость об отвозных за границу закавказских товаров по Закав-' казскому карантинному таможенному округу. Зап. Кавк. отд. Русск. геогр. общ., III : 308—311.
А в о н и м. 1884—1885. Численность народонаселения больших административных отделов Кавказского края в 1882 году и приблизительная плотность населения этих отделов. Изв. Кавк. отд. Русск. геогр. общ., VIII : 96—108.
Анон и м. 1902. Падение ледников с Гимрайхоха. Изв. Кавк. отд. Русск. геогр. общ., XV, 2—5 : 205—210.
Анучин Д. Н. 1884. Отчет о поездке в Дагестан летом 1882 г. Изв. Русск. геогр. общ.,. 4 : 357—449.
Анфимов Н. В. 1935. Новые данные к истории Азиатского Боспора. Сов. археол., VII : 258—267.
Анфимов Н. В. 1951. Раскопки Семибратнего городища. Кратк. сообщ. Инет, ист. матер, культуры, XXXVII : 238—244.
Апостолов ^!. Я. 1898. Климатическое значение лесов в связи с облесением Кубанской области. Кубанск. сб., IV : 1—16. .
Аристов Н. 1866. Промышленность древней Руси. СПб : I—VI-J—1—321.
Артамонов М. И. 1937. Раскопки курганов в долине р. Маныч в 1935 г. Сов. археол., IV : 93—133.
Артамонов М. И. 1952. Белая Вежа. Сов. археол., XVI : 42—76.
Архангельский А. Д. 1934. Геологическое строение СССР. Л.—М., 2-е изд.: 1—223.
Архангельский А. Д., Н. М. Страхов. 1938. Геологическое строение и история Черного моря. М.—Л. : 5—226.
Асланян А. Т. 1950. Стратиграфия вулканогенных пород Приереванского района (Армянская ССР). Докл. АН АрмССР, XI, 5 : 145—150.
Байер н Ф. 1871. О древних сооружениях на Кавказе. Сб. сведений о Кавказе, 1 : 298—325.
Баранов В. И. 1948. Этапы развития флоры и растительности СССР в третичном периоде. Уч. зап. Казанск. гос. унив. им. В. И. Ульянова-Ленина, 108, 3, 7 : 1—135.
Баранов В. И. 1952. О чем говорят песчаники Камышина и пески Ергеней. Сталинград : 1—46.
[Бар. бар о И.]. 1836. Путешествие в Тану Иосафата Барбаро, венецианского дворянина. Библиотека иностр, писателей о России. СПб, 1 : I—XV1+1—156.
Батурин В. П. 1931. Физико-географические условия века продуктивной толщи Тр. Азерб. нефт. иссл. инет., 1 : 52—96.
Башкиров А. С. 1931. Искусство Дагестана, М. : 1—117, табл. 1—107.
Белл. 1776. Белевы путешествия через Россию в разные асиятские земли, а именно: в Испаган, в Пекин, в Дербент, в Константинополь. Перевел с французского М. Попов. СПб., ч. I : 1—250; ч. II : 244; ч. III : 1—246.
Белоусов В. В. 1939. Большой Кавказ. Опытные геотектонические исследования. Тр. Центр, научно-иссл. геол.-разв. инет., 126.
Берг Л. С. 1934. Уровень Каспийского моря за историческое время. Пробл. физ. географии, Изд. АН СССР, 1 : 11—65.
Берг Л. С. 1938. Основы климатологии. Учпедгиз, Л. : 1—427.
Берг Л. С. 1947. Климат и жизнь. Географгиз, М.—Л., 1 : 1—356.
Берг Л. С. 1950. Некоторые соображения о послеледниковых изменениях климата и о лесостспье. Вопр. геогр., 23 : 57—84.
Берг Л. С. 1952. Географические зоны Советского Союза, II. М.-—Л. : 1—510.
Бердзенишвили Н. А., А. Джавахишвили, С. Н. Джана ши а. 1946. История Грузии с древнейших времен до начала XIX века, 1. Госизд. ГрузССР, Тбилиси : 1—454.
^Берцелиус С. Л. (Нальчагаров). 1927. Верхнесарматские отложения Эльдара.
’ Изв. общ. обслед. и изуч. Азерб., 5, Баку : 181—193.
Бламберг И. Ф. 1853. Статистическое обозрение Персии, составленное подполковником И. Ф. Бламбергом в 1841 г. Зап. Русск. геогр. общ., XII, СПб. : 1—305.
Богачев Б. В. 1918. Очерки географии Всевеликого войска Донского. Ростов : 1—202.
Богачев В. В. 1928. Урмийское и Ванское озера. Изв. Азерб. гос. унив., отд. естествозн. и мед., VII : 165—205.
Богачев В. В. 1932. Геологические экскурсии в окрестностях Баку. Баку : 3—85.
.ш
Литература
В о г а ч ев В. В. 4938. Миоцен Закавказья. Тр. Азерб. фил. АН СССР, X, 44, Баку: 1—46.
Б огд^а и В. С., Б. А. Шуманов. 1925. Закубанские плавни в почвенно-ботаническом и мелиоративном отношениях. Тр. Кубанск. инет., 3 : 15—76.
Б он л а н Г. В. 1832. Описание Украины. СПб. : 1—179.
* Боровка Г. И. 1922. Бронзовый олень из Ульского кургана. Изв. Акад. инет, матер, культуры, II : 193—203.
Б р о н е в с к и й С. 1823. Новейшие географические и исторические известия о Кавказе. М., 1 : I—XXXI+1—352; 2 : I—IX+1—465.
Б уза-нд Ф. 1953. История Армении Фавстоса Бузанда. Изд. АН АрмССР, Ереван : > V—XVI+1—237.
Бурчак-Абрамович Н. О., Н. К. Верещагин. 1947. Священная роща в Кабристане. Докл. АН АзербССР, III, 4 : 168—170.
Бурчак-Абрамович Н. И., Н.К. Верещагин. 1950. Древние можжевельники (Juniperus polycarpos С. Koch) в Нахичеванской АССР. Изв. АН АзербССР, 12 : 33—37.
Буш Н. А. 1900. Описание и главнейшие результаты третьего путешествия по северо-западному Кавказу в 1899 г. Изв. Русск. геогр. общ., 36 : 227—298.
Буш Н. А. 1935. Ботанико-географический очерк Кавказа. М.—Л. : 5—105.
Буш Н. А., Е. А. Б у ш. 1936. Растительный покров восточной Юго-Осетии и его динамика. Тр. Совета по изуч. производ. сил, сер. Закавказ., XVIII : 1—264. - В а и д о в Р. М. 1952. Археологические работы в Мингечауре в 1950 г. Кратк. сообщ. J Инет. ист. матер, культуры, XLVI : 86—100.
5 Варданянц Л. А. 1933а. Материалы по геоморфологии Большого Кавказа, I. Центральный и Западный Кавказ. Изв. Гос. геогр. общ., LXV, 2 : 116—139.. Варданянц Л. А. 19336. Материалы по геоморфологии Большого Кавказа. II. Большой Кавказ. Изв. Гос. геогр. общ., LXV, 3 : 187—211.
Варданянц Л. А. 1935. Метеорологический фактор в развитии оледенения. Изв. 4 Гос., геогр. общ., 67, 4 : 460—470.
Варданяна Л. А. 1939. К истории развития рельефа Центрального Кавказа. Тр. Сов. секции Междун. ассоц. по изуч. четверт. пер. Европы, IV : 95—98.
Варданянц Л. А. 1948. Постплиоценовая история Кавказско-Черноморско-Каспийской области. Ереван : 1—184.
В а р е н ц о в М. Н. 1933. Геологическая история Таманского полуострова в после-третичное время. Тр. Второй Междун. конф, ассоц. по изуч. четвертичн. периода Европы, 3 : 88—101.
Васильев Я. К. 1898. Значение рек Кубанской области в лесном хозяйстве. Кубанск. сб., IV : 1—30.
Вассоевич Н. Б. 1928. Палеонтологические заметки по плиоценовым и после-третичным отложениям Таманского полуострова. Изв. Геол, комит., 47, 6 : 711— 731.
Вассоевич Н. Б. 1929. Капустина балка. Таманский полуостров. Азерб. нефт. хоз., 8—9 : 16—37.
В а х у шт и Царевич. 1904. География Грузии. Перевод и редакция М. Г. Джана-швили. Зап. Кавк. отд. Русск. геогр. общ., XXIV, 5, Тифлис : I—XLIX4-1—241.
Вейденбаум Е. 1878. Священные рощи и деревья у кавказских народов. Изв. Кавк. отд. Русск. геогр. общ., V, 3 : 153—179.
Вейденбаум Е. 1879. Кавказские пещеры. Пятый археол. съезд в Тифлисе, М. : 55—57.
Верещагин Н. К. и Н. О. Бурчак-Абрамович. 1948. Рисунки на скалах юго-восточного Кабристана. Изв. Гос. геогр. общ., LXXX, 5 : 507—518.
[Веселовский Н. И.]. 1901. Раскопки Н. И. Веселовского. Кубанская обл. Отчет Археол. комисс. за 1898 г., СПб. : 1—65.
[ Веселовский Н. И.]. 1903. Раскопки Н. И. Веселовского. Кубанская обл. Отчет Археол. комисс. за 1901 г., СПб. : 66—89.
[Веселовский Н. И.]. 1907. Раскопки Н. И. Веселовского. Кубанская обл. Отчет Археол. комисс. за 1904 г. СПб. : 85—97.
[Веселовский Н. И.]. 1910. Раскопки И. И. Веселовского. Кубанская обл. Отчет Археол. комисс. за 1907 г. СПб. : 85—91.
[В и ж е н е р Б.]. 1890. Извлечение из записок Блеза до-Виженер (1573). Мемуары, относящиеся к истории Южн. России. 1, Киев : 59—-88.
Винда В. 1910. Майкопские залежи нефти в связи с нефтепромышленностью Кубанской области, ч. 1. Екатеринодар : 1—404.
Вирхов Р. 1884. Замечания о Кавказе, состоянии его в доисторическое время, об его народах и промышленности последних. Изв. Кавк. отд. Русск. геогр. общ., ' VIII, 2.
Прочая литература
6JS
Воейков А. И. 1881. Климатические условия ледниковых явлений настоящих и прошедших. Зап. СПб. минерал, общ., II сер., ч. 16 : 21—90.
Воейков А. И. 1894- Воздействие человека на природу. Землеведение, 11 : 1—32.
Воларович П. и Н. Леднев. 1913—4929. Детальная геологическая карта
Апшеронского полуострова. Кирмаку—Бинагадинский район. Масштаб 1 : ’ 8400, Л.
Волков Ф. 1899. Палеолитическая стоянка на Черноморье (ст. Ильская, к тогу от Новороссийска). Киевская старина, 6 : 167—168. : f
Вульф Е. В. 1944. Историческая география растений. История флор земного шара.
М.—Л. : 1—545.
В я з и г и н А. 1929. Памятник первобытной религии на Северном Кавказе — древнее осетинское святилище Реком. Результаты археол. эксп. А. А. Миллера. Революция и горец, 5 (7), Ростов/Дон : 75—76.
Гайдукевич В. Ф. 1948. Боспорское царство. М.—Л. : 1—622.
Галстян Б. Я. 1930. Основные черты географии почв ССР Армении. Закавк. краеведческ. сб., сер. А., естествозн., 1, Тбилиси : 248—265. • t
Гамбарян П. П. 1934. Ыурнусское местонахождение диатомита. Сб. научно-иссл. трудов НИИС при СНК ССР Армении, 1, Ереван : 5—35. ;
Ган К. Ф. 1884, 1894. Известия древних греческих и римских писателей о Кавказе. Тифлис, 1884, 1, IV : 1—1248; 1890, 2, IX : 1—224.
Гату ев С. А. 1932. Обзор третичных отложений восточного Предкавказья. Тр. Геол, инет., II : 1—35.
Г а т у е в С. А. 1947, Новейшие движения земной коры в области Восточного Предкавказья. Природа, 3 : 47—48.
Гвоздецкий Н. А. 1954. Физическая география Кавказа, 1. Изд. Моск. унив. : 1—208.
Герасимов И. II. 1936. О значении эпейрогенических движений в развитии рельефа Прикаспийской и Западно-сибирской низменности. Изв. Гос. геогр. общ. 5 : 611—619.
Герасимов И. П. 1937. Каспийское море в четвертичный период. Тр. Сов. секции Междун. ассоц. по изуч. четвертичн. периода Европы, III : 5—29.
Герасимов И. II. иК. К. М а р ко в. 1939. Четвертичная геология. М. : 1—361.
Геродот. 1888. История в девяти книгах. Перевод, предисловие и указатель Ф. Г. Мищенко. М. I : 1—639; II : I—CLXVIII-H—394.
Герц К. 1870. Археологическая топография Таманского полуострова. М. : 1—128., Гзелишвили И. А. 1953. К вопросу о стратиграфии «Сагварджиле». Сообщ. АН ГрузССР, XIV, 9 : 669—675.
Гмелин С. Г. 1771, 1777, 1785. Путешествие по России для исследования трех царств естества, СПб. 1, 1771 : 1—272; 2, 1777 : 1—361; 3, 1785 : 1—737.
Гниловской В. Г. 1955. Клухорский район. Матер, по изуч. Ставропольского края, 7. Ставропольск. краевой музеи и общ. краевед. : 61—82.
Гниловской В. Г., А. С. Егоров. 1955. Геоморфологические условия района Косякинского карьера. Матер, по изуч. Ставропольского края, 7. Ставропольск. краевой муз. и общ. краевед. : 305—312.
Голубятников Д-В. 1925. Продуктивная толща Апшеронского полуострова. Азерб. нефт. хоз., 8—9.
Горецкий Г. И. 1948. Новые стоянки конца неолита в эпохи бронзы на террасах нижнего Дона и Маныча как геологические документы. Изв. Всесоюзн. геогр. общ., 80, 5 : 535—540.
Городцов В. А. 1935. О результатах археологических исследований Елизаветинского городища и могильника в 1934 г. Сов. этногр., 3 : 71—76.
Городцов В. А. 1936. Елизаветинское городище и сопровождающие его могильники по раскопкам 1935 года. Сов. археол., 1 : 171 — 187.
Городцов В. А. 1940. Ильская палеолитическая стоянка по раскопкам 1937 года.
Бюлл. комисс. по изуч. четвертичн. периода, 6—7 : 89—92.
Городцов В. А. 1941. Результаты исследования Ильской палеолитической стоянки Матер, и исслед. по археол. СССР, 2 : 7—26.
Грен А. И. 1895. Краткий очерк истории Кавказского перешейка, 1. Языческий период. Киев : 1 —123.
Григоров Н. 1892. Село Татев Зангезурского уезда Елизаветпольской губернии. Сб. матер, для описан, мести, и племен Кавказа, XIII : 59—125.
Григорьев С. Г. 1913. Долина окрестностей Кисловодска. Сб. в честь 70-летнего юбилея проф. Д. Н. Анучина. М. : 77—106.
Громов В. И. 1940а. Итоги изучения геологических условий нахождения палеолита на Кавказе и его значение для четвертичной стратиграфии. Бюлл. Комисс.
» ho изуч. четвертичн. периода, 6—7 : 93—96.
616
Л итература
Гро м о в В. И. 19406. Материалы к изучению террас реки Терека между г. Орджоникидзе и Моздоком. Тр. Инет. геол, наук, 33, геол. сер. 10 : 17—35.
Громов В. И. 1950а. Геологический возраст палеолита на территории СССР. Матер, по четвертичн. периоду СССР, 2 : 90—106.
Громов В. И. 19506. О верхней границе третичного периода. Матер, по четвертичн. периоду СССР, 2 : 5—12.
Гроссгейм А. А. 1930. Очерк растительного покрова Закавказья. Тифлис : 1—38..
Гроссгейм А. А. 1932. Очерк растительности Кура-Араксинской низменности.
Матер, к общ. схеме использ. водн. ресур. Кура-Аракс. басе. 4, Баку : 57—125.
Гроссгейм А. А. 1936. Анализ флоры Кавказа. Тр. Ботан. инет. Азерб. фил. АН СССР, 1 : 1—257.
Гроссгейм А. А. 1946. Растительные ресурсы Кавказа. Изд, АН АзербССР, Баку: 1—671.
Гроссгейм А. А. 1948. Растительный покров Кавказа. Изд. Моск. общ. испыт. природы, М. : 3—265.
Губкин И. М. 1914. Заметка о возрасте слоев с Elasmotherium и Elephas на Таманском полуострове. Изв. Акад, наук, сер. VI, VIII, 9, СПб. : 587—590.
Гуммель Я. И. 1948. Некоторые памятники раннебронзовой эпохи Азербайджана. Кратк, сообщ. Инет. ист. матер, культуры, XX : 15—28.
Г уме ль Я. И. и А. А. Яценко-Хмелевский. 1941. Древесная растительность города Ганджи в эпоху Низами Ганджеви, XII в. н. э. Сообщ. АН ГрузССР, II, 8 : 733—740.
[Гюль дев шт едт]. 1879. Дневник путешествия в Южную Россию академика Гильденштедта в 1773—1774 г. Перевод М. Шугунова. Зап. Одесск. общ. истории и древностей, XI : 180—228.
Де ген-Ковалевский Б. Е. 1935. Отчет о работах. Работы на строительстве Баксанской гидроэлектростанции. Изв. Акад. ист. матер, культуры, 110 :11—28.
Дементьев К. 1888. Высыхание Ставропольской губ. Изв. Кавк. отд. Русск. геогр. общ., 265—287.
Деминский Ф. А. 1901. Некоторые сведения о Кабристанском полицейском участке Шемахинского уезда Бакинской губернии. Зап. Кавк. отд. Русск. геогр. общ., XXII, 2 : 1—70.
Демченко М. А. 1947. Отступание ледников в бассейне реки Терек. Изв. Все-союзн. геогр. общ., 5 : 590—592.
Джанашия Н. 1915. Религиозные верования абхазов. Христианский восток,. IV, 1 : 72—112.
Джафар-Заде И. М. 1946. Элементы археологической культуры древней Му-гани (оружие и керамика). Изв. АН АзербССР, 9 : 21—51.
Динник Н. Я. 1884а. Горы и ущелья Терской области. Зап. Кавк. отд. Русск. геогр. общ., ХШ, 1 : 1—48.
Динник Н. Я. 18846. Горы и ущелья Кубанской области. Зап. Кавк. отд. Русск, геогр. общ., ХШ, 1 : 307—363-
Динник Н. Я. 1884в. Осетия и верховья Риона. Зап. Кавк. отд. Русск. геогр. общ., ХШ, 1 : 49—100.
Динник Н. Я. 1890а. Путешествие по Дигории. Зап. Кавк. отд. Русск. геогр. XIV, 1 : 1—61.
общ.,
Д и н н и к Н. Я. 18906. Поездка в Балкарию в 1887 году. Зап. Кавк. отд. Русск.
геогр. общ., XIV, 1 : 199—247.
Ди н н и к Н. Я. 1890в. Современные древние ледники Кавказа. Зап. Кавк. отд.
Русск. геогр. общ., XIV, 1 : 282—416.
Динник Н. Я. 1893а. Путешествие по Пшавии и Тушетии. Зап. Кавк. отд. Русск. геогр. общ., XV : 91—147.
Дин ник Н. Я. 18936. Путешествие по Зап. Осетии. Зап. Кавк. отд. Русск. геогр. общ., XV : 51—90.
Динник Н. Я. 1911. Лесная зона северо-западного Кавказа. Тр. Ставропольск. общ. изуч. Сев.-Кавк, края, 1 : 15—27.
Динник Н. Я. 1912. Путешествие по Закатальскому округу и Дагестану. Изв.
Кавк. отд. Русск. геогр. общ., XXI, 2 : 1—60.
Долгушин А. А. 1924. Леса Закавказья и их эксплуатация. Тр. Закавказск. научн. ассоц. при Зак. Цик, Тифлис : 1—225.
Домбровский Б. С. 1914. Заметка о геологических условиях нахождения костей ископаемых млекопитающих в местности Эльдара (Тифлисской губернии). Тр. Геол. муз. АН, VIII, 14 ; 231—238.
Дорн Б. 1875. Каспий. Прилож. к XXVI т. Зап. Акад, наук : 1—LVI+1—718.
Думитрашко Н. В. 1949. О древнем оледенении Малого Кавказа. Тр. Инет, геогр., 43 : 33—52.
Прочая литература
617
:Дюбуа. деМонпере Ф. 1937. Путешествие вокруг Кавказа. Изд. Инет, абхаз* ской культуры, 1, 6, Сухуми : 1—175.
*Е ф и м е н к о П. И. 1953. Первобытное общество. Л. : 1—658.
Е фи менко П. П. и Н. А. Береговая. 1941. Палеолитические местонахо-- ждения СССР. Матер, и исслед. по археол. СССР, 2 : 254—290.
;Жиж\(енко Б. П., В. А. Колесников, А. Г. Э б е р з и н. 1940. Неоген - СССР. Стратиграфия СССР, XII. Изд. АН СССР, М.—Л. : 1—677.
(Жирмунский А. М. 1936. Новые опыты синхронизации четвертичных отложений ! Западной и Восточной Европы. Матер, по четвертичн. периоду СССР : 46—62. (Жуков М. М. 1939. Миграция дельт рек Волги, Кумы и Урала за послеледниковой i время. Тр. Сов. секции Междун. ассоц. по изуч. четвертичн. периода Европы, [ IV : 22—25.
Жуков М. М. 1945. Плиоценовая и четвертичная история севера Прикаспийской впадины. Пробл. Зап. Казахстана, II, Изд. АН СССР, М.—Л. : 1—234.
Заклинская Е. Д. 1953. К вопросу об основных этапах в развитии кайнозой-? ской флоры юга Европейской части СССР на основании данных спорово-пыльце-• вых анализов. Докл. АН СССР, LXXXIX, 5 : 913—917.
Ваклинский Н. С. 1931. Леса Абхазии:!—210.
Замятнин С. Н. 1934. Итоги последних исследований Ильского палеолитического местонахождения. Тр. Второй Междун. конф, по изуч. четвертичн. пе-> риода Европы, 5 : 207—219.
Вамятнин С. Н. 1935. Новые данные по палеолиту Закавказья. Сов. этногр. 2 : .116—123.
Вамятнин С. Н. 1937а. Палеолит Абхазии. Тр. Инет, абхазской культуры, X : 1—54.
Вамятнин С. Н. 19376. Пещерные навесы Мгвимеви, близ Чиатуры (Грузия).
Сов. археол., 3 : 57—76.
Замятнин С. Н. 1940. Навалишинская и Ахштырская пещеры на Черноморском ! побережье Кавказа. Бюлл. Комисс. по изуч. четвертичн. периода, 6—7 : 100— ; 101. .
Вамятнин С. Н.1947. Находки нижнего палеолита в Армении. Изв. АН АрмССР, ? общ. науки, 1 : 15—26.
Замятнин С. Н. 1950а. О первоначальном заселении пещер. Кратк. сообщ.
: Инет. ист. матер, культуры, XXXI : 55—63.
Замятнин С. И. 19506. Изучение палеолитического периода на Кавказе за 1936— 1948 гг. Матер, по четвертичн. периоду СССР, 2 : 127—139.
Замятнин С. Н. 1957. Палеолит западного Закавказья. Сб. Музея антропол. и этногр., XVII : 432—499.
3 а р и д з е Г. М. и Н. Ф. Т а т р и ш в и л и. 1948. О возрасте Цалкинского лавового комплекса. Докл. АН СССР, LIX, 1 : 111—113.
Захаров С. А. 1913. О почвенных областях и зонах Кавказа. Сб. в честь 70-летнего юбилея проф. Д. Н. Анучина, М. : 557—588.
Захаров С. А. 1934. Опыт классификации почв Закавказья. Тр. почв. секц. Груз, фил. АН СССР, 1 : 1—92.
Захаров С. А. 1935. Борьба леса и степи на Кавказе (по данным почвенных исследований). Почвоведение, 4 : 501—549.
Зедельмейер О. М. 1927. Распространение торфяных болот и сфагновых мхов на Кавказе. Торфяное дело, 7 : 153—156.
Зунтуриди И. Г. 1931. К вопросу о вековых колебаниях земной коры в Закавказье. Закавк. краеведческ. сб., 1 : 203—212.
II. К. 1948. Хроника научной жизни. Научная сессия по археологии Закавказья. Вести, древней истории, 3 : 169—181.
Л в а н о в с к и й А. А. 1911. По Закавказью. Матер, по археол. Кавказа, 6, VIII : 1—194.
Псссен А. А. 1931. Северокавказские сосуды с изображением оленя. Сообщ. Акад, ист. матер, культуры, 2 .* 14—18.
Псссен А. А. 1935. Отчет о работах, XI. Работы на Сулаке. Изв. Акад. ист. матер, культуры, 110 : 29—39.
Псссен А. А. 1940. Проблема изучения археологических памятников Колхидской I низменности в целях характеристики современных (послеледниковых) геол о-
I гических процессов. Бюлл. Комисс. по изуч. четвертичн. периода, 6—7 : F 107—108.
। И е с с е н А. А. и П и о т р о в с к и й Б. Б. 1940. Моздокский могильник. Археол.. L эксп. Гос. Эрмитажа, 1 : 1—53.
Ильинский А. П. 1937. Растительность земного шара. Изд. АН СССР, М.—Л. ; 1 1—458. .
$18
Лите ратура
К аганкатва ци Моисей. 1861. История Агван. СПб.: 1—371.
Лаландадзе А. Н. 1948. Периодизация памятников Самтавро. Вести, древней истории, 3 : 171—172.
Каландадзе А. Н. и Д. Н. Т у’ш а б р а ми шв и л и. 1955. Новые раскопки в пещере Гварджилас-Клде. Кратк. сообщ. Инет, археол., АН УССР, 4 : 155— 157.
<Каллистов д. п. 1949. Очерки по истории северного Причерноморья античной эпохи. Л. : 1—284.
Канделаки Н. и Г. Дзоценидзе. 1937. Карстовые пещеры: Цхалтубо и Сатаплия. Запов. и памяти, природы ГрузССР, 1, Тифлис : 124—135.
Караулов Н. А. 1901. Сведения арабских писателей о Кавказе, Армении и Азербайджане. Ал. Истархий. Книга путей и царств. Сб. матер, для описан, мести, и племен Кавказа, XXIX, Тифлис : 1—73.
Карта растительности Европейской части СССР, 1950, 1 : 2500. Под редакцией Е. М. Лавренко, М.—Л.
.К а с пи ев Б. Ф. 1939. Палеорека! в западном Ставрополье. Тр. Ворошиловск. гос. пед. инет., 1 : 75—90.
Каспиев Б. Ф. 1940. К плиоценовой истории Ставрополья. Природа, 1 : 93—94.
Каспиев Б. Ф. 1941. Древние реки западного склона Ставропольской возвышенности. Тр. Ворошиловск. гос. пед. инет., III : 95—110.
Кверфельдт Э.К. 1940. Черты реализма в рисунках на тканях и коврах времени сефевидов. Гос. Эрмитаж, Тр. Отд. Востока, III : 263—273.
Кецховели Н. 1942. Прошлое прибрежных лесов восточной Грузии и к вопросу об их восстановлении. Тр. Груз. с.-х. инет., XVIII : 29—32.
.Киладзе Н. В. 1944. Палеолитические находки в Мгвимеви. Вести, гос. муз. Грузии, ХП-В : 279—290 (на груз, языке).
Киладзе Н. В. 1953. Многослойный археологический памятник «Сагварджиле». Сообщ. АН ГрузССР, XIV, 9 : 561—567.
.Кириллов М. 1915. Действующие горные потоки в Нухинском лесничестве Елизаветпольской губернии и лес, как естественный фактор борьбы с ними. Леей. журн., 8—9 : 1251—1273.
Клопотовский Б. А. 1947. Почвенно-географический очерк Армении. Изв. АН АрмССР, сер. естеств. науки, 7 : 3—26.
Клопотовский Б. А. 1948. Ахалцихская котловина — узел основных ландшафтов Закавказья. Тр. Второго Всесоюзн. геогр. съезда, 1 : 356—367.
Книпович Т. Н. 1949. Танаис. М.—Л. : 1—149.
Князев В. 1946. Тебердинский государственный заповедник. Тбилиси : 1—61.
Кобылина М. М. 1951. Раскопки Фанагории. Кратк. сообщ. Инет. ист. матер, культуры, XXXVII : 232—237.
Ковалевский С. А. 1933. Лик Каспия. Тр. Геол.-Развед. конторы Азнефти, 2, Баку : 1—129.
Ковалевский С. А. 1936. Континентальные толщи Аджиноура. Азнефтеиздат, Баку—М. : 3—180.
Колаковский А. А. 1947а. Фитоландшафты Абхазии и история их развития (тезисы диссерт.), АН ГрузССР, Тбилиси : 1—28.
Колаковский А. А. 19476. К вопросу о происхождении флоры Колхиды в связи с историей флоры Ангариды и Средиземноморской фитогеографической области. Сообщ. АН ГрузССР, VIII, 3 : 151—159.
Колаковский А. А. 1951. Предварительные данные о находке плиоценовой флоры в Западной Грузии. Ботан. журн., 4 : 408—410.
Комаров А. В. 1879. Пещеры и древние могилы в Дагестане. Пятый археол. съезд в Тифлисе : 432—439.
Котов Ф. А. 1852. О ходу в Персидское царство и из Персиды в Турскую землю и в Индию и в Урмуз, где корабли приходят. Временник Моск. общ. ист. и древн. российских, кн. 14, М. : 1—22.
Краснов А. Н. 1893—1894. Кавказские цепи гор, параллельные Главному хребту и их роль в группировке лесной и степной флоры Западного Кавказа. Тр. Общ. испыт. природы при Харьковск. унив., XXVIII : 71—80.
Кричевский Е. 10. и Круглов А. П. 1941. Неолитическое поселение близ г. Нальчика. Матер, и исслед. по археол. СССР, 3 : 51.
К р и ш т о ф о в и ч А. Н. 1930. Основные черты развития третичной флоры Азии. Изв. Главн. ботан. сада СССР : 391—402.
Крон гельм К. 1873. О сталактитовой пещере близ Сухума. Изв. Кавк. отд. Русск. геогр. общ., II : 38—39.
Кру г л о в А. П. 1946а. Северо-восточный Кавказ во II—I тысячелетии до н. э. Кратк. сообщ. Инет. ист. матер, культуры, XIII : 130—133.
Прочая литература
61&
t
i
й
с.
i.
а-
ш
су
ГЗ.
в».
гве ли.
зв.
нд-
-61. гер.
[ИИ, дат, 1ТИЯ вязи 1СТИ. овой
сеол.
эмлю ст. и ребту Общ.
близ Азии. отд.
Н. 3.
Круглов А. П. 19466. Культовые места горного Дагестана. Кратк. сообщ. Инет, ист. матер, культуры, XII : 31—40.
Круглов А. П., Б. Б. Пиотровский, Г. В. Подгаецкий. 1941. Могильник в Нальчике. Матер, и исслед. по археол. СССР, 3 : 67.
Круковский Стефан. 1916. Пещера Гварджилас-Клде в Ргани. Изв. Кавк. » музея, X, 3 : 253—259.
Крупнов Е. И. 1946. Краткий очерк археологии Кабардинской АССР. Кабгос-издат, Нальчик : 1—47.
Крупнов Е. И. 1947. Северокавказская археологическая экспедиция. Кратк. сообщ. Инет. ист. матер, культуры, XVII : 99—104.
Крупнов Е. И. 1948. Археологические памятники верховьев реки Терека и бассейна реки Сунжи. Тр. Гос. ист. муз., Археол. сб., XVII : 7 : 55.
Крупнов Е. И. 1949. О поселениях скифского времени на Северном Кавказе. Кратк. сообщ. Инет. ист. матер, культуры, XXIV : 27—41.
I р ы м с к и й А. Е. 1934. Страницы из истории Северного, или Кавказского, Азербайджана, Кабала. Сб. в честь акад. С. Ф. Ольденбурга, Л : 289—305.
! сенофонт. 1896. Поход (Анабазис). Перевод В. А. Вроблевского. СПб. : 1—61.
; у д р я в ц е в Н. А. 1932. Геологические исследования в междуречье А лаза ни и Куры. Тр. Нефт. геол.-разв. инет. сер. Б, 32 : 1—52.
Кудрявцев Н. А. 1933. О возрасте Мирзаанской (Ширакской) толщи. Нефт. хоз., 4 : 215—219.
Кузнецов Н. И. 1890. Путешествие по Кавказу летом 1890 г. Изв. Русск. геогр. общ., XXVI : 413—432.
Кузнецов Н. И. 1896. Наши степи. Научи, обозрение, 7 : 194—-203.
Кузнецов Н. И. 1900. О причинах безлесия Армении. Изв. Русск. геогр. общ., 36 : 576—578.
К у з н е ц о в Н. И. 1909. Принципы деления Кавказа на ботанико-географические провинции. Зап. Акад, наук^ сер. VIII, XXIV, I : III—IV+1 —174.
(Кузнецов Н. И.1910. Нагорный Дагестан и значение его в истории развития флоры Кавказа. Изв. Русск. геогр. общ., XLVI : 213—260.
Кузнецов С. С. 1923. Геологический очерк Ставропольской губернии : 1—66. Кузнецов С. С. 1941. Вопросы геоморфологии Закавказья. Геология СССР, X : 529—546.
J Кутепов Н. 1896. Естественные и бытовые условия древней русской охоты. 1 Природа и охота, 3 : 21—48.
| К у ф т и п Б. А. 1939. Материалы к археологии Колхиды, I, Тбилиси : 1—359.
в К у ф т и н Б. А. 1940. К вопросу о ранних стадиях бронзовой культуры на терри-
1 тории Грузии. Кратк. сообщ. Инет. ист. матер, культуры, VIII : 5—35.
Ку фт ин Б. А. 1941. Археологические раскопки в Триалети, I. Опыт периодиза-
ции памятников. Тбилиси : 1—231.
Л а в р е н к о Е. М. 1938. История флоры и растительности СССР по данным современного распределения растений. Растительность СССР, 1 : 235—296.
.Л а в р е н к о Е. М. 1939. Вопрос о причинах безлесия степей как проблема исторической ботанической географии. Сб. «Академия наук СССР Президенту АН СССР академику В. Л. Комарову к 70-летию со дня рождения» : 486—516.
F Л а в р е н к о Е. М. 1958. О положении лесной части Кавказа в системе ботанико-гео-I графического районирования Палеарктики. Бот. журн., XLIII, 9 : 1237—1253. : Л’а в р о в Л. И. 1952. Развитие земледелия на северо-западном Кавказе с древнейших ; * времен до середины XVIII в. Матер, по ист. земледелия СССР, Изд. АН СССР, М.—Л., сб. 1 : 179—225.
Лал аян Е. А. 1929. Археологические раскопки в Новобаязетском уезде ССР Армении. Русск. антропол. журн., XVII, 3—4 : 59—63.
л мберти А. 1913. Описание Колхиды, называемой теперь Мингрелией (1654 г.) Перевод К. Ф. Гана. Сб. матер, для описан, местн. и племен Кавказа, 43 : 1—232.
Латынин Б. А. 1935. К вопросу об истории ирригации. Сб. «Академия наук СССР академику Н. Я, Марру», XLV : 697—708.
Латышев В. В. 1893, 1904. Известия древних писателей греческих и латинских о Скифии и Кавказе. СПб. 1893, I, 1—3 : I—VIII-f-1—9^6; 1904, II, 1, 2 : 1—454.
Латышев В. В. 1947, 1948. Известия древних писателей о Скифии и Кавказе. Вести, древней истории, 1947, I, 1—4 : 263—316; 249—332; 235—315; 171—289; 1948, II, 1—4 : 224—315; 215—314; 218—330; 226—298.
Левинсон-Лессинг Ф. Ю. 1928. Армянское вулканическое нагорье. Природа, 5 : 429—446.
Л е г к о б ы т о в В. С. 1836. Обозрение российских владений за Кавказом в статистическом, этнографическом и финансовом отношении. СПб. I : 1—399; II : 1—401; III : 1—392; IV : 1—401.
620
Л ите рату ра
Леонтьев Г. С. 1938. «Дождевые тени» за Скалистым хребтом Центрального Кавказа. Изв. Гос. геогр. общ, 2 : 272—279.
Леонтьев Л. Н. 1949. Тектоническое строение и история геотектонического развития Малого Кавказа. Бюлл. Моск. общ. испыт. природы, отд. геол., XXIV (4) : 3—37.
Леонтьев О. К. и П. В. Ф е до ров. 1953. К истории Каспийского моря в поздне-и послехвалынское время. Изв. АН СССР, сер. геогр., 4 : 64—74.
Литвин М. 1890. О нравах татар, литовцев и московитян. Мемуары, относящиеся к ист. Южн. России, I, Киев : 1—58.
-Лопатин Г. В. 1949. Эрозия и сток наносов в Европейской части СССР и Северном Кавказе. Изв. Всесоюзн. геогр. общ., 81, 5 : 473—483.
Лукин А. Л. 1941. Материалы по археологии бзыбской Абхазии. Тр. Отд. ист. первобытн. культуры, I : 18—97.
Любин В. П. 1953. Каменный век Юго-Осетии (в связи с некоторыми вопросами нижнего палеолита Закавказья и стран Передней Азии). Автореф. АН СССР,. Инет. ист. матер, культуры, Л. : 3—21.
Любин В. П. 1959. Высокогорная пещерная стоянка Кударо I (Юго-0сетия). Изв. Всес. геогр. общ. 91, 2 : 173—183.
[Люк Жан]. 1879. Описание перекопских и ногайских татар, черкесов, мингрелов и грузин Жана де Люка, монаха Доминиканского ордена (1625). Перевод П. Юрченко. Зап. Одесск. общ. ист. и древн. XI : 473—493.
Л ю л ь е. 1870. Верования, религиозные обряды и предрассудки у черкес. Зад. Кавк. отд. Русск. геогр. общ., V : 125.
Л я й е л ь Ч. 1864. Геологические доказательства древности человека с некоторыми замечаниями о теориях происхождения видов. СПб. : 1—512.
Магакьян А. К. 1948. Остатки лесов в Сисианском районе АрмССР. Изв. АН АрмССР, 1 : 2—19.
Маевский И. А. 1902а. Условия орошения и колонизации на Мугани и на Кура-Араксинской равнине вообще. Изв. Кавк. отд. Русск. геогр. общ., XV, 9 : 64—167.
Маевский И. А. 19026. Городища и курганы Кура-Араксинской равнины. Изв. Кавк. отд. Русск. геогр. общ., XV, 2—5 : 57—64.
М а з а р о в и ч А. Н. 1946. К вопросу о стратиграфии антропогена. Бюлл. Моск, общ. испыт. природы, отд. геол., XXI (4) : 13—24.
Мазарович А. Н. 1950. Принципы стратиграфии четвертичных отложений. Матер. по четвертичн. периоду СССР, 2 : 30—36.
Макалатия С. И. 1935. Культовые места и связанные с ними ритуальные обряды у грузин-мохевцев. Сб. «Академия паук СССР академику Н. Я. Марру», XLV : 517—522.
Макаров В. Н. 1940. Заповедники СССР. Сельхозгиз, М. : 3—158.
Малеев В. П. 1938. Растительность причерноморских стран (Эвксинской провинции) и ее происхождение и связи. Тр. Бот. инет. АН СССР, сер. III, 4 : 136—252.
Малеев В. П. 1939а. О происхождении грушевых лесов северо-западного Кавказа. Сов. ботан., 5 (3) : 50—54.
Малеев В. П. 19396. О следах ксеротермического периода на северо-западном Кавказе. Сов. ботан., 4 : 68—75.
Малеев В. П. 1940. О растительности полян предгорий северо-западного Кавказа. Сов. ботан., 2 : 30—47.
Малеев В. П. 1941. Третичные реликты во флоре западного Кавказа и основные этапы четвертичной истории его флоры и растительности. Матер, по ист. флоры и растительности СССР, 1 : 61—144.
Малеев В. П. 1946. Основные этапы развития растительности Средиземноморья в четвертичный период в связи с историей растительности юга СССР. Тр. Инет, геогр., 37 : 321—323.
Малей н А. 1911. История монголов Иоганна де Плано-Карпини и путешествие в восточные страны Вильгельма де Рубрука. СПб. : VII—XV1+1—224.
М а м е д а л и е в Ю. Г. и С. 3. К а и л а и. 1948. Исследования битума из бинагадинского кладбища четвертичной фауны и флоры. Изв. АН АзербССР, 6 : 7—20.
М а н и е в и ч Е. Д. 1949. Гребень и фиала из кургана Солохи. Сов. археол., XIII : 217—239.
Марков Е. Л. 1939. Охотничьи поверья и песни сванов Абхазии. Монадире, 9 : 15—16.
Марковин В. И. 1953. Наскальные изображения в Дагестане. Изв. Всесоюзн. геогр. общ., 85, 2 : 209—219.
Маркович В. В. 1897. В лесах Ичкерии. Воспоминания лесничего о чеченском лесе. Леей, журн., 6 : 975—1079.
Прочая литература
621
Маруашвили Л. И. 1938а. О древнем оледенении Малого Кавказа. Природа, 7—8 : 21.
Маруашвили Л. И. 19386. Пещера Абласкира — замечательное карстовое образование в Абхазии. Природа, 10 : 117—120.
Маруашвили Л. И. 1946. Зуртакетская палеолитическая стоянка в Южной ’ Грузии и ее геологическое значение. Природа, 12 : 56—62.
Маруашвили Л, И. 1949. Пещеры Южной Грузии. Природа, 3 : 55—58.
Маруашвили Л. И. 1952. Палеогеография четвертичных образований Закавказья. Матер, по четвертичн. периоду СССР, 3 : 168—187.
Маруашвили Л. И. 1956. Целесообразность пересмотра существующих представлений о палеогеографических условиях ледникового времени на Кавказе. Изд. АН ГрузССР, Тбилиси : 3—129.
Марш Г. 1866. Человек и природа или о влиянии человека на изменение физико-географических условий природы. Перев. Н. А. Неведомского, СПб. : 1—587.
Мастан-Заде А. 1939. Битуминозные отложения Апшеронского полуострова, Изв. Азерб. фил. АН СССР, 6.
Махатадзе Л. Б. 1957. Дубравы Армении. Ботан. инет. АН АрмССР, Ереван : 1—326.
Медведев Я.С. 1915. Растительность Кавказа. Тр. Тифлисск. ботан. сада, XXVII, 1 : 1—108.
Медведев Я. С. 1931. Из недавнего фитогеологического прошлого восточного Закавказья. Закавк. краеведческ. сб., 1 : 64—72.
Мелик-Саркисян С. А. 1897. Муганская степь. Естественноисторический и сельскохозяйственный очерк. СПб. : 1—65.
Мелик-Шахназаров Е. 1898. Селение Арпеваник Зангезурского уезда Елисаветпольской губернии. Сб. матер, для описан, местн. и племен Кавказа, XXV : 1—58.
Миланкович М. 1939. Математическая климатология и астрономическая теория колебаний климата. Гос. объед. научно-технич. изд., М.—Л. : 3—207.
Милановский Е. 1935. Плиоценовые и четвертичные отложения Сызранского района. Тр. Комисс. по изуч. четвертичн. периода, 4,2 : 175—222.
Миллер А. А. 1922. Изображение собаки в древностях Кавказа. Изв. Росс. Акад, ист. матер, культуры, II : 287—324.
Мир ч и н к Г. Ф. 1928. Четвертичная история Черного моря. Природа, 6 : 606.
Мирчи н к Г. Ф. 1936а. О четвертичном орогенезе и эпейрогенезе на территории СССР. Матер, по четвертичн. периоду СССР : 121—124.
Мирч инк Г. Ф. 19366. Корреляция континентальных четвертичных отложений Русской равнины и соответствующих отложений Кавказа и Понто-Каспия. Матер, по четвертичн. периоду СССР : 10—38.
Мирчи нк М. Ф. 1934. Обзор геологического строения Апшеронского полуострова. Тр. Азерб. отд. Закавк. фил. АН СССР, VI : 5—39.
Михайловская О. Н. 1934. Четвертичные террасы Абхазии. Тр. Первого Всесоюзн. геогр. съезда, 11—18 апреля 1933 г., 3 : 82.
Михеев А. А. 1926. Флора Азербайджана. Растительные сообщества междуречной равнины рек Куры, Поры и Алазани. Изв. Общ. обсл. изуч. Азербайдж., № 3, Баку: 8—56.
К М и щ е н к о П. И. 1923. Остатки лесов в степном правобережном районе Кубани, к Тр. Кубанск. с.-х. инет. I, 1, Краснодар : 114—123.
ж Мищенко П. И. 1928. К истории леса и степи на Кубани. Дневник Всесоюзн.
В съезда ботаников в 1928 г. в Ленинграде : 119—120.
{Муратов М. В. 1951. История Черноморского бассейна в связи с развитием окру--/кающих его областей. Бюлл. Моск. общ. испыт. природы, отд. геол., XXVI I (1) : 7—34.
F Муратов М. В. и Н. А. Г в о з д е ц к и й. 1948. Наблюдения над современными ' геоморфологическими процессами в бассейне реки Хасаут и Эшкакои (Северный f Кавказ). Бюлл. Комисс. по изуч. четвертичн. периода, 12 : 101 — 107.
Н Мчедлишвили II. А. 1951. К палеографии Кавказа в чокракском веке в И связи с данными палеоботаники. Докл. АП СССР, 78, 5: 1013—1016.
Мчедлишвили П. А. 1954а. Еще раз о возрасте годердзекой флоры. Бюлл.
Ж Моск. общ. испыт. природы, отд. геол., 29, 1 : 21—33.
Мчедлишвили П. А. 19546. Новые данные о понтических растительных остатках
I Грузии. Докл. АН СССР, 96, 1 : 185—187. , .
Н. Б. 1861. Озеро Гокча. Газета «Кавказ», 61.
® Н а л и в к и н Д. В. 1956. Учение о фациях. Географические условия образования « осадков. Изд. АН СССР, М.—Л. 1 : 1—534; II : 1—388.
Низами Ганджеви. 1940. Искандер Паме. Изд. Азерб. фил. АН СССР, Баку: 1—391.
$22
Л ите ра/пу ра
.Никитин П. А. 1933. Четвертичные флоры низового Поволжья. Тр. Комисс. по изуч. четвертичн. периода, III : 65—120.
Николаев Н. И. 1937. О возрасте четвертичной волжской фауны млекопитающих. Бюлл. Моск. общ. испыт. природы, отд. геол., XV (6) : 487—514.
Николаев Н. И. 1941. О четвертичных тектонических движениях и возрасте рельефа Центрального Кавказа и Предкавказья. Докл. АН СССР, 30, 1 : 47—50.
Николаев Н. И. 1947. О нижней границе антропогена. Бюлл. Комисс. по изуч. четвертичн. периода, 10 : 77—80.
Николаев Н. И. 1949. Новейшая тектоника СССР. Тр. Комисс. по изуч. четвертичн. периода, VIII : 1—296.
Николаев Н. И. 1950. Значение неотектоники в установлении нижней границы четвертичной системы и ее стратиграфического подразделения. Матер, по четвертичн. периоду СССР, 2 : 13—20.
Николаев Н. И. 1953. Стратиграфия четвертичных отложений Прикаспийской низменности и Нижнего Поволжья. Сб. «Стратигр. четвертичн. отлож. и новейшая тектоника Прикаспийск. низм.», Инет, геогр. АН СССР. Комисс. по изуч. четвертичн. периода : 5—40.
Ниорадзе 1933. Палеолитический человек из пещеры Девис-Хврели. Тр. Муз. Грузии, VI : I—IV+1—ПО (на груз, языке).
Ниорадзе Г. К. 1934. Палеолит Грузии. Тр. Второй Междун. конф, ассоц. по изуч. четвертичн. периода Европы, V : 219—229.
Ниорадзе Г. К. 1953. Человек каменного века в пещере Сакажиа. Тбилиси: 1—88 (на груз, языке).
Новопокровский И. В. 1925. Растительность Северо-Кавказского края. Матер, к перспект. плану восстанови, и развития сельск. хоз. Сев.-Кавк, края, I, II : 1—27.
О л е а р и й Адам. 1870. Подробное описание путешествия голштинского посольства в Московию и Персию в 1633, 1636, 1639 гг. Перевод П. Барсова, М.:1—XIV+ 1—1038, 1—32+1—X.
Оленич-Гнененко А. П. 1955. В горах Кавказа. Ростов : 1—158.
О р б е л и И. А. и К. В. Тренер. 1935. Сасанидский металл. М.—Л. : 1—85.
Освальд Ф. 1915—1916. К истории тектонического развития Армянского нагорья. Перевод А. И. Шишкиной. Зап. Кавк. отд. Русск. геогр. общ., XXIX, 2 : 1—77.
Осипов Г. 1898. Селение Чайкенд, Елисаветпольского уезда той же губернии. Сб. матер, для описан, местн. и племен Кавказа, XXV : 59—136.
Очерк сельского и лесного хозяйства Ленкоранского уезда, Бакинской губернии. 1914 : 1—231.
П а в л о в А. П. 1925. Неогеновые и послетретичные отложения южной и восточной Европы. Мемуары Геол. отд. Общ. любит, естеств., [антропол. и этногр., 5 : 1—125.
Павлов А. П. 1936. Геологическая история европейских земель и морей в связи с историей ископаемого человека. Изд. АН СССР, М.—Л. : 1—343.
П а ли би н И. В. 1913. К вопросу о посттретичной флоре Северного Кавказа.. Тр. Тифлисск. Ботан. сада, XII, 2 : 159—168.
П алибин И. В. 1936. Этапы развития флоры Прикаспийских стран со времени мелового периода. М.—Л. : 1—60.
Паничкина М. 3. 1948. К вопросу о верхнем палеолите в Армении. Изв. АН АрмССР, 7, общ. науки : 67—79.
П аничкина М. 3. 1950а. Древнепалеолитическая стоянка Сатани-Дар в Армении. Кратк. сообщ. Инет. ист. матер, культуры, XXXV : 66—73.
Паничкина М. 3. 19506. Палеолит Армении. Изд. Эрмитажа, Л. : 1— 115.
Паничкина М. 3. 1951. Аширабадское мустьерское местонахождение в Армении. Кратк. сообщ. Инет. ист. матер, культуры, XXXVI : 76—86.
Паничкина М. 3. 1952. К вопросу о и а значении шеллнеких орудий (по материалам Сатани-Дара). Кратк. сообщ. Инет. ист. матер, культуры, XLXI : 19—30.
Пантюхов И. И. 1892. Ахалкалакский уезд. Тр. Кавк. общ. сельск. хоз.. 11—12 : 693—720.
П а с с е к Т. С. и Б. А. Латынин. 1926. Очерк доистории Северного Азербайджана. Изв. общ. обслед. и изуч. Азербайджана, 3 : 112—157.
Паффенгольц К. Н. 1934. Бассейн озера Гокча (Севан). Тр. Всесоюзн. геол.-разв. объед. НКТП СССР, 219 : 105.
Паффенгольц К. Н. 1948. Геология Армении. М.—Л. : 1—332.
Прочая литература
623
аффен гольц К. Н. 1950. О происхождении озер Севан (Армения), Ван (Анатолия) и Урмия (Иран). Изв., АН СССР, сер. геол., 1 : 125—138.
аффенгольц К. Н. 1951. К стратиграфии вулканических толщ Джавахет-ского нагорья. Тр. Инет. геол, и минерал. АН ГрузССР : 351—369.
Пахомов Е. А. 1923. Краткий курс истории Азербайджана. Баку: 1—48.
П е*т р о в В. А. 1939. Растительные остатки закированного слоя Бинагадов. Изв. Азерб. фил. АН СССР, 6 : 100—107.
Петрушевский И. П. 1949. Очерки по истории феодальных отношений в Азербайджане и Армении. Изд. ЛГУ, Л. : 1—380.
Пид он лич ко И. Г. 1946. О ледниковом периоде, 1. Киев : 1—170.
Пидопличко И. Г. 1952. Новый метод определения геологического возраста ископаемых костей четвертичной системы. Киев : 1—89.
Пидопличко И. Г. иП. С. Макеев. 1952. О климатах и ландшафтах прошлого, 1. Киев, 1—85.
Пиотровский Б. Б. 1944. История и культура Урарту. Ереван : IX—ХП-ф-1—364.
Пиотровский Б. Б. 1947. Археологическое изучение древнейшего Закавказья. Вести. Древн. истории : 167—187.
Пиотровский Б. Б. 1949. Археология Закавказья. Л. : 1—131.
Пиотровский Б. Б. 1950. Кармир-блур. Результаты раскопок 1939—1949. Ереван : 1—98.
Погребова Н. Н. 1950. К вопросу о скифском зверином стиле. Кратк. сообщ. Инет. ист. матер, культуры, XXXIV : 129—141.
Покровский М. В. иН. В. Анфимов. 1937. Карта древнейших поселений и могильников Прикубанья. Сов. археол., IV : 265—279.
Полное собрание законов Российской империи с 1649 года. 1830. X, 1737—1739 : 1—995.
Полынов Б. Б. 1926, 1927. Пески Донской области, их почвы и ландшафты. Тр. Почв. инет. им. Докучаева, 1926, I : 3—198; 1927, II : 1—198.
Поляков И. С. 1882. Дневник археологических работ, проведенных в Закавказском крае осенью 1879 г. Пятый археол. съезд в Тифлисе. Протоколы под-гот. комит. : 157—218.
П о п к о И. 1880. Терские казаки I. Гребенское войско. СПб. : 1—516.
Попов В. И. 1950. Фациальное развитие осадков горных склонов и подгорных, пустынных равнин. Матер, по четвертичн. периоду СССР, 2 : 192—205.
Попов Г. И. 1955. История Манычского проливав связи со стратиграфией Черноморских и каспийских четвертичных отношений. Бюлл. Моск. общ. ист. прир., Отд. геол., XXX, 2 : 31—49.
Попов Н. К. иМ. С. Гагошидзе. 1940. Обзор и оценка существующих методов борьбы с селями и наводнениями в Алазанской и Автаранской долинах. Селевые явления и борьба с ними. Тбилиси : 27—52.
Потто В. А. 1912. Два века терского казачества (1577—1801). Владикавказ, I : 1—115; II : 1—247.
Православлев П. А. 1930. Современные движения земной коры в Понто-* Каспийской области. Тр. III Всесоюзн. съезда геологов, 20—26 сентября 1928 г., 1, Ташкент : 123—178.
Православлев П. А. 1932. К гидрологии Прикубанской степной равнины. Тр. Всесоюзн. геол.-разв. объед., 188 : 1—71.
Православлев П. А. и О. М. Аншелес. 1930. Вулканический пепел из лёссовидных суглинков правого побережья реки Кубани около станицы Темиж-бекской. Тр. Ленингр. общ. естествоиспыт. 60, 1 : 149—159.
Прилипко Л. И. 1954. Лесная растительность Азербайджана. Баку : 1—485.
Пчел ин а Е. Г. 1940. Армянские памятники на территории Азербайджанской!. ССР. Гос. Эрмитаж, Тр. Отд. Востока, III : 243—256.
Пыльцов Н. Н. 1904. О внезапных наводнениях и селях в Нухинском уезде Елизаветпольской губернии. Изв. Кавк. отд. Русск. геогр. общ., XVII, 4 : 244—282.
Радде Г. И. 1866. Путешествие в Мингрельских Альпах и в трех их верхних долинах. Зап. Кавк. отд. Русск. геогр. общ., VII : 112—113.
Радде Г. И. 1901. Основные черты растительного мира на Кавказе. Зап. Кавк, отд. Русск. геогр. общ., XXII, 3 : 1—199.
Раушенбах В. М. 1952. Новый метод определения возраста археологических* памятников и геологических отложений. Природа, 2 : 102—106.
Рашид-Ад-Дин. 1946. Сборник летописей. Перевод с персидского А. К. Арендса.
III, Изд. АН СССР, М.—Л. : 1—340.
Рейнгард А. Л. 1931. Подразделение четвертичного ледникового периода*
$24
Литература
в Альпах и на Кавказе по новым данным. Изв. Гос. геогр. общ., LXIII, 1 : 105— 117.
Рейнгард А. Л. 1932а. О происхождении исполинских конгломератов Кабардинского хребта в Северном Кавказе. Тр. Комисс. по изуч. четвертичн. периода 1 : 13—23.
Рейнгард А. Л. 19326. Несколько слов о хронологической связи между оледенениями Кавказа и каспийскими трансгрессиями. Зап. Росс, минерал, общ. LXI, 1 : 151—158.
Рейнгард А. Л. 1933. О террасах Кубанской долины у Баталпашинска (Кавказ). Тр. Комисс. по изуч. четвертичн. периода. Ill, 1 : 121—125.
Рейнгард А. Л. 1936а. Основные черты четвертичной истории Предкавказ-ской равнины. Западная часть (работы 1933 и 1934 гг.). Центр, научно-иссл. геол.-развед. инет.
Р ейн г ар д А. Л. 19366. Четвертичные отложения восточного Предкавказья. Проол. сов. геол., М., VI, 10 : 919.
Рейнгард А. Л. 1937. К проблеме стратиграфии ледникового периода Кавказа.
Тр. Сов. секции Междун. ассоц. по изуч. четвертичн. периода Европы, 1 ; 9—3°.
Рейнгард А. Л. 1939. К вопросу о послетретичных тектонических движениях на Кавказе. Тр. Сов. секции Междун. ассоц. по изуч. четвертичн. периода Европы, IV : 25—28.
Рейнгард А. Л. 1941. Четвертичная система. Геология СССР, X, Закавказье: 5—614.
Рейнгард А. Л. 1947а. Четвертичная система. Общий обзор. Геология СССР, IX. Северный Кавказ: 329—331.
Рейнгард А. Л. 19476. Четвертичная система. Континентальные отложения. Геология СССР, IX, Северный Кавказ : 332—343.
JP е с л е р Э.]. 1896. Раскопки Э. Реслера. Отчет Археол. комисс. за 1896 г. СПб.
Ровинский. 1809. Хозяйственное описание Астраханской и Кавказской губерний по гражданскому и естественному их состоянию, СПб. : 1—527.
Роговский П. А. 1928. К истории лесной растительности на правом берегу реки Кубани в зоне степей.Изв. Кубанск. пед. инет., 1 : 126-—164.
Роговский П. А. 1938. К истории флоры Северного Кавказа. Тр. Краснодарок, пед. инет., VII .111—122.
Рождаев П. 1925. Основные черты организации крестьянского хозяйства яа Северном Кавказе. Природные условия Северокавказского, края : 14.
Розенберг Ф. А. 1922. Персидская миниатюра конца XVI в. Работа Али Риза-и Абоси. Изв. Росс. Акад. ист. матер, культуры, II : 177—190-
Р о н о в А. Б. 1949. История осадконакопления и колебательных движений Европейской части СССР. Тр. Геогр. инет., 3 (130) ; 1—390.
Россиков К. Н. 1886—1888. Поездка в юго-западную часть южной Чечни и в западный Дагестан. Изв. Кавк. отд. Русск. геогр. общ., IX : 99—103.
Россиков К. 1893. Усыхание озер на северном склоне Кавказского хребта. Зап. Кавк. отд. русск. геогр. общ., XV : 189—225.
Ростовцев М. И. 1918. Эллинство и иранство на юге России. Пгр.: V—VII-^ 1 — 190.
Рощин И. И. 1928. Борьба с селевыми потоками. Изв. Тифлисск. политеха, инет., III : 132—160.
Русов А. А. 1879. Отчет о летних и осенних археологических работах (1880) в южном Дагестане. Пятый археол. съезд в Тифлисе : 503—621.
Руставели Шота. 1937. Витязь в тигровой шкуре. М.—Л. : VII—X4-1—274.
Рябинин А. Н. 1913. Геологические исследования в Ширакской степи и ее окрестностях. Тр. Геол, комит., 93 : 1—73.
Сардарян С. А. 1954. Палеолит в Армении. Изд. АН АрмССР, Ереван : 1— 182.
Сатунин К. А. 1910. Экскурсия в пещеры Сухумского округа. Пещеры великаны Абласкира и Адзаба. Изв. Кавк. отд. Русск. геогр. общ., XXI, 1.
Саушкин Ю. Г. 1946. Культурный ландшафт. Вонр. геогр., 1 : 97—106.
Свириденко П. А. 1925. Главнейшие этапы развития дела защиты растений на Северном Кавказе. Сев.-Кавк, краевед, зем. уиравл., Краев, станц. защ. от вредит., сер. А, 8, Ростов : 1 —16.
С в и щ е в с кий Д. 1939. Разрушение морского берега у г. Батуми как явление общее для восточного побережья Черного моря. Изв. Геогр. общ., 5 : 658— 689.
Северный Кавказ после районирования (итоги и выводы), 1925, Ростов н/Дону I : 1—541; II : 1—423. ’
Прочая литература
625
Сизов В- 1889. Восточное побережье Черного моря. Археологические экскурсии. Матер, по археол. Кавказа, II, М. : 1—182.
Йиницын И. В. 1948. Памятники пред скифской эпохи в степях Нижнего Поволжья. Сов. археол., X : 143—160*
Си н с кая Е. Н. 1933. Основные черты эволюции лесной растительности Кавказа *в связи с историей видов. Ботан. журн: СССР, XVIII, 5, 6 : 370—406.
Синская Е. Н. 1939. Что такое центр видообразования в свете учения Ч. Дарвина. Природа, 11 : 51—59.
Скальковский А. 1885. История Новой Сечи или последнего Коша Запорожского, I—III. Одесса : 1—330.
Скворцов А. Ф. 1950. Об эрозии почв в бассейне реки Риони. Сообщения АН ГрузССР, XI, 10 : 643—649.
Смирнов К.Ф. 1952. Основные этапы развития меотосарматской культуры Среднего Прикубанья. Кратк. сообщ. Инет. ист. матер, культуры, XLVI : 3—18.
Смирнов Я. И. 1909. Восточное серебро. СПб. : 1—18, табл. СХХХ.
Снегирев И. Л. 1937. Древний Восток. Атлас по древней истории Египта, Передней Азии, Индии и Китая. Соцэкгиз, Л. : 1—59, текст 1—259 (атлас).
Соболев Д. 1938. К геоморфологии окрестностей Кисловодска. Природа, 3: 112—120.
Собрание путешествий к татарам и другим восточным народам в XIII,XIV и XV столетиях. I. Плано-Карпини. II. Асцелин. Перевод Д. Языкова, СПб., 1825: 1—263.
Соймонов Ф. И. 1763. Описание Каспийского моря и чиненных на оном российских завоеваний яко часть истории Петра Великого. Ежемесячные сочинения и известия о ученых делах. Генварь 1763 года. Акад, наук, СПб. : 1—35, 99—123; 195—246, 291—320, 387—432, 483—506, 3—35, 99—119, 195—233, 291—334, 387—400.
Соколов Н. А. 1904. К истории причерноморских степей с конца третичного периода. Почвоведение, VI, 2, 3 : 105—124 и 198—220.
Солнцев 3. Я. 1940. Рубки в горных лесах Северного Кавказа. Лесн. хоз., 39—18.
Соловьев Л. Н. 1939. 3неолитическое селище у Очемчирского порта в Абхазии. Матер, по ист. Абхазии, 1, Сухуми : 5—55.
,С о с н о в с к и й Д. И. 1915. Процессы исчезновения лесов в ближайших окрестностях Тифлиса. Изв. Кавк. отд. Русск. геогр. общ., 23, 1 : 37—51.
Стащук Н. И. 1952. Заселение Ставрополья в конце XVIII века и в первой половине XIX века. Матер, по изучен. Ставропольск. края, 4 : 137—173.
Степанов Н. А. 1934. Возобновительные рубки в буковых лесах Северного Кавказа. Ростов/Дону : 1—71.
Страбон. 1879. География в семнадцати книгах. Перевод Ф. Г. Мищенко. М. : t V—XXV+856+I—CCLXXVII.
Страже в В. И. 1926, Бронзовая культура в Абхазии. Изв. Абхазского научн. общ., IV : 106—123.
Сукачев В. Н. 1936. Основные черты развития растительности СССР во время плейстоцена. Матер, по четвертичн. периоду СССР : 62—89.
Султанов Р. Г. 1947. Геологические условия залегания костеносного слоя в Бинагадинском и Кирмакинском «кладбищах» Апшерона. Изв. АН АзербССР, 6 : 60—68.
Сысоев В. М. 1898. Краткий археологический очерк Кубанской области и Черноморской губернии. Ку банек, сб., IV, Екатеринодар : 1—14.
Сысоев В. М. 1925. Краткий очерк истории Азербайджана. Баку : 1 — 143.
Сысоев В. М. и В. А. Щербина. 1898. Краткий указатель археологических естественно исторических и этнографическо-промышленных вещей и предметов, имеющихся при Кубанском областном статистическом комитете. Ку банек, сб., IV, Екатеринодар : 1—43.
Тан ф и л ь е в Г. И. 1896. Доисторические степи Европейской России. Землеведение : 73—92.
Тахтаджян Армен. 1941. Ботанико-географические провинции и округа Армении. Тр. Ботан. инет. Арм. фил. АП СССР, 2 : 7—14S.
Толстой И. иН. Кондаков. 1889. Русские древности в памятниках искусства, II. Древности скифо-сарматские. СПб. : 1 —160.
Троицкий Н. А. 1927. Остатки лесов в Ахалкалакском уезде. Вести. Тифлисск. Ботан. сада, нов. сер., 3—4; 83—89.
Троицкий Н. А. 1931. «Священная роща» осетин в верховьях Арагвы. Закавк. краеведчсск. сб., 1 : 20—27.
40 Н. К. Верещагин
626
Литература
Троицкий II. А. 1934. Влияние деятельности человека на растительность в бассейне верхнего течения реки Бамбак (Армения). Тр. Зоотехн. инет., 1, Эривань : 105—143.
Троицкий Н. А. 1939.' Остатки лесов в верхнем течении реки Дзорагет. Тр. Ереванск. гос. унив., IX.
Тума Джонов И. И. 19401 Бук и буковые леса в нагорном Дагестане. Тр. Тбнлисск. ботан. инет. Груз. фил. АН СССР, VII : 57—83.
Тумаджанов 11. II. 1947. Лесная растительность долины Теберды в свете послеледниковой истории развития фи то ландшафтов. Тр. Тбилисск. Ботан. инет. Груз. фил. АН СССР, XI : 4—106.
Турутанова-Кетова А. И. 1933. Некоторые данные к новейшей ископаемой флоре Армении. Тр. Геол. инет. АН СССР, Ill : 209—215.
Т у т к о в с к и й II. А. 1909. Ископаемые пустыни северного полушария. Прилож. к журн. «Землеведение» : 313.
Уварова П. С. 1900. Могильники Северного Кавказа. Матер, по археол. Кавказа, VIII, М. : 1 — 381.
Уварова II. С. 1902. Археология. Коллекции Кавк, муз., V, Тифлис : 1—231.
У с а м а И б н Мунк ы з. 1922—1923. Книга назидания. XII век. М. : 1— . 207.
Ф а р м а к о в с к и й Б. В. 1914. Архаический период в России. Матер, по археол. России, 34 : 15—78.
Ф а р м а к о в с к и й М. В. 1922. Горит из кургана Солохи. Изв. Росс. Акад, ист. матер, культуры, II : 23—48.
Фел и ц и н Е. Д. 1884. Краткий очерк истории заселения Кубанской области, с картой. Изв. Кавк. отд. Русск. геогр. общ., VIII, 1 : 250—284.
Федоров А. А. 1952. История высокогорной флоры Кавказа в четвертичное время. Матер, по четвертичн. периоду СССР, 3 : 49—86.
Ф и г у р о вс к и й И. В. 1916. Деление Кавказа на физико-географические области и районы. Изв. Кавк. отд. Русск. геогр. общ., XXIV, 2 : 145.
•Ф и г у р о в с к й й И. В. 1923. Климаты Кавказа. Изв. Кавк. отд. Русск. геогр общ., XIX, 1.
Филиппов Н. М. 1890. Об изменении уровня Каспийского моря. Зап. Русск. геогр. общ., XX, 2 : 1—112.
Флеров А. Ф. 1931. Песчаные ландшафты Черноморско-Азовского побережья Кавказа, их происхождение и развитие. Изв. Гос. геогр. общ. LXIII : 21—43.
Ф л е ров А. Ф. и В. Н. Б а л а н д и н. 1931. Степи Северо-Кавказского края. Изд. «Сев. Кавказ», Ростов н/Дону: 1—127.
'Формозов А. А. 1952. Нижнепалеолитические местонахождения Прикубанья (по данным разведок 1950 г.). Кратк. сообщ. Инет. ист. матер, культуры, XLVI:31—41.
Фортунатов Б. 1933. О генеральном плане реконструкции промысловой фауны в Европейской части СССР и Украины. Природа и социалист, хоз., 6 : 90—107.
ФюронР. 1955. Введение в геологию и гидрогеологию Турции. Изд. иностр, литер., М.—Л. : 1—142.
X а р к е в и ч С. С. 1954. Роль четвертичного эпейрогенс за в формировании высокогорной флоры Большого Кавказа. Ботан. журн., 39, 4 : 498—514.
• X о р с п с к и й Моисей. 1893. История Армении. Перевод Н. О. Эмина. М. : V-XXIV+1 — 323.
Цейпор Ф. Е. 1936. Палеобиология и климаты прошлого. Пробл. палеонтол I, М. : 199—216.
Ц ер ет е л и Д. В. 1954. К палеографии и истории развития рельефа Алазансксв долины в четвертичном периоде. Сообщ. АН ГрузССР, XV, № 7 : 437—444.
Цилоссани Н. О. 1879. Дневник раскопок, веденных в южном Дагестане в 1870 г. Пятый археол. съезд в Тифлисе : 456—473.
Ч е р д ы н це в В. В. 1955. Об изотопном составе радиоэлементов в природных объектах в связи с вопросами геохронологии. Тр. Третьей сессии комисс. по спред, абс. возраста геол, формаций (25—27 марта 1954). М. : 175—233.
Шавров И. Н. 1913. Персидское побережье Каспийского моря, его производи-;
тельность и торговля. Зап. Кавк. отд. Русск. геогр. общ., XXVI, 10:1—93. i Шанце р К. В. 1940. Условия залегания и геологическая датировка абхазского] палеолита. Бюлл. Комисс. по изуч. четвертичн. периода, 6—7 : 97—100. i
Шан не р Е. В. 1951. Аллювий равнинных рек умеренного пояса и его значение j для познания закономерностей строения и формирования аллювиальных свит.! Тр. 14нет. геол, наук, 135, геол. сер. (55) : 1 — 274.
л Шафранов П. 1885. Образование Ри опекой низменности в течение исторического периода. Сб. сведений о Кутаисск. губ., 1, Кутаис : 1—64.
Прочая литература
627
к.
в-
11.
)Л-ад тп, ное
об-эгр.
!5КЬЯ —43.
:рая.
)ИКу-:уры.
|ауны |-107. io стр.
: X -
|Ьтол., ilHCKCM I—444. гестапо
родных Исе. но вз.
3 ВОДИ-
ГО лил [аачеппе |х свит, г' ^ЯССКОГО
Швецов М. С. 1928. Геологическое строение западной оконечности Кабардинского хребта. Тр. Научно-иссл. нефт. инет., 3: 1—75.
Ш е л к о в н и к о в А. Б. 1907. Поездка в Зуванд в июле 1906 г. Изв. Кавк, муз., 2, 3:171 — 242.
Полковников А. Б. 1909. Nelumbo nucifera Gaertn. на разливах Аракса на ' Мугани. Изв. Кавк, муз., IV, 3 : 157—172.
ill е л к о в н и к о в А. Б. 1929. Облесснность берегов оз. Севан. Сб. «Бассейн оз. Севан (Гокча)». Паучн. результ. эксп. 1927. Изд. АН СССР и Упр. водн. .хоз. Арм., Л. : 457—466.
• и л ьд е р В. 1892. Боржомский звериный отвод. СПб. : 1—20.
i и р а к ц а А. 1918. Вопросы и решения Анании Ширакца. Перевод И. Орбели, Пгр. : 1—80.
I и ф ф е р.с Е. В. 1953. Растительность Северного Кавказа и его природные кормовые угодья. Изд. АН СССР, М.—Л. : 1—399.
1 лепне.в Н. И. 1947. Эпейрогенические колебания в Закавказье и уровни морей Черного и Каспийского. Изв. АН АзербССР, 2 : 109—113.
I о п е н. И. 1852. Исторический памятник состояния Армянской области в эпоху ее присоединения к Российской империи. СПб. : 1—1231.
I т е й н. 1879. О пещерах и могилах в Дагестане. Пятый археол. съезд в Тифлисе : 473—482.
[тукенберг И. Ф. 1858а. (1860). Статистическое описание Ставропольской губернии с землею черноморских казаков. Статист, труды И. Ф. Штукенберга, 1 : 1—41.
[тукенберг И. Ф. 18586. (1860). Статистическое описание Закавказского края. Статист, труды И. Ф. Штукенберга, 2:3—58.
Ц е р б и н а Ф. 1913. История Кубанского Казачьего Войска. Екатсринодар, I: 1—700; II : 1—848.
i Щукин И. С. 1925. Следы сухой послеледниковой эпохи на Северном Кавказе.
| Землеведение, XXVI, 1 : 47—71.
к Щукин И. и А. Щукина. 1927. Очерки Армянского нагорья. Землеведение, 1 , XXIX, !—2, § 5—50.
В Эбер з и н А. Г. .1939. Тектонические движения Таманского полуострова за чет- верти чпый период. Тр. Сов. секции Междун. ассоц. по изуч. четвертичн. периода
Ж Европы, IV : 28—29.
К 10 л кин а О. Н. 1948. Вопросы древней истории на сессии АН СССр. Вести. К древн. истории, 2 : 160—164.
.Юрковский Н. 1861. Несколько слов о главных причинах истребления лесов • за Кавказом. Газета «Кавказ» : 88.
Я. К. В. 1879. Лесоистребление на Кавказе и его последствия. Газета «Кавказ» : 139—143. -
Я ков лев С. А. 1914. Почвы и грунты по линии Армавир-Туапсинской-железной дороги. Изд. Бюро по почвовед, и землед. при Уч. комитете Гл. упр. землеустр.
«и землед., сообщ. XV, СПб. : 1—332. ;
Ярошенко Г.Д. 1940. К истории высокогорной растительности Кавказа. Изв. Арм. фил. АН СССР, 4—5 : 223—229.
Ярошенко Г. Д. 1945. Динамика развития лесной растительности Северной Армении за последние 300 лет. Докл. АН АрмССР, IIJ, 5 : 151-^154.
Ярошенко П. Д. 1941. О причинах безлесия южной Армении. Изв. Арм. фил.
tAH СССР, 2 : 49—56. ;
Ярошенко П. Д. 1956. Смены растительного покрова Закавказья. Изд. АН
. СССР, М.—Л. : 1—240ч .:
Я ц е н к о - X м е л е в с к и й А. А. и Г. В. Канделаки. 1940. Древесные угли из археологических раскопок в Дабна Гоми (Западная Грузия). Сообщ. Груз. фил. АН СССР, 1,5: 353—360.
Яценко-Хмелевский А. А. и Г. В. Канделаки. 1941а. Древесные угли из раскопок Б ешта итонской циклопической крепости (Цалка). Сообщ. Груз. фил. АП СССР, II : 441—448.
Яцеяко-Хмелевский и Г. В. Канделаки. 19416. Эльдарская сосна в окрестностях г. Ганджи в XII в. н. э. Сообщ. V. Сообщ. АП ГрузССР, II, 6 : 540—550.
В а с 11 о f с п-Е с h t А. 1937. Bildicho Darstellung des Risenhirschcs aus vorge-schichtJicher and gcschichtlicher. Zoitschr. filr Saugetiorkunde, I : 81—88.
1 Bibers t ein F. S. 1800. Bcschrcibung der Lander zwischen den Fiusscn Terek und Kur am Caspischen Моего. Frankfurt am Main : 1—211.
Bia и с k о n h о r n M. 1901. Ncues zur Geologie und Palaontologie Aegyptens, IV Pl iocan und Quartar. Zeitschr. deutsch. Geolog. Gescllsch., LI II : 453—457.
40*
-.' 628 .
Л ише рату ра
Blanckenhorn М. 1910. Das Klima der Quartarperiode in Syrien, Palastina : und Aegypten. Die Veranderungen des Klimas. Stockholm : 1—427.
Blanckenhorn M. 1921—1922. Die Steinzeit Palastina-Syriens und Nord-africas. Das Land der Bibel. Leipzig, 1921, I : 1—48; II : 1—26; 1922, III : 1—45.
Bossert H. T. 1942. Altanatolien. Die Altestenkulturen des Mittelmeereises: 11—320.
Brown T. B. 1951. Excavations in Azerbaijan 1948. London : I—XIV4-1—279. Chardin. 1686, 1711, 1735. Voyage en Perse et autres lieux de 1’Orient. London, 1686, 1—11 : 1—254; Amsterdam, III, 1711:1—334; 1735, IV : 1—324.
Clarke E. D. 1810. Travels in various countries of Europe, Asia and Africa, I. Russia, Tartary and Turkey. London : 1—759.
Coon C. S. 1951. Cave explorations in Iran. 1949. Philad., Univ. Museum., Univ, of Pennsylvania : 1—125.
Coon C. S. 1952. Excavations in Hotu cave, Iran, a preliminary report. Proc, of the American Philosoph. Soc., 96 (3) : 231—249.
Dubois de Montp ere u x F. 1839. Voyage autour du Caucase, chez les Tsher-kesses et les Abkhases, en Colchide, en Armenie et en Crimee, II. Paris : 1—199.
Garrod D. A. 1930. The Palaeolithic of Southern Kurdistan. Excavations in the caves of Zarsi and Hazar-Merd. Bull. Amer. School Prehist. Research, 6 : 8—33.
Garrod D. A. and Bate D. M. A. 1937. The Stone Age of Mount Carmel. Excavations at Wady el Mughara. Oxford, I : XII+250, 55.
Geiger W. 1882. Ostiranische Kultur im Altertum, Erlangen : 3—520.
Keller Otto. 1909. Die Antike Tierwelt. Erster Band Saugetiere. Leipzig: 1—434.
К о r n e r O. 1930. Die homerische Tierwelt. Munchen : 1—100.
О s t e n H. H. 1929—1930. Explorations in Hittit Asia Minor. Seasons of 1927—1929, 1—2. Chicago : 1—147.
Oswald F. 1910. Zur tektonischen Entwicklungsgeschichte des armenischen Hoch-landes. Petermanns Geograph. Mitteil., 56, Gotha, I : 8—14; II : 69—74; III: 126—131.
О z a n s о у Fkret. 1955. Sur les gisements continentaux et les Mammiferes du Neogene et du Villafranchien d*Ancara (Turquie). C. R. Acad. Sci. Paris, 240, 9 : 992—994.
Penck A. 1939. Palaolitische und Geologische Ghronologie. Zeitschr. der deutsch. Geol. Gesellsch., H. 1, Bd 91 : 57—64.
Pfannenstiel Max. 1941. Die altsteinzeitlichen Kulturen Anatoliens. Istanbu-ler Forsch., 15, Berlin : 1—50.
P i 11 a r d E. 1929. Decouverte do la civilisation paleolitique en Asie Mineure. Arch, suisses d’Anthr. gener., V, 2 : 135—165.
Sauter M. R. 1948. Prehistoire de la Mediterranee paleolitique-mesolithique. Paris : 1—184.
Schaeffer C. F. A. 1948. Stratigraphie comparee et chronologie de ГAsie Occi-dentale. London : 1—668.
Simpson G. C. 1934. World climate during the quarternary period. Quarterly Journal of the Royal Meteorol. Soc., 60, 257 : 425—479.
Soergel W. 1925. Die Glioderung und absolute Zeitrechnung des Eiszeitalters, Berlin. : 1—133.
Soergel W. 1941. Der Klimacharakter des Mammuts. Palaeontologische Zeitschrift, 22, 3—4 : 29.
Stahl A. F. 1907. Geologische Beobachtungen in Zentral- und Nord west Persien. Peterm. Mitt., 53, VIII : 169—177; IX : 205—214.
Z e u n e r F. 1935. The Pleistocene Chronology of Central Europe. Geol. Mag., LXXII, 854 : 350—376.
Zeuner F. E. 1953. Dating the past. An introduction to geochronology. London: 1—516.
Приложение 1
ДИНАМИКА ДОБЫЧИ И ЗАГОТОВОК ШКУР ПУШНЫХ ЗВЕРЕЙ
НА КАВКАЗСКОМ ПЕРЕШЕЙКЕ
с 1925 по 1955 гг.
(графики 1—23)
«
ц h
11
i \
i t
/ i
Шокал । 1
-------Предкавказье I \
------j? Закавказье } *
....... Дагестан * I
—I--!--1-“--1--1--1™
27 29 31 33
* *—— -t — . *.. ,, - J —J , * J _ Ь * - 4— »—— __ 1 „Ji , |_„ t
35 37 39 1 47 43 45 47 45 1 57 53 55
1940г 1950г.
1930г
632
П риложение 1
Графики 3—6
633
* Лис и и О
--------зокоЬнозье
25 27 29 ' 31 33 35 37 39 1 91 93 95 97 99 1 51 53 55
ШОг ‘ W02 1950?
63^<
Приложение 1
47
43
1940г.
И
45 47
1950г
25 27 33 35 37 39 ' 41
1930 г. 19402,
Енотовидная собака
—- Предкавказье — Закавказье
45
49 ‘ 51 S3 55
1950г.
Меддедь
Предкавказье Закавказье
Графики —10
635
<---L_< < I i - i ill t i i i i I ! 1_i ।___i___I-—t--1--1--1-1---1-1--1
/5 21 29 1 31 33 35 37 39 ! ol 43 45 97 99 1 51 53 55
' 1930г 1940 г 1950 г
636
Приложение 1
1930г. 1940г. 1950г
638
Приложение 1
Рысь
I I I !
I
-------Предкабказье ----------- Армения t 1 320
-------Занобмзье --------------Грузия q I *
-------- верный кабказ ........ Дагестан !( р 1 *
--------Азербайджан \ * « 1 /
JO £f df jj jo jz jy V'l ч/ yy 07 63 oo
1930г
19М)г
1950г
Графики 15—18
639
Заяц-русак
1330г 1940г 1950г
640
Приложение 1
Кобказска* Uc --и
-------Азербайджан
------- Грузия
ЮЗОг. 1940г. 1950г
I
Графики 19—22
Ш
2890357
41 Н. К. Верещагин
64%
Приложение 1. График 23
Приложение 2
КА РТЫ РА СПРОСТРА НЕНИЯ ЗВЕРЕЙ КАВКАЗА
(карты 1—97)
41*
Карты 1—2
$45
ОСТОВ
БАТУМИ
Х/ о f
СУХУМИ
АСТРАХАНЬ
МАХАЧКАЛА
К А РТА 2
КРОТЫ
• Talpa orientalis Ogn
* Talpa caucasica Sat э Историческая эпоха (^Объединенный ареал
♦ Talpa caucasica ognevi
Tp Strop.
Вероятный ареал Talpa cf caucasica Sat ф Верхний плейстоцен • 100 шкурок
Talpa sp.
▲ Нижний плиоцен
200км
ВЫХУХОЛЬ
Desmans moschata L.
» Коллекционные экземпляры
Desmans aff moschata L Верхний плейстоцен Desmans sp
д Нижний плиоцен
646
Приложение 2
Карты 3—6
647
Приложение 2
fiaрты 7—10
649
650
Приложение 2
Карты 11—14
651
$52
Приложение 2
Карты 15—18
653
38
КАРТА 17 ПОЗДНИЙ КОЖАН
Vespertiho serotinus Schr
КОЖАН ОГНЕВА
Vespertilio ognevi Bobr
СЕВЕРНЫЙ КОЖАНОК
♦ Vespertilio nilssonii Keys, et Blas
ШИРОКОУХИЙ СКЛАДЧАТОГУБ м Tadarida taenictis Raf.
654
Приложение 2
Карты 19—22
655
Г
200 км
АСТРАХАНЬ
РОСТОВ*
о
БИЛИС
ЕРЕВАН
@ —
.Х-о® 40V
СУХУМИ
БАТУМИ
МАХАЧКАЛА
КАРТА 21
КОРСАК
Vulpes corsac L Коллекционные экземпляры / шкурка ЕШШ.нэ
Vulpes aff corsac L о Средний плейстоцен ( С Вероятный ареал В плейстоцене
ПОЛОСАТАЯ ГИЕНА
Hyaena hyaena. L
Добытые экземпляры * Начало XX в © Валорен
АКУ
ПЕЩЕРНАЯ ГИЕНА
Crocuta crocuta spelaea Coldf
ф Верхний плейстоцен
® Средний плейстоцен Crocuta sp.
▲ Плиоцен
ЗЛЬДАРСКАЯ ГИЕНА
Crocuta еЫаПса V вод д Верхний миоцен
£56
Приложение 2
№
Карты
657 -
42 Н. К. Верещагин
658
Приложение 2
Карты 27—30
•с I h
660
П риложение 2
$61
Карты 31—34
I
662
Приложение 2
Карты 43—43
667
4 Xr-iWV ' '
668
Приложение 2
Карты 47—50
669 '
. 670
Приложение 2
Карты 51—54
671
«72
Приложение 2
К
Карты 55—58
I-
43 Н. К. Верещагин
673
674
Приложение 2
Нарты 59—62
675
43*
676
Приложение 2
Карты 63—66
677
Приложение 2
•678
Карты 67—70
679
680
Приложение 2
Нарты 71—74
68Т
46
РОСТОВ
КРАС
ОРДЖОНИКИДЗЕ
СУХУМИ
ТБИЛИСИ
БАТУМИ
ЕРЕВАН
$ & рм
?
* £
ПЛОСКОЛОБЫЙ СЛОН
Elephas of. plaru frons Bale
▲ Верхний плиоцен Elephas sp
А Плиоцен
50 0 50 too ISO 200км
АСТРАХАНЬ
МАХАЧКАЛА
КАРТА 74
ЮЖНЫЙ СЛОН Elephas (ArchidisKOdon) meridionalis Mesli e Нижний плейстоцен о Верхний плиоцен
ФАНАГОРИЙСКИЙ СЛОН
Еlephas (Phanaporoloxodonl mammontoides garull ф Верхний плиоцен
БАКУ
-
Приложение 2
683
Карты 75—78
684
П риложение 2
Карты 79—82
685
686
Приложение 2
IBIF
687
Карты 83—86
<688
Приложение 2
Карты 87—90
689
44 Н. К. Верещагин
1кя№гН!ЯЙНи1КНст1ННН8*доЯга1*Яя^ II wl
690
Приложение 2
Карты 91—94
691
40
АСТРАХАНЬ
РОСТОВ
^СТАВРОПОЛЬ
.КРАС
СУХУМИ
ЕРЕВАН
БАТУМИ
МАХАЧКАЛА
БИНАГАДИНСКИЙ ТУР
Bos ma st an -zadei Burts ch. e Средний плейстоцен Bos sp.
о Верхний плиоцен
УРМИЙСКИЙ ТУР
Urmia! os azerbaidzanicus Burtsch.
А Верхний миоцен
'• 50 200км
КАРТА 93
ПЕРВОБЫТНЫЙ ТУР
Bos primigenius BoJ. г (s.lato)
• II тысячелетие до н.з.
о !олоцен_
® Верхний плейстоцен ф Средний плейстоцен i
БАКУ
? б
44*
692
Приложение 2
Карты <95—<97 693
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ МЛЕКОПИТАЮЩИХ1
амбликастор 53
амфицион 73 ‘
антилопа 46, 47, 58, 69, 73
1 антракотерий 73
! анхитерий 31, 32, 73
аргали — см. баран
архар — см. баран , *
ацератерий 73
байбак — см. сурок
баран 109, 117, 119, 121, 125, 127, 163, 373—374*—375*—376, 690+
барс 127, 144, 170, 275—276*—277*, . 403, 405, 661+
барсук 127, 135, 144, 148, 153,400, 402*, . 637°, 660'
— марагинский 660+ -i
— сехендский 660+
бегемот 160 / .
беличьи 285—293
белка 58, 571, 692* алтайская 571*
— кавказская 470, 640°, 664+
обыкновенная 692*, см. также алтайская
белозубка белобрюхая 647+
—. длиннохвостая 143, 154, 390, 647*
— малая 647*
— малютка 647+
— персидская 647*
— серая 647*
бенаратерий 73
бизон 77, 91, 93, 96, 101
бобр 47, 53, 58, 118, 519
— амбликастор 665*
— болотный 525, 538,* 539*, 573, 642°, 692*
./ речной 82, 117, 123, 293—294* —
•,f 297, 665+
— таманский 665+
—- трогонтерий 73, 665+ ч
бобровые 293—297 \
бородавочник 52 ; /
бурозубка 143, 389
— кавказская 646+
— малая 646+ * J
— обыкновенная 646*
бык 61, 93, 100, 131, 136, 139, 152, 160,
163, 383*
бык Мастана 156
верблюд 58, 69, 77, 100, 131, 159, 161 — гигантский 685*
— Кноблоха 164, 685* ,
— куяльникский 685*
— одногорбый 163, 222, 685*
вечерница гигантская 651* ,
— Лейслера 651* >
— рыжая 651+
волк 99, 104, 117, 118, 122, 127, 135, 136, 139*, 144, 148, 153, 158, 159, 256, 391, 632°, 654+
— красный 653 +
— таманский 73, 654+ >
выдра 82, 143, 144, 637°, 660+ : j
выдро-тюлень 69 i
выхухоль 53, 254, 645* !
газель 45, 46, 57, 73
гепард 144, 148, 154, 280*—281*—282*— 283, 519, 663+
гигантское непарнопалое 35*
гиена 40*, 62, 73, 129, 135, 136, 139, 159, 260—263
— пещерная 125, 148, 153, 158, 261, 655+ — полосатая 144, 261—262*—263, 655+ — пятнистая 144
— эльдарская 655+ гиеновые 260—263 гиппарион 39, 42, 44, 46, 47, 58, 73, 77 гиппопотам 58, см. также бегемот горностай 658+
грызуны 27, 44, 58, 106, 110, 120, 136, 137, 140, 148, 149, 162, 164, 187, 207, 233, 285—327, 405, 470, 473, 529, 564*, 566, 692+
дельфин 69
джейран 57, 82, 171, 189, 354—357, 480*, 535*, 688+
дикобраз 58, 121, 122, 125, 127, 148, - 471, 681+
— бипагадипский — см. дикобраз Виноградова
1 * — отмечены рисунки; ° — графики заготовок шкур; + — карты распространения. Сводные списки антропогеновых видов помещены на стр. 234—245 и 498—511; в указателе эти страницы не отмечены.
^*й^«втаи«а»^л«
Указатель русских названий
695
— Виноградова 149, 154, 681+
— индийский 149
дикороцерус 32 .
динотерий 42 f .
длиннокрыл 650+ 1
ежовые 250 . -
емуранчик 480*, 669+
енотовидная собака 525*, 572, 634°, 691*
енот-полоскун 526, 572*, 691+
еж обыкновенный 143, 219*, 250, 388, 645+
ёж ушастый 143, 154, 191, 251, 388, 645+
жвачные 62 . <
жирафа 39, 42, 44—46, 73
жирафиды 48
зайцеобразные 58, 140, 148, 149, 233, 283—285, 405, 469, 473, 692+
зайчик земляной 668+
заяц 58, 139, 148, 191, 405
заяц-беляк 125
заяц-русак 149, 283—285, 405, 469, 639°, 664+
заячьи 283—285 >
землеройка 250
зубр 77, 93, 95, 96, 102, 103, 109, 112, ИЗ, 117—122, 125, 197*, 378+, 379*, 381, 448*, -449*, 450, 519
— длиннорогий 91, 100
— кавказский 376*—380, 690+
— первобытный 100, 110, 376—380, 514, 690+
— таманский 73, 690+
зубробизон 575, 692+
иктитерий 46
кабан 82, 96, 98, 103, 108, 109, 118, 119, 122, 123, 137, 152, 154, 334—338, 435, 463*, 684+
кабан апшеронский 684+
— таманский 684+ . . . .
кит 26
— беззубый 37 : .
— древний 26, 42 ...... ..
китообразные 26
кожан двухцветный 652+
— Огнева 653+
— поздний 653+
кожанок Бобринского 652+
кожанок северный 653+
козел 88, 98, 104, 108, 112, 117—121, 125, 127, 160, 212, 220*, 371*
— безоаровый 370—371—372*—373,
478*, 689+
— восточнокавказский 203*, 206*, 363— 364*—367*—368*—370*, 445, 475, 689+
— западнокавказский 363—365*—370, 445, 475, 689+
— кавказский 47, 112, 118, 121, 122, 128, 443
козлорог 39, 42, 73
копытные 26, 27*, 30, 40*, 41*, 55*, 78, 106, 108, 109, 117, 120—122, 126,
127, 134, 137, 140, 152, 163, 164, 194—196, 202—205, 209, 535
корсак 144, 148, 153, 160, 259, 399*,-400*, 633°, 655+
косуля 73, 102, 109, 119, 121—123, 125, 351—353, 354, 687+
косуля сибирская 687+
кот камышовый 144, 279, 480*, 663+
— лесной 122, 143, 144, 278, 405, 662+
— пятнистый 148, 154, 663+
кошачьи 270—283 i.. . - -
кошка 104, 148 ;
— дикая 638° 1 '
креодонты 26 . ‘ ;
кролик 537*, 692+
крот 143, 251—252—253*—254, 388, 631°, 645+
— кавказский 388, 389, 405 "
кротовые 251
крыса 57
— водяная — см. полёвка водяная
— серая 142, 299—300*—301, 551*, 670+
— черная 297—298*—299, 470,551*, 670+ куница 46, 122, 148, 160, 402, 635°
— белодушка 143, 144, 267, 404*, 556*, 656+
— лесная 143, 144, 268—269—270, 402, 404*, 657+ /
куньи 266—270
кулан 189, 331—332*—333, 474, 519, 683+
кутора водяная 646*
лань 164, 345, 570, 685+ ' ’ t
— европейская 692+ ’
ласка 143, 144, 659+
ластоногие 69
лев 270—272*—273*—274, 519, 661+
леопард — см. барс
летучая мышь 105, 106, 528
лисица 57, 73, 99, 122, 127, 139, 148, 153, 158, 258, 391—392*—394*—395— 396—397—398*, 632°, 633°, 654+
— Хоменко 654+
— черносеребристая 691+
лось 104, 109, 122, 123, 197, 348*, 349*, 440, 479*, 519
— кавказский 180, 347—351, 352*, 686+ лофиомерикс 28
лошадиные 44, 327—333
лошадь 48, 53, 61, 69, 77, 94, 96, 100, 101, 112, 113, 117—122, 129, 131, 134—136, 139, 151, 153, 159, 163,'164. 171, 432*—434, 474, 514, 684+
— нижнечетвертичная 684*
— Стенопа 66, 69, 73, 93, 161, 684+
— четвертичного периода 327—329— 330*—331
макак 167, 233
мамонт 91—95, 101—103, 122, 125, 128, 419*, 420, 425—427, 682+
манул 663+
мастодонт 32, 42, 46, 52, 66, 71, 77, 415
— болотный 30, 73
— длиннорылый 32, 73 . .
696 Указатель русских названий
мастодонт овернский 69, 73 махайрод — см. саблезубый тигр медведь 105, 107, ИЗ, 122, 136, 144, 153, 265*, 400—401*—402*,. 634°
— бурый 105, 108, 117, 143, 160, 264— 266, 399, 656+
— большой пещерный 264*
— малый пещерный 91, 263, 655+
— пещерный 104—112, 117—119, 125, 127, 148, 160, 514, 655+
медвежьи 263—266
мезопитек 45
микромерикс 32
морская корова 26
муфлон 478*
— арменийский 373—376, 690+
мышиные 297—327
мышовка кавказская 95, 667+
— лесная 471, 667+
I — южная 667+
мышь домовая 408, 470, 551*, 671+
— желтогорлая 409, 471, 672+
. — кавказская желтопятная 409, 410, 470, 672+
— лесная 121, 149, 409—411, 470, 556*, 564*, 673+
— малоазийская 409, 470, 477*, 672+ мышь-малютка 471, 672+
мышь полевая 301—303, 471, 479*, 671+
>• — степная 148, 149, 564*, 671+
насекомоядные 58, 120, 140, 143, 164, 185, 233, 250—255, 388, 465, 529
некромит 69
непарнокопытные 41, 44, 46, 68, 69, 151, 244, 327—333, 430, 475
нетопырь-карлик 651+
— Куля 652+
— Натузиуса 652+
— Сави 652+
порка 143, 404, 479*, 636°, 659+
— американская 570, 572, 691+
носорог 31, 32, 39, 42—46, 52, 53,58, 61, 122, 129, 132*, 136, 137, 139, 151, 154, 159, 430
— безрогий 44
— бинагадинский 683+
— волосатый 102, 125, 430, 683+
— восточный 683+
— двурогий 73
— кавказский 683+
— Мерка 92, 161, 164, 683+
— этрусский 61, 77, 683+
ночница Бехштейна 649+
— Наттерера 649+
— остроухая 649ь
— трехцветная 649+
— усатая 649+
нутрия — см. бобр болотный
обезьяна 58, 127, 233
— человекообразная 41
овцебык 98, 125
олень 44, 48, 53, 57, 58, 61, 66, 69, 73, 91, 94, 96, 101, 102, 108, 117—122,
134—136, 152, 162, 171, 197*, 435— 440
— благородный 57, 105, 131, 159, 164, 338—345, 435—347*—438*—439, 514, 686+
— гигантский 94, 103, 153, 164, 346, 685+
— пятнистый 570, 575, 692+
— северный 98, 125
— сложнорогий 73, 685+
оленьи 338—354
ондатра 525, 526*, 692+
осёл 122, 135, 151, 154, 163
— плейстоценовый 683*
паранхитерий 31, 73
парнокопытные 27*, 35*, 41, 44, 46, 58, 152, 199, 244, 333—383, 434, 475, 692+
пасюк — см. серая крыса
перевязка 144, 154, 401, 657+
песец 98, 125
пеструшка степная 323, 676+
песчанка 74, 82, 139
— араздаянская 676+
— большая 187, 675+
— Виноградова 471, 478*, 564*, 676+
— краснохвостая 137, 148, 149, 154, 307, 411, 471, 675+
— малоазийская 305—306, 471, 478*, 564*, 676+
— персидская 306—307, 471, 478*, 564*, 675+
— полуденная 675+
— тамарисковая 675+
пищуха 170
— арменийская 469, 664+
— рыжеватая 664+ •
— степная 664+
платибелодон 30—34
подковонос 105 - '
— Блазиуса 648+
— большой 648+
— малый 648+
— Мегели 648+
— южный 648+
полёвка 139, 148, 154, 411
— апшеронская 148, 149, 413, 679+
— водяная 82, 153, 326, 411, 412, 470, 641°, 677+
— — длиннохвостая 180, 322, 470, 477*, 679+
— — кавказская 184, 317—318*—319* — 320, 413, 470, 477*, 678+
— кустарниковая 148, 181, 187, 315— 317, 413, 470, 477*, 558*, 679+
— малоазийская 320—322, 413, 471, 678*
— общественная — см. полёвка степная
— обыкновенная 148, 149, 153, 312— 315, 414, 471, 564*, 680+
— прометеева 47, 121, 122, 323—326, 324*, 411, 470, 477*, 558*, 677+
— рыжая 323, 471, 479*, 680+
— серая — см. полёвка обыкновенная
— снеговая 88, 187, 412, 564*
— степная 80, 148, 149, 154, 191, 219*, 308—312, 415, 564*, 680+
полорогие 44, 48, 101, 354—383
Указатель русских названий
697
f полчок 470, 641°, 666+
приматы 44
примитивные копытные 26
( псовые 255—260
росомаха 266, 401, 403, 658+
рукокрылые 233, 390, 467
рысь 143, 144, 160, 277, 405, 638°, 622+
сайга 82, 96, 98, 136, 152, 153, 156, 158, 160, 187, 357—360, 440—441*—442*— 443, 479*, 687+, 692+
сайгак — см. сайга
свиные 334—338
свинья 44, 53, 62
свинья гигантская 73
семантор 69
сеноставка 58
серна 98, 108, 117, 118, 121, 125, 127, 160, 360*—362*—363, 688+
— кавказская 475
складчатогуб широкоухий 653
скунс 570, 572, 691+
слепушонка 137, 148
— горная 149, 154, 471, 478, 677 +
— обыкновенная 479*, 677+
! слепыш гигантский 669+
; — горный 471, 478*, 669+
i — обыкновенный 58, 187, 669+
слон 53, 61, 62, 78, 91, 94, 99, 100, 103, 161, 415—418, 420—429
— армянский 74
— Бюста 92, 417
— древний 682+
— плосколобый 66, 77, 416, 681+
— фанагорийский 681+
► — трогонтериевый 91—94, 100, 102, 103,
161, 164, 417, 420, 427, 682+
— южный 69, 73, 77, 91, 94, 416, 417*, 420, 428, 681+
соня лесная 137, 149, 470, 666+
стегодон 71
сурок 127, 161, 179, 664+
— алтайский 573—575*, 692+
— кавказский 664+
суслик 80, 640°
— европейский — см, суслик малоазийский
— малоазийский 292, 406, 407, 471, 478*, 665+
— малый 191, 285—292, 406, 407*, 479*, 665+
схизотерий 28
тапир 52, 53, 73
тарпан 519, 684+
тигр 274, 275*, 519, 661+
— саблезубый 46, 47
тигролев 104, 117, 119, 125, 144, 148г
153, 270*, 661+
тр агул иды 44, 73
тур бинагадинский 691+
— карликовый 163
— первобытный 100, 122, 156, 158, 160, 164, 380—382*, 450, 519, 691+
— урмийский 691+
тушканчик 82, 137, 139, 148, 406
— большой 149, 153, 160, 408, 479*, 668+
— горный 149, 154
— малоазийский 407, 471, 668+
— малый 149, 407, 667+
— мохноногий 669+
тюлень 28, 37, 42, 51, 62, 69, 189
удабнопитек — см. человекообразная обезьяна
ушан 650+
халикотерий 73
хищники 26, 51, 80, 95, 117, 139
хищные 32*, 40*, 41*, 44—46, 54*, 58, 106, 109, 117, 120—122, 126, 127, 140, 143, 144, 148, 159, 185, 233, 255—283, 391, 469, 534
хоботные 44, 46, 47, 54*, 58, 68, 415, 691+ хомяк 73, 80, 104, 105, 122, 127, 139, 408 — дагестанский 154
— древнекавказский 674+
— малоазийский 47, 112, 121, 122, 125, 303—305, 408, 471, 478*, 673+
— обыкновенный 110, 471, 479*, 674+
— равнинный 149
— черноватый 148, 149, 191
Хомячок мышевидный 471, 478*, 564*, 675+
— серый 137, 148, 149, 154, 408, 471, 564*, 674+
хорёк степной 479*, 636°, 658+
цетотерий 28, 37
шакал 144, 148, 170, 255, 480*, 631°, 653+
широкоушка азиатская 650+
— европейская 650+
эласмотерий 77, 100, 129, 159, 431
— кавказский 683+
— сибирский 683+
эотрагус 32
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ МЛЕКОПИТАЮЩИХ
Aceratherium 41, 43
— filholi 27* ’ ‘
— incisivum 52 • -
— persia 44
— transcaucasicum 41*
— zernovi 44
Achtiaria 41
— borissiakii 39
— expectans 44
Acinonyx jubatus 141, 145*, 154, 280— 283, 469, 663+
Alactagulus acontion 473, 668+
Alces alces 105, 108*, 109, 117, 118, 440, 475, 686+
— — caucasicus 180, 202, 205, 347, 686+
Alcicephalus neumayri 45
Allactaga 127, 406
— elater 146*, 219, 230, 408, 473, 530, 667+
— — dzhafarovi 141
— jacuius 408, 473, 668+
— jacuius bogatchevi 141, 146*, 153, 408, 668+
— williamsi 141, 154, 219, 227, 228, 230, 408, 473, 530, 550, 668+
— — dzafarovi 146*, 668+
Ambelodon fricki 32
Amblycastor caucasicus 52, 665+
Amphicyon 31, 32*
Anancus arvernensis 52, 54*, 71, см. также Mastodon arvernensis
Anchitherium aurelianense 31 .
Anthracotherium 27
Antilopinae 45, 47
Apodemus 409
— agrarius 149, 301, 473, 544, 671+
— flavicollis 149, 410*, 473, 672+
— fulvipectus 409, 410*, 473, 550, 672+
— mystacinus 409, 473, 672+
— sylvaticus 141, 149, 153, 182, 183, 185, 193, 206, 208, 218, 219, 225, 228, 230, 410*, 473, 530, 544, 547, 550, 566, 673+
— — arianus 534
Archeoceti 26
Archidiskodon — cm. Elephas
Artiodactyla 27, 39, 41, 43, 44, 47, 48, 52, 57, 59, 60, 61, 63, 69, 71, 74, 96, 123, 127, 141, 193, 233, 333—383, 434, 474, 476, 530, 547, 550, 692+
Arvicola 411, 677+
— terrestris 111*, 141, 149, 153, 183, 185, 186*, 195, 227, 228, 230, 326, 411, 474, 534, 547, 550, 677+
— — cubanensis 412
— — kuruschi 412
— — persicus 412
— — tanaiticus 412
Barbastella barbastellus 467, 650+
— darj el ingens is 467, 650+
Benaratherium callistrati 27, 27*
Bison 61, 63, 67*, 71, 77, 100, 101, 118, 448*, 690+
— bonasus 122, 152, 211, 475, 692+
— — bonasus 448*, 449*
— — caucasicus 202, 205, 448*, 449*, 376, 690+
— longicornis 448*, 449*
— priscus 96, 97*, 99, 109, 111, 112, 115*—117, 119, 124*, 127, 161, 163, 376, 448*, 449*, 690+
— — deminutus 92, 96, 103, 448*
— — longicornis 91, 92
— — schoetensacki 63, 77, 93, 100, 376, 448*, 449*
— sp. (tamanensis) 448*, 449*, 690+
Bos 59, 61, 91, 100, 118, 131, 161
— mastan-zadei 141, 152*, 154, 380,
691+
— minutus 163, 380
— primigenius 100, 152, 160, 161, 380, 475, 691+
— taurus fossilis 100
— trochoceros 162*, 380
Bovidae 41, 71, 354—383
Bubalis 69
Calomyscus bailwardi 230, 473, 675+
Camelopardalis 39
— attic a 45 . . •
— parva 42 ' . ' ’
Camelus 59, 131
— dromedarius 163, 223*, 685+
— knoblochi 161, 685+
— kujalnikensis 60, 63
Canidae 52, 54*, 255—260
Canis 60, 99, 141, 144, 160, 654+
— aureus 144,155, 170, 210, 212, 227, 255 — 256, 469, 530, 547, 550, 653+
— lupus 60, 96,97*, 105, 108*, 109, 115*, 117, 118, 119, 122, 127, 144, 153, 177,
У казателъ латинских названий
699
179, 208, 227, 256—258, 469, 530, 547, 550, 654+ • ;
— — apsheronicus 141, 145*, 654+
— tamanensis 63, 67*, 654+
‘Capra 105, 127
aegagrus 225, 227, 370, 475, 689+
— caucasica 99, 109, 111*, 112, 115*, 117—118—119, 122, 124*, 127, 193, 194*, 205, 209*, 211, 212, 444*, 475, 689+
— — caucasica 446*
— — severtzovi 446*
— cyl indr icornis 202, 205, 212, 443, 444, 446*, 475, 689+
— dinniki 444
— severtzovi 444 ;
Capreolus 101
— capreolus 108*, 109, 117, 118, 122, 127, 130*, 177,183,190,193, 202, 205, 208— 209*— 210, 217, 351, 475, 547, 550, 687+
— — pygargUS 687+ .
Caprovinae 119
Carnivora 31, 39, 41, 44, 52, 57, 59, 60, 63, 74, 123, 127, 141, 233, 255—283, 391, 468, 476, 530,534,547,550,691+
Castor fiber 117, 211, 293, 473, 665+
— tamanensis 63, 67*, 665+
Castoridae 47, 293—297
Cervidae 41, 47, 55*, 63, 71, 101, 338—354 Cervus 57, 59,- 60, 61, 66, 71, 91, 161, 686+
— elaphus 60, 93, 96, 97*, 99, 108*, 109, ill*, 115*—118, 122, 124*, 127, 131, 152, 153, 162, 175, 177, 181, 183, 190, 193—194*, 195, 208, 210, 217, 222, 225, 227, 338., 437*—439*, 475, 550, 686+
— — binagadensis 141
— — maral 161, 202, 205, 438
— nippon 575, 692+
— pliotarandoides 685+
Cetacea 31, 39, 43 .
Cetotherium 37, 39, 43 .
— meyeri 28
— priscum 43 .
Chalicotheriidae 31
Chalicotherium pentelici 44 = .
Chionomys — cm. Microtus
Chiroptera 123, 127, 233, 390, 466, 476, 533, 534
Citellus citellus 292, 473, 665+
— pygmaeus 149, 191, 193, 194*, 285, 473, 665+
— — boeinii 406
— — musicus 406
— — planicola 406
— — satunini 406
Clethrionomys glareolus 323, 474, 680^ Cricetus 53*, 408, 674+
— cricetus 77, 99, 105—108*—409, 111*, 182, 183, 185, 186*, 190, 406, 473, 674+
Cricetulus migratorius 154, 182, 183, 185, 206, 218, 219, 225, 227, 228, 230, 231, 473, 530, 533, 534, 566, 674+
— — argiropuloi 141, 164*, 674+ vp.4
— Crocidura 52
— lasia 466, 674+
— leucodon 143, 228, 389, 466, 530, 533, 534,. 550, 647+
— russula 141, 143, 154, 218, 219, 225, 227, 228, 230, 466, 550, 647+
— — caspica 390
— — giildenstaedti 389, 390, 530, 532— 533—534, 544
— — monacha 390, 547
— suaveolens 466, 647+
— zarudnyi 466, 647+
Crocuta 129, 655+
— eldarica 39, 41*, 655+
— spelaea 96, 97*, 99, 127, 141, 144,145*, 153, 160, 261, 655*
Cuon 127, 653+ C' у : . 4
Dama 685+
— dama 692+ .
— mesopotamica 162*, 170*, 345
Delphinus 26, 39, 210,
— delphis 69, 175
Desmana 52, 53*, 250, 645+
— moschata 254—255, 466, 645+
Dicerorhinae 32
Dicerorhinus 43, см. также Rhinoceros — caucasicus 31, 683+ — etruscus — cm. Rhinoceros etruscus — orientalis 52, 56*, 683+
Dicrocerus elegans 31
— salomeae 41
Dinocyon thenardi 52, 54*
Dinotherium 41, 52
— giganteum 39, 42, 48
Dipus sagitta 473, 669+
Dyromys nitedula 141, 149, 185, 218, 219, 228, 473, 530, 544, 547, 550, 666+
Elasmotherium 60
— caucasicum 63, 77, 431 -
— sibiricum 100
Elephas 74, 91, 681+
— antiquus 59, 63, 68, 101, 415, 682+
— armeniacus 74, 161
— lyrodon 59
— mammontoides 681+
— meridionalis 59—63, 67*—71, 74, 78, 93, 94, 161, 415, 423*, 681+
— primigenius 91, 92, 94, 96, 97*, 99, 101, 103, 104, 128, 162, 418, 421*, 682+
— trogontherii 63, 69, 77, 78, 91—94,102, 103, 160, 161, 415, 417, 422*, 682+
— — primigenius 93
— wusti 92, 417
Ellobius lutescens 141, 146*, 149, 154, 227, 228, 230, 231, 473, 677+
— talpinus 149, 193, 194*, 473, 677+ Eostylocerus 41
Eotragus martinianus 31, 36*
Equidae 327—333
Equus 61, 71, 91, 100, 101, 129, 160, 163 ~ caballus 96, 97*, 112, 115*, 117, 119, 131, 141, 150, 151*, 153, 161, 162, 163, 211, 327, 433*, 43, 475, 684+
, 700
Указатель латинских названий
Equus caballus gmelini 151, 433*, 434, 475,. 684+
— — latipes 433*, 434
— hemionus 151, 155, 188, 189*, 190, 215, 227, 331, 475, 683+
— hidruntinus 96, 97*, 117, 141, 150*, 151, 154, 683+
— przewalskii 433*, 434
— stenonis 59—61, 69, 70*, 74, 77, 100, 161, 432, 684+
— siissenbornensis 63, 67*, 77, 94, 129, 432, 684+
Erinaceidae 250
Erinaceus europaeus 122, 141—143, 153, 182, 183, 212, 219, 227, 228, 230, 250, 388, 466, 530, 547, 550, 645+
— — rumanicus 645*
Eucladocerus 61, 63, 67*, 69, 70*, 71, 94
Eucladocerus pliotarandoides 60—62*
Felidae 270—283
Felis 109
— chaus 144, 155, 279, 469, 530, 550, 663+
— issiodorensis 52, 54*
— lybica 141, 144, 145*, 154, 225, 227, 469, 663+
—lynx 115*, 117, 144, 155, 175, 195, 277, 469, 550, 662+
— — orientalis 405
— silvestris 99, 111*, 115, 144, 183, 211, 278, .547, 550, 662*
— — caucasicus 405 ' ‘
Gazella 39—44, 47, 52, 57, 63, 170*
— brevicornis 45
— gaudry 45
— subgutturosa 100, 152, 155, 188, 189*, 214, 215, 225, 354, 475, 530, 688+
Gerbillus 74
Giraffidae 41, 47, 52
Glis glis 183, 212, 473, 544, 547, 550, 666+
Gulo gulo 122, 124*, 127, 266, 658+
Halitherium schinzi 26
Helicophora rotundocornis 45
Helladotherium 39
— gaudry 45
Hemiechinus auritus 141—143, 154, 182, 183, 191, 218, 219, 225, 230, 231, 251, 388, 466, 530, 533, 645"
Hipparion 41—43, 47, 52, 55*, 60, 69, 74 — crassum 69
— gracile 38, 39, 41, 43, 44, 47, 52, 55*
— mediterranean! 44
Hippopotamidae 31, 35*
Hippopotamus 160
Hyaena 31, 32*, 40*, 41, 59, 60
— eximia 39
— hyaena 155, 261—263, 469, 655+
— striata 129, 144
Hyaenidae 260—263
Hypsodontus miocenicus 31, 36*
Hystricidae 41
Hystrix 127, 130*, 681+
— hirsutirostris 117
— leucura 474, 681+
— vinogradovi 141, 146*, 149, 154, 681
Iniops 26
— caucasicum 26
Insectivora 52, 123, 141, 233, 250—255,.. 388, 465, 476, 530, 533, 534, 547, 550
Ictitherium tauricum 44
Kubanochoerus robustus 31, 36*
Lagomorpha 52, 74, 123, 141, 233, 283— 285,405,469,473, 476, 530, 534,547,. 550,692+
Lagurus lagurus 323, 473, 676+ Leporidae 283—285
Lepus 52, 53*, 74, 101, 664+
— europaeus 96, 99, 127, 149, 154, 175,. 177, 179, 182, 183, 185, 186*, 191, 193, 195, 206, 208, 209*, 212, 214, 224*, 227, 228, 230, 231, 283—285,. 473, 530, 547, 550, 664+
— — caucasicus 405
— — cyrensis 405, 534
— — gureevi 141, 146*, 664+
Listriodon 43
Lophiomeryx benarensis 27
Lutra lutra 144, 177, 469, 547, 550, 660+' Lutreola lutreola 144, 659* — — turovi 404 — vison 691+
Macaca 127, 130*
Machairodus 47, 60
Marmota 127, 130*, 160, 664+
— baibacina 573, 692+
— bobac 179, 664+
Martes 108*, 109, 111*, 115*, 193, 402:
— foina 117, 127, 144, 155, 224*, 225,. 227, 267, 469, 550, 656+
— — nehringi 404
— martes 268—270, 469, 657+
Mastodontidae 71
Mastodon 31, 39, 41, 60
— arvernensis 59, 66, 69, см. также Anancus
— borsoni 48
— longirostris 39 • ' .
— pentelici 39, 44
— tapiroides 48
Megaceros 103, 160, 161
— euryceros 63, 94, 96, 97*, 99, 109,. 141, 152, 153, 346, 685+
— hibernicus 100
Meles 59, 660+
— meles 99, 115*, 117, 127, 144, 153, 160,, 177, 179, 183, 193, 208, 209*, 222, 227, 469, 530, 547, 550, 660+
— — minor 141, 145*, 402,* 660+
— — urartuorum 660+
— maraghanus 660+
— polaki 660+
Mephitis mephitis 691+
Meriones 225, 228, 473, 676+
— blackleri 230, 305, 473, 534, 676«-
— erythrourus 154, 218, 219, 473, 530,. 533, 675+
Указатель латинских названий
701
— — intermedins 141, 146*, 411, 675+ — meriodianus 473, 675+
— persicus 227, 230, 231, 306, 473, 675+
— tamariscinus 473, 675+
— vinogradovi 473, 676v
Mesocricetus 47, 408
— auratus 112, 118, 119, 122, 124*, 127, 130*, 154, 182, 183, 185, 195, 227, 228, 230, 231, 303, 473, 673"
— — nigriculus 186*, 191, 408
— — planicola 141, 146*, 408, 673
— — raddei 408, 673+
Micromeryx flourensianus 31
Micromys minutus 149, 182, 473, 672+ Microtinae 208
Microtus 411
— apscheronicus 141, 149, 413, 679+ arvalis 141, 149, 153, 182, 183, 185, 206, 208, 230, 231, 312, 414, 474, 550, 680+
— — gudauricus 414
— — macrocranius 414
— — mystacinus 414
— — transcaucasicus 414
— gud 127, 185, 317, 474, 678+
— - gud 413
— — Ighesicus 413
— — nenjukovi 413
— guentheri 314
— majori 127, 182, 183, 208, 228, 315, 474, 547, 679+ '
— nivalis 227, 228, 413, 320, 474, 678+
— — dementievi 413
— — loginovi 413
— — satunini 413
— roberti 115*, 180, 322, 474, 679+
socialis 141, 149, 154, 191, 218, 219, 227—231, 308—312, 415, 474, 530, 532—534, 544, 550, 680+
-----schidlovskii 309
Microzeuglodon 26
— caucasicum 26
Miniopterus schreibersii 230, 467, 650+
Muridae 71, 96, 127, 297—327
Mus 52, 671 +
— musculus 141, 149, 153, 182, 183 185, 206, 208, 218, 219, 225, 230, 408, 473, 547, 566, 671+
— — abbotti 532, 550
— — forraosovi 544, 671+
— — liortulanus 671 +
— — musculus 530, 532—534
-----tataricus 530, 532—534, 544, 550, 566, 671+
Mustela erminea 469, 658+
— filholi 74
— nivalis 127, 144, 155, 183, 185, 186*, 206, 208, 227, 230, 469, 530, 534, 547, 550, 659+
— — caucasica 122, 182, 228, 404
— — dinniki 182, 404
Mustelidae 266—270
Myocastor coypus 525, 547, 692+
Myotis 390
— bechsteinii 467, 649+
— emarginatus 467, 649+
— mystacinus 467, 528, 534, 649+
— nattereri 467, 649+
— oxygnathus 227, 228, 467, 649+
Necromites nestoris 69
Neomys 127, 389
— halcaricus 289
— daghestanicus 389
— fodiens 228, 389, 466, 646+
— leptodactylus 389
— schelkovikovi 389
Nyctalus leisleri 467, 651+
— noctula 467, 528, 651+
— siculus 467, 651+
Nyctereutes procyonoides 524, 691+
Ochotona 171, 227, 228, 473, 664+
— antiqua 52, 664+
— pusilia 664+
— rufescens 664+
Oiceros atropatanes 45
— boulei 45
— rothi 45
Ondatra zibethica 525, 692+
Orictolagus cuniculus 692+
Orycteropus sp. 31
— gaudryi 44
Otocolobus manul 469, 663+
Ovis 127, 195
— ammon 119, 127, 130*, 141, 152*, 154, 373, 690+
— gmelini 163, 195*, 225, 227, 373, 475, 690+
— ophion 109
Palaeocricetus caucasicus 31, 674+ Palaeoloxodon 416
Palaeomastodon 32
Palaeoryx longicephalus 48 '*
— pallasi 45
Panthera 63, 67*, 661+
— leo 144, 155, 270—274, 469, 661+ — pardus 127, 130*, 144, 155, 171, 193, 194*, 211, 275—277, 403, 469, 661+
— spelaea 99, 104, 115*, 117, 119, 127, 141, 144, 145*, 153, 270, 661+
— tigris 274, 469, 661+
Paracamelus 77
— gigas 69, 71*, 100, 685+ — kujalnikensis 685+ Parahippus 32
Paranchitherium karpinskii 31, 34+
Paratragocerus caucasicus 31, 36* Parelephas — см. Elephas Perissodactyla 27, 31, 39, 41, 43, 44, 47, 52, 59, 60, 61, 63, 69, 71, 74, 123, 141, 233, 327—333, 430, 474, 476.
Phanagoroloxodonmammontoides 63, 415, 681 +
Phiomia 32
Phoca 28, 39, 47
— caspica 62, 69, 188, 215
— pontica 43 -
— vindobonensis 28
Phocidae 37
Phronetragus arknethensis 41
702
Указателъ латинских названий
Pinnipedia 39, 43
Pitymys 315, 562, см. Microtus majori
Platy'belodon danovi 31, 33*
Plecotus auritus 467, 650+ ..
Pliocervus 52
Primates 39, 41, 44, 233
Proboscidae 31, 39, 41, 44, 47, 48, 52, 59—63, 69, 71, 233, 415
Procapreolus 52, 55*, 687+
Ргосуоп lotor 526, 550, 691+ ' ‘ ;
Prometheomys 47, 411
— schaposchnicovi 122, 124*, 127, 323, 473, 677+
Propotamochoerus provincialis 52, 55*
Protoryx carolinae 45
Protragelaphus scozesi 45 c
Pseudalces 52
Pu tori us 74
— eversmanni 144, 183, 185, 469, 658+ — putorius 469, 658+ .....
Rattus norvegicus 155, 182, 183, 185, 186*, 206, 212, 219, 299, 473, 530, 533, 547, 566, 670+
— rattus 208, 209*, 297, 473, 530, 547, 550, 670+
Rhinoceros 43, 60, 71, 127, 130*, 161 — antiquitatis 92
— binagadensis 141, 150, 151*, 154, 430, 431*, 683+
— etruscus 61, 63, 71, 74, 160, 430, 683+
— mercki 74, 92, 129, 161, 430, 683+
— morgani 44
— schleiermacheri 32
— tichorhinus 102, 430, 431*, 683+
Rhinocerotidae 38, 47, 52, 56*
Rhinolophidae 390
Rhinolophus blasii 467, 648+
— euryaJe 467, 648+
— ferrum-equinum 109, 467, 528, 648+
— hipposideros 467, 528, 648+
— mehelyi 467, 648+
Rhombomys opimus 187, 473, 675+
Rodentia 27, 31, 41, 52, 59, 60, 63, 71, 74, 123, 141, 233, 285—327, 405, 470, 473, 476, 530, 533, 534, 547, 550, 692+
Rupicapra rupicapra 108*, 111*, 115*, 117, 118, 122, 193, 475
-----caucasica 202, 205 , 211, 360, 688+
Saiga borealis 441*, 443
— prisca — cm. S. tatarica
— tatarica 96, 97*, 100, 357, 441*—443, 475, 687+, 692+
— — binagadensis 141, 152*, 153, 441*, 443
— — mongolica 442+
Schisotherium chucua 27 *, 27
Scirtopoda telum 473, 669+
Sciuridae 285—293
Sciurus anomalus 473, 550, 664+ '•
— vulgaris 692+
— — altaicus 570 :
Semantor macrurus 69
Sicista betulina 473, 667+ ' :
— caucasica 96, 97*, 473, 667+
— subtilis 473, 667b . ' ; < -
Sorex 250 38Q
— araneus 185, 208, 466, 547,. 550, 646+^
— minutus 228, 389, 466, 646+
— raddei 389, 466, 646+
Spalax giganteus 187, 473, 669+
— leucodon 473, 669+
— microplitalmus 77, 182, 183, 185, 473 r 669+ . ...
Spelaearctos rossicus 91
Stegodon 71
Steneofiber 52, 53*, 293,665+
Strepsicerotini 63
Suidae 334—338
Suncus etruscus 466, 647+
Sus 41, 43, 47, 60
— apscheronicus 141, 152*, 154, 334?. 435, 436*, 684+
— erymanthius 39, 44
— scrota 96, 108*, 109, 111*, 116—118, 122, 127, 152, 175, 177, 183, 188, 193, 195, 202, 205, 208, 210, 222, 334, 435, 475, 530, 547, 550, 684+
— — attila 435, 436*
— tamanensis 63, 67*, 334, 435, 436* r 684*
— vittatus 434
Tadarida taeniotis 467, 653+
Talpa 52, 53*, 127, 250, 388, 645+
— coeca 547
— caucasica 122, 124*, 182, 183,. 186* 251, 466, 645*
— — caucasica 389
— — ognevi 388, 389, 645+
— orientalis 251, 466, 645+
— — orientalis 389
— — talyschensis 388, 389
— — transcaucasica 388, 389
Talpidae 251—254
Tamanalces 67*, 686+
— caucasicus 63
Tapirus arvernensis 52, 55*
Tragelaphus 63
— hontom-schindleri 45
Tragocerus 39, 40*, 41, 44, 74
— Feskevitschi 39, 41, 41*, 44
— rugosifron 45
Trogontherium cuvier i 59, 60, 63, 67*
71, 665+
Tubulidentata 31, 44
Udabnopithecus garedziensis 41
Urmiabos azerbaidzanicus 45, 691+
Ursavus 31, 263
Ursidae 263—266
Ursus 60, 263
— arctos 109, 111, 117, 118, 144, 153, 183, 188, 189*, 208, 209*, 211, 222, 227, 264—266, 402*, 469, 550, 656*
— arctos binagadensis 141, 145*, 4O2*„ 656+
— — caucasicus 122, 399, 401*, 402*
— — meridionalis 122, 402*
— arvernensis 52, 54*
Ука&ателъ латинских названий
703'
— kamiensis 400—402*
— karmalkiensis 401*, 402*
— rossicus 263, 655*
— spelaeus 96, 99, 105, 108*—112, 115*, 117-119, 122, 127, 130*, 264, 655+
Vespertilio 390
— bobrinskii 467, 652+
— kuhlii 467, 528, 533, 652+ т ; ... ,
— murinus 467, 652+
— nathusii 467, 528, 652+
— nilssonii 467, 653+
— ognevi 467, 653+
— pipistrellus 467, 528, 534, 651+
— savii 467, 652+
— serotinus 467, 528, 533, 653+
Vormela peregusna 141, 145*, 154, 183, 227, 469, 530, 657+
Vulpes 99
— corsac 141, 145*, 153, 259—260, 399*,. 469, 655+
— fulvus 691+
— khomenkoi 57, 391, 654+
— vulpes 108*, 109, 115*, 117, 122, 127,. 144, 153, 179, 183, 188—190, 193,„ 206, 215, 222, 224*, 227, 258, 469y 530, 547, 550, 654+
— — alpherakyi 141, 145*, 392,. 654т
— — alticola 392
— — kurdistanica 392
Zeuglodon 26
Предисловие........................................................ 3
Введение........................................................... 5
Часть I. ПАЛЕОНТОЛОГИЧЕСКОЕ И АРХЕОЛОГИЧЕСКОЕ ОБОСНОВАНИЕ ИСТОРИИ ФОРМИРОВАНИЯ ФАУНЫ
Глава I. Развитие ландшафтов и фаун млекопитающих Кавказа в третичном периоде..................................................... 25
Глава II. Развитие ландшафтов и фаунистических комплексов Кавказа в четвертичном периоде ................................................ 76
Часть IL АНАЛИЗ ПРОИСХОЖДЕНИЯ четвертичных млекопитающих КАВКАЗА В СВЯЗИ С ИХ РАСПРОСТРАНЕНИЕМ, ЭКОЛОГИЕЙ И МОРФОГЕНЕЗОМ
Глава III. Обзор прежнего и современного распространения и численности некоторых млекопитающих................................. 249
Глава IV. Материалы по стратиграфической и географической изменяемости кавказских млекопитающих четвертичного периода ......... 387
Часть Ш. ТЕРИОГЕОГРАФИЧЕСКОЕ РАЙОНИРОВАНИЕ КАВКАЗСКОГО ПЕРЕШЕЙКА И ПУТИ НОВЕЙШЕЙ ЭВОЛЮЦИИ ТЕРИОФАУНЫ
Глава V. Анализ голоценовой териофауны и схема районирования .... 455
Гл а в а VI. Изменения экологических группировок и ареалов млекопитающих под влиянием деятельности человека в различных зонах........ 514
Заключение ...................................................... 580
Литература....................................................... 583
Зоологическая литература.................................... 583
Прочая литература .......................................... 612
Приложение 1. Динамика добычи и заготовок шкур пушных зверей на
Кавказском перешейке с 1925 по 1955 гг. (графики 1—23)...... 629
Приложение 2. Карты распространения зверей Кавказа (карты 1—97) 643
Указатель русских названий млекопитающих ........................ 694
Указатель латинских названий млекопитающих....................... 698
НИКОЛАЙ КУЗЬМИЧ ВЕРЕЩАГИН. МЛЕКОПИТАЮЩИЕ КАВКАЗА Утверждено к печати Зоологическим институтом Академии наук СССР
Редактор издательства Г. И. Козлова. Технический редактор А. В. Смирнова Корректоры Р. Г. Гершинская и Н. М. Медведева
Сдано в набор 29/V 1959 г. Подписано к печати 28/IX 1959 г. РИСО АН СССР № 1-118В. Формат бумаги 70 X lOSVie- Бум. л. 22. Печ. л. 44 = 60.28 усл. печ. л. + 3 внл. Уч.-изд. л. 59.84
3 вил. Изд. № 836. Тип. зак. № 197, М-45575. Тираж 1500.
Цена 44 р. 40 к.
Ленинградское отд. Издательства АН СССР. Ленинград, В-164, Менделеевская лин., д. 1
i-я тип. Издательства АН СССР. Ленинград, В-34, 9 линия, д. 12
I
ИСПРАВЛЕНИЯ И О П Е Ч А Т К И
Страница Строка Н опечатано Должно быть
20 26 сверху стратиграфических с тр а тит р а фи ч е с к и х
68 27 » развита развития
115 12 » по всем сборам обоих по всем сборам приведены
коллекций приведены
118 3 снизу масштаб а ми масштабами
127 Табл., боковик, Масаса р. Масаса sp.
4 сверху
154 24 снизу к роду Pity ту s к подроду Pitymys
155 6 сверху Capra primigenia Frass Capra priniigenia Fraas
155 19 » Р anther aleo Panthera leo
168 18 » Адзми Азии
180 6 » Mircrotus M icrotus
244 18 снизу протянижеи протяжении
265 8 сверху галеценовых голоценовых
337 21 » одинако однако
349 3 » «Зоогеографии» «Зоографии»
410 Подпись к рис. желтопятой желтонятной
2 снизу
420 Табл., боковик, в Пине в ПИНе
5 снизу
432 9 сверху однополости однопалости
437 6 » ландшафта и зон ландшафтных зон
442 3 снизу популляция популяция
452 17 сверху описанные случаи описанные примеры
553 26 снизу мумшулы мушмулы
559 16 сверху стен скал
559 17 » синатронпая синантропная
Н. К. Верещагин. Млекопитающие Кавказ.
. Необходима, шов).
человека дают IX горизонтов смены челове-й и районов, ие палеолити-иные геологи-, необходимо, ческому слою связи с неред-онты больше одок остатков штически тип ла и простей-азатели лишь тичным видам эннее приме-ращение с ко-прежних ис-сохранности я вполне воз-одразделений нь обеднения нерализацию, j свежем раз-наличию за-а для опреде-[ного периода ли использо-юводившиеся в Армении, ной Осетии, пл на карте
р, спиртовых очисленными ческом музее ии Академии ии Академии ии Академии гьской и Ба-гсло пунктов коллекцион-ости и аренда рственных жазье за 25-
ли уточнены
42
шуляцию лисиц
48
52“
5V
56’
46
идарту 40-х годов _ слегка темно-бурь1м •членений окрашены пей Азии эти лисицы рацию — темпоокра-
РОСТОВ
Терек
352
г. Чугуш
СУХУМИ
3704
19
3738
5х6 7 8^9
шобурым особям л®
4 5'6~7“~8 9 10
* С
Ъ Я о о
'Т Славянская
50 100 150 200 км
* СталинириА^
О 10* (
Тбилиси
76
РАСПРЕДЕЛЕНИЕ ТИПОВ ОКРАСКИ ЛИСИЦ ПО ЗАГОТОВКАМ 1945 Г
(в процентах) I
1 Гемно-рыжая
2 сыжая
з Светло-рыжая
4 Рыже-серая t
5 Серо-рыжая
6 Темно-серая с рыжеваты ремнем ? Светло-серая срыхеваты/ремнем
8 Грязно-серая
9 Светло-серо-рыжая 9 Заготовительные лишня
ьче предкавказски » кавказских исключ ая, на ощупь шелк -гзио-буровато-рьк*-ОД именем V-
(лее к югу, веР0Я™вд ЦИИ. Из высокогории п центрального Д карты aPea^°B pgcS пения типов <шр гь учтена возра^
хутор Веселый
КРАСНОДАР
МАТЕРИАЛЫ КРЯЖЕВАНИЯ ШКУР ЛИСИЦЫ на ростовской пушной БАЗЕ
заготовкам 1945-1946 гг.
исследованных шнур Пункты приемки пушнины
Сальск
Арзгир
12 3 4 5 6 7-8' л-^
з Новолабинская
Ланчхуты
Л’
БАТУМИ
О» 10
о.Чечень
Моздок
Буйнакск
7 8-9
г.Алагез # А оз. Севан
Ереван/ I
4'5 6^7 8 9J0 г.Арарат
Нахичивань
ВЫХОД КРЯЖЕЙ ПО ГЛУБИННЫМ ПУНКТАМ (в процентах)
1 Украинский
2 Донской
3 Кубанский
4 Северокавказский красный
5 Северокавказский нрасно--серый
6 Закавказский красный
7 Закавказский красно-серый
8 Закавказский серый
9 Караганка
10 Ереванский
11 Сиводушка и крестовка О Лисица без кряжа
Дербент 'О’
3 4 5 6,7-8 9 10
о.Артема
о
Г-лиг/ О.ЖиЛОЙ ЬАНУ
о-Булла о.Свиной
4 ЭвОЛЮЦии
СТЫНЬ
песчанок)
пустынь MpWnv [ТЛ лгттолг™,.-
востоку, на устье выходя к Каспию фешейке с преде. ™ — Хвалынском тми отложениями ?с чано-глинистую 1 песками, а югопослеледниковой -восточная треть шсгрессиях Кастой полупустыни бугристые пески лиманному побе-сли, перемежаю-ъгшепиях.
предста вителями составляют виды
Ца участка идет
убиквисты, зани-льпые кавказцы, кабан заселяют > сами существеп-
эщие (II группа) ушастым ежом, мячком, степной him, образующим j древних средне-кие (VII и IX новых регрессии палым сусликом, В историческую
о уцелела сайга-придают, I Ч* ше, чем в ИР®"
I, по-впдямо^’ ,з Средней Аз»"-ды VIII ГРУ"“‘Л’ 1И лишь Ш""аЛ
46
48
Восточноевропейский округ Преднавказский район С К Северокаспийский округ
Терсно- Нумский участок
СК
РОСТОВ
ПК
г. Чугуш
г. Эльбрус
КАВ
рцОН
Кр
г.Алагез *
оз. Севан
ЕРЕВАН
ПА
BE
Кума
тк
Рис. 195.
о.Чечень
МАХАЧКАЛА
о
БАТУМИ
BE
ПК
ГК ДГ ПА Ар НТ СВ Кр
ТК Терсно- Нумский участок КАВ Кавказский округ
Гирканский район
Дагестанский район Переднеазиатский округ Араксинский участок Нагорно-Талышсний участок Севанский район Нуринсний район
о.Артема
О
БАНУ оЖилой
• о.Булла
'О.Свиной
С«3
ЗООГЕОГРАФИЧЕСКОЕ районирование КАВКАЗСКОГО перешейка Mammalia
200 нм