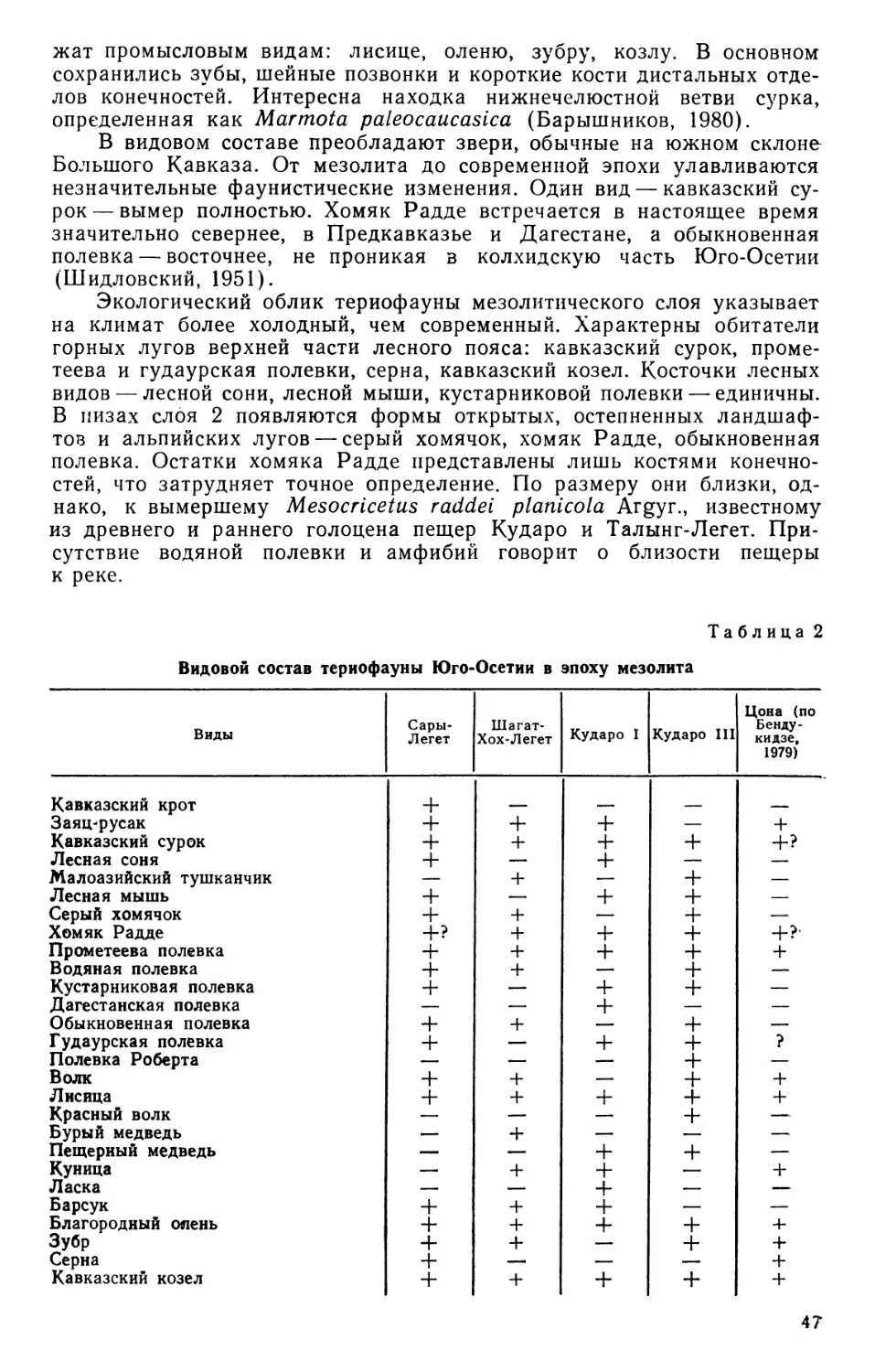
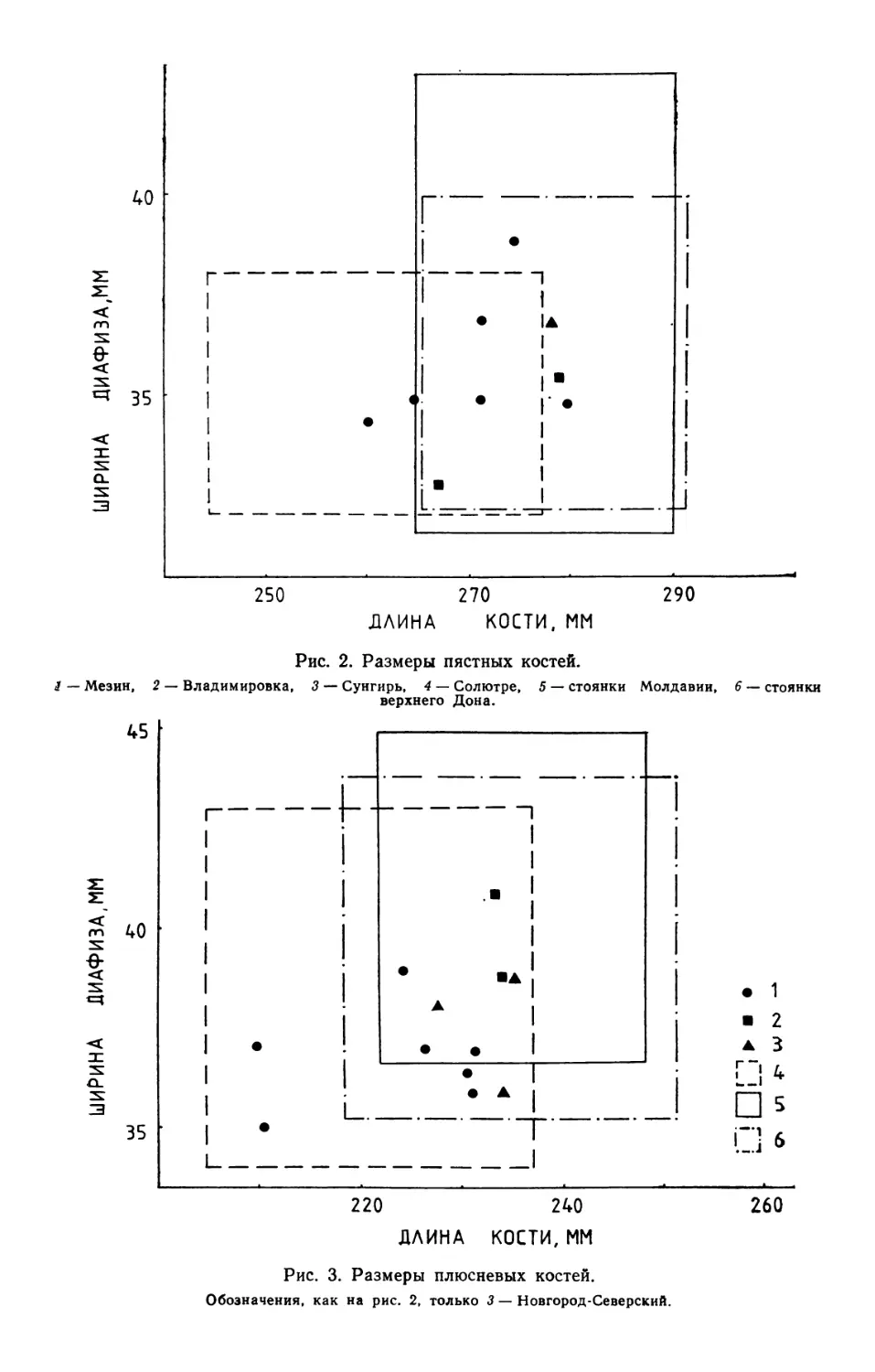
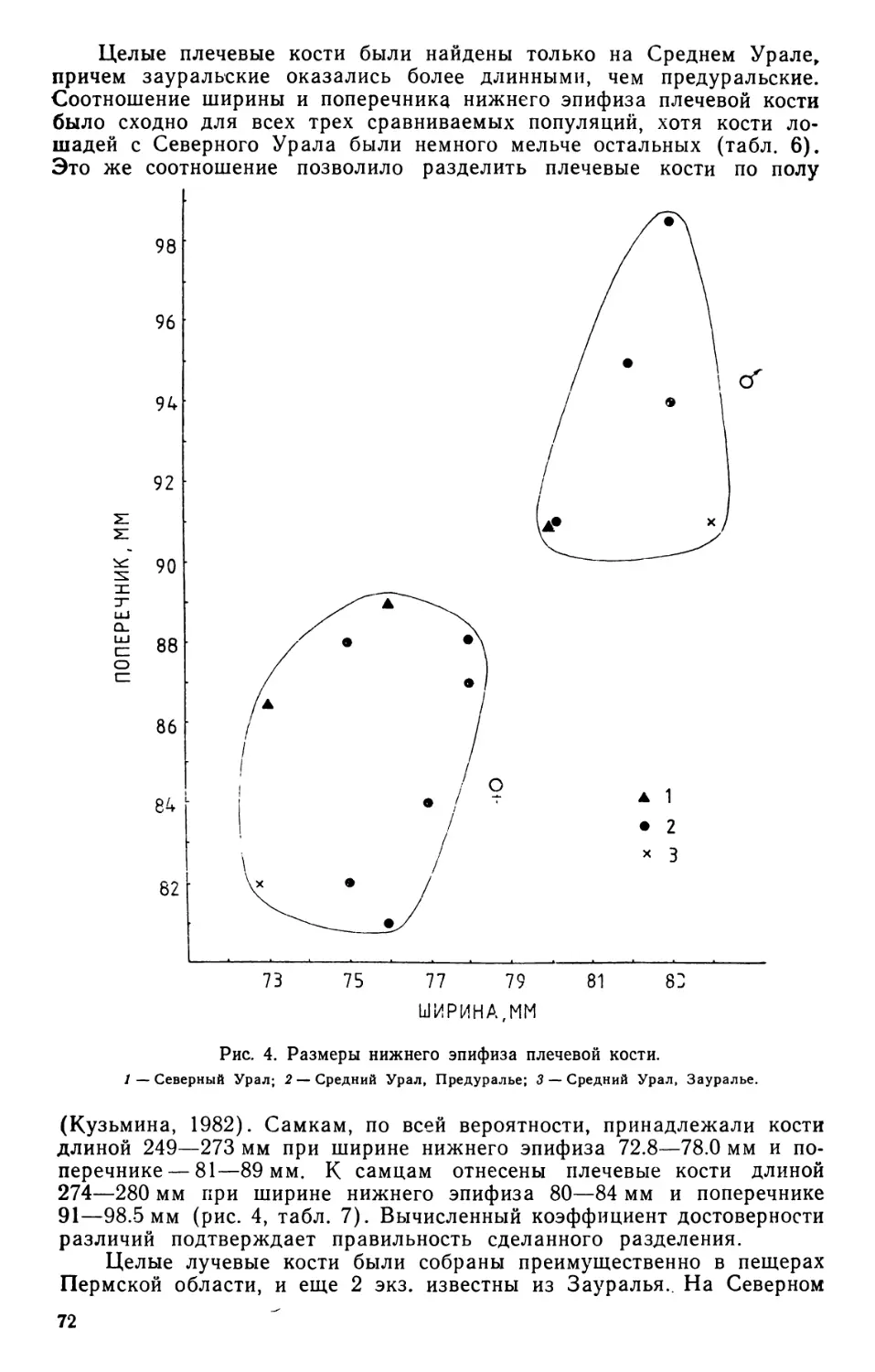
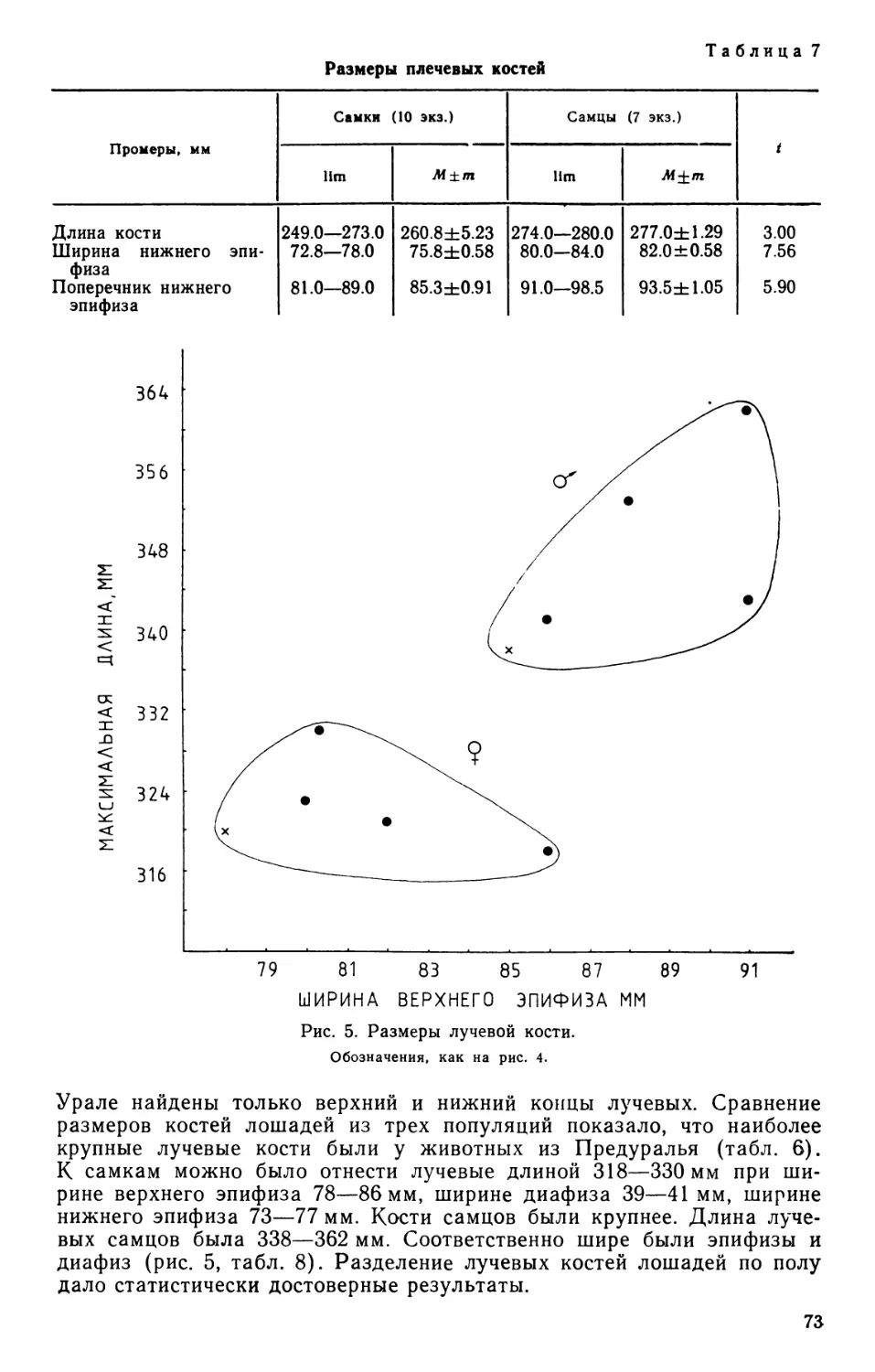
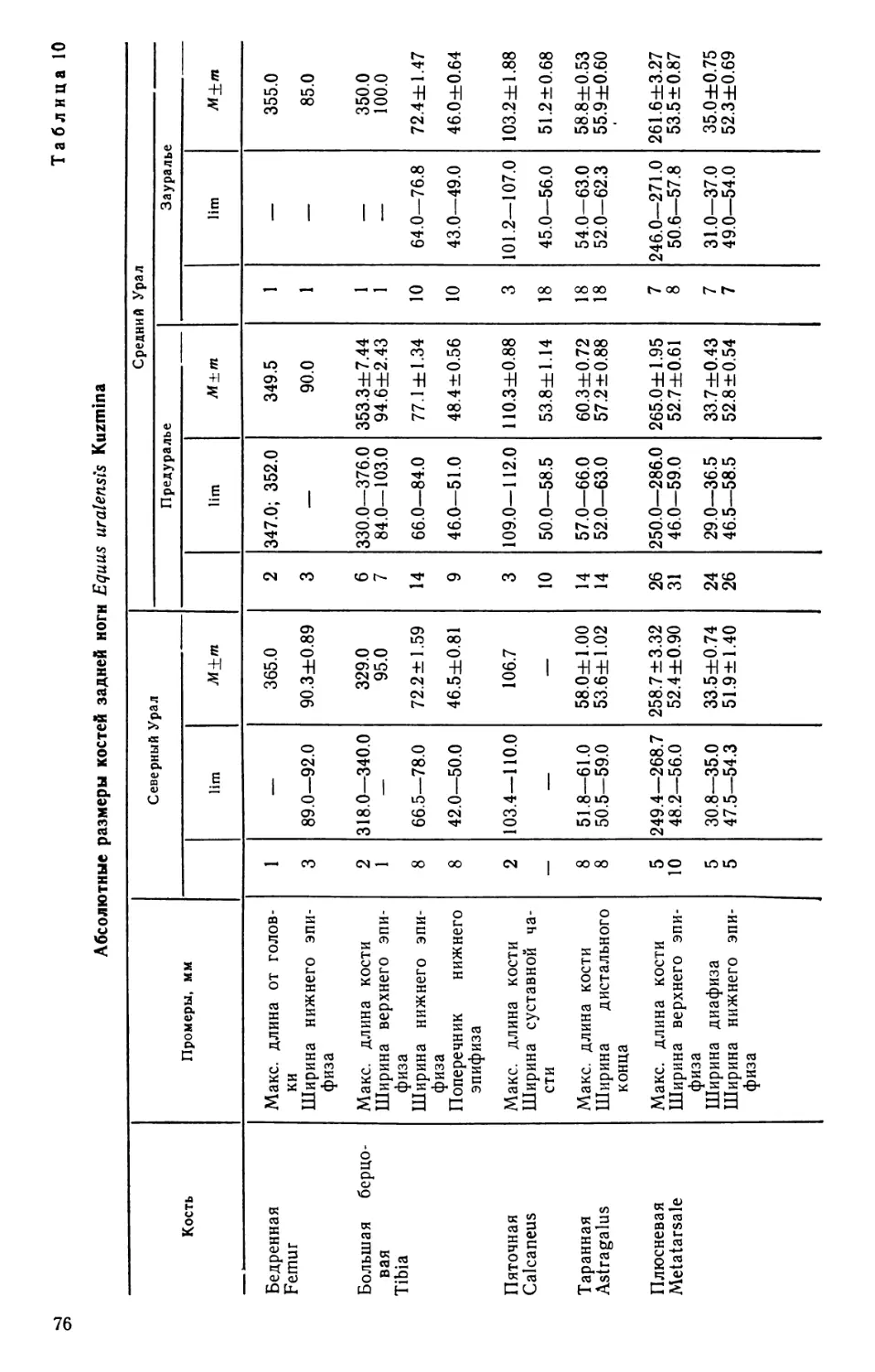
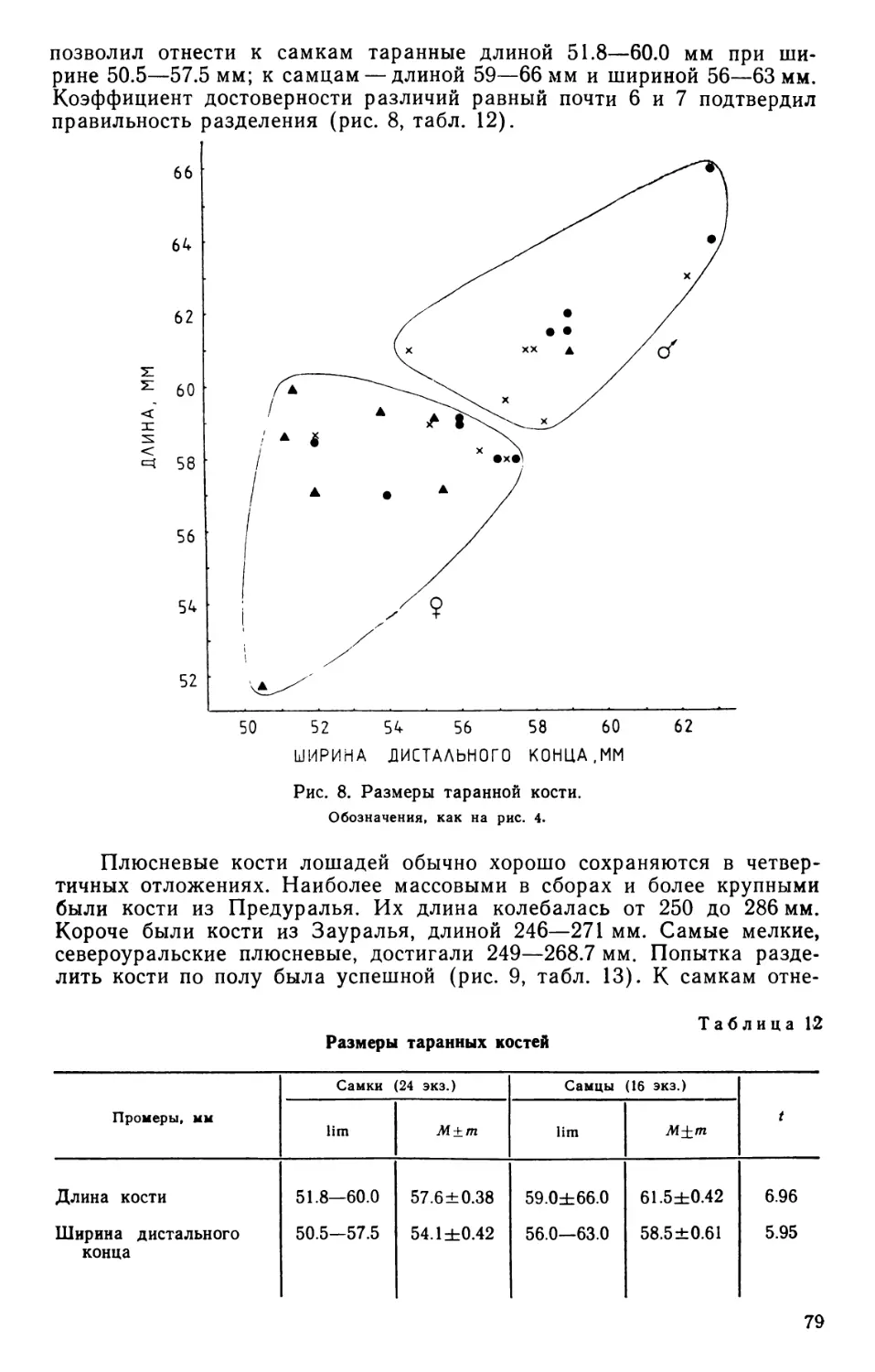
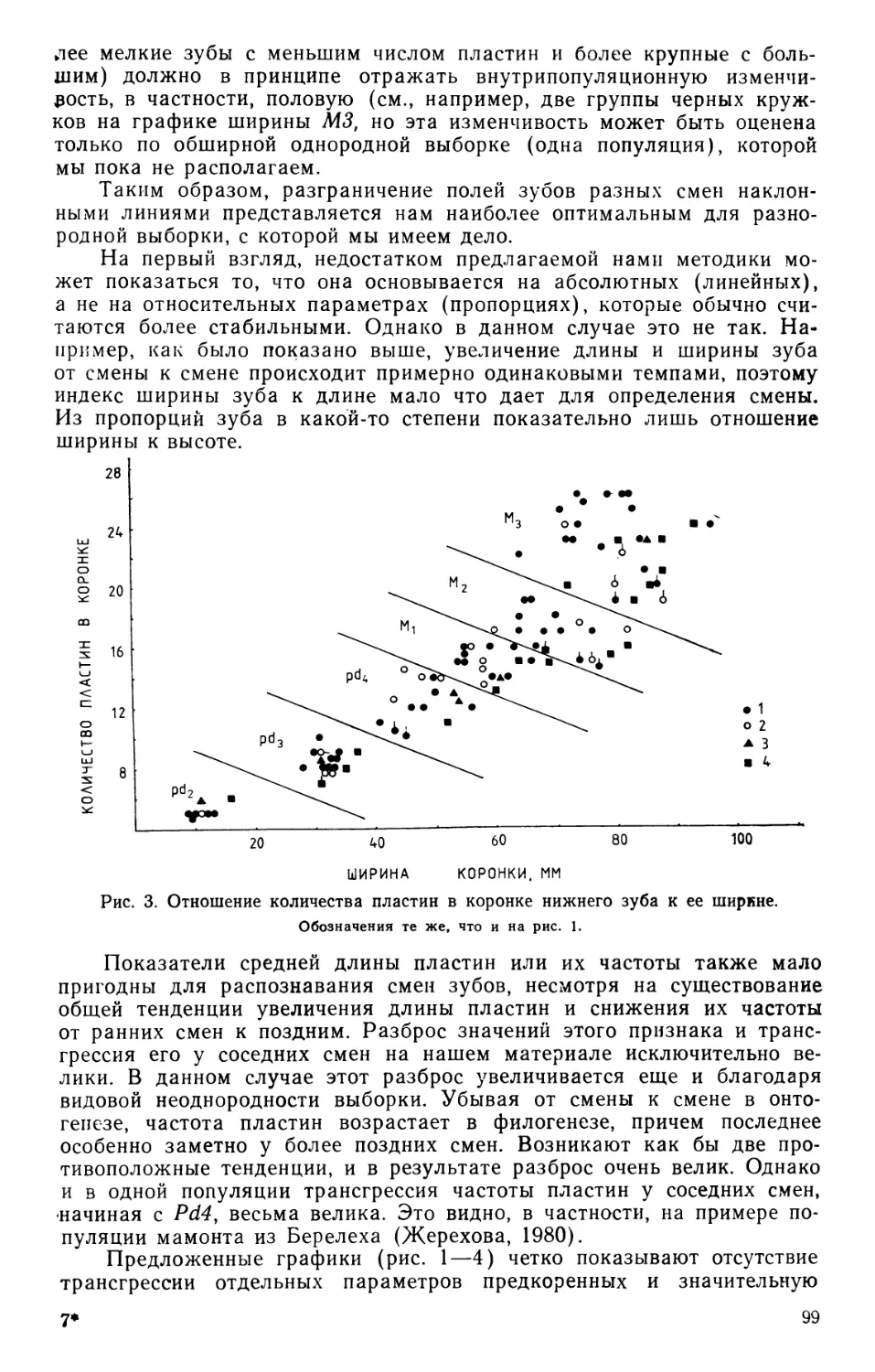
/
































































































































Author: Верещагин Н.К. Кузьмина И.Е.
Tags: палеонтология биология млекопитающие
ISBN: 0206-0477
Year: 1985




Text
ISSN 0206-0477
АКАДЕМИЯ НАУК СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
МЛЕКОПИТАЮЩИЕ
СЕВЕРНОЙ ЕВРАЗИИ
В ЧЕТВЕРТИЧНОМ ПЕРИОДЕ
АКАДЕМИЯ НАУК СССР
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА
Том 131
МЛЕКОПИТАЮЩИЕ
СЕВЕРНОЙ ЕВРАЗИИ
В ЧЕТВЕРТИЧНОМ ПЕРИОДЕ
Под редакцией
Н. К. Верещагина и И. Е. Кузьминой
ЛЕНИНГРАД
1985
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE
Vol. 131
MAMMALS OF NORTHERN EURASIA
DURING QUATERNARY
Главный редактор
директор Зоологического института АН СССР
О. А. Скарлато
Редакционная коллегия:
Я. И. Старобогатов (редактор серии), Ю. С. Балашов, Л. Я. Боркин,
И, С. Даревский, В. А. Заславский, И. М. Кержнер, В. А. Тряпицин,
И. М. Фокин, С. Я. Цалолихин, В. В, Хлебович
Рецензенты:
Р. С. Полякова, Л. И. Хозацкий
Сборник содержит статьи, в которых рассматриваются причины вымирания мле-
копитающих в четвертичном периоде, вопросы систематики и морфологии мамонтов
и некоторых видов мамонтовой фауны, новые находки мезолитических животных,
описания предметов изобразительного искусства, выполненные позднепалеолитическими
художниками. Книга представляет интерес для зоологов, палеозоологов, геологов,
палеогеографов и археологов.
2002000000—005
М----------------Без объявления © Зоологический институт АН СССР, 1985
055(02)3—85
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1985, ТОМ 131
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, 1985, VOL. 131
USSR ACADEMY OF SCIENCES
ВЫМИРАНИЕ МЛЕКОПИТАЮЩИХ
В ЧЕТВЕРТИЧНОМ ПЕРИОДЕ СЕВЕРНОЙ ЕВРАЗИИ
Н. К. Верещагин, Г. Ф. Барышников
Зоологический институт АН СССР, Ленинград
N К. Vereshchagin, G. F Baryshnikov. Extinction of mammals during Quaternary
in northern Eurasia
Предисловие
Угасание филогенетических ветвей и видов животных — процесс
сложный, зависящий от многих биологических и физических факторов.
В методическом плане палеонтолог, занимающийся проблемой выми-
рания, оперирует прежде всего голыми фактами присутствия или от-
сутствия ископаемых остатков видов на различных стратиграфических
уровнях. Далее он должен быть уверен в прямом генетическом родстве
систематических форм на воссоздаваемых филогенетических ветвях.
Кроме того, для написания реальной истории вымирания четвертичных
млекопитающих исследователе должен учитывать тафономические пред-
посылки и использовать факты смежных наук — геологии, палеогеогра-
фии, археологии. Необходимо помнить, что само обнаружение наиболее
поздних остатков исчезнувшего вида свидетельство не окончательного
вымирания, а еще относительно высокой численности его популяций
(Ефремов, 1950). Только правильная интерпретация морфологических,
палеозоогеографических и тафономических данных может помочь найти
объяснение фактов вымирания и понять их сущность. Для фантазий
и абстрактных рассуждений здесь не должно быть места.
В разработках объяснения причин и темпов вымирания четвертич-
ных млекопитающих в настоящее время существуют четыре основных
направления:
1. Исследование влияний смен природных (экологических) факто-
ров среды — климата, погодных условий, характера грунтов, естествен-
ных изменений ландшафтов и биотопов.
2. Исследование пластичности в морфологических структурах и фи-
зиологических адаптациях у вымерших и вымирающих видов, ее недо-
статочности при инадаптивной эволюции.
3. Исследование влияния хищников, эпизоотий и биоценотических
перестроек.
4. Изучение истребительной деятельности человека, действующей
непосредственно или через изменение среды обитания животных.
В теоретическом плане можно предполагать, что на ранних этапах
четвертичного периода преобладало влияние первых трех групп факто-
ров, а на поздних — четвертой. Очевидно также,, что наиболее интен-
сивное вымирание происходило в результате сложных комбинаций на-
званных причин. Мы уделили здесь внимание лишь первому и четвер-
тому направлениям.
Колоссальная широтная и долготная протяженность Северной
Евразии, разнообразие ее климатов и ландшафтов создавали возмож-
3
ность плейстоценовым млекопитающим, вымирая на одних участках,
сохраняться на других. Это усложняет общую картину их угасания, но
позволяет реалистично оценивать влияние природных факторов и дея-
тельности человека для судеб отдельных видов зверей и их комплексов.
Иными словами, привлечение фактов из области изменения ареалов
четвертичных млекопитающих полезно для обсуждения причин выми-
рания.
Типы развития ареалов, их динамичность разнообразны, но в основ-
ном сводятся к первоначальной экспансии — быстрому расселению вида
из очага формирования, последующей консервации ареала пли его уга-
сания, и к вторичным вспышкам распространения из реликтовых участ-
ков. Такие реликты, как правило, процветают, если в их судьбу не вме-
шивается деятельность человека. Вымирание бывает обычно связано
на первых этапах с быстрым или медленным сокращением ареалов
видов и численности их популяций. Сплошные ранее области обитания
могут дробиться при этом на обособленные участки. В ряде случаев
такие факты позволяют легче понять и объяснить причинность вымира-
ния, особенно если дело заключается в коренных или частичных пре-
образованиях сред обитания — ландшафтных зон и биотопов.
Последовательное сокращение ареала под влиянием природных
процессов часто совпадает с путями предыдущего расселения вида
с его первоначальной родины. Характерный пример такого частичного
вымирания на западных участках области своего распространения
дают центральноазиатские степные звери: желтая пеструшка, корсак,
кулан и сайга. Их ареалы стремительно сокращались с запада на во-
сток на протяжении последних веков нашей эры под влиянием мало-
изученных экологических факторов, но явно усиленных в XIX и XX ве-
ках деятельностью человека.
В данной работе мы ограничиваемся рассмотрением судеб терио-
фауны только на заключительных этапах четвертичного периода,
а именно, для позднего плейстоцена и голоцена. Факты и причины вы-
мирания зверей в раннем и среднем плейстоцене мало понятны и пока
трудно объяснимы.
Эволюция видового состава млекопитающих
Северной Евразии в четвертичном периоде
В настоящее время известно около 530 видов наземных млекопи-
тающих, населявших в четвертичное время (включая виллафранк)
Европу и азиатскую часть СССР. По отрядам они распределены сле-
дующим образом:
Насекомоядные Insectivora 35
Рукокрылые Chiroptera 30
Приматы Primates 8
Зайцеобразные Lagomorpha 30
Грызуны Rodentia 220
Хищные Carnivora 75
Хоботные Proboscidea 14
Непарнокопытные Perissodactyla 28
Парнокопытные Artiodactyla 100
Для эпохи позднего плейстоцена — вюрма (висконсина) —и голо
цена на территории СССР известны остатки до 184 видов наземны!
зверей и из них до 30 вымерших.
4
По отрядам они распределяются следующим образом:
Отряд Число известных форм Из них вымерших
Insectivora 6
Chiroptera 10 —
Primates 1 1
Lagomorpha 8 2
Rodentia <83 8
Carnivora 38 5
Proboscidea 1 1
Perissodactyla 6 4
Artiodactyia 31 9
Итого । 184 30
Следует оговориться, что отнесение некоторых видов в категорию
вымерших встречает известные трудности. Они обусловлены отсут-
ствием четких представлений о прямом генетическом родстве форм
в ряде филумов и наличием проблемы одомашнивания. Так, например,
в отряде хищных волжский волк {Canis volgensis М. PavL), наиболее
вероятный предок домашней собаки, передал часть генов собачьим по-
родам.
Недостаточно ясен вопрос с пещерной гиеной рода Crocuta и пе-
щерным львом рода Panthera, которые, возможно, являются лишь хо-
лодоустойчивыми северными подвидами ныне живущих африканоазиат-
ских тропических видов (об этом см. далее).
Некоторые плейстоценовые зайцеобразные и грызуны, описанные
как самостоятельные виды, могут быть на самом деле лишь ближай-
шими предками современных форм. В частности, это относится к дон-
скому зайцу {Lepus tanaiticus Gureev) — вероятному предку современ-
ного беляка (или его побочной ветви?), и к некоторым сусликам.
В отряде непарнопалых к вымершим формам отнесена плейстоценовая
лошадь (Equus ferus Bodd., s. lato), хотя один из ее подвидов (видов)
дал несомненно начало процветающим породам домашних лошадей.
Аналогично в отряде парнокопытных первобытного тура {Bos primi-
genius Boj.) мы считаем условно вымершим, поскольку его гены на-
верняка имеются в современном крупном рогатом скоте.
Учитывая обширность территории Советского Союза, видовой спи-
сок зверей, остатки которых известны в ископаемом состоянии, дается
по крупным регионам (табл. 1).
Далее сделана попытка оценки факторов вымирания для некото-
рых видов млекопитающих на основе их iеографического распростра-
нения, морфологических и экологических особенностей и датировок
местонахождений.
Обзор некоторых вымерших в плейстоцене млекопитающих
Мамонт — Mammuthus pritnigenius Blumenbach, 1799
История мамонтов в Северной Евразии известна теперь с ниж-
него плейстоцена, в слоях которого впервые обнаруживаются тонко-
эмалевые зубы крупных слонов. Возможно, впрочем, что они принад-
лежат архидискодонам (род Archidiskodon), конвергентно прпобревшим
«мамонтовые» признаки в суровых условиях ранних оледенений северо-
восточной Сибири.
5
Таблица 1
Видовой состав зверей четвертичного периода, известных по
ископаемым остаткам в СССР *
Отряды и виды Русская равнина и Крым Кавказ Средняя Азия Сибирь Приморье
Inseclivora
Erinaceus europaeus L. Г пг — г —
Erinaceus amurensis Schrenk — — — — пг
Desmana rnoschata L. пг — — — —
Talpa caucasica Satunin — пг — — —
Sorex sibirica Dukelsky — — — пг —
Crocidura russula Giildenstaedt — пг — — —
Chiroptera
Rhinolophus mehelyi Matschic — пг — — —
Rhinolophus ferrumequinum Schreber г пг г — —
Myotis blythi Tomes г пг г г —
Myotis natter er i Kuhl г пг г г г
Nyctalus noctula Schreber пг г г г —
Nyctalus lasiopterus Schreber пг г г — —
Eptesicus serotinus Schreber пг г г — —
Eptesicus nilssoni Keyserling et Blasius пг г г пг —
Vespertilio murinus L. пг пг г г г
Minioplerus schreibersi Kuhl г пг г — г
Primates
+Macaca sp. — ?п — — —
Lagomorpha
-{-Lepus tanaiticus Gureev п — п —
Lepus timidus L. ?г — — ?г —
Lepus tolai Pallas — — пг ?г —
Lepus europaeus Pallas пг пг г г —
Ochotona daurica Pallas — — — пг —
Ochotona alpina Pallas — — — пг —
-{-Ochotona azerica Gadziev et Aliev — п — — —
Ochotona pusilia Pallas пг ?п г — —
Rodentia
Pteroniys volans L. г — — ?г —
Sciurus vulgaris L. пг — — г г
Sciurus anomalus Gmeiin — пг — — —
Tamias sibiricus Laxmann г — — г г
Spermophilus undulatus Pallas — — г пг —
+Spermophilus glactalis Vinogradov — — — п —
Spermophilus relictus Kashkarov — — пг — —
-{-Spermophilus severskensis I. Gromov п — — — —
Spermophilus suslicus Gdldenstaedt пг — — — —
+ Spermophilus musicoides I. Gromov п — — — —
Spermophilus musicus Menetrie — пг — — —
Spermophilus pygmaeus Pallas пг г г — —
+ Spermophilus superciliosus Каир п — ? ? —
Spermophilus fulvus Lichtenstein пг — г — —
Spermophilus erythrogenys Brandt — — г пг —
Marmota bobac MOller пг —. г г —
+ Marmota paleocaucasica Baryshnikov —, пг — — —
Marmota marmota L. п — — — —
* П — поздний плейстоцен, Г — голоцен.
6
п зодолжен и е табл. I
к 2 s к 4)
Я к - к
Отряды и виды Sga х CQ Я X X < S СП X S о S X
>» CQ cu ax X О С
Marmota baibacina Kastschenko Г ПГ
Marmota sibirica Radde — ПГ
Marmota camlschatica Pallas ПГ
Marmota caudata Geoffroy — — ПГ
Castor fiber L. ПГ ПГ ПГ г
Castor canadensis Kuhe — ?
-\-Hystrix vinogradovi Argyropulo П П? __
Hystrix leucura Sykes — ПГ ПГ
Dryomys nitedula Pallas г ПГ Г
Glis glis L. г ПГ
Sicista subtilis Pallas ПГ г г
Sicista caucasica Vinogradov — ?Г —
Allactaga jaculus Pallas ПГ — г г
Allactaga elater Lichtenstein ПГ Г г
Allactaga williamsi Thomas .— ПГ
Pygerethmus platyurus Lichtenstein п г
Alactcigulus pygmaeus Pallas ПГ г г
Dipus sagitta Pallas ПГ Г г г
SciriopodaJelum Lichtenstein ПГ Г г г
Paradipus ctenodactylus Vinogradov — ПГ
N annospalax leucodon Nordmann ?г —
Spalax microphtalmus Giildenstaedt ?г ПГ
Apodemus sylvaticus L. ПГ ПГ г г
Apodemus flavicollis Melchior ?г Г
Mas musculus L. ПГ Г г г г
Rattus rattoides Hodgson — ПГ
Nesokia indie a Gray ПГ
Ellobius lutescens Thomas ПГ
Ellobius talpinus Pallas ПГ ?ПГ г
Ellobius tancrei Blasius ПГ
Allocricetulus eversmanni Brandt ПГ г г
Tschcrskia albipes Ognev ПГ
+ Cricetulus argyropuloi I. Gromov — п
Cricetulus tnigralorius Pallas ПГ ПГ г г
Mesocricetus raddei Nehring ПГ
Cricetus cricetus L. ПГ ПГ г ПГ
Merionus erythrourus Gray Г ПГ
Merionus meridianus Pailas ПГ Г г
Rombomys opitnus Lichtenstein Г ПГ
Myospalax myospalax Laxmann — — ПГ
Myospalax aspalax Pallas ПГ
Prometheomys scliaposchnikovi Satunin ПГ
Clethriononiys rufocanus Sunderval г ПГ г
Clethrionornys glareolus Schreber ПГ Г ПГ
Clethriononiys rutilus Pallas г ПГ г
Lagurus lagurus Pallas ПГ г ПГ
Eolagurus luteus Eversmann п г ПГ
+ Dicrostonyx quilielmi Sanford п п
Dicrostonyx torquatus Pallas ?г ?г
Lemmus sibiricus Kerr ПГ ПГ
Myopus schisticolor Lilljeborg г г
Arvicola terrestris L. ПГ ПГ г
Pitymys subterraneus Selys-Longchamps ПГ
Pitymys majori Thomas ПГ
Pitymys daghestanicus Schidlovskii ПГ
Microtus gregalis Pallas ПГ г ПГ
Microtus socialis Pallas ?г ПГ г
Microtus fortis Biichner ПГ г
Microtus maximoviezii Schrenk г П
Microtus oeconomus Pallas ПГ г ПГ
Microtus agrestis L. ПГ — ПГ —
7
Продолжение табл. 1
Отряды и виды Русская равнина и Крым Кавказ Средняя Азия Сибирь Приморье
Microius arvalis Pallas s. 1. ПГ ПГ — ПГ
Microtus transcaspicus Satunin — — ПГ — —
Lasiopodomys brandti Radde — — — ПГ —
Chionomys gud Satunin — ПГ — — —
Carnivora
Nyctereutes procyonoides Gray — — — — ПГ
Canis lupus L. ПГ ПГ ПГ ПГ ПГ
Canis aureus L. г Г ?г — —
+Canis volgensis M. Pavlova ПГ — — — —
Alopex lagopus L. ПГ — — ПГ —
Vulpes vulpes L. ПГ ПГ ПГ ПГ ПГ
Vulpes corsac L. ПГ Г г г —
Cuon alpinus Pallas г ПГ г ПГ ПГ
Selenarctos thibetanus G. Cuvier — 7 — — ?г
Ursus arctos L. ПГ ПГ г ПГ ПГ
+ Ursus spelaeus Rosenmiiller et Heinroth п ПГ ? — —
+ Ursus rossicus Borissiak п п — п ——
Thalarctos maritimus Phipps — — —. ПГ —
Martes zibellina L. ?г — — ПГ ПГ
Martes martes L. ПГ ПГ — — —
Martes foina Erxleben г ПГ г — —
Martes flavigula Boddaert — — — ПГ
Gulo gulo L. ПГ п — ПГ ПГ
Mustela erminea L. г г г ПГ ?г
Mustela nivalis L. ПГ ПГ г ПГ г
Mustela sibirica Pallas г — — ПГ ПГ
Mustela altaica Pallas — — г ?г г
Putorius eversmanni Lesson ПГ г г ПГ —
Vormela peregusna Guldenstaedt г ПГ г — —
Meles meles L. ПГ ПГ г ПГ ПГ
Lutra lutra L. ПГ Г г ПГ ПГ
+ Crocuta spelaea Goldfuss п п п п
Hyaena hyaena L. — г ПГ — —
Panthera leo L. г г — — —
4-Panthera spelaea Goldfuss п п п п п
Panthera tigris L. — г г — ПГ
Panthera pardus L. ПГ ПГ — ПГ
Uncia uncia Schreber — — ПГ г —
Felis silvestris Schreber ПГ ПГ — — —
Felis libyca Forster г г ПГ — —
Fellis euptilura Elliot — — — — ?г
Felis chaus Guldenstaedt ПГ ПГ г — —
Lynx lynx L. ПГ ПГ г г ПГ
Proboscidea
-^Mainmuthus primigenius Blumenbach п п ? п п
Perissodactyla
+ Equus ferus Boddaert s. lato * ПГ ПГ ПГ ПГ ПГ
Equus przewalskii Poliakov — — ?г —
+Equus hydruntinus Regalia ПГ п 7 — —
Equus hemionus Pallas г г ПГ ПГ —
+ Dicerorhinus kirchbergensis Jaeger — п — — —
-FCoelodonta antiquitaiis Blumenbach ПГ ? п п п
* Включая Е. latipes V. Gromova, Е. uralensis Kuzmi Е. lenensis Russanov, E. gmeli
Antonins.
8
П родолжениетабл. 1
55? к
к сх
a я £ я к S о
Отряды и виды Q H-. CX О S* о СО к ®' ХО я 5 5
>» co CX cxs X О.С о О О, С
Artiodactyla Sus scrofa L. ПГ пг пг Г ПГ
+ Camelus knoblochi Nehring ПГ ? пг — —
Moschus moschiferus L. — — — Г пг
Dama mesopotamica Brooke — п? — — —
Cervus nippon Temminck — — —. — пг
Cervus elaphus L. ПГ пг пг пг пг
Capreolus capreolus L. ПГ пг — — —
Capreolus pygargus Pallas П г пг пг пг
+ Megaloceros giganteus Blumenbach П п п п —
Alces alces L. ПГ пг — пг пг
Rangifer tarandus L. ПГ — — пг ?
4- Bos primigenius Bojanus ПГ п пг п ?
+ Poephagus baikalensis N. Verestchagin — — — II ?
+ Bison priscus Bojanus П п п пг п
Bison bonasus L. г ?г — — —
+Spirocerus kiakhtensis M. Pavlova — — — п —
Gazella subgutturosa Giildenstaedt г г пг — —
Procapra gutturosa Pallas — — — пг —
+ Parabubalis capricornis V. Gromova — — — п —
Saiga tatarica L. пг пг пг II —
Ovibos moschatus Zimmermann п — — пг —
Naemorhedus caudatus Miln-Edwards — — — — пг
Rupicapra rupicapra L. — пг — — —
Capra aegagrus Erxleben — пг г — —
Capra sibirica Pallas — — пг пг —
Capra caucasica Giildenstaedt et Pallas — пг — — —
+ Capra priska Woldrich — п — — —
Ovis orientalis Gmelin — пг — — —
Ovis vignei Bl у th — — пг — —
Ovis ammon L. ? п г пг —
Ovis nivicola Eschscholz — — — пг —
В СССР рядом ведущих палеонтологов и геологов принята обосно-
ванная и удобная для стратиграфических целей гипотеза происхожде-
ния мамонтов (род Mammuthus) от последовательного ряда форм архи-
дискодонтных слонов: Громова (Archidiskodon gromovi Garutt et Ale-
xeeva), южного (A. meridionalis Nesti) и таманского (A. tamanensis
Dubrovo) из верхнего виллафранка (хапровский и таманский фаунисти-
ческие комплексы) с промежуточной формой трогонтериевого слона
(A. trogontherii Pohlig), характерного для раннего плейстоцена (тирас-
польского комплекса) (Громов, 1948; Дуброво, 1964; Громова, 1965;
Алексеева, 1977; Гарутт, 1981).
По мнению Э. Тениуса (Thenius, 1980) и Н. К. Верещагина пра-
вильнее, однако, считать, что разделение архидискодонов и мамонтов
произошло на границе позднего плиоцена, а мамонтов и близкого
к ним азиатского слона (Elephas maximus L.) — в нижнем плейсто-
цене.
Формирование холодовыносливого вида мамонта произошло, оче-
видно, на территории северо-восточной Азии. В среднем плейстоцене
он был уже широко распространен по всей перигляциальной области
Евразии. Впрочем, крупных мамонтов из хазарской фауны Восточной
Европы иногда рассматривают как особый вид Mammuihus chosaricus
Dubrovo.
9
Наибольшего развития ареал Л1. primigenius достиг в рисс-вюрме
и особенно в вюрме (валдае), когда мамонты были распространены
от берегов Атлантического до Тихого океанов и далее через Аляску
в Северной Америке. На севере они жили за Полярным кругом у гра-
ниц Арктического бассейна, а на юге — до пустынь Средней Азии и
Монголии. В Европе проникали до Испании, южной оконечности Ита-
лии и Закавказья.
Остатки мамонтов обычны в слоях мустьерских стоянок Крыма
(Чокурча) и Северного Кавказа (Ильская, Даховская), но на стоян-
ках позднего палеолита они здесь уже не встречаются. Южная гра-
ница ареала вида, видимо, отступила к северу.
Исследования советских биологов, палеогеографов и геологов
внесли теперь отчетливые представления в биологию и экологию ма-
монтов. Эти слоны были хорошо приспособлены на севере к сухому,
холодному и резко континентальному климату. Об этом свидетель-
ствует мощный меховой покров, обросший шерстью хобот, уши в 5—6
раз меньшие по площади, чем у азиатского, и в 10—12 раз меньше,
чем у африканского слона, короткий волосатый хвост, обильные жиро-
вые отложения под кожей.
Материалы по питанию основаны на конкретных исследованиях
содержимого желудков. Мамонты летом питались преимущественно
травой (луговыми злаками, осоками, пушицей), концевыми побегами
кустарников (пв, берез, ольхи). Они обдирали бивнями кору деревьев —
ивы, чозении, лиственницы. Зимой, при замерзании водопоев и отсут-
ствии снега, мамонты могли скоблить бивнями грунтовый лед на вер-
тикальных стенках обрывов или выламывать его из морозобойных
трещин. Это, как кажется, подтверждается характерной боковой стер-
тостью концов бивней с молодого возраста и случаями поломок бивней
при таких усилиях. Фактических данных о зимнем питании зверей пока
нет, но, очевидно, его основу составляли также подсохшая трава и
побеги лиственных кустарников и хвойных: лиственницы, сосны, пихты.
Мамонты совершали, вероятно, длительные кочевки по долинам
рек меридианного направления, особенно в случае сильных снегопадов
и засух. В долинах и поймах рек они чаще всего гибли при внезапных
наводнениях, разливах и при переходах по непрочному, припорошенному
снегом льду озер и рек.
Туши мамонтов захоронялись под оползнями береговых склонов,
в растопленных солнцем потоках мерзлых суглинков, в толщах песка
и ила речных потоков и старых русел, а также в дельтовых участках и,
замерзая в осадках, сохранялись тысячелетиями. Редкие, но идеаль-
ные случаи захоронения замороженных трупов бывали при попадании
животных в коварные промоины в грунтовом льде, промытые в ледо-
вой толще небольшими ручейками. Сохранение мерзлых туш и вообще
мягких тканей мамонтов до наших дней было возможным лишь при
наличии в эпоху их жизни многолетней мерзлоты, при зимних темпе-
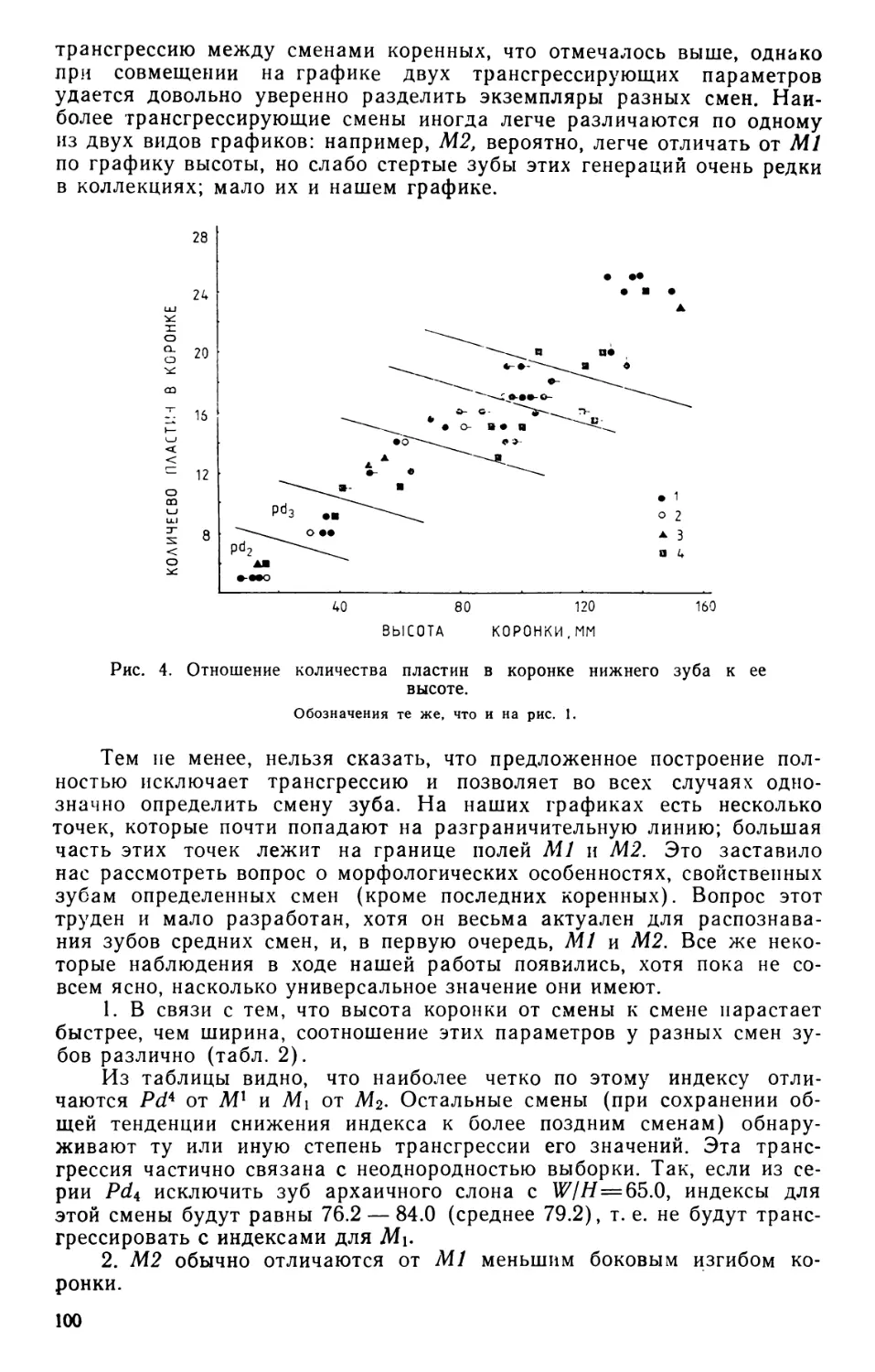
ратурах 60—80°С ниже нуля (рис. 1—3, вклейка).
На северо-востоке Сибири, в Якутии мерзлые туши мамонтов захо-
ронялись также в надувах лессовой пыли, формировавшей покровную
сартанскую толщу суглинков. Это доказывается наличием мощных ле-
довых жил, уходящих на глубину в 30—40 м и обжимающих столбы
пылеватого грунта с костями или частями разорванных тел мамонтов.
Такие факты, подтверждающие холодовыносливость мамонтов, имеют
принципиальное значение для правильного объяснения условий их
жизни, гибели и вымирания.
Наиболее загадочно в истории мамонтов — малое количество их
10
остатков древнее 50—60 тыс. лет, т. е. за пределами последней ледни-
ковой эпохи (вюрма, висконсина), а также и за возможностями радио-
углеродного датирования. Между тем, по тафономической теории усло-
вия для захоронения их костяков должны были быть обильны именно
в рисс-вюрмское (днепровско-валдайское) межледниковье, когда оттаи-
вание грунтов, эрозия и перестройка озерно-речной сети были наи-
большими.
В проблеме плейстоценовых вымираний наибольшее внимание
всегда уделялось мамонтам. При отсутствии реальных фактов, экологи-
ческих и тафономических наблюдений логические выводы подменялись
схоластическими дискуссиями или фантастическими домыслами на про-
тяжении почти двух столетий.
С первых печатных статей о мамонте статского советника и уче-
ного В. Н. Татищева (Татищев, Гмелин, 1780) в ходу было представ-
ление о том, что мамонты жили в более теплом климате, подобно аф-
риканским и азиатским слонам, а погибали от охлаждения его. Это
представление устойчиво бытует и до сих пор.
Для объяснения массовой гибели и консервации скелетов и мерз-
лых туш мамонтов, как правило, прибегали и прибегают к гипотезе
потопов в различных ее вариациях. Обычно речь идет о переносе туш
мамонтов разлившимися сибирскими реками с юге на север (Мидден-
дорф, 1860). Реже привлекается для этого гигантская волна, якобы
прокатившаяся через Гималаи в результате вулканических взрывов
(Pallas, 1773) или от удара метеорита в области Тихого океана. Ги-
потеза гибели мамонтов от гигантских наводнений была подробно из-
ложена в обширном обзоре природы захоронений животных Г Говор-
том (Howorth, 1887). По обилию примеров различных типов захоро-
нений в четвертичных осадках это сочинение столетней давности не
имело равных.
Исследователи, не знакомые с природой захоронений трупных и
костных остатков, считают также удовлетворительным объяснением
гибели зверей и сохранности их мерзлых туш быстрое охлаждение
климата Сибири в результате захвата части атмосферы ворвавшимся
в нее астероидом.
Первые научные представления о жизни мамонтов и отношении их
мерзлых трупов к грунтовым льдам Арктики были заложены работами
Русской полярной экспедиции под начальством Э. Толля. Они создали
основу для заключений о причинах вымирания наших северных сло-
нов. Сам Э. Толль (1897) связывал гибель этих гигантов с раздробле-
нием существовавшего в прошлом между Азией и Америкой обшир-
ного Арктического материка, в результате чего климат стал менее кон-
тинентальным, но охладился, при этом исчезли богатые пастбища. По-
холоданием и развитием зоны тайги объяснял гибель мамонтовой
фауны северо-востока Сибири палеонтолог и геолог И. Д. Черский
(1891).
Позднее обзоры причин вымирания мамонтов публиковали в на-
чале XX века палеонтолог М. В. Павлова (1924), журналист Б. Дигби
(Digby, 1926), побывавший в начале века в Якутии; препаратор
Е. Пфиценмайер (Pfizenmayer, 1926), проводивший раскопки Березов-
ского мамонта; географ И. Толмачев (Tolmachoff, 1929) и другие.
В основном дело сводилось к соображениям о роли истребления ма-
монтов человеком, о значении физических смен среды обитания или
чрезмерного развития бивней.
Туманное и путаное объяснение гибели мамонтов от холода дал
на основе своей ледниковой теории капитан Е. С. Гернет (1930: 2-е
11
изд., 1981). Геолог В. И. Громов (1948) объяснял гибель мамонтов
похолоданием климата в вюрме. Геофизик М. И. Будыко (1967) ука-
зал на эту же причину, добавив математический расчет такой гибели
в результате низкого уровня размножения северных слонов и истреби-
тельного влияния первобытных охотников.
Для территории Украины и юга Русской равнины палеонтолог
И. Г. Пидопличко (1969) считал несомненным истребление обитавших
там мамонтов палеолитическими охотниками. Проводя параллели с охо-
тами африканских негров, он указывал, что при поголовье мамонтов
в Восточной Европе в полумиллиона голов, их могли истребить в па-
леолите в течение одного тысячелетия. Так же считала Н. Л. Корниец
(1962).
Ихтиолог и палеогеограф Г У. Линдберг (1972) пытался объяс-
нить гибель мамонтов подъемом уровня Мирового океана, в результате
чего животные якобы гибли на Русской равнине от перелива вод
Балтики в Черное море, а на Новосибирских островах от голода, отре-
занные морем от материка.
Палеогеограф А. А. Величко (1973) подробно и убедительно на-
рисовал картину формирования гиперзоны продуктивной тундро-степи,
обитаемой мамонтовой фауной, и обосновал причину разрушения ма-
монтового комплекса и вымирания самих мамонтов под влиянием
ландшафтных смен — размораживания грунтов, заболачивания боль-
ших пространств, развития лесов на границе плейстоцена и голоцена.
Зоолог Н. К. Верещагин (1971, 1979) считает основной причиной
вымирания мамонтов кардинальную перестройку климатов и ландшаф-
тов Северной Евразии в конце последней ледниковой эпохи. Однако
в южных районах Восточной Европы и Сибири могло преобладать и
истребительное влияние первобытного человека.
По мнению палеогеографа и мерзлотоведа С. В. Томирдиаро (1977)
гибель мамонтов и мамонтовой фауны на северо-востоке Сибири была
обусловлена в конце вюрма (валдая) «экологической катастрофой»,
вызванной потеплением климата, повлекшим за собой разморажива-
ние Полярного бассейна и вытаивание грунтовых льдов. Кормная
тундростепь превращалась в заозеренную, болотистую, моховую тундру.
В современной арктической тундре Евразии с ее плотным, переду-
ваемым ветрами снегом мамонты не могли бы существовать.
Географ Д. Д. Квасов (1977) также считает одной из причин ги-
бели мамонтов и мамонтовой фауны послеледниковое потепление
климата и наступившее его увлажнение в связи с образованием об-
ширных водоемов — приледниковых озер.
Наконец, появилось и сочинение Г Краузе (Krause, 1977), в кото-
ром сравниваются некоторые морфологические черты мамонтов: шерст-
ный покров, строение ушей, жировые отложения и т. д., с такими же
чертами других северных животных. В результате с прямолинейной
логикой человека, не знакомого с палеогеографическими фактами, за-
является, что холодоустойчивость мамонтов только «научная фикция»
и что эти звери, жившие в тепле, погибли от стужи. Подобные рас-
суждения, однако, не новы и только подтверждают цепкость наивных
представлений XVIII—XIX века.
Антропогенным воздействиям на популяции мамонтов в Евразии
неоднократно уделялось внимание палеобиологами, особенно в связи
с исследованием палеолитических стоянок. В Восточной Европе и Си-
бири были обнаружены массовые «кладбища» костей мамонтов и зага-
дочные сооружения из их черепов и костей. Обычно эти сооружения
рассматриваются как развалины основания хижин (Пидопличко, 1969),
12
но некоторые из них могли иметь культовое назначение или использо-
ваться в качестве музыкальных установок (Бибиков, 1981). Для проб-
лемы вымирания мамонтов эти завалы костей интересны тем, что мо-
гут указывать на существование массовых активных охот. Однако
прямых фактов для этого пока не существует. На костях не обнару-
жены, например, следы ударов орудий нападения.
В то же время имеются многочисленные актуалистические наблю-
дения, как первичные «кладбища» животных образуются без участия
человека за счет концентрации трупов утонувших зверей в староречьях
и заводях (Верещагин, 1972). Полностью мацерированные кости ма-
монтов могли переноситься из таких обсохших природных скоплений
палеолитическими охотниками на высокие террасы для устройства со-
оружений различного назначения. Отнюдь не отрицая вероятность су-
ществования активных охот на мамонтов при помощи копий, отрав-
ленных наконечников дротиков и стрел, загона в естественные ловушки
и искусственные ямы, на непрочный лед и даже в вязкое болото, сле-
дует указать, что бесспорных материалов по этому поводу до сих пор нет.
Природные массовые захоронения мамонтовых костей — «мамонто-
вые кладбища» первого порядка (in situ) залегают в водных осадках
в долинах рек, в озерных котловинах. Вторичные захоронения, обра-
зованные с участием первобытных людей, залегают также в долинах
рек и по краям озерных котловин, но чаще всего в лёссах и лёссовид-
ных суглинках высоких террас, как в Восточной Европе, так и в Си-
бири (рис. 9, вклейка).
Учитывая грандиозность ареала мамонтов и разнообразие природ-
ных условий в его пределах, нельзя утверждать, что эти звери вымерли
от одной природной причины, скажем, от потепления и увлажнения
климата, вызвавшего экологическую катастрофу в виде замены ланд-
шафта холодной тундростепи заозеренной тундрой и тайгой. Стооон-
ники именно этого взгляда прибегают для пояснения исчезновения
мамонтов в зоне древних и современных степей южной Сибири к воз-
действию позднепалеолитического и более многочисленного здесь нео-
литического человека.
Такие же недостатки таятся и в пояснениях наличия «эволюцион-
ного тупика», ибо мы пока не знаем негативных черт в морфологиче-
ской и физиологической организации мамонтов по сравнению с живу-
щими слонами, которые могли бы дать объяснение факту их выми-
рания.
Пока для разумных объяснений остается только оперирование с по-
следними хронологическими датировками. Для Европы и Сибири они
таковы (Berglund а. oth., 1976; Орлова, 1979; I Вереща! ин, 1982):
Швеция Западная Европа Локарп 13 360±95 (Lu-796)
Франция 13 13 Ла Комбье 13 090±120 (Lu-796 2) 260±110 (Lu-865) 390±300 (Ly-433)
Швейцария Пра Роде 12 270±210 (Ly-877)
Европейская часть СССР Кунда, Эстонская ССР 9 780±260 (ТА-12)
Костенки 2, Воронеж- 11 ская обл. Тимоновка 1, Брянская 12 обл. Юдиново, Брянская обл. 13 13 Авдеево, Курская обл. 13 000±200 (ГИН-93) 200±300 (ИГАН-82) 650±200 (ЛУ-153) 830±850 (ЛУ-103) 900 ±200 (ИГАН-78)
13
Сибирь
Юрибей, Гыданский п-ов
р. Ниж. Таймыра,
п-ов Таймыр
оз. Таймыр
р. Мамонтовая, Таймыр
Северная Земля
Берелех, Якутская АССР
9 600+300
(ВСЕГИНГЕО)
9 730+100 (МГУ-763)
10 000+70 (ЛУ-1153)
9 670±60 (ГИН-1828)*
10 300+100
(ГИН-1828 т. к.)
Яр Березовский, р. Ир-
тыш
11 140+180
12 100 ± 80
12 780+80
11 450+250
11 500+60
10 370±90
12 000+130
12 850+110
13 700+80
12 860±90
(ГИН-3067)
(ГИН-1783)
(ГИН-2677)
(Т-297)
(ЛУ-610)
(СОАН-327)
(ЛУ-149)
(ЛУ-1055)
(МАГ-114)
(СО АН-1283)
К ним близки наиболее поздние сроки существования мамонтов
в Северной Америке (Kurten and Anderson, 1980) — 10370±160 (Lost
Chicken Cr.) для M. primigenius; 11 280±350 (Union Pacific), 11 260±
±360 (Lehner Ranch), 11 230±340 (Murray Springs), U160±500 (Do-
mebo) для M. jeffersonii Osborn. Наряду с другими сходными датами
угасания плейстоценовых видов в Северном полушарии, они могут слу-
жить основой для теоретического поиска причин вымирания нашего
волосатого слона.
С угасанием мамонтов вымер и их своеобразный желудочный овод
Cobboldia russanovi Grunin, 1973 — один из трех видов оводов, извест-
ных для хоботных.
Большой пещерный медведь —
Ursus spelaeus Rosenmuller et Heinroth, 1794
История пещерных медведей прослеживается с раннего плейсто-
цена, когда у европейского Ursus etruscus Cuvier начинают проявляться
спелеоидные черты, что привело затем, через промежуточную форму
U. deningeri Reich., к образованию в среднем плейстоцене — риссе
U. spelaeus (Erdbrink, 1953; Громова, 1965). Родиной большого пещер-
ного медведя следует считать карстовые области средней Европы, по-
бережье Средиземного моря. Ареал вида простирался на север до юж-
ной Англии, Бельгии, южной Польши и включал Урал, Крым и Кавказ.
Каждый карстовый район обладал своей популяцией пещерных мед-
ведей, которые, видимо, были мало склонны к миграциям. Изолиро-
ванная равнинными степями небольшая колония этих животных оби-
тала, например, на Жигулевской возвышенности (Самарская лука)
в Среднем Поволжье. Пещерные медведи Кавказа также были изоли-
рованы и эволюционировали низкими темпами, сохраняя до конца
плейстоцена архаичные признаки, свойственные U. deningeri (Барыш-
ников, Дедкова, 1978).
Морфологические черты U. spelaeus'. гигантский рост и вес (до
1000 кг), тупобугорчатые и уплощенные коренные, вздутый, выпуклый
лоб, мощный сагиттальный гребень, узкое отверстие хоан, укороченная
голень — свидетельства его малой подвижности и преимущественной
растениеядности, несомненно превышавшей таковую современного бу-
рого медведя (U. arctos L.). В экологическом смысле этот зверь был
тесно связан с пещерами для жилья и вывода молодых, что оказалось
* Данные Л. Д. Сулержицкого (Геологический институт АН СССР, Москва).
14
роковым для вида в конце вюрма. Основной тафономический тип ме-
стонахождения его остатков — пещерные отложения. Остатков медве-
дей из аллювия, лёсса и покровных суглинков практически не известно.
Палеолитические племена настойчиво охотились на пещерных мед-
ведей, и в Западной Европе жизнь человека в значительной степени
обязана этому зверю. В ашельских слоях стоянки Кударо 1 на Кавказе
найдены остатки не менее чем от 200 особей медведей.
Вымирание вида произошло, вероятно, вместе с мамонтом и носо-
рогом в позднем вюрме (мадлене), так как на стоянках мезолита Рус-
ской равнины и Урала его остатков уже не находят.
Предложено много гипотез для объяснения причин вымирания пе-
щерных медведей. Их обзор дан Б. Куртеном (Kurten, 1965, 1976) и
Л. К. Габунией (1969). Наиболее ходовые — дегенеративное вырожде-
ние как следствие инбридинга в изолированных популяциях, несовер-
шенство морфологических приспособлений к потреблению раститель-
ной! пищи вследствие инадаптивного характера эволюции и, наконец,
истребление у пещер палеолитическими охотниками. Л. К. Габуния, на-
ряду с Л. Ш. Давиташвили (1969), придает еще значение конкуренции
с бурыми медведями, якобы вытеснявшими пещерных, но это принятие
следствия за причину. Всеядный бурый зверь мог отчасти конкуриро-
вать с травоядным пещерным, но не был причиной его вымирания.
Кроме того, бурых медведей в плейстоцене было мало.
Нами подмечено роковое значение тяготения этих зверей к пеще-
рам. В конце ледниковой эпохи некоторые пещеры, регулярно заселяв-
шиеся медведями, оказывались коварными ловушками при быстрых
оттепелях и подъемах воды в речках и ручьях. Такой смертельной ло-
вушкой оказывались, например, на протяжении сотен лет подряд пе-
щеры Урала в верховьях Камы и Печоры (Верещагин, 1982).
Несколько дольше, чем в других районах, пещерные медведи про-
жили на Западном Кавказе и в верховьях притоков Риони (Кударо 2).
Из тамошних пещер известны кости пещерных медведей с хорошо со-
хранившимся коллагеном.
Основным фактором вымирания этого вида была все же смена
сухого и континентального климата на влажный, что резко ухудшило
микроклиматический режим их пещерных убежищ, сделало их сырыми,
некомфортными.
Другой близкий вид — малый пещерный медведь Ursus rossicus
Borissiak—морфологически еще более специализированный, обитал
в Восточной Европе и Западной Сибири и вне областей карста, т. е. был
менее связан с пещерами. Тем не менее, он вымер подобно своему ги-
гантскому собрату, что подтверждает несовершенство морфофизиологи-
ческой организации подрода Spelaearctos,
Пещерная гиена — Crocuta spelaea Goldfuss, 1823
Из плиоценовых и четвертичных отложений Старого Света описаны
остатки различных видов гйен двух основных филогенетических ветвей,
представленных в современной фауне родами Crocuta и Hyaena. В плей-
стоцене Западной Европы встречались представители обоих родов. По-
лосатая гиена (Hyaena hyaena L.) исчезла здесь лишь на протяжении
плейстоцена, а пещерная — в его конце (Thenius, 1980).
Черепные и скелетные признаки пещерной гиены очень сходны
с таковыми пятнистой гиены (Crocuta crocuta Erxl.) Африки. Поэтому
некоторые палеонтологи нередко считают ее северным, пещерным под-
15
Рис. 4. Нижняя челюсть пещерного медведя из Воронцовской пещеры на Кавказе,
голоценового типа сохранности. Краснодарский музей.
видом типичной африканской крокуты. Это утверждение не доказано,
спорно.
В Европе С. spelaea появилась в интергляциале кромер, а на тер-
ритории СССР — в составе тираспольской фауны. При этом ранние
формы (например, мелкая гиена из Зюссендорфа) по ряду признаков
стоят ближе к современной пятнистой, чем к пещерной (Soergel, 1936).
Пещерная гиена — высокоспециализированная форма с очень мас-
сивными костедробящими коренными зубами и слабыми клыками. Уро-
вень развития ее прогрессивных и регрессивных признаков выше, чем
у современной крокуты. Неуклюжее на вид сложение и приподнятость
передней части тела были характерны у этого зверя, как и у пещер-
ных медведей.
Ареал пещерной гиены занимал всю Европу, субтропические и
средние широты Северной Азии, в Сибири примерно до 56° с. ш. В арк-
тическую зону она, по-видимому, не проникала, не поднималась, оче-
видно, и высоко в горы, так как на высокогорных стоянках Кавказа
(Кударо, Цона) ее остатков не найдено.
Если концепция единого африкано-евразийского вида правдива, то
произошло не вымирание, а лишь катастрофическое сокращение север-
ной евразийской части ареала. Аналогичная ситуация, возможно, суще-
ствует с пещерным львом (об этом далее).
В слоях многих пещер Европы известны массовые скопления ко-
стей пещерной гиены, а также ее копролиты и кости жертв. Так, в пе-
щере Киркдаль в Англии найдены многочисленные остатки гиен всех
возрастов — от новорожденных до старых, а в пещере Торневтон соб-
рано более 20 тысяч зубов этого вида (Kurten, 1968). В СССР кости
гиен обычны в пещерах Алтая («Логово Гиены») и Приморья («Гео-
графического общества») (Оводов, 1980). В пещерах звери устраивали
свои логова, а иногда и гибли, проваливаясь в карстовые колодцы
(Агеева и др., 1978).
Подобно африканской пятнистой гиене, пещерная питалась преиму-
щественно трупами крупных копытных и завершала таким образом по-
следнюю стадию использования «вторичной биомассы» до уничтожения
ее личинками насекомых и трупоядными бактериями. Возможно, она
охотилась также на грызунов, детенышей копытных и мамонтов.
Сокращение численности и ареала пещерной гиены шло парал-
лельно исчезновению стад степных и лесостепных копытных при забо-
лачивании равнин и развитии таежной зоны на границе плейстоцена
и голоцена. Палеолитические охотники добывали гиен лишь попутно,
как и современные нимроды Африки, так как обломки костей, черепов,
зубов изредка попадаются в слоях стоянок, особенно датируемых солю-
трейской — мадленской культурой. Ни в одной из мезолитических и
неолитических стоянок в пределах СССР остатков этого вида не об-
наружено.
Причины вымирания пещерной гиены некоторые палеозоологи ви-
дят также в чрезмерной специализации и в антропическом факторе —
резком уменьшении числа диких копытных в связи с зарождением зем-
леделия и скотоводства (Пидопличко, 1951).
Пещерный лев — Panthera spelaea Goldfuss, 1810
История этой гигантской кошки спорна и загадочна. В Европе она
известна с кромерского межледниковья — фауна Мосбаха. Часть евро-
пейских палеонтологов (Рябинин, 1919; Hemmer, 1967; Kurten, 1968)
считала и считает пещерного льва лишь подвидом современного афри-
2 Млекопитающие -
17
канского Pantera leo L. Сходство скелетных признаков этих зверейг
столь далеко разобщенных географически и хронологически, действи-
тельно значительное, но далеко не абсолютное. Обнаружена большая
морфологическая специализация в кошачьем типе, чем у современного
льва (Верещагин, 1971).
Концепция единого вида подразумевает существование колоссаль-
ного ареала от мыса Доброй Надежды на юге Африки до Скандина-
вии, Таймыра, Берингии в Евразии, а в Северной Америке — до Кали-
форнии и Мексики. Аналогий такому ареалу нет среди видов наземной
мировой териофауны.
Другие ученые (Goldfuss, 1810; Leidy, 1853; Громова, 1932) счи-
тали, что пещерный лев был особым видом. Кроме холодовыносли-
вости и нередко крупных размеров, он обладал признаками черепа и
скелета, сочетающими черты льва и тигра Р. tigris L. Его ареал был
ограничен северной половиной Евразии, включая Европу с Британскими
островами, Крым, Кавказ, Сибирь, Приморье, Берингию, а далее
Аляску и всю южную половину Северной Америки, где обитал близ-
кий подвид — Р. spelaea atrox Leidy.
По образу жизни и морфологии пещерный лев был более сходен
со львом, чем с тигром. В горах он пользовался пещерами в качестве
убежищ, но на равнинах обходился и без них. Питание лошадьми, оле-
нями, бизонами, овцебыками, сайгаками, а в Америке — лошадьми, би-
зонами, верблюдами, гигантскими ленивцами обусловило его полную*
зависимость от состояния численности названных травоядных.
Вымирание пещерных львов последовало, очевидно, непосред-
ственно вслед за резким падением численности популяций плейстоце-
новых копытных в конце вюрма (валдая-висконсина). Интересно, что-
это произошло, по-видимому, синхронно в Евразии и Северной Аме-
рике около 10 тыс. лет от наших дней.
Естественно, что сторонники единого африкано-голарктического
вида льва должны признавать лишь вымирание североевразийской и
североамериканской плейстоценовых популяций этого замечательного
зверя. В Европе лев африканского типа дольше всего продержался
по палеонтологическим данным в северном Причерноморье (до III тыс.
до н.э.; Бибикова, 1973), а на Балканском полуострове даже до на-
чала нашей эры, если верить широко известному свидетельству Геро-
дота о нападении здесь львов на караван верблюдов Ксеркса.
Волосатый носорог — Coelodonta antiquitatis Blumenbach, 1799
Этот характерный «спутник» мамонта, по-видимому, сформиро-
вался к раннему плейстоцену в степях Монголии, Забайкалья и Север-
ного Китая, откуда происходит предковая раннечетвертичная форма
С. tologoijensis Beljaeva (Тологой на р. Селенга), а также наиболь-
шее количество его ископаемых остатков, датируемых преимущественно
средним и верхним плейстоценом.
В Западной Европе волосатый носорог известен с мпнделя (Фран-
кенхаузен, Борнхаузен), причем коренные зубы его менее специализи-
рованы, чем у более поздних форм (Sickenberg, 1962). Многочислен-
ным волосатый носорог становится в риссе. На Русской равнине его
черепа встречаются наряду с более редкими черепами носорога Мерка
(Dicerorhinus kirchbergensis Jaeger) в аллювии Допа, Волги, датируе-
мом второй половиной среднего плейстоцена.
Наибольшее распространение волосатого носорога приходится на
вюрм (валдай), когда он широко расселился по всей Северной Евра-
18
зии, проник до побережья Северного моря на западе, до Италии, Се-
верного Кавказа, Киргизии и Таджикистана на юге, а на северо-во-
стоке Сибири до бассейнов Колымы и Анадыря и побережья Охотского
моря. Что послужило преградой его расселения в Новый Свет по Бе-
рингийской суше в вюрме (висконсине) в континентальную фазу, пока
неясно.
Ряд морфологических и экологических черт — низкая посадка и на-
клон головы, мощный меховой покров, питание травой (судя по содер-
жимому кишечника Чурапчинского экземпляра из Якутии), свидетель-
ствуют об обитании этого носорога в ландшафте холодной тундростепи.
Здесь намечается парадоксальное экологическое сходство евразийского
тундростепного носорога с белым носорогом африканских саванн.
Рис. 5. Время вымирания гигантского оленя в Евразии.
Обычно считается, что этот вид вымер одновременно с мамонтом
или даже раньше, причем в Сибири раньше, чем в Западной Европе.
Между тем, определяя остаточный коллаген в костях волосатых носо-
рогов из покровных суглинков Украины, И. Г. Пидопличко (1952) по-
лучал иногда очень поздние датировки, порядка 0.8—1 тысячелетия
до наших дней. «Свежие» образцы костей носорогов с отчетливым за-
пахом коллагена имеются также в коллекциях Казанского универси-
тета. Иные материалы из разных мест, датированные Си, указывают
даты порядка 12—14 тыс. лет до наших дней (рис. 6, 7).
В позднем палеолите на волосатого носорога энергично охотились.
В слоях стоянок на Русской равнине его кости составляют от 1,5 до
3% костей животных промысловой группы (Верещагин, 1979), а на
2* 19
стоянках Предбайкалья — до 3—4% (Ермолова, 1978). В степном За-
байкалье остатки этих зверей особенно обильны как в природных за-
хоронениях, так и на палеолитических стоянках. Способы охоты на
носорогов в палеолите и влияние ее на вымирание вида мало изучены.
Активная охота с дротиками и стрелами изображена на стене грота
Коломбьер (Франция). Вероятно, существовала и пассивная — при по-
мощи «волчьих ям», вырытых на тропах.
Существенно отметить, что рисунков волосатою носорога и его
костей пока не известно из слоев стоянок мезолитической и неолити-
ческой эпох. Таким образом, для объяснения причин вымирания вида
в конце плейстоцена и начале голоцена остается обсуждение морфо-
физиологических несоответствий организма носорога сменившейся среде
обитания в начале голоценовой эпохи. Аналогия с судьбой мамонта
и причинность вымирания были довольно велики.
Гигантский олень — Megaloceros giganteus Blumenbach, 1803
История большерогих оленей трибы Megacerini известна в Европе
с виллафранка, в СССР — с эпохи хапровского териокомплекса (Ба-
рышников, 1981). Их эволюция шла по линии увеличения размеров
тела, усложнения рогов и развития их лопаты.
Родиной гигантского оленя была, вероятно, Европа. Ранние пред-
ставители рода — Megaloceros giganteus aniecedes Вегск. из миндель-
рисса Германии (Штейнгейм)—имели рога с широкой лопатой, но
направленные назад, позволявшие им жить в лесу. В среднем плейсто-
цене (риссе) стал обычен крупный М. giganteus с широко раскинутыми
в стороны рогами (Громова, 1965). Его остатки обильны в хазарских
слоях долины Волги, где на отмелях они составляют от 2 до 8% всего
костного материала (Верещагин, 1959). Карликовые формы большеро-
гих оленей известны с юга Италии и островов Средиземного моря
(Bonfiglio, 1978).
Ареал гигантского оленя занимал всю Европу, на севере до Скан-
динавии и Тиманского кряжа, Северную Африку, Кавказ, Казахстан,
Сибирь, на севере до Тюмени, и до Забайкалья (Кяхга) на востоке.
Остатки вида обычны на мустьерских памятниках Русской рав-
нины: Старые Дуруиторы (Молдавия)—2,0% костей промысловых
видов (Давид, 1980), Кормань IV (Украина)—2,2 (Татаринов, 1977);
Северного Кавказа: Ильская — 1,3 (Верещагин, 1959), Баракаевская —
12,0 (Барышников, 1979) и многочисленны в Крыму: Шайтан-Коба —
39,4 (Громов, 1948). В Закавказье они редки — 0,07% в Ахштырской
пещере (Верещагин, 1959).
Переразвитость рогов самцов гигантского оленя, достигавших до
3,7 м в размахе, дает основание считать, что этот олень жил и питался
в открытых ландшафтах по долинам рек на луговых полянах, чере-
дующихся с перелесками. Это подтверждается и сравнением с лесным
зверем — лосем. У гигантского оленя глазницы были крупнее, зубы
гипсодонтнее, нижняя челюсть массивнее.
К позднему валдаю (вюрму) ареал гигантского оленя резко со-
кратился (рис. 5). Он исчез в Сибири, Казахстане и на Кавказе. Са-
мые поздние находки в Сибири датируются началом позднего палео-
лита (Верхнеленская гора; Ермолова, 1978). На Русской равнине они
редки. Так, в верхнепалеолитических слоях стоянок Кормань IV на
Днестре кости этого оленя составляют всего 0,1% остатков промысло-
вых видов (Татаринов, 1977), а на стоянке Костенки VIII на Дону —
20
0,2 (Верещагин, Кузьмина, 1977). Более обычен он был в Крыму — до
7,8% остатков травоядных млекопитающих (Бибикова, Белан, 1979).
На стоянках Западной Европы эпохи ориньяка и мадлена кости
гигантского оленя единичны. Во Франции он исчез в аллерёде (Вои-
chud, 1965), а на севере ФРГ встречался еще в эпоху предбореала
(Guenther, 1960). Возможно, что в Ирландии этот олень дожил до на-
шей эры (Mitchell, Parkes, 1949) и был уничтожен средневековыми
англосаксами.
В восточной Европе вымирание вида произошло, вероятно, в конце
плейстоцена, так как ни в одной мезолитической и неолитической
стоянке его костей не обнаружено (Паавер, 1965; Верещагин, 1979).
Указания на нахождение костей оленя голоценовой сохранности на'
Украине (Тарасовка Днепропетровской области; Пидопличко, 1951) и
в Сибири (Камышлов; Черский, 1891) требуют подтверждения.
Причины вымирания вида неясны. Указывается обычно на колос-
сальные энергетические затраты для роста исполинских рогов самцов,
малую плодовитость и относительную теплолюбивость, сокращение
в вюрме (валдае) площади мезофитных лугов, истребление древними
охотниками.
Первобытный бизон — Bison priscus Bojanus, 1827
Происхождение бизонов связывается с югом Азии, так как в плио-
ценовых отложениях Индии (Сивалики) и Китая обнаружены их древ-
нейшие остатки (подрод Eobison). В СССР остатки мелкого В. (ЕоЫ-
son) tamanensis N. Ver. найдены в верхнеплиоценовых-нижнеплейсто-
ценовых слоях берегов Азовского моря в таманском териокомплексе.
Из раннего плейстоцена Европы и Северной Азии известны более
крупные В. voigtstedtensis Fischer (Германия, Англия) и В. schoetensa-
cki Freud., причем ареал второго вида, которого К. К. Флёров (1979)
считает лесным жителем, прослежен на восток до р. Вилюй в Якутии.
В это время бизоны уже проникли в Америку.
В раннем плейстоцене, вероятно, происходит разделение бизонов
на степную и лесную ветви (Hilzheimer, 1918; Флёров, 1979). Остатки
лесных короткорогих форм в верхнечетвертичных слоях редки. На Кав-
казе раздробленные кости бизонов известны из слоев стоянок Кударо
в составе лесных фаун с ашельской эпохи до голоцена (Верещагин,
Барышников, 1980).
Значительно больше материалов по эволюции степного В. priscus.
В хазарской фауне среднего плейстоцена Поволжья и Казахстана он
был представлен огромным В. р. gigas Flerow, расстояние между кон-
цами рогов которого достигало 2 м. В Сибири и на Аляске в это время
встречался В. р. crassicornis Rich., приспособленный к обитанию в хо-
лодном климате северных лесостепей, о чем говорит большая ширина
и объем его носовой полости (Флёров, 1979).
Обширные пространства позднеплейстоценовых перигляциальных
степей Европы и Западной Сибири населял В. р. mediator Hilzh.,
а тундростепей Восточной Сибири, Аляски и Канады — В. р, occidenta-
l's Lucas. Проникновение в арктические широты — на Гыданский и
Таймырский полуострова и на Берингийскую сушу — было осуществ-
лено именно степными формами бизонов, обитателями открытых остеп-
ненных ландшафтов. Это подтверждено палеогеографическими мате-
риалами и сравнением экологии бизонов с экологией и распростране-
нием более мезофильных быков — туров.
До исторической эпохи в Голарктике уцелевают четыре географи-
22
чески изолированных формы, различные ио экологии и морфологии:
это лесной зубр Bison bonasus L. — обитатель лесов Европы и Кав-
каза; измельчавший потомок В. priscus subsp., сохранившийся местами
в степях юга Русской равнины (бассейн Дона) до VIII—XIII вв. нашей
эры (Верещагин, 1971), а в остепненной долине Ангары и в Прибай-
калье— до VI—VII вв. (Ермолова, 1978); лесной бизон Bison priscus
athabascae Rhoads, населявший редколесья и светлохвойную тайгу
северо-востока Сибири и тайгу Канады и уцелевший до современности
только в Канаде; бизон прерий Северной Америки В. bison L., процве-
тавший там до прихода европейцев.
В целом картина эволюции адаптаций зубров и бизонов к лесному
и степному образу жизни на протяжении четвертичного периода
сложна. Сложна и картина эволюции их ареалов и численности под
влиянием природных и антропогенных факторов. В настоящее время
известно, что зубры (и бизоны) не переносят длительного снегового
покрова глубже 40 см и поэтому перспективы расселения зверей, раз-
множаемых в заповедниках и лесопарках, у нас в СССР ограничены.
Судя по современным наблюдениям над тысячным поголовьем зубро-
бизонов (гибридных зубров) в Кавказском заповеднике в 60—70 годах
нашего столетия, становится ясным, что адаптация к лесным и степ-
ным условиям у зубров и бизонов не абсолютны. Гибридные особи
отлично прижились в лесных условиях Западного Кавказа, подни-
маются летом и на горные луга альпийского пояса, но в предгорную
степь не выходят.
Грандиозный ареал первобытных бизонов в плейстоцене Евразии
отчетливо показывает, что причиной его сильнейшей деградации в голо-
цене могла быть только природная экологическая катастрофа. Такой
катастрофой могло быть потепление и увлажнение климата в север-
ных частях ареала, вызвавшее размораживание плейстоценовой тундро-
степи и глубокоснежье. Уцелевшие в лесах и степях Европы, Кавказа
и в степях Сибири популяции зубров и бизонов были почти выбиты —
уничтожены человеком к началу XX века. Детальный обзор такого
истребления дан В. Г Гептнером (1961) и С. В. Кириковым (1969).
Овцебык — Ovibos moschatus Zimmermann, 1780
Многочисленный в плейстоцене овцебык интересен для проблемы
вымирания тем, что дожил местами до наших дней. Это позволяет су-
дить о его палеоэкологии на актуалистической основе.
Родиной овцебыка была, вероятно, северо-восточная Сибирь. От-
сюда известен ряд предковых форм из рода Praeovibos (Шер, 1971),
описанного ранее из раннего плейстоцена Европы (Франкенхаузен).
Наиболее древние остатки настоящего Ovibos известны из миндель-
ского гравия Зюссенборна в Германии (Soergel, 1942). Ряд отличий
в черепе плейстоценовых евразийских овцебыков от современных аме-
риканских дает основание некоторым зоологам для выделения их
в особый вид О. pallantis Н. Smith, 1827 (Ryziewicz, 1955), что, однако,
не бесспорно. На протяжении плейстоцена наблюдается изменение
в пропорциях метаподий овцебыков: они делаются короче и массивнее
(Kahlke, 1963), приближаясь к массивности метаподий современных
экземпляров.
В среднем и позднем плейстоцене ареал овцебыка занимал почти
всю Европу и Северную Азию. В Западной Европе он доходил на юге
до Дордони (Эйзи) во Франции, юго-востока Венгрии и до Добруджи
в Румынии и даже, видимо, в наиболее холодные периоды, до Испании
23
(Арбреда; Estevez, 1979). На Русской равнине южная граница про-
ходила на широте Киева и Волгограда. В Сибири она шла севернее,
так как на палеолитических стоянках Прибайкалья его остатки отсут-
ствуют (Ермолова, 1978). «Следы» северной границы уходят на Тай-
мыр и в шельф Полярного бассейна у Новосибирских островов.
Современные овцебыки живут стадами обычно в 20—30 голов,
среди которых всего 2—3 взрослых быка. Летом самцы держатся от-
дельно. В позднеплейстоценовых слоях Европы остатки самцов к сам-
кам составляют 7 1, что по мнению В. Зоргеля (Soergel, 1942), гово-
рит о гибели в прошлом от хищников и первобытных охотников пре-
имущественно быков, живших одиночками, изгнанными из стада в ходе
турнирных боев.
Теперь стало известно, что овцебык не выносит плотного снегового
покрова глубже 30 см и мало способен к тебеневке и далеким мигра-
циям. Зато он переносит жестокие морозы до —50° С и ветер на остро-
вах Канадского архипелага и побережья северо-востока Гренландии и
довольствуется скудной травянистой и кустарниковой растительностью
каменистой тундры.
Опыты интродукции канадских и аляскинских овцебыков в СССР —
на южных склонах хребта Бырранга на Таймыре и на острове Вран-
геля, проводившиеся с 70-х годов, показали, что они могут жить лишь
на немногих участках современной арктической тундры Евразии.
С окончанием ледниковой эпохи — позднего валдая (вюрма III) —
ареал овцебыка, вероятно, стремительно отступал к северу, и дольше
всего небольшая популяция задержалась на Таймыре (рис. 8, вклейка).
Наиболее поздние датировки по С14 поверхностно лежавших черепов и
роговых футляров овцебыков известны с северного Таймыра: 3800±200;
2900±95 (Верещагин, 1971); 2920±50 (ГИН-2945). В Европе наиболее
поздние находки приурочены к позднеледниковым отложениям Скан-
динавии, например, череп из Эстерсунда в Швеции имеет возраст около
9 тыс. лет (Borgen, 1979).
Основной ограничивающий фактор в пределах современной тундро-
вой зоны Сибири — это, безусловно, плотный снеговой покров, препят-
ствующий кормежке и вызвавший в послеледниковье вымирание овце-
быка во всей Евразии. Истребительная деятельность человека обусло-
вила сильное сокращение ареала вида в Северной Америке за послед-
ние столетия.
Подводя некоторые итоги нашего краткого обзора, можно утвер-
ждать, что, во-первых, плейстоценовые виды зверей вымирали не одно-
временно, часть видов как бы отступала, стягивалась в своеобразные
рефугии, доживая там свой век. Во-вторых, палеонтологические факты
и материалы почти не дают оснований говорить о том. что у рассмо-
тренных видов были какие-то дефекты в морфофизиологической орга-
низации, которые обусловили их угасание.
О локальных причинах вымирания и местных сдвигах
ареалов млекопитающих в плейстоцене и голоцене
Факты современного частичного вымирания млекопитающих отчет-
ливо показывают значение внешних факторов среды для их существо-
вания и распространения. В качестве наглядных примеров приведем
пульсации ареалов мезофильных и относительно ксерофильных видов.
Кавказский крот (Talpa caucasica Satun.) в западном Предкав-
казье распространен по долинам рек и на предгорной равнине, где
в засушливые годы периодически вымирает на обширных просгран-
24
ствах. Факторы вымирания: резкое сокращение биомассы почвенных
беспозвоночных и, вероятно, «ухудшение» микроклиматических условий
обитания в норах — сильный прогрев, повышенная сухость. Иссушение
и остепнение здешних пространств вследствие сельскохозяйственного
освоения и вырубки приречных лесов может привести и к полному вы-
теснению вида из равнинных участков.
В результате иссушения луговых пространств в послеледниковую
эпоху кроты сохранились на реликтовых участках в мезофильных био-
топах в окружении степных и даже полупустынных группировок расте-
ний и животных. Такой замечательный пример кротового рефугиума
имеется в Караязском дубовом лесу долины Куры восточнее Тбилиси
(Верещагин, 1959). Реликтовыми оказались участки обитания зверьков
на окруженной сухими степями Ставропольской возвышенности и на
альпийских лугах Армянского нагорья.
Сходный с кротом пример пульсации границ ареала показывает
относительно ксерофильный вид — общественная полевка (Microtus so-
cialis Pall.). В полупустыне и степях восточного Закавказья (Азер-
байджан) этот зверек процветает в пределах выпадения осадков от 350
до 800 мм в год. В результате ряда засушливых зимних сезонов соз-
даются стрессовые ситуации, и полевка вымирает на больших площа-
дях, причем стремительное (в течение одного-двух сезонов) отступание
границы ее ареала в предгорья Кавказа западнее г. Баку происходит
на расстояние 100—120 км.
Подобное вымирание в годовом цикле может перерастать в много-
летнее и вековое, а осложненное иными, не климатическими факто-
рами, вести к вымиранию вида. По-видимому, к подобным примерам
следует отнести постепенное исчезновение группировки степных грызу-
нов и мелких хищных в прикаспийских степях на протяжении послед-
них веков нашей эры.
А. Н. Формозов (1938) описал вымирание в середине XIX — начале
XX века малой пищухи (Ochotona pusiila Pail.), желтой пеструшки
(Eolagurus luteus Eversm.), большой песчанки (Phombomys opimus
Licht.), слепушонки (Ellobius talpinus Pall.) и гигантского слепыша
(Spalax giganteus Nehr.) в степях и полупустынях северного Прикас-
пия. В середине нашего столетия стали известны новые факты исчез-
новения в волго-донских и предкавказских степях корсака (Vulpes cor-
sac L.) и степного хоря (Putorius eversmanni Lesson.) (Верещагин,
1959). В данном случае решающее значение имело, возможно, истреб-
ление ядами и газами малых сусликов как основной пищи и довольно
интенсивный промысел упомянутых хищников.
Наглядна также эволюция ареала сайги (Saiga tatarica L.), имею-
щей устойчивые адаптации к резко континентальному климату, степ-
ным и пустынным ландшафтам, питанию злаками и полынями и к бы-
строму бегу по твердым грунтам. Родиной вида могут считаться остеп-
ненные плоскогорья Центральной Азии, где встречается родственная
антилопа оронго (Pantholops hodgsoni Abel). В позднем плейстоцене
сайгаки были распространены от Англии на западе до Аляски на во-
стоке и от Новосибирских островов на севере до среднеазиатских
пустынь на юге. Возможно, впрочем, что арктические степи Сибири
населял другой плейстоценовый вид (Saiga borealis Tscherskyi).
В историческую эпоху ареал этой антилопы стремительно отсту-
пал на восток, но в средние века сайгаки жили во множестве в степях
бассейна Днестра, Днепра и, вероятно, еще встречались в пуштах
Венгрии западнее Карпат. К 20-м годам нашего века только несколько
сот животных уцелело в степях Калмыкии — междуречье Волги и Дона,
25
в Приаральских Каракумах и в западной части Бет-Пак-Далы. При та-
ком падении численности полностью вымер особый вид подкожного
овода (Pallasiomyia antilopum), сохранившийся лишь на изолирован-
ной популяции монгольских сайгаков (Грунин, 1957).
Основным фактором вымирания сайги в Сибири было исчезнове-
ние плейстоценовых тундровостепных ландшафтов и расширение таеж-
ных. На Русской равнине и в Казахстане в XIX—начале XX в. оно
было связано с непосредственным истреблением зверей кочевыми ско-
товодами и земледельцами, особенно близ водопоев. Последующий
быстрый рост численности и восстановление ареала вида в 30—40 годы
объяснялись главным образом обезлюдением степи и полупустыни,
а также попытками государственной охраны — полным запретом охоты.
Современная численность этой антилопы в Казахстане достигает в от-
дельные годы 1800—2000 тысяч голов, что объясняется регулированием
промысла и истреблением волков стрельбой с самолетов.
В целом же создается отчетливое представление, что в позднем
плейстоцене и голоцене степная группировка зверей центральноазиат-
ского корня, включающая хищных (корсак, степной хорь), грызунов
(желтая пеструшка, степной сурок — байбак), зайцеобразных (малая
пищуха), копытных (кулан, тарпан, сайга), непрерывно отступала
с конца плейстоцена из Западной Европы, а в последние века новой
эры и из Восточной на родину — в степи Казахстана и Монголии,
как бы повторяя свой путь плейстоценовой экспансии в обратном на-
правлении.
Широко известный факт отступления мезофильных видов грызу-
нов с иссушенных и остепненных в послеледниковую эпоху равнин
в горы иллюстрируется в Европе и Азии многими примерами. В полу-
пустыне Добруджи по правобережью Дуная румынские палеонтологи
обнаружили при раскопке пещеры Адам остатки снеговой полевки
(Chionomys nivalis Martins), обитающей ныне в Южных Карпатах.
В Предкавказье близкая к ней гудаурская полевка (Chionomys gud
Satun.), населяющая высокогорья Кавказа, уцелела в Пятигорье на
горе Развалка на высотах 500—600 м над уровнем моря, в участке все-
сезонной мерзлоты в россыпях тешенитовых скал.
Хромосомные виды обыкновенной полевки (Microtus arvalis Pall.,
М. subarvalis Meyer, Orlov et Skholl), широко распространенные по
мезофильным лугам Русской равнины и в высоко! орье Кавказа, на юж-
ном пределе ареала имеют пятнистый ареал по альпийским лугам
горных хребтов, поэтому есть основание утверждать, что подобные изо-
ляции произошли в результате вымирания зверьков в послеледниковое
время на опустыненных днищах и склонах межгорных долин.
Вообще мобильность границ ареалов мезофильных и ксерофиль-
ных мелких млекопитающих на протяжении плейстоцена и голоцена
была очень велика. Это отчетливо прослежено в Восточном Закавказье
в связи с открытием Бинагадинской среднеплейстоценовой фауны в би-
тумах Апшеронского полуострова.
Из Закавказья в голоцене (?) отступили в Предкавказье ареалы
корсака и сайги, а на юг, в иранские плоскогорья — ареалы горной
слепушонки, дикобраза и муфлона. Сдвиги ареалов других местных
видов были менее значительны. Так, граница ареала обыкновенной
полевки отступила к северо-западу на 120—130 км, а золотистого хо-
мяка (Mesocricetus raddei Nehr.), уцелевшего в Дагестане, — на 200—
250 км.
Иные факторы обусловили большие колебания площадей и границ
распространения речного бобра (Castor fiber L.). На северо-востоке
26
СССР известны местонахождения следов деятельности бобров в виде
ископаемых плотин, огрызков ветвей и стволов деревьев. На реках
Енисей, Алдан, Пенжина эти места отстоят на многие тысячи кило-
метров от реликтовых колоний бобров, сохранившихся, например,
в Туве и у восточных подножий Урала по рекам Конда и Сосьва
(Граве, 1931; Скалой, 1951).
В историческое время бобры были малочисленны в Сибири в пре-
делах многолетней (вековой) мерзлоты, которая препятствует устрой-
ству устойчивых нор в берегах и обусловливает сильное промерзание
водоемов и бобровых хижин, особенно при образовании наледей. Про-
никновение бобров далеко на север Сибири было, очевидно, возмож-
ным в эпохи потеплений и сильного размораживания грунтов, т. е.
исчезновении вековой мерзлоты. По нашим представлениям, уцелев-
шие к XX столетию от перепромысла реликтовые колонии бобров
в бассейне Иртыша и других мест Сибири образовались в послеледни-
ковую эпоху — не более чем 8—9 тыс. лет тому назад, поэтому их «або-
ригенность» относительна. Аборигенность более древняя (плио-плейсто-
ценовая) была свойственна колониям бобров, существовавшим в Пе-
редней Азии, Казахстане и существующей пока в 1Монголии на р. Бу-
луган.
К XX веку бобровые колонии в пределах России уцелели либо во
владениях монастырей в европейской части, либо под охраной тузем-
ных шаманов, т. е. также на религиозной, культовой основе. Крупные
государственные мероприятия по расселению (реакклиматизации) боб-
ров, предпринятые в 50—60 годах в Европейской части СССР и южной
Сибири, увенчались полным успехом и отчетливо показали, что основ-
ной причиной исчезновения бобров в историческое время на огромных
пространствах Восточной Европы, Кавказа, Передней и Средней Азии
и Сибири было истребление их человеком в погоне за ценной шкурой
и бобровой струей. Впрочем, в южных районах страны большое значе-
ние имело истребление скотоводами приречных, галерейных лесов —
кормовой базы наших замечательных грызунов.
Климатические и ландшафтные изменения
на границе плейстоцена и голоцена
как причина вымирания мамонтовой группировки
Как уже было сказано выше, в XVIII, XIX и начале XX в. боль-
шинство европейских ученых считало, что климат и среда обитания
мамонтовой фауны в Сибири были теплее и резко отличались от совре-
менных. Зоолог Ф. Брандт (1866, с. 3) справедливо писал, что скудная
растительность нынешних тундр не была бы в состоянии прокормить
гигантских мамонтов, носорогов, бизонов, а наличие в прошлом пыш-
ного растительного покрова в Арктике он объяснял более теплым кли-
матом.
В первой половине XX в. мнения разделились. Были высказаны
соображения о температурных условиях, близких современным. Орни-
толог А. Я. Тугаринов (1928, 1934) считал, что тундра в эпоху жизни
в ней пещерного льва и сайги была «не той, что сейчас», представляла
собой «открытый ландшафт при значительной ксеротермичности кли-
мата, достаточно холодного, с малым количеством зимних осадков»
(1928, с. 699). Существовала, по его мнению, сумма факторов в виде
увеличения влажности, наступления затем засушливого периода и но-
вого увеличения влажности, которые повлекли отступание ареалов ло-
шадей, верблюдов, сайгаков с восточно-сибирских степных языков
27
в область сплошных степей и пустынь (1934, с. 63—64). Мамонт, носо-
рог, быки и пещерный медведь якобы вымерли при условиях далеко
не резкого изменения климата, «при этом не в сторону ухудшения»,
«очевидно они обитали на грани возможного жизненного оптимума и
достаточно было лишь весьма незначительною изменения в условиях
существования, чтобы наступил кризис».
Превосходный орнитолог и палеозоолог П. В. Серебровский (1935),
по-видимому, признавал существование в вюрме (валдае, висконсине)
примерно тех же ландшафтных зон в пределах северной Евразии, что
и ныне, однако одновременно он признавал и существование гран-
диозного европейского ледника.
Палеонтолог И. Г Пидопличко (1951, 1969) весьма последова-
тельно утверждал, что мамонты могли бы отлично жить на территории
современной Украины, подчеркивая тем самым отсутствие резких кли-
матических и ландшафтных различий между двумя эпохами и значе-
ние этого фактора для вымирания плейстоценовых зверей. Гляциолог
Д. Дайсон (1966, с. 138) прямо подтверждал сходство ландшафтных
сред плейстоценовой и современной с точки зрения обитания мамонтов.
Столь категорические выводы делались на основе косвенных фак-
тов и умозрительных соображений, так как детальных палеоклимати-
ческих исследований еще не было. Иные факты вскрылись работами
многочисленных отечественных палеогеографов, мерзлотоведов и гео-
28
морфологов в арктической и субарктической зонах и в области былого
плейстоценового развития многолетней мерзлоты.
Наиболее солидны результаты исследований природных процессов
в плейстоцене, полученные А. А. Величко (1973, 1982) и С. В. Томир-
диаро (1977, 1980). На основе геоморфологических наблюдений на
Русской равнине и на северо-востоке Сибири, палеогеографических со-
поставлений оба исследователя реконструировали экологическую об-
становку эпохи мамонта в период максимума похолодания в вюрме
(валдае) как очень суровую с резкоконтинентальным климатом, обусло-
вившим развитие многолетней мерзлоты на глубину до 1,5 км и обра-
зование мощных ледовых жил-стен подземного льда на глубину до
40 м, обжимавших земляные столбы. Южная граница мерзлоты продви-
галась в Европе до 46—48° с. ш. Были характерны низкие зимние тем-
пературы, составлявшие, судя по палеоботаническим данным, на ши-
роте южной Англии, Белоруссии и Среднерусской возвышенности около
—30° С (Величко, 1982). Летние температуры снижались не столь су-
щественно. Количество осадков в перигляциальной зоне не превышало
250—300 мм в год.
Вместо зональных смешанных и широколиственных лесов в Европе
был распространен перигляциальный тип растительности, занимавший
широкую полосу между Скандинавским ледниковым щитом на севере
и Альпийским ледником на юге (Гричук, 1982). На юге Русской рав-
нины и на среднедунайских низменностях его сменяли перигляциаль-
•ные степи.
Малоснежье зимой и развитие пышного травяного покрова на твер-
дых, ссыхающихся при обильной инсоляции летом грунтах позволяли
жить лошадям, бизонам, сайгакам на огромных пространствах Север-
ной Евразии. Для границы плейстоцена и голоцена характерно наличие
резких климатических колебаний с короткой периодичностью: потепле-
ние беллиг (12400—12000 лет назад), похолодание средний дриас
(12000—11800), потепление аллерёд (11800—11000), похолодание позд-
ний дриас (11000—10300) и т. д., имевших решающее значение для
жизни плейстоценовых видов животных. Именно в эти даты происхо-
дила массовая гибель мамонтов и их «спутников» в арктической зоне
(рис. 6, 7), о чем свидетельствуют сотни тысяч костей разрозненных ске-
летов и единичных мерзлых туш, захороненных в сартанской толще
(поздний висконсин) северной Якутии и Таймыра. Судя по современ-
ным примерам массовой гибели диких и домашних копытных в степях
Казахстана (Слудский, 1963), такая гибель зверей в конце плейстоцена
объясняется скорее всего появлением частых снежных бурь (бура-
нов) зимой и преобразованием плейстоценовой кормной тундростепи
в заболоченную и заозеренную тундру.
В субарктических широтах в это время на открытых пространствах
шло быстрое наступление таежных и смешанных лесов, обусловившее
развитие лесной фауны.
Наиболее убедительным доказательством коренных ландшафтных
изменений на границе плейстоцена и голоцена (с точки зрения палео-
зоолога) является смена степного облика териофауны на лесной на
Русской равнине, Северном Урале (Кузьмина, 1971), в Сибири и даже
в Приморье.
Вымирание зверей под влиянием человека
Истребительное влияние человека на промысловые виды животных
(тем более, на «вредные») в век техноцена общеизвестно. Оно нашло
29
яркое отражение в многочисленных природоохранных законах, меро-
приятиях, книгах и инструкциях по промыслу и истреблению и не нуж-
дается в доказательствах. Иное положение существовало в эпоху палео-
лита, когда численность первобытного населения была не велика,
а технические средства, потребности и экономические связи держались
на низком уровне.
Первобытные орды раннего и позднего палеолита оставили много-
численные следы своего воздействия на популяции промысловых зве-
рей, птиц и рыб в Западной и Восточной Европе, на Кавказе, в Сред-
ней Азии, Сибири и на Дальнем Востоке. Это сотни тысяч осколков
искусственно разбитых костей в слоях открытых и пещерных палеоли-
тических, мезолитических и неолитических стоянок. Видовой состав
зверей, добывавшихся первобытными охотниками, в сущности пол-
ностью совпадает со списками определенного костного материала по
крупным позвоночным. Сюда входят хищные, крупные грызуны, зайце-
образные, непарно- и парнокопытные, хоботные и изредка даже при-
маты. Обзоры способов охотничьего промысла в палеолите и неолите
Европы опубликованы В. Зоргелем (Soergel, 1922), К. Линднером
(Lindner, 1937) и встречаются во многих археологических и этногра-
фических сочинениях.
Для территории Советского Союза первая палеозоологическая
сводка о видовом составе остатков животных, обнаруженных на палео-
литических стоянках, дана В. И. Громовым (1948), а специальные па-
леоохотничьи обзоры — С. А. Семеновым (1968) и Н. К. Верещагиным
(1971). По этим работам известно, что на Русской равнине, Кавказе,
в Средней Азии, Сибири и Приморье человек добывал и осваивал для
пищи и технических целей 1 вид обезьяны, 28 — хищных, 2 — хоботных,
5 — зайцеобразных, 5 — грызунов, 4 — непарнопалых и 23—парнопа-
лых, а всего до 67—70 промысловых видов наземных млекопитающих.
Отчетливо видны региональные особенности, связанные с зоогео-
графическими чертами и изученностью территорий. Так, например,
наибольшее разнообразие добываемых хищных и копытных было на
Русской равнине и Кавказе, наименьшее — в Средней Азии. Однако
материалы эти расширяются и уточняются. Основная закономерность —
использование местных, подручных ресурсов и при этом наиболее
массовых, многочисленных. Попытки объективной оценки значимости
отдельных видов зверей в рационах древних охотников затруднены
различной тафономической природой захоронений их остатков. Кости
крупных животных (мамонтов и копытных) лучше и легче сохраняются
в слоях отложений и учитываются в большей степени, чем кости сред-
них и мелких по величине хищных и грызунов. Нет и удовлетвори-
тельной оценки величины древних мясных рационов. Такие расчеты
сильно расходятся. По этнографическим сведениям известно, что со-
временные туземцы Сибири — ненцы, долгане, юкагиры, якуты, эвенки
и эвенны — способны съедать от 2 до 5 кг и более мяса северного
оленя в сутки.
Ф. Моуэт (1963) приводил суточную норму у современных эскимо-
сов Канады в 2кг мяса карибу (северного оленя). Для человека
позднего палеолита Восточной Европы советскими исследователями
принимались явно заниженные нормы: С. Н. Бибиков (1967)—600г
в сутки; И. Г Пидопличко (1969) — 800—1000 г. Эти нормы, очевидно,
сильно разнились летом и зимой. Летом поедалось много растительного
корма: корневищ рогоза, тростника, дудок зонтичных, плодов и ягод,
орехов, съедобных трав и даже коры деревьев, например, луба ивы.
И. Г. Пидопличко (1969, с. 153) считал, что в летние и осенние ме-
30
сяцы Уз всей потребляемой пищи составляли растительные продукты,
рыба и мясо мелких зверей и птиц. При расчете на весь год расти-
тельная пища, по его мнению, составляла вместе с мясом мелких жи-
вотных 74 годового рациона, а мясо мамонтов — 3/4. Судя по материа-
лам Костенковских (Донских) палеолитических стоянок, основу пита-
ния древних поселенцев составляли в большинстве случаев лошади и
зайцы, а не мамонты (Верещагин, Кузьмина, 1977).
Учитывая суровость климата северной Евразии эпохи палеолита
и краткость вегетационного периода, Н. К. Верещагин (1971) прини-
мал среднегодовую норму потребления мяса в 2 кг в сутки на чело-
века. При такой норме населению позднего палеолита численностью
в 15 000 человек (цифра явно заниженная), обитавших в южной поло-
вине Русской равнины, требовалось в год до 10 500 т мяса. Это коли-
чество могли дать до 1000 убитых полувзрослых мамонтов (убойный
вес до 1000 кг), до 60 000 лошадей (убойный вес до 100 кг) и до
10 000 бизонов (убойный вес до 300 кг). Первобытным охотникам, оче-
видно, требовались напряженные круглогодичные усилия, если учесть
трудности охоты с примитивными орудиями на крупных стадных и
одиночных животных. Тем не менее эти цифры прямо указывают на
весьма мощный пресс палеолитического человека на животное насе-
ление равнин. Впрочем, имеется также основание подозревать, что
древние обитатели Северной Евразии нередко использовали для еды
трупы утонувших и замерзших мамонтов и копытных. На реке Берелёх
в Северной Якутии такие факты установлены при раскопке береговых
байджерахов * в 1980 г. на известном грандиозном «кладбище» мамон-
тов, образовавшемся около 13 тыс. лет тому назад.
В ряде случаев первобытные охотники Северной Евразии без-
условно пользовались остатками добычи стайных хищников — пещер-
ных львов и волков, как это делали недавно аборигены Африки. Такой
двойной пресс хищников и расселявшихся человеческих племен мог
губительно сказываться на популяциях мамонтов и копытных в позд-
нем плейстоцене.
Непостоянство удач на первобытных охотах и миграции мамонтов
и копытных должны были безусловно развивать способы заготовки
мяса впрок, прежде всего при помощи вяления и сушки на солнце и
ветру и сохранения в мороженом виде зимой, а за счет мерзлоты
грунтов — и летом.
В послепалеолитическую эпоху, при зарождении животноводства
и земледелия пресс на промысловых животных продолжал увеличи-
ваться, благодаря совершенствованию технических приемов добычи
диких животных, одомашниванию предков собаки и лошади.
Именно собака и лошадь создали предпосылки для разведки, по-
иска и поимки крупной и мелкой добычи — пушных, мясных и мясо-
сальных зверей. О видовом составе добываемых животных, об изобре-
тении и совершенствовании лука, гарпунов, рогатин и их применении
на охотах у различных племен в неолитическую эпоху и в век бронзы
свидетельствуют обильные рисунки (петроглифы) на скалах — писа-
ницы, обнаруженные и описанные этнографами на огромной террито-
рии от Скандинавии до Камчатки и от побережий Средиземного моря
до Тихого океана (Kuhn, 1956; Савватсев, 1970; Верещагин и Бурчак-
/Хбрамович, 1948; Мариковский, 1953; Окладников, 1959; Диков, 1971;
Формозов, 1969; Дэвлет, 1980 и др.).
* Байджсрах—(якут.), столб и бугор грунта, обжатого жилами грунтового
льда при полигональном растрескивании поверхности тундры.
31
Обильные костные остатки крупных диких животных — медведей,
кабанов, лосей — обнаруживаются в слоях неолитических свайных по-
селений в Восточной Прибалтике. Удельный вес в добыче морских
зверей — дельфинов и ластоногих — оказывается значительным в при-
морских поселениях (Паавер, 1965; Верещагин, Русаков, 1979). Сход-
ная картина наблюдается и на стоянках эпохи неолита и бронзы по
побережьям морей Дальнего Востока и Приморья.
Нет сомнения, что именно неолитические племена Восточной
Европы и Сибири добивали последних, уцелевших кое-где мамонтов,
овцебыков, винторогих антилоп, забайкальских бубалов, байкальских
яков. Сарматским и скифским племенам еще достались в степях Во-
сточной Европы и Южной Сибири почти нетронутые табуны и стада
тарпанов, куланов, сайгаков, первобытных туров и бизонов.
О грандиозных облавных охотах («паганеях», «джаркэ») конницы
монголов и татар на степных копытных — бизонов, благородных оленей,
сайгаков — имеются сведения у средневековых летописцев (Рашид-Ад-
Дин, 1946; Кириков, 1959). Такие охоты подтверждены, например,
обильными костными материалами из слоев VIII—XIII вв. хазарского
городища и крепости Саркел на Дону (Верещагин, 1971) и превосход-
ными остеологическими исследованиями В. И. Цалкина (1962), В. И. Би-
биковой (1953), Н. Г. Тимченко (1972) по серин раскопанных архео-
логами городищ на Русской равнине в середине нашего столетия.
При всем разнообразии материалов учесть значение охотничьего про-
мысла на протяжении нашей эры в истреблении отдельных видов зве-
рей чрезвычайно трудно. Общей тенденцией на протяжении веков яв-
ляется лишь уменьшение процента остатков диких видов по сравнению
с возрастающим обилием домашних. По учетам многотысячных сборов
костей при раскопках средневековых поселений Приднепровья (Украи-
на) кости диких животных (кабанов, куланов, косуль, оленей, лосей,
сайгаков, бизонов и туров) составляли от 0,4 до 10,6%, а по числу
особей — от 5,0 до 29% от таких же чисел домашних животных (Тим-
ченко, 1972).
Широко известные факты сокращения областей распространения
крупных млекопитающих и даже их полного исчезновения в резуль-
тате интенсивного охотничьего промысла, вырубки лесов, распашки
целинных земель можно иллюстрировать судьбой первобытного тура
и крупных кошек.
Ископаемые остатки первобытного тура (Bos priinigenius Bojanus,
1827) появляются впервые в нижнечетвертичных отложениях Европы,
Малой Азии и Кавказа. Уже тогда это были очень крупные животные.
Плейстоценовый ареал тура был обширен, но значительно меньше, чем
у бизонов. Кроме Европы, остатки этих быков известны в Передней
Азии, Предкавказье и Закавказье, Средней Азии, южной Сибири до
Прибайкалья. В течение среднего и позднего плейстоцена звери
остаются довольно крупными, но к концу периода и в голоцене распа-
даются на крупную и мелкую формы. В эпоху палеолита тур был одо-
машнен, причем для этого использовались, видимо, обе формы.
В экологическом смысле туры были более мезофильны и тепло-
любивы, чем степные бизоны, и держались по долинам и поймам рек
(особенно при высыхании, выгорании под солнцем водораздельных
пространств) и не поднимались высоко в горы. В средневековых пущах
Литвы и Польши они придерживались сплошных смешанных лесов,
даже сырых и заболоченных, питаясь там (частично) веточным кормом
(Гептнер и др., 1961).
На Украине кости тура присутствуют на стоянках эпохи неолита,
32
энеолита, бронзы и вплоть до X—XI вв. (Бибикова, 1953), на Кавказе
имеются его изображения на скалах мезолитического и неолитического
времени. В Передней и Средней Азии звери встречались до века
бронзы в приречных зарослях и тростниках Ефрата, Амударьи и Сыр-
дарьи. В связи с вырубанием лесов туры к XV в. исчезли из Западной
и Средней Европы, дольше всего (до начала XVIII в.), сохраняясь
в пущах (охраняемых феодалами лесах) на территории современных
Польши и Белоруссии.
Плейстоценовые остатки тигра (Panthera ligris L.) найдены в Ин-
дии, Китае и на юге Приморского края. В начале XX в. тигр встре-
чался в пределах СССР в Закавказье, по тугаям среднеазиатских рек,
в Приморье и по долине Амура. Заходы одиночных зверей отмечены
для юга Западной Сибири и Якутии (Гентнер, Слудский, 1972). На Се-
верном Кавказе зверь исчез, видимо, в XII в. Прямое преследование
и развитие сельского хозяйства Закавказья привели к тому, что уже
в 20-х годах нашего столетия лишь отдельные особи заходили сюда
из Ирана. В Средней Азии тигры жили вплоть до 50-х годов нашего
века, когда уничтожение тугаев, истребление кабана и охота привели
к их полному истреблению. Численность вида неуклонно снижалась
в Приморье, где к началу 80-х годов было около 180—200 особей, на-
ходящихся под охраной (Животченко, 1981).
Ареал гепарда (Acinonyx jubatus Schreb.) в середине века включал
равнины восточного Закавказья, где он продержался, возможно, до
XVIII в. (Верещагин, 1959). В нашем столетии этот зверь населял
в СССР пустынные районы Туркмении и Узбекистана. В связи с умень-
шением численности джейранов и прямым истреблением гепард стал
здесь редок и к настоящему времени, вероятно, полностью исчез.
К этим примерам можно добавить широко известное истребление
стеллеровой коровы (Hydrodamalis gigas Zimm.) у побережья Коман-
дорских островов в 1768 г.; исчезновение тарпана (Equus gmelini
Anton.) и кулана (Е. hemionus Pall.) в середине XIX в. в степях юга
Русской равнины; уничтожение в 20-х годах нашего века последних
кавказских зубров (Bison bonasus caucasicus Satun.) и другие данные,
дополняющие картину обеднения видового состава фауны (рис. 10,
вклейка).
Быстрое исчезновение на планете огромных культурных ценностей
в виде генетического фонда диких животных заставили биологов при-
нимать все более решительные меры по его сохранению в виде изда-
ния Всемирной и республиканских «Красных книг» и разработке зако-
нов об охране животного мира.
Сопоставляя темпы вымирания (уничтожения) животных под
влиянием человека в эпохи палеолита, неолита и ранних металлических
культур с современностью, мы должны признать, что в век техноцена
они увеличились, ускорились необычайно.
Кроме непосредственного и видимого косвенного уничтожения че-
ловеком ценных и вредных млекопитающих, теперь стали широко из-
вестны факторы сильного влияния хищников и эпизоотий, а также
различного рода стрессовых ситуаций, действующих через разрушение
структуры популяций. Повинен в этом бывает также и человек с его
перепромыслом — недопустимой промысловой нагрузкой на популяции^
В охотоведческой практике нередко встречаются положения, когда
рост стада диких копытных или хищников бывает резко замедлен или
вообще отсутствует в результате нарушения оптимальной структуры
стада. Преимущественный убой крупных особей, выбивание самцов
3 Млекопитающие 33
.(для трофеев!) разрушает возможность нормального воспроизводства,,
что ведет популяции к истощению. Подобные же результаты разруше-
ния структуры популяции бывают следствием влияния хищников и
эпизоотий, неурожаев кормовых растений и т. п.
Заключение
Изучение фактических материалов по географии и экологии вы-
мерших и вымирающих млекопитающих четвертичеого периода в пре-
делах СССР отчетливо показало, что существовало четыре группы
факторов разрушения мамонтовой группировки и исчезновения с лица
земли ряда видов.
1. Изменение среды обитания, включая сезонные и вековые коле-
бания климата, влекущие изменения ландшафтов и биотопов. Сюда же
можно отнести тектонические поднятия суши и колебания уровня Ми-
рового океана, не только менявших циркуляцию воздушных масс, но
и ослаблявших давление естественного отбора в условиях островной
и полуостровной изоляции.
2. Утеря видами зверей резистентности, способности достаточно
быстро адаптироваться в процессе эволюции к новым абиотическим
и биотическим условиям среды. Речь идет о чрезмерной специализации
и инадаптивном характере эволюции.
3. Нарушение структуры популяций под влиянием внешних фак-
торов— более прогрессивных конкурентов, хищников, паразитов, эпи-
зоотий, стрессовых ситуаций.
4. Прямое истребительное влияние человека и косвенное воздей-
ствие его хозяйственной деятельности на промысловые и непромысло-
вые виды млекопитающих.
Принимая в качестве постулатов все названные группы факторов,,
следует сделать попытку оценки их удельного веса во времени и про-
странстве, т. е. с учетом этапов и географических зон.
Приведенные выше примеры быстрого (молниеносного в геологи-
ческом смысле) сокращения ареалов и численности непромысловых
видов млекопитающих Северной Евразии на протяжении четвертичного
периода и в наши дни указывают на главенствующее значение в их
вымирании внешних, абиотических факторов среды. Почти полное вы-
мирание крупных зверей мамонтовой группировки, способных к даль-
ним миграциям, в пределах тундровой и таежной зон и частичное их
сохранение в зонах лесостепей и степей также подтверждает решаю-
щее и абсолютное доминирование климатических и ландшафтных из-
менений для жизни и смерти и этой группы видов.
Лучшим доказательством сказанному является повсеместная смена
«степного» по облику фаунистического позднеплейстоценового ком-
плекса на лесной и таежный на огромных пространствах высоких и
умеренных широт северного полушария. В данном случае не было
ни вытеснения примитивных видов более прогрессивными, ни наруше-
ния популяционных структур, ни влияния человека. Изменение среды
обитания было столь резко и велико, что морфофизиологическая эво-
люция «не поспевала» за этими изменениями.
Что касается истребительной деятельности палеолитического и со-
временного человека, которому нередко отводят решающее значение
в вымирании четвертичных гигантов — мамонтов, носорогов, пещерных
медведей, то его влияние на популяции млекопитающих неуклонно
росло, но определяющим стало лишь в последние столетия. Кроме того,
роль человека в разных географических зонах была не одинакова,.
34
значительно возрастая в древнейших очагах цивилизации Средиземно-
морья, Передней и Средней Азии и Китая и уменьшаясь до минимума
в пределах полярных пустынь.
Значение остальных факторов вымирания было, очевидно, второ-
степенным, лишь в той или иной степени способствовавшим сокраще-
нию ареалов и численности видов под воздействием климатических и
ландшафтных изменений или антропогенного пресса.
ЛИТЕРАТУРА
Агеева Е. А., Гричан Ю. В., Оводов Н. Д. Плейстоценовая гиена в каменной ло-
вушке.— Природа, 1978, № 12, с. 114—115.
Алексеева Л. И. Териофауна раннего антропогена Восточной Европы. — Тр. Геол,
ин-та АН СССР, 1977, вып. 300, 214 с.
Барышников Г. Ф. Териофауна и позднеплейстоценовые ландшафты Горного При-
кубанья.— Вести. ЛГУ, 1979, № 12, с. 53—62.
Барышников Г Ф. Отряд Artiodactyla Owen, 1848 — Парнопалые. — В кн.: Каталог
млекопитающих СССР (плиоцен — современность). М., Наука, 1981, с. 343—408.
Барышников Г Ф., Дедкова И. И. Пещерные медведи Большого Кавказа. — Тр.
Зоол. ин-та АН СССР, 1978, т. 75, с. 69—77.
Бибиков С. Н. Опыт палеоэкономического моделирования в археологии. — В кн.: Тез.
докл. Всесоюзн. сессии, посвящ. итогам археол. и этнограф, исслед. Кишинев,
1966.
Бибиков С. Н. Древнейший музыкальный комплекс из костей мамонта. Киев, Нау-
кова думка, 1981, 108 с.
Бибикова В. И. Фауна раннетрипольского поселения Лука-Врублевицкая. — Матер,
и иссл. по археол. СССР, 1953, № 38, с. 411—458.
Бибикова В. И, Костные остатки льва из энеолитических поселений северо-западного
Причерноморья. — Вести, зоологии, 1973, № 1, с. 57—63.
Бибикова В. И., Белан Н. Г Локальные варианты и группировки позднепалеолити-
ческого териокомплекса юго-восточной Европы. — Бюл. Моск, о-ва испыт. при-
роды, отд. биол., 1979, т. 84, вып. 3, с. 3—14.
Брандт Ф. Ф. Об ископаемом носороге. — Натуралист, 1865, № 15, с. 1—8.
Будыко М. И. О причинах вымирания некоторых животных в конце плейстоцена.—
Изв. АН СССР, сер. геогр., 1967, № 2, с. 28—36.
Величко А. А. Природный процесс в плейстоцене. М., Наука, 1973, 256 с.
Величко А. А. Основные особенности последнего климатического макроцикла и со-
временное состояние природной среды. — В кн.: Палеогеография Европы за по-
следние сто тысяч лет (атлас-монография). М., Наука, 1982, с. 131—143.
Верещагин Н. К. Млекопитающие Кавказа. История формирования фауны. М. — Л.,
изд-во АН СССР, 1959, 703 с.
Верещагин Н. К Пещерный лев и его история в Голарктике и в пределах СССР. —
Тр. Зоол. ин-та АН СССР, 1971, т. 49, с. 123—199.
Верещагин Н. К. Охоты первобытного человека и вымирание плейстоценовых млеко-
питающих в СССР —Тр. Зоол. ин-та АН СССР, 1971, т. 49, с. 200—232.
Верещагин И. К. О происхождении мамонтовых «кладбищ». — Природная обстановка
и фауна прошлого, 1972, вып. 6, Киев, с. 131—148.
Верещагин Н. К. Почему вымерли мамонты. Л., Наука, 1979, 196 с.
Верещагин Н. К. Новая гыданская (юрибейская) находка мамонта. — Вести, зооло-
гии, 1982, № 3, с. 32—38.
Верещагин И. К. Кизеловская пещера — ловушка зверей на Среднем Урале. — Тр..
Зоол. ин-та АН СССР, 1982, т. 111, с. 37—44.
Верещагин Н. К., Барышников Г Ф. Остатки млекопитающих в восточной галерее*
пещеры Кударо I. — В. кн.: Кударские пещерные палеолитические стоянки в Юго-
Осетии. М., Наука, 1980, с. 51—62.
Верещагин Н. К., Бу рчак-Абрамович Н. О. Рисунки на скалах юго-восточного Каб-
ристана. — Изв. Гос. геогр. о-ва, 1948, т. 80, № 5, с. 507—518.
Верещагин Н. К., Кузьмина И. Е. Остатки млекопитающих нз палеолитических стоя-
нок на Дону и верхней Десне. — Тр. Зоол. ин-та АН СССР, 1977, т. 72, с. 77—
ПО.
Верещагин Н. К, Русаков О. Н. Копытные северо- запада СССР. Л., Наука, 1979,
309 с.
Габуния Л. К. Вымирание древних рептилий и млекопитающих. Тбилиси, Мецние-
реба, 1969, 236 с.
Гарутт В. Е. Отряд Proboscidea liliger, 1811. — Хоботные. — В кн.: Каталог млеко-
питающих СССР (плиоцен — современность). Л., Наука, 1981, с. 304—318.
Гептнер В. Г Насимович А. А., Банников А. Г Млекопитающие Советского Союза.
3* 35-
Т. 1, Парнокопытные и непарнокопытные, М., Высшая школа, 1961, 776 с.
Гептнер В. Г., Слудский А. А. Млекопитающие Советского Союза. Т. 2, ч. 2, Хищные
(гиены и кошки). М., Высшая школа, 1972, 552 с.
Гернет Е. С. Ледяные лишаи. М., Наука, 1981, 145 с.
Граве Г. Л. Речной бобр в пределах СССР и его хозяйственное значение. — Тр. по
лесн. опытн. делу, 1931, т. 14, с. 76—144.
Гричук В. П. Растительность Европы в позднем плейстоцене. — В кн.: Палеогеогра-
фия Европы за последние сто тысяч лет (атлас-монография). М., Наука, 1982,
с. 92—109.
Г ромов В. И. Палеонтологическое и археологическое обоснование стратиграфии кон-
тинентальных отложений четвертичного периода на территории СССР (млеко-
питающие, палеолит). — Тр. Ин-та геол, наук, 1948, т. 64, геол, сер., № 17, 520с.
Громова Вера. Новые материалы по четвертичной фауне Поволжья и по истории
млекопитающих Восточной Европы и Северной Азии вообще. — Тр. Комисе, по
изуч. четв. периода, 1932, вып. 2, с. 69—180.
Громова Вера. Краткий обзор четвертичных млекопитающих Европы (опыт сопостав-
ления). М., Наука, 1965, 144 с.
Грунин К. Я. Носоглоточные овода (Oestridae). — Фауна СССР Насекомые двукры-
лые, т. 19, вып. 3, М., изд-во АН СССР, 1957, 147 с.
Грунин К. Я. Первая находка личинок желудочного овода мамонта — Cobboldia
(Mamontia, subgen. n./russanovi, sp. n./Diptera, Gasterophilidae). — Энтомол. обозр.,
1973, т. 52, вып. 1, с. 228—233.
Давид А. И. Териофауна плейстоцена Молдавии. Кишинев, Штиинца, 1980, 188 с.
Давиташвили Л. Ш. Причины вымирания организмов. М., Наука, 1969, 440 с.
Дайсон Д. Л. В мире льда. Л., Гидрометеоиздат, 1966, 230 с.
Диков Н. Н. Наскальные загадки древней Чукотки. М., Наука, 1971, 128 с.
Дуброво И. А. Слоны рода Archidiskodon на территории СССР — Палеонтол. жури.,
1964, № 3, с. 82—94.
Дэвлет М. А. Петроглифы Мугур-Саргола. М., Наука, 1980, 269 с.
Ермолова Н. М. Териофауна долины Ангары в позднем антропогене. Новосибирск,
Наука, 1978, 223 с.
Ефремов И. А. Тафономия и геологическая летопись. 1. — Тр. Палеонтол. ин-та АН
СССР, 1950, т. 24, 176 с.
Животченко В. П. Амурский тигр. Успехи в восстановлении вида. — Природа, 1981,
№ 1, с. 91—97.
Квасов Д. Д. Увеличение влажности климата на границе плейстоцена и голоцена —
одна из причин вымирания мамонтов. — Тр. Зоол. ин-та АН СССР, 1977, т. 73,
с. 71—77.
Кириков С. В. Изменения животного мира в природных зонах СССР 1. Степная
зона и лесостепь. М., изд-во АН СССР, 1959, 174 с.
Кириков С. В. Изменения животного мира в природных зонах СССР 2. Лесная
зона и лесотундра. М., изд-во АН СССР, 1960, 155 с.
Корниец Н. Л. Про причини вымирания мамонта на территории Украши. Киев,
Наукова думка, 1962, с. 93—168.
Кузьмина И. Е. Формирование териофауны Северного Урала в позднем антропо-
гене.— Тр. Зоол. ин-та АН СССР, 1971, т. 49, с. 44—122.
Линдберг Г. У. Крупные колебания уровня океана в четвертичный период. Л., Наука,
1972, 447 с.
Мариковский П. И. Способы и объекты охоты по мотивам наскальных рисунков Чу-
лакских гор (Казахская ССР). — Зоол. журн., 1953, т. 32, № 6, с. 1064—1074.
Миддендорф А. Путешествие на север и восток Сибири, ч. I, СПб., 1860, 490 с.
Моуэт Ф. Люди оленьего края (пер. с англ.). М., Изд-во иностр, лит., 1963, 313 с.
Оводов Н. Д. Пещерные местонахождения остатков млекопитающих Сибири и Даль-
него Востока (краткий обзор). — В кн.: Край Дальнего Востока и Сибири. Вла-
дивосток, 1980, с. 154—163.
Окладников А. Н. Шишкинские писаницы. Иркутск, Иркутск, книжн. изд., 1959, 209 с.
Орлова Л. А. Радиоуглеродный возраст ископаемых остатков мамонта на террито-
рии СССР. — Изв. Сиб. отд. АН СССР, сер. обществ, наук, 1979, № 6/2, с. 89—97
Паавер К. Л. Формирование териофауны и изменчивость млекопитающих Прибалтики
в голоцене. Тарту, изд-во АН Эстонской ССР, 1965, 494 с.
Павлова М. В. Причины вымирания животных в прошедшие геологические эпохи—
В кн.: Современные проблемы естествознания. М. — Пгр., Госиздат, 1924, с. 5—88.
Пидопличко И. Г. О ледниковом периоде. 2. Киев, изд-во АН УССР, 1951, 262 с.
Пидопличко И. Г Новый метод определения геологического возраста ископаемых
Костей четвертичной системы. Киев, изд-во АН УССР, 1951, 89 с.
Пидопличко И. Г Позднепалеолитические жилища из костей мамонта на Украине.
Киев, Наукова Думка, 1969, 162 с.
36
Рашид-Ад Дин. Сборник летописей (пер. с персид.). Т. 3, М.— Л., изд-во АН СССР
1946, 340 с.
Рябинин А. Е. Ископаемые львы Урала и Поволжья. — Тр. Геол, ком., 1919, т. 168,
24 с.
Савватеев Ю. А. Залавруга. Археологические памятники низовья р. Выг. I. Петро-
глифы. Л., Наука, 1970, 441 с.
Семенов С. А. Развитие первобытной техники в каменном веке. Л., Наука, 1968,
359 с.
Серебровский П. В. История животного мира СССР Л., Леноблиздат, 1935, 127 с.
Скалой В. Н. Речные бобры Северной Азии. — Матер, к позн. фауны и флоры СССР,
нов. сер., отд. зоол., 1951, вып. 25, 208 с.
Слудский А. А. Джуты в евразийских степях и пустынях. — Тр. Ин-та зоол. АН Ка-
захССР, т. 20, с. 1—88.
Соколов В. Е. О прошлых ареалах копытных Казахстана по наскальным изображе-
ниям.— Бюл. Моск, о-ва испыт. природы, 1964, т. 69, вып. 1, с. 113—117.
Татаринов К. А. Фауна позвоночных стоянки Кормань IV В кн.: Многослойная
палеолитическая стоянка Кормань IV. М., Наука, 1977, с. 112—118.
Татищев В. Н., Гмелин И. Г О костях, которые из земли выкапываются. — Истор.,
генеал. и географ, прим, в Ведомостях, 1730, № 80—83 и 88—93; с. 319—336,
363—372.
Тимченко Н. Г. К истории охоты и животноводства в Киевской Руси. Киев, Наукова
думка, 1972, 204 с.
Толлъ Э. В. Ископаемые ледники Новосибирских островов и их отношение к трупам
мамонтов и к ледниковому периоду. — Зап. Русск. географ, о-ва, 1897, т. 32, № 1,
139 с.
Томирдиаро С. В. Изменение физико-географической обстановки на равнинах северо-
востока Азии на границе плейстоцен — голоцен как основная причина вымирания
териофауны мамонтового комплекса. — Тр. Зоол. ин-та АН СССР, 1977, т. 73,
с. 64—72.
Томирдиаро С. В. Лёссово-ледовая формация Восточной Сибири в позднем плейсто-
цене и голоцене. М., Наука, 1980, 184 с.
Тугаринов А. Я. О происхождении арктической фауны. — Природа, 1929, № 7—8,
с. 653—680.
Тугаринов А. Я. Опыт истории арктической фауны Евразии. — Тр. II конф. Ассоц.
по изуч. четвертичн. пер., 1934, вып. 5, с. 55—65.
Формозов А. А. Очерки по первобытному искусству. М., Наука, 1969, 253 с.
Формозов А. Н. К вопросу о вымирании некоторых степных грызунов в позднечет-
вертичное и историческое время. — Зоол. журн., 1938, т. 17, вып. 2, с. 260—272.
Флеров К. К. Систематика и эволюция. — В кн.: Зубр. Морфология, систематика,
эволюция, экология. М., Наука, 1979, с. 9—127.
Цалкин В. И. К истории животноводства и охоты в Восточной Европе. — Матер, и
исслед. по археол. СССР, 1962, № 104, М., 130 с.
Черский И. Описание коллекций послетретичных млекопитающих животных, собран-
ных новосибирскою экспедициею 1885—1886 гг. — Прил. к 65 тому «Записок
Акад, наук», 1891, т. 1, 706 с.
Шер А. В. Млекопитающие и стратиграфия плейстоцена крайнего северо-востока
СССР и Северной Америки. М., Наука, 1971, 312 с.
Berglund В. Е., Hakansson S. and Lagerlund Е. Radiocarbondated mammoth (Mam-
muthus primigenius Blumenbach) finds in South Sweden. — Boreas, 1976, vol. 5,
p. 177—191.
Bonfiglio L. Resti di Cervide (Megaceros) dell’Eutireeniano di Bovetto (RC). — Quater-
naria, 1978, t. 20. p. 87—108.
Borgen U. Ett fynd av fossil myskoxe i Jamtland och nagot om myskoxarnas biologi
och historia. — Fauna och flora (Sver.), 1979, vol. 74, N 1, p. 1—12.
Bouchud J. Le Cervus megaceros dans la sud et sud-ouest de la France. — Israel J.
Zool., 1965, vol. 14, N 1/4, p. 24—37.
Digby B. The Mammoth and Mammoth-Hunting in North-East Siberia. London, H. F.
a. G. Witherby, 1926. 224 p.
Erdbrink D. P. A review of fossil and recent bears of the Old World, with remarks
on their phylogeny based upon their dentition. Proefschrift, Deventer, 1953. 597 p.
Estevez J. Primer hallazgo del buey almizclado (Ovibos moschatus Zimmermann) en
el pleistocene peninsular. — Acta geol. hisp., 1978 (1979), vol. 13, N 2, p. 59—60.
Goldfuss G. Osteologische Beitrage zur Kenntnis verschiedener Saugethiere der Vor-
welt. IV Uber den Schadel des Hohlenlowen. — Nova Acta Phys. Acad. Caesarea
Leopold. Carol., Nat. Currosorum. Halle, 1821, Bd 10, H. 2, S. 489—494.
Guenther E. W. Funde des Riesenhirsches in Schleswig-Holstein und ihre zeitliche
Einordnung. — Steinzeitfragen der Alten und neuen Welt. Zotz-Festschrift. Bonn,
1960, S. 201—206.
37
Hemmer H. Fossilbelege zur Verbreitung und Artgeschichte des Lowen, Panthera leo
(Linne, 1758), mit Gabbild. — Saugethierkundliche Mitteilungen, 15, S. 289—300.
Hilzheimer M. Dritter Beitrage zur Kenntnis der Bisonten. — Arch. Naturgesch., Abt. A.,
1918, Bd 84, H. 6, S. 44—87.
Howorth H. The mammoth and the flood. London, Sampson Low, Marston. Searle, Ri-
vington, 1887, 464 p.
Kahlke H. D. Ovibos aus den Kiesen von Siissenborn. Ein Beitrag zur Systematik und
Phylogenie der Ovibovinen und zur Stratigraphie des Pleistozans. — Geologie, 1963,
Jg. 12, N 8, S. 942—972.
Krause H. The mammoth — in Ice and Snow? Stuttgart, 1977. 107 p.
Kuhn H. The Rock Pictures of Europe. London. 1956, 266 p.
Kurten B. Life and Death of the pleistocene Cave Bear. — Acta zool. Fennica, 1958,
t. 95, p. 1—59.
КиНёп В. Pleistocene mammals of Europe. The World Naturalist. London, Weidenfeld
and Nicolson, 1968, VIII+317 p.
Kurten B. The Cave Bear Story. New York, Columbia Univ. Press, 1976, 163 p.
Kurten B., Anderson E. Pleistocene mammals of North America. New York, Columbia
University Press, 1980, 443 p.
Leidy Y. Description of an extinct species of American lion: Felis atrox. — Trans. Amer.
Phil. Soc., 1853, vol. 5, N 10, p. 319—321.
Lindner К Die Jagd der Vorzeit. Bd. 1. Berlin und Leipzig, 1937. 435 p.
Mitchell G. F., Parkes H. M. The Giant Deer in Ireland. — Proc. Royal Irish. Acad.,
1949, vol. 52, sect. B., N 7, p. 291—314.
Pallas P. S. De reliquiis animalium exoticorum per Asian borealem repertis complemen-
tum. — Nov. Comm. Acad. Sci. Petrop., 1773, t. 17, p. 576—606.
Pfizenmayer N. W. Mammutleichen und Urweltmenschen in Nordost Sibirien. Leipzig,
1926, 341 S.
Ryzievicz Z. Systematic place of the fossil musk-ox from the Eurasien diluvium. — Prace
Wroclawsk. towar. naukow., ser. B., 1955, N. 49, 73 S.
Sickenberg O. Die Saugetierreste aus den elsterzeitlichen Kiesen (Pleistozan) von Born-
hausen am Harz. — Geol. Jahrb., 1962, Bd. 79, S. 707—736.
Soergel W. Die Jagd der Vorzeit. Jena, 1922, 149 S.
Soergel W. Hyaena brevirostris Aymard und Hyaena ex aff. crocuta Erxl. aus den Kie-
sen von Siissenborn. — Z. Deutsch. Geol. Ges.. 1936, Bd 88, S. 525—539.
Soergel W. Die Verbreitung des diluvialen Moschusochsen in Mitteleuropa. — Beitr.
Geol. Thiiringen, 1942, Bd 7, S. 75—95.
Thenius E. Grundziige der Faunen- und Verbreitungsgeschichte des Saugetiere. Eine
historische Tiergeographie. Jena, VEB Gustav Fischer Verlag, 1980, 375 S.
Tolmachoff J. P. The carcasses of the mammoth and rhinoceros found in the frozen
ground of Siberia. — Trans. Amer. Philos. Soc. Nov. Ser., vol. 23, part 1, 1929,
p. 1—74.
SUMMARY
Possible causes of destruction of mammoth fauna and extinction of some of its
representatives are discussed. Regarded as a significant factor in this process are va-
riations of species habitat due to paleolandscape and paleoclimate changes on the
boundary of Pleistocene and Holocene. The influence of exterminatory human activity
was high in the few regions only and became decisive only during historical time. Other
factors, i. e. extreme specialization, inadaptive evolution etc., were of secondary impor-
tance.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1985, ТОМ 131
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, 1985, VOL. 131
USSR ACADEMY OF SCIENCES
ВРЕМЯ И ПУТИ РАССЕЛЕНИЯ ВЕРБЛЮДОВ
В ВОСТОЧНОМ ПОЛУШАРИИ
Л. И. Алексеева
Геологический институт АН СССР, Москва
L. /. Alexeeva Period and routs of migration of camels in eastern hemisphere
Вымершие мозоленогие, остатки которых известны из многих позд-
некайнозойских местонахождений Евразии и Африки, являются ими-
грантамн в Старый свет. Самые древние находки остатков верблюдов
встречены на территории юга Восточной Европы в отложениях раннего
понта. В свете современных хроностратиграфических представлений
комплекс фауны млекопитающих южнорусского понта примерно отве-
чает интервалу ниже 5 млн. лет (Aguirre и др., 1974, с. 162). Это по-
зволяет утверждать, что первое появление верблюдов в Евразии про-
изошло в предвиллафранкское время. Следует отметить, что пока не
очень четко установлено, что считать нижним виллафранком и какой
рубеж принять за его нижнюю границу. Несмотря на то, что имеются
некоторые расхождения относительно даты начала виллафранка, можно
видеть, что его основание лежит где-то около 4 млн. лет. Так, во Фран-
ции большинством исследователей нижняя граница проводится между
отложениями с фауной Этуэр и отложениями с фауной Перпиньяна
и Монпелье (Heintz и Piveteau, 1968), т. е. это примерно на уровне
3,5 млн. лет (Бут, 1972). Абсолютные датировки, полученные в резуль-
тате изучения континентальных отложений долины Омо, позволяет по-
местить основание виллафранка в Африке на рубеж 4 млн. лет (Агат-
bourg, 1959). В коллективной работе итальянских исследователей
(Ambrosetti, Azzaroli, Bonadonna и Follieri, 1972) граница нижнего
виллафранка опущена даже немного ниже 4 млн. лет.
Появившись на территории юга Восточной Европы в понте, вер-
блюды прочно вошли в состав фауны зоны северного Причерноморья
и продолжали там обитать в течение всего эоплейстоцена (время су-
ществования фауны руссильонско — виллафранкского типа). Пока
точно не установлено, к какому роду относятся костные остатки самых
древних мозоленогих Евразии. Высказаны предположения (Свистун,
1971, Аубекерова, 1974), что они принадлежат роду Gigantocamelus,
известному до сих пор только на территории Северной Америки. Что
касается верблюдов эоплейстоцена, то все их остатки относятся к роду
Paracarnelus, получившему в раннечетвертичное время широчайшее
распространение. Ареал рода Paracarnelus протягивался от Китая и
Монголии на запад, захватывая Южную Сибирь, Казахстан, Среднюю
Азию и Восточную Европу с прилегащими районами. Западные точки
ареала зафиксированы на территории Румынии и Венгрии, самые юж-
ные— в зоне Предгималайского прогиба (Индия и Пакистан). На тер-
ритории СССР род Paracarnelus был представлен 4 видами (alexejevi,
kujalnensis, praebactrianus, gigas). Единичные находки последних вер-
блюдов этого рода встречены в отложениях времени существования
тираспольского комплекса (нижний плейстоцен). В следующем, син-
39
гильском комплексе (начало среднего плейстоцена) на территории Евро-
пейской части СССР появляются верблюды рода Cainelus (Алексеева,.
1974). Окончательное исчезновение верблюдов с территории Восточной
Европы произошло в начале периода развития максимального оледе-
нения, т. е. в период существования хазарского фаунистического ком-
плекса.
Самые древние находки верблюдов на африканском континенте
известны в Тунисе, в местонахождении Ишкёль. По мнению ряда
исследователей, это местонахождение является нижневиллафранкским
(Arambourg, 1969; Coppens, 1971). Согласно данным Ж. Жеже (Jaeger,
1971), фауна Ишкёль отвечает несколько более древней эпохе, пред-
шествуя фауне таких местонахождений, как Ним, Сэт и Руссильон
( = Серра-ан-Ваке). Это утверждение дает право предположить, чта
время проникновения верблюдов на территорию Северной Африки
уходит в предвиллафранкскую эпоху. Местонахождения остатков вер-
блюдов виллафранкского времени известны в более восточных райо-
нах, прилегающих к Северной Африке. Camelus sp. указан в составе
фауны местонахождения Убедийя в долине Иордана (Haas, 1963).
В последние годы появились сведения о присутствии верблюдов в внл-
лафранке Турции. В краткой заметке О. Зикенберга и X. Тобина (Si-
ckenberg u. Tobien, 1971) указаны Camelidae, найденные там на терри-
тории Анатолии в местонахождениях фауны виллафранкского типа
вместе с последними гиппарионами, с жирафами рода Macedonotherium,
с быками рода Leptobos, с оленями, газелями и другими представите-
лями развитой виллафранкской фауны.
Самые южные находки остатков мозоленогих на африканском:
континенте известны из района оз. Чад (Gautier, 1966) и на террито-
рии Эфиопии в долине Омо (Coppens и Howell, 1974). Согласно на-
ходкам в Омо, появление верблюдов в Центральной Африке фикси-
руется интервалом 1,8—1,3 млн. лет (верхний виллафранк средиземно-
морской зоны).
Интересно отметить, что просмотр списков фауны млекопитающих
североафриканских местонахождений показал, что верблюды там не
обитали в это время. Их вторичное появление в Северной Африке от-
мечается только в среднем плейстоцене (450—300 тыс. лет, миндель и
миндель — рисе). Остатки верблюдов этого времени встречены в Ал-
жире, в фауне Тернифина. Затем верблюд найден в атере Северного
Судана (Gautier, 1966). К сожалению, пока не выяснен вопрос о род-
ственных связях древних форм африканских верблюдов. Л. Готье
(1966), основываясь на абсолютных размерах остатков, отнес северо-
суданскую форму верблюда к Camelus thomasi, описанному из ашель-
ских слоев Алжира. Кроме того, этим же исследователем было уста-
новлено, что северосуданский верблюд родствен верблюдам-бактринам.
На этом основании вероятнее всего построено его утверждение, что вер-
блюд из района оз. Чад близ Коро-Торо также родствен С. thomasi
и, следовательно, бактриану. Однако исследователи фауны Омо (Cop-
pens и Howell, 1974) проявили осторожность и указали в списке фауны
центральноафриканскую форму верблюда только как Camelus sp. Не
исключено, что виллафранкский верблюд Африки был представлен ро-
дом Paracamelus, широко распространенным в это время в Евразии,
а верблюды рода Camelus попали в Африку в поствиллафранкское
время где-то в минделе или миндель — риссе. Р. Кок (Coque, 1962)
связывает с поствиллафранкской эпохой появление в Африке целого-
ряда евразийских иммигрантов. Материалы местонахождения Аин-эль-
Геттер (Тунис) позволяют видеть, что в это время туда проникли быки.
40
рода Bos, Dicerorhinus mercki и Camelus dromedarius, и, как уже от-
мечалось выше, именно с поствиллафранкским временем связано по-
явление верблюдов рода Camelus на территории Восточной Европы.
Итак, суммируя имеющиеся данные по местонахождениям остат-
ков мозоленогих в Старом свете, можно предположить, что миграция
этой группы имела две волны. Первая проявилась в предвиллафранк-
ское время. Затем, широко расселившись, эта группа животных на
протяжении всего виллафранка занимала одно из доминирующих мест
в составе фаун виллафранкского типа. Все остатки верблюдов из евра-
зийских местонахождений руссильонской и виллафранкской фаун при-
надлежат роду Paracamelus, поэтому есть некоторое основание пола-
гать, что африканские находки верблюдов виллафранкского времени
тоже принадлежат роду Paracamelus. Вторая волна миграции мозоле-
ногих, обеспечившая широкое распространение верблюдов рода Came-
lus, связана с поствиллафранкским временем. До сих пор степень
родственных взаимоотношений родов Paracamelus и Camelus точно не
установлена. По мнению Я. И. Хавесона (1954), род Camelus близко-
родствен Р. gigas, основная площадь ареала которого связана с азиат-
ской частью Старого света. Вероятнее всего формирование рода Ca-
melus произошло на территории Азии. Оттуда верблюды этого рода
распространились в поствиллафранкское время на запад до Восточной
Европы и проникли также через Закавказье и Ближний Восток на тер-
риторию Северной Африки.
ЛИТЕРАТУРА
Алексеева Л. И. Верблюды Северного Причерноморья и Предкавказья. — В кн.: Мле-
копитающие позднего кайнозоя юго-запада СССР Кишинев, Штиинца, 1974,
с. 53—77.
Аубекерова П. А. Новый гигантский верблюд из плиоценовых отложений Текесской
впадины. — Матер, по истории фауны и флоры Казахстана, т. VI, изд-во Наука
КазССР, 1974, с. 77—82.
Бут П. Абсолютный возраст вулканогенных формаций Оверни и Веле и хронология
четвертичной фауны млекопитающих Европы. — В кн.: Геология и фауна ниж-
него и среднего плейстоцена Европы. М., Наука, 1972, с. 7—24.
Свистун В. И. Новые находки остатков верблюдов (Tylopoda, Camelidae) в отложе-
ниях понта юга Европейской части СССР. — Вест, зоологии, 1971, № 1, с. 64—68.
Хавесон Я. И. Третичные верблюды Восточного полушария (род Paracamelus). — Тр.
Палеонт. ин-та АН СССР, 1954, т. XVII, вып. 2, с. 100—162.
Aguirre Е., Т Alberdi, L. Thaler, N. Lopes et R. Bustos. Murcia — Granada. Libro-Guida
Coloq. Intern, sombre biostrat. cont. neogeno sup. у cuatern. inf., Madrid, 1974,
168 p.
Ambrosetti P.} A. Azzaroli, F. F. Bonadonna et M. Follieri. A scheme of pleistocene
chronology for the tyrrhenian side of Central Italy. — Boll. Soc. Geol. It., 1972,
t. 91, p. 169—184.
Arambourg C. Les correlations paleontologiques et chronologiques entre le Pleistocene
inferieur de 1’Europe et de 1’Afrique. — C. R. Soc. Geol. Fr., 1961, N 3, p. 92—93.
Coppens Y. Les Vertebres villafranchiens de Tunisie: gisements nouveaux significa-
tion.— C. R. Acad. Sci. Paris, 1971, t. 278, Ser. D, p. 51—54.
Coppens Y. a. F. C. Howell. Les Faunes de Mammiferes fossiles des formations plio-
pleistocenes de 1’Omo en Ethiopie (Proboscidea, Perissodactyla, Artiodactyla).—
C. R. Acad. Sc. Paris, 1974, t. 278, Ser. D, p. 2275—2278.
Coque R. La Tunisie presaharienne (Etude geomorphologiques), Paris, 1962, 475 p.
Gautier A. Camelus thomasi from the Northern Sudan and its bearing on the relation-
ship C. thomasi — C. bactrianus. — Journ. Paleont., 1966, vol. 40, N 6, p. 1368—1382.
Haas G. Preliminary remarks on the early Quaternary faunal Assamblage from Ubei-
diya, Jordan valley. — South Afr. Journ. Sci., 1963, vol. 59, N 3, p. 73—76.
Heintz E. et J. Piveteau. Principaux resultats systematiques et biostratigraphiques de
1’etude des Cervides villafranchiens de France et d’Espagne. — C. R. Acad. Sci.
Paris, 1968, t. 266, Ser. D, p. 2184—2186.
Jaeger J.-J. Les Micromammiferes du “Villafranchien” inferieur du lac Ichkeul (Tuni-
sia): donnes stratigraphiques et biostratigraphiques nouvelles. — C. R. Acad. Sci.
Paris, 1971, t. 273, Ser. D, p. 562—565.
Sickenberg О. a. H. Tobien. New Neogene and Lower Quaternary vertebrate faunas
in Turkey. — News!. Stratigr., Leiden, 1971, vol. 1, N 3, p. 51—61.
SUMMARY
There were two stages in distribution of Tylopoda in the Old World. The first
occured in previllafranchien time. All remains of camels from Eurasian localities of russi-
lion and villafrank fauna belonged to genus Paracarnelus. The second wave of migra-
tion is related to a wide distribution of camelys of genus Camelus. The exact relation-
ship between genera Paracarnelus and Camelus has not been established et. Genus
Camelus is most likely to have been formed on the territory of Asia, from where they
spread westwards up to Eastern Europe and penetrated also through Transcaucasia
and Near East into North Africa.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1985, ТОМ 131
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, 1985, VOL. 131
USSR ACADEMY OF SCIENCES
САРЫ-ЛЕГЕТ—НОВАЯ МЕЗОЛИТИЧЕСКАЯ СТОЯНКА
В ЮГО-ОСЕТИИ
Г. Ф. Барышников, Д. А. Чистяков
Зоологический институт АН СССР, Институт археологии АН СССР (ЛО), Ленинград
G. F. Baryshnikov, D. А. Т Chistyakov. Sary-Leget — a new Mesolitic site in southern
Osetia (Caucasus)
В 1977 г. Кавказской палеолитической экспедицией Ленинградского
отделения Института археологии АН СССР (руководитель В. П. Лю-
,бин) была обнаружена новая мезолитическая пещерная стоянка Сары-
Легет. Как и другие известные юго-осетинские пещерные памятники
каменного века (Кударо 1—5, Цона, Шагат-Хох-Легет), она находится
в северо-западной, колхидской части Джавского района Юго-Осетин-
ской АО Грузинской ССР. Географически — это участок южного склона
центральной части Большого Кавказа, расположенный на востоке
Нижне-Рачинской межгорной впадины и обрамленный многочислен-
ными хребтами и их отрогами (Любин, 1977).
Пещера лежит на правом борту глубокого (свыше 300 м) и узкого
ущелья р. Грамулы, левого притока Джорджори (бассейн р. Риони).
Сама речка имеет ширину всего около 3 м, ширина же поймы с отме-
лями достигает 50 м. Ущелье сложено известняками, порфиритами
и сланцами. Близ устья реки, над долиной выступают огромные скалы
известняков, представляющие собой по мнению Н. Б. Вассоевича «экзо-
тические глыбы турона, появившиеся тут в результате надвига» (Пра-
солов, Соколов, 1931, с. 403—404). В одной из этих скал (в невысоком,
пятидесятиметровом, отдельно стоящем, сильно изъеденном карстом
известняковом останце) и расположена пещера Сары-Легет* (рис. 1,
вклейка). Она находится в 250 м от шоссейной дороги Цхинвали — Они
и в 4,5 км на юго-восток от кударского пещерного комплекса. Ее отно-
сительная высота 15 м, абсолютная — около 1540—1560 м над уровнем
моря.
Пещера (общая площадь около 30 м2) состоит из двух камер (за-
падной — 8 м2 и восточной — 16 м2), соединенных коридором, ширина
которого 1.2 м, длина 3.6, высота 1.8. Она имеет два входа и одно, не-
доступное для человека, отверстие-лаз с восточной стороны скалы
(рис. 2). В настоящее время попасть в пещеру можно только через
аркообразный вход в восточную камеру (ширина входа 2.3 м, высота
1.6 м). Перед ним имеется небольшая предвходовая площадка (6.5 м2),
р двух сторон ограниченная обрывом к реке. Экспозиция входов —
юго-западная.
Большая часть пещеры (около 20 м2) покрыта глинистыми отло-
жениями; на небольшом участке (6 м2) западной камеры обнажен
скальный пол. Наиболее высокий участок скального дна выступает
в центральной части этой камеры; отсюда он плавно понижается в сто-
* В буквальном переводе с осетинского «Сары-Легет» означает «пещера в го-
лове». Пещера получила такое название потому, что скала напоминает по очерта-
ниям голову барана.
43
рону западной стены пещеры и в сторону соединительного коридора.
Несмотря на то, что стены, потолок и, судя по обнаженному
участку, скальное дно очень сильно затронуты карстовыми процессами,
пещера относительно сухая и удобная для кратковременного пребыва-
ния небольших групп людей.
В августе 1977 г. в восточной камере, в 1.5 м от капельной линии,
на участке, примыкающем к соединительному коридору (рис. 2), был
заложен разведывательный шурф 1.6Х1.3м, который выявил следую-
щую стратиграфию пещерных слоев* (рис. 3).
Слой 1. Сильно гумусированный, рыхлый темно-коричневый су-
глинок с незначительным количеством мелкого (размером до 3—4 см)
щебня. Встречены угли и малочисленные нефоссилизованные кости
животных. Мощность слоя 0.04—0.12 м. Контакт с нижележащим слоем
четкий.
1— >иурф 1977 г., 2 — капельная линия, 3 — сквозные трещины в стенах и
узкие лазы, 4 — обрывистый край предвходовой площадки.
Слой 2. Светло-коричневый суглинок, насыщен более крупным
(до 5 см) угловатым щебнем. На поперечном (Л — В) разрезе и при-
мыкающем к нему северном участке разреза В — С зафиксировано
большое количество угловатых крупных обломков известняка и, преиму-
щественно разбитых, крупных галек и валунов. Целые валуны дости-
гают размеров 29—37 см. Гальки и валуны принесены человеком в пе-
щеру с реки и, видимо, здесь же разбиты друг о друга. Анализ облом-
ков и осколков валунов не оставляет сомнений в том, что они не пред-
назначались и не были использованы в качестве орудий труда. Назна-
чение их и характер локализации совместно с крупными обломками
известняка в северо-западной и, главным образом, в западной части
шурфа установить без дополнительных раскопок не представляется воз-
можным.
* Характер отложений, зафиксированный на поперечном (Л — В) и продольном
(В — С) разрезах дается суммарно.
44
A
О 0.5M
i______________________i
Рис. 3. Разрез рыхлых четвертичных отложений пещеры Сары-Легет по линии А—В — В—С.
/ — гумусный слой 1, 2 — впускные ямы, 3 — приблизительная граница слоев 2 и 3 на разрезе В—С, 4 — скальный пол пещеры,
5 — кремневые пластинки, 6 — отщеп, 7 — номера слоев.
В верхах слоя, в 15—25 см от дневной поверхности современного*
пола пещеры, встречено шесть кремневых изделий мезолитического
облика. К этому же слою приурочены обломки костей крупных млеко-
питающих и большая часть обнаруженных в пещере остатков грызунов.
На поперечном разрезе А — В слой 2 частично залегает непосред-
ственно на скальном полу. Мощность слоя колеблется от 0.26 до 0.62 м.
Слой 3. Светло-серый (сизоватый) суглинок, плотный, с немного-
численным мелким (3—4 см) угловатым щебнем. Залегает непосред-
ственно на скальном полу пещеры. Остатков материальной культуры
не встречено. Мощность слоя до 0.30 м.
Слои 1 и 2 нарушены тремя поздними впускными ямами, одна
из которых—очажная — имеет диаметр 70 см, глубину 23 см. Назначе-
ние двух других непонятно, так как никаких культурных остатков, за
исключением древесных углей, в них не встречено.
Из отложений пещеры извлечено свыше 450 костных остатков по-
звоночных животных — млекопитающих, птиц и земноводных. Среди
млекопитающих определено 20 видов, преимущественно грызунов
(табл. 1). Обломки костей крупных форм немногочисленны, принадле-
Таблица 1
Видовой состав и число костных остатков позвоночных из пещеры Сары-Легет
Г о л о ц e н
Виды M e з о л ] я т Всего
1 2 3 Шурф
a 6 B
Млекопитающие — Mammalia 9 89 104 90 10 73 375
Кавказский крот — Talpa caucasica Satun. — 4 — 1 — 1 6
Землеройка-белозубка — Crocidura sp. — — — — — 1 1
Рукокрылые — Chiroptera indet. — — — 2 — 1 3
Заяц-русак — Lepus europaeus Pall. — 2 — — 1 3 6
Кавказский сурок — Marmota paleocaucasica — 8 4 — — 8 20
Baryschn. Лесная соня — Dryomys niiedula Pall. 1 1 2
Лесная мышь — Apodetnus sylvaticus L. — — — 1 — — 1
Серый хомячок — Cricetulus migratorius — — 1 — — — 1
Pall. Хомяк Радде — Mesocricetus conf, raddei 1 6 2 9
Nehr. Прометеева полевка — Prometheomys scha- 11 12 10 7 40
poschnikovi Satun. Водяная полевка — Arvicola terrestris L. — 1 1 2
Кустарниковая полевка — Pitymys majori — 1 1 — — — 2
Thom. Обыкновенная полевка—Mier ot us arvalis 3 3
L. s. 1. Гудаурская полевка — Chionomys gud 3 2 5
Satun. Полевковые — Microtinae indet. 8 56 72 58 8 40 242
Волк — Canis lupus L. — __ 1 1 — 2
Лисица — Vulpes vulpes L. — — 2 4 — 6 12
Барсук — Meles tneles L. — — 2 1 1 2 6
Благородный олень — Cervus elaphus L. 1 1 — 2 — — 4
Зубр — Bison bonasus L. — — 1 — — — 1
Серна — Rupicapra rupicapra L. — — 1 — — — 1
Кавказский козел — Capra caucasica Giild. — 1 4 — — — 5
et Pall.
Птицы — Aves — 3 9 2 — 3 17
Земноводные — Amphibia — 26 12 5 2 12. 57
46
жат промысловым видам: лисице, оленю, зубру, козлу. В основном
сохранились зубы, шейные позвонки и короткие кости дистальных отде-
лов конечностей. Интересна находка нижнечелюстной ветви сурка,
определенная как Marmota paleocaucasica (Барышников, 1980).
В видовом составе преобладают звери, обычные на южном склоне
Большого Кавказа. От мезолита до современной эпохи улавливаются
незначительные фаунистические изменения. Один вид — кавказский су-
рок— вымер полностью. Хомяк Радде встречается в настоящее время
значительно севернее, в Предкавказье и Дагестане, а обыкновенная
полевка — восточнее, не проникая в колхидскую часть Юго-Осетии
(Шидловский, 1951).
Экологический облик териофауны мезолитического слоя указывает
на климат более холодный, чем современный. Характерны обитатели
горных лугов верхней части лесного пояса: кавказский сурок, проме-
теева и гудаурская полевки, серна, кавказский козел. Косточки лесных
видов — лесной сони, лесной мыши, кустарниковой полевки — единичны.
В низах слоя 2 появляются формы открытых, остепненных ландшаф-
тов и альпийских лугов — серый хомячок, хомяк Радде, обыкновенная
полевка. Остатки хомяка Радде представлены лишь костями конечно-
стей, что затрудняет точное определение. По размеру они близки, од-
нако, к вымершему Mesocricetus raddei planicola Argyr., известному
из древнего и раннего голоцена пещер Кударо и Талынг-Легет. При-
сутствие водяной полевки и амфибий говорит о близости пещеры
к реке.
Таблица 2
Видовой состав териофауны Юго-Осетии в эпоху мезолита
Виды Сары- Легет Шагат- Хох-Легет Кударо I Кударо III Цона (по Бенду- кидзе, 1979)
Кавказский крот +
Заяц-русак + + + — +
Кавказский сурок + + + + +?
Лесная соня + — + — —
Малоазийский тушканчик — + — + —
Лесная мышь + — 4- + —
Серый хомячок + 4- — + —
Хомяк Радде +? + + + +?•
Прометеева полевка + 4- + + +
Водяная полевка + + — + —
Кустарниковая полевка + — + + —
Дагестанская полевка — — + — —
Обыкновенная полевка + 4- — + —
Гудаурская полевка + — + + ?
Полевка Роберта — — — + —
Волк + 4- — + 4-
Лисица + 4- 4- + 4-
Красный волк — — — + —
Бурый медведь — 4- — — —-
Пещерный медведь — + + —
Куница 4- + — 4-
Ласка — — + — ——
Барсук + 4- + — —
Благородный олень + 4- + + 4-
Зубр + 4- — + 4-
Серна + —— — — 4-
Кавказский козел + 4- + + 4-
47
Сопоставление териофауны Сары-Легет со списками видов из дру-
гих мезолитических памятников Юго-Осетии (табл. 2) показывает их
значительное сходство, говорящее, очевидно, о стратиграфической (вре-
менной) близости. Для всех стоянок этой эпохи характерны кавказский
сурок, хомяк Радде, прометеева и обыкновенная полевка. Отсутствие
в Сары-Легет остатков малоазийского тушканчика и пещерного мед-
ведя объясняется либо ее более молодым возрастом, либо чисто тафо-
номическими причинами.
Рис. 4. Мезолитические кремневые изделия из пещеры Сары-Легет.
Встреченные в слоях пещеры каменные изделия (рис. 4) — пять
призматических пластинок и небольшой отщеп — изготовлены из не
местного турон-сенонского (имеретинского?) кремня розового (2 экз.),
светло-серого (3 экз.) и голубоватого (1 экз.) цвета.*
Кремни не имеют намеренной вторичной обработки, за исключе-
нием одной розовой кремневой пластинки, у которой одним ударом
усечена ударная площадка и на правом крае у обломанного дисталь-
ного конца имеется небольшая выемка (рис. 4,4).
* При обследовании пещеры в 1980 г. на возвышающемся скальном полу
в центре западной камеры был найден еще один отщеп из светлого кремня (рис. 4,/).
48
Несмотря на немногочисленность и типологическую невыражен-
ность, кремневые изделия из пещеры Сары-Легет могут быть предва-
рительно соотнесены и синхронизированы с материалами из слоя 2
пещеры Кударо III (Любин, Левковская, 1972; Любин и др., 1978),
слоя 3 пещеры Цона (Каландадзе, 1965), слоя 4 пещеры Кведи (Га-
буния, Церетели, 1977) и, учитывая данные анализа териофауны этих
стоянок, датированы мезолитическим временем.*
Археологические и палеонтологические данные позволяют, по-види-
мому, рассматривать пещеру Сары-Легет как кратковременный лагерь
мезолитических охотников.
ЛИТЕРАТУРА
Барышников Г Ф. Сурок в палеолите Кавказа. — Тр. Зоол. ин-та АН СССР, 1980,
т. 93, с. 50—59.
Бендукидзе О. Г Голоценовая фауна позвоночных Грузии. Тбилиси, Мецниереба,
1979, 122 с.
Габуния М. К., Церетели Л. Д. Мезолит Грузии — Кр. сообщ. Ин-та археологии
АН СССР, 1977, № 149, с. 34—41.
Каландадзе А. Н. Цонская пещера и ее культура. — В кн.: Пещеры Грузии, Тбилиси,
1965, вып. 3, с. 32—36.
Любин В. П. Мустьерские культуры Кавказа. Л., Наука, 1977, 224 с.
Любин В. П., Левковская Г. М. Пещера Кударо III (Юго-Осетия).— Матер, и
исслед. по археол. СССР, 1972, № 185, с. 25—40.
Любин В. П., Селиванова Н. Б., Барышников Г Ф., Левковская Г М. Пещера Ку-
даро III. — В кн.: Археология и палеогеография раннего палеолита Крыма и
Кавказа. М., Наука, 1978, с. 87—95.
Прасолов Л. И., Соколов Н. Н. Почвенно-географический очерк Юго-Осетии. — Тр.
Совета по изуч. произвол, сил, сер. Закавказск., 1931, вып. 2, с. 341—465.
Шидловский М. В. Грызуны Юго-Осетии. — Тр. Зоол. ин-та АН ГССР, 1951, т. 10,
с. 187—222.
SUMMARY
Archaeological and paleontological materials from the new cave site Sary-Leget
(Dzhav Region of South-Osetian Autonomous Region of Georgian SSR) allow to re-
late them to and synchronize with materials of West Georgian mesolithic sites (Ku-
daro I, layer 2; Kudaro III, layer 2; Tsona, layer 3; Kvedi, layer 4). 20 species of
mammals have been identified including Caucasian marmot, Raddi hamster, fox, bison,
Caucasian goat. The new site represents a transitory hunting camp and can be dated
as mesolithic.
* Археологические материалы из пещеры Шагат Хох-Легет еще не изданы.
4 Млекопитающие
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1985, ТОМ 131
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, 1985, VOL. 131
USSR ACADEMY OF SCIENCES
ПОЗДНЕПЛЕЙСТОЦЕНОВАЯ ШИРОКОПАЛАЯ ЛОШАДЬ
БАССЕЙНА ДЕСНЫ
Н. Г. Белан
Институт зоологии АН УкрССР, Киев
N. G. Belan. Late Pleistocene wide-toed horse from Desna basin
В последние годы плейстоценовые лошади привлекли внимание-
ряда исследователей (Громова, 1949; Musil, 1962; Radulesco, Samson^
1962; Sikenberg, 1962; Nobis, 1971; Давид, 1974; Samson, 1975; Кузь-
мина, 1980 и др.), однако до сих пор остается много спорных и нере-
шенных вопросов, удовлетворительно не разработана систематика этой
группы, номенклатура позднеплейстопеновых лошадей.
Материалы из Мезина в разное время послужили основой для
описания двух видов позднеплейстоценовых лошадей Восточной Европы.
В. И. Громова на материалах из средне- и позднепалеолитических
стоянок (в том числе из Мезина) описала Equus caballus tatipes (Гро-
мова, 1949)*. Г. Нобис на материалах только из Мезина (фрагменты
верхних и нижних челюстей) выделил новый подвид — мезинскую ло-
шадь Е. ferus ferus, обитавшую в позднем плейстоцене Восточной
Европы (Nobis, 1971), сближая ее с лошадью солютре (Е. ferus solu-
treensis Nobis) Западной Европы. Подвид talipes исследователь отнес
к Е. retnagensis Skorkowski ( = £'. germanicus Nehring), считая ее ти-
пичной лошадью среднего палеолита (мустье) Евразии.
В основу данного исследования были положены значительно уве-
личившиеся коллекции из позднепалеолитической стоянки Мезин Ко-
рейского района Черниговской области. Кроме того, привлекались ма-
териалы из ряда других местонахождений Черниговской области. Раз-
меры костей даны в мм, индекты — в %, высота в холке — в см.
Череп
В коллекции часто встречаются фрагменты черепов, изолирован-
ные зубы молодых животных. Возраст определялся по срокам проре-
зывания у позднеспелых пород (Ellenberger-Baum, 1943; Климов, 1955).
Возраст изолированных зубов устанавливался по высоте коронки (No-
bis, 1971). Всего было просмотрено 20 фрагментов черепа и верхних
изолированных зубов и 13 нижних челюстей и зубов. Значительную
часть добытых лошадей составляли молодые и полувзрослые особи
(45.4%).
В коллекции из Мезина хранятся 22 фрагмента мозговой и лице-
вой части черепа (табл. 1), к сожалению, целых черепов нет.
Лучше других сохранился череп 5-571 (рис. 1, вклейка), однако
и он сильно разрушен: почти полностью отсутствует крыша черепа, на
нижней стороне сохранился правый зубной ряд, на левой стороне от-
* В последнее время преобладает мнение, что название caballus не должно
употребляться для обозначения диких лошадей, так как оно было дано К. Линнеем
для домашней лошади.
50
Таблица 1
Размеры черепов лошадей позднего палеолита (мм, % %)
Промер Мезин, 1956 Табурище, 1956
5-571 5-573 абс. 5-565 абс. 5-567* абс. 34-2886
абс. %% абс. %%
1. Основная длина 498 100 (520) — — 493.0 100
2. Теменная длина 554 111.2 — — — ±540 109.5
3. Премолярная основная длина 367 73.7 ±387 — — 367.5 74.5
4. От prosthion до заднего края неба 269 54.0 — 270 — 266.5 54.1
5. От заднего края неба до basion 227 46.6 240 — — 227.5 46.1
6. От prosthion до ближайшей точ- ки края глазницы — — — — — 327 66.3
7. То же до наиболее удаленной точки заднего края глазницы ±390 78.3 — — — 385 78.1
8. От переднего края Р2 до бли- жайшей точки края глазницы — — — — — 193.5 39.2
9. От acrocranion до ближайшей точки заднего края глазницы ±210 43.7 — — — 195 39.5
10. От basion до сошниковой вы- резки 122 24.5 — — — 123.5 25.1
11. От сошниковой вырезки до заднего края неба 108 21.7 — — — 104 21.1
12. Предкоренная длина от Р2 133.5 26.8 — 130 — 128 26.0
13. Длина зубного ряда Р2—М3 ±183 36.7 ±190 184 ±176 186 37.7
14. Длина верхнечелюстного бугра 33 6.6 29.5 26.0 38.0 31.5 6.4
15. Длина лицевого гребня ±185 + 37.1 — — — 185 37.5
16. Анатомическая мозговая ось — — — — — 177 35.9
17. Анатомическая лицевая ось — — — — — 384 77.9
18. Ширина лба у заднего края глаз- ниц — — — — — 207.5 42.1
19. Ширина у наружных краев сус- тавных ямок — — — — — 200 40.6
20. Ширина на уровне передних краев глазниц — — — — — 166 33.7
21. Ширина между передними точ- ками лицевых гребней 150 30.1 — — — 147 29.8
22. Наибольшая ширина челюсти у наружных краев альвеол 129 25.9 — — — 125 25.4
23. Ширина мозговой коробки наи- большая — — 119 — 122 113 22.9
24. То же в заглазничнвм сужении — — 96 — 93.5 87 17.6
25. Резцовая ширина — — 79.5 80 76.0 76 15.4
26. Больший поперечник глазниц — — — — — 65 13.2
27. Малый поперечник глазниц — — — — — 56.5 11.5
30. Затылочная высота большая 103.5 20.8 103.5 — 103 103 20.9
31. Затылочная высота малая 65 13.0 64 — 68 62 12.6
32. Высота черепа позади М3 — — — — — ±127 25.8
35. Наименьшая ширина хоанальной вырезки 40.5 8.1 — — — 41 8.3
* 5-567 + 5-568+5-570 — записаны
под одним номером 5-567.
сутствует резцовая часть, диастема, ряд Ртх— Рт2. Череп принадле-
жал молодой особи, постоянный Рт3 еще не затронут стиранием, Рт4
еще не сменился, М3 прорезывается, зацеп уже постоянный, окраек —
молочный. Имеется так называемый волчий зуб (Рт1). Череп принад-
лежал животному в возрасте 3—4 лет, имеет средние размеры (основ-
ная длина около 500 мм, общая длина — 554 мм).
Судя по относительной длине от prosthion до наиболее удаленной
точки заднего края глазницы (78.3%), индексу положения глазницы
(185.7%), лицевой отдел был сравнительно длинным. Предкоренной
отдел средней длины, зубы довольно крупные. Верхнечелюстной бугор
короткий как абсолютно (33.0 мм), так и относительно, по отношению
к основной длине он составляет 6.6%, а к длине зубного ряда— 18.1%.
Лицевой гребень длинный (37.1% основной длины), передний его край
на уровне середины Рт*. Судя по наибольшей ширине челюсти (26%),
лицевая часть черепа средней ширины. Небо укороченное (индекс по-
ложения нёба 118.5%, задний край его на уровне Л12). Однако все эти
признаки, по-видимому, связаны с молодым возрастом животного. Сош-
никовый индекс невысокий—112.9%. Хоанальная вырезка средней
ширины. Судя по основной длине черепа, животное попадает в верх-
ние пределы средних лошадей по классификации Витта (1952). По
коэффициентам Кизевальтера и Неринга (цит. по Громовой, 1949), вы-
сота в холке должна быть несколько выше: по теменной длине от 140
до 166 см, а по основной — от 137 до 166 см, в среднем 149.5 см.
Очень трудно провести сравнение рассматриваемого нами черепа
№ 5-571 с черепами других позднепалеолитических лошадей, так как,
во-первых, череп принадлежал полувзрослому животному (у взрослого
животного пропорции его могут быть несколько иными), а во-вторых,
в литературе известны описания и размеры не более десяти частично
разрушенных черепов позднеплейстоценовых лошадей Европы (Гро-
мова, 1949; Давид, 1974; Кузьмина, 1975, 1980). По индивидуальному
возрасту мезинский череп близок к черепу из Фёбена (цит. по Громо-
вой, 1949), однако мезинский череп намного мельче (основная длина
фёбенского — 540 мм). По-видимому, у мезинского черепа более корот-
кий лицевой отдел, значительно короче верхнечелюстной бугор. Оба че-
репа близки по относительной длине зубного ряда. Мезинский череп так-
же несколько короче черепа из Ремагена, отнесенного к Е. germatiicus
Nehr., у которого основная длина 528 мм. Череп из Ремагена более
длинномордый (индекс положения глазницы 193,3%), имеет более ко-
роткий зубной ряд (32,6% основной длины), более длинный верхнече-
люстной бугор. Близки эти черепа по предкоренной длине, и также, ве-
роятно, по ширине лицевого отдела, затылочной высоте. Череп из Ме-
зина отличается и от черепа из Шуссенрида (цит. по Громовой, 1949):
последний мельче (основная длина 480 мм), более короткомордый,
с более коротким зубным рядом (34%) и более широким лбом. Череп
из Мезина, возможно, был мельче черепа из стоянки Старые Дуруи-
торы (Давид, 1974), у которого восстановленная основная длина
должна была составлять 540 мм. Мезинский череп имел более длинную
лицевую часть и предкоренную длину, более короткий верхнечелюст-
ной бугор, более длинный лицевой гребень, более широкую хоаналь-
ную вырезку. Видимо, рассматриваемые черепа близки по относитель-
ной ширине лицевой части черепа, относительной длине зубного ряда.
Исследуемый череп также был мельче сильно разрушенного черепа
из Костенок (Кузьмина, 1980), вероятно, имел более узкий лоб и моз-
говую коробку; черепа были близки по относительной длине зубного
ряда и ширине хоанальной вырезки.
52
От черепа № 5-573 сохранилась мозговая часть с правым зубным
рядом. Очевидно, она принадлежала животному в возрасте 5—6 лет.
Вершина нёба на уровне задней половины М2, передний край лицевого
гребня на уровне середины Р4. Данный череп был крупнее вышеопи-
санного черепа № 5-571. По премолярной длине (около 387 мм) и длине
от заднего края нёба до basion (240,0 мм) основная длина должна
была достигать примерно 520 мм. Верхнечелюстной бугор короткий.
Зубной ряд довольно длинный — около 190 мм, по отношению к пре-
молярной длине составляет 49,2% и уступает значению этого индекса
на черепе № 5-571.
Череп № 5-565 — сохранилась лицевая часть, принадлежавшая же-
ребцу в возрасте 7—8 лет. Задний край нёба расположен на уровне
последней трети М2, лицевой гребень — на уровне переднего края Л41.
Резцовая часть широкая. Наблюдается сильная вогнутость верхнече-
люстной кости в диастемной части. Верхнечелюстной бугор короткий
и выражен слабо, достигает в длину 28,5 мм, индекс—15,5%. Судя по
фотографии и размерам, правая ветвь верхней челюсти именно этого
черепа была выделена Нобисом (Nobis, 1971, taf. 4, tabl. XXXII) как
голотип Equus ferus ferus.
Кроме описанных выше, в коллекции из Мезина имеются еще
остатки черепов худшей сохранности. Среди них свод черепа с заты-
лочной частью № 5-567, левая верхняя челюсть № 5-568, правая верх-
няя челюсть № 5-569, резцовая часть со стертыми клыками 5-570, по-
видимому, принадлежали одному черепу старого животного с широкой
мозговой коробкой, затылком средней высоты и средней длины верхне-
челюстным бугром.
По размерам к черепу № 5-571 близок череп 34-2886 из аллю-
виального местонахождения позднеплейстоценового возраста у с. Табу-
рища (г. Орджоникидзе) Днепропетровской области. Этот череп отли-
чается хорошей сохранностью и принадлежал кобыле в возрасте около
4 лет. Череп из Табурища сходен в черепом 5-571 по основной длине
(табл. 1), по длине от prosthion до заднего края нёба, передней глаз-
ной линии, от basion до сошниковой вырезки, длине зубного ряда,
длине верхнечелюстного бугра, лицевого гребня, ширине лицевой части,
затылочной высоте. В отличие от черепа из Мезина, череп из Табу-
рища принадлежал к группе длинномордых * лошадей, относительная
ширина глазницы на нем достигает 197,4%. Длинномордость в основ-
ном обусловлена укороченным мозговым отделом (анатомическая моз-
говая ось составляет 35,9%, задняя глазная линия — 39,6%). Лоб до-
вольно узкий, наибольшая ширина составляет 42,1% основной длины,
ширина у наружных краев суставных ямок — 40,6%, лобно-основной
указатель 237,6%. По градации Громовой (1949, ч. II), череп из Табу-
рища относится к группе узколобых лошадей, хотя лоб был и несколько
шире, чем у лошади из Ремагена и хозарских лошадей Среднего По-
волжья. Резцовая часть широкая (15,4%), однако уже, чем у лошади
из Ремагена.
Рассмотрев все фрагменты черепа из Мезина и череп из Табу-
рища, можно сделать некоторые выводы. Позднеплейстоценовая ло-
шадь бассейнов Десны и Днепра обладала черепом средней длины
со сравнительно длинным лицевым отделом, по-видимому, была узко-
лобой, мозговая коробка довольно широкая, также широкой была и
резцовая часть, зубы относительно крупные, верхнечелюстной бугор
короткий.
Отмечается некоторое сходство с черепами Е. missi М. Pavlova
(Громова, 1949) по индексу положения нёба, относительной наибольшей
53
ширине челюсти, относительной высоте затылка, относительной ширине
хоанальной вырезки, однако у позднепалеолитических лошадей были
относительно крупные зубы, более длинный лицевой гребень, значи-
тельно короче верхнелюстной бугор, более широкая резцовая часть.
Отмечается также некоторое сходство с черепами Е. chosaricus, раз-
меры которых приведены Громовой (1949). Они близки, по-видимому,
в общих размерах, по длине верхнечелюстного бугра, резцовой ширине,
ширине мозговой коробки, наименьшей ширине хоанальной вырезки.
По сравнению с Е. uralensis (Кузьмина, 1975) широкопалая ло-
шадь Украины была более длинномордой, с относительно более длин-
ным зубным рядом, более широким лбом.
Верхние коренные зубы позднепалеолитической лошади бассейна
Десны довольно крупные, эмаль относительно толстая, складчатость
средняя, так на задней стенке передней марки и на передней стенке
задней марки 1—3 складочки. Протокон длинный, сильно вытянут
вдоль оси зуба, внутренний край его часто волнистый. Наружные
стенки пара- и метакона глубоко вогнуты, мезостиль большей частью
раздвоенный или расширенный на премолярах, на молярах закруглен-
ный, нераздвоенный. Размеры зубов приведены в табл. 2.
По размерам зубов, длине протокона и рисунку эмали зубы позд-
неплейстоценовой лошади Десны существенно не отличаются от зубов
из других позднепалеолитических памятников, однако они обладают
относительно более длинным протоконом.
Таблица 2
Размеры верхних коренных зубов позднепалеолитической лошади Десны
Признак 11m м •_/п
1. Длина ряда Рт2—А43, альвеолярная 4 ±176 ±190 183,25 —
2. То же по коронкам 6 173,0—186,5 179,91 —
3. Длина ряда Рт2—Рт\ альвеолярная 1 99,0 — —
4. То же по коронкам 5 97,5—106,0 101,30 —
5. Длина ряда ЛП—М3, альвеолярная 4 78,5—89,0 82,50 —
6. То же по коронкам 8 77,5—91,0 82,31 —
7. Молярно-премолярный индекс (6 : 4) 4 82,5—85,8 84,25 —
8. Длина Рт2 7 40,0—43,5 42,14 0,51
9. Ширина Рт2 7 25,5—30,0 27,43 0,60
10. Длина протокона Рт2 7 9,0—12,5 11,00 0,42
11. Индекс длины протокона Рт2 7 22,5—28,5 26,03 0,46
12. Длина Рт3 И 28,5—34,0 30,82 0,47
13. Ширина Рт3 И 28,0—31,0 29,59 0,33
14. Длина протокона Рт3 11 13,5—16,0 14,73 0,24
15. Индекс длины портокона Рт3 11 45,1—50,0 47,65 9,46
16. Длина Рт4 12 25,0—33,5 29,75 0,64
17. Ширина Рт4 12 27,0—32,5 29,92 0,48
18. Длина протокона Рт4 12 13,0—17,0 14,79 0,35
19. Индекс длины протокона Рт4 12 40,6—54,8 50,91 1,22
20. Длина М1 15 23,5—28,5 25,57 0,45
21. Ширина М1 15 27,0—31,5 28,80 0,38
22. Длина протокона М1 15 13,5—15,5 14,57 0,20
23. Индекс длины протокона М1 15 54,0—63,8 57,16 0,87
24. Длина М2 15 24,0—30,0 26,47 0,55
25. Ширина М2 14 26,5—32,0 28,61 0,36
26. Длина протокона М2 15 13,5—16,5 15,27 0,21
27. Индекс длины протокона М2 15 53,3—62,5 57,82 0,83
28. Длина М3 16 27,0—31,5 29,38 0,36
29. Ширина М3 16 23,0—28,5 25,06 0,36
30. Длина протокона М3 16 14,0—18,0 15,69 0,24
31. Индекс длины протокона М3 16 50,0—63,2 53,50 0,82
54
Нижние челюсти представлены 7 экз. из Мезина. Все они повреж-
дены, 4 из них принадлежали молодым животным.
Нижняя челюсть 5-566 — правая горизонтальная ветвь с полным
зубным рядом и симфизальной частью. По степени стирания коренных
зубов, резцов челюсть принадлежала жеребцу в возрасте примерно
8 лет. Горизонтальная ветвь относительно высокая, индекс ее высоты
перед Рт2— 38,1%, перед Mi — 46,4%. Наибольшая выпуклость ниж-
него края находится на уровне — Рт4. Нижний край ветви несколь-
ко волнистый. Подбородочная вырезка средней ширины. Симфизальная
часть довольно массивная, наименьшая ширина ее — 52 мм, резцовая
часть широкая — 70 мм. Нижнечелюстное отверстие крупное. Симфиз и
.диастема средней длины, зубы крупные. Обращает внимание складча-
тость эмали, на передней стенке задней долинки Рт4— 3 складочки.
Челюсть 5-582 — фрагмент правой горизонтальной ветви с рядом
Рт4 — М3. По степени стертости зубной поверхности челюсть принад-
лежала жеребцу в возрасте около 10 лет. Очень глубокая выемка
двойной петли.
Кроме рассмотренных нижних челюстей, имеются еще фрагменты
челюстей молодых животных и изолированные зубы (табл. 3). Разме-
ры зубов колеблются в значительных пределах. Например, длина Рт2
варьирует от 33,5 до 37,5мм, Рт3 — Рт4 — 26,5—34,5, АЛ — М2 — 25,5—
34,5 мм. Двойная петля наиболее короткая на Рт2 и Л13, относительно
«самая длинная на Рт3. Двойная петля кабаллоидная, складчатость
эмали средняя, за исключением зубов челюсти 5-566. Передняя петля
(метаконид) большей частью округлая, средняя (метастилид)—более
вытянутая, субтреугольной формы. Выемка двойной петли, как правило,
неглубокая, за исключением 5-582. Задняя петля (энтоконид) округ-
лая. Талонид короткий и широкий. Нижняя стенка задней долинки
прямая или волнистая. Наружная долинка, как правило, на молярах
достигает (или даже заходит) в шейку двойной петки, на премолярах
не достигает ее. На молочных премолярах наружная долинка глубоко
заходит в шейку двойной петли. Почти всегда имеется шпора.
Среди просмотренных нижних зубов не обнаружили ни один, у ко-
торого была бы такая глубокая и длинная наружная долинка, как это
•отмечено Радулеско и Самсоном (Radulesco, Samson, 1962) у описан-
ной ими Е. scythicus.
Кости посткраниального скелета
В коллекции из Мезина имеются все части скелета лошади, из
других стоянок Подесенья — изолированные зубы, отдельные кости и
их фрагменты. Большая часть костей была разбита, однако целыми
оказались следующие кости конечностей: 3 плечевые, 3 лучевые,
8 пястных, 2 большеберцовые, 7 плюсневых; кроме того, небольшие
серии таранных и пяточных костей (по 12 экз.), 25 первых фаланг.
Размеры костей посткраниального скелета приведены в табл. 4, сюда
же включены данные Громовой (1949).
Таранные кости представлены серией из 12 экз., которые можно
было измерить. Как правило, внутренняя длина их меньше наиболь-
шей ширины (только в одном случае они равны), в среднем внутрен-
няя длина составляет 63,66 мм, а наибольшая ширина кости — 68,68 мм.
Также широки и нижние суставные поверхности: отношение их ширины
к внутренней длине кости колеблется от 84,2 до 96,8%. Нижний гре-
бень наружного блока опускается низко, нижний конец внутреннего
‘блока или слегка загибается внутрь, или прямой. Наружный край
55
Таблица 3
Размеры нижней челюсти лошади из Мезина
Признак 11m м
1. Высота горизонтальной ветви позади М3 1 108.5 —
2. Высота ветви перед Ah 3 72.0—84.5 80.2
3. Высота ветви перед Рт2 4 57.5—71.0 65.8
4. Наибольшая толщина ветви 3 29.0—30.0 29.5
5. Длина симфиза 3 81.0—85.0 82.5
6. Длина диастемы 3 73.5—75.5 74.6
7. Индекс длины симфиза к длине диастемы (6:5) 3 10.7—11.5 11.0
8. Длина зубного ряда Рт2—М3 2 178.0— 181.0 179.5
9. по коронкам 2 170.0— 174.5 172.2
10. Рт2—Рт4 по альвеолам 5 94.5—103.0 97.8
11. „по коронкам 6 89.5—103.5 95.7
12. All—А1з по альвеолам 2 82.0—91.0 86.5
13. „ по коронкам 2 79.5—85.5 82.5
14. Индекс (2 : 8) 1 46.4 —
15. (3:8) 1 38.1 —
16. (12:10) 2 86.3—96.2 91.2
17. (13:11) 2 87.3—95.2 91.2
18. Резцовая ширина 3 67.5—70.0 68.8
19. Наименьшая ширина челюсти 3 43.5—50.5 47.1
20. Индекс (18:6) 3 89.4—95.5 92.3
21. Длина Рт2 7 33.5-37.5 35.2
22. Ширина Рт2 7 15.0—19.0 17.2
23. Индекс ширины Рт2 7 40.0—56.7 48.1
24. Длина двойной петли 6 15.0—19.5 17.0
25. Индекс двойной петли 6 49.0—54.9 47.8
26. Длина Рт3—Рт4 12 26.5—34.5 30.25
27. Ширина Рт3—Рт4 12 16.0—20.5 18.3
28. Индекс ширины Рт3—Рт4 12 46.3—75.4 60.6
29. Длина двойной петли 11 16.5—19.0 17.5
30. Индекс двойной петли 11 49.2—65.5 58.1
31. Длина М}—М2 13 25.5—34.5 29.6
32. Ширина Л1!—М2 13 15.5—21.5 16.5
33. Индекс ширины Mi— М2 13 45.5—82.6 61.4
34. Длина двойной петли 13 13.0-17.0 14.9
35. Индекс двойной петли 13 38.2—58.1 50.8
36. Длина М3 2 30.5—32.0 31.25
37. Ширина М3 2 16.0—17.0 16.5
38. Индекс ширины М3 2 52.4—53.1 52.7
39. Длина двойной петли М3 2 13.0—15.0 14.0
40. Индекс двойной петли 2 42.6—46.8 44.7
41. Длина талонида М3 1 6.0 —
42. Индекс талонида М3 1 19.7
55
Таблица *
Размеры и индексы костей позднеплейстоценовой широкопалой лошади Десны
Признак 11m м ±/п С
Лопатка
Ширина через шишковидный бугор 6 94.5—99.5 97.08 0.79 1.77 1.8
Ширина суставной поверхности 6 58.0—63.0 60.83 1.02 2.29 3.8
Поперечник суставной поверхности Плечевая 6 45.0—51.0 49.00 1.03 2.30 47
Наибольшая длина 3 298-306 303.00 1.94 3.87 1.28
Ширина верхнего конца 5 87.5—101 94.9 2.78 5.55 5.8
Ширина нижнего конца 8 81.0—93.0 86.31 1.50 3.98 4.61
Ширина нижнего суставного блока 9 73.0—85.5 78.89 1.36 3.86 4.9
Поперечник нижнего эпифиза Лучевая 7 84.5—95.0 90.14 1.37 3.62 4.0
Наибольшая длина 3 325.0— 336.0 332 4.31 6.08 1.8
Ширина верхнего эпифиза 6 84.5—95.0 88.33 1.77 3.96 4.5
Ширина нижнего эпифиза 5 77.0—85.0 80.60 2.05 4.1 5.1
Ширина нижней суставной поверх- ности Пястная 6 63.5—71.0 66.25 1.49 3.34 5.0
Наибольшая длина 8 210.0— 231.5 225.65 2.94 7.80 3.4
Ширина верхнего эпифиза 9 51.0—56.5 54.32 0.75 2.14 3.7
Ширина нижнего эпифиза 8 50.7—53.5 52.37 0.40 1.07 2.4
Ширина диафиза 8 35.0—39.0 37.37 0.44 1.18 3.1
Обхват диафиза I фаланга передней конечности 8 96.0—104.0 102.0 1.06 2.82 2.7
Наибольшая длина 10 83.0—91.5 87.40 0.91 2.72 3.1
Длина по средней линии передней поверхности 10 75.5—82.0 79.6 0.64 1.92 2.4
Ширина верхнего эпифиза 10 53.0—62.5 58.6 0.85 2.54 4.3
Ширина нижнего эпифиза 10 45.5—50.0 47.4 0.43 1.28 2.7
Ширина диафиза 10 36.0—41.0 38.80 0.47 1.40 3.6
Индекс ширины диафиза Бедренная 10 41.8—48.5 44.20 0.55 1.66 3.8
Ширина нижнего эпифиза Большая берцовая 5 93.0—99.0 96.4 1.11 2.22 2.5
Наибольшая длина 2 361.0; 361.5 — — —
Ширина верхнего эпифиза 7 97.0—105.0 101.64 1.02 2.51 2.53
Ширина нижнего эпифиза Таранная 11 74.0—86.5 79.96 1.05 3.33 4.1
Наибольшая длина 12 59.0—68.5 63.66 0.87 2.90 4.5
Ширина нижней суставной поверх- ности Пяточная 12 50.5—60.0 55.08 1.03 3.43 6.1
Наибольшая длина 12 107.0— 123.5 116.18 1.24 4.12 3.5
Наибольшая ширина 12 52.0—59.0 56.32 0.62 2.08 3.7
57
Продолжение таблицы 4
Признак 11m м с
Плюсневая
Наибольшая длина 7 260.5— 280.0 271.71 2.59 6.35 2.3
Ширина верхнего эпифиза 7 53.0—61.5 56.86 1.26 3.08 5.4
Ширина нижнего эпифиза 7 50.5—58.0 54.54 1.18 2.89 5.29
Ширина диафиза 7 34.5—39.0 36.07 0.66 1.62 4.5
Обхват диафиза 6 102.0— 119.0 108.17 2.73 6.11 5.6
I фаланга задней конечности
Наибольшая длина 15 81.0—91.0 86.86 0.66 2.49 2.8
Длина по средней линии передней 15 75.0—85.0 78.46 0.69 2.58 3.3
поверхности 0.54
Ширина верхнего эпифиза 15 58.0—67.0 61.74 2.02 3.3
Ширина нижнего эпифиза 15 45.5—53.0 48.00 0.58 2.19 4.5
Ширина диафиза 15 36.0—41.0 38.46 0.37 1.38 3.6
Индекс ширины диафиза 15 41.6—46.5 44.26 0.43 1.62 3.6
II фаланга
Наибольшая длина 6 44.0—52.0 48.58 1.38 3.08 6.34
Длина по средней линии 6 37.0—42.5 39.08 0.96 2.16 5.5
Ширина верхнего эпифиза 6 48.0—61.0 54.75 1.85 4.15 7.6
Ширина нижнего эпифиза 6 45.0—54.5 50.92 0.41 0.91 1.8
Ширина диафиза 6 41.0—49.0 46.58 0.78 1.75 3.8
нижней суставной поверхности у некоторых таранных костей образует
выступ, у других он почти не заметен. Мускульный бугор развит хо-
рошо и сильно выступает внутрь.
Все пяточные кости имеют типичное строение, однако, как отме-
чала Громова (1949, ч. 1, с. Г69), пяточная № 2663 обладает очень
узким телом, относительная ширина кости составляет 45,5%, а попе-
речник только 43,8%—минимальное значение для украинских и мол-
давских лошадей позднего плейстоцена. Наиболее крупная пяточная
длиной 123.5 мм перекрывает максимальное значение для молдавских
лошадей мустьерского возраста, имевших максимальные размеры (Да-
вид, 1974).
В типологии лошадей большое значение имеют метаподии. В Ме-
зине были найдены 8 целых пястных костей (рис. 2). Дляина их ко-
леблется от 210 до 232 мм. 6 пястных костей располагаются в соседних
классах: 225—230—235 мм, сюда же оносятся 2 пястные из позднепа-
леолитических слоев Владимировки Кировоградской области длиной
в 234 и 235,5 мм. Однако в Мезине были найдены 2 экз. очень корот-
ких пястных (6-631 и 5-609) от разных особей, достигавших в длину
210 и 211 мм. Возникает вопрос, принадлежат ли эти две кости широко-
палой лошади или относятся к другой, более мелкой форме? Размах
вариации (у) мезинской выборки небольшой — 22 мм, он значительно
меньше, чем у широкопалой лошафи Дона (по Кузьминой, 1980, и = 33)
и лошади Солютре (Nobis, 1971, и = 32,5 мм). Статистическая обработка
по методу Ашмарина с соавторами (1971) показала, что пястные дли-
ной 210, 211мм входят в мезинскую совокупность. По-видимому, мел-
кие особи из Мезина принадлежали той же форме позднепалеолити-
ческой лошади, что и крупные. Однако вопрос этот окончательно не
ясен: не исключена возможность, что могли одновременно существовать
мелкая и крупная формы позднеплейстоценовых лошадей (Sikenberg,
1962).
58
ШИРИНА ДИАФИЗА,ММ | ШИРИНА ДИАФИЗА,ММ
40
250 270 290
ДЛИНА КОСТИ, ММ
Рис. 2. Размеры пястных костей.
2 — Владимировка, 3 — Сунгирь, 4 — Солютре, 5 — стоянки Молдавии, 6 — стоянки
верхнего Дона.
220 240
260
ДЛИНА КОСТИ, ММ
Рис. 3. Размеры плюсневых костей.
Обозначения, как на рис. 2, только 3 — Новгород-Северский.
Все пястные кости из Мезина по относительной ширине диафиза
принадлежат к группе широконогих лошадей по градации В. И. Гро-
мовой, ширина их диафиза составляла 15,6—17,7% наибольшей длины.
По относительной ширине эпифизов кость 5-630 (длиной 231,5 мм) по-
падает в группу средних лошадей, находясь в ее максимальных преде-
лах. Из морфологических особенностей нужно отметить сильное раз-
витие боковых метаподиев, особенно rntcir. они широкие, спускаются
почти на 3Л длины основной пястной кости. Дистальная депрессия хо-
рошо видна на 5 костях, слабо развита на одной кости. Гребень ниж-
него валика выступает' резко, индекс выступания 78,8—85,6%. Пять
пястных в надсуставных буграх уже, чем в суставе, у трех эти при-
знаки равны.
В коллекции имеются 6 целых плюсневых костей из Мезина и 1
из Новгород-Северского (рис. 3). Длина их колебалась в пределах
20 мм — от 260,5 до 280 мм. Кости массивные, по относительно ширине
верхнего и нижнего концов все плюсневые относятся к группе широко-
ногих лошадей. По относительной ширине диафиза 2 кости — к группе
средненогих, остальные — к широконогим лошадям. 5 плюсневых в су-
ставе были шире, чем в надсуставных буграх. На задней поверхности
кости в дистальном отделе заметна депрессия на трех костях, на
остальных она выражена слабо. Гребень нижнего валика развит хо-
рошо, индекс выступания — 77,7—82,3%.
Первые фаланги встречаются часто, их размеры приведены
в табл. 4. Разделение фаланг на передние и задние проведено по ра-
боте Браунера (1916), В. И. Громовой (1963) и путем сравнения с фа-
лангами современных лошадей. Передние фаланги в среднем несколько
длиннее задних, на задних фалангах верхний конец несколько шире
и массивнее. Надсуставные бугры на задних фалангах выступают
резче; так, индекс ширины в надсуставных буграх на передних фалан-
гах колебался от 55,2 до 60,4%, в среднем составляя 56,44%; на зад-
них соответственно от 55,5% до 63,6, в среднем — 58,4%. У большей
части I фаланг треугольная ямка, расположенная на задней поверх-
ности, средней глубины. Шероховатые полосы треугольного поля на
задней поверхности опускаются на 2/3 длины, боковые гребни поля хо-
рошо выражены, на некоторых фалангах у вершины треугольника
имеется вздутие. Ямка на задней поверхности нижнего конца у боль-
шинства фаланг выражена четко, а у некоторых слабее.
Среди копытных фаланг можно было измерить две от передней
конечности и одну от задней. Передние были типичными для широко-
палой лошади: длина вдоль передней поверхности — 59 и 61 мм, наи-
большая ширина — 96,5 и 99,0 мм, длина опорного края по кривой — 207
и 217 мм. По градации Громовой (1949, ч. II) они относятся к группе
лошадей с широким копытом. Задняя фаланга была мельче и уже,
длина составляла 55 мм, ширина 80 мм, длина опорного края — 165 мм.
Этот промер не достигает минимальных пределов для позднепалеолити-
ческих лошадей (Громова, 1949, Давид, 1974); возможно, это связано
с молодым возрастом особи. По пропорциям фаланга принадлежала
лошади, относящейся к группе лошадей с широкими копытами.
Как было указано выше, в нашем распоряжении было 23 кости
конечностей и 1 череп, по которым можно восстановить рост лошадей,
обитавших в позднем плейстоцене в среднем течении Десны. Высота
в холке, рассчитанная по коэффициентам Кизевальтера (цит. по Гро-
мовой, 1949) и по Витту (1952) колебалась от 128 до 152 см, в сред-
нем 140—141 см. Более 2/з составляли средние по росту животные, мел-
кие и крупные лошади не отмечены. Примечательно то, что средние
60
Таблица 5
Высота в холке широкопалой лошади бассейна Десны
Высота в холке lim М & ±/71 с
По Кизевальтеру 129,0—149,6 140,76 0,91 4,38 3,1
По Витту 128,0—152,0 140,33 1,01 4,96 3,5
показатели, вычисленные двумя способами, оказались очень близкими;
характеристика вариационных рядов выглядит следующим образом
(табл. 5).
Полученные нами показатели мы смогли сравнить с опубликован-
ными данными из ряда позднепалеолитических памятников Европы,
давших обильный материал: Солютре, Франция, первая половина позд-
него палеолита (Nobis, 1971)*; стоянки Молдавии, поздний палеолит
(Давид, 1980); стоянки на Дону — Костёнки и др., первая половина
позднего палеолита (Кузьмина, 1980).
Нобис (1971), подчеркивая большое сходство между лошадьми
Солютре и Мезина, отнес их к одному виду Е. ferus Boddaert, назвав
первых Е. f. solutreensis, вторых — Е. f. ferus. В нашем распоряжении
было значительно больше остатков мезинской лошади, особенно костей
посткраниального скелета. Сравнение с данными Нобиса по Солютре
дало несколько иные результаты. По абсолютным размерам большин-
ства признаков кости мезинской лошади были крупнее: по длине мета-
подиев, таранной, пяточной, I фаланг, ширине верхнего и нижнего эпи-
физов метаподиев, нижнего конца берцовой, ширине пяточной различия
статистически достоверны. Однако по относительной ширине кости
этих лошадей или были сходны, или даже несколько стройнее у ме-
зинской; так, у последней статистически достоверно ниже среднее зна-
чение относительной ширины диафиза пястной кости и I фаланги зад-
ней конечности. По высоте в холке лошадь Десны статистически досто-
верно была выше лошади Солютре. Так, по коэффициентам Витта
в среднем высота в холке мезинской лошади составляла 140,33± 1,01 см,
малорослых было 12,5%, средних — 70,8% и 16,7% рослых животных.
В Солютре лошадь в среднем на 3,77 см ниже и достигала в холке
136,56±0,42 см, 1,4% относились к мелким, 4,5%—малорослым,
50,7% —средним и только 2,9% —рослым лошадям.
Сравнение наших материалов с данными Давида (1980) по широ-
копалой лошади позднего палеолита Молдавии показывает, что ло-
шадь Десны была мельче. У нее были короче метаподии, I фаланги,
несколько уже эпифизы и диафиз пястной кости, I фаланг, нижний
эпифиз и диафиз плюсневой кости. Однако обе эти лошади, по-види-
мому, были сходны в пропорциях костей.
Наиболее крупными (по промерам И. Е. Кузьминой, 1980) из из-
вестных нам позднеплейстоценовых лошадей были лошади Дона Ко-
стенковских стоянок, из слоев, датируемых ориньяк-солютрейской эпо-
хой. У лошади Десны достоверно короче были лучевая, пястная, плюс-
невая, I передняя фаланга (/ = 2,97—4,3), и в среднем она была на
5 см ниже. Также достоверно уже у деснянской лошади были эпифизы
и диафиз пястной кости и I передней фаланги, нижний суставной блок
плечевой кости (/ = 2,44 — 3,4). В то же время у названных лошадей
* Статистическая обработка данных измерений, приведенных Г. Нобисом, про-
ведена автором данной статьи.
61
были близки по ширине верхний и нижний эпифиз лучевой кости, ниж-
ний эпифиз бедренной, берцовой кости, эпифизы и диафиз плюсневой
кости, I задней фаланги, по длине — таранная и пяточная кости. Эти
две формы были сходны в пропорциях костей; например, относитель-
ная ширина диафиза пястной кости у лошади бассейна Десны колеба-
лась от 15,6 до 17,7%, в среднем составляя 16,48±0,27%, у лошади
Дона соответственно— 14,8—18,3, М = 16,6%.
_Таким образом, в Европейской части СССР в позднем плейстоцене
наиболее мелкими размерами обладали широкопалые лошади, обитав-
шие в мадлене, в бассейне Десны; несколько крупнее их были лошади
Молдавии и наиболее крупными — лошади Дона ориньяк-солютрей-
ской эпохи (рис. 2 и 3).
В позднем плейстоцене Европейской части СССР, кроме Урала,
обитала широкопалая лошадь Е. latipes, представленная разными
группами. В течение этого отрезка времени под влиянием быстро из-
меняющихся условий она могла быть крупнее в более благоприятных,
или мельче в менее благоприятных условиях. Например, на Дону
в ориньяк-солютрейскую эпоху лошади были многочисленны (Вере-
щагин, Кузьмина, 1977), а на Десне в мадлене они были мельче и реже
встречались (Бибикова, Белан, 1979). Все эти лошади имели массивные
кости конечностей, широкие копыта, длинный протокон на верхних
зубах.
Сопоставление с лошадьми других частей Европы затруднено, так
как разными авторами в позднем плейстоцене описано много форм,
сходных или тождественных. Например, лошадь в Солютре Прат (Prat,
цит. по Samson, 1975) назвал Е. с. gallicus, Нобис (1971) эту же ло-
шадь назвал Е. ferus solutreensis. Мы вполне согласны с Самсоном
(Samson, 1975), что последнее название неудачно, так как, во-первых,
Боддаерт (Boddaert, 1785) название Е. ferus предложил для рецентных
лошадей. Во-вторых, в отношении этого названия должна действовать
ст. 23в Международного кодекса зоологической номенклатуры о забы-
тых названиях. В последней сводке по лошадям Румынии Самсон
(Samson, 1975) установил 5 форм лошадей в позднем вюрме, среди
которых Е. transilvanicus он сближает с Е. latipes.
Трудно согласиться с разделением Нобисом (1971) позднеплейсто-
ценовых лошадей на 2 вида: Е. reinagensis (-=Е. germanicus) для сред-
него палеолита и Е. ferus для позднего палеолита. По размерам костей
молдавские и донские позднепалеолитические лошади должны отно-
ситься к Е. remagensis. На основании размеров метаподиев из Влади-
мировки, Сунгиря и др., ошибочно датируя их мезолитическим време-
нем, Нобис делает вывод об увеличении размеров лошадей в голоцене
и о непрерывной линии развития, называя их Е. ferus gmelini. Однако
материалы из Сунгиря и Владимировки датируются поздним плейсто-
ценом и хорошо укладываются в пределы вариации Е. talipes (рис. 2,
3). Как показали исследования серии костей лошади из позднемезоли-
тической стоянки у с. Мирное Одесской области и зубов из мезолити-
ческого слоя Владимировки (Белан, Бибикова, 1980), голоценовая
лошадь отличается от позднеплейстоценовой и авторы считают ее
тарпаном Е. gmelini.
Выводы
В бассейне Десны во второй половине позднего палеолита (мад-
лене) обитала широкопалая лошадь Е. latipes. Она имела череп сред-
ней длины со сравнительно длинным лицевым отделом, по-видимому,
62
узким, лбом, довольно широкой мозговой коробкой, широкой резцовой
частью, коротким верхнечелюстным бугром и относительно крупными
зубами с длинным протоконом. Кости конечностей средней длины, мас-
сивные, копыта широкие. По высоте в холке эти лошади относились
-к группам малорослых, средних и рослых лошадей (128—152 см),
в среднем рост составлял 140—141 см.
Лошадь Десны была несколько мельче позднеплейстоценовых ло-
шадей Дона и Молдавии, живших в более благоприятных условиях.
ЛИТЕРАТУРА
Ашмарин И. П., Васильев Н. Н., Амбросов В. А. Быстрые методы статистической
обработки и планирование экспериментов. Л., изд-во ЛГУ, 1971, 78 с.
Бибикова В. И., Белан Н. Г. Локальные варианты и группировки позднепалеолити-
ческого териокомплекса Юго-Восточной Европы. — Бюл. МОИП, отд. биол., 1979,
т. 84, в. 3, с. 3—14.
Бибикова В. И., Белан Н. Г. Раннеголоценовый тарпан на территории Северо-Запад-
ного Причерноморья. — Вести, зоологии, 1981, № 3, с. 21—26.
Браунер А. А. Материалы к познанию домашних животных России. I. Лошадь кур-
ганных погребений. Одесса, 1916, 184 с.
Вангенгейм Э. А. Описание остатков лошади Equus caballus cf. laubachensis Freu-
denberg из верхнепалеолитической стоянки Сунгирь. — Тр. геол, ин-та АН СССР,
1966, вып. 162, с. 118—135.
Верещагин Н. К., Кузьмина И. Е. Остатки млекопитающих из палеолитических стоя-
нок на Дону и верхней Десне. — Тр. Зоол. ин-та АН СССР, 1977, т. 72, с. 77—ПО.
Витт В. О. Лошади Пазырыкских курганов. — Сов. археология, 1952, т. 16, с. 165—
205.
Громова Вера. История лошадей (рода Equus) в Старом Свете. — Тр. Палеонт. ин-та,
1949, т. XVII, в. 1, 374 с.; в. 2, 163 с.
Громова Вера. О скелете тарпана (Equus caballus gmelini Ant.) и других современ-
ных диких лошадей (часть II). — Тр. Моск, о-ва испыт. природы, 1963, т. X,
с. 10—61.
Давид А. И. Остатки Equus caballus latipes Grom, в Молдавии. — В кн.: Млекопи-
тающие позднего кайнозоя юго-запада СССР. Кишинев, Штиинца, 1974, с. 3—53.
Давид А. И. Териофауна плейстоцена Молдавии. Кишинев, Штиинца, 1980, 188 с.
Климов А. Ф. Анатомия домашних животных. М., Гос. изд-во сельхоз. литературы,
1955, т. I, 576 с.
Кузьмина И. Е. Некоторые данные о млекопитающих Среднего Урала в позднем
плейстоцене. — Бюл. Комис, по изуч. четверт. периода, 1975, в. 43, с. 63—77.
Кузьмина И. Е. Позднеплейстоценовая широкопалая лошадь верхнего Дона. — Тр.
Зоол. ин-та АН СССР, 1980, т. 93, с. 91—118.
Boddaert Р. Elenchus Animalium 1. Rotterdam, 1784, s. 159—160.
Ellenberger-Baum. Handbuch der vergleichenden Anatomie der Haustiere. Berlin, Sprin-
ger-Verlag, 1943, 1170 s.
Musil R. Die Hohle „Sveduv Stul” ein typisher Hohlenhyanenhorst. — In: Die Erfor-
schung der Hohle Sveduv Stul 1953—1955 — Anthropos, 1962, i3, N. S. 5, s. 97—
260.
Nobis G. Vom Wieldpferd zum Hauspferd. — Fundamenta Reihe B. Koln Bohlau Ver-
lag, 1971, Bd 6, 96 s.
Radulesco C. S. und Samson P. Quelques observations zur «Equus scythicus» — Zoolo-
gischer Anzeiger. 1962, Bd. 169, s. 347—355.
Samson P. Les Equides fossiles de Romanic — Geologica Romana, Roma, 1975, vol. 14,
p. 165—352.
Sikenberg O. Uber die Gro e der pleistozanen Pferde der Caballus — Gruppe in Europa
und Nordasien — Eiszeitalter und Gegenwart, 1962, Bd. 12, s. 99—124.
SUMMARY
A study of horses bone remains from sites Mezin, Novgorod-Seversky, Chulatov
of Chernigov Region has shown that Equus latipes inhabited in the region of Desna
in the socond half of Late Pleistocene. It had a scull of median length with a relatively
long facial region, apparently narrow forehead, rather wide brain case and relatively
large teeth with a long protocone. Limb bones are of middle length, massive hoofs are
wide. By height in the withers the horses of Desna belonged to groups of dwarfish,
medium height and tall horses 128—152 cm), their average height being 140—141 cm.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1985, ТОМ 131
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, 1985, VOL. 131
USSR ACADEMY OF SCIENCES
УРАЛЬСКАЯ ПОЗДНЕПЛЕЙСТОЦЕНОВАЯ ЛОШАДЬ
И. Е. Кузьмина
Зоологический институт АН СССР, Ленинград
I. Е. Kuzmina. Urals late Pleistocene horse
Введение
В результате геологических исследований и археологических рас-
копок, проведенных преимущественно в пещерах Северного и Среднего
Урала, за последние 20 Лет собран обширный палеонтологический ма-
териал, насчитывающий около 31 тыс. костей 35 видов млекопитающих.
Сборы были сделаны на Северном Урале сотрудниками Коми филиала
АН СССР Б. И. Гуслицером и В. И. Канивцом в 1960—1965 гг.; на
западных склонах Среднего Урала — сотрудникОхМ Института археоло-
гии АН СССР О. Н. Бадером и членом Пермского отделения Геогра-
фического общества СССР Е. П. Близнецовым в 1965—1978 гг.; на во-
сточных склонах Среднего Урала — археологом Уральского государ-
ственного университета В. Т. Петриным и зоологом Института эколо-
гии растений и животных Уральского научного центра АН СССР
Н. Г Смирновым в 1974 г. Я принимала участие в раскопках Мед-
вежьей пещеры на Северном Урале в 1961 г., на стоянке Бызовая на
Средней Печоре в 1965 г., в гроте Столбовой на р. Усьва, в гротах
Близнецова «Черные кости» на р. Чаньва в бассейне Камы на Среднем
Урале в 1968 г. В 1978 г. обследовала грот Ладейный близ г. Алек-
сандровска Пермской области, а в 1980 г. обработала палеонтологи-
ческие сборы Геологического музея в Свердловске и в краеведческих
музеях Свердловска, Нижнего Тагила, Серова, Краснотурьинска, Кар-
пинска, Североуральска Среднего Зауралья.
Основная масса ископаемых остатков хранится в Зоологическом
институте АН СССР в Ленинграде. Среди них обычны костные об-
ломки волка, песца, пещерных медведей — большого и малого, донского
зайца, степной пищухи, копытного лемминга, мамонта, уральской ло-
шади, шерстистого носорога, северного оленя, овцебыка и бизона. Ви-
довой состав и относительная численность видов изменялись с севера
на юг и с запада на восток. На Северном Урале было больше песца,
донского зайца, копытного лемминга, северного оленя и овцебыка, на
Среднем Урале — большого и малого пещерных медведей, светлого
хоря, барсука, сурка, суслика, мамонта, лошади, шерстистого носо-
рога, сайги и бизона. При этом пещерные медведи, светлый хорь, бар-
сук, сурок, суслик и шерстистый носорог чаще встречались на запад-
ных склонах Среднего Урала, а мамонт, лошадь, сайга и бизон —
на восточных склонах. Таким образом, на Северном и Среднем Урале
в позднем плейстоцене преобладали виды открытых биотопов тундры,
степи и лесостепи, приспособленные к холодному и сухому климату.
Однако Зауралье, по-видимому, было несколько более континентальным
и сухим, чем Предуралье, и в нем было еще меньше лесных стаций
обитания, чем на западных склонах Урала. Причем, на Северном Урале
64
найдено 548 костей лошади, на западных склонах Среднего Урала —
592 кости, на восточных склонах —577 костей. В процентном отноше-
нии это составляло на Северном Урале 2.2%, в Предуралье—13.7%»
в Зауралье — 33.7% от общего числа ископаемых костей для каждого
региона в отдельности (Кузьмина, 1971, 1975, 1982).
Первоначальными исследованиями было установлено, что поздне-
плейстоценовая лошадь Северного Урала достоверно отличалась от ши-
рокопалой лошади Русской равнины (Кузьмина, 1971). Позже, после
находки и изучения черепа лошади из пещеры «Черные кости» в Перм-
ской области, уральская лошадь была выделена в самостоятельный
подвид Equus caballus uralensis Kuzmina (1975). Сравнение промеров
черепа и костей скелета ленской лошади с аналогичными промерами
уральской лошади показало их несомненные отличия (Кузьмина, 1977).
При составлении Каталога млекопитающих СССР (1981) проведено
некоторое упорядочение в подроде настоящих лошадей, и несколько
подвидов были повышены до вида, в частности уральская лошадь.
Перейдем к более подробному рассмотрению внешнего вида и устрой-
ства скелета уральской лошади.
Экстерьер
В позднем плейстоцене на Урале обитали лошади средних разме-
ров. Высота их в холке, вычисленная по методике Витта (1952), коле-
балась от 125 до 150 см, а в среднем была 139 см. Большой размах
крайних значений роста связан, вероятно, с половыми отличиями жи-
вотных (табл. 1). Жеребец, основная длина черепа которого была
517 мм, в холке достигал, по-видимому, 150 см.
Таблица 1
Высота в холке уральских лошадей (по методике Витта, 1952)
Кость Кол-во экз. Длина кости, мм Высота в холке, см
lim м lim м
Череп, основная длина 1 517 150
Лучевая 11 312—362 333 128—150 137
Пястная 28 201—228 216 125—140 133
Берцовая 9 318—376 348 125—148 135
Плюсневая Площадь копыта и копытной 38 фаланги ; 246—286 у некоторых 266 видов 128—150 Табл рода Equus 139 и ц а 2
Копыто, см2 Копытная фаланга, см2
Виды переднее передняя задняя
Е. latipes — 6 43.0 40.9
Е. uralensis — — — 10 42.3 37.6
Е. lenensis 1 121 115 9 39.6 36.1
Судить о пропорциях туловища уральской лошади можно лишь
косвенно, сравнивая ее с другими видами лошадей. Так, ширина диа-
физа пястной кости, отнесенная к основной длине черепа у Equus ura-
lensis составляла 6.96%, у сибирской Е. lenensis— 7.07%, у широко-
5 Млекопитающие
65
палой лошади Русской равнины Е. talipes — 7.80%. Площадь копытной
фаланги передней ноги уральской лошади была 42.3 см2, задней ноги —
37.6 см2. По этому признаку уральская лошадь занимала промежуточ-
ное положение между широкопалой лошадью и ленской (табл. 2).
Таким образом, если ленскую и широкопалую лошадей можно было
считать мелким и крупным «тяжеловозами», то уральскую лошадь
следует относить к лошадям более легкой конституции, сухим и под-
вижным (Кузьмина, 1975, 1980; Верещагин, Лазарев, 1977).
Череп
В 1968 г. во время археологических раскопок под руководством
О. Н. Бадера, в пещере «Черные кости», на берегу небольшой р. Анюши,
относящейся к бассейну Чаньвы, на западных склонах Среднего Урала
был найден череп лошади сильно поврежденный при раскопках. Осе-
вой череп взрослой особи оказался без значительной части теменной
кости, дистальной части носовых, наружной части левой верхнечелюст-
ной, задней части небной кости. Отсутствовали также решетчатая
кость, сошник, крыловидная и передняя часть клиновидной кости
(рис. I,вклейка). Тем не менее можно было сделать наиболее важные
промеры черепа (табл. 3).
Таблица 3
Промеры черепа ^(мм) уральской лошади Equus uralensis Kuzmina
Основная длина 517.0 Ширина заглазничного суже- 84.0
Кбндилобазальная длина 543.0 ния
Длина морды от prostion до 330.0 Ширина в суставных отрост- 207.0
ближайшей точки края ках
глазницы Наибольший поперечник 67.0
Длина вернхего зубного ряда 175.5 глазницы
Длина диастемы 103.0 Высота позади М3 118.0
Ширина лба у заднего края 214.0 Мин. высота черепа в основ- 104.0
глазниц ной кости
Череп принадлежал жеребцу в возрасте около 8 лет, что установ-
лено по методике Корневен и Лесбр (1932). Все швы на черепе срос-
лись, клыки хорошо развиты и начали стираться, резцы в профиль
изгибаются плавной дугой, резцы в верхней челюсти с хорошо разви-
той чашечкой. Он отличался узостью лба и мозговой коробки и при-
ближался по этим промерам к черепу арабской лошади (Кузьмина,
1975).
В краеведческом музее Свердловска была найдена затылочная
часть черепа лошади позднеплейстоценового возраста без точного ме-
стонахождения. Сохранившиеся скуловые отростки позволили измерить
скуловую ширину. Она равнялась 198 мм, ширина между концами слу-
ховых трубок была 113 мм, заглазничное сужение — 85 мм.
Кроме того, найдено еще пять фрагментов черепа с полностью со-
хранившимся верхним рядом коренных. Размеры верхнего зубного
ряда уральских лошадей и Предуралья составляли 172—178 мм, в сред-
нем 175.6 мм. В двух случаях сохранилась альвеола Рт[, и тогда длина
верхнего ряда коренных составила 172—184 мм, в среднем 178 мм.
Длина 2 экз. из Зауралья была 164—170 мм, в среднем 167 мм. Свя-
зать изменения размеров верхнего зубного ряда с полом не удалось.
Зубы
Предкоренные зубы были крупнее коренных. Парастиль и мезо-
стиль сильно выступали за контур зуба, Рт3 и Рт4 слегка сплюснуты
€6
и имеют небольшую выемку. Передняя марка в нижнем левом углу
имеет глубокую складочку, вдающуюся в переднюю марку и почти
достигающую передней наружной лопасти — паракона. Этот признак
прослеживается на всех предкоренным и коренных, за исключением
Рт2 и М3. На передней стенке задней марки центральная складочка
сильно развита, так что она отчленяет нижнюю часть задней марки
в виде крестообразной ямки, особенно на предкоренных. Внутренняя
долинка имеет сильно развитую шпору, особенно ярко выраженную
на предкоренных зубах (рис. 2, вклейка). Протокон на коренных зубах
узкий, плоский и длинный, на предкоренных Рт3 и Рт4 несколько
раздут в задней половине. Относительная длина протокона равномерно
увеличивается от Рт2 к Л13 (табл. 4).
Нижние челюсти представлены полными правой и левой ветвями
от одной особи и еще пятью фрагментами от взрослых особей. Не-
сколько челюстей сохранилось от жеребят разного возраста: на ста-
дии трех молочных коренных и стадии прорезания первого и второго
коренных. У взрослых длина челюсти от сочленовного отростка до
альвеолы центрального резца составляла 430 и 445 мм, длина ряда
коренных (п=5) изменялась от 165 до 180 и в среднем была 176.2 мм.
Длина диастемы от коренных до резцов была 100 мм, у жеребца —
от коренных до клыков — 78 мм. Высота челюсти в сочленовном от-
ростке составляла (п=3) 239—256 мм; между Рт4 и М\ в четырех
случаях была 79—84 мм, в среднем 81.5 мм; в диастеме высота была
50 мм. Ширина резцовой части была 66 мм.
Эмаль коренных зубов уральской лошади тоньше, чем у широко-
палой лошади Equus latipes. Для уральской лошади характерно, что
передняя часть петли — метаконид — на предкоренных зубах напоми-
нает равносторонний треугольник с закруглениями на вершинах, на
коренных — округлой формы. Метастилид почти треугольной формы
с заостренной вершиной. Наружная долинка хорошо развита и глу-
боко вдается в шейку двойной петли (особенно на коренных зубах),
причем шпора на ней не развита. На предкоренных зубах наружная
долинка менее длинная, только приближается к шейке двойной петли
и шпоры на ней нет. Размеры постоянных зубов нижней челюсти пред-
ставлены в табл. 5. Талонид последнего коренного зуба при слабой
степени стирания был всего 6 мм длиной, что составляло 17—18%
длины зуба. Иногда он был крестообразной формы (рис. 3, вклейка).
У средне- и сильно стертых Л43 он достигал 7—10 мм, что соответство-
вало 20.9—28.7% длины зуба, т. е. талонид последнего коренного зуба
уральской лошади был относительно короче талонида широкопалой
лошади.
Кости конечностей
Кости конечностей уральской лошади частично уже описаны (Кузь-
мина, 1971, 1975), особенно для животных, обитавших на Северном
Урале и на западных склонах Среднего Урала, в Предуралье. Теперь
можно провести сравнение, добавив данные о зауральских лошадях —
с восточных склонов Среднего Урала. Всего для этого измерено
342 кости по схеме, принятой ранее (Кузьмина, 1971).
Лопатки лошадей немногочисленны в сборах. В каждой из срав-
ниваемых групп оказалось по 3 экз. Из них наиболее крупными по
всем промерам оказались лопатки зауральских лошадей, затСм мельче
были лопатки североуральских животных и самыми мелкими — из
пермской популяции (табл. 6).
5*
67
3
Таблица 4
Размеры постоянных зубов верхней челюсти
Р/п2 Рт3 Рт1
Промеры, Jim М т lim М :• т lim М±т
Длина И 27.5—40.0 37.3+1.06 17 26.2—32.2 29.9+0.38 22 24.6—32.8 29.4+0.39
Ширина 11 23.2—28.5 25.2+0.47 17 25.5-31.8 28.6+0.33 22 23.6—29.5 27.3+0.38
Длина протокона — — — 17 10.6—15.0 12.8+0.27 22 12.0-15.4 13.5+0.23
Относительная длина протокона, % — — — 17 34.8—53.4 42.8+1.10 22 37.8—52.1 46.0 + 0.73
М' ЛР ЛР
Промеры, lim М±т lim М-^т lim М ± т
Длина 10 23.6-28.2 26.6+0.45 15 22.3—28.5 25.5+0.37 15 24.2-32.2 29.3+0.52
Ширина 10 28.0—29.0 28.5+0.29 15 25.6-29.2 27.1+0.28 14 22.5-26.0 24.5+0.25
Длина протокона 10 12.5—14.2 13.4+0.18 15 11.6—15.2 13.4+0.29 14 13.6—17.3 15.4+0.34
Относительная длина протокона, % 10 48.9—51.1 50.5 + 0.35 15 44.6—59.5 52.7+1.10 14 44.6—66.1 53.0+1.58
Таблица 5
Размеры постоянных зубов нижней челюсти
Промеры, мм Рт2 Рт9 Pmk
lim М±т lim Af +/72 lim M±m
Длина 10 30.6-34.5 32.8+0.39 4 26.8—29.0 28.1+0.48 5 25.5—28.2 26.9±0.48
Ширина 10 15.2-18.2 16.8±0.32 4 16.7—20.0 18.7 + 0.70 5 17.5—20.5 19.0±0.55
Промеры, мм Aft м2 Af3
lim Al ±т lim Al - m lim Af + m
Длинна 7 24.0-26.4 24.9+0.34 1 24.8—27.7 25.8+0.37 14 31.0-36.5 33.4+0.47
Ширина 7 17.0-19.3 17.8 + 0.34 1 15.6-18.2 16.9+0.42 14 14.0—16.3 15.2+0.21
Длина талонида — — — — — —* 12 7.0—10.0 8.4+0.34
Относительная длина талонида, % о — — — — — — 12 20.9—28.7 25.2+0.72
о
Таблица 6
Абсолютные размеры костей передней ноги Equus uralensis Kuzmina
Кость Промеры, мм Северный Урал Средний Урал
Предуралье Зауралье
п lim М z т п | lim М±т п | lim М±т
Лопатка Scapula Ширина через шишко- видный бугор 3 85.0—95.0 90.3zE2.91 3 87.0—92.0 89.0zEl.52 3 96.0—103.0 100.0±2.08
Ширина суставной по- 3 47.3—50.0 48.8±0.79 3 54.0—58.0 55.3zEl.33 3 59.0—64.0 61.0± 1.52
верхности Поперечник суставной поверхности 3 54.5—61.5 59.0±2.25 3 47.0—48.0 47.5±0.29 3 52.0—55.0 53.3±0.88
Плечевая Humerus Макс, длина от голов- ки — — — 5 249.0—276.0 265.6±5.47 3 265.0—280.0 274.3 ±4.70
Ширина нижнего эпи- физа Поперечник нижнего эпифиза 3 73.0-80.0 76.3±2.03 10 75.0—83.0 78.7±0.99 5 72.0—84.0 77.6±2.40
3 86.5—91.0 88.8zEl.30 10 81.0-98.5 88.8zEl.82 4 82.0—94.0 88.1 ±2.69
Лучевая Макс, длина кости — — — 9 312.0—362.0 333.7zE5.71 2 320.0; 338.0 329.0
Radius Ширина верхнего эпи- физа Ширина диафиза 1 — 84.0 8 80.0-91.0 85.9± 1.41 4 78.0—87.2 83.0±1.99
— — — 7 39.0—44.5 41.8±0.78 3 39.0-41.0 40.0±0.57
Ширина нижнего эпи- физа 1 — 80.0 8 73.0—82.0 76.8±0.99 3 73.0—77.0 74.3± 1.22
Пястная Макс, длина кости 6 212.0—224.0 217.6± 1.75 9 201.0—228.0 217.3zE3.18 13 208.0—224.0 214.8±1.41
Metacarpale Ширина верхнего эпи- физа Ширина диафиза 7 49.0—55.3 52.2±0.84 9 49.0—55.0 52.3±0.65 93 48.0—54.5 50.9±0.60
6 31.5-38.5 36.0±1.03 8 33.0-39.0 36.0±0.76 12 34.0—39.0 35.8 + 0.56
Ширина нижнего эпи- физа 6 47.5—54.0 51.4zE0.95 9 49.0—55.5 51.lzE0.72 13 47.5-53.6 50.4±0.44
Промеры,
Кость
Северный Урал
п lim
Первая фаланга Сагиттальная длина 8 73.0—86.2 79.5 ±1.45
Phalanx I по переднему краю Ширина верхнего эпи- 8 54.0—62.0 58.5±0.88
физа Ширина диафиза 8 38.0—41.3 39.2±0.39
Ширина нижнего эпи- 8 45.6—49.3 47.9±0.52
физа
Вторая фаланга Сагиттальная длина по 5 36.0—40.0 38.3±0.86
Phalanx II переднему краю Ширина верхнего эпи- 6 52.2—58.5 55.9±0.93
физа Ширина диафиза 6 45.8—50.3 48.4±0.77
Ширина нижнего эпи- 4 49.3—53.5 52.0±0.94
физа
Третья фаланга Сагиттальная длина по 4 51.0—60.0 54.7±2.04
Phalanx III переднему краю Ширина суставной по- 4 48.5—54.6 52.5± 1.38
верхности Поперечник суставной 4 26.2—31.5 29.4± 1.20
поверхности Макс, ширина копыта 4 92.5-100.0 95.6 ±1.57
Продолжение табл. 6
Средний Урал
Предуралье Зауралье
п lim | М -т п Пт М±т
10 73.0—81.0 76.4±0.71 15 72.0—83.6 77.3±1.93
10 56.0—61.0 58.4±0.44 13 55.0—61.2 57.8±0.56
10 36.5—39.5 38.0±0.28 15 37.0—43.0 38.9±0.43
10 44.0—49.0 46.6 + 0.58 15 45.0—52.0 47.8±0.52
7 35.5—42.0 37.5±0.88 9 36.0—39.8 37.9±0.44
7 53.0—58.0 55.5±0.78 9 53.5—58.2 55.9±0.56
7 45.0-52.0 48.6±0.99 9 46.5—52.0 49.0±0.66
7 48.2—56.5 52.6± 1.13 8 51.0-54.2 52.3±0.40
7 47.5—60.0 53.3±1.60 2 55.0; 58.0 56.5
7 50.0—56.0 52.4±0.84 2 51.0; 52.0 51.5
7 26.0—30.0 28.4±0.58 2 26.0; 27.0 26.5
6 91.0-102.0 94.7 ±1.66 2 94.0; 95.0 94.5
Целые плечевые кости были найдены только на Среднем Урале,
причем зауральские оказались более длинными, чем предуральские.
Соотношение ширины и поперечника нижнего эпифиза плечевой кости
было сходно для всех трех сравниваемых популяций, хотя кости ло-
шадей с Северного Урала были немного мельче остальных (табл. 6).
Это же соотношение позволило разделить плечевые кости по полу
73 75 77 79 81 83
ШИРИНА,ММ
Рис. 4. Размеры нижнего эпифиза плечевой кости.
1 — Северный Урал; 2 — Средний Урал, Предуралье; 3 — Средний Урал, Зауралье.
(Кузьмина, 1982). Самкам, по всей вероятности, принадлежали кости
длиной 249—273 мм при ширине нижнего эпифиза 72.8—78.0 мм и по-
перечнике— 81—89 мм. К самцам отнесены плечевые кости длиной
274—280 мм при ширине нижнего эпифиза 80—84 мм и поперечнике
91—98.5 мм (рис. 4, табл. 7). Вычисленный коэффициент достоверности
различий подтверждает правильность сделанного разделения.
Целые лучевые кости были собраны преимущественно в пещерах
Пермской области, и еще 2 экз. известны из Зауралья.. На Северном
72
Таблица 7
Размеры плечевых костей
Промеры, мм Самки (10 экз.) Самцы (7 экз.) t
lim М ±т 11m М
Длина кости 249.0—273.0 260.8±5.23 274.0—280.0 277.0±1.29 3.00
Ширина нижнего эпи- физа 72.8—78.0 75.8±0.58 80.0-84.0 82.0 ±0.58 7.56
Поперечник нижнего эпифиза 81.0—89.0 85.3±0.91 91.0—98.5 93.5±1.05 5.90
Рис. 5. Размеры лучевой кости.
Обозначения, как на рис. 4.
Урале найдены только верхний и нижний концы лучевых. Сравнение
размеров костей лошадей из трех популяций показало, что наиболее
крупные лучевые кости были у животных из Предуралья (табл. 6).
К самкам можно было отнести лучевые длиной 318—330 мм при ши-
рине верхнего эпифиза 78—86 мм, ширине диафиза 39—41 мм, ширине
нижнего эпифиза 73—77 мм. Кости самцов были крупнее. Длина луче-
вых самцов была 338—362 мм. Соответственно шире были эпифизы и
диафиз (рис. 5, табл. 8). Разделение лучевых костей лошадей по полу
дало статистически достоверные результаты.
73
Таблица 8
Размеры лучевых костей
Промеры, мм Самки (5 экз.) Самцы (5 экз.) t
lim М ±т lim Af ±/п
Длина кости 318.0—330.0 322.4zfc2.06 338.0—362.0 347.4±4.43 5.12
Ширина верхнего эпи- физа Ширина диафиза 78.0—86.0 81.9±1.37 85.0—91.0 88.2±1.24 3.41
39.0—41.0 40.2±0.48 40.0—44.5 42.6±0.83 2.50
Ширина нижнего эпи- физа 73.0—77.0 74.6±0.99 75.0—82.0 78.1 ± 1.16 2.29
Таблица 9
Размеры пястных костей
Промеры, мм Самки (19 экз.) Самцы (9 экз.) t
lim М±т lim М±т
Длина кости 201.0—218.0 212.6zfcl.02 221.0—228.0 223.8±0.92 8.18
Ширина верхнего эпифиза 48.0—54.5 50.8zfc0.46 49.0—55.5 53.2±0.58 3.24
Ширина диафиза 31.5—39.0 35.5zfc0.43 33.0—39.0 36.9±0.74 1.65
Ширина нижнего эпифиза 47.5—53.6 50.4 ±0.40 49.0—55.5 51.8 ±0.72 1.71
Локтевая кость из Предуралья имела в длину 83 мм. Ширина лок-
тевого отростка равнялась 58 мм. По-видимому, она принадлежала
самке.
При сравнении размеров пястной кости лошадей Северного Урала,
Предуралья и Зауралья кажется, что особи северной популяции ближе
к Предуральским, а зауральские мельче (табл. 6). Если же учесть
половой состав попавших в выборки костей животных, окажется, что
из северной популяции рассматриваются 4 2 и 2d, в предуральской —
4 9 и 5 d, а в зауральской—112 и 2d (рис. 6). Таким образом, из-
мельчание зауральских животных может быть кажущимся, так как
в выборке оказалось больше самок, чем самцов.
К самкам были отнесены пястные кости длиной 201—218 мм при
ширине верхнего эпифиза 48.0—54.5 мм. К самцам — длиной 221—
228 мм и шириной верхнего эпифиза 49.0—55.5 мм. Эти размеры дали
статистически достоверные отличия (/ равен 8.18 и 3.24 соответ-
ственно). Различия в ширине диафиза и ширине нижнего эпифиза не
достигли статистически достоверного уровня (табл. 9). Таким образом,
у самок наиболее крупными были особи из североуральской и заураль-
ской популяции, а у самцов — предуральские.
Фаланги передней и задней конечности рассмотрим вместе, после
трубчатых костей задней конечности.
Целые бедренные кости были очень малочисленны в сборах — на
Северном Урале и в Зауралье найдено всего по одной кости и две
в Предуралье. Наиболее крупный экземпляр длиной 365 мм оказался
с Северного Урала, мельче — длиной 355 мм — был из Зауралья,
74
а Предуральские кости были еще короче: 347 и 352 мм (табл. 10).
Однако выборки слишком малы, чтобы судить о географической и по-
ловой изменчивости.
Рис. 6. Размеры пястной кости.
Обозначения, как на рис. 4.
Большая берцовая кость сохранилась целой чаще, чем бедренная,
особенно нижний конец кости. Самые мелкие кости были найдены на
Северном Урале, несколько крупнее в Зауралье, и самые крупные —
в Предуралье (табл. 10). По соотношению ширины и толщины ниж-
него эпифиза большой берцовой кости можно разделить найденные
ископаемые остатки по полу (рис. 7). Тогда получается, что самкам
принадлежали кости шириной 64.0—75.2 мм при толщине 42.0—48.5 мм,
а самцам — шириной 74—84 мм и толщиной 47—51мм. Коэффициент
достоверности различий более 6 подтвердил правильность разделения
(табл. 11).
Целые пяточные кости с приросшим верхним эпифизом встреча-
лись изредка. На Северном Урале найдено всего 2 экз., на Среднем
Урале — по 3 экз. в Предуралье и Зауралье (табл. 10). Чаще встреча-
Таблица 10
Абсолютные размеры костей задней ногн Equus uralensis Kuzmina
Северный Урал Средний Урал
Кость Промеры, мм Предуралье Зауралье
lim М±т lim М±т lim Л4±т
Бедренная Femur Макс, длина от голов- ки Ширина нижнего эпи- физа 1 3 89.0—92.0 365.0 90.3±0.89 2 3 347.0; 352.0 349.5 90.0 1 1 — 355.0 85.0
Большая берцо- вая Tibia Макс, длина кости Ширина верхнего эпи- физа Ширина нижнего эпи- физа Поперечник нижнего эпифиза 2 1 8 318.0—340.0 66.5—78.0 329.0 95.0 72.2 ±1.59 6 7 14 330.0—376.0 84.0—103.0 66.0—84.0 353.3±7.44 94.6±2.43 77.1 ±1.34 1 1 10 64.0-76.8 350.0 100.0 72.4± 1.47
8 42.0—50.0 46.5±0.81 9 46.0-51.0 48.4±0.56 10 43.0—49.0 46.0±0.64
Пяточная Calcaneus Макс, длина кости Ширина суставной ча- сти 2 103.4—110.0 106.7 3 10 109.0-112.0 50.0—58.5 110.3±0.88 53.8±1.14 3 18 101.2—107.0 45.0—56.0 103.24=1.88 51.2±0.68
Таранная Astragalus Макс, длина кости Ширина дистального конца 8 8 51.8—61.0 50.5-59.0 58.0±1.00 53.6± 1.02 14 14 57.0—66.0 52.0—63.0 60.3±0.72 57.2 ±0.88 18 18 54.0-63.0 52.0-62.3 58.8±0.53 55.9±0.60
Плюсневая Metatarsale Макс, длина кости Ширина верхнего эпи- физа Ширина диафиза Ширина нижнего эпи- физа 5 10 249.4—268.7 48.2—56.0 258.7±3.32 52.4±0.90 26 31 250.0—286.0 46.0-59.0 265.0± 1.95 52.7±0.61 7 8 246.0—271.0 50.6—57.8 261.64=3.27 53.5±0.87
5 5 30.8—35.0 47.5—54.3 33.5±0.74 51.9± 1.40 24 26 29.0—36.5 46.5—58.5 33.7±0.43 52.8 ±0.54 7 7 31.0—37.0 49.0—54.0 35.0±0.75 52.3±0.69
Кость Промеры, мм Северный Урал
lim Af±m
Первая фаланга Сагиттальная длина по 12 68.8—78.5 72.7+0.85
Phalanx I переднему краю Ширина верхнего эпи- 12 50.5—59.3 54.4+0.81
физа Ширина диафиза Ширина нижнего эпи- 12 12 34.2-36.6 42.8-45.4 35.4+0.23 43.9 + 0.25
физа
Вторая фаланга Сагиттальная длина по 3 38.8—41.5 40.4+0.83
Phalanx II переднему краю о 54.2—57.5 55.3+1.08
Ширина верхнего эпи- о
физа Ширина диафиза 3 о 46.0—47.8 49.0—52.2 46.6+0.60 50.7 + 0.93
Ширина нижнего эпи- о
физа
Третья фаланга Сагиттальная длина по 4 51.5-59.5 55.6+1.65
Phalanx III переднему краю л 45.5—51.5 48.8+1.36
Ширина суставной по- 4
верхности Поперечник суставной 4 26.0—28.3 27.0 + 0.57
поверхности Макс, ширина копыта 2 73.0; 80.0 76.5
______Продолжение табл. 10
Средний Урал
Предуралье Зауралье
lim М + т lim М т
6 71.8—810 75.9+1.43 10 70.0—81.0 76.4+1.16
6 53.5-59.0 56.9±0.91 9 54.8—63.0 57.1+0.94
6 34.0—36.5 35.2h-0.47 10 33.0—37.5 35.7+0.43
6 43.5—47.5 45.4±0.60 9 42.5-46.5 44.4+0.43
5 38.0—40.0 38.8+0.37 8 35.5—40.3 38.1+0.54
5 51.0—56.0 53.7+0.86 8 47.5—54.0 52.2 + 0.74
5 42.0—47.0 45.0+0.89 8 40.5-46.5 43.5+0.66
5 48.0—50.5 49.4 + 0.48 8 43.0-51.0 48.1+0.91
6 53.0—57.0 54.8+0.87 4 50.0-57.5 55.1 + 1.76
6 44.5—52.5 49.0+1.18 5 49.0—53.0 51.0+0.70
6 26.0—28.2 27.5+0.65 5 26.0—29.0 27.5+0.63
6 80.0—89.0 86.8+1.53 4 80.0—87.0 82.9+1.50
лись пяточные кости с поврежденными (погрызенными) или не при-
росшими верхними эпифизами. Наиболее крупные пяточные кости
найдены в Предуралье, длиной 109—112 мм при ширине суставной
части 50.0—58.5 мм. Самыми мелкими были кости из Зауралья — дли-
ной 101.2—107 мм и шириной 45—56 мм, североуральские занимали
промежуточное положение. Связать с полом разные размеры костей
оказалось затруднительным.
ПОПЕРЕЧНИК НИЖНЕГО ЭПИФИЗА, ММ
Рис. 7. Размеры нижнего эпифиза большой берцовой кости.
Обозначения, как на рис. 4.
Таранные кости обычны в палеонтологических сборах. В трех срав-
ниваемых популяциях наиболее мелкие кости оказались на Северном
Урале, немного крупнее — в Зауралье, и самые крупные были найдены
в Предуралье (табл. 10). Анализ соотношения длины и ширины кости
Таблица 11
Размеры больших берцовых костей
Промеры, мм Самки Самцы t
п lim М±т п lim М ± т
Макс, длина кости 2 330; 340 335.0 4 347—376 356.8±6.58 3.31
Ширина верхнего эпифиза Ширина нижнего эпифиза Поперечник ниж- него эпифиза 2 95; 96 95.5 3 97—103 100.0 ±0.82 5.48
17 64.0—75.2 70.6±0.81 11 74—84 78.6±0.92 6.56
17 42.0—48.5 45.7±0.42 11 47—51 49.1 ±0.35 6.30
78
позволил отнести к самкам таранные длиной 51.8—60.0 мм при ши-
рине 50.5—57.5 мм; к самцам — длиной 59—66 мм и шириной 56—63 мм.
Коэффициент достоверности различий равный почти 6 и 7 подтвердил
правильность разделения (рис. 8, табл. 12).
ШИРИНА ДИСТАЛЬНОГО КОНЦА,ММ
Рис. 8. Размеры таранной кости.
Обозначения, как на рис. 4.
Плюсневые кости лошадей обычно хорошо сохраняются в четвер-
тичных отложениях. Наиболее массовыми в сборах и более крупными
были кости из Предуралья. Их длина колебалась от 250 до 286 мм.
Короче были кости из Зауралья, длиной 246—271 мм. Самые мелкие,
североуральские плюсневые, достигали 249—268.7 мм. Попытка разде-
лить кости по полу была успешной (рис. 9, табл. 13). К самкам отне-
Размеры таранных костей
Таблица 12
Промеры, мм Самки (24 экз.) Самцы (16 экз.) t
lim М±т lim М±т
Длина кости 51.8—60.0 57.6+0.38 59.0±66.0 61.5±0.42 6.96
Ширина дистального конца 50.5-57.5 54.1 ±0.42 56.0—63.0 58.5±0.61 5.95
79
сены кости длиной 246.0—262.2 мм при ширине верхнего эпифиза 47.0—
55.5 мм. К самцам — плюсневые длиной 263—286 мм при ширине верх-
него эпифиза 50—59 мм. Коэффициент достоверности различий был
очень высок, особенно для длины костей. Ширина верхнего и нижнего
эпифизов также показала статистически достоверные отличия.
47 49 51 53 55 57 59
ШИРИНА ВЕРХНЕГО ЭПИФИЗА,ММ
Рис. 9. Размеры плюсневой кости.
Обозначения, как на рис. 4.
Фаланги лошади обычны в палеонтологических сборах на Урале.
Разделить первые фаланги на передние и задние помогла форма костей
и их размеры. Первые фаланги передней ноги имеют параллельные
стенки диафиза, тогда как у фаланг задней ноги они сужаются к сере-
дине диафиза, поэтому у первых фаланг задних ног ширина диафиза
в середине наименьшая. На задней стенке первой фаланги передней
конечности У-образная шероховатость составляет две трети длины
кости. Она опускается своей вершиной значительно ниже середины
80
Таблица 13
Размеры плюсневых костей
Промеры, мм Самки (19 экз.) Самцы (19 экз.) t
lim Л1 ±т lim М^т
Макс, длина кости 246.0—262.2 255.8±0.94 263.0—286.0 271.4±1.45 9.02
Ширина верхнего эпи- физа 47.0—55.5 51.4±0.55 50.0—59.0 54.7±0.52 4.40
Ширина диафиза 29.0—37.0 33.0±0.56 32.0—36.5 34.8±0.31 2.86
Ширина нижнего эпи- физа 46.5—55.0 51.1+0.54 49.0—58.5 53.9 ±0.49 3.84
Рис. 10. Размеры первой фаланги передней (•) и задней (X) ноги.
кости, тогда как на костях задних ног она чуть больше половины, не-
значительно опускаясь ниже середины кости.
Первые фаланги передней ноги у самок достигали размеров 72.0—
77.5 мм в длину при ширине диафиза 36.5 —40.0 мм, у самцов 75.0—
86.2 мм в длину при ширине диафиза 38—43 мм. Различия статистиче-
ски достоверны (рис. 10, табл. 14). Установлено, что на Северном
Урале собраны первые фаланги передних ног 2$ и 6d, в Предуралье —
7 $ и 4 в, в Зауралье — 9$ и 6 <?, поэтому средние значения размеров
кости различных географических популяций Урала можно сравнивать
6 Млекопитающие
81
Таблица 14
Размеры первой фаланги передней ноги
Промеры, мм Самки (18 экз.) Самцы (16 экз.) t
lim М±т Пт М±т
Сагиттальная длина по переднему краю 72.0—77.5 74.9±0.35 75.0—86.2 80.5±0.68 7.37
Ширина верхнего эпи- физа 54.0—60.3 57.4±0.42 55.0—62.0 59.0dz0.50 2.42
Ширина диафиза 36.5—40.0 37.9i0.23 38.0—43.0 39.6dz0.35 4.15
Ширина верхнего эпи- физа 44.0—49.0 46.6±0.36 45.0—52.0 48.4±0.47 3.15
Таблица 15
Размеры первой фаланги задней ноги
Промеры, мм Самки (13 экз.) Самцы (15 экз.) t
lim М + т lim М±т
Сагиттальная длина по переднему краю 68.8—73.4 71.2dz0.37 75.0—81.0 77.7+0.52 10.16
Ширина верхнего эпи- физа 50.0—59.3 54.8dz0.80 53.5—63.0 56.9zb0.71 1.98
Ширина диафиза 34.2—36.6 35.5dz0.22 33.0—37.5 35.4dz0.35 0.24
Ширина нижнего эпи- физа 42.8-47.5 44.3±0.35 42.5—46.5 44.5 ±0.34 0.41
параллельно, отдельно для самцов и самок. Такое сравнение показало,
что больше всего сходства у самцов Северного Урала и Зауралья.
Первые фаланги этих животных выше и стройнее. У лошадей из Пред-
уралья эти кости ниже и шире. Такая же закономерность проявилась
при сравнении костей самок этих же популяций.
Первые фаланги задней конечности отличались от передних не
только формой, но и размерами. Сравнение было проведено отдельно
между самками и самцами. Оно показало, что у самок длина первых
фаланг передней конечности выше, чем у этих же костей задних ног
в среднем на 3.7 мм (/=7.25). У самцов в этом промере разница
меньше. Она достигает 2.8 мм (/ = 3.29). Наибольшие различия отме-
чены в ширине диафиза. У самок она составляла 2.4 мм, у самцов
4.2мм (/=7.5 и 8.57 соответственно). Сравнение верхнего и нижнего
эпифизов показало, что больше различий в ширине нижнего эпифиза,
коэффициент достоверности различий достигал у самок 4.6, у самцов
6.72. Тем самым статистически подтверждена правильность рис. 10.
Первые фаланги задней ноги у самок достигали длины 68.8—
73.4 мм при ширине диафиза 34.2—36.6 мм, у самцов 75—81 мм длиной
и 33.0—37.5 мм ширины диафиза. Различия были достоверны (/=10.16)
82
Таблица 16
Размеры второй фаланги передней ноги
Промеры, мм Самки (15 экз.) Самцы (7 экз.) t
lim М + т 11m М±т
Сагиттальная длина по переднему краю 35.5-39.0 36.8+0.27 38.2—42.0 39.8+0.44 5.77
Ширина верхнего эпи- физа 52.2—57.5 54.8+0.40 57.0—58.5 57.7+0.23 6.30
Ширина диафиза 45.0—50.3 47.7+0.41 49.6—52.0 50.9+0.41 5.52
Ширина нижнего эпи- физа 48.2—56.0 51.8+0.54 52.0—56.5 53.7+0.79 1.98
Рис. 11. Размеры второй фаланги передней (•) и задней (X) ноги.
только для длины первой фаланги. Остальные размеры кости были
сходны у самцов и самок (рис. 10, табл. 15).
Отличать вторые фаланги передних и задних ног не трудно, так
как они различны по форме. Вторые фаланги передних ног относи-
тельно низкие и широкие, а задних ног выше и уже (рис. 11). Вторые
фаланги передних ног у самцов и самок отличаются размерами. К сам-
кам отнесены кости длиной 35.5—39.0 мм при ширине диафиза 45.0—
50.3 мм, к самцам длиной 38.2—42.0 мм при ширине диафиза 49.6—
52.0 мм. Различия достигли статистически достоверного уровня не
только по этим промерам, но и ширине верхнего эпифиза (табл. 16).
На Северном Урале найдены вторые фаланги 4$ и 2d, в Пред-
уралье 5$ и 2<J, в Зауралье 6$ и 3d. Во всех популяциях количество
самок вдвое превышало число самцов. Сравнение средних размеров
показало, что более высокими и узкими были вторые фаланги перед-
них ;ног на Северном Урале и в Зауралье. В Предуралье они были
короче и шире (табл. 6).
6*
83
Таблица 17
Размеры второй фаланги задней ноги
Промеры, Самки (12 экз.) Самцы (4 экз.) t
lim М ± т lim М +т
Сагиттальная длина по переднему краю 35.5-39.0 38.1 ±0.34 40.0—41.5 40.7+0.34 5.42
Ширина верхнего эпи- физа 47.5—56.0 52.8 + 0.64 52.0—57.4 54.6±1.14 1.37
Ширина диафиза 40.5—47.0 44.3+0.59 41.8—47.8 45.4+1.27 0.78
Ширина нижнего эпи- физа 43.0—51.0 48.8+0.66 46.2—52.2 49.5+1.27 0.49
ШИРИНА СУСТАВНОЙ ПОВЕРХНОСТЯМИ
Рис. 12. Размеры третьей фаланги передней (•) и задней (X) ноги.
Вторые фаланги задних ног у самцов и самок отличаются разме-
рами. К самкам отнесены кости длиной 35.5—39.0 мм при ширине диа-
физа 40.5—47.0 мм, к самцам длиной 40.0—41.5 мм при ширине диа-
физа 41.8—47.8 мм (рис. И, табл. 17). Половой состав сравниваемых
выборок был следующий: на Северном Урале найдены вторые фаланги
задних ног 1$ и 2d; в Предуралье 4 $ и 1 d, в Зауралье 7? и 1 <5.
Сравнение размеров этих костей самцов показало, что самыми мел-
84
кими, низкими и узкими были кости из Зауралья. Средние размеры
костей самок трех сравниваемых популяций выявили ту же закономер-
ность.
Третьи фаланги передних ног отличались от таких же костей зад-
них ног тем, что они выше и шире (рис. 12, 13, вклейка; табл. 6, 10).
К самкам отнесены фаланги высотой 50.0—55.6 мм при ширине копыта
91—96 мм, к самцам высотой 58—60 мм при ширине копыта 98—102 мм.
Материал невелик, чтобы сделать выводы о географической изменчи-
вости размеров этой кости, однако самые узкие третьи фаланги были
из Зауралья.
Из третьих фаланг задних ног к самкам отнесены кости высотой
47.5—53 мм при ширине копыта 73—92.5 мм, к самцам — высотой
55.0—59.5 мм при ширине копыта 80—95 мм. Самыми широкими эти
кости были в Предуралье, а на Северном Урале и в Зауралье уже.
Географическая изменчивость и распространение
Сравнение размеров костей конечностей уральской лошади из трех
«популяций» показало, что в позднем плейстоцене на Урале обитал
хорошо охарактеризованный морфологически, особый вид уральской
лошади, отличавшийся от широкопалой и ленской лошадей. Географи-
ческая изменчивость уральской лошади проявилась в укрупнении боль-
шинства костей скелета у животных из Предуралья. Североуральские
и, в особенности, зауральские лошади имели более узкие кости конеч-
ностей. Вероятнее всего, это связано с обитанием в более сухом, кон-
тинентальном климате. Более крупные размеры лошадей Предуралья
и более широкие фаланги этих животных свидетельствуют, что они
обитали в более мягком, влажном климате, на менее твердом грунте,
который связан с большей облесенностью местности. Это подтверждает
сделанные ранее выводы, полученных при сравнении видового состава
и относительной численности млекопитающих на западных и восточ-
ных склонах Среднего Урала (Кузьмина, 1982). Отличия лошадей из
Предуралья и Зауралья достигали, по-видимому, подвидового уровня.
Итак, ископаемые остатки уральской лошади известны пока только
из позднеплейстоценовых отложений Северного и Среднего Урала.
На север она была распространена до 65° с. ш. по Средней Печоре
(стоянка Бызовая), на юг до 55° с. ш. в Каслинском районе Челябин-
ской области (грот Зотинский) и Сухоложском районе Свердловской
области (грот Безымянный). На западе предположительно можно счи-
тать, что она была распространена до Вятки, на восток по Зауралью,
ограниченному реками Обью и Тоболом (рис. 14).
Систематическое положение
В настоящее время систематическое положение уральской лошади
таково: отр. Perissodactyla
сем. Equidae
род Equus Linnaeus, 1758
Е. (Equus) uralensis Kuzmina, 1975
Диагноз. Лошадь высотой в холке 125—150см, легкой, «сухой»
конституции, с костями конечностей более узкими, чем у широкопалой
лошади. Кости конечностей ленской лошади короче. Череп длинный,
узкий, низкий с узким лбом и мозговой коробкой. Основная длина че-
репа 517мм, скуловая ширина 198мм (38.3%), ширина между кон-
85
ПЛЕЧЕВАЯ ЛУЧЕВАЯ
ПЯСТНАЯ БЕДРЕННАЯ БОЛЬШАЯ ПЛЮСНЕВАЯ
БЕРЦОВАЯ
Рис.
15. Отличия в размерах
1—Equus latipes,
крупных костей конечностей трёх видов
лошадей.
2 — Е. uralensis, 3 — Е. lenensis.
иами слуховых трубок 113 мм (21.8%), ширина у заднего края глазниц
214 мм (41.5%), ширина заглазничного сужения 84 мм (16.3%), высота
позади М3—118 мм (22.8%), длина верхнего зубного ряда 176 мм
(34%), нижнего— 165—180мм (34%) основной длины черепа. Эмаль
зубов средней толщины и складчатости. Относительная длина прото-
кона на Л41 48.9 мм — 51.1%, на М2 44.6 мм — 59.5%, на Af3 44.6 —
66.1% длины зуба. Длина пястных костей 201—228 мм при ширине
диафиза 31—39 мм. Длина плюсневых костей 246—286 мм при ширине
диафиза 29—37 мм. Ширина третьей фаланги передней ноги 91—102 мм,
задней ноги 73—95 мм. От позднеплейстоценовых видов лошадей Е. 1а-
tipes и Е. lenensis отличается длинным, узким и низким черепом, вы-
сокими и узкими костями конечностей (табл. 6, 10; рис. 15).
Голотип. Осевой череп (колл. Зоологического института АН
СССР, Ленинград, № 29724), пещера «Черные кости» Пермской обла-
сти, раскопки О. Н. Бадера, Е. П. Близнецова и И. Е. Кузьминой
в 1968 г.
Материал. Нижние челюсти и кости конечностей более 1500экз.
в коллекциях Зоологического института АН СССР.
Геологический возраст. Поздний плейстоцен.
Распространение. По предгорьям Северного и Среднего
Урала от 55° с. ш. до 65° с. ш. На западе, возможно, ограничен тече-
нием р. Вятки, на востоке Обью и Тоболом.
Выводы
Уральская лошадь — Equus uralensis Kuzmina, 1975 — узко рас-
пространенный вид, обитавший на Урале в позднем плейстоцене, вхо-
дил в состав мамонтового териокомплекса. При дальнейшем изучении
вымерших лошадей Евразии возможно расширение ареала и выясне-
ния происхождения вида, а также выделение двух подвидов.
В позднем плейстоцене на Урале эта лошадь обитала в лесостеп-
ных ландшафтах, особенно в Предуралье, а на Северном Урале и в За-
уралье— в более открытых местообитаниях. По внешнему виду — круп-
ное животное, высотой в холке 125—150 см, легкого склада, с высо-
кими, стройными конечностями, копытами более узкими, чем у широко-
палой лошади. Половой диморфизм выражен в размерах и пропор-
циях костей конечностей. Количество взрослых самок в табунах пре-
вышало количество самцов в 2 и более раза. В популяциях представ-
лены все возрастные группы от жеребят до старых особей. Большое
число раздробленных костей говорит о том, что в жизни палеолитиче-
ских охотников Урала лошадь играла важную роль как источник пищи.
ЛИТЕРАТУРА
Верещагин Н. К., Лазарев П. А. Описание частей трупа и скелетных остатков селе-
риканской лошади. — Тр. Зоол. ин-та АН СССР, 1977, т. 63, с. 85—185.
Витт В. О. Лошади Пазырыкских курганов. — Сов. археология, 1952, № 14, с. 165—
205.
Каталог млекопитающих СССР, плиоцен — современность. Л., Наука, 1981, 456 с.
Корневен и Лесбр. Распознавание возраста по зубахМ и производным эпителия.
М. — Л., изд-во сельхоз. и колхозн. лит., 1932, 213 с.
Кузьмина И. Е. Формирование териофауны Северного Урала в позднем антропо-
гене.— Тр. Зоол. ин-та АН СССР, 1971, т. 49, с. 44—122.
Кузьмина И. Е. Некоторые данные о млекопитающих Среднего Урала в позднем
плейстоцене. — Бюл. Комис, по изуч. четвертичн. периода. М., 1975, т. 43, с. 63—77.
Кузьмина И. Е. О происхождении и истории териофауны Сибирской Арктики.—Тр.
Зоол. ин-та АН СССР, 1977, т. 63, с. 18—55.
87
Кузьмина И. Е. Позднеплейстоценовая широкопалая лошадь Верхнего Дона. — Тр.
Зоол. ин-та АН СССР, 1980, т. 93, с. 91—118.
Кузьмина И. Е. Видовой состав и относительная численность млекопитающих Сред-
него Урала в позднем плейстоцене. — Тр. Зоол. ин-та АН СССР, 1982, т. 111>
с. 44—48.
Кузьмина И. Е. Определение пола у млекопитающих по ископаемым костным остат-
кам.— III съезд Всес. Териол. о-ва, тез. докл. М., 1982, т. 1, с. 43—44.
Лазарев П. А. Антропогеновые лошади Якутии. М., Наука, 1980, 190 с.
SUMMARY
Urals Late Pleistocene horse Equus uralensis Kuzmina, 1975, is a species with nar-
row distribution range, that inhabited in the Urals during the Late Pleistocene and was
a part of mammoth teriocomplex. A further study of extinct horses of Eurasia may
extend its area and clearify origin of the species, also perhaps distinguish two sub-
species.
In the Late Pleistocene in the Urals this horse was inhabiting in forest steppe land-
scapes, particularly in Preuralia, and in more exposed habitats of the Northern Urals
and Transuralia. In its appearance it is a tall animal, its height in the withess being
125—150 cm, slight, with high slender limbs, hoofs narrower than of Equus latipis.
Sexual dimorphism is expressed in sizes and proportions of limb bones. The number
of adult females in herds was 2 and more times higher than of males. All age groups
from colts to old individuals are represented in the populations. The large amount of
splintered bones is the evidence of the important role of hoise as a source of food
for paleolithic hunters.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1985, ТОМ 131
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, 1985, VOL. 131
USSR ACADEMY OF SCIENCES
МЕЗОЛИТИЧЕСКАЯ КОКШАРОВСКО-ЮРЬИНСКАЯ СТОЯНКА
В СРЕДНЕМ ЗАУРАЛЬЕ
Ю. Б. Сериков, И. Е. Кузьмина
Нижнетагильский краеведческий музей, Зоологический институт АН СССР,
Ленинград
Ju. В. Serikov, I. Е. Kuzmina. Mesolitic Koksharovsko-Yurinskaya site in Middle Trans-
uralia
Кокшаровско-Юрьинская стоянка находится на восточном берегу
Кокшаровского торфяника в 70 м к западу от деревни Кокшарово
Верхнесалдинского района Свердловской области (рис. 1). Памятник
открыт в 1979 г. Нижнетагильским археологическим отрядом Ураль-
ского государственного университета под руководством Ю. Б. Сери-
кова и исследован в 1980 г. Стоянка располагается на низком корен-
ном берегу торфяника, в северной части которого находится зарастаю-
щее озеро. Раскопками вскрыто 132 м2 площади. Характер памятника
довольно сложный. Стоянка состоит из двух частей — береговой и при-
брежной. На берегу культурные остатки ранней бронзы и неолита за-
легают сразу под дерном, в черном иловатом слое мощностью до 40 см.
В прибрежной части изделия раннего железа и поздней бронзы зале-
гают в торфе, на глубине соответственно 60—70 см и 80—100 см (Се-
риков, 1980). Под торфом в слое темно-серого песка залегают куль-
турные остатки ранней бронзы, энеолита, неолита и мезолита. Мощ-
ность этого слоя достигает 50—60 см. Еще ниже, в слое галечника и
светло-серого песка, залегают материалы только мезолитического вре-
мени. Раскопки дали 804 каменных и 186 костяных изделия (рис. 2).
Из каменных изделий можно отметить нуклеусы, скребки, резцы,
резчики, микропластинки с ретушью. Интересны находки в этом слое
шлифованных тёсел, их обломков и шлифовальных плит. Из костяных
орудий на стоянке найдены игловидные, биконические и вкладышевые
наконечники стрел, ретушеры, обломки ножей, лощила, проколки, гар-
пун, рыболовные грузила и некоторые другие предметы. Ближайшие
аналоги всему комплексу находок встречаются в материалах Шигир-
ского торфяника (Толмачев, 1914). Там в конце прошлого века в про-
цессе золотодобычи собраны тысячи костяных изделий. Была
выделена даже шигирская культура. Правда, разные исследователи
относили ее к различным археологическим эпохам от бронзового века
(Дмитриев, 1951) до мезолита (Раушенбах, 1956). Наши первые рас-
копки на Кокшаровском торфянике неоспоримо доказывают, что боль-
шая часть шигирских костяных изделий, игловидных и биконических
наконечников стрел, гарпунов, вкладышевых орудий датируется мезо-
литическим временем. На Кокшаровско-Юрьинской стоянке подобные
костяные изделия залегают в четких стратифицированных слоях
в сочетании с кремневым инвентарем, который имеет широкие анало-
гии в мезолитических памятниках Среднего Зауралья, таких как
Выйка II, «Серый камень» на Горбуновском торфянике, Крутяки I и др.
Кроме костяных и каменных изделий, в раскопе обнаружены кости
млекопитающих, птиц и рыб (см. таблицу). В более древних отложе-
89
ниях, датируемых мезолитом определены кости волка, бурого медведя,
бобра, северного оленя и лося, причем кости лося наиболее многочис-
ленные, от двух особей, взрослого и молодого. В слое с примесью
гальки чаще всего встречались кости волка (16 экз.), бобра (76 экз.)
и лося (93 экз.). Найдены также остатки трех видов куньих — куницы,
барсука и черного хоря. Наибольший интерес представляют кости со-
бак. К ней отнесены две левых ветви нижних челюстей и один обломок
правой, верхний эпифиз лучевой кости, метаподия, хвостовой позвонок
и обломок зуба. Значит, они представляют остатки как минимум двух
особей. Лучше всего сохранилась левая ветвь нижней челюсти с предко-
ренными зубами и предпоследним коренным. У нее немного обломаны
угловой и сочленовный отростки, венечного нет совсем (рис. 3, вклейка).
Длина зубного ряда без резцов была равна 100 мм, у другого экзем-
пляра— 101мм, длина Рт2— 8.9 мм, ширина 5 мм, длина Рт3—
11.6 мм, ширина 5.7 мм, длина Рт4—13 мм, ширина 6.4 мм. Альвеола
All была в длину 22 м при ширине 7.3 мм. Длина М2— 9.4 мм, ширина
6.9 мм. Альвеола 7И3— в длину 5 мм, в ширину 3.5 мм. Высота челюсти
между Рт± и М\ была 26.5 мм, за Л13 — 30 мм. Характерная кривизна
зубного ряда при рассматривании сверху и малые размеры предкорен-
ных зубов показывают, что эта челюсть принадлежала собаке, а не
волку (Кузьмина, Сериков, 1982).
Среди костных остатков, датируемых мезолитом и годных для из-
мерений, была и нижняя челюсть бобра. Длина зубного ряда ее была
равна 37 мм. Плечевая кость бобра сохранилась без верхнего эпифиза,
ширина нижнего эпифиза ее равнялась 30 мм.
В отложениях темно-серого песка, датируемых неолитом, были
собраны остатки волка, бурого медведя, черного хоря, барсука, бобра,
полевок, северного оленя, лося, птиц и рыб. Больше всего было костей
лося, из них преобладали зубы и их мелкие обломки. Годной для из-
мерений оказалась единственная таранная кость. Ее длина была 78 мм,
ширина нижнего эпифиза 49.3 мм.
SO
Рис. 2. Каменные и костяные изделия, найденные на Кокшаровско-Юрьинской стоянке.
Видовой состав и количество костных остатков животных
на Кокшаровско-Юрьинской стоянке
Раскопки Ю. Б. Серикова в 1979, 1980 гг.
Виды Галечник, погребенный торф светлый песок, мезолит Темно-серый песок, неолит Торф, бронза
косте й особей костей особей костей особей
Дикие Волк — Canis lupus L. 16 2 14 3 1 1
Бурый медведь — Ursus arctos 5 1 7 1 — —
L. Черный хорь — Mustela puto- 8 1 2 1 — —
rius L. Куница — Martes martes L. 2 1 2 1 — —
Барсук — Meles meles L. 1 1 5 1 — —
Бобр — Castor fiber L. 76 7 32 5 — —
Полевки — Microtinae 22 — 32 — — —
Лось—Alces alces L. 93 6 336 12 11 2
Северный олень — Rangifer ta- 4 1 24 2 7 1
randus L. Птицы — Aves 86 — 54 — 4
Рыбы — Pisces — — 5 — — —
Домашние Собака — Canis familiaris L. 8 2 — — —
Всего
костей
31
12
10
4
6
108
54
440
35
144
5
особей
2
2
12
20
4
6
2
2
8
2
В самом верхнем слое торфа, датируемом бронзой, найдены
остатки волка, северного оленя, лося и 4 кости птиц, ближе не опре-
деленные.
В целом полученные при раскопках костные остатки млекопитаю-
щих свидетельствуют, что в окрестностях Кокшаровско-Юрьинской
стоянки на протяжении голоцена существовава тайга, где жили типично
лесные обитатели — лось, северный олень, бобр, волк и бурый мед-
ведь. Жители этой стоянки занимались преимущественно охотой на
зверей и птиц и рыбной ловлей. Костные остатки собак по меньшей
мере от двух особей подтверждают, что мезолитические охотники уже
имели четвероногих помощников.
ЛИТЕРАТУРА
Дмитриев П. А. Шигирская культура на восточном склоне Урала. — Матер, и исслед.
по археологии СССР, 1951, № 21, с. 28—93.
Кузьмина И. Е., Сериков Ю. Б. Новые находки голоценовых млекопитающих в Сред-
нем Зауралье. — В кн.: Экология горных млекопитающих. Свердловск, УНЦ АН
СССР, 1982, с. 60—61.
Раушенбах В. М. Среднее Зауралье в эпоху неолита и бронзы. — Тр. Гос. истор. му-
зея, 1956, вып. 29, с. 103—104.
Сериков Ю. Б. Работы Нижнетагильского отряда. — Археол. открытия. 1979. М.,
Наука, 1980, с. 171—172.
Толмачев В. Я. Древности Восточного Урала. — Зап. Уральск, о-ва любит, естеств.,
1914, т. 34, вып. 8, с. 151—266.
SUMMARY
In 1979 members of Nizhnetagil expedition of the Urals University discovered
Koksharovsko-Yurinskaya site. Analysis of archeological findings has shown that the site
was inhabited from Mesolithic till Bronze Age. The chief occupation of the inhabitants
of the site was hunting taiga animals and birds. The most common prey were elks,
reindeers, beavers, wolves, brown bears and birds. The hunters used dogs.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1985, ТОМ 131
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, 1985, VOL. 131
USSR ACADEMY OF SCIENCES
О МЕТОДИКЕ ОПРЕДЕЛЕНИЯ ГЕНЕРАЦИЙ
КОРЕННЫХ ЗУБОВ ВЫМЕРШИХ СЛОНОВ
А. В. Шер, В. Е. Гарутт
Палеонтологический институт АН СССР, Москва-
Зоологический институт АН СССР, Ленинград
А. V Sher, V Е. Garutt. On the method of identification of molar generations in ele-
phants
Зубная система хоботных Proboscidea характеризуется многими
своеобразными чертами строения по сравнению с другими группами
млекопитающих. Она является довольно сильно редуцированной, и эта
редукция выражена наиболее ярко у слонов Elephantidae — самых про-
грессивных представителей отряда.
В ротовой полости слонов одновременно функционируют два верх-
них резца — dentes incisivi — (I), образующие бивни, и четыре жева-
тельных зуба, по одному в каждой половине верхней и нижней челю-
сти. Постоянные предкоренные зубы (премоляры)—dentes praemola-
res (Р)—у слонов редуцированы; в виде рудиментарных остатков
они сохранились лишь у наиболее древних, архаичных видов (Гарутт,
1977). У всех остальных слонов функционируют лишь молочные пре-
моляры— dentes praemolares decidui (Pd) и коренные (моляры) —
dentes molares (М).
Коренные зубы слонов сменные. В процессе пережевывания пищи
они постепенно стираются, выпадают и замещаются последующими,
формирующимися в полостях челюстей. В периоды замены одного зуба
другим в ротовой полости слона могут одновременно функционировать
до восьми жевательных зубов: по два зуба в каждой половине челюсти
наверху и внизу. Подобные смены зубов происходят в течение жизни
слона пять раз. И только у самых архаичных форм слонов, у которых
в онтогенезе сохранялись постоянные предкоренные, смена зубов имела
место шесть или даже семь раз.
Зубы разных генераций отличаются друг от друга размерами,
числом пластин и их частотой, т. е. теми параметрами, которые весьма
важны для определения эволюционного положения вымерших форм,
поэтому при изучении слонов точное определение генерации каждого
зуба играет первостепенную роль. Общепризнано, что наиболее легко
диагностируются зубы двух первых молочных генераций — Pd2 и Pd3,
а также самый последний заднекоренной — М3. Труднее отличать по-
следний предкоренной — Pd4 от первого коренного — Ml, первый ко-
ренной от второго коренного — М2. С проблемой диагностики смен
зубов сталкивается каждый исследователь, обрабатывающий любую
серию ископаемых зубов слонов, поэтому многие авторы рассматри-
вали этот вопрос в методическом аспекте (Pohlig, 1888; Pavlow, 1910;
Корниец, 1959; Дуброво, 1960; Щеглова, 1961; Гарутт и Форонова,
1976; Гарутт, 1977; Жерехова, 1977; Урбанас, 1980 и другие). Тем не
менее, общепризнанная и надежная методика диагностики пока отсут-
ствует, в результате чего возникают затруднения в определении смен
93
зубов и разногласия между исследователями ископаемого материала
по слонам. Более того, отдельные авторы считают невозможным уве-
ренное разделение Pd4 и Ml даже на однородных сериях из одного
местонахождения (Урбанас, 1980). Очевидно, что при неправильном
определении генерации зуба будут неоправданно расширяться пределы
изменчивости показателей, важных для диагностики видов, а также не-
верно оцениваться возрастной состав популяций.
Всего нами исследовано более 300 зубов разных смен. Материал
хранится в коллекциях Палеонтологического, Зоологического и Геоло-
гического институтов Академии наук СССР. Авторы признательны
руководству и сотрудникам этих учреждений за предоставление воз-
можности изучения этого материала. Руководствуясь изложенными
выше требованиями, мы стремились не включать в выборку зубы, пара-
метры которых не могли быть точно определены. При необходимости
включения таких зубов (например, когда в челюсти имеются зубы двух
рмен и один из них значительно стерт), величина параметра фиксиро-
валась как минимальная и его обозначение на графиках сопровожда-
лось специальным значком.
После такой выбраковки в нашей серии осталось 238 зубов, в том
числе 155 зубов ранних и средних смен (Pd2 — М2). Основу этой вы-
борки составляли зубы мамонтов Matnmuthus primigenius из позднего
плейстоцена Восточной Сибири, в том числе большая серия из Бере-
лехского «кладбища». Вторая часть изученной коллекции — зубы древ-
них слонов с мамонтоидными признаками из северо-восточной Якутии
(конец плиоцена — начало плейстоцена), систематическое описание ко-
торых подготовлено авторами. В эту же группу нами включен опуб-
ликованный ранее (Гарутт, 1966) материал по молочным зубам из
местонахождения Тологой. Наконец, для сравнения мы включили в вы-
борку некоторое количество зубов мамонтов из позднего плейстоцена
Европейской части Союза, в основном из позднепалеолитических стоя-
нок (Авдееве, Юдиново). Сводка включенного в выборку материала
приведена в табл. 1.
Таблица 1
Состав выборки зубов слонов разных смен
Время и место находок ₽d2 Pd3 м} М2
Поздний плейстоцен Сибири (в основном, Восточной) Берелехское «кладбище» Поздний плейстоцен Европейской части Союза Древние слоны Восточной Сибири 2 9 7 9 9 31
5 2 10 7 8 9 9 8 И 6 18 7
1 0 3 1 4 2 4 1 4 0 4 0
1 1 1 2 2 3 1 1 № О 1 14
1 2 1 3 ; з 8
Всего по сменам 5 19 21 19 17 52
8 16 15 17 18 31
Примечание. В числителе — количество верхних зубов, в знаменателе ниж-
них.
Многие исследователи справедливо считают, что важнейшим клю-
чем к распознаванию зубов соседних смен служат остатки особей, по-
гибщих в то время, когда у них в каждой ветви челюсти функциониро-
94
рало одновременно по два зуба. В распоряжении авторов имелось 19
нижних и 10 верхних челюстей с зубами двух смен. Челюсти были
вскрыты для получения точных параметров зубов и послужили важ-
ными опорными точками на предлагаемых ниже графиках.
Общая тенденция изменения параметров зуба от смены к смене
в онтогенезе хорошо известна: это — возрастание размеров коронки
и увеличение числа пластин с одновременным снижением их частоты.
Параметры заднекоренных зубов соседних смен перекрываются, причем
трансгрессия иногда весьма значительна. Молочные переднекоренные
зубы почти по всем параметрам практически не имеют трансгрессии.
По имеющимся в нашем распоряжении нижним челюстям мамонта
с зубами двух соседних смен мы попытались, хотя бы весьма прибли-
зительно, оценить соответствие между размерами коронок и общими
размерами животного, приняв за показатель последних длину нижней
челюсти.* Оказалось, что с начала функционирования Pd3 до начала
стирания Ml длина нижней челюсти возрастает примерно в два раза,
а от начала стирания Ml до появления М3 — в полтора-два раза. При
етом длина Ml больше таковой Pd3 в среднем в два с половиной раза,
а М3 длиннее Ml в полтора-два раза. Таким образом, на первой из на-
званных стадий «рост» зубов ют смены к смене несколько опережает
по темпу рост челюсти, а на второй они выравниваются по темпу. Это
опережение как бы компенсирует относительную укороченность перед-
некоренных зубов: если длина сформированного Pd3 примерно равна
половине длины челюсти или чуть меньше ее, то длина М3 составляет
около двух третей нижней челюсти.
Молочные предкоренные увеличиваются в размере от смены
к смене скачками, коренные — более постепенно. Это хорошо
иллюстрируют цифры, полученные нами в результате анализа сводных
таблиц В. Мальо (Maglio, 1973) для мамонта М. primigenius и южного
слона Archidiskodon tneridionalis, суммирующих промеры большого
числа зубов разных смен. Длина Pd3 в среднем больше, чем длина Pd2
в 2:6 раза, a Pd4 длиннее Pd3 в среднем в 1.9 раза. В каждой из
остальных смен (Pd4 — Ml, Ml—М2 и М2 — М3) каждый последую-
щий зуб длиннее предыдущего в 1,3 раза, т. е. в среднем на 30%.
Близкие цифры получаются по сводной таблице В. Е. Гарутта и
И. В. Фороновой (1976) для мамонта.
Перечисленные выше признаки неравномерности развития зубов
в онтогенезе легко объясняются при знакомстве с их наиболее раннимй
стадиями. На рентгенограмме черепа детеныша мамонта из Киргиляха
(Верещагин, 1981) отчетливо видно, что в челюстях у семимесячного
животного присутствуют уже все молочные предкоренные зубы, при-
чем Pd2 и Pd3 функционируют, а у Pd4 оформлены все пластины и пе-
редние уже начинают срастаться у основания. Раннее заложение всех
цредкоренных зубов, каждый из которых намного крупнее предыду-
щего, является, по-видимому, тем механизмом, который обеспечивает
резерв зубной системы при увеличении общих размеров животного
в онтогенезе. Отсюда следует, что размеры коронок молочных зубов
в меньшей степени подвержены колебаниям в зависимости от размеров
растущей особи.
В отличие от предкоренных, коренные закладываются более посте-
пенно, процесс их формирования и последующего функционирования
* За длину нижней челюсти мы в данном случае приняли расстояние от перед-
него конца альвеолы зуба до заднего края челюсти параллельно жевательной по-
верхности (Дуброво, 1960).
95
значительно растянут во времени. Так, по аналогии с ныне живущими
слонами (Гарутт, 1977 и др.), если последний предкоренной зуб (Pd4)
функционирует 5—7 лет, то первый коренной (Ml) «работает» уже 10—
12 лет, второй (М2) — более 20 и третий (М3) более 30 лет. Если же
учесть время формирования коронки зуба до его прорезывания, то
жизнь каждого заднекоренного зуба еще дольше. За это долгое время
каждый зуб проходит свой «онтогенез». Процесс этот изучен еще недо-
статочно. Можно предполагать зависимость размеров моляров от об-
щих размеров животного, которые продолжают увеличиваться, по край-
ней мере, до появления М3, а следовательно, большую изменчивость
размерных характеристик коренных зубов по сравнению с предкорен-
ными.
Указанные обстоятельства — меньший коэффициент возрастания
размеров коренных зубов от смены к смене и большая степень их ин-
дивидуальной изменчивости — объясняют значительно более широкую
трансгрессию признаков у соседних смен заднекоренных по сравнению
с предкоренными.
Из всех параметров зубов разных смен наиболее независимым
от особенностей онтогенеза, генетически обусловленным признаком яв-
ляется полное число пластин в коронке. Хотя этот признак также под-
вержен индивидуальной изменчивости в пределах каждого вида, но
в меньшей степени, чем размерные величины. Из последних наиболее
изменчива длина зуба, кроме того, истинное значение длины можно
измерить только на зубах, почти не затронутых стиранием, а такие
Зубы весьма редки в коллекциях. Стирание переднего отдела коронки
начинается еще тогда, когда задняя часть коронки, скрытая в аль-
веоле, полностью не сформирована, т. е. пластины этого отдела еще
слабо связаны между собой цементом. Первой стирается передне-верх-
няя площадка.* При промерах незначительно стертых зубов (показа-
тели стертости 1 и 2 по Гарутту и Фороновой, 1976) отсутствием этой
части обычно пренебрегают, но она часто довольно сильно выступает
вперед, и измеренная без ее учета длина коронки будет занижена.
Еще более грубая ошибка в измерении длины происходит в тех слу-
чаях, когда коронки с уже уничтоженной стиранием передней секцией
пластин (в районе первого корня) принимаются за целые. Подобную
ошибку рассматривает в недавней работе И. А. Дуброво (Dubrovo,
1981).
Менее изменчива высота коронки зуба (Н), если понимать под ней
полную высоту самой большой пластины. Однако на практике чаще
измеряется наиболее высокая из сохранившихся пластин, не затрону-
тых стиранием, а таковая далеко не всегда оказывается самой высо-
кой пластиной в коронке данного зуба.
Наименее изменчива максимальная ширина зуба. Она сравни-
тельно медленно, но неуклонно возрастает от смены к смене. По весьма
приближенным оценкам по тем же таблицам В. Мальо (1973), В. Е. Га-
рутта и И. В. Фороновой (1976) видно, что закономерность изменения
ширины зуба от смены к смене примерно такая же, как и длины.
Резко, в 2.5—3 раза ширина Pd3 превышает ширину Pd2, Pd4 шире,
чем Pd3 примерно в 1.5 раза; в следующих сменах этот коэффициент
более или менее постоянен и колеблется по данным ряда авторов
у различных видов слонов в среднем от 1.2 до 1.3, т. е. каждый корен-
* Для простоты изложения и у верхних, и у нижних зубов мы условно назы-
ваем «нижней» прикорневую часть коронки, а «верхней» — ту, где расположена жева-
тельная поверхность.
£6
дой зуб шире предыдущего в среднем на 20—30%. Меньшая изменчи-
вость ширины коронки определенной смены (по сравнению с другими
размерами) отражается в меньшей трансгрессии зубов соседних смен
до данному признаку. Это отмечали некоторые исследователи и ранее
(Щеглова, 1961; Жерехова, 1977).
Нам представляется вполне закономерным, что трансгрессия раз-
меров коронки зубов соседних смен незначительна у предкоренных зу-
0ов и выражена более сильно у коренных, и что наиболее устойчивыми
параметрами являются число пластин и ширина коронки и наиболее
изменчивым — ее длина. Тем не менее даже и эти устойчивые признаки
имеют трансгрессию, поэтому, как справедливо отмечает В. В. Щег-
лова (1961), для установления порядкового номера (смены) зуба не-
обходимо пользоваться комплекссом признаков.
В процессе обработки нашей коллекции мы пришли к выводу, что
наиболее целесообразно строить сопряженные графики наименее транс-
грессирующих показателей: 1—числа пластин в коронке и ее ширины,
2—числа пластин в коронке и ее высоты. Данные графики для нашей
коллекции (отдельно для верхних и нижних зубов) приведены на
рис. 1—4. Значками помечены экземпляры, для которых оба пара-
метра поддаются точному определению или надежной реконструкции.
Значками со стрелками — экземпляры, для которых известно точное
значение одного параметра (по оси, перпендикулярной направлению
стрелки), а второй не поддается точному определению из-за стертости
коронки и для него приведена минимальная оценка. В этом случае
ртрелка показывает направление, по которому на графике должна ле-
жать точка, соответствующая первоначальной величине параметра.
После нанесения всех точек на графики мы провели разграничи-
тельные линии, отделяющие поля зубов каждой смены. При этом мы
руководствовались положением на графике опорных точек, принад-
лежность которых к определенной смене не вызывала сомнений, в пер-
вую очередь — экземпляров с двумя зубами разных смен в одной че-
люсти. Вначале проводилась линия, разделяющая поля с минимальным
разрывом, после чего обычно не составляло труда провести парал-
лельно ей остальные разграничительные линии.
Понятно, что положение разграничительных линий имеет чисто
эмпирический характер и определяется выбранным масштабом и кон-
кретными точками на графике, поэтому угол наклона этих линий
в значительной степени случайный. Особенно это относится к пред-
коренным зубам, где распределение точек более кучное, а разрыв ме-
жду соседними сменами большой. Так, между полями Pd2 и Pd3 раз-
граничительную линию можно было бы провести и вертикально, и
горизонтально. Однако в целом проведение наклонных разграничитель-
ных линий показалось нам в данном случае более адекватно отражаю-
щим характер распределения материала, чем нанесение традиционных
для скаттер-диаграмм овалов. Это связано в первую очередь с тем,
что наша выборка неоднородна не только по половому, но и по систе-
матическому признаку. Легко видеть, что в косых полях наших гра-
фиков точки, соответствующие зубам древних слонов, тяготеют
д нижней правой части поля. Это объясняется тем, что при несколько
более крупных линейных размерах эти зубы имеют меньшее полное
число пластин. С другой стороны, зубы мамонтов из Берелеха в целом
отклоняются в левую верхнюю часть полей, что отражает морфоло-
гическое своеобразие этой одной из наиболее поздних популяций ма-
7 Млекопитающие
97
зонтов (относительно мелкие размеры при довольно высоком полном:
числе пластин). Особенно заметны эти различия на М2 и М3, где раз-
мах изменчивости параметров зубов максимален. Таким образом, рас-
пределение точек по диагонали, параллельной разграничительным ли-
Рис. 1. Отношение количества пластин в коронке верхнего зуба к ее ширине.
1 — мамонты позднего плейстоцена Сибири (в основном Восточной), 2 — мамонты Берелехского
«кладбища» (р. Индигирка), 3 — мамонты позднего плейстоцена Европейской части СССР (позд-
непалеолитические стоянки Авдеево, Юдиново и др.), 4 — древние слоны с мамонтоидными призна-
ками из Восточной Сибири (олерская свита северо-восточной Якутии, поздний плиоцен — ранний,
плейстоцен; Тологой, ранний плейстоцен).
Рис. 2. Отношение количества пластин в коронке верхнего зуба к ее высоте.
Обозначения те же, что и на рис. 1.
$шям, соответствует общей тенденции увеличения числа пластин при
ртносительном сокращении линейных размеров зубов, и, следовательно^
в той или иной степени отражает эволюционную изменчивость в пре-
делах нашей выборки. Распределение точек по другой диагонали (бо-
98
лее мелкие зубы с меньшим числом пластин и более крупные с боль-
шим) должно в принципе отражать внутрипопуляционную изменчи-
вость, в частности, половую (см., например, две группы черных круж-
ков на графике ширины М3, но эта изменчивость может быть оценена
только по обширной однородной выборке (одна популяция), которой
мы пока не располагаем.
Таким образом, разграничение полей зубов разных смен наклон-
ными линиями представляется нам наиболее оптимальным для разно-
родной выборки, с которой мы имеем дело.
На первый взгляд, недостатком предлагаемой нами методики мо-
жет показаться то, что она основывается на абсолютных (линейных),
а не на относительных параметрах (пропорциях), которые обычно счи-
таются более стабильными. Однако в данном случае это не так. На-
пример, как было показано выше, увеличение длины и ширины зуба
от смены к смене происходит примерно одинаковыми темпами, поэтому
индекс ширины зуба к длине мало что дает для определения смены.
Из пропорций зуба в какой-то степени показательно лишь отношение
ШИРИНА КОРОНКИ, мм
Рис. 3. Отношение количества пластин в коронке нижнего зуба к ее ширине.
Обозначения те же, что и на рис. 1.
Показатели средней длины пластин или их частоты также мало
пригодны для распознавания смен зубов, несмотря на существование
общей тенденции увеличения длины пластин и снижения их частоты
от ранних смен к поздним. Разброс значений этого признака и транс-
грессия его у соседних смен на нашем материале исключительно ве-
лики. В данном случае этот разброс увеличивается еще и благодаря
видовой неоднородности выборки. Убывая от смены к смене в онто-
генезе, частота пластин возрастает в филогенезе, причем последнее
особенно заметно у более поздних смен. Возникают как бы две про-
тивоположные тенденции, и в результате разброс очень велик. Однако
и в одной популяции трансгрессия частоты пластин у соседних смен,
начиная с Pd4, весьма велика. Это видно, в частности, на примере по-
пуляции мамонта из Берелеха (Жерехова, 1980).
Предложенные графики (рис. 1—4) четко показывают отсутствие
трансгрессии отдельных параметров предкоренных и значительную
7*
99
трансгрессию между сменами коренных, что отмечалось выше, однако
при совмещении на графике двух трансгрессирующих параметров
удается довольно уверенно разделить экземпляры разных смен. Наи-
более трансгрессирующие смены иногда легче различаются по одному
из двух видов графиков: например, М2, вероятно, легче отличать от Ml
по графику высоты, но слабо стертые зубы этих генераций очень редки
в коллекциях; мало их и нашем графике.
Рис. 4. Отношение количества пластин в коронке нижнего зуба к ее
высоте.
Обозначения те же, что и на рис. 1.
Тем не менее, нельзя сказать, что предложенное построение пол-
ностью исключает трансгрессию и позволяет во всех случаях одно-
значно определить смену зуба. На наших графиках есть несколько
точек, которые почти попадают на разграничительную линию; большая
часть этих точек лежит на границе полей Ml и М2. Это заставило
нас рассмотреть вопрос о морфологических особенностях, свойственных
зубам определенных смен (кроме последних коренных). Вопрос этот
труден и мало разработан, хотя он весьма актуален для распознава-
ния зубов средних смен, и, в первую очередь, Ml и М2. Все же неко-
торые наблюдения в ходе нашей работы появились, хотя пока не со-
всем ясно, насколько универсальное значение они имеют.
1. В связи с тем, что высота коронки от смены к смене нарастает
быстрее, чем ширина, соотношение этих параметров у разных смен зу-
бов различно (табл. 2).
Из таблицы видно, что наиболее четко по этому индексу отли-
чаются Pd* от Л41 и Mi от М2. Остальные смены (при сохранении об-
щей тенденции снижения индекса к более поздним сменам) обнару-
живают ту или иную степень трансгрессии его значений. Эта транс-
грессия частично связана с неоднородностью выборки. Так, если из се-
рии Pd± исключить зуб архаичного слона с №7/7=65.0, индексы для
этой смены будут равны 76.2 — 84.0 (среднее 79.2), т. е. не будут транс-
грессировать с индексами для Л4ь
2. М2 обычно отличаются от Ml меньшим боковым изгибом ко-
ронки.
100
3. М2 отличаются от предыдущей смены относительно более ранней
закладкой и последующим мощным разрастанием очень широкого зад-
него корня. Хотя на М2, как и на остальных зубах, первым заклады-
вается передний корень, задний быстро его обгоняет. На Ml разраста-
ние заднего корня происходит, видимо, позднее. По нашим наблюде-
ниям, первые коренные зубы (особенно нижние) отделены в челюсти
от формирующихся вторых сплошной прокладкой костной ткани. Та-
кая перегородка существует и между М2 и М3 на ранних стадиях фор-
мирования последнего, однако создается впечатление, что М3 относи-
тельно раньше начинает «давить» на М2, yl давление это более мощное,
что и приводит к своеобразному строению заднего конца вторых ко-
ренных зубов.
Таблица 2
Отношение ширины коронки
к ее полной высоте у разных смен зубов, WIH, %
3 убы lim м
(V X Pd4 9 65.4—94.5 78.0
X X м1 6 53.5—67.7 58.4
(V 0 м2 8 50.7—61.7 .54.9
(V X Pd4 4 65.0-84.0 75.7
X £ м, 7 65.0-72.0 68.8
X X М2 4 61.0—64.0 62.4
4. Вероятно, в связи с последним обстоятельством и тот факт, что
пластины М2 имеют субпараллельные края или даже несколько рас-
ширяются книзу, особенно в задних 2/3 коронки. Форма же пластин Ml
(как и Pd4) ближе к овальной (сужение книзу), поэтому у первых
коренных зубов максимальная ширина коронки отмечается близко
к середине ее высоты, а у вторых—в основании, в задней трети ко-
ронки.
Предлагаемые графики помогли решить стоявшую перед нами
практическую задачу — распознавание смен зубов в коллекции из се-
веро-восточной Якутии, с которой мы работали. Мы не склонны прида-
вать предложенным графикам значение универсального шаблона для
определения смен зубов, однако практическое использование их при не-
которых условиях, по-видимому, возможно.
Главным условием является точность определения параметров
зуба, в первую очередь, полного числа пластин и ширины коронки.
В большинстве случаев этого достаточно. В отдельных затруднитель-
ных случаях, когда на графике ширины зуб попадает на границу полей
двух смен, следует воспользоваться графиком высоты (если ее можно
определить), индексом WIH или морфологическими признаками. Если
полное число пластин не поддается уверенному определению, смену
зуба можно установить по ширине или высоте зуба лишь с той или
иной степенью приближенности.
Мы умышленно не приводим крайних значений диапазона измене-
ний параметров зубов разных смен, так как такие цифры можно да-
вать лишь для однородных выборок. Вместе с тем отметим, что даже
по нашей неоднородной выборке мы получили значительно более узкие
101
диапазоны для отдельных смен, чем приводимые иногда в литературе.
Так например, И. Е. Жерехова (1977) для М2 берелехских мамонтов
приводит диапазон изменения полного числа пластин 12—20. По на-
шим наблюдениям, достоверные значения полного числа пластин на
этой серии устанавливаются лишь в пределах 17—19 пластин, а по всей
нашей неоднородной выборке—15—19 пластин. Полное же число пла-
стин, равное 12, может иметь только последний предкоренной. Несо-
мненно, это ошибка, связанная либо с тем, что сохранившееся число
пластин принималось за полное, либо с неправильным определением
смены зуба.
Сравнение данных, полученных нами при повторном изучении бе-
релехской серии, с данными, приводимыми И. Е. Жереховой (1977),
показало значительные расхождения почти во всех параметрах зубов.
Характерно, что эти расхождения носят систематический характер.
Так, минимальные значения полного числа пластин Pd4, Ml и М2
у И. Е. Жереховой сильно занижены (в подсчет включались зубы, у ко-
торых часть пластин уже уничтожена стиранием). Резко занижены
минимальные значения высоты коронки у Pd4 и Ml (измерялись стер-
тые зубы). В данных по ширине коронки, напротив, заметно завы-
шены максимальные ее значения для Pd4 — М2, что, вероятно, связано
с тем, что в серию по данной смене ошибочно включались стертые
зубы следующей, более поздней смены, из-за занижения их числа
пластин.
Естественно, что ошибки в определении смен зубов и оценке их
параметров не могли не повлиять на обоснованность выводов автора
о возрастном и половом составе берелехской популяции. Примеры
подобных ошибок, приводящих к необоснованному расширению диапа-
зона изменений параметров определенных смен зубов, можно найти
и в работах других авторов. Это еще раз показывает необходимость
более внимательного отношения к определениям параметров исследуе-
мых зубов.
Кратко остановимся на выводах, вытекающих из приведенных гра-
фиков.
1. Предлагаемый метод парных графиков сопряженных парамет-
ров позволяет более надежно распознавать смены зубов, чем ранее
предлагавшиеся методики.
2. Графики четко демонстрируют относительно слабую трансгрес-
сию признаков у предкоренных зубов и более сильно выраженную
у коренных. Наибольшая трансгрессия параметров отмечается между
Ml и М2.
3. Наибольший размах вариации всех параметров, превосходящий
в несколько раз изменчивость зубов остальных смен, обнаруживают
последние коренные — М3. Из остальных зубов наибольший диапазон
изменчивости оказывается у последних предкоренных Pd4.
4. При составлении графиков обязательным условием является
тщательность и точность промеров зубов и определения полного числа
пластин с учетом талонов. Практикуемое обычно измерение максималь-
ной ширины зуба, заключенного в челюсти, часто дает ошибку, выхо-
дящую за пределы допустимой для данного метода.
5. При включении в выборку зубов разных видов слонов разброс
точек на графике возрастает от ранних смен к поздним.
102
ЛИТЕРАТУРА
Верещагин Н. К. Морфометрическое описание мамонтенка. — В кн/. Магаданский ма-
монтенок. Л., Наука, 1981, с. 52—80.
Гарутт В. Е. Сем. Elephantidae Gray. 1821. — Тр. Геолог, ин-та АН СССР, 1УЬ6,
т. 152, с. 47—58.
Гарутт В. Е. Зубная система слонов в онтогенезе и филогенезе.—Тр. Зоол. ин-та
АН СССР, 1977, т. 73, с. 3—36.
Гарутт В. Е., Форонова И. В. Исследование зубов вымерших слонов. Методические
рекомендации. Новосибирск, Ин-т геол, и геофиз. СО АН СССР, 1976, с. 1—36.
Дуброво И. А. Древние слоны СССР. — Тр. Палеонтол. ин-та АН СССР, 1960, т. 85,
с. 1—78.
Дуброво И. A. (Dubrovo /. A.) Die Fossilen Elefanten Japans. Quartapalaontologie.—
В. 4., Berlin, 1981, S. 49—84.
Жерехова И. E. Описание и измерения зубов мамонтов Берелеха. — Тр. Зоол. ин-та
АН СССР, 1977, т. 72, с. 50^—58.
Корниец Н. Л. Досл1дження шдив!дуального вшу мамонт!в. — Доповш АН УРСР,
1959, № 7, с. 780—784.
Павлова М. (Pavlow Marie). Les Elephants fossiles de la Russie. — Nouveaux Memoi-
res de la Soc. Imperiale des Natur de Moscou, 1910, t. XVII, p. 1—56.
Урбанас E. В. Зубы мамонта из позднепалеолитических стоянок села Костенки Во-
ронежской области. — Тр. Зоол. ин-та АН СССР, 1980, т. 93, с. 81—90.
Щеглова В. В. К вопросу о различиях в коренных зубах разного порядкового но-
мера у мамонта. — Докл. АН БССР, 1961, т. 5, с. 465—469.
Maglio V Origin and evolution of the Elephantidae. — Trans. Amer. Philosophical Soc.
New., ser. 63, 1973, vol. 3, 149 p.
Pohlig H. Dentition und Kranologie des Elephas antiquus Falc. mit Beitragen iiber Ele-
phas primigenius Blum, und Elephas meridionalis Nesti. — Nova Acta Leop. Carol.,
1888, Bd. LIII, h. 1, 279 S.
SUMMARY
Identifying generations of elephant teeth scientists are often faced with difficul-
ties because of transgression of numerical indices of features in crowns of teeth of pre-
vious or subsequent generations. The difficulties are enhanced, since it is sometimes
impossible to establish primary parameters of crowns that have been partly erased.
The authors describe methods for reconstruction of such crowns.
To make identification of teeth generations in elephants more exact the authors
suggest to use several least transgressing features, instead of one, and to construct
for that purpose paired graphs of conjugate parameters.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1985, ТОМ 131
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, 1985, VOL. 131
USSR ACADEMY OF SCIENCES
ПРОИСХОЖДЕНИЕ И СТРАТИГРАФИЯ
БЕРЕЛЕХСКОГО «КЛАДБИЩА» МАМОНТОВ
Н. К. Верещагин, В. В. Украинцева
Зоологический институт АН СССР, Ботанический институт АН СССР, Ленинград
N. К. Vereshchagin, V. V. Ukraintseva. The origin and stratigraphy of Berelyokh mam-
month “burial ground”
Введение
Массовое захоронение трупных остатков мамонтов и некоторых
их «спутников» на р. Берелех — левом притоке Индигирки, располо-
жено под 71° с. ш. и 145° в. д. (от Гринвича) под северной границей
лесотундры. Оно изучалось геологами, палеонтологами и археологами
в 1970, 1971, 1972, 1980 гг. (Верещагин, 1971, 1972, 1974, 1977, 1979,
1981; Ложкин, 1977; Медведев и Воронова, 1977). Костеносный слой
залегал там на глубине 3.5—4.5 м от бровки высокой (12 м) террасы
левого берега Берелеха и на высоте 7—8 м над меженным уровнем
реки в урочище Угамыт. В 70-х годах этот слой прослеживался на
протяжении 180 м склона берега. В участке его нижнего по течению
реки конца на глубине 1.5 и 2.5 м залегал культурный слой поздне-
палеолитической стоянки (Верещагин и Мочанов, 1972; Мочанов, 1977).
Выше по течению от головного участка костеносного слоя на 300—350 м
по срезанному краю едомы хорошо выделяются в рельефе останцы —
байджерахи — с пылевидными супесями и суглинками. Они возвы-
шаются там над обрывом берега на 20—22 м от меженного уровня
реки и, обдуваемые ветром, служат летом пристанищем лосей, спасаю-
щихся от гнуса и комаров. Над костеносным участком полигоны и
байджерахи в 1970 г. намечались в виде квадратов и пологих бугров,
оползавших по склону, заросшему крупными злаками и сложноцвет-
ными. К 1980 г., в результате раскопок археологов в 1971 г. и новых
размывов склона мотопомпой, проведенных в 1972 г. Б. С. Русановым
и П. А. Лазаревым, мощные оползни быстро обрезали берег на ши-
рину 10—15 м и повели к образованию полигонов, байджерахов и ко-
ротких овражков непосредственно на плато, над изучавшимся участком
яра. Диаметр полигонов достигал здесь в 1980 г. 22 м, а мощность
(ширина) ледовых жил — до 2 м. Всего по простиранию костеносного
участка выявилось 5 байджерахов, разделенных короткими крутыми
овражками. Пылеватые серые лессовидные супеси и суглинки останцов
обжатого грунта в этом участке оказались в общем сходны с суглин-
ками выше лежащих байджерахов Угамытской едомы.
Видовой состав и число учтенных нами в 1970, 1971 и 1980 гг. ко-
стей и особей зверей в участке «кладбища» оказался следующим*
* Число костей, вымытых ^оползнями, течением и размывами Б. С. Русанова
в 1972—1979 гг. и вновь замытых в донных отложениях Берелеха, неизвестно.
104
Видовой состав Число костей Число особей Процент особей
Мамонт 8830 156 88.6
Росомаха 84-1 труп 5 2.8
Пещерный лев 1 1 0.56
Волосатый носорог 1 1 0.56
Лошадь Черского 19 5 2.84
Северный олень 34 5 2.84
Первобытный бизон 11 3 1.70
Белая куропатка 1 труп
Итого 8904 176 100.0
Кроме остатков млекопитающих и трупика белой куропатки, в ко-
стеносном слое обнаружены экскременты грызунов (леммингов и узко-
черепных полевок) и чешуя сибирской плотвы, а также обильные
остатки жуков-долгоносиков, жужелиц, листогрызов и навозников
(Медведев и Воронова, 1977), свидетельствующие о преобладании
тундровых, тундрово-степных и степных видов при полном отсутствии
таежных.
Над костеносным горизонтом на глубине 2.0 м от бровки залегал
полуметровый слой с обилием ветвей и корней кустарников — ольхи,
ивы, а выше на глубине 1.5 м — слоистая толща, переполненная расти-
тельным детритом, мощностью в 0.5—0.8 м. Датировка бивня мамонта
с глубины 3.5 м по радиокарбону С14 оказалась 12 240±160 лет
(ЛУ-149); обрезки кожи и связок из того же слоя, датированные в Ма-
гадане, дали 13 700±400 лет. Слой с ветвями и корнями кустарников
был датирован по древесине соответственно в 11.830± 110 лет (ЛУ-147)
и в 10 440=100 лет и И 870±60 лет (НИИ ДВНЦ АН СССР).
Описания геологического разреза Берелехского яра геологи не
опубликовали. Их заключение сводилось к тому, что массовая гибель
мамонтов и формирование природного костеносного горизонта прохо-
дили в «няпанскую стадию сартанского оледенения» в условиях откры-
того тундрового ландшафта, а формирование «горизонта ископаемых
почв» (наш слой с ветвями и корнями кустарников) соответствует кли-
матическим зонам беллинг и аллерёд — с затемным потеплением около
12 000 лет тому на^ад (Ложкин, 1977). Описание разреза было опубли-
ковано нами (Верещагин, 1977).
Тафономическая ситуация и природа образования Берелехского
«кладбища» представлялась нам в начале 70-х годов в виде существо-
вания старицы праБерелеха — пойменного озера, куда сносились трупы
животных (Верещагин, 1977). Возможно, что и палеолитические охот-
ники принимали активное участие в гибели некоторой части мамонтов,
загоняя их в трясину.
В культурном слое позднепалеолитической стоянки, датированном
«дюктайской культурой» позднего палеолита (около 10 000 лет тому
назад), было собрано археологами в 1971 г. 1046 костных фрагментов,
принадлежащих мамонту, волку, зайцу-беляку, лошади, северному
оленю, белой куропатке, гусю-гуменнику и рыбе. Основой питания
древних охотников были заяц-беляк и белая куропатка, а остатки ма-
монтов (в виде осколков разбитых бивней и трубчатых костей) проис-
ходили, вероятно, от трупов этого зверя из природного «кладбища» и
по типу сохранности были несколько древнее костей волка и зайца.
105
По участку палеолитической стоянки, раскопанной в 1971 г.,
Ю. А. Мочанов (1977) составил такое описание разреза (в сокращен-
ном виде):
«1. Дерн мощностью 3—5 см.
2. Желтовато-коричневая комковатая льдистая супесь с верти-
кально-волнистыми затеками голубовато-серой супеси. ЛАощность пачки
40—50 см. От подошвы этой супеси на глубину 5 м уходят ледяные
жилы.. .
3. Горизонтально-слоистая пачка, состоящая из прослоек серовато-
коричневой супеси толщиной в 0.1—0.2 см и прослоек разнозернистого
песка толщиной в 0.1—0.5 см. Мощность пачки — 4—4.5 м, и она пред-
ставляет пойменную фацию аллювия, обогащенную детритом. На глу-
бине 2.5 м расколотые кости мамонтов и других животных, разнообраз-
ные каменные изделия. Датировки культурного слоя по древесине дали
цифры: 10 660±90 лет (ЛЕ-998); 12 930±80 лет (ГИН-1021) ; и 13 420±
200 лет (ИМ-152).
4. Горизонтально-слоистая пачка, состоящая из прослоек серовато-
голубой супеси мощностью 1—Зсм и желтоватых разнозернистых пес-
ков мощностью 0.5—5 см. Видимая мощность — 7—7.5 м. Отдельные
прослойки супеси обогащены растительным детритом. Датировка по
образцу детрита из слоя, лежащего на 20 см выше меженного уровня
р. Берелех — 42 000 лет (ЛЕ-1112). По-видимому, это — размытые от-
ложения фации аллювия 20-метровой террасы, в которую вложена
12-метровая терраса».
Это описание в общих чертах подтверждает наше заключение
о том, что древние берелехцы жили на затопляемой сезонными павод-
ками пойменной террасе праБерелеха и использовали трупы погибших
в реке и старице животных. Возможно, они принимали и активное
участие в их гибели, т. е. загоняли зверей на непрочный лед или на
подмытую водой сплавину пойменного озера.
Ю. А. Мочанов явно пытался зачем-то удревнить слой стоянки,
синхронизировать ее с основным костеносным горизонтом, хотя должно
быть понятно, что палеоохотники не могли жить в болотной жиже и
озере, а устраивали стойбище на берегу. Возможно, что археолог имел
дело со сползшим по склону участком, так как в 1970 г. я находил ору-
дия и разбитые кости на глубине 1.5 м от бровки.
С 29 июля по 9 августа 1980 г. отряд Зоологического института
АН СССР в составе Н. К. Верещагина, А. Н. Николаева, В. В. Про-
кофьевой, А. В. Прокофьева (Институт цитологии АН СССР),
К. С. Ефремова (Институт геологии ЯкФАН) вновь обследовал Бере-
лехское «кладбище» (рис. вклейка). Были собраны вымытые тече-
нием за прошлые годы и вытаявшие кости и перевезены затем на пра-
вый берег. Мамонтовых костей оказалось 399 (в том числе от подсос-
ных— 10, подростковых — 206, взрослых—170, старых—13; примерно
от 12 особей), лошадиных — 7, оленьих — 3, бизоньих — 3, росо-
машьих— 1. Это было в 9 раз меньше, чем в 1970 г. и в два раза
меньше, чем в 1971 г.
Головной участок костеносного слоя, лишенный защиты в виде
сползавших по склону костей, оказался размыт, и на его месте обра-
зовался затопленный водой котлован диаметром в 18 м. Ниже по тече-
нию береговой склон стал менее крутым, но вместо пологих попереч-
ных ложбин образовались резко выраженные, глубиною в 5—6 м
овражки, разделяющие столбы байджерахов. На протяжении 100—
120 м костеносного слоя образовалось 5 выраженных байджерахов с со-
ответствующими бортовыми остатками полигонов (рис. 2).
106
Рис. 2. Расположение байджерахов над участком костеносного слоя в 1980 г.
Треугольником отмечена палеолитическая стоянка.
Посредине костеносного слоя, т. е. на 50—60 м ниже по течению
от головной точки, была произведена зачистка двух смежных байдже-
рахов с шириной геологической канавки в 1 м на глубину в 5—6 м и
отобрано 8 образцов грунта на пыльцу и механический состав. Разрезы
обоих байджерахов дали в общем сходную картину. Приводим описа-
ние разреза по байджераху № 3 (рис. 7, вклейка).
Прослойки торфа в профиле берегового (байджерахового) разреза
оказались наклонены на 3—4° в речную сторону. Это, вероятно, указы-
вает на то, что именно здесь располагался участок берега бывшего за-
растающего водоема (рис. 3). Подтверждением этому является и зале-
гание костей мамонта на глубине в 2.5—3.0 м, т. е. на метр с лишним
выше основного костеносного слоя, размывавшегося в 1970—1971 гг.
Профиль глубин современного русла Берелеха по промерам 1980 г.
показан на рис. 3.
Из новых палеонтологических находок 1980 г. интересен обломок
таза молодого мамонта с правильными насечками (рис. 4, вклейка).
Глубина и мощность, м Состав и включения
0—0.2
0.2—0.5
0.5—1.0
1.0—1.5
1.5—1.7
1.7—2.0
2.0—2.5
2.5—3.0
3.0—3.7
3.7—4.0
4.0—4.5
4.5—5.0
5.0—12.0
12.0
Торфяно-дерновый слой, бурый с коричневыми
примазками
Сизый суглинок, мерзлота на глубине 52 см
Бурая торфяная прослойка толщиной 5 см, об-
разец № 7
Темно-бурый и сизый суглинок с торфяными
прослойками, обломок таза небольшого ма-
монта с насечками — следами счета, образец
№ 6
Торфяная коричневая прослойка 0.1 м
Бурый суглинок с торфом
Сизый суглинок с линзой коричневого торфа
толщиной 15 см, образец № 5
Сизый суглинок с волнистыми прослойками бу-
рого торфа, обломок бивня диаметром в 50 мм,
р3, обломок ребра и обломок таза эмбриона
мамонта, образец № 4
Сизый суглинок с включениями бурого торфа и
небольшой охристой линзой, образец № 3
Суглинок, перемешанный с бурым торфом; об-
разец № 2
Сизый суглинок с прослойкой бурого торфа тол-
щиной до 25 см
Сизый суглинок с небольшими торфяными при-
мазками, образец № 1
Суглинок, перекрытый оползнем темно-бурого
грунта с костями мамонтов
Меженный уровень Берелеха
107
ДРЕВНЯЯ СТАРИЦА
С УТОНУВШИМИ МАМОНТАМИ
современный
ЛАНДШАФТ ЛЕСОТУНДРЫ
Рис. 3. Схема формирования «кладбища» мамонтов. Профиль по меридиану.
Его залегание на глубине 1.5 м от бровки дает новое свидетельство
того, что кости мамонтов переносились древними охотниками с уровня
3.5—4.0 м, т. е. со дна водоема — на берег, где вновь захоронялись
в отложениях аллювия и ветровых надувах пыли. Разница с уровнем
культурного слоя стоянки, изученной Мочановым (1977), в 1 м гово-
рит о том, что, возможно, существовало два культурных слоя, либо
(что более вероятно) рельеф берега водоема в стародавние времена
повышался в северо-западном направлении, т. е. к головному участку
«кладбища». Следует также еще напомнить, что при накоплении под-
земного льда столбы обжатого грунта полигонов испытывают, кроме
деформаций, тем большее поднятие, чем больше создавалась мощность
ледовых жил.
Кроме орнаментированного обломка таза, найденного в третьем
байджерахе, на береговой отмывке у воды, против раскопа археологов,
был поднят великолепный листовидный на-
конечник копья из зеленоватого окремне-
лого сланца длиной 69 мм. (рис. 5).
Из описанной пятиметровой толщи на
палинологический анализ было отобрано
8 образцов. Все образцы (за исключением
двух, отобранных в интервале 4—5 м) ока-
зались насыщенными пыльцой и спорами.
Состав палинологических спектров ис-
следованной толщи отложений (рис. 6)
свидетельствует, что растительность в рай-
оне берелехского «кладбища» мамонтов
претерпевала неоднократные изменения.
По соотношению пыльцы деревьев, кустар-
ников, трав и спор мхов в составе спект-
ров исследованных образцов пыльцевая
Рис. 5. Листовидный наконеч-
ник копья. Находка 1980 г.
диаграмма подразделяется на четыре эта-
па в истории растительности и климата
этого региона.
Этап I (гл. 3.90—3.60 м). Охарактеризован спектрами двух образ-
цов: образца № 3, отобранного с глубины 3.9 м из расчистки третьего
байджераха и образца № За с той же глубины, но из головного участка
костеносного горизонта. Пыльца древесных пород, кустарников и ку-
старничков представлена здесь почти в равных количествах, составляя
соответственно 21.0% и 23.0%; на долю пыльцы травянистых растений
приходится 36%. Споры мхов BryalesF Sphagnum составляют 20%.
В группе пыльцы древесных растений доминирует пыльца березы дре-
вовидной Betula sp. ex. sect. Albae 45.6%. Пыльца березы кустарниковой
Belula sp. ex. sect. Fruticosa составляет 12.0%, на долю пыльцы кустар-
ничковых берез Betula exilis, В. sp. приходится 33.7%• Пыльца ив Sa-
lix sp. составляет 6%. Отмечены единичные пыльцевые зерна листвен-
ницы Larix sp., и ольховника Alnus fruticosa. В группе пыльцы травя-
нистых растений господствует пыльца злаковых Роасеае (43.0%) и
осоковых Сурегасеае (22.7%). Пыльца камнеломок Saxifraga spp. состав-
ляет 9.3%, полыней Artemisia sp. — 8.0%. Отмечены единичные пыль-
цевые зерна — Sparganium sp., Ranunculus sp., Sanguisorba officinalis,
Gentiana tennela, Stellaria sp. В группе споровых растений доминируют
споры не менее 4 видов гипновых мхов, на долю которых приходится
89.4%; отмечены единичные споры Sphagnum, а также Sellaginela si-
birica.
109
С учетом того, что пыльца лиственницы сохраняется плохо (Вась-
ковский, 1957; Украинцева, 1980), а пыльца лиственных пород, и в част-
ности берез, на большие расстояния не переносится, можно полагать,
что в эпоху формирования толщи осадков в интервале глубин 3.9—3.6 м
на исследованной территории получали распространение лиственнично-
березовые заболоченные леса с подлеском из кустарниковых и кустар-
ничковых берез. Ольховников, ивняков было мало. Современная север-
ная граница древовидной березы Betula pendula находится на менее чем
в 500 км южнее местонахождения Берелехского «кладбища» мамонтов,
и это дает основание считать, что в эпоху формирования костеносного-
Рис. 6. Спорово-пыльцевая диаграмма отложений II надпойменной терассы р. Берелех.
а — пыльца деревьев, б — пыльца кустарников и кустарничков, в — пыльца трав, г — споры мхов
и папоротников, д — почвенный слой.
слоя климат района исследования был теплее современного. Радиокар-
боновые даты— 11 870±60 лет и 11 830 ±110 лет, полученные по кор-
ням и ветвям, отобранным из горизонта, лежащего стратиграфически
выше (Ложкин, 1977), позволяют считать, что этот этап в развитии
растительности соответствует, вероятно, по времени кокоревскому по-
теплению Сибири (Кинд, 1973) или аллерёду Европы (Нейштадт, 1971),
когда почти по всей территории Средней Европы получали распростра-
нение преимущественно сосновые и березовые леса. Это потепление
отчетливо прослеживается и на палеотемпературной кривой Дангарда.
Этап II (гл. 3.50—2.50м). Господствует пыльца травянистых ра-
стений Роасеае, Сурегосеа и споры гипновых мхов Bryales, составляю-
щие соответственно 50.4—54.5% и 31.5—36.6%. На долю пыльцы кус-
тарников и кустарничков приходится лишь 13.5—13.8%. Пыльца
деревьев представлена лишь единичными зернами лиственницы Larix
cf. gmelini. Состав охарактеризованных выше спектров и ход кривых
основных компонентов (рис. *5) свидетельствуют, что формирование
толщи отложений в интервале глубин 3.50—2.50 м происходило в усло-
виях резкого похолодания климата, что обусловливало сокращение
лесных формаций и исчезновение из их состава древовидной березы.
Широкое развитие в этот период получали кустарничково-травянистые
ПО
и кустарничково-моховые сообщества как на ллакорах, так и в доли-
нах рек и у приозерных понижений. Лиственничники, близкие или ана-
логичные по своему составу современным, если и были распространены
в эту эпоху, то лишь по долинам рек.
Этап III (гл. 2.40—1.20 м). Охарактеризован спектром одного
образца, отобранного на глубине 1.5 м. В составе его преобладает
пыльца травянистых растений, составляющая 44.5%; на долю пыльцы
кустарников и кустарничков приходится 24.9%; споры мхов, папорот-
ников и хвощей составляют в сумме 27.0%; пыльца деревьев Larix sp.,
Betula sp. ex sect. Albae (в том числе и несомненно дальнезаносные
зерна Pinus sibirica, Р sylvestris) составляет лишь 4.6%. Состав
спектра свидетельствует о широком развитии в эту эпоху кустарничко-
вых моховых заболоченных тундр, злаково-осоковых сообществ по бе-
регам рек и водоемов. В составе растительности вновь увеличивается
значение лиственницы, распространенной в долине реки.
Этап IV (гл. 1.0м). Господствующим типом растительности ста-
новятся кустарничковые Betula exilis, Betula sp. ex. Nanae и кочкарные
Cassiope tetragona сообщества заболоченных тундр, что было обуслов-
лено общим похолоданием климата. О последнем свидетельствует и
обедненный видовой состав пыльцы.
В группе травянистых растений преобладает пыльца верескоцвет-
ных (67%), мало злаков (до 8%) и осоковых (до 14%). Много спор
сфагновых (61.4%) и гипновых мхов (33%).
В результате палинологического анализа Берелехского разреза вы-
ясняется, что после длительного существования безлесных ландшафтов
вследствие потепления климата, около 12 тыс. лет тому назад (этап I)
произошло появление лиственично-березовых и березовых лесов, соот-
ветствующее кокоревскому потеплению Сибири и беллингу Европы.
Затем последовало новое похолодание (этап II), соответствующее сред-
нему дриасу Европы и тукрик Северной Америки. На этапе III насту-
пило новое потепление, вероятно, аналогичное Таймырскому и алле-
реду Европы. Последующее похолодание на этапе IV может быть со-
поставлено с норильской стадией Сибири, поздним Висконсином Аме-
рики и верхним дриасом Европы.
Заключение
Палеонтологические, тафономические, археологические и геоморфо-
логические исследования и наблюдения по Берелехскому Мамонтову
«кладбищу» приводят к следующим выводам.
1. Современная река Берелех в историческую эпоху перерезала и
размыла на протяжении 180 м отложения слабопроточного озера типа
пойменной старицы. Эти отложения бурого цвета, переполненные расти-
тельным детритом, составляли верхнюю треть второй 12-метровой над-
пойменной террасы. Основной костеносный горизонт залегал до размыва
толщи на глубине 3.5—4 м от бровки и на высоте 7—8 м над межен-
ным уровнем реки. На глубине 2 м от бровки залегал слой с обилием
корней и ветвей ив и ольховника.
2. Накопление озерно-аллювиальных (озерно-старичных) отложе-
ний в участке мамонтов «кладбища» происходило в позднем плейсто-
цене (сартане) и в послеледниковое время. Костеносный горизонт дати-
ровался по костям и коже мамонта в пределах 13700—12400 лет,
а^верхний слой с корнями кустарников—11830—10440 лет.
111
3. Из склоновых отложений и со дна реки за три года размыва
было собрано в общей сложности 8904 кости. 8830 костей от 156 осо-
бей принадлежали мамонтам, остальные 74 кости — единичным особям
росомах, пещерных львов, волосатых носорогов, лошадей, северных оле-
ней и первобытных бизонов. Преимущественная гибель полувзрослых
мамонтов и самок требует объяснения с этологических позиций. Про-
валы под непрочный лед или в сплавину наиболее вероятны.
4. Полевые учеты и сборы вымываемых из берегового склона ко-
стей в 1970, 1971 и 1980 гг. показали, что «кладбище» уже истощилось,
т. е. оказалось перемытым. Это подтвердили и наши раскопки 1980 г.
двух байджерахов против центрального участка «кладбища», которые
составляли уже краевую (прибрежную) зону водоема. Костеносный
слой (3.5—4.0 м) там выклинивался, а единичные кости мамонтов
со следами воздействия первобытного человека залегали на глубинах
2.5 и 1.5 м, что указывало на перенос их со дна на берег водоема
(рис. 5).
5. Слой палеолитической стоянки с разбитыми бивнями мамонтов,
костями зайцев, северных оленей, волков и белых куропаток находился
в 1970 г. на глубине 1.5 м от бровки (по Ю. А. Мочанову, 1977 — на
глубине 2.5 м). Основой питания древних берелехцев были зайцы, куро-
патки и северные олени.
6. Палинологический анализ образцов грунта из краевого участка
водоема показал, что образование Мамонтова «кладбища» происходило
в условиях безлесных ландшафтов травянистых и кустарничково-тра-
вянистых тундр с осоково-злаковыми лугами в пойменных понижениях.
Безлесие подтверждается и отсутствием остатков лесных зверей и лес-
ных насекомых.
7. Образование верхней части разреза с глубины 2.5 м от бровки
(или 9.5 м от меженного уровня р. Берелех) происходило на этапе раз-
вития ландшафта лесотундры и ерниковых тундр. В это время «клад-
бище» посещалось первобытными охотниками. Северо-якутские ма-
монты были уже близки к вымиранию.
ЛИТЕРАТУРА
Васъковский А. П. Краткий очерк растительности, климата и хронологии в верховьях
рек Колымы, Индигирки и на побережье Охотского моря. — В кн.: Ледниковый
период на территории европейской части СССР и Сибири. М. 1959, с. 511—545.
Верещагин Н. К. Берелехское «кладбище» мамонтов. — Тр. Зоол. ин-та АН СССР,
1977, т. 72, с. 5—50.
Верещагин Н. К. Почему вымерли мамонты. Л., Наука, 1979, 193 с.
Кинд Н. В. Хронология позднего антропогена по радиометрическим данным. — В кн.:
Стратиграфия. Палеонтология. М., Наука, 1973, с. 5—49.
Ложкин А. В. Условия обитания берелехской популяции мамонтов. — Тр. Зоол. ин-та,
1977, т. 72, с. 67—68.
Медведев Л. Н., Воронова Н. В. Колеоптерологический анализ геологических разре-
зов мамонтовых «кладбищ» в Северной Якутии. — Тр. Зоол. ин-та, 1977, т. 72,
с. 72—77.
Мочанов Ю. А. Древнейшие этапы заселения человеком северо восточной Азии. Ново-
сибирск, Наука, 1977, 263 с.
Нейштадт М. И. О нижней границе голоцена. — В кн.: Палинология голоцена. М.,
Наука, 1971, с. 7—13.
Соколов С. Я., Связева О. А., Кубли В. А. Ареалы деревьев и кустарников СССР.
Т. I, Л., Наука, 1977, с. 163.
Украинцева В. В. Природная среда и условия гибели мамонтенка. — В кн.: Магадан-
ский мамонтенок. Л., Наука, 1980, с. 254—264.
112
SUMMARY
Taphonomic observations of 1970, 1971, and 1980 have shown that Berelyokh «bu-
rial ground» was formed within the period of 14—12 thousand years before present.
The animals died in the valley of the ancient Berelyokh and their carcasses were car-
ried to the ancient river and burried there beneath a layer of silty sediments: 8830
bones from 156 mammothes and 74 bones of solitary gluttons, cave lions, rhinoceroses
horses, reindeers, protobisons have been studied. Predominant in the epoch of life and
death of mammothes in that region were treeless landscapes. Buried close to the «bu-
rial ground» 1.5 and 2.5 m beneath the ancient river-bank is a cultural layer of a paleo-
lithic site containing bones of hares, reindeers, wolves, willow ptarmigans, and imple-
ments of silicon argilite and mammoth tusks.
8 Млекопитающие
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1985, ТОМ 131
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, 1985, VOL. 131
USSR ACADEMY OF SCIENCES
ГРАВИРОВАННЫЕ ИЗОБРАЖЕНИЯ ЖИВОТНЫХ
В ГМЕЛИНСКОЙ ПОЗДНЕПАЛЕОЛИТИЧЕСКОЙ СТОЯНКЕ
(КОСТЕНКИ 21)
Н. Д. Праслов
Институт археологии АН СССР (ЛО), Ленинград
N. D. Praslov. Engravect animals in Gmelin Late Paleolitic site (Kostenki 21)
На территории нашей страны известно довольно большое количе-
ство предметов палеолитического искусства. Наряду с многочислен-
ными украшениями в виде подвесок и разнообразными изделиями
с орнаментом, найдены сюжетные изображения человека и различных
зверей, главным образом в объемной скульптуре (Абрамова, 1962).
В этих огромных материалах поражает исключительная редкость сю-
жетных гравюр. Помимо великолепного силуэтного изображения ма-
монта и змей в Мальте, к ним можно добавить своеобразный рисунок
мамонта на бивне из Берелеха (Бадер, Флинт, 1977), тонкий рисунок
антилопы на ребре мамонта и несколько фрагментов с гравировкой
на камне и кости в верхнем слое Костенок I (Ефименко, 1958), а также
миниатюрное изображение оленя на ребре мамонта в Тимоновской
стоянке на Десне (Грехова, 1977). Сложная композиция фантастиче-
ской птицы(?), выгравированной на бивне мамонта в Кирилловской
палеолитической стоянке на Днепре до сих пор остается уникальной
и не поддается достоверной дешифровке. Единичным и изолированным
на большой территории южнорусских степей остается гравированное
антропоморфное изображение на лощиле из рога оленя на Муралов-
ской стоянке в Приазовье (Праслов, Филиппов, 1967). Поэтому не-
сомненный интерес представляют две новые гравюры, найденные в ниж-
нем культурном слое Костенок 21.
Гмелинская стоянка (или Костенки 21) расположена на северной
окраине с. Костенки в долине Дона на низкой надпойменной террасе.
Дон здесь, после крутого поворота по широкой пойме, постоянно вре-
зается в правый коренной берег и ежегодно разрушает в среднем около
1 м плейстоценовых отложений. В двадцатые годы нашего столетия
основные размывы весенними водами приходились на тот участок, где
в долину впадает мощный ручей, вытекающий из Попова лога. В 1923 г.
в размытом береговом обрыве на правом борту ручья была открыта
позднепалеолитическая стоянка Костенки 3. Впоследствии Дон здесь
уничтожил довольно значительную площадку первой надпойменной
террасы, разрушив полностью позднепалеолитическое поселение. Основ-
ное направление размыва берега стало постепенно смещаться вниз по
течению, разрушая новые участки хорошо выраженной площадки пер-
вой надпойменной террасы. В 1956 г. примерно в 200 м от Костенок 3
ниже по течению Дона была выявлена линза культурных остатков,
совершенно отличных от Костенок 3 как по составу каменных изделий,
так и по условиям залегания. Это новое палеолитическое поселение
114
получило название Гмелинской стоянки и порядковый номер Ко-
стенки 21.
Раскопки этой стоянки не были плановыми и всегда имели харак-
тер спасательных работ. Так было в 1956—1958, I960, 1964, 1971 и
1976—1979 гг. Вскрывались участки с высокой концентрацией культур-
ных остатков, отложившихся, как выяснилось, на местах жилых соору-
жений. Особенно значительная площадь была вскрыта в 1971 г. Тогда
же удалось установить, что Гмелинская стоянка представляла собой
огромное по площади поселение, состоящее из легких шалашеобразных
хижин, располагавшихся вдоль берега Дона на расстоянии 10—12 м
друг от друга на протяжении около 200 м.
Культурный слой приурочен к слабо выраженной ископаемой
почве, сформировавшейся на пойменном аллювии в тот период, когда
уровень Дона резко понизился, и терраса как бы вышла из пойменного
состояния, т. е. перестала заливаться весенними водами. В это же
время произошло и снижение активности склоновых процессов, что спо-
собствовало накоплению гумуса и других органических веществ, т. е.
формированию почвы.
В отличие от других палеолитических памятников Костенок, куль-
турный слой Гмелинской стоянки характеризуется большим количе-
ством каменных изделий и сравнительно малым числом костных остат-
ков животных. По определению И. Е. Кузьминой, здесь встречены
остатки мамонта, лошади, бизона, северного оленя, волка, песца, зайца
и сурка. Преобладают костные остатки зайца и мамонта, причем, абсо-
лютное большинство костей — в виде обломков и осколков. В пределах
жилых площадок целых костей практически нет, да и фрагменты несут
на себе явные следы обработки или иные признаки искусственного
раскалывания, но даже и таких искусственных осколков костей встре-
чено очень мало. Это говорит о том, что палеолитические обитатели
Гмелинской стоянки поддерживали свои жилища (да и соседние уча-
стки) в определенном порядке и чистоте. Здесь нет изобилия так назы-
ваемых кухонных отбросов, на особую заботу о чистоте в жилищах
указывает и наличие каменной вымостки из плиток девонского извест-
няка около одного из жилищ на северном участке. Каменные изделия,
собранные внутри жилищ, представлены в основном законченными
формами орудий. Процент их по отношению к отбросам производства
очень высок.
В тридцатые — пятидесятые годы на южной окраине тогда еще
не открытой Гмелинской стоянки, из суглинка на высоте 3.5—4 м над
уровнем Дона (т. е. с уровня культурного слоя) вымывалось большое
количество крупных целых костей мамонта. Большинство исследовате-
лей склонялось к тому, что где-то именно здесь производил свои рас-
копки С. Г. Гмелин. Мне кажется, что это скопление крупных костей
действительно связано с нижним культурным слоем Гмелинской
стоянки. По-видимому, обитатели стоянки сбрасывали сюда основную
массу костей, поэтому становится понятным, почему на площади посе-
ления их так мало и представлены они только технологическими отхо-
дами.
Совершенно иную картину представляют остатки огромного в плане
комплекса, расположенного на южной окраине поселения. Здесь нет
такой четкой локализации находок, как на местах жилых сооружений.
Огромное количество расщепленного кремня позволяет восстановить
всю технологию обработки орудий. Зольные пятна расплывчаты, четких
очагов нет. Повсюду много мелких костных осколков, расщепленных
пластин бивня мамонта, начатых обработкой, или сломанных ребер
8Ф
115
лошади и бизона. Среди кремневых орудий также много сломавшихся
в процессе обработки. Особенно хорошо это можно проследить на на-
конечниках стрел с боковой выемкой. По-видимому, здесь была свое-
образная площадка для изготовления каменных и костяных орудий,
а также предметов искусства. Именно в этом комплексе собрано больше
всего изделий из кости: серия овальных подвесок из бивня мамонта
(причем, на разных стадиях обработки), рукоять кинжала, вырезанная
из ребра лошади или бизона, «жезл» или выпрямитель для стрел, из-
готовленный из бивня, и т. д. Именно в этом комплексе на квадрате
Л-21 в 1971 г. и было найдено первое гравированное изображение жи-
вотного.
Каменный диск, на котором выполнена гравюра, представляет со-
бой своеобразную «лепешку» из слабо сцементированного мелкозерни-
стого песчаника. Он имеет в плане форму неправильного круга или
слегка асимметричного овала размерами 101X91 мм, плоско-выпуклого
в сечении. Максимальная толщина—16 мм. Одна сторона его совер-
шенно ровная, гладкая, искусственно отшлифована. Неровные края
также сглажены, скорее всего, в процессе использования. Гравюра вы-
полнена на выпуклой стороне (рис. i, вклейка). Ее неровный естествен-
ный рельеф позволил первобытному художнику сделать не просто
силуэтное изображение, а использовать наиболее выпуклую часть
для показа массивности тела животного.
На поверхности диска изображение занимает большую площадь,,
практически всю центральную выпуклую часть. Контур животного по-
казан различными по ширине и глубине линиями, прочерченными
кремневым орудием.
При анализе данной гравюры следует учитывать, что она выпол-
нена на песчанике. Песчаник является твердым абразивом, поэтому
провести линии на нем довольно трудно. Этим, по-моему, объясняется
то, что силуэт зверя изображен не столь пластичными линиями. По-
жалуй, наиболее четкой изогнутой линией показано лишь брюхо. Эта
линия асимметрична в сечении. Ее верхний край более прямой и более
отвесный, от него поверхность процарапанной канавки постепенно
повышается, сливаясь с общим массивом. В наиболее глубокой части
линия врезана в поверхность песчаника на 1 мм. Примерно на такую же
глубину врезана и линия, показывающая контур спины. Она углублена
также примерно на I мм, имеет ширину 2—2,5 мм, менее асимметрична
в сечении, чем брюшная линия.
На линии контура спины хорошо видно, что она состоит как бы
из трех прямолинейных отрезков, соединяющихся на изгибах в холке
(переход к голове) и у хвоста. Жесткий твердый песчаник вынуждал
первобытного мастера прилагать большие усилия при нанесении ли-
ний. При этом режущий инструмент иногда срывался, и на поверх-
ности диска оставались лишние царапины явно случайного характера.
Это видно хорошо на кончике хвоста: режущий инструмент ближе
к краю диска скользил, отклоняясь от прямой основной линии.
Тело изображенного зверя оконтурено достаточно четко. Хорошо
прослеживаются линии спины, хвоста, брюха, передней и задней ноги.
К сожалению, из-за повреждения этой стороны песчаникового диска
плохо «читается» голова, особенно ее нижняя часть. Хорошо просле-
живается переход от холки к круглому лбу, и дальше слабо выражен-
ная изогнутая линия уходит на край диска. Сравнительно неплохо
выражен переход от передней ноги к шее. Но далее восстановлению
линий мешает выпавший кусочек песчаника. Это очень затрудняет ре-
шение вопроса о том, какой же зверь в данном случае изображен.
116
Подробно проанализировав общие контуры и пропорции, невольно
лриходишь к выводу о том, что изображен крупный зверь с массивным
корпусом и довольно массивными ногами. Такие пропорции характерны
для быков и мамонта. По массивности корпуса можно было бы пред-
положить изображение носорога, однако у носорогов очень короткие
ноги. Наблюдательные первобытные охотники обычно четко переда-
вали соотношение пропорций различных частей тела животного, причем
иногда даже подчеркивая те или иные особенности. У зверя из Косте-
нок 21 ноги крупные, массивные, высота их примерно около двух пя-
тых общей высоты зверя в холке.
Общая длина зверя не сильно превышает его высоту: соотношение
длины и высоты примерно 7 : 5. Такая пропорция характерна для всех
слонов, в том числе и для мамонта. В какой-то мере эти пропорции
характерны и для зубров, однако у зубров более тонкие ноги, да и иной
силуэт морды. На песчаниковом диске Костенок 21 хорошо виден плав-
ный переход линии от крутого лба к краю диска, и под этой линией
имеется еще одна слабо выраженная слегка изогнутая линия, воз-
можно, изображающая бивень. Все это заставляет склониться к мне-
нию, что в данном случае изображен мамонт. Это мнение не бесспорно.
Осмотр данного изображения разными специалистами, в том числе и
такими крупными зоологами, как Н. К. Верещагин, В. Е. Гарутт,
И. Е. Кузьмина и многими другими, не позволил придти к единому
выводу. Однако все сходятся на том, что изображен крупный зверь
не меньше зубра.
Одной из особенностей данного изображения являются тонкие ли-
нии, перечеркивающие переднюю и заднюю ноги. На передней ноге
насчитывается 7 таких линий, на задней — 5 длинных и 2 коротких
черточки. Две длинных тонких линии нанесены выше брюха, одна —
ниже. Они как бы веером расходятся от передней ноги. Эти линии
не имеют отношения к изображению самого животного и, несомненно,
имеют глубокую семантику, отражающую определенную идею перво-
бытных обитателей Гмелинской стоянки. Эта идея выражена и на вто-
рой гравюре, найденной в Гмелинской стоянке 15 июня 1982 г., при-
мерно в 20 м севернее первой.
На второй гравюре совершенно отчетливо прослеживается схема-
тичное контурное изображение мамонта (рис. 2, вклейка). Оно выпол-
нено на ровной пришлифованной поверхности расколотой пополам
сланцевой гальки тончайшими линиями и штрихами. Площадь изобра-
жения 24X20 мм. Хорошо виден длинный опушенный хобот, контуры
головы и спины, бивней нет. Рисунок выполнен уверенной рукой тон-
чайшими плавными линиями. Только на спине имеются следы двойной
линии. Голова отчленена от туловища двумя короткими линиями. Из
них одна как бы продолжает округлую линию спины, вторая — прямая,
идет от линии лба и затем почти под прямым углом опускается вниз.
Необычно массивно изображена двумя параллельными линиями
тумбообразная передняя нога, перечеркнутая четырьмя поперечными
короткими штрихами. Рядом с ногой сделано несколько коротких вер-
тикальных штрихов, изображающих, несомненно, длинную шерсть
на брюхе. Здесь же прослеживаются несколько дополнительных штри-
хов, сохранившихся от абразивной подправки этой поверхности и учтен-
ных первобытным художником при работе над изображением.
Мамонт, выгравированный на плоской поверхности сланцевой
гальки, обращен головой влево, точно так же, как и на песчаниковом
диске. Слева, чуть выше и впереди, ближе к краю плоскости, просле-
живается тонкая изогнутая линия, которая как будто бы не связана
117
с изображением. Вместе с тем, она как бы продолжает контур спины
и, возможно, является случайной, оставшейся от первоначального на-
броска рисунка, композиционно не вписавшегося в контуры гальки.
Оба рисунка, найденные в Гмелинской стоянке, имеют радиоугле-
родный возраст около 22 тыс. лет (Праслов, Иванова, 1982). Они
исполнены схематично, в реалистической манере, силуэтно, и свиде-
тельствуют о высоком общем уровне культуры обитателей этого посе-
ления. Их стилистический анализ и семантика не могут быть поняты
без привлечения больших материалов по западно-европейскому палео-
литическому искусству, однако это не входит в задачу данного сооб-
щения.
ЛИТЕРАТУРА
Абрамова 3. А. Палеолитическое искусство на территории СССР. — Свод археологи-
ческих источников, вып. А 4—3, 1962, 84 с.
Бадер О. И., Флинт В. Е. Гравировка на бивне мамонта с Берелёха. — Тр. Зоол.
ин-та АН СССР, 1977, т. 72, с. 68—72.
Грехова Л. В. Обработанная кость Тимоновской стоянки. — Б кн.: Проблемы палео-
лита Восточной и Центральной Европы. Л., Наука, 1977, с. 90.
Ефименко П. П. Костенки 1. М. — Л.; изд-во АН СССР, 1958, 408 с.
Праслов Н. Д., Иванова М. А. Костенки 21 (Гмелинская стоянка). — В кн.: Палео-
лит Костенковско-Борщевского района на Дону. 1879—1979. Л., Наука, 1982,
с. 209.
Праслов Н. Д., Филиппов А. К. Первая находка палеолитического искусства в южно-
русских степях. — Краткие сообщ. ин-та археологии АН СССР, вып. Ill, М., 1967,
с/ 24—30.
SUMMARY
Paleolithic plot engravings are extremely rare on the territory of the Russian Plain.
In 1971 excavations of Gmelin late paleolithic site in Kostenki have revealed a sand-
stone disc flatly convex with one side polished and the other convex side covered with
engraving of a big animal. The author regards it as an engraving of a mammoth.
The second engraving of a mammoth was found on the polished side of a cleaved slaty
pebble. It was found in 1982. Both the engravings have been performed in schematic
realistic silhouette style. Age of the site is about 22 thousand years. The findings show
a high cultural level of its late paleolithic inhabitants.
РЕФЕРАТЫ
УДА 551.791 :575.79
Вымирание млекопитающих в четвертичном периоде Северной Евразии. Вереща-
гин Н. К., Барышников Г. Ф. — В кн.: Млекопитающие Северной Евразии в чет-
вертичном периоде. Л., Зоол. ин-т АН СССР, 1985, с. 3—38. (Тр. Зоол ин-та АН
СССР, т. 131).
Анализируются причины разрушения мамонтовой фауны и вымирания ее отдельных
представителей. Решающим фактором в этом процессе признаются изменения среды
обитания видов, обусловленные палеоландшафтными и палеоклиматическими сменами
на границе плейстоцена и голоцена. Влияние истребительной деятельности человека
имело региональный характер и стало решающим лишь в историческое время. Зна-
чение остальных факторов (чрезмерной специализации, инадаптивной эволюции и т. д.)
было второстепенным. Библ. 115 назв., табл. 1, ил. 10.
УДК 569.735
Время и пути расселения верблюдов в восточном полушарии. Алексеева Л. И. —
В кн.: Млекопитающие Северной Евразии в четвертичном периоде. Л., Зоол. ин-т
АН СССР, 1985, с. 39—42. (Тр. Зоол. ин-та АН СССР, т. 131).
Расселение мозоленогих в Старом свете происходило в два этапа. Первый был в пред-
виллафранкское время. Все остатки верблюдов из Евразийских местонахождений
руссильонской и виллафранкской фауны принадлежали роду Paracarnelus. Вторая
волна миграции связана с широким расселением верблюдов рода Camelus. Степень
родственных взаимоотношений родов Paracarnelus и Camelus точно не установлена.
Вероятнее всего, формирование рода Camelus произошло на территории Азии. Оттуда
они распространились на запад до Восточной Европы и проникли также через За-
кавказье и Ближний восток в Северную Африку. Библ. 16 назв.
УДК 551.795 + 56(119)(479.225)
Сары-Легет — новая мезолитическая стоянка в Юго-Осетии. Барышников Г. Ф.,
Чистяков Д. А. — В кн.: Млекопитающие Северной Евразии в четвертичном пе-
риоде. Л., Зоол. ин-т АН СССР, 1985, с. 43—49. (Тр. Зоол. ин-та АН СССР, т. 131).
Археологические и палеонтологические материалы из новой пещерной стоянки Сары-
Легет (Джавский р-н Юго-Осетинской АО ГССР) позволяют соотнести и синхронизи-
ровать их с материалами мезолитических стоянок западной Грузии (Кударо I,
слой 2; Кударо III, слой 2; Цона, слой 3; Кведи, слой 4). Определено 20 видов мле-
копитающих, в том числе кавказский сурок, хомяк Радде, лисица, зубр, кавказский
козел. Новый памятник представляет собой кратковременный охотничий лагерь и мо-
жет быть датирован мезолитическим временем. Библ. 9 назв., табл. 2, ил. 4.
УДК 569.723(477.51)
Позднеплейстоценовая широкопалая лошадь бассейна Десны. Белан Н. Г. — В кн.:
Млекопитающие Северной Евразии в четвертичном периоде. Л., Зоол. ин-т АН СССР,
1985, с. 50—6'3. (Тр. Зоол. ин-та АН СССР, т. 131).
В результате изучения костных остатков лошади из стоянок Мезин, Новгород-Север-
ский, Чулатов Черниговской области установлено, что на Десне во II половине позд-
него плейстоцена обитала широкопалая лошадь Equus latipes. Она имела череп сред-
ней длины со сравнительно длинным лицевым отделом, по-видимому, узким лбом, до-
вольно широкой мозговой коробкой, широкой резцовой частью, коротким верхне-
челюстным бугром и относительно крупными зубами с длинным протоконом. Кости
конечностей средней длины, массивные, копыта широкие. По высоте в холке лошади
Десны относились к группам малорослых, средних и рослых лошадей (128—152 см),
в среднем рост составлял 140—141 см. Библ. 21 назв., табл. 4, ил. 3.
119
УДК 599.61.72.73.74
Уральская позднеплейстоценовая лошадь. Кузьмина И. Е. — В кн.: Млекопитаю-
щие Северной Евразии в четвертичном периЪде. Л., Зоол. ин-т АН СССР, 1985,
с. 64—88. (Тр. Зоол. ин-та АН СССР, т. 131).
Уральская лошадь — Equus uralensis Kuzmina, 1975 — узко распространенный вид,
обитавший на Урале в позднем плейстоцене, входил в состав мамонтового териоком-
плекса. При дальнейшем изучении вымерших лошадей Евразии возможно расширение
ареала и выяснение происхождения вида, а также выделение двух подвидов.
В позднем плейстоцене на Урале эта лошадь обитала в лесостепных ландшаф-
тах особенно в Предуралье, а на Северном Урале и в Зауралье — в более открытых
местообитаниях. По внешнему виду — крупное животное, высотой в холке 125—150 см,
легкого склада, с высокими стройными конечностями, копытами более узкими, чем
у широкопалой лошади. Половой диморфизм выражен в размерах и пропорциях
костей конечностей. Количество взрослых самок в табунах превышало количество-
самцов в 2 и более раза. В популяциях представлены все возрастные группы от же-
ребят до старых особей. Большое число раздробленных костей говорит о том, что
в жизни палеолитических охотников Урала лошадь играла важную роль как источ-
ник пищи. Библ. 9 назв., табл. 17, ил. 15.
УДК 569.614
Мезолитическая Кокшаровско-Юрьинская стоянка в Среднем Зауралье. Сери-
ков Ю. Б., Кузьмина И. Е. — В кн.: Млекопитающие Северной Евразий в чет-
вертичном периоде. Л., Зоол. ин-т АН СССР, 1985, с. 89—92. (Тр. Зоол. ин-та АН
СССР, т. 131).
Работами Нижнетагильского отряда экспедиции Уральского университета в 1979 г.
открыта Кокшаровско-Юрьинская стоянка. Анализ археологических находок показал,
что стоянка была обитаема от мезолита до бронзы. Жители стоянки занимались пре-
имущественно охотой на таежных зверей и птиц. Наиболее частой добычей были лоси,
северные олени, бобры, волки, бурые медведи, птицы. Охотники имели помощников —
собак. Библ. 5 назв., табл. 1, ил. 3.
УДК 569.614
О методике определения генераций коренных зубов слонов. Ш е р А. В., Г а -
рутт В. Е. — В кн.: Млекопитающие Северной Евразии в четвертичном периоде. Л.,
Зоол. ин-т АН СССР, 1985, с. 93—103. (Тр. Зоол. ин-та АН СССР, т. 131).
При определении генераций (смен) зубов слонов у исследователей часто возникают
затруднения, связанные с трансгрессией цифровых показателей признаков в коронках
зубов соседних смен. Трудности в определении усиливаются еще за счет того, что
не всегда можно установить первоначальные параметры коронок, подвергнувшихся
частичному стиранию. Авторами излагается методика реконструкций подобных коро-
нок. Для более надежного установления генераций зубов слонов авторы предлагают
пользоваться не одним признаком, а несколькими наименее трансгрессирующими, и
с этой целью строить парные графики сопряженных параметров. Библ. 13 назв.,
табл. 2, ил. 2.
УДК 569.614
Происхождение и стратиграфия Берелехского «кладбища» мамонтов. В е р щ а -
гин Н. К., Украинцева В. В. — В кн.: Млекопитающие Северной Евразии в чет-
вертичном периоде. Л., Зоол. ин-т АН СССР, 1985, с. 104—ИЗ. (Тр. Зоол. ин-та
АН СССР, т. 131).
Тафономические наблюдения в 1970, 1971 и 1980 гг. показали, что Бсрелехское «клад-
бище» образовалось в хронологическом отрезке 14—12 тыс. лет до наших дней. Ги-
бель животных происходила в долине реки — праБерелехе, а их трупы сносились
в староречье, где и захоронялись под толщей иловатых осадков. Было исследовано
8830 костей от 156 особей мамонтов и 74 кости от единичных особей росомах, пе-
щерных львов, носорогов, лошадей, северных оленей, первобытных бизонов. В эпоху
жизни и гибели мамонтов в районе преобладали безлесные ландшафты. По соседству
с «кладбищем», на глубине 1.5 и 2,5 м от бровки, захоронен культурный слой палео-
литической стоянки с костями зайцев, северных оленей, волков, белых куропаток и
орудиями из окремнелого аргилита и бивней мамонтов. Библ. 10 назв., ил. 7.
120
УДК 599.61 (470.324.571.66)
Гравированные изображения животных в Гмелинской позднепалеолитической стоянке
(Костенки 21). Праслов Н. Д. — В кн.: Млекопитающие Северной Евразии в четвер-
тичном периоде. Л., Зоол. ин-т АН СССР, 1985, с. 114—118. (Тр. Зоол. ин-та АН
СССР, т. 131).
На территории Русской равнины сюжетные гравюры эпохи палеолита исключительно
редки. В 1971 г. при раскопках Гмелинской позднепалеолитической стоянки в Костен-
ках найден песчаниковый плоско-выпуклый диск, одна сторона которого отшлифована,
а на второй, выпуклой, выгравировано изображение крупного зверя. Автор видит
в нем изображение мамонта. Второе изображение мамонта выгравировано на отшли-
фованной плоскости расколотой сланцевой гальки. Оно найдено в 1982 г. Оба изо-
бражения выполнены в схематично-реалистической силуаэтноп манере. Возраст
стоянки около 22 тыс. лет. Находки свидетельствуют о высоком уровне культуры
позднепалеолитических обитателей стоянки. Библ. 6 назв., ил. 2.
СОДЕРЖАНИЕ стр.
Верещагин Н, К., Барышников Г. Ф. Вымирание млекопитающих в четвертич-
ном периоде Северной Евразии . . . . 3
Алексеева Л, И. Время и пути расселения верблюдов в восточном полушарии 39
Барышников Г, Ф., Чистяков Д. А. Сары-Легет — новая мезолитическая стояцка
в Юго-Осетии ... 43
Белан Н. Г. Позднеплейстоценовая широкопалая лошадь бассейна Десны 50
Кузьмина И. Е. Уральская позднеплейстоценовая лошадь ... 64
Сериков Ю. Б., Кузьмина И. Е. Мезолитическая Кокшаровско-Юрьинская
стоянка в Среднем Зауралье . . . . . 89
Шер А. В., Гарутт В. Е. О методике определения генерации коренных зубов
слонов................. ... ... . 93
Верещагин, Н. К-, Украинцева В. В. Происхождение н стратиграфия Берелёх-
ского «кладбища» мамонтов.............................. .... . 104
Праслов Н. Д. Гравированные изображения животных в Гмелинской поздне-
палеолитической стоянке (Костенки 21) 114
Рефераты 119
CONTENTS page
Vereshchagin N K., Baryshnikov G. F. Extinction of mammals during Quaternary
in northern Eurasia . . . . . . ,x. 3
Alexeeva L. I. Period and routs of migration of camels in eastern hemisphere 39
Baryshnikov G. F., Chistyakov D. A. Sary-Leget — a new Mesolitic site in
southern Osetia (Caucasus) ... 43
Belan N. G. Late Pleistocene wide-toed horse from Desna 50
Kuzmina I. E. Urals late Pleistocene horse .... 64
Serikov Ju. B., Kuzmina I. E. Mesolitic Koksharovsko-Yurinskaya site in Middle
Transurals . . . ............................. . . 89
Sher A. V., Garutt V. E. On the method of identification of molars generations
in elephants.................................. . . . . .93
Vereshchagin N K-, Ukraintseva V. V. The origin and stratigraphy of Berelyokh
mammoth „burial ground” . .... ............... 104
Praslov N. D. Engravect animals in Gmelin Late Paleolitic site (Kostenki 21) 114
Abstracts (in Russian) 119
ЗООЛОГИЧЕСКИМ ИНСТИТУТОМ АН СССР были выпущены
следующие книги по четвертичным млекопитающим:
Мамонтовая фауна Русской равнины и Восточной Сибири.
Труды Зоол. ин-та АН СССР, 1977, т. 72. Цена 1 руб. (тираж распродан).
Мамонтовая фауна и среда ее обитания в антропогене СССР.
Труды Зоол. ин-та АН СССР, 1977, т. 73. Цена 1 руб.
Млекопитающие Восточной Европы в антропогене.
Труды Зоол. ин-та АН СССР, 1980, т. 93. Цена 1 руб.
Мамонтовая фауна азиатской части СССР.
Труды Зоол. ин-та АН СССР, 1982, т. 111. Цена 1 руб.
Эти книги можно заказать наложенным платежом по адресу:
199034, Ленинград, Университетская наб., 1
Зоологический институт АН СССР,
Отделение истории териофауны
МЛЕКОПИТАЮЩИЕ СЕВЕРНОЙ ЕВРАЗИИ
В ЧЕТВЕРТИЧНОМ ПЕРИОДЕ
Труды Зоологического института АН СССР
Том 131
Утверждено к печати
редакционно-издательским советом
Зоологического института АН СССР
План 1985 г.
Редактор Т. А. Асанович
Художник С. Е. Станкевич
Сдано в набор 04.02.85. Подписано к печати 24.06.85. М-27920. Формат 70X100/16.
Бумага типографская № 1. Гарнитура литературная. Печать высокая. Объем 7,75 п.
л.-Н п. л. вкл., усл.-печ. л. 10,85, уч.-изд. л. 10. Тираж 800 экз. Заказ № 379.
Цена 1 р. 20 к.
Зоологический институт АН СССР, 199034, Ленинград, Университетская наб., 1.
Типография № 2 Ленуприздата, 191104, Ленинград, Литейный пр., 55.
К статье И. К. Верещагина и Г ,Ф. Барышникова, стр. 3—38
Рис. 1. Голова Хатангского мамонта.
Фото Н. К. Верещагина, 1978 г.
К статье И. К- Верещагина и Г. Ф. Барышникова, стр. 3—38
юсм
Рис. 2. Череп одного из последних мамонтов Берелеха
с выломанными бивнями.
К статье Н. К. Верещагина и Г Ф. Барышникова, стр. 3—38
Рис. 3. Остатки туши Юрибейской мамонтихи.
Фото Н. К- Верещагина, 1979 г.
К статье И. К. Верещагина и Г Ф. Барышникова, стр. 3—38
Рис. 8. Череп одного из поздних овцебыков с северного Таймыра.
Рис. 9. Учет костей на берелехском «кладбище» мамонтов.
Фото А. В. Ложкина. 1970 г.
К статье И. К. Верещагина и Г Ф. Барышникова, стр. 3—38
Рис. 10. Черепа последних зубров на Центральном Кавказе. Пещера Дигоризед.
Фото Н. К- Верещагина, 1947 г.
К статье Г. Ф. Барышникова и Д. А. Чистякова, стр. 43—49
Рис. 1. Вид на пещеру Сары-Легет с западной стороны.
Фото Г Ф. Барышникова, 1977 г.
К статье И. Г Белан, стр. 50—63
Рис. 1. Череп молодого жеребца широкопалой лошади из Мезина, № 5-571.
К статье И. Е. Кузьминой, стр. 64—88
Рис. 1. Череп уральской лошади Equus uralensis Kuzmina, 1975. Позд-
ний плейстоцен, пещера «Черные кости» Пермской области, колл. ЗИН
АН СССР, № 29724, голотип.
а — сверху, б — снизу, в — сбоку.
К статье И. Е. Кузьминой, стр. 64—88
Рис. 2. Зубы верхней челюсти.
а — из Предуралья; б — из Зауралья.
К статье И. Е. Кузьминой, стр. 64—88
Рис. 3. Зубы нижней челюсти.
сбоку; б, г — сверху, из Предуралья; в — сверху, из Зауралья.
К статье И. Е. Кузьминой, стр. 64—88
а
Рис. 13. Фаланги.
а — первая передняя, б — третья передняя, в — третья задняя.
К статье Ю. Б. Серикова и И. Е. Кузьминой, стр. 89—92
О 5СМ
Рис. 3. Нижняя челюсть собакй.
Вид сверху, несколько увеличен.
К статье Н. К. Верещагина и В. В. Украинцевой, стр. 104—113
Рис. 1. Общий вид Берелехского яра с юго-востока.
Фото Н. К- Верещагина, 1980 г.
Рис. 4. Первобытная «бухгалтерия». Насечки на обломке таза мамонтенка.
К статье И. К. Верещагина и В. В. Украинцевой, стр. 104—113
Рис. 7. Расчистка байджераха № 1.
Белый треугольник — место находки таза с насечками.
К статье Н. Д. Праслова, стр. 114—118.
Рис. 1. Гмелинская стоянка. Изображение крупного зверя на
песчаниковом диске (прорисовка и фото автора).
К статье Н. Д. Праслова, стр. 114—118
Рис. 2. Гмелинская стоянка. Прорисовка и фотография
изображения мамонта на сланце (увеличено).